instituto de pesquisas jardim botânico do rio de janeiro...
TRANSCRIPT

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
Dinâmica de populações da espécie exótica invasora Artocarpus heterophyllus L. (Moraceae)
no Parque Nacional da Tijuca - Rio de Janeiro
Rodolfo Cesar Real de Abreu
2008

II
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Dinâmica de populações da espécie exótica invasora Artocarpus heterophyllus L. (Moraceae)
no Parque Nacional da Tijuca - Rio de Janeiro
Rodolfo Cesar Real de Abreu
Dissertação apresentada ao Programa de Pós-Graduação em Botânica, Escola Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Botânica. Orientador: Dr. Pablo José Francisco Pena Rodrigues Co – orientador: Dr. Renato Soares Armelin
Rio de Janeiro 2008

III
Dinâmica de populações da espécie exótica invasora
Artocarpus heterophyllus L. (Moraceae) no Parque Nacional da Tijuca – Rio de Janeiro
Rodolfo Cesar Real de Abreu Dissertação submetida ao corpo docente da Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - JBRJ, como parte dos requisitos necessários para a obtenção do grau de Mestre.
Banca Examinadora: Dr. Pablo José F.P. Rodrigues (Orientador – JBRJ)
________________________________________
Dra. Giselda Durigan (USP – São Carlos)
________________________________________
Dra. Tânia Sampaio Pereira (JBRJ)
_ _______________________________________
Dr. Lucas Del Bianco Faria (LNCC – RJ)
________________________________________
em 31/03/2008.
Rio de Janeiro 2008

IV
Abreu, Rodolfo Cesar Real de.
A162d Dinâmica de populações da espécie invasora Artocarpus heterophyllus L. (Moraceae) no Parque Nacional da Tijuca – Rio de Janeiro / Rodolfo Cesar Real de Abreu. – Rio de Janeiro, 2008.
xiv, 77 f. : il. Dissertação (Mestrado) – Instituto de Pesquisas Jardim Botânico
do Rio de Janeiro/Escola Nacional de Botânica Tropical, 2008. Orientador: Pablo José Francisco Pena Rodrigues. Bibliografia. 1. Artocarpus heterophyllus. 2. Modelos matriciais. 3. Espécie
invasora. 4. Dinâmica transiente. 5. Mata Atlântica. 6. Parque Nacional da Tijuca (RJ). 7. Rio de Janeiro (Estado). I.Título. II. Escola Nacional de Botânica Tropical.
CDD 583.45098153

V
À minha família.

VI
AGRADECIMENTOS:
Muitas pessoas contribuíram para o desenvolvimento deste trabalho, deixo aqui
registrado meus sinceros agradecimentos:
À minha família por todo apoio e incentivo.
Ao amigo e orientador Dr. Pablo J. F. Pena Rodrigues por todas as conversas e
conselhos nestes anos de convivência.
Ao professor Dr. Renato Soares Armelin pela imprescindível co-orientação.
À Dra. Giselda Durigan por aceitar o convite em compor a banca examinadora.
À Dra. Tânia Sampaio Pereira por aceitar o convite em compor a banca examinadora.
Ao Programa Mata Atlântica pelo apoio logístico através da PETROBRAS.
Aos professores Dr. Rogério Ribeiro (PUC-RIO), Dr. Sérgio Cardoso (JBRJ), Dr.
Lucas Faria (LNCC) e Dra. Cláudia Barros (JBRJ) pelas valiosas sugestões nas disciplinas de
seminários.
Aos professores Dr. Flavio Antonio Maës dos Santos (UNICAMP) e Dr. Renato
Soares Armelin pelo curso de modelagem matricial.
Ao IBAMA do Parque Nacional da Tijuca por informações a respeito da espécie no
local.
Aos funcionários da secretaria da ENBT e da biblioteca Barbosa Rodrigues pelo pronto
atendimento e simpatia.
À todos os pesquisadores que procurei com dúvidas sobre o trabalho.
Aos amigos da mesma área que ajudaram nos trabalhos de campo e contribuíram com
idéias: Letícia Melo, Izar Aximoff, Rafael Pinto, Guilherme Malvar, Mariela Figueredo,
Jerônimo Sansevero, Mariana Iguatemy, Raul Lins e Talita Reis.

VII
Aos amigos de outras áreas: minha irmã Geisa e Igor Bonomo, pela ajuda no campo
(“nesta floresta cheia de perigos!”).
Aos amigos que me acolheram durante a disciplina na UNICAMP: Juliana Furlaneto,
André Gil e Dudu Meirelles.
Aos alunos das turmas de mestrado e doutorado da ENBT pelas agradáveis trocas e
divertida convivência dentro e fora da sala de aula.
À todos do laboratório: Pablo (o chefe), Jerônimo, Mariana, Pablinho, Mariela, Talita,
Monique, Andrea, Carol, Lilian e Raul, pelas trocas, momentos de descontração,
recomendações bibliográficas e incentivos.
À Mariela Figueredo pela cuidadosa revisão do texto final.
À Miguel de Moraes pela revisão dos ‘abstracts’.
Agradecimentos especiais à Talita Reis pelo apoio durante diversas fases da elaboração
do texto, pela leitura crítica de todas as versões, sugestões de formatação, idéias e envio de
referências bibliográficas. Agradeço também pela convivência e incentivo.
Aos amigos Marcos Franco e Izidro Souza pela troca de idéias.
Aos amigos Miguel de Moraes, Alex, Belinha, Mari e Dum pelos divertidos treinos.
Aos amigos que eventualmente perguntaram sobre o andamento do trabalho com as
jaqueiras.
Enfim, agradeço à todos aqueles que porventura eu possa ter esquecido.

VIII
RESUMO
Espécies invasoras são vistas como uma ameaça à saúde dos ecossistemas. Ações
como a remoção destas espécies estão se tornando essenciais para a manutenção da
biodiversidade. Neste contexto, Artocarpus heterophyllus Lamk. (Moraceae), conhecida como
jaqueira, é uma espécie arbórea nativa do sudeste asiático historicamente introduzida no
Parque Nacional da Tijuca (PNT). Atualmente, compõe a comunidade vegetal do PNT com
pronunciada monodominância em alguns trechos, onde desenvolveu interações com a fauna
local. Este trabalho procurou gerar informações sobre o estágio atual de desenvolvimento da
população de jaqueiras no PNT através do estudo da demografia da espécie por meio de
modelagem matricial. Desta forma, o trabalho considerou as duas fases de dinâmica
populacional da espécie. Na fase assintótica, foi possível realizar a comparação com outros
trabalhos e detectar padrões de colonização desta exótica na Mata Atlântica do PNT. Análises
retrospectivas e prospectivas foram realizadas com base nas matrizes anuais obtidas no
período 2005 – 2008. Na fase transiente foram feitas análises prospectivas de sensibilidades e
de elasticidades, além de simulações do manejo da espécie. Os resultados indicaram que as
populações de jaqueiras se expandem menos de 1% ao ano com duração da fase transiente de
179 anos. Os indivíduos permanecem por longos períodos nas classes de diâmetros, com
baixas taxas de transição e mortalidade. O manejo com intervenções de até 5% voltado para as
sobrevivências nas classes de JUVENIS 1 e ADULTOS 1 e 2, além reduzir a taxa de
crescimento populacional da espécie, é suficiente para que a população entre em declínio.
Estas pequenas intervenções quando feitas no longo-prazo podem mitigar os impactos sobre a
biota local.

IX
ABSTRACT
Invasive species are a major threat to healthy ecosystems. Actions like removing this
kind of species are becoming essential to maintain biodiversity. In this way, Artocarpus
heterophyllus Lamk. (Moraceae), commonly known as jackfruit, is a woody tree native from
Southeast Asia historically introduced at Parque Nacional da Tijuca (PNT). Nowadays, it
belongs to the plant community of PNT with a substantial dominance at some regions where
local fauna interactions are developed. This work aims to generate data about the present
development stage of jackfruit population at PNT. The population demography was studied
through matrix models and two kinds of population dynamics were considered. On asymptotic
dynamic the colonization patterns were detected and compared to other Atlantic Forest
species. Retrospective and prospective analyses were developed based on annual matrices
from 2005 to 2008. On transient dynamic, prospective sensitivities, elasticities analysis and
management simulations were conducted. Results indicates that jackfruit populations have a
small growth expanding less than 1% a year and it would stay for 179 years in transient
dynamic. Stases on diameter classes are high, with small rates of individual growth and
mortality. Environmental management interventions up to 5% on survival rates on classes
JUVENILES 1, and ADULTS 1 and 2 are the best options to generate the aimed effect of
population decline. Small and long-term interventions mitigate impacts on local organisms.

X
SUMÁRIO
LISTA DE FIGURAS ............................................................................................................ XII
LISTA DE TABELAS .......................................................................................................... XIV
INTRODUÇÃO GERAL .......................................................................................................... 1
REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................................... 6
CAPÍTULO 1 – DINÂMICA DE POPULAÇÕES DA ESPÉCIE EXÓTICA INVASORA
ARTOCARPUS HETEROPHYLLUS (LAMK.) NA MATA ATLÂNTICA ..................... 9
RESUMO .................................................................................................................... 10
ABSTRACT ................................................................................................................ 11
INTRODUÇÃO ........................................................................................................... 12
MATERIAIS E MÉTODOS ....................................................................................... 15
RESULTADOS ............................................................................................................ 27
DISCUSSÃO ............................................................................................................... 38
CONCLUSÕES ........................................................................................................... 44
REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................... 45
CAPÍTULO 2 – SIMULAÇÕES DE MANEJO ATRAVÉS DA MODELAGEM
POPULACIONAL DE UMA ÁRVORE INVASORA NO PARQUE NACIONAL DA
TIJUCA – RJ .......................................................................................................................... 50
RESUMO ..................................................................................................................... 51
ABSTRACT ................................................................................................................. 52
INTRODUÇÃO ........................................................................................................... 53

XI
MATERIAIS E MÉTODOS ........................................................................................ 56
RESULTADOS ............................................................................................................ 63
DISCUSSÃO ............................................................................................................... 67
CONCLUSÕES ........................................................................................................... 72
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................ 73

XII
LISTA DE FIGURAS:
Figura 1.1 – Imagem aérea da localização do Parque Nacional da Tijuca na cidade do Rio de Janeiro. Cortesia IBAMA PNT. ……………………………………………….......……….... 16 Figura 1.2 – Exemplo da alta produção de frutos por planta adulta de A. heterophyllus no PNT........................................................................................................................................... 18 Figura 1.3 – Exemplo de fruto da jaqueira em fase de desenvolvimento ............................... 18 Figura 1.4 – Início da emergência de plântulas após a decomposição do fruto no chão da floresta ...................................................................................................................................... 18 Figura 1.5 – Detalhe da fase inicial de germinação................................................................. 18 Figura 1.6 – Representação das taxas vitais na matriz de estágios At (Lefkovitch 1965). Onde, S: probabilidade de um indivíduo permanecer na mesma classe durante t (t = 1 ano); G: probabilidade do indivíduo passar para classe seguinte em t; R: probabilidade de retração de indivíduos para classe anterior; F: fecundidade de cada classe reprodutiva.............................23 Figura 1.7 – Histograma de classes de diâmetros (DAP em cm) das sub-populações de jaqueiras no Parque Nacional da Tijuca – RJ no ano de 2005...................................................28 Figura 1.8 – Matrizes de estágios (Lefkovitch 1965) das taxas de transição de A. heterophyllus no PNT durante o período de estudo (2005 – 2008). 2006 = dinâmica 2005 – 2006; 2007 = dinâmica 2006 – 2007; 2008 = dinâmica 2007 – 2008; ANOS = matriz sumário dos anos de estudo (2005 – 2008). Os valores entre parênteses indicam o número de parcelas utilizadas na construção da matriz.............................................................................................30 Figura 1.9 – Representação gráfica da matriz de estágios do ciclo de vida de A. heterophyllus no PNT – RJ, no período 2005 – 2008. Em vermelho a fecundidade, em preto o crescimento e em azul a permanência em uma mesma classe..........................................................................32 Figura 1.10 – Representação das sensibilidades reais do ciclo de vida de A. heterophyllus no PNT – RJ no período 2005 – 2007. Em vermelho a fecundidade, em preto o crescimento e em azul a permanência em uma mesma classe................................................................................34 Figura 1.11 – Representação das elasticidades do ciclo de vida de A. heterophyllus no PNT – RJ no período 2005 – 2007. Em vermelho a fecundidade, em preto o crescimento e em azul a permanência em uma mesma classe..........................................................................................35 Figura 1.12 – Contribuições dos estágios da história de vida para o � populacional de jaqueiras no PNT – RJ, nos anos de 2006 a 2008......................................................................36 Figura 1.13 – Contribuições anuais das taxas demográficas para o � populacional de jaqueiras no PNT - RJ. Nos anos de 2006 a 2008. ...................................................................................37

XIII
Figura 2.1 – Matriz de estágios (Lefkovitch 1965) de A. heterophyllus no PNT para o período 2005 – 2008. ..............................................................................................................................63 Figura 2.2 – Simulações da intensidade de manejo por Taxa Vital. Onde: s – sobreviência (permanências + crescimentos) dos indivíduos nas classes; g – crescimento para classe seguinte; f – fecundidade (entrada de novos indivíduos DAP � 5cm). ...................................65 Figura 2.3 – Projeções das elasticidades das taxas vitais no tempo. De t+1 até t+178: fase de dinâmica transiente; t assintótico: tempo > 178 anos fase de dinâmica assintótica; s: sobrevivência; g: crescimento; f: fecundidade. ........................................................................ 66

XIV
LISTA DE TABELAS:
Tabela 1.1 – Sumário de parâmetros utilizados para caracterizar a estrutura da comunidade nas parcelas estudadas no Parque Nacional da Tijuca em 2005. Onde: AB total = área basal de toda a comunidade; AB jaq = área basal ocupada pelas jaqueiras na comunidade; AB outros = área basal ocupada por outros indivíduos excetuando-se as jaqueiras; N total = número de indivíduos da camunidade; N jaq = número de indivíduos de jaqueiras na comunidade............27 Tabela 1.2 – Taxas de transição e taxa de crescimento populacional (�) da dinâmica anual da população de jaqueiras no Parque Nacional da Tijuca – RJ, no período 2005 – 2008. Tx Trans = taxa de transição, onde S: permanência na classe, G: crescimento, passagem para a classe seguinte, F: fecundidades, os numerais indicam as respectivas classes; elem. = elemento da matriz (elementos não listados apresentaram valor zero); 2006 = dinâmica 2005 – 2006; 2007 = dinâmica 2006 – 2007; 2008 = dinâmica 2007 – 2008; ANOS = matriz sumário dos anos de estudo (2005 – 2008). Os valores entre parênteses indicam o número de parcelas utilizadas na construção da matriz. � = autovalor dominante da matriz, que corresponde a taxa finita de crescimento populacional; ICs = intervalos de confiança calculados para o �. ........................29 Tabela 2.1 – Taxas vitais de Artocarpus heterophyllus no PNT. Crescimentos estão incluídos nas sobrevivências. ...................................................................................................................63 Tabela 2.2 – Sensibilidades da estrutura populacional de A. heterophyllus em relação às taxas vitais na fase de dinâmica transiente em t+1. J1 – Juvenis 1; J2 – Juvenis 2; PR – Pré-Reprodutivos; A1 – Adultos 1; A2 – Adultos 2; Taxas vitais: s = sobrevivência, g = crescimento, f = fecundidade. .................................................................................................. 64 Tabela 2.3 – Elasticidades da estrutura populacional de A. heterophyllus em relação às taxas vitais na fase de dinâmica transiente em t+1. J1 – Juvenis 1; J2 – Juvenis 2; PR – Pré-Reprodutivos; A1 – Adultos 1; A2 – Adultos 2; Taxas vitais: s = sobrevivência, g = crescimento, f = fecundidade. ...................................................................................................64

1
INTRODUÇÃO GERAL
Invasões biológicas vêm sendo debatidas no mundo há pelo menos cinco décadas
(Elton 1958), enquanto no Brasil ainda são relativamente pouco estudadas. O tema esteve
inicialmente em evidência em 1992 durante a Conferência das Nações Unidas sobre Meio
Ambiente (Rio 92), quando foi elaborado o texto da Convenção sobre Diversidade Biológica -
CDB, que define como um dos itens prioritários (8º artigo) o cuidado com espécies exóticas e
os problemas que estas podem causar à biodiversidade (o decreto legislativo nº 2 de 1994
aprova a CDB). Ainda assim, pouco tem sido feito para conter o avanço de tais espécies no
Brasil e o primeiro Simpósio científico de âmbito nacional que tratou exclusivamente do tema
foi realizado recentemente, em 2005 (MMA 2005). Em um país de dimensões continentais
como o Brasil, representante da maior biodiversidade do mundo, a presença de tais espécies
não deve ser negligenciada (Myers et al. 2000). Uma vez que a redução da biodiversidade
mundial está relacionada com a destruição direta de habitats e com as invasões biológicas
(Simberloff 2003), respectivamente, ambos impactos amplamente experimentados pelos
ecossistemas brasileiros (e.g. Pivello et al. 1999; Scariot 2001). Além disso, as modificações
dos habitats naturais tais como a fragmentação e demais impactos relacionados, podem
favorecer a disseminação de espécies exóticas (Williamson 1996). A Mata Atlântica, por
exemplo, com 92,4% de sua área original desmatada e atualmente disposta em fragmentos,
está teoricamente dentre os ecossistemas mais vulneráveis (Myers et al. 2000; Morellato &
Haddad 2000). Outros ecossistemas brasileiros tais como o Cerrado (Pivello et al. 1999;
Rodrigues-da-Silva & Filgueiras 2003) e a Floresta Amazônica (Scariot 2001), também estão
sujeitos a este tipo de degradação (Myers et al. 2000).
Elton (1958), no trabalho: ‘The Ecology of invasions by animals and plants’, descreve
algumas modificações que espécies invasoras podem causar ao colonizar novos ambientes.

2
Porém, somente a partir da década de 1980 começaram a surgir publicações sobre o assunto,
culminando em 1999 com o surgimento de uma revista especializada: “Biological Invasions”.
No entanto, esta área da ecologia é relativamente nova (Richardson et al. 2000; Davis et al.
2001; Colautti & MacIsaac 2004) e ainda pouco explorada. É considerada como uma
disciplina à parte da ecologia, quando na realidade deveria ser tratada de forma integrada, uma
vez que aborda temas como colonização de habitats, dinâmica de clareiras, dinâmica de
metapopulações, pragas, ervas-daninhas e restauração ecológica, por exemplo (Davis et al.
2001). Um dos problemas relacionados está na incorporação de termos e conceitos por
cientistas, sejam de formação acadêmica similar ou diferente (e.g. Ciências agrárias,
ambientais, biológicas, florestais, veterinárias, dentre outras). Por exemplo, o termo invasão é
aplicado por ecólogos para espécies nativas ou exóticas quando tratam de processos de
sucessão ecológica (Johnstone 1986). Sendo assim, tentativas de uniformização da
terminologia associada às espécies exóticas e invasoras são propostas em alguns trabalhos
(e.g. Richardson et al. 2000). Esta rápida emergência da ecologia de invasões no mundo e a
identificação dos diversos problemas causados têm aumentado a demanda de estudos sobre
tais espécies.
De maneira geral, as ciências naturais classificam as espécies exóticas como aquelas
espécies não-nativas do ambiente em questão. Tais espécies podem ser introduzidas
intencionalmente para suprir demandas ou acidentalmente, podendo gerar grandes prejuízos
ambientais e econômicos (Pimentel et al. 2001). Nos tempos atuais, acredita-se que todas as
regiões do globo foram afetadas pela introdução de espécies exóticas, desde o continente mais
biodiverso até a ilha oceânica mais isolada (Cassey et al. 2005). Esta transposição de barreiras
biogeográficas pelas espécies ocorria de maneira natural e lenta, porém, ultimamente ocorre de
forma cada vez mais acelerada inclusive com espécies que seriam incapazes de ultrapassar tais

3
barreiras (Richardson et al. 2000; Pimentel et al. 2001). As modificações realizadas pelas
espécies exóticas invasoras (EEIs) nos novos ambientes podem provocar a extinção de
diversas espécies nativas, sendo comparadas aos eventos de extinções em massa ocorridos em
períodos geológicos (Cassey et al. 2005). Estas “invasões em massa” provocadas pelo homem
(Cassey et al. 2005) são capazes de provocar homogeneizações de biotas locais (McKinney &
Lockwood 1999). Todavia, estes são casos extremos e ressalta-se que a magnitude dos
problemas gerados pelas invasões biológicas irá variar em função da espécie exótica
introduzida, do tempo decorrido após a introdução e do ambiente em questão (Williamson
1996).
Neste contexto, ações como a remoção de plantas e animais não-nativos e o controle de
predadores e parasitas que ameaçam espécies raras estão se tornando ferramentas essenciais
para a manutenção da biodiversidade local e global (Wilcove & Chen 1998). Porém, o
controle e a remoção de espécies exóticas e invasoras são extremamente difíceis e
significativamente mais onerosos do que a prevenção da entrada destas (Mack 2000;
Wittenberg & Cock 2001). Uma vez que bilhões de dólares são gastos anualmente com tais
espécies (Williamson 1996; Pimentel et al. 2001), países como África do Sul, Austrália,
Brasil, Estados Unidos, Índia e Reino Unido, possuem legislações específicas para prevenir a
entrada de organismos estrangeiros (Pimentel et al. 2001). No Brasil, as invasões biológicas
podem ser estudadas com relativo sucesso, pois as introduções de espécies exóticas no país
são relativamente recentes, tendo sido iniciadas a partir do período colonial (Ferrão 1993) com
espécies conhecidas (e.g. Floresta da Tijuca, Chaves et al. 1966).
Neste contexto, em meados de 1800 no Parque Nacional da Tijuca (PNT) - RJ ocorreu
a introdução da espécie exótica Artocarpus heterophyllus Lamk. (Moraceae), conhecida como
jaqueira (Chaves et al. 1966). O PNT está inserido em zona urbana do município do Rio de

4
Janeiro (22 55’-23 00’S e 43 11’-43 19’W) e possui aproximadamente 4.000 ha de Mata
Atlântica em distintos graus de preservação (IBAMA 2004). Nas áreas mais preservadas, as
árvores emergentes podem atingir 25 m e o dossel varia em torno de 20 m de altura, onde
podem ser encontrados representantes das famílias Leguminosae, Sapotaceae, Vochysiaceae,
Bombacaceae, Euphorbiaceae, Meliaceae, Lauraceae, Lecythidaceae, Moraceae,
Melastomataceae, dentre outras. No sub-bosque as famílias mais representativas são Palmae,
Cyatheaceae (samambaias gigantes), Rubiaceae, Myrtaceae e Piperaceae. Apresenta ainda
trepadeiras e uma grande diversidade de espécies epífitas (Chaves et al. 1966). Além disso, em
alguns lugares pode ser observada uma dominância em estrutura e biomassa da espécie
arbórea exótica Artocarpus heterophyllus (jaqueira): MORACEAE (Abreu 2005; Cunha et al.
2006). Desta forma, o PNT foi escolhido como área de estudo.
Este trabalho pretende gerar informações sobre o estágio atual de desenvolvimento da
população de jaqueiras no PNT e para isto, foi estudada a demografia da espécie visando a
descrição dos padrões populacionais por meio de modelagem matricial. Para a análise
matricial de populações estruturadas dois períodos distintos da fase da dinâmica populacional
podem ser detectados (Hastings 2004). O período de dinâmica transiente, em que a população
ainda não atingiu a estabilidade, e o período de dinâmica assintótica, onde tal estabilidade é
alcançada. Portanto, a identificação da fase da dinâmica em que a população estudada se
encontra pode ser fundamental na determinação dos fatores que irão reger o comportamento
populacional (Hastings 2004). Sendo assim, o presente trabalho foi dividido em dois capítulos.
No capítulo 1 foi assumida a premissa de que a população encontrava-se em fase de dinâmica
assintótica. Desta forma, foi possível realizar a comparação com outros trabalhos e a detecção
de padrões de colonização da espécie exótica A. heterophyllus na Mata Atlântica do PNT.
Análises retrospectivas e prospectivas foram realizadas com base nas matrizes anuais obtidas

5
no período 2005 – 2008. No capítulo 2, assumiu-se a fase de dinâmica transiente da
população. Foram feitas análises de sensibilidades, análises de elasticidades e simulações do
manejo da espécie. Através destas análises, procurou-se sugerir medidas eficazes para o
manejo da espécie exótica.

6
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, R.C.R. 2005. Estrutura de populações de uma espécie exótica na Mata Atlântica: o
caso da potencial invasora Artocarpus heterophyllus L. (jaqueira) no Parque Nacional da
Tijuca – RJ. Monografia de Bacharelado em Ciências Biológicas. Universidade Federal do
Estado do Rio de Janeiro – UNIRIO.
Cassey, P.; Blackburn, T.M.; Duncan, R.P. & Chown, S.L. 2005. Concerning invasive species:
reply to Brown and Sax. Austral Ecol. 30: 475-480.
Chaves, C.M.; Martins, H.F.; Carauta, J.P.P.; Lanna-Sobrinho, J.P.; Vianna, M.C. & Silva,
S.A.F. 1966. Arboreto Carioca 3. Centro de Conservação da Natureza – Rio de Janeiro –
Brasil – 28p +il.
Cunha, A.; Vieira, M.V. & Grelle, C.E.V. 2006. Preliminary observations on habitat, support
use and diet in two non-native primates in an urban Atlantic forest fragment: The capuchin
monkey (Cebus sp.) and the common marmoset (Callithrix jacchus) in the Tijuca forest,
Rio de Janeiro. Urban Ecosystems 9(4): 351-359.
Coulatti, R.I. & MacIsaac, H.J. 2004. A neutral terminology to define ‘invasive’ species.
Diversity and Distributions 10:135-141.
Davis, M.A.; Thompson, K. & Grime, J.P. 2001. Charles S. Elton and the dissociation of
invasion ecology from the rest of ecology. Diversity and Distributions 7: 97-102.
Elton, C.S. 1958. The ecology of invasions by animals and plants. Methuen and Co, London.
Ferrão, J.E.M. 1993. A aventura das plantas e os descobrimentos portugueses. 2ª ed., Lisboa,
Instituto de Investigaçäo Científica Tropical.
Hastings, A. 2004. Transients: the key to long-term ecological understanding? Trends in
Ecology and Evolution 19:39-45.

7
IBAMA, 2004. Proposta de ampliação e correção de limites do Parque Nacional da Tijuca.
Gestão compartilhada IBAMA e Prefeitura da Cidade do Rio de Janeiro.
Johnstone, I.M. 1986. Plant invasion windows: a time-based classification of invasion
potential. Biological Reviews 61: 369-394.
Mack, R.N. 2000. Cultivation fosters plant naturalization by reducing environmental
stochasticity. Biological Invasions 2:111-122.
McKinney, M.L. & Lockwood, J.L. 1999. Biotic homogenization: a few winners replacing
many losers in the next mass extinction. Trends in Ecology and Evolution 14(11): 450-
453.
Morellato, L.P.C. & Haddad, C.F.B. 2000. Introduction: The Brazilian Atlantic Forest.
Biotropica 32(4b): 786-792.
Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B. & Kent, J. 2000.
Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
MMA, 2005. Espécies Exóticas Invasoras: Situação Brasileira. Disponível em:
http://www.mma.gov.br/invasoras/ acessado em janeiro de 2006.
Pimentel, D.; McNair, S.; Janecka, J.; Wightman, J.; Simmonds, C.; O'Connel, C.; Wong, E.;
Russel, L.; Zern, J.; Aquino, T. & Tsomondo, T. 2001. Economic and environmental
threats of alien plant, and microbe invasions. Agriculture, Ecosystems & Environment
84:1-20.
Pivello, V.R.; Shida, C.N. & Meirelles, S.T. 1999. Alien grasses in Brazilian savannas: a
threat to the biodiversity. Biodiversity and Conservation 8:1281-1294.
Richardson, D.M.; Pysek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D. & West, C.J.
2000. Naturalization and invasion of alien plants: concepts and definitions. Diversity and
Distributions 6: 93-107.

8
Rodrigues-da-Silva, R. & Filgueiras, T.S. 2003. Gramíneas (Poaceae) da área de relevante
interesse ecológico (ARIE) “Santuário de Vida Silvestre do Riacho Fundo”, Distrito
Federal, Brasil. Acta Botanica Brasilica 17(3): 467-486.
Scariot, A. 2001. Weedy and secondary palm species in central Amazonian Forest fragments.
Acta Botanica Brasilica 15(2): 271-280.
Simberloff, D. 2003. Confronting introduced species: a form of xenophobia? Biological
Invasions 5:179-192.
Wilcove, D.S. & Chen, L.Y. 1998. Management Costs for Endangered Species. Conservation
Biology 12(6):1405-1407.
Williamson, M. 1996. Biological Invasions. Chapman & Hall. UK.
Wittenberg, R. & Cock, M.J.W. 2001. Especies exóticas invasoras: Una guía sobre las mejores
prácticas de prevención y gestión. CAB Internacional, Wallingford, Oxon, Reino Unido,
xvii – 228.

9
CAPÍTULO 1
DINÂMICA DE POPULAÇÕES DA ESPÉCIE EXÓTICA INVASORA
ARTOCARPUS HETEROPHYLLUS LAMK. (MORACEAE) NA MATA ATLÂNTICA

10
RESUMO
Artocarpus heterophyllus Lamk. (Moraceae) é uma espécie arbórea nativa do
sudeste asiático. Conhecida como jaqueira, esta espécie provavelmente foi introduzida no
Parque Nacional da Tijuca – RJ (PNT) há cerca de 150 anos. Para identificar os processos
ou estágios do ciclo de vida que influenciam a população, um estudo demográfico foi
conduzido e um modelo matricial foi elaborado para caracterizar a dinâmica populacional
de A. heterophyllus no PNT no período 2005 – 2008. A estrutura populacional, através do
padrão “J reverso”, atestou a capacidade de regeneração da espécie exótica no local. A
matriz de transições indicou que os indivíduos permanecem por longos períodos nas
classes de diâmetros, um padrão recorrente para populações de árvores tropicais de
estágios sucessionais avançados. Foi observado que as jaqueiras apresentam características
populacionais similares às espécies nativas de Mata Atlântica. O autovalor dominante (λ)
de 1,0021, exibiu a tendência anual de expansão populacional. Verificou-se que a
população encontra-se em fase de dinâmica transiente e que levaria 179 anos para alcançar
a estrutura estádio-estável. As sensibilidades indicaram que as taxas mais sensíveis às
variações no λ foram as permanências nas classes reprodutivas e a transição de ADULTOS
1 para ADULTOS 2. Já as elasticidades destacaram a importância das permanências de
ADULTOS 2. Análises de Resposta da Tabela de Vida (LTREs) indicaram que as histórias
de vida (fecundidades, crescimentos e permanências) e as taxas demográficas (classes de
diâmetros) contribuíram de forma distinta nos diferentes anos. Foi possível concluir que a
exótica A. heterophyllus apresentou-se efetivamente estabelecida na Mata Atlântica,
caracterizando-se como espécie invasora deste ambiente.
Palavras-chave: Artocarpus heterophyllus; jaqueira; modelos matriciais; taxa de
crescimento assintótico; sensibilidades; elasticidades; LTRE; espécie invasora; Mata
Atlântica; Parque Nacional da Tijuca; Brasil;

11
ABSTRACT
Artocarpus heterophyllus Lamk. (Moraceae) is a native woody tree from Southeast
Asia. Commonly known as ‘jaqueira’ (Portuguese to jackfruit), this species was probably
brought to Parque Nacional da Tijuca – RJ (PNT) 150 years ago. To identify the processes
or life-cycle stages that can influence population behavior, a demographic matrix model
was built to characterize population dynamics of ‘jaqueiras’ from PNT between 2005 and
2008. Population structure represented by a ‘reversed J’ distribution shows that the exotic
species regenerates well, locally. Transition matrix indicates high stasis on diameter
classes, a well-know pattern to woody tropical trees on advanced succession stages. The
exotic species shows population characters with similarities to the native ones from
Atlantic Forest. The dominant eigenvalue (λ) of 1.0021 shows population tendency to a
small growth by year, indicating that the population is currently in transient dynamics,
taking 179 years to reach stable-stage distribuction. Sensitivities analyses indicates that
stasis rates and transition rates from ADULTS 1 to ADULTS 2 are the main factors
influencing λ. Nevertheless, elasticities analyses already shows valuable influences of rates
from ADULTS 2 class. Life Table Response Experiments (LTRE) indicates that life
history (fertility, growth ans stasis) and demographic rates (diameter classes) contributed in
different ways on distinct years. These results supports the conclusion that the exotic tree
A. heterophyllus is stablished at the Atlantic forest as an invasive species of local
environment.
Key-words: Artocarpus heterophyllus, jackfruit, matrix models; assintotic growth rate;
sensitivities; elasticities; LTRE; invasive species; Atlantic Forest; Tijuca National Park;
Brasil;

12
INTRODUÇÃO
Artocarpus heterophyllus Lamk. (jaqueira) é uma espécie arbórea nativa do sudeste
asiático (Chaves et al. 1967; Ferrão 1993). A espécie foi introduzida no Parque Nacional
da Tijuca – RJ (PNT) há aproximadamente 150 anos através de um reflorestamento
conduzido por Archer (Bandeira 1993). O PNT está inserido em área urbana do município
do Rio de Janeiro onde esta espécie exótica está bem estabelecida e amplamente
distribuída. O porte de alguns indivíduos nas sub-populações locais aparentemente indica
que estes foram introduzidos na época do reflorestamento supracitado, atestando a boa
aclimatação da espécie às novas condições do ecossistema invadido. Neste, diversos
fatores potencialmente colaboram para a habilidade competitiva da espécie e de acordo
com Cronk & Fuller (1995), o clima semelhante ao do habitat de origem pode facilitar a
adaptação das exóticas ao novo ambiente. Atualmente, tais sub-populações distribuem-se
de maneira agregada em escala de paisagem e formam em alguns locais densas florestas
monodominantes, onde indivíduos da espécie ocupam todos os estratos da mata (Abreu
2005). Além disso, interagem com a fauna local favorecendo apenas os animais capazes de
utilizarem seus frutos, inclusive alguns primatas exóticos (Cunha et al. 2006). Um dos
efeitos ecológicos conhecidos provocado por populações de espécies exóticas é o
crescimento em densas concentrações, excluindo outras espécies (Wiliamson 1996),
portanto, na medida em que se expande localmente e ocupa o lugar de espécies nativas,
esta espécie exótica altera a biodiversidade local.
Em populações estruturadas, expansões ou retrações populacionais podem ser
detectadas atráves de modelos matriciais. Nestes, a taxa de crescimento finito da
população, o lambda (λ), é usada para quantificar o crescimento populacional e também
como ponto de partida para realização de outras análises (Caswell 2001). Neste sentido,
apesar dos modelos matriciais auxiliarem na descrição da estrutura populacional e nos

13
fatores decorrentes desta (e.g. as taxas de transição entre classes ou estágios), estes ainda
são sub-utilizados no Brasil. Em parte porque para árvores tropicais, a detecção de padrões
populacionais temporais consistentes torna-se difícil devido ao ciclo de vida longo destas
espécies e à dificuldade em realizar estudos de longo prazo (Mantovani 2003). Como
exemplo, poucas espécies nativas de Mata Atlântica têm a estrutura populacional descrita
através desta ferramenta, destacando-se as arbóreas: Miconia cinnamomifolia D.C. Naudin
(Melastomataceae, Pereira 1998), Schefflera angustissima (March.) Frodin (Araliaceae) e
Andira anthelmia (Vell.) March. (Fabaceae, Armelin 2005); a arbustivo-arbórea: Psycotria
suterella Müll. Arg. (Rubiaceae, Bertani 2006); e as palmeiras: Euterpe edulis Mart.
(Silva-Mattos et al. 1999) e Attalea humilis Mart. ex. Spreng. (Souza & Martins, 2004).
Tal escassez de estudos é agravada quando consideram-se as espécies vegetais exóticas,
especialmente as arbóreas, como no caso de A. heterophyllus.
Espécies exóticas podem tornar-se invasoras caso proliferem continuamente nos
novos ambientes, causando diversos problemas à biota local e favorecendo outras espécies
exóticas (Cronk & Fuller 1995; Williamson 1996). A modelagem matemática permite a
detecção de padrões de invasão de espécies exóticas (Neubert & Caswell 2000). Os
modelos matriciais permitem a aplicação de Análises de Sensibilidades e de Análises de
Elasticidades. Através destas, podem ser identificadas e quantificadas as causas que
contribuem negativa ou positivamente para o fitness ou taxa de crescimento finito (λ) da
população (Fox & Gurevitch 2000; Armelin 2005; Caswell 2007). As sensibilidades
quantificam mudanças absolutas na população (Fox & Gurevitch 2000), e as elasticidades
são derivadas das análises de sensibilidade e quantificam proporcionalmente a contribuição
do λ para cada taxa de transição da matriz, sendo estas últimas indicadas para estudos
comparativos entre populações (Fox & Gurevitch 2000; Armelin 2005). Além disso,
elasticidades também podem ser utilizadas para subsidiar a aplicação do manejo em

14
populações estruturadas (e.g. Armelin 2005). Este seria um dos passos para a
decomposição da estrutura populacional e indicação das classes de tamanhos, ou estágios
da história de vida, a serem manejados com eficácia (Morris & Doak 2002). A partir destas
análises o objetivo pode ser o aumento ou a redução do valor de λ, tratando da conservação
ou do manejo de espécies, pois λ < 1 indica retração populacional, λ = 1 estabilidade e λ >
1 indica expansão da população (Caswell 2001).
Desta forma, o estudo da ecologia da espécie exótica em questão pode auxiliar a
detectar padrões, como por exemplo, a taxa de crescimento populacional. A partir das
informações geradas pelos modelos matriciais e da comparação com padrões de espécies
nativas torna-se possível inferir se A. heterophyllus comporta-se como espécie invasora no
ambiente de Mata Atlântica estudado. Neste contexto, este trabalho objetivou: 1) descrever
o ciclo de vida da espécie no PNT no período de 2005 a 2008, 2) verificar se a população
de jaqueiras no PNT está em expansão, 3) verificar os parâmetros responsáveis por
alterações no crescimento populacional, e 4) verificar a maneira como as taxas
demográficas e histórias de vida contribuíram para a taxa de crescimento populacional nos
anos de estudo.

15
MATERIAIS E MÉTODOS
Área de Estudo
O clima do PNT é classificado como Tropical úmido, e de acordo com a
classificação de Köppen, nas regiões mais altas é do tipo Cf e nas regiões mais baixas tipo
Am. A precipitação anual varia de 1300 mm a pouco mais de 3000 mm, com precipitações
abundantes observadas inclusive no inverno (Mattos et al. 1970) e chuvas torrenciais
associadas a temperaturas quentes nos meses de verão (dez-fev) (Coelho-Netto et al.
2007). Litossolos, cambissolos e latossolos prevalecem na porção superior e solos
podozolicos na porção inferior (Coelho-Netto et al. 2007).
O PNT (Figura 1.1) é considerado uma das maiores florestas urbanas do mundo
sendo procurado por muitos turistas e pela população local como área de lazer (IBAMA
2004). Porém, o Rio de Janeiro é o segundo centro econômico e segunda cidade mais
populosa do Brasil, com aproximadamente 6.000.000 de habitantes (IBGE 2006), o que faz
com que o PNT experimente alguns impactos como o crescimento urbano desordenado em
suas regiões limítrofes (Coelho-Netto et al. 2007), a poluição atmosférica (Azevedo et al.
1999) e a presença de espécies não-nativas de fauna (Cunha et al. 2006) e flora (Bandeira
1993), introduzidas de forma acidental ou intencional.
A espécie
Artocarpus heterophyllus Lamarck, popularmente chamada de jaqueira é uma
espécie originária do Sudeste Asiático (Chaves et al. 1967; Ferrão 1993). A introdução
desta espécie no Brasil ocorreu durante o período colonial, onde para cumprir instruções
Régias, os Vice-Reis da Índia deveriam enviar periodicamente para Portugal (metrópole),
África e Brasil (colônias), plantas de interesse para tentar introduzi-las nestas novas
regiões (Ferrão 1993). No ano de 1682, no estado da Bahia, já havia 11 jaqueiras de
procedência desconhecida. Porém, o primeiro registro de envio para o Brasil data de

16
Figu
ra 1
.1 –
Imag
em a
érea
da
loca
lizaç
ão d
o Pa
rque
Nac
iona
l da
Tiju
ca n
a ci
dade
do
Rio
de
Jane
iro.
Cor
tesi
a IB
AM
A P
NT.

17
janeiro de 1683 (Ferrão 1993). As recém-chegadas sementes e mudas eram enviadas para
um horto na Bahia para que fossem aclimatadas e posteriormente disseminadas para outras
regiões do País (Dean 2002). O Rio de Janeiro, em 1803, já possuía exemplares
aparentemente naturalizados (Chaves et al. 1967).
A jaqueira é considerada uma árvore de grande porte, atinge mais de 10 m de
altura, o tronco pode ultrapassar 1 m de diâmetro, é monóica e exibe caulifloria (Chaves et
al. 1967; Crane et al. 2002). Pode ser polinizada por insetos ou pelo vento, resiste bem a
estresses climáticos e em condições ideais de cultivo, pode frutificar em 3 ou 4 anos (Crane
et al. 2002). No Brasil, uma jaqueira bem desenvolvida chega a produzir até 100 frutos por
ano (Figuras 1.2 e 1.3), alguns destes chegando a pesar mais de 30 quilos (Chaves et al.
1967). Indivíduos da espécie frutificam assincronicamente durante todo o ano, com grande
parte dos mesmos frutificando preferencialmente nos meses de verão (dez-fev). Além
disso, os frutos são consumidos pela população humana e pela fauna de mamíferos, como
roedores e primatas (Cunha et al. 2006). Observações de campo indicaram alta germinação
de sementes (ex. Figuras 1.4 e 1.5).
Atualmente, sua distribuição abrange praticamente todo território Nacional com
certa amplitude de habitats, podendo ser observada colonizando áreas abertas e áreas de
mata, ambas associadas a ambientes antrópicos (Carauta & Diaz 2002). No Rio de Janeiro,
coloniza densamente áreas de borda de Mata Atlântica sendo encontrada em Unidades de
Conservação tais como Parque Nacional da Tijuca e as Reservas Biológicas Poço das
Antas, União e Tinguá (Abreu 2005).

18
No habitat original
Nativa do sudeste asiático, Artocarpus heterophyllus é típica de estágios
sucessionais avançados e cresce naturalmente até 1300 m de altitude no sub-bosque da
Floresta Tropical Úmida Monodominante, todavia a espécie é considerada rara (até 1
Figura 1.2 – Exemplo da alta produção de frutos por planta adulta de A. heterophyllus no PNT.
Figura 1.4 – Início da emergência de plântulas após a decomposição do fruto no chão da floresta.
Figura 1.5 – Detalhe da fase inicial de germinação.
Figura 1.3 – Exemplo de fruto da jaqueira em fase de desenvolvimento.

19
indivíduo por ha) neste ambiente, onde poucas espécies dominam os diferentes estratos da
mata (Chittibabu & Parthasarathy 2000; Bhuyan et al. 2003). É tolerante a sombra, porém
necessita de um pouco de luz e espaço para seu desenvolvimento durante os estágios
iniciais de vida, germinando melhor em clareiras. Os frutos amadurecem entre julho e
agosto e o número de sementes varia em tamanho e massa (1,5 – 14 g), entretanto, as
sementes viáveis variam de 4 a 14g de massa, e dentre seus dispersores são mencionados
roedores, macacos e porcos selvagens (Khan 2004). As sementes possuem taxa de
predação relativamente alta (Sodhi et al. 2003) e germinam ainda na estação chuvosa,
durante o verão (maio-setembro), onde ocorrem 85% da precipitação anual (Chittibabu &
Parthasarathy 2000; Bhuyan et al. 2003). Desta forma, as sementes germinam e suas
plântulas se estabelecem, tornando-se capazes de tolerar bem a estação seca (dezembro-
fevereiro, Chittibabu & Parthasarathy 2000; Bhuyan et al. 2003).
Amostragem das sub-populações de A. heterophyllus no PNT
Em 2001 foi realizado um estudo piloto em Unidades de Conservação do Rio de
Janeiro: Reserva Biológica União, Reserva Biológica do Tinguá e Parque Nacional da
Tijuca, visando definir a forma e o tamanho adequado das parcelas e as áreas prioritárias
para a realização de estudos com as populações da espécie exótica (Abreu 2005). Após este
estudo, em 2004, optou-se pela marcação de 20 parcelas permanentes em sub-populações
de jaqueiras distribuídas pelo PNT. Foram marcadas parcelas circulares com 10 m de raio
(314 m2 cada) totalizando uma área amostral de 0,628 ha. Nestas parcelas todos os
indivíduos da comunidade vegetal com diâmetro a altura do peito (DAP) ≥ 5 cm foram
marcados e mensurados. Indivíduos com troncos múltiplos visivelmente interligados foram
considerados um único indivíduo. Nestes casos, cada tronco foi medido separadamente e
posteriormente somaram-se suas áreas basais, e da área basal resultante, obteve-se um

20
único diâmetro virtual ≥ 5cm. Todos os indivíduos foram mapeados dentro das sub-
populações e foi verificada a presença de estruturas reprodutivas.
Distribuição de Freqüências
As classes de diâmetros utilizadas foram estabelecidas arbitrariamente de acordo
com critérios biológicos observados no campo (e.g., presença de estruturas reprodutivas e
tipo de casca do tronco) e objetivando futuras comparações com outros estudos sobre
populações de espécies arbóreas e comunidades. Histogramas de freqüências foram
elaborados utilizando-se o programa STATISTICA 6.0. Os demais gráficos e tabelas foram
elaborados no programa Excel.
A classe JUVENIS 1 foi constituída por arvoretas de pequeno a médio porte com
DAP variando de 5 a 9,99 cm, e copa pouco desenvolvida. Os indivíduos da classe
JUVENIS 2 caracterizaram-se por árvores de pequeno a médio porte com DAP variando
de 10 a 14,99 cm, ainda com copa proporcionalmente pouco desenvolvida. A classe de
PRÉ-REPRODUTIVOS reuniu indivíduos de 15 a 24,99 cm de DAP, sendo composta por
grandes árvores imaturas (não reprodutivas) com copa relativamente ampla. Os indivíduos
reprodutivos foram divididos nas classes ADULTOS 1 e 2. Os ADULTOS 1, com DAP
variando de 25 a 44,99 cm, apresentaram tronco liso e copa ampla. A classe ADULTOS 2
englobou indivíduos com diâmetro igual ou superior a 45 cm que, em geral, apresentavam
fendilhamentos na casca e copa ampla.
Dinâmica de populações
Todos os indivíduos foram marcados e mensurados no período de novembro de
2004 a janeiro de 2005 e tiveram o DAP remensurado anualmente nos meses de janeiro de
2006 e 2007, e dezembro de 2007, que daqui em diante será referido como remedição de

21
2008. Indivíduos que obedeciam ao critério de inclusão adotado, ou seja, atingiram DAP
de 5 cm, foram marcados e incluídos na amostragem durante os recensos.
Excepcionalmente no ano de 2006, por problemas de impossibilidade de acesso à algumas
áreas de estudo, apenas 10 parcelas foram remensuradas, entretanto, nos outros anos todas
as 20 parcelas foram consideradas.
Ciclo de Vida
Para a elaboração das matrizes de transição das dinâmicas de 2006 – 2007,
respectivamente primeiro e segundo ano da dinâmica populacional, foram consideradas
apenas as 10 parcelas dos sítios que foram remensurados anualmente. Para a dinâmica de
2007 – 2008, todas as 20 parcelas da amostra foram consideradas. A partir daí, com base
nos 3 anos de remedições foi construída a matriz de transição média A. Ao tratar o ciclo de
vida como uma cadeia de Markov, assume-se que eventos fututros dependem apenas do
presente, e não do passado, a partir daí é possível determinar a duração das fases deste
ciclo e outros parâmetros relacionados com a idade da planta, mesmo que o ciclo de vida
não tenha sido classificado por idade (Armelin 2005).
Matrizes de transição – Foi construída uma matriz de transição At, quadrada e de
ordem i x j, com i = j e representando o número de classes da população. A matriz de
transição contém as probabilidades de transição de uma classe para outra no período t
(Caswell 2001). Para o cálculo dos parâmetros dos ciclos de vida desejados foram
utilizados os valores médios dos dados anuais de 2006 a 2008. Foram construídas,
portanto, matrizes de transição anuais médias (t = 1 ano).
Considera-se j como a classe de origem da planta no início do período t, e i como a
classe de destino dessa mesma planta no final deste período. As entradas relativas às

22
transições foram calculadas dividindo-se o número de plantas em ij pelo total de plantas
em j.
Uma vez que este trabalho não contemplou classes de diâmetros menores do que 5
cm, as entradas ij relativas a fecundidade foram estimadas a partir do número de recrutas
que entravam na amostragem em cada recenso. Assumiu-se não ser possível relacionar o
indivíduo recruta à planta-mãe e que a espécie estudada, por frutificar durante quase todo o
ano, tem reprodução em fluxo contínuo. Desta forma, o cálculo da fecundidade procedeu-
se pela fórmula:
F = √sj . fj . √s0 (Morris & Doak 2002), onde:
F = fecundidade;
sj = probabilidade de um indivíduo da classe j (ADULTO 1 ou 2, neste caso) sobreviver
por metade do tempo do próximo recenso, ou seja, por seis meses (t = 1 ano);
fj = número de recrutas por adulto da classe;
s0 = probabilidade de um recruta sobreviver por metade do tempo do próximo recenso;
Após a obtenção da fecundidade total para cada ano, para se obter a fecundidade
por classe de adultos, multiplicou-se F pelo peso arbitrário dado às mesmas, peso de 0,4
para ADULTOS 1, por serem adultos menores e 0,6 para ADULTOS 2, por estes serem
maiores e aparentemente frutificarem mais.
Considerou-se que a população estava em fase de dinâmica assintótica. Foram
calculados o autovalor dominante e os autovetores esquerdo (v) e direito (w) associados ao
λ (autovalor dominante). O autovalor dominante da matriz de Lefkovitch (ou matriz de
estágios) expressa a taxa de crescimento assintótico da população, representado por λ
(lambda). O autovetor direito (w) associado a λ indica proporcionalmente a contribuição de
cada classe para o λ quando a população atinge a estrutura-estádio-estável (EEE), caso as

23
condições ambientais mantenham-se constantes. O autovetor esquerdo (v) associado ao
lambda, contém os valores reprodutivos de cada classe. Valores reprodutivos podem ser
definidos como a contribuição futura esperada que um indivíduo de uma classe específica
fará ao crescimento populacional (Morris & Doak 2002).
Foi calculado o tempo que a população atual levaria para atingir a estrutura-estádio-
estável, para isto, foi utilizado o ‘power method’ proposto por Caswell (2001). Neste, a
população é projetada em t → infinito e observa-se a partir de quando ocorre a
estabilização do autovalor dominante, ou seja, a partir de que ano a população atingirá a
fase assintótica. Todos os cálculos foram realizados no programa MATLAB®.
S11 R12 0 F14 F15G21 S22 R23 0 00 G32 S33 R34 00 0 G43 S44 R450 0 0 G54 S55
At =
S11 R12 0 F14 F15G21 S22 R23 0 00 G32 S33 R34 00 0 G43 S44 R450 0 0 G54 S55
S11 R12 0 F14 F15G21 S22 R23 0 00 G32 S33 R34 00 0 G43 S44 R450 0 0 G54 S55
At =
AJUSTES – As informações aqui obtidas em 3 anos de dinâmica não foram
suficientes para parametrizar o modelo de ciclo de vida das jaqueiras, pois eventos como a
mortalidade de indivíduos ADULTOS 2 não puderam ser detectados durante o período de
estudo. A mortalidade na última classe é um evento necessário para fechar o ciclo de vida
da espécie estudada, portanto, assim como em outros trabalhos (Caswell 2001, Armelin
2005) foi incluída no modelo a mortalidade artificial de um indivíduo da última classe no
último ano de estudo (2008).
Figura 1.6 – Representação das taxas vitais na matriz de estágios At (Lefkovitch 1965). Onde, S: probabilidade de um indivíduo permanecer na mesma classe durante o período t (t = 1 ano); G: probabilidade do indivíduo passar para classe seguinte em t; R: probabilidade de retração de indivíduos para classe anterior; F: fecundidade de cada classe reprodutiva.

24
A taxa de mortalidade anual de ADULTOS 2 foi estimada dividindo-se 1
(mortalidade suposta) pelo total de indivíduos da classe no último recenso, o resultado é
gerado com uma superestimativa e para corrigir tal viés, o valor encontrado é ajustado para
o período total da estimação ‘t(final) + 1’, aplicando-se a fórmula (R. Armelin, comunicação
pessoal):
M = [1+(m/n)]1/(tfinal+1) – 1 , onde:
M = mortalidade corrigida;
m = mortalidade suposta (1 indivíduo);
n = número de indivíduos na classe no último recenso;
tfinal = tempo total de dinâmica.
Sensibilidades e Elasticidades da fase assintótica
Por ser derivado da matriz de transição At, o λ (medida de ‘fitness’ populacional) é
sensível a variações nas taxas demográficas que compõem a matriz. E dois derivativos de
λ, a sensibilidade e a elasticidade, podem ser utilizados para avaliar os impactos das
mudanças destas taxas.
sij = ∂λ1 / ∂aij onde:
sij = sensibilidades = derivada parcial de λ1 em relação à aij;
As sensibilidades sij quantificam mudanças absolutas em λ, resultante de uma
mudança absoluta infinitesimal na matriz de transição aij (de Kroon et al. 2000), sendo
indicadas para ações de manejo (Fox & Gurevitch 2000; Armelin 2005).
A análise de elasticidade é uma ferramenta utilizada para decompor a taxa de
crescimento populacional nas contribuições feitas pelas transições do ciclo de vida.
Matematicamente, a elasticidade eij quantifica a mudança proporcional no λ resultante de

25
uma mudança proporcional infinitesimal na matriz de transição aij e é calculada a partir da
sensibilidade.
eij = (aij / λ1)* sij onde:
eij = elasticidade = derivada da sensibilidade de λ1 em relação à aij;
Elasticidades então, comparam efeitos relativos em λ com as mesmas mudanças
relativas nos valores dos parâmetros demográficos. Uma característica importante é que
todas elasticidades de uma matriz de transição somam uma unidade, o que permite a
comparação da importância relativa das diferentes taxas vitais (e.g., sobrevivência,
crescimento e reprodução) dentro do ciclo de vida, sendo mais indicadas para estudos
comparativos entre populações (Fox & Gurevitch 2000; Armelin 2005). Desta forma por
exemplo, elasticidades de diferentes transições podem ser somadas quando desejado (de
Kroon et al. 2000).
Análise Retrospectiva – LTRE (Life Table Response Experiments)
LTREs foram utilizadas para avaliar o impacto real de aij ocorridos em λ, ou seja,
para investigar a importância das contribuições da história de vida e das taxas
demográficas, ocorridas no passado, e a maneira como estas influenciaram o λ
populacional atual. Desta forma:
λ(t+1) ≈ λ(t) + ∑ij (aij(t+1) - aij
(t) ) ∂λ/∂aij │A*
A(t) = matriz de referência = matriz do tempo ‘t’ adotada como controle;
A(t+1) = matriz de tratamento = matriz do tempo ‘t+1’;
∂λ/∂aij │A* = sij ; sensibilidades extraídas da matriz A*;
A* = (A(t+1) + A(t))/2 = matriz média entre tratamento e controle;

26
Duas LTREs foram feitas, uma adotando a matriz de 2006 como referência e a de
2007 como tratamento, para verificar as mudanças ocorridas no período 2006 – 2007, e
outra adotando o ano de 2007 como referência e 2008 como tratamento, para estudo do
período 2007-2008. A LTRE mostra os mecanismos demográficos que influenciaram a
variação no desempenho da população ao longo dos anos de estudo, ou seja, podem ser
observadas as contribuições para o ∆λ (λ2 – λ1 e/ou λ3 – λ2), e estas contribuições, quando
somadas, resultam no ∆λ observado.
Uma vez que o objetivo desta análise é a decomposição e posterior comparação das
variações em λ entre os anos, para que não fossem geradas estimativas enviesadas, foram
utilizadas as matrizes que contemplaram 10 parcelas e não foram incluídas mortalidades
artificiais na última classe do último ano.

27
RESULTADOS
As vinte parcelas amostradas dentro das sub-populações de jaqueiras no PNT,
exibiram desde 40% até 100% da comunidade local composta por indivíduos de A.
heterophyllus. Um total de 558 indivíduos foi amostrado em 2005 e a área basal média de
jaqueiras foi de 1,649 ± 0,762 m².parcela-1 com número aproximado de 30
indivíduos.parcela-1 (Tabela 1.1).
Tabela 1.1 – Sumário de parâmetros utilizados para caracterizar a estrutura da comunidade nas parcelas estudadas no Parque Nacional da Tijuca em 2005. Onde: AB total = área basal de toda a comunidade; AB jaq = área basal ocupada pelas jaqueiras na comunidade; AB outros = área basal ocupada por outros indivíduos excetuando-se as jaqueiras; N total = número de indivíduos da camunidade; N jaq = número de indivíduos de jaqueiras na comunidade. AB total AB jaq AB outros N total N jaq Total 42,982 32,987 9,995 695 558 Média ± DP 2,149 ± 0,787 1,649 ± 0,762 0,952 ± 2,115 34,75 ± 9,5 27,9 ± 11,5
A estrutura da população de jaqueiras no PNT no ano de 2005 (Figura 1.7), exibiu o
padrão de distribuição log-linear assimétrica para a direita, conhecida como “J reverso”. A
maior parte dos indivíduos (40%) encontrou-se na primeira classe de tamanho e as classes
de adultos reprodutivos somaram 29% dos indivíduos da população.

28
Ciclo de Vida
O ciclo de vida de A. heterophyllus foi observado para cada ano na tabela 1.2 e
representado na figura 1.8 na forma da matriz de estágios. Para a população de jaqueiras no
PNT, o ‘fitness’ populacional (λ) nos anos de 2006, 2007 e 2008 apresentou valores de
1,0074, 1,0100 e 0,9990, respectivamente. O valor de λ = 1,0021 foi observado na matriz
média.
A taxa de mortalidade corrigida, estimada para complementar o modelo do ciclo de
vida da espécie foi de 0,42 % para a última classe.
Figura 1.7 – Histograma de classes de diâmetros (DAP em cm) das sub-populações de jaqueiras no Parque Nacional da Tijuca – RJ no ano de 2005.

29
Tx Trans elem. 2006(10) 2007(10) 2008 (10) 2008 (20) ANOS
S1 a11 0.9659 0.9659 0.9551 0.9309 0.9466 S2 a22 0.9167 0.9167 0.9714 0.9529 0.9363 S3 a33 0.8974 0.9714 0.9730 0.9885 0.9627 S4 a44 0.9787 0.9796 0.9796 0.9717 0.9752 S5 a55 1 1 1 0.9831* 0.9909* G1 a21 0.0341 0.0227 0.0337 0.0369 0.0331 G2 a32 0.0833 0.0833 0.0286 0.0353 0.0573 G3 a43 0.0769 0.0286 0.0270 0.0115 0.0311 G4 a54 0.0213 0.0204 0.0204 0.0189 0.0198 F1 a14 0.0102 0.0221 0.0209 0.0215 0.0181 F2 a15 0.0152 0.0331 0.0314 0.0323 0.0269
λ 1.0074 1.0100 1.0096 0.9989 1.0021 ICs (1.0039 - 1.0104) (1.0000 - 1.0165) (1.0000 - 1.0157) (0.9770 - 1.0002) (0.9926 - 1.0079)
*mortalidade artificial de 1 indivíduo inserida na última classe
Tabela 1.2 – Taxas de transição e taxa de crescimento populacional (λ) da dinâmica anual da população de jaqueiras no Parque Nacional da Tijuca – RJ, no período 2005 – 2008. Tx Trans = taxa de transição, onde S: permanência na classe, G: crescimento, passagem para a classe seguinte, F: fecundidades, os numerais indicam as respectivas classes; elem. = elemento da matriz (elementos não listados apresentaram valor zero); 2006 = dinâmica 2005 – 2006; 2007 = dinâmica 2006 – 2007; 2008 = dinâmica 2007 – 2008; ANOS = matriz sumário dos anos de estudo (2005 – 2008). Os valores entre parênteses indicam o número de parcelas utilizadas na construção da matriz. λ = autovalor dominante da matriz, que corresponde a taxa finita de crescimento populacional; ICs = intervalos de confiança calculados para o λ.
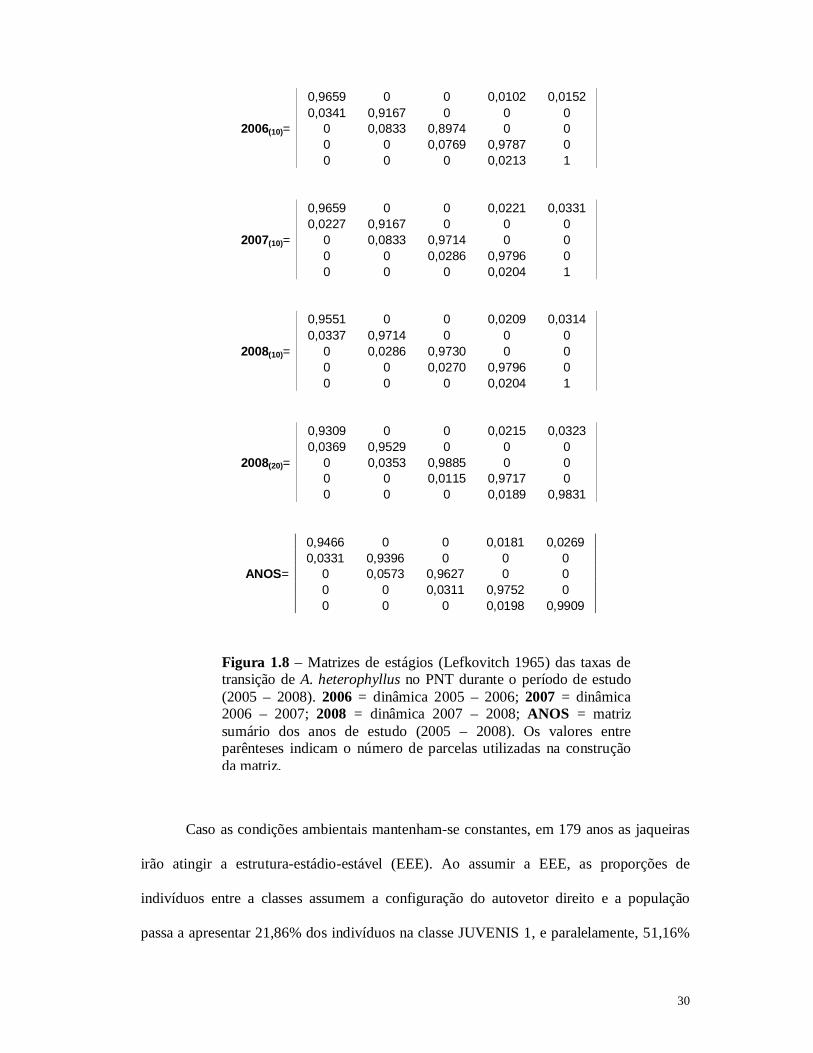
30
0,9659 0 0 0,0102 0,0152 0,0341 0,9167 0 0 0 2006(10)= 0 0,0833 0,8974 0 0 0 0 0,0769 0,9787 0 0 0 0 0,0213 1
0,9659 0 0 0,0221 0,0331 0,0227 0,9167 0 0 0 2007(10)= 0 0,0833 0,9714 0 0 0 0 0,0286 0,9796 0 0 0 0 0,0204 1
0,9551 0 0 0,0209 0,0314 0,0337 0,9714 0 0 0 2008(10)= 0 0,0286 0,9730 0 0 0 0 0,0270 0,9796 0 0 0 0 0,0204 1
0,9309 0 0 0,0215 0,0323 0,0369 0,9529 0 0 0 2008(20)= 0 0,0353 0,9885 0 0 0 0 0,0115 0,9717 0 0 0 0 0,0189 0,9831
0,9466 0 0 0,0181 0,0269 0,0331 0,9396 0 0 0 ANOS= 0 0,0573 0,9627 0 0 0 0 0,0311 0,9752 0 0 0 0 0,0198 0,9909
Caso as condições ambientais mantenham-se constantes, em 179 anos as jaqueiras
irão atingir a estrutura-estádio-estável (EEE). Ao assumir a EEE, as proporções de
indivíduos entre a classes assumem a configuração do autovetor direito e a população
passa a apresentar 21,86% dos indivíduos na classe JUVENIS 1, e paralelamente, 51,16%
Figura 1.8 – Matrizes de estágios (Lefkovitch 1965) das taxas de transição de A. heterophyllus no PNT durante o período de estudo (2005 – 2008). 2006 = dinâmica 2005 – 2006; 2007 = dinâmica 2006 – 2007; 2008 = dinâmica 2007 – 2008; ANOS = matriz sumário dos anos de estudo (2005 – 2008). Os valores entre parênteses indicam o número de parcelas utilizadas na construção da matriz.

31
dos indivíduos nas classes reprodutivas, com 18,49% na classe ADULTOS 1 e 32,67% em
ADULTOS 2. O autovetor direito (w) associado a λ é representado a seguir:
As contribuições dos valores reprodutivos (v) são crescentes nas classes não-
reprodutivas e muito similares nas classes reprodutivas (duas últimas), em torno de 25,5%
cada uma, indicando que indivíduos reprodutivos contribuem mais para o crescimento
populacional da espécie, conforme esperado.
.
No ciclo de vida da espécie sumarizado para o período de um ano, a permanência
de indivíduos nas classes é indicada pelas setas azuis, a passagem para a classe superior
pelas setas pretas e a fecundidade é representada em vermelho (Figura 1.9). Em JUVENIS
1 observou-se que 94,66% dos indivíduos permaneceram na mesma classe e 3,31% destes
passaram para a classe superior anualmente. Os indivíduos JUVENIS 2, em relação as
demais classes, apresentaram a menor taxa de permanência (93,63%) e o mais rápido
recrutamento para a classe superior, com 5,73% dos indivíduos passando anualmente para
a classe de PRÉ-REPRODUTIVOS. Esta última, apresentou a taxa de recrutamento para a
classe seguinte de 3,11% e permanenceram na mesma classe 96,27% dos indivíduos. Para
ADULTOS 1, a permanência foi de 97,52% com aproximadamente 2% dos indivíduos
0,2186 0,1099 w = 0,1599 0,1849 0,3267
0,1057 0,1776 v = 0,2039 0,2585 0,2543

32
passando para a classe superior anualmente. A permanência na última classe (ADULTOS
2) é a mais alta apresentando o valor de 99,09 %.
JUVENIS 15 – 9,9 cm
JUVENIS 210 – 14,9 cm
PRÉ-REPRODUTIVOS15 – 24,9 cm
ADULTOS 125 – 44,9 cm
ADULTOS 2> 45 cm
0,0331
0,0573
0,0311
0,0198
0,9466
0,9363
0,9752
0,9909
0,9627
0,02
69
0,01
81
Limitações do modelo - Uma vez que modelos ecológicos exibem algumas limitações que
podem ser corrigidas no longo prazo (Caswell 2001), as matrizes utilizadas para a
elaboração do ciclo de vida da espécie (período de 2005 - 2008) precisaram ser ajustadas.
Desta forma, o modelo trabalhado para as jaqueiras no período estudado exibiu duas
limitações: a primeira delas foi a ausência de mortalidade na última classe. Para solucionar
este problema mortalidades artificiais foram inseridas (1 indivíduo na matriz do último
ano) na classe ADULTOS 2 com a finalidade de encerrar o ciclo de vida da espécie (vide
materiais e métodos). Como segunda limitação, o inicío do ciclo de vida deu-se a partir do
Figura 1.9 – Representação gráfica da matriz de estágios do ciclo de vida de A. heterophyllus no PNT – RJ, no período 2005 – 2008. Em vermelho a fecundidade, em preto o crescimento e em azul a permanência em uma mesma classe.

33
critério de inclusão adotado (DAP ≥ 5), podendo resultar em um aumento na velocidade da
dinâmica populacional (R. Armelin, comunicação pessoal), ou seja, pode ocorrer uma
superestimação no valor de lambda. O início do ciclo de vida ideal seria a partir da classe
de SEMENTES, onde o valor de lambda seria estimado adequadamente (Caswell 2001).
Sensibilidades e Elasticidades
A análise de sensibilidade (Figura 1.10) indicou que as taxas que mais
influenciaram o λ populacional foram principalmente as permanências (setas azuis) nas
classes reprodutivas (ADULTOS 1 e 2), a transição (setas pretas) de ADULTOS 1 para
ADULTOS 2, e a fecundidade (setas vermelhas) dos ADULTOS 2. Em seguida,
influenciaram o λ populacional a transição de PRÉ-REPRODUTIVOS para ADULTOS 1 e
a transição de JUVENIS 1 para JUVENIS 2. As menores contribuições ao crescimento da
população deveram-se tanto à permanência quanto à transição dos indivíduos JUVENIS 2.
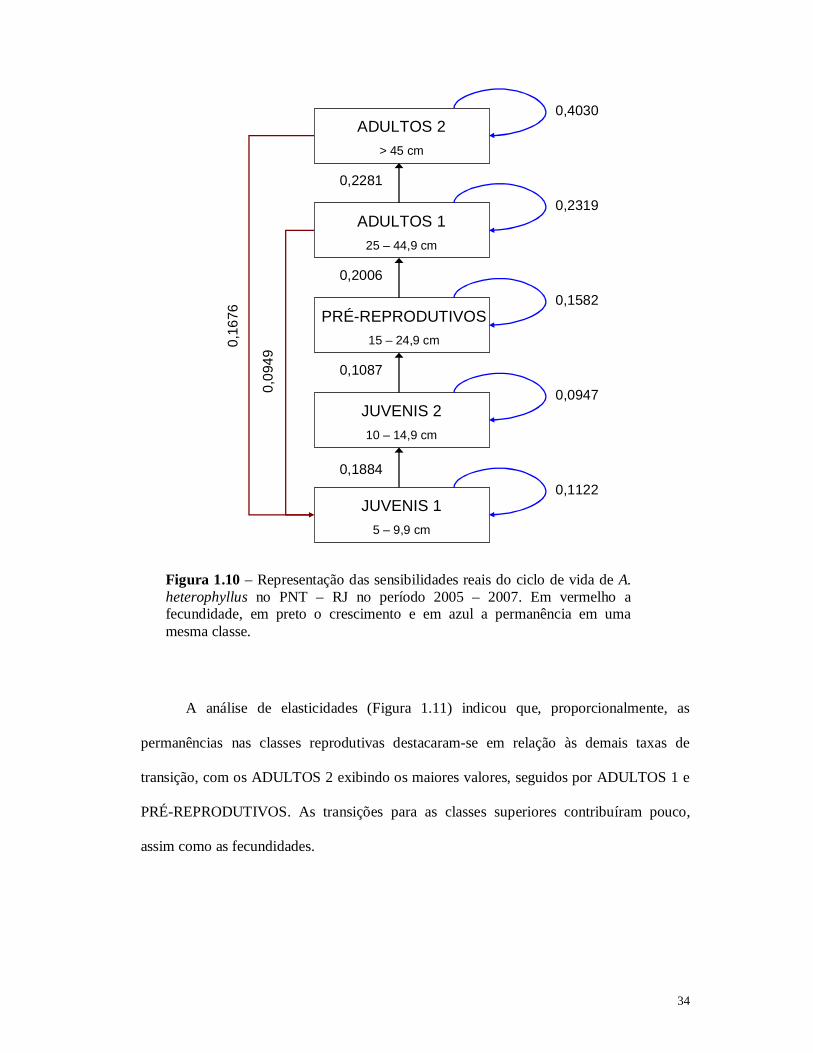
34
JUVENIS 210 – 14,9 cm
PRÉ-REPRODUTIVOS15 – 24,9 cm
ADULTOS 125 – 44,9 cm
ADULTOS 2> 45 cm
0,1884
0,1087
0,2006
0,2281
0,09
49
0,16
76
0,1122
0,0947
0,2319
0,4030
0,1582
JUVENIS 15 – 9,9 cm
A análise de elasticidades (Figura 1.11) indicou que, proporcionalmente, as
permanências nas classes reprodutivas destacaram-se em relação às demais taxas de
transição, com os ADULTOS 2 exibindo os maiores valores, seguidos por ADULTOS 1 e
PRÉ-REPRODUTIVOS. As transições para as classes superiores contribuíram pouco,
assim como as fecundidades.
Figura 1.10 – Representação das sensibilidades reais do ciclo de vida de A. heterophyllus no PNT – RJ no período 2005 – 2007. Em vermelho a fecundidade, em preto o crescimento e em azul a permanência em uma mesma classe.

35
JUVENIS 15 – 9,9 cm
JUVENIS 210 – 14,9 cm
PRÉ-REPRODUTIVOS15 – 24,9 cm
ADULTOS 125 – 44,9 cm
ADULTOS 2> 45 cm
0,0062
0,0062
0,0062
0,0045
0,00
170,00
45
0,1060
0,0885
0,2257
0,3985
0,1520
Experimentos de Resposta da Tabela de Vida - Life Table Response Experiments (LTRE)
A variação do crescimento populacional pôde ser avaliada em relação aos estágios
das histórias de vida dos indivíduos e/ou em relação às taxas demográficas. As matrizes
anuais utilizadas na comparação estão representadas na tabela 1.2 e na figura 1.8 . Em
relação aos estágios das histórias de vida, aqueles que influenciaram positivamente o ∆λ de
2006 para 2007, ou seja, contribuíram para o crescimento populacional, foram
respectivamente: a passagem de indivíduos para as classes superiores (crescimentos) e o
Figura 1.11 – Representação das elasticidades do ciclo de vida de A. heterophyllus no PNT – RJ no período 2005 – 2007. Em vermelho a fecundidade, em preto o crescimento e em azul a permanência em uma mesma classe.
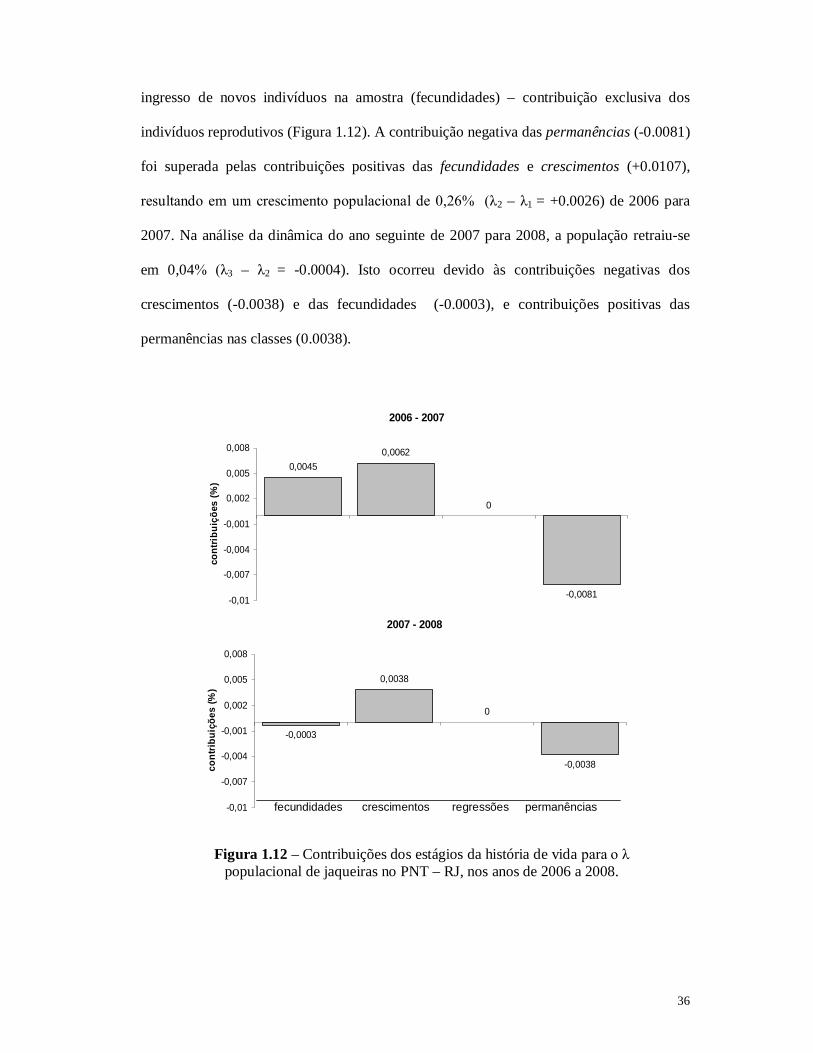
36
ingresso de novos indivíduos na amostra (fecundidades) – contribuição exclusiva dos
indivíduos reprodutivos (Figura 1.12). A contribuição negativa das permanências (-0.0081)
foi superada pelas contribuições positivas das fecundidades e crescimentos (+0.0107),
resultando em um crescimento populacional de 0,26% (λ2 – λ1 = +0.0026) de 2006 para
2007. Na análise da dinâmica do ano seguinte de 2007 para 2008, a população retraiu-se
em 0,04% (λ3 – λ2 = -0.0004). Isto ocorreu devido às contribuições negativas dos
crescimentos (-0.0038) e das fecundidades (-0.0003), e contribuições positivas das
permanências nas classes (0.0038).
2006 - 2007
0,00450,0062
0
-0,0081-0,01
-0,007
-0,004
-0,001
0,002
0,005
0,008
cont
ribui
ções
(%)
2007 - 2008
-0,0003
0,0038
0
-0,0038
-0,01
-0,007
-0,004
-0,001
0,002
0,005
0,008
cont
ribui
ções
(%)
fecundidades crescimentos regressões permanências
Figura 1.12 – Contribuições dos estágios da história de vida para o λ populacional de jaqueiras no PNT – RJ, nos anos de 2006 a 2008.

37
As taxas demográficas (Figura 1.13) indicaram que de 2006 para 2007, somente os
JUVENIS 1 contribuíram negativamente para o ∆λ, os JUVENIS 2 não influenciaram, e
PRÉ-REPRODUTIVOS, ADULTOS 1 e ADULTOS 2 contribuíram positivamente. As
contribuições foram crescentes, com destaque para os indivíduos reprodutivos. No ano
seguinte, de 2007 para 2008, a retração da população em 0,04% ocorreu devido a
contribuição negativa de todas as classes, exceto de JUVENIS 1, que contribuiu
positivamente com 0,1%.
2007 - 2008
0,001
-0,0009-0,0001 -0,0001 -0,0002
-0,007
-0,004
-0,001
0,002
0,005
cont
ribui
ções
(%)
JUVENIS 1 JUVENIS 2 PRÉ-REPRO ADULTOS 1 ADULTOS 2
2006 - 2007
-0,0024
0 0,00040,001
0,0035
-0,007
-0,004
-0,001
0,002
0,005
cont
ribui
ções
(%)
Figura 1.13 – Contribuições anuais das taxas demográficas para o λ populacional de jaqueiras no PNT - RJ. Nos anos de 2006 a 2008.

38
DISCUSSÃO
A espécie exótica estudada apresenta intensa regeneração no PNT (Figura 1.2 e
1.5), onde estabeleceu-se com sucesso passados pelo menos 150 anos de sua introdução. O
bom crescimento em clareiras, a tolerância à sombra e a colonização do sub-bosque são
características da espécie no habitat de origem (Khan 2004). Conforme observado em
campo, esta espécie coloniza densamente regiões de borda de mata e a ação antropogênica
é um dos fatores que auxilia sua propagação. Além disso, também foi observado que em
regiões de encosta, devido à dispersão barocórica, há uma tendência da espécie de se
propagar em sentido descendente. Fatores climáticos podem favorecer a propagação de
Artocarpus heterophyllus na Mata Atlântica, pois no habitat original a espécie encontra-se
submetida ao clima de monção com estação seca pronunciada (Chittibabu & Parthasarathy
2000; Bhuyan et al. 2003). Já na Mata Atlântica, o clima tropical e as precipitações
abundantes durante todo o ano parecem não limitar a reprodução da espécie, de modo que
na população podem ser observados indivíduos frutificando assincronicamente durante
todo o ano, com picos de frutificação no verão. Isto induz a regeneração da espécie e
permite assumir que a reprodução é contínua.
A espécie é típica de florestas tropicais monodominantes onde poucas espécies
dominam os diversos estratos da mata, porém, é considerada rara neste ambiente e possui
distribuição aleatória com poucos indivíduos por hectare (Chittibabu & Parthasarathy
2000; Bhuyan et al. 2003). Desta forma, A. heterophyllus pode encontrar facilidades ao
colonizar um ambiente como a Mata Atlântica. Por exemplo, na Mata Atlântica não há
espécies com altos valores de dominância no estrato superior da mata (Rodrigues 2004).
Sendo assim, uma espécie que co-evoluiu em um ambiente extremamente competitivo
pode expressar sua alta capacidade de crescimento neste novo habitat, passando a dominar
o novo local colonizado e exibindo distribuição gregária em escala regional (Abreu 2005).

39
Ciclo de vida
Em populações de espécies arbóreas que ocupam o dossel de florestas tropicais
Silvertown et al. (1993) observaram elevadas permanências. Conforme verificado para A.
heterophyllus a maior parte dos indivíduos da população (de 93,6% a 99,1%) permaneceu
na mesma classe no período de um ano (t). Estas plantas possuem crescimento lento devido
ao investimento em tecidos lenhosos e lignificados que resultam em uma madeira de
qualidade superior às plantas de crescimento rápido, capazes de atingir o dossel da mata
nas fases iniciais de sucessão ecológica, necessitando de luz direta para o desenvolvimento
(Tilman 1988; Alvarez-Buylla 1994). Há indícios de que as jaqueiras cresçam rapidamente
até atingir a maturidade pois as mudanças de classe foram mais notáveis na passagem de
JUVENIS 2 para PRÉ-REPRODUTIVOS (5,73%) e destas, para ADULTOS 1 (3,11%).
Uma vez que as classes de tamanho aqui utilizadas foram expressas em DAPs, neste
trabalho não foram observadas regressões entre as classes, corroborando o padrão
encontrado por Silvertown et al. (1996) para espécies arbóreas.
As taxas mais altas de permanências foram observadas nas classes reprodutivas, o
que era esperado, pois além das amplitudes destas classes (pelo menos 19,9 cm) serem
maiores do que as demais, na última classe de indivíduos não há transição para classes
superiores e esta taxa de permanência só se altera devido a eventuais mortalidades
ocorridas. Além disso, não foram observadas mortalidades para ADULTOS 2. Desta
forma, a mortalidade estimada de 0,46% para a espécie exótica é inferior aos valores
esperados para mortalidade de árvores de dossel de Mata Atlântica, que estão entre 1 e
2,5% (R. Armelin, comunicação pessoal), atestando a alta adaptação de A. heterophyllus
ao ambiente de Mata Atlântica. Em espécies arbóreas, a taxa de crescimento populacional
depende principalmente da sobrevivência dos adultos estabelecidos (Silvertown et al.
1996). Neste trabalho, as altas permanências encontradas são reflexos da sobrevivência e

40
do lento crescimento em diâmetro típico de árvores tropicais não-pioneiras (Primack
1985).
A população de jaqueiras do PNT apresentou taxa de crescimento populacional λ =
1,0021, não diferindo significativamente de 1, valor que indica estabilidade populacional
(Caswell 2001). Isto demonstra que a população desta espécie exótica está efetivamente
estabelecida em ambiente de Mata Atlântica, e espécies exóticas estabelecidas podem ser
consideradas espécies invasoras (Richardson et al. 2000). Para outras espécies arbóreas de
florestas tropicais, os valores de λ também não diferiram significativamente de 1, como em
Cecropia obtusifolia no México λ = 1,0137 (Alvarez-Buylla 1994). Na Mata Atlântica
brasileira, este padrão de estabilidade populacional se repete, conforme encontrado para as
árvores nativas Miconia cinnamomifolia, com λ variando de 0,8887 à 1,0159 (Pereira
1998), Scheflera angustissima, λ = 1,0004 e Andira anthelmia, λ = 0,9832 (Armelin 2005)
e para a arbustivo-arbórea Psychotria suterella, λ = 0,94 a 0,97, tanto em fragmentos
quanto em mata contínua (Bertani 2006). Isto pode indicar que a população de A.
heterophyllus estudada apresenta um padrão de crescimento similar às populações de
espécies nativas de Mata Atlântica, o que pode ser considerado um indício da adaptação da
espécie a este ambiente.
Sensibilidades e Elasticidades
Sensibilidades e elasticidades são usadas para prever o impacto que variações nas
taxas de transição podem causar ao λ, e enquadram-se na categoria de análises prospectivas
(Caswell 2000; Zuidema & Franco 2001). Dependendo das taxas vitais consideradas
variações no λ podem apresentar correlações com variações nas sensibilidades ou
elasticidades (Zuidema & Franco 2001). De Kroon et al. (2000), apontam outras diferenças
entre sensibilidades e elasticidades, onde as sensibilidades estimam os efeitos no λ caso

41
ocorram variações absolutas nos valores dos parâmetros, mesmo que sejam parâmetros
medidos em escalas diferentes (i.e. crescimentos/permanências x fecundidades). Pequenas
mudanças nos elementos da matriz que apresentam altas sensibilidades, resultam em
grandes alterações no λ (de Kroon et al. 1986). Neste contexto, as sensibilidades calculadas
para A. heterophyllus indicam que pequenas alterações em qualquer uma das taxas geradas
pelos ADULTOS 2 podem afetar bastante o ‘fitness’ populacional da espécie, assim como
as permanências de ADULTOS 1, e a entrada e saída de indivíduos nesta classe. Estas
informações podem servir para subsidiar um manejo adequado da espécie, pois para
reduzir o ‘fitness’ populacional desta exótica estabelecida no PNT podem ser tomadas
medidas que evitem a entrada de indivíduos nas classes reprodutivas como a criação de um
“gargalo” no recrutamento para a classe reprodutiva; a remoção de ADULTOS 1 e/ou 2; a
redução da fecundidade da espécie (expressa na forma de novos indivíduos com DAP >
5cm); ou ainda a implementação simultânea destas medidas.
No entanto, as sensibilidades não medem a contribuição ao λ, e neste caso, devem
ser usadas as sensibilidades relativas (de Kroon et al. 1986), também conhecidas como
elasticidades. Segundo o teorema de Euller (Mesterton-Gibbons 1993), os valores das
elasticidades de uma matriz de projeção somam uma unidade (de Kroon et al. 1986;
Silvertown et al. 1996), e por serem proporcionais permitem a comparação entre diferentes
espécies. Porém, deve-se tomar cuidado ao fazer estas comparações, já que o número de
estágios escolhidos (biológica ou matematicamente) para elaboração do ciclo de vida pode
influenciar nas contribuições das taxas de fecundidades, permanências e crescimentos
observadas nas elasticidades (Enright et al. 1995). Neste contexto, conforme observado
para as elasticidades das jaqueiras no PNT, as grandes contribuições das permanências nas
classes são típicas de populações com baixas taxas de crescimento (de Kroon et al. 2000).

42
As elasticidades das transições são as mesmas para todos os estágios não
reprodutivos e nunca aumentam nas classes reprodutivas, além disso, as transições entre
classes são sempre maiores do que fecundidades. Entretanto, esta é uma característica
inerente às propriedades matemáticas das matrizes de estágio com mais de uma classe
reprodutiva e sem reprodução clonal (Matos & Silva Matos 1998). Logo, os padrões aqui
encontrados são semelhantes a outros estudos com matrizes deste tipo para árvores
tropicais, como por exemplo, para Scheflera angustissima (Araliaceae) em Mata Atlântica
de São Paulo (Armelin 2005) e para Cecropia obtusifolia (Cecropiaceae) em floresta
tropical no México (Alvarez-Buylla 1994). Assim como para a espécie arbustivo-arbórea
Psychotria suterella (Rubiaceae) em Mata Atlântica de São Paulo, com matrizes de
estágios que incluem reprodução clonal e por sementes, Bertani (2006) também encontrou
que as permanências nas classes são responsáveis pelas maiores elasticidades. Segundo
Matos & Silva Matos (1998), estas propriedades têm conseqüências importantes no estudo
de sistemas biológicos, pois as elasticidades das fecundidades sempre serão menores do
que a maior transição entre classes, desta forma, as sensibilidades seriam
preferencialmente indicadas para comparar a influência de mudanças nos parâmetros
demográficos (Souza & Martins 2004).
LTREs – Experimentos de resposta da tabela de vida
As variações no λ não especificam as contribuições de cada classe ou taxa vital ao
incremento ou retração populacional, para isso podem ser utilizadas LTREs, que permitem
a comparação de taxas vitais mesmo que medidas em escalas diferentes (Caswell 1996).
Neste sentido, o pequeno incremento populacional observado de 2006 para 2007 (0,26%)
ocorreu principalmente devido às contribuições positivas dos crescimentos (0,0062) e
fecundidades (0,0045). Isto não foi observado de 2007 para 2008, onde houve uma

43
pequena retração da população (0,04%), pois as fecundidades contribuíram negativamente
(-0,0003) e os valores de crescimentos e permanências foram simétricos. Grandes
variações temporais nas taxas de fecundidade foram reportadas por Bruna & Oli (2005) em
cinco anos de estudos para a herbácea Heliconia acuminata na Amazônia. Em relação às
taxas demográficas, de 2006 para 2007, todas as classes contribuíram positivamente,
exceto JUVENIS 1 (-0,24%) e o oposto foi observado na comparação de 2007 para 2008,
onde apenas os JUVENIS 1 contribuíram positivamente (+0,1%). Apesar de reprodutivos
contribuírem positivamente com a fecundidade, estes, da mesma forma que os indivíduos
das demais classes, contribuem negativamente para o ∆λ ao permanecerem na mesma
classe, ou seja, indivíduos que não crescem não contribuem positivamente para o λ
populacional. Por outro lado, esta constatação aplica-se somente aos ADULTOS 1, pois
apenas estes podem crescer e mudar de classe uma vez que os ADULTOS 2 compõem a
última classe, estes não recrutam para classes superiores. Embora tenha ocorrido uma
pequena retração da população em 2008, as permanências nas classes foram menores. De
maneira geral, incrementos ou retrações do ‘fitness’ populacional (λ) devem-se a
compensações dos valores de maior magnitude, positivos ou negativos, sobre valores de
menor magnitude.

44
CONCLUSÕES
De acordo com os modelos matriciais estudados é possível concluir que Artocarpus
heterophyllus apresenta características de espécie invasora no ambiente de Mata Atlântica.
O ciclo de vida da espécie é típico de populações de árvores que apresentam baixas taxas
de crescimento, onde indivíduos permanecem alguns anos na mesma classe. A forma como
a população está estruturada após pelo menos 150 anos de introdução no Parque Nacional
da Tijuca indica um pequeno crescimento populacional anual e corrobora tal conclusão.
Adicionalmente, a atuação do homem como veículo dispersor devido aos frutos
comestíveis, também atrativos para a fauna de mamíferos, e as similaridades do ambiente
invadido com o habitat original são facilidades encontradas pela espécie na Mata Atlântica.
Ainda que a população de jaqueiras cresça em média menos de 1% ao ano, os padrões de
dinâmica populacional encontrados indicam que esta espécie exótica se comporta de
maneira similar às espécies nativas de Mata Atlântica, exibindo sucesso de estabelecimento
neste habitat.
Durante os anos de estudo, verificaram-se pequenas retrações e expansões
populacionais, consequentemente, as taxas demográficas e histórias de vida dos indivíduos
contribuíram de forma distinta nos diferentes anos. A emergência de novos padrões
populacionais capazes de preencher eventuais lacunas está condicionada ao
acompanhamento da dinâmica da população em longo-prazo.

45
REFERÊNCIAS BIBLIOGRÁFICAS
Abreu, R.C.R. 2005. Estrutura de populações de uma espécie exótica na Mata Atlântica: o
caso da potencial invasora Artocarpus heterophyllus L. (jaqueira) no Parque Nacional
da Tijuca – RJ. Monografia de Bacharelado em Ciências Biológicas. Universidade
Federal do Estado do Rio de Janeiro – UNIRIO.
Alvarez-Buylla, E.R. 1994. Density Dependence and Patch Dynamics in Tropical Rain
Forests: Matrix Models and Applications to a Tree Species. The American Naturalist
143(1):155-191.
Armelin, R.S. 2005. As dinâmicas de Schefflera angustissima (March.) Frodin (Araliaceae)
e de Andira anthelmia (Vell.) March. (Fabaceae) na reserva de Morro Grande, São
Paulo. Tese de Doutorado. Universidade de São Paulo, Instituto de Biociências.
Azevedo, D.A., Moreira, L.O., Siqueira D.S. 1999. Composition of extractable organic
matter in aerosols from urban areas of Rio de Janeiro city, Brazil. Atmospheric
Environment 33:4987-5001.
Bandeira, C.M. 1993. Parque Nacional da Tijuca. Markom Books, São Paulo.
Bertani, D.F. 2006. Ecologia de populações de Psychotria suterella Müll. Arg. (Rubiaceae)
em uma paisagem fragmentada de Mata Atlântica. Tese de Doutorado em Biologia
Vegetal. Universidade Estadual de Campinas – UNICAMP, São Paulo, Brasil.
Bhuyan, P.; Khan, M.L. & Tripathi, R.S. 2003. Tree diversity and population structure in
disturbed and human-impacted stands of tropical wet evergreen forest in Arunachal
Pradesh, Eastern Himalayas, India. Biodiversity and Conservation. 12: 1753-1773.
Bruna, E.M. & Oli, M.K. 2005. Demographic Effects of Habitat Fragmentation on a
Tropical Herb: Life-Table Response Experiments. Ecology 86: 1816-1824
Carauta, J.P.P. & Diaz, B.E. 2002. Figueiras no Brasil. Editora UFRJ, Rio de Janeiro, RJ.

46
Caswell, H. 1996. Analysis of life table response experiments II. Alternative
parameterizations for size- and stage-structured models. Ecological Modelling 88: 73-82
_______ 2000. Prospective and Retrospective perturbation analyses: their roles in
Conservation Biology. Ecology 81: 619-627.
_______ 2001. Matrix population models: construction, analysis and interpretation. 1st ed.
Sunderland, Massachussets, Sinauer Assoc. Inc.
_______ 2007. Sensivity analysis of transient population dynamics. Ecology Letters 10: 1-
15
Chaves, C.M.; Martins, H.F.; Carauta, J.P.P.; Lanna-Sobrinho, J.P.; Vianna, M.C. & Silva,
S.A.F. 1967. Arboreto Carioca 3. Centro de Conservação da Natureza – Rio de Janeiro
– Brasil – 28p +il.
Chittibabu, C.V. & Parthasarathy, N. 2000. Attenuated tree species diversity in human-
impacted tropical evergreen forest sites at Kolli hills, Eastern Ghats, India. Biodiversity
and Conservation 9: 1493–1519.
Coelho-Netto, A.L.; Avelar, A.S.; Fernandes, M.C. & Lacerda, W.A. 2007. Landslide
susceptibility in a mountainous geoecosystem, Tijuca Massif, Rio de Janeiro: the role of
morphometric subsivision of the terrain. Geomorphology 87:120-131.
Crane, J.H.; Balerdi, C.F. & Campbell, R.J. 2002. The Jackfruit (Artocarpus heterophyllus
Lam.) in Florida. University of Florida IFAS Extension. Fact Sheet HS-882.
http://edis.ifas.ufl.edu
Cronk, Q.C.B. & Fuller, J.L. 1995. Plant Invaders. Chapman & Hall, London, UK.
Cunha, A.; Vieira, M.V. & Grelle, C.E.V. 2006. Preliminary observations on habitat,
support use and diet in two non-native primates in an urban Atlantic forest fragment:
The capuchin monkey (Cebus sp.) and the common marmoset (Callithrix jacchus) in the
Tijuca forest, Rio de Janeiro. Urban Ecosystems 9(4): 351-359.

47
de Kroon, H.; Plaisier, A.; van Groenendael, J. & Caswell, H. 1986. Elasticity: the relative
contribution of demographic parameters to population growth rate. Ecology 67: 1427-
1431.
de Kroon, H.; Groenendael, J. van; Ehrlén, J. 2000. Elasticities: A review of methods and
model limitations. Ecology 81:607-618.
Dean, W. 2002. A Ferro e Fogo: a história e a devastação da Mata Atlântica brasileira.
Companhia das Letras, São Paulo.
Enright, N.J., Franco, M. & Silvertown, J. 1995. Comparing plant life histories using
elasticity analysis: the importance of life span and the number of life-cycle stages.
Oecologia 104: 79-84.
Ferrão, J.E.M. 1993. A aventura das plantas e os descobrimentos portugueses. 2ª ed.,
Lisboa, Instituto de Investigaçäo Científica Tropical.
Fox, G.A. & Gurevitch, J. 2000. Population numbers count: Tools for near-term
demographic analysis. The American Naturalist 156:242-256.
IBAMA, 2004. Proposta de ampliação e correção de limites do Parque Nacional da Tijuca.
Gestão compartilhada IBAMA e Prefeitura da Cidade do Rio de Janeiro.
IBGE, 2006. http://www.ibge.gov.br/cidadesat/topwindow.htm?1 - acessado em junho de
2006.
Khan, M.L. 2004. Effects of seed mass on seedling success in Artocarpus heterophyllus L.,
a tropical tree species of north-east India Acta Oecologica 25 : 103-110.
Lefkovitch, L.P. 1965. The study of population growth in organisms grouped by stages.
Biometrics 21:1-18.
Mantovani, W. 2003. Ecologia da floresta pluvial Atlântica. In: Vanda Carneiro de
Claudino Sales. (orgs). Ecossistemas brasileiros: manejo e conservação. Fortaleza, p.
265-278.

48
Matos, M.B. de & Silva-Matos, D.M. 1998. Mathematical constraints on transition matrix
elasticity analysis. Journal of Ecology 86: 706-708
Mattos, C.C.L.V. de; Mattos, M. D. L. V. de & Laroche R.C. 1970. Aspectos do Clima e
da Flora do Parque Nacional da Tijuca. Brasil Florestal 1: 5-12.
Mesterton-Gibbons, M. 1993. Why demographic elasticities sum to one: a postscript to de
Kroon et al. Ecology 74: 2467-2468.
Morris, W.F. & Doak, D.F. 2002. Quantitative Conservation Biology: Theory and Practice
of Population Viability Analysis. Sinauer Associates Inc., 480 pp.
Neubert, M.G. & Caswell, H. 2000. Demography and Dispersal: calculation and sensitivity
analysis of invasion speed for structured populations. Ecology 81: 1613-1628.
Pereira, T.S. 1998. Ecologia de Miconia cinnamomifolia (DC.) Naudin - Jacatirão na
sucessão secundária da Mata Atlântica. Tese de Doutorado. Universidade de São Paulo
– USP.
Primack, R.B.; Ashton, P.S.; Chai, P. & Lee, H.S. 1985. Growth rates and population
structure of Moraceae Trees in Sarawak, East Malaysia. Ecology 66: 577-588.
Richardson, D.M.; Pysek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D. & West, C.J.
2000. Naturalization and invasion of alien plants: concepts and definitions. Diversity
and Distributions 6: 93-107.
Rodrigues, P.J.F.P. 2004. A vegetação da Reserva Biológica União e os efeitos de borda na
Mata Atlântica fragmentada. Tese de Doutorado. Universidade Estadual do Norte
Fluminense – UENF. Campos dos Goytacazes, RJ.
Silva-Mattos, D.M.; Freckleton, R.P. & Watkinson, A.R. 1999. The role of density
dependence in the population dynamics of a tropical palm. Ecology 80:2635-2650.

49
Silvertown, J.; Franco, M. & Menges, E. 1996. Interpretation of Elasticity Matrices as an
Aid to the Managment of Plant Populations for Conservation. Conservation Biology 10:
591-597.
__________; Franco, M.; Pisanty, I. & Mendoza, A. 1993. Comparative plant demography
- relative importance of life-cicle components to the finite rate of increase in woody and
herbaceous perennials. Journal of Ecology 81: 465-476.
Sodhi, N.S.; Peh, K.S.H.; Lee, T.M.; Turner, I.M.; Tan, H.T.W.; Prawiradilaga, D.M. &
Darjono, 2003. Artificial nest and seed predation experiments on tropical southeast
Asian islands. Biodiversity and Conservation 12: 2415-2433.
Souza, A.F. & Martins, F.R. 2004. Population structure and dynamics of a neotropical
palm in fire-impacted fragments of the Brazilian Atlantic Forest. Biodiversity and
Conservation 13: 1611-1632
Williamson, M. 1996. Biological Invasions. Chapman & Hall. UK.
Tilman, D. 1988. Plant strategies and the structure of plant communities. Princeton, New
Jersey. Princeton University Press.
Zuidema, P.A. & Franco, M. 2001. Integrating vital rate variability into perturbation
analysis: an evaluation for matrix population models of six plant species. Journal of
Ecology 89: 995-1005

50
CAPÍTULO 2
SIMULAÇÕES DE MANEJO ATRAVÉS DA MODELAGEM POPULACIONAL DE
UMA ÁRVORE INVASORA NO PARQUE NACIONAL DA TIJUCA – RJ

51
RESUMO
Artocarpus heterophyllus Lamk (Moraceae), uma espécie arbórea nativa do sudeste
asiático, atualmente é considerada espécie invasora no Parque Nacional da Tijuca (PNT),
região de Mata Atlântica do Rio de Janeiro. A dominância da espécie invasora em alguns
trechos, como no caso da jaqueira, e a conseqüente substituição da comunidade nativa
apresentam evidências suficientes para justificar o manejo da espécie. Atualmente em fase de
dinâmica transiente, a população de jaqueiras no PNT atingiria a fase assintótica somente
daqui a 179 anos. Desta forma, este estudo analisou a fase transiente desta invasora
historicamente estabelecida na região. Foram sugeridas formas de manejo considerando-se os
possíveis impactos causados nas comunidades de fauna e flora locais. Para isto, foram
caracterizados o ciclo de vida e as taxas vitais da espécie. Na análise de perturbação, as
sensibilidades das taxas vitais indicaram que um manejo voltado para sobrevivência e
crescimento de JUVENIS 1 e de ADULTOS 1 seria bastante eficiente na redução do λ
populacional. Já as análises de elasticidades indicaram que as melhores opções de manejo
seriam baseadas nas taxas de sobrevivência, destas classes e dos ADULTOS 2. As simulações
do manejo indicaram que pequenas intervenções seriam suficientes para que a população
entrasse em declínio, reduzindo-se em até 5% as sobrevivências das classes, principalmente
nas classes reprodutivas. O manejo feito de forma gradativa e no longo prazo mostrou-se mais
eficiente do que altas intervenções. Desta forma, intervir demasiadamente seria desnecessário
e poderia resultar no desperdício de recursos, além de dificultar a colonização do ambiente por
espécies nativas.
Palavras-chave: Artocarpus heterophyllus, dinâmica transiente, sensibilidades, elasticidades,
manejo, espécie invasora, Parque Nacional da Tijuca, Mata Atlântica.

52
ABSTRACT
Artocarpus heterophyllus Lamk (Moraceae) is a woody tree native from Southeast
Asia and nowadays is considered as an invasive species at the Atlantic Forest from Parque
Nacional da Tijuca – RJ (PNT). The species dominates forest places affecting ecosystem
composition. This population is currently on transient dynamic phase and would take 179
years to reach stable-stage structure. In this way, this work focuses on transient dynamics from
A. heterophyllus population. Environmental management actions that take in account the
impact over local fauna and flora communities have been suggested. For this species life cycle
and vital rates were characterized. On perturbation analysis, sensitivities from vital rates
shows that the best options are the management of survival and growth rates from
JUVENILES 1 and ADULTS 1. Elasticities analysis indicates best management options
regarding survival rates are the classes cited above and ADULTS 2 class. Management
simulations support that small interventions are better than big ones and reductions up to 5%
on survival rates are enough to population declines. Gradual and long-term management
shows better results than intense management. In this sense, too many interventions are
unnecessary and can expense resources besides representing an environmental barrier for
colonization by native plant species.
Key–words: Artocarpus heterophyllus, transient dynamics, sensitivities, elasticities,
environmental management, invasive specie, Parque Nacional da Tijuca, Atlantic Forest.

53
INTRODUÇÃO
Artocarpus heterophyllus Lamk (Moraceae) atualmente é considerada espécie invasora
no Parque Nacional da Tijuca (PNT), uma região de Mata Atlântica do Rio de Janeiro (ver
capítulo 1). A espécie foi introduzida historicamente na cidade do Rio de Janeiro por volta do
ano de 1811 (Atala et al. 1966), e na área do atual PNT em meados de 1800, há
aproximadamente 150 anos (Bandeira 1993) provavelmente a partir do reflorestamento
realizado por Manuel Gomes Archer e Thomas Nogueira da Gama nas encostas da então
Floresta da Tijuca e da Floresta das Paineiras (Pádua & Coimbra-Filho 1979). Estes locais
haviam sido desmatados para o cultivo do café (Coffea arabica L.) e neste reflorestamento
foram utilizadas espécies nativas e exóticas consorciadas (Bandeira 1993; Dean 2002; Ferrão
1993; Pádua & Coimbra-Filho 1979). Outro fator que contribuiu para a introdução da espécie
no Brasil foi a política da Metrópole portuguesa ao incentivar a troca de espécies entre as
Colônias, uma vez que já era conhecido o fato de que a produtividade poderia ser maior em
regiões introduzidas (Ferrão 1993).
Atualmente no PNT são observados locais com monodominância de jaqueiras em
todos os estratos da mata, desde plântulas até adultos reprodutivos, com sub-populações que
gradativamente se expandem e crescem suprimindo espécies da flora nativa (ver capítulo 1). O
sucesso na colonização deste novo ambiente permitiu que a espécie desenvolvesse complexas
interações ecológicas com a fauna local, constituindo inclusive uma importante fonte de
recursos para mamíferos (Cunha et al. 2006). Por outro lado, a alteração promovida e a
possível exclusão competitiva da espécie invasora sobre as espécies da flora nativa também
podem afetar indireta e negativamente outros animais nativos comprometendo a
biodiversidade local. Adicionalmente, é provável que as raízes destas árvores desempenhem
papel importante na contenção de horizontes de solos (Rogério Ribeiro comunicação pessoal).

54
Espécies exóticas invasoras são reconhecidas mundialmente como espécies que
causam problemas aos ecossistemas que colonizam (Cronk & Fuller 1995; Williamson 1996).
Desta forma, intervenções para a retirada destas espécies dos habitats invadidos estão se
tornando ferramentas essenciais para manutenção da biodiversidade local e global (Wilcove &
Chen 1998). Como medida assessória ao desenvolvimento de programas de controle destas
espécies são aplicados modelos matemáticos (DeWalt 2006; Parker 2000; Shea & Kelly 1998).
Estes têm exercido papel fundamental na previsibilidade das respostas dos organismos, e até
mesmo dos ecossistemas, ao tipo de manejo aplicado (Caswell 1989). Mediante simples
mudanças nos objetivos a serem alcançados, os mesmos modelos utilizados na conservação de
espécies ameaçadas de extinção podem ser utilizados no manejo de espécies exóticas (Caswell
1989; 2007; Morris & Doak 2002). Por exemplo, para conservar uma espécie em extinção o
objetivo almejado seria uma forma de aumentar a taxa de crescimento finito da população
(representado por lambda – λ), em contraste, para controlar uma espécie exótica, um manejo
que vise a redução do λ deve ser aplicado (Morris & Doak 2002). Neste contexto, modelos
matriciais podem ser utilizados para estes fins, pois permitem a aplicação de análises
prospectivas (Armelin 2005; Caswell 2000; 2007; de Kroon et al. 1986; de Kroon et al. 2000;
Fox & Gurevitch 2000), ou seja, de projeções teóricas da intervenção feita na população em
tempo futuro pré-determinado.
As análises de modelos matriciais de populações estruturadas comumente são
elaboradas assumindo-se que a população atingiu a fase de dinâmica assintótica (Caswell 2007;
Fox & Gurevitch 2000; Yearsley 2004), o que implica em assumir que a população tornou-se
estável. A fase assintótica é uma fase estacionária onde a população atingiu o equilíbrio. Isto
não indica que o crescimento populacional cessou, mas que o autovalor dominante (λ) tornou-
se constante e desta forma, as proporções obtidas entre as classes da variável-estado são

55
mantidas (Morris & Doak 2002). Porém, populações de arvóres tropicais podem levar anos
para atingir a fase assintótica, como por exemplo o período de 256 anos, encontrado para a
palmeira Astrocaryum mexicanum em floresta tropical no México (Piñero et al. 1984). A fase
que precede tal dinâmica é conhecida como fase de dinâmica transiente. Desta forma, a
dinâmica transiente e a dinâmica assintótica possuem comportamentos distintos que requerem
análises diferentes (Fox & Gurevitch 2000). A população de jaqueiras no PNT encontra-se
atualmente em fase de dinâmica transiente e os modelos matriciais determinísticos apontaram
um período de 179 anos até esta população atingir a fase assintótica (ver capítulo 1). Logo, é
recomendado que as ações de manejo priorizem o período de dinâmica transiente da espécie.
Este estudo tem como objetivo principal a detecção da(s) forma(s) de manejo mais
indicada(s) para esta espécie exótica, que atualmente compõe a comunidade do PNT. E
procurou responder às seguintes questões: 1) Qual a(s) taxa(s) vital(is) que mais infuencia(m)
o crescimento populacional? 2) Ao simular um manejo da espécie, em que taxas vitais são
notadas as maiores mudanças? 3) Qual o nível de intervenção necessário para um manejo
efetivo? E a partir de que intensidade as intervenções tornam-se pouco eficazes e passam a ser
desperdício de recursos?

56
MATERIAIS E MÉTODOS
Área de Estudo
O clima do PNT é classificado como Tropical úmido, e de acordo com a classificação
de Köppen, nas regiões mais altas é do tipo Cf e nas regiões mais baixas tipo Am. A
precipitação anual varia de 1300 a pouco mais de 3000 mm, com precipitações abundantes
observadas inclusive no inverno (Mattos et al. 1976) e chuvas torrenciais associadas a
temperaturas quentes nos meses de verão (dez-fev) (Coelho-Netto et al. 2007). Litossolos,
cambissolos e latossolos prevalecem na porção superior e solos podozolicos na porção inferior
(Coelho-Netto et al. 2007).
O PNT é considerado uma das maiores florestas urbanas do mundo sendo procurado
por muitos turistas e pela população local como área de lazer (IBAMA 2004). Porém, o Rio de
Janeiro é o segundo centro econômico e segunda cidade mais populosa do Brasil, com
aproximadamente 6.000.000 de habitantes (IBGE 2006), o que faz com que o PNT
experimente alguns impactos como o crescimento urbano desordenado em suas regiões
limítrofes (Coelho-Netto et al. 2007), a poluição atmosférica (Azevedo et al. 1999) e a
presença de espécies não-nativas de fauna (Cunha et al. 2006) e flora (Bandeira 1993),
introduzidas de forma acidental ou intencional.
A espécie
Artocarpus heterophyllus Lamarck, popularmente chamada de jaqueira é uma espécie
originária do Sudeste Asiático (Chaves et al. 1969; Ferrão 1993). A introdução desta espécie
no Brasil ocorreu durante o período colonial, onde para cumprir instruções Régias, os Vice-
Reis da Índia deveriam enviar periodicamente para Portugal (metrópole), África e Brasil
(colônias), plantas de interesse para tentar introduzi-las nestas novas regiões (Ferrão 1993). No

57
ano de 1682, no estado da Bahia, já havia 11 jaqueiras de procedência desconhecida. Porém, o
primeiro registro de envio para o Brasil data de janeiro de 1683 (Ferrão 1993). As recém-
chegadas sementes e mudas eram enviadas para um horto na Bahia para que fossem
aclimatadas e posteriormente disseminadas para outras regiões do País (Dean 2002). O Rio de
Janeiro, em 1803, já possuía exemplares aparentemente naturalizados (Chaves et al. 1969).
Considerada uma árvore de grande porte, atinge mais de 10 m de altura, seu tronco
pode ultrapassar 1 m de diâmetro, é monóica e exibe caulifloria (Chaves et al. 1969; Crane et
al. 2002). Podem ser polinizadas por insetos ou pelo vento, resistem bem a estresses
climáticos e em condições ideais de cultivo, podem frutificar em 3 ou 4 anos (Crane et al.
2002). No Brasil, uma jaqueira bem desenvolvida chega a produzir até 100 frutos por ano,
alguns destes chegando a pesar mais de 30 quilos (Chaves et al. 1969). Indivíduos da espécie
frutificam assincronicamente durante todo o ano, com grande parte dos indivíduos frutificando
preferencialmente nos meses de verão (dez-fev). Além disso, os frutos são consumidos pela
população humana e pela fauna de mamíferos, como roedores e primatas (Cunha et al. 2006).
Atualmente sua distribuição abrange praticamente todo território Nacional, com certa
amplitude de habitats, podendo ser observada colonizando áreas abertas e áreas de mata,
ambas associadas a ambientes antrópicos (Carauta & Diaz 2002). No Rio de Janeiro, coloniza
densamente áreas de borda de Mata Atlântica sendo encontrada em algumas Unidades de
Conservação tais como Parque Nacional da Tijuca e as Reservas Biológicas de Poço das Antas,
da União e do Tinguá (Abreu 2005).
No habitat original
Nativa do sudeste asiático, A. heterophyllus é típica de estágios sucessionais avançados
e cresce naturalmente até 1300 m de altitude no sub-bosque da Floresta Tropical Úmida

58
Monodominante, todavia a espécie é considerada rara (até 1 indivíduo por ha) neste ambiente,
onde poucas espécies dominam os diferentes estratos da mata (Bhuyan et al. 2003; Chittibabu
& Parthasarathy 2000). É tolerante a sombra, porém necessita de um pouco de luz e espaço
para seu desenvolvimento durante os estágios iniciais de vida, germinando melhor em
clareiras. Os frutos amadurecem entre julho e agosto e o número de sementes varia em
tamanho e massa (1,5 – 14 g), entretanto, as sementes viáveis variam de 4 a 14g de massa, e
dentre seus dispersores são mencionados roedores, macacos e porcos selvagens (Khan 2004).
As sementes possuem taxa de predação relativamente alta (Sodhi et al. 2003) e germinam
ainda na estação chuvosa, durante o verão (maio-setembro), onde ocorre 85% da precipitação
anual (Bhuyan et al. 2003; Chittibabu & Parthasarathy 2000). Desta forma, as sementes
germinam e suas plântulas se estabelecem, tornando-se capazes de tolerar bem a estação seca
[dezembro – fevereiro] (Bhuyan et al. 2003; Chittibabu & Parthasarathy 2000).
Amostragem das sub-populações de A. heterophyllus no PNT
Em 2001 foi realizado um estudo piloto nas Unidades de Conservação do Rio de
Janeiro: Reserva Biológica União, Tinguá e Parque Nacional da Tijuca, visando definir a
forma e o tamanho adequado das parcelas e as áreas prioritárias para a realização de estudos
com as populações da espécie exótica (Abreu 2005). Após este estudo, em 2004, optou-se pela
marcação de 20 parcelas permanentes em sub-populações de jaqueiras distribuídas pelo PNT.
Foram marcadas parcelas circulares com 10 m de raio (314 m2 cada) totalizando uma área
amostral de 0,628 ha. Nestas parcelas todos os indivíduos da comunidade vegetal com
diâmetro a altura do peito (DAP) ≥ 5 cm foram marcados e mensurados. Indivíduos com
troncos múltiplos visivelmente interligados foram considerados um único indivíduo. Nestes
casos, cada tronco foi medido separadamente e posteriormente somaram-se suas áreas basais,

59
e da área basal resultante, obteve-se um único diâmetro virtual. Todos os indivíduos foram
mapeados dentro das sub-populações e a presença de estruturas reprodutivas foi verificada.
Todos os indivíduos foram marcados e mensurados no período de novembro de 2004 a
janeiro de 2005 e tiveram o DAP remensurado anualmente nos meses de janeiro de 2006 e
2007, e dezembro de 2007, que daqui em diante será referido como remedição de 2008.
Indivíduos que obedeciam ao critério de inclusão adotado, ou seja, atingiram DAP de 5 cm,
foram marcados e incluídos na amostragem durante os recensos. Excepcionalmente no ano de
2006, por problemas de impossibilidade de acesso à algumas áreas de estudo, apenas 10
parcelas foram remensuradas, nos outros anos todas as 20 parcelas foram consideradas.
Modelo Matricial
Matrizes de transição – Foi construída uma matriz de transição At, quadrada e de
ordem i x j, com i = j e representando o número de classes da população. A matriz de transição
contém as probabilidades de transição de uma classe para outra no período t (Caswell 1989),
neste caso t = 1 ano. Para o cálculo dos parâmetros dos ciclos de vida desejados foram
utilizados os valores médios dos dados anuais de 2006 a 2008. Considera-se j como a classe
de origem da planta no início do período t e i a classe de destino dessa mesma planta no final
deste período. As entradas relativas às transições foram calculadas dividindo-se o número de
plantas em ij pelo total de plantas em j.
Uma vez que este trabalho não contemplou classes de diâmetros menores do que 5 cm,
as entradas ij relativas a fecundidade foram estimadas a partir do número de recrutas que
entravam na amostragem em cada recenso. Assumiu-se não ser possível relacionar o indivíduo
recruta à planta-mãe e que a espécie estudada, por frutificar durante quase todo o ano, tem

60
reprodução em fluxo contínuo. Desta forma, o cálculo da fecundidade procedeu-se pela
fórmula:
F = √sj . fj . √s0 (Morris & Doak 2002), onde:
F = fecundidade;
sj = probabilidade de um indivíduo da classe j (ADULTO 1 ou 2, neste caso), sobreviver por
metade do tempo do próximo recenso, ou seja, por seis meses (t = 1 ano);
fj = número de recrutas por adulto da classe;
s0 = probabilidade de um recruta sobreviver por metade do tempo do próximo recenso;
Após a obtenção da fecundidade, para se obter a fecundidade por classe de adultos,
multiplicou-se F pelo peso arbitrário dado às mesmas, peso de 0,4 para ADULTOS 1, por
serem adultos menores e 0,6 para ADULTOS 2, por estes serem maiores e aparentemente
frutificarem mais.
Foi calculado o autovalor dominante da matriz de Lefkovitch (ou matriz de estágios)
que pode ser considerado uma medida de ‘fitness’ da população e expressa a taxa de
crescimento, representado por λ (lambda) e seu respectivo intevalo de confiança com base em
reamostragens aleatórias (bootstrap das histórias de vida) ao nível de 5% de significância.
AJUSTES – Eventos como a mortalidade de indivíduos ADULTOS 2 não foram
detectados durante o período de estudo. Uma vez que a mortalidade na última classe é um
evento necessário para fechar o ciclo de vida da espécie, foi incluída no modelo a mortalidade
artificial de um indivíduo da última classe no último ano de estudo (2008) assim como em
outros trabalhos (Armelin 2005; Caswell 1989).

61
Taxas vitais
As taxas vitais foram extraídas a partir da matriz de transição A. As sobrevivências
(“s”) equivaleram às somas das colunas por classe (permanências + transições para classes
superiores), excetuando-se as fecundidades. Já os crescimentos, corresponderam a parcela de
indivíduos que passaram a classes superiores divididos pela sobrevivência total da classe. A
fecundidade foi a mesma descrita na construção da matriz (ver tópico “modelo matricial”).
Sensibilidades e Elasticidades da fase transiente
As análises da fase assintótica consideram apenas os autovalores e autovetores
dominantes, já as análises da fase transiente trabalham com todos os autovalores e autovetores
obtidos nos cálculos dos modelos matriciais, não apenas com os dominantes, pois durante a
transiência é importante a contribuição de todos os autovalores e autovetores (Fox &
Gurevitch 2000). Desta maneira, as análises de sensibilidades e as de elasticidades são
baseadas nas contribuições de todos os autovalores e autovetores obtidos a partir da matriz A
(Caswell 2007; Fox & Gurevitch 2000; Yearsley 2004).
Etapas dos cálculos das sensibilidades e elasticidades na fase transiente podem ser
observadas no trabalho de Caswell (2007), onde também são fornecidos os algoritmos do
programa MATLAB® para realização destes. Os algoritmos aqui utilizados foram adaptados
para espécie estudada (Renato Armelin comunicação pessoal) e podem ser obtidos mediante
requisição ao autor.

62
Simulações do manejo
Para verificar as taxas vitais que mais influenciaram a taxa de crescimento da
população, foram feitas análises prospectivas com simulações da retirada de indivíduos da
população, desta forma, ao suprimir as taxas vitais reduz-se o número de indivíduos. Para isto,
cada taxa de transição da matriz A foi recalibrada individualmente com a retirada de
indivíduos nas proporções desejadas e, posteriormente, o λ foi recalculado. Desta forma foi
possível estimar a maneira como cada taxa vital influencia o comportamento do λ.
Projeções temporais das Elasticidades
A partir das elasticidades obtidas em relação às taxas vitais foram selecionadas as
projeções temporais t+1, t+5, t+10 e a fase assintótica. Tais projeções foram
proporcionalizadas, em valores percentuais, para que pudessem ser plotadas na mesma escala.

63
RESULTADOS
Ciclo de vida
A matriz de estágios das jaqueiras no PNT exibiu a alta permanência dos indivíduos
nas classes no período de 1 ano (Figura 2.1). O crescimento para classes superiores variou em
torno de 2 a 6%. As fecundidades não foram expressas em unidades percentuais, portanto não
puderam ser comparadas com permanências e crescimentos por estarem em escalas diferentes.
O autovalor dominante obtido foi λ=1,0021 com intervalo de confiança (0,9926 – 1,0079).
0,9466 0 0 0,0181 0,0269 0,0331 0,9396 0 0 0
A = 0 0,0573 0,9627 0 0 0 0 0,0311 0,9752 0 0 0 0 0,0198 0,9909
Taxas vitais
Ao longo do ciclo de vida, as jaqueiras apresentaram altas taxas de sobrevivência e
baixas taxas de crescimento. Todas as sobrevivências foram superiores a aproximadamente
98%. A classe JUVENIS 2 exibiu o maior crescimento para classe superior (Tabela 2.1).
Tabela 2.1 – Taxas vitais de Artocarpus heterophyllus no PNT. Crescimentos estão incluídos nas sobrevivências.
JUVENIS 1 JUVENIS 2 PRÉ-REP. ADULTOS 1 ADULTOS 2 Sobrevivência 0,9797 0,9969 0,9938 0,9950 0,9909 Crescimento 0,0338 0,0575 0,0313 0,0199 -
Figura 2.1 – Matriz de estágios (Lefkovitch 1965) de A. heterophyllus no PNT para o período 2005 – 2008.

64
Sensibilidades e Elasticidades da fase transiente
As sensibilidades da estrutura populacional em t+1 indicaram que pequenas mudanças
nas taxas vitais associadas respectivamente aos JUVENIS 1 (s1 e g21) e aos ADULTOS 1
podem causar grandes variações no λ populacional (Tabela 2.2).
Tabela 2.2 – Sensibilidades da estrutura populacional de A. heterophyllus em relação às taxas vitais na fase de dinâmica transiente em 2008 – 2009 (t+1). J1 – Juvenis 1; J2 – Juvenis 2; PR – Pré-Reprodutivos; A1 – Adultos 1; A2 – Adultos 2; Taxas vitais: s = sobrevivência, g = crescimento, f = fecundidade.
Ao proporcionalizar as sensibilidades, as sobrevivências foram as maiores
responsáveis por alterações no λ (Tabela 2.3). Tratando-se destas elasticidades, destacaram-se
as contribuições dos JUVENIS 1 (s1), ADULTOS 2 (s5) e ADULTOS 1 (s4).
Tabela 2.3 – Elasticidades da estrutura populacional de A. heterophyllus em relação às taxas vitais na fase de dinâmica transiente em 2008 – 2009 (t+1). J1 – Juvenis 1; J2 – Juvenis 2; PR – Pré-Reprodutivos; A1 – Adultos 1; A2 – Adultos 2; Taxas vitais: s = sobrevivência, g = crescimento, f = fecundidade. T+1 s1 s2 s3 s4 s5 g1 g2 g3 g4 f
J 1 1,0665 0,0833 -0,0344 0,1666 J 2 0,0825 0,9175 0,0825 -0,0560 PR 0,0550 0,945 0,0550 -0,0305 A 1 0,0255 0,9745 0,0255 -0,0198 A2 0,0347 0,9653 0,0347 0,0000
t +1 s1 s2 s3 s4 s5 g21 g32 g43 g54 f
J 1 227,427 17,561 -212,595 76,655 J 2 7,335 80,113 212,595 -84,737 PR 4,888 84,277 84,737 -86,461 A 1 2,723 103,891 86,461 -105,47 A 2 2,109 59,000 105,47 0,000

65
Simulações do manejo
As taxas vitas relativas às sobrevivências dos indivíduos nas classes (“s”) foram as que
mais influenciaram na redução do fitness populacional, já as transições para classes superiores
(“g”) e a fecundidade (“f”) pouco influenciaram a variação do λ (Figura 2.2). De maneira geral,
partindo-se do λ atual de 1,0021, a simulação da intensidade do manejo revelou que pequenas
intervenções podem ser mais eficientes do que grandes intervenções. Tratando-se da
fecundidade (f) e do crescimento até a primeira classe reprodutiva (g21, g32 e g43), as
intervenções só começam a reduzir o λ para valores abaixo de 1 quando são removidos
aproximadamente 35% dos indivíduos. Quando considerado apenas o crescimento para a
última classe reprodutiva (g54), o efeito desejado da redução do λ abaixo de 1 surge somente
depois da retirada de ≈ 40% dos indivíduos. Tratando-se das sobrevivências nas classes (s1 –
s5) grandes efeitos são notados com a redução de apenas 5% dos indivíduos.
Figura 2.2 – Simulações das reduções nas taxas vitais. Onde: s – sobreviência (permanências + crescimentos) dos indivíduos nas classes; g – crescimento para classe seguinte; f – fecundidade (entrada de novos indivíduos DAP ≥ 5cm);
0,9900
0,9920
0,9940
0,9960
0,9980
1,0000
1,0020
1,0040
0% 20% 40% 60% 80% 100% Reduções nas taxas vitais
Lam
bd
a (λ)
s1 s2 s3 s4 s5 g(21;32;43) g54 f
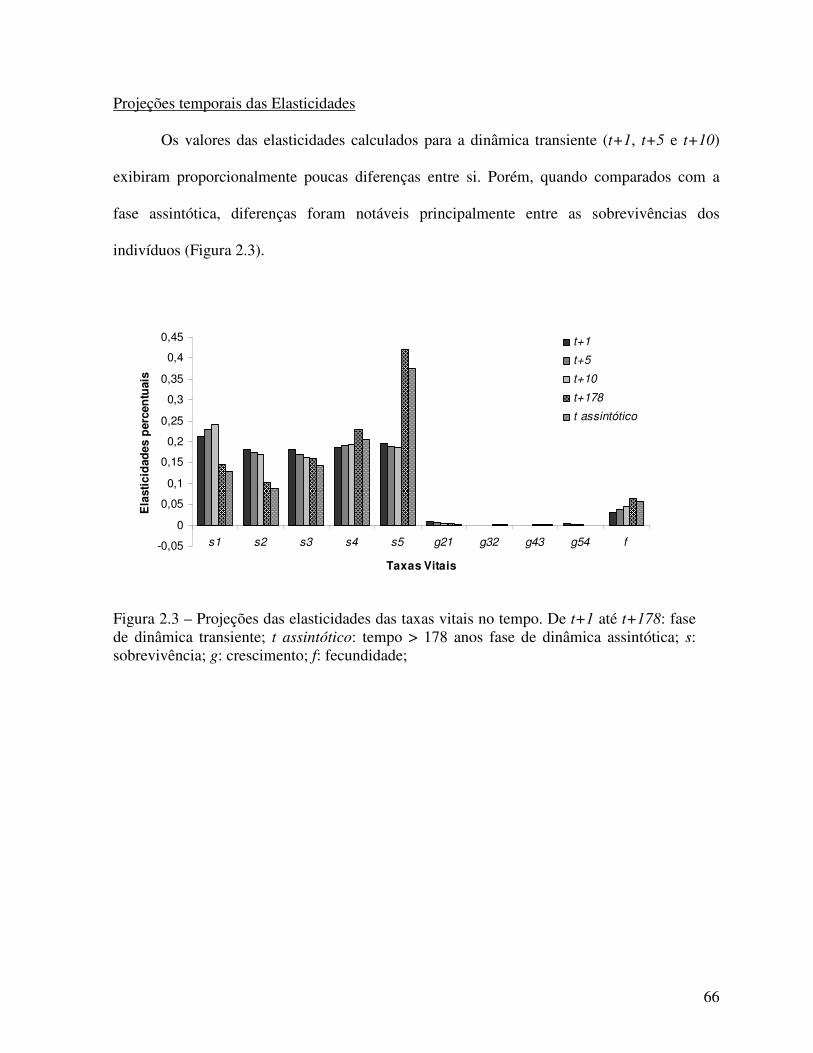
66
Projeções temporais das Elasticidades
Os valores das elasticidades calculados para a dinâmica transiente (t+1, t+5 e t+10)
exibiram proporcionalmente poucas diferenças entre si. Porém, quando comparados com a
fase assintótica, diferenças foram notáveis principalmente entre as sobrevivências dos
indivíduos (Figura 2.3).
-0,05
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,45
s1 s2 s3 s4 s5 g21 g32 g43 g54 f
Taxas Vitais
Ela
sti
cid
ad
es p
erc
en
tuais
t+1
t+5
t+10
t+178
t assintótico
Figura 2.3 – Projeções das elasticidades das taxas vitais no tempo. De t+1 até t+178: fase de dinâmica transiente; t assintótico: tempo > 178 anos fase de dinâmica assintótica; s: sobrevivência; g: crescimento; f: fecundidade;

67
DISCUSSÃO
De acordo com o Sistema Nacional de Unidades de Conservação (SNUC), o PNT está
categorizado como uma Unidade de Proteção Integral. Estas unidades têm como principal
objetivo a preservação da flora e fauna nativas do ecossistema em questão, neste caso a Mata
Atlântica. Conforme mencionado, a invasão e a conseqüente substituição da comunidade
nativa apresentam evidências suficientes para justificar o manejo de espécies capazes de
dominar a comunidade local (Blossey 1999). Porém quando os efeitos diretos provocados
pelos invasores não são notados, o controle não costuma ser considerado uma alternativa
(Courchamp et al. 1999). Desta maneira, a forma gradativa como as populações de jaqueiras
colonizam o ambiente, e o lento crescimento típico de espécies arbóreas (Silvertown et al.
1993), podem fazer com que os efeitos provocados por estas plantas no ecossistema demorem
a ser percebidos.
Neste aspecto, De Wit et al. (2001) analisaram distintas possibilidades de manejo para
a espécie invasora Acacia mearnsii De Wild. na África do Sul e verificaram que a opção de
não manejar mostra-se, em médio e longo-prazos, insustentável para a natureza. Ao contrastar
esta opção com diferentes técnicas de manejo, combinadas ou separadas, resultados benéficos
foram pelo menos quatro vezes maiores do que os custos (De Wit et al. 2001). Porém, o
sucesso de um programa de manejo de espécies invasoras depende essencialmente de
planejamento e do conhecimento ecológico acerca da espécie-alvo, uma vez que a utilização
de técnicas de controle inadequadas ou aplicadas por um período menor do que o necessário
pode comprometer a credibilidade destes programas, tornando-os extremamente ineficientes e
onerosos (Starfinger et al. 2003; Zavaleta et al. 2001). Além disso, como fator agravante, pode
ocorrer a re-invasão de locais previamente manejados (Panetta & Sparkes 2001).

68
Tratando da necessidade de manejo das jaqueiras no PNT, na análise de perturbação,
as sensibilidades da fase transiente da estrutura populacional em t+1 indicaram que um
manejo voltado para a classe de JUVENIS 1 e/ou de ADULTOS 1 produziria grandes
alterações no crescimento populacional, pois as sensibilidades das taxas vitais relativas à
sobrevivência e ao crescimento destas classes apresentaram altos valores (Tabela 2.2). Já as
análises de elasticidades indicaram que as melhores opções de manejos seriam baseadas nas
taxas de sobrevivência, principalmente para os JUVENIS 1, ADULTOS 2 e ADULTOS 1,
nesta ordem (Tabela 2.3). As elasticidades indicaram também que um manejo baseado apenas
nas fecundidades, como por exemplo, com enfoque apenas na remoção de frutos ou na
remoção dos JUVENIS 1 que entram anualmente na amostra (como no modelo estudado),
além de ser dispendioso em diversos sentidos (mão-de-obra, veículos especiais, recursos
financeiros) seria pouco eficaz na redução do λ. Ou seja, os custos econômicos seriam
superiores aos ganhos ambientais.
As simulações do manejo indicaram que pequenas intervenções foram mais eficientes
do que grandes esforços de controle, ou seja, a redução em até 30% das sobrevivências dos
indivíduos de cada classe, produz reduções no λ similares à retirada, por exemplo, de 80% dos
indivíduos. Além disso, uma intervenção de até 5% mostrou-se bastante eficaz (Figura 2.2).
Isto ocorre porque cada taxa vital possui uma relação particular com a taxa de crescimento
assintótico da população e tais relações não são lineares (Morris & Doak 2002). Conforme
indicado pelas simulações, as taxas que mais influenciaram a redução do λ foram as
sobreviências nas classes, principalmente nas classes reprodutivas. Porém o manejo destas
classes deve ser feito com cautela uma vez que a retirada de ADULTOS implica em uma
diminuição quase imediata da disponibilidade de frutos para a fauna local, enfatizando
portanto, a importância do manejo ser feito gradativamente e no longo prazo.

69
De acordo com as condições iniciais da população estudada e com a duração da
respectiva fase transiente, o resultado de análises de sensibilidades e de elasticidades baseados
na fase assintótica pode diferir dos resultados das mesmas análises feitas durante a fase
transiente (Caswell 2007; Fox & Gurevitch 2000; Yearsley 2004). Segundo Fox & Gurevitch
(2000), estas fases possuem comportamentos distintos, logo, requerem análises diferentes.
Neste sentido, a escala temporal em que o manejo será aplicado também revela-se importante,
pois eventuais atrasos nas intervenções podem exigir que novas análises sejam realizadas antes
da implementação do manejo (Yearsley 2004). O exemplo das elasticidades da fase transiente
em t+1, t+5, t+10 e t assintótico corrobora tal observação (Figura 2.3).
Os resultados dos modelos matemáticos dos tipos de manejo desejados podem ser
previstos em simulações (ex. figura 2.2) e estas podem ser realizadas considerando o manejo
de taxas vitais isoladas, como no presente trabalho, ou a combinação de taxas relacionadas ou
não (Morris & Doak 2002). Como exemplos de taxas vitais intimamente relacionadas estão as
taxas de sobrevivências de ADULTOS e as taxas de fecundidade. A retirada de ADULTOS
implica necessariamente em uma pequena redução da fecundidade e isto pode potencializar a
redução do λ em médio e longo-prazos, uma vez que estes são responsáveis pela produção dos
novos indivíduos e consequentemente pela manutenção da população no tempo. Ou seja,
biologicamente uma pequena mudança na fecundidade é matematicamente equivalente a uma
grande mudança na sobrevivência (Morris & Doak 2002).
O controle das populações de invasoras em geral é mais fácil onde as espécies estão
em baixas densidades ou em regiões limítrofes das populações (Ehrenfeld 1999). A contenção
do avanço populacional da invasora pode também favorecer as espécies nativas que são
suprimidas na competição por recursos (Wittenberg & Cock 2001). Sendo assim, para as
jaqueiras no PNT recomenda-se que o controle das populações também seja feito atingindo

70
principalmente indivíduos nas periferias das sub-populações. Por outro lado, uma vez que a
declividade no PNT é muito variável (Coelho-Netto et al. 2007) e considerando-se fatores
como a barocoria da espécie invasora (Chaves et al. 1967) e a distribuição gregária das sub-
populações (Abreu 2005), uma forma de otimizar o esforço de controle seria priorizar o
manejo de indivíduos que estão acima da encosta, pois há uma tendência dos frutos se
propagarem em sentido descendente do relevo (ver capítulo 1).
Independentemente dos indicadores a serem considerados se faz necessário que as
opções de manejo sejam realistas e viáveis (Armelin 2005). Neste sentido, a redução em até
5% das sobrevivências, principalmente nas classes JUVENIS 1 e ADULTOS 1 e 2, mostra-se
suficiente para que a população seja controlada e entre em declínio. Tal declínio pode ser
potencializado caso reduções como esta ocorram simultaneamente nas três classes. Em casos
como este, onde as exóticas causam perdas na biodiversidade, os efeitos podem ser observados
diretamente na forma de interações ecológicas (Roemer et al. 2002). Portanto, manejando-se
com pequenas intervenções e em longo-prazo, os locais não estariam sujeitos ao abandono da
fauna associada às jaqueiras devido à escassez de comida ou abrigo (Zavaleta et al. 2001).
A resolução de questões relativas à conservação de espécies através do manejo mostra-
se muitas vezes mais complexa do que a simples remediação da causa primária (Courchamp et
al. 2003), como por exemplo, o foco apenas na remoção dos invasores. Diversos fatores
devem ser considerados, como as novas interações ecológicas desenvolvidas (Courchamp et al.
1999; 2000) e a forma como a remoção da espécie afetaria o ambiente físico local. De acordo
com Courchamp et al. (2003) a remoção de espécies pode ser emocionalmente difícil,
politicamente desagradável e legalmente desafiadora. Segundo Myers et al. (2000), seis
exigências devem ser atendidas para que seja obtido sucesso em Programas de Manejo: 1) os
recursos devem ser suficientes para o financiamento do programa até sua conclusão; 2) o

71
indivíduo ou instituição responsável deve ter autoridade para tomada de decisões sobre
interesses individuais ou governamentais conflitantes com o programa; 3) a biologia do
organismo alvo deve permitir que ele seja suscetível aos procedimentos de controle; 4) a re-
invasão deve ser prevenida; 5) a espécie-alvo deve ser detectável em baixas densidades; 6)
após o controle da espécie, deve ser realizada a restauração ou o manejo da comunidade ou
ecossistema (Myers et al. 2000). Neste sentido, é fundamental que os programas de remoção
de espécies invasoras sejam muito bem planejados.

72
CONCLUSÕES
As interações ecológicas desenvolvidas por A. heterophyllus no novo ambiente devem
ser consideradas ao se manejar esta árvore invasora. Baseado nas simulações e análises
realizadas, o manejo feito de forma gradativa e no longo prazo mostrou-se mais eficiente do
que intervenções radicais. Intervenções pequenas e gradativas com reduções das taxas vitais
em até 5% ao ano, por possuírem custo menor do que grandes intervenções (>50%) tornam-se
economicamente viáveis. O manejo de forma gradativa permite também a recomposição do
ambiente através da recolonização natural de espécies nativas, preservando o ambiente
florestal. Sendo assim, intervir demasiadamente seria desnecessário e poderia resultar no
desperdício de recursos, além de dificultar colonização do ambiente pelas espécies nativas.
Especificamente para a população de jaqueiras do PNT, as taxas vitais que mais
influenciaram o crescimento populacional foram as elevadas sobrevivências (todas superiores
a 98%) dos indivíduos nas classes. Desta forma, as simulações indicaram que as reduções
destas sobrevivências foram mais efetivas nas classes de JUVENIS 1, ADULTOS 1 e 2.
Os dados demográficos e as questões relativas às interações com outras espécies aqui
aplicados, foram específicos para a população de jaqueiras do PNT. O modelo de manejo
visou mitigar o impacto que a retirada de uma espécie invasora estabelecida historicamente
pode causar à biota de uma Unidade de Conservação com alta diversidade de espécies.
Portanto, a aplicação do manejo da espécie em outras localidades está sujeita a estudos
populacionais locais e a consideração de fatores como a recomposição do ambiente nativo.

73
REFERÊNCIAS BIBLIOGRÁFICAS:
Abreu, R.C.R. 2005. Estrutura de populações de uma espécie exótica na Mata Atlântica: o
caso da potencial invasora Artocarpus heterophyllus L. (jaqueira) no Parque Nacional da
Tijuca – RJ. In, Monografia de Bacharelado em Ciências Biológicas. Universidade Federal
do Estado do Rio de Janeiro (UNIRIO), Departamento de Ciências Naturais.
Armelin, R.S. 2005. As dinâmicas de Schefflera angustissima (March.) Frodin (Araliaceae) e
de Andira anthelmia (Vell.) March. (Fabaceae) na reserva de Morro Grande, São Paulo. In,
Instituto de Biociências. Departamento de Ecologia. Universidade de São Paulo - USP,
São Paulo, p. 212.
Atala, F.; Bandeira, C.M.; Martins, H.M.; Coimbra-Filho, A.F.; Chaves, C.M.; Tamara, R.;
Carauta, J.P.P.; Silveira, E.K.P. & Vianna, M.C. (eds.). 1966. Floresta da Tijuca. Centro de
Conservação da Natureza, Rio de Janeiro 152p.
Azevedo, D.A.; Moreira, L.O. & Siqueira, D.S.D.; 1999. Composition of extractable organic
matter in aerosols from urban areas of Rio de Janeiro city, Brazil. Atmospheric
Environment 33: 4987-5001.
Bandeira, C.M. 1993. Parque Nacional da Tijuca. Markom Books, São Paulo.
Bhuyan, P.; Khan, M.L. & Tripathi, R.S. 2003. Tree diversity and population structure in
disturbed and human-impacted stands of tropical wet evergreen forest in Arunachal
Pradesh, Eastern Himalayas, India. Biodiversity and Conservation 12: 1753-1773.
Blossey, B. 1999. Before, during and after: the neeed for long-term monitoring in invasive
plant species management. Biological Invasions 1: 301-311.
Carauta, J.P.P. & Diaz, B.E. 2002. Figueiras no Brasil. Editora UFRJ, Rio de Janeiro, RJ.
Caswell, H. 1989. Matrix population models: construction, analysis and interpretation. Sianuer
Associates, Sunderland.

74
Caswell, H. 2000. Prospective and Retrospective perturbation analyses: their roles in
Conservation Biology. Ecology 81: 619-627.
Caswell, H. 2007. Sensivity analysis of transient population dynamics. Ecology Letters 10: 1-
15.
Chaves, C.M.; Martins, H.F.; Carauta, J.P.P.; Lanna-Sobrinho, J.P.; Vianna, M.C. & Silva,
S.A.F. (eds.). 1967. Arboreto Carioca 3. Centro de Conservação da Natureza, Rio de
Janeiro.
Chaves, C.M.; Martins, H.F.; Carauta, J.P.P.; Lanna-Sobrinho, J.P.; Vianna, M.C. & Silva,
S.A.F. (eds.). 1969. Arboreto Carioca 4. Centro de Pesquisas Florestais e Conservação da
Natureza, Rio de Janeiro.
Chittibabu, C.V. & Parthasarathy, N. 2000. Attenuated tree species diversity in human-
impacted tropical evergreen forest sites at Kolli hills, Eastern Ghats, India. Biodiversity
and Conservation 9: 1493-1519.
Coelho-Netto, A.L.; Avelar, A.S.; Fernandes, M.C. & Lacerda, W.A. 2007. Landslide
susceptibility in a montainous geoecosystem, Tijuca Massif, Rio de Janeiro: The role of
morphometric subdivision of the terrain. Geomorphology 87: 120-131.
Courchamp, F.; Langlais, M. & Sugihara, G., 1999. Control of rabbits to protect island from
cat predation. Biological Conservation 89: 219-225.
Courchamp, F.; Langlais, M. & Sugihara, G. 2000. Rabbits killing birds: modelling the
hyperpredation process. Journal of Animal Ecology 69: 154-164.
Courchamp, F.; Woodroffe, R. & Roemer, G. 2003. Removing Protected Populations to Save
Endangered Species. Science 302: 1532.

75
Crane, J.H.; Balerdi, C.F. & Campbell, R.J. 2002. The Jackfruit (Artocarpus heterophyllus
Lam.) in Florida. University of Florida IFAS Extension. Fact Sheet HS-882.
http://edis.ifas.ufl.edu.
Cronk, Q.C.B. & Fuller, J.L. 1995. Plant Invaders: The threat to natural ecosystems. Chapman
& Hall, London, UK.
Cunha, A.A.; Vieira, M.V. & Grelle, C.E. 2006. Preliminary observations on habitat, support
use and diet in two non-native primates in an urban Atlantic forest fragment: The capuchin
monkey (Cebus sp.) and the commom marmoset (Callithrix jacchus) in the Tijuca forest,
Rio de Janeiro. Urban Ecosystems 9: 351-359.
de Kroon, H.; Plaisier, A.; van Groenendael, J. & Caswell, H., 1986. Elasticity: the relative
contribution of demographic parameters to population growth rate. Ecology 67: 1427-1431.
de Kroon, H.; van Groenendael, J. & Ehrlén, J. 2000. Elasticities: a Review of methods and
model limitations. Ecology 81: 607-608.
De Wit, M.P.; Crookes, D.J. & van Wilgren, B.W. 2001. Conflicts of interest in environmental
management: estimating the costs and benefits of a tree invasion. Biological Invasions 3:
167-178.
Dean, W. 2002. A ferro e fogo: a história e a devastação da Mata Atlântica brasileira.
Companhia das Letras, São Paulo.
DeWalt, S.J. 2006. Population dynamics and potential for biological control of an exotic
invasive shrub in Hawaiian rainforests. Biological Invasions 8: 1145-1158.
Ehrenfeld, J.G. 1999. Structure and dynamics of populations of Japanese barberry (Berberis
thunbergii DC.) in deciduous forests of New Jersey. Biological Invasions 1: 203-213.
Ferrão, J.E.M. 1993. A aventura das plantas e os descobrimentos portugueses. Instituto de
Investigação Científica Tropical, Lisboa.

76
Fox, G.A. & Gurevitch, J. 2000. Population Numbers Count: Tools for Near-Term
Demographic Analysis. The American Naturalist 156: 242-256.
IBAMA, 2004. Proposta de ampliação e correção de limites do Parque Nacional da Tijuca. In.
Gestão compartilhada IBAMA e Prefeitura da Cidade do Rio de Janeiro.
IBGE, 2006. http://www.ibge.gov.br/cidadesat/topwindow.htm?1 - acessado em junho de
2006.
Khan, M.L. 2004. Effects of seed mass on seedling success in Artocarpus heterophyllus L., a
tropical tree species of north-east India. Acta Oecologica 25: 103-110.
Mattos, C.C.L.V.D.; Mattos, M.D.L.V.D. & Laroche, R.C. 1976. Aspectos do clima e da flora
do Parque Nacional da Tijuca. Brasil Florestal 7.
Morris, W.F. & Doak, D.F. 2002. Quantitative Conservation Biology: Theory and Practice of
Population Viability Analysis. Sinauer Associates Inc.
Myers, J.H.; Simberloff, D.; Kuris, A.M. & Carey, J.R. 2000. Eradication revisited: dealing
with exotic species. Trends in Ecology and Evolution 15: 316-320.
Pádua, M.T.J. & Coimbra-Filho, A.F. 1979. Os Parques Nacionais e as Reservas Biológicas
do Brasil. Instituto Brasileiro de Desenvolvimento Florestal, Brasília.
Panetta, F.D. & Sparkes, E.C. 2001. Reinvasion of a riparian forest community by an animal-
dispersed tree following control measures. Biological Invasions 3: 75-88.
Parker, I.M. 2000. Invasion dynamics of Cytisus scoparius: a matrix model approach
Ecological Applications 10: 726-743.
Piñero, D., Martinez-Ramos, M. & Sarukhan, J. 1984. A population model of Astrocaryum
mexicanum and a sensivity analysis of its finite rate of increase. Journal of Ecology 72:
977-991.

77
Roemer, G.W.; Josh, C.D. & Courchamp, F. 2002. Golden eagles, feral pigs, and insular
carnivores: How exotic species turn native predators in prey. PNAS 99: 791-796.
Shea, K. & Kelly, D. 1998. Estimating biocontrol agent impact with matrix models: Carduus
nutans in New Zeland Ecological Applications 8: 824-832.
Silvertown, J.; Franco, M.; Pisanty, I. & Mendoza, A. 1993. Comparative plant demography -
relative importance of life-cicle components to the finite rate of increase in woody and
herbaceous perennials. Journal of Ecology 81: 465-476.
Sodhi, N.S.; Peh, K.S.H.; Lee, T.M.; Turner, I.M.; Tan, H.T.W.; Prawiradilaga, D.M., Darjono,
2003. Artificial nest and seed predation experiments on tropical southeast Asian islands.
Biodiversity and Conservation 12: 2415-2433.
Starfinger, U.; Kowarik, I.; Rode, M. & Schepker, H. 2003. From desirable ornamental plant
to pest to accepted addition to the flora? - the perception of an alien tree species through
the centuries. Biological Invasions 5: 323-335.
Wilcove, D.S. & Chen, L.Y. 1998. Management Costs for Endangered Species. Conservation
Biology 12: 1405-1407.
Williamson, M. 1996. Biological Invasions. Chapman & Hall, London, UK.
Wittenberg, R. & Cock, M.J.W. (eds). 2001. Espécies exóticas invasoras: Una guía sobre las
mejores prácticas de prevención y gestión. CAB International, Wallingford, Oxon, Reino
Unido.
Yearsley, J.M. 2004. Transient population dynamics and short-term sensivity analysis of
matrix population models. Ecological Modelling 177: 245-258.
Zavaleta, E.S.; Hobbs, R.J. & Mooney, H.A. 2001. Viewing invasive species removal in a
whole-ecosystem context. Trends in Ecology and Evolution 16: 454-459.