aspectos imunolÓgicos e mÉtodos diagnÓsticos de -...
TRANSCRIPT

i
UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
SEMINÁRIOS APLICADOS
ASPECTOS IMUNOLÓGICOS E MÉTODOS DIAGNÓSTICOS de
Salmonella SP. APLICADOS À AVICULTURA
Denizard André de Abreu Delfino
Orientadora: Valéria de Sá Jayme – UFG
GOIÂNIA
2013

ii
DENIZARD ANDRÉ DE ABREU DELFINO
ASPECTOS IMUNOLÓGICOS E MÉTODOS DIAGNÓSTICOS de
Salmonella SP. APLICADOS À AVICULTURA
Seminário apresentado junto à
disciplina Seminários Aplicados do
curso de Doutorado em Ciência
Animal da Escola de Veterinária e
Zootecnia da Universidade Federal
de Goiás.
Área de concentração:
Sanidade Animal
Linha de pesquisa:
Epidemiologia das doenças infecciosas
Orientadora:
Prof. Dr. Valéria de Sá Jayme – UFG
Comitê de orientação:
Prof. Dr. Maria Auxiliadora Andrade – UFG
Prof. Dr. Guido Fontgalland Coelho Linhares – UFG
GOIÂNIA
2013

iii
SUMÁRIO
1. INTRODUÇÃO ................................................................................................... 4
2. REVISÃO DA LITERATURA .............................................................................. 6
2.1- Salmonella ...................................................................................................... 6
2.2 DIAGNÓSTICO .............................................................................................. 10
2.2.1 Diagnóstico bacteriológico convencional ..................................................... 11
2.2.2- Diagnóstico sorológico ............................................................................... 13
2.2.2.1Aspectos imunológicos das aves .............................................................. 13
2.2.3- Soroaglutinação rápida (SAR) .................................................................... 16
2.2.4 Soroaglutinação Lenta (SAL) ...................................................................... 18
2.2.5 – Teste Imunoenzimático (ELISA) ............................................................... 18
2.2.6 Diagnóstico molecular ................................................................................. 19
2.2.6.1 Fatores de virulência ................................................................................ 19
3. CONSIDERAÇÕES FINAIS ............................................................................. 27
REFERÊNCIAS .................................................................................................... 29

4
1. INTRODUÇÃO
Salmonella sp. continua a ser um agente patogênico de distribuição
mundial, com importância epidemiológica tanto do ponto de vista humano quanto
animal, uma vez que está relacionada com contaminações nas indústrias de
alimentos, e tem sido isolada e identificada com frequência como agente
etiológico de surtos de doenças veiculadas por alimentos e, consequentemente,
muito pesquisada (BEAM et al., 2013).
As enfermidades atribuídas à bactéria Salmonella, com exceção da
febre tifóide, não são de notificação compulsória, o que dificulta estimar dados da
real incidência dos casos. Porém, estima-se que a incidência seja bastante
elevada entre a população mundial (CRUMP et al., 2004).
A incidência de salmonelose no homem continua aumentando em
várias partes do mundo, mesmo com todo desenvolvimento tecnológico utilizado
na produção de alimentos e a adoção de melhores práticas higiênico-sanitárias. A
preocupação com a qualidade dos alimentos envolve não só os riscos de
veiculação de enfermidades para o consumidor, mas também perdas econômicas
em virtude das alterações microbianas ocorridas nos alimentos. Os surtos de
salmonelose humana podem ter um custo bastante elevado, pois devem ser
computadas as despesas médicas e a diminuição da produtividade (GOMES,
2008).
A vigilância de doenças de origem alimentar é complexa devido a
vários fatores, dentre os quais se destaca a falta de notificação de casos, em
razão do serviço médico muitas vezes não ser procurado. Embora a doença
possa ser severa ou até mesmo serem reportados frequentemente casos fatais, é
mais comum casos com sintomatologia moderada. Além disso, muitos patógenos
são transmitidos não só por meio dos alimentos, mas também através da água ou
de pessoa para pessoa. Essa variedade de forma de transmissão também
dificulta o diagnóstico (BARROW, et al., 2010).
Salmonella sp., Campylobacter spp. e Listeria spp. estão entre os
principais agentes causadores de enfermidades transmitidas por alimentos no
homem, além de representarem risco para a sanidade avícola. As aves são os
principais carreadores de Salmonella sp., determinando alta taxa de mortalidade,
especialmente em aves jovens e o estado de portador, dependendo do sorovar,

5
da dose infectante, da susceptibilidade da espécie, da idade e do estado imune
do hospedeiro (CHAPPEL, et al., 2009).
O diagnóstico da salmonelose nas aves pode servir como um meio de
reduzir a infecção em humanos. Para apoiar o diagnóstico, são importantes
informações epidemiológicas e clínicas, incluindo histórico, sinais clínicos, lesões
e mortalidade (RODRIGUES, 2011; CHA, et al., 2013).
Por todas essas questões, técnicas que permitam um diagnóstico
preciso e eficiente são cada vez mais necessárias para minimizar ou evitar riscos
à saúde humana, como surtos de infecções, principalmente devido às tóxico
infecções alimentares, e à produção animal, em especial quanto à avicultura
industrial, devido às restrições comerciais.
Considerando a importância do patógeno, buscou-se fazer uma revisão
sobre Salmonella sp., abordando aspectos relacionados ao microrganismo,
causas alimentares, ao sistema imunológico das aves de produção e aos métodos
de diagnóstico do patógeno.

6
2. REVISÃO DA LITERATURA
2.1- Salmonella
O gênero Salmonella recebeu este nome em função do seu
descobridor Daniel Elmer Salmon (1850–1914). São bactérias em forma de
bastonetes, que fazem parte da família Enterobacteriaceae, não formam esporos,
apresentam coloração negativa em Gram e são formadas principalmente por
sorovares que apresentam motilidade, sendo que as variantes imóveis incluem
Salmonella enterica sorovar Gallinarum e Salmonella enterica sorovar Pullorum
(BARROW, et al., 2010).
O patôgeno caracteriza-se por ser um bacilo curto, aeróbio e anaeróbio
facultativo, desprovido de cápsula, não formador de esporos imóvel ou móvel com
flagelos peritríquios. Cresce numa variação de temperatura de 5oC – 45oC,
entretanto, a temperatura ótima para seu crescimento é de aproximadamente
37oC. Cresce muito bem em pH entre 4–9, sendo 7 o pH ideal. Inclui
microrganismos fermentadores de L-rhamnose, L-arabinose, D-sorbitol, D-manitol,
D-manose, D-xilose, maltose e trehalose, porém, não fermentam lactose e
sacarose (GAST, 2008).
Os antígenos na camada de lipopolissacarídeos (LPS) facilitam o
contato direto da bactéria com o ambiente, contribuindo na interação do agente
com o hospedeiro, isso faz com que Salmonella sp. possa resistir em diferentes
locais e condições, como fezes, poeira, ambientes secos, acidez do estômago,
lúmen do intestino, espaço extracelular dos tecidos do hospedeiro e dentro de
macrófagos (SUO et al.,2010).
O gênero Salmonella é dividido em duas espécies: Salmonella bongori
que possui 23 sorovares e Salmonella enterica que se divide em seis
subespécies: Salmonella enterica subespécie enterica, Salmonella enterica
subespécie salamae, Salmonella enterica subespécie arizonae, Salmonella
enterica subespécie diarizonae, Salmonella enterica subespécie houtenae e
Salmonella enterica subespécie indica, sendo que a espécie enterica está
relacionada com a salmonelose transmitida ao homem, portanto de importância
em saúde pública (GUIBOURDENCHE et al., 2010).

7
O outro grupo é composto por um pequeno número de sorovares que
causam doenças sistêmicas restritas a determinadas espécies, semelhantes à
febre tifóide, tais como: Salmonella enterica sorovar Typhi em seres humanos,
Salmonella enterica sorovar Choleraesuis em suínos, Salmonella enterica sorovar
Dublin em bovinos, Salmonella enterica sorovar Pullorum e Salmonella enterica
sorovar Gallinarum em aves (BARROW et al., 1994; WIGLEY et al., 2001;
BERCHIERI JR. et al., 2010).
O número de sorovares isolados inclui mais de 2.625 sorotipos, entre
os quais 1.367 pertencem à subespécie enterica. Estes estão amplamente
difundidos na natureza, sendo capazes de infectar o trato intestinal de animais,
tanto de sangue frio quanto de sangue quente, entre eles o homem (SUO et al.,
2010). Os componentes deste grupo são os principais responsáveis pela
contaminação de ovos e de carcaças durante o processamento de abate
(BERCHIERI JR. et al., 2010).
Para a distinção dos sorovares, são utilizados antissoros que reagem
com antígenos presentes na célula bacteriana. Existem três diferentes tipos de
antígenos que podem identificar os sorovares de Salmonella sp.: antígeno
somático (O), antígeno flagelar (H) e o antígeno capsular de virulência (Vi)
(FRANCO & LANDGRAF, 1996).
O antígeno somático O apresenta-se na porção mais externa da
parede bacteriana e corresponde à parte antigênica da camada de
lipopolissacarídeos (LPS) presentes na membrana celular de todas as bactérias
Gram negativas como Salmonella sp. (REEVES et al., 1996; SELANDER et al.,
2002). Já os antígenos H da fase um são homólogos a outras bactérias entéricas,
enquanto antígenos H de fase dois são específicos do gênero Salmonella
(BARROW, 2010).
Ainda assim, segundo a Organização Mundial da Saúde (OMS)
Salmonella sp. é o agente bacteriano mais frequentemente envolvido em casos
de doenças transmitidas por alimentos em todo o mundo. O agente é
normalmente transmitido ao homem por meio de alimentos de origem animal,
como carne, ovos e leite (BORSOI, et al., 2011).
Surtos de salmonelose, portanto, têm sido descritos com relativa
frequência, em diversas regiões. Em trabalho conduzido no Rio Grande do Sul, foi

8
verificada a ocorrência de surtos de salmonelose em 99 relatórios de
investigação, correspondendo a 74,7%, dos registros, sendo 11,4% associados
ao consumo de carne de aves (NADVORNY et al., 2004). No Estado do Rio de
Janeiro, em estudo de 53 surtos que acometeram 461 pessoas, Salmonella sp. foi
responsável por 7% dos casos, atingindo 15,8% dos indivíduos, inclusive com um
óbito (FERNANDEZ et al., 2001). Nos Estados Unidos, ZHAO et al. (2001)
relataram a ocorrência de 1,4 milhões de casos notificados de salmonelose em
seres humanos.
Destaca-se que os produtos avícolas tem sido responsáveis por surtos
humanos e têm se tornado fontes de infecções frequentes. Assim, tanto a
presença quanto a disseminação de Salmonella sp. nos alimentos representam
uma relevante preocupação na indústria avícola, uma vez que podem determinar
decréscimo no consumo de carne de aves, representando, assim, uma ameaça
ao comércio avícola nacional e internacional (IKUNO et al., 2004).
A bactéria é rotineiramente detectada em amostras clínicas de
alimentos e de meio ambiente, utilizando-se cultivo microbiológico, depois de ser
realizado um enriquecimento seletivo que possibilita posterior sorotipagem
(GAST, 2003).
Em pesquisa com diversos alimentos enviados para um laboratório na
cidade de São Paulo, SP, nas 140 amostras isoladas de Salmonella sp., havia 17
sorotipos com S. Enteritidis presente em dez dos 12 tipos de alimentos
pesquisados, sendo a carne de frango “in natura” o alimento em que mais se
obteve isolados, 77,1% (LÍRIO et al., 1997). Em trabalho com cortes de frango
(coxa e peito) verificaram-se, de um total de 15 amostras, quatro positivas para
Salmonella sp. (26,7%) (GONÇALVES et al., 1998). Já em carcaças de frango,
CASON et al. (1997) detectaram uma frequência de Salmonella sp. de 94%,
enquanto SANTOS et al. (2000), avaliando 150 carcaças de frango congeladas,
identificaram 32% das amostras positivas.
No Brasil, existem muitos estudos de prevalência e frequência de
isolamento de Salmonella spp. em produtos de origem avícola. Um estudo
realizado em 1996 pesquisou este micro-organismo em 45 cortes (15 peitos, 15
asas e 15 coxas) oriundos do comércio de Jaboticabal (SP). Dos cortes
examinados, 35% estavam contaminados, sendo 46,6% de asas, 40% de peito e
9
20% de coxas. Neste estudo foram identificados 10 sorovares de Salmonella sp.,
sendo que o sorovar mais prevalente foi S. Enteritidis (COSTA et al.,1997).
No período de 1996 a 1997, 32% das amostras de carcaças de frango
analisadas em São Paulo apresentaram a bactéria (SANTOS et. al, 2000).
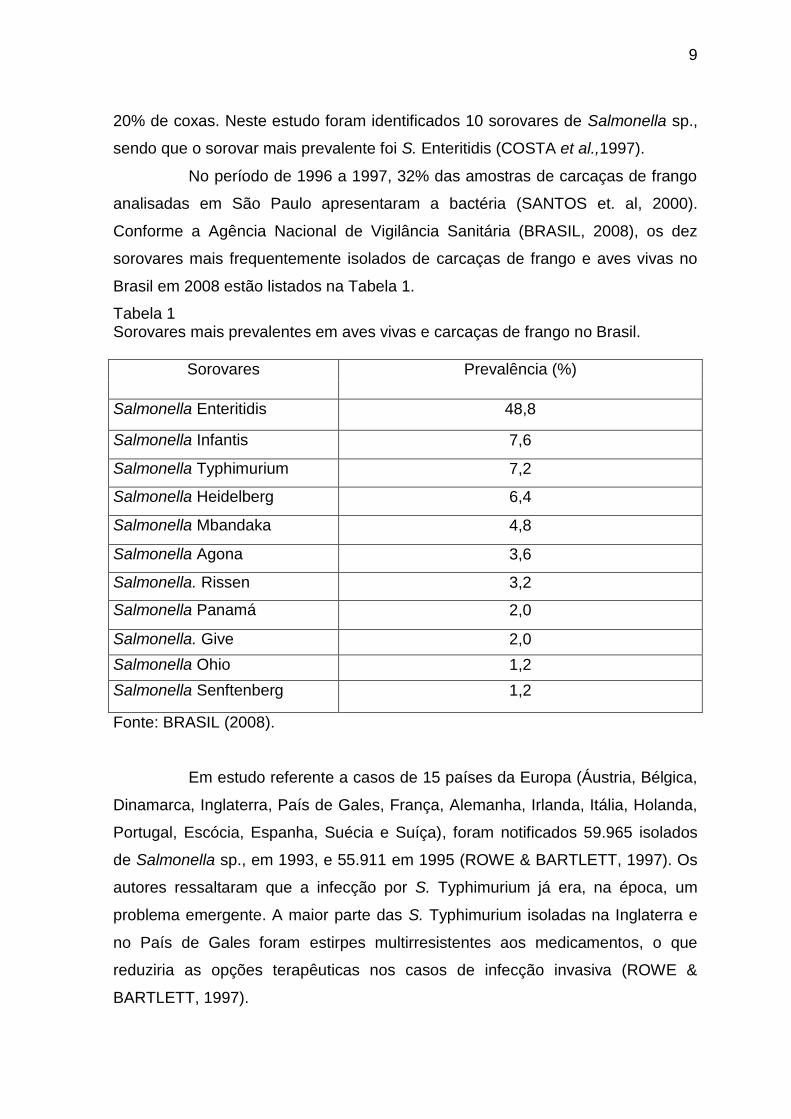
Conforme a Agência Nacional de Vigilância Sanitária (BRASIL, 2008), os dez
sorovares mais frequentemente isolados de carcaças de frango e aves vivas no
Brasil em 2008 estão listados na Tabela 1.
Tabela 1 Sorovares mais prevalentes em aves vivas e carcaças de frango no Brasil.
Sorovares Prevalência (%)
Salmonella Enteritidis 48,8
Salmonella Infantis 7,6
Salmonella Typhimurium 7,2
Salmonella Heidelberg 6,4
Salmonella Mbandaka 4,8
Salmonella Agona 3,6
Salmonella. Rissen 3,2
Salmonella Panamá 2,0
Salmonella. Give 2,0
Salmonella Ohio 1,2
Salmonella Senftenberg 1,2
Fonte: BRASIL (2008).
Em estudo referente a casos de 15 países da Europa (Áustria, Bélgica,
Dinamarca, Inglaterra, País de Gales, França, Alemanha, Irlanda, Itália, Holanda,
Portugal, Escócia, Espanha, Suécia e Suíça), foram notificados 59.965 isolados
de Salmonella sp., em 1993, e 55.911 em 1995 (ROWE & BARTLETT, 1997). Os
autores ressaltaram que a infecção por S. Typhimurium já era, na época, um
problema emergente. A maior parte das S. Typhimurium isoladas na Inglaterra e
no País de Gales foram estirpes multirresistentes aos medicamentos, o que
reduziria as opções terapêuticas nos casos de infecção invasiva (ROWE &
BARTLETT, 1997).

10
No tocante a questão de resistência, o uso de antibióticos como
promotores de crescimento tem gerado muita polêmica, tanto com relação às
preocupações com a saúde animal, pela perda da eficiência ao longo do tempo,
quanto com a saúde pública, pelo desenvolvimento de cepas resistentes
(CALIXTO et al., 2002).
O surgimento de cepas resistentes de Salmonella sp. é comum e
agravado com o uso indiscriminado de antibióticos em rações animais,
principalmente, os promotores de crescimento. Por isso, rações avícolas têm sido
relatadas como um elo de grande importância no ciclo epidemiológico da
salmonelose aviária, agravando assim, as infecções alimentares causadas por
estas bactérias (REIS et al., 1995).
Portanto, pesquisas diversas têm mostrado a importância do patógeno
em alimentos de origem animal, particularmente aviária, destacando-se que aves
de corte são potenciais carreadoras de patógenos em abatedouros, açougues e
cozinhas industriais e domésticas (GLYNN et al., 2006).
2.2 DIAGNÓSTICO
Embora a salmonella tenha uma multiplicação rápida, mais de 72 horas
são necessárias para sua cultura e sorotipagem (WOODWARD & KIRWAN,
1996). A técnica PCR e outras técnicas moleculares constituem uma ferramenta
de grande utilidade no diagnóstico microbiológico por propiciarem um resultado
rápido e com alto grau de especificidade (WOODWARD & KIRWAN, 1996).
STONE et al. (1994), além dos fatores supracitados, ressaltaram ainda o fato de
que o uso das várias técnicas de PCR propiciam um aumento do número de
amostras positivas e, ao mesmo tempo, reduzido número de falsos negativos,
além de permitirem o processamento de grande número de amostras em um curto
período de tempo.
A avaliação clínica e a microbiologia convencional em conjunto com
técnicas capazes de detectar infecções inaparentes tais como testes sorológicos,
Imunositoquímica e diagnóstico molecular, são ferramentas utilizadas para auxiliar
no diagnóstico (MORENO et al., 2009).
Para diagnóstico por isolamento bacteriano de aves com sinais clínicos
da doença ou suspeitas sorológicas, é comum a utilização de suabes de órgãos

11
ou de fragmentos colhidos com material estéril. Na monitoria de lotes, são mais
usadas as coletas através de suabes de cloaca, suabes de arrasto, cama, fezes
frescas, conteúdo de comedouros e água, e no caso de reprodutoras e poedeiras
comerciais, ovos e embriões também podem ser cultivados (BERCHIERI JÚNIOR
& FREITAS NETO, 2009).
Já o diagnóstico sorológico, que mede uma resposta sorológica do
organismo frente a um antígeno específico, tem grande importância na medicina
aviária, pois permite o diagnóstico de doenças e o monitoramento do plantel
avícola (MUNOZ, 2005).
Destaca-se que além da tipificação sorológica, os sorovares de
Salmonella sp. podem ser classificados pelas técnicas fenotípicas e genotípicas.
Dentre as fenotípicas, as mais importantes são: biotipagem relacionada com
reações bioquímicas; fagotipagem baseada no emprego de bacteriófagos
específicos e reações antígeno-anticorpo, que se baseiam na presença ou
ausência de atividades metabólicas ou biológicas expressas pelo microrganismo
(FARBER, 1996).
Os métodos genotípicos se baseiam na reação em cadeia da
polimerase (PCR), que é uma técnica muito específica, podendo ser analisados
DNA cromossomais ou plasmidiais, possibilitando a designação do perfil
bacteriano a partir da utilização de genes conhecidos e únicos para determinada
espécie (DOUBLET et al., 2008).
Segundo VELILLA & TERZOLO (2009), as técnicas de caracterização
genotípicas são adequadas para identificar diferentes sorovares de Salmonella
sp. mediante a detecção de variações na sequência do DNA, sendo mais
vantajosas que as fenotípicas, uma vez que todas as bactérias podem ter seu
DNA extraído e, assim, a capacidade discriminatória dos métodos genotípicos é
maior que nos fenotípicos
Esses citados métodos de diagnóstico serão, pela sua importância e
aplicabilidade na medicina aviária, incluindo na salmonelose, descritos
sequencialmente.
2.2.1 Diagnóstico bacteriológico convencional

12
Segundo a Portaria nº 126 de 03/11/1995 da Secretaria de Defesa
Agropecuária do Ministério da Agricultura Pecuária e Abastecimento (MAPA), que
aprova as normas de Credenciamento e Monitoramento de Laboratórios para
diagnóstico da Pulorose e Tifo Aviário, o diagnóstico da Salmonella sp. deve ser
realizado por método bacteriológico tradicional, com o isolamento e identificação
do agente (BRASIL, 1994).
O diagnóstico bacteriológico é o teste gold standard para detecção de
Salmonella sp. nas fezes dos animais, no entanto, para o processamento das
amostras por essa técnica é necessário um prazo aproximado de uma semana.
No r PCR as amostras são processadas em um prazo aproximado de 24 horas,
incluindo o período de enriquecimento bacteriano. As técnicas moleculares vêm
sendo usadas com sucesso na detecção de Salmonella sp., inclusive dos
sorotipos específicos (DILMAGHANI et al., 2011).
A microbiologia convencional utiliza para o diagnóstico: isolamento,
identificação bioquímica e caracterização antigênica. Para a identificação final da
presença de Salmonella sp. as fases de pré-enriquecimento, enriquecimento
seletivo, isolamento em meio sólido, seleção de colônias suspeitas e sorologia
são necessárias (BRASIL, 2003).
Nas etapas de detecção, os testes bioquímicos utilizados apresentam
características que permitem, de acordo com a mesma fonte, observar, quando
há crescimento bacteriano, mudanças de pH e alterações de coloração dos meios
de cultura. O meio Sulphur Indol Motility (SIM) determina se os micro-organismos
têm motilidade ou não, dependendo do sorovar, sem motilidade (negativo para
Salmonella Gallinarum ou Salmonella Pullorum). Na presença de Salmonella sp. o
meio Triple Sugar Iron (TSI) apresenta bisel alcalino com produção de gás
positiva ou negativa e o meio Lysine Iron Agar (LIA) se mostra alcalino na base e
com ou sem presença de gás H2S. O caldo uréia apresenta reação de urease-
negativa para a presença de Salmonella sp. A interpretação dos resultados dos
testes é realizada após a incubação a 37º C por 24 horas (DILMAGHANI et al.,
2011).
Sequencialmente, as colônias que apresentarem a leitura dos testes
bioquímicos preliminares compatíveis com Salmonella sp. passam pela
caracterização bioquímica definitiva com testes para utilização do citrato e

13
malonato, transformação da fenilalanina, descarboxilação da lisina e ornitina,
fermentação de carboidratos, prova de Voges-Proskauer e prova do vermelho de
metila. Para confirmação do perfil esperado para Salmonella sp., prossegue-se a
caracterização antigênica com soro antissomático O polivalente de Salmonella sp,
sendo a identificação final de sorovar realizada pelos laboratórios credenciados
ou de referência (BRASIL, 1994).
Essa metodologia tradicional ou microbiologia convencional para
detecção de Salmonella sp., requer no mínimo quatro dias para obtenção de
resultado negativo. Assim, nem sempre são os mais adequados frente a
determinadas necessidades. As indústrias de alimentos, por exemplo, requerem
resultados em menor tempo para reduzir a estocagem de produtos e custos
associados, assim, métodos rápidos para detecção de Salmonella sp., por
exemplo, o ensaio imunoenzimático – ELISA, são utilizados com frequência por
causa do seu benefício de detecção rápida e eficiente (FLOWERS et al., 1988).
Assim, uma abordagem sucinta sobre testes sorológicos será
sequencialmente procedida.
2.2.2- Diagnóstico sorológico
2.2.2.1Aspectos imunológicos das aves
A eliminação do agente depende da resposta humoral e da resposta
imune local no intestino, que se desenvolvem após o nascimento, sendo que a
imunidade local parece ser mais efetiva na salmonelose sistêmica (BARROW, et
al., 2010).
O sistema imune das aves é constituído de órgãos linfóides centrais,
bursa de Fabricius, timo e órgãos linfóides periféricos que são o baço, glândula de
Harder e os tecidos linfóides associados ao sistema digestório (GALT) e sistema
respiratório (BALT). A bursa é um apêndice da cloaca, estrutura exclusiva das
aves, sendo o local de desenvolvimento dos linfócitos B e no timo, estrutura
localizada na região cervical, onde ocorre o desenvolvimento de linfócitos T.
Durante a fase embrionária e logo após a eclosão, os linfócitos B e T migram dos

14
órgãos linfóides centrais para as regiões linfóides periféricas (GOMES, 2008;
MONTASSIER, 2009).
Acrescenta-se ainda, que os tecidos linfóides do sistema digestório
(GALT) compreendem as placas de Peyer, encontradas ao longo do trato
intestinal, as tonsilas cecais e a Bursa de Fabricius. Estes órgãos, captam
antígenos presentes na luz intestinal que vão estimular células responsáveis pelo
desenvolvimento da imunidade geral e específica, denominadas células T e
células B precursoras de IgA, responsáveis por bloquear os receptores,
diminuindo a quantidade de bactérias na luz intestinal (BARROW, 2010; GAST et
al., 2011).
A primeira linha do sistema de defesa dos vertebrados é a resposta
imune inata, constituída de barreiras físicas e células fagocitárias, sendo que nas
aves as principais são os macrófagos e heterófilos. A imunidade adquirida ou
adaptativa é subdividida em imunidade passiva, pela qual ocorre transferência de
anticorpos de uma ave imunizada para outra (matriz-pinto), ou imunidade ativa,
que é induzida pela exposição da ave a antígenos. Uma vez identificada a
presença do antígeno, são desencadeadas as respostas humoral e celular. A
resposta imune inata pode influenciar o caráter da imunidade adquirida, que por
sua vez, possui ramos efetores que se utilizam de mecanismos da inata como os
fagócitos (ABBAS & LICHTMAN, 2005).
De acordo com MORGULES (2005), a Bursa de Fabricius controla a
imunidade humoral, atuando na secreção de anticorpos pelos linfócitos B. Os
anticorpos interagem e neutralizam o patógeno permitindo a digestão, lise e morte
celular. Por outro lado, a resposta imune celular envolve a interação entre células
diferentes montando uma resposta especifica ao antígeno invasor. Os macrófagos
além de detectar, fagocitar, digerir e apresentar fragmentos do patógeno aos
linfócitos T, secretam citocinas, como a IL-1, capaz de alterar a atividade dos
linfócitos T, e IL-2 indutoras da proliferação clonal de células que responderão
especificamente ao antígeno.
As salmonelas virulentas, segundo TERZOLO (2011), possuem um
sistema denominado PhoP/PhoQ, capaz de regular genes que permitem a
adaptação do microrganismo nas condições do ambiente interno dos órgãos das
aves. Este sistema PhoQ é um regulador da expressão de aproximadamente 40

15
genes envolvidos na sobrevivência da bactéria no interior dos macrófagos, na
resistência a peptídeos antimicrobianos e no crescimento em quantidades
limitadas de Mg2+. Cepas mutantes com supressão do sistema PhoP têm elevado
nível de atenuação aliada à perfeita capacidade de colonização do intestino.
BERCHIERI JÚNIOR & FREITAS NETO (2009) relataram que
infecções por Salmonella ocorrem principalmente por via oral. Na superfície apical
do epitélio intestinal, este microrganismo induz a ruptura e o afastamento das
microvilosidades, provocando endocitose. As bactérias são fagocitadas pelos
macrófagos e por fagócitos polimorfonucleares, permanecendo nos fagossomos
nos quais se multiplicam e estabelecem a infecção sistêmica. Ao penetrar no
epitélio intestinal, estimulam receptores das células intestinais “toll-like receptor 5”
(TLR5), que desencadeiam a produção de IL-1, IL-6, IL-8, que são citocinas pró-
inflamatórias. A ação dos macrófagos e heterófilos, juntamente com a secreção
de IL-1, IL-6, IL-8, configura a resposta imune inata, que auxilia na prevenção da
infecção sistêmica e estimula a resposta imunidade ativa (celular e humoral).
A IL-1 ativa macrófagos e células T. Em macrófagos do fígado há a
produção de IL-18 que estimula as células Th-1 (T helper) a produzirem INF-gama
e também estimula a produção de IL-2, iniciando a resposta imune celular. O
INF-gama ativa MHC II, que apresentam antígenos bacterianos aos linfócitos T,
que irão destruir células infectadas. IL-2 ativam macrófagos e induzem a
multiplicação de linfócitos T e B (BERCHIERI JÚNIOR & FREITAS NETO, 2009).
Além disso, bactérias do gênero Salmonella são capazes de estimular
resposta imunológica via complexo de histocompatibilidade (MHC) de classes I e
II, ao atingirem as células M da mucosa intestinal. Células T CD8+ e T CD4+ são
as células que reconhecem os antígenos apresentados pelo MHC I e II,
respectivamente (WEISS & KRUSCH, 2001).
Estudos desenvolvidos por VAN IMMERSSEL et al. (2005),
confirmaram que após a invasão de Salmonella sp. no epitélio intestinal, a
bactéria é capaz de se multiplicar intracelularmente, causar inflamação da parede
intestinal e promover a migração de macrófagos dos vasos para a lâmina própria,
realizando a fagocitose. Mas, diferente do que acontece com outros patógenos,
as substâncias antimicrobianas não são capazes de eliminar a bactéria. Com isso,

16
os macrófagos migram de volta para o sangue, disseminando o agente para os
órgãos internos, estabelecendo, assim, a doença sistêmica.
Uma observação importante no estudo de respostas imunes do
organismo das aves à salmonela, é que no caso das infecções por Salmonella
Enteritidis, quando este sorovar está presente num lote de aves, outros
normalmente encontrados, desaparecem. Podendo ressaltar que a infecção por
Salmonella Enteritidis possui um excelente mecanismo de resistência as outras
salmonelas em aves (CHA et al., 2013).
A sorologia é uma metodologia laboratorial que visa o estudo e a
mensuração das reações antígeno-anticorpo através do soro, após a exposição
do hospedeiro a um determinado agente estranho, ou seja, a sorologia mede uma
resposta específica do organismo frente a um antígeno específico. Consiste em
detectar e quantificar a presença de anticorpos para um determinado agente. Esta
técnica é a melhor opção para diagnosticar uma infecção, uma vez que o agente
infeccioso tende a ser eliminado com a evolução do quadro. Apenas anticorpos
permanecem e servem como prova de que a infecção ocorreu. Portanto, a
sorologia representa um método indireto para o diagnóstico de infecções
(MAIJALA, et al., 2005).
Na avicultura, um programa de monitoramento sorológico tem como
objetivo reduzir os riscos de infecções em uma população específica e aumentar
o controle sanitário dos plantéis avícolas. Para tal, faz-se necessário desenvolver
e implementar normas e procedimentos rígidos em todos os segmentos da
produção por meio de programas de biossegurança (RODRIGUES, 2011)..
Especificamente para salmoneloses aviárias, Salmonella enterica
sorovar Gallinarum e Salmonella enterica sorovar Pullorum podem ser detectadas
sorologicamente pelo uso de testes de aglutinação em tubo, aglutinação rápida ou
teste de micro aglutinação (ZANELLA, 2007), os quais serão abordados a seguir:
2.2.3- Soroaglutinação rápida (SAR)
A SAR é uma prova simples para monitorar o estado de saúde de
plantéis avícolas, dotada de grande sensibilidade para a detecção dos anticorpos,
porém, requer especificidade quanto à identificação do agente. Assim, deve-se

17
usar provas mais específicas para suporte de um diagnóstico mais preciso
(RODRIGUES, 2011).
A realização de provas de Soroaglutinação rápida (SAR) com
antígenos polivalentes O (somático) utilizado para tirar aves reagentes para
Salmonella Pullorum e Salmonella Gallinarum, pode ser útil para reconhecer aves
infectadas por diferentes espécies de salmonelas do grupo paratifóife,
particularmente Salmonella Enteritidis (OLIVEIRA, 2004).
A descrição original do teste, usando soro, foi realizada por RUNNELS
em 1927. Posteriormente, BUNYEA, et al. (1929) fizeram uma adaptação da
prova usando uma gota de sangue total e uma gota de suspensão salina densa
de Salmonella Pullorum viva como antígeno.
Em 1931, SCHAFFER publicou um trabalho alertando sobre o risco do
uso de antígeno vivo na realização do teste de SAR em granjas e no mesmo
trabalho descreve um novo antígeno produzido com Salmonella Pullorum
inativada com formol e corado com cristal violeta.
O uso do teste de aglutinação rápida tem sido novamente enfatizado
devido ao reaparecimento de Salmonella Pullorum em operações integradas de
frangos de corte e devido à possibilidade de detecção de lotes infectados por
Salmonella Enteritidis. O teste é, entretanto, sujeito a alterações nos resultados.
Podem ser encontrados resultados falso-positivos, pelo teste apresentar
especificidade inadequada, e falso-negativos, em consequência de sua baixa
sensibilidade (BERCHIERI JÚNIOR & FREITAS NETO, 2009).
A Soroaglutinação é um teste de triagem que detecta imunoglobulinas
M e G, sendo IgM a principal. A concentração de proteína/g.cm3 detectada é
0,05. Este teste é qualitativo, assim, determina a positividade e negatividade de
animais, também é considerado teste de ligação secundária, o qual se processa
em dois estágios. O primeiro estágio é a interação entre antígeno e anticorpo,
uma reação que não é intensamente afetada pela temperatura, mas pode ser
revertida por força iônica alta ou um pH baixo. O segundo estágio é determinado
pelo estado físico do antígeno. Portanto, se o s anticorpos combinam-se com
antígenos solúveis em solução sob condições apropriadas, os complexos
precipitam-se. Se os antígenos são particulados – por exemplo, bactérias ou
eritrócitos, então, eles aglutinam-se. (MONTASSIER, 2009).

18
Apesar da larga utilização do teste de SAR na triagem das aves, o
achado de aves com sorologia positiva não pode ser interpretado como infecção,
sendo sempre recomendada a cultura bacteriológica, para identificação do
organismo infectante (OLIVEIRA et al., 2004).
2.2.4 Soroaglutinação Lenta (SAL)
O teste é utilizado em várias provas diagnósticas. Geralmente tomam-
se diluições seriadas do soro, sobre as mesmas adiciona-se uma suspensão do
antígeno padronizado. Após um período de incubação em temperatura adequada,
procede-se a leitura, verificando-se a formação do aglutinado no fundo do tubo ou
da microplaca (MUNOZ, 2005).
A salmonelose também pode ser detectada por teste de
Soroaglutinação lenta em tubos e ELISA. O ensaio imunoenzimático, ELISA,
detecta portadores de resposta sorológica ao agente da pulorose e do tifo aviário.
Oliveira et al, (2004) verificaram que o ELISA é sensível para detectar resposta
sorológica para Salmonella Gallinarum e Salmonella Pullorum.
Os testes de microaglutinação e soroaglutinação macroscópica em
tubos (prova lenta) também detectam anticorpos contra S. Enteritidis, embora com
variação de sensibilidade entre elas (MONTASSIER, 2009).
2.2.5 – Teste Imunoenzimático (ELISA)
Salmonella Enteritidis é um patógeno importante da avicultura e tem
sido isolada de frangos de corte, matrizes e lotes de aves de postura comercial. A
identificação bacteriológica de aves positivas é difícil devido à excreção
intermitente da bactéria. A presença de anticorpos nem sempre determina uma
infecção, mas indica exposição prévia. A análise por imunoabsorção ligada a
enzimas tem demonstrado ser útil na detecção de anticorpos contra salmonela
aviária e em especial no controle em larga escala de lotes para verificar a
presença de infecção causada por Salmonella Enteritidis (BURKHOLDER, et al.,
2008).

19
O kit pode ser usado como método de triagem inicial para detecção de
anticorpos contra Salmonella Enteritidis. Entretanto, uma vez que o ELISA para
Salmonella Enteritidis baseia-se nos antígenos flagelares G e M, podem gerar
potencialmente resultados falso-positivos. Portanto, os resultados positivos da
detecção devem sempre ser confirmados por bacteriológicos convencionais
(ABBAS & LICHTMAN, 2005).
2.2.6 Diagnóstico molecular
2.2.6.1 Fatores de virulência
Os micro-organismos patogênicos se distinguem de outros da mesma
espécie, por possuírem e expressarem genes que codificam os fatores de
virulência e que conferem à bactéria a habilidade de causar doença no seu
hospedeiro. Estes fatores propiciam a invasão e colonização das células do
hospedeiro pelo micro-organismo, levando à ocorrência de uma série de eventos
que levam ao aparecimento da doença. Estes fatores podem ser mecanismos de
invasão, que interfiram na resposta imune do hospedeiro ou mecanismos de
resistência a antimicrobianos (VIEIRA, 2009).
De acordo com VAN ASTEN & VAN DICK (2005), os genes
codificadores dos fatores de virulência podem estar alojados no próprio
cromossoma do patógeno, como, por exemplo, nas ilhas cromossomais,
denominadas ilhas de patogenicidade ou em elementos genéticos transmissíveis
que são os plasmídeos, bacteriófagos e transposons.
Assim como em outros microrganismos patogênicos, as bactérias do
gênero Salmonella possuem fimbrias ou pili, que são estruturas protéicas,
dispostas em forma helicoidal e responsáveis pela aderência as superfícies,
permanência no ambiente em que se encontram e formação de biofilmes
(GIBSON et al., 2007).
Antes de invadir uma célula do hospedeiro, a salmonela deve encontrar
e se aderir a um ou mais tipos de células intestinais. Geneticamente, em
Salmonella enterica sorovar Typhimurium são descritos quatro tipos diferentes de
fimbrias, que são as fimbrias do tipo I (FIM), as fimbrias codificadas por
plasmídeos (PE), fimbria polar longa (LPF) e as fimbrias agregativas (AGF).

20
Postula-se ainda, que cada tipo de fimbria tem tropismo especifico para o tipo
celular que será colonizado e que participam diretamente no processo de
recrutamento de leucócitos para o sitio de infecção (VIEIRA, 2009).
Em Salmonella enterica sorovar Enteritidis foram descritos pelo menos
cinco tipos diferentes de fimbrias: SEF14, SEF17, SEF21, fimbria tipo 3 e BFP
(bundle-forming pili). A fimbria SEF14 é codificada por um gene de 5,3Kb que foi
estratificado, com o auxilio da enzima de restrição Hind III, em uma porção de
3,9Kb. Após sequenciamento genético, este fragmento foi subdividido em três
genes: SEFA, SEFB E SEFC. Estes genes são responsáveis pela estrutura e
expressão da fimbria SEF14, destacando-se que a identificação de suas
sequências nucleotídicas constitui a primeira sequência de um operon fimbrial
importante com relação à capacidade invasiva deste agente. Entretanto, o gene
SEFA codifica apenas uma fimbria, enquanto que sefB e sefC codificam proteínas
similares às proteínas acessórias de Escherichia coli e Klebsiella pneumoniae. As
transcrições de sefB e sefC se iniciam nas regiões de sefA, portanto, não se
expressam na ausência de sefA (CLOUTHIER et al., 1993).
Posteriormente, estudo desenvolvido por COLLINGHAN &
WOODWARD (2001) permitiu a identificação de mais dois genes no operon
codificadores de fimbrias sefD e sefE. Foi constatada a presença de genes sefA e
sefD em 100% dos isolados de Salmonella enterica sorovar Dublin e Salmonella
enterica sorovar Enteritidis, entretanto, apenas em 38,9% das 18 cepas de
Salmonella enterica sorovar Pullorum encontrou-se o gene sefD (ZHU et al.,
2010).
NAUGHTON et al. (2001) analisaram a contagem de Salmonella sp.
em estômago e intestino de ratos e observaram que as bactérias que
expressaram as fimbrias SEF21, estavam em maior número nas primeiras horas
após a inoculação. Para estes pesquisadores, a fimbria SEF21 é fator
considerável de virulência nas fases iniciais de infecção por esse patógeno.
Em pesquisa desenvolvida por SALEHI et al. (2011), foram
identificados genes fimbriais codificadores de SEF14, SEF17 e SEF21 em 100%
de Salmonella enterica sorovar Enteritidis isoladas em 45 amostras de fezes de
diarréia de humanos e em 25 de excretas de aves assintomáticas. Os autores
concluíram que, embora a presença de genes fimbriais seja essencial para a

21
aderência e colonização da bactéria no epitélio intestinal do hospedeiro,
isoladamente os mesmos não são capazes de determinar infecção clinica.
Portanto, outros fatores de virulência como, por exemplo, endotoxinas, resistência
à ação de fagócitos e genes plasmidiais, são indispensáveis para instalação da
enfermidade.
As ilhas de patogenicidade são extensas regiões do cromossoma, com
variação de 10.000 até 200.000 pares de base, que apresentam características
próprias, de grande instabilidade e que têm pelo menos um gene associado à
virulência. Dentre as características específicas das ilhas de patogenicidade,
destaca-se a elevada quantidade de guanidina e citosina em relação ao restante
do sequenciamento do cromossoma, que geralmente se encontra associada a
genes codificadores de RNA transportador (VIEIRA, 2009).
Em Salmonella sp. são denominadas “Salmonella patogenicity islands”
(SPI), sendo que neste gênero foram descritas mais de dez ilhas de
patogenicidade diferentes (SALEHI et al., 2011), sendo cinco delas em sorovares
de interesse veterinário (SPI-1, SPI-2, SPI-3, SPI-4, SPI-5) (BERCHIERI JÚNIOR
& FREITAS NETO, 2009).
A sobrevivência do microrganismo no hospedeiro e a manifestação da
fase sistêmica da doença são processos estabelecidos pelas ilhas SPI-2, SPI-3 e
SPI-4. Já a SPI-5 codifica fatores de virulência relacionados à inflamação e
secreção de cloretos, caracterizando a fase entérica da doença (MARCUS et al.,
2000). A SPI-1 está presente em todos os sorovares de Salmonella sp. e contém
genes que são indispensáveis para a invasão do patógeno em células não
fagocíticas, como as células do epitélio intestinal: inv, spa e hil (HACKER et al.,
1997).
A SPI-1 contém genes responsáveis pela codificação do sistema de
secreção tipo III (SSTT), o qual favorece a entrada de proteínas denominadas
“efetoras” no interior da célula do hospedeiro. Por meio deste mecanismo, as
bactérias exportam proteínas com auxílio de uma espécie de seringa molecular.
Uma vez dentro da célula alvo, as proteínas efetoras interagem com proteínas do
hospedeiro, desencadeando algumas sequências de reações químicas capazes
de configurar diversas alterações celulares, tais como, escape do sistema de
defesa, mudanças no citoesqueleto e morte celular (MOTA et al., 2005).

22
A ilha de patogenicidade denominada SPI-2, promove a ativação de
outro tipo de SSTT, que estabelece o trânsito de proteínas bacterianas no meio
intracelular no momento em que a bactéria está dentro dos fagossomos. Genes
pertencentes a SPI-2, provocam a inativação da enzima NADPH-oxidase,
impedindo a morte bacteriana por intermediários reativos de oxigênio e,
consequentemente, criando ambiente satisfatório à sobrevivência do patógeno
(BERCHIERI JÚNIOR & FREITAS NETO, 2009).
De acordo com GALAN et al. (1992), o operon invABC em diversos
sorovares de Salmonella sp., desempenha função importante na invasão de
células epiteliais. O gene invA é o primeiro do operon, estando situado na mesma
unidade transcricional dos genes invB e invC, sendo que invD está alojado em
outra unidade.
Destaca-se que WHANG et al. (2009) afirmaram que o gene invA
parece estar conservado em todos os sorovares de Salmonella sp. sendo eleito
para detecção desta bactéria pela técnica da Reação em Cadeia da Polimerase
(PCR).
Em estudo realizado por PAVLOVA et al. (2011), foram infectados
macrófagos alveolares de suínos com Salmonella enterica sorovar Typhimurium e
Salmonella enterica sorovar Enteritidis e suas estirpes mutantes que foram
desprovidas da SPI-1. Os pesquisadores observaram que os mutantes de ambos
sorovares foram cerca de cinco vezes menos eficientes no processo de invasão
do hospedeiro. Destacaram, entretanto, que apesar desta condição, os
macrófagos infectados por estas bactérias responderam de forma satisfatória à
infecção pelo aumento da expressão de citocinas inflamatórias como, por
exemplo, IL-1b, IL-8 E IL-23. Concluíram ainda que a SPI-1, além de auxiliar na
invasão celular é responsável pela supressão da expressão de citocinas pelos
macrófagos.
Os plasmídeos constituem elementos genéticos extracromossomais
que se replicam de maneira independente, carreiam informações genéticas
adicionais e podem ser perdidos ou adquiridos de forma espontânea (ARBEIT,
1999), sendo que, segundo ROTGER & CASADÉSUS (1999), a maioria dos
sorovares de Salmonella enterica subspécie enterica não possui plasmídeos.
Contudo, estão presentes nos sorovares frequentemente associados a infecções

23
humanas e de animais de criação, como Salmonella enterica sorovar Enteritidis,
Salmonella enterica sorovar Pullorum, Salmonella enterica sorovar Gallinarum,
Salmonella enterica sorovar Dublin, Salmonella enterica sorovar Typhimurium e
Salmonella enterica sorovar Abortus-ovis.
Acentua-se que os plasmídeos apresentam tamanhos variados de
acordo com o sorovar de Salmonella sp., no entanto, existe uma região
cromossômica, de 7,8Kb, que se mantém conservada nos diversos sorovares do
gênero. Esta região foi sequenciada e foi determinada a presença de cinco genes
denominados spv (Salmonella Plasmid Virulence): spvA, spvB, spvC, spvD, spvR.
Os genes spvA, spvB, spvC, spvD são considerados genes estruturais, mas
somente são ativados mediante a presença do produto codificado pelo spvR, que
é produzido na fase estacionária de crescimento. Bactérias patogênicas
portadoras de operon spv expressam fatores de virulência que contribuem para
sua sobrevivência em ambientes desfavoráveis como, por exemplo, privação de
nutrientes, pH ácido e altas temperaturas (VALONE et al., 1993).
As técnicas associadas à biologia molecular e à engenharia genética
progrediram muito desde a década de 50, quando o DNA teve sua estrutura
descrita. Os métodos genotípicos se baseiam na reação em cadeia da polimerase
(PCR), que é uma técnica muito específica, podendo ser analisados DNA
cromossomais ou plasmidiais, possibilitando a designação do perfil bacteriano a
partir da utilização de genes conhecidos e únicos para determinada espécie
(DOUBLET et al., 2008).
Assim, o desenvolvimento da PCR representou um avanço tecnológico,
possibilitando diagnóstico rápido, rentável e automatizado. Além destas citadas
vantagens, as técnicas de amplificação do ácido nucléico apresentam, segundo
MORENO et al. (2009), outras como a utilização de amostras clínicas com
pequenas quantidades de microrganismos, elevadas sensibilidade e
especificidade e segurança no diagnóstico.
Nesse contexto, WEI et al. (2013) destacaram que a PCR apresenta
capacidade de detectar um gene especifico de um microrganismo isolado ou em
grupo. Além desta, há a possibilidade de detectar a presença de um patógeno
sem a necessidade de cultivo e isolamento de cultura pura e detecção de grupos

24
taxonômicos amplificados, como no caso das provas de região alvo da molécula
de RNA ribossômico com níveis diferentes de variabilidade.
Deve ser registrado que para aplicação da técnica de PCR, o Comitê
Internacional de Normatização da Europa (European Committee for
Standardization) estabeleceu protocolos de PCR para detecção de patógenos.
Um método de PCR padronizado deve cumprir alguns critérios como precisão de
análise e de diagnóstico, alta probabilidade de detecção, baixa contaminação e
protocolos que sejam acessíveis e de fácil interpretação (MALORNY et al., 2004).
Destaca-se que um ponto crítico da técnica da PCR é a seleção das
sequências que flanqueiam os segmentos de DNA que se pretende amplificar,
sendo que a especificidade do método de diagnóstico molecular está vinculada à
precisão dos iniciadores em desempenhar esta tarefa. No caso de Salmonella sp.
a região escolhida deve ser comum à maioria das cepas, deve codificar proteínas
importantes na patogenicidade e não deve apresentar homologia com outros
patógenos, evitando assim resultados falso-positivos (STONE et al., 1994). O
conjunto de genes invABCDE distingue entre Salmonella sp. e outras bactérias
como, por exemplo, Escherichia coli, pela hibridização com sonda específicas ou
amplificação de DNA, uma vez que estão presentes em uma série de sorovares
de Salmonella sp. e ausentes na região correspondente de outros patógenos
(HEID, et al., 2004).
Existe uma variedade de iniciadores utilizados para detectar
Salmonella sp.: agfA, IS200, hin, H-li, iagAB, spvR, viaB, mkfA, ompC, oriC,
sendo a principal característica a especificidade. Por exemplo, o gene invA está
presente nas duas espécies de Salmonella, mas o gene invAB somente em
alguns sorovares de Salmonella enterica (BAUNLER et al., 1997).
A PCR em Tempo Real permite a quantificação das amostras
amplificadas, é de grande relevância para diagnósticos de patógenos e doenças
genéticas. Dentre as vantagens, desta técnica em relação à PCR qualitativa estão
a facilidade na quantificação, maior sensibilidade, maior precisão,
reprodutibilidade e acurácia, velocidade na análise, melhor controle de qualidade
no processo e menor risco de contaminação (HEID, et al., 2004).
A reação em cadeia de polimerização em tempo real combina a
metodologia de PCR convencional com um mecanismo de detecção e

25
quantificação por fluorescência. A metodologia, por não mais requerer a detecção
em gel de eletroforese necessário na análise da PCR, permite que os processos
de amplificação, detecção e quantificação de DNA, sejam realizados em uma
única etapa, agilizando a obtenção de resultados e diminuindo o risco de
contaminação da amostra e dando maior precisão. A possibilidade de monitorar a
PCR em tempo real ( PCR) revolucionou, pois, o processo de quantificação de
fragmentos de DNA e RNA e na aplicação em diagnósticos, como a detecção de
patógenos ou doenças, como a salmonelose (MULLIS et al., 1990).
A r PCR realiza a quantificação destes ácidos nucléicos de maneira
precisa e com maior reprodutibilidade, porque determina valores durante a fase
exponencial da reação. O ponto que detecta o ciclo na qual a reação atinge o
limiar da fase exponencial é denominado de Cycle Threshold (CT). Este ponto
permite a quantificação exata e reprodutível baseado na fluorescência (HEID, et
al., 2004).
A r PCR requer uma plataforma de instrumentação que contém um
termociclador com sistema ótico para a excitação da fluorescência e na coleção
da emissão e um computador com um software para aquisição de dados e análise
final da reação. Estas máquinas diferem na capacidade da amostra (96-poços
padrão, processamento de poucas amostras ou requerem tubos capilares de vidro
especializados), no método da excitação (lasers ou fontes claras do espectro
largo com filtros ajustáveis) e na sensibilidade total. Há também diferenças nos
softwares para o processamento dos dados (HEID, et al., 2004).
A emissão dos compostos fluorescentes gera um sinal que aumenta na
proporção direta da quantidade de produto da PCR. Sendo assim, os valores da
fluorescência são gravados durante cada ciclo e representam a quantidade de
produto amplificado. Os compostos fluorescentes mais utilizados são o SYBR®
Green e TaqMan®. (MULLIS et al., 1990), que serão sucintamente enfocados.
O SYBR® Green se liga entre a fita dupla de DNA e com a excitação
da luz emitida pelo sistema ótico do termociclador, emite uma fluorescência verde.
As vantagens da utilização do SYBR® Green são: baixo custo, facilidade no uso e
sensibilidade. A desvantagem é a ligação em todo DNA fita dupla que surge
durante a reação, incluindo os dímeros dos iniciadores e outros produtos
inespecíficos, podendo superestimar a concentração do fragmento alvo. O

26
SYBR® Green não ligado ao DNA exibe uma fluorescência muito pequena (HEID,
et al., 2004).
.Entretanto, a fluorescência é realçada quando ligado na fita dupla do
DNA. No começo da amplificação, a mistura da reação contém o DNA
desnaturado, os iniciadores e o SYBR® Green. As moléculas não-ligadas do
SYBR® Green apresentam fluorescência fraca produzindo um sinal mínimo sendo
este subtraído durante a análise de computador. Após o reconhecimento dos
iniciadores, algumas moléculas do SYBR® Green podem ligar-se na fita dupla
previamente formada. Durante a polimerização catalisada pela enzima Taq DNA
polimerase, as moléculas do SYBR® Green vão se ligando ao DNA recentemente
sintetizado. Assim, a reação é monitorada continuamente e um aumento da
fluorescência é observado em tempo real. No ciclo seguinte, na etapa de
desnaturação do DNA, as moléculas do SYBR® Green são liberadas e há queda
no sinal da fluorescência. A detecção da fluorescência no fim da etapa de
extensão de cada ciclo da PCR permite monitorar a quantidade crescente de DNA
amplificado (VITZTHUM et al., 1999).
Já TaqMan® é uma sonda (fragmento de DNA marcado usado para
hibridizar outra molécula de DNA) utilizada para detectar sequências específicas
nos fragmentos de DNA amplificados na PCR. Esta sonda apresenta em uma
extremidade um fluoróforo, e na outra extremidade um quencher (molécula que
aceita energia do fluoróforo na forma de luz e a dissipa na forma de luz ou calor).
Os produtos da reação são detectados pela fluorescência gerada após a atividade
exonuclease 5'—>3' da Taq DNA polimerase. Durante a PCR em tempo real a
sonda TaqMan® hibridiza com a sequência da fita simples de DNA complementar
alvo para a amplificação.
No processo da amplificação, a sonda TaqMan® é degradada devido à
atividade exonuclease 5'—>3' da Taq DNA polimerase, separando o quencher da
molécula fluorescente durante a extensão. A separação do fluoróforo do quencher
resulta em um aumento da intensidade da fluorescência. Assim, durante o
processo de amplificação a emissão de luz é aumentada de forma exponencial.
Esse aumento da fluorescência ocorre apenas quando a sonda hibridiza e quando
a amplificação da sequência alvo é estabelecida (HEID et al., 1996). A reação

27
com a TaqMan® é considerada um método sensível para determinar a presença
ou ausência de sequências específicas (VITZTHUM et al., 1999).
Registre-se que esse sistema de quantificação em tempo real, além da
aplicação direta na análise de sequência de patógenos como a Salmonella sp.,
dentre outros agentes virais, bacterianas ou de protozoários a partir de várias
fontes, possibilita a identificação de alelos em DNA genômico, a análise de
patógenos em produtos transgênicos e em alimentos, sendo esta última também
importante para a bactéria aqui abordada (ZHU, et al., 2010).
Muitos estudos vêm sendo conduzidos empregando a técnica de PCR.
Dentre eles, pode-se citar o de MALORNY et al ( 2004), que visando avaliar
amostras de alimentos contaminadas com Salmonella sp. avaliaram 110 amostras
de alimentos pela bacteriologia convencional e pelo PCR em tempo real. Os
primers e a sonda eram específicas para um locus altamente conservado
composto por cinco genes no operon de Salmonella sp. denominado ttr. Até
então, esta região não teria sido relatada para detecção desta bactéria. Os genes
ttrA, ttrB e ttrC são responsáveis pela codificação de proteína tetrationato
reductase. Todas as amostras analisadas, tanto pela microbiologia como pelo
diagnóstico molecular foram positivas, entretanto, a PCR em tempo real (q-PCR),
foi concluída em 24h enquanto que a bacteriologia demorou cinco dias, além de
demonstrar alta seletividade, precisão e probabilidade de detecção
(SHIVAPRASAD & BARROW, 2008).
Destaca-se que a técnica de PCR utilizada nesse referido estudo, a de
PCR em tempo real, é empregada principalmente, segundo GLYNN et al. ( 2006),
devido à característica qualitativa da PCR tradicional, que a torna limitante nos
estudos quantitativos.
3. CONSIDERAÇÕES FINAIS
As infecções causadas por Salmonella sp. são de grande relevância,
tanto na Saúde Pública como na sanidade avícola, uma vez que podem afetar
homens e aves e causar grandes perdas na produção avícola, com impactos
sanitários e econômicos.

28
Assim, o controle da salmonelose nos sistemas de criação é
fundamental, dependendo, dentre outros fatores, do conhecimento da
epidemiologia, patogênese e das características do patógeno, principalmente
aquelas relacionadas à virulência e transmissão.
Estudos do grau de infecção das aves e do ambiente de criação de
uma determinada região devem ser conduzidos para que sejam tomadas
decisões a respeito do monitoramento e controle da bactéria, bem como a
utilização de algumas ferramentas como, por exemplo, a vacinação.
Outro aspecto importante e atual refere-se à resistência antimicrobiana,
pois o uso indiscriminado de antibióticos pode levar à seleção de cepas
resistentes e estas podem estar presentes nos alimentos de origem animal e
causar graves infecções em seres humanos.
Portanto, há necessidade de ampliação de pesquisas na área que
permitam não apenas mensurar a circulação do patógeno na avicultura, mas,
dentre outros aspectos, identificar seus principais fatores de risco e avaliar e
desenvolver técnicas de diagnóstico efetivas visando à sua prevenção e controle,
minimizando os impactos da salmonelose tanto no setor avícola quanto para a
saúde humana.
A redução de tais impactos representa um dos maiores desafios para a
avicultura atualmente, pois evitar a ocorrência de doenças transmissíveis aos
seres humanos, garantir a saúde e o desempenho das aves bem como atender às
demandas comerciais são aspectos indissociáveis, destacando-se a crescente
relevância da segurança alimentar, que deve ser aplicada tanto para alimentos
destinados à exportação quanto para abastecimento do mercado interno.

29
REFERÊNCIAS
1. ABBAS, A.K.; LICHTMAN, A.H. Propriedades gerais das respostas imunológicas. In: Imunologia Celular e Molecular. Rio de Janeiro. ELSEVIER. 5aed. 580p. 2005.
2. BARROW, P. A.; HUGGINS, M. B.; LOVELL, M. A. Host Specificity of Salmonella Infection in Chickens and Mice Is Expressed In Vivo Primarily at the Level of the Reticuloendothelial System, Infection and Immunity, Washington, v. 62, n. 10, p. 4602-4610, 1994.
3. BARROW, P. A.; JONES, M.A.; THOMSON, N. Salmonella. In: GYLES, C.L.; PRESCOTT, J.F.; SONGER, G.; THOEN, C.O. Pathogenesis of bacterial infections in animals. 4 ed. Iowa: Blackwell Publishing, 2010. Cap. 14, p 231-257.
4. BAUNLER, J. A.; HEFFRON, F.; REISSBRODT, R. Rapid detection of Salmonella enterica with primers specific for iroB. Journal of Clinical Microbiology, p. 1224-1230, 1997
5. BERCHIERI JÚNIOR, A.; FREITAS NETO, O.C.; Salmoneloses. In: Doenças das Aves. Campinas, São Paulo. FACTA. 2aed. p.435-456. 2009
6. BERCHIERI JR., A.; MURPHY, C. K.; MARSTON, K.; BARROW, P. A. Observations on the persistence and vertical transmission of Salmonella enterica sorovares Pullorum and Gallinarum in chickens: effect of bacterial and host genetic background. Avian Pathology, Huntingdon, v. 30, n. 3, p. 221 - 231, 2010.
7. BEAM, A.; GARBER, L.; SAKUGAWA, J.; KOPRAL, C. Salmonella awareness and related management practices in U.S. urban backyard chicken flocks. Prev Vet Med, v. n. p. 2013.
8. BRASIL, Ministério da Agricultura, do Abastecimento e da Reforma Agrária Programa Nacional de Sanidade Avícola.Atos Legais. Portaria nº 193 de 19 de setembro de 1994. Diário Oficial da República Federativa do Brasil, Poder Executivo, Brasília – DF, 22 de setembro de 1994. Seção 1, p14309-14312.
9. BRASIL. Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa Agropecuária Instrução Normativa nº 62, de 26 de agosto de 2003. Oficializa os Métodos Analíticos Oficiais para Análises microbiológicas para Controle de Produtos de Origem Animal e Água. Diário Oficial da União, Brasília, DF, 18 de set. 2003. Seção 1, p.14.
10. BRASIL. Programa Nacional de Monitoramento da Prevalência e da Resistência Bacteriana em Frangos. Agência Nacional de Vigilância Sanitária (ANVISA), Brasília, DF, 2008.186p.

30
11. BURKHOLDER, K.M.; THOMPSON, K.L.; EINSTEIN, M.E.; APPLEGATE,T.J.; PATTERSON, J.A. Influence of Stressors on Normal Intestinal Microbiota, Intestinal Morphology,and Susceptibility to Salmonella Enteritidis Colonization in Broilers. Poultry Science, Champaign, v.87, n. 9, p. 1734-1741, 2008.
12. BUNYEA, H., HALL, W.J., DORSET, M. A. A simplified agglutination test for Pullorum disease. Journal American Veterinary Medical Association, v. 28, n.4, p. 408-410, 1929.
13. BORSOI, A., L. R. SANTOS, L. B. RODRIGUES, H. L. S. MORAES, C. T. P. SALLE, V. P. N. Behavior of Salmonella Heidelberg and Salmonella Enteritidis strains following Broiler Chick inoculation: Evaluation of cecal morphometry, liver and cecum bacterial counts and fecal excretion patterns. Brazilian Journal of Microbiology. 42:266-273. 2011.
14. CALIXTO, A. E. R.; SERAFINI, A. B.; KIPNIS, A.; ANDRÉ, M. C. D. P. B. Prevalência de Salmonella e ocorrência de cepas resistentes a antimicrobianos em insumos de rações para aves produzidos por um matadouro-frigorífico com fiscalização permanente, em Goiânia, GO. Higiene Alimentar, São Paulo, v. 16, n. 101, p. 56-62, 2002.
15. CASON, J. A.; BAILEY, J. S.; STERN, N. J.; WHITTEMORE, A. D., COX, N. A. Relationship between aerobic bacteria, Salmonellae, and Campylobacter on broiler carcasses. Poultry Science, Champaign, v. 76, n. 8, p. 1037-1041, 1997.
16. CHA, S.Y.; KANG M.; YOON, R.H.; PARK, C. K.; MOON, O. K.; JANG, H.K.Comp Immunol Microbiol Infect Dis. 2013 Sep;36(5):473-9. doi: 10.1016/j.cimid.2013.03.004. Epub Apr 22. 2013.
17. CHAPPEL, L,; KAISER, P.; BARROW, P.; B, JONES, M. A.; JOHNSTON, C.; WIGLEY, P.The immunobiology of avian systemic salmonellosis, Veterinary Immunology and Immunopathology, Amsterdam, v. 128, n. 1-3, p. 53-59, 2009.
18. CRUMP, J.A.; LUBY, S.P.; MINTZ, E.D. The global burden of typhoid fever. Bulletin of the Word Health Organization. v.82, n.5, p.346-353, 2004.
19. CLOUTHIER, S.C.; COLISSON, S.K.; LIPPERT, D.; AUSIO, J.; WHITE, A.P.; KAY, W.W. Characterization of three fimbrial genes, sefABC, of Salmonella Enteritidis. Journal of Bacteriology. v.175, n.9, p.2523-2533. 1993.
20. COSTA, F.N.; ROSSI JUNIOR, O.D.; NADER FILHO, A.; TAVECHIO, A.T. Sorovares de Salmonella isolados de carcaças e frangos obtidos na indústria e no comércio em Jaboticabal, Estado de São Paulo, em 1996. Revista Brasileira de Ciência Veterinária,v.4, n.3, 1997. p.97-100
21. COLLINGHAN, R.J.; WOODWARD, M.J. The SEF14 fimbrial antigen of Salmonella enterica serovar Enteritidis is encoded within a pathogenicity islet. Veterinary Microbiology. v.80, p.235-245. 2001.

31
22. DILMAGHANI, M.; AHMADI, M; ZAHRAEI-SALEHI, T.; TALEBI, T. Detection of Samonella enteric Serovar Typhimurium from avians using Multiplex-PCR. Veterinary Research Forum, v.2, n.3, p. 157-165, 2011.
23. DOUBLET, B.; DOUARD, G.; TARGANT, H.; MEUNIER, D.; MADEC, J.Y.; CLOECKAERT, A. Antibiotic marker modifications of lambda red and FLP helper plasmids, pKD46 and pCP20, for inactivation of chromosomal genes using PCR products in multidrugresistant strains. Journal of Microbiological Methods. v.75, n.2, p.359-361. 2008.
24. FARBER, J. M. An introduction to the hows and whys of molecular typing. Journal Food Protection. v. 59, n.10, p. 1091-1101, 1996.
25. FERNANDEZ, A. T.; FORTES, M. L. M.; ALEXANDRE, M. H. S.; BASTOS, C. S. P.; VIANNA, E. P. L. Ocorrência de surtos de doenças transmitidas por alimentos na cidade do Rio de Janeiro. 2001 Disponível em: <http://www.unigranrio.br/veterinaria/surtosnacidade_rj.doc>. Acesso em: 25 set. 2013.
26. FRANCO, B. D. G. M; LANDGRAF. M. T. D. Microbiologia dos Alimentos. São Paulo, ed. Atheneu, p. 27-171, 1996.
27. FLOWERS, R.S.; KLATT, M.J.; KEELAN, S.L. Visualimmunoassay for detection of Salmonella in foods: collaborative study. Journal of the Association of Official Analytical Chemists, v.71, n.5, p.973-980, 1988
28. GALAN, J. E.; GINICHIO, C.; COSTEAS, P. Molecular and functional characterization of the Salmonella invasion gene invA: homology of invA to members of a new protein family. Journal Bacteriology, v. 174, p. 4338-4340, 1992.
29. GAST, R. F. Paratyphoid infections. In. SAIF, Y. M. Diseases of Poultry. 11 ed. Ames. Iowa University Press, p. 583-599, 2003.
30. GAST, R.K. Salmonella infections – Paratyphoid infections. In: Disease of Poultry. 12aed. Iowa. p.636-665. 2008.
31. GAST, R.K.; GURAYA, R.G.; GUARD, J.; HOLT, P.S. Frequency and magnitude of internal organ colonization following exposure of laying hens to different oral doses of Salmonella Enteritidis. International Journal of Poultry Science, Pakistan v. 10, n. 4, p. 325 - 331, 2011.
32. GIBSON, D.L.; WHITE, A.P.; RAJOTTEI, C.M.; KAY, W.W. agfC and agfE facilitate extracellular thin aggregative fimbriae synthesis in Salmonella Enteritidis. Microbiology. v.153, p.1131-1140. 2007.
33. GUIBOURDENCHE, M.; ROGGENTIN, P.; MIKOLEIT, M.; FIELDS, P.I.; BOCKEMÜHL, J.; GRIMONT, P.A.D.; WEILL, F.X. Supplement 2003 e 2007 (no47) to the White-Kauffmann – LeMinor scheme. Research in Microbiology. n.61, p.26-29. 2010.

32
34. GLYNN, B.; LAHIFF, S.; WERNECKE, M.; BARRY, T.; SMITH, T.J.; MAHER, M. Current and emerging molecular diagnostic technologies applicable to bacterial food safety. International Journal Dairy Technology. v.59, n.2, p.126-139. 2006.
35. GONÇALVES, P. M. R.; FRANCO, R. M.; ZAMBORLINI, L. C. Enumeração de enterococos e coliformes fecais, pesquisa de Salmonella e indicação presuntiva de Proteus, em cortes e miúdos de frango congelados. Higiene Alimentar, São Paulo, v. 12, n. 53, p. 42-47, 1998.
36. GOMES, M. J. P. ENTEROBACTERIACEAS (Salmonella spp). BACTERIOLOGIA DA FAVET – UFRGS. Laboratorio de Analises clinicas veterinarias. Microbiologia Clinica, 2008. Disponivel em: www.ufrgs.br/labacvet. Acessado em: 28/09/2013.
37. HACKER, J.; BLUM-OEHLER, G.; MÜHLDORFER, I.; TSCHÄPE, H. Pathogenicity islands of virulent bacteria: structure, function and impact on microbial evolution. Molecular Microbiology. n.23, n.6, p.1089-1097. 1997.
38. HEID, C.A, STEVENS, J., LIVAK, K.J. & WILLIAMS, P.M. Real time quantitative PCR. Genome Res. 6(10):986-94, 1996.
39. IKUNO, A. A.; KANASHIRO, A. M. I.; KIYOTA, S.; CASTRO, A. G. M.; FERREIRA, V. C. A. Multiplex PCR for accurate diagnosis of poultry infection by using Salmonella invA, sefA, spvC genes sequences as molecular markers. Arquivos do Instituto Biológico, São Paulo, v. 71, p. 265-267, 2004.
40. LÍRIO, V. S.; SILVA, E. A.; STEFORI, S.; CAMARGO, D.; RECCO, E. A. P.; MALUF, Y.; MIYAZAWA, T. T.; NEVES, D. V. A.; OLIVEIRA, V. M. R. Freqüência de 17 sorotipos de Salmonella isolados de alimentos. Higiene Alimentar, São Paulo, v. 12, n. 55, p. 36-42, 1997.
41. MAIJALA, R.; RANTA, J.; SEUNA, E. The efficiency of the Fininish Salmonella Control Programme. Food Control, Reading, v. 16, n. 8, p 669-675, 2005.
42. MARCUS, S.L.; BRUMELL, J.H.; PFEIFER, G., FINLAY, B.B. Salmonella pathogenicity islans: big virulence in small packages. Microbes Infection. v.2, p.145-156, 2000.
43. MONTASSIER, H. J. Fisiologia do sistema imune. In: Doença das Aves. Campinas, São Paulo. FACTA. 2aed. p.391-434. 2009.
44. MARLONY,B.; PACCASSONI, E.; FACH, P.; BUNGE, C.; MARTIN, A.; HELMUTH, R. DIAGNOSTIC Real-Time PCR for detection of Salmonella in food. Applied and Environmental Microbiology. v.70, n.2, p.7046-7052, 2004.
45. MORENO, A. M. Técnicas moleculares de diagnóstico. In: Patologia Aviária, Manole, Barueri – SP, p. 413-427, 2009.
46. MORGULES, M.S.F.A. Imunomoduladores. In: Farmacologia Aplicada a Avicultura. São Paulo. Roca. p.249-264, 2005.

33
47. MOTA, L.J.; SORG, I.; CORNELIS, G.R.. Type III secretion: the bactéria-eukaryotic express. Fems Microbiology Letters. v. 252, p.1-10, 2005.
48. MUNOZ, R. Sorologia como ferramenta de monitoria e diagnóstico. Conferência Apico; 2005. Santos, São Paulo.Brasil.
49. MULLIS, K.B. Target amplification for DNA analysis by the polymerase chain reaction. Ann Biol Clin 48(8): 579-82, 1990.
50. NAUGHTON, P.J.; GRANT, G.; SOJKA, M. BARDOCZ, S.; THORNS, C.J.; PUSZTAI, A. Survival and various compartments of the rat gastrointestinal tract in vivo. Journal of Medical Microbiology. v.50, p.1049-1054, 2001.
51. NADVORNY, A.; FIGUEIREDO, D. M. S. SCHMIDT, V. Ocorrência de Salmonella sp. em surtos de doenças transmitidas por alimentos no Rio Grande do Sul em 2000. Acta Scientiae Veterinariae, Porto Alegre, v. 32, n. 1, p. 47- 51, 2004.
52. PAVLOVA, B.; VOLF, J.; ONDRACKOVA, P.; MATIASOVIC, J.; STEPANOVA, H.; CRHANOVA, M.; KARASOVA, D.; FALDYNA, M.; RYCHLIK, I. SPI-1-encoded type III secretion system of Salmonella enterica is required for the suppression of porcine alveolar macrophage cytokine expression. Veterinary Research. v.42, n.16, p.1-7, 2011.
53. OLIVEIRA, G.H.; BERCHIERI, J.A.; MONTASSIER, H.J.; FERNANDES, A.C.; Assessment of serological response of chicken to Salmonella Galinarum e Salmonella Pullorum by ELISA. Rev. Bras. Cien. Avic, v.6,n2, p.111-115, 2004.
54. REEVES, P.R.; HOBBS, M.; VALVANO, M.A.; SKURNIK, M.; WHITFIELD, C.; COPLIN, D.; KIDO, N.; KLENA, J.; MASKELL, D.; RAETZ, C.R.H.; RICK, P.D. Bacterial polysaccharide synthesis and gene nomenclature. Trends in Microbiology, Oxford, v. 4, n. 12, p. 495-503, 1996.
55. REIS, R. B.; KRUGER, C. S.; MACIEL, M. S. Salmonella spp. em produtos cárneos comercializados no município de Cuiabá-MT. Avaliação da metodologia de pesquisa. Modelos de resistência a drogas antimicrobianas. Ciência e Tecnologia, Rio de Janeiro, v. 15, n. 1, p. 74-78, 1995.
56. RODRIGUES, D.P. Perspectivas atuais e falhas no diagnóstico antigênico de Salmonella spp.: importância no reconhecimento dos sorovares circulantes, emergentes e exóticos. In: Simpósio Internacional sobre Salmonelose Aviária. Rio de Janeiro. 2011
57. ROWE, B.; BARTLETT, C. R. L. Salmonella enteritidis e Salmonella typhimurium na Europa Ocidental, de 1993 a 1995: relatório da vigilância da Salm-Net. Euro Surveillance Monthly, Oslo, v. 2, n. 2, p. 4-6, 1997.
58. ROTGER, R.; CASADÉUS, J. The virulence plasmids of Salmonella. International Microbiology. v.2, p.177-184, 1999.

34
59. RUNNELS, R. A. An application of the rapid method agglutination test in the diagnosis of Bacillary White Diarrhoea Infection. Journal of the American Veterinary Medical Association, v. 70, p. 660-667, 1927
60. SANTOS, D. M. S. S.; BERCHIERI JR., A.; FERNANDES, S. A.; TAVECHIO, A. T.; AMARAL, L. A. Salmonella em carcaças de frango congeladas. Pesquisa Veterinária Brasileira, Rio de Janeiro, v. 20, n. 1, p. 39-42, 2000.
61. SALEHI, T.Z.; MADADGAR, O.; NASERLI, S.; FASAEI, B.N.; GHAFARI, M.M.; TAMAI, I.A. Detection of sef14, sef17, sef21 fimbrial virulence genes of Salmonella Enteritidis by multiplex PCR. Journal of Animal and Veterinary Advances. v.10, n.11, p.1421-1426, 2011.
62. SELANDER, R.K.; LI, J.; NELSON, K. Evolutionary genetics of Salmonella enterica. In: NEIDHARDI, F.C.; CURTISS, R.; INGRAHAM, J.L.; LIN, E.C. C.; LOW, K. B.; SOUZA, E.R.N.; CARVALHO, E.P.; DIONÍZIO, F.L. Estudo da presença de Salmonella sp. em poedeiras submetidas à muda forçada. Ciência Agrotécnica, Lavras, v.26, n.1, p.140-147, 2002.
63. SCHAFFER, J. M. Stained antigen for the rapide whole blood test for Pullorum disease. Journal of the American Veterinary Medical Association, v. 79, p. 236-240, 1931 64. SHIVAPRASAD, H.L.; BARROW, P.A. Salmonella infections – Pullorum disease and fowl typhoid. In: Disease of Poultry. 12aed. Iowa. p.620-536. 2008.
65. SUO, B.; HE, Y.; TU, S. I.; SHI, X. A multiplex real-time polymerase chain reaction for simultaneous detection of salmonella sp., escherichia coli o157, and listeria monocytogenes in meat products. Foodborne Pathogens and Disease, v. 7, n. 6, p. 619-628, 2010.
66. STONE, G. G.; OBERST, R. D.; HAYS, M. P.; McVEY, S.; GALLAND, J.; CURTSS, R.; KELLY, S. M.; CHEMGAPPA, M. Detection of S. Typhimurium from rectal swabs of experimentally infected beagles by short cultivation and PCR hybridization. Journal of Clinical Microbiology, v. 33, n. 5, p. 1292-1295, 1994.
67. WIGLEY, P.; BERCHIERI JR., A.; PAGE, K. L.; SMITH, A. L.; BARROW, P. A. Salmonella enterica Sorovar Pullorum Persists in Splenic Macrophages and in the Reproductive Tract during Persistent, Disease Free Carriage in Chickens. Infection and Immunity, Washington, v. 69, n. 12, p. 7873 - 7879, 2001.
68. WEI, B.; CHA, S. Y.; KANG, M.; PARK, I. J.; MOON, O. K.; PARK, C. K.; JANG, H. K. Development and application of a multiplex PCR assay for rapid detection of 4 major bacterial pathogens in ducks. Poult Sci, v. 92, n. 5, p. 1164-1170, 2013.
69. WEISS, S.; KRUSCH, S. Bacteria-mediated transfer of eukaryotic expression plasmids into mammalian host cells. Biol. Chem., Berlin, v. 382, p. 533-541, 2001.

35
70. TERZOLO, H.R. Estudio bacteriológico de las salmmonelosis de las aves (S. pullorum, S. gallinarum, S. Enteritidis y S. Typhimurium) em La América Latina. In: Simpósio Internacional sobre Salmonelose Aviária. Rio de Janeiro. 2011.
71. VAN IMMERSEEL, F.; METHNER, U,; RYCHLIK, I.; NAGY, B.; VEGE, P.; MARTIN, G.; FOSTER, N.; DUCATELLE, R.; BARROW, P. A. Vaccination and early protection against non-host-specific Salmonella serotypes in poultry exploitation of innate immunity and microbial activity. Epidemiology Infectious, v. 133, n6, p. 959-978, 2005.
72. VALONE, S.E.; CHIKAM, G.K.; MÜLLER, V.L. Stress induction of the virulence proteins (SpvA, B, C) from native plasmid pSDL2 of Salmonella Dublin. Infection and Immunity. v.61, n.2, p.707-713, 1993.
73. VAN ASTEN, A.J.A.M.; VAN DICK, J.E. Distribuition of “classic” virulence factors among Salmonella spp. Fems Immunology and Medical Microbiology. v. 44, n.3, p.251-259, 2005.
74. VIEIRA, M.A.M. Ilhas de patogenicidade. In: O Mundo da Saúde. São Paulo. v.33, n.4, p.406-414, 2009.
75. VELILLA, A.V.; TERZOLO, H.R. Biología molecular aplicada a La detección y caracterización de Salmonella enterica. In: XXI Congresso Latinoamericano de Avicultura. Cuba. 2009. Veterinary Microbiology. v. 133, p.328-334, 2009.
76. VITZTHUM, F; GEIGER, G; BISSWANGER, H.; BRUNNER, H.; BERNHAGEN, J. A. Quantitative Fluorescence-Based Microplate Assay for the Utraviolet Transilluminator Gel imaging System. Anais Biochem. 276(1).59-64, 1999.
77. WHANG, Y.P.; LIA,L.; SHENA, J.Z.; YANGB, F.J.; WU, Y.W. Quinolone-resistance in Salmonella is associated with decreased mRNA expression of virulence genes invA and avrA growth and intracellular invasion and survival. 2009.
78. WOODWARD, M. J; KIRWAN, S. E. S. Detection of Salmonella enteritidis in eggs by the polymerase chain reaction. Veterinary Record, London, v. 138, p. 411-413, 1996.
79. ZANELLA, A. Poultry disease manual: characteristics and control of infections. 1.ed. Bologna: Tipoarte, 2007. 93. p.
80. ZHAO, C.; GE, B.; VILLENA, J.; SUDLER, R.; YEH, E.; ZHAO, S.; WHITE, D. G.; WAGNER, D.; MENG, J. Prevalence of Campylobacter spp., Escherichia coli, and Salmonella serovars in retail chicken, turkey, pork, and beef from the greater Washington, D. C., area. Applied Environmental Microbiology, Baltimore, v. 67, n. 12, p. 5431-5436, 2001.
81. ZHU, C.H.; WU, J.; CHEN, W.W.; HASSAN, H.M.; ZHU, G.Q. Difference and variation of the sef14 operon gene clusters in Salmonella Pullorum. Journal of Basic Microbiology. v. 50, n.1, p.S120-S123, 2010.