instituto de pesquisas jardim botânico do rio de janeiro escola...
TRANSCRIPT

Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Programa de Pós-graduação Stricto Sensu
Dissertação de Mestrado
Anatomia foliar, ontogenia das estruturas secretoras foliares
e composição do óleo essencial de Myrsine coriacea (Sw.)
R.Br. ex Roem. & Shult. e Myrsine venosa A.DC.
(Myrsinoideae – Primulaceae)
Bruna Nunes de Luna
Rio de Janeiro
2013

ii
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro
Escola Nacional de Botânica Tropical
Programa de Pós-graduação Stricto Sensu
Anatomia foliar, ontogenia das estruturas secretoras foliares
e composição do óleo essencial de Myrsine coriacea (Sw.)
R.Br. ex Roem. & Shult. e Myrsine venosa A.DC.
(Myrsinoideae – Primulaceae)
Bruna Nunes de Luna
Rio de Janeiro
2013
Dissertação apresentada ao Programa de
Pós-Graduação em Botânica, Escola
Nacional de Botânica Tropical, do
Instituto de Pesquisas Jardim Botânico
do Rio de Janeiro, como parte dos
requisitos necessários para a obtenção
do título de Mestre em Botânica.
Orientadora: Claudia Franca
Barros

iii
Título: Anatomia foliar, ontogenia das estruturas secretoras foliares e
composição do óleo essencial de Myrsine coriacea (Sw.) R.Br. ex Roem.
& Shult. e Myrsine venosa A.DC. (Myrsinoideae – Primulaceae)
Bruna Nunes de Luna
Dissertação submetida ao corpo docente da Escola Nacional de Botânica Tropical,
Instituto de Pesquisas Jardim Botânico do Rio de Janeiro - JBRJ, como parte dos
requisitos necessários para a obtenção do grau de Mestre.
Aprovada por:
Profª. Drª Claudia Franca Barros (Orientadora) _________________________
Profª. Dra Helena Regina Pinto Lima _________________________
Profª. Dra Silvia Rodrigues Machado _________________________
Profª. Dra Maura da Cunha _________________________
Profª. Dra Cecília Gonçalves Costa _________________________
em 26/02/ 2013
Rio de Janeiro
2013

iv
Luna, Bruna Nunes de.
L961o Anatomia foliar, ontogenia das estruturas secretoras foliares e
composição do óleo essencial de Myrsine coriacea (Sw.) R.Br. ex Roem.
& Shult. e Myrsine venosa A.DC. (Myrsinoideae – Primulaceae) / Bruna
Nunes de Luna. – Rio de Janeiro, 2013.
xiii, 65f. : il. ; 28 cm.
Dissertação (mestrado) – Instituto de Pesquisas Jardim Botânico do
Rio de Janeiro / Escola Nacional de Botânica Tropical, 2013.
Orientadora: Claudia Franca Barros
Bibliografia.
1.Primulaceae. 2.Anatomia vegetal. 3.Cavidades secretoras. 4.
Tricomas glandulares. 5.Testes histoquímicos. 6. Óleos essenciais. 7.
Terpenóides. I. Título. II. Escola Nacional de Botânica Tropical.
CDD 583.672

v
Dedicatória
Aos meus amores:
Ilma, Jose Carlos, Lucas e Luiz Felipe

vi
Agradecimentos
À CAPES, pelo auxílio financeiro.
À minha orientadora, Profa. Dr
a. Claudia Franca Barros, pelos ensinamentos, incentivo, apoio
constante, confiança e amizade ao longo dessa trajetória.
À Profa. Dr
a. Maria de Fátima Freitas, pela sugestão dessas belas espécies com as quais
trabalhamos nesta dissertação, pelo auxílio com as coletas, e por toda a parceria, dedicação e
amizade.
À Profa. Dr
a. Alice Sato, pela confiança e acolhida em seu laboratório e pela parceria na análise
dos óleos essenciais.
À Anna Carina Antunes e Defaveri, Msc., pela imensa ajuda na análise e interpretação dos
resultados dos óleos essenciais, pelo apoio e amizade.
Ao Prof. Dr. Humberto Bizzo, pela análise dos óleos essenciais.
Ao Prof. Dr. Emílio de Castro Miguel, pelo auxílio na observação e análise dos materiais para
microscopia eletrônica, por todos os ensinamentos sobre microscopia e por todo apoio e
confiança.
À Profa. Dr
a. Denise Espellet Klein, da UNIRIO, pelo aceite da orientação do estágio em
docência, por todas as informações, ensinamentos e dicas preciosas, que contribuíram para minha
formação profissional.
À Aline Machado, Msc., pelo apoio com as coletas.
Aos técnicos do Laboratório de Botânica Estrutural, Elaine Zózimo e Rogério Figueiredo, por
todo apoio e auxílio prestado. E aos companheiros de laboratório, Felipe, Raquel, Monique e
Alessandra.
Aos amigos Tahysa Mota Macedo e Warlen Costa, pelo apoio nas coletas, pelas trocas de
experiências, incentivo, conversas, risadas e amizade. À Simone Petrucci e Fernanda Masullo que
viveram comigo esta etapa final, trazendo alegria com os papos do almoço e do final do dia,
pelas dicas técnicas que deixaram o trabalho mais bonito.

vii
Às amigas do PBV – UFRJ, Aline Carvalho, Aline Vieira e Anna Carina Defaveri, pelos
momentos de descontração, amizade e apoio.
Aos colegas e amigos do curso de pós-graduação da ENBT, pelas dicas que aprimoraram o
projeto inicial e pelo apoio.
Aos docentes da ENBT, que contribuíram com sugestões e críticas construtivas, essenciais para o
desenvolvimento deste trabalho.
Aos meus grandes amigos, Lais Elias, Thaiany Alves, Talita Jimenez e Bernardo Nunes, que me
permitem ver além das minhas capacidades, em vocês eu deposito toda minha confiança, amizade
e lealdade.
Aos Legattos, que compõem uma parte essencial da minha vida e proporcionam os momentos
musicais mais profissionais, descontraídos e importantíssimos para manter meu equilíbrio entre a
razão e a emoção.
À minha incrível família, Ilma Nunes de Luna, Jose Carlos de Luna e Lucas Nunes de Luna, por
acreditar e dar sentido às minhas decisões, com todo apoio e amor.
Ao meu amor, Luiz Felipe Machado, por ser o meu maior incentivador e amigo. Por todo o
apoio, carinho e paciência que teve comigo durante essa trajetória.
Ao meu querido Deus, que realiza cada sonho meu da maneira mais perfeita e delicada, em seu
tempo. Obrigada por ser o centro da minha vida.

viii
“O começo de todas as ciências é o espanto das coisas serem o que são”
Aristóteles

ix
Sumário
Lista de figuras ............................................................................................................................................... x
Lista de tabelas ............................................................................................................................................. xi
Resumo ........................................................................................................................................................ xii
Abstract ...................................................................................................................................................... xiii
1 - Introdução ............................................................................................................................................... 2
1.1- Primulaceae ............................................................................................................................... 2
1.2- Estruturas secretoras em plantas ................................................................................................ 3
2 - Objetivos ................................................................................................................................................ 10
3 - Materiais e métodos .............................................................................................................................. 12
3.1- Espécies selecionadas .............................................................................................................. 12
3.2- Microscopia óptica ................................................................................................................... 14
3.3 - Microscopia de epifluorescência ............................................................................................. 15
3.4 - Microscopia confocal .............................................................................................................. 15
3.5 - Microscopia eletrônica de varredura ....................................................................................... 15
3.6 - Testes histoquímicos ............................................................................................................... 16
3.7 - Extração dos óleos essenciais .................................................................................................. 16
4 - Resultados .............................................................................................................................................. 20
4.1 - Anatomia das folhas expandidas ............................................................................................. 20
4.1.1 - Pecíolo ..................................................................................................................... 20
4.1.2 - Lâmina foliar ........................................................................................................... 20
4.2 - Natureza química da secreção ................................................................................................. 25
4.3 - Ontogênese das estruturas secretoras ...................................................................................... 28
4.3.1 - Tricomas peltados .................................................................................................... 28
4.3.2 - Tricoma glandular pluricelular ................................................................................ 28
4.3.3 - Cavidades e ductos .................................................................................................. 33
4.4 - Óleos essenciais ...................................................................................................................... 36
5 - Discussão ................................................................................................................................................ 44
6 – Conclusões ............................................................................................................................................. 52
7 – Referências bibliográficas .................................................................................................................... 55

x
Lista de figuras
Pag.
Figura 1 Exemplos de estruturas secretoras vegetais internas e externas............................................. 4
Figura 2 Resumo das funções dos metabólitos secundários................................................................. 5
Figura 3 Hábito de Myrsine coriacea................................................................................................... 12
Figura 4 Hábito de Myrsine venosa...................................................................................................... 13
Figura 5 Mapa de distribuição das espécies......................................................................................... 13
Figura 6 Primórdios foliares de Myrsine coriacea e M. venosa........................................................... 14
Figura 7 Aparelho de Clevenger........................................................................................................... 17
Figura 8 Aspectos da anatomia do pecíolo de Myrsine coriacea e M. venosa..................................... 22
Figura 9 Aspectos da anatomia foliar de Myrsine coriacea e M. venosa............................................. 24
Figura 10 Testes histoquímicos em M. coriacea.................................................................................... 26
Figura 11 Testes histoquímicos em M. venosa...................................................................................... 27
Figura 12 Ontogenia dos tricomas glandulares peltados das folhas de Myrsine coriacea e M.venosa.. 30
Figura 13 Ontogenia dos tricomas glandulares pluricelulares das folhas de M. coriacea..................... 32
Figura 14 Ontogenia das cavidades secretoras das folhas de M. coriacea............................................. 34
Figura 15 Ontogenia das cavidades secretoras das folhas de M. venosa................................................ 35
Figura 16 Coloração do óleo essencial extraído das folhas secas de M. coriacea e M. venosa............. 36
Figura 17 Porcentagem dos componentes majoritários do óleo essencial de M. coriacea e M. venosa 37
Figura 18 Fórmula estrutural dos componentes majoritários do óleo essencial de M. coriacea............ 38
Figura 19 Fórmula estrutural dos componentes majoritários do óleo essencial de M. venosa............... 38

xi
Lista de Tabelas
Tabela 1 Testes histoquímicos realizados e substâncias detectadas...................................... 16
Tabela 2 Diferenças anatômicas observadas entre Myrsine coriacea e M. venosa............... 22
Tabela 3 Testes histoquímicos realizados nas folhas adultas e primórdios foliares de
Myrsine coriacea e M. venosa, com os componentes analisados e os respectivos
reagentes utilizados. (+) - presente; (-) – ausente................................................... 24
Tabela 4 Composição do óleo essencial das folhas secas de Myrsine coriacea................... 39
Tabela 5 Composição do óleo essencial das folhas secas de Myrsine venosa...................... 40

xii
RESUMO
Luna, B.N: Anatomia foliar, ontogenia das estruturas secretoras foliares e composição do
óleo essencial de Myrsine coriacea (Sw.) R. Br. ex Roem. & Shult. e Myrsine venosa A.DC.
(Myrsinoideae – Primulaceae). 2013. Dissertação (Mestrado). Escola Nacional de Botânica
Tropical - Instituto de Pesquisas do Jardim Botânico do Rio de Janeiro.
A família Primulaceae (Ordem Ericales) apresenta como uma de suas principais características a presença
de cavidades secretoras encontradas em diferentes órgãos da planta (vegetativos e reprodutivos). Estas
estruturas secretam substâncias resiníferas e outras conhecidas como hidroquixibenzoquinonas, exclusivas
desta família. Primulaceae apresenta um clado monofilético formado pelos gêneros Primula, Lysimachia e
Myrsine, que pode ser subdividido em Primuloideae, circunscrevendo Primula, gênero sem cavidades
secretoras e Myrsinoideae, que engloba Lysimachia e Myrsine, gêneros com espécies que apresentam
cavidades secretoras. As espécies selecionadas como objeto de estudo do presente trabalho pertencem ao
gênero Myrsine, amplamente distribuído no Brasil e popularmente conhecido como capororoca. Estudos
recentes têm indicado o potencial farmacológico do exsudado produzido por espécies de Myrsine, mas
pouco se conhece a respeito da estrutura que o produz. O objetivo deste trabalho foi caracterizar a
anatomia foliar e elucidar o processo de formação das estruturas secretoras em Myrsine coriacea e M.
venosa, bem como analisar a composição da fração volátil (óleos essenciais) da secreção produzidas por
essas estruturas. Para as análises anatômicas e de ontogênese, foram utilizados métodos de análises
anatômicas e histoquímicas e para a análise do óleo essencial, métodos de cromatografia gasosa e
espectrometria de massa. Os tricomas glandulares, idioblastos, cavidades e ductos foram observados em
ambas as espécies, sendo essas últimas estruturas secretoras descritas pela primeira vez em M. venosa. Os
tricomas glandulares, localizados na lâmina foliar e pecíolo, são do tipo peltado, originam-se de células
protodérmicas e diferenciam-se antes do completo desenvolvimento foliar. M. coriacea também apresenta
tricomas pluricelulares glandulares dispersos por toda a lâmina foliar, nervura mediana e pecíolo. As
cavidades e ductos secretores são oriundos de células do meristema fundamental, e resultam de um
processo de formação esquizógeno. Os componentes principais do óleo essencial de M. coriacea foram o
β – elemeno, germacreno-D, biciclogermacreno e T-muurolol, enquanto os principais constituintes do óleo
de M. venosa foram γ – muuroleno, biciclogermacreno, δ – cadineno e β – cariofileno.
Palavras-chave: anatomia foliar, ontogênese de estruturas secretoras, cavidades secretoras, tricomas
glandulares, testes histoquímicos, óleo essencial, terpenóides.

xiii
ABSTRACT
Luna, B.N. Leaf anatomy, ontogenesis of leaves secretory structures and essential oil
composition from Myrsine coriacea (Sw.) R.Br. ex Roem. & Shult and M. venosa A. DC.
(Myrsinoideae – Primulaceae). 2013. Master thesis. Escola Nacional de Botânica Tropical -
Instituto de Pesquisas do Jardim Botânico do Rio de Janeiro.
Primulaceae (Ericales) can be recognized, among other characteristics, by the presence of secretory
cavities. These structures produce a resiniferous substance and others known as hidroxybenzoquinones.
Primulaceae presents a monophyletic clade formed by Primula, Lysimachia and Myrsine, which can be
subdivided in two subfamilies: Primuloideae, composed by Primula, genus without secretory cavities and
Myrsinoideae, with Lysimachia and Myrsine, both presenting secretory cavities. Selected species for this
study belongs to Myrsine, widely distributed in Brazil, popularly known as “capororoca”. In the present
study we aimed to elucidate the formation of the secretory structures found in the leaves of Myrsine
coriacea and M. venosa and the composition of the volatile fraction from the secretion produced by these
structures. To elucidate leaves secretory structures development and morphology, there were perfomed
anatomical and histochemical analyses and for the essential oil analyses, methods of gas chromatography
and mass spectrometry were used. Leaf anatomical analyses showed characters common to both species,
such as dorsiventral mesophyll, uniseriate epidermis and anisocytic stomata. Secretory structures found in
M. coriacea and M. venosa are glandular trichomes, idioblasts, and secretory cavities and ducts. Glandular
trichomes can be peltate type, found in both species, or pluricelular type, found only in M. coriacea.
Trichomes are initiated by the enlargement of protodermal cells, followed by their division and are
completely formed before the leaf expansion. Secretory cavities and ducts are formed by schizogeny from
ground meristem cells. Principal components from M. coriacea essential oil were β-elemene, germacrene-
D, bicyclogermacrene and T-muurolol, while the major components of M. venosa essential oil were, γ-
muurolene, bicyclogermacrene, δ-cadinene and β-caryophyllene.
Key-words: leaf anatomy, ontogenesis of secretory structures, histochemical tests, essential oil,
terpenóides, secretory cavities, glandular trichomes, essential oil, sesquiterpenes.
1

2
1 – Introdução
1.1 – Primulaceae
Primulaceae, subordinada à ordem Ericales (APG III 2009), apresenta distribuição
pantropical. Seus representantes, cerca de 2.500 espécies, apresentam porte arbóreo, arbustivo ou
herbáceo. A monofilia desta família é sustentada, dentre outros caracteres, pela presença de
cavidades ou canais secretores, característica ausente em alguns táxons, e pela placentação
central-livre com um eixo espesso, geralmente globoso. O clado monofilético Primula +
Lysimachia + Myrsine pode ser subdividido em dois subclados: Primuloideae, constituído por
plantas herbáceas sem cavidades secretoras, e Myrsinoideae, que compreende os gêneros que
apresentam cavidades secretoras internas (Judd et al. 2009). A literatura acerca da anatomia dos
representantes da família caracterizam as cavidades secretoras como sendo oriundas de um
processo de formação esquizógeno e que secretam material resinífero, com coloração amarelada
ou alaranjada (Lersten 1986; Metcalfe & Chalk 1950; Otegui et al. 1998a; 1998b).
As estruturas secretoras em Primulaceae produzem, principalmente, compostos derivados
de substâncias quimicamente conhecidas como hidroxibenzoquinonas, que variam de composição
de acordo com a espécie, permitindo sua utilização como caráter quimiotaxonômico para
distinção genérica dos representantes de Myrsinoideae (Ogawa & Natori 1968; Otegui et al.
1998b).
O gênero Myrsine L. (= Rapanea), circunscrito no subclado Myrsinoideae, possui cerca
de 300 espécies de distribuição pantropical (Freitas 2003), dentre as quais 26 ocorrem no Brasil
(Freitas 2012). Os representantes de Myrsine, popularmente conhecidos como “capororoca” ou
“pororoca”, são utilizados na medicina popular para o tratamento de doenças que acometem
animais e seres humanos. A infusão das folhas de M. coriacea (Sw.) R. Br. ex Roem & Schult.,
por exemplo, segundo Bueno et al.(2005), é usada para tratar infecções de garganta causadas pela
caxumba.
Diferentes substâncias com atividade biológica foram extraídos e identificados em
algumas espécies de Myrsine. Baccarin et al. (2011) desenvolveram, a partir do extrato
hidroetanólico de M. coriacea, um método eficaz de isolamento do ácido mirsinóico B. O
interesse nessa substância deve-se à sua comprovada ação antinociceptiva (Hess et al. 2010),
sendo, portanto, precursora potencial para o desenvolvimento de analgésicos. Os ácidos

3
mirsinóicos B, C, E e F apresentam potencial atividade anti-inflamatória (Hirota et al. 2002;
Makabi et al. 2003), os ácidos terpeno-benzóicos em Myrsine seguinii H. Lév, possuem ação
anti-inflamatória (Mizushina et al. 2000), o extrato etanólico de Myrsine africana L., apresenta
potencial ação anti-inflamatória e anti-espasmódica (Ahmad et al. 2011; Azam et al. 2011). Não
há referências acerca da presença ou composição de óleos essenciais em Myrsine.
Destacam-se também produtos secretados por outros representantes de Primulaceae, como
as saponinas triterpênicas isoladas em Ardisia Sw. e Maesa Forssk. (Huang et al. 2000; Koike et
al. 2001), que apresentam ação anti-leishmaniose (Germonprez et al. 2004; Vermeersch et al.
2009); as saponinas e flavonóides em Anagallis L., com efeito anti-microbiano e anti-
inflamatório (López et al. 2011), o extrato etanólico de Lysimachia ramosa Wall. ex Duby, com
ação anti-helmíntica (Challam et al. 2010); e as alquilbenzoquinonas em Ardisia kivuensis Taton,
que se revelaram efetivas no combate à linhagens de células de câncer (Ndonsta et al. 2011).
Apesar da importância taxonômica e farmacêutica, o estudo de estruturas secretoras em
Primulaceae é escasso. Otegui & Maldonado (1998) elencaram características anatômicas foliares
diagnósticas, incluindo tricomas glandulares, para espécies de Myrsine da América do Sul.
Otegui et al. (1998b) analisaram as cavidades e tecidos secretores, exceto tricomas, observando
os seus aspectos ultraestruturais associados à secreção de hidroxibenzoquinonas. Os mesmos
autores atentam para a necessidade de investigação em outras espécies de Primulaceae para a
elucidação do processo de formação das cavidades secretoras. Acerca do desenvolvimento das
cavidades secretoras, Guttemberg (1928), descreve o processo em Lysimachia vulgaris L. e o
classifica como esquizolisígeno. Entretanto, Lersten (1986) reavalia este processo de formação
em Lysimachia nummularia L., chegando à conclusão de que as cavidades secretoras são
esquizógenas.
1.2 - Estruturas secretoras em plantas
Estruturas secretoras são encontradas em diversas famílias vegetais (Angiospermas e
Gimnospermas), que muitas vezes não são filogeneticamente relacionadas (Metcalfe & Chalk
1957). Muitas apresentam morfologia semelhante, mas secretam substâncias e desempenham
funções distintas (Mauseth 1988). As classificações utilizadas são as propostas por Fahn (1988),
que relacionam as estruturas secretoras à sua função e ao material secretado. As estruturas
secretoras localizadas na superfície dos órgãos vegetais são classificadas como estruturas
externas (p.ex. tricomas), e a secreção é do tipo exógena, uma vez que é exsudada para o exterior

4
do órgão vegetal. As estruturas localizadas internamente nos diferentes órgãos vegetais são
chamadas internas (p. ex. cavidades, ductos, laticíferos), e liberam secreção do tipo endógena, a
qual é armazenada dentro da própria célula secretora ou em espaços intercelulares (Kisser 1958;
Fahn 1984) (Fig. 1).
Figura 1 – Exemplos de estruturas secretoras vegetais internas e externas. Fonte: Fahn (1988); Evert
(2006); Konno (2011); Milani et al.(2012).
A variabilidade morfológica reflete a diversidade de metabólitos secretados por estes
tecidos. Substâncias da mesma natureza química podem ser secretadas por estruturas distintas,
por exemplo, tricomas glandulares e cavidades secretoras como sítios de produção de óleos
essenciais. A secreção pode não sofrer muitas modificações ao longo do processo secretor, p.ex.
em hidatódios, estruturas que secretam água e solutos provenientes do tecido xilemático, ou pode
ser inteiramente sintetizada por células ou tecidos secretores (Fahn 1979). Neste último caso de
secreção de substâncias complexas, podem-se citar as resinas, gomas, mucilagens, óleos e látex
(Langenheim 2003). Esses metabólitos secundários, produtos do metabolismo vegetal,

5
desempenham diferentes funções no organismo. Algumas das principais funções estão
sumarizadas na Figura 2.
Figura 2 – Resumo das funções dos metabólitos secundários. Adaptado de Wink (2010)
Uma das principais categorias de metabólitos produzidos pelas estruturas secretoras são
os óleos essenciais, formados principalmente por componentes terpênicos. Os terpenos, por sua
vez, são formados por unidades de isoprenos (cadeias de cinco carbonos - C5). Em se tratando de
óleos essenciais, as classes de terpenos que os constituem são os monoterpenos (que possuem
duas cadeias de isoprenos - C10, p. ex. linalol, lavandulol, citronelal) e os sesquiterpenos (três
cadeias de isoprenos - C15, p. ex. β-cariofileno, curcumeno) (Bakkali et al. 2008).
Os óleos essenciais podem ser sintetizados em todos os órgãos das plantas (flores, folhas,
caule e raízes) e armazenados em células ou estruturas específicas, como as estruturas secretoras

6
citadas anteriormente. A composição e produção dos óleos essenciais são reguladas por fatores
fisiológicos, bioquímicos, metabólicos e genéticos (Sangwan et al. 2001).
Estima-se que sejam conhecidos cerca de 3.000 óleos essenciais e destes, 300 são usados
na indústria alimentícia ou de fragrâncias. Além desse conhecido uso dos óleos essenciais, eles
podem servir como estratégia de defesa das plantas contra organismos patógenos ou fitófagos,
devido às suas propriedades antimicrobianas (Bassolé & Juliani 2012). Ainda podem atuar
atraindo animais polinizadores e dispersores de semente (Langeheim 2003).
Em todas as 32 ordens reconhecidas de eudicotiledôneas (APG III 2009) estão
circunscritas espécies, famílias ou clados que apresentam algum tipo de estrutura secretora
(Bottega & Corsi 2000; Cicarelli et al. 2001; Corsi & Biasci 1998; Cury & Appezzato-da-Glória
2009; Gomes & Lombardi 2010; Kalachanis & Psaras 2005; Kim & Mahlberg 2000; Klein et al.
2004; Liang et al. 2009; Otegui & Maldonado 1998; Rocha et al. 2011; Teixeira & Rocha 2009;
Thadeo et al. 2009). Sabe-se, portanto, que um mesmo tipo de estrutura ocorre em ordens
distintas, que não apresentam relação filogenética direta. Como, por exemplo, as cavidades
secretoras encontradas em Copaifera langsdorfii Desf. e C. trapezifolia Hayne (Leguminosae-
Caesalpinoideae- Fabales, respectivamente, Rodrigues et al. 2011b e Milani et al. 2012), e em
Myrsine laetevirens (Mez) Arechav (Primulaceae-Myrsinoideae- Ericales, Otegui et al. 1998).
Em ambos os casos o processo de formação das cavidades e a morfologia dessas estruturas são
semelhantes, entretanto Fabales é classificada hierarquicamente no cladograma das relações
filogenéticas entre as Rosídeas, enquanto Ericales está agrupada entre as Asterídeas (Judd et al
2009).
As principais abordagens utilizadas na investigação de estruturas secretoras são a
morfologia, o processo de diferenciação e desenvolvimento, o tipo de substância secretada e a
dinâmica do processo secretor (Fahn 1979; 1988). Acerca da ontogenia pode-se destacar o
trabalho de Moura et al. (2005), com as células secretoras das folhas de Lantana camara L.; o de
Benicci & Tani (2004), com as cavidades secretoras em Citrus limon (L.) Osbeck e C. sinensis
(L.) Osbeck; de Milani et al. (2012), sobre as glândulas de oleorresina em Copaifera trapezifolia,
o de Rodrigues et al. (2011a), que discorre sobre a ontogenia das cavidades secretoras de óleo em
Pterodon pubescens (Benth.) Benth., considerando a participação da bainha de células
parenquimáticas durante esse processo e o de Rodrigues et al. (2011b) sobre o sistema secretor de
óleo-resinas em Copaifera langsdorfii.

7
São várias as famílias cujas espécies são modelo para estudo da morfologia e
funcionamento das estruturas secretoras e natureza das substâncias por elas produzidas. Podem-se
destacar os seguintes trabalhos: Wist & Davis (2006) sobre a anatomia e a ultraestrutura do
nectário floral de Echinacea purpurea (L.) Moench (Asteraceae); Fahn & Shimony (2001), sobre
a ultraestrutura das células secretoras de duas espécies de Fagonia L. (Zygophyllaceae); Fahn
(2000), sobre a estrutura e função das células secretoras de diferentes espécies; Bottega & Corsi
(2000), sobre a estrutura e função de tricomas glandulares do cálice de Rosmarinus offcinalis L.
(Labiatae); Knight et al. (2001), sobre as glândulas de óleo do fruto de Citrus sinensis;
Combrinck et al. (2007), sobre a morfologia e histoquímica de tricomas secretores em Lippia
scaberrima Sond. (Verbenaceae); Miguel et al. (2010) sobre os coléteres de espécies de Bathysa
C. Presl. (Rubiaceae); Klein et al. (2004), sobre coléteres de Simira Aubl. (Rubiaceae); Liang et
al. (2009), sobre a ação sequencial das pectinases e celulases durante o processo de formação das
cavidades em frutos de Citrus L.; Xu et. al (2011), sobre o desenvolvimento de células secretoras
e células cristalíferas em Eichornia crassipes (Mart.) Solms; Christodoulakis et al. (2011), sobre
a estrutura foliar e histoquímica de Ecballium elaterium (L.) A. Rich..
Poucos, entretanto, são os estudos conduzidos concernentes à anatomia do
desenvolvimento das estruturas secretoras. Na maior compilação de dados sobre estruturas
secretoras, Fahn (1979) reúne algumas considerações evolutivas acerca de nectários, ductos e
tricomas glandulares, baseadas principalmente no trabalho de Denissova (1975). A hipótese
considera que as estruturas secretoras originaram-se de células parenquimáticas não
especializadas, e, a partir das modificações evolutivas, surgiram progressivamente: (i) tecidos
secretores endógenos; (ii) tecidos secretores endógenos esquizógenos, com acúmulo de material
secretado; (iii) tecidos secretores com lúmen esquizolisógeno e (iv) estruturas glandulares
exógenas (tricomas glandulares). Portanto, acredita-se que no curso da evolução, as estruturas
secretoras inicalmente desenvolveram-se no interior dos tecidos vegetais, e em eventos
posteriores surgiram as cavidades e os ductos secretores e, finalmente, os tricomas glandulares.
Esta diversidade morfológica e de localização das estruturas secretoras as tornam importantes em
estudos filogenéticos (Rudall et al. 2003). Contudo, não se sabe como a evolução tem atuado na
formação de estruturas com alto grau de diferenciação morfológica.
Cavidades secretoras são estruturas constituídas pelas células epiteliais e pelo lúmen
formado pelo arranjo destas células. Em algumas espécies, além das células epiteliais, há uma
bainha de células parenquimáticas. A secreção, que é produzida pelas células epiteliais, é

8
secretada para fora da célula e armazenada no lúmen da estrutura, é quimicamente diversa. Sua
natureza é majoritariamente lipofílica, mas pode apresentar variações na composição de acordo
com a família, gênero ou espécie. As funções destas estruturas nas plantas são, em geral,
atribuídas à defesa química ou mecânica dos órgãos onde estão presentes (Fahn 1979). Cavidades
secretoras ocorrem em diversas famílias, a exemplo de Asteraceae (Russin et al. 1988),
Hypericaceae (Curtis & Lersten 1990; Ciccarelli et al. 2001), Myrtaceae (Kalachanis & Psaras
2005), Rutaceae (Turner 1986), Salicaceae (Thadeo et al. 2009) e Primulaceae, incluindo
Myrsinaceae e Theophrastaceae (Metcalfe & Chalk 1950; Otegui & Maldonado 1998).
As cavidades secretoras podem ser formadas a partir de três processos de
desenvolvimento: (i) a esquizogenia, através da qual a lamela média das células epiteliais é
desfeita, ocorrendo rearranjo das mesmas, resultando na formação de um espaço intercelular, o
lúmen, encontrada, por exemplo, nas famílias Hypericaceae (Curtis & Lersten 1990),
Polygonaceae (Curtis & Lersten 1994) e Leguminosae (Teixeira & Rocha 2009); (ii) lisigenia,
através da qual há morte celular para a formação do lúmen; e (iii) esquizolisigenia, uma
combinação dos processos anteriores, ocorrendo morte celular seguida de rearranjo espacial das
células epiteliais, ocorre, por exemplo, nas famílias, Myrtaceae (Cicarelli et al. 2008), Asteraceae
(Cury & Appezzato da Glória 2009) e Rutaceae (Chen & Wu 2010). A categoria de
desenvolvimento por lisigenia tem sido redescutida por Turner (1999) e Turner et al. (1998),
onde são apresentadas evidências que demonstram a interferência de artefatos, produzidos
durante o processamento do material, na interpretação do tipo de desenvolvimento da estrutura
secretora. Desta forma, atualmente acredita-se que nenhuma estrutura desenvolva-se apenas por
lisogenia, mas por processos combinados de esquizogenia e lisogenia.
Tricomas são estruturas resultantes da diferenciação de células da protoderme (Fahn
1988). São amplamente distribuídos entre as plantas, com ocorrência reportada em mais de 60
famílias de angiospermas (Metcalfe & Chalk 1957). Tais estruturas possuem grande variedade
morfológica e funcional (Evert 2006; Fahn 1988; Metcalfe & Chalk 1957; Wagner 1991). O
crescimento e divisão diferencial das células da protoderme são os responsáveis pela origem de
tricomas com diferentes graus de organização (Castro et al. 2009). Alguns dos tipos morfológicos
de tricomas mais comuns são as papilas (pequenas projeções de células epidérmicas), os tricomas
simples ou unicelulares, os tricomas estrelados (com forma de estrela), os peltados, que
apresentam placa discoide (cabeça pluricelular), e os tricomas dendríticos ou ramificados. Esta
diversidade morfológica dos tricomas, juntamente com outras características anatômicas que

9
apresentam, como as diferenças na espessura da parede, espessamento da cutícula, tipos celulares
encontrados e o tipo de conteúdo secretado são características taxonômicas comumente utilizadas
na descrição de táxons (Evert 2006).
Tricomas glandulares são sítios de produção de metabólitos secundários, como taninos,
mucilagem, resinas e óleos essenciais. Estas substâncias participam na defesa do organismo
contra insetos fitófagos ou organismos patógenos (Croteau & Johnson 1984; Fahn 1988), uma
vez que apresentam propriedades repelentes, a partir do contato de quimiorreceptores presentes
nos patógenos/insetos e na substância secretada (Wagner 1991). Em plantas carnívoras, diferentes
tipos tricomas glandulares são responsáveis pela produção da secreção para apreensão
(substâncias mucilaginosas) e digestão (enzimas proteolíticas) das presas. Atuam também na
regulação da temperatura do vegetal, aumentam a reflexão de luz, auxiliam a diminuição da perda
de água, podem secretar íons e poluentes, ou têm função de atrair polinizadores (Wagner 1991).
Além disso, muitos dos produtos secretados pelos tricomas glandulares apresentam importância
econômica, sendo utilizados na produção de fármacos, cosméticos e alimentos. Em Varronia
curassavica Jacq. (= Cordia verbenacea DC.), por exemplo, os tricomas são os sítios de
produção do óleo essencial, que é extraído e empregado na confecção de fármacos utilizados no
tratamento de mialgias.
Diante do exposto, considerando a importância das estruturas secretoras à família
Primulaceae sob os diferentes aspectos supracitados, a saber: filogenético, taxonômico e
medicinal, o presente trabalho objetivou esclarecer o processo de desenvolvimento das estruturas
secretoras e caracterizar quimicamente a fração volátil da secreção por elas produzida, a fim de
ampliar o conhecimento acerca dos aspectos morfológicos e químicos particulares dessas
estruturas.

10
2 - Objetivos
Geral
Caracterizar a anatomia foliar de Myrsine coriacea e Myrsine venosa, o desenvolvimento
das estruturas secretoras foliares e a composição química da secreção.
Específicos
Descrever a anatomia foliar de Myrsine coriacea e M. venosa, indicando caracteres
anatômicos diagnósticos;
Caracterizar as diferentes estruturas secretoras;
Descrever o processo de formação das cavidades secretoras nas folhas de Myrsine
coriacea e M. venosa, em diversos estágios de desenvolvimento foliar;
Caracterizar o processo de formação dos tricomas glandulares em Myrsine coriacea e M.
venosa, em diversos estágios de desenvolvimento foliar;
Comparar a ontogênese das estruturas secretoras de Myrsine coriacea e M. venosa com as
descritas para espécies de outros gêneros da família Primulaceae;
Analisar a composição química das estruturas secretoras através de testes histoquímicos;
Analisar a composição dos óleos essenciais das folhas secas nessas duas espécies;

11

12
3 - Materiais e métodos
3.1 - Espécies selecionadas
As espécies selecionadas foram Myrsine coriacea (Sw.) R.Br. ex Roem. (= Rapanea
ferruginea Mez) (Fig. 3) e M. venosa A. DC (= Rapanea venosa (Mez) A.DC.) (Fig. 4). Myrsine
coriacea é uma espécie de ampla distribuição encontrada em diversas formações florestais do
Brasil (Fig. 5A). São arbustos a arvoretas, com altura entre 3-10m. É considerada pioneira nos
estágios sucessionais e distribui-se preferencialmente em regiões abertas e na orla da mata. As
folhas e ramos terminais são cobertos total ou parcialmente por tricomas ramificados, de
colocação ferrugínea. Myrsine venosa é endêmica da flora brasileira (Fig. 5B) com ocorrências
reportadas em formações vegetais de campos rupestres e restinga, e também em regiões
sombreadas de mata. São arbustos de 3-5m de altura. As folhas apresentam na face abaxial
cavidades secretoras hialinas, que se tornam nigrescentes após a desidratação (Freitas & Carrijo
2008).
Ápices caulinares e folhas expandidas do 4o
e 5o
nós de 5 indivíduos adultos de M. venosa
foram coletados na Floresta da Tijuca – Mirante do Excelsior (22°55' e 23°00' S, 43°12'e 43°19'
W) (Figura 4), Rio de Janeiro (Brasil), no período de Maio de 2011 a fevereiro de 2012 e os de
M. coriacea coletados no Morro das Camarinhas (APA Andorinhas – Ouro Preto, Minas Gerais)
e no arboreto do Jardim Botânico do Rio de Janeiro.
Figura 3 - Myrsine coriacea. A - Hábito de um indivíduo localizado no arboreto do Jardim Botânico do Rio de
Janeiro. B - Detalhe dos ramos com folhas adultas. Caule e pecíolos cobertos por tricomas ferrugíneos. C - Detalhe
13
do pecíolo em folhas jovens, evidenciando grande densidade de tricomas cobrindo essa estrutura. D - Primórdios
foliares (■) circundados por folhas jovens (●).
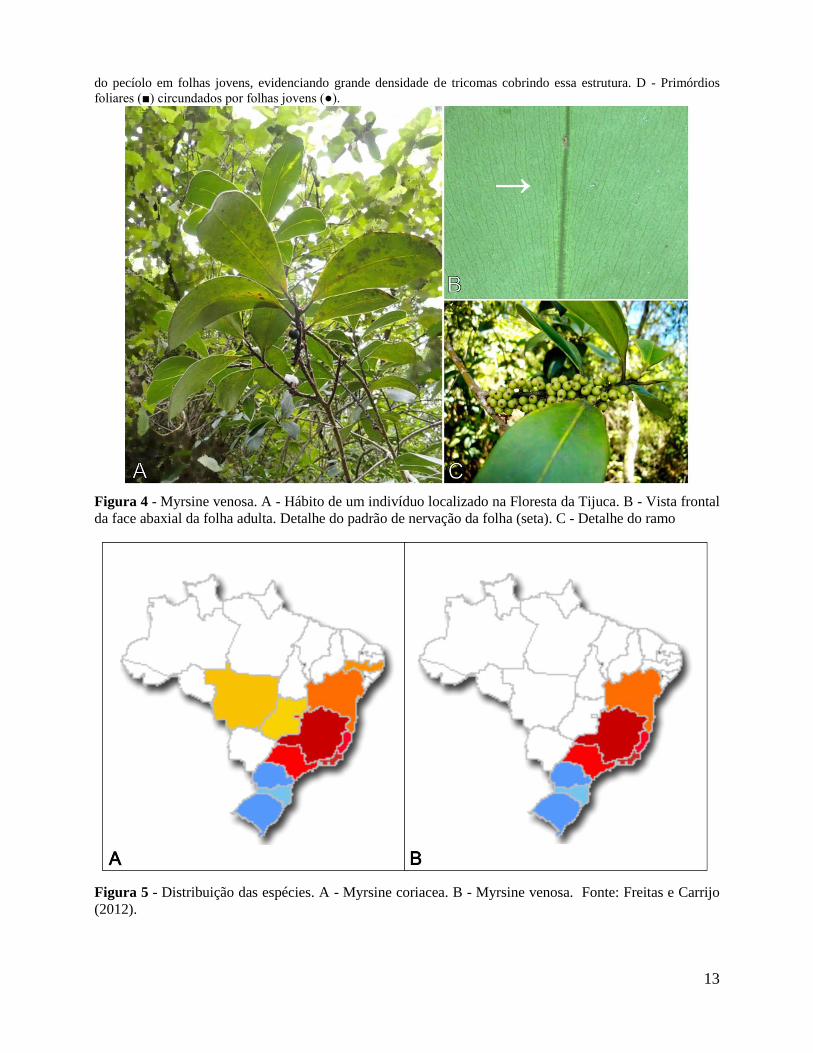
Figura 4 - Myrsine venosa. A - Hábito de um indivíduo localizado na Floresta da Tijuca. B - Vista frontal
da face abaxial da folha adulta. Detalhe do padrão de nervação da folha (seta). C - Detalhe do ramo
Figura 5 - Distribuição das espécies. A - Myrsine coriacea. B - Myrsine venosa. Fonte: Freitas e Carrijo
(2012).

14
3.2 - Microscopia óptica de campo claro
Fragmentos do terço médio de folhas completamente expandidas com cerca de 3 mm²,
meristemas apicais e primórdios foliares em diferentes estágios de desenvolvimento foram
fixados em uma solução aquosa de glutaraldeído 2,5 %, paraformaldeído 4,0 % em tampão
cacodilato 0,05 M a 7,2 pH por 48 horas (Klein 2004). Os primórdios foliares arranjam-se
alternadamente na gema caulinar, esta organização, no caule desenvolvido, corresponde à
filotaxia alterna das folhas. A fim de que se identificassem as fases de desenvolvimento das
estruturas secretoras, cada primórdio foi dissecado da gema apical e fixado e seccionado
separadamente. Estabeleceram-se, desta forma, 4 fases de desenvolvimento foliar – Fase 1: 1,5
mm; Fase 2: 3 mm; Fase 3: 6 mm; Fase 4: 12 mm (Fig. 6A e 6B), que correspondem a diferentes
estágios de desenvolvimento das estruturas secretoras.
Posteriormente, os fragmentos foram lavados em água destilada, desidratados em série
etanólica crescente e incluídos em Historresina (Leica®). As secções (4-6 µm de espessura),
obtidas em micrótomo rotativo, foram coradas com azul de toluidina 0,01 % em tampão fosfato.
A epiderme foi dissociada para a análise das estruturas epidérmicas. Nesta etapa, secções
do terço médio das folhas fixadas foram levadas à estufa por 48h, ou até o total clareamento do
material, numa solução de peróxido de hidrogênio e ácido acético (1:1) (Franklin 1945). As
amostras foram posteriormente coradas em safranina aquosa 1%.
As lâminas foram observadas e descritas ao microscópio Olympus BX 50. As imagens
foram obtidas com a câmera digital Coolsnap acoplada ao mesmo microscópio utilizando o
software Image Pro-Plus (4.0 versão para Windows).
Figura 6 - Primórdios foliares de Myrsine coriacea (A) e M. venosa (B). Gradação do mais interno
(extrema esquerda) ao mais externo (extrema direita).

15
3.3 - Microscopia de epifluorescência
Para a observação em microscopia de epifluorescência, foram utilizadas a epiderme
dissociada e secções transversais de folhas em diferentes estágios de desenvolvimento. O material
foi fixado e processado como descrito para microscopia óptica de campo claro. Para a observação
da cutícula e parede celular, a epiderme dissociada foi exposta, no escuro, aos fluorocromos
Auramina O 0,05% (Filtro de excitação (Ex) 450-480 nm/Filtro de emissão (Em) 515 nm) e
Calcofluor (Ex360-370 nm/ Em420), e para a observação do material genético nuclear, secções
transversais das folhas foram expostas ao flurocromo DAPI (Ex360-370 nm/ Em420). As lâminas
foram observadas e descritas ao microscópio Olympus BX 50. As imagens foram obtidas com a
câmera digital Coolsnap acoplada ao mesmo microscópio utilizando o software Image Pro-Plus
(4.0 versão para Windows).
3.4 - Microscopia confocal
Para observação dos tricomas em diferenciação, foi utilizada a epiderme dissociada de
folhas em diferentes estágios de desenvolvimento, fixadas e processadas como descrito para
microscopia óptica de campo claro. O material foi observado ao microscópio Confocal de
Varredura a Laser Leica TCS SPE. Utilizou-se o canal de laser de 405nm para excitação do
fluorocromo Auramina O 0,01% (Heslop-Harrison 1977) na emissão de 428 nm visando a
detecção de compostos lipídicos. As imagens foram capturadas pela aquisição direta com “Z
step” variando entre 1-1,5 μm, que gerou um conjunto de 40 seções ópticas, através do software
LAS AF LITE 2.6.0. (Leica Microsystems).
3.5 - Microscopia eletrônica de varredura
Para análises micromorfológicas, o material foi fixado como descrito para microscopia
óptica de campo claro, pós-fixado em uma solução de tetróxido de ósmio 1 M em tampão
cacodilato 0,05 M por 3 horas (Klein et al. 2004). Depois deste período realizou-se a lavagem no
mesmo tampão (2x por 30 minutos), seguido da desidratação em série cetônica crescente (30-
100% - 1 hora).
Após a desidratação, o material foi submetido ao ponto crítico no equipamento Leica EM
CPD030. Em seguida, os fragmentos foram aderidos à fita adesiva de carbono em suportes
específicos para microsopia eletrônica de varredura, e recobertos por ouro em aparelho Emitech
K550X. As análises foram realizadas no microscópio eletrônico de varredura Zeiss EVO 040.

16
3.6 - Testes histoquímicos
Para detectar a natureza química dos componentes da secreção, foram realizados testes
histoquímicos em folhas jovens e adultas, recém-coletadas e cortadas à mão livre. Os reagentes
utilizados, a substância por eles detectada e as respectivas referências estão listados na Tabela 1.
Os controles foram realizados seguindo as referências mencionadas.
Tabela 1 – Testes histoquímicos realizados, com as substâncias por eles detectadas e as respectivas
referências.
3.7 - Extração e análise do Óleo Essencial
A extração do óleo essencial de M. coriacea e M. venosa foi realizada por hidrodestilação
em aparelho de Clevenger no Laboratório de Cultura de Tecidos Vegetais (LCTV/UNIRIO) (Fig.
7). Cerca de 60g de folhas secas foram rasuradas e inseridas em balão de fundo redondo com 1L
de capacidade. Ao conteúdo no balão foram adicionados 600mL de água destilada e iniciada a
extração, por três horas. Em seguida, o óleo essencial teve seu volume mensurado em µL. Ao
óleo, ainda na coluna graduada do aparelho de Clevenger, foram acrescentados 2,0 mL de
diclorometano (CH2Cl2). A solução de óleo essencial em diclorometano foi recolhida juntamente
com o solvente e o hidrolato e armazenado em freezer, a aproximadamente -14oC, até a análise. O
rendimento do processo de extração do óleo essencial (µL/mg) foi calculado dividindo-se o
Substância detectada Teste Referência
Amido Lugol Sass (1951)
Substâncias lipofílicas
Sudan III Johansen (1940)
Sudan IV Johansen (1940)
Sudan Black B Pearse (1980)
Sudan Red 7B Brundrett et al. (1991)
Lipídios ácidos Sulfato azul do Nilo Cain (1947)
Óleos essenciais e óleoresinas Reagente de NADI David & Carde (1964)
Terpenóides com grupo
carbonilo
2,4-Dinitrofenilhidrazina Ganter & Jollés (1969)
Borracha Oil red O Jayabalan & Shah (1986)
Compostos fenólicos gerais Cloreto de ferro III Johansen (1940)
Taninos Vanilina clorídrica Mace & Howell (1974)
Lignina Floroglucinol clorídrico Sass (1951)
Flavonóides Hidróxido de potássio 5% Costa (1982)

17
volume de óleo essencial obtido (µL) pela massa fresca de folhas (mg) da qual o óleo essencial
foi extraído.
A composição química do óleo essencial foi determinada por cromatografia em fase
gasosa com a colaboração do Dr. Humberto Ribeiro Bizzo, no Laboratório de Óleos Essenciais
da Empresa Brasileira de Pesquisa Agropecuária (EMBRAPA). Foi utilizado cromatógrafo
Perkin-Elmer Autosystem, com detector por ionização de chama (CG-DIC) e coluna capilar
apolar HP-5MS (fase estacionária de 5% difenil 95% dimetilpolisiloxano; 30 m de comprimento;
0,25 mm de diâmetro interno; 0,25 µm de espessura de filme), sendo hidrogênio o gás de arraste
(1mL min-1
a 40°C). Foi injetado um volume de 0,5µl da amostra de óleo essencial em solução de
diclorometano no modo sem divisão de amostra (splitless) para M. coriacea e com divisão de
amostra (split) para M. venosa, com taxa de divisão 1:20. O injetor foi mantido em 250oC e o
detector em 280oC. A programação de temperatura do cromatógrafo foi a seguinte: 60-240
oC,
com taxa de aquecimento de 3 oC min
-1, e isotérmico (240
oC) por dez minutos.
Para a análise por cromatografia em fase gasosa acoplada à espectrometria de massas
(CG-EM) foi utilizado cromatógrafo Agilent 6890 acoplado a um detector por espectrometria de
massas Agilent 5973. Foi utilizada uma coluna capilar HP-5MS. O gás de arraste utilizado foi
hélio (1mL min-1
), sendo a ionização eletrônica (feixe de 70eV). A programação de temperatura
foi 60-240oC, com taxa de aquecimento de 3
oC min
-1, e isotérmico por dez minutos, sendo a
temperatura do injetor a mesma utilizada para as análises com CG-DIC.
Figura 7 - Aparelho de Clevenger esquematizado. A – Condensador; B – Balão volumétrico, com a
amostra e água destilada; C – Manta de aquecimento; D – Isopor com água gelada, que permite o
resfriamento do condensador.

18
O índice de retenção linear (IRL) foi calculado através da seguinte fórmula:
A identificação dos componentes do óleo essencial foi realizada por comparação dos
espectros de massas das amostras com os espectros de massas da biblioteca Wiley 6ª edição e por
comparação dos índices de retenção de Kovats calculados e os índices de retenção linear
calculados (IKC e IRLC) com os compilados por Adams (2007).
O índice de retenção de Kovats (IK) foi calculado utilizando-se os dados de tempo de
retenção (TR) em minutos de uma série homóloga de hidrocarbonetos alifáticos saturados (C7-
C26) nas mesmas condições das análises cromatográficas com o DIC através da seguinte
fórmula:
na qual TRn = tempo de retenção da molécula;
TRn-1 = tempo de retenção do alcano anterior à molécula;
TRn+1 = tempo de retenção do alcano posterior à molécula;
n = número de átomos de carbono do alcano anterior à molécula.

19

20
4 - Resultados
4.1 - Anatomia das folhas expandidas
4.1.1 - Pecíolo
Em secção transversal, o pecíolo nas duas espécies é revestido por epiderme
uniestratificada, constituído por células arredondas ou retangulares, que apresentam parede
periclinal externa com cutícula espessa (Fig. 8A). Neste tecido são observados tricomas
pluricelulares glandulares ramificados somente em Myrsine coriacea (Fig. 8B). O colênquima
angular subepidérmico, em ambas as espécies, é formado por 4-5 camadas de células (Fig. 8A).
Adjacente ao colênquima encontra-se o tecido parenquimático cujas células têm formato circular
(Fig. 8C). Nas células do parênquima e do colênquima são encontrados grãos de amido (Fig. 8C).
No parênquima também são observados idioblastos contendo cristais em forma de drusas (Fig.
8D), e idioblastos mucilaginosos distribuídos pelo tecido. (Fig. 8E).
Em ambas as espécies, cavidades secretoras são encontradas imersas na região do
colênquima, parênquima cortical e medula do pecíolo. Em M. venosa também ocorrem canais
secretores nestas porções. As cavidades e canais secretores, em secção transversal, têm formato
circular ou elíptico, e são formados por epitélio secretor bem delimitado, e lúmen conspícuo (Fig.
8F, 8G). Em geral, M. coriacea encerra uma cavidade na região interna do sistema vascular,
enquanto em M. venosa observam-se 2 ou mais cavidades/ductos na porção interna do sistema
vascular (Fig.8F, 8G). Em ambas as espécies, o sistema vascular é colateral e arranja-se em forma
de arco na porção distal do pecíolo, enquanto na porção proximal, o sistema vascular é fechado.
O sistema vascular é circundado por uma bainha de fibras não lignificadas (Fig. 8F e 8G).
4.1.2 - Lâmina Foliar
Ambas as espécies apresentam folhas hipoestomáticas, com estômatos do tipo anisocítico
(Fig. 9A) localizados no mesmo nível das demais células epidérmicas, e epiderme
uniestratificada (Fig. 9B). Em vista frontal, as células comuns, nas faces abaxial e adaxial da
epiderme, têm paredes anticlinais sinuosas em M. venosa (Fig. 9C) e retas em M. coriacea (Fig.
9D). Na face abaxial das paredes periclinais externas das células comuns da epiderme, em ambas
as espécies, observam-se estrias epicuticulares organizadas paralelamente ao maior eixo das
células (Fig. 9E). A face adaxial é irregular em M. coriacea (Fig. 9F) e estriada em M. venosa
(Fig. 9H).

21
Em secção transversal, as células comuns da epiderme têm formato poligonal, e são
recobertas por cutícula fina (Fig. 9B), estas células podem secretar conteúdo fenólico, observado
a partir na reação ao cloreto férrico (Fig. 9G). Em M. venosa, geralmente, as células da epiderme
adaxial são mais largas no sentido periclinal (Fig. 9B). O conteúdo vacuolar destas células é
denso, visivelmente marcado pelo corante azul de toluidina, indicando a presença de conteúdo
mucilaginoso. Tricomas glandulares peltados ocorrem nas duas faces da epiderme (Fig. 9D, 9E,
9F e 9H). Em M. coriacea, também ocorrem tricomas do tipo pluricelular ramificado Os tricomas
pluricelulares ramificados são formados por um número variável de células (Fig. 9D, 9F e 9I). Os
tricomas peltados constituem-se de uma célula basal, uma célula do pedúnculo, e cerca de 10
células da cabeça em M. coriacea (Fig. 9J) e 30 em M. venosa (Fig. 9K). As células do pedúnculo
apresentam as paredes anticlinais com forte espessamento cuticular. Esse tipo de tricoma é
observado em regiões de depressão da epiderme (Fig. 9E, 9H e 9L).
O mesofilo é dorsiventral, o parênquima paliçádico apresenta uma camada de células e o
parênquima lacunoso é formado por cerca de sete estratos celulares. Bem como as células
epidérmicas, as células do parênquima paliçádico podem secretar compostos fenólicos. Ao longo
do mesofilo observam-se cavidades secretoras distribuídas esparsamente nas porções do terço
médio, ápice e bordo (Fig. 9M). Em M. venosa, também são observados ductos secretores, que se
diferenciam das cavidades por apresentar, em vista frontal, lúmen com maior extensão (Fig. 9N).
No mesofilo, em ambas as espécies, também são encontrados idioblastos contendo drusas.
As principais diferenças acerca da anatomia foliar em M. coriacea e M. venosa estão
listadas na Tabela 2.
Tabela 2 – Características anatômicas distintivas observadas nas folhas de Myrsine coriacea e M. venosa.
(+) = presente; (-) = ausente.
Myrsine coriacea Myrsine venosa
Cavidades secretoras na medula do pecíolo 1 >2
Tricomas glandulares pluricelulares + -
Número de células da cabeça – Tricomas
peltados
10 30
Coloração da secreção nas cavidades secretoras Amarela Incolor
Ductos secretores - +
Parede anticlinal das células epidérmicas na
face adaxial (VF)
Reta Sinuosa
Epiderme com células alongadas no sentido
periclinal (ST)
- +
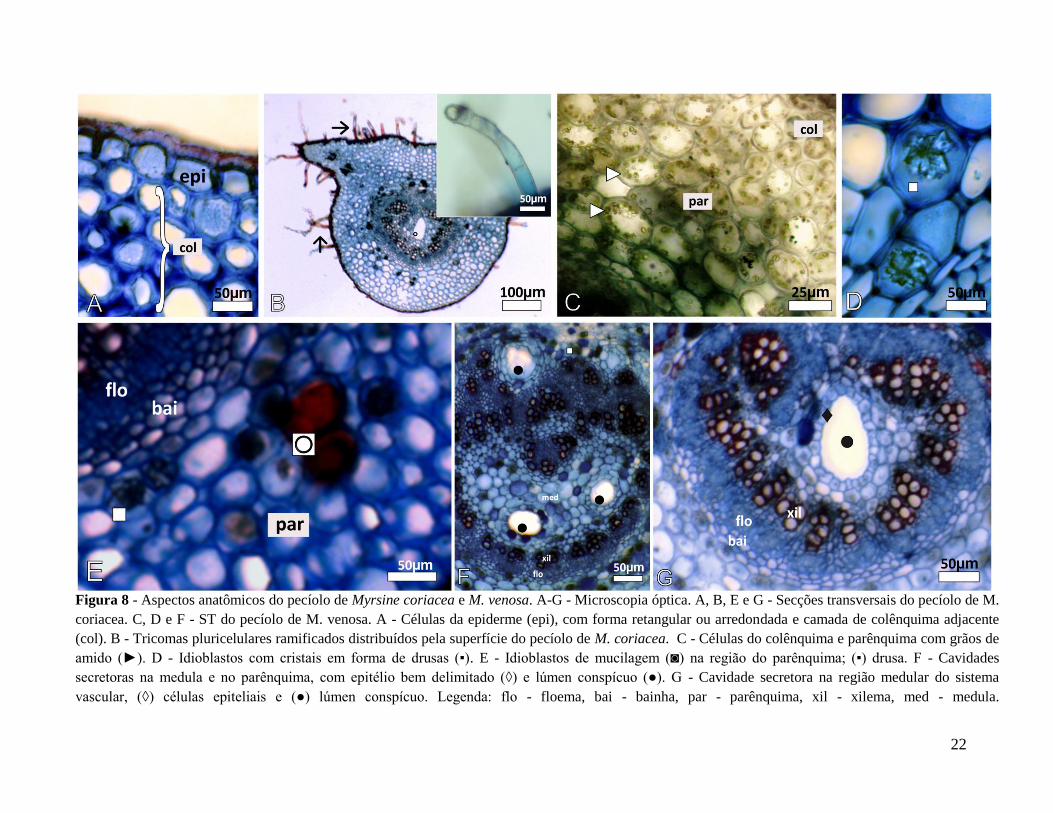
22
Figura 8 - Aspectos anatômicos do pecíolo de Myrsine coriacea e M. venosa. A-G - Microscopia óptica. A, B, E e G - Secções transversais do pecíolo de M.
coriacea. C, D e F - ST do pecíolo de M. venosa. A - Células da epiderme (epi), com forma retangular ou arredondada e camada de colênquima adjacente
(col). B - Tricomas pluricelulares ramificados distribuídos pela superfície do pecíolo de M. coriacea. C - Células do colênquima e parênquima com grãos de
amido (►). D - Idioblastos com cristais em forma de drusas (▪). E - Idioblastos de mucilagem (◙) na região do parênquima; (▪) drusa. F - Cavidades
secretoras na medula e no parênquima, com epitélio bem delimitado (◊) e lúmen conspícuo (●). G - Cavidade secretora na região medular do sistema
vascular, (◊) células epiteliais e (●) lúmen conspícuo. Legenda: flo - floema, bai - bainha, par - parênquima, xil - xilema, med - medula.

23
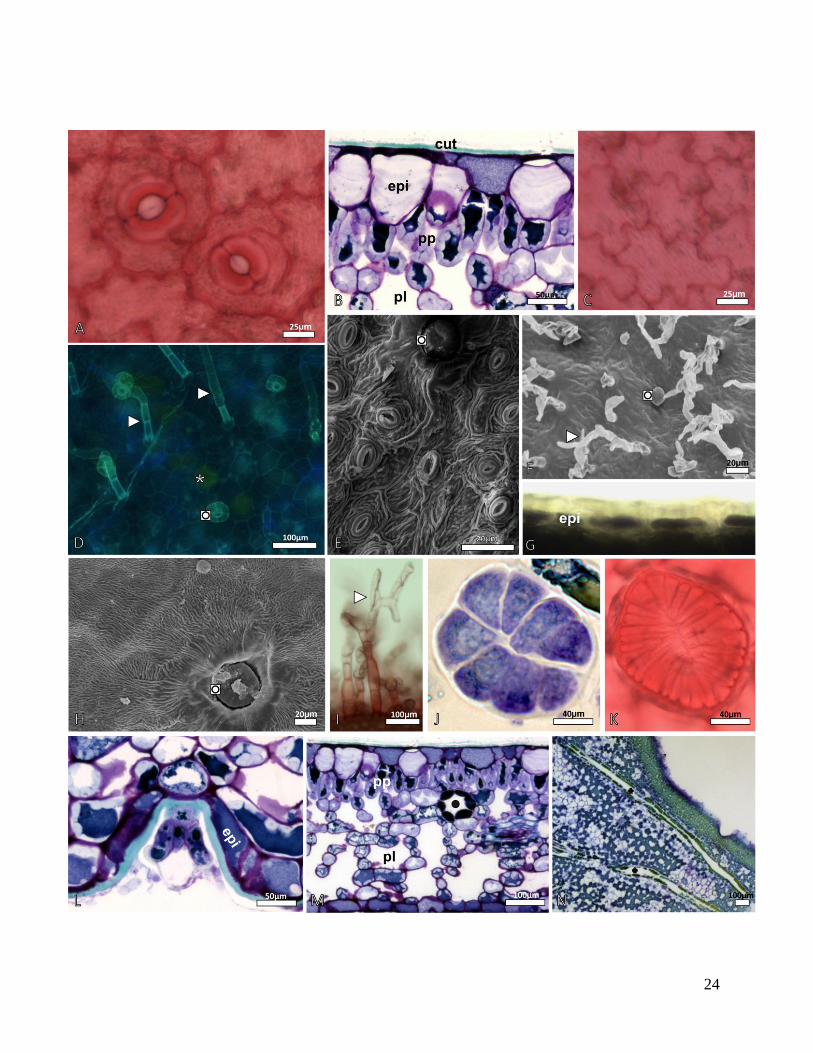
Figura 9 - Aspectos da anatomia foliar de Myrsine coriacea e M. venosa. A-D, H-N -
Microscopia óptica (MO), D - Microscopia de fluorescência (MF) (fluorocromos - auramina e
calcofluor), E-G - Microscopia eletrônica de varredura (MEV). A - Vista frontal da face abaxial
da epiderme de M. venosa, evidenciando os estômatos do tipo anisocíticos. B - ST do mesofilo de
M. venosa. Epiderme uniestratificada, com células volumosoas alongadas periclinalmente. C -
VF da epiderme adaxial de M. venosa destacando as paredes anticlinais sinuosas das células
comuns da epiderme. D - VF da epiderme adaxial de M. coriacea, evidenciando as células
comuns da epiderme com paredes anticlinais retas. Notar a presença de tricomas pluricelulares
(►), tricomas peltados (◙) e cavidades secretoras imersa no mesofilo (*). E - VF da epiderme
abaxial de M. venosa, onde se observam estriações epicuticulares na parede periclinal externa das
células da epiderme; (◙) tricoma peltado. F - VF da epiderme adaxial de M. coriacea, com
superfície epicuticular lisa. G - VF da epiderme adaxial de M. venosa, evidenciando superfície
epicuticular estriada. H - ST do mesofilo de M. coriacea, reação positiva das células epidérmicas
ao cloreto férrico, indicando a presença de compostos fenólicos. I - ST do mesofilo de M.
coriacea, com detalhe do tricoma pluricelular com ramificações (►). J e K - VF do mesofilo de
M. coriacea e M. venosa, respectivamente, com células da cabeça do tricoma peltado. L - ST
mesofilo M. venosa, com tricoma peltado localizado em região de depressão da epiderme. M - ST
do mesofilo de M. venosa, (●) cavidade secretora. N - Secção paradérmica (SP) do mesofilo de
M. venosa, evidenciando ductos secretores (♣). Legenda: cut-cutícula; epi-epiderme; pp-
parênquima paliçádico; pl-parênquima lacunoso.
24
25
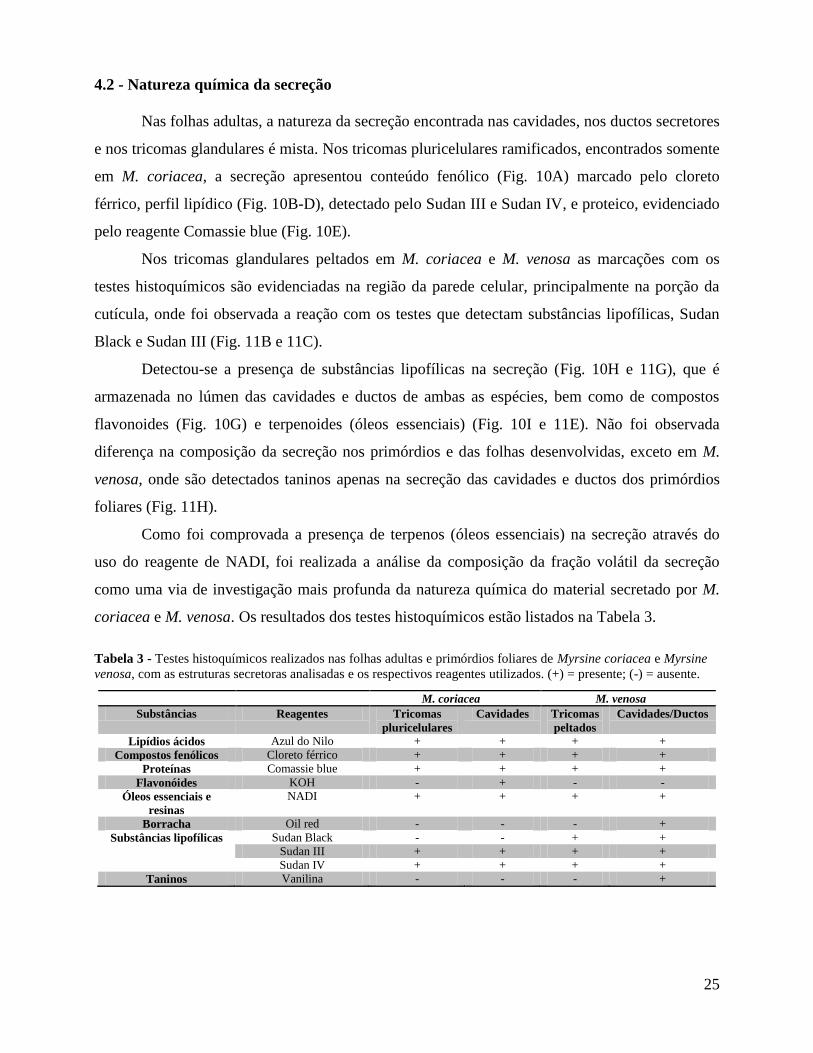
4.2 - Natureza química da secreção
Nas folhas adultas, a natureza da secreção encontrada nas cavidades, nos ductos secretores
e nos tricomas glandulares é mista. Nos tricomas pluricelulares ramificados, encontrados somente
em M. coriacea, a secreção apresentou conteúdo fenólico (Fig. 10A) marcado pelo cloreto
férrico, perfil lipídico (Fig. 10B-D), detectado pelo Sudan III e Sudan IV, e proteico, evidenciado
pelo reagente Comassie blue (Fig. 10E).
Nos tricomas glandulares peltados em M. coriacea e M. venosa as marcações com os
testes histoquímicos são evidenciadas na região da parede celular, principalmente na porção da
cutícula, onde foi observada a reação com os testes que detectam substâncias lipofílicas, Sudan
Black e Sudan III (Fig. 11B e 11C).
Detectou-se a presença de substâncias lipofílicas na secreção (Fig. 10H e 11G), que é
armazenada no lúmen das cavidades e ductos de ambas as espécies, bem como de compostos
flavonoides (Fig. 10G) e terpenoides (óleos essenciais) (Fig. 10I e 11E). Não foi observada
diferença na composição da secreção nos primórdios e das folhas desenvolvidas, exceto em M.
venosa, onde são detectados taninos apenas na secreção das cavidades e ductos dos primórdios
foliares (Fig. 11H).
Como foi comprovada a presença de terpenos (óleos essenciais) na secreção através do
uso do reagente de NADI, foi realizada a análise da composição da fração volátil da secreção
como uma via de investigação mais profunda da natureza química do material secretado por M.
coriacea e M. venosa. Os resultados dos testes histoquímicos estão listados na Tabela 3.
Tabela 3 - Testes histoquímicos realizados nas folhas adultas e primórdios foliares de Myrsine coriacea e Myrsine
venosa, com as estruturas secretoras analisadas e os respectivos reagentes utilizados. (+) = presente; (-) = ausente.
M. coriacea M. venosa
Substâncias Reagentes Tricomas
pluricelulares
Cavidades Tricomas
peltados
Cavidades/Ductos
Lipídios ácidos Azul do Nilo + + + +
Compostos fenólicos Cloreto férrico + + + +
Proteínas Comassie blue + + + +
Flavonóides KOH - + - -
Óleos essenciais e
resinas
NADI + + + +
Borracha Oil red - - - +
Substâncias lipofílicas Sudan Black - - + +
Sudan III + + + +
Sudan IV + + + +
Taninos Vanilina - - - +
26
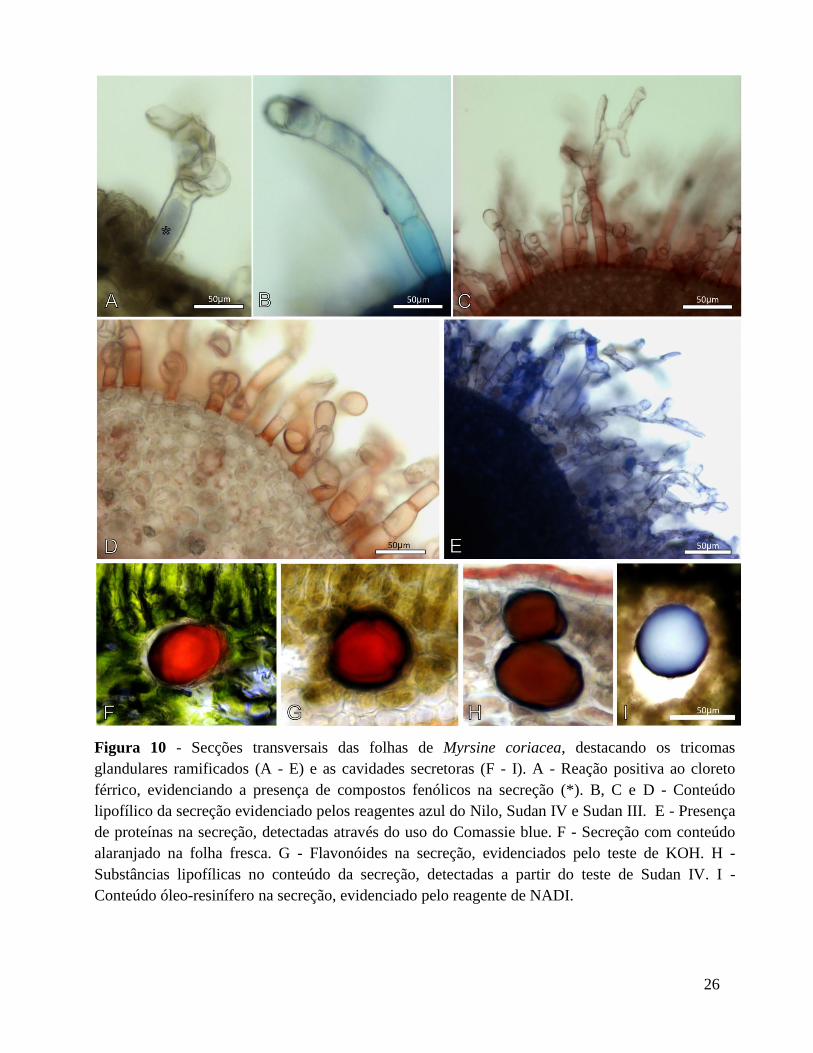
Figura 10 - Secções transversais das folhas de Myrsine coriacea, destacando os tricomas
glandulares ramificados (A - E) e as cavidades secretoras (F - I). A - Reação positiva ao cloreto
férrico, evidenciando a presença de compostos fenólicos na secreção (*). B, C e D - Conteúdo
lipofílico da secreção evidenciado pelos reagentes azul do Nilo, Sudan IV e Sudan III. E - Presença
de proteínas na secreção, detectadas através do uso do Comassie blue. F - Secreção com conteúdo
alaranjado na folha fresca. G - Flavonóides na secreção, evidenciados pelo teste de KOH. H -
Substâncias lipofílicas no conteúdo da secreção, detectadas a partir do teste de Sudan IV. I -
Conteúdo óleo-resinífero na secreção, evidenciado pelo reagente de NADI.
27
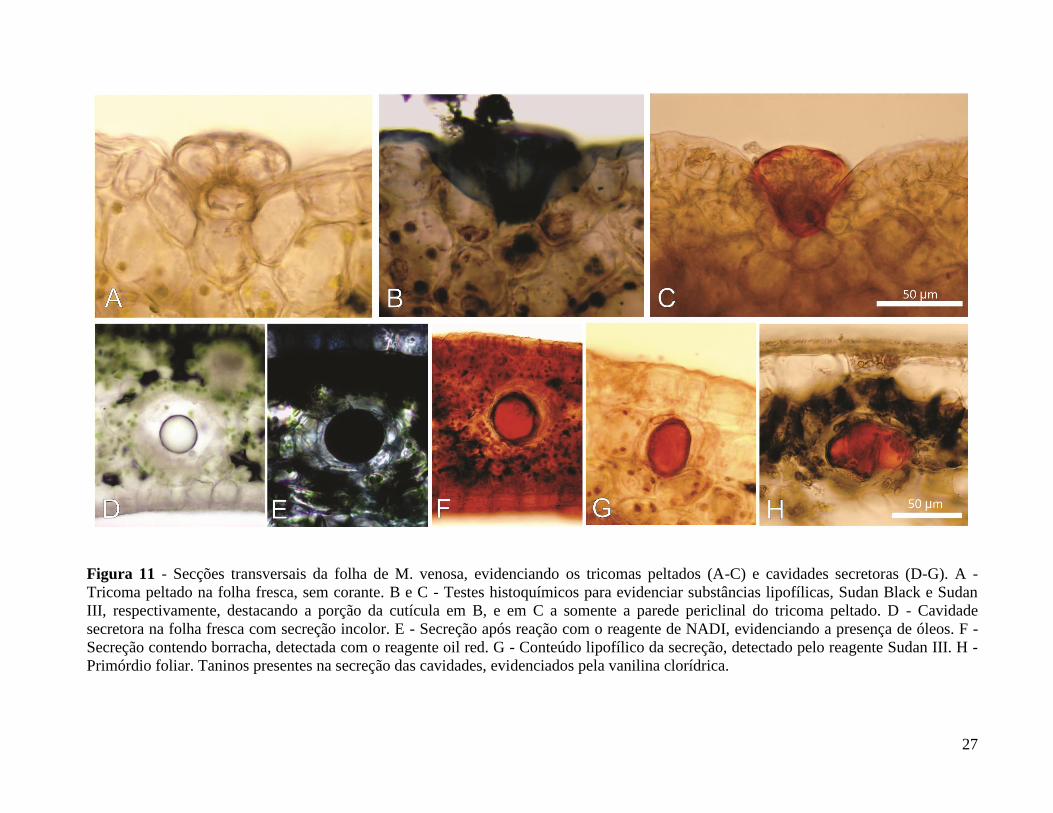
Figura 11 - Secções transversais da folha de M. venosa, evidenciando os tricomas peltados (A-C) e cavidades secretoras (D-G). A -
Tricoma peltado na folha fresca, sem corante. B e C - Testes histoquímicos para evidenciar substâncias lipofílicas, Sudan Black e Sudan
III, respectivamente, destacando a porção da cutícula em B, e em C a somente a parede periclinal do tricoma peltado. D - Cavidade
secretora na folha fresca com secreção incolor. E - Secreção após reação com o reagente de NADI, evidenciando a presença de óleos. F -
Secreção contendo borracha, detectada com o reagente oil red. G - Conteúdo lipofílico da secreção, detectado pelo reagente Sudan III. H -
Primórdio foliar. Taninos presentes na secreção das cavidades, evidenciados pela vanilina clorídrica.

28
4.3 - Ontogênese das estruturas secretoras
4.3.1 - Tricomas peltados
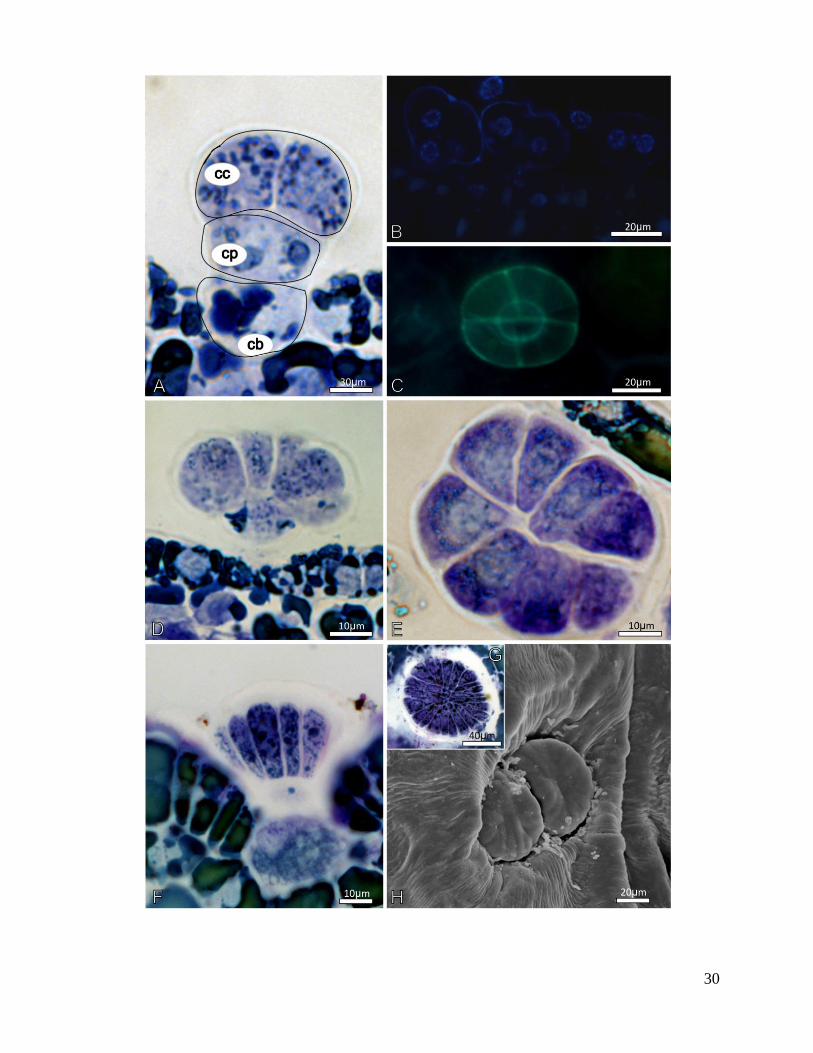
A sequência de eventos da ontogenia dos tricomas acontece nas fases iniciais do
desenvolvimento foliar, que corresponde às fases 1 a 3 do desenvolvimento do primórdio, após
essa etapa os tricomas peltados estão totalmente diferenciados.
O desenvolvimento dos tricomas peltados, observados em M. coriacea e em M. venosa,
incia-se com a expansão de uma célula protodérmica e sua posterior divisão periclinal, formando
a célula basal e a apical (estas etapas ocorrem quando o primórdio foliar atinge 1,5 mm,
correspondente à fase 1). A célula apical, por sua vez, sofre nova divisão periclinal, formando a
célula secretora da cabeça (Fig. 12A e 12B). A nova divisão ocorre no sentido anticlinal, de
forma simétrica, dando origem a duas células da cabeça (Fig. 12A e 12B). Cada nova célula sofre
seguidas divisões anticlinais, até resultar na estrutura completamente desenvolvida, com 10
células em M. coriacea e 30 células em M. venosa (Fig. 12C e 12D). Os tricomas atingem o
estágio completo de desenvolvimento nos primórdios com cerca de 6 mm, fase 3.
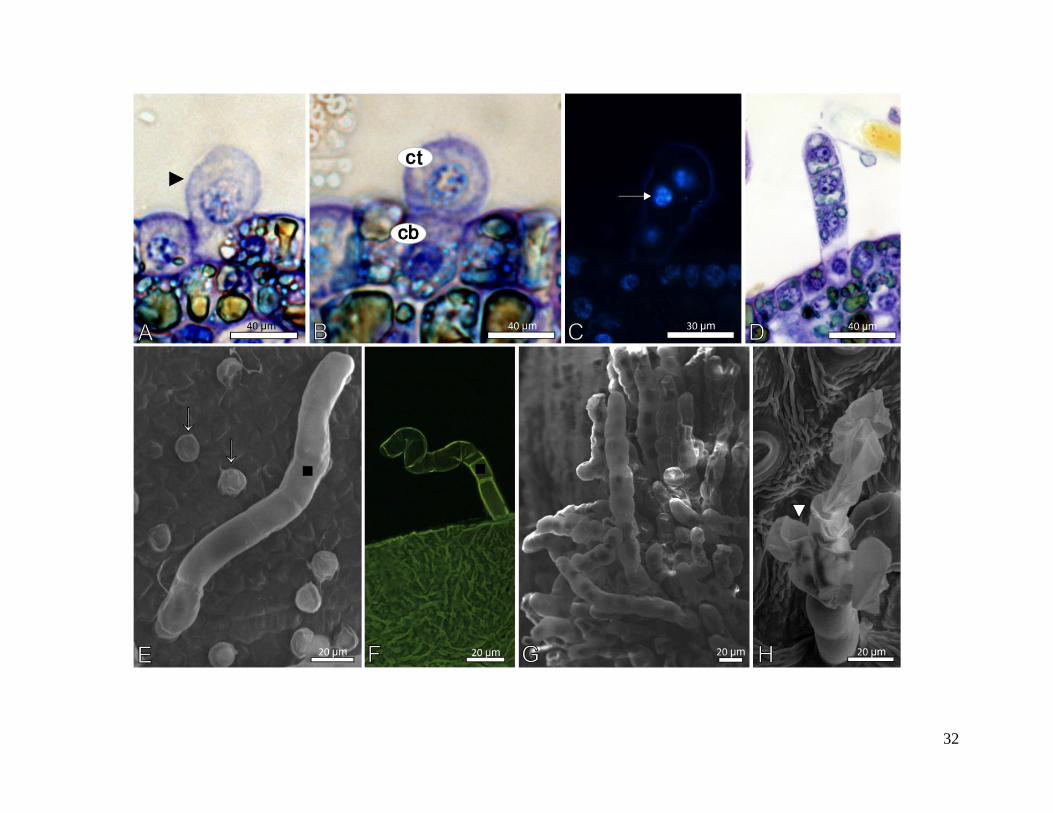
4.3.2 - Tricomas pluricelulares
A ontogenia dos tricomas pluricelulares, encontrados somente em M. coriacea, inicia-se
com a expansão da célula inicial, de origem protodérmica (Fig. 13A) e posterior divisão
periclinal (Fig. 13B). Esta divisão resulta na formação de uma célula basal e em outra apical,
ambas bastante vacuoladas e com núcleo volumoso (Fig. 13B). A célula apical sofre seguidas
divisões periclinais (Fig. 13C, 13D, 13E), formando um longo tricoma pluricelular com formato
cilíndrico (Fig. 13F, 13G). Ao final de sua diferenciação é possível observar cada célula que
forma o tricoma. Em alguns tricomas, células da porção apical podem sofrer divisões anticlinais,
formando uma estrutura ramificada (Fig. 13H).

29
Figura 12 - Ontogenia dos tricomas glandulares peltados das folhas de Myrsine coriacea e
Myrsine venosa. A - Células iniciais do tricoma peltado, que se originam a partir de uma célula
da protoderme. Inicialmente, a célula precursora do tricoma divide-se periclinalmente, formando
a célula da base (cb) e a célula do pedúnculo (cp). Posteriormente a essa divisão, a célula do
pedúnculo sofre nova divisão periclinal, formando a célula da cabeça (cc). Esta, por sua vez,
sofrerá divisão anticlinal. B - Tricomas peltados em estágio inicial, com o fluorocromo DAPI,
que evidencia o material genético nuclear. Microscopia de epifluorescência. C - Visão frontal do
primórdio foliar, evidenciando o tricoma peltado com quatro células da cabeça. Microscopia de
epifluorescência. D - Secção transversal do primórdio foliar. Nesta fase, as células da cabeça do
tricoma sofrem novas divisões anticlinais. E - Vista frontal do tricoma peltado, após as divisões
anticlinais das células da cabeça. F - Secção transversal da folha desenvolvida, com o tricoma
peltado localizado em região de depressão da epiderme. G - Vista frontal da epiderme de M.
venosa, evidenciando tricoma peltado com cerca de 30 células da cabeça. H - Superfície foliar de
M. venosa, evidenciando tricomas peltados em região de depressão da epiderme. Microscopia
eletrônica de varredura. A, D, E e G - Microscopia óptica.
30

31
Figura 13 - Ontogenia dos tricomas glandulares pluricelulares das folhas de Myrsine coriacea. A
- Célula protodérmica do primórdio foliar em expansão (►). B - Células iniciais do tricoma
pluricelular, após a divisão periclinal da célula protodérmica, que resulta na formação de uma
célula da base (cb) e uma célula do tricoma (ct). C - Tricoma pluricelular, após divisão anticlinal
da célula do tricoma. Células com núcleo evidenciado (seta) pelo fluorocromo DAPI, que detecta
material genético nuclear. Microscopia de epifluorescência. D - Vista transversal do tricoma, com
as células apresentando pequenos vacúolos, que são densamente preenchidos. E - Superfície do
primórdio foliar evidenciando tricomas pluricelulares em diferentes fases de desenvolvimento:
em estágio inicial (seta) e tricoma pluricelular desenvolvido (▪). F - Tricoma pluricelular com
Auramina como fluorocromo. G - Superfície do primórdio densamente preenchida por tricomas
pluricelulares. H - Superfície da folha adulta, evidenciando tricoma pluricelular após a divisão
periclinal de células subapicais, formando uma estrutura ramificada (►). A, B e D - Microscopia
óptica. E, G e H - Microscopia eletrônica de varredura. F - Microscopia confocal de varredura a
laser.
32

33
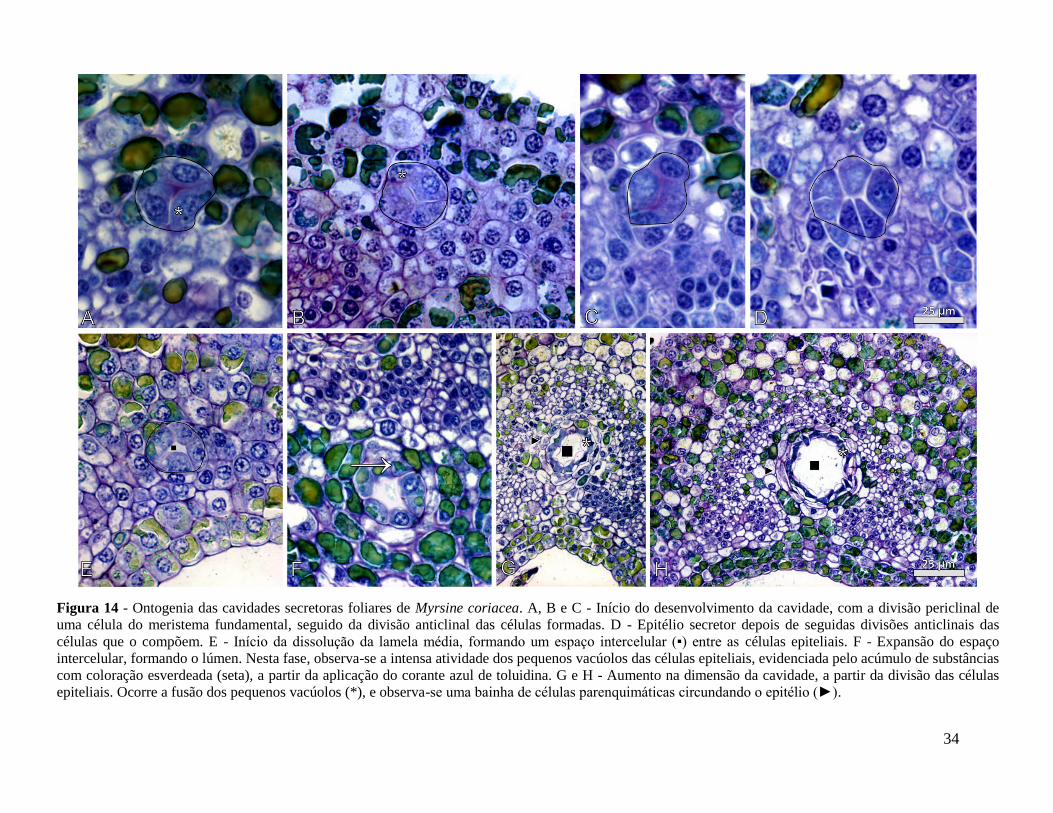
4.3.3 - Cavidades e ductos secretores
As cavidades secretoras, em ambas as espécies, e os ductos secretores, em M. venosa, são
formados anteriormente à diferenciação das células do mesofilo. No primórdio foliar com 1,5 mm
(fase 1), o mesofilo é composto pelas células do meristema fundamental, caracterizadas pela
presença de núcleo proeminente, esférico e volumoso (Fig. 15B). O processo de ontogenia dessas
estruturas é semelhante em ambas as espécies.
Quando a folha atinge o estágio de 3 mm, fase 2, uma célula do meristema fundamental
divide-se no plano periclinal; subsequentemente, as células formadas sofrem divisão no plano
anticlinal, resultando em um arranjo de quatro células (Fig. 14A-D; 15A-C). Estas são facilmente
distintas das demais células do tecido pelo arranjo em roseta, e pela presença de pequenos
vacúolos densamente preenchidos encontrados no citoplasma. Novas divisões anticlinais ocorrem
nestas células e quando o arranjo atinge cerca de 6-8 células, inicia-se a dissolução da lamela
média entre elas, promovendo o seu afastamento (Fig. 14E e 15D). Na medida em que novas
divisões celulares ocorrem, a distância entre as células aumenta e forma-se um grande espaço
intercelular, o lúmen (Fig. 14F-H; 15E, 15F). Estas células que circundam e delimitam o lúmen
constituem as células epiteliais, ou epitélio secretor. As células epiteliais variam em tamanho e
formato, podendo ser circulares, tabulares ou de formato irregular (Fig. 14B, D e H). Após o
desenvolvimento completo das cavidades e ductos secretores, o núcleo dessas células deixa de ser
visível e o vacúolo é preenchido por substâncias de natureza fenólica (Fig. 14H e 17F). Por vezes,
as células que circundam o epitélio secretor arranjam-se de maneira a formar uma bainha
parenquimática (Fig. 14H e 15F). A ontogenia das cavidades não ocorre de maneira simultânea
nos primórdios foliares, sendo observadas cavidades em diferentes estágios de desenvolvimento
numa mesma fase do primórdio (15G).
Os ductos secretores são estruturas lineares e destacam-se das cavidades por apresentarem
o lúmen da estrutura com maior extensão (Fig. 15J). Ao final do desenvolvimento do primórdio e
expansão foliar, as cavidades e ductos distribuem-se por todo o mesofilo, nervura mediana e
pecíolo (Fig. 15H). O processo secretor promovido pelas células epiteliais se dá somente a partir
do completo desenvolvimento da cavidade, que corresponde às fases de folha jovem e
completamente expandidas. As células epiteliais secretam as substâncias e as enviam de forma
não destrutiva, ou seja, através de um processo merócrino, ao lúmen. A secreção tem cor
alaranjada em M. coriacea, e é incolor em M. venosa.
34
Figura 14 - Ontogenia das cavidades secretoras foliares de Myrsine coriacea. A, B e C - Início do desenvolvimento da cavidade, com a divisão periclinal de
uma célula do meristema fundamental, seguido da divisão anticlinal das células formadas. D - Epitélio secretor depois de seguidas divisões anticlinais das
células que o compõem. E - Início da dissolução da lamela média, formando um espaço intercelular (▪) entre as células epiteliais. F - Expansão do espaço
intercelular, formando o lúmen. Nesta fase, observa-se a intensa atividade dos pequenos vacúolos das células epiteliais, evidenciada pelo acúmulo de substâncias
com coloração esverdeada (seta), a partir da aplicação do corante azul de toluidina. G e H - Aumento na dimensão da cavidade, a partir da divisão das células
epiteliais. Ocorre a fusão dos pequenos vacúolos (*), e observa-se uma bainha de células parenquimáticas circundando o epitélio (►).
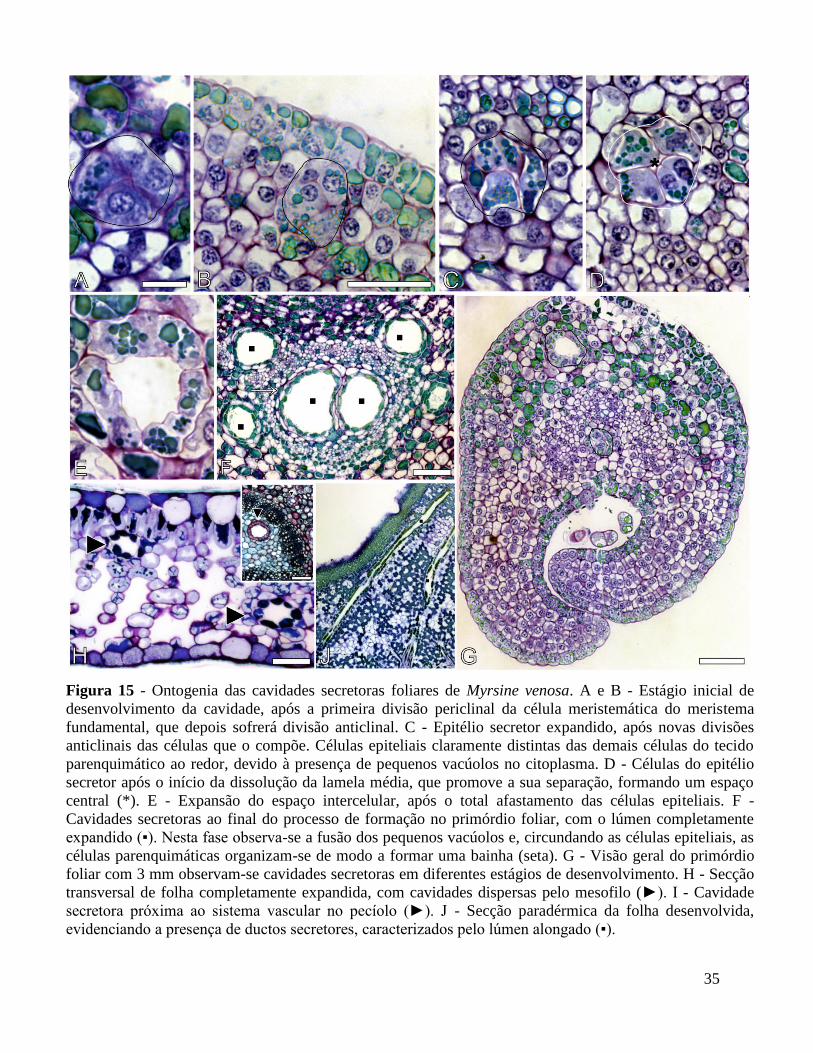
35
Figura 15 - Ontogenia das cavidades secretoras foliares de Myrsine venosa. A e B - Estágio inicial de
desenvolvimento da cavidade, após a primeira divisão periclinal da célula meristemática do meristema
fundamental, que depois sofrerá divisão anticlinal. C - Epitélio secretor expandido, após novas divisões
anticlinais das células que o compõe. Células epiteliais claramente distintas das demais células do tecido
parenquimático ao redor, devido à presença de pequenos vacúolos no citoplasma. D - Células do epitélio
secretor após o início da dissolução da lamela média, que promove a sua separação, formando um espaço
central (*). E - Expansão do espaço intercelular, após o total afastamento das células epiteliais. F -
Cavidades secretoras ao final do processo de formação no primórdio foliar, com o lúmen completamente
expandido (▪). Nesta fase observa-se a fusão dos pequenos vacúolos e, circundando as células epiteliais, as
células parenquimáticas organizam-se de modo a formar uma bainha (seta). G - Visão geral do primórdio
foliar com 3 mm observam-se cavidades secretoras em diferentes estágios de desenvolvimento. H - Secção
transversal de folha completamente expandida, com cavidades dispersas pelo mesofilo (►). I - Cavidade
secretora próxima ao sistema vascular no pecíolo (►). J - Secção paradérmica da folha desenvolvida,
evidenciando a presença de ductos secretores, caracterizados pelo lúmen alongado (▪).

36
4.4 - Óleos essenciais
O rendimento do óleo essencial extraído de M. coriacea foi de 1,16µL/g e de 1,22µL/g
em M. venosa. A coloração do óleo essencial, extraído das folhas secas de ambas as espécies, é
diferente da coloração da secreção observada nas cavidades secretoras nas folhas frescas,
enquanto em M. coriacea o óleo essencial extraído é amarelado (Fig. 16A), a secreção nas
cavidades é alaranjada. Em M. venosa o óleo essencial extraído apresenta coloração esverdeada
(Fig. 16B), enquanto a secreção nas cavidades/ductos é incolor.
Figura 16 – Coloração do óleo essencial extraído de A. Myrsine coriacea; B. M. venosa.
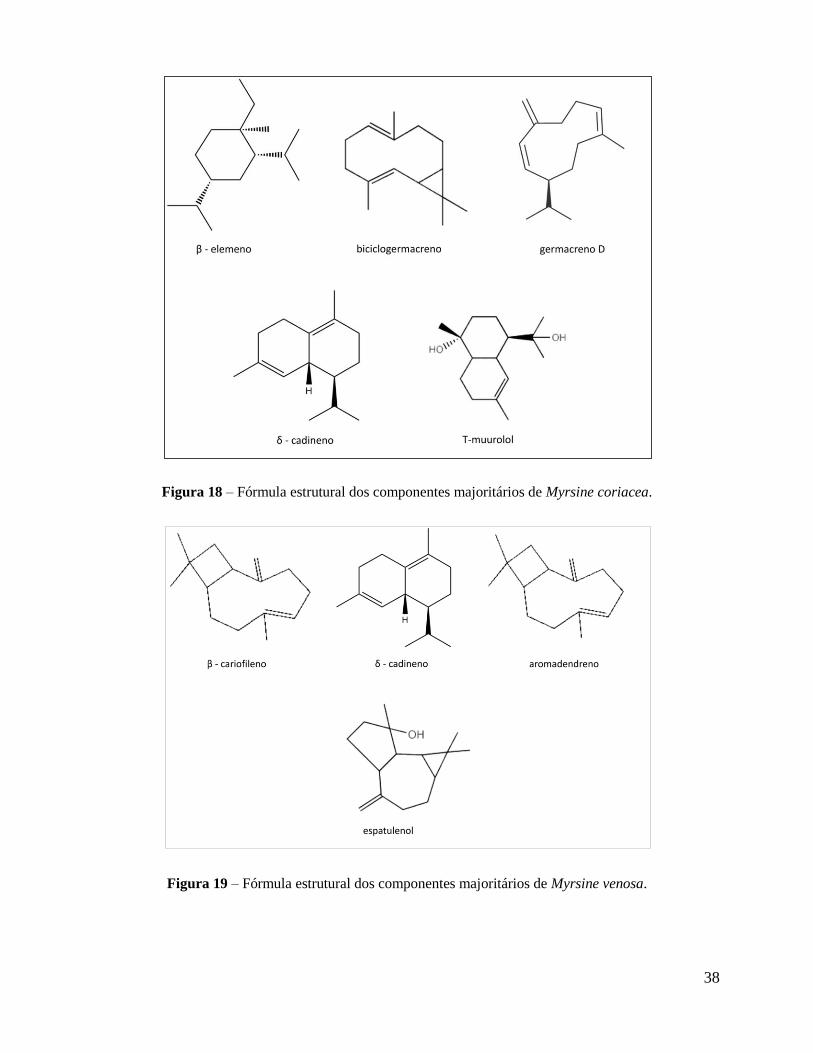
Os constituintes identificados no óleo essencial das folhas secas de M. coriacea e M.
venosa estão listados nas Tabelas 4 e 5, respectivamente. Do óleo essencial de M. coriacea
identificaram-se 22 componentes, que corresponderam a 89,2% do volume do óleo essencial
obtido. Os componentes majoritários foram: β-elemeno (18,74%), biciclogermacreno (11,6%),
germacreno D (9,12%), T-muurolol (7,48%).
De M. venosa, foram identificados 32 componentes, que representaram 77,5% do volume
total do óleo essencial obtido. Os componentes majoritários foram: β-cariofileno (9,78), δ-
cadineno (6,22%), aromadendreno (5,1%) e espatulenol (4,73%). As fórmulas estruturais dos
componentes majoritários do óleo essencial de M. coriacea e M. venosa são apresentadas nas
Figuras 18 e 19, respectivamente. A maior parte dos constituintes presentes no óleo essencial
apresenta cadeia cíclica.
A maior fração dos óleos essenciais das duas espécies é constituída por sesquiterpenos
hidrocarbonetos, seguido por sesquiterpenos oxigenados (alcoóis), como observado na Figura 17

37
e nas Tabelas 4 e 5. Ambas as espécies apresentam porcentagem baixa (inferior a 20%) de
monoterpenos, além disso, M. coriacea apresenta pequena porcentagem de ácido carboxílico e
éster.
Além dos componentes identificados do óleo essencial de M. coriacea e M. venosa,
respectivamente, as Tabelas 4 e 5 ainda apresentam o tempo de retenção, o índice de retenção de
Kovats calculado, o índice de retenção de Kovats segundo Adams (2007), o índice de retenção
linear calculado, o índice de retenção linear ou índice aritmético tabelado segundo Adams (2007)
e a área relativa de cada componente do óleo. Essa tabela também informa o número de
referência CAS (Chemical Abstract Service), a fórmula molecular e o peso molecular
aproximado.
Figura 17 - Porcentagem dos componentes identificados do óleo essencial extraído das folhas secas de
Myrsine coriacea e Myrsine venosa, respectivamente. Notar a predominância de sesquiterpenos no óleo
analisado de ambas as espécies. HC - hidrocarbonetos; -OH - álcoois; =O – cetona; Outros - éster e ácido
carboxílico; ni - não identificados.
0
20
40
60
80
100
Myrsine coriacea
Monoterpenos (-OH)
Sesquiterpenos (HC)
Sesquiterpenos (-OH)
Outros
ni
0
20
40
60
80
100
Myrsine venosa
Monoterpenos (HC)
Monoterpenos (-OH)
Sesquiterpenos (HC)
Sesquiterpenos (-OH)
Sesquiterpenos (=O)
ni
38
Figura 18 – Fórmula estrutural dos componentes majoritários de Myrsine coriacea.
Figura 19 – Fórmula estrutural dos componentes majoritários de Myrsine venosa.
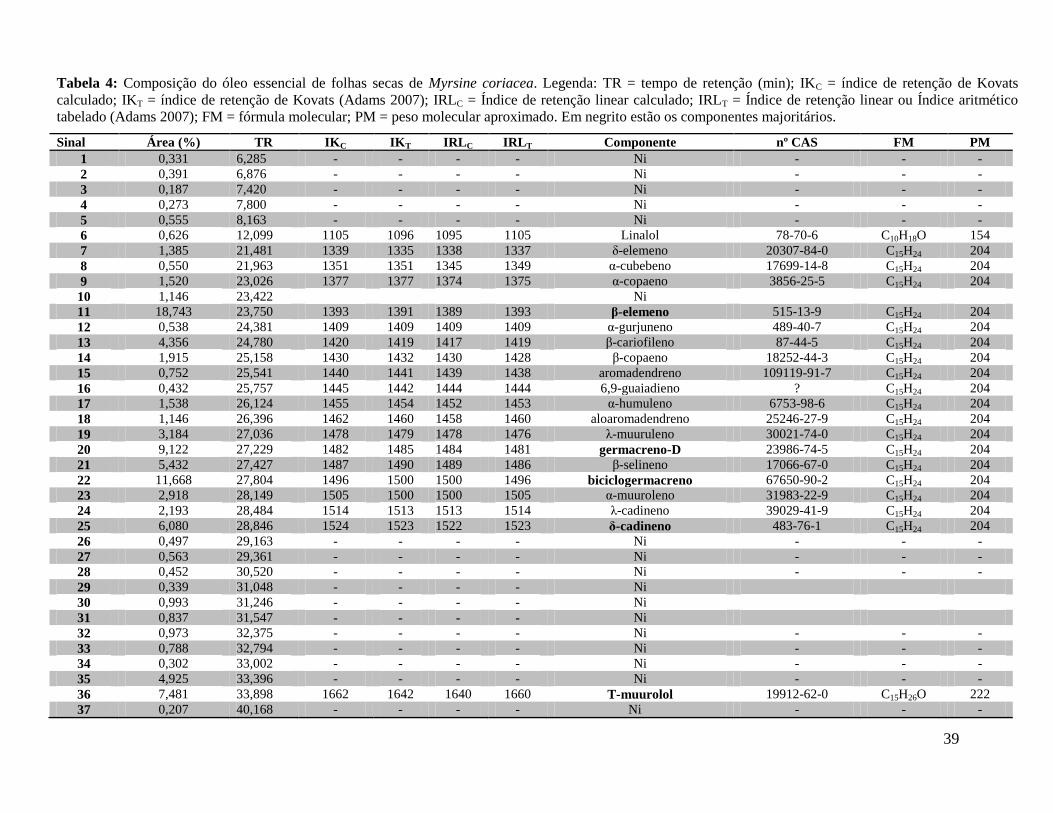
39
Tabela 4: Composição do óleo essencial de folhas secas de Myrsine coriacea. Legenda: TR = tempo de retenção (min); IKC = índice de retenção de Kovats
calculado; IKT = índice de retenção de Kovats (Adams 2007); IRLC = Índice de retenção linear calculado; IRLT = Índice de retenção linear ou Índice aritmético
tabelado (Adams 2007); FM = fórmula molecular; PM = peso molecular aproximado. Em negrito estão os componentes majoritários.
Sinal Área (%) TR IKC IKT IRLC IRLT Componente nº CAS FM PM
1 0,331 6,285 - - - - Ni - - -
2 0,391 6,876 - - - - Ni - - -
3 0,187 7,420 - - - - Ni - - -
4 0,273 7,800 - - - - Ni - - -
5 0,555 8,163 - - - - Ni - - -
6 0,626 12,099 1105 1096 1095 1105 Linalol 78-70-6 C10H18O 154
7 1,385 21,481 1339 1335 1338 1337 δ-elemeno 20307-84-0 C15H24 204
8 0,550 21,963 1351 1351 1345 1349 α-cubebeno 17699-14-8 C15H24 204
9 1,520 23,026 1377 1377 1374 1375 α-copaeno 3856-25-5 C15H24 204
10 1,146 23,422 Ni
11 18,743 23,750 1393 1391 1389 1393 β-elemeno 515-13-9 C15H24 204
12 0,538 24,381 1409 1409 1409 1409 α-gurjuneno 489-40-7 C15H24 204
13 4,356 24,780 1420 1419 1417 1419 β-cariofileno 87-44-5 C15H24 204
14 1,915 25,158 1430 1432 1430 1428 β-copaeno 18252-44-3 C15H24 204
15 0,752 25,541 1440 1441 1439 1438 aromadendreno 109119-91-7 C15H24 204
16 0,432 25,757 1445 1442 1444 1444 6,9-guaiadieno ? C15H24 204
17 1,538 26,124 1455 1454 1452 1453 α-humuleno 6753-98-6 C15H24 204
18 1,146 26,396 1462 1460 1458 1460 aloaromadendreno 25246-27-9 C15H24 204
19 3,184 27,036 1478 1479 1478 1476 λ-muuruleno 30021-74-0 C15H24 204
20 9,122 27,229 1482 1485 1484 1481 germacreno-D 23986-74-5 C15H24 204
21 5,432 27,427 1487 1490 1489 1486 β-selineno 17066-67-0 C15H24 204
22 11,668 27,804 1496 1500 1500 1496 biciclogermacreno 67650-90-2 C15H24 204
23 2,918 28,149 1505 1500 1500 1505 α-muuroleno 31983-22-9 C15H24 204
24 2,193 28,484 1514 1513 1513 1514 λ-cadineno 39029-41-9 C15H24 204
25 6,080 28,846 1524 1523 1522 1523 δ-cadineno 483-76-1 C15H24 204
26 0,497 29,163 - - - - Ni - - -
27 0,563 29,361 - - - - Ni - - -
28 0,452 30,520 - - - - Ni - - -
29 0,339 31,048 - - - - Ni
30 0,993 31,246 - - - - Ni
31 0,837 31,547 - - - - Ni
32 0,973 32,375 - - - - Ni - - -
33 0,788 32,794 - - - - Ni - - -
34 0,302 33,002 - - - - Ni - - -
35 4,925 33,396 - - - - Ni - - -
36 7,481 33,898 1662 1642 1640 1660 T-muurolol 19912-62-0 C15H26O 222
37 0,207 40,168 - - - - Ni - - -

40
38 2,713 44,779 1991 1960 1959 1990 ácido palmítico 57-10-3 C16H32O2 256
39 0,618 47,916 2095 2085 2095 2095 metil linoleato 112-63-0 C19H34O2 294
40 0,396 48,699 - Ni
41 0,254 58,810 - Ni
42 0,680 64,447 - Ni
Total 86,00%
Monoterpenos Total 1,0%
(OH) 1,0%
Sesquiterpenos Total 85,9%
(HC) 73,5%
(OH) 12,4%
Outros 3,3%
Tabela 5: Composição do óleo essencial de folhas secas de Myrsine venosa. Legenda: TR = tempo de retenção (min); IKC = índice de retenção de Kovats
calculado; IKT = índice de retenção de Kovats (Adams 2007); IRLC = Índice de retenção linear calculado; IRLT = Índice de retenção linear ou Índice aritmético
tabelado (Adams 2007); FM = fórmula molecular; PM = peso molecular aproximado. Em negrito estão os componentes majoritários.
Sinal Área (%) TR IKC IKT IRLC IRLT Componente nº CAS FM PM
1 0,066 5,036 - - - - ni - - -
2 3,125 5,192 937 939 932 932 α-pineno 80-56-8 C10H16 136
3 1,436 6,178 980 979 976 974 β-pineno 127-91-3 C10H16 136
4 0,753 6,503 992 991 990 988 mirceno 127-35-3 C10H16 136
5 0,162 7,216 - - - - ni - - -
6 1,199 7,562 1031 1029 1027 1024 limoneno 138-86-3 C10H16 136
7 0,584 8,482 1061 1060 1056 1054 γ-terpineno 99-85-4 C10H16 136
8 0,353 9,451 - - - - ni - - -
9 0,097 9,846 1089 1089 1087 1086 terpinoleno 586-62-9 C10H16 136
10 0,081 10,436 - - - - ni - - -
11 0,054 10,603 - - - - ni - - -
12 0,047 10,784 - - - - ni - - -
13 0,148 11,228 - - - - ni - - -
14 0,194 12,116 - - - - ni - - -
15 0,606 12,661 1179 1177 1176 1174 4-terpineol 562-74-3 C10H18O 154
16 0,255 13,169 - - - - ni - - -
17 0,180 13,400 - - - - ni - - -
18 0,073 16,443 - - - - ni - - -
19 0,081 18,710 - - - - ni - - -
20 1,211 18,985 - - - - ni - - -
21 1,052 19,460 1338 1338 1336 1335 δ-elemeno 20307-84-0 C15H2 204
22 0,051 20,038 1350 1351 1348 1345 α-cubebeno 17699-14-8 C15H2 204
23 0,269 20,296 1371 1375 1369 1373 α-ylangeno 14912-44-8 C15H2 204
24 1,203 20,477 1375 1377 1373 1374 α-copaeno 3856-25-5 C15H2 204
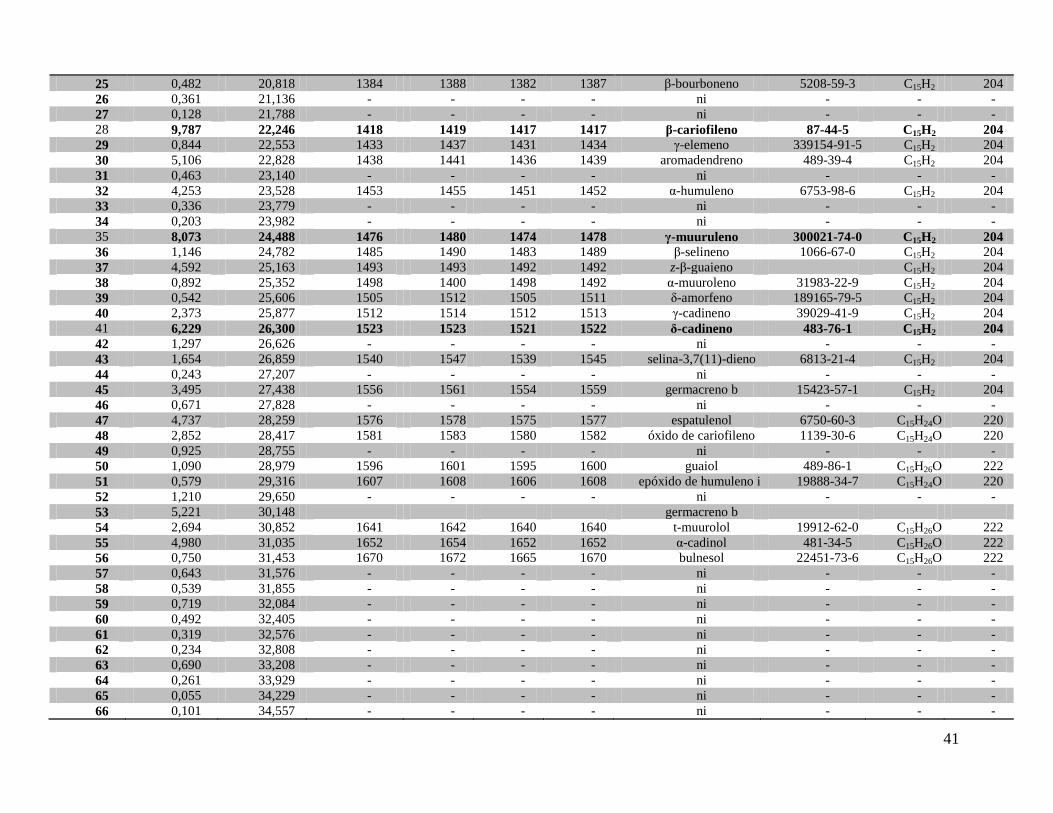
41
25 0,482 20,818 1384 1388 1382 1387 β-bourboneno 5208-59-3 C15H2 204
26 0,361 21,136 - - - - ni - - -
27 0,128 21,788 - - - - ni - - -
28 9,787 22,246 1418 1419 1417 1417 β-cariofileno 87-44-5 C15H2 204
29 0,844 22,553 1433 1437 1431 1434 γ-elemeno 339154-91-5 C15H2 204
30 5,106 22,828 1438 1441 1436 1439 aromadendreno 489-39-4 C15H2 204
31 0,463 23,140 - - - - ni - - -
32 4,253 23,528 1453 1455 1451 1452 α-humuleno 6753-98-6 C15H2 204
33 0,336 23,779 - - - - ni - - -
34 0,203 23,982 - - - - ni - - -
35 8,073 24,488 1476 1480 1474 1478 γ-muuruleno 300021-74-0 C15H2 204
36 1,146 24,782 1485 1490 1483 1489 β-selineno 1066-67-0 C15H2 204
37 4,592 25,163 1493 1493 1492 1492 z-β-guaieno C15H2 204
38 0,892 25,352 1498 1400 1498 1492 α-muuroleno 31983-22-9 C15H2 204
39 0,542 25,606 1505 1512 1505 1511 δ-amorfeno 189165-79-5 C15H2 204
40 2,373 25,877 1512 1514 1512 1513 γ-cadineno 39029-41-9 C15H2 204
41 6,229 26,300 1523 1523 1521 1522 δ-cadineno 483-76-1 C15H2 204
42 1,297 26,626 - - - - ni - - -
43 1,654 26,859 1540 1547 1539 1545 selina-3,7(11)-dieno 6813-21-4 C15H2 204
44 0,243 27,207 - - - - ni - - -
45 3,495 27,438 1556 1561 1554 1559 germacreno b 15423-57-1 C15H2 204
46 0,671 27,828 - - - - ni - - -
47 4,737 28,259 1576 1578 1575 1577 espatulenol 6750-60-3 C15H24O 220
48 2,852 28,417 1581 1583 1580 1582 óxido de cariofileno 1139-30-6 C15H24O 220
49 0,925 28,755 - - - - ni - - -
50 1,090 28,979 1596 1601 1595 1600 guaiol 489-86-1 C15H26O 222
51 0,579 29,316 1607 1608 1606 1608 epóxido de humuleno ii 19888-34-7 C15H24O 220
52 1,210 29,650 - - - - ni - - -
53 5,221 30,148 germacreno b
54 2,694 30,852 1641 1642 1640 1640 t-muurolol 19912-62-0 C15H26O 222
55 4,980 31,035 1652 1654 1652 1652 α-cadinol 481-34-5 C15H26O 222
56 0,750 31,453 1670 1672 1665 1670 bulnesol 22451-73-6 C15H26O 222
57 0,643 31,576 - - - - ni - - -
58 0,539 31,855 - - - - ni - - -
59 0,719 32,084 - - - - ni - - -
60 0,492 32,405 - - - - ni - - -
61 0,319 32,576 - - - - ni - - -
62 0,234 32,808 - - - - ni - - -
63 0,690 33,208 - - - - ni - - -
64 0,261 33,929 - - - - ni - - -
65 0,055 34,229 - - - - ni - - -
66 0,101 34,557 - - - - ni - - -
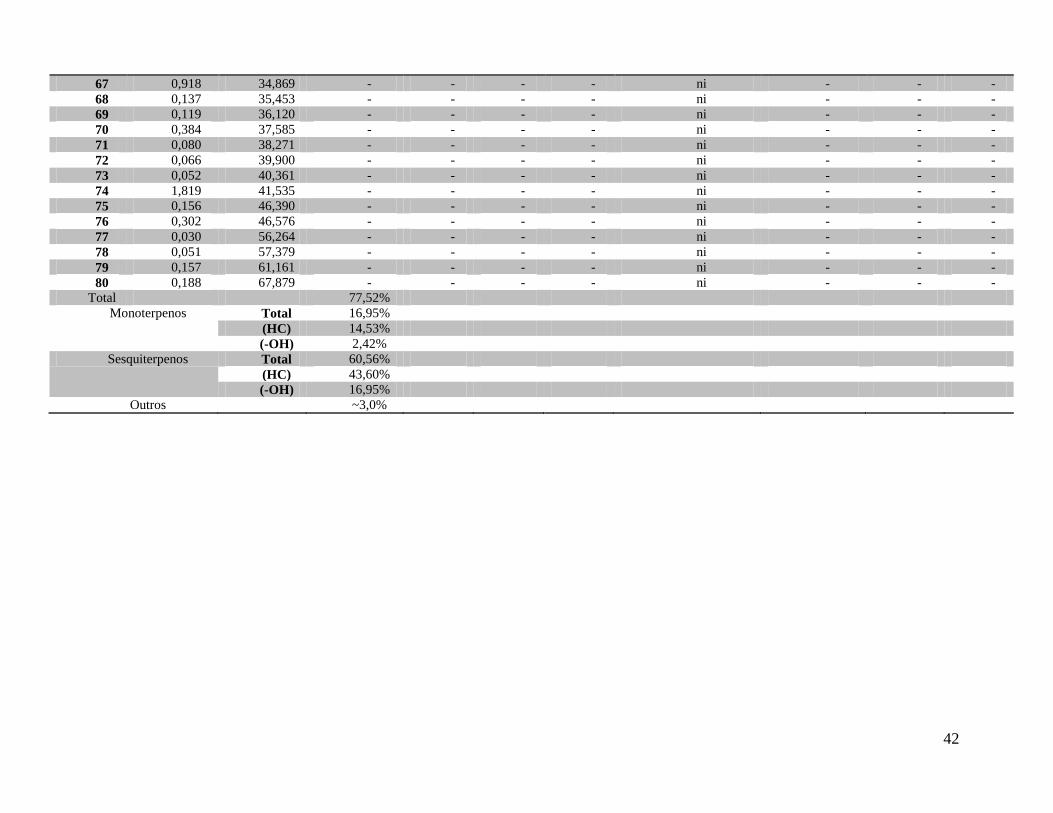
42
67 0,918 34,869 - - - - ni - - -
68 0,137 35,453 - - - - ni - - -
69 0,119 36,120 - - - - ni - - -
70 0,384 37,585 - - - - ni - - -
71 0,080 38,271 - - - - ni - - -
72 0,066 39,900 - - - - ni - - -
73 0,052 40,361 - - - - ni - - -
74 1,819 41,535 - - - - ni - - -
75 0,156 46,390 - - - - ni - - -
76 0,302 46,576 - - - - ni - - -
77 0,030 56,264 - - - - ni - - -
78 0,051 57,379 - - - - ni - - -
79 0,157 61,161 - - - - ni - - -
80 0,188 67,879 - - - - ni - - -
Total 77,52%
Monoterpenos Total 16,95%
(HC) 14,53%
(-OH) 2,42%
Sesquiterpenos Total 60,56%
(HC) 43,60%
(-OH) 16,95%
Outros ~3,0%

43

44
5 - Discussão
Muitos autores têm utilizado aspectos da anatomia foliar para a segregação de espécies e
táxons, como por exemplo: Diane et al. (2003), em espécies de Heliotropiaceae; Matias et al.
(2007), em espécies de Echinodorus Rich. (Alismataceae); e Oliveira et al.(2011), em
Campomanesia Ruiz & Pav. (Myrtaceae). Myrsine coriacea e M. venosa apresentam
características taxonômicas comuns a outras espécies de Primulaceae - Myrsinoideae (Metcalfe &
Chalk 1979; Otegui & Maldonado 1998; Gostin et al. 2011, Luna & Barros, dados não
publicados), como a epiderme uniestratificada com estômatos anisocíticos localizados no mesmo
nível das demais células epidérmicas, mesofilo dorsiventral com cavidades secretoras e
idioblastos contendo drusas.
A ornamentação epicuticular é uma das características epidérmicas mais informativas à
taxonomia em folhas (p.ex: Solereder 1908; Metcalfe & Chalk 1979, Barthlott et al. 1998), e tem
sido utilizada na identificação de espécies, por exemplo, em Trifolium L. – Fabaceae (Zoric et al.
2009) e em Posoqueria Aubl. – Rubiaceae (Arruda et al. 2010). M. venosa apresenta estrias
epicuticulares na em ambas as faces da epiderme, enquanto M. coriacea apresenta epiderme
abaxial estriada e epiderme adaxial irregular. Análises do relevo epicuticular em outras espécies
de Myrsine podem fornecer características relevantes à taxonomia do gênero. Outros caracteres
epidérmicos distintivos observados em M. coriacea e M. venosa foram o contorno das células
epidérmicas em vista frontal, que é sinuoso em M. venosa e liso em M. coriacea e a forma das
células epidérmicas em secção transversal, que são periclinalmente alongadas em M. venosa em
relação às de M. coriacea. Em semelhante análise, Freitas (2003) utilizou caracteres anatômicos
das folhas como recurso adicional à taxonomia de espécies de Myrsine do sul e sudeste do Brasil,
observando diferenças significativas, principalmente, em relação à ornamentação da cutícula na
epiderme foliar. Luna et al. (dados não publicados) identificaram caracteres da anatomia foliar
diagnósticos para a segregação de quatro espécies de Stylogyne A.DC., gênero neotropical de
Primulaceae (Myrsinoideae), dentre os quais destacam-se a forma do sistema vascular no
pecíolo, a coloração e forma das cavidades secretoras, e a ornamentação cuticular.
Diferentes tipos de estruturas secretoras são encontrados entre as espécies de Primulaceae,
como os tricomas glandulares, cavidades secretoras e hidatódios (Grosse, 1908; Solereder, 1908;
Metcalfe &Chalk 1950; Lersten 1977). As estruturas secretoras encontradas em Myrsine coriacea
são os tricomas pluricelulares ramificados, os tricomas peltados e as cavidades secretoras e em

45
M. venosa são as cavidades e ductos secretores e os tricomas peltados. As características gerais
das estruturas secretoras, como a micromorfologia e a localização, têm sido utilizadas por muitos
autores em outras famílias como características diagnósticas úteis, por exemplo, em
Lonchocarpus – Fabaceae (Teixeira et al. 2000) e em Pilocarpus Vahl. – Rutaceae (Muntoreanu
et al. 2011), nos quais as cavidades secretoras são uma das características utilizadas na
segregação das espécies, e em Stachys L. – Lamiaceae (Salmaki et al. 2009), no qual os aspectos
anatômicos dos tricomas são úteis à delimitação das espécies.
A diversidade morfológica e funcional das estruturas secretoras presentes nas plantas é
objeto de estudo recorrente, no que concerne à anatomia e à química de produtos naturais. A
organização estrutural das células e tecidos dessas estruturas permite sua utilização como
atributos importantes para a classificação de famílias, gêneros ou espécies. Em Primulaceae, por
exemplo, a presença de cavidades secretoras e a natureza da secreção, no clado Myrsinoideae, são
uma das principais características distintivas que o separa dos demais clados da família (Judd et
al. 2008).
Evert (2006) refere que em alguns gêneros de Primulaceae - Ardisia, Lysimachia e
Myrsine - a secreção é produzida por células parenquimáticas que secretam o material em
espaços intercelulares comuns. Essa característica não foi observada nas espécies de Myrsine
analisadas no presente trabalho: M. coriacea apresenta cavidades secretoras, e M. venosa contém
cavidades e ductos secretores. Bem como, não foram observados em Stylogyne depauperata Mez,
S. pauciflora Mez, S. sordida Mez e S. warmingii Mez (Luna et al. dados não publicados). Em
ambas as espécies descritas no presente trabalho, essas estruturas são formadas por epitélio
secretor bem delimitado, desenvolvido a partir de divisões anticlinais e periclinais de células do
meristema fundamental do primórdio foliar, oriundo de um processo de desenvolvimento
esquizógeno, e são estas células que secretam o material em espaços intercelulares formados
durante a ontogênese da estrutura, o lúmen. Semelhante desenvolvimento também foi observado
nas cavidades secretoras foliares de Stylogyne depauperata (Primulaceae) (Luna & Barros, dados
não publicados).
As estruturas secretoras internas das folhas em Myrsine são observadas na forma de
pontuações translúcidas, alaranjadas ou nigrescentes, quando observadas contra a luz (Freitas &
Carrijo 2008). Em M. coriacea essa observação nem sempre é possível, em geral, somente nas
folhas mais jovens observam-se as estruturas com conteúdo alaranjado. Em M. venosa,
entretanto, observam-se estruturas hialinas por todo o limbo foliar. As análises anatômicas

46
revelaram que essas estruturas, vistas a olho nu, correspondem a cavidades secretoras em M.
coriacea e à cavidades e ductos secretores em M. venosa, presentes em todo o mesofilo e no
pecíolo de ambas as espécies. A observação de ductos secretores nas folhas de M. venosa é a
primeira ocorrência relatada para a família, para a qual só haviam sido reportadas cavidades
secretoras (Metcalfe & Chalk 1950, Otegui & Maldonado 1998). De acordo com Fahn (1979),
cavidades e ductos secretores diferenciam-se pelo tamanho do lúmen da estrutura. Cavidades
secretoras apresentam formato globoso ou elipsoide e lúmen reduzido em comprimento, se
comparadas aos ductos secretores, que possuem lúmen bastante alongado.
A origem das cavidades secretoras em Primulaceae foi descrita por Solereder (1908),
Guttemberg (1928) e Lersten (1986) em Lysimachia. Guttemberg (1928) afirma que a ontogenia
das cavidades secretoras foliares de Lysimachia vulgaris ocorre de forma esquizolisígena. No
entanto, Lersten (1986), analisando as cavidades secretoras foliares em uma espécie congênere,
L. nummularia, atestou que o trabalho de Guttemberg (1928) apresentou um resultado com a
interferência de artefatos produzidos durante o processamento do material, concluindo que as
cavidades em L. nummularia têm origem esquizógena. Exemplos de diferentes interpretações
dadas a respeito do desenvolvimento de estruturas em um mesmo gênero são apresentados por
Turner et al. (1998), ao analisar a ontogenia das cavidades esquizolisígenas de Citrus limon
(Rutaceae), espécie cuja origem das cavidades secretoras já foi discutida por diferentes autores
(Chen & Wu 2010; Knight et al. 2001; Turner 1999) observam evidências de que artefatos, como
a má fixação dos tecidos e a interferência dos meios de montagem, podem interferir na
interpretação dos resultados.
A origem esquizógena das cavidades secretoras foliares em M. coriacea e M. venosa está
em concordância com os resultados de Lersten (1986) em Lysimachia nummularia, em que o
lúmen secretor das cavidades é formado a partir da separação das células epiteliais, em função da
dissolução da lamela média. Este processo também é observado em outras famílias, como em
Asteraceae (Appezzato-da-Glória et al. 2008), Hypericaceae (Curtis & Lersten 1990; Ciccarelli et
al. 2001), Leguminosae (Teixeira & Rocha 2009; Rodrigues & Machado 2009; Rodrigues et al.
2011a, 2011b; Milani et al. 2012) e Rubiaceae (Vieira et al. 2001). O compartilhamento do
processo de desenvolvimento de estruturas presentes em grupos filogeneticamente distintos pode
significar que, estas estruturas são caracteres homoplásicos, ou seja, são comuns a diferentes
espécies, mas ausentes no ancestral comum a elas; ou são homologias, características
compartilhadas por espécies, que são encontradas no ancestral comum a elas (Ridley 2006). Para

47
que se conheça conclusivamente a importância das estruturas secretoras num contexto evolutivo
faz-se necessária a utilização de estudos que abordem a morfologia, ontogenia e filogenia de
maneira integrada (Schonenberger et al. 2005).
O surgimento das cavidades e ductos nas duas espécies se dá a partir das células do
meristema fundamental, semelhante ao descrito por Lersten (1986) em Lysimachia nummularia,
cujas cavidades iniciam-se a partir de células do mesofilo. O padrão de divisão celular observado
apresenta semelhanças com o relatado por Milani et al. (2012) em Copaifera trapezifolia
(Leguminoseae), onde a célula inicial divide-se periclinalmente, e posteriormente as células
resultantes sofrem divisões anticlinais até que o epitélio secretor seja completamente formado.
As cavidades e ductos das espécies analisadas podem apresentar uma bainha de células
parenquimáticas envolvendo as células epiteliais. A bainha, nessas estruturas, é muitas vezes
relacionada à substituição das células epiteliais quando ocorre a liberação da secreção de maneira
holócrina, ou seja, as células epiteliais morrem e liberam a secreção, assim, os limites da
cavidade ou ducto é garantido pelas células da bainha (Fahn 1979). Observação semelhante foi
feita por Machado & Carmello-Guerreiro (2001) e Rodrigues et al. (2011), em Schinus
terebinthifolia Raddi e Copaifera langsdorfii, respectivamente. Neste trabalho, entretanto, a
bainha não parece desempenhar essa função, uma vez que a liberação da secreção ocorre de
maneira merócrina, isto é, em que não há morte das células epiteliais.
No clado Myrsinoideae, a secreção produzida pelas células epiteliais, e encerrada no
lúmen das cavidades secretoras, foi descrita por Metcalfe & Chalk (1950) como de natureza
resinífera e coloração amarelada. No entanto, estas características não foram padrão neste
trabalho, visto que, apesar de a secreção ter coloração entre tons de vermelho e amarelo em M.
coriacea, a secreção é composta por uma mistura complexa de substâncias lipofílicas, óleos
essenciais e fenóis. Em M. venosa, é incolor tornando-se alaranjada (por um processo de
oxidação) após o seccionamento da folha e tem natureza lipofílica, sendo igualmente composta
por óleo-resina. A mudança na coloração da secreção é relatada em outras famílias, como em
Piperaceae (Pothomorphe umbellata), por Marinho et al. (2011). A complexidade de substâncias
secretadas pelas cavidades e outras estruturas secretoras é comumente relatada em outras famílias
vegetais (Anacardiaceae – Sant’Anna Santos et al. 2006; Flacourtiaceae – Thadeo et al. 2009;
Lythraceae – Lusa & Bona 2011; Piperaceae – Marinho et al. 2011).
Tricomas são estruturas que apresentam grande variabilidade morfológica, podendo ser
classificados em unicelulares ou pluricelulares, simples ou compostos, secretores ou não secretores

48
(Behnke 1984). Esta diversidade morfológica é um atributo taxonômico importante empregado na
delimitação de alguns grupos, como Rondeletia L. - Rubiaceae (Kocsis et al. 2004) e Trifolium -
Leguminosae (Zoric et al. 2009). Em Primulaceae, Otegui & Maldonado (1998), analisando
espécies de Myrsine e Fico et al. (2007), analisando espécies de Primula L., utilizaram
características dos tricomas glandulares foliares para a distinção das espécies. Apesar de haver
semelhança morfológica entre os tricomas de M. coriacea e M. venosa, o número de células da
cabeça dos tricomas peltados é uma característica distintiva observada entre as duas espécies,
enquanto em M. coriacea a cabeça é formada por cerca de 10 células, em M. venosa são cerca de
30 células que formam essas estruturas. A presença de tricomas pluricelulares ramificados
somente em M. coriacea é também uma característica distintiva em relação à M. venosa.
De origem protodérmica, os tricomas peltados em M. coriacea e M. venosa e os
pluricelulares ramificados em M. coriacea, têm origem ainda nos primeiros estágios de
desenvolvimento dos primórdios foliares. A presença de tricomas glandulares em fases iniciais
do desenvolvimento, provavelmente está associada à defesa do órgão vegetal, e quando esta
função não é mais necessária, a estrutura sofre senescência e por isso não é mais observada em
folhas completamente expandidas.
A ontogenia dos tricomas peltados, segue o padrão semelhante ao observado por Turner et
al. (2000) em Mentha piperita L., onde uma célula protodérmica inicial se expande, formando
uma protuberância, que posteriormente se divide. Após o completo desenvolvimento do tricoma
peltado, a parede anticlinal da célula basal sofre maior deposição de cutina. Esta especialização,
provavelmente tem como função prevenir a passagem das substâncias via apoplasto, restringindo
sua passagem via simplasto, além de fornecer suporte estrutural para toda a estrutura, como
também é discutido por Ascensão & Pais (1998) em Leonotis leonurus (L.) R. Br. (Lamiaceae),
por Sacchetti et al. (1999) em Calceolaria adscendens Lidl. (Scrophulariaceae), e por Gersbach
(2002) em Prostanthera ovalifolia R. Br. (Lamiaceae). Em relação à ontogenia dos tricomas
pluricelulares ramificados de M. coriacea não foram encontradas referências de ocorrência para
outras espécies de Primulaceae ou de outras famílias.
A liberação da secreção pelos tricomas glandulares carece de maiores informações. A
secreção não foi observada no espaço subcuticular do tricoma peltado, sugerindo que: (i) a
secreção é volátil, sendo liberada através de microporos na cutícula; (ii) o processamento do
material leva ao rompimento da cutícula, acarretando a saída da secreção. De fato, a liberação da
secreção é um assunto que suscita debates, principalmente pela complexidade da passagem de

49
substâncias através de uma barreira física, como a parede celular (Miguel et al. 2010). Kim &
Mahlberg (1991) e Kim & Mahlberg (2000), analisando tricomas glandulares de espécies de
Cannabaceae, reportam a presença de vesículas secretoras que carreiam o material produzido nas
organelas das células do disco (ou células da cabeça) e chegam à cavidade secretora (ou espaço
subcuticular). A partir de então, o material pode ser volatilizado e liberado sem injuriar a
cutícula, ou só é liberado a partir do rompimento da mesma. No entanto, esse mecanismo de
liberação permanece desconhecido para a maioria das espécies providas de tricomas glandulares.
Quanto à atuação ecológica desses tricomas, não foi possível obter a análise química de
cada tipo de tricoma separadamente, mas a análise histoquímica, com o emprego de reagentes
específicos, evidenciou a presença de uma mistura complexa de substâncias, como compostos
fenólicos e substâncias lipofílicas, nos tricomas peltados de ambas as espécies e a presença de
proteínas, lipídos e compostos fenólicos na secreção dos tricomas pluricelulares ramificados
exclusivos à M. coriacea. A secreção destes compostos age, provavelmente, como primeira
barreira de defesa das folhas.
De modo geral, presume-se que a presença das estruturas secretoras nos representantes da
família Primulaceae é uma estratégia adaptativa de defesa das plantas, especialmente pelos
produtos secretados detectados neste trabalho, como compostos fenólicos e terpenos. Contudo, a
evolução dessas estruturas, tratando-se tanto da morfologia, quanto da secreção, dentro da família
permanece sem solução e é um assunto relevante a ser analisado no futuro, visto que poderá
prover informações sobre a importância dessas estruturas à família, num contexto evolutivo. Em
semelhante abordagem, Chauveau et al. (2011) realizaram análises filogenéticas com base em
dados moleculares em espécies de Sisyrinchium L. (Iridaceae), puderam traçar uma hipótese a
respeito da origem evolutiva dos tricomas encontrados nas flores de 85 espécies do gênero.
Concluíram que o surgimento dos tricomas secretores de óleo contribuiu para a diversificação do
gênero.
Langenheim (2003) destaca a importância das estruturas secretoras como sítios de
produção dos compostos de origem vegetal que são amplamente utilizados pelo homem, entre
eles, óleos essenciais, resinas, alcaloides, mucilagem, látex, gomas e muitos outros compostos
derivados do metabolismo secundário das plantas. A prospecção dos produtos naturais de
espécies de Primulaceae tem resultado na descoberta de substâncias com amplo espectro de
atividades biológicas e farmacológicas: antinociceptiva (Hess et al. 2010), anti-inflamatória
(Ahmad 2011), antileishmaniose (Vermeersch et al. 2009), anti-helmíntica (Challam et al. 2010)

50
e antioxidante (Sumino et al. 2002), por exemplo. Análises recentes em Myrsine coriacea
(=Rapanea ferruginea) permitiram o isolamento do ácido myrsinóico B, a partir de extratos
hidroetanólicos do caule (Baccarin et al. 2011). Este ácido é utilizado na indústria farmacêutica
por suas propriedades anti-inflamatórias e analgésicas (Hess et al. 2010). Sobre a composição e
aplicação dos óleos essenciais, não há referências para Primulaceae, apesar de muitas
propriedades de reconhecida importância e aplicabilidade desses metabólitos secundários
extraídos de outras famílias serem reportadas, por suas propriedades antimicrobianas, anti-
fúngicas, antioxidantes, anti-nociceptivas, diuréticas em espécies de Alismataceae (Pimenta et al.
2006), Asteraceae (Buyisile 2009; ’Cavar et al. 2012; Heinrich et al. 2002; Radulovíc et al. 2012;
Tolouee et al. 2010), Lamiaceae (Lange et al. 2000; Gersbach 2002), Poaceae (Dar et al. 2011) e
muitas outras famílias.
Os óleos essenciais exercem um importante papel na proteção das plantas contra
patógenos ou organismos fitófagos. Além disso, são amplamente explorados nas indústrias
farmacêutica, alimentícia, cosmética e de perfumes (Bakkali et al. 2008). A análise dos óleos
essenciais das folhas de M. venosa revelou a presença de β-cariofileno, γ-muuruleno e δ-
cadineno, como componentes majoritários, enquanto em M. coriacea foram o β-elemeno, α-
selineno e α-cadinol. Como esta é uma análise inédita, preconizou-se a identificação dos
constituintes do óleo essencial, contudo, ensaios posteriores são necessários para a elucidação das
propriedades do óleo.
Pesquisadores brasileiros desenvolveram o fitofármaco Acheflan – Ache®, a partir do
óleo essencial da erva-baleeira (Varronia curassavica - Boraginaceae), que apresenta
propriedades anti-inflamatórias, sendo indicada no tratamento de mialgias. Sua atividade
farmacológica se dá pela presença do constituinte ativo alfa-humuleno, cujas propriedades
lipofílicas e hidrofílicas permitem sua absorção cutânea. Para exercer a atividade desejada, a
empresa Ache® padronizou a composição no óleo essencial do alfa-humuleno em 2,3% - 2,9%.
Em Myrsine venosa a composição de alfa-humuleno no óleo essencial foi de 2,7% - 4,3%,
indicando possível propriedade anti-inflamatória do óleo essencial dessa espécie.
A diferença significativa observada na composição dos óleos entre as duas espécies pode
se dar devido à influência de fatores abióticos, sazonais ou pode ser uma característica inerente
às espécies. Neste estudo, os estágios de coleta foram padronizados e o local de ocorrência das
mesmas tem características bastante homogêneas, assim sendo, acredita-se que esta composição
reflita uma característica própria de cada espécie.

51
A secreção também pode desempenhar papéis ecológicos importantes ao vegetal. As
propriedades atrativas das substâncias voláteis liberadas por osmóforos nas flores (Pansarin et al.
2009), ou a oferta de substâncias nutritivas por nectários (Fahn & Shimony 2001), para a atração
de polinizadores, a proteção das gemas através da secreção dos coléteres estipulares (Klein et al.
2004), todas são estratégias adaptativas das plantas, que se beneficiam dos produtos do seu
metabolismo secundário. A secreção de compostos fenólicos e substâncias lipofílicas em M.
coriacea e M. venosa podem ter papel na proteção dos primórdios foliares e folhas jovens durante
o processo de expansão foliar, contra patógenos e organismos fitófagos.
Os testes histoquímicos revelaram a presença de taninos na secreção das cavidades e
ductos presentes nos primórdios de M. venosa. Taninos são um grupo de compostos fenólicos
produzidos a partir do metabolismo secundário das plantas (Rhoades & Cates 1976). Esses
compostos podem assumir papel importante na defesa do vegetal, uma vez que uma de suas
propriedades é a de reduzir a capacidade de digestão. Sendo assim, acreditamos que a presença
destes metabólitos em uma estrutura em desenvolvimento é uma das estratégias protetoras da
planta. Nas folhas desenvolvidas das espécies estudadas, as estruturas secretoras secretam um
misto de substâncias como, compostos fenólicos, proteínas e substâncias lipofílicas, que também
podem atuar na defesa do órgão vegetal (Fahn 1988).
Sumarizando, as análises realizadas neste trabalho permitiram a caracterização anatômica
das folhas de M. coriacea e M. venosa, através das quais foram indicados caracteres anatômicos
distintivos entre as espécies, também foram classificadas as etapas ontogenéticas pelas quais as
estruturas secretoras foliares – cavidades, tricomas peltados e pluricelulares ramificados em M.
coriacea e cavidades, ductos e tricomas peltados em M. venosa – percorrem até atingirem o
completo desenvolvimento, e foram identificados os constituintes químicos da secreção
produzida por essas estruturas.

52
6 – Conclusões
As principais características anatômicas distintivas entre M. coriacea e M. venosa são: o
relevo epicuticular na face adaxial, o contorno das paredes anticlinais das células
epidérmicas em vista frontal, a presença de tricomas pluricelulares ramificados somente
em M. coriacea, a diferença entre o número de células da cabeça dos tricomas peltados
nas duas espécies; a presença de ductos seceretores em M. venosa; a coloração da
secreção das cavidades secretoras;
Alguns dos caracteres utilizados para diferenciar M. coriacea e M. venosa, a saber, a
ornamentação do relevo epicuticular e as características acerca das estruturas secretoras
(tricomas e cavidades), são semelhantes aos utilizados por outros autores em Myrsine e
em espécies de Stylogyne;
As estruturas secretoras foliares observadas em Myrsine coriacea são os tricomas
peltados, os tricomas pluricelulares ramificados e as cavidades secretoras, e as de M.
venosa são as cavidades e ductos secretores e os tricomas peltados;
Os tricomas pluricelulares ramificados em M. coriacea são estruturas glandulares e
correspondem aos tricomas ferrugíneos observados nas folhas à olho nu;
A observação de ductos secretores em M. venosa é a primeira ocorrência relatada para a
família;

53
Os ductos e as cavidades secretoras em M. venosa são oriundos do mesmo processo de
desenvolvimento;
As cavidades secretoras, em ambas as espécies e os ductos secretores em M. venosa são
formados a partir de um processo esquizógeno;
A diferença observada da coloração dos óleos essenciais para a coloração da secreção nas
cavidades secretoras atesta a complexidade química da secreção, evidenciada pelos testes
histoquímicos, através dos quais foi possível concluir que a secreção é composta por
substâncias lipídicas, fenólicas e proteicas;
O óleo essencial das espécies analisadas apresentou diferenças em relação aos
componentes majoritários identificados. Em M. coriacea os constituintes majoritários
foram β-elemeno, germacreno-D, biciclogermacreno e T-muurolol, enquanto os de M.
venosa foram γ-muuroleno, biciclogermacreno, δ-cadineno e β-cariofileno;
O α-humuleno, constituintes do óleo essencial presente em menor porcentagem, pode
apresentar propriedade anti-inflamatória, tal como observado em outras espécies
comercialmente exploradas pela indústria farmacêutica.

54

55
7 – Referências bibliográficas
Adams, R. P. 2007. Identification of essential oils components by gas chromatography mass
spectroscopy. 4th
, Allured Pubishing Corporation, Carol Stream, Illinois, USA, 804 pp.
Ahmad, B., Azam, S., Bashir, S., Hussain, F. & Chaudhary, M. I. 2011. Insecticidal, brine shrimp
cytotoxicity, antifungal and nitric oxide free radical scavenging activities of the aerial parts
of Myrsine africana L. African Journal of Biotechnology 10(8), 1448-1453.
APG III. 2009. An update of the Angiosperm Phylogeny Group classification for the orders and
families of flowering plants: APG III. Botanical Journal of the Linnean Society 161: 105–
121.
Appezzato-da-Glória B., Hayashi A.H., Cury G., Misaki-Soares M.K., Rocha R. 2008.
Occurrence of secretory structures in underground systems of seven Asteraceae species.
Botanical Journal of the Linnean Society 157, 789–796.
Arruda, R. C. O., Gomes, D. M. S., Magalhães, M. L., Azevedo, A. C. & Gomes, M. 2010. Leaf
anatomy and micromorphology of six Posoqueria Aublet species (Rubiaceae). Rodriguesia
61: 505-518.
Ascensão, L. & Pais, M. S. 1998. The Leaf Capitate Trichomes of Leonotis leonurus:
Histochemistry, Ultrastructure and Secretion. Annals of Botany 81, 263–271.
Ascensão, L., Mota, L. & Castro, M. De M. 1999. Glandular Trichomes on the Leaves and
Flowers of Plectranthus ornatus: Morphology, Distribution and Histochemistry. Annals of
Botany 84 (4), 437-447.
Azam, S., Bashir, S. & Ahmad, B. 2011. Anti-spasmodic action of crude methanolic extract and a
new compound isolated from the aerial parts of Myrsine Africana. BMC Complementary
and Alternative Medicine 11, 55.
Baccarin, T., Muceneeki, R. S., Bresolin, T. M. B., Yunes, R. A., Malheiros, A., Lucinda-Silva,
R. M. 2011. Development and validation of an HPLC-PDA method for the determination
of myrsinoic acid B in the extracts of Rapanea ferruginea Mez. Talanta 85, 1221– 1224.
Bakkali, F., Averbeck, S., Averbeck, D. & Idaomar, M. 2008. Biological effects of essential oils
– A review. Food and Chemical Toxicology 46(2), 446-475.
Barthlott, W., Neinhuis, C., Cutler, D., Ditsch, F., Meusel, I., Theisen, I. & Wilhelmi, H. 1998.
Classification and terminology of plant epicuticular waxes. Botanical Journal of the Linnean
Society 126: 227-236.

56
Bassolé, I.H., Juliani, H.R. 2012. Essential oils in combination and their antimicrobial properties.
Molecules. 17(4), 3989-4006.
Behnke, H.D. (1984) Plant trichomes - structure and ultrastructure: general terminology,
taxonomic applications, and aspects of trichome-bacteria interaction in leaf tips of
Discorea. In: Rodriguez, E.; Healey, P.L. e Metha, I. (1984) Biology and chemistry of plant
trichomes. New York: Plenum Press, 95-112.
Bennici, A., Tani, C. 2004. Anatomical and ultrastructural study of the secretory cavity
development of Citrus sinensis and Citrus limon: Evaluation of schizolysigenous ontogeny.
Flora 199, 464– 475.
Bottega, S., Corsi, G., 2000. Structure, secretion and possible functions of calyx glandular hairs
of Rosmarinus officinalis L. (Labiatae). Botanical Journal of the Linnean Society 132, 325
–335.
Brundrett, M.C., Kendrick, B., Peterson, C.A. 1991. Efficient lipid staining in plant material with
Sudan Red 7B or Fluoral Yellow 088 in polyethylene glycol-glycerol. Biotecnic &
Histochemistry 66, 111-116.
Bueno N.R., Oliveira R. C., Brito R. C, Pott A., Pott V.J., Scheidt G.N. & da Silva Batista M.
2005. Medicinal plants used by the Kaiowá and Guaraní indigenous populations in the
Caarapó Reserve, Mato Grosso do Sul, Brazil. Acta Botanica Brasilica 19 (1), 39-44.
Burt, S. 2004. Essential oils: their antibacterial properties and potential applications in foods - a
review. International Journal of Food Microbiology 94, 223-253.
Buyisile, M. 2009. Morphological and chemical composition of the essential oil of the leaf of
Schistostephium heptalobium. African Journal of Biotechnology 8(8), 1509-1519.
Cain, A.J. 1947. The use of Nile blue in the examination of lipids. Scientific Monthly 88, 383-
392.
Castro, E.M., Pereira, F.J. & Paiva, R. 2009. Histologia vegetal: estrutura e função de órgãos
vegetativos. Lavras: UFLA. 234 p.
Challam, M., Roy, B., Tandon V. 2010. Effect of Lysimachia ramosa (Primulaceae) on helminth
parasites: Motility, mortality and scanning electron microscopic observations on surface
topography. Veterinary Parasitology 169, 214–218
Chauveau, O., Eggers, L., Raquin, C. et al. 2011 Evolution of oil-producing trichomes in
Sisyrinchium (Iridaceae): insights from the first comprehensive phylogenetic analysis of
the genus. Annals of Botany 107, 1287–1312.

57
Chen, Y. & Wu H. 2010. Programmed cell death involved in the schizolysigenous formation of
the secretory cavity in Citrus sinensis L. (Osbeck). Chinese Science Bulletin 55(20): 2160–
2168.
Christodoulakis, N. S., Kollia, K., and Fasseas, C. 2011. Leaf structure and histochemistry of
Ecballium elaterium (L.) A. Rich. (squirting cucumber). Flora 206, 191-197.
Ciccarelli, D., Andreucci, A. C. & Pagni, A. M. 2001. Translucent glands and secretory canals in
Hypericum perforatum L. (Hypericaceae): morphological, anatomical and histochemical
studies during the course of ontogenesis. Annals of Botany 88 (4), 637-644.
Combrinck, S., Du Plooy, G. W., McCrindle, R. I., & Botha, B. M. 2007. Morphology and
histochemistry of the glandular trichomes of Lippia scaberrima (Verbenaceae). Annals of
Botany 99(6), 1111-1119.
Corsi, G. & Biasci, D. 1998. Secretory Structures and Localization of Alkaloids in Conium
maculatum L. (Apiaceae). Annals of Botany 81: 157–162.
Costa, A. F. 1982. Farmacognosia. 2. ed. Lisboa: Fundação Calouste Gulbenkian. v. 3. 1032 p.
Croteau, R. & Johnson, M.A. 1984. Biosynthesis of terpenoids in glandular trichomes. In:
Rodriguez E, Healey PL, Metha J, eds. Biology and chemistry of plant trichomes. New
York: Plenum Press, 133–185.Curtis, J. D. & Lersten, N. R. 1990.
Curtis, J.D. & Lersten, N.R. 1990. Internal secretory structures in Hypericum (Clusiaceae): H.
perforatum L. and H. balnearicum L. New Phytologist 114, 571–580.
Cury, G. & Appezzato-da-Glória, B. 2009. Internal secretory spaces in thickened underground
systems of Asteraceae species. Australian Journal of Botany 57: 229–239.
´Cavar, S., Maksimovic, M., Vidic, D., Paric, A. 2012. Chemical composition and antioxidant
and antimicrobial activity of essential oil of Artemisia annua L. from Bosnia. Industrial
Crops and Products 37, 479– 485.
Dar, M. Y., Shah, W. A., Rather, M. A., Qurishi, Y., Hamidc, A., Qurishi, M. A. 2011. Chemical
composition, in vitro cytotoxic and antioxidant activities of the essential oil and major
constituents of Cymbopogon jawarancusa (Kashmir). Food Chemistry 129, 1606–1611.
David, R., Carde, J.P., 1964. Coloration différentielle dês inclusions lipidique et ter- peniques des
pseudophylles du pine maritime aumoyen du reactif Nadi. Comptes Rendus Aca. Sci. Paris
257, 1338–1340.
Denissova G. A. 1975. Attempt of classification of terpenoid-containing conceptacles in plants.
(In Russian.) - Bot. Zh. SSSR. 60: 1698-1706.

58
Diane, N, Jacob, C. & Hilger, H.H. 2003. Leaf anatomy and foliar trichomes in Heliotropiaceae
and their systematic relevance. Flora 198: 468–485.
Evert, R. F. 2006. Esau’s Plant anatomy : meristems, cells, and tissues of the plant body : their
structure, function, and development. Rev. ed. of: Plant anatomy / Katherine Esau. 2nd. ed.
1965.
Fahn, A. & Shimony C. 2001. Nectary structure and ultrastructure of unisexual flowers of
Ecballium elaterium (L.) A. Rich. (Cucurbitaceae) and their presumptive pollinators.
Annals of Botany 87, 27–33.
______. 1979. Secretory tissues in plants. London, Academic Press.
______ 1984. Secretory tissues and factors influencing their development. Phyton 28(1), 13-26.
______ 1988. Secretory tissues in vascular plants. New Phytologist 108, 229-257.
______. 2000. Structure and function of secretory cells. Advances in Botanical Research 31, 37–
75.
Fahn, A. & Shimony, C., 1998. Ultrastructure and Secretion of the Secretory Cells of Two
Species of Fagonia L. (Zygophyllaceae). Annals of Botany 81, 557-565.
Fico, G., Rodondi, G., Flamini, G., Passarella, D., Tomè, F., 2007. Comparative phytochemical
and morphological analyses of three Italian Primula species. Phytochemistry 68, 1683–
1691.
Franklin, G.L. 1945. Preparation of thin sections of synthetic resins and wood-resin composites,
and a new macerating method for wood. Nature 155(3924): 51.
Freitas, M. F. & Carrijo, T. T. 2008. A família Myrsinaceae nos contrafortes do maciço da Tijuca
e entorno do Jardim Botânico do Rio de Janeiro. Rodriguesia 59(4), 813-828.
Freitas, M. F. 2003. Estudos taxonômicos em espécies de Myrsine (Myrsinaceae) das regiões
sudeste e sul do Brasil. Tese de Doutorado, Unicamp, Campinas, SP.
Freitas, M.F., 2012. Myrsine in Lista de Espécies da Flora do Brasil. Jardim Botânico do Rio de
Janeiro. (http://floradobrasil.jbrj.gov.br/2012/FB010223).
Ganter, P. & Jollés, G. 1969. Histologie normale et pathologique. Vols I e II. Gauthier – Villars.
Paris.
Germonprez, N., Van Puyvelde, L., Maes, L., Van Tri, M. & de Kimpe, N. 2004. New
pentacyclic triterpenes saponins with strong anti-leishmanial activity from the leaves of
Maesa balansae. Tetrahedron 60, 219–228.

59
Gersbach, P.V., 2002. The essential oil secretory structures of Prostanthera ovalifolis
(Lamiaceae). Annals of Botany 89, 255–260.
Gifford, E.M. & Foster, A.S. 1989. Morphology and evolution of vascular plants. 3rd ed.
Freeman and Co., New York.
Gomes, S. M. A. & Lombardi, J. A. 2010. Leaf anatomy as a contribution to the taxonomy of
Salacioideae N.Halle´ ex Thorne & Reveal (Celastraceae). Plant Systematic and Evolution
289:13–33.
Gostin I. N, Zamfirache, M. M. & Ştefan M. 2011. Anatomical and micromorphological aspects
at Ardisia crenata Sims. Analele ştiinţifice ale Universităţii “Alexandru Ioan Cuza” din Iaşi
Biologie vegetală 57 (1).
Gould, S. J. 1977. Ontogeny and Phylogeny. Cambridge MA: Harvard Univ. Press
Grosse, A. 1908. Anatomisch-systematische Untersuchungen der Myrsinaceen. Botanische
Jahrbücher für Systematik 41(96): 1-46.
Guttenberg, H. V. 1928. Die Harzdrusen von Lysimachia vulgaris L. Planta 1, 232-238.
Harbome, J. B. 1984. Phytochemical methods, ed. 2. - Chapman & Hall, London, New York.
Heinrich G, Pfeifhofer HW, Stabentheiner E, Sawidis T. 2002. Glandular hairs of Sigesbeckia
jorullensis Kunth (Asteraceae): morphology, histochemistry and composition of essential
oil. Annals of Botany 89, 459–469.
Heslop-Harrison, Y. 1977. The pollen–stigma interaction: pollen- tube penetration in Crocus.
Annals of Botany, 41: 913–922.
Hess, S., Padoani, C., Scorteganha, L. C., Holzmann, I., Malheiros, A., Yunes, R. A., Delle
Monache, F., & De Souza, M. M. (2010). Assessment of mechanisms involved in
antinociception caused by myrsinoic acid B. Biological Pharmaceutical Bulletin 33, 209-
215.
Hirota, M., Miyakazi, S., Minakuchi, T., Takagi, T. & Shibota, H. 2002. Myrsinoic acids B, C
and F, anti-inflammatory compounds from Myrsine seguinii. Biosci. Biotechonol.
Biochem. 66 (3), 655-659.
Huang J, Ogihara Y, Zhang H, Shimizu N, Takeda T. 2000. Triterpenoid saponins from Ardisia
mamillata. Phytochemistry 54(8), 817-22.
Jayabalan M. & Shah, J.J. 1986. Histochemical techniques to localize rubber in guayule
(Parthenium argentatum Gray). Stain Technol. 61(5), 303-8.
Johansen, D.A. 1940. Plant Microtechnique. McGraw-Hill, New York.

60
Judd, W.S., Campbell, C.S., Kellog, E.A., Stevens, P.F. & Donoghue, M.J. 2008. Sistemática
Vegetal Um Enfoque Filogenético. 3a Edição, ARTMED, Porto Alegre.
Kalachanis, D. & Psaras, G.K. 2005. Structure and development of the secretory cavities of
Myrtus communis leaves. Biologia Plantarum 49(1), 105-110.
Kim, E.S. & Mahlberg, P. G. 1991. Secretory Cavity Development in Glandular Trichomes of
Cannabis sativa L . (Cannabaceae). American Journal of Botany 78(2), 220-229.
Kim, E.S. & Mahlberg, P. G. 2000. Early development of the secretory cavity of peltate glands in
Humulus lupus L. (Cannabaceae). Molecules and cells 10(5), 487-492.
Kisser, J. G. 1958. Die Ausscheidung von atherischen Glen und Harzen. Handbuch der
Pflanzenphysiol 10, 91-131.
Klein, D. E., Gomes, V. M., Silva-Neto, S. J., Da Cunha, M., 2004. The structure of colleters in
several species of Simira (Rubiaceae). Annals of Botany 94, 733 - 740.
Knight, T. G.; Klieber, A. & Sedgley, M. 2001. The relationship between oil gland and fruit
development in Washington Navel Orange (Citrus sinensis L. Osbeck). Annals of Botany
88, 1039–1047.
Kocsis, M.; Darók, J. & Borhidi, A. 2004. Comparative leaf anatomy and morphology of some
neotropical Rondeletia (Rubiaceae) species. Plant Systematics and Evolution 218(1-4):
205-218.
Koike, K., Jia, Z., & Nikaido, T. 2001. New Triterpenoid Saponins from Maesa tenera. Chem.
Pharm. Bull. 49(6), 758-761.
Lange, B. M., Wildung, M. R., Stauber, E. J., Sanchez, C., Pouchnik, D., Croteau, R. 2000.
Probing essential oil biosynthesis and secretion by functional evaluation of expressed
sequence tags from mint glandular trichomes. Proceedings of the National Academy of
Sciences of the United States of America 97(6), 2934-2939.
Langenheim, J.H. 2003. Plant resins. Timber, Cambridge, p 586.
Lersten, N. R. 1977. Trichome forms in Ardisia (Myrsinaceae) in relation to the bacterial leaf
nodule symbiosis. Botanical Journal of the Linnean Society 75: 229–244.
Lersten, N.R. 1986. Re-investigation of secretory cavity development in Lysimachia
(Primulaceae). The New Phytologist 102, 193-197.
Liang, S., Haiyang Wang, H., Yang, M., Wu, H. 2009 Sequential actions of pectinases and
cellulases during secretory cavity formation in Citrus fruits. Trees 23, 19–27.

61
López, V., Jäger, A. K., Akerreta, S., Cavero, R. Y., Calvo, M.I. 2011. Pharmacological
properties of Anagallis arvensis L. (“scarlet pimpernel”) and Anagallis foemina Mill. (“blue
pimpernel”) traditionally used as wound healing remedies in Navarra (Spain). Journal of
Ethnopharmacology 134(3), 1014-1017.
Lusa, M. G. & Bona, C. 2011. Caracterização morfoanatômica e histoquímica de Cuphea
carthagenensis (Jacq.) J.f. Macbr. (Lythraceae). Acta botanica brasilica 25(2), 517-527.
Mace, M.E. & Howell, C.R. 1974. Histochemistry and indentification of condensed tannin
precursor in roots of cotton seedlings. Canadian Journal of Botany 52, 2423-2426.
Machado, S.R. & Carmello-Guerreiro, S.M. 2001. Estrutura e desenvolvimento de canais
secretores em frutos de Schinus terebinthifolius Raddi (Anacardiaceae). Acta Botanica
Brasilica 15, 189-195.
Machado, S.R. & Gregório, E.A. 2001. Zinc iodide-osmium tetroxide (ZIO) reactive sites in the
extrafloral nectary of Citharexylum mirianthum Cham. (Verbenaceae). Tissue & Cell 33(1),
72-77.
Machado, S.R., Gregório, E.A., Yanagizawa Y. & Carmello, S.M. 1995. Ultrastructural aspects
of the peltate glandular trichomes of the gynoecium in Zeyheria digitalis (Vell.) Hoehne
(Bignoniaceae). Brazilian Journal of Botany 18, 197–205.
Makabe H, Miyazaki S, Kamo T, Hirota M. 2003. Myrsinoic acid E an anti-inflammatory
compound from Myrsine seguinii. Bioscience, Biotechnology and Biochemistry 67, 2038-
2041.
Marinho, C. R., Zacaro, A. A. & Ventrella, M. C. 2011. Secretory cells in Piper umbellatum
(Piperaceae) leaves: a new example for the development of idioblasts. Flora 206, 1052-
1062.
Matias, L.Q., Soares, A. & Scatena, V. L. 2007. Systematic consideration of petiole anatomy of
species of Echinodorus Richard (Alismataceae) from north-eastern Brazil. Flora 202: 395–
402.
Mauseth, J.D. 1988. Plant Anatomy, Benjamin/Cummings
Metcalfe, C. R. & Chalk. 1950, Anatomy of the dicotyledons. Vol. II. Oxford, Oxford Claredon
Press.
Metcalfe, C.R. & Chalk, L. 1957. Anatomy of the dicotyledons. Oxford: Caredon Press.

62
Miguel, E. C.; Klein, D. E. ; De Oliveira, M. A. ; Da Cunha, M. 2010. Ultrastructure of secretory
and senescence phase in colleters of Bathysa gymnocarpa K. Schum. and B. stipulata
(Vell.) C, Presl. (Rubiaceae). Revista Brasileira de Botânica 33, 455-466.
Milani, J. M., Rocha, J. F., Teixeira, S. P. 2012. Oleoresin glands in copaíba (Copaifera
trapezifolia Hayne: Leguminosae), a Brazilian rainforest tree. Trees 26 (3), 769-775.
Mizushina, Y., Miyazaki, S., Ohta, K., Hirota, M., and Sakaguchi, K. 2000. Novel anti-
inflammatory compounds from Myrsine seguinii, terpeno-benzoic acids, are inhibitors of
mammalian DNA polymerase. Biochim. Biophys. Acta General Subjects 1475, 1–4.
Moura, M.Z.D., Dos Santos Isaias, R.M. & Soares, G.L.G. 2005. Ontogenesis of internal
secretory cells in leaves of Lantana camara (Verbenaceae). Botanical Journal of the
Linnean Society 148, 427–431.
Muntereanu, T. G., Cruz, R. S., Melo-de-Pina, G. F. 2011. Comparative leaf anatomy and
morphology of some neotropical Rutaceae: Pilocarpus Vahl and related genera. Plant
systematic and evolution 296: 87-99.
Ndonsta, B.L.; Tatsimo, J.S.N.; Csupor, D.; Forgo, P.; Berkecz, R.; Berenyi, A.; Tene, M.;
Molnar, J.; Zupko, I.; Hohmann, J.; Tane, P. 2011. Alkylbenzoquinones with
antiproliferative effect against human cancer cell lines from stem of Ardisia kivuensis.
Phytochemistry Letters 4(3), 227-230.
Ogawa, H. & Natori, S. 1968. Hydroxybenzoquinones from Myrsinaceae plants II – Distribution
among Myrsinaceae plants in Japan. Phytochemistry 7, 773-782.
Oliveira, M. I. U., Funch, L. S., Ribeiro dos Santos, F. A., Landrum, L. R. 2011. Aplicação de
caracteres morfoanatômicos foliares na taxonomia de Campomanesia Ruiz & Pavon
(Myrtaceae). Acta botanica brasillica 25(2): 455-465.
Otegui M., Maldonado S. 1998. Morfologia foliar de las especies de Myrsine L. (Myrsinaceae)
del Cono Sur de America del Sur. Candollea 53, 349-363.
Otegui, M., Lima, C., Maldonado, S. & Lederkremer R, M. 1998a. Histological and Chemical
Characterization of Myrsine laetevirens Seed. International Journal of Plant Sciences
159(5), 762-772.
Otegui, M.S., Gaspar, M. L., Maldonado, S., Varetti, E. L. & Pollero, R. 1998b. Studies on
tissues associated to hydroxybenzoquinone secretion in Myrsine laetevirens (Myrsinaceae).
Nordic Journal of Botany 18, 447-459.

63
Pearse, A.G.E. 1980. Histochemistry theoretical and applied. Vol. II, 4th edition. Longman
Group Limited.Reynolds, E. S., 1963. The use of lead citrate at high pH as an electron-
opaque stain in electron microscopy. Journal of Cell Biology 17, 208-212.
Pimenta, D. S., Figueiredo, M. R. & Kaplan, M. A. C. 2006. Essential oil from two populations
of Echinodorus grandiflorus (Cham. & Schltdl.) Micheli (Chapéu de couro) Anais da
Academia Brasileira de Ciências 78(4), 623-628.
Radulovic´, N. S., Dekic´, M. S., Rand-elovic´, P. J., Stojanovic´, N. M., Zarubica, A. R.,
Stojanovic´-Radic, Z. Z. 2012. Toxic essential oils: Anxiolytic, antinociceptive and
antimicrobial properties of the yarrow Achillea umbellata Sibth. et Sm. (Asteraceae)
volatiles. Food and Chemical Toxicology 50, 2016–2026.
Rhoades, D.F., Cates, R.G., 1976. Toward a general theory of plant anti-herbivore chemistry.
Recent Advances in Phytochemistry 10, 168–213.
Rocha, J. F., Pimentel, R.R. & Machado, S.R. 2011. Estruturas secretoras de mucilagem em
Hibiscus pernambucensis Arruda (Malvaceae): distribuição, caracterização
morfoanatômica e histoquímica. Acta Botanica Brasilica 25(4): 751-763.
Rodrigues, T. M. & Machado, S. R. 2009. Developmental and structural features of secretory
canals in root and shoot wood of Copaifera langsdorffii Desf. (Leguminosae–
Caesalpinioideae) Trees 23, 1013–1018.
Rodrigues, T. M., Teixeira, S. D. P., & Machado, S. R. 2011b. The oleoresin secretory system in
seedlings and adult plants of copaíba (Copaifera langsdorffii Desf., Leguminosae–
Caesalpinioideae). Flora 206(6), 585-594.
Rodrigues, T.M., Santos, D.C. & Machado, S.R. 2011a. The role of the parenchyma sheath and
PCD during the development of oil cavities in Pterodon pubescens (Leguminosae-
Papilionoideae). Comptes Rendus Biologies 334(7), 535-543.
Rudall, P.J. & Bateman, R.M. 2003. Evolutionary change in flowers and inflorescences: evidence
from naturally occurring terata. Trends in Plant Science 8(2): 76−82.
Russin, W. A., Uchytil, T. F., Feistner, G. & Durbin, R. D. 1988. Developmental Changes in
Content of Foliar Secretory Cavities of Tagetes erecta (Asteraceae). American Journal of
Botany 75(12), 1787-1793.
Sacchetti, G., Romagnoli, C., Nicoletti, M., Di Fabio, A., Bruni, A. & Poli, F. 1999. Glandular
trichomes of Calceolaria adscendens Lidl. (Scrophulariaceae): Histochemistry,
Development and Ultrastructure. Annals of Botany 83(1), 87-92.

64
Salmaki, Sh., Zarre, Z., Jamzad, C. & Bräuchler. 2009. Trichome micromorphology of Iranian
Stachys (Lamiaceae) with emphasis on its systematic implication. Flora 204: 371–381.
Sangwan, N.S., Farooqi, A.H.A., Shabih, F., Sangwan, R.S., 2001. Regulation of essential oil
production in plants. Plant Growth Regul. 24, 3–21.
Sant’ Anna – Santos, B. F., Thadeo, M., Meira, R. M. S. A. & Ascensão, L. 2006. Anatomia e
histoquímica das estruturas secretoras do caule de Spondias dulcis Forst. F.
(Anacardiaceae). Revista Árvore 30(3), 481-489.
Sass, J.E. 1951. Botanical microtechnique. 2nd
ed. Iowa: State College Press. 228 p
Sass, J.E. 1958. Elements of botanical microthecnique. New York- London, Mcgraw-Hill Book
Company Inc. 222p.
Schönenberger, J., Anderberg, A.A., Sytsma, K.J., 2005. Molecular phylogenetics and patterns of
floral evolution in the Ericales. Int. J. Plant Sci. 166, 265–288.
Simões, C.M.O. & Spitzer, V. Óleos voláteis. In: SIMOES, C.M.O.; SCHENKEL, E.P.;
GOSMANN, G. (Eds). Farmacognosia: da planta ao medicamento. 2. ed. Porto Alegre:
Universidade Federal do Rio Grande do Sul; Universidade Federal de Santa Catarina, 2000.
p.394-412.
Solereder, H. 1908. Systematic anatomy of dicotyledons. A handbook for laboratories of pure
applied Botany. Translated by L.A. Boodle & F.G. Fritsch, 2.v., Clarendon press, Oxford.
Sumino, M., Sekine, T., Ruangrungsi, N., Igarashi K., & Ikegami, F. 2002. Ardisiphenols and
other antioxidant principles from the fruits of Ardisia colorata. Chemical Pharmacological
Bulletin 50(11), 1484-1487.
Teixeira, S. P. & Rocha, J. F. 2009. Oil glands in the Neotropical genus Dahlstedtia Malme
(Leguminosae, Papilionoideae, Millettieae). Revista Brasileira de Botânica 32, 33-40.
Teixeira, S. P., Castro M. M. & Tozzi. A. M. G. A. 2000. Secretory cavities and pellucid dots in
leaflets of Lonchocarpus (Leguminosae, Papilionoideae, Millettieae). Plant Systematics and
Evolution 221(1-2): 61-68.
Thadeo, M., Meira, R. M. S. A., Azevedo, A. A., & Araújo, J. M. D. 2009. Anatomia e
histoquímica das estruturas secretoras da folha de Casearia decandra Jacq. (Salicaceae).
Revista Brasileira de Botânica, 32(2), 329-338.
Tolouee, M., Alinezhad, S., Saberi, R., Eslamifar, A., Zad, S. J., Jaimand,K. 2010.Effect of
Matricaria chamomilla L. flower essential oil on the growth and ultrastructure of
Aspergillus niger vanTieghem. International Journal of Food Microbiology 139,127-133.

65
Turner, G. W. 1986. Comparative development of secretory cavities in the tribes Amorpheae and
Psoraleae (Leguminosae: Papilionoideae) American Journal of Botany 73(8), 1178-1192.
Turner, G. W. 1999. A Brief History of the Lysigenous Gland Hypothesis. Botanical Review. 65,
76-88.
Turner, G. W., A. M. Berry & E. M. Gifford. 1998. Schizogenous secretory cavities of Citrus
limon (L.) Burm. f. and a reevaluation of the lysigenous gland concept. International
Journal of PIant Science 159, 75-88.
Turner, G. W., Gershenzon, J., and Croteau, R. B. 2000. Development of peltate glandular
trichomes of peppermint. Plant Physiology 124, 665-680.
Vermeesch M., Foubert K., da Luz R. I., Puyvelde L.V., Piters L., Cos P. & Mães L. 2009.
Selective antileishmania activity of 13,28-epoxy-oleanane and related triterpene saponins
from the plant families Myrsinaceae, Primulaceae, Aceraceae e Icacinaceae. Phytotherapy
Research 23, 1404-14.
Vieira, R. C., Delprete, P. G., Leitão, G. G., Leitão, S. G. 2001. Anatomical and chemical
analyses of leaf secretory cavities of Rustia Formosa (Rubiaceae). American Journal of
Botany 88, 2151 -2156.
Vitalini S., Flamini G., Valaguzza A., Rodondi G., Iriti M. & Fico G. 2011. Primula spectabilis
Tratt. aerial parts: morphology, volatile compounds and flavonoids. Phytochemistry 72(11-
12), 1371-1378.
Wagner, G. J. 1991. Secreting Glandular Trichomes: More than Just Hairs. Plant Physiology 96,
675-679.
Wink, M. 2010. Functions and biotechnology of plant secondary metabolites: Functions of plant
secondary metabolites and their exploitation in biotechnology. Annual plant reviews, 2nd.
ed. v. 39.
Wist, T. J., & Davis, A. R. 2006. Floral Nectar Production and Nectary Anatomy and
Ultrastructure of Echinacea purpurea (Asteraceae). Annals of Botany 97, 177-193.
Xu, G. X., Tan, C., Wei, X. J., Gao, X. Y., and Zheng, H. Q. 2011. Development of secretory
cells and crystal cells in Eichhornia crassipes ramet shoot apex. Protoplasma 248, 257-266.
Zoric, L., Merkulov, L., Lukovic, J., Boza, P. & Polic, D. 2009. Leaf epidermal characteristics of
Trifolium L. species from Serbia and Montenegro. Flora 204(3): 198-209.