apostila curso de inverno 2005
TRANSCRIPT

2º curso de inverno tópicos em
fisiologia comparativa
11 a 29 jul 2005
departamento de fisiologia instituto de biociências - usp http://www.ib.usp.br/cursodeinverno

2o curso de inverno – tópicos em fisiologia comparativa
comissão organizadora
Ana Paula Canel Bluhm Andreas Betz Daniel Carneiro Carrettiero Denise Loli Eduardo Koji Tamura Gláucia Jansen Da Re Lopes James Fernando Malta da Silva Renata Brandt Nunes Rodrigo Pavão Roseli Barbosa alunos de pós-graduação do departamento de fisiologia – ib/usp
Orientador:
Profa. Dra. Maria Aparecida Visconti
Apoio south american society for comparative physiology and biochemistry pró-reitoria de cultura e extensão comissão de pós-graduação instituto de biociências
i
2o curso de inverno – tópicos em fisiologia comparativa
CRONOGRAMA
Data 8 às 12 h 14 às 18 h
11/07/05
Aula inaugural
Profa. Dra. Maria Aparecida Visconti
Módulo I
Fisiologia celular Daniel Carneiro Carrettiero
Eduardo Koji Tamura Lidiana Duarte de Almeida Paula
12/07/05
Módulo I
Fundamentos básicos de técnicas em cultura celular
Daniel Carneiro Carrettiero Eduardo Tamura
Lidiana Duarte de Almeida Paula
Módulo I
Célula pigmentar: teórica e prática Ana Paula Canel Bluhm
Gláucia Jansen Da Re Lopes Fernanda Pizão Farhat
Leonardo Henrique Ribeiro Graciani de LimaRoseli Barbosa
13/07/05
Módulo I
Metabolismo em parasitas tripanossomatídeos: descrição de alvos
quimioterápicos Profa. Dra. Lucile Maria Floeter-Winter
Marcos Gonzaga dos Santos Ariel Mariano Silber
Maria Fernanda Laranjeira da Silva
Módulo I
Mecanismos de osmorregulação em animais - parte I
James Fernando Malta da Silva
14/07/05
Módulo I
Relógio biológico em mamíferos: mecanismos moleculares e controle da
ritmicidade interna do organismo Cíntia Etsuko Yamashita
Pedro Augusto Carlos Magno Fernandes
Módulo II
Neuroanatomia básica Emerson Ferraz Coelho
Prática de neuroanatomia
Andreas Betz Daniel Carneiro Carrettiero
Emerson Ferraz Coelho Jéssica Ruivo Maximino
Regiane Xavier de Moraes
15/07/05
Módulo II Variabilidade cardíaca e modelamento
matemático de sistemas biológicos Breno Teixeira Santos
Módulo II As toxinas de anêmonas do mar como
ferramentas para entender a fisiologia de tecidos, órgãos e sistemas.
André Junqueira Zaharenko
ii
2o curso de inverno – tópicos em fisiologia comparativa
18/07/05
Módulo II Evolução do sistema nervoso
Adriano Alonso Pereira da Cunha Rodrigo Pavão
Memória em Aves
Silvia Maria Matsuda
Módulo II Mecanismos centrais do controle
cardiovascular Jéssica Ruivo Maximino Video: Reação de alerta
19/07/05
Módulo III Comportamento e fisiologia de formigas
Pedro Leite Ribeiro
Módulo III Ecofisiologia de lagartos
Renata Brandt Nunes
Metabolismo energético da vocalização em anfíbios anuros do gênero Scinax: “O canto da
perereca” Adne Abbud Righi
20/07/05
Módulo III Substâncias de organismos marinhos que podem causar acidentes e intoxicações
Cynthia Grazielle Martins Delboni Jeanete Lopes Naves
Módulo III Consciência como expressão do
funcionamento do sistema nervoso Adriano Alonso Pereira da Cunha
Exercício físico, hipertensão arterial e comportamentos: interações, efeitos e
benefícios Regiane Xavier de Moraes
21/07/05
Módulo III Ritmicidade biológica: conceitos e
aplicações em metabolismo energético Denise Loli
Módulo III Mecanismo de osmoregulação em animais -
parte II James Fernando Malta da Silva
22/07/05
Módulo III Ritmos circadianos e funções de
aprendizagem e memória Luiz Fernando Lopes do Espírito Santo
Fisiologia do sistema respiratório em
ambientes aquáticos Renato Massaaki Honji
Módulo III
Definição dos laboratórios para estágio
25/07/05 Estágio em laboratório Estágio em laboratório 26/07/05 Estágio em laboratório Estágio em laboratório 27/07/05 Estágio em laboratório Estágio em laboratório 28/07/05 Estágio em laboratório Estágio em laboratório
29/07/05 Apresentação dos resultados obtidos durante o estágio
Apresentação dos resultados obtidos durante o estágio
iii

2o curso de inverno – tópicos em fisiologia comparativa
INDICE
FISIOLOGIA CELULAR.................................................................................................................1
FUNDAMENTOS BÁSICOS DE TÉCNICAS EM CULTURA CELULAR...............................5
CÉLULA PIGMENTAR ...................................................................................................................8
METABOLISMO EM PARASITAS TRIPANOSOMATÍDEOS: DESCRIÇÃO DE ALVOS
QUIMIOTERÁPICOS ....................................................................................................................11
MECANISMOS DE OSMORREGULAÇÃO EM ANIMAIS.....................................................15
RELÓGIO BIOLÓGICO DE MAMÍFEROS: MECANISMOS MOLECULARES E
CONTROLE DA RITMICIDADE INTERNA DO ORGANISMO............................................26
NEUROANATOMIA BÁSICA ......................................................................................................28
VARIABILIDADE CARDÍACA E MODELAMENTO MATEMÁTICO DE SISTEMAS
BIOLÓGICOS..................................................................................................................................33
AS TOXINAS DE ANÊMONAS DO MAR COMO FERRAMENTAS PARA ENTENDER A
FISIOLOGIA DE TECIDOS, ÓRGÃOS E SISTEMAS..............................................................38
EVOLUÇÃO DO SISTEMA NERVOSO......................................................................................43
MEMÓRIA EM AVES....................................................................................................................45
MECANISMOS CENTRAIS DO CONTROLE CARDIOVASCULAR....................................49
COMPORTAMENTO E FISIOLOGIA DE FORMIGAS ..........................................................53
ECOFISIOLOGIA DE LAGARTOS.............................................................................................57
METABOLISMO ENERGÉTICO DA VOCALIZAÇÃO EM ANFÍBIOS ANUROS DO
GÊNERO SCINAX: “O CANTO DA PERERECA” ...................................................................60
CONSCIÊNCIA COMO EXPRESSÃO DO FUNCIONAMENTO DO SISTEMA NERVOSO
............................................................................................................................................................62
EXERCÍCIO FÍSICO, HIPERTENSÃO ARTERIAL E COMPORTAMENTOS:
INTERAÇÕES, EFEITOS E BENEFÍCIOS ................................................................................64
SUBSTÂNCIAS DE ORGANISMOS MARINHOS QUE PODEM CAUSAR ACIDENTES E
INTOXICAÇÕES ............................................................................................................................68
RITMICIDADE BIOLÓGICA: CONCEITOS E APLICAÇÕES EM METABOLISMO
ENERGÉTICO ................................................................................................................................71
RITMOS CIRCADIANOS E FUNÇÕES DE APRENDIZAGEM E MEMÓRIA....................75
FISIOLOGIA DO SISTEMA RESPIRATÓRIO EM AMBIENTES AQUÁTICOS................78
ANEXO............................................................................................................................................. 81
iv

2o curso de inverno – tópicos em fisiologia comparativa
FISIOLOGIA CELULAR
Daniel Carneiro Carrettiero
Eduardo Koji Tamura
Lidiana Duarte de Almeida Paula
EVOLUÇÃO E COMPARTIMENTALIZAÇÃO CELULAR
Todos os organismos, e todas as células que os constituem, acredita-se, descendem de um
ancestral celular comum e surgiram através do processo de evolução por seleção natural. Mas como
um aglomerado de matéria poderia ter originado a primeira célula?
Uma pequena parte da grande energia liberada no ambiente primitivo através das radiações e
descargas elétricas de altíssimas voltagens teria se acumulado nas substâncias através de ligações
químicas originando, assim, as primeiras moléculas precursoras da vida: os polipeptídeos e os
polinucleotídeos. Entre estas moléculas estava uma, muito parecida com o RNA que conhecemos
hoje, a qual acredita-se que desencadeou todos os processos que levariam a formação da vida. Um
dos passos importantíssimos que possibilitaria esta transformação foi o desenvolvimento dos
catalisadores que ajudariam no processo de replicação. Um segundo passo importante foi o
confinamento destas moléculas que se auto-replicavam em um compartimento individualizado.
Desta forma surgiu a primeira célula. Esta célula primitiva seria parecida com o que conhecemos
hoje por célula procariota.Tipos distintos de células se desenvolveram e a associação de alguns
daria origem às então conhecidas células eucariontes animais.
A complexidade de reações internas envolvidas na comunicação com o ambiente se tornou tão
grande que foi possível o aparecimento de estruturas especializadas como o Reticulo
Endoplasmático, o Complexo de Golgi, as vesículas, os centríolos e o citoesqueleto.
No processo de divisão celular algumas células se mantiveram unidas possibilitando o
aprimoramento da comunicação celular e o surgimento da divisão de trabalho originando, assim, os
primeiros organismos multicelulares. Hoje em dia, em um organismo vertebrado, existem mais de
200 tipos celulares diferentes.
1

2o curso de inverno – tópicos em fisiologia comparativa
SINALIZAÇÃO CELULAR
Para o desenvolvimento e funcionamento normal do organismo, tanto em reposta ao ambiente
externo, como exemplo uma lesão tecidual causada por alguma injúria, ou para o próprio controle
fisiológico como crescimento e diferenciação celular, o organismo necessita se adaptar e provocar
efeitos diversos que variam de acordo com a necessidade. As células executam funções e
características que são coordenadas por uma maquinaria interna do organismo, que envia sinais para
outras células que respondem de modo especifico, ou seja, as próprias células se encarregam de
gerar e/ou propagar o sinal.
Um dos constituintes necessários para a sinalização e comunicação celular é a presença de
moléculas sinalizadoras, que podem ser as mais variadas moléculas possíveis, como hormônios,
aminoácidos, proteínas e gases. A comunicação entre a célula sinalizadora e a célula alvo pode ser
através de diferentes modos: sinalização endócrina, parácrina, autócrina, sináptica e dependente de
contato.
Para o reconhecimento dessas moléculas sinalizadoras e para a propagação e ampliação do
sinal dentro da célula são necessárias proteínas, conhecidas como receptores que são classificadas
em cinco tipos: receptores acoplados a proteína G, canais iônicos, associados a enzimas, nucleares e
órfãos. Esses receptores atuam de forma distinta, mas têm como função propagar o sinal e
principalmente direcionar o efeito, visto que células diferentes podem responder de maneira similar
e a mesma célula também pode responder de diferentes maneiras dependendo do estímulo, ou seja,
depende da molécula sinalizadora e dos receptores específicos a essas moléculas, que direcionam o
sinal para resultar no efeito apropriado.
Após a ativação dos receptores, o sinal é propagado para uma cascata de reações que incluem
outras moléculas, principalmente enzimas, que controlam o sinal adequadamente dentro da célula,
essas enzimas podem ser ativadas ou inibidas por moléculas conhecidas como segundos
mensageiros que são produzidos após a ativação de receptores de membrana, podemos citar como
exemplo o AMPc, GMPc, DAG, IP3. Outro mecanismo importante, para controle do
comportamento celular é o processo que consiste na adição ou remoção em proteínas celulares de
grupamentos fosfato (PO43-).
O conjunto de reações que compõem todo o processo de sinalização, resulta finalmente no
efeito final da célula que pode ser na transcrição de genes específicos e/ou em reações morfológicas
e fisiológicas da célula.
2

2o curso de inverno – tópicos em fisiologia comparativa
CICLO CELULAR
Basicamente, o ciclo celular é o programa para o crescimento e divisão (proliferação) celular.
Existem 4 fases no ciclo celular: G1 (e G0), S, G2 e M. A fase G1 é caracterizada por expressão de
genes e síntese de proteínas. Isto permite a célula crescer e produzir todas as proteínas necessárias
para a síntese de DNA. Durante a fase S, a célula replica seu DNA, permitindo que a célula divida-
se em duas células filhas, cada uma delas com uma cópia completa de DNA. Mas, antes que a
célula possa fazer isto, ela necessita entrar na terceira fase do ciclo celular: a fase G2. Na fase G2, a
célula novamente cresce e sintetiza proteínas permitindo a ela o processo de divisão. Completada
esta fase a célula finalmente entra na quarta fase do ciclo celular: a fase final M. Nesta fase a célula
passa por um processo denominada citocinese, originando 2 células filhas. O ciclo celular está
completo. As células agora, têm duas opções: podem começar outro ciclo entrando na fase G1 ou
podem tornar-se quiescentes entrando em G0.
Na maioria das células há vários pontos de checagem, nos quais as células podem ficar retidas
se eventos prévios não forem inteiramente completados.
APOPTOSE
Por definição Apoptose ou Morte Celular Programada é um tipo de "autodestruição celular"
que requer energia e síntese protéica para a sua execução. Está relacionado com a homeostase na
regulação fisiológica do tamanho dos tecidos, exercendo um papel oposto ao da mitose.
Fisiologicamente, esse suicídio celular ocorre no desenvolvimento embrionário, na organogênese,
na renovação de células epiteliais e hematopoiéticas, na involução cíclica dos órgãos reprodutivos
da mulher, na atrofia induzida pela remoção de fatores de crescimento ou hormônios, na involução
de alguns órgãos e ainda na regressão de tumores. Portanto consiste em um tipo de morte
programada, desejável e necessária que participa na formação dos órgãos e que persiste em alguns
sistemas adultos como a pele e o sistema imunológico.
3

2o curso de inverno – tópicos em fisiologia comparativa
Bibliografia:
Molecular Biology of the cell. 4.ed. Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts.K. &
Walter. P. Garland Science, 2002
Lenhinger Principles od Biochemistry, 3.ed. Nelson. D.L. & Cox. M. M. Worth Publishers, 2000.
http://fai.unne.edu.ar/biologia/cel_euca/celula3.htm
4

2o curso de inverno – tópicos em fisiologia comparativa
FUNDAMENTOS BÁSICOS DE TÉCNICAS EM CULTURA CELULAR
Daniel Carneiro Carrettiero
Eduardo Koji Tamura
Lidiana Duarte de Almeida Paula
Dadas as condições apropriadas, a maior parte das células vegetais e animais poderão viver,
multiplicar-se e até mesmo expressar propriedades diferenciadas em uma placa de cultura de
tecidos. As células podem ser observadas sob o microscópio ou analisadas bioquimicamente, e os
efeitos da adição ou remoção de moléculas específicas, tais como hormônios ou fatores de
crescimento podem ser explorados. Experimentos com células oriundas de cultura são, às vezes,
ditos como tendo sido conduzidos in vitro para contrastá-los daqueles experimentos com
organismos intactos, os quais são referidos como conduzidos in vivo.
O cultivo de células in vitro possibilita uma vasta aplicação nas áreas de: pesquisa, industria e
médica, todas com fins comerciais. O avanço na área de pesquisa com células tronco promete
avanços significativos no tratamento de diversas doenças.
O uso de células in vitro possibilita o estudo direto de respostas celulares específicas, ou seja,
sem grandes interferências dos inúmeros sinais provenientes do organismo. Atualmente é uma
grande ferramenta para o conhecimento dos mais diversos sistemas desde organismos procariotos
até os grandes grupos de vertebrados.
O estudo com células em cultura vem sendo desenvolvida e aprimorada desde o século XIX,
alguns dos principais estudos foram:
1885 – Wilheum Roux demonstrou que existia a possibilidade de manter células de
embriões de galinha in vitro em uma solução fisiológica.
1907 – Ross Harrison manteve por semanas fragmentos da crista neural de embriões de sapo
utilizando linfa coagulada como meio de cultura.
1910 – Rous utilizando extratos de células tumorais de galinha filtrado induziu a formação
de tumor, posteriormente foi demonstrada a presença de um oncovírus nestes extratos (vírus
do sarcoma de Rous).
1911 – Lewis & Lewis desenvolveram o primeiro meio de cultura líquido
1912 – Alexis Carrel manteve uma cultura de fibroblastos derivados do coração de galinha
por tempo indeterminado em cultura.
1948 – Earle e colaboradores desenvolveram células clonais a partir de células isoladas.
5

2o curso de inverno – tópicos em fisiologia comparativa
1952 - George Gey e colaboradores estabeleceram a primeira linhagem contínua de células
conhecida como HeLa,
1954 – Levi-Montalcini e colaboradores descobriram a existência de fatores de crescimento.
1954 – Abercrombie e colaboradores descrevera o fenômeno de inibição por contato.
1961 – Hatflick & Moorhead demonstraram que os fibroblastos humanos possuem um
número finito de divisões celulares.
1965 – Ham desenvolveu o primeiro meio de cultura sem soro.
1975 – Köhler & Milstein desenvolveram uma linhagem de células híbrida (hibridoma) para
a produção de anticorpos monoclonais.
1977 – Wigler & Axel descreveram um método eficiente para a introdução de genes de
mamíferos em células cultivadas.
1986 – Martin & Evans e colaboradores isolaram células embrionárias pluripotentes de
camundongos.
1998 – Thompson & Gearhart isolaram células tronco humanas.
Existem três tipos básicos de cultura celular:
Células normais (cultura primária): retiradas diretamente de um organismo para utilização em
experimentos.
Células estabelecidas: retiradas de um determinado tecido e cultivadas em laboratório,
possuem inibição por contato e apresentam características próximas do tecido de origem.
Células transformadas (linhagens celulares): retiradas de tumores ou imortalizadas através de
tratamentos físicos, químicos ou modificações genéticas, são muito utilizadas por poderem ser
mantidas indefinidamente em cultura, porém podem apresentar perda das características do
tecido de origem.
As características de células cultivadas in vitro são diretamente relacionadas com o ambiente
em que estão inseridas e um dos fatores que influenciam é o substrato, ou seja, o local em que as
células aderem, os substratos mais conhecidos são: polímeros sintéticos, como poliestireno e teflon;
polímeros biodegradáveis, formados por géis de colágeno ou outras biomoléculas, e polímeros
híbridos, contendo proteínas da matriz extracelular e peptídeos bioativos. Outros fatores que
influenciam a fisiologia das células cultivadas são os gases presentes no ambiente, principalmente o
oxigênio (O2) e o dióxido de carbono (CO2), a maioria das células necessitam somente da pressão
atmosférica de O2, a concentração de CO2 varia de acordo com o tipo celular, as células
normalmente são enriquecidas com até 10% CO2, mas como o CO2 é convertido em gás carbônico
quando dissolvido, deve se adicionar agentes tamponantes na solução, como bicarbonato de sódio
ou HEPES.
6

2o curso de inverno – tópicos em fisiologia comparativa
Inicialmente os meios de culturas utilizados continham misturas complexas, como extratos de
embriões ou linfas, mas para eliminação de fatores desconhecidos que poderiam influenciar na
preparação, diversos meios de culturas foram criados contendo quantidades especificas dos
ingredientes, com alguns componentes básicos como água, íons, fontes de energia (ex:
carboidratos), fontes de nitrogênio (aminoácidos), vitaminas, antibióticos e indicadores de pH.
Os meios de cultura são separados em complexos e quimicamente definidos, os meios
complexos são suplementados com soros (ex: soro fetal bovino), e os quimicamente definidos não
são suplementados com soro, mas por um conjunto de fatores de proteínas, fatores de crescimento e
hormônios. A temperatura que as culturas de células devem ser mantidas, depende da temperatura
corpórea do animal de onde foram extraídas, portanto nas culturas de células de animais de sangue
quente a temperatura deve ser de 37oC.
Bibliografia Básica:
Culture of Animal Cells: A Manual of Basic Techiques, 2. ed. Freshney. R.I. Jonh Wiley &
Sons, 1994.
Cd interativo do livro Molecular Biology of the cell. 4.ed. Alberts, B., Johnson, A., Lewis, J.,
Raff, M., Roberts.K. & Walter. P. Garland Science, 2002
http://www.cnice.mecd.es/eos/MaterialesEducativos/mem2001/biologia/ayuda/thesaurus.htm
7

2o curso de inverno – tópicos em fisiologia comparativa
CÉLULA PIGMENTAR
Ana Paula Canel Bluhm
Fernanda Pizão Farhat
Gláucia Jansen Da Re Lopes
Leonardo Henrique Ribeiro Graciani de Lima
Roseli Barbosa
A maioria dos animais apresenta cores variadas com as mais incríveis combinações e nuances.
Cor é uma propriedade física de certas substâncias cujas moléculas absorvem luz em certos
comprimentos de onda e, portanto, transmitem o comprimento de onda complementar ao absorvido.
Estas substâncias são denominadas pigmentos e há uma grande variedade deles presente na
natureza.
Os grânulos de pigmento são produzidos e armazenados nas células pigmentares ou
cromatóforos, que são células especializadas, com muitas projeções citoplasmáticas, que lhe
conferem aspecto estrelado. Nos vertebrados, são originárias da crista neural e migram para
diversas regiões, apresentando cores variadas, sendo classificadas conforme a natureza química dos
pigmentos que armazenam:
Melanóforos: pretos ou pardos, contêm grânulos de melanina (melanossomos);
Eritróforos: vermelhos, contêm diferentes proporções de pigmentos carotenóides e
pteridínicos (eritrossomos);
Xantóforos: amarelos, contêm pigmentos pteridínicos e carotenóides em proporções
variadas (xantossomos);
Leucóforos: brancos, contêm grânulos de purinas (leucossomos);
Iridóforos: iridescentes cores metálicas contêm purinas depositadas em finas placas
cristalinas.
Em aves e mamíferos a diversidade de cromatóforos foi perdida e o único tipo de célula
pigmentar presente é o melanócito, célula que sintetiza e armazena melanina.
8

2o curso de inverno – tópicos em fisiologia comparativa
O padrão de cor e a presença de listras e/ou manchas são determinados geneticamente, mas
alguns animais podem mudar de cor em resposta a estímulos ambientais, como temperatura
(termorregulação), cor do ambiente (favorecendo caça ou escape), iluminação, presença de outros
animais (atração de presas ou parceiro sexual), estações do ano e ciclos de dia/noite, permitindo sua
adaptação a uma nova situação. Alguns animais podem sofrer mudanças de cor de forma rápida
(mudança de cor fisiológica) ou de forma lenta (mudança de cor morfológica). A mudança na
coloração dos animais é, na maioria das vezes, regulada pela ação de hormônios ou
neurotransmissores.
A mudança de cor fisiológica é uma propriedade de alguns invertebrados, como cefalópodes e
crustáceos e dos vertebrados pecilotérmicos. São adaptações rápidas, que se completam em minutos
ou segundos, não estão presentes em aves e mamíferos, que apresentam apenas uma lenta migração
de grânulos de melanina para os queratinócitos vizinhos, penas e pêlos, alem da mudança de cor
morfológica.
Já a mudança de cor morfológica, que pode ocorrer em todos os animais, é decorrente de
alterações na quantidade ou no tipo de pigmento dentro das células pigmentares. Pode ocorrer como
uma resposta direta da célula pigmentar à luz ou à temperatura, como é o caso da resposta de
melanócitos humanos à luz UV (bronzeamento). São mudanças extremamente lentas, que podem
levar dias ou semanas, mas são mudanças duradouras.
Os seres humanos também possuem melanócitos na pele. A cor de nossa pele não depende do
número de melanócitos presente em uma determinada área, já que esta proporção é semelhante em
todas as raças. O que varia é a taxa de produção ou o tipo de melanina presente, determinados
geneticamente. Quando somos expostos à luz solar, a irradiação UV estimula a síntese de melanina
nos melanócitos, processo conhecido popularmente como bronzeamento. Este processo tem a
importante função de proteger o DNA de nossas células contra danos causados pela radiação
ultravioleta. Como este processo é muito lento, recomenda-se a exposição gradual ao sol, para
permitir a síntese de melanina em quantidade adequada. O câncer de pele é uma doença comum,
causada por uma exposição excessiva à luz solar.
Existem diversas patologias associadas à pigmentação, que podem ser causadas por
mecanismos fisiológicos deficientes: albinismo, vitiligo, hiperpigmentação, hipopigmentação,
melanose, lentigo, manchas diversas.
9

2o curso de inverno – tópicos em fisiologia comparativa
Bibliografia
Visconti, M.A. Regulação hormonal da célula pigmentar de vertebrados – Tese (Livre-Docência) –
Departamento de Fisiologia/Instituto de Biociências-USP, 1999.
Benabou, M.H.P. Modulação hormonal de células de eritroforoma da linhagem GEM 81 por α-
MSH e MCH – Tese (Mestrado em Fisiologia) – Departamento de Fisiologia/Instituto de
Biociências-USP, 1999.
Molecular Biology of the cell. 4.ed. Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts.K. &
Walter. P. Garland Science, 2002.
http://fisio.ib.usp.br/labpig/
www.pubmed.com
10

2o curso de inverno – tópicos em fisiologia comparativa
METABOLISMO EM PARASITAS TRIPANOSOMATÍDEOS: DESCRIÇÃO DE ALVOS
QUIMIOTERÁPICOS
Ariel Mariano Silber
Maria Fernanda Laranjeira da Silva
Marcos Gonzaga dos Santos
Lucile Maria Floeter-Winter
Os protozoários da família Trypanosomatidae apresentam dois gêneros, Trypanosoma e
Leishmania, com relevância na saúde pública, uma vez que são agentes de graves enfermidades
parasitárias.
Os protozoários do gênero Leishmania, formam um grupo de organismos que apresentam dois
hospedeiros obrigatórios em seu ciclo de desenvolvimento: um flebotomíneo e um mamífero Neste
ciclo o homem aparece acidentalmente, e essa infeção pode resultar um complexo quadro de
desdobramentos clínicos, denominados genericamente como leishmaniose. As leishmanioses são
consideradas pelo Tropical Diseases Research Programme Organização Mundial de
Saúde(TDR/OMS), uma das seis doenças de maior importância em Saúde Pública eleitas para o
desenvolvimento desse programa. Entre as protozooses, ocupam o segundo lugar, sendo superadas
apenas pela malária.
Do ponto de vista biológico, organismos do gênero Leishmania apresentam características
interessantes. São parasitas intracelulares obrigatórios de células do sistema fagocítico mononuclear
de um grande número de hospedeiros vertebrados, os quais adquirem a infecção, na quase totalidade
das ocasiões, através do contato com o vetor flebotomíneo (Zuckerman & Lainson, 1977). Nos
mamíferos, a principal célula parasitada é o macrófago. O interessante é que essa célula apresenta
diversos mecanismos microbicidas: enzimas lisossômicas, intermediários reativos de oxigênio e
nitrogênio e mediadores derivados de lipídeos. Contudo o parasita é capaz, por diversas estratégias,
algumas conhecidas, de escapar desses mecanismos microbicidas e sobreviver no ambiente hostil.
A arginase é uma enzima integrante do ciclo da uréia (Krebs-Henseleit) nos animais
uricotélicos, e é expressa em alguns tripanosomatídeos, entre eles Leishmania. A arginase utiliza
como substrato L-arginina, produzindo L-ornitina e uréia. Inicialmente seu papel funcional foi
associado apenas aos processos metabólicos envolvidos nessa interconversão arginina-ornitina-
11

2o curso de inverno – tópicos em fisiologia comparativa
citrulina. Além disso, a expressão específica é uma das características utilizadas na identificação de
organismos da família (Camargo, 1999).
Nos últimos anos, uma série de trabalhos vem sendo publicada relatando as propriedades de
uma outra enzima, a óxido nítrico sintase induzida (iNOS) que, como a arginase, utiliza a L-
arginina como substrato, produzindo citrulina e óxido nítrico (Nathan e Xie, 1994, Mori & Gotoh,
2000), e como a arginase é expressa em macrófagos. A produção de NO é uma importante resposta
microbicida dos macrófagos. Nessas células, a enzima iNOS é dependente de L-arginina, assim a
arginase pode atuar negativamente na regulação dos níveis de NO produzidos, consumindo o
substrato da iNOS (Boucher e cols., 1999). Recentemente, foi demonstrado que a presença de um
inibidor de arginase, a Nω-hydroxyl-L-arginina, diminui a capacidade de L. major em estabelecer a
infecção em macrófagos (Iniesta et al.., 2001).
Com base nestes resultados, postulamos que um dos papéis funcionais da arginase de
Leishmania estaria relacionado com a sobrevivência do parasita no interior dos macrófagos,
competindo com a iNOS pelo mesmo substrato.
Nosso laboratório tem-se dedicado à caracterização do segmento gênico que codifica a
arginase de Leishmania, assim como seu produto de transcrição (da Silva et al., 2002). As
informações geradas por essa caracterização permitiram a construção de um transfectante com
apenas uma cópia do gene nocauteado e esse organismo mutante foi utilizado em ensaios de
infecção in vitro, com macrófagos da linhagem J774. Os resultados dos experimentos apontam para
uma baixa taxa na infectividade de macrófagos por L. (L.) amazonensis com uma cópia do gene de
arginase nocauteado. Isso parece indicar que a arginase possa ser importante para a virulência e
viabilidade do parasita na célula hospedeira. Estudos complementares mostraram que a diminuição
da síntese de ornitina, também é responsável por uma diminuição na taxa de proliferação dos
parasitas, uma vez que esse composto é precursor na síntese de poliaminas e portanto essencial para
a replicação do DNA.
A doença de Chagas é um importante problema de saúde pública na América Latina, onde se
estima que existem aproximadamente 16 - 18 milhões de pessoas infectadas na América Latina e
sul dos Estados Unidos (http://www.who.int/ctd/chagas/disease.htm).
O T. cruzi apresenta um ciclo de vida complexo alternando entre hospedeiros mamíferos e
inseto vetor, um hematófago da família Triatominae. Durante esse ciclo, o T. cruzi passa por
distintos estádios evolutivos. Dois deles estão presentes no inseto vetor: tripomastigota metacíclico
e epimastigota. Outros dois estão presentes no hospedeiro mamífero: tripomastigota e amastigota.
No interior do hospedeiro mamífero, o T. cruzi deve obrigatoriamente invadir as células para poder
se diferenciar para formas replicativas e estabelecer a infecção (Brener, 1973).
12

2o curso de inverno – tópicos em fisiologia comparativa
Pouco se sabe sobre os eventos que acontecem a partir da diferenciação das formas invasivas
no interior das células hospedeiras. Uma incógnita a respeito das características dessas formas
intracelulares é como sustentam o seu crescimento dentro da célula e quais são os fatores que
disparam a diferenciação entre os estágios. Nosso grupo abordou esse problema analisando o
transporte de prolina e glicose ao longo do ciclo intracelular de T. cruzi (Silber et al. 2002). Entre os
resultados obtidos observou-se que esses parasitas não são capazes de consumir glicose durante a
fase intracelular do ciclo de vida, já que não expressam o transportador correspondente. Porém, eles
têm altos níveis de prolina intracelular, que é consumida ao longo desse processo. É interessante
ressaltar que quando a prolina intracelular é consumida o parasita deve importá-la do ambiente, no
caso, o citoplasma da célula hospedeira. Nesse sentido, se observou também que a diferenciação
entre as formas intracelulares é um processo dependente do funcionamento do metabolismo e
transporte de prolina (Tonelli et al. 2004). Esses dados e outros nos levaram a propor que as
enzimas do metabolismo de aminoácidos, são alvos relevantes para a identificação de compostos
líderes para o desenho de novas drogas com perspectivas terapêuticas para a doença de Chagas
(Silber et al. 2005).
Bibliografia
Boucher JL, Moali C & Tenu JP (1999) - Nitric oxide biosynthesis, nitric oxide synthase inhibitors
and arginase competition for L-arginine utilization; Cellular and Molecular Life Sciences 55:
1015-1028,.
Brener Z. (1973) Biology of Trypanosoma cruzi. Ann. Rev. Microbiol. 27, 347.
Camargo EP (1999) - Phytomonas and other trypanosomatid parasites of plants and fruit. Adv.
Parasitol., 42: 29-112.
da Silva ER, Moreno TM, Pioker FC, Silva CHTP & Floeter-Winter LM (2002) - Genomic
organisation and transcription characterisation of the gene encoding Leishmania (Leishmania)
amazonensis arginase and its protein structure prediction. Int. J. Parasitol.,32: 727-737.
Iniesta V, Gómez-Nieto LC, Corraliza I (2001) - The inhibition of arginase by N�-hydroxy-L-
arginine controls the growth of Leishmania inside macrophage. J. Exp. Med. 193, 777-83.
Lainson R, ShawJJ, Silveira FT, Braga RR, Ryan L, Póvoa MM & Ishikawa EAY (1986) - A
Leishmania e as leishmanioses. - Instituto EvandroChagas - 50 anos de contribuição à
Ciências Biológicas e à Medicina Tropical.
Mori M & Gotoh T (2000) - Regulation of Nitric Oxide Production by Arginine Metabolic
Enzymes. BBRC, 275:715-719.
13

2o curso de inverno – tópicos em fisiologia comparativa
Nathan C & Xie QW (1994) - Nitric oxide synthases: roles, tolls, and controls. Cell. 23;78(6):915-
918.
Silber, A. M.; Tonelli, R. R.; Martinelli, M.; Colli, W.; Alves, M. J. (2002) Active transport of L-
proline in Trypanosoma cruzi. J. Eukaryot. Microbiol. 49, 441.
Silber, A. M.; Colli, W.; Ulrich, H.; Alves, M. J.; Pereira, C. A. (2005) Amino acid metabolic routes
in Trypanosoma cruzi: possible therapeutic targets against Chagas' disease.Curr. Drug Targets
Infect. Disord. 5, 53.
Tonelli, R. R.; Silber, A. M.; Almeida-de-Faria, M.; Hirata, I. Y.; Colli, W.; Alves, M. J. (2004) L-
proline is essential for the intracellular differentiation of Trypanosoma cruzi. Cell. Microbiol.
6, 733.
Zuckerman A & Lainson R (1997) - Leishmania. In Parasitic Protozoa, Vol I. (J. P. Kreier, Ed.).
Academic Press Inc: New York.
14

2o curso de inverno – tópicos em fisiologia comparativa
MECANISMOS DE OSMORREGULAÇÃO EM ANIMAIS.
James Fernando Malta da Silva
COMENTÁRIOS GERAIS
Adaptado de R. Gilles (1979) por
Luis Alberto Valotta
James Fernando Malta da Silva
A vida na Terra é dependente de água. Os seres vivos são sistemas bioquímicos altamente
sofisticados organizados em torno das propriedades desta molécula. A água constitui-se no principal
meio onde as interações moleculares indispensáveis à vida ocorrem. Também se constitui no
veículo que transporta as moléculas para diferentes locais onde essas interações podem ocorrer.
Além dos muitos compostos orgânicos encontrados como solutos nos seres vivos, os íons
inorgânicos também são de fundamental importância: participam como cofatores em muitas reações
enzimáticas; formam os gradientes químicos os quais podem atuar como estoques de energia
potencial; e influenciam na permeabilidade das membranas biológicas a outros solutos. Os diversos
solutos encontrados nas células vão, por outro lado, influenciar a mobilidade osmótica da água e,
portanto, irão desempenhar um papel proeminente na manutenção da arquitetura celular. Além
disso, muitos dos sistemas enzimáticos que controlam as interações químicas características da vida
estão localizadas em estruturas altamente organizadas. Isto aponta a importância da manutenção da
estrutura celular e volume nas reações as quais envolvem estas enzimas como catalisadores.
Basicamente, as células podem ser vistas como máquinas químicas extremamente complexas
nas quais a localização e a concentração de várias espécies moleculares interatuantes devem ser
precisamente controladas no sentidos de manter atividade ótima. Em tal contexto, o controle e a
manutenção do volume celular podem ser considerados requisitos essenciais à vida. Além disso, o
problema da regulação de volume celular é um dos elementos cruciais na conquista de diferentes
biótopos e no estabelecimento de organismos em ambientes aquáticos com flutuações de
salinidade. De acordo, a vida foi originada em algum tipo de oceano e a capacidade de controlar o
volume celular é um dos principais pré-requisitos para a invasão de outros tipos de habitats como
os ambientes de água doce e terrestre. Os organismos que habitam este meio desenvolveram
adaptações osmóticas específicas habilitando a sua manutenção em suas comunidades. Há várias
maneiras através das quais o problema da manutenção do volume celular pode ser resolvido. O
15

2o curso de inverno – tópicos em fisiologia comparativa
organismo pode isolar-se completamente do meio externo, evitando dessa forma o ganho ou a
perda de água. Esta solução não foi mantida por um grande número de espécies ao longo da
evolução. Trocas com o meio externo são necessárias para satisfazer as necessidades celulares.
Alguns esporos bacterianos podem sobreviver por longos períodos com um conteúdo baixo de água
e sem trocas com o seu meio ambiente; nesta situação, entretanto, seus processos vitais são
essencialmente suspensos. Na maioria dos organismos, a água atravessa a membrana celular por
difusão em resposta a gradientes osmóticos. Há duas maneiras de evitar mudanças no volume
celular enquanto mantém-se a possibilidade de trocas entre o fluído intracelular e o meio ambiente.
O primeiro método consiste no controle da Concentração Osmótica (CO) do fluído intracelular em
relação a eventuais modificações do meio externo. O segundo método implica no controle da CO
do fluído que circunda as células em quaisquer condições externas. A última solução foi adotada
por diversos eucariotos e foi denominada por Florkin (1962), de a “regulação anisosmótica
extracelular”. Embora a existência de um fluído extracelular diferente do meio externo foi
observada precocemente na evolução, a efetiva regulação deste meio (os fluídos corpóreos) é um
atributo de apenas alguns grupos zoológicos altamente evoluídos. Pode ser encontrado em alguns
vermes e moluscos, mas, essencialmente, ocorre em artrópodes e em vertebrados. Além disso,
muitos dessas espécies são incapazes de manter o estado osmótico de seu sangue quando a CO do
ambiente varia.
Os mais eficientes reguladores anisosmóticos formam a categoria denominada dos assim
chamados animais homeostáticos; essas espécies podem manter a CO do seu sangue estacionária
independente das condições externas. Além de alguns crustáceos e peixes, representantes deste
grupo são encontrados entre répteis, aves, e mamíferos. Os íons inorgânicos Na+ e Cl- são
predominantes como efetor osmótico sanguíneo na maioria dos reguladores anisosmóticos. Uréia é
usada por alguns vertebrados inferiores. Este composto orgânico é encontrado essencialmente em
ciclostomados e em elasmobrânquios, mas também tem um papel em vários anfíbios e répteis.
Há apenas alguns animais homeostáticos. Em todos as outras espécies, as células têm que,
algumas vezes, se defrontar com importantes mudanças na CO de seu meio ambiente. Além disso,
os eficientes mecanismos de controle da CO sangüínea que atuam em espécies homeostáticas
podem estar encobertos sob certas condições ou podem apresentar uma certa demora em
responder a uma nova situação. Isto aponta para a importância dos mecanismos de controle
osmótico do fluído intracelular na manutenção do volume celular.
Na maioria das espécies de animais eucarióticas, os fluídos intra e extracelulares são mantidos
próximos da condição isosmóticas. Os mecanismos implicados neste processo foram denominados
por Florkin (1962) como mecanismos de “regulação isosmótica intracelular”. Trabalham para
manter o equilíbrio osmótico apesar da presença de solutos aniônicos não-difusíveis no interior das
16

2o curso de inverno – tópicos em fisiologia comparativa
células; estas partículas geram uma pressão osmótica a qual, de outra forma, iria induzir o
inchamento e a lise das células de animais por possuírem membranas facilmente distensíveis. Além
disso, estes mecanismos são de fundamental importância na resposta regulatória de volume que
estas células são capazes de desenvolver após mudanças na CO de seu meio ambiente. Em todos os
tecidos e células estudados até o momento, eles implicam no controle ativo do montante de vários
efetores osmóticos intracelulares entre os quais os íons inorgânicos Na+, K+ e Cl- e os aminoácidos
livres possuem um papel proeminente.
Os mecanismos de regulação isosmótica intracelular foram encontrados em tecidos e células
de muitas espécies de vários grupos zoológicos incluindo protozoários, invertebrados, e
vertebrados. É importante notar que muitos desses organismos ou não possuem, ou possuem de
forma muito frágil, a capacidade de regulação anisosmótica extracelular. Pode-se, portanto, concluir
que a regulação isosmótica em nível celular um processo mais primitivo o qual apareceu
precocemente e persistiu ao longo da evolução. Processos de regulação anisosmóticos devem ter
sido adquiridos posteriormente, adicionando às espécies que o possuíam um novo leque de
possibilidades.
Bibliografia Geral
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. & Watson, J.D. (1994) Molecular Biology of
The Cell (3rd edition). Garland Publishing Inc.
Florkin, M. (1962) La regulation isosmotique intracellulaire chez les inevertebrés marins
euryhalins. Bull. Acad. R. Belg. Cl. Sci., 48, 687-694.
Gilles, R. (Editor) (1979) Mechanisms of osmoregulation in animals: maintenance of cell volume
(1st edition) John Wiley & Sons.
Gupta, B. L., Moreton, R. B., Oschman, J. L. & Wall, B. J. (1977) Transport of ions and water in
animals (1st edition). Academic Press.
Schmidt-Nielsen, K. (1997) Animal Physiology : Adaptation and Environment (1st edition)
Cambridge University Press.
Stone, G., Johnston, I. A.& Willmer, P. J. (2000) Environmental Physiology of Animals (2nd
edition). Blackwell Science Inc.
Strange, K. (Editor) (1994) Cellular and Molecular Physiology of Cell Volume Regulation (1st
edition). CRC Press.
Withers, P. C. (1992) Comparative Animal Physiology (1st edition). Harcourt Brace.
17

2o curso de inverno – tópicos em fisiologia comparativa
EXERCÍCIO TEÓRICO-PRÁTICO
BALANÇO OSMÓTICO EM AMBIENTES MARINHO, DE ÁGUA DOCE E XÉRICO
Elaborado por
Prof. Dr. Luiz Carlos Salomão
Introdução
Nos animais aquáticos, especialmente nos animais marinhos e estuarinos, as variações da
salinidade do meio podem resultar em variações nas concentrações iônica e osmótica do sangue e
fluídos extracelulares. Animais marinos hiposmóticos estão sujeitos ao efluxo de água e influxo de
íons, contrariamente aos animais de água doce, que por serem hiperosmóticos, estão sujeitos ao
influxo de água e efluxo de íons, alterando a Concentração Osmótica (CO) do sangue e líquidos
tissulares.
Há dois padrões básicos de resposta dos animais a tais variações na salinidade, isto é,
osmoconformação e osmorregulação. No primeiro caso, a CO do sangue, hemolinfa ou líquidos
extracelulares varia linearmente com a variação da CO do meio. No segundo caso, a CO se mantém
constante apesar das variações na salinidade do meio. Entre estes dois casos extremos,
osmorregulação e osmoconformação, ocorrem respostas intermediárias. Para se saber o padrão de
resposta osmótica, em laboratório, geralmente submetem-se os animais a meios de diferentes
salinidades, ou seja, de composições iônicas diferentes e determinam-se as concentrações iônicas e
a CO do sangue destes animais nestas diferentes condições experimentais.
Já no ambiente terrestre, em que a grande vantagem é a maior disponibilidade de oxigênio, o
balanço hídrico é de outra natureza e, muitas vezes, é obtido tanto por ajustes fisiológicos como
comportamentais. Tal é que se observa, por exemplo, no banco hidromineral do rato canguru
Dipodomys merriami. Entre os mamíferos, 40% das espécies pertence a Ordem dos roedores, a mais
numerosa. Distribue-se por todo planeta, mas principalmente na América do Sul. Adaptaram-se aos
diferentes ambientes, das regiões polares ao equador, das montanhas as praias e do deserto aos
pântanos. A maioria dos roedores é terrestre, mas alguns são arborícolas ou semi-aquáticos. A
maior parte dos roedores é de pequeno porte, isto é, de 10 a 20 cm de comprimento e de 50 a 500 g
de massa corporal. Alguns, no entanto, como uma espécie de porco espinho (Hystrix cristata) do
norte da África chega a atingir massas corpóreas de 20-50 kg. A presença dos roedores tem
relevante papel ecológico por serem a principal fonte de alimento para aves e mamíferos carnívoros,
havendo uma relação bem estabelecida entre o tamanho da população de roedores e suas presas. A
18

2o curso de inverno – tópicos em fisiologia comparativa
relação com os homens não se limita à destruição da agricultura ou à transmissão de doenças. São
úteis como animais de laboratório, por consumirem certos insetos e por propiciarem o arejamento
do solo cavando galerias subterrâneas.
Entre os mamíferos, são os roedores que ocuparam os mais diferentes ambientes com relação
à disponibilidade à água. Estão presentes nos desertos mais áridos onde a água não está disponível,
mas que também se tornou dispensável para eles. Nesse sentido, deve-se ressaltar as pesquisas de
Schmidt-Nielsen (1964) sobre os hábitos e a fisiologia renal do rato canguru que vive numa região
tão inóspita, quanto à disponibilidade de água, que poucas outras espécies lhe fazem companhia.
Respostas osmóticas em Perna perna
A Tab. 1 apresenta resultados em experimentos realizados com o molusco bivalve Perna
perna. Os mexilhões Perna perna foram coletados em costões nas proximidades de São Sebastião e
foram transferidos para tanques de cimento amianto e mantidos em água de mesma salinidade do
local de coleta, isto é, 1000 mOsm/kg H2O por cerca de 24 h. A seguir foram distribuídos em
tanques de cimento amianto contendo água do mar diluída com água destilada, obtendo-se, assim,
as diferentes salinidades experimentais. Em cada salinidade experimental foram colocados
mexilhões com cunha entre as valvas e sem cunha. A hemolinfa dos animais com cunha foi coletada
após 6 h de exposição aos diferentes meios, tempo previamente determinado considerando ser este
período o suficiente para as trocas osmo-iônicas. Nos animais sem cunha, as amostras foram obtidas
após 24 h. O objetivo das cunhas era o de manter a livre exposição das partes moles do animal aos
meios experimentais. Manteve-se arejamento contínuo durante todo o experimento.
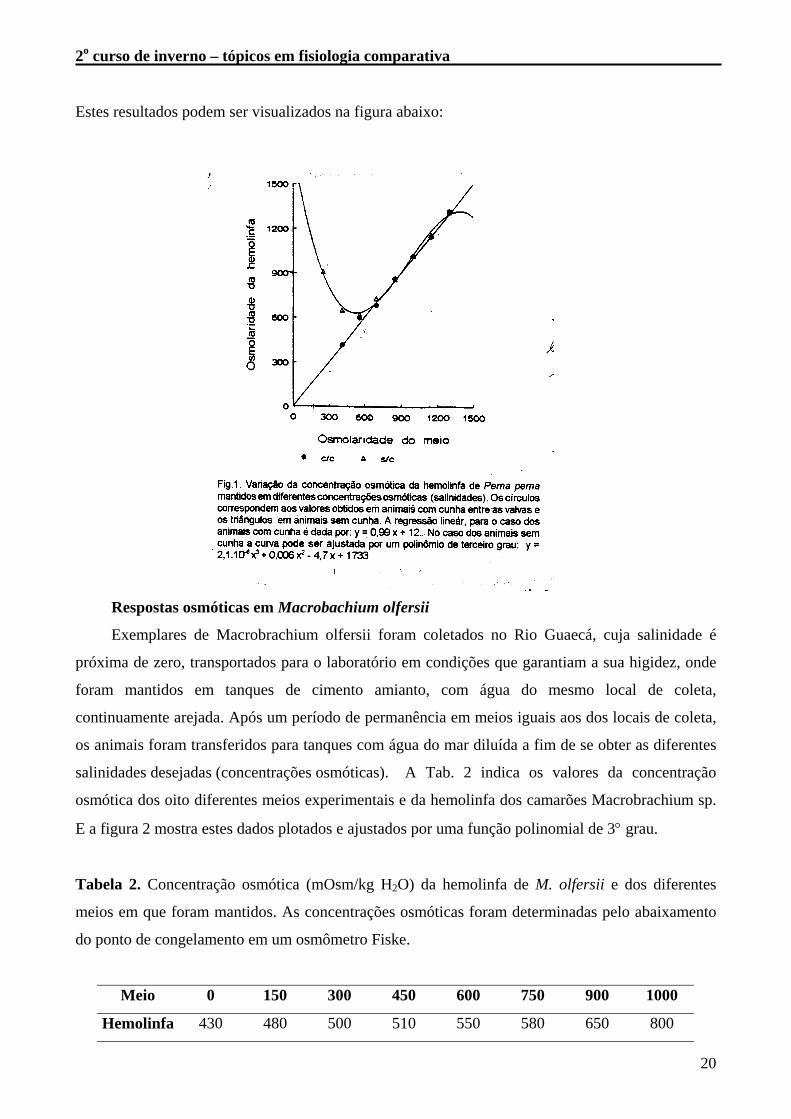
Tabela 1. Concentração osmótica da hemolinfa de Perna perna, com cunha e sem cunha mantidos
em diferentes salinidades. Valores em mOsm/kg H2O. As concentrações osmóticas foram
determinadas pelo abaixamento do ponto de congelamento conforme descrito por Salomão (Bolm
Fisiol. Animal, Univ. S. Paulo, 4: 143-152, 1980).
Meio 250 410 560 700 850 1000 1150 1300
c/ cunha 417 598 685 864 1016 1150 1320
s/ cunha 910 650 620 730 860 1020 1160 1315
19
2o curso de inverno – tópicos em fisiologia comparativa
Estes resultados podem ser visualizados na figura abaixo:
Respostas osmóticas em Macrobachium olfersii
Exemplares de Macrobrachium olfersii foram coletados no Rio Guaecá, cuja salinidade é
próxima de zero, transportados para o laboratório em condições que garantiam a sua higidez, onde
foram mantidos em tanques de cimento amianto, com água do mesmo local de coleta,
continuamente arejada. Após um período de permanência em meios iguais aos dos locais de coleta,
os animais foram transferidos para tanques com água do mar diluída a fim de se obter as diferentes
salinidades desejadas (concentrações osmóticas). A Tab. 2 indica os valores da concentração
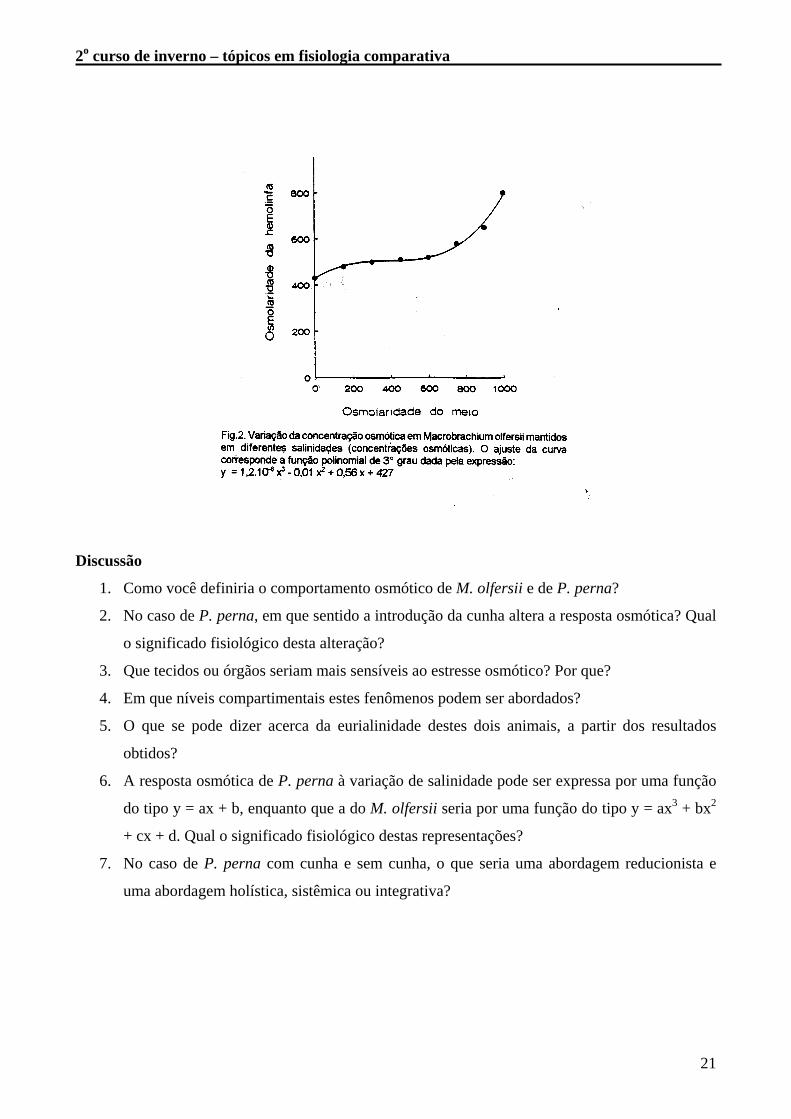
osmótica dos oito diferentes meios experimentais e da hemolinfa dos camarões Macrobrachium sp.
E a figura 2 mostra estes dados plotados e ajustados por uma função polinomial de 3° grau.
Tabela 2. Concentração osmótica (mOsm/kg H2O) da hemolinfa de M. olfersii e dos diferentes
meios em que foram mantidos. As concentrações osmóticas foram determinadas pelo abaixamento
do ponto de congelamento em um osmômetro Fiske.
Meio 0 150 300 450 600 750 900 1000
Hemolinfa 430 480 500 510 550 580 650 800
20
2o curso de inverno – tópicos em fisiologia comparativa
Discussão
1. Como você definiria o comportamento osmótico de M. olfersii e de P. perna?
2. No caso de P. perna, em que sentido a introdução da cunha altera a resposta osmótica? Qual
o significado fisiológico desta alteração?
3. Que tecidos ou órgãos seriam mais sensíveis ao estresse osmótico? Por que?
4. Em que níveis compartimentais estes fenômenos podem ser abordados?
5. O que se pode dizer acerca da eurialinidade destes dois animais, a partir dos resultados
obtidos?
6. A resposta osmótica de P. perna à variação de salinidade pode ser expressa por uma função
do tipo y = ax + b, enquanto que a do M. olfersii seria por uma função do tipo y = ax3 + bx2
+ cx + d. Qual o significado fisiológico destas representações?
7. No caso de P. perna com cunha e sem cunha, o que seria uma abordagem reducionista e
uma abordagem holística, sistêmica ou integrativa?
21
2o curso de inverno – tópicos em fisiologia comparativa
Osmorregulação no rato canguru
O balanço hidromineral no rato canguru se torna crítico em razão do ambiente inóspito em que
vive fazendo-o depender apenas da pouca água contida nos alimentos e da água metabólica.
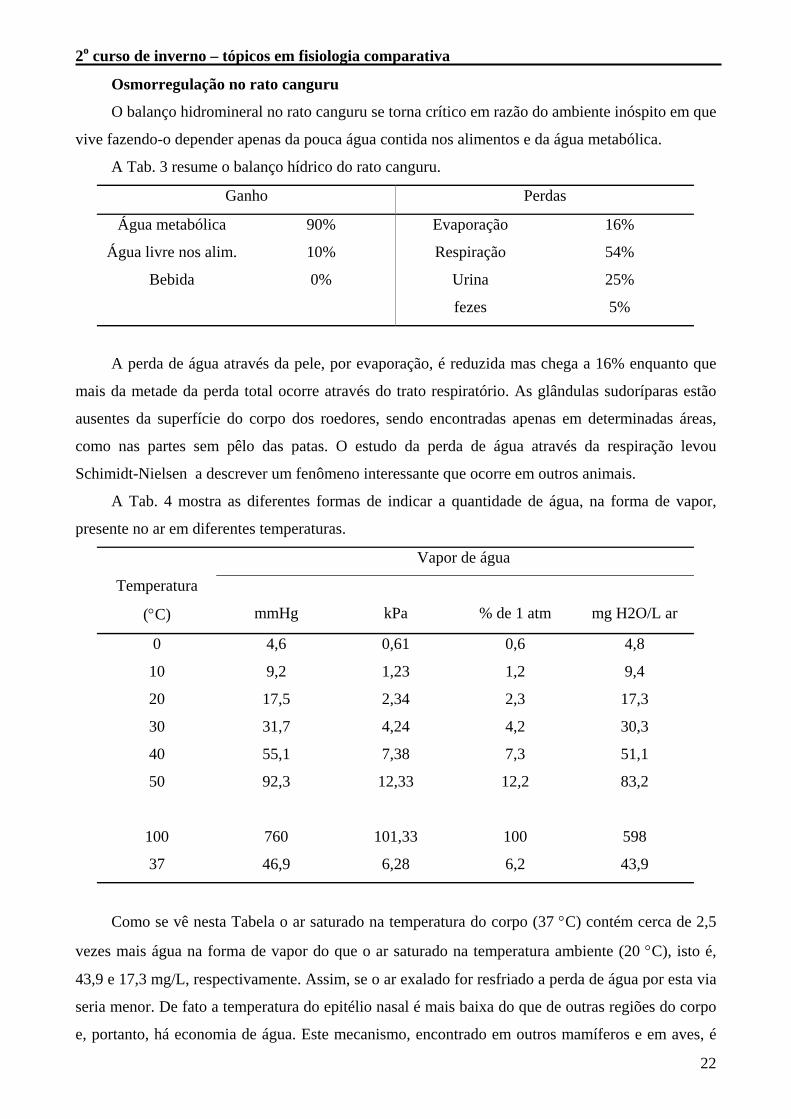
A Tab. 3 resume o balanço hídrico do rato canguru.
Ganho Perdas
Água metabólica 90% Evaporação 16%
Água livre nos alim. 10% Respiração 54%
Bebida 0% Urina 25%
fezes 5%
A perda de água através da pele, por evaporação, é reduzida mas chega a 16% enquanto que
mais da metade da perda total ocorre através do trato respiratório. As glândulas sudoríparas estão
ausentes da superfície do corpo dos roedores, sendo encontradas apenas em determinadas áreas,
como nas partes sem pêlo das patas. O estudo da perda de água através da respiração levou
Schimidt-Nielsen a descrever um fenômeno interessante que ocorre em outros animais.
A Tab. 4 mostra as diferentes formas de indicar a quantidade de água, na forma de vapor,
presente no ar em diferentes temperaturas.
Vapor de água
Temperatura
(°C) mmHg kPa % de 1 atm mg H2O/L ar
0 4,6 0,61 0,6 4,8
10 9,2 1,23 1,2 9,4
20 17,5 2,34 2,3 17,3
30 31,7 4,24 4,2 30,3
40 55,1 7,38 7,3 51,1
50 92,3 12,33 12,2 83,2
100 760 101,33 100 598
37 46,9 6,28 6,2 43,9
Como se vê nesta Tabela o ar saturado na temperatura do corpo (37 °C) contém cerca de 2,5
vezes mais água na forma de vapor do que o ar saturado na temperatura ambiente (20 °C), isto é,
43,9 e 17,3 mg/L, respectivamente. Assim, se o ar exalado for resfriado a perda de água por esta via
seria menor. De fato a temperatura do epitélio nasal é mais baixa do que de outras regiões do corpo
e, portanto, há economia de água. Este mecanismo, encontrado em outros mamíferos e em aves, é
22
2o curso de inverno – tópicos em fisiologia comparativa
denominado de mecanismo de contra-corrente nasal. No homem a temperatura do ar exalado está
próxima daquela do corpo. Logo, não há economia de água.
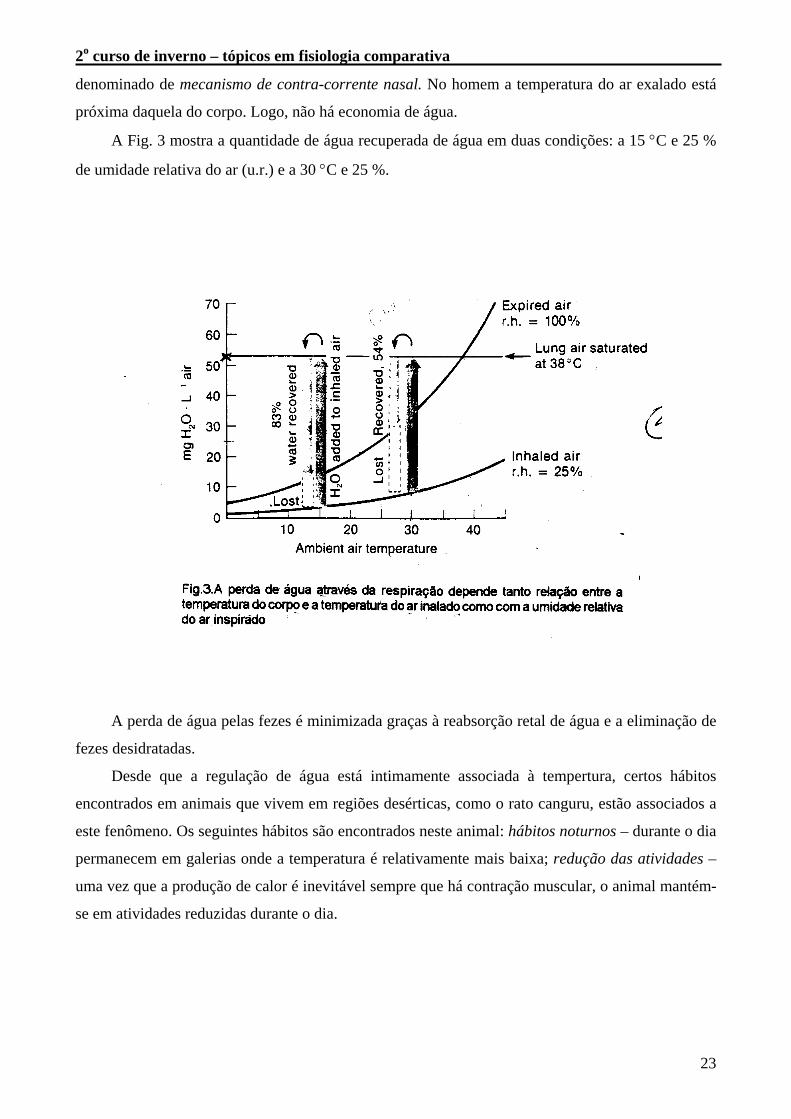
A Fig. 3 mostra a quantidade de água recuperada de água em duas condições: a 15 °C e 25 %
de umidade relativa do ar (u.r.) e a 30 °C e 25 %.
A perda de água pelas fezes é minimizada graças à reabsorção retal de água e a eliminação de
fezes desidratadas.
Desde que a regulação de água está intimamente associada à tempertura, certos hábitos
encontrados em animais que vivem em regiões desérticas, como o rato canguru, estão associados a
este fenômeno. Os seguintes hábitos são encontrados neste animal: hábitos noturnos – durante o dia
permanecem em galerias onde a temperatura é relativamente mais baixa; redução das atividades –
uma vez que a produção de calor é inevitável sempre que há contração muscular, o animal mantém-
se em atividades reduzidas durante o dia.
23

2o curso de inverno – tópicos em fisiologia comparativa
A Fig. 4 resume as estratégias utilizadas pelo rato canguru para sobreviver num ambiente de
grande restrição hídrica.
Produção de urina concentrada
Esta talvez seja a mais importante adaptação fisiológica do rato canguru. A concentração
osmótica da urina deste animal é superior a 6000 mOsm/kg H2O. É um valor elevado, embora
valores superiores a 9000 mOsm/kg H2O possam ser considerados em outros roedores de regiões
desérticas. (Lembre-se que a concentração osmótica da urina humana varia de cerca de 60 a 1200
mOsm/kg H2O) Rim capaz de produzir urina mais concentrada que o plasma só é encontrada em
ammíferos e aves.
O “truque simples”, como diz Schimidt-Nielsen, para a produção de urina concentrada reside
num fenômeno conhecido com “efeito multiplicador de contra-corrente”. Esquemas deste fenômeno
são encontrados em praticamente todos os livros de fisiologia. No entanto, valeria a pena ressaltar
que os elementos essenciais deste mecanismos são: (1) alça de Henle longa; (2) fluxo em sentido
contrário nos dois ramos da alça; (3) transporte ativo; (4) um ramo que reabsorve ativamente soluto
deve ser impermeável à água.
24

2o curso de inverno – tópicos em fisiologia comparativa
Discussão
1. Que relação há entre umidade relativa do ar e o balanço hídrico do rato canguru?
2. Que relação há entre a temperatura ambiente, balanço hídrico e temperatura corporal?
3. Em que o mecanismo de contra-corrente nasal difere do mecanismo multiplicador de contra-
corrente encontrado no rim?
4. Em que se assemelha a coriza observada no ser humano nos dias frios com aquele observado
nos animais com focinho frio?
5. Aves e mamíferos são capazes de produzir urina concentrada. Por que?
Bibliografia
Fyhn, H. J. Rodents. (1979) In: Maloiy, G. M. O. (Editor). Comparative Physiology of
Osmorregulation in Animal. London, Academic Press, v. 2
Salomão, L.C. & Luneta, J.E. (1989) The effects of salinity changes on the osmotic and ionic
concentrations in the hemolymph of Perna perna (Mollusca: Bivalvia). Bol. Fisiol. Anim., 13:
29-38.
Schmidit-Nielsen, K. How animals work (1st edition). Cambridge University Press. 1988
Stucchi-Zucchi, A. & Salomão, L. C. (2000) Effect of osmo-ionic concentration on the compound
action potential of the cerebro-visceral connective of Perna perna (Mollusca: Bivalvia).
Comp. Biochem. Physiol., 101(A): 109-112.
25

2o curso de inverno – tópicos em fisiologia comparativa
RELÓGIO BIOLÓGICO DE MAMÍFEROS: MECANISMOS MOLECULARES E
CONTROLE DA RITMICIDADE INTERNA DO ORGANISMO
Cíntia Etsuko Yamashita
Pedro Augusto Carlos Magno Fernandes
Aspectos Abordados:
O intuito desta aula será passar um pouco da evolução dos relógios biológicos ao longo das
espécies até chegarmos aos mamíferos onde o estudo será aprofundado. Discutiremos os
mecanismos moleculares que controlam a ritmicidade interna do relógio central e como ele é
sincronizado por fatores externos como, por exemplo, o ciclo claro/escuro ambiental.
O relógio biológico central transmite as informações rítmicas ambientais para diversos
osciladores internos que, por sua vez, promovem respostas fisiológicas que se refletem em padrões
comportamentais específicos. Para ilustrar este processo tomarei por base o controle do relógio
sobre a produção rítmica dos glicocorticóides e da melatonina. Aproveitando para fazer um gancho
com a minha área de pesquisa que consiste na investigação da inter-relação entre as glândulas
pineal e adrenal durante um processo inflamatório crônico.
Introdução:
Os seres vivos, na sua forma mais simples, como os seres unicelulares, até sua forma mais
complexa, como os vertebrados, são estruturados no tempo e no espaço. A maioria dos parâmetros
bioquímicos, fisiológicos e comportamentais dos organismos apresenta flutuações diárias que
persistem sob condições constantes, indicando que osciladores endógenos foram incorporados aos
sistemas em resposta às variações do ciclo claro-escuro ambiental (Menaker et al., 1997).
No centro dos sistemas que controlam e regulam os ritmos circadianos dos vertebrados estão
três estruturas que se interconectam num “eixo circadiano central”, que são: os núcleos
supraquiasmáticos (NSQ), a retina e as glândulas pineal e adrenal (Menaker et al., 1997). Estes
otimizam as atividades básicas diárias, a vida reprodutiva e algumas respostas sazonais.
O sistema mínimo requerido para adaptação às variações ambientais seria um relógio
endógeno, independente do meio ambiente, um sistema sensor que pudesse detectar as alterações
rítmicas do meio ambiente e um ou mais elementos sincronizadores, que teriam como função o
26

2o curso de inverno – tópicos em fisiologia comparativa
ajuste do meio interno e das funções vitais às variações ambientais. Recentemente, foram descritos
os mecanismos moleculares do relógio central (NSQ) e de relógios biológicos locais, chamados de
servo relógios (Reppert e Weaver, 2002). Também é conhecida a forma como a retina é capaz de
detectar luz e enviar a informação fótica para os NSQ (Provencio et al., 2000) e o papel de diversos
marcadores internos como, por exemplo, o da melatonina – hormônio produzido e liberado pela
glândula pineal- como marcador do escuro e o dos glicocorticóides -produzidos ritmicamente pela
glândula adrenal- como antecipadores do período de atividade, agindo portanto sobre o ciclo
vigília/sono dos indivíduos.
Bibliografia:
Cronobiologia:Princípios e Aplicações; Organizadores: Nelson Marques e Luiz Menna-Barreto;
editora Fiocruz, 2003.
Menaker, M., Moreira, L.F., Tosini, G. Evolution of circadian organization in vertebrates. Braz. J.
Med. Biol. Res., 30: 305-313, 1997.
Reppert, S.M. & Weaver, D.R. Coordination of circadian timing in mammals. Nature 418: 935-941,
2002.
Provencio, I; Rodriguez, I.R.; Jiang, G.; Hayes, W.P.; Moreira, E.F. & Rollag, M.D. A novel human
opsin in the inner retina. J. Neurosc. 20: 600-605, 2000.
Lang, V., Sizonenko, PC. Melatonin and human adreno-cortical function. In: Miles, A., Philbrick,
D.R.S. Thompson, C (eds) Melatonin, clinical perspectives. Oxford University Press, pp 62-
78, 1988.
Sites Recomendados:
www.nature.com
www.pubmed.com
27

2o curso de inverno – tópicos em fisiologia comparativa
NEUROANATOMIA BÁSICA
Emerson Ferraz Coelho
Divisões do Sistema Nervoso:
Divisão do Sistema Nervoso com base em critérios anatômicos:
Sistema Nervoso Central:
Neuro eixo: Encéfalo (cérebro, cerebelo e tronco encefálico) e medula
Sistema Nervoso Periférico:
Nervos cranianos e espinais, gânglios, terminações nervosas sensitivas e motoras.
Divisão do Sistema Nervoso com base em critérios embriológicos
Prosencéfalo, mesencéfalo e rombencéfalo.
Divisão do Sistema Nervoso com base em critérios funcionais:
Sistema Nervoso da Vida de Relação (Somático).
Sistema Nervoso da Vida Vegetativa (Visceral).
Bibliografia: Neuroanatomia Funcional, Angelo Machado.
28

2o curso de inverno – tópicos em fisiologia comparativa
ROTEIRO DE AULA PRÁTICA
NEUROANATOMIA
Andreas Betz
Daniel Carneiro Carrettiero
Emerson Ferraz Coelho
Jéssica Ruivo Maximino
Regiane Xavier de Moraes
OBJETIVO: Apresentar o Sistema Nervoso (SN) e suas possíveis divisões didáticas.
O SN é um todo. Sua divisão em partes tem um significado exclusivamente didático, pois várias
delas estão intimamente relacionadas do ponto de vista morfológico e funcional. O SN pode ser
divido levando-se em conta critérios anatômicos, funcionais e embriológicos.
DIVISÃO DO SISTEMA NERVOSO COM BASE EM CRITÉRIOS ANATÔMICOS
Sistema Nervoso
Sistema NervosoCentral
Sistema NervosoPeriférico
Encéfalo
Medula espinhal
CérebroCerebeloTronco Encefálico
MesencéfaloPonteBulbo
NervosGângliosTerminações Nervosas
EspinhaisCranianos
Sistema Nervoso
Sistema NervosoCentral
Sistema NervosoPeriférico
Encéfalo
Medula espinhal
CérebroCerebeloTronco Encefálico
MesencéfaloPonteBulbo
NervosGângliosTerminações Nervosas
EspinhaisCranianos
DIVISÃO DO SISTEMA NERVOSO COM BASE EM CRITÉRIOS FUNCIONAIS
Sistema NervosoSomático
Sistema NervosoVisceral
AferenteEferente
AferenteEferente
SimpáticoParassimpático
Sistema NervosoSomático
Sistema NervosoVisceral
AferenteEferente
AferenteEferente
SimpáticoParassimpático
DIVISÃO DO SISTEMA NERVOSO COM BASE EM CRITÉRIOS
Prosencéfalo
Mesencéfalo
Rombencéfalo
TelencéfaloDiencéfalo
MetencéfaloMielencéfalo
Cérebro
Mesencéfalo
Rombencéfalo
Prosencéfalo
Mesencéfalo
Rombencéfalo
TelencéfaloDiencéfalo
MetencéfaloMielencéfalo
Cérebro
Mesencéfalo
Rombencéfalo
29
2o curso de inverno – tópicos em fisiologia comparativa
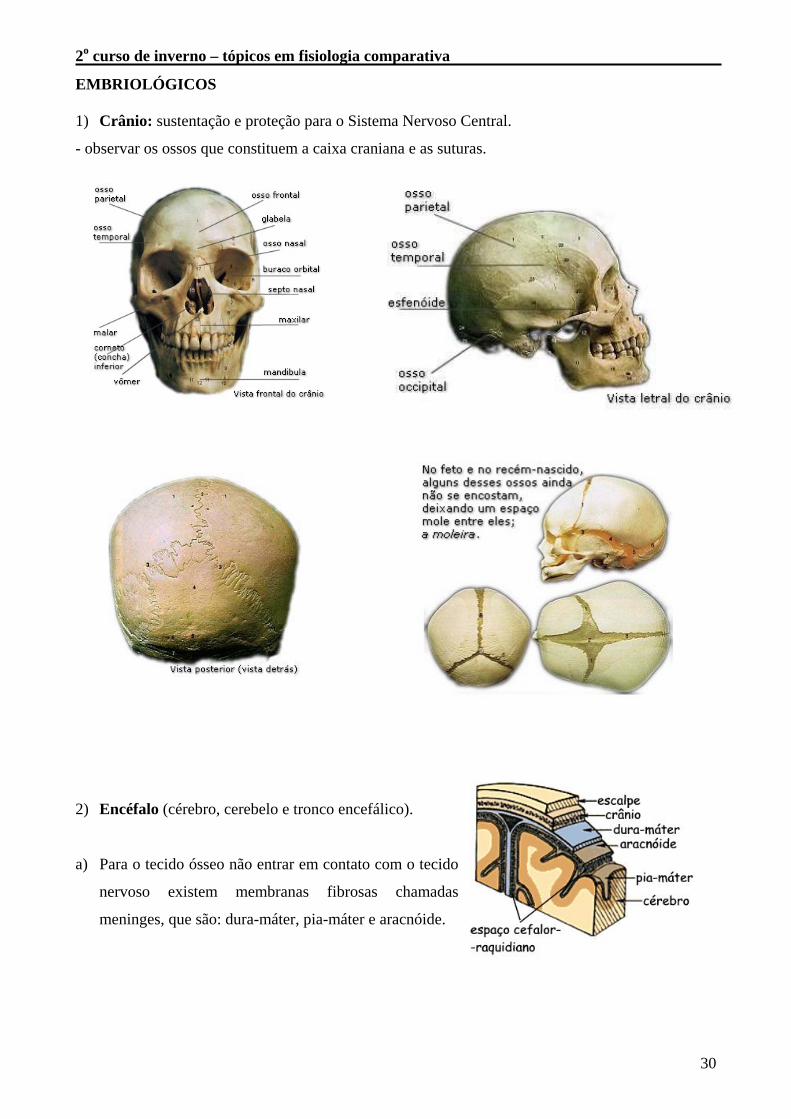
EMBRIOLÓGICOS 1) Crânio: sustentação e proteção para o Sistema Nervoso Central.
- observar os ossos que constituem a caixa craniana e as suturas.
2) Encéfalo (cérebro, cerebelo e tronco encefálico).
a) Para o tecido ósseo não entrar em contato com o tecido
nervoso existem membranas fibrosas chamadas
meninges, que são: dura-máter, pia-máter e aracnóide.
30
2o curso de inverno – tópicos em fisiologia comparativa
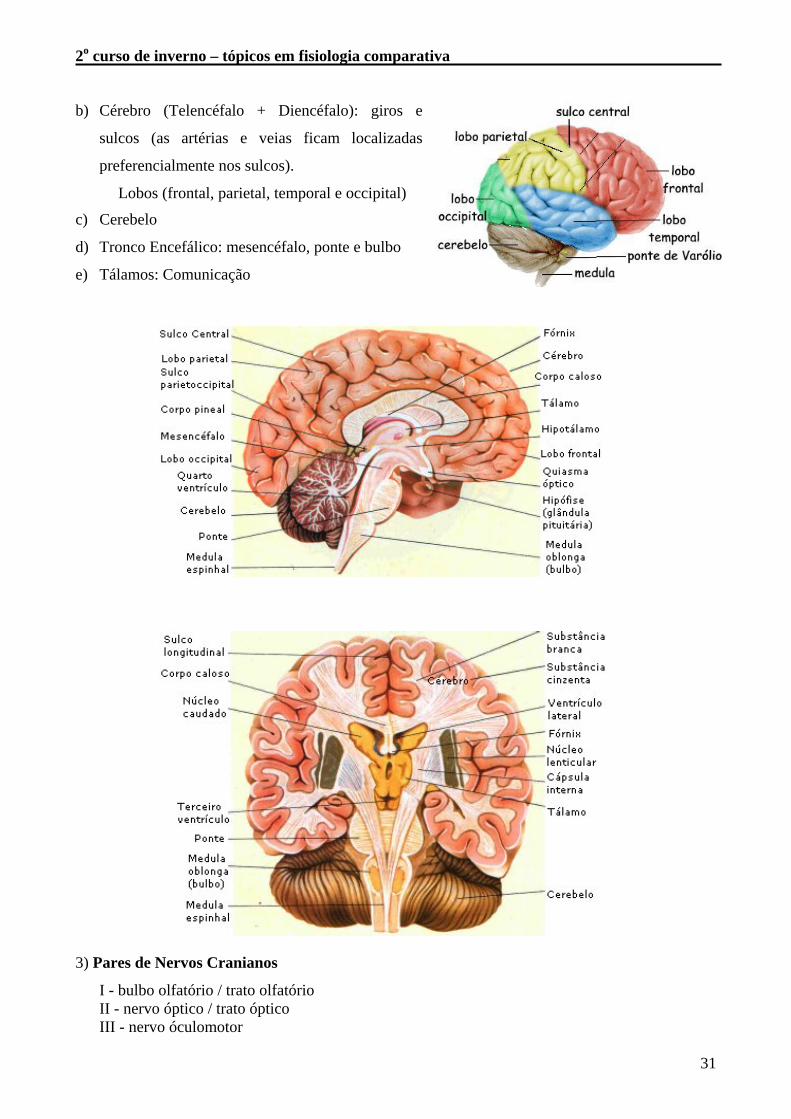
b) Cérebro (Telencéfalo + Diencéfalo): giros e
sulcos (as artérias e veias ficam localizadas
preferencialmente nos sulcos).
Lobos (frontal, parietal, temporal e occipital)
c) Cerebelo
d) Tronco Encefálico: mesencéfalo, ponte e bulbo
e) Tálamos: Comunicação
3) Pares de Nervos Cranianos
I - bulbo olfatório / trato olfatório II - nervo óptico / trato óptico III - nervo óculomotor
31
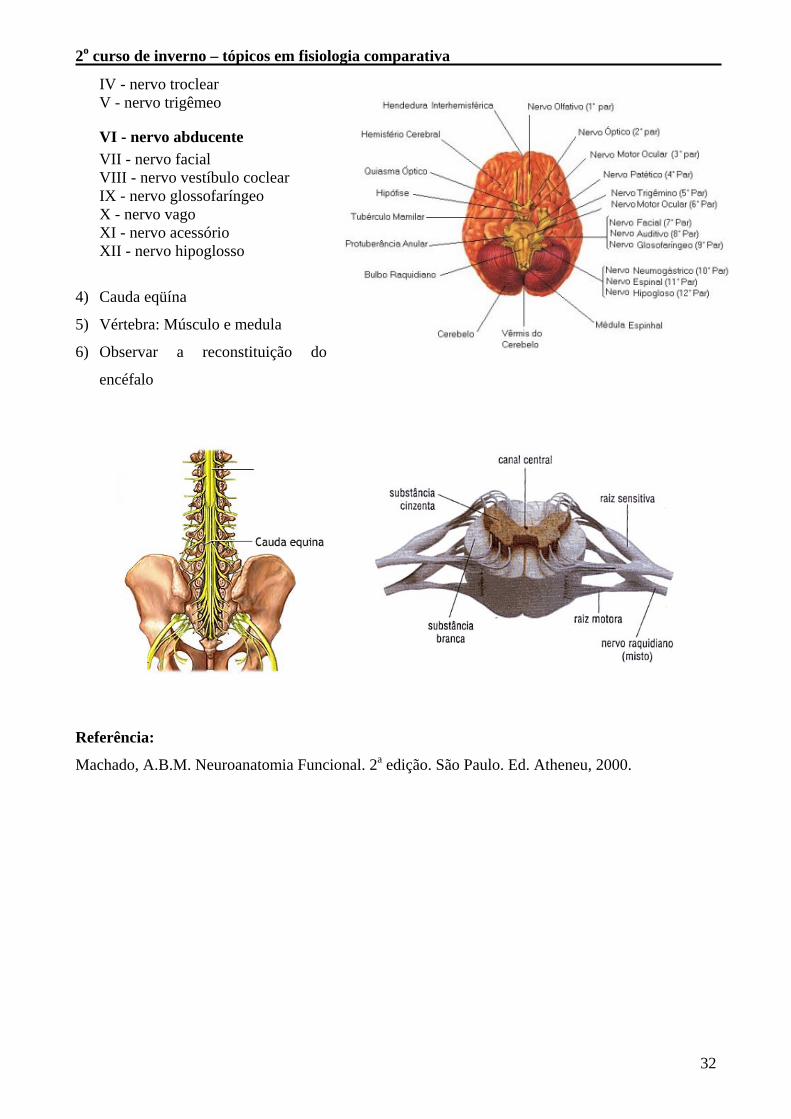
2o curso de inverno – tópicos em fisiologia comparativa
IV - nervo troclear V - nervo trigêmeo
VI - nervo abducente VII - nervo facial VIII - nervo vestíbulo coclear IX - nervo glossofaríngeo X - nervo vago XI - nervo acessório XII - nervo hipoglosso
4) Cauda eqüína
5) Vértebra: Músculo e medula
6) Observar a reconstituição do
encéfalo
Referência:
Machado, A.B.M. Neuroanatomia Funcional. 2a edição. São Paulo. Ed. Atheneu, 2000.
32

2o curso de inverno – tópicos em fisiologia comparativa
VARIABILIDADE CARDÍACA E MODELAMENTO MATEMÁTICO DE SISTEMAS
BIOLÓGICOS
Breno Teixeira Santos
Variabilidade Cardíaca
Cada vez mais acumulam-se fatos na literatura indicando que diversos sinais biológicos
possuem altos níveis de variabilidade associada. Essa, por sua vez, parece indicar condições
saudáveis de funcionamento nos diversos órgãos como o cérebro e o coração.
O sistema cardiovascular está intimamente associado à demanda metabólica dos organismos.
Esse sistema se encontra sob controle nervoso e hormonal visando à adequação de fluxo aos órgãos
nas mais diversas situações de vida. Em mamíferos, o resultado desse controle aparece na forma de
uma intensa variabilidade na freqüência cardíaca, variabilidade essa que é independente da escala
temporal de análise. Quando o organismo se encontra exposto a situações adversas, como doenças
ou exercício extenuante, há, aparentemente, perda dessa variabilidade.
Visão geral do sistema cardiovascular
O sistema cardiovascular é o responsável pela manutenção de fluxo convectivo de matéria e
calor em praticamente todos os metazoários. Particularmente em mamíferos e outros vertebrados,
existe um órgão, o coração, responsável por gerar entalpia no sistema vascular, mantendo fluxo aos
órgãos. O funcionamento do coração como fonte de entalpia, é, portanto, controlado de acordo com
as necessidades do organismo de uma forma geral e de tecidos específicos em situações
particulares, como, por exemplo, o aumento do aporte sangüíneo ao trato digestivo quando da
alimentação (e.g., Secor, 2003). A manutenção da entalpia no sistema é, basicamente, em função da
manutenção da pressão intravascular, que, na sua porção das grandes artérias é a pressão arterial
(P.A.). Como o sangue será distribuído aos órgãos a partir das artérias e como é nessa porção do
sistema que o coração coloca pressão, o controle adequado da perfusão tecidual, no nível do
organismo, se dá através da manutenção da P.A. em níveis compatíveis com a demanda.
A pressão arterial é mantida através da combinação de duas variáveis principais: (1) débito
cardíaco (Q ), e (2) resistência periférica (Rp) (Milnor, 1997). Entretanto, além desses dois
componentes principais, podemos colocar a viscosidade sangüínea, turbulência nos ductos e outras
variáveis menos importantes como concorrentes no valor de P.A. a cada instante de tempo. O débito
cardíaco é resultado do quanto o coração ejeta a cada contração, i.e., o volume sistólico (VS), e do
&
33

2o curso de inverno – tópicos em fisiologia comparativa
número de contrações por unidade de tempo, i.e., a freqüência cardíaca (f). Já a resistência
periférica é resultado, basicamente, do raio, comprimento e complacência dos ductos. Assim, a
diferença de energia entre os pontos do sistema é o que causa o fluxo, e o “formato” geral abaixo se
aplica ao sistema cardiovascular, numa visão global do processo de circulação (Hoppensteadt &
Peskin, 2002):
P.A. = Rp (1a) Q&
e
= VS . f (1b) Q&
O controle da pressão arterial em mamíferos foi objeto de numerosos e extensos estudos. De
uma maneira geral, os estímulos oriundos do sistema nervoso autônomo simpático tendem a elevar
a pressão arterial, porém isso depende do local de ação (e.g., vasodilatação no território muscular
por receptores α-adrenérgicos). Mais diretamente, no miocárdio, agem os dois sistemas autônomos.
A inervação simpática aumenta a força de contração e a freqüência cardíaca, basicamente devido à
liberação de epinefrina. A epinefrina aumenta, ainda, a velocidade de condução do impulso elétrico
nos tecidos especializados (Milnor, 1997). Por outro lado, a acetilcolina, liberada nos terminais
vagais da inervação parassimpática, diminui a freqüência de disparos do marca-passo cardíaco e
velocidade de condução, além de levar a um decréscimo na força de contração.
A força de contração é dependente de características intrínsecas da musculatura cardíaca, mas,
também, do grau de estiramento prévio dessa musculatura. Exatamente como nos demais músculos
estriados, o cardíaco tem sua força de contração ligada ao deslizamento dos filamentos de miosina
sobre a actina devido à formação de pontes cruzadas (Guyton & Hall, 2000). No miocárdio, o grau
de estiramento prévio à contração é decorrente do grau de enchimento ventricular, ou seja, do
volume ao final da diástole. O aumento do volume diastólico final, até um certo limite, aumenta,
assim, o grau de estiramento das fibras miocárdicas, e leva, com isso, a um aumento na força de
contração (Lei de Frank-Starling, e.g., Milnor, 1997).
O volume sistólico, ou seja, o volume ejetado a cada contração, encontra-se diretamente
relacionado à força de contração. Quando há elevação da freqüência cardíaca, como na estimulação
simpática, ocorre uma diminuição do tempo para enchimento ventricular devido ao encurtamento
do tempo de diástole e o inverso ocorre com o estímulo parassimpático. Logo, segundo a equação
(1b) e o discutido acima para o efeito da pré-carga, paradoxalmente, um aumento de freqüência
pode ser acompanhado por uma diminuição de débito cardíaco e vice-versa.
34

2o curso de inverno – tópicos em fisiologia comparativa
Como apresentado anteriormente, o controle da pressão arterial é de suma importância para a
manutenção de fluxo adequado aos órgãos; e agem, diretamente sobre o miocárdio, os sistemas
nervosos autônomos responsáveis por grande parte desse controle. A análise da freqüência cardíaca
revela uma grande variabilidade dessa ao longo do tempo. Independentemente da escala (e.g.,
segundo, minutos, horas, etc.), detecta-se uma constante modificação do número de batimentos
cardíacos por unidade de tempo (Goldberger et al., 1990). Apesar de que essa variação poderia estar
refletindo somente fenômenos locais decorrentes da relação pré-carga e volume sistólico (como
explicado acima, Lei de Frank-Starling), acumulam-se evidências ao longo das últimas décadas a
respeito do papel do controle nervoso gerando esse fenômeno. Assim, aceita-se, hoje em dia, que a
variabilidade de freqüência indica o funcionamento normal e adequado do órgão, como fonte de
entalpia, e do sistema cardiovascular, como fonte de fluxo sangüíneo (Goldberger, 1996).
Tem-se, como contra-exemplo, a perda da variação de freqüência quando o organismo
encontra-se fora de sua condição fisiológica. A insuficiência cardíaca congestiva de grau avançado
(Poon & Merrill, 1997), a isquemia miocárdica (Goldberger, 1996), a displasia broncopulmonar em
infantes pré-termo (Patzak et al., 1997), o período que antecede a fibrilação ventricular (Goldberger
et al, 1988), a apnéia obstrutiva do sono (Trzebski & Smietanowski, 2001), são todas situações nas
quais se evidenciou a perda de variabilidade. Mesmo em experimentos mais simples, pode-se
detectar uma maior estabilidade da freqüência cardíaca em decorrência de situações não-fisiológicas
(ver Figura 1). Assim, a avaliação em tempo real da variância da freqüência cardíaca é um valioso
elemento de diagnóstico com antecedência de condições não normais do sistema cardiovascular.
Mais ainda, condições de estresse para o sistema vêm sendo abordadas através da observação dos
componentes de variância presentes no espectro do sinal (e.g., Yeragania et al., 2003).
Figura 1: Intervalo <RR> (freqüência cardíaca) sob diferentes condições: ventilação normal
(traçado verde, porções oscilatórias) e em apnéia com glote aberta (traçado verde, porções sem
oscilação). Note que nos trechos 1, 3, 5 e 7, correspondendo a períodos de ventilação normal, que o
<RR> varia bastante e de forma aleatória. Nos demais trechos, onde foi imposta a condição
estressante, o sinal varia consideravelmente menos (Andrade e Santos, 2003).
35

2o curso de inverno – tópicos em fisiologia comparativa
Modelamento Matemático
O modelamento matemático de sistemas biológicos é algo já bastante difundido em algumas
áreas da biologia como ecologia e fisiologia. Essa aula tem por objetivo demonstrar a utilidade,
importância e aplicabilidade dessa ferramenta dentro do estudo da biologia e, mais especificamente,
na fisiologia.
Mas afinal o que é um modelo? Uma boa definição é a apresentada no preâmbulo do capítulo
4 – Modelling, do livro Biomechanics of the Musculo-Skeletal System de Niggs e Herzog:
“Modelagem, uma tentativa de representar a realidade, é freqüentemente utilizada quando o
entendimento de um certo fenômeno se torna difícil. Um modelo é uma ferramenta poderosa para
aumentar o entendimento sobre um mecanismo e tem, portanto, sido muito aplicada em situações do
dia-a-dia e/ou em situações de pesquisa... Modelagem, freqüentemente combinada com dados
experimentais, torna-se uma poderosa ferramenta científica. ”
Normalmente, embora nem sempre, um modelo é uma representação simplificada da
realidade, específica para um determinado fim. Um modelo pode ser criado a partir de dois
paradigmas distintos: dedução ou indução. No primeiro, a partir de conhecimento sobre o sistema,
opera-se sobre princípios gerais (normalmente físicos e químicos) e deduz-se analiticamente uma
solução única. A segunda forma baseia-se em uma coleção de dados experimentais e, por indução,
tenta-se explicar os dados observados, o não leva a uma solução única.
Qual a melhor solução a adotar? Resposta: não há uma melhor. Sua escolha depende do nível
de conhecimento que se tem sobre o sistema, se é possível com os métodos conhecidos deduzir algo
ou ainda, se é factível obter dados experimentais.
Independente da escolha deve-se sempre ter em mente que um modelo possui basicamente
dois objetivos:
1- Aumentar o conhecimento e o “insight” sobre a realidade e
2- Estimar ou predizer variáveis de interesse.
Modelos podem ser: analíticos, semi-analíticos, modelos de caixa preta, ou conceituais.
Modelos analíticos são deduzidos matematicamente com base em conhecimento real ou especulado
e possui provas determinísticas. Os semi-analíticos diferem dos puramente analíticos pelo fato de
serem tão complicados que impossibilitam a dedução total – nesses casos pode ser possível chegar a
níveis de dedução intermediários. Modelos de caixa preta, ou de regressão, buscam funções de
36

2o curso de inverno – tópicos em fisiologia comparativa
transferência que mapeiem um conjunto de entradas em saídas respectivas. Por fim os conceituais
são hipóteses baseadas em conhecimento real ou dados experimentais.
Finalizaremos a aula com alguns exemplos de modelagem e análise matemática aplicadas ao
estudo de variabilidade cardíaca.
Biliografia
Andrade, F. R. & Santos, B. T. (2003)Comparação dos efeitos de três diferentes tipos de apnéia
voluntária na freqüência cardíaca. 6º Semana Temática da Biologia.
Goldberger, A. L. (1996) Non-linear dynamics for clinicians: chaos theory, fractals, and complexity
at the bedside. The Lancet 347: 1312-1314.
Goldberger, A. L., Rigney, D. R., Mietus, J., Antman, E. M. & Greenwald, S. (1998) Non-linear
dynamics in sudden cardiac death syndrome: heart-rate oscillations and bifurcations.
Experientia 44: 983-987.
Goldberger, A., Rigney, D. & West, B. J. (1990) Chaos and Fractals in Human Physiology. Sci.
Amer. Fevereiro: 35-41.
Guyton, A.C. & Hall, J. E. (2000) Textbook of Medical Physiology, 10a edição W B Saunders.
Hoppensteadt, F. C. & Peskin, C. S. (2002) Modeling and Simulation in Medicine and the Life
Sciences 2a edição. Springer, Nova York.
Milnor, W. R. (1997)Cardiovascular Physiology. Oxford University Press.
Secor, S. M. (2003)Gastric function and its contribution to the postprandial metabolic response of
the Burmese python Python molurus. J. Exp. Biol. 206:1621-1630.
Patzak, A., Schluter, B., Orlow, W., Mrowka, R., Gerhardt, D., Schubert, E., Presson, P. B.,
Barschdorff, D. & Trowitzsch, E. (1997) Linear and nonlinear properties of heart rate control
in infants at risk. Am. J. Physiol. Regulatory 273, R540-R547.
Poon, C. & Merrill, C. (1997) Decreased of cardiac chaos in congestive heart failure. Nature 389:
492-495.
Trzebski, A. & Smietanowski, M. (2001) Non-linear dynamics of cardiovascular system in humans
exposed to repetitive apneas modeling obstrutive sleep apnea: aggregates time series data
analysis. Anatomic Neuroscience: Basic and Clinical, 90, pg. 106-115.
37

2o curso de inverno – tópicos em fisiologia comparativa
AS TOXINAS DE ANÊMONAS DO MAR COMO FERRAMENTAS PARA ENTENDER A
FISIOLOGIA DE TECIDOS, ÓRGÃOS E SISTEMAS
André Junqueira Zaharenko.
As anêmonas do mar são animais que pertencem ao filo Cnidaria e a classe Anthozoa. Todas
as anêmonas, assim como os celenterados em geral, possuem estruturas celulares microscópicas,
similares a arpões, denominadas de nematocistos, responsáveis pelo papel de paralisar presas e
também atuam na defesa dos animais. Estas estruturas contêm potentes neurotoxinas paralisantes
que agem sobre crustáceos e peixes pelo simples contato com esses animais. Essa estratégia é de
vital importância para os mais distintos animais, desde esses cnidários, passando por moluscos
predadores, escorpiões, aranhas e até serpentes.
Todos esses tipos de animais produzem toxinas protéicas e peptídicas em glândulas
especializadas e as injetam, a partir de estruturas também especializadas na inoculação, como
ferrões, presas e, no caso dos cnidários, nematocistos. Durante muitos anos os cientistas em geral
se debruçaram em tentativas de purificar e elucidar os mecanismos de ação das toxinas, ainda que
por décadas as limitações tecnológicas e operacionais dificultassem essa tarefa.
Um dos aspectos mais importantes que devemos nos focar, quando vamos tentar investigar
quais são os possíveis mecanismos de ação e alvos das toxinas, é exatamente qual o tipo de animal
predado pelo nosso objeto de estudo. As anêmonas, por exemplo, predam em geral peixes e
crustáceos planctônicos. Essas presas, por sua vez, são de rápida locomoção e poderiam facilmente
escapar de um predador que também não fosse rápido na captura.
Quando pensamos nos ambientes ocupados pelas anêmonas, logo devemos nos ater ao fato de
que esses animais são sésseis, ou seja, vivem fixos em um substrato rochoso e basicamente não se
movem. Se as presas das anêmonas são animais de rápida locomoção, as toxinas direcionadas a sua
captura devem ter um efeito o mais efetivo e letal possível. Seguindo nessa linha de raciocínio, os
alvos mais conhecidos onde as toxinas anêmonas atuam são: na condução nervosa e na contração
muscular das presas. Toxinas que bloqueiam a neurotransmissão, levando a paralisia seguida de
morte, são as moléculas mais investigadas nesse grupo de animais até hoje.
Remetendo aos conhecimentos básicos de fisiologia e biologia celular, sabemos que um
potencial de ação (PA) é evocado basicamente pela mudança de voltagem do interior de uma célula
nervosa através da entrada de íons sódio (despolarização) e a saída de íons potássio na mesma. Este
38

2o curso de inverno – tópicos em fisiologia comparativa
último processo é um pouco mais tardio e permite o restabelecimento da voltagem inicial no interior
da célula (repolarização), o chamado potencial de membrana. A seqüência desses eventos,
controlados refinadamente pelos canais para Na+ e K+ dependentes de voltagem, leva a transmissão
do impulso elétrico, com consequências fisiológicas importantes para a comunicação neuronal e a
contração muscular. Para uma revisão completa, a leitura do capítulo 11 (páginas 523-547) do livro
MOLECULAR BIOLOGY OF THE CELL (Alberts et al., 3a e 4a edições; 1994, 2002) é bastante
recomendada.
Durante a passagem de um PA de um neurônio a outro, temos no neurônio pré-sináptico,
durante a despolarização, a abertura de canais para Ca2+ dependentes de voltagem próximos ao
botão sináptico. Quando se abrem, esses canais permitem o influxo de Ca2+ que provoca,
subsequentemente, a alteração da concentração intracelular desse íon e a ativação de maquinarias
secretórias de neurotransmissores. Vesículas contendo neurotransmissores são liberadas na fenda
sináptica e estes ligam-se aos respectivos receptores pós-sinápticos, propagando o PA ou
desencadeando eventos secundários importantes para a fisiologia celular. A contração muscular
desencadeia-se da mesma maneira, apenas diferindo na sequência de eventos pós-sinápticos
desencadeadas pela ligação da acetilcolina (no caso de mamíferos, por exemplo) ou do glutamato
(no caso de crustáceos e invertebrados) nos respectivos receptores das fendas sinápticas. É
importante ressaltarmos que nesse caso não há um neurônio pós-sináptico e sim uma musculatura
inervada por um neurônio pré-sináptico.
Essa revisão de conceitos básicos é importante quando nos deparamos com os mecanismos de
ação de neurotoxinas de anêmonas e de outros animais peçonhentos. A maioria dos peptídeos de
anêmonas descritos e estudados age em canais para Na+ ou para K+ dependentes de voltagem.
Retardam o processo de inativação dos canais de Na+ e bloqueiam os canais de K+, produzindo um
influxo enorme de íons Na+ e uma diminuição drástica da saída de íons K+ nas células. Isso faz com
que o PA tenha sua amplitude e duração aumentadas dramaticamente.
Para a fisiologia celular, a liberação de neurotransmissores passa a ser tremenda, levando a
contrações musculares que não cessam, paralisando um animal que tenha sofrido injeções diretas
dessas toxinas em seus tecidos.
Existem, atualmente, cerca de 10 subtipos de canais de Na+ dependentes de voltagem (os
chamados, Nav) distribuídos nos sistemas nervosos central e periférico, tecido cardíaco, medula
espinhal e músculo esquelético. São proteínas transmembranares que diferem levemente em termos
de sequências primárias e parâmetros cinéticos.
Como nosso grupo de pesquisa vem trabalhando há longos anos com neurotoxinas de
anêmonas do mar, recentemente publicamos um trabalho no qual a comparação dos efeitos de 3
39

2o curso de inverno – tópicos em fisiologia comparativa
toxinas praticamente idênticas difere pouco dependendo dos subtipos de Nav ensaiados (Oliveira et
al., 2004). Ou seja, isso mostra que a atuação das moléculas em seus sítios de ligação é sutilmente
modulada por alguns aminoácidos em suas estruturas. Dependendo do subtipo de Nav ensaiado,
havia efeitos preferenciais ou não de cada uma das toxinas. Durante a aula expositiva os resultados
serão apresentados e discutidos com os alunos. A leitura do trabalho de Oliveira et al.(2004) também
é requerida.
Conforme os diferentes tipos de toxinas são purificados e caracterizados, cada vez mais essas
moléculas são empregadas como ferramentas farmacológicas, para em laboratório induzirem seus
efeitos e ajudarem os cientistas a investigar diferentes aspectos da fisiologia.
Muitas companhias farmacêuticas revendem toxinas com esse propósito, a preços
elevadíssimos. Acessando o site www.alomone.com e clicando em Ion Channel Modulators e
Neurotoxins, podemos ver a lista de diferentes toxinas com seus respectivos preços.
Embora companhias farmacêuticas forneçam toxinas como substâncias para pesquisa básica,
muitas delas vêm investindo milhões de dólares no desenvolvimento de fármacos a partir dessas
moléculas. Como exemplo, existem peptídeos que bloqueiam especificamente canais de K+
dependentes de voltagem expressos em linfócitos-T. Esse tipo de bloqueio leva a uma supressão do
sistema imune e, consequentemente, esse tipo de toxina torna-se altamente atrativo como um
remédio para tratar artrite reumatóide e rejeição a órgãos transplantados. Na revisão apresentada
por Chandy et al., 2001, os alunos podem acompanhar os avanços recentes nesse sentido.
Finalizando, queremos mostrar que a partir de protótipos naturais pode-se obter substâncias
altamente eficazes para o estudo da fisiologia e que sirvam também como modelos para o
desenvolvimento de medicamentos.
REFERÊNCIAS:
ALBERTS, B. et al. 1994. Molecular Biology of the Cell. Garland Publishing. New York. 3rd
edition. 1294p.
CHANDY, K.G., CAHALAN, M., PENNINGTON, M., NORTON, R. ., WULFF, H., GUTMAN,
G.A. 2001. Potassium channels in T lymphocytes: toxins to therapeutic immunosupressants.
Toxicon. 39: 1269-1276.
OLIVEIRA, J. S., REDAELLI, E., ZAHARENKO, A. J., CASSULINI, R. R., KONNO, K., CURIA,
G., PIMENTA, D.C., FREITAS, J. C., CLARE, J. J., WANKE, E. 2004. Binding of sea
anemone toxins to Nav 1.1-1.6 Sodium Channels: Unexpected Contributions from Differences
in the IV/ S3-S4 Outer Loop. Journal of Biological Chemistry. 279 (32): 33323-33335.
40

2o curso de inverno – tópicos em fisiologia comparativa
ROTEIRO DE AULA PRÁTICA
Durante a aula prática, os alunos acompanharão procedimentos de fracionamento de peçonhas
totais pela técnica cromatográfica de gel-filtração. Basicamente, a metodologia consistirá conforme
descrito abaixo:
Purificação da peçonha por gel-filtração em gel de Sephadex G-50 e estimativa do conteúdo
protéico:
O fracionamento da peçonha liofilizada de anêmona será realizado através de uma coluna
(1,9cm X 131cm) empacotada com gel Sephadex G-50 (Pharmacia LKB- Biotechnology, Uppsala,
Sweden), para se obter as frações neurotóxicas e hemolíticas já detectadas neste tipo de
fracionamento. A peçonha previamente centrifugada e liofilizada (cerca de 1-2g de material seco;
200mg de proteína) será dissolvida em 10-20mL de tampão acetato de amônio 0,1M, pH 7,0 e
aplicada ao topo da coluna, equilibrada e eluída por gravidade com o mesmo tampão. Cerca de 60
frações de 10mL cada uma foram coletadas, agrupadas e liofilizadas.
A absorbância de cada fração, uma medida relativa da quantidade de proteína contida em
cada uma, será registrada diretamente na saída da coluna, através da passagem do efluente por um
detector de UV (Spectra/ ChromTM Flow Thru UV Monitor com unidade óptica de 280nm e UV
Monitor Controller, Spectrum, Austria) antes de entrar no coletor de frações. A informação, captada
pelo detector, é integrada e registrada permanentemente em papel (Spectra/ Chrom TM 1 Channel
Recorder, Spectrum, Áustria). O registro direto na saída da coluna permite o acompanhamento
constante do processo de filtração em gel, desta maneira, otimizando o processo de coleta.
Para estimar-se a quantidade de proteína presente na peçonha e nas frações obtidas, durante
esta etapa e nas subseqüentes, será empregado um “kit” que se baseia no método de dosagem do
ácido bicinconínico (BCA) seguindo-se o protocolo do manual do fabricante (Pierce, Rockford,
USA). Utiliza-se albumina sérica bovina como padrão.
Purificação por cromatografia de fase reversa (RP-HPLC) da fração neurotóxica (FR III):
Esta etapa será realizada no Laboratório Especial de Espectrometria de Massa (LEEM) do
Centro de Toxinologia Aplicada (CAT/CEPID- FAPESP) do Instituto Butantan.
O conjunto da fração neurotóxica (FR III) obtida de cromatografias por filtração em gel vai ser
ressuspendido em água Milli-Q (Millipore Inc.) e injetado em um sistema de purificação Shimadzu
de HPLC constituído por um detector UV-VIS SPD-10A VP, bombas LC-10AD VP e um sistema
controlador SCL-10A VP (Shimadzu Corp., Japan). As amostras são então cromatografadas em uma
coluna C-18 de fase reversa ODS (4.6 x 150 mm, 5µm; Hi-Q™) com um gradiente linear de 10 a
41

2o curso de inverno – tópicos em fisiologia comparativa
60% do solvente “B” composto de acetonitrila + 0,1% de ácido trifluoroacético (CH3CN / 0,1%TFA)
com fluxo de 1,0 mL/min durante 40min, através de monitoramento em UV 214 nm. O solvente “A”
é composto de 0,1% de TFA em água Milli-Q.
Todos os picos obtidos são coletados manualmente e individualmente ou agrupados em frações
para serem posteriormente submetidos a espectrometria de massa e ensaios biológicos.
Monitoramento de atividade neurotóxica das amostras em nervo sensorial de crustáceo.
Técnica de “sucrose-gap”:
A preparação será realizada utilizando-se nervos sensoriais de crustáceos decápodos braquiúros
da espécie Callinectes danae (siri azul), coletados no canal de São Sebastião. Testaremos o efeito de
neurotoxinas isoladas de peçonhas de anêmonas sobre o potencial de ação axonal de nervo de siri.
O procedimento consiste na separação do 2o ou 3o par de patas provocando-se autotomia por
compressão do artículo proximal (base-ísquio). Como o nervo ocupa uma posição aproximadamente
central no pereiópodo, o mesmo é exposto até o dáctilo, removendo-se um a um os artículos por
secção das membranas artrodiais, apódemas musculares e separação dos côndilos articulares.
Técnica de “sucrose-gap” - Essa técnica consiste no isolamento elétrico de uma área
superficial do nervo, na região entre os eletrodos de registro, através de lavagens sucessivas com
uma solução de sacarose 1M. São feitas 10 lavagens com sacarose a fim de remover íons da
superfície do nervo, deixando a área eletricamente isolada, impedindo a passagem de corrente na
superfície do nervo. O potencial de membrana pode ser medido de uma câmara contendo KCl para
outra com salina. Assim, eletrodos antes e após a barreira de sacarose registram tanto a voltagem
produzida pela corrente iônica que passa através da membrana e no interior das fibras nervosas
durante o potencial de ação, como o potencial de repouso. A câmara de acrílico que é utilizada no
experimento para estimulação e registro dos potenciais de ação contém 8 sulcos escavados, unidos
centralmente por um sulco onde se encontra apoiado o nervo e, para isolar um sulco do outro
adiciona-se vaselina. Os eletrodos de estimulação se encontram nos sulcos 1 (positivo) e 2
(negativo), enquanto que os de registro nos sulcos 5 e 8, separados por dois sulcos contendo sacarose
1M para isolamento. Os cinco primeiros sulcos são preenchidos com solução fisiológica de C. danae
e o último com solução de KCl isosmótica (0,46M). Para amplificação dos sinais obtidos utilizamos
um pré-amplificador modelo CP511AC (Grass Instruments Co., Warwick, USA). Os dados serão
adquiridos através de um microcomputador tipo PC e processados através do programa WinWCP
(Strathclyde Electrophysiology Software, University of Strathclyde, Glasgow, UK
http://www.strath.ac.uk/Departments/PhysPharm/ses.htm).
42

2o curso de inverno – tópicos em fisiologia comparativa
EVOLUÇÃO DO SISTEMA NERVOSO
Adriano Alonso
Rodrigo Pavão
Dado que todos os organismos compartilham um ancestral comum, porque eles diferem tanto
em relação a suas capacidades de sentir, lembrar e responder ao mundo que o cerca? Como
surgiram as habilidades para pensar e sentir? De que maneira diferimos de outros animais em
relação a essas capacidades? As resposta para essas perguntas dependem crucialmente do
entendimento de como o sistema nervoso evoluiu.
A evolução estaria ocorrendo de forma natural e não determinada, pois não sabemos qual será
a característica que acrescentará valor adaptativo aos indivíduos, ou seja, quais características serão
filogeneticamente conservadas. Podemos comparar a evolução a gotas sucessivas de água que
percorrem caminhos diferentes ao serem jogadas em uma rocha – não se pode prever qual caminho
será percorrido nem onde irão cair, devido à mudanças na umidade, vento etc. Cada caminho
diferente, nessa analogia, seria uma linhagem que pode ter originado os animais atuais, mostrando
toda a diversidade de serem que existem hoje.
Cada organismo tem feições neuroanatômicas distintas que, em última instância, refletem sua
relação com o meio. O estudo evolutivo do sistema nervoso é feito essencialmente através de
evidências indiretas. Isso porque o sistema nervoso não é fossilizável e o comportamento de seres
extintos pode apenas ser inferido. As características do sistema nervoso desses seres, como a
conformação e complexidade dos circuitos e conexões sinápticas são inacessíveis; o que é
observável é o volume da caixa craniana e as impressões nela gravadas. Assim, o estudo do sitema
nervoso e do comportamento baseiam-se na premissa de que capacidades funcionais de sistemas
filogeneticamente mais antigos são refletidas em animais originários dessas linhagens primitivas.
Vale a pena lembrar que essa concepção sobre animais primitivos e derivados, que parece
estar intimamente relacionada com a capacidade de processar informação e de agir favoravelmente
para benefício individual, não pode ser diretamente interpretada como superioridade. Um pequeno
sistema nervoso pode ser mais vantajoso do que uma grande, dependendo das circunstâncias
envolvidas. Um encéfalo grande e capaz de lidar com muita informação tem gasto energético maior,
ampla necessidade de oxigênio. Limitações a esses fatores podem beneficiar animais com encéfalos
pequenos e mais econômicos.
Dois princípios nortearam a configuração dos sistemas nervosos dos grandes grupos de
animais: (1) a capacidade de integrar mais informação sensorial e motora e (2) o sistema ter
43

2o curso de inverno – tópicos em fisiologia comparativa
dimensões reduzidas, com menor número de neurônios envolvidos. O resultado da atuação conjunta
desses fatores resultou em sistemas eficientes, capazes de processar informações diversas e de gerar
comportamentos complexos.
Neste módulo discutiremos a evolução do sistema nervoso através de uma abordagem
multidisciplinar envolvendo dinâmica dos genes reguladores do desenvolvimento, história
geológica da Terra, neuroanatomia e ecologia comportamental. As capacidades sensoriais, motoras
e cognitivas de animais de diferentes níveis da escala filogenética e as estruturas neuroanatômicas
relacionadas serão apresentadas conjuntamente. Uma vez descritos e comparados os sistemas
nervosos desses diferentes grupos, serão apresentadas as teorias mais consistentes sobre seus
padrões evolutivos.
Bibliografia
Allman J. M. (1999). Evolving Brains. Scientific American Library
Anderson P A V (1989). Evolution of the First Nervous Systems. Plenum Press.
Bullock T H (1977). Introduction to Nervous Systems. W H Freeman and Company
Dethier V G & Stellar E (1973). Comportamento Animal – textos básicos de biologia moderna.
Editora Edgard Clucher Ltda, Editora da Universidade de São Paulo.
Maturana H R & Varela F J (1988). The Tree of Knowledge: The Biological Roots of Human
Understanding. New Science Library Shambhala.
Sarnat, B H & Netsky M G (1981). Evolution of Nervous System. Oxford University Press.
Sites
http://www.ib.usp.br/~rpavao
44

2o curso de inverno – tópicos em fisiologia comparativa
MEMÓRIA EM AVES
Sylvia Maria Matsuda
O que é memória? Segundo Tulving (1995, p. 751) memória no sistema biológico sempre
requer aprendizagem (aquisição da informação) e aprendizagem implica em retenção (memória)
desta informação. Memória seria então a retenção de informações adquiridas. Mas como verificar se
um organismo realmente reteve dada informação?
Segundo Xavier (1993, p. 62) memória pode ser definida como a capacidade do organismo de
alterar o seu comportamento em função de experiências anteriores, uma vez que, dada uma
informação, é possível averiguar através de testes se esta informação fora retida ou não dependendo
do comportamento-resposta do indivíduo. Em geral, testes de memória apresentam, num primeiro
momento, certos estímulos a um indivíduo e depois de passado algum tempo é perguntado
(verbalmente ou não) ao indivíduo sobre tais estímulos.
Em seres humanos, é relativamente fácil verificar se uma dada informação foi guardada ou
não, podemos simplesmente perguntar ou, em habilidades não verbais, podemos realizar testes de
desempenho, se o desempenho melhora a partir de informações anteriores, podemos dizer que
houve aprendizagem e conseqüentemente retenção na memória.
Mas, e em animais não-humanos, como fazer testes que verifiquem sua memória? Seguindo o
mesmo princípio que nos seres humanos, mas como não podemos perguntar, realizamos testes que
analisem seu desempenho. Em geral testes de memória em animais envolvem a manipulação de
alimento (recompensa) e do local de oferecimento deste alimento Em aves, por exemplo, são
comuns testes de memória espacial, que seria a memória sobre a disposição de objetos, alimentos,
outros indivíduos, e da sua própria localização num determinado espaço.
Um teste simples de memória espacial seria oferecer, num primeiro momento, em dois locais
distintos, um recipiente com alimento e outro sem alimento e deixar o animal investigar o local até
achar o alimento e consumí-lo. Num segundo momento, oferecer os dois recipientes novamente,
agora sem alimento em ambos (a fim de evitar pistas olfativas) e verificar para qual se dirigem
primeiramente. Podemos tanto treiná-los a procurar no mesmo recipiente ou no oposto, dependendo
do tipo de pergunta que estamos fazendo.
Testes de memória em animais são importantes na construção de modelos que podem, dentro
de certos limites, ser extrapolados para humanos. Ao contrário dos humanos, testes com animais
permitem a manipulação invasiva, possibilitando, por exemplo, que o experimentador, através de
45

2o curso de inverno – tópicos em fisiologia comparativa
uma lesão em determinada região encefálica, verifique possíveis perdas de habilidade mnemônica e
consiga estabelecer uma relação entre esse dado substrato neural com alguma função de memória.
Ou ainda, testes de memória são muito utilizados para comparar o comportamento de diferentes
espécies ou grupos animais frente a um mesmo problema e, a partir dos resultados, extrapolar qual o
seu papel na ecologia do animal.
Por exemplo, estudos com diferentes espécies de pássaros das famílias Corvidae, Sittidae e
Paridae, mostraram que espécies estocadoras de alimento, ou seja, que coletam e estocam seus
alimentos em diferentes locais para depois serem consumidos, apresentam, no geral, melhor
desempenho e maior taxa de aprendizagem em testes de memória espacial do que espécies não-
estocadoras (ver revisão em Krebs, et al., 1996). Além disso, observou-se que o hipocampo* de aves
estocadoras de alimento é relativamente maior, proporcionalmente a todo o encéfalo e tamanho
corpóreo, em relação às espécies não estocadoras (Clayton, 1998; Healy, 1996; Krebs et al., 1996).
Sugere-se que esse aumento hipocampal em pássaros estocadores evoluiu em resposta a um
aumento na demanda cognitiva que deve acompanhar o comportamento de estocagem (Clayton,
1998) uma vez que, para estocar e reaver seus alimentos esses pássaros contariam com uma
memória e um mapa espacial apurados do ambiente.
Os pombos, por serem aves de fácil reprodução e manuseio e serem biologicamente bem
conhecidos, em geral são utilizados em estudos que envolvem o mapeamento de substratos neurais
e suas funções. Por exemplo, através de experimentos com pombos, hoje é consenso que o
hipocampo das aves é homólogo ao hipocampo dos mamíferos, apesar de diferirem
neuroanatomicamente e fisiologicamente, parecem possuir a mesma origem e função. (Striedter,
2002)
Uma outra família de aves bastante estudada é a dos beija-flores. Por serem aves
extremamente especializadas e possuírem uma das maiores taxas metabólicas dentre os endotermos,
são altamente dependentes da disponibilidade alimentar do meio. Acredita-se, portanto, que uma
memória espacial apurada seja um importante componente para seu sucesso alimentar, uma vez que
estes se beneficiariam se guardassem informações sobre os diferentes locais de plantas com flores e
fossem capazes de retornar no dia seguinte ou na estação seguinte quando voltarão a florescer, ou
até no mesmo dia, quando da reposição do nectar.
*substrato neural consensualmente considerado importante na aquisição e consolidação de
memória de longa duração (Clayton, 1998); o hipocampo seria responsável pela combinação de
estímulos elementares na construção de uma única representação do ambiente (Carlson, 1994) e,
dentre outras funções, a formação hipocampal forneceria para o restante do sistema nervoso um
mapa espacial do ambiente (O’Keefe & Nadel, 1978; Santos, 1999).
46

2o curso de inverno – tópicos em fisiologia comparativa
De fato, observou-se que, beija-flores das espécies Archilocus colubris e Selasphorus rufus,
aprendem locais de bebedouros idênticos, reforçados e não reforçados, em diferentes padrões de
distribuição (Miller & Miller, 1971; Brown & Gass, 1993; Hurly, 1995; Sutherland & Gass, 1995).
Ainda que, S. rufus é capaz de lembrar e evitar locais de bebedouros recentemente visitados e
utilizar sua altura para lembrar de sua localização (Henderson et al., 2001). E estudos envolvendo
diferentes espécies de beija-flores - Calypte anna, A. colubris e S. rufus - demonstraram que eles:
(1) respondem preferencialmente à disposição espacial em detrimento às pistas intrínsecas dos
bebedouros, como cor ou padrão de cores (Collias & Collias, 1968; Hurly & Healy, 1996; Miller &
Miller, 1971; Miller et al., 1985); (2) aprendem a utilizar a cor dos bebedouros para encontrar o
reforço (Brown & Gass, 1993; Collias & Collias, 1968; Miller & Miller, 1971; Miller et al., 1985)
e; (3) aprendem mais rapidamente tarefas espaciais do que tarefas que envolvem pistas visuais
(Brown & Gass, 1993; Miller et al., 1985, Healy & Hurly, 1998). Sugere-se que, na natureza, os
beija-flores se utilizariam tanto de pistas intrínsecas quanto de pistas espaciais durante seu
forrageamento. Pistas intrínsecas, como cor ou padrões de cores das flores, para a procura e
generalização de novas fontes de alimentares e, se utilizariam de pistas espaciais na discriminação
entre flores recentemente visitadas e para visitas subseqüentes depois de passado seu tempo de
renovação de néctar (Brown & Gass, 1993; Hurly and Healy, 1996; Miller et al.,1985).
Pudemos verificar que experimentos de memória em animais e, mais especificamente em
aves, são importantes tanto para a comparação comportamental entre as diferentes espécies e táxons
quanto para a criação e consolidação de modelos de memória que ajudam no entendimento dos
nossos próprios mecanismos de formação e evolução da memória.
Bibliografia
Brown, G. S. & Gass, C. L. (1993) Spatial association learning by hummingbirds. Animal
Behaviour 46: 487-497.
Carlson, N. R. (1994) Physiology of Behavior. 5ed., 704p., Ally and Bacon, Boston, USA.
Clayton, N. S. (1998) Memory and the hippocampus in food-storing birds: a comparative approach.
Neuropharmacology 37:441-452.
Collias, N. E. & Collias, E. C. (1968) Anna’s hummingbirds trained to select different colors in
feeding. The Condor 70: 273-274.
Healy, S. D. (1996) Ecological specialization in the avian brain. In Neuroethological Studies of
Cognitive and Perceptual Processes, eds. Moss, C. F. & Shettleworth. S. J. USA: Westview
Press, Inc., a Division of HarperCollins Publishers, Inc. 330p.
47

2o curso de inverno – tópicos em fisiologia comparativa
Healy, S. D. & Hurly, T. A. (1998) Rufous hummingbirds’ (Selasphorus rufus) memory for
flowers: patterns or actual spatial locations? Journal of Experimental Psychology: Animal
Behavior Processes 24 (4): 396-404.
Henderson, J., Hurly, T. A. & Healy, S. D. (2001) Rufous hummingbirds’ memory for flower
location. Animal Behaviour 61: 981-986.
Hurly, T. A. (1995) Spatial memory in rufous hummingbirds: memory for rewarded and non-
rewarded sites. Animal Behaviour 51(1): 177-183.
Hurly, T. A. & Healy, S. D. (1996) Memory for flowers in rufous hummingbird: location or visual
cues? Animal Behaviour 51: 1149-1157.
Krebs, J. H., Clayton, N. S., Healy, S. D., Cristol, D. A., Patel, S. N. & Jolliffe, A. R. (1996) The
ecology of the avian brain: food-storing memory and the hippocampus. IBIS 138: 34-46.
Miller, R. S. & Miller, R. E. (1971) Feeding activity and color preference of Ruby-throated
hummingbirds. Condor 73: 309-313.
Miller, R. S., Tamm, S., Sutherland, G. D. & Gass, C. L. (1985) Cues for orienting in hummingbird
foraging: color and position. Canadian Journal of Zoology, 63: 18-21.
O’Keefe, J & Nadel, L. (1978) The hippocampus as a cognitive map. Oxford: Clarendon Press.
Santos, A. M. G. (1999) Aprendizagem e memória no Labirinto Aquático de Morris. In Técnicas
para o Estudo do sistema Nervoso. Org. Xavier, G. F. Ed. Plêiade, São Paulo.
Sutherland, G. D. & Gass, C. L. (1995) Learning and remembering of spatial patterns by
hummingbirds. Animal Behaviour 50:1237-1286.
Striedter, G. F. (2002) Brain homology and function: an uneasy alliance. Brain Research Bulletin
57(3/4): 239-242.
Tulving, E. (1983) Ecphoric processes in episodic memory. Philosophical Transactions of the Royal
Society of London Series B- Biological Sciences 302(1110): 361-371.
Xavier, G. F. (1993) Modularidade da memória e o sistema nervoso. Psicologia USP 4:61-115.
48

2o curso de inverno – tópicos em fisiologia comparativa
MECANISMOS CENTRAIS DO CONTROLE CARDIOVASCULAR
Jessica Ruivo Maximino
1. INTRODUÇÃO
A perfusão tecidual é garantida pela manutenção da força motriz da circulação em níveis
adequados e razoavelmente constantes ao longo de toda a vida dos indivíduos, estejam eles em
repouso ou desenvolvendo diferentes atividades comportamentais. Os níveis de pressão arterial
(PA), gerados pela atividade cardíaca e vascular são controlados por diversos sistemas inter-
relacionados que realizam funções específicas.
Assim, conhece-se alguns mecanismos de ajuste da PA:
- Mecanismo de controle a curto-prazo da PA (Mecanismos neuro-humorais)
- Mecanismos de controle a longo-prazo da PA (Mecanismos de fluidos
corporais)
2. CONTROLE DA PRESSÃO ARTERIAL PELO SISTEMA NERVOSO CENTRAL
O Sistema Nervoso Central (SNC) tem um papel importante na regulação do sistema
cardiovascular, por controlar tanto a atividade do sistema nervoso autonômico quanto a liberação
de fatores hormonais circulantes. O SNC modifica agudamente a PA e os batimentos cardíacos,
facilitando a homeostase e as respostas apropriadas ao meio ambiente (WYSS et al., 1990).
Assim, tem um papel fundamental na regulação a curto-prazo da PA.
O controle a curto-prazo da PA é realizado particularmente pelo reflexo barorreceptor, o
qual é iniciado em terminações nervosas localizadas no seio carotídeo e no arco aórtico, as quais
captam alterações no estiramento da parede arterial (KUMADA et al., 1990).
Sabemos que o SNC contribui diretamente em algumas formas de hipertensão (WYSS et al.,
1990) e algumas áreas encefálicas parecem ter uma grande importância na regulação
cardiovascular.
O núcleo do trato solitário (NTS) está localizado na porção dorsal da medula oblonga está
envolvido na recepção e integração de múltiplos processos viscerosensoriais, incluindo o
49

2o curso de inverno – tópicos em fisiologia comparativa
controle cardiovascular, respiratório, gustatório, hepático e renal (LAWRENCE & JARROT, 1996).
Ele é o principal núcleo de integração dos impulsos provenientes dos barorreceptores localizados
no seio carotídeo e no arco aórtico, dos quimiorreceptores dos corpos carotídeos e das aferências
provenientes do coração, pulmão e de outras vísceras (MIURA & REIS, 1971; LIPSKI et al., 1975 ;
CIRIELLO & CALARESU, 1981).
O NTS influencia o controle cardiovascular através de suas projeções para núcleos
medulares (OTAKE, 1993), área postrema (SAPER et al.,1983), locus coeruleus (LC) e núcleos
hipotalâmicos, como o núcleo paraventricular do hipotálamo (PVN) (SAWCHENKO & SWANSON,
1982).
Assim, além do NTS outros centros bulbares estão envolvidos no controle cardiovascular,
como neurônios da porção caudal (CVL) e da porção rostral (RVL) da medula ventrolateral
(VLM), áreas da formação reticular nesta região do SNC. A CVL recebe projeções diretas do
NTS que, por sua vez, envia projeções inibitórias à RVL. A RVL envia projeções para os
neurônios pré-ganglionares simpáticos na coluna intermédio lateral da medula espinhal
exercendo dessa forma, efeito modulatório sobre o tono do sistema nervoso simpático (revisado
por DAMPNEY, 1994). Essas projeções constituem o barorreflexo, assim, a circuitaria do
barorreflexo é ativada em decorrência da variação da PA.
O NTS é rico em variedade e quantidade de neurotransmissores, sendo que a participação
destes no controle neural da PA vem sendo muito discutida (VAN GIERSBERGEN et al., 1992;
LAWRENCE & JARROT, 1996). Mais de trinta neurotransmissores bem como seus respectivos
receptores são descritos no NTS, sendo que o papel funcional de cada um deles ainda não foi
adequadamente demonstrado. Além disso, é possível que interações entre sistemas de
neurotransmissão neste núcleo ampliem a capacidade de modulação das respostas que acontecem
após determinado estímulo pressórico.
O LC, localizado na ponte também tem participação relevante nas respostas reflexas
autonômicas e neuroendócrinas em decorrência de alterações da PA. O LC envia projeções para
diversas áreas do encéfalo e medula espinhal (FOOTE et al., 1983), recebendo aferências
principalmente da medula oblonga (ASTON-JONES et al., 1986).
Outro núcleo importante no controle da PA é o PVN, localizado bilateralmente ao terceiro
ventrículo, o PVN recebe e envia projeções diretas para o NTS podendo modular o processamento
bulbar do controle cardiovascular (SAWCHENKO & SWANSON, 1982). A estimulação deste núcleo
pode causar tanto efeito pressor quanto depressor. O efeito difere para cada órgão alvo. O fato de
existir projeções diretas do PVN para a coluna intermédio lateral e do NTS para o PVN, pode-se
relacionar esse circuito com o barorreflexo (SAWCHENKO & SWANSON, 1982).
50

2o curso de inverno – tópicos em fisiologia comparativa
No SNC diversos mediadores químicos participam do controle/modulação da PA, tais
como as catecolaminas, o neuropeptídeo Y (NPY), a angiotensina II (Ang II), o gaba, o
glutamato, a vasopressina entre outros.
Referências Bibliográficas:
Aston-Jones, G.; Ennis, M.; Pieribone, V.A.; Nickell, W.T. & Shipley, M.T. (1986) The brain
nucleus locus coeruleus: restricted afferent control of a broad efferent network. Science ,
7;234 (4777):734-737.
Ciriello, J. & Calaresu, F.R. (1981) Projections from buffer nerves to the nucleus of the solitary
tract: an anatomical and electrophysiological study in the cat. J. Auton. Nerv. Syst., 3(2-4):
299-310.
Dampney, R.A.L. (1994) Functional organization of central pathways regulating the cardiovascular
system. Physiol. Rev., 74(2): 323-364.
Foote, S.L.; Bloom, F.E. & Aston-Jones,G. (1983) Nucleus locus coeruleus: new evidence of
anatomical and physiological specificity. Physiol. Rev.,63(3): 844-914.
Kumada, M.; Terui, N. & Kuwaki, T.(1990) Arterial baroreceptor reflex: its central and peripheral
neural mechanisms. Prog. Neurobiol., 35(5):331-61.
Lawrence, A.J. & Jarrott, B. (1996) Neurochemical modulation of cardiovascular control in the
nucleus tractus solitarius. Prog. Neurobiol., 48(1): 21-53.
Lipski, J., McAllen, R.M. & Spyer, K.M. (1975) The sinus nerve and baroreceptor input to the
medulla of the cat. J. Physiol., 251(1): 61-78.
Miura, M. & Reis, D.J. (1971) The paramedian reticular nucleus: a site of inhibitory interaction
between projections from fastigial nucleus and carotid sinus nerve acting on blood pressure. J.
Physiol., 216(2): 441-460.
Otake, K.; Nakamura, Y. & Ezure, K. (1993) Projections from the commissural subnucleus of the
solitary tract onto catecholamine cell groups of the ventrolateral medulla. Neurosci. Lett.,
12;149(2):213-216.
Saper, C.B.; Reis, D.J. & Joh, T. (1983) Medullary catecholamine inputs to the anteroventral third
ventricular cardiovascular regulatory region in the rat. Neurosci. Lett.,11;42(3):285-291.
Sawchenko, P.E. & Swanson, L.W. (1982) Immunohistochemical identification of neurons in the
paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in
the rat. J. Comp. Neurol., 1;205(3):260-272.
51

2o curso de inverno – tópicos em fisiologia comparativa
van Giersbergen, P.L.; Palkovits, M. & De Jong, W. (1992) Involvement of neurotransmitters in the
nucleus tractus solitarii in cardiovascular regulation. Physiol. Rev., 72(3):789-824.
Wyss, J.M.; Oparil, S. & Chen, Yui-Chen. (1990) The role of the central nervous system in
hypertension. In: Hypertension: Pathophysiology, Diagnosis and Management, edited by J. H.
Laragh and B.M. Brenner. New York: Raven, p.679-701.
Site interessante:
www.pubmed.com
52

2o curso de inverno – tópicos em fisiologia comparativa
COMPORTAMENTO E FISIOLOGIA DE FORMIGAS
Pedro Leite Ribeiro
As sociedades dos insetos são conhecidas por se constituírem de indivíduos especializados em
diferentes tarefas. Em certas espécies de formigas, esta especialização está relacionada com o
aparecimento de diferenças surpreendentes no tamanho entre indivíduos de uma mesma colônia,
geralmente associadas a tarefas diferentes (Bonabeu, 1997). Temos, por exemplo, em colônias do
gênero Atta formigas cuja largura da cápsula cefálica pode variar de 0,8mm até 3,0mm.
Simplificando, podemos subdividir estes diferentes tamanhos em quatro grupos, de acordo com suas
funções. Assim, temos as jardineiras e “babás” que são as menores operárias da colônia e têm a
função de cuidar das hifas e das formas imaturas. Já as generalistas, de cápsula cefálica com
aproximadamente 1,4mm são responsáveis por vários tipos de atividades dentro do ninho, como a
preparação de vegetais, que deve ser feita antes da incorporação dos mesmos à esponja, descarte de
lixo e reconstrução das esponjas de fungo. Temos ainda a não menos importante casta das
forrageadoras com 2,2mm de cápsula cefálica, essas formigas exploram o meio externo ao ninho e
são responsáveis pela coleta de vegetação. Por último, temos as defensoras ou soldados, com
cápsula cefálica de 3,0mm, como o próprio nome sugere são elas as responsáveis pela defesa do
ninho contra invasores, principalmente de outras formigas. (Adam e Ratnieks, 2001; Roces e
Holldobler, 1994; Andrade et al., 2002; Carvalho, 1972; Wilson, 1980; Wilson, 1971). A divisão de
tarefas não é completamente rígida, sendo que a colaboração entre os indivíduos de diferentes
castas propicia o surgimento de padrões de comportamentos característicos de um
“superorganismo”, favorecendo a colônia de uma forma global (Hölldobler e Wilson 1984). A
manutenção do equilíbrio homeostático de uma colônia requer uma rápida percepção de fatores
desestabilizadores externos ou internos e uma comunicação que possibilite a reorganizaçao na
direção do equilíbrio.
Além da flexibilidade na troca de tarefas entre as diferentes castas de formigas operárias
variações do meio ambiente promovem comportamentos que conduzem ao reequilíbrio do
microambiente da colônia. Kleineidam e Roces em 2000, observaram em ninhos de formigas
cortadeiras Atta vollenveideri diferentes comportamentos em função da idade da colônia. Os ninhos
de colônias grandes, maduras, asseguram um bom micro-clima para o crescimento do fungo. Essas
colônias constroem montes de terra ao redor das entradas do ninho cuja arquitetura promove a
ventilação preservando a concentração de CO2 em baixos níveis. Além disso, esses montes de terra
53

2o curso de inverno – tópicos em fisiologia comparativa
impedem a entrada da enxurrada nos olheiros. As colônias jovens, pequenas e que ainda não
construíram montículos protetores se vêem obrigadas a fecharem todas as saídas do ninho para
protegê-lo do excesso de chuva. Quando fecham as entradas a concentração de CO2 aumenta
rapidamente, a taxa de respiração da colônia reduz-se, a respiração do fungo simbiótico também é
reduzida e o crescimento da colônia fica comprometido. Contudo, através do fechamento, as
operárias impedem a inundação e garantem a continuidade da colônia.
Freqüentemente a homeostase de uma colônia envolve processos complexos de
comportamentos onde aspectos individuais e sociais interagem para suprir, com maiores benefícios,
as necessidades energéticas da colônia (Roces, 2002). Em colônias de Atta cephalotes, a primeira
formiga forrageadora que encontra uma nova fonte de alimento, corta um fragmento de folha bem
menor do que cortaria numa situação de forrageamento bem estabelecida. Dessa forma, essa
primeira formiga diminui o tempo de corte e chega mais rapidamente ao ninho onde passará as
informações da fonte para suas irmãs. Assim, a coleta total para a colônia aumenta através do
recrutamento embora a primeira forrageadora tenha prejudicado sua performance. Num primeiro
momento, o recrutamento torna-se mais importante que a atividade individual de corte. Essa
capacidade individual de distinguir as diferentes situações de forrageamento possibilita maior
rapidez no reequilibrio energético da colônia (Roces, 1993). A velocidade com que as operárias
modificam seu comportamento em resposta aos estímulos externos varia de acordo com o tipo de
comportamento e o contexto. O grau de flexibilidade pode variar com o tamanho e a idade da
colônia, as operárias de colônias jovens parecem mais versáteis, mais flexíveis em suas tarefas, mas
a colônia como um todo é menos homeostática. Uma vez que a proporção entre as castas é muito
mais frágil em colônias jovens, e os mecanismos de suporte, como a mudança de tarefas não é
100% eficiente (Lenoir, 1979a; Gordon,1987).
Todo processo de homeostase de uma colônia que envolve novas organizações e mudanças de
comportamentos, individuais ou sociais, depende de um sofisticado mecanismo de comunicação
química. As intercomunicações ocorrem através de emanações de substâncias químicas
(feromônios) exaladas de diferentes partes do corpo das formigas e percebíveis por suas irmãns,
através das antenas que são órgãos de percepção química. Esses avisos químicos têm, de acordo
com as substâncias desprendidas e circunstâncias do momento, funções diferentes como: alarme de
invasão, chamados para os cuidados com a prole, recrutamento para a busca de alimentos e muitas
outras ainda não bem conhecidas (Hölldobler e Wilson, 1990). Além de se comunicar através de
feromônios as formigas são capazes de perceber diversas variações ambientais entre elas as de
temperatura e umidade. Uma vez identificadas essas variações as operárias podem modificar o seu
comportamento. Desta maneira, a colônia, aparentemente, reconhece a existência de um problema e
passa a se comportar de forma a tentar corrigi-lo. (Roces e Kleineidam, 2000).
54

2o curso de inverno – tópicos em fisiologia comparativa
Nesta aula nos basearemos na descrição e discussão de experimentos realizados em nosso
laboratório. Estes experimentos mostram a plasticidade comportamental que formigas Atta sexdens
rubropilosa podem apresentar em diversas situações.
Referências:
Adam, G.H. and Ratnieks F.L.W. 2001. Task partitioning, division of labour and nest
compartmentalisation collectively isolate hazardous waste in the leafcutting ant Atta
cephalotes. Behav. Ecol. Sociobiol. 49: 387-392.
Andrade, A.P.P., L.C. Forti, A.A. Moreira, M.A.C. Boareto, V.M. Ramos, C.A.O. Matos.
Sociobiology vol. 40, nº 2, 2002.
Autuori, M. 1941. Contribuição para o conhecimento da saúva. (Atta ssp – Hymenoptera
Formicidae) I- Evolução do sauveiro Atta sexdens rubropilosa Forel, 1908.
Autuori, M. 1942. Contribuição para o conhecimento da saúva (Atta ssp. Hymenoptera).
(Formicidae) III- Excavação de um sauveiro (Atta sexdens rubropilosa. Forel, 1908).Arq. Inst.
Biol. 13: 137-148.
Autuori, M. 1947. Contribuição para o conhecimento da saúva (Atta ssp. –
HymenopteraFormicidae) IV- O sauveiro depois da 1º revoada ( Atta sexdens rubropilosa,
Forel, 1988). Arq. Inst. Biol. 18: 39-70.
Bergmeier, T. e al. 1948. Combate à saúva. Biologia- Formicidas- Extintores- Cuiabanas Ed. Chac.
e Quint. S. Paulo, nº 42, 35.
Bonabeau E., Theraulaz G., Deneubourg J.L., Aron, S., Camazine S., 1997. Self-organization in
social insects. Trends in Ecology and Evolution. Vol 12, nº 5, 188-193.
Carvalho, A.M.A. 1972. Alguns dados sobre a divisão do trabalho entre operárias de Att sexdens
rubropilosa, Forel, 1908 em colônias iniciais mantidas em laboratório. Tese de doutorado,
Instituto de Psicologia, Universidade de São Paulo, São Paulo.
Carvalho, J. H. 1935. Ligeiras notas sobre o combate à saúva. Min. Agric., Dept. Nac. da Prod.
Vegetal Serv. Defesa Sanitária Vegetal. Publicação nº 3 18 pp.
Cherret, J.M. 1968. The foraging behaviour of Atta Cephalotes ( l.). ( HymenopteraFormicidae). I.
Foraging pattern and plant species attacked in tropical rain forest.
Journal of Animal Ecology, 37: 387-402.
Detrain, C and J.M. Pasteels 1992. Caste polyethism and collective defense in the ant,
Pheidole pallidula: the outcome of quantitative differences in recruitment. Behav. Ecol.
Sociobiol. 29:405-412.
Gordon, D.M. 1999. Ants at work: How an Insect Society is Organized. The Free Press,
55

2o curso de inverno – tópicos em fisiologia comparativa
Simon and Schuster, de Nova York, USA.
Haines, B. L.1978. Element and energy flows through colonies of the leaf-cutting ant,Atta
colombica in Panama. Biotropica, 10 (4): 270-277.
Hölldobler, B. e E.O. Wilson 1990. The Ants. The Belknap Press of Harvard University Press,
Cambridge, Massachusetts 732 pp.
Hölldobler, B. and F.O. Wilson 1994. Journey to the ants. Library of Congress Cataloging- in-
Publication data. USA. 228 pp.
Kerr, W. 1961. Acasalamento de rainhas com vários machos em duas espécies da tribo Attini (
Hymenoptera, Formicoidea). Rev. Brasil. Biol. 21(1): 45-48
Mariconi, F.A.M. 1970. As saúvas. Editora Agronómica Ceres, São Paulo, Brasil. 167 pp.
Mariconi, F.A.M., E. Berti Filho, L.R. Fontes.1996. Anais do Simpósio sobre Formigas
Cortadeiras dos Países do Mercosul. Fundação de Estudos Agrários Luis de Queiroz-
Piracicaba, São Paulo, Brasil. 139 pp.
Mueller U.G. e W.T. Wcislo ; 1998. Nesting biology of the fungus-growing ant Cyphomyrmex
longiscapus Weber (Attini, formicidae).
Oliveira Filho, M.L. 1934. Combate a saúva. Bol. Agric. 35: 541-610.
Ribeiro, P.L. e C.A. Navas 2004. A study on the infuence of relative humidity in the selection of a
garbage disposal location in Atta sexdens rubropilosa, Forel, 1908 (Hymenoptera-
Formicidae) (submetido).
Roces F. e Nuñez J. 1995. Thermal Sensitivity During Brood Care in Workers of Two
Camponotus Ant Species: Circadian Variation and Its Ecological Correlates. J. Insect.
Physiol. Vol 41, nº 8, pp. 659-669. 1995.
Roces, F. and Hölldobler B. 1994. Leaf density and a trade-off between load-size selection and
recruitment behavior in the ant Atta cephalotes. Oecologia 97: 1-8.
Roces, F e C. Kleineidam, 2000. Humidity preference for fungus culturing by workers of the leaf-
cutting ant Atta sexdens rubropilosa . Insectes soc. 47: 348-350.
Stahel, G. and D.C. Geijskes 1940. Observations about temperature moisture in Atta-nests. Rev.
Entomol. 11: 766-775.
Wilson, E.O.1971. The Insects Societies The Belknap of Harvard University Press.
Wilson, E.O. 1980a. Caste and Division of Labour in Leaf-cutter Ants (Hymenoptera: Formicidae:
Atta) Behavioural Ecology and Sociobiology. vol 7, 143-156.
56

2o curso de inverno – tópicos em fisiologia comparativa
ECOFISIOLOGIA DE LAGARTOS
Renata Brandt Nunes
A ecofisiologia pode ser interpretada como o estudo de como os organismos funcionam
e respondem a mudanças em seus ambientes naturais. Neste contexto, a aula tratará principalmente
das implicações fisiológicas das mudanças de temperatura nos Squamata, com ênfase nos lagartos,
pois é bastante profunda a influência deste fator na vida destes animais. Será importante recordar a
terminologia relacionada à regulação da temperatura, como os conceitos de endotermo e ectotermo
assim como pecilotermo e homeotermo; e ainda heliotermos, tigmotermos; a diferença entre ser
termorregulador e termoconformador; o que é a temperatura preferencial e conceitos relacionados
aos limites de temperatura de atividade.
A aula em si será baseada na discussão sobre os mecanismos morfológicos e fisiológicos
da termorregulação e a relação com aspectos da história natural dos lagartos no contexto metabólico
e dependente de temperatura, como distribuição e seleção de microhabitat; atividade diária e
sazonal; comportamento; alimentação, digestão e dieta; reprodução; infecção e balanço hídrico.
Sugestões de Leitura
Bennet AF (1980). The thermal dependence of lizard behaviour. Animal Behaviour, 28: 752-762.
Deeming DC & Fergusson MWJ (1991) Physiological effects of incubation temperature on
embryonic development in reptiles and birds. In Egg incubation: its effects on embryonic
development in birds and reptiles, eds D. C. Deeming & M. W. J. Fergusson, 147-171.
Cambridge: Cambridge University Press.
Espinoza RE & Tracy CR (1997) Thermal biology, metabolism and hibernation. In: The biology,
husbandry and health care of reptiles. Ackerman LJ ed. Volume 1, Biology of Reptiles.
T.F.H. Publication,, Neptune City, NJ.
Huey RB (1982). Temperature, Physiology, and the Ecology of Reptiles. In Biology of the Reptilia
vol 12. Gans C. et al. eds, Academic Press.
McNab BK (2002). The Physiological Ecology of Vertebrates: a view from energetics. Cornell
University Press.
57

2o curso de inverno – tópicos em fisiologia comparativa
Packard GC (1991). The physiological and ecological importance of water to embryos of oviparous
reptiles. In Egg incubation: its effects on embryonic development in birds and reptiles, eds D.
C. Deeming & M. W. J. Fergusson, 213-228. Cambridge: Cambridge University Press
Pough FH (1980). The advantages of ectothermy for tetrapods. American Naturalist 115:92-112.
Pough FH & Gans C (1982). The vocabulary of reptilian thermoregulation. In Biology of the
Reptilia vol 12. Gans C. et al. eds, Academic Press.
Pough HF, Andrews RM, Cadle JE, Crump ML, Savitzky AH, Wells KD (1998). Herpetology. New
Jersey: Prentice – Hall, Inc.
Links Interessantes
Páginas de pesquisadores, com informações sobre as linhas de pesquisa e links para outras páginas
de herpetologia.
http://www.ib.usp.br/~rbrandt - Minha página pessoal
http://fisio.ib.usp.br/ecofis - Carlos A. Navas
http://rydberg.biology.colostate.edu/faculty/profile.php?name=Packard
http://lamar.colostate.edu/~packard/ – Gary Packard
http://faculty.washington.edu/hueyrb/index.html - Laboratory of Evolutionary Physiology, dirigido
por Raymond B. Huey. Dispõe ainda de conselhos para estudantes de pós-graduação ou
interessados em fazer pós-graduação
http://uts.cc.utexas.edu/~varanus/ - Pianka´s Lab Page. Página do Laboratório do Eric Pianka.
http://www.omnh.ou.edu/personnel/herpetology/vitt/ - Laurie J. Vitt
http://compphys.bio.uci.edu/bennett/bennett.htm – Albert F. Bennet
http://oeb.indstate.edu/faculty/Angilletta.htm – Michael J. Angilletta Jr.
Cursos de Ecofisiologia na Internet
http://wildlife.wisc.edu/courses/401/ - University of Wisconsin – Madison
http://www.eeb.uconn.edu/courses/eeb296/ - University of Connecticut
58

2o curso de inverno – tópicos em fisiologia comparativa
http://bioweb.wku.edu/faculty/Crawford/526.html – Western Kentucky University
Informações gerais sobre lagartos
http://tolweb.org/tree?group=Diapsida&contgroup=Amniota – Tree of Life, com cladogramas e
discussões sobre filogenia de todos os grupos de seres vivos. Este link direciona para o grupo dos
diapsida.
http://www.ucmp.berkeley.edu/diapsids/diapsids.html – UCPM Berkeley, cladogramas e todo tipo
de informação biológica (incluindo fósseis) sobre a diversidade da vida, este link direciona para o
grupo dos diapsida
59

2o curso de inverno – tópicos em fisiologia comparativa
METABOLISMO ENERGÉTICO DA VOCALIZAÇÃO EM ANFÍBIOS ANUROS DO
GÊNERO SCINAX: “O CANTO DA PERERECA”
Adne Abbud Righi
A comunicação vocal em anfíbios anuros é de grande importância dado que torna possível o
reconhecimento entre indivíduos da mesma espécie, além de ter papel central na atração de fêmeas,
na competição entre machos e na defesa de territórios. A atividade vocal é um exercício realizado
através de contrações dos músculos do tronco, cujas características principais são retículo
sarcoplasmático bem desenvolvido, grande número de mitocôndrias por unidade de volume e alta
densidade de capilares sangüíneos nas espécies mais ativas, assim indicando a existência de uma
ampla demanda de oxigênio durante o exercício. De acordo com tais características, muitos
pesquisadores acreditam que a intensa atividade vocal em anfíbios anuros seja mantida
essencialmente pelo metabolismo aeróbio. Porém, outros autores sugerem que o uso localizado da
glicólise independente de oxigênio pode ter um papel importante na vocalização em espécies cuja
massa muscular envolvida é relativamente pequena. Dessa forma, tal estratégia levaria à rápida
síntese de ATP e lactato nas células destes tecidos, como resultado de um aumento repentino do
fluxo glicolítico.
Com base no exposto anteriormente, o intuito desta aula será fazer uma análise integrada a
respeito das modificações bioquímicas e fisiológicas associadas à manutenção da atividade vocal.
Para tal estudo será utilizado como modelo a espécie Scinax gr. perpusillus, que exibe diferentes
padrões e níveis de atividades.
Sugestões de Leitura:
Bennett, A.F. & Licht, P. (1973) Relative contributions of anaerobic and aerobic energy production
during activity in Amphibia. J. Comp. Physiol. 87:351-360.
Carvalho, J.E. (2004) Correlações entre as capacidades metabólicas energéticas, o ambiente térmico
e a atividade em anfíbios anuros do gênero Scinax (Anura / Hylidae). Tese de Doutorado.
Instituto de Biociências da USP, São Paulo.
Cummings, J.W. (1979) Physiological and biochemical adaptations to training in Rana pipiens. J.
Comp. Physiol. 134: 345-350.
Gatten Jr., R.E. (1985) The uses of anaerobiosis by Amphibians and Reptiles. Amer. Zool. 25: 945-
954.
60

2o curso de inverno – tópicos em fisiologia comparativa
Gleeson, T. T. (1991) Patterns of metabolic recovery from exercise in amphibians and repitailes. J.
Exp. Biol. 160:187-207.
Gleeson, T. T. (1996) Post – exercise lactate metabolism: a comparative review of sites, pathways
and
regulation. Annu. Rev. Physiol. 58: 565-581.
Grafe, T.U. (1996) Energetics of vocalization in the African reed frog (Hyperolius marmoratus).
Comp. Biochem. Physiol. 114A: 235-243.
Grafe, T.U. and Thein, J. (2001) Energetics of calling and metabolic substrate use during prolonged
exercise in the European treefrog Hyla arborea. J. Comp. Physiol. 171B: 69-76.
Oliveira, F. B. (2002) Comportamento Vocal de Scinax gr. perpusillus (Anura, Hylidae):
Correlações Fisiológicas e Ecológicas da Variação Intraespecífica. Dissertação de Mestrado.
Instituto de Biociências da USP, São Paulo.
Pough, F.H. and Gatten Jr., R.E. (1984) The use of anaerobic metabolism by frogs in a breeding
chorus. Comp. Biochem. Physiol. 78A: 337-340.
Pough, F.H.; Magnusson, W.E., Ryan, M.J. Wells, K.D. and Taigen, T.L. (1992) Behavioral
energetics. In: Ferder, M.E. and Burggren, W.M. (eds.) Enviromental Physiology of the
Amphibians. The Univ. Chicago Press. pp. 395-436.
Taigen, T.L. & Pough, F.H. (1985) Metabolic Correlates of Anuran Behaviour. Amer. Zool. 25:
987-
997.
Taigen, T.L., Wells, K.D. & Marsh, R.L. (1985) The enzymatic basis of high metabolic rates in
calling frogs. Physiol. Zool. 58 (6): 719-726.
Wells, K.D. & Taigen, T.L. (1984) Reproductive behaviour and aerobic capacities of American
toads
(Bufo americanus): Is behaviour constrained by physiology? Herpetologica 40: 292-298.
Wells, K.D. (2001) The energetics of calling in frogs. In: Ryan, M.J. (ed.) Anuran Communication.
pp.
45-60. Smithsonian Inst. Press.
61

2o curso de inverno – tópicos em fisiologia comparativa
CONSCIÊNCIA COMO EXPRESSÃO DO FUNCIONAMENTO DO SISTEMA NERVOSO
Adriano Alonso
Pessoas de várias formações diferentes já abordaram o tema consciência, tais como psicólogos
e filósofos. O livro de Marcel e Bisiach (1992) é uma coletânea de artigos que têm algumas dessas
maneiras de se analisar o tema, assim como o volume 8 da revista Psicologia USP (1997).
Uma forma de se abordar seria aquela que trata da consciência como uma entidade mística, a
consciência seria parte da essência de ser humano, temos um corpo que é ligado a uma consciência.
Essa forma seria uma derivação do conceito de alma. Essa abordagem impossibilita um estudo
científico sobre a consciência, já que traz consigo explicações místicas, metafísicas ou teológicas.
Uma segunda abordagem costuma se perguntar sobre a experiência que temos no mundo,
tentam explicar o motivo que essa experiência é dessa forma e, portanto, está ligada aos sentidos.
Traz uma subjetividade que também não é bem-vinda, pois embora os seres humanos percebam o
mundo através dos mesmos sentidos, cada um possui uma experiência única do mundo em que
vivemos. O que eu sinto ao tomar um copo de vinho é influenciado por muitos fatores, pela minha
experiência com vinhos e pelas condições atmosféricas, só para citar 2 exemplos. Essa experiência,
quando vivida por outra pessoa, será diferente da experiência que tive, já que estou em outro corpo e
vivencio outras experiências.
Já uma terceira abordagem, mais recente, assume que consciência seria um fenômeno
biológico como qualquer outro e por isso possa ser estudada, podemos investigar sua função e seu
funcionamento, como fazemos com qualquer outro fenômeno biológico complexo (Gazzaniga,
2002). Para esses autores, a consciência seria fruto da evolução do sistema nervoso. Uma explicação
satisfatória sobre consciência deve conter explicações sobre a continuidade evolutiva. Assim como
deve refletir a organização do sistema nervoso, que é funcionalmente modular, ou seja, cada módulo
é responsável por uma função, como por exemplo, o córtex visual primário . Podemos observar
vários comportamentos que indicam que os vários módulos do sistema nervoso contribuem para uma
experiência única do mundo em que vivemos, como o fenômeno da “visão-às-cegas”, em que
pacientes com lesões no córtex estriado reagem a estímulos visuais embora, quando questionados,
dizem que não estão “vendo nada”. Ao mesmo tempo em que um objeto tem características de
diversas modalidades sensoriais como visuais e tácteis, por exemplo, e temos uma única sensação
desse objeto, os vários módulos do sistema nervoso também proporcionariam uma única experiência
consciente.
62

2o curso de inverno – tópicos em fisiologia comparativa
A consciência seria formada pelo funcionamento desses diversos módulos que processam
informações específicas e, portanto não está localizada em nenhum local específico do sistema
nervoso. Essa última forma de olhar para a consciência possibilita que ela seja estudada de forma
científica, explica vários comportamentos e não tem a necessidade ser de complementada por
nenhum outro tipo de explicação mística ou metafísica.
Bibliografia
Campos, A.; Santos, A.M.G. & Xavier, G.F. (1997). A consciência como fruto da evolução e do
funcionamento do sistema nervoso. Psicologia USP, São Paulo, v.8, n.2, p.181-226. Instituto
de Psicologia - USP.
Gazzaniga, M.S.; Ivry, R.B. & Magnun, G.R. (2002). Consciousness. In Cognitive Neuroscience.
Segunda Edição, Cap. 18, p. 325-333.
Marcel, A.J.; & Bisiach, E. (1992). Consciousness in Contemporary Science. Oxford University
Press.
Psicologia USP, São Paulo, v.8, n.2, p.181-226. Instituto de Psicologia - USP.
63

2o curso de inverno – tópicos em fisiologia comparativa
EXERCÍCIO FÍSICO, HIPERTENSÃO ARTERIAL E COMPORTAMENTOS:
INTERAÇÕES, EFEITOS E BENEFÍCIOS
Regiane Xavier de Moraes
Neste tópico será sucintamente abordado a relação do exercício físico com a hipertensão
arterial, além de atualidades e tendências em pesquisa na área da Fisiologia do Exercício e doenças
cardiovasculares. Além disso, será também elucidada a importância do treinamento físico na
alteração de características e hábitos comportamentais.
O sedentarismo pode contribuir para o aparecimento e/ou agravamento de doenças
cardiovasculares como a hipertensão arterial. De acordo com 3° Consenso Brasileiro de
Hipertensão (1998), 15 a 20% da população brasileira está acometida. Atualmente 90 a 95% dos
idosos são hipertensos.
Durante os últimos tempos, o exercício físico, bem como as suas implicações e conseqüências,
tem sido extensamente estudado por cientistas de todo o mundo. Usualmente, os exercícios,
aeróbicos e/ou de resistência, mais recomendados e utilizados são a caminhada e a corrida, a
natação, a musculação e o ciclismo. Em animais, as metodologias normalmente utilizadas são a
roda de corrida espontânea, além da corrida induzida em esteiras rolantes adaptadas e a natação
forçada. No entanto, rodas de corrida espontânea são os métodos de treinamento animal mais
indicados para o estudo de parâmetros fisiológicos por não causar estresse e injúria aos animais. As
pesquisas buscam compreender as ações do exercício no organismo, quais os mecanismos centrais e
periféricos que as norteiam e, principalmente, quais os benefícios que poucas horas de mudança na
rotina diária podem causar tanto para uma pessoa ou animal saudável como para os acometidos por
patologias.
A realização do exercício físico provoca uma série de respostas fisiológicas nos diversos
sistemas corporais, em particular no cardiovascular e nervoso. Objetivando manter a homeostasia
celular, diante do aumento das necessidades metabólicas, há incremento do débito cardíaco,
redistribuição do fluxo e aumento da perfusão sanguínea para a musculatura em atividade.
Sabe-se que exercícios físicos regulares, quando são adequadamente prescritos, e de baixa
intensidade podem provocar alterações autonômicas importantes que influenciam o sistema
cardiovascular. Dentre estas, a atenuação da hipertensão arterial tanto em humanos quanto em ratos
espontaneamente hipertensos (SHR). A atividade física contribui para a melhora do controle
64

2o curso de inverno – tópicos em fisiologia comparativa
barorreflexo e para a redução de 8 a 11 mmHg da pressão arterial sistólica e diastólica,
respectivamente, em indivíduos hipertensos (Hagberg et al., 2000). Estudos mostram que a
diminuição da pressão arterial deve-se à diminuição do débito cardíaco que está associado à
diminuição da frequência cardíaca pós exercício (bradicardia de repouso) (Véras-Silva et. al., 1997).
Entretanto, alguns autores propõem que exercícios crônicos provocam queda na resistência vascular
sistêmica e, consequentemente na redução da pressão arterial (Nelson et al.,1986). O treinamento
físico normaliza o tônus simpático, que controla a frequência cardíaca em SHRs (Gava et al.,1995)
e diminui a atividade nervosa simpática em humanos, ou seja, estes resultados sugerem que a
atividade física pode modular a atividade nervosa simpática para o coração e vasos periféricos,
explicando, em partes, a queda pressórica.
Modulações específicas da frequência cardíaca durante o exercício constituem um mecanismo
muito preciso de manutenção do suprimento do fluxo sanguíneo para o cérebro, coração, pele e
músculos em atividade.
Alguns neurotransmissores possuem importantes funções que garantem condições necessárias
para a realização da atividade física. Dentre estes estão a vasopressina (AVP) e a ocitocina (OT). A
AVP e a OT são produzidas em neurônios magnocelulares do Núcleo Paraventricular do
Hipotálamo (PVN) que envia e recebe projeções do Núcleo do Trato Solitário (NTS). O PVN e o
NTS são importantes centros de controle cardiovascular (Michelini e Morris, 1999).
A AVP facilita a resposta taquicárdica durante a atividade física. Contraditoriamente, a OT
diminui a taquicardia e contribui para a bradicardia. Desta forma, estes neurotransmissores possuem
efeitos específicos e opostos no controle da frequência cardíaca. Este balanço entre o estímulo
excitatório (AVP) e inibitório (OT) provê a eficiência do ajuste fisiológico requerido
momentaneamente, já que a taquicardia é necessária para suprir a maior demanda de fluxo
sanguíneo e a maior taxa metabólica da musculatura em atividade durante o exercício. Assim, no
NTS de indivíduos treinados, a AVP e OT atuam como moduladores da frequência cardíaca durante
a atividade física por potencializar ou moderar, respectivamente, a taquicardia (Michelini, 2001).
É importante enfatizar que as vias vasopressinérgicas e ocitocinérgicas do tronco encefálico
não são os únicos mecanismos centrais envolvidos na gênese da taquicardia. Assim, projeções
descendentes vasopressinérgicas e ocitocinérgicas do PVN para o NTS são parte do mecanismo
central de modulação do reflexo barorreceptor no controle da frequência cardíaca durante o
exercício e outras condições ambientais (Michelini, 2001).
Podem ser observadas ainda outras alterações cardiovasculares decorrentes do treinamento
físico tais como a hipertrofia cardíaca. Exercícios aeróbicos, por meio do aumento de volume
sanguíneo, podem estimular adaptações na morfologia cardíaca, metabolismo energético e funções.
65

2o curso de inverno – tópicos em fisiologia comparativa
Estes podem produzir hipertrofia cardíaca excêntrica, na qual o aumento da massa ventricular é
proporcional ao aumento da câmara cardíaca (Frohlic et al., 1992). Trata-se de uma resposta
fisiológica e compensatória fundamental para suportar o aumento da carga de trabalho. Estas
alterações estruturais, morfo-funcionais e metabólicas do coração induzidas pelo exercício resultam
em maior volume de ejeção sistólica, que torna-se mais vigorosa, e maior esvaziamento ventricular.
Entretanto, a hipertrofia cardíaca pode se instalar em resposta a certos estados patológicos
crônicos como e hipertensão arterial. Na hipertrofia concêntrica o aumento da massa ventricular não
é proporcional ao aumento da câmara cardíaca. Desta forma o trabalho cardíaco é feito contra uma
excessiva resistência ao fluxo sanguíneo. O coração hipertrofiado pode falhar e tornar-se incapaz,
em casos mais graves, de prover o fluxo sanguíneo normal para o indivíduo hipertenso.
Vários são os fatores desencadeantes da hipertensão arterial. Dentre eles, o excesso de peso, a
alimentação rica em gordura e sal e pobre em frutas, verduras e legumes, o tabagismo, o alcoolismo
e os fatores genéticos. Outros fatores importantes são os relacionados aos comportamentos e à
capacidade de reação em diversas situações cotidianas. Assim, os comportamentos que atualmente
acometem quase a totalidade das pessoas entre crianças e adultos, como o estresse e a ansiedade,
podem desencadear ou acentuar o estado hipertensivo. A ansiedade, o estresse e a hiperatividade
são acentuadas características comportamentais de SHRs. Interessantemente, foi observado que
SHRs efetivamente treinados em rodas de corrida espontânea apresentam diminuição do
medo/ansiedade, estresse e hiperatividade (Moraes, R.X. in dissertação de mestrado, 2005).
Em suma, o exercício físico crônico de intensidade baixa a moderada possui implicações
clínicas importantes já que pode reduzir ou mesmo abolir a necessidade de uso de medicamentos
anti-hipertensivos, diminuindo, desta forma, o custo do tratamento, extinguindo efeitos colaterais e
principalmente promovendo melhora do quadro clínico de indivíduos hipertensos. Assim, pode ser
tido como uma importante conduta não farmacológica no tratamento e controle da hipertensão
arterial. Somam-se à estas descobertas o fato de que o treinamento físico colabora não só para a
manutenção e conquista da saúde cardiovascular, mas, também para a saúde mental, já que atua
beneficamente na redução de comportamentos maléficos e deletérios que prejudicam os sistemas
corporais e podem levar a graves patologias psicossomáticas. Finalmente, uma mudança na rotina
diária para o desenvolvimento da prática de esportes, além de ser prazeroso e desestressante,
colabora para a melhora na qualidade de vida bem como para a manutenção e ganho da saúde física
e mental de indivíduos hipertensos e saudáveis.
66

2o curso de inverno – tópicos em fisiologia comparativa
REFERÊNCIAS BIBLIOGRÁFICAS CITADAS
Frohlic, E.D., et al. (1992) The Heart in Hypertension. N. England. J. Med. 327: 998-1008. Gava, N.S.; Véras-Silva, A.S.; Negrão, C.E.;et al. (1995) Low-Intensit Exercise Training Attenuates
Cardiac β-adrenergic Tone During Exercise in Sponatneously Hypertensive Rats.
Hypertension. 26 (2): 1129-33.
Hagberg, J.M.; Park, J.J.; Brown, M.D. (2000) The Role of Exercise Training in the Treatment of
Hypertension: an Update. Sports Med, 30: 193–206.
Michelini, L.C. (2001) Oxytocin in the NTS - a New Modulator of Cardiovascular Control During
Exercise. Annals New York Acad. Sci. 940: 206-20.
Michelini, L.C.; Morris, M. (1999) Endogenous Vasopressin Modulates the Cardiovascular
Responses to Exercise. Annals New York Acad. Sci. 897:198-221.
Moraes, R.X. Análise da Expressão Gênica da Tirosina Hidroxilase e Vasopressina em Áreas do
Sistema Nervoso Central de Ratos Espontaneamente Hipertensos Submetidos a Treinamento
Físico Espontâneo. Dissertação de mestrado, 2005. Biblioteca da Biologia: LC: QP 361.5.
Nelson, L.; Jennings, G.L.; Esler, M.D.; et al. (1986) Effect of Changing Levels of Physical
Activity on Blood-pressure and Haemodynamics in Essential Hypertension. Lancet. 2: 473–
76.
Véras-Silva, A.S.; Mattos, K.C.; Gava, N.S.; et al. (1997) Low-intensity Exercise Training
Decreases Cardiac Output and Hypertension in Spontaneously Hypertensive Rats. Am J
Physiol: Heart Circ Physiol. 273(6 Pt2): H2627-H2631.
Referências Bibliográficas Sugeridas
Dufloth, D.L.; Morris, M.; Michelini, L.C. (1997) Modulation of Exercise Tachycardia by
Vasopressin in the Nucleus Tractus Solitarii. Am.J.Physiol. 273:R1271-R82.
Kramer, J.M.; Beatty, J.A.; Plower,E.D.; Waldrop, T.G. (2002) Exercise and Hypertension: a model
for Central Neural Plasticity. Clin. Exp. Pharmacol. Physiol. Jan; 29(1-2): 122-6.
Kramer, J.M.; Plowey,E.D.; Beatty,J.A.; Little,H.R.; Waldrop,T.G. (2000) Hypothalamus,
Hypertension and Exercise. Brain Res Bull. 53(1):77-85.
Silva, G.J.J.; Brum,P.C.; Negrão,C.E.; Krieger,E.M. (1997) Acute and Chronic Effects of Exercise
on Baroreflexes in Spontaneously Hypertensive Rats. Hypertension. v.30(3).
Turner, D.L. (1991) Cardiovascular and Respiratory Control Mechanisms During Exercise: an
Integrated View. J.Exp.Biol. 160:309-340.
67

2o curso de inverno – tópicos em fisiologia comparativa
SUBSTÂNCIAS DE ORGANISMOS MARINHOS QUE PODEM CAUSAR ACIDENTES E
INTOXICAÇÕES
Cynthia Grazielle Martins Delboni
Jeanete Lopes Naves
Em todos os níveis da escala filogenética animal, existem vários exemplos de ataque, defesa
e outros comportamentos que dependem de substâncias repelentes, paralisantes ou de outras ações
farmacológicas. Durante os milhões de anos de evolução os organismos desenvolveram um
refinamento dessas substâncias para diversas funções, a captura de presas e as defesas químicas em
geral. A produção e a alta diversidade de toxinas têm estimulado outros animais a desenvolver
técnicas de sobrevivência a estes compostos. Rápida excreção ou acúmulo destas substâncias em
compartimentos do corpo, tais como tecido de gordura ou cutícula estão entre os mecanismos
desenvolvidos para combater o efeito letal dos compostos.
Para o homem se envolver em acidentes ou intoxicações por qualquer substância produzida
por organismos vivos, é necessário que esta exerça alguma influência química em um ou mais
constituintes das suas células a fim de produzir uma resposta farmacológica. Em outras palavras, é
necessário que as moléculas dessas substâncias fiquem muito próximas das moléculas celulares para
que o funcionamento destas seja alterado.
Popularmente chamamos de veneno toda substância química, ou mistura de substâncias
químicas, que provoca intoxicação ou a morte com baixas doses, como também reservado, segundo
alguns autores, especificamente para designar substâncias provenientes de animais, nos quais teriam
importantes funções de autodefesa ou de predação, como é o caso de veneno de cobra, de abelha,
etc. Entretanto existe diferentes terminologia para as diferentes substâncias que podem causar
algum dano a um sistema biológico – veneno, peçonha, toxina.
Venenos (em inglês ‘poisons’) são produtos metabólicos produzidos ou estocados em órgãos
que, em condições naturais, afetam o organismo que os ingere e podem também atuar, de modo
artificial, por via parenteral. Animais venenosos produzem substâncias tóxicas em glândulas ou
tecidos, mas eles podem também adquirir compostos do ambiente, de outros animais, plantas ou
microrganismos. Eles os estocam ou acumulam em seu corpo ou em algum órgão particular e fazem
uso passivo destes compostos. Envenenamento geralmente é o resultado da ingestão do animal pelo
seu predador, tais como os humanos. Ex: dinoflagelado marinho Garbierdiscus toxicus e aqueles
que adquirem suas toxinas através da cadeia trófica.
68

2o curso de inverno – tópicos em fisiologia comparativa
Peçonhas (em inglês ‘venoms’) são originadas em glândulas especializadas associadas a
dutos excretores e possuindo ou não, estruturas inoculadoras; a maioria dessas substâncias contém
proteínas que são facilmente inativadas pela digestão. Animais peçonhentos utilizam à peçonha para
aquisição de presas, que pode incluir uma pré-digestão, e como defesa contra predadores,
obviamente a mais importante função de interação com os humanos. Ex: Peixe escorpião e
cnidários.
Peçonhas e venenos são compostos que são deletérios para outro organismo em certa
dosagem, interferindo em importantes atividades fisiológicas de plantas, animais ou humanos,
causando envenenamento. Peçonhas e venenos raramente são substâncias puras, na maioria dos
casos representam misturas de numerosos compostos, que podem ser tóxicos ou ter outra atividade.
Peçonhas consistem principalmente de peptídeos ou proteínas. Venenos são geralmente produtos ou
metabólicos secundários produzidos por microrganismos, plantas ou animais.
Toxinas são sempre de origem natural, são substâncias quimicamente bem definidas, puras e
homogêneas.
Uso terapêutico
Para que uma droga seja de algum modo útil tanto como instrumento terapêutico quanto
cientifico, ela deve agir seletivamente em determinadas células e tecidos, ou seja, deve mostrar alto
grau de especificidade de ligação ao sítio.
Para isso são necessários muitos estudos, principalmente de químicos e biólogos. Muitas
vezes, no final, a pesquisa não leva a nenhum produto de interesse farmacológico. Diferente do que
ocorreu com o ácido okadaico que se tornou uma importante ferramenta de estudo de mecanismos
moleculares. Ele foi primeiro isolado de esponjas marinhas e depois reconhecido como um
constituinte da microalga Prorocentrum lima.
Toxicologia x toxinologia
A disciplina científica ligada à peçonhas, venenos e toxinas de plantas e animais é
denominada de toxinologia, e pode ser considerado um ramo da toxicologia. Entretanto, toxinologia
inclui mais do que química e modo de ação das toxinas. Explica também, a biologia da peçonha ou
veneno produzido pelo organismo, a estrutura e função do aparato de peçonha, o uso da peçonha ou
veneno, e papel ecológico deles.
Toxinas no ambiente marinho
69

2o curso de inverno – tópicos em fisiologia comparativa
A vida no mar tem sua complexidade e diversidade bem observada em recifes de corais
tropicais. Neste ambiente, organismos de todas as classes e ordens vivem juntos expostos a uma
enorme pressão de competição intra e inter-específica. Essa luta por espaço em ambientes
consolidados, defesa contra competidores de espaço e predadores, proteção contra infecção por
bactéria e fungos são através do uso de produtos tóxicos.
“Todas as substâncias são venenos; não há nenhuma que não seja veneno. A dose correta
diferencia o veneno do remédio.”
Referências bibliográficas
Freitas, J.C. (1991). Nomenclatural em toxinologia. Relações com a comunicação química entre
organismos e propriedades biológicas das toxinas. Mem Inst Butantan 53 (2), 191-195.
Halstead, B.W. (1988). Poisonous and venomous marine animals of the world. 2ª ed. The Darwin
Press, INC. 1168 p.
Halstead, B.W., Auerbach, P.S., Campbell, D. (1990). A colour atlas of dangerous marine animals.
Wolf Medical Publications Ltd. 192 p.
Mebs, D. (2002). Venomous and Poisonous Animals: A Handbook for Biologists, Toxicologists
and Toxinologists, Physicians and Pharmacists. CRC Press. 360 p.
Scheuer, P.J. (1991). Drugs from the sea. Chemistry and Industry 8, 276-279.
Rang, H.P., Dale, M.M., Ritter, J.M.(1997). Farmacologia. 3ª ed. Rio de Janeiro: Editora Guanabara
Koogan, 692 p.
Oga, S. (2003). Fundamentos de Toxicologia. 2ª ed. São Paulo: Atheneu Editora, 474 p.
70

2o curso de inverno – tópicos em fisiologia comparativa
RITMICIDADE BIOLÓGICA: CONCEITOS E APLICAÇÕES EM METABOLISMO
ENERGÉTICO
Denise Loli
INTRODUÇÃO
Os ritmos biológicos, que estão presentes em todos seres vivos, parecem ter-se originado
desde os primórdios da vida. A ritmicidade encontrada em animais e plantas não é apenas expressão
reflexa dos eventos temporais do meio externo, mas possui um caráter endógeno, sendo
determinado geneticamente (Cipolla-Neto et al 1988). Halberg apud Withrow (1959), introduziu o
termo circadiano (latim circa: em torno de; dies: dia), para caracterizar os ritmos com períodos em
torno de 24 horas, quando o organismo é mantido em condições constantes.
Os ritmos circadianos são gerados endogenamente por um oscilador ou relógio biológico e sua
expressão independe do ambiente. Um ritmo biológico pode persistir em condições constantes por
muitos dias ou mesmo anos. Endogenamente, estes osciladores permitem a um organismo
antecipar-se a eventos no ambiente, antes mesmo que eles aconteçam. (Dunlap et al., 2004). O ritmo
obtido em condições naturais (exemplo, fotoperíodo natural) é denominado diário.
CONCEITOS IMPORTANTES EM ESTUDOS DE RITMOS BIOLÓGICOS
As condições ambientais constantes constituem-se de um ambiente completamente livre de
oscilações de iluminação, temperatura, ruídos, umidade relativa, chegada de alimento e de outras
variáveis que possam ser percebidas pela espécie em estudo. Estas condições constantes são
denominadas "de livre-curso". A característica endógena de um ritmo só se expressa em condições
de livre-curso e o período assim medido é chamado de "período do ritmo em livre-curso",
representado pela letra grega T (tau) (Marques et al, 1997).
O tau é específico de cada espécie e varia, no entanto, dependendo das características das
condições constantes (exemplo: iluminação) (Aschoff, 1960). Diversas condições podem abolir a
expressão rítmica e são chamadas "não permissivas" (Pittendrigh, 1960). O mecanismo pelo qual a
expressão do ritmo é afetada é conhecido como "mascaramento" (Aschoff, 1960). Quando exposto
ao ciclo ambiental, o tau (T) do ritmo endógeno torna-se igual ao do ciclo e tem suas fases acertadas
por ele. A este processo deu-se o nome "arrastamento" (Pittendrigh, 1960) e ao ciclo ambiental
arrastador, Zeitgeber, neologismo alemão criado por Jurgen Aschoff, que significa "doador de
tempo" (Aschoff, 1960).
71

2o curso de inverno – tópicos em fisiologia comparativa
Três propriedades importantes caracterizam um ritmo biológico: 1) a endogenicidade 2) a
possibilidade do ritmo endógeno ser arrastado por um Zeitgeber e 3) a compensação de T a
variações da temperatura ambiente (Almeida, 2004).
APLICAÇÕES EM METABOLISMO ENERGÉTICO
Os estudos relativos a variações diárias na taxa metabólica tem sido estudados em diversos
animais. Em seres humanos, por exemplo, Ravussin et al (1986) estudaram as variações diárias da
taxa metabólica em 177 indivíduos, considerando a massa de gordura do corpo, determinada por
densitometria, e a atividade física espontânea.
Diversos insetos foram estudados sob o ponto de vista do ritmo circadiano da taxa respiratória,
como por exemplo por Banks et al. (1975) que pesquisaram o ritmo circadiano da taxa respiratória
de duas espécies de baratas Blaberus, observando um ritmo circadiano endógeno. Estudos da
ritmicidade metabólica de insetos Tribolium confusum (Coleoptera) e Camponotus rufipes
(Hymenoptera, Formicidae) indicaram uma flutuação rítmica da taxa respiratória sob condições de
ciclo normal claro/escuro 12:12 para as duas espécies.
Proni et al (2004) investigaram a taxa respiratória de abelhas operárias isoladas de
Nannotrigona (Scaptotrigona) postica, em fotoperíodo invertido (dia=escuro / noite=claro) por um
período de 24 horas, mostrando um ritmo circadiano metabólico invertido que provavelmente indica
a existência de um ritmo exógeno que aparece como uma resposta imediata às mudanças no
ambiente. Em operárias isoladas de Tetragonisca e Trigona um padrão definido de ritmo
circadiano metabólico foi observado, apresentando um maior consumo de oxigênio durante o dia,
com um pico entre 10h e 15h. À noite o consumo diminuiu, alcançando os níveis mínimos entre 2h
e 3h (Proni & Macieira, 2004).
AULA PRÁTICA
OBJETIVOS:
Demonstração de metodologias para coleta e análise de dados, em abelhas sem ferrão
Melipona quadrifasciata (mandaçaia), de variações diárias de taxa metabólica e termorregulação
individual e colonial. Discutir sobre as possibilidades de análises de dados de séries temporais
MATERIAIS E MÉTODOS
72

2o curso de inverno – tópicos em fisiologia comparativa
Para cálculo de taxa metabólica, é utilizado sistema de respirometria, podendo-se utilizar a
colônia inteira e/ou operárias isoladas.
Para estudos de termorregulação individual, utiliza-se termômetro infravermelho (medição
de temperatura torácica), em temperatura ambiente constante. Para a termorregulação colonial,
utilizam-se sensores ligados a carregadores de dados de temperatura (Hobo e Stoway), em
diferentes regiões da colônia de abelhas. Pode-se utilizar a colônia aberta (forrageiras saem da
colônia) ou colônia fechada em condições constantes de iluminação e temperatura.
BIBLIOGRAFIA
Cipolla-Neto, J.; N. Marques &, L.S. Menna-Barreto. 1988. Introdução ao estudo da
cronobiologia. São Paulo, Icone Edusp.
Withrow, R.B. 1959. Photoperiodism and Related Phenomena in Plants and Animals.
Washington, American Association for the Advancement of Science.
Dunlap, J. C.; Loros, J. J.; DeCoursey, P.J. (2004) Chronobiology: biological timekeeping.
Massachusetts: Sinauer.
Marques, M. D.; Golombek, D.; Moreno, C. Adaptação Temporal. In: Marques, N. &
Menna-Barreto, L. (1997) Cronobiologia: princípios e aplicações. EDUSP, São Paulo.
Aschoff, J. (1960) Exogenous and endogenous components in circadian rhythms. Cold Sping
Harbor Symp. Quant. Biol. 25: 11-28.
Pittendrigh, C.S. (1960) Circadian rhythms and circadian organization of living systems. Cold
Spring Harbor Symposium on Quantitative Biology 25: 159-84.
Almeida, G.F. (2004) Estudo de componentes rítmicos detectados na colônia de
Frieseomelitta varia (Hymenoptera: Apidae: Meliponinae). Dissert de Mestrado, FFCLRP-USP.
Ravussin, E.; Lillioja, S.; Anderson, T et al. (1986) Determinants of 24-hour energy
expenditure in man. J Clin Invest 78: 1568-78.
Banks, W.M.; A.S. Bruce. & H.T. Peart. 1975. The effects of temperature, sex and circadian
rhythm on the oxygen consumption in two species of cokroaches. Comparative Biochemistry
Physiology, Great Britain, 52 (A): 223-227.
Proni, E.A.; Macieira, O.J.D.; Morias, A.A.; Tristão, F.S.M. (2004) Circadian rhythm of the
respiratory rate of workers of Nannotrigona (Scaptotrigona) postica (Hymenoptera: Apidae:
Meliponinae) in inverted photoperiodic conditions during springtime. Proceedings of VI Encontro
sobre abelhas, Ribeirão Preto.
73

2o curso de inverno – tópicos em fisiologia comparativa
Proni, E. & Macieira, OJ.D. (2004) Ritmo circadiano da taxa respiratória de Tetragonisca
angustula fiebrigi (Schwarz), T. a. angustula (Latreille) e Trigona spinipes (Fabricius)
(Hymenoptera, Apidae, Meliponinae). Rev. Bras. Zool.21(4).
74

2o curso de inverno – tópicos em fisiologia comparativa
RITMOS CIRCADIANOS E FUNÇÕES DE APRENDIZAGEM E MEMÓRIA
Luiz Fernando Lopes do Espírito Santo
Os sistemas biológicos que evoluíram na Terra estiveram, desde a sua origem, expostos a
variações cíclicas do ambiente. Após o aparecimento de sistemas capazes de lidar com essas
oscilações ambientais, a seleção natural levou à manutenção desses sistemas.
São evidentes as vantagens adaptativas proporcionadas por sistemas que controlam ritmos
biológicos.. Num ambiente cujo funcionamento é cíclico, é possível prever a natureza das alterações
ambientais que ocorrerão. Organismos equipados com sistemas de temporização podem antecipar
essa ocorrência e iniciar os ajustes fisiológicos necessários para lidar com essa alteração ambiental
antes mesmo que ela ocorra. Vantagens adaptativas desse tipo levaram à seleção dos sistemas
controladores de ritmos biológicos e sua importância é ressaltada pelo fato de eles estarem presentes
em praticamente todos os grupos de animais.
Nos últimos anos se intensificou o interesse por identificar o papel de ritmos circadianos em
processos de aprendizagem e memória. Em mamíferos uma das estruturas responsáveis pelo
controle da ritmicidade circadiana é o conjunto formado pelos núcleos supraquiasmáticos (NSQ),
considerados os principais relógios (ou osciladores) circadianos. Nesse circuitaria neural os tratos
retino-hipotalâmicos transmitem informação temporal do ambiente externo, especificamente
informações sobre os ciclos claro-escuro (CE), de tal forma a permitir a sincronização da
ritmicidade endógena dos NSQ à ciclicidade ambiental. O sistema circadiano possui ainda diversas
vias eferentes, tanto neurais como humorais, que transmitem a informação sobre o tempo, gerada
nos NSQ e sincronizada ao meio externo, para o restante do organismo.
Aprendizagem e memória, envolvem diferentes estruturas do sistema nervoso. Definimos
aprendizagem como os processos subjacentes que levam à mudança de comportamento em
função de uma experiência prévia. Uma forma interessante de ilustrar a complexidade da
interação entre aprendizagem, memória e processos temporais é lembrar que o própio sistema
circadiano é capaz de "aprender" uma resposta. Em outras palavras, o funcionamento do sistema
circadiano pode ser modulado através de processos de aprendizagem e memória.
Do ponto de vista da circuitaria neural, as estruturas relacionadas com a temporização
circadiana e as estruturas envolvidas em processos de aprendizagem e memória mantém intimas
conexões, sugerindo que processos de aprendizagem e memória também são modulados pelo
funcionamento do sistema circadiano.
75

2o curso de inverno – tópicos em fisiologia comparativa
A maioria dos trabalhos que tentam investigar este assunto procura avaliar a eficiência da
aquisição e arquivamento de informação em função da hora do dia. Os resultados são ainda
escassos e inconsistentes. A aprendizagem do tipo espaço-temporal é particularmente sensível à
influência do sistema circadiano. Neste tipo de aprendizagem o organismo não raro estabelece uma
associação entre um evento biologicamente significante (e.g., disponibilidade de alimento) e uma
localização espacial em determinada hora do dia. Estes estudos podem resultar numa melhor
compreensão sobre a interação entre os sistemas de memória e o sistema circadiano na produção do
comportamento adaptativo.
APRENDIZAGEM ESPAÇO-TEMPORAL
Nos últimos anos diversos estudos têm utilizado animais em tarefas de aprendizagem
espaço-temporal para determinar sua capacidade de estabelecer associações entre a
disponibilidade de alimento num local determinado e numa determinada hora do dia. Vários
experimentos demonstraram que animais manifestam um bom desempenho numa tarefa de
aprendizagem com desafios espaço-temporais. O que é preciso ser demonstrado é que esta
estratégia se baseia em mecanismos de temporização circadianos, e conseqüentemente a
participação dos núcleos supraquiasmáticos nestes processos.
Eventos biologicamente significantes acontecem nas dimensões espacial e temporal.
Também o sistema cognitivo de vertebrados muito provavelmente evoluiu de tal forma a permitir a
percepção das regularidades espaço-temporais significantes, e usar essa informação para organizar
suas atividades eficientemente. De fato, diversos vertebrados aproveitam fontes de alimentos que
manifestam padrões de disponibilidade espaço-temporais. Estudos de campo têm mostrado que
alguns predadores ajustam seu horário de caça ao pico de atividade de sua pressa predileta
(Rijnsdorp et al., 1981). Por exemplo, caçadores de ostras antecipam os ritmos da maré associados à
disponibilidade de moluscos (Daan & Koene, 1981). Porém, nesses estudos não se demonstrou
inequivocamente que os animais estabelecem uma associação espaço-temporal, ou se simplesmente
respondem aos estímulos ambientais.
É provável que mamíferos possam fazer uso de múltiplas estratégias (além da mensuração
circadiana) para desempenhar tarefas de tipo espaço-temporal e que o uso de uma ou outra
estratégia pode variar em função das condições experimentais. Possivelmente, em determinadas
situações, mamíferos utilizam uma combinação de duas ou mais estratégias simultaneamente para
lidar com as demandas ambientais (Carr et al., 1999b).
76

2o curso de inverno – tópicos em fisiologia comparativa
REFERÊNCIAS BIBLIOGRÁFICAS
Abe H., Sugimoto S. (1987) Food-anticipatory response to restricted food access based on the
pigeon’s biological clock. Animal Learning & Behavior, 15:353-359.
Carr J.A.R., Tan A.O., Wilkie D.M. (1999) Further evidence that rats use ordinal timing in a daily
time-place task. Behavioural Processes, 48:35-48.
Carr J.A.R., Wilkie D.M. (1999) Rats are reluctant to use circadian timing in a daily time-place
task. Behavioural Processes, 44:287-299.
Daan S., Koene P. (1981) On the timing of foraging flights by oystercatchers, Haematopus
ostralegus, on tidal muflats. Netherlands Journal of Sea Research, 15:1-22.
Marques N. e Menna-Barreto L. (org), (1997) Cronobiologia: princípios e aplicações Editora da
Universidade de São Paulo.
Mistllberger R.E., de Groot M.H.M., Bossert J.M., Marchant E.G. (1996) Discrimination of
circadian phase in intact and suprachiasmatic nuclei-ablated rats. Brain Res., 739:12-18.
Mistlberger, R.E. (1990) Circadian pitfalls in experimental designs employing food restriction.
Psychobiology, 18(1):23-29.
Rijnsdorp A., Daan S., Dijkstra C. (1981) Hunting in the kestrel, Falco tinnunculus, and the
adaptive significance of daily habits. Oeceologia 50:391-406.
Saksida, L.M. & Wilkie, D.M. (1994) Time-of-day discrimination by pigeons, Columba livia.
Animal Learning and Behavior, 22(2):142-154.
77

2o curso de inverno – tópicos em fisiologia comparativa
FISIOLOGIA DO SISTEMA RESPIRATÓRIO EM AMBIENTES AQUÁTICOS
Renato Massaaki Honji
Quando nos referimos aos habitantes encontrados no ambiente aquático, lembramos
rapidamente dos teleósteos (peixes ósseos), que são o mais numeroso e diverso grupo de
vertebrados. Os peixes representam aproximadamente 50% dos vertebrados, englobando cerca de
28.000 espécies viventes que ocupam ambientes aquáticos mais diversos, ocorrendo desde as altas
latitudes até as fossas submarinas dos oceanos. Essa diversidade de espécies viventes também
apresenta uma grande variedade morfológica, fisiológica e adaptações comportamentais dentre
outras características. Levando-se em consideração apenas à fisiologia da respiração em peixes,
podemos encontrar os seguintes tipos de respiração: branquial, aérea (facultativa e obrigatória) e
pulmonar.
A maioria das espécies de peixes apresenta respiração branquial. As brânquias são geralmente
ventiladas com um fluxo unidirecional de água, no qual a simples abertura da boca e do opérculo,
adicionado ao deslocamento do animal na água, faz com que haja um fluxo em uma única direção
(peixes migradores como os atuns, albacora entre outros). A grande maioria dos peixes apresenta
musculatura esquelética na cavidade bucal e opercular, que mantém o bombeamento ativo da água
nas brânquias, mantendo assim um suplemento regular de O2. As brânquias dos peixes consistem
geralmente de quatro arcos branquiais e desses arcos estendem-se duas fileiras de filamento
branquiais, dos quais, cada filamento possui várias lamelas que são estruturas achatadas e
densamente enfileiradas onde ocorrem as trocas gasosas. Conforme a água flui entre essas lamelas
em uma direção, o fluxo sanguíneo flui em direção oposta, esse tipo de fluxo é denominado
contracorrente. Desta forma, quando o sangue está saindo das lamelas, o mesmo encontra a água
cujo oxigênio ainda não foi removido e conforme a água passa entre as lamelas, ela encontra o
sangue com uma pressão de oxigênio sempre abaixo e, portanto continua liberando mais oxigênio.
Desta forma depois de passar pelas brânquias, a água pode ter perdido mais ou menos 80 a 90% de
seu conteúdo de oxigênio.
Além das brânquias, muitas espécies de peixes apresentam estruturas capazes de realizar
trocas gasosas como, por exemplo: vesícula gasosa, intestino, estômago, esôfago entre outras
estruturas. Esses animais são chamados de peixes com respiração aérea.
A maioria das espécies de peixes com respiração aérea habita ambientes aquáticos no qual, em
algum período do dia ou estação, a concentração de oxigênio é muito baixa ou em ambientes
hipóxicos, ou seja, são locais no qual o nível de oxigênio é reduzido. Esses peixes responderão à
diminuição da concentração de oxigênio na água, nadando até a superfície para sorver uma bolha de
78

2o curso de inverno – tópicos em fisiologia comparativa
ar pela boca, o que resulta num melhoramento no suprimento de oxigênio. A respiração aérea pode
ser facultativa ou obrigatória, ou seja, se o ambiente não estiver hipóxico, o animal consegue retirar
da água toda quantidade de oxigênio necessária para a sua manutenção, apenas bombeando a água
através das brânquias. Quando o ambiente estiver hipóxico, essas espécies retiram uma parte do
oxigênio necessário para a sua manutenção da atmosfera (respiração aérea facultativa). Os rios da
Amazônia são um bom exemplo de ambiente aquático, no qual se observa uma variação de
concentração de oxigênio durante a estação de seca, sendo que muitos Siluriformes apresentam esse
tipo de respiração aérea facultativa. Os peixes com respiração aérea obrigatória são aquelas espécies
que necessitam subir até a superfície para respirar ar atmosférico, no qual sorvem uma bolha de ar e
o oxigênio é absorvido através das estruturas relacionadas acima. Quem disse que peixe não morre
afogado!!! Neste caso se o peixe com respiração aérea obrigatória for impedido de subir até a
superfície, ele morre afogado sim.
Existem apenas três espécies de peixes com respiração pulmonar, uma espécie australiana
(Neoceratodus), outra espécie africana (Protopterus) e uma sul-americana (Lepidosiren).
Protopterus e Lepidosiren vivem em águas paradas e em lagos, quando a falta de chuvas pode
ocasionar o ressecamento total do seu habitat. Essas espécies estivam até a próxima estação
chuvosa, quando elas saem dos seus casulos que estavam enterrados na lama. A espécie australiana
habita rios e corpos de água lênticos, no qual, também estivam em períodos de seca.
Os animais endotérmicos como, por exemplo, as aves e mamíferos que possuem uma elevada
taxa metabólica, necessitam de um maior suprimento de oxigênio se comparados com os animais
ectotérmicos (peixes, anfíbios e répteis). Conforme avançamos na escala filogenética em direção a
esses animais endotérmicos, observa-se que os pulmões tornam-se cada vez mais complexos. Os
pulmões de mamíferos são amplamente subdivididos em sacos denominados alvéolos que
aumentam enormemente a área de superfície disponível para as trocas gasosas. Essa enorme área de
troca é essencial para a alta taxa de transferência de oxigênio necessária para manter essa elevada
taxa metabólica que esses animais endotérmicos apresentam. Em geral os pulmões servem
principalmente para a respiração aérea e as brânquias para a respiração aquática, é claro que existem
exceções. Apesar do ar atmosférico conter mais oxigênio, as brânquias são em geral inadequadas
para a respiração aérea, pois na água o peso das brânquias é facilmente sustentado e no ar, as
mesmas não conseguem sustentar o seu próprio peso. Por isso que os peixes morrem se
permanecerem por muito tempo fora da água (asfixia).
A renovação de ar nos pulmões dos mamíferos deve ser constante, devido a grande taxa
metabólica. A ventilação nos pulmões é do tipo bidirecional, no qual o ar entra e sai do sistema
respiratório por bombeamento ativo. É importante salientar que o sistema respiratório nas aves é
mais complexo se comparado com os mamíferos.
79

2o curso de inverno – tópicos em fisiologia comparativa
Neste contexto, como os organismos com respiração pulmonar conseguem permanecer
submersos sem o acesso ao oxigênio atmosférico durante o mergulho? Algumas espécies de aves,
mamíferos e répteis marinhos e outras dulciaquícolas, permanecem grande parte de seu ciclo diário
submersos, sem acesso ao oxigênio atmosférico. Esses animais apresentam ajustes fisiológicos
cardiovasculares e respiratórios associados durante o período de mergulho. Normalmente essas
espécies permanecem durante o mergulho voluntário, em torno de 10 a 20 minutos, no entanto
existem alguns casos extremos como a foca Weddell que é capaz de permanecer submersa por
períodos de até 70 minutos. Os ajustes fisiológicos ao mergulho apresentado por esses animais,
evidenciando a integração dos sistemas cardiovasculares e respiratórios será discutido durante a
apresentação.
Referências Bibliográficas Recomendadas:
Butler, P. J. 2001. Diving beyond the limits. NewPhysiol. Sci. 16: 222-227.
Graham, J. B. 1997. Air-Breathing Fishes. Evolution, Diversity and Adaptation. Academic
Press.299p.
Moyle, P. B. & Cech, J. J. 1996. Fishes. An Introduction to Ichthyology. Prentice Hall, Upper
Saddle River. New Jersey. 590p.
Randall, D.; Burggren, W. & French, K. 2000. Fisiologia Animal. Mecanismos e Adaptações.
Guanabara Koogan. 729p.
Schmidt-Nielsen, K. 2002. Fisiologia Animal. Adaptação e Meio Ambiente. Santos, Livraria e
Editora. São Paulo – SP. 600p.
Val, A. L.; Almeida-Val, V. M. & Randall, D. J. 1996. Physiology and biochemistry of the fishes of
the amazon. Inpa-Manaus. 402p.
Williams, T. M.; Davis, R. W.; Fuiman, L. A.; Francis, J.; Le Boeuf, B. J.; Homing, M.;
Calambokidis, J. & Croll, D. A. 2000. Sink or swim: strategies for cost-efficient diving by
marine mammals. Science 288: 133-136.
Withers, P. C. 1992. Comparative Animal Physiology. Fort Worth. Saunders Publ. 949p.
80

2o curso de inverno – tópicos em fisiologia comparativa
ANEXO
Relação de ministrantes e laboratórios
Renata Brandt Nunes: [email protected]
Adne Abbud Righi: [email protected]
Pedro Leite Ribeiro: [email protected]
•Laboratório de Ecofisiologia Teórica e Fisiologia Evolutiva - Prof. Dr. Carlos Arturo Navas
Iannini Tel: (11)3091-7609
Jessica Ruivo Maximino: [email protected]
Regiane Xavier de Moraes: [email protected]
Emerson Ferraz Coelho: [email protected]
Daniel Carneiro Carrettiero: [email protected]
Andreas Betz: [email protected]
•Laboratório de Neurotransmissão e Modulação Neural da Pressão Arterial - Profa. Dra. Débora
Rejane Fior Chadi
Tel: (11)3091-7608
Profa. Dra. Lucile Maria Floeter-Winter: [email protected]
Marcos Gonzaga dos Santos: [email protected]
Maria Fernanda Laranjeira da Silva: [email protected]
Ariel Mariano Silber: [email protected]
•Laboratório de Fisiologia de Tripanosomatídeos - Profa. Dra. Lucile Maria Floeter-Winter
Tel: (11)3091-7503
Adriano Alonso Pereira da Cunha: [email protected]
Rodrigo Pavão: [email protected]
Luiz Fernando Lopes do Espírito Santo: [email protected]
Sylvia Maria Matsuda: [email protected]
•Laboratório de Neurociências e Comportamento – Prof. Dr. Gilberto Fernando Xavier
Tel: (11)3091-7504
81

2o curso de inverno – tópicos em fisiologia comparativa
Eduardo Koji Tamura: [email protected]
Pedro Augusto Carlos Magno Fernandes: [email protected]
Cíntia Etsuko Yamashita: [email protected]
Lidiana Duarte de Almeida Paula: [email protected]
•Laboratório de Cronofarmacologia - Profa. Dra. Regina Pekelmann Markus
Tel: (11)3091-7612
James Fernando Malta da Silva: [email protected]
•Laboratório de Osmo-regulação e Transporte Transmembrânico - Prof. Dr. Luis Carlos Salomão
Tel: (11)3091-7518
Profa. Dra. Maria Aparecida Visconti: [email protected]
Roseli Barbosa: [email protected]
Ana Paula Canel Bluhm: [email protected]
Gláucia Jansen Da Re Lopes: [email protected]
Leonardo Henrique Ribeiro Graciani de Lima: [email protected]
Fernanda Pizão Farhat: [email protected]
•Laboratório de Fisiologia Comparativa da Pigmentação – Profa. Dra. Maria Aparecida Visconti e
Prof. Dra. Ana Maria de Lauro Castrucci
Tel: (11)3091-7610
Denise Loli: [email protected] / [email protected]
•Laboratório de Fisiologia Respiratória Comparativa e Metabolismo Energético – Prof. Dr. José
Eduardo Pereira Wilken Bicudo
Tel: (11)3091-7607
Breno Teixeira Santos: [email protected]
•Laboratório de Fisiologia Teórica – Prof. Dr. José Guilherme Chaui Berlinck
Tel: (11)3091-7479
Cynthia Grazielle Martins Delboni: [email protected]
•Laboratório de Biologia Celular de Invertebrados Marinhos – Prof. Dr. Márcio Reis
Custódio
Tel: (11)3091-7611
82

2o curso de inverno – tópicos em fisiologia comparativa
Jeanete Lopes Naves: [email protected]
André Junqueira Zaharenko: [email protected]
•Laboratório de Toxicologia e Produtos Naturais - Prof. Dr. José Carlos de Freitas
Tel: (11)3091-7522
Renato Massaaki Honji: [email protected] / [email protected]
•Laboratório de Fisiologia Respiratória e Metabolismo Energético – Profa. Dra. Silvia Cristina
Ribeiro de Souza
Tel: (11)3091-7479
Laboratório de Metabolismo e Reprodução de Peixes do Núcleo de Ciências Ambientais da
Universidade de Mogi das Cruzes – Profa. Dra. Renata Guimarães Moreira
83