universidade ceuma - uniceuma prÓ-reitoria de pÓs ... costa... · manifestaram efeito citotóxico...
TRANSCRIPT

0
UNIVERSIDADE CEUMA - UNICEUMA
PRÓ-REITORIA DE PÓS-GRADUAÇÃO, PESQUISA E EXTENSÃO
MESTRADO EM BIOLOGIA PARASITÁRIA
FRANCYELLE COSTA MORAES
Estudo da patogenicidade de isolados de Burkholderia sp e Stenotrophomonas
maltophillia em amostras clínicas
SÃO LUÍS
2014

1
FRANCYELLE COSTA MORAES
Estudo da patogenicidade de isolados de Burkholderia sp e Stenotrophomonas
maltophillia em amostras clínicas
Dissertação apresentada ao Programa de
Mestrado em Biologia Parasitária da
Universidade CEUMA, linha de pesquisa em
Patogenicidade Celular e Molecular de Micro-
organismos, para obtenção do título de Mestre.
Orientadora:
Profa. Dr
a. Patricia de Maria Silva Figueirêdo
Co-orientador:
Prof. Dr. Valério Monteiro Neto
SÃO LUÍS
2014

2
FRANCYELLE COSTA MORAES
Estudo da patogenicidade de isolados de Burkholderia sp e Stenotrophomonas
maltophillia em amostras clínicas
A Comissão julgadora da Defesa do Trabalho Final de Mestrado em
Biologia Parasitária, em sessão pública realizada no dia / / , considerou a
candidata:
( ) APROVADO ( ) REPROVADO
1. Examinador ________________________________________________
2. Examinador ________________________________________________
3. Examinador ________________________________________________
4. Presidente (Orientador)________________________________________

3
LISTA DE FIGURAS
Figura 1 - Distribuição das amostras de Burkholderia sp e S. maltophillia.por
amostra clínica.................................................................................................................
37
Figura 2- Perfil de susceptibilidade antimicrobiana dos isolados de Burkholderia
sp......................................................................................................................................
39
Figura3-Perfil de susceptibilidade antimicrobiana dos isolados de Stenotrophomonas
maltophillia......................................................................................................................
42
Figura 4- Microscopia óptica (100X) da adesão, invasão e pontos de vacuolização
em células HEp-2 (A) e Pneumócitos A549 (B).............................................................
49
Figura 5- Placa de Ágar Vermelho Congo. Amostras produtoras de cápsula tiveram
crescimento de colônias enegrecidas e as não produtoras tiveram crescimento de
colônias vermelhas.........................................................................................................
54
Figura 6- Produção das exoenzimas: A) Halos de degradação da protease em ágar
Skim Milk 10%; C) Hidrólise de gelatinase em ágar gelatina 12%...............................
60
Figura 7- Atividade hemolítica: A) Halo de hemólise em ágar sangue humano tipo B,
5%; B) Halo de hemólise em ágar sangue de carneiro 5%..............................................
66

4
LISTA DE TABELAS
Tabela 1- Frequencia absoluta e relativa de resistência intermédiária aos
antimicrobianos por Burkholderia sp................................................................................
40
Tabela 2- Correlação entre a capacidade hidrofóbica e os sítios de isolamento das
estirpes de S. maltophillia.................................................................................................
46
Tabela 3- Correlação entre a capacidade hidrofóbica e os sítios de isolamento das
estirpes de Burkholderia sp...............................................................................................
46
Tabela 4- Capacidade de adesão das amostras clínicas de Burkholderia sp às
superfícies abióticas e células humanas.............................................................................
48
Tabela 5- Capacidade de adesão das amostras clínicas de S. maltophillia às
superfícies
abióticas.............................................................................................................................
51
Tabela 6- Correlação entre hidrofobicidade e formação de biofilme diante das cepas de
Burkholderia sp..................................................................................................................
57
Tabela 7- Correlação entre hidrofobicidade e formação de biofilme diante das cepas de
Burkholderia sp............................................................................................................
57
Tabela 8- Atividae enzimática e hemolítica de amostras clínicas de S. maltophillia
correlacionando os sítios de isolamento............................................................................
59
Tabela 9- Atividae enzimática e hemolítica de amostras clínicas de Burkholderia sp
correlacionando os sítios de isolamento............................................................................
62
.

5
LISTA DE ABREVIATURAS
ATCC - American Type Culture Collection
BHI - Brain Heart Infusion
CVC - Catéter Venoso Central
EDTA - Etilenodiamina-tetra ácido acético
ELISA - Enzyme-linked Immunosorbent Assay
et al – et alli, e outros
ITU - Infecção do Trato Urinário
NCCLS - Clinical and Laboratory Standards Institute
PBS – tampão salina fosfato (phosphate buffered saline)
PCR – Reação em cadeia de polimerase (Polimerase Chain Reaction)
pH – potencial hidrogeniônico
μg - microgramas
μl – microlitros
rpm – rotações por minuto
ºC – graus Celsius ou graus centígrados

6
RESUMO
As bactérias Burkholderia sp e S. maltophillia constituem bacilos gram-negativos
que estão associadas à elevada prevalência de infecções nosocomiais, aumento da taxa de
morbimortalidade e também aos altos custos de materiais hospitalares e recursos humanos das
unidades de saúde. Neste estudo foram avaliadas 14 estirpes de Burkholderia sp e 18 de S.
maltophillia isoladas de amostras clínicas em laboratórios de São Luís-MA. A elucidação dos
fatores de virulência destas espécies é de suma importância para a produção científica acerca
da patogenicidade bacteriana. Os isolados foram analisados quanto as suas propriedades de
virulência: resistência antimicrobiana, expressão de ESBL, hidrofobicidade, formação de
biofilme, adesão em materiais inertes e células, potencial citotóxico, atividade enzimática
(amilase, DNAse, elastase, fosfolipase, gelatinase, lipase, proteinase) e hemolítica. A maioria
(57,4%), das amostras de Burkholderia sp foram isoladas de sangue, 75% foram resistentes
ao sulfametoxazol/trimetropim, 64% foram produtoras de ESBL, demonstraram elevada
capacidade de adesão nos mais variados substratos testados, 78,5% formaram biofilme, não
manifestaram efeito citotóxico com o sobrenadante, apresentaram hemólise apenas em ágar
sangue e, diante de todos os eritrócitos. Com exceção de amilase e elastase produziram todas
as enzimas pesquisadas. As cepas de S. maltophillia foram mais isoladas de amostras de
secreção traqueal (70,58%), revelaram baixa resistência aos antibióticos avaliados, 63%
expressaram ESBL, adesão ao vidro e a formação de biofilme foi constatada em todas as
cepas, revelaram-se capaz de aderir também ao látex e inox, não produziram lipase,
fosfolipase e amilase, mas produziram as outras enzimas em percentuais elevados, mostraram-
se capaz de hemolisar as diferentes hemácias testadas.Ressalta-se que nenhuma das espécies
foi capaz de causar hemólise em meio líquido. Os resultados sugerem que os fatores de
virulência destas espécies podem ocasionar danos teciduais e possivelmente agravamento do
quadro clínico dos pacientes acometidos por infecções causadas por tais patógenos.
.
Palavras-chave: Patogenicidade, Fatores de Virulência; Burkholderia sp; S. maltophillia.

7
ABSTRACT
Bacteria of the genus Burkholderia sp and S. maltophillia are gram-negative bacilli that are
associated with high prevalence of nosocomial infections, increased morbidity and mortality
rates as well as the high cost of hospital supplies and human resources in health units. This
study included 14 strains of Burkholderia sp and 18 S. maltophillia isolated from clinical
specimens in laboratories of Sao Luis. The elucidation of the virulence factors of these species
is of paramount importance to the scientific production about the bacterial pathogenicity. The
isolates were analyzed for their virulence properties: antimicrobial resistance, ESBL
expression, hydrophobicity, hemagglutination, biofilm formation, adhesion to inert materials
and cells, cytotoxic, enzyme (amylase, DNase, elastase, phospholipase, gelatinase, lipase ,
proteinase), and hemolytic. The vast majority (57.4%) samples of Burkholderia sp were
isolated from blood, 75% showed resistance to sulfamethoxazole / trimethoprim, 64%
produced ESBL showed high adhesion capacity in various substrates tested, 78.5 % formed
biofilm, expressed no cytotoxic effect with the supernatant showed hemolysis on blood agar
before all erythrocytes, except for amylase and elastase produced all the enzymes studied. The
strains of S. maltophillia were more isolated from tracheal samples (70.58%), showed low
resistance to antibiotics tested, 63% expressed ESBL produced biofilm, showed 100%
adhesion to glass, proved able to also adhere to latex and steel did not produce lipase,
phospholipase and amylase, but other enzymes produced in high percentages, as well as other
species, were able to hemolyze erythrocytes testadas.Os different results suggest that the
virulence factors of these species can cause the tissue damage and possibly worsening of
clinical symptoms of patients with infections caused by these pathogens.
Keywords: Pathogenicity, Virulence Factors, Burkholderia sp, S. maltophillia.

8
SUMÁRIO
1 REVISÃO BIBLIOGRÁFICA ........................................................................................... 12
1.1 BACILOS GRAM-NEGATIVOS NÃO FERMENTADORES ...................................................... 12
1.2 BURKHOLDERIA SP ............................................................................................................ 13
1.3 STENOTROPHOMONAS MALTOPHILLIA ................................................................................ 16
1.4 RESISTÊNCIA AOS ANTIBIÓTICOS ..................................................................................... 18
1.5 ADESÃO E FORMAÇÃO DE BIOFILME ................................................................................. 21
1.6 DETERMINANTES DE PATOGENICIDADE ............................................................................ 23
2 OBJETIVOS ........................................................................................................................ 26
2.1 GERAL ............................................................................................................................. 26
2.2 ESPECÍFICOS .................................................................................................................... 26
3 METODOLOGIA ................................................................................................................ 27
3.1 AMOSTRAS BACTERIANAS ................................................................................................ 27
3.2 RESISTÊNCIA ANTIMICROBIANA ....................................................................................... 27
3.3 IDENTIFICAÇÃO DE CEPAS PRODUTORAS DE ESBL ...................................................... 28
3.3.1 Detecção fenotípica de beta-lactamase ................................................................... 28
3.4 CAPACIDADE HIDROFÓBICA ............................................................................................ 28
3.5 CAPACIDADE DE ADESÃO A SUPERFÍCIES INERTES E CÉLULAS HUMANAS .................... 28
3.5.1 Adesão ao vidro ....................................................................................................... 28
3.5.2 Adesão ao látex siliconizado e aço inox .................................................................. 29
3.5.3 Adesão a células humanas ....................................................................................... 29
3.5.3.1 Cultivo e preparo das monocamadas celulares ............................................... 29
3.5.3.2 Realização do teste de adesão .......................................................................... 30
3.6 PRODUÇÃO DE BIOFILME ................................................................................................. 30
3.6.1 Avaliação da formação de cápsula ......................................................................... 30
3.6.2 Análise da indução de biofilme por especrotofotometria ........................................ 31
3.7 DETECÇÃO DE CITOTOXINA ........................................................................................... 31
3.7.1 Preparo do sobrenadante das amostras bacterianas .............................................. 31
3.7.2 Teste de citotoxidade ............................................................................................... 32
3.8 ANÁLISE ENZIMÁTICA ................................................................................................... 32
3.8.1 Amilase .................................................................................................................... 32
38.2 DNase ....................................................................................................................... 32
3.8.3 Elastase .................................................................................................................... 33
3.8.4 Fosfolipase .............................................................................................................. 33
3.8.5 Gelatinase ................................................................................................................ 33
3.8.6 Lipase ...................................................................................................................... 33
3.8.7 Proteinase ................................................................................................................ 34
3.9 PRODUÇÃO DE HEMOLISINAS ......................................................................................... 34
3.9.1 Atividade hemolítica em meio sólido ....................................................................... 34
3.9.2. Atividade hemolítica com o pellet e sobrenadante bacteriano (teste em
microplaca) ....................................................................................................................... 34
3.10 ANÁLISES ESTATÍSTICAS .............................................................................................. 35
4 RESULTADOS E DISCUSSÃO ........................................................................................ 35
4.1 AMOSTRAS E SÍTIOS DE ORIGEM ....................................................................................... 35
4.2 SUSCEPTIBILIDADE AOS ANTIMICROBIANOS ..................................................................... 38
4.3 EXPRESSÃO DAS ESBL FENOTIPICAMENTE ...................................................................... 43
4.4 DETERMINANTES DE ADESÃO ........................................................................................... 43

9
5.4.1 CAPACIDADE HIDROFÓBICA .......................................................................................... 43
5.5 ADESÃO A MATERIAIS INERTES E CÉLULAS HUMANAS ..................................................... 46
5.6 PRODUÇÃO DE EXOENZIMAS E HEMOLISINAS ................................................................... 56
REFERÊNCIAS ..................................................................................................................... 67

10
1 INTRODUÇÃO
Autoridades e profissionais de saúde buscam a solução para uma problemática
antiga, que desde o século XIX quando surgiram os hospitais perdura nas unidades de saúde, a
Infecção Hospitalar (IH) (MOURA et al., 2008). No cenário atual constata-se uma
revolucionária expansão tecnológica nos procedimentos rotineiros da assistência a saúde, que
embora aumente as chances de sobrevida dos pacientes paralelamente torna-os suscetíveis a
adquirir infecções e eleva os custos hospitalares (TURRINI & SANTO, 2002).
A aquisição de tais afecções está relacionada a vários fatores: condições fisiológicas
inerentes a cada enfermo, procedimentos invasivos que são significantes portas de entrada
para micro-organismos oportunistas (que por vezes são cepas resistentes), internação
prolongada, entre outros (ANDRADE E ANGERAMI, 1999; TURRINI, 2000;
HINRICHSEN, 2004). Nesta conjuntura o Ministério da Saúde através da Portaria N°
2.616/98, conceituou IH como “aquela adquirida após a admissão do paciente e que se
manifeste durante a internação ou após a alta, quando puder ser relacionada com a internação
ou procedimentos hospitalares”.
Hinrichsen (2004) estima que entre os agentes etiológicos das infecções nosocomiais
as bactérias são as mais prevalentes com cerca de 90% dos episódios avaliados, em seguida
encontram-se os fungos com 9% e vírus, protozoários e helmintos apresentando 1% dos
casos. Entre os grupos bacterianos envolvidos nestes processos infecciosos, os bacilos gram-
negativos não fermentadores (BGN-NF) representam uma parcela significativa entre os
responsáveis pela etiologia destes processos (FROTA, 1998). Os bacilos Gram negativos
classificados como não fermentadores são assim designados por não utilizarem as vias
fermentativas para a obtençaão de energia, para tanto necessitam de oxigênio para a
degradação de carboidratos através da via oxidativa. Além disso, não formam esporos, são
aeróbios estritos e ubíquos, estando amplamente distribuídos no meio ambiente: animais,
vegetais, água, solo e seres humanos de onde são isolados geralmente de amostras clínicas e
estão associados às infecções oportunistas em vários sítios anatômicos (KONEMAN et al.,
2008).

11
No ambiente hospitalar as Unidades de Terapia Intensiva (UTI´s), de hemodiálise e
de imunodeprimidos são os locais mais afetados por estes agentes, que vêm adquirindo uma
maior importância clínica devido aos seus mecanismos de resistência aos antibióticos e
desinfetantes (PÉREZ-MIRAVETE, 2001; LARANJEIRA, 2010). Uma pesquisa realizada
em um hospital universitário na Argentina envolvendo 84 pacientes com bacteremia
hospitalar por bacilos gram negativos (BGN), constatou que Acinetobacter baumanii,
Burkholderia sp e Stenotrophomonas maltophillia (bacilos gram negativos não fermentadores
– BGNNF) foram isolados com maior incidência, manifestando elevada resistência aos
antibióticos testados e foram mais frequentes nas amostras de pacientes que evoluíram a óbito
(LIZASO et al., 2008).
Burkholderiam spp e S. maltophillia constituem gêneros bacterianos que estão
amplamente distribuídos em habitats naturais e antrópicos. Recentemente, essas bactérias
foram reconhecidas como ameaça aos pacientes hospitalizados, em especial aos doentes
imunodeprimidos. Um outro problema evidenciado em tais micro-organismos é a resistência
intrínseca a uma grande maioria dos antibióticos disponíveis clinicamente, incluindo
aminoglicosídeos, cefalosporinas e β-lactâmicos ( KONEMAN et al., 2008).
Stenotrophomonas maltophilia é considerado um patógeno emergente, que exibe
resistência intrínseca à maioria dos antimicrobianos, e adquirida aos poucos disponíveis
contra ele possivelmente, em virtude do emprego de fármacos cada vez mais potentes
(ALMEIDA et al., 2005) . Apresenta-se em grande diversidade de ambientes, aumentado
assim, as possíveis fontes de contaminação. No gênero Burkholderia as espécies que
possuem enorme relevância no contexto clínico é a Burkholderia pseudomallei, responsável
pela etiologia da melioidose, uma doença infecciosa cuja taxa de mortalidade é de 95%
quando não tratada (KONEMAN et al., 2008) e as estirpes do Complexo Burkholderia
cepacia .
Entre os fatores de virulência destes micro-organismos, estão: formação de biofilme,
adesão, exotoxina A (inibe a síntese de proteínas com lesão celular e semelhante a toxina
diftérica), exotoxina S (inibe a síntese protéica), neuramidase (auxilia a adesão da bactéria
pois destrói muco), fosfolipase C (lise de membranas plasmáticas), elastase (cliva inibidores
de proteases leucocitárias e componentes do complemento), leucocidina (toxina leucotóxica) e

12
resistência múltipla a antibióticos por produzir vários plasmídios R (KONEMAN, 2008;
SOUSA et al., 2011).
Analisando a relevância destes bacilos no ambiente nosocomial como potenciais
agentes infecciosos especialmente entre pacientes imunodeprimidos, torna-se imprescindível
um estudo que objetiva estudar os fatores de virulência e de estirpes de Burkholderia sp e S.
maltophillia.
2 REVISÃO BIBLIOGRÁFICA
2.1 Bacilos Gram-Negativos Não Fermentadores
Os bacilos Gram-negativos não fermentadores (BGN-NF) compõem um grupo muito
diversificado, são estritamente aeróbios, não esporulados e se caracterizam pelo fato de serem
incapazes de utilizar carboidratos como fonte de energia por meio de fermentação,
degradando-os pela via oxidativa (KONEMAN et al., 2001; MENEZES et al., 2004).
Os BGNNF por não usarem a via fermentativa da glicose geralmente usam a via
Entner- Doudoroff para obter ATP através da glicose, oxidando-a. As bactérias que realizam
esse processo conseguem menos energia sob a forma de ATP do que as bactérias que realizam
a via de Embden-Meyerhof; por outro lado, essas bactérias conseguem outra forma de
coenzima reduzida NADPH. Esta é a coenzima usada preferencialmente em vias anabólicas,
fornecendo elétrons e íons de hidrogênio para os processos de biossíntese celular
(KONEMAN et al., 2001; ANVISA, 2004; VERMELHO et al., 2008).
A partir da década de 70 as infecções por BGNNF tornaram-se cada vez mais
prevalentes em unidades de saúde, tendo como principais representantes Pseudomonas
aeruginosa, Acinetobacter spp, Stenotrophomonas maltophillia e Burkholderia cepacia
(KONEMAN et al., 2001;FALAGAS e KOPTERIDES 2006; CEZÁRIO et al., 2009). Essas
bactérias estão relacionados predominantemente com infecções hospitalares (FIGUEIREDO-
MENDES, 2005; NOUÉR, 2007). A capacidade de sobrevivência desses patógenos
possibilita adaptação a condições adversas, sendo necessário apenas um local úmido para sua
supervivência e viabilidade por diversos meses. Os BGNNF têm boa afinidade com a água,
por se tratar de um grupo de micro-organismos de natureza ubiquitária, presente em várias

13
condições ambientais. Quando encontrados em ambientes nosocomiais podem ser
considerados patógenos em potencial, em função da sua habilidade em se multiplicar, higiene
precária e condutas terapêuticas inadequadas (uso indiscriminado de antimicrobianos de
amplo espectro), o que contribui para aumentar o estado de morbimortalidade dos pacientes
imunodeprimidos colocando suas vidas em risco (LARANJEIRA et al., 2010; SOARES,
2005).
Além disso, Stenotrophomonas maltophillia e Burkholderia cepacia são exemplos de
não fermentadores que vêm apresentando sensibilidade diminuída a um grande número de
fármacos (SADER et al., 2001). Almodóvar et al. (2007) realizaram um estudo com o
objetivo de constatar a ocorrência de BGNNF de glicose em 97 amostras de água tratada para
diálise e 27 amostras de dialisatos, analisadas entre junho de 2005 e dezembro de 2006. Nos
resultados, detectou-se 29,6% de BGNNF nas amostras de dialisatos e 49,5% nas amostras de
água tratada, sendo mais frequente estirpes do Complexo Burkholderia cepacia (59,0%),
seguido de Stenotrophomonas maltophillia (13,1%).
2.2 Burkholderia sp
O gênero Burkholderia pertence à divisão das β-proteobactérias, inclui cerca de 40
espécies descritas e é formado por bacilos Gram-negativos não fermentadores (BGN-NF),
móveis, aeróbios e não esporulados (BERRIATUA et al., 2001; SAVOIA, ZUCCA, 2007;
MAHENTHIRALINGAM, BALDWIN, DOWSON, 2008). Essas bactérias exibem uma
enorme versatilidade metabólica, permitindo sua adaptação a vários ambientes (CHIARINI
etal, 2006). Podem ocupar uma diversidade ampla de nichos ecológicos naturais ou artificiais,
podendo ser isoladas em solos, plantas, animais, humanos e ambiente nosocomial, onde se
revelou um emergente patógeno oportunista (GOLDMAN et al 1986; COENYE E
VANDAMME, 2003).
Algumas espécies são alvos de interesse agrícola e biotecnológico de indústrias que
almejam a biorremediação e o biocontrole (SAVOIA, ZUCCA, 2001;
MAHENTHIRALINGAM, BALDWIN, DOWSON, 2008). Seu potencial como agente
promotor de crescimento em plantações de diversas culturas, retrata uma opção válida para
uma diversidade de pesticidas e fertilizantes químicos e também como agentes de

14
biorremediação, por serem capazes de degradar compostos existentes em alguns pesticiadas e
herbicidas utilizados rotineiramente (TABACCHIONI et al., 2002; KILBANE et al., 1983).
Entretanto, a sua capacidade de causar infecções em seres humanos também colocou algumas
dessas espécies no grupo de patógenos humanos oportunistas, atingindo principalmente
pacientes com fibrose cística- FC, pacientes com doença granulomatosa crônica (DGC),
imunodeprimidos e internos em tratamentos intensivos (BERRIATUA et al., 2001).
Em 1950, William Burkholder descreveu uma bactéria que causava podridão em
aliáceas denominando-a de cepacia, nome derivado de cebola (Allium cepa) que
posteriormente ficou conhecida como Pseudomonas cepacia, um patógeno de plantas (GIL,
2001). Também neste ano, este microorganismo foi descrito como um patógeno humano que
causava endocardite bacteriana. Em 1971, identificou-se mais uma doença causada por tal
agente, o “pé podre” evidenciado em tropas norte-americanas em formação no pântano, em
exercício na Flórida (DAVIES; RUBIN, 2007).
Com o advento da biologia molecular foi possível a realização da análise taxonômica
molecular de cepas dessa espécie de Pseudomonas, ocasionando a descoberta do gênero
Burkholderia, sendo a espécie B. cepacia considerada espécie tipo deste novo gênero
(YABUUCHI et al., 1992; MAHENTHIRALINGAM; BALDWIN; DOWSON, 2008).
Apesar da similaridade fenotípica dos isolados dessa espécie tipo, existem várias diferenças
genéticas que foram evidenciadas em análises de hibridização de DNA, ocasionando a
diferenciação de isolados anteriormente classificados como de uma única espécie em várias
espécies filogeneticamente associadas (VANDAMME et al., 1997). A heterogenicidade dos
grupos identificados acarretaram na denominação do complexo Burkholderia cepacia (CBc),
composto inicialmente por 9 espécies: B. cepacia, B.multivorans, B. cenocepacia, B. stabilis,
B. vietnamiensis, B. dolosa, B. ambifaria, B. anthina e B. pyrrocinia (VANDAMME et al.,
2002; COENYE et al., 2001; VERMIS et al., 2004; COENYE, VANDAMME, 2001).
Atualmente a posição taxonômica de clustres gênicos de recA isolados do CBc através de
taxonomia polifásica revelou as espécies: B. ubonensis, B. latens, B.diffusa, B. arboris, B.
seminalis e B. metallica, B. contaminans e B.lata totalizando, até o momento, 17 espécies
dentro do complexo (VANLAERE et al., 2008).
Cepas do CBc destacam-se entre as demais espécies do gênero por estarem
frequentemente associadas à infecções hospitalares (IHs). O potencial de adaptação,
resistência e disseminação desses micro-organismos estão relacionados aos surtos de

15
contaminação de água para hemodiálise, antissépticos, desinfetantes, gel de ECG, soluções
anestésicas entre outros medicamentos, possibilitando a contaminação cruzada e
consequentemente a própria infecção nosocomial (COENYE et al 2001).
Na década de 80 demonstrou-se a capacidade de transmissão entre pacientes dentro
e fora da unidade de saúde (SPEERT et al., 2002; HAUSER et al., 2011). De acordo com a
literatura a transmissão entre pacientes com fibrose cística e sem fibrose cística tem sido
associada com doenças graves e óbito, causando uma maior preocupação no ambiente de
saúde (DAVIES; RUBIN, 2007). Dentre as principais infecções ocasionadas por essas
bactérias destaca-se: as infecções do trato respiratório em pacientes com FC e DGC, infecção
do trato urinário (ITUs) em pacientes cateterizados, septicemia associada ao uso de cateter
intravascular, entre outras infecções oportunistas (MURRAY et al., 2003).
Em infecções da corrente sanguínea-ICS, cerca de 90% estão associadas a um cateter
vascular (SIEGMAN-IGRA et al., 1997) e representa um terço de todas as infecções
associadas à assistência à saúde e com elevadas taxas de morbimortalidade (CHRISTOFF et
al., 2010). Estima-se que a taxa de letalidade tem sido de 10 a 20% dos casos de ICS
(SIEGMAN-IGRA et al., 1997).
Após a colonização inicial pelo CBc o curso clínico é variável. No entanto, a grande
maioria dos pacientes apresenta rápido declínio da função pulmonar evoluindo ao óbito em
consequência da falência cardiorrespiratória. Ocasionalmente podem ocorrer quadros mais
graves com alta taxa de mortalidade, em consequência a doença progressiva, invasiva, com
célere decadência pulmonar, progredindo para um tipo de bacteremia manifestada por
pneumonia necrotizante progressiva, conhecida como "Síndrome Cepacia” (GOVAN E
DERETIC 1996; VANDAMME et al. 1997; COENYE et al. 2001; LI PUMA 2010).
O tratamento de infecções causadas por esses patógenos torna-se difícil devido a sua
alta resistência à grande maioria dos antibióticos de uso clínico. Este problema é complicado
pelo desenvolvimento de resistência durante terapia e também pelo desenvolvimento de
resistência cruzada a outros antimicrobianos (WIGFIELD et al., 2002). O tratamento
adequado ainda é questionado, sendo que a utilização da ceftazidima, da sulfametoxazol-
trimetoprim, cloranfenicol ou tetraciclina já foi considerada útil. Quando é admissível, a
terapia com dois antibióticos sinérgicos é ideal (HODSON, 1995).

16
2.3 Stenotrophomonas maltophillia
Ao longo do tempo a taxonomia de Stenotrophomonas maltophilia foi bastante
variável, pertencendo esta bactéria originalmente ao gênero Bacillus A, descrito em 1887 por
Booker, como o agente responsável por uma diarréia que ocorria no verão. Desde então,
outros autores sugeriram sua inclusão em novos gêneros e/ou espécies. Desta foram, a mesma
foi mencionada como Bacillus bookeri, Bacterium bookeri, Alcaligenes bookeri. Atualmente,
está classificada em um único gênero, Stenotrophomonas, composto por uma única espécie,
Stenotrophomonas maltophilia, anteriormente denominado Xanthomonas maltophilia ou
Pseudomonas maltophilia (DENTON e KEER, 1998; PALLERONI e BRADBURY, 1993).
Morfologicamente as células de S. maltophillia são bacilos Gram-negativos, lineares
ou ligeiramente curvados de 0,5-1,5 μm de comprimento. Podem ocorrer individualmente ou
em pares, são móveis por meio de flagelos polares. As propriedades bioquímicas incluem: a
respiração aeróbia estrita, testes negativos para oxidase e positivos a catalase e esculina, a não
fermentação de açúcares e a oxidação da maltose. Ao mesmo tempo, apresentam crescimento
ótimo à temperatura de 35ºC em ágar Mac-Conkey (MARGESIN E SCHINNER, 1991;
MURRAY et al., 2003 ).
S. maltophillia pode ser encontrada em múltiplos ambientes e regiões geográficas,
incluindo até mesmo a Antártica (VAZQUEZ et al., 1995), ocupando nichos ecológicos
diferentes e fontes variadas de água tais como poços, lagos de reservatórios municipais, rios e
até água utilizada na hemodiálise e na indústria farmacêutica. Também podem ser isolados a
partir do solo, leite cru, ovos, peixe congelado e carcaça de animais. No ambiente nosocomial,
essa bactéria já foi isolada de água de torneira (HOEFEL et al., 2005), pias, respiradores,
cateteres de sucção, monitores de pressão arterial, máquina produtora de gelo (DENTON e
KEER, 1998; ROBIN e JANDA, 1996), equipamento de diálise (FLAHERTY et al., 1993),
soluções desinfetantes (MUKHOPADHYAY et al., 2003), e, ocasionalmente, das mãos de
profissionais de saúde (GUO et al., 2005). Pesquisas evidenciaram como microambiente para
essa espécie outras fontes secas e pobres de nutrientes como, curativos de algodão e placas de
petri (HIRAI et al., 1991). Atualmente, foram caracterizadas outras fontes de isolamento
inesperadas, como sêmen e embriões bovinos congelados (BIELANSKY et al., 2003).

17
Contudo, S. maltophillia está associada a uma variedade cada vez maior de
síndromes clínicas, sendo considerada um agente patogênico oportunista. Colonização e
infecção hospitalar são os casos mais prevalentes. Esta linhagem é capaz de causar
bacteremia, septicemia, endocardite, mastoidite, conjuntivite, meningite, feridas pós-
operatórias, abcessos, infecções do trato urinário e pneumonia (FUJITA et al., 1996; JULVE
et al., 1998). A frequência de isolados clínicos de S. maltophillia descrita em diversos
hospitais, abrangendo pacientes colonizados e infectados varia de 2 a 37,7 casos por 10.000
altas (DEL TORO et al., 2002; DENTON e KEER, 1998). Tan et al., (2008) observou em um
hospital terciário de Taiwan que a taxa de infecção por 10.000 admissões era de 5,3 em 1999
e aumentou para 9,8 em 2004. No Brasil, uma pesquisa realizada no Hospital das Clínicas da
FMUSP entre os anos de 1999 e 2006, evidenciou que a incidência de infecção de corrente
sanguínea por S. maltophillia variou de 0,12 a 1,0 (média de 0,55) por 1.000 pacientes-dia
(GIRÃO et al., 2008).
Embora S. maltophillia seja considerada um patógeno nosocomial emergente
(BERGOGNE-BEREZIN,1993; FANG e MADINGER 1996), esta bactéria também pode
ocasionar infecções na comunidade (HEATH e CURRIE, 1995). Em um estudo de Laing et
al. (1995), analisou 63 pacientes de três hospitais de cuidados agudos, descreveu que 15
(23,8%) casos de infecção e/ou colonização por S. maltophillia foram contraídas na
comunidade. Em outro estudo, de 109 amostras 26 (24%) foram consideradas como tendo
sido adquirida na comunidade (YAO et al., 1995).
Outro fato importante é o aumento na incidência de S maltophilia associada à
pneumonia no paciente portador de fibrose cística (CANTÓN et al., 2003; MOORE et al.,
2003), atualmente essa bactéria é considerada o principal agente etiológico desta problemática
(STEINKAMP et al., 2005). Embora recente e ainda poucos estudos disponíveis, cabe
ressaltar também à importância clínica desse microorganismo para os pacientes infectados
pelo HIV, já que foi possível associá-lo a esse grupo de pacientes (MANFREDI e CHIODO,
2002; CALZA et al., 2003). A taxa de mortalidade é considerada um forte indicador do
potencial patogênico de qualquer bactéria, com as estirpes de S. maltophillia esse indicador é
em torno de 40%, e vêm sendo associada à sua presença entre indivíduos gravemente
debilitados ou imunodeprimidos (APISARNTHANARAK et al., 2003; TSAI et al., 2003 ).
Recentemente, a literatura evidenciou que pacientes com queimaduras também
apresentaram alta incidência de bacteremia por S. maltophillia, a taxa de mortalidade nesses

18
casos foi de 30,77%, e está relacionada ao tamanho da área do corpo atingida pela queimadura
e do tempo prolongado de hospitalização (TSAI et al., 2006).
Os fatores de risco associados às infecções por S. maltophillia geralmente estão
vinculados aos procedimentos realizados no ambiente nosocomial, como o uso de cateteres
intravasculares, utilização prolongada de tubo endotraqueal, de nebulizadores e de ventilação
mecânica e, até mesmo, o uso de lentes de contato (DENTON e KEER, 1998).
2.4 Resistência aos antibióticos
Bactérias resistentes a antibióticos que ocasionam infecções associadas a assistência
a saúde representam um desafio contínuo e crescente para as unidades de saúde, tanto na
terapêutica clínica de pacientes quanto na prevenção da transmissão cruzada (HIDRON,
2008). A quantidade de infecções originadas por patógenos multirresistentes tem aumentado
expressivamente na última década. Tempo de internação prolongado, aumento dos custos,
mortalidade elevada e redução das opções terapêuticas fazem da prevenção e controle de
bactérias multirresistentes uma ação prioritária (WEINTROB et al., 2010).
As estruturas biológicas que definem tal fenótipo se estabelecem por uma
combinação de determinantes intrínsecos ou adquiridos, compreendendo a produção de
enzimas inativadoras de fármacos, a redução na permeabilidade da membrana externa, a
expressão de um sistema de efluxo multidrogas (BARRY et al., 1985), bem como a
transferência de genes de resistência (BARUCHEL et al., 1986), mediada por plasmídeos
(BATES et al., 1993; . BAUERNFEIND, 1993; BELCOUR et al., 1993).
Mecanismos de resistência de espécies de Burkholderia incluem permeabilidade
seletiva na parede celular, alteração de alvos intracelulares das drogas, deterioração
enzimática ou inativação de drogas ou o efluxo ativo de antibióticos (WIGFIELD et al.,
2002). Burkholderia cepacia é intrinsecamente resistente aos antibióticos antipseudomonas:
colistina e polimixina (DE SOYZA; CORRIS, 2003).
S. maltophillia é um micro-organismo multirresistente aos antimicrobianos clássicos,
incluindo ß-lactâmicos, quinolonas e aminoglicosídeos (BALTIMORE et al., 1990; BANDIN
et al., 1996 ). Adicionalmente, S. maltophillia, especialmente no ambiente nosocomial, é

19
considerada reservatório de genes móveis para ß-lactamases de amplo espectro, o que atribui
capacidade aumentada de transmissão de resistência e, portanto, maior importância
epidemiológica (BENEZRA et al., 1988 ). A resistência aos antibióticos ß- lactâmicos é
bastante elaborada, pois acontece via produção de enzimas cujo gene codificador tem origem
distinta (cromossomal, plasmidial ou nos transposons) e ainda por um mecanismo de efluxo
multidrogas (BAUERNFEIND, 1993).
Em adição à resistência intrínseca, espécies de Burkholderia podem também adquirir
resistência durante a terapia devido à produção de betalactamases, auto-regulação de
antibióticos em bombas de efluxo e alteração de alvos de antibióticos. A resistência induzida é
um problema particular em pacientes com fibrose cística, que frequentemente recebem
terapias múltiplas e prolongadas de antibióticos (DAVIES; RUBIN, 2007). Esses patógenos
são muito resistentes aos antimicrobianos por possuir um genoma extremamente flexível que
é capaz de sofrer várias mutações, adaptando-se às mais distintas condições (MOORE;
BANCOCK, 1986). Por vezes, essas bactérias demonstram susceptibilidade aos fármacos in
vitro, mas clinicamente são resistentes à antibioticoterapia. Ressalta-se que a resistência à
classe dos ß- lactâmicos e inibidores de ß- lactamase é crescente. O sequenciamento do
genoma dessas bactérias revelou a presença de genes de ß- lactamase de classe A, C e D, logo
a terapêutica necessita da realização de testes de sensibilidade sempre (LEELARASAMEE,
2004).
Os aminoglicosídeos mostram atividade ínfima contra S. maltophillia em razão de
alterações na conformação da membrana externa, dependentes de temperatura, e também à
presença de enzimas modificadoras dessas drogas (DENTON e KEER 1998). A recente
demonstração da transferência gênica entre micro-organismos Gram-positivos e S.
maltophillia pode também explicar esse fenótipo de sensibilidade reduzida (ALONSO et al.,
2000).
A resistência intrínseca aos aminoglicosídeos também é observada entre membros do
CBc (MOORE; BANCOCK, 1986). Esses organismos são resistentes a esses antibióticos
devido ao incomum componente lipopolissacarídeo de suas membranas celulares. Em
pesquisa realizada observou-se que a maioria de 366 cepas estudadas foram resistentes à
ciprofloxacina, amicacina, gentamicina, tobramicina, cefuroxima, cefotaxima, imipenem,
cloranfenicol, tetraciclina e sulfas e mais de 77% foram sensíveis à ceftazidima, 34%
piperacilina/tazobactam e meropenem. A migração de sequências de inserção dentro do

20
cromossomo de Burkholderia cepacia pode afetar a expressão de genes que modulam a
resistência antimicrobiana. Logo, isso poderia explicar porque cepas com o mesmo ribotipo,
porém com diferentes antibiogramas, podem ser isolados do mesmo paciente (LEDSON;
GALLAGHER; CORKHILL, 1998).
A eficácia clínica das quinolonas, assim como a suscetibilidade in vitro de S.
maltophillia a esses antibióticos, é questionável. A partir do ácido nalidíxico, no mínimo
(GALES et al., 2001) fluoroquinolonas são largamente utilizadas no tratamento contra micro-
organismos multirresistentes, entretanto, nos últimos anos, foram isoladas cepas de S.
maltophillia resistentes a alguns desses agentes (WEISS et al., 2000). Em geral, são dois os
mecanismos para essa resistência: a) modificações nas proteínas de membrana externa
modificando a permeabilidade aos fármacos e b) superexpressão das proteínas do sistema de
efluxo (RIBERA et al., 2002).
Resistência a múltiplos antibióticos tem sido repetidamente relatada; em um estudo
foi encontrada multirresistência em 48% (27/56) das cepas. Um total de 57% (17/30) de
bacteriemias por gram-negativos foram devidas às bactérias resistentes a múltiplos
antibióticos; estavam incluídos 50% de Pseudomonas aeruginosa, 100% de Burkholderia
cepacia e 50% de Klebsiella pneumoniae isoladas. Quando pacientes com Burkholderia
cepacia foram excluídos, a proporção de pacientes com fibrose cística que tinham bacteriemia
por bactérias gram-negativas resistentes a múltiplos antibióticos foi análoga a pacientes com
fibrose não-cística (HUSAIN et al., 2006).
Visto que várias classes de antibióticos não fornecem resultados eficientes, tanto nos
ensaios in vitro como in vivo, a escolha no tratamento das infecções por S. maltophillia e
Burkholderia sp é a associação entre o sulfametoxazol e o trimetroprim (BETRIU et al., 2001;
MERMEL et al., 2001;FRITSCHE et al., 2005). Entretanto, algumas pesquisas mostraram
taxas de 2 a 10% de estirpes de S. maltophillia resistentes a esses quimioterápicos. Do mesmo
modo, outras combinações mostraram- se eficazes nos testes in vitro envolvendo as seguintes
drogas: ticarcilina/ácido clavulânico cefalosporinas, aztreonam, fluorquinolonas, azitromicina,
claritromicina, colistina e rifampicina ( POULOS et al., 1995; GIAMARELLOS-
BOURBOULIS et al., 2002).
Ao avaliar o perfil de sensibilidade in vitro de isolados clínicos do CBc Chaves
(2010), revelou que 83,3% das cepas foram sensíveis ao meropenem e 61,1% ao

21
sulfametoxazol-trimetoprim. Tais evidências limitam ainda mais o valor terapêutico
individual de cada um desses fármacos diante de infecções causadas por tais bactérias
(ALMEIDA et al., 2005) .
2.5 Adesão e formação de biofilme
Sabe-se que os micro-organismos aderem às células hospedeiras para evitar sua
destruição e a maioria destes são posteriormente internalizados para encontrar um ambiente
apropriado para sobrevivência e replicação, distante das defesas imunológicas do hospedeiro
(PIZARRO-CERDÁ & COSSART, 2006). A adesão bacteriana tem sido descrita como o
equilíbrio de interações físico-químicas atrativas e repulsivas entre as bactérias e superfícies.
A natureza adesiva das bactérias é devido a vários recursos de sua membrana externa, como
pili, proteínas e lipopolissacarídeos (LPS) em razão disto as respostas das células do
hospedeiro ao processo de adesão é modificável de acordo com o tipo de bactéria e a célula
(CAMESANO & LOGAN, 2000; WALKER et al.,2004;TRABULSI & ALTERTHUM,
2008).
Todos os tipos de adesão celular são intercedidos por adesinas, proteínas de
transmembrana com uma propriedade extracelular envolvida no reconhecimento e interação
ligante, e um propriedade intracelular que traduz sinais para a reorganização do citoesqueleto
da célula e outros eventos. As adesinas são transdutores mecânicos e sua ativação desencadeia
rearranjos de actina importantes, geralmente mediados pelas famílias Rho e Ras de pequenas
GTPases. Existem vários tipos de adesinas, de acordo com o tipo de bactéria (PEREZ-
MORENO et al., 2003; TRABULSI & ALTERTHUM, 2008).
Em uma pesquisa de aderência de B.pseudomallei à células da faringe, inoculou-se
em camundongos por via intrabronquial, bactérias desta espécie e verificou-se cargas
bacterianas significativas nos pulmões, o que indica que B. pseudomallei tem a capacidade de
aderir ao trato respiratório in vivo ( AHMED et al., 1999). Garcia et al., (2000) comprovou
adesão de células HEp-2 por S. maltophillia, o que sugere que esta espécie também pode
aderir a outras células epiteliais. Esta evidência pode explicar a presença de S. maltophillia
nas células das vias respiratórias. A maioria dos isolados desta bactéria foi obtida de infecções
nosocomiais, que em vários casos, os pacientes estavam em uso de dispositivos médicos.

22
As alterações mucocutâneas ocasionadas por procedimentos invasivos tais como
intubação ou uso de sondas, cateteres, próteses valvulares e etc., podem impor risco
aumentado de sobrevivência e multiplicação de cepas de S. maltophillia (CANDEL et al.,
2002). O passo inicial da infecção por espécies de Burkholderia envolve a colonização do
trato respiratório superior, que evolui para o trato inferior. A desenvoltura da bactéria para
formar biofilmes tem sido relacionada com sua capacidade para causar doença no hospedeiro
humano (CUNHA et al., 2004).Alguns autores ao investigar surtos de endocardite por S.
maltophillia sugerem que as possíveis fontes de infecções são os dispositivos de acesso
vascular e/ou as válvulas cardíacas em casos de cirurgias (GUTIERREZ et al., 1996).
A adesão bacteriana à superfície de um biomaterial é um processo em que a bactéria
se liga firmemente à superfície e pode ser descrita como um processo de duas fases, sendo a
primeira, a fase física, instantânea e reversível e a segunda, a fase molecular e celular,
dependente do tempo e irreversível (AN et al., 1998). Tais características resultam por fim na
formação de microcolônias embebidas em uma matriz de polímeros extracelulares
(exopolissacarídeos - EPS), o biofilme (POULSEN et al. 1999; BOARI et al. 2009). Este tipo
de disposição é vantajosa a qualquer espécie de micro-organismo, por ocasionar proteção
contra adversidades como desidratação, colonização por bacteriófagos e resistência a
antimicrobianos (HIRAI, 1991; VICTOR et al., 1994).
Em casos de bacteremia por S. maltophillia os dispositivos intravasculares são
importantes determinantes na patogênese desta bactéria, (MUDER et al., 1996; )tem sido
sugerido que estes dispositivos podem ser o fonte primária desta manifestação clínica. Por
exemplo, Murder e seus colaboladores (1998) informam que, em uma análise com 91
pacientes com bacteremia por S. maltophillia, 84% destes indivíduos usavam um cateter
venoso central. Adicionalmente, Douce et al. (2008), expuseram investigação de um surto em
setembro de 2006, após 4 pacientes com cateter venoso central terem sido identificados com
bacteriemia pelo CBc.
O processo patogênico da infecção associada ao cateter está relacionado com a
migração dos micro-organismos da pele no local de inserção cutânea do cateter e, com a
colonização da ponta do cateter, a qual é o caminho mais comum de infecção para os cateteres
inseridos perifericamente e cateteres de curta duração (DONLAN, COSTERTON, 2002)

23
O agravamento de infecções por Burkholderia cepacia é devido em parte à sua
formação de biofilmes, que são geralmente resistentes aos antibióticos. A iniciação de um
biofilme é depende do pili do microorganismo e flagelos usados para fixação. A maturação do
biofilme está relacionada com a produção do exopolissacarídeo cepaciano que é conhecido
por estabilizar a arquitetura tridimensional do biofilme além de, está intimamente relacionado
a resistência aos antibióticos (CUAHA et al., 2004).
2.6 Determinantes de patogenicidade
Stenotrophomonas maltophillia e Burkholderia sp são importantes patógenos em
pacientes hospitalizados, principalmente aqueles com antibioticoterapia de espectro estendido
e com fibrose cística -FC (MARQUES, 2011). Pouco se sabe sobre os fatores de virulência
desses micro-organismos (DENTON e KEER, 1998; BERNHARDT et al., 2003), a falta de
distinção entre colonização e infecção gerou a crença de que S. maltophillia é um organismo
de patogenicidade limitada (DENTON e KEER, 1998).
Esse ponto de vista está sendo reconsiderado em algumas pesquisas, até
recentemente, S. maltophillia foi considerada uma bactéria incomum entre os isolados
clínicos, fato que já não ocorre atualmente, já que este patógeno vem apresentando uma
prevalência cada vez maior nos diagnósticos de laboratórios de microbiologia (DENTON e
KEER, 1998). Já se sabe que são várias as manifestações clínicas causadas por S.
maltophillia, a bacteremia é considerada uma das manifestações mais comuns (ELTING et al.,
1990; KHARDORI et al., 1991 ;JANG et al., 1992; MUDER et al., 1996) e sua frequência
vêm aumentando (KRCMERY et al., 1996). Essa manifestação pode ser secundária a uma
infecção pulmonar, urinária ou gastrointestinal (AOUN et al., 1992 ), ainda que na maioria
das vezes não é possível determinar a porta de entrada, Kollef et al. (1995) considerou o
isolamento de S. maltophillia como de “alto-risco”, sendo este um dos importantes
responsáveis pela mortalidade em pacientes com pneumonia associada à ventilação mecânica.
A percentagem de mortalidade em pacientes neutropênicos com pneumonia é de 40%
(KHARDORI et al., 1991).
O prognóstico ruim dos pacientes acometidos por tais manifestações vêm sendo
associado a produção de uma multiplicidade de enzimas extracelulares que podem exercer um

24
papel importante na patogenicidade bacteriana. Elas podem ultrapassar as barreiras de defesa
do hospedeiro, além de ocasionar a proliferação bacteriana fornecendo os próprios peptídeos e
aminoácidos das proteínas de membrana às bactérias (ZAVASCKI, 2003; JAFFAR-
BANDJEE et al., 1995).As exoenzimas evidenciadas em S. maltophillia incluem: DNase,
RNase, fibrinolisina, lipases, hialuronidase, proteases e elastase, (DE OLIVEIRA-GARCIA et
al., 2003; YAMAMURA et al., 2002; WINDHORST et al., 2002 ). Para averiguar tal
associação, estudiosos relataram um caso de ectima gangrenoso em um paciente com
leucemia que adquiriu bacteremia por S. maltophillia. A escolha ocorreu porque a produção
de protease e elastase bacteriana é considerada vital na patogênese de casos de ectima
gangrenoso associado com septicemia por P. aeruginosa. Assim, eles analisaram o isolado de
S. maltophillia para capacidade de produzir estas enzimas e evidenciaram que este patógeno
era um bom produtar de protease e elastase (BOTTONE et al., 1986 ).
Para as estirpes de Burkholderia sp acredita-se que fatores pulmonares do hospedeiro
tais como presença de cable pilli, além da capacidade de algumas cepas replicarem
intracelularmente, e produzirem enzimas extracelulares contribuem para a aptidão de
persistência de infecção em pacientes com fibrose cística (SAJJAN et al., 2001). Os
mecanismos que estabelecem a patogênese das cepas do CBc envolvem múltiplos fatores de
virulência como a produção de hemolisinas, enzimas (proteases e lípases), formação de
biofilme, capacidade de adesão, resistência ao soro humano e aos antibióticos (GOVAN et al.
1996).
Além disso, outros fatores são considerados determinantes para os processos
infecciosos e/ou colonização, como é o caso de adesinas associadas a pilus, flagelo,
lipopolissacárido (LPS, constituinte da parede das células das bactérias gram negativas) e
outros exopolissacárideos (SALDÍAS, et al., 2009a, SAJJAN, et al., 2006), sistemas de
secreção do tipo II, III, IV e VI (MCKEON, et al. 2010a, SAJJAN, et al., 2008), sideróforos,
catalases e superóxido dismutases, proteases, lipases (SAJJAN, et al., 2006) e sistema de
“quorum sensing” (um sistema de resposta a estímulos, regulando a expressão de expressão
de genes de acordo com a densidade populacional) (SOKOL, et al. 2003).
Embora pouco se saiba sobre a lipase, foi provado que desempenham um papel na
invasão de células epiteliais do pulmão. Metaloproteases também desempenham um papel
importante na virulência de "Burkholderia cepacia" no tecido pulmonar (CALLAGHAN e
MCCLEAN, 2009).

25
Há relato de seis pacientes com sérias infecções causadas por um gel de ultra-
sonografia contaminado com Burkholderia cepacia, sendo isoladas do sangue dos pacientes e
eram idênticas às isoladas no gel de ultra-som, demonstrando habilidade da bactéria em
degradar conservantes utilizados em cosméticos e produtos industriais e farmacêuticos
(encontrados em cremes, pastas, produtos de beleza, cola, gorduras e óleos) in vitro, mediante
enzimas específicas (CASHMAN; WARSHAW, 2005). Espécies pertencentes ao CBc,
manifestam vários mecanismos clássicos de virulência, incluindo exoenzimas, hemolisinas e
endotoxinas. Pacientes infectados por uma mesma estirpe também pode demonstrar uma
variação na evolução clínica, fato que aponta a resposta imune do paciente como fator
decisivo no combate às infecções (BERNHARDT et al., 2003).

26
3 OBJETIVOS
3.1 Geral
Caracterizar os fatores de virulência de Burkholderia sp e Stenotrophomonas
maltophillia isoladas de amostras clínicas que possam ser importantes na elucidação de seus
mecanismos de patogenicidade.
3.2 Específicos
Conhecer o perfil de susceptibilidade aos antibióticos de uso clínico;
Analisar a expressão das ESBL fenotipicamente;
Conferir a aptidão hidrofóbica;
Avaliar a capacidade de adesão a materiais inertes e células humanas;
Constatar produção de cápsula, formação de biofilme
Verificar atividade citotóxica;
Detectar fenotipicamente a produção de exoenzimas e hemolisinas;

27
4 METODOLOGIA
4.1 Amostras bacterianas
Foram avaliadas 14 cepas de Burkholderia sp e 18 de Stenotrophomonas maltophillia
isoladas de diferentes amostras clínicas. Todas as cepas foram identificadas pelo método
automatizado Vitek 2 (bioMérieux)® em laboratórios particulares de São Luís-MA e
gentilmente cedidas ao Laboratório de Microbiologia Médica da Universidade CEUMA-
UNICEUMA. Posteriormente, realizou-se o reisolamento das amostras em Ágar Sangue (5%)
pela técnica de semeadura em superfície para certificação da pureza das amostras
(KONEMAN et al. 2008). Para o estoque, as cepas foram mantidas em caldo BHI (Brain
Heart Infusion), acrescido de 15% de glicerol a - 20 ºC (SAMBROOK; FRTSCH;
MANIATIS, 2002).
A presente pesquisa foi aprovada pelo Comitê de Ética em Pesquisa do
UNICEUMA, segundo emissão dos pareceres: 000348/11 de acordo com as normas da
Resolução CNS 196/96.
4.2 Resistência antimicrobiana
O perfil de resistência das amostras bacterianas estudadas foi estabelecido pelo
método automatizado Vitek 2 (bioMérieux)®, nos laboratórios particulares que doaram as
cepas ao Laboratório de Microbiologia Médica da Universidade CEUMA-UNICEUMA. Os
antibióticos utilizados diante do gênero Burkholderia sp foram: ceftazidima, meropenem,
minociclina e sulfametaxazol/trimetropim. Para a espécie Stenotrophomonas maltophillia
utilizou-se: minociclina, levofloxacino e sulfametaxazol/trimetropim.

28
4.3 Identificação de cepas produtoras de ESBL
4.3.1 Detecção fenotípica de beta-lactamase
Todas as amostras foram submetidas ao método do disco aproximação para detecção
da produção de ESBL. Para cada estirpe foi preparada uma suspensão bacteriana com solução
salina 0,9% e turbidez correspondente a 0,5 da escala de McFarland (aproximadamente 3 x
108 células/ml). Posteriormente, as suspensões foram homogeneizadas e semeadas em placas
de ágar Müeller-Hinton (Merck). Discos de aztreonam (30 µg), cefotaxima (30 µg),
ceftazidima (30 µg) e cefepime (30 µg), foram colocados a uma distância de 20 mm do disco
de amoxicilina/ácido clavulânico (30 µg). As amostras foram consideradas produtoras de
ESBL mediante a formação de uma zona irregular de inibição (“ghost-zone”) entre o disco
composto e o disco de uma das drogas betalactâmicas (CLSI, 2008).
4.4 Capacidade Hidrofóbica
As amostras de Burkholderia sp e S. maltophillia foram semeadas em Ágar BHI por
24 horas a 37°C, posteriormente colônias foram suspensas em concentrações crescentes de
sulfato de amônio de 0,5 M, 1,0 M, 1,5 M, 2,0 M, 2,5M e 3,0 M. Após este momento,
observou-se a formação de grumos em até dois minutos indicando resultado positivo
(SCHMIDT et al., 1998). Sendo que a formação de grumos em concentrações até 1,5 M de
sulfato de amônio indica hidrofobicidade forte, a partir de 1,5 até 2,5M, moderada e acima de
2,5M fraca (LOCATELLI et al. 2004).
4.5 Capacidade de adesão a superfícies inertes e células humanas
4.5.1 Adesão ao vidro
Utilizou-se a metodologia de Garcia et al. (2002), com algumas modificações.
Lamínulas redondas de vidro foram colocadas em poços de microplacas (24 poços) contendo
BHI. Posteriormente, inoculou-se 100μL da suspensão bacteriana que havia crescido em BHI,
centrifugada e previamente lavada em PBS até adquirir compração igual a escala de 0,5
MacFarland. Depois, a placa foi lavada 3 vezes com PBS (pH 7,2) e as lamínulas coradas pela

29
técnica de Gram. A microplaca foi lavada novamente com PBS (pH 7,2), as lamínulas
retiradas e fixadas com Entellan® em lâminas para serem examinadas por microscopia de luz.
4.5.2 Adesão ao látex siliconizado e aço inox
No presente trabalho, foram utilizados fragmentos de látex siliconizado (cateter
urinário, Solidor®) medindo 0,5 mm de diâmetro e fragmentos de agulha medindo 1 cm. Os
fragmentos foram transferidos para tubos de ensaio, contendo 5 mL de caldo BHI e
esterilizados em autoclave por 15 minutos a 120°C para posteriormente serem utilizados. As
suspensões bacterianas foram preparadas a partir de inóculos previamente crescidos em Ágar
Muller Hinton, de onde se retirou colônias bacterianas e transferiu-se para tubos com salina
0,9% até adquirirem a turbidez 0,5 da escala de MacFarland. Em seguida, retirou-se 0,1ml de
cada suspensão preparada e inoculou-se nos tubos contendo BHI + látex siliconizado e BHI +
inox. Os tubos foram incubados em estufa bacteriológica a 37°C por 3 h e, a seguir, retirados,
lavados em solução fisiológica 0,9%, transferidos para 5mL de solução fisiológica
esterilizada.
Em seguida, foi realizada uma intensa homogeneização em aparelho Vortex durante
30 segundos. Depois, foram retiradas alíquotas de 0,1ml da suspensão obtida que foram
semeadas em duplicata em Ágar Muller Hinton, e as placas foram incubadas a 37ªC/ 24h,
sendo, então, determinado o número de unidades formadoras de colônia por mililitro
(UFC/mL) de Burkholderia sp e S. maltophillia recuperada de cada fragmento avaliado
(GOMES et al. 2009 com adequações).
Os tubos controles negativos não receberam inóculo bacteriano, mas passaram pelo
mesmo tratamento que as amostras bacterianas.
4.5.3 Adesão a células humanas
4.5.3.1 Cultivo e preparo das monocamadas celulares
As células HEp-2 (carcinoma epidermóide de laringe), HEla (carcinoma do colo do
útero), Vero (rim de macaco verde africano) e Pnemócitos A549 foram descongeladas em
banho-maria a 37º C e transferidas para uma garrafa de cultura de células contendo meio

30
MEM (Meio Mínimo Essencial de Eagle modificado / Cultilab), suplementado com 10% de
soro fetal bovino (SFB / Cultilab) e 1% de solução de antibióticos contendo penicilina (1000
U /mL; Sigma) e estreptomicina (250 µg / mL; Sigma). Os frascos de cultura foram incubados
em atmosfera de 5% de CO2 e 37º C/ 48 h, até a formação do tapete celular. Após este período
o meio foi descartado, sendo adicionado à cultura celular uma solução de ATV (Associação
Tripsina-Versene / Cultilab) para o descolamento do tapete. As células foram ressuspensas no
meio MEM, acrescido de 10% de soro fetal bovino (SFB) e 1% de antibióticos, para um
número estimado de 2,5 x células / mL. A suspensão de células foi distribuída em microplacas
de 24 cavidades, contendo lamínulas redondas, em volumes de 1mL por cavidade. As placas
foram incubadas a 37 º C em atmosfera de 5% de CO2 por 48 h ou até que a monocamada
atingisse a semi-confluência.
4.5.3.2 Realização do teste de adesão
Um inóculo de 40μL das amostras de Burkholderia sp e S. maltophillia cultivadas
em BHI por 24h a 37º C, centrifugada e lavada em PBS até adquirir compração igual a escala
de 0,5 MacFarland foi adicionado em placas de 24 orifícios, contendo monocamadas celulares
semi-confluentes de células Vero e HEp-2 contidas em MEM acrescido de SFB a 2% na
ausência ou presença de D-manose a 2%. Após 2h de incubação a 37º C em atmosfera de 5%
de CO2 (período de infecção), as placas foram lavadas com PBS e adicionado ao meio 1mL
de MEM acrescido de SFB com e sem manose. Após 3h (período de multiplicação), cada uma
das placas foram lavadas com PBS e as células foram fixadas com metanol por um período
mínimo de 10 minutos. As preparações foram coradas com May-Grunwald / Giemsa e
analisadas quanto ao padrão de aderência em microscópio óptico (Nikon) (ZARANZA et al.,
2012).
4.6 Produção de Biofilme
4.6.1 Avaliação da formação de cápsula
A produção de cápsula como fator presuntivo para formação de biofilme foi
realizada pelo método de semeadura em Ágar Vermelho Congo (AVC), descrito por Freeman,
Falkiner e Keane (1989), com poucas adaptações. Posteriormente à semeadura em AVC,
observou-se a coloração apresentada pelas colônias. As amostras foram consideradas

31
produtoras de cápsula apenas quando apresentaram coloração preta.
4.6.2 Análise da indução de biofilme por especrotofotometria
Primeiramente, as amostras bacterianas em pesquisa foram cultivadas em BHI,
incubadas a 37°C/24 horas. As mesmas foram padronizadas por espectrofotometria, deixando
todas com densidade óptica igual (Do) a 0,7. Logo após, uma porção de cada cultivo foi
diluída a 1:40 em meio BHI. Cada amostra foi adicionada em microplacas com 96 poços com
fundo em U, em volume de 200 μL/cavidade, depois de preenchidas as placas foram
novamente incubadas a 37°C por 24 horas. Após o tempo de incubação, as placas foram
lavadas por três vezes com tampão salina fosfato e deixadas a temperatura ambiente.
Adicionou-se às placas 200μL/cavidade de Cristal Violeta, que foram incubadas por 15
minutos a temperatura ambiente. Posteriormente a este procedimento, cada poço foi lavado
com água destilada por três vezes, e deixadas secar em temperatura ambiente. As placas
coradas com cristal violeta foram submetidas a espectrofotometria com filtro de 540nm para
aferir as respectivas absorbâncias (Ab) de cada poço. Baseando-se nas densidades ópticas
produzidas pelos isolados (D.o.i) e tomando-se por base o controle negativo (D.o.c), os
isolados foram classificados nas seguintes categorias (STEPANOVIC et al., 2004):
Não produtor: D.o.i < D.o.c
Produtor fraco: D.o.c < D.o.i
Produtor moderado: (2x D.o.c) < D.o.i ≤ (4 X D.o.c)
Produtor forte: (4 X D.o.c) < D.o.i
4.7 Detecção de citotoxina
4.7.1 Preparo do sobrenadante das amostras bacterianas
As amostras de Burkholderia sp e S. maltophillia foram cultivadas em 3mL de BHI,
incubadas a 37º C/24 h. Após a incubação, as bacterianas foram centrifugadas (3600
rpm/10min) e seu sobrenadante foi coletado. Após a obtenção do sobrenadante, o mesmo foi
centrifugado por mais 3 vezes e filtrado, para a retirada total das células bacterianas e, em
seguida inoculado nos poços da microplaca para realização do experimento.

32
4.7.2 Teste de citotoxidade
As placas de cultura das linhagens de célula Vero e HEp-2 foram preparadas como
descrito no item 4.5.3.1, e utilizadas para detecção de citotoxinas existentes nos sobrenadantes
de cultura das amostras de Burkholderia sp e S. maltophillia (item 4.8.1). Alíquotas de 100μL
dos sobrenadantes foram adicionadas em duplicata, em diluições seriadas na razão 2. As
placas foram incubadas em estufa a 37º C em atmosfera de 5% de CO2 e a leitura dos
resultados foi realizada em 18h, 24h e 72h após a incubação, com auxílio de um microscópio
invertido (Axiovert – Zeiss). Foi utilizado como controle positivo a solução de fenol a 2%
com PBS, como controle negativo utilizou-se as células das linhagens citadas anteriormente e
meios de cultura utilizados, insentos de qualquer amostra bacteriana, para confirmar pureza .
(KONOWALCHUK; SPEIRS; STAVRIC, 1977).
4.8 Análise Enzimática
Para verificar a atividade enzimática dos isolados de Burkholderia sp e S.
maltophillia foram analisadas a produção das seguintes enzimas extracelulares:
4.8.1 Amilase
O ágar amilase foi preparado com 2g de amido de milho para cada 1000 ml de Ágar
BHI. Após a preparação e esterilização as placas foram semeadas por picada central,
incubadas a 37°C por 24-48h. Adicionou-se solução de lugol à placa e observou-se a
ocorrência de halos amarelos ao redor do crescimento bacteriano (BUZZINI E MARTINI et
al. 2002).
4.8.2 DNase
As amostras de S. maltophillia foram semeadas por picada central em Ágar DNase
com Azul de Toluidina (Himedia), incubadas a 37°C por 24-48h. Posteriormente, observou-se
um halo translúcido em torno do inóculo e a produção de uma reação rosa após a adição de
ácido hidroclorídrico à placa para constatar a positividade do teste (WILSON, 1977).

33
4.8.3 Elastase
Preparou-se uma solução com 2% de Ágar bacteriológico acrescido de 1% de
Elastina (INLAB) diluídas em tampão Tris-HCL (hidroximetil) aminometano 0,03M pH 8,0.
As amostras previamente crescidas em caldo BHI foram inoculadas pontualmente em placas
de Petri com o meio de cultura descrito anteriormente e incubadas por 48 horas a 37° C
durante 5 dias em temperatura ambiente. A atividade de elastase foi determinada pela
observação de zonas claras ao redor do local de inoculação, em decorrência da ação da
enzima (ZARANZA et al., 2012).
4.8.4 Fosfolipase
A fosfolipase foi detectada utilizando 10 ml de “egg yolk” 50% para cada 90 ml de
Ágar Muller-Hinton estéril. Com o auxílio da alça de platina, as amostras foram semeadas e a
positividade da prova foi observada a partir da presença de zona clara ao redor do inóculo
após 24-48h de incubação a 37° C (FIGUEIRÊDO et al., 2006 )
4.8.5 Gelatinase
Primeiramente, as cepas foram inoculadas em BHI e após a incubação a 37°C
durante 24 horas, foram semeadas em tubos (por profundidade) em uma solução de gelatina a
12%, preparada em tampão fosfato pH 7,4 e também em placas de Ágar Muller Hinton
contendo 5% de gelatina. Após incubação a 37°C por 24 horas, observou-se se ocorreu a
liquefação da gelatina (ZARANZA et al., 2012).
4.8.6 Lipase
Para a detecção da atividade lipolítica foi utilizado o método descrito por Edberg,
Gallo e Kontnick (1996) com modificações. As amostras bacterianas previamente cultivadas
em Ágar Nutriente a 37°C/ 24 h, foram semeadas por picada em placas com ágar BHI
suplementado com 1% de Tween-80 e também em placas com Ágar Muller Hinton com 1%
de azeite de oliva (MESSIAS et al., 2011). As culturas foram incubadas à temperatura
ambiente e avaliadas diariamente durante 3 dias. O aspecto turvo ao redor do inóculo foi
considerado como teste positivo.

34
4.8.7 Proteinase
A atividade proteolítica foi positiva quando verificada zona clara ao redor dos
inóculos realizados em placas de Ágar Muller Hinton (Himedia) acrescido de 10% de “Skim
Milk” (HIMEDIA) (FLACH, 2006, com algumas modificações).
4.9 Produção de hemolisinas
4.9.1 Atividade hemolítica em meio sólido
As amostras de Burkholderia sp e S. maltophillia foram cultivadas em caldo BHI
(Brain Hearth Infusion – Oxoid) durante 24 h/ 37º C. Após esse período, semeou-se as
bactérias por picada, em placas de Ágar Mueller Hinton com 5% de sangue de carneiro,
cavalo e humano (A, B e O). As placas foram incubadas a 37º C durante 3 dias (72 h), a cada
24 h verificou-se a presença ou não de halos de hemólise notificando em qual tempo
observou-se a positividade do teste (GARCIA et al., 2002). Zonas de hemólise, parcial ou
total, puderam ser visualizadas ao redor das colônias ou abaixo delas em casos de amostras
hemolíticas.
4.9.2. Atividade hemolítica com o pellet e sobrenadante bacteriano (teste em microplaca)
Para a execução deste teste, utilizou-se a metodologia descrita por Bhakdi et al.
(1986), com algumas modificações. Os isolados foram semeados e incubadas a 37 ºC/ 24h,
em: caldo BHI puro, BHI + cloreto de cálcio (CaCl2- 10 mM), BHI + ácido etileno diamino
di-orto- hidroxifenil acético (12,5 μg/ml EDDA), BHI + ácido etileno diamina tetra acético
(10 mM de EDTA ) e BHI + cloreto férrico (FeCl3 - 10 mM) (BEUTIN et al. 1988).
Posteriormente, as amostras foram centrifugadas três vezes (3600 rpm/15min), o
sobrenadante e o “pellet” coletados separadamente. Em microplacas de 96 poços de fundo
“U” alíquotas de 100 μL de suspensão de hemácias de carneiro a 1%, antecipadamente
lavadas 3 vezes em PBS (pH 7,4) foram adicionadas a 100 μL do sobrenadante e também a
100 μL do “pellet” (com a turbidez equiparada a escala 0,5 de Mac Farland). Em seguida, as
microplacas foram incubadas em estufa a 37 °C por 1h, e depois analisadas visualmente para
a ocorrência ou não da hemólise. A formação de um “botão” de hemácias no fundo do(s)
poço(s) da microplaca indicou o resultado negativo para hemólise e, a ausência desse “botão”

35
foi considerado positivo.
Como controle positivo de hemólise adicionou-se água destilada a solução de
hemácia e, para o controle negativo reservou-se os 5 meios descritos anteriormente,
adicionado a solução de hemácias.
4.10 Análises estatísticas
No processamento eletrônico dos dados foi utilizado o Programa Biostat 5.0. Os
testes estatísticos empregados foram Coeficiente de Contigência C, Q de Cochran, Exato de
Fisher e McNemar para a análise dos dados. Em todos os testes o nível de significância
aplicado foi de 5%.
5 RESULTADOS E DISCUSSÃO
5.1 Amostras e sítios de origem
As estirpes de Burkholderia sp e S. maltophillia analisadas nesta pesquisa, foram
isoladas de amostras clínicas de distintos sítios anatômicos (Figura 1).
Entre os isolados de Burkholderia sp, a amostra mais frenquente foi proveniente de
sangue, correspondendo a 57,14%. A presença desta bactéria em amostras advindas de
sangue também foi evidenciada por Lizaso et al (2008), que ao identificarem os agentes
etiológicos de bacteremias encontraram Burkholderia sp em 28% das hemoculturas
pesquisadas.
A litertura também relata divergências, Douce et al (2008) ressaltam que
Burkholderia cepacia é uma causa incomum de infecções de corrente sanguínea (ICS).
Surtos hospitalares de pequenos focos envolvendo pacientes sem fibrose cística, têm sido
relacionado à contaminação de fonte comum, tais como: reservatórios de água, desinfetantes,
soluções antissépticas, anestésicos, soluções e medicamentos intravenosos (HOLMES;

36
GOVAN; GOLDESTEIN, 1998; ÁLVAREZ-LERMA et al., 2008). Tal contaminação pode
ser intrínseca, acontecendo durante o preparo de tais soluções ou extrínseca, ocorrendo por
exemplo, durante a manipulação de vias multidoses (DIAS et al., 2008).
Figura 1. Distribuição das amostras de Burkholderia sp e S. maltophillia.por amostra clínica
Em um surto de infecção grave originada pelo Complexo Burkholderia cepacia em
uma unidade de terapia intensiva multidisciplinar da Espanha, um hidratante corporal
contaminado serviu como reservatório e origem da infecção. A eliminação do reservatório foi
associado com a erradicação da Burkholderia cepacia do hospital (ÁLVAREZ-LERMA et al.,
2008).
Já em hospital universitário, de 250 leitos no Líbano, Nasser et al. (2004) relataram
uma epidemia de ICS, causada por Burkholderia cepacia relacionada a cateter. Ocorreram
411 episódios de ICS por tal micro-organismo, acometendo 361 pacientes no Hospital Saint
George. Os fatores de risco para este tipo de infecção relacionada a cateter, variam de acordo
com o tipo de cateter, localização do sítio de inserção e a duração de permanência do mesmo,
tamanho do hospital, unidade ou setor de prestação de serviço, (MERMEL et al., 2001).
Este fato deve ser ressaltado, tendo em vista que possivelmente o percentual elevado
de amostras sanguíneas contaminadas por Burkholderia sp encontrado no presente estudo,
14,28%
57,14%
7,14%
21,42%
70,58%
11,76% 11,76% 5,88%
0
10
20
30
40
50
60
70
80
Secreção
traqueal
Sangue Swab nasal Urina Não informado
Burkholderia sp S. maltophillia

37
pode estar associado ao uso de cateteres ou dispositivos intravasculares, uma vez que se trata
de amostras clínicas oriundas de pacientes hospitalizados.
Diversos estudos acerca de contaminação de cateteres venosos centrais de curta
permanência, evidenciaram que bacilos gram-negativos (BGN´s) poderiam ser a causa de 22 a
54% das ICS´s associadas. Outros estudos demonstraram que os BGN´s causaram 26 a 55%
de ICS relacionada a cateter de longa permanência. Além disso, as infusões contaminadas são
uma importante fonte de bacteriemias por esses bacilos, pois uma vez introduzidos, podem se
proliferar em tais fluidos (RAAD, 2004).
Ao analisarmos os isolados de S. maltophillia verificou-se que esta bactéria
apresentou maior frequência entre as amostras de secreção traqueal, 70,58%. Estudiosos
relatam que o aparelho respiratório é considerado o sítio anatômico mais comum de
isolamento de S. maltophillia em pacientes hospitalizados, representando a origem de 56 a
69% dos isolados (DAVIN-REGLI et al., 1996; GERNER-SMIDT et al., 1995), comprovando
o percentual citado anteriormente.
A presença deste patógeno neste tipo de amostra clínica pode estar associada à
ventilação mecânica, traqueostomia, a exposição prévia a antibióticos de largo espectro, ao
uso de equipamento de vias respiratórias, tais como nebulizadores (ELTING et al., 1990;
KHARDORI et al., 1991; KOLLEF et al., 1995; LAING et al., 1995). A literatura diz que
algumas condições pré-existentes do pulmão, como a doença pulmonar obstrutiva crônica,
fibrose cística, ou obstrução brônquica de muitos pacientes, são consideradas fatores
determinantes na instalação de S. maltophillia (LAING et al., 1995).
Klick e Du Moulin (1976) relataram casos em uma unidade de terapia intensiva no
qual um analisador de oxigênio contaminado com S. maltophillia foi a fonte de disseminação.
Korn et al (1996), também descreveram um surto associado ao tubos endotraqueais de
ventiladores mecânicos. Turner-Hubbarde e colaboradores (1996), isolaram S. maltophillia a
partir de amostras broncoalveolar de broncoscópios
O envolvimento de cepas de S. maltophillia com o trato respiratório está
significativamente associado ao aumento da mortalidade (MORRISON et al., 1986). Kollef et
al. (1995), relataram que o isolamento de um patógeno de "alto-risco", tal como S.
maltophillia foi o mais importante fator de mortalidade em casos de pneumonia associada à

38
ventilação mecânica. A taxa de mortalidade em pacientes neutropênicos com pneumonia é de
40% (KHARDORI et al.,1991).
5.2 Susceptibilidade aos antimicrobianos
A análise da resistência aos antibióticos entre amostras bacterianas de origem
hospitalar é um instrumento de suma importância, pois ajuda a escolher a antibioticoterapia
mais adequada às infecções por bactérias resistentes (MARRA et al., 2006).
Entre as amostras de Burkholderia sp avalidas no presente trabalho, observamos
sensibilidade em 80% das cepas frente ao meropenem e 100% frente à ceftazidima e
minociclina (Figura 2), demonstrando assim, que esses antibióticos são boas escolhas na
terapêutica de infecções causadas por tais agentes.
Figura 2- Perfil de susceptibilidade aos antimicrobianos dos isolados de Burkholderia sp.
Os resultados aqui expostos, evidenciaram percentuais maiores de sensibilidade
frente a ceftazidima e meropenem quando comparados a pesquisa realizada com 366 cepas
clínicas de Burkholderia sp. Em tal pesquisa, constatou-se 77% de sensibilidade à
ceftazidima, piperacilina/tazobactam e meropenem. Entretanto, evidenciou-se resistência à
100%
80%
25%
100%
20%
75%
Ceftazidima Meropenem Sulf/Trim Minociclina
Susceptibilidade aos antimicrobianos de isolados de
Burkholderia sp
Sensível Intermediário Resistente

39
ciprofloxacina, amicacina, gentamicina, tobramicina, cefuroxima, cefotaxima, imipenem,
cloranfenicol, tetraciclina e sulfas (LEDSON; GALLAGHER; CORKHILL, 1998).
As cepas aqui pesquisadas também apresentaram elevada taxa de resistência frente
ao sulfametaxazol/trimetropim (75%) e sensibilidade intermediária ao meropenem (20%).
Fato que causa preocupação, já que estas são duas das poucas drogas de escolha para o
tratamento de infecções por Burkholderia sp e em alguns casos já não mostram-se capaz de
inibir a ação desta.
Os mecanismos envolvidos na resistência ao sulfametaxazol/trimetropim ainda não
estão bem elucidados, mas acredita-se que a obtenção de integrons de classe 1, contendo o
gene sul1 na sua extremidade 3 não traduzda, favorece esse perfil, bem como a capacidade de
formar biofilme que, apesar de não ser, de fato, um mecanismo de resitência, é capaz de
dificultar a atividade da droga sobre a bactéria (TRAUB et al., 1998).
Segundo alguns estudos, estipes de Burkholderia sp são intrinsecamente resistentes
aos aminoglicosídeos; a taxa de resistência aos beta-lactâmicos é geralmente elevada. Quanto
aos carbapenêmicos, o meropenem tem boa atividade in vitro, 90% das cepas são inibidas
com uma concentração inibitória mínima de 8mcg/ml (BURNS; SAIMAN, 1999). Tal
percentil de sensibilidade é mais elevado que o encontrado neste trabalho, sugerindo que as
estirpes clínnicas avaliadas estejam expressando possíveis fatores de resistência, o que
possibilita a diminuição da sensibilidade ao referido antibiótico.
Ressalta-se, que as únicas cepas que manifestaram sensibilidade intermediária ao
meropenem, foram as oriundas de secreção traqueal (Tabela 1). Deste modo, sugere-se que as
cepas isoladas da traqueia apresentam maior probabilidade de terem resistência intermediária
ao meropenem quando comparadas as cepas isoladas do sangue ( valor de p=0.005).
Tabela 1- Frequência absoluta e relativa de resistência intermediária aos
antimicrobianos por Burkholderia sp
Sítio
Anatômico
Meropenem
(%)
Ceftazidima
(%)
Sulf/trim
(%)
Minociclina
(%)
Traqueal 4 (80) 0 0 0
Sangue 0 0 0 0

40
Cabe esclarecer que a resistência ao meropenem já é um problema real, Sader et al.,
(2001), verificaram o perfil de sensibilidade aos antibióticos de bactérias isoladas do trato
respiratório e, um dos resultados evidenciados foi a resistência aos carbapenêmicos
(imipenem e meropenem) por BGNF´s.
Recentemente, Dentini et al.,(2011), realizaram pesquisa com 283 pacientes com
fibrose cística atendidos no ambulatório de Pediatria do HC/UNICAMP e, destes 11,3 %
tiveram suas amostras de secreção de orofaringe e de escarro positivas para o Complexo
Burkholderia cepacea. Dentre as estirpes isoladas apenas 38,06% mostraram-se sensíveis à
todos os antimicrobianos testados, os mesmos que foram avaliados no presente estudo.
Entretanto, 54,48% mostraram-se resistentes ao sulfametoxazol-trimetropim, esse percentual
de resistência superior a 50%, mostra que sobram cada vez menos alternativas terapêuticas
para combater infecções por tal micro-organismo.
Em análises de amostras de escarro de pacientes com fibrose cística infectados com
Pseudomonas aeruginosa e Burkholderia cepacia, evidenciou-se a presença de N-
acylhomoserine lactones (AHL). As AHL são responsáveis por controlar a expressão dos
genes de virulência bacteriana e também estão envolvidas no estímulo a maturação de
biofilmes resistentes a antibióticos (WARD et al., 2003), podendo assim explicar em parte, a
elevada taxa de resistência entre os isoldados de secreção traqueal.
Desigualmente ao que foi exposto anteriormente, Chaves (2010) analisou o perfil de
susceptibilidade “in vitro” e constatou que 61,1% das cepas do Complexo Burkholderia
cepacia foram sensíveis ao sulfametoxazol-trimetoprim.
Diferenças nas taxas de sensibilidade aos antimicrobianos também foram descritas
em trabalho realizado por González-Saldaña et al., (2008), onde a resistência a
sulfametaxazol/trimetropim e a tendência a sensibilidade intermediária ao meropenem
também foi constatada. Além da resistência a sulfametoxazol/trimetropim, também há a
possibilidade de intolerância a essa droga, tornando-se fundamental a analise in vitro de
outras, que alternativamente possam ser utilizadas no tratamento das infecções por esse agente
(RODRIGUES et al., 2011).
Os resultados obtidos corroboram a literatura ao relatar que o tratamento de
infecções pelo Complexo Burkholderia cepacia torna-se difícil devido a sua resistência à
maioria dos antibióticos disponíveis clinicamente. Este problema é complicado pelo

41
desenvolvimento de resistência durante terapia e o desenvolvimento de resistência cruzada a
outros antimicrobianos (WIGFIELD et al., 2002).
Quanto ao perfil de susceptibilidade das estirpes de S. maltophillia, evidenciou-se
altas taxas de sensibilidade aos antibióticos utilizados : 83,3% a minociclina, 77,77%
levofloxacino e 88,8% ao sulfametaxazol/trimetropim (Figura 3).
Figura 3- Perfil de susceptibilidade aos antimicrobianos dos isolados de S. maltophillia.
Garcia et al. (2002) também relataram alta taxa de sensibilidade ao
sulfametaxazol/trimetropim, sendo este ainda, uma boa sugestão para o tratamento de
infecções causadas por tal bactéria.
A resistência a sulfametaxazol/trimetropim observada em 5,5% das amostras
(oriundas de secreção traqueal), apesar de não ser expressiva e não ter demonstrado aumento
significativo nos últimos anos, pode dificultar a decisão terapêutica empírica nas infecções
graves, visto que a resistência a essa droga pode estar associada a resistência a outros
antibióticos considerados ativos contra esse patógeno (PAEZ et al., 2008).
Ainda existem poucos trabalhos que verificam a resistência de isolados clínicos de S.
maltophillia para a minociclina. No entanto, esses estudos mostram alta porcentagem de
sensibilidade a essa droga, variando entre 97% e 100% (MILATOVIC et al., 2003 BETRIU et
al., 2002).
Cantón et al., (2003) analisaram 47 estirpes de S. maltophillia em pacientes com FC
e mostraram percentagem de resistência menor que 1% para minociclina. Hu et al., (2011) ao
77,77%
88,88%
16,66%
5,5% 5,5%
Minociclina Levofloxacina Sulf/Trim
Susceptibilidade antimicrobiana dos isolados de S. maltophillia
Sensível Resistente
83,33

42
avaliarem concentrações inbitórias em 102 estirpes mostraram uma sensibilidade a
minociclina de 74,5%. Contudo, a taxa de sensibilidade a miniciclina exposta no gráfico
acima, encontra-se em conformidade com a literatura.
Para Paez (2011), que obteve taxa de 100% de sensibilidade a minociclina o que
poderia explicar tal fato seria a não comercialização da droga no Brasil, a sua
indisponibilidade por via intravenosa, a falta de estudos clínicos com esse medicamento e o
seu uso limitado na prática clínica.
Ao avaliar a sensibilidade de levofloxacino através da microdiluição em caldo,
diante de 1584 amostras de S. maltophillia isoladas ao redor do mundo, Farrel et al., (2010)
evidenciaram uma sensibilidade de 83,4%.
Gesu et al., (2003) compararam a atividade de levofloxacino e ciprofloxacino, em
124 cepas analisadas e obtiveram sensibilidade de 85,5% e 58,9% respectivamente. Tais
pesquisas corroboram em parte com as taxas encontradas, onde constatou-se 77,77% de
sensibilidade frente ao levofloxacino.
As diferenças de sensibilidade aos antimicrobianos que foram evidenciadas, sugerem
que as unidades de assistência a saúde não usam um protocolo único de administração de
antibióticos para o tratamento de infecções causadas por estirpes de Burkholderia sp e S.
maltophillia. Tal medida parece contribuir indiretamente para a resistência antimicrobiana
múltipla. Já que, cada unidade de saúde utiliza diferentes antibióticos, muitas vezes
empiricamente, sem realização de testes de sensibilidade para identificar a melhor escolha
afim de combater a infecção. Isso pode contribuir para que as estirpes bacterianas tenham
contato com maior número de antibióticos e ao longo do tempo desenvolvam mecanismos de
resistência a esses.
Deste modo, pôde-se concordar com a literatura ao relatar que Burkholderia sp e S.
maltophillia apresentam resistência a várias classes de antibióticos. Principalmente às drogas
de interesse clínico, seja por meio da obtenção de genes de outras espécies ou por mutações
na membrana plasmática bacteriana (ZHANG et al., 2000;WIGFIELD et al., 2002).

43
5.3 Expressão das ESBL fenotipicamente
Alguns isolados produtores de ESBL apresentam completa resistência aos
antibióticos beta–lactâmicos de amplo espectro, expressando essas enzimas nos testes
fenotípicos, embora muitos isolados não expressem esse fenótipo, mesmo possuindo os genes
codificadores dessas enzimas (PATTERSON et al., 2001).
Fenotipicamente obtve-se positividade em 63% das cepas de S. maltophillia e 64%
das cepas de Burkholderia sp para ESBL. Não foi encontrado na literatura trabalhos que
tenham realizado este teste de expressão fenotípica diante de tais micro-organismos. Mas com
os dados obtidos, pode-se confirmar a produção da enzima beta-lactamase por estas duas
espécies.
A resitência aos beta-lactâmicos é considerada bastante complexa pois ocorre via
produção de enzimas cujo gene codificador tem origem distinta (cromossomal, plasmidial ou
nos transposons) e ainda por um mecanismo de efluxo multidrogas (ZHANG et al., 2000).
Duas ß lactamases têm origem cromossomal: L1, uma carbapenemase dependente de zinco e
L2 uma cefalosporinase inibida pelo ácido clavulânico. Um plasmídeo R de 5,6 quilobases
(kb) confere fenótipo resistente frente à penicilina e à cefazolina, além de outro, de 200 kb,
que também possui genes codificadores das â lactamases L1 e L2. Adicionalmente, S.
maltophillia, sobretudo no ambiente hospitalar, é considerada reservatório de genes móveis
para ß-lactamases de amplo espectro, o que confere capacidade aumentada de transmissão de
resistência e, portanto, maior relevância epidemiológica (DENTON e KEER, 1998).
5.3 Determinantes de adesão
5.3.1 Capacidade hidrofóbica
Burkholderia sp e S. maltophillia estão entre os bacilos Gram-negativos não
fermentadores (BGN-NF) mais envolvidos a surtos de infecções hospitalares (CAETANO et
al, 2011). Tal evidência pode estar associada a capacidade de adesão a superfícies bióticas ou
abióticas (PEETERS et al. 2008; ROSE et al. 2009).

44
Para constatar a capacidade de adesão dos isolados em estudo, iniciou-se a pesquisa
avaliando a capacidade hidrofóbica de cada cepa, já que alguns estudiosos têm comprovado
que a hidrofobicidade da superfície celular bacteriana é um dos fatores que contribuem no
processo de adesão bacteriana aos diferentes materiais (CAPELLO; GUGLIELMINO, 2006).
Assim, quanto mais hidrofóbica for a superfície celular bacteriana, possivelmente maior será
sua capacidade de formar biofilmes já que, as características físico-químicas da superfície
podem desempenhar uma forte influência sobre a adesão bacteriana, as quais aderem com
mais facilidade às superfícies hidrofóbicas (ex: plástico) do que às hidrofílicas (ex:vidro ou
metais) (RODRIGUES et al., 2009).
A capacidade hidrofóbica das bactérias aqui avaliadas foi determinada por meio de
agregação bacteriana em concentrações crescentes de sulfato de amônio (LOCATELI et al.,
2004). De acordo com a literatura, durante a fase exponencial de crescimento as bactérias
tendem a se tornar mais hidrofóbicas e formar grumos, aderindo umas às outras ou a
superfícies presentes no meio de cultura (VAN LOOSDRECHT et al., 1987).
Deste modo, constatou-se que do total de cepas de S. maltophillia 6 (33,3%),
apresentaram hidrofobicidade do tipo fraca, 3 (16,6%) moderada e 9 (50%) forte. Percentuais
de capacidade hidrofóbica parecidos a esses também foram obtidos entre os isolados de
Burkholderia sp, na ordem: 4 (30,7%) fracos, 3 (23,07%) moderados e 6 (46,15%) fortes. A
capacidade hidrofóbica de ambas as estirpes bacterianas apresentaram forte capacidade
hidrofóbica e quando comparada com o sítio de isolamento das bactérias não apresentou
significância estatística (Tabelas 2 e 3).
Em estudo desenvolvido por Locateli et al. (2004), foi constatado a capacidade
hidrofóbica moderada das culturas de P. aeruginosa e S. aureus, enquanto que a outra espécie
avaliada, S. epidermidis não formou grumos na concentração mais elevada de 3M,
apresentando uma característica hidrofílica diante do teste realizado.

45
Tabela 2- Correlação entre a capacidade hidrofóbica e os sítios de isolamento das estirpes de
Stenotrophomonas maltophillia.
Hidrofobicidade Sítio Anatômico Coeficiente de
Contigência C
Valor de p
Outros Traqueia
Fraco 3 (25%) 3 (50%) 0,43 0,12
Moderado 1 (8,3%) 2 (33,3%)
Forte 8 (66,7%) 1 (16,7%)
Tabela 3- Correlação entre a capacidade hidrofóbica e os sítios de isolamento das estirpes de
Burkholderia sp.
Hidrofobicidade Sítio Anatômico Coeficiente de
Contigência C
Valor de p
Sangue Traqueia
Fraco 2 (14,2%) 2 (14,2%) 0,37 0,36
Moderado 3 (21,4%) 0
Forte 4 (28,5%) 2 (14,2%)
De Rossi Passerini et al. (2007) fizeram um trabalho semelhante ao nosso para
averiguar a capacidade hidrofóbica de amostras clínicas de S. maltophillia e, observaram que
doze das treze (92,3%) cepas avaliadas apresentaram hidrofobiciadade moderada a forte,
constatando assim um percentual mais elevado que o obtido no presente trabalho. Já Pompilio
et al. (2008), encontraram 11 (27,5%) das estirpes de S. maltophillia com hidrofobicidade
moderada, taxa mais compatível com este estudo.
Na literatura, raros são os estudos que avaliam a hidrofobicidade dos isolados de
Burkholderia sp, não foi encontrado trabalhos que tenham realizado tal teste com amostras
clínicas. Chakraborty et al. (2010) pesquisaram a hidrofobicidade de 3 estirpes
biodegradantes: Burkholderia cepacia ( ES1 ) e duas linhagens de Burkholderia multivorans
( NG1 e HN1 ), onde foi possível constatar que o aumento da hidrofobicidade bacteriana
resultou em aumento da adesão ao vidro.

46
Pensa-se que a superfície de uma célula hidrofóbica pode ser uma vantagem para as
bactérias patogênicas “in vivo”, pela promoção de interações com as células hospedeiras
(ONG et al. de 2004 ; KESPICHAYAWATTANA et al. de 2004 ; BRETT . et al , 1998 ).
5.3 Adesão a materiais inertes e células humanas
As infecções associadas aos materiais médico-hospitalares resultam da adesão
bacteriana à superfície de um biomaterial (KATSIKOGIANNI et al., 2004). Bactérias
patogênicas possuem uma variedade de mecanismos de adesão e invasão celular, que permite
explorar uma grande quantidade de componentes da superfície da célula hospedeira e
possibilitam a ocupação de nichos diferentes dentro do corpo humano (SANTOS, 2011).
Assim, a adesão é considerada um pré-requisito para a patogênese de quase todas as doenças
infecciosas (HULTGREN et al., 1996).
Entre os bastonetes Gram- negativos não fermentadores (BGNNF) Pseudomonas
aeruginosa, S. maltophilia e Burkholderia cepacia são comumente associados a bacteremias
(REIS, 2010), já que podem colonizar equipamentos hospitalares e secreções humanas
(SENOL, 2004).
Todos os isolados bacterianos avaliados neste estudo foram submetidos aos testes de
adesão a superfícies abióticas, utilizando: lamínula de vidro, aço inoxidável e látex
siliconizado. As estipes de Burkholderia sp também foram submetidas ao teste de adesão as
células humanas HEp-2 e Pneumócitos A549, enquanto que as estirpes de S. maltophillia
foram avaliadas apenas na presença da linhagem de Pneumócitos A549.
A capacidade de adesão das amostras clínicas de Burkholderia sp foi significante e
apresentaram elevados índices de positividade para adesão, tanto às superfícies abióticas
quanto as linhagens das células humanas avaliadas (Tabela 4). Idependentemente do sítio de
isolamento, todas as cepas aderiram ao vidro. Entre as estirpes provenientes de sangue, a taxa
de adesão ao inox e ao látex foi a mesma, 88,9 %. Já as amostras oriundas da traqueia
aderiram um pouco menos ao inox, 80% e, 100% ao látex. Comparando a adesão às células
humanas observou-se que diante das células HEp-2, os percentis variaram de 60% a 66,7%
entre os isolados da traqueia e os procedentes de sangue, respectivamente. A adesão aos

47
Penumócitos ocorreu em apenas 40% das amostras de traqueia e 33,3% das amostras
sanguíneas.
Tabela 4- Capacidade de adesão das amostras clínicas de Burkholderia sp à superfícies abióticas e
células humanas.
Sítio
Anatômico
Abiótico Células Q de
Cochran
Valor de
p
Vidro Inox Látex HEp-2 Pneumócito
Sangue 9 (100%) 8 (88.9%) 8 (88.9%) 6(66.7%) 3 (33.3%) 33,57 <0.0001
Traqueia 5 (100%) 4 (80%) 5 (100%) 3 (60%) 2 (40%)
Estatisticamente, as cepas de de Burkholderia sp têm menos propensão de aderir aos
pneumócitos. Não há diferença na adesão das cepas segundo o sítio anatômico (Valor de p
>0.05). Não há diferença na adesão das cepas segundo o sítio anatômico, valor de p >
0.05.Assim, a adesão das cepas de Burkholderia sp independe o sítio anatômico na qual foi
isolada, portanto independentemente se ela é isolada de amostras sanguíneas ou de traqueia a
probabilidade de aderência é a mesma.
Gelsuu et al. (2012), também evidenciaram a adesão de B. cepacia a lamínulas de
vidro. Além disso, constataram que a presença de substratos como alginato, ajudam a adesão
com consequente formação de biofilme nas lamínulas (Gelsuu et al., 2012). No trabalho de
Nornberg (2009), com micro-organismos psicotróficos isolados de leite bovino, a autora
encontrou contagens elevadas de colônias de B. cepacia em substrato de aço inox indicando
que esta cepa tem alta capacidade de aderir, confirmando os achados evidenciados no
presente estudo. O percentual baixo de adesão aos Pneumócitos em relação a outra linhagem,
pode ser devido ao elevado índice de isolados que apresentarem efeito citotóxico. Foram
observadas alterações morfológicas, pontos de vacuolização citoplasmática, invasão e
destruição completa do tapete de células o que tornou inviável a visualização da adesão, caso
esta tenha ocorrido (Fugura 4). Tais dados demonstram que as cepas avaliadas tiveram maior
afinidade com os Pneumócitos, justificando os altos índices de colonização e infecções
pulmonares.

48
Figura 4- Microscopia óptica (100 X) da adesão, invasão e pontos de vacuolização em células HEp-2(A) e
Pneumócitos A549 (B).

49
Apesar de ter sido observado por microscopia óptica o efeito citotóxico de algumas
cepas de Burkholderia sp, ao avaliarmos a citotoxicidade do sobrenadante das culturas
bacterianas em células Vero e HEp-2 não constatamos nenhum efeito diante das células
avaliadas.
Adesão e subsequente invasão a células de mamíferos têm sido implicadas como
passos essenciais na patogênese de muitas bactérias intracelulares facultativas como a B.
pseudomallei (TOMICH et al, 2002; COWAN et al, 2000; HAAGER et al, 2003). Os
mecanismos envolvidos na invasão de células incluem a fagocitose, a endocitose mediada por
bactérias e por proteínas de membrana específicos (por exemplo: invasina, proteína de adesão
extracelular (PAE), flagelina e adesina (CHEN et al, 2001; TOMICH et al, 2002; HAGGAR
et al, 2003; INGLIS et al, 2003).
Shahin e Proll (2004) também relataram o potencial de adesão e invasão por cepas de
B. pseudomallei em pequenas células epiteliais humanas das vias respiratórias, SAEC
(Human small airway epithelial cells/ SAEC).
Martin e Mohr (2000) evidenciaram que um virulento e altamente transmissível
isolado clínico de B. cepacia, estirpe J2315, é capaz de entrar, sobreviver e replicar
intracelularmente em culturas de macrófagos humanos e células epiteliais pulmonares,
confirmando em parte as evidências aqui encontradas. Também observaram que um isolado
ambiental de B. cepacia, estirpe J2540, é capaz de entrar em macrófagos cultivados e células
epiteliais com frequências semelhantes às da estirpe clínica. No entanto, em contraste com a
estirpe clínica, o isolado ambiental de B. cepacia é incapaz de sobreviver ou replicar
intracelularmente.
Semelhante aos dados obtidos no presente trabalho, Brown et al. (2002), relatam
que o fenótipo de B. pseudomallei aderente foi capaz de aderir de forma eficiente para
diversas linhagens de células epiteliais, entre elas HEp-2. Vale ressaltar também, que neste
mesmo estudo o teste de adesão realizado a 30° C com as culturas bacterianas em fase
estacionária foi considerado fatores decisivos na capacidade de adesão, diante das linhagens
celulares avaliadas (BROWN et al. 2002). Para Shahin e Proll (2004), existem casos de
bactérias invasoras que são dependentes da temperatura, para expressar algum gene que
codifica proteína ou receptor específico de membrana que são necessárias para aderir e
invadir diferentes tipos de células hospedeiras.

50
Quanto aos isolados de S. maltophillia, o percentual de adesão ao vidro também foi
alto. Todos os isolados aderiram ao referido material com exceção dos advindos da traqueia,
que tiveram um percentil de 92,3% de adesão. Entretanto, as taxas de aderência ao inox e ao
látex foram baixas. Das estirpes advindas da traqueia apenas 15,4% e 30,8% aderiram ao
inox e ao látex, respectivamente. Dos isolados procedentes de outros sítios anatômicos apenas
uma amostra proveniente de sangue aderiu ao inox e a adesão ao látex foi observada em
somente duas amostras, uma oriunda de sangue e outra de swab nasal (Tabela 5).
Tabela 5- Capacidade de adesão das amostras clínicas de S. maltophillia às superfícies
abióticas
Quando comparamos a capacidade de adesão entre as superfícies abióticas avaliadas,
pôde-se observar que as estirpes de S. maltophillia aqui testadas mostraram maior afinidade
ao vidro. Importante ressaltar que a literatura relata que as bactérias aderem com mais
facilidade às superfícies plásticas, consideradas hidrofóbicas, que ao vidro, considerado
hidrofílico (RODRIGUES et al., 2009).
Entretanto, estudos sugerem que a hidrofobicidade da superfície dos materiais pode
não desempenhar papel determinante no processo inicial de adesão bacteriana e que outros
fatores podem ser fundamentais para o processo, como a rugosidade da superfície (GRENHO,
2010).
Oliveira-Garcia et al. (2003) evidenciaram a presença de fímbrias em S.
maltophillia, que agem como pontes entre as bactérias que aderem às superfícies inertes e
células epiteliais cultivadas. A adesão a superfícies e materiais inertes deve ser estudada para
melhor entender a patogênese de infecções hospitalares (CHRISTENSEN et al. 1995).
Corroborando em parte com dados aqui obtidos, Jucker et al. (1996), descreveu a
aderência ao teflon e ao plástico por S. maltophillia isolada de cateter urinário de um
Sítio Anatômico Abiótico
Q de
Cochran
Valor de
p
Vidro Inox Látex
Outros 5 (100%) 1 (20%) 2 (40%) 23,28 <0.0001
Traqueia 12 (92.3%) 2 (15.4%) 4 (30.8%)

51
paciente com infecção urinária. O autor demonstrou que o isolado de S. maltophillia
carregado positivamente favoreceu a sua adesão consideravelmente.
Semelhante aos resultados evidenciados, Reis (2010) constatou a adesão ao inox e ao
PVC (Polyvinyl chloride) por isolados clínicos de P. aeruginosa, S. maltophillia e B. cepacia.
Ademais, Garcia et al., (2002) também conferiram a adesão a células HEp-2, plástico e vidro
por amostras clínicas de S. maltophillia. Supõe-se que este micro-organismo também pode
aderir a outras células epiteliais, conforme dados aqui obtidos, adesão aos Pneumócitos por
45% dos isolados de S. maltophillia. Isto pode explicar a presença de S. maltophillia em
amostras oriundas das vias respiratórias (GARCIA et al., 2002).
Hollanda (2001) e Di Martino et al., (2002) descrevem que a adesão em células HEp-
2 por P. aeruginosa ocorre devido ao reconhecimento de receptores nas células epiteliais e
isso contribui na patogênese pulmonar deste patógeno em pacientes com fibrose cística, nos
quais causa uma infecção pulmonar crônica. Esse mecanismo pode ser o mesmo utilizado
pelas estirpes de S. maltophillia e Burkholderia sp.
Contudo, a variação da capacidade de adesão pode está relacionada às características
da superfície de adesão (Jackson et al.,2002). Na literatura, as opiniões variam em relação ao
efeito das características da superfície na adesão bacteriana. Alguns pesquisadores relatam
que há uma correlação positiva entre adesão e aumento da rugosidade, enquanto outros
pesquisadores não reportam nenhuma correlação entre a habilidade da bactéria aderir e
irregularidades ou rugosidade das superfícies. Este aparente conflito está relacionado com o
nível de rugosidade estudada, com as espécies bacterianas estudadas, os parâmetros físico-
químicos da superfície e o método usado para detectar a bactéria na superfície (FLINT et al.,
2000).
5.3.2 Produção de cápsula e formação de biofilme
Os micro-organismos tem a capacidade de se atrair ou repelir a superfícies bióticas
ou abióticas, isso depende de diferentes interações não específicas, especialmente por
interações hidrofóbicas (DOYLE, 2000; BRIANDET et al., 2001; COSTA et al., 2006). A
hidrofobicidade modula tanto a adesão quanto a formação de biofilme, contudo, outros fatores
como: a presença de fímbrias, flagelos e cápsula, também estão envolvidos (POMPILIO et al.,
2008).

52
Muitos trabalhos têm estudado a produção de cápsula para estabelecer sua relação
com adesão bacteriana a biomateriais (LOCATELI et al., 2004). Em estudo realizado para
avaliar a aderência de cepas de S. epidermidis a diferentes tipos de LIO (lentes intra-oculares),
concluiu-se que isolados produtores de cápsula foram mais aderentes do que os cápsulas-
negativos (GARCÍA-SÁENZ et al., 2000).
A produção de cápsula foi observada como teste presuntivo para formação de
biofilme dos isolados em estudo. Assim, evidenciamos a produção de cápsula em Ágar
Vermelho Congo (Figura 5) em 7 (38%) e 6 (42%) das amostras de S. maltophillia e
Burkholderia sp, respectivamente. Dentre as produtoras de cápsula, 5 (71,4%) das amostras
de S. maltophillia foram isoladas de secreção traqueal, enquanto que, 5 (83,3%) das estirpes
de Burkholderia sp foram provenientes de sangue. Tal difereneça pode ser explicada pela
predominância de estirpes de S. maltophillia em amostras de secreção traqueal e de
Burkholderia sp em amostras sanguíneas.
Em pesquisa realizada com amostras clínicas de S. maltophillia advindas do Hospital
de Clínicas de José de San Martín, Argentina, os pesquisadores encontraram 100% de
positividade para produção de cápsula (DE ROSSI PASSERINI et al. (2007), percentual
acima do obtido. A habilidade desta bactéria em aderir fortemente e formar biofilme em
superfícies plásticas (DI BONAVENTURA et al., 2004), favorece a contaminação de
materiais hospitalares (DENTON & KEER 1998) e consequentemente a contaminação
cruzada.

53
Figura 5 – Placa de Ágar Vermelho Congo. Amostras produtoras de cápsula tiveram crescimento de
colônias enegrecidas e não produtoras tiveram crescimento de colônias vermelhas.

54
Estudos que caracterizaram alguns tipos de polissacarídeos capsulares existentes no
genoma de B. pseudomallei, constataram que a expressão da cápsula está dependente do
ambiente particular, podendo alterar ou não a hidrofobiciade celular. Fato este que indica que
B. pseudomallei produz estas cápsulas para promover uma vantagem de sobrevivência ou no
hospedeiro ou no ambiente (SHAUNA et al., 2010).
Sarkar et al., (2007) relataram que B. pseudomallei possui um agrupamento de genes
que codificam diferentes tipos de polissacarídeo capsular, os quais desempenham um papel
na virulência desta espécie. Pouco se sabe sobre a função do polissacarídeo capsular, mas o
mesmo pode proporcionar resistência à fagocitose por redução dos níveis de complemento C3
(RECKSEIDLER-ZENTENO et al., 2005).
B. thailandensis não produz polissacarídeo capsular, e já foi relatado que a
capacidade de B. pseudomallei de aderir e invadir as células epiteliais humanas é
significativamente mais elevada do que a de B. thailandensis (ONG et al. de 2004;
KESPICHAYAWATTANA et al. de 2004 ; BRETT . et al , 1998 ). Fato este que pode
comprovar a importância da cápsula na capacidade de adesão de uma determinada estirpe
produtora deste polissacarídeo capsular.
Acredita-se que a formação do biofilme está intimamente relacionada com a
produção de cápsula, composta principalmente de polissacarídeo, muito embora o tipo de
ambiente seja considerado um dos fatores preponderantes para o desenvolvimento do biofilme
(STEINBERGER & HOLDEN, 2004).
Biofilmes são notoriamente difíceis de erradicar e são uma fonte de muitas infecções
crônicas. De acordo com o Instituto Nacional de Saúde, biofilmes são clinicamente
importante, representando mais de 80% das infecções microbianas do corpo. Um biofilme
maduro pode tolerar os antibióticos, em concentrações de 10-1000 vezes mais do que os
necessários para matar as bactérias. Bactérias em biofilmes são resistentes à fagocitose,
tornando biofilmes extremamente difíceis de erradicar a partir de hospedeiros vivos (LEWIS,
2001).
Ao avaliarmos a formação de biofilme por espectofotometria, verificou-se que as
cepas de Burkholderia sp obtiveram a seguinte classificação: 67% produziram biofilme
moderadamente, enquanto que 33% produziram fracamente. Entre os isolados de S.
maltophillia 16,6% foram produtores moderados e 83,3% produtores fracos. Nenhuma das

55
duas espécies avaliadas evidenciaram estirpes produtotas fortes de biofilme. Entretanto, a
capacidade de formar biofilme independentemente da classificação, por si só já condiz com
um fator de virulência bacteriana e de risco aos pacientes expostos a tais micro-organismos.
Assim como Reis et al. (2010), foi possível constatar a formação de biofilme em
poliestireno por cepas de B. cepacia e S. maltophillia. Desta forma, torna-se evidente que a
produção de biofilme nas superfícies inertes acarreta na persistência dos patógenos no
ambiente e aumenta o risco de infecções.
Nornberg (2009), ao avaliar uma estirpe de B. cepacia capaz de aderir ao inox,
verificou que a mesma mostrou-se altamente hábil para formar biofilme em microplacas,
conforme evidenciado no presente trabalho. Os valores de absorvância apresentaram-se
baixos, entretanto, foram observados a 4°C, sugerindo que esta bactéria pode aderir e formar
biofilmes mesmo a temperaturas de refrigeração, mostrando, capacidade semelhante de
bactérias como Pseudomonas putida, Proteus vulgaris e Citrobacter freundii que ocasionam a
deterioração de alimentos. Importante ressalvar que não foi encontrado estudos que tenham
classificado o tipo de biofilme produzido por estirpes clínicas de B. cepacia e S. maltophillia,a
literatura apenas relata a formação ou não deste fator de virulência.
Bactérias em biofilmes colonizam uma grande variedade de dispositivos médicos,
como cateteres, marcapassos cardíacos artificiais, válvulas cardíacas protéticas e aparelhos
ortopédicos, e estão associados a várias doenças humanas, como a endocardite, infecções de
feridas e queimaduras, otite média crônica com efusão e fibrose cística ( COSTERTON et al. ,
1999 ).
Fazendo a correlação entre as variáveis hidrofobicidade e biofilme (Tabelas 6 e 7),
estatisticamente não houve significância frente as duas espécies bacterianas avaliadas.
Embora, todos os isolados tenham apresentado capacidade hidrofóbica e de produção de
biofilme. Dados esses sugerem a presença de outros fatores como: fímbrias, flagelos, adesinas
não fimbriais (ZARANZA, 2011) e cápsula (DE ROSSI PASSERINI et al., 2007), possam
estar relacionados com a adesão das espécies em questão. A produção de cápsula foi
observada em algumas cepas, o que pode corroborar em partes com os referidos autores.

56
Tabela 6- Correlação entre hidrofobicidade e formação de biofilme diante das cepas clínicas de
Burkholderia sp.
Hidrofobicidade Biofilme Coeficiente de
Contigência C
Valor de
P
Sim Não
Fraco 2 (14,2%) 2 (14,2%) 3,74 0,15
Moderado 2 (14,2%) 0
Forte 1 (7,14%) 4 (28,5%)
Tabela 7- Correlação entre hidrofobicidade e formação de biofilme diante das cepas clínicas de
Stenotrophomonas maltophillia.
Hidrofobicidade Biofilme Coeficiente de
Contigência C
Valor de
p
Sim Não
Fraco 3 (16,7%) 3 (16,7%) 0,33 0,31
Moderado 0 3 (16,7%)
Forte 4 (22,2%) 5 (27,8%)
Na literatura não foram encontradas referências que tenham realizado correlação
entre esses mecanismos de virulência em amostras de Burkholderia sp.
Diferentemente, uma correlação positiva foi observada por Pompilio et al., (2004),
entre adesão - formação de biofilme e hidrofobicidade da superfície celular, sugerindo pela
primeira vez que a hidrofobicidade modula tanto adesão e formação de biofilmes em
superfícies de poliestireno frente a culturas de S. maltophillia. Esse mesmo estudo também
mostrou que a capacidade de formação de biofilme de maneira semelhante, pode ser
encontrada em estirpes exibindo diferentes valores de hidrofobicidade, sugerindo, assim, que
a formação de biofilmes em S. maltophillia é regulado por fatores diferentes (POMPILIO et
al., 2004).
5.4 Produção de exoenzimas e hemolisinas
As infecções bacterianas são consideradas complexas, por constituírem um processo
multifatorial que envolve fatores inerentes ao patógeno e ao hospedeiro. A habilidade em

57
produzir enzimas hidrolíticas e hemolisinas é considerada um importante fator de virulência, a
interação entre esses fatores colabora para o desenvolvimento de doenças causadas por
determinadas bactérias (HOLANDA, 2001).
A produção de enzimas extracelulares por Burkholderia sp e S. maltophillia também
tem fundamental importância para o desenvolvimento de processos infecciosos, sendo já
conhecidos de alguns estudiosos os seguintes fatores de virulência: produção de DNAse,
RNAse, fibrinolisina, hemolisinas, lipases, hialuronidases, proteases e elastases (ROBIN et al.
1996; DENTON e KERR 1998; GARCIA et al 2002; SEGONDS et al. 2006; SHAGINIAN et
al. 2007).
Quanto as cepas de S. maltophillia não evidenciou-se atividade amidolítica,
fosfolipídica e lipolítica (Tabela 08). Sugerindo que essas exoenzimas não fazem parte do
padrão de virulência dos isolados bacterianos aqui avaliados. No entanto, foi constatada a
produção de proteinase, DNAse e gelatinase (12%) em percentuais elevados, principlamente
entre os isolados de secreção traqueal, que manifestaram maior propensão a produzir tais
exoenzimas. Ressalta-se, que a maioria das amostras de S. maltophillia foram isoladas de
secreção traqueal, portanto, este fato pode esclarecer os percentis elevados diante da produção
das referidas enzimas.
Estatisticamente, a correlação entre a produção de exoenzimas e os sítios de
isolamento das cepas, não apresentou diferença. Mas, avaliando as estipes entre si quanto a
produção dessas enzimas obteve-se valor de p < 0,0001 (Q de Cochran), demonstrando assim
significância. Deste modo, as amostras clínicas de S. maltophillia aqui estudadas, produzem
exoenzimas que podem desempenhar um papel importante em sua patogênese.
Outro resultado que merece atenção é a atividade gelatinolítica, que foi observada
apenas quando utilizou-se o meio com 12% de gelatina. Essa concentração é 2,4 vezes maior
que a concentração empregada na outra metodologia, que também foi testada mas não houve
positividade frente ao referido procedimento. A explicação para tal acontecimento pode ser a
própria composição dos meios utilizados: gelatina 5% foi preparado em Ágar Muller Hinton,
considerado um meio de cultura nutritivo e, gelatina 12% teve como base apenas tampão
salina fosfato (PBS). Deste modo, o meio de concentração elevada oferecia exclusivamente a
própria gelatina como fonte de nutrientes e, por uma questão de sobrevivência as bactérias
semeadas tiveram que produzir gelatinase para metabolizar o único nutriente ofertado.

58
Tabela 8- Atividade enzimática e hemolítica de amostras clínicas de S. maltophillia correlacionando os
sítios de isolamento .
Virulência Sítio anatômico n (%) Valor de p
Outros Traqueal
Atividade Enzimática Proteinasea 1 (20%) 9 (69.2%) ns
Lipase
b 0 0 ns
Dnase
a 5 (100%) 7 (53.8%) ns
Fosfolipase
b 0 0 ns
Amilase
b 0 0 ns
Gelatinase 5%
b 0 0 ns
Gelatinase 12%
a 5 (100%) 9 (69.2%) ns
Hemólise Tipo A1 3 (60%) 8 (61.5%) ns
Tipo B
1 4 (80%) 8 (61.5%) ns
Tipo O
1 4 (80%) 9 (69.2%) ns
Carneiro
2 1 (20%) 4 (30.8%) ns
Cavalo1 4 (80%) 7 (53.8%) ns
Letras e número diferentes indicam diferença significativa entre a produção de atividade enzimáticas nas cepas
analisadas. Portanto as cepas apresentam uma menor atividade lipolítica, fosfolipidíca, amidolítica e gelatinolítica
(5%). As cepas apresentam menor capacidade hemolítica no sangue de carneiro.
Travassos et al. (2004) ao verificarem a produção de exoenzimas em 39 isolados de
S. maltophillia, constatou que 97,6% das cepas produziram DNAse e gelatinase, percentis
acima dos resultados aqui obtidos : 66,6% e 77,7 % respectivamente. Na Figura 06 observa-
se os halos de degradação de fosfolipídeos, proteínas e gelatina.Ainda neste mesmo estudo,
encontraram 100% de positividade para a expressão de fosfolipase, lipase e protease. Destas,
a protease foi a única enzima que foi observada pela presente pesquisa, 55,5% de isolados
produtores.
Ao avaliar a hidrólise de protease, Garcia et al. (2002) também constataram estirpes
produtoras de tal exoenzima, fato que corrobora com os dados já citados. A produção de
proteases exerce um papel expressivo na patogênese bacteriana, participando da invasão
celular, de danos tecidual e evasão da defesa do hospedeiro (Travis et al. 1995 ). As
divergências observadas entre os resultados aqui obtidos e os dados da literatura podem ser
devido à origem diferente de substrato e meios de cultura ou relacionados com o potencial
particular exoenzimático de cada cepa infectante.

59
Figura 6- Produção das exoenzimas: A) Halos de degradação da protease em ágar Skim Milk 10%;
B)Hidrólise de gelatina em ágar gelatina a 12% .
A
B

60
Quanto a atividade hemolítica das cepas clínicas de S. maltophillia, foi possível
observar que houve hemólise em todos os tipos de sangue avaliados. Entretanto, os isolados
clínicos apresentaram menor atividade hemolítica no sangue de carneiro (apenas 27,7% dos
isolados), sugerindo que as hemolisinas possuem variados padrões de ação, dependente do
tipo de eritrócito testado (Tabela 8). O halo de hemólise só foi possível ser visualizado com
precisão após 72h de incubação.
Assim como citado anteriormente com as exoenzimas, estatisticamente não houve
significância ao correlacionar a produção de hemolisinas e os sítios de isolamento. Ou seja,
idependentemente do sítio de isolamento, S. maltophillia causa hemólise. Ao avaliarmos
apenas a produção de hemolisinas pelas estirpes, evidenciou-se Q de Cochran : 21,8, valor de
p 0,0002.
Garcia et al. (2002), obtiveram resultados semelhantes aos conseguidos, a atividade
hemolítica dos isolados de S. maltophillia também mostrou uma variedade de padrões, sendo
o tipo de sangue, a temperatura e o tempo de incubação condicionantes para a positividade do
teste. A maior parte dos isolados expressou atividade hemolítica somente após 96 horas a 30 º
C e 37 º C. Após 48h a 30 º C, apenas 4% das cepas apresentaram hemólise em ágar sangue
humano e de 2% em sangue de coelho. Estes dados corroboram em parte com os resultados
aqui obtidos, uma vez que precisou-se de 72h de incubação a 37°C para ocorrer a hemólise
nos eritrócitos analisados. Sugerindo assim, que o tempo mínimo de incubação para que esses
micro-organismos consigam produzir este fator de virulência, é 72h.
Tais evidências podem refletir a virulência de S. maltophillia e explicar os casos de
bacteremias causados por este patógeno que ocasiona alto índice de mortalidade (WU et al.
2006).
Diante das estirpes de Burkholderia sp, observou-se a produção de proteinase, lípase,
DNAse, fosfolipase (verificar halo de degradação de fosfolipídeos na Figura 7) e gelatinase,
apenas a amilase e elastase que não foram produzidas (Tabela 9). Dentre as enzimas
produzidas, a lipase e a gelatinase (no meio com 12% de gelatina) foram as que apresentaram
menores percentis de positividade. A atividade lipolítica só foi evidenciada em 2 (22,2%) dos
isolados oriundos de sangue. No caso da atividade gelatinolítica, ocorreu divergência nos
valores encontrados frente às duas metodologias utilizadas. O teste que utiliza 5% de gelatina
revelou positividade em 8 (88,9%) dos isolados provenientes de sangue e 2 (40%) advindos
de secreção traqueal. Mas, diante da metodologia que utiliza 12% de gelatina e PBS, apenas

61
um isolado de secreção traqueal foi capaz de degradar a gelatina presente no meio. Entendeu-
se que as estirpes avaliadas só conseguem expressar tal atividade diante de baixos percentuais
de gelatina.
Tabela 9- Atividade enzimática e hemolítica de amostras clínicas de Burkholderia sp
correlacionando os sítios de isolamento.
Os resultados de Arcuri et al. (2008) demonstraram que nenhuma B. cepacia
causou proteólise, embora tenham apresentado atividade lipolítica em todas as temperaturas.
È importante ressaltar que essa mesma pesquisa realizou os referidos testes em temperaturas
de refrigeração (4°C, 7ºC, 10ºC e 21°C), diferentemente da nossa metodologia, que empregou
uma temperatura superior (37°C), e obtivemos percentuais elevados de proteinase: 77% dos
isolados advindos do sangue e 60% da traqueia, e baixa atividade lipolítica, o que pode
explicar a discordância com nossos dados.
Virulência Sítio anatômico n (%) Valor de p
Sangue Traqueal
Atividade
Enzimática Proteinasea
7 ( 77.8) 3 (60) ns
Lipaseb
2 (22.2) 0 ns
Dnasea
6 (66.7) 3 (60) ns
Fosfolipasea
7 ( 77.8) 0 0,01
Amilaseb
0 0 ns
Elastaseb 0 0 ns
Gelatinase 5%a
8 (88.9%) 2 (40%) ns
Gelatinase 12%b
0 1 (20%) ns
Hemólise Tipo A1
8 (88.9) 4 (80) ns
Tipo B2
1 (11.1) 0 ns
Tipo O1
6 (66.7) 1 (20) ns
Carneiro1
7 ( 77.8) 1 (20) ns
Cavalo1
6 (66.7) 1 (20) ns
Letras e número diferentes indicam diferença significativa entre a produção de atividade
enzimáticas nas cepas analisadas. Portanto as cepas apresentam menos atividade lipolítica,
amilase, elastase e gelatinase a 12%. As cepas apresentam menor capacidade hemolítica no
sangue do tipo B.

62
Figura 7- Halos de degradação de fosfolipídeos em Àgar Muller Hinton acrescido Egg Yolk.

63
O'Grady et al.(2010), relatou que Burkholderia cenocepacia possui uma proteína
específica que influencia a regulação de genes responsáveis pela expressão de protease e
lipase. Encontramos cepas produtoras dessas duas enzimas, embora somente 2 isolados
provenientes de sangue apresentaram atividade lipolítica. Além disso, foi constatado por esse
mesmo autor, que a temperatura influencia a expressão desta proteína, a 37°C os níveis de
expressão foram melhores que a 29°C.A atividade enzimática de alguns micro-organismos é
espécie especifica e depende da temperatura de armazenamento (ALATOSSAVA &
ALATOSSAVA, 2006) confirmando nossa hipótese.
Num trabalho desenvolvido por Pilatz e seus colaboradores (2006), com 9 cepas de
Burkholderia pseudomallei, 88,8% apresentaram atividade proteolítica e 100% foram
produtoras de fosfolipase, semelhante aos nossos resultados, em que também encontramos
altas porcentagens de produção dessas duas enzimas: 71,4% e 50%, concomitantemente.
Igualmente em Burkholderia pseudomallei foi detectada atividade colagenolítica/
gelatinolítca, sugerindo que este micro-organismo expressa uma colagenase/gelatinase
durante o seu crescimento. Esta proteína pode ser considerada um fator importante com
potencial papel na virulência deste micro-organismo (RAINBOW et al., 2004). Do mesmo
modo, a produção de gelatinase foi evidenciada em percentuais superiores a 80%.
Nossos resultados sugerem que a expressão de tantas exoenzimas pode ter sido a
causa da destruição do tapete celular durante o teste de adesão às células humanas, onde
algumas cepas manifestaram tal efeito.
De acordo com trabalhos realizados com P. aeruginosa, as exoenzimas colaboram
para a destruição tecidual, interferem com a resposta imunológica do hospedeiro e facilitam a
disseminação sendo assim, capazes de aprimorar os mecanismos de virulência envolvidos na
destruição de lipídeos e proteínas de membrana e imunoglobulinas (EDBERG et al 1996;
KVITKO, 2010). Desta forma, podemos idealizar o processo de patogenicidade de
Burkholderia sp e S. maltophillia sendo similar ao da P. aeruginosa, por fazerem parte do
mesmo grupo bacteriano, Bacilos Gram-negativos não fermentaores e por possuírem alguns
fatores de virulência em comum.
As proteinases como as hemolisinas e exotoxinas se destacam como fator de
virulência extracelular de Burkholderia sp, pois facilitam o rompimento da integridade
epitelial durante a colonização dos tecidos possuindo potencial imunogênico (SILVA, 2010).
A
C

64
Além disso, outras enzimas podem auxiliar no processo de colonização destacando-se as
elastases, fosfolipase C, neuramidase, exoenzima S e lectina (JAFFAR- BANDJEE et
al.,1995).
Quanto a atividade hemolítica as estipes de Burkholderia sp não tiveram boa
afinidade diante do sangue tipo B (Q de Cochran = 20, valor de p = 0.0004). Mas, diante de
todos os outros tipos sanguíneos, os percentuais de hemólise foram altos, superiores a 60%.
Na figura 8 é possível verificarmos os halos de hemólise em diferentes tipos de sangue.
No entanto, não foi constatada atividade hemolítica em meio líquido, sugerindo que
essa atividade parece ser dependente do método de detecção e/ou a hemolisina pode ser parte
constituinte da célula bacteriana e precisa do contato com os eritrócitos para se expressar.
Hemolisinas representam um grupo de toxinas bacterianas que, variavelmente de
muitas outras toxinas, não são internalizadas pelas células agindo como agentes ativos de
membranas levando à lise e morte celular, além de sua característica principal de lisar
eritrócitos (ROWE; WELK, 1994).
Em pesquisa realizada por Corbett et al. (2003), a hemolisina de B. cepacia em
baixas concentrações foi capaz de induzir a degradação nucleossomal consistente com a
apoptose em neutrófilos humanos e macrófagos de rato. A exposição dos neutrófilos humanos
a concentrações mais elevadas de toxina resultou no aumento da atividade da desgranulação
de neutrófilos.
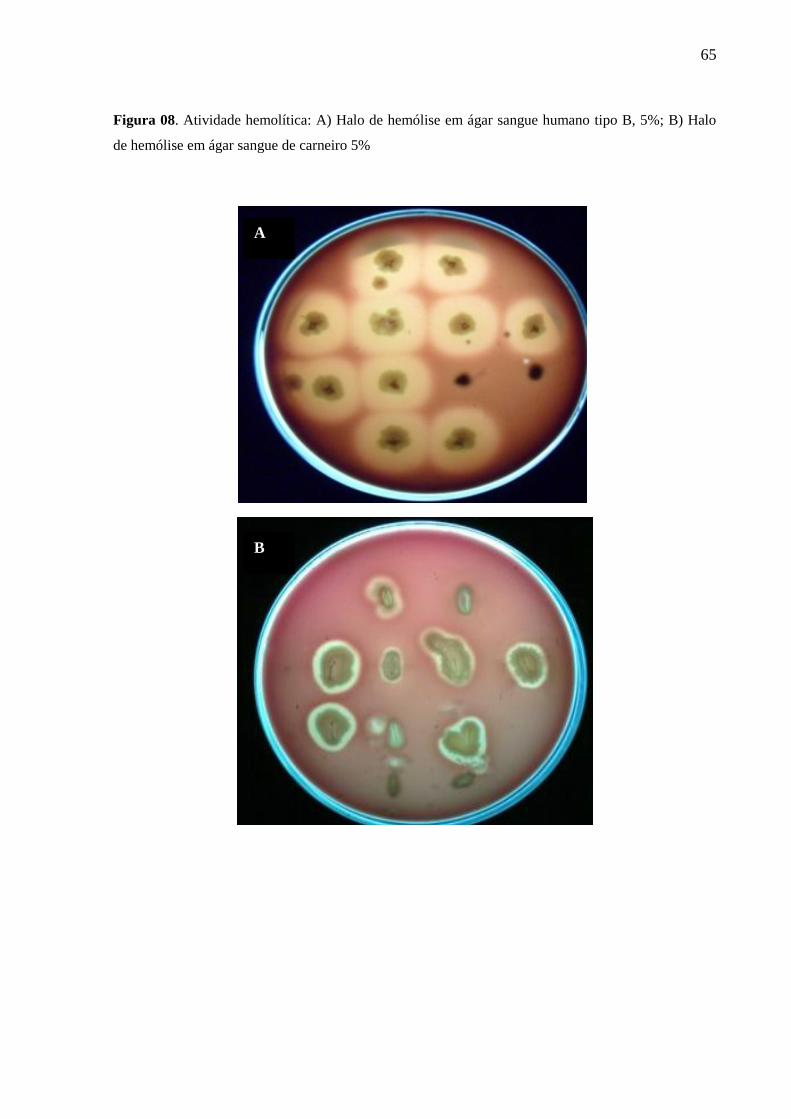
65
Figura 08. Atividade hemolítica: A) Halo de hemólise em ágar sangue humano tipo B, 5%; B) Halo
de hemólise em ágar sangue de carneiro 5%
B
A

66
6 Conclusão
De acordo com os resultados obtidos observou-se que:
1. Os isolados de Burkholderia sp foram mais isolados em amostras de sangue enquanto que
S. maltophillia foi isolada com mais frequência em secreção traqueal;
2. A resistência a sulfametaxazol-trimetropim diante das cepas de Burkholderia sp evidencia
uma problema na terapêutica a ser adotada em casos de infecções por esse patógeno;
3. A baixa taxa de resistência a minociclina, levofloxacino e sulfametaxazol/trimetropim
pode implicar em dificuldade de tratamento de infecções causadas por S. maltophillia no
futuro;
4. A presença de ESBL foi detectada em mais de 60% de ambas as espécies, demonstrando a
resit~encia antimicrobiana acentuada de ambas as espécies.
5. As estirpes apresentaram aptidão hidrofóbica, produção de cápsula, formação de biofilme e
capacidade de aderir em diferentes superfícies e células humanas o que pode favorecer o
processo patogênico desses micro-organismos;
6. Nenhuma das bactérias estudadas produziu amilase nem elastase. Entretanto a produção de
outras exoenzimas e hemolisinas foi evidenciada, fato que pode explicar o efeito
citotóxico observado com a cultura pura de algumas estirpes.
Considerando os itens 1, 2, 3, 4, 5 e 6 , concluo que as espécies estudadas podem ser
consideradas importantes patógenos no ambiente nosocomial, por apresentarem diversos
fatores de virulência que podem condicionar graves infecções.

67
REFERÊNCIAS
Alonso A, Sanchez P, Martinez JL. Stenotrophomonas maltophilia D457R contains a cluster
of genes from gram-positive bacteria involved in antibiotic and heavy metal resistance.
Antimicrob Agents Chemother 2000;44(7):1778-82.
ANDRADE, D., ANGERAMI, E.L., PADOVANI, C.R. Condição microbiológica dos leitos
hospitalares antes e depois de sua limpeza. Rev. Saúde Pública 2000;34(2):163-9.
ANDREU, A. et al. Urovirulence determinants in Escherichia coli strain causing prostatitis.
Journal Infection Diseases. v.176, 1997.
Antimicrobial Surveillance Program (1997-1999). Clin Infect Dis 2001;32 Suppl 2:S104-13.
Balandreau J, Viallard V, Cournoyer B, Coenye T, Laevens S,Vandamme P (2001)
Burkholderia cepacia genomovar III is acommon plant associated bacterium. Appl Environ
Microbiol
Baltimore, R. S., and H. B. Jenson. 1990. Puncture wound osteochondritis of the foot caused
by Pseudomonas maltophilia. Pediatr. Infect. Dis. J. 9:143–144.
Bandin, I., P. Heinen, L. L. Brown, A. E. Toranzo. 1996. Comparison of different ELISA kits
for detecting Renibacterium salmoninarum. Bull. Eur. Assoc. Fish Pathol. 16:19–22.
Barry, A. L., R. J. Fass, J. P. Anhalt, H. C. Neu, C. Thornsberry, R. C. Tilton, B. G. Painter,
and J. A. Washington II. 1985. Ciprofloxacin disk susceptibility tests: interpretive zone size
standards for 5-microgram disks. J. Clin. Microbiol. 21:880–883.

68
Baruchel, A., O. Hartmann, A. Andremont, and C. Tancrede. 1986. Severe gram-negative
infections in neutropenic children cured by imipenem/cilastatin in combination with an
aminoglycoside. J. Antimicrob. Chemother. 18(Suppl. E):167–173.
Bates, J. H., G. D. Campbell, A. L. Barron, G. A. McCracken, P. N. Morgan, E. B. Moses,
and C. M. Davis. 1992. Microbial etiology of acute pneumonia in hospitalised patients. Chest
100:1005–1012.
Bauernfeind, A. 1993. Comparative in-vitro activities of the new quinolone, Bay y 3118, and
ciprofloxacin, sparfloxacin, tosufloxacin, CI 960 and CI 990. J. Antimicrob. Chemother.
31:505–522.
Bauernfeind, A., R. M. Bertele, K. Harms, G. Horl, R. Jungwirth, C. Petermuller, B.
Przyklenk, and C. Weisslein-Pfister. 1987. Qualitative and quantitative microbiological
analysis of sputa of 102 patients with cystic fibrosis. Infection 15:270–277.
Belcour, B., D. Courtois, C. Ehret, and V. Petiard. 1993. Rapid production of irones by
maturation of orris rhizomes by two bacterial strains. Phytopathology 34:1313–1315.
Benezra, D., T. E. Kiehn, J. W. M. Gold, A. E. Brown, A. D. M. Turnbull, and D. Armstrong.
1988. Prospective study of infections in indwelling central venous catheters using quantitative
blood cultures. Am. J. Med. 85: 495–498.
Betriu C, Sánchez A, Palau ML, Gómez M, Picazo JJ. Antibiotic resistance surveillance of
Stenotrophomonas maltophilia, 1993-1999. J Antimicrob Chemother 2001;48(1):152-4.
CHRISTOFF, J.; TOLENTINO, J.; MAWDSLEY, E.; MATUSHEK, S.Optimizing empirical
antimicrobial therapy for infection due to gram-negative pathogens in the intensive care unit:
utility of a combination antibiogram. Infection Control and Hospital Epidemiology, Chicago,
v 31, n3, p 249-255, Mar. 2010.J Clin Microbiol. 1986 Dec;24(6):995-7.
DAVIES, J. C.; RUBIN, B. K. Emerging and unusual gram-negative infections in cystic
fibrosis. Seminars in Respiratory and Critical Care Medicine, New York, v. 28, n. 3, p. 312-
321, July 2007.
DE SOYZA, A.; CORRIS, P. A. Lung transplantation and the Burkholderia cepacia Complex.
The Journal of Heart and Lung Transplantation, Newcastle, v. 22 , n. 9, p. 954-958, Sept.
2003.

69
Denton M, Kerr KG. Microbiological and clinical aspects of infection associated with
Stenotrophomonas maltophilia. Clin Microbiol Rev 1998;11(1):57-80.
EVANS Jr., D.J., EVANS, D.G., YOUNG, L.S., PITT, J. Hemagglutination typing of
Escherichia coli: definition of seen hemogglutination types. Journal Clinical Microbiology.
v.12, p.235-242, 1980.
FREITAS, Felipe Teixeira de Mello. Fatores de risco associados à aquisição de neumonia por
Stenotrophomonas maltophilia na Unidade de terapia intensiva do instituto de Infectologia
Emílio Ribas no período de março a novembro de 2007: investigação de um surto. 2008. 37f.
Monografia (conclusão de curso) – Instituto de Infectologia Emílio Ribas. São Paulo.
FREITAS, M. R.; COSTA, R. M. M.; SILVA, E. F.; DINIZ, S. R. D.; NETO, F. V. A.;
MEDEIROS, W. D. A.; PASCOAL, W. F. B.; MELO, C. R. Surto de Burkholderia cepacia
em pacientes cirúrgicos. Revista Paraense de Medicina, Belém, v. 21, n. 4, p. 77-77, dez.
2007.
Fritsche TR, Sader HS, Stilwell MG, Dowzicky MJ, Jones RN Antimicrobial activity of
tigecycline tested against organisms causing community- acquired respiratory tract infection
and nosocomial pneumonia. Diagn Microbiol Infect Dis 2005;52(3):187-93
FROTA, Cristiane Cunha; MOREIRA, José Luciano Bezerra. Frequency of Nonfermentative
Gram-Negative Bacilli Isolated from Clinical Materials of Pacients at Universidade Federal
do Ceará Hospital Complex - Brazil. Rev. Microbiol., São Paulo, v. 29, n. 3, Sept. 1998
Gales AC, Jones RN, Forward KR, Linares J, Sader HS, Verhoerf J. Emerging importance of
multidrug-resistant Acinetobacter species and Stenotrophomonas maltophilia as pathogens in
seriously ill patient: geographic patterns, epidemiological features, and trends in the SENTRY
Goldman, D. A., and J. D. Klinger. 1986. Pseudomonas cepacia: biology, mechanisms of
virulence, epidemiology. J. Pediatr. 108:806–812.
HINRICHSEN, S.L. Biossegurança e controle de infecções: risco sanitário hospitalar. Rio
de Janeiro: Medsi, 2004, p. 273-281.
KONEMAN, E.W. et al. Diagnóstico Microbiológico: Textos e Atlas Colorido. 6ed. São
Paulo: MEDSI, 2008.

70
L. Chiarini, A. Bevivino, C. Dalmastri, S. Tabacchioni, and P. Visca, “Burkholderia cepacia
complex species: health hazards and biotechnological potential,” Trends in Microbiology, vol.
14, no. 6, pp. 277–286, 2006.
LARANJEIRA, Vani Dos Santos et al . Pesquisa de Acinetobacter sp e Pseudomonas
aeruginosa produtores de metalo-β-lactamase em hospital de emergência de Porto Alegre,
Estado do Rio Grande do Sul, Brasil. Rev. Soc. Bras. Med. Trop., Uberaba, v. 43, n.
4, Aug. 2010 .
LEDSON, M. J.; GALLAGHER, M. J.; CORKHILL, J. E. Chronic Burkholderia cepacia
bronchiectasis in a non cystic fibrosis individual. Thorax, London, v. 53, n. 5, p. 430-432,
May 1998.
Leelarasamee, A. Recent development in melioidosis. Curr. Opin. Infec. Dis. V 17, p 131-
136, 2004
LIZASO, Diego et al . Epidemiología y factores de riesgo de mortalidad de las bacteriemias
intrahospitalarias por bacilos gramnegativos. Rev. chil. infectol., Santiago, v. 25, n.
5, oct. 2008 .
MOURA, Maria Eliete Batista et al . Infecção hospitalar: estudo de prevalência em um
hospital público de ensino. Rev. bras. enferm., Brasília, v. 60, n. 4, Aug. 2007
Peeters E., Nelis HJ, Coenye T. 2008. Avaliação da eficácia de processos de desinfecção
contra biofilmes Burkholderia cenocepacia . J. Hosp. Infect . 70 :361-368
PELEG, Anton Y.; SEIFERT, Harald; PATERSON, David L.. Acinetobacter baumannii:
Emergence of a Successful Pathogen. Clin Microbiol Rev. 2008 Jul;21(3):538-82.
PÉREZ-MIRAVETE, Adolfo; JIMÉNEZ-TAPIA, Yolanda y ALCÁZAR-LÓPEZ, Virginia.
Los generos Pseudomonas, Xanthomonas y similares dentro de la patología de un hospital
pediátrico. Bioquimia [en línea] 2001, vol. 26 [citado 2011-03-23].
Pseudomonas maltophilia exoenzyme activity as correlate in pathogenesis of ecthyma
gangrenosum. Bottone EJ, Reitano M, Janda JM, Troy K, Cuttner J.
Ribera A, Domenech-Sanchez A, Ruiz J, Benedi VJ, Jimenez de Anta MT, Vila J. Mutations
in gyrA and parC QRDRs are not relevant for quinolone resistance in epidemiological

71
unrelated Stenotrophomonas maltophilia clinical isolates. Microb Drug Resist 2002;8(4):245-
51.
Rose H., A. Baldwin, Dowson CG, Mahenthiralingam E. 2009. susceptibilidade Biocida do
complexo Burkholderia cepacia . J. Antimicrob. Chemoter . 63 :502-510.
SCALETSKY, I.C.A., SILVA, M.L.M., TRABULSI, L.R. Distinctive patterns of adherence
of Escherichia coli to HeLa cells. Infection Immunology, v.2, p.534-536, 1984.
Segonds, C., T. Heulin, N. Marty, and G. Chabanon. 1999. Differentiation of Burkholderia
species by PCR-restriction fragment length polymorphism analysis of the 16S rRNA gene and
application to cystic fibrosis isolates. J. Clin. Microbiol. 37:2201–2208.
SIEGMAN-IGRA, Y.; ANGLIN A. M.; SHAPIRO, D. E.; ADAL, K. A.; STRAIN, B. A.;
FARR, B. M. Diagnosis of vascular catheter-related bloodstream infection:a meta-analysis.
Journal of Clinical Microbiology, Washington, v. 35, n. 4, p. 928-936, Apr. 1997.
SOUSA,Sílvia A.;RAMOS,Christian G.; LEITÃO, Jorge H. “Burkholderia cepacia Complex:
Emerging Multihost Pathogens Equipped with a Wide Range of Virulence Factors and
Determinants,” International Journal of Microbiology, vol. 2011, Article ID 607575, 9
pages, 2011. doi:10.1155/2011/607575
STEPANOVIC, S.; CIRKOVICK, I.; RANIN, L.; SVABIC-VLAHOVIC, M. Biofilm
formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl.
Microbiol. v. 38, p. 428-432, 2004.
TURRINI, R. N. T. Percepção das Enfermeiras sobre fatores de risco para a infecção
hospitalar. Rev.Esc.Enf.USP, v. 34, n. 2, p. 174-84, jun. 2000.
TURRINI, Ruth N.T.; SANTO, Augusto H.. Infecção hospitalar e causas múltiplas de morte.
J. Pediatr. (Rio J.), Porto Alegre, v. 78, n. 6, Dec. 2002 .
WEINTROB, A. C.; ROEDIGER, M. P.; BARBER, M.; SUMMERS, A.; FIEBERG, A. M.;
DUNN, J.; SELDON, V.; LEACH, F.; HUANG, X.; NIKOLICH, M. P.; WORTMANN, G.
W. Natural history of colonization with gram-negative multidrug-resistant organisms among
hospitalized patients. Infection Control and Hospital Epidemiology, Chicago, v. 31, n. 4, p.
330-337, Apr. 2010.

72
Weiss K, Restieri C, De Carolis E, Laverdière M, Guay H. Comparative activity of new
quinolones against 326 clinical isolates of Stenotrophomonas maltophilia. J Antimicrob
Chemother 2000;45(3):363-5.
WIGFIELD, S. M.; RIGG, G. P.; KAVARI, M.; WEBB, A. K.; MATTHEWS, R. C.;
BURNIE, J. P. Identification of na immunodominant drug efflux pump in Burkholderia
cepacia. Journal of Antimicrobial Chemotherapy, Oxford, v. 49, n. 4, p. 619-624, Apr. 2002.