plasticidade sináptica à curto prazo na junção ...livros01.livrosgratis.com.br/cp154350.pdf ·...
TRANSCRIPT

PRISCILA ELISA SILVEIRA
Plasticidade sináptica à curto prazo na junção neuromuscular: papel de diferentes mecanismos de depuração de cálcio
e do receptor de IP3.
Belo Horizonte 2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

PRISCILA ELISA SILVEIRA
Plasticidade sináptica à curto prazo na junção neuromuscular: papel de diferentes mecanismos de depuração de cálcio
e do receptor de IP3.
Tese submetida ao departamento de
Fisiologia e Biofísica do Instituto de
Ciências Biológicas na Universidade
Federal de Minas Gerais, como requisito
parcial para obtenção do grau de Doutor
em Ciências.
Orientador: Profa. Dra. Lígia Araújo Naves Kushmerick
Co-orientador: Prof. Dr. Christopher Kushmerick
D.Sci. Jordi Molgó

Este trabalho foi realizado no Laboratório de Eletrofisiologia Celular (ELETROCEL) do
Departamento de Fisiologia e Biofísica – Instituto de Ciências Biológicas da Universidade
Federal de Minas Gerais e no Laboratoire de Neurobiologie Cellulaire et Moleculaire do
Centre National de la Recherche Scientifique em Gif-sur-Yvette/França.

Dedico este trabalho aos meus pais, José Roberto e
Vera Lúcia, pelo amor incondicional e por serem os
grandes alicerces da minha vida.

AGRADECIMENTOS
Agradeço a Deus, primeiramente, fonte de amor e vida.
À CAPES por proporcionar o desenvolvimento deste trabalho.
À Dra. Lígia Araújo Naves Kuschmerick e ao Dr. Christopher Kuschmerick, pela confiança,
aprendizado e orientação na realização deste trabalho.
Aos meus irmãos Fernando e Leandro pelo apoio e companheirismo constantes.
Ao Antônio por todo amor, carinho e respeito.
À Rose pela amizade e cumplicidade.
Aos amigos e colegas do Laboratório Eletrocel (Miguel José Lopes, Sílvia Guatimosim, Éder,
Ricardo, Adriano, Naiara, Luciana, Verônica, Fabíola, Pedro, Mariana, Fabrício, Marcinha,
Enéas, Aline e Amanda) pelo convívio e aprendizado.
À equipe do Dr. Jordi Molgó (Evelyne Benoit, Roland Bournaud, Sabine de la Porte, Patricia
Villeneuve, Sébastien Schlumberger, Agnese Bontadi, Emmanuelle Girard, Klara Rozman,
Cesare Colassante, Enrique Jaimovich, Romulo Araoz e Nadine Garrido) que tão bem me
receberam e pelo crescimento científico e pessoal.
Às amizades da UFMG que permanecem, Kátia, Lúcio, Viviane, Ana Paula Araújo, Ana
Paula Côrrea, Juliana, Laura, Samuel, Daniel, Adaliene, Luciana, Maira e Patrícia.
Aos professores do Departamento de Fisiologia e Biofísica e Farmacologia, principalmente à
Dra. Adelina Martha dos Reis, pelo apoio e confiança em meu trabalho.
À Celinha pelo carinho e disposição em ajudar sempre que necessário.
Aos amigos, Ana Cristina, Janice, Gisele, Flávia, Fábio, Rebeca, Monique, Uriatan, Rodrigo
Sírio, Priscilla, Rodrigo Cardoso, Ivna e Charles, que souberam compreender a minha
ausência e torcem por mim.

Aos colegas e amigos residentes da Maison du Brésil em Paris. Vocês foram essenciais nos
momentos mais difíceis e nos mais alegres também. Amizade e saudade toda a vida.
Aos que colocaram pedras no meu caminho e que me deram exemplos do que não ser.
“Pedras no caminho? Guardo todas, um dia vou construir um castelo...” (Fernando Pessoa)
Ao meu instrumento de pesquisa, as rãs. Sem elas nada disso teria sido possível. Com elas
pude perceber as semelhanças e diferenças entre nós, seres humanos, e os animais, e
acima de tudo aprender a respeitá-los.

“A sabedoria não se transmite, é preciso que nós
a descubramos fazendo uma caminhada que
ninguém pode fazer em nosso lugar e que
ninguém nos pode evitar, porque a sabedoria é
uma maneira de ver as coisas.”
(MARCEL PROUST)

LISTA DE ABREVIATURAS
2-APB - 2-Aminoethyl diphenyl borate
ACh – acetilcolina
AChE – enzima acetilcolinesterase
BAPTA, AM - 1,2-Bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid
tetrakis(acetoxymethyl ester)
Ca+2 – íon cálcio
CaCl2 – cloreto de cálcio
CCCP – carbonilcianeto-3-clorofenilhidrazona
CEDA, SE – (5-(and-6)-carboxyeosin diacetate, succinimidyl ester)
CGP 31157 - 7-Chloro-5-(2-chlorophenyl)-1,5-dihydro-4,1-benzothiaze pin-2(3H)-one
CICR – liberação de cálcio induzida pelo cálcio
CPA – ácido ciclopiazônico
DAG – diacilglicerol
DMSO – dimetilsulfóxido
EGTA - Ethylene glycol tetraacetic acid
EPP – potencial de placa motora
EPP1 – potencial de placa motora seguido do 1º estímulo
EPP2 – potencial de placa motora seguido do 2º estímulo
fMEPPs – frequência dos potenciais de placa motora miniatura
F – facilitação
Gq/11 – proteína G heteromérica
HEPES - 4-(2-Hydroxyethyl)piperazine-1-ethanesulfonic acid, N-(2-Hydroxyethyl)piperazine-
N′-(2-ethanesulfonic acid)

ICICR – liberação de cálcio induzida pelo inositol trifosfato
IP3 – inositol trifosfato
IP3R – receptor de inositol trifosfato
JNM – junção neuromuscular
KCl – cloreto de potássio
LiCl – cloreto de lítio
LTP – potenciação de longa duração
M1 – receptor muscarínico do tipo 1
M2 – receptor muscarínico do tipo 2
MCU – uniporter mitocondrial
MeOH – metanol
MEPP – potencial de placa em miniatura
Mg+2 – íon magnésio
MgCl2 – cloreto de magnésio
Na+ – íon sódio
NaCl – cloreto de sódio
NaOH – hidróxido de sódio
NCX – trocador sódio cálcio
NCXm – trocador sódio cálcio mitocondrial
PbTx-3 - brevetoxina
pH – potencial hidrogeniônico
PKC – proteína quinase do tipo C
PLC – fosfolipase do tipo C
PMCA – cálcio ATPase da membrana plasmática

PMR1/ATP 2 C1 – cálcio ATPase do tipo P
RE – retículo endoplasmático
Ru360 - (μ)[(HCO2)(NH3)4Ru]2OCl3
Ry - rianodina
RyR – receptor de rianodina
SERCA- cálcio ATPase do retículo saco-endoplasmático
SNARE – soluble N-ethylmaleimide factor attachment protein receptors
SNC – sistema nervoso central
Tg – tapsigargina
U-73122 - 1-[6-((17b-3-Methoxyestra-1,3,5(10)-trien-17-yl)amino)hexyl]-1H-pyrrole-2,5-dione
V – Volt

LISTA DE FIGURAS
FIGURA 1 – Animal após destruição do sistema nervoso central..........................................33
FIGURA 2 – Foto mostrando a mesa onde eram realizados os experimentos em JNM........34
FIGURA 3 – Organograma do sistema para a aquisição dos dados.....................................35
FIGURA 4 – Registro da diferença de potencial medida fora e dentro de uma
célula......................................................................................................................................36
FIGURA 5 – Registro dos potenciais de placa motora evocados na presença de 6 µM de d-
tubocurarina............................................................................................................................38
FIGURA 6 – Registro dos potenciais de placa motora espontâneos.....................................38
FIGURA 7 – Potenciais de placa motora registrados com pulsos pareados nos diferentes
intervalos entre os pulsos, na presença de curare.................................................................40
FIGURA 8 – Facilitação por pulsos pareados com intervalo de 10 ms na presença de curare.
Projeção do primeiro EPP para o cálculo da amplitude do segundo EPP.............................40
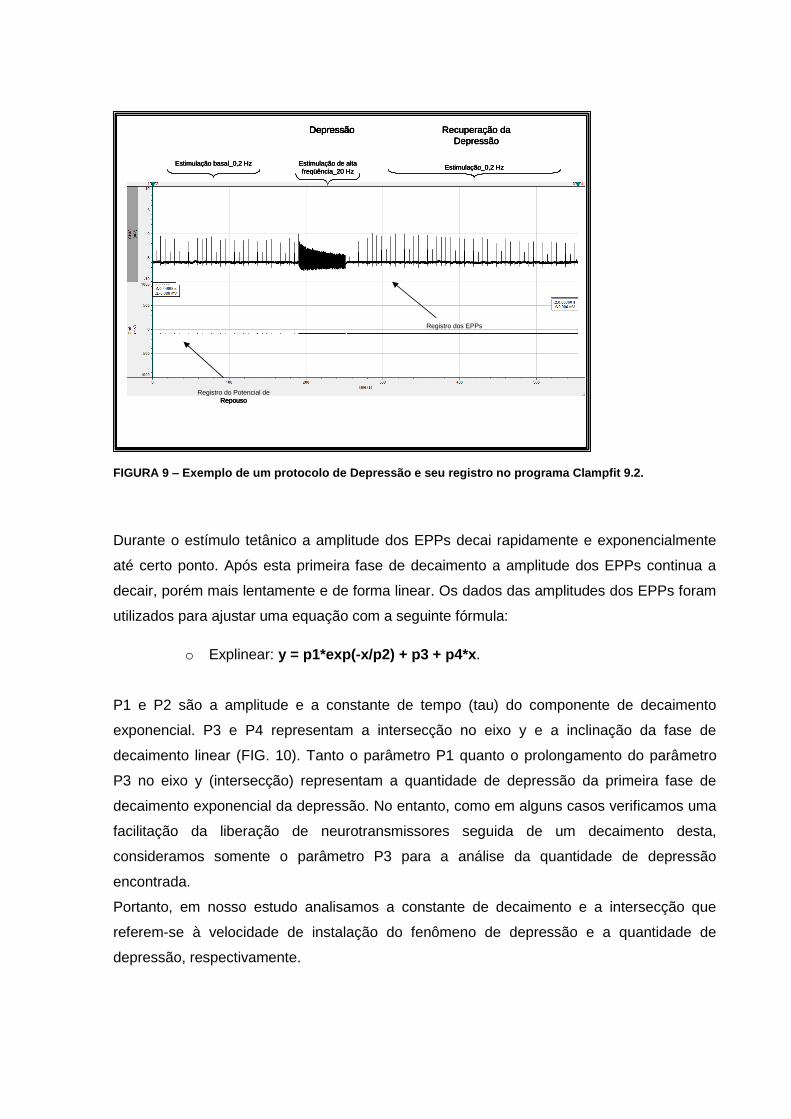
FIGURA 9 – Exemplo de um protocolo de Depressão e seu registro no programa Clampfit
9.2...........................................................................................................................................42
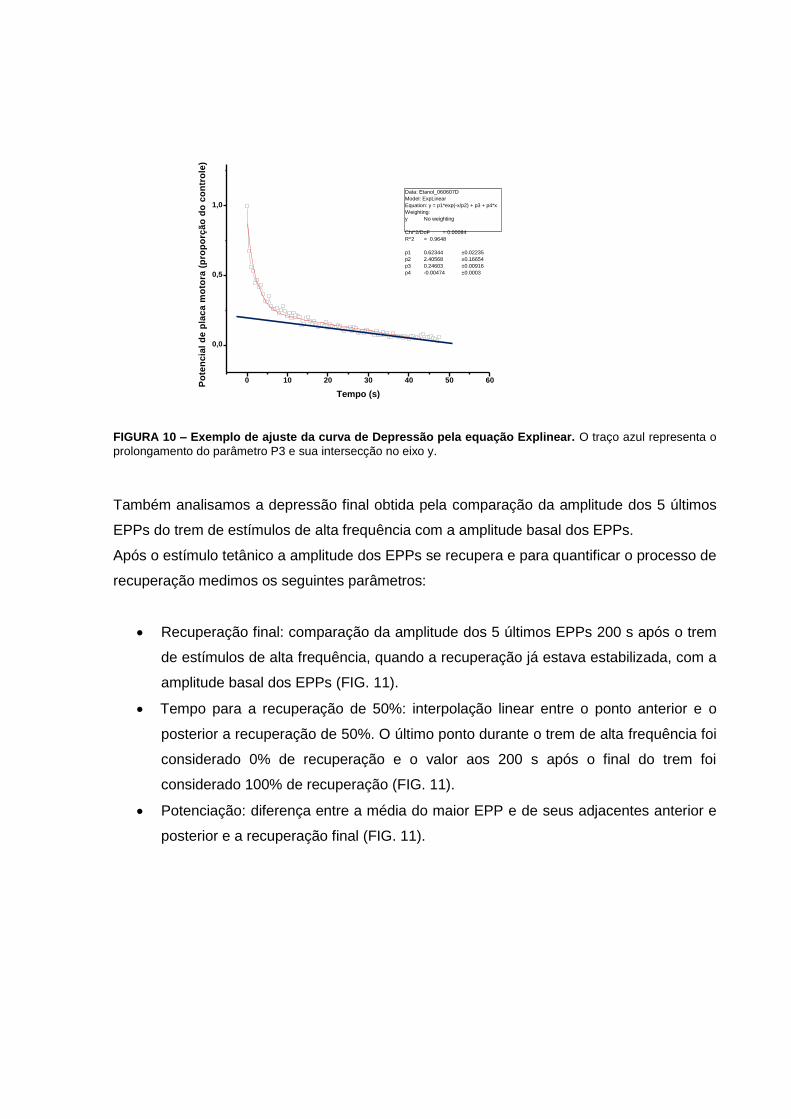
FIGURA 10 – Exemplo de ajuste da curva de Depressão pela equação Explinear...............43
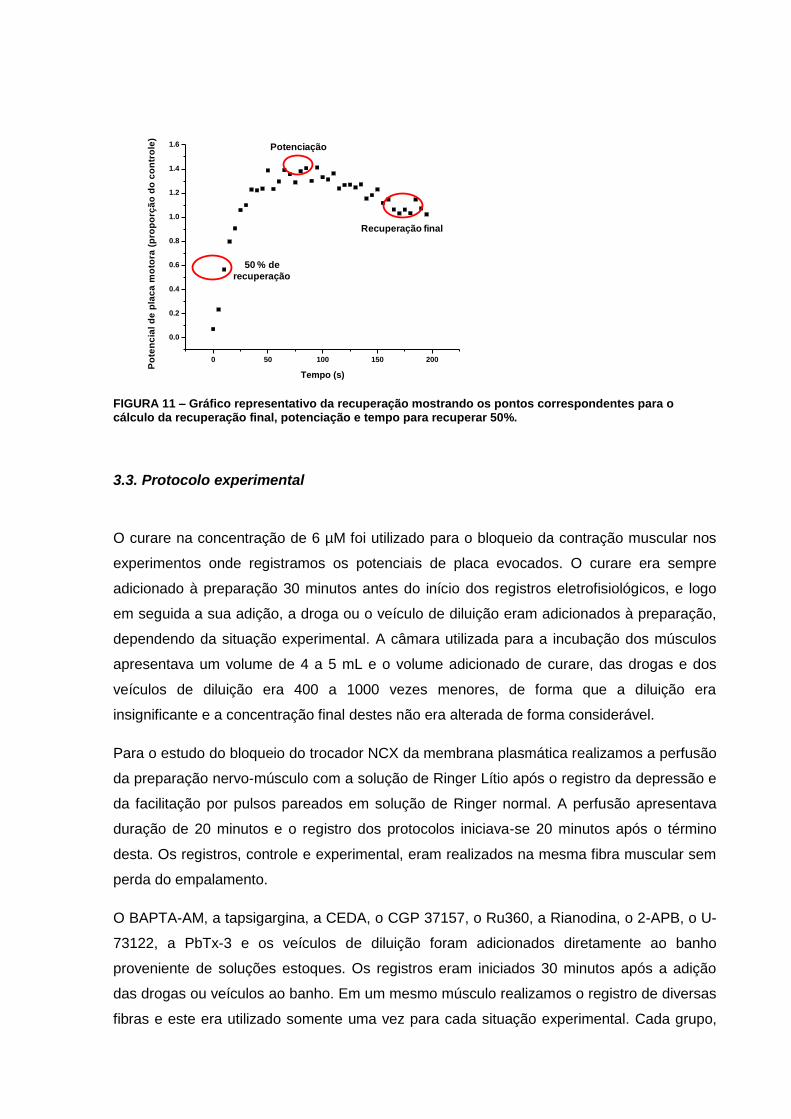
FIGURA 11 – Gráfico representativo da recuperação mostrando os pontos correspondentes
para o cálculo da recuperação final, potenciação e tempo para recuperar
50%.........................................................................................................................................44
FIGURA 12 - Organograma da situação experimental...........................................................45
FIGURA 13 - Efeito do BAPTA-AM no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................49
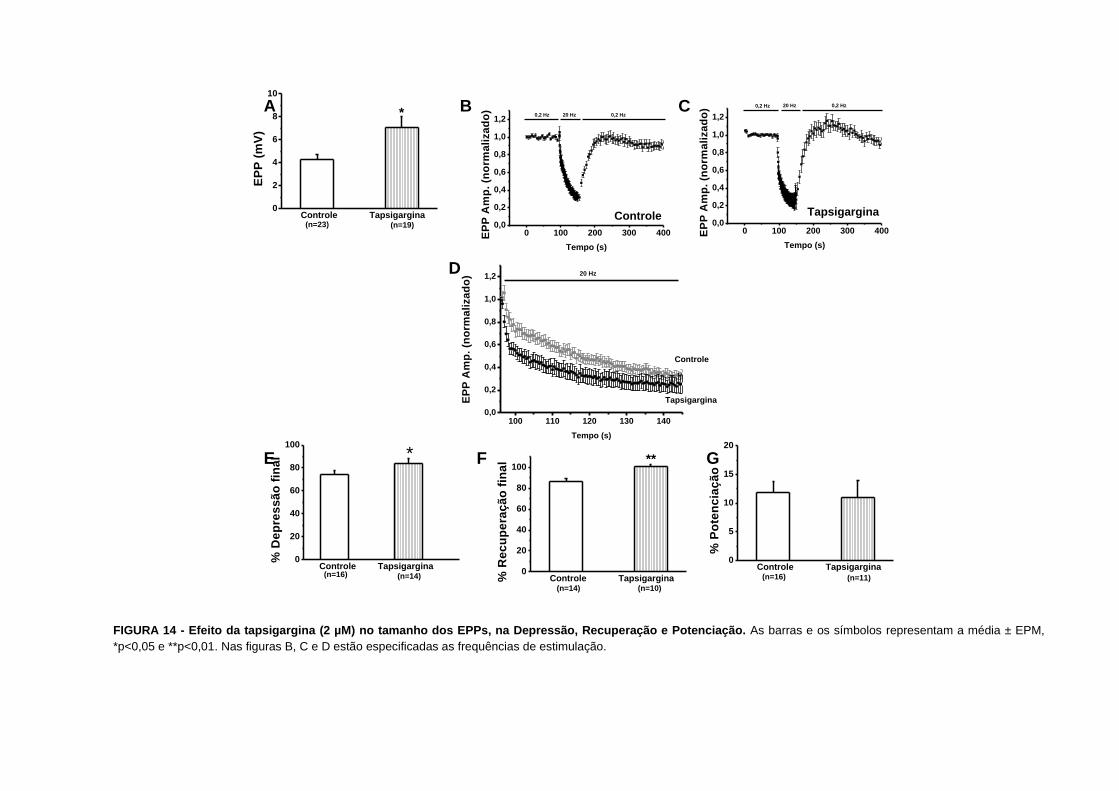
FIGURA 14 - Efeito da tapsigargina no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................51
FIGURA 15 - Efeito do CEDA no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................54

FIGURA 16 - Efeito do CGP no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................57
FIGURA 17 - Efeito do Ru360 no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................60
FIGURA 18 - Efeito do Ringer Lítio no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................63
FIGURA 19 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio
intracelular na depressão final................................................................................................65
FIGURA 20 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio
intracelular na depressão (análise da intersecção no eixo y).................................................66
FIGURA 21 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio
intracelular na constante de decaimento da
depressão...............................................................................................................................67
FIGURA 22 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio
intracelular na recuperação final.............................................................................................68
FIGURA 23 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio
intracelular na potenciação.....................................................................................................69
FIGURA 24 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio
intracelular na facilitação por pulsos pareados.......................................................................70
FIGURA 25 – Efeito da PbTx-3 na frequência dos potenciais de placa motora miniatura.....71
FIGURA 26 – Potenciais de placa motora miniatura representativos registrados em solução
de Ringer 0 Ca+2 + 2,5 mM EGTA (A) e após 30 minutos de incubação com 50 nM de PbTx-
3 (B)........................................................................................................................................72
FIGURA 27 – Efeito da TTX sobre o aumento da fMEPPs ocasionado pela PbTx-3............72
FIGURA 28 – Efeito da tapsigargina sobre o aumento da fMEPPs ocasionado pela PbTx-3-
................................................................................................................................................73
FIGURA 29 – Efeito do 2-APB sobre o aumento da fMEPPs ocasionado pela PbTx-3.........74
FIGURA 30 – Efeito do U-73122 sobre o aumento da fMEPPs ocasionado pela PbTx-3.....74

FIGURA 31 - Efeito da Rianodina no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................76
FIGURA 32 - Efeito do bloqueio do receptor de rianodina pela rianodina na Facilitação por
pulsos pareados.....................................................................................................................77
FIGURA 33 - Efeito do 2-APB no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................79
FIGURA 34 - Efeito do bloqueio do receptor de IP3 pelo 2-APB na Facilitação por pulsos
pareados.................................................................................................................................80
FIGURA 35 - Efeito do U-73122 no tamanho dos EPPs, na Depressão, Recuperação e
Potenciação............................................................................................................................82
FIGURA 36 – Efeito do bloqueio da fosfolipase C na facilitação por pulsos pareados..........83

LISTA DE TABELAS
TABELA I - Soluções nutridoras............................................................................................32
TABELA II – Facilitação por pulsos pareados – Controle x Tapsigargina.............................52
TABELA III – Facilitação por pulsos pareados – Controle x CEDA.......................................55
TABELA IV – Facilitação por pulsos pareados – Controle x CGP.........................................58
TABELA V – Facilitação por pulsos pareados – Controle x Ru360.......................................61
TABELA VI – Facilitação por pulsos pareados – Controle x Ringer Lítio..............................64
TABELA VII – Facilitação por pulsos pareados – Controle x Rianodina...............................77
TABELA VIII – Facilitação por pulsos pareados – Controle x 2-APB....................................80
TABELA IX – Facilitação por pulsos pareados – Controle x U-73122...................................83

RESUMO
Mecanismos de depuração do cálcio são importantes para a homeostase celular, mas sua
importância na plasticidade sináptica à curto prazo não está completamente esclarecida.
Nós comparamos a contribuição dos diferentes mecanismos de depuração do cálcio na
depressão sináptica e na facilitação por pulsos pareados. O retículo endoplasmático (RE) é
considerado um estoque de cálcio intracelular. O cálcio liberado pelo RE pode ocorrer pela
estimulação dos receptores de rianodina (RyR) ou pelas receptores de inositol trifosfato
(IP3R). No intuito de esclarecer sobre sua presença na junção neuromuscular e sua
importância na plasticidade sináptica, nós investigamos o mecanismo de ação da
brevetoxina-3 (PbTx-3), uma toxina conhecida por aumentar a concentração de IP3 e liberar
cálcio intracelular via IP3Rs. Nós também estudamos o papel dos IP3Rs e dos RyRs em
duas formas de plasticidade sináptica à curto prazo na junção neuromuscular: depressão e
facilitação por pulsos pareados.
Nosso estudo foi realizado na preparação nervo-músculo cutâneo peitoral de rãs. Utilizamos
o registro intracelular para medir a liberação espontânea, a depressão e a facilitação por
pulsos pareados.
Verificamos que o BAPTA-AM diminui a depressão, evidenciando a importância do cálcio
intracelular neste fenômeno. O bloqueio da Ca+2-ATPase do retículo endoplasmático
(SERCA) aumentou a depressão e a facilitação por pulsos pareados, entretanto o bloqueio
dos outros mecanismos de depuração do cálcio (Ca+2-ATPase da membrana plasmática,
uniporter e trocador Na+/Ca+2 mitocondriais) não apresentou alterações nestes parâmetros.
Estes resultados sugerem uma importante contribuição da SERCA na depuração de cálcio
nestes fenômenos de plasticidade sináptica à curto prazo. O bloqueio do trocador Na+/Ca+2
da membrana plasmática aumentou a depressão e diminuiu a facilitação por pulsos
pareados. A interpretação da substituição do sódio extracelular pelo lítio não é conclusiva,
pois o lítio pode alterar o potencial de ação pré-sináptico.
Nos experimentos com a PbTx-3 verificamos que esta toxina aumenta a liberação
espontânea. A tetrodotoxina e a tapsigargina impedem tal efeito, indicando sua relação com
a entrada de sódio e a liberação de cálcio de estoques intracelulares. Verificamos o
envolvimento do IP3R e da fosfolipase C (PLC) através da utilização de dois antagonistas: o
2-APB, um bloqueador do IP3R e o U-73122, um inibidor da PLC. O 2-APB e o U-73122
inibiram o aumento da liberação espontânea. Nossos resultados sugerem que a PbTx-3
promove a liberação de cálcio de estoques intracelulares sensíveis ao IP3 no terminal pré-

sináptico ocasionando aumento da liberação espontânea e este efeito depende da diferença
de potencial elétrico.
No estudo do papel dos IP3Rs na plasticidade sináptica à curto prazo verificamos que o 2-
APB diminui a depressão, mas não apresenta efeito na facilitação por pulsos pareados. A
PLC e os RyRs estão envolvidos na depressão sináptica e na facilitação por pulsos
pareados, visto que o U-73122 e a rianodina diminuem a intensidade destes fenômenos.
Nossos resultados sugerem o envolvimento da PLC e dos RyRs na depressão sináptica e
na facilitação por pulsos pareados, enquanto que o IP3R está envolvido somente na
depressão sináptica.
Nosso estudo esclarece sobre o mecanismo de ação da PbTx-3 e mostra evidências sobre a
presença do IP3R na junção neuromuscular e de seu papel na plasticidade sináptica à curto
prazo. Com relação aos mecanismos de depuração do cálcio nós sugerimos um maior
envolvimento da SERCA na depressão sináptica e na facilitação por pulsos pareados.
Palavras-chaves: junção neuromuscular, acetilcolina, depuração de cálcio, receptor de
inositol trifosfato e plasticidade sináptica.

SUMMARY
Mechanisms of calcium clearance are important for calcium homeostasis, but their role in
short-term plasticity is incompletely understood. We compared the contribution of different
mechanisms of calcium clearance in synaptic depression and paired pulse facilitation. The
endoplasmic reticulum (ER) is a well-characterized intracellular store of calcium. Calcium
release from the ER can be evoked by stimulation of the ryanodine receptor (RyR) or the
inositol (1,4,5)-trisphosphate receptor (IP3R). To clarify the presence of IP3Rs at the
neuromuscular junction and its role in synaptic plasticity we investigated the mechanism of
action of brevetoxin-3 (PbTx-3), a toxin known generate IP3 and release calcium from the ER
by activating IP3Rs. We also searched for the role of IP3Rs and RyR in two forms of short-
term plasticity at the neuromuscular junction: depression and paired pulse facilitation.
Our studies were carried out on cutaneous pectoris nerve-muscle preparations from frogs.
We recorded spontaneous release, paired pulse facilitation and synaptic depression using
intracellular recording.
We show that treating the neuromuscular junction with BAPTA-AM decreased depression.
These data indicate that intracellular calcium is important for synaptic depression. Blockage
of the endoplasmic reticulum Ca2+-ATPase (SERCA) increased depression and paired pulse
facilitation, whereas blockage of others mechanisms of calcium clearance (plasma
membrane Ca2+-ATPase, mitochondrial Ca2+ uniporter and mitochondrial Na+-Ca2+
exchanger) was without effect on these parameters. These results suggest an important and
major involvement of SERCA in calcium clearance during these phenomena of short-term
synaptic plasticity. Blockage of plasma membrane Na+-Ca2+ exchanger by substituting Li+ for
Na+ increased depression and decreased paired pulse facilitation. However the interpretation
of the effect of Li+ substitution is not straightforward, because it may also affect the
presynaptic action potential.
In addition, we show that PbTx-3 markedly enhanced spontaneous quantal transmitter
release. Tetrodotoxin and thapsigargin prevented these effects, indicating that it is related to
Na+ entry and to calcium release from intracellular stores. We verified the involvement of
IP3R and phospholipase C (PLC) by using two antagonists: 2-APB, a blocker of IP3R and U-
73122, a PLC inhibitor. Both of the treatments blocked the effect of PbTx-3 on spontaneous
quantal transmitter release sugesting PbTx-3 elicits Ca2+ release from the IP3 sensitive
intracellular Ca2+ stores of the presynaptic nerve terminal leading to an enhancement of
spontaneous transmitter release and that this effect depends on voltage.

In order to evaluate the involvement of IP3Rs in short-term plasticity we used 2-APB. Our
results show that 2-APB decreased depression, but had no effect on paired pulse facilitation.
PLC and RyRs are involved on synaptic depression and paired pulse facilitation. Our results
show that U-73122 and rianodine decreased depression and paired pulse facilitation. These
results suggest an involvement of PLC and RyRs in synaptic depression and facilitation.
IP3Rs are involved only in synaptic depression.
Our study clarifies the mechanism of action of PbTx-3, shows additional evidence for the
presence of IP3R at the neuromuscular junction and its role in short-term synaptic plasticity.
Concerning the mechanism of calcium clearance we suggest a major involvement of SERCA
in synaptic depression and paired pulse facilitation.
Keywords: neuromuscular junction, acetylcholine, calcium clearance, inositol (1,4,5)-
trisphosphate receptor and synaptic plasticity.

SUMÁRIO
I. INTRODUÇÃO 21
1.1. JUNÇÃO NEUROMUSCULAR 21
1.2. NEUROTRANSMISSÃO COLINÉRGICA 23
1.3. PLASTICIDADE SINÁPTICA 24
II. OBJETIVOS 29
III. MATERIAIS E MÉTODOS 30
3.1. MATERIAIS 30
3.1.1. ANIMAIS 30
3.1.2. EQUIPAMENTOS 30
3.1.3. DROGAS E REAGENTES 31
3.1.4. SOLUÇÕES 32
3.2. MÉTODOS 33
3.2.1. PREPARAÇÃO NERVO-MÚSCULO 33
3.2.2. SISTEMA DE AQUISIÇÃO DE DADOS 34
3.2.3. REGISTROS ELETROFISIOLÓGICOS 36
3.2.4. PROTOCOLOS DE PLASTICIDADE SINÁPTICA À CURTO PRAZO 39
3.3. PROTOCOLO EXPERIMENTAL 44
3.4. ANÁLISE ESTATÍSTICA 45
IV. RESULTADOS 47
4.1. INFLUÊNCIA DA CONCENTRAÇÃO INTRACELULAR DE CÁLCIO SOBRE A RESPOSTA AO
ESTÍMULO TETÂNICO. 47
4.2.1. BLOQUEIO DA CÁLCIO-ATPASE DO RETÍCULO ENDOPLASMÁTICO 50
4.2.2. BLOQUEIO DA CÁLCIO-ATPASE DA MEMBRANA PLASMÁTICA 53
4.2.3. BLOQUEIO DO TROCADOR NA+/CA
+2 DA MITOCÔNDRIA 56
4.2.4. BLOQUEIO DO UNIPORTER MITOCONDRIAL 59
4.2.5. BLOQUEIO DO TROCADOR NA+/CA
+2 DA MEMBRANA PLASMÁTICA 62
4.3. IMPORTÂNCIA DO RECEPTOR DE IP3 DO TERMINAL PRÉ-SINÁPTICO NO AUMENTO DA ENTRADA
DE SÓDIO, NA DEPRESSÃO E NA FACILITAÇÃO POR PULSOS PAREADOS. 71
4.3.1. EFEITO DA BREVETOXINA NA LIBERAÇÃO ESPONTÂNEA 71
4.3.2. BLOQUEIO DO RECEPTOR DE RIANODINA 75
4.3.3. BLOQUEIO DO RECEPTOR DE IP3 78
4.3.4. BLOQUEIO DA FOSFOLIPASE C 81
V. DISCUSSÃO 84

5.1. ANÁLISE DA LIBERAÇÃO DE NEUROTRANSMISSORES NA JUNÇÃO NEUROMUSCULAR 84
5.2. PLASTICIDADE SINÁPTICA 86
5.2.1. DEPRESSÃO SINÁPTICA 86
5.2.2. FACILITAÇÃO POR PULSOS PAREADOS 88
5.3. EFEITO DOS DIFERENTES MECANISMOS DE DEPURAÇÃO DO CÁLCIO INTRACELULAR SOBRE A
DEPRESSÃO E A FACILITAÇÃO POR PULSOS PAREADOS 89
5.4. EFEITO DA BREVETOXINA NA LIBERAÇÃO ESPONTÂNEA 95
5.5. EFEITO DOS RECEPTORES DE RIANODINA E DE IP3 SOBRE A DEPRESSÃO E A FACILITAÇÃO
POR PULSOS PAREADOS 97
VI. CONCLUSÃO 101
VII. REFERÊNCIAS BIBLIOGRÁFICAS 102
ANEXO I – PROTOCOLOS EXPERIMENTAIS 117
ANEXO II – ARTIGOS EM PROCESSO DE ELABORAÇÃO E SUBMISSÃO 122

I. INTRODUÇÃO
1.1. Junção neuromuscular
A junção neuromuscular (JNM), também denominada de placa motora ou de junção
mioneural, é a região de comunicação especializada onde sinais são transmitidos de um
neurônio a uma célula muscular (KATZ, 1966). Em virtude da grande dimensão da fibra
muscular e da acessibilidade, a JNM foi a primeira sinapse química de vertebrados a ser
bem caracterizada, e onde as bases da teoria quântica da liberação de neurotransmissores
foram estabelecidas (KATZ, 1966; VAN DER KLOOT & MOLGÓ, 1994).
Muitas das propriedades funcionais da JNM de rã foram elucidadas inicialmente por B. Katz
e colaboradores na Inglaterra. Seus estudos produziram as bases do conhecimento sobre a
transmissão química (DELL CASTILLO & KATZ, 1953, 1954a, b; FATT & KATZ, 1952; KATZ
& MILEDI, 1965, 1967). Estudos realizados nesta sinapse servem de suporte para o estudo
da neurotransmissão colinérgica no Sistema Nervoso Central (SNC) e em outras espécies
(VAN DER KLOOT & MOLGÓ, 1994). Além disso, vários mecanismos de modulação da
neurotransmissão descobertos na junção neuromuscular foram verificados posteriormente
no SNC, reforçando que a JNM é um modelo adequado para estudar a neurotransmissão
colinérgica (WOOD & SLATER, 2001).
As fibras musculares são inervadas por motoneurônios mielinizados que se originam do
corno anterior da medula espinhal (RUFF, 2003). As sinapses químicas da JNM apresentam
três estruturas distintas e interligadas: o terminal pré-sináptico, a fenda sináptica e o aparato
de recepção pós-sináptico. O terminal pré-sináptico da JNM contém vesículas, mitocôndrias
e retículo endoplasmático (BIRKS et al., 1960; WESTRUM & GRAY, 1986). O axônio motor
dá origem em sua porção distal a um conjunto de ramos terminais não mielinizados de 1,5
μm de diâmetro que percorrem sulcos rasos na superfície da fibra muscular por extensões
de aproximadamente 100 μm (BIRKS et al., 1960; DE ROBERTIS & BENNETT, 1955;
DULHUNTY & FRANZINI-ARMSTRONG, 1975; KATZ, 1966). Esta região é recoberta por
um tipo de célula glial denominada célula de Schwann. Estudos recentes sugerem que estas
células são capazes de modular a neurotransmissão (AULD & ROBITAILLE, 2003;
CASTONGUAY & ROBITAILLE, 2001; ROBITAILLE, 1998). Ao longo do terminal pré-
sináptico encontram-se as zonas ativas identificadas pela concentração de vesículas
sinápticas de aproximadamente 50 nm de diâmetro que apresentam acetilcolina (ACh) em

seu interior (JAHN & SUDHOF, 1994). O número de vesículas presentes em cada JNM é de
aproximadamente 3 x 106 (BIRKS et al., 1960; DE ROBERTIS & BENNET, 1955). A zona
ativa é uma região especializada na liberação dos neurotransmissores para a fenda
sináptica, pois apresenta uma maquinaria especializada para tal secreção. O grande número
de mitocôndrias no terminal nervoso supre a demanda metabólica da síntese e liberação de
neurotransmissores (BIRKS et al., 1960). Ainda compondo o terminal pré-sináptico estão
presentes os canais para cálcio sensíveis à voltagem, dispostos em fileiras e associados à
zona ativa. A disposição dos canais para cálcio nas zonas ativas permite um rápido aumento
na concentração intracelular de cálcio de cerca de 100 a 1000 nM nas regiões do terminal
nervoso onde a fusão de vesículas ocorre (AUGUSTINE et al., 1991; ROBITAILLE et al.,
1990; SMITH & AUGUSTINE, 1988; ZHAI & BELLEN, 2004).
O terminal pós-sináptico constitui a superfície da fibra muscular, recoberta por uma
membrana basal, e é especializado na resposta rápida e específica à liberação de
neurotransmissores pelo terminal pré-sináptico. A terminação axônica e a célula muscular
são separadas pela fenda sináptica, que apresentam espessura de 50 a 100 nm (WOOD &
SLATER, 2001) e onde existe um material amorfo, rico em carboidratos, e também a enzima
acetilcolinesterase (AChE) (BIRKS et al., 1960; WOOD & SLATER, 2001). Cada vesícula
sináptica libera cerca de 10.000 moléculas de ACh na fenda sináptica (KUFFLER &
YOSHIKAMI, 1975; MARTIN, 1966; MILEDI et al., 1983). Um potencial de ação no terminal
nervoso ocasiona a fusão de 50 a 300 vesículas sinápticas (KATZ & MILEDI, 1979). A AChE
na lâmina basal da membrana pós-sináptica e na fenda sináptica hidrolisa a ACh na fenda
sináptica. Grande parte da ACh liberada na fenda sináptica é hidrolisada antes de alcançar
os receptores nicotínicos. A concentração da AChE é de aproximadamente 3.000
moléculas/µm² na membrana pós-sináptica (MCMAHAN et al., 1978), que é cerca de 5 a 8
vezes menor que a concentração de receptores para a ACh (LAND et al., 1981). O terminal
pós-sináptico apresenta um grande número de receptores nicotínicos. A concentração de
receptores nicotínicos na placa motora é de cerca de 15.000 a 20.000 receptores/µm2
(LAND et al., 1981; RUFF, 1986). O arranjo dos receptores permite a detecção rápida e
eficiente da ACh liberada durante a exocitose (HALL & SANES, 1993).
Portanto, a estrutura da junção neuromuscular possibilita a integração entre o sistema
nervoso e as células musculares, constituindo também o principal modelo experimental para
estudo das funções sinápticas colinérgicas em virtude de sua simplicidade morfológica, de
suas dimensões amplas e de sua acessibilidade quando comparada, por exemplo, com uma
sinapse entre neurônios no sistema nervoso central (SANES & LICHTMAN, 2001).

1.2. Neurotransmissão colinérgica
A transmissão sináptica na JNM envolve a liberação de neurotransmissores presentes nas
vesículas sinápticas e a ativação dos receptores pós-sinápticos. Para que tal fenômeno
ocorra é necessária a despolarização do terminal pré-sináptico, abertura dos canais de
cálcio sensíveis à voltagem e influxo do íon cálcio (PARNAS et al., 2000). Além dos canais
para cálcio, estão presentes canais para potássio no terminal nervoso, incluindo o canal de
potássio ativado por cálcio. Há evidências de que estes canais são componentes das zonas
ativas (ROBITAILLE et al., 1993a, b) e que estão envolvidos em limitar a duração da
despolarização do terminal nervoso, e assim limitar a entrada de cálcio e a liberação do
transmissor.
A ACh presente em uma única vesícula é frequentemente referida como um quantum de
transmissor. A exocitose espontânea de vesículas ocorre na maioria dos terminais nervosos
já estudados. Na JNM esta liberação espontânea origina pequenas despolarizações de
cerca de 0,5 mV na membrana da fibra muscular, sendo denominadas de potenciais de
placa motora em miniatura (MEPPs, do inglês miniature end plate potential). A frequência de
ocorrência destes potenciais em miniatura é geralmente é de 1 a 10 por segundo (VAN DER
KLOOT, 1991). O potencial de placa motora em miniatura, também denominado de
potencial de placa espontâneo, apresenta o mesmo formato dos potenciais de placa motora
ou potenciais evocados (EPP, do inglês end plate potential), sendo que a amplitude dos
potenciais evocados, quando apresenta valores até 6 a 8 mV, é geralmente um valor
múltiplo dos potenciais espontâneos. Além disso, ambos respondem de forma equivalente
quando inibidos por curare. (DEL CASTILLO & KATZ, 1954b; FATT & KATZ, 1952; MARTIN,
1966). Estas evidências proporcionaram a formulação da teoria quântica da liberação de
neurotransmissores.
No motoneurônio em condições fisiológicas, o impulso nervoso causa a liberação de 20 a
200 quanta, dependendo da espécie, em uma fração de milissegundos. Na JNM de rã a
duração da corrente de placa motora (EPC) é de 2 ms (VAN DER KLOOT, 1991; WOOD &
SLATER, 2001) e o tempo entre a liberação do neurotransmissor e sua resposta é de 0,5 ms
(BALDO et al., 1986; BARRET & STEVENS, 1972). Os quanta atuam em conjunto para
originar uma grande despolarização na região da JNM, o potencial de placa motora. O
número médio de quanta liberado por um impulso nervoso em uma junção neuromuscular é
conhecido como o conteúdo quântico do potencial de placa motora. O conteúdo quântico
pode ser determinado eletrofisiologicamente, e é comumente utilizado na mensuração da
liberação evocada do transmissor (WOOD & SLATER, 2001).

A ACh liberada na fenda sináptica se difunde por esta estrutura, e caso alcance a superfície
externa da membrana plasmática muscular da placa motora, liga-se a uma proteína
receptora específica denominada de receptor nicotínico (um canal para cátions dependente
de ligante). A combinação de duas moléculas de ACh com sua proteína receptora leva à
abertura deste canal, originando a despolarização denominada de potencial de placa. A
despolarização é transitória porque a ação de ACh é interrompida por sua hidrólise pela
acetilcolinesterase e pela alteração conformacional do receptor nicotínico (KUFFLER &
YOSHIKAMI, 1975; LINDER et al., 1984).
A JNM apresenta um fator de segurança alto, de tal maneira que um potencial de ação pré-
sináptico tem sempre como resposta um potencial de ação pós-sináptico. Entretanto, em
certas doenças neuromusculares, como na miastenia gravis, essa margem de segurança
está reduzida (ANTOZZI, 2003; VICENT, 2002). Como a JNM representa o último elo entre
o SNC e o desencadeamento da atividade motora, doenças que afetam sua função
apresentam sérias consequências clínicas (MCCONVILLE & VICENT, 2002). Portanto, é de
interesse entender os mecanismos que controlam a liberação de acetilcolina nesta sinapse.
1.3. Plasticidade sináptica
A plasticidade sináptica consiste na habilidade do Sistema Nervoso de alterar a função em
reposta a mudanças nas aferências em situações fisiológicas e patológicas. Nestes casos, é
verificada que a margem de segurança está alterada, pois em situações de estimulação
repetitiva verifica-se o aumento ou a diminuição da liberação de neurotransmissores, o que
ocasiona alteração da eficácia sináptica e consequentemente alteração da força muscular,
caso este fenômeno ocorra na JNM. A plasticidade sináptica foi descoberta por técnicas de
eletrofisiologia, sendo o registro eletrofisiológico das respostas sinápticas (potenciais ou
correntes sinápticas) capaz de verificar se a função sináptica foi modificada. Estudos dos
fenômenos de plasticidade auxiliam no esclarecimento da fisiologia neuromuscular e na
elucidação da etiologia de distúrbios que apresentam eficácia sináptica alterada como
ocorre em doenças neurodegenerativas e também no desenvolvimento de novos e mais
eficazes tratamentos (MCCONVILLE & VICENT, 2002).
De acordo com a duração do fenômeno a plasticidade sináptica pode ser caracterizada
como de curto ou de longo prazo. A plasticidade sináptica de curto prazo apresenta duração
de milissegundos a poucos minutos e a de longo prazo apresenta duração de minutos,
horas, dias ou até a vida inteira, sendo que neste caso ocorre síntese de proteínas e

alterações morfológicas (BLISS, 1990; LINDEN & CONNOR, 1995; PAN & ZUCKER, 2009;
ZUCKER & REGEHR, 2002). Neste trabalho consideramos somente a plasticidade sináptica
à curto prazo por esta estar mais diretamente relacionada à alterações no terminal pré-
sináptico da JNM.
A plasticidade sináptica à curto prazo na JNM é comumente classificada como depressão,
facilitação, aumentação e potenciação de acordo com o estímulo e duração deste para sua
geração e consequente alteração na liberação de neurotransmissores (aumento ou redução)
(MAGLEBY & ZENGEL, 1976b, 1982; ZUCKER, 1989).
A depressão pode ser definida como a diminuição da liberação de neurotransmissores
durante a estimulação repetida, sendo uma consequência da estimulação nervosa em
soluções de cálcio normal. A magnitude da depressão está diretamente relacionada à
quantidade de ACh liberada pelo terminal nervoso (TAKEUCHI, 1958; THIES, 1965). Alguns
estudos sugerem que a depleção dos pools de ACh está associada à depressão
neuromuscular (OTSUKA et al., 1962). O terminal nervoso contém uma determinada
quantidade de quantas disponíveis para a liberação imediata, pool de vesículas prontamente
liberáveis. Parte deste estoque é liberado a cada potencial de ação. Se os potenciais de
ação ocorrem mais frequentemente do que o repreenchimento do estoque, a liberação
quântica por estímulo diminui (LILEY & NORTH, 1953).
Diferentemente do que ocorre na depressão, nos processos de facilitação, aumentação e
potenciação a estimulação nervosa repetitiva ocasiona o aumento da liberação de ACh
(MAGLEBY & ZENGEL, 1975a,b).
A facilitação é mais facilmente visualizada quando o terminal nervoso é estimulado duas
vezes, sendo o intervalo entre os pulsos de milissegundos ou dezenas de milissegundos.
Neste caso, o segundo EPP apresenta maior amplitude e é designado como “facilitado”. À
medida que o intervalo entre os pulsos aumenta a facilitação diminui. A facilitação pode
ocorrer pelo aumento na concentração de uma ou mais substâncias seguidas do potencial
de ação. Katz e Miledi (1968) sugerem que esta substância seja o íon cálcio. Parte do cálcio
que entra no terminal após o primeiro pulso é depurado lentamente, portanto a entrada de
cálcio após o segundo potencial de ação é adicionada ao cálcio residual já presente,
aumentando a liberação de neurotransmissores (BITTNER & SCHATZ, 1981; DODGE &
RAHAMIMOFF, 1967; STOCKBRIDGE & HINES, 1982).
A aumentação diferencia-se da facilitação porque apresenta um decaimento mais lento. Sua
constante de decaimento, 7 segundos, é intermediária entre as constantes da facilitação e
da potenciação (MAGLEBY & ZENGEL, 1976a, b).

A potenciação consiste no aumento do número de quantas liberados por um potencial de
ação, assim como também um aumento da liberação espontânea. Pode desenvolver-se
após um único estímulo nervoso, mas é mais aparente após a estimulação de alta
frequência e em situações de baixa concentração de cálcio (MAGLEBY & ZENGEL, 1975a).
A constante de decaimento encontra-se entre 6 segundos a 2 minutos de acordo com as
condições de estimulação (MAGLEBY & ZENGEL, 1975b).
O cálcio é o principal segundo mensageiro envolvido nos processos de neurotransmissão e
plasticidade sináptica. Como vários tipos celulares os neurônios apresentam fontes de cálcio
intra e extracelulares e as utilizam para a liberação de neurotransmissores (BERRIDGE,
1998). Em condições basais existe um equilíbrio entre os mecanismos de influxo e efluxo de
cálcio que mantêm sua concentração citosólica baixa entre 10 a 100 nM. Quando ocorre a
despolarização da membrana celular e estimulação das células por despolarização do
potencial de membrana, transmissores químicos ou estímulos mecânicos como pressão ou
estiramento, ocorre um aumento na concentração intracelular de cálcio que pode alcançar 1
M ou mais, dependendo do tipo celular. O aumento da concentração citoplasmática é
necessário para que o cálcio exerça suas funções (CARAFOLI & BRINI, 2000). Os influxos
de cálcio, originários de porções restritas da membrana plasmática, fazem com que o
aumento da concentração deste íon possa ser transitório e delimitado a pequenos volumes
do citoplasma. Isso ocorre devido à pequena mobilidade do cálcio no citoplasma e à alta
eficiência de vários sistemas sequestradores que restauram rapidamente a concentração do
cálcio aos níveis normais, sem que o restante do citoplasma seja influenciado (BERRIDGE
et al., 2003., DODGE & RAHAMIMOFF, 1967).
No terminal pré-sináptico da JNM o retículo endoplasmático (RE) é a principal organela
responsável por estocar e liberar o cálcio. O íon cálcio pode ser liberado pelos receptores de
rianodina (RyR) e pelos receptores de inositol trifosfato (IP3R) presentes nesta organela
(BERRIDGE, 1998; COLLIN et al., 2005; FILL & COPELLO, 2002; NOWYCKY & THOMAS,
2002; PAREKH, 2003).
Os mecanismos de remoção do cálcio citoplasmático são importantes para a manutenção
da atividade neuronal (GWAG et al., 1999; JANH et al., 2006). A extrusão do cálcio para o
meio extracelular pode ser realizada por duas famílias de proteínas: o trocador Na+/Ca+2
(NCX) e a cálcio-ATPase da membrana plasmática (PMCA). Além disso, o cálcio intracelular
pode ser removido do citoplasma por uma variedade de tampões, bombas e transportadores
específicos de organelas. A recaptação para o RE é regulada por uma família de cálcio-
ATPases de retículo sarco-endoplasmático (SERCA) (BERRIDGE et al., 2003). A
recaptação pela mitocôndria é mediada pelo uniporter mitocondrial (MCU), já a recaptação

pelo complexo de Golgi é mediada pela cálcio-ATPase do tipo P (PMR1/ATP2C1)
(NOWYCKY & THOMAS, 2002).
A etiologia de algumas doenças neuromusculares está relacionada ao comprometimento na
depuração do cálcio intracelular (GUATTEO et al., 2007). Em outras, a sinalização do cálcio,
envolvendo os RyR e IP3R é que está comprometida (CHEN et al., 2008; GAILLY, 2002).
Este fato reforça a importância de estudos da sinalização do cálcio intracelular na JNM.
Existem vários estudos que correlacionam o cálcio de organelas intracelulares e a
neurotransmissão (BERRIDGE et al., 2003). Com relação aos mecanismos de depuração do
cálcio intracelular já se sabe quais apresentam um papel fisiológico na neurotransmissão e
na plasticidade sináptica de diversas sinapses, como citado acima. Jensen et al. (2007)
verificaram que a PMCA é importante na cinética da depuração pré-sináptica de cálcio em
terminais excitatórios da região CA3 do hipocampo. O bloqueio da PMCA aumenta a
facilitação por pulsos pareados, além de aumentar a frequência das correntes pós-sinápticas
excitatórias miniaturas sem alterar o tamanho destas. Castonguay e Robitaille (2001)
verificaram que os estoques intracelulares de cálcio na JNM de rã, mais especificadamente
o RE, atuam como um mecanismo de depuração do cálcio para limitar a duração e a
liberação de neurotransmissores. Bennett et al. (2007) desenvolveram um modelo
matemático do sequestro de cálcio e o aplicaram às observações experimentais dos
transientes de cálcio na JNM em diferentes protocolos de estimulação seguidos do bloqueio
de diferentes vias de depuração de cálcio. O modelo matemático foi desenvolvido levando-
se em consideração parâmetros de diferentes sinapses, pois nem todos os parâmetros
relacionados à depuração do cálcio na junção neuromuscular são conhecidos. Verificou-se
que diferentes mecanismos de sequestro de cálcio apresentam contribuições diferenciadas.
No entanto, não existem trabalhos que quantifiquem eletrofisiologicamente e comparem a
importância de diferentes mecanismos de depuração em fenônemos de plasticidade à curto
prazo na JNM.
Nos neurônios do SNC, principalmente no hipocampo e no cerebelo, a plasticidade sináptica
é mediada pela liberação de cálcio induzida pelo cálcio (CICR, do inglês Ca2+-induced Ca2+
release) e pela liberação de cálcio dependente de inositol trifosfato (IP3) (CARTER et al.,
2002; EMPTAGE et al., 2001; INOUE et al., 1998; LAURI et al., 2003). Alguns trabalhos
evidenciam que nas junções neuromusculares de rã e de camundongo de que a liberação
de cálcio de estoques intracelulares pode modular a liberação de neurotransmissores
(BRAILOIU & MIYAMOTO, 2000; BRAILOIU et al., 2003; DROPIC et al., 2005; HACHISUKA
et al., 2007; KUBOTA et al., 2005; NARITA et al., 2000; PIRIZ et al., 2003). Com relação aos
RyR, seu papel na neurotransmissão, na plasticidade sináptica e sua presença no terminal

pré-sináptico já estão bem esclarecidos (KUBOTA et al., 2005; NARITA et al., 1998, 2000).
No entanto, existem somente evidências indiretas da presença do IP3R no terminal pré-
sináptico da JNM, como o trabalho de Brailoiu e Miyamoto (2000), onde foi verificada que a
admistração de IP3 ocasiona um aumento do conteúdo e do tamanho quântico.
Posteriormente este mesmo grupo de pesquisadores verificaram que a modulação da
liberação de neurotransmissores pela serotonina na JNM de lagostina envolve estoques de
cálcio intracelulares sensíveis à rianodina e ao IP3 (DROPIC, et al., 2005). Yang et al. (2001)
verificaram que neurotrofinas do tipo 3 induzem potenciação da transmissão sináptica na
junção neuromuscular de girino e este efeito está relacionado à ativação dos IP3Rs. No
entanto, não há estudos que identifiquem a presença do IP3R no terminal pré-sináptico e
nem esclareçam seu papel em fenômenos de plasticidade sináptica à curto prazo na JNM.
Tendo em vista as evidências da correlação de doenças neuromusculares e sinalização de
cálcio intracelular, a inexistência de estudos eletrofisiológicos que quantifiquem e comparem
a participação de diferentes mecanismos de depuração do cálcio na plasticidade sináptica à
curto prazo e do papel dos IP3R nestes fenômenos, neste trabalho nós nos propusemos a
aprofundar o estudo da regulação do cálcio intracelular em alguns fenômenos de
plasticidade à curto prazo. Para a realização deste estudo utilizamos os registros
eletrofisiológicos, considerados ferramentas úteis e fidedignas para avaliar informações de
forma qualitativa e quantitativa da liberação de neurotransmissores.
A plasticidade sináptica é um fenômeno complexo e amplo. Escolhemos verificar nossos
objetivos em fenômenos de plasticidade sináptica à curto prazo, pois esta apresenta funções
fisiológicas importantes como a ativação de sinapses silenciosas e a indução da plasticidade
sináptica à longo prazo (MUKHAMEDYAROV, et al., 2009). Os mecanismos envolvidos nas
alterações das respostas sinápticas não estão completamente elucidados. O esclarecimento
dos mecanismos fisiológicos envolvidos é importante e deve ser realizado com diferentes
técnicas nas diversas sinapses, uma vez que mecanismos distintos podem estar envolvidos
na neurotransmissão.

II. OBJETIVOS
Verificar a importância dos diferentes mecanismos de depuração do cálcio
intracelular em fenômenos de plasticidade sináptica à curto prazo. Os mecanismos
de depuração estudados foram:
Cálcio-ATPase do retículo endoplasmático;
Cálcio-ATPase da membrana plasmática;
Trocador sódio-cálcio mitocondrial;
Uniporter mitocondrial;
Trocador sódio-cálcio da membrana plasmática.
Verificar a presença do receptor de inositol trifosfato através da utilização da
brevetoxina do tipo 3, uma toxina cujo efeito está relacionado ao aumento da
concentração intracelular de inositol trifosfato e à liberação de cálcio via ativação dos
receptores deste segundo mensageiro.
Verificar a importância do receptor de rianodina do terminal pré-sináptico na
depressão e na facilitação por pulsos pareados.
Verificar a importância do receptor de inositol trifosfato do terminal pré-sináptico na
depressão e na facilitação por pulsos pareados.

III. MATERIAIS E MÉTODOS
3.1. Materiais
3.1.1. Animais
Realizamos experimentos com rãs (Lithobates catesbeianus e Rana temporaria) de ambos
os sexos, pesando entre 20 e 70 g. Os animais foram criados em ranário e mantidos no
laboratório de Eletrofisiologia Celular (ICB-UFMG) e no Laboratoire de Neurobiologie
Cellulaire et Moleculaire (CNRS-Gif sur Yvette/França), em tanque com água corrente.
Nosso estudo foi aprovado pelos comitês de éticas em experimentação animal do Brasil
(CETEA/UFMG) (protocolo n° 164/2007) e da França.
3.1.2. Equipamentos
3.1.2.1. Sistema para aquisição de dados:
Estimulador Grass SD9 ou S48 (Astro-Med Inc., EUA);
Ociloscópio (Tektronix, Beaverton, EUA);
Micromanipulador (World Precision Instruments, EUA);
Microscópio esteroscópio PGH Rundfunk – Fernsehen (Carl Zeiss, Alemanha);
Amplificador I – Dagan 8500 (Dagan Co., Minneapolis, EUA) ou Axoclamp-2A (Axon
Instruments, EUA);
Amplificador II – Ectron 750 (Ectron Co., EUA) ou Grass AC/DC strain gage (Astro-
Med Inc., W. Warnick, EUA);
Conversor A/D Labmaster (Axon Instruments, EUA);
Microcomputador contendo o software SES (Strathclyde Electrophysiology Software -
University of Strathclyde, Escócia);

3.1.2.2. Material cirúrgico: tesouras, pinças anatômicas e alfinetes para insetos;
3.1.2.3. Câmara de acrílico contendo Sylgard® (EUA) em sua base;
3.1.2.4. Mesa antivibratória TMC (EUA);
3.1.2.5. Estirador de Microeletrodos modelo PN-30 (Narishige; Japão) ou DKI modelo
700 C (Tujunga, CA, EUA);
3.1.2.6. Balanças Eletrônicas (OHAUS – presicion standard, EUA e BEL engineering
Mark 210 A, EUA);
3.1.2.7. Medidor de pH – Handylab 1 (SCHOTT – Alemanha);
3.1.2.8. Capilares GC150F-15 (Clark Electromedical Instruments – Inglaterra);
3.1.2.9. Pipetadores automáticos (Daigger, Finlândia; Boeco, Alemanha; Oxford
Benchmate, Japão).
3.1.3. Drogas e reagentes
1-[6-((17b-3-Methoxyestra-1,3,5(10)-trien-17-yl)amino)hexyl]-1H-pyrrole-2,5-dione
(U-73122) (Calbiochem – EUA).
2-Aminoethyl diphenyl borate (2-APB) (Sigma-Aldrich – França).
7-Chloro-5-(2-chlorophenyl)-1,5-dihydro-4,1-benzothiaze pin-2(3H)-one (CGP
37157) (Tocris, EUA).
BAPTA, AM (1,2-Bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid
tetrakis(acetoxymethyl ester) (Molecular Probes – EUA).
Brevetoxina (PbTx-3) (Latoxan, França).
CEDA, SE (5-(and-6)-carboxyeosin diacetate, succinimidyl ester) (Invitrogen,
Molecular Probes – EUA).
d-tubocurarina (Sigma-Aldrich - EUA).
(μ)[(HCO2)(NH3)4Ru]2OCl3 (Ru360) (Calbiochem – EUA).
Tapsigargina (Tg) (ALOMONE LABS - Jerusalém, ISRAEL).
Tetrodotoxina (TTX) Sigma (Sigma-Aldrich, França)
Rianodina (Ry) (Sigma-Aldrich – França).
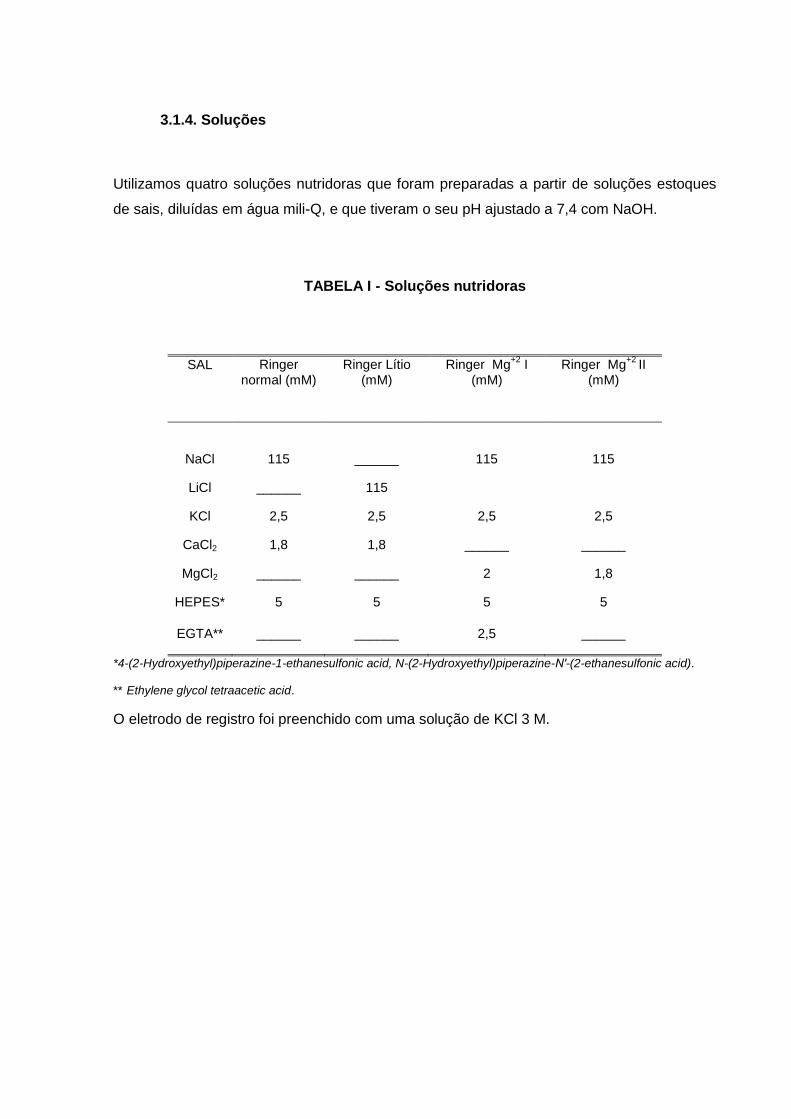
3.1.4. Soluções
Utilizamos quatro soluções nutridoras que foram preparadas a partir de soluções estoques
de sais, diluídas em água mili-Q, e que tiveram o seu pH ajustado a 7,4 com NaOH.
TABELA I - Soluções nutridoras
SAL Ringer normal (mM)
Ringer Lítio (mM)
Ringer Mg+2
I (mM)
Ringer Mg+2
II (mM)
NaCl 115 ______ 115 115
LiCl ______ 115
KCl 2,5 2,5 2,5 2,5
CaCl2 1,8 1,8 ______ ______
MgCl2 ______ ______ 2 1,8
HEPES* 5 5 5 5
EGTA** ______ ______ 2,5 ______
*4-(2-Hydroxyethyl)piperazine-1-ethanesulfonic acid, N-(2-Hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid).
** Ethylene glycol tetraacetic acid.
O eletrodo de registro foi preenchido com uma solução de KCl 3 M.
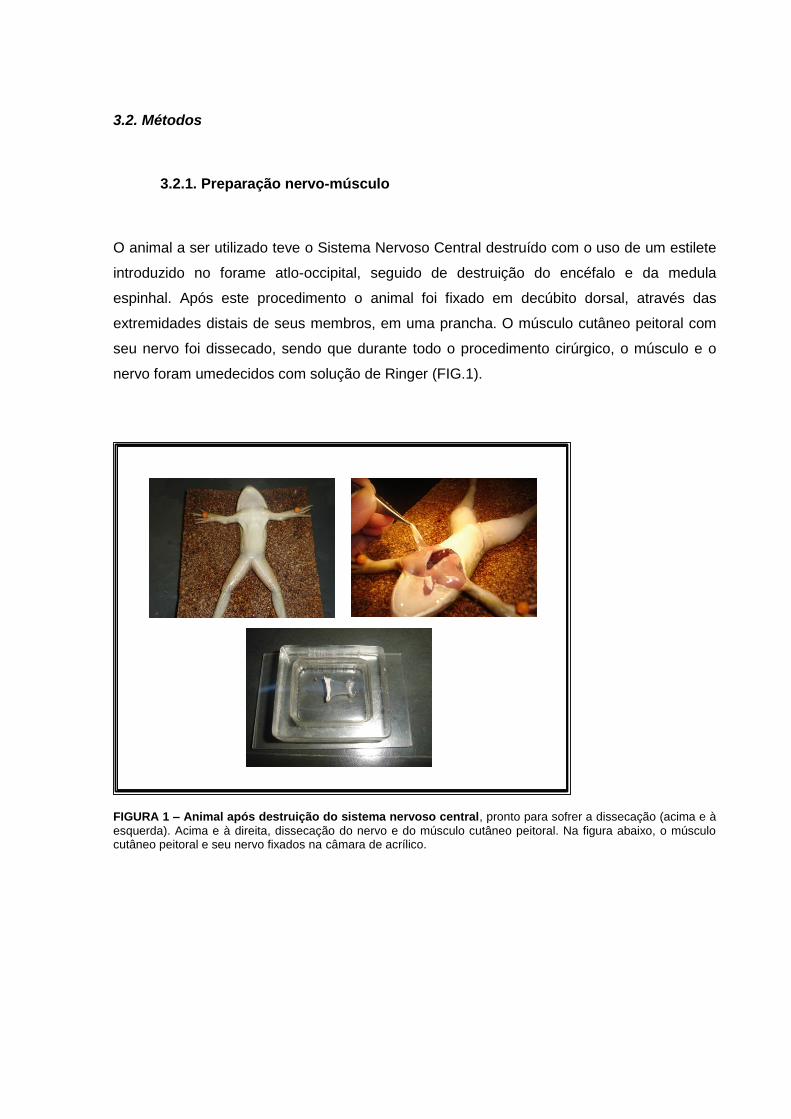
3.2. Métodos
3.2.1. Preparação nervo-músculo
O animal a ser utilizado teve o Sistema Nervoso Central destruído com o uso de um estilete
introduzido no forame atlo-occipital, seguido de destruição do encéfalo e da medula
espinhal. Após este procedimento o animal foi fixado em decúbito dorsal, através das
extremidades distais de seus membros, em uma prancha. O músculo cutâneo peitoral com
seu nervo foi dissecado, sendo que durante todo o procedimento cirúrgico, o músculo e o
nervo foram umedecidos com solução de Ringer (FIG.1).
FIGURA 1 – Animal após destruição do sistema nervoso central, pronto para sofrer a dissecação (acima e à
esquerda). Acima e à direita, dissecação do nervo e do músculo cutâneo peitoral. Na figura abaixo, o músculo cutâneo peitoral e seu nervo fixados na câmara de acrílico.

3.2.2. Sistema de aquisição de dados
O sistema de aquisição de dados era montado em cima de uma mesa antivibratória (TMC,
EUA), que sustentava a preparação neuromuscular. Sobre esta mesa havia um “eletrodo de
sucção” sustentado por uma haste e conectado a uma seringa (5 ml), a qual sofria uma leve
pressão negativa para se obter a interiorização do nervo. Na mesa também se encontrava
um micromanipulador, que sustentava e conectava o prendedor de microeletrodos,
permitindo sua aproximação lenta e precisa da junção neuromuscular (FIG. 2).
FIGURA 2 – Foto mostrando a mesa onde eram realizados os experimentos em JNM.
A diferença de potencial transmembrana era captada pelo eletrodo de registro em relação
ao eletrodo de aterramento presente na solução. O sinal era enviado ao pré-amplificador,
que o enviava ao amplificador I, que apresentava duas saídas, ambas com um ganho de
10X e filtragem passa-baixa de 10 KHz: a) uma que era filtrada (passa-alta – 0,1 Hz) para
remover o potencial de repouso antes de ser amplificada 50 a 1000X (cinquenta a mil
vezes) pelo amplificador II; b) a segunda saída dirigia-se direto ao osciloscópio para a
visualização do sinal (FIG. 3). A digitilização foi realizada por um conversor A/D Lab Master
com frequência de amostragem de 20 KHz.
Eletrodo de
referência
Eletrodo de registro
Eletrodo estimulado
rr Mesa
antivibratória
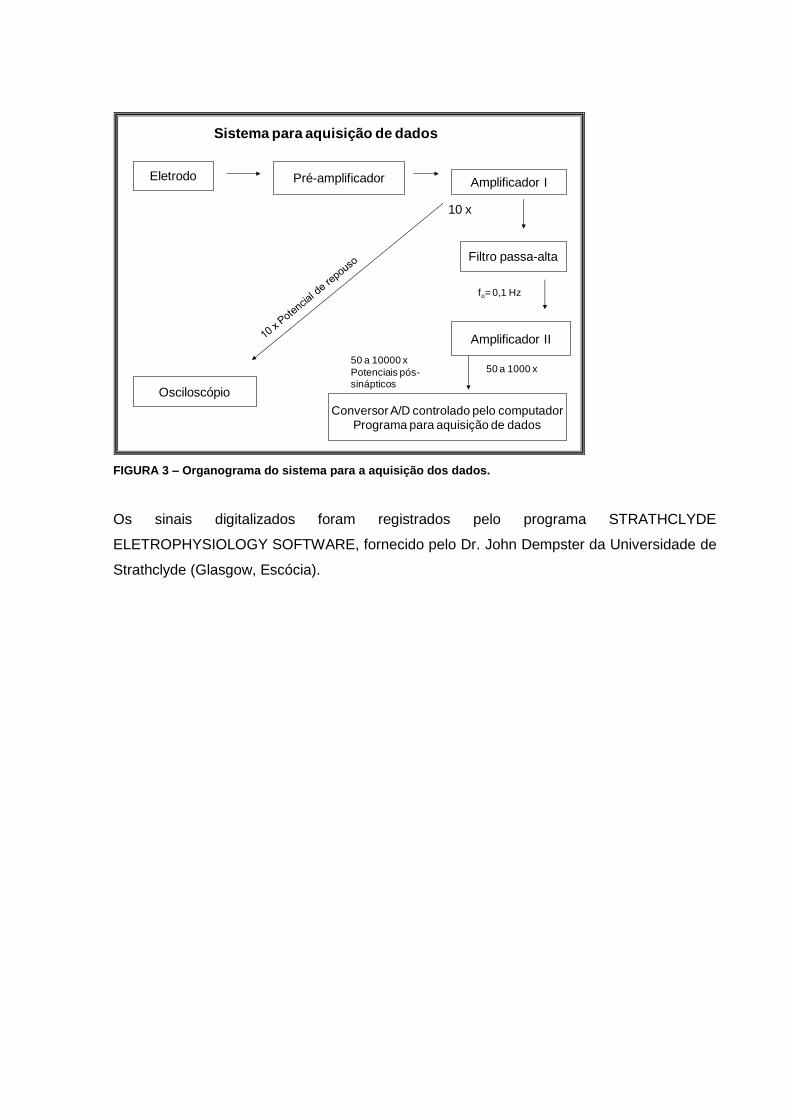
Eletrodo Pré-amplificador Amplificador I
Filtro passa-alta
Amplificador II
Conversor A/D controlado pelo computador
Programa para aquisição de dados
Osciloscópio
50 a 1000 x
10 x
Sistema para aquisição de dados
50 a 10000 x
Potenciais pós-sinápticos
fo= 0,1 Hz
FIGURA 3 – Organograma do sistema para a aquisição dos dados.
Os sinais digitalizados foram registrados pelo programa STRATHCLYDE
ELETROPHYSIOLOGY SOFTWARE, fornecido pelo Dr. John Dempster da Universidade de
Strathclyde (Glasgow, Escócia).

3.2.3. Registros eletrofisiológicos
3.2.3.1. Medida dos potenciais de placa
Empalamento
Amplificador de voltagem e
osciloscópio
Fibra muscular
Eletrodo
extracelular
Vm
(m
V)
+ 60
0
- 90
Potencial de repouso
Tempo (ms)
Eletrodo
de registroEmpalamento
Amplificador de voltagem e
osciloscópio
Fibra muscular
Eletrodo
extracelular
Vm
(m
V)
+ 60
0
- 90
Potencial de repouso
Tempo (ms)
Empalamento
Amplificador de voltagem e
osciloscópio
Fibra muscular
Eletrodo
extracelular
Vm
(m
V)
+ 60
0
- 90
Potencial de repouso
Tempo (ms)
Eletrodo
de registro
FIGURA 4 – Registro da diferença de potencial medida fora e dentro de uma célula.
O microeletrodo intracelular foi utilizado para registrar os potencias de placa motora.
Mensuramos as variações do potencial de membrana da fibra muscular na região da placa
motora. O registro foi realizado pela diferença de potencial do eletrodo de referência
(denominado zero ou aterramento) em relação ao eletrodo de registro. Quando ambos estão
fora da célula, nenhuma diferença de potencial é registrada. Assim que o eletrodo de
registro é inserido na célula o osciloscópio mostra a mudança de potencial, ou seja, o
potencial de repouso da membrana. (FIG. 4).
Inicialmente realizamos o registro do potencial de membrana, através do empalamento da
célula muscular monitorada no osciloscópio. O empalamento era aceito quando uma
deflexão brusca de pelo menos -70 mV ocorria. No entanto, os registros eram realizados em
células que apresentavam um potencial de membrana entre -80 a -90 mV. Como o registro
dos potenciais ocorria em fibras que apresentavam diferentes potenciais de repouso
corrigimos a amplitude destes potenciais para -90 mV (KATZ & THESLEFF, 1957; VAN DER
KLOOT & COHEN, 1984) considerando o potencial de reversão de -15 mV (TAKEUCHI &
TAKEUCHI, 1960). A amplitude dos potenciais também foi corrigida pela somação não-
linear (MARTIN, 1955).

A localização do empalamento com relação à placa motora foi avaliada de acordo com os
potenciais obtidos. Os potenciais de placa eram observados, primeiramente, no osciloscópio
e depois na tela do microcomputador. Estes devem apresentar baixo tempo de subida (1
ms) e amplitude elevada, pois quanto mais longe da junção menor seria o sinal, ou seja, o
sinal é inversamente proporcional à distância. Deste modo o registro era realizado o mais
próximo possível da placa motora com o mínimo de perda eletrotônica e com uma proporção
sinal/ruído adequada, permitindo a visualização clara e total do potencial de placa evocado
sem gerar dúvidas durante sua análise.
Os microeletrodos de borosilicato, utilizados no registro intracelular, eram confeccionados
em um estirador de pipetas modelo PN-30 (Narishige) ou DKI modelo 700 C. Os
microeletrodos eram preenchidos com KCl (3M) e inseridos dentro da célula muscular na
região das junções neuromusculares. Estes apresentavam um diâmetro pequeno (< 1 μm) e
resistência de 5 a 15 MΩ.
3.2.3.2. Medidas da liberação evocada
Para o estudo de potenciais de placa evocados, o nervo foi sugado por um eletrodo
conectado ao estimulador. Inicialmente determinamos o limiar de excitação muscular
(tensão elétrica mínima para a qual ocorre contração muscular). Os registros dos potenciais
de placa evocados foram realizados com estímulos supralimiares (3 a 5 vezes o limiar) para
garantir a excitação da fibra pré-sináptica estudada (FIG. 5).
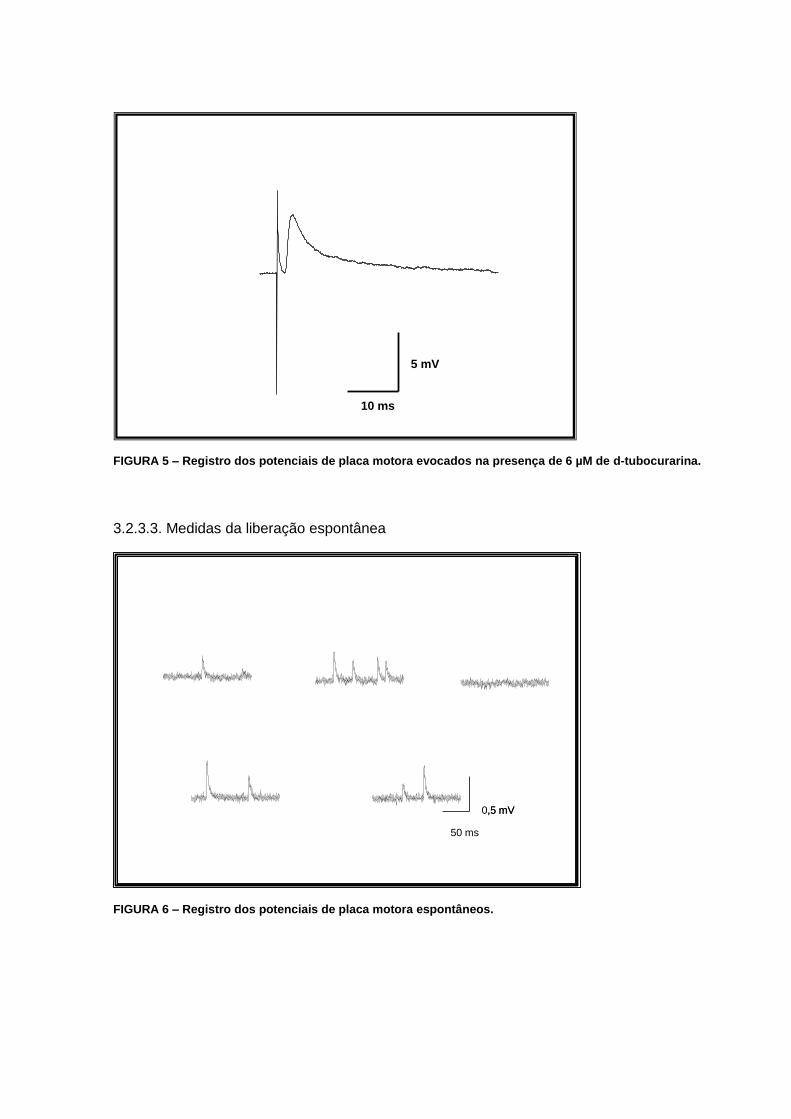
10 ms
5 mV
FIGURA 5 – Registro dos potenciais de placa motora evocados na presença de 6 µM de d-tubocurarina.
3.2.3.3. Medidas da liberação espontânea
0,5 mV
50 ms
0,5 mV
50 ms
FIGURA 6 – Registro dos potenciais de placa motora espontâneos.

A liberação espontânea foi medida através do empalamento da célula muscular e registro
dos potencias de placa motora em miniatura (MEPPs), antes e após o tratamento com a
droga. Foram registrados 100 (cem) MEPPs e sua frequência foi calculada de acordo com o
intervalo de tempo (FIG. 6).
Para o registro dos potenciais espontâneos não havia a necessidade de preservação do
nervo. Nos experimentos realizados com brevetoxina do tipo 3 utilizamos solução de Ringer
livre de Ca+2 com 2 mM Mg+2 e 2 mM EGTA.
3.2.4. Protocolos de Plasticidade Sináptica à curto prazo
Para o estudo de dois fenômenos de plasticidade sináptica à curto prazo utilizamos o
protocolo de facilitação por pulsos pareados e o de depressão. Nos protocolos utilizados
realizamos a mensuração da amplitude dos potenciais de placa evocados.
3.2.4.1. Facilitação por pulsos pareados
No protocolo da facilitação registramos 35 (trinta e cinco) pulsos pareados com frequência
de 0,2 Hz, duração de pulso de 100 μs e intervalo entre os pulsos que variavam entre 5 - 60
ms (FIG. 7). Em alguns casos, quando o intervalo entre os pulsos era de 15 ms ou menos, o
potencial de placa motora gerado pelo primeiro estímulo ultrapassava o potencial de placa
motora gerado pelo segundo estímulo. Nestes casos a amplitude do segundo potencial de
placa motora era calculada do seu pico até a projeção do primeiro potencial de placa motora
(FIG. 8).

FIGURA 7 – Potenciais de placa motora registrados com pulsos pareados nos diferentes intervalos entre os pulsos, na presença de curare. Potencial de repouso –92 mV.
FIGURA 8 – Facilitação por pulsos pareados com intervalo de 10 ms na presença de curare. Projeção do primeiro EPP para o cálculo da amplitude do segundo EPP. Potencial de repouso –92 mV.

A facilitação foi calculada pela seguinte fórmula:
o F = (EPP2 –EPP1) / EPP1, onde
EPP1: média da amplitude dos dez últimos EPPs seguidos do 1º estímulo,
EPP2: média da amplitude dos dez últimos EPPs seguidos do 2º estímulo.
Portanto, a facilitação era definida como o aumento na média da amplitude do potencial de
placa motora seguido do segundo estímulo em relação a média da amplitude do potencial
seguido do primeiro estímulo.
3.2.4.2. Depressão
No protocolo de depressão registramos os potenciais de placa motora durante a estimulação
basal com uma frequência de 0,2 Hz e duração de pulso de 100 µs por três minutos. Em
seguida estimulamos o terminal nervoso a uma frequência de 20 Hz durante um minuto para
o desencadeamento da depressão. Após o término do trem de alta frequência retornamos a
estimulação inicial de 0,2 Hz por mais cinco minutos para o registro da recuperação da
depressão (FIG. 9). Nos gráficos em que expressamos a amplitude dos potenciais de placa
motora durante as estimulações em função do tempo, plotamos somente a amplitude dos
vinte últimos potenciais antes da estimulação de alta frequência e a amplitude de todos os
potenciais de placa motora foi normalizada com a média destes últimos vinte potenciais
durante a estimulação basal de 0,2 Hz.
Estimulação basal_0,2 Hz Estimulação de alta
freqüência_20 HzEstimulação_0,2 Hz
Registro do Potencial de
Repouso
Registro dos EPPs
Depressão Recuperação da
Depressão
Depressão
Estimulação basal_0,2 Hz Estimulação de alta
freqüência_20 HzEstimulação_0,2 Hz
Registro do Potencial de
Repouso
Registro dos EPPs
Depressão Recuperação da
Depressão
Estimulação basal_0,2 Hz Estimulação de alta
freqüência_20 HzEstimulação_0,2 Hz
Registro do Potencial de
Repouso
Registro dos EPPs
Depressão Recuperação da
Depressão
Depressão
FIGURA 9 – Exemplo de um protocolo de Depressão e seu registro no programa Clampfit 9.2.
Durante o estímulo tetânico a amplitude dos EPPs decai rapidamente e exponencialmente
até certo ponto. Após esta primeira fase de decaimento a amplitude dos EPPs continua a
decair, porém mais lentamente e de forma linear. Os dados das amplitudes dos EPPs foram
utilizados para ajustar uma equação com a seguinte fórmula:
o Explinear: y = p1*exp(-x/p2) + p3 + p4*x.
P1 e P2 são a amplitude e a constante de tempo (tau) do componente de decaimento
exponencial. P3 e P4 representam a intersecção no eixo y e a inclinação da fase de
decaimento linear (FIG. 10). Tanto o parâmetro P1 quanto o prolongamento do parâmetro
P3 no eixo y (intersecção) representam a quantidade de depressão da primeira fase de
decaimento exponencial da depressão. No entanto, como em alguns casos verificamos uma
facilitação da liberação de neurotransmissores seguida de um decaimento desta,
consideramos somente o parâmetro P3 para a análise da quantidade de depressão
encontrada.
Portanto, em nosso estudo analisamos a constante de decaimento e a intersecção que
referem-se à velocidade de instalação do fenômeno de depressão e a quantidade de
depressão, respectivamente.
0 10 20 30 40 50 60
0,0
0,5
1,0
Data: Etanol_060607D
Model: ExpLinear
Equation: y = p1*exp(-x/p2) + p3 + p4*x
Weighting:
y No weighting
Chi^2/DoF = 0.00084
R^2 = 0.9648
p1 0.62344 ±0.02235
p2 2.40568 ±0.16654
p3 0.24603 ±0.00916
p4 -0.00474 ±0.0003
Tempo (s)
Po
ten
cia
l d
e p
lac
a m
oto
ra (
pro
po
rçã
o d
o c
on
tro
le)
FIGURA 10 – Exemplo de ajuste da curva de Depressão pela equação Explinear. O traço azul representa o
prolongamento do parâmetro P3 e sua intersecção no eixo y.
Também analisamos a depressão final obtida pela comparação da amplitude dos 5 últimos
EPPs do trem de estímulos de alta frequência com a amplitude basal dos EPPs.
Após o estímulo tetânico a amplitude dos EPPs se recupera e para quantificar o processo de
recuperação medimos os seguintes parâmetros:
Recuperação final: comparação da amplitude dos 5 últimos EPPs 200 s após o trem
de estímulos de alta frequência, quando a recuperação já estava estabilizada, com a
amplitude basal dos EPPs (FIG. 11).
Tempo para a recuperação de 50%: interpolação linear entre o ponto anterior e o
posterior a recuperação de 50%. O último ponto durante o trem de alta frequência foi
considerado 0% de recuperação e o valor aos 200 s após o final do trem foi
considerado 100% de recuperação (FIG. 11).
Potenciação: diferença entre a média do maior EPP e de seus adjacentes anterior e
posterior e a recuperação final (FIG. 11).
0 50 100 150 200
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6P
ote
nc
ial
de
pla
ca
mo
tora
(p
rop
orç
ão
do
co
ntr
ole
)
Tempo (s)
Recuperação final
Potenciação
50 % de
recuperação
FIGURA 11 – Gráfico representativo da recuperação mostrando os pontos correspondentes para o cálculo da recuperação final, potenciação e tempo para recuperar 50%.
3.3. Protocolo experimental
O curare na concentração de 6 µM foi utilizado para o bloqueio da contração muscular nos
experimentos onde registramos os potenciais de placa evocados. O curare era sempre
adicionado à preparação 30 minutos antes do início dos registros eletrofisiológicos, e logo
em seguida a sua adição, a droga ou o veículo de diluição eram adicionados à preparação,
dependendo da situação experimental. A câmara utilizada para a incubação dos músculos
apresentava um volume de 4 a 5 mL e o volume adicionado de curare, das drogas e dos
veículos de diluição era 400 a 1000 vezes menores, de forma que a diluição era
insignificante e a concentração final destes não era alterada de forma considerável.
Para o estudo do bloqueio do trocador NCX da membrana plasmática realizamos a perfusão
da preparação nervo-músculo com a solução de Ringer Lítio após o registro da depressão e
da facilitação por pulsos pareados em solução de Ringer normal. A perfusão apresentava
duração de 20 minutos e o registro dos protocolos iniciava-se 20 minutos após o término
desta. Os registros, controle e experimental, eram realizados na mesma fibra muscular sem
perda do empalamento.
O BAPTA-AM, a tapsigargina, a CEDA, o CGP 37157, o Ru360, a Rianodina, o 2-APB, o U-
73122, a PbTx-3 e os veículos de diluição foram adicionados diretamente ao banho
proveniente de soluções estoques. Os registros eram iniciados 30 minutos após a adição
das drogas ou veículos ao banho. Em um mesmo músculo realizamos o registro de diversas
fibras e este era utilizado somente uma vez para cada situação experimental. Cada grupo,

controle e experimental, apresentava 3 a 5 músculos diferentes, sendo realizado o registro
de 2 a 7 fibras em cada.
Os registros da frequencia dos MEPPs, da facilitação por pulsos pareados, da depressão e
da recuperação da depressão foram realizados após o tratamento com as drogas, com os
veículos de diluição ou com as diferentes soluções nutridoras. Os experimentos com os
veículos de diluição das drogas e os realizados em solução de Ringer normal fizeram parte
dos nossos grupos controles.
Os experimentos controles e experimentais eram realizados de forma intercalada e na
mesma época do ano no intuito de reduzir a variabilidade.
Segue abaixo um esquema representativo dos protocolos (FIG. 12). A descrição de cada
protocolo encontra-se no Anexo I.
FIGURA 12 - Organograma da situação experimental.
3.4. Análise estatística
O teste de normalidade Shapiro-Wilk foi aplicado para observar se as variáveis estudadas
possuíam distribuição normal. As variáveis que não exibiam distribuição normal foram
submetidas à transformação logarítmica (log 10). Utilizamos o teste t de Student não
pareado para comparação entre os grupos experimentais e seus respectivos controles. O
teste t de Student pareado foi utilizado para a comparação dos grupos controles e

experimentais quando a preparação foi perfundida com solução de Ringer lítio. No entanto,
em alguns casos utilizamos o teste t de Student não pareado como na ocorrência da perda
de algum experimento da amostra. A análise One-Way ANOVA seguida do post-test de
Newman-Keuls foi utilizada para a comparação dos diferentes grupos experimentais quando
a preparação foi tratada com PbTx-3. A análise Two-Way ANOVA seguida do post-test de
Bonferroni foi utilizada para a análise da facilitação por pulsos pareados. Diferenças foram
consideradas significativas ao nível de p < 0,05. Os resultados foram expressos como média
erro padrão da média (EPM) e o n significa o número de fibras registradas. Os resultados
dos dados que não apresentaram distribuição normal e que foram submetidos à
transformação logarítmica foram expressos como média desvio padrão da média (DP).

IV. RESULTADOS
4.1. Influência da concentração intracelular de cálcio sobre a resposta ao estímulo
tetânico.
Durante uma estimulação nervosa prolongada frequentemente se observa diminuição da
resposta pós-sináptica, fenômeno conhecido como depressão. Este fenômeno tem origens
pré e/ou pós sinápticas. Com o intuito de verificar a variação da depressão em diferentes
concentrações de cálcio, nós utilizamos o BAPTA-AM, um quelante de cálcio intracelular, na
concentração de 100 µM. Estes experimentos tiveram como objetivo avaliar o quanto da
depressão obtida com nosso protocolo é de origem pré-sináptica. Nossa hipótese de
trabalho foi que concentrações elevadas de cálcio intracelular levam à maior depressão em
decorrência de uma maior liberação de neurotransmissores. Nossos resultados mostram
que o BAPTA-AM diminuiu a amplitude dos EPPs antes da tetania de 2,20,2 mV (n=18)
para 1,30,1 mV (n=16), p=0,0003 (FIG. 13A). Nós também observamos que a depressão
foi menor nas preparações tratadas com BAPTA-AM. A depressão final diminuiu de 97±2%
(n=15) para 61±4% (n=15), p=2x10-8 (FIG. 13E e 19). Estes experimentos sugerem que
nosso protocolo é sensível a variações da concentração intracelular de cálcio.
Estudos anteriores mostraram que a facilitação é dependente da concentração de cálcio
intracelular (MUKHAMEDYAROV et al., 2009; TANABE & KIJIMA, 1992; VAN DER KOOT &
MOLGÓ, 1993).
Com relação à recuperação da depressão e a potenciação também verificamos que estes
fenômenos são sensíveis a concentração de cálcio intracelular. Nossos resultados mostram
que o BAPTA-AM aumentou a intensidade e a velocidade da recuperação e da potenciação.
A recuperação final aumentou de 922% (n=17) para 11511% (n=15), p=0,04 (FIG. 13F e
22) e o tempo necessário para a recuperação diminuiu de 26,5±2,2 s (n=15) para 12,6±3,1 s
(n=12), p=0,0009. Já a potenciação aumentou de 102% (n=16) para 277% (n=15), p=0,02
(FIG. 13G e 23).

Em nossos experimentos subsequentes nós utilizamos a depressão sináptica como
indicador da concentração de cálcio intracelular, para verificarmos a participação de
diferentes sistemas de depuração do cálcio intracelular. Nós também pesquisamos o efeito
do bloqueio dos diferentes sistemas sobre a facilitação por pulsos pareados.

Controle BAPTA-AM 0,0
0,5
1,0
1,5
2,0
2,5
(n=16)
EP
P (
mV
)
***
(n=18)
Controle BAPTA-AM 0
20
40
60
80
100
(n=13)(n=15)
% D
ep
res
sã
o f
ina
l
***
Controle BAPTA-AM 0
5
10
15
20
25
30
35
40
(n=15)(n=16)
% P
ote
nc
iaç
ão
*
Controle BAPTA-AM 0
20
40
60
80
100
120
(n=15)(n=17)
% R
ec
up
era
çã
o f
ina
l *
0 100 200 300 400 5000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6 0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
BAPTA-AM
0,2 Hz20 Hz
0 100 200 300 400 5000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,60,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
BAPTA-AM
Controle
20 Hz
A B C
D
E F G
FIGURA 13 - Efeito do BAPTA-AM (100 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ± EPM,
*p<0,05 e ***p<0,001. Nas figuras B, C e D estão especificadas as frequências de estimulação.

4.2. Importância dos diferentes mecanismos de depuração do cálcio intracelular em
fenômenos de plasticidade sináptica à curto prazo.
4.2.1. Bloqueio da Cálcio-ATPase do retículo endoplasmático
A tapsigargina é um bloqueador específico da cálcio-ATPase do retículo endoplasmático
(SAGARA & INESI, 1991; THASTRUP et al., 1990) e nos a utilizamos na concentração de 2
µM para verificarmos a contribuição de tal mecanismo de depuração do cálcio intracelular
nos fenômenos de depressão e facilitação por pulsos pareados. O controle foi realizado com
o veículo de diluição etanol a 0,028%.
Nossos resultados mostram que a tapsigargina aumentou a amplitude dos EPPs antes da
tetania de 4,30,5 mV (n=23) para 7,10,9 mV (n=19), p=0,01 (FIG. 14A). Nós também
observamos que a depressão foi maior nas preparações tratadas com tapsigargina. A
depressão final aumentou de 76±3% (n=15) para 87±4% (n=13), p=0,03 (FIG. 14E e 19).
Nossos resultados também indicam uma maior depressão verificada por uma intersecção
mais baixa no eixo y de 0,550,04 (n=15 a 16) para 0,390,05 (n=11 a 12), p=0,02 (FIG. 20).
O tempo para a instalação da depressão diminuiu de 5,91,5 (n=15) para 2,60,4 (n=13),
p=0,04 (FIG. 21). Com relação à recuperação final, verificamos que esta aumentou de
863% (n=14) para 1002% (n=10), p=0,002 (FIG. 14F e 22).
Nós também observamos um aumento na facilitação por pulsos pareados quando o intervalo
entre os pulsos foi de 5 a 15 ms (FIG. 24A e tabela II).
Controle Tapsigargina0
20
40
60
80
100
(n=14)(n=16)
% D
ep
res
sã
o f
ina
l *
Controle Tapsigargina 0
2
4
6
8
10
(n=19)
EP
P (
mV
)
*
(n=23)
Controle Tapsigargina 0
5
10
15
20
(n=11)(n=16)
% P
ote
nc
iaç
ão
Controle Tapsigargina 0
20
40
60
80
100
(n=10)(n=14)
% R
ec
up
era
çã
o f
ina
l **
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,20,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Tapsigargina
0,2 Hz 20 HzA B C
D
E F G
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
Tapsigargina
20 Hz
FIGURA 14 - Efeito da tapsigargina (2 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ± EPM,
*p<0,05 e **p<0,01. Nas figuras B, C e D estão especificadas as frequências de estimulação.

TABELA II – Facilitação por pulsos pareados – Controle x Tapsigargina
Intervalo entre os pulsos (ms)
Controle
(n = 16)
2 µM Tapsigargina
(n = 6 a 9)
P
5 1,52 0,06 1,96 0,15** 0,004
10 1,06 0,04 1,38 0,06 ** 0,006
15 0,87 0,05 1,07 0,06 * 0,02
20 0,78 0,04 0,86 0,05 0,28
30 0,65 0,04 0,76 0,05 0,14
40 0,53 0,04 0,58 0,04 0,47
50 0,39 0,03 0,49 0,04 0,09
60 0,30 0,03 0,36 0,06 0,22

4.2.2. Bloqueio da Cálcio-ATPase da membrana plasmática
No intuito de verificarmos a contribuição da Ca+2 ATPase da membrana plasmática como
mecanismo de depuração nos fenômenos de depressão e facilitação por pulsos pareados,
utilizamos a CEDA na concentração de 20 µM para bloqueá-la. O controle foi realizado com
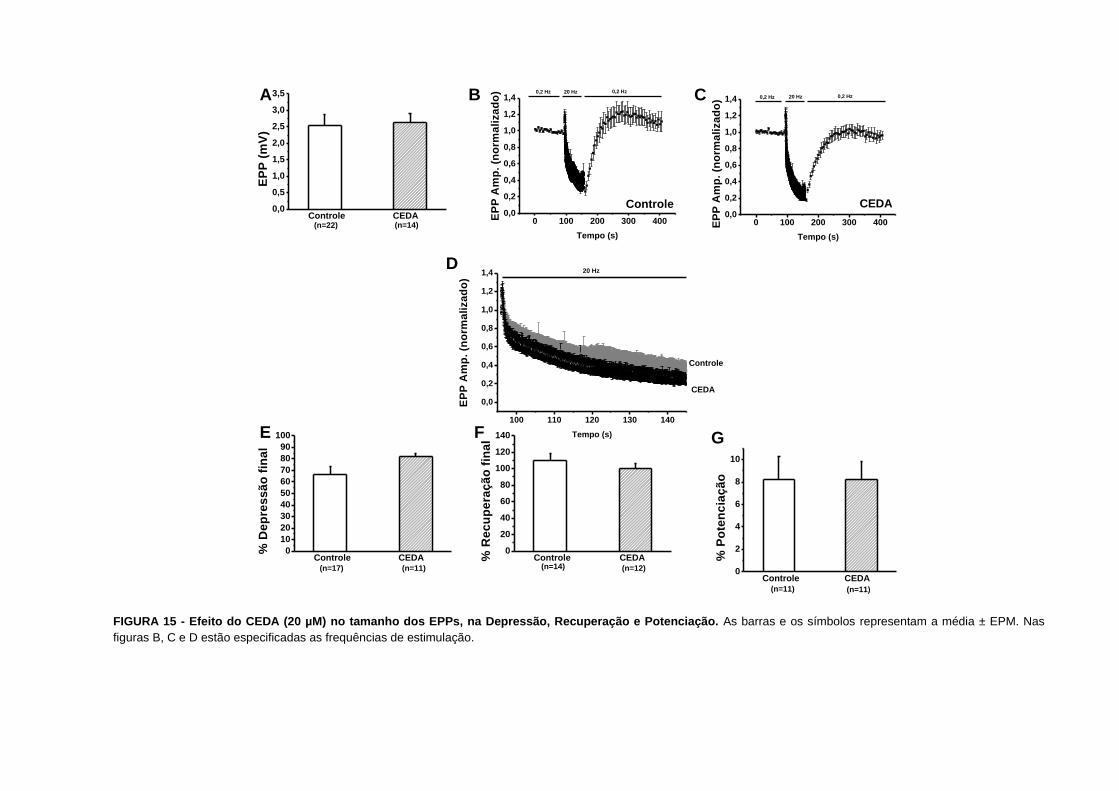
o veículo de diluição DMSO a 0,17%.
Nossos resultados não evidenciam nenhuma diferença nos parâmetros estudados na
depressão e nem na facilitação por pulsos pareados.
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4 0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
CEDA
0,2 Hz 20 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,40,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz20 Hz
Controle CEDA 0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
(n=14)
EP
P (
mV
)
(n=22)
Controle CEDA 0
10
20
30
40
50
60
70
80
90
100
(n=11)(n=17)
% D
ep
res
sã
o f
ina
l
Controle CEDA 0
2
4
6
8
10
(n=11)(n=11)
% P
ote
nc
iaç
ão
Controle CEDA0
20
40
60
80
100
120
140
(n=12)(n=14)
% R
ec
up
era
çã
o f
ina
l
100 110 120 130 140
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
CEDA
Controle
20 Hz
A B C
D
E F G
FIGURA 15 - Efeito do CEDA (20 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ± EPM. Nas
figuras B, C e D estão especificadas as frequências de estimulação.
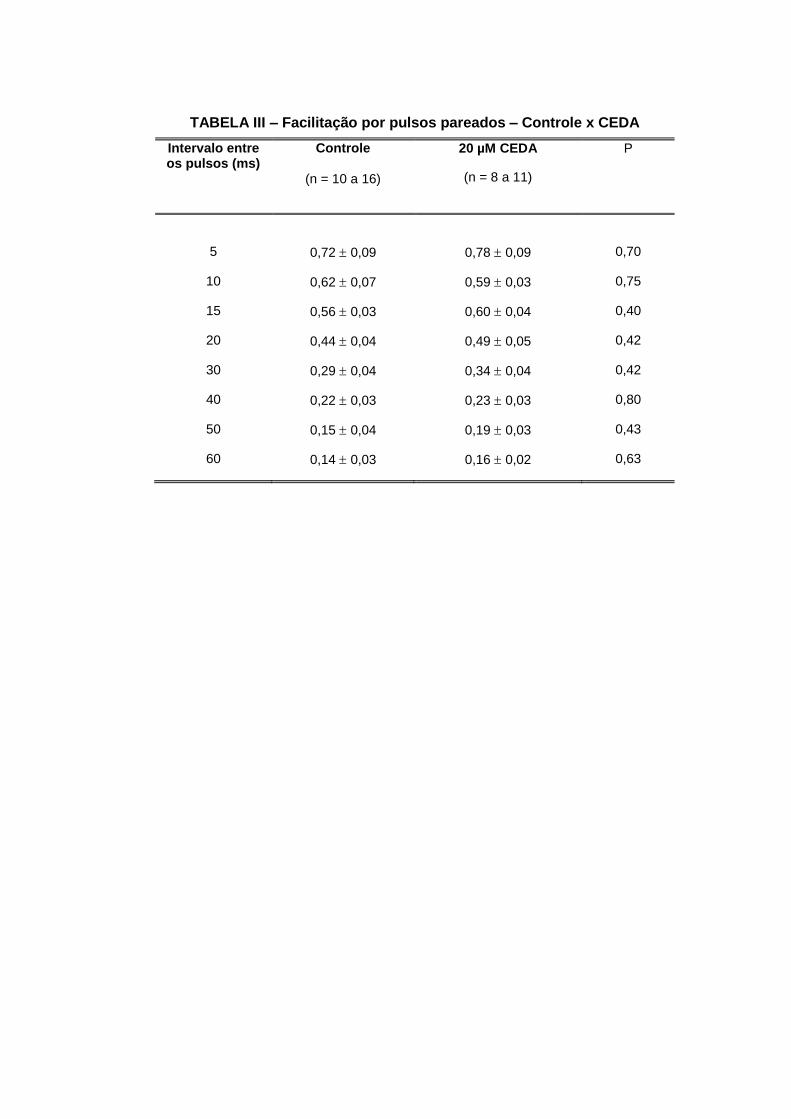
TABELA III – Facilitação por pulsos pareados – Controle x CEDA
Intervalo entre os pulsos (ms)
Controle
(n = 10 a 16)
20 µM CEDA
(n = 8 a 11)
P
5 0,72 0,09 0,78 0,09 0,70
10 0,62 0,07 0,59 0,03 0,75
15 0,56 0,03 0,60 0,04 0,40
20 0,44 0,04 0,49 0,05 0,42
30 0,29 0,04 0,34 0,04 0,42
40 0,22 0,03 0,23 0,03 0,80
50 0,15 0,04 0,19 0,03 0,43
60 0,14 0,03 0,16 0,02 0,63

4.2.3. Bloqueio do trocador Na+/Ca+2 da mitocôndria
O trocador Na+/Ca+2 mitocondrial (NCXm) tem o papel de extrudar o cálcio recaptado pela
mitocôndria (GARCÍA-CHACÓN et al., 2006; WHITE & REYNOLDS, 1997; ZHONG et al.,
2001), mas em situações de aumento da concentração intracelular de cálcio, ele pode atuar
de modo reverso e recaptar o cálcio citoplasmático (GRIFFITHS, 1999; SMETS et al., 2004).
Portanto, decidimos verificar o papel deste trocador como mecanismo de depuração do
cálcio intracelular e para isso utilizamos o bloqueador CGP 37157 na concentração de 50
µM (COX et al., 1993). O controle foi realizado com o veículo de diluição DMSO a 0,05%.
Nossos resultados indicam uma maior depressão verificada por uma intersecção mais baixa
no eixo y de 0,530,07 (n=14 a 15) para 0,320,03 (n=8 a 10), p=0,03 (FIG. 20) e também
um aumento no tempo para a instalação desta de 7,41,4 s (n=14 a 15) para 15,94,5 s
(n=8 a 10), p=0,04 (FIG. 21). Entretanto, um resultado mais condizente com a função do
trocador NCXm refere-se à recuperação e a potenciação. Nossos resultados mostram um
aumento no tempo da recuperação de 17,2±1,7 s (n=18) para 22,3±1,6 s (n=17), p=0,03.
Verificamos também uma diminuição na potenciação de 133% (n=18) para 51% (n=18),
p=0,004 (FIG. 16G e 23).
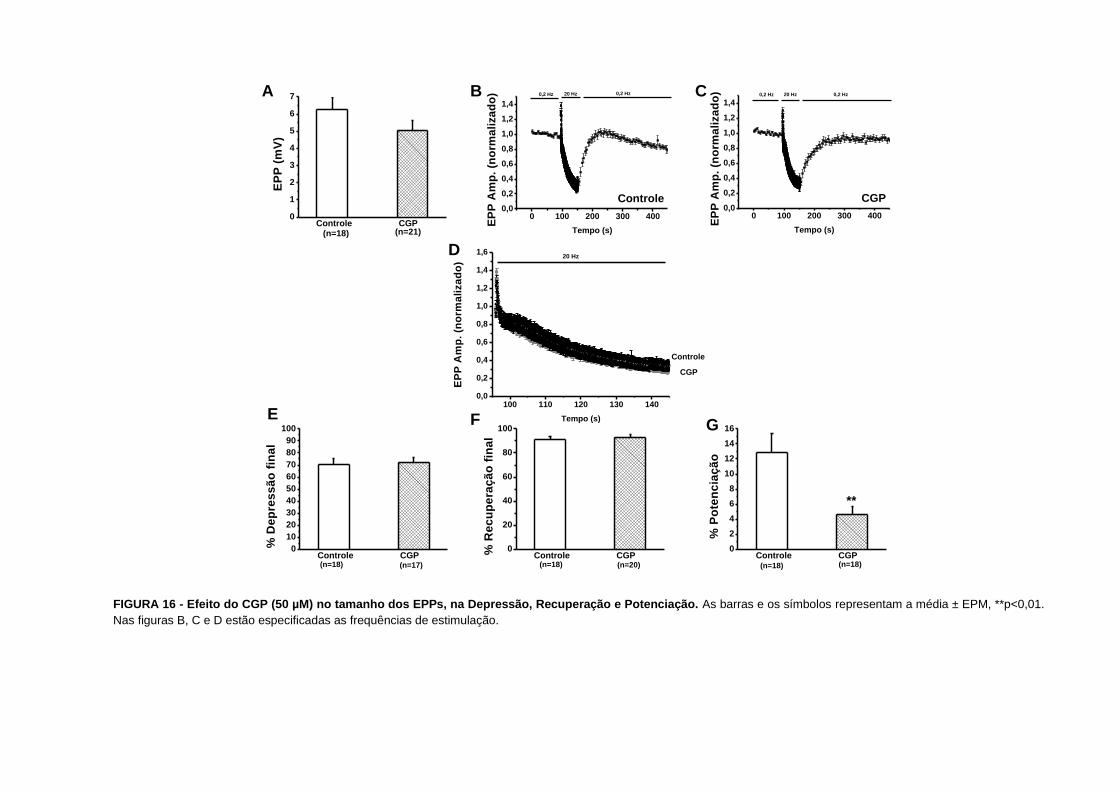
Controle CGP 0
10
20
30
40
50
60
70
80
90
100
(n=17)
% D
ep
res
sã
o f
ina
l
(n=18)Controle CGP
0
2
4
6
8
10
12
14
16
(n=18)
% P
ote
nc
iaç
ão
**
(n=18)
Controle CGP 0
20
40
60
80
100
(n=20)
% R
ec
up
era
çã
o f
ina
l
(n=18)
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,40,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
CGP
0,2 Hz 20 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
CGP
20 Hz
Controle CGP 0
1
2
3
4
5
6
7
(n=21)
EP
P (
mV
)
(n=18)
A B C
D
E F G
FIGURA 16 - Efeito do CGP (50 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ± EPM, **p<0,01.
Nas figuras B, C e D estão especificadas as frequências de estimulação.
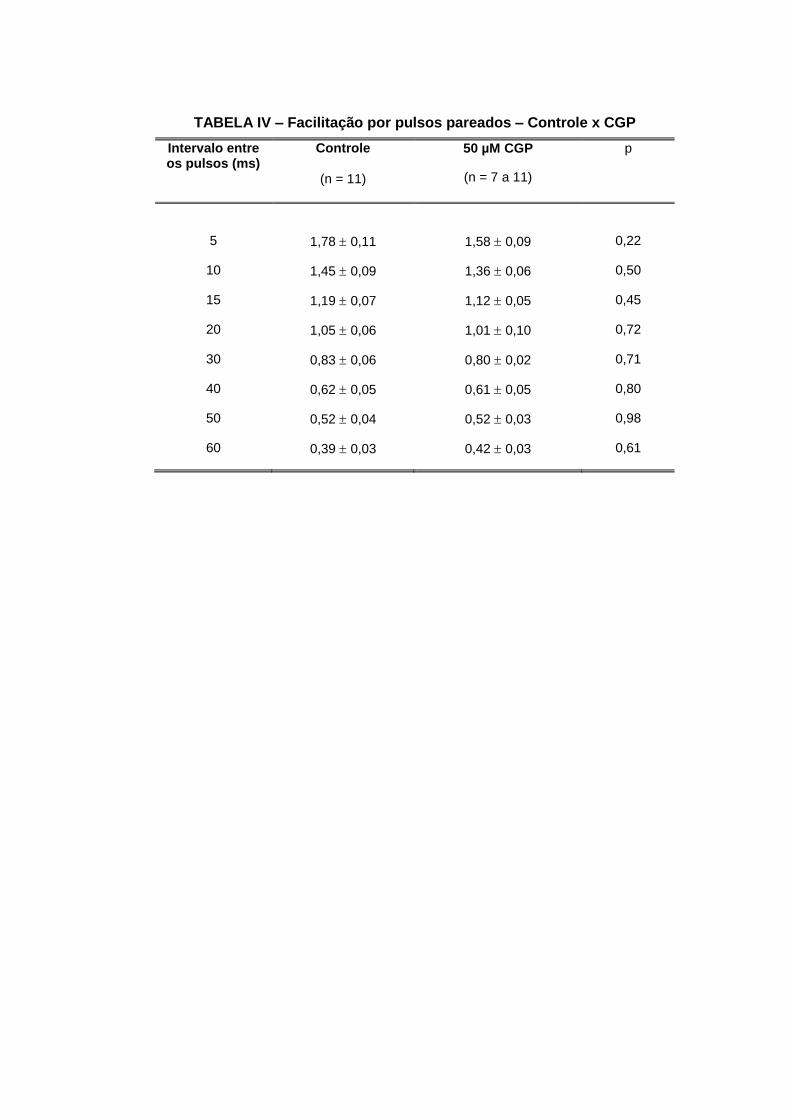
TABELA IV – Facilitação por pulsos pareados – Controle x CGP
Intervalo entre os pulsos (ms)
Controle
(n = 11)
50 µM CGP
(n = 7 a 11)
p
5 1,78 0,11 1,58 0,09 0,22
10 1,45 0,09 1,36 0,06 0,50
15 1,19 0,07 1,12 0,05 0,45
20 1,05 0,06 1,01 0,10 0,72
30 0,83 0,06 0,80 0,02 0,71
40 0,62 0,05 0,61 0,05 0,80
50 0,52 0,04 0,52 0,03 0,98
60 0,39 0,03 0,42 0,03 0,61

4.2.4. Bloqueio do uniporter mitocondrial
No intuito de verificarmos a contribuição do uniporter mitocondrial como mecanismo de
depuração nos fenômenos de depressão e facilitação por pulsos pareados, utilizamos o
bloqueador Ru360 na concentração de 10 µM para bloqueá-lo (MATLIB et al., 1998). O
controle foi realizado com o veículo de diluição, água deoxigenada no mesmo volume
utilizado do Ru360, 10 µl.
Com relação aos parâmetros analisados na depressão verificamos que as preparações
tratadas com o Ru360 apresentaram uma diminuição da depressão verificada pela
intersecção maior no eixo y de 0,450,07 (n=7 a 9) para 0,640,04 (n=9 a 12), p=0,03 (FIG.
20).
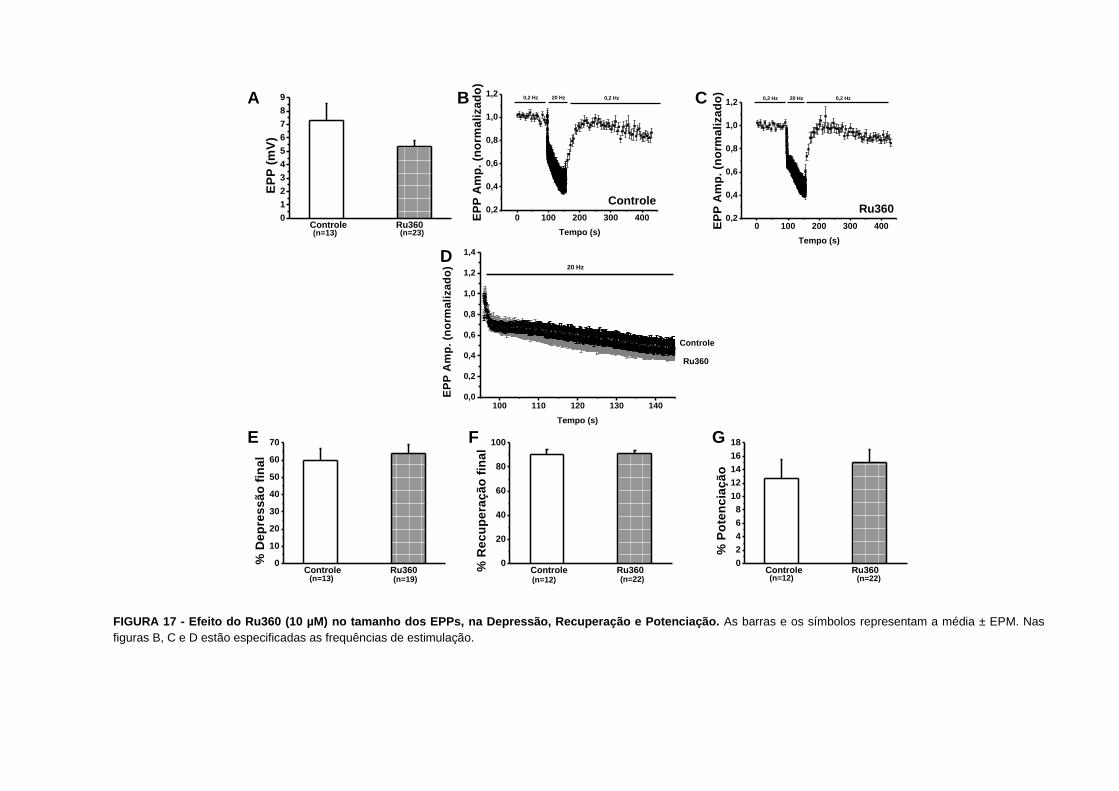
Controle Ru360 0
10
20
30
40
50
60
70
(n=19)
% D
ep
res
sã
o f
ina
l
(n=13)Controle Ru360
0
2
4
6
8
10
12
14
16
18
(n=22)
% P
ote
nc
iaç
ão
(n=12)Controle Ru360
0
20
40
60
80
100
(n=22)
% R
ec
up
era
çã
o f
ina
l
(n=12)
0 100 200 300 4000,2
0,4
0,6
0,8
1,0
1,20,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
0 100 200 300 4000,2
0,4
0,6
0,8
1,0
1,20,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Ru360
0,2 Hz 20 Hz
Controle Ru360 0
1
2
3
4
5
6
7
8
9
(n=23)
EP
P (
mV
)
(n=13)
A B C
D
E F G
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
Ru360
20 Hz
FIGURA 17 - Efeito do Ru360 (10 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ± EPM. Nas
figuras B, C e D estão especificadas as frequências de estimulação.
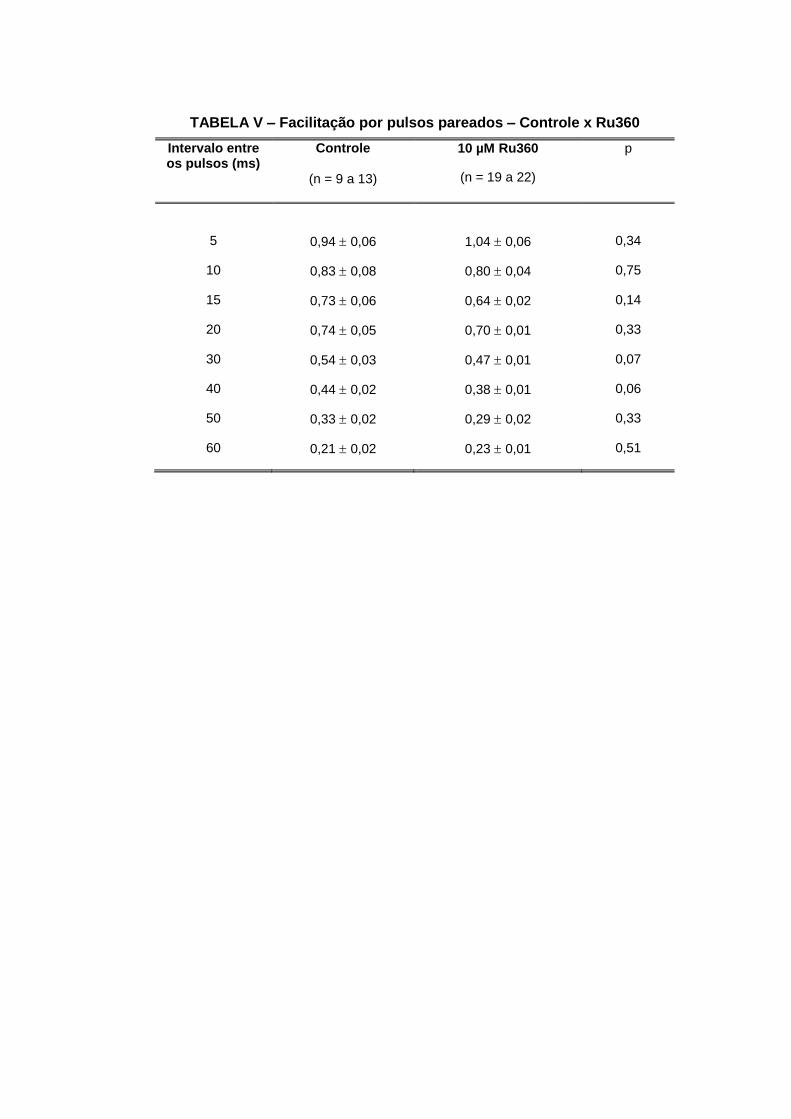
TABELA V – Facilitação por pulsos pareados – Controle x Ru360
Intervalo entre os pulsos (ms)
Controle
(n = 9 a 13)
10 µM Ru360
(n = 19 a 22)
p
5 0,94 0,06 1,04 0,06 0,34
10 0,83 0,08 0,80 0,04 0,75
15 0,73 0,06 0,64 0,02 0,14
20 0,74 0,05 0,70 0,01 0,33
30 0,54 0,03 0,47 0,01 0,07
40 0,44 0,02 0,38 0,01 0,06
50 0,33 0,02 0,29 0,02 0,33
60 0,21 0,02 0,23 0,01 0,51

4.2.5. Bloqueio do trocador Na+/Ca+2 da membrana plasmática
O trocador Na+/Ca+2 da membrana plasmática pode ser bloqueado através da utilização de
uma solução extracelular, onde o íon sódio é substituído pelo lítio (HERMONI et al., 1987).
Nossos resultados mostram que as preparações incubadas com Ringer lítio apresentaram
uma diminuição na amplitude dos EPPs durante o estímulo de baixa frequência de 4,20,5
mV (n=14) para 2,80,4 mV (n=14), p=0,002 (FIG.18A). Nós também observamos um
aumento na depressão final de 727% (n=11) para 805% (n=11), p=0,01 (FIG.18E e 19) e
uma diminuição na potenciação de 163% (n=9) para 62% (n=8), p=0,02 (FIG.18G e 23).
Com relação à facilitação por pulsos pareados verificamos uma menor facilitação por pulsos
pareados quando o intervalo entre os pulsos foi de 5-50 ms (FIG. 24E e tabela VI).
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,20,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Lítio
0,2 Hz 20 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,20,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
Controle Lítio 0
10
20
30
40
50
60
70
80
90
(n=11)
% D
ep
res
sã
o f
ina
l
*
(n=11)
Controle Lítio 0
1
2
3
4
5
(n=14)
EP
P (
mV
)
**
(n=14)
Controle Lítio 0
5
10
15
20
25
(n=9)
% P
ote
nc
iaç
ão
*
(n=9)Controle Lítio
0
20
40
60
80
100
120
(n=9)
% R
ec
up
era
çã
o f
ina
l
(n=7)
A B C
D
E F G
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
Lítio
20 Hz
FIGURA 18 - Efeito do Ringer Lítio no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os pontos representam a média ± EPM, *p<0,05 e
**p<0,01. Nas figuras B, C e D estão especificadas as frequências de estimulação.
TABELA VI – Facilitação por pulsos pareados – Controle x Ringer Lítio
Intervalo entre os pulsos (ms)
Controle
(n = 10)
Ringer Lítio
(n = 10)
p
5 1,29 0,11 0,66 0,11 *** 0,0008
10 0,95 0,11 0,45 0,05 ** 0,001
15 0,88 0,06 0,49 0,09 ** 0,001
20 0,72 0,09 0,47 0,09 * 0,01
30 0,64 0,05 0,36 0,06 ** 0,001
40 0,44 0,05 0,21 0,04 *** 0,0004
50 0,34 0,05 0,18 0,05 ** 0,002
60 0,25 0,03 0,17 0,06 0,23
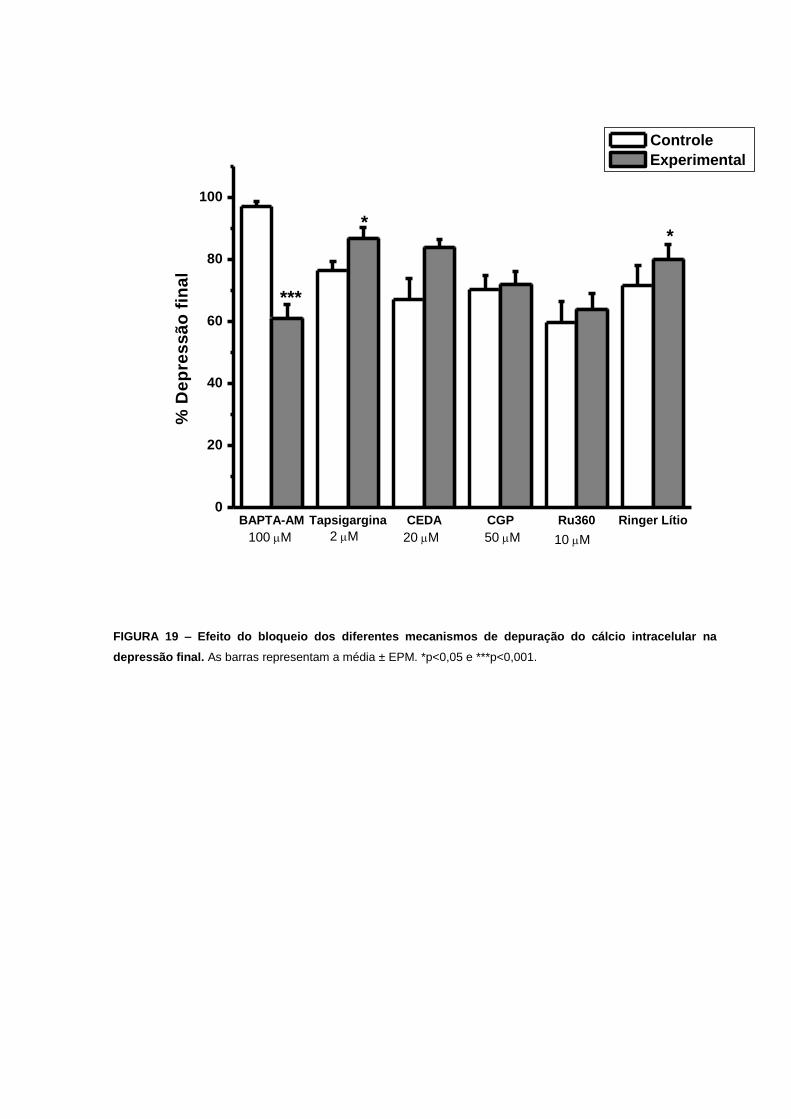
BAPTA-AM Tapsigargina CEDA CGP Ru360 Ringer Lítio0
20
40
60
80
100
10 M50 M20 M2 M
*
% D
ep
res
sã
o f
ina
l
Controle
Experimental
***
*
100 M
FIGURA 19 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio intracelular na
depressão final. As barras representam a média ± EPM. *p<0,05 e ***p<0,001.
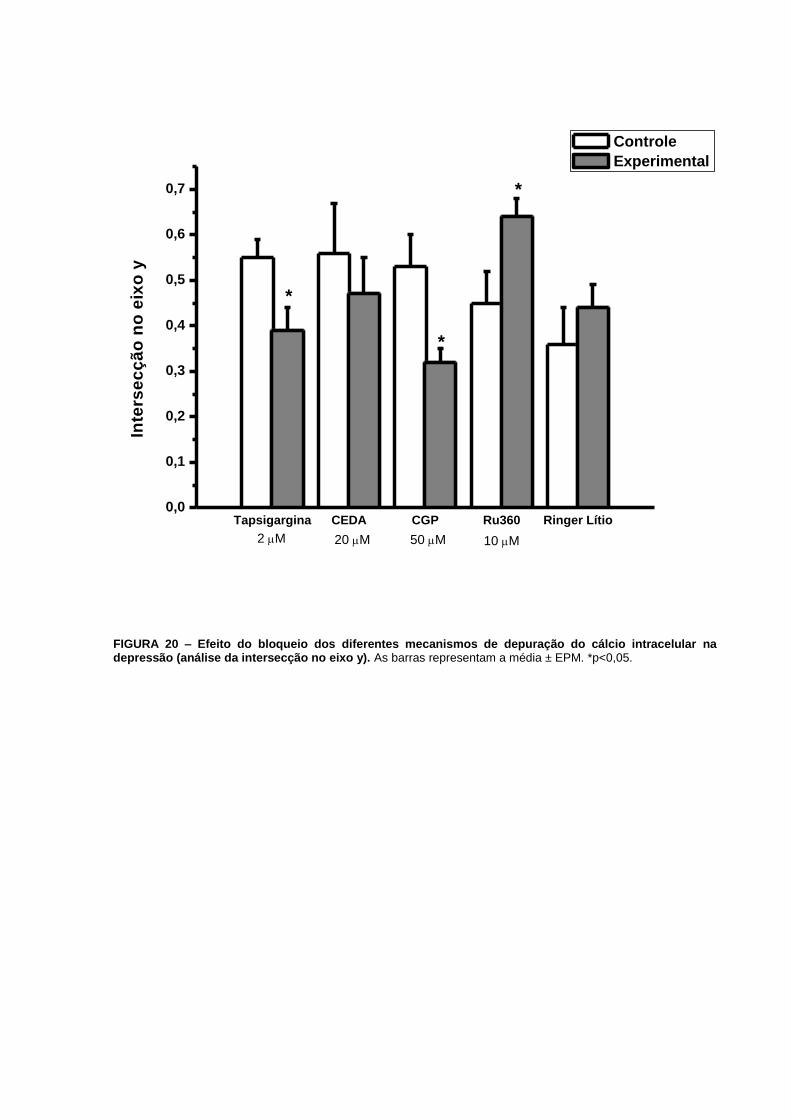
Tapsigargina CEDA CGP Ru360 Ringer Lítio0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
10 M50 M20 M
*
*
Inte
rse
cç
ão
no
eix
o y
Controle
Experimental
*
2 M
FIGURA 20 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio intracelular na depressão (análise da intersecção no eixo y). As barras representam a média ± EPM. *p<0,05.

Tapsigargina CEDA CGP Ru360 Ringer Lítio0
5
10
15
20
25
*
50 M 10 M20 M2 M
Co
ns
tan
te d
e d
ec
aim
en
to
Controle
Experimental
*
FIGURA 21 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio intracelular na constante de decaimento da depressão. As barras representam a média ± EPM. *p<0,05.
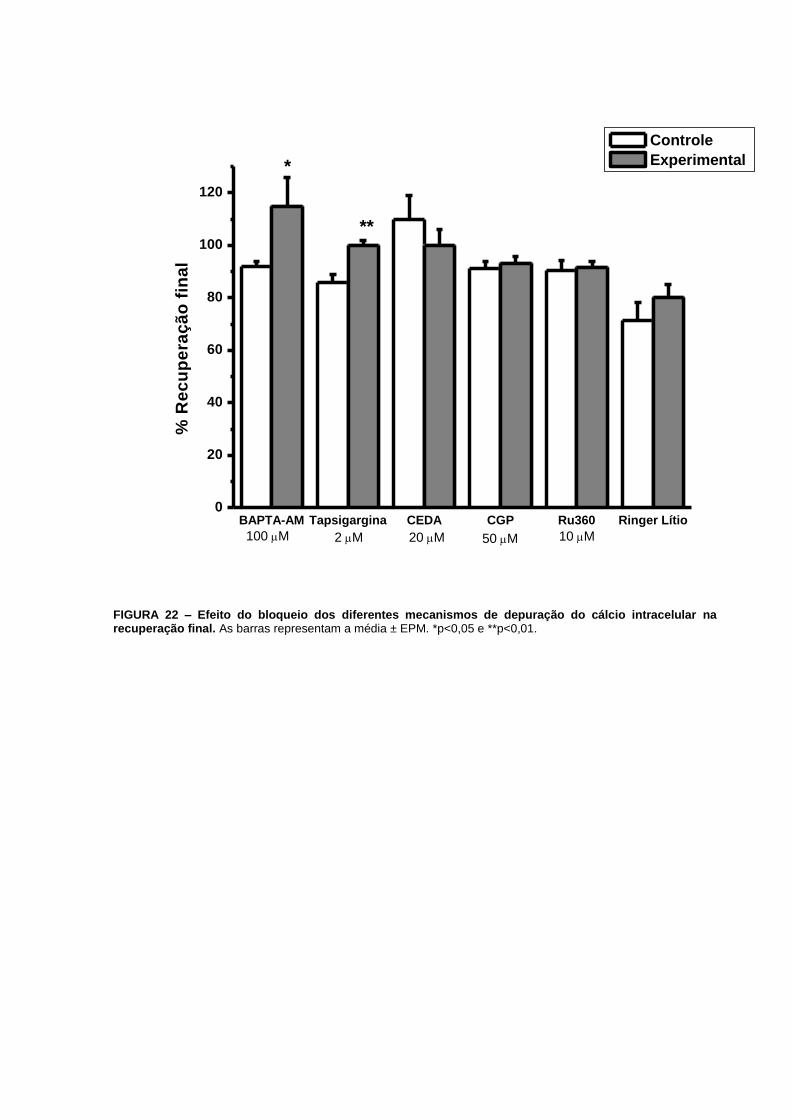
BAPTA-AM Tapsigargina CEDA CGP Ru360 Ringer Lítio0
20
40
60
80
100
120
50 M2 M 20 M 10 M
% R
ec
up
era
çã
o f
ina
l
Controle
Experimental*
**
100 M
FIGURA 22 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio intracelular na recuperação final. As barras representam a média ± EPM. *p<0,05 e **p<0,01.
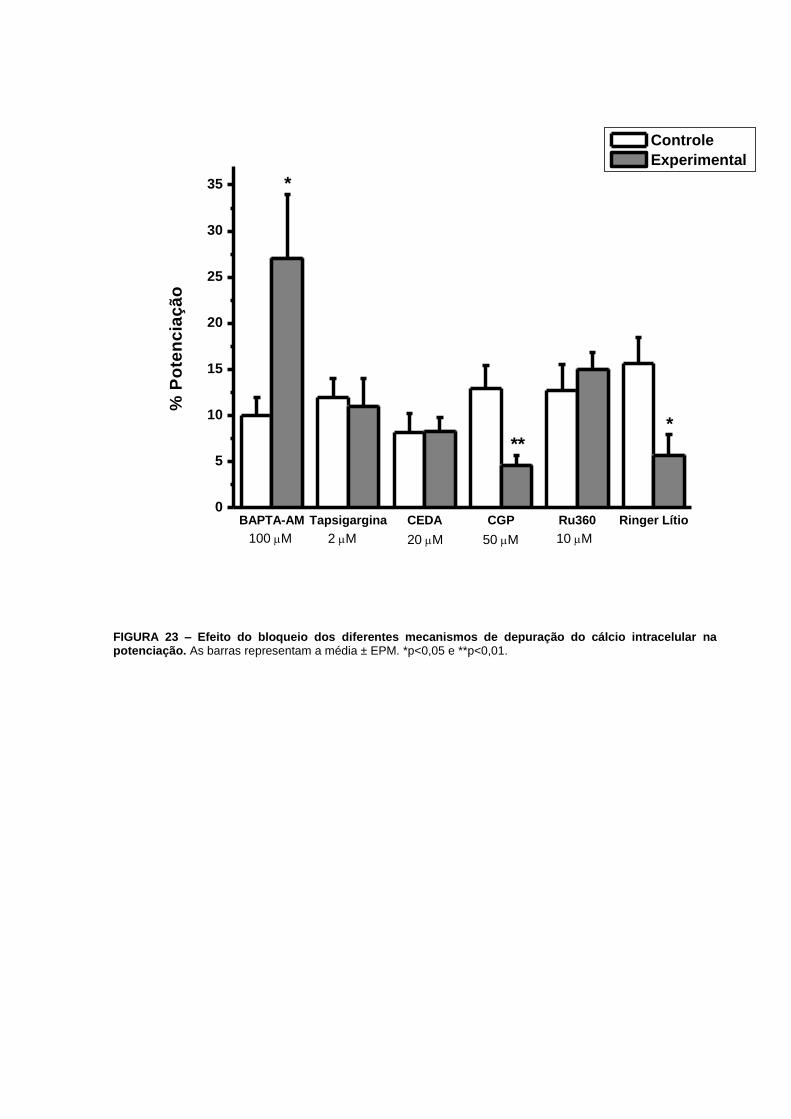
BAPTA-AM Tapsigargina CEDA CGP Ru360 Ringer Lítio0
5
10
15
20
25
30
35
50 M 10 M20 M2 M
% P
ote
nc
iaç
ão
Controle
Experimental
***
*
100 M
FIGURA 23 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio intracelular na potenciação. As barras representam a média ± EPM. *p<0,05 e **p<0,01.

0 10 20 30 40 50 600,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
*
**
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 6 a 9)
2 M Tapsigargina (n = 16)
**
0 10 20 30 40 50 600,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
2,0
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 11)
50 M CGP (n = 7 a 11)
0 10 20 30 40 50 600,0
0,2
0,4
0,6
0,8
1,0
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 10 a 16)
20 M CEDA (n = 8 a 11)
0 10 20 30 40 50 600,0
0,2
0,4
0,6
0,8
1,0
1,2
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 9 a 13)
10 M Ru360 (n = 19 a 22)
0 10 20 30 40 50 600,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
*****
**
****
***
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 10)
Ringer Lítio (n = 10)
*
A B
C D E
FIGURA 24 – Efeito do bloqueio dos diferentes mecanismos de depuração do cálcio intracelular na facilitação por pulsos pareados. Os símbolos representam a
média ± EPM, *p<0,05, **p<0,01 e ***p<0,001.
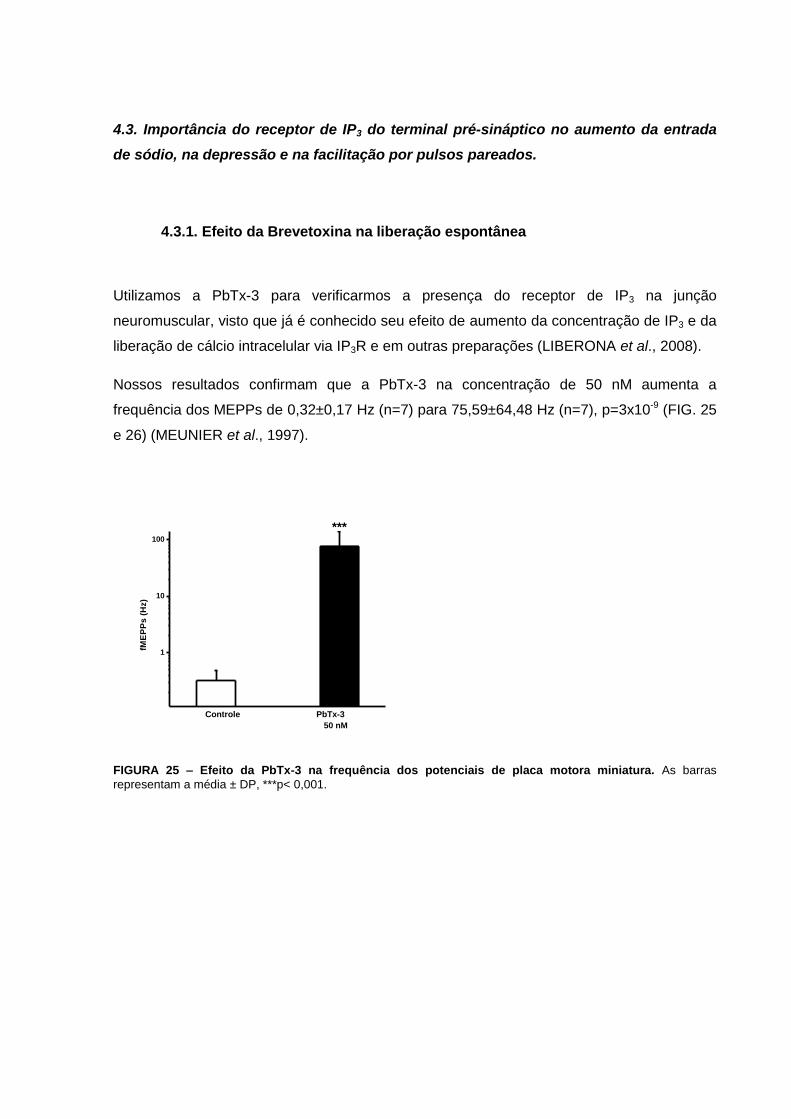
4.3. Importância do receptor de IP3 do terminal pré-sináptico no aumento da entrada
de sódio, na depressão e na facilitação por pulsos pareados.
4.3.1. Efeito da Brevetoxina na liberação espontânea
Utilizamos a PbTx-3 para verificarmos a presença do receptor de IP3 na junção
neuromuscular, visto que já é conhecido seu efeito de aumento da concentração de IP3 e da
liberação de cálcio intracelular via IP3R e em outras preparações (LIBERONA et al., 2008).
Nossos resultados confirmam que a PbTx-3 na concentração de 50 nM aumenta a
frequência dos MEPPs de 0,32±0,17 Hz (n=7) para 75,59±64,48 Hz (n=7), p=3x10-9 (FIG. 25
e 26) (MEUNIER et al., 1997).
Controle PbTx-3
1
10
100
fME
PP
s (
Hz)
***
50 nM
FIGURA 25 – Efeito da PbTx-3 na frequência dos potenciais de placa motora miniatura. As barras
representam a média ± DP, ***p< 0,001.
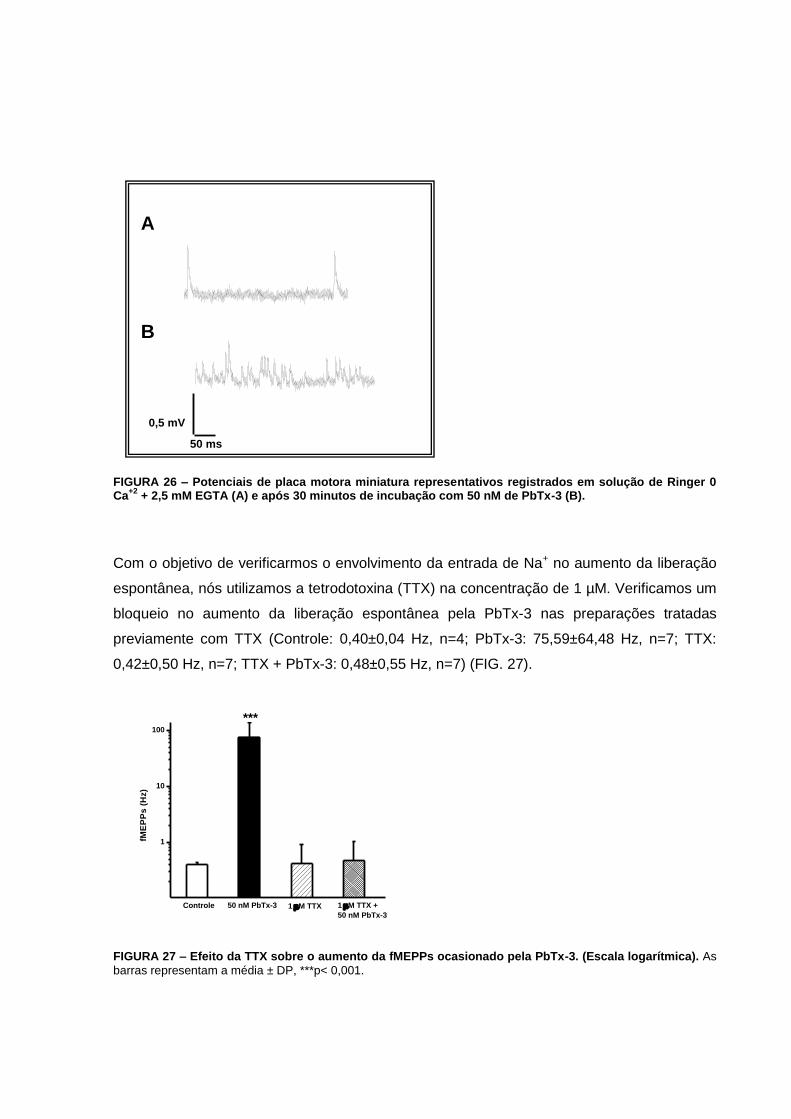
0,5 mV
50 ms
A
B
FIGURA 26 – Potenciais de placa motora miniatura representativos registrados em solução de Ringer 0 Ca
+2 + 2,5 mM EGTA (A) e após 30 minutos de incubação com 50 nM de PbTx-3 (B).
Com o objetivo de verificarmos o envolvimento da entrada de Na+ no aumento da liberação
espontânea, nós utilizamos a tetrodotoxina (TTX) na concentração de 1 µM. Verificamos um
bloqueio no aumento da liberação espontânea pela PbTx-3 nas preparações tratadas
previamente com TTX (Controle: 0,40±0,04 Hz, n=4; PbTx-3: 75,59±64,48 Hz, n=7; TTX:
0,42±0,50 Hz, n=7; TTX + PbTx-3: 0,48±0,55 Hz, n=7) (FIG. 27).
1
10
100
1 M TTX +
50 nM PbTx-3
fME
PP
s (
Hz)
***
Controle 50 nM PbTx-3 1 M TTX
FIGURA 27 – Efeito da TTX sobre o aumento da fMEPPs ocasionado pela PbTx-3. (Escala logarítmica). As
barras representam a média ± DP, ***p< 0,001.
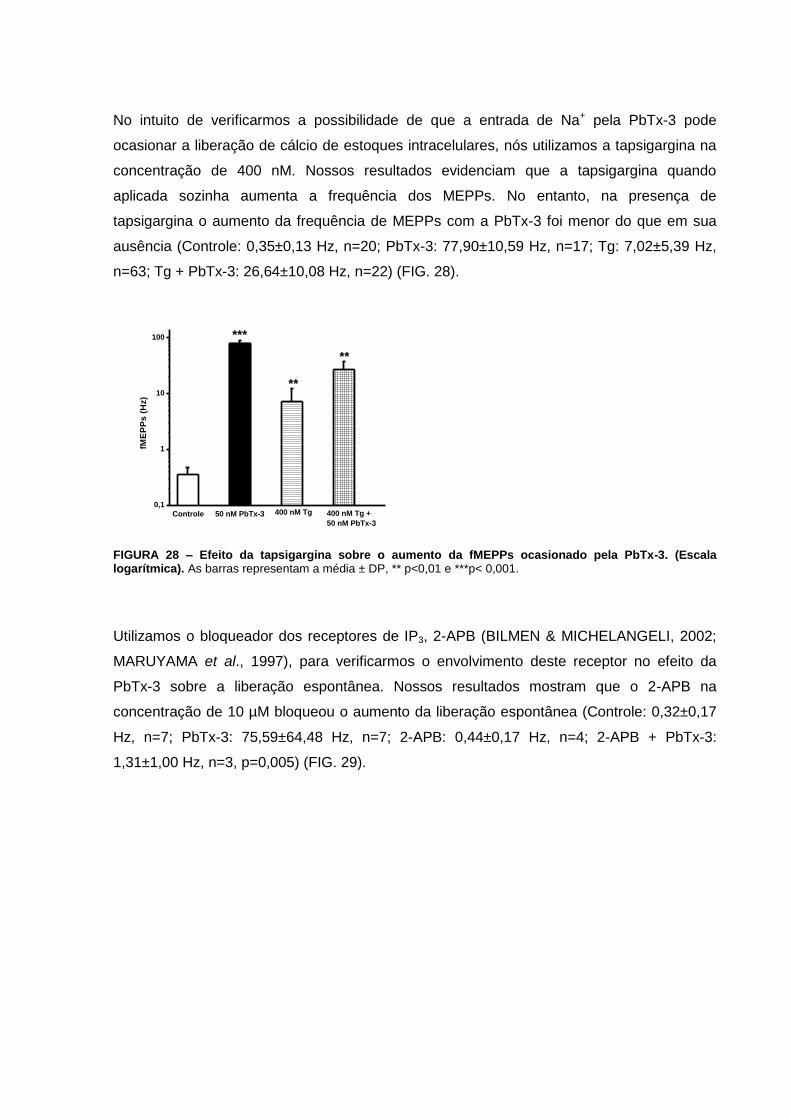
No intuito de verificarmos a possibilidade de que a entrada de Na+ pela PbTx-3 pode
ocasionar a liberação de cálcio de estoques intracelulares, nós utilizamos a tapsigargina na
concentração de 400 nM. Nossos resultados evidenciam que a tapsigargina quando
aplicada sozinha aumenta a frequência dos MEPPs. No entanto, na presença de
tapsigargina o aumento da frequência de MEPPs com a PbTx-3 foi menor do que em sua
ausência (Controle: 0,35±0,13 Hz, n=20; PbTx-3: 77,90±10,59 Hz, n=17; Tg: 7,02±5,39 Hz,
n=63; Tg + PbTx-3: 26,64±10,08 Hz, n=22) (FIG. 28).
0,1
1
10
100
**
**
fME
PP
s (
Hz)
***
Controle 50 nM PbTx-3 400 nM Tg 400 nM Tg +
50 nM PbTx-3
FIGURA 28 – Efeito da tapsigargina sobre o aumento da fMEPPs ocasionado pela PbTx-3. (Escala logarítmica). As barras representam a média ± DP, ** p<0,01 e ***p< 0,001.
Utilizamos o bloqueador dos receptores de IP3, 2-APB (BILMEN & MICHELANGELI, 2002;
MARUYAMA et al., 1997), para verificarmos o envolvimento deste receptor no efeito da
PbTx-3 sobre a liberação espontânea. Nossos resultados mostram que o 2-APB na
concentração de 10 µM bloqueou o aumento da liberação espontânea (Controle: 0,32±0,17
Hz, n=7; PbTx-3: 75,59±64,48 Hz, n=7; 2-APB: 0,44±0,17 Hz, n=4; 2-APB + PbTx-3:
1,31±1,00 Hz, n=3, p=0,005) (FIG. 29).
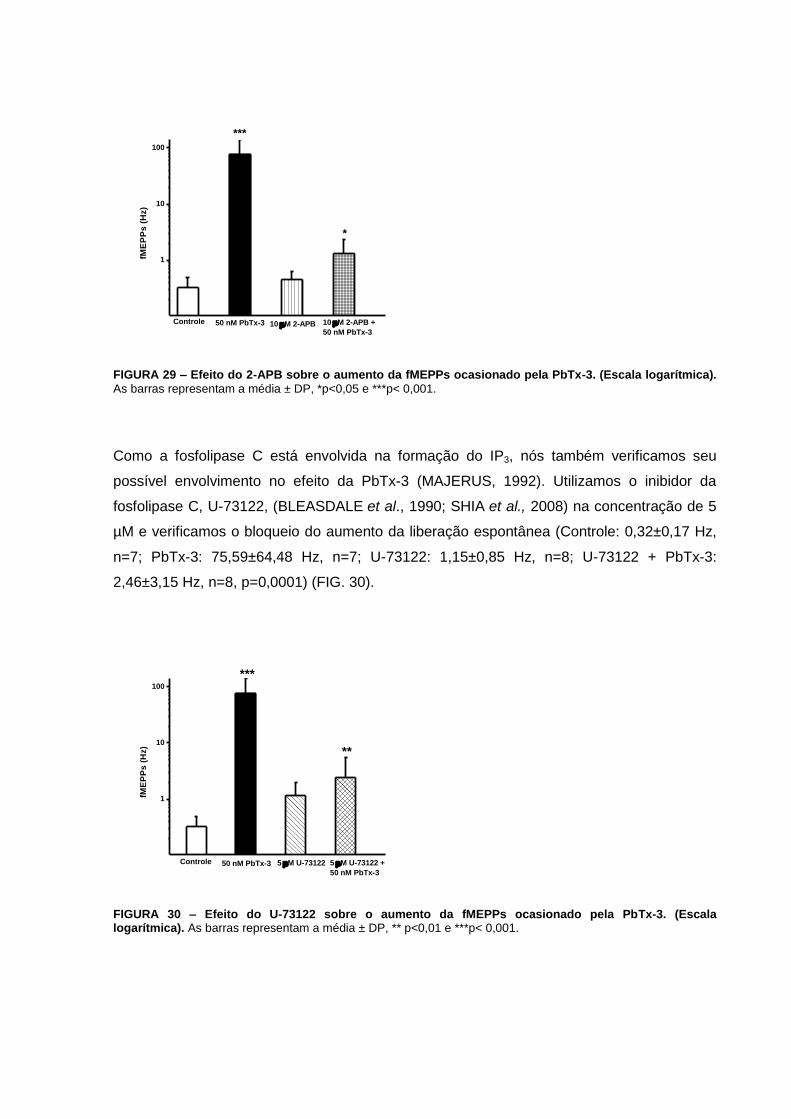
1
10
100
*
***
fME
PP
s (
Hz)
Controle 50 nM PbTx-3 10 M 2-APB 10 M 2-APB +
50 nM PbTx-3
FIGURA 29 – Efeito do 2-APB sobre o aumento da fMEPPs ocasionado pela PbTx-3. (Escala logarítmica).
As barras representam a média ± DP, *p<0,05 e ***p< 0,001.
Como a fosfolipase C está envolvida na formação do IP3, nós também verificamos seu
possível envolvimento no efeito da PbTx-3 (MAJERUS, 1992). Utilizamos o inibidor da
fosfolipase C, U-73122, (BLEASDALE et al., 1990; SHIA et al., 2008) na concentração de 5
µM e verificamos o bloqueio do aumento da liberação espontânea (Controle: 0,32±0,17 Hz,
n=7; PbTx-3: 75,59±64,48 Hz, n=7; U-73122: 1,15±0,85 Hz, n=8; U-73122 + PbTx-3:
2,46±3,15 Hz, n=8, p=0,0001) (FIG. 30).
1
10
100
**
fME
PP
s (
Hz)
***
Controle 50 nM PbTx-3 5 M U-73122 5 M U-73122 +
50 nM PbTx-3
FIGURA 30 – Efeito do U-73122 sobre o aumento da fMEPPs ocasionado pela PbTx-3. (Escala logarítmica). As barras representam a média ± DP, ** p<0,01 e ***p< 0,001.

4.3.2. Bloqueio do receptor de Rianodina
No intuito de verificarmos a contribuição do receptor de rianodina nos fenômenos de
depressão e facilitação por pulsos pareados, utilizamos a rianodina na concentração de 10
µM para bloqueá-lo (MEISSNER, 1986; OGAWA, 1994). O controle foi realizado com o
veículo de diluição etanol a 0,1%.
Nossos resultados mostram uma diminuição da depressão final de 783% (n=12) para
419% (n=8), p=0,0003 (FIG. 31E). Também verificamos uma diminuição da intensidade da
depressão pela intersecção mais alta no eixo y de 0,490,04 (n=12) para 0,700,04 (n=6),
p=0,01 e um aumento no tempo de sua instalação de 4,21,1 s (n=12) para 14,16,0 s
(n=6), p=0,03.
Com relação à facilitação por pulsos pareados verificamos uma diminuição da facilitação
quando o intervalo entre os estímulos foi de 5-15 ms (FIG. 32 e tabela VII).
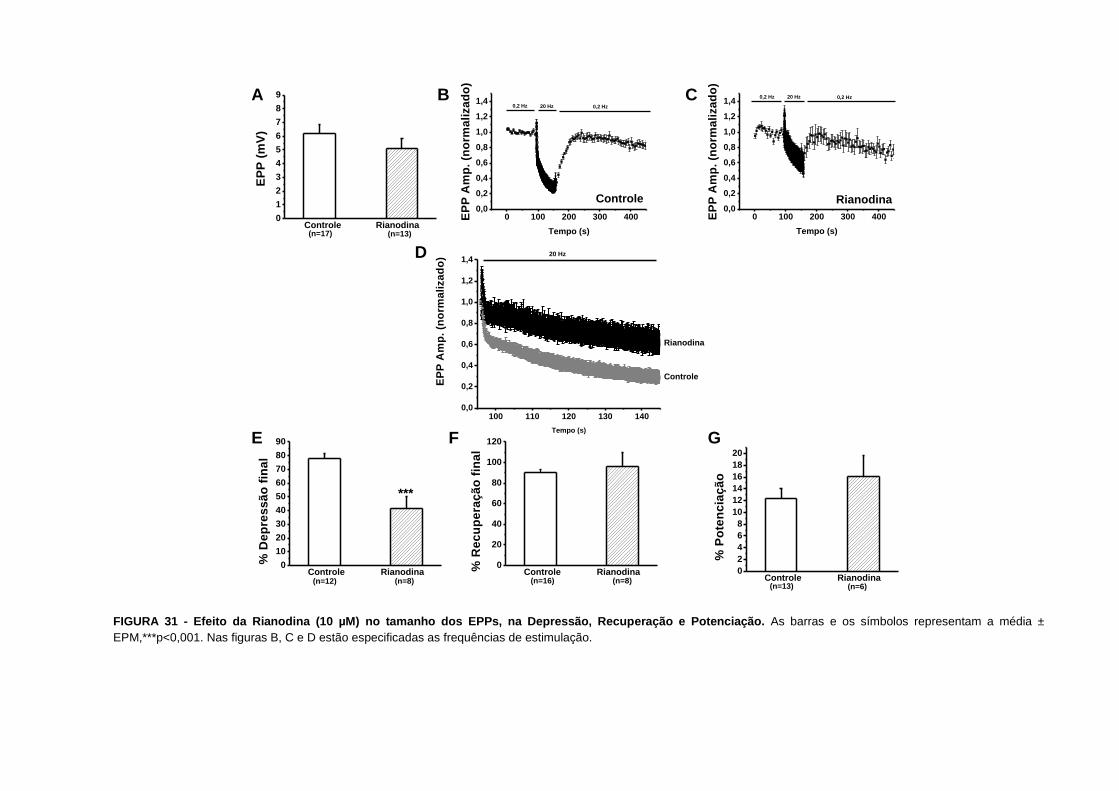
Controle Rianodina 0
1
2
3
4
5
6
7
8
9
(n=13)
EP
P (
mV
)
(n=17)
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,40,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,40,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Rianodina
0,2 Hz 20 Hz
Controle Rianodina 0
10
20
30
40
50
60
70
80
90
(n=12)
% D
ep
res
sã
o f
ina
l
***
(n=8) Controle Rianodina 0
2
4
6
8
10
12
14
16
18
20
(n=6)(n=13)
% P
ote
nc
iaç
ão
Controle Rianodina 0
20
40
60
80
100
120
(n=8)(n=16)
% R
ec
up
era
çã
o f
ina
l
A B C
E
D
F G
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
EP
P A
mp
. (n
orm
ali
za
do
)Tempo (s)
Rianodina
Controle
20 Hz
FIGURA 31 - Efeito da Rianodina (10 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ±
EPM,***p<0,001. Nas figuras B, C e D estão especificadas as frequências de estimulação.
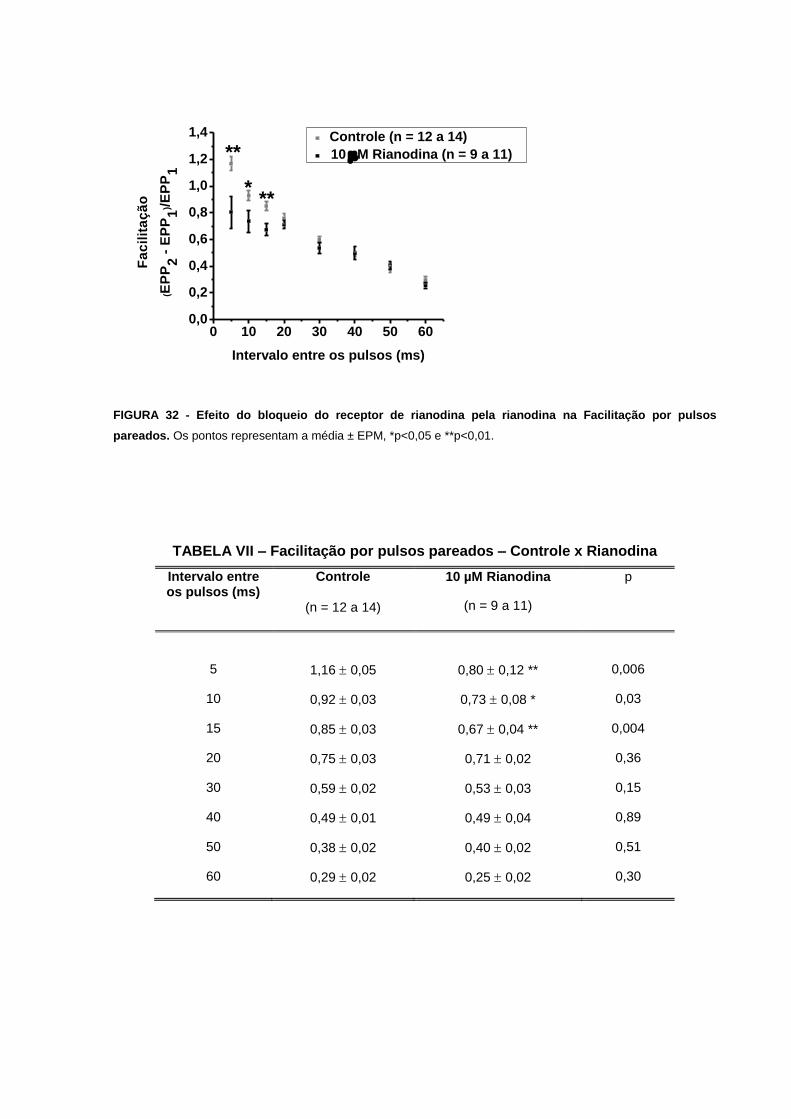
0 10 20 30 40 50 600,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
***
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 12 a 14)
10 M Rianodina (n = 9 a 11)**
FIGURA 32 - Efeito do bloqueio do receptor de rianodina pela rianodina na Facilitação por pulsos
pareados. Os pontos representam a média ± EPM, *p<0,05 e **p<0,01.
TABELA VII – Facilitação por pulsos pareados – Controle x Rianodina
Intervalo entre os pulsos (ms)
Controle
(n = 12 a 14)
10 µM Rianodina
(n = 9 a 11)
p
5 1,16 0,05 0,80 0,12 ** 0,006
10 0,92 0,03 0,73 0,08 * 0,03
15 0,85 0,03 0,67 0,04 ** 0,004
20 0,75 0,03 0,71 0,02 0,36
30 0,59 0,02 0,53 0,03 0,15
40 0,49 0,01 0,49 0,04 0,89
50 0,38 0,02 0,40 0,02 0,51
60 0,29 0,02 0,25 0,02 0,30

4.3.3. Bloqueio do receptor de IP3
No intuito de verificarmos a contribuição do receptor de IP3 nos fenômenos de depressão e
facilitação por pulsos pareados, utilizamos o 2-APB na concentração de 10 µM para
bloqueá-lo (BILMEN & MICHELANGELI, 2002; MARUYAMA et al., 1997). O controle foi
realizado com o veículo de diluição metanol a 0,002%. Apesar do fato do 2-APB inibir a
Ca+2-ATPase do retículo endoplasmático e de também ser um bloqueador da entrada de
cálcio operada por estoques (BOOTMAN et al., 2002), utilizamos uma concentração que não
apresentou efeito sobre a frequência (FIG. 29) e a amplitude (dado não mostrado) dos
MEPPs.
Nossos resultados mostram uma diminuição da depressão final de 923% (n=14) para
677% (n=16), p=0,002 (FIG. 33E). Verificamos também um aumento no tempo de
instalação da depressão de 0,90,2 s (n=10) para 3,50,9 s (n=13 a 15), p=0,02.
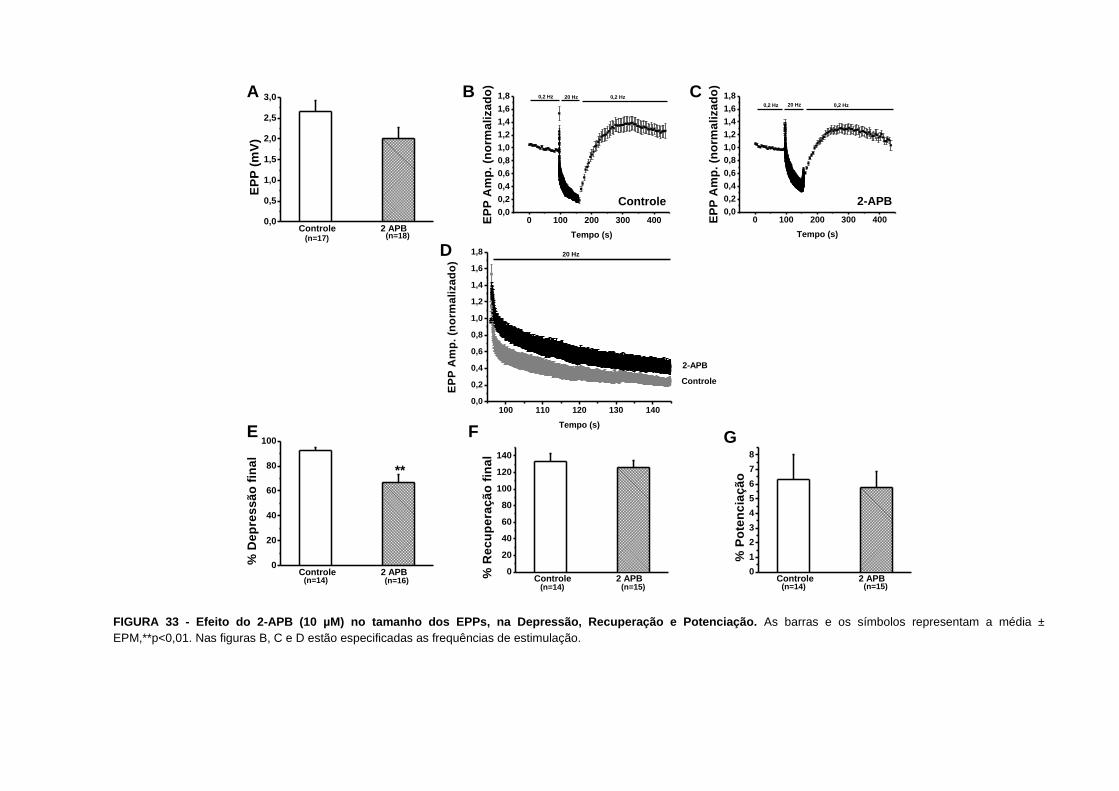
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,80,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
2-APB
20 Hz0,2 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8 0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
Controle 2 APB 0
20
40
60
80
100
(n=16)
% D
ep
res
sã
o f
ina
l
**
(n=14)
Controle 2 APB 0,0
0,5
1,0
1,5
2,0
2,5
3,0
(n=18)
EP
P (
mV
)
(n=17)
Controle 2 APB 0
1
2
3
4
5
6
7
8
(n=15)
% P
ote
nc
iaç
ão
(n=14)Controle 2 APB
0
20
40
60
80
100
120
140
(n=15)
% R
ec
up
era
çã
o f
ina
l
(n=14)
A B C
D
E F G
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
1,8
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
2-APB
Controle
20 Hz
FIGURA 33 - Efeito do 2-APB (10 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ±
EPM,**p<0,01. Nas figuras B, C e D estão especificadas as frequências de estimulação.
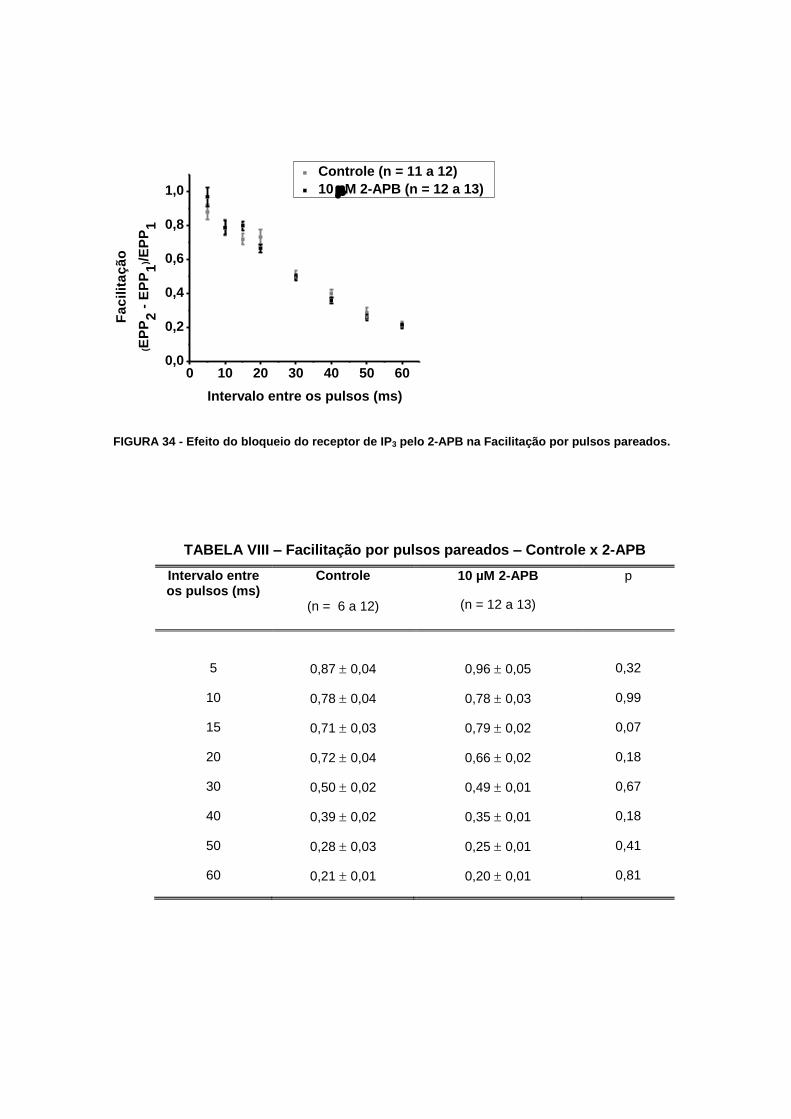
0 10 20 30 40 50 600,0
0,2
0,4
0,6
0,8
1,0
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalo entre os pulsos (ms)
Controle (n = 11 a 12)
10 M 2-APB (n = 12 a 13)
FIGURA 34 - Efeito do bloqueio do receptor de IP3 pelo 2-APB na Facilitação por pulsos pareados.
TABELA VIII – Facilitação por pulsos pareados – Controle x 2-APB
Intervalo entre os pulsos (ms)
Controle
(n = 6 a 12)
10 µM 2-APB
(n = 12 a 13)
p
5 0,87 0,04 0,96 0,05 0,32
10 0,78 0,04 0,78 0,03 0,99
15 0,71 0,03 0,79 0,02 0,07
20 0,72 0,04 0,66 0,02 0,18
30 0,50 0,02 0,49 0,01 0,67
40 0,39 0,02 0,35 0,01 0,18
50 0,28 0,03 0,25 0,01 0,41
60 0,21 0,01 0,20 0,01 0,81

4.3.4. Bloqueio da fosfolipase C
A formação de IP3 requer a ativação da fosfolipase C (MAJERUS, 1992). Portanto,
utilizamos o inibidor da fosfolipase C, U-73122, (BLEASDALE et al.,1990; SHIA et al.,2008)
na concentração de 5 µM no intuito de verificarmos seu papel nos fenômenos de
plasticidade sináptica à curto prazo, depressão e facilitação por pulsos pareados. O controle
foi realizado com o veículo de diluição DMSO a 0,05%.
Nossos resultados mostram uma diminuição na amplitude dos EPPs antes da tetania de
3,90,6 mV (n=22) para 1,90,2 mV (n=18), p=0,002 (FIG. 35A). Também verificamos uma
diminuição da depressão final de 884% (n=22) para 726% (n=18), p=0,02 (FIG. 35E) e no
tempo de sua instalação de 1,50,2 s (n=16) para 0,90,1 s (n=23), p=0,004. Com relação à
recuperação evidenciamos um aumento na recuperação final de 942% (n=22) para
1197% (n=17), p=0,0002 (FIG. 35F) e uma diminuição no tempo de sua instalação de
25,7±2,0 s (n=17) para 17,6±1,9 s (n=15), p=0,006.
A análise da facilitação por pulsos pareados revelou uma diminuição na facilitação quando o
intervalo entre os pulsos foi de 5-30 ms (FIG. 36 e tabela IX).
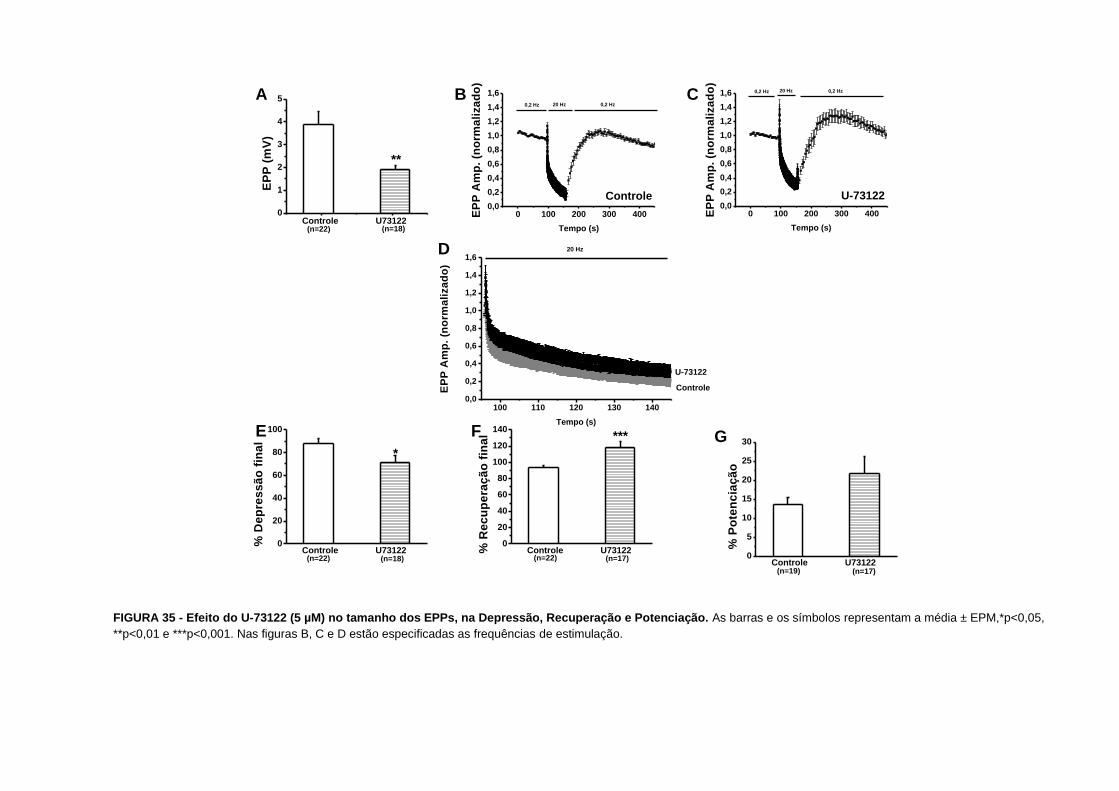
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
Controle
0,2 Hz 20 Hz
0 100 200 300 4000,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6 0,2 Hz
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
U-73122
0,2 Hz 20 Hz
Controle U73122 0
20
40
60
80
100
(n=18)
% D
ep
res
sã
o f
ina
l
*
(n=22)
Controle U73122 0
1
2
3
4
5
(n=18)
EP
P (
mV
)
**
(n=22)
Controle U73122 0
20
40
60
80
100
120
140
(n=17)(n=22)
% R
ec
up
era
çã
o f
ina
l ***
Controle U73122 0
5
10
15
20
25
30
(n=17)
% P
ote
nc
iaç
ão
(n=19)
A B C
E F
D
G
100 110 120 130 1400,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
1,6
EP
P A
mp
. (n
orm
ali
za
do
)
Tempo (s)
U-73122
Controle
20 Hz
FIGURA 35 - Efeito do U-73122 (5 µM) no tamanho dos EPPs, na Depressão, Recuperação e Potenciação. As barras e os símbolos representam a média ± EPM,*p<0,05,
**p<0,01 e ***p<0,001. Nas figuras B, C e D estão especificadas as frequências de estimulação.
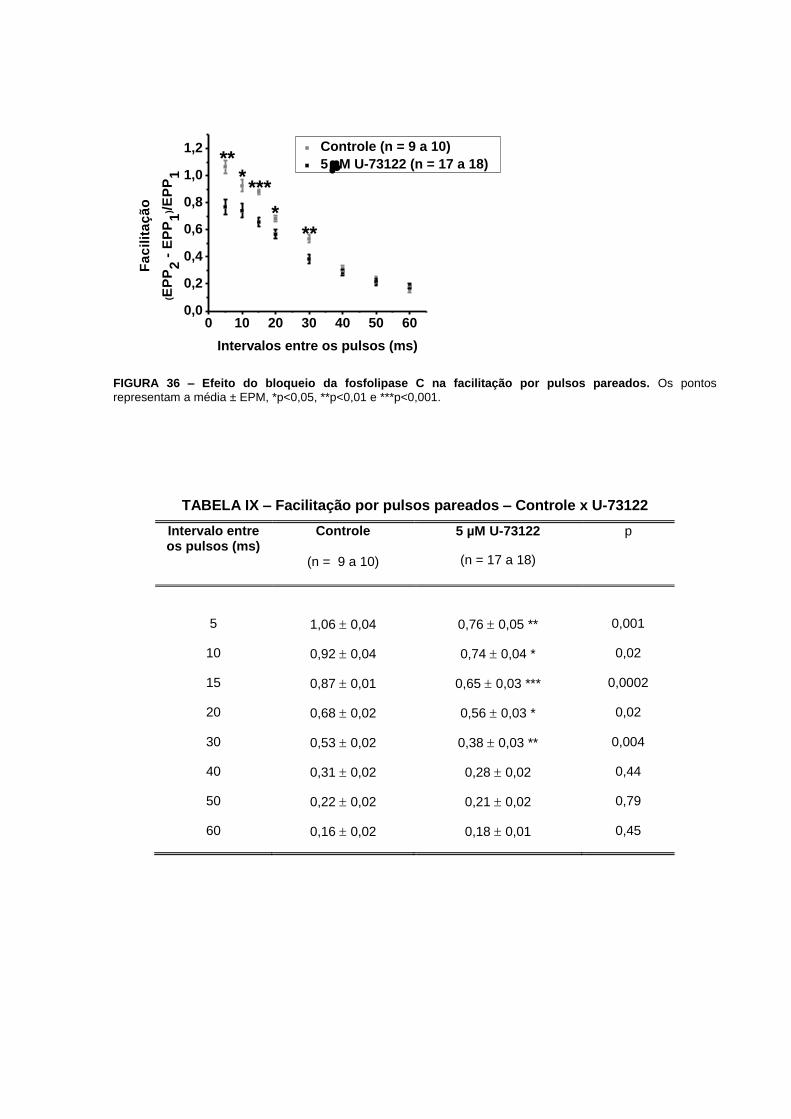
0 10 20 30 40 50 600,0
0,2
0,4
0,6
0,8
1,0
1,2
**
*
*
***
Fa
cil
ita
çã
o
(EP
P2
- E
PP
1)/E
PP
1
Intervalos entre os pulsos (ms)
Controle (n = 9 a 10)
5 M U-73122 (n = 17 a 18)**
FIGURA 36 – Efeito do bloqueio da fosfolipase C na facilitação por pulsos pareados. Os pontos
representam a média ± EPM, *p<0,05, **p<0,01 e ***p<0,001.
TABELA IX – Facilitação por pulsos pareados – Controle x U-73122
Intervalo entre os pulsos (ms)
Controle
(n = 9 a 10)
5 µM U-73122
(n = 17 a 18)
p
5 1,06 0,04 0,76 0,05 ** 0,001
10 0,92 0,04 0,74 0,04 * 0,02
15 0,87 0,01 0,65 0,03 *** 0,0002
20 0,68 0,02 0,56 0,03 * 0,02
30 0,53 0,02 0,38 0,03 ** 0,004
40 0,31 0,02 0,28 0,02 0,44
50 0,22 0,02 0,21 0,02 0,79
60 0,16 0,02 0,18 0,01 0,45

V. DISCUSSÃO
5.1. Análise da liberação de neurotransmissores na junção neuromuscular
A presença de receptores nicotínicos na região pós-sináptica da junção neuromuscular torna
esta região um detector sensível ao neurotransmissor acetilcolina. Portanto, a liberação de
acetilcolina é facilmente detectada por registros intracelulares na fibra muscular (DEL
CASTILLO & KATZ, 1957). Assim, o registro da liberação espontânea e evocada na junção
neuromuscular permite o estudo da liberação de neurotransmissores.
A liberação de acetilcolina na junção neuromuscular pode ocorrer de duas formas:
espontânea ou evocada por um estímulo nervoso. Registros eletrofisiológicos possibilitam a
medição de tais formas de liberação através de técnicas que utilizam eletrodos intra e
extracelulares (FATT & KATZ, 1951, 1952).
O registro eletrofisiológico dos potenciais sinápticos evocados requer a adoção de medidas
para bloquear a contração muscular. Como tínhamos o intuito de estudar os mecanismos de
depressão e facilitação por pulsos pareados em condições fisiológicas, descartamos a
possibilidade da utilização de baixa concentração de cálcio e alta concentração de
magnésio. Outras alternativas para a realização do bloqueio da contração muscular são a
fibra cortada ou cut fiber, a utilização de formamida ou curare. O método de cut fiber
consiste na dilaceração das fibras musculares com um estilete de forma a bloquear a
geração e a condução do potencial de ação na fibra muscular. As propriedades elétricas da
membrana da fibra muscular são alteradas e verificadas por uma grande despolarização do
potencial de membrana (BARSTAD, 1962; BARSTAD & LILLEHEIL, 1968; DODSON &
FORSYTHE, 2004; SABATINI & REGEHR, 1999). A utilização da formamida consiste na
destruição dos túbulos T da membrana da fibra muscular durante a mudança da preparação
nervo-músculo de uma solução hipertônica de formamida para uma solução isosmótica
normal. Este tratamento também altera as propriedades elétricas da membrana da fibra
muscular e como consequência verifica-se também uma grande despolarização do potencial
de membrana (DODSON & FORSYTHE, 2004; ESCALONA DE MOTTA et al., 1982a, b;
HERRERA, 1984; SABATINI & REGEHR, 1999).
Considerando que o curare não despolariza a membrana da fibra muscular, em nosso
estudo nós o utilizamos para realizar o bloqueio da contração muscular. Esta prática tem a

vantagem de produzir menores potenciais sinápticos evocados, o que minimiza o problema
da correção de somação não linear (MARTIN, 1955). O curare, além do efeito pós-sináptico
de bloqueio do receptor nicotínico, também pode apresentar efeitos pré-sinápticos, como a
interferência no processo de liberação de transmissores em situações de alta frequência de
estimulação (GIBB & MARSHALL, 1984; GLAVINOVÍC, 1979; MAGLEBY et al., 1981). No
entanto, mesmo podendo interferir no processo de liberação de neurotransmissores
verificamos que nosso protocolo para o estudo da depressão sináptica era adequado, pois
em uma situação de estimulação de alta frequência evidenciamos que a depressão
apresentava origem pré-sináptica e relacionada a uma alteração na concentração de cálcio
intracelular. Com relação à facilitação por pulsos pareados existem evidências de que as
preparações tratadas com curare apresentam uma maior facilitação com menores intervalos
entre os pulsos (MATZNER et al., 1988), entretanto, nós estudamos a facilitação nas
condições controles e experimentais nas mesmas concentrações de curare. Nós
observamos um efeito de diminuição dos EPPs com o tempo, mesmo em baixas frequências
de estimulação nervosa (0,2 Hz), e este efeito se estabilizava por volta do terceiro pulso. No
intuito de impedir a interferência de tal efeito em nossas medições, nós analisamos somente
os 10 últimos pulsos dos 35 pulsos aplicados ao terminal nervoso. Além disso, a facilitação
por pulsos pareados encontrada em nosso estudo apresenta um decaimento de natureza
exponencial também encontrado em outros estudos (ATLURI & REGEHR, 1996; MAGLEBY
& ZENGEL, 1982) e uma maior facilitação à menores intervalos entre os pulsos
(MUKHAMEDYAROV, et al., 2006, 2009).
Todos os experimentos, controles e experimentais, foram realizados nas mesmas condições
experimentais. Além disso, o curare continua a ser uma ferramenta muito utilizada para o
bloqueio da contração muscular em diversos estudos eletrofisiológicos (BENNETT et al.,
2007; HACHISUKA, et al., 2007; KUBOTA et al., 2005). Portanto nossas condições
experimentais estavam adequadas para justificar a metodologia de análise empregada.

5.2. Plasticidade sináptica
Inicialmente nosso objetivo consistia na análise da depressão final e da facilitação por
pulsos pareados. No entanto, no decorrer do estudo e das análises verificamos a
possibilidade do estudo de outros parâmetros, como a recuperação final e a potenciação.
Estes parâmetros estão relacionados aos fenômenos de plasticidade estudados e
contribuem para o entendimento de tais fenômenos.
Os mecanismos envolvidos na transmissão sináptica no sistema nervoso periférico são
relevantes para a compreensão de como os circuitos neurais no SNC são continuamente
alterados (PERSONIUS & BALICE-GORDON, 2000). Informações provenientes de outros
tecidos e sistemas esclarecem sobre o papel da liberação de cálcio intracelular nos
neurônios. Na JNM a plasticidade sináptica à curto prazo contribui para o aumento da
margem de segurança que está relacionada ao controle de movimento (MUKAMEDYAROV
et al., 2009; WOOD & SLATER, 1997, 2001).
5.2.1. Depressão sináptica
A estimulação nervosa repetida em situações de concentração normal de cálcio pode
ocasionar a diminuição dos potenciais pós-sinápticos, sendo comumente denominada de
depressão sináptica. O mecanismo de depressão pode estar relacionado à diminuição da
liberação pré-sináptica de transmissores e/ou à diminuição da sensibilidade dos receptores
pós-sinápticos ao transmissor. Na JNM de rã uma diminuição na liberação de
neurotransmissores está envolvida na depressão sináptica (BETZ, 1970; DEL CASTILLO &
KATZ, 1954b; VAN DER KLOOT & MOLGÓ, 1994; WU & BETZ, 1998; ZUCKER, 1989) e a
sensibilidade dos receptores pós-sinápticos medida através da amplitude dos MEPPs não
está alterada (DEL CASTILLO & KATZ, 1954b; ZENGEL & SOSA, 1994). A diminuição na
liberação de transmissores pode ser causada pela depleção do pool de vesículas
prontamente liberáveis, ou por uma diminuição na probabilidade de liberação. Em alguns
casos, a intensidade da depressão pode ser diminuída com a redução da quantidade de
transmissores liberados, como ocorre nas situações de baixo cálcio. Este fato sugere que
durante a depressão ocorre a depleção do pool de vesículas prontamente liberáveis
(ZUCKER, 1989). A diminuição na probabilidade de liberação pode ser causada por uma
diminuição no influxo pré-sináptico de cálcio seguido de um potencial de ação ou uma
modulação direta da maquinaria de cálcio intracelular (BETZ, 1970; OTSUKA et al., 1962).

Takeuchi e Takeuchi (1962) mostraram que o potencial de ação pré-sináptico na JNM de
lagostina não está alterado durante a depressão. Portanto, o mecanismo da depressão
parece não estar relacionado às alterações no potencial de ação, o que exclui a
possibilidade de um influxo reduzido de cálcio ser devido à variações no potencial de ação
(BETZ, 1970). Entretanto, pode ser que o influxo de cálcio seja diminuído durante a
depressão por bloqueio dos canais de cálcio durante este processo. Existem também relatos
de diminuição da liberação de neurotransmissores por ativação de receptores nicotínicos
pré-sinápticos e bloqueio de canais para cálcio (VAN DER KLOOT et al., 1997) e este
mecanismo pode estar ativado durante a depressão.
Verificamos que as preparações tratadas com o quelante de cálcio intracelular, BAPTA-AM,
apresentaram EPPs menores. Este resultado sugere uma menor liberação de
neurotransmissores em uma situação de menor concentração intracelular de cálcio. Nossos
resultados evidenciaram uma menor depressão nestas preparações, o que sugere que o
aumento na concentração intracelular de cálcio está relacionado à depleção de vesículas
sinápticas e à depressão sináptica. Nosso resultado sugere que o mecanismo da depressão
sináptica está relacionado a um aumento na concentração intracelular de cálcio rápido e
localizado próximo dos canais de cálcio presentes no terminal pré-sináptico. Esta hipótese é
corroborada por estudos anteriores que verificaram que a micro injeção de EGTA, um
quelante mais lento de cálcio, em preparações nervo músculo não apresentaram alteração
na depressão sináptica (SWANDULLA et al., 1991).
O estudo do efeito do BAPTA-AM sobre a depressão também nos permitiu verificar que a
recuperação da depressão e a potenciação pós-tetânica também são fenômenos sensíveis
às variações da concentração intracelular de cálcio. Na situação de menor concentração
intracelular de cálcio verificamos que tanto a recuperação como a potenciação foram
maiores. A recuperação e a potenciação são fenômenos correlacionados e mais facilmente
visualizados em situações de baixa concentração de cálcio. Depressão e potenciação
apresentam o mesmo curso temporal (KALKSTEIN & MAGLEBY, 2004; MAGLEBY &
ZENGEL, 1982), mas em situações de baixa concentração de cálcio a potenciação torna-se
mais evidente. Verificamos também que a recuperação ocorreu mais rapidamente. Portanto,
tanto a intensidade, quanto a velocidade da recuperação dependem da concentração do
cálcio intracelular.
Assim, nossos resultados demonstram que a intensidade dos fenômenos de depressão,
recuperação e potenciação são sensíveis as variações da concentração de cálcio
intracelular.

Nossos resultados demonstram que nosso protocolo de depressão é adequado para o
estudo deste fenômeno de plasticidade sináptica, uma vez que ele reflete uma alteração
pré-sináptica, ou seja, uma alteração na concentração intracelular de cálcio e consequente
alteração na liberação de neurotransmissores.
5.2.2. Facilitação por pulsos pareados
Com relação à facilitação por pulsos pareados já foi demonstrado que este fenômeno de
plasticidade sináptica à curto prazo também é sensível às variações na concentração
intracelular de cálcio (MUKHAMEDYAROV et al., 2009; TANABE & KIJIMA, 1992; VAN DER
KOOT & MOLGÓ, 1993). Apesar de a facilitação ser um dos fenômenos de plasticidade
mais estudados e frequentemente relacionada ao cálcio residual, os mecanismos envolvidos
neste processo não estão completamente esclarecidos. A hipótese do cálcio residual propõe
que parte do cálcio que entra no terminal após o primeiro pulso é depurado lentamente,
portanto a entrada de cálcio após o segundo potencial de ação é adicionada ao cálcio
residual já presente, aumentando a liberação de neurotransmissores (BITTNER & SCHATZ,
1981; DODGE & RAHAMIMOFF, 1967; STOCKBRIDGE & HINES, 1982). A liberação de
neurotransmissores requer um aumento na concentração de cálcio de cerca de 50-200 µM
(ZEFIROV & CHERANOV, 2000). Llinás et al. (1992) demonstraram que a concentração de
cálcio intracelular nos microdomínios, onde ocorre a liberação de neurotransmissores, é de
aproximadamente 200-300 µM. No entanto, a concentração residual de cálcio no terminal
pré-sináptico, mesmo após uma atividade rítmica de alta frequência e prolongada não
ultrapassa mais que 3-5 µM (ANGLESON & BETZ, 2001). Portanto, a hipótese de cálcio
residual não pode ser a única responsável por tal processo. Até o momento não existem
dados concretos sobre a natureza específica da facilitação. Provavelmente, mecanismos
distintos com diferentes cinéticas e importância podem estar relacionados.
Em nosso estudo, além de quantificarmos através dos registros eletrofisiológicos a
importância dos diferentes mecanismos de depuração do cálcio intracelular em uma mesma
preparação e verificarmos o papel dos receptores de IP3 nos fenômenos de plasticidade
sináptica à curto prazo, depressão e facilitação por pulsos pareados, também buscamos
elucidar os mecanismos envolvidos nestes fenômenos de plasticidade.

5.3. Efeito dos diferentes mecanismos de depuração do cálcio intracelular sobre a
depressão e a facilitação por pulsos pareados
Tendo em vista nossos resultados de diminuição da intensidade da depressão em uma
situação de menor concentração de cálcio intracelular ao incubarmos as preparações com o
BAPTA-AM, nós utilizamos a variação na intensidade de depressão como indicativo da
concentração de cálcio nos microdomínios de liberação de neurotransmissores. Desta
forma, avaliamos a contribuição dos diferentes sistemas depuradores de cálcio durante a
depressão. Nós também verificamos a participação destes sistemas durante estimulações
menos intensas capazes de produzir facilitação. Os primeiros experimentos objetivaram
verificar a contribuição das Ca+2-ATPases do RE (SERCA) e da membrana plasmática
(PMCA). As Ca+2-ATPases pertencem a família de ATPases do tipo P, caracterizadas pela
conservação temporária da energia do ATP na forma de uma enzima fosforilada
intermediária (PEDERSEN & CARAFOLI, 1987). As Ca+2-ATPases apresentam alta
afinidade ao cálcio, mas o transporte é realizado mais lentamente em comparação aos
trocadores Na+/Ca+2 (CARAFOLI, 1991; DIPOLO & BEAUGE, 1983; LEHOSTSKY, 1995).
A contribuição da SERCA foi verificada através da utilização da tapsigargina, um inibidor
não competitivo desta classe de enzima (ROGERS, et al., 1995). Nas células não
musculares a isoforma mais encontrada é a do tipo 2b e 3 (BRINI & CARAFOLI, 2009;
CARAFOLI & BRINI, 2000). A tapsigargina tem a capacidade de prender a bomba no estado
inativo de forma irreversível, sendo a inibição maior em solução livre de cálcio e na presença
de EGTA. Nesta situação o cálcio não pode ser recaptado para o RE, permanecendo por
mais tempo no citoplasma (SAGARA & INESI, 1991). Após o tratamento da preparação
nervo-músculo com tapsigargina verificamos um aumento na amplitude dos EPPs. Este
resultado sugere uma maior liberação de neurotransmissores na situação de maior
concentração de cálcio intracelular. Nossos resultados também evidenciam uma maior
depressão, condizente com a depleção das vesículas sinápticas devido à maior
concentração intracelular de cálcio. A maior recuperação final evidenciada pode ser
justificada por uma liberação lenta do cálcio pela mitocôndria após o término do estímulo
tetânico (TANG & ZUCKER, 1997; VERSTREKEN, 2005). Não verificamos efeito na
potenciação, provavelmente, pelo fato deste fenômeno ser mais facilmente visualizado em
situações de menor concentração de cálcio, no entanto, recuperação e potenciação são
fenômenos correlacionados (KALKSTEIN & MAGLEBY, 2004; MAGLEBY & ZENGEL,
1982).

Com relação à facilitação por pulsos pareados, verificamos uma maior facilitação quando o
intervalo entre os pulsos foi de 5 a 15 ms. Este resultado evidencia a importância deste
mecanismo de depuração de cálcio intracelular no fenômeno de facilitação e indica a
possível localização do RE próximo das zonas ativas, como já demonstrado
morfologicamente por Westrum & Gray (1986). O efeito significativo somente nos menores
intervalos entre os pulsos sugere a participação da SERCA na primeira fase da facilitação,
onde os mecanismos envolvidos encontram-se próximos dos locais de liberação de
neurotransmissores (MUKHAMEDYAROV et al., 2009). Além disto, este resultado sugere
que a SERCA é um mecanismo de depuração rápido quando ocorrem variações na
concentração intracelular de cálcio.
O segundo mecanismo de depuração do cálcio intracelular estudado foi a Ca+2-ATPase da
membrana plasmática (PMCA). No tecido nervoso as isoformas do tipo 1, 3 e 4 são as mais
encontradas (BRINI & CARAFOLI, 2009). Diferentemente da SERCA, não existe um inibidor
específico da PMCA como a tapsigargina é para a SERCA. No entanto, a CEDA é a
ferramenta farmacológica mais frequentemente utilizada e considerada mais adequada para
o bloqueio (BASSANI, et al., 1995; CHOI & EISNER, 1999; GATTO et al., 1995).
Nossos resultados não evidenciaram nenhum efeito sobre os parâmetros analisados na
depressão e nem na facilitação por pulsos pareados, o que sugere que a PMCA como
mecanismo de depuração do cálcio intracelular não apresenta uma importância significativa
nesta preparação. De forma contrária, Jensen et al. (2007) verificaram que o bloqueio da
PMCA com CEDA aumentou a frequência das correntes excitatórias pós-sinápticas e a
facilitação por pulsos pareados na região hipocampal CA3 de ratos. Neste mesmo trabalho
também foi realizado o bloqueio da SERCA com ácido ciclopiazônico (CPA) e nenhum efeito
foi encontrado. Outros estudos na JNM de rã (BENNET, et al., 2007; SUZUKI, et al., 2002) e
de lagostina (LIN, et al., 2005) também não verificaram uma contribuição importante da
PMCA no sequestro de cálcio citoplasmático. Portanto, diferentes preparações podem
apresentar mecanismos distintos para o controle da concentração intracelular de cálcio.
Outra organela importante para a homeostase do cálcio intracelular é a mitocôndria. A
mitocôndria apresenta em sua membrana dois sistemas de transporte que contribuem para
o controle da concentração intracelular de cálcio. O trocador Na+/Ca+2 (NCXm) e o uniporter
mitocondriais (MCU) (CARAFOLI, 1987). O NCXm opera de forma eletroneutra, mas pode
operar em alguns casos eletrogenicamente na direção do efluxo de Ca+2, ou seja, trocando
mais de dois íons Na+ para cada íon Ca+2 (CROMPTON et al., 1976). Em situações de
grande aumento da concentração intracelular de cálcio o NCXm pode atuar de modo reverso

(GRIFFITHS, 1999; SMETS et al., 2004). Portanto, resolvemos também investigar o papel
deste trocador no sequestro do cálcio intracelular.
Um dos fatores limitantes para o esclarecimento do papel do NCXm na fisiologia celular é a
ausência de informação sobre sua estrutura molecular. Nenhum gene do NCXm foi
encontrado no genoma mitocondrial (ANDERSON et al., 1981; BARRELL et al., 1980), o que
sugere que o NCXm é codificado por um gene nuclear, transferido para o citoplasma e
posteriomente para a mitocôndria. O mesmo acontece para o uniporter mitocondrial.
Utilizamos o bloqueador CGP37157 (COX et al., 1993) e verificamos que a intensidade da
depressão é maior e sua instalação é mais lenta. Este resultado sugere a atuação do NCXm
como mecanismo depurador do cálcio citoplasmático em situações de aumento de sua
concentração intracelular. Entretanto, sua cinética é diferenciada, pois a instalação da
depressão é mais lenta, o que indica que este mecanismo de depuração pode ser utilizado
mais tardiamente durante a depressão. Sugerimos que a SERCA seja o principal
mecanismo de depuração pela maior intensidade de depressão apresentada e instalação
mais rápida. No entanto, devemos considerar o fato do CGP também ativar a liberação de
cálcio (HERNÁNDEZ-SANMIGUEL et al., 2006) e reduzir o repreenchimento do RE
(ARNAUDEAU et al., 2001; MALLI et al., 2005). Portanto, a maior depressão final observada
pode estar relacionada à liberação de cálcio pelo RE e não à inibição do trocador NCXm ao
atuar de forma reversa e como depurador de cálcio intracelular.
O bloqueio do NCXm também apresentou efeitos sobre a recuperação e a potenciação pós-
tetânica. Verificamos uma recuperação mais lenta e uma menor potenciação, resultado este
mais condizente com a função do trocador. O bloqueio do trocador impede a extrusão de
cálcio pela mitocôndria após o estimulo tetânico, o que acaba por influenciar a recuperação
e a potenciação pós-tetânica. Nossos resultados assemelham-se com resultados de
trabalhos anteriores em outras preparações que demonstraram que a mitocôndria é capaz
de sequestrar o cálcio que entra no terminal nervoso durante a estimulação repetida numa
frequência que pode exceder 10-20 Hz (GARCÍA-CHACON et al., 2006). O sequestro de
cálcio pela mitocôndria contribui para a liberação de neurotransmissores, de forma a
minimizar a depleção de vesículas sinápticas após a tetania (DAVID & BARRETT, 2003).
Além disso, Tong (2007) verificou na JNM de drosófila um aumento significativo na
densidade de mitocôndrias nas sinapses estimuladas tetanicamente, enquanto que nas
sinapses não estimuladas, a densidade desta organela não foi alterada. Este rearranjo é
considerado essencial para a potenciação pós-tetânica.

No que se refere à facilitação por pulsos pareados, não verificamos nenhum efeito. Portanto,
sugere-se que o NCXm não atua como um mecanismo de depuração neste fenômeno de
plasticidade à curto prazo.
Outro sistema de transporte de cálcio presente na membrana mitocondrial é o uniporter. O
MCU consiste em um canal iônico altamente seletivo presente na membrana interna da
mitocôndria que permite a recaptação do cálcio citoplasmático para o interior desta
organela. O cálcio atravessa a membrana mitocondrial interna graças ao gradiente de força
representado pelo potencial transmembrana negativo (RIZZUTO et al., 2000, 2004). O
Ru360 foi utilizado em nosso estudo para verificar a participação do MCU como mecanismo
depurador do cálcio intracelular na depressão e facilitação por pulsos pareados (MATLIB et
al., 1998; ZAZUETA et al., 1999). Verificamos que as preparações nervo-músculo tratadas
com o Ru360 apresentaram uma menor intensidade de depressão. Hernandez-Guijo et al.,
(2001) demonstraram em células cromafins que o bloqueio da recaptação de cálcio pela
mitocôndria aumenta a inibição das correntes de cálcio dependentes de voltagem. Já foi
demonstrado que o Ru360 produz uma redução nos transientes de cálcio induzidos por
estimulação na JNM de lagarto. Este efeito decorre da redução da entrada de cálcio pela
inibição dos canais de cálcio presentes na membrana plasmática, principalmente os do tipo
N (DAVID et al., 1997; 1999). Portanto, nossa observação mostrando que houve uma
depressão de menor intensidade sugere que o Ru360 tenha bloqueado os canais de cálcio
voltagem-dependentes. A entrada reduzida de cálcio leva a uma menor liberação de
neurotransmissores e consequentemente uma depressão de menor intensidade como
verificamos. A análise dos demais parâmetros da depressão e a análise da facilitação por
pulsos pareados não apresentaram nenhuma diferença, o que sugere que este mecanismo
de depuração do cálcio intracelular não deve desempenhar um papel importante nestes
fenômenos de plasticidade sináptica à curto prazo na junção neuromuscular. No entanto, o
efeito do Ru360 sobre os canais de cálcio pode ter comprometido a análise do papel do
MCU.
A membrana plasmática também apresenta um trocador Na+/Ca+2 capaz de extrudar o
cálcio. Este sistema de transporte pertence a uma família de proteínas que transporta três
íons sódio para cada íon cálcio bidirecionalmente através da membrana plasmática
(CARAFOLI et al., 2001). O transporte é considerado eletrogênico (LAGNADO &
MCNAUGHTON, 1990). O gradiente eletrogênico para o transporte realizado pelo trocador é
o gradiente de concentração do sódio, do cálcio e o potencial de membrana (ANNUNZIATO
et al., 2004; HINATA & KIMURA, 2004). O tecido nervoso apresenta praticamente todas as
isoformas deste trocador (LI et al., 2000; THURNEYSEN et al., 2002).

Na ausência de um inibidor específico do NCX da membrana plasmática, realizamos o
bloqueio deste através da substituição da solução de Ringer normal pela solução de Ringer
lítio. Na ausência de sódio no meio extracelular o cálcio não pode ser extrudado pelo
trocador, permanecendo por mais tempo no citoplasma (HERMONI et al., 1987).
Verificamos que as preparações incubadas com a solução de Ringer lítio apresentaram
EPPs menores, maior depressão final, menor potenciação e menor facilitação quando o
intervalo entre os pulsos foi de 5 a 50 ms. Nossos resultados podem refletir uma
contribuição deste mecanismo de depuração na depressão e facilitação por pulsos
pareados, no entanto alguns aspectos devem ser esclarecidos antes de sugerirmos uma
participação. Caso o trocador atue realmente como um mecanismo de depuração nesta
preparação, seu bloqueio desencadearia um aumento na concentração intracelular de cálcio
e possíveis alterações nos fenômenos estudados. Estas alterações poderiam refletir um
aumento de intensidade da depressão e da facilitação por pulsos pareados. No entanto, os
efeitos do lítio sobre o conteúdo quântico e a liberação espontânea encontrados na literatura
são contraditórios. Crawford (1975) e Brainsteanu & Volle (1975) verificaram um aumento do
conteúdo quântico e da frequência dos MEPPs, enquanto que Benoit et al., (1973) e
Onodera & Yamakawa (1966) evidenciaram uma diminuição do tamanho e do conteúdo
quântico. Nossos resultados assemelham-se mais ao efeito inibitório do lítio na liberação de
neurotransmissores. Assim, a substituição do Ringer normal pelo Ringer lítio não foi uma
ferramenta adequada para bloquearmos o NCX da membrana plasmática. EPPs com
menores amplitudes, menor potenciação e facilitação por pulsos pareados são resultados
que podem refletir uma menor liberação de neurotransmissores. Alguns trabalhos
evidenciam que o lítio pode diminuir a velocidade de condução do potencial de ação e
reduzir a excitabilidade do terminal nervoso, bloqueando a neurotransmissão (ORTIZ &
JUNGE, 1978). Além disso, a permeabilidade dos receptores nicotínicos ao lítio é reduzida
quando comparada ao sódio (WATANABE & NARAHASHI, 1979). Estes efeitos podem
justificar os resultados encontrados, inclusive a maior depressão. Nesta situação a
depressão pode não estar relacionada à depleção de vesículas sinápticas decorrente de um
aumento na concentração intracelular de cálcio, mas sim a um bloqueio da
neurotransmissão (KELLY, 1968) ou a uma diminuição da sensibilidade do canal da placa
motora na presença no lítio (WATANABE & NARAHASHI, 1979).
Portanto, dentre os mecanismos de depuração estudados verificamos que somente a
SERCA apresenta uma contribuição nos fenômenos de depressão e facilitação por pulsos
pareados e talvez o NCXm na depressão. A ausência de efeito na depressão e na facilitação
por pulsos pareados quando realizamos o bloqueio da PMCA e do uniporter mitocondrial

sugere que estes mecanismos não apresentam importância significativa na depuração de
cálcio intracelular na plasticidade sináptica à curto prazo na JNM.
O bloqueio do NCX da membrana plasmática através da substituição do Ringer normal pelo
Ringer lítio não foi uma ferramenta adequada, mas podemos considerar que a presença de
íons sódio no meio extracelular é essencial para a neurotransmissão (SABATINI &
REGEHR, 1999). Apesar de não podermos verificar adequadamente o papel do NCX da
membrana plasmática como mecanismo de depuração do cálcio intracelular na depressão e
na facilitação por pulsos pareados, dados da literatura sugerem que este trocador não é o
principal participante da regulação da liberação de neurotransmissores dependente de
cálcio. Os NCX da membrana plasmática encontram-se concentrados no terminal nervoso,
mas não estão presentes nas zonas ativas onde as vesículas sinápticas estão posicionadas
e os canais de cálcio estão localizados (JUHASOVA et al., 2000; LUTHER et al., 1992).
Nossos resultados sugerem uma importante contribuição da mitocôndria no fenômeno de
potenciação como já demonstrado em estudos anteriores (DAVID & BARRETT, 2003;
GARCÍA-CHACON et al., 2006; TONG, 2007). Ao bloquearmos a SERCA verificamos uma
maior recuperação final. A recuperação e a potenciação são fenômenos correlacionados e
como já mencionados podem estar relacionados com a liberação lenta de cálcio pela
mitocôndria. Quando realizamos o bloqueio do NCXm verificamos uma menor potenciação e
uma recuperação mais lenta. Estes resultados confirmam a importância da mitocôndria na
potenciação pós-tetânica.
Podemos fazer uma correlação dos nossos resultados com os resultados obtidos por
Bennett et al., (2007). Neste trabalho foi verificado que diferentes mecanismos de depuração
do cálcio intracelular apresentam diferentes contribuições nos fenômenos de plasticidade
sináptica. A facilitação a qual o autor se refere consiste no aumento da concentração
intracelular de cálcio seguida de uma estimulação de alta frequência e curta duração. No
entanto, a mensuração da variação da concentração intracelular de cálcio não é uma
medida direta de alteração nos fenômenos de plasticidade como o registro eletrofisiológico
realizado em nosso trabalho. Também foi verificada uma contribuição importante do RE e da
mitocôndria como mecanismos de depuração. Com relação à PMCA, foi verificada uma
pequena contribuição deste mecanismo no sequestro de cálcio citoplasmático. Para verificar
a contribuição da mitocôndria foi utilizado o CCCP, um desacoplador metabólico da
mitocôndria que dissipa o gradiente de prótons na membrana mitocondrial interna, liberando
cálcio para o citoplasma (PENG, 1998). No entanto, esta ferramenta não é específica para
nenhum transportador ou trocador presente na mitocôndria. Portanto, nosso trabalho
mostra-se mais adequado para o estudo da participação de diferentes mecanismos de

depuração do cálcio intracelular em fenômenos de plasticidade sináptica à curto prazo, por
utilizar o registro eletrofisiológico e bloqueadores direcionados.
O efeito predominante do bloqueio da SERCA na depressão e facilitação por pulsos
pareados sugere que este mecanismo de depuração apresenta maior importância. O RE
parece estar mais envolvido no ajuste fino da concentração citosólica de cálcio. Já a
mitocôndria apresenta um papel secundário nestes fenômenos. A recaptação de cálcio pela
mitocôndria é de baixa afinidade, apesar de sua grande capacidade de estocagem deste íon
(VERCESI et al., 1978). A afinidade da mitocôndria ao cálcio é insuficiente para que esta
apresente um papel importante na regulação do cálcio intracelular quando a concentração
intracelular de cálcio é sub-µM (CROMPTON et al., 1976). O RE atua como um estoque
imediato de cálcio, enquanto a mitocôndria parece estar mais envolvida na potenciação pós-
tetânica.
5.4. Efeito da brevetoxina na liberação espontânea
Neurotoxinas são ferramentas importantes para o estudo da fisiologia das sinapses.
Brevetoxinas são poliéteres cíclicos lipossolúveis produzidos pelos dinoflagelados
Ptychodiscus brevis (BADEN, 1989). Esta toxina apresenta efeitos nas fibras nervosas
mielinizadas e nas junções neuromusculares de vertebrados. O alvo molecular desta toxina
é a quinta alça transmembrânica do canal de sódio, uma proteína essencial para a
excitabilidade celular (GAWLEY et al., 1992; MATTEI et al., 1999; POLI et al., 1986). A
PbTx-3 em concentrações nanomolares aumenta a permeabilidade ao sódio em células
excitáveis, e consequentemente afeta vários mecanismos sódio-dependentes nas células
nervosas. Esta ação é atribuída a uma modificação dos canais de sódio, que permanecem
permanentemente abertos, inclusive em potenciais de repouso da membrana, ao invés de
permanecerem no estado fechado. Como consequência, as brevetoxinas causam uma
despolarização do potencial de membrana, e consequentemente um aumento da liberação
espontânea na JNM (ATCHISON et al., 1986).
A PbTx-3 foi utilizada para identificarmos indiretamente a presença do receptor de IP3 na
junção neuromuscular, visto que o efeito da PbTx-3 de aumento da concentração de IP3 e
aumento da liberação de cálcio intracelular via IP3R em outras preparações já é conhecido
(LIBERONA et al., 2008).

A concentração utilizada de PbTx-3 foi 50 nM, pois estudos anteriores demonstraram que
esta concentração permite o monitoramento do potencial de repouso, apesar da
despolarização da membrana das fibras musculares de cerca de 8 - 18 mV (MEUNIER,
1997). Nossos resultados confirmam que a entrada de sódio pelos canais de sódio sensíveis
à TTX está envolvida no aumento da liberação espontânea produzida pela PbTx-3
(MEUNIER, 1997).
No intuito de esclarecer se o mecanismo de ação da PbTx-3 envolve a liberação de cálcio
de estoques sensíveis à tapsigargina utilizamos a tapsigargina. A tapsigargina produziu um
aumento na frequência dos MEPPs, o que sugere que o bloqueio da Ca+2-ATPase é capaz
de aumentar a concentração citosólica de cálcio. Este efeito está relacionado ao aumento
dos EPPs na presença de tapsigargina já demonstrado por Castonguay e Robitaille (2001).
A tapsigargina pode induzir o aumento da concentração citosólica de cálcio, que ocorre pela
liberação de cálcio dos estoques sensíveis à IP3 sem a hidrólise de fosfolipídios de inositol
(JACKSON et al., 1988). O aumento da frequência dos MEPPs quando realizamos a
incubação da preparação com PbTx-3 e tapsigargina foi menor do que na ausência de
tapsigargina. Portanto, a PbTx-3 aumenta a liberação espontânea pela liberação de cálcio
de estoques intracelulares sensíveis à tapsigargina.
Estudos anteriores demonstraram que o mecanismo de ação da PbTx-3 não envolve a
liberação de cálcio da mitocôndria ou dos receptores de rianodina (LIBERONA et al., 2008).
O aumento na frequência dos MEPPs pela PbTx-3 não é bloqueado por agentes
farmacológicos conhecidos por impedir a liberação de cálcio de estoques intracelulares,
como o TMB-8, heparina e dantrolene (MEUNIER, 1997). Entretanto, a utilização de alguns
agentes, como a heparina, pode não ser a mais adequada. A heparina bloqueia os IP3Rs,
mas apresenta uma lenta difusão na célula (NILSSON et al., 1988; WARRIER, 2005), além
de apresentar outros efeitos como o desacoplamento de proteínas G e a ativação de
receptores de rianodina (BEZPROZVANNY, 1993; DASSO & TAYLOR, 1991; EHRLICH et
al., 1994). Portanto, para investigar se o mecanismo de ação da PbTx-3 está relacionado à
ativação dos IP3Rs é necessária a utilização de agentes farmacológicos mais específicos.
Nossos resultados demonstram que o 2-APB e o U-73122 bloqueiam o efeito da PbTx-3 de
aumento da liberação espontânea. Apesar do 2-APB também poder inibir a Ca+2-ATPase do
RE e de bloquear a entrada de cálcio operada por estoques, utilizamos uma concentração
que não apresentou efeito na frequência e na amplitude dos MEPPs (dado não mostrado)
(BOOTMAN et al., 2002). Esta mesma concentração foi utilizada nos experimentos para
estudo da plasticidade sináptica à curto prazo. Portanto, concluímos que a PbTx-3 aumenta
a frequencia dos MEPPs pela ativação da fosfolipase C (PLC) e dos IP3Rs.

A ativação da PLC e a consequente ativação dos IP3Rs pode estar relacionada à vários
mecanismos. O grande influxo de sódio pode ativar proteínas G, as vias de sinalização da
PLC e do fosfatidil inositol 3-quinase (ELTIT et al., 2004, 2006). Gusovsky e Daly (1988)
sugere que o aumento da concentração intracelular de sódio pode ativar diretamente a PLC.
A ativação da PLC ocasiona a formação de segundos mensageiros como o diacilglicerol
(DAG) e o IP3. O IP3 é capaz de desencadear a liberação de cálcio de estoques
intracelulares, mais especificamente do RE. O DAG juntamente com o cálcio estimula a
proteína quinase C (PKC) e esta está envolvida na fosforilação de componentes do
complexo SNARE, possibilitanto o acesso das vesículas aos locais de liberação na
membrana plasmática (CATTERALL, 1999; MORGAN et al., 2005; VAUGHAN et al., 1998).
A PLC também é capaz de modular a liberação de neurotransmissores pela fosforilação de
substratos de proteínas envolvidas no recrutamento de vesículas sinápticas e no processo
de exocitose (BROWN & SIHRA, 2008; GILMAN, 1987).
Portanto, a entrada de sódio e ativação de cascatas intracelulares envolvidas na liberação
de neurotransmissores proporcionam o aumento da liberação espontânea. Nossos
resultados, além de auxiliarem na elucidação do mecanismo de ação da PbTx-3, sugerem a
presença do IP3R e da PLC e o importante papel destes na modulação da liberação de
neurotransmissores na JNM.
5.5. Efeito dos receptores de rianodina e de IP3 sobre a depressão e a facilitação por
pulsos pareados
O esclarecimento do papel do IP3R na plasticidade sináptica à curto prazo na JNM é um dos
objetivos de nosso trabalho, no entanto, também verificamos o papel do RyR no intuito de
realizarmos uma comparação destes dois canais liberadores de cálcio presentes na mesma
organela.
No terminal pré-sináptico da JNM de rã a principal isoforma encontrada do RyR é a do tipo 3
(KUBOTA et al., 2005). Com o bloqueio dos RyRs verificamos uma depressão menor e de
instalação mais lenta. Com relação à facilitação por pulsos pareados evidenciamos uma
menor facilitação quando o intervalo entre os pulsos foi de 5-15 ms. Portanto, nossos
resultados sugerem uma importante contribuição do cálcio liberado pelos receptores de
rianodina nestes fenômenos de plasticidade sináptica à curto prazo.

Narita et al., (1998, 2000) demonstraram que o mecanismo de CICR na JNM de rã necessita
da entrada de cálcio através da estimulação tetânica. Os RyRs localizam-se próximos dos
canais de cálcio presentes no terminal pré-sináptico e amplificam a exocitose evocada por
impulsos e a plasticidade sináptica (KUBOTA et al., 2005; NARITA et al., 2000).
Posteriormente, este mesmo grupo de pesquisadores demonstrou que a CICR ocorre em
resposta à entrada de cálcio, na ausência de estimulação tetânica (KUBOTA et al., 2005).
Em nosso protocolo de estimulação tetânica verificamos depressão sináptica em contraste
com os resultados encontrados por Narita et al., (1998, 2000) que observaram potenciação.
Narita et al., (1998) não observaram alteração na facilitação após a estimulação tetânica de
curta duração, porém, a aumentação e a potenciação aumentaram após a estimulação
tetânica de longa duração. Realizamos nosso estudo em condições de conteúdo quântico
normal (1,8 mM Ca+2). Nestas condições a depressão é observada imediatamente após a
estimulação tetânica, ao invés de pronunciada aumentação e potenciação, comumente
encontradas em situações de baixo cálcio (KALKSTEIN & MAGLEBY, 2004). Além disso, a
facilitação referida no trabalho de Narita et al. (1998, 2000), não é a facilitação por pulsos
pareados que estudamos, e sim o aumento da amplitude dos EPPs. Neste caso, para uma
melhor comparação de efeito na facilitação a amplitude dos MEPPs também deve ser
considerada.
Nosso resultado de menor facilitação por pulsos pareados quando o intervalo entre os
pulsos foi de 5-15 ms está de acordo com o rápido aumento na concentração intracelular de
cálcio observada após o disparo do potencial de ação (LLINÁS et al., 1995a, b), com a
localização dos RyRs próxima dos canais de cálcio no terminal pré-sináptico (KUBOTA et
al., 2005; NARITA et al., 2000) e com a ocorrência da CICR mesmo na ausência de
estimulação tetânica (KUBOTA et al., 2005).
As três isoformas dos IP3Rs são expressas na maioria dos tipo celulares, o que sugere a
importância deste receptor em vários processos fisiológicos (DE SMEDT et al., 1997;
FUJINO et al., 1995, TAYLOR et al., 1999). No tecido nervoso a isoforma mais comumente
encontrada é a do tipo 1 (FURUICHI et al., 1993; NAKAGAWA et al., 1991), entretanto, não
se sabe qual isoforma é expressa no terminal pré-sináptico das junções neuromusculares.
A sinalização do cálcio intracelular mediada pelos IP3Rs é importante para a
neurotransmissão e a plasticidade sináptica, visto que a formação de IP3 pode ocorrer
seguida a ativação de receptores presentes na membrana plasmática por
neurotransmissores (COHEN et al., 1998; NISHIZAKI & MORI, 1998; POWER & SAH,
2002). A liberação de cálcio dependente do IP3 (ICICR, do inglês IP3-dependent calcium-
induced calcium release) é um mecanismo comumente encontrado nos neurônios piramidais

do hipocampo, no neocórtex e nas células de Purkinje do córtex cerebelar
(BEZPROZVANNY et al., 1991; FINCH & AUGUSTINE, 1998; NAKAMURA et al., 2000;
TAKECHI et al., 1998). As preparações tratadas com 2-APB apresentaram uma depressão
menor e de instalação mais lenta. Estes dados sugerem que os IP3Rs são ativados durante
estimulação intensa. No entanto, não verificamos nenhum efeito na facilitação por pulsos
pareados. Nossos resultados sugerem a participação dos IP3Rs somente na depressão
sináptica, sendo esta a primeira evidência da participação dos IP3Rs na plasticidade
sináptica à curto prazo na JNM.
Como a PLC está envolvida na formação do IP3, também verificamos seu envolvimento na
depressão sináptica e na facilitação por pulsos pareados. Verificamos que as preparações
tratadas com o inibidor da PLC, U-73122, apresentaram EPPs menores. Este resultado
sugere uma menor liberação de neurotransmissores em uma situação de menor
concentração intracelular de cálcio. Verificamos também uma depressão menor e de
instalação mais rápida. Com relação à recuperação evidenciamos que esta foi maior e mais
rápida. Portanto, sugerimos que a ativação da fosfolipase C é importante para este
fenômeno de plasticidade sináptica à curto prazo. Nossos resultados sugerem que o
provável efeito estimulatório nos IP3Rs durante estimulação prolongada seja mediado pela
fosfolipase C. Este efeito pode ser devido ao aumento na concentração intracelular de sódio
(ELTIT et al., 2004, 2006, GUSOVSKY & DALY, 1988).
Com relação à facilitação por pulsos pareados verificamos uma menor facilitação quando o
intervalo entre os pulsos foi de 5-30 ms, o que sugere também uma importante contribuição
da ativação da fosfolipase C neste fenômeno de plasticidade sináptica à curto prazo.
Entretanto, como nós não observamos alteração na facilitação por pulso pareados por
bloqueio de IP3Rs, o efeito da fosfolipase C na facilitação não deve ser por ativação destes
receptores. Outros alvos da fosfolipase C devem ser investigados.
Nossos resultados evidenciam uma importante contribuição dos IP3Rs e da PLC na
plasticidade sináptica à curto prazo na JNM. Durante a estimulação de alta frequencia do
protocolo de depressão também ocorre uma grande entrada de íons sódio juntamente com o
cálcio. Somente alguns aspectos foram considerados sobre o envolvimento da via de
sinalização do IP3 na neurotransmissão quando discutimos o efeito da PbTx-3 na liberação
espontânea. O diacilglicerol, formado pela ativação da fosfolipase C, juntamente com o
cálcio estimula a proteína quinase C (PKC). A PKC é capaz de modular canais de cálcio
sensíveis à voltagem e consequentemente alterar o influxo pré-sináptico de cálcio
(CATTERALL, 1999; VAUGHAN et al., 1998). A ativação da cascata de sinalização
intracelular do IP3 pode ocorrer com a ativação de diferentes receptores presentes na

membrana celular. Nas células piramidais do hipocampo, a liberação de cálcio de estoques
sensíveis à IP3 e o aumento da potenciação de longo prazo (LTP, do inglês long term
potentiation) são desencadeadas pela ativação de receptores glutamatérgicos, que muito
provavelmente atuam via cascatas intracelulares similares às ativadas por receptores
muscarínicos (COHEN et al., 1998). A ativação de receptores purinérgicos e adrenérgicos
também estão relacionadas com a produção de IP3 (NISHIZAKI & MORI, 1998).
A produção de IP3 induzida pela ativação dos receptores muscarínicos é um dos principais
mecanismos de liberação de cálcio do RE nos neurônios piramidais (POWER & SAH, 2002).
Na sinapse colinérgica da JNM de rã receptores muscarínicos regulam a liberação de ACh.
Dois subtipos de receptores estão envolvidos neste processo. O subtipo 1 (M1) regula o
aumento da liberação de ACh, enquanto que o subtipo 2 (M2) está envolvido na inibição da
liberação (REN & HARTY, 1994; SLUTSKY et al., 1999; VANNUCCHI & PEPEU,1995). O
receptor muscarínico M1 está acoplado à PLC e via proteína G heteromérica (Gq/11)
hidrolisa fosfolipídeos de membrana, produzindo IP3 e DAG (BARDO et al., 2006; ROSE &
KONNERTH, 2001). Portanto, a ativação do receptor muscarínico pré-sináptico na JNM pela
maior quantidade de ACh liberada durante a facilitação por pulsos pareados e a depressão
pode ativar a via de sinalização da PLC, produzir IP3 e DAG e proporcionar modificações na
eficácia sináptica.
Outra alternativa para a ativação da PLC consiste na fosforilação e ativação desta proteína
por proteínas quinases dependentes do cálcio ativadas durante a estimulação tetânica
(KOYANO et al., 1985).
Além dos efeitos da ativação dos RyRs e dos IP3Rs por seus agonistas específicos,
evidências recentes sugerem que o IP3 pode mobilizar cálcio via RyRs e provavelmente de
forma mais eficiente (LOUVET & COLLIN, 2005).
Portanto, verificamos uma importante contribuição dos IP3Rs e da PLC em fenômenos de
plasticidade sináptica à curto prazo. A PLC parece apresentar efeitos variados, pois
verificamos efeito tanto na depressão sináptica quanto na facilitação por pulsos pareados,
muito provavelmente por sua ativação estar relacionada à formação e ativação de outros
componentes, como a PKC e o DAG, também envolvidos na sinalização de cálcio
intracelular.

VI. CONCLUSÃO
Nosso estudo demonstra que a Ca+2-ATPase presente no RE apresenta um importante
papel na depuração do cálcio intracelular na plasticidade sináptica à curto prazo na JNM.
Estudos anteriores já haviam demonstrado tal importância (BENNETT et al., 2007), no
entanto, em nosso estudo realizamos a mensuração da eficácia sináptica através do registro
eletrofisiológico, que é considerado adequado na detecção de alterações quantitativas e
qualitativas da função sináptica. Além disso, o efeito da tapsigargina na facilitação por
pulsos pareados é novo e contribuiu para confirmar o papel primordial da facilitação no
mecanismo de depuração do cálcio intracelular e da presença do RE próximo dos locais de
liberação de neurotransmissores.
Nossos resultados confirmam evidências de estudos anteriores do papel da mitocôndria no
fenômeno de potenciação pós-tetânica (DAVID & BARRETT, 2003). Também sugerimos a
provável importância da mitocôndria no fenômeno de depressão sináptica.
Com relação aos IP3Rs demonstramos indiretamente a presença destes receptores no
terminal pré-sináptico da JNM e sua importância na modulação da liberação de
neurotransmissores através da utilização de um bloqueador para este receptor e para a
enzima PLC envolvida na sua sinalização nos fenômeno de depressão e facilitação por
pulsos pareados. Além disso, estudamos o mecanismo de ação da brevetoxina-3. Nossos
resultados são a primeira evidência da importância do IP3R em fenômenos de plasticidade
sináptica à curto prazo na JNM. Portanto, podemos considerar que os terminais pré-
sinápticos não possuem somente estoques intracelulares de cálcio sensíveis ao IP3, mas
também toda a maquinaria necessária, como receptores metabotrópicos e a enzima
fosfolipase C, para gerar o IP3.
Também evidenciamos que os mecanismos da depressão sináptica e da facilitação por
pulsos pareados estão relacionados à liberação de cálcio de estoques intracelulares
sensíveis ao IP3 e a ativação da fosfolipase C.
Por fim, sugerimos que estes resultados possam contribuir, pelo menos em parte, para um
maior entendimento da plasticidade sináptica na JNM. Além disso, estes resultados podem
ajudar no esclarecimento da plasticidade sináptica de outros sistemas colinérgicos, como no
sistema nervoso central.

VII. REFERÊNCIAS BIBLIOGRÁFICAS
Anderson, S., Bankier, A.T., Barrell, B.G., de Bruijn, M.H., Coulson, A.R., Drouin, J., Eperon, I.C., Nierlich, D.P., Roe, B.A., Sanger, F., Schreier, P.H., Smith, A.J., Staden, R., Young, I.G. (1981). "Sequence and organization of the human mitochondrial genome." Nature 290(5806): 457-465. Angleson, J. K., Betz, W.J. (2001). "Intraterminal Ca2+ and spontaneous transmitter release at the frog neuromuscular junction." Journal of Neurophysiology 85(1): 287-294. Annunziato, L., Pignataro, G., Di Renzo, G.F. (2004). "Pharmacology of brain Na+/Ca2+ exchanger: from molecular biology to therapeutic perspectives." Pharmacological Reviews 56(4): 633-654. Antozzi, C. (2003). "Myastenia gravis and myasthenic syndrome." Neurological Sciences 24: S260-S263. Arnaudeau, S., Kelley, W.L., Walsh, J.V., Demaurex, N. (2001). "Mitochondria recycle Ca2+ to the endoplasmic reticulum and prevent the depletion of neighboring endoplasmic reticulum regions." Journal of Biological Chemistry 276: 29430-29439. Atchison, W. D., Luke, V.S., Narahashi, T., Vogel, S.M. (1986). "Nerve membrane sodium channels as the target site of brevetoxins at neuromuscular junctions." British Journal of Pharmacology 89(4): 731-738. Atluri, P. P., Regehr, W.G. (1996). "Determinants of the time course of facilitation at the granule cell to Purkinje cell synapse." Journal of Neuroscience 16(18): 5661-5671. Augustine, G. J., Adler, E.M., Charlton, M.P. (1991). "The calcium signal for transmitter secreation from presynaptic nerve terminals." Annals of the New York Academy of Sciences 635: 365-381. Auld, D., Robitaille, R. (2003). "Perisynaptic schwann cells at the neuromuscular junction: nerve- and activity-dependent contributions to synaptic efficacy, plasticity and reinnervation." The Neuroscientist 9(2): 144-157. Baden, D. G. (1989). "Brevetoxins: unique polyether dinoflagellate toxins." FASEB Journal 3: 1807-1817. Baldo, G. J., Cohen, I.S., Van der Kloot, W. (1986). "Estimating the time course of evoked quantal release at the frog neuromuscular junction using end-plate current latencies." Journal of Physiology 374: 503-513. Bardo, S., Cavazzini, M.G., Emptage, N. (2005). "The role of the endoplasmic reticulum Ca2+ store in the plasticity of central neurons." TRENDS in Pharmacological Sciences 27(2): 78-84. Barrell, B. G., Anderson, S., Bankier, A.T., de Bruijn, M.H., Chen, E., Coulson, A.R., Drouin, J., Eperon, I.C., Nierlich, D.P., Roe, B.A., Sanger, F., Schreier, P.H., Smith, A.J., Staden, R., Young, I.G. (1980). "Different pattern of codon recognition by mammalian mitochondrial tRNAs." Proceedings of the National Academy of Sciences of United States of America 77(6): 3164-3166.

Barstad, J. A. B. (1962). "Presynaptic effect of the neurotransmitter." Experimentia 18: 579-580. Barstad, J. A. B., Lilleheil, G. (1968). "Transversally cut diaphragm preparation from the rat." Archives internationales de pharmacodynamie et the thérapie 175: 373-390. Bassani, R. A., Bassani, J.W.M., Bers, D.M. (1995). "Relaxation in ferret ventricular myocites: role of the sarcolemmal Ca ATPase." European Journal of Physiology 430: 573-578. Bennett, M. R., Farnell, L., Gibson, W.G., Dickens, P. (2007). "Mechanisms of calcium sequestration during facilitation at actives zones of an amphibian neuromuscular junction." Journal of Theoretical Biology 247: 230-241. Benoit, P. N., Audibert-Benoit, M.l., Peyrot, M. (1973). "Importance des effets presynaptiques dans le blocage de la jonction neuromusculaire de grenouille par substitution du lithium au sodium dans le milieu de survie." Archives Italiennes de Biologie 3: 323-335. Berridge, M. J. (1998). "Neuronal Calcium Signaling." Neuron 21: 13-26. Berridge, M. J., Bootman, M.D., Roderick, H.L. (2003). "Calcium signalling: dynamics, homeostasis and remodelling." Nature Reviews: Molecular Cell Biology 4: 517-529. Betz, W. J. (1970). "Depression of transmitter release at the neuromuscular junction of the frog." Journal of Physiology 206: 629-644. Bezprozvanny, I., Watras, J., Ehrlich., B.E. (1991). "Bell-shaped calcium-response curves of Ins(1,4,5)P3- and calcium-gated channels from endoplasmic reticulum of cerebellum." Nature 351: 751-754. Bezprozvanny, I. B., Ondrias, K., Kaftan, E., Stoyanovsky, D.A., Ehrlich, B.E. (1993). "Activation of the calcium release channel (ryanodine receptor) by heparin and other polyanions is calcium dependent." Molecular Biology of the Cell 4(3): 347-352. Bilmen, J. G., Michelangeli, F. (2002). "Inhibition of the type 1 inositol 1,4,5-trisphosphate receptor by 2-aminoethoxydiphenylborate." Cellular Signalling 14(11): 955-60. Birks, R., Huxley, H.E., Katz, B. (1960). "The fine structure of the neuromuscular junction of the frog." Journal of Physiology 150: 134-144. Bittner, G. D., Schatz, R.A. (1981). "An examination of the residual calcium theory for facilitation of transmitter release." Brain Research 210: 431-436. Bleasdale, J. E., Thakur,N.R., Gremban, R.S., Bundy, G.L., Fitzpatrick, F.A., Smith, R.J., Bunting, S. (1990). "Selective inhibition of receptor-coupled phospholipase C-dependent processes in human platelets and polymorphonuclear neutrophils." Journal of Pharmacology and Experimental Therapeutics 255(2): 756-768.

Bliss, T. V. (1990). "Long-term potentiation." Science 249(4972): 973. Bootman, M. D., Collins, T.J., Mackenzie, L., Roderick, H.L., Berridge M.J., Peppiatt C.M. (2002). "2-aminoethoxydiphenyl borate (2-APB) is a reliable blocker of store-operated Ca2+ entry but an inconsistent inhibitor of InsP3-induced Ca2+ release." FASEB Journal 16(10): 1145-50. Brailoiu, E., Miyamoto, M.D. (2000). "Inositol trisphosphate and cyclic adenosine diphosphate-ribose increase quantal transmitter release at frog motor nerve terminals: possible involvement of smooth endoplasmic reticulum." Neuroscience 95(4): 927-931. Brailoiu, E., Miyamoto, M.D., Dun, N.J. (2003). "Inositol derivatives modulate spontaneous transmitter release at the frog neuromuscular junction." Neuropharmacology 45(5): 691-701. Branisteanu, D. D., Volle, R.L. (1975). "Modification by lithium of transmitter release at the neuromuscular junction of the frog." Journal of Pharmacology and Experimental Therapeutics 194: 362-372. Brini, M., Carafoli, E. (2009). "Calcium pumps in health and disease." Physiological Reviews 89: 1341-1378. Brown, D. A., Sihra, T.S. (2008). "Presynaptic signaling by heterotrimeric G-proteins." Handbook of Experimental Pharmacology 184: 207-260. Carafoli, E. (1987). "Intracellular calcium homeostasis." Annual Review of Biochemistry 56: 395-433. Carafoli, E. (1991). "The calcium ATPase of the plasma membrane." Annual Review of Physiology 53: 531-547. Carafoli, E., Brini, M. (2000). "Calcium pumps: structural basis for and mechanism of calcium transmembrane transport." Current Opinion in Chemical Biology 4(2): 152-161. Carafoli, E., Santella, L., Branca, D., Brini, M. (2001). "Generation, control, and processing of cellular calcium signals." Critical Reviews in Biochemistry and Molecular Biology 36(2): 107-260. Carter, A. G., Vogt, K.E., Foster, K.A., Regehr, W.G. (2002). "Assessing the role of calcium-induced calcium release in short-term presynaptic plasticity at excitatory central synapses." Journal of Neuroscience 22(1): 21-28. Castonguay, A., Robitaille, R. (2001). "Differential regulation of transmitter release by presynaptic and glial Ca2+ internal stores at the neuromuscular synapse." Journal of Neuroscience 21(6): 1911-1922. Catterall, W. A. (1999). "Interactions of presynaptic Ca2+ channels and snare proteins in neurotransmitter release." Annals of the New York Academy of Sciences 868: 144-159. Chen, X., Tang, T-S., Tu, H., Nelson, O., Pook, M., Hammer, R., Nukina, N., Bezprozvanny, I. (2008). "Deranged calcium signaling and neurodegeneration in spinocerebellar ataxia type 3." Journal of Neuroscience 28(48): 12713-12724. Choi, H. S., Einer, D.A. (1999). "The role of sarcolemmal Ca2+-ATPase in the regulation of resting calcium concentration in rat ventricular myocytes." Journal of Physiology 515(1): 109-

118. Cohen, A. S., Raymond, C.R., Abraham, W.C. (1998). "Priming of long-term potentiation induced by activation of metabotropic glutamate receptors coupled to phospholipase C." Hippocampus 8: 160-170. Collin, T., Marty, A., Llano, I. (2005). "Presynaptic calcium stores and synaptic transmission." Current Opinion in Neurobiology 15: 275-281. Cox, D. A., Conforti, L., Sperelakis, N., Matlib, M.A. (1993). "A role for the mitochondrial Na+-Ca2+ exchanger in the regulation of oxidative phosphorylation in isolated heart mitochondria." Journal of Biological Chemistry 268: 938-947. Crawford, A. C. (1975). "Lithium ions and the release of transmitter at the frog neuromuscular junction." Journal of Physiology 246: 109-142. Crompton, M., Capano, M., Carafoli, E. (1976). "The sodium induced efflux of calcium from heart mitochondria. A possible mechanism for the regulation of mitochondrial calcium." European Journal of Biochemistry 69(2): 453-462. Dasso, L. L., Taylor, C.W. (1991). "Heparin and other polyanions uncouple alpha 1-adrenoceptors from G-proteins." Biochemical Journal 280(3): 791-795. David, G., Barrett, J.N., Barrett, E.F. (1997). "Stimulation-induced changes in [Ca2+] in lizard motor nerve terminals." Journal of Physiology 504(1): 83-96. David, G. (1999). "Mitochondrial clearance of cytosolic Ca2+ in stimulated lizard motor nerve terminals procees without progressive elevation of mitochondrial matrix [Ca2+]." Journal of Neuroscience 19(17): 7495-7506. David, G., Barrett, E.F. (2003). "Mitochondrial Ca2+ uptake prevents desynchonization of quantal release and minimizes depletion during repetitive stimulation of mouse motor nerve terminals." Journal of Physiology 548(2): 425-438. De Robertis, E. D., Bennet, H.S. (1955). "Some features of the submicroscopic morphology of synapses in frog and earthworm." The Journal of Biophysical and Biochemical Cytology 1: 47-58. De Smedt, H., Missiaen, L., Parys, J.B., Henning, R.H., Sienaert, I., Vanlingen, S., Gijsens, A., Himpens, B., Casteels, R. (1997). "Isoform diversity of the inositol trisphosphate receptor in cell types of mouse origin." Biochemical Journal 322(Pt.2): 575-583. Del Castillo, J., Katz, B. (1953). "Statistical aspects of transmision at single nerve muscle junction." Journal of Physiology 120(4): 32. Del Castillo, J., Katz, B. (1954a). "The effect of magnesium on the activity of motor nerve endings." Journal of Physiology 124: 553-559. Del Castillo, J., Katz, B. (1954b). "Quantal components of the end plate potential." Journal of Physiology 124: 560-573. Del Castillo, J., Katz, B. (1957). "Interaction at end-plate receptors between different choline derivatives." Proceedings of the Royal Society of London. Series B, containing papers of a Biological character 146(924): 369-381.

Dipolo, R., Beauge, L. (1983). "The calcium pump and sodium-calcium exchange in squid axons." Annual Review of Physiology 45: 313-324. Dodge, F. A. J., Rahamimoff, R. (1967). "Co-operative action a calcium ions in transmitter release at the neuromuscular junction." Journal of Physiology 193(2): 419-432. Dodson, P. D., Forsythe, I.D. (2004). "Presynaptic K+ channels: electrifying regulators of synaptic terminal excitability." Trends in Neurosciences 27(4): 210-217. Dropic, A. J., Brailoiu, E., Cooper, R.L. (2005). "Presynaptic mechanism of action by 5-HT in nerve terminals: Possible involvement of ryanodine and IP3 sensitive Ca2+ stores." Comparative Biochemistry and Physiology 142: 335-361. Dulhunty, A. F., Franzini-Armstrong, C. (1975). "The relative contribution of the folds and caveolae to the surface membrane of frog skeletal muscle fibres at different sarcomere lengths." Journal of Physiology 250: 513-539. Ehrlich, B. E., Kaftan, E., Bezprozvannaya, S., Bezprozvanny, I. (1994). "The pharmacology of intracellular Ca(2+)-release channels." Trends in Pharmacological Sciences 15(5): 145-149. Eltit, J. M., Hidalgo, J., Liberona, J.L., Jaimovich, E. (2004). "Slow calcium signals after tetanic electrical stimulation in skeletal myotubes." Biophysical Journal 86(5): 3042-3051. Eltit, J. M., García, A.A., Hidalgo, J., Liberona, J.L., Chiong, M., Lavandero, S., Maldonado, E., Jaimovich, E. (2006). "Membrane electrical activity elicits inositol 1,4,5-trisphosphate-dependent slow Ca2+ signals through a Gbetagamma/phosphatidylinositol 3-kinase gamma pathway in skeletal myotubes." Journal of Biological Chemistry 281(17): 12143-12154. Emptage, N. J., Reid, C.A., Fine, A. (2001). "Calcium stores in hippocampal synaptic boutons mediate short-term plasticity, store-operated Ca2+ entry, and spontaneous transmitter release." Neuron 29: 197-208. Escalona de Motta, G., Cordova, F., de Leon, M., del Castillo, J. (1982a). "Mechanim of the excitation-contraction uncoupling of frog skeletal muscle by formamide." Journal of Neuroscience Research 7: 163-178. Escalona de Motta, G., Smith, D.S., Cayer, M., del Castillo, J. (1982b). "Mechanism of excitation-contraction uncoupling of frog skeletal muscle by formamide." Biol Bull Marine Biol Lab Woods Hole Mass 163: 276-286. Fatt, P., Katz, B. (1951). "An analysis of the end-plate potential recorded with an intra-cellular electrode." Journal of Physiology 115: 320-370. Fatt, P., Katz, B. (1952). "Spontaneous subtreshold activity at motor nerve endings." Journal of Physiology 117: 109-128. Fill, M., Copello, J.A. (2002). "Ryanodine receptor calcium release channels." Physiological Reviews 82: 893-922. Finch, E. A., Augustine, G.J. (1998). "Local calcium signalling by inositol-1,4,5-trisphosphate in Purkinje cell dendrites." Nature 396: 753-756. Fujino, I., Yamada, N., Miyawaki, A., Hasegawa, M., Furuichi, T., Mikoshiba, K. (1995). "Differential expression of type 2 and type 3 inositol 1,4,5-trisphosphate receptor mRNAs in

various mouse tissues: in situ hybridization study." Cell and tissue research 280(2): 201-210. Furuichi, T., Simon-Chazottes, D., Fujino, I., Yamada, N., Hasegawa, M., Miyawaki, A., Yoshikawa, S., Guénet, J.L., Mikoshiba, K. (1993). "Widespread expression of inositol 1,4,5-trisphosphate receptor type 1 gene (Insp3r1) in the mouse central nervous system." Receptors channels 1(1): 11-24. Gailly, P. (2002). "New aspects of calcium signaling in skeletal muscle cells: implications in Duchenne muscular dystrophy." Biochimica et Biophysica Acta 1600: 38-44. García-Chacón, L. E. G., Nguyen, K.T., David, G., Barret, E.F. (2006). "Extrusion of Ca2+ from mouse motor terminal mitochondria via a Na+-Ca2+ exchanger increases post-tetanic evoked release." Journal of Physiology 574(3): 663-675. Gatto, C., Hale, C.C., Xu, W., Milanick, M.A. (1995). "Eosin, a potent inhibitor of the plasma membrane Ca pump, does not inhibit the cardiac Na-Ca exchanger." Biochemistry 34: 965-972. Gawley, R. E., Rein, K.S., Kinoshita, M., Baden, D.G. (1992). "Binding of brevetoxins and ciguatoxin to the voltage-sensitive sodium channel and conformational analysis of brevetoxin B." Toxicon 30(7): 780-785. Gibb, A. J., Marshall J.G. (1984). "Pre- and post-junctional effects of tubocurarine and other nicotinic antagonists during repetitive stimulation in rat." Journal of Physiology 351: 275-297. Gilman, A. G. (1987). "G proteins: transducers of receptor-generated signals." Annual Review of Biochemistry 56: 615-649. Glavinovic, M. I. (1979). "Presynaptic action of curare." Journal of Physiology 290: 499-506. Griffiths, E. J. (1999). "Reversal of mitochondrial Na/Ca exchange during metabolic inhibition in rat cardiomyocytes." FEBS Letters 453: 400-404. Guatteo, E., Carunchio, I., Pieri, M., Albo, F., Canu, N., Mercuri, M.B., Zona, C. (2007). "Altered calcium homeostasis in motor neurons following AMPA receptor but not voltage-dependent calcium channels activation in a genetic model of amyotrophic lateral sclerosis." Neurobiology of Disease 28: 90-100. Gusovsky, F., Daly, J.W. (1988). "Formation of second messengers in response to activation of ion channels in excitable cells." Cellular and Molecular Neurobiology 82(5): 157-169. Gwag, B. J., Canzoniero, L.M., Sensi, S.L., Demaro, J.A., Koh, J.Y., Goldberg, M.P., Jacquin, M., Choi, D.W. (1999). "Calcium ionophores can induce either apoptosis or necrosis in cultured cortical neurons." Neuroscience 90(4): 1339-1348. Hachisuka, J., Soga-Sakakibara, S., Kubota, M., Narita, K., Kuba, K. (2007). "Enhancement of Ca2+-induced Ca2+ release by cyclic ADP-ribose in frog motor nerve terminals." Neuroscience 146: 123-134. Hall, Z. W., Sanes, J. R. (1993). "Synaptic structure and development: the neuromuscular junction." Cell/Neuron 72(10): 99-121. Hermoni, M., Barzilai, A., Rahamimoff, H. (1987). "Modulation of the Na+-Ca2+ antiport by its ionic environment: the effect of lithium." Israel Journal of Medical Sciences 23(1-2): 44-48. Hernandez-Guijo J.M., M.-F., V.E., Ruiz-Nuno, A., Villarroya, M., Garcia, A.G., Gandia, L.

(2001). "Calcium-dependent inhibition of L, N, and P/Q Ca2+ channels in chromaffin cells: role of mitochondria." Jounal of Neuroscience 21(8): 2553-2560. Hernandez-SanMiguel, E., Vay, L., Santo-Domingo, J., Lobatón, CD., Moreno, A., Montero, M., Alvarez, J. (2006). "The mitochondrial Na+/Ca2+ exchanger plays a key role in the control of cytosolic Ca2+ oscillations." Cell Calcium 40: 53-61. Herrera, A. A. (1984). "Polyneuronal innervation and quantal transmitter release in formamide-treated frog sartorius muscles." Journal of Physiology 355: 267-280. Hinata, M., Kimura, J. (2004). "Forefront of Na+/Ca2+ exchanger studies: stoichiometry of cardiac Na+/Ca2+ exchanger; 3:1 or 4:1?" Journal of Pharmacological Science 96(1): 15-18. Inoue, T., Kato, K., Kohda, K., Mikoshiba, K. (1998). "Type 1 inositol 1,4,5-trisphosphate receptor is required for induction of long-term depression in cerebellar Purkinje neurons." Journal of Neuroscience 18(14): 5366-5373. Jackson, T. R., Patterson, S.I., Thastrup, O., Hanley, M.R. (1988). "A novel tumor promoter, thapsigargin, transiently increases cytoplasmic free Ca2+ without generation of inositol phosphates in NG115-401L neuronal cells." Biochemical Journal 253(1): 81-86. Jahn, R., Südhof, T.C. (1994). "Synaptic vesicles and exocytocis." Annual Review of Neuroscience 17: 219-246. Jahn, K., Grosskreutz, J., Haastert, K., Ziegler, E., Schlesinger, F., Grothe, C., Dengler, R., Bufler, J. (2006). "Temporospatial coupling of networked synaptic activation of AMPA-type glutamate receptor channels and calcium transients in cultured motoneurons." Neuroscience 142(4): 1019-1029. Jensen, T. P., Filoteo, A.G., Knopfel, T., Empson, R.M. (2007). "Presynaptic plasma membrane Ca2+-ATPase isoform 2a regulates excitatory synaptic transmission in rat hippocampal CA3." Journal of Physiology 579(1): 85-99. Juhazsova, M., Church, P., Blaustein, M.P., Stanley, E.F. (2000). "Location of calcium transporters at presynaptic terminals." European Journal of Neuroscience 12: 839-846. Kalkstein, J. M., Magleby, K.L. (2004). "Augmentation increases vesicular release probability in the presence of masking depression at the frog neuromuscular junction." Journal of Neuroscience 24(50): 11391-11403. Katz, B., Thesleff, S. (1957). "On the factors wich determine the amplitude of the miniature end-plate potential." Journal of Physiology (London) 138: 267-278. Katz, B., Miledi, R. (1965). "The effect of calcium on acetylcholine release from motor nerve terminals." Proceedings of the Royal Society of London. Series B, containing papers of a Biological character 161: 496-503. Katz, B. (1966). Nerve, Muscle and Synapse., McGraw-Hill Book Company. Katz, B., Miledi, R. (1967). "Ionic requirements of synaptic transmitter release." Nature 215: 651. Katz, B., Miledi, R. (1968). "The role of calcium in neuromuscular facilitation." Journal of Physiology 195: 481-492.

Katz, B., Miledi, R. (1979). "Estimates of quantal content during chemical potentiation of transmitter release." Proceedings of the Royal Society of London 205: 369-378. Kelly, J. S. (1968). "The antagonism of Ca2+ by Na+ and other monovalent ions at the frog neuromuscular junction." Quartely journal of experimental physiology and cognate medical science 53: 239-249. Koyano, K., Kuba, K., Minota, S. (1985). "Long-term potentiation of transmitter release induced by repetitive presynaptic activities in bull-frog sympathetic ganglia." Journal of Physiology 359: 219-233. Kubota, M., Narita, K., Murayama, T., Suzuki, S., Soga, S., Usukura, J., Ogawa, Y., Kuba, K. (2005). "Type-3 ryanodine receptor involved in Ca2+-induced Ca2+ release and transmitter exocytosis at frog motor nerve terminals." Cell Calcium 38: 557-567. Kuffler, S. W., Yoshikami, D. (1975). "The number of transmitter molecules in a quantum: an estimate from iontophoretic application of acetylcholine at the neuromuscular synapse." Journal of Physiology 251: 465-482. Lagnado, L., McNaughton, P.A. (1990). "Electrogenic properties of the Na:Ca exchange." Journal of Membrane Biology 113: 177-191. Land, B. R., Salpeter, E.E., Salpeter, M.M. (1981). "Kinetic parameters for acetylcholine interaction in intact neuromuscular junction." Proceedings of the National Academy of Sciences 78: 7200-7204. Lauri, S. E., Bortolotto, Z.A., Nistico, R., Bleakman, D., Ornstein,P.L., Lodge, D., Isaac; J.T., Collingridge, G.L. (2003). "A role for Ca2+ stores in kainate receptor-dependent synaptic facilitation and LTP at mossy fiber synapses in the hippocampus." Neuron 39: 327-341. Lehostsky, J. (1995). "Plasma membrane Ca2+ pump functional specialization in the brain. Complex of isoform expression and regulation by effectors." Molecular and Chemical Neuropathology 25: 175-187. Li, L., Guerini, D., Carafoli, E. (2000). "Calcineurin controls the transcription of Na+/Ca2+ exchanger isoforms in developing cerebellar neurons." Journal of Biological Chemistry 275(27): 20903-20910. Liberona, J. L., Cárdenas, J.C., Reyes, R., Hidalgo, J., Molgó, J., Jaimovich, E. (2008). "Sodium-dependent action potentials induced by brevetoxin-3 trigger both IP3 increase and intracellular Ca2+ release in rat skeletal myotubes." Cell Calcium 44(3): 289-297. Liley, A. W., North, K.A.K. (1953). "An electrical investigation of the effects of repetitive stimulation on mammalian neuromuscular junction." Journal of Neurophysiology 16: 509-527. Lin, J.-W., Fu, Q., Allana, T. (2005). "Probing the endogenous Ca2+ at the presynaptic terminals of the crayfish neuromuscular junction." Journal of Neurophysiology 99: 377-386. Linden, D. J., Connor, J.A. (1995). "Long-term synaptic depression." Annual Review of Neuroscience 18: 319-357. Linder, T. M., Pennefather, P., Quastel, D.M. (1984). "The time course of miniature endplate currents and its modification by receptor blockade and ethanol." Journal of General Physiology 83: 435-468.

Llinás, R., Sugimori, M., Silver, R.B. (1992). "Microdomains of high calcium concentration in a presynaptic terminal." Science 256(5057): 677-679. Llinás, R., Sugimori, M; Silver., R.B. (1995a). "The concept of calcium concentration microdomains in synaptic transmission." Neuropharmacology 34(11): 1443-1451. Llinás, R., Sugimori, M., Silver., R.B. (1995b). "Time resolved calcium microdomains and synaptic transmission." Journal of Physiology - Paris(89): 2. Louvet, L., Collin, T. (2005). "Involvement of ryanodine receptors in IP3-mediated calcium signalling in neurons. A modelling approach." Neuroscience Letters 381(1-2): 149-153. Luther, P. W., Yip, R. K., Bloch, R.J., Ambesi, A., Lindenmayer, G.E., Blaustein, M.P. (1992). "Presynaptic Localization of Sodium/Calcium Exchangers in Neuromuscular Preparations." Jounal of Neuroscience 12(12): 4898-4904. Magleby, K. L., Zengel, J.E. (1975a). "A dual effect of repetitive stimulation on post-tetanic potentiation of transmitter release at the frog neuromuscular junction." Journal of Physiology 245: 163-182. Magleby, K. L., Zengel, J.E. (1975b). "A quantitative description of tetanic and post-tetanic potentiation of transmitter release at the frog neuromuscular junction." Journal of Physiology 245: 183-208. Magleby, K. L., Zengel, J.E. (1976a). "Augmentation: a process that acts to increase transmitter release at the frog neuromuscular junction." Journal of Physiology 257: 449-470. Magleby, K. L., Zengel, J.E. (1976b). "Long-term changes in augmentation, potentiation and depression of transmitter release as a function of repeated activity at the frog neuromuscular junction." Journal of Physiology 257: 471-494. Magleby, K. L., Palotta. B.S., Terrar, D. (1981). "The effect of d-tubocurarine on neuromuscular transmission during repetitive stimulation in the rat, mouse and frog." Journal of Physiology 312: 97-113. Magleby, K. L., Zengel, J.E. (1982). "A quantitative description of stimulation-induced changes in transmitter release at the frog neuromuscular junction." Journal of General Physiology 80: 613-638. Majerus, P. W. (1992). "Inositol phosphate biochemistry." Annual Review of Biochemistry 61: 225-250. Malli, R., Frieden, M., Trenker, M., Graier, W.F. (2005). "The role of mitochondria for Ca2+ refilling of the endoplasmic reticulum." Journal of Biological Chemistry 280: 12114-12122. Martin, A. R. (1955). "A further study of the statistical composition of the end plate potential." Journal of Physiology 130: 114-122. Martin, A. R. (1966). "Quantal nature of synaptic transmission." Physiological Reviews 46: 51-66. Maruyama, T., Kanaji, T., Nakade, S., Kanno, T., Mikoshiba, K. (1997). "2APB, 2-aminoethoxydiphenyl borate, a membrane-penetrable modulator of Ins(1,4,5)P3-induced Ca2+ release." Journal of Biochemistry 122(3): 498-505.

Matlib, M. A., Zhou, Z., Knight, S., Ahmed, S., Choi, K.M., Krause-Bauer, J., Phillips, R., Altschuld, R., Katsube, Y., Sperelakis, N., Bers, D.M. (1998). "Oxigen-bridged dinuclear ruthenium amine complex specifically inhibits Ca2+ uptake into mitochondria in vitro and in situ in single cardiac myocytes." Journal of Biological Chemistry 273(17): 10223-10231. Mattei, C., Dechraoui, M-Y., Molgó, J., Meunier, F.A., Legrand, A-M., Benoit, E. (1999). "Neurotoxins targetting receptor site 5 of voltage-depende sodium channels increase the nodal volume of myelinated axons." Journal of Neuroscience Research 55: 666-674. Matzner, H., Parnas, H., Parnas, I. (1988). "Presynaptic effects of d-tubocurarine on neurotransmitter release at the neuromuscular junction of the frog." Journal of Physiology 398: 109-121. Mcconville, J., Vincent, A. (2002). "Diseases of the neuromuscular junction." Current Opinion in Pharmacology 2(3): 296-301. McMahan, U. J., Sanes, J.R., Marshall, L.M. (1978). "Cholinesterase is associated with the basal lamina at the neuromuscular junction." Nature 271: 172-174. Meissner, G. (1986). "Ryanodine activation and inhibition of the Ca2+ release channel of sarcoplasmic reticulum." Journal of Biological Chemistry 261(14): 6300-6306. Meunier, F. A., Colassante, C., Molgó, J. (1997). "Sodium-dependent increase in quantal secreation induced by Brevetoxin-3 in Ca2+-free medium is associated with depletion of synaptic vesicles and swelling of motor nerve terminals in situ." Neuroscience 78(3): 883-893. Miledi, R., Molenaar, P.C., Polak, R.L. (1983). "Electrophysiology and chemical determination of acetylcholine release at the frog neuromuscular junction." Journal of Physiology 334: 245-254. Morgan, A., Burgoyne, R.D., Barclay, J.W., Craig, T.J., Prescott, G.R., Ciufo, L.F., Evans, G.J., Graham, M.E. (2005). "Regulation of exocytosis by protein kinase C." Biochemical Society transactions 33(6): 1341-1344. Mukhamedyarov, M. A., Zefirov, A.L., Palotás, A. (2006). "Paired-pulse facilitation of transmitter release at different levels of extracellular calcium concentration." Neurochemical Research 31: 1055-1058. Mukhamedyarov, M. A., Grishin; S.N., Zefirov, A.L., Palotás, A. (2009). "The mechanisms of multi-component paired-pulse facilitation of neurotransmitter release at the frog neuromuscular junction." European Journal of Physiology 458(3): 563-70. Nakagawa, T., Shiota, C., Okano, H., Mikoshiba, K. (1991). "Differential localization of alternative spliced transcripts encoding inositol 1,4,5-trisphosphate receptors in mouse cerebellum and hippocampus: in situ hybridization study." Journal of Neurochemistry 57(5): 1807-1810. Nakamura, T., Nakamura, K., Lasser-Ross, N., Barbara, J.G., Sandler, V.M., Ross, W.N. (2000). "Inositol 1,4,5-trisphosphate (IP3)-mediated Ca2+ release evoked by metabotropic agonists and backpropagating action potentials in hippocampal CA1 pyramidal neurons." Journal of Neuroscience 20: 8365-8376. Narita, K., Akita, T., Osanai, M., Shirasaki, T., Kijima, H., Kuba, K. (1998). "A Ca2+ -induced Ca2+ release mechanism involved in asynchronous exocytosis at frog motor nerve terminals."

Journal of General Physiology 112: 593-609. Narita, K., Akita, T., Hachisuka, J., Huang, S-M., Ochi, K., Kuba, K. (2000). "Functional coupling of Ca2+ channels to ryanodine receptors at presynaptic terminals: amplification of exocytosis and plasticity." Journal of General Physiology 115: 519-532. Nilsson, T., Zwiller, J., Boynton, A.L., Berggren, P.O. (1988). "Heparin inhibits IP3-induced Ca2+ release in permeabilized pancreatic beta-cells." FEBS Letters 229(1): 211-214. Nishizaki, T., Mori, M. (1998). "Diverse signal transduction pathways mediated by endogenous P2 receptors in cultured rat cerebral cortical neurons." Journal of Neurophysiology 79: 2513-2521. Nowycky, M. C., Thomas, A.P. (2002). "Intracellular calcium signaling." Journal of Cell Science 115(19): 3715-3716. Ogawa, Y. (1994). "Role of ryanodine receptors." Critical Reviews in Biochemistry and Molecular Biology 29(4): 229-274. Onodera, K., Yamakawa, K. (1966). "The effects of lithium on the neuromuscular junction of the frog." Japanese Journal of Physiology 16(5): 541-550. Ortiz, C. L., Junge, D. (1978). "Depressant action of lithium at the crayfish neuromuscular junction: pre- and postsynaptic effects." Journal of Experimental Biology 75: 171-187. Otsuka, M., Endo, M., Nonamura, Y. (1962). "Presynaptic nature of neuromuscular depression." Japanese Journal of Physiology 12: 573-584. Pan, B., Zucker, R.S. (2009). "A general model of synaptic transmission and short-term plasticity." Neuron 62: 539-554. Parekh, A. B. (2003). "Store-operated Ca2+ entry: dynamic interplay between endoplasmic reticulum, mitochondria and plasma membrane." Journal of Physiology 547(2): 333-348. Parnas, H., Segel, L., Dudel, J., Parnas, I. (2000). "Autoreceptors, membrane potential and the regulation of transmitter release." Trends in Neurosciences 23: 60-68. Pedersen, P., Carafoli, E. (1987). "Ion motive ATPases. I. Ubiquity, properties, and significance to cell function." Trends in Biochemical Sciences 12: 146-150. Peng, Y. Y. (1998). "Effects of mitochondrion on calcium transients at intact presynaptic terminals depend on frequency of nerve firing." Journal of Neurophysiology 80(1): 186-195. Personious, K. E., Balice-Gordon, R.J. (2000). "Activity-dependent editing of neuromuscular synaptic connections." Brain Research Bulletin 53(5): 513-522. Piriz, J., Rosato Siri, M.D., Pagani, R., Uchitel, O.D. (2003). "Nifedipine-mediated mobilization of intracellular calcium stores increases spontaneous neurotransmitter release at neonatal rat motor nerve terminals." Journal of Pharmacology and Experimental Therapeutics 306: 658-663. Poli, M. A., Mende, T.J., Baden, D.G. (1986). "Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptossomes." Molecular Pharmacology 30: 129-135.

Power, J. M., Sah, P. (2002). "Nuclear calcium signaling evoked by cholinergic stimulation in hippocampal CA1 pyramidal neurons." Journal of Neuroscience 22: 3454-3462. Ren, J., Harty, R.F. (1994). "Presynaptic muscarinic receptors modulate acetylcholine release from rat antral mucosal/submucosal nerves." Digestive diseases and sciences 39(5): 1099-1106. Rizzuto, R., Bernardi, P., Pozzan, T. (2000). "Mitochondria as all-round players of the calcium game." Journal of Physiology 529(1): 37-47. Rizzuto, R., Duchen, M.R., Pozzan, T. (2004). "Flirting in little space: the ER/mitochondria Ca2+ liaison." Science's STKE : signal transduction knowledge environment 215: re1. Robitaille, R., Adler, E. M., Charlton, M. P. (1990). "Strategic localization of calcium channels at transmitter release sites of frog neuromuscular synapses." Neuron 5: 773-779. Robitaille, R., Adler, E.M., Charton, M.P. (1993a). "Calcium channels and calcium gated potassium channels at the fog neuromuscular junction." Journal of Physiology 87: 15-24. Robitaille, R., Garcia, M.L., Kaczorowski, G.J., Charlton, M.P. (1993b). "Functional colocalization of calcium and calcium-gated potassium channels in control of transmitter release." Neuron 11: 645-655. Robitaille, R. (1998). "Modulation of synaptic efficacy and synaptic depression by glial cells at the frog neuromuscular junction." Neuron 21: 847-855. Rogers, T. B., Inesi, G., Wade, R., Lederer, W.J. (1995). "Use of thapsigargin to study Ca2+ homeostasis in cardiac cells." Bioscience reports 15(5): 341-349. Rose, C. R., Konnerth, A. (2001). "Stores not just for storage, intracellular calcium release and synaptic plasticity." Neuron 31: 519-522. Ruff, R. L. (1986). "Ionic channels: II. Voltage- and agonist-gated and agonist-modified channel properties ans structure." Muscle Nerve 9(9): 767-786. Ruff, R. L. (2003). "Neurophysiology of the neuromuscular junction: overview." Annals of the New York Academy of Sciences 998: 1-10. Sabatini, B. L., Regehr, W.G. (1999). "Timing of synaptic transmission." Annual Review of Physiology 61: 521-542. Sagara, Y., Inesi, G. (1991). "Inhibition of the sarcoplasmic reticulum Ca2+ transport ATPase by thapsigargin at subnanomolar concentrations." Journal of Biological Chemistry 266(21): 13503-13506. Sanes, J. R., Lichtman, J. W. (2001). "Induction, assembly, maturation and maintenance of a postsynaptic apparatus." Nature Reviews/Neuroscience 2: 791-805. Shia, T.-J. S., Liu, S-X.L., Hammarbergb, H., Watanabec, M., Xua,Z-Q.D., Hökfelt, T. (2008). " Phospholipase Cß3 in mouse and human dorsal root ganglia and spinal cord is a possible target for treatment of neuropathic pain." Proceedings of the National Academy of Sciences 105(50): 20004-20008. Slutsky, I., Parnas, H., Parnas, I. (1999). "Presynaptic effects of muscarine on ACh release at the frog neuromuscular junction." Journal of Physiology 514(3): 769-782.

Smets, I., Caplanusi, A., Despa,S., Molnar, Z., Radu, M., vandeVen, M., Ameloot, M., Steels, P. (2004). "Ca2+ uptake in mitochondria occurs via the reverse action of the Na+/Ca2+ exchanger in metabolically inhibited MDCK cells." American Journal of Physiology - Renal Physiology 286: F784-F794. Smith, S. J., Augustine, G.J. (1988). "Calcium ions, active zones, and synaptic transmitter release." Trends in Neurosciences 10: 458-464. Stockbridge, N., Hines, M. (1982). "A re-examination of the residual calcium theory for facilitation of transmnitter release." Brain Research 250: 378-380. Suzuki, S., Osanai, M., Mitsumoto, N., Akita, T., Narita, K., Kijima, H., Kuba, K. (2002). "Ca2+-dependent Ca2+ clearance via mitochondrial uptake and plasmalemmal extrusion in frog motor nerve terminals." Journal of Neurophysiology 87: 1816-1823. Swandulla, D., Hans, M., Zipser, K., Augustine, G.J. (1991). "Role of residual calcium in synaptic depression and posttetanic potentiation: fast and slow calcium signaling in nerve terminals." Neuron 7(6): 915-926. Takechi, H., Eilers, J., Konnerth, A. (1998). "A new class of synaptic response involving calcium release in dendritic spines." Nature 396: 757-760. Takeuchi, A. (1958). "The long-lasting depression in neuromuscular transmission of frog." Japanese Journal of Physiology 8: 102-113. Takeuchi, A., Takeuchi, N. (1960). "On the permeability of end-plate membrane during the action of transmitter." Journal of Physiology 154: 52-67. Takeuchi, A., Takeuchi, N. (1962). "Electrical changes in pre- and postsynaptic axons of the giant synapse of Loligo." Journal of General Physiology 45: 1181-1193. Tanabe, N., Kijima, H. (1992). "Ca2+-dependent and -independent components of transmitter release at the frog neuromuscular junction." Journal of Physiology 455: 271-289. Tang, Y.-G., Zucker, R.S. (1997). "Mitochondrial involvement in post-tetanic potentiation of synaptic transmission." Neuron 18: 483-491. Taylor, C. W., Genazzani, A.A., Morris, S.A. (1999). "Expression of inositol trisphosphate receptors." Cell Calcium 26(6): 237-251. Thastrup, O., Cullen, P.J., Drøbak, B.K., Hanley, M.R., Dawson, A.P. (1990). "Thapsigargin, a tumor promoter, discharges intracellular Ca2+ stores by specific inhibition of the endoplasmic reticulum Ca2+-ATPase." Proceedings of the National Academy of Sciences 87(7): 2466-2470. Thies, R. E. (1965). "Neuromuscular depression and the apparent depletion of transmitter in mammalian muscle." Journal of Neurophysiology 28: 427-442. Thurneysen, T., Nicoll, D.A., Philipson, K.D., Porzig, H. (2002). "Immunohistochemical detection of the sodium-calcium exchanger in rat hippocampus cultures using subtype-specific antibodies." Annals of the New York Academy of Sciences 976: 367-375. Tong, J. J. (2007). "Mitochondrial delivery is essential for synaptic potentiation." The Biological Bulletin 212: 169-175.

Van der Kloot, W., Cohen, I.S. (1984). "End-plate potentials in a model muscle fiber: Corrections for the effect of membrane potential on currents and channel lifetimes." Biophysical Journal 45: 905-912. Van Der Kloot, W. (1991). "The regulation of quantal size." Progress in Neurobiology 36: 93-130. Van der Kloot, W., Molgó, J. (1993). "Facilitation and delayed release at about 0º C at the frog neuromuscular junction: effects of calcium chelators, calcium transport inhibitors and okadaic acid." Journal of Neurophysiology 69(3): 717-729. Van der Kloot, W., Molgó, J. (1994). "Quantal acetylcholine release at the vertebrate neuromuscular junction." Physiological Reviews 74: 899-991. Van Der Kloot, W., Molgó, J., Naves, L.A. (1997). "Cholinergic agonists decrease quantal output at the frog neuromuscular junction by targeting a calcium channel blocked by omega-conotoxin." Pflügers Archiv - European Journal of Physiology 434(6): 735-741. Vannucchi, M. G., Pepeu, G. (1995). "Muscarinic receptor modulation of acetylcholine release from rat cerebral cortex and hippocampus." Neuroscience Letters 190: 53-56. Vaughan, P. F., Walker, J.H., Peers, C. (1998). "The regulation of neurotransmitter secretion by protein kinase C." Molecular Neurobiology 18(2): 125-155. Vercesi, A., Reynafarje, B., Lehningerg, A.L. (1978). "Stoichiometry of H+ ejection and Ca2+ uptake coupled to electron transport in rat heart mitochondria." Journal of Biological Chemistry 253(18): 6379-6385. Verstreken, P., Ly, C.V., Venken, K.J., Koh, T.W., Zhou, Y., Bellen, H.J. (2005). "Synaptic mitochondria are critical for mobilization of reserve pool vesicles at Drosophila neuromuscular junctions." Neuron 47(3): 365-378. Vicent, A. (2002). "Unravelling the pathogenesis of myasthenia gravis." Nature Reviews/Immunology 2: 797-804. Warrier, A., Borges, S., Dalcino, D., Walters, C., Wilson, M. (2005). "Calcium from internal stores triggers GABA release from retinal amacrine cells." Journal of Neurophysiology 94(6): 4196-4208. Watanabe, S., Narahashi, T. (1979). "Cation selectivity of acetylcholine-activated ionic channel of frog end plate." Journal of General Physiology 74(5): 615-628. Westrum, L. E., Gray, E.G. (1986). "New observations on the substructure of the active zone of brain synapses and motor endplates." Proceedings of the Royal Society B: Biological Sciences 229(1254): 29-38. White, R. J., Reynolds, I.J. (1997). "Mitochondria accumulate Ca2+ following intense glutamate stimulation of cultured rat forebrain neurones." Journal of Physiology 498(1): 31-47. Wood, S. J., Slater, C.R. (1997). "The contribution of postsynaptic folds to the safety factor for neuromuscular transmission in rat fast- and slow-twitch muscles." Journal of Physiology 500: 165-176.

Wood, S. J., Slater, C.R. (2001). "Safety factor at the neuromuscular junction." Progress in Neurobiology 64: 393-429. Wu, L.-G., Betz, W, J. (1998). "Kinetcs os synaptic depression and vesicle recycling after tetanic stimulation of frog motor nerve terminals." Biophysical Journal 74: 3003-3009. Yang, F., He, X., Feng, L., Mizuno, K., Liu, X-W., Russel, J., Xiong, W-C., Lu, B. (2001). "PI-3 kinase and IP3 are both necessary and sufficient to mediate NT3-induced synaptic potentiation." Nature Neuroscience 4(1): 19-28. Zazueta, C., Sosa-Torres, M.E., Correa, F., Garza-Ortiz, A. (1999). "Inhibitory properties of ruthenium amine complexes on mitichondrial calcium uptake." Journal of Bioenergetics and Biomembranes 31(6): 551-557. Zefirov, A. L., Cheranov, S. (2000). "The molecular mechanisms of quantum mediator secretion in the synapse." Usp Fiziol Nauk 31(3): 3-22. Zengel, J. E., Sosa, M.A. (1994). "Changes in MEPP frequency during depression of evoked release at the frog neuromuscular junction." Journal of Physiology 477(2): 267-277. Zhai, R. G., Bellen, H.J. (2004). "The architecture of the active zone in the presynaptic nerve terminal." Physiology 19(262-270). Zhong, N., Beaumont, V., Zucker, R.S. (2001). "Roles of mitochondrial and reverse mode Na+/Ca2+ exchange and the plasmalemma Ca2+ ATPase in post-tetanic potentiation at crayfish neuromuscular junction." Journal of Neuroscience 21(24): 9598-9607. Zucker, R. S. (1989). "Short-term synaptic plasticity." Annual Review of Neuroscience 12: 13-31. Zucker, R. S., Regehr, W.G. (2002). "Short-term synaptic plasticity." Annual Review of Physiology 64: 355-405.

ANEXO I – Protocolos experimentais
1) Protocolo do efeito do BAPTA-AM na Depressão.
Ringer 0 mM Ca+2 + 1,8 mM Mg+2
+ 100 M BAPTA-AM / 0,05% DMSO
Ringer 1,8 mM Ca+2
+ 6 M curare
Registro da Depressão
120 minutos
30 minutos
2) Protocolo de bloqueio da Ca+2-ATPase do retículo endoplasmático.
Ringer 0 mM Ca+2 + 2,5 mM
EGTA
+ 2 M Tapsigargina / 0,028%
etanol
Ringer 1,8 mM Ca+2
+ 2 M Tapsigargina / 0,028% etanol + 6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
30 minutos
30 minutos

3) Protocolo de bloqueio da Ca+2-ATPase da membrana plasmática.
Ringer Normal
+ 20 M CEDA / 0,17% DMSO
Ringer Normal
+ 6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
40 minutos
30 minutos
Lavagem
4) Protocolo de bloqueio do trocador Na+/Ca+2 pelo CGP 31157.
Ringer Normal
+ 50 M CGP / 0,05% DMSO
6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
30 minutos

5) Protocolo de bloqueio do uniporter mitocondrial pelo Ru360.
Ringer Normal
+ 10 M Ru360 / Água deoxigenada
6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
30 minutos
6) Protocolo de bloqueio do trocador Na+/Ca+2 pela substituição do Ringer normal pelo
Ringer Lítio.
Ringer Normal
6 M curare
Registro da Depressão e da Facilitação por pulsos pareados
30 minutos
Perfusão_Ringer Lítio
(20 min.) 6 M curare
Registro da Depressão e da Facilitação por pulsos pareados
20 minutos
Obs.: Resgistros foram realizados
na mesma fibra sem perda do
empalamento.

7) Protocolos dos experimentos realizados com PbTx-3.
Ringer 0 Ca+2 + 2,5 mM EGTA
Registro da fMEPPs
60 minutos
Adição de 50 nM PbTx-3 à
preparação
Registro da fMEPPs
60 minutos
Adição de TTX, Tapsigargina, 2-APB
ou U-73122
Registro da fMEPPs
30 minutos
Registro da fMEPPs
Ringer 0 Ca+2 + 2,5 mM EGTA
30 minutos
30 minutos
Adição de 50 nM PbTx-3
30 minutos
Registro da fMEPPs
A) B)
Obs.: Resgistros foram realizados
na mesma fibra sem perda do
empalamento.
8) Protocolo de bloqueio do receptor de rianodina pela rianodina.
Ringer Normal
+ 10 M Ry / 0,1% EtOH
6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
30 minutos

9) Protocolo de bloqueio do receptor de IP3 pelo 2-APB.
Ringer Normal
+ 10 M 2-APB / 0,002% MeOH
6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
30 minutos
10) Protocolo de bloqueio da fosfolipase C pelo U-73122.
Ringer Normal
+ 5 M U-73122 / 0,05% DMSO
6 M curare
Registro da Depressão e
da Facilitação por pulsos
pareados
30 minutos

ANEXO II – Artigos em processo de elaboração e submissão
1) Artigo em processo de submissão
Extracellular calcium-independent increase in quantal secretion induced by brevetoxin-3 is
related to IP3 receptor and phospholipase C.
Priscila E. Silveira1,2, Emmanuelle Girard1, Ligia A. Naves2 and Jordi Molgό1
1 CNRS, Institut de Neurobiologie Alfred Fessard – FRC2118, Laboratoire de Neurobiologie
Cellulaire et Moléculaire – UPR9040, Gif sur Yvette, France.
2 Laboratório de Eletrofisiologia Celular, Departamento de Fisiologia e Biofísica, ICB-UFMG,
Belo Horizonte, MG, Brazil.
Revista pretendida: Toxicon.
2) Artigo em processo de elaboração
Synaptic plasticity at the neuromuscular junction: the role of IP3 and Ryanodine receptors.
Priscila E. Silveira 1,2, Christopher Kushmerick 1, Jordi Molgό 2 and Ligia A. Naves 1
1 Laboratório de Eletrofisiologia Celular, Departamento de Fisiologia e Biofísica, ICB-UFMG,
Belo Horizonte, MG, Brazil;
2 CNRS, Institut de Neurobiologie Alfred Fessard – FRC2118, Laboratoire de Neurobiologie
Cellulaire et Moléculaire – UPR9040, Gif sur Yvette, France.
Revista pretendida: Neuroscience Letters ou Neuroreport.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo