bioquímica i - ucbweb.castelobranco.brucbweb.castelobranco.br/webcaf/arquivos/bioquimica_l.pdf ·...
TRANSCRIPT

Rio de Janeiro / 2008
TODOS OS DIREITOS RESERVADOS À
UNIVERSIDADE CASTELO BRANCO
VICE-REITORIA DE ENSINO DE GRADUAÇÃO E CORPO DISCENTE
COORDENAÇÃO DE EDUCAÇÃO A DISTÂNCIA
BIOQUÍMICA I

Un3b Universidade Castelo Branco
Bioquímica I / Universidade Castelo Branco. – Rio de Janeiro: UCB, 2008. - 64 p.: il.
ISBN 978-85-86912-74-0
1. Ensino a Distância. 2. Título.
CDD – 371.39
UNIVERSIDADE CASTELO BRANCO
Todos os direitos reservados à Universidade Castelo Branco - UCB
Nenhuma parte deste material poderá ser reproduzida, armazenada ou transmitida de qualquer forma ou por quaisquer meios - eletrônico, mecânico, fotocópia ou gravação, sem autorização da Universidade Castelo Branco - UCB.
Universidade Castelo Branco - UCBAvenida Santa Cruz, 1.631Rio de Janeiro - RJ21710-250 Tel. (21) 2406-7700 Fax (21) 2401-9696www.castelobranco.br

Responsáveis Pela Produção do Material InstrucionalResponsáveis Pela Produção do Material Instrucional
Coordenadora de Educação a DistânciaCoordenadora de Educação a DistânciaProf.ª Ziléa Baptista Nespoli
Coordenador do Curso de GraduaçãoCoordenador do Curso de GraduaçãoMaurício Magalhães - Ciências Biológicas
ConteudistaConteudistaAndré Sias das Neves
Supervisor do Centro Editorial – CEDISupervisor do Centro Editorial – CEDIJoselmo Botelho


Apresentação
Prezado(a) Aluno(a): É com grande satisfação que o(a) recebemos como integrante do corpo discente de nossos cursos de graduação,
na certeza de estarmos contribuindo para sua formação acadêmica e, conseqüentemente, propiciando oportunidade para melhoria de seu desempenho profi ssional. Nossos funcionários e nosso corpo docente esperam retribuir a sua escolha, reafi rmando o compromisso desta Instituição com a qualidade, por meio de uma estrutura aberta e criativa, centrada nos princípios de melhoria contínua.
Esperamos que este instrucional seja-lhe de grande ajuda e contribua para ampliar o horizonte do seu conhe-cimento teórico e para o aperfeiçoamento da sua prática pedagógica.
Seja bem-vindo(a)!Paulo Alcantara Gomes
Reitor


Orientações para o Auto-Estudo
O presente instrucional está dividido em oito unidades programáticas, cada uma com objetivos defi nidos e conteúdos selecionados criteriosamente pelos Professores Conteudistas para que os referidos objetivos sejam atingidos com êxito.
Os conteúdos programáticos das unidades são apresentados sob a forma de leituras, tarefas e atividades com-plementares.
As Unidades 1, 2, 3 e 4 correspondem aos conteúdos que serão avaliados em A1.
Na A2 poderão ser objeto de avaliação os conteúdos das oito unidades.
Havendo a necessidade de uma avaliação extra (A3 ou A4), esta obrigatoriamente será composta por todo o conteúdo de todas as Unidades Programáticas.
A carga horária do material instrucional para o auto-estudo que você está recebendo agora, juntamente com os horários destinados aos encontros com o Professor Orientador da disciplina, equivale a 60 horas-aula, que você administrará de acordo com a sua disponibilidade, respeitando-se, naturalmente, as datas dos encontros presenciais programados pelo Professor Orientador e as datas das avaliações do seu curso.
Bons Estudos!

Dicas para o Auto-Estudo
1 - Você terá total autonomia para escolher a melhor hora para estudar. Porém, seja disciplinado. Procure reservar sempre os mesmos horários para o estudo.
2 - Organize seu ambiente de estudo. Reserve todo o material necessário. Evite interrupções.
3 - Não deixe para estudar na última hora.
4 - Não acumule dúvidas. Anote-as e entre em contato com seu monitor.
5 - Não pule etapas.
6 - Faça todas as tarefas propostas.
7 - Não falte aos encontros presenciais. Eles são importantes para o melhor aproveitamento da disciplina.
8 - Não relegue a um segundo plano as atividades complementares e a auto-avaliação.
9 - Não hesite em começar de novo.

SUMÁRIO
Quadro-síntese do conteúdo programático ................................................................................................. 11Contextualização da disciplina ................................................................................................................... 12
UNIDADE I
COMPOSIÇÃO QUÍMICA DOS SERES VIVOS
1.1 – Água ................................................................................................................................................... 131.2 – Produto iônico da água ....................................................................................................................... 141.3 – Conceitos de ácidos e bases ............................................................................................................... 141.4 – pH ....................................................................................................................................................... 14 1.5 – Tampão biológico ............................................................................................................................... 151.6 – Íons minerais e osmolaridade ............................................................................................................. 161.7 – Aminoácidos ...................................................................................................................................... 171.8 – Proteínas ............................................................................................................................................. 171.9 – Vitaminas e coenzimas ....................................................................................................................... 20
UNIDADE II
ENZIMOLOGIA
2.1 – Conceito ............................................................................................................................................. 242.2 – Regulação da atividade enzimática .................................................................................................... 252.3 – Cinética enzimática ............................................................................................................................ 25
UNIDADE III
ENERGÉTICA BIOQUÍMICA
3.1 – Princípios de termodinâmica .............................................................................................................. 283.2 – Papel de aceptores de energia ............................................................................................................ 28
UNIDADE IV
METABOLISMO DE CARBOIDRATOS
4.1 – Estrutura geral .................................................................................................................................... 304.2 – Respiração celular e fermentação ...................................................................................................... 304.3 – Fotossíntese ........................................................................................................................................ 344.4 – Shunt das pentoses ............................................................................................................................. 374.5 – Glicogenólise e glicogênese ............................................................................................................... 384.6 – Gliconeogênese .................................................................................................................................. 39

UNIDADE V
METABOLISMO DE LIPÍDIOS
5.1 – Estrutura geral .................................................................................................................................... 415.2 – Metabolismo de lipídios ..................................................................................................................... 415.3 – Biossíntese de esteróides .................................................................................................................... 425.4 – Prostaglandinas, prostaciclinas e leucotrienos ................................................................................... 43
UNIDADE VI
METABOLISMO DE AMINOÁCIDOS E PROTEÍNAS
6.1 – Ciclo da uréia ..................................................................................................................................... 446.2 – Biossíntese de aminoácidos naturais .................................................................................................. 45
UNIDADE VII
REGULAÇÃO ENDÓCRINA DO METABOLISMO
7.1 – Insulina ............................................................................................................................................... 467.2 – Glucagon e adrenalina ........................................................................................................................ 46
UNIDADE VIII
METABOLISMO DE PURINAS E PIRIMIDINAS
8.1 – Bases nitrogenadas ............................................................................................................................. 508.2 – Biossíntese de nucleotídios ................................................................................................................ 508.3 – Metabolismo de nucleotídios ............................................................................................................. 518.4 – Importância biológica dos nucleotídios ............................................................................................. 52
Glossário ..................................................................................................................................................... 55Gabarito ....................................................................................................................................................... 57Referências bibliográfi cas ........................................................................................................................... 61
11
I – COMPOSIÇÃO QUÍMICA DOS SERES VIVOS1.1 – Água1.2 – Produto iônico da água1.3 – Conceitos de ácidos e bases1.4 – pH1.5 – Tampão biológico1.6 – Íons minerais e osmolaridade1.7 – Aminoácidos1.8 – Proteínas1.9 – Vitaminas e coenzimas
II – ENZIMOLOGIA2.1 – Conceito2.2 – Regulação da atividade enzimática2.3 – Cinética enzimática
III – ENERGÉTICA BIOQUÍMICA3.1 – Princípios de termodinâmica3.2 – Papel de aceptores de energia
IV – METABOLISMO DE CARBOIDRATOS4.1 – Estrutura geral4.2 – Respiração celular e fermentação4.3 – Fotossíntese4.4 – Shunt das pentoses4.5 – Glicogenólise e glicogênese4.6 – Gliconeogênese
V – METABOLISMO DE LIPÍDIOS5.1 – Estrutura geral5.2 – Metabolismo de lipídios5.3 – Biossíntese de esteróides5.4 – Prostaglandinas, prostaciclinas e leucotrienos
VI – METABOLISMO DE AMINOÁCIDOS E PRO-TEÍNAS6.1 – Ciclo da uréia6.2 – Biossíntese de aminoácidos naturais
VII – REGULAÇÃO ENDÓCRINA DO METABO-LISMO7.1 – Insulina7.2 – Glucagon e adrenalina
VIII – METABOLISMO DE PURINAS E PIRIMIDI-NAS8.1 – Bases nitrogenadas8.2 – Biossíntese de nucleotídios8.3 – Metabolismo de nucleotídios8.4 – Importância biológica dos nucleotídios
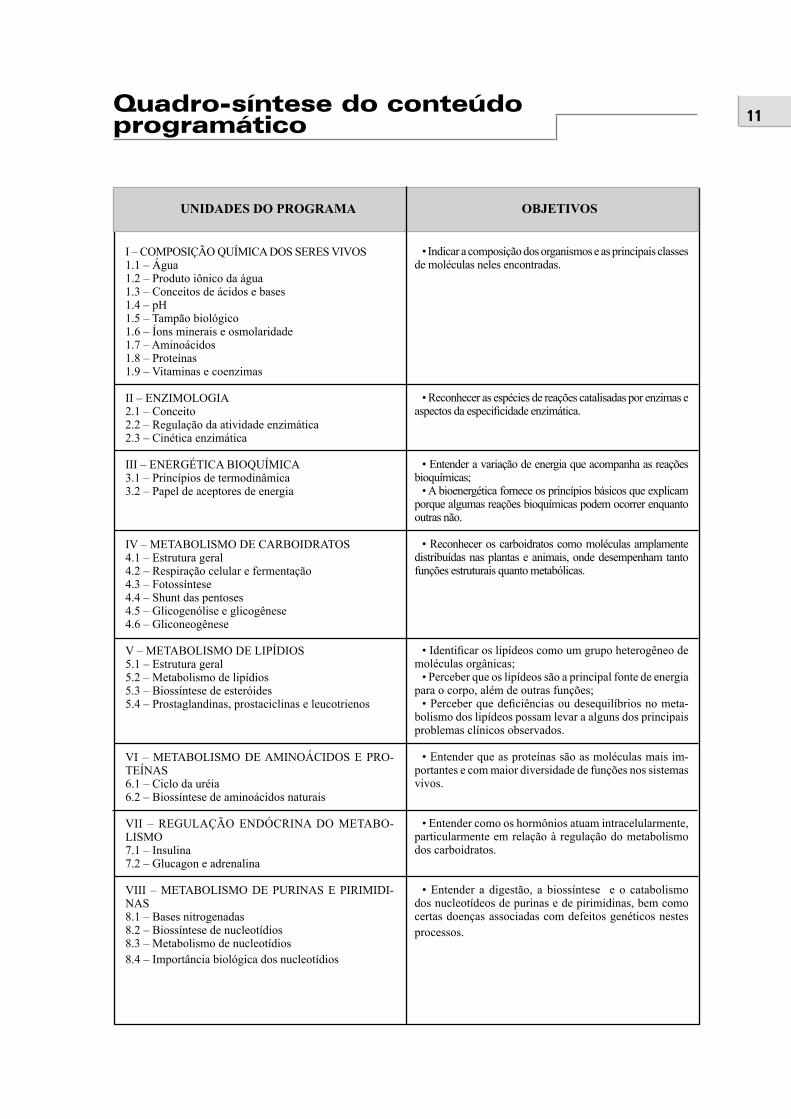
UNIDADES DO PROGRAMA OBJETIVOS
• Indicar a composição dos organismos e as principais classes de moléculas neles encontradas.
• Reconhecer as espécies de reações catalisadas por enzimas e aspectos da especifi cidade enzimática.
• Entender a variação de energia que acompanha as reações bioquímicas;
• A bioenergética fornece os princípios básicos que explicam porque algumas reações bioquímicas podem ocorrer enquanto outras não.
• Reconhecer os carboidratos como moléculas amplamente distribuídas nas plantas e animais, onde desempenham tanto funções estruturais quanto metabólicas.
• Identifi car os lipídeos como um grupo heterogêneo de moléculas orgânicas;
• Perceber que os lipídeos são a principal fonte de energia para o corpo, além de outras funções;
• Perceber que defi ciências ou desequilíbrios no meta-bolismo dos lipídeos possam levar a alguns dos principais problemas clínicos observados.
• Entender que as proteínas são as moléculas mais im-portantes e com maior diversidade de funções nos sistemas vivos.
• Entender como os hormônios atuam intracelularmente, particularmente em relação à regulação do metabolismo dos carboidratos.
• Entender a digestão, a biossíntese e o catabolismo dos nucleotídeos de purinas e de pirimidinas, bem como certas doenças associadas com defeitos genéticos nestes processos.
Quadro-síntese do conteúdo programático

12 Contextualização da Disciplina
Não se sabe exatamente quando a Bioquímica começou a emergir como disciplina independente. Na Alemanha se estabeleceu o primeiro departamento de Química Fisiológica e o primeiro periódico de bioquímica. Hoje, existem várias dezenas de periódicos, cada qual dando enfoque maior a determinada área da bioquímica.
É importante ressaltar a enorme infl uência que a bioquímica vem exercendo em todas as ciências biológicas, tornando-se uma ciência básica por excelência.
A bioquímica se constitui numa tentativa de descrever vida em termos químicos. Não se limitando, no entanto, a um simples relato das análises da matéria viva e das várias reações através das quais os componentes da vida são metabolizados, mas a maneira pela qual as reações biológicas são controladas.
Considerando que a vida na célula depende de milhares de reações químicas diferentes, cada uma catalisada por enzima específi ca, é de grande interesse saber como estão elas sincronizadas. A investigação dos mecanismos reguladores constitui um dos capítulos mais atraentes da bioquímica.
Espera-se ainda que, à medida que os conhecimentos básicos de bioquímica se tornem maiores, seja possível dirimir questões como a da transformação de células cancerosas e de diversas outras doenças.

13UNIDADE I
COMPOSIÇÃO QUÍMICA DOS SERES VIVOS
1.1- Água
A água é a substância mais abundante nos sistemas vi-vos, perfazendo 70% ou mais da massa da maioria dos organismos. A água permeia todas as porções de todas as células e é o meio no qual ocorrem o transporte de nutrientes, as reações metabólicas catalisadas enzima-ticamente e a transferência de energia química. Todos os aspectos da estrutura celular e de suas funções são adaptados às propriedades físicas e químicas da água. A molécula de água e seus produtos de ionização, H+ e OH-, infl uenciam profundamente a estrutura, a auto-montagem e as propriedades de todos os componentes celulares, incluindo as enzimas e outras proteínas, ácidos nucléicos e lipídios.
As pontes de hidrogênio entre as moléculas de água fornecem as forças coesivas que fazem da água um líquido à temperatura ambiente e que favorecem o or-denamento extremo das moléculas de água cristalizada (gelo). As biomoléculas polares dissolvem-se facilmen-te na água porque elas podem substituir as interações moleculares água-água energeticamente favoráveis por interações ainda mais favoráveis, como água-soluto. Diferentemente, as biomoléculas não polares interfe-rem com as interações água-água e são muito pouco solúveis na mesma água. Estas moléculas não-polares tendem, quando em solução aquosa, a agregarem-se de forma a minimizar os efeitos energeticamente desfa-voráveis provocados por sua presença.
Quando comparada com a maioria dos outros líquidos comuns, a água tem ponto de fusão, ponto de ebulição e calor de vaporização maiores que todos. Estas pro-priedades incomuns da água são uma conseqüência da existência de forças de atração fortes entre moléculas de água adjacentes, e estas forças dão à água líquida uma grande coesão interna.
As biomoléculas polares, porém não carregadas, como os açúcares, dissolvem-se facilmente na água devido ao efeito estabilizador das numerosas pontes de hidrogênio que elas formam entre os grupos hidroxila e o oxigênio do grupo carbonila do açúcar de um lado, e as moléculas polares da água de outro. Os alcoóis, aldeídos e cetonas formam, todos, pontes de hidrogênio com a água, como também o fazem compostos con-
tendo ligações N - H, por isso, as moléculas contendo tais grupos tendem a ser solúveis na água.
A água é um solvente polar. Ela dissolve facilmente a maioria das biomoléculas, as quais são, geralmen-te, compostos carregados eletricamente ou polares; compostos que se dissolvem facilmente na água são hidrofílicos. Em contraste, solventes não-polares, como o clorofórmio e o benzeno, são solventes muito pobres para as biomoléculas polares, mas dissolvem facilmente as biomoléculas não-polares como os lipídios e as ceras.
A água dissolve sais como o NaCl por hidratação e estabilização dos íons sódio e cloreto, enfraquecendo suas interações eletrostáticas e, desta forma, contra-pondo-se à tendência deles a se associarem, formando uma rede cristalina.
Os gases biologicamente importantes CO2, O2 e N2 são não-polares. Esses gases são muito pouco solúveis em água. Por isso, alguns organismos têm proteínas solúveis na água que são transportadoras de O2 (he-moglobina e mioglobina) e que facilitam o transporte do mesmo. Em solução aquosa, o dióxido de carbono forma ácido carbônico (H2CO3) e este é transportado na sua forma dissociada.
Outros dois gases, NH3 e H2S, também têm funções biológicas em alguns organismos. Estes, entretanto, são polares e se dissolvem facilmente na água.
Compostos anfi páticos têm regiões que são polares (ou carregadas) e regiões que são não-polares. Quando compostos anfi páticos são misturados com a água, as duas regiões da molécula do soluto experimentam tendências confl itantes; a região hidrofílica, polar ou carregada, interage favoravelmente com o solvente que tende a dissolver a molécula, entretanto a região hidrofóbica não-polar tem a tendência oposta, ou seja, a de evitar o contato com a água.
Muitas biomoléculas são anfi páticas, proteínas, pigmen-tos, algumas vitaminas, esteróides e fosfolipídios de mem-brana têm regiões superfi ciais polares e não-polares.

141.2 - Produto Iônico da Água
Embora muitas das propriedades solventes da água possam ser explicadas em função das características da molécula de H2O eletricamente não-carregada, é também necessário levar em consideração o pequeno grau de ionização da água em íons (H+) e íons hidroxila (OH-). Quando ácidos ou bases fracos são dissolvidos na água,
eles podem produzir H+ por ionização (se forem ácidos) ou consumir H+ ao serem protonizados (se forem bases); esses processos são também governados pelas respecti-vas constantes de equilíbrio. A concentração total do íon hidrogênio originário de todas as fontes é experimental-mente mensurável; ela é expressa como pH da solução.
1.3 - Conceitos de Ácidos e Bases
Os ácidos são compostos moleculares que, quando em solução aquosa, se ionizam, formando uma solu-ção eletrolítica, sendo que o cátion é sempre o hidro-gênio. Essa é a defi nição de Arrhenius para ácidos.
De modo geral, os ácidos têm sabor azedo, como pode ser observado quando se experimenta o limão (ácido cítrico) ou o vinagre (ácido acético). Embora esses dois ácidos sejam orgânicos, essa propriedade também está presente nos ácidos inorgânicos.
Na dissociação, o cátion é sempre hidrogênio, mas o ânion pode ser composto de átomos de mais de um elemento. Veja os exemplos abaixo:
Solução de ácido clorídrico – HCl HCl → H+ + Cl-
Solução de ácido sulfúrico – H2SO4 H2SO4 → 2 H+ + SO4
-2
Solução de ácido fosfórico – H3PO4 H3PO4 → 3 H+ + PO4
-3
Por formarem uma solução eletrolítica, em solução aquosa, os ácidos conduzem eletricidade.
As bases, também chamadas de hidróxidos, são com-
postos químicos que se ionizam na água, como os áci-dos; e o ânion formado é sempre OH-, conhecido como radical hidroxila. O cátion, em geral, é um metal.
As bases têm gosto adstringente (que produz constri-ção, “amarra” a boca), como banana verde ou caju.
Veja alguns exemplos:
Hidróxido de sódio (soda cáustica) – NaOH NaOH → Na+ + OH-
Hidróxido de cálcio – Ca(OH)2 Ca(OH)2 → Ca2+ + 2 OH-
Hidróxido de potássio – KOH KOH → K+ + OH-
Assim como os ácidos, as bases formam soluções ele-trolíticas, por isso conduzem corrente elétrica. Soluções básicas também são chamadas de soluções alcalinas.
A grandeza que indica o quanto uma solução é ácida ou alcalina chama-se pH. Envolve números que vão de 1 a 14. Soluções com pH inferior a 7 são ácidas, e com pH superior a 7 são alcalinas. Soluções neutras, como a água, têm pH 7. Quanto menor o pH de um ácido, mais forte ele é. Quanto maior o pH de uma base, mais forte ela é.
1.4 - pH
As moléculas de água têm uma pequena tendência para ionizar-se reversivelmente e liberar o íon hidro-gênio e o íon hidroxila, dando o equilíbrio: H2O ↔ Η++ΟΗ−. Esta ionização reversível é crucial para o papel da água nas funções celulares. Quando as con-centrações de H+ e OH- são exatamente iguais, como na água pura, a solução é dita estar em pH neutro.
A escala de pH designa a concentração de H+ e OH-. O produto iônico da água é a base para a escala de pH. O valor 7,0 para o pH de uma solução precisamente neutra não é um número escolhido arbitrariamente; ele deriva do valor absoluto do produto iônico da água a 25oC. As soluções que têm pH maior que 7 são alcalinas ou básicas; a concentração de OH- é maior que aquela de H+; por outro lado, soluções que têm pH menor do que 7 são ácidas.
Figura 1

15As medidas de pH são um dos procedimentos mais
importantes e mais freqüentes na prática bioquímica. O pH afeta a atividade e a estrutura das macromoléculas biológicas, como a atividade catalítica das enzimas. As medidas do pH do sangue e da urina são comumente empregadas no diagnóstico de doenças. O pH do plasma sanguíneo de pessoas com diabetes severa, por exemplo, é freqüentemente menor que o valor normal de 7,4; esta
condição é chamada de acidose. Em outros estados pa-tológicos, o pH do sangue é maior que o normal e esta condição é conhecida como alcalose.
Quase todos os processos biológicos são dependentes do pH; uma pequena variação no pH do meio produz uma grande variação na velocidade da maioria dos processos biológicos que se desenvolvem neste mesmo meio.
1.5 - Tampão Biológico
As células e os organismos matam um pH citosólico constante e específi co, geralmente próximo de pH 7,0, o que mantém as biomoléculas em seu estado iônico ótimo. Em organismos multicelulares, o pH dos fl ui-dos extracelulares (sangue, por exemplo) é também estreitamente regulado. A constância do pH é conse-guida primariamente através da existência de tampões biológicos; estes são misturas de ácidos fracos e suas bases conjugadas. O tamponamento biológico pode ser ilustrado pela descrição do funcionamento dos sistemas tampões fosfato e carbonato em seres humanos.
Tampões são substâncias que em solução aquosa dão a estas soluções a propriedade de resistir às va-riações do seu pH quando às mesmas são adicionadas quantidades relativamente pequenas de ácido (H+) ou base (OH-).
O plasma sangüíneo é tamponado, em parte, pelo siste-ma tampão bicarbonato que consiste de ácido carbônico (H2CO3) como doador de prótons e do (HCO3
- ) como receptor de próton H2CO3 ↔ H++HCO3
- . O pH do sistema tampão bicarbonato depende da
concentração do H2CO3 e do H2CO3- , os compostos
que doam e recebem os prótons, respectivamente. A concentração de H2CO3, por sua vez, depende da con-centração do CO2 dissolvido, o qual depende da con-centração ou da pressão parcial do CO2 na fase gasosa; desta forma, o pH do tampão bicarbonato exposto a uma fase gasosa é, em última análise, determinado pela concentração de HCO3
- na fase aquosa e pela pressão parcial de CO2 na fase gasosa.
Nos animais com pulmões, o sistema tampão bicar-bonato é um sistema tampão fi siológico efetivo em va-lores de pH próximos a 7,4 porque o H2CO3 do plasma sanguíneo está em equilíbrio com um reservatório de CO2 (g) de grande capacidade localizado nos espaços aéreos dos pulmões. Este sistema tampão envolve três equilíbrios reversíveis entre o CO2 gasoso nos pulmões e o bicarbonato HCO3
- no plasma sanguíneo. Quando H+ é adicionado ao sangue, como ocorre quando ele atravessa os tecidos, a reação 1 desloca-se para um novo equilíbrio, no qual a concentração de H2CO3 está aumentada. Isto aumenta a concentração de CO2 (d) no sangue (reação 2), o que faz com que a pressão
de CO2 (g) nos espaços aéreos pulmonares aumente (reação 3); o CO2 extra é expirado.
Por sua vez, quando adicionamos ao plasma sangüíneo ocorrem os eventos opostos: a concentração de H+ diminui, provocando a dissociação do H2CO3 em H+ e HCO3
- , isto causa uma maior dissolução de CO2 (g) no plasma sangüíneo. A velocidade respiratória, quer dizer, a velocidade com que o CO2 é inalado e expi-rado, pode ajustar rapidamente esses equilíbrios com a fi nalidade de manter o pH do sangue praticamente constante.
O pH do sangue humano tem um valor próximo a 7,40. Sempre que os mecanismos reguladores do pH falham ou são sobrepujados, como pode acontecer em casos de diabetes grave, não-controlada, quando uma superprodução metabólica de ácidos causa acidose, o pH do sangue pode cair para 6,80 ou menos, provocan-do danos celulares irreparáveis e, por conseqüência, a morte. Em outras doenças o pH pode elevar-se a níveis mortais. Embora muitos aspectos de estrutura celular e de seu funcionamento sejam infl uenciados pelo pH, é a atividade catalítica das enzimas que é especialmente sensível. As enzimas mostram, tipicamente, atividade catalítica máxima em um valor de pH característico, chamado pH ótimo. Em cada um dos lados do pH ótimo a atividade enzimática declina rapidamente. Assim, uma pequena variação do pH pode provocar uma grande diferença na velocidade de algumas rea-ções cruciais catalisadas enzimaticamente. O controle biológico do pH das células e dos fl uidos corporais é, portanto, de importância central em todos os aspectos das atividades celulares e do metabolismo.

16
Figura 2
Os organismos têm se adaptado efetivamente aos seus ambientes aquosos e têm desenvolvido maneiras de explorar as propriedades incomuns da água. O alto calor específi co da água (a energia calórica necessária para aumentar de 1ºC a temperatura de um grama de água) é útil para as células e organismos porque possibilita que a água atue como “tampão de calor”, permitindo que a temperatura de um organismo permaneça relativamente constante, mesmo que a temperatura do ar fl utue e que calor seja gerado em
quantidades relativamente grandes como um produto colateral do metabolismo. Além disso, alguns verte-brados exploram o alto valor do calor de vaporização da água usando (e, portanto, perdendo) o excesso de calor corporal para evaporar o suor. O alto grau de coesão interna da água líquida, devido às pontes de hidrogênio, é explorado pelas plantas como um meio de, durante o processo de transpiração, transportar nutrientes dissolvidos das raízes até as folhas. Mesmo a densidade mais baixa do gelo que a da água líquida tem conseqüências biológicas importantes nos ciclos vitais de muitos organismos aquáticos. Poços e lagoas congelam da superfície para o fundo, e a camada de gelo na superfície isola a água imediatamente abaixo dela do ar muito mais frio, impedindo que a lagoa, e os organismos que nela vivem, congelem totalmente. De fundamental importância para todos os organismos vivos é o fato de que muitas das propriedades físicas e biológicas das macromoléculas celulares, particular-mente das proteínas e dos ácidos nucléicos, derivam da sua interação com as moléculas de água do meio ambiente em que se encontram. A infl uência da água no curso da evolução biológica tem sido profunda e determinante. Se alguma forma de vida evoluiu em alguma outra parte do universo, é improvável que ela se assemelhe àquelas da Terra, a menos que seu lugar de origem também seja um local no qual existe grande quantidade de água líquida como um solvente acessível.
1.6 - Íons Minerais e Osmolaridade
Os eletrólitos mais importantes encontrados nos fl uidos biológicos são Na+, K+, Ca++, Mg++ e H+, entre os cátions, e Cl-, fosfatos, bicarbonatos e ácidos orgâ-nicos, entre os ânions.
A apresentação da distribuição dos eletrólitos em compartimentos baseia-se no fato de que os cátions não são capazes de atravessar as membranas celulares, senão lentamente, já que só a água passa livremente, de um compartimento para outro. Assim, os ajustes osmóticos entre os compartimentos fazem-se à custa dos movimentos livres da água.
A composição do líquido intersticial é próxima do plasma sanguíneo, exceto quanto às proteínas, que existem em pequena quantidade. No sangue, a dis-tribuição dos cátions e ânions é diferente, mas suas cargas elétricas estão em equilíbrio, isto é, a soma dos ânions é igual à dos cátions, quando são expressos em miliequivalente por litro.
O sódio, na concentração de 142 mEq/litro, e os de-mais (K+, Ca++, Mg++) perfazem o total de 153 mEq/litro de fl uido extracelular; o ânion principal é o cloreto, acompanhado de bicarbonato, fosfato, sulfato, ânions de ácidos orgânicos (piruvato e outros).
Osmolaridade (Efeito Gibbs-Donnan)
Quando duas soluções de concentrações diferentes estão separados por uma membrana, o movimento dos líquidos faz-se do ambiente menos concentrado para o mais concentrado. Esse é o processo da os-mose, e as substâncias dissolvidas exercem pressão osmótica, que motiva a passagem dos líquidos. A pressão osmótica é função do número de partícu-las dissolvidas e não depende nem do peso, nem da valência, e nem da carga elétrica das mesmas. O equilíbrio osmótico faz-se pela entrada e saída de água entre os compartimentos, podendo os sais acompanhar o movimento da água.
Os animais superiores têm a prioridade de manter constante, dentro de certos limites, sua composição em eletrólitos e a percentagem de água do meio interno. A existência de um meio interno foi estabelecida por Claude Bernard, que lançou a “constância ou a fi xi-dez do meio interno é a própria condição da vida”. A manutenção dessa constância foi denominada de ho-meostase, e depende das forças osmóticas dos líquidos biológicos e da sua regulação por meio de glândulas endócrinas e pela excreção renal.

171.7 - Aminoácidos
Os aminoácidos têm características estruturais comuns. Todos os 20 aminoácidos encontrados nas proteínas têm um grupo carboxila e um grupo amino ligados ao mesmo átomo de carbono (o carbono α).
Eles diferem um dos outros através de suas cadeias laterais ou grupos R, os quais variam em estrutura, tamanho e carga, e infl uenciam a solubilidade do ami-noácido em água. Os aminoácidos em soluções aquosas estão ionizados e podem agir como ácidos ou bases.
Aproximadamente 300 aminoácidos adicionais foram en-contrados nas células e têm uma grande variedade de funções, mas eles nunca aparecem em proteínas. A ornitina e a citrulina merecem uma nota especial, porque são intermediárias impor-tantes na biossíntese da arginina e no ciclo da uréia.
Os peptídeos que ocorrem biologicamente variam muito de tamanho, desde moléculas pequenas contendo dois ou três aminoácidos até grandes macromoléculas contendo milhares de aminoácidos.
Duas moléculas de aminoácidos podem ser unidas covalen-temente através de uma ligação amida substituída, chamada ligação peptídica, para formar um dipeptídio. Tal ligação é formada por remoção dos elementos da água de um grupo α-carboxila de um aminoácido e do grupo α-amino de outro.
R H R
H N CH C OH + H N CH COO
O
1 2
+ 3
-
R H R
H N CH C N CH COO
O
1 2
+ 3
-
H O2
Três aminoácidos podem ser reunidos por duas liga-ções peptídicas para formar um tripeptídio, da mesma maneira, os aminoácidos podem ser reunidos para formar tetra e pentapeptídios. Quando um pequeno número de aminoácidos é reunido desta forma, a es-trutura é chamada de oligopeptídio e quando muitos aminoácidos são reunidos, o produto é chamado de polipeptídio. As proteínas podem ter milhares de unidades de aminoácidos. Embora os termos “pro-teína” e “polipeptídios” possam ser, algumas vezes, intercambiáveis.
As ligações peptídicas podem ser hidrolisadas por aquecimento tanto com ácido forte quanto com base forte, para liberar os aminoácidos, para liberar os aminoácidos unidos por elas.
As ligações peptídicas podem também ser hidroli-sadas por determinadas enzimas chamadas proteases. Elas são enzimas proteolíticas e são encontradas em todas as células e tecidos, onde elas degradam proteínas que se tornaram desnecessárias ou danifi -cadas, além de ajudarem na digestão dos alimentos protéicos.
Existem muitos oligopeptídios e polipeptídios pequenos que ocorrem naturalmente, possuem atividades biológicas importantes e pronunciadas e, por isso, exercem seus efeitos em concentrações muito pequenas. Por exemplo, um certo número de hormônios de vertebrados são polipeptídios pe-quenos. O hormônio insulina contém duas cadeias polipeptídicas, uma com 30 resíduos de aminoácidos e a outra com 21. Outros hormônios polipeptídios são o glucagon, um hormônio pancreático de 29 resíduos que tem ação oposta àquela da insulina, e a corticotrofi na, um hormônio com 39 resíduos de aminoácidos, secretado pela hipófi se anterior e que estimula o córtex adrenal.
1.8 - Proteínas
Quase tudo que ocorre nas células envolve uma ou mais proteínas. As proteínas fornecem a estrutura, catalisam as reações celulares e executam milhares de outras tarefas. O papel central ocupado por elas é evidenciado no fato de que a informação genética é, em última instância, expressa como proteínas. Para cada proteína existe um segmento de DNA (um gene) que guarda a informação, especifi cando sua seqüência de aminoácidos. Em uma célula típica existem milhares de diferentes tipos de proteínas, cada
uma delas codifi cada por um gene e, cada uma delas, executando uma função específi ca. As proteínas estão entre as macromoléculas biológicas mais abundantes e também são extremamente versáteis em suas funções.
Funções biológicas das proteínas
1. Enzimas – o grupo de proteínas mais variado e mais altamente especializado é aquele cujos compo-

18nentes exibem atividade catalítica – as enzimas. São virtualmente catalisadas por enzimas todas as reações químicas nas quais participam as biomoléculas orgâ-nicas das células. Muitos milhares de enzimas dife-rentes, cada uma capaz de catalisar um tipo de reação química diferente, foram descobertos em diferentes organismos.
2. Proteínas transportadoras – proteínas transpor-tadoras existentes no plasma sanguíneo ligam-se a íons ou a moléculas específi cas os quais são transportados de um órgão para outro. A hemoglobina dos eritrócitos liga-se ao oxigênio à medida que o sangue atravessa os pulmões, transporta-os até os tecidos periféricos e, aí, libera-o para que possa participar da oxidação dos nutrientes, com concomitante liberação de energia. O plasma sanguíneo também contém lipoproteínas que transportam lipídios do fígado para outros órgãos. Outros tipos de proteínas de transporte estão presentes nas membranas plasmáticas e nas membranas intra-celulares de todos os organismos; elas estão aptas a ligarem-se, por exemplo, à glicose, aos aminoácidos ou às outras substâncias e transportá-las através dessas membranas.
3. Proteínas nutrientes e de armazenamento – as sementes de muitas plantas armazenam proteínas nu-trientes necessárias para a germinação e o crescimento do broto. Exemplos particularmente bem estudados são as proteínas das sementes do trigo, milho e arroz. A ovoalbumina, a principal proteína da clara do ovo, e a caseína, a principal proteína do leite, são outros exemplos de proteínas nutrientes. A ferritina encon-trada em algumas bactérias e em tecidos animais e vegetais armazena átomos de ferro.
4. Proteínas contráteis ou de motilidade – algumas proteínas habilitam células e organismos com a capa-cidade de contraírem-se, de mudarem de forma, ou de se deslocarem no meio ambiente. A actina e a miosina funcionam no sistema contrátil do músculo esquelé-tico e também em muitas células não musculares. A tubulina é a proteína com a qual os microtúbulos são construídos. Os microtúbulos agem de forma concen-trada com a proteína dineína nos cílios e fl agelos para propelir as células.
a) Proteínas estruturais – muitas proteínas servem como fi lamentos de suporte, cabos ou lâminas para fornecer proteção ou resistência a estruturas biológicas. O principal componente das cartilagens e dos tendões é a proteína fi brosa colágeno, a qual tem alta resistên-cia à tensão. O couro é quase que colágeno puro. Os ligamentos contêm elastina, uma proteína estrutural capaz de distender-se em duas dimensões. O cabelo, as unhas e as penas consistem principalmente da proteína resistente e insolúvel denominada queratina. O maior componente das fi bras da seda e da teia das aranhas é
a fi broína. Os ligamentos “em dobradiça” das asas de certos insetos são feitos de resilina, uma proteína que tem propriedades elásticas próximas da perfeição.
5. Proteínas de defesa – muitas proteínas defendem os organismos contra a invasão de outras espécies ou os protegem de ferimentos. As imunoglobulinas ou anticorpos, proteínas especializadas sintetizadas pelos linfócitos dos vertebrados, podem reconhecer e preci-pitar, ou neutralizar, invasores como bactérias, vírus ou proteínas estranhas oriundas de outras espécies. O fi brinogênio e a trombina são proteínas que participam da coagulação do sangue que previne a perda de sangue quando o sistema vascular é lesado. Venenos de ser-pente, toxinas bacterianas e proteínas vegetais tóxicas, como a ricina, também parecem ter funções defensivas. Algumas destas proteínas, incluindo o fi brinogênio, a trombina e alguns venenos também são enzimas.
6. Proteínas reguladoras – algumas proteínas aju-dam a regular a atividade celular ou fi siológica. Entre elas estão muitos hormônios. Alguns exemplos incluem a insulina, a qual regula o metabolismo dos açúcares e o hormônio do crescimento da hipófi se. A resposta celular a muitos sinais hormonais é freqüentemente regulada por uma classe de proteínas que se ligam ao GTP e são chamadas proteínas G (o GTP é estreita-mente relacionado ao ATP com a guanina substituindo a adenina). Outras proteínas reguladoras ligam-se ao DNA e regulam a biossíntese de enzimas e das molé-culas de RNA envolvidas na divisão celular, tanto em procariotos como em eucariotos.
7. Outras proteínas – existem numerosas outras proteínas cujas funções podem ser ditas exóticas e de difícil classifi cação. A monelina, uma proteína de uma planta africana, tem um sabor intensamente doce. Ela está sendo estudada como um adoçante não-tóxico e quase sem calorias para uso humano. O plasma san-guíneo de alguns peixes da Antártica contém proteínas anticoagulantes, as quais protegem do congelamento o sangue destes animais.
É extraordinário que todas estas proteínas, com suas propriedades e funções tão diferentes, sejam constru-ídas com o mesmo grupo de 20 aminoácidos.
Algumas proteínas contêm grupos químicos dife-rentes dos aminoácidos. Muitas proteínas, como as enzimas ribonuclease e quimotripsina, contêm apenas aminoácidos e nenhum outro grupo químico; elas são consideradas proteínas simples. Entretanto, algumas proteínas contêm componentes químicos em adição aos aminoácidos; elas são chamadas proteínas conjugadas. A parte não-aminoácido de uma proteína conjugada é geralmente chamada de seu grupo prostético. As prote-ínas conjugadas são classifi cadas com base na natureza química dos seus grupos prostéticos; por exemplo:

19lipoproteínas contêm lipídios, glicoproteínas contêm moléculas de açúcares e metaloproteínas contêm um metal específi co. Um certo número de proteínas contém mais do que um grupo prostético. Geralmente o grupo prostético desempenha um papel importante na função biológica da proteína.
Proteínas Conjugadas
Grupo prostético
Lipídios
Carboidratos
Grupo fosfato
Heme(ferro- porfi rina)
Nucleotídeos de fl avina
Ferro
Zinco
Cálcio
Molibdênio
Cobre
Classe
Lipoproteínas
Glicoproteínas
Fosfoproteínas
Heme-proteínas
Flavoproteínas
Metaloproteína
Exemplo
Β1-lipoproteína do sangue
ImunoglobulinaG
Caseína do leite
Hemoglobina
Succinato desidrogenase
Ferritina
Álcool desidrogenase
Calmodulina
Dinitrogenase
Plastocianina
Função de uma proteína depende da sua seqüência de aminoácidos
A bactéria E. coli produz perto de 3.000 proteínas diferen-tes. Um ser humano produz de 50.000 a 100.000 proteínas diferentes. Em ambos os casos, cada tipo separado de proteína tem uma estrutura única e esta estrutura confere a ela uma função única. Mais de 1.400 doenças genéticas hu-manas têm sido identifi cadas como resultantes da produção de proteínas defeituosas. Talvez um terço dessas proteínas defeituosas assim o são, porque um único aminoácido da seqüência foi mudado; portanto, se a estrutura primária é alterada, a função da proteína também pode mudar.
A seqüência de aminoácidos pode ser deduzida da seqüência de bases no DNA
O desenvolvimento rápido dos métodos de seqüen-ciamento do DNA, a elucidação do código genético
e o desenvolvimento de técnicas para o isolamento de genes tornam possível deduzir a seqüência de um polipeptídio pela determinação da seqüência de nucleotídeos em seu gene. As duas técnicas são com-plementares.
Existem quatro níveis na arquitetura das proteínas:
1. Estrutura primária – inclui todas as ligações covalentes entre os aminoácidos que compõem uma proteína e é defi nida pela seqüência dos aminoácidos unidos por ligações peptídicas e pela localização das pontes dissulfeto. O arranjo espacial relativo dos ami-noácidos não é especifi cado.
2. Estrutura secundária – refere-se aos arranjos regulares e recorrentes no espaço de resíduos de ami-noácidos adjacentes em uma cadeia polipeptídica.
3. Estrutura terciária – refere-se ao relacionamento espacial entre todos os aminoácidos em um polipep-tídio.
4. Estrutura quaternária – especifi ca a relação espacial dos polipeptídios, ou subunidades, no interior de uma dada proteína.

20
As proteínas perdem a estrutura e a função, quan-do desnaturadas
A maneira de demonstrar a importância da estrutura específi ca das proteínas para a função biológica que exercem é alterar esta estrutura e determinar o efeito que isto causa nesta função. Uma alteração extrema é a perda total da sua estrutura tridimensional, um processo chamado desnaturação. Este é o processo familiar que ocorre quando um ovo é cozido. A clara do ovo, a qual contém a proteína solúvel albumina do ovo, ou ovoalbumina, coagula pelo aquecimento para formar uma substância branca e sólida. Esta substância não redissolverá, quando resfriada, para reproduzir a solução límpida de proteína que era,
Figura 3
antes do aquecimento, a clara do ovo original. O aquecimento da albumina do ovo produziu, portanto, uma mudança irreversível. Este efeito do calor ocorre em, virtualmente, todas as proteínas globulares, in-dependentemente do seu tamanho ou da sua função biológica, embora, a temperatura precisa, na qual o processo ocorre, possa variar e o seu efeito nem sempre será irreversível. Algumas proteínas globu-lares desnaturadas pelo calor, extremos de pH, ou reagentes desnaturantes, recuperarão a sua estrutura nativa e sua atividade biológica, um processo chama-do renaturação. Quando estes agentes são retirados da solução em que as proteínas se encontram, esta mesma solução retorna às condições nas quais a conformação protéica nativa é possível e estável. A mudança na estrutura produzida pela desnaturação é quase invariavelmente associada à perda de função. Isto é uma conseqüência esperada do princípio de que a estrutura tridimensional específi ca das proteínas é crítica para o exercício de suas funções.
As proteínas podem ser desnaturadas não somente pelo aquecimento, mas também por valores extremos de pH, por alguns solventes orgânicos miscíveis com a água, como o etanol e a acetona, por algumas subs-tâncias em solução como a uréia, ou por exposição da proteína a substâncias detergentes.
1.9 - Vitaminas e Coenzimas
Vitaminas são compostos orgânicos não-relacionados qui-micamente, que não podem ser sintetizados por humanos e, portanto, devem ser suprimidos pela dieta. Nove vitaminas (ácido fólico, cobalamina, ácido ascórbico, piridoxina, tiamina, niacina, ribofl avina, biotina e ácido pantotênico) são classifi cados como hidrossolúveis, enquanto quatro vitaminas (vitaminas A, D, K e E) são ditas lipossolúveis. As vitaminas são requeridas para a execução de funções celulares específi cas. Por exemplo, muitas das vitaminas hidrossolúveis são precursores de coenzimas para as enzi-mas do metabolismo intermediário. Em contraste com as vitaminas hidrossolúveis, somente uma vitamina liposso-lúvel (vitamina K) tem função de coenzima.
Ácido Fólico
O ácido fólico (ou folato), o qual desempenha um pa-pel chave no metabolismo dos grupos de um carbono, é essencial para a biossíntese de vários compostos.
Cobalina (Vitamina B12)
A vitamina B12 é necessária em humanos para duas reações enzimáticas essenciais: a síntese de metionina e a isomerização da metilmalonil-CoA, que é produ-
zida durante a degradação de alguns aminoácidos e de ácidos graxos com número impar de átomos de carbonos. Quando a vitamina é defi ciente, ácidos graxos anormais acumulam-se e são incorporados nas membranas celulares, incluindo as do sistema nervoso. Isso pode contribuir para algumas das manifestações neurológicas da defi ciência da vitamina B12.
Ácido Ascórbico (Vitamina C)
A forma ativa da vitamina C é o ácido ascórbico. A principal função do ascorbato é como agente redutor em diversas reações diferentes. A vitamina C tem um papel muito bem-documentado como coenzima nas reações de hidroxilação, como por exemplo na hidroxilação dos resíduos prolil- e lisil- do colágeno. A vitamina C é, dessa forma, necessária para a manu-tenção normal do tecido conectivo, assim como para recompor tecidos danifi cados. A vitamina C também facilita a absorção do ferro da dieta no intestino.
Piridoxina (Vitamina B6)
Vitamina B6 é um termo coletivo para piridoxina, pi-ridoxal e piridoxamina, todos derivados da piridina. A

21piridoxina ocorre principalmente nas plantas, enquanto o piridoxal e a piridoxamina são encontrados em ali-mentos obtidos de animais. Todos os três compostos podem servir como precursores da coenzima biologi-camente ativa, o piridoxal-fosfato. O piridoxal-fosfato funciona como uma coenzima para um grande número de enzimas, particularmente aquelas que catalisam reações envolvendo aminoácidos.
Tiamina (Vitamina B1)
O pirofosfato de tiamina (TPP) é a forma biologica-mente ativa da vitamina, formada pela transferência do grupo pirofosfato da ATP para a tiamina. O piro-fosfato de tiamina serve como coenzima na formação ou na degradação de α-cetóis pela transcetolase e na descarboxilação oxidativa dos α-cetoácidos. A beri-béri é uma grave síndrome de defi ciência de tiamina observada em áreas onde o arroz polido é o principal componente da dieta.
Niacina
Niacina, ou ácido nicotínico, é um derivado subs-tituído da piridina. As formas biologicamente ativas da coenzima são nicotinamida-adenina-dinucleotídeo (NAD+) e seu derivado fosforilado, nicotinamida-adenina-dinucleotídeo-fosfato (NADP+). O NAD+ e o NADP+ servem como coenzimas nas reações de oxidação-redução nas quais a coenzima sofre redução do anel piridina, pela incorporação de um íon hidreto (átomo de hidrogênio + um elétron). As formas redu-zidas do NAD+ e do NADP+ são NADH e NADPH, respectivamente.
Riboflavina (Vitamina B2)
As duas formas biologicamente ativas são fl avina mononucleotídeo (FMN) e fl avina adenina dinucleo-tídeo (FAD), formadas pela transferência de um AMP do ATP para FMN. O FMN e o FAD são capazes de aceitar reversivelmente dois átomos de hidrogênio, formando FMNH2 ou FADH2. O FMN e o FAD são fortemente ligados – algumas vezes covalentemente – a fl avoenzimas que catalisam a oxidação ou a redução de um substrato.
Biotina
A biotina é uma coenzima nas reações de carboxila-ção, nas quais ela serve como carregador do dióxido de carbono ativado. A biotina liga-se covalentemente ao grupo ε-amino de resíduos de lisina nas enzimas dependentes de biotina. A defi ciência de biotina não ocorre naturalmente porque a vitamina está amplamen-te distribuída nos alimentos. Além disso, uma grande
porcentagem da biotina necessária para os humanos é suprimida por bactérias intestinais. Entretanto, a adição de claras de ovos cruas à dieta como uma fonte de proteína induz sintomas de defi ciência de biotina, isto é, dermatite, glossite, perda de apetite e náusea. As claras de ovos cruas contém uma glicoproteína, a avidina, a qual se liga fortemente à biotina e impede a sua absorção a partir do intestino. No entanto, com uma dieta normal, estima-se que seriam necessários 20 ovos por dia para induzir uma síndrome da defi ciência. Assim, a inclusão ocasional de ovos crus à dieta não leva à defi ciência de biotina.
Ácido Pantotênico
O ácido pantotênico é um componente da coenzima A, a qual atua na transferência de grupos acila. A coen-zima A contém um grupo tiol que transporta compostos acila como ésteres de tiol ativados. Exemplos de tais estruturas são a succinil-CoA, a acil-CoA e a acetil-CoA. O ácido pantotênico é também um componente da sintetase dos ácidos graxos. Os ovos, o fígado e as leveduras são as mais importantes fontes de ácido pantotênico, embora a vitamina seja amplamente distribuída.
Vitamina A
Os retinóides, uma família de moléculas relacionadas ao retinol (vitamina A), são essenciais para a visão, a reprodução, o crescimento e a manutenção dos tecidos epiteliais. O ácido retinóico, derivado da oxidação do retinol da dieta, medeia a maioria das ações dos reti-nóides, exceto para a visão, a qual depende do retinal, o derivado aldeídico do retinol. O ácido retinóico liga-se com alta afi nidade a proteínas receptoras específi cas, no núcleo de células-alvo, tais como as células epi-teliais. O complexo ativado receptor-ácido retinóico interage com a cromatina nuclear, estimulando a síntese de RNA retinóide-específi co, resultando na produção de proteínas específi cas, as quais medeiam várias funções fi siológicas. Por exemplo, os retinóides controlam a expressão do gene da queratina na maior parte dos tecidos epiteliais do corpo. As proteínas receptoras específi cas do ácido retinóico são parte da superfamília dos reguladores transcricionais que incluem os hormônios esteróides e da tireóide, e o 1,25-diidroxicolecalciferol, os quais funcionam de maneira similar.
Vitamina D
As vitaminas D são um grupo de esteróides que apresentam uma função do tipo hormonal. A molécula ativa, 1,25-diidroxicolecalciferol (1,25-diOH-D3), liga-se a proteínas receptoras intracelulares. O complexo 1,25-diOH-D3-receptor interage com o DNA no núcleo

22de células-alvo de modo semelhante à vitamina A e estimula seletivamente ou reprime de modo específi -co a transcrição gênica. A ação mais proeminente do 1,25-diOH-D3 é a regulação dos níveis plasmáticos de cálcio e fósforo.
Vitamina K
O principal papel da vitamina K é exercido na modifi cação pós-traducional de vários fatores de coagulação sangüínea, quando essa vitamina serve como uma coenzima na carboxilação de certos resíduos de ácido glutâmico presentes nessas pro-teínas. A vitamina K existe em diversas formas. Por
Exercícios de Auto-Avaliação
1. A fenilcetonúria é uma doença que resulta de um defeito na enzima fenilalanina hidroxilase, que participa do catabolismo do aminoácido fenilalanina. A falta de hidroxilase produz o acúmulo de fenilalanina que, por transaminação, forma ácido fenilpirúvico. Quando em excesso, o ácido fenilpirúvico provoca retardamento mental severo. Por outro lado, o portador desse defeito enzimático pode ter uma vida normal desde que o defeito seja diagnosticado imediatamente após o nascimento e que sua dieta seja controlada. A fenilcetonúria é tão comum que mesmo nas latas de refrigerantes dietéticos existe o aviso:
“Este produto contém fenilalanina!”.
Qual o principal cuidado a tomar com a dieta alimentar de um portador desse defeito enzimático? Por quê?
2. O gato siamês é um animal de rara beleza, pois a pelagem de seu corpo é clara com extremidades – orelhas, focinho, pés e cauda – pretas. A presença do pigmento que dá a cor negra a essas extremidades é o resultado da atividade de uma enzima que fi ca inativada acima de 34°C. Explique por que esses animais têm a pelagem negra nas extremidades do corpo.
3. O gene A é responsável pela produção do polipeptídeo X. Seu alelo a não produz o polipeptídeo X. Assim, indivíduos de genótipos AA ou Aa produzem o polipeptídeo X, que está ausente nos indivíduos aa. Os dois gráfi cos, I e II, referem-se à velocidade de formação de um determinado produto (VFP), em mg/hora, em dois indivíduos da mesma espécie, quando suas temperaturas variam.
Sabendo que a velocidade de formação do produto (VFP) está relacionada à presença ou ausência do poli-peptídeo X, responda:
a) Qual dos gráfi cos se refere a indivíduo AA ou Aa e qual se refere a indivíduo aa?
b) Pelos dados dos gráfi cos, qual seria a função mais provável do polipeptídeo X no processo de formação do produto? Como você explicaria o comportamento da curva no gráfi co correspondente ao indivíduo AA ou Aa?
4. Quais são os principais sinais que surgem no organismo humano como resultado da falta das seguintes vitaminas: B1, B12‚ e K, respectivamente.
5. Dê exemplos de alimentos que possuem grande quantidade de água.
exemplo, nas plantas ela existe como fi loquinona (ou vitamina K1) e nas bactérias da fl ora intestinal como menaquinona (ou vitamina K2). Para a terapia, está disponível um derivado sintético da vitamina K, a menadiona.
Vitamina E
As vitaminas E consistem em oito tocoferóis de ocor-rência natural, dos quais o α-tocoferol é o mais ativo. A principal função da vitamina E é como antioxidante na prevenção da oxidação não-enzimática de componentes celulares (por exemplo, ácidos graxos poliinsaturados) pelo oxigênio molecular e por radicais livres.

236. A vitamina A, conhecida como beta caroteno e retinol, é importante para o bom funcionamento da visão e
para a integridade da pele. Cite um alimento de origem animal e um de origem vegetal ricos nesta vitamina.
7. Qual é a vitamina cuja falta acarreta o escorbuto, avitaminose que se manifesta como sangramento nas gengivas, queda dos dentes e rachaduras na pele?
8. Por que a vitamina D (esteróide) é importante para o crescimento?
9. Uma prática corriqueira na preparação de comida é colocar um pouco de “leite” de mamão ou suco de abacaxi para amaciar a carne. Hoje em dia, os supermercados já vendem um amaciante de carne industrializado.
a) Explique o amaciamento da carne promovido pelo componente presente no mamão, no abacaxi ou no ama-ciante industrializado e compare esse processo com a digestão.
b) Se o amaciante, natural ou industrializado, for adicionado durante o cozimento, qual será o efeito sobre a carne? Por quê?
10. É muito comum que mulheres apresentem um quadro de anemia durante a gravidez. As mulheres anêmi-cas queixam-se de cansaço constante, além de uma acentuada “falta de ar”. Essa condição, em geral, pode ser tratada por meio da ingestão de sais de ferro, ou de uma dieta rica em ferro. Explique de que forma a dose extra de ferro alivia os sintomas de falta de ar.
11. Em condições normais, nem todo o gás oxigênio transportado pelo sangue é liberado nos tecidos corporais; um pouco dele continua retido nas moléculas de hemoglobina. No entanto, um aumento da temperatura ou uma queda do pH faz com que a hemoglobina libere uma quantidade adicional de gás oxigênio.
a) Explique a relação entre atividade muscular e aumento de temperatura.
b) Explique a relação entre atividade muscular e queda de pH.
c) Explique de que maneira o comportamento da hemoglobina, descrito no texto, pode ser benéfi co para músculos em atividade intensa.
Atividades Complementares
Depois de ler a Unidade I, sublinhe os termos que você não conhece pesquisando sobre eles.

24 UNIDADE II
ENZIMOLOGIA
2.1 - Conceito
Praticamente todas as reações no corpo são mediadas por enzimas, as quais são proteínas catalisadoras que aumentam a velocidade das reações, sem sofrerem alterações no processo global. Dentre as muitas reações biológicas que são ener-gicamente possíveis, as enzimas seletivamente canalizam reatantes (chamados substratos) para rotas úteis. As enzimas direcionam, assim, todos os eventos metabólicos.
Os nomes de enzimas mais comumente usados têm o sufi xo “-ase” adicionado ao nome do substrato da reação (por exemplo, glicosidase, urease, sacarase) ou à descri-ção da ação realizada (por exemplo, lactato-desidrogenase e adenilato-ciclase). Algumas enzimas mantêm seu nome trivial original, o qual não tem qualquer associação com a reação enzimática, por exemplo, tripsina e pepsina.
As enzimas são catalisadores protéicos que aumentam a velocidade de uma reação química e não são consumidos durante a reação que catalisam. Alguns tipos de RNA po-dem atuar como enzimas, geralmente catalisando a quebra e a síntese de ligações fosfo-diéster. Os RNAs com atividade catalítica são chamados ribozimas e são encontrados com muito menos freqüência que as proteínas catalisadoras.
As moléculas de enzimas contêm uma região específi ca formando uma fenda que é chamada sítio ativo. O sítio ativo contém cadeias laterais de aminoácidos, as quais criam uma superfície tridimensional complementar ao substra-to. O sítio ativo liga o substrato, formando um complexo enzima-substrato (ES). O complexo ES é convertido em enzima-produto (EP), o qual subseqüentemente se dissocia em enzima e produto. As enzimas são altamente especí-fi cas, interagindo com um ou alguns poucos substratos e catalisando apenas um tipo de reação química.
Praticamente todas as reações têm uma barreira de energia separando os reatantes dos produtos. Essa barreira, denominada energia livre de ativação, é a diferença entre a energia dos reatantes e aquela de um intermediário de alta energia, que ocorre durante a for-mação do produto. Por exemplo, a fi gura abaixo mostra as alterações na energia durante a conversão de uma molécula do reatante A no produto B, passando pelo estado de transição (intermediário de alta energia).
Energia livre de ativação: O pico de energia é a
diferença na energia livre entre os reatantes e T, onde um intermediário rico em energia é formado durante a conversão do reatante em produto. Devido à grande energia de ativação, as velocidades das reações quími-cas não-catalisadas são freqüentemente lentas.
Velocidade da reação: Para as moléculas reagirem, devem conter energia sufi ciente para superar a barreira de energia do estado de transição. Na ausência de uma enzima, somente uma pequena proporção da popula-ção de moléculas pode possuir energia sufi ciente para atingir o estado de transição entre reatante e produto. A velocidade da reação é determinada pelo número dessas moléculas “energizadas”. Em geral, quanto menor a energia livre de ativação, mais moléculas têm energia sufi ciente para superar o estado de transição e, assim, mais rápida é a velocidade da reação.
Fatores que afetam a velocidade da reação
As diferentes enzimas mostram diferentes respostas às alterações de concentração de substrato, tempera-tura e pH.
Concentração do substrato: A velocidade de uma reação (v) é o número de moléculas de substrato convertidas em produto por unidade de tempo. A velocidade de uma reação catalisada por enzima au-menta conforme a concentração do substrato, até uma velocidade máxima (Vmax) ser atingida.
Figura 4 Figura 5

25Temperatura: A velocidade de reação aumenta com
a temperatura, até um pico de velocidade ser atingido. Esse aumento é devido ao aumento do número de moléculas com energia sufi ciente para atravessar a bar-reira de energia e formar os produtos da reação. Uma elevação maior da temperatura resulta em redução na velocidade de reação, como resultado da desnaturação da enzima, induzida pela temperatura.
Figura 6 Figura 7
pH: Valores extremos de pH também podem le-var à desnaturação da enzima, pois a estrutura da molécula protéica cataliticamente ativa depende do caráter iônico das cadeias laterais dos aminoácidos. O pH no qual a atividade máxima da enzima é atin-gida difere para cada enzima e, geralmente, reflete a [H+] na qual a enzima funciona no organismo.
2.2 - Regulação da Atividade Enzimática
A regulação da velocidade das reações enzimá-ticas é essencial para o organismo coordenar seus numerosos processos metabólicos. As velocidades da maioria das enzimas respondem a mudanças na concentração dos substratos, pois, o nível intracelular de muitos dos substratos se encontra na faixa do KM (constante de Michaelis). Dessa forma, um aumento na concentração do substrato é refl etido no aumento da velocidade de reação, o que tende a fazer a con-centração do substrato retornar ao valor normal. Além disso, algumas enzimas com funções reguladoras especializadas respondem a efetores alostéricos ou a modifi cações covalentes, ou ainda, possuem a velo-cidade de sua síntese alterada quando as condições fi siológicas são alteradas.
Sítios alostéricos de ligação: As enzimas alostéricas são reguladas por moléculas chamadas efetores (tam-bém chamados de modifi cadores ou moduladores), os
quais ligam-se de forma não-covalente a outro sítio que não o sítio catalítico.
Regulação de enzimas por modifi cação covalente:Muitas enzimas podem ser reguladas pela adição ou pela remoção de grupos fosfato de resíduos específi cos. A fosforilação de proteínas é reconhecida como uma das principais formas pelas quais os processos celulares são regulados.
Indução a repressão da síntese de enzimas: Os me-canismos reguladores descritos previamente modifi cam a atividade de moléculas enzimáticas existentes. Entre-tanto, as células também podem regular a quantidade de enzima presente – em geral alterando a velocidade da síntese da enzima. O aumento (indução) ou a diminuição (repressão) da síntese da enzima leva a uma alteração nas população total de sítios ativos (nesse caso, a efi ciência das moléculas existentes na enzima não é afetada).
2.3 - Cinética Enzimática
Michaelis e Menten propuseram um modelo sim-ples, que explica a maioria das características das reações catalisadas por enzimas. Nesse modelo, a enzima combina-se reversivelmente com o substrato, formando um complexo ES que, subseqüentemente, degrada-se em produto, regenerando a enzima livre. O modelo, envolvendo uma molécula de substrato, é representado a seguir:
K1 K2 E + S ↔ ES → E + P K-1
onde S é o substrato E é a enzima ES é o complexo enzima-substrato K1, K - 1 e K2 são as constantes de velocidade
Equação de Michaelis-Menten
A equação de Michaelis-Menten descreve como a velo-cidade da reação varia com a concentração do substrato:
VO = VMAX [S] Km + [S]

26Onde VO = velocidade inicial de reação VMAX = velocidade máxima KM = constante de Michaelis = (K- 1 + K2 )/ K 1 [S] = concentração de substrato
Ao derivar-se a equação de velocidade de Michaelis-Menten, são feitas as considerações a seguir.
1. Concentrações relativas de E e S: A concentração de substrato ([S]) é muito maior do que a concentração da enzima ([E]), de modo que a porcentagem de substra-to ligado à enzima em qualquer tempo é pequena.
2. Hipótese do estado de equilíbrio: A [ES] não varia com
o tempo hipótese do estado de equilíbrio), isto é, a velocidade de formação de ES é igual àquela da degradação de ES (para E + S e para E + P). Em geral, um intermediário em uma série de reações é dito estar em estado de equilíbrio quando sua ve-locidade de síntese é igual a sua velocidade de degradação.
3. Velocidade inicial: Somente as velocidades iniciais da reação (VO) são utilizadas na análise das reações enzimáticas. Isso signifi ca que a velocidade de reação é medida assim que a enzima e o substrato são misturados. Nesse momento, a concentração de produto é muito pequena e, assim sendo, a velocidade de reação inversa de P para S pode ser ignorada.
Conclusões importantes sobre a cinética de Mi-chaelis-Menten
1. Características do KM. KM - a constante de Mi-chaelis – é característico de uma enzima e de determinado substrato seu, e refl ete a afi nidade da enzima para aquele substrato. O KM é numericamente igual à concentração do substrato na qual a velocidade da reação é igual a ½ VMAX.. O KM não varia com a concentração da enzima.
Figura 8
a. Km baixo – Um Km numericamente pequeno refl ete uma alta afi nidade da enzima pelo substrato, pois uma baixa concentração de substrato é neces-sária para atingir a metade da saturação da enzima – isto é, atingir a velocidade que é ½ Vmax (fi gura ao lado).
b. Km alto – Um Km numericamente grande (ele-vado) refl ete uma baixa afi nidade da enzima pelo substrato, pois é necessária uma alta concentração de substrato para atingir a metade da saturação da enzima.
2. Relação entre a velocidade e a concentração da enzima – a velocidade da reação é diretamente proporcional à concentração da enzima em qual-quer concentração de substrato. Por exemplo, se a concentração da enzima é reduzida pela metade, a velocidade inicial da reação (VO), assim como Vmax, são reduzidas à metade da velocidade ori-ginal.
3. Ordem de reação – quando a [S] é muito menor que o Km, a velocidade da reação é aproximadamen-te proporcional à concentração do substrato. A velo-cidade da reação é então dita de primeira ordem com relação ao substrato. Quando a [S] é muito maior do que o Km, a velocidade é constante e igual à VMAX. A velocidade da reação, nesse caso, é independente da concentração de substrato e é dita de ordem zero em relação à concentração de substrato.
Figura 9
Exercícios de Auto-Avaliação
1. As estatinas, por seu grande êxito na prevenção da doença coronariana, estão entre os medicamentos mais prescritos no mundo. Essas substâncias atuam sobre a enzima que regula a síntese de colesterol pelo fígado, denominada, simplifi cadamente, de HMG-CoA redutase.

27Para testar a efi ciência de vários derivados de estatinas, utilizou-se uma preparação de HMG-CoA redutase
isolada de tecido hepático. A velocidade de reação dessa preparação enzimática foi medida em função de con-centrações crescentes de seu substrato HMG-CoA, na ausência e na presença de uma concentração fi xa de três derivados de estatina. Nesses experimentos, o pH, a temperatura, a concentração da enzima e a concentração dos co-fatores necessários foram sempre mantidos constantes. O gráfi co a seguir representa os resultados en-contrados; a curva 1 foi obtida na ausência de estatinas.
a) Nomeie o tipo de mecanismo de ação das estatinas sobre a enzima HMG-CoA redutase hepática e justifi que sua resposta.
b) Aponte uma substância sintetizada a partir do colesterol em nosso organismo, não caracterizada como hormônio, e sua respectiva função.
2. A glicoquinase e a hexoquinase são duas enzimas que reagem com o mesmo substrato, a glicose. Ambas são enzimas intracelulares que fosforilam a glicose formando glicose 6-fosfato (G6P). Dependendo da enzima produtora, a G6P pode ou ser degradada na via da glicólise para gerar energia ou então ser usada para síntese de glicogênio.
A glicólise ocorre nos tecidos em geral e a síntese de glicogênio ocorre principalmente no fígado. A síntese do glicogênio somente acontece quando existe excesso de glicose no sangue. Essa é uma forma de armazenar esse açúcar.
Observe a fi gura a seguir, que apresenta as velocidades de reação dessas duas enzimas em função da concen-tração da glicose. Níveis normais de glicose no sangue estão ao redor de 4mM.
Qual das duas enzimas gera G6P para síntese de glicogênio hepático? Justifi que sua resposta.
Atividades Complementares
Depois de ler a Unidade II, sublinhe os termos que você não conhece pesquisando sobre eles.

28 UNIDADE III
ENERGÉTICA BIOQUÍMICA
3.1 - Princípios de Termodinâmica
A bioenergética descreve a transferência e a utilização da energia em sistemas biológicos. Ela utiliza algumas idéias básicas da termodinâmica, em especial o con-ceito de energia livre. Mudanças na energia livre (ΔG) fornecem uma medida da possibilidade, em termos energéticos, de que uma reação química ocorra e nos permitem, portanto, prever se uma reação ou processo pode acontecer.
O sinal de ΔG prediz o sentido da reação
A variação na energia livre, ΔG, pode ser utilizada para predizer o sentido de uma reação em condições de pressão e temperatura constantes. Considere a reação: A ↔ B.
1. ΔG negativo. Se ΔG é um valor negativo, há uma perda líquida de energia, e a reação anda espontane-amente no sentido em que está escrita, ou seja, A é convertido em B. A reação é dita exergônica.
2. ΔG positivo. Se ΔG é um valor positivo, há ganho líquido de energia e a reação não anda espontaneamente de B para A. A reação é dita endergônica, e alguma energia deve ser adicionada ao sistema para fazer com que a reação ande de A para B.
3. ΔG igual a zero. Se ΔG = 0, os reatantes estão em equilíbrio (quando uma reação ocorre esponta-neamente – ou seja, alguma energia livre está sendo perdida – a reação então continua, até que ΔG atinja o zero e o equilíbrio seja estabelecido).
ΔG de reações no sentido direto e inverso
A energia livre de uma reação (A B) no sentido direto (aquele em que está escrita) é de igual magni-tude, mas de sinal oposto àquela da reação no sentido inverso (B A). Por exemplo, se o ΔG da reação no sentido direto é -5.000 cal/mol, então o ΔG da reação no sentido inverso é +5.000 cal/mol.
3.2 - Papel de Aceptores de Energia
ATP como um Carregador de Energia
Reações ou processos que apresentam ΔG muito maior que zero, como íons movendo-se contra um gradiente de concentração através de uma mem-brana celular, podem ocorrer pelo acoplamento do movimento endergônico dos íons com um segundo processo espontâneo que apresente um ΔG bastante negativo, como a hidrólise de trifosfato de adenosina (ATP). A fi gura abaixo mostra um modelo mecânico de acoplamento de energia. Uma engrenagem à qual está amarrado um peso gira espontaneamente no sentido de alcançar o estado de menor energia, nesse caso com o peso na posição mais baixa (fi gura A). O movimento contrário (fi gura B) é energeticamente desfavorecido e não ocorre espontaneamente. A fi gura C mostra que o movimento energeticamente favorável de uma engrenagem pode ser utilizado para girar uma segunda engrenagem em um sentido para o qual ela não giraria espontaneamente. O exemplo mais simples de acoplamento energético em reações biológicas ocorre quando as reações que requerem energia e as reações que produzem energia compartilham um intermediário comum.
Figura 10

29O ATP consiste em uma molécula de adenosina
(adenina + ribose) à qual estão ligados três grupos de fosfato. Se um fosfato for removido, será produzido o difosfato de adenosina (ADP); se dois fosfatos forem removidos, teremos como resultado monofosfato de
adenosina (AMP). A energia livre padrão para a hidró-lise do ATP, ΔG0, é aproximadamente -7.300 cal/mol para cada um dos dois grupos fosfato terminais. Em função desse ΔG0 grande e negativo, o ATP é denomi-nado um composto fosfatado de alta energia.

30 UNIDADE IV
METABOLISMO DE CARBOIDRATOS
4.1 - Estrutura Geral
Os carboidratos são as moléculas orgânicas mais abundantes na natureza. Eles possuem uma grande variedade de funções, as quais incluem o forneci-mento de uma fração signifi cativa da energia na dieta da maioria dos organismos e a atuação como uma forma de armazenamento de energia no corpo e como componentes da membrana celular, mediando algumas formas de comunicação intracelular. Os carboidratos também servem como componentes estruturais de muitos organismos, incluindo a pa-rede celular de bactérias, o exoesqueleto de muitos insetos e as fi bras de celulose das plantas. A fórmula empírica para muitos dos carboidratos mais simples é (CH2O)n, daí o nome “hidratos de carbono”.
Os monossacarídeos (açucares simples) podem ser classifi cados de acordo com o número de átomos de carbono que contêm. Exemplos de alguns monossa-carídeos comumente encontrados em humanos estão listados na fi gura A. Os carboidratos com um aldeído como seu grupo funcional mais oxidado são deno-minados aldoses, enquanto aqueles com um grupo cetona como seu grupo funcional mais oxidado são chamados cetoses (fi gura B). Por exemplo, o glice-
raldeído é uma aldose, enquanto a diidroxiacetona é uma cetose. Os carboidratos que apresentam um grupo carbonila livre recebem o sufi xo “-ose”. (As cetoses [com algumas exceções, como a frutose] recebem duas letras adicionais no seu sufi xo; “-ulose”, como xilulose). Os monossacarídeos podem ligar-se por ligações glicosídi-cas, criando estruturas maiores. Os dissacarídeos contêm duas unidades de monossacarídeos, os oligossacarídeos contêm cerca de 3 a 12 unidades de monossacarídeos e os polissacarídeos contêm mais de 12 unidades de mo-nossacarídeos, podendo chegar a centenas de unidades de açucares em sua estrutura.
4.2 - Respiração Celular e Fermentação
Respiração Aeróbica
O metabolismo oxidativo nas mitocôndrias é alimentado não só pelo piruvato produzido a partir da glicólise de açúcares no citosol, mas também a partir de ácidos graxos.
A B
Figura 11

31
Figura 12
O piruvato e os ácidos graxos são seletivamente transportados do citosol para a matriz mitocondrial onde são quebrados a grupamentos acetílicos de dois carbonos da acetil coenzima A (acetil CoA); o grupa-mento acetílico é então introduzido no ciclo do ácido cítrico para futura degradação, e o processo termina com a passagem dos elétrons de alta energia derivados do acetil para a cadeia respiratória.
Para assegurar um suprimento contínuo de combus-tível para o metabolismo oxidativo, as células animais armazenam ácidos graxos, na forma de gorduras, e glicose, na forma de glicogênio.
O glicogênio é um polímero de glicose grande e ramifi cado contido em grânulos no citoplasma; a sua síntese e degradação são altamente reguladas de acordo com a necessidade. Quando tal necessidade aumenta, as células degradam o glicogênio para liberar glicose 1-fosfato, a qual é então submetida à glicólise.
As reações de glicólise convertem a molécula de glicose de seis carbonos (e açúcares relacionados) a duas moléculas de piruvato de três carbonos.
Figura 13
O piruvato é transportado do citoplasma para a matriz mitocondrial, onde ele encontra um comple-xo multienzimático gigante, o complexo piruvato desidrogenase. Este complexo – que contém múl-tiplas cópias de três enzimas, cinco coenzimas e duas proteínas regulatórias – rapidamente converte o piruvato em acetil CoA, liberando CO2 como produto lateral.
O ciclo do ácido cítrico responde por cerca de dois terços da oxidação total dos compostos de carbono da maioria das células, e os seus produtos fi nais são CO2 e elétrons de alta energia, os quais passam via NADH e FADH2, para a cadeia respiratória. O CO2 é liberado pelas células como produto residual desprezível, enquanto os elétrons de alta energia são movidos ao longo da cadeia respiratória e fi nalmente combinados com o O2 para produzir H2O.

32O ciclo do ácido cítrico se inicia quando a acetil CoA,
formada a partir de ácidos graxos ou piruvato, reage com o composto de quatro carbonos oxalacetato para produzir o ácido cítrico de seis carbonos, de onde o nome do ciclo é derivado. A seguir, como resultado de sete reações seqüenciais mediadas por enzimas, dois átomos de carbono são removidos na forma de
CO2 e o oxalacetato é regenerado. Cada volta do ciclo produz duas moléculas de CO2 a partir dos dois átomos de carbono que entraram nos ciclos prévios. Porém, o resultado líquido, no que concerne o grupamento ace-tílico da acetil CoA, é CH3COOH (como acetil CoA) + 2H2O + 3NAD+ + FAD ligado à proteína → 2CO2 + 3H+ + 3NADH + FADH2 ligado à proteína.
Figura 14
A mais importante contribuição do ciclo do ácido cítrico para o metabolismo é a extração de elétrons de alta energia durante a oxidação dos dois átomos de carbono do acetil para o CO2. Estes elétrons, tran-sitoriamente sustentados pelo NADH e pelo FADH2, são rapidamente transferidos à cadeia respiratória na membrana mitocondrial interna.
Embora o ciclo do ácido cítrico seja parte do meta-bolismo aeróbico, nenhuma das reações que levam à produção de NADH e FADH2 faz uso direto do oxigênio molecular; somente nas reações catabólicas fi nais, que ocorrem na membrana mitocondrial interna, é que o oxigênio é consumido diretamente. Nos estágios iniciais da oxidação, aproximadamente toda a energia disponível
pela queima de carboidratos, gorduras e outros produtos alimentares é primeiramente salva na forma de elétrons de alta energia removidos dos substratos por NAD+ e FAD. Estes elétrons, carregados por NADH e FADH2, são então combinados com o oxigênio molecular através da cadeia respiratória. Devido ao fato da grande quanti-dade de energia liberada ser empregada pelas enzimas da membrana interna para a conversão de ADP + Pi o termo fosforilação oxidativa é usado para descrever esta última série de reações.
De acordo com a hipótese quimiosmótica, os inter-mediários químicos de alta energia são substituídos por uma ligação entre os processos químicos (“quimi”) e os processos de transporte (“osmótico”- do grego osmos,

33empurrar) por isso, acoplamento quimiosmótico. À medida que os elétrons de alta energia, derivados dos hidrogênios de NADH e FADH2, são transportados pela cadeia respiratória na membrana mitocondrial interna, a energia liberada pelas suas passagens de uma molécula carreadora para a próxima é utilizada para bombear prótons (H+) da matriz mitocondrial para o
espaço intermembranas, através da membrana interna. Isto gera um gradiente eletroquímico de prótons através da membrana mitocondrial interna, e o refl uxo de H+ a favor desse gradiente é, por sua vez, utilizado para dirigir a enzima ATP sintase ligada à membrana, a qual catalisa a conversão de ADP +Pi em ATP, completando o processo de fosforilação oxidativa.
Figura 15
A reação energeticamente favorável H2 + ½ O2 → H2O é realizada em várias pequenas etapas, de tal forma que grande parte da energia liberada pode ser convertida em uma forma estocável de energia, ao in-vés de ser perdida para o meio ambiente como calor.
O processo de transporte de elétrons inicia quando o íon hidreto é removido do NADH, para regenerar NAD+, e é convertido em um próton e dois elétrons (H: -→ H+ + 2e-).
Os dois elétrons são passados para o primeiro de uma série de mais de 15 carreadores de elétrons diferentes da cadeia respiratória. Os elétrons iniciam com ener-gias muito altas e gradativamente as perdem, à medida que fl uem ao longo da cadeia. Na sua maior parte, os elétrons passam de um átomo metal para outro, sendo que cada átomo encontra-se fortemente ligado a uma molécula protéica que altera a afi nidade eletrônica do átomo de metal.
É importante salientar que as várias proteínas en-volvidas estão agrupadas em três grandes complexos enzimáticos respiratórios. Cada complexo da cadeia tem uma afi nidade maior para elétrons do que o seu predecessor, e os elétrons fl uem seqüencialmente de um complexo para o outro até que sejam fi nalmente
transferidos para o oxigênio, o qual tem, entre os componentes da cadeia, a mais alta afi nidade pelos elétrons.
A fosforilação oxidativa só é viável graças à íntima associação entre os carreadores de elétrons e moléculas protéicas. As proteínas guiam os elétrons através da cadeia respiratória, de tal forma que estes se movem seqüencialmente de um complexo enzimático para outro.
À medida que um elétron de alta energia passa ao longo da cadeia transportadora de elétrons, parte da energia liberada é utilizada para direcionar três complexos enzimáticos respiratórios que bombeiam H+ para fora da matriz mitocondrial. O resultante gra-diente eletroquímico de prótons, através da membrana interna, dirige H+ de volta através da ATP sintase, um complexo protéico transmembrana que utiliza a energia do fl uxo de H+ para sintetizar ATP a partir de ADP e Pi na matriz.
Devido ao fato do oxigênio ter grande afi nidade por elétrons, uma grande quantidade de energia livre é liberada quando ele é reduzido para formar água. Portanto, a evolução da respiração celular, onde o O2 é convertido em água, permitiu aos organismos pro-

34cessar muito mais energia do que poderia ser obtida pelo metabolismo anaeróbico.
Fermentação
No processo de fermentação, o ATP é fabricado por um evento de fosforilação que utiliza energia liberada quando uma molécula orgânica rica em hidrogênios, tal como a glicose, é parcialmente oxidada. Sem o O2 para servir como aceptor fi nal de elétrons, os elétrons perdidos pela molécula orgâ-nica oxidada devem ser transferidos (via NADH ou NADPH) para uma molécula orgânica diferente (ou para uma porção diferente da mesma molécula), a qual, conseqüentemente, se torna mais reduzida. Ao fi nal do processo de fermentação, uma (ou mais) das moléculas orgânicas produzidas é excretada no meio como produto fi nal do metabolismo; outras, tal como o piruvato, são retidas na célula para biossíntese.
Os produtos fi nais excretados são diferentes para cada organismo, mas tendem a ser ácidos orgânicos (compostos de carbono que contêm um grupamento COOH). Em células bacterianas, entre os mais impor-tantes desses produtos estão o ácido lático (que também
é acumulado na glicólise anaeróbica de mamíferos) e os ácidos fórmico, acético, propiônico, butírico e succínico. Duas rotas de fermentação presentes em bactérias de hoje estão ilustradas abaixo.
4.3 - Fotossíntese
As muitas reações que ocorrem durante a fotossíntese podem ser agrupadas em duas grandes categorias. Nas reações fotossintetizantes de transferência de elétrons (também chamadas de reações da “fase clara” ou da “fase luminosa”), a energia derivada da luz solar energiza um elétron da clorofi la, capacitando-o a se mover através de uma cadeia transportadora de elétrons da membrana tila-cóide de uma maneira muito semelhante ao movimento de elétrons na cadeia respiratória das mitocôndrias. A clorofi la obtém os seus elétrons da água com a liberação de O2. Durante o processo de transporte de elétrons, H+ é bombeado através da membrana tilacóide e a força próton-motriz resultante direciona a síntese de ATP no estroma. Como fase fi nal desta série de reações, os elétrons de alta energia são carregados (juntamente com H+) no NADP+, convertendo-o em NADPH. Todas estas reações estão confi nadas ao cloroplasto. Nas reações de fi xação do carbono (também chamadas de reações da fase escura), o ATP e o NADPH produzidos pelas reações fo-tossintetizantes de transferência de elétrons servem como fonte de energia e como força redutora, respectivamente, para dirigir a conversão de CO2 em carboidratos. As reações de fi xação do carbono, que iniciam no estroma cloroplástico e continuam no citosol, produzem sacarose nas folhas do vegetal; deste local, o açúcar é exportado para outros tecidos como fonte da moléculas orgânicas e energia para o crescimento.
Complexos multiprotéicos, chamados de fotossiste-mas, catalisam a conversão da energia luminosa cap-turada em moléculas excitadas de clorofi la em formas de energia utilizável.
O complexo da antena é importante para a captação da luz. Nos cloroplastos, ele consiste de um aglomerado de várias centenas de moléculas de clorofi la unidas por proteínas que as sustentam fi rmemente sobre a membrana tilacóide.
Figura 16
Figura 17

35Fase Clara
A fotossíntese em plantas e cianobactérias produz ATP e NADPH diretamente por um processo de duas etapas, chamado fotofosforilação acíclica. Como dois fotossistemas em série são utilizados para energizar um elétron, o elétron pode ser transferido por todo o caminho entre a água e o NADPH. Na medida em que os elétrons de alta energia passam através dos fotossistemas, acoplados para gerar NADPH, parte da sua energia é desviada para a síntese de ATP.
No primeiro dos dois fotossistemas – chamado de fotos-sistemas II por razões históricas –, os oxigênios de duas moléculas de água ligam-se a um agrupamento de átomos de manganês de uma enzima pouco conhecida, capaz de romper a molécula de água, o que permite que os elétrons sejam removidos um de cada vez para preencher os espaços criados pela luz nas moléculas de clorofi la do centro de reação. Logo que os quatro elétrons das duas moléculas de água tenham sido removidos (o que requer quatro quanta de energia luminosa), O2 é liberado; o fotossistema II, portanto, catalisa a reação 2H2O + 4 prótons → 4H+ + 4e- + O2.
Figura 18
As quinonas passam os seus elétrons a uma bomba de H+ chamada complexo b6-ƒ. O complexo b6-f bombeia H+ para o espaço tilacóide, através da membrana tilacói-de (ou para o citosol, através da membrana plasmática em cianobactérias), e o gradiente eletroquímico resul-tante dirige a síntese de ATP pela ATP sintase. O aceptor fi nal de elétrons, nesta cadeia transportadora de elétrons, é o segundo fotossistema (fotossistema Ι), que aceita um elétron no espaço criado pela luz em uma molécula de clorofi la do seu centro de reação. Cada elétron que entra no fotossistema I é excitado em um nível muito alto de energia, o que permite que este seja transferido para o centro de ferro-enxofre da ferredoxina e, então, ao NADP+ para gerar NADPH; esta última etapa envolve a captura de um H+ do meio.
Para produzir ATP extra, os cloroplastos de algumas espécies vegetais podem converter o fotossistema I para um modo cíclico, de forma que produza ATP, ao
invés de NADPH. Neste processo, chamado de fotofos-forilação cíclica, os elétrons de alta energia do fotossis-tema I são transferidos de volta para o complexo b6-f, ao invés de seguirem o seu caminho para o NADP+, e o elétron é então reciclado para o fotossistema I com uma baixa energia. O único resultado líquido, além da conversão de parte da energia luminosa em calor, é que H+ é bombeado através da membrana tilacóide pelo complexo b6-f para aumentar, assim, o gradiente eletroquímico de prótons que propulsiona a síntese de ATP.
Em resumo, a fotofosforilação cíclica envolve so-mente o fotossistema I e produz ATP sem formação de NADPH ou O2. Portanto, as atividades relativas dos fl uxos cíclico e acíclico de elétrons podem determinar quanto de energia luminosa é convertida em força redutora (NADPH) e quanto é convertida nas altas energias das ligações de fosfato (ATP).

36Fase Escura
Na reação central de fi xação do carbono, na qual um átomo de carbono inorgânico é convertido em carbono orgânico, o CO2 da atmosfera combina-se com o com-posto ribulose 1,5- bifosfato de cinco carbonos mais água para resultar em duas moléculas do composto 3- fosfoglicerato de três carbonos. Esta reação de “fi -xação do carbono”, descoberta em 1948, é catalisada no estroma cloroplástico por uma grande enzima de ribulose bifosfato carboxilase.
Porém, para produzir um suprimento sufi ciente de ribulose 1,5- bifosfato, é necessário uma série de rea-ções que consomem grandes quantidades de NADPH e ATP. Três moléculas de CO2 são fi xadas pela ribulose bifosfato carboxilase para produzir 6 moléculas de 3- fosfoglicerato (contendo 6x3=18 átomos de carbono no total: 3 do CO2 e 15 da ribulose 1,5- bifosfato). Os 18 átomos de carbono, então, sofrem um ciclo de reações que regeneram as 3 moléculas de ribulose 1,5 bifosfato utilizadas na etapa inicial de fi xação do carbono (3x5 = 15 átomos de carbono).
Isso deixa uma molécula de gliceraldeído 3- fosfato (3 átomos de carbono) como ganho líquido. Neste ciclo de fi xação do carbono (ou ciclo de Calvin-Benson), três moléculas de ATP e duas moléculas de NADPH
são consumidas para cada molécula de CO2 contida em carboidrato. A reação simplifi cada é:
3CO2 + 9ATP + 6NADPH + água → gliceraldeído 3-fosfato + 8Pi + 9ADP + 6NADP+
O gliceraldeído 3-fosfato produzido nos cloroplastos pelo ciclo de fi xação do carbono é um açúcar de três carbonos que serve como intermediário central na glicólise. Grande parte dele é exportado para o citosol, onde pode ser convertido em frutose 6-fosfato e glicose 1-fosfato, pela inversão de várias reações da glicose. A glicose 1-fosfato é então convertida no nucleotídeo-açúcar UDP- glicose, e este combina-se com a glicose 6-fosfato para formar sacarose-fosfato, o precursor imediato do dissacarídeo sacarose. A sacarose é a principal forma de transportar açúcar entre as células vegetais; assim como a glicose é transportada no san-gue dos animais, fornecendo o carboidrato necessário pelo resto do vegetal.
A maior parte do gliceraldeído 3-fosfato que perma-nece no cloroplasto é convertido em amido no estroma. Assim como o glicogênio em células animais, o amido é um grande polímero de glicose que serve como car-boidrato de reserva. À noite, o amido é quebrado para auxiliar no suprimento das necessidades metabólicas do vegetal.
Figura 19

374.4 - Shunt das Pentoses
A via das pentoses-fosfato ocorre no citosol da célula. Nenhum ATP é consumido ou produzido diretamente no ciclo. O carbono 1 da lactose-6-fos-fato é liberado na forma de CO2, e dois NADPHS são produzidos para cada molécula de glicose-6-fosfato que entra na parte oxidante da via. A velocidade e o sentido das reações reversíveis da via são determina-dos pela oferta e demanda de intermediários do ciclo. A via proporciona a maior quantidade de NADPH do organismo, sendo que esse nucleotídio tem como função agir como redutor bioquímico. A via produz também a ribose-5-fosfato, necessária para a biossín-tese de nucleotídeos, e proporciona um mecanismo para o uso metabólico de açúcares de 5 carbonos,
obtidos da dieta ou da degradação de carboidratos estruturais do organismo.
A coenzima NADP+ difere da coenzima NAD+ apenas pela presença de um grupo fosfato (- PO2-4) em uma das unidades da ribose. Essa mudança aparentemente pequena permite que o NADP+ interaja com as enzimas específi cas para NADP+, que cumprem papéis únicos na célula.
O NADPH pode ser considerado uma molécula de alta energia, da mesma forma que o NADH. Contudo, os elétrons do NADPH são destinados à biossíntese redutora, em vez da transferência para o oxigênio, como é o caso dos elétrons do NADH.
Figura 20
O peróxido de hidrogênio é um membro da família das espécies reativas de oxigênio, formado a partir da redução parcial do oxigênio molecular. Esses compostos são formados continuadamente como subprodutos do metabolismo aeróbico, por meio de reações com drogas e toxinas do ambiente e quando o nível de antioxidantes é reduzido, situações que criam condições para o estresse oxidativo. Os intermediários altamente reativos de oxigênio podem causar danos
químicos graves ao DNA, às proteínas e aos lipídios insaturados, podendo levar à morte celular. Essas espécies reativas de oxigênio tem sido implicadas em uma série de processos patológicos, entre os quais câncer, doenças infl amatórias e envelhecimento. A célula possui diversos mecanismos protetores, que minimizam o potencial tóxico desses compostos.
Deficiência da Glicose-6-fosfato-desidrogenase
Figura 21

38A defi ciência da glicose-6-p-desidrogenase (G6PD) é uma
doença hereditária que se caracteriza por anemia hemolítica, causada pela incapacidade de destoxifi car agentes oxidantes. A defi ciência na G6PD é a mais comum anormalidade enzi-mática que produz doenças em seres humanos, afetando mais de 200 milhões de indivíduos no mundo inteiro. A sua mais alta incidência está no Oriente Médio, na África tropical e na Ásia, além de partes do mediterrânio. A defi ciência de G6PD é ligada ao cromossomo X. A vida de muitos indivíduos com defi ciência de G6PD é, de certo modo, reduzida por compli-cações resultantes da hemolise crônica. Esse efeito negativo da defi ciência de G6PD foi contrabalançado, ao longo da evolução, por uma vantagem na sobrevivência – uma maior resistência à malária falcípara, apresentada por mulheres portadoras da mutação (o traço falciforme e a β-talassemia menor também conferem resistência).
A diminuição da G6PD prejudica a capacidade da célula de formar NADPH. Isso resulta em uma
queda na destoxificação celular de radicais livres e peróxidos, formados no interior da célula. A oxidação leva à desnaturação de proteínas, que formam massas insolúveis (chamadas de corpos de HEINZ) que se associam às membranas do eritrócito. A oxidação adicional das proteínas das membranas faz com que os eritrócitos se tornem rígidos e não deformáveis, sendo removidos da circulação por macrófagos no baço e no fígado. Embora a deficiência da G6PD ocorra em todas as células do indivíduo afetado, ela é mais grave nos eritrócitos, onde a via das pentoses-fosfato corresponde a única forma de gerar NADPH. Outros tecidos têm fontes alternativas para a pro-dução de NADPH. Os eritrócitos não tem núcleo ou ribossomos e não pode renovar seu suprimento da enzima, sendo assim os eritrócitos são espe-cialmente vulneráveis a variantes da enzima com estabilidade reduzida.
4.5 - Glicogenólise e Glicogênese
A glicose é fonte preferencial de energia para o encéfalo e fornece energia necessária para células com poucas ou nenhuma mitocôndria, como os eritrócitos maduros, ela também é essencial como fonte de energia para o múscu-lo em exercício. A glicose sangüínea pode ser obtida de três fontes principais: dieta, degradação do glicogênio e Gliconeogênese. A ingestão da glicose e de seus precur-sores é esporádica e, dependendo do tipo de alimentação, nem sempre representa uma fonte segura de glicose para o sangue. Em contraste, a gliconeogênese pode fornecer uma síntese sustentada de glicose, mas é um tanto lenta para responder a uma redução no nível sangüíneo dessa substância. Sendo assim, o corpo desenvolveu mecanismos para armazenar um suprimento de glicose em uma forma rapidamente mobilizável, o glicogênio. Na ausência de uma fonte de glicose na alimentação, esse composto é rapida-mente liberado a partir do glicogênio hepático e renal.
Os principais estoques de glicogênio no corpo se en-contram nos músculos esqueléticos e no fígado, embora a maioria das outras células armazena pequenas quantida-des para uso próprio. A função do glicogênio muscular é servir como reserva de combustível para a síntese de ATP durante a contração muscular. A função do glicogênio hepático é manter a concentração de glicose sangüínea, especialmente durante o início do jejum.
Síntese de Glicogênio (Glicogênese ou Gli-cogenogênese)
O glicogênio é sintetizado a partir das moléculas de α–D–glicose. O processo ocorre no citosol e requer
energia fornecida pelo ATP (para a fosforilação da glicose) e trifosfato de uridina (UTP).
Degradação do Glicogênio (Glicogenólise)
A via de degradação, que mobiliza o glicogênio armazenado no fígado e no músculo esquelético, não é o inverso das reações de síntese. Em vez disso, um conjunto de enzimas citosólicas diferentes é necessá-rio. Os passos para a degradação do glicogênio são: encurtamento de cadeias, remoção das ramifi cações, conversão de glicose-1-fosfato em glicose-6-fosfato e degradação lisossômica do glicogênio.
Regulação da Síntese e da Degradação do Glicogênio
A síntese e a degradação do glicogênio são firme-mente reguladas. No fígado, a síntese do glicogênio é acelerada quando o corpo está bem alimentado, enquanto a degradação do glicogênio é acelerada em períodos de jejum. No músculo esquelético, a degradação do glicogênio ocorre durante o exercí-cio e a síntese começa assim que o músculo entra novamente em descanso. A regulação da síntese e da degradação do glicogênio ocorre em dois níveis. Em primeiro lugar, a glicogênio-síntase e a glico-gênio-fosforilase são controladas alostericamente. Em segundo, as vias de síntese e de degradação do glicogênio são reguladas hormonalmente.

394.6 - Gliconeogênese
Alguns tecidos como o encéfalo, os eritrócitos, a medula renal, o cristalino e a córnea, os testículos e o músculo em exercício, requerem um suprimento contínuo de glicose como combustível metabólico. O glicogênio hepático pode satisfazer essas necessidades por apenas 10 a 18 horas na ausência de ingestão de carboidratos. Durante um jejum prolongado os depósi-tos de glicogênio hepático são depletados. E a glicose é formada a partir de precursores como o lactato, o piruvato, o glicerol e os α-cetoácidos (catabolismo de aminoácidos). Durante um jejum de uma noite, cerca de 90% da gliconeogênese ocorre no fígado, com os rins fornecendo 10% das moléculas de glicose recém-sintetizadas. Durante o jejum prolongado, no entanto, os rins contribuem com aproximadamente 40% da produção total de glicose.
Substratos para a gliconeogênese são moléculas que podem ser usadas na produção líquida de glicose. In-cluem todos os intermediários da glicólise e do ciclo
do ácido cítrico, glicerol, lactato e α-cetoácidos, são os mais importantes precursores gliconeogênicos.
O glicerol é liberado durante a hidrólise de triacil-gliceróis, no tecido adiposo, e é levado ao fígado pelo sangue. O glicerol é fosforilado pela glicerol-cinase, resultando em glicerol-fosfato, que é oxidado pela glicerol-fosfato-desidrogenase, produzindo diidro-xiacetona-fosfato, um intermediário da glicólise (os adipócitos não podem fosforilar o glicerol, pois não apresentam a glicerol-cinase).
O lactato é liberado no sangue pelo músculo esque-lético em exercício e pelas células que não possuem mitocondrias, como os eritrócitos. Esse lactato é captado pelo fígado e reconvertido em glicose, que é liberada de volta para a circulação.
Os aminoácidos obtidos pela hidrólise de proteínas teciduais são fontes de glicose no jejum.
Exercícios de Auto-Avaliação
1. Algumas etapas metabólicas encontradas na citoplasma das células hepáticas de mamíferos.
Cite as duas etapas, dentre as representadas, que são estimuladas pela ação da insulina.
2. O esquema abaixo representa um processo bioquímico utilizado na fabricação de pães, vinhos, cervejas e outros produtos de grande importância para o ser humano.
a) Que processo bioquímico está representado no esquema?b) Qual o papel desse processo no funcionamento das células que são capazes de realizá-lo?c) Em células musculares é responsável a ocorrência desse processo bioquímico? Explique.
3. Existem seres vivos, ou mesmo células de um organismo, que são chamados de anaeróbicos facultativos. Estes respiram aerobicamente enquanto há oxigênio disponível. No entanto, se o oxigênio faltar, esses seres ou essas células podem degradar a glicose anaerobicamente, realizando a fermentação. Pergunta-se:
a) Na fermentação, o consumo de glicose é maior ou menor do que o usado no processo aeróbico?b) Justifi que sua resposta.

404. Os açúcares complexos, resultantes da união de muitos monossacarídeos, são denominados polissacarídeos.a) Cite dois polissacarídeos de reserva energética, sendo um de origem animal e outro de origem vegetal.b) Indique um órgão animal e um órgão vegetal, onde cada um destes açúcares pode ser encontrado.
5. Analise os gráfi cos seguintes.
a) Considerando P, Q e Z, qual deles corresponde a água, a carboidratos e a fi bras?b) Com base no gráfi co da semente, explique sucintamente qual a vantagem adaptativa de se apresentar tal
proporção de carboidratos, lipídios, proteínas e água na composição de seus tecidos.
Atividades Complementares
Depois de ler a Unidade IV, sublinhe os termos que você não conhece pesquisando sobre eles.

41UNIDADE V
METABOLISMO DE LIPÍDIOS
5.1 - Estrutura Geral
Os lipídios biológicos constituem um grupo de com-postos que, apesar de quimicamente diferentes entre si, exibem a sua insolubilidade em água como característica defi nidora e comum a todos. As funções biológicas dos diferentes lipídios são igualmente diversas. Em muitos organismos, óleos e gorduras são as formas principais de armazenar energia, enquanto os fosfolipídios e os esteróis representam perto de metade da massa das mem-branas biológicas. Outros lipídios, embora presentes em quantidades relativamente pequenas, desempenham papéis cruciais como cofatores enzimáticos, transporta-dores de elétrons, pigmentos que absorvem radiações luminosas, âncoras hidrofóbicas, agentes emulsifi cantes, hormônios e mensageiros intracelulares.
Óleos e gorduras, empregados quase universalmente como formas de armazenamento de energia nos orga-nismos vivos, são compostos, do ponto de vista químico, altamente reduzidos e derivados dos ácidos graxos.
Os ácidos graxos são ácidos carboxílicos com cadeias hidrocarbonadas de 4 a 36 átomos de carbono. Em alguns ácidos graxos esta cadeia é totalmente saturada (não contém duplas ligações) e não é ramifi cada; outros contêm uma ou mais duplas ligações.
Os lipídios mais simples constituídos de ácidos gra-xos são os triacilgliceróis, também referidos como
triglicerídios, gorduras ou gorduras neutras. Os tria-cilgliceróis são moléculas hidrofóbicas e não-pola-res, essencialmente insolúveis em água. Na maioria das células eucarióticas os triacilgliceróis formam uma fase separada de gotículas microscópicas ole-osas no citosol aquoso, servindo como depósitos de combustível metabólico. Nos animais vertebrados as células especializadas chamadas adipócitos, ou células gordurosas, armazenam grandes quantidades de triacilgliceróis como gotas de gordura, as quais preenchem quase toda a célula. Os triacilgliceróis também são armazenados nas sementes de muitos tipos de plantas, fornecendo energia e precursores biossintéticos quando da germinação da semente.
Como combustíveis estocados os triacilgliceróis têm duas vantagens signifi cativas sobre os polissa-carídios, como o glicogênio e o amido. Os átomos de carbono dos ácidos graxos são quimicamente mais reduzidos que aqueles dos açúcares e, por-tanto, a oxidação dos triacilgliceróis libera, grama por grama, uma quantidade de energia mais do que duas vezes maior que a liberada pelos carboidratos. Como os triacilgliceróis são hidrofóbicos e por isso desidratados, o organismo que transporta gordura como combustível não tem que carregar o peso extra da água de hidratação que está sempre armazenada aos polissacarídios armazenados.
5.2 - Metabolismo de Lipídios
Os ácidos graxos existem no organismo na forma livre e também como moléculas mais complexas, tais como os triacilglicerol. Em todos os tecidos, existem níveis baixos de ácidos graxos livres, entretanto, algumas vezes, pode-se encontrar quantidades substanciais no plasma, em especial durante o jejum. Ácidos graxos livres no plasma estão circulando a partir da origem (triacilgliceróis do tecido adiposo ou das lipoprote-ínas da circulação) para o sítio de consumo (outros tecidos).
Estrutura
Um ácido graxo consiste em uma cadeia hidrofóbica de hidrocarboneto com um grupo carboxila terminal que tem afi nidade com a água, conferindo ao ácido graxo natureza anfi pática (apresenta uma região hidro-
fóbica e outra hidrofílica). Nos ácidos graxos de cadeia longa, a porção hidrofóbica é predominante. Essas moléculas são altamente insolúveis em água e preci-sam ser transportadas pela circulação associadas às proteínas. Mais de 90% dos ácidos graxos encontrados no plasma estão em forma de ésteres de ácidos graxos (triacilgliceróis, ésteres de colesterol e fosfolipídeos), contidos nas partículas de lipoproteínas da circulação. Ácidos graxos não-esterifi cados são transportados na circulação associados com a albumina.

42Síntese
Uma grande proporção de ácidos graxos usados pelo organismo é suprida pela dieta. Carboidratos, proteínas e outras moléculas, quando obtidas da dieta em excesso em relação às necessidades desses compostos, podem ser convertidos em ácidos graxos, que são armazenados como triacilgliceróis. Em humanos, a síntese dos ácidos graxos ocorre principalmente no fígado e nas glândulas mamárias em lactação e, em menor extensão, no tecido adiposo. O processo incorpora carbonos a partir da acetil-CoA na cadeia de ácido graxo em crescimento, usando ATP e NADPH.
Armazenamento
Mono-, Di - e triacilgliceróis consistem em uma, duas ou três moléculas de ácidos graxos esterifi cando uma molécula de glicerol (se uma molécula de acilglicerol é sólida à temperatura ambiente, é chamada de “gordu-ra”, se é líquida, é chamada de “óleo”). Como os TAGS são fracamente solúveis em água e não formam micelas estáveis, eles coalescem dentro dos adipócitos, forman-do gotas oleosas. Essas gotas lipídicas citosólicas são a maior reserva energética do organismo.
No tecido adiposo, o TAG é armazenado no citosol das células. Ele serve como um “depósito de gordura”, prontamente mobilizado como combustível quando o organismo necessitar. Pouco TAG é armazenado no fígado. Por outro lado, muitos TAGS são exportados,
agrupados com ésteres de colesterol, fosfolipídeos e proteínas para formar partículas lipoprotéicas cha-madas de lipoproteínas de muito baixa densidade (VLDLS). A VLDL nascente é secretada para o sangue, onde amadurece e funciona entregando lipídeos endó-genos para os tecidos periféricos.
Corpos cetônicos: um combustível alternativo para as células
A mitocôndria do fígado tem a capacidade de con-verter acetil-CoA proveniente da β-oxidação de ácidos graxos em corpos cetônicos. Os compostos classifi -cados como corpos cêtonicos são o acetoacetato, o 3-hidroxibutirato e a acetona (um produto colateral não-metabolizável). O acetoacetato e o 3-hidroxibuti-rato são transportados pelo sangue aos tecidos perifé-ricos. Ali, eles podem ser convertidos novamente em acetil-CoA, que é oxidada no ciclo do ácido cítrico. Os corpos cetônicos são importantes fontes de energia para os tecidos periféricos, porque: 1) são solúveis em meio aquoso e não necessitam ser incorporados a lipoproteínas ou transportados pela albumina, como outros lipídeos; 2) são produzidos no fígado durante períodos em que a quantidade de acetil-CoA excede a capacidade oxidativa do fígado e 3) são usados pelos tecidos extra-hepáticos, como os músculos esquelético e cardíaco e o córtex renal, em quantidade proporcional a sua concentração no sangue. Mesmo o cérebro pode usar corpos cetônicos como fonte de energia, se os níveis sangüíneos aumentarem sufi cientemente (isso é importante durante o jejum prolongado).
5.3 - Biossíntese de Esteróides
O colesterol é o esteróide característico dos tecidos animais e desempenha várias funções essenciais no organismo. Por exemplo, o colesterol é componente de todas as membranas celulares, modulando sua fl uidez e, em tecidos especializados, e o precursor de ácidos bi-liares, hormônios esteróides e da vitamina D. Portanto, é muito importante para o organismo manter um supri-mento contínuo de colesterol. Um complexo sistema de transporte, biossíntese e mecanismos regulatórios está envolvido no atendimento dessa necessidade. O fígado desempenha um papel central na regulação da homeostasia do colesterol.
A maior parte do colesterol plasmático está na forma esterifi cada, que é uma molécula ainda mais hidrofóbi-ca que o colesterol livre. Os ésteres de colesterol não são encontrados nas membranas celulares, existindo, normalmente, em pequena quantidade na maioria das células. Devido a sua hidrofobicidade, o colesterol e seus ésteres são transportados associados a proteínas, como componentes das lipoproteínas, ou são solubili-zados por fosfolipídeos e sais biliares na bile.
O colesterol é sintetizado por praticamente todos os tecidos humanos, embora fígado, intestino, córtex adrenal e os tecidos reprodutivos, incluindo ovários, testículos e placenta, sejam os maiores contribuintes do “conjunto” de colesterol. A síntese acontece no citoplasma, sendo as enzimas encontradas no citosol e nas membranas do retículo endoplasmático.
Em humanos, a estrutura cíclica do colesterol não pode ser degradada até CO2 e H2O. O núcleo esterol é eliminado intacto pela conversão em ácidos e sais biliares, que são excretados nas fezes, e pela secreção de colesterol na Bile, a qual o transporta até o intestino para eliminação. Bactérias intestinais podem modifi car o colesterol antes da excreção.
Hormônios Esteróides
O colesterol é o precursor de todas as classes de hormônios esteróides: glicocorticóides (por exemplo, cortisol), mineralocorticóides (por exemplo, aldoste-rona) e hormônios sexuais – andrógenos, estrógenos e

43progestágenos (glicocorticóides e mineralocorticóides são coletivamente chamados de corticosteróides). A síntese e a secreção do cortisol, da aldosterona e dos andrógenos ocorrem no córtex adrenal, a síntese dos estrógenos, nos ovários e na placenta e da testosterona, nos testículos. Os hormônios esteróides são transpor-tados pelo sangue de seus sítios de síntese até o local de ação. Devido a sua hidrofobicidade, eles devem
5.4 - Prostaglandinas, Prostaciclinas e Leucotrienos
As prostaglandinas (PGS) e os compostos relaciona-dos tromboxanos (TXS) e leucotrienos (LTS) são coleti-vamente conhecidos como eicosanóides, refl etindo sua origem a partir de ácidos graxos poliinsaturados com 20 carbonos. São compostos extremamente potentes, que provocam uma ampla faixa de respostas fi sioló-gicas e patológicas. Embora tenham sido comparados com hormônios em termos de suas ações, os eicosa-nóides diferem dos hormônios verdadeiros porque são produzidos em quantidade muito pequenas por quase
ser complexados com uma proteína plasmática. A albumina plasmática pode atuar como transportador inespecífi co, por exemplo, para a aldosterona. Entre-tanto, proteínas plasmáticas específi cas para trans-portar esteróides ligam os esteróides mais fortemente que a albumina, como a globulina transportadora de corticosteróides (transcortina) e a proteína ligadora de hormônios sexuais.
todos os tecidos em vez de serem produzidos por um determinado tecido ou uma glândula especializada. Eles também atuam mais localmente, e não em alvos distantes, atingidos por meio da corrente circulatória, como fazem os hormônios. Os eicosanóides não são armazenados e têm uma meia-vida extremamente curta, sendo rapidamente metabolizados a compostos inativos no seu local de síntese. Suas ações biológicas são mediadas por receptores nas membranas plasmática e nuclear, que variam nos diferentes tecidos.
Exercícios de Auto-Avaliação
1. Com relação às substâncias químicas dos seres vivos resolva os itens a seguir:a) Qual é a forma de armazenamento dos carboidratos nos tecidos animais e vegetais, respectivamente?b) Qual é a unidade monomérica dos ácidos nucléicos?c) Em qual tipo de lipídeo são classifi cados os óleos e gorduras?d) Cite um dos aspectos que permite distinguir as diversas proteínas.
2. Os esteróides são lipídios bem diferentes dos glicerídeos e das ceras, apresentando uma estrutura composta por quatro anéis de átomos de carbono interligados. O colesterol é um dos esteróides mais conhecidos, devido à sua associação com as doenças cardiovasculares. No entanto, este composto é muito importante para o homem, uma vez que desempenha uma série de funções. Com base no que estudou, diga:
a) Duas principais funções do colesterol.b) Duas origens do colesterol sangüíneo.
3. Os lipídios têm papel importante na estocagem de energia, estrutura de membranas celulares, visão, controle hormonal, entre outros. São exemplos de lipídios: fosfolipídios, esteróides e carotenóides.
a) Como o organismo humano obtém os carotenóides? Que relação têm com a visão?b) A quais das funções citadas no texto acima os esteróides estão relacionados? Cite um esteróide importante
para uma dessas funções.c) Cite um local de estocagem de lipídios em animais e um em vegetais.
Atividades Complementares
Depois de ler a Unidade V, sublinhe os termos que você não conhece pesquisando sobre eles.

44 UNIDADE VI
METABOLISMO DE AMINOÁCIDOS E PROTEÍNAS
6.1 - Ciclo da Uréia
A uréia é a principal forma de eliminação dos grupos amino oriundos dos aminoácidos e perfaz cerca de 90% dos com-ponentes nitrogenados da urina. Um átomo de nitrogênio da molécula de uréia é fornecido por NH3 livre e o outro pelo aspartato. O carbono e o oxigênio da uréia são derivados do CO2. A uréia é produzida pelo fígado e então transportada pelo sangue até os rins, para ser excretada na urina.
Formação da citrulina As duas primeiras reações que levam à síntese de uréia ocorre na mitocôndria, enquanto as demais enzimas do ciclo estão localizadas no citosol. A ornitina e a citrulina são aminoácidos básicos, que participam do ciclo da uréia.
Clivagem da arginina, resultando em ornitina e uréia A arginase cliva a arginina em ornitina e uréia. Essa enzima ocorre quase que exclusiva-mente no fígado.
Destino da uréia A uréia sai do fígado por di-fusão e é transportada no sangue até os rins, onde é fi ltrada e excretada na urina. Parte da uréia difunde do sangue para o intestino, onde é clivada em CO2 e NH3 pela urease bacteriana. Essa amônia é parcial-mente perdida nas fezes e parcialmente reabsorvida para o sangue.
Figura 22

456.2 - Biossíntese de Aminoácidos Naturais
O catabolismo dos aminoácidos encontrados nas proteínas envolve a remoção dos grupos amino, seguindo-se a quebra dos esqueletos carbonados resultantes. Essas vias conver-gem para formar sete produtos intermediários: oxalacetato, α–cetoglutarato, piruvato, fumarato, succinil-CoA, acetil-Coa e acetoacetil-CoA. Esses produtos entram diretamente nas vias do metabolismo intermediário, resultando na síntese de glicose ou lipídeos, ou na produção de energia livre por sua oxidação a CO2 e H2O no ciclo do ácido cítrico. Os aminoácidos não–es-senciais podem ser sintetizados em quantidades suficientes a partir de intermediários do metabolismo ou, como no caso da cisteína e da tirosina, a partir de aminoácidos essenciais. Em contraste, os aminoácidos essenciais não podem ser sintetizados (ou produzidos em quantidades suficientes) pelo organismo e, portanto, devem ser obtidos a partir da dieta, afim de que ocorra uma síntese protéica normal. Defeitos genéticos nas vias do metabolismo de aminoácidos podem causar doenças graves.
Os aminoácidos podem ser classifi cados em glico-gênicos e cetogênicos, em função do intermediário produzido durante seu catabolismo.
Os aminoácidos cujo catabolismo produz piruvato ou um dos intermediários do ciclo do ácido cítrico são denominados glico-gênicos. Esses intermediários são substratos para a gliconeogê-nese e podem, portanto, originar a formação líquida de glicose ou de glicogênio no fígado e de glicogênio no músculo.
Figura 23
Os aminoácidos cujo catabolismo produz aceto-acetato ou um dos seus precursores (acetil-CoA ou acetoacetil-CoA) são denominados cetogêni-cos. Leucina e lisina são os únicos aminoácidos exclusivamente cetogênicos encontrados nas proteínas. Seus esqueletos carbonados não são substratos para a gliconeogênese e não podem, portanto, originar a produção líquida de glicose ou de glicogênio no fígado, ou de glicogênio no músculo.
As vias pelas quais os aminoácidos são cataboli-zados são divididas, por conveniência, de acordo com o intermediário (ou intermediários) produzido a partir de um determinado aminoácido.
Aminoácidos que produzem oxalacetato
A asparagina é hidrolisada pela asparaginase, produzindo amônia e aspartato. Algumas células leucêmicas de divisão rápida são incapazes de sin-tetizar asparagina em quantidade sufi ciente para seu crescimento. Isso torna a asparagina um aminoácido essencial para aquelas células, as quais captarão asparagina do sangue. A asparaginase, que hidrolisa a asparagina dando aspartato, pode ser administrada sistematicamente para tratar pacientes com leucemia. A asparaginase diminui o nível de asparagina no plasma e, assim, priva as células cancerosas de um nutriente essencial.
Aminoácidos que produzem fumarato
1. Fenilalanina e tirosina A hidroxilação da fenilalanina leva à formação de tirosina. O metabolismo da fenilalanina e da tirosina con-fluem, levando por fim à formação de fumarato e acetoacetato. A fenilalanina e a tirosina são, portanto, aminoácidos tanto glicogênicos quanto cetogênicos.
2. Deficiências herdadas Deficiências herda-
das nas enzimas do metabolismo da fenilalanina e da tirosina levam às doenças fenilcetonúria e al-captonúria, e a condição denominada albinismo.

46 UNIDADE VII
REGULAÇÃO ENDÓCRINA DO METABOLISMO
Quatro tecidos principais exercem função dominante no metabolismo energético: fígado, tecido adiposo, músculo e encéfalo. Esses tecidos contêm conjuntos exclusivos de enzimas, de forma que cada órgão é especializado no estoque, no uso e na formação de combustíveis específi cos. Esses tecidos não funcionam isoladamente, ao contrário, eles formam uma comuni-dade, na qual um tecido pode fornecer substrato a outro,
7.1 - Insulina
A insulina é um hormônio polipeptídico produzido pelas células β das ilhotas pancreáticas (langerhans) – grupos de células que fazem parte da porção endócrina do pâncre-as. As ilhotas pancreáticas compreendem somente cerca de 1 a 2% do total de células pancreáticas. A insulina é um dos mais importantes hormônios que coordenam a utilização de combustíveis pelos tecidos. Seus efeitos metabólicos são anabólicos, favorecendo, por exemplo, a síntese de glicogênio, de triacilgliceróis e de proteínas.
Efeitos Metabólicos da Insulina
1. Efeitos sobre o metabolismo de carboidra-tos Os efeitos da insulina no metabolismo da glicose são mais proeminentes em três tecidos: fígado, músculo e tecido adiposo. No fígado, a insulina diminui a produção de glicose por inibir a gliconeogênese e a degradação de glicogênio. No músculo e no fígado, a insulina aumenta a síntese de glicogênio. No músculo e no tecido adiposo, a insulina aumenta a captação de glicose por au-
ou processar compostos produzidos por outros órgãos. A comunicação entre os tecidos é mediada pelo sistema nervoso, pela disponibilidade de substratos circulantes e pela variação nos níveis de hormônios plasmáticos. A integração do metabolismo energético é controlada principalmente pelas ações de dois hormônios: a insu-lina e o glucagon, com as catecolaminas adrenalina e noradrenalina exercendo uma função de apoio.
mentar o número de transportadores de glicose na membrana celular. Assim, administração intrave-nosa de insulina causa uma diminuição imediata na concentração de glicose no sangue.
2. Efeitos sobre o metabolismo de lipídeos O tecido adiposo responde dentro de minutos à administração de insulina, a qual causa uma importante redução na libera-ção de ácidos graxos:
a. Diminuição na degradação de triacilgli-ceróis A insulina diminui os níveis de ácidos graxos circulantes por inibir a atividade da lipase sensível a hormônio no tecido adiposo.
b. Aumento na síntese de triacilgliceróis A insu-lina aumenta o transporte e o metabolismo da glicose nos adipócitos, fornecendo o substrato glicerol-3-fosfa-to para a síntese de triacilgliceróis. A insulina também aumenta a atividade da lipase lipoprotéica no tecido adiposo, por aumentar a síntese da enzima, fornecendo, assim, ácidos graxos para esterifi cação.
7.2 - Glucagon e Adrenalina
O glucagon é um hormônio polipeptídico secretado pelas células α das ilhotas pancreáticas (Langerhans). O glucagon, juntamente com a adrenalina, o cortisol e o hormônio do crescimento (os “hormônios contra-reguladores”), se opõe a muitas das ações da insulina. Em especial, o glucagon age na manutenção dos níveis de glicose sangüínea, pela ativação da glicogenólise e da gliconeogênese hepáticas.
Efeitos Metabólicos do Glucagon
1. Efeitos sobre o metabolismo de carboidratos A administração intravenosa de glucagon leva a um aumento imediato na glicemia. Isso resulta de um aumento na de-
gradação do glicogênio hepático (não muscular) e de um aumento da gliconeogênese.
2. Efeitos sobre o metabolismo de lipídeos O glucagon favorece a oxidação hepática de ácidos graxos e a subse-qüente formação de corpos cetônicos a partir de acetil-CoA. O efeito lipolítico do glucagon no tecido adiposo é mínimo em humanos.
3. Efeitos sobre o metabolismo protéico O glucagon aumenta a captação de aminoácidos pelo fí-gado, resultando em aumento na disponibilidade de esqueletos carbonados para a gliconeogênese. Como conseqüência, os níveis plasmáticos de aminoácidos estão diminuídos.

47Adrenalina
Níveis elevados de adrenalina circulante produzida pela medula adrenal ou de noradrenalina produzida pela intervenção simpática do pâncreas, ou de ambas, estimulam a liberação de glucagon. Assim, durante períodos de estresse, trauma ou exercício intenso, os níveis elevados de adrenalina podem impedir o efeito dos substratos circulantes sobre as células α. Nessas situações – independentemente da concentração de glicose no sangue – os níveis de glucagon se elevam em antecipação ao aumento na utilização de glicose. Em contraste, os níveis de insulina são reduzidos.
O glucagon e a adrenalina são os hormônios mais im-portantes na regulação aguda e a curto prazo de glicemia. O glucagon estimula a glicogenólise e a gliconeogênese hepáticas. A adrenalina promove a glicogenólise e a li-pólise, inibe a secreção de insulina e inibe a captação de glicose mediada por insulina nos tecidos periféricos. A adrenalina normalmente não é essencial para combater a hipoglicemia, mas pode assumir um papel crítico quando a secreção do glucagon está defi ciente, por exemplo, nos estágios tardios do diabetes melito tipo I (dependente de insulina). A prevenção ou correção da hipoglicemia falha quando a secreção de ambos, adrenalina e gluca-gon, estão defi cientes.
Exercícios de Auto-Avaliação
1. Os seguintes eventos ocorrem em células das Ilhotas de Langerhans do pâncreas:I. Secreção de hormônio no sangue.II. Produção de proteína pelos ribossomos.III. Transporte de proteína ao aparelho de Golgi.a) Em que ordem esses eventos ocorrem?b) Que hormônios são produzidos nessas células?
2. Uma conquista recente no campo da biotecnologia é o uso de bactérias para a produção de proteína animal de interesse comercial. Por exemplo, hoje já estão sendo comercializadas insulina e somatotrofi na (ou soma-tropina) humanas produzidas por bactérias.
a) Em que locais do corpo humano são produzidas essas proteínas e qual é a principal função de cada uma delas no organismo?
b) Explique sucintamente o processo por meio do qual se modifi cam bactérias para que elas passem a produzir proteínas humanas.
3. Uma jovem que sempre foi saudável chegou a um hospital em estado de coma. O histórico da paciente revelou que ela recebera erroneamente injeção de uma dose excessiva de insulina.
a) Por que a injeção de insulina induziu o coma na jovem?b) A insulina é normalmente administrada a pacientes com disfunção de que órgão? Qual é a doença causada
pela defi ciência de insulina?
4. Qual é o papel da insulina, hormônio produzido nas Ilhotas de Langerhans do pâncreas?
5. O que é diabetes mellitus? Qual sua causa?
6. O esquema abaixo representa a ação de alguns hormônios na captação ou na liberação de glicose pela célula hepática.

48Cite:a) um hormônio que atua como mostrado em A e a molécula que, após decomposta, resulta no liberação da glicose.b) um hormônio que atua como mostrado em B e a alteração no sangue que estimula a secreção deste hormônio
pela glândula endócrina.
7. Já no início do século passado, demonstrava-se, experimentalmente, que a retirada do pâncreas alterava o metabolismo dos glicídios em animais, provocando hiperglicemia não-reversível, mesmo com a administração de extratos integrais pancreáticos.
Os cientistas Banting e Best realizaram, em 1921, uma experiência que consistiu em obstruir o duto excretor principal do pâncreas de um cão. Tal manobra destrói a parte exócrina do órgão, mas não altera as ilhotas pan-creáticas responsáveis pela atividade endócrina dessa glândula.
Semanas após, os cientistas retiraram o pâncreas, assim degenerado, e injetaram seu extrato integral em um outro cão pancreatectomizado, medindo suas alterações glicêmicas ao longo de três dias.
No gráfi co abaixo, elaborado pelos próprios cientistas, as setas indicam os momentos das injeções.
Observe que o extrato de pâncreas de uma das injeções foi previamente incubado com suco pancreático.
Variação da glicemia de cão pancreatectomizado após injções de extrato de pâncreas degenerado
(Adaptado de BARRINGTON, E. J. W. “The chemical basis of physiological regulation”. Glenview: Scott, Foresman and Company, 1968.)
(*) extrato de pâncreas degenerado previamente incubado com suco pancreático.
a) Explique as causas das alterações da glicemia notadas no cão após as injeções de extrato de pâncreas e a injeção de extrato de pâncreas previamente incubado com suco pancreático.
b) Indique a conseqüência da ação do hormônio pancreático envolvido neste experimento, tanto sobre a síntese quanto sobre a degradação de gorduras.
8. A diabete é uma doença que resulta da falta de produção da insulina, um hormônio produzido no pâncreas. A insulina, um hormônio protéico, é necessária para o transporte da glicose para o interior das células, onde é eventualmente catabolizada.
Freqüentemente utiliza-se o teste de tolerância à glicose para diagnosticar a diabete. Neste teste, o paciente ingere uma solução açucarada e, a intervalos regulares de tempo, é medida a concentração de glicose sangüínea. As curvas a seguir mostram o teste realizado em um indivíduo normal e outro diabético.

49a) Qual das curvas representa o indivíduo diabético, A ou B? Justifi que sua resposta.b) Por que este hormônio é necessariamente injetado na corrente sangüínea e não administrado por via oral?
9. Até recentemente, a terapia para os diabéticos dependentes de insulina (DDI) dependia da injeção de doses de insulina suína, que possui uma estrutura muito parecida com a insulina humana. Um problema associado com essa terapia era usar a dose correta, pois o tratamento crônico obrigava os diabéticos a aplicar doses crescentes da insulina suína, para compensar o aumento da reação do organismo contra o hormônio.
Atualmente, com as técnicas de engenharia genética, é possível obter insulina humana para o tratamento dos DDI. No entanto, para os DDI que mudaram da insulina suína para a insulina humana, doses menores do hormônio foram sufi cientes.
Explique por que são administradas doses menores de insulina humana em relação à insulina suína.
10. Em menos de uma hora, Magali tomou um litro de sorvete de chocolate. Sua mãe repreendeu-a e a proibiu de tomar sorvete por um mês. Revoltada, Magali resolveu fazer greve de fome e passou todo o dia seguinte sem se alimentar. Explique como os hormônios do pâncreas atuaram para manter a concentração de glicose sangüínea em níveis constantes, quando Magali:
a) consumiu o sorvete.b) fez greve de fome.
11. Em 1920, F. Banting e C. Best, na Universidade de Toronto, obtiveram a cura de cães que apresentavam altos níveis de glicose no sangue, tratando-os com o extrato de uma glândula. Indique o hormônio e a glândula envolvidos no tratamento dos cães.
Atividades Complementares
Depois de ler a Unidade VII, sublinhe os termos que você não conhece pesquisando sobre eles.
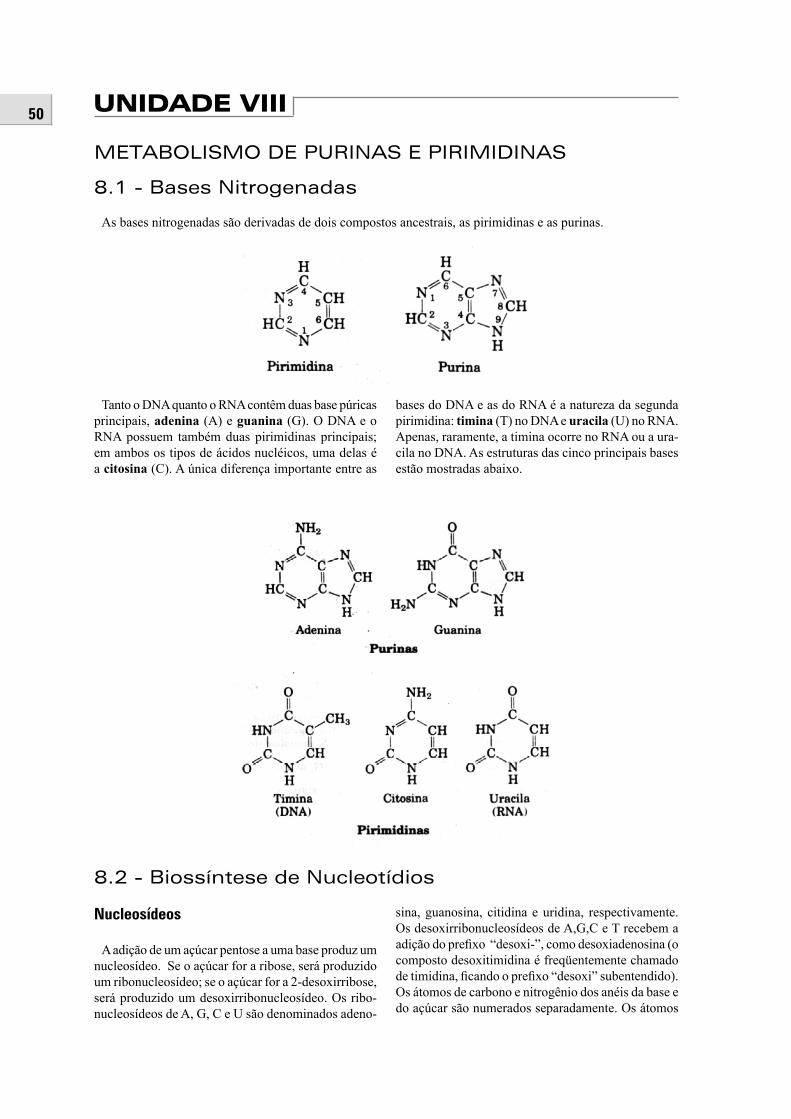
50 UNIDADE VIII
METABOLISMO DE PURINAS E PIRIMIDINAS
8.1 - Bases Nitrogenadas
As bases nitrogenadas são derivadas de dois compostos ancestrais, as pirimidinas e as purinas.
Tanto o DNA quanto o RNA contêm duas base púricas principais, adenina (A) e guanina (G). O DNA e o RNA possuem também duas pirimidinas principais; em ambos os tipos de ácidos nucléicos, uma delas é a citosina (C). A única diferença importante entre as
bases do DNA e as do RNA é a natureza da segunda pirimidina: timina (T) no DNA e uracila (U) no RNA. Apenas, raramente, a timina ocorre no RNA ou a ura-cila no DNA. As estruturas das cinco principais bases estão mostradas abaixo.
8.2 - Biossíntese de Nucleotídios
Nucleosídeos
A adição de um açúcar pentose a uma base produz um nucleosídeo. Se o açúcar for a ribose, será produzido um ribonucleosídeo; se o açúcar for a 2-desoxirribose, será produzido um desoxirribonucleosídeo. Os ribo-nucleosídeos de A, G, C e U são denominados adeno-
sina, guanosina, citidina e uridina, respectivamente. Os desoxirribonucleosídeos de A,G,C e T recebem a adição do prefi xo “desoxi-”, como desoxiadenosina (o composto desoxitimidina é freqüentemente chamado de timidina, fi cando o prefi xo “desoxi” subentendido). Os átomos de carbono e nitrogênio dos anéis da base e do açúcar são numerados separadamente. Os átomos

51dos anéis das bases são numerados de 1 a 6 nas piri-midas e de 1 a 9 nas purinas, enquanto os carbonos da pentose são numerados de 1’ a 5’. Dessa forma,
quando nos referimos ao carbono 5’ de um nucleosídeo (ou nucleotídeo), estamos especifi cando um átomo de carbono presente na pentose, e não na base.
Figura 24
8.3 - Metabolismo de Nucleotídios
A degradação dos ácidos nucléicos da dieta ocorre no intestino delgado, onde um grupo de enzimas pancreá-ticas hidrolisa os nucleotídeos a nucleosídeos e bases livres. Dentro das células, os nucleotídeos púricos são seqüencialmente degradados por enzimas específi cas, sendo que o produto fi nal da via é o ácido úrico. Os mamíferos, com exceção dos primatas, oxidam a seguir o ácido úrico a alantoína, que, em alguns animais que não os mamíferos, pode ser seqüencialmente degradada a uréia ou amônia.
As ribonucleases e as desoxirribonucleases, secretadas pelo pâncreas, hidrolisam o RNA e o DNA, produzindo principalmente oligonucleotídeos. Os oligonucleotídeos são, a seguir, hidrolisados pelas fosfodiesterase pancre-áticas, produzindo uma mistura de mononucleotídeos
3’ e 5’. Uma família de nucleotidases remove hidro-liticamente os grupos fosfato, liberando nucleosídeos que podem ser absorvidos pelas células da mucosa intestinal ou podem ser degradados em bases livres, antes da absorção. As purinas e as pirimidinas da dieta não são usadas em larga escala para a síntese de ácidos nucléicos teciduais. Em vez disso, as purinas da dieta são geralmente convertidas em ácido úrico pelas células da mucosa intestinal. A maior parte do ácido úrico entra no sangue e é excretada pela urina. Por esta razão, indi-víduos com tendência a desenvolver gota devem ser cau-telosos quanto ao consumo de alimentos que contenham altas quantidades de ácidos nucléicos, como miúdos de animais, anchovas, sardinhas ou leguminosas, como o feijão. O restante das purinas da dieta é metabolizado pela microbiota intestinal.
Nucleotídeos
Nucleotídeos são ésteres mono, di ou trifosfatados dos nucleosídeos. O primeiro grupo fosfato é ligado por uma ligação éster à hidroxila 5’ da pentose. Se um grupo fosfato é ligado no carbono 5’ da pentose, a estrutura será um nucleosídeo monofosfato (NMP), como AMP ou CMP. Se um segundo ou terceiro fos-fato forem adicionados ao mesmo nucleosídeo, serão
formados um nucleosídeo difosfato (por exemplo, ADP) ou trifosfato (por exemplo ATP). O segundo e o terceiro fosfato são ligados ao nucleotídeo por ligação de “alta energia”. Os grupos fosfato são responsáveis pelas cargas negativas associadas aos nucleotídeos e também são o motivo pelo qual o DNA e o RNA são chamados “ácidos nucléicos”.
Figura 25

52
Exercícios de Auto-Avaliação
1. Os ácidos nucléicos são moléculas formadas pelo encadeamento de um grande número de unidades chamadas nucleotídeos. Cada nucleotídeo é formado por uma base nitrogenada, uma pentose e um radical fosfato.
Em relação às substâncias químicas que formam os nucleotídeos, considere as assertivas:I. Existem cinco tipos principais de bases nitrogenadas: adenina (A), guanina (G), citosina (C), timina (T) e
uracila (U).II. A adenina e a guanina são bases pirimídicas por possuírem em comum um anel da substância conhecida
como pirimidina.III. O açúcar presente nos ácidos nucléicos pode ser a ribose ou desoxirribose.IV. O RNA aparece associado à proteína nos cromossomos, possuindo fi lamento de nucleotídeos duplo.Indique o(s) iten(s) correto(s) e justifi que.
2. Os organismos vivos requerem energia para o crescimento e manutenção do seu metabolismo. Moléculas orgânicas como a apresentada no esquema a seguir conservam a energia que é utilizada para a biossíntese dos componentes celulares, a partir de precursores simples. Analise a fi gura e descreva a composição molecular deste importante transportador de energia.
3. Considerando que na fi gura tem-se uma representação plana de um segmento da molécula de DNA, analise as proposições a seguir.
8.4 - Importância Biológica dos Nucleotídios
Os ribonucleosídeos e os desoxirribonucleosídeos fosfatados (nucleotídeos) são essenciais para todas a células. Sem eles, nem o DNA nem o RNA pode-riam ser produzidos e, dessa forma, as proteínas não poderiam ser sintetizadas, nem as células poderiam proliferar. Os nucleotídeos também são utilizados como carreadores de intermediários ativados na síntese de alguns carboidratos, lipídeos e proteínas e são componentes estruturais de várias coenzimas essenciais, como, por exemplo, a coenzima A, o FAD, o NAD+ e o NADP+. Nucleotídeos, tais como o AMP cíclico (AMPC) e o GMP cíclico (GMPC),
também são utilizados como segundos mensageiros em cascata de sinalização celular. Além disso, os nucleotídeos desempenham um papel importante como “moedas” de energia na célula. Finalmente, os nucleotídeos são importantes compostos reguladores para muitas das rotas do metabolismo intermediário, inibindo ou ativando enzimas-chave. As bases púricas e pirimídicas encontradas em nucleotídeos podem ser sintetizadas de novo ou podem ser obtidas por vias de salvação, as quais permitem a reutilização das bases pré-formadas resultantes do metabolismo normal da célula ou da dieta.

531) Um nucleotídeo é formado por um grupo fosfato (I), uma molécula do açúcar desoxirribose (II) e uma
molécula de base nitrogenada.2) Um nucleotídeo com Timina (T) em uma cadeia pareia com um nucleotídeo com Adenina (A) em outra
cadeia.3) Um nucleotídeo com Guanina (G) em uma cadeia pareia com um nucleotídeo com Citosina (C) em outra
cadeia. 4) Pontes de hidrogênio se estabelecem entre as bases nitrogenadas T e A e entre as bases nitrogenadas C e G. Qual(is) informação(ões) está(ão) correta(s)?
Atividades Complementares
Depois de ler a Unidade VIII, sublinhe os termos que você não conhece pesquisando sobre eles.

54
Se você:
1) concluiu o estudo deste guia;2) participou dos encontros;3) fez contato com seu tutor;4) realizou as atividades previstas;
Então, você está preparado para as ava-liações.
Parabéns!

55Glossário
Acetil – grupo químico derivado do ácido acético. Grupos acetil são importantes no metabolismo e freqüen-temente são adicionados a proteínas como modifi cações covalentes.
Acetil-CoA – pequena molécula hidrossolúvel que transporta os grupos acetil nas células. Consiste do grupo acetil ligado à coenzima A (CoA) por uma ligação tioéster facilmente hidrolisável.
Ácido graxo – composto como o ácido palmítico que possui um ácido carboxílico ligado a uma longa cadeia de hidrocarboneto. Usado como uma importante fonte de energia durante o metabolismo, e o ponto de partida para a síntese de fosfolipídeos.
Ácido nucléico – RNA ou DNA; consiste de uma cadeia de nucleotídeos unidos por ligações fosfodiéster.Adrenalina (epinefrina) – hormônio liberado pelas células “chromaffi ns” (na glândula adrenal) e por alguns
neurônios em resposta ao estresse. Produz uma resposta “lute ou fuja”, que inclui aumento dos batimentos cardíacos e do nível de açúcar no sangue.
Aeróbico – descreve um processo que necessita ou que ocorre na presença de oxigênio gasoso (O2).Álcool – molécula orgânica polar que contém um grupo hidroxila funcional (-OH) ligado a um átomo de
carbono não pertencente a um anel aromático. Um exemplo é o álcool etílico.Alteração da energia livre (DG) – alteração da energia livre durante uma reação: resultado da energia livre
das moléculas produzidas subtraído da energia livre das moléculas que iniciaram a reação. Um valor maior e negativo de DG indica que a reação tem uma forte tendência de ocorrer.
Amido – polissacarídeo composto exclusivamente por unidades de glicose, usado como um estoque de energia nas células vegetais.
Anaeróbico – descreve uma célula, organismo, ou processo metabólico que funciona na ausência de ar, ou, mais precisamente, na ausência de oxigênio molecular.
Anfi pátia – que possui regiões hidrofílicas e hidrofóbicas, como um fosfolipídio ou uma molécula de-tergente.
ATP (5’-trifosfato de adenosina) – nucreosídeo trifosfatado composto de adenina, ribose e três grupos fosfato, é o principal carreador celular de energia química. Os grupos fosfato terminais são altamente reativos e sua hidrólise, ou transferência a outra molécula, liberam uma grande quantidade de energia livre.
Carboidrato – termo geral para designar açúcares e compostos relacionados contendo carbono, hidrogênio e oxigênio, geralmente com a fórmula empírica ((CH2O)n.
Catabolismo – termo geral utilizado para as reações catalisadas por enzimas em uma célula, na qual molécu-las complexas são degradadas a moléculas mais simples com liberação de energia. Os produtos intermediários destas reações são algumas vezes chamados de catabólitos.
Catalisador – substância que acelera uma reação química sem sofrer alterações. Enzimas são proteínas ca-talíticas.
Coenzima – pequena molécula fortemente associada com uma enzima que participa na reação catalisada pela enzima, freqüentemente pela formação de uma ligação covalente temporária ao substrato. Exemplos incluem a biotina, NAD+ e coenzima A.
Cofator – coenzima ou íon inorgânico que é necessária para a atividade de uma enzima.Colesterol – molécula lipídica com uma estrutura característica de quatro anéis esteróides, componente im-
portante das membranas plasmáticas de células animais.Enzima – proteína que catalisa uma reação química específi ca.Fosfolipídio – a principal categoria de moléculas lipídicas utilizadas na construção de membranas biológicas.
Normalmente composta de dois ácidos graxos ligados por um fosfato de glicerol a um grupo polar, um dentre uma variedade de tipos.
Glicogênio – polissacarídeo composto exclusivamente por unidades de glicose, usados para armazenar energia nas células animais. Grandes grânulos de glicogênio são especialmente abundantes nas células dos músculos e fígado.
Glicólise – via metabólica universal no citosol, na qual açúcares são parcialmente degradados com produção de ATP. Literalmente, “divisão do açúcar”.
Glicoproteína – qualquer proteína contendo uma ou mais cadeias de oligossacarídeo ligadas covalen-temente.
Glicose - açúcar de seis carbonos com papel fundamental no metabolismo das células vivas. Armazenada na forma polimérica como glicogênio nas células animais, e como amido nas células vegetais.
Ligação Iônica - ligação entre dois átomos, um com carga positiva e outro com carga negativa. Um dos tipos de ligação não covalente.

56Ligação não covalente – ligação química na qual, ao contrário da ligação covalente, não há compartilhamento
de elétrons. Ligações não covalentes são relativamente fracas, mas podem ser somadas, produzindo uma inte-ração forte e altamente específi ca entre moléculas.
Ligação peptídica – ligação química entre o grupo carbonila de um aminoácido e o amino grupo de um se-gundo aminoácido – uma forma especial de ligação amídica.
Lipase – enzima que catalisa a clivagem dos ácidos graxos da molécula de glicerol em um triglicerídeo.Lipídio – molécula orgânica, insolúvel em água, que se dissolve prontamente em solventes orgânicos apolares.
Uma classe, os fosfolipídios, formam a base estrutural das membranas biológicas.Metabolismo – a soma total de processos químicos que ocorrem em uma célula viva.NAD+ (Nicotina Adenina Dinucleotídeo) – coenzima que participa em uma reação de oxidação, aceitando
um íon hidreto (H-) de uma molécula doadora. O NADH formado é um transportador de elétrons importante para a oxidação fosforilativa.
NADP+ (Nicotina Adenina Dinucleotídeo Fosfato) – coenzima muito relacionada ao NAD+ que é amplamente utilizada em vias biossintéticas, ao invés de vias catabólicas.
Oxidação (verbo oxidar) – perda da densidade de elétrons de um átomo, como ocorre durante a adição de oxigênio a uma molécula ou quando um hidrogênio é removido. Contrário de redução.
pH – medida comum da acidez de uma solução: “p” refere-se à potência de 10. “H”, a hidrogênio. É defi nido como o logaritmo negativo da concentração dos íons hidrogênio em moles por litro (M). Então, pH 3 (10-3 M H+) é ácido e pH 9 (10-9 M H+) é alcalino.
Polímero – grande molécula formada por uma série de ligações covalentes, que unem várias unidades idênticas ou semelhantes (monômeros).
Polipeptídeo – polímero linear composto por múltiplos aminoácidos. Proteínas são grandes polipeptídeos, e os dois termos podem ser usados como sinônimos.
Proteína – o principal componente macromolecular das células. Um polímero linear de aminoácidos unidos por ligações peptídicas em uma seqüência específi ca.
Respiração – termo genérico para qualquer processo em uma célula, na qual a incorporação de moléculas de O2 é acoplada à produção de CO2.
Sítio ativo – região da superfície de uma enzima na qual a molécula de substrato deve ligar-se para sofrer uma reação catalítica.

57Gabarito
Unidade I
1. Evitar a ingestão de alimentos que contenham o aminoácido fenilalanina pois os “fenilcetonúricos” são incapazes de metabolizar essa substância e correm risco de apresentar graves distúrbios metabólicos com con-seqüências irreversíveis.
2. As extremidades do corpo perdem calor para o meio ambiente com mais facilidade e costumam, portanto, apresentar uma temperatura inferior à do restante do corpo. Como a enzima só é ativa abaixo de 34°C, a síntese do pigmento que confere cor negra só ocorrerá nas extremidades do corpo.
3.a) O gráfi co I refere-se a um indivíduo AA ou Aa, capazes de produzir o polipeptídeo. O gráfi co II representa
a formação da substância no indivíduo aa.b) O polipeptídeo X é uma enzima. A análise do gráfi co I revela que a velocidade da formação do produto é
dependente da temperatura, o que indica tratar-se de uma reação catalisada.
4. A vitaminose B2 é Beribéri.A vitaminose B12 – Anemia perniciosa.Avitaminose K é a difi culdade de coagulação sangüínea e hemorragias.
5. Leite, sopas, refrigerantes, frutas etc.
6. Gema de ovo, ova de peixe, cenoura, tomate, beterraba etc.
7. Vitamina C (ácido ascórbico).
8. Auxilia a absorção de cálcio no intestino.
9.a) Os amaciantes naturais e industrializados contêm proteases, enzimas relacionadas com a hidrólise das
proteínas fi brosas que “endurecem” a carne. No corpo humano, a digestão das proteínas da carne tem início na cavidade gástrica, por ação da enzima pepsina. Prossegue no duodeno, onde atua a tripsina presente no suco pancreático e é fi nalizada pela atividade das peptidases existentes no suco entérico.
b) O cozimento causará a desnaturação das enzimas presentes nos amaciantes. Desta forma, a carne não sofrerá qualquer efeito, pois as enzimas desnaturadas não poderão desempenhar seu papel como catalisadores biológicos.
10. A mulher grávida possui uma demanda maior de oxigênio devido à presença do feto. Uma dieta rica em ferro aumenta a disponibilidade do complexo ferro-hemoglobina e portanto, permite o transporte de mais oxi-gênio, o que reduz a sensação de “falta de ar”.
11.a) Quanto mais intensa for a atividade muscular maior será a taxa respiratória e, conseqüentemente, maior
será a liberação de calor.b) A respiração celular aeróbia intensifi cada na atividade muscular resulta no aumento da produção de CO,
este combina-se com a água produzindo ácido carbônico. Este fato reduz o pH do meio facilitando a liberação do oxigênio pela hemoglobina.
c) Em atividade muscular intensa há maior produção de calor e diminuição do pH. Maior quantidade de oxigênio é liberado pela hemoglobina o que resulta em maior disponibilidade de ATP necessário à contração muscular.
Unidade II
1. a) Inibição competitiva.Na inibição enzimática do tipo competitivo, o inibidor, mantido em concentração constante, exerce seu efeito
com maior intensidade em concentrações baixas de substrato. Com o aumento da concentração do substrato,

58devido ao efeito competitivo, a inibição tende a diminuir. Dessa forma, em excesso de substrato, a velocidade máxima de reação é a mesma na ausência ou na presença do inibidor.
b) Uma dentre as substâncias e respectiva função:- sais biliares – emulsifi cação de gorduras durante a digestão.- vitamina D (Dƒ) – metabolismo do cálcio e desenvolvimento do tecido ósseo.
2. A hexoquinase possui uma grande afi nidade pela glicose, ou seja, ela atinge a velocidade máxima com uma concentração muito pequena de glicose. A glicoquinase exibe uma afi nidade bem menor pois somente atinge sua velocidade máxima em concentrações bem mais altas do substrato. Logo, a enzima que contribui para a formação de glicogênio hepático é a glicoquinase, pois esta somente produz G6P com máxima efi ciência quando há excesso de glicose no sangue.
Unidade IV
1. A insulina, hormônio produzido pelo pâncreas, estimula a glicogenogênese e glicólise.
2.a) Respiração celular anaeróbia ou fermentação.b) Produção de energia química (ATP) a partir de compostos orgânicos.c) Fibras musculares esqueléticas submetidas a esforços podem entrar em débito de oxigênio. Nesta situação,
a produção energética ocorre anaerobicamente através da fermentação láctica. O ácido láctico, subproduto deste processo, é tóxico para os músculos e causa a fadiga muscular.
3.a) Na fermentação, o consumo de glicose é maior.b) O rendimento energético da fermentação é menor do que a produção energética obtida na respiração aeróbica.
4. a) Polissacarídeo de reserva animal: Glicogênio. Polissacarídeo de reserva vegetal: Amido.b) Glicogênio é armazenado nos músculos esqueléticos e no fígado. Amido pode ser armazenado na raiz
(mandioca), no caule (batata-inglesa) e nas sementes (milho).
5. a) Q - água P - carboidratos Z - fi bras.b) Durante a germinação ocorre utilização de água e ativação enzimática. Carboidratos e lipídios fornecem
energia para a multiplicação e as proteínas fornecem nitrogênio para formação dos ácidos nucléicos.
Unidade V
1. a) Animais: glicogênio Vegetais: amidob) Nucleotídeos.c) Glicerídeos.d) Estrutura terciária Grupo prostético
2. a) As duas principais funções do colesterol são: participar da composição estrutural das membranas dos animais
e ser precursor de hormônios sexuais (estrógenos, andrógenos e progesterona).b) O colesterol sangüíneo tem origem endógena ou exógena (proveniente da dieta).
3.a) Cenoura, tomate, mamão e outros vegetais são ricos em carotenóides. A vitamina A é essencial para a síntese
dos pigmentos visuais.

59b) Controle hormonal. Testosterona, estrógenos e progesterona são exemplos.c) Animais: tecido conjuntivo adiposo subcutâneo. Vegetais: sementes.
Unidade VII
1.a) II - III - Ib) Insulina e glucagon.
2.a) A insulina é hormônio produzido nas Ilhotas pancreáticas e é responsável pela manutenção da taxa glicêmica
constante. A somatropina é produzida na adenohipófi se e é responsável pelo crescimento ósseo e muscular.b) Segmentos de DNA (genes) contendo as informações para a produção destas proteínas são enxertados em
bactérias em laboratório. As bactérias “transgênicas” passam então a produzir estas substâncias.
3. a) A injeção de insulina causou um quadro grave de hipoglicemia que conduziu a jovem ao coma.b) Insulina é administrada a pacientes que apresentam disfunção na porção endócrina do pâncreas (Ilhotas
pancreáticas). A falta deste hormônio causa a diabete melito.
4. Regulação da glicemia, ou seja, da taxa de glicose na corrente sangüínea. A insulina remove o excesso de glicose do sangue promovendo seu armazenamento no fígado, na forma de glicogênio.
5. Hiperglicemia causada por defi ciência de insulina, hormônio produzido pelas ilhotas pancreáticas.
6. a) Glucagon ou Adrenalina (epinefrina).Glicogêniob) InsulinaAumento da taxa de glicose sangüínea circulante.
7.a) Após as injeções de extrato de pâncreas degenerado, a glicemia foi mantida baixa algum tempo, por ação
da insulina. Quando, porém, foi injetado extrato de pâncreas degenerado pré-incubado com suco pancreático, a insulina, sendo um hormônio polipeptídico, foi degradada pela ação das enzimas proteolíticas deste suco, não havendo resposta hipoglicêmica.
b) Aumento da síntese e diminuição da degradação de gorduras.
8.a) Curva A porque a taxa de glicose no sangue permanece alta com o passar do tempo.b) A insulina é uma proteína e seria digerida pelas enzimas proteolíticas do suco gástrico e pancreático, antes
de ser absorvida.
9. Mesmo exibindo uma discreta diferença estrutural, a insulina suína não é reconhecida como uma molécula própria do organismo humano e, portanto, induzia a formação de anticorpos. Assim, nos doentes crônicos, parte da dose injetada era neutralizada pelos anticorpos, o que os obrigava a aumentar a dose gradualmente. No entanto, ao mudar para a insulina humana era necessário diminuir a dose, já na ausência de anticorpos bloqueadores, era possível administrar a dose fi siológica do hormônio.
10.a) Ao consumir o sorvete, rico em glicose, o pâncreas de Magali secretou a insulina, hormônio hipoglice-
miante.b) Na greve de fome haverá secreção do glucagon pelas células do pâncreas. Esse hormônio estimula a quebra
do glicogênio hepático, ou seja, apresenta função hiperglicemiante.
11. Insulina, hormônio produzido pela Ilhotas de Langerhans do Pâncreas.

60Unidade VIII
1. Os itens I e III estão corretos, pois as bases nitrogenadas encontradas nos ácidos nucléicos são: DNA, ade-nina, timina, guanina e citosina e no RNA, adenina uracila, guanina e citosina.
2. 1- Base Nitrogenada: Adenina.2- Açúcar: Ribose.3- Constituinte inorgânico: Fosfato.4- Energia de ligação: Alta.5- Ácido nucléico: Ribonucléico.
3. 1, 2, 3 e 4.

61Referências Bibliográficas
ALBERTS, Bruce. Biologia Molecular da Célula. 4. ed. Porto Alegre: Artmed, 2004.LULLMANN, Heinz. Farmacologia: texto e atlas. 4. ed. Porto Alegre: Artmed, 2004.MURRAY, Robert K. Harper: Bioquímica. 8. ed. São Paulo: Atheneu, 1998.NELSON, D. L. & COX, LEHNINGER, M. Princípios de bioquímica. 3. ed. São Paulo: Sarvier, 2002.PETER H. Raven. Biologia Vegetal. 6. ed. Rio de Janeiro: Guanabara Koogan, 2001.STRYER, L.; TYMOCZKO, J.L. & BERG, J.M. Bioquímica. 5. ed. Rio de Janeiro: Guanabara Koogan, 2004.VILLELA, Gilberto Guimarães. Bioquímica. 4. ed. Rio de Janeiro: Guanabara Koogan, 1984.VOET, Donald; VOET, Judith & PRATT, Charlotte. Fundamentos de bioquímica. Porto Alegre: Artes Médicas, 2000.
Fonte das Figuras
Figura 1: Disponível em: http://www.springway.com.br/diversos/escala_ph.gifFigura 2: VILLELA, Gilberto Guimarães. Bioquímica. Rio de Janeiro: Guanabara Koogan, [s/d].Figura 3: Disponível em: http://profs.ccems.pt/OlgaFranco/10ano/biomoleculas.htmFigura 4 a 10 e 20 a 25: HARVEY, Richard A. Bioquímica ilustrada. Porto Alegre: Artmed, [s/d].Figura 11: Disponível em: http://www.geocities.com/capecanaveral/launchpad/9071/cetoses.gifFigura 12 e 16: ALBERTS, Bruce. Biologia molecular da célula. Porto Alegre: Artmed, [s/d].Figura 13 a 15 e 17 a 19: RAVEN, Peter H. Biologia vegetal. Rio de Janeiro: Guanabara Koogan, [s/d].


