instituto de pesquisas jardim botânico do rio de janeiro...
TRANSCRIPT

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
Análise dos efeitos de borda sobre a composição, dinâmica e
estrutura da comunidade arbórea na Mata Atlântica da
Reserva Biológica do Tinguá - RJ
Mariela Figueredo Simões de Jesus
2009

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro Escola Nacional de Botânica Tropical
Análise dos efeitos de borda sobre a composição, dinâmica e estrutura da comunidade arbórea na Mata Atlântica da
Reserva Biológica do Tinguá - RJ
Mariela Figueredo Simões de Jesus
Dissertação apresentada ao Programa de Pós-Graduação em Botânica, Escola Nacional de Botânica Tropical, do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro, como parte dos requisitos necessários para a obtenção do título de Mestre em Botânica.
Orientador: Dr. Pablo J. F. Pena Rodrigues
Rio de Janeiro 2009

iii
Análise dos efeitos de borda sobre a composição, dinâmica e estrutura da comunidade arbórea na Mata Atlântica da Reserva Biológica do Tinguá-RJ
Mariela Figueredo Simões de Jesus Dissertação submetida ao corpo docente da Escola Nacional de Botânica Tropical, Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - JBRJ, como parte dos requisitos necessários para a obtenção do grau de Mestre.
Aprovada por: Prof. Dr. Pablo J. F. Pena Rodrigues (Orientador) ____________________ Prof. Dra. Gislene Ganade ______________________ Prof. Dra. Marcia C. M. Marques ______________________ em 30/04/2009.
Escola Nacional de Botânica Tropical-JBRJ 2009

iv
Jesus, Mariela Figueredo Simões de.
J58a Análise dos efeitos de borda sobre a composição, dinâmica e estrutura da comunidade arbórea na Mata Atlântica da Reserva Biológica do Tinguá - RJ / Mariela Figueredo Simões de Jesus. – Rio de Janeiro, 2009.
ix, 149 f. : il. Dissertação (Mestrado) – Instituto de Pesquisas Jardim Botânico do Rio
de Janeiro/Escola Nacional de Botânica Tropical, 2009. Orientador: Pablo José Francisco Pena Rodrigues Bibliografia. 1. Fragmentação. 2. Mata Atlântica. 3. Efeitos de borda. 4. Comunidade
arbórea. 5. Reserva Biológica do Tinguá (RJ). 5. Rio de Janeiro (RJ). I. Título. II. Escola Nacional de Botânica Tropical.
CDD 577.35

v
AGRADECIMENTOS Agradeço, primeiramente, ao Prof. Dr. Pablo José Francisco Pena Rodrigues por ter aceitado
me orientar, pelo apoio e amizade nestes dois anos de convivência e principalmente pela
confiança depositada em mim.
A Dra. Marcia C. M. Marques, a Dra. Gislene Ganade e ao Dr. Haroldo Lima por terem
aceitado participar da banca.
Aos avaliadores convidados das disciplinas Seminários I e II, Dr. Leandro Freitas e Dra
Alexandra Pires, pelas sugestões e questionamentos.
Aos colegas do Laboratório de Ecologia Vegetal: Jerônimo Sansevero, Pablo Pietro, Talita
Reis, Monique Gabriel, Rodolfo Abreu e Mariana Iguatemy que me receberam tão bem. Pelo
apoio, pelas boas conversas e ensinamentos, e pelos momentos de descontração e boas
risadas.
Aos que me ajudaram no campo, Talita, Jerônimo e Raul, em especial a Lilian e Deborah
pela grande colaboração em todas as etapas, pricipalmente na identificação das plantas.
A todos os taxonomistas que colaboraram na etapa mais árdua do trabalho, a identificação
das plantas: Sebastião Neto, Marcelo Souza, João Marcelo Braga, Adriana Lobão, Ronaldo
Marquete, Arline Souza, Ariane Peixoto, Elsie Guimarães, Tatiana Carrijo, Maria de Fátima
Freitas, Alexandre Christo, José Fernando Baumgratz, Massimo Bovini, Marcus Coelho, Ana
Joffily (UFF), Genise Vieira (UFRRJ), Maria Silva Ferrucci (UNNE-CONICET), Anderson
Alves-Araújo (UFPE) e Marcelo Vianna (Museu). Em especial, ao Haroldo Lima e
Alexandre Quinet pela imprescindível ajuda e paciência.
A FAPERJ pela bolsa de pesquisa concedida, que foi de extrema importância para a minha
dedicação exclusiva ao trabalho e para minha permanência no Rio.
A Petrobras pelo apoio financeiro através do Programa Mata Atlântica.
A todos os funcionários da ENBT e do JBRJ, em especial a Janúzia, Kátia, Luciano e a
guarda Edimar.
Aos meus colegas de turma, por compartilharmos nossas dúvidas, frustrações e sucessos a
cada etapa do trabalho cumprida, mesmo em rápidas conversas pelos corredores.
Aos técnicos de campo (“mateiros”) Seu Walter e Adilson Pintor por suas importantíssimas
colaborações e conhecimentos.

vi
Ao IBAMA e ao chefe da Reserva Biológica do Tinguá, Luís Henrique dos Santos Teixeira,
por nos cederem alojamento e facilitarem o bom andamento do trabalho.
A Dona Terezinha e a Colônia de Férias dos Aeroviários, em especial ao Jaime e a Umbelina,
por nos cederem alojamentos e boas refeições ao final dos longos dias de trabalho.
A todos os meus amigos que mesmo entendendo muito pouco do meu trabalho sempre
mostraram interesse e me apoiaram durante toda a trajetória.
Em especial aos meus pais, Aldírio Simões (in memorian) e Maristela de Figueredo, que
sempre priorizaram a minha educação, pelo total apoio e confiança nas minhas escolhas
mesmo de longe. Amo vocês!
E ao meu companheiro de vida, João Hildebrando Borges Jr, por todo o amor, apoio,
dedicação e principalmente paciência durante todos esses anos de convivência.
A todos, MUITO OBRIGADA!

vii
RESUMO Os processos de fragmentação e perda de habitat, decorrentes de desmatamentos,
representam uma das maiores ameaças a biodiversidade em florestas tropicais, assim como
na Mata Atlântica. Com a fragmentação é criada uma borda onde ocorrem diversas alterações
bióticas e abióticas, chamadas efeitos de borda. Mesmo a fragmentação interna, onde a
matriz é circundada pelo remanescente florestal, pode causar alterações semelhantes no
ecossistema. Neste contexto, este estudo analisou os efeitos de borda de aberturas lineares de
dossel de diferentes idades sobre a comunidade arbórea na Reserva Biológica do Tinguá-RJ.
Para isso, a comunidade arbórea (DAP≥5 cm) foi avaliada em parcelas permanentes no
período de 2003 a 2008, a partir da medição anual de diâmetro, além da altura e coleta de
material botânico. Com esses dados foi possível comparar a estrutura, composição e
dinâmica da vegetação entre bordas de diferentes idades (B-200 e B-30; aprox. 200 e 30
anos, respectivamente) e interiores preservados (controles: INT-1 e INT-2). Foi verificado
que a comunidade está sujeita a efeitos de borda e que a intensidade destes varia com a idade
da borda. A borda B-30 diferiu principalmente de INT-1 quanto a estrutura e composição
apresentando maior densidade total, densidade de perfilhadas, maior representatividade de
árvores de estágios sucessionais iniciais e espécies raras com pequena área basal e menores
alturas, além da perda de biomassa. Tais diferenças foram associadas a maior
heterogeneidade ambiental decorrente da maior freqüência de aberturas de clareiras na borda.
A análise das classes de tamanhos mostrou que as árvores pequenas (10cm≤DAP≤30cm) são
as principais responsáveis por estas alterações e refletem modificações pretéritas nos
processos sucessionais que teriam iniciado há 30 anos com o surgimento da borda, onde o
crescimento destas teria sido favorecido. Os efeitos de borda sobre a dinâmica foram menos
intensos e podem estar relacionados ao curto período avaliado neste estudo (4 anos) ou a
heterogeneidade dos sítios. Por outro lado, o longo tempo decorrido desde o surgimento da
borda B-200 teria induzido um tamponamento bastante avançado nesta borda, com o
crescimento de vegetação secundária e diminuição das modificações microclimáticas, além
do favorecimento de um equilíbrio semelhante ao interior florestal com o retorno de espécies
secundárias tardias a comunidade.

viii
ABSTRACT The processes of fragmentation and habitat lost due the deforestation represent the greatest
threat to biodiversity in tropical forests, as well as Atlantic Forest. With the fragmentation, a
forest edge is created and occur several biotic and abiotic changes, called edge effects. Even
the internal fragmentation, where the habitat matrix is surrounded by forest remnant, can
cause similar changes in the ecosystem. In this context, this work assessed the edge effects of
linear canopy openings of different ages on the tree community of an Atlantic forest reserve.
For this, the tree community (DBH≥5 cm) was evaluated in permanents plots (total area: 1,17
ha) in the period 2003 to 2008, from the annual measurement of diameter, height and collect
of botanical material. Thus, it was possible to compare the structure, composition and
dynamic of vegetation among edges of different ages (B-200 and B-30; approx. 200 and 30
years old, respectively) and preserved interiors (controls: INT-1 and INT-2). It was found
that the community is subject to edge effects and the intensity of these varies with the age of
the edge. The edge B-30 differed mainly from INT-1 as the structure and composition with
higher density, density of tillering, greater representation of early successional trees and rare
species with low basal area and lower heights, beyond the loss of biomass. Such differences
were associated with greater environmental heterogeneity caused by the higher frequency of
gap openings at the edge. The size classes analysis has shown that the small trees (10 cm≤
DBH≤ 30 cm) are primarily responsible for these changes and reflect changes in the past
successional processes that would have started 30 years ago with the appearance of the edge,
where the growth of these would have been favored. The edge effects on the dynamic were
less intense and may be related to the short evaluation period in this study (4 years) or to the
heterogeneity of sites. Moreover, the long time since the appearance of edge B-200 would
have caused a buffer well advanced in this edge, with the growth of secondary vegetation and
reduction of microclimatic changes, as well as to favoring a balance similar to the forest
interior with the return of the late secondary species to the community.

ix
SUMÁRIO
Análise dos efeitos de borda sobre a composição, dinâmica e estrutura da comunidade arbórea na Mata Atlântica da Reserva Biológica do Tinguá-RJ. Introdução ........................................................................................................... 1 Materiais e métodos .......................................................................................... 14 Resultados ......................................................................................................... 22 Discussão .......................................................................................................... 51 Conclusões ........................................................................................................ 66 Perspectivas ...................................................................................................... 68 Referências bibliográficas ................................................................................ 87 Anexos ............................................................................................................ 111

1
INTRODUÇÃO
O processo de desmatamento e as consequentes fragmentação e perda de habitat
têm sido as maiores ameaças à biodiversidade em todos os ecossistemas terrestres,
principalmente em florestas tropicais (Whitmore 1990; Whitmore 1997; Mittermeier et al.
1998; Brooks et al. 2002). Dentro deste cenário, o bioma Mata Atlântica que atualmente
abriga cerca de 60% da população brasileira sofre processos de devastação, reflexo da
ocupação territorial e exploração desordenada dos recursos naturais, com ritmo acelerado nas
últimas três décadas (Dean 1996; Pinto et al. 2006). Este histórico de destruição reduziu a
Mata Atlântica em remanescentes florestais de variados tamanhos que correspondem juntos a
cerca de 7% de sua cobertura original (Rizzini 1997; Morelatto & Haddad 2000),
enquadrando-a como um hotspot para a conservação da biodiversidade devido ao alto grau de
endemismo e seus níveis de ameaça (Myers et al. 2000). Isto porque, originalmente, este
bioma cobria uma área de 1,3 milhões de km2 do território brasileiro, estendendo-se do Rio
Grande do Norte até o Rio Grande do Sul por mais de 3.300 km (Morelatto & Haddad 2000).
Os sucessivos impactos resultantes de diferentes ciclos de exploração, da concentração da
população humana e dos maiores núcleos urbanos e industriais levaram a uma drástica
redução na cobertura vegetal natural, que resultou numa paisagem atualmente fragmentada
(Fonseca 1985; Dean 1996).
A disposição desses remanescentes florestais em fragmentos predominantemente
pequenos resulta em alterações severas nos ecossistemas que compõem a Mata Atlântica,
especialmente pela perda e fragmentação de habitats (Tabarelli et al. 1999) que alteram a
quantidade e a qualidade dos habitats disponíveis. Eventos sequenciais e/ou concomitantes
ocorrem em uma floresta a partir da supressão da vegetação (por corte ou fogo) e criação dos

2
limites do fragmento que modificam diversos processos ecológicos (Laurance & Bierregaard
Jr 1997). Com a diminuição da área florestal inicial preservada é criada uma borda artificial
resultante da interação entre dois sistemas adjacentes distintos que constituem os limites
externos da floresta remanescente e do novo ambiente criado, a “matriz” (Saunders et al.
1991). Na borda florestal ocorre uma sequência de eventos abióticos e bióticos que interagem
de maneira complexa e expõem os organismos remanescentes a novas condições de entorno,
chamados “efeitos de borda”, que dependem basicamente das características do fragmento e
do habitat da matriz (Murcia 1995; Laurance & Bierregaard Jr 1997; Mesquita et al. 1999;
Harper et al. 2005). Vale ressaltar que estas bordas antrópicas se diferenciam de ecótonos
naturais devido principalmente ao grau de contraste entre os dois habitats (Rodrigues &
Nascimento 2006), que é muito maior imediatamente após o isolamento de um fragmento.
Dependendo do grau de isolamento do fragmento e das características da matriz (tipo de
vegetação, extensão), esta pode exercer grande influência no fragmento levando a degradação
do remanescente florestal (Saunders et al. 1991; Gascon et al. 2000; Laurance et al. 2000;
Harper et al. 2005).
Em muitos casos, a fragmentação não causa o isolamento total do habitat
circundado por uma matriz e, no entanto, pode acarretar alterações semelhantes no
ecossistema. A fragmentação interna é ainda muito pouco estudada apesar de ser bastante
comum na paisagem e se caracteriza principalmente por aberturas lineares na floresta
decorrentes da construção de rodovias (Pohlman et al. 2007), linhas de transmissão de
energia (Goosem 1997; Rodrigues 2004; Pohlman et al. 2007), instalação de oleodutos ou
gasodutos (Rodrigues 2004; Reis 2008), que atravessam extensas áreas florestadas. As
consequências destas aberturas lineares de dossel ainda não estão bem elucidadas, mas é

3
possível que nestes casos, o efeito da borda ocorra com maior intensidade que o efeito do
isolamento sobre as comunidades de fragmentos ou até mesmo, de grandes remanescentes, já
que as faixas desmatadas são circundadas pelo habitat natural e possuem duas vezes sua
extensão constituindo duas bordas, permitindo, desta forma, que os efeitos de borda penetrem
nos dois lados da abertura linear (Goosem 1997; Rodrigues 2004; Pohlman et al. 2007; Reis
2008). Todavia, muitos estudos têm apresentado que as características físicas do fragmento,
como forma, tamanho e isolamento de outras áreas, além das características e atividades
praticadas na matriz, são de extrema importância nos mecanismos de alterações do
fragmento, principalmente na borda adjacente a matriz impactante (Laurance & Yensen
1991; Nascimento et al. 2006; Rodrigues & Nascimento 2006). Sob esta perspectiva, também
é esperado que a fragmentação interna, como a abertura linear de dossel, cause modificações
com intensidades menores nas bordas do fragmento, já que este está disposto de maneira
diferente na paisagem, com a matriz circundada pelo remanescente. Desta forma, as
alterações no fragmento remanescente causadas pela matriz estariam diretamente
relacionadas ao tamanho da abertura de dossel e a distância entre as bordas deste fragmento.
Dentro do contexto dos efeitos de borda, a primeira resposta após a criação de uma
borda é a formação de um gradiente de condições microclimáticas borda-interior, com maior
incidência de luz lateral e velocidade de vento próximo à borda do fragmento (Lovejoy et al.
1986; Kapos 1989; Williams-Linera 1990, Murcia 1995, Laurance 1997; Harper et al. 2005;
Pohlman et al. 2007). Estas primeiras alterações podem gerar um aumento na temperatura do
solo e do ar e no déficit de pressão de vapor, além da redução na umidade do solo e do ar
(Kapos et al. 1997; Newmark 2005; Pohlman et al. 2007). Estas modificações
microclimáticas causarão estresses fisiológicos principalmente nas árvores próximas a borda,

4
respostas comuns a dessecação do ambiente que foi modificado causando maior queda de
folhas e até mesmo a morte destas por senescência (Ferreira & Laurance 1997; Sizer &
Tanner 1999; Laurance et al. 2000). Além disso, o aumento na intensidade de ventos laterais,
oriundos da matriz, leva à maior frequência na queda de árvores grandes e galhos (Ferreira &
Laurance 1997; D’Angelo et al. 2004) e, consequentemente, no número de clareiras abertas,
permitindo a chegada de maior intensidade luminosa no sub-bosque da floresta (Nascimento
& Laurance 2004). Com isso, nestes locais, ocorre um favorecimento da germinação de
sementes e uma aceleração no recrutamento de plântulas e árvores de espécies adaptadas a
maior incidência de luz e de crescimento rápido, caracterizadas como pioneiras (Williams-
Linera 1990; Laurance et al. 1998b; Sizer & Tanner 1999; Laurance et al. 2006; Nascimento
et al. 2006), além das espécies tolerantes à sombra que também se beneficiam de maiores
intensidades luminosas (Hubbell & Foster 1987). Isto marca o início de uma série de
modificações nos processos ecológicos na borda e que tende a diminuir em direção ao
interior florestal mais preservado (Laurance & Bierregaard Jr 1997).
Em florestas tropicais preservadas, a dinâmica natural da comunidade arbórea
ocorre a uma taxa moderada e aproximadamente constante com a substituição dos indivíduos
mortos por novos e tem fundamental importância para a manutenção da diversidade biológica
e da maior parte dos processos ecológicos (Liberman et al. 1985; Turner 2001; Townsend et
al. 2006). Porém, a intensidade e frequência com que os distúrbios ocorrem em uma floresta
irão determinar a riqueza de espécies no local e a integridade e heterogeneidade dos
ambientes (Begon 2006). Em comunidades caracterizadas por distúrbios muito frequentes ou
por distúrbios muito raros, é favorecida a dominância de poucas espécies mais competitivas e
de poucas espécies características de estágios sucessionais iniciais, respectivamente. Com

5
isso, de acordo com a hipótese do distúrbio intermediário, é esperado que comunidades
apresentando taxas de distúrbios moderadas tendam a uma maior riqueza de espécies
(Townsend et al. 2006).
Dentro desta abordagem, a presença de bordas em fragmentos ou remanescentes
florestais modificaria a intensidade dos distúrbios, podendo acelerar a dinâmica dos
processos naturais e causar um desequilíbrio no ecossistema. Entretanto, tais efeitos podem
ser entendidos como multicausais e dependentes do foco de estudo, como, espécie,
população, comunidade ou a interação entre elas, e da escala espacial e temporal da
observação (Debinski & Holt 2000; Rodrigues & Nascimento 2006). De acordo com o
conhecimento atual sobre fragmentação e efeitos de borda sobre as comunidades vegetais,
estas modificações causariam alterações em sua dinâmica, estrutura, composição florística e
seus processos ecológicos, como polinização, dispersão e herbivoria (Lovejoy et al. 1986;
Laurance & Yensen 1991; Bierregaard Jr. et al. 1992; Murcia 1995; Oliveira-Filho et al.
1997; Laurance et al. 2002; Scariot et al. 2003; Harper et al. 2005; Oliveira et al. 2004;
Rodrigues 2004; Iguatemy 2008; Pietro 2008; Reis 2008), podendo levar à extinção de
espécies (Cerqueira et al. 2003; Saunders et al. 1991; Laurance & Bierregaard Jr 1997),
principalmente raras e/ou endêmicas, além daquelas com maior requerimento de área e que
respondem rapidamente à dinâmica de fragmentação.
Identificar e interpretar tais modificações nas comunidades e suas interações
causadas pelo processo de fragmentação florestal pode ser bastante complexo e somente nas
últimas três décadas tornou-se foco principal de pesquisas em regiões tropicais, já que
anteriormente o entendimento destes processos era restrito a florestas temperadas. Entretanto,
os maiores esforços e resultados no Brasil se restringem as pesquisas de longo-prazo em

6
fragmentos experimentais de diferentes tamanhos na Floresta Amazônica, pelo PDBFF
(Projeto Dinâmica Biológica de Fragmentos Florestais) que já mostra padrões e processos
florestais bastante consistentes acerca dos efeitos de área e efeitos de borda sobre as
comunidades vegetais e animais dos fragmentos (Bierregaard Jr. et al. 1992; Laurance &
Bierregaard Jr. 1997; Bierregaard Jr. et al. 2001). Estes estudos acerca dos efeitos da
fragmentação florestal foram iniciados pautados na Teoria de Biogeografia de Ilhas (sensu
MacArthur & Wilson 1967 apud Begon et al. 2006), que relacionava a perda de espécies
como uma consequência da redução e do isolamento de habitats. Atualmente, diante dos
diversos estudos sobre fragmentação é possível observar que estes processos são muito mais
complexos do que a teoria propunha, e que os ecossistemas e as comunidades vão responder
a fragmentação de formas distintas (Gascon et al. 2000; Bissonette & Storch 2002).
As primeiras alterações reportadas podem ser detectadas na estrutura florestal já nos
primeiros anos (de 1 a 5 anos) após a fragmentação e criação da borda, com uma hiper-
dinamização das comunidades vegetais. Com o aumento da dessecação e mortalidade na
borda, a densidade de árvores grandes tende a diminuir, há um aumento no estoque de
serapilheira (Didham 1998; Lovejoy et al. 1986, Laurance et al. 1998a; Laurance et al. 2000;
Oliveira-Filho et al. 1997; Ferreira & Laurance 1997) ao mesmo tempo em que ocorre uma
mudança na taxa de recrutamento com o crescimento de árvores menores de sub-bosque
florestal, devido ao aumento na frequência de clareiras, aumentando sua densidade,
principalmente dentro dos 15 a 60 metros próximos à borda (Kapos 1989; Williams-Linera
1990, Oliveira-Filho et al. 1997; Laurance et al. 1998b; Laurance et al. 2002; Laurance et al.
2006; Nascimento et al. 2006). Esta mudança estrutural altera a distribuição de tamanhos das
árvores, com a substituição de árvores maiores por árvores menores, gerando uma perda de

7
biomassa total que diminui a capacidade de estoque de carbono armazenado pela comunidade
na borda (Nascimento & Laurance 2004). A mudança da vegetação na borda é similar a
florestas de crescimento secundário (Breugel et al. 2007), causando assim alterações em
outras comunidades associadas, como a fauna (Murcia 1995). A maior intensidade luminosa
pode causar flutuações na abundância de animais, aumentando o desempenho de animais
favorecidos por maior luminosidade nas bordas florestais em detrimento de outros, causando
um desequilíbrio nas estruturas tróficas associadas e nos processos de polinização, herbivoria
e dispersão (Laurance et al. 2001a; Laurance et al. 2002; Pires 2006).
Entretanto, com o aumento da idade da borda, estas mudanças na estrutura da
vegetação tendem a diminuir, mas não desaparecem totalmente (D’Angelo et al. 2004,
Oliveira-Filho et al. 1997). As diferenças entre interiores protegidos e bordas mais antigas
tornam-se mais sutis, com a proliferação de lianas e vegetação secundária que submetem
estas bordas a um processo de “tamponamento” ao longo do tempo, tornando-as menos
permeáveis a entrada de luz lateral e vento, diminuindo assim a dessecação (Camargo &
Kapos 1995; Kapos et al. 1997; Mesquita et al. 1999). Isto porque as espécies arbóreas que
não morrem sofrem aclimatação nestes novos habitats criados na borda (Laurance et al.
2001b), tornando-se mais abundantes e constituindo maior heterogeneidade a comunidade.
O tempo necessário para o restabelecimento estrutural e dinâmico inicial da
vegetação nas bordas de fragmentos e seus processos ecológicos, similar ao interior
preservado, ainda é muito discutido, já que a heterogeneidade e as peculiaridades de cada
ambiente são bastante complexas, podendo direcionar a evolução dos processos ecológicos
de maneiras distintas, e são dependentes do tamanho, forma, isolamento e qualidade
ambiental do fragmento, da qualidade da matriz e a que tipo de manejo está sujeita (Murcia

8
1995; Jules & Shahani 2003; Hill & Curran 2005; Nascimento et al. 2006; Rodrigues &
Nascimento 2006).
Justificativa
Estudos sobre a composição, estrutura e dinâmica de comunidades arbóreas são
utilizados com frequência para verificar os impactos causados pelos efeitos de borda,
principalmente na Floresta Amazônica (Ferreira & Laurance 1997; Laurance et al. 1998a;
Laurance et al. 2002; Harper et al. 2005; Nascimento et al. 2006; Laurance et al. 2006;
Laurance et al. 2007), e na Mata Atlântica, apesar de crescentes (Oliveira et al. 1997; Viana
et al. 1997; Rodrigues 2004; Oliveira et al. 2004; Oliveira-Filho et al. 2007; Pietro 2008)
ainda são escassos diante da importância desse bioma.
Considerando que 98,6% dos remanescentes de Mata Atlântica protegidos em
Unidades de Conservação (UC) no Corredor da Serra do Mar são menores que 100 hectares
(Pinto et al. 2006), a Reserva Biologia do Tinguá, que constitui uma UC de
aproximadamente 25.000 hectares, atualmente tem grande importância para a conservação e
o entendimento acerca dos processos ecológicos deste bioma. Pelo fato desta região estar
experimentando um processo de fragmentação interna pela implantação de duas aberturas
lineares de dossel de idades diferentes (30 e 200 anos, aproximadamente), que
potencialmente submetem este fragmento a efeitos de borda com distintas magnitudes, este
estudo tem como principal objetivo avaliar esses efeitos sobre a composição, estrutura e
dinâmica da comunidade arbórea.
Neste contexto, foram utilizadas parcelas permanentes como unidades amostrais
para avaliar os efeitos de borda de aberturas lineares de dossel de diferentes idades sobre a
comunidade arbórea num trecho de Mata Atlântica montana no estado do Rio de Janeiro. As

9
seguintes hipóteses foram testadas neste estudo: 1) A composição florística, a estrutura e a
dinâmica da comunidade arbórea nas localidades de borda são diferentes do interior florestal;
2) A vegetação arbórea na borda mais recente (30 anos) é diferente da vegetação na borda
mais antiga (200 anos), sendo esta última mais semelhante ao interior florestal.
Área de estudo
O estudo foi realizado na Reserva Biológica do Tinguá (ReBio do Tinguá) que
constitui um dos maiores e mais bem preservados remanescentes de Mata Atlântica do
Estado do Rio de Janeiro. A reserva foi criada em 1989 pelo decreto n 97.780 e atualmente
abrange uma área de aproximadamente 25.000 hectares e 150 km de perímetro, englobando
parte dos municípios de Nova Iguaçu (55,14%), Duque de Caxias (37,44%), Petrópolis
(4,26%) e Miguel Pereira (3,16%). Está situada entre as coordenadas 2222’20” e 2245’00”
de latitude S e 4340’00” e 4305’40” de longitude W (IBAMA 2006) (Figura 1).
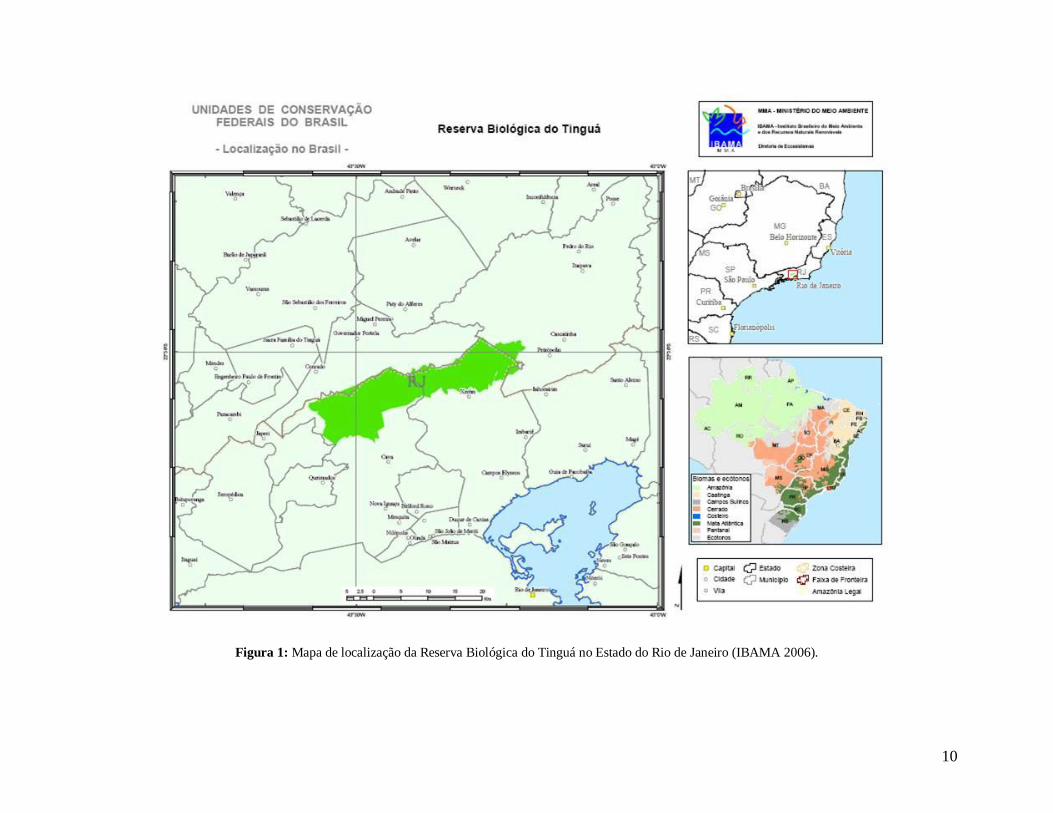
10
Figura 1: Mapa de localização da Reserva Biológica do Tinguá no Estado do Rio de Janeiro (IBAMA 2006).

11
O clima é do tipo Cwb, segundo Köppen, com verões frescos e chuvas típicas da
estação, correspondendo ao tropical de altitude, sendo que nos pontos mais altos a estação
seca é pouco pronunciada. A temperatura média anual varia entre 13 e 23C, e a pluviosidade
média fica entre 1500 e 2600 mm com regime de distribuição periódica e precipitação
máxima nos meses de dezembro e fevereiro (IBAMA 2006). O relevo, de uma forma geral, é
bastante acidentado com escarpas sulcadas por rios torrenciais. Destaca-se topograficamente,
na escarpa da Serra do Mar, o Maciço do Tinguá, que consiste em uma unidade montanhosa
imponente, bastante elevada, alcançando 1.600 m de altitude (IBAMA 2006). Os solos são
do tipo Cambissolos, Latossolos e Podzólicos, freqüentemente associados entre si e a solos
Litólicos com afloramentos de rochas (IBAMA 2006).
A vegetação local é predominantemente florestal, com a exceção de formações
campestres pouco acessíveis como no topo do Pico do Tinguá, onde predominam
afloramentos rochosos e solos muito secos. A classificação fitogeográfica é Floresta
Ombrófila Densa (IBGE 1992), ou Floresta Pluvial Tropical (Rizzini 1997). Na
caracterização fisionômico-florística desta cobertura vegetal, utilizando a terminologia
empregada por Veloso et al. (1991), foram reconhecidas as quatro subformações: Floresta
Submontana, Floresta Montana, Floresta Altomontana e Campos de Altitude (Relatório JBRJ
2002), que se encontram em bom estado de preservação devido, principalmente, à presença
de nascentes que abastecem o sistema de águas do Estado, correspondendo atualmente a uma
das únicas porções ainda florestadas da Baixada Fluminense (JBRJ 2002). Logo, trata-se de
uma unidade de conservação estratégica tanto sob o ponto de vista da sua importância
socioeconômica, quanto do ponto de vista ecológico, sendo crucial à preservação do Bioma
Mata Atlântica.

12
Apesar do bom estado de conservação, é possível observar algumas atividades
conflitantes dentro dos limites da ReBio, como a exploração de palmito (Euterpe edulis
Mart.) (Figura 2), a ação constante de caçadores e a visitação intensa da população que utiliza
a área para lazer, principalmente nos locais de mananciais hídricos do entorno da ReBio,
além da presença de oleodutos subterrâneos localizados em dois grandes corredores
desmatados que cortam a área da Reserva (Figura 3).
Figura 2: Troncos de palmitos (Euterpe edulis Mart.) cortados na Reserva Biológica do Tinguá-RJ.
Aberturas lineares de dossel
A área da Reserva Biológica do Tinguá é atravessada por dois dutos subterrâneos da
Petrobras, chamados ORBEL1 e ORBEL2 (Oleodutos Rio-Belo Horizonte 1 e 2,
respectivamente) que transferem produtos refinados da Refinaria de Duque de Caxias para
Belo Horizonte. Estão localizados sob faixas desmatadas de aproximadamente 20 m de
largura, podendo chegar a 40 m em alguns trechos, e possuem idades diferentes (Figura 3).
ORBEL1 foi implantado na década de 60 sob uma antiga estrada - Estrada do Comércio -
construída no Brasil Império e que viabilizava o transporte de ouro de Vila Velha ao porto
(Rodrigues 1996), datada do começo do século XIX, constituindo uma borda de mata de
quase 200 anos. Já ORBEL2 foi implantado mais tarde, no início da década de 80, e

13
corresponde a uma borda bem mais recente de aproximadamente 30 anos, já que a faixa foi
aberta na mata para a instalação deste oleoduto (Souza 2003).
Figura 3: Aspecto geral dos corredores desmatados onde se encontram os oleodutos subterrâneos ORBEL1 (A) e ORBEL2 (B) na Reserva Biológica do Tinguá-RJ.
Os corredores, onde os oleodutos se encontram, são monitorados e sofrem manejo
periódico por funcionários da Petrobras que capinam o local impedindo o crescimento de
vegetação lenhosa, já que seus sistemas radiculares podem perfurar e causar danos aos dutos
enterrados. Com isso, a vegetação nestes locais é predominantemente de porte herbáceo, com
o estabelecimento da espécie exótica Hedychium coronarium J. König (Zingiberaceae; lírio-
do-brejo) em grande parte na localidade ORBEL1, e a mata adjacente a esses limites (nas
bordas) é caracterizada pela vegetação arbustivo-arbórea, muito semelhante
fisionomicamente aos interiores florestais, apesar de se encontrar em estágios sucessionais
menos avançados devido ao fato de estar sujeita a efeitos de borda locais.
A B 200 anos 30 anos

14
A partir da presença destas bordas, do grau de periculosidade dos oleodutos que
transportam produtos inflamáveis e dos possíveis efeitos deletérios que as aberturas lineares
de dossel poderiam estar exercendo sobre a vegetação da Reserva Biológica do Tinguá,
fenômeno conhecido como “efeitos de borda”, viu-se a necessidade do conhecimento e
monitoramento constante da comunidade arbórea e de plântulas nestes locais a partir do
estabelecimento de parcelas permanentes, comparando áreas de bordas de diferentes idades
(aproximadamente 30 e 200 anos) com áreas de interior preservado (controle). Este projeto
teve início no ano de 2003 com a alocação das primeiras parcelas experimentais pela equipe
do Laboratório de Ecologia Vegetal do Jardim Botânico do Rio de Janeiro (JBRJ), tendo
como responsável o Dr. Pablo José Francisco Pena Rodrigues (Rodrigues et al. 2008; Reis
2008). Os dados da comunidade arbórea de 2003 a 2006, analisados neste trabalho, foram
retirados do banco de dados do projeto, sem a participação da referida autora no trabalho de
campo, o que passou a ocorrer nos anos 2007 e 2008.
Para este trabalho, convencionou-se nomear ORBEL1 de localidade B-200
(referente a uma borda mais antiga de aproximadamente 200 anos) e ORBEL2 de localidade
B-30 (referente a uma borda mais recente de aproximadamente 30 anos).
MATERIAIS E MÉTODOS
As primeiras parcelas permanentes foram estabelecidas no ano de 2003, totalizando
24 parcelas (10 m x 30 m cada; área amostrada total: 0,72 hectares), sendo 9 na borda B-200,
5 na borda mais recente B-30 e 10 parcelas “controle” distribuídas no interior florestal,
localidade INT-1, mais preservado e próximo a B-200, porém situadas a mais de 400 m de
qualquer borda e sem vestígios de perturbação recente. Em 2004, o desenho amostral foi

15
ampliado totalizando 10 parcelas (10 m x 30 m) na borda B-200, 10 parcelas na borda B-30,
e 10 parcelas no INT-1. As 30 parcelas totalizavam uma área de 0,9 ha. As parcelas
localizadas nas bordas foram alocadas longitudinalmente às bordas, distantes 5 m, no
máximo, do limite externo destas (Figura 4).
No ano de 2007 o desenho amostral foi ampliado novamente com a marcação de
mais 9 parcelas no interior florestal, nomeado localidade INT-2, próximas a B-30, porém
localizadas a mais de 400 m de distância de qualquer borda. Totalizando assim 1,17 hectares
de área estudada (Tabela 1). Todas as parcelas foram georeferenciadas para a confecção de
mapas e uma melhor visualização da distribuição das mesmas (Figura 5). As duas aberturas
lineares de dossel (B-200 e B-30) estão distantes, no mínimo (nos pontos mais próximos), 4
kilômetros entre si.
Figura 4: Esquema de alocação da parcela permanente (10 x 30 metros) na borda da mata na Reserva Biológica do Tinguá-RJ.

16
Figura 5: Visão geral da localização das parcelas nos anos de 2007 e 2008 nas localidades: borda B-200 (vermelho) e interior INT-1 (amarelo) (A); e nas localidades: borda B-30 (verde) e interior INT-2 (azul) (B) na Reserva Biológica do Tinguá-RJ.
A
B B
B-200 INT-1
B-30 INT-2

17
Todas as parcelas foram marcadas de forma aleatória e apresentavam fisionomia
florestal. Porém, para se obter uma padronização das amostragens, foram evitados topos de
morros, vales e a proximidade com grandes clareiras. A área amostrada se localizava na faixa
altitudinal de 500 a 1000 m, caracterizada pela formação de Floresta Montana (Veloso et al.
1991). Este tipo florestal ocupa a maior parte da área da Rebio do Tinguá, se encontra em
excelente estado de preservação e é muito diversificada, apresentando grande variação
estrutural, característica em grande parte influenciada pela rica rede de riachos entrecortando
as encostas, pelos diferentes níveis de declividade e pela presença de afloramentos rochosos
dispersos em muitos locais (JBRJ 2002).
Na busca de uma análise comparativa temporal da composição florística e estrutural
da comunidade arbórea, desde 2003, todas as árvores (indivíduos mortos e vivos, incluindo
palmeiras) com DAP maior ou igual a 5 cm (DAP: diâmetro à altura do peito a 1,30 m do
solo ou 0,50 m acima das raízes tabulares) localizadas dentro das parcelas foram marcadas
com placas numeradas (Figura 6), tiveram seu diâmetro medido anualmente e material
botânico coletado (fértil e/ou estéril) para posterior identificação (até espécie ou morfo-
espécie), com exceção de Euterpe edulis Mart. que era facilmente identificada no campo. O
material coletado foi herborizado e armazenado no JBRJ. Nos meses de julho a agosto dos
anos 2003, 2004, 2005, 2006, 2007 e 2008, todos os indivíduos sobreviventes e mortos foram
remedidos, e os novos que atingiram o critério de inclusão (DAP≥5 cm), foram incluídos na
amostragem como recrutas. Indivíduos com troncos perfilhados (tronco múltiplo) foram
incluídos na amostragem quando a soma das áreas de cada tronco constituiu um diâmetro
virtual maior ou igual a 5 cm (critério de inclusão). Todas as árvores também foram

18
estimadas quanto à altura total no ano de 2007, com a utilização de vara de medição de
altura.
Figura 6: Árvore marcada com placa numerada dentro da parcela permanente do estudo na Reserva Biológica do Tinguá-RJ.
Análise dos dados
A comunidade arbórea foi analisada como um todo (DAP≥5 cm), e para as classes
de tamanho: arvoretas (5 cm≤DAP<10 cm) e árvores (DAP≥10 cm), para facilitar a
comparação com estudos que avaliaram apenas este último grupo; e árvores pequenas (5
cm≤DAP≤30 cm) e árvores grandes (DAP>30 cm), com o objetivo de obter grupos mais
homogêneos.
Os dados de diâmetro, família e espécie para cada planta coletada e identificada no
ano de 2008 foram exportados para o Programa Mata Nativa 2 (CIENTEC 2006) para os
cálculos dos parâmetros fitossociológicos de todas as árvores e suas classes de tamanho
supracitadas, considerando a comunidade arbórea como um todo e as localidades

19
separadamente. Os parâmetros calculados para cada espécie foram: N (número de
indivíduos), NP (número de parcelas onde a espécie está presente), AB (área basal), DR
(densidade relativa), DoR (dominância relativa), VC (valor de cobertura) e VI (valor de
importância). Para estimar a diversidade para a comunidade e suas classes nas localidades,
foram calculados o índice de diversidade de Shannon (H’) e equabilidade de Pielou (J).
Também foi analisada a ordenação das parcelas e localidades para todas as árvores e para as
classes, através da análise não-métrica de escalas multidimensionais (NMDS) a partir do
Programa XLSTAT (Addinsoft 2009). A matriz de similaridade florística utilizada para a
NMDS foi calculada a partir da distância de Bray-Curtis com a utilização do Software MVSP
3.1 (Kovach 1999). Para as comparações da estrutura da comunidade arbórea foram
calculadas, para as diferentes localidades (B-200, B-30, INT-1 e INT-2) e para cada ano
(2003, 2004, 2005, 2006, 2007 e 2008), os seguintes parâmetros: densidade e área basal de
indivíduos vivos e mortos e densidade média de indivíduos perfilhados por parcela, além da
estrutura de tamanho (diâmetro e altura).
Para comparar a dinâmica da comunidade arbórea, foram calculadas para cada
localidade as taxas médias anuais de mortalidade (Sheil et al. 1995) (a), de recrutamento
(Sheil et al. 2000) (b) e de turnover (substituição de árvores mortas por recrutas) (Phillips et
al. 1994) (c), a partir dos valores de cada parcela para os intervalos entre os anos: 2003-2004,
2004-2005, 2005-2006, 2006-2007, 2007-2008 e 2004-2008, conforme se segue:
a) m = 1 – (N1 / N0) 1/t
Onde,
N0 = número de árvores vivas no início do intervalo;
N1 = N0 menos o número de árvores mortas no intervalo;

20
t = intervalo de tempo, em anos, entre os dois levantamentos.
b) r = 1 – (1 – nr/Nt) 1/t
Onde,
nr = número de recrutas no intervalo;
Nt = número de árvores vivas no final do intervalo;
t = intervalo de tempo, em anos, entre os dois levantamentos.
c) turnover = (m + r) / 2
Onde,
m = taxa de mortalidade;
r = taxa de recrutamento;
Os valores das taxas de mortalidade, recrutamento (m e r) e turnover foram
multiplicados por 100 para a transformação em porcentagem após a análise estatística.
A ocorrência da palmeira Euterpe edulis Mart. também foi analisada separadamente
devido a sua alta ocorrência no local de estudo, por constituir um importante componente na
estrutura e dinâmica de florestas tropicais e por seus frutos representarem recurso importante
para a fauna local. Além disso, assim como em todas as florestas tropicais do mundo, as
palmeiras são bastante utilizadas pelos humanos, e no caso desta espécie, principalmente para
a obtenção de seu meristema apical como alimento (“palmito”).
As comparações entre as localidades para densidade, área basal, mortos em pé,
perfilhados, densidade de Euterpe edulis, taxas de mortalidade, recrutamento, turnover e
índice de diversidade de Shannon foram feitas através de análise de variância One-way
ANOVAs. Já as distribuições diamétricas (DAP) e de alturas foram comparadas através de
ANOVA aninhada (em que as árvores estão aninhadas em localidades). Em ambas, foram

21
aplicados testes posteriores do tipo Tukey (com HSD para número de parcelas igual e
Unequal N HSD para número de parcelas diferente, dependendo do ano de estudo), a 5% de
significância. Os dados que não apresentaram normalidade e homogeneidade de variâncias, a
partir dos testes de Shapiro-Wilk e de Levene, respectivamente, foram transformados
utilizando log10 (x+1) (Zar 1999). Nas comparações dos parâmetros que não atenderam as
premissas de normalidade e homocedastidade, foram utilizados testes não-paramétricos de
Kruskal-Wallis com testes posteriores do tipo Mann-Whitney, a 5% de significância. Todos
os testes estatísticos foram realizados a partir do programa Statistica 6.0 (StatSoft Inc. 2001).
Em decorrência das diferentes áreas amostradas em cada localidade nos diferentes
anos de estudo, variando de 0,15 ha a 0,30 ha, todos os resultados apresentados foram
extrapolados para 1 hectare através de “regra de três”, facilitando a comparação entre os
anos.

22
RESULTADOS
Florística e estrutura da comunidade arbórea da ReBio do Tinguá
Nas 39 parcelas analisadas neste estudo (área total de 1,17 ha) foram encontradas
441 espécies (245 espécies e 196 morfo-espécies) subordinadas a 158 gêneros e 64 famílias.
Do total de 2378 indivíduos arbóreos identificados, apenas 34 permaneceram indeterminados
e foram distribuídos em 22 morfo-espécies (Anexo 1). As famílias mais bem representadas
em número de indivíduos foram: Myrtaceae (481), Arecaceae (363), Nyctaginaceae (194),
Rubiaceae (162) e Lauraceae (142).
As famílias mais ricas em espécies foram Myrtaceae (101), Lauraceae (38),
Rubiaceae (35), Fabaceae (31), Sapotaceae (21) e Euphorbiaceae (18). Apesar da grande
representatividade em abundância das famílias Arecaceae e Nyctaginaceae, estas
apresentaram baixíssima riqueza devido à dominância das espécies Euterpe edulis (344
indivíduos), presente em todas as parcelas, e Guapira opposita (194 indivíduos), ausente em
apenas 3 parcelas, que foram as espécies mais abundantes e perfizeram juntas 29% do total
de indivíduos (Figura 7).
Das 441 espécies, 43 foram consideradas comuns e apresentaram de 10 a 50
indivíduos (9,7% do total de espécies), 209 espécies ocasionais com 2 a 9 indivíduos (47,3%)
e 189 espécies (42,5%) apresentaram padrão de raridade com apenas 1 indivíduo cada.

23
Figura 7: Distribuição do número total de indivíduos por espécie para a comunidade arbórea (DAP≥5 cm) no ano de 2008 na Reserva Biológica do Tinguá-RJ.
A riqueza média de espécies por parcela foi de 33 espécies (dp= ±7), com mínima
de 19 e máxima de 47 e a comunidade apresentou índice de diversidade de Shannon (H’)
total de 4,9 nats e equabilidade (J) de 0,81.
As principais espécies (e morfo-espécies) com os maiores valores de importância
(VI) em 2008 foram: Euterpe edulis, Guapira opposita, Malpighiaceae sp4, Cupania
oblongifolia, Gomidesia spectabilis, Ocotea dispersa e Alchornea triplinervia (Anexo 2). A
espécie E. edulis apresentou o maior VI devido principalmente à alta densidade relativa
(DR=14,5%), já que obteve a terceira maior dominância relativa (DoR=3,41%). Já G.
opposita apresentou a maior DoR (9,7%) e alta DR (8,2%), seguida de Malpighiaceae sp4
com DoR igual a 5,3%, que compensou sua baixa DR (1,05%) no cálculo do VI (Anexo 2).
Composição florística das localidades
As localidades de borda (B-200 e B-30) e interior (INT-1 e INT-2) apresentaram
grandes semelhanças entre as dez famílias mais representativas em número de indivíduos
344 194
55 45
39 34 32
27 26 26
0 50 100 150 200 250 300 350 400
Euterpe edulis Mart. Guapira opposita (Vell.) Reitz Gomidesia spectabilis O.Berg
Cupania oblongifolia Mart. Psychotria glaziovii Müll. Arg.
Myrceugenia myrcioides (Cambess.) O.Berg Ocotea dispersa (Nees) Mez
Cabralea canjerana (Vell.) Mart. Myrtaceae sp16
Myrcia tenuivenosa Kiaersk.
número de indivíduos

24
(Figura 8). A família Myrtaceae foi a mais abundante nas localidades B-200, B-30 e INT-2,
Arecaceae a mais abundante em INT-1, representada principalmente pela espécie dominante
Euterpe edulis; Nyctaginaceae (apenas Guapira opposita), que esteve entre as três famílias
mais representativas em B-200, B-30 e INT-1, além de Lauraceae e Fabaceae. Apesar de os
indivíduos de “palmitos” serem bastante abundantes em todas as localidades, em B-30 esta
foi menor, refletindo possivelmente ao corte seletivo nos anos anteriores (resultados
mostrados mais abaixo). As famílias Rubiaceae e Sapotaceae estavam entre as mais
abundantes em B-30, INT-1 e INT-2, Euphorbiaceae e Sapindaceae nas localidades B-200 e
INT-1 e Melastomataceae e Chrysobalanaceae em B-30 e INT-2. Monimiaceae e Salicaceae
apenas foram mais abundantes em B-200 e Annonaceae em INT-2.
B-200
162
62
58
44
37
36
35
22
21
15
0 20 40 60 80 100 120 140 160 180
Myrtaceae
Areaceae
Nyctaginaceae
Lauraceae
Euphorbiaceae
Sapindaceae
Meliaceae
Monimiaceae
Fabaceae
Salicaceae
número de indivíduos
A
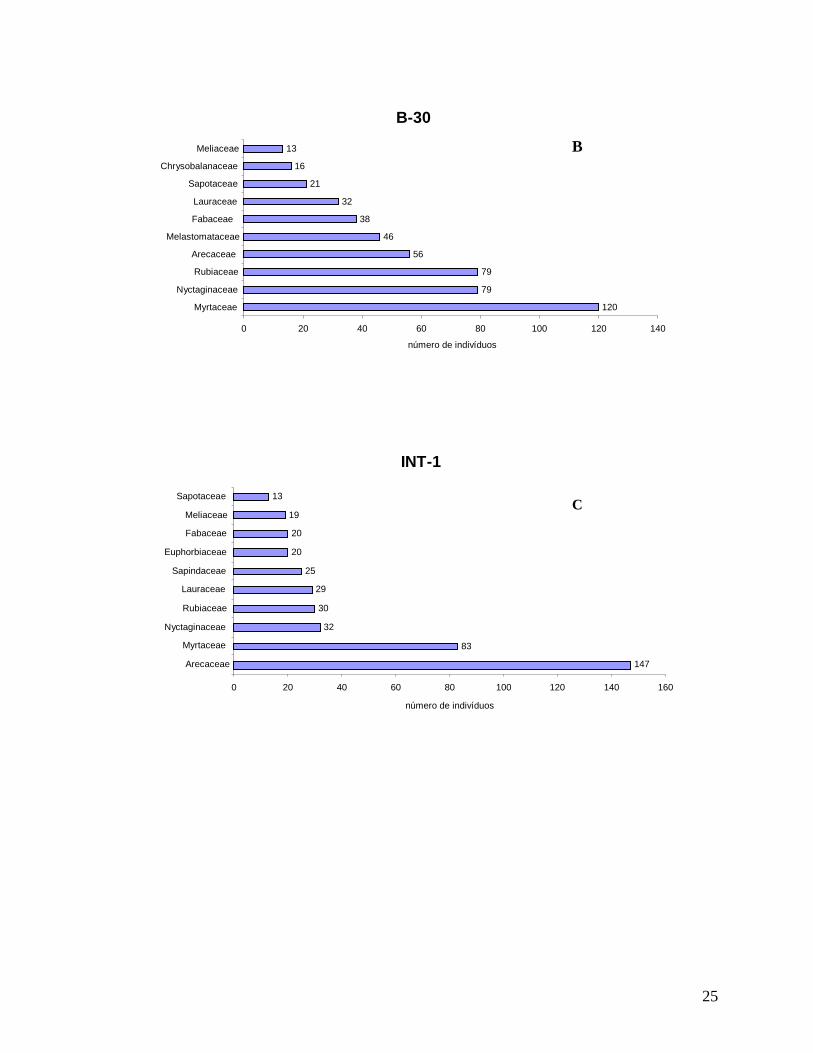
25
B-30
120
79
79
56
46
38
32
21
16
13
0 20 40 60 80 100 120 140
Myrtaceae
Nyctaginaceae
Rubiaceae
Arecaceae
Melastomataceae
Fabaceae
Lauraceae
Sapotaceae
Chrysobalanaceae
Meliaceae
número de indivíduos
INT-1
147 83
32 30 29
25 20 20 19
13
0 20 40 60 80 100 120 140 160 Arecaceae
Myrtaceae Nyctaginaceae
Rubiaceae Lauraceae
Sapindaceae Euphorbiaceae
Fabaceae Meliaceae
Sapotaceae
número de indivíduos
B
C

26
Figura 8: Distribuição do número de indivíduos por família para a comunidade arbórea (DAP≥5 cm) nas localidades (A) B-200; (B) B-30; (C) INT-1 e (D) INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
As famílias com maiores riquezas de espécies em todas as localidades foram
Myrtaceae, Rubiaceae, Lauraceae e Fabaceae (Figura 9). As espécies Euterpe edulis e
Guapira opposita foram as mais representativas em número de indivíduos em todas as
localidades, enquanto as outras espécies mais abundantes variaram de acordo com a
localidade.
INT-2
116 98
42 37 37
28 25
21 19
18
0 20 40 60 80 100 120 140 Myrtaceae Arecaceae Rubiaceae Lauraceae
Sapotaceae Chrysobalanaceae
Nyctaginaceae Melastomataceae
Annonaceae Fabaceae
número de indivíduos
D
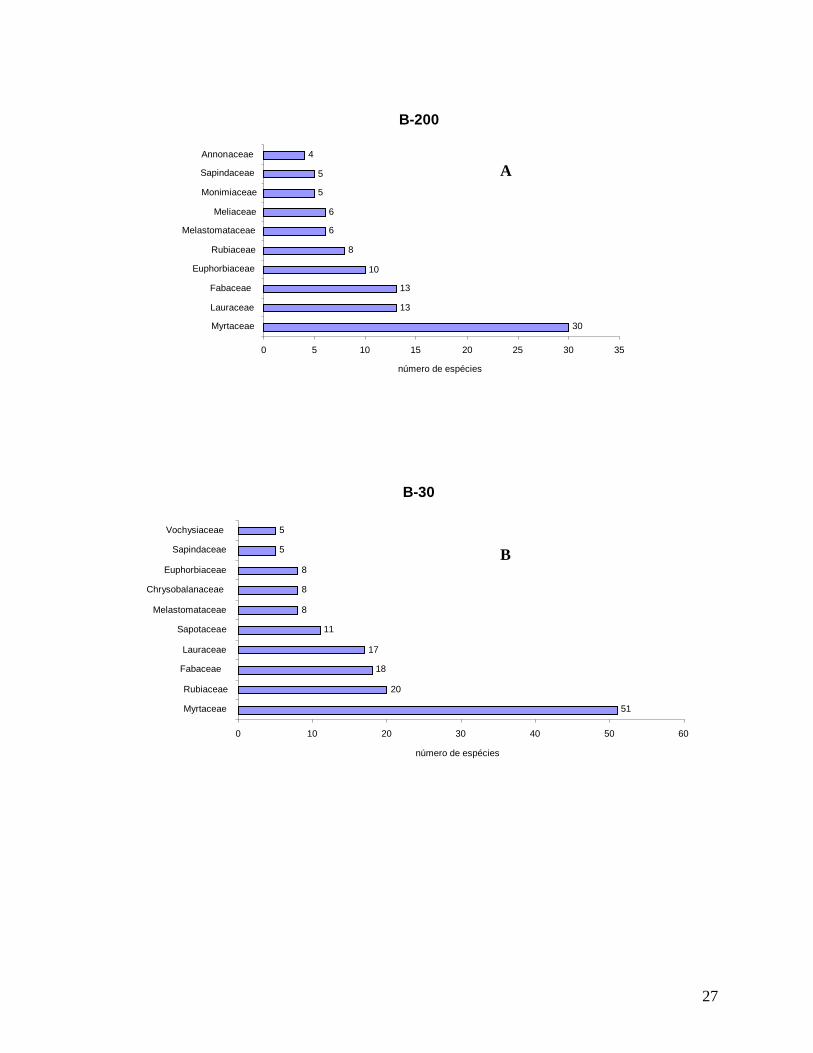
27
B-200
30 13 13
10 8
6 6
5 5
4
0 5 10 15 20 25 30 35 Myrtaceae Lauraceae Fabaceae
Euphorbiaceae Rubiaceae
Melastomataceae Meliaceae
Monimiaceae Sapindaceae Annonaceae
número de espécies
B-30
51
20
18
17
11
8
8
8
5
5
0 10 20 30 40 50 60
Myrtaceae
Rubiaceae
Fabaceae
Lauraceae
Sapotaceae
Melastomataceae
Chrysobalanaceae
Euphorbiaceae
Sapindaceae
Vochysiaceae
número de espécies
A
B
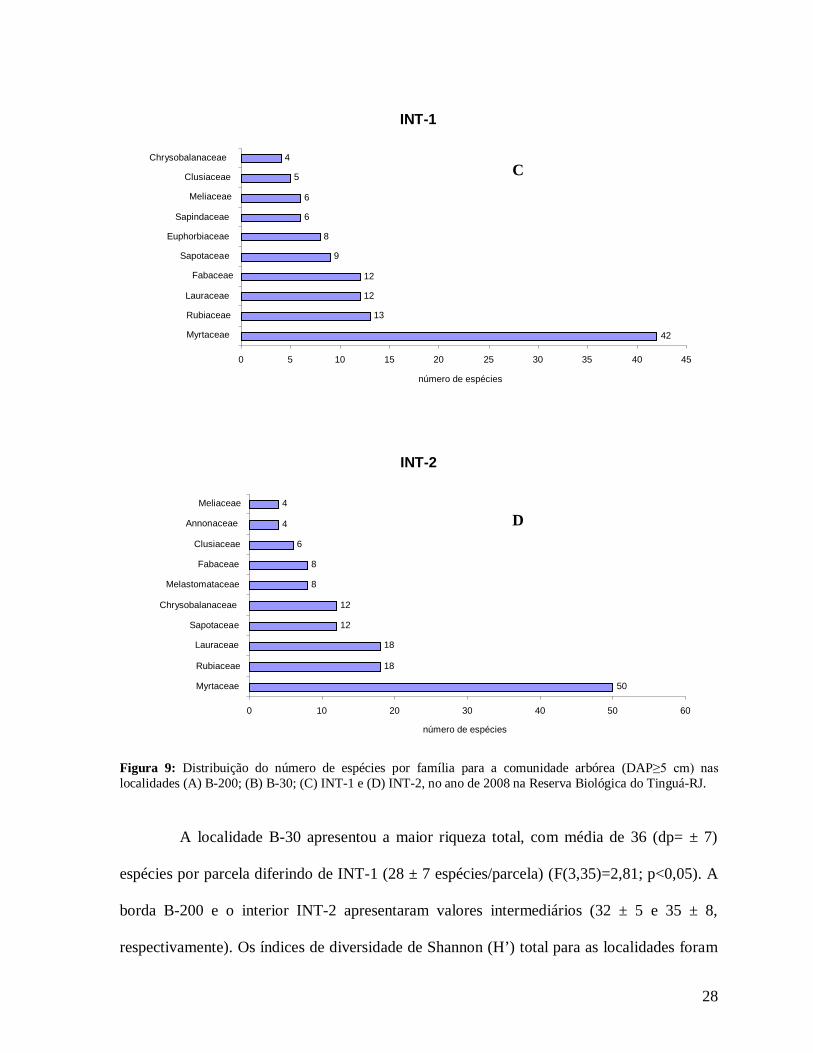
28
Figura 9: Distribuição do número de espécies por família para a comunidade arbórea (DAP≥5 cm) nas localidades (A) B-200; (B) B-30; (C) INT-1 e (D) INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
A localidade B-30 apresentou a maior riqueza total, com média de 36 (dp= ± 7)
espécies por parcela diferindo de INT-1 (28 ± 7 espécies/parcela) (F(3,35)=2,81; p<0,05). A
borda B-200 e o interior INT-2 apresentaram valores intermediários (32 ± 5 e 35 ± 8,
respectivamente). Os índices de diversidade de Shannon (H’) total para as localidades foram
INT-1
42
13
12
12
9
8
6
6
5
4
0 5 10 15 20 25 30 35 40 45
Myrtaceae
Rubiaceae
Lauraceae
Fabaceae
Sapotaceae
Euphorbiaceae
Sapindaceae
Meliaceae
Clusiaceae
Chrysobalanaceae
número de espécies
INT-2
50
18
18
12
12
8
8
6
4
4
0 10 20 30 40 50 60
Myrtaceae
Rubiaceae
Lauraceae
Sapotaceae
Chrysobalanaceae
Melastomataceae
Fabaceae
Clusiaceae
Annonaceae
Meliaceae
número de espécies
C
D

29
bastante similares entre si, com maior valor para B-30 (H’=4,7), seguida de INT-2 (H’=4,4),
B-200 (H’=4,2) e INT-1 (H’=4,1). Já a média da diversidade de Shannon por parcela na
localidade B-30 diferiu apenas de INT-1 (H’=3,2 ± 0,27 e H’=2,8 ± 0,41, respectivamente;
B-200, H’=3,1 ± 0,27 e INT-2, H’=3,2 ± 0,38).
Todas as localidades compartilharam apenas 23 espécies (5,2% do total de
espécies). Os interiores (INT-1 e INT-2) compartilharam entre si 66 espécies (15%) e as
bordas (B-200 e B-30) 71 espécies (16%). A localidade B-30 apresentou 78 espécies
exclusivas (17%), seguida de INT-2 com 74 (17%), B-200 com 53 (12%) e INT-1 com 51
(12%). As dez espécies mais abundantes nas quatro localidades diferiram parcialmente.
Euterpe edulis e Guapira opposita foram as mais abundantes em todas as localidades (Figura
10), assim como para a comunidade como um todo, já citado anteriormente. Na localidade B-
200, das dez espécies mais abundantes que ocorreram com mais de 10 indivíduos, destacam-
se ainda as Myrtaceae: Gomidesia spectabilis (44) e Myrceugenia myrcioides (30), como a
terceira e quarta espécies em maior abundância, respectivamente. De maneira diferente, B-30
apresentou 6 espécies com mais de 10 indivíduos, destacando-se a Rubiaceae: Psychotria
glaziovii (26) e a Melastomataceae: Miconia cubatanensis (15), INT-1 com 4 espécies,
destacando-se a Sapindaceae: Cupania oblongifolia (15) e a Rubiaceae: Bathysa gymnocarpa
(11) e INT-2 com 6 espécies, destacando-se a Annonaceae: Duguetia microphylla (16) e a
Chrysobalanaceae: Licania sp1 (12).
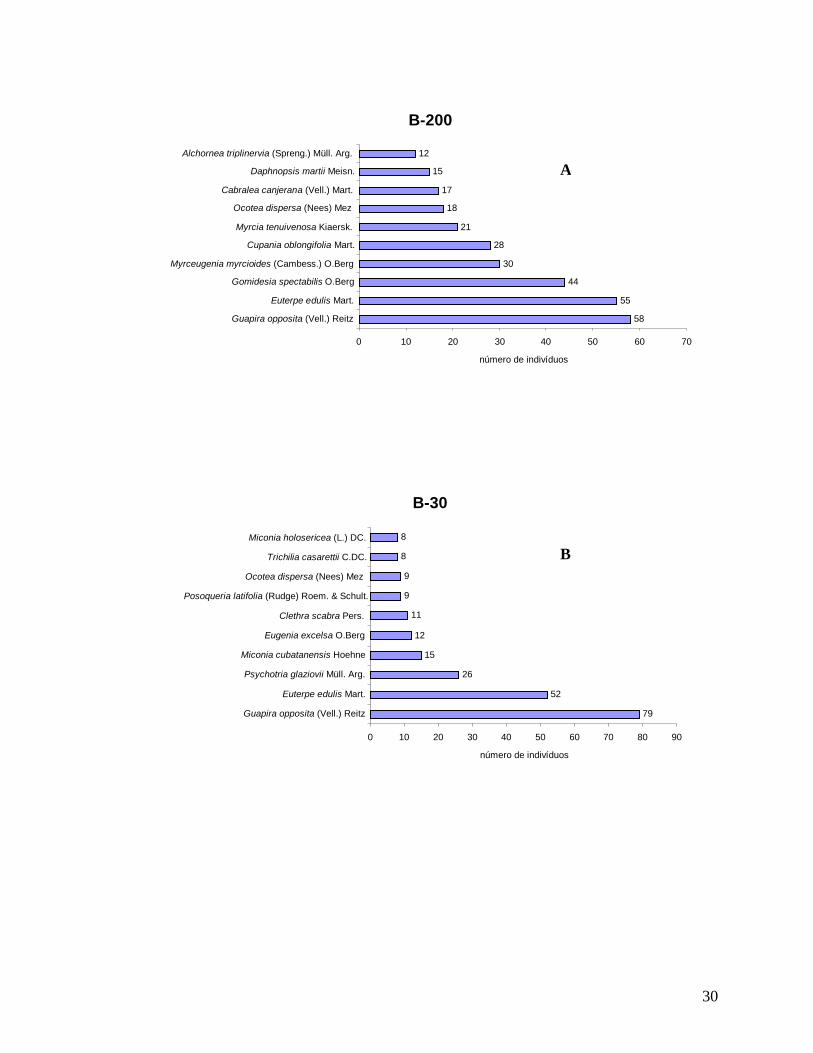
30
B-200
58 55
44 30
28 21
18 17
15 12
0 10 20 30 40 50 60 70 Guapira opposita (Vell.) Reitz
Euterpe edulis Mart. Gomidesia spectabilis O.Berg
Myrceugenia myrcioides (Cambess.) O.Berg Cupania oblongifolia Mart.
Myrcia tenuivenosa Kiaersk. Ocotea dispersa (Nees) Mez
Cabralea canjerana (Vell.) Mart. Daphnopsis martii Meisn.
Alchornea triplinervia (Spreng.) Müll. Arg.
número de indivíduos
B-30
79 52
26 15
12 11
9 9
8 8
0 10 20 30 40 50 60 70 80 90 Guapira opposita (Vell.) Reitz
Euterpe edulis Mart. Psychotria glaziovii Müll. Arg.
Miconia cubatanensis Hoehne Eugenia excelsa O.Berg
Clethra scabra Pers. Posoqueria latifolia (Rudge) Roem. & Schult.
Ocotea dispersa (Nees) Mez Trichilia casarettii C.DC.
Miconia holosericea (L.) DC.
número de indivíduos
B
A
31
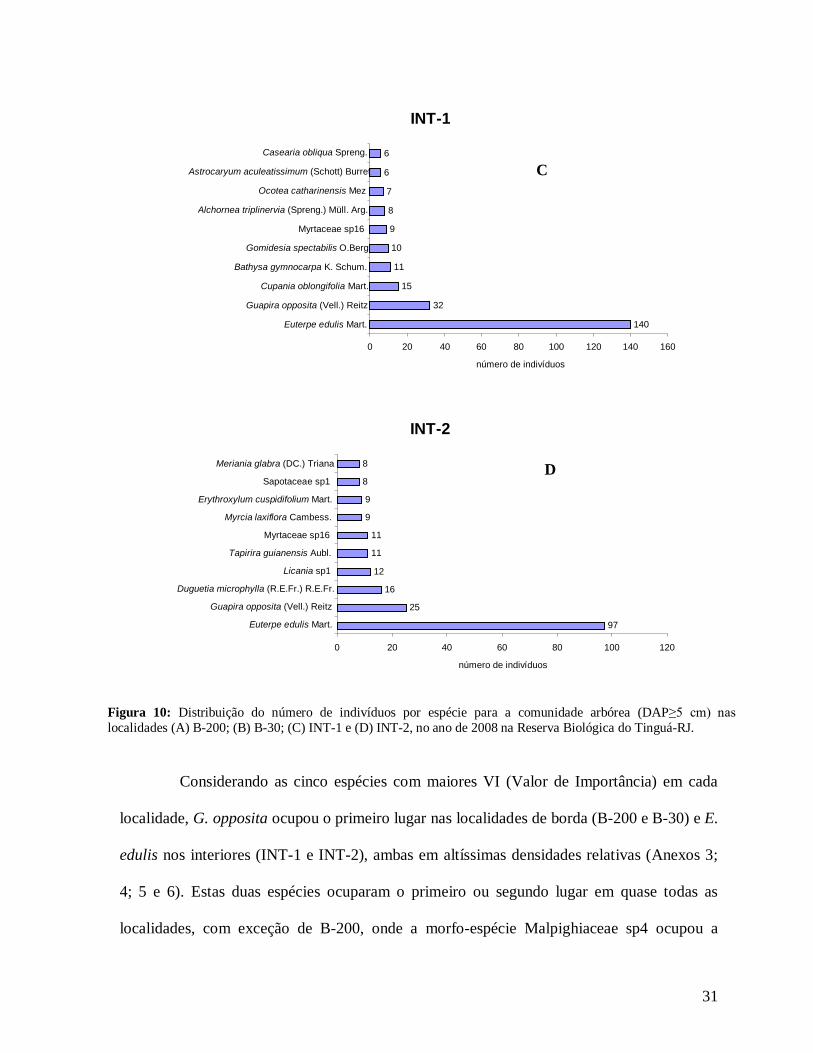
Figura 10: Distribuição do número de indivíduos por espécie para a comunidade arbórea (DAP≥5 cm) nas localidades (A) B-200; (B) B-30; (C) INT-1 e (D) INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
Considerando as cinco espécies com maiores VI (Valor de Importância) em cada
localidade, G. opposita ocupou o primeiro lugar nas localidades de borda (B-200 e B-30) e E.
edulis nos interiores (INT-1 e INT-2), ambas em altíssimas densidades relativas (Anexos 3;
4; 5 e 6). Estas duas espécies ocuparam o primeiro ou segundo lugar em quase todas as
localidades, com exceção de B-200, onde a morfo-espécie Malpighiaceae sp4 ocupou a
INT-1
140 32
15 11 10 9 8 7 6 6
0 20 40 60 80 100 120 140 160 Euterpe edulis Mart.
Guapira opposita (Vell.) Reitz Cupania oblongifolia Mart.
Bathysa gymnocarpa K. Schum. Gomidesia spectabilis O.Berg
Myrtaceae sp16 Alchornea triplinervia (Spreng.) Müll. Arg.
Ocotea catharinensis Mez Astrocaryum aculeatissimum (Schott) Burret
Casearia obliqua Spreng.
número de indivíduos
INT-2
97 25
16 12 11 11
9 9 8 8
0 20 40 60 80 100 120 Euterpe edulis Mart.
Guapira opposita (Vell.) Reitz Duguetia microphylla (R.E.Fr.) R.E.Fr.
Licania sp1 Tapirira guianensis Aubl.
Myrtaceae sp16 Myrcia laxiflora Cambess.
Erythroxylum cuspidifolium Mart. Sapotaceae sp1
Meriania glabra (DC.) Triana
número de indivíduos
D
C

32
segunda posição, apesar da baixa DR, que foi compensada por sua maior DoR levando-a ao
segundo maior VC (Valor de Cobertura) (Anexo 3). Euterpe edulis, apesar de ocorrer em
altas densidade e frequência, muito similar a G. opposita, constituiu baixíssima DoR,
diminuindo sua posição com relação ao VI. Na borda B-30 destacam-se a espécie Psychotria
glaziovii com alta densidade e baixa DoR, e as espécies Copaifera lucens e Posoqueria
latifolia (4° e 5° lugares de maior VI), com baixíssimas densidades e grandes DoR (Anexo
4).
Na localidade INT-1, Schefflera sp1 com apenas 3 indivíduos apresentou grande
DoR (grande área basal), ocupando o 4° lugar de maior VI (Anexo 5) e em INT-2, Ocotea
catharinensis em baixíssima abundância ocupou a 3° posição devido seus indivíduos
apresentarem grande AB total (Anexo 6). Destaca-se também nesta localidade (INT-2) a
morfo-espécie Chrysobalanaceae sp3 que com apenas 1 indivíduo ocupou o 9° lugar de
maior VI, com DoR superior a DoR da espécie E. edulis (mais alto valor de VI) (Anexo 6).
As classes arvoretas e árvores pequenas das localidades apresentaram muitas
semelhanças florísticas. Considerando as cinco famílias mais abundantes em número de
indivíduos, destacam-se Myrtaceae e Arecaceae presentes em todas as localidades, sendo
Myrtaceae a família mais rica em espécies. Nestas classes de menores tamanhos,
Nyctaginaceae aparece também entre as mais abundantes em quase todas as localidades, com
exceção de INT-2, onde aparece exclusivamente a família Sapotaceae entre as cinco, e
Lauraceae no grupo das arvoretas em B-200 e no grupo das árvores pequenas nas
localidades B-200, INT-1 e INT-2. As espécies Euterpe edulis e Guapira opposita aparecem
como as mais abundantes, presentes em todas as localidades (Anexos 7 e 9). Apesar de
Myrtaceae apresentar maior riqueza, algumas espécies tem maior representatividade em

33
abundância, como Gomidesia spectabilis nas localidades B-200 e INT-1, Myrceugenia
myrcioides em B-200 e Eugenia excelsa em B-30. As espécies Psychotria glaziovii
(Rubiaceae) e Miconia cubatanensis (Melastomataceae) foram muito abundantes em B-30
nas duas classes, exclusivamente (Anexos 7 e 9). Da mesma forma Cupania oblongifolia
(Sapindaceae) em B-200 e INT-1 apenas na classe das árvores pequenas (Anexo 9).
Na classe das árvores, Myrtaceae também aparece em todas as localidades como a
mais abundante e mais rica em espécies. Porém, na classe das árvores grandes ocorre apenas
em B-30 e INT-1. Na medida em que as árvores dos interiores (INT-1 e INT-2) são
consideradas, a família Nyctaginaceae perde sua representatividade em abundância,
caracterizados assim por maior densidade de indivíduos menores. Arecaceae aparece como
uma das mais abundantes em todas as localidades, na classe das árvores, porém metade do
total de indivíduos de E. edulis em B-30 apresentou diâmetro maior que 10 cm, diferente das
outras localidades em que a maioria dos indivíduos pertence a classe das arvoretas (5
cm≤DAP<10 cm). Já a família Lauraceae aparece como uma das mais abundantes e mais
ricas em todas as localidades, tanto no grupo das árvores quanto das árvores grandes,
destacando-se Ocotea dispersa como uma das mais abundantes nas duas localidades de borda
no primeiro grupo (Anexo 8) e Ocotea catharinensis como a mais abundante no interior INT-
2 no segundo grupo (Anexo 10). A família Sapindaceae está entre as mais abundantes e mais
ricas apenas em B-200 e INT-1, representada pela espécie Cupania oblongifolia, com
distribuição agregada (Anexo 8).
A partir da análise de ordenação foi possível avaliar a similaridade florística entre
as parcelas analisadas na procura de um padrão geral para as localidades. Na análise de todas
as árvores (DAP≥5 cm) foi possível observar que as parcelas das diferentes localidades
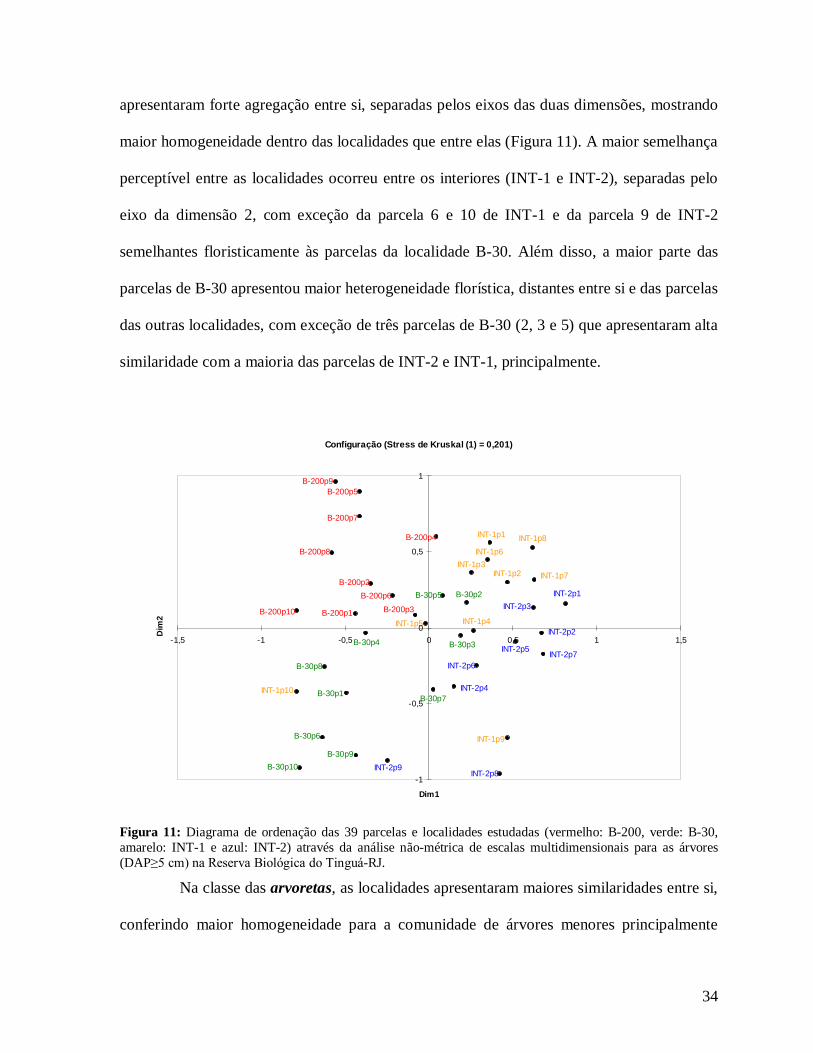
34
apresentaram forte agregação entre si, separadas pelos eixos das duas dimensões, mostrando
maior homogeneidade dentro das localidades que entre elas (Figura 11). A maior semelhança
perceptível entre as localidades ocorreu entre os interiores (INT-1 e INT-2), separadas pelo
eixo da dimensão 2, com exceção da parcela 6 e 10 de INT-1 e da parcela 9 de INT-2
semelhantes floristicamente às parcelas da localidade B-30. Além disso, a maior parte das
parcelas de B-30 apresentou maior heterogeneidade florística, distantes entre si e das parcelas
das outras localidades, com exceção de três parcelas de B-30 (2, 3 e 5) que apresentaram alta
similaridade com a maioria das parcelas de INT-2 e INT-1, principalmente.
Configuração (Stress de Kruskal (1) = 0,201)
B-200p1
B-200p2
B-200p3
B-200p4
B-200p5
B-200p6
B-200p7
B-200p8
B-200p9
B-200p10
B-30p1
B-30p2
B-30p3B-30p4
B-30p5
B-30p6
B-30p7
B-30p8
B-30p9B-30p10
INT-1p1
INT-1p2INT-1p3
INT-1p4INT-1p5
INT-1p6
INT-1p7
INT-1p8
INT-1p9
INT-1p10
INT-2p1
INT-2p2
INT-2p3
INT-2p4
INT-2p5
INT-2p6INT-2p7
INT-2p8INT-2p9
-1
-0,5
0
0,5
1
-1,5 -1 -0,5 0 0,5 1 1,5
Dim1
Dim
2
Figura 11: Diagrama de ordenação das 39 parcelas e localidades estudadas (vermelho: B-200, verde: B-30, amarelo: INT-1 e azul: INT-2) através da análise não-métrica de escalas multidimensionais para as árvores (DAP≥5 cm) na Reserva Biológica do Tinguá-RJ.
Na classe das arvoretas, as localidades apresentaram maiores similaridades entre si,
conferindo maior homogeneidade para a comunidade de árvores menores principalmente
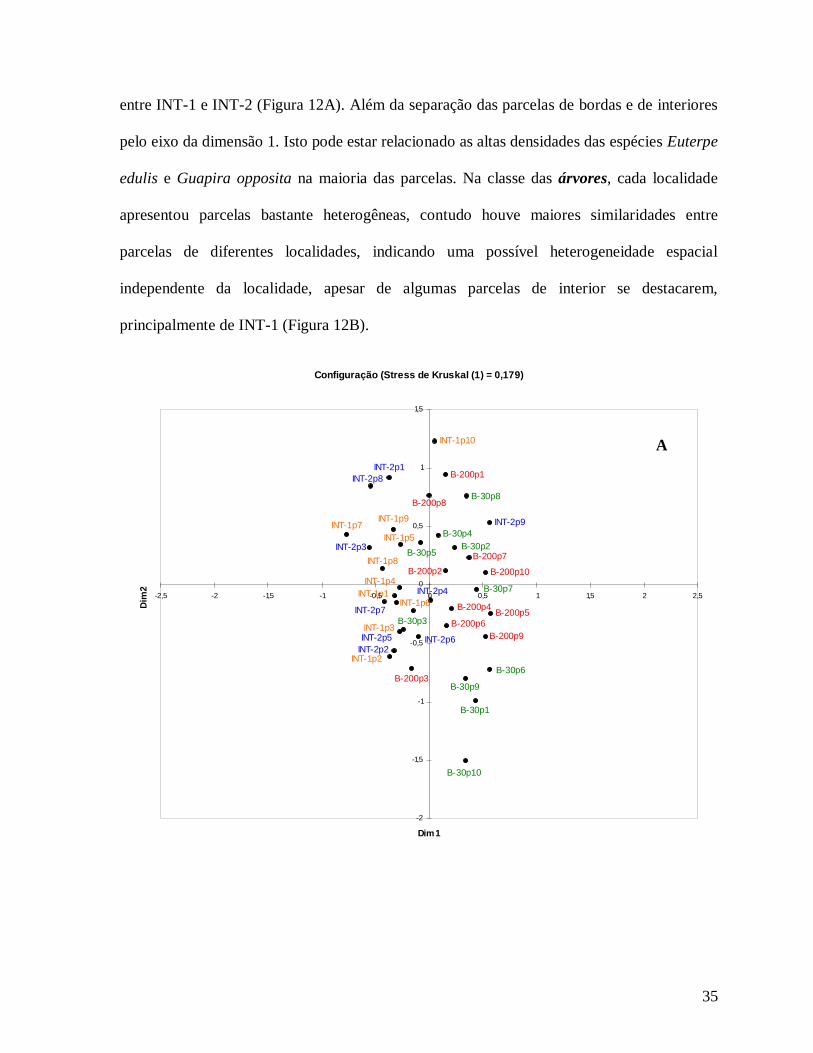
35
entre INT-1 e INT-2 (Figura 12A). Além da separação das parcelas de bordas e de interiores
pelo eixo da dimensão 1. Isto pode estar relacionado as altas densidades das espécies Euterpe
edulis e Guapira opposita na maioria das parcelas. Na classe das árvores, cada localidade
apresentou parcelas bastante heterogêneas, contudo houve maiores similaridades entre
parcelas de diferentes localidades, indicando uma possível heterogeneidade espacial
independente da localidade, apesar de algumas parcelas de interior se destacarem,
principalmente de INT-1 (Figura 12B).
Configuração (Stress de Kruskal (1) = 0,179)
B-200p1
B-200p2
B-200p3
B-200p4 B-200p5B-200p6
B-200p7
B-200p8
B-200p9
B-200p10
B-30p1
B-30p2
B-30p3
B-30p4
B-30p5
B-30p6
B-30p7
B-30p8
B-30p9
B-30p10
INT-1p1
INT-1p2
INT-1p3
INT-1p4
INT-1p5
INT-1p6
INT-1p7
INT-1p8
INT-1p9
INT-1p10
INT-2p1
INT-2p2
INT-2p3
INT-2p4
INT-2p5 INT-2p6
INT-2p7
INT-2p8
INT-2p9
-2
-1,5
-1
-0,5
0
0,5
1
1,5
-2,5 -2 -1,5 -1 -0,5 0 0,5 1 1,5 2 2,5
Dim1
Dim
2
A
36
Configuração (Stress de Kruskal (1) = 0,217)
INT-2p9
INT-2p8
INT-2p7
INT-2p6
INT-2p5
INT-2p4
INT-2p3
INT-2p2
INT-2p1
INT-1p10
INT-1p9
INT-1p8
INT-1p7
INT-1p6
INT-1p5
INT-1p4
INT-1p3
INT-1p2
INT-1p1
B-30p10
B-30p9
B-30p8
B-30p7B-30p6B-30p5
B-30p4
B-30p3
B-30p2
B-30p1
B-200p10B-200p9
B-200p8
B-200p7
B-200p6
B-200p5
B-200p4
B-200p3
B-200p2
B-200p1
-1
-0,5
0
0,5
1
1,5
-2 -1,5 -1 -0,5 0 0,5 1 1,5
Dim1
Dim
2
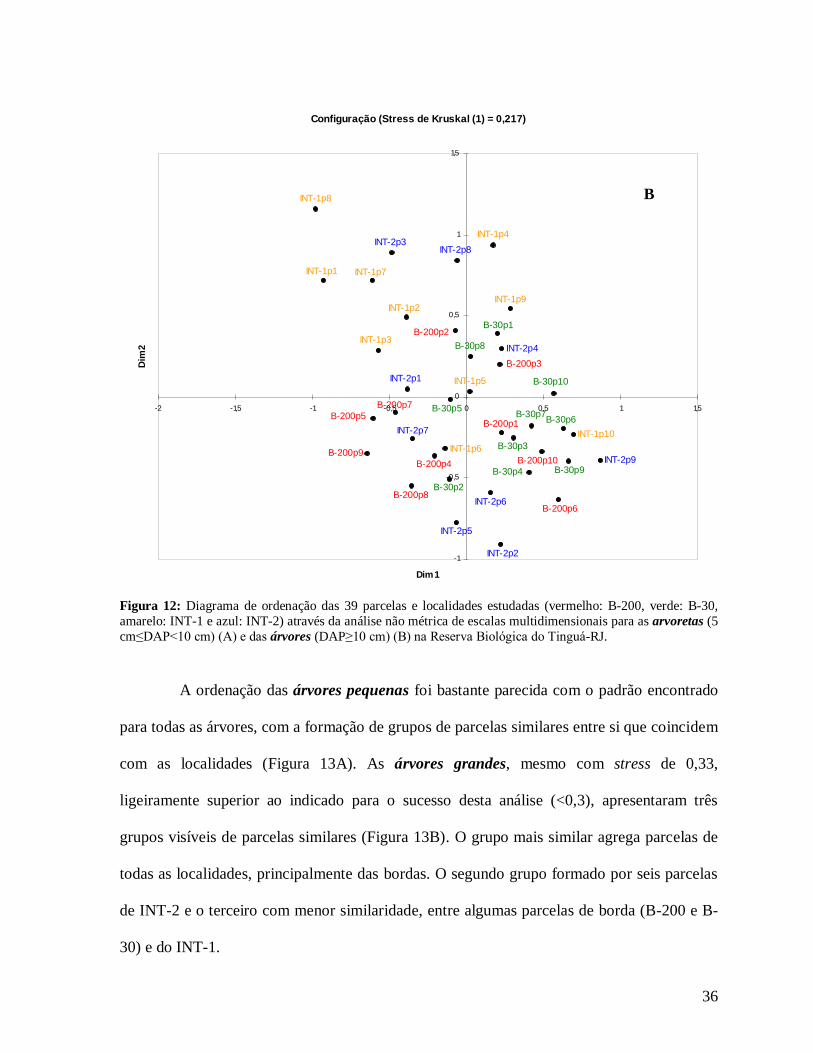
Figura 12: Diagrama de ordenação das 39 parcelas e localidades estudadas (vermelho: B-200, verde: B-30, amarelo: INT-1 e azul: INT-2) através da análise não métrica de escalas multidimensionais para as arvoretas (5 cm≤DAP<10 cm) (A) e das árvores (DAP≥10 cm) (B) na Reserva Biológica do Tinguá-RJ.
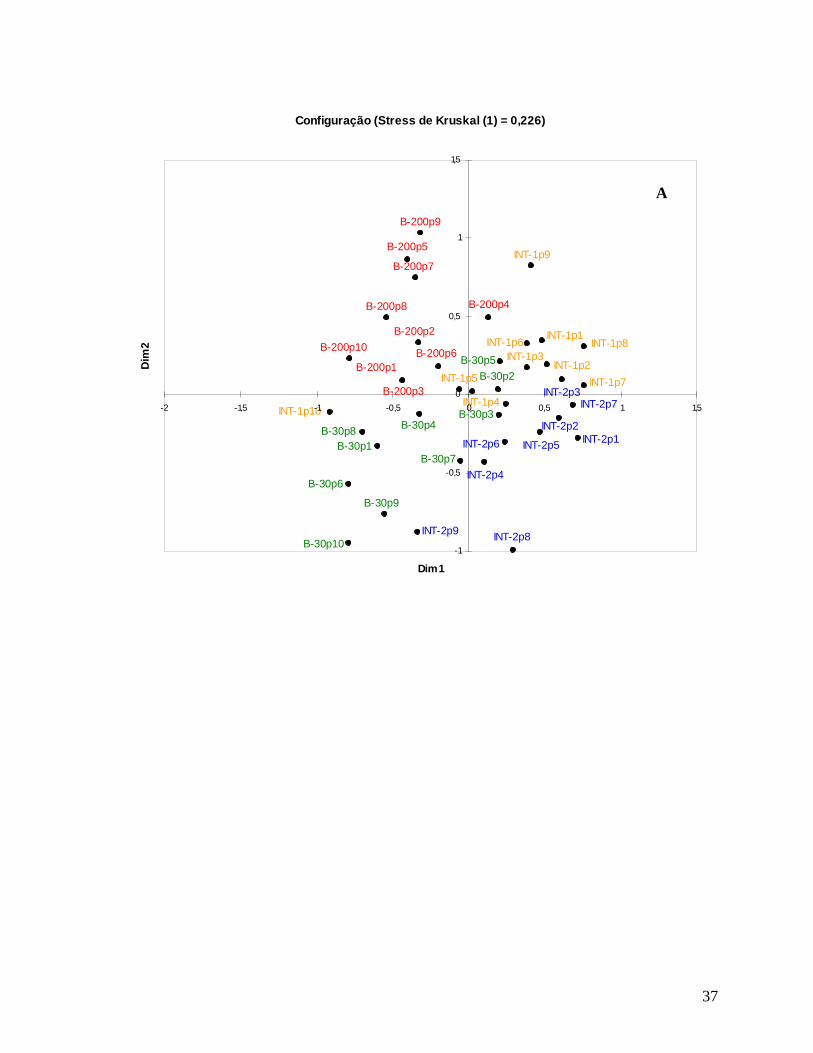
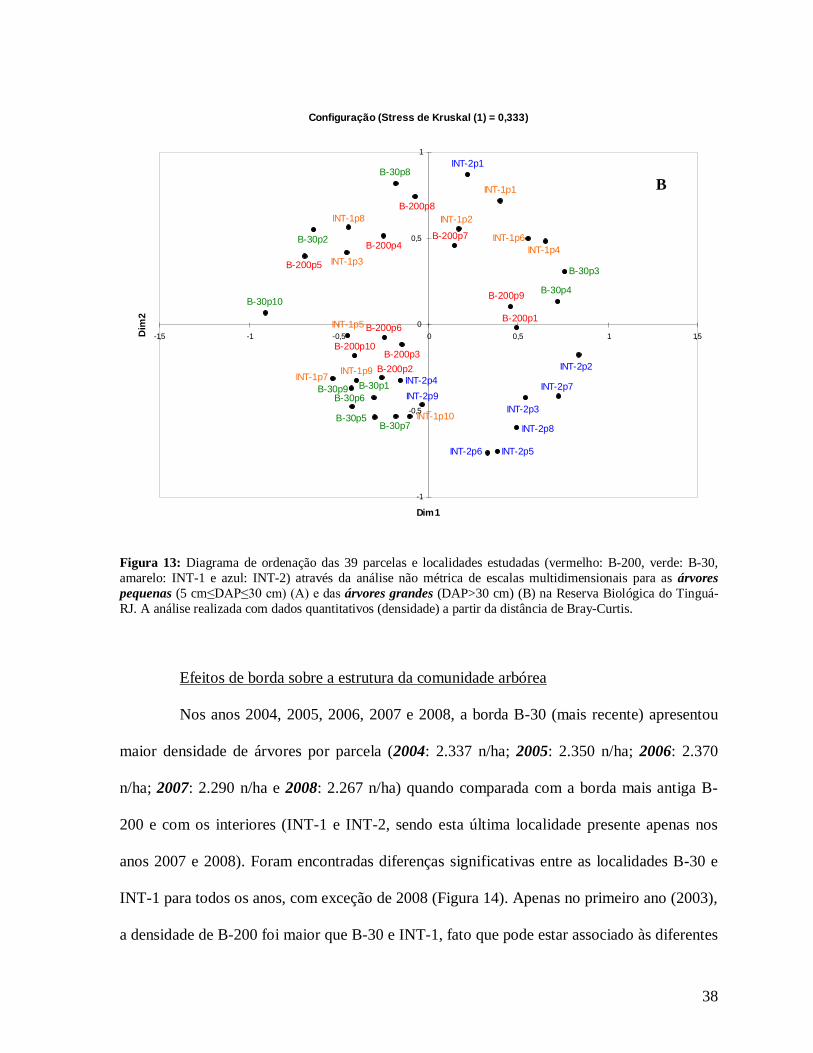
A ordenação das árvores pequenas foi bastante parecida com o padrão encontrado
para todas as árvores, com a formação de grupos de parcelas similares entre si que coincidem
com as localidades (Figura 13A). As árvores grandes, mesmo com stress de 0,33,
ligeiramente superior ao indicado para o sucesso desta análise (<0,3), apresentaram três
grupos visíveis de parcelas similares (Figura 13B). O grupo mais similar agrega parcelas de
todas as localidades, principalmente das bordas. O segundo grupo formado por seis parcelas
de INT-2 e o terceiro com menor similaridade, entre algumas parcelas de borda (B-200 e B-
30) e do INT-1.
B
37
Configuração (Stress de Kruskal (1) = 0,226)
INT-2p9 INT-2p8
INT-2p7
INT-2p6 INT-2p5
INT-2p4
INT-2p3
INT-2p2INT-2p1
INT-1p10
INT-1p9
INT-1p8
INT-1p7
INT-1p6
INT-1p5
INT-1p4
INT-1p3INT-1p2
INT-1p1
B-30p10
B-30p9
B-30p8
B-30p7
B-30p6
B-30p5
B-30p4B-30p3
B-30p2
B-30p1
B-200p10
B-200p9
B-200p8
B-200p7
B-200p6
B-200p5
B-200p4
B-200p3
B-200p2
B-200p1
-1
-0,5
0
0,5
1
1,5
-2 -1,5 -1 -0,5 0 0,5 1 1,5
Dim1
Dim
2
A
38
Configuração (Stress de Kruskal (1) = 0,333)
B-200p1
B-200p2B-200p3
B-200p4
B-200p5
B-200p6
B-200p7
B-200p8
B-200p9
B-200p10
B-30p1
B-30p2
B-30p3
B-30p4
B-30p5
B-30p6
B-30p7
B-30p8
B-30p9
B-30p10
INT-1p1
INT-1p2
INT-1p3INT-1p4
INT-1p5
INT-1p6
INT-1p7
INT-1p8
INT-1p9
INT-1p10
INT-2p1
INT-2p2
INT-2p3
INT-2p4
INT-2p5INT-2p6
INT-2p7
INT-2p8
INT-2p9
-1
-0,5
0
0,5
1
-1,5 -1 -0,5 0 0,5 1 1,5
Dim1
Dim
2
Figura 13: Diagrama de ordenação das 39 parcelas e localidades estudadas (vermelho: B-200, verde: B-30, amarelo: INT-1 e azul: INT-2) através da análise não métrica de escalas multidimensionais para as árvores pequenas (5 cm≤DAP≤30 cm) (A) e das árvores grandes (DAP>30 cm) (B) na Reserva Biológica do Tinguá-RJ. A análise realizada com dados quantitativos (densidade) a partir da distância de Bray-Curtis.
Efeitos de borda sobre a estrutura da comunidade arbórea
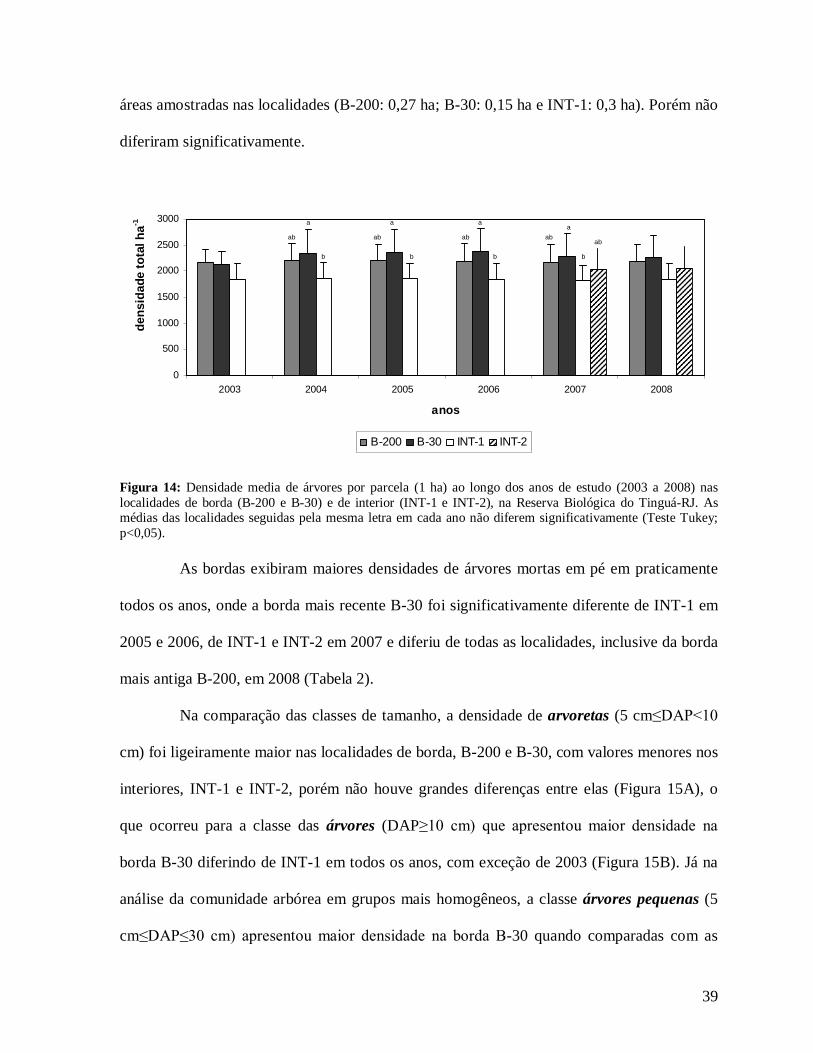
Nos anos 2004, 2005, 2006, 2007 e 2008, a borda B-30 (mais recente) apresentou
maior densidade de árvores por parcela (2004: 2.337 n/ha; 2005: 2.350 n/ha; 2006: 2.370
n/ha; 2007: 2.290 n/ha e 2008: 2.267 n/ha) quando comparada com a borda mais antiga B-
200 e com os interiores (INT-1 e INT-2, sendo esta última localidade presente apenas nos
anos 2007 e 2008). Foram encontradas diferenças significativas entre as localidades B-30 e
INT-1 para todos os anos, com exceção de 2008 (Figura 14). Apenas no primeiro ano (2003),
a densidade de B-200 foi maior que B-30 e INT-1, fato que pode estar associado às diferentes
B
39
áreas amostradas nas localidades (B-200: 0,27 ha; B-30: 0,15 ha e INT-1: 0,3 ha). Porém não
diferiram significativamente.
Figura 14: Densidade media de árvores por parcela (1 ha) ao longo dos anos de estudo (2003 a 2008) nas localidades de borda (B-200 e B-30) e de interior (INT-1 e INT-2), na Reserva Biológica do Tinguá-RJ. As médias das localidades seguidas pela mesma letra em cada ano não diferem significativamente (Teste Tukey; p<0,05).
As bordas exibiram maiores densidades de árvores mortas em pé em praticamente
todos os anos, onde a borda mais recente B-30 foi significativamente diferente de INT-1 em
2005 e 2006, de INT-1 e INT-2 em 2007 e diferiu de todas as localidades, inclusive da borda
mais antiga B-200, em 2008 (Tabela 2).
Na comparação das classes de tamanho, a densidade de arvoretas (5 cm≤DAP<10
cm) foi ligeiramente maior nas localidades de borda, B-200 e B-30, com valores menores nos
interiores, INT-1 e INT-2, porém não houve grandes diferenças entre elas (Figura 15A), o
que ocorreu para a classe das árvores (DAP≥10 cm) que apresentou maior densidade na
borda B-30 diferindo de INT-1 em todos os anos, com exceção de 2003 (Figura 15B). Já na
análise da comunidade arbórea em grupos mais homogêneos, a classe árvores pequenas (5
cm≤DAP≤30 cm) apresentou maior densidade na borda B-30 quando comparadas com as
0
500
1000
1500
2000
2500
3000
2003 2004 2005 2006 2007 2008
anos
dens
idad
e ha
-1
B-200 B-30 INT-1 INT-2
ab
a
b
ab
a
b
ab a
a
ab
b b
ab
dens
idad
e to
tal h
a-1
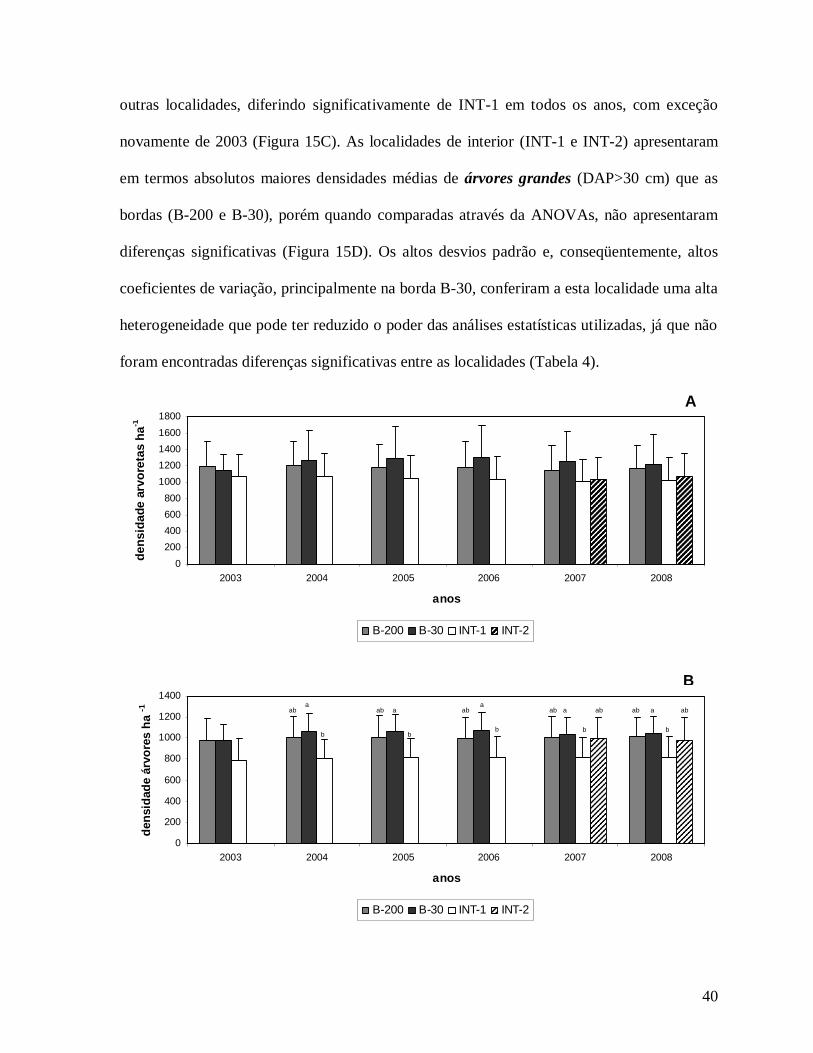
40
outras localidades, diferindo significativamente de INT-1 em todos os anos, com exceção
novamente de 2003 (Figura 15C). As localidades de interior (INT-1 e INT-2) apresentaram
em termos absolutos maiores densidades médias de árvores grandes (DAP>30 cm) que as
bordas (B-200 e B-30), porém quando comparadas através da ANOVAs, não apresentaram
diferenças significativas (Figura 15D). Os altos desvios padrão e, conseqüentemente, altos
coeficientes de variação, principalmente na borda B-30, conferiram a esta localidade uma alta
heterogeneidade que pode ter reduzido o poder das análises estatísticas utilizadas, já que não
foram encontradas diferenças significativas entre as localidades (Tabela 4).
0200400600800
10001200140016001800
2003 2004 2005 2006 2007 2008
anos
dens
idad
e ar
vore
tas
ha-1
B-200 B-30 INT-1 INT-2
dens
idad
e ar
vore
tas
ha-1
A
0
200
400
600
800
1000
1200
1400
2003 2004 2005 2006 2007 2008
anos
dens
idad
e ár
vore
s ha
-1
B-200 B-30 INT-1 INT-2
ab a
b
ab a
b b
ab a
a ab ab
b
ab ab
b
a
B
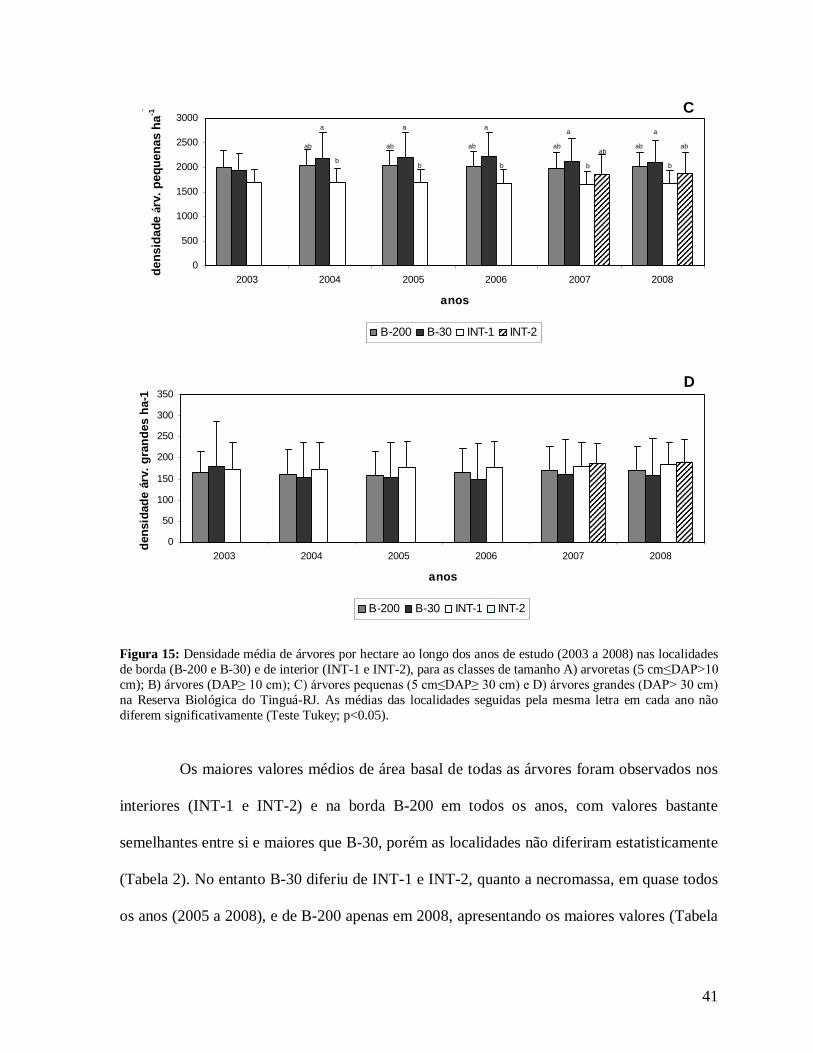
41
Figura 15: Densidade média de árvores por hectare ao longo dos anos de estudo (2003 a 2008) nas localidades de borda (B-200 e B-30) e de interior (INT-1 e INT-2), para as classes de tamanho A) arvoretas (5 cm≤DAP>10 cm); B) árvores (DAP≥ 10 cm); C) árvores pequenas (5 cm≤DAP≥ 30 cm) e D) árvores grandes (DAP> 30 cm) na Reserva Biológica do Tinguá-RJ. As médias das localidades seguidas pela mesma letra em cada ano não diferem significativamente (Teste Tukey; p<0.05).
Os maiores valores médios de área basal de todas as árvores foram observados nos
interiores (INT-1 e INT-2) e na borda B-200 em todos os anos, com valores bastante
semelhantes entre si e maiores que B-30, porém as localidades não diferiram estatisticamente
(Tabela 2). No entanto B-30 diferiu de INT-1 e INT-2, quanto a necromassa, em quase todos
os anos (2005 a 2008), e de B-200 apenas em 2008, apresentando os maiores valores (Tabela
0
500
1000
1500
2000
2500
3000
2003 2004 2005 2006 2007 2008
anos
dens
idad
e ár
v. p
eque
nas
ha-
1
B-200 B-30 INT-1 INT-2
a a a a
a
ab ab ab ab ab
ab ab
b b b b b
dens
idad
e ár
v. p
eque
nas
ha-1
C
0
50
100
150
200
250
300
350
2003 2004 2005 2006 2007 2008
anos
dens
idad
e ár
v. g
rand
es h
a-1
B-200 B-30 INT-1 INT-2
D

42
2). Analisando os valores de área basal das classes de tamanho, arvoretas e árvores, as
localidades não contrastaram entre si, apesar de as arvoretas apresentarem valores de
biomassa ligeiramente maiores na localidade B-30 e as árvores nas localidades INT-1, INT-2
e B-200, em todos os anos de estudo (Tabela 3). Entretanto, os valores médios de área basal
das árvores pequenas foram significativamente maiores em B-30 em relação ao interior INT-
1, em 2004, 2005 e 2006. A área basal das árvores grandes não diferiu entre as localidades,
devido provavelmente aos altos coeficientes de variação (CV) na borda B-30 (Tabela 4).
Distribuição de tamanhos
Todas as localidades apresentaram distribuição de diâmetros na forma de J reverso
em todos os anos (Figura 16). Comparando estas localidades, a borda B-30 apresentou
maiores densidades de árvores nas três primeiras classes de diâmetro (DAP = 5 a 39,9 cm)
(Figura 16), constituindo, em média, 98% dos indivíduos amostrados em quase todos os
anos, com exceção do ano 2003 (Figura 17). Neste, a borda mais antiga B-200 apresentou
maior densidade nas duas primeiras classes, fato que pode estar relacionado à menor área
amostrada em B-30 (0,15 ha), que pode ter subestimado a densidade nesta localidade.
INT-1 apresentou os menores valores nas três primeiras classes, apesar de esta
localidade ter exibido maiores porcentagens de indivíduos na primeira classe em todos os
anos (Figura 17). Além disso, foram verificadas maiores densidades de árvores grandes
(DAP≥40 cm), em todos os anos, nas localidades de interior, INT-1 e INT-2, e B-200 e
baixos valores na borda mais recente (Figura 16).
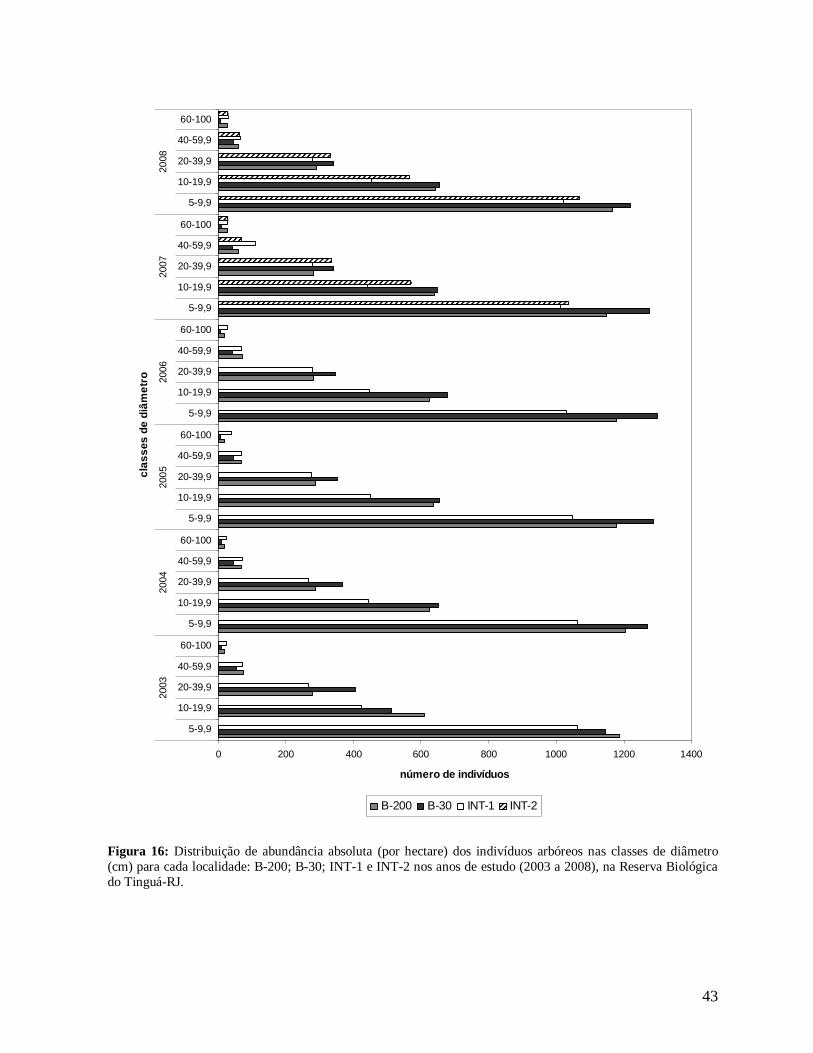
43
0 200 400 600 800 1000 1200 1400
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
2003
2004
2005
2006
2007
2008
clas
ses
de d
iâm
etro
número de indivíduos
B-200 B-30 INT-1 INT-2
Figura 16: Distribuição de abundância absoluta (por hectare) dos indivíduos arbóreos nas classes de diâmetro (cm) para cada localidade: B-200; B-30; INT-1 e INT-2 nos anos de estudo (2003 a 2008), na Reserva Biológica do Tinguá-RJ.
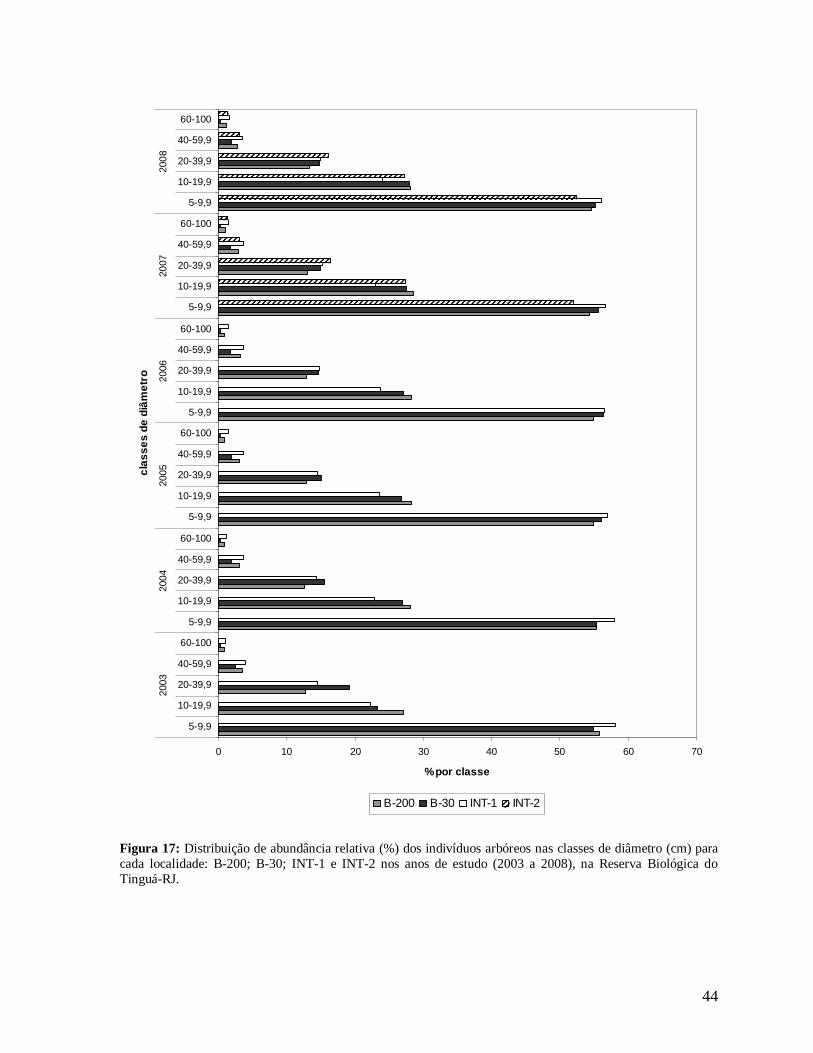
44
0 10 20 30 40 50 60 70
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
5-9,9
10-19,9
20-39,9
40-59,9
60-100
2003
2004
2005
2006
2007
2008
clas
ses
de d
iâm
etro
% por classe
B-200 B-30 INT-1 INT-2
Figura 17: Distribuição de abundância relativa (%) dos indivíduos arbóreos nas classes de diâmetro (cm) para cada localidade: B-200; B-30; INT-1 e INT-2 nos anos de estudo (2003 a 2008), na Reserva Biológica do Tinguá-RJ.
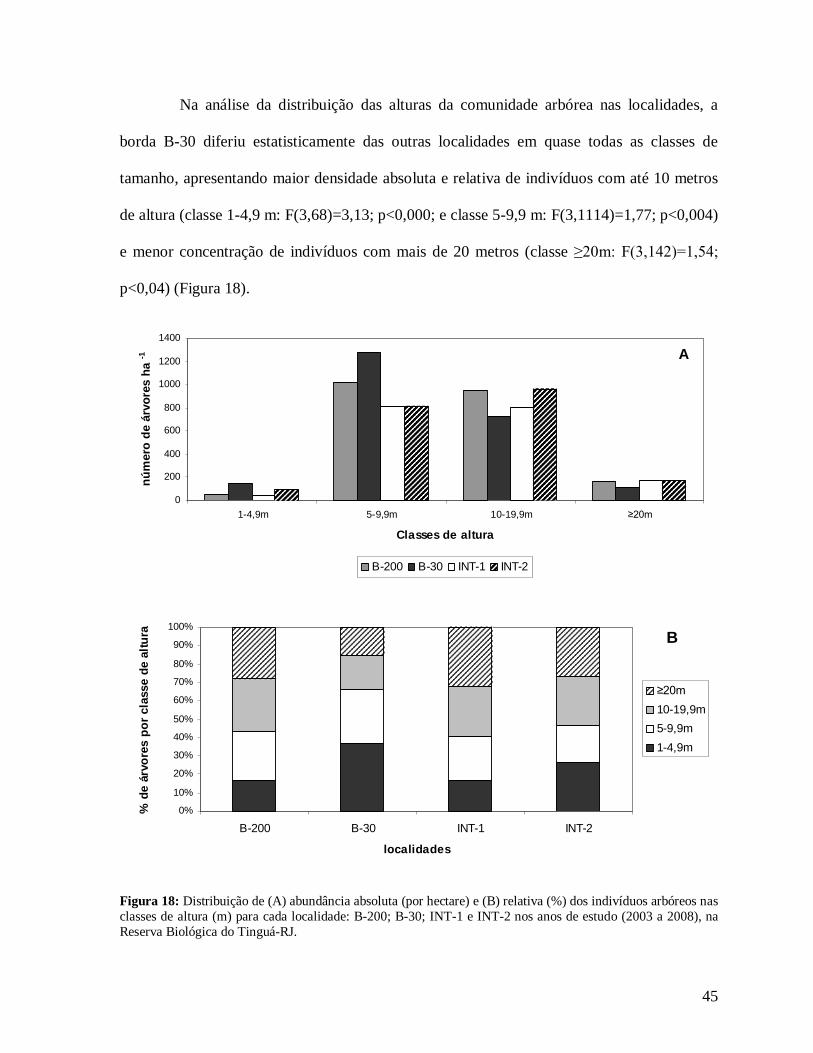
45
Na análise da distribuição das alturas da comunidade arbórea nas localidades, a
borda B-30 diferiu estatisticamente das outras localidades em quase todas as classes de
tamanho, apresentando maior densidade absoluta e relativa de indivíduos com até 10 metros
de altura (classe 1-4,9 m: F(3,68)=3,13; p<0,000; e classe 5-9,9 m: F(3,1114)=1,77; p<0,004)
e menor concentração de indivíduos com mais de 20 metros (classe ≥20m: F(3,142)=1,54;
p<0,04) (Figura 18).
0
200
400
600
800
1000
1200
1400
1-4,9m 5-9,9m 10-19,9m ≥20m
Classes de altura
núm
ero
de á
rvor
es h
a-1
B-200 B-30 INT-1 INT-2
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
B-200 B-30 INT-1 INT-2
localidades
% d
e ár
vore
s po
r cla
sse
de a
ltura
≥20m10-19,9m5-9,9m1-4,9m
Figura 18: Distribuição de (A) abundância absoluta (por hectare) e (B) relativa (%) dos indivíduos arbóreos nas classes de altura (m) para cada localidade: B-200; B-30; INT-1 e INT-2 nos anos de estudo (2003 a 2008), na Reserva Biológica do Tinguá-RJ.
A
B
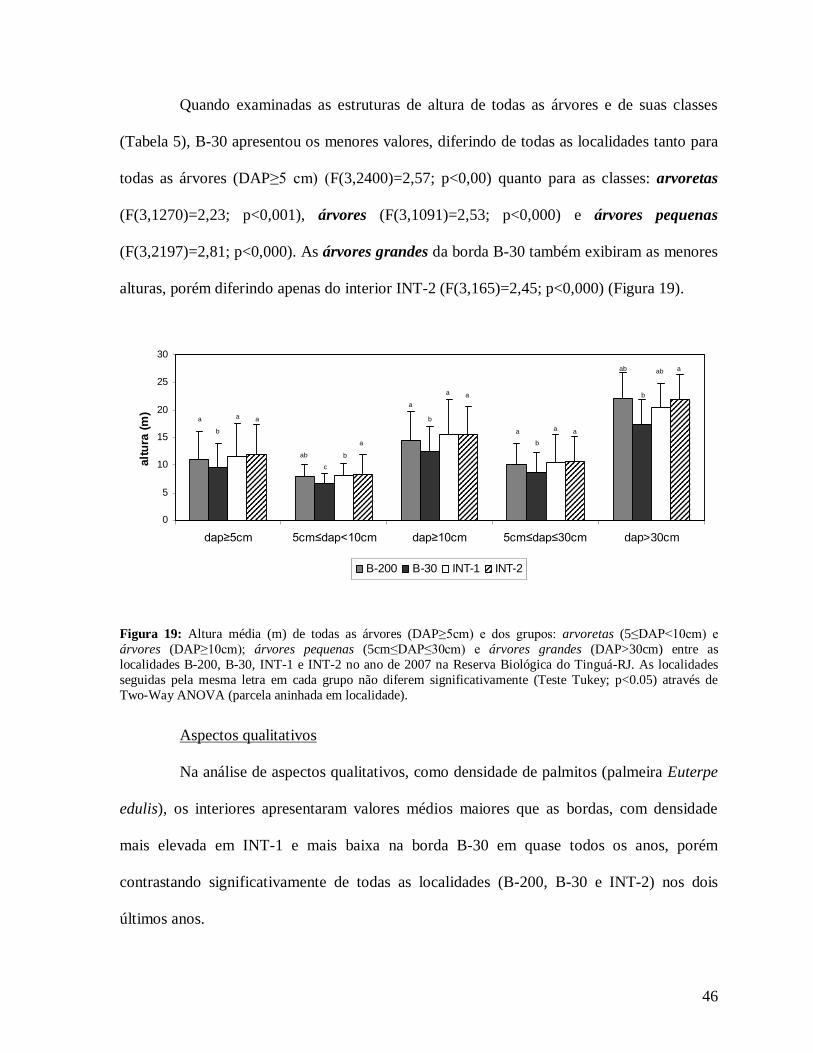
46
0
5
10
15
20
25
30
dap≥5cm 5cm≤dap<10cm dap≥10cm 5cm≤dap≤30cm dap>30cm
altu
ra (m
)
B-200 B-30 INT-1 INT-2
Quando examinadas as estruturas de altura de todas as árvores e de suas classes
(Tabela 5), B-30 apresentou os menores valores, diferindo de todas as localidades tanto para
todas as árvores (DAP≥5 cm) (F(3,2400)=2,57; p<0,00) quanto para as classes: arvoretas
(F(3,1270)=2,23; p<0,001), árvores (F(3,1091)=2,53; p<0,000) e árvores pequenas
(F(3,2197)=2,81; p<0,000). As árvores grandes da borda B-30 também exibiram as menores
alturas, porém diferindo apenas do interior INT-2 (F(3,165)=2,45; p<0,000) (Figura 19).
Figura 19: Altura média (m) de todas as árvores (DAP≥5cm) e dos grupos: arvoretas (5≤DAP<10cm) e árvores (DAP≥10cm); árvores pequenas (5cm≤DAP≤30cm) e árvores grandes (DAP>30cm) entre as localidades B-200, B-30, INT-1 e INT-2 no ano de 2007 na Reserva Biológica do Tinguá-RJ. As localidades seguidas pela mesma letra em cada grupo não diferem significativamente (Teste Tukey; p<0.05) através de Two-Way ANOVA (parcela aninhada em localidade).
Aspectos qualitativos
Na análise de aspectos qualitativos, como densidade de palmitos (palmeira Euterpe
edulis), os interiores apresentaram valores médios maiores que as bordas, com densidade
mais elevada em INT-1 e mais baixa na borda B-30 em quase todos os anos, porém
contrastando significativamente de todas as localidades (B-200, B-30 e INT-2) nos dois
últimos anos.
a a a
ab
c
b
a
b
a a
a
b
b
a a a
a ab ab
b

47
Já a densidade de árvores com tronco múltiplo (perfilhadas) foi mais elevada em B-
30, seguida de B-200, INT-1 e INT-2, diferindo estatisticamente dos interiores em
praticamente todos os anos, com exceção de 2003 (Tabela 5).
Dinâmica da Comunidade
Foi verificado um padrão geral na dinâmica das árvores para todas as localidades
(B-200, B-30, INT-1 e INT-2) e em todos os intervalos de estudo (2003-2004, 2004-2005,
2005-2006, 2006-2007, 2007-2008 e 2004-2008), com maiores densidades e dinâmica mais
acelerada nas menores classes de tamanho, com altos valores de recrutas (ingressos), de
indivíduos mortos e de indivíduos vivos que caíram nos intervalos, além de muitos
indivíduos mudando de classe de tamanho. De forma diferente, os indivíduos das classes
maiores se caracterizaram por uma dinâmica muito mais lenta, como pouca mudança de
classe e baixos valores de indivíduos mortos e caídos nos intervalos (Tabela 6, 7, 8 e 9).
Nos intervalos 2003-2004, 2004-2005, 2005-2006 e 2006-2007 foi observada maior
dinâmica da comunidade na borda mais recente B-30, com altos valores de recrutas e de
indivíduos mortos (Tabela 7), principalmente, e maior estabilidade nas localidades INT-1 e
B-200 (Tabela 6 e 8). No último intervalo, 2007-2008, com a inclusão de INT-2, este padrão
ainda se manteve parcialmente (Tabela 9), com maiores valores de mortos na localidade B-30
(Tabela 7). Apesar disso, a outra localidade de borda (B-200, mais antiga), apresentou maior
estabilidade (Tabela 6), seguida pelos interiores, INT-1 (Tabela 8) e INT-2 (Tabela 9), que
apresentaram dinâmicas bastante semelhantes entre si com maiores valores de recrutas.
Avaliando a dinâmica desta comunidade num intervalo de tempo maior (2004-2008; quatro
anos), foi verificado a manutenção deste mesmo padrão, com uma pronunciada aceleração da

48
dinâmica na borda mais recente B-30 (Tabela 7), e maior estabilidade em INT-1 e B-200
(Tabela 8 e 6, respectivamente).
Considerando as mudanças nas densidades de árvores, não foi encontrado nenhum
padrão claro entre as localidades, com aumentos e diminuições nestes valores de acordo com
os intervalos estudados de 1 ano. Porém, no intervalo de 4 anos, foi observado um
decréscimo no número total de indivíduos em todas as localidades (B-200: -0,76%; B-30: -
3,00% e INT-1: -1,25%), sendo a maior diminuição na borda mais recente. No grupo das
arvoretas (5 cm ≤ DAP < 10 cm), estes decréscimos foram mantidos com maior diminuição
no interior (B-200: -3,05%; B-30: -3,94% e INT-1: -4,08%), diferentemente dos grupos das
árvores (DAP ≥ 10 cm), árvores pequenas (5 cm ≤ DAP ≤ 30 cm) e árvores grandes (DAP
> 30 cm) que tiveram acréscimos na borda mais antiga e no interior (ou permaneceram
estáveis). Já a borda mais recente apresentou decréscimos no número de indivíduos nestes
grupos (B-200: 2,00%; B-30: -1,88% e INT-1: 2,49%; B-200: 1,01%; B-30: -1,09% e INT-1:
1,02%; B-200: 0%; B-30: -2,08% e INT-1: 1,85%, respectivamente).
Com relação aos valores de área basal nas localidades no intervalo de 4 anos, de
maneira geral, as localidades B-200 e INT-1 apresentaram incrementos que ocorreram,
principalmente, devido ao aumento na biomassa dos indivíduos maiores (DAP > 30 cm).
Todavia, na borda B-30 houve um decréscimo para todas as árvores e também para quase
todos os grupos de tamanho (arvoretas, árvores e árvores pequenas), com exceção das
árvores grandes que apresentaram um acréscimo na área basal, porém aproximadamente sete
vezes menor que nas outras localidades (todas as árvores: B-200: 4,77%; B-30: -2,02%;
INT-1: 4,80%; arvoretas: B-200 e INT-1: 0%; B-30: -0,93%; árvores: B-200: 5,79; B-30: -

49
1,57%, INT-1: 5,07%; árvores pequenas: B-200: -0,53%; B-30: -5,32%; INT-1: -0,16%;
árvores grandes: B-200: 8,97%; B-30: 1,69%; INT-1: 7,69%).
Em geral, a borda B-30 apresentou maiores taxas de mortalidade, recrutamento e
turnover em todos os intervalos estudados, e as localidades B-200, INT-1 e INT-2, esta
última localidade no intervalo 2007-2008, apresentaram taxas bastante semelhantes entre si.
Apesar disso, a borda mais recente B-30 diferiu marginalmente de B-200 e INT-1 apenas no
intervalo 2004-2005 (H(2,30)=5,99; p=0,05). No intervalo de 4 anos, a taxa de mortalidade
anual em B-30 foi superior à taxa de recrutamento, sendo que as outras localidades (B-200 e
INT-1) apresentaram o oposto, isto é, taxas de recrutamentos maiores que as taxas de
mortalidade (Tabela 10). Além disso, B-200 e INT-1 apresentaram valores muito
semelhantes entre mortos (mortos em pé e morte por queda) e recrutas neste mesmo intervalo
(2004-2008) (Tabela 6; 8), mostrando uma dinâmica bastante estável. Entretanto, em B-30, a
densidade de mortos superou a de recrutas (Tabela 7), provavelmente como resultado das
taxas de mortalidade e recrutamento citadas acima.
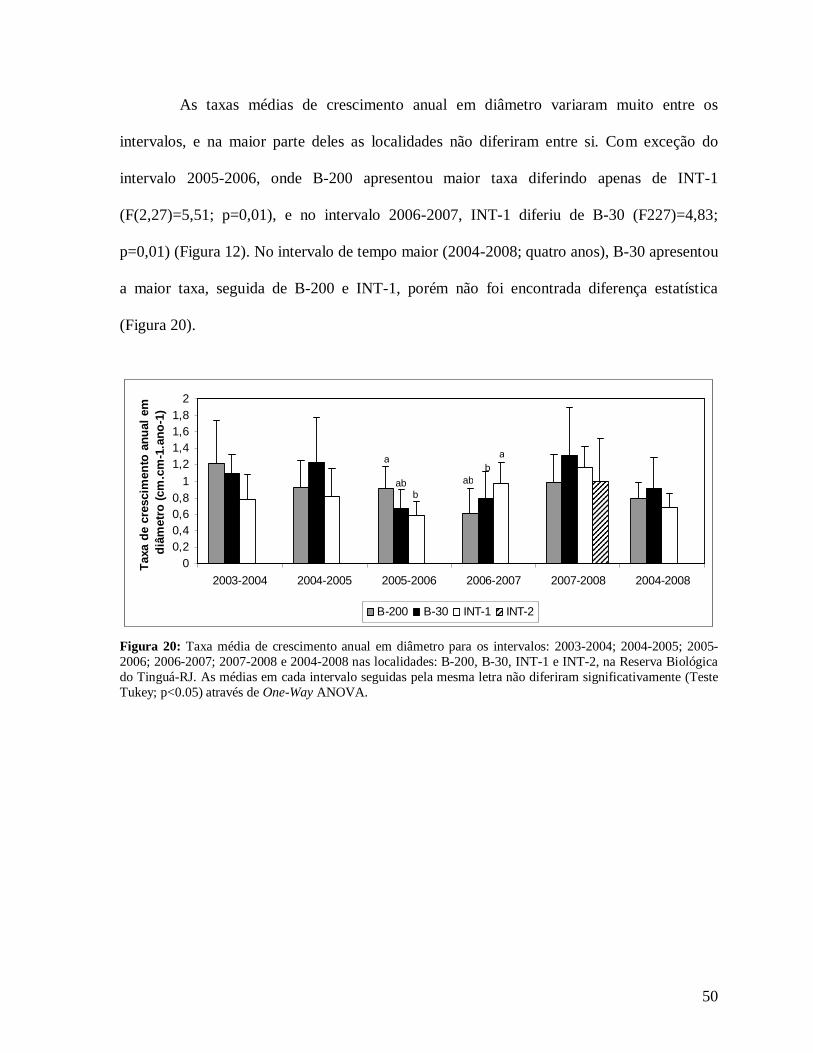
50
As taxas médias de crescimento anual em diâmetro variaram muito entre os
intervalos, e na maior parte deles as localidades não diferiram entre si. Com exceção do
intervalo 2005-2006, onde B-200 apresentou maior taxa diferindo apenas de INT-1
(F(2,27)=5,51; p=0,01), e no intervalo 2006-2007, INT-1 diferiu de B-30 (F227)=4,83;
p=0,01) (Figura 12). No intervalo de tempo maior (2004-2008; quatro anos), B-30 apresentou
a maior taxa, seguida de B-200 e INT-1, porém não foi encontrada diferença estatística
(Figura 20).
Figura 20: Taxa média de crescimento anual em diâmetro para os intervalos: 2003-2004; 2004-2005; 2005-2006; 2006-2007; 2007-2008 e 2004-2008 nas localidades: B-200, B-30, INT-1 e INT-2, na Reserva Biológica do Tinguá-RJ. As médias em cada intervalo seguidas pela mesma letra não diferiram significativamente (Teste Tukey; p<0.05) através de One-Way ANOVA.
00,20,40,60,8
11,21,41,61,8
2
2003-2004 2004-2005 2005-2006 2006-2007 2007-2008 2004-2008
Taxa
de
cres
cim
ento
anu
al e
m
diâm
etro
(cm
.cm
-1.a
no-1
)
B-200 B-30 INT-1 INT-2
a
ab b
ab b
a

51
DISCUSSÃO
Composição florística
A altíssima riqueza local encontrada para a comunidade arbórea neste estudo (441
espécies e morfo-espécies; 377 espécies por hectare) coloca a Reserva Biológica do Tinguá
como uma das regiões mais ricas da Mata Atlântica, quando comparada a outros trechos
deste bioma, mesmo considerando os distintos critérios de inclusão, tamanhos das áreas e
esforços amostrais (Rodrigues 1996; Guedes-Bruni et al. 1997; Thomaz & Monteiro 1997;
Kurtz & Araújo 2000; Silva & Nascimento 2001; Moreno et al. 2003; Rodrigues 2004;
Carvalho 2005; Garcia 2007) (Tabela 11).
O dossel da mata na ReBio do Tinguá foi dominado por muitas espécies raras
(apenas 1 indivíduo na área amostrada). Esta altíssima riqueza de espécies raras pode estar
relacionada a grande heterogeneidade do ambiente, que possivelmente é potencializado pela
presença das aberturas lineares de dossel de diferentes idades, constituindo quatro bordas,
que atravessam o remanescente florestal, elevando ainda mais a riqueza e diversidade total
(441 espécies; H’= 4,9). Alguns estudos na Mata Atlântica já relacionaram o padrão de
grande diversidade, mostrando altos índices de diversidade de Shannon, a alta densidade de
espécies raras (Guedes-Bruni et al. 1997; Rodrigues 2004). Além disso, como ocorre em
algumas florestas tropicais (e.g. Kurtz & Araújo 2000; Moreno et al. 2003; Rodrigues 2004),
a estrutura da comunidade arbórea da ReBio do Tinguá exibe um grande número de espécies
com baixos valores acerca dos parâmetros fitossociológicos (DR, FR e DoR), atribuídos
principalmente as estas espécies raras e ocasionais (com 1 a 9 indivíduos), e poucas espécies
abundantes mostrando grande contribuição, seja por sua alta densidade e frequência ou
grande área basal total.

52
De forma semelhante a outros estudos em remanescentes e fragmentos de Mata
Atlântica, a família Myrtaceae também foi considerada a mais representativa em número de
indivíduos e/ou espécies na comunidade arbustiva-arbórea (Rodrigues 1996; Guedes-Bruni et
al. 1997; Kurtz & Araújo 2000; Oliveira-Filho & Fontes 2000; Rodrigues 2004; Pietro 2008)
ou ficou entre as três famílias com maior riqueza (Silva & Nascimento 2001; Moreno et al.
2003; Garcia 2007). De acordo com Mori et al. (1983), esta família representa elemento
importante na estrutura da comunidade arbórea da Mata Atlântica, oferecendo fração
representativa de recursos alimentares a fauna. Este estudo comprova a grande importância
das Myrtaceae na comunidade, ao mesmo tempo em que confirma a lacuna acerca de seu
conhecimento taxonômico pelo grande número de morfo-espécies encontradas, as quais
muitas possivelmente representam espécies novas (Anexo 1).
Outras famílias relevantes na ReBio do Tinguá, como Fabaceae, Euphorbiaceae,
Rubiaceae, Sapindaceae e Meliaceae também aparecem entre as mais importantes em outros
estudos, principalmente na Mata Atlântica no estado do Rio de Janeiro (Rodrigues 1996;
Guedes-Bruni et al. 1997; Kurtz & Araújo 2000; Silva & Nascimento 2001; Moreno et al.
2003; Rodrigues 2004; Carvalho 2005; Garcia 2007). Fabaceae apareceu como a família
mais rica na Mata do Carvão (RJ), na ReBio Santa Cândida (MG) e na Região de Imbaú
(RJ), considerados locais com alto grau de perturbação (ex.: ambiente fragmentado), e em um
remanescente relativamente bem preservado na Região de Imbé (RJ) (Silva & Nascimento
2001; Moreno et al. 2003; Carvalho 2005; Garcia 2007), ocupando a 4a posição em riqueza
neste estudo. Rodrigues (1996), num estudo na ReBio do Tinguá, encontrou apenas seis das
dez famílias mais ricas encontradas neste estudo (Myrtaceae, Lauraceae, Fabaceae,
Euphorbiaceae, Sapotaceae e Melastomataceae), porém com critério de inclusão mais amplo

53
(DAP≥2,5 cm) que favoreceu a quantificação de grande riqueza em Piperaceae, característica
de estratos inferiores.
Apesar da grande riqueza de Myrtaceae, suas espécies foram pouco abundantes
assim como as espécies de Lauraceae, a segunda família mais rica. No entanto, Myrtaceae
dividiu a dominância da comunidade arbórea com Arecaceae, sendo a primeira atribuída a
muitas espécies com pouca abundância, e a segunda a uma única espécie, Euterpe edulis, que
apresentou quase o dobro de indivíduos da segunda espécie mais abundante, Guapira
opposita. Segundo Mantovani (1993) e Guedes-Bruni et al. (1997), E. edulis pode ser
categorizada como espécie clímax e servir como indicadora de floresta em bom estado de
preservação. De forma semelhante, G. opposita, apesar de mostrar grande plasticidade
fenotípica e se desenvolver bem em diversos ecossistemas, é bastante freqüente
principalmente em associações clímax (Reitz 1981). Desta forma, a grande
representatividade destas espécies, além da alta riqueza de espécies raras, indicam o bom
estado de conservação em que se encontra a vegetação da ReBio do Tinguá.
Efeitos de borda sobre a estrutura e composição
A borda mais recente (B-30), de aproximadamente 30 anos, exibiu em geral maior
densidade total de árvores e diferiu principalmente de INT-1, nos anos 2004 a 2007. A maior
parte dos estudos em fragmentos florestais também relata maior densidade média nas bordas
diferentes dos interiores preservados (Laurance & Bierregaard Jr 1997; Oliveira-Filho et al.
1997; Laurance et al. 2002; Rodrigues 2004). Na análise das classes de tamanho, ficou
associada à maior densidade de árvores pequenas (5 cm≤DAP≥30 cm) principalmente
árvores com DAP de 10 a 30 cm, na localidade B-30. O efeito da densidade das arvoretas (5
cm≤DAP≥10 cm) na classe de árvores pequenas entre estas duas localidades (B-30 e INT-1)

54
foi desconsiderado, já que em sua análise separadamente não foi encontrada diferença
estatística, indicando que a comunidade arbórea atualmente não apresenta dificuldades de
regeneração. Contudo, tais árvores (10 cm≤DAP≥30 cm) pertenceram, em sua maioria, a
espécies características de estágios sucessionais iniciais e apresentaram padrões de
distribuição distintos na borda, como Miconia cubatanensis e Clethra scabra (espécies
restritas a apenas 1 parcela), Psychotria glaziovii, Eugenia excelsa e Miconia holosericea
(espécies presentes na maioria das parcelas), além de Guapira opposita, que se desenvolve
bem em ambientes distintos, todas alcançando o diâmetro máximo de 35 cm na amostragem.
De forma semelhante, Nascimento & Laurance (2006) encontraram maior densidade de
arvoretas e árvores (DAP≥1 cm) de espécies pioneiras em locais próximos à borda de
fragmentos (13 a 17 anos) comparados a floresta contínua e interiores florestais (>300 m da
borda), principalmente para a classe de 10 a 20 cm de diâmetro, onde estas apresentaram
densidade três vezes maior que os interiores. Neste estudo as bordas das aberturas lineares
não diferiram dos interiores em relação a densidade de arvoretas, diferentemente das
arvoretas da borda e do interior em um fragmento de 150 anos na Mata Atlântica (Oliveira-
Filho et al. 2007). Além deste resultado diferente encontrado, vale ressaltar que a idade do
fragmento estudado (150 anos; Oliveira-Filho et al. 2007) é muito semelhante a da borda B-
200 (200 anos) deste estudo, e no entanto apresenta estrutura distinta do interior, se
assemelhando nem mesmo a borda mais recente (B-30), considerando que as arvoretas desta
localidade não diferiram estruturalmente dos interiores (INT-1 e INT-2). Esta sugerida
homogeneidade estrutural entre as localidades para as arvoretas na ReBio do Tinguá pode ter
sido induzida pelos mais altos valores de CV obtidos para a borda B-30, que mostram mais
uma vez a heterogeneidade dos sítios (parcelas) desta localidade, com diferentes parcelas

55
apresentando tanto altíssimas densidades de arvoretas quanto baixíssimas densidades. De
forma semelhante, outros estudos mostram esta heterogeneidade dos descritores analisados o
que diminui o poder das análises estatísticas utilizadas, camuflando as diferenças estruturais
entre os locais estudados (Rodrigues 2004; Nascimento & Laurance 2006). Tal
heterogeneidade pode ser explicada pelas diferentes intensidades dos efeitos de borda ao
longo das bordas (Rodrigues 2004), ou até mesmo pela variabilidade das características
intrínsecas locais, muito comuns em florestas tropicais, como diferentes tipos de solos, de
nutrientes e posição topográfica dos sítios (Clark & Clark 2000).
A análise da composição florística das localidades também mostrou uma maior
abundância de espécies raras com pequena área basal total, incrementando ainda mais a
abundância das árvores pequenas (5 cm≤DAP≥ 30 cm). Este grupo, na borda mais recente,
também diferiu da localidade INT-1 em relação à área basal em alguns anos (2004, 2005 e
2006), onde B-30 agregou valores mais altos, associados as altíssimas densidades médias
nestes anos. A maior frequência de clareiras associada a maior mortalidade na borda (Ferreira
& Laurance 1997), decorrente da maior queda de árvores e galhos, oferece uma
diversificação de habitats que favorece o estabelecimento de diversas espécies arbóreas
(Nascimento & Laurance 2006). Espécies raras pioneiras e secundárias de estratos mais
baixos da floresta são as principais favorecidas por grandes clareiras (Denslow 1987; Hubbell
& Foster 1987; Gomes et al. 2003). Como não é de se esperar que clareiras sejam abertas
com alta frequência em florestas relativamente preservadas e em equilíbrio (Swaine et al.
1987), como os interiores florestais da ReBio do Tinguá, a maior frequência destas em
bordas favorece o estabelecimento de espécies raras adaptadas a maior intensidade de luz,

56
além da ocorrência de outras espécies comuns, como principalmente pioneiras e ocasionais
que podem ocorrer tanto em bordas como interiores (Rodrigues 2004).
Segundo a “hipótese do distúrbio intermediário” a heterogeneidade de nichos
oferecidos decorrentes de distúrbios a muitas espécies diferentes tende a aumentar a riqueza
no local (Townsend et al. 2006). Como não foram mensuradas a quantidade e a frequência
das clareiras abertas neste estudo, não é possível afirmar que as bordas apresentam mais
clareiras e que estas são formadas com maior frequência que nos interiores. Porém,
analisando a comunidade de plântulas nas mesmas localidades na ReBio do Tinguá (B-200,
B-30 e INT-1) em 2008, Reis (2008) encontrou maior abertura de dossel na borda mais
recente (B-30) diferindo do INT-1, além da maior heterogeneidade espacial. Da mesma
forma, Iguatemy (2008), comparando bordas de idades diferentes (30 e 50 anos) numa
Floresta Atlântica de baixada no Rio de Janeiro, também encontrou maior abertura na borda
mais recente (30 anos). Teoricamente sabe-se que cada espécie possui maior aptidão a locais
específicos que ofereçam os recursos necessários ao seu desenvolvimento, e que seu sucesso
depende de suas tolerâncias fisiológicas e sua habilidade competitiva que delimitam seu
padrão de ocupação (Begon et al. 2006; Olifiers & Cerqueira 2006). No caso específico das
espécies raras, a ocupação espacial de mais de um indivíduo de mesma espécie num mesmo
microhabitat pode ser limitado por fatores externos, como a ação de predadores de sementes
específicos, herbívoros ou patógenos que impediriam o estabelecimento de juvenis próximos
ao adulto da mesma espécie (Modelo de Janzen-Connel) (Clark & Clark 1984). Estas
interações seriam um dos principais fatores responsáveis pela manutenção da alta diversidade
de espécies arbóreas em florestas tropicais. Dentro deste contexto, mesmo reconhecendo os
limites do desenho experimental deste estudo para identificar o padrão de distribuição das

57
espécies, pode-se inferir que a maior riqueza de espécies da localidade B-30 pode estar
diretamente relacionada a alta diversidade de microhabitats oferecidos, a alta densidade e ao
efeito da mínima distância crítica das espécies raras (Clark & Clark 1984; Crawley 1997;
Townsend et al. 2006).
A distribuição diamétrica encontrada em todas as localidades foi semelhante ao que
ocorre em comunidades naturais (aspecto de J-reverso), que garante que o processo dinâmico
florestal se perpetue, com a morte de indivíduos grandes, geralmente por senescência, dando
lugar ao desenvolvimento de indivíduos jovens (Swaine et al. 1987). Foi encontrada maior
representatividade de árvores mais baixas na borda B-30 em todas as classes analisadas, fato
que parece estar relacionado a dominância de espécies de estágios sucessionais iniciais
pertencentes aos estrato arbóreo mais baixo da floresta, com até 10 metros de altura, como já
foi relatado, que no processo dinâmico florestal substituem as árvores maiores que morrem
(B-30 também apresentou maior necromassa que as outras localidades), além de indivíduos
juvenis de espécies de estágios sucessionais tardios. Outros estudos têm relatado maior
densidade de árvores com alturas médias menores em áreas perturbadas ou bordas de
fragmentos (Oliveira-Filho et al. 1997; Rodrigues 2004). Entretanto, em ambientes com
menores intensidades luminosas, como em interiores de mata mais fechada, as árvores
tendem a direcionar sua energia para o crescimento em altura, tentando alcançar estratos
superiores onde o aproveitamento da luz é maior (Crawley 1997), o que pode ter reforçado
esta diferença entre as localidades.
Pode-se ainda inferir que as árvores maiores nesta borda (B-30), que agregam
maiores biomassas, estão sendo substituídas por estes novos indivíduos em densidade
suficiente para a manutenção da biomassa total da comunidade, já que não foram encontradas

58
diferenças significativas entre a área basal total de todas as árvores e dentro das classes das
diferentes localidades. Esta manutenção da biomassa total para a comunidade arbórea foi
encontrada para uma floresta tropical bem preservada na Costa Rica (Lieberman &
Lieberman 1987), que num intervalo de 13 anos praticamente não variou, com o
recrutamento e crescimento das árvores substituindo as perdas por mortalidade.
Em termos proporcionais, INT-1 apresentou maior porcentagem de árvores na
primeira classe de diâmetro (arvoretas: 5 cm≤DAP<10 cm) mas não diferiu das outras
localidades em relação à densidade média. Isto está associado a população de Euterpe edulis,
onde aproximadamente 91% dos indivíduos desta espécie amostrados nesta localidade
pertencem a classe das arvoretas (5 cm≤DAP>10 cm), representando 43% do total de
indivíduos de todas as espécies desta classe. A dificuldade oferecida pelos interiores
florestais de remanescentes ou mesmo fragmentos grandes aos caçadores e cortadores de
palmito, atribuída a distância a ser percorrida e até mesmo a topografia do relevo, favorece a
preservação destas áreas e o equilíbrio natural das populações de Euterpe edulis, colocando-a
como a espécie dominante, e onde o estrato regenerante é mantido pela dispersão dos frutos
de indivíduos adultos em abundância (T. Reis, comunicação pessoal). Na ReBio do Tinguá,
os indivíduos com diâmetros na faixa de 5 a 9,9 cm são muito abundantes na população
correspondendo a classe com as maiores taxas reprodutivas e coincidindo ainda com o
tamanho mínimo preferencial para o corte e extração do palmito pelos cortadores. Segundo
Baez & Balslev (2007), as palmeiras (Arecaceae) representam elemento florístico e ecológico
importante em florestas tropicais e suas diversas espécies respondem de maneiras distintas as
modificações nas condições ambientais que ocorrem em bordas de fragmentos. Dentro deste
contexto, as menores densidades médias de E. edulis nas bordas podem estar associadas a

59
diferentes motivos: a) aos níveis de distúrbio do habitat, que no caso dos efeitos de borda, a
densidade de palmitos diminuiria com a proximidade da borda e quanto mais recente esta
fosse. Porém, neste caso, a densidade foi muito semelhante nas duas bordas mostrando que a
idade da borda pouco influi na sua ocorrência; b) e principalmente, pela facilidade de acesso
dos “palmiteiros” nas bordas, devido à proximidade, diminuindo consideravelmente a
densidade e manutenção destas populações nestes locais, e consequentemente, afetando a
fauna associada a esta espécie já que esta é responsável por oferecer grande parte dos
recursos alimentares. Além disso, assim como a maior parte das espécies de palmeiras, sua
menor população em bordas pode influenciar criticamente a estrutura e fisionomia de
fragmentos florestais (Baez & Balslev 2007), como ocorreu neste estudo.
A maior riqueza de espécies arbóreas na borda mais recente, associada
possivelmente a maior abundância encontrada no local, pode estar relacionada a grande
heterogeneidade de ambientes que constitui um mosaico de condições abióticas amostrados
nesta localidade, conferindo assim a possibilidade de estabelecimento de diferentes espécies
em nichos distintos e espacialmente próximos entre si (Denslow 1995). Isto porque, de fato,
esta borda apresentou tanto sítios com menor estrutura arbórea e grande riqueza quanto sítios
com maior estrutura e menor riqueza. De maneira semelhante, Rodrigues (2004) também
encontrou grande variação nos padrões florísticos e estruturais em bordas de fragmentos de
Mata Atlântica de baixada, indicando que os efeitos de borda estão distribuídos de forma
assimétrica ao longo das bordas.
Por outro lado, a borda mais antiga (B-200) apresentou parâmetros com valores
intermediários entre B-30 e os interiores (INT-1 e INT-2), assemelhando-se mais a estes
últimos. Tais fatos sugerem que a borda mais recente esteja sujeita a efeitos de borda mais

60
intensos que a borda mais antiga, e que processos sucessionais de extrema importância teriam
ocorrido na comunidade próxima à borda num período entre 30 e 200 anos alterando sua
estrutura no sentido de alcançar maior equilíbrio semelhante ao interior florestal, tais como o
crescimento de vegetação secundária (Kapos 1989; Kapos et al. 1997; Mesquita et al. 1999)
e o retorno de espécies de estágios sucessionais tardios e de crescimento mais lento a
comunidade. De acordo com alguns estudos, esta influência da idade da borda sobre a
magnitude e extensão dos efeitos de borda pode estar associada às modificações nas
condições microclimáticas dentro de remanescentes florestais criadas pela borda, que
desaparecem ou tornam-se mais complexas com sua idade (Kapos 1989; Williams-Linera
1990; Matlack 1994). A estrutura da vegetação próxima à borda é alterada, logo após a
criação desta, onde muitas árvores perdem suas folhas e morrem em pé, aparentemente
porque ocorrem mudanças abruptas na intensidade de luz, temperatura e umidade nestes
locais que excedem suas tolerâncias fisiológicas (Laurance et al. 2002). Com isso, ocorre um
aumento nos distúrbios do dossel, como conseqüência do aumento na mortalidade e queda de
árvores maiores devido também à maior intensidade de ventos (Lovejoy et al. 1986;
Laurance 1997; Laurance et al. 2000). Todavia, com o tempo estas modificações abióticas na
matriz, neste caso as aberturas lineares de dossel, seriam diminuídas e tornar-se-iam mais
uniformes (Williams-Linera 1990), podendo levar a um aumento de espécies tolerantes à
seca ou a aclimatização fisiológica de plantas próximas às bordas (Laurance et al. 2002) que
substituiriam espacialmente as espécies pioneiras e secundárias ao final de seu ciclo de vida
mais curto. O longo tempo decorrido desde a abertura da Estrada do Comércio (há
aproximadamente 200 anos), aqui chamada borda B-200, pode ter induzido um
tamponamento bastante avançado e favorecido o crescimento em altura e diâmetro de

61
espécies secundárias tardias, que voltariam a ocupar o alto dossel florestal, aumentando a
biomassa total. Consequentemente, o cenário atual mostra uma borda sob efeitos de borda tão
tênues que a tornam semelhante estruturalmente ao interior florestal.
A alta ocorrência de árvores perfilhadas na borda mais recente sugere que a borda
de 30 anos oferece condições mais adversas (como maior velocidade do vento, altas
temperaturas e maior dessecamento no ar e no solo) ao desenvolvimento das plantas, e fica
evidente o efeito da idade da borda, onde a mais antiga exibe menor densidade de troncos
múltiplos, porém apresentando valores intermediários entre B-30 e os interiores (INT-1 e
INT-2). Apesar da necromassa no solo não ter sido quantificada neste estudo, a partir da
medição de galhos e troncos quebrados, é possível inferir que os troncos múltiplos de árvores
(perfilhos) em B-30 estão mais propícios a morte e quebra, aumentando a intensidade de
clareiras na borda, e seus tamanhos estão diretamente relacionados ao tamanho dos galhos
(Laurance et al. 1998a; Sizer et al. 2000).
Efeitos de borda sobre a dinâmica
A comunidade arbórea na borda mais recente (B-30) apresentou dinâmica
levemente mais acelerada que as outras localidades (B-200 e INT-1) no período de estudo
(2004 a 2008), caracterizado pela mais alta mortalidade principalmente dos indivíduos com
até 20 cm de diâmetro. Apesar do fato de a densidade total de árvores em B-30 ter diminuído
a uma taxa mais alta que nas outras localidades neste período, esta continuou mantendo os
maiores valores médios em todos os anos, diferindo de INT-1. A separação em classes
mostra que esta diminuição em B-30 ocorre principalmente para as árvores maiores de 10 cm
(DAP). De maneira diferente, nas outras localidades (B-200 e INT-1), a densidade de todas
as classes aumentou ao longo dos anos, com exceção das arvoretas que experienciam uma

62
diminuição em todas as localidades, inclusive B-30. É provável que o total de arvoretas
mortas somado as arvoretas que cresceram e mudaram de classe (para DAP≥10 cm) esteja
superando a densidade de recrutas, porém com a manutenção de sua biomassa total nesta
classe. Desta forma, o crescimento das árvores da classe regenerante estaria suprindo esta
perda de biomassa. É esperado que as árvores de algumas espécies pioneiras cresçam a uma
taxa maior nas menores classes de tamanho, em busca de maior aproveitamento da luz, e que
esta taxa diminua na medida em que alcançam tamanhos maiores (Clark & Clark 1999).
No entanto, a diminuição no número total de árvores em B-30 está levando a perda
de biomassa total, ao mesmo tempo em que as outras localidades (B-200 e INT-1) estão
apresentando incrementos com valores próximos a 5%. Novamente, as árvores com DAP
maior que 10 cm estão sofrendo as maiores modificações estruturais com perda de biomassa,
principalmente das árvores com DAP de 10 a 30 cm na borda mais recente. Nascimento &
Laurance (2004) encontraram alterações significativas na quantidade e distribuição de
biomassa acima do solo em fragmentos florestais amazônicos, principalmente entre bordas e
interiores. No entanto, a diminuição de biomassa nas bordas destes fragmentos foi
relacionada a alta mortalidade de árvores grandes nestes locais (DAP>60 cm), resposta
comum da vegetação nos primeiros anos após a fragmentação.
Desta maneira, estes resultados sugerem que as árvores com tamanhos medianos
(com DAP variando de 10 a 30 cm) correspondem a fração principal das modificações na
estrutura e dinâmica da comunidade arbórea na borda mais recente (30 anos), tornando-a
diferente estruturalmente das outras localidades. O cenário atual nesta localidade reflete
modificações pretéritas nos processos sucessionais, que podem ter iniciado logo após a
criação da borda, há aproximadamente 30 anos, quando a vegetação desta borda passou a

63
experimentar novas condições microclimáticas externas, como aumento na dessecação, na
morte e queda de grandes árvores. Alguns estudos em fragmentos recentes na floresta
amazônica mostraram que o aumento na morte e danos em árvores com diâmetro maior que
10 cm está relacionado as alterações microclimáticas próximo a borda (Ferreira & Laurance
1997; Laurance et al. 1998a). A morte destas árvores em B-30, logo após a criação da borda,
teria estimulado tanto a germinação de plântulas quanto o crescimento de pequenas árvores
de crescimento rápido, que são beneficiadas pela maior intensidade luminosa no sub-bosque
em clareiras recém-formadas (Sizer & Tanner 1999; Laurance et al. 1998b; Laurance et al.
2006). Reis (2008) analisando a abertura do dossel e a quantidade de radiação direta e difusa
que chega a comunidade de plântulas nas mesmas localidades deste estudo (B-200, B-30 e
INT-1) no ano de 2008, encontrou maior radiação, além da maior abertura de dossel, na
borda mais recente B-30. Neste contexto, considerando elevada radiação solar na borda B-30
desde sua criação, as espécies encontradas neste levantamento podem ter tido seu
crescimento favorecido nestas condições pretéritas, há aproximadamente 30 anos com a
abertura linear para a instalação do oleoduto, e atualmente constituem este grupo de árvores
com 10 a 30 cm de diâmetro, que podem ainda estar sendo favorecidas. Contudo, neste
mesmo estudo (Reis 2008) foi observado que os efeitos de borda não atuam mais sobre a
densidade de plântulas nas bordas da ReBio do Tinguá, atribuindo isso ao processo de
tamponamento que protege a comunidade dos fatores externos, estando, desta forma,
atravessando um período de retorno as condições estruturais iniciais semelhantes ao interior
preservado. Segundo Matlack (1994), as modificações na estrutura vegetal próximo a borda
de fragmentos tendem a permanecer mesmo algumas décadas após a criação da borda, no
caso da ReBio do Tinguá, apenas para a comunidade arbórea na borda mais recente.

64
As taxas de mortalidade e recrutamento e o equilíbrio dinâmico entre elas encontrados
na localidade B-200, semelhante à INT-1, e até mesmo em B-30, na maioria dos intervalos
estudados, correspondem ao esperado para florestas tropicais maduras, onde estas não
ultrapassam 3% (Lieberman et al. 1985; Phillips et al. 1996; Laurance et al. 2004; Lewis et
al. 2004; Saiter 2007). Em fragmentos amazônicos que variaram de 14 a 18 anos de idade,
foi encontrada baixa incidência de árvores mortas em pé na borda que foi relacionada a
redução na intensidade das alterações microclimáticas na borda (D’Angelo et al. 2004) ao
longo dos anos. Esta redução teria sido causada pela proliferação de lianas e vegetação
secundária (Camargo & Kapos 1995), e pelo aumento de árvores tolerantes a seca próximas
as bordas (Laurance et al. 2001b). Além do possível efeito do tamponamento induzido pela
idade da borda (B-30), o fato de não terem sido encontradas diferenças entre as taxas de
mortalidade e entre as taxas de recrutamento das localidades de borda e interior, na maioria
dos anos estudados, pode estar relacionado aos altos desvios padrão destas médias que
mostram a heterogeneidade dos sítios. Apenas no intervalo de quatro anos (2004-2008), todas
as parcelas apresentaram taxas de mortalidade diferentes de zero. E mesmo assim, foram
muito heterogêneas dentro de todas as localidades, diminuindo o poder das análises
estatísticas.
No entanto, as maiores taxas de mortalidade, recrutamento e turnover na borda mais
recente (B-30) no intervalo 2004-2005, diferente de B-200 e INT-1, mostram que o maior
dinamismo na comunidade arbórea em decorrência da proximidade da borda pode continuar
ocorrendo em diferentes escalas espaciais e temporais, porém com taxas menores, não apenas
logo após a criação da borda, mas pode também se perpetuar por alguns anos, como
mostrado aqui, até 30 anos. Além disso, florestas úmidas podem apresentar pulsos cíclicos

65
mais dinâmicos, com maiores taxas de mortalidade e/ou recrutamento relacionadas a
flutuações climáticas (Phillips et al. 1994) ou, neste caso, a perturbações permanentes como a
presença da borda. De acordo com os resultados deste estudo, esta mudança na dinâmica da
borda mais recente está influenciando uma mudança na estrutura florestal e tem relevante
efeito sobre a composição e diversidade florestal desta localidade.
Oliveira-Filho et al. (2007) comparando a dinâmica entre borda e interior de um
remanescente florestal de Mata Atlântica (MG) num intervalo de 5 anos, encontraram uma
redução na densidade e aumento na área basal da comunidade arbórea tanto na borda de
aproximadamente 150 anos quanto no interior. Esse padrão foi similar ao encontrado para
borda B-200 e INT-1, no intervalo de 4 anos, apesar de o primeiro estudo ter relacionado tal
fato as taxas de mortalidade superiores as taxas de recrutamento tanto na borda quanto no
interior florestal. Além disso, contabilizou maior densidade de mortos que recrutas no
interior, diferente deste estudo. No entanto, estas duas localidades (B-200 e INT-1)
apresentaram alguns indivíduos de Euterpe edulis Mart. cortados em algumas parcelas
durante o período de monitoramento, que somados ao mortos refletiriam esta diminuição nas
densidades.

66
CONCLUSÕES
Os resultados florísticos encontrados para a comunidade arbórea na Reserva Biológica
do Tinguá mostram que este remanescente se encontra em alto grau de preservação, num
estágio sucessional bastante avançado e que abriga grande diversidade de espécies arbóreas.
Dado o tamanho de sua área total (26.000 ha) e a pequena área total amostrada (1,17 ha),
pode-se dizer que este remanescente apresenta fração importante de plantas ainda
desconhecidas pela ciência para a Mata Atlântica.
A comunidade está sujeita aos efeitos de borda e a intensidade destes varia não
apenas com a proximidade da matriz, mas também com o tempo de existência das bordas
(idade). Isto porque as duas aberturas lineares de dossel (adjacentes às bordas B-200 e B-30)
exibem larguras e tipos de matriz bastante semelhantes (vegetação herbácea) o que excluiria
o efeito da característica da matriz sobre a vegetação diferenciada nas bordas.
É possível inferir que os efeitos de borda causados pela presença das duas aberturas
lineares ocorreram com grande intensidade em algum momento, provavelmente nos
primeiros anos após o surgimento destas. Tais efeitos de borda pretéritos teriam acelerado a
dinâmica da comunidade, com altas taxas de mortalidade e recrutamento, que refletiu no
cenário atual de estrutura e composição arbórea modificada na borda mais recente em relação
ao interior.
Ficou constatado que os efeitos de borda estão afetando a estrutura e composição em B-
30, porém o efeito sobre a dinâmica tem sido pouco intenso pelo menos nos últimos quatro
anos (2004-2008). Este intervalo pode ter sido relativamente curto para perceber mudanças
mais elucidativas na comunidade, já que a aceleração da dinâmica pode ocorrer em pulsos
cíclicos dependentes de alterações climáticas, como períodos menos chuvosos.

67
As diferenças estruturais e de dinâmica da comunidade arbórea entre a borda mais
recente de 30 anos e o interior indicam que estas aberturas lineares de dossel, apesar de
parecerem menos impactantes que matrizes que circundam fragmentos (na fragmentação
“clássica”) são, de forma semelhante, causadoras de modificações no habitat da borda e suas
comunidades relacionadas. No entanto, a semelhança estrutural e dinâmica da comunidade
entre a borda mais antiga (aprox. 200 anos) e o interior sugere que as modificações causadas
no habitat da borda pela fragmentação podem se estender por algumas décadas, no caso da
outra borda do estudo, até 30 anos, ou até períodos mais longos, porém podem não ocorrer
indefinitivamente, como alguns autores já propuseram (Matlack 1994; Oliveira-Filho et al.
1997; D’Angelo et al. 2004). Todavia, o cenário encontrado para esta borda mais antiga pode
também estar relacionado ao tamanho e impacto da abertura linear de dossel na floresta, que
apresenta largura média de 30 metros, e seria assim muito menos impactante que outro tipo
de matriz. Entretanto, os efeitos da borda mais recente na vegetação mostram o contrário, que
a pequena largura da abertura linear de dossel é suficiente para causar tais alterações.

68
PERSPECTIVAS
- O monitoramento constante da comunidade arbórea nas bordas dos oleodutos é de
extrema importância para a percepção de padrões mais elucidativos quanto as modificações
que os efeitos de borda causam na vegetação. O acompanhamento das mudanças estruturais e
dinâmicas, assim como dos processos sucessionais principalmente na borda mais recente
ajudará a identificar em qual estágio sucessional esta fração da vegetação se encontra e como
esta evolui em intervalos maiores de tempo.
- A avaliação de alguns parâmetros microclimáticos, tais como, temperatura e umidade
no solo e no ar, e velocidade de vento nas bordas e nos interiores, além de quantificação dos
nutrientes e da necromassa caída no solo e seu tempo de decomposição, correlacionando com
dados de declividade e orientação de cada sítio, poderão ajudar na elucidação destes
processos e explicar a grande heterogeneidade dos dados encontrados neste estudo.
- A busca constante de material botânico fértil dos indivíduos amostrados ajudará na
identificação precisa das plantas e mostrará a real representatividade da flora da ReBio do
Tinguá na diversidade da Mata Atlântica.
- A identificação dos novos recrutas e o monitoramento da mortalidade dos indivíduos
sobreviventes oferecerão subsídios para a caracterização da dinâmica das espécies.
- A futura amostragem da comunidade arbustiva (1 cm≤DAP<5 cm) e a identificação
de seus indivíduos, representarão, junto aos dados existentes da comunidade de plântulas e da
comunidade arbórea, um avanço no entendimento dos processos ecológicos de florestas
tropicais, e de que forma e com que intensidade os efeitos de borda alteram a vegetação.
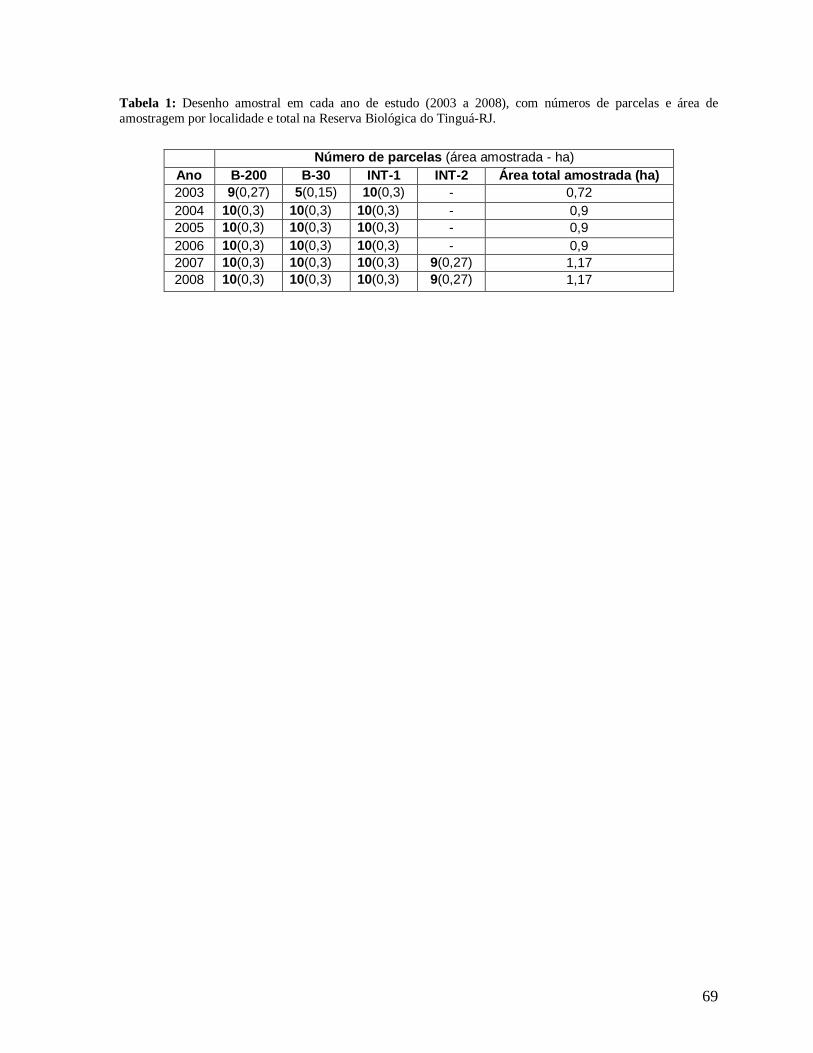
69
Tabela 1: Desenho amostral em cada ano de estudo (2003 a 2008), com números de parcelas e área de amostragem por localidade e total na Reserva Biológica do Tinguá-RJ.
Número de parcelas (área amostrada - ha) Ano B-200 B-30 INT-1 INT-2 Área total amostrada (ha) 2003 9(0,27) 5(0,15) 10(0,3) - 0,72 2004 10(0,3) 10(0,3) 10(0,3) - 0,9 2005 10(0,3) 10(0,3) 10(0,3) - 0,9 2006 10(0,3) 10(0,3) 10(0,3) - 0,9 2007 10(0,3) 10(0,3) 10(0,3) 9(0,27) 1,17 2008 10(0,3) 10(0,3) 10(0,3) 9(0,27) 1,17
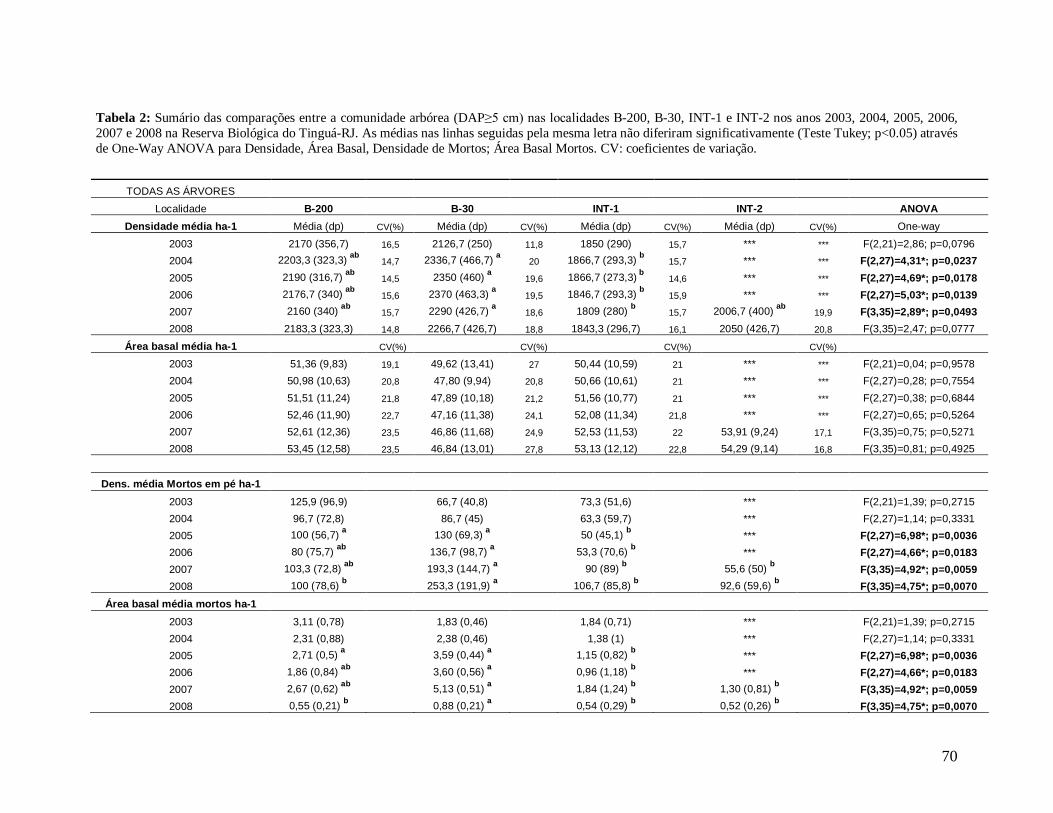
70
Tabela 2: Sumário das comparações entre a comunidade arbórea (DAP≥5 cm) nas localidades B-200, B-30, INT-1 e INT-2 nos anos 2003, 2004, 2005, 2006, 2007 e 2008 na Reserva Biológica do Tinguá-RJ. As médias nas linhas seguidas pela mesma letra não diferiram significativamente (Teste Tukey; p<0.05) através de One-Way ANOVA para Densidade, Área Basal, Densidade de Mortos; Área Basal Mortos. CV: coeficientes de variação.
TODAS AS ÁRVORES
Localidade B-200 B-30 INT-1 INT-2 ANOVA Densidade média ha-1 Média (dp) CV(%) Média (dp) CV(%) Média (dp) CV(%) Média (dp) CV(%) One-way
2003 2170 (356,7) 16,5 2126,7 (250) 11,8 1850 (290) 15,7 *** *** F(2,21)=2,86; p=0,0796 2004 2203,3 (323,3) ab 14,7 2336,7 (466,7) a 20 1866,7 (293,3) b 15,7 *** *** F(2,27)=4,31*; p=0,0237 2005 2190 (316,7) ab 14,5 2350 (460) a 19,6 1866,7 (273,3) b 14,6 *** *** F(2,27)=4,69*; p=0,0178 2006 2176,7 (340) ab 15,6 2370 (463,3) a 19,5 1846,7 (293,3) b 15,9 *** *** F(2,27)=5,03*; p=0,0139 2007 2160 (340) ab 15,7 2290 (426,7) a 18,6 1809 (280) b 15,7 2006,7 (400) ab 19,9 F(3,35)=2,89*; p=0,0493 2008 2183,3 (323,3) 14,8 2266,7 (426,7) 18,8 1843,3 (296,7) 16,1 2050 (426,7) 20,8 F(3,35)=2,47; p=0,0777
Área basal média ha-1 CV(%) CV(%) CV(%) CV(%)
2003 51,36 (9,83) 19,1 49,62 (13,41) 27 50,44 (10,59) 21 *** *** F(2,21)=0,04; p=0,9578 2004 50,98 (10,63) 20,8 47,80 (9,94) 20,8 50,66 (10,61) 21 *** *** F(2,27)=0,28; p=0,7554 2005 51,51 (11,24) 21,8 47,89 (10,18) 21,2 51,56 (10,77) 21 *** *** F(2,27)=0,38; p=0,6844 2006 52,46 (11,90) 22,7 47,16 (11,38) 24,1 52,08 (11,34) 21,8 *** *** F(2,27)=0,65; p=0,5264 2007 52,61 (12,36) 23,5 46,86 (11,68) 24,9 52,53 (11,53) 22 53,91 (9,24) 17,1 F(3,35)=0,75; p=0,5271 2008 53,45 (12,58) 23,5 46,84 (13,01) 27,8 53,13 (12,12) 22,8 54,29 (9,14) 16,8 F(3,35)=0,81; p=0,4925
Dens. média Mortos em pé ha-1
2003 125,9 (96,9) 66,7 (40,8) 73,3 (51,6) *** F(2,21)=1,39; p=0,2715 2004 96,7 (72,8) 86,7 (45) 63,3 (59,7) *** F(2,27)=1,14; p=0,3331 2005 100 (56,7) a 130 (69,3) a 50 (45,1) b *** F(2,27)=6,98*; p=0,0036 2006 80 (75,7) ab 136,7 (98,7) a 53,3 (70,6) b *** F(2,27)=4,66*; p=0,0183 2007 103,3 (72,8) ab 193,3 (144,7) a 90 (89) b 55,6 (50) b F(3,35)=4,92*; p=0,0059 2008 100 (78,6) b 253,3 (191,9) a 106,7 (85,8) b 92,6 (59,6) b F(3,35)=4,75*; p=0,0070
Área basal média mortos ha-1
2003 3,11 (0,78) 1,83 (0,46) 1,84 (0,71) *** F(2,21)=1,39; p=0,2715 2004 2,31 (0,88) 2,38 (0,46) 1,38 (1) *** F(2,27)=1,14; p=0,3331 2005 2,71 (0,5) a 3,59 (0,44) a 1,15 (0,82) b *** F(2,27)=6,98*; p=0,0036 2006 1,86 (0,84) ab 3,60 (0,56) a 0,96 (1,18) b *** F(2,27)=4,66*; p=0,0183 2007 2,67 (0,62) ab 5,13 (0,51) a 1,84 (1,24) b 1,30 (0,81) b F(3,35)=4,92*; p=0,0059 2008 0,55 (0,21) b 0,88 (0,21) a 0,54 (0,29) b 0,52 (0,26) b F(3,35)=4,75*; p=0,0070
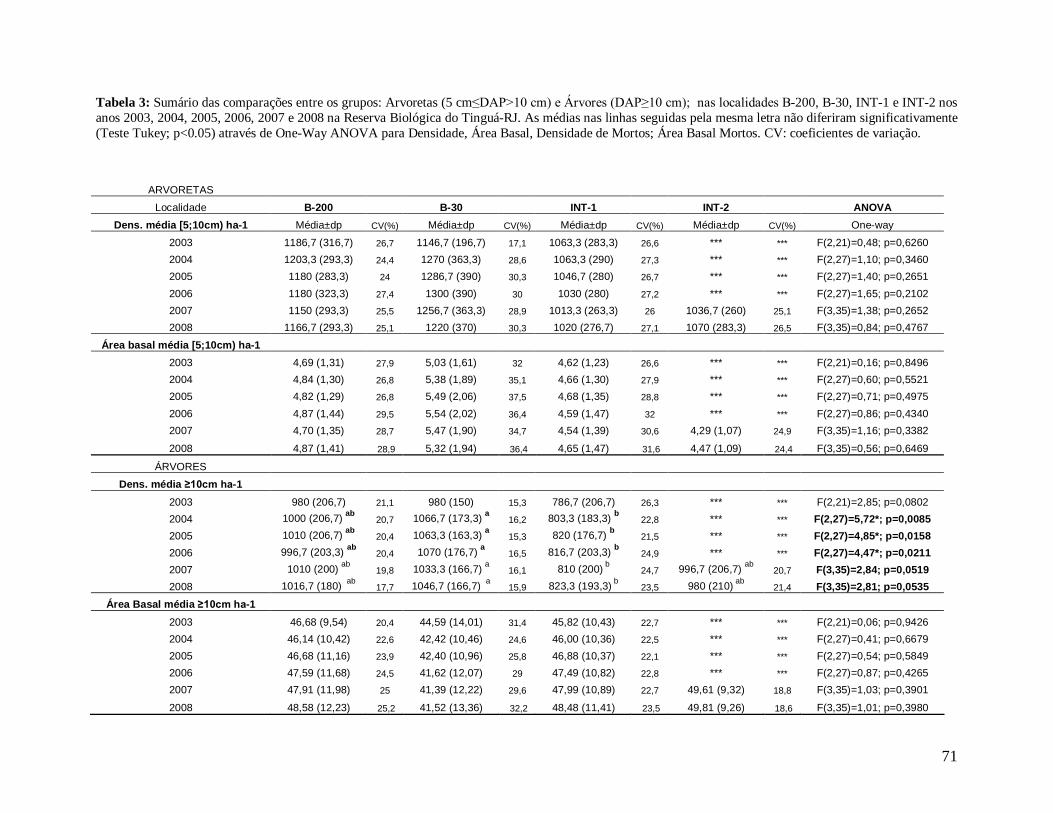
71
Tabela 3: Sumário das comparações entre os grupos: Arvoretas (5 cm≤DAP>10 cm) e Árvores (DAP≥10 cm); nas localidades B-200, B-30, INT-1 e INT-2 nos anos 2003, 2004, 2005, 2006, 2007 e 2008 na Reserva Biológica do Tinguá-RJ. As médias nas linhas seguidas pela mesma letra não diferiram significativamente (Teste Tukey; p<0.05) através de One-Way ANOVA para Densidade, Área Basal, Densidade de Mortos; Área Basal Mortos. CV: coeficientes de variação.
ARVORETAS
Localidade B-200 B-30 INT-1 INT-2 ANOVA Dens. média [5;10cm) ha-1 Média±dp CV(%) Média±dp CV(%) Média±dp CV(%) Média±dp CV(%) One-way
2003 1186,7 (316,7) 26,7 1146,7 (196,7) 17,1 1063,3 (283,3) 26,6 *** *** F(2,21)=0,48; p=0,6260 2004 1203,3 (293,3) 24,4 1270 (363,3) 28,6 1063,3 (290) 27,3 *** *** F(2,27)=1,10; p=0,3460 2005 1180 (283,3) 24 1286,7 (390) 30,3 1046,7 (280) 26,7 *** *** F(2,27)=1,40; p=0,2651 2006 1180 (323,3) 27,4 1300 (390) 30 1030 (280) 27,2 *** *** F(2,27)=1,65; p=0,2102 2007 1150 (293,3) 25,5 1256,7 (363,3) 28,9 1013,3 (263,3) 26 1036,7 (260) 25,1 F(3,35)=1,38; p=0,2652 2008 1166,7 (293,3) 25,1 1220 (370) 30,3 1020 (276,7) 27,1 1070 (283,3) 26,5 F(3,35)=0,84; p=0,4767
Área basal média [5;10cm) ha-1
2003 4,69 (1,31) 27,9 5,03 (1,61) 32 4,62 (1,23) 26,6 *** *** F(2,21)=0,16; p=0,8496 2004 4,84 (1,30) 26,8 5,38 (1,89) 35,1 4,66 (1,30) 27,9 *** *** F(2,27)=0,60; p=0,5521 2005 4,82 (1,29) 26,8 5,49 (2,06) 37,5 4,68 (1,35) 28,8 *** *** F(2,27)=0,71; p=0,4975 2006 4,87 (1,44) 29,5 5,54 (2,02) 36,4 4,59 (1,47) 32 *** *** F(2,27)=0,86; p=0,4340 2007 4,70 (1,35) 28,7 5,47 (1,90) 34,7 4,54 (1,39) 30,6 4,29 (1,07) 24,9 F(3,35)=1,16; p=0,3382
2008 4,87 (1,41) 28,9 5,32 (1,94) 36,4 4,65 (1,47) 31,6 4,47 (1,09) 24,4 F(3,35)=0,56; p=0,6469
ÁRVORES
Dens. média ≥10cm ha-1
2003 980 (206,7) 21,1 980 (150) 15,3 786,7 (206,7) 26,3 *** *** F(2,21)=2,85; p=0,0802 2004 1000 (206,7) ab 20,7 1066,7 (173,3) a 16,2 803,3 (183,3) b 22,8 *** *** F(2,27)=5,72*; p=0,0085 2005 1010 (206,7) ab 20,4 1063,3 (163,3) a 15,3 820 (176,7) b 21,5 *** *** F(2,27)=4,85*; p=0,0158 2006 996,7 (203,3) ab 20,4 1070 (176,7) a 16,5 816,7 (203,3) b 24,9 *** *** F(2,27)=4,47*; p=0,0211 2007 1010 (200) ab 19,8 1033,3 (166,7) a 16,1 810 (200) b 24,7 996,7 (206,7) ab 20,7 F(3,35)=2,84; p=0,0519 2008 1016,7 (180) ab 17,7 1046,7 (166,7) a 15,9 823,3 (193,3) b 23,5 980 (210) ab 21,4 F(3,35)=2,81; p=0,0535
Área Basal média ≥10cm ha-1
2003 46,68 (9,54) 20,4 44,59 (14,01) 31,4 45,82 (10,43) 22,7 *** *** F(2,21)=0,06; p=0,9426 2004 46,14 (10,42) 22,6 42,42 (10,46) 24,6 46,00 (10,36) 22,5 *** *** F(2,27)=0,41; p=0,6679 2005 46,68 (11,16) 23,9 42,40 (10,96) 25,8 46,88 (10,37) 22,1 *** *** F(2,27)=0,54; p=0,5849 2006 47,59 (11,68) 24,5 41,62 (12,07) 29 47,49 (10,82) 22,8 *** *** F(2,27)=0,87; p=0,4265 2007 47,91 (11,98) 25 41,39 (12,22) 29,6 47,99 (10,89) 22,7 49,61 (9,32) 18,8 F(3,35)=1,03; p=0,3901
2008 48,58 (12,23) 25,2 41,52 (13,36) 32,2 48,48 (11,41) 23,5 49,81 (9,26) 18,6 F(3,35)=1,01; p=0,3980
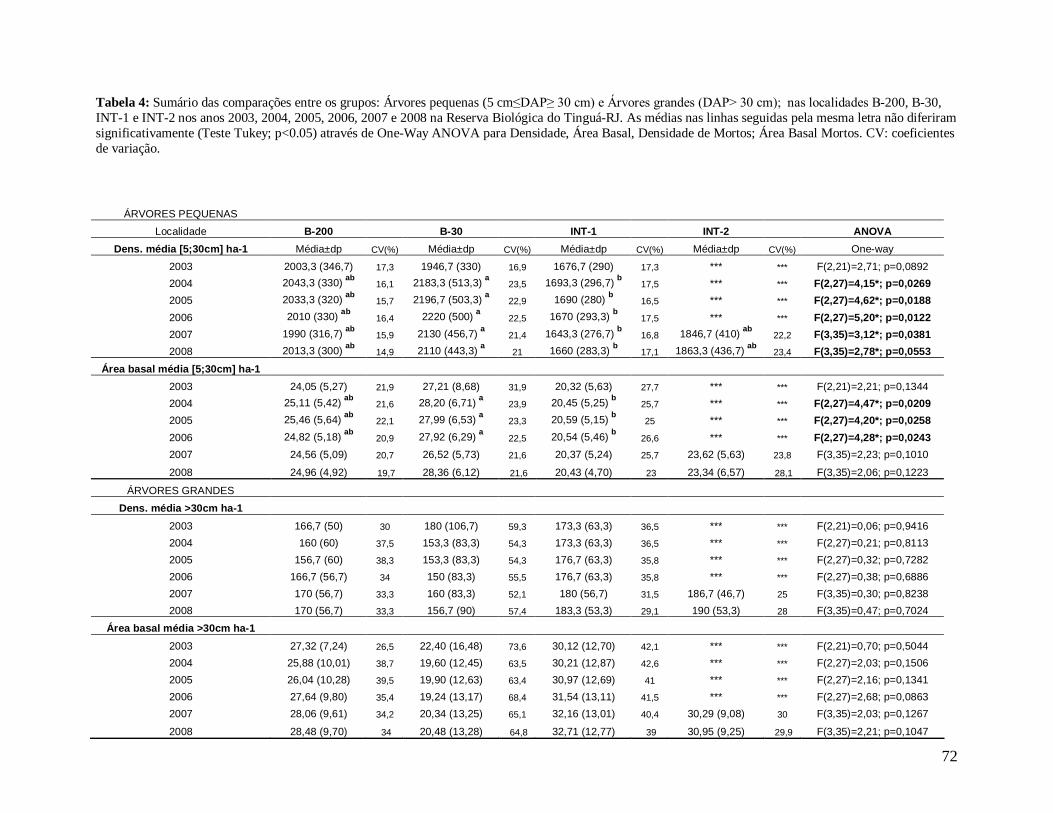
72
Tabela 4: Sumário das comparações entre os grupos: Árvores pequenas (5 cm≤DAP≥ 30 cm) e Árvores grandes (DAP> 30 cm); nas localidades B-200, B-30, INT-1 e INT-2 nos anos 2003, 2004, 2005, 2006, 2007 e 2008 na Reserva Biológica do Tinguá-RJ. As médias nas linhas seguidas pela mesma letra não diferiram significativamente (Teste Tukey; p<0.05) através de One-Way ANOVA para Densidade, Área Basal, Densidade de Mortos; Área Basal Mortos. CV: coeficientes de variação.
ÁRVORES PEQUENAS
Localidade B-200 B-30 INT-1 INT-2 ANOVA Dens. média [5;30cm] ha-1 Média±dp CV(%) Média±dp CV(%) Média±dp CV(%) Média±dp CV(%) One-way
2003 2003,3 (346,7) 17,3 1946,7 (330) 16,9 1676,7 (290) 17,3 *** *** F(2,21)=2,71; p=0,0892 2004 2043,3 (330) ab 16,1 2183,3 (513,3) a 23,5 1693,3 (296,7) b 17,5 *** *** F(2,27)=4,15*; p=0,0269 2005 2033,3 (320) ab 15,7 2196,7 (503,3) a 22,9 1690 (280) b 16,5 *** *** F(2,27)=4,62*; p=0,0188 2006 2010 (330) ab 16,4 2220 (500) a 22,5 1670 (293,3) b 17,5 *** *** F(2,27)=5,20*; p=0,0122 2007 1990 (316,7) ab 15,9 2130 (456,7) a 21,4 1643,3 (276,7) b 16,8 1846,7 (410) ab 22,2 F(3,35)=3,12*; p=0,0381 2008 2013,3 (300) ab 14,9 2110 (443,3) a 21 1660 (283,3) b 17,1 1863,3 (436,7) ab 23,4 F(3,35)=2,78*; p=0,0553
Área basal média [5;30cm] ha-1
2003 24,05 (5,27) 21,9 27,21 (8,68) 31,9 20,32 (5,63) 27,7 *** *** F(2,21)=2,21; p=0,1344 2004 25,11 (5,42) ab 21,6 28,20 (6,71) a 23,9 20,45 (5,25) b 25,7 *** *** F(2,27)=4,47*; p=0,0209 2005 25,46 (5,64) ab 22,1 27,99 (6,53) a 23,3 20,59 (5,15) b 25 *** *** F(2,27)=4,20*; p=0,0258 2006 24,82 (5,18) ab 20,9 27,92 (6,29) a 22,5 20,54 (5,46) b 26,6 *** *** F(2,27)=4,28*; p=0,0243 2007 24,56 (5,09) 20,7 26,52 (5,73) 21,6 20,37 (5,24) 25,7 23,62 (5,63) 23,8 F(3,35)=2,23; p=0,1010
2008 24,96 (4,92) 19,7 28,36 (6,12) 21,6 20,43 (4,70) 23 23,34 (6,57) 28,1 F(3,35)=2,06; p=0,1223
ÁRVORES GRANDES
Dens. média >30cm ha-1
2003 166,7 (50) 30 180 (106,7) 59,3 173,3 (63,3) 36,5 *** *** F(2,21)=0,06; p=0,9416 2004 160 (60) 37,5 153,3 (83,3) 54,3 173,3 (63,3) 36,5 *** *** F(2,27)=0,21; p=0,8113 2005 156,7 (60) 38,3 153,3 (83,3) 54,3 176,7 (63,3) 35,8 *** *** F(2,27)=0,32; p=0,7282 2006 166,7 (56,7) 34 150 (83,3) 55,5 176,7 (63,3) 35,8 *** *** F(2,27)=0,38; p=0,6886 2007 170 (56,7) 33,3 160 (83,3) 52,1 180 (56,7) 31,5 186,7 (46,7) 25 F(3,35)=0,30; p=0,8238 2008 170 (56,7) 33,3 156,7 (90) 57,4 183,3 (53,3) 29,1 190 (53,3) 28 F(3,35)=0,47; p=0,7024
Área basal média >30cm ha-1
2003 27,32 (7,24) 26,5 22,40 (16,48) 73,6 30,12 (12,70) 42,1 *** *** F(2,21)=0,70; p=0,5044 2004 25,88 (10,01) 38,7 19,60 (12,45) 63,5 30,21 (12,87) 42,6 *** *** F(2,27)=2,03; p=0,1506 2005 26,04 (10,28) 39,5 19,90 (12,63) 63,4 30,97 (12,69) 41 *** *** F(2,27)=2,16; p=0,1341 2006 27,64 (9,80) 35,4 19,24 (13,17) 68,4 31,54 (13,11) 41,5 *** *** F(2,27)=2,68; p=0,0863 2007 28,06 (9,61) 34,2 20,34 (13,25) 65,1 32,16 (13,01) 40,4 30,29 (9,08) 30 F(3,35)=2,03; p=0,1267
2008 28,48 (9,70) 34 20,48 (13,28) 64,8 32,71 (12,77) 39 30,95 (9,25) 29,9 F(3,35)=2,21; p=0,1047
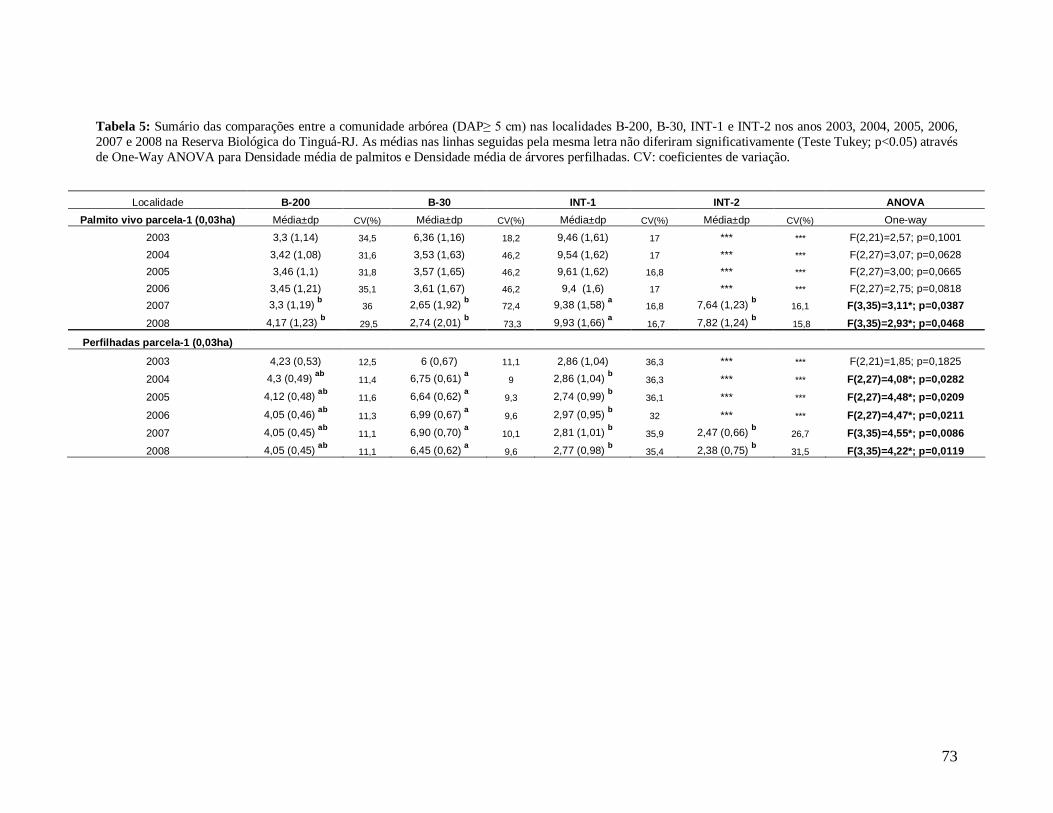
73
Tabela 5: Sumário das comparações entre a comunidade arbórea (DAP≥ 5 cm) nas localidades B-200, B-30, INT-1 e INT-2 nos anos 2003, 2004, 2005, 2006, 2007 e 2008 na Reserva Biológica do Tinguá-RJ. As médias nas linhas seguidas pela mesma letra não diferiram significativamente (Teste Tukey; p<0.05) através de One-Way ANOVA para Densidade média de palmitos e Densidade média de árvores perfilhadas. CV: coeficientes de variação.
Localidade B-200 B-30 INT-1 INT-2 ANOVA Palmito vivo parcela-1 (0,03ha) Média±dp CV(%) Média±dp CV(%) Média±dp CV(%) Média±dp CV(%) One-way
2003 3,3 (1,14) 34,5 6,36 (1,16) 18,2 9,46 (1,61) 17 *** *** F(2,21)=2,57; p=0,1001 2004 3,42 (1,08) 31,6 3,53 (1,63) 46,2 9,54 (1,62) 17 *** *** F(2,27)=3,07; p=0,0628 2005 3,46 (1,1) 31,8 3,57 (1,65) 46,2 9,61 (1,62) 16,8 *** *** F(2,27)=3,00; p=0,0665 2006 3,45 (1,21) 35,1 3,61 (1,67) 46,2 9,4 (1,6) 17 *** *** F(2,27)=2,75; p=0,0818 2007 3,3 (1,19) b 36 2,65 (1,92) b 72,4 9,38 (1,58) a 16,8 7,64 (1,23) b 16,1 F(3,35)=3,11*; p=0,0387 2008 4,17 (1,23) b 29,5 2,74 (2,01) b 73,3 9,93 (1,66) a 16,7 7,82 (1,24) b 15,8 F(3,35)=2,93*; p=0,0468
Perfilhadas parcela-1 (0,03ha)
2003 4,23 (0,53) 12,5 6 (0,67) 11,1 2,86 (1,04) 36,3 *** *** F(2,21)=1,85; p=0,1825
2004 4,3 (0,49) ab 11,4 6,75 (0,61) a 9 2,86 (1,04) b 36,3 *** *** F(2,27)=4,08*; p=0,0282 2005 4,12 (0,48) ab 11,6 6,64 (0,62) a 9,3 2,74 (0,99) b 36,1 *** *** F(2,27)=4,48*; p=0,0209 2006 4,05 (0,46) ab 11,3 6,99 (0,67) a 9,6 2,97 (0,95) b 32 *** *** F(2,27)=4,47*; p=0,0211 2007 4,05 (0,45) ab 11,1 6,90 (0,70) a 10,1 2,81 (1,01) b 35,9 2,47 (0,66) b 26,7 F(3,35)=4,55*; p=0,0086 2008 4,05 (0,45) ab 11,1 6,45 (0,62) a 9,6 2,77 (0,98) b 35,4 2,38 (0,75) b 31,5 F(3,35)=4,22*; p=0,0119
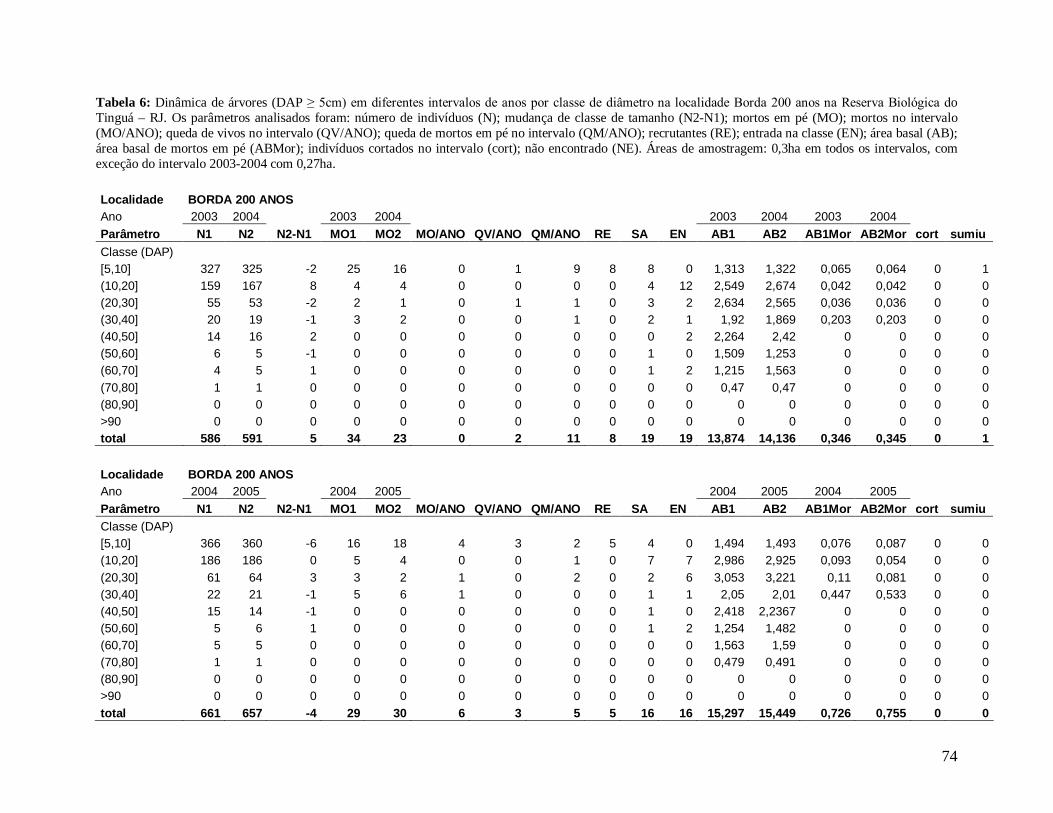
74
Tabela 6: Dinâmica de árvores (DAP ≥ 5cm) em diferentes intervalos de anos por classe de diâmetro na localidade Borda 200 anos na Reserva Biológica do Tinguá – RJ. Os parâmetros analisados foram: número de indivíduos (N); mudança de classe de tamanho (N2-N1); mortos em pé (MO); mortos no intervalo (MO/ANO); queda de vivos no intervalo (QV/ANO); queda de mortos em pé no intervalo (QM/ANO); recrutantes (RE); entrada na classe (EN); área basal (AB); área basal de mortos em pé (ABMor); indivíduos cortados no intervalo (cort); não encontrado (NE). Áreas de amostragem: 0,3ha em todos os intervalos, com exceção do intervalo 2003-2004 com 0,27ha. Localidade BORDA 200 ANOS Ano 2003 2004 2003 2004 2003 2004 2003 2004 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 327 325 -2 25 16 0 1 9 8 8 0 1,313 1,322 0,065 0,064 0 1 (10,20] 159 167 8 4 4 0 0 0 0 4 12 2,549 2,674 0,042 0,042 0 0 (20,30] 55 53 -2 2 1 0 1 1 0 3 2 2,634 2,565 0,036 0,036 0 0 (30,40] 20 19 -1 3 2 0 0 1 0 2 1 1,92 1,869 0,203 0,203 0 0 (40,50] 14 16 2 0 0 0 0 0 0 0 2 2,264 2,42 0 0 0 0 (50,60] 6 5 -1 0 0 0 0 0 0 1 0 1,509 1,253 0 0 0 0 (60,70] 4 5 1 0 0 0 0 0 0 1 2 1,215 1,563 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,47 0,47 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 586 591 5 34 23 0 2 11 8 19 19 13,874 14,136 0,346 0,345 0 1
Localidade BORDA 200 ANOS Ano 2004 2005 2004 2005 2004 2005 2004 2005 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 366 360 -6 16 18 4 3 2 5 4 0 1,494 1,493 0,076 0,087 0 0 (10,20] 186 186 0 5 4 0 0 1 0 7 7 2,986 2,925 0,093 0,054 0 0 (20,30] 61 64 3 3 2 1 0 2 0 2 6 3,053 3,221 0,11 0,081 0 0 (30,40] 22 21 -1 5 6 1 0 0 0 1 1 2,05 2,01 0,447 0,533 0 0 (40,50] 15 14 -1 0 0 0 0 0 0 1 0 2,418 2,2367 0 0 0 0 (50,60] 5 6 1 0 0 0 0 0 0 1 2 1,254 1,482 0 0 0 0 (60,70] 5 5 0 0 0 0 0 0 0 0 0 1,563 1,59 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,479 0,491 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 661 657 -4 29 30 6 3 5 5 16 16 15,297 15,449 0,726 0,755 0 0
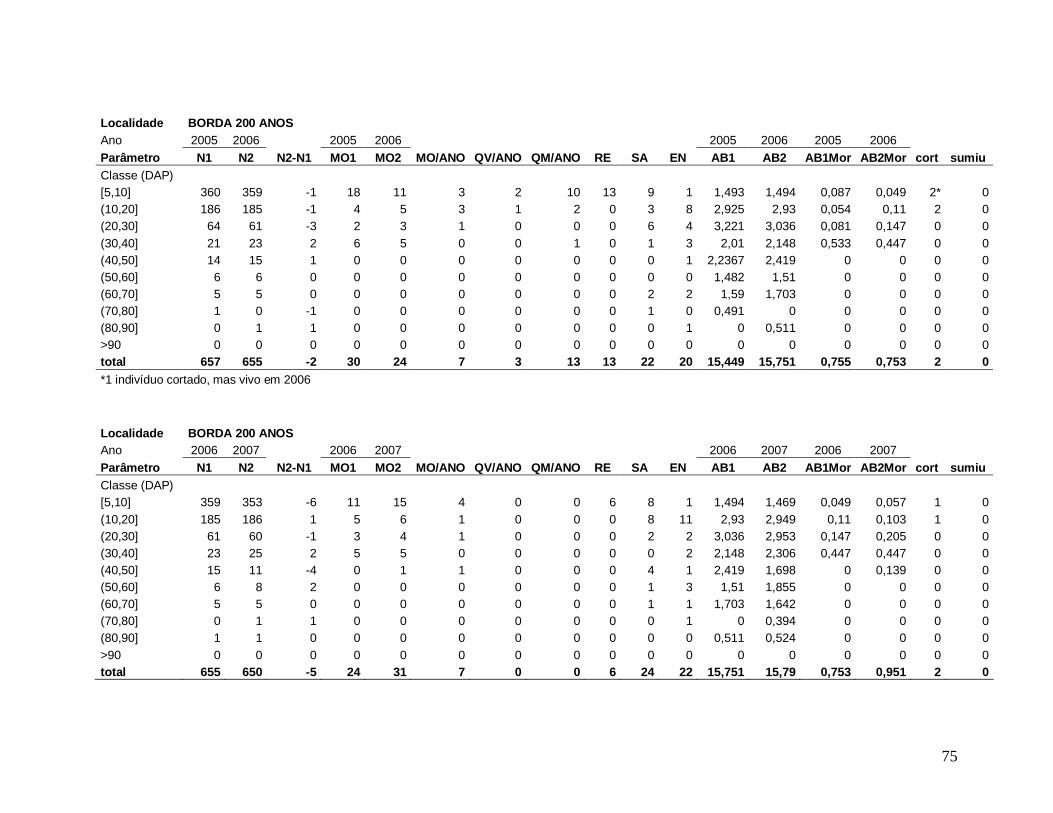
75
Localidade BORDA 200 ANOS Ano 2005 2006 2005 2006 2005 2006 2005 2006 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 360 359 -1 18 11 3 2 10 13 9 1 1,493 1,494 0,087 0,049 2* 0 (10,20] 186 185 -1 4 5 3 1 2 0 3 8 2,925 2,93 0,054 0,11 2 0 (20,30] 64 61 -3 2 3 1 0 0 0 6 4 3,221 3,036 0,081 0,147 0 0 (30,40] 21 23 2 6 5 0 0 1 0 1 3 2,01 2,148 0,533 0,447 0 0 (40,50] 14 15 1 0 0 0 0 0 0 0 1 2,2367 2,419 0 0 0 0 (50,60] 6 6 0 0 0 0 0 0 0 0 0 1,482 1,51 0 0 0 0 (60,70] 5 5 0 0 0 0 0 0 0 2 2 1,59 1,703 0 0 0 0 (70,80] 1 0 -1 0 0 0 0 0 0 1 0 0,491 0 0 0 0 0 (80,90] 0 1 1 0 0 0 0 0 0 0 1 0 0,511 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 657 655 -2 30 24 7 3 13 13 22 20 15,449 15,751 0,755 0,753 2 0 *1 indivíduo cortado, mas vivo em 2006
Localidade BORDA 200 ANOS Ano 2006 2007 2006 2007 2006 2007 2006 2007 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 359 353 -6 11 15 4 0 0 6 8 1 1,494 1,469 0,049 0,057 1 0 (10,20] 185 186 1 5 6 1 0 0 0 8 11 2,93 2,949 0,11 0,103 1 0 (20,30] 61 60 -1 3 4 1 0 0 0 2 2 3,036 2,953 0,147 0,205 0 0 (30,40] 23 25 2 5 5 0 0 0 0 0 2 2,148 2,306 0,447 0,447 0 0 (40,50] 15 11 -4 0 1 1 0 0 0 4 1 2,419 1,698 0 0,139 0 0 (50,60] 6 8 2 0 0 0 0 0 0 1 3 1,51 1,855 0 0 0 0 (60,70] 5 5 0 0 0 0 0 0 0 1 1 1,703 1,642 0 0 0 0 (70,80] 0 1 1 0 0 0 0 0 0 0 1 0 0,394 0 0 0 0 (80,90] 1 1 0 0 0 0 0 0 0 0 0 0,511 0,524 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 655 650 -5 24 31 7 0 0 6 24 22 15,751 15,79 0,753 0,951 2 0
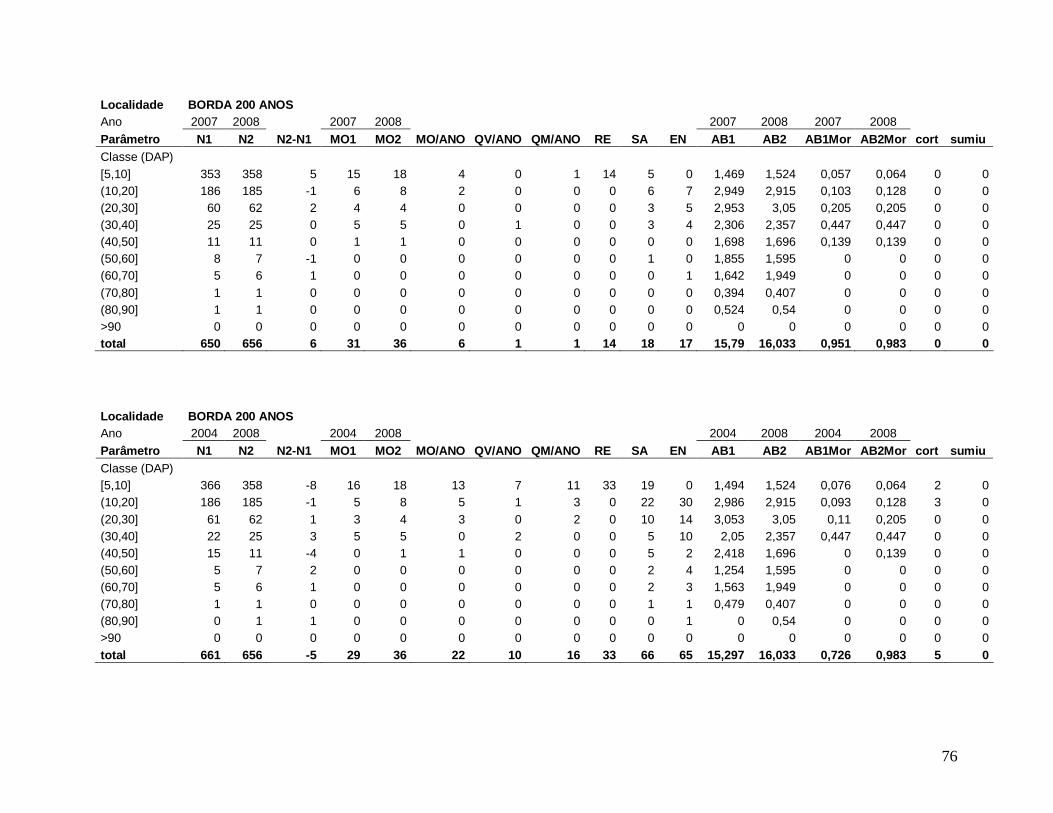
76
Localidade BORDA 200 ANOS Ano 2007 2008 2007 2008 2007 2008 2007 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 353 358 5 15 18 4 0 1 14 5 0 1,469 1,524 0,057 0,064 0 0 (10,20] 186 185 -1 6 8 2 0 0 0 6 7 2,949 2,915 0,103 0,128 0 0 (20,30] 60 62 2 4 4 0 0 0 0 3 5 2,953 3,05 0,205 0,205 0 0 (30,40] 25 25 0 5 5 0 1 0 0 3 4 2,306 2,357 0,447 0,447 0 0 (40,50] 11 11 0 1 1 0 0 0 0 0 0 1,698 1,696 0,139 0,139 0 0 (50,60] 8 7 -1 0 0 0 0 0 0 1 0 1,855 1,595 0 0 0 0 (60,70] 5 6 1 0 0 0 0 0 0 0 1 1,642 1,949 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,394 0,407 0 0 0 0 (80,90] 1 1 0 0 0 0 0 0 0 0 0 0,524 0,54 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 650 656 6 31 36 6 1 1 14 18 17 15,79 16,033 0,951 0,983 0 0
Localidade BORDA 200 ANOS Ano 2004 2008 2004 2008 2004 2008 2004 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 366 358 -8 16 18 13 7 11 33 19 0 1,494 1,524 0,076 0,064 2 0 (10,20] 186 185 -1 5 8 5 1 3 0 22 30 2,986 2,915 0,093 0,128 3 0 (20,30] 61 62 1 3 4 3 0 2 0 10 14 3,053 3,05 0,11 0,205 0 0 (30,40] 22 25 3 5 5 0 2 0 0 5 10 2,05 2,357 0,447 0,447 0 0 (40,50] 15 11 -4 0 1 1 0 0 0 5 2 2,418 1,696 0 0,139 0 0 (50,60] 5 7 2 0 0 0 0 0 0 2 4 1,254 1,595 0 0 0 0 (60,70] 5 6 1 0 0 0 0 0 0 2 3 1,563 1,949 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 1 1 0,479 0,407 0 0 0 0 (80,90] 0 1 1 0 0 0 0 0 0 0 1 0 0,54 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 661 656 -5 29 36 22 10 16 33 66 65 15,297 16,033 0,726 0,983 5 0
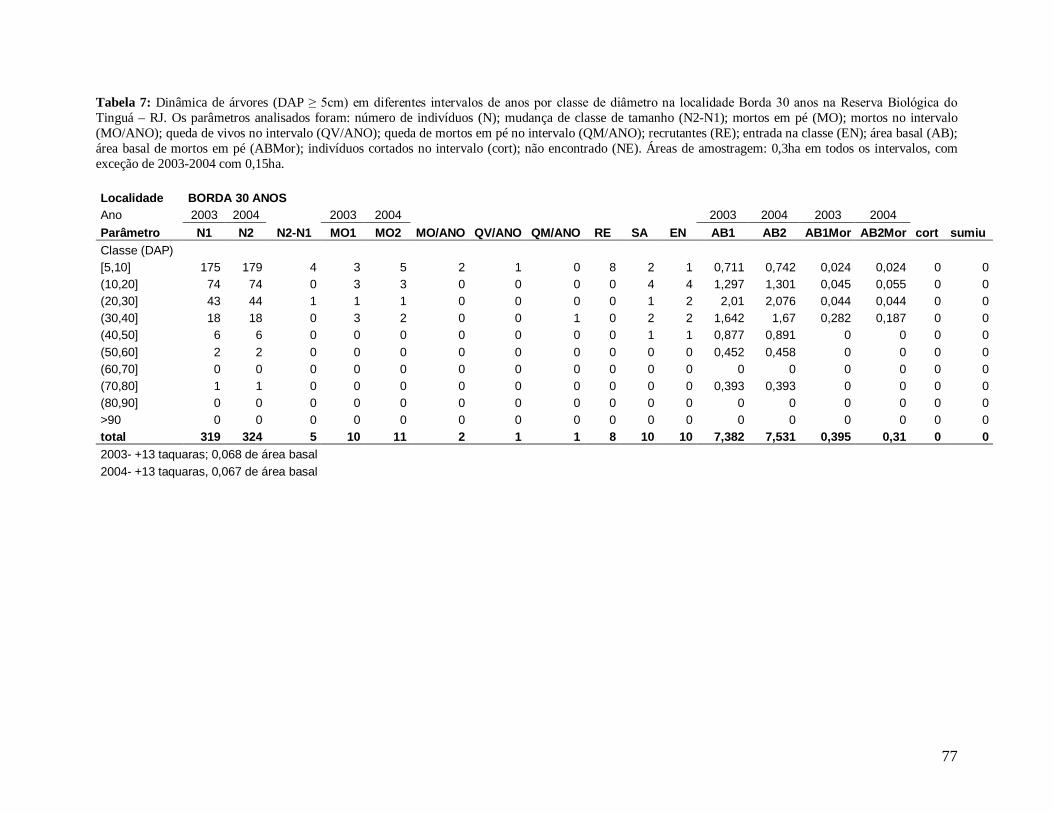
77
Tabela 7: Dinâmica de árvores (DAP ≥ 5cm) em diferentes intervalos de anos por classe de diâmetro na localidade Borda 30 anos na Reserva Biológica do Tinguá – RJ. Os parâmetros analisados foram: número de indivíduos (N); mudança de classe de tamanho (N2-N1); mortos em pé (MO); mortos no intervalo (MO/ANO); queda de vivos no intervalo (QV/ANO); queda de mortos em pé no intervalo (QM/ANO); recrutantes (RE); entrada na classe (EN); área basal (AB); área basal de mortos em pé (ABMor); indivíduos cortados no intervalo (cort); não encontrado (NE). Áreas de amostragem: 0,3ha em todos os intervalos, com exceção de 2003-2004 com 0,15ha. Localidade BORDA 30 ANOS Ano 2003 2004 2003 2004 2003 2004 2003 2004 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 175 179 4 3 5 2 1 0 8 2 1 0,711 0,742 0,024 0,024 0 0 (10,20] 74 74 0 3 3 0 0 0 0 4 4 1,297 1,301 0,045 0,055 0 0 (20,30] 43 44 1 1 1 0 0 0 0 1 2 2,01 2,076 0,044 0,044 0 0 (30,40] 18 18 0 3 2 0 0 1 0 2 2 1,642 1,67 0,282 0,187 0 0 (40,50] 6 6 0 0 0 0 0 0 0 1 1 0,877 0,891 0 0 0 0 (50,60] 2 2 0 0 0 0 0 0 0 0 0 0,452 0,458 0 0 0 0 (60,70] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,393 0,393 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 319 324 5 10 11 2 1 1 8 10 10 7,382 7,531 0,395 0,31 0 0 2003- +13 taquaras; 0,068 de área basal 2004- +13 taquaras, 0,067 de área basal
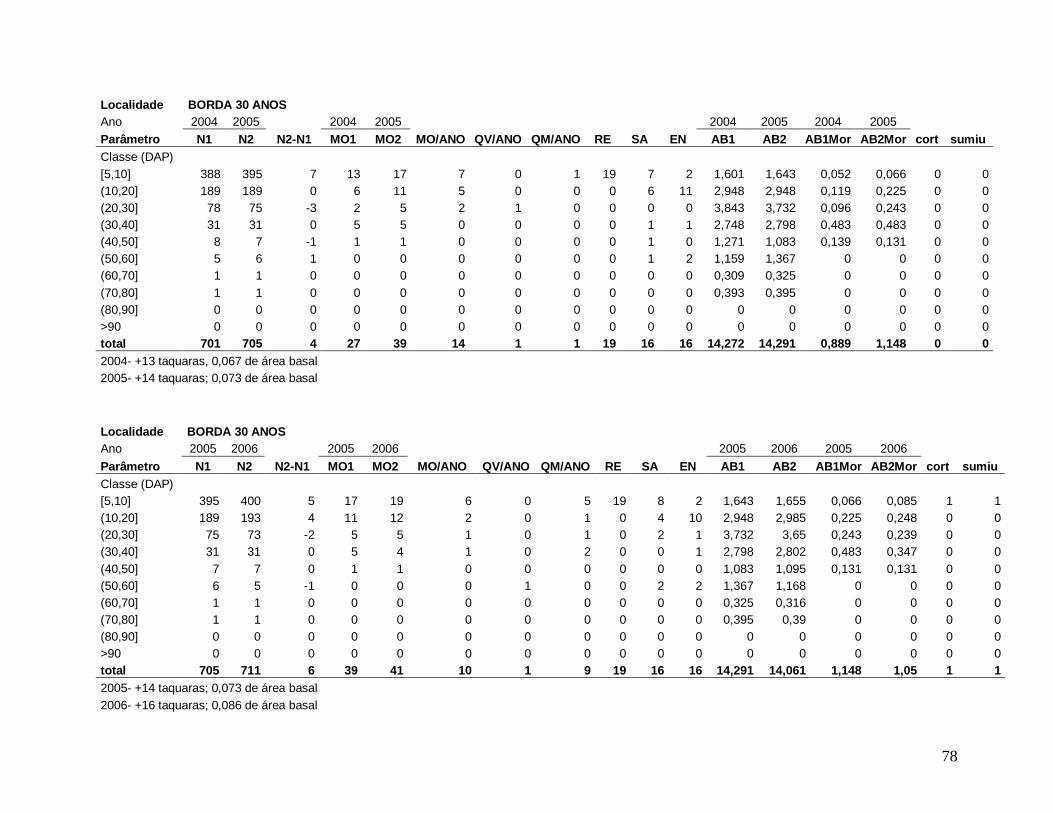
78
Localidade BORDA 30 ANOS Ano 2004 2005 2004 2005 2004 2005 2004 2005 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 388 395 7 13 17 7 0 1 19 7 2 1,601 1,643 0,052 0,066 0 0 (10,20] 189 189 0 6 11 5 0 0 0 6 11 2,948 2,948 0,119 0,225 0 0 (20,30] 78 75 -3 2 5 2 1 0 0 0 0 3,843 3,732 0,096 0,243 0 0 (30,40] 31 31 0 5 5 0 0 0 0 1 1 2,748 2,798 0,483 0,483 0 0 (40,50] 8 7 -1 1 1 0 0 0 0 1 0 1,271 1,083 0,139 0,131 0 0 (50,60] 5 6 1 0 0 0 0 0 0 1 2 1,159 1,367 0 0 0 0 (60,70] 1 1 0 0 0 0 0 0 0 0 0 0,309 0,325 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,393 0,395 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 701 705 4 27 39 14 1 1 19 16 16 14,272 14,291 0,889 1,148 0 0 2004- +13 taquaras, 0,067 de área basal 2005- +14 taquaras; 0,073 de área basal
Localidade BORDA 30 ANOS Ano 2005 2006 2005 2006 2005 2006 2005 2006 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 395 400 5 17 19 6 0 5 19 8 2 1,643 1,655 0,066 0,085 1 1 (10,20] 189 193 4 11 12 2 0 1 0 4 10 2,948 2,985 0,225 0,248 0 0 (20,30] 75 73 -2 5 5 1 0 1 0 2 1 3,732 3,65 0,243 0,239 0 0 (30,40] 31 31 0 5 4 1 0 2 0 0 1 2,798 2,802 0,483 0,347 0 0 (40,50] 7 7 0 1 1 0 0 0 0 0 0 1,083 1,095 0,131 0,131 0 0 (50,60] 6 5 -1 0 0 0 1 0 0 2 2 1,367 1,168 0 0 0 0 (60,70] 1 1 0 0 0 0 0 0 0 0 0 0,325 0,316 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,395 0,39 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 705 711 6 39 41 10 1 9 19 16 16 14,291 14,061 1,148 1,05 1 1 2005- +14 taquaras; 0,073 de área basal 2006- +16 taquaras; 0,086 de área basal
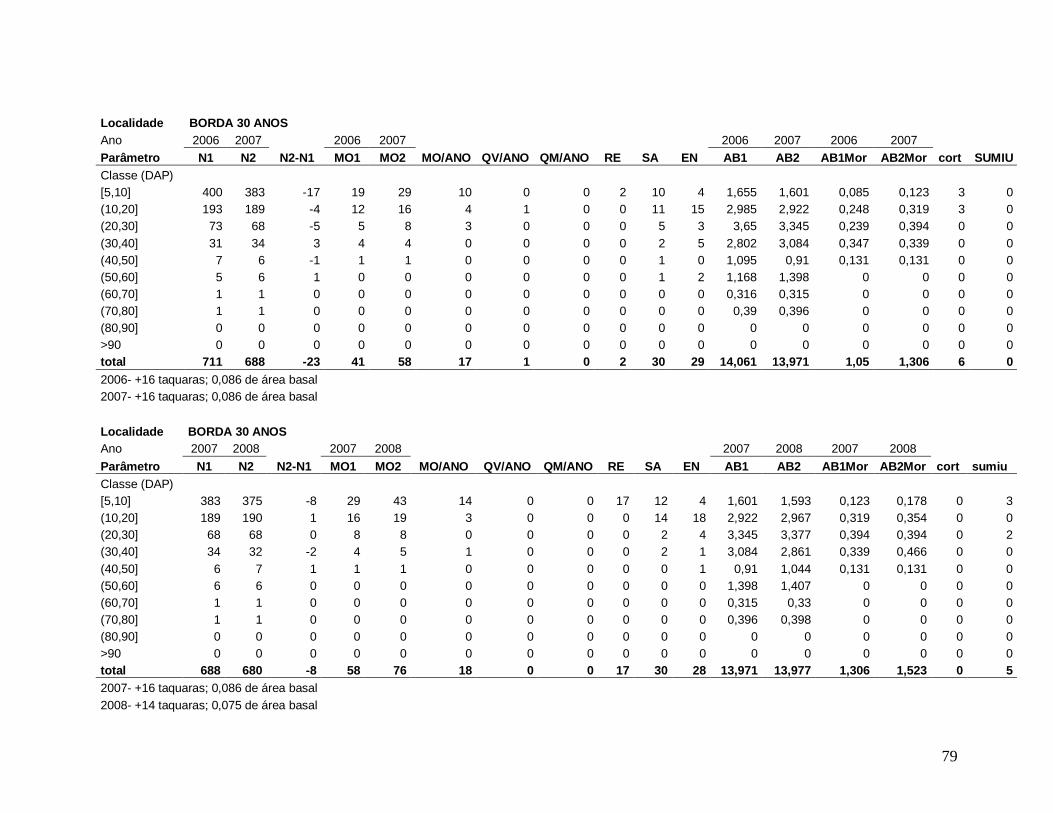
79
Localidade BORDA 30 ANOS Ano 2006 2007 2006 2007 2006 2007 2006 2007 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort SUMIU Classe (DAP) [5,10] 400 383 -17 19 29 10 0 0 2 10 4 1,655 1,601 0,085 0,123 3 0 (10,20] 193 189 -4 12 16 4 1 0 0 11 15 2,985 2,922 0,248 0,319 3 0 (20,30] 73 68 -5 5 8 3 0 0 0 5 3 3,65 3,345 0,239 0,394 0 0 (30,40] 31 34 3 4 4 0 0 0 0 2 5 2,802 3,084 0,347 0,339 0 0 (40,50] 7 6 -1 1 1 0 0 0 0 1 0 1,095 0,91 0,131 0,131 0 0 (50,60] 5 6 1 0 0 0 0 0 0 1 2 1,168 1,398 0 0 0 0 (60,70] 1 1 0 0 0 0 0 0 0 0 0 0,316 0,315 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,39 0,396 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 711 688 -23 41 58 17 1 0 2 30 29 14,061 13,971 1,05 1,306 6 0 2006- +16 taquaras; 0,086 de área basal 2007- +16 taquaras; 0,086 de área basal
Localidade BORDA 30 ANOS Ano 2007 2008 2007 2008 2007 2008 2007 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 383 375 -8 29 43 14 0 0 17 12 4 1,601 1,593 0,123 0,178 0 3 (10,20] 189 190 1 16 19 3 0 0 0 14 18 2,922 2,967 0,319 0,354 0 0 (20,30] 68 68 0 8 8 0 0 0 0 2 4 3,345 3,377 0,394 0,394 0 2 (30,40] 34 32 -2 4 5 1 0 0 0 2 1 3,084 2,861 0,339 0,466 0 0 (40,50] 6 7 1 1 1 0 0 0 0 0 1 0,91 1,044 0,131 0,131 0 0 (50,60] 6 6 0 0 0 0 0 0 0 0 0 1,398 1,407 0 0 0 0 (60,70] 1 1 0 0 0 0 0 0 0 0 0 0,315 0,33 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,396 0,398 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 688 680 -8 58 76 18 0 0 17 30 28 13,971 13,977 1,306 1,523 0 5 2007- +16 taquaras; 0,086 de área basal 2008- +14 taquaras; 0,075 de área basal
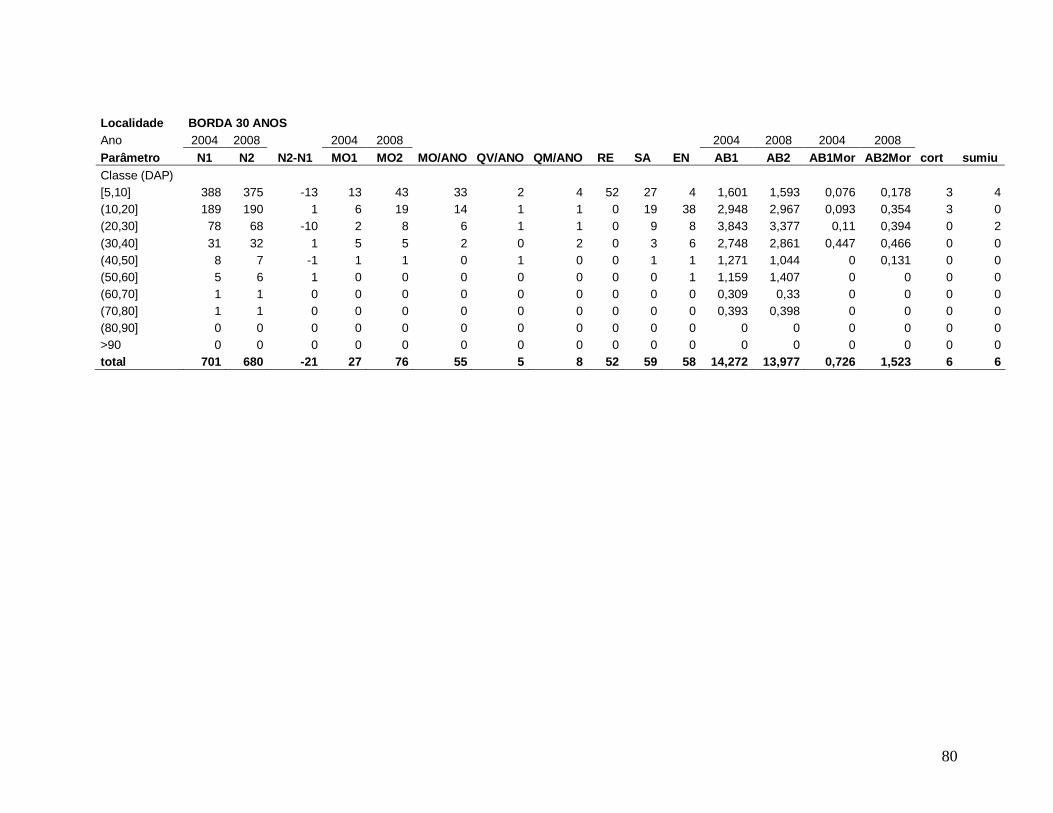
80
Localidade BORDA 30 ANOS Ano 2004 2008 2004 2008 2004 2008 2004 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 388 375 -13 13 43 33 2 4 52 27 4 1,601 1,593 0,076 0,178 3 4 (10,20] 189 190 1 6 19 14 1 1 0 19 38 2,948 2,967 0,093 0,354 3 0 (20,30] 78 68 -10 2 8 6 1 1 0 9 8 3,843 3,377 0,11 0,394 0 2 (30,40] 31 32 1 5 5 2 0 2 0 3 6 2,748 2,861 0,447 0,466 0 0 (40,50] 8 7 -1 1 1 0 1 0 0 1 1 1,271 1,044 0 0,131 0 0 (50,60] 5 6 1 0 0 0 0 0 0 0 1 1,159 1,407 0 0 0 0 (60,70] 1 1 0 0 0 0 0 0 0 0 0 0,309 0,33 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,393 0,398 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 701 680 -21 27 76 55 5 8 52 59 58 14,272 13,977 0,726 1,523 6 6
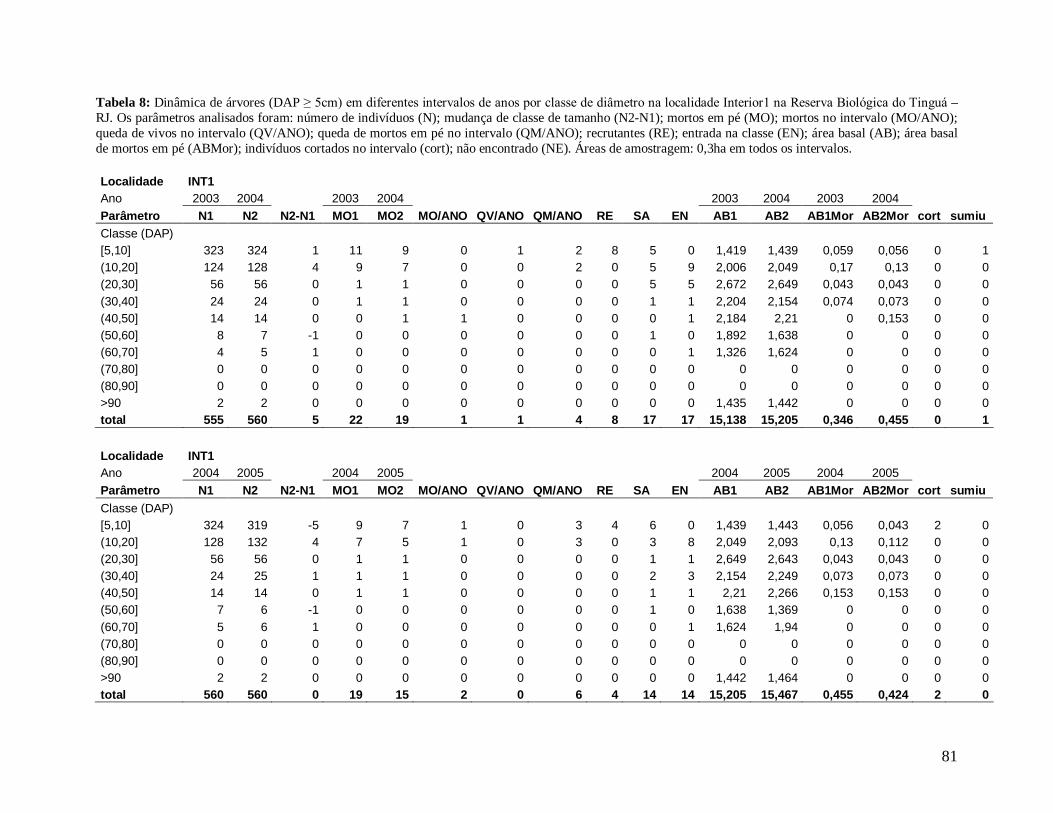
81
Tabela 8: Dinâmica de árvores (DAP ≥ 5cm) em diferentes intervalos de anos por classe de diâmetro na localidade Interior1 na Reserva Biológica do Tinguá – RJ. Os parâmetros analisados foram: número de indivíduos (N); mudança de classe de tamanho (N2-N1); mortos em pé (MO); mortos no intervalo (MO/ANO); queda de vivos no intervalo (QV/ANO); queda de mortos em pé no intervalo (QM/ANO); recrutantes (RE); entrada na classe (EN); área basal (AB); área basal de mortos em pé (ABMor); indivíduos cortados no intervalo (cort); não encontrado (NE). Áreas de amostragem: 0,3ha em todos os intervalos. Localidade INT1 Ano 2003 2004 2003 2004 2003 2004 2003 2004 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 323 324 1 11 9 0 1 2 8 5 0 1,419 1,439 0,059 0,056 0 1 (10,20] 124 128 4 9 7 0 0 2 0 5 9 2,006 2,049 0,17 0,13 0 0 (20,30] 56 56 0 1 1 0 0 0 0 5 5 2,672 2,649 0,043 0,043 0 0 (30,40] 24 24 0 1 1 0 0 0 0 1 1 2,204 2,154 0,074 0,073 0 0 (40,50] 14 14 0 0 1 1 0 0 0 0 1 2,184 2,21 0 0,153 0 0 (50,60] 8 7 -1 0 0 0 0 0 0 1 0 1,892 1,638 0 0 0 0 (60,70] 4 5 1 0 0 0 0 0 0 0 1 1,326 1,624 0 0 0 0 (70,80] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 2 2 0 0 0 0 0 0 0 0 0 1,435 1,442 0 0 0 0 total 555 560 5 22 19 1 1 4 8 17 17 15,138 15,205 0,346 0,455 0 1
Localidade INT1 Ano 2004 2005 2004 2005 2004 2005 2004 2005 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 324 319 -5 9 7 1 0 3 4 6 0 1,439 1,443 0,056 0,043 2 0 (10,20] 128 132 4 7 5 1 0 3 0 3 8 2,049 2,093 0,13 0,112 0 0 (20,30] 56 56 0 1 1 0 0 0 0 1 1 2,649 2,643 0,043 0,043 0 0 (30,40] 24 25 1 1 1 0 0 0 0 2 3 2,154 2,249 0,073 0,073 0 0 (40,50] 14 14 0 1 1 0 0 0 0 1 1 2,21 2,266 0,153 0,153 0 0 (50,60] 7 6 -1 0 0 0 0 0 0 1 0 1,638 1,369 0 0 0 0 (60,70] 5 6 1 0 0 0 0 0 0 0 1 1,624 1,94 0 0 0 0 (70,80] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 2 2 0 0 0 0 0 0 0 0 0 1,442 1,464 0 0 0 0 total 560 560 0 19 15 2 0 6 4 14 14 15,205 15,467 0,455 0,424 2 0
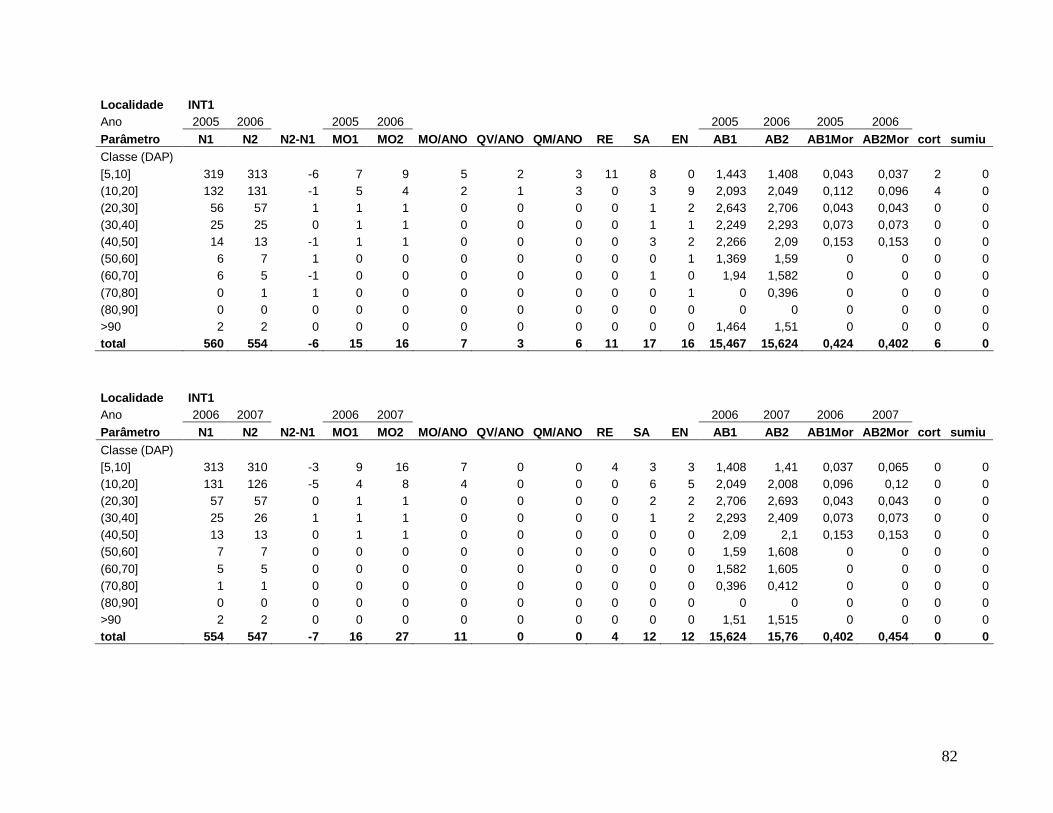
82
Localidade INT1 Ano 2005 2006 2005 2006 2005 2006 2005 2006 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 319 313 -6 7 9 5 2 3 11 8 0 1,443 1,408 0,043 0,037 2 0 (10,20] 132 131 -1 5 4 2 1 3 0 3 9 2,093 2,049 0,112 0,096 4 0 (20,30] 56 57 1 1 1 0 0 0 0 1 2 2,643 2,706 0,043 0,043 0 0 (30,40] 25 25 0 1 1 0 0 0 0 1 1 2,249 2,293 0,073 0,073 0 0 (40,50] 14 13 -1 1 1 0 0 0 0 3 2 2,266 2,09 0,153 0,153 0 0 (50,60] 6 7 1 0 0 0 0 0 0 0 1 1,369 1,59 0 0 0 0 (60,70] 6 5 -1 0 0 0 0 0 0 1 0 1,94 1,582 0 0 0 0 (70,80] 0 1 1 0 0 0 0 0 0 0 1 0 0,396 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 2 2 0 0 0 0 0 0 0 0 0 1,464 1,51 0 0 0 0 total 560 554 -6 15 16 7 3 6 11 17 16 15,467 15,624 0,424 0,402 6 0
Localidade INT1 Ano 2006 2007 2006 2007 2006 2007 2006 2007 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 313 310 -3 9 16 7 0 0 4 3 3 1,408 1,41 0,037 0,065 0 0 (10,20] 131 126 -5 4 8 4 0 0 0 6 5 2,049 2,008 0,096 0,12 0 0 (20,30] 57 57 0 1 1 0 0 0 0 2 2 2,706 2,693 0,043 0,043 0 0 (30,40] 25 26 1 1 1 0 0 0 0 1 2 2,293 2,409 0,073 0,073 0 0 (40,50] 13 13 0 1 1 0 0 0 0 0 0 2,09 2,1 0,153 0,153 0 0 (50,60] 7 7 0 0 0 0 0 0 0 0 0 1,59 1,608 0 0 0 0 (60,70] 5 5 0 0 0 0 0 0 0 0 0 1,582 1,605 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,396 0,412 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 2 2 0 0 0 0 0 0 0 0 0 1,51 1,515 0 0 0 0 total 554 547 -7 16 27 11 0 0 4 12 12 15,624 15,76 0,402 0,454 0 0
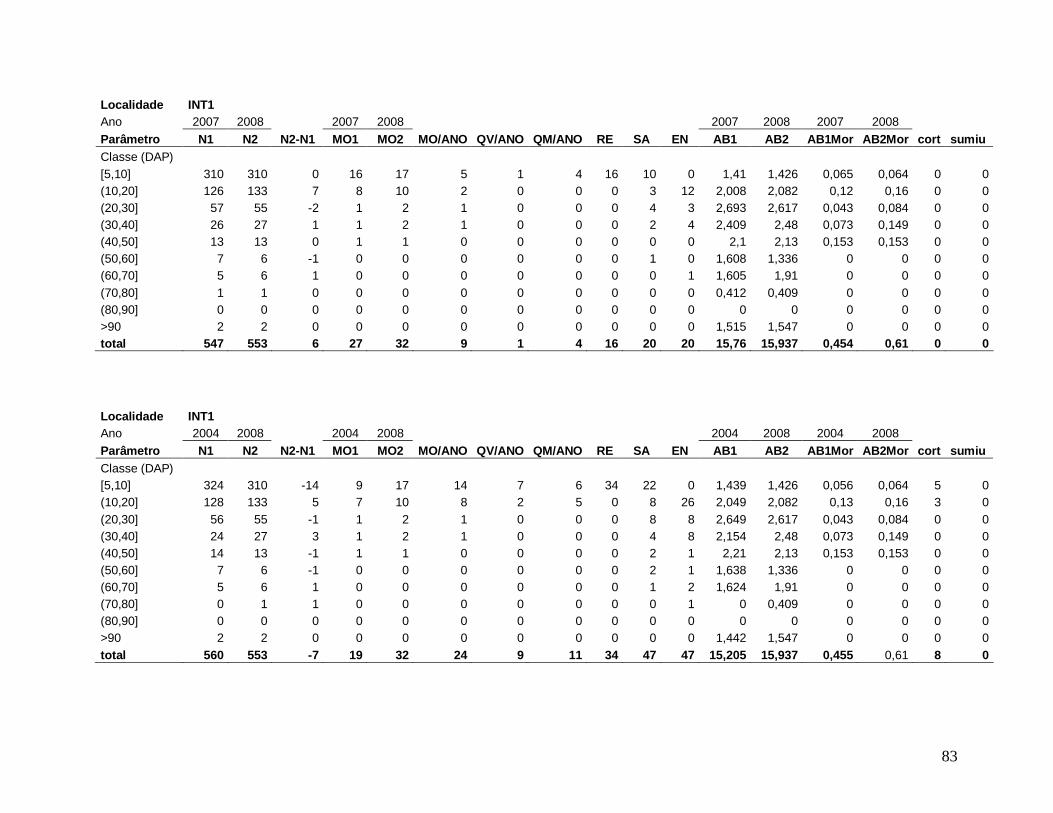
83
Localidade INT1 Ano 2007 2008 2007 2008 2007 2008 2007 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 310 310 0 16 17 5 1 4 16 10 0 1,41 1,426 0,065 0,064 0 0 (10,20] 126 133 7 8 10 2 0 0 0 3 12 2,008 2,082 0,12 0,16 0 0 (20,30] 57 55 -2 1 2 1 0 0 0 4 3 2,693 2,617 0,043 0,084 0 0 (30,40] 26 27 1 1 2 1 0 0 0 2 4 2,409 2,48 0,073 0,149 0 0 (40,50] 13 13 0 1 1 0 0 0 0 0 0 2,1 2,13 0,153 0,153 0 0 (50,60] 7 6 -1 0 0 0 0 0 0 1 0 1,608 1,336 0 0 0 0 (60,70] 5 6 1 0 0 0 0 0 0 0 1 1,605 1,91 0 0 0 0 (70,80] 1 1 0 0 0 0 0 0 0 0 0 0,412 0,409 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 2 2 0 0 0 0 0 0 0 0 0 1,515 1,547 0 0 0 0 total 547 553 6 27 32 9 1 4 16 20 20 15,76 15,937 0,454 0,61 0 0
Localidade INT1 Ano 2004 2008 2004 2008 2004 2008 2004 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1 AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 324 310 -14 9 17 14 7 6 34 22 0 1,439 1,426 0,056 0,064 5 0 (10,20] 128 133 5 7 10 8 2 5 0 8 26 2,049 2,082 0,13 0,16 3 0 (20,30] 56 55 -1 1 2 1 0 0 0 8 8 2,649 2,617 0,043 0,084 0 0 (30,40] 24 27 3 1 2 1 0 0 0 4 8 2,154 2,48 0,073 0,149 0 0 (40,50] 14 13 -1 1 1 0 0 0 0 2 1 2,21 2,13 0,153 0,153 0 0 (50,60] 7 6 -1 0 0 0 0 0 0 2 1 1,638 1,336 0 0 0 0 (60,70] 5 6 1 0 0 0 0 0 0 1 2 1,624 1,91 0 0 0 0 (70,80] 0 1 1 0 0 0 0 0 0 0 1 0 0,409 0 0 0 0 (80,90] 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 >90 2 2 0 0 0 0 0 0 0 0 0 1,442 1,547 0 0 0 0 total 560 553 -7 19 32 24 9 11 34 47 47 15,205 15,937 0,455 0,61 8 0
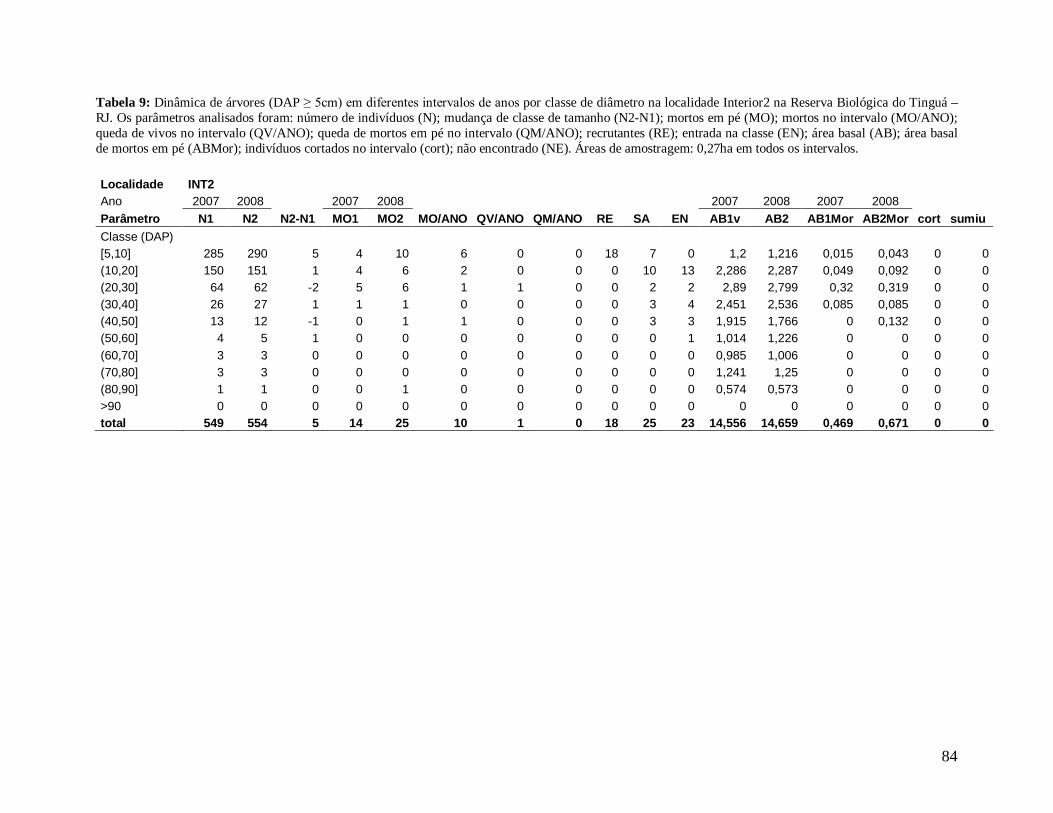
84
Tabela 9: Dinâmica de árvores (DAP ≥ 5cm) em diferentes intervalos de anos por classe de diâmetro na localidade Interior2 na Reserva Biológica do Tinguá – RJ. Os parâmetros analisados foram: número de indivíduos (N); mudança de classe de tamanho (N2-N1); mortos em pé (MO); mortos no intervalo (MO/ANO); queda de vivos no intervalo (QV/ANO); queda de mortos em pé no intervalo (QM/ANO); recrutantes (RE); entrada na classe (EN); área basal (AB); área basal de mortos em pé (ABMor); indivíduos cortados no intervalo (cort); não encontrado (NE). Áreas de amostragem: 0,27ha em todos os intervalos. Localidade INT2 Ano 2007 2008 2007 2008 2007 2008 2007 2008 Parâmetro N1 N2 N2-N1 MO1 MO2 MO/ANO QV/ANO QM/ANO RE SA EN AB1v AB2 AB1Mor AB2Mor cort sumiu Classe (DAP) [5,10] 285 290 5 4 10 6 0 0 18 7 0 1,2 1,216 0,015 0,043 0 0 (10,20] 150 151 1 4 6 2 0 0 0 10 13 2,286 2,287 0,049 0,092 0 0 (20,30] 64 62 -2 5 6 1 1 0 0 2 2 2,89 2,799 0,32 0,319 0 0 (30,40] 26 27 1 1 1 0 0 0 0 3 4 2,451 2,536 0,085 0,085 0 0 (40,50] 13 12 -1 0 1 1 0 0 0 3 3 1,915 1,766 0 0,132 0 0 (50,60] 4 5 1 0 0 0 0 0 0 0 1 1,014 1,226 0 0 0 0 (60,70] 3 3 0 0 0 0 0 0 0 0 0 0,985 1,006 0 0 0 0 (70,80] 3 3 0 0 0 0 0 0 0 0 0 1,241 1,25 0 0 0 0 (80,90] 1 1 0 0 1 0 0 0 0 0 0 0,574 0,573 0 0 0 0 >90 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 total 549 554 5 14 25 10 1 0 18 25 23 14,556 14,659 0,469 0,671 0 0
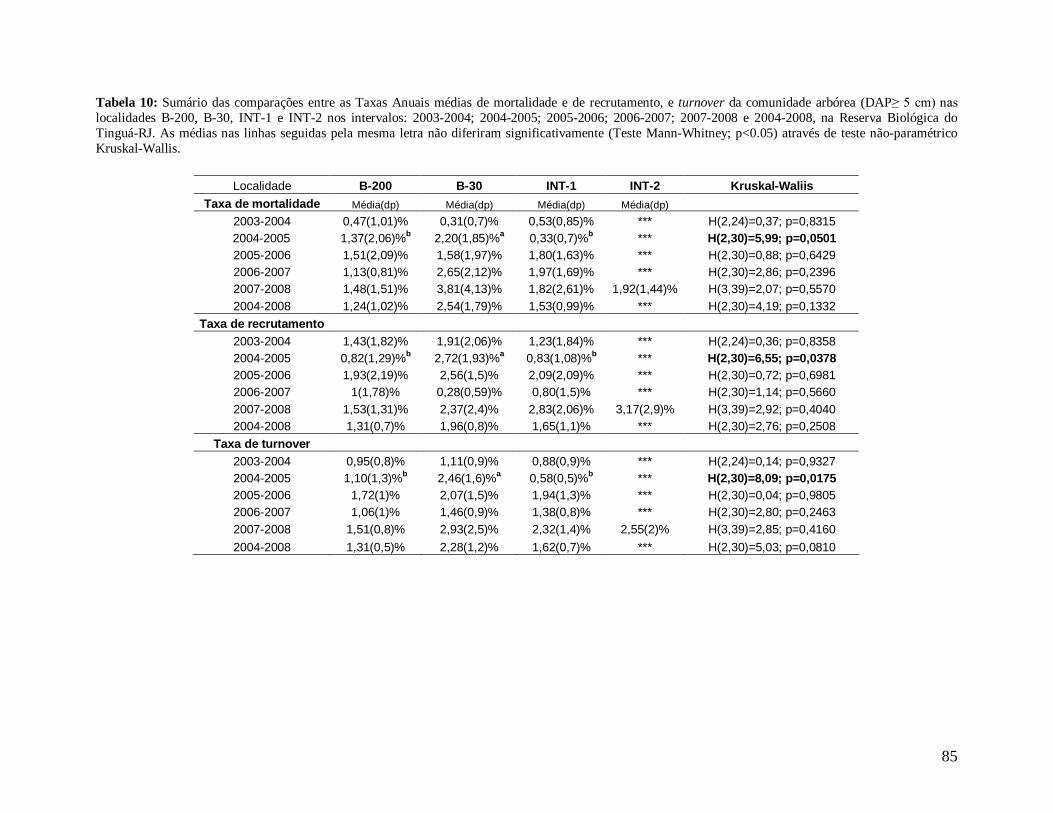
85
Tabela 10: Sumário das comparações entre as Taxas Anuais médias de mortalidade e de recrutamento, e turnover da comunidade arbórea (DAP≥ 5 cm) nas localidades B-200, B-30, INT-1 e INT-2 nos intervalos: 2003-2004; 2004-2005; 2005-2006; 2006-2007; 2007-2008 e 2004-2008, na Reserva Biológica do Tinguá-RJ. As médias nas linhas seguidas pela mesma letra não diferiram significativamente (Teste Mann-Whitney; p<0.05) através de teste não-paramétrico Kruskal-Wallis.
Localidade B-200 B-30 INT-1 INT-2 Kruskal-Waliis Taxa de mortalidade Média(dp) Média(dp) Média(dp) Média(dp)
2003-2004 0,47(1,01)% 0,31(0,7)% 0,53(0,85)% *** H(2,24)=0,37; p=0,8315 2004-2005 1,37(2,06)%b 2,20(1,85)%a 0,33(0,7)%b *** H(2,30)=5,99; p=0,0501 2005-2006 1,51(2,09)% 1,58(1,97)% 1,80(1,63)% *** H(2,30)=0,88; p=0,6429 2006-2007 1,13(0,81)% 2,65(2,12)% 1,97(1,69)% *** H(2,30)=2,86; p=0,2396 2007-2008 1,48(1,51)% 3,81(4,13)% 1,82(2,61)% 1,92(1,44)% H(3,39)=2,07; p=0,5570 2004-2008 1,24(1,02)% 2,54(1,79)% 1,53(0,99)% *** H(2,30)=4,19; p=0,1332
Taxa de recrutamento 2003-2004 1,43(1,82)% 1,91(2,06)% 1,23(1,84)% *** H(2,24)=0,36; p=0,8358 2004-2005 0,82(1,29)%b 2,72(1,93)%a 0,83(1,08)%b *** H(2,30)=6,55; p=0,0378 2005-2006 1,93(2,19)% 2,56(1,5)% 2,09(2,09)% *** H(2,30)=0,72; p=0,6981 2006-2007 1(1,78)% 0,28(0,59)% 0,80(1,5)% *** H(2,30)=1,14; p=0,5660 2007-2008 1,53(1,31)% 2,37(2,4)% 2,83(2,06)% 3,17(2,9)% H(3,39)=2,92; p=0,4040 2004-2008 1,31(0,7)% 1,96(0,8)% 1,65(1,1)% *** H(2,30)=2,76; p=0,2508
Taxa de turnover 2003-2004 0,95(0,8)% 1,11(0,9)% 0,88(0,9)% *** H(2,24)=0,14; p=0,9327 2004-2005 1,10(1,3)%b 2,46(1,6)%a 0,58(0,5)%b *** H(2,30)=8,09; p=0,0175 2005-2006 1,72(1)% 2,07(1,5)% 1,94(1,3)% *** H(2,30)=0,04; p=0,9805 2006-2007 1,06(1)% 1,46(0,9)% 1,38(0,8)% *** H(2,30)=2,80; p=0,2463 2007-2008 1,51(0,8)% 2,93(2,5)% 2,32(1,4)% 2,55(2)% H(3,39)=2,85; p=0,4160 2004-2008 1,31(0,5)% 2,28(1,2)% 1,62(0,7)% *** H(2,30)=5,03; p=0,0810
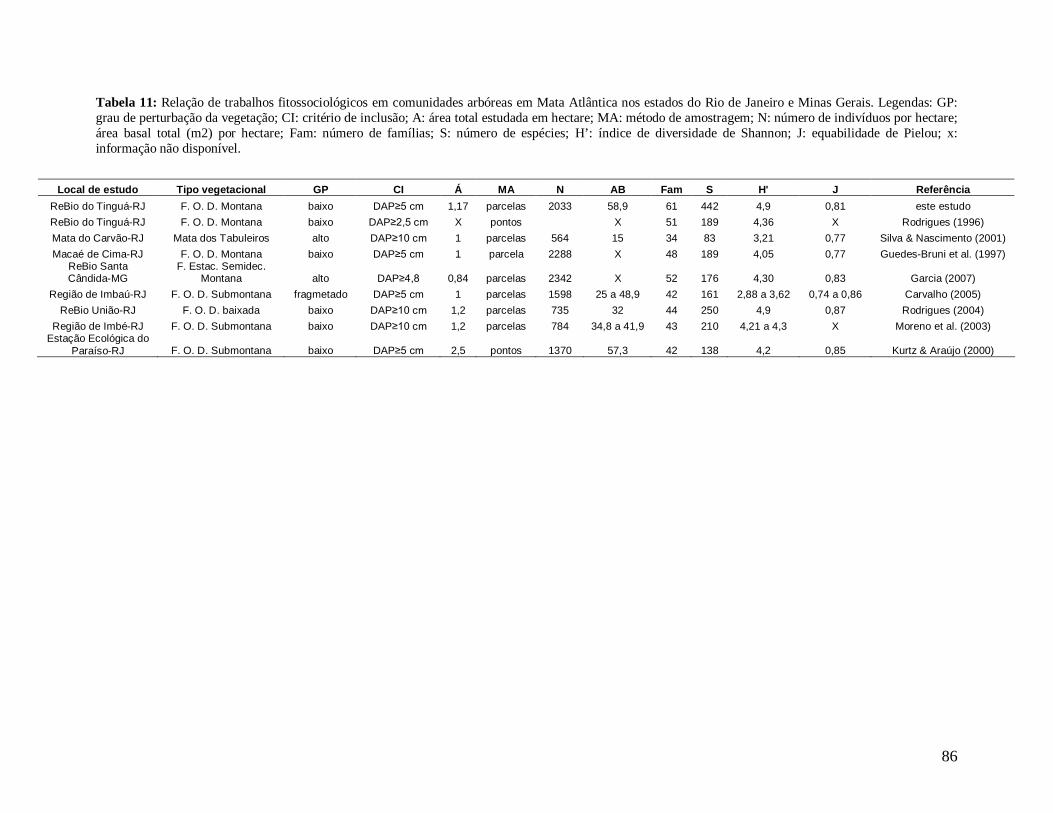
86
Tabela 11: Relação de trabalhos fitossociológicos em comunidades arbóreas em Mata Atlântica nos estados do Rio de Janeiro e Minas Gerais. Legendas: GP: grau de perturbação da vegetação; CI: critério de inclusão; A: área total estudada em hectare; MA: método de amostragem; N: número de indivíduos por hectare; área basal total (m2) por hectare; Fam: número de famílias; S: número de espécies; H’: índice de diversidade de Shannon; J: equabilidade de Pielou; x: informação não disponível.
Local de estudo Tipo vegetacional GP CI Á MA N AB Fam S H' J Referência ReBio do Tinguá-RJ F. O. D. Montana baixo DAP≥5 cm 1,17 parcelas 2033 58,9 61 442 4,9 0,81 este estudo ReBio do Tinguá-RJ F. O. D. Montana baixo DAP≥2,5 cm X pontos X 51 189 4,36 X Rodrigues (1996) Mata do Carvão-RJ Mata dos Tabuleiros alto DAP≥10 cm 1 parcelas 564 15 34 83 3,21 0,77 Silva & Nascimento (2001) Macaé de Cima-RJ F. O. D. Montana baixo DAP≥5 cm 1 parcela 2288 X 48 189 4,05 0,77 Guedes-Bruni et al. (1997)
ReBio Santa Cândida-MG
F. Estac. Semidec. Montana alto DAP≥4,8 0,84 parcelas 2342 X 52 176 4,30 0,83 Garcia (2007)
Região de Imbaú-RJ F. O. D. Submontana fragmetado DAP≥5 cm 1 parcelas 1598 25 a 48,9 42 161 2,88 a 3,62 0,74 a 0,86 Carvalho (2005) ReBio União-RJ F. O. D. baixada baixo DAP≥10 cm 1,2 parcelas 735 32 44 250 4,9 0,87 Rodrigues (2004)
Região de Imbé-RJ F. O. D. Submontana baixo DAP≥10 cm 1,2 parcelas 784 34,8 a 41,9 43 210 4,21 a 4,3 X Moreno et al. (2003) Estação Ecológica do
Paraíso-RJ F. O. D. Submontana baixo DAP≥5 cm 2,5 pontos 1370 57,3 42 138 4,2 0,85 Kurtz & Araújo (2000)

87
REFERÊNCIAS BIBLIOGRÁFICAS
Addinsoft. 2009. XLSTAT Versão 2009 – Statistical software for MS Excel. Disponível em
www.xlstat.com. Acessado em fevereiro de 2009.
Baez, S. & Balslev, H. 2007. Efeitos de borda on palm diversity in rain forests fragments in
western Ecuador. Biodiversity and Conservation 16: 2201-2211.
Begon, M.; Townsend, C. R. & Harper, J. L. 2006. Ecology: from individuals to ecosystems.
4a edição. Blackwell Publishing, Cambridge.
Bierregaard Jr., R. O.; Lovejoy, T. E.; Kapos, V.; Santos, A. A. & Hutchings, R W. 1992.
The biological dynamics of tropical rainforest fragments: a prospective comparison of
fragments and continuous forest. BioScience 42(11): 859-866.
Bierregaard Jr., R. O.; Gascon, C.; Lovejoy, T. E. & Mesquita, R. C. G. 2001. Lessons from
Amazonia: the ecology and conservation of a fragmented forest. Yale University Press,
London.
Bissonette, J. A. & Storch, I. 2002. Fragmentation: Is the message clear? Conservation
Ecology 6 (2): 14 (on line).
Breugel, M. V.; Bongers, F. & Martínez-Ramos, M. 2007. Species dynamics during early
secondary forest succession: recruitment, mortality and species turnover. Biotropica
35(5): 610-619.
Brooks, T. M.; Mittermeier, R. A.; Mittermeier, C. G.; Fonseca, G. A. B. da; Rylands, A. B.;
Konstant, W. R.; Flick, P.; Pilgram, J.; Olfield, S.; Magin, G. & Hilton-Taylor, C. 2002.
Habitat loss and extinction in the hotspots of biodiversity. Conservation Biology 16:
909-923.
Camargo, J. L. & Kapos, V. 1995. Complex adge effects on soil moisture and microclimate
in Central Amazonian forest. Journal of Tropical Ecology 11: 205-221.
Carvalho, F. A. 2005. Efeitos da fragmentação florestal na florística e estrutura da Mata
Atlântica submontana da região de Imbaú, município de Silva Jrdim, RJ. Dissertação
de Mestrado, Universidade Estadual do Norte Fluminense, Campos de Goytacazes (RJ),
128p.

88
Cerqueira, R.; Brant, A.; Nascimento, M. T.; Pardini, R. 2003. Fragmentação: Alguns
conceitos. In: Rambaldi, D. M. & Oliveira, D. A. S. (Org.). Fragmentação de
ecossistemas: causas, efeitos sobre a biodiversidade e recomendações de políticas
públicas. CID, Brasília, p. 23-40.
CIENTEC. 2006. Software Mata Nativa 2: Sistema para análise fitossociológica, elaboração
de inventários e planos de manejo de florestas nativas. Viçosa (MG), Cientec.
Clark, D. A. & Clark, D. B. 1984. Spacing dynamics of a tropical rain forest tree: evaluation
of the Janzen-Connel model. The American Naturalist 124(6): 769-788.
Clark, D. A. & Clark, D. B. 1999. Assessing the growth of tropical rain forest trees: issues
for forest modeling and management. Ecological Applications 9(3): 981-997.
Clark, D. B. & Clark, D. A. 2000. Landscape-scale variation in forest structure and biomass
in a tropical rain forest. Forest Ecology and Management 137: 185-198.
Crawley, M. J. (Ed.). 1997. Plant ecology. 2a edição, Blackweel Science, London.
D’Angelo, S. A.; Andrade, A. C. S.; Laurance, S. G.; Laurance, W. F. & Mesquita, R. C. G.
2004. Inferred causes of tree mortality in fragmented and intact Amazonian forests.
Journal of Tropical Ecology 20: 243-246.
Dean, W. 1996. A ferro e fogo: a história e a devastação da Mata Atlântica brasileira. Cia
das Letras, São Paulo.
Debinski, D. M. & Holt, R. D. 2000. A survey and overview of habitat fragmentation
experiments. Conservation Biology 14: 342-355.
Denslow, J. S. 1987 Tropical rainforest gaps and tree species diversity. Annual Review of
Ecology and Systematics 18: 431-451.
Denslow, J. S. 1995. Disturbance and diversity in tropical rain forests: the density effect.
Ecological Applications 5(4): 962-968.
Didham, R. K. 1998. Altered leaf-litter decomposition rates in tropical forest fragments.
Oecologia 116: 397-406.
Ferreira, L. V. & Laurance, W. F. 1997. Effects of forest fragmentation on mortality and
damage of selected trees in Central Amazonia. Conservation Biology 11(3): 797-801.

89
Fonseca, G. A. B. 1985. The vanishing brazilian atlantic forest. Biological Conservation
34(1): 17-43.
Garcia, P. O. 2007. Estrutura e composição do estrato arbóreo em diferentes trechos da
Reserva Biológica Municipal Santa Cândida, Juiz de Fora-MG. Dissertação de
Mestrado, Universidade Federal de Juiz de Fora (MG), 91p.
Gascon, C.; Williamson, G. B. & Fonseca, G. A. B. 2000. Receding edges and vanishing
fragments. Science 288: 1356-1358.
Gomes, E. P. C.; Mantovani, W & Kageyama, P. Y. 2003. Mortality and recruitment of tree
in a secundary montane rain forest in southeastern Brazil. Brazilian Journal of Biology
63(1): 47-60.
Goosem, M. 1997. Internal fragmentation: The effects of roads, highways, and powerline
clearings on movements and mortality of Rainforest Vertebrates. In: Laurance, W. F. &
Bierregaard-Jr., R. O. (Eds.) Tropical Forest Remnants: ecology, management and
conservation of fragmented communities. Chicago University Press, Chicago, p. 241-
255.
Guedes-Bruni, R. R.; Pessoa, S. V. A. & Kurtz, B. C. 1997. Florística e estrutura do
componente arbustivo-arbóreo de um trecho preservado de floresta montana na Reserva
Ecológica de Macaé de Cima. In: Lima, H. C. & Guedes-Bruni, R. R. (Eds.). Serra de
Macaé de Cima: diversidade florística e conservação em Mata Atlântica. Jardim
Botânico do Rio de Janeiro, Rio de Janeiro, p. 127-145.
Harper, K. A.; Macdonald, S. E.; Burton, P. J.; Chen, J.; Brosofske, K. D.; Saunders, S. C.;
Euskirchen, E. S.; Roberts, D.; Jaiteh, M. S. & Esseen, P. 2005. Edge influence on
forest structure and composition in fragmented landscapes. Conservation Biology
19(3): 768-782.
Hill, J. L. & Curran, P. J. 2005 Fragment shape and tree species composition in tropical
forests: a landscape level investigation. African Journal of Ecology 43: 35-43.
Hubbell, S. P. & Foster, R. B. 1987. La estructura espacial en gran escala de un bosque
neotropical. Revista de Biologia Tropical 35: 7-22.

90
IBAMA. 2006. Plano de Manejo da Reserva Biológica do Tinguá, RJ. MMA/IBAMA
disponível em http://www.ibama.gov.br. Acessado em junho de 2008.
IBGE. 1992. Manual Técnico da Vegetação Brasileira. IBGE, Rio de Janeiro.
Iguatemy, M. A. 2008. Os efeitos de borda sobre a dinâmica da estrutura da comunidade de
plântulas na Mata Atlântica do centro norte do Estado do Rio de Janeiro. Dissertação de
Mestrado. Universidade Estadual de Campinas-UNICAMP, Campinas, SP, 90p.
JBRJ- Instituto de Pesquisas Jardim Botânico do Rio de Janeiro. 2002. Relatório
técnicocientífico - Projeto Tinguá.
Jules, E. S. & Shahani, P. 2003. A broader ecological context to habitat fragmentation: Why
matrix habitat is more important than we thought. Journal of Vegetation Science 14:
459-464.
Kapos, V. 1989. Effects of isolation on the water status of Forest patches in the Brazilian
Amazon. Journal Tropical Ecology 5: 173-185.
Kapos, V.; Wandelli, E.; Camargo, J. L. & Ganade, G. 1997. Edge related changes in
enviroment and plant responses due to Forest fragmentation in Central Amazonia. In:
Laurance, W. F. & Bierregaard-Jr, R. O. (Eds.). Tropical Forest Remnants: ecology,
management and conservation of fragmented communities. Chicago University Press,
Chicago, p. 33-44.
Kovach, W. L. 1999. MVSP – A multi-variate statistical package-Vesrão 3.1. Kovach
Computing Services. Disponível em www.exetersoftware.com. Acessado em fevereiro
de 2009.
Kurtz, B. C. & Araújo, D. S. D. 2000. Composição florística e estrutura do componente
arbóreo de um trecho de Mata Atlântica na Estação Ecológica Estadual do Paraíso,
Cachoeiras de Macacu, Rio de Janeiro, Brasil. Rodriguésia 51: 69-112.
Laurance, W. F. & Yensen, E. 1991. Predicting the impacts of edge effects in fragmented
habitats. Biological Conservation 55: 77-92.
Laurance, W. F. 1997. Hyper-disturbed parks: edge effects and the ecology of isolated rain
forest reserves in tropical Australia. In: Laurance, W. F. & Bierregaard Jr., R. O. (Eds.).

91
Tropical Forest Remnants: ecology, management and conservation of fragmented
communities. Chicago University Press, Chicago, p. 71-84.
Laurance, W. F. & Bierregaard Jr, R. O. (Eds.). 1997. Tropical Forest Remnants: ecology,
management and conservation of fragmented communities. Chicago University Press,
Chicago, 616p.
Laurance, W. F.; Ferreira, L. V.; Rankin de Merona, J. M. & Laurance, S. G. 1998a. Rain
forest fragmentation and the dynamics of amazonian tree communities. Ecology 79(6):
2032-2040.
Laurance, W. F.; Ferreira, L. V.; Rankin de Merona, J. M. & Laurance, S. G. 1998b. Effects
of forest fragmentation on recruitment patterns in Amazonian tree communities.
Conservation Biology 12: 460-464.
Laurance, W. F.; Delamônica, P.; Laurance, S. G.; Vasconcelos, H. L. & Lovejoy, T. E.
2000. Rainforest fragmentation kills big trees. Nature 404: 836.
Laurance, W. F.; Didham, R. K. & Power, M. E. 2001a. Ecological boundaries: a search for
synthesis. Trends in Ecology and Evolution 16(2): 70-71.
Laurance, W. F.; Williamson, G. B.; Delamonica, P.; Oliveira, A.; Gascon, C.; Lovejoy, T. &
Pohl, L. 2001b. Effects of a strong drought on Amazonian forest fragments and edges.
Journal of Tropical Ecology 17: 771-785.
Laurance, W. F.; Lovejoy, T. E.; Vasconcelos, H. L.; Bruna, E. M.; Didham, R. K.; Stouffer,
P. C.; Gascon, C.; Bierregaard Jr., R. O; Laurance, S. G. & Sampaio, E. 2002.
Ecosystem decay of Amazonian Forest fragments: a 22-year investigation.
Conservation Biology 16(3): 605-618.
Laurance, W. F.; Oliveira, A. A.; Laurance, S. G.; Condit, R.; Nascimento, H. E. M.;
Sanchez-Thorin, A. C.; Lovejoy, T. E.; Andrade, A.; D’Angelo, S.; Ribeiro, J. E. &
Dick, C. W. 2004. Pervasive alteration of tree communities in undisturbed Amazonian
forest. Nature 428: 171-175.
Laurance, W. F.; Nascimento, H. E. M.; Laurance, S. G.; Andrade, A. C.; Fearnside, P. M.;
Ribeiro, J. E. L. & Capretz, R. L. 2006. Rain forest fragmentation and the proliferation
of successional trees. Ecology 87(2): 469-482.

92
Laurance, W. F.; Nascimento H. E. M.; Laurance, S. G.; Andrade, A.; Ewers, R. M.; Harms,
K. E.; Luizão, R. C. C. & Ribeiro, J. E. 2007. Habitat fragmentation, variable edge
effects, and the landscape-divergence hypothesis. Plos One 10(1017): 1-8.
Lewis, S. L.; Phillips, O. L.; Baker, T. R.; Lloyd, J.; Malhi, Y.; Almeida, S.; Higuchi, N.;
Laurance, W. F.; Neill, D. A.; Silva, J. N. M.; Terborgh, J. W.; Lezama, A. T.;
Martínez, R. V.; Brown, S.; Chave, J.; Kuebler, C.; Vargas P. N. & Vincet, B. 2004.
Concerted changes in tropical forest estructure and dynamics: evidence from 50 South-
American long-term plots. Philosophical Transactions of Royal Society B 359: 421-
436.
Lieberman, D.; Lieberman, M.; Peralta, R. & Hartshorn, G. S. 1985. Mortality patterns and
stand turnover rates in a wet tropical forest in Costa Rica. Journal of Ecology 73: 915-
924.
Lieberman, D. & Lieberman, M. 1987. Forest tree growth and dynamics at La Selva, Costa
Rica (1969-1982). Journal of Tropical Ecology 3: 347-358.
Lovejoy, T. E.; Bierregaard Jr, R. O.; Rylands, A. B.; Malcon, J. R.; Quintela, C. E.; Harpe,
L. H.; Brown-Jr, K. S.; Powell, A. H.; Powell, G. V. N.; Schubart, H. O. R. & Hays, M.
B. 1986. Edge and other effects of isolation on Amazon forest fragments. In: Soulé, M.
E. (Eds.) Conservation Biology: The science of scarcity and diversity. Sinauer
Associates, Sunderland, p. 257-285.
Mantovani, W. 1993. Estrutura e dinâmica da floresta atlântica na Juréia, Iguape, SP. Tese
de livre docência, Universidade de São Paulo-USP, 128p.
Matlack, G. R. 1994. Vegetation dynamics of the forest edge - trends in space and
successional time. Journal of Ecology 82: 113-123.
Mesquita, R. C. G.; Delamônica, P. & Laurance, W. F. 1999. Effects of surrouding
vegetation on edge-related tree mortality in Amazonian forest fragments. Biological
Conservation 91(1): 129-134.
Mittermeier, R. A.; Myers, N.; Thompsen, J. G.; Fonseca, G. A. B. da; Olivieri, S. 1998.
Biodiversity hotspots and major wilderness areas: approaches to setting conservation
priorities. Conservation Biology 12: 516-520.

93
Morellato, L. P. C. & Haddad, C. F. B. 2000. Introduction: The Brazilian Atlantic forest.
Biotropica 32: 786-792.
Moreno, M. R.; Nascimento, M. T. & Kurtz, B. C. 2003. Estrutura e composição forística do
estrato arbóreo em duas zonas altitudinais na Mata Atlântica de encosta da região de
Imbé, RJ. Acta Botanica Brasilica 17(3): 371-386.
Mori, S. A.; Boom, B. M.; Carvalho, A. M. & Santos, T. S. 1983. Ecological importance of
Myrtaceae in an eastern brazilian wet forest. Biotropica 15(1): 68-70.
Murcia, C. 1995. Edge Effects in Fragmented Forests: implications for conservation. Tree
10(2): 58-62.
Myers, N.; Mittermeier, R. A.; Mittermeier, C. G.; Fonseca, G. A. B. & Kents, J. 2000.
Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
Nascimento, E. M. & Laurance, W. F. 2004. Biomass dynamics in amazonian forest
fragments. Ecological Applications 14(4): 127-138.
Nascimento, H. E. M.; Andrade, A. C. S.; Camargo, J. L. C.; Laurance, W. F.; Laurance, S.
G. & Ribeiro, J. E. L. 2006. Effects of the surrounding matrix on tree recruitment in
Amazonian forest fragments. Conservation Biology 20(3): 853-860.
Nascimento, E. M. & Laurance, W. F. 2006. Efeitos de área e de borda sobre a estrutura
florestal em fragmentos de floresta de terra-firme após 13-17 anos de isolamento. Acta
Amazônica 36(2): 183-192.
Newmark, W. 2005. Diel variation in the difference in air temperature between the forest
edge and interior in the Usambara Mountains, Tanzania. African Journal of Ecology 43:
177-180.
Olifiers, N. & Cerqueira, R. 2006. Fragmentação de habitats: efeitos históricos e ecológicos.
In: Rocha, C. F. D.; Bergallo, H. G.; Sluys, M. V. & Alves, M. A. S. (Eds.). Biologia
da conservação: essências. RiMa, São Carlos, p. 261-279.
Oliveira, M. A.; Grillo, A. S. & Tabarelli, M. 2004. Forest edges in the Brazilian Atlantic
forest: drastic changes in tree species assemblages. Oryx 38: 4.

94
Oliveira-Filho, A. T.; Mello, J. M, de; Scolforo, J. R. S. 1997. Effects of past disturbance and
edges on tree community structure and dynamics within a fragment of tropical
semideciduous forest in south-eastern Brazil over five-year period (1987-1992). Plant
Ecology 131: 45-66.
Oliveira-Filho, A. T. & Fontes, M. A. L. 2000. Patterns of floristic differentiation among
atlantic forests in southeastern Brazil and the influence of climate. Biotropica 32(4b):
793-810.
Oliveira-Filho, A. T.; Carvalho, W. A. C.; Machado, E. L. M.; Higuchi, P.; Appolinário, V.;
Castro, G. C.; Silva, A. C.; Santos, R. M.; Borges, L. F.; Corrêa, B. S. & Alves, J. M.
2007. Dinâmica da comunidade e populações arbóreas da borda e interior de um
remanescente florestal na Serra da Mantiqueira, Minas Gerais, em um intervalo de
cinco anos (1999-2004). Revista Brasileira de Botânica 30(1): 149-161.
Phillips, O. L. 1996. Long-term environmental change in tropical forests: increasing tree
turnover. Environmental Conservation 23: 235-248.
Phillips, O. L.; Hall, P.; Gentry, A. H.; Sawyer, S. A. & Vásquez, R. 1994. Dynamics and
species richness of tropical rain forests. Proceedings of National Academy of Science
91: 2805-2809.
Pietro, P. V. 2008. Efeitos de borda sobre o sub-bosque da Mata Atlântica de terras baixas
na Reserva Biológica União-RJ. Dissertação de Mestrado. Escola Nacional de Botânica
Tropical – JBRJ, Rio de Janeiro, 123p.
Pinto, P. P.; Bedê, L.; Paese, A.; Fonseca, M.; Paglia, A. & Lamas, I. 2006. Mata Atlântica
brasileira: os desafios para conservação da biodiversidade de um hotspot mundial. In:
Rocha, C. F. D.; Bergallo, H. G.; Sluys, M. V. & Alves, M. A. S. (Eds.). Biologia da
conservação: essências. RiMa, São Carlos, p. 91-118.
Pires, A. S.; Fernandez, F. A. S. & Barros, C. S. 2006. Vivendo em um mundo empedaços:
efeitos da fragmentação florestal sobre comunidades e populações animais. In: Rocha,
C. F. D.; Bergallo, H. G.; Sluys, M. V. & Alves, M. A. S. (Eds.). Biologia da
conservação: essências. RiMa, São Carlos, p. 231-260.

95
Pohlman, C.; Turton, S. M. & Goosem, M. 2007. Edge effects of linear canopy openings on
Tropical Rain Forest understory microclimate. Biotropica 39(1): 62-71.
Reis, T. S. 2008. Os efeitos de borda na estrutura e dinâmica da comunidade de plântulas
em um trecho de Mata Atlântica do sudeste brasileiro. Dissertação de Mestrado.
Universidade Federal do Rio de Janeiro-UFRJ, Rio de Janeiro, 179p.
Reitz, R. 1981. Nictagináceas. In: Reitz, R. (Ed.). Flora Ilustrada Catarinense, Itajaí.
Rizzini, C. T. 1997. Tratado de Fitogeografia do Brasil: aspectos ecológicos, sociológicos e
florísticos. Âmbito Cultural Edições, Rio de janeiro.
Rodrigues, H. C. 1996. Composição florística e fitossociológica de um trecho de Mata
Atlântica na Reserva Biológica do Tinguá, Nova Iguaçu, RJ. Dissertação de Mestrado.
Museu Nacional da Universidade Federal do Rio de Janeiro, Rio de Janeiro, 77p.
Rodrigues, P. J. F. P. & Nascimento, M. T. 2006. Fragmentação florestal: breves
considerações teóricas sobre efeitos de borda. Rodriguésia 57(1): 63-74.
Rodrigues, P. J. F. P. 2004. A vegetação da Reserva Biológica União e os efeitos de borda na
Mata Atlântica fragmentada. Tese de Doutorado. Universidade Estadual do Norte
Fluminense-UENF, Campo de Goytacazes, 153p.
Rodrigues, P. J. F. P.; Iguatemy, M. A.; Prieto, P. V.; Reis, T. S.; Sansevero, J. B. B. &
Simões-Jesus, M. F. 2008. As pesquisas no longo prazo em parcelas permanentes do
Jardim Botânico do Rio de Janeiro. In: Sanquetta, C. R. (Ed.). Experiências de
monitoramento no Bioma Mata Atlântica com uso de parcelas permanentes (meio
digital-CD). Editora C. R. Sanquetta, Curitiba, p. 47-80.
Saiter, F. Z. 2007. Dinâmica da comunidade e distribuição geográfica de espécies do
componente arbóreo de uma floresta pluvial do sudeste do Brasil. Dissertação de
Mestrado, Universidade Federal do Rio de Janeiro-UFRJ, Rio de Janeiro, 108p.
Saunders, D. A.; Hobbs, R. J. & Margules, C. R. 1991. Biological consequences of
ecosystem fragmentation: a review. Conservation Biology 5: 18-32.
Scariot, A.; Freitas, S. R.; Mariano-Neto, E.; Nascimento, M. T.; Oliveira, L. C.; Sanaiotti,
T.; Sevilha, A. C. & Villela, D. 2003. Vegetação e flora. In: Rambaldi, D. M. &

96
Oliveira, D. A. S. (Org.). Fragmentação de ecossistemas. Causas, e efeitos sobre a
biodiversidade e recomendações de políticas públicas. CID, Brasília, p. 103-123.
Sheil, D.; Burslen, D. & Alder, D. 1995. The interpretation and misinterpretation of mortality
rate measures. Journal of Ecology 83: 331-333.
Sheil, D.; Jennings, S. & Savill, P. 2000. Long term permanent plot observation of vegetation
dynamics in Budongo, a ungandan rain forest. Journal of Tropical Ecology 16: 765-
800.
Silva, G. C. & Nascimento, M. T. 2001. Fitossociologia de um remanescente de mata sobre
de tabuleiros no norte do estado do Rio de Janeiro (Mata do Carvão). Revista Brasileira
de Botânica 24(1): 51-62.
Sizer, N. & Tanner, E. V. J. 1999. Responses of wood plant seedlings to edge formation in a
lowland tropical rainforest, Amazonia. Biological Conservation 91: 135-142.
Sizer, N. C.; Tanner, E. V. J. & Ferraz, I. D. K. 2000. Edge effects on litterfall mass and
nutrient concentrations in forest fragments in central Amazonia. Journal of Tropical
Ecology 16: 853-863.
Souza, J. C. 2003. Reserva Biológica do Tinguá: Discutindo o processo de co-gestão a partir
de uma iniciativa local. Dissertação de Mestrado. Escola Nacional de Ciências
Estatísticas, ENCE – IBGE, Rio de Janeiro, 234p.
StatSoft Inc. 2001. STATISTICA (data analysis software system), version 6.
www.statsoft.com. Acessado em fevereiro de 2008.
Swaine, M. D.; Liebermam, D. & Putz, F. E. 1987. The dynamics of tree populations in
tropical forest: a review. Journal of Tropical Ecology 3: 359-366.
Tabarelli, M.; Mantovani, W. & Peres, C. A. 1999. Effects of habitat fragmentation on plant
guild structure in the montane Atlantic Forest of Southeastern Brazil. Biological
Conservation 91: 119-127.
Thomaz, L. D. & Monteiro, R. 1997. Composição florística da Mata Atlântica de enconsta da
Estação Biológica de Santa Lúcia, município de Santa Tereza-ES. Boletim do Museu de
Biologia Prof. Mello Leitão (Nova Série) 7: 3-48.

97
Townsend, C. R.; Begon, M. & Harper, J. L. 2006. Fundamentos em ecologia. Artmed, Porto
Alegre.
Turner, I. M. 2001. The ecology of trees in the Tropical Rain Forest. Cambridge University
Press, New York.
Veloso, H. P.; Rangel-Filho, A. L. R. & Lima, J. C. A. 1991. Classificação da vegetação
brasileira, adaptada a um sistema universal. IBGE, Rio de Janeiro.
Viana, V. M.; Tabanez, A. A. J. & Batista, J. L. 1997. Dynamics and restoration of forst
fragments in the brazilian atlantic moist forest. In: Laurance, W. F. & Bierregaard Jr, R.
O. (Eds.). Tropical Forest Remnants: ecology, management and conservation of
fragmented communities. Chicago University Press, Chicago, p. 351-365.
Whitmore, T. C. 1990. An introduction to tropical rain forests. Blackwell, London.
Whitmore, T. C. 1997. Tropical forest disturbance, disappearance, and species loss. In:
Laurance, W. F. & Bierregaard, R. O. (Eds.). Tropical forest remnants. The University
of Chicago Press, Chicago & London, p. 3-13.
Williams-Linera, G. 1990. Vegetation structure and environmental conditions of forest edges
in Panama. Journal of Ecology 78: 356-373.
Zar, J. 1999. Biostatistical Analysis. 4a edição. Prentice Hall, Englewood Cliffs.

98
ANEXOS

99
Anexo 1: Listagem das espécies encontradas nas 39 parcelas (10x30m) alocadas em trecho de floresta montana Reserva Biológica do Tinguá (RJ), ordenadas em ordem alfabética de famílias.
Família Espécie Anacardiaceae Astronium sp1 Tapirira guianensis Aubl. Annonaceae Annona cacans Warm. Annona dolabripetala Raddi Duguetia microphylla (R.E.Fr.) R.E.Fr. Guatteria australis A. St.-Hil. Guatteria campestris R.E.Fr. Unonopsis sp1 Xylopia brasiliensis Spreng. Apocynaceae Apocynaceae sp1 Aquifoliaceae Ilex cf. congonhinha Loes. Ilex cf. dumosa Reissek Araliaceae Schefflera cf. angustissimus Marchal

100
Schefflera cf. anomalus Taub. Schefflera sp1 Schefflera sp2 Araliaceae sp1 Arecaceae Astrocaryum aculeatissimum (Schott) Burret Attalea dubia (Mart.) Burret Euterpe edulis Mart. Geonoma sp1 Polyandrococos caudescens (Mart.) Barb. Rodr. Asteraceae Critoniopsis stellata (Spreng) H. Rob. Asteraceae sp1 Vernonanthura discolor (Spreng.) H. Rob. Bignoniaceae Jacaranda puberula Cham. Jacaranda sp1 Bignoniaceae sp1 Bignoniaceae sp2 Tabebuia sp1 Boraginaceae Cordia sellowiana Cham. Burseraceae Protium widigrenii Engl. Caricaceae Jacaratia spinosa (Aubl.) A.DC. Celastraceae Maytenus sp1 Chloranthaceae Hedyosmum brasiliense Miq. Chrysobalanaceae Hirtella hebeclada Moric. ex DC. Hirtella sp1 Licania sp1 Licania sp2 Chrysobalanaceae sp1 Chrysobalanaceae sp2 Chrysobalanaceae sp3 Chrysobalanaceae sp4 Chrysobalanaceae sp5 Chrysobalanaceae sp6

101
Chrysobalanaceae sp7 Chrysobalanaceae sp8 Chrysobalanaceae sp9 Chrysobalanaceae sp10 Clethraceae Clethra scabra Pers. Clusiaceae Chrysochlamys saldanhae (Engl.) Oliveira-Filho Clusia lanceolata Cambess. Garcinia brasiliensis Mart. Garcinia sp1 Garcinia sp2 Clusiaceae sp1 Clusiaceae sp2 Clusiaceae sp3 Clusiaceae sp4 Clusiaceae sp5 Tovomitopsis sp1 Combretaceae Terminalia januariensis DC. Cunoniaceae Lamanonia ternata Vell. Dichapetalaceae Stephanopodium organense (Rizzini) Prance Ebenaceae Ebenaceae sp1 Ebenaceae sp2 Elaeocarpaceae Sloanea guianensis (Aubl.) Benth. Erythroxylaceae Erythroxylum cuspidifolium Mart. Erythroxylum pulchrum A.St.-Hil. Euphorbiaceae Actinostemon cf. verticillatus (Klotzsch) Baill. Actinostemon cf. klotzschii (Didr.) Pax Actinostemon concolor (Spreng.) Müll. Arg. Actinostemon sp1 Alchornea glandulosa Poepp. & Endl. Alchornea triplinervia (Spreng.) Müll. Arg. Hieronyma alchorneoides Allemão Pachystroma sp1

102
Pachystroma sp2 Pera glabrata (Schott) Poepp. ex Baill. Pera leandri Baill. Pera sp1 Pogonophora schomburgkiana Miers Sebastiania brasiliensis Spreng. Senefeldera verticillata (Vell.) Croizat Euphorbiaceae sp1 Euphorbiaceae sp2 Euphorbiaceae sp3 Fabaceae – Caesalpinioideae Copaifera lucens Dwyer Copaifera trapezifolia Hayne Hymenaea courbaril L. Moldenhawera polysperma (Vell.) Stellfeld Peltogyne angustiflora Ducke Senna multijuga (Rich.) H. S. Irwin & Barneby Tachigali paratyensis (Vell.) H.C. Lima Tachigali pilgeriana (Harms) Oliveira-Filho Tachigali urbaniana L.G. Silva & H.C. de Lima Fabaceae – Mimosoideae Inga capitata Desv. Inga lanceifolia Benth. Inga marginata Willd. Inga mendoncaei Harms Inga tenuis (Vell.) Mart. Piptadenia gonoacantha (Mart.) J.F.Macbr. Pseudopiptadenia inaequalis (Benth.) Rauschert Stryphnodendron polyphyllum Mart. Fabaceae – Papilionoideae Andira cf. ormosioides Benth. Balizia pedicellaris (DC.) Barneby & J.W.Grimes Dahlstedtia pinnata (Benth.) Malme Dalbergia foliolosa Benth. Machaerium nyctitans (Vell.) Benth. Machaerium sp1 Machaerium sp2 Machaerium sp3 Myrocarpus frondosus Allemão Ormosia fastigiata Tul. Pterocarpus rohrii Vahl Swartzia myrtifolia Sm. Swartzia sp1 Zollernia ilicifolia (Brongn.) Vogel Hypericaceae Vismia martiana Reichardt Lacistemataceae

103
Lacistema pubescens Mart. Lauraceae Aiouea saligna Meisn. Aniba firmula (Nees & Mart.) Mez Beilschmiedia angustifolia Kosterm. Cinnamomum riedelianum Kosterm. Cinnamomum sp1 Cinnamomum sp2 Cryptocarya micrantha Meisn. Cryptocarya saligna Mez Cryptocarya sp1 Cryptocarya sp2 Endlicheria paniculata (Spreng.) J.F.Macbr. Mezilaurus navaluim (Allemão) Taub. ex Mez Nectandra membranacea (Sw.) Griseb. Nectandra sp1 Ocotea aciphylla (Nees) Mez Ocotea aff. vaccinioides (Meisn.) Mez Ocotea bicolor Vattimo Ocotea catharinensis Mez Ocotea diospyrifolia (Meisn.) Mez Ocotea dispersa (Nees) Mez Ocotea divaricata (Nees) Mez Ocotea domatiata Mez Ocotea elegans Mez Ocotea glaziovii Mez Ocotea lancifolia (Schott) Mez Ocotea nectandrifolia Mez Ocotea odorifera (Vell.) Rohwer Ocotea schottii (Meisn.) Mez Ocotea sp1 Ocotea sp2 Ocotea sp3 Ocotea teleiandra (Meins.) Mez Ocotea villosa Kosterm. Persea sp1 Persea willdenowii Kosterm. Rhodostemonodaphne macrocalyx (Meisn.) Rohwer ex Madriñán Urbanodendron bahiense (Meisn.) Rohwer Urbanodendron verrucosum (Nees) Mez Lecythidaceae Lecythis sp1 Loganiaceae Strychnos acuta Progel Lythraceae Lafoensia vandelliana Cham. & Schltdl.

104
Magnoliaceae Magnolia ovata (A.St.-Hil.) Spreng. Malpighiaceae Byrsonima sp1 Malpighiaceae sp1 Malpighiaceae sp2 Malpighiaceae sp3 Malpighiaceae sp4 Malvaceae Ceiba erianthos (Cav.) K. Schum. Eriotheca pentaphylla (Vell.) A. Robyns Melastomataceae Meriania glabra (DC.) Triana Miconia brasiliensis (Spreng.) Triana Miconia budlejoides Triana Miconia cf. eichleri Cogn. Miconia chartacea Triana Miconia cubatanensis Hoehne Miconia holosericea (L.) DC. Miconia pusilliflora (DC.) Triana Miconia sp1 Mouriri sp1 Tibouchina arborea (Gardner) Cogn. Tibouchina estrellensis (Raddi) Cogn. Meliaceae Cabralea canjerana (Vell.) Mart. Cedrela sp1 Guarea guidonia (L.) Sleumer Guarea macrophylla Vahl Guarea sp1 Trichilia casarettii C.DC. Trichilia lepidota Mart. Monimiaceae Mollinedia aff. fruticulosa Perkins Mollinedia elegans Tul. Mollinedia lamprophylla Perkins Mollinedia longifolia Tul. Mollinedia oligantha Perkins Mollinedia salicifolia Perkins Mollinedia schottiana (Spreng.) Perkins Mollinedia stenophylla Perkins Mollinedia triflora (Spreng.) Tul. Moraceae

105
Sorocea cf. guilleminiana Gaudich. Moraceae sp1 Moraceae sp2 Myristicaceae Virola bicuhyba (Schott.) Warb. Virola sp1 Myrsinaceae
Myrsine hermogenesii (Jung-Mendaçolli & Bernacci) M.F. Freitas & L.S. Kinoshita
Myrsine parvula (Mez) Otegui Myrsine squarrosa (Mez) M.F. Freitas & L.S. Kinoshita Stylogyne lhotzkyana (A.DC.) Mez Myrtaceae Calyptranthes cf. brasiliensis Spreng. Calyptranthes cf. grandifolia O.Berg Calyptranthes cf. concinna DC. Calyptranthes lucida Mart. ex DC. Calyptranthes strigipes O.Berg Campomanesia guaviroba (DC.) Kiaersk. Eugenia cf. copacabanensis Kiaersk. Eugenia cf. macrosperma DC. Eugenia cf. punicifolia (Kunth) DC. Eugenia cf. rotundifolia Casar. Eugenia cf. tinguyensis Cambess. Eugenia excelsa O.Berg Eugenia florida DC. Eugenia macahensis O.Berg Eugenia microcarpa O.Berg Eugenia neoglomerata Sobral Eugenia prasina O.Berg Eugenia pseudodichasiantha Kiaersk. Eugenia rostrata O.Berg Eugenia cf. speciosa Cambess. Eugenia splendens (Sw.) DC Eugenia sulcata Spring ex Martius Eugenia villae-novae Kiaersk. Eugenia xanthoxyloides Cambess. Gomidesia spectabilis O.Berg Marlierea cf. strigipes O. Berg Marlierea excoriata Mart. Marlierea obscura O.Berg Marlierea silvatica (Gardner) Kiaersk. Marlierea sp1 Marlierea tomentosa Cambess. Myrceugenia miersiana (Gardner) D. Legrand & Kausel Myrceugenia myrcioides (Cambess.) O.Berg Myrcia acuminatissima O.Berg Myrcia cf. guianensis (Aubl.) DC.

106
Myrcia laxiflora Cambess. Myrcia multiflora (Lam.) DC. Myrcia pubipetala Miq. Myrcia splendens (Sw.) DC. Myrcia tenuivenosa Kiaersk. Myrcia tijucensis Kiaersk. Myrciaria cf. disticha O.Berg Myrciaria tenella (DC.) O.Berg Myrtaceae sp1 Myrtaceae sp2 Myrtaceae sp3 Myrtaceae sp4 Myrtaceae sp5 Myrtaceae sp6 Myrtaceae sp7 Myrtaceae sp8 Myrtaceae sp9 Myrtaceae sp10 Myrtaceae sp11 Myrtaceae sp12 Myrtaceae sp13 Myrtaceae sp14 Myrtaceae sp15 Myrtaceae sp16 Myrtaceae sp17 Myrtaceae sp18 Myrtaceae sp19 Myrtaceae sp20 Myrtaceae sp21 Myrtaceae sp22 Myrtaceae sp23 Myrtaceae sp24 Myrtaceae sp25 Myrtaceae sp26 Myrtaceae sp27 Myrtaceae sp28 Myrtaceae sp29 Myrtaceae sp30 Myrtaceae sp31 Myrtaceae sp32 Myrtaceae sp33 Myrtaceae sp34 Myrtaceae sp35 Myrtaceae sp36 Myrtaceae sp37 Myrtaceae sp38 Myrtaceae sp39 Myrtaceae sp40 Myrtaceae sp41 Myrtaceae sp42

107
Myrtaceae sp43 Myrtaceae sp44 Myrtaceae sp45 Myrtaceae sp46 Myrtaceae sp47 Myrtaceae sp48 Myrtaceae sp49 Myrtaceae sp50 Myrtaceae sp51 Myrtaceae sp52 Myrtaceae sp53 Neomitranthes glomerata (D. Legrand) D. Legrand Plinia cf. rivularis (Cambess.) Rotman Plinia edulis (Vell.) Sobral Psidium cattleianum Sabine Siphoneugena densiflora O.Berg Nyctaginaceae Guapira opposita (Vell.) Reitz Ochnaceae Ouratea sp1 Ochnaceae sp1 Olacaceae Heisteria sp1 Heisteria sp2 Heisteria sp3 Phyllanthaceae Margaritaria nobilis L. f. Savia dictyocarpa Müll. Arg. Picramniaceae Picramnia glazioviana Engl. Polygonaceae Coccoloba sp1 Proteaceae Roupala sp1 Roupala sp2 Roupala sp3 Roupala sp4 Proteaceae sp1 Quiinaceae Quiina glazovii Engl. Rhamnaceae

108
Rhamnaceae sp1 Rosaceae Prunus brasiliensis Dietrich Rubiaceae Alibertia concolor (Cham.) K. Schum. Alibertia sp1 Alibertia sp2 Alseis floribunda Schott Amaioua intermedia Mart. Bathysa australis (A.St.-Hil.) Benth. & Hook. f. Bathysa cuspidata (St.-Hil.) Hook. f. Bathysa gymnocarpa K. Schum. Bathysa stipulata (Vell.) J. Presl Coussarea nodosa (Benth.) Müll. Arg. Faramea cf. coerulea (Nees & Mart.) DC. Faramea pachyantha Müll. Arg. Guettarda viburnoides Cahm. & Schltdl. Ixora schottiana Müll. Arg. Posoqueria acutifolia Mart. Posoqueria latifolia (Rudge) Roem. & Schult. Psychotria glaziovii Müll. Arg. Psychotria nuda (Chan. & Schltdl.) Wawra Psychotria pubigera Schltdl. Psychotria sp1 Psychotria sp2 Psychotria sp3 Psychotria sp4 Psychotria sp5 Psychotria suterella Müll. Arg. Psychotria vellosiana Benth. Rudgea recurva Müll. Arg. Rudgea sp1 Rudgea sp2 Rudgea vellerea Müll. Arg. Rustia formosa Klotzsch Simira pikia (K. Schum.) Steyerm. Rubiaceae sp1 Rubiaceae sp2 Tocoyena sellowiana (Cham. & Schltdl.) K. Schum. Rutaceae Almeidea sp1 Zanthoxylum rhoifolium Lam. Salicaceae Casearia aff. arborea (Rich.) Urb. Casearia aff. pauciflora Cambess Casearia obliqua Spreng.

109
Casearia sylvestris Sw. Sapindaceae Allophyllus melanophloeus Radlk. Allophylus edulis (A.St.-Hil., Cambess. & A.Juss.) Radlk. Cupania concolor Radlk. Cupania furfuracea Radlk. Cupania oblongifolia Mart. Cupania schizoneura Radlk. Matayba cf. grandis Radlk. Matayba sp1 Sapotaceae Chrysophyllum flexuosum Mart. Chrysophyllum sp1 Chrysophyllum sp2 Diploon cuspidatum (Hoehne) Cronquist Ecclinusa ramiflora Mart. Manilkara sp1 Manilkara sp2 Manilkara sp3 Micropholis cf. compta Pierre Micropholis sp1 Micropholis sp2 Pouteria bapeba T.D. Penn Pouteria caimito (Ruiz & Pav.) Radlk. Pouteria cf. guianensis Aubl. Pouteria gardneri (Mart. & Miq.) Baehni Pouteria glomerata (Miq.) Radlk. Sarcaulus cf. brasiliensis (A. DC.) Eyma Sapotaceae sp1 Sapotaceae sp2 Sapotaceae sp3 Sapotaceae sp4 Simaroubaceae Simarouba amara Aubl. Siparunaceae Siparuna brasiliensis (Spreng.) A.DC. Solanaceae Metternichia princeps Mik. Solanum leucodendron Sendtn. Solanum swartzianum Roem. & Schult. Solanaceae sp1 Symplocaceae Symplocos sp1

110
Theaceae Gordonia fruticosa (Schrad.) H. Keng Thymeliaceae Daphnopsis martii Meisn. Lamiaceae Aegiphila sellowiana Cham. Vitex sp1 Urticaceae Cecropia sp1 Cecropia sp2 Cussapoa microcarpa (Schott) Rizzini Vochysiaceae Vochysiaceae sp1 Vochysiaceae sp2 Vochysiaceae sp3 Vochysiaceae sp4 Vochysiaceae sp5 Vochysiaceae sp6 Vochysiaceae sp7 Vochysiaceae sp8 Vochysiaceae sp9 Vochysia sp1 Indeterminadas Indeterminada sp1 Indeterminada sp2 Indeterminada sp3 Indeterminada sp4 Indeterminada sp5 Indeterminada sp6 Indeterminada sp7 Indeterminada sp8 Indeterminada sp9 Indeterminada sp10 Indeterminada sp11 Indeterminada sp12 Indeterminada sp13 Indeterminada sp14 Indeterminada sp15 Indeterminada sp16 Indeterminada sp17 Indeterminada sp18 Indeterminada sp19 Indeterminada sp20 Indeterminada sp21 Indeterminada sp22
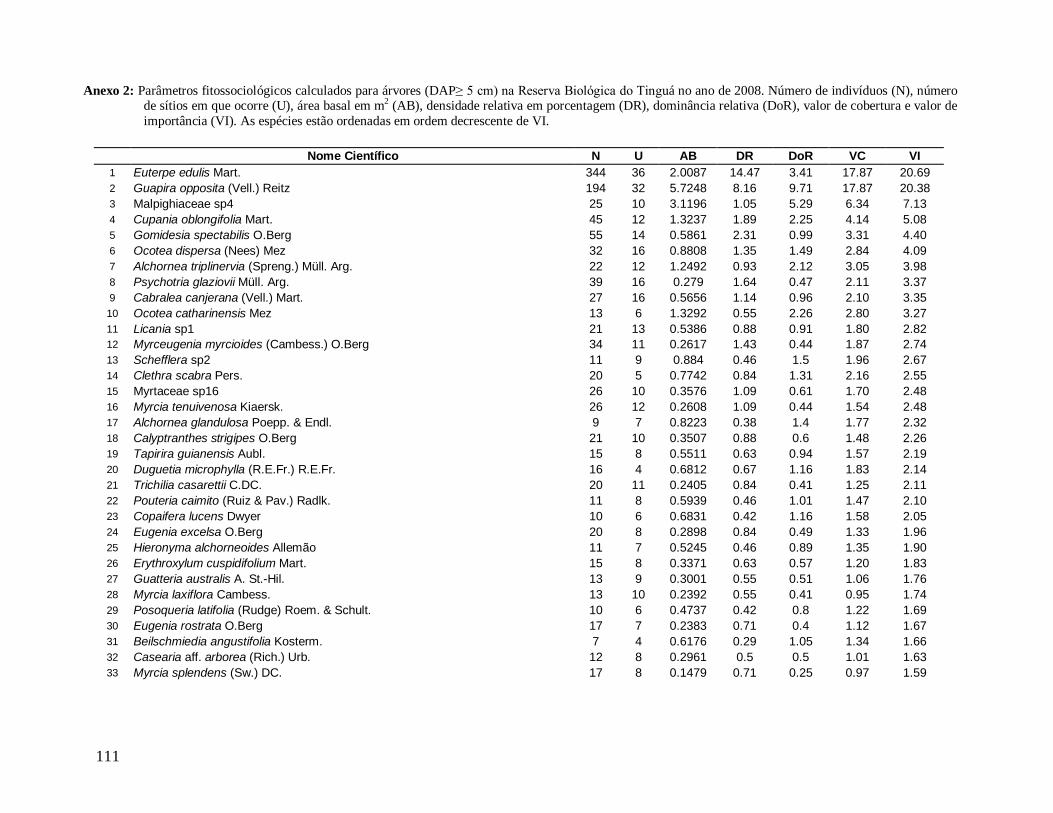
111
Anexo 2: Parâmetros fitossociológicos calculados para árvores (DAP≥ 5 cm) na Reserva Biológica do Tinguá no ano de 2008. Número de indivíduos (N), número de sítios em que ocorre (U), área basal em m2 (AB), densidade relativa em porcentagem (DR), dominância relativa (DoR), valor de cobertura e valor de importância (VI). As espécies estão ordenadas em ordem decrescente de VI.
Nome Científico N U AB DR DoR VC VI 1 Euterpe edulis Mart. 344 36 2.0087 14.47 3.41 17.87 20.69 2 Guapira opposita (Vell.) Reitz 194 32 5.7248 8.16 9.71 17.87 20.38 3 Malpighiaceae sp4 25 10 3.1196 1.05 5.29 6.34 7.13 4 Cupania oblongifolia Mart. 45 12 1.3237 1.89 2.25 4.14 5.08 5 Gomidesia spectabilis O.Berg 55 14 0.5861 2.31 0.99 3.31 4.40 6 Ocotea dispersa (Nees) Mez 32 16 0.8808 1.35 1.49 2.84 4.09 7 Alchornea triplinervia (Spreng.) Müll. Arg. 22 12 1.2492 0.93 2.12 3.05 3.98 8 Psychotria glaziovii Müll. Arg. 39 16 0.279 1.64 0.47 2.11 3.37 9 Cabralea canjerana (Vell.) Mart. 27 16 0.5656 1.14 0.96 2.10 3.35 10 Ocotea catharinensis Mez 13 6 1.3292 0.55 2.26 2.80 3.27 11 Licania sp1 21 13 0.5386 0.88 0.91 1.80 2.82 12 Myrceugenia myrcioides (Cambess.) O.Berg 34 11 0.2617 1.43 0.44 1.87 2.74 13 Schefflera sp2 11 9 0.884 0.46 1.5 1.96 2.67 14 Clethra scabra Pers. 20 5 0.7742 0.84 1.31 2.16 2.55 15 Myrtaceae sp16 26 10 0.3576 1.09 0.61 1.70 2.48 16 Myrcia tenuivenosa Kiaersk. 26 12 0.2608 1.09 0.44 1.54 2.48 17 Alchornea glandulosa Poepp. & Endl. 9 7 0.8223 0.38 1.4 1.77 2.32 18 Calyptranthes strigipes O.Berg 21 10 0.3507 0.88 0.6 1.48 2.26 19 Tapirira guianensis Aubl. 15 8 0.5511 0.63 0.94 1.57 2.19 20 Duguetia microphylla (R.E.Fr.) R.E.Fr. 16 4 0.6812 0.67 1.16 1.83 2.14 21 Trichilia casarettii C.DC. 20 11 0.2405 0.84 0.41 1.25 2.11 22 Pouteria caimito (Ruiz & Pav.) Radlk. 11 8 0.5939 0.46 1.01 1.47 2.10 23 Copaifera lucens Dwyer 10 6 0.6831 0.42 1.16 1.58 2.05 24 Eugenia excelsa O.Berg 20 8 0.2898 0.84 0.49 1.33 1.96 25 Hieronyma alchorneoides Allemão 11 7 0.5245 0.46 0.89 1.35 1.90 26 Erythroxylum cuspidifolium Mart. 15 8 0.3371 0.63 0.57 1.20 1.83 27 Guatteria australis A. St.-Hil. 13 9 0.3001 0.55 0.51 1.06 1.76 28 Myrcia laxiflora Cambess. 13 10 0.2392 0.55 0.41 0.95 1.74 29 Posoqueria latifolia (Rudge) Roem. & Schult. 10 6 0.4737 0.42 0.8 1.22 1.69 30 Eugenia rostrata O.Berg 17 7 0.2383 0.71 0.4 1.12 1.67 31 Beilschmiedia angustifolia Kosterm. 7 4 0.6176 0.29 1.05 1.34 1.66 32 Casearia aff. arborea (Rich.) Urb. 12 8 0.2961 0.5 0.5 1.01 1.63 33 Myrcia splendens (Sw.) DC. 17 8 0.1479 0.71 0.25 0.97 1.59
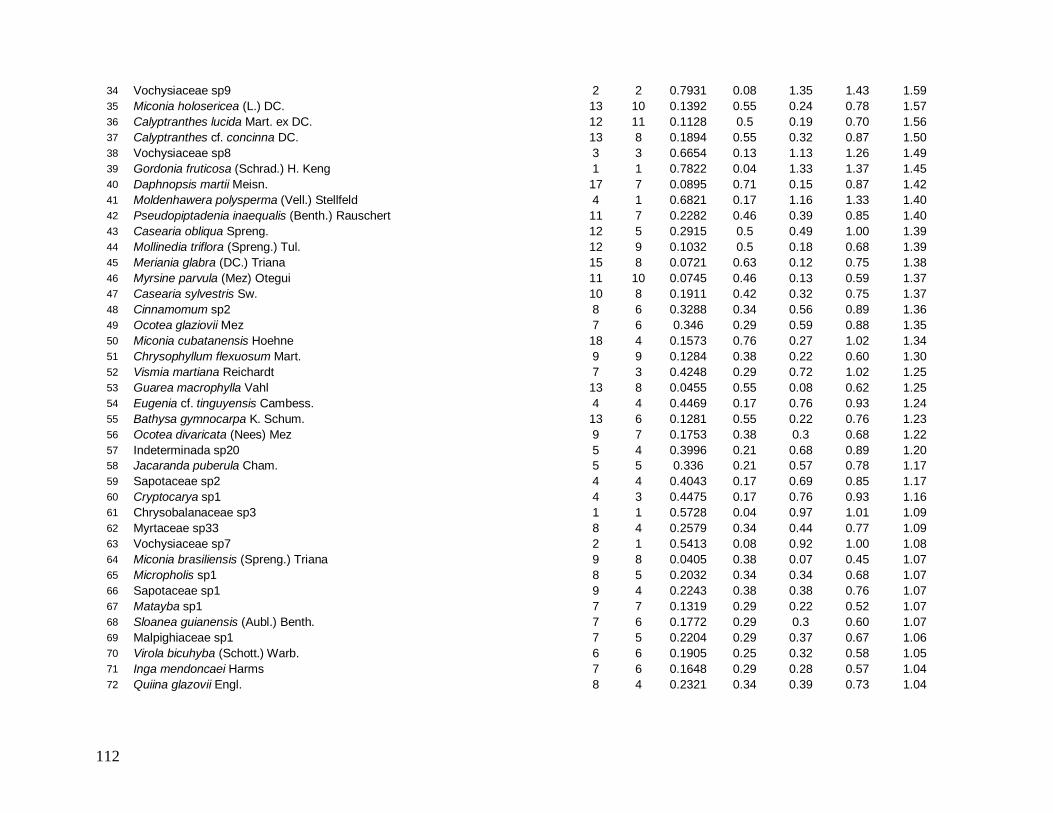
112
34 Vochysiaceae sp9 2 2 0.7931 0.08 1.35 1.43 1.59 35 Miconia holosericea (L.) DC. 13 10 0.1392 0.55 0.24 0.78 1.57 36 Calyptranthes lucida Mart. ex DC. 12 11 0.1128 0.5 0.19 0.70 1.56 37 Calyptranthes cf. concinna DC. 13 8 0.1894 0.55 0.32 0.87 1.50 38 Vochysiaceae sp8 3 3 0.6654 0.13 1.13 1.26 1.49 39 Gordonia fruticosa (Schrad.) H. Keng 1 1 0.7822 0.04 1.33 1.37 1.45 40 Daphnopsis martii Meisn. 17 7 0.0895 0.71 0.15 0.87 1.42 41 Moldenhawera polysperma (Vell.) Stellfeld 4 1 0.6821 0.17 1.16 1.33 1.40 42 Pseudopiptadenia inaequalis (Benth.) Rauschert 11 7 0.2282 0.46 0.39 0.85 1.40 43 Casearia obliqua Spreng. 12 5 0.2915 0.5 0.49 1.00 1.39 44 Mollinedia triflora (Spreng.) Tul. 12 9 0.1032 0.5 0.18 0.68 1.39 45 Meriania glabra (DC.) Triana 15 8 0.0721 0.63 0.12 0.75 1.38 46 Myrsine parvula (Mez) Otegui 11 10 0.0745 0.46 0.13 0.59 1.37 47 Casearia sylvestris Sw. 10 8 0.1911 0.42 0.32 0.75 1.37 48 Cinnamomum sp2 8 6 0.3288 0.34 0.56 0.89 1.36 49 Ocotea glaziovii Mez 7 6 0.346 0.29 0.59 0.88 1.35 50 Miconia cubatanensis Hoehne 18 4 0.1573 0.76 0.27 1.02 1.34 51 Chrysophyllum flexuosum Mart. 9 9 0.1284 0.38 0.22 0.60 1.30 52 Vismia martiana Reichardt 7 3 0.4248 0.29 0.72 1.02 1.25 53 Guarea macrophylla Vahl 13 8 0.0455 0.55 0.08 0.62 1.25 54 Eugenia cf. tinguyensis Cambess. 4 4 0.4469 0.17 0.76 0.93 1.24 55 Bathysa gymnocarpa K. Schum. 13 6 0.1281 0.55 0.22 0.76 1.23 56 Ocotea divaricata (Nees) Mez 9 7 0.1753 0.38 0.3 0.68 1.22 57 Indeterminada sp20 5 4 0.3996 0.21 0.68 0.89 1.20 58 Jacaranda puberula Cham. 5 5 0.336 0.21 0.57 0.78 1.17 59 Sapotaceae sp2 4 4 0.4043 0.17 0.69 0.85 1.17 60 Cryptocarya sp1 4 3 0.4475 0.17 0.76 0.93 1.16 61 Chrysobalanaceae sp3 1 1 0.5728 0.04 0.97 1.01 1.09 62 Myrtaceae sp33 8 4 0.2579 0.34 0.44 0.77 1.09 63 Vochysiaceae sp7 2 1 0.5413 0.08 0.92 1.00 1.08 64 Miconia brasiliensis (Spreng.) Triana 9 8 0.0405 0.38 0.07 0.45 1.07 65 Micropholis sp1 8 5 0.2032 0.34 0.34 0.68 1.07 66 Sapotaceae sp1 9 4 0.2243 0.38 0.38 0.76 1.07 67 Matayba sp1 7 7 0.1319 0.29 0.22 0.52 1.07 68 Sloanea guianensis (Aubl.) Benth. 7 6 0.1772 0.29 0.3 0.60 1.07 69 Malpighiaceae sp1 7 5 0.2204 0.29 0.37 0.67 1.06 70 Virola bicuhyba (Schott.) Warb. 6 6 0.1905 0.25 0.32 0.58 1.05 71 Inga mendoncaei Harms 7 6 0.1648 0.29 0.28 0.57 1.04 72 Quiina glazovii Engl. 8 4 0.2321 0.34 0.39 0.73 1.04
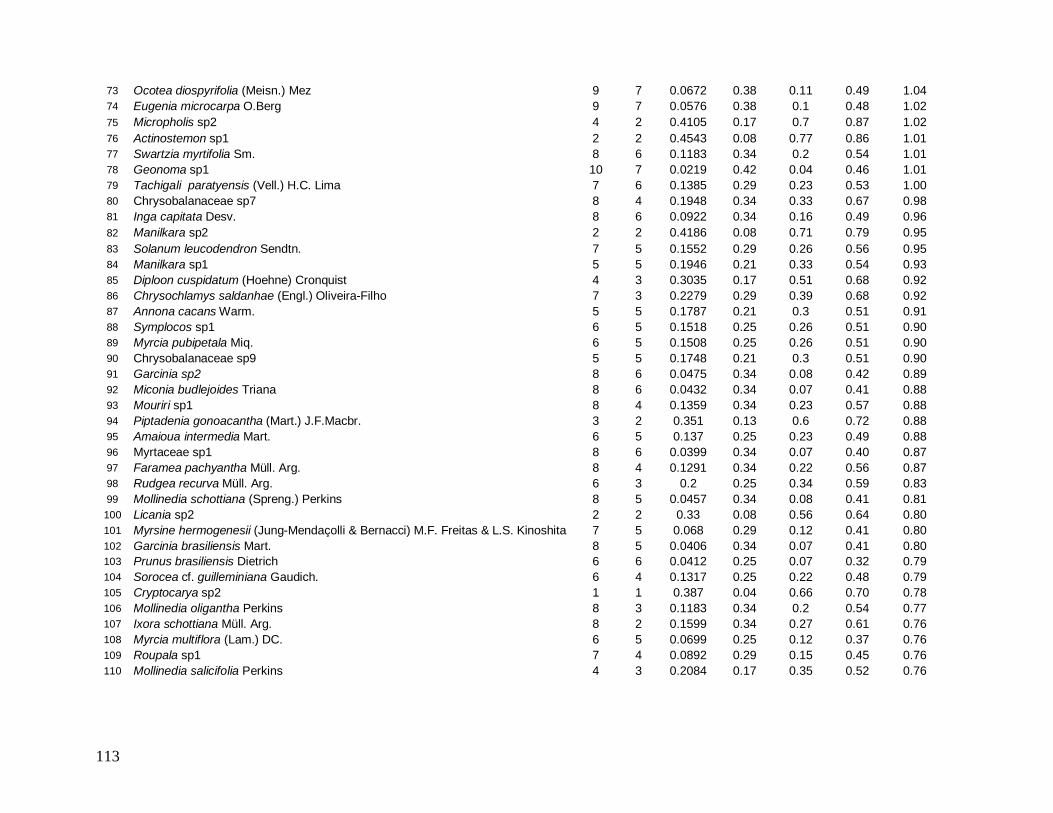
113
73 Ocotea diospyrifolia (Meisn.) Mez 9 7 0.0672 0.38 0.11 0.49 1.04 74 Eugenia microcarpa O.Berg 9 7 0.0576 0.38 0.1 0.48 1.02 75 Micropholis sp2 4 2 0.4105 0.17 0.7 0.87 1.02 76 Actinostemon sp1 2 2 0.4543 0.08 0.77 0.86 1.01 77 Swartzia myrtifolia Sm. 8 6 0.1183 0.34 0.2 0.54 1.01 78 Geonoma sp1 10 7 0.0219 0.42 0.04 0.46 1.01 79 Tachigali paratyensis (Vell.) H.C. Lima 7 6 0.1385 0.29 0.23 0.53 1.00 80 Chrysobalanaceae sp7 8 4 0.1948 0.34 0.33 0.67 0.98 81 Inga capitata Desv. 8 6 0.0922 0.34 0.16 0.49 0.96 82 Manilkara sp2 2 2 0.4186 0.08 0.71 0.79 0.95 83 Solanum leucodendron Sendtn. 7 5 0.1552 0.29 0.26 0.56 0.95 84 Manilkara sp1 5 5 0.1946 0.21 0.33 0.54 0.93 85 Diploon cuspidatum (Hoehne) Cronquist 4 3 0.3035 0.17 0.51 0.68 0.92 86 Chrysochlamys saldanhae (Engl.) Oliveira-Filho 7 3 0.2279 0.29 0.39 0.68 0.92 87 Annona cacans Warm. 5 5 0.1787 0.21 0.3 0.51 0.91 88 Symplocos sp1 6 5 0.1518 0.25 0.26 0.51 0.90 89 Myrcia pubipetala Miq. 6 5 0.1508 0.25 0.26 0.51 0.90 90 Chrysobalanaceae sp9 5 5 0.1748 0.21 0.3 0.51 0.90 91 Garcinia sp2 8 6 0.0475 0.34 0.08 0.42 0.89 92 Miconia budlejoides Triana 8 6 0.0432 0.34 0.07 0.41 0.88 93 Mouriri sp1 8 4 0.1359 0.34 0.23 0.57 0.88 94 Piptadenia gonoacantha (Mart.) J.F.Macbr. 3 2 0.351 0.13 0.6 0.72 0.88 95 Amaioua intermedia Mart. 6 5 0.137 0.25 0.23 0.49 0.88 96 Myrtaceae sp1 8 6 0.0399 0.34 0.07 0.40 0.87 97 Faramea pachyantha Müll. Arg. 8 4 0.1291 0.34 0.22 0.56 0.87 98 Rudgea recurva Müll. Arg. 6 3 0.2 0.25 0.34 0.59 0.83 99 Mollinedia schottiana (Spreng.) Perkins 8 5 0.0457 0.34 0.08 0.41 0.81 100 Licania sp2 2 2 0.33 0.08 0.56 0.64 0.80 101 Myrsine hermogenesii (Jung-Mendaçolli & Bernacci) M.F. Freitas & L.S. Kinoshita 7 5 0.068 0.29 0.12 0.41 0.80 102 Garcinia brasiliensis Mart. 8 5 0.0406 0.34 0.07 0.41 0.80 103 Prunus brasiliensis Dietrich 6 6 0.0412 0.25 0.07 0.32 0.79 104 Sorocea cf. guilleminiana Gaudich. 6 4 0.1317 0.25 0.22 0.48 0.79 105 Cryptocarya sp2 1 1 0.387 0.04 0.66 0.70 0.78 106 Mollinedia oligantha Perkins 8 3 0.1183 0.34 0.2 0.54 0.77 107 Ixora schottiana Müll. Arg. 8 2 0.1599 0.34 0.27 0.61 0.76 108 Myrcia multiflora (Lam.) DC. 6 5 0.0699 0.25 0.12 0.37 0.76 109 Roupala sp1 7 4 0.0892 0.29 0.15 0.45 0.76 110 Mollinedia salicifolia Perkins 4 3 0.2084 0.17 0.35 0.52 0.76
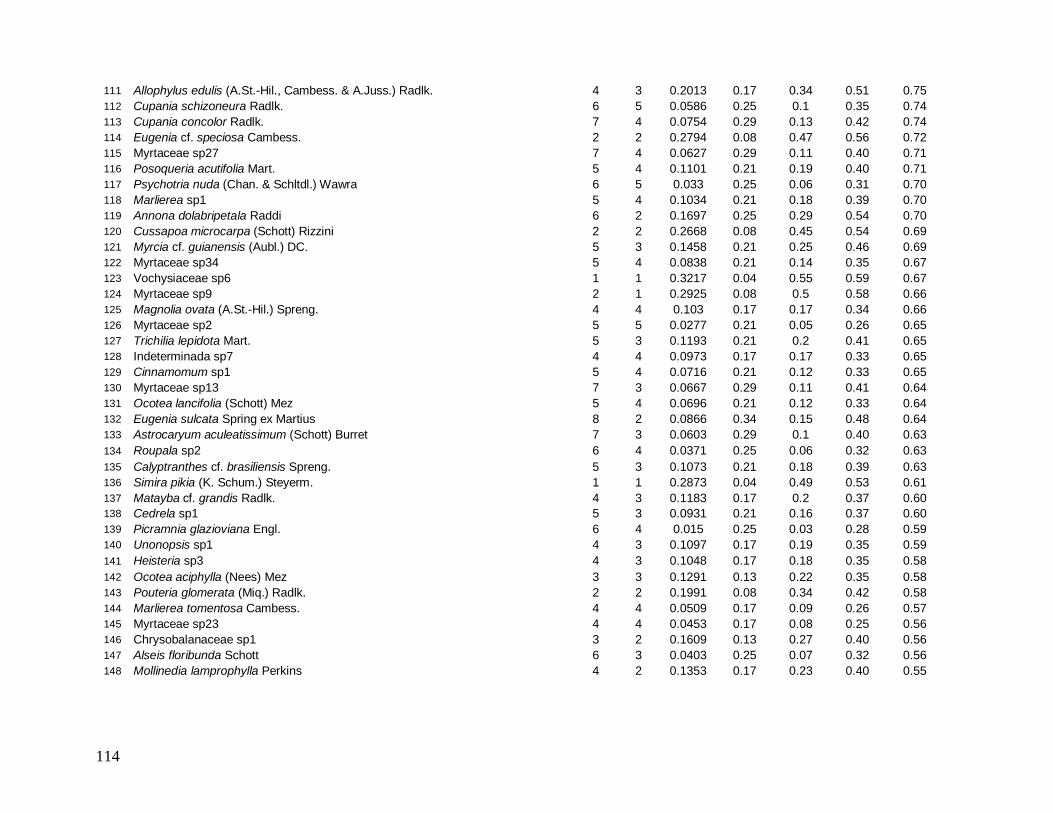
114
111 Allophylus edulis (A.St.-Hil., Cambess. & A.Juss.) Radlk. 4 3 0.2013 0.17 0.34 0.51 0.75 112 Cupania schizoneura Radlk. 6 5 0.0586 0.25 0.1 0.35 0.74 113 Cupania concolor Radlk. 7 4 0.0754 0.29 0.13 0.42 0.74 114 Eugenia cf. speciosa Cambess. 2 2 0.2794 0.08 0.47 0.56 0.72 115 Myrtaceae sp27 7 4 0.0627 0.29 0.11 0.40 0.71 116 Posoqueria acutifolia Mart. 5 4 0.1101 0.21 0.19 0.40 0.71 117 Psychotria nuda (Chan. & Schltdl.) Wawra 6 5 0.033 0.25 0.06 0.31 0.70 118 Marlierea sp1 5 4 0.1034 0.21 0.18 0.39 0.70 119 Annona dolabripetala Raddi 6 2 0.1697 0.25 0.29 0.54 0.70 120 Cussapoa microcarpa (Schott) Rizzini 2 2 0.2668 0.08 0.45 0.54 0.69 121 Myrcia cf. guianensis (Aubl.) DC. 5 3 0.1458 0.21 0.25 0.46 0.69 122 Myrtaceae sp34 5 4 0.0838 0.21 0.14 0.35 0.67 123 Vochysiaceae sp6 1 1 0.3217 0.04 0.55 0.59 0.67 124 Myrtaceae sp9 2 1 0.2925 0.08 0.5 0.58 0.66 125 Magnolia ovata (A.St.-Hil.) Spreng. 4 4 0.103 0.17 0.17 0.34 0.66 126 Myrtaceae sp2 5 5 0.0277 0.21 0.05 0.26 0.65 127 Trichilia lepidota Mart. 5 3 0.1193 0.21 0.2 0.41 0.65 128 Indeterminada sp7 4 4 0.0973 0.17 0.17 0.33 0.65 129 Cinnamomum sp1 5 4 0.0716 0.21 0.12 0.33 0.65 130 Myrtaceae sp13 7 3 0.0667 0.29 0.11 0.41 0.64 131 Ocotea lancifolia (Schott) Mez 5 4 0.0696 0.21 0.12 0.33 0.64 132 Eugenia sulcata Spring ex Martius 8 2 0.0866 0.34 0.15 0.48 0.64 133 Astrocaryum aculeatissimum (Schott) Burret 7 3 0.0603 0.29 0.1 0.40 0.63 134 Roupala sp2 6 4 0.0371 0.25 0.06 0.32 0.63 135 Calyptranthes cf. brasiliensis Spreng. 5 3 0.1073 0.21 0.18 0.39 0.63 136 Simira pikia (K. Schum.) Steyerm. 1 1 0.2873 0.04 0.49 0.53 0.61 137 Matayba cf. grandis Radlk. 4 3 0.1183 0.17 0.2 0.37 0.60 138 Cedrela sp1 5 3 0.0931 0.21 0.16 0.37 0.60 139 Picramnia glazioviana Engl. 6 4 0.015 0.25 0.03 0.28 0.59 140 Unonopsis sp1 4 3 0.1097 0.17 0.19 0.35 0.59 141 Heisteria sp3 4 3 0.1048 0.17 0.18 0.35 0.58 142 Ocotea aciphylla (Nees) Mez 3 3 0.1291 0.13 0.22 0.35 0.58 143 Pouteria glomerata (Miq.) Radlk. 2 2 0.1991 0.08 0.34 0.42 0.58 144 Marlierea tomentosa Cambess. 4 4 0.0509 0.17 0.09 0.26 0.57 145 Myrtaceae sp23 4 4 0.0453 0.17 0.08 0.25 0.56 146 Chrysobalanaceae sp1 3 2 0.1609 0.13 0.27 0.40 0.56 147 Alseis floribunda Schott 6 3 0.0403 0.25 0.07 0.32 0.56 148 Mollinedia lamprophylla Perkins 4 2 0.1353 0.17 0.23 0.40 0.55
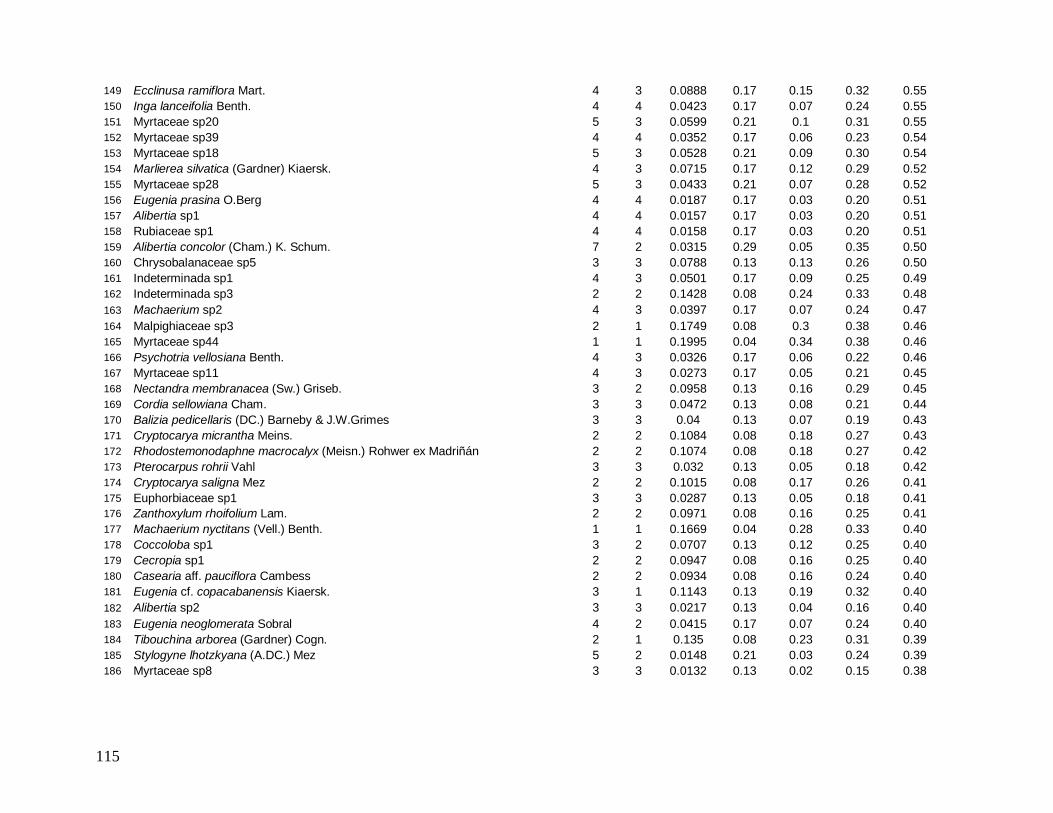
115
149 Ecclinusa ramiflora Mart. 4 3 0.0888 0.17 0.15 0.32 0.55 150 Inga lanceifolia Benth. 4 4 0.0423 0.17 0.07 0.24 0.55 151 Myrtaceae sp20 5 3 0.0599 0.21 0.1 0.31 0.55 152 Myrtaceae sp39 4 4 0.0352 0.17 0.06 0.23 0.54 153 Myrtaceae sp18 5 3 0.0528 0.21 0.09 0.30 0.54 154 Marlierea silvatica (Gardner) Kiaersk. 4 3 0.0715 0.17 0.12 0.29 0.52 155 Myrtaceae sp28 5 3 0.0433 0.21 0.07 0.28 0.52 156 Eugenia prasina O.Berg 4 4 0.0187 0.17 0.03 0.20 0.51 157 Alibertia sp1 4 4 0.0157 0.17 0.03 0.20 0.51 158 Rubiaceae sp1 4 4 0.0158 0.17 0.03 0.20 0.51 159 Alibertia concolor (Cham.) K. Schum. 7 2 0.0315 0.29 0.05 0.35 0.50 160 Chrysobalanaceae sp5 3 3 0.0788 0.13 0.13 0.26 0.50 161 Indeterminada sp1 4 3 0.0501 0.17 0.09 0.25 0.49 162 Indeterminada sp3 2 2 0.1428 0.08 0.24 0.33 0.48 163 Machaerium sp2 4 3 0.0397 0.17 0.07 0.24 0.47 164 Malpighiaceae sp3 2 1 0.1749 0.08 0.3 0.38 0.46 165 Myrtaceae sp44 1 1 0.1995 0.04 0.34 0.38 0.46 166 Psychotria vellosiana Benth. 4 3 0.0326 0.17 0.06 0.22 0.46 167 Myrtaceae sp11 4 3 0.0273 0.17 0.05 0.21 0.45 168 Nectandra membranacea (Sw.) Griseb. 3 2 0.0958 0.13 0.16 0.29 0.45 169 Cordia sellowiana Cham. 3 3 0.0472 0.13 0.08 0.21 0.44 170 Balizia pedicellaris (DC.) Barneby & J.W.Grimes 3 3 0.04 0.13 0.07 0.19 0.43 171 Cryptocarya micrantha Meins. 2 2 0.1084 0.08 0.18 0.27 0.43 172 Rhodostemonodaphne macrocalyx (Meisn.) Rohwer ex Madriñán 2 2 0.1074 0.08 0.18 0.27 0.42 173 Pterocarpus rohrii Vahl 3 3 0.032 0.13 0.05 0.18 0.42 174 Cryptocarya saligna Mez 2 2 0.1015 0.08 0.17 0.26 0.41 175 Euphorbiaceae sp1 3 3 0.0287 0.13 0.05 0.18 0.41 176 Zanthoxylum rhoifolium Lam. 2 2 0.0971 0.08 0.16 0.25 0.41 177 Machaerium nyctitans (Vell.) Benth. 1 1 0.1669 0.04 0.28 0.33 0.40 178 Coccoloba sp1 3 2 0.0707 0.13 0.12 0.25 0.40 179 Cecropia sp1 2 2 0.0947 0.08 0.16 0.25 0.40 180 Casearia aff. pauciflora Cambess 2 2 0.0934 0.08 0.16 0.24 0.40 181 Eugenia cf. copacabanensis Kiaersk. 3 1 0.1143 0.13 0.19 0.32 0.40 182 Alibertia sp2 3 3 0.0217 0.13 0.04 0.16 0.40 183 Eugenia neoglomerata Sobral 4 2 0.0415 0.17 0.07 0.24 0.40 184 Tibouchina arborea (Gardner) Cogn. 2 1 0.135 0.08 0.23 0.31 0.39 185 Stylogyne lhotzkyana (A.DC.) Mez 5 2 0.0148 0.21 0.03 0.24 0.39 186 Myrtaceae sp8 3 3 0.0132 0.13 0.02 0.15 0.38
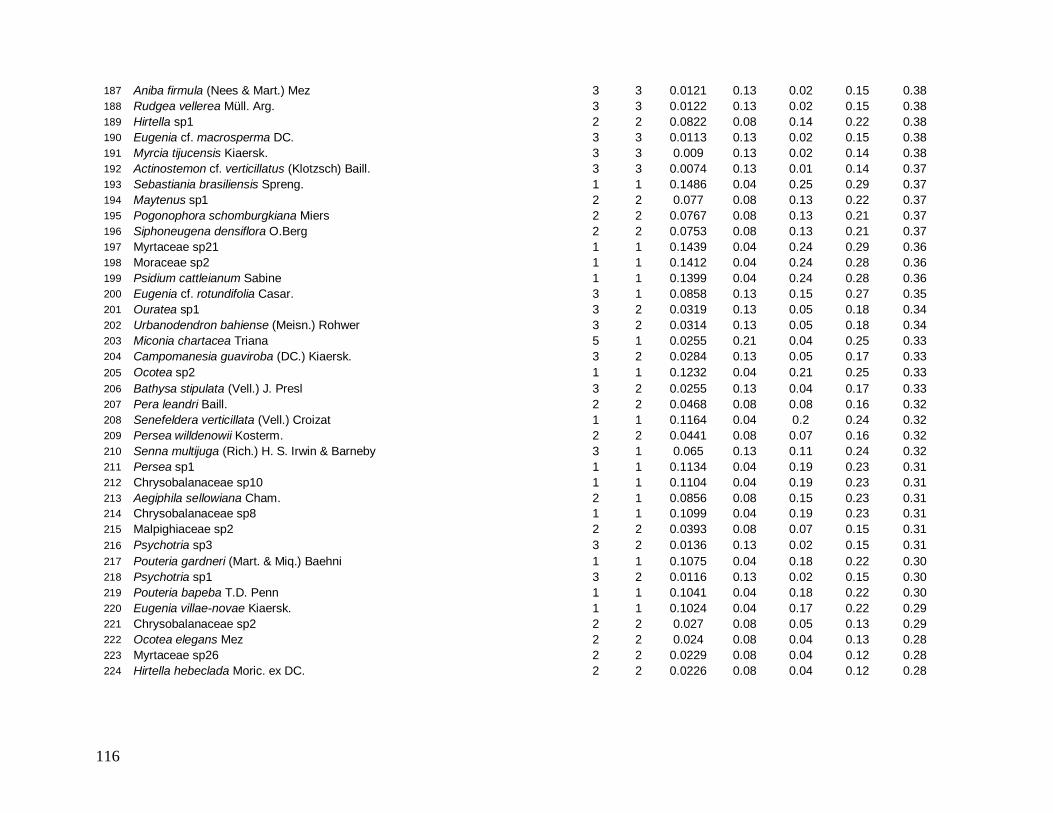
116
187 Aniba firmula (Nees & Mart.) Mez 3 3 0.0121 0.13 0.02 0.15 0.38 188 Rudgea vellerea Müll. Arg. 3 3 0.0122 0.13 0.02 0.15 0.38 189 Hirtella sp1 2 2 0.0822 0.08 0.14 0.22 0.38 190 Eugenia cf. macrosperma DC. 3 3 0.0113 0.13 0.02 0.15 0.38 191 Myrcia tijucensis Kiaersk. 3 3 0.009 0.13 0.02 0.14 0.38 192 Actinostemon cf. verticillatus (Klotzsch) Baill. 3 3 0.0074 0.13 0.01 0.14 0.37 193 Sebastiania brasiliensis Spreng. 1 1 0.1486 0.04 0.25 0.29 0.37 194 Maytenus sp1 2 2 0.077 0.08 0.13 0.22 0.37 195 Pogonophora schomburgkiana Miers 2 2 0.0767 0.08 0.13 0.21 0.37 196 Siphoneugena densiflora O.Berg 2 2 0.0753 0.08 0.13 0.21 0.37 197 Myrtaceae sp21 1 1 0.1439 0.04 0.24 0.29 0.36 198 Moraceae sp2 1 1 0.1412 0.04 0.24 0.28 0.36 199 Psidium cattleianum Sabine 1 1 0.1399 0.04 0.24 0.28 0.36 200 Eugenia cf. rotundifolia Casar. 3 1 0.0858 0.13 0.15 0.27 0.35 201 Ouratea sp1 3 2 0.0319 0.13 0.05 0.18 0.34 202 Urbanodendron bahiense (Meisn.) Rohwer 3 2 0.0314 0.13 0.05 0.18 0.34 203 Miconia chartacea Triana 5 1 0.0255 0.21 0.04 0.25 0.33 204 Campomanesia guaviroba (DC.) Kiaersk. 3 2 0.0284 0.13 0.05 0.17 0.33 205 Ocotea sp2 1 1 0.1232 0.04 0.21 0.25 0.33 206 Bathysa stipulata (Vell.) J. Presl 3 2 0.0255 0.13 0.04 0.17 0.33 207 Pera leandri Baill. 2 2 0.0468 0.08 0.08 0.16 0.32 208 Senefeldera verticillata (Vell.) Croizat 1 1 0.1164 0.04 0.2 0.24 0.32 209 Persea willdenowii Kosterm. 2 2 0.0441 0.08 0.07 0.16 0.32 210 Senna multijuga (Rich.) H. S. Irwin & Barneby 3 1 0.065 0.13 0.11 0.24 0.32 211 Persea sp1 1 1 0.1134 0.04 0.19 0.23 0.31 212 Chrysobalanaceae sp10 1 1 0.1104 0.04 0.19 0.23 0.31 213 Aegiphila sellowiana Cham. 2 1 0.0856 0.08 0.15 0.23 0.31 214 Chrysobalanaceae sp8 1 1 0.1099 0.04 0.19 0.23 0.31 215 Malpighiaceae sp2 2 2 0.0393 0.08 0.07 0.15 0.31 216 Psychotria sp3 3 2 0.0136 0.13 0.02 0.15 0.31 217 Pouteria gardneri (Mart. & Miq.) Baehni 1 1 0.1075 0.04 0.18 0.22 0.30 218 Psychotria sp1 3 2 0.0116 0.13 0.02 0.15 0.30 219 Pouteria bapeba T.D. Penn 1 1 0.1041 0.04 0.18 0.22 0.30 220 Eugenia villae-novae Kiaersk. 1 1 0.1024 0.04 0.17 0.22 0.29 221 Chrysobalanaceae sp2 2 2 0.027 0.08 0.05 0.13 0.29 222 Ocotea elegans Mez 2 2 0.024 0.08 0.04 0.13 0.28 223 Myrtaceae sp26 2 2 0.0229 0.08 0.04 0.12 0.28 224 Hirtella hebeclada Moric. ex DC. 2 2 0.0226 0.08 0.04 0.12 0.28
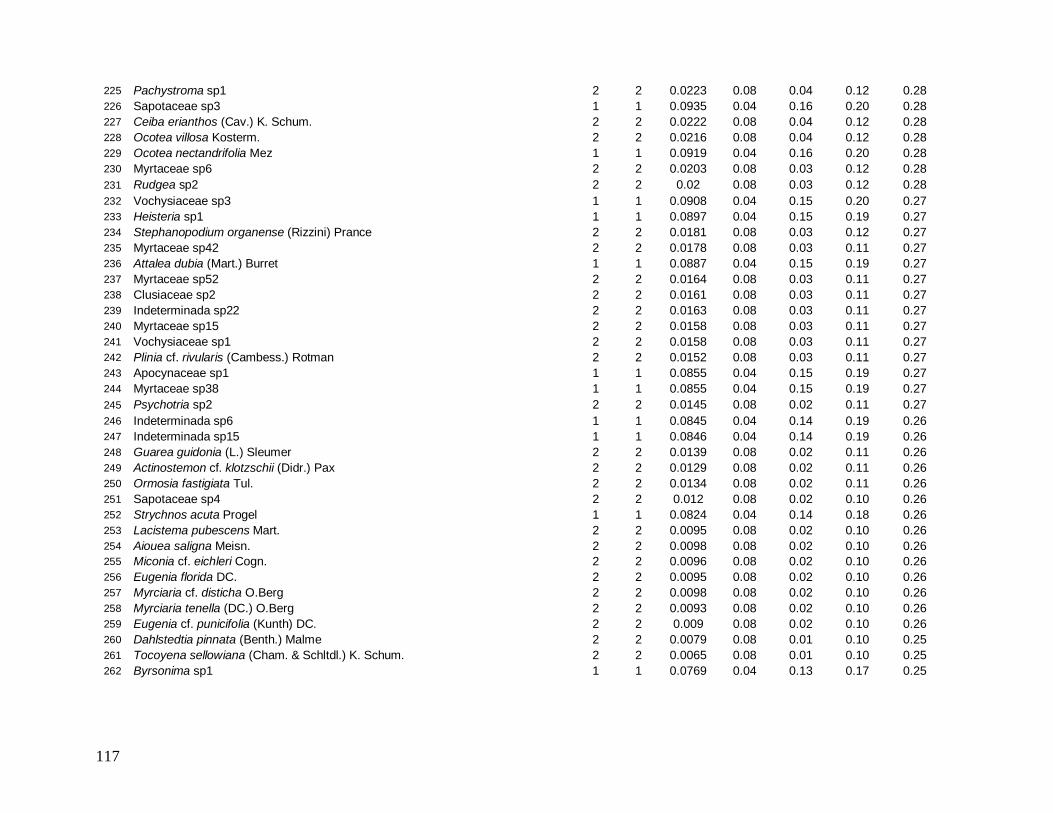
117
225 Pachystroma sp1 2 2 0.0223 0.08 0.04 0.12 0.28 226 Sapotaceae sp3 1 1 0.0935 0.04 0.16 0.20 0.28 227 Ceiba erianthos (Cav.) K. Schum. 2 2 0.0222 0.08 0.04 0.12 0.28 228 Ocotea villosa Kosterm. 2 2 0.0216 0.08 0.04 0.12 0.28 229 Ocotea nectandrifolia Mez 1 1 0.0919 0.04 0.16 0.20 0.28 230 Myrtaceae sp6 2 2 0.0203 0.08 0.03 0.12 0.28 231 Rudgea sp2 2 2 0.02 0.08 0.03 0.12 0.28 232 Vochysiaceae sp3 1 1 0.0908 0.04 0.15 0.20 0.27 233 Heisteria sp1 1 1 0.0897 0.04 0.15 0.19 0.27 234 Stephanopodium organense (Rizzini) Prance 2 2 0.0181 0.08 0.03 0.12 0.27 235 Myrtaceae sp42 2 2 0.0178 0.08 0.03 0.11 0.27 236 Attalea dubia (Mart.) Burret 1 1 0.0887 0.04 0.15 0.19 0.27 237 Myrtaceae sp52 2 2 0.0164 0.08 0.03 0.11 0.27 238 Clusiaceae sp2 2 2 0.0161 0.08 0.03 0.11 0.27 239 Indeterminada sp22 2 2 0.0163 0.08 0.03 0.11 0.27 240 Myrtaceae sp15 2 2 0.0158 0.08 0.03 0.11 0.27 241 Vochysiaceae sp1 2 2 0.0158 0.08 0.03 0.11 0.27 242 Plinia cf. rivularis (Cambess.) Rotman 2 2 0.0152 0.08 0.03 0.11 0.27 243 Apocynaceae sp1 1 1 0.0855 0.04 0.15 0.19 0.27 244 Myrtaceae sp38 1 1 0.0855 0.04 0.15 0.19 0.27 245 Psychotria sp2 2 2 0.0145 0.08 0.02 0.11 0.27 246 Indeterminada sp6 1 1 0.0845 0.04 0.14 0.19 0.26 247 Indeterminada sp15 1 1 0.0846 0.04 0.14 0.19 0.26 248 Guarea guidonia (L.) Sleumer 2 2 0.0139 0.08 0.02 0.11 0.26 249 Actinostemon cf. klotzschii (Didr.) Pax 2 2 0.0129 0.08 0.02 0.11 0.26 250 Ormosia fastigiata Tul. 2 2 0.0134 0.08 0.02 0.11 0.26 251 Sapotaceae sp4 2 2 0.012 0.08 0.02 0.10 0.26 252 Strychnos acuta Progel 1 1 0.0824 0.04 0.14 0.18 0.26 253 Lacistema pubescens Mart. 2 2 0.0095 0.08 0.02 0.10 0.26 254 Aiouea saligna Meisn. 2 2 0.0098 0.08 0.02 0.10 0.26 255 Miconia cf. eichleri Cogn. 2 2 0.0096 0.08 0.02 0.10 0.26 256 Eugenia florida DC. 2 2 0.0095 0.08 0.02 0.10 0.26 257 Myrciaria cf. disticha O.Berg 2 2 0.0098 0.08 0.02 0.10 0.26 258 Myrciaria tenella (DC.) O.Berg 2 2 0.0093 0.08 0.02 0.10 0.26 259 Eugenia cf. punicifolia (Kunth) DC. 2 2 0.009 0.08 0.02 0.10 0.26 260 Dahlstedtia pinnata (Benth.) Malme 2 2 0.0079 0.08 0.01 0.10 0.25 261 Tocoyena sellowiana (Cham. & Schltdl.) K. Schum. 2 2 0.0065 0.08 0.01 0.10 0.25 262 Byrsonima sp1 1 1 0.0769 0.04 0.13 0.17 0.25
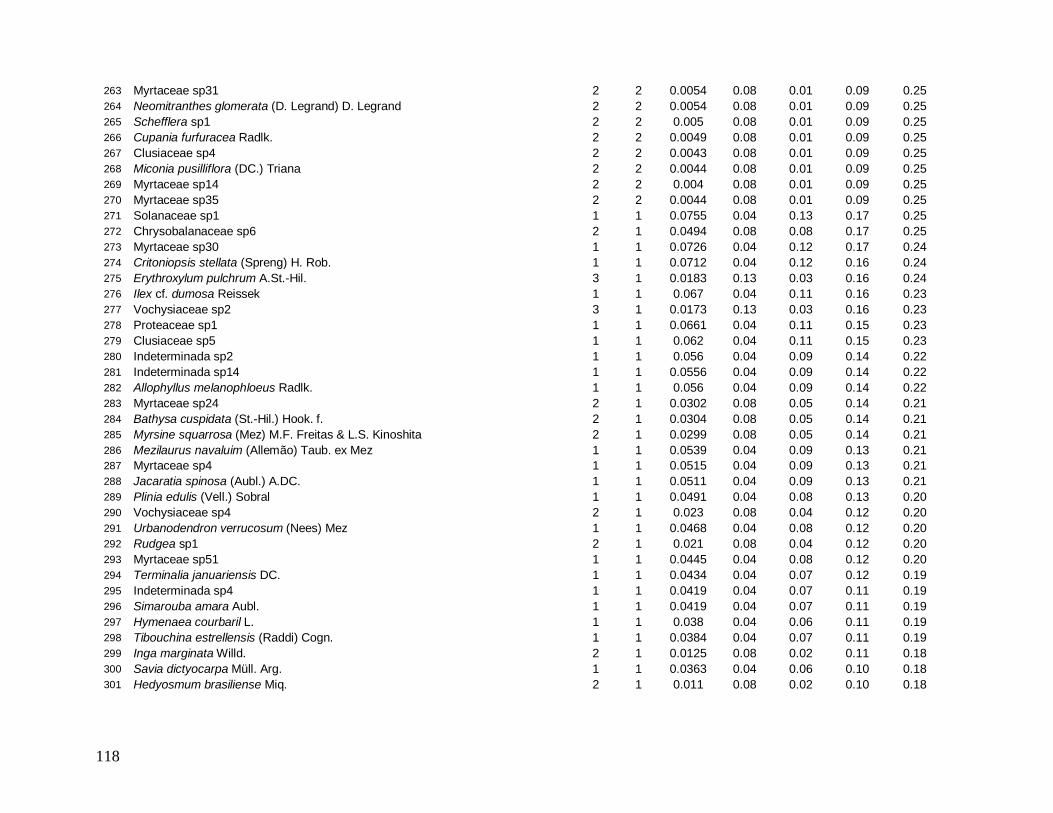
118
263 Myrtaceae sp31 2 2 0.0054 0.08 0.01 0.09 0.25 264 Neomitranthes glomerata (D. Legrand) D. Legrand 2 2 0.0054 0.08 0.01 0.09 0.25 265 Schefflera sp1 2 2 0.005 0.08 0.01 0.09 0.25 266 Cupania furfuracea Radlk. 2 2 0.0049 0.08 0.01 0.09 0.25 267 Clusiaceae sp4 2 2 0.0043 0.08 0.01 0.09 0.25 268 Miconia pusilliflora (DC.) Triana 2 2 0.0044 0.08 0.01 0.09 0.25 269 Myrtaceae sp14 2 2 0.004 0.08 0.01 0.09 0.25 270 Myrtaceae sp35 2 2 0.0044 0.08 0.01 0.09 0.25 271 Solanaceae sp1 1 1 0.0755 0.04 0.13 0.17 0.25 272 Chrysobalanaceae sp6 2 1 0.0494 0.08 0.08 0.17 0.25 273 Myrtaceae sp30 1 1 0.0726 0.04 0.12 0.17 0.24 274 Critoniopsis stellata (Spreng) H. Rob. 1 1 0.0712 0.04 0.12 0.16 0.24 275 Erythroxylum pulchrum A.St.-Hil. 3 1 0.0183 0.13 0.03 0.16 0.24 276 Ilex cf. dumosa Reissek 1 1 0.067 0.04 0.11 0.16 0.23 277 Vochysiaceae sp2 3 1 0.0173 0.13 0.03 0.16 0.23 278 Proteaceae sp1 1 1 0.0661 0.04 0.11 0.15 0.23 279 Clusiaceae sp5 1 1 0.062 0.04 0.11 0.15 0.23 280 Indeterminada sp2 1 1 0.056 0.04 0.09 0.14 0.22 281 Indeterminada sp14 1 1 0.0556 0.04 0.09 0.14 0.22 282 Allophyllus melanophloeus Radlk. 1 1 0.056 0.04 0.09 0.14 0.22 283 Myrtaceae sp24 2 1 0.0302 0.08 0.05 0.14 0.21 284 Bathysa cuspidata (St.-Hil.) Hook. f. 2 1 0.0304 0.08 0.05 0.14 0.21 285 Myrsine squarrosa (Mez) M.F. Freitas & L.S. Kinoshita 2 1 0.0299 0.08 0.05 0.14 0.21 286 Mezilaurus navaluim (Allemão) Taub. ex Mez 1 1 0.0539 0.04 0.09 0.13 0.21 287 Myrtaceae sp4 1 1 0.0515 0.04 0.09 0.13 0.21 288 Jacaratia spinosa (Aubl.) A.DC. 1 1 0.0511 0.04 0.09 0.13 0.21 289 Plinia edulis (Vell.) Sobral 1 1 0.0491 0.04 0.08 0.13 0.20 290 Vochysiaceae sp4 2 1 0.023 0.08 0.04 0.12 0.20 291 Urbanodendron verrucosum (Nees) Mez 1 1 0.0468 0.04 0.08 0.12 0.20 292 Rudgea sp1 2 1 0.021 0.08 0.04 0.12 0.20 293 Myrtaceae sp51 1 1 0.0445 0.04 0.08 0.12 0.20 294 Terminalia januariensis DC. 1 1 0.0434 0.04 0.07 0.12 0.19 295 Indeterminada sp4 1 1 0.0419 0.04 0.07 0.11 0.19 296 Simarouba amara Aubl. 1 1 0.0419 0.04 0.07 0.11 0.19 297 Hymenaea courbaril L. 1 1 0.038 0.04 0.06 0.11 0.19 298 Tibouchina estrellensis (Raddi) Cogn. 1 1 0.0384 0.04 0.07 0.11 0.19 299 Inga marginata Willd. 2 1 0.0125 0.08 0.02 0.11 0.18 300 Savia dictyocarpa Müll. Arg. 1 1 0.0363 0.04 0.06 0.10 0.18 301 Hedyosmum brasiliense Miq. 2 1 0.011 0.08 0.02 0.10 0.18
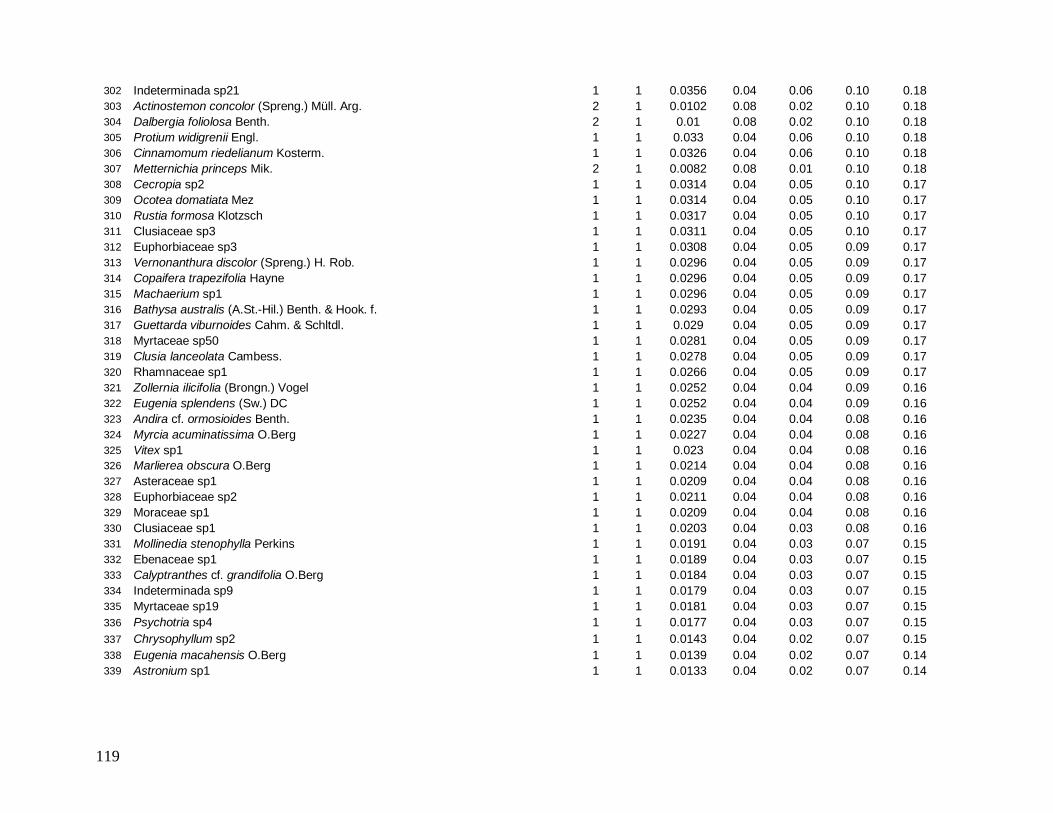
119
302 Indeterminada sp21 1 1 0.0356 0.04 0.06 0.10 0.18 303 Actinostemon concolor (Spreng.) Müll. Arg. 2 1 0.0102 0.08 0.02 0.10 0.18 304 Dalbergia foliolosa Benth. 2 1 0.01 0.08 0.02 0.10 0.18 305 Protium widigrenii Engl. 1 1 0.033 0.04 0.06 0.10 0.18 306 Cinnamomum riedelianum Kosterm. 1 1 0.0326 0.04 0.06 0.10 0.18 307 Metternichia princeps Mik. 2 1 0.0082 0.08 0.01 0.10 0.18 308 Cecropia sp2 1 1 0.0314 0.04 0.05 0.10 0.17 309 Ocotea domatiata Mez 1 1 0.0314 0.04 0.05 0.10 0.17 310 Rustia formosa Klotzsch 1 1 0.0317 0.04 0.05 0.10 0.17 311 Clusiaceae sp3 1 1 0.0311 0.04 0.05 0.10 0.17 312 Euphorbiaceae sp3 1 1 0.0308 0.04 0.05 0.09 0.17 313 Vernonanthura discolor (Spreng.) H. Rob. 1 1 0.0296 0.04 0.05 0.09 0.17 314 Copaifera trapezifolia Hayne 1 1 0.0296 0.04 0.05 0.09 0.17 315 Machaerium sp1 1 1 0.0296 0.04 0.05 0.09 0.17 316 Bathysa australis (A.St.-Hil.) Benth. & Hook. f. 1 1 0.0293 0.04 0.05 0.09 0.17 317 Guettarda viburnoides Cahm. & Schltdl. 1 1 0.029 0.04 0.05 0.09 0.17 318 Myrtaceae sp50 1 1 0.0281 0.04 0.05 0.09 0.17 319 Clusia lanceolata Cambess. 1 1 0.0278 0.04 0.05 0.09 0.17 320 Rhamnaceae sp1 1 1 0.0266 0.04 0.05 0.09 0.17 321 Zollernia ilicifolia (Brongn.) Vogel 1 1 0.0252 0.04 0.04 0.09 0.16 322 Eugenia splendens (Sw.) DC 1 1 0.0252 0.04 0.04 0.09 0.16 323 Andira cf. ormosioides Benth. 1 1 0.0235 0.04 0.04 0.08 0.16 324 Myrcia acuminatissima O.Berg 1 1 0.0227 0.04 0.04 0.08 0.16 325 Vitex sp1 1 1 0.023 0.04 0.04 0.08 0.16 326 Marlierea obscura O.Berg 1 1 0.0214 0.04 0.04 0.08 0.16 327 Asteraceae sp1 1 1 0.0209 0.04 0.04 0.08 0.16 328 Euphorbiaceae sp2 1 1 0.0211 0.04 0.04 0.08 0.16 329 Moraceae sp1 1 1 0.0209 0.04 0.04 0.08 0.16 330 Clusiaceae sp1 1 1 0.0203 0.04 0.03 0.08 0.16 331 Mollinedia stenophylla Perkins 1 1 0.0191 0.04 0.03 0.07 0.15 332 Ebenaceae sp1 1 1 0.0189 0.04 0.03 0.07 0.15 333 Calyptranthes cf. grandifolia O.Berg 1 1 0.0184 0.04 0.03 0.07 0.15 334 Indeterminada sp9 1 1 0.0179 0.04 0.03 0.07 0.15 335 Myrtaceae sp19 1 1 0.0181 0.04 0.03 0.07 0.15 336 Psychotria sp4 1 1 0.0177 0.04 0.03 0.07 0.15 337 Chrysophyllum sp2 1 1 0.0143 0.04 0.02 0.07 0.15 338 Eugenia macahensis O.Berg 1 1 0.0139 0.04 0.02 0.07 0.14 339 Astronium sp1 1 1 0.0133 0.04 0.02 0.07 0.14
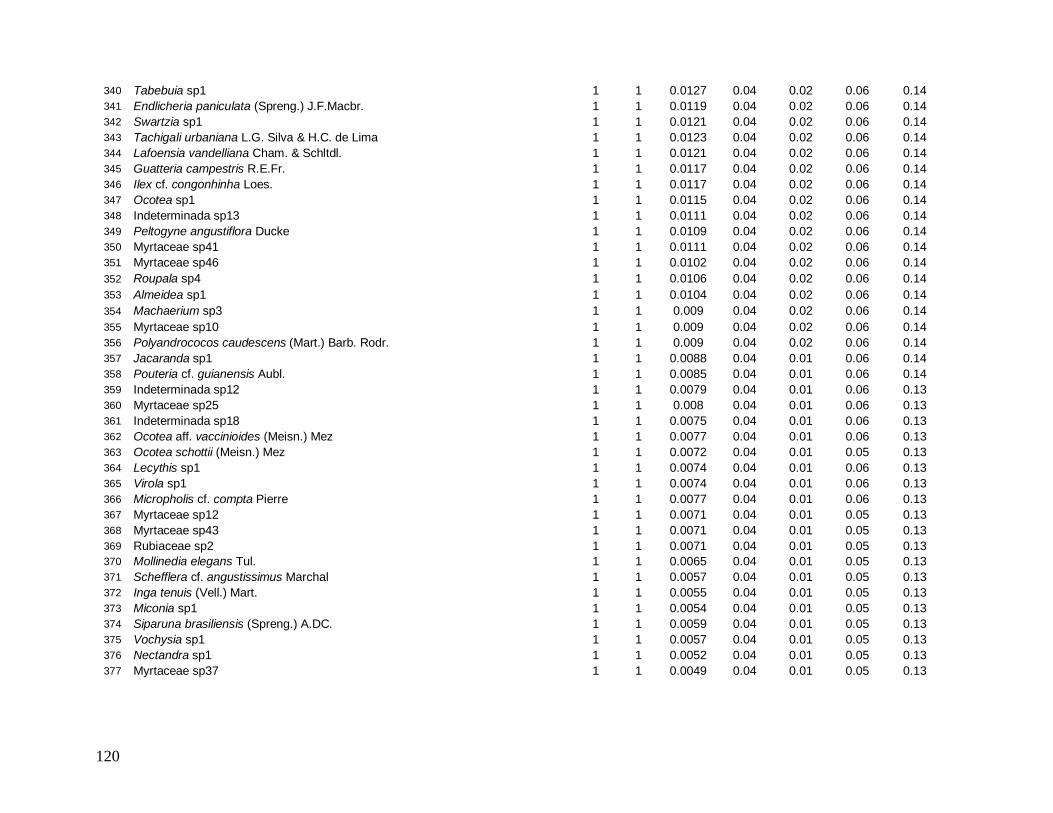
120
340 Tabebuia sp1 1 1 0.0127 0.04 0.02 0.06 0.14 341 Endlicheria paniculata (Spreng.) J.F.Macbr. 1 1 0.0119 0.04 0.02 0.06 0.14 342 Swartzia sp1 1 1 0.0121 0.04 0.02 0.06 0.14 343 Tachigali urbaniana L.G. Silva & H.C. de Lima 1 1 0.0123 0.04 0.02 0.06 0.14 344 Lafoensia vandelliana Cham. & Schltdl. 1 1 0.0121 0.04 0.02 0.06 0.14 345 Guatteria campestris R.E.Fr. 1 1 0.0117 0.04 0.02 0.06 0.14 346 Ilex cf. congonhinha Loes. 1 1 0.0117 0.04 0.02 0.06 0.14 347 Ocotea sp1 1 1 0.0115 0.04 0.02 0.06 0.14 348 Indeterminada sp13 1 1 0.0111 0.04 0.02 0.06 0.14 349 Peltogyne angustiflora Ducke 1 1 0.0109 0.04 0.02 0.06 0.14 350 Myrtaceae sp41 1 1 0.0111 0.04 0.02 0.06 0.14 351 Myrtaceae sp46 1 1 0.0102 0.04 0.02 0.06 0.14 352 Roupala sp4 1 1 0.0106 0.04 0.02 0.06 0.14 353 Almeidea sp1 1 1 0.0104 0.04 0.02 0.06 0.14 354 Machaerium sp3 1 1 0.009 0.04 0.02 0.06 0.14 355 Myrtaceae sp10 1 1 0.009 0.04 0.02 0.06 0.14 356 Polyandrococos caudescens (Mart.) Barb. Rodr. 1 1 0.009 0.04 0.02 0.06 0.14 357 Jacaranda sp1 1 1 0.0088 0.04 0.01 0.06 0.14 358 Pouteria cf. guianensis Aubl. 1 1 0.0085 0.04 0.01 0.06 0.14 359 Indeterminada sp12 1 1 0.0079 0.04 0.01 0.06 0.13 360 Myrtaceae sp25 1 1 0.008 0.04 0.01 0.06 0.13 361 Indeterminada sp18 1 1 0.0075 0.04 0.01 0.06 0.13 362 Ocotea aff. vaccinioides (Meisn.) Mez 1 1 0.0077 0.04 0.01 0.06 0.13 363 Ocotea schottii (Meisn.) Mez 1 1 0.0072 0.04 0.01 0.05 0.13 364 Lecythis sp1 1 1 0.0074 0.04 0.01 0.06 0.13 365 Virola sp1 1 1 0.0074 0.04 0.01 0.06 0.13 366 Micropholis cf. compta Pierre 1 1 0.0077 0.04 0.01 0.06 0.13 367 Myrtaceae sp12 1 1 0.0071 0.04 0.01 0.05 0.13 368 Myrtaceae sp43 1 1 0.0071 0.04 0.01 0.05 0.13 369 Rubiaceae sp2 1 1 0.0071 0.04 0.01 0.05 0.13 370 Mollinedia elegans Tul. 1 1 0.0065 0.04 0.01 0.05 0.13 371 Schefflera cf. angustissimus Marchal 1 1 0.0057 0.04 0.01 0.05 0.13 372 Inga tenuis (Vell.) Mart. 1 1 0.0055 0.04 0.01 0.05 0.13 373 Miconia sp1 1 1 0.0054 0.04 0.01 0.05 0.13 374 Siparuna brasiliensis (Spreng.) A.DC. 1 1 0.0059 0.04 0.01 0.05 0.13 375 Vochysia sp1 1 1 0.0057 0.04 0.01 0.05 0.13 376 Nectandra sp1 1 1 0.0052 0.04 0.01 0.05 0.13 377 Myrtaceae sp37 1 1 0.0049 0.04 0.01 0.05 0.13
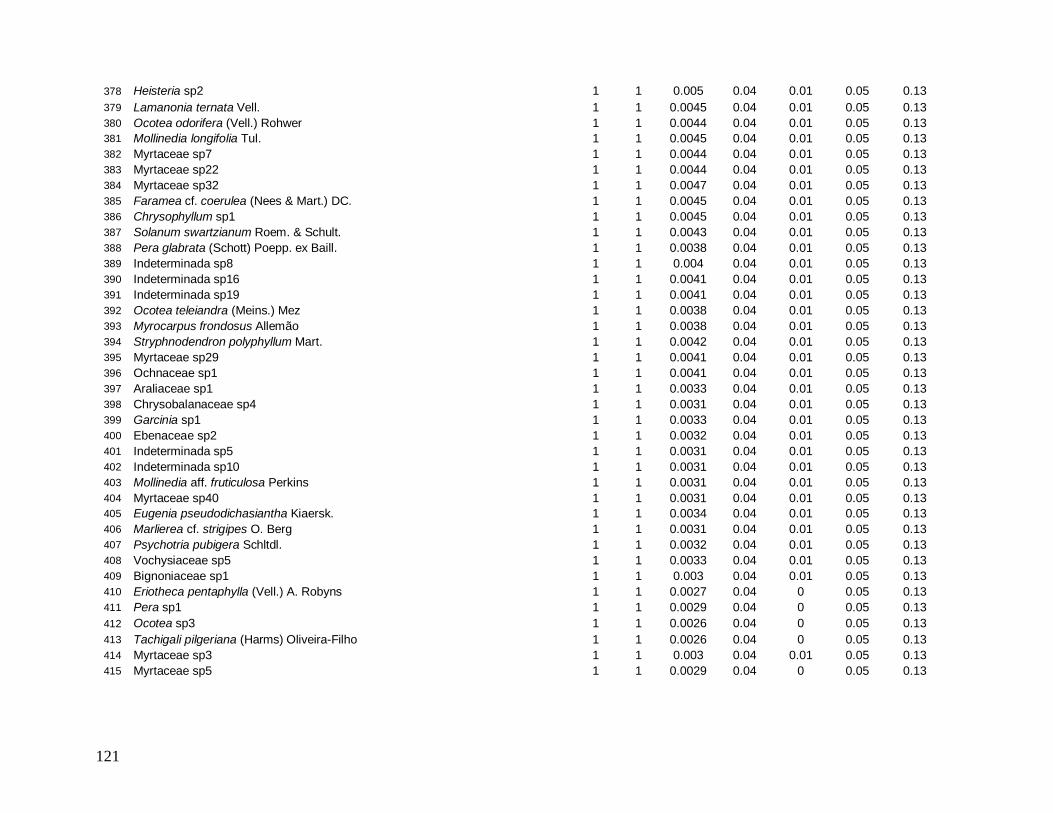
121
378 Heisteria sp2 1 1 0.005 0.04 0.01 0.05 0.13 379 Lamanonia ternata Vell. 1 1 0.0045 0.04 0.01 0.05 0.13 380 Ocotea odorifera (Vell.) Rohwer 1 1 0.0044 0.04 0.01 0.05 0.13 381 Mollinedia longifolia Tul. 1 1 0.0045 0.04 0.01 0.05 0.13 382 Myrtaceae sp7 1 1 0.0044 0.04 0.01 0.05 0.13 383 Myrtaceae sp22 1 1 0.0044 0.04 0.01 0.05 0.13 384 Myrtaceae sp32 1 1 0.0047 0.04 0.01 0.05 0.13 385 Faramea cf. coerulea (Nees & Mart.) DC. 1 1 0.0045 0.04 0.01 0.05 0.13 386 Chrysophyllum sp1 1 1 0.0045 0.04 0.01 0.05 0.13 387 Solanum swartzianum Roem. & Schult. 1 1 0.0043 0.04 0.01 0.05 0.13 388 Pera glabrata (Schott) Poepp. ex Baill. 1 1 0.0038 0.04 0.01 0.05 0.13 389 Indeterminada sp8 1 1 0.004 0.04 0.01 0.05 0.13 390 Indeterminada sp16 1 1 0.0041 0.04 0.01 0.05 0.13 391 Indeterminada sp19 1 1 0.0041 0.04 0.01 0.05 0.13 392 Ocotea teleiandra (Meins.) Mez 1 1 0.0038 0.04 0.01 0.05 0.13 393 Myrocarpus frondosus Allemão 1 1 0.0038 0.04 0.01 0.05 0.13 394 Stryphnodendron polyphyllum Mart. 1 1 0.0042 0.04 0.01 0.05 0.13 395 Myrtaceae sp29 1 1 0.0041 0.04 0.01 0.05 0.13 396 Ochnaceae sp1 1 1 0.0041 0.04 0.01 0.05 0.13 397 Araliaceae sp1 1 1 0.0033 0.04 0.01 0.05 0.13 398 Chrysobalanaceae sp4 1 1 0.0031 0.04 0.01 0.05 0.13 399 Garcinia sp1 1 1 0.0033 0.04 0.01 0.05 0.13 400 Ebenaceae sp2 1 1 0.0032 0.04 0.01 0.05 0.13 401 Indeterminada sp5 1 1 0.0031 0.04 0.01 0.05 0.13 402 Indeterminada sp10 1 1 0.0031 0.04 0.01 0.05 0.13 403 Mollinedia aff. fruticulosa Perkins 1 1 0.0031 0.04 0.01 0.05 0.13 404 Myrtaceae sp40 1 1 0.0031 0.04 0.01 0.05 0.13 405 Eugenia pseudodichasiantha Kiaersk. 1 1 0.0034 0.04 0.01 0.05 0.13 406 Marlierea cf. strigipes O. Berg 1 1 0.0031 0.04 0.01 0.05 0.13 407 Psychotria pubigera Schltdl. 1 1 0.0032 0.04 0.01 0.05 0.13 408 Vochysiaceae sp5 1 1 0.0033 0.04 0.01 0.05 0.13 409 Bignoniaceae sp1 1 1 0.003 0.04 0.01 0.05 0.13 410 Eriotheca pentaphylla (Vell.) A. Robyns 1 1 0.0027 0.04 0 0.05 0.13 411 Pera sp1 1 1 0.0029 0.04 0 0.05 0.13 412 Ocotea sp3 1 1 0.0026 0.04 0 0.05 0.13 413 Tachigali pilgeriana (Harms) Oliveira-Filho 1 1 0.0026 0.04 0 0.05 0.13 414 Myrtaceae sp3 1 1 0.003 0.04 0.01 0.05 0.13 415 Myrtaceae sp5 1 1 0.0029 0.04 0 0.05 0.13
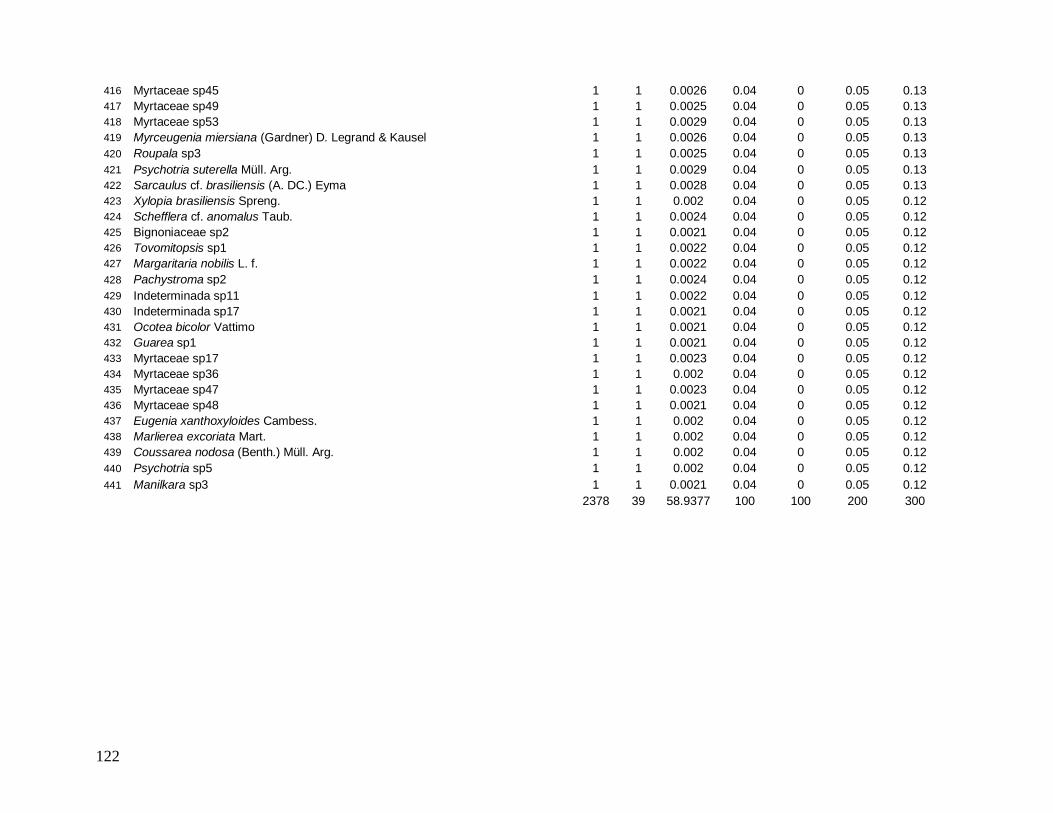
122
416 Myrtaceae sp45 1 1 0.0026 0.04 0 0.05 0.13 417 Myrtaceae sp49 1 1 0.0025 0.04 0 0.05 0.13 418 Myrtaceae sp53 1 1 0.0029 0.04 0 0.05 0.13 419 Myrceugenia miersiana (Gardner) D. Legrand & Kausel 1 1 0.0026 0.04 0 0.05 0.13 420 Roupala sp3 1 1 0.0025 0.04 0 0.05 0.13 421 Psychotria suterella Müll. Arg. 1 1 0.0029 0.04 0 0.05 0.13 422 Sarcaulus cf. brasiliensis (A. DC.) Eyma 1 1 0.0028 0.04 0 0.05 0.13 423 Xylopia brasiliensis Spreng. 1 1 0.002 0.04 0 0.05 0.12 424 Schefflera cf. anomalus Taub. 1 1 0.0024 0.04 0 0.05 0.12 425 Bignoniaceae sp2 1 1 0.0021 0.04 0 0.05 0.12 426 Tovomitopsis sp1 1 1 0.0022 0.04 0 0.05 0.12 427 Margaritaria nobilis L. f. 1 1 0.0022 0.04 0 0.05 0.12 428 Pachystroma sp2 1 1 0.0024 0.04 0 0.05 0.12 429 Indeterminada sp11 1 1 0.0022 0.04 0 0.05 0.12 430 Indeterminada sp17 1 1 0.0021 0.04 0 0.05 0.12 431 Ocotea bicolor Vattimo 1 1 0.0021 0.04 0 0.05 0.12 432 Guarea sp1 1 1 0.0021 0.04 0 0.05 0.12 433 Myrtaceae sp17 1 1 0.0023 0.04 0 0.05 0.12 434 Myrtaceae sp36 1 1 0.002 0.04 0 0.05 0.12 435 Myrtaceae sp47 1 1 0.0023 0.04 0 0.05 0.12 436 Myrtaceae sp48 1 1 0.0021 0.04 0 0.05 0.12 437 Eugenia xanthoxyloides Cambess. 1 1 0.002 0.04 0 0.05 0.12 438 Marlierea excoriata Mart. 1 1 0.002 0.04 0 0.05 0.12 439 Coussarea nodosa (Benth.) Müll. Arg. 1 1 0.002 0.04 0 0.05 0.12 440 Psychotria sp5 1 1 0.002 0.04 0 0.05 0.12 441 Manilkara sp3 1 1 0.0021 0.04 0 0.05 0.12
2378 39 58.9377 100 100 200 300
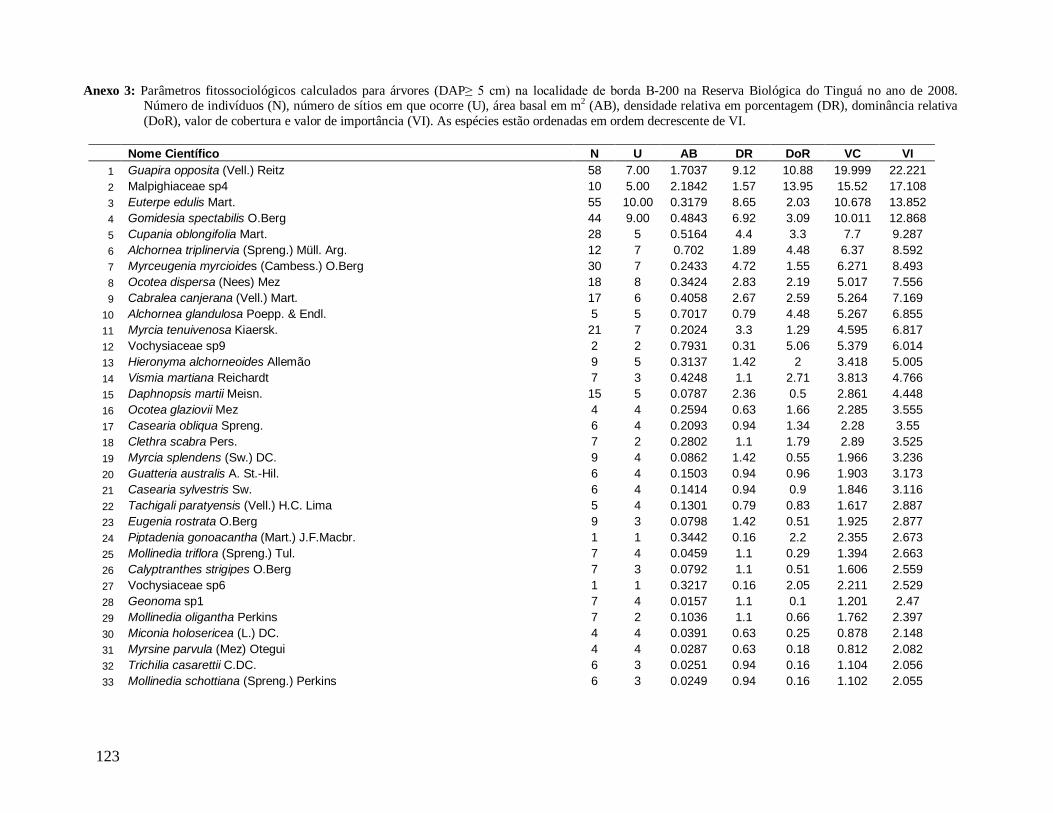
123
Anexo 3: Parâmetros fitossociológicos calculados para árvores (DAP≥ 5 cm) na localidade de borda B-200 na Reserva Biológica do Tinguá no ano de 2008. Número de indivíduos (N), número de sítios em que ocorre (U), área basal em m2 (AB), densidade relativa em porcentagem (DR), dominância relativa (DoR), valor de cobertura e valor de importância (VI). As espécies estão ordenadas em ordem decrescente de VI.
Nome Científico N U AB DR DoR VC VI
1 Guapira opposita (Vell.) Reitz 58 7.00 1.7037 9.12 10.88 19.999 22.221 2 Malpighiaceae sp4 10 5.00 2.1842 1.57 13.95 15.52 17.108 3 Euterpe edulis Mart. 55 10.00 0.3179 8.65 2.03 10.678 13.852 4 Gomidesia spectabilis O.Berg 44 9.00 0.4843 6.92 3.09 10.011 12.868 5 Cupania oblongifolia Mart. 28 5 0.5164 4.4 3.3 7.7 9.287 6 Alchornea triplinervia (Spreng.) Müll. Arg. 12 7 0.702 1.89 4.48 6.37 8.592 7 Myrceugenia myrcioides (Cambess.) O.Berg 30 7 0.2433 4.72 1.55 6.271 8.493 8 Ocotea dispersa (Nees) Mez 18 8 0.3424 2.83 2.19 5.017 7.556 9 Cabralea canjerana (Vell.) Mart. 17 6 0.4058 2.67 2.59 5.264 7.169
10 Alchornea glandulosa Poepp. & Endl. 5 5 0.7017 0.79 4.48 5.267 6.855 11 Myrcia tenuivenosa Kiaersk. 21 7 0.2024 3.3 1.29 4.595 6.817 12 Vochysiaceae sp9 2 2 0.7931 0.31 5.06 5.379 6.014 13 Hieronyma alchorneoides Allemão 9 5 0.3137 1.42 2 3.418 5.005 14 Vismia martiana Reichardt 7 3 0.4248 1.1 2.71 3.813 4.766 15 Daphnopsis martii Meisn. 15 5 0.0787 2.36 0.5 2.861 4.448 16 Ocotea glaziovii Mez 4 4 0.2594 0.63 1.66 2.285 3.555 17 Casearia obliqua Spreng. 6 4 0.2093 0.94 1.34 2.28 3.55 18 Clethra scabra Pers. 7 2 0.2802 1.1 1.79 2.89 3.525 19 Myrcia splendens (Sw.) DC. 9 4 0.0862 1.42 0.55 1.966 3.236 20 Guatteria australis A. St.-Hil. 6 4 0.1503 0.94 0.96 1.903 3.173 21 Casearia sylvestris Sw. 6 4 0.1414 0.94 0.9 1.846 3.116 22 Tachigali paratyensis (Vell.) H.C. Lima 5 4 0.1301 0.79 0.83 1.617 2.887 23 Eugenia rostrata O.Berg 9 3 0.0798 1.42 0.51 1.925 2.877 24 Piptadenia gonoacantha (Mart.) J.F.Macbr. 1 1 0.3442 0.16 2.2 2.355 2.673 25 Mollinedia triflora (Spreng.) Tul. 7 4 0.0459 1.1 0.29 1.394 2.663 26 Calyptranthes strigipes O.Berg 7 3 0.0792 1.1 0.51 1.606 2.559 27 Vochysiaceae sp6 1 1 0.3217 0.16 2.05 2.211 2.529 28 Geonoma sp1 7 4 0.0157 1.1 0.1 1.201 2.47 29 Mollinedia oligantha Perkins 7 2 0.1036 1.1 0.66 1.762 2.397 30 Miconia holosericea (L.) DC. 4 4 0.0391 0.63 0.25 0.878 2.148 31 Myrsine parvula (Mez) Otegui 4 4 0.0287 0.63 0.18 0.812 2.082 32 Trichilia casarettii C.DC. 6 3 0.0251 0.94 0.16 1.104 2.056 33 Mollinedia schottiana (Spreng.) Perkins 6 3 0.0249 0.94 0.16 1.102 2.055
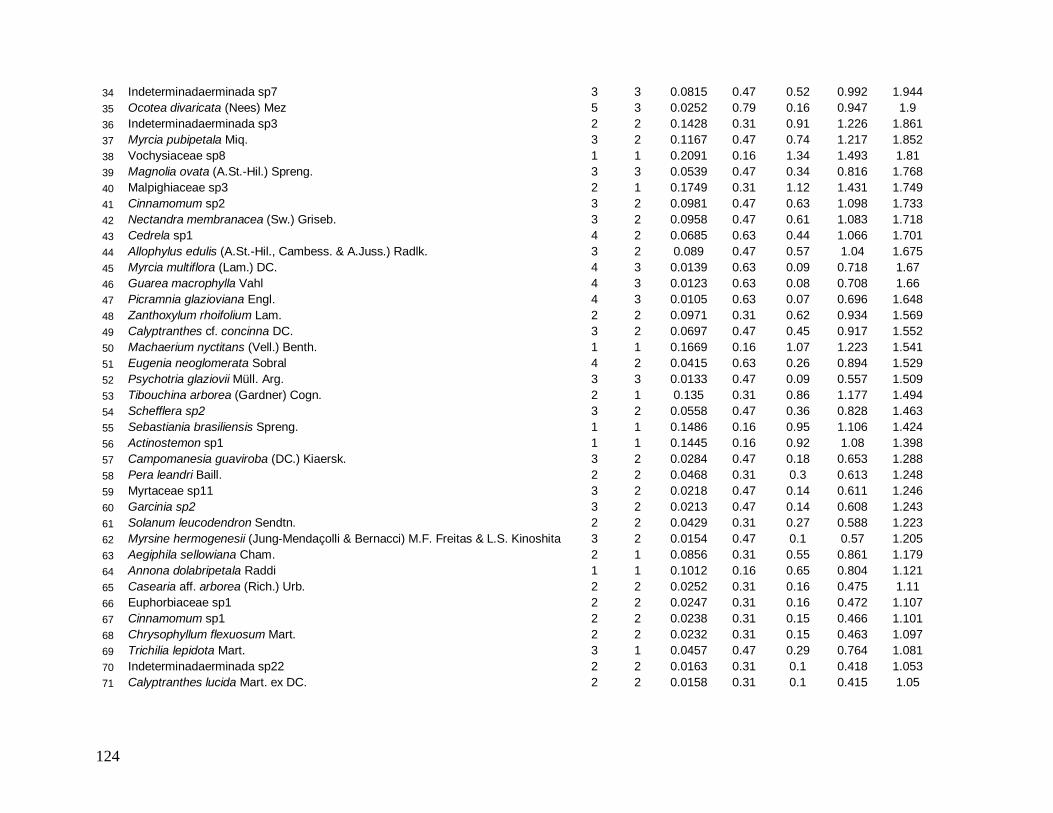
124
34 Indeterminadaerminada sp7 3 3 0.0815 0.47 0.52 0.992 1.944 35 Ocotea divaricata (Nees) Mez 5 3 0.0252 0.79 0.16 0.947 1.9 36 Indeterminadaerminada sp3 2 2 0.1428 0.31 0.91 1.226 1.861 37 Myrcia pubipetala Miq. 3 2 0.1167 0.47 0.74 1.217 1.852 38 Vochysiaceae sp8 1 1 0.2091 0.16 1.34 1.493 1.81 39 Magnolia ovata (A.St.-Hil.) Spreng. 3 3 0.0539 0.47 0.34 0.816 1.768 40 Malpighiaceae sp3 2 1 0.1749 0.31 1.12 1.431 1.749 41 Cinnamomum sp2 3 2 0.0981 0.47 0.63 1.098 1.733 42 Nectandra membranacea (Sw.) Griseb. 3 2 0.0958 0.47 0.61 1.083 1.718 43 Cedrela sp1 4 2 0.0685 0.63 0.44 1.066 1.701 44 Allophylus edulis (A.St.-Hil., Cambess. & A.Juss.) Radlk. 3 2 0.089 0.47 0.57 1.04 1.675 45 Myrcia multiflora (Lam.) DC. 4 3 0.0139 0.63 0.09 0.718 1.67 46 Guarea macrophylla Vahl 4 3 0.0123 0.63 0.08 0.708 1.66 47 Picramnia glazioviana Engl. 4 3 0.0105 0.63 0.07 0.696 1.648 48 Zanthoxylum rhoifolium Lam. 2 2 0.0971 0.31 0.62 0.934 1.569 49 Calyptranthes cf. concinna DC. 3 2 0.0697 0.47 0.45 0.917 1.552 50 Machaerium nyctitans (Vell.) Benth. 1 1 0.1669 0.16 1.07 1.223 1.541 51 Eugenia neoglomerata Sobral 4 2 0.0415 0.63 0.26 0.894 1.529 52 Psychotria glaziovii Müll. Arg. 3 3 0.0133 0.47 0.09 0.557 1.509 53 Tibouchina arborea (Gardner) Cogn. 2 1 0.135 0.31 0.86 1.177 1.494 54 Schefflera sp2 3 2 0.0558 0.47 0.36 0.828 1.463 55 Sebastiania brasiliensis Spreng. 1 1 0.1486 0.16 0.95 1.106 1.424 56 Actinostemon sp1 1 1 0.1445 0.16 0.92 1.08 1.398 57 Campomanesia guaviroba (DC.) Kiaersk. 3 2 0.0284 0.47 0.18 0.653 1.288 58 Pera leandri Baill. 2 2 0.0468 0.31 0.3 0.613 1.248 59 Myrtaceae sp11 3 2 0.0218 0.47 0.14 0.611 1.246 60 Garcinia sp2 3 2 0.0213 0.47 0.14 0.608 1.243 61 Solanum leucodendron Sendtn. 2 2 0.0429 0.31 0.27 0.588 1.223 62 Myrsine hermogenesii (Jung-Mendaçolli & Bernacci) M.F. Freitas & L.S. Kinoshita 3 2 0.0154 0.47 0.1 0.57 1.205 63 Aegiphila sellowiana Cham. 2 1 0.0856 0.31 0.55 0.861 1.179 64 Annona dolabripetala Raddi 1 1 0.1012 0.16 0.65 0.804 1.121 65 Casearia aff. arborea (Rich.) Urb. 2 2 0.0252 0.31 0.16 0.475 1.11 66 Euphorbiaceae sp1 2 2 0.0247 0.31 0.16 0.472 1.107 67 Cinnamomum sp1 2 2 0.0238 0.31 0.15 0.466 1.101 68 Chrysophyllum flexuosum Mart. 2 2 0.0232 0.31 0.15 0.463 1.097 69 Trichilia lepidota Mart. 3 1 0.0457 0.47 0.29 0.764 1.081 70 Indeterminadaerminada sp22 2 2 0.0163 0.31 0.1 0.418 1.053 71 Calyptranthes lucida Mart. ex DC. 2 2 0.0158 0.31 0.1 0.415 1.05
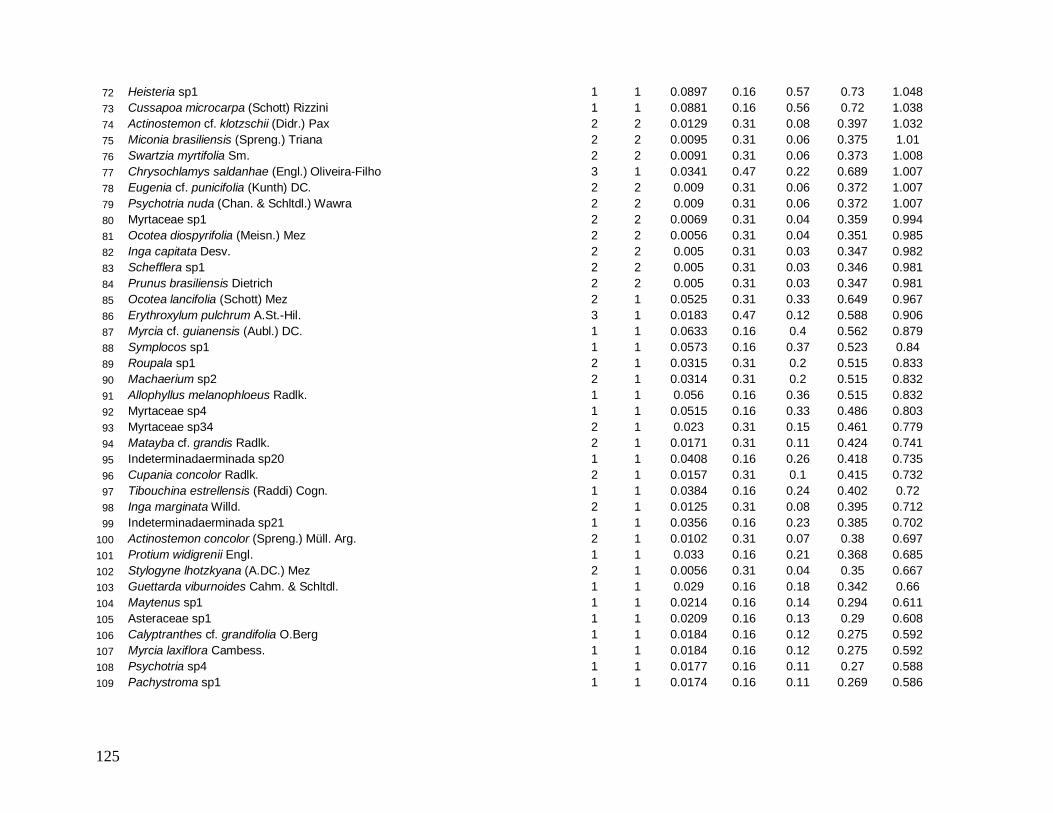
125
72 Heisteria sp1 1 1 0.0897 0.16 0.57 0.73 1.048 73 Cussapoa microcarpa (Schott) Rizzini 1 1 0.0881 0.16 0.56 0.72 1.038 74 Actinostemon cf. klotzschii (Didr.) Pax 2 2 0.0129 0.31 0.08 0.397 1.032 75 Miconia brasiliensis (Spreng.) Triana 2 2 0.0095 0.31 0.06 0.375 1.01 76 Swartzia myrtifolia Sm. 2 2 0.0091 0.31 0.06 0.373 1.008 77 Chrysochlamys saldanhae (Engl.) Oliveira-Filho 3 1 0.0341 0.47 0.22 0.689 1.007 78 Eugenia cf. punicifolia (Kunth) DC. 2 2 0.009 0.31 0.06 0.372 1.007 79 Psychotria nuda (Chan. & Schltdl.) Wawra 2 2 0.009 0.31 0.06 0.372 1.007 80 Myrtaceae sp1 2 2 0.0069 0.31 0.04 0.359 0.994 81 Ocotea diospyrifolia (Meisn.) Mez 2 2 0.0056 0.31 0.04 0.351 0.985 82 Inga capitata Desv. 2 2 0.005 0.31 0.03 0.347 0.982 83 Schefflera sp1 2 2 0.005 0.31 0.03 0.346 0.981 84 Prunus brasiliensis Dietrich 2 2 0.005 0.31 0.03 0.347 0.981 85 Ocotea lancifolia (Schott) Mez 2 1 0.0525 0.31 0.33 0.649 0.967 86 Erythroxylum pulchrum A.St.-Hil. 3 1 0.0183 0.47 0.12 0.588 0.906 87 Myrcia cf. guianensis (Aubl.) DC. 1 1 0.0633 0.16 0.4 0.562 0.879 88 Symplocos sp1 1 1 0.0573 0.16 0.37 0.523 0.84 89 Roupala sp1 2 1 0.0315 0.31 0.2 0.515 0.833 90 Machaerium sp2 2 1 0.0314 0.31 0.2 0.515 0.832 91 Allophyllus melanophloeus Radlk. 1 1 0.056 0.16 0.36 0.515 0.832 92 Myrtaceae sp4 1 1 0.0515 0.16 0.33 0.486 0.803 93 Myrtaceae sp34 2 1 0.023 0.31 0.15 0.461 0.779 94 Matayba cf. grandis Radlk. 2 1 0.0171 0.31 0.11 0.424 0.741 95 Indeterminadaerminada sp20 1 1 0.0408 0.16 0.26 0.418 0.735 96 Cupania concolor Radlk. 2 1 0.0157 0.31 0.1 0.415 0.732 97 Tibouchina estrellensis (Raddi) Cogn. 1 1 0.0384 0.16 0.24 0.402 0.72 98 Inga marginata Willd. 2 1 0.0125 0.31 0.08 0.395 0.712 99 Indeterminadaerminada sp21 1 1 0.0356 0.16 0.23 0.385 0.702
100 Actinostemon concolor (Spreng.) Müll. Arg. 2 1 0.0102 0.31 0.07 0.38 0.697 101 Protium widigrenii Engl. 1 1 0.033 0.16 0.21 0.368 0.685 102 Stylogyne lhotzkyana (A.DC.) Mez 2 1 0.0056 0.31 0.04 0.35 0.667 103 Guettarda viburnoides Cahm. & Schltdl. 1 1 0.029 0.16 0.18 0.342 0.66 104 Maytenus sp1 1 1 0.0214 0.16 0.14 0.294 0.611 105 Asteraceae sp1 1 1 0.0209 0.16 0.13 0.29 0.608 106 Calyptranthes cf. grandifolia O.Berg 1 1 0.0184 0.16 0.12 0.275 0.592 107 Myrcia laxiflora Cambess. 1 1 0.0184 0.16 0.12 0.275 0.592 108 Psychotria sp4 1 1 0.0177 0.16 0.11 0.27 0.588 109 Pachystroma sp1 1 1 0.0174 0.16 0.11 0.269 0.586
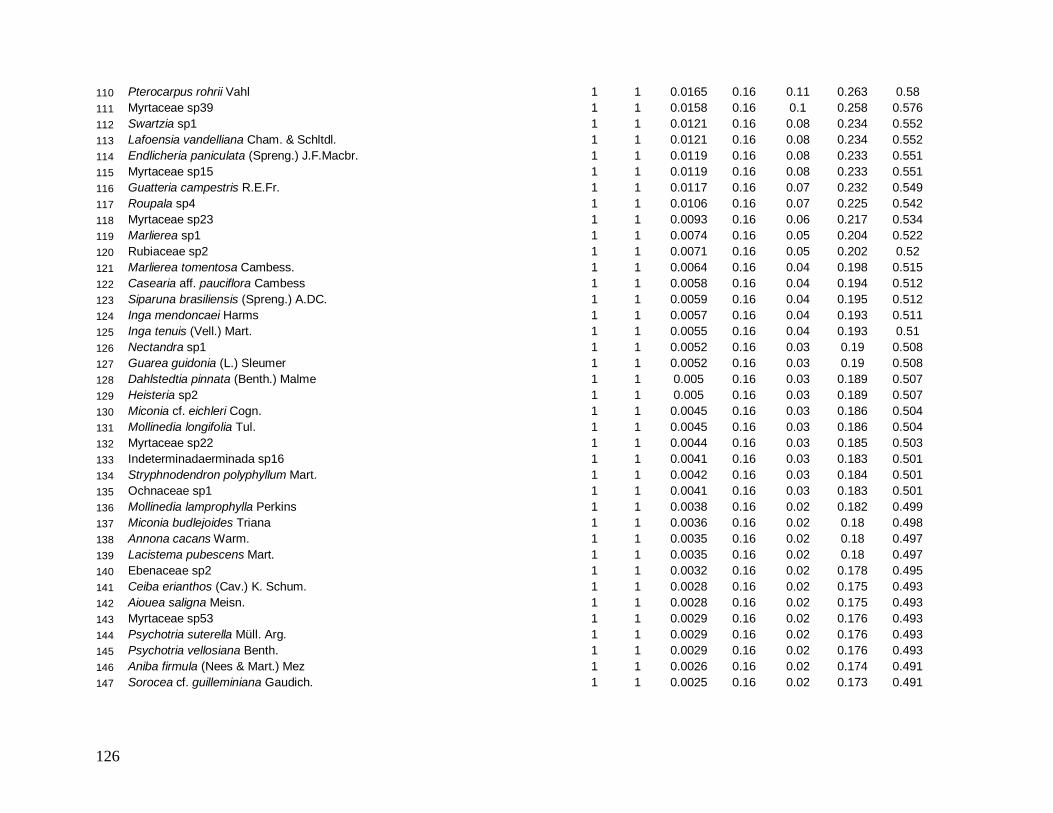
126
110 Pterocarpus rohrii Vahl 1 1 0.0165 0.16 0.11 0.263 0.58 111 Myrtaceae sp39 1 1 0.0158 0.16 0.1 0.258 0.576 112 Swartzia sp1 1 1 0.0121 0.16 0.08 0.234 0.552 113 Lafoensia vandelliana Cham. & Schltdl. 1 1 0.0121 0.16 0.08 0.234 0.552 114 Endlicheria paniculata (Spreng.) J.F.Macbr. 1 1 0.0119 0.16 0.08 0.233 0.551 115 Myrtaceae sp15 1 1 0.0119 0.16 0.08 0.233 0.551 116 Guatteria campestris R.E.Fr. 1 1 0.0117 0.16 0.07 0.232 0.549 117 Roupala sp4 1 1 0.0106 0.16 0.07 0.225 0.542 118 Myrtaceae sp23 1 1 0.0093 0.16 0.06 0.217 0.534 119 Marlierea sp1 1 1 0.0074 0.16 0.05 0.204 0.522 120 Rubiaceae sp2 1 1 0.0071 0.16 0.05 0.202 0.52 121 Marlierea tomentosa Cambess. 1 1 0.0064 0.16 0.04 0.198 0.515 122 Casearia aff. pauciflora Cambess 1 1 0.0058 0.16 0.04 0.194 0.512 123 Siparuna brasiliensis (Spreng.) A.DC. 1 1 0.0059 0.16 0.04 0.195 0.512 124 Inga mendoncaei Harms 1 1 0.0057 0.16 0.04 0.193 0.511 125 Inga tenuis (Vell.) Mart. 1 1 0.0055 0.16 0.04 0.193 0.51 126 Nectandra sp1 1 1 0.0052 0.16 0.03 0.19 0.508 127 Guarea guidonia (L.) Sleumer 1 1 0.0052 0.16 0.03 0.19 0.508 128 Dahlstedtia pinnata (Benth.) Malme 1 1 0.005 0.16 0.03 0.189 0.507 129 Heisteria sp2 1 1 0.005 0.16 0.03 0.189 0.507 130 Miconia cf. eichleri Cogn. 1 1 0.0045 0.16 0.03 0.186 0.504 131 Mollinedia longifolia Tul. 1 1 0.0045 0.16 0.03 0.186 0.504 132 Myrtaceae sp22 1 1 0.0044 0.16 0.03 0.185 0.503 133 Indeterminadaerminada sp16 1 1 0.0041 0.16 0.03 0.183 0.501 134 Stryphnodendron polyphyllum Mart. 1 1 0.0042 0.16 0.03 0.184 0.501 135 Ochnaceae sp1 1 1 0.0041 0.16 0.03 0.183 0.501 136 Mollinedia lamprophylla Perkins 1 1 0.0038 0.16 0.02 0.182 0.499 137 Miconia budlejoides Triana 1 1 0.0036 0.16 0.02 0.18 0.498 138 Annona cacans Warm. 1 1 0.0035 0.16 0.02 0.18 0.497 139 Lacistema pubescens Mart. 1 1 0.0035 0.16 0.02 0.18 0.497 140 Ebenaceae sp2 1 1 0.0032 0.16 0.02 0.178 0.495 141 Ceiba erianthos (Cav.) K. Schum. 1 1 0.0028 0.16 0.02 0.175 0.493 142 Aiouea saligna Meisn. 1 1 0.0028 0.16 0.02 0.175 0.493 143 Myrtaceae sp53 1 1 0.0029 0.16 0.02 0.176 0.493 144 Psychotria suterella Müll. Arg. 1 1 0.0029 0.16 0.02 0.176 0.493 145 Psychotria vellosiana Benth. 1 1 0.0029 0.16 0.02 0.176 0.493 146 Aniba firmula (Nees & Mart.) Mez 1 1 0.0026 0.16 0.02 0.174 0.491 147 Sorocea cf. guilleminiana Gaudich. 1 1 0.0025 0.16 0.02 0.173 0.491
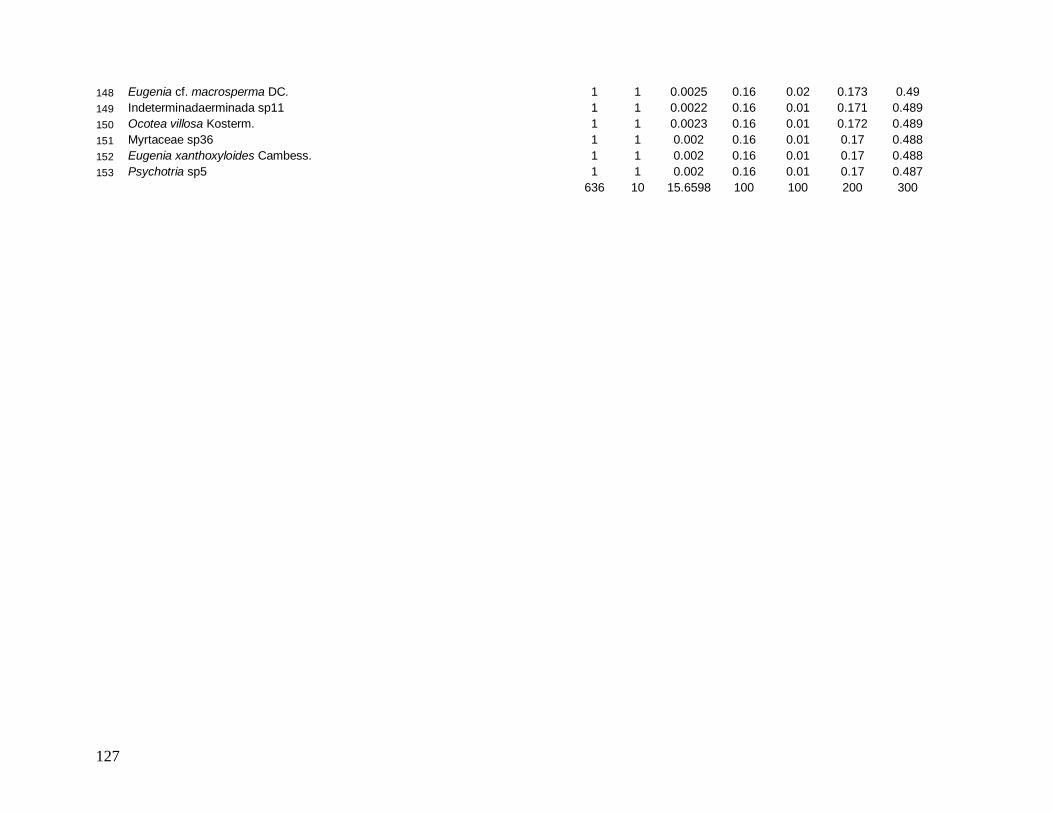
127
148 Eugenia cf. macrosperma DC. 1 1 0.0025 0.16 0.02 0.173 0.49 149 Indeterminadaerminada sp11 1 1 0.0022 0.16 0.01 0.171 0.489 150 Ocotea villosa Kosterm. 1 1 0.0023 0.16 0.01 0.172 0.489 151 Myrtaceae sp36 1 1 0.002 0.16 0.01 0.17 0.488 152 Eugenia xanthoxyloides Cambess. 1 1 0.002 0.16 0.01 0.17 0.488 153 Psychotria sp5 1 1 0.002 0.16 0.01 0.17 0.487 636 10 15.6598 100 100 200 300
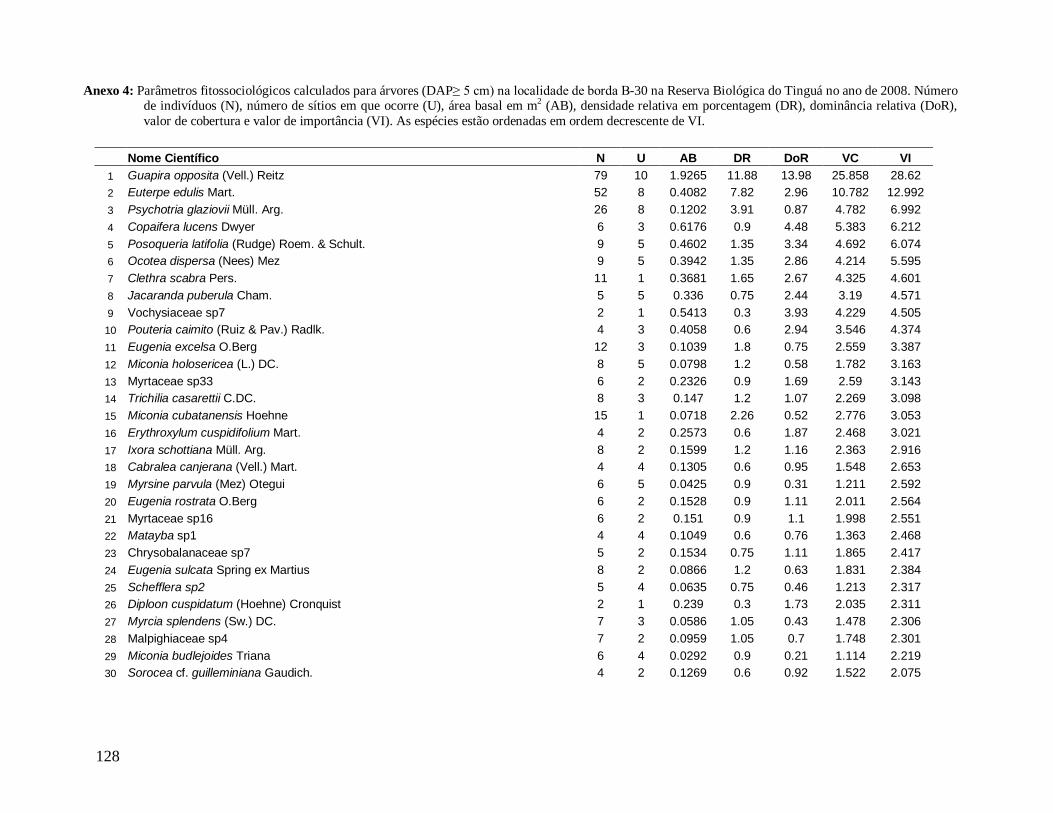
128
Anexo 4: Parâmetros fitossociológicos calculados para árvores (DAP≥ 5 cm) na localidade de borda B-30 na Reserva Biológica do Tinguá no ano de 2008. Número de indivíduos (N), número de sítios em que ocorre (U), área basal em m2 (AB), densidade relativa em porcentagem (DR), dominância relativa (DoR), valor de cobertura e valor de importância (VI). As espécies estão ordenadas em ordem decrescente de VI.
Nome Científico N U AB DR DoR VC VI 1 Guapira opposita (Vell.) Reitz 79 10 1.9265 11.88 13.98 25.858 28.62 2 Euterpe edulis Mart. 52 8 0.4082 7.82 2.96 10.782 12.992 3 Psychotria glaziovii Müll. Arg. 26 8 0.1202 3.91 0.87 4.782 6.992 4 Copaifera lucens Dwyer 6 3 0.6176 0.9 4.48 5.383 6.212 5 Posoqueria latifolia (Rudge) Roem. & Schult. 9 5 0.4602 1.35 3.34 4.692 6.074 6 Ocotea dispersa (Nees) Mez 9 5 0.3942 1.35 2.86 4.214 5.595 7 Clethra scabra Pers. 11 1 0.3681 1.65 2.67 4.325 4.601 8 Jacaranda puberula Cham. 5 5 0.336 0.75 2.44 3.19 4.571 9 Vochysiaceae sp7 2 1 0.5413 0.3 3.93 4.229 4.505 10 Pouteria caimito (Ruiz & Pav.) Radlk. 4 3 0.4058 0.6 2.94 3.546 4.374 11 Eugenia excelsa O.Berg 12 3 0.1039 1.8 0.75 2.559 3.387 12 Miconia holosericea (L.) DC. 8 5 0.0798 1.2 0.58 1.782 3.163 13 Myrtaceae sp33 6 2 0.2326 0.9 1.69 2.59 3.143 14 Trichilia casarettii C.DC. 8 3 0.147 1.2 1.07 2.269 3.098 15 Miconia cubatanensis Hoehne 15 1 0.0718 2.26 0.52 2.776 3.053 16 Erythroxylum cuspidifolium Mart. 4 2 0.2573 0.6 1.87 2.468 3.021 17 Ixora schottiana Müll. Arg. 8 2 0.1599 1.2 1.16 2.363 2.916 18 Cabralea canjerana (Vell.) Mart. 4 4 0.1305 0.6 0.95 1.548 2.653 19 Myrsine parvula (Mez) Otegui 6 5 0.0425 0.9 0.31 1.211 2.592 20 Eugenia rostrata O.Berg 6 2 0.1528 0.9 1.11 2.011 2.564 21 Myrtaceae sp16 6 2 0.151 0.9 1.1 1.998 2.551 22 Matayba sp1 4 4 0.1049 0.6 0.76 1.363 2.468 23 Chrysobalanaceae sp7 5 2 0.1534 0.75 1.11 1.865 2.417 24 Eugenia sulcata Spring ex Martius 8 2 0.0866 1.2 0.63 1.831 2.384 25 Schefflera sp2 5 4 0.0635 0.75 0.46 1.213 2.317 26 Diploon cuspidatum (Hoehne) Cronquist 2 1 0.239 0.3 1.73 2.035 2.311 27 Myrcia splendens (Sw.) DC. 7 3 0.0586 1.05 0.43 1.478 2.306 28 Malpighiaceae sp4 7 2 0.0959 1.05 0.7 1.748 2.301 29 Miconia budlejoides Triana 6 4 0.0292 0.9 0.21 1.114 2.219 30 Sorocea cf. guilleminiana Gaudich. 4 2 0.1269 0.6 0.92 1.522 2.075
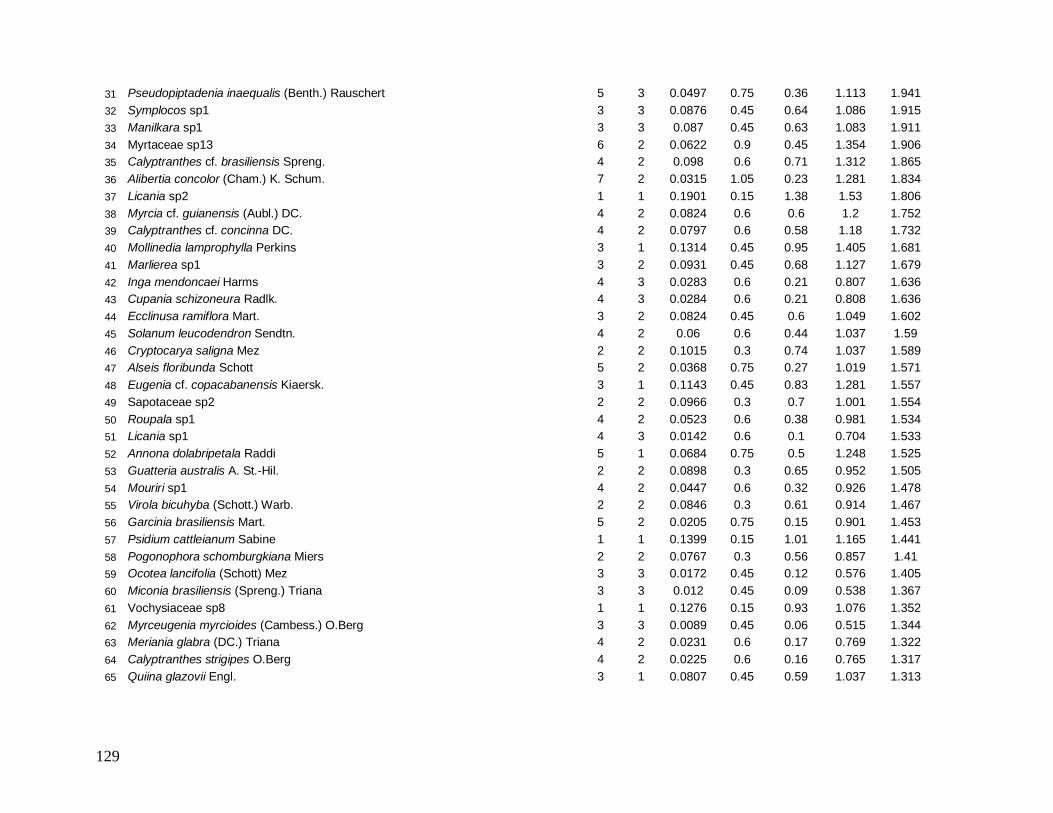
129
31 Pseudopiptadenia inaequalis (Benth.) Rauschert 5 3 0.0497 0.75 0.36 1.113 1.941 32 Symplocos sp1 3 3 0.0876 0.45 0.64 1.086 1.915 33 Manilkara sp1 3 3 0.087 0.45 0.63 1.083 1.911 34 Myrtaceae sp13 6 2 0.0622 0.9 0.45 1.354 1.906 35 Calyptranthes cf. brasiliensis Spreng. 4 2 0.098 0.6 0.71 1.312 1.865 36 Alibertia concolor (Cham.) K. Schum. 7 2 0.0315 1.05 0.23 1.281 1.834 37 Licania sp2 1 1 0.1901 0.15 1.38 1.53 1.806 38 Myrcia cf. guianensis (Aubl.) DC. 4 2 0.0824 0.6 0.6 1.2 1.752 39 Calyptranthes cf. concinna DC. 4 2 0.0797 0.6 0.58 1.18 1.732 40 Mollinedia lamprophylla Perkins 3 1 0.1314 0.45 0.95 1.405 1.681 41 Marlierea sp1 3 2 0.0931 0.45 0.68 1.127 1.679 42 Inga mendoncaei Harms 4 3 0.0283 0.6 0.21 0.807 1.636 43 Cupania schizoneura Radlk. 4 3 0.0284 0.6 0.21 0.808 1.636 44 Ecclinusa ramiflora Mart. 3 2 0.0824 0.45 0.6 1.049 1.602 45 Solanum leucodendron Sendtn. 4 2 0.06 0.6 0.44 1.037 1.59 46 Cryptocarya saligna Mez 2 2 0.1015 0.3 0.74 1.037 1.589 47 Alseis floribunda Schott 5 2 0.0368 0.75 0.27 1.019 1.571 48 Eugenia cf. copacabanensis Kiaersk. 3 1 0.1143 0.45 0.83 1.281 1.557 49 Sapotaceae sp2 2 2 0.0966 0.3 0.7 1.001 1.554 50 Roupala sp1 4 2 0.0523 0.6 0.38 0.981 1.534 51 Licania sp1 4 3 0.0142 0.6 0.1 0.704 1.533 52 Annona dolabripetala Raddi 5 1 0.0684 0.75 0.5 1.248 1.525 53 Guatteria australis A. St.-Hil. 2 2 0.0898 0.3 0.65 0.952 1.505 54 Mouriri sp1 4 2 0.0447 0.6 0.32 0.926 1.478 55 Virola bicuhyba (Schott.) Warb. 2 2 0.0846 0.3 0.61 0.914 1.467 56 Garcinia brasiliensis Mart. 5 2 0.0205 0.75 0.15 0.901 1.453 57 Psidium cattleianum Sabine 1 1 0.1399 0.15 1.01 1.165 1.441 58 Pogonophora schomburgkiana Miers 2 2 0.0767 0.3 0.56 0.857 1.41 59 Ocotea lancifolia (Schott) Mez 3 3 0.0172 0.45 0.12 0.576 1.405 60 Miconia brasiliensis (Spreng.) Triana 3 3 0.012 0.45 0.09 0.538 1.367 61 Vochysiaceae sp8 1 1 0.1276 0.15 0.93 1.076 1.352 62 Myrceugenia myrcioides (Cambess.) O.Berg 3 3 0.0089 0.45 0.06 0.515 1.344 63 Meriania glabra (DC.) Triana 4 2 0.0231 0.6 0.17 0.769 1.322 64 Calyptranthes strigipes O.Berg 4 2 0.0225 0.6 0.16 0.765 1.317 65 Quiina glazovii Engl. 3 1 0.0807 0.45 0.59 1.037 1.313
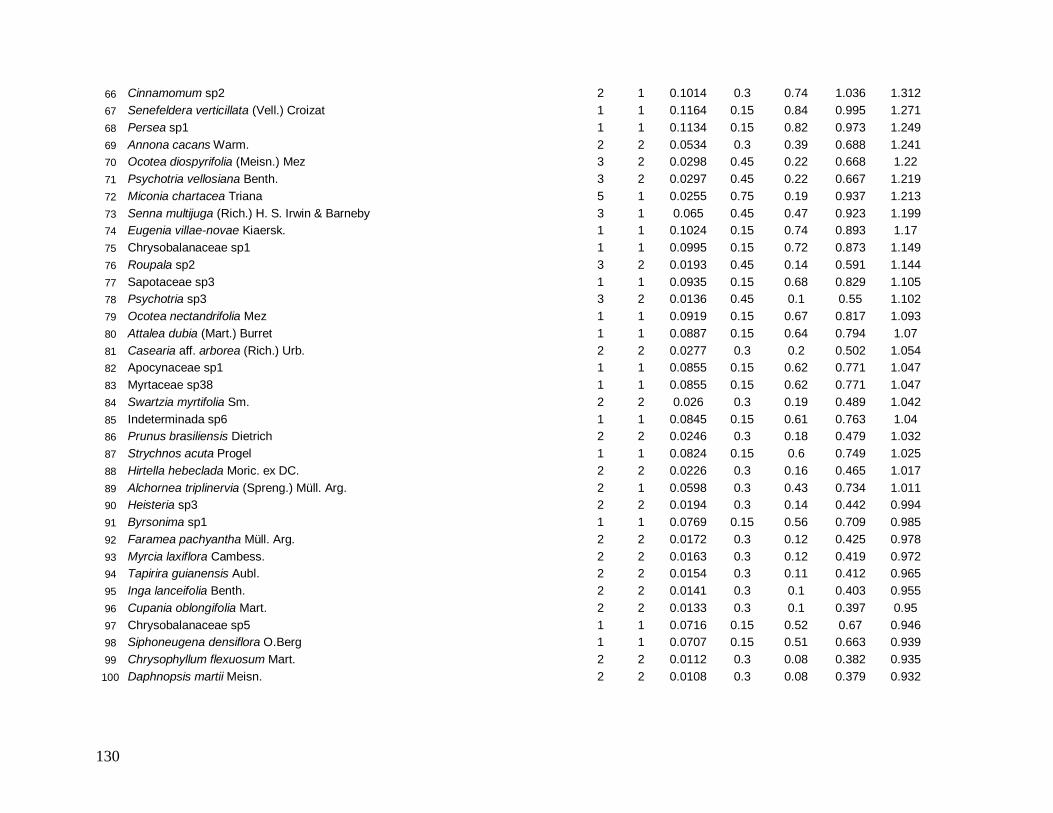
130
66 Cinnamomum sp2 2 1 0.1014 0.3 0.74 1.036 1.312 67 Senefeldera verticillata (Vell.) Croizat 1 1 0.1164 0.15 0.84 0.995 1.271 68 Persea sp1 1 1 0.1134 0.15 0.82 0.973 1.249 69 Annona cacans Warm. 2 2 0.0534 0.3 0.39 0.688 1.241 70 Ocotea diospyrifolia (Meisn.) Mez 3 2 0.0298 0.45 0.22 0.668 1.22 71 Psychotria vellosiana Benth. 3 2 0.0297 0.45 0.22 0.667 1.219 72 Miconia chartacea Triana 5 1 0.0255 0.75 0.19 0.937 1.213 73 Senna multijuga (Rich.) H. S. Irwin & Barneby 3 1 0.065 0.45 0.47 0.923 1.199 74 Eugenia villae-novae Kiaersk. 1 1 0.1024 0.15 0.74 0.893 1.17 75 Chrysobalanaceae sp1 1 1 0.0995 0.15 0.72 0.873 1.149 76 Roupala sp2 3 2 0.0193 0.45 0.14 0.591 1.144 77 Sapotaceae sp3 1 1 0.0935 0.15 0.68 0.829 1.105 78 Psychotria sp3 3 2 0.0136 0.45 0.1 0.55 1.102 79 Ocotea nectandrifolia Mez 1 1 0.0919 0.15 0.67 0.817 1.093 80 Attalea dubia (Mart.) Burret 1 1 0.0887 0.15 0.64 0.794 1.07 81 Casearia aff. arborea (Rich.) Urb. 2 2 0.0277 0.3 0.2 0.502 1.054 82 Apocynaceae sp1 1 1 0.0855 0.15 0.62 0.771 1.047 83 Myrtaceae sp38 1 1 0.0855 0.15 0.62 0.771 1.047 84 Swartzia myrtifolia Sm. 2 2 0.026 0.3 0.19 0.489 1.042 85 Indeterminada sp6 1 1 0.0845 0.15 0.61 0.763 1.04 86 Prunus brasiliensis Dietrich 2 2 0.0246 0.3 0.18 0.479 1.032 87 Strychnos acuta Progel 1 1 0.0824 0.15 0.6 0.749 1.025 88 Hirtella hebeclada Moric. ex DC. 2 2 0.0226 0.3 0.16 0.465 1.017 89 Alchornea triplinervia (Spreng.) Müll. Arg. 2 1 0.0598 0.3 0.43 0.734 1.011 90 Heisteria sp3 2 2 0.0194 0.3 0.14 0.442 0.994 91 Byrsonima sp1 1 1 0.0769 0.15 0.56 0.709 0.985 92 Faramea pachyantha Müll. Arg. 2 2 0.0172 0.3 0.12 0.425 0.978 93 Myrcia laxiflora Cambess. 2 2 0.0163 0.3 0.12 0.419 0.972 94 Tapirira guianensis Aubl. 2 2 0.0154 0.3 0.11 0.412 0.965 95 Inga lanceifolia Benth. 2 2 0.0141 0.3 0.1 0.403 0.955 96 Cupania oblongifolia Mart. 2 2 0.0133 0.3 0.1 0.397 0.95 97 Chrysobalanaceae sp5 1 1 0.0716 0.15 0.52 0.67 0.946 98 Siphoneugena densiflora O.Berg 1 1 0.0707 0.15 0.51 0.663 0.939 99 Chrysophyllum flexuosum Mart. 2 2 0.0112 0.3 0.08 0.382 0.935
100 Daphnopsis martii Meisn. 2 2 0.0108 0.3 0.08 0.379 0.932
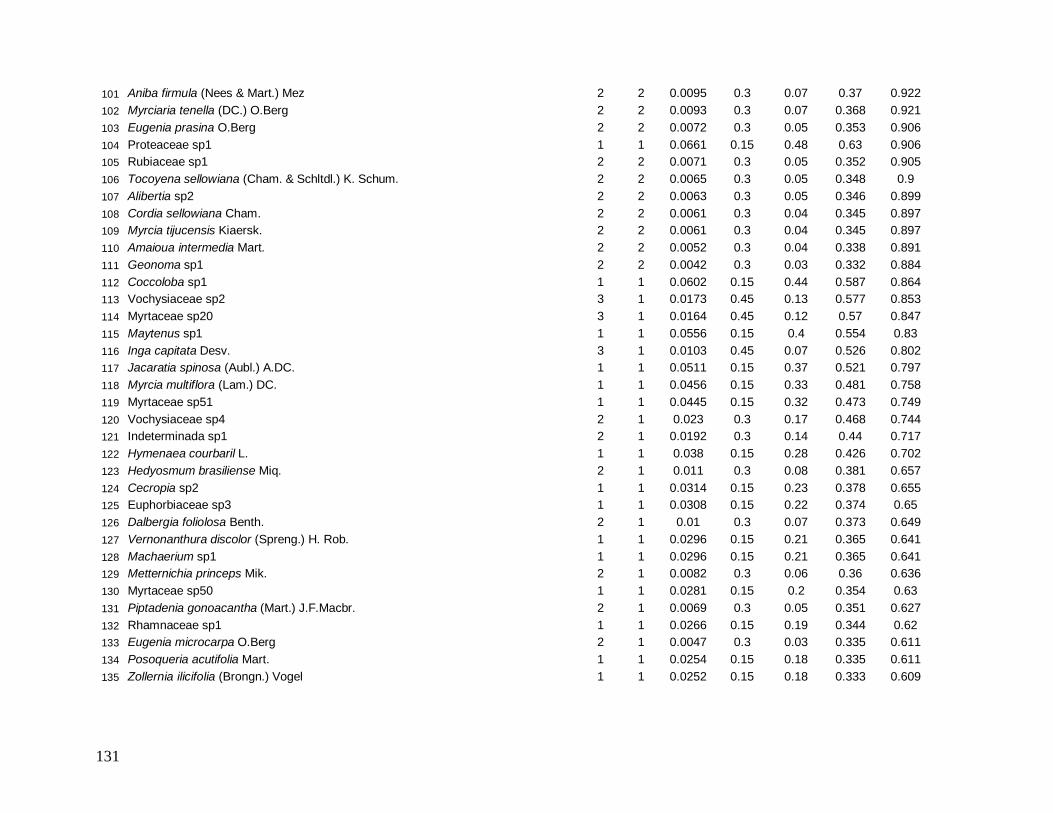
131
101 Aniba firmula (Nees & Mart.) Mez 2 2 0.0095 0.3 0.07 0.37 0.922 102 Myrciaria tenella (DC.) O.Berg 2 2 0.0093 0.3 0.07 0.368 0.921 103 Eugenia prasina O.Berg 2 2 0.0072 0.3 0.05 0.353 0.906 104 Proteaceae sp1 1 1 0.0661 0.15 0.48 0.63 0.906 105 Rubiaceae sp1 2 2 0.0071 0.3 0.05 0.352 0.905 106 Tocoyena sellowiana (Cham. & Schltdl.) K. Schum. 2 2 0.0065 0.3 0.05 0.348 0.9 107 Alibertia sp2 2 2 0.0063 0.3 0.05 0.346 0.899 108 Cordia sellowiana Cham. 2 2 0.0061 0.3 0.04 0.345 0.897 109 Myrcia tijucensis Kiaersk. 2 2 0.0061 0.3 0.04 0.345 0.897 110 Amaioua intermedia Mart. 2 2 0.0052 0.3 0.04 0.338 0.891 111 Geonoma sp1 2 2 0.0042 0.3 0.03 0.332 0.884 112 Coccoloba sp1 1 1 0.0602 0.15 0.44 0.587 0.864 113 Vochysiaceae sp2 3 1 0.0173 0.45 0.13 0.577 0.853 114 Myrtaceae sp20 3 1 0.0164 0.45 0.12 0.57 0.847 115 Maytenus sp1 1 1 0.0556 0.15 0.4 0.554 0.83 116 Inga capitata Desv. 3 1 0.0103 0.45 0.07 0.526 0.802 117 Jacaratia spinosa (Aubl.) A.DC. 1 1 0.0511 0.15 0.37 0.521 0.797 118 Myrcia multiflora (Lam.) DC. 1 1 0.0456 0.15 0.33 0.481 0.758 119 Myrtaceae sp51 1 1 0.0445 0.15 0.32 0.473 0.749 120 Vochysiaceae sp4 2 1 0.023 0.3 0.17 0.468 0.744 121 Indeterminada sp1 2 1 0.0192 0.3 0.14 0.44 0.717 122 Hymenaea courbaril L. 1 1 0.038 0.15 0.28 0.426 0.702 123 Hedyosmum brasiliense Miq. 2 1 0.011 0.3 0.08 0.381 0.657 124 Cecropia sp2 1 1 0.0314 0.15 0.23 0.378 0.655 125 Euphorbiaceae sp3 1 1 0.0308 0.15 0.22 0.374 0.65 126 Dalbergia foliolosa Benth. 2 1 0.01 0.3 0.07 0.373 0.649 127 Vernonanthura discolor (Spreng.) H. Rob. 1 1 0.0296 0.15 0.21 0.365 0.641 128 Machaerium sp1 1 1 0.0296 0.15 0.21 0.365 0.641 129 Metternichia princeps Mik. 2 1 0.0082 0.3 0.06 0.36 0.636 130 Myrtaceae sp50 1 1 0.0281 0.15 0.2 0.354 0.63 131 Piptadenia gonoacantha (Mart.) J.F.Macbr. 2 1 0.0069 0.3 0.05 0.351 0.627 132 Rhamnaceae sp1 1 1 0.0266 0.15 0.19 0.344 0.62 133 Eugenia microcarpa O.Berg 2 1 0.0047 0.3 0.03 0.335 0.611 134 Posoqueria acutifolia Mart. 1 1 0.0254 0.15 0.18 0.335 0.611 135 Zollernia ilicifolia (Brongn.) Vogel 1 1 0.0252 0.15 0.18 0.333 0.609
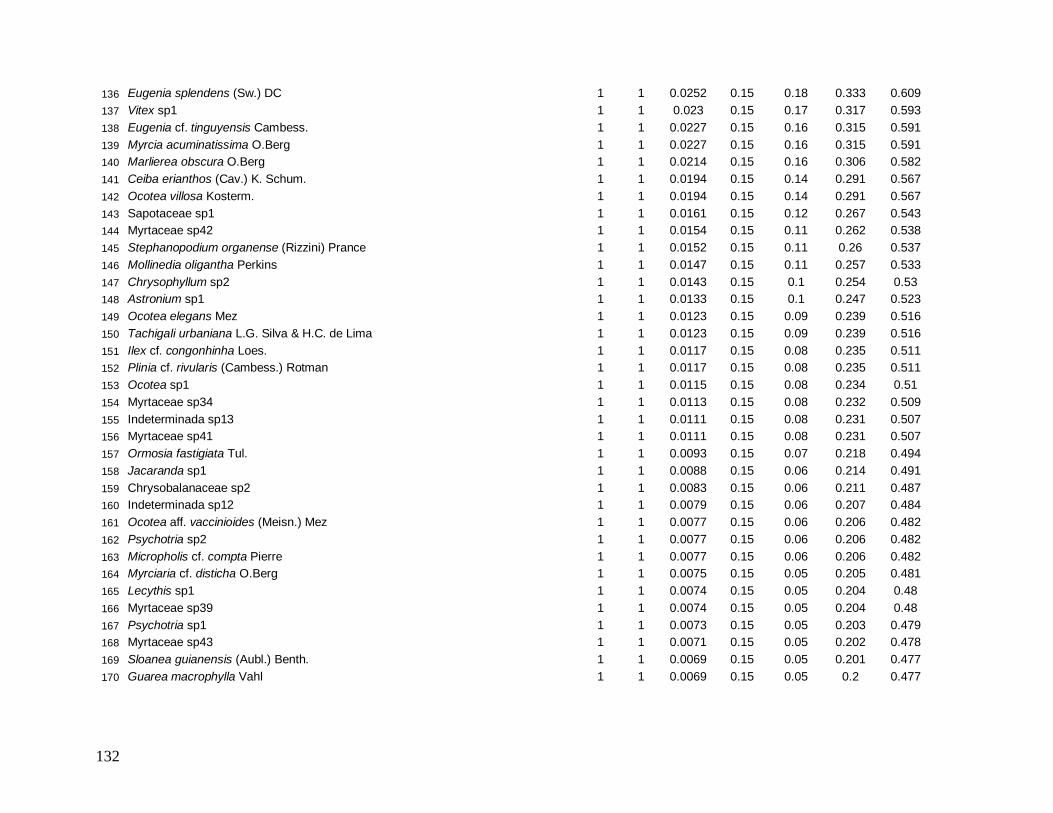
132
136 Eugenia splendens (Sw.) DC 1 1 0.0252 0.15 0.18 0.333 0.609 137 Vitex sp1 1 1 0.023 0.15 0.17 0.317 0.593 138 Eugenia cf. tinguyensis Cambess. 1 1 0.0227 0.15 0.16 0.315 0.591 139 Myrcia acuminatissima O.Berg 1 1 0.0227 0.15 0.16 0.315 0.591 140 Marlierea obscura O.Berg 1 1 0.0214 0.15 0.16 0.306 0.582 141 Ceiba erianthos (Cav.) K. Schum. 1 1 0.0194 0.15 0.14 0.291 0.567 142 Ocotea villosa Kosterm. 1 1 0.0194 0.15 0.14 0.291 0.567 143 Sapotaceae sp1 1 1 0.0161 0.15 0.12 0.267 0.543 144 Myrtaceae sp42 1 1 0.0154 0.15 0.11 0.262 0.538 145 Stephanopodium organense (Rizzini) Prance 1 1 0.0152 0.15 0.11 0.26 0.537 146 Mollinedia oligantha Perkins 1 1 0.0147 0.15 0.11 0.257 0.533 147 Chrysophyllum sp2 1 1 0.0143 0.15 0.1 0.254 0.53 148 Astronium sp1 1 1 0.0133 0.15 0.1 0.247 0.523 149 Ocotea elegans Mez 1 1 0.0123 0.15 0.09 0.239 0.516 150 Tachigali urbaniana L.G. Silva & H.C. de Lima 1 1 0.0123 0.15 0.09 0.239 0.516 151 Ilex cf. congonhinha Loes. 1 1 0.0117 0.15 0.08 0.235 0.511 152 Plinia cf. rivularis (Cambess.) Rotman 1 1 0.0117 0.15 0.08 0.235 0.511 153 Ocotea sp1 1 1 0.0115 0.15 0.08 0.234 0.51 154 Myrtaceae sp34 1 1 0.0113 0.15 0.08 0.232 0.509 155 Indeterminada sp13 1 1 0.0111 0.15 0.08 0.231 0.507 156 Myrtaceae sp41 1 1 0.0111 0.15 0.08 0.231 0.507 157 Ormosia fastigiata Tul. 1 1 0.0093 0.15 0.07 0.218 0.494 158 Jacaranda sp1 1 1 0.0088 0.15 0.06 0.214 0.491 159 Chrysobalanaceae sp2 1 1 0.0083 0.15 0.06 0.211 0.487 160 Indeterminada sp12 1 1 0.0079 0.15 0.06 0.207 0.484 161 Ocotea aff. vaccinioides (Meisn.) Mez 1 1 0.0077 0.15 0.06 0.206 0.482 162 Psychotria sp2 1 1 0.0077 0.15 0.06 0.206 0.482 163 Micropholis cf. compta Pierre 1 1 0.0077 0.15 0.06 0.206 0.482 164 Myrciaria cf. disticha O.Berg 1 1 0.0075 0.15 0.05 0.205 0.481 165 Lecythis sp1 1 1 0.0074 0.15 0.05 0.204 0.48 166 Myrtaceae sp39 1 1 0.0074 0.15 0.05 0.204 0.48 167 Psychotria sp1 1 1 0.0073 0.15 0.05 0.203 0.479 168 Myrtaceae sp43 1 1 0.0071 0.15 0.05 0.202 0.478 169 Sloanea guianensis (Aubl.) Benth. 1 1 0.0069 0.15 0.05 0.201 0.477 170 Guarea macrophylla Vahl 1 1 0.0069 0.15 0.05 0.2 0.477
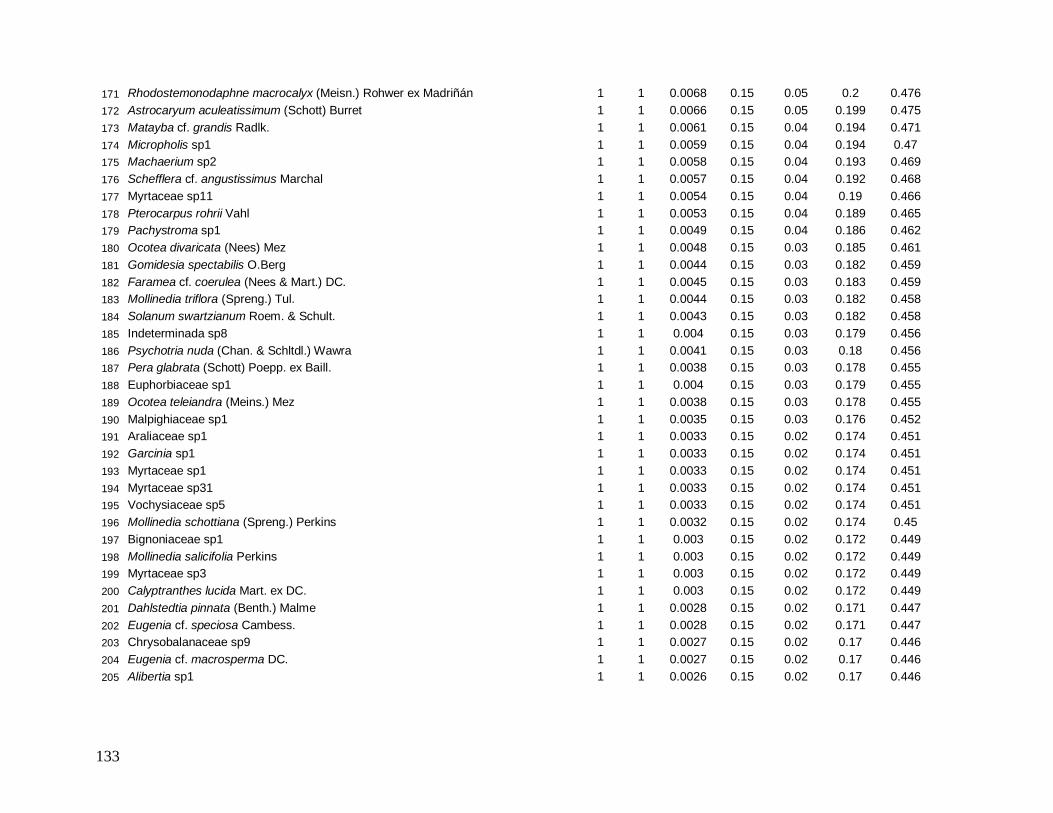
133
171 Rhodostemonodaphne macrocalyx (Meisn.) Rohwer ex Madriñán 1 1 0.0068 0.15 0.05 0.2 0.476 172 Astrocaryum aculeatissimum (Schott) Burret 1 1 0.0066 0.15 0.05 0.199 0.475 173 Matayba cf. grandis Radlk. 1 1 0.0061 0.15 0.04 0.194 0.471 174 Micropholis sp1 1 1 0.0059 0.15 0.04 0.194 0.47 175 Machaerium sp2 1 1 0.0058 0.15 0.04 0.193 0.469 176 Schefflera cf. angustissimus Marchal 1 1 0.0057 0.15 0.04 0.192 0.468 177 Myrtaceae sp11 1 1 0.0054 0.15 0.04 0.19 0.466 178 Pterocarpus rohrii Vahl 1 1 0.0053 0.15 0.04 0.189 0.465 179 Pachystroma sp1 1 1 0.0049 0.15 0.04 0.186 0.462 180 Ocotea divaricata (Nees) Mez 1 1 0.0048 0.15 0.03 0.185 0.461 181 Gomidesia spectabilis O.Berg 1 1 0.0044 0.15 0.03 0.182 0.459 182 Faramea cf. coerulea (Nees & Mart.) DC. 1 1 0.0045 0.15 0.03 0.183 0.459 183 Mollinedia triflora (Spreng.) Tul. 1 1 0.0044 0.15 0.03 0.182 0.458 184 Solanum swartzianum Roem. & Schult. 1 1 0.0043 0.15 0.03 0.182 0.458 185 Indeterminada sp8 1 1 0.004 0.15 0.03 0.179 0.456 186 Psychotria nuda (Chan. & Schltdl.) Wawra 1 1 0.0041 0.15 0.03 0.18 0.456 187 Pera glabrata (Schott) Poepp. ex Baill. 1 1 0.0038 0.15 0.03 0.178 0.455 188 Euphorbiaceae sp1 1 1 0.004 0.15 0.03 0.179 0.455 189 Ocotea teleiandra (Meins.) Mez 1 1 0.0038 0.15 0.03 0.178 0.455 190 Malpighiaceae sp1 1 1 0.0035 0.15 0.03 0.176 0.452 191 Araliaceae sp1 1 1 0.0033 0.15 0.02 0.174 0.451 192 Garcinia sp1 1 1 0.0033 0.15 0.02 0.174 0.451 193 Myrtaceae sp1 1 1 0.0033 0.15 0.02 0.174 0.451 194 Myrtaceae sp31 1 1 0.0033 0.15 0.02 0.174 0.451 195 Vochysiaceae sp5 1 1 0.0033 0.15 0.02 0.174 0.451 196 Mollinedia schottiana (Spreng.) Perkins 1 1 0.0032 0.15 0.02 0.174 0.45 197 Bignoniaceae sp1 1 1 0.003 0.15 0.02 0.172 0.449 198 Mollinedia salicifolia Perkins 1 1 0.003 0.15 0.02 0.172 0.449 199 Myrtaceae sp3 1 1 0.003 0.15 0.02 0.172 0.449 200 Calyptranthes lucida Mart. ex DC. 1 1 0.003 0.15 0.02 0.172 0.449 201 Dahlstedtia pinnata (Benth.) Malme 1 1 0.0028 0.15 0.02 0.171 0.447 202 Eugenia cf. speciosa Cambess. 1 1 0.0028 0.15 0.02 0.171 0.447 203 Chrysobalanaceae sp9 1 1 0.0027 0.15 0.02 0.17 0.446 204 Eugenia cf. macrosperma DC. 1 1 0.0027 0.15 0.02 0.17 0.446 205 Alibertia sp1 1 1 0.0026 0.15 0.02 0.17 0.446
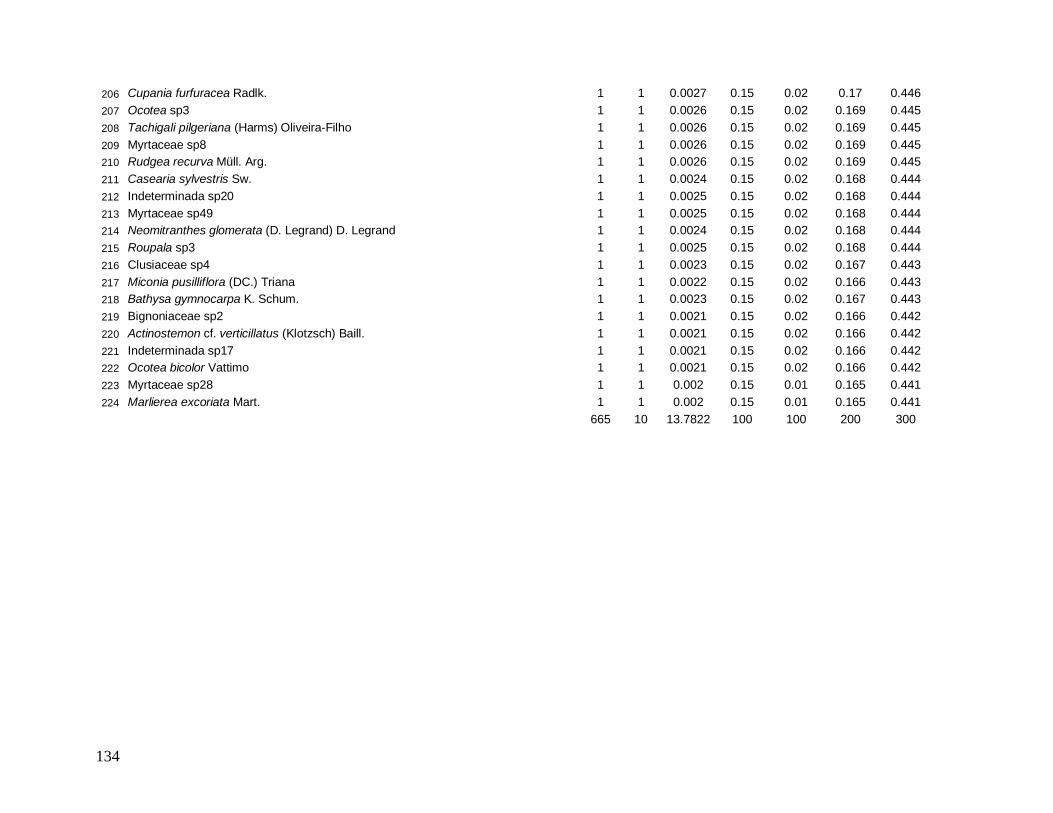
134
206 Cupania furfuracea Radlk. 1 1 0.0027 0.15 0.02 0.17 0.446 207 Ocotea sp3 1 1 0.0026 0.15 0.02 0.169 0.445 208 Tachigali pilgeriana (Harms) Oliveira-Filho 1 1 0.0026 0.15 0.02 0.169 0.445 209 Myrtaceae sp8 1 1 0.0026 0.15 0.02 0.169 0.445 210 Rudgea recurva Müll. Arg. 1 1 0.0026 0.15 0.02 0.169 0.445 211 Casearia sylvestris Sw. 1 1 0.0024 0.15 0.02 0.168 0.444 212 Indeterminada sp20 1 1 0.0025 0.15 0.02 0.168 0.444 213 Myrtaceae sp49 1 1 0.0025 0.15 0.02 0.168 0.444 214 Neomitranthes glomerata (D. Legrand) D. Legrand 1 1 0.0024 0.15 0.02 0.168 0.444 215 Roupala sp3 1 1 0.0025 0.15 0.02 0.168 0.444 216 Clusiaceae sp4 1 1 0.0023 0.15 0.02 0.167 0.443 217 Miconia pusilliflora (DC.) Triana 1 1 0.0022 0.15 0.02 0.166 0.443 218 Bathysa gymnocarpa K. Schum. 1 1 0.0023 0.15 0.02 0.167 0.443 219 Bignoniaceae sp2 1 1 0.0021 0.15 0.02 0.166 0.442 220 Actinostemon cf. verticillatus (Klotzsch) Baill. 1 1 0.0021 0.15 0.02 0.166 0.442 221 Indeterminada sp17 1 1 0.0021 0.15 0.02 0.166 0.442 222 Ocotea bicolor Vattimo 1 1 0.0021 0.15 0.02 0.166 0.442 223 Myrtaceae sp28 1 1 0.002 0.15 0.01 0.165 0.441 224 Marlierea excoriata Mart. 1 1 0.002 0.15 0.01 0.165 0.441
665 10 13.7822 100 100 200 300
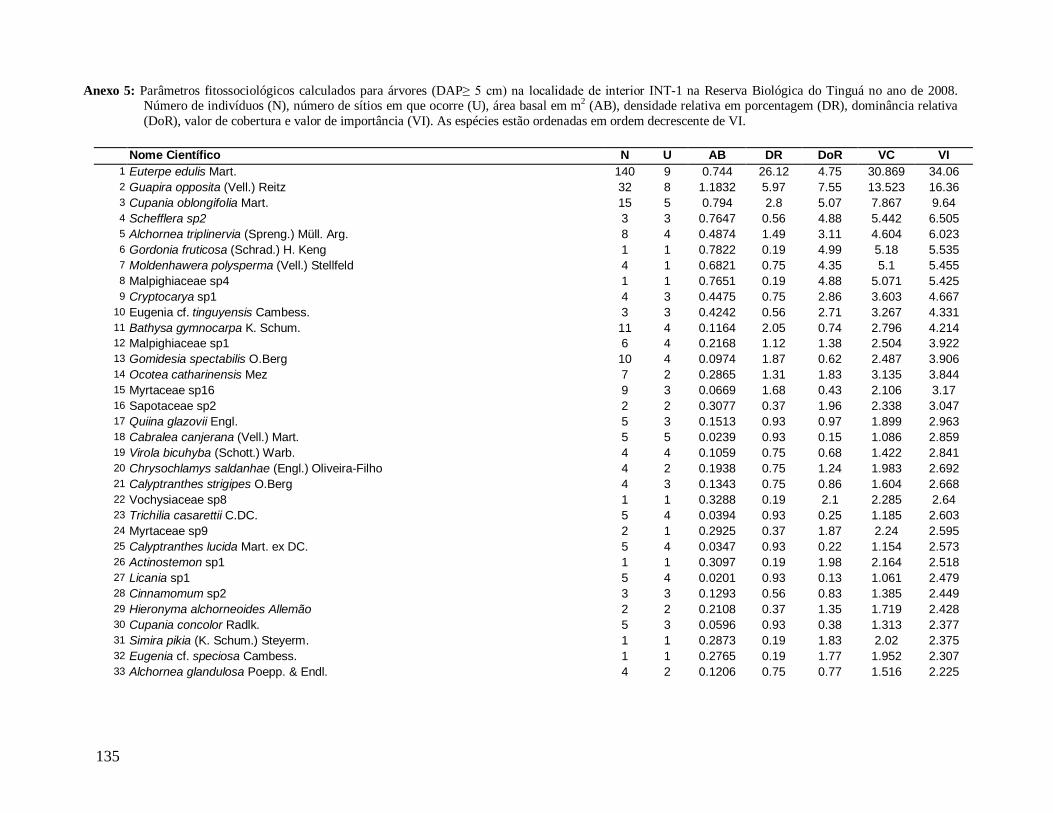
135
Anexo 5: Parâmetros fitossociológicos calculados para árvores (DAP≥ 5 cm) na localidade de interior INT-1 na Reserva Biológica do Tinguá no ano de 2008. Número de indivíduos (N), número de sítios em que ocorre (U), área basal em m2 (AB), densidade relativa em porcentagem (DR), dominância relativa (DoR), valor de cobertura e valor de importância (VI). As espécies estão ordenadas em ordem decrescente de VI.
Nome Científico N U AB DR DoR VC VI
1 Euterpe edulis Mart. 140 9 0.744 26.12 4.75 30.869 34.06 2 Guapira opposita (Vell.) Reitz 32 8 1.1832 5.97 7.55 13.523 16.36 3 Cupania oblongifolia Mart. 15 5 0.794 2.8 5.07 7.867 9.64 4 Schefflera sp2 3 3 0.7647 0.56 4.88 5.442 6.505 5 Alchornea triplinervia (Spreng.) Müll. Arg. 8 4 0.4874 1.49 3.11 4.604 6.023 6 Gordonia fruticosa (Schrad.) H. Keng 1 1 0.7822 0.19 4.99 5.18 5.535 7 Moldenhawera polysperma (Vell.) Stellfeld 4 1 0.6821 0.75 4.35 5.1 5.455 8 Malpighiaceae sp4 1 1 0.7651 0.19 4.88 5.071 5.425 9 Cryptocarya sp1 4 3 0.4475 0.75 2.86 3.603 4.667
10 Eugenia cf. tinguyensis Cambess. 3 3 0.4242 0.56 2.71 3.267 4.331 11 Bathysa gymnocarpa K. Schum. 11 4 0.1164 2.05 0.74 2.796 4.214 12 Malpighiaceae sp1 6 4 0.2168 1.12 1.38 2.504 3.922 13 Gomidesia spectabilis O.Berg 10 4 0.0974 1.87 0.62 2.487 3.906 14 Ocotea catharinensis Mez 7 2 0.2865 1.31 1.83 3.135 3.844 15 Myrtaceae sp16 9 3 0.0669 1.68 0.43 2.106 3.17 16 Sapotaceae sp2 2 2 0.3077 0.37 1.96 2.338 3.047 17 Quiina glazovii Engl. 5 3 0.1513 0.93 0.97 1.899 2.963 18 Cabralea canjerana (Vell.) Mart. 5 5 0.0239 0.93 0.15 1.086 2.859 19 Virola bicuhyba (Schott.) Warb. 4 4 0.1059 0.75 0.68 1.422 2.841 20 Chrysochlamys saldanhae (Engl.) Oliveira-Filho 4 2 0.1938 0.75 1.24 1.983 2.692 21 Calyptranthes strigipes O.Berg 4 3 0.1343 0.75 0.86 1.604 2.668 22 Vochysiaceae sp8 1 1 0.3288 0.19 2.1 2.285 2.64 23 Trichilia casarettii C.DC. 5 4 0.0394 0.93 0.25 1.185 2.603 24 Myrtaceae sp9 2 1 0.2925 0.37 1.87 2.24 2.595 25 Calyptranthes lucida Mart. ex DC. 5 4 0.0347 0.93 0.22 1.154 2.573 26 Actinostemon sp1 1 1 0.3097 0.19 1.98 2.164 2.518 27 Licania sp1 5 4 0.0201 0.93 0.13 1.061 2.479 28 Cinnamomum sp2 3 3 0.1293 0.56 0.83 1.385 2.449 29 Hieronyma alchorneoides Allemão 2 2 0.2108 0.37 1.35 1.719 2.428 30 Cupania concolor Radlk. 5 3 0.0596 0.93 0.38 1.313 2.377 31 Simira pikia (K. Schum.) Steyerm. 1 1 0.2873 0.19 1.83 2.02 2.375 32 Eugenia cf. speciosa Cambess. 1 1 0.2765 0.19 1.77 1.952 2.307 33 Alchornea glandulosa Poepp. & Endl. 4 2 0.1206 0.75 0.77 1.516 2.225
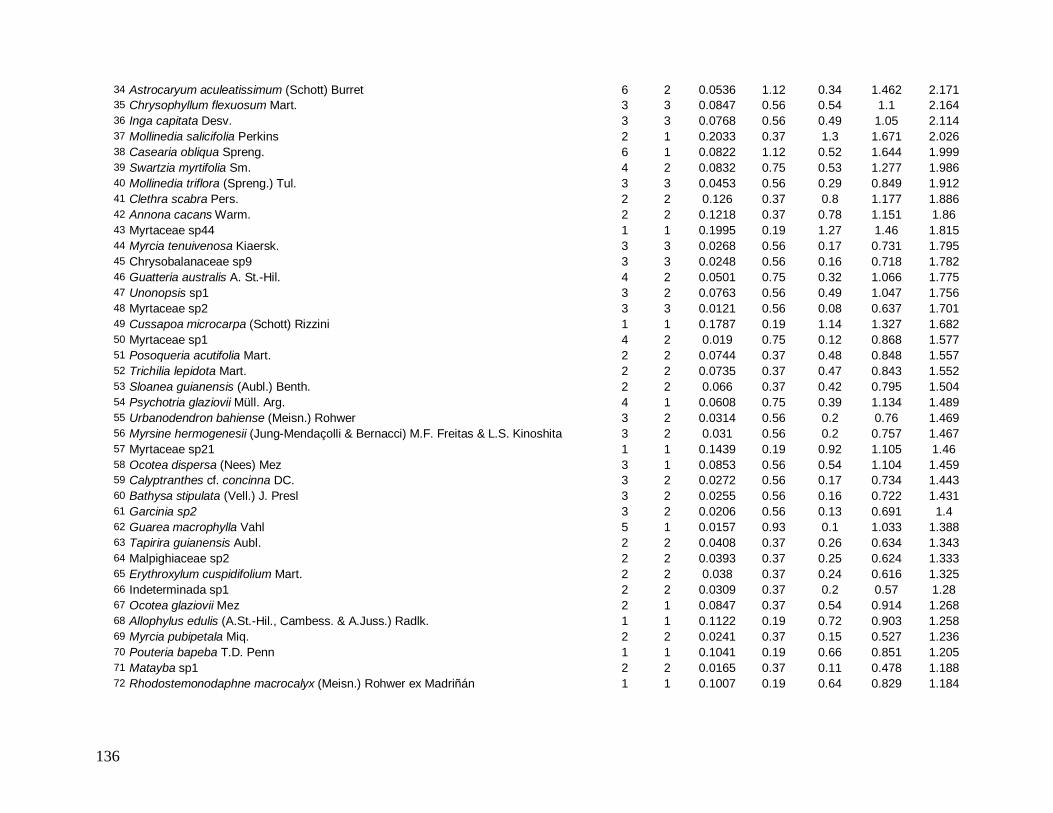
136
34 Astrocaryum aculeatissimum (Schott) Burret 6 2 0.0536 1.12 0.34 1.462 2.171 35 Chrysophyllum flexuosum Mart. 3 3 0.0847 0.56 0.54 1.1 2.164 36 Inga capitata Desv. 3 3 0.0768 0.56 0.49 1.05 2.114 37 Mollinedia salicifolia Perkins 2 1 0.2033 0.37 1.3 1.671 2.026 38 Casearia obliqua Spreng. 6 1 0.0822 1.12 0.52 1.644 1.999 39 Swartzia myrtifolia Sm. 4 2 0.0832 0.75 0.53 1.277 1.986 40 Mollinedia triflora (Spreng.) Tul. 3 3 0.0453 0.56 0.29 0.849 1.912 41 Clethra scabra Pers. 2 2 0.126 0.37 0.8 1.177 1.886 42 Annona cacans Warm. 2 2 0.1218 0.37 0.78 1.151 1.86 43 Myrtaceae sp44 1 1 0.1995 0.19 1.27 1.46 1.815 44 Myrcia tenuivenosa Kiaersk. 3 3 0.0268 0.56 0.17 0.731 1.795 45 Chrysobalanaceae sp9 3 3 0.0248 0.56 0.16 0.718 1.782 46 Guatteria australis A. St.-Hil. 4 2 0.0501 0.75 0.32 1.066 1.775 47 Unonopsis sp1 3 2 0.0763 0.56 0.49 1.047 1.756 48 Myrtaceae sp2 3 3 0.0121 0.56 0.08 0.637 1.701 49 Cussapoa microcarpa (Schott) Rizzini 1 1 0.1787 0.19 1.14 1.327 1.682 50 Myrtaceae sp1 4 2 0.019 0.75 0.12 0.868 1.577 51 Posoqueria acutifolia Mart. 2 2 0.0744 0.37 0.48 0.848 1.557 52 Trichilia lepidota Mart. 2 2 0.0735 0.37 0.47 0.843 1.552 53 Sloanea guianensis (Aubl.) Benth. 2 2 0.066 0.37 0.42 0.795 1.504 54 Psychotria glaziovii Müll. Arg. 4 1 0.0608 0.75 0.39 1.134 1.489 55 Urbanodendron bahiense (Meisn.) Rohwer 3 2 0.0314 0.56 0.2 0.76 1.469 56 Myrsine hermogenesii (Jung-Mendaçolli & Bernacci) M.F. Freitas & L.S. Kinoshita 3 2 0.031 0.56 0.2 0.757 1.467 57 Myrtaceae sp21 1 1 0.1439 0.19 0.92 1.105 1.46 58 Ocotea dispersa (Nees) Mez 3 1 0.0853 0.56 0.54 1.104 1.459 59 Calyptranthes cf. concinna DC. 3 2 0.0272 0.56 0.17 0.734 1.443 60 Bathysa stipulata (Vell.) J. Presl 3 2 0.0255 0.56 0.16 0.722 1.431 61 Garcinia sp2 3 2 0.0206 0.56 0.13 0.691 1.4 62 Guarea macrophylla Vahl 5 1 0.0157 0.93 0.1 1.033 1.388 63 Tapirira guianensis Aubl. 2 2 0.0408 0.37 0.26 0.634 1.343 64 Malpighiaceae sp2 2 2 0.0393 0.37 0.25 0.624 1.333 65 Erythroxylum cuspidifolium Mart. 2 2 0.038 0.37 0.24 0.616 1.325 66 Indeterminada sp1 2 2 0.0309 0.37 0.2 0.57 1.28 67 Ocotea glaziovii Mez 2 1 0.0847 0.37 0.54 0.914 1.268 68 Allophylus edulis (A.St.-Hil., Cambess. & A.Juss.) Radlk. 1 1 0.1122 0.19 0.72 0.903 1.258 69 Myrcia pubipetala Miq. 2 2 0.0241 0.37 0.15 0.527 1.236 70 Pouteria bapeba T.D. Penn 1 1 0.1041 0.19 0.66 0.851 1.205 71 Matayba sp1 2 2 0.0165 0.37 0.11 0.478 1.188 72 Rhodostemonodaphne macrocalyx (Meisn.) Rohwer ex Madriñán 1 1 0.1007 0.19 0.64 0.829 1.184
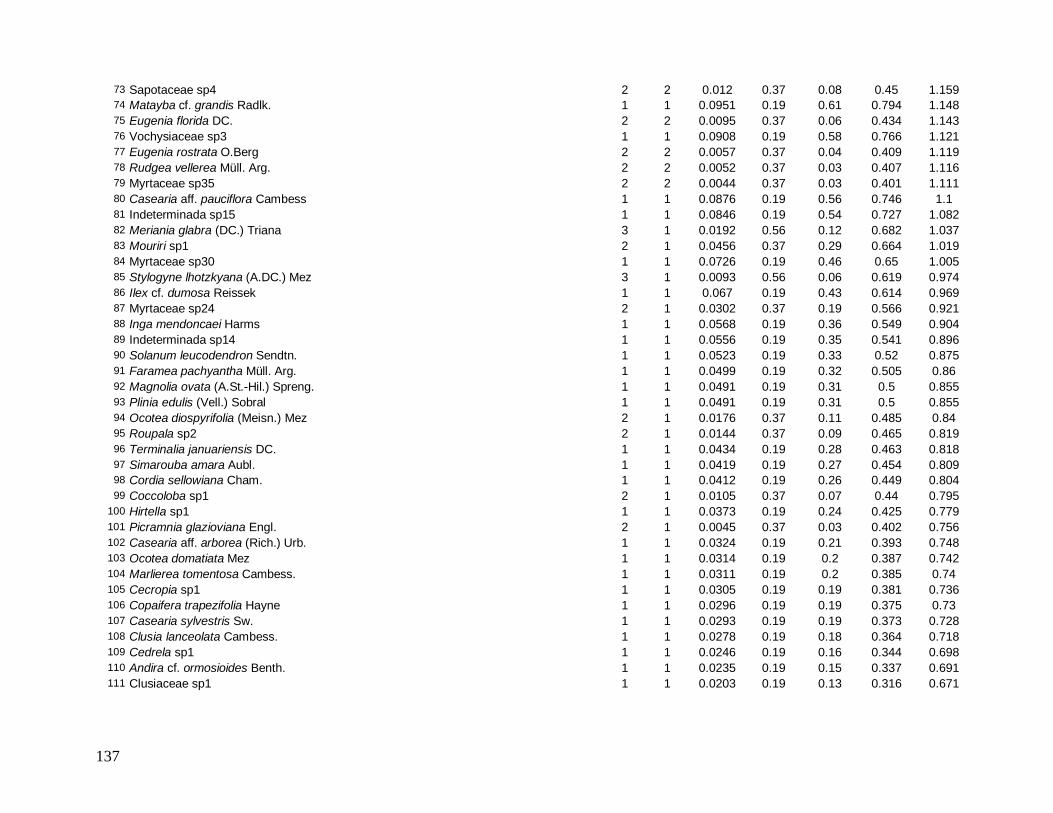
137
73 Sapotaceae sp4 2 2 0.012 0.37 0.08 0.45 1.159 74 Matayba cf. grandis Radlk. 1 1 0.0951 0.19 0.61 0.794 1.148 75 Eugenia florida DC. 2 2 0.0095 0.37 0.06 0.434 1.143 76 Vochysiaceae sp3 1 1 0.0908 0.19 0.58 0.766 1.121 77 Eugenia rostrata O.Berg 2 2 0.0057 0.37 0.04 0.409 1.119 78 Rudgea vellerea Müll. Arg. 2 2 0.0052 0.37 0.03 0.407 1.116 79 Myrtaceae sp35 2 2 0.0044 0.37 0.03 0.401 1.111 80 Casearia aff. pauciflora Cambess 1 1 0.0876 0.19 0.56 0.746 1.1 81 Indeterminada sp15 1 1 0.0846 0.19 0.54 0.727 1.082 82 Meriania glabra (DC.) Triana 3 1 0.0192 0.56 0.12 0.682 1.037 83 Mouriri sp1 2 1 0.0456 0.37 0.29 0.664 1.019 84 Myrtaceae sp30 1 1 0.0726 0.19 0.46 0.65 1.005 85 Stylogyne lhotzkyana (A.DC.) Mez 3 1 0.0093 0.56 0.06 0.619 0.974 86 Ilex cf. dumosa Reissek 1 1 0.067 0.19 0.43 0.614 0.969 87 Myrtaceae sp24 2 1 0.0302 0.37 0.19 0.566 0.921 88 Inga mendoncaei Harms 1 1 0.0568 0.19 0.36 0.549 0.904 89 Indeterminada sp14 1 1 0.0556 0.19 0.35 0.541 0.896 90 Solanum leucodendron Sendtn. 1 1 0.0523 0.19 0.33 0.52 0.875 91 Faramea pachyantha Müll. Arg. 1 1 0.0499 0.19 0.32 0.505 0.86 92 Magnolia ovata (A.St.-Hil.) Spreng. 1 1 0.0491 0.19 0.31 0.5 0.855 93 Plinia edulis (Vell.) Sobral 1 1 0.0491 0.19 0.31 0.5 0.855 94 Ocotea diospyrifolia (Meisn.) Mez 2 1 0.0176 0.37 0.11 0.485 0.84 95 Roupala sp2 2 1 0.0144 0.37 0.09 0.465 0.819 96 Terminalia januariensis DC. 1 1 0.0434 0.19 0.28 0.463 0.818 97 Simarouba amara Aubl. 1 1 0.0419 0.19 0.27 0.454 0.809 98 Cordia sellowiana Cham. 1 1 0.0412 0.19 0.26 0.449 0.804 99 Coccoloba sp1 2 1 0.0105 0.37 0.07 0.44 0.795
100 Hirtella sp1 1 1 0.0373 0.19 0.24 0.425 0.779 101 Picramnia glazioviana Engl. 2 1 0.0045 0.37 0.03 0.402 0.756 102 Casearia aff. arborea (Rich.) Urb. 1 1 0.0324 0.19 0.21 0.393 0.748 103 Ocotea domatiata Mez 1 1 0.0314 0.19 0.2 0.387 0.742 104 Marlierea tomentosa Cambess. 1 1 0.0311 0.19 0.2 0.385 0.74 105 Cecropia sp1 1 1 0.0305 0.19 0.19 0.381 0.736 106 Copaifera trapezifolia Hayne 1 1 0.0296 0.19 0.19 0.375 0.73 107 Casearia sylvestris Sw. 1 1 0.0293 0.19 0.19 0.373 0.728 108 Clusia lanceolata Cambess. 1 1 0.0278 0.19 0.18 0.364 0.718 109 Cedrela sp1 1 1 0.0246 0.19 0.16 0.344 0.698 110 Andira cf. ormosioides Benth. 1 1 0.0235 0.19 0.15 0.337 0.691 111 Clusiaceae sp1 1 1 0.0203 0.19 0.13 0.316 0.671
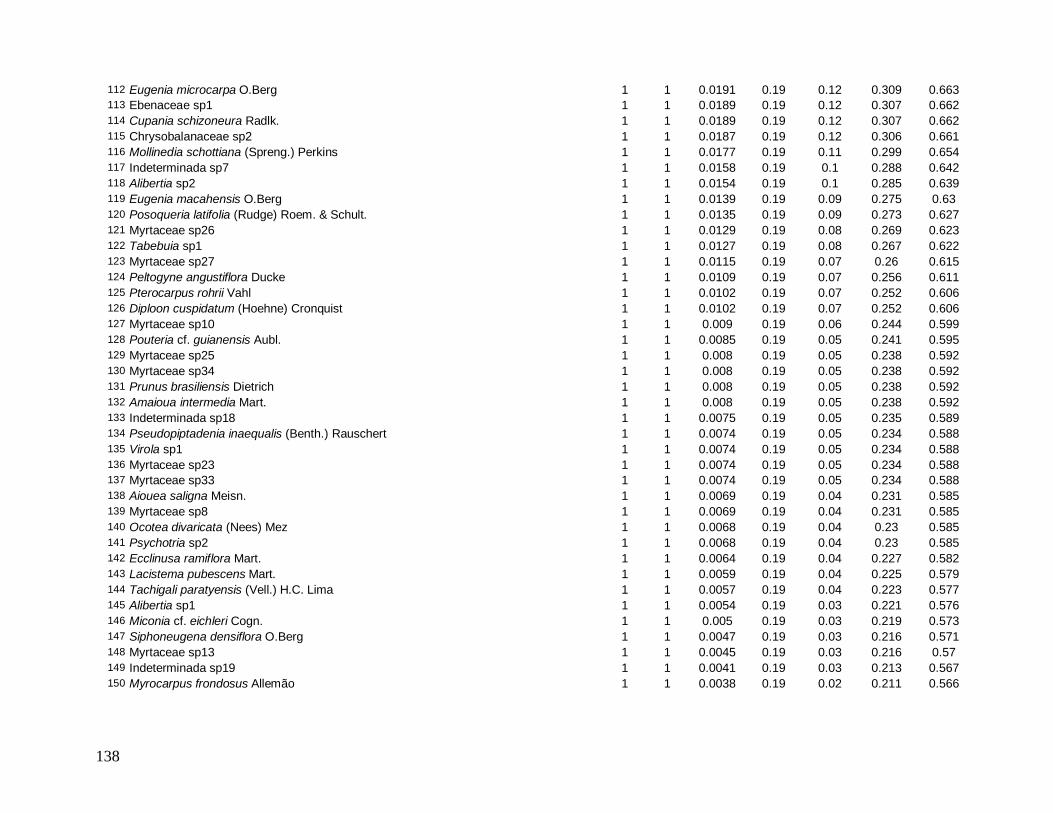
138
112 Eugenia microcarpa O.Berg 1 1 0.0191 0.19 0.12 0.309 0.663 113 Ebenaceae sp1 1 1 0.0189 0.19 0.12 0.307 0.662 114 Cupania schizoneura Radlk. 1 1 0.0189 0.19 0.12 0.307 0.662 115 Chrysobalanaceae sp2 1 1 0.0187 0.19 0.12 0.306 0.661 116 Mollinedia schottiana (Spreng.) Perkins 1 1 0.0177 0.19 0.11 0.299 0.654 117 Indeterminada sp7 1 1 0.0158 0.19 0.1 0.288 0.642 118 Alibertia sp2 1 1 0.0154 0.19 0.1 0.285 0.639 119 Eugenia macahensis O.Berg 1 1 0.0139 0.19 0.09 0.275 0.63 120 Posoqueria latifolia (Rudge) Roem. & Schult. 1 1 0.0135 0.19 0.09 0.273 0.627 121 Myrtaceae sp26 1 1 0.0129 0.19 0.08 0.269 0.623 122 Tabebuia sp1 1 1 0.0127 0.19 0.08 0.267 0.622 123 Myrtaceae sp27 1 1 0.0115 0.19 0.07 0.26 0.615 124 Peltogyne angustiflora Ducke 1 1 0.0109 0.19 0.07 0.256 0.611 125 Pterocarpus rohrii Vahl 1 1 0.0102 0.19 0.07 0.252 0.606 126 Diploon cuspidatum (Hoehne) Cronquist 1 1 0.0102 0.19 0.07 0.252 0.606 127 Myrtaceae sp10 1 1 0.009 0.19 0.06 0.244 0.599 128 Pouteria cf. guianensis Aubl. 1 1 0.0085 0.19 0.05 0.241 0.595 129 Myrtaceae sp25 1 1 0.008 0.19 0.05 0.238 0.592 130 Myrtaceae sp34 1 1 0.008 0.19 0.05 0.238 0.592 131 Prunus brasiliensis Dietrich 1 1 0.008 0.19 0.05 0.238 0.592 132 Amaioua intermedia Mart. 1 1 0.008 0.19 0.05 0.238 0.592 133 Indeterminada sp18 1 1 0.0075 0.19 0.05 0.235 0.589 134 Pseudopiptadenia inaequalis (Benth.) Rauschert 1 1 0.0074 0.19 0.05 0.234 0.588 135 Virola sp1 1 1 0.0074 0.19 0.05 0.234 0.588 136 Myrtaceae sp23 1 1 0.0074 0.19 0.05 0.234 0.588 137 Myrtaceae sp33 1 1 0.0074 0.19 0.05 0.234 0.588 138 Aiouea saligna Meisn. 1 1 0.0069 0.19 0.04 0.231 0.585 139 Myrtaceae sp8 1 1 0.0069 0.19 0.04 0.231 0.585 140 Ocotea divaricata (Nees) Mez 1 1 0.0068 0.19 0.04 0.23 0.585 141 Psychotria sp2 1 1 0.0068 0.19 0.04 0.23 0.585 142 Ecclinusa ramiflora Mart. 1 1 0.0064 0.19 0.04 0.227 0.582 143 Lacistema pubescens Mart. 1 1 0.0059 0.19 0.04 0.225 0.579 144 Tachigali paratyensis (Vell.) H.C. Lima 1 1 0.0057 0.19 0.04 0.223 0.577 145 Alibertia sp1 1 1 0.0054 0.19 0.03 0.221 0.576 146 Miconia cf. eichleri Cogn. 1 1 0.005 0.19 0.03 0.219 0.573 147 Siphoneugena densiflora O.Berg 1 1 0.0047 0.19 0.03 0.216 0.571 148 Myrtaceae sp13 1 1 0.0045 0.19 0.03 0.216 0.57 149 Indeterminada sp19 1 1 0.0041 0.19 0.03 0.213 0.567 150 Myrocarpus frondosus Allemão 1 1 0.0038 0.19 0.02 0.211 0.566
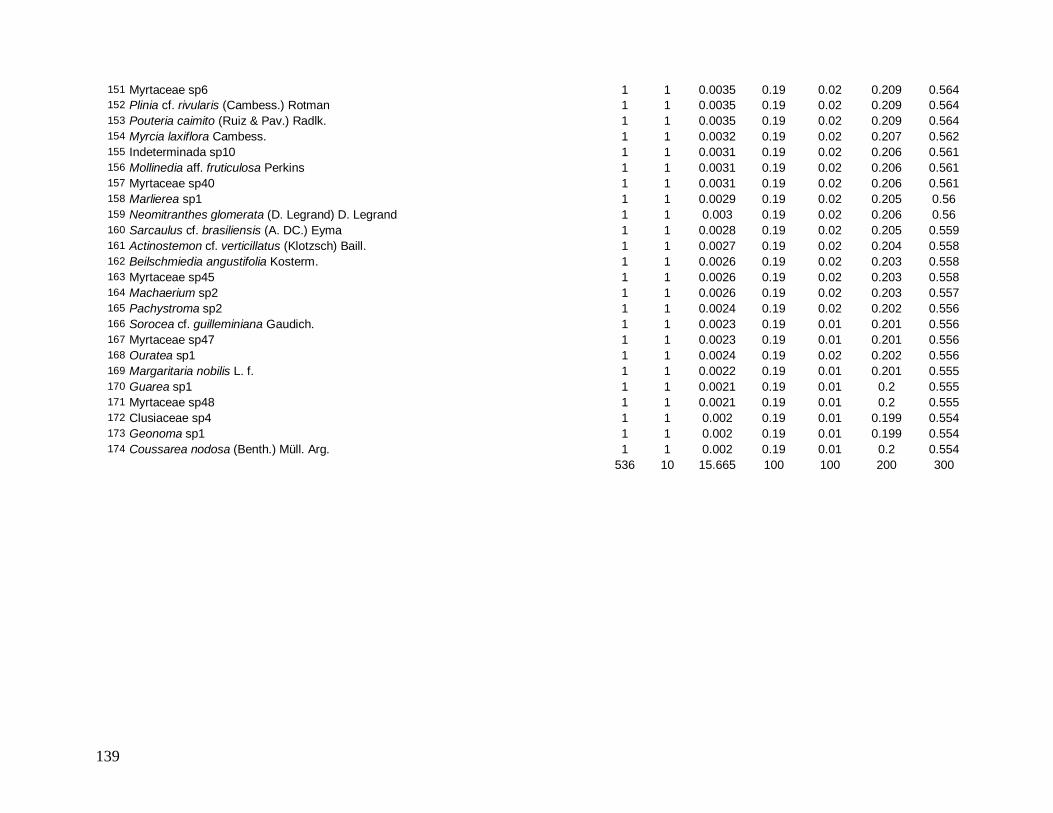
139
151 Myrtaceae sp6 1 1 0.0035 0.19 0.02 0.209 0.564 152 Plinia cf. rivularis (Cambess.) Rotman 1 1 0.0035 0.19 0.02 0.209 0.564 153 Pouteria caimito (Ruiz & Pav.) Radlk. 1 1 0.0035 0.19 0.02 0.209 0.564 154 Myrcia laxiflora Cambess. 1 1 0.0032 0.19 0.02 0.207 0.562 155 Indeterminada sp10 1 1 0.0031 0.19 0.02 0.206 0.561 156 Mollinedia aff. fruticulosa Perkins 1 1 0.0031 0.19 0.02 0.206 0.561 157 Myrtaceae sp40 1 1 0.0031 0.19 0.02 0.206 0.561 158 Marlierea sp1 1 1 0.0029 0.19 0.02 0.205 0.56 159 Neomitranthes glomerata (D. Legrand) D. Legrand 1 1 0.003 0.19 0.02 0.206 0.56 160 Sarcaulus cf. brasiliensis (A. DC.) Eyma 1 1 0.0028 0.19 0.02 0.205 0.559 161 Actinostemon cf. verticillatus (Klotzsch) Baill. 1 1 0.0027 0.19 0.02 0.204 0.558 162 Beilschmiedia angustifolia Kosterm. 1 1 0.0026 0.19 0.02 0.203 0.558 163 Myrtaceae sp45 1 1 0.0026 0.19 0.02 0.203 0.558 164 Machaerium sp2 1 1 0.0026 0.19 0.02 0.203 0.557 165 Pachystroma sp2 1 1 0.0024 0.19 0.02 0.202 0.556 166 Sorocea cf. guilleminiana Gaudich. 1 1 0.0023 0.19 0.01 0.201 0.556 167 Myrtaceae sp47 1 1 0.0023 0.19 0.01 0.201 0.556 168 Ouratea sp1 1 1 0.0024 0.19 0.02 0.202 0.556 169 Margaritaria nobilis L. f. 1 1 0.0022 0.19 0.01 0.201 0.555 170 Guarea sp1 1 1 0.0021 0.19 0.01 0.2 0.555 171 Myrtaceae sp48 1 1 0.0021 0.19 0.01 0.2 0.555 172 Clusiaceae sp4 1 1 0.002 0.19 0.01 0.199 0.554 173 Geonoma sp1 1 1 0.002 0.19 0.01 0.199 0.554 174 Coussarea nodosa (Benth.) Müll. Arg. 1 1 0.002 0.19 0.01 0.2 0.554
536 10 15.665 100 100 200 300
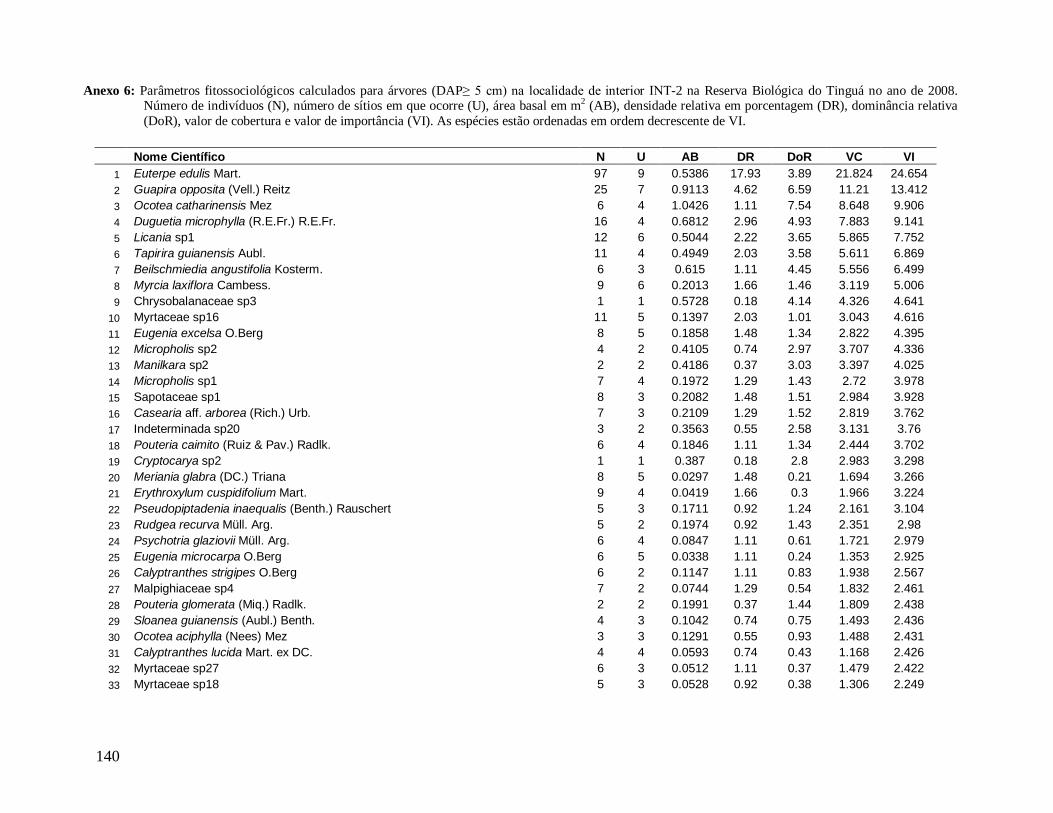
140
Anexo 6: Parâmetros fitossociológicos calculados para árvores (DAP≥ 5 cm) na localidade de interior INT-2 na Reserva Biológica do Tinguá no ano de 2008. Número de indivíduos (N), número de sítios em que ocorre (U), área basal em m2 (AB), densidade relativa em porcentagem (DR), dominância relativa (DoR), valor de cobertura e valor de importância (VI). As espécies estão ordenadas em ordem decrescente de VI.
Nome Científico N U AB DR DoR VC VI
1 Euterpe edulis Mart. 97 9 0.5386 17.93 3.89 21.824 24.654 2 Guapira opposita (Vell.) Reitz 25 7 0.9113 4.62 6.59 11.21 13.412 3 Ocotea catharinensis Mez 6 4 1.0426 1.11 7.54 8.648 9.906 4 Duguetia microphylla (R.E.Fr.) R.E.Fr. 16 4 0.6812 2.96 4.93 7.883 9.141 5 Licania sp1 12 6 0.5044 2.22 3.65 5.865 7.752 6 Tapirira guianensis Aubl. 11 4 0.4949 2.03 3.58 5.611 6.869 7 Beilschmiedia angustifolia Kosterm. 6 3 0.615 1.11 4.45 5.556 6.499 8 Myrcia laxiflora Cambess. 9 6 0.2013 1.66 1.46 3.119 5.006 9 Chrysobalanaceae sp3 1 1 0.5728 0.18 4.14 4.326 4.641
10 Myrtaceae sp16 11 5 0.1397 2.03 1.01 3.043 4.616 11 Eugenia excelsa O.Berg 8 5 0.1858 1.48 1.34 2.822 4.395 12 Micropholis sp2 4 2 0.4105 0.74 2.97 3.707 4.336 13 Manilkara sp2 2 2 0.4186 0.37 3.03 3.397 4.025 14 Micropholis sp1 7 4 0.1972 1.29 1.43 2.72 3.978 15 Sapotaceae sp1 8 3 0.2082 1.48 1.51 2.984 3.928 16 Casearia aff. arborea (Rich.) Urb. 7 3 0.2109 1.29 1.52 2.819 3.762 17 Indeterminada sp20 3 2 0.3563 0.55 2.58 3.131 3.76 18 Pouteria caimito (Ruiz & Pav.) Radlk. 6 4 0.1846 1.11 1.34 2.444 3.702 19 Cryptocarya sp2 1 1 0.387 0.18 2.8 2.983 3.298 20 Meriania glabra (DC.) Triana 8 5 0.0297 1.48 0.21 1.694 3.266 21 Erythroxylum cuspidifolium Mart. 9 4 0.0419 1.66 0.3 1.966 3.224 22 Pseudopiptadenia inaequalis (Benth.) Rauschert 5 3 0.1711 0.92 1.24 2.161 3.104 23 Rudgea recurva Müll. Arg. 5 2 0.1974 0.92 1.43 2.351 2.98 24 Psychotria glaziovii Müll. Arg. 6 4 0.0847 1.11 0.61 1.721 2.979 25 Eugenia microcarpa O.Berg 6 5 0.0338 1.11 0.24 1.353 2.925 26 Calyptranthes strigipes O.Berg 6 2 0.1147 1.11 0.83 1.938 2.567 27 Malpighiaceae sp4 7 2 0.0744 1.29 0.54 1.832 2.461 28 Pouteria glomerata (Miq.) Radlk. 2 2 0.1991 0.37 1.44 1.809 2.438 29 Sloanea guianensis (Aubl.) Benth. 4 3 0.1042 0.74 0.75 1.493 2.436 30 Ocotea aciphylla (Nees) Mez 3 3 0.1291 0.55 0.93 1.488 2.431 31 Calyptranthes lucida Mart. ex DC. 4 4 0.0593 0.74 0.43 1.168 2.426 32 Myrtaceae sp27 6 3 0.0512 1.11 0.37 1.479 2.422 33 Myrtaceae sp18 5 3 0.0528 0.92 0.38 1.306 2.249
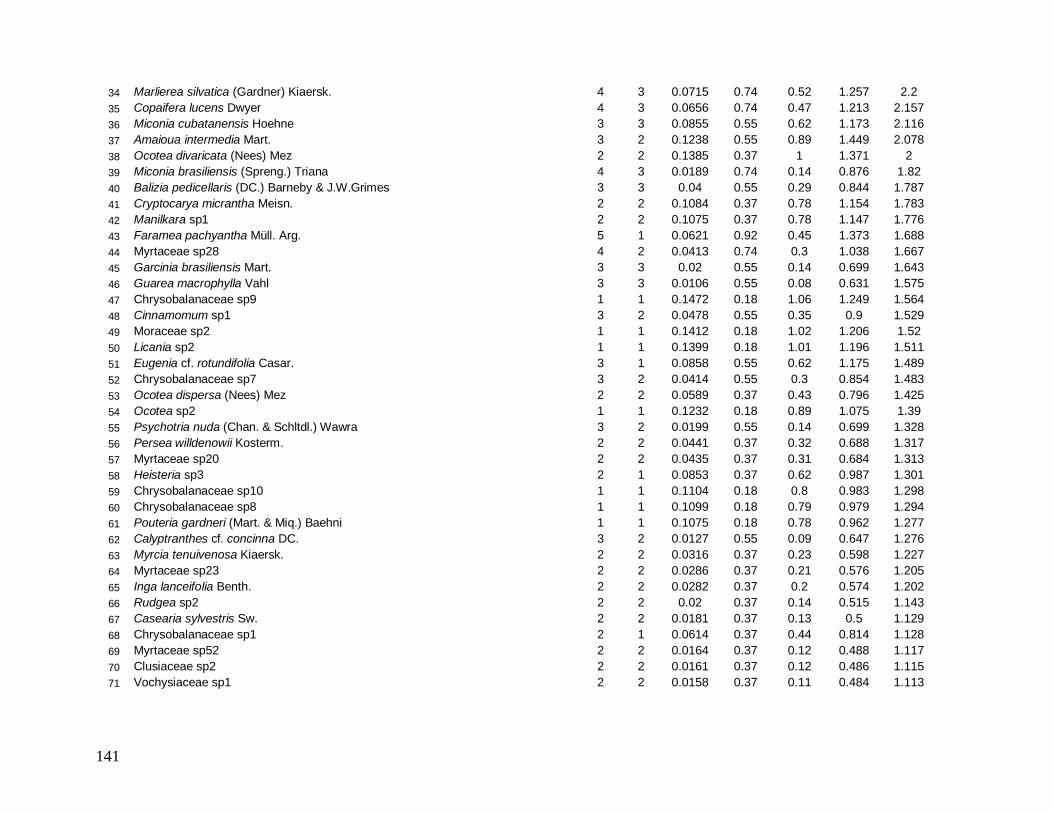
141
34 Marlierea silvatica (Gardner) Kiaersk. 4 3 0.0715 0.74 0.52 1.257 2.2 35 Copaifera lucens Dwyer 4 3 0.0656 0.74 0.47 1.213 2.157 36 Miconia cubatanensis Hoehne 3 3 0.0855 0.55 0.62 1.173 2.116 37 Amaioua intermedia Mart. 3 2 0.1238 0.55 0.89 1.449 2.078 38 Ocotea divaricata (Nees) Mez 2 2 0.1385 0.37 1 1.371 2 39 Miconia brasiliensis (Spreng.) Triana 4 3 0.0189 0.74 0.14 0.876 1.82 40 Balizia pedicellaris (DC.) Barneby & J.W.Grimes 3 3 0.04 0.55 0.29 0.844 1.787 41 Cryptocarya micrantha Meisn. 2 2 0.1084 0.37 0.78 1.154 1.783 42 Manilkara sp1 2 2 0.1075 0.37 0.78 1.147 1.776 43 Faramea pachyantha Müll. Arg. 5 1 0.0621 0.92 0.45 1.373 1.688 44 Myrtaceae sp28 4 2 0.0413 0.74 0.3 1.038 1.667 45 Garcinia brasiliensis Mart. 3 3 0.02 0.55 0.14 0.699 1.643 46 Guarea macrophylla Vahl 3 3 0.0106 0.55 0.08 0.631 1.575 47 Chrysobalanaceae sp9 1 1 0.1472 0.18 1.06 1.249 1.564 48 Cinnamomum sp1 3 2 0.0478 0.55 0.35 0.9 1.529 49 Moraceae sp2 1 1 0.1412 0.18 1.02 1.206 1.52 50 Licania sp2 1 1 0.1399 0.18 1.01 1.196 1.511 51 Eugenia cf. rotundifolia Casar. 3 1 0.0858 0.55 0.62 1.175 1.489 52 Chrysobalanaceae sp7 3 2 0.0414 0.55 0.3 0.854 1.483 53 Ocotea dispersa (Nees) Mez 2 2 0.0589 0.37 0.43 0.796 1.425 54 Ocotea sp2 1 1 0.1232 0.18 0.89 1.075 1.39 55 Psychotria nuda (Chan. & Schltdl.) Wawra 3 2 0.0199 0.55 0.14 0.699 1.328 56 Persea willdenowii Kosterm. 2 2 0.0441 0.37 0.32 0.688 1.317 57 Myrtaceae sp20 2 2 0.0435 0.37 0.31 0.684 1.313 58 Heisteria sp3 2 1 0.0853 0.37 0.62 0.987 1.301 59 Chrysobalanaceae sp10 1 1 0.1104 0.18 0.8 0.983 1.298 60 Chrysobalanaceae sp8 1 1 0.1099 0.18 0.79 0.979 1.294 61 Pouteria gardneri (Mart. & Miq.) Baehni 1 1 0.1075 0.18 0.78 0.962 1.277 62 Calyptranthes cf. concinna DC. 3 2 0.0127 0.55 0.09 0.647 1.276 63 Myrcia tenuivenosa Kiaersk. 2 2 0.0316 0.37 0.23 0.598 1.227 64 Myrtaceae sp23 2 2 0.0286 0.37 0.21 0.576 1.205 65 Inga lanceifolia Benth. 2 2 0.0282 0.37 0.2 0.574 1.202 66 Rudgea sp2 2 2 0.02 0.37 0.14 0.515 1.143 67 Casearia sylvestris Sw. 2 2 0.0181 0.37 0.13 0.5 1.129 68 Chrysobalanaceae sp1 2 1 0.0614 0.37 0.44 0.814 1.128 69 Myrtaceae sp52 2 2 0.0164 0.37 0.12 0.488 1.117 70 Clusiaceae sp2 2 2 0.0161 0.37 0.12 0.486 1.115 71 Vochysiaceae sp1 2 2 0.0158 0.37 0.11 0.484 1.113
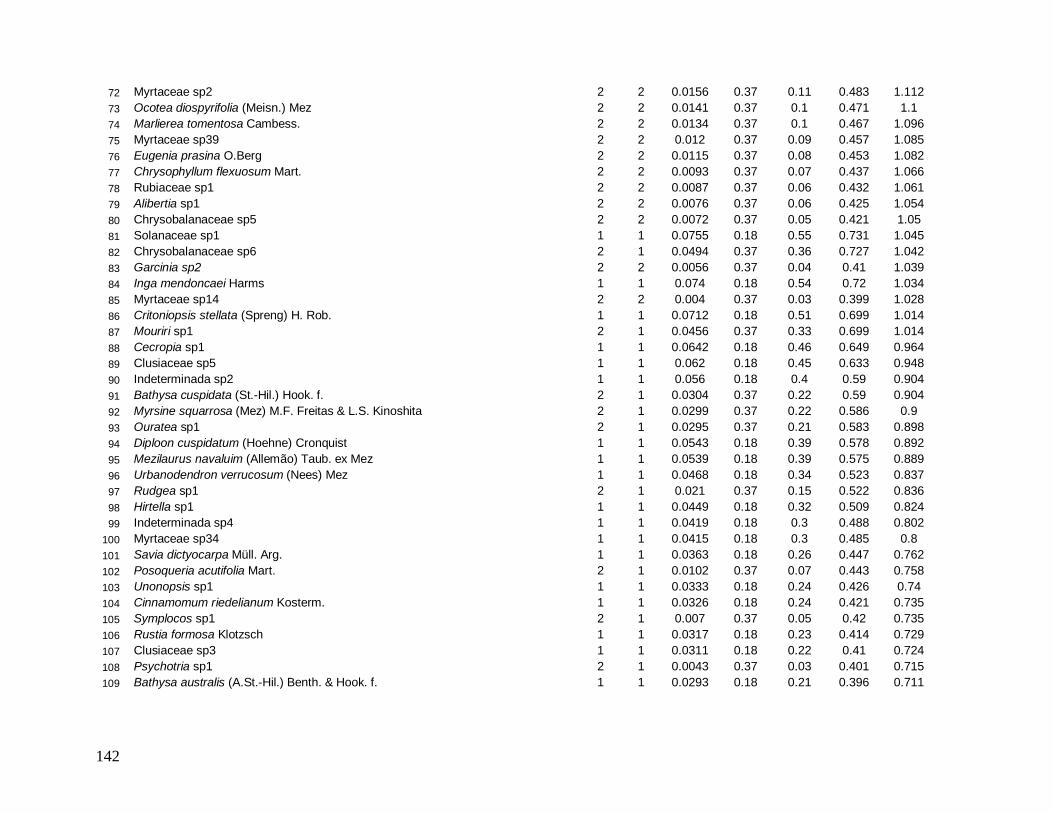
142
72 Myrtaceae sp2 2 2 0.0156 0.37 0.11 0.483 1.112 73 Ocotea diospyrifolia (Meisn.) Mez 2 2 0.0141 0.37 0.1 0.471 1.1 74 Marlierea tomentosa Cambess. 2 2 0.0134 0.37 0.1 0.467 1.096 75 Myrtaceae sp39 2 2 0.012 0.37 0.09 0.457 1.085 76 Eugenia prasina O.Berg 2 2 0.0115 0.37 0.08 0.453 1.082 77 Chrysophyllum flexuosum Mart. 2 2 0.0093 0.37 0.07 0.437 1.066 78 Rubiaceae sp1 2 2 0.0087 0.37 0.06 0.432 1.061 79 Alibertia sp1 2 2 0.0076 0.37 0.06 0.425 1.054 80 Chrysobalanaceae sp5 2 2 0.0072 0.37 0.05 0.421 1.05 81 Solanaceae sp1 1 1 0.0755 0.18 0.55 0.731 1.045 82 Chrysobalanaceae sp6 2 1 0.0494 0.37 0.36 0.727 1.042 83 Garcinia sp2 2 2 0.0056 0.37 0.04 0.41 1.039 84 Inga mendoncaei Harms 1 1 0.074 0.18 0.54 0.72 1.034 85 Myrtaceae sp14 2 2 0.004 0.37 0.03 0.399 1.028 86 Critoniopsis stellata (Spreng) H. Rob. 1 1 0.0712 0.18 0.51 0.699 1.014 87 Mouriri sp1 2 1 0.0456 0.37 0.33 0.699 1.014 88 Cecropia sp1 1 1 0.0642 0.18 0.46 0.649 0.964 89 Clusiaceae sp5 1 1 0.062 0.18 0.45 0.633 0.948 90 Indeterminada sp2 1 1 0.056 0.18 0.4 0.59 0.904 91 Bathysa cuspidata (St.-Hil.) Hook. f. 2 1 0.0304 0.37 0.22 0.59 0.904 92 Myrsine squarrosa (Mez) M.F. Freitas & L.S. Kinoshita 2 1 0.0299 0.37 0.22 0.586 0.9 93 Ouratea sp1 2 1 0.0295 0.37 0.21 0.583 0.898 94 Diploon cuspidatum (Hoehne) Cronquist 1 1 0.0543 0.18 0.39 0.578 0.892 95 Mezilaurus navaluim (Allemão) Taub. ex Mez 1 1 0.0539 0.18 0.39 0.575 0.889 96 Urbanodendron verrucosum (Nees) Mez 1 1 0.0468 0.18 0.34 0.523 0.837 97 Rudgea sp1 2 1 0.021 0.37 0.15 0.522 0.836 98 Hirtella sp1 1 1 0.0449 0.18 0.32 0.509 0.824 99 Indeterminada sp4 1 1 0.0419 0.18 0.3 0.488 0.802
100 Myrtaceae sp34 1 1 0.0415 0.18 0.3 0.485 0.8 101 Savia dictyocarpa Müll. Arg. 1 1 0.0363 0.18 0.26 0.447 0.762 102 Posoqueria acutifolia Mart. 2 1 0.0102 0.37 0.07 0.443 0.758 103 Unonopsis sp1 1 1 0.0333 0.18 0.24 0.426 0.74 104 Cinnamomum riedelianum Kosterm. 1 1 0.0326 0.18 0.24 0.421 0.735 105 Symplocos sp1 2 1 0.007 0.37 0.05 0.42 0.735 106 Rustia formosa Klotzsch 1 1 0.0317 0.18 0.23 0.414 0.729 107 Clusiaceae sp3 1 1 0.0311 0.18 0.22 0.41 0.724 108 Psychotria sp1 2 1 0.0043 0.37 0.03 0.401 0.715 109 Bathysa australis (A.St.-Hil.) Benth. & Hook. f. 1 1 0.0293 0.18 0.21 0.396 0.711
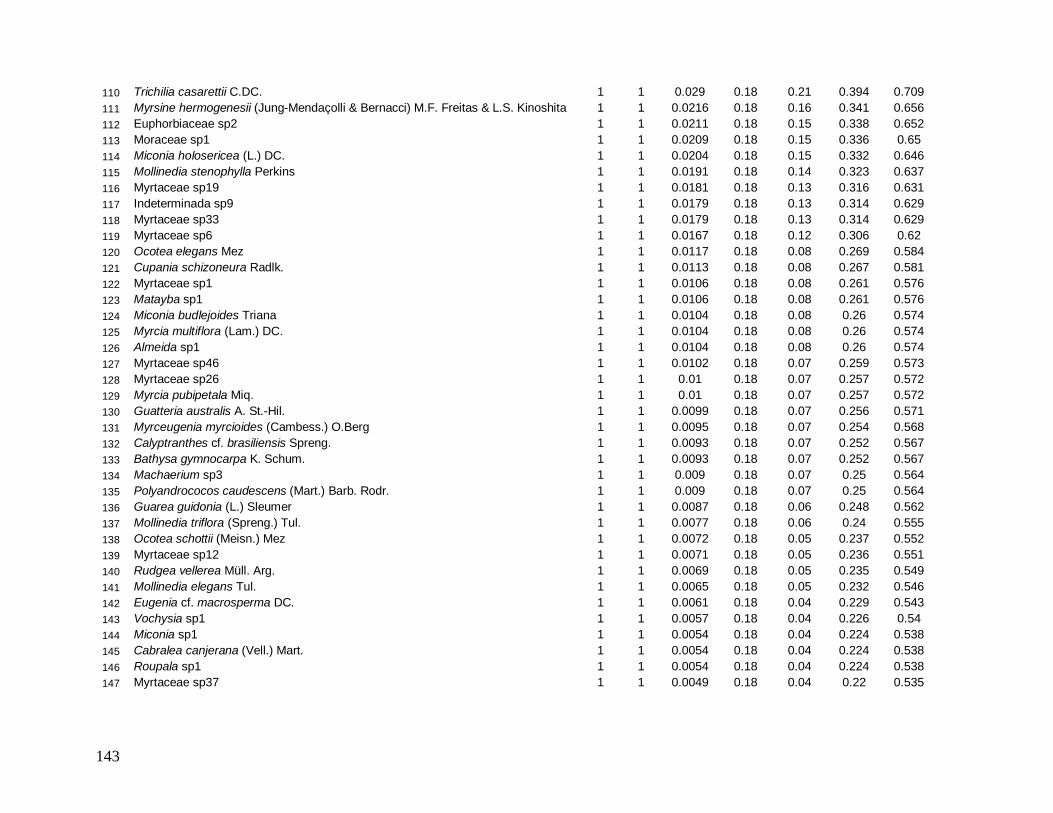
143
110 Trichilia casarettii C.DC. 1 1 0.029 0.18 0.21 0.394 0.709 111 Myrsine hermogenesii (Jung-Mendaçolli & Bernacci) M.F. Freitas & L.S. Kinoshita 1 1 0.0216 0.18 0.16 0.341 0.656 112 Euphorbiaceae sp2 1 1 0.0211 0.18 0.15 0.338 0.652 113 Moraceae sp1 1 1 0.0209 0.18 0.15 0.336 0.65 114 Miconia holosericea (L.) DC. 1 1 0.0204 0.18 0.15 0.332 0.646 115 Mollinedia stenophylla Perkins 1 1 0.0191 0.18 0.14 0.323 0.637 116 Myrtaceae sp19 1 1 0.0181 0.18 0.13 0.316 0.631 117 Indeterminada sp9 1 1 0.0179 0.18 0.13 0.314 0.629 118 Myrtaceae sp33 1 1 0.0179 0.18 0.13 0.314 0.629 119 Myrtaceae sp6 1 1 0.0167 0.18 0.12 0.306 0.62 120 Ocotea elegans Mez 1 1 0.0117 0.18 0.08 0.269 0.584 121 Cupania schizoneura Radlk. 1 1 0.0113 0.18 0.08 0.267 0.581 122 Myrtaceae sp1 1 1 0.0106 0.18 0.08 0.261 0.576 123 Matayba sp1 1 1 0.0106 0.18 0.08 0.261 0.576 124 Miconia budlejoides Triana 1 1 0.0104 0.18 0.08 0.26 0.574 125 Myrcia multiflora (Lam.) DC. 1 1 0.0104 0.18 0.08 0.26 0.574 126 Almeida sp1 1 1 0.0104 0.18 0.08 0.26 0.574 127 Myrtaceae sp46 1 1 0.0102 0.18 0.07 0.259 0.573 128 Myrtaceae sp26 1 1 0.01 0.18 0.07 0.257 0.572 129 Myrcia pubipetala Miq. 1 1 0.01 0.18 0.07 0.257 0.572 130 Guatteria australis A. St.-Hil. 1 1 0.0099 0.18 0.07 0.256 0.571 131 Myrceugenia myrcioides (Cambess.) O.Berg 1 1 0.0095 0.18 0.07 0.254 0.568 132 Calyptranthes cf. brasiliensis Spreng. 1 1 0.0093 0.18 0.07 0.252 0.567 133 Bathysa gymnocarpa K. Schum. 1 1 0.0093 0.18 0.07 0.252 0.567 134 Machaerium sp3 1 1 0.009 0.18 0.07 0.25 0.564 135 Polyandrococos caudescens (Mart.) Barb. Rodr. 1 1 0.009 0.18 0.07 0.25 0.564 136 Guarea guidonia (L.) Sleumer 1 1 0.0087 0.18 0.06 0.248 0.562 137 Mollinedia triflora (Spreng.) Tul. 1 1 0.0077 0.18 0.06 0.24 0.555 138 Ocotea schottii (Meisn.) Mez 1 1 0.0072 0.18 0.05 0.237 0.552 139 Myrtaceae sp12 1 1 0.0071 0.18 0.05 0.236 0.551 140 Rudgea vellerea Müll. Arg. 1 1 0.0069 0.18 0.05 0.235 0.549 141 Mollinedia elegans Tul. 1 1 0.0065 0.18 0.05 0.232 0.546 142 Eugenia cf. macrosperma DC. 1 1 0.0061 0.18 0.04 0.229 0.543 143 Vochysia sp1 1 1 0.0057 0.18 0.04 0.226 0.54 144 Miconia sp1 1 1 0.0054 0.18 0.04 0.224 0.538 145 Cabralea canjerana (Vell.) Mart. 1 1 0.0054 0.18 0.04 0.224 0.538 146 Roupala sp1 1 1 0.0054 0.18 0.04 0.224 0.538 147 Myrtaceae sp37 1 1 0.0049 0.18 0.04 0.22 0.535
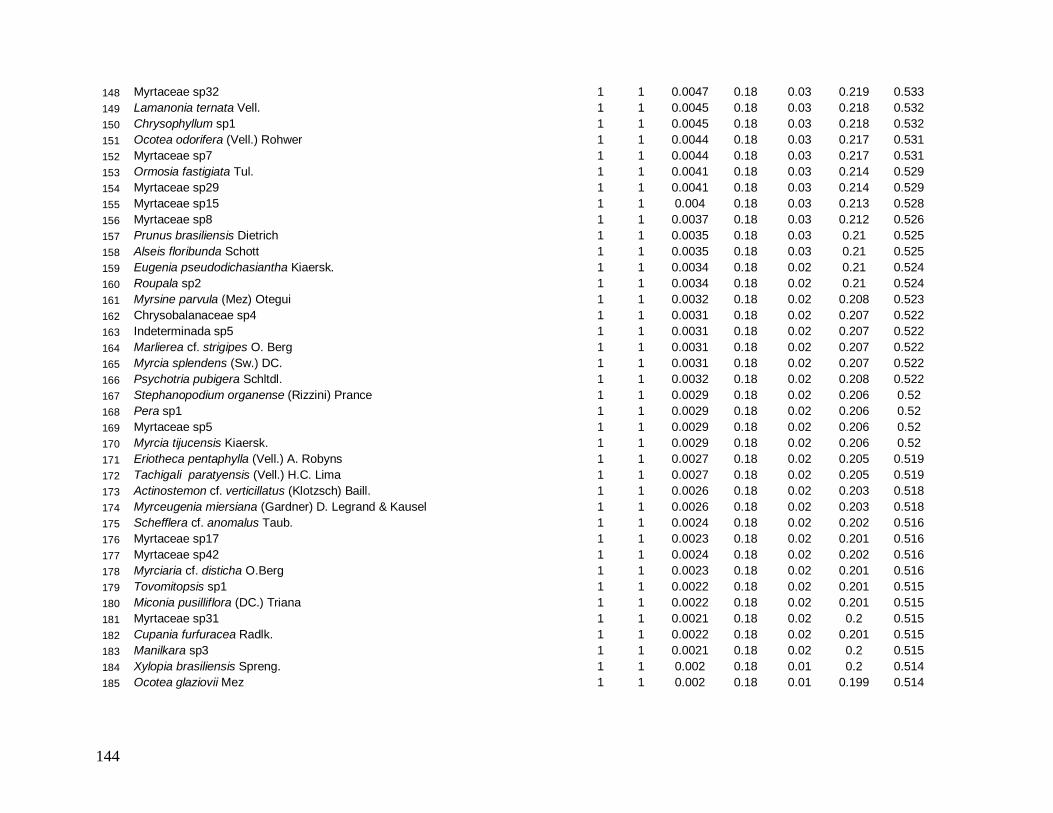
144
148 Myrtaceae sp32 1 1 0.0047 0.18 0.03 0.219 0.533 149 Lamanonia ternata Vell. 1 1 0.0045 0.18 0.03 0.218 0.532 150 Chrysophyllum sp1 1 1 0.0045 0.18 0.03 0.218 0.532 151 Ocotea odorifera (Vell.) Rohwer 1 1 0.0044 0.18 0.03 0.217 0.531 152 Myrtaceae sp7 1 1 0.0044 0.18 0.03 0.217 0.531 153 Ormosia fastigiata Tul. 1 1 0.0041 0.18 0.03 0.214 0.529 154 Myrtaceae sp29 1 1 0.0041 0.18 0.03 0.214 0.529 155 Myrtaceae sp15 1 1 0.004 0.18 0.03 0.213 0.528 156 Myrtaceae sp8 1 1 0.0037 0.18 0.03 0.212 0.526 157 Prunus brasiliensis Dietrich 1 1 0.0035 0.18 0.03 0.21 0.525 158 Alseis floribunda Schott 1 1 0.0035 0.18 0.03 0.21 0.525 159 Eugenia pseudodichasiantha Kiaersk. 1 1 0.0034 0.18 0.02 0.21 0.524 160 Roupala sp2 1 1 0.0034 0.18 0.02 0.21 0.524 161 Myrsine parvula (Mez) Otegui 1 1 0.0032 0.18 0.02 0.208 0.523 162 Chrysobalanaceae sp4 1 1 0.0031 0.18 0.02 0.207 0.522 163 Indeterminada sp5 1 1 0.0031 0.18 0.02 0.207 0.522 164 Marlierea cf. strigipes O. Berg 1 1 0.0031 0.18 0.02 0.207 0.522 165 Myrcia splendens (Sw.) DC. 1 1 0.0031 0.18 0.02 0.207 0.522 166 Psychotria pubigera Schltdl. 1 1 0.0032 0.18 0.02 0.208 0.522 167 Stephanopodium organense (Rizzini) Prance 1 1 0.0029 0.18 0.02 0.206 0.52 168 Pera sp1 1 1 0.0029 0.18 0.02 0.206 0.52 169 Myrtaceae sp5 1 1 0.0029 0.18 0.02 0.206 0.52 170 Myrcia tijucensis Kiaersk. 1 1 0.0029 0.18 0.02 0.206 0.52 171 Eriotheca pentaphylla (Vell.) A. Robyns 1 1 0.0027 0.18 0.02 0.205 0.519 172 Tachigali paratyensis (Vell.) H.C. Lima 1 1 0.0027 0.18 0.02 0.205 0.519 173 Actinostemon cf. verticillatus (Klotzsch) Baill. 1 1 0.0026 0.18 0.02 0.203 0.518 174 Myrceugenia miersiana (Gardner) D. Legrand & Kausel 1 1 0.0026 0.18 0.02 0.203 0.518 175 Schefflera cf. anomalus Taub. 1 1 0.0024 0.18 0.02 0.202 0.516 176 Myrtaceae sp17 1 1 0.0023 0.18 0.02 0.201 0.516 177 Myrtaceae sp42 1 1 0.0024 0.18 0.02 0.202 0.516 178 Myrciaria cf. disticha O.Berg 1 1 0.0023 0.18 0.02 0.201 0.516 179 Tovomitopsis sp1 1 1 0.0022 0.18 0.02 0.201 0.515 180 Miconia pusilliflora (DC.) Triana 1 1 0.0022 0.18 0.02 0.201 0.515 181 Myrtaceae sp31 1 1 0.0021 0.18 0.02 0.2 0.515 182 Cupania furfuracea Radlk. 1 1 0.0022 0.18 0.02 0.201 0.515 183 Manilkara sp3 1 1 0.0021 0.18 0.02 0.2 0.515 184 Xylopia brasiliensis Spreng. 1 1 0.002 0.18 0.01 0.2 0.514 185 Ocotea glaziovii Mez 1 1 0.002 0.18 0.01 0.199 0.514

145
186 Mollinedia salicifolia Perkins 1 1 0.002 0.18 0.01 0.2 0.514 541 9 13.8307 100 100 200 300
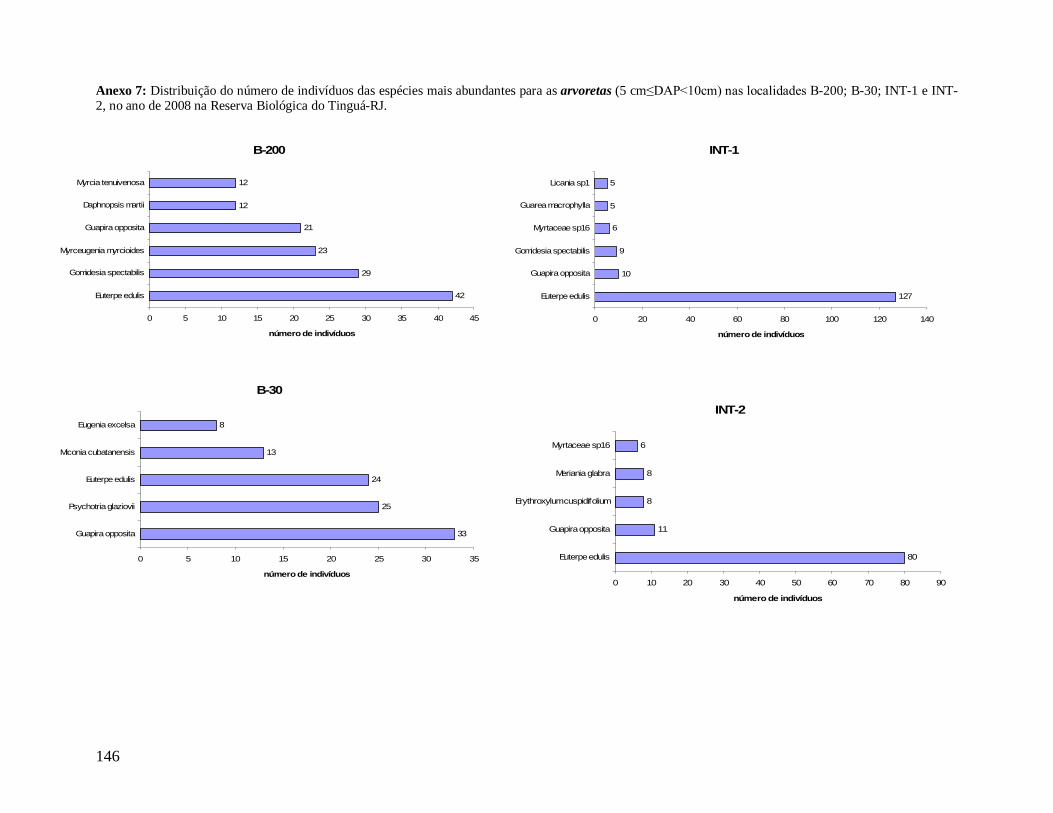
146
Anexo 7: Distribuição do número de indivíduos das espécies mais abundantes para as arvoretas (5 cm≤DAP<10cm) nas localidades B-200; B-30; INT-1 e INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
B-200
42
29
23
21
12
12
0 5 10 15 20 25 30 35 40 45
Euterpe edulis
Gomidesia spectabilis
Myrceugenia myrcioides
Guapira opposita
Daphnopsis martii
Myrcia tenuivenosa
número de indivíduos
B-30
33
25
24
13
8
0 5 10 15 20 25 30 35
Guapira opposita
Psychotria glaziovii
Euterpe edulis
Miconia cubatanensis
Eugenia excelsa
número de indivíduos
INT-1
127
10
9
6
5
5
0 20 40 60 80 100 120 140
Euterpe edulis
Guapira opposita
Gomidesia spectabilis
Myrtaceae sp16
Guarea macrophylla
Licania sp1
número de indivíduos
INT-2
80
11
8
8
6
0 10 20 30 40 50 60 70 80 90
Euterpe edulis
Guapira opposita
Erythroxylum cuspidifolium
Meriania glabra
Myrtaceae sp16
número de indivíduos
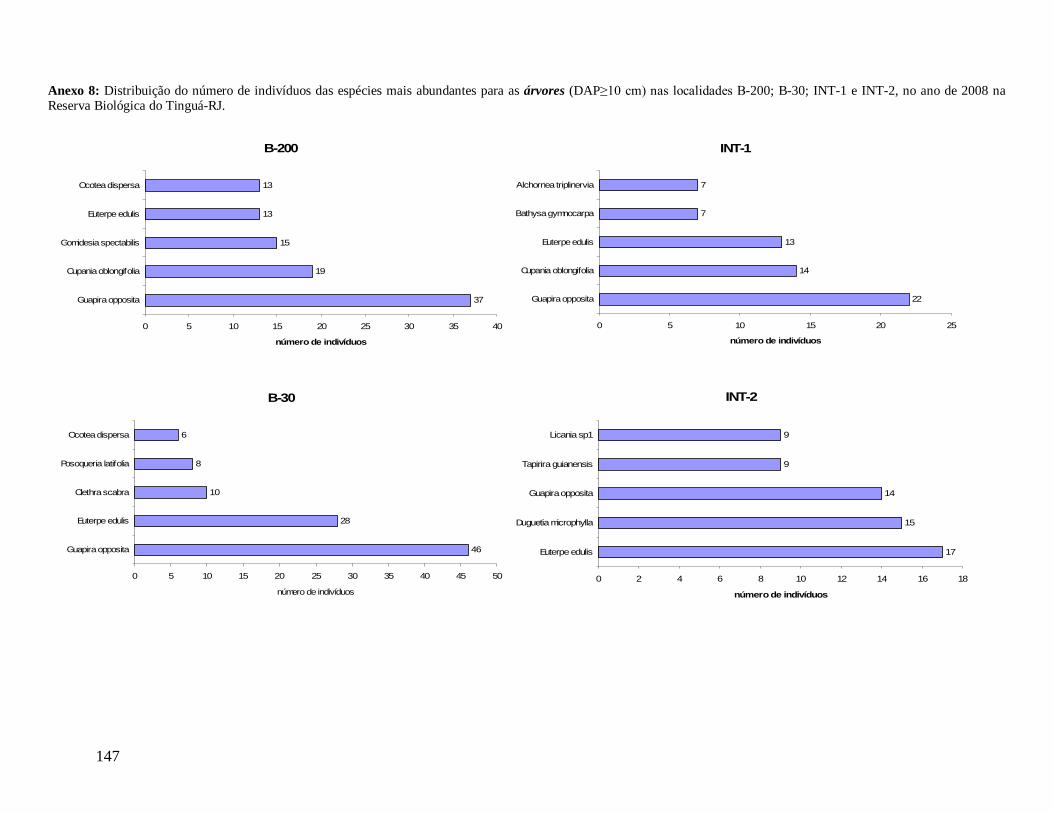
147
Anexo 8: Distribuição do número de indivíduos das espécies mais abundantes para as árvores (DAP≥10 cm) nas localidades B-200; B-30; INT-1 e INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
B-200
37
19
15
13
13
0 5 10 15 20 25 30 35 40
Guapira opposita
Cupania oblongifolia
Gomidesia spectabilis
Euterpe edulis
Ocotea dispersa
número de indivíduos
B-30
46
28
10
8
6
0 5 10 15 20 25 30 35 40 45 50
Guapira opposita
Euterpe edulis
Clethra scabra
Posoqueria latifolia
Ocotea dispersa
número de indivíduos
INT-1
22
14
13
7
7
0 5 10 15 20 25
Guapira opposita
Cupania oblongifolia
Euterpe edulis
Bathysa gymnocarpa
Alchornea triplinervia
número de indivíduos
INT-2
17
15
14
9
9
0 2 4 6 8 10 12 14 16 18
Euterpe edulis
Duguetia microphylla
Guapira opposita
Tapirira guianensis
Licania sp1
número de indivíduos
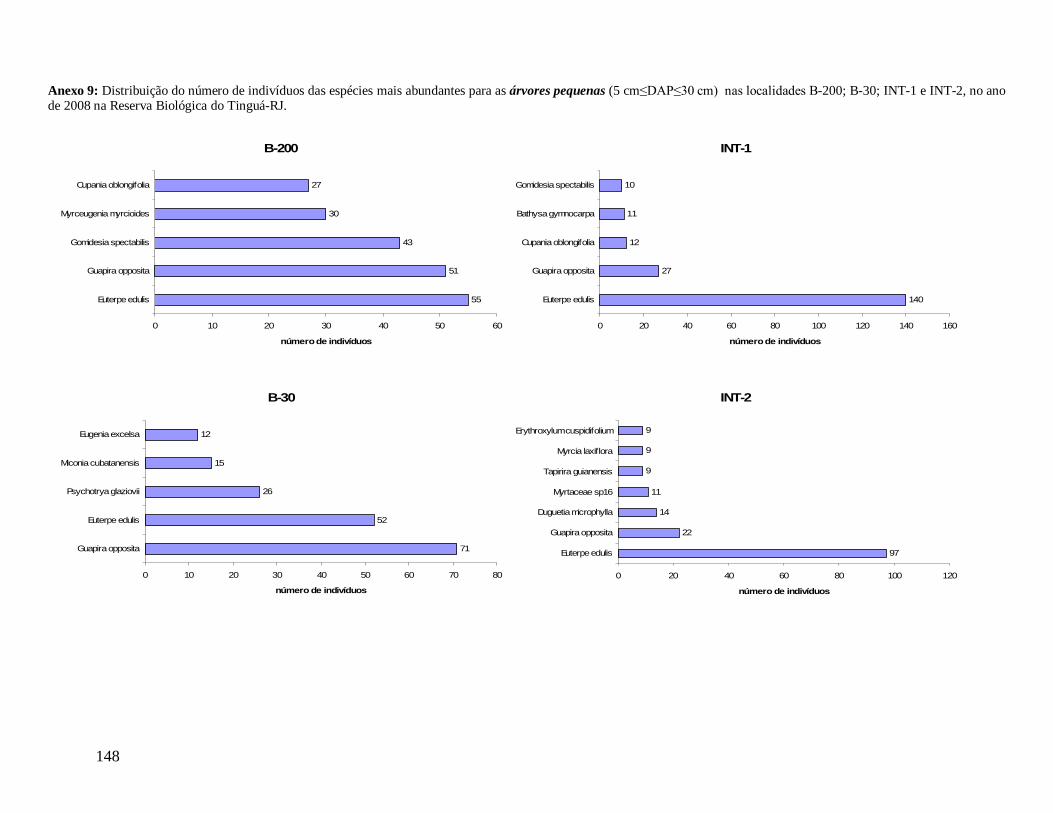
148
Anexo 9: Distribuição do número de indivíduos das espécies mais abundantes para as árvores pequenas (5 cm≤DAP≤30 cm) nas localidades B-200; B-30; INT-1 e INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
B-200
55
51
43
30
27
0 10 20 30 40 50 60
Euterpe edulis
Guapira opposita
Gomidesia spectabilis
Myrceugenia myrcioides
Cupania oblongifolia
número de indivíduos
B-30
71
52
26
15
12
0 10 20 30 40 50 60 70 80
Guapira opposita
Euterpe edulis
Psychotrya glaziovii
Miconia cubatanensis
Eugenia excelsa
número de indivíduos
INT-1
140
27
12
11
10
0 20 40 60 80 100 120 140 160
Euterpe edulis
Guapira opposita
Cupania oblongifolia
Bathysa gymnocarpa
Gomidesia spectabilis
número de indivíduos
INT-2
97
22
14
11
9
9
9
0 20 40 60 80 100 120
Euterpe edulis
Guapira opposita
Duguetia microphylla
Myrtaceae sp16
Tapirira guianensis
Myrcia laxiflora
Erythroxylum cuspidifolium
número de indivíduos
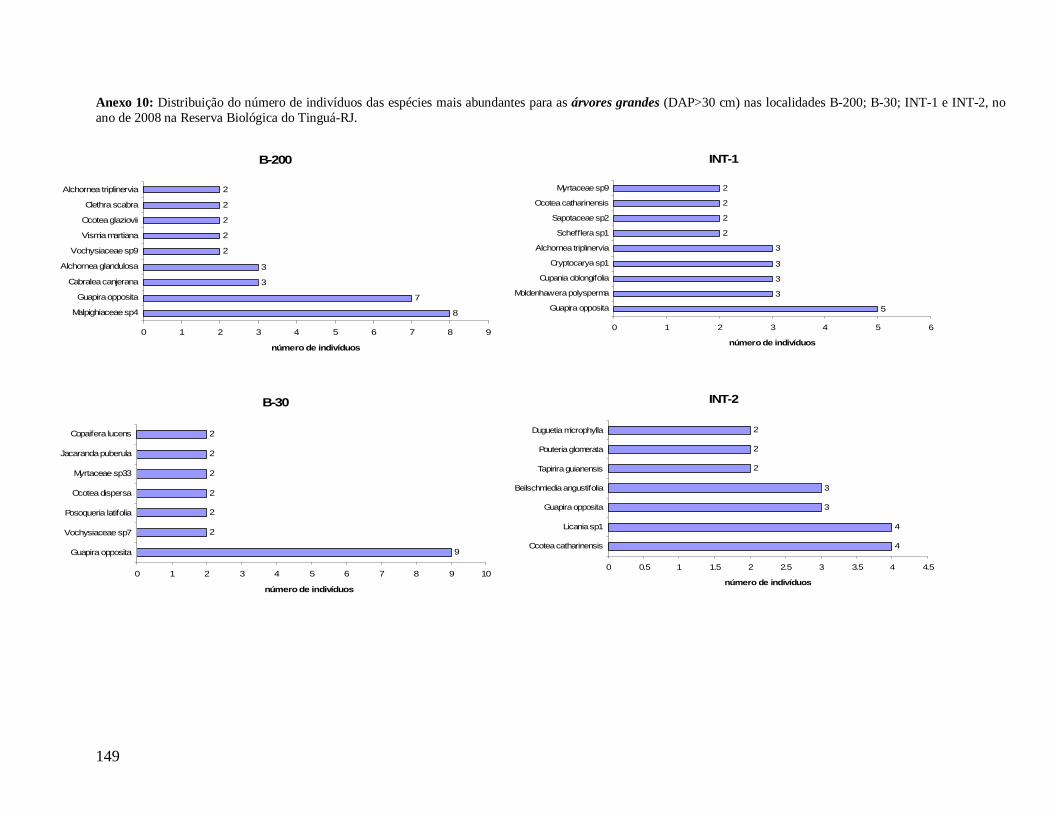
149
Anexo 10: Distribuição do número de indivíduos das espécies mais abundantes para as árvores grandes (DAP>30 cm) nas localidades B-200; B-30; INT-1 e INT-2, no ano de 2008 na Reserva Biológica do Tinguá-RJ.
B-200
8
7
3
3
2
2
2
2
2
0 1 2 3 4 5 6 7 8 9
Malpighiaceae sp4
Guapira opposita
Cabralea canjerana
Alchornea glandulosa
Vochysiaceae sp9
Vismia martiana
Ocotea glaziovii
Clethra scabra
Alchornea triplinervia
número de indivíduos
B-30
9
2
2
2
2
2
2
0 1 2 3 4 5 6 7 8 9 10
Guapira opposita
Vochysiaceae sp7
Posoqueria latifolia
Ocotea dispersa
Myrtaceae sp33
Jacaranda puberula
Copaifera lucens
número de indivíduos
INT-1
5
3
3
3
3
2
2
2
2
0 1 2 3 4 5 6
Guapira opposita
Moldenhawera polysperma
Cupania oblongifolia
Cryptocarya sp1
Alchornea triplinervia
Schefflera sp1
Sapotaceae sp2
Ocotea catharinensis
Myrtaceae sp9
número de indivíduos
INT-2
4
4
3
3
2
2
2
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5
Ocotea catharinensis
Licania sp1
Guapira opposita
Beilschmiedia angustifolia
Tapirira guianensis
Pouteria glomerata
Duguetia microphylla
número de indivíduos