universidade de sÃo paulo - usp€¦ · aprender muito mais com ele. ao carlos mendonça pela...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
Faculdade de Ciências Farmacêuticas
Programa de Pós-Graduação em Tecnologia Bioquímico-Farmacêutica
Área de Tecnologia de Fermentações
Produção de L-asparaginase pela levedura Leucosporidium muscorum
CRM 1648 isolada de sedimento marinho coletado na Península Antártica
Rominne Karla Barros Freire
Tese para obtenção do Título de DOUTORA
Orientador: Prof. Dr. Adalberto Pessoa Jr.
Coorientador: Prof. Dr. Aldo Tonso
São Paulo, 2019

UNIVERSIDADE DE SÃO PAULO
Faculdade de Ciências Farmacêuticas
Programa de Pós-Graduação em Tecnologia Bioquímico-Farmacêutica
Área de Tecnologia de Fermentações
Produção de L-asparaginase pela levedura Leucosporidium muscorum
CRM 1648 isolada de sedimento marinho coletado na Península Antártica
Rominne Karla Barros Freire
Versão Corrigida da Tese conforme resolução CoPGr 6018 de 13/10/2011. Original
encontra-se disponível no Serviço de Pós-Graduação da FCF/USP.
Tese para obtenção do Título de DOUTORA
Orientador: Prof. Dr. Adalberto Pessoa Jr.
Coorientador: Prof. Dr. Aldo Tonso
São Paulo, 2019

Rominne Karla Barros Freire
Produção de L-asparaginase pela levedura Leucosporidium muscorum
CRM 1648 isolada de sedimento marinho coletado na Península Antártica
Comissão Julgadora
da
Tese para obtenção do título de DOUTORA
__________________________________
Prof. Dr. Orientador/Presidente
__________________________________
1° Examinador
__________________________________
2° Examinador
__________________________________
3° Examinador
__________________________________
4° Examinador
São Paulo, ___ de ___________ de 2019.


Epígrafe
Como ligar 9 pontos utilizando apenas 4 linhas retas?
Quando olhamos a figura e temos um olhar mais rígido, tendenciamos a limitar-nos à
imagem do quadrado formado pelos pontos. Esse olhar fixo, limitado, faz a gente não
alcançar a solução do problema que é, justamente, OLHAR PARA FORA, sair dos
limites, ir além. Para fazer Ciência é preciso “sair da caixa”, para fazer Ciência é preciso
“pensar vivo” e o conhecimento vivo é aquele que nunca se fecha, pois o final
representa sempre o começo de um novo pensar.
Rominne Freire
(trecho do portfólio desenvolvido como parte do curso livre de Pedagogia Waldorf, em
Fevereiro de 2017).
“A coisa mais bela que podemos experimentar é o mistério.
Essa é a fonte de toda a arte e ciências verdadeiras”.
(Albert Einstein).
1
2
3
4

Aos meus pais e irmãos,
que sempre me apoiaram nas minhas escolhas.

Agradecimentos
Aos meus pais, Lana e Wilson, pelo carinho e por terem me apoiado e me incentivado
durante toda a minha trajetória.
Aos meus irmãos (Radha e Endrigo), sobrinhas e sobrinho (Cecília, Clarice e Dante),
cunhado e cunhada (Gilles e Aline), pelo carinho e apoio emocional.
Ao meu orientador Prof. Adalberto por toda a paciência, disponibilidade e por ter
aceitado a orientação desta tese.
Ao meu coorientador Prof. Aldo, sempre muito atencioso e disposto a passar horas com
seus alunos em longas conversas e aprendizados. Gratidão por ter aceitado o convite e
por ter compreendido e apoiado as minhas escolhas, em especial pela orientação nos
ensaios em biorreator.
Ao Prof. Dr. Paul Long, por ter me recebido em seu grupo de pesquisa, no Instituto de
Ciências Farmacêuticas do King’s College, em Londres. Eu ainda não estava
oficialmente matriculada no doutorado, mas foi uma experiência enriquecedora, da
qual sou grata pela indicação do prof. Adalberto.
Ao prof. Dr. Felipe Rebello, profa. Dra. Gisele Monteiro, profa. Dra. Carlota Rangel,
Profa. Dra. Lara Sette e Prof. Dr. João Carlos pela disponibilidade em me auxiliar e
sanar dúvidas com relação a este trabalho. Em especial a Profa. Lara Sette e seu aluno
Fábio Inforsato, por ceder as amostras de leveduras utilizadas neste trabalho e as
informações acerca da coleta, e ao prof. Felipe Rebello pela disponibilidade em me
orientar a respeito das análises estatísticas utilizadas neste trabalho.
Ao prof. Marco Antonio Stephano que participou do meu processo de entrada no
doutorado, pelas sugestões, conversas nos corredores da Faculdade de Ciências
Farmacêutica e pela oportunidade de organizar junto com ele e outros colegas o Curso
de Inverno da FCF/USP nos anos de 2015 e 2017.

Aos amigos de bancada (e da vida) que me apoiaram, em especial a João Santos,
Joãozinho, um irmão querido, que sempre me acolheu e esteve do meu lado ao longo de
todo esse processo, pela orientação nas tentativas de purificação da enzima, pelas
correções dos meus textos e incentivo para que eu não desistisse dessa jornada. Ao
Ignácio Moguel, pela paciência e por ter me ajudado nas muitas tentativas para a
purificação da enzima. Ao Rafael Ferraro, pela oportunidade de ensiná-lo e, assim,
aprender muito mais com ele. Ao Carlos Mendonça pela disponibilidade e atenção na
leitura e correção dos meus textos.
Por todas as conversas de incentivo, em especial a minha mãe, Lana (mãe, amiga,
companheira em todos os momentos), a César Diaz, João Santos e ao prof. Aldo, que
não me deixaram desistir.
A toda equipe (alunos, professores e técnicos) dos laboratórios de Biotecnologia,
Nanobio e Biomol, aos novos e aos que já passaram por aqui, em especial à Larissa
Bentim, Alexsandra Apolinário e Luciana Bueno pela amizade e apoio; à Letícia
Parizotto pelo auxílio nos ensaios em biorreator; ao Tales Costa-Silva e ao Juan
Flores-Santos pela parceria no artigo COSTA-SILVA et al., 2018b.
Ao apoio financeiro da CAPES e FAPESP (que apoia o projeto temático no qual este
trabalho está inserido).
A todos os que estiveram presentes na minha vida nesse período (2014-2019), aos que
ficaram e aos que se foram, aos que me bem quiseram e àqueles que me mal quiseram,
pois, como já disse a Monja Coen em uma de suas palestras, os nossos “inimigos” são
os nossos maiores amigos, pois são eles que apontam nossos erros, nossas falhas e
fraquezas, são o espelho que mostram as nossas sombras e, assim, impulsionam o nosso
crescimento. Por isso, gratidão a todos!

Sumário
Lista de Figuras........................................................................................................ 10
Lista de Tabelas........................................................................................................ 13
Lista de Equações..................................................................................................... 14
Lista de abreviações................................................................................................. 15
RESUMO................................................................................................................. 16
ABSTRACT............................................................................................................. 17
Introdução e Justificativa.......................................................................................... 18
I. REVISÃO BIBLIOGRÁFICA............................................................................. 21
1.1.L-ASNase como agente quimioterápico............................................................. 21
1.2. Estrutura e mecanismo de ação........................................................................ 23
1.3.Leucemia Linfoblástica Aguda.......................................................................... 25
1.4. Tratamento de LLA............................................................................................ 26
1.5. Outras aplicações da enzima L-ASNase........................................................... 29
1.6. Em busca de alternativas às L-ASNases convencionais................................... 30
1.7. Produção de L-ASNase em biorreator.............................................................. 33
1.8. Leveduras da região Antártica.......................................................................... 35
1.8.1. Interesse biotecnológico................................................................................. 36
1.8.2. Leucosporidium muscorum............................................................................ 40
II. OBJETIVOS........................................................................................................ 42
2.1. Objetivo Geral................................................................................................... 42
2.2. Objetivos Específicos........................................................................................ 42
III. MATERIAIS E MÉTODOS.............................................................................. 43
3.1. Leveduras isoladas da Antártica....................................................................... 43
3.2. Preservação das linhagens................................................................................ 45
3.3. Meios de cultivo................................................................................................. 45
3.4. Condições de cultivo em frascos agitados (shaker).......................................... 46
3.5. Seleção dos isolados produtores de L-ASNase................................................. 46
3.6. Análise univariada de fontes de carbono.......................................................... 48
3.7. Análise univariada de fontes de nitrogênio....................................................... 48
3.8. Avaliação da interação de fontes nutricionais por delineamento fatorial
completo para melhoramento do crescimento e produção de L-
ASNase...................................................................................................................... 49
3.9. Análise univariada das condições de cultivo: temperatura, pH inicial,
concentração celular inicial (X0) e concentração de água do mar no
meio.......................................................................................................................... 50
3.10. Ensaios em biorreator..................................................................................... 51
3.10.1. Preparo do biorreator.................................................................................... 51
3.10.2. Ensaios em biorreator: condições de cultivo e oxigênio
dissolvido.................................................................................................................. 52
3.11. Metodologias Analíticas.................................................................................. 55
3.11.1.Monitoramento dopH.................................................................................... 55
3.11.2. Monitoramento do crescimento celular........................................................ 56
3.11.3. Dosagem de atividade enzimática................................................................ 57
3.11.4. Dosagem de proteínas totais......................................................................... 59
3.11.5. Dosagem de sacarose.................................................................................... 59
3.11.6. Dosagem de prolina...................................................................................... 60
3.11.7. Cálculo dos parâmetros cinéticos................................................................. 60
IV. RESULTADOS E DISCUSSÃO....................................................................... 61

4.1. Triagem de leveduras adaptadas ao frio produtoras de L-ASNase (avaliação
qualitativa)............................................................................................................... 61
4.2. Perfil de produção de L-ASNase pelas leveduras selecionadas (avaliação
quantitativa) e seleção do melhor produtor............................................................. 65
4.3. Triagem e seleção dos fatores nutricionais....................................................... 67
4.3.1. Avaliação de fontes de carbono ..................................................................... 68
4.3.2. Avaliação de fontes de nitrogênio.................................................................. 70
4.4. Otimização dos fatores nutricionais por delineamento fatorial
completo................................................................................................................... 73
4.5. Análise univariada das variáveis temperatura (°C), concentração inicial de
células (X0), pH inicial e concentração de água do mar na composição do meio
de cultivo.................................................................................................................. 79
4.6. Cultivos em biorreator e avaliação do oxigênio dissolvido............................. 84
4.6.1. Ensaio 1: sem controle de oxigênio dissolvido (OD < 20%)......................... 84
4.6.2. Ensaio 2: sem controle de oxigênio dissolvido (OD > 20%)......................... 87
4.6.3. Ensaio 3: oxigênio dissolvido controlado em 80%........................................ 90
4.6.4. Ensaio 4: oxigênio dissolvido controlado em 20%........................................ 93
4.6.5. Avaliação comparativa dos ensaios em biorreator......................................... 95
V. CONCLUSÕES................................................................................................... 100
VI. PERSPECTIVAS FUTURAS........................................................................... 102
VII. REFERÊNCIAS.............................................................................................. 103
APÊNDICES.......................................................................................................... 117
apêndice A................................................................................................................ 118
apêndice B................................................................................................................ 122
apêndice C................................................................................................................ 122
apêndice D................................................................................................................ 126
apêndice E................................................................................................................ 127
apêndice F................................................................................................................. 132
apêndice G................................................................................................................ 133
apêndice H................................................................................................................ 136
apêndice I................................................................................................................. 137

Lista de Figuras
Figura 1 – Número de publicações registradas na plataforma PUBMED do
NCBI que faz referência ao termo “asparaginase” desde 1949 até os dias atuais.
Pesquisa realizada em fevereiro de 2019................................................................. 22
Figura 2 – Mecanismo de ação da enzima L-ASNase. Fonte: adaptado de
AVRAMIS, 2012........................................................................................................ 23
Figura 3 –A atividade enzimática em relação a temperatura. A atividade das
enzimas psicrófilas (círculo vazio) α-amilase (gráfico esquerdo) e celulase
(gráfico direito) e seus correspondentes mesófilos (círculo cheio). Fonte:
Adaptado de FELLER, 2013.................................................................................... 38
Figura 4 – Imagem de células em microscópio e morfologia da colônia de
linhagem de L. muscorum. Fonte: http://www.westerdijkinstitute.nl/collections,
acesso em 12/12/18.................................................................................................. 41
Figura 5 – Mapa da Baia do Almirantado, localizada na Ilha do Rei George,
Península Antártica. Os números no mapa da Baia do Almirantado (A) indicam
os lugares onde as amostras foram coletadas. 1. Refúgio 2; 2. EACF (Estação
Antártica Comandante Ferraz, base brasileira de pesquisa); 3. Botany Point.
Fontes: (A) grupo de pesquisa da Profa. Dra. Lara Sette; (B) acesso no Google
Maps; e (C) adaptada de ANDRADE et al., 2015.................................................... 44
Figura 6 – Distribuição de cada cepa e dos brancos (amostras sem
microrganismo) em placas de 96 poços para pré-inóculo e produção..................... 47
Figura 7 – Biorreator BIOSTAT® B-MO em uso para cultivo de L.
muscorumCRM 1648 e produção de L-ASNase..................................................... 51
Figura 8 – Representação do painel de controle do biorreator BIOSTAT®
B-MO
e todos os parâmetros possíveis de controle: agitação, temperatura, pH, OD,
vazão de ar puro de entrada (1) e vazão de O2 puro de entrada (2), este último
controlado por pulsos de abertura (% de tempo de abertura) da válvula de O2 (3).
No início do ensaio 4 a conFiguração foi modificada, conforme representado no
círculo pontilhado................................................................................................... 55
Figura 9 – Valores de densidade óptica em função de valores de massa seca
celular, sendo o R2 = 0,987.......................................................................................
56
Figura 10 – Reações químicas no método do ácido L-aspartil-β-hidroxâmicode
quantificação de L-ASNase. Fonte: adaptada de MAGRI et al., 2018.................... 57
Figura 11 – Sequência de passos para a execução do teste de atividade pelo
método do ácido L-aspartil-β-hidroxâmico.............................................................. 58
Figura 12 – Triagem de produtores de L-ASNase entre 150 isolados da Antártica
pelo método do ácido L-aspartil-β-hidroxâmico. A intensidade da cor vermelha
indica maior ou menor produção da enzima............................................................ 62
Figura 13 – Resultado do cultivo dos isolados produtores de L-ASNase em meio
CDM com azul de bromotimol. Da esquerda para a direita: meio não inoculado,
meio sem fonte de nitrogênio, isolados 2, 26, 1, 16, 86, 94 e 85............................. 63
Figura 14 – Morfologia das colônias dos isolados produtores................................ 64
Figura 15 – Curva de crescimento e produção de L-ASNase por 7 isolados da
Antartica .................................................................................................................. 66
Figura 16 – Crescimento celular (A) e produção de L-ASNase (B) pela linhagem
L. muscorum CRM 1648 em meio Czapek Dox com adição de diferentes fontes
de carbono............................................................................................................... 69

Figura 17 – Crescimento celular (A) e produção de L-ASNase (B) pela linhagem
L. muscorum CRM 1648 em meio Czapek Dox com diferentes fontes de
nitrogênio.................................................................................................................. 71
Figura 18 – Gráfico da superfície de resposta em 3D de MSC (Y1) e atividade
de L-ASNase (Y2) em função da concentração de sacarose (X1), extrato de
levedura (X2)e prolina (X3). Valores fixados: X1 = 17, X2 = 15 e X3 = 16......... 78
Figura 19 – Efeito do meio Czapek Dox no crescimento de L. muscorum CRM
1648 e produção de L-ASNase após o delineamento experimental. CD: meio
Czapek Dox inicial, com glicose e prolina; CDM: meio Czapek Dox melhorado
neste trabalho, contendo prolina (20 g.L-1
), extrato de levedura (15 g.L-1
) e
sacarose (20 g.L-1
).................................................................................................... 79
Figura 20 – Avaliação do crescimento de L. muscorum CRM 1648 em meio
CDM por análise de DO e massa seca celular, bem como de Ln(msc), quando X0
= 0,5 g.L-1
(A), X0 = 1,0 g.L-1
(B) e X0 = 1,5 g.L-1
(C). msc: massa seca celular....... 80
Figura 21 – Curva comparativa de crescimento de L. muscorum CRM 1648 e
produção de L-ASNase quanto a concentração celular inicial (X0)......................... 81
Figura 22 – Análise dos dados de inclinação das curvas de crescimento de L.
muscorum CRM 1648 (A) e produção de L-ASNase (B) por regressão linear em
cada nível de X0........................................................................................................ 81
Figura 23 – Curva de crescimento de L. muscorum CRM 1648 e produção de L-
ASNase em diferentes temperaturas......................................................................... 82
Figura 24 – Curva de crescimento de L. muscorum CRM 1648 e produção de L-
ASNase em diferentes valores de pH inicial do meio CDM.................................... 83
Figura 25 – Curva de crescimento de L. muscorum CRM 1648 e produção de L-
ASNase em diferentes concentrações de água do mar no meio CDM..................... 84
Figura 26 – Perfis das variáveis pH, temperatura (°C), agitação (rpm) e volumes
adicionados de ácido (HCl 20%) e base (NaOH 20%) registradas pelo biorreator
durante o Ensaio 1.................................................................................................... 84
Figura 27 – Perfis das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 1
em biorreator............................................................................................................ 85
Figura 28 – Perfil das variáveis crescimento (DO e MSC), pH e produção de L-
ASNase no Ensaio 1 em biorreator.......................................................................... 86
Figura 29 – Perfil das variáveis produção de CO2 (CER) e Ln de CER na fase
exponencial (A) e crescimento celular (massa seca celular, [X]) mais ln(X) e
ln(X) na fase exponencial (ln(X)exp) no Ensaio 1................................................... 86
Figura 30 – Perfis das variáveis pH, temperatura (°C) e agitação (rpm)
registradas pelo biorreator durante o Ensaio 2......................................................... 87
Figura 31 – Perfis das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 2
em biorreator............................................................................................................ 88
Figura 32 – Perfil de crescimento [X], consumo de sacarose e de prolina [S] e
produção de L-ASNase no Ensaio 2 em biorreator.................................................. 89
Figura 33 – Perfil das variáveis crescimento celular (massa seca celular, [X]),
ln(X) e ln(X) na fase exponencial (ln(X)exp) no Ensaio 2...................................... 89
Figura 34 – Medida de pH do meio CDM e atividade de L-ASNase ao longo do
cultivo de L. muscorum CRM 1648 em biorreator no ensaio 2............................... 90
Figura 35 – Perfis das variáveis pH, temperatura (°C) e agitação (rpm)
registradas pelo biorreator durante o ensaio 3......................................................... 91
Figura 36 – Perfis das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 3
em biorreator............................................................................................................ 91
Figura 37 – Perfil de crescimento [X], consumo de sacarose e prolina [S] e 92

produção de L-ASNase no Ensaio 3 em biorreator.................................................
Figura 38 – Perfil das variáveis crescimento celular (massa seca celular, [X]),
ln(X) e ln(X) na fase exponencial (ln(X)exp) no Ensaio 3...................................... 92
Figura 39 – Perfis das variáveis pH, temperatura (°C) e agitação (rpm)
registradas pelo biorreator no Ensaio 4.................................................................... 93
Figura 40 – Perfis das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 4
em biorreator. O valor médio de OD foi de 25,1 %................................................. 94
Figura 41 –Perfil de crescimento [X], consumo de sacarose e de prolina [S] e
produção de L-ASNase no Ensaio 4 em biorreator.................................................. 95
Figura 42 – Perfil das variáveis crescimento celular (massa seca celular, [X]),
ln(X) e ln(X) na fase exponencial (ln(X)exp) no Ensaio 4...................................... 95
Figura 43 – Comparação do perfil de OD em cada ensaio...................................... 97
Figura 44 – Comparação do perfil de crescimento (A) e de produção de L-
ASNase (B) nas condições de OD avaliadas em biorreator..................................... 97
Figura 45 – Perfil comparativo da produtividade em células Px (A) e em produto
L-ASNase Pp (B) nos ensaios 1-4 em biorreator..................................................... 98
Figura 46 – Perfil comparativo do consumo dos substratos sacarose (A) e
prolina (B) nos ensaios 2-4 em biorreator................................................................ 99

Lista de Tabelas
Tabela 1 –Propriedades das L-ASNases disponíveis comercialmente. Fontes: de
Abud, 2005 e Narta et al., 2007................................................................................... 27
Tabela 2 – Referências encontradas na literatura relacionadas à produção de L-
ASNase por espécies de leveduras............................................................................... 32
Tabela 3 – Informações sobre a espécie, material de origem e local de coleta dos
isolados identificados neste trabalho como produtores de L-ASNase......................... 43
Tabela 4 – Meios de cultivo utilizados para a preservação, manutenção e
crescimento de leveduras isoladas da Antártica........................................................... 46
Tabela 5 – Fontes de carbono e respectivas concentrações (C) em g.L-1
de cada
fonte avaliada no meio CD para crescimento da levedura alvo e produção de L-
ASNase......................................................................................................................... 48
Tabela 6 – Fontes de nitrogênio e respectivas concentrações (C) em g.L-1
de cada
fonte avaliada no meio CD para crescimento da levedura e produção de L-
ASNase......................................................................................................................... 49
Tabela 7 – Fatores e níveis avaliados em delineamento fatorial completo (I) para
melhoramento do crescimento de L. muscorum CRM 1648 e produção de L-
ASNase......................................................................................................................... 49
Tabela 8 – Fatores e níveis avaliados em delineamento fatorial completo (II) para
melhoramento do crescimento de L. muscorum CRM 1648 e produção de L-
ASNase......................................................................................................................... 50
Tabela 9 – Matriz do planejamento fatorial completo(I) e suas respectivas
respostas para a avaliação do crescimento de L. muscorumCRM 1648 e produção
de L-ASNase................................................................................................................ 74
Tabela 10 –Análise de Variância (ANOVA) por regressão de superfície de
resposta de MSC (Y1) e atividade de L-ASNase (Y2) em função da concentração
de sacarose (X1), extrato de levedura (X2) e prolina (X3) – delineamento
experimental I.............................................................................................................. 75
Tabela 11 – Matriz do planejamento fatorial completo (II)e suas respectivas
respostas para a avaliação do crescimento de L. muscorumCRM 1648 e produção
de L-ASNase................................................................................................................ 76
Tabela 12 –Análise de Variância (ANOVA) por regressão de superfície de
resposta de MSC (Y1) e atividade de L-ASNase (Y2) em função da concentração
de sacarose (X1), extrato de levedura (X2) e prolina (X3) – delineamento
experimental II............................................................................................................. 77

Lista de Equações
Equação 1 – Cálculo do volume de suspensão celular para inoculação do meio de
cultivo........................................................................................................................... 46
Equação 2 – Cálculo de predição da resposta massa seca celular no delineamento
experimental................................................................................................................. 50
Equação 3 – Cálculo de predição da resposta produção de L-ASNase no
delineamento experimental.......................................................................................... 50
Equação 4 – Cálculo da taxa de captação de O2 dissolvido (OUR)............................ 52
Equação 5 – Cálculo da taxa de produção de CO2 (CER).......................................... 52
Equação 6 – Cálculo do coeficiente respiratório (RQ)............................................... 52
Equação 7 – Cálculo do fluxo de ar de saída no biorreator (Φs)................................ 53
Equação 8 – Cálculo de massa seca celular (em gramas por litro)............................. 56
Equação 9 – Cálculo da relação entre DO e MSC...................................................... 56
Equação 10 – Cálculo da velocidade específica de crescimento (µx)......................... 60
Equação 11 – Cálculo da Integral da equação 10....................................................... 60
Equação 12 – Cálculo da produtividade em células................................................... 60
Equação 13 – Cálculo da produtividade em produto (L-ASNase)............................. 60

Lista de abreviações
L-ASNase –L-asparaginase
Asn – asparagina
LLA – leucemia linfoblástica aguda
ASNS – asparagina sintetase
MS – Ministério da Saúde
PI – ponto isolelétrico
FDA – Food and Drug administration
OFAT –one-factor at a time
DoE – Delineamento Experimental
CCD – Central Composite Design
OD – oxigênio dissolvido
DO – densidade óptica
CD – Czapek-Dox
CDM – Czapek-Dox Modificado
EACF – Estação Antártica Comandante Ferraz
CRM – Central de Recursos Microbianos da UNESP
vvm – volume de gás por volume de meio por minuto
LpM – litro de gás por litro de meio por minuto
AHA –-hidroxamato aspártico
GOD/POD– glicose oxidase/peroxidase
Eq – equação
UV – ultravioleta
TCA – ácido tricloroacético
RCN – repressão catabólica pelo nitrogênio
FN – fonte de nitrogênio
FC – fonte de carbono
MSC – massa seca celular
DEC – digestivo enzimático de caseína
RQ – coeficiente respiratório
OUR –taxa de captação de O2 dissolvido
CER –taxa de produção de CO2

RESUMO
FREIRE, R.K.B. Produção de L-asparaginase pela levedura Leucosporidium muscorum
CRM 1648 isolada de sedimento marinho coletado na Península Antártica. 2019. 147f.
Tese (Doutorado) – Faculdade de Ciências Farmacêuticas, Universidade de São Paulo,
São Paulo, 2019.
L-asparaginase (L-ASNase) é uma enzima com propriedades interessantes para a
indústria médica, farmacêutica e de alimentos, que tem recebido atenção especial,
inclusive no Brasil, por fazer parte do protocolo de tratamento de distúrbios
linfoproliferativos, como a leucemia linfoblástica aguda (LLA). No mercado desde a
década de 1970, as enzimas de origem bacteriana enfrentam algumas limitações por
provocarem reações adversas graves em quase 80% dos pacientes em tratamento. Nesse
contexto, L-ASNases provenientes de leveduras se destacam como alternativa, por
serem mais próximas às congêneres humanas. A Antártica ainda é um ambiente pouco
explorado, com grande diversidade de microrganismos com potencial para a produção
de moléculas biológicas de interesse industrial. Nesse contexto, 150 leveduras isoladas
de amostras de sedimento marinho coletadas na Península Antártica como parte do
projeto MICROSFERA (PROANTAR/CNPq) foram avaliadas para a produção de L-
ASNase. A triagem resultou em 9 isolados produtores, dos quais 7 pertencem ao gênero
Leucosporidium. A linhagem L. muscorum CRM 1648 foi a que produziu mais enzima
(540 U.L-1
), com maior produtividade (5,6 U.L-1
.h-1
) e, por isso, foi alvo deste estudo. A
análise univariada de fontes de carbono e nitrogênio indicou maior crescimento desse
microrganismo e produção de L-ASNase em meio CD com extrato de levedura, prolina
e sacarose. Ureia, cloreto de amônio e sulfato de amônio resultaram em baixa ou
nenhuma produção da enzima, sugerindo que a metabolização de fontes de nitrogênio
por essa linhagem está sob a influência do fenômeno de repressão catabólica pelo
nitrogênio (RCN). Dois delineamentos experimentais do tipo fatorial completo
resultaram em um aumento de 10 vezes na produção e produtividade da enzima (4582,5
U.L-1
e 63,6 U.L-1
.h-1
, respectivamente). A análise univariada da concentração inicial de
inóculo (X0), pH inicial do meio, temperatura e adição de água do mar mostrou que a
melhor condição para a produção foi: pH = 5,5 ou 6,5, cultivo a 15°C com adição de
água do mar (25-50% m/v). A variável X0 não foi significativa nas concentrações
avaliadas. Cultivos em biorreator (batelada) foram conduzidos em quatro diferentes
níveis de oxigênio dissolvido (OD): (1) OD não controlado e abaixo de 20%, (2) OD
não controlado e acima de 20%, (3) OD controlado em 80% e (4) OD controlado em
20%. Os resultados mostraram que OD é fator limitante para o crescimento de L.
muscorum CRM 1648 e produção de L-ASNase por essa levedura e deve ser mantido
acima de 35% para maior produção da enzima.Neste trabalho, a composição do meio e
condições de cultivo foram estabelecidas para favorecer a produção de uma nova L-
ASNase livre de atividade glutaminásica por levedura adaptada ao frio, abrindo espaço
para novos estudos acerca de seu potencial antileucêmico e possível uso como
alternativa às enzimas já existentes no mercado no tratamento de LLA.
Palavras-chaves: L-asparaginase; Leucosporidium muscorum; leveduraadaptada ao
frio; leucemia linfoblástica aguda; Antártica; biorreator; delineamento experimental;
oxigênio dissolvido.

ABSTRACT
FREIRE, R.K.B. L-asparaginase production by the yeast Leucosporidium muscorum
CRM 1648 isolated from marine sediments collected in Antarctic Peninsula. 2019. 147f.
Tese (Doutorado) – Faculdade de Ciências Farmacêuticas, Universidade de São Paulo,
São Paulo, 2019.
L-asparaginase (L-ASNase) is an enzyme with interesting properties for medical,
pharmaceutical and food industry, which has received special consideration, especially
in Brazil, for being part of lymphoproliferative disorders treatment, such as acute
lymphoblastic leukemia (ALL). Bacterial enzymes are on the market since the 1970s
and face some limitations related to theirserious adverse reactions that reach almost
80% of all patients in treatment. In this context, L-ASNases from yeasts are highlighted
as important alternative to bacterial enzymes, due to the closerphylogeny to human
congeners. Antarctic environment has much to be explored, with a vast diversity of
microorganisms with potential to produce biomolecules with industrial interest. A total
of 150 yeasts isolated from Antarctic marine sediments as part of MICROSFERA
project (PROANTAR/CNPq) were evaluated for L-ASNase production. The screening
resulted in 9 producers, 7 species from the genus Leucosporidium. L. muscorum CRM
1648 was the strain that yielded the highest L-ASNase activity (540 U.L-1
) and
volumetric productivity (5.6 U.L-1
.h-1
). Carbon and Nitrogen sources were evaluated by
a method of one-factor at a time (OFAT). From the gather results, sucrose, yeast extract
and proline resulted in a maximal growth and highest enzyme production.The absence
or low production of L-ASNase in medium with urea, ammonium chloride and
ammonium sulfate suggests the presence of nitrogen catabolic repression (NCR).
Carbon and nitrogen concentration were evaluated by full factorial design and yielded
about ten times higher enzyme and volumetric productivity (4582.5 U.L-1
and 63.6 U.L-
1.h
-1, respectively). Initial inoculum concentration (X0), initial pH, temperature and
concentration of seawater in the culture were evaluated by OFAT analysis and the best
condition for L-ASNase production was: pH = 5.5 or 6.5, at 15 °C with addition of
seawater (25-50 wt%). X0 was not considered a significant variable. Bioreactor assays
(in batch regime) were performed in four different dissolved oxygen (DO) levels: (1)
without DO control (DO remained under 20%), (2) without DO control (DO remained
above 20%), (3) DO controlled at 80%, and (4) DO controlled at 20%.The results
showed that DO is a key factor for growth of L. muscorum CRM 1648 and production
of L-ASNase by this yeast and should be maintained above 35% for higher production
of this enzyme.At this work, the medium and culture conditions were established to
support the production of a novel glutaminase-free L-ASNase by a cold adapted yeast,
opening a new path for further studies regarding its antileukemic potential and possible
use as an alternative for ALL treatment.
Keywords: L-asparaginase; Leucosporidium muscorum; cold-adapted yeast; acute
lymphoblastic leukemia; Antarctica; bioreactor; experimental design; dissolved oxygen.

18
INTRODUÇÃO E JUSTIFICATIVA
Produção de L-asparaginase no Brasil: uma necessidade urgente
Desde a década de 70, quando a enzima L-asparaginase (L-ASNase) de
Escherichia coli foi extraída, purificada e introduzida no protocolo de tratamento de
pacientes com leucemia linfoblástica aguda (LLA), a taxa de sobrevida aumentou
sensivelmente e hoje é em torno de 80-90% (PIETERS et al., 2011; KATO; MANABE,
2018). Combinada com esteróides, vincristina e, às vezes, antraciclina nas etapas
iniciais do tratamento, essa enzima se tornou um produto terapêutico de grande
importância para a erradicação completa das células leucêmicas (KATO; MANABE,
2018), ainda que apresente limitações quanto à estabilidade no soro e possa causar
hipersensibilidade em 3-78% dos pacientes (apud NARTA et al., 2007).
Mais recentemente, outras L-ASNases de origem bacteriana foram lançadas no
mercado, uma forma peguilada proveniente da forma nativa de E. coli e outra nativa,
proveniente da espécie Erwinia chrysanthemi (reclassificada como Dickeya dadantii, de
acordo com GRENIER et al 2006). No Brasil, no entanto, a única formulação liberada
para tratamento de LLA pela ANVISA é a nativa de E. coli. Além disso, o Brasil não é
um país de fabricação de biofármacos, muito pelo contrário, costuma importar esse tipo
de medicamento. Por depender da importação de L-ASNase, em 2013 o país sofreu
problemas no seu abastecimento pela empresa fornecedora Bagó, que descontinuou a
comercialização de Elspar®, fórmula de L-ASNase inicialmente fabricada pela Merck
Sharp & Dohme (MSD) e, posteriormente, pela Lundbeck INC e OSO
Biopharmaceutical (MINISTÉRIO DA SAÚDE, 2017).
De acordo com a Recomendação n°10/2013 do Ministério Público Federal, a
ausência de L-ASNase nos protocolos de tratamento no Brasil é um problema de saúde
pública e representaria um retrocesso de pelo menos 20 anos no tratamento de LLA
nesse país, principalmente porque dados da época (2012-2013) do INCA (Instituto
Nacional do Câncer) previam 11.500 novos casos/ano de câncer em crianças, dos quais
25% seriam de LLA e 15% de Linfoma Não Hodgkin (MINISTÉRIO PÚBLICO
FEDERAL, Recomendação n°10/2013). Embora dados atuais do INCA mostrem que no
Brasil, de 2000 a 2005, a sobrevida relativa em cinco anos de leucemia infantil tenha
resultado em 70% (INCA, 2016), na falta de L-ASNase cerca de 5 mil crianças e
adolescentes no Brasil teriam sua chance de cura comprometida (MINISTÉRIO
PÚBLICO FEDERAL, Recomendação n°10/2013).

19
Por ser a L-ASNase tão importante no protocolo de tratamento de LLA,
associada ao fato de que o Brasil ainda não produzir biofármacos e de que a enzima
Elspar® (formulação nativa de E. coli) é, dentre as formulações comerciais, aquela que
mais provoca reações imunológicas, existe uma urgente necessidade da sua produção
nacional, bem como a pesquisa de fontes alternativas dessa enzima. Por isso, desde
2013 há uma demanda do Ministério da Saúde (MS) para que o Brasil comece a
produzir L-ASNase. Recentemente, o MS adquiriu uma alternativa mais barata da
enzima, de nome comercial Leuginase®, cuja eficácia foi contestada por profissionais da
área da saúde, gerando polêmica que foi parar na mídia brasileira e resultou em sua
retirada do mercado (Fantástico, Rede Glogo, 2017). Desde então há risco de
desabastecimento dos hospitais que pode vir a comprometer o tratamento de pacientes
com LLA.
Essas questões motivaram o projeto temático (FAPESP 2013/08617-7)
Produção de L-asparaginase extracelular: da bioprospecção à engenharia de um
biofármaco antileucêmico, que propõe incentivar a pesquisa nessa área, com a
finalidade de buscar diferentes fontes produtoras dessa enzima em sua forma nativa,
bem como utilizar de ferramentas de engenharia genética para melhorar os biofármacos
que já estão no mercado ao reduzir as repostas imunológicas e, portanto, efeitos
colaterais associados ao tratamento com as L-ASNases convencionais.
Em razão de suas condições inóspitas, a Antártica tem se revelado um ambiente
bastante propício para a descoberta de novas biomoléculas com características
interessantes para a indústria. Por isso, esse ambiente foi um dos escolhidos para
rastrear microrganismos produtores de L-ASNase e este é o campo de pesquisa onde se
encaixa este trabalho.
O presente trabalho focou na seleção de uma levedura produtora de L-ASNase,
melhoramento do meio e condições de cultivo em frascos agitados e avaliação da
demanda de oxigênio dissolvido (OD) em biorreator, visto que o entendimento da
fisiologia do microrganismo produtor é essencial e compõe as primeiras etapas em um
processo industrial que visa à produção de produtos biológicos. Optou-se por explorar
apenas leveduras porque todas as L-ASNases disponíveis no mercado são de origem
bacteriana e existe a expectativa de que a enzima de origem fúngica, por sofrer
modificações pós-traducionais, possa resultar em um tratamento com menor resposta
imunológica e, logo, com mais qualidade de vida para os pacientes com LLA.
Conscientemente, espera-se que este trabalho não seja o fim, mas apenas o começo de

20
novas pesquisas tanto sobre a levedura Leucosporidium muscorum e seu potencial
biotecnológico, como acerca da enzima L-ASNase produzida por esse microrganismo.

21
I. REVISÃO BIBLIOGRÁFICA
1.1. L-ASNase como agente quimioterápico
L-asparaginase, L-asparagina amidohidrolase (EC 3.5.1.1), é um importante
agente quimioterápico utilizado para o tratamento de uma infinidade de desordens
linfoproliferativas e linfomas, principalmente de leucemia linfoblástica aguda (LLA) em
crianças e jovens (NARTA et al., 2007). Clementi (1922) foi o primeiro a observar
atividade elevada dessa enzima no soro de porquinhos da índia (Cavia porcellus). Anos
mais tarde, Kidd (1953) reportaria a regressão de linfosarcomas em ratos tratados
também com o soro desse animal. Em 1961, Broome relacionaria o estudo de Kidd com
a observação feita por Clementi, concluindo que a enzima L-ASNase seria o agente
responsável pela regressão de linfosarcomas (apud NARTA et al., 2007).
A extração de L-ASNase, em quantidades suficientes para a produção industrial,
a partir do soro de porquinhos da índia envolve uma operação complicada, de forma que
outras fontes produtoras da enzima foram buscadas. Mashburn e Wriston (1964), bem
como Campbell e Mashburn (1969) reportaram a purificação de L-ASNase de
Escherichia coli e demonstraram que esta tinha potencial tumoricida similar à enzima
produzida por porquinhos da índia, com a vantagem de ser produzida mais facilmente e
por um custo mais baixo (apud NARTA et al., 2007). Dessa forma, a obtenção da
enzima a partir de sistemas mais simples, como bactérias, possibilitou a produção em
larga escala para os primeiros estudos pré-clínicos e clínicos. Desde então, L-ASNases
de diversas fontes têm sido estudadas, desde aquelas produzidas por microrganismos
procariotos como Escherichia coli (CEDAR; SCHWARTZ, 1968), Erwinia carotovora
(ROTH et al., 2013) e Pseudomonas aeruginosa (EL-BESSOUMY et al., 2004), até
organismos eucariotos, como Aspergillus terreus, roedores, primatas e plantas
(KHUMAR; SOBHA, 2012).
Uma pesquisa na plataforma PUBMED do National Center for Biotechnology
Information (NCBI) realizada dia 26 de dezembro de 2018 mostra o registro de
publicações que fazem referência a palavra “asparaginase”. Desde 1949 até os dias
atuais são quase 5.000 publicações, as quais se mostram expressivas nos primeiros anos
da década de 70, época na qual a enzima nativa de E. coli foi lançada no mercado. O
tema tornou-se evidente novamente desde 2012, como mostra a Figura 1, período no
qual o Ministério da Saúde começou a anunciar futuros problemas no abastecimento de
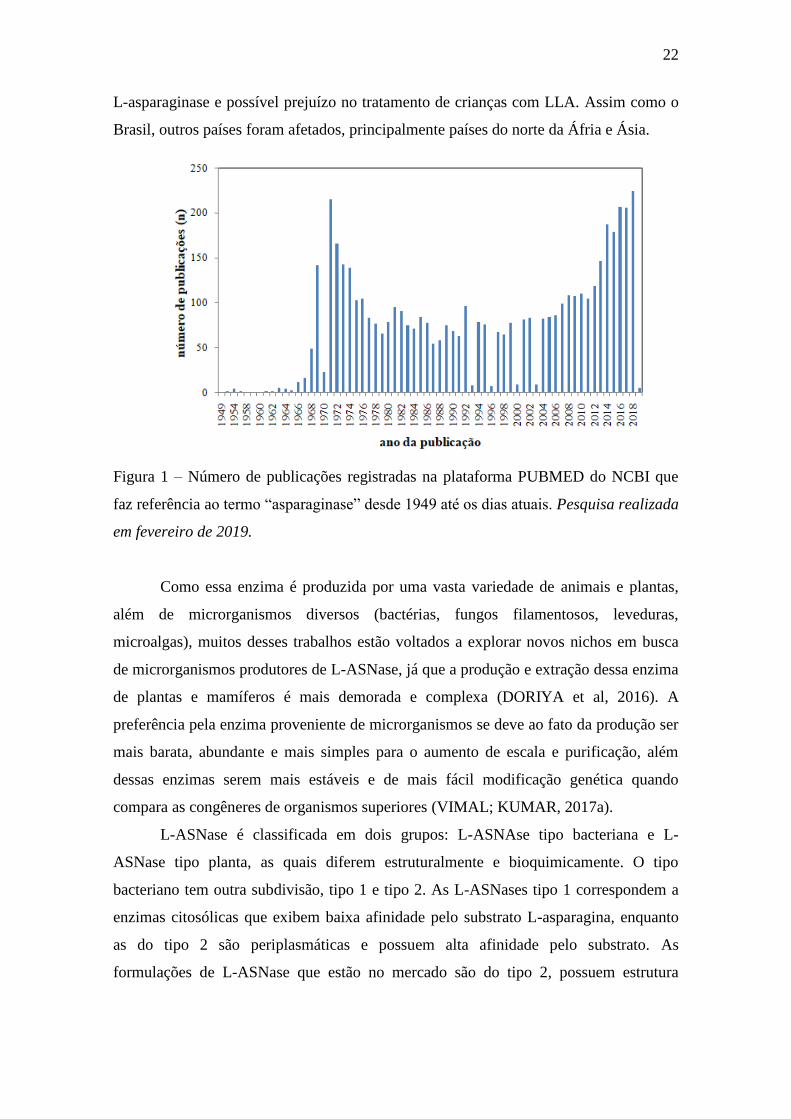
22
L-asparaginase e possível prejuízo no tratamento de crianças com LLA. Assim como o
Brasil, outros países foram afetados, principalmente países do norte da Áfria e Ásia.
Figura 1 – Número de publicações registradas na plataforma PUBMED do NCBI que
faz referência ao termo “asparaginase” desde 1949 até os dias atuais. Pesquisa realizada
em fevereiro de 2019.
Como essa enzima é produzida por uma vasta variedade de animais e plantas,
além de microrganismos diversos (bactérias, fungos filamentosos, leveduras,
microalgas), muitos desses trabalhos estão voltados a explorar novos nichos em busca
de microrganismos produtores de L-ASNase, já que a produção e extração dessa enzima
de plantas e mamíferos é mais demorada e complexa (DORIYA et al, 2016). A
preferência pela enzima proveniente de microrganismos se deve ao fato da produção ser
mais barata, abundante e mais simples para o aumento de escala e purificação, além
dessas enzimas serem mais estáveis e de mais fácil modificação genética quando
compara as congêneres de organismos superiores (VIMAL; KUMAR, 2017a).
L-ASNase é classificada em dois grupos: L-ASNAse tipo bacteriana e L-
ASNase tipo planta, as quais diferem estruturalmente e bioquimicamente. O tipo
bacteriano tem outra subdivisão, tipo 1 e tipo 2. As L-ASNases tipo 1 correspondem a
enzimas citosólicas que exibem baixa afinidade pelo substrato L-asparagina, enquanto
as do tipo 2 são periplasmáticas e possuem alta afinidade pelo substrato. As
formulações de L-ASNase que estão no mercado são do tipo 2, possuem estrutura

23
tetramérica composta por quatro subunidades idênticas, cada qual constituída por 326
resíduos de aminoácidos (DORIYA et al, 2016).
1.2. Estrutura e mecanismo de ação
A enzima L-ASNase age hidrolisando o grupamento amida da cadeia lateral do
aminoácido, não essencial, L-asparagina (Asn), levando à formação de ácido aspártico e
amônia (AVRAMIS, 2012), como mostra a Figura 2.
Figura 2 – Mecanismo de ação da enzima L-ASNase. Fonte: adaptado de AVRAMIS,
2012.
O aspartato resultante dessa reação sofre transaminação com α-cetoglutarato,
gerando glutamato e oxaloacetato que, em células de mamíferos, é convertido em
malato no citosol, sendo transportado para a matriz mitocondrial pelo transportador
malato-α-cetoglutarato. Em bactérias, o oxaloacetato produzido na reação de
transaminação pode ser utilizado diretamente no ciclo do ácido cítrico para a geração de
energia na forma de potencial redutor (NELSON; COX, 2014).
Em células humanas normais, a asparagina é um aminoácido não essencial, já
que pode ser sintetizada a partir de outros aminoácidos (como glutamato, glutamina e
aspartato), através de transaminases e asparagina sintetase (MARINI et al., 2017). O
modo de ação da enzima L-ASNase consiste no fato das células leucêmicas
necessitarem de elevadas quantidades do aminoácido L-asparagina para a sua
proliferação, além de não o sintetizarem em quantidades suficientes em razão de baixa
ou nenhuma capacidade de produção de asparagina sintetase (NARTA et al., 2007;
RAETZ; SALZER, 2010; KUMAR; SOBHA, 2012). Assim, para satisfazer a elevada
demanda desse aminoácido, as células leucêmicas precisariam de asparagina fornecida

24
pela dieta ou proveniente de sua biossíntese, que é limitada (KUMAR; SOBHA, 2012).
Dessa forma, a ação antineoplásica efetiva dessa enzima tem como base a necessidade
nutricional dos linfoblastos leucêmicos, uma vez que deprimem a asparagina presente
no soro, fator essencial para a síntese de proteínas, DNA e RNA, resultando em morte
celular. Uma vantagem desse tratamento é que as células saudáveis não sofrem com os
efeitos diretos das L-ASNases por serem capazes de produzir normalmente o
aminoácido L-asparagina a partir de outros aminoácidos, como já mencionado.
A enzima L-ASNase também é capaz de converter L-glutamina em ácido
glutâmico e amônia, ainda que essa reação ocorra em menor velocidade e mais baixa
afinidade pelo substrato (CHAN et al., 2014). Uma questão ainda pouco abordada e
esclarecida é se a atividade glutaminásica da enzima L-ASNase seria prejudicial ao
tratamento de LLA, já que aparentemente esta estaria associada a alguns efeitos
adversos (neurotoxicidade, principalmente), ou se essa atividade atuaria juntamente com
a atividade asparaginásica no combate a esse tipo de leucemia. Há indícios na literatura
de que glutaminases microbianas também possuem atividade antitumoral e foi reportada
a regressão completa de leucemia em camundongos tratados com L-ASNase em
condições na qual a L-glutamina circulante foi depletada (apud WADE et al., 1971). De
acordo com COVINI et al. (2012), a hidrólise de glutamina pela L-ASNase contribuiria
com os efeitos tóxicos no tratamento de LLA mas, por outro lado, seria necessária para
sustentar a depleção do aminoácido asparagina, contribuindo significativamente para a
atividade antineoplásica da enzima.
CHAN et al. (2014) avaliaram o efeito da ausência de atividade glutaminásica na
L-ASNase de E. coli comercial (Elspar®). Eles observaram que a enzima modificada
por mutagênese sítio-específica (mutante Q59L) conseguia matar as células leucêmicas
não produtoras de asparagina sintetase (ASNS negativas), mas que não tinha atividade
alguma sobre as células cancerígenas produtoras dessa enzima (ASNS positivas). Esse
resultado sugere que a ausência de atividade glutaminásica em variantes da enzima L-
ASNase poderia alcançar melhor resultado terapêutico em células leucêmicas ASNS
negativas e talvez possa ser interessante em alguns casos, já que supostamente reduziria
a toxicidade no tratamento com L-ASNase. Por outro lado, a atividade glutaminásica
continua sendo essencial para matar as células leucêmicas produtoras de asparagina
sintetase (ASNS positivas) (CHAN et al., 2014). Faltam na literatura estudos adicionais
que corroborem e se aprofundem mais nesse assunto.

25
1.3. Leucemia Linfoblástica Aguda
O termo leucemia refere-se a um grupo de doenças complexas e diferentes entre
si que afetam a produção dos glóbulos brancos (HAMERSCHLAK, 2010). Pode ser
classificada como linfoide ou mieloide, de acordo com o tipo celular afetado, e quanto
ao tempo de desenvolvimento, podendo ser de forma aguda ou crônica. A forma aguda é
caracterizada pelo aumento rápido de células imaturas no sangue, fazendo com que a
medula óssea seja incapaz de produzir células sanguíneas saudáveis (INCA, 2016). A
leucemia corresponde a aproximadamente 1/3 dos casos de câncer em crianças de 0-14
anos em todo o mundo, sendo a LLA o tipo mais freqüente (STELIAROVA-FOUCHER
et al., 2017).
De acordo com dados do Instituto Nacional do Câncer (INCA, 2018), no Brasil,
para o biênio de 2018-19 estimam-se 5.840 casos novos de leucemia em homens e
4.860 em mulheres, aproximadamente 7% de aumento em relação a estimativa para os
anos 2016-17 (INCA, 2016). Os valores atuais correspondem a um risco estimado de
5,75 casos novos a cada 100 mil homens e 4,56 para cada 100 mil mulheres. Sem
considerar os tumores de pele não melanoma, a leucemia em homens é o quinto tipo de
câncer mais frequente na Região Norte (4,17/100 mil), na Região Nordeste ocupa a
oitava posição (4,90/100 mil), nas Regiões Sudeste (5,79/100 mil) e Sul (8,67/100 mil)
ocupa a décima posição e na Região Centro Oeste (4,88/100 mil) está na décima
primeira posição. Para as mulheres, é a sexto tipo de câncer mais frequente na Região
Norte (3,29/100 mil), o nono tipo mais frequente na Região Sul (6,50/100 mil), ocupa a
décima posição na Região Nordeste (3,66/100 mil) e a décima primeira nas Regiões
Sudeste (4,86/100 mil) e Centro-Oeste (3,93/100 mil).
A leucemia linfoblástica aguda (LLA) resulta na produção descontrolada de
blastos de características linfoides e no bloqueio da produção normal de glóbulos
vermelhos, brancos e plaquetas. É o tipo de câncer mais comum em crianças,
representando, aproximadamente, 25% dos cânceres diagnosticados na população com
menos de 15 anos (EGLER et al., 2016). Seu pico de incidência ocorre entre os 2 e 5
anos de idade, sendo quatro vezes mais freqüente que a leucemia mieloide aguda
(http://bvsms.saude.gov.br/bvs/publicacoes/inca, acesso em 22/10/2018). Nos Estados
Unidos, dos 4.000 casos de LLA diagnosticados anualmente, dois terços são em
crianças e adolescentes (LOPES et al., 2017). Nessa faixa etária, a melhora pode ocorrer

26
em até 80% dos casos com uma taxa de sobrevivência de 90% a depender do acesso ou
não ao tratamento.
Desde a inserção de L-ASNase no tratamento de LLA, há mais de 40 anos,
houve aumento da sobrevida dos pacientes de <10% a >80% (AVRAMIS;
PANOSYAN, 2005). Apesar desse avanço, ainda se observa recidiva em 10-25% dos
casos, principalmente após longos períodos de tratamento. No caso de pacientes na fase
da adolescência e adultos jovens, entre 15-39 anos e diagnosticados com LLA, o
tratamento não tem sido tão favorável, embora a literatura sugira aumento substancial
da sobrevida de jovens adultos tratados com protocolos similares aos pediátricos,
inclusive com L-ASNase (EGLER et al., 2016).
1.4. Tratamento de LLA
O tratamento completo da LLA deve considerar a idade do paciente, a
imunofenotipagem, a citogenética, a contagem inicial de glóbulos, as condições clínicas
e o envolvimento ou não do sistema nervoso, testículos e gânglios, e é realizado com
quimioterapia. Os pacientes necessitam ser tratados assim que o diagnóstico é
confirmado e o objetivo inicial é a remissão com restauração da produção normal de
glóbulos vermelhos, brancos e plaquetas (HAMERSCHLAK, 2010).
No Brasil, o tratamento da LLA também se dá com a combinação de várias
drogas. É importante a escolha adequada do melhor esquema de tratamento e sua
sequência para garantir as melhores chances de cura aos pacientes. A escolha do
esquema quimioterápico deve ser feita com base na idade, quadro clínico, resultados
laboratoriais e resposta ao tratamento inicial. A presença de fatores prognósticos
desfavoráveis ou recidiva de doença deve dirigir a abordagem do paciente para
tratamentos mais agressivos, incluindo o transplante de medula óssea nas suas diversas
modalidades (HAMERSCHLAK, 2010).
O tratamento de LLA consiste em quatro fases ou blocos, cada uma composta
por um coquetel de medicamentos, a fim de evitar resistência ao tratamento. São elas
indução da remissão, consolidação, reindução e manutenção ou continuação, com uma
duração total de tratamento de 2 a 3,5 anos, com terapia intensiva após os primeiros 6 a
9 meses (EGLER et al., 2016). O primeiro bloco de quimioterapia dura de 4 a 6
semanas e tem como objetivo induzir a remissão completa do câncer, a qual pode
ocorrer em até 90% dos casos (COOPER et al., 2015). O tratamento nessa fase inclui,
dentre outras drogas, L-ASNase que é também utilizada na segunda fase do tratamento

27
(consolidação), com duração de 4 a 8 meses e que serve de terapia preventiva para
eliminar doenças subclínicas do sistema nervoso central (EGLER et al., 2016). Esse
medicamento, que é um biofármaco, desde que começou a ser utilizado há
aproximadamente 50 anos, permanece como um dos mais importantes agentes no
tratamento de LLA (MARINI et al., 2017). Sua importância pode ser estimada em
termos de seu valor de mercado, cuja contribuição fica em torno de 40 % em relação ao
total de enzimas que são produzidas atualmente para uso médico (VIMAL; KUMAR,
2017a). Foi estabelecido por estudos em macacos Rhesus e humanos que o nível de
atividade enzimática de L-ASNase deve ser ≥ 0,1 U/mL para adequada depleção do
aminoácido asparagina (MARINI et al., 2017).
Atualmente três formas de L-ASNase estão disponíveis no mercado para
tratamento de LLA: a forma nativa obtida da linhagem bacteriana Escherichia coli K12,
de nome comercial Elspar®, Merck & CO (JAFFE et al., 1971); a sua forma peguilada
(PEG-ASNase), que possui uma ligação covalente com metoxi-polietileno glicol, m-
PEG (PASUT; VERONESE, 2012), comerciamente denominada Oncaspar® (Enzon
Pharmaceuticals, Inc.); e a L-asparaginase obtida da linhagem bacteriana Erwinia
chrysanthemi, denominada crisantaspase e comercialmente chamada de Erwinase®,
EUSA Pharma Inc (KEEGAN et al., 2011). As diferentes formulações apresentam
diferenças em relação à atividade, afinidade pelo substrato (Km), peso molecular, tempo
de meia vida, ponto isoelétrico (PI), resultando em asparaginases com diferentes
propriedades, toxicidade e eficácia (Tabela 1).
Tabela 1 – Propriedades das L-ASNases disponíveis comercialmente. Fontes: ABUD,
2005 e NARTA et al., 2007.
E. coli Erwinia chrysanthemi
nativa Nativa peguilada
atividade específica (U.mg-1
) 280-400 280-400 650-700
KM (µM)-L-asparaginase 12 12 12
KM (µM)-L-glutaminase 3000 3000 1400
massa molecular (kDa) 141 145 138
ponto isolelétrico (PI) 5 5 8,7
tempo de meia-vida (dias) 1,24 ± 0,17 5,73 ± 3,24 0,65 ± 0,13

28
Dois mecanismos de resistência ao tratamento com L-ASNase têm sido
propostos. O primeiro refere-se ao aumento dos níveis de produção de asparagina
sintetase pelas células leucêmicas, de forma que elas conseguem ser auto-suficientes na
produção do aminoácido Asn; o outro mecanismo seria a produção de anticorpos anti-
asparaginase que neutralizariam a enzima impedindo a sua ação enzimática (EL-
GHONEMY, 2014).
Aprovada pela FDA (do inglês, Food and Drug Administration) em 1978, a
forma nativa de E. coli é a mais imunogênica (RIZZARI et al., 2013). Sua degradação
pelas proteases lisossomais catepsina B e asparaginil endopeptidase resulta na
inativação e exposição de epítopos, com consequente ativação da resposta imune
(OFFMAN et al., 2011). Essa resposta pode ser sintomática, com sinais de
hipersensibilidade clínica, ou assintomática, quando está associada com o aparecimento
de anticorpos neutralizantes, efeito conhecido por “inativação silenciosa”, resultando
em declínio rápido dos níveis de atividade enzimática (COVINI et al., 2012). Um
estudo com 154 crianças com LLA mostrou que, após 29 dias do início do tratamento,
aproximadamente 35% das crianças tratadas com L-ASNase de E. coli desenvolveram
anticorpos anti-asparaginase, dos quais 56% apresentaram sintomas clínicos e 44%
apresentaram inativação silenciosa (RIZZARI et al., 2013). A estimativa é que pelo
menos 10% dos pacientes tratados com L-ASNase desenvolvem inativação silenciosa
durante a terapia (MARINI et al., 2017).
A forma peguilada foi aprovada pelo FDA em fevereiro de 1994 como a segunda
linha de tratamento para pacientes com LLA sensibilizados pela formulação nativa de E.
coli (PIETERS et al., 2011). Essa formulação reduz a imunogenicidade da proteína,
aumentando sua estabilidade no plasma, tem meia vida mais longa e pode ser
administrada a doses mais baixas e intervalos mais longos (apud EL-GHONEMY,
2014).
Por outro lado, além de também induzir a inativação silenciosa, é comprovada a
existência de reação-cruzada entre anticorpos desenvolvidos nos pacientes previamente
tratados com L-ASNase nativa e PEG-ASNase, não sendo recomendada neste caso a
mudança de uma para outra (PIETERS et al., 2011).
A alternativa quando há reação a alguma das formulações de E. coli é substituí-
la por crisantaspase, aprovada em 2011 pelo FDA para utilização em segunda ou
terceira linha de tratamento de LLA (KEEGAN et al., 2011). Embora a crisantaspase
não apresente reação-cruzada com os outros dois tipos, ainda pode induzir reações

29
alérgicas em um terço dos pacientes (PIETERS et al., 2011; RIZZARI et al., 2013).
Além disso, apresenta meia vida biológica menor e maior neurotoxicidade do que as
outras asparaginases de E. coli (OFFMAN et al., 2011). De acordo com a literatura, os
efeitos neurotóxicos são atribuídos à atividade de glutaminase, que é de
aproximadamente 2% para E. coli e de 10% para E. chrysanthemi (PIETERS et al.,
2011; AVRAMIS, 2012).
Dessa forma, a imunogenicidade continua sendo uma problemática na utilização
de L-ASNase no protocolo de tratamento de LLA. Atribuída à forma tetramérica da
enzima e, sobretudo, à presença de agregados nas formulações, reflete um sistema de
produção ineficiente que tem prejudicado a aprovação dessas preparações (PIETERS et
al., 2008). Por essa razão, a elucidação das propriedades dessa enzima, bem como a
busca por novas fontes e condições otimizadas de cultivo, poderão contribuir para o
desenvolvimento de um biofármaco antileucêmico que não possua as limitações
presentes nas formulações já existentes no mercado.
1.5. Outras aplicações da enzima L-ASNase
Apesar da enzima L-ASNase ser mais conhecida e explorada como agente
quimioterápico, possui outras propriedades interessantes para a indústria médica e
veterinária que, inclui o tratamento de doenças infecciosas, autoimunes, câncer em cães
e gatos (VIMAL; KUMAR, 2017a).
Bactérias patogênicas do gênero Streptococcus, por exemplo, durante aderência
a células hospedeiras, libera toxinas (estreptolisinas) que induzem o estresse celular e,
por consequência, o aumento dos níveis de asparagina sintetase e produção do
aminoácido asparagina, que está envolvida com a expressão aumentada de genes da
proliferação dessa bactéria (BARUCH et al., 2014). L-ASNase age bloqueando a
proliferação do patógeno, de forma que esse estudo sugere o uso terapêutico dessa
enzima contra esse tipo de infecção (VIMAL; KUMAR, 2017a; BARUCH et al., 2014).
Outra área de aplicação para essa molécula, cujo interesse tem sido crescente, é
na indústria de alimentos, como agente mitigante da acrilamida (VIMAL; KUMAR,
2017a; ALAM et al., 2018). Alguns alimentos ricos em amido quando fritos ou
aquecidos em temperatura elevada (acima de 120°C) liberam acrilamida. Ela é
resultante da reação de Maillard que ocorre entre os grupos carbonil dos açúcares
redutores e o grupamento amino do aminoácido asparagina, que estão presentes nesses
alimentos (VIMAL; KUMAR, 2017a; ALAM et al., 2018; QESHMI et al., 2018). O uso

30
da asparagina como agente mitigante da acrilamida é importante porque a acrilamida é
neurotóxica e classificada como agente carcinogênico (MEDEIROS et al., 2012). Há
duas marcas de L-ASNase no mercado lançadas com esse propósito, para atender a
indústria de alimentos: Acrylaway® e PreventASe®. Alimentos industrializados como
biscoitos, salgadinhos, batata frita, cafés, cereais e torradas quando pré-tratados com
Acrylaway® apresentaram redução de 50 a 90% dos níveis de acrilamida (VIMAL;
KUMAR, 2017a). No entanto, existe demanda do mercado para buscar novas fontes de
L-ASNase, com propriedades desejáveis para a indústria de alimentos, como
estabilidade em ampla variedade de pH e temperaturas, elevada taxa de conversão de
substrato, baixo Km (elevada especificidade pelo substrato) e alto Kcat (elevada
eficiência catalítica) para reduzir o tempo dessa etapa no processo industrial (VIMAL;
KUMAR, 2017a).
1.6. Em busca de alternativas às L-ASNases convencionais
Estratégias têm sido desenvolvidas com o intuito de melhorar a estabilidade das
L-ASNases, aumentar a meia vida e reduzir a imunogenicidade dessas enzimas.
Inicialmente, foram propostas modificações químicas como forma de ocultar epítopos
expostos na estrutura proteica dessa enzima, imunologicamente ativos e capazes de
estimular as reações de hipersensibilidade (NARTA et al., 2007). A proposta mais
conhecida nesse sentido resultou no desenvolvimento da forma peguilada da enzima
(ABUCHOWSKI et al., 1979). Outra alternativa seria eliminar os epítopos
imunodominantes por mutagênese sítio-específica. OFFMAN et al. (2011), através de
estudos de modelagem e dinâmica molecular com L-ASNase nativa de E. coli,
mostraram que a substituição de um único aminoácido pode bloquear a clivagem dessa
enzima por proteases presentes no soro, mantendo ou melhorando a sua atividade
enzimática.
Além do melhoramento das L-ASNases já existentes no mercado, muito estudos
têm apostado na busca por outras fontes da enzima. Um exemplo são as L-ASNases
produzidas por bactérias termofílicas, as quais possuem a vantagem de serem mais
estáveis em temperaturas elevadas (PRITSA; KYRIAKIDIS, 2001; BANSAL et al.,
2012); produzidas por bactérias do gênero Rhizobium (HUERTA-SAQUERO et al.,
2013); Yersinia pseudotubercullosis (POKROVSKAYA et al., 2012); actinobactérias
(KHAMNA et al., 2009; MEENA et al., 2015b; SAXENA et al., 2015); Erwinia spp.
(KOTZIA; LABROU, 2007); Pectobacterium spp. (KUMAR et al., 2010); linhagens de

31
Bacillus (ALRUMMAN et al., 2019; ERVA et al., 2018; GHOLAMIAN et al., 2013;
HYMAVATHI et al., 2010); Pseudomonas aeruginosa (GECKIL et al., 2006) e
Zymomonas mobilis (EINSFELDT, 2014).
Embora as L-ASNases bacterianas continuem sendo as mais estudadas, alguns
trabalhos têm apontado o interesse por fontes eucarióticas (leveduras e fungos
filamentosos) em razão dessas células realizarem diversas modificações pós-
traducionais (glicosilação, fosforilação, acilação, metilação, formação de pontes
dissulfeto) que podem estar relacionadas à estabilidade proteica e ao mascaramento de
sítios imunogênicos. Dessa forma, a busca por fontes eucarióticas de L-ASNase poderia
resultar em uma enzima que produzisse menos efeitos colaterais em humanos
(SHRIVASTAVA et al., 2015).
Microrganismos eucarióticos como fungos filamentosos do gênero Aspergillus,
Penicillium e Fusarium, são comumente reportados na literatura como produtores de L-
ASNase (da ROCHA et al., 2019; CACHUMBA et al., 2019; COSTA-SILVA et al.,
2018a; VALA et al., 2018; KUMAR; SOBHA, 2012), além de Trichosporon asashii
(ASHOK et al., 2019) e espécies endofíticas (SILVA et al., 2018). Também já foi
registrada a produção de L-ASNase por microalgas das espécies Spirulina maxima (El
BAKY; El BAROTY, 2016) e Chlorella vulgaris (EBRAHIMINEZHAD et al., 2014).
Há poucos trabalhos abordando a pesquisa de L-ASNases produzidas por
leveduras. KRISHNAPURA et al. (2015) citam em seu artigo de revisão alguns
trabalhos nesse sentido, de produção de L-ASNase por Rhodosporidium toruloides,
Candida guilliermondii e S. cerevisiae, todos eles trabalhos muito antigos da década de
80. RAMAKRISHNAN; JOSEPH (1996) reportaram a caracterização de uma L-
ASNase de Rhodosporidium toruloides, sugerindo que esta seria um homodímero (e não
um tetrâmero, como são as de origem bacteriana), com uma subunidade de massa
molecular de 87 kDa, e kM 1,43 mM para asparagina e 6,45 mM para glutamina, com
atividade ótima em pH 6,35 e temperatura de 37ºC (RAMAKRISHNAN; JOSEPH,
1996). Esse trabalho traz a possibilidade de que novas estruturas moleculares da enzima,
menores que os seus congêneres bacterianos, possam ser menos imunogênicos e, por
isso, uma alternativa interessante no tratamento de LLA.
LOPES et al. (2017) também relatam em seu artigo de revisão a produção de L-
ASNase pelos mesmos gêneros já citados aqui, em publicações também antigas. Uma
busca no banco de dados do PUBMED (acesso em Janeiro de 2019) mostrou as
seguintes referências, mais recentes, sobre a produção de L-ASNase por leveduras

32
(Tabela 2): produção heteróloga de ASP3 de S. cerevisiaepor P. pastoris (FERRARA et
al., 2006; 2010; GIRÃO et al., 2016), produção de L-ASNase em biorreator por
linhagem de Candida utilis (MOMENI et al., 2015) e a produção dessa enzima pela
levedura Yarrowia lipolytica (DARVISHI et al., 2019). A tese de MOGUEL (2018) traz
resultados sobre a produção de L-ASNase de linhagem de L. scotti, levedura isolada de
sedimentos marinhos coletados na Antártica. Alguns métodos de rompimento celular
foram avaliados utilizando a linhagem L. muscorum CRM 1648, alvo do presente
trabalho, mostrando que os métodos mecânicos, principalmente rompimento por pérolas
de vidro e sonicação, foram mais eficientes para a recuperação de L-ASNase (COSTA-
SILVA et al., 2018b). Esses e todos os trabalhos que envolvem o tema estão
referenciados na Tabela 2 a seguir.
Tabela 2 – Referências encontradas na literatura relacionadas à produção de L-ASNase
por espécies de leveduras.
Taxonomia Referências
Candida utilis KIL et al., 1995; MOMENI et al., 2015
Candida guilliermondii STEPANYAN; DAVTYAN, 1988
Pichia polymorpha FODA et al., 1980
Rhodosporidium toruloides RAMAKRISHNAN; JOSEPH, 1996
Rhodotorula sp. FODA et al., 1980
Saccharomyces cerevisiae FERRARA et al., 2006 e 2010; GIRÃO et al., 2014 e 2016
Leucosporidium scotti MOGUEL, 2018
Leucosporidium muscorum COSTA-SILVA et al., 2018b
Yarrowia lipolytica DARVISHI et al., 2019
Portanto, ainda há muito a ser explorado entre as leveduras que, por serem
organismos eucarióticos e apresentarem as vantagens já citadas anteriormente em
relação a sistemas procarióticos, apresentam grande perspectiva de uso para a produção
de biofármacos, como a L-ASNase. Dessa forma, a pesquisa desses microrganismos em
diversos nichos traz uma perspectiva para encontrar uma L-ASNase com ação
antileucêmica e, ao mesmo tempo, menos imunogênica que as congêneres disponíveis
no mercado.

33
1.7. Produção de L-ASNase em biorreator
A produção de L-ASNase é feita principalmente por cultivo submerso (em meio
líquido), embora também haja relatos na literatura de produção em meio sólido por
fungos filamentosos do gênero Aspergillus. (CACHUMBA et al., 2019; DUTTA et al.,
2015) e Trichoderma viride (ELSHAFEI; EL-GHONEMY, 2015), pela bactéria
Myroides gitamensis (PRASAD TALLURI et al., 2019) e actinobactérias do gênero
Streptomyces (BASHA et al., 2009). As vantagens da produção submersa é que esta é
bem estabelecida, a manipulação dos componentes do meio é mais simples, não requer
pré-tratamento do substrato, além do controle dos parâmetros físicos (pH, OD e
temperatura) e purificação do produto serem realizados mais facilmente do que na
fermentação em estado sólido (VIMAL; KUMAR, 2017a).
A otimização do processo de produção tem papel importante na indústria
biotecnológica por melhorar os rendimentos e produtividade do processo sendo,
portanto, decisiva para o sucesso comercial do produto (KUMAR et al., 2017). O
melhoramento do processo é feito por meio da manipulação dos nutrientes e das
condições do processo (pH, temperatura, tempo de cultivo, concentração inicial do
inóculo, agitação) utilizando metodologias de análises estatísticas.
Uma das técnicas mais simples e mais utilizada é a análise univariada (OFAT,
do inglês one-factor at a time), na qual é avaliado um fator por vez, pela variação dos
níveis do fator em estudo (FUKUDA et al., 2018). Essa metodologia, por não conseguir
avaliar a interação entre os fatores, costuma ser associada a técnicas de Delineamento
Experimental (DoE) que se divide em: técnicas de rastreamento dos fatores (variáveis
independentes – x) que são ou não significativos para as respostas avaliadas (variáveis
dependentes – y), como por exemplo o estudo fatorial completo de dois níveis ou
fracionado e Plackett-Burmann; e técnicas de otimização, como Box-Behnken,
delineamento composto central (CCD, do inglês Central Composite Design) e fatorial
completo de 3 níveis (FUKUDA et al., 2018). Como mostra o Apêndice A, as análises
mais utilizadas para o melhoramento do processo de produção de L-ASNase de
diferentes origens têm sido até então a análise univariada, o CCD e o fatorial completo
e/ou fracionado.
A otimização do meio e condições de cultivo enfrentam alguns limites, pois
ainda é possível melhorar a produtividade e o rendimento do processo caso aumente a
capacidade de produção (volume de meio) e crie condições apropriadas de oxigenação

34
em um sistema que permita o controle e monitoramento dos parâmetros principais,
como a aeração, agitação, pH e o oxigênio dissolvido. Por essa razão, a etapa seguinte
ao melhoramento dos parâmetros nutricionais e de cultivo em frascos agitados é o
cultivo em biorreator. O tipo de processo mais utilizado para cultivo em biorreator e
produção de L-ASNase é a batelada (cultivo descontínuo) seguida pela batelada
alimentada (descontínuo alimentado), como sugere o Apêndice A.
Ao analisar as publicações nos últimos 5 anos envolvendo o termo em inglês
“asparaginase production” (Apêndice A), realizada no banco de dados do PUBMED e
no Google Acadêmico, observa-se que os grupos que mais tem publicado estão
localizados no Brasil e, principalmente, na Ásia e Sudoeste da África (Índia, Irã e
Egito). Bactérias e fungos filamentosos são os grupos de microrganismos que mais têm
sido explorados para a produção dessa enzima. Além disso, é possível encontrar
organismos produtores de L-ASNase em nichos diversos ao redor do mundo, desde
solo, rizosfera a ambiente marinho. As fontes de carbono e de nitrogênio utilizadas para
produção dessa enzima variam muito de acordo com o microrganismo utilizado na
produção, mas parece existir uma tendência maior no uso de glicose como fonte de
carbono e asparagina como fonte de nitrogênio (Apêndice A).
A bactéria Pectobacterium carotovorum foi detectada como produtora de uma
L-ASNase livre de atividade glutaminásica e cultivada em biorreator com 1,5 litros de
volume útil, por 12 horas, em cultivo descontínuo e descontínuo alimentado, utilizando
um meio com glicose e asparagina como fontes principais de carbono e nitrogênio, pH
controlado em 8,6, a 30°C, com cascata de agitação variando de 160 a 600 rpm, aeração
de 1,5 vvm e OD de 20% da saturação com ar. Nessas condições, a bactéria foi capaz de
produzir, aproximadamente, 40 U.mL-1
da enzima, valor duas vezes maior do que o
obtido nos cultivos em batelada (KUMAR et al., 2017).
Fungo filamentoso da espécie Aspergillus terreus foi cultivado em meio Czapek-
Dox contendo glicose e prolina como fontes de carbono e nitrogênio em cultivo
descontínuo com 5 litros de volume útil, pH 9,5, sendo cultivado a 35°C e aeração de
0,5 vvm, obtendo uma produção final aproximada de 14 U.g-1
de massa seca celular
(COSTA-SILVA et al., 2018a).
A levedura Yarrowia lipolytica foi avaliada quanto a produção de L-ASNase, em
um processo descontínuo, utilizando meio Czapek-Dox, pH inicial igual a 7,0, com
glicose e asparagina como fontes de carbono e nitrogênio. Essa levedura foi cultivada
por 24 horas em um biorreator de 5 litros a 29°C, 500 rpm e 1 vvm de aeração,

35
produzindo 210 U.mL-1
da enzima, valor 12 vezes maior do que o obtido em frascos
agitados (DARVISHI et al., 2019).
A levedura Candida utilis foi cultivada em biorreator (cultivo descontínuo) com
melado de beterraba e extrato de levedura como fontes de carbono e nitrogênio, pH
ajustado para 7,0 e temperatura de 30°C. A produção de L-ASNase foi máxima e igual a
245,6 U.mL-1
com 20 horas de cultivo sob agitação controlada em 300 rpm e aeração de
1,25 vvm (MOMENI et al., 2015).
Mais raramente, a produção de L-ASNase foi feita em cultivo contínuo. Bactéria
da espécie Zymomonas mobilis foi cultivada em biorreator com 0,3 litros úteis de meio
contendo sacarose, asparagina e extrato de levedura como fontes de carbono e
nitrogênio, a 30°C, em modo descontínuo até atingir a fase exponencial, quando o
sistema passou a ser alimentado continuamente. A maior produção (117 U.L-1
) ocorreu
em uma taxa de diluição igual a 0,20 h-1
(MENEGAT et al., 2016).
Os métodos de quantificação da enzima são variados e os mais utilizados,
segundo o Apêndice A, é o Método de Nessler, seguido pelo método de L-aspartil-β-
hidroxâmico, este último sendo utilizado com bem menos freqüência do que o primeiro.
O problema do uso do método de Nessler é que, por quantificar indiretamente a enzima
L-ASNase através da amônia que é liberada da reação de hidrólise da asparagina, pode
superestimar a atividade enzimática em razão da presença de interferentes no caldo de
cultivo (proteínas que, ao serem metabolizadas, liberam amônia). Os resultados de
produção da enzima são expressos em diferentes unidades, em unidades totais da
enzima (U), unidades por volume de cultivo (U.mL-1
ou U.L-1
), unidades por massa seca
de células (U.g-1
ou U.mg-1
) ou unidades da enzima por mg de proteínas totais (U.mg-1
).
Essas diferenças acabam dificultando a análise comparativa da produção de L-ASNase
entre os diversos microrganismos estudados.
1.8. Leveduras da região Antártica
As leveduras são microrganismos eucarióticos que habitam quase todos os
ambientes terrestres, inclusive ambientes onde ficam expostas permanentemente a
temperaturas abaixo de 5 °C (CARRASCO et al., 2016). Mais de 80% da biosfera da
Terra é permanentemente ou periodicamente exposta a essas temperaturas baixas, o que
incluem mares profundos (90% dos oceanos apresentam temperaturas < 5 °C), desertos
gelados e regiões glaciais, como o ambiente Ártico e Antártico (BUZZINI et al., 2012;
TSUJI et al., 2013).

36
O primeiro registro de vida microbiana em um ambiente frio data de 400 anos
AC, em escritos do filósofo grego Aristóteles que mostravam a descrição do que depois
descobriram ser um organismo fotossintético que deixava a neve avermelhada
(DEMING, 2009). Apenas em 1887 o cientista germânico Forster descreveria a
capacidade de uma bactéria reproduzir-se a 0 °C e em 1902 seria cunhada a
terminologia psicrófilo por Schmidt-Nielsen para descrever microrganismos com tal
comportamento (DEMING, 2009).
A definição clássica utilizada para classificar organismos adaptados ao frio se
baseia nos parâmetros cinéticos de crescimento em diferentes temperaturas, de acordo
com o qual o termo psicrófilo é reservado a organismos que podem crescer em
temperatura abaixo ou igual a 0°C e cujas temperaturas ótima e máxima para
crescimento são ≤ 15°C e ≤ 20°C, respectivamente. Aqueles que podem crescer a 0 °C
e cuja máxima temperatura de crescimento pode estar acima de 20 °C são denominados
psicrotolerantes ou psicrotróficos (MORITA, 1975).
Embora até hoje esses termos sejam ainda os mais utilizados, foi proposto o uso
do termo estenopsicrófilo para o grupo que inclui os psicrófilos estritos ou obrigatórios,
e euripsicrófilo, para o grupo que inclui os organismos psicrófilos ou psicrotróficos
facultativos (WANG et al., 2017). Essa definição leva em consideração que, em
ambientes frios, o que diferencia um grupo do outro não é a temperatura ótima e
máxima de crescimento, mas sim a extensão da faixa de temperatura em que o
microrganismo é capaz de crescer (RUSSELL, 2006). A maioria dos microrganismos
recuperada de amostras das regiões Ártica e Antártica são psicrófilos euritérmicos,
justamente porque a temperatura do solo nessas regiões pode sofrer grandes variações,
de 30 a 40 °C (RUSSELL, 2006).
1.8.1. Interesse biotecnológico
A região Antártica, situada no polar austral, é a porção mais ao sul da Terra, com
uma área correspondente a 14 milhões Km2, o que faz da Antártica o quinto maior
continente do mundo (TSUJI et al., 2013; SHIVAJI; PRASAD, 2009).
Aproximadamente 98% dessa região da Terra é coberta por gelo e neve, com
temperaturas na região costeira que variam de 5 a -35 °C (TSUJI et al., 2013).
O estudo de microrganismos provenientes da Antártica tem atraído o interesse de
pesquisadores desde 1960 em razão das condições climáticas severas, onde predominam
o frio extremo, ciclos de congelamento/descongelamento, alta salinidade, exposições

37
excessivas a luz UV alternadas com longos períodos de escuridão e baixa
disponibilidade de nutrientes (apud DUARTE et al., 2013; apud BUZZINI et al., 2012;
di MENNA, 1958). Desde então, espécies de bactérias, leveduras e fungos filamentosos
tem sido isoladas de diversos habitats na Antártica, como solo, gelo, liquens,
macroalgas, esponjas, sedimento marinho, água do mar, plantas briófitas e
angiospermas (Deschampsia antarctica e Colobanthus quitensis) e biofilmes (VAZ et
al., 2011; DUARTE et al., 2013; TSUJI et al., 2013; DUARTE et al., 2016;
CARRASCO et al., 2016; MARTORELL et al., 2017; INFORSATO, 2017; WENTZEL
et al., 2018; FERREIRA et al., 2019). Foram identificadas aproximadamente 70
espécies de leveduras provenientes da Antártica, sendo 13 do filo Ascomycota e 57 do
filo Basidiomycota (BUZZINI et al., 2012). Aparentemente 90% das leveduras isoladas
na Antártica e em regiões de clima frio pertencem ao filo Basidiomycota, sendo que
desse grupo espécies dos gêneros Cryptococcus, Candida, Rhodotorula e Mrakia são
mais comumente reportadas (CARRASCO et al., 2012; MARTORELL et al., 2017;
DUARTE et al., 2018).
Estudos têm mostrado que a capacidade de microrganismos crescerem em
temperaturas tão baixas e condições tão adversas estaria associada a ajustes estruturais e
fisiológicos, adaptações evolutivas para amenizar a redução das reações químicas
devido às baixas temperaturas (de GARCIA; van BROOCK, 2016; FELLER, 2013;
D’AMICO et al., 2002). Entre os mecanismos de adaptação estão a modificação de
lipídeos de membrana, com maior presença de ácidos graxos insaturados ou
poliinsaturados, que aumenta a fluidez da membrana; a síntese de proteínas termo-
protetoras (proteínas de choque-frio) e de crioprotetores, reduzindo a presença de
cristais de gelo no citoplasma; e desenvolvimento de mecanismos de proteção contra
espécies reativas de oxigênio (FELLER, 2013; de GARCIA; van BROOCK, 2016). A
produção de enzimas hidrolíticas extracelulares, por exemplo, seria uma forma de
adaptação já que, com essas enzimas é possível aproveitar as fontes de carbono
disponíveis no ambiente, contribuindo para o reciclo de nutrientes e mineralização da
matéria orgânica (apud BUZZINI et al., 2012; apud CARRASCO et al., 2016). Além
disso, também é relatada a produção de outras enzimas de adaptação ao frio e proteínas
anticongelantes, que controlam a formação de cristais de gelo, evitando danos físicos na
estrutura da célula (TSUJI et al., 2013).
O efeito da temperatura sobre as reações químicas é descrito pela equação de
Arrhenius k = Ae-Ea/RT
, onde k é a constante da reação, A é uma constante para cada

38
sistema, relacionada à colisão das moléculas, EA é a energia de ativação (energia
mínima requerida para que uma reação se inicie), R é a constante universal dos gases e
T é a temperatura absoluta em Kelvin. Por essa reação percebe-se que qualquer redução
na temperatura provoca uma redução exponencial na velocidade de reação. Para superar
esse efeito, os psicrófilos e psicrotolerantes sintetizam enzimas com maior eficiência
catalítica, pelo aumento do seu poder catalítico (kcat) ou da redução de sua afinidade
pelo substrato (Km) ou ainda pela mudança em ambos os parâmetros (D’AMICO et al.,
2002; FELLER, 2013), como mostra a Figura 3.
Figura 3 – A atividade enzimática em relação a temperatura. A atividade das enzimas
psicrófilas (círculo vazio) α- amilase (gráfico esquerdo) e celulase (gráfico direito) e
seus correspondentes mesófilos (círculo cheio). Fonte: Adaptado de Feller (2013).
O aumento do poder catalítico dessas enzimas pode ser até 10 vezes maior a
baixas (e moderadas) temperaturas do que por seus homólogos mesófilos (FELLER;
GERDAY, 2003). Isso é possível em razão dessas enzimas apresentarem uma estrutura
mais flexível no sítio catalítico, que permite fazer e desfazer interações não covalentes,
tornando-as mais ou menos ativas (FELLER, 2013). Essas propriedades fazem com que
as enzimas adaptadas ao frio sejam altamente promissoras do ponto de vista
biotecnológico, pois aceleram o tempo de processos realizados a temperaturas
baixas/moderadas, reduzem a concentração enzimática requerida, resultam em alta
produtividade nas reações com componentes termo-sensíveis, previnem transformações
químicas indesejáveis e a perda de compostos voláteis e ainda diminuem os gastos com
energia elétrica (MARGESIN et al., 2003).

39
A produção de enzimas psicrófilas – amilases, lipases, proteases, celulases,
pectinases, β-gluconidades, esterases – tem sido reportada em leveduras e diferem dos
similares produzidos por outros microrganismos (HAMID et al., 2014; CARRASCO et
al., 2016). Amilases ativas no frio e celulases possuem alto potencial de aplicação em
processos que requerem baixas temperaturas, como os requeridos na indústria de
alimentos, biocombustíveis e detergentes (apud CARRASCO et al., 2016).
A triagem de microrganismos isolados a partir de amostras coletadas nas Ilhas
Shetland do Sul, na Antártica, produtores de lipases, proteases e xilanases, revelou a
presença de 21 espécies de leveduras dos filos ascomicota e basidiomicota, entre elas as
mais abundantes foram Candida glaebosa, Cryptococcus victoriae, Meyerozyma
(Pichia) guilliermondii, Rhodotorula mucilaginosa e R. laryngis (DUARTE et al.,
2013). Entre as leveduras avaliadas nesse estudo, 46,4%, 37,1% e 14,4% foram capazes
de produzir lipases (a 15 °C), xilanases (a 15 °C) e proteases (a 25 °C), respectivamente.
Uma protease ácida produzida pela levedura isolada de ambiente marinho na
Antártica, Rhodotorula muscilaginosa, foi purificada e caracterizada (LARIO et al.,
2015). Essa enzima apresentou características interessantes como ótima atividade
catalítica em pH 5,0 e temperatura de 50 °C, além de ser estável em altas concentrações
salinas.
Um total de 226 microrganismos (60 fungos filamentosos e 166 leveduras)
isolados de amostras de sedimento marinho, coletadas próxima a Estação Antártica
Comandante Ferraz (EACF), estação brasileira localizada na Ilha Rei George (Antártica
marítimica) foram avaliados quanto a produção de lipases e proteases. A levedura da
espécie Metschnikowia australis foi a mais abundante na região e a que apresentou
maior atividade lipolítica (0,88 U.mL-1
), enquanto o fungo filamentoso do gênero
Pseudogymnoascus apresentou a maior atividade proteolítica (6,21 U.mL-1
) entre os
isolados avaliados (INFORSATO, 2017).
Enzimas pectinolíticas são utilizadas para a degradação de compostos de pectina
nas indústrias de processamento de frutas e vegetais (apud HAMID et al., 2014). Essas
enzimas produzidas por leveduras psicrófilas Candida capitatum e R. muscilaginosa
exibiram 50-80% de sua atividade catalítica ótima em pH 3-5 e temperaturas de 6 e 12
°C, propriedades estas interessantes para a indústria de vinhos e clarificação a baixas
temperaturas (SAHAY et al., 2013). Além disso, pectinases psicrófilas têm potencial
para manter valores nutricionais, sabor e características sensoriais, propriedades
importantes na indústria de alimentos (NAKAGAWA et al., 2002).

40
Uma linhagem de Leucosporidium scotti, também isolada da Península
Antartica, foi avaliada e mostrou capacidade para a produção de L-ASNase e de
acumular lipídios mono e poliinsaturados, ao ser cultivada em substratos de baixo custo,
como sacarose e glicerol (MOGUEL, 2018). Seu cultivo em biorreator resultou em
produtividade de 36,9 U.L-1
.h-1
dessa enzima em condições otimizadas de cultivo (15
°C, X0: 5 g.L-1
; pH 7,0; 48 h; kLa: 89-92 h-1
). Esse trabalho apontou que L-ASNases
advindas desse gênero de levedura tem potencial terapêutico, com uma eficiência
catalítica próxima a da congênere de E. coli e mais elevada que a congênere de S.
cerevisiae, indicando que as L-ASNases de espécies de Leucosporidium sp. possam
apresentar propriedades interessantes para a indústria farmacêutica e de alimentos.
1.8.2. Leucosporidium muscorum
Do reino Fungi, filo Basidiomicota e família Leucosporidiales, a espécie
Leucosporidium muscorum já teve outras denominações, entre elas: Candida muscorum,
Azymocandida muscorum, Rhodotorula muscorum, Leucosporidiella muscorum e
Vanrija muscorum (www.mycobank.org, acesso em: 10/01/19). Foi primeiramente
isolada de solo de floresta na Ilha de Chatham, localizada aproximadamente a 800 km a
Leste da Ilha Sul de Nova Zelândia, também denominada Ilha Te Waipounamu (di
MENNA, 1958). Também foi registrado o isolamento dessa espécie de levedura em
pequenas poças no glacial de Midre Lovénbreen, localizado em Spitsbergen, território
ártico da Noruega (PATHAN et al., 2010); de musgos em decomposição do tipo
Sphagnum, coletados na Nova Zelândia (SAMPAIO et al., 2003; GARCÍA et al., 2015);
de esponja marinha nas proximidades da Baia Fildes, na Ilha do Rei George (LAICH et
al., 2014); e de sedimentos marinhos próximo as bases brasileira e peruana na Baia do
Almirantado (INFORSATO, 2017), local de onde vieram as amostras utilizadas neste
trabalho.
Quando cultivada em agar Sabouraud, apresenta colônia cremosa e de coloração
creme ou amarelada, célula ovóide ou cilíndrica (2,5-4) x (7-19) µ (di MENNA, 1958).
Essa espécie foi avaliada em diferentes temperaturas (25, 30, 35, 37, 40, 42 e 45°C),
mas não houve crescimento em temperatura acima de 25 °C
(http://www.westerdijkinstitute.nl/collections, acesso em 12/11/18). Outro trabalho
mostrou que essa levedura é capaz de crescer a 4, 22 e 25 °C, mas não a 30 °C
(PATHAN et al., 2010). O mesmo trabalho mostra que L. muscorum consegue crescer
em meio com alta concentração de sais (≤ 4,0 % de NaCl, m/v) e foi capaz de produzir

41
urease, pectinase e amilase, quando cultivadas a 8°C e 22°C, mas não produziu
proteases, celulases, nem lipases nas condições avaliadas (PATHAN et al., 2010). Por
outro lado INFORSATO (2017) avaliou 170 isolados da Antártica e observou que um
isolado da espécie L. muscorum foi aquele que mais produziu protease.
Aparentemente, a espécie L. muscorum não realiza fermentação e assimila as
seguintes fontes de nutrientes: D-glicose, L-sorbose, D-xilose, D-arabinose, sacarose,
maltose, trealose, celobiose, rafinose, glicerol, manitol, D-gluconato, succinato, citrato,
nitrato, nitrito, cadaverina, lisina, creatinina, D-galactose, D-glucosamina, D-ribose e
lactose (KURTZMAN; FELL, 1998). De acordo com di MENNA (1958), a assimilação
de galactose e lactose é mais fraca. Parte desses testes foi avaliado e confirmado em
outro trabalho (LAICH et al., 2014). O seu principal mecanismo de adaptação ao frio
parece estar associado a elevada concentração de ácidos graxos insaturados presentes
em sua membrana celular (PATHAN et al., 2010; GARCÍA et al., 2015).
Figura 4 – Imagem de células em microscópio e morfologia da colônia de linhagem de
L. muscorum. Fonte: http://www.westerdijkinstitute.nl/collections, acesso em 12/12/18.

42
II. OBJETIVOS
2.1. Objetivo Geral
Avaliar a capacidade de crescimento e produção de L-ASNase por leveduras
isoladas de amostras de sedimentos marinhos coletados na Península Antártica,
especialmente pela linhagem Leucosporidium muscorum CRM 1648, em condições
diversas de cultivo em frascos agitados e em biorreator.
2.2. Objetivos específicos
2.2.1 Rastrear e selecionar leveduras produtoras de L-ASNase isoladas na
Antártica (seleção qualitativa).
2.2.2 Avaliar a produção de L-ASNase por esses microrganismos e selecionar o
melhor produtor (seleção quantitativa).
2.2.3 Avaliar as melhores fontes de carbono e de nitrogênio para o crescimento e
produção da enzima pela levedura L. muscorum CRM 1648.
2.2.4 Avaliar por delineamento experimental a melhor combinação entre as
fontes de carbono e nitrogênio selecionadas na etapa anterior para o melhoramento da
produção de L-ASNase.
2.2.5 Avaliar as melhores condições de cultivo (concentração inicial do inóculo,
pH inicial, temperatura e presença de água do mar) para a produção de L-ASNase e
crescimento de L. muscorum CRM 1648.
2.2.6 Avaliar em biorreator (cultivo descontínuo) o crescimento de L. muscorum
CRM 1648 e a produção de L-ASNase em situações de baixa e alta disponibilidade de
oxigênio dissolvido.

43
III. MATERIAIS E MÉTODOS
3.1. Leveduras isoladas de sedimento marinho na Antártica
As linhagens de leveduras utilizadas neste trabalho foram isoladas de amostras
de sedimentos marinhos coletadas na fase IV da OPERANTAR XXXIII, em Janeiro de
2015, pelo grupo de pesquisa da Profa. Dra. Lara Sette (UNESP, Rio Claro-SP), no
âmbito do projeto MICROSFERA, edital PROANTAR/CNPq, coordenado pela prof.
Dra. Vivian Pellizari, IO/USP, São Paulo-SP (INFORSATO, 2017). A coleta foi
realizada na Baia do Almirantado, na Ilha Rei George, localizada na Península Antártica
(Figura 5). Neste trabalho foram utilizados 150 isolados, cujas fontes e locais de coleta
estão listados na Tabela 3 e apontados na Figura 5.
Tabela 3 – Informações sobre a espécie, material de origem e local de coleta dos
isolados identificados neste trabalho como produtores de L-ASNase.
código do
isolado espécies material de origem local de coleta (dado GPS)
1 Leucosporidium
muscorum
sedimento marinho
(zona entre marés)
Refugio 2
(62˚ 04.341'S 58˚ 25.233'W)
2 L. muscorum sedimento marinho Botany Point
(62˚ 05.734'S 58˚ 19.919'W)
16 L. muscorum sedimento marinho
(zona entre marés)
Refugio 2
(62˚ 04.341'S 58˚ 25.233'W)
26 L. muscorum sedimento marinho
(zona entre marés)
Refugio 2
(62˚ 04.341'S 58˚ 25.233'W)
85 L. muscorum sedimento marinho
(zona entre marés)
Refugio 2
(62˚ 04.341'S 58˚ 25.233'W)
86 L. muscorum sedimento marinho Refugio 2
(62˚ 04.373'S 58˚ 25.335'W)
94 L. muscorum sedimento marinho
(zona entre marés)
Refugio 2
(62˚ 04.341'S 58˚ 25.233'W)
74 Glaciozyma antarctica sedimento marinho EACF
(62˚ 05.130'S 58˚ 23.356'W)
17 G. antarctica sedimento marinho EACF
(62˚ 05.130'S 58˚ 23.356'W)
EACF: Estação Antártica Comandante Ferraz, base brasileira de pesquisa.
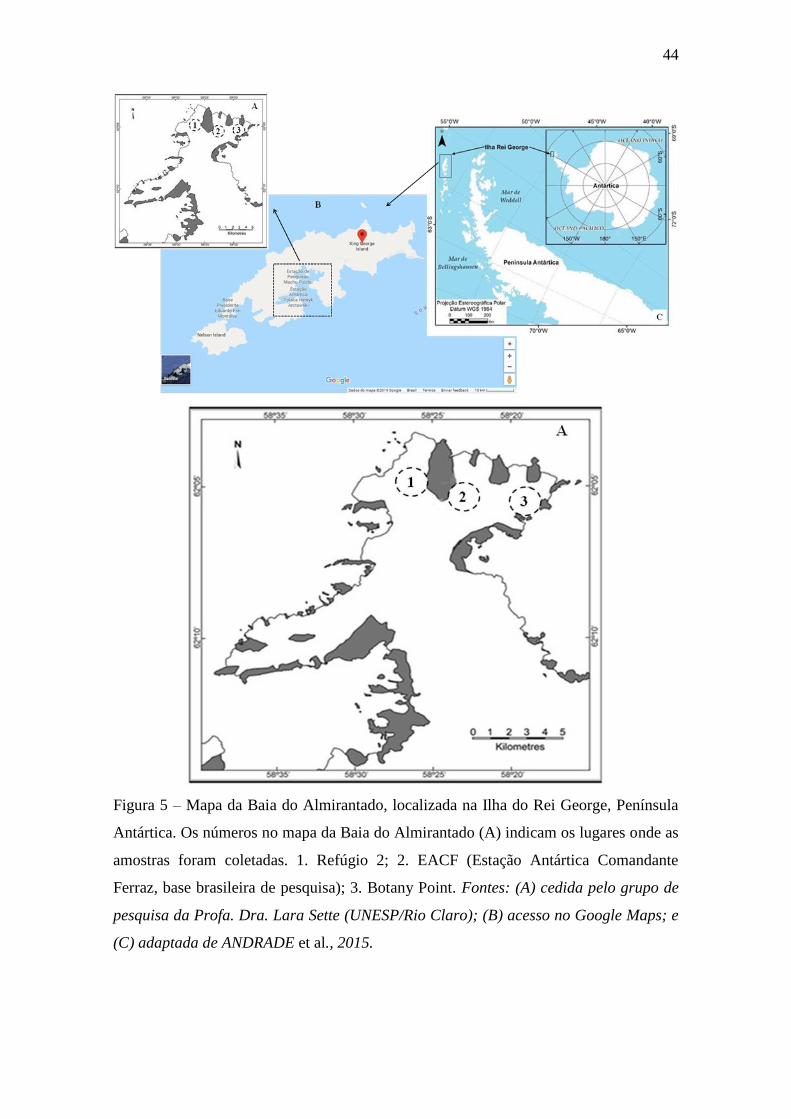
44
Figura 5 – Mapa da Baia do Almirantado, localizada na Ilha do Rei George, Península
Antártica. Os números no mapa da Baia do Almirantado (A) indicam os lugares onde as
amostras foram coletadas. 1. Refúgio 2; 2. EACF (Estação Antártica Comandante
Ferraz, base brasileira de pesquisa); 3. Botany Point. Fontes: (A) cedida pelo grupo de
pesquisa da Profa. Dra. Lara Sette (UNESP/Rio Claro); (B) acesso no Google Maps; e
(C) adaptada de ANDRADE et al., 2015.

45
3.2. Preservação das linhagens
As linhagens foram preservadas por ultracongelamento em freezer a -80 °C,
conforme a literatura (HUNTER-CEVERA; BELT, 1996; KIRSOP; DOYLE, 1991).
Para cada isolado, uma colônia foi semeada em meio sólido inclinado ágar Sabouraud e,
após 72 horas de crescimento em estufa a 15 °C, foi adicionada a cada tubo solução de
glicerol (10% v/v). As células foram ressuspendidas nessa solução e a suspensão foi
aliquotada em microtubos de 1,5 mL. Por fim, as alíquotas foram pré-resfriadas por 20
minutos a - 20°C, tendo-se o cuidado de evitar o congelamento da suspensão e,
consequentemente, a formação de cristais que possam danificar as células do
microrganismo. Esse resfriamento tem a função de permitir que o crioprotetor envolva
inteiramente as células, aumentando a sua ação de proteção. Após esse tempo de pré-
resfriamento, os microtubos foram mantidos em ultrafreezer (- 80°C).
3.3. Meios de cultivo
Neste trabalho foi utilizado caldo Sabouraud para preservação e ativação dos
isolados; ágar Sabouraud para a manutenção das linhagens; e meio Czapek Dox (CD)
(SAXENA; SINHA, 1981), para crescimento e produção de L-ASNase (Tabela 4). Este
último meio também serviu como base para a avaliação de diversas fontes de carbono e
nitrogênio. Os componentes dos meios foram esterilizados em autoclave (120 °C, 15
minutos). Os elementos traços do meio CD foram preparados a parte em uma solução
concentrada 50 vezes, esterilizados e mantido resfriado. Dados da literatura (SAXENA;
SINHA, 1981) mostram que a asparagina é utilizada para substituir o aminoácido
prolina no meio CD, mas optou-se por utilizar este último em razão desta fonte de
nitrogênio ter se mostrado melhor para a produção de L-ASNase em testes realizados
pelo grupo (MOGUEL, 2018; COSTA-SILVA et al., 2018a). Após o preparo do meio, o
pH foi ajustado para 5,5, utilizando-se soluções de NaOH e HCl (0,5 M cada).
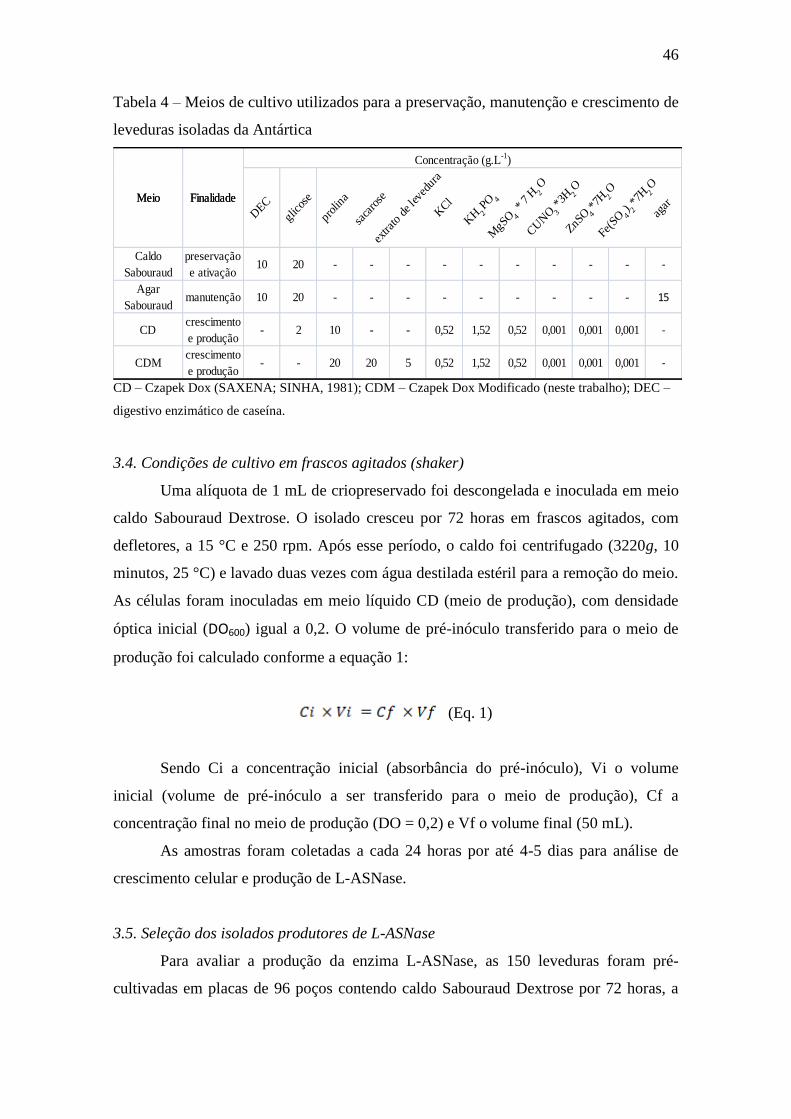
46
Tabela 4 – Meios de cultivo utilizados para a preservação, manutenção e crescimento de
leveduras isoladas da Antártica
Meio Finalidade
Concentração (g.L-1
)
DEC
glic
ose
prol
ina
saca
rose
extra
to d
e le
vedu
ra
KCl
KH 2
PO4
MgS
O 4 *
7 H 2
O
CU
NO 3
*3H 2
O
ZnSO 4
*7H 2
O
Fe(S
O 4) 2
*7H 2
O
agar
Meio Finalidade
Caldo
Sabouraud
preservação
e ativação10 20 - - - - - - - - - -
Agar
Sabouraudmanutenção 10 20 - - - - - - - - - 15
CDcrescimento
e produção- 2 10 - - 0,52 1,52 0,52 0,001 0,001 0,001 -
CDMcrescimento
e produção- - 20 20 5 0,52 1,52 0,52 0,001 0,001 0,001 -
CD – Czapek Dox (SAXENA; SINHA, 1981); CDM – Czapek Dox Modificado (neste trabalho); DEC –
digestivo enzimático de caseína.
3.4. Condições de cultivo em frascos agitados (shaker)
Uma alíquota de 1 mL de criopreservado foi descongelada e inoculada em meio
caldo Sabouraud Dextrose. O isolado cresceu por 72 horas em frascos agitados, com
defletores, a 15 °C e 250 rpm. Após esse período, o caldo foi centrifugado (3220g, 10
minutos, 25 °C) e lavado duas vezes com água destilada estéril para a remoção do meio.
As células foram inoculadas em meio líquido CD (meio de produção), com densidade
óptica inicial (DO600) igual a 0,2. O volume de pré-inóculo transferido para o meio de
produção foi calculado conforme a equação 1:
(Eq. 1)
Sendo Ci a concentração inicial (absorbância do pré-inóculo), Vi o volume
inicial (volume de pré-inóculo a ser transferido para o meio de produção), Cf a
concentração final no meio de produção (DO = 0,2) e Vf o volume final (50 mL).
As amostras foram coletadas a cada 24 horas por até 4-5 dias para análise de
crescimento celular e produção de L-ASNase.
3.5. Seleção dos isolados produtores de L-ASNase
Para avaliar a produção da enzima L-ASNase, as 150 leveduras foram pré-
cultivadas em placas de 96 poços contendo caldo Sabouraud Dextrose por 72 horas, a

47
15 °C e sob agitação de 300 rpm. As placas foram seladas com BreathSeal, adesivo que
permite trocas gasosas com o meio externo sem que haja contaminação. Palitos estéreis
foram utilizados para inocular cada isolado, um a um, por poço, conforme distribuição
apresentada na Figura 6.
Figura 6 – Distribuição de cada cepa e dos brancos (amostras sem microrganismo) em
placas de 96 poços para pré-inóculo e produção.
Foram denominadas “Branco” (B) as amostras contendo apenas o meio de
cultivo estéril, para controle de contaminação das placas. Após 72 horas de crescimento,
uma alíquota de cada isolado foi transferida para outra placa (obedecendo a ordem
estabelecida na Figura 6) contendo meio CD. Os isolados foram então cultivados por
mais 120 horas, a 15 °C e agitação de 800 rpm. Após esse período, cada isolado foi
avaliado qualitativamente para a produção da enzima L-ASNase pelo método
colorimétrico L-aspartil-β-hidroxâmico (ver metodologias analíticas, item 3.11.3). O
critério de seleção foi a intensidade da coloração vermelha, após revelação com cloreto
férrico.
As leveduras identificadas como produtoras nesta etapa foram cultivadas em
volumes maiores (50 mL), utilizando para isso frascos de 250 mL com defletores.
Foram feitos dois blocos, em um deles foi adicionado azul de bromotimol (0,007%)
para inferir a produção de L-ASNase através da mudança de pH e, consequentemente,
da coloração do meio de cultivo, que se torna azul quando básico. Para isso, foi utilizada
a metodologia proposta por MAHAJAN et al. (2013). O azul de bromotimol é um
indicador de pH que em meio ácido, como é o caso do meio CD (pH = 5,5), é
alaranjado, enquanto em meio básico, torna-se azul. Como a produção de L-ASNase e
consequente degradação de asparagina resulta em amônia e, portanto, aumento do pH, a

48
mudança de coloração dos meios para a cor azul é indicativo da produção da enzima de
interesse. Meios CD não inoculados e sem fonte de nitrogênio (prolina) foram utilizados
como controle.
O outro bloco foi cultivado conforme já descrito (item 3.4). Alíquotas foram
retiradas de cada frasco a cada 24 horas durante 5 dias para avaliação quantitativa do
crescimento e da produção de L-ASNase. A partir dessa fase, foi escolhido um isolado
para dar continuidade ao trabalho e o critério de escolha foi a produção da enzima alvo.
O melhor produtor foi taxonomicamente identificado como pertencente a espécie
Leucosporidium muscorum e depositado na Central de Recursos Microbianos da
UNESP (CRM-UNESP) sob o código de acesso CRM 1648.
3.6. Análise univariada de fontes de carbono
Sete fontes de carbono (glicerol, sacarose, xilose, extrato de levedura, sorbitol,
frutose, além de glicose) foram avaliadas na composição do meio CDM com o intuito
de observar qual desses nutrientes teria impacto mais positivo sobre o crescimento da
levedura e produção de L-ASNase. Os demais componentes do meio foram mantidos
nas mesmas concentrações já descritas (Tabela 4). As fontes de carbono e concentrações
utilizadas estão descritas na Tabela 5. A prolina foi mantida como fonte de nitrogênio
em todas as análises. O cultivo foi feito em triplicata, conforme já descrito (item 3.4).
Tabela 5 – Fontes de carbono e respectivas concentrações (C) em g.L-1
de cada fonte
avaliada no meio CD para crescimento da levedura alvo e produção de L-ASNase.
Fonte C (g.L-1
)
1 glicose 2,0
2 glicerol 2,0
3 sacarose 2,0
4 xilose 2,0
5 extrato de levedura 2,0
6 sorbitol 2,0
7 frutose 2,0
3.7. Análise univariada de fontes de nitrogênio
Este ensaio foi feito como o ensaio anterior, mas manteve-se a glicose como
fonte de carbono, variando a fonte de nitrogênio. Foram avaliadas 9 fontes de

49
nitrogênio: asparagina, glutamina, cloreto de amônio, sulfato de amônio, ureia,
casaminoácidos, acetato de amônio, extrato de levedura e prolina (Tabela 6).
Tabela 6 – Fontes de nitrogênio e respectivas concentrações (C) em g.L-1
de cada fonte
avaliada no meio CD para crescimento da levedura e produção de L-ASNase.
Fonte C (g.L-1
)
1 asparagina 10,0
2 glutamina 10,0
3 prolina 10,0
4 cloreto de amônio 10,0
5 sulfato de amônio 10,0
6 ureia 10,0
7 casaminoácidos 10,0
8 acetato de amônio 10,0
9 extrato de levedura 10,0
3.8. Avaliação da interação de fontes nutricionais por delineamento fatorial completo
para melhoramento do crescimento e produção de L-ASNase
A fim de avaliar as possíveis interações e efeitos sinérgicos entre as variáveis
que melhor favoreçam o crescimento da levedura selecionada e produção da enzima L-
ASNase, foi utilizado um planejamento experimental do tipo fatorial completo 33 com
27 ensaios e 4 replicações no ponto central. Os dados foram gerados e analisados pelo
programa Minitab® 17, sendo X1 (sacarose), X2 (extrato de levedura) e X3 (prolina) as
variáveis independentes selecionadas nas análises univariadas anteriores, e avaliadas em
três níveis (-1, 0, +1), conforme Tabela 7.
Tabela 7 – Fatores e níveis avaliados em delineamento fatorial completo (I) para
melhoramento do crescimento de L. muscorum CRM 1648 e produção de L-ASNase.
Fatores níveis (g.L
-1)
-1 0 1
X1 5 10 15
X2 0 5 10
X3 5 10 15

50
Com base nos resultados do delineamento anterior, foi estruturado um segundo
planejamento experimental, também do tipo fatorial completo 33, para a maximização
das respostas. Neste novo planejamento, o ponto central do planejamento anterior
passou a ocupar o nível -1 (Tabela 8). Os experimentos 1, 2, 4, 5, 10, 11, 13 e 14 não
foram repetidos nesta etapa em razão de terem sido realizados na etapa anterior. Foram
realizadas 4 repetições no ponto central.
Tabela 8 – Fatores e níveis avaliados em delineamento fatorial completo (II) para
melhoramento do crescimento de L. muscorum CRM 1648 e produção de L-ASNase.
Fatores níveis (g.L
-1)
-1 0 1
X1 10 15 20
X2 5 10 15
X3 10 15 20
As respostas avaliadas Y1 e Y2, massa seca celular (g.L-1
) e produção de L-
ASNase (U.L-1
), respectivamente, foram analisadas pelas seguintes equações, que
incluem todos os termos de interações, sem considerar a significância.
3.9. Análise univariada das condições de cultivo: temperatura, pH inicial,
concentração celular inicial (X0) e concentração de água do mar no meio
A levedura L. muscorum CRM 1648 foi pré-cultivada em caldo Sabouraud
dextrose e transferida para o meio de produção, conforme já descrito (item 3.4). Czapek
Dox modificado (CDM) foi utilizado como meio de produção. Foram avaliados em
blocos separados as seguintes variáveis: temperatura, pH inicial do meio, concentração
de água do mar e a concentração inicial do inóculo (X0), com a finalidade de avaliar o
efeito de cada uma dessas variáveis sobre o crescimento da levedura e produção de L-
ASNase. As temperaturas avaliadas foram: 9, 12, 15, 18, 21 e 30°C; pH inicial: 6,5, 5,5

51
e 4,5; concentração de água do mar: 0, 25 e 50% do volume total de meio (50 mL); e
X0: 0,5; 1,0 e 1,5 g.L-1
. Cada condição foi feita em triplicata.
3.10. Ensaios em biorreator
3.10.1. Preparo do biorreator
Todos os experimentos em biorreator foram realizados no modelo BIOSTAT®
B-MO (Figura 7), do fabricante Sartorius, equipado com sonda de temperatura, pH e de
O2 dissolvido (OD). Ao equipamento está acoplado também um analisador de gases (O2
e CO2), que permite detectar as porcentagens dos gases no gás de saída da dorna.
Assim, é possível ter a informação a respeito do O2 consumido e CO2 produzido ao
longo do cultivo, além de permitir a medição, regulação e avaliação online de
temperatura, aeração, agitação, volume de meio ou qualquer substrato, ácido, base ou
antiespumante que seja adicionado ao cultivo, pH e OD no meio. Os parâmetros e
variáveis foram registradas a cada minuto pela unidade de controle.
Figura 7 – Biorreator BIOSTAT®
B-MO em uso para cultivo de L. muscorum CRM
1648 e produção de L-ASNase.
A sonda de pH foi calibrada antes da esterilização da dorna, utilizando dois
pontos de calibração: uma solução tampão onde pH = 7,0, seguida de outra solução
tampão (pH = 4,0). Após montagem e esterilização do biorreator (121 °C, 1 atm, 30
minutos), o sensor de O2 foi conectado ao equipamento e polarizado por no mínimo 3
horas nas condições de máxima agitação e aeração a ser utilizada no experimento. Após
esse período, a sonda de O2 foi calibrada também por uma calibração de dois pontos. O

52
ponto 0 (0% de OD) foi obtido pela desconexão do conector da sonda de O2 com a
unidade de controle, enquanto o ponto 100 (100% de OD) foi obtido pela reconexão
desta mesma sonda com a unidade de controle, estando o meio de cultivo saturado com
ar.
3.10.2. Ensaios em biorreator: condições de cultivo e oxigênio dissolvido
Para os cultivos em biorreator foi utilizado o meio CDM. O pré-inóculo foi feito
como já descrito (item 3.4). As células foram centrifugadas (3220g, 10 minutos, 25 °C),
ressuspendidas em meio CDM e uma alíquota dessa suspensão celular foi transferida
para um frasco de 250 mL contendo 100 mL de meio CDM estéril, conforme equação 1
(item 3.4). Esse volume foi inoculado na dorna de forma asséptica, sendo X0 igual a 1
g.L-1
e Vf igual a 1 litro (volume útil do biorreator).
Durante o cultivo microbiano, o oxigênio gasoso foi transferido para o meio de
cultivo através de ar aspergido, mas em algumas situações foi necessária a adição de
oxigênio puro. O balanço gasoso foi calculado de acordo com a relação de transferência
de O2 para o meio de cultivo (OUR, do inglês Oxygen Uptake Rate) e produção de CO2
(CER, do inglês Carbon Dioxide Evolution Rate), através das medidas de fração molar
gasosa que são obtidas pelo analisador de gases acoplado a saída de ar do biorreator. A
relação do consumo de O2 e produção de CO2 é denominada Coeficiente Respiratório
(RQ, do inglês Respiratory Coefficient). Dessa forma, o balanço gasoso foi feito
conforme as equações:
Onde,
RQ: coeficiente respiratório
OUR: velocidade de consumo de O2 dissolvido (mmol O2.L-1
.h-1
)
CER: velocidade de produção de CO2 (mmol CO2.L-1
.h-1
)
QO2: velocidade específica de respiração (mmol O2.g-1
.h-1
)

53
X: concentração de células na dorna (g.L-1
)
V: volume de meio de cultivo (L)
: fluxo de gás (mmol.h-1
) de entrada (Φe) ou saída (Φs).
y: Fração molar de entrada (ye) ou saída (ys) de O2 (Eq. 4) ou CO2 (Eq. 5)
O fluxo de ar de saída (Φs) foi calculado supondo que o fluxo de gases inertes,
basicamente nitrogênio, não varia na entrada e saída de ar. Dessa forma, Φs foi
calculado conforme equação a seguir:
Sendo que, no caso do gás de entrada ser ar, a expressão (1 – yO2e – yCO2e)
corresponde a 0,7901.
No cultivo descontínuo, durante a fase Lag (fase em que as células ainda estão
em adaptação, ou seja, X = X0 e não há reprodução celular) e na fase de transição da
fase Lag para a fase de crescimento exponencial, o valor de X é muito baixo e há pouco
consumo de oxigênio pelas células, ou seja, QO2 é mínimo. Durante o cultivo, X
aumenta e o mesmo ocorre com QO2 (aumenta a demanda respiratória por célula), que
atinge o seu valor máximo na fase exponencial, quando a velocidade específica de
crescimento (µ) é máxima.
Em razão do sistema de transferência de O2 ser muito importante em um sistema
aeróbico de cultivo, como é o caso do empregado neste trabalho, foi avaliado qual seria
a demanda de oxigênio dissolvido (OD) pela levedura L. muscorum CRM 1648 que
permitiria maior crescimento e produção da enzima L-ASNase, visto que não foram
encontrados trabalhos na literatura que tragam informações a respeito da fisiologia
desse microrganismo. As condições avaliadas foram: Ensaio 1 - sem controle de OD
(OD < 20%), Ensaio 2 - sem controle de OD (OD > 20%), Ensaio 3 – OD controlado
em 80% e Ensaio 4 – OD controlado em 20%. Em todos os ensaios foi adicionado
previamente no meio de produção 2 mL de antiespumante.
O Ensaio 1 (OD não controlado e < 20%) foi realizado a 15 °C, 500 rpm e
controle do pH através da adição de HCl (20%) e NaOH (20%) para mantê-lo igual a
5,5. Neste ensaio, a entrada de ar (1) foi ativada, com aeração de 1 LpM (litro de ar por
litro de meio por minuto), e a entrada de O2 puro (2) foi mantida desligada (Figura 8).

54
Os outros ensaios em biorreator (ensaios 2, 3 e 4) também foram conduzidos na mesma
temperatura, com algumas mudanças na agitação e aeração, e sem controle do pH (sem
adição de ácido e base) porque foi observado após o ensaio 1 que este parâmetro
poderia interferir na produção de L-ASNase.
O Ensaio 2 (OD não controlado e > 20%) foi conduzido com cascata de agitação
variando de 200 a 1200 rpm e aeração com ar puro e igual a 1 LpM (entrada de O2 puro
desligada, conforme ensaio anterior). No entanto, como o aumento da agitação para o
máximo programado (1200 rpm) não foi suficiente para evitar a queda de OD ao longo
do tempo, a aeração foi alterada para 1,5 LpM após 20 horas de cultivo. Os parâmetros
de agitação e aeração utilizados neste ensaio não foram capazes de controlar OD, mas
este se manteve acima de 20%.
O Ensaio 3 (OD controlado em 80%) foi conduzido com agitação constante de
600 rpm, aeração de 1 LpM com ar puro e cascata de adição de oxigênio (O2) puro à
linha de ar variando de 0 a 60% (porcentagem do tempo de abertura da válvula de O2).
Nesse ensaio, as entradas 1 e 2 estão ligadas, permitindo o incremento do ar com O2
puro (Figura 8). Para controlar OD, o equipamento foi programado para abrir a válvula
de O2 (3) sempre que OD < 80%, de forma que o ar puro seria incrementado com esse
gás de acordo com o consumo de O2 por L. muscorum CRM 1648.
O Ensaio 4 (OD controlado em 20%) foi conduzido de forma que OD se
mantivesse em torno de 20%. Para isso, no início, gás nitrogênio foi conectado à entrada
de ar (1) e à entrada de O2 puro foi conectado ar puro (2). No entanto, por problemas na
fonte de nitrogênio no início do cultivo (nas primeiras 6 horas) e na tentativa de manter
os níveis de OD dentro do planejado, a entrada de N2 foi desligada, mantendo-se apenas
pulsos de ar puro (entrada 2 ligada e entrada 1 desligada), com a abertura da válvula (3)
controlada em 0-20% (Figura 8). Quando OD atingiu um valor próximo a 20%, a
configuração do sistema foi restabelecida para o original, conforme Figura 8 (Ar na
entrada de ar e O2 na entrada de O2). Dessa forma, entradas 1 e 2 foram mantidas
ligadas, com aeração de 1LpM de ar puro e cascata de O2 puro, com abertura da válvula
controlada em 0-30%. A cascata seria acionada sempre que OD < 20%. A agitação foi
mantida em 600 rpm durante todo o ensaio. Os limites da cascata de O2 puro precisaram
ser ajustados ao longo de todo o cultivo nos ensaios 3 e 4, de acordo com o crescimento
da levedura e consumo de O2.
Em todos os ensaios, foram retiradas amostras de, aproximadamente, 12-15 mL
a cada 3, 6 ou 12 horas, a depender da fase de cultivo (no início a freqüência de coleta

55
foi maior) para a medição das seguintes variáveis: densidade óptica (DO) e massa seca
celular para avaliar o crescimento; medida de pH offline para a confirmação dos valores
obtidos de forma online; avaliação do consumo dos substratos prolina e sacarose; e
atividade enzimática de L-ASNase. Os valores das variáveis dos processos referentes
aos quatro ensaios estão registrados nos Apêndices B a I.
Figura 8 – Representação do painel de controle do biorreator BIOSTAT® B-MO e todos
os parâmetros possíveis de controle: agitação, temperatura, pH, OD, vazão de ar puro de
entrada (1) e vazão de O2 puro de entrada (2), este último controlado por pulsos de
abertura (% de tempo de abertura) da válvula de O2 (3). No início do ensaio 4 a
configuração foi modificada, conforme representado no círculo pontilhado.
3.11.Metodologias Analíticas
3.11.1.Monitoramento do pH
O pH foi medido em pHmetro de bancada (KASVI, modelo K39-1014B). O
equipamento foi calibrado antes das medições utilizando dois pontos de calibração: uma
solução tampão com pH = 7,0 e outra solução tampão com pH = 4,0.

56
3.11.2.Monitoramento do crescimento celular
O crescimento celular foi avaliado por densidade óptica e gravimetria. A medida
de densidade óptica foi realizada em comprimento de onda (λ) igual a 600 nm em
espectrofotômetro Expectra Max Plus 384. Em todas as análises utilizou-se como
“branco” o próprio meio de cultivo sem células ou água (para as amostras diluídas em
água destilada).
A concentração celular também foi avaliada pela massa seca celular
(gravimetria) e expressa em g.L-1
. Amostras de 1 a 5 mL foram centrifugadas (3220g x
5 min) em tubo previamente pesado, o sobrenadante foi removido e reservado (para as
medidas de substrato e pH), as células lavadas (duas vezes) e centrifugadas novamente.
A água foi removida com cuidado e as células foram secadas em estufa em temperatura
acima de 60 °C. O valor da concentração de biomassa em gramas por litro foi calculado
conforme equação a seguir:
(Eq. 8)
Onde,
P1 – peso do tubo vazio (g)
P2 – peso tubo com a biomassa seca (g)
V – volume de cultivo centrifugado (L)
A relação entre DO e massa seca celular (X), apresentada na Figura 9, foi
estabelecida pela equação:
(Eq. 9)
Figura 9 – Valores de densidade óptica em função de valores de massa seca celular,
sendo o R2 = 0,987.
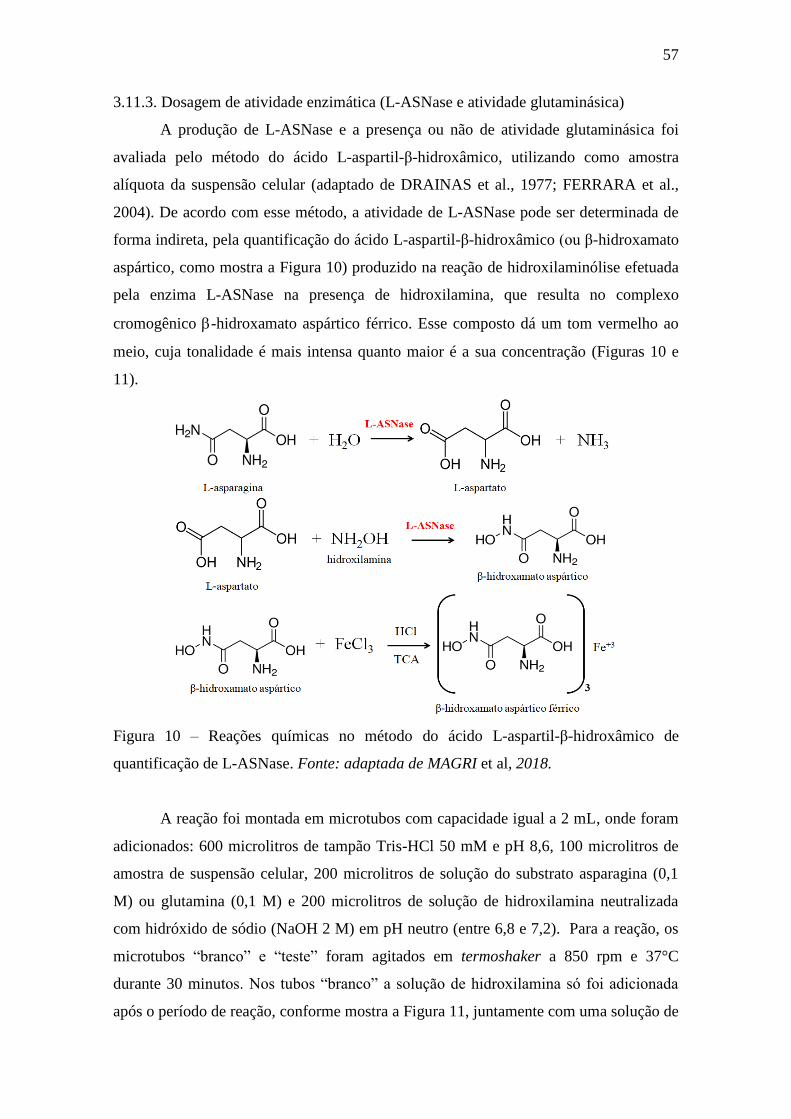
57
3.11.3. Dosagem de atividade enzimática (L-ASNase e atividade glutaminásica)
A produção de L-ASNase e a presença ou não de atividade glutaminásica foi
avaliada pelo método do ácido L-aspartil-β-hidroxâmico, utilizando como amostra
alíquota da suspensão celular (adaptado de DRAINAS et al., 1977; FERRARA et al.,
2004). De acordo com esse método, a atividade de L-ASNase pode ser determinada de
forma indireta, pela quantificação do ácido L-aspartil-β-hidroxâmico (ou β-hidroxamato
aspártico, como mostra a Figura 10) produzido na reação de hidroxilaminólise efetuada
pela enzima L-ASNase na presença de hidroxilamina, que resulta no complexo
cromogênico -hidroxamato aspártico férrico. Esse composto dá um tom vermelho ao
meio, cuja tonalidade é mais intensa quanto maior é a sua concentração (Figuras 10 e
11).
Figura 10 – Reações químicas no método do ácido L-aspartil-β-hidroxâmico de
quantificação de L-ASNase. Fonte: adaptada de MAGRI et al, 2018.
A reação foi montada em microtubos com capacidade igual a 2 mL, onde foram
adicionados: 600 microlitros de tampão Tris-HCl 50 mM e pH 8,6, 100 microlitros de
amostra de suspensão celular, 200 microlitros de solução do substrato asparagina (0,1
M) ou glutamina (0,1 M) e 200 microlitros de solução de hidroxilamina neutralizada
com hidróxido de sódio (NaOH 2 M) em pH neutro (entre 6,8 e 7,2). Para a reação, os
microtubos “branco” e “teste” foram agitados em termoshaker a 850 rpm e 37°C
durante 30 minutos. Nos tubos “branco” a solução de hidroxilamina só foi adicionada
após o período de reação, conforme mostra a Figura 11, juntamente com uma solução de

58
TCA e cloreto férrico (100 g.L-1
FeCl3, 50 g.L-1
TCA em uma solução de HCl a 0,66M),
que também foi adicionada nos tubos “teste”. Os microtubos foram centrifugados 3220g
por 5 minutos e 1 mL do sobrenadante foi utilizado para a medição, que foi feita em
espectrofotômetro Expectra Max Plus 384, em comprimento de onda (λ) igual a 500
nanômetros (nm).
Figura 11 – Sequência de passos para a execução do teste de atividade pelo método do
ácido L-aspartil-β-hidroxâmico.
*Solução de hidroxilamina (2M) neutralizada com solução de NaOH (2M).
**10 % (p/v) FeCl3, mais 5 % (p/v) ácido tricloroacético (TCA) em solução de HCl (0,66M).
Uma unidade de atividade (L-ASNase ou atividade glutaminásica) foi definida
como a quantidade de enzima capaz de produzir 1 µmol de β-hidroxamato aspártico ou
1 µmol de ácido γ-hidroxamato glutâmico (no caso de a glutamina ser utilizada como
substrato) por minuto de tempo e por volume de amostra (MOGUEL, 2018). O valor de
produção de L-ASNase foi calculado em U.L-1
(unidade de enzima por volume de
meio). A atividade total (U) foi calculada multiplicando o valor de atividade em U.L-1
pelo volume total (v) do meio de produção.

59
3.11.4. Dosagem de sacarose
Para a dosagem de sacarose, foi utilizado o método colorimétrico GOD/invertase
(TEIXEIRA et al., 2012). Nesse método, foi adicionado 85 µL de água destilada, 5 µL
de amostra (sobrenadante das amostras coletadas nos experimentos em biorreator) e 10
µL de invertase por poço de uma placa de 96 poços. A invertase foi diluída em água até
atingir a concentração de 8000 U.mL-1
. A reação ocorreu a 55°C por 10 minutos. Nessa
etapa, a invertase atua catalisando a quebra de sacarose, formando glicose e frutose.
Após esse período, 200 µL de GOD/POD (sigla em inglês para glicose
oxidase/peroxidase) foi adicionado em cada poço e a placa foi encubada por mais 15
minutos a 37 °C. Nessa etapa, a enzima GOD catalisa a reação de oxidação de uma
molécula de glicose formando peróxido (H2O2) e ácido glucurônico. A molécula de
H2O2por sua vez reage com 4-aminoantipirina e fenol, sob a ação catalisadora da
peroxidase, através de uma reação oxidativa de acoplamento formando uma
antipirilquinonimina vermelha, cuja intensidade da cor é proporcional a concentração de
glicose na amostra e no padrão (e, portanto, proporcional a concentração de sacarose). A
leitura foi feita em espectrofotômetro Expectra Max Plus 384 a 490 nm. A curva padrão
foi construída com sacarose diluída em água nas concentrações de 0,0, 0,05, 0,1, 0,15,
0,20 e 0,25% (g/100 mL), faixa onde mostrou linearidade. Cada análise foi feita em
duplicata.
3.11.5. Dosagem de prolina
A análise do consumo de prolina foi feita colorimetricamente como descrito por
ABRAHAM et al. (2010). Em microtubos de 2 mL foi adicionado 100 µL de amostra,
100 µLde ácido sulfosalicílico (solução a 3%), 200 µL de ácido acético glacial e 200 µL
de nihidrina ácida (1,25 g de nihidrina em 30 mL de ácido acético glacial e 20 mL de
ácido ortofosfórico a 6M). Os microtubos foram incubados a 96 °C por 1 hora e depois
desse período colocados em gelo para a adição de 1 mL de tolueno com o intuito de
parar a reação. As amostras foram agitadas em vortex por 20 segundos e a fase orgânica
(fase topo) foi coletada. A leitura foi feita em cubeta de vidro, no espectrofotômetro
Expectra Max Plus 384 a 520 nm.

60
3.11.6. Cálculo dos parâmetros cinéticos
Os resultados obtidos nos ensaios em biorreator referentes ao crescimento celular,
formação do produto (L-ASNase) e consumo de substratos (sacarose e prolina) foram
colocados em gráficos para a determinação dos seguintes parâmetros cinéticos:
a) velocidade específica de crescimento (µx)
Foi calculada nos ensaios em frascos agitados e biorreator a partir de dados de
biomassa. Foram utilizados os pontos da fase logarítmica de crescimento, onde a
velocidade específica de crescimento é constante e máxima (µx = µmax) (SCHMIDELL
et al., 2001).
Supondo µx constante, o valor de µmax foi calculado pela inclinação da reta
tangente resultante da representação gráfica do logaritmo natural (Ln) de X em função
do tempo. Essa relação é estabelecida pelas equações:
(Eq. 10)
Quando µx é constante, a integral da equação acima resulta em:
(Eq. 11)
b) Produtividade
O desempenho do processo de produção de células e produto (L-ASNase) foi
avaliado pela produtividade ao longo do cultivo, respectivamente, Px e Pp, calculada
conforme as equações 15 e 16, sendo o sufixo f = final e i = inicial.
(Eq. 12)
(Eq. 13)

61
IV. RESULTADOS E DISCUSSÃO
A região Antártica é conhecida pelas suas condições inóspitas, apresentando o clima
mais frio e seco da terra, além de alta salinidade, baixa precipitação e disponibilidade de
nutrientes, ciclos alternados de congelamento e descongelamento e alta irradiação UV
alternada por longos períodos de escuridão (apud DUARTE et al., 2013). Apesar do
ambiente adverso, essa região tem se revelado uma fonte de numerosos microrganismos
psicrófilos e psicrotolerantes, denominados de forma generalizada de microrganismos
adaptados ao frio, com grande capacidade metabólica para produzir uma diversidade de
substâncias de interesse industrial. Por essa razão tem sido cada vez mais comum a
triagem de biomoléculas em microrganismos isolados dessa região, inclusive em
leveduras (BUZZINI et al., 2012; DUARTE et al., 2013; LARIO et al., 2015;
CARRASCO et al., 2016).
4.1. Triagem de leveduras adaptadas ao frio produtoras de L-ASNase (avaliação
qualitativa)
A triagem de microrganismos, principalmente leveduras e fungos filamentosos,
produtores de L-ASNase normalmente é feito em meio sólido Czapek Dox (CD)
suplementado com o indicador vermelho de fenol na concentração de 0,009% (m/v) ou
azul de bromotimol a 0,007% (m/v) (MAHAJAN et al., 2013). A avaliação qualitativa
da produção de L-ASNase em meio sólido, no entanto, não foi empregada neste
trabalho porque foi observado por outros pesquisadores desse grupo de pesquisa que a
habilidade de certos fungos de produzirem L-ASNase em meio sólido pode ser diferente
da habilidade de produção em meio líquido. Essa observação também foi reportada por
LAPMAK et al. (2010).
Foram cultivadas em caldo CD, 150 leveduras de amostras isoladas de
sedimentos marinhos na Península Antártica, provenientes das regiões Refúgio 2, EACF
e Botany Point (Figura 5), em busca de produtores da enzima L-ASNase. Após 120
horas de cultivo, cada amostra foi avaliada quanto a presença dessa enzima na
suspensão celular. Nesse momento o objetivo não era fazer uma avaliação quantitativa,
mas identificar qualitativamente os produtores, conforme mostrado na Figura 12. Foi
considerado produtor da enzima aquele que assumiu a coloração vermelha no meio de
reação após a adição de cloreto férrico/TCA/HCl. Como existe relação entre a
intensidade da coloração vermelha e a produção do complexo L-aspartil-β-hidroxamato

62
com íon férrico, as amostras com coloração amarelo-alaranjada foram consideradas
negativas para a produção de L-ASNase. Os isolados 17 (placa II, B6-7) e 74 (placa I,
E7) aparentemente produziram menos que os demais (coloração rosa claro), porém os
isolados 85 (placa II, G3) e 94 (placa II, D5) foram aqueles que visualmente produziram
mais enzima, pois atingiram uma coloração vinho, bem mais escura que os demais. Esse
resultado foi confirmado posteriormente, com o cultivo em frascos agitados em maior
volume útil (50 mL), no qual foi possível avaliar a produção de L-ASNase ao longo do
tempo. Os isolados 17 e 74 foram descartados na etapa de cultivo em frascos devido a
contaminações sucessivas com fungos filamentosos.
Figura 12 – Triagem de produtores de L-ASNase entre 150 isolados da Antártica pelo
método do ácido L-aspartil-β-hidroxâmico. A maior intensidade da cor vermelha indica
maior produção da enzima.
Os isolados identificados nesta etapa como produtores da enzima alvo foram
cultivados em frascos agitados contendo meio líquido CD e azul de bromotimol
(0,007%), de acordo com a metodologia proposta por MAHAJANet al. (2013). Um
frasco com meio estéril e outro com meio CD sem prolina (fonte de nitrogênio) foram
utilizados como controle. A produção da enzima foi avaliada visualmente pela mudança

63
de coloração do meio, que se tornou azul após 72 horas de cultivo. Como azul de
bromotimol é um indicador de pH, ficando laranja em meio ácido e azul quando em
meio básico, a intensidade da cor azul indicou maior produção de L-ASNase (a reação
catalisada por esta enzima resulta na produção de amônia, que torna o meio básico). O
resultado deste cultivo (Figura 13) confirma maior produção de L-ASNase pelos
isolados 85 e 94.
Figura 13 – Resultado do cultivo dos isolados produtores de L-ASNase em meio CDM
com azul de bromotimol. Da esquerda para a direita: meio não inoculado, meio sem
fonte de nitrogênio, isolados 2, 26, 1, 16, 86, 94 e 85.
Os resultados do seqüenciamento de marcadores taxonômicos e análises
filogenéticas (INFORSATO, 2017) revelaram que os isolados 17 e 74 pertencem a
mesma espécie Glaciozyma antarctica e que os outros isolados produtores pertencem a
espécie Leucosporidium muscorum.
A morfologia das colônias dos isolados produtores (Figura 14) confirma que
todos eles apresentaram colônias de cor creme, com superfície lisa, aspecto cremoso e
textura mucoide. Nenhum desses isolados produziram pigmento em meio ágar
Sabouraud. A morfologia das colônias mostrada na Figura 14 corrobora com as
características morfológicas já descritas para a espécie tipo L. muscorum (di MENNA,
1958).
Nessa etapa do trabalho, 9 isolados (6%) foram identificados como potenciais
produtores de L-ASNase, sendo que desse total 7 (77,8%) pertencem a espécie L.
muscorum. Espécies da ordem Leucosporidiales reconhecidas como potenciais fontes
de enzimas extracelulares adaptadas ao frio e de interesse industrial tem sido isoladas
predominantemente de ambientes frios e reportadas como leveduras psicrotolerantes
(VAZ et al., 2011). O isolamento de leveduras de regiões diversas na Antártica resultou
em 89 isolados dentre os quais dois eram da espécie L. muscorum, um proveniente da
rizosfera de uma planta nativa da Antártica (Deschampsia antarctica) e o outro de

64
sedimento marinho próximo a estação Machu Picchu, na Baia do Almirantado (VAZ et
al., 2011), muito próximo ao local onde foram isoladas as leveduras utilizadas neste
trabalho.
Pode-se supor, portanto, que a espécie L. muscorum é característica de regiões
polares, de clima mais frio,por ter sido isolada até então apenasde regiões mais
próximas aos pólos (di MENNA, 1958; SAMPAIO et al., 2003; PATHAN et al., 2010;
GARCÍA et al., 2015), uma vez que não foram encontrados registros
dessemicrorganismo em regiões tropicais. Por ser essa espécie filogeneticamente
próxima a leveduras do gênero Glaciozyma spp., e dos gêneros Rhodotorula spp. e
Rhodosporidium spp. (WANG et al., 2015), que já foram identificados como produtores
de L-ASNase (RAMAKRISHNAN; JOSEPH, 1996; FODA et al., 1980; IMADA et al.,
1973), associado ao fato de que outra espécie do gênero Leucosporidium sp. também foi
identificada como produtora dessa enzima (MOGUEL, 2018),é possível haver alguma
correlação entre esse grupo de leveduras e a produção de L-ASNase, que precisa ser
melhor investigada.
Figura 14 –Morfologia das colônias dos isolados produtores.
Já foi demonstrado que a espécie L. muscorum é capaz de produzir diversas
enzimas, incluindo proteases, amilases, pectinases, urease (INFORSATO, 2017;
PATHAN et al., 2010), mas não foi identificada a produção de L-ASNase por essa

65
levedura, sendo este trabalho o primeiro a trazer essa informação. É válido registrar que
os isolados 85 e 94 são provenientes de uma mesma região (Refúgio 2, entre marés) e,
portanto, poderiam tratar da mesma linhagem, sugerindo que mesmo uma mesma
linhagem pode trazer especificidades quanto a produção de biomoléculas. O mesmo
pode-se dizer dos isolados 1, 16 e 26, também isolados de uma mesma região (tabela 3).
4.2. Perfil de produção de L-ASNase pelas leveduras selecionadas (avaliação
quantitativa) e seleção do melhor produtor
As leveduras selecionadas na etapa anterior (no total 7 isolados: 1, 2, 16, 26, 85,
86 e 94) foram cultivadas em volume maior de meio (50 mL), utilizando frascos do tipo
Erlenmeyer com defletores e 250 mL de volume total. O objetivo desta etapa foi avaliar
a produção de L-ASNase por cada isolado ao longo do cultivo e selecionar um deles,
com maior capacidade de produção da enzima, para dar continuidade às etapas
subsequentes.
O perfil de crescimento foi muito similar entre eles, mas os isolados 85 e 94
produziram 2 vezes mais do que os demais (Figura 15), com produtividade média igual
a 4,8 e 5,6 U.L-1
.h-1
em 96 horas de cultivo, respectivamente.
Como os resultados do teste quantitativo foi bem similar ao obtido
qualitativamente nas placas de 96 poços, é possível concluir que esse método de triagem
foi eficiente na seleção de microrganismos produtores de L-ASNase e vale ser
valorizado no presente trabalho, já que o uso desse tipo de metodologia para a avaliação
da produção de L-ASNase não foi previamente encontrado na literatura consultada. A
triagem normalmente é realizada em meio sólido (MAHAJAN et al., 2013) e isso tem se
mostrado um problema quando o cultivo é feito em meio líquido, pois nem sempre
aqueles que se mostram produtores em meio sólido conseguem manter a produção em
meio submerso, como já havia sido notado em nosso grupo de pesquisa (dados não
publicados).
O isolado 94 foi identificado como sendo da espécie Leucosporidium muscorum
e depositado na Central de Recursos Microbianos da UNESP (CRM-UNESP), com o
número de acesso CRM 1648. Nessa estapa do trabalho a linhagem L. muscorum CRM
1648 foi selecionada e se tornou o microrganismo alvo deste trabalho. Esse isolado foi o
escolhido porque, apesar de ter apresentado um perfil de produção de L-ASNase muito
semelhante ao do isolado 85 até aproximadamente 85 horas de cultivo, atingiu maior
produção eprodutividade após esse período (Figura 15).

66
Figura 15 – Curva de crescimento e produção de L-ASNase por sete idolados da
Antartica.
Legenda:
Também foi avaliada apresença de atividade glutaminásica na suspensão celular
e de atividade de L-ASNase no sobrenadante. O resultado das análises indicou ausência
de atividade glutaminásica e de enzima extracelular para os 7 isolados analisados.
A avaliação de 108 cepas de Rhodotorula spp., entre as espécie R. flava, R.
rubra, R. macerans, mostrou que 30 delas apresentaram atividade de L-ASNase, 8 delas
apresentaram atividade de L-ASNase e glutaminase e que nenhuma apresentou apenas
atividade glutaminásica (IMADA et al., 1973). Linhagem de Leucosporidium scotti
produtora de L-ASNase apresentou atividade glutaminásica baixa, próxima a zero
(MOGUEL, 2018). Como já comentado anteriormente, a ausência de atividade
glutaminásica pode ter um efeito positivo no tratamento de pacientes com LLA, uma
vez que a hidrólise de glutamina por L-ASNase contribuiria com os efeitos neurotóxicos
no tratamento desse tipo de leucemia (COVINI et al., 2012), além disso, a ausência de
atividade glutaminásica em algumas variantes de L-ASNase teria melhor resultado
terapêutico em células leucêmicas ASNS negativas (CHAN et al., 2014).
A produção extracelular de L-ASNase foi identificada para leveduras da espécie
Rhodotorula rosa (ARIMA et al., 1972) e R. rubra (IMADA et al., 1973). Mais
frequentemente, a enzima produzida por leveduras parece ficar alojada no periplasma da
célula (MOGUEL, 2018; FERRARA et al., 2004). Dessa forma, é possível que a
enzima L-ASNase produzida pela levedura L. muscorum CRM 1648 seja do tipo
periplasmática.

67
4.3.Triagem e seleção de fatores nutricionais
A triagem e seleção dos componentes do meio de cultivo são importantes para
tornar o processo de produção economicamente viável, principalmente porque essa
composição é específica para cada organismo produtor (GHOLAMIAN et al., 2013).
Dessa forma, não há parâmetros físicos nem nutricionais estabelecidos para a máxima
produção de L-ASNase pelas diferentes fontes microbianas (ELSHAFEI; EL-
GHONEMY, 2015). Em estudos prévios, a produção de L-ASNase pela bactéria
Serratia marcescens foi maior na presença de extrato de levedura, lactose e frutose, mas
reduziu na presença de glicose (HEINEMANN; HOWARD,1969). Por outro lado, a
produção dessa enzima pela bactéria Erwinia aroideae foi melhorada na presença de
lactose, mas reprimida quando na presença de glicose (LIU; ZAJIC, 1971). Em
mutantes de S. marcescens, nomeados 933 e WF, a produção de L-ASNase foi
estimulada na presença de glicose e sacarose e inibida por lactose, respectivamente, mas
em ambos mutantes a produção dessa enzima foi estimulada por extrato de levedura
(SUKUMARAN et al., 1979).
De acordo com a literatura (SCHIMIDELL et al., 2001), algumas características
gerais devem ser consideradas ao compor o meio de cultivo: 1. Este deve ser o mais
barato possível, embora, no caso de produtos de alto valor agregado como um
biofármaco, o custo do meio não impacte tanto no valor final do produto; 2. Atender às
necessidades nutricionais do microrganismo; 3. Auxiliar no controle do processo, por
exemplo, evitando variações drásticas do pH ou formação excessiva de espuma; 4. Não
dificultar a recuperação do produto; 5. Ter composição razoavelmente fixa para não
prejudicar a manutenção do controle de qualidade do processo; 6. Não dificultar o
tratamento final do efluente (SCHIMIDELL et al., 2001). A princípio, foi levado em
conta principalmente o item 2, a fim de avaliar quais fontes de carbono e nitrogênio
mais favorecem o crescimento de L. muscorum CRM 1648 e a produção de L-ASNase
por essa levedura.
Os microrganismos utilizam como fonte de carbono e, frequentemente, também
como fonte de energia, diversos açúcares, entre eles: glicose, sacarose, frutose, ou ainda
polissacarídeos, como o amido e a celulose (SCHIMIDELL et al., 2001). Como fonte de
nitrogênio são frequentemente utilizados sais, como o sulfato de amônio ((NH4)2SO4), o
fosfato de amônio ((NH4)2HPO4), aminoácidos e uréia. Como fontes de fósforo são
utilizadas comumente os sais de fosfato (fosfato de sódio, potássio, amônio etc). Ainda
necessita-se adicionar outros elementos, entre minerais como Na (sódio), Ca (cálcio),

68
Mg (magnésio) e elementos traços como Mn (manganês), Cu (cobre), Fe (ferro), Zn
(Zinco), entre outros a depender da composição média do microrganismo usado. Para
leveduras, a composição média de carbono é de 48% da massa seca celular, podendo
variar de 46-52%, enquanto a composição média de nitrogênio é de 7,5 % (6 a 8,5%) da
massa seca celular (OKAFOR, 2007).
A produção de enzimas pode ser influenciada por diferentes fatores incluindo
pH, salinidade, disponibilidade de oxigênio e fontes de nutrientes. O estudo das
variáveis em um bioprocesso pode ser feito em um procedimento experimental do tipo
OFAT, análise univariada, que envolve o estudo de uma variável por vez,fixando-se as
demais variáveis envolvidas no processo (FUKUDA et al., 2018).Esse foi o
procedimento experimental utilizado para avaliar o efeito de 7 diferentes fontes de
carbono (FC) e 9 fontes de nitrogênio (FN) sobre o crescimento celular e produção de
L-ASNase pela levedura L. muscorum 1648. Algumas fontes de nitrogênio, como
prolina, extrato de levedura, glutamina são também fontes de carbono, mas foram
utilizadas neste trabalho como fonte única de nitrogênio na composição do meio CD e,
por essa razão, foram classificadas dessa forma.O extrato de levedura foi a única fonte
avaliada nas duas situações, ora como fonte de carbono, ora como fonte de nitrogênio.
4.3.1.Avaliação de fontes de carbono
No meio de cultivo, as fontes de carbono são utilizadas para melhorar o
crescimento e, subsequentemente, melhorar a produção de metabólitos primários, por
exemplo, enzimas (KALYANASUNDARAM et al., 2015). Nesta etapa, glicose,
glicerol, sacarose, xilose, extrato de levedura, sorbitol e frutose foram utilizados na
concentração inicial de 2 g.L-1
, mantendo-se os outros componentes do meio CD nas
suas respectivas concentrações, conforme materiais e métodos. Essas fontes de carbono
foram escolhidas por estarem disponíveis no laboratório e porque são frequentemente
citadas na literatura em experimentos de otimização do meio de cultivo para a produção
de L-ASNase por diferentes tipos de microrganismos (GHOLAMIAN et al., 2013;
KALYANASUNDARAM et al., 2015).
De acordo com os resultados (Figura 16), o extrato de levedura foi a fonte de
carbono que proporcionou maior crescimento de L. muscorum CRM 1648 nas primeiras
72 horas de cultivo (DO = 13,2 ± 1,0), três vezes maior quando comparado ao meio CD
com glicose (DO = 3,7 ± 0,17). Embora a quantidade de carbono seja similar em todos
os meios, além de carbono, o extrato de levedura é fonte adicional de outros nutrientes,
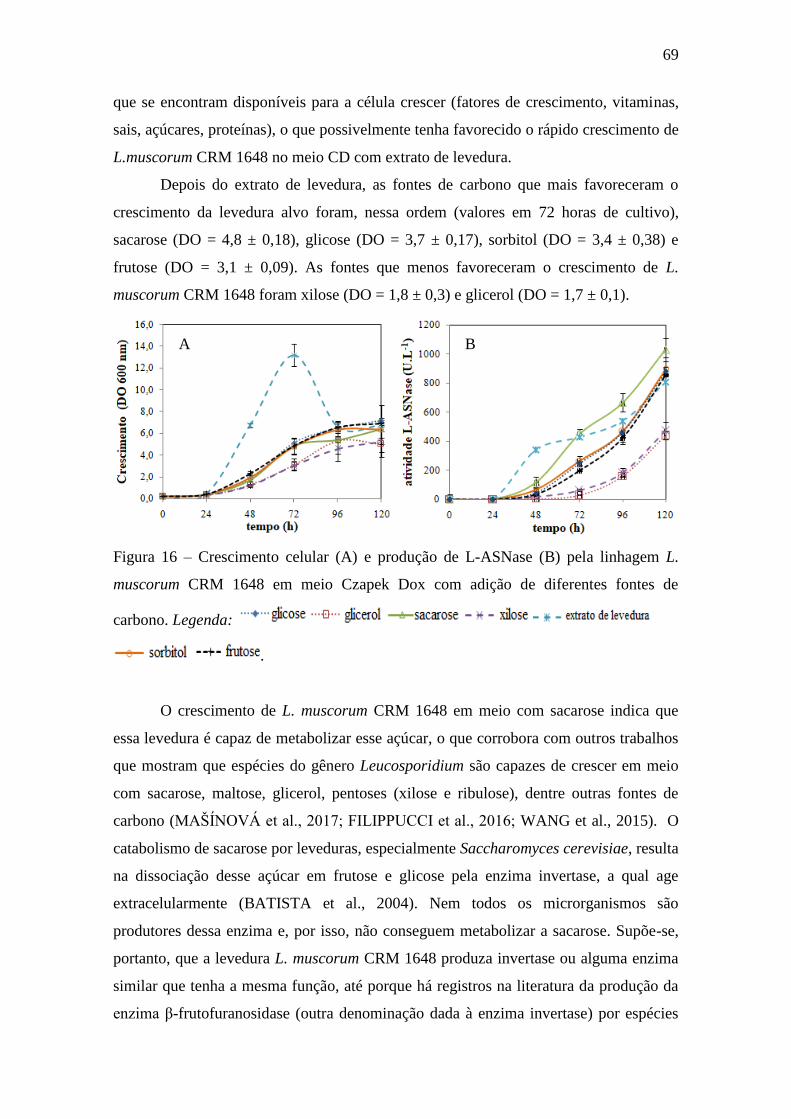
69
que se encontram disponíveis para a célula crescer (fatores de crescimento, vitaminas,
sais, açúcares, proteínas), o que possivelmente tenha favorecido o rápido crescimento de
L.muscorum CRM 1648 no meio CD com extrato de levedura.
Depois do extrato de levedura, as fontes de carbono que mais favoreceram o
crescimento da levedura alvo foram, nessa ordem (valores em 72 horas de cultivo),
sacarose (DO = 4,8 ± 0,18), glicose (DO = 3,7 ± 0,17), sorbitol (DO = 3,4 ± 0,38) e
frutose (DO = 3,1 ± 0,09). As fontes que menos favoreceram o crescimento de L.
muscorum CRM 1648 foram xilose (DO = 1,8 ± 0,3) e glicerol (DO = 1,7 ± 0,1).
Figura 16 – Crescimento celular (A) e produção de L-ASNase (B) pela linhagem L.
muscorum CRM 1648 em meio Czapek Dox com adição de diferentes fontes de
carbono. Legenda:
.
O crescimento de L. muscorum CRM 1648 em meio com sacarose indica que
essa levedura é capaz de metabolizar esse açúcar, o que corrobora com outros trabalhos
que mostram que espécies do gênero Leucosporidium são capazes de crescer em meio
com sacarose, maltose, glicerol, pentoses (xilose e ribulose), dentre outras fontes de
carbono (MAŠÍNOVÁ et al., 2017; FILIPPUCCI et al., 2016; WANG et al., 2015). O
catabolismo de sacarose por leveduras, especialmente Saccharomyces cerevisiae, resulta
na dissociação desse açúcar em frutose e glicose pela enzima invertase, a qual age
extracelularmente (BATISTA et al., 2004). Nem todos os microrganismos são
produtores dessa enzima e, por isso, não conseguem metabolizar a sacarose. Supõe-se,
portanto, que a levedura L. muscorum CRM 1648 produza invertase ou alguma enzima
similar que tenha a mesma função, até porque há registros na literatura da produção da
enzima β-frutofuranosidase (outra denominação dada à enzima invertase) por espécies
A B

70
de leveduras taxonomicamente próximas ao gênero Leucosporidium como, por
exemplo, a espécie Rhodotorula dairenenses (GUTIÉRREZ-ALONSO et al., 2009) e R.
glutinis (RUBIO et al., 2002).
Sacarose foi a fonte de carbono que resultou em maior produção de L-ASNase
(1028,3 ± 81,4 U.L-1
) no final do cultivo (após 120 horas), seguida por sorbitol (895,8 ±
77,5 U.L-1
), glicose (878,8 ± 32,8 U.L-1
), frutose (856,5 ± 9,2 U.L-1
) e extrato de
levedura (806,4 ± 19,7 U.L-1
). Meio CD com xilose ou glicerol resultou em uma
produção pelo menos duas vezes menor (471,7 ± 59,7 U.L-1
e 439,6 ± 28,9 U.L-1
,
respectivamente) em relação ao meio com sacarose.
Linhagem de Leucosporidium scotti isolada de amostra da Península Antártica
foi avaliada em meio CD com diferentes fontes de carbono e nitrogênio e, dentre as
fontes de carbono consideradas no estudo, sacarose seguida por glicerol foram as que
mais favoreceram tanto o crescimento como a produção de L-ASNase por essa levedura
(MOGUEL, 2018). Resultado similar foi obtido por linhagem de Leucosporidium
creatinivorum, que mostrou crescimento também em meio com xilose e maltose, mas
não foi capaz de crescer em galactose e celobiose (FILIPPUCCI et al., 2016). Em outro
estudo, L. muscorum foi capaz de assimilar sorbose, xilose, lactose e rafinose (LAICH
et al., 2014). Sacarose e glicerol não foram avaliadas nesse trabalho.
Como conclusão desta etapa, sacarose foi a fonte de carbono que mais favoreceu
a produção de L-ASNase e, juntamente com o extrato de levedura, a que mais
contribuiu para o crescimento de L. muscorum CRM 1648.
4.3.2.Avaliação de fontes de nitrogênio
O nitrogênio (orgânico e inorgânico) é metabolizado para produzir aminoácidos,
ácidos nucléicos, proteínas e componentes da parede celular. Juntamente com o
carbono, a fonte de nitrogênio (FN) auxilia no crescimento e maior produção de enzimas e,
dependendo do tipo de fonte de carbono adicionada no meio, podem proporcionar maior ou
menor produção de L-ASNase (KALYANASUNDARAM et al., 2015).
De acordo com o gráfico de crescimento (Figura 17), L. muscorum CRM 1648
não foi capaz de crescer em meio com ureia ou com cloreto de amônio. Por outro lado,
o crescimento foi mais acelerado em meio CD com extrato de levedura (DO = 6,4 ±
0,12 em 75 horas de cultivo). Meio CD com prolina resultou em maior valor de DO no
final do cultivo (DO = 8,2 ± 0,14). Dessa forma, as fontes de nitrogênio que
proporcionaram maior crescimento foram, nessa ordem, prolina, casaminoácidos e
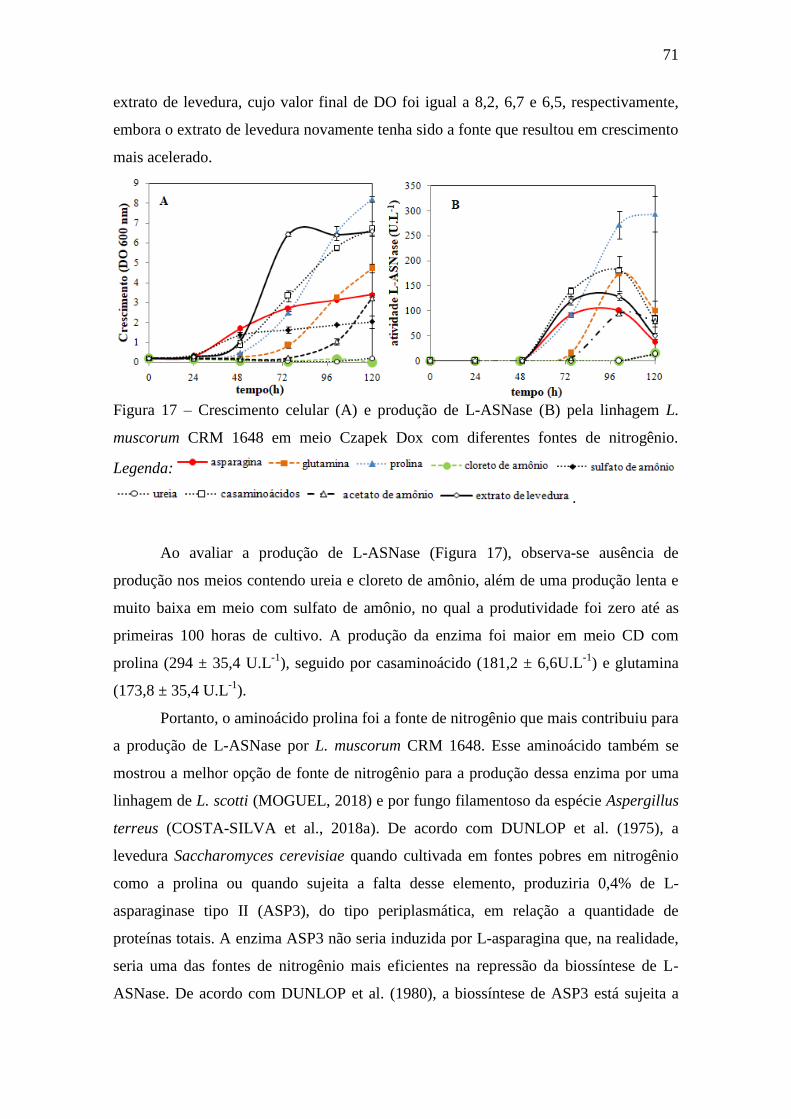
71
extrato de levedura, cujo valor final de DO foi igual a 8,2, 6,7 e 6,5, respectivamente,
embora o extrato de levedura novamente tenha sido a fonte que resultou em crescimento
mais acelerado.
Figura 17 – Crescimento celular (A) e produção de L-ASNase (B) pela linhagem L.
muscorum CRM 1648 em meio Czapek Dox com diferentes fontes de nitrogênio.
Legenda:
.
Ao avaliar a produção de L-ASNase (Figura 17), observa-se ausência de
produção nos meios contendo ureia e cloreto de amônio, além de uma produção lenta e
muito baixa em meio com sulfato de amônio, no qual a produtividade foi zero até as
primeiras 100 horas de cultivo. A produção da enzima foi maior em meio CD com
prolina (294 ± 35,4 U.L-1
), seguido por casaminoácido (181,2 ± 6,6U.L-1
) e glutamina
(173,8 ± 35,4 U.L-1
).
Portanto, o aminoácido prolina foi a fonte de nitrogênio que mais contribuiu para
a produção de L-ASNase por L. muscorum CRM 1648. Esse aminoácido também se
mostrou a melhor opção de fonte de nitrogênio para a produção dessa enzima por uma
linhagem de L. scotti (MOGUEL, 2018) e por fungo filamentoso da espécie Aspergillus
terreus (COSTA-SILVA et al., 2018a). De acordo com DUNLOP et al. (1975), a
levedura Saccharomyces cerevisiae quando cultivada em fontes pobres em nitrogênio
como a prolina ou quando sujeita a falta desse elemento, produziria 0,4% de L-
asparaginase tipo II (ASP3), do tipo periplasmática, em relação a quantidade de
proteínas totais. A enzima ASP3 não seria induzida por L-asparagina que, na realidade,
seria uma das fontes de nitrogênio mais eficientes na repressão da biossíntese de L-
ASNase. De acordo com DUNLOP et al. (1980), a biossíntese de ASP3 está sujeita a

72
forte repressão catabólica por uma variedade de compostos de nitrogênio, fenômeno
denominado de repressão catabólica pelo nitrogênio (RCN).
SARQUIS et al. (2004) avaliaram a produção de L-ASNase por espécies de
fungos do gênero Aspergillus spp. em diferentes FN, inclusive prolina, e também
observaram que esse aminoácido era aquele que mais favorecia a produção de L-
ASNase pela espécie A. terreus e que glutamina e ureia foram as fontes que resultaram
em menor produção da enzima, indicando que a metabolização de fontes de nitrogênio
por esses fungos filamentosos sofre influência do fenômeno de RCN (SARQUIS et al.,
2004).
Embora microrganismos consigam utilizar uma grande variedade de fontes de
nitrogênio, o crescimento irá variar conforme o tipo de FN disponível no meio. Por essa
razão, a fim de selecionar a melhor FN, os microrganismos desenvolvem mecanismos
de regulação que permitem o uso de nutrientes que melhor suportem o seu crescimento
(SANCHEZ; DEMAIN, 2002). Amônia, glutamina e asparagina são reconhecidas como
fontes de nitrogênio mais facilmente metabolizáveis ou mais ricas. A ausência de fontes
ricas em nitrogênio resulta na liberação a nível transcricional das vias de catabolismo de
fontes de N consideradas não preferenciais ou “pobres” (prolina, ureia, glutamato), o
que caracteriza o fenômeno de repressão catabólica pelo nitrogênio (HOLFMAN-
BANG, 1999). Segundo a literatura (MAGAZANIK; KAIZER, 2002), as leveduras ao
crescerem nessas fontes de nitrogênio aumentam a expressão de enzimas envolvidas na
síntese de glutamato e glutamina e, por consequência, aumentam a atividade de
permeases responsáveis pela captação de aminoácidos a serem utilizados como fonte de
N.
Segundo ARIMA et al (1972), os fungos e as leveduras mostram modos de
regulação de L-ASNase similar aos das células vegetais, onde a produção extracelular
da enzima é estimulada por uma carência de nitrogênio. Há na literatura estudos sobre o
mecanismo de RCN na levedura S. cerevisiae (DUNLOP et al., 1980; OLIVEIRA,
1999; HOFMAN-BANG, 1999). OLIVEIRA (1999) cita que a produção de ASP3 seria
induzida pela falta de nitrogênio e não pela presença de asparagina. MOGUEL (2018)
também sugere que este fenômeno ocorra em linhagem de L. scotti.
Com base nessas informações, a ausência de atividade no meio com ureia,
sulfato de amônio e cloreto de amônio, observada quando L. muscorum CRM 1648 foi
cultivada nessas fontes de nitrogênio, muito provavelmente é consequência do
fenômeno de RCN. A presença de atividade após 120 horas de cultivo nos meios com

73
essas fontes de nitrogênio indica que esta foi consumida (falta de nitrogênio, do inglês
nitrogen starvation), desreprimindo o catabolismo de fontes de N. Além disso,
explicaria também o fato da prolina, que é uma das FN mais pobres, ser aquela que mais
favorece a produção de L-ASNase.
Como conclusão desta etapa, o aminoácido prolina foi a FN que mais favoreceu
a produção de L-ASNase e, juntamente com o extrato de levedura, a que mais
contribuiu para o crescimento de L. muscorum CRM 1648. Sacarose, extrato de
levedura e prolina foram, portanto, as fontes de carbono e nitrogênio que mais
contribuíram para o crescimento dessa levedura e produção da enzima e, por isso, foram
selecionadas para compor o meio Czapek-Dox Modificado (CDM), que foi otimizado
na etapa a seguir.
4.4. Otimização dos fatores nutricionais por delineamento fatorial completo
As combinações e resultados obtidos no delineamento experimental (I) estão
dispostos na Tabela 9. Os resultados experimentais referentes aos pontos centrais para
ambas as respostas (crescimento e produção) foram muito próximos, indicando
confiabilidade nos resultados.
O delineamento experimental mostrou que a produção da enzima tende a
aumentar com o aumento da concentração de prolina e sacarose (15 g.L-1
cada), porém
em nível intermediário de extrato de levedura (5 g.L-1
), como mostra o experimento 26
(3658,3 U.L-1
de L-ASNase), que teve a maior produção de L-ASNase (Tabela 9). A
análise estatística desse ensaio foi realizada por Análise de Variância (ANOVA) para
ambas as respostas, sendo significativos os valores de p< 0,1 (Tabela 10).
Os resultados de ANOVA (Tabela 10) mostram que as concentrações de
sacarose, extrato de levedura e prolina têm relação direta com o crescimento e produção
da enzima, uma vez que os termos lineares desses três fatores foram significativos (p<
0,1) para as duas respostas. Ao analisar a Soma dos Quadrados, a resposta crescimento
sofre maior influência do fator sacarose (SQ = 167,58), seguida pelo fator extrato de
levedura (SQ = 35,77) e, em menor grau, pela prolina (SQ = 8,9). Esses fatores têm
menor influência sobre a produção de L-ASNase, mas também existe um efeito sobre
essa resposta, que é mais afetada pela sacarose (SQ = 7,78) e em menor grau pelos
outros dois fatores (SQ aproximadamente igual a 0,7 para ambos). Os termos
quadráticos não influenciaram nenhuma das respostas, mas as interações foram
significativas, exceto a interação de sacarose ou de extrato de levedura com prolina, que

74
não foi significativa para o crescimento de L. muscorum CRM 1648 (p>0,1). Os valores
de R2 e R
2ajustado estão bem próximo a 100%, indicando que os resultados estão bem
ajustados com o modelo.
Tabela 9 – Matriz do planejamento fatorial completo (I) e suas respectivas respostas
para a avaliação do crescimento de L. muscorum CRM 1648 e produção de L-ASNase.
Exp.
Fatores (g.L-1
) X (g.L-1
) L-ASNase (U.L-1
)
sacarose ext. de
levedura prolina experimental previsto experimental previsto
1 5 0 5 2,36 1,72 911,79 797,25
2 5 5 5 4,13 4,94 430,83 1075,00
3 5 10 5 5,13 6,15 681,52 408,75
4 5 0 10 3,23 4,62 1463,32 1450,50
5 5 5 10 7,65 7,84 2150,42 1924,25
6 5 10 10 8,88 9,05 1996,29 1454,00
7 5 0 15 3,80 5,89 1377,9 1149,75
8 5 5 15 7,82 9,11 1184,77 1819,50
9 5 10 15 9,68 10,33 1210,77 1545,25
10 5 0 5 5,74 1,72 367,69 797,25
11 10 5 5 9,35 10,55 623,96 974,00
12 10 10 5 11,98 12,53 870,94 536,50
13 10 0 10 8,74 9,46 1996,29 1525,00
14 10 5 10 11,88 13,45 2098,42 2227,50
15 10 10 10 13,14 15,43 1327,76 1986,00
16 10 0 15 9,15 10,74 1485,61 1628,50
17 10 5 15 12,4 14,73 3240,48 2527,00
18 10 10 15 14,44 16,71 2813,37 2481,50
19 15 0 5 7,99 8,69 397,40 137,75
20 15 5 5 12,02 13,45 809,66 873,00
21 15 10 5 15,12 16,19 882,08 664,25
22 15 0 10 8,65 11,59 1374,19 1599,50
23 15 5 10 14,41 16,34 2896,94 2530,75
24 15 10 10 16,94 19,09 1819,87 2518,00
25 15 0 15 8,53 12,87 1541,32 2107,25
26 15 5 15 15,01 17,62 3658,31 3234,50
27 15 10 15 16,61 20,37 3491,18 3417,75
PC1 10 5 10 11,86 13,45 2330,55 2227,50
PC2 10 5 10 11,93 13,45 2135,56 2227,50
PC3 10 5 10 11,75 13,45 2525,53 2227,50
PC4 10 5 10 11,47 13,45 2107,71 2227,50
Dados do 4° dia de cultivo
MSC – massa seca celular
Exp – experimento; PC – ponto central.
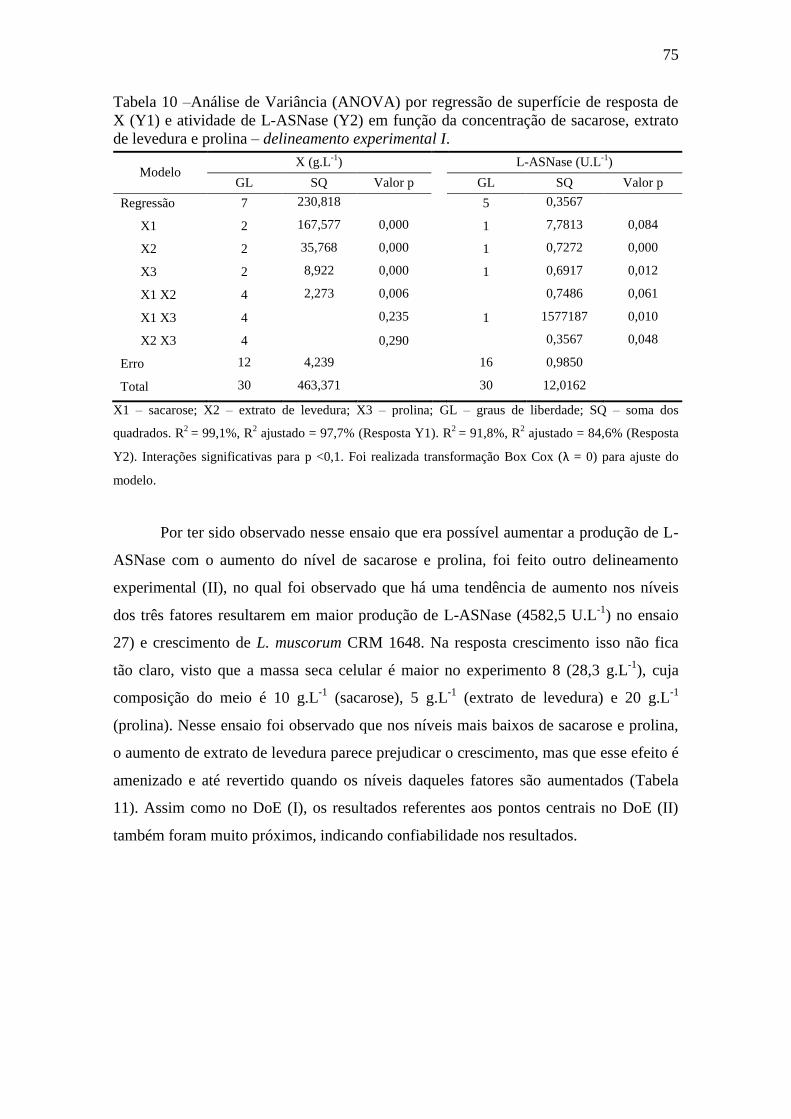
75
Tabela 10 –Análise de Variância (ANOVA) por regressão de superfície de resposta de
X (Y1) e atividade de L-ASNase (Y2) em função da concentração de sacarose, extrato
de levedura e prolina – delineamento experimental I.
Modelo X (g.L
-1) L-ASNase (U.L
-1)
GL SQ Valor p GL SQ Valor p
Regressão 7 230,818 5 0,3567
X1 2 167,577 0,000 1 7,7813 0,084
X2 2 35,768 0,000 1 0,7272 0,000
X3 2 8,922 0,000 1 0,6917 0,012
X1 X2 4 2,273 0,006 0,7486 0,061
X1 X3 4 0,235 1 1577187 0,010
X2 X3 4 0,290 0,3567 0,048
Erro 12 4,239 16 0,9850
Total 30 463,371 30 12,0162
X1 – sacarose; X2 – extrato de levedura; X3 – prolina; GL – graus de liberdade; SQ – soma dos
quadrados. R2
= 99,1%, R2 ajustado = 97,7% (Resposta Y1). R
2 = 91,8%, R
2 ajustado = 84,6% (Resposta
Y2). Interações significativas para p <0,1. Foi realizada transformação Box Cox (λ = 0) para ajuste do
modelo.
Por ter sido observado nesse ensaio que era possível aumentar a produção de L-
ASNase com o aumento do nível de sacarose e prolina, foi feito outro delineamento
experimental (II), no qual foi observado que há uma tendência de aumento nos níveis
dos três fatores resultarem em maior produção de L-ASNase (4582,5 U.L-1
) no ensaio
27) e crescimento de L. muscorum CRM 1648. Na resposta crescimento isso não fica
tão claro, visto que a massa seca celular é maior no experimento 8 (28,3 g.L-1
), cuja
composição do meio é 10 g.L-1
(sacarose), 5 g.L-1
(extrato de levedura) e 20 g.L-1
(prolina). Nesse ensaio foi observado que nos níveis mais baixos de sacarose e prolina,
o aumento de extrato de levedura parece prejudicar o crescimento, mas que esse efeito é
amenizado e até revertido quando os níveis daqueles fatores são aumentados (Tabela
11). Assim como no DoE (I), os resultados referentes aos pontos centrais no DoE (II)
também foram muito próximos, indicando confiabilidade nos resultados.
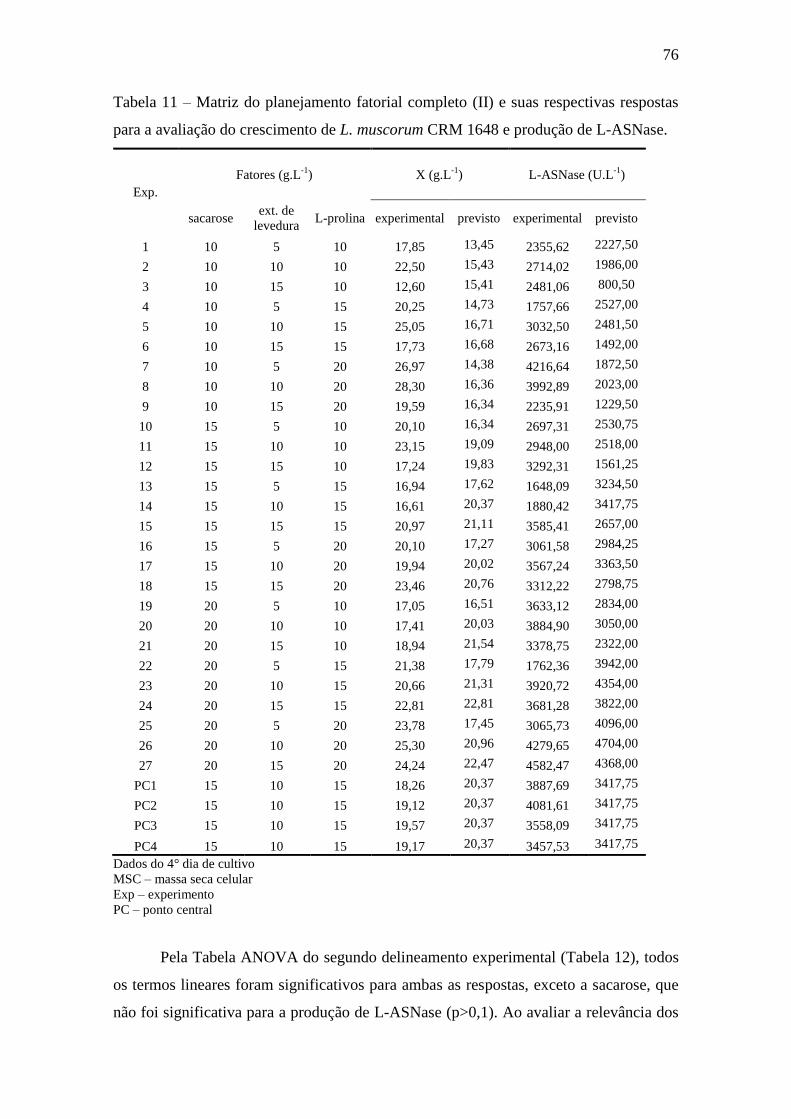
76
Tabela 11 – Matriz do planejamento fatorial completo (II) e suas respectivas respostas
para a avaliação do crescimento de L. muscorum CRM 1648 e produção de L-ASNase.
Exp.
Fatores (g.L-1
) X (g.L-1
) L-ASNase (U.L-1
)
sacarose ext. de
levedura L-prolina experimental previsto experimental previsto
1 10 5 10 17,85 13,45 2355,62 2227,50
2 10 10 10 22,50 15,43 2714,02 1986,00
3 10 15 10 12,60 15,41 2481,06 800,50
4 10 5 15 20,25 14,73 1757,66 2527,00
5 10 10 15 25,05 16,71 3032,50 2481,50
6 10 15 15 17,73 16,68 2673,16 1492,00
7 10 5 20 26,97 14,38 4216,64 1872,50
8 10 10 20 28,30 16,36 3992,89 2023,00
9 10 15 20 19,59 16,34 2235,91 1229,50
10 15 5 10 20,10 16,34 2697,31 2530,75
11 15 10 10 23,15 19,09 2948,00 2518,00
12 15 15 10 17,24 19,83 3292,31 1561,25
13 15 5 15 16,94 17,62 1648,09 3234,50
14 15 10 15 16,61 20,37 1880,42 3417,75
15 15 15 15 20,97 21,11 3585,41 2657,00
16 15 5 20 20,10 17,27 3061,58 2984,25
17 15 10 20 19,94 20,02 3567,24 3363,50
18 15 15 20 23,46 20,76 3312,22 2798,75
19 20 5 10 17,05 16,51 3633,12 2834,00
20 20 10 10 17,41 20,03 3884,90 3050,00
21 20 15 10 18,94 21,54 3378,75 2322,00
22 20 5 15 21,38 17,79 1762,36 3942,00
23 20 10 15 20,66 21,31 3920,72 4354,00
24 20 15 15 22,81 22,81 3681,28 3822,00
25 20 5 20 23,78 17,45 3065,73 4096,00
26 20 10 20 25,30 20,96 4279,65 4704,00
27 20 15 20 24,24 22,47 4582,47 4368,00
PC1 15 10 15 18,26 20,37 3887,69 3417,75
PC2 15 10 15 19,12 20,37 4081,61 3417,75
PC3 15 10 15 19,57 20,37 3558,09 3417,75
PC4 15 10 15 19,17 20,37 3457,53 3417,75
Dados do 4° dia de cultivo
MSC – massa seca celular
Exp – experimento
PC – ponto central
Pela Tabela ANOVA do segundo delineamento experimental (Tabela 12), todos
os termos lineares foram significativos para ambas as respostas, exceto a sacarose, que
não foi significativa para a produção de L-ASNase (p>0,1). Ao avaliar a relevância dos
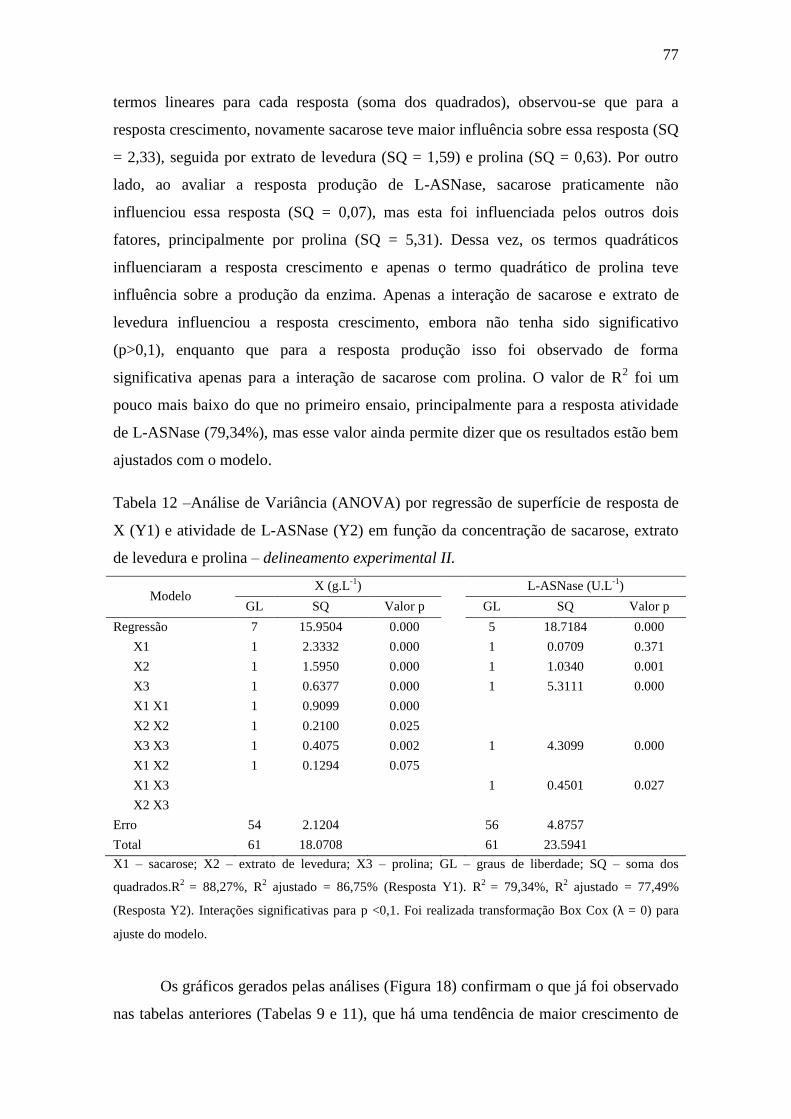
77
termos lineares para cada resposta (soma dos quadrados), observou-se que para a
resposta crescimento, novamente sacarose teve maior influência sobre essa resposta (SQ
= 2,33), seguida por extrato de levedura (SQ = 1,59) e prolina (SQ = 0,63). Por outro
lado, ao avaliar a resposta produção de L-ASNase, sacarose praticamente não
influenciou essa resposta (SQ = 0,07), mas esta foi influenciada pelos outros dois
fatores, principalmente por prolina (SQ = 5,31). Dessa vez, os termos quadráticos
influenciaram a resposta crescimento e apenas o termo quadrático de prolina teve
influência sobre a produção da enzima. Apenas a interação de sacarose e extrato de
levedura influenciou a resposta crescimento, embora não tenha sido significativo
(p>0,1), enquanto que para a resposta produção isso foi observado de forma
significativa apenas para a interação de sacarose com prolina. O valor de R2 foi um
pouco mais baixo do que no primeiro ensaio, principalmente para a resposta atividade
de L-ASNase (79,34%), mas esse valor ainda permite dizer que os resultados estão bem
ajustados com o modelo.
Tabela 12 –Análise de Variância (ANOVA) por regressão de superfície de resposta de
X (Y1) e atividade de L-ASNase (Y2) em função da concentração de sacarose, extrato
de levedura e prolina – delineamento experimental II.
Modelo X (g.L
-1) L-ASNase (U.L
-1)
GL SQ Valor p GL SQ Valor p
Regressão 7 15.9504 0.000 5 18.7184 0.000
X1 1 2.3332 0.000 1 0.0709 0.371
X2 1 1.5950 0.000 1 1.0340 0.001
X3 1 0.6377 0.000 1 5.3111 0.000
X1 X1 1 0.9099 0.000
X2 X2 1 0.2100 0.025
X3 X3 1 0.4075 0.002 1 4.3099 0.000
X1 X2 1 0.1294 0.075
X1 X3 1 0.4501 0.027
X2 X3
Erro 54 2.1204 56 4.8757
Total 61 18.0708 61 23.5941
X1 – sacarose; X2 – extrato de levedura; X3 – prolina; GL – graus de liberdade; SQ – soma dos
quadrados.R2
= 88,27%, R2 ajustado = 86,75% (Resposta Y1). R
2 = 79,34%, R
2 ajustado = 77,49%
(Resposta Y2). Interações significativas para p <0,1. Foi realizada transformação Box Cox (λ = 0) para
ajuste do modelo.
Os gráficos gerados pelas análises (Figura 18) confirmam o que já foi observado
nas tabelas anteriores (Tabelas 9 e 11), que há uma tendência de maior crescimento de

78
L. muscorum CRM1648 nos níveis mais elevados dos três fatores. Na produção de L-
ASNase por essa levedura, observa-se um comportamento sinérgico entre a sacarose e o
extrato de levedura, indicando uma maior produção quanto maior é o nível de cada um
desses nutrientes, sugerindo inclusive a possibilidade de melhorar ainda mais a
produção ao extrapolar os níveis desses dois fatores. Por outro lado, ao avaliar o
sinergismo entre prolina e sacarose ou prolina e extrato de levedura, observa-se que os
fatores sacarose e extrato de levedura praticamente não influenciam a resposta produção
e que esta depende quase que exclusivamente do aumento de prolina no meio, indicando
a grande relevância desse fator para a produção de L-ASNase por L. muscorum CRM
1648.
Figura 18. Gráfico da superfície de resposta em 3D de MSC (Y1) e atividade de L-
ASNase (Y2) em função da concentração de sacarose (X1), extrato de levedura (X2), e
prolina (X3). Valores fixados: X1 = 17, X2 = 15 e X3 = 16.
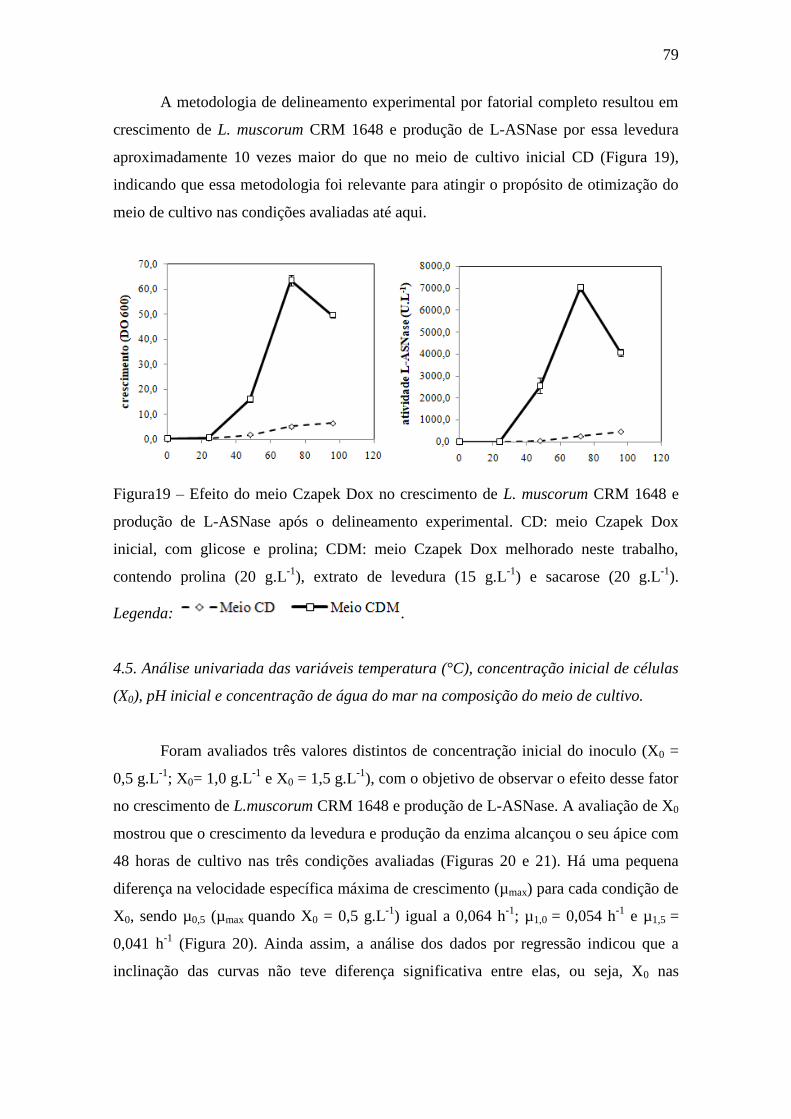
79
A metodologia de delineamento experimental por fatorial completo resultou em
crescimento de L. muscorum CRM 1648 e produção de L-ASNase por essa levedura
aproximadamente 10 vezes maior do que no meio de cultivo inicial CD (Figura 19),
indicando que essa metodologia foi relevante para atingir o propósito de otimização do
meio de cultivo nas condições avaliadas até aqui.
Figura19 – Efeito do meio Czapek Dox no crescimento de L. muscorum CRM 1648 e
produção de L-ASNase após o delineamento experimental. CD: meio Czapek Dox
inicial, com glicose e prolina; CDM: meio Czapek Dox melhorado neste trabalho,
contendo prolina (20 g.L-1
), extrato de levedura (15 g.L-1
) e sacarose (20 g.L-1
).
Legenda: .
4.5. Análise univariada das variáveis temperatura (°C), concentração inicial de células
(X0), pH inicial e concentração de água do mar na composição do meio de cultivo.
Foram avaliados três valores distintos de concentração inicial do inoculo (X0 =
0,5 g.L-1
; X0= 1,0 g.L-1
e X0 = 1,5 g.L-1
), com o objetivo de observar o efeito desse fator
no crescimento de L.muscorum CRM 1648 e produção de L-ASNase. A avaliação de X0
mostrou que o crescimento da levedura e produção da enzima alcançou o seu ápice com
48 horas de cultivo nas três condições avaliadas (Figuras 20 e 21). Há uma pequena
diferença na velocidade específica máxima de crescimento (µmax) para cada condição de
X0, sendo µ0,5 (µmax quando X0 = 0,5 g.L-1
) igual a 0,064 h-1
; µ1,0 = 0,054 h-1
e µ1,5 =
0,041 h-1
(Figura 20). Ainda assim, a análise dos dados por regressão indicou que a
inclinação das curvas não teve diferença significativa entre elas, ou seja, X0 nas
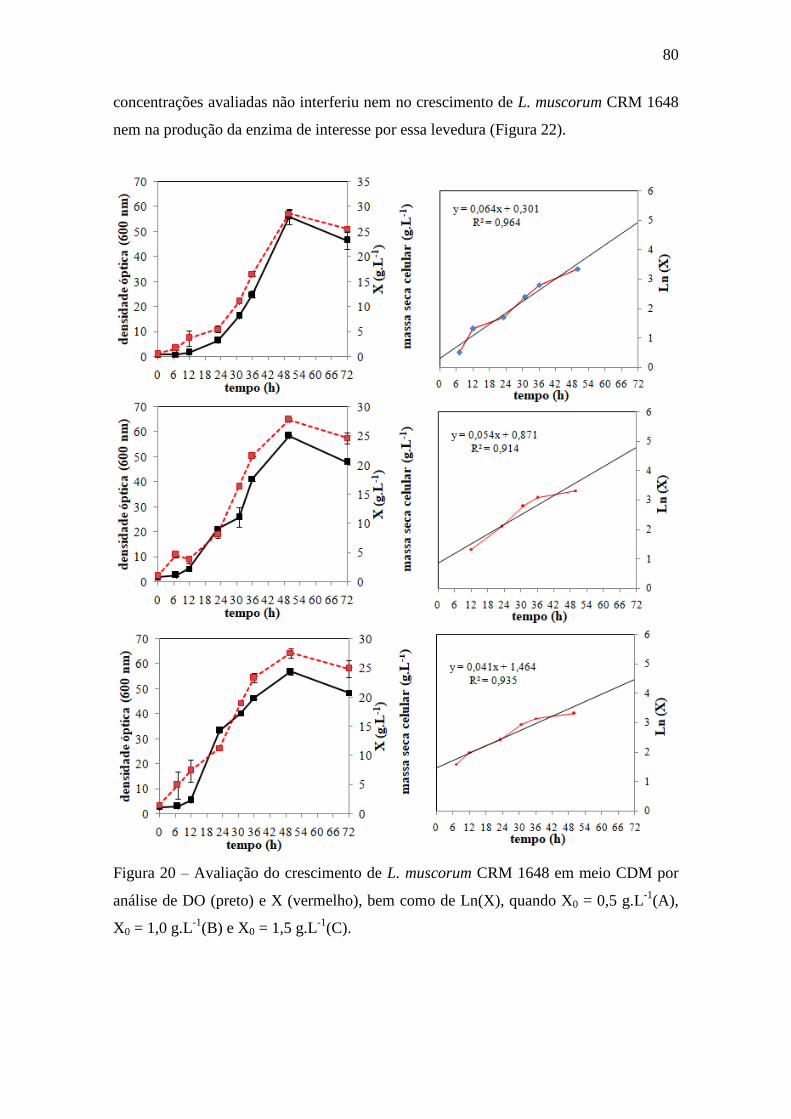
80
concentrações avaliadas não interferiu nem no crescimento de L. muscorum CRM 1648
nem na produção da enzima de interesse por essa levedura (Figura 22).
Figura 20 – Avaliação do crescimento de L. muscorum CRM 1648 em meio CDM por
análise de DO (preto) e X (vermelho), bem como de Ln(X), quando X0 = 0,5 g.L-1
(A),
X0 = 1,0 g.L-1
(B) e X0 = 1,5 g.L-1
(C).
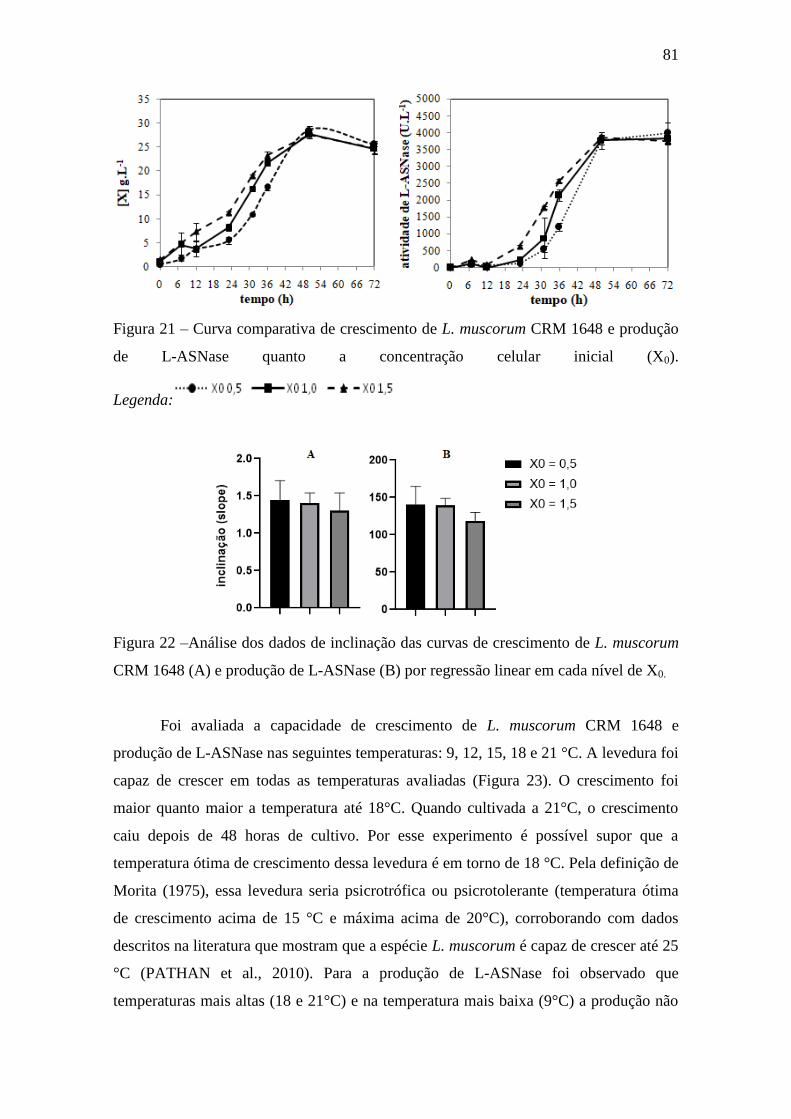
81
Figura 21 – Curva comparativa de crescimento de L. muscorum CRM 1648 e produção
de L-ASNase quanto a concentração celular inicial (X0).
Legenda:
Figura 22 –Análise dos dados de inclinação das curvas de crescimento de L. muscorum
CRM 1648 (A) e produção de L-ASNase (B) por regressão linear em cada nível de X0.
Foi avaliada a capacidade de crescimento de L. muscorum CRM 1648 e
produção de L-ASNase nas seguintes temperaturas: 9, 12, 15, 18 e 21 °C. A levedura foi
capaz de crescer em todas as temperaturas avaliadas (Figura 23). O crescimento foi
maior quanto maior a temperatura até 18°C. Quando cultivada a 21°C, o crescimento
caiu depois de 48 horas de cultivo. Por esse experimento é possível supor que a
temperatura ótima de crescimento dessa levedura é em torno de 18 °C. Pela definição de
Morita (1975), essa levedura seria psicrotrófica ou psicrotolerante (temperatura ótima
de crescimento acima de 15 °C e máxima acima de 20°C), corroborando com dados
descritos na literatura que mostram que a espécie L. muscorum é capaz de crescer até 25
°C (PATHAN et al., 2010). Para a produção de L-ASNase foi observado que
temperaturas mais altas (18 e 21°C) e na temperatura mais baixa (9°C) a produção não
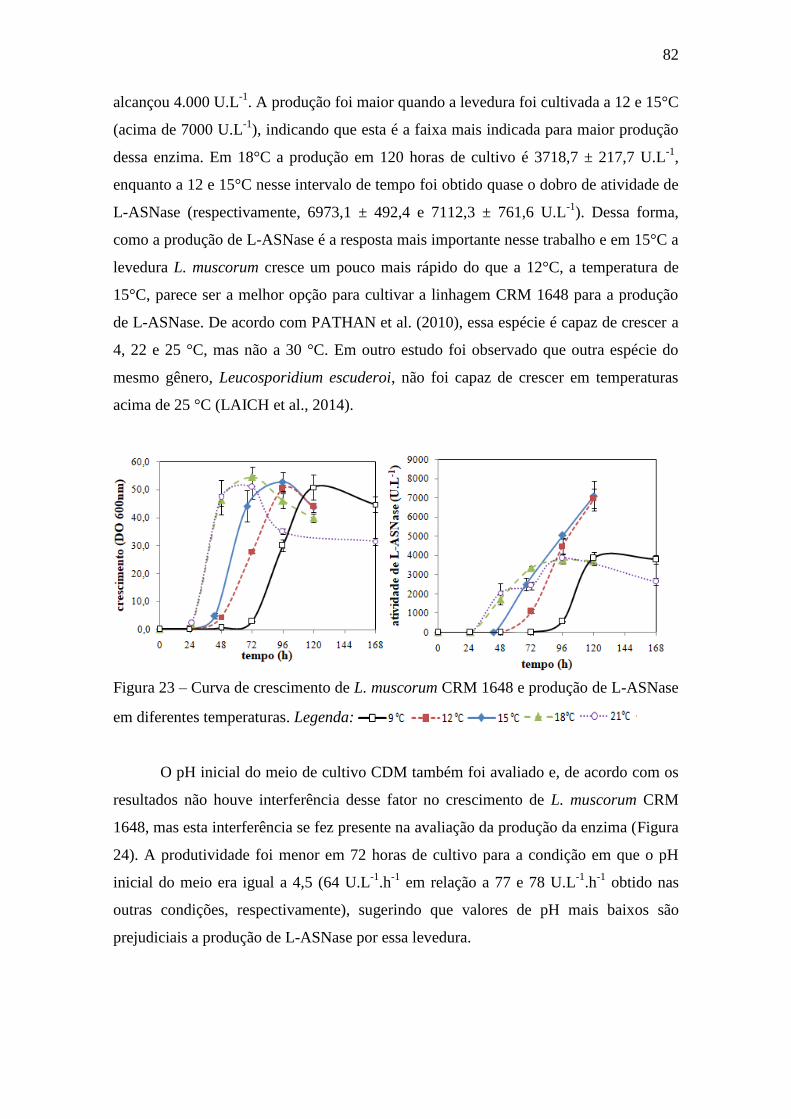
82
alcançou 4.000 U.L-1
. A produção foi maior quando a levedura foi cultivada a 12 e 15°C
(acima de 7000 U.L-1
), indicando que esta é a faixa mais indicada para maior produção
dessa enzima. Em 18°C a produção em 120 horas de cultivo é 3718,7 ± 217,7 U.L-1
,
enquanto a 12 e 15°C nesse intervalo de tempo foi obtido quase o dobro de atividade de
L-ASNase (respectivamente, 6973,1 ± 492,4 e 7112,3 ± 761,6 U.L-1
). Dessa forma,
como a produção de L-ASNase é a resposta mais importante nesse trabalho e em 15°C a
levedura L. muscorum cresce um pouco mais rápido do que a 12°C, a temperatura de
15°C, parece ser a melhor opção para cultivar a linhagem CRM 1648 para a produção
de L-ASNase. De acordo com PATHAN et al. (2010), essa espécie é capaz de crescer a
4, 22 e 25 °C, mas não a 30 °C. Em outro estudo foi observado que outra espécie do
mesmo gênero, Leucosporidium escuderoi, não foi capaz de crescer em temperaturas
acima de 25 °C (LAICH et al., 2014).
Figura 23 – Curva de crescimento de L. muscorum CRM 1648 e produção de L-ASNase
em diferentes temperaturas. Legenda:
O pH inicial do meio de cultivo CDM também foi avaliado e, de acordo com os
resultados não houve interferência desse fator no crescimento de L. muscorum CRM
1648, mas esta interferência se fez presente na avaliação da produção da enzima (Figura
24). A produtividade foi menor em 72 horas de cultivo para a condição em que o pH
inicial do meio era igual a 4,5 (64 U.L-1
.h-1
em relação a 77 e 78 U.L-1
.h-1
obtido nas
outras condições, respectivamente), sugerindo que valores de pH mais baixos são
prejudiciais a produção de L-ASNase por essa levedura.
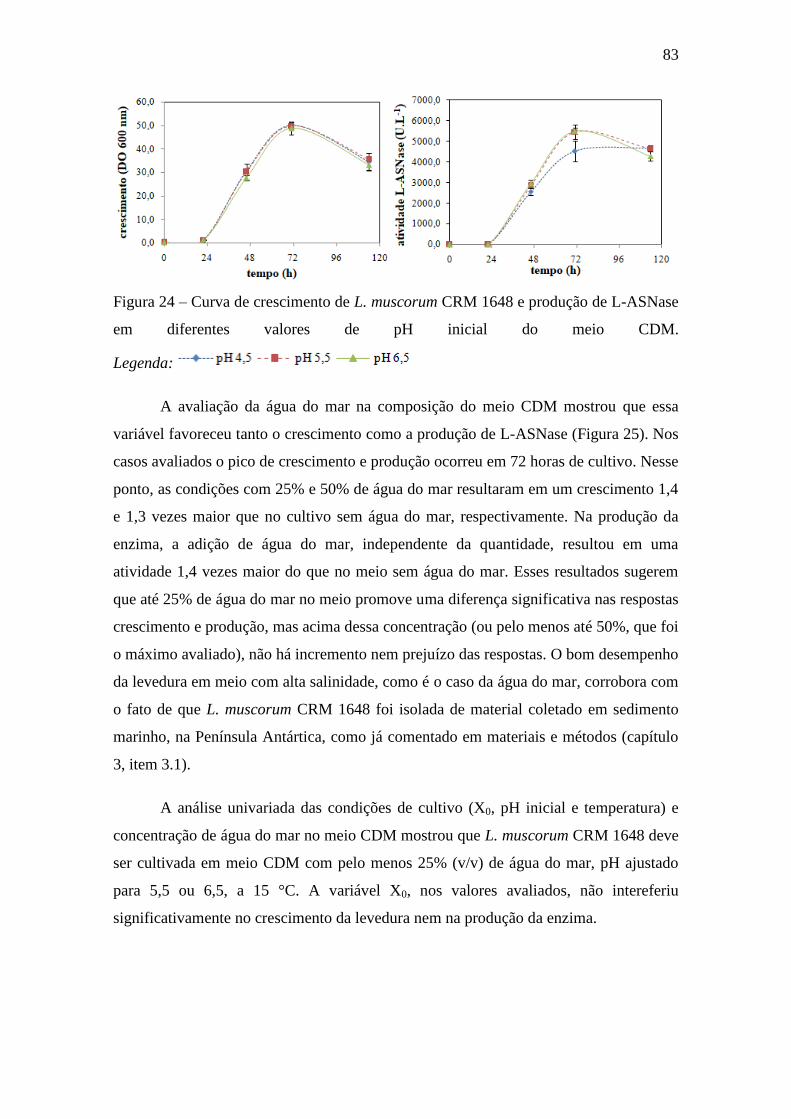
83
Figura 24 – Curva de crescimento de L. muscorum CRM 1648 e produção de L-ASNase
em diferentes valores de pH inicial do meio CDM.
Legenda:
A avaliação da água do mar na composição do meio CDM mostrou que essa
variável favoreceu tanto o crescimento como a produção de L-ASNase (Figura 25). Nos
casos avaliados o pico de crescimento e produção ocorreu em 72 horas de cultivo. Nesse
ponto, as condições com 25% e 50% de água do mar resultaram em um crescimento 1,4
e 1,3 vezes maior que no cultivo sem água do mar, respectivamente. Na produção da
enzima, a adição de água do mar, independente da quantidade, resultou em uma
atividade 1,4 vezes maior do que no meio sem água do mar. Esses resultados sugerem
que até 25% de água do mar no meio promove uma diferença significativa nas respostas
crescimento e produção, mas acima dessa concentração (ou pelo menos até 50%, que foi
o máximo avaliado), não há incremento nem prejuízo das respostas. O bom desempenho
da levedura em meio com alta salinidade, como é o caso da água do mar, corrobora com
o fato de que L. muscorum CRM 1648 foi isolada de material coletado em sedimento
marinho, na Península Antártica, como já comentado em materiais e métodos (capítulo
3, item 3.1).
A análise univariada das condições de cultivo (X0, pH inicial e temperatura) e
concentração de água do mar no meio CDM mostrou que L. muscorum CRM 1648 deve
ser cultivada em meio CDM com pelo menos 25% (v/v) de água do mar, pH ajustado
para 5,5 ou 6,5, a 15 °C. A variável X0, nos valores avaliados, não intereferiu
significativamente no crescimento da levedura nem na produção da enzima.
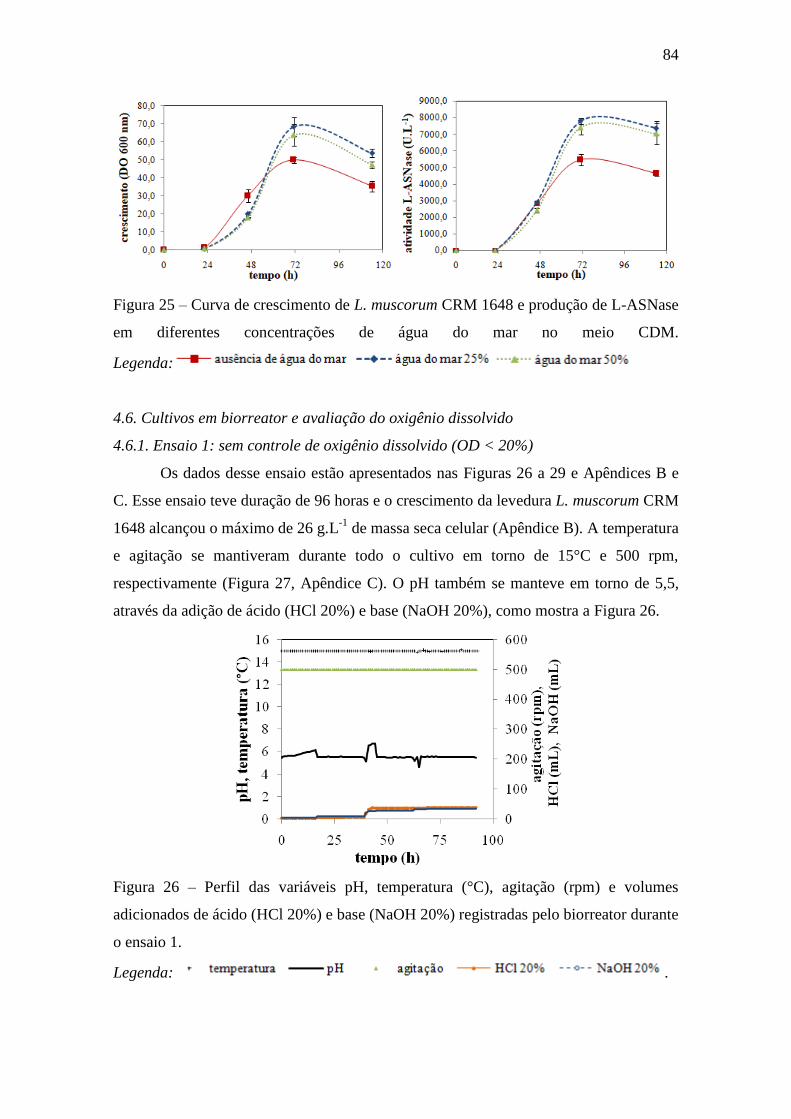
84
Figura 25 – Curva de crescimento de L. muscorum CRM 1648 e produção de L-ASNase
em diferentes concentrações de água do mar no meio CDM.
Legenda:
4.6. Cultivos em biorreator e avaliação do oxigênio dissolvido
4.6.1. Ensaio 1: sem controle de oxigênio dissolvido (OD < 20%)
Os dados desse ensaio estão apresentados nas Figuras 26 a 29 e Apêndices B e
C. Esse ensaio teve duração de 96 horas e o crescimento da levedura L. muscorum CRM
1648 alcançou o máximo de 26 g.L-1
de massa seca celular (Apêndice B). A temperatura
e agitação se mantiveram durante todo o cultivo em torno de 15°C e 500 rpm,
respectivamente (Figura 27, Apêndice C). O pH também se manteve em torno de 5,5,
através da adição de ácido (HCl 20%) e base (NaOH 20%), como mostra a Figura 26.
Figura 26 – Perfil das variáveis pH, temperatura (°C), agitação (rpm) e volumes
adicionados de ácido (HCl 20%) e base (NaOH 20%) registradas pelo biorreator durante
o ensaio 1.
Legenda: .
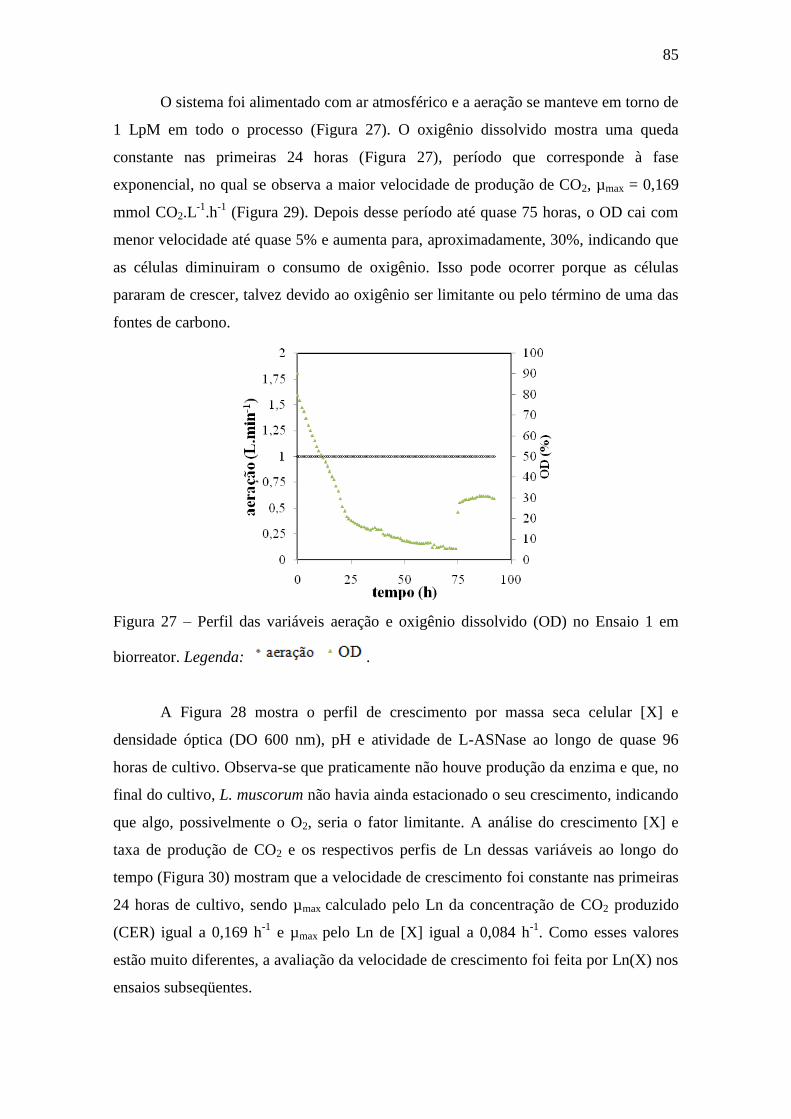
85
O sistema foi alimentado com ar atmosférico e a aeração se manteve em torno de
1 LpM em todo o processo (Figura 27). O oxigênio dissolvido mostra uma queda
constante nas primeiras 24 horas (Figura 27), período que corresponde à fase
exponencial, no qual se observa a maior velocidade de produção de CO2, µmax = 0,169
mmol CO2.L-1
.h-1
(Figura 29). Depois desse período até quase 75 horas, o OD cai com
menor velocidade até quase 5% e aumenta para, aproximadamente, 30%, indicando que
as células diminuiram o consumo de oxigênio. Isso pode ocorrer porque as células
pararam de crescer, talvez devido ao oxigênio ser limitante ou pelo término de uma das
fontes de carbono.
Figura 27 – Perfil das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 1 em
biorreator. Legenda: .
A Figura 28 mostra o perfil de crescimento por massa seca celular [X] e
densidade óptica (DO 600 nm), pH e atividade de L-ASNase ao longo de quase 96
horas de cultivo. Observa-se que praticamente não houve produção da enzima e que, no
final do cultivo, L. muscorum não havia ainda estacionado o seu crescimento, indicando
que algo, possivelmente o O2, seria o fator limitante. A análise do crescimento [X] e
taxa de produção de CO2 e os respectivos perfis de Ln dessas variáveis ao longo do
tempo (Figura 30) mostram que a velocidade de crescimento foi constante nas primeiras
24 horas de cultivo, sendo µmax calculado pelo Ln da concentração de CO2 produzido
(CER) igual a 0,169 h-1
e µmax pelo Ln de [X] igual a 0,084 h-1
. Como esses valores
estão muito diferentes, a avaliação da velocidade de crescimento foi feita por Ln(X) nos
ensaios subseqüentes.

86
Figura 28 – Perfil das variáveis crescimento (DO e MSC), pH e produção de L-
ASNaseno ensaio 1 em biorreator.
Legenda: .
Figura 29 – Perfil das variáveis produção de CO2 (CER) e Ln de CER na fase
exponencial (A) e crescimento celular (massa seca celular, [X]) mais ln(X) e ln(X) na
fase exponencial (ln(X)exp) no Ensaio 1. Legenda:
(A)
(B)
Como já havia sido observado nos cultivos em frascos agitados que o pH tende a
aumentar bastante quando há a produção de L-ASNase, variando de um pH inicial igual
a 5,5 até, aproximadamente, 8,5 no final do cultivo (dados não mostrados), havia nessa
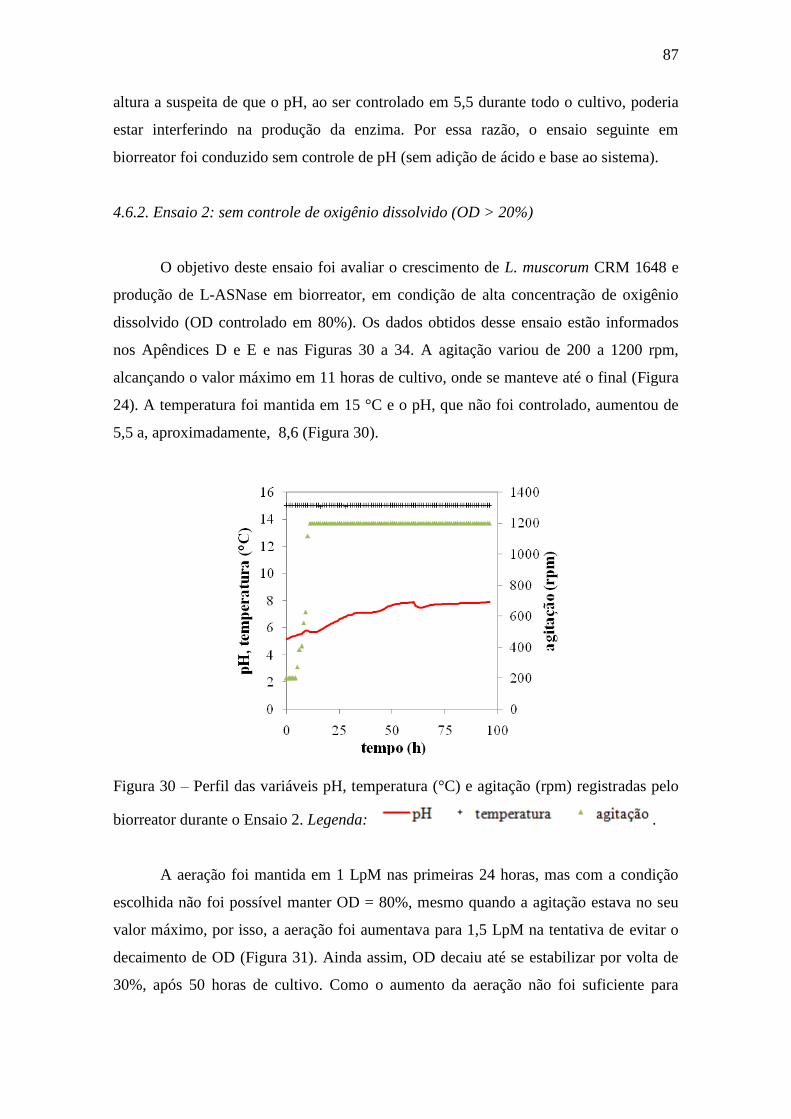
87
altura a suspeita de que o pH, ao ser controlado em 5,5 durante todo o cultivo, poderia
estar interferindo na produção da enzima. Por essa razão, o ensaio seguinte em
biorreator foi conduzido sem controle de pH (sem adição de ácido e base ao sistema).
4.6.2. Ensaio 2: sem controle de oxigênio dissolvido (OD > 20%)
O objetivo deste ensaio foi avaliar o crescimento de L. muscorum CRM 1648 e
produção de L-ASNase em biorreator, em condição de alta concentração de oxigênio
dissolvido (OD controlado em 80%). Os dados obtidos desse ensaio estão informados
nos Apêndices D e E e nas Figuras 30 a 34. A agitação variou de 200 a 1200 rpm,
alcançando o valor máximo em 11 horas de cultivo, onde se manteve até o final (Figura
24). A temperatura foi mantida em 15 °C e o pH, que não foi controlado, aumentou de
5,5 a, aproximadamente, 8,6 (Figura 30).
Figura 30 – Perfil das variáveis pH, temperatura (°C) e agitação (rpm) registradas pelo
biorreator durante o Ensaio 2. Legenda: .
A aeração foi mantida em 1 LpM nas primeiras 24 horas, mas com a condição
escolhida não foi possível manter OD = 80%, mesmo quando a agitação estava no seu
valor máximo, por isso, a aeração foi aumentava para 1,5 LpM na tentativa de evitar o
decaimento de OD (Figura 31). Ainda assim, OD decaiu até se estabilizar por volta de
30%, após 50 horas de cultivo. Como o aumento da aeração não foi suficiente para

88
manter OD em 80% e o aumento da agitação poderia provocar o cisalhamento da célula,
a adição de oxigênio puro foi a alternativa adotada no ensaio seguinte (ensaio 3).
Figura 31 – Perfil das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 2 em
biorreator. Legenda: .
A Figura 32 mostra o perfil de crescimento [X], produção de L-ASNase e
consumo dos substratos sacarose e prolina. Dessa vez, foi detectada produção de L-
ASNase. A produtividade em células e em produto é máxima por volta de 47 horas de
cultivo e, respectivamente, igual a 0,58 g.L-1
.h-1
e 75,4 U.L-1
.h-1
(Apêndice D). Nessa
condição e intervalo de tempo L. muscorum CRM 1648 produziu, aproximadamente, 27
g.L-1
de massa seca celular e 3.500 U.L-1
de L-ASNase (Apêndice D). A velocidade
específica de crescimento é constante e máxima nesse intervalo de tempo (Figura 33),
sendo µmax de [X] igual a 0,085 h-1
. A produção parece estar associada ao crescimento,
pois se observa que o consumo dos substratos e produção de células e produtos ocorrem
simultaneamente e, quando acabam os dois substratos, por volta de 47 horas de cultivo,
não se observa mais crescimento nem produção da enzima (Figura 32). Os substratos
são consumidos ao mesmo tempo, embora o consumo de sacarose seja um pouco mais
rápido que o consumo de prolina.

89
Figura 32 – Perfil de crescimento [X], consumo de sacarose e de prolina [S] e produção
de L-ASNase no Ensaio 2 em biorreator. Legenda:
Figura 33 – Perfil das variáveis crescimento celular (massa seca celular, [X]), ln(X) e
ln(X) na fase exponencial (ln(X)exp) no Ensaio 2.
Legenda: .
Como nesse experimento não houve controle de pH, foi observado que houve
produção e que existe uma correlação entre o aumento dessa variável e aprodução de L-
ASNase (Figura 34). Não é possível dizer que o aumento do pH ao longo do cultivo está
associado a produção da enzima L-ASNase, mas sugere que o seu monitoramento

90
poderia ser utilizado na indústria como método simples, rápido e de baixo custo para
monitorar a produção da enzima por L. muscorum CRM 1648.
Figura 34 – Medida de pH do meio CDM e atividade de L-ASNase ao longo do cultivo
de L. muscorum CRM 1648 em biorreator no Ensaio 2. Legenda:
4.6.3. Ensaio 3: oxigênio dissolvido controlado em 80%
Os resultados desse ensaio estão apresentados nas Figuras 35 a 38 e Apêndices F
e G. Como as condições estabelecidas não foram suficientes para manter OD em torno
de 80 %, foi feito outro ensaio no qual a agitação foi mantida em 600 rpm e foi
adicionado oxigênio puro. A temperatura foi mantida a 15°C e o pH aumentou ao longo
do cultivo de 5,5 até aproximadamente 8,5 (Figura 35, Apêndice F).
A aeração foi mantida em 1 LpM e, embora OD tenha oscilado bastante ao longo
do tempo, mostrou um perfil bem diferente do apresentado nos dois ensaios anteriores,
não houve uma queda constante, mas oscilações em torno do valor médio de 85 %
(Figura 36). A manutenção de OD foi feita com a adição de oxigênio puro na corrente
de vazão fixa de ar e esse procedimento requer atenção 24 horas e modificação
constante da porcentagem de fechamento e abertura da válvula, que dar acesso a
mangueira de O2 puro (valvO2). A válvula só foi aberta a partir de 15 horas de cultivo,
aproximadamente, quando L. muscorum CRM 1648 começou a crescer
exponencialmente (Figuras 37-38). Até 50 horas de cultivo foi observada muita
oscilação na abertura e fechamento da válvula de O2 e conseqüente variação de OD no
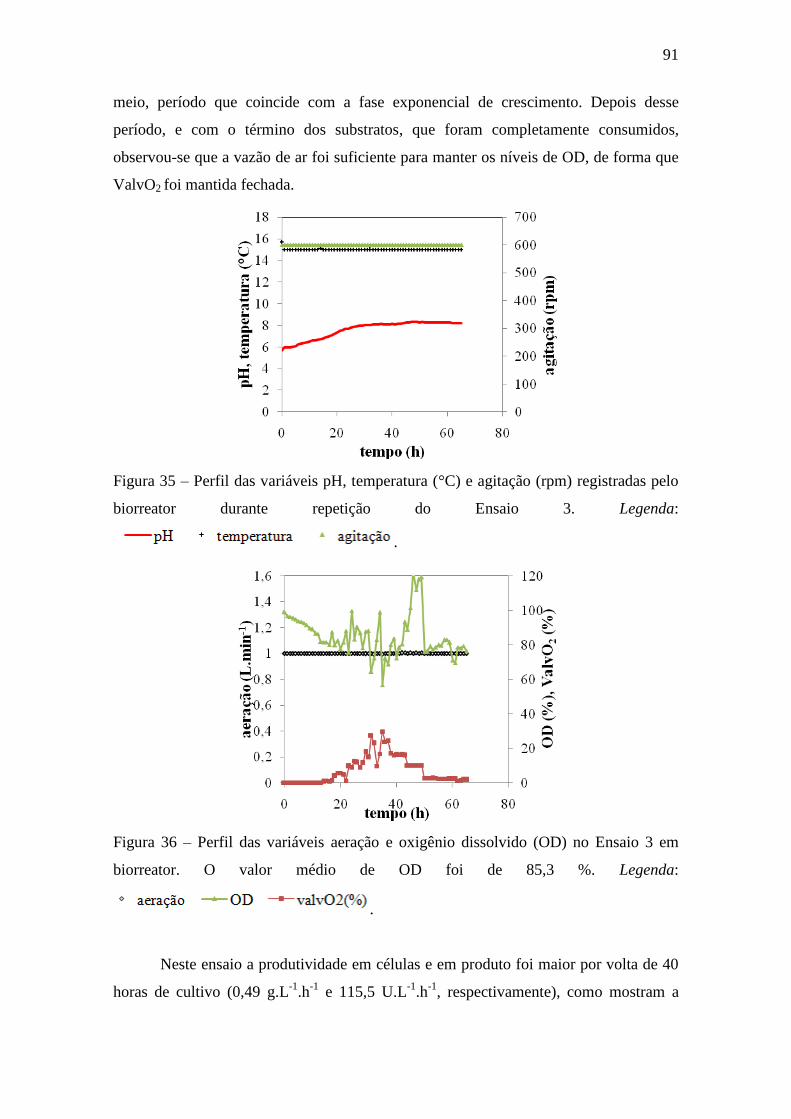
91
meio, período que coincide com a fase exponencial de crescimento. Depois desse
período, e com o término dos substratos, que foram completamente consumidos,
observou-se que a vazão de ar foi suficiente para manter os níveis de OD, de forma que
ValvO2 foi mantida fechada.
Figura 35 – Perfil das variáveis pH, temperatura (°C) e agitação (rpm) registradas pelo
biorreator durante repetição do Ensaio 3. Legenda:
.
Figura 36 – Perfil das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 3 em
biorreator. O valor médio de OD foi de 85,3 %. Legenda:
.
Neste ensaio a produtividade em células e em produto foi maior por volta de 40
horas de cultivo (0,49 g.L-1
.h-1
e 115,5 U.L-1
.h-1
, respectivamente), como mostram a

92
Figura 37 e o APÊNDICE F. Nesse intervalo, foram produzidas, aproximadamente, 20
g.L-1
de células e 4700 U.L-1
de L-ASNase. Também foi observado que o consumo,
crescimento e produção da enzima ocorreram concomitantemente. Os substratos foram
consumidos completamente, mas a sacarose foi consumida primeiro, após 35 horas de
cultivo, enquanto a concentração de prolina só zerou em 50 horas (Figura 37). A
velocidade específica máxima de crescimento foi maior neste ensaio, igual a 0,11 h-1
(Figura 38).
Figura 37 – Perfil de crescimento [X], consumo de sacarose e prolina [S] e produção de
L-ASNase no Ensaio 3 em biorreator. Legenda:
.
Figura 38 – Perfil das variáveis crescimento celular (massa seca celular, [X]), ln(X) e
ln(X) na fase exponencial (ln(X)exp) no Ensaio 3. Legenda:
.

93
4.6.4. Ensaio 4: oxigênio dissolvido controlado em 20%
O objetivo deste ensaio foi avaliar o crescimento de L. muscorum CRM 1648 e
produção de L-ASNase em biorreator, em condição de baixa concentração de oxigênio
dissolvido (OD controlado em 20%). Os dados obtidos desse ensaio estão informados
nos Apêndices H e I e nas Figuras 39 a 42. A agitação foi mantida em 600 rpm e a
temperatura foi controlada em 15 °C durante todo o cultivo. O pH aumentou ao longo
do cultivo como nos ensaios anteriores, chegando a 8,6 no final (Figura 39).
Figura 39 – Perfil das variáveis pH, temperatura (°C) e agitação (rpm) registradas pelo
biorreator no ensaio 4. Legenda: .
Este ensaio foi montado de forma que nitrogênio puro era injetado na entrada de
ar do biorreator e era alimentado com pulsos de ar puro na entrada de O2, conforme
descrito em materiais e métodos (item 3.10.2). Isso foi feito para que OD do meio caísse
rapidamente para 20%. No entanto, após 6 horas de cultivo, a mangueira de nitrogênio
rompeu e esvaziou a fonte de nitrogênio. Por isso, a entrada de nitrogênio foi fechada e
OD caiu continuamente durante as primeiras 15 horas de cultivo até, aproximadamente,
20%, quando pequenos pulsos de ar começaram a ser injetados no meio provocando
oscilações, inicialmente sutis, de OD (Figura 40). Essas oscilações se tornaram maiores
quando L. muscorum CRM 1648 começou a crescer exponencialmente. Quando os
pulsos de ar não eram mais capazes de manter OD em torno de 20% e este começou a
cair abaixo desse valor, a entrada de ar foi aberta (aeração do sistema igual a 1 LpM a
partir de 43 horas de cultivo) e o sistema foi alimentado com ar puro (nessa etapa, o ar
puro foi ligado na entrada de ar e O2 puro na entrada de O2). No intervalo de tempo de

94
43-53 horas de cultivo o sistema foi alimentado com pulsos de O2 puro para tentar
manter OD = 20% e sustentar o crescimento das células, que ainda estavam em fase
exponencial (Figuras 41 e 42). O sistema deixou de ser alimentado com O2 puro depois
de 53 horas, pois as células pararam de crescer a partir desse ponto, já que nesse
momento as células passaram a respirar menos, de forma que havia menor necessidade
de OD no meio. Embora OD tenha oscilado, o valor médio dessa variável ao longo do
cultivo foi de 25,1% (Figura40).
Figura 40 – Perfil das variáveis aeração e oxigênio dissolvido (OD) no Ensaio 4 em
biorreator. O valor médio de OD foi de 25,1 %.
Legenda: .
No final do crescimento exponencial, com aproximadamente 52 horas de
cultivo, foi obtida a maior produtividade em células e em produto (0,51 g.L-1
.h-1
e 97,5
U.L-1
.h-1
, respectivamente) deste ensaio, como mostra o Apêndice H. Nesse intervalo,
foram produzidas, aproximadamente, 27 g.L-1
de células e 5116 U.L-1
de L-ASNase.
Novamente foi observado que o consumo, crescimento e produção da enzima ocorreram
concomitantemente, mas dessa vez, a prolina não foi consumida em sua totalidade, ao
contrário da sacarose, que em 43 horas já não foi mais detectada no meio de cultivo
(Figura 41). Este ensaio resultou em menor valor de velocidade específica máxima de
crescimento, igual a 0,07 h-1
(Figura 42).

95
Figura 41 – Perfil de crescimento [X], consumo de sacarose e de prolina [S] e produção
de L-ASNase no Ensaio 4 em biorreator. Legenda:
.
Figura 42 – Perfil das variáveis crescimento celular (massa seca celular, [X]), ln(X) e
ln(X) na fase exponencial (ln(X)exp) no Ensaio 4. Legenda:
.
4.6.5. Avaliação comparativa dos ensaios em biorreator
Ao avaliar comparativamente a relação de OD no meio de cultivo dos quatro
ensaios em biorreator (Figura 43) com o crescimento de L. muscorum CRM 1648
(Figura 44 A), foi observado crescimento mais rápido nos ensaios 2 e 3, nos quais OD

96
se manteve bem acima de 20%, quando comparado aos ensaios 1 e 4, nos quais OD se
manteve em torno de 20% e até abaixo desse valor, chegando a 5% em alguns
momentos. Além disso, a curva de crescimento no Ensaio 1 começa a se distanciar das
curvas dos outros três ensaios após 40 horas de cultivo, quando OD nesse ensaio já está
muito próximo a 10% (Apêndice C). Outra observação é que, no Ensaio 1 as células de
L. muscorum CRM 1648 ficaram expostas por, aproximadamente, 12 horas a um valor
de OD em torno de 5% (de 64 a 75 horas de cultivo, como mostra o Apêndice C),
enquanto no Ensaio 4 OD se manteve em 20%, havendo oscilações nesse valor que fez
OD cair em torno de 5% por 2 horas (entre 36 e 37 horas de cultivo), e outra oscilação
logo em seguida (próximo a 50 horas), quando OD subiu acima de 100% (Figura 43,
Apêndice I). Essa diferença, aparentemente, fez com que a levedura desacelerasse seu
crescimento no Ensaio 1, mas continuasse a crescer progressivamente no ensaio 4. O
valor final de X nos 4 ensaios foi semelhante e igual a, aproximadamente, 26 g. L-1
, no
entanto observa-se uma grande diferença no comportamento das curvas nos Ensaios 2, 3
e 4 em relação ao Ensaio 1 (Figura 44 A). Dessa forma, OD demonstrou ser uma
variável crítica por interferir na velocidade de crescimento da levedura, embora a
produção de células no final do cultivo tenha sido igual nas quatro condições.
A análise da relação de OD com a síntese de produto mostrou que essa variável
também foi crítica para a produção da enzima L-ASNase pela levedura L. muscorum
CRM 1648. No Ensaio 3 a produção foi maior e mais rápida possivelmente porque
nesse ensaio OD se manteve sempre elevado, controlado em 80%. Nos Ensaios 2 e 4,
nos quais OD se manteve acima de 20%, a produção foi bastante similar até
aproximadamente 50 horas de cultivo, quando no Ensaio 4 ocorre o pico de OD, que
extrapola 100%. Nesse ponto, observa-se que a enzima continua sendo produzida no
Ensaio 4, onde as células por alguns instantes (50 a 53 horas de cultivo, como mostra o
Apêndice I) são expostas a concentrações elevadas de OD. Em contrapartida, no Ensaio
2, a partir desse ponto, OD cai progressivamente de, aproximadamente, 35% a 30%
(Apêndice E). Esse comportamento sugere que não só valores elevados de OD são
favoráveis para a produção de L-ASNase, como também valores abaixo de 35% já
começam a prejudicar a sua produção.
A curva de produção de L-ASNase no Ensaio 1 não foi adicionada a Figura 44 B
porque esse resultado teve influência de outro fator, o pH, que interferiu com a análise
de OD nesse ensaio. Ainda assim, como existe correlação entre o crescimento de L.
muscorum CRM 1648 e produção de L-ASNase por essa levedura, é possível supor que,

97
a produção da enzima seria mais baixa no Ensaio 1 em relação aos outros três ensaios,
já que OD se mostrou limitante tanto para o crescimento de L. muscorum CRM 1648
como para a produção de L-ASNase por esse microrganismo.
Figura 43 – Comparação do perfil de OD em cada ensaio. Legenda: Ensaio 1: triângulo
azul vazio; Ensaio 2: círculo vermelho vazio; Ensaio 3: quadrado verde cheio; Ensaio 4:
losango roxo cheio.
Figura 44 – Comparação do perfil de crescimento (A) e de produção de L-ASNase (B)
nas condições de OD avaliadas em biorreator. Legenda: Ensaio 1: triângulo azul; Ensaio
2: círculo vermelho; Ensaio 3: quadrado verde; Ensaio 4: losango roxo.
A produtividade em células (Px) foi maior entre 40 e 50 horas de cultivo (Figura
45) nos ensaios 2 (0,58 g.L-1
.h-1
) e 3 (0,50 g.L-1
.h-1
), que tiveram níveis mais elevados

98
de OD no meio, em relação aos ensaios 4 (0,51 g.L-1
.h-1
) e, principalmente, em relação
ao ensaio 1 (0,34 g.L-1
.h-1
).
A produtividade em produto L-ASNase (Pp) também foi maior nesse mesmo
intervalo de tempo, entre 40 e 50 horas, nos ensaios 2 (75 U.L-1
.h-1
) e 3 (116 U.L-1
.h-1
),
confirmando que existe correlação entre o crescimento de L.muscorum CRM 1648 e a
produção da enzima. O valor de Pp do Ensaio 2 chega no seu máximo em
aproximadamente 50 horas de cultivo quando começa a cair, o que possivelmente está
relacionado a queda contínua de OD para valores abaixo de 35%. Pelo perfil de Px e Pp
do Ensaio 2 é possível supor que a manutenção de valores mais elevados de OD é mais
importante para a produção de L-ASNase do que para o crescimento da levedura, visto
que a queda de OD abaixo de 35% teve um efeito mais negativo sobre Pp do que sobre
Px. Como o controle e manutenção do pH prejudicou a produção de L-ASNase no
Ensaio 1, o perfil de Pp desse ensaio não foi incluído na Figura 45.
Figura 45 – Perfil comparativo da produtividade em células Px (A) e em produto L-
ASNase Pp (B) nos Ensaios 1-4 em biorreator. Legenda: Ensaio 1: triângulo azul;
Ensaio 2: círculo vermelho; Ensaio 3: quadrado verde; Ensaio 4: losango roxo.
Quanto ao perfil de consumo dos substratos (Figura 46), embora L. muscorum
CRM 1648 tenha consumido sacarose mais rapidamente do que prolina em todas as
condições de OD avaliadas, o consumo desses dois nutrientes foi mais rápido em
condição de maior disponibilidade de oxigênio no meio, ou seja, nos ensaios 2, 3 e 4,
respectivamente, mostrando que OD é importante por interferir na velocidade de
consumo das fontes de carbono (e nitrogênio, no caso de prolina) e, consequentemente,
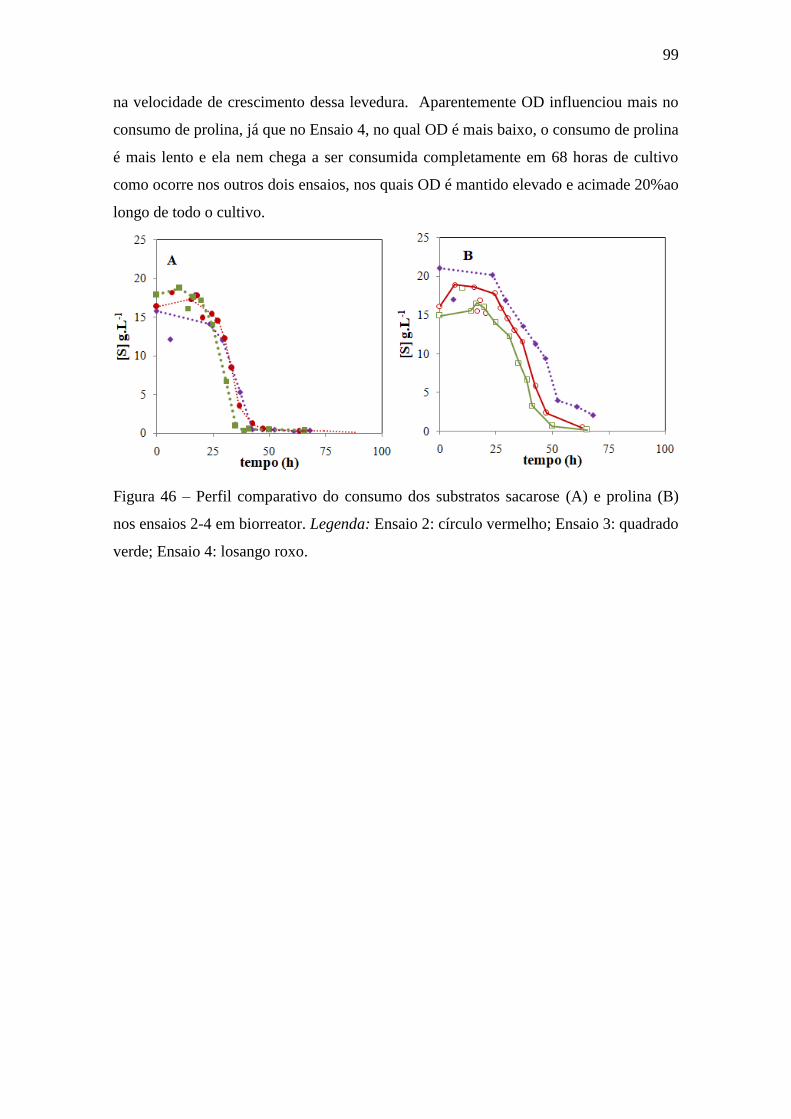
99
na velocidade de crescimento dessa levedura. Aparentemente OD influenciou mais no
consumo de prolina, já que no Ensaio 4, no qual OD é mais baixo, o consumo de prolina
é mais lento e ela nem chega a ser consumida completamente em 68 horas de cultivo
como ocorre nos outros dois ensaios, nos quais OD é mantido elevado e acimade 20%ao
longo de todo o cultivo.
Figura 46 – Perfil comparativo do consumo dos substratos sacarose (A) e prolina (B)
nos ensaios 2-4 em biorreator. Legenda: Ensaio 2: círculo vermelho; Ensaio 3: quadrado
verde; Ensaio 4: losango roxo.

100
V. CONCLUSÕES
Este trabalho explorou o potencial de um ambiente extremo (Antártica
marítima), ao investigar novas fontes microbianas produtoras da enzima L-ASNase que,
dentre outras aplicações, possui papel chave no tratamento da leucemia linfoblástica
aguda. Dos 150 isolados pesquisados, 6% produziu a enzima e desses quase 80% era da
espécie Leucosporidium muscorum, sugerindo que não há muitos produtores de L-
ASNase nesse ambiente (ou pelo menos não foi detectado), mas que os poucos que tem
em sua maioria pertencem ao gênero Leuscosporidium.
De acordo com os resultados apresentados, a linhagem L. muscorum CRM 1648
foi capaz de produzir uma L-ASNase sem atividade glutaminásica em maior quantidade
do que os demais isolados, tornando-se alvo deste trabalho. Ao estudar as condições de
cultivo e substratos (fontes de carbono e nitrogênio) para melhor crescimento e
produção de L-ASNase foi possível conhecer um pouco das características dessa
levedura, que cresceu melhor em meio CD com extrato de levedura e produziu mais L-
ASNase com sacarose como fonte de carbono e prolina como fonte de nitrogênio. A
preferência dessa linhagem por fontes pobres de nitrogênio como a prolina sugere a
presença do fenômeno de Repressão Catabólica pelo Nitrogênio, que ocorre em outras
espécies de levedura, como Saccharomyces cerevisiae.
O estudo da temperatura de cultivo indicou que a temperatura ótima de
crescimento dessa levedura é 18°C, mas que a produção da enzima é mais favorecida
quando o cultivo é realizado a 15°C. A condição de cultivo que mais favoreceu a
produção de L-ASNase por L. muscorum CRM 1648 foi o cultivo a 15°C, pH inicial
igual a 5,5 ou 6,5 com adição de pelo menos 25% de água do mar. A concentração
inicial do inoculo (X0), por outro lado, não influenciou significativamente nenhuma das
respostas.
Os cultivos em biorreator com diferentes ofertas de oxigênio dissolvido às
células mostraram que o crescimento da levedura e, em especial, a produção da enzima
são favorecidos quando OD é mantido elevado, em torno de 80%.
Embora seja necessário purificar e caracterizar a enzima produzida por L.
muscorum CRM 1648 este trabalho mostrou que esse microrganismo é produtor de uma
L-ASNase livre de atividade glutaminásica, característica que pode ser interessante para
o uso dessa enzima no tratamento de LLA.

101
Além disso, todo o trabalho referente ao melhoramento do meio e das condições
de cultivo em biorreator resultou em valores aproximadamente 10 vezes mais elevados
de produção de L-ASNase e produtividade (4582 U.L-1
e 63,6 U.L-1
.h-1
,
respectivamente) quando comparado aos valores obtidos antes do delineamento
experimental.
Essa etapa (upstream) é de suma importância para tornar o processo viável
industrialmente, visto que a baixa produtividade pode tornar o processo muito caro e
inviabilizá-lo, ainda que o produto seja de alto valor agregado, como é o caso de uma
enzima de uso clínico. Somado a isso, não foram encontrados relatos de produção de L-
ASNase por essa levedura e o que tem acerca do metabolismo e crescimento é muito
escasso e superficial. As informações que resultam desse trabalho são, portanto, inéditas
e importantes para o desenvolvimento de um processo produtivo que vise à produção de
L-ASNase pela levedura de origem antártica L. muscorum CRM 1648.

102
VI. PERSPECTIVAS FUTURAS
Embora este trabalho tenha sido relevante no que concerne ao estudo da fase
upstream, no melhoramento das condições de cultivo para a produção de L-ASNase,
pela levedura adaptada ao frio L. muscorum CRM 1648, se faz necessário purificar a
enzima, caracterizá-la e avaliá-la quanto ao potencial antileucêmico para poder afirmar
seu uso no tratamento de LLA.
Esses estudos foram iniciados e parte dele resultou no artigo COSTA-SILVA et
al., 2018b, no qual colaborei, mas encontramos algumas dificuldades na recuperação da
enzima nas etapas finais de purificação. Foram avaliados alguns sistemas de duas fases
aquosas, além de cromatografia de troca iônica e por exclusão molecular, até então sem
sucesso. É preciso, portanto, avaliar outros sistemas de purificação e testar alguns
inibidores de proteases para reduzir a degradação do produto ao longo do processo.
Outro ponto interessante seria explorar a produção de outros potenciais produtos
de interesse industrial por L. muscorum CRM 1648, como por exemplo, lipídios mono e
poliinsaturados, visto que há relatos na literatura que confirmam a produção desse tipo
de lipídio por leveduras do gênero Leucosporidium, que pode ser interessante para a
indústria de biodiesel e de alimentos.

103
VII. REFERÊNCIAS
ABDELRAZEK, N.A. et al. Experimental and bioinformatics studyfor production of l‑asparaginase from
Bacilluslicheniformis: a promising enzyme for medicalapplication. AMB Express, v. 9, p. 39 -55, 2019.
ABRAHAM, E.; SZABADOS, L.; ERDEL, L. Methods for determination of proline in plants. Methods in
Molecular Biology, v. 639, p. 317-331, 2010.
ABUCHOWSKI A. et al. Treatment of L5178Y tumor bearing BDf mice with a non-immunogenic L-
glutaminase asparaginase.Cancer Treat Rep, v. 63, p.1127–9, 1979.
ABUD, A. K. de S. Estudo do controle de qualidade do processo de produção de L-asparaginase por
zymomonas mobilis. 2005. 243f. Tese (Doutorado) – Engenharia Química, Universidade Federal do Rio
de Janeiro, Rio de Janeiro, RJ.
ANDRADE, A.M. Influência da radiação solar na distribuição superficial da vegetação na Península
Fildes, Antártica Marítima. In: XVII SIMPÓSIO BRASILEIRO DE SENSORIAMENTO REMOTO.
João Pessoa, PB: SBSR. 2015.
ASHOK, A. et al. Microbes Producing L-asparaginase free of glutaminase and urease isolated from extreme
locations ofantarctic soil and moss. Scientific Reports, v. 9, n. 1, p. 1423-1432.
ALAM, S. et al. Asparaginase conjugated magnetic nanoparticles used for reducing acrylamide formation in
food model system. Bioresource Technology, v. 269, p. 121-126, 2018.
ALRUMMAN, S.A. et al. Production and anticancer activityof an L-Asparaginase from
Bacilluslicheniformis isolated from the RedSea, Saudi Arabia. Scientific Reports, v. 9, n. 1, p. 3756-
3770, 2019.
ARIMA, K., et al. Production of extracellular L-asparaginases by microorganisms. Agricultural and
Biological Chemistry, v.36, p.356-361, 1972.
AVRAMIS, V.I. Asparaginases: Biochemical Pharmacologyand Modes of Drug Resistance. Anticancer
Research, vol. 32, p. 2423-2438, 2012.
AVRAMIS, V.I.; PANOSYAN, E.H. Pharmacokinetic/pharmacodynamic relationships of asparaginase
formulations: the past, the present and recommendations for the future.Clinical pharmacokinetics, v. 44,
n. 4, p. 367-93, 2005.

104
BADOEI-DALFARD, A. L-asparaginase production in the pseudomonas pseudoalcaligenes strain JHS-71
isolated from Jooshan Hot-spring. Molecular Biology Research Communications, v. 5, n. 1, p. 1-10,
2016.
BANSAL, S. et al. Hyperthermophilic asparaginase mutants with enhanced substrate affinity and
antineoplastic activity: structural insights on their mechanism of action. FASEB Journal, v. p. 1161-
1171, 2012.
BARUCH, M. et al. An extracellular bacterial pathogen modulates host metabolism to regulate its
ownsensing and proliferation. Cell, v.156, p. 97–108, 2014.
BATISTA, A.S. Sucrose fermentation by Saccharomyces cerevisiae lacking hexose transport.Journal
Molecular Microbiology and Biotechnology,v. 8, n.1, p.26-33, 2004.
BUZZINI, P. et al. Psychrophilic yeasts from worldwide glacial habitats: diversity,adaptation strategies and
biotechnological potential. FEMS Microbiol Ecol., v. 82, p. 217-241, 2012.
CACHUMBA, J.J.M. et al. Extracellular L-asparaginase production in solidstatefermentation by using
sugarcane bagasse assupport material. Preparative Biochemistry and Biotechnology, 2019.
https://doi.org/10.1080/10826068.2019.1566152.
CARVALHO, J.C.M. et al. Tecnologia de Fermentações. In: VITOLO, M. Biotecnologia Farmacêutica:
Aspectos sobre aplicação industrial. São Paulo, SP: Blucher, 2015. Cap.3. p.144-145.
CARRASCO, M. et al. Screening and characterization of amylase and cellulase activities in psychrotolerant
yeasts. BMC Microbiology, v.16, p.21-29, 2016.
CEDAR, H.; SCHWARTZ, J.H. Production of L-asparaginase II by Escherichia coli. Journal of
bacteriology, v. 96, n. 6, p. 2043-2048, 1968.
CHAN, W.K.. et al. The glutaminase activity of L-asparaginase is not required for anticancer activity against
ASNS-negative cells. BLOOD, v.123, n. 23, 2014.
COOPER, S.L.; BROWN, P. A. Treatment of Pediatric Acute Lymphoblastic Leukemia. Pediatr Clin North
Am.,v. 62, n. 1, p. 61–73, 2015.
COSTA-SILVA, T.A. et al. Optimization of culture conditions and benchscaleproduction of anticancer
enzyme L-asparaginaseby submerged fermentation from Aspergillus terreus CCT 7693. Preparative
Biochemistry and Biotechnology, v. 49, n. 1, p. 95-104, 2018a.

105
COSTA-SILVA, T.A. et al. Microbial cell disruption methods for efficient release of enzyme L-asparaginase.
Preparative Biochemistry and Biotechnology, v. 48, n. 8, p. 707-717, 2018b.
COVINI, D. et al. Expanding Targets for a Metabolic Therapy of Cancer: L-Asparaginase. Recent Patents
on Anti-Cancer Drug Discovery, v. 7, p. 4-13, 2012.
D’AMICO, S. et al. Molecular basis of cold adaptation. Phil. Trans. R. Soc. Lond. B, v. 357, p. 917–925,
2002.
DARVISHI, F.; FARAJI, N.; SHAMSI, F. Production and structural modeling of a novel asparaginase
inYarrowia lipolytica. International Journal of Biological Macromolecules, v. 125, p. 955-961, 2019.
DEMING, J.W. Extremophiles: Cold environments. In: Schaechter, M. (ed). The Desk Encyclopedia of
Microbiology. Oxford: Elsevier, 2009. p. 483-494.
DESHPANDE, N.; CHOUBEY, P.; AGASHE, M. Studies on optimization of growth parameters forL-
asparaginase production by Streptomyces ginsengisoli. The Scientific World Journal, 2014. doi:
10.1155/2014/895167.
di MENNA, M.E. Yeasts from Antarctica. Journal of General Microbiology, v.23, p. 295–300, 1960.
DIAS, F.F.G.; SATO, H.H. Sequential optimization strategyformaximum L-asparaginase productionfrom
Aspergillus oryzae CCT 3940. Biocatalysis and Agricultural Biotechnology, v. 6, p. 33-39, 2016.
DORIYA, K.; JOSE, N.; GOWDA, M.; KUMAR, D.S. Solid state fermentation Vs submerged fermentation
for the production of L-asparaginase. Advances in Food and Nutrition Research, v. 78, p. 115-135,
2016.
DRAINAS, C. et al. Aspartic hydroxamate resistance and asparaginase regulationin the fungus Aspergillus
nidulans. Journal of General Microbiology, v. 98, p. 493-501, 1977.
DUARTE, A.W.F. et al. Taxonomic assessment and enzymes production by yeast isolates from marine and
terrestrial Antartic samples. Extremophiles, v. 17, n. 6, p. 1023-1035, 2013.
DUARTE, A.W.F. et al. Yeasts from macroalgae and lichens that inhabit theSouth Shetland Islands,
Antarctica. Environmental Microbiology Reports, v. 8, n. 5, p. 874-885, 2016.
DUARTE, A.W.F. et al. Cold-adapted enzymes produced by fungi fromterrestrial and marine Antarctic
environments. Critical Reviews in Biotechnology, v. 38, n. 4, p. 600-619, 2018.

106
DUNLOP, P.C. et al. Nitrogen Catabolite Repression of Asparaginase II in Saccharomyces cerevisiae.
Journal of Bacteriology, v. 142, n. 1, p. 422426, 1980.
DUTTA, S.; GHOSH, S.; PRAMANIK, S. L-asparaginase and L-glutaminase from Aspergillus fumigatus
WL002: production and some physicochemical properties. Applied Biochemistry and Microbiology, v.
51, n. 4, p. 425-431, 2015.
EBRAHIMINEZHAD, A. et al. Chlorella vulgaris - a novel microalgal source forL-asparaginase production.
Biocatalysis and Agricultural Biotechnology, v. 3,p. 214–217, 2014.
EGLER, R.A., AHUJA, S.P., MATLOUB, Y. L-asparaginase in the treatment of patients with acute
lymphoblastic leukemia. Journal of Pharmacology and Pharmacotherapeutics, v. 7, n. 2, p. 62-71,
2016.
EINSFELDT, K. Desenvolvimento de uma nova L-asparaginase recombinante de Zymomonas mobilis
para aplicação como biofármaco. 2014. 154f. Tese (Doutorado) – Engenharia Química, COPPE,
Universidade Federal do Rio de Janeiro, Rio de Janeiro, RJ.
El BAKY, H.H.A; El BAROTY, G.S. Optimization of growth conditions for purification and production of
L-Asparaginase by Spirulina maxima. Evidence-Based Complementary and Alternative Medicine,
2016. doi: 10.1155/2016/1785938.
EL-BESSOUMY, A.A.; SARHAN, M.; MANSOUR, J. Production, isolation, and purification of L-
asparaginase from Pseudomonas Aeruginosa 50071 using solid-state fermentation. Journal of
Biochemistry and Molecular Biology, v. 37, n. 4, p. 387-393, 2004.
EBRAHIMINEZHAD, A. et al. Chlorella vulgaris, a novel microalgal source for L-asparaginase production.
Biocatalysis and Agricultural Biotechnology, v. 3, p. 214-217, 2014.
EL-GHONEMY, D.H.E. Microbial Amidases and their Industrial Applications: A Review. Journal of
Medical Microbiology & Diagnosis,v.4, n.1, p.173-178, 2014.
EL-NAGGAR, N.E.A. Extracellular production of the oncolytic enzyme, L-asparaginase, by newly isolated
Streptomyces sp. strain NEAE-95 as potential microbialcell factories: optimization of culture conditions
using responsesurface methodology. Current Pharmaceutical Biotechnology, v. 16, p. 162-178, 2015.
EL-NAGGAR, N.E.A. et al. Purification, characterization and immunogenicity assessment of
glutaminasefree L-asparaginase from Streptomyces brollosae NEAE-115. BMC Phamacology and
Toxicology, v. 19, p. 51-66, 2018.

107
ELSHAFEI, A.M.; EL-GHONEMY, D.H. Screening and media optimization for enhancing L-asparaginase
production, an anticancer agent, from different filamentous fungi in solid state fermentation. British
Biotecnology Journal, v. 9, n. 3, p. 1-15, 2015.
ERVA, R.R. et al. Optimization of L-asparaginase production from novel Enterobacter sp., by
submergedfermentation using response surface methodology. Preparative Biochemistry and
Biotechnology, v. 47, n. 3, p. 219-228, 2017.
ERVA, R.R.; VENKATESWARULU, T.C.; PAGALA, B. Multi level statistical optimization of L-
asparaginase from Bacillus subtilis VUVD001. 3 Biotech, v. 8, n. 1, p. 1-8, 2018.
FANTÁSTICO, Rede Globo. Ministério da Saúde anuncia que não compra mais leuginase da China.
Disponível em:<http://g1.globo.com/fantastico/noticia/2017/07/ministerio-da-saude-anuncia-que-nao-
compra-mais-leuginase-da-china.html>, acesso em 25/05/2017.
FARAG, A.M.; HASSAN, S.W.; BELTAGY, E.A.; EL-SHENAWY, M.A. Optimization of production of
anti-tumorL-asparaginase by free and immobilized marine Aspergillus terreus. Egyptian Journal of
Aquatic Research, v. 41, p. 295–302, 2015.
FELLER, G.; GERDAY, C. Psychrophilic enzymes: hot topics in cold adaptation. Nature. v.1, p.200-208,
2003.
FELLER, G. Psychrophilic Enzymes: from folding tofunction and biotechnology. Scientifica, v. 2013, p.1-
28, 2013.
FENG, Y. et al. Enhanced extracellular production of L-asparaginase from Bacillus subtilis 168 by B.
subtilis WB600 through a combined strategy. Applied Microbiology and Biotechnology, v. 101, v. 4, p.
1509-1520, 2017.
FERRARA, M. A. et al.Asparaginase production by a recombinant Pichia pastoris strain harbouring
Saccharomyces cerevisiae ASP3 gene. Enzyme and Microbial Technology, v. 39, p. 1457-1463, 2006.
FERRARA M.N., SEVERINO, N.M.B., VALENTE, R.H., PERALES, J., BON, E.P.S. High-yield extraction
of periplasmic asparaginase produced by recombinant Pichia pastoris harbouring the Saccharomyces
cerevisiae ASP3 gene. EnzymeandMicrobial Technology, v. 47, p. 71-76, 2010.
FERREIRA, E.M.S. et al. Taxonomy and richness of yeasts associated with angiosperms,bryophytes, and
meltwater biofilms collected in the AntarcticPeninsula. Extremophiles, v. 23, p. 151-159, 2019.

108
FILIPPUCCI, S. et al. Study of Holtermanniella wattica, Leucosporidium creatinivorum, Naganishia
adeliensis, Solicoccozyma aeria, and Solicoccozyma terricola for their lipogenicaptitude from different
carbon sources. Biotechnology for Biofuels, v. 9, p. 259-273, 2016.
FODA, M.S; ZEDAN, H.H.; HASHEM, S.A. Formation and properties of L-glutaminase and L-asparaginase
activities in Pichia polymorpha. Acta Microbiologica Polonica, v.29, p.343-352, 1980.
FUKUDA, I.M. et al. Design of experiments (DoE) applied to pharmaceutical and analytical Quality by
Design (QbD). Brazilian Journal of Pharmaceutical Sciences, v. 54, p. 1-16, 2018.
de GARCÍA, V. et al. Sex in the cold: taxonomic reorganization of psychrotolerant yeasts in the order
Leucosporidiales. FEMS Yeast Research, v. 15, n. 4, p. 1-12, 2015.
de GARCIA, V.; van BROOK, M.R. Cold-Adapted Yeasts from Patagonia Argentina. International
Journal of Biodiversity, v. 12, n. 1, p. 1-10, 2016.
GECKIL, H. et al. Effect of Vitreoscilla hemoglobin on production of a chemotherapeutic enzyme, L-
asparaginase, by Pseudomonas aeruginosa. Biotechnology Journal, v. 1, p. 203–8, 2006.
GIRÃO, L.F.C. et al. Saccharomyces cerevisiae asparaginase II, a potential antileukemic drug: Purification
and characterization of the enzyme expressed in Pichia pastoris. BMC Proceedings, v. 8 (Suppl 4), p78-
79. 2014.
GIRÃO, L.F.C. et al. Saccharomyces cerevisiae asparaginase II, a potential antileukemic drug: Purification
and characterization of the enzyme expressed in Pichia pastoris. Protein Expression and Purfication, v.
120, p. 118-125, 2016.
GHOLAMIAN, S. et al. Optimization of culture media for L-Asparaginase production by newly isolated
bacteria, Bacillus sp. GH51, Microbiology, v. 82, n. 6, p. 856- 863, 2013.
GONÇALVES, A.B. et al. Fungal production of the anti-leukemic enzyme L-asparaginase:from screening to
medium development. Acta Scientiarum, v. 38, n. 4, p. 387-394, 2016.
GUTIÉRREZ-ALONSO, P. et al. Biochemical characterization of a β-fructofuranosidase from Rhodotorula
dairenensis with transfructosylating activity. FEMS Yeast Res, v. 9, p. 768–773, 2009.
HAMERSCHLAK, N. Bone marrow transplantation for acute leukemias in Brazil. Where are we going?
Revista Brasileira de Hematologia e Hemoterapia, v. 32, n. 2, p. 97, 2010.

109
HAMID, B. et al. Psychrophilic yeasts and their biotechnological applications - A review. African Journal
of Biotechnology, v. 12, n. 22, p. 2188-2197, 2014.
HEINEMANN, B.; HOWARD, A.J. Influence of dissolved oxygen levels on production of L-asparaginase
and prodigiosin by Serratia marcescens.Applied Microbiology, v. 18, p. 550–554, 1969.
HOFMAN-BANG, J. Nitrogen Catabolite Repression in Saccharomyces cerevisiae. Molecular
Biotechnology, v. 12, 1999.
HUERTA-SAQUERO, A. et al. Rhizobium etli asparaginase II: an alternative for acute lymphoblastic
leukemia (ALL) treatment. Bioengineered, v. 4, n. 1, p. 30-36, 2013.
HUNTER-CEVERA, J. C.; BELT, A. MaintenanceCultures for Biotechnology and Industry. London:
Academic Press, 1996.
HYMAVATHI, M. et al. Impact of Carbon and Nitrogen Sources on L-Asparaginase Production by Isolated
Bacillus circulans (MTCC 8574):Application of Saturated Plackett-Burman Design. Chemical and
Biochemical Engineering, v.4, p. 473-480, 2010.
IMADA, A. et al. Asparaginase and Glutaminase Activities of Micro-organisms. Journal of General
Microbiology, v.76, p. 85-99, 1973.
INFORSATO, F.J. Fungos de sedimentos marinhos da Antártica: diversidade e prospecção de
enzimas.2017. 138f. Dissertação (Mestrado) – Instituto de Biociências, Unersidade Estadual Paulista, Rio
Claro, São Paulo, SP.
INSTITUTO NACIONAL DO CÂNCER. Estimativa 2016: incidência de câncer no Brasil. Rio de Janeiro:
INCA, 2016.
INSTITUTO NACIONAL DO CÂNCER. Estimativa 2018: incidência de câncer no Brasil.Rio de Janeiro:
INCA, 2018.
JAFFE, N. et al. L-Asparaginase in the Treatment of Neoplastic Diseases. Cancer Research,v. 31, p. 942-
949, 1971.
KEEGAN, P. Erwinase (Erwinia L-asparaginase) is indicated as a component of a multi agent
chemotherapeutic regimen for the treatment of patients with Acute lymphoblastic leukemia (ALL) who
have developed hypersensitivity to E. coli derived asparaginase. Division Director Summary Review,
22p, 2011.

110
KALYANASUNDARAM, I. et al. Production, purification and characterisation of extracellular L-
asparaginase from salt marsh fungal endophytes. World Journal of Pharmacy and Pharmaceutical
Sciences, v. 4, n. 3, p. 663-677, 2015.
KHAMNA, S., YOKOTA, A., LUMYONG, S. L-Asparaginase production by actinomycetes isolated from
someThai medicinal plant rhizosphere soils. IJIB, v. 6, n. 1, p. 22-26, 2009.
KIL, J.O; KIM, G.N.; PARK, I. Extraction of extracellular L-asparaginase from Candida utilis. Bioscience,
Biotechnology, and Biochemistry, v..59, p.749-750. 1995.
KIRSOP, B.E.; DOYLE, A. Maintenance of Microorganisms and Cell Cultures - A Manual of
Laboratory Methods. 2nd ed., London: Academic Press, 1991.
KOTZIA, G.A.; LABROU N.E. L-asparaginase from Erwinia chrysanthemi 3937: cloning, expression and
characterization. Journal of Biotechnology, v.127, p.657–69, 2007.
KRISHNAPURA, P. R., BELUR, P.D., SUBRAMANYA, S. A critical review on properties and applications
of microbial L- asparaginases. Critical Reviews in Microbiology, p. 1-18, 2015.
KRISHNAPURA, P. R., BELUR, P.D. Isolation and screening of endophytes from the rhizomes of some
Zingiberaceae plants for L-asparaginase production. Preparative Biochemistry and Biotechnology, v.
46, n. 3, p. 281-287, 2016.
KUMAR, S. et al.Batch and fed-batch bioreactor studies for the enhanced production of glutaminase-free
Lasparaginase from Pectobacterium carotovorumMTCC 1428. Preparative Biochemistry and
Biotechnology, v. 47, n. 1, p. 74-80, 2017.
KUMAR S., VENKATA D.V., PAKSHIRAJAN K. Localization and production of novel L-asparaginase
from Pectobacterium carotovorum MTCC 1428. Proc Biochem, v. 45, p.223–9, 2010.
KUMAR, D.S.; SOBHA, K. L-asparaginase from microbes: a comprehensive review. Advances in
Bioresearch, v. 3, n. 4, 2012.
KURTZMAN, C.P.; FELL, J.W. The yeasts: a taxonomic study. Amsterdam: Elsevier, 1998.
LAICH, F.; CHÁVEZ, R.; VACA, I. Leucosporidium escuderoi f.a., sp. nov., a basidiomycetousyeast
associated with an Antarctic marine sponge. Antonie van Leeuwenhoek, v. 105, p. 593-601, 2014.
LAPMAK, K. et al. L-Asparaginase production by Bipolaris sp. BR438 isolated from Brown Rice in
Thailand. Chiang Mai Journal of Science, v. 37, n. 1, p. 160-164, 2010.

111
LARIO, L.D. et al. Production, purification, and characterization of anextracellular acid protease from the
marineAntarctic yeast Rhodotorula mucilaginosa L7. Fungal Biology, v. 119, p.1129-1136, 2015.
LIU, F.S; ZAJIC, J.E. L-Asparaginase synthesis by Erwinia aroideae, Applied Microbiology, v. 23,p. 667–
668, 1971.
LOPES, A. M. et al. Therapeutic L-asparaginase: upstream, downstream and beyond. Critical Reviews in
Biotechnology, p. 1-18, 2017.
MAGAZANIK, B.; KAISER, C.A. Nitrogen regulation in Saccharomyces cerevisiae. Gene, v. 290, p. 1-18,
2002.
MAHAJAN, R. A rapid, efficient and sensitive plate assay for detection and screening of L-asparaginase-
producing microorganisms, Federation of European MicrobiologicalSocieties Microbiological
Letters, v. 341, p. 122-126, 2013.
MARGESIN, R. et al. Cold adapted Microorganisms: Adaptationstrategies and biotechnological potential.
Encyclopedia of Environmental Microbiology, p.871-885,2003.
MARINI, B.L. et al. Catalyzing improvements in ALL therapy with asparaginase. Blood Reviews, v. 31, p.
328-338, 2017.
MARTORELL, M.M. et al. Bioprospection of cold-adapted yeasts with biotechnological potential from
Antarctica. Jornal of Basic Microbiology, v. 57, p. 504-516, 2017.
MAŠÍNOVÁ, T. et al. Libkindia masarykiana gen. et sp. nov., Yurkovia mendeliana gen. et sp. nov. and
Leucosporidium krtinense f.a. sp. nov., isolated from temperate forest soils. International Journal of
Systematic and Evolutionary Microbiology, v. 67, p. 902-908, 2017.
MEDEIROS, V.R.; MESTDAGH, F.; de MEULENAER, B. Acrylamide formation in fried potato products
present and future, a critical review on mitigation strategies. Food Chemistry. v, 133, n. 4, p. 1138-1154,
2012.
MEENA, B. et al. Novel glutaminase free L-asparaginase from Nocardiopsis alba NIOT-VKMA08:
production, optimization, functionaland molecular characterization. Bioprocess and Biosystems
Engineering, v. 38, n. 2, p. 373-388, 2015a.

112
MEENA, B. et al. L-Asparaginase from Streptomyces griseus NIOT-VKMA29: optimization of process
variables using factorial designs and molecular characterization of L-asparaginase gene. Scientific
Reports, v. 5, 2015b.
MENEGAT, F.B. et al. Optimization of asparaginase production from Zymomonas mobilis by continuous
fermentation. Acta Scientiarum, v. 38, n. 2, p. 163-168, 2016.
MINISTÉRIO PÚBLICO FEDERAL. PROCURADORIA DA REPÚBLICA EM SÃO PAULO.
Recomendação nº 10/2013, de 5 de Abril de 2013, São Paulo.
MINISTÉRIO DA SAÚDE, Esclarecimentos L-asparaginase. Disponível
em:<http://portalarquivos2.saude.gov.br/images/pdf/2017/junho/22/2.%20b%20-%20L
ASPARAGINASE_CIT_22_06_2017.pdf>, acesso em 01/01/2019.
MOGUEL, I.S. Production of L-asparaginase of pharmaceutical interest from yeasts isolated from the
Antarctic continent. 2018. 114f. Tese (Doutorado) – Tecnologia Bioquímico-Farmacêutica, Faculdade
de Ciências Farmacêuticas, Universidade de São Paulo, São Paulo, SP.
MOMENI, V.; ALEMZADEH, I.; VOSOUGHI, M. Enhancement of L-asparaginase production by Candida
utilis in a 13 L fermenter and its purification. International Journal of Engineering, v. 28, n. 8, p. 1134-
1139, 2015.
MORITA, R.Y. Psychrophilic bacteria. Bacteriological Reviews, v. 39, n. 2, p. 144-167, 1975.
MOSTAFA, E.E.; EL-DEEN, A.M.N.; AWAD, H.M.; MAWAD, M.A. Partial characterization of L-
asparaginase-producing by Streptomyces sp. SAH1_CWMSG isolated from Rizosphere soil in Egypt.
Journal of Chemical and Pharmaceutical Research, v. 8, n. 4, p. 1290-1307, 2016.
NARAGANI, K.; MUVVA, V. Optimum conditions for L-asparaginase production by Rhodococcus
erythropolis VLK-12 isolated from marine habitats. European Journal of Biotechnology and
Bioscience, v. 4, n. 10, p. 1-5, 2016.
NAKAGAWA, T. et al. Cold-activepectinolytic activity of psychrophilic basidiomycetous yeast
Cystofilobasidium capitatum strain PPY-1. Journal of Bioscience and Bioengineering, v 94, p.175-177,
2002.
NARTA, U.K..;KANWAR, S.S.; AZMI, W. Pharmacological and clinical evaluation of l-asparaginase in the
treatment of leukemia. Critical Reviews in Oncology/Hematology, v. 61, p. 208–221, 2007.
NELSON, D.L.; COX, M.M. Princípios de bioquímica de Lehninger. Porto Alegre:Artmed, 2014.

113
OKAFOR, N. Modern Industrial Microbiology and Biotechnology. Estados Unidos: Science Publishers,
2007.
OFFMAN, M.N. et al.Rational engineering of L-asparaginase reveals importance of dual activity for cancer
cell toxicity. Blood Journal, v. 117, p. 1614-1621, 2011.
OLIVEIRA, E. M. et al. L-Asparaginase II of Saccharomyces cerevisiaeActivity profile during growth using
an ure2 mutant P40-3C and a P40-3C+URE2p strain. Applied Biochemistry and Biotechnology, v. 77,
n.1, p.311-316, 1999.
PASUT, G.; VERONESE, F. M. State of the art in PEGylation: The great versatility achieved after forty
years of research. Journal of Controlled Release, v. 161, p. 461-472, 2012.
PATHAN, A.A.K. et al. Diversity of yeasts from puddles in the vicinity of MidreLove´nbreen Glacier, Arctic
and bioprospecting for enzymesand fatty acids. Current Microbiology, v. 60, p. 307-314, 2010.
PIETERS, R. et al. L-asparaginase treatment in acute lymphoblastic leukemia: a focus on Erwinia
asparaginase. Cancer, v. 117, p. 238-249, 2011.
POKROVSKAYA, M.V. et al. Cloning, expression and characterization of the recombinant Yersinia
pseudotuberculosis L-asparaginase. Protein Expression and Purification, v. 82, P. 150-154, 2012.
PRITSA, A.A.; KYRIAKIDIS, D.A. L-asparaginase of Thermusthermophilus: purification, properties and
identification of essentialamino acids for its catalytic activity. Molecular and Cellular Biochemistry, v.
216, p. 93–101, 2001.
PRASAD TALLURI, V.S.S.L.; LANKA, S.S.; RAJAGOPAL SALADI, V. Statistical Optimization of
Process Parameters by Central Composite Design (CCD) for an enhanced Production of L-asparaginase
by Myroides gitamensisBSH-3, a novel species. Avicenna Journal of Medical Biotechnology, v. 11, n.
1, p. 59-66, 2019.
QESHMI, F.I. et al. Marine microbial L-asparaginase: Biochemistry, molecular approaches and applications
in tumor therapy and in food industry. Microbiological Research, v. 208, p. 99-112, 2018.
RAETZ, E.A., SALZER, W.L. Tolerability and Efficacy of L-Asparaginase Therapy in Pediatric Patients
With Acute Lymphoblastic Leukemia. Medical Progress, v. 32, n. 7, p. 554-563, 2010.

114
RAMAKRISHNAN, M.S.; JOSEPH, R. Characterization of an extracellular asparaginase of Rhodosporidium
toruloides CBS14 exhibiting unique physicochemical properties. Canadian Journal of Microbiology.
v.42, n.4, p.316-325, 1996.
RIZZARI, C. et al. Optimizing asparaginase therapy for acute lymphoblastic leukemia. Co-oncology, v.25, p.
S1-S9, 2013.
da ROCHA, W.R.V et al. Screening and optimizing fermentation production of L-asparaginase by
Aspergillus terreus strain S-18 isolated from the Brazilian Caatinga Biome. Journal of Applied
Microbiology, 2019. doi: 10.1111/jam.14221.
ROTH, J.E.S.N. et al. Recombinant Erwinia carotovora L-asparaginase II production in Escherichia coli fed-
batch cultures. Brazilian Journal of Chemical Engineering, v. 30, n. 2, p. 245-256, 2013.
RUBIO, M.C.; RUNCO R.; NAVARRO A.R. Invertase from a strain of Rhodotorula glutinis.
Phytochemistry,v.61, p. 605–609, 2002.
RUSSELL, N.J. Antarctic micro-organisms: coming in from the cold. Culture, Oxoid, v. 27, n. 2, 2006.
SAHAY, S. et al. Evaluation of pectinolytic activities for oenological uses from psychrotrophic yeasts.
Letters in Applied Microbiology, v. 57, p.115-121, 2013.
SAMPAIO, JP. Et al. Taxonomic studies in the Microbotryomycetidae: Leucosporidium golubevii sp. nov.,
Leucosporidiella gen. nov. and the new orders Leucosporidiales and Sporidiobolales. Mycological
Progress, v. 2, p. 53-68, 2003.
SANCHEZ, S.; DEMAIN, A.L. Metabolic regulation of fermentation process. Enzyme and Microbial
Technology, v. 31, p. 895–906, 2002.
SANTOS, J.A. Potencial biotecnológico de fungos marinhos eantárticos da central de recursos
microbianos da UNESP (CRM-UNESP). 2015. 90f. Dissertação (Mestrado) – Instituto de Biociências,
Universidade Estadual Paulista, São Paulo, SP.
SARQUIS, M.I.M. et al. Production of L-asparaginase by Filamentous Fungi. Memórias do Instituto
Oswaldo Cruz, v. 99, n.5, p. 489-492, 2004.
SAXENA, R.K.; SINHA, U. L-asparaginase and glutaminase activities in the culture filtrates of Aspergillus
nidulans. Current Science, v. 50, p. 218-219, 1981.

115
SAXENA, A.; UPADHYAY, R.; KANGO, N. Isolation and identification of actinomycetes for production of
novel extracellular glutaminase free L-asparaginase. Indian Journal of Experimental Biology, v. 53, p.
786-793, 2015.
SCHIMIDELL, W. et al. Biotecnologia Industrial: Engenharia Bioquímica. São Paulo, SP:Blucher, 2001.
SHAKAMBARI, G. et al. Agro waste utilization for cost-effective production of L‑Asparaginase by
Pseudomonas plecoglossicida RS1 with anticancer and acrylamide mitigation potential. ACS Omega, v.
2, p. 8108-8117, 2017.
SHARAFI, Z. et al. Screening for Type II L-Asparaginases: Lessons from the Genus Halomonas. Iranian
Journal of Pharmaceutical Research, v.16, n. 4, p. 1565-1573, 2017.
SHIVAJI, S.; PRASAD, G.S. Antarctic yeasts: biodiversity and potential applications. In:
SATYANARAYANA, T.; KUNZE, G (ed(s)). Yeast biotechnology: diversity and applications.
Amsterdam: Springer, 2009, p 3-18.
SHRIVASTAVA, A. et al. Recent developments in L-asparaginase discovery and its potential as anticancer
agent. Critical Reviews in Oncology/Hematology, v. 100, p. 1-10, 2015.
SILVA, L.F. Penicillium and Talaromyces endophytes from Tillandsia catimbauensis, a bromeliad endemic
in the Brazilian tropical dry forest, and their potential for L-asparaginase production. World Journal of
Microbiology and Biotechnology, v. 34, p.162-174, 2018.
SMITH, P.K. et al. Measurement of protein using bicinchoninic acid. Analytical Biochemistry, v. 150, p.76-
85, 1985.
STELIAROVA-FOUCHER, E. et al. International incidence of childhood cancer, 2001–10: a population-
based registry study. Lancet Oncology, v. 18, p. 719-731, 2017.
STEPANYAN, K.R.; DAVTYAN, M.A. Some questions of thermostability of L-asparaginase of yeast
Candida guilliermondii. BiologicheskiiZhurnal Armenii, v.41, p.599-603, 1988.
SUKUMARAN, C.P.; SINGH, D.V.; MAHADEVAN, P.R. Synthesis of L-asparaginase by Serratia
marcescens (Nima). Journal of Biosciences, v. 3, p. 263–269, 1979.
TEIXEIRA, A.I. et al. Development of a method to quantify sucrose in soybean grains. Food Chemistry, v.
130, p. 1134-1136, 2012.

116
TSUJI, M. et al. Cold adaptation of fungi obtained from soil and lake sedimentin the Skarvsnes ice-free area,
Antarctica. FEMS Microbiology Letters, v. 346, p. 121-130, 2013.
VALA, A.K. et al. Characterization of L-asparaginase from marine-derived Aspergillus niger AKV-MKBU,
its antiproliferative activity and bench scale production using industrial waste. International Journal of
Biological Macromolecules, v. 108, p. 41-46, 2018.
VAZ, A.B.M. et al. The diversity, extracellular enzymatic activities and photoprotective compounds of yeasts
isolated in Antarctica. Brazilian Journal of Microbiology, v. 42, p. 937-947, 2011.
VIMAL, A.; KUMAR, A. Biotechnological production and practical application of L-asparaginase enzyme.
Biotechnology and Genetic Engeneering Reviews, v. 33, n. 1, p. 40 – 61, 2017a.
VIMAL, A.; KUMAR, A. In vitro screening and in silico validation revealed key microbes for higher
production of significant therapeutic enzyme L-asparaginase. Enzyme and Microbial Technology, v. 98,
p. 9 – 17, 2017b.
WANG, Q. M. et al. Living strategy of cold-adapted fungi with the reference to several representative
species. Mycology, v. 8, n. 3, p. 178-188, 2017.
WANG, Q. M. et al.Phylogenetic classification of yeasts and related taxa within Puccinio mycotina. Studies
in Mycology, v. 81, p. 149-189, 2015.
WADE, H.E.; ROBINSON, H.K.; PHILLIPS, B.W. Asparaginase and Glutaminase Activities of Bacteria.
Journal of General Microbiology, v.69, n. 2, p. 299-31, 1971.
WENTZEL, L.C.P. et al. Fungi from Admiralty Bay (King George Island, Antarctica) soils and marine
sediments. Microbial Ecology, v. 77, n. 1, p. 12-24, 2018.

117
APÊNDICES

118
APÊNDICE A. Dados comparativos sobre publicações envolvendo a produção de L-ASNase por bactérias, fungos filamentosos, leveduras e
microalgas nos últimos 5 anos de acordo com dados do PUBMED e Google Acadêmico (acesso: março de 2019).
espécie origem
condições de cultivo indicador de produção de L-
ASNase
análise da
atividade
análise
estatística referência país
cultivo FC FN pH T°C agitação
(rpm) aeração (vvm)
tempo (dias)
Bacillus licheniformis solo ferment. submersa glicose cloreto de
amônio 7,4 39,6 200
sem
aeração 1 8,0 U.mL-1.h-1 Nessler CCD
ABDELRAZEK et al.
(2019) Egito
Bacillus licheniformis costa do Mar
Vermelho ferment. submersa glicose
sulfato de
amônio 6,5 37 250
sem
aeração 3 11,74 U.mL-1 Nessler univariada
ALRUMMAN et al.
(2019)
Arábia
Saudita
Myroides gitamensis solo de matadouro fermentação em
estado sólido farelo de trigo
extrato de levedura
7,5 37 120 sem
aeração 2 296,0 U Nessler
univariada; CCD
PRASAD TALLURI et al. (2019)
Índia
Bacillus subtilis coleção de cultura ferment. submersa glicose asparagina 8,3 35 200 sem
aeração 6 2,9 U.mL-1 Nessler
univariada;
ANNs ERVA et al. (2018) Índia
Streptomyces
brollosae solo ferment. submersa glicose asparagina 7 30 150
sem
aeração 7 795,0 U Nessler ausente
EL-NAGGAR et al.
(2018) Egito
Pseudomonas plecoglossicida
ambiente marinho ferment. submersa cascas de
cebola e alho asparagina ND 37 estático
sem aeração
1 1,1 U.mL-1 Nessler CCD SHAKAMBARI et
al. (2017) Índia
Halomonas sp. ambiente halofílico ferment. submersa glicose asparagina 7 30 180 sem
aeração 1 6,2 U.mL-1 Nessler CCD
SHARAFI et al.
(2017) Irã
Pectobacterium carotovorum
coleção de cultura
ferment. submersa
(descontínuo e desc.
alimentado)
glicose asparagina e ext.
levedura 8,5 30 160-600 1,5-2,0 0,5 40,0 U.mL-1 Nessler ausente KUMAR et al. (2017) Índia
Enterobacter sp. coleção de cultura ferment. submersa citrato de sódio cloreto de
amônio 6 33 200
sem
aeração 1,5 18,35 U.mL-1 Nessler
univariada;
CCD ERVA et al. (2017) Índia

119
espécie origem
condições de cultivo indicador de
produção de
L-ASNase
análise de L-ASNase
análise estatística
referência país
cultivo FC FN pH T°C agitação
(rpm)
aeração
(vvm)
tempo
(dias)
pseudomonas
pseudoalcaligenes fontes termais
fermentação
submersa glicose asparagina ND 30 160
sem
aeração 2 240,0 U.mL-1 Nessler ausente
BADOEI-
DALFARD (2016) Irã
Bacillus subtilis coleção de cultura
fermentação
submersa (descontínuo e
desc. alimentado)
sacarose asparagina ND 37 600-1000 1,8 2 407,6 U.mL-1 Nessler ausente FENG et al. (2016) China
Zymomonas mobilis coleção de cultura fermentação
submersa
(contínuo)
sacarose extrato de levedura e
asparagina
ND 30 ND ND 1 117,4 U.L-1 Nessler fatorial
completo
MENEGAT et al.
(2016) Brasil
Streptomyces ghanensis
solo fermentação
submersa glicerol
sulfato de amônio
7 28 200 sem
aeração 5 106, 9 U Nessler univariada
MOSTAFA et al. (2016)
Egito
Rhodococcus erythropolis
sedimento marinho fermentação
submersa glicerol
extrato de levedura
7 30 ND sem
aeração 3 10,9 U Nessler univariada
NARAGANI; MUVVA (2016)
Índia
Streptomyces
phaeochromogenes solo
fermentação
submersa amido asparagina 7,2 28 150
sem
aeração 2 19,2 U.mL-1 Nessler ausente
SAXENA et al.
(2015) Índia
Streptomyces parvus
solo fermentação
submersa glicose, amido
asparagina,
extrato de
levedura
7 30 200 sem
aeração 8 92,4 U.mL-1 Nessler
Plackett
Burman; Box-
Behnken
EL-NAGGAR et al. (2015)
Egito
Streptomyces olivaceus
solo fermentação
submersa glicose asparagina 6,8 35 200
sem aeração
3 68,6 U.mL-1 Nessler
Plackett-
Burman;
CCD
EL-NAGGAR et al. (2015)
Egito
Streptomyces griseus sedimento marinho fermentação
submersa amido
extrato de
levedura 8 35 estático
sem
aeração 6 57,8 U.mL-1 Nessler
Box-
Behnken
MEENA et al.
(2015b) Índia
Nocardiopsis alba sedimento marinho fermentação
submersa maltose
extrato de
levedura 8 30 125
sem
aeração 6 18,5 U.mL-1 Nessler univariada
MEENA et al.
(2015a) Índia
Streptomyces
ginsengisoli sedimento de rios
fermentação
submersa glicose peptona 8 30 120
sem
aeração 5 3,23 U.mL-1 Nessler univariada
DESHPANDE et al.
(2014) Índia
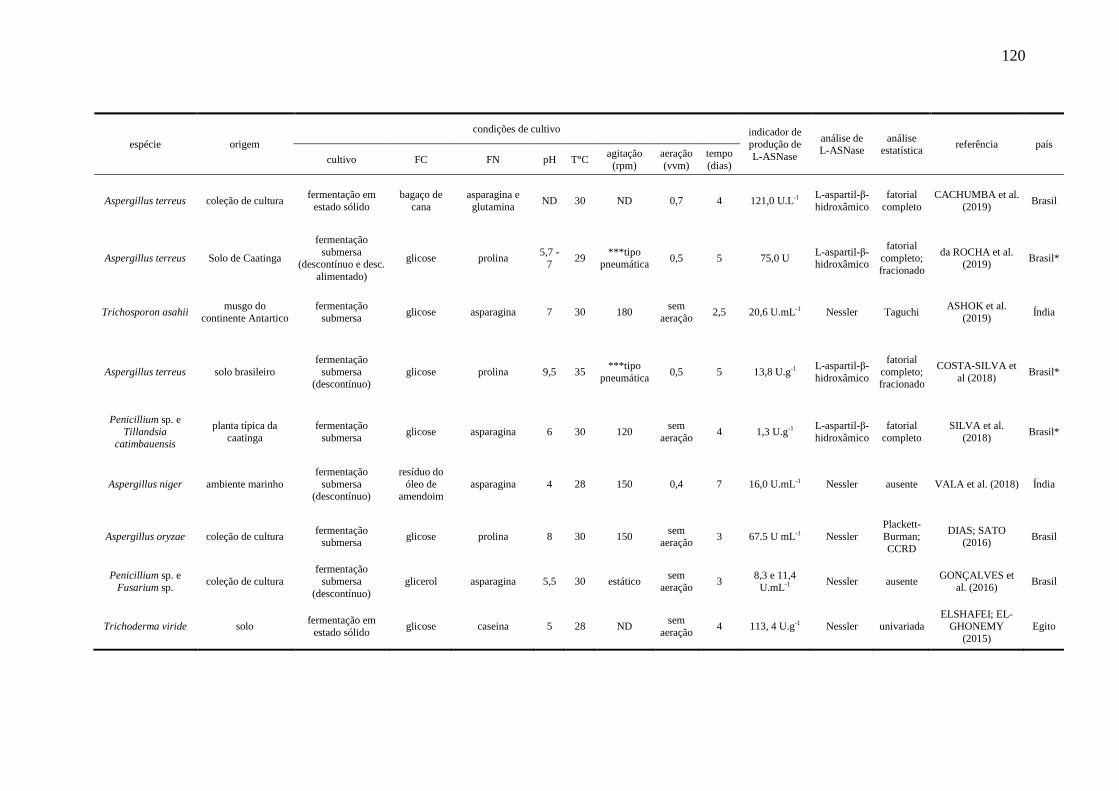
120
espécie origem
condições de cultivo indicador de
produção de
L-ASNase
análise de L-ASNase
análise estatística
referência país
cultivo FC FN pH T°C agitação
(rpm)
aeração
(vvm)
tempo
(dias)
Aspergillus terreus coleção de cultura fermentação em
estado sólido
bagaço de
cana
asparagina e
glutamina ND 30 ND 0,7 4 121,0 U.L-1
L-aspartil-β-
hidroxâmico
fatorial
completo
CACHUMBA et al.
(2019) Brasil
Aspergillus terreus Solo de Caatinga
fermentação
submersa (descontínuo e desc.
alimentado)
glicose prolina 5,7 -
7 29
***tipo pneumática
0,5 5 75,0 U L-aspartil-β-hidroxâmico
fatorial
completo;
fracionado
da ROCHA et al. (2019)
Brasil*
Trichosporon asahii musgo do
continente Antartico
fermentação
submersa glicose asparagina 7 30 180
sem
aeração 2,5 20,6 U.mL-1 Nessler Taguchi
ASHOK et al.
(2019) Índia
Aspergillus terreus solo brasileiro
fermentação
submersa (descontínuo)
glicose prolina 9,5 35 ***tipo
pneumática 0,5 5 13,8 U.g-1
L-aspartil-β-
hidroxâmico
fatorial
completo; fracionado
COSTA-SILVA et
al (2018) Brasil*
Penicillium sp. e
Tillandsia catimbauensis
planta típica da
caatinga
fermentação
submersa glicose asparagina 6 30 120
sem
aeração 4 1,3 U.g-1
L-aspartil-β-
hidroxâmico
fatorial
completo
SILVA et al.
(2018) Brasil*
Aspergillus niger ambiente marinho
fermentação
submersa (descontínuo)
resíduo do
óleo de amendoim
asparagina 4 28 150 0,4 7 16,0 U.mL-1 Nessler ausente VALA et al. (2018) Índia
Aspergillus oryzae coleção de cultura fermentação
submersa glicose prolina 8 30 150
sem aeração
3 67.5 U mL-1 Nessler
Plackett-
Burman;
CCRD
DIAS; SATO (2016)
Brasil
Penicillium sp. e
Fusarium sp. coleção de cultura
fermentação
submersa
(descontínuo)
glicerol asparagina 5,5 30 estático sem
aeração 3
8,3 e 11,4
U.mL-1 Nessler ausente
GONÇALVES et
al. (2016) Brasil
Trichoderma viride solo fermentação em
estado sólido glicose caseina 5 28 ND
sem
aeração 4 113, 4 U.g-1 Nessler univariada
ELSHAFEI; EL-GHONEMY
(2015)
Egito
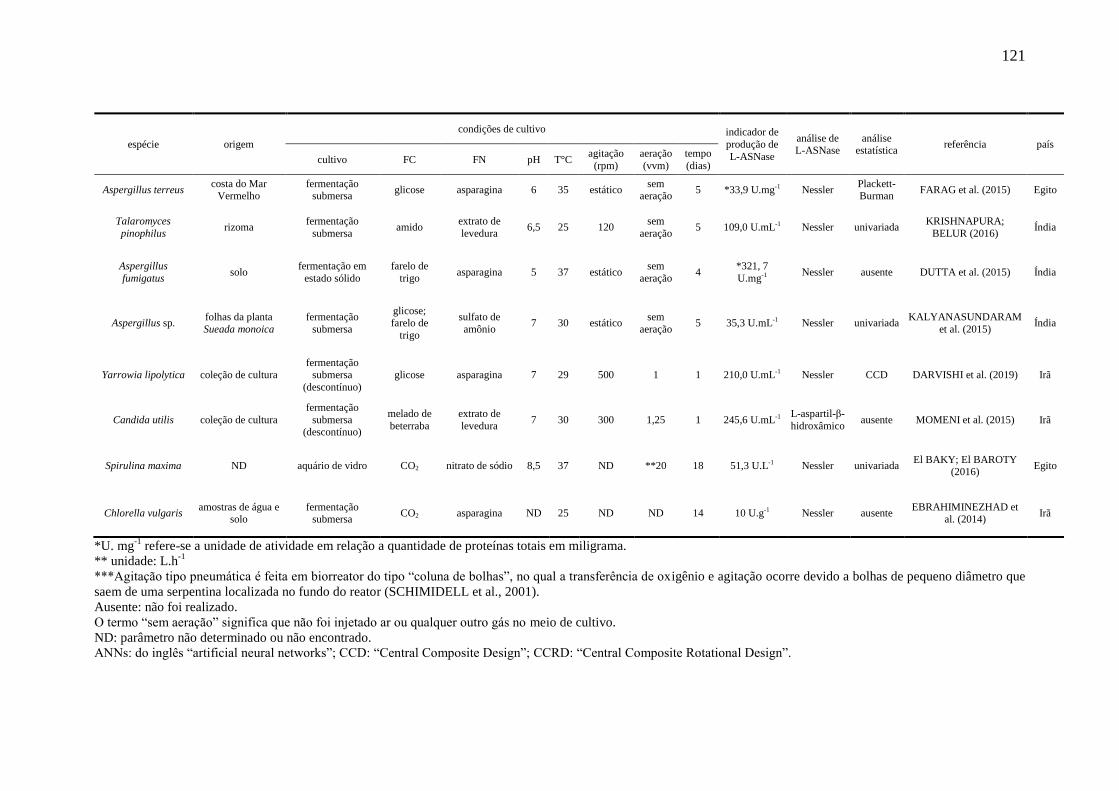
121
espécie origem
condições de cultivo indicador de
produção de
L-ASNase
análise de L-ASNase
análise estatística
referência país
cultivo FC FN pH T°C agitação
(rpm)
aeração
(vvm)
tempo
(dias)
Aspergillus terreus costa do Mar
Vermelho fermentação
submersa glicose asparagina 6 35 estático
sem aeração
5 *33,9 U.mg-1 Nessler Plackett- Burman
FARAG et al. (2015) Egito
Talaromyces
pinophilus rizoma
fermentação
submersa amido
extrato de
levedura 6,5 25 120
sem
aeração 5 109,0 U.mL-1 Nessler univariada
KRISHNAPURA;
BELUR (2016) Índia
Aspergillus
fumigatus solo
fermentação em
estado sólido
farelo de
trigo asparagina 5 37 estático
sem
aeração 4
*321, 7
U.mg-1 Nessler ausente DUTTA et al. (2015) Índia
Aspergillus sp. folhas da planta
Sueada monoica
fermentação
submersa
glicose; farelo de
trigo
sulfato de
amônio 7 30 estático
sem
aeração 5 35,3 U.mL-1 Nessler univariada
KALYANASUNDARAM
et al. (2015) Índia
Yarrowia lipolytica coleção de cultura fermentação
submersa
(descontínuo)
glicose asparagina 7 29 500 1 1 210,0 U.mL-1 Nessler CCD DARVISHI et al. (2019) Irã
Candida utilis coleção de cultura
fermentação
submersa (descontínuo)
melado de
beterraba
extrato de
levedura 7 30 300 1,25 1 245,6 U.mL-1
L-aspartil-β-
hidroxâmico ausente MOMENI et al. (2015) Irã
Spirulina maxima ND aquário de vidro CO2 nitrato de sódio 8,5 37 ND **20 18 51,3 U.L-1 Nessler univariada El BAKY; El BAROTY
(2016) Egito
Chlorella vulgaris amostras de água e
solo fermentação
submersa CO2 asparagina ND 25 ND ND 14 10 U.g-1 Nessler ausente
EBRAHIMINEZHAD et al. (2014)
Irã
*U. mg-1
refere-se a unidade de atividade em relação a quantidade de proteínas totais em miligrama.
** unidade: L.h-1
***Agitação tipo pneumática é feita em biorreator do tipo “coluna de bolhas”, no qual a transferência de oxigênio e agitação ocorre devido a bolhas de pequeno diâmetro que
saem de uma serpentina localizada no fundo do reator (SCHIMIDELL et al., 2001).
Ausente: não foi realizado.
O termo “sem aeração” significa que não foi injetado ar ou qualquer outro gás no meio de cultivo.
ND: parâmetro não determinado ou não encontrado.
ANNs: do inglês “artificial neural networks”; CCD: “Central Composite Design”; CCRD: “Central Composite Rotational Design”.
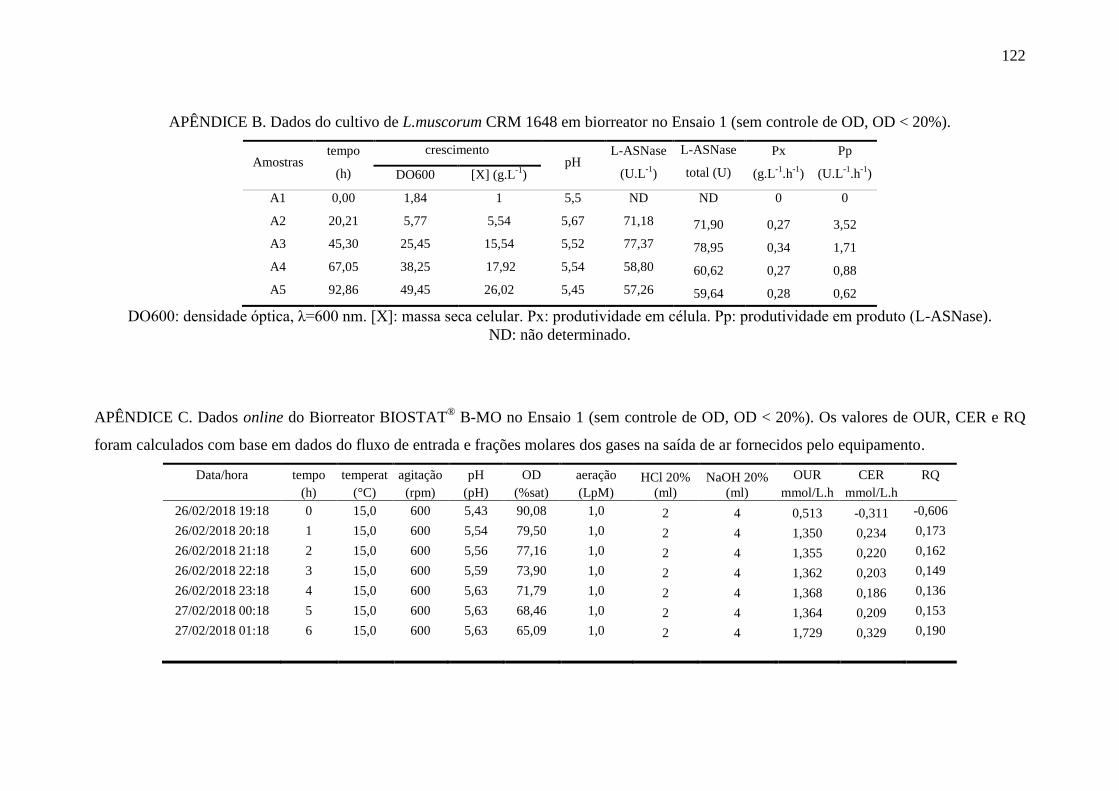
122
APÊNDICE B. Dados do cultivo de L.muscorum CRM 1648 em biorreator no Ensaio 1 (sem controle de OD, OD < 20%).
Amostras tempo
(h)
crescimento pH
L-ASNase
(U.L-1
)
L-ASNase
total (U)
Px
(g.L-1
.h-1
)
Pp
(U.L-1
.h-1
) DO600 [X] (g.L-1
)
A1 0,00 1,84 1 5,5 ND ND 0 0
A2 20,21 5,77 5,54 5,67 71,18 71,90 0,27 3,52
A3 45,30 25,45 15,54 5,52 77,37 78,95 0,34 1,71
A4 67,05 38,25 17,92 5,54 58,80 60,62 0,27 0,88
A5 92,86 49,45 26,02 5,45 57,26 59,64 0,28 0,62
DO600: densidade óptica, λ=600 nm. [X]: massa seca celular. Px: produtividade em célula. Pp: produtividade em produto (L-ASNase).
ND: não determinado.
APÊNDICE C. Dados online do Biorreator BIOSTAT® B-MO no Ensaio 1 (sem controle de OD, OD < 20%). Os valores de OUR, CER e RQ
foram calculados com base em dados do fluxo de entrada e frações molares dos gases na saída de ar fornecidos pelo equipamento.
Data/hora tempo temperat agitação pH OD aeração HCl 20% NaOH 20% OUR CER RQ
(h) (°C) (rpm) (pH) (%sat) (LpM) (ml) (ml) mmol/L.h mmol/L.h
26/02/2018 19:18 0 15,0 600 5,43 90,08 1,0 2 4 0,513 -0,311 -0,606
26/02/2018 20:18 1 15,0 600 5,54 79,50 1,0 2 4 1,350 0,234 0,173
26/02/2018 21:18 2 15,0 600 5,56 77,16 1,0 2 4 1,355 0,220 0,162
26/02/2018 22:18 3 15,0 600 5,59 73,90 1,0 2 4 1,362 0,203 0,149
26/02/2018 23:18 4 15,0 600 5,63 71,79 1,0 2 4 1,368 0,186 0,136
27/02/2018 00:18 5 15,0 600 5,63 68,46 1,0 2 4 1,364 0,209 0,153
27/02/2018 01:18 6 15,0 600 5,63 65,09 1,0 2 4 1,729 0,329 0,190

123
Data/hora tempo temperat agitação pH OD aeração HCl 20% NaOH 20% OUR CER RQ
(h) (°C) (rpm) (pH) (%sat) (LpM) (ml) (ml) mmol/L.h mmol/L.h
27/02/2018 02:18 7 15,0 600 5,64 62,60 1,0 2 4 2,092 0,500 0,239
27/02/2018 03:18 8 15,0 600 5,68 60,19 1,0 2 4 2,636 0,657 0,249
27/02/2018 04:18 9 15,0 600 5,73 57,66 1,0 2 4 3,039 0,929 0,306
27/02/2018 05:18 10 15,0 600 5,79 55,04 1,0 2 4 3,142 1,025 0,326
27/02/2018 06:18 11 15,0 600 5,85 52,71 1,0 2 4 3,102 1,205 0,388
27/02/2018 07:18 12 15,0 600 5,90 50,89 1,0 2 4 3,467 1,341 0,387
27/02/2018 08:18 13 15,0 600 5,95 49,45 1,0 2 4 3,011 1,589 0,528
27/02/2018 09:18 14 15,0 600 6,01 47,39 1,0 2 4 3,372 1,745 0,517
27/02/2018 10:18 15 15,0 600 6,06 45,21 1,0 2 4 3,131 2,126 0,679
27/02/2018 11:18 16 15,0 600 6,12 42,72 1,0 2 4 2,878 2,505 0,870
27/02/2018 12:18 17 15,0 600 6,18 40,28 1,0 2 4 2,775 2,887 1,040
27/02/2018 13:18 18 15,0 600 5,54 38,77 1,0 4 9 3,376 3,376 1,000
27/02/2018 14:18 19 15,0 600 5,55 35,81 1,0 4 9 3,313 3,877 1,170
27/02/2018 15:18 20 15,0 600 5,54 33,33 1,0 4 9 4,098 4,499 1,098
27/02/2018 16:18 21 15,0 600 5,55 29,54 1,0 5 9 3,959 5,081 1,283
27/02/2018 17:18 22 15,0 600 5,54 25,92 1,0 5 9 5,407 5,712 1,056
27/02/2018 18:18 23 15,0 600 5,55 23,56 1,0 5 9 6,521 6,135 0,941
27/02/2018 19:18 24 15,0 600 5,55 20,86 1,0 5 9 7,005 6,690 0,955
27/02/2018 20:18 25 15,0 600 5,54 19,85 1,0 5 9 8,274 6,963 0,842
27/02/2018 21:18 26 15,0 600 5,52 19,07 1,0 5 9 8,448 7,418 0,878
27/02/2018 22:18 27 15,0 600 5,54 18,50 1,0 5 9 8,839 7,520 0,851
27/02/2018 23:18 28 15,0 600 5,53 17,75 1,0 5 9 8,826 7,657 0,868
28/02/2018 00:18 29 15,0 600 5,55 17,30 1,0 5 9 9,264 7,835 0,846
28/02/2018 01:18 30 15,0 600 5,53 16,61 1,0 5 9 9,349 8,103 0,867
28/02/2018 02:18 31 15,0 600 5,55 16,10 1,0 6 9 9,662 8,523 0,882

124
Data/hora tempo temperat agitação pH OD aeração HCl 20% NaOH 20% OUR CER RQ
(h) (°C) (rpm) (pH) (%sat) (LpM) (ml) (ml) mmol/L.h mmol/L.h
28/02/2018 03:18 32 15,0 600 5,53 15,89 1,0 6 9 9,645 8,682 0,900
28/02/2018 04:18 33 15,0 600 5,55 15,43 1,0 6 9 9,991 8,935 0,894
28/02/2018 05:18 34 15,0 600 5,53 14,90 1,0 6 9 10,556 9,212 0,873
28/02/2018 06:18 35 15,0 600 5,55 14,45 1,0 6 9 10,670 9,520 0,892
28/02/2018 07:18 36 15,0 600 5,53 15,10 1,0 6 9 8,913 9,728 1,091
28/02/2018 08:18 37 15,0 600 5,51 15,59 1,0 6 9 7,500 9,819 1,309
28/02/2018 09:18 38 15,0 600 5,52 14,86 1,0 6 9 7,981 9,947 1,246
28/02/2018 10:18 39 15,0 600 5,55 14,70 1,0 6 9 7,800 9,959 1,277
28/02/2018 11:18 40 15,0 600 5,48 14,70 1,0 6 9 10,045 12,085 1,203
28/02/2018 12:18 41 15,0 600 5,12 12,52 1,0 16,48 21,97 8,804 12,601 1,431
28/02/2018 13:18 42 14,9 600 6,56 11,87 1,0 32,48 25,97 6,513 9,453 1,451
28/02/2018 14:18 43 15,0 600 6,62 12,33 1,0 34,48 25,97 7,987 10,229 1,281
28/02/2018 15:18 44 15,0 600 6,70 11,97 1,0 35,48 25,97 5,997 9,948 1,659
28/02/2018 16:18 45 15,0 600 6,74 11,21 1,0 36,48 25,97 6,544 9,507 1,453
28/02/2018 17:18 46 15,0 600 5,53 11,05 1,0 37,48 26,97 8,083 10,069 1,246
28/02/2018 18:18 47 15,0 600 5,52 10,74 1,0 37,48 26,97 8,696 10,465 1,204
28/02/2018 19:18 48 15,0 600 5,51 10,70 1,0 37,48 26,97 9,402 10,732 1,141
28/02/2018 20:18 49 15,0 600 5,52 10,26 1,0 37,48 26,97 9,447 10,363 1,097
28/02/2018 21:18 50 15,0 600 5,50 9,56 1,0 37,48 26,97 9,577 10,168 1,062
28/02/2018 22:18 51 15,0 600 5,49 9,23 1,0 37,48 26,97 9,490 10,050 1,059
28/02/2018 23:18 52 15,0 600 5,45 9,01 1,0 37,48 26,97 9,633 10,127 1,051
01/03/2018 00:18 53 15,0 600 5,47 8,85 1,0 37,48 26,97 9,404 9,928 1,056
01/03/2018 01:18 54 15,0 600 5,50 8,64 1,0 37,48 26,97 9,410 9,713 1,032
01/03/2018 02:18 55 15,0 600 5,53 8,39 1,0 37,48 26,97 9,120 9,305 1,020
01/03/2018 03:18 56 15,0 600 5,47 8,31 1,0 37,48 26,97 9,485 9,331 0,984

125
Data/hora tempo temperat agitação pH OD aeração HCl 20% NaOH 20% OUR CER RQ
(h) (°C) (rpm) (pH) (%sat) (LpM) (ml) (ml) mmol/L.h mmol/L.h
01/03/2018 04:18 57 15,0 600 5,50 8,11 1,0 37,48 26,97 9,529 9,243 0,970
01/03/2018 05:18 58 15,0 600 5,46 7,91 1,0 37,48 26,97 9,550 9,116 0,955
01/03/2018 06:18 59 15,0 600 5,46 7,96 1,0 37,48 26,97 9,569 9,205 0,962
01/03/2018 07:18 60 15,0 600 5,54 8,04 1,0 37,48 26,97 9,100 9,173 1,008
01/03/2018 08:18 61 15,0 600 5,55 8,24 1,0 37,48 26,97 8,569 9,112 1,063
01/03/2018 09:18 62 15,0 600 5,53 8,11 1,0 37,48 26,97 6,958 8,809 1,266
01/03/2018 10:18 63 15,0 600 5,46 8,16 1,0 37,48 26,97 7,009 8,604 1,228
01/03/2018 11:18 64 14,9 600 5,18 6,02 1,0 37,48 32,97 9,630 10,873 1,129
01/03/2018 12:18 65 15,0 600 5,55 7,25 1,0 37,48 32,97 9,747 10,949 1,123
01/03/2018 13:18 66 15,0 600 4,61 6,13 1,0 37,48 32,97 11,105 13,560 1,221
01/03/2018 14:18 67 15,0 600 5,56 6,07 1,0 37,48 32,97 13,288 12,694 0,955
01/03/2018 15:18 68 15,0 600 5,54 6,40 1,0 37,48 32,97 11,062 10,976 0,992
01/03/2018 16:18 69 15,0 600 5,51 6,71 1,0 37,48 32,97 15,126 14,196 0,939
01/03/2018 17:18 70 14,9 600 5,56 5,54 1,0 38,48 33,77 17,984 14,095 0,784
01/03/2018 18:18 71 15,0 600 5,51 5,60 1,0 38,48 33,77 16,205 14,529 0,897
01/03/2018 19:18 72 15,0 600 5,56 5,67 1,0 38,48 33,77 16,781 14,949 0,891
01/03/2018 20:18 73 15,0 600 5,53 5,60 1,0 38,48 33,77 16,856 14,915 0,885
01/03/2018 21:18 74 15,0 600 5,56 5,61 1,0 38,48 33,77 18,323 14,846 0,810
01/03/2018 22:18 75 15,0 600 5,51 5,55 1,0 38,48 33,77 18,417 14,877 0,808
01/03/2018 23:18 76 14,9 600 5,54 23,01 1,0 38,48 33,77 7,796 6,733 0,864
02/03/2018 00:18 77 15,0 600 5,54 27,62 1,0 38,48 33,77 5,746 2,843 0,495
02/03/2018 01:18 78 15,0 600 5,53 28,41 1,0 38,48 33,77 5,384 2,258 0,419
02/03/2018 02:18 79 15,0 600 5,51 28,87 1,0 38,48 33,77 5,259 1,890 0,359
02/03/2018 03:18 80 15,0 600 5,54 29,18 1,0 38,48 33,77 5,347 1,598 0,299
02/03/2018 04:18 81 15,0 600 5,52 29,33 1,0 38,48 33,77 4,732 1,385 0,293
02/03/2018 05:18 82 15,0 600 5,55 29,53 1,0 38,48 33,77 4,959 1,207 0,243
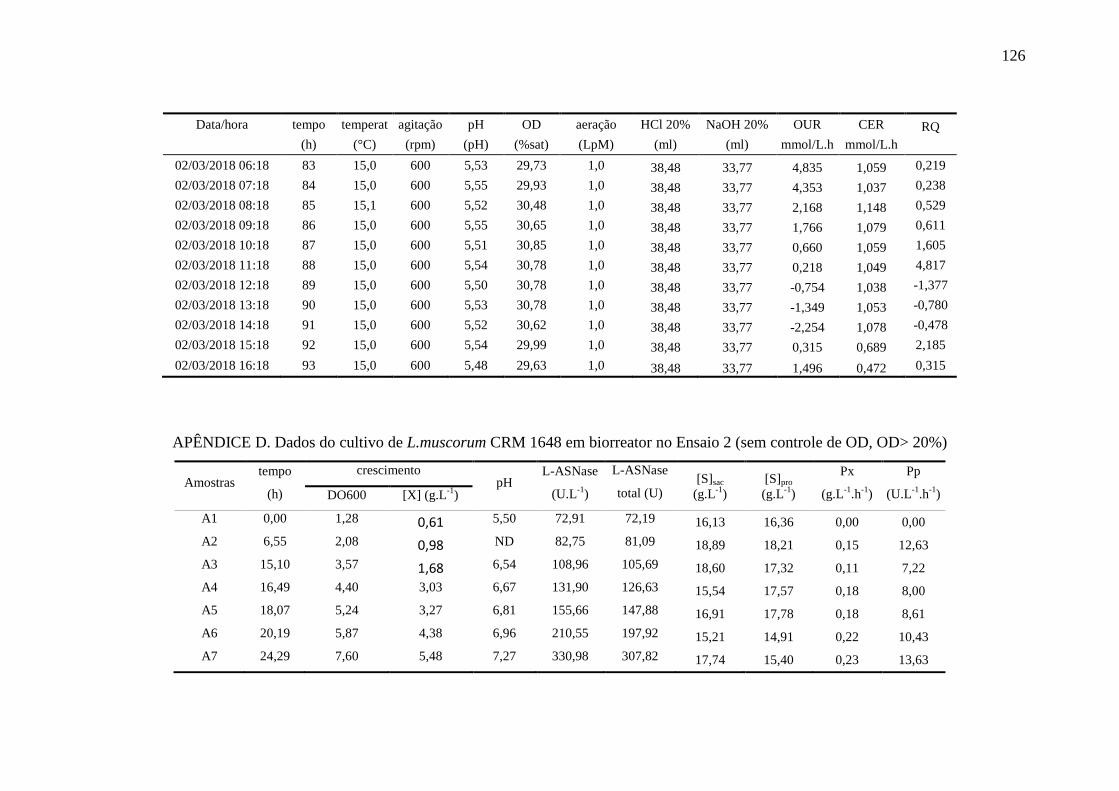
126
Data/hora tempo temperat agitação pH OD aeração HCl 20% NaOH 20% OUR CER RQ
(h) (°C) (rpm) (pH) (%sat) (LpM) (ml) (ml) mmol/L.h mmol/L.h
02/03/2018 06:18 83 15,0 600 5,53 29,73 1,0 38,48 33,77 4,835 1,059 0,219
02/03/2018 07:18 84 15,0 600 5,55 29,93 1,0 38,48 33,77 4,353 1,037 0,238
02/03/2018 08:18 85 15,1 600 5,52 30,48 1,0 38,48 33,77 2,168 1,148 0,529
02/03/2018 09:18 86 15,0 600 5,55 30,65 1,0 38,48 33,77 1,766 1,079 0,611
02/03/2018 10:18 87 15,0 600 5,51 30,85 1,0 38,48 33,77 0,660 1,059 1,605
02/03/2018 11:18 88 15,0 600 5,54 30,78 1,0 38,48 33,77 0,218 1,049 4,817
02/03/2018 12:18 89 15,0 600 5,50 30,78 1,0 38,48 33,77 -0,754 1,038 -1,377
02/03/2018 13:18 90 15,0 600 5,53 30,78 1,0 38,48 33,77 -1,349 1,053 -0,780
02/03/2018 14:18 91 15,0 600 5,52 30,62 1,0 38,48 33,77 -2,254 1,078 -0,478
02/03/2018 15:18 92 15,0 600 5,54 29,99 1,0 38,48 33,77 0,315 0,689 2,185
02/03/2018 16:18 93 15,0 600 5,48 29,63 1,0 38,48 33,77 1,496 0,472 0,315
APÊNDICE D. Dados do cultivo de L.muscorum CRM 1648 em biorreator no Ensaio 2 (sem controle de OD, OD> 20%)
Amostras tempo
(h)
crescimento pH
L-ASNase
(U.L-1
)
L-ASNase
total (U) [S]sac
(g.L-1
)
[S]pro
(g.L-1
)
Px
(g.L-1
.h-1
)
Pp
(U.L-1
.h-1
) DO600 [X] (g.L-1
)
A1 0,00 1,28 0,61 5,50 72,91 72,19 16,13 16,36 0,00 0,00
A2 6,55 2,08 0,98 ND 82,75 81,09 18,89 18,21 0,15 12,63
A3 15,10 3,57 1,68 6,54 108,96 105,69 18,60 17,32 0,11 7,22
A4 16,49 4,40 3,03 6,67 131,90 126,63 15,54 17,57 0,18 8,00
A5 18,07 5,24 3,27 6,81 155,66 147,88 16,91 17,78 0,18 8,61
A6 20,19 5,87 4,38 6,96 210,55 197,92 15,21 14,91 0,22 10,43
A7 24,29 7,60 5,48 7,27 330,98 307,82 17,74 15,40 0,23 13,63
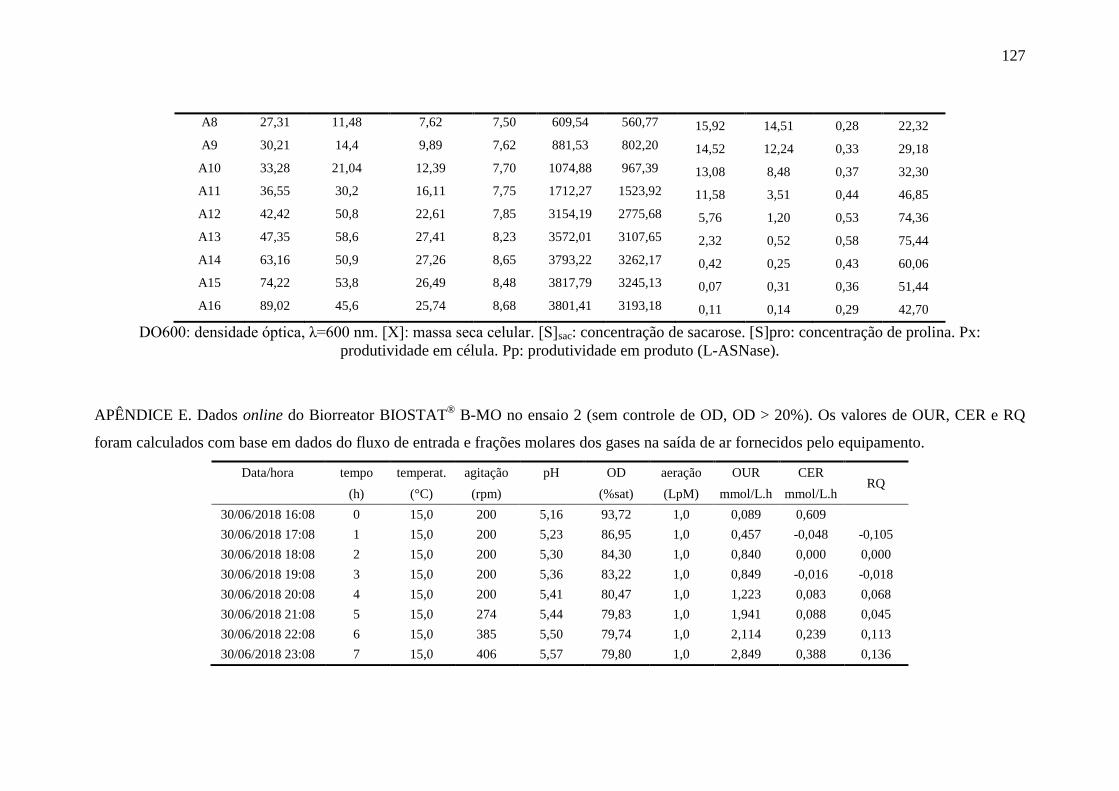
127
A8 27,31 11,48 7,62 7,50 609,54 560,77 15,92 14,51 0,28 22,32
A9 30,21 14,4 9,89 7,62 881,53 802,20 14,52 12,24 0,33 29,18
A10 33,28 21,04 12,39 7,70 1074,88 967,39 13,08 8,48 0,37 32,30
A11 36,55 30,2 16,11 7,75 1712,27 1523,92 11,58 3,51 0,44 46,85
A12 42,42 50,8 22,61 7,85 3154,19 2775,68 5,76 1,20 0,53 74,36
A13 47,35 58,6 27,41 8,23 3572,01 3107,65 2,32 0,52 0,58 75,44
A14 63,16 50,9 27,26 8,65 3793,22 3262,17 0,42 0,25 0,43 60,06
A15 74,22 53,8 26,49 8,48 3817,79 3245,13 0,07 0,31 0,36 51,44
A16 89,02 45,6 25,74 8,68 3801,41 3193,18 0,11 0,14 0,29 42,70
DO600: densidade óptica, λ=600 nm. [X]: massa seca celular. [S]sac: concentração de sacarose. [S]pro: concentração de prolina. Px:
produtividade em célula. Pp: produtividade em produto (L-ASNase).
APÊNDICE E. Dados online do Biorreator BIOSTAT® B-MO no ensaio 2 (sem controle de OD, OD > 20%). Os valores de OUR, CER e RQ
foram calculados com base em dados do fluxo de entrada e frações molares dos gases na saída de ar fornecidos pelo equipamento.
Data/hora tempo temperat. agitação pH OD aeração OUR CER RQ
(h) (°C) (rpm) (%sat) (LpM) mmol/L.h mmol/L.h
30/06/2018 16:08 0 15,0 200 5,16 93,72 1,0 0,089 0,609
30/06/2018 17:08 1 15,0 200 5,23 86,95 1,0 0,457 -0,048 -0,105
30/06/2018 18:08 2 15,0 200 5,30 84,30 1,0 0,840 0,000 0,000
30/06/2018 19:08 3 15,0 200 5,36 83,22 1,0 0,849 -0,016 -0,018
30/06/2018 20:08 4 15,0 200 5,41 80,47 1,0 1,223 0,083 0,068
30/06/2018 21:08 5 15,0 274 5,44 79,83 1,0 1,941 0,088 0,045
30/06/2018 22:08 6 15,0 385 5,50 79,74 1,0 2,114 0,239 0,113
30/06/2018 23:08 7 15,0 406 5,57 79,80 1,0 2,849 0,388 0,136

128
Data/hora tempo temperat. agitação pH OD aeração OUR CER RQ
(h) (°C) (rpm) (%sat) (LpM) mmol/L.h mmol/L.h
01/07/2018 00:08 8 15,0 555 5,70 79,72 1,0 2,737 0,563 0,206
01/07/2018 01:08 9 15,0 626 5,79 79,35 1,0 3,118 0,694 0,223
01/07/2018 02:08 10 15,0 1116 5,81 78,89 1,0 3,822 0,893 0,234
01/07/2018 03:08 11 15,0 1200 5,70 79,53 1,0 3,686 0,982 0,266
01/07/2018 04:08 12 15,0 1200 5,69 77,40 1,0 4,196 1,098 0,262
01/07/2018 05:08 13 15,0 1200 5,70 74,57 1,0 4,550 1,332 0,293
01/07/2018 06:08 14 15,0 1200 5,72 71,95 1,0 4,812 1,634 0,340
01/07/2018 07:08 15 15,0 1200 5,75 70,52 1,0 5,084 1,874 0,369
01/07/2018 08:08 16 14,9 1200 5,82 69,84 1,0 5,333 2,214 0,415
01/07/2018 09:08 17 15,0 1200 5,93 67,25 1,0 5,247 2,644 0,504
01/07/2018 10:08 18 15,0 1200 6,04 65,30 1,0 5,168 3,056 0,591
01/07/2018 11:08 19 15,0 1200 6,13 63,43 1,0 5,565 3,628 0,652
01/07/2018 12:08 20 15,0 1200 6,22 61,89 1,0 5,498 4,139 0,753
01/07/2018 13:08 21 15,0 1200 6,30 61,53 1,0 6,253 4,800 0,768
01/07/2018 14:08 22 15,0 1200 6,38 60,84 1,5 6,109 5,373 0,880
01/07/2018 15:08 23 15,0 1200 6,44 59,57 1,5 6,257 6,207 0,992
01/07/2018 16:08 24 15,0 1200 6,52 58,18 1,5 6,556 6,883 1,050
01/07/2018 17:08 25 15,0 1200 6,62 55,32 1,5 7,553 7,331 0,971
01/07/2018 18:08 26 15,0 1200 6,71 52,36 1,5 9,807 8,107 0,827
01/07/2018 19:08 27 15,0 1200 6,77 50,60 1,5 11,308 8,920 0,789
01/07/2018 20:08 28 14,9 1200 6,84 49,01 1,5 13,366 9,729 0,728
01/07/2018 21:08 29 15,0 1200 6,91 47,58 1,5 13,959 10,694 0,766
01/07/2018 22:08 30 15,0 1200 6,94 46,62 1,5 15,352 12,030 0,784
01/07/2018 23:08 31 15,0 1200 6,95 45,46 1,5 16,033 12,861 0,802
02/07/2018 00:08 32 15,0 1200 7,04 44,02 1,5 17,682 14,141 0,800
02/07/2018 01:08 33 15,0 1200 7,09 43,01 1,5 18,110 14,991 0,828

129
Data/hora tempo temperat. agitação pH OD aeração OUR CER RQ
(h) (°C) (rpm) (%sat) (LpM) mmol/L.h mmol/L.h
02/07/2018 02:08 34 15,0 1200 7,11 41,91 1,5 18,890 15,664 0,829
02/07/2018 03:08 35 15,0 1200 7,10 40,86 1,5 21,233 16,972 0,799
02/07/2018 04:08 36 15,0 1200 7,09 39,87 1,5 21,847 18,405 0,842
02/07/2018 05:08 37 15,0 1200 7,11 39,16 1,5 21,846 18,930 0,867
02/07/2018 06:08 38 15,0 1200 7,12 37,89 1,5 22,720 20,022 0,881
02/07/2018 07:08 39 15,0 1200 7,12 37,19 1,5 25,060 20,867 0,833
02/07/2018 08:08 40 15,0 1200 7,13 36,95 1,5 24,776 21,440 0,865
02/07/2018 09:08 41 15,0 1200 7,14 36,97 1,5 25,051 22,226 0,887
02/07/2018 10:08 42 15,0 1200 7,17 36,89 1,5 24,959 22,889 0,917
02/07/2018 11:08 43 15,0 1200 7,21 36,89 1,5 24,187 23,068 0,954
02/07/2018 12:08 44 15,0 1200 7,25 36,43 1,5 22,777 23,196 1,018
02/07/2018 13:08 45 15,0 1200 7,30 36,32 1,5 22,270 22,743 1,021
02/07/2018 14:08 46 15,0 1200 7,39 36,66 1,5 18,769 17,029 0,907
02/07/2018 15:08 47 15,0 1200 7,50 37,36 1,5 14,218 12,959 0,911
02/07/2018 16:08 48 15,0 1200 7,58 37,00 1,5 12,221 10,961 0,897
02/07/2018 17:08 49 15,0 1200 7,63 35,84 1,5 11,547 9,778 0,847
02/07/2018 18:08 50 15,0 1200 7,67 34,84 1,5 11,837 8,944 0,756
02/07/2018 19:08 51 15,0 1200 7,71 34,44 1,5 12,047 8,399 0,697
02/07/2018 20:08 52 15,0 1200 7,75 33,98 1,5 11,451 7,837 0,684
02/07/2018 21:08 53 15,0 1200 7,78 33,51 1,5 11,675 7,521 0,644
02/07/2018 22:08 54 15,0 1200 7,80 33,16 1,5 11,797 7,320 0,621
02/07/2018 23:08 55 15,0 1200 7,81 32,71 1,5 11,908 7,193 0,604
03/07/2018 00:08 56 15,0 1200 7,82 32,34 1,5 12,076 7,127 0,590
03/07/2018 01:08 57 15,0 1200 7,83 31,99 1,5 12,284 7,021 0,572
03/07/2018 02:08 58 15,0 1200 7,84 31,58 1,5 12,187 6,978 0,573
03/07/2018 03:08 59 15,0 1200 7,87 31,29 1,5 12,328 6,741 0,547

130
Data/hora
tempo temperat. agitação pH
OD aeração OUR CER RQ
(h) (°C) (rpm) (%sat) (LpM) mmol/L.h mmol/L.h
03/07/2018 04:08 60 15,0 1200 7,92 31,00 1,5 13,310 7,324 0,550
03/07/2018 05:08 61 15,0 1200 7,61 31,00 1,5 13,162 7,207 0,548
03/07/2018 06:08 62 15,0 1200 7,51 30,94 1,5 13,324 6,819 0,512
03/07/2018 07:08 63 15,0 1200 7,47 30,76 1,5 13,230 6,673 0,504
03/07/2018 08:08 64 15,0 1200 7,48 30,71 1,5 13,010 5,654 0,435
03/07/2018 09:08 65 15,0 1200 7,53 30,76 1,5 11,079 5,687 0,513
03/07/2018 10:08 66 15,0 1200 7,59 30,82 1,5 9,948 5,651 0,568
03/07/2018 11:08 67 15,0 1200 7,64 30,71 1,5 9,195 5,682 0,618
03/07/2018 12:08 68 15,0 1200 7,67 30,45 1,5 9,381 5,180 0,552
03/07/2018 13:08 69 15,0 1200 7,69 30,12 1,5 9,745 5,055 0,519
03/07/2018 14:08 70 15,0 1200 7,71 29,89 1,5 9,148 5,038 0,551
03/07/2018 15:08 71 15,0 1200 7,72 29,60 1,5 9,667 4,837 0,500
03/07/2018 16:08 72 15,0 1200 7,73 29,42 1,5 9,777 4,645 0,475
03/07/2018 17:08 73 15,0 1200 7,74 29,07 1,5 10,750 4,377 0,407
03/07/2018 18:08 74 15,0 1200 7,75 28,84 1,5 12,101 4,347 0,359
03/07/2018 19:08 75 15,0 1200 7,75 28,66 1,5 10,985 4,034 0,367
03/07/2018 20:08 76 15,0 1200 7,75 28,43 1,5 12,150 3,889 0,320
03/07/2018 21:08 77 15,0 1200 7,76 28,31 1,5 12,488 3,771 0,302
03/07/2018 22:08 78 15,0 1200 7,76 28,25 1,5 13,684 3,622 0,265
03/07/2018 23:08 79 15,0 1200 7,76 28,14 1,5 12,759 3,377 0,265
04/07/2018 00:08 80 15,0 1200 7,77 28,08 1,5 12,892 3,178 0,246
04/07/2018 01:08 81 15,0 1200 7,78 28,02 1,5 12,967 3,238 0,250
04/07/2018 02:08 82 15,0 1200 7,79 27,90 1,5 9,686 3,009 0,311
04/07/2018 03:08 83 15,0 1200 7,80 27,85 1,5 12,793 2,854 0,223
04/07/2018 04:08 84 15,0 1200 7,81 27,85 1,5 11,960 2,945 0,246
04/07/2018 05:08 85 15,0 1200 7,81 27,81 1,5 12,126 2,624 0,216
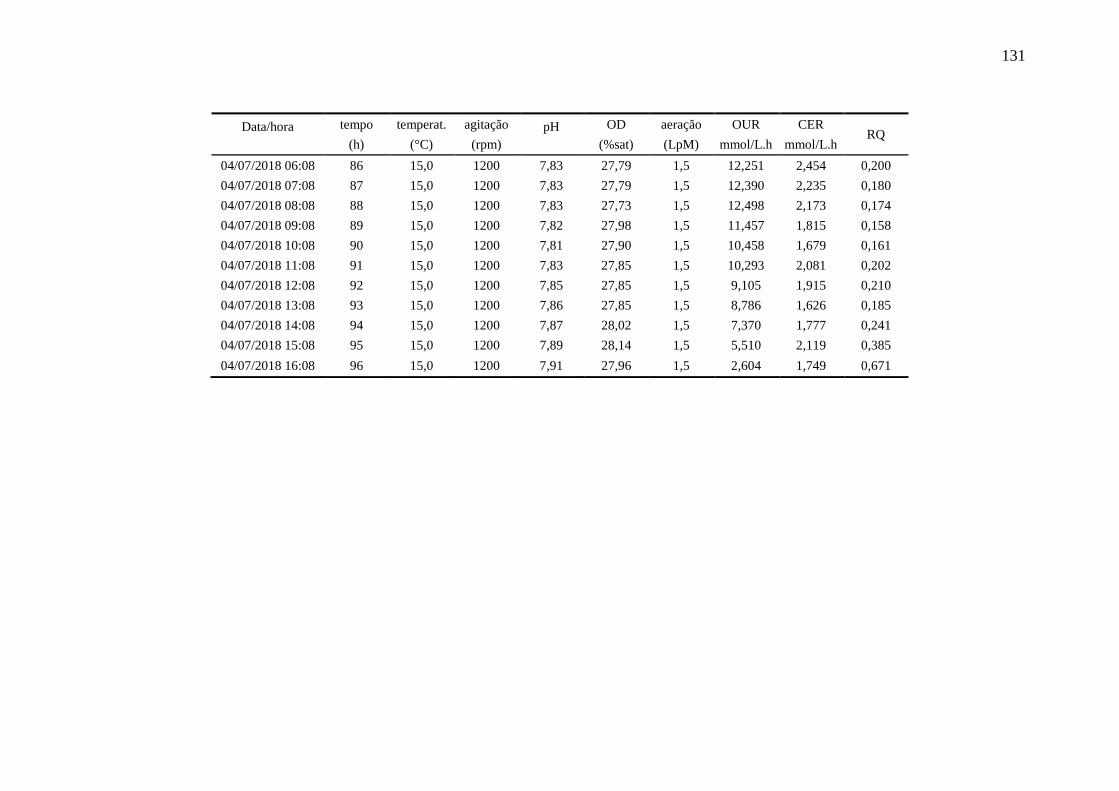
131
Data/hora
tempo temperat. agitação pH
OD aeração OUR CER RQ
(h) (°C) (rpm) (%sat) (LpM) mmol/L.h mmol/L.h
04/07/2018 06:08 86 15,0 1200 7,83 27,79 1,5 12,251 2,454 0,200
04/07/2018 07:08 87 15,0 1200 7,83 27,79 1,5 12,390 2,235 0,180
04/07/2018 08:08 88 15,0 1200 7,83 27,73 1,5 12,498 2,173 0,174
04/07/2018 09:08 89 15,0 1200 7,82 27,98 1,5 11,457 1,815 0,158
04/07/2018 10:08 90 15,0 1200 7,81 27,90 1,5 10,458 1,679 0,161
04/07/2018 11:08 91 15,0 1200 7,83 27,85 1,5 10,293 2,081 0,202
04/07/2018 12:08 92 15,0 1200 7,85 27,85 1,5 9,105 1,915 0,210
04/07/2018 13:08 93 15,0 1200 7,86 27,85 1,5 8,786 1,626 0,185
04/07/2018 14:08 94 15,0 1200 7,87 28,02 1,5 7,370 1,777 0,241
04/07/2018 15:08 95 15,0 1200 7,89 28,14 1,5 5,510 2,119 0,385
04/07/2018 16:08 96 15,0 1200 7,91 27,96 1,5 2,604 1,749 0,671
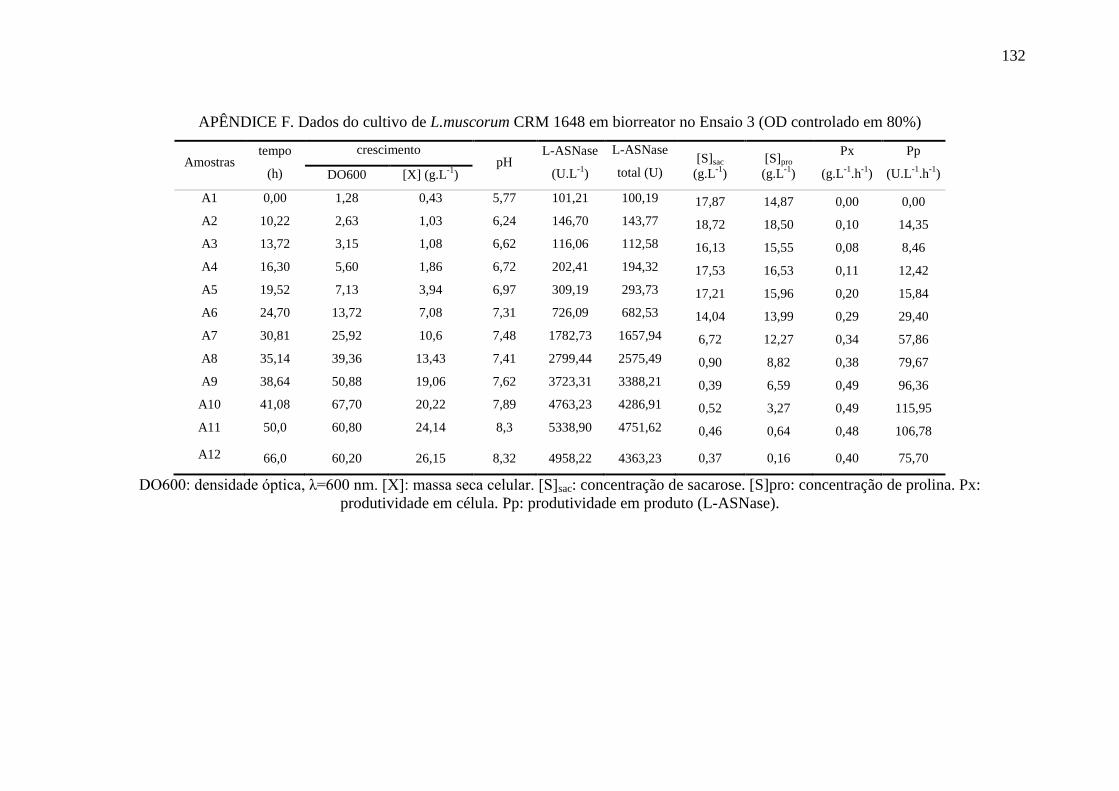
132
APÊNDICE F. Dados do cultivo de L.muscorum CRM 1648 em biorreator no Ensaio 3 (OD controlado em 80%)
Amostras tempo
(h)
crescimento pH
L-ASNase
(U.L-1
)
L-ASNase
total (U) [S]sac
(g.L-1
)
[S]pro
(g.L-1
)
Px
(g.L-1
.h-1
)
Pp
(U.L-1
.h-1
) DO600 [X] (g.L-1
)
A1 0,00 1,28 0,43 5,77 101,21 100,19 17,87 14,87 0,00 0,00
A2 10,22 2,63 1,03 6,24 146,70 143,77 18,72 18,50 0,10 14,35
A3 13,72 3,15 1,08 6,62 116,06 112,58 16,13 15,55 0,08 8,46
A4 16,30 5,60 1,86 6,72 202,41 194,32 17,53 16,53 0,11 12,42
A5 19,52 7,13 3,94 6,97 309,19 293,73 17,21 15,96 0,20 15,84
A6 24,70 13,72 7,08 7,31 726,09 682,53 14,04 13,99 0,29 29,40
A7 30,81 25,92 10,6 7,48 1782,73 1657,94 6,72 12,27 0,34 57,86
A8 35,14 39,36 13,43 7,41 2799,44 2575,49 0,90 8,82 0,38 79,67
A9 38,64 50,88 19,06 7,62 3723,31 3388,21 0,39 6,59 0,49 96,36
A10 41,08 67,70 20,22 7,89 4763,23 4286,91 0,52 3,27 0,49 115,95
A11 50,0 60,80 24,14 8,3 5338,90 4751,62 0,46 0,64 0,48 106,78
A12 66,0 60,20 26,15 8,32 4958,22 4363,23 0,37 0,16 0,40 75,70
DO600: densidade óptica, λ=600 nm. [X]: massa seca celular. [S]sac: concentração de sacarose. [S]pro: concentração de prolina. Px:
produtividade em célula. Pp: produtividade em produto (L-ASNase).
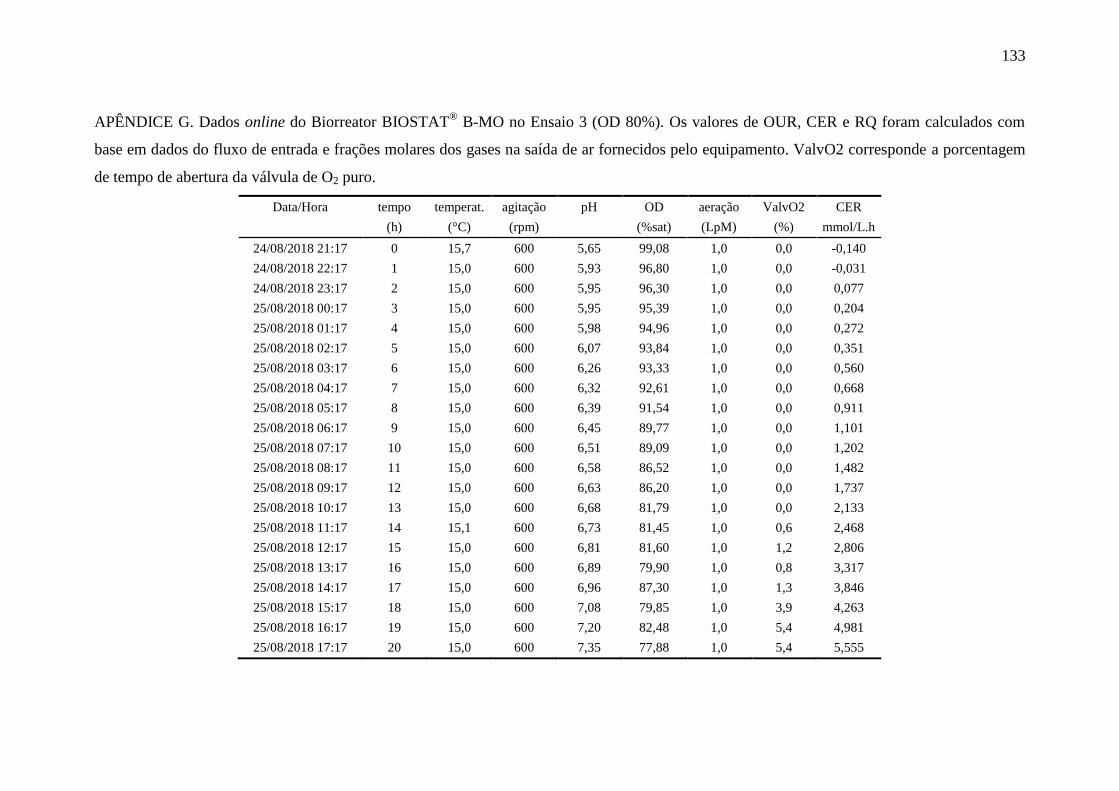
133
APÊNDICE G. Dados online do Biorreator BIOSTAT® B-MO no Ensaio 3 (OD 80%). Os valores de OUR, CER e RQ foram calculados com
base em dados do fluxo de entrada e frações molares dos gases na saída de ar fornecidos pelo equipamento. ValvO2 corresponde a porcentagem
de tempo de abertura da válvula de O2 puro.
Data/Hora tempo temperat. agitação pH OD aeração ValvO2 CER
(h) (°C) (rpm)
(%sat) (LpM) (%) mmol/L.h
24/08/2018 21:17 0 15,7 600 5,65 99,08 1,0 0,0 -0,140
24/08/2018 22:17 1 15,0 600 5,93 96,80 1,0 0,0 -0,031
24/08/2018 23:17 2 15,0 600 5,95 96,30 1,0 0,0 0,077
25/08/2018 00:17 3 15,0 600 5,95 95,39 1,0 0,0 0,204
25/08/2018 01:17 4 15,0 600 5,98 94,96 1,0 0,0 0,272
25/08/2018 02:17 5 15,0 600 6,07 93,84 1,0 0,0 0,351
25/08/2018 03:17 6 15,0 600 6,26 93,33 1,0 0,0 0,560
25/08/2018 04:17 7 15,0 600 6,32 92,61 1,0 0,0 0,668
25/08/2018 05:17 8 15,0 600 6,39 91,54 1,0 0,0 0,911
25/08/2018 06:17 9 15,0 600 6,45 89,77 1,0 0,0 1,101
25/08/2018 07:17 10 15,0 600 6,51 89,09 1,0 0,0 1,202
25/08/2018 08:17 11 15,0 600 6,58 86,52 1,0 0,0 1,482
25/08/2018 09:17 12 15,0 600 6,63 86,20 1,0 0,0 1,737
25/08/2018 10:17 13 15,0 600 6,68 81,79 1,0 0,0 2,133
25/08/2018 11:17 14 15,1 600 6,73 81,45 1,0 0,6 2,468
25/08/2018 12:17 15 15,0 600 6,81 81,60 1,0 1,2 2,806
25/08/2018 13:17 16 15,0 600 6,89 79,90 1,0 0,8 3,317
25/08/2018 14:17 17 15,0 600 6,96 87,30 1,0 1,3 3,846
25/08/2018 15:17 18 15,0 600 7,08 79,85 1,0 3,9 4,263
25/08/2018 16:17 19 15,0 600 7,20 82,48 1,0 5,4 4,981
25/08/2018 17:17 20 15,0 600 7,35 77,88 1,0 5,4 5,555
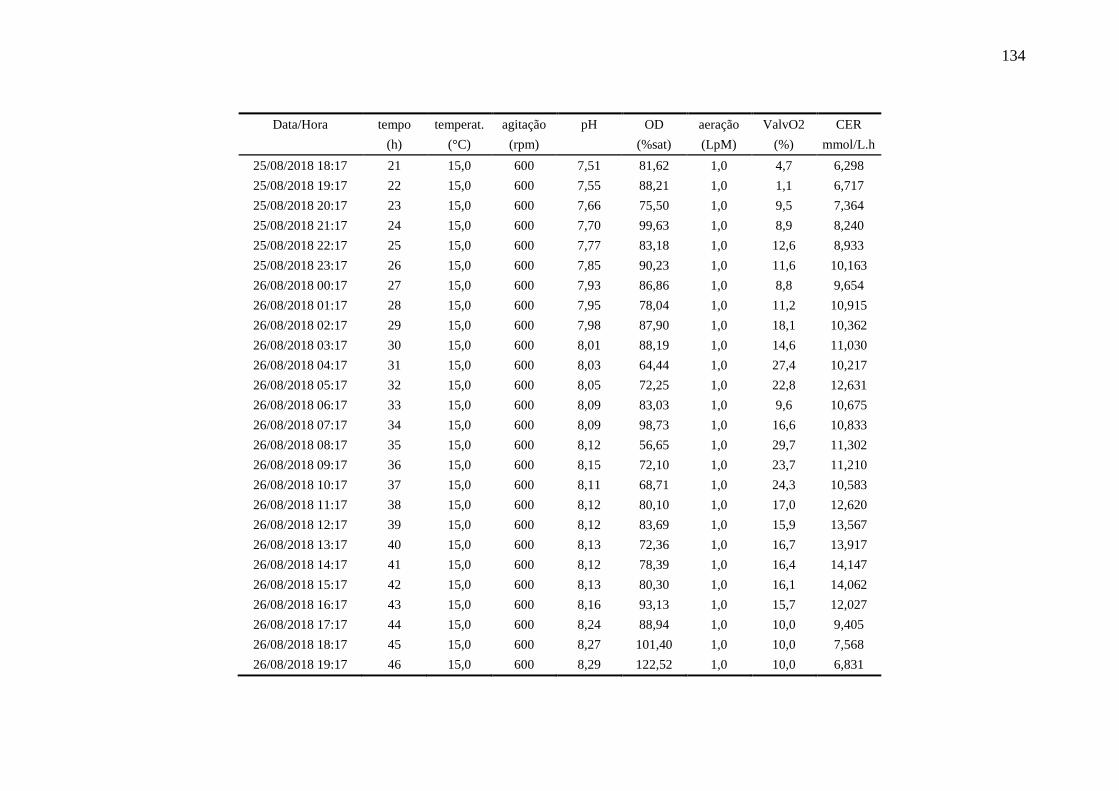
134
Data/Hora tempo temperat. agitação pH OD aeração ValvO2 CER
(h) (°C) (rpm)
(%sat) (LpM) (%) mmol/L.h
25/08/2018 18:17 21 15,0 600 7,51 81,62 1,0 4,7 6,298
25/08/2018 19:17 22 15,0 600 7,55 88,21 1,0 1,1 6,717
25/08/2018 20:17 23 15,0 600 7,66 75,50 1,0 9,5 7,364
25/08/2018 21:17 24 15,0 600 7,70 99,63 1,0 8,9 8,240
25/08/2018 22:17 25 15,0 600 7,77 83,18 1,0 12,6 8,933
25/08/2018 23:17 26 15,0 600 7,85 90,23 1,0 11,6 10,163
26/08/2018 00:17 27 15,0 600 7,93 86,86 1,0 8,8 9,654
26/08/2018 01:17 28 15,0 600 7,95 78,04 1,0 11,2 10,915
26/08/2018 02:17 29 15,0 600 7,98 87,90 1,0 18,1 10,362
26/08/2018 03:17 30 15,0 600 8,01 88,19 1,0 14,6 11,030
26/08/2018 04:17 31 15,0 600 8,03 64,44 1,0 27,4 10,217
26/08/2018 05:17 32 15,0 600 8,05 72,25 1,0 22,8 12,631
26/08/2018 06:17 33 15,0 600 8,09 83,03 1,0 9,6 10,675
26/08/2018 07:17 34 15,0 600 8,09 98,73 1,0 16,6 10,833
26/08/2018 08:17 35 15,0 600 8,12 56,65 1,0 29,7 11,302
26/08/2018 09:17 36 15,0 600 8,15 72,10 1,0 23,7 11,210
26/08/2018 10:17 37 15,0 600 8,11 68,71 1,0 24,3 10,583
26/08/2018 11:17 38 15,0 600 8,12 80,10 1,0 17,0 12,620
26/08/2018 12:17 39 15,0 600 8,12 83,69 1,0 15,9 13,567
26/08/2018 13:17 40 15,0 600 8,13 72,36 1,0 16,7 13,917
26/08/2018 14:17 41 15,0 600 8,12 78,39 1,0 16,4 14,147
26/08/2018 15:17 42 15,0 600 8,13 80,30 1,0 16,1 14,062
26/08/2018 16:17 43 15,0 600 8,16 93,13 1,0 15,7 12,027
26/08/2018 17:17 44 15,0 600 8,24 88,94 1,0 10,0 9,405
26/08/2018 18:17 45 15,0 600 8,27 101,40 1,0 10,0 7,568
26/08/2018 19:17 46 15,0 600 8,29 122,52 1,0 10,0 6,831
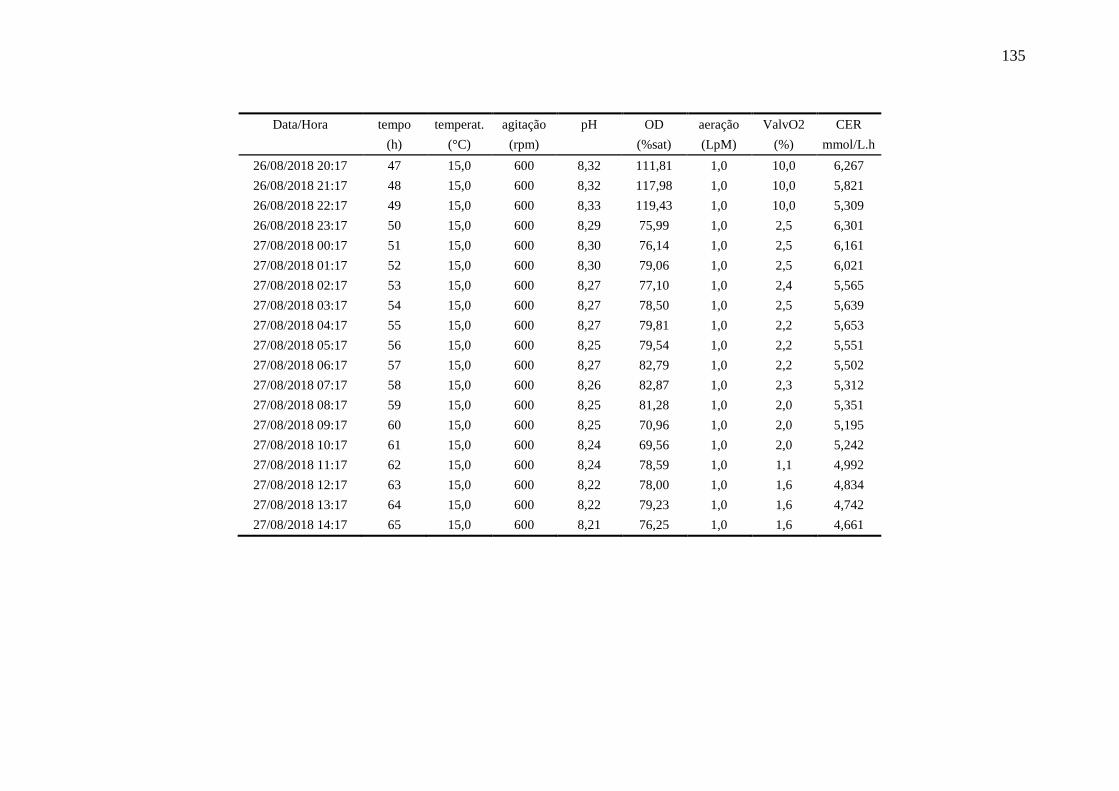
135
Data/Hora tempo temperat. agitação pH OD aeração ValvO2 CER
(h) (°C) (rpm)
(%sat) (LpM) (%) mmol/L.h
26/08/2018 20:17 47 15,0 600 8,32 111,81 1,0 10,0 6,267
26/08/2018 21:17 48 15,0 600 8,32 117,98 1,0 10,0 5,821
26/08/2018 22:17 49 15,0 600 8,33 119,43 1,0 10,0 5,309
26/08/2018 23:17 50 15,0 600 8,29 75,99 1,0 2,5 6,301
27/08/2018 00:17 51 15,0 600 8,30 76,14 1,0 2,5 6,161
27/08/2018 01:17 52 15,0 600 8,30 79,06 1,0 2,5 6,021
27/08/2018 02:17 53 15,0 600 8,27 77,10 1,0 2,4 5,565
27/08/2018 03:17 54 15,0 600 8,27 78,50 1,0 2,5 5,639
27/08/2018 04:17 55 15,0 600 8,27 79,81 1,0 2,2 5,653
27/08/2018 05:17 56 15,0 600 8,25 79,54 1,0 2,2 5,551
27/08/2018 06:17 57 15,0 600 8,27 82,79 1,0 2,2 5,502
27/08/2018 07:17 58 15,0 600 8,26 82,87 1,0 2,3 5,312
27/08/2018 08:17 59 15,0 600 8,25 81,28 1,0 2,0 5,351
27/08/2018 09:17 60 15,0 600 8,25 70,96 1,0 2,0 5,195
27/08/2018 10:17 61 15,0 600 8,24 69,56 1,0 2,0 5,242
27/08/2018 11:17 62 15,0 600 8,24 78,59 1,0 1,1 4,992
27/08/2018 12:17 63 15,0 600 8,22 78,00 1,0 1,6 4,834
27/08/2018 13:17 64 15,0 600 8,22 79,23 1,0 1,6 4,742
27/08/2018 14:17 65 15,0 600 8,21 76,25 1,0 1,6 4,661
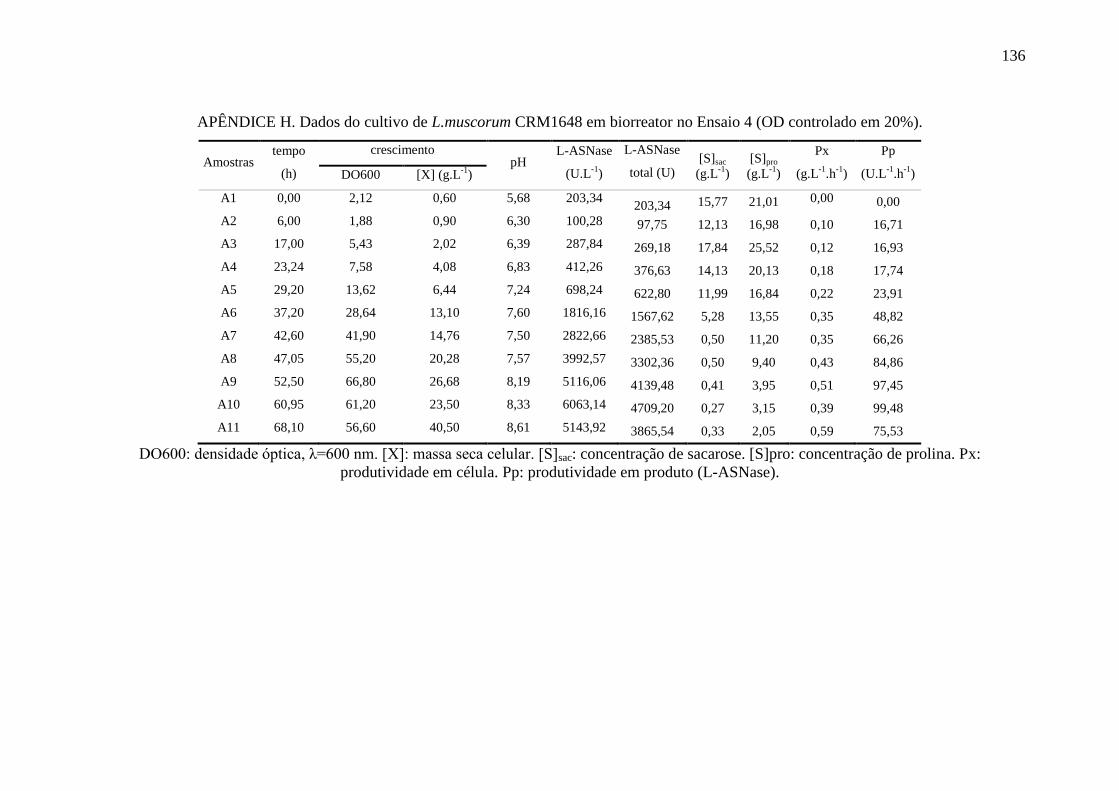
136
APÊNDICE H. Dados do cultivo de L.muscorum CRM1648 em biorreator no Ensaio 4 (OD controlado em 20%).
Amostras tempo
(h)
crescimento pH
L-ASNase
(U.L-1
)
L-ASNase
total (U) [S]sac
(g.L-1
)
[S]pro
(g.L-1
)
Px
(g.L-1
.h-1
)
Pp
(U.L-1
.h-1
) DO600 [X] (g.L-1
)
A1 0,00 2,12 0,60 5,68 203,34 203,34 15,77 21,01 0,00 0,00
A2 6,00 1,88 0,90 6,30 100,28 97,75 12,13 16,98 0,10 16,71
A3 17,00 5,43 2,02 6,39 287,84 269,18 17,84 25,52 0,12 16,93
A4 23,24 7,58 4,08 6,83 412,26 376,63 14,13 20,13 0,18 17,74
A5 29,20 13,62 6,44 7,24 698,24 622,80 11,99 16,84 0,22 23,91
A6 37,20 28,64 13,10 7,60 1816,16 1567,62 5,28 13,55 0,35 48,82
A7 42,60 41,90 14,76 7,50 2822,66 2385,53 0,50 11,20 0,35 66,26
A8 47,05 55,20 20,28 7,57 3992,57 3302,36 0,50 9,40 0,43 84,86
A9 52,50 66,80 26,68 8,19 5116,06 4139,48 0,41 3,95 0,51 97,45
A10 60,95 61,20 23,50 8,33 6063,14 4709,20 0,27 3,15 0,39 99,48
A11 68,10 56,60 40,50 8,61 5143,92 3865,54 0,33 2,05 0,59 75,53
DO600: densidade óptica, λ=600 nm. [X]: massa seca celular. [S]sac: concentração de sacarose. [S]pro: concentração de prolina. Px:
produtividade em célula. Pp: produtividade em produto (L-ASNase).
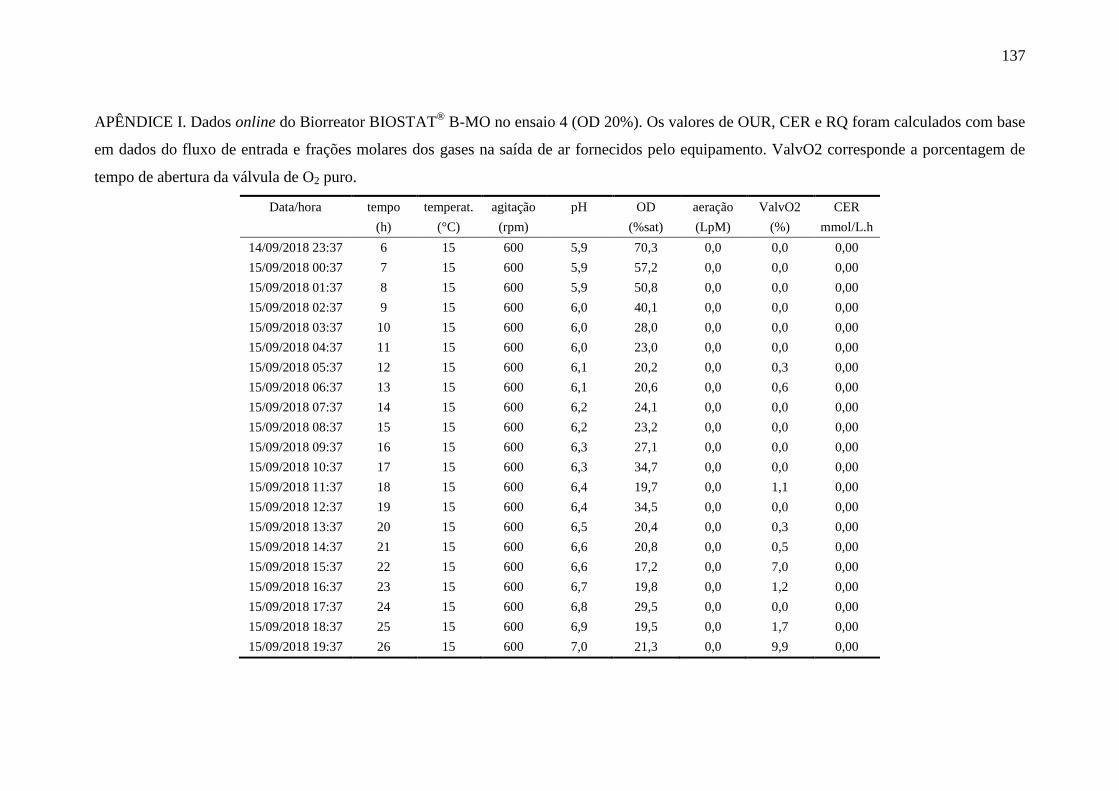
137
APÊNDICE I. Dados online do Biorreator BIOSTAT® B-MO no ensaio 4 (OD 20%). Os valores de OUR, CER e RQ foram calculados com base
em dados do fluxo de entrada e frações molares dos gases na saída de ar fornecidos pelo equipamento. ValvO2 corresponde a porcentagem de
tempo de abertura da válvula de O2 puro.
Data/hora tempo temperat. agitação pH OD aeração ValvO2 CER
(h) (°C) (rpm) (%sat) (LpM) (%) mmol/L.h
14/09/2018 23:37 6 15 600 5,9 70,3 0,0 0,0 0,00
15/09/2018 00:37 7 15 600 5,9 57,2 0,0 0,0 0,00
15/09/2018 01:37 8 15 600 5,9 50,8 0,0 0,0 0,00
15/09/2018 02:37 9 15 600 6,0 40,1 0,0 0,0 0,00
15/09/2018 03:37 10 15 600 6,0 28,0 0,0 0,0 0,00
15/09/2018 04:37 11 15 600 6,0 23,0 0,0 0,0 0,00
15/09/2018 05:37 12 15 600 6,1 20,2 0,0 0,3 0,00
15/09/2018 06:37 13 15 600 6,1 20,6 0,0 0,6 0,00
15/09/2018 07:37 14 15 600 6,2 24,1 0,0 0,0 0,00
15/09/2018 08:37 15 15 600 6,2 23,2 0,0 0,0 0,00
15/09/2018 09:37 16 15 600 6,3 27,1 0,0 0,0 0,00
15/09/2018 10:37 17 15 600 6,3 34,7 0,0 0,0 0,00
15/09/2018 11:37 18 15 600 6,4 19,7 0,0 1,1 0,00
15/09/2018 12:37 19 15 600 6,4 34,5 0,0 0,0 0,00
15/09/2018 13:37 20 15 600 6,5 20,4 0,0 0,3 0,00
15/09/2018 14:37 21 15 600 6,6 20,8 0,0 0,5 0,00
15/09/2018 15:37 22 15 600 6,6 17,2 0,0 7,0 0,00
15/09/2018 16:37 23 15 600 6,7 19,8 0,0 1,2 0,00
15/09/2018 17:37 24 15 600 6,8 29,5 0,0 0,0 0,00
15/09/2018 18:37 25 15 600 6,9 19,5 0,0 1,7 0,00
15/09/2018 19:37 26 15 600 7,0 21,3 0,0 9,9 0,00
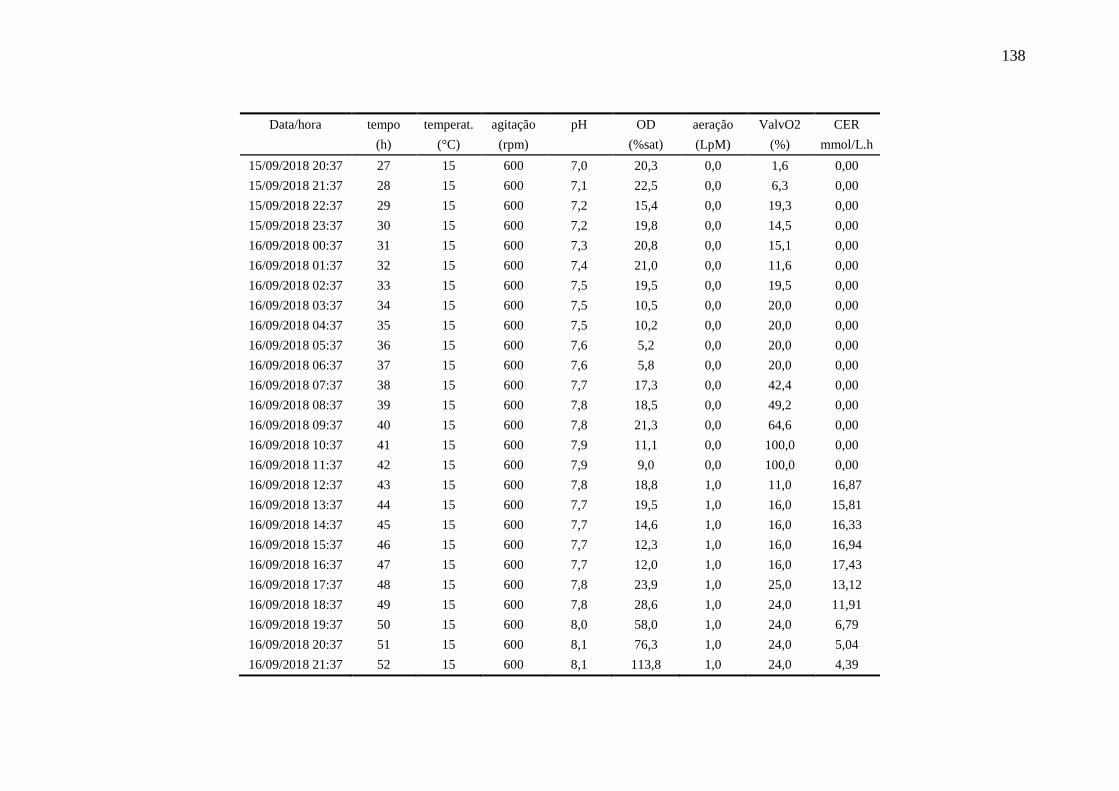
138
Data/hora tempo temperat. agitação pH OD aeração ValvO2 CER
(h) (°C) (rpm) (%sat) (LpM) (%) mmol/L.h
15/09/2018 20:37 27 15 600 7,0 20,3 0,0 1,6 0,00
15/09/2018 21:37 28 15 600 7,1 22,5 0,0 6,3 0,00
15/09/2018 22:37 29 15 600 7,2 15,4 0,0 19,3 0,00
15/09/2018 23:37 30 15 600 7,2 19,8 0,0 14,5 0,00
16/09/2018 00:37 31 15 600 7,3 20,8 0,0 15,1 0,00
16/09/2018 01:37 32 15 600 7,4 21,0 0,0 11,6 0,00
16/09/2018 02:37 33 15 600 7,5 19,5 0,0 19,5 0,00
16/09/2018 03:37 34 15 600 7,5 10,5 0,0 20,0 0,00
16/09/2018 04:37 35 15 600 7,5 10,2 0,0 20,0 0,00
16/09/2018 05:37 36 15 600 7,6 5,2 0,0 20,0 0,00
16/09/2018 06:37 37 15 600 7,6 5,8 0,0 20,0 0,00
16/09/2018 07:37 38 15 600 7,7 17,3 0,0 42,4 0,00
16/09/2018 08:37 39 15 600 7,8 18,5 0,0 49,2 0,00
16/09/2018 09:37 40 15 600 7,8 21,3 0,0 64,6 0,00
16/09/2018 10:37 41 15 600 7,9 11,1 0,0 100,0 0,00
16/09/2018 11:37 42 15 600 7,9 9,0 0,0 100,0 0,00
16/09/2018 12:37 43 15 600 7,8 18,8 1,0 11,0 16,87
16/09/2018 13:37 44 15 600 7,7 19,5 1,0 16,0 15,81
16/09/2018 14:37 45 15 600 7,7 14,6 1,0 16,0 16,33
16/09/2018 15:37 46 15 600 7,7 12,3 1,0 16,0 16,94
16/09/2018 16:37 47 15 600 7,7 12,0 1,0 16,0 17,43
16/09/2018 17:37 48 15 600 7,8 23,9 1,0 25,0 13,12
16/09/2018 18:37 49 15 600 7,8 28,6 1,0 24,0 11,91
16/09/2018 19:37 50 15 600 8,0 58,0 1,0 24,0 6,79
16/09/2018 20:37 51 15 600 8,1 76,3 1,0 24,0 5,04
16/09/2018 21:37 52 15 600 8,1 113,8 1,0 24,0 4,39

139
Data/hora tempo temperat. agitação pH OD aeração ValvO2 CER
(h) (°C) (rpm) (%sat) (LpM) (%) mmol/L.h
16/09/2018 22:37 53 15 600 8,1 117,2 1,0 24,0 4,07
16/09/2018 23:37 54 15 600 8,1 17,8 1,0 1,0 6,96
17/09/2018 00:37 55 15 600 8,1 16,2 1,0 1,0 6,94
17/09/2018 01:37 56 15 600 8,1 11,5 1,0 1,0 6,64
17/09/2018 02:37 57 15 600 8,1 11,1 1,0 1,0 6,51
17/09/2018 03:37 58 15 600 8,2 9,4 1,0 1,0 6,33
17/09/2018 04:37 59 15 600 8,2 8,8 1,0 1,0 6,32
17/09/2018 05:37 60 15 600 8,2 8,4 1,0 1,0 6,24
17/09/2018 06:37 61 15 600 8,2 8,3 1,0 1,0 6,21
17/09/2018 07:37 62 15 600 8,2 8,5 1,0 1,0 6,21
17/09/2018 08:37 63 15 600 8,3 18,8 1,0 4,0 6,08
17/09/2018 09:37 64 15 600 8,3 20,0 1,0 3,8 5,91
17/09/2018 10:37 65 15 600 8,3 19,9 1,0 3,9 5,92
17/09/2018 11:37 66 15 600 8,3 19,4 1,0 4,0 5,86
17/09/2018 12:37 67 15 600 8,3 19,7 1,0 3,7 5,78
17/09/2018 13:37 68 15 600 8,3 21,8 1,0 3,0 5,75
17/09/2018 14:37 69 15 600 8,3 23,5 1,0 3,0 5,65

140