alterações imunológicas e epigenéticas em células natural...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
PÓS-GRADUAÇÃO EM IMUNOLOGIA BÁSICA E APLICADA
LUANA SILVA SOARES
Alterações imunológicas e epigenéticas em células natural killer de pacientes infectados pelo HIV-1
Ribeirão Preto 2016

LUANA SILVA SOARES
Alterações imunológicas e epigenéticas em células natural killer
de pacientes infectados pelo HIV-1
Tese apresentada à Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo para obtenção do título de Doutora em Ciências. Área de concentração: Imunologia Básica e Aplicada. Orientadora: Prof.a Dr.a Fabiani Gai Frantz
Ribeirão Preto 2016

AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL DESTE
TRABALHO, POR QUALQUER MEIO CONVENCIONAL OU ELETRÔNICO, PARA
FINS DE ESTUDO E PESQUISA, DESDE QUE CITADA A FONTE.
Catalogação da publicação
Faculdade de Medicina de Ribeirão Preto
Universidade de São Paulo
Soares, Luana Silva
Alterações imunológicas e epigenéticas em células natural killer de pacientes infectados pelo HIV-1. Ribeirão Preto, 2016.
113 p. : il. ;; 30 cm
Tese de Doutorado, apresentada à Faculdade de Medicina de Ribeirão Preto/USP. Área de concentração: Imunologia Básica e Aplicada.
Orientador: Frantz, Fabiani Gai.
1. Imunidade Inata 2. HIV/AIDS. 3. Metilação Global. 4. Células natural killer.

SOARES, L. S. Alterações imunológicas e epigenéticas em células natural killer de pacientes infectados pelo HIV-1. Tese de Doutorado apresentada à Faculdade de Medicina de Ribeirão Preto – Universidade de São Paulo para obtenção do título de Doutora em Ciências.
Aprovada em: ___/___/___
Banca Examinadora
Prof. Dr. ______________________________________ Instituição: ___________________
Julgamento: _____________________ Assinatura: __________________________________
Prof. Dr. ______________________________________ Instituição: ___________________
Julgamento: _____________________ Assinatura: __________________________________
Prof. Dr. ______________________________________ Instituição: ___________________
Julgamento: _____________________ Assinatura: __________________________________
Prof. Dr. ______________________________________ Instituição: ___________________
Julgamento: _____________________ Assinatura: __________________________________
Prof. Dr. ______________________________________ Instituição: ___________________
Julgamento: _____________________ Assinatura: __________________________________

Dedico este trabalho aos meus queridos pais Marcos e Roselene Soares pelo amor
incondicional, pela força nos momentos em que mais precisei, sobretudo, por sonharem
comigo um sonho que eu não achava possível de ser realizado.

AGRADECIMENTOS
À minha orientadora Profa. Dra. Fabiani Gai Frantz, pela oportunidade e confiança para a realização deste trabalho. Muito obrigada pelos ensinamentos, científicos e pessoais, e sobretudo, por contribuir na minha formação. Agradeço imensamente todo o apoio e crescimento que me foi proporcionado!
À Profa. Dra. Lúcia Faccioli, pelos ensinamentos e oportunidades de discussões científicas, assim como pelo compartilhamento da infra-estrutura de seu laboratório.
Ao Prof. Dr. Celio Lopes Silva, pelo auxílio desde o meu mestrado, ao disponibilizar a infra-estrutura de seu laboratório para realização dos experimentos de clonagem, assim como sua excelente equipe técnica representada pela Aninha, Iza e Wendy, que não medem esforços para te ajudar e compartilhar conhecimento.
Ao Prof. Dr. Valdes Roberto Bollela, por intermediar nosso acesso à Unidade Especial de Tratamento de Doenças Infecciosas (UETDI) do Hospital das Clínicas de Ribeirão Preto e estar sempre disposto a ajudar no que fosse preciso durante todo o desenvolvimento do projeto.
À Dra. Fernanda Guioti Puga, por auxiliar na abordagem dos pacientes atendidos no Centro de Saúde Escola da FMRP/USP.
À Profa. Dra. Cleni Marzocchi Machado, por disponibilizar a infra-estrutura de seu laboratório para realização dos experimentos de quimioluminescência.
À Profa. Dra. Silvana Giuliatti, pela atenção e auxílio nas análises de bioinformática.
Aos meus queridos pais e à minha irmã de coração Joana, por estarem sempre presentes em minha vida independente da distância, sobretudo, por me darem força e acreditarem em mim mais do que eu mesma. Amor é muito pouco perto do que sinto por vocês!
A todos os outros membros da minha família, pelo apoio e carinho em todos os momentos, assim como pela alegria dos nossos reencontros.
Ao Thiago Malardo (Pamonha!), por todos esses anos, que não foram poucos, de companheirismo, alegrias, conquistas, trapalhadas e sempre com muito carinho e amor envolvidos. Por me dar uma segunda família em Ribeirão Preto que não mediram esforços para que eu sempre me sentisse em casa. Muito obrigada também, Valéria, Euvaldo e Vanessa!
Aos queridos amigos do LIIP e LIME, por compartilharem seu conhecimento, seja nas discussões científicas e/ou no dia a dia da bancada. Por fazerem as, muitas vezes, incansáveis horas de trabalho mais divertidas e produtivas. Agradeço especialmente ao Leonardo Galvão, Morgana Prado, Karina Zoccal, Francisco Silva e Mouzarllem Barros.
Às amigas Fabiana Zambuzi (Fabiii), Priscilla Cardoso (Priii) e Verônica Brauer (Veee), por todo auxílio direto e/ou indireto para a realização deste trabalho. Pela amizade, risadas, e conversas infinitas sobre a vida. Obrigada por me aguentarem e por sempre compartilharem das minhas gordices.

À Milena Sobral Espíndola, pela amizade e companhia de bancada, viagens e congressos. Pelos ensinamentos científicos, inclusive aqueles relacionados à escolha de um bom vinho!
Ao Murilo Soares, pela atenção e auxílio nas análises epigenéticas relacionadas à metilação.
À equipe técnica do laboratório, Caroline Fontanari, Thaísa Araújo, Alyne Galvão, Carlos Sorgi e Fabiana Morais, pelo apoio técnico essencial para manutenção do laboratório e realização dos experimentos.
À Wendy, meu anjo da guarda desde que cheguei em Ribeirão Preto, por estar sempre presente e disposta a ajudar. Por todo carinho e preocupação, além dos conselhos científicos e pessoais. Pelo exemplo de profissional e, sobretudo, pelo auxílio nos experimentos de clonagem. Muito obrigada pela oportunidade de conviver com você e o Fernando! Vocês são muito especiais para mim!
À Natália de Paula, querida amiga sempre disposta a me ouvir e a me ajudar, seja cientificamente ou pessoalmente. Pela amizade durante todos esses anos, e com certeza pela pipoca doce e o amendoim! Brincadeiras à parte, obrigada por ser essa inspiração de menina/mulher batalhadora e perseverante.
Às meninas do Interpós, pelos ensinamentos futebolísticos, amizade e horas de diversão, sem as quais não conseguimos recuperar as energias para mais um dia de labuta.
Aos funcionários da UETDI, por todo auxílio durante a abordagem dos pacientes e coleta de informações para a realização do estudo, em especial ao Chico.
A todos os doadores de sangue, especialmente aos pacientes HIV-1+, que aceitaram participar do trabalho e que depositaram em nós parte de suas esperanças de um futuro melhor.
À Faculdade de Ciências Farmacêuticas de Ribeirão Preto e à Faculdade de Medicina de Ribeirão Preto, por fornecerem a infra-estrutura para a realização deste trabalho.
Ao Programa de Pós-Graduação em Imunologia Básica e Aplicada, pela oportunidade de crescimento científico e pessoal proporcionada pela troca de conhecimentos. Agradeço também à secretária do programa, Ana Cristine, pelo auxílio, principalmente nas questões burocráticas.
À FAPESP, pelo auxílio financeiro fundamental para realização deste trabalho (Processos FAPESP n° 2011/24026-3 e n°2011/12199-0).

“In examining disease, we gain wisdom about anatomy and physiology and biology. In examining the person with disease, we gain wisdom about life."
Oliver Sacks

SOARES, L.S.
RESUMO
SOARES, L. S. Alterações imunológicas e epigenéticas em células natural killer de pacientes infectados pelo HIV-1. 2016. 113f. Tese (Doutorado) – Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 2016. A infecção pelo HIV caracteriza-se como doença inflamatória crônica e está associada a diversas disfunções do sistema imune, as quais aumentam a suscetibilidade dos indivíduos infectados a infecções oportunistas. Dentre os componentes da imunidade inata, as células natural killer possuem importante função de defesa contra os vírus, no entanto seu fenótipo e função estão alterados durante a infecção pelo HIV. Dessa forma, o objetivo deste estudo foi avaliar as mudanças fenotípicas e funcionais das células NK em pacientes infectados pelo HIV-1, sob tratamento antirretroviral ou não, e a relação destas alterações com possíveis modificações epigenéticas em genes do sistema imune nestas células. O perfil inflamatório sistêmico foi avaliado por meio da dosagem de mediadores plasmáticos, sendo que pacientes HIV-1+ apresentaram maior ativação imunológica, sobretudo os virgens de tratamento, o que foi constatado pelo aumento de moléculas pró-inflamatórias, principalmente: TNF-α, IP-10, sCD14 e sCD163. Ao avaliarmos especificamente a população de células NK, observou-se diminuição do número de células circulantes nos indivíduos infectados, tratados ou não, assim como alteração da proporção das subpopulações destas células. A expansão característica das células NK não funcionais foi observada tanto em pacientes não tratados, quanto naqueles sob tratamento antirretroviral, embora apenas nos indivíduos sob tratamento, houve recuperação parcial da função citotóxica celular. O fenótipo das células NK do grupo HIV-1+ sugeriu desenvolvimento do estado de exaustão celular, com poucas células desse grupo expressando os receptores característicos de NK como o NKp46 e alguns receptores de inibição (KIR2DL1, KIR2DL2/L3 e KIR2DL1), assim como pela inibição da função citotóxica e redução na produção de citocinas. Em contrapartida, as células NK do grupo TARV demonstraram maior estado de ativação verificado pelos altos níveis de produção de citocinas inflamatórias como TNF-α, e que poderia estar relacionado também ao maior tempo de infecção desses pacientes. Aliado a isso, as células NK dos indivíduos HIV-1+ sob tratamento apresentaram aumento da produção de espécies reativas de oxigênio, indicativo do estado de imunosenescência nessas células. Considerando as disfunções observadas, investigou-se a possível associação com modificações epigenéticas nessas células. A porcentagem de metilação global do DNA de células NK dos pacientes HIV-1+ sob estágio avançado da doença (AIDS) estava aumentada, embora o perfil de metilação do gene KIR2DL2 permaneceu inalterado entre os grupos. A frequências das enzimas de ativação e repressão da transcrição envolvidas nas mudanças epigenéticas também não demonstraram modulação evidente. Este é um dos primeiros estudos a demonstrar a ocorrência de modificações epigenéticas em células NK de pacientes HIV-1+, ainda que sutis. Além disso, os resultados aqui apresentados sugerem exaustão das células NK dos indivíduos recentemente infectados em contraste com possível estado de imunosenescência nos pacientes HIV-1+ sob tratamento antirretroviral e com maior tempo de infecção. Outros marcadores devem ser investigados para confirmação dessa hipótese e melhor compreensão da patogênese do HIV, com o intuito de fornecer informações e novas estratégias que possam contribuir para o reestabelecimento da homeostase das células do sistema imune, como as células natural killer. Palavras-chave: imunidade inata, HIV/AIDS, metilação global, células natural killer.

SOARES, L.S.
ABSTRACT
SOARES, L. S. Immunological and epigenetic alterations in natural killer cells from patients infected with HIV-1. 2016. 113f. Tese (Doutorado) – Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, 2016. HIV infection is characterized as an inflammatory chronic disease and is associated to many dysfunctions of the immune system, which increase the susceptibility of infected individuals to opportunistic infections. Among the innate immunity components, natural killer cells have an important function of defense against viruses, however their function and phenotype are disturbed during HIV infection. In this way, the aim of this study was to evaluate phenotypic and functional changes in natural killer cells from HIV-1 infected patients, with or without antiretroviral treatment, and correlate these changes to epigenetic alterations in genes of the immune system on these cells. The systemic inflammatory profile was evaluated by quantifying the plasmatic mediators, wherein HIV-1+ patients showed higher immunologic activation, mainly the treatment-naïve patients, which was observed by the increase of pro-inflammatory molecules levels, mainly TNF-α, IP-10, sCD14 e sCD163. When we specifically evaluated the NK cell population, it was verified a reduction in the numbers of circulating NK cells from infected individuals, treated or not, as well as changing in the subpopulation proportions of these cells. The characteristic expansion of the non-functional NK cells was observed both in the treatment-naïve patients as well in the treated ones, although only in the treated HIV-1+ patients, there was a partial recovery of the cytotoxic function. The NK cell phenotype from HIV-1+ group suggested an exhaustion state of these cells, with fewer cells from this group expressing the characteristic receptors of NK such as NKp46 and some inhibitory receptors (KIR2DL1, KIR2DL2/L3 e KIR2DL1), as well as by the reduced cytotoxic function and cytokine production. In contrast, NK cells from TARV group showed higher activation state that was verified by the elevated levels of inflammatory cytokines production such as TNF-α, and that could also be related to the long-term infection on these patients. Besides this, NK cells from HIV-1 treated individuals exhibited increased production of oxygen reactive species, indicative of immunosenescence in these cells. Considering the observed dysfunctions, we investigated the possible association with epigenetic modifications in these cells. The percentage of global DNA methylation of NK cells from HIV-1+ patients on advanced stage of disease (AIDS) was increased, although the methylation profile of the KIR2DL2 gene remained unchanged between the groups. The frequency of enzymes of activation and repression of transcription involved in epigenetic changes also showed no apparent modulation. This is one of the first studies to show the occurrence of epigenetic modifications on NK cells from HIV-1+ patients, although these changes were subtle. Besides this, the results showed here suggested exhaustion of NK cells from recently infected individuals, in contrast to the potential immunosenescence state of HIV-1+ patients under antiretroviral treatment and with long-term infection. Other markers should be investigated in order to confirm this hypothesis, as well as contributing to the understanding of HIV pathogenesis, with the aim of providing information and new strategies which may help in the recovery of the immune cells homeostasis, such as natural killer cells. Key words: innate immunity, HIV/AIDS, global methylation, natural killer cells.

SOARES, L.S.
SUMÁRIO
1. Introdução .......................................................................................................................... 13 1.1. Histórico, etiologia e epidemiologia da infecção por HIV ............................................... 14 1.2. Patogênese do HIV ........................................................................................................... 16 1.3. Disfunções associadas às células natural killer durante a infecção por HIV .................... 18 1.4. Correlação entre mecanismos epigenéticos e doenças ...................................................... 21 2. Objetivos ............................................................................................................................. 24 2.1. Objetivo Geral ................................................................................................................... 25 2.2. Estratégias utilizadas para alcançar o objetivo ................................................................. 25 3. Delineamento Experimental .............................................................................................. 26 4. Material e Métodos ............................................................................................................ 28 4.1. Casuística .......................................................................................................................... 29 4.2. Obtenção de células mononucleares do sangue periférico ............................................... 29 4.3. Ensaio imunoenzimático para quantificação das moléculas CD14 e CD163 solúveis no
plasma .................................................................................................................................. 30 4.4. Quantificação de citocinas e quimiocinas no plasma ....................................................... 30 4.5. Análise fenotípica das células NK e suas subpopulações por citometria de fluxo ........... 31 4.6. Separação das células NK por esferas magnéticas ........................................................... 31 4.7. Análise fenotípica das células NK por PCR em Tempo Real ........................................... 32 4.8. Avaliação funcional das células NK por ensaio de citotoxicidade ................................... 33 4.8.1. Cultivo das células alvo ................................................................................................. 33 4.8.2. Ensaio de citotoxicidade ................................................................................................ 33 4.8.3. Quantificação de citocinas e quimiocinas em sobrenadante de cultura ......................... 33 4.8.4. Quantificação da produção de espécies reativas de oxigênio por quimioluminescência34 4.9. Avaliação de mudanças epigenéticas relacionadas às células NK .................................... 34 4.9.1. Determinação do padrão de metilação global do DNA cromossômico ......................... 34 4.9.2. Avaliação do padrão de metilação gene-específico ....................................................... 34 4.9.2.1. Análise da sequência genômica referente ao gene KIR2DL2 ..................................... 35 4.9.2.2. Quantificação da metilação do gene KIR2DL2 após modificação por bissulfito ....... 36 4.9.3. Quantificação da expressão de enzimas que promovem as alterações epigenéticas ..... 38 4.9.4. Imunoprecipitação da cromatina .................................................................................... 38 4.10. Análise estatística ............................................................................................................ 40 5. Resultados ........................................................................................................................... 41 5.1. Características da população em estudo ........................................................................... 42 5.2. Análise de moléculas relacionadas à progressão da doença ............................................. 44 5.2.1. Quantificação das moléculas CD14 e CD163 solúveis no plasma ................................ 44 5.2.2. Quantificação de citocinas e quimiocinas do plasma .................................................... 46 5.2.3. Correlação entre os marcadores de ativação do plasma ................................................. 49 5.3. Avaliação fenotípica das células NK e suas subpopulações ............................................. 53 5.3.1. Determinação da frequência de células NK e a distribuição de suas subpopulações .... 53 5.3.2. Análise da expressão dos receptores de ativação e inibição das células NK ................. 57 5.4. Avaliação funcional das células NK ................................................................................. 62 5.4.1. Análise da citotoxicidade das células NK purificadas ................................................... 62 5.4.2. Quantificação das citocinas e quimiocinas no sobrenadante do ensaio de citotoxicidade
.............................................................................................................................................. 64 5.4.3. Correlações entre os parâmetros que caracterizam as disfunções das células NK nos
indivíduos HIV-1+ ............................................................................................................... 68 5.4.4. Determinação da produção de granzima B pelas células NK ........................................ 74

SOARES, L.S.
5.4.5. Quantificação da produção de espécies reativas de oxigênio pelas células NK ............ 76 5.5. Caracterização das modificações epigenéticas relacionadas às disfunções das células NK
.............................................................................................................................................. 78 5.5.1. Determinação do padrão de metilação global do DNA das células NK ........................ 78 5.5.2. Avaliação do padrão de metilação específico do gene KIR2DL2/L3 ............................ 80 5.5.3. Análise da frequência de enzimas que promovem as modificações epigenéticas ......... 85 5.5.4. Investigação de modificações epigenéticas associadas às histonas em genes da resposta
imune de células NK ............................................................................................................ 90 6. Discussão ............................................................................................................................. 93 7. Conclusões ........................................................................................................................ 104 8. Referências Bibliográficas ............................................................................................... 106 APÊNDICE ........................................................................................................................... 114 APÊNDICE A – FIGURAS SUPLEMENTARES ................................................................ 115 ANEXOS ............................................................................................................................... 130 ANEXO I – MANUSCRITO PARA PUBLICAÇÃO .......................................................... 131 ANEXO II – Aprovação do Comitê de Ética em Pesquisa para Seres Humanos .................. 150 ANEXO III – Aprovação do Centro de Saúde Escola para coleta de amostras .................... 151 ANEXO IV – Termo de Consentimento Livre e Esclarecido – Indivíduos Controles .......... 152 ANEXO V – Termo de Consentimento Livre e Esclarecido – Pacientes HIV-1+ ................ 154

SOARES, L.S.
13
1. Introdução

SOARES, L.S. Introdução
14
1. Introdução
1.1. Histórico, etiologia e epidemiologia da infecção por HIV
A Síndrome da Imunodeficiência Adquirida (AIDS do inglês Acquired immune
deficiency syndrome) foi primeiramente reconhecida como doença no início dos anos 80 [1] e
desde então, diversos grupos de pesquisa têm se dedicado ao estudo dessa patologia em busca
de sua prevenção e cura efetivas. Os primeiros casos de AIDS foram descritos em 1981 em
homens homossexuais com infecção pulmonar rara por Pneumocystis carinii [2]. Em seguida,
surgiram relatos de casos de sarcoma de Kaposi que também foram associados à AIDS [3].
Nesse cenário, em 1982, o Centro de Controle e Prevenção de Doenças (CDC) dos Estados
Unidos passou a definir os casos de AIDS a partir da “ocorrência de pelo menos uma doença
preditiva de um defeito na imunidade celular e de causa desconhecida para diminuição da
resistência a essa doença. Tais doenças incluíam pneumonia por Pneumocystis carinii,
sarcoma de Kaposi e outras infecções oportunistas” [1].
De junho de 1981 a setembro de 1982, aproximadamente 75% dos casos de AIDS
ocorreram entre homens homossexuais ou bissexuais e havia evidências de possível
transmissão sexual, levando essa síndrome a ser inicialmente chamada de GRID, do inglês
Gay-related immune deficiency [4]. Essa denominação alimentou grande parte do preconceito
e estigma social que perduram, embora de forma mais discreta, até os dias atuais.
Considerando que a falta de informação pode induzir a atitudes e julgamentos errôneos, a
evolução do conhecimento sobre a AIDS foi e é de extrema importância no contexto da saúde
mundial. Até então não havia sido isolado o agente etiológico responsável pela síndrome, até
que em 1983, dois principais grupos de pesquisadores (liderados por Luc Montagnier e Robert
Gallo) descreveram o isolamento de um novo retrovírus, o qual foi posteriormente definido
como agente causador da AIDS e denominado HIV, do inglês Human Immunodeficiency
Virus [5-7]. O grupo do pesquisador francês Luc Montagnier inicialmente nomeou o vírus de
LAV, do inglês lymphadenopathy-associated virus [7], e a ele foi creditado o primeiro relato
verdadeiro do isolamento desse vírus, rendendo a ele e à Françoise Barré-Sinoussi,
pesquisadora de seu grupo, o prêmio Nobel de Fisiologia ou Medicina de 2008.
A caracterização molecular do HIV ocorreu em 1985 e forneceu a base para o estudo da
diversidade, origem e evolução desse vírus [8-10]. O HIV é um vírus pertencente ao gênero
Lentivirus, da família Retroviridae. Existem duas linhagens principais de vírus conhecidas por
causar a doença em humanos, o HIV do tipo 1 e 2, no entanto HIV-1 é descrito como mais

SOARES, L.S. Introdução
15
comum e mais patogênico. Acredita-se que o HIV-1 tenha surgido na espécie humana após a
manipulação da carne de chimpanzé (Pan troglodytes troglodytes) infectada com o vírus da
imunodeficiência símia (SIV) por caçadores selvagens. Esse evento teria ocorrido entre 1900
e 1930 próximo ao sudeste de Camarões, na região ocidental da África Central [11, 12]. Por
outro lado, o HIV-2 teria se originado do SIV proveniente do macaco verde africano “sooty
mangabey” (Cercocebus atys), o qual era comumente caçado na África ocidental [13].
Outro fato importante na história da AIDS/HIV foi a aprovação do AZT (zidovudina)
como primeiro antirretroviral licenciado para uso em humanos em 1987 [14]. A partir de
então, surgiram diversos medicamentos antirretrovirais que auxiliaram na contenção da
expansão da infecção, mas sem levar à cura [15]. Vale ressaltar que o Brasil foi um dos países
pioneiros na acessibilidade ao tratamento após promulgação da Lei 9.313, em 1996, que
garantiu o direito e o acesso gratuito à terapia antirretroviral a todos os indivíduos portadores
do HIV [16].
Apesar de haver tratamento disponível que auxilia no controle do desenvolvimento da
doença, a cura ainda é um assunto muito discutido. O primeiro relato de cura da infecção por
HIV ocorreu em 2008 e é conhecido como o “paciente de Berlim” [17]. Embora existam
outros possíveis casos, estes são denominados de cura funcional, ou seja, supressão da
replicação viral na ausência de tratamento, entretanto, sem a completa eliminação do
reservatório viral [18, 19]. Enquanto diversos grupos de pesquisa têm se empenhado na busca
da cura e prevenção efetivas, tem-se falado bastante em profilaxia pré e pós exposição como
alternativas para frear o avanço da epidemia da AIDS no mundo [20].
Apesar de diversos esforços, o Programa Conjunto das Nações Unidas sobre HIV/AIDS
(UNAIDS) relatou a incidência de 2 milhões de casos de infecção por HIV em 2014, e a
morte de 1,2 milhões de pessoas em decorrência da AIDS. Estima-se que existam
aproximadamente 36,9 milhões de pessoas com HIV no mundo, sendo a maior prevalência na
África Subsaariana (aproximadamente 25,8 milhões de pessoas) [21]. No Brasil, desde o
início da epidemia de AIDS até junho de 2015, foram registrados em torno de 798 mil casos,
sendo a maior concentração nas regiões Sudeste (53,8%) e Sul (20%). Por outro lado, a região
Sudeste é a única a apresentar tendência de queda no número de casos nos últimos dez anos,
enquanto as regiões Norte, Nordeste e Centro-Oeste ainda demonstram crescimento
significativo. Quanto à mortalidade por AIDS, até dezembro de 2014, havia sido reportado
cerca de 291 mil óbitos em todo o Brasil [22].
Em 2014, foram definidas três metas para o monitoramento clínico das pessoas com
HIV como uma das principais ferramentas para acompanhamento da evolução da epidemia.

SOARES, L.S. Introdução
16
Essas metas ficaram conhecidas como 90-90-90, cujo objetivo é atingir, até 2020, o
diagnóstico de no mínimo 90% das pessoas infectadas por HIV, sendo que dessas pessoas,
90% deveriam estar em tratamento antirretroviral, e 90% dessas últimas estariam em
supressão viral. Essas metas também são denominadas de cascata de cuidado contínuo e no
Brasil, estimou-se que, ao fim de 2014, 83% das pessoas com HIV tinham sido
diagnosticadas, e dessas, 52% estavam em tratamento. Das pessoas sob tratamento
antirretroviral, 46% apresentavam supressão viral por pelo menos seis meses após início da
terapia [22]. Esses dados sugerem um cenário relativamente positivo quanto ao controle da
doença no Brasil, embora não seja o ideal. Para tanto, faz-se necessário a conscientização da
população e adoção de política públicas, aliados a estudos cada vez mais detalhados visando a
maior compreensão da patogênese do HIV.
1.2. Patogênese do HIV
O curso da infecção por HIV apresenta variações entre as pessoas, no entanto, a maioria
delas desenvolvem doenças oportunistas que são utilizadas como indicativo do quadro de
imunodeficiência [23]. A principal via de transmissão do vírus é a sexual, podendo também
ocorrer a transmissão vertical, por compartilhamento de seringa ou agulha contaminada,
transfusão sanguínea e em menor número, por acidente com material perfuro-cortante não
esterilizado [24]. Acredita-se que as primeiras células a serem infectadas sejam os linfócitos T
CD4+ de memória residentes, mas também pode ocorrer infecção de outras células como os
monócitos e macrófagos, que podem servir como reservatórios virais; e células dendríticas,
que podem transportar o vírus e promover a infecção de outras células nos linfonodos
drenantes [25]. A entrada do vírus na célula ocorre após ligação da proteína viral gp120 com a
molécula CD4, seguida da interação entre a subunidade gp41 com os correceptores CCR5 ou
CXCR4 presentes na membrana das células alvo, propiciando assim a fusão do envelope viral
à membrana celular. No interior da célula, o RNA viral é transformado em DNA pela
transcriptase reversa do vírus e a partir da maquinaria celular são geradas novas partículas
virais com potencial de infectar outras células [26].
A partir de estudos realizados em pacientes, observou-se que aproximadamente dez dias
após a transmissão, o RNA viral passa a ser detectado no plasma e em seguida, o vírus
alcança os linfonodos drenantes, onde pode infectar outras células T, assim como células
dendríticas e células B. Dentro de 21 a 28 dias após a infecção, observa-se a maior carga viral

SOARES, L.S. Introdução
17
no sangue, sendo que neste período podem aparecer os primeiros sintomas associados à
doença e ocorre redução significativa das células T CD4+ presentes na mucosa [27].
A infecção por HIV caracteriza-se não somente pelos altos níveis de replicação viral e
depleção de linfócitos T CD4+ de forma sistêmica, mas também pela disfunção generalizada
do sistema imune correlacionada a um estado de inflamação crônica que está fortemente
associado à progressão do paciente à AIDS. Esse perfil inflamatório permanece, mesmo que
em níveis menores, nos pacientes que estão sob tratamento antirretroviral evidenciando a
participação de outros fatores na indução desse perfil [28, 29]. Em 2006, Brenchley [30] e
colaboradores demonstraram a ocorrência do fenômeno de translocação microbiana devido à
quebra da integridade da mucosa gastrintestinal, que foi proporcionada pela depleção maciça
de linfócitos T CD4+ naquela região após a infecção pelo HIV. Dessa forma, esse fenômeno
amplifica a inflamação sistêmica observada nos pacientes HIV-1+, sendo sugerido alguns
marcadores da fase aguda da infecção, como elevados níveis plasmáticos de IP-10, IL-10 e
TNF-α, os quais poderiam ser considerados indicadores da progressão rápida da doença, em
paralelo à contagem de linfócitos T CD4+ e carga viral [31].
Em pacientes com boa adesão ao tratamento antirretroviral, é possível controlar a
replicação viral e recuperar a contagem de linfócitos T CD4+, no entanto, por não existir cura
efetiva, o paciente deve tomar a medicação por toda a vida. Além disso, a resposta imune
nestes indivíduos não é completamente restaurada e tem-se observado aumento da ocorrência
de co-morbidades não relacionadas à infecção por HIV como: doenças cardiovasculares,
neurocognitivas, osteoporose, doenças relacionadas ao fígado e rim, assim como alguns tipos
de câncer. Considerando que muitas dessas patologias ocorrem principalmente em indivíduos
idosos, tem-se especulado que a infecção por HIV possa promover de alguma forma o avanço
do envelhecimento natural [32].
Cada célula possui funções específicas na resposta imune ao HIV e, na maioria das
vezes, suas funções tornam-se alteradas como influência direta e/ou indireta do vírus. A
maioria das células descritas por desempenhar funções protetoras ao início da infecção por
HIV, como as células dendríticas e células T CD8+, passam a exibir disfunções com o
decorrer da infecção dificultando o controle da replicação viral e contribuindo para o
desenvolvimento de infecção com caráter sistêmico [33, 34]. Estes fatores associados à
redução significativa de linfócitos T CD4+ tornam os indivíduos infectados por HIV
suscetíveis a diversas infecções oportunistas que dependem da imunidade celular para seu
controle efetivo [35].
Diante desse painel, outras células do sistema imune passaram a ser estudadas no

SOARES, L.S. Introdução
18
contexto da infecção por HIV, visto que as principais células associadas à imunidade
adaptativa (linfócitos T CD4+ e T CD8+) estão com suas funções diminuídas, o que dificulta o
controle efetivo da infecção por HIV, assim como de infecções oportunistas. Células
classicamente caracterizadas como sendo da imunidade inata têm sido alvo de vários estudos
por serem rapidamente ativadas e tornarem-se funcionais logo ao início das infecções [36].
Além disso, recentemente tem sido demonstrado um fenótipo de memória associado à função
destas células, especificamente das células NK [37], caracterizando-as como importante alvo
de estudo na infecção por HIV.
1.3. Disfunções associadas às células natural killer durante a infecção por HIV
As células NK foram primeiramente identificadas em 1975 e descritas como linfócitos
grandes e granulares com citotoxicidade natural contra células tumorais [38]. Atualmente, são
classificadas como células inatas linfóides pertencentes ao grupo 1, com base na expressão
dos fatores de transcrição: T-box, EOMES e T-bet; e na produção de IFN-γ [39]. Além de
atuarem na defesa contra tumores, as células NK constituem importante mecanismo de
proteção a infecções, principalmente aquelas causadas por vírus [40].
Em humanos, as células NK representam de 5 a 15% dos linfócitos encontrados no
sangue periférico e são fenotipicamente caracterizadas como CD3-CD56+ [41]. Também
expressam a molécula CD16 (receptor para a porção Fc dos anticorpos) e conforme a
expressão de CD16 e CD56, as células NK são subdivididas em subpopulações
funcionalmente distintas. A subpopulação CD56dimCD16+ corresponde a 90% das células NK
encontradas no sangue periférico e baço e possui função predominantemente citotóxica.
Enquanto as células NK CD56brightCD16- estão presentes em maior quantidade nos linfonodos
e estão envolvidas na produção de citocinas como IFN-γ, entre outras [34].
A função efetora desempenhada pelas células NK depende sobretudo do balanço dos
sinais advindos de seus receptores de ativação e inibição (KAR – Killer Activation Receptor e
KIR – Killer Inhibtory Receptor). Os receptores de ativação reconhecem moléculas que
passam a ser expressas nas células sob condições de estresse, assim como substâncias
derivadas dos patógenos. Enquanto os receptores de inibição interagem com as moléculas de
MHC de classe I, sendo que essa interação além de estar envolvida no processo de ativação
das células NK, também é importante durante o desenvolvimento dessa população celular [42,
43].

SOARES, L.S. Introdução
19
Dentre as principais funções efetoras das células NK está a lise de células alvo
infectadas antes mesmo do recrutamento das células da imunidade adaptativa [44], destacando
a importância dessas células na proteção do hospedeiro durante a infecção por HIV, assim
como em infecções de caráter oportunista. No entanto, ao longo dos anos têm sido descritas
algumas disfunções das células NK em pacientes infectados pelo HIV, como a observação de
que havia diminuição dos mecanismos de lise celular mediada por essas células conforme a
doença progredia [45-47]. Em 2003, Portales et al. [48] reportaram a diminuição das reservas
de perforina e granzima nas células NK de indivíduos HIV-1+, o que poderia explicar a
diminuição da citotoxicidade celular observada nestes pacientes. Além disso, a viremia
persistente em alguns indivíduos tem sido associada à expressão anormal de vários receptores
nas células NK, como a menor expressão dos receptores de citotoxicidade natural (NCRs),
entre eles: NKp46 e NKp30. Esse fato pode estar associado à diminuição dos mecanismos
citotóxicos desencadeados pelas células NK e consequente redução da eliminação de células
infectadas in vivo [49].
Por outro lado, a maioria dos trabalhos demonstram aumento da expressão dos
receptores inibitórios na superfície das células NK de indivíduos infectados pelo HIV-1, entre
eles: KIR2DL1, KIR2DL2 e KIR3DL1, principalmente nos indivíduos com carga viral
detectável [50, 51]. No entanto, os resultados quanto à expressão desses receptores nas células
NK variam de um estudo para outro conforme o estágio da doença e a subpopulação
analisada. Em resumo, a maior expressão dos receptores inibitórios e menor dos de ativação,
durante a infecção por HIV-1, contribuem para um estado de supressão das células NK,
comprometendo a resposta imune destes indivíduos.
Outra função alterada em células NK de pacientes HIV-1+ é a produção de citocinas e
quimiocinas. Em indivíduos com carga viral detectável, observou-se menor produção de IFN-
γ, TNF-α e GM-CSF pelas células NK, influenciando principalmente na maturação e
migração de células dendríticas [52]. Dessa forma, há diminuição da diferenciação da resposta
imune para o padrão Th1 e a interação entre essas duas populações celulares é comprometida.
Além disso, células NK possuem importante função de eliminar células dendríticas imaturas,
o que também diminui durante a infecção pelo HIV-1, causando o acúmulo destas células
imaturas nos indivíduos infectados [34]. Em relação às quimiocinas, a produção de CCL3,
CCL4 e CCL5 pelas células NK auxilia no bloqueio da entrada de vírus com tropismo para o
correceptor CCR5. A menor produção dessas quimiocinas também contribui para o aumento
da replicação viral, evidenciando outra importante função das células NK durante a infecção
pelo HIV-1 [53].

SOARES, L.S. Introdução
20
Um dos primeiros estudos a reportarem mudanças fenotípicas nas células NK durante a
infecção pelo HIV-1 utilizando a técnica de citometria de fluxo foi publicado em 1995 [54].
Dentre os principais resultados estava o relato da diminuição das células NK CD56+CD16+
citotóxicas e a expansão daquelas CD56-CD16+. Esse último fenótipo foi posteriormente
atribuído a células NK denominadas não funcionais, caracterizadas pela expressão de altos
níveis de receptores inibitórios, menor produção de citocinas e por possuir defeitos na lise de
células alvo infectadas [52]. Acredita-se que grande parte das disfunções associadas às células
NK durante a infecção pelo HIV-1 seja devido à expansão dessa subpopulação não funcional.
Em resumo, o vírus da imunodeficiência humana está envolvido na indução de diversas
alterações fenotípicas e funcionais das células NK. Esses efeitos podem ser desencadeados
mediante a ligação direta do vírus aos receptores de quimiocinas, ou como resultado da
ativação generalizada do sistema imune pelo vírus, ou ainda após infecção direta das células
NK [55]. Esta última possibilidade tem sido descrita em um subtipo de células NK, que além
de expressar os receptores CXCR4 e CCR5, passa também a expressar o receptor CD4,
propiciando assim a entrada do vírus na célula NK [56]. Quando no interior das células, em
geral, o vírus é capaz de modular a expressão das moléculas de HLA, diminuindo
especificamente a expressão dos HLA-A e B, mas não dos HLA-C e E. Dessa forma, a célula
infectada escapa dos mecanismos efetores dos linfócitos T CD8+ direcionados aos epítopos
restritos aos HLA-A/B, assim como da morte mediada pelas células NK, pois estas células
expressam receptores inibitórios que reconhecem os HLA-C/E. Este mecanismo de evasão é
desencadeado pela proteína Nef (fator negativo) do vírus, que também está envolvida na
diminuição da expressão de ligantes dos receptores de NK (NKG2D) como MICA, ULBP1 e
ULBP2 nas células infectadas [57, 58].
Alguns estudos têm descrito a associação entre a expressão de certos receptores de NK
e moléculas de HLA com o maior controle da replicação viral e, portanto, menor progressão
da doença desencadeada por HIV [59, 60]. Alter et al. [61] reportaram, em 2009, a expansão
de células NK KIR3DS1+ e KIR3DL1+ durante a infecção de indivíduos com HIV-1 e que
expressavam o HLA-Bw4-80I. Essa observação estava relacionada à supressão da replicação
viral em células alvo que expressavam essa molécula de HLA. Os receptores do tipo KIR
constituem um grupo de moléculas altamente polimórficas, sendo que alguns destes genes
encontram-se silenciados, enquanto outros não. Durante o desenvolvimento e maturação das
células NK ocorrem modificações em nível de expressão gênica, denominadas mudanças
epigenéticas, como a demetilação dos genes KIR para que ocorra sua expressão nas células
NK [62]. Considerando que na infecção por HIV são observadas modificações na expressão

SOARES, L.S. Introdução
21
de diversos receptores do tipo KIR, assim como de outras moléculas, sugere-se que mudanças
epigenéticas possam estar envolvidas nesse processo.
Num estudo conduzido com vietnamitas usuários de drogas e que permaneciam
soronegativos mesmo após vários anos de exposição ao HIV, verificou-se maior capacidade
efetora das células NK quanto a sua atividade citolítica e produção de quimiocinas e citocinas
do que em indivíduos HIV soropositivos [63]. E uma das hipóteses levantadas quanto a esta
atividade aumentada das células NK vista em indivíduos expostos, mas não infectados, foi a
possível associação com as características genéticas do indivíduo que favorecem a expressão
de certos receptores e moléculas nas células NK. Essas observações também foram feitas em
indivíduos HIV-1+ considerados progressores lentos e naqueles denominados controladores
de elite [51]. Considerando que muitas das alterações descritas até aqui podem ser resultado
também de modificações epigenéticas, justifica-se a importância do estudo destes
mecanismos na infecção por HIV, especificamente em relação às alterações funcionais
desencadeadas como consequência destas modificações nas células NK.
1.4. Correlação entre mecanismos epigenéticos e doenças
O termo epigenética refere-se aos mecanismos que controlam a expressão gênica, sem
modificar a sequência de DNA dos organismos. As modificações epigenéticas são
potencialmente herdáveis e a importância do seu estudo tem sido associada à observação de
que muitas doenças se desenvolvem após a ocorrência de mudanças epigenéticas errôneas. Os
mecanismos epigenéticos incluem modificações químicas no DNA e/ou nas suas histonas
associadas, resultando na mudança da acessibilidade física do DNA aos fatores
transcricionais. Tais modificações podem ser agrupadas em quatro principais categorias:
metilação do DNA, alterações na histona, posicionamento dos nucleossomos, e a regulação do
DNA por RNAs não codificantes (Ex.: microRNAs) também é considerada um mecanismo
epigenético [64, 65].
Os mecanismos envolvidos na regulação epigenética dos produtos gênicos podem ser
generalizados como ativador transcricional ou silenciador gênico. Sabe-se, por exemplo, que a
acetilação da histona 4 (AcH4) ou a tri-metilação da lisina 4 da histona 3 (H3K4me3)
geralmente aumentam o acesso ao DNA e estão envolvidas com ativação gênica [66],
enquanto a di- ou tri-metilação da lisina 27 da histona 3 (H3K27me2 ou me3) fecham o DNA
resultando em silenciamento gênico [67]. Dentre as enzimas envolvidas em tais modificações
estão as histona-acetiltransferases (HATs), histona-metiltransferases (HMTs), histona-

SOARES, L.S. Introdução
22
quinases (HKs), entre outras [68]. Já os eventos de metilação do DNA e a regulação pelos
microRNAS estão relacionados principalmente ao silenciamento gênico [65].
Em doenças auto-imunes, câncer e desordens neurológicas já foram relatadas várias
alterações epigenéticas, as quais foram associadas ao aparecimento ou agravamento das
doenças [65]. Além disso, estudos recentes demonstraram o envolvimento de mudanças
epigenéticas no direcionamento do fenótipo de macrófagos, assim como na função de células
dendríticas durante a sepse [69-71].
Até 2011, ainda eram poucos os trabalhos publicados com o assunto epigenética e a
relação patógeno versus hospedeiro [65]. Em relação ao patógeno, existem várias evidências
do envolvimento dos mecanismos epigenéticos para sua manutenção e sobrevivência no
hospedeiro. Por exemplo, a acetilação de histonas em Toxoplasma gondii está relacionada à
mudança entre os estágios replicativo e não replicativo desse patógeno [72]. O controle
epigenético também tem sido associado à expressão de fatores de virulência em protozoários
(Ex.: Plasmodium falciparum) e em bactérias (Ex.: Samonella enterica) [73, 74]. Essas
observações são de grande importância, pois sugerem novas possibilidades de intervenções
terapêuticas em doenças infecciosas.
As modificações epigenéticas também podem ocorrer no hospedeiro como
consequência da infecção por determinado patógeno. Durante a infecção por Mycobacterium
tuberculosis, a expressão de genes induzidos por IFN-γ é reprimida via acetilação de histonas
nos macrófagos alveolares, o que se correlaciona ao desenvolvimento da doença crônica em
alguns pacientes [75]. Interessantemente, na infecção pelo vírus Influenza, a proteína viral
NS1 possui uma sequência de aminoácidos similar à cauda da histona H3 de humanos. Dessa
forma, essa proteína “sequestra” o fator de transcrição hPAF1 do hospedeiro, suprimindo a
produção de proteínas antivirais e consequentemente a resposta imune contra o vírus [76].
No contexto da infecção por HIV-1, já foram realizados estudos caracterizando o
epigenoma das principais células alvo durante a infecção (células T CD4+, monócitos e
macrófagos). Vale ressaltar que o vírus da imunodeficiência humana foi o primeiro patógeno
a ser descrito como indutor de mudanças epigenéticas nas células hospedeiras. Sabe-se que as
proteínas Nef e Tat induzem aumento da enzima DNA metiltransferase 1 (DNMT1),
favorecendo a metilação do DNA e consequente repressão gênica [77].
Os mecanismos de regulação epigenética mais estudados na infecção por HIV-1 são
aqueles relacionados à indução e manutenção da latência viral. Essa latência é mantida em
parte pela atividade da enzima histona deacetilase (HDAC), envolvida na remoção de grupos
acetil das histonas, afetando dessa forma a expressão gênica [78]. Diante desse conhecimento,

SOARES, L.S. Introdução
23
há alguns anos sugeriu-se a utilização do medicamento vorinostat como aliado ao tratamento
convencional contra o HIV. O vorinostat é um inibidor de HDAC utilizado na terapia de
alguns tipos de câncer. Essa sugestão de tratamento recebeu o nome de “shock and kill”, pois
com o emprego do vorinostat ocorreria a produção de novas partículas virais que seriam
retiradas da latência e as drogas antirretrovirais atuariam eliminando tais vírus [79]. Essa
estratégia abriu novas perspectivas de tratamento da infecção por HIV aplicando os
conhecimentos adquiridos na área de epigenética.
Diante desse painel, consideramos que o estudo das alterações epigenéticas em genes
relacionados à resposta imune efetora das células NK pode ter um papel chave na elucidação
dos mecanismos pelos quais o HIV influencia na função dessas células, além de fornecer
subsídios para novas intervenções terapêuticas.

SOARES, L.S.
24
2. Objetivos

SOARES, L.S. Objetivos
25
2. Objetivos
2.1. Objetivo Geral
Avaliar as mudanças fenotípicas e funcionais das células NK em pacientes infectados
pelo HIV-1, sob tratamento antirretroviral ou não, e a relação destas alterações com possíveis
modificações epigenéticas em genes do sistema imune.
2.2. Estratégias utilizadas para alcançar o objetivo
a) Caracterização fenotípica das células NK de pacientes HIV-1+, em tratamento
antirretroviral ou não, por meio da comparação da expressão de receptores específicos e
distribuição das subpopulações celulares em relação aos indivíduos controle;
b) Avaliação da função efetora de células NK de pacientes HIV-1+, em tratamento
antirretroviral ou não, a partir de ensaios funcionais in vitro e quantificação da produção de
moléculas solúveis (citocinas, quimiocinas, granzima B e espécies reativas de oxigênio);
c) Análise do padrão de metilação global e específico de genes relacionados aos receptores
das células NK de pacientes HIV-1+, em tratamento antirretroviral ou não;
d) Determinação da expressão de enzimas relacionadas à ativação e repressão gênica em
células NK de pacientes HIV-1+, em tratamento antirretroviral ou não;
e) Análise das modificações de histonas associadas à ativação ou repressão dos genes
relacionados à resposta imune das células NK.

SOARES, L.S.
26
3. Delineamento Experimental
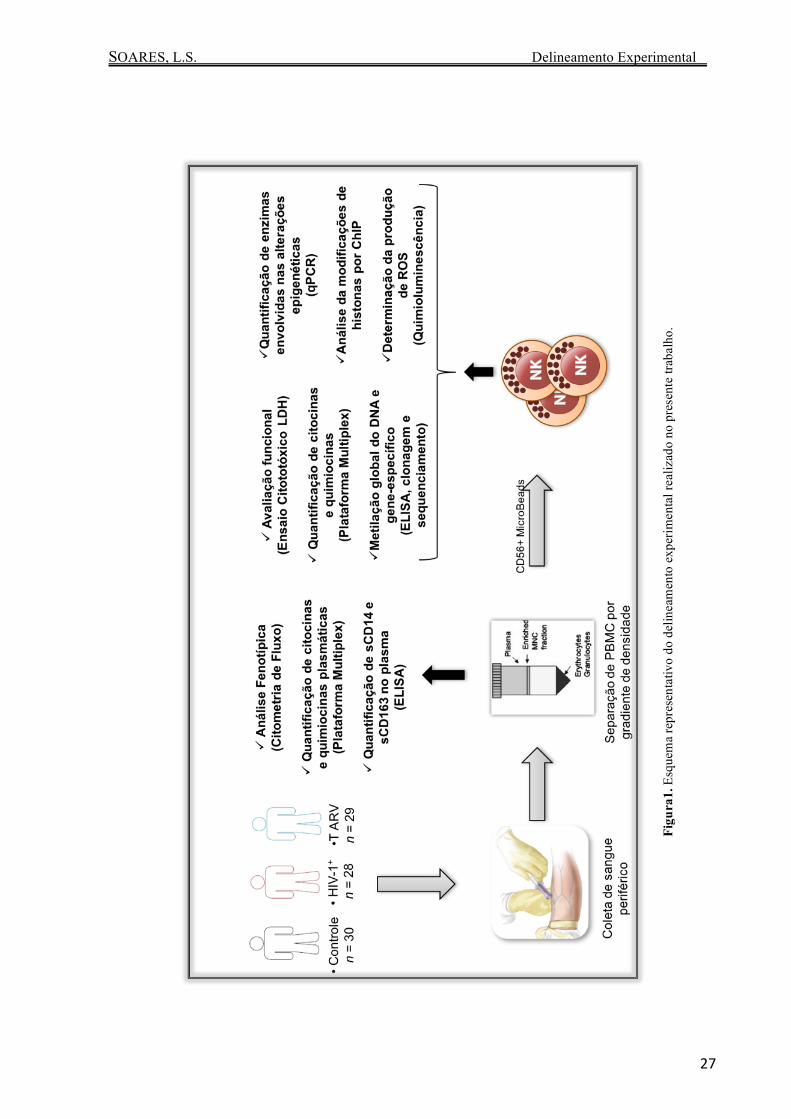
SOARES, L.S. Delineamento Experimental
27
Figu
ra1.
Esq
uem
a re
pres
enta
tivo
do d
elin
eam
ento
exp
erim
enta
l rea
lizad
o no
pre
sent
e tra
balh
o.

SOARES, L.S.
28
4. Material e Métodos

SOARES, L.S. Material e Métodos
29
4.Material e Métodos
4.1. Casuística
O projeto foi aprovado pelo Comitê de Ética em Pesquisa do Hospital das Clínicas da
Faculdade de Medicina de Ribeirão Preto da Universidade de São Paulo em reunião realizada
em 13/08/2012 (Processo HCRP n° 9818/2012).
A coleta das amostras do grupo experimental foi realizada a partir de pacientes
soropositivos para HIV-1, sob tratamento antirretroviral ou não, atendidos na Unidade do
Hospital das Clínicas de Ribeirão Preto da Faculdade de Medicina de Ribeirão Preto (HC-
FMRP-USP). Como critérios de inclusão, foram selecionados pacientes HIV-1+ virgens de
tratamento (n=28) e pacientes HIV-1+ em uso regular da terapia antirretroviral por no mínimo
6 meses (n=29). E como critérios de exclusão, foram excluídos do estudo todos os pacientes
menores de 18 anos e maiores de 65 anos de idade, assim como os que não consentiram em
participar da pesquisa.
Os pacientes foram abordados no momento da consulta pelo clínico e convidados a
participar voluntariamente do estudo, registrando a sua autorização por meio da assinatura do
termo de consentimento livre e esclarecido aprovado pelo Comitê de Ética e Pesquisa do HC-
FMRP-USP (Anexo V).
Como controle, obtivemos amostras de sangue de indivíduos com sorologia negativa
para HIV (n=30) com idade entre 18 e 65 anos atendidos regularmente no Hemocentro de
Ribeirão Preto e concordantes com o termo de consentimento livre e esclarecido aprovado
pelo Comitê de Ética e Pesquisa do HC-FMRP-USP (Anexo IV).
4.2. Obtenção de células mononucleares do sangue periférico
Foram coletadas amostras de sangue total (40 mL cada) dos indivíduos via punção
venosa em tubo contendo heparina sódica (Vacutainer®; Becton, Dickinson and Company).
Em seguida, procedeu-se à separação do plasma após centrifugação a 400 x g por 10 min a
20°C. Homogeneizou-se o sangue e diluiu-se em tampão fosfato salina (PBS) pH 7,4 na
proporção de 1:2. As amostras de sangue diluídas foram aplicadas cuidadosamente sobre a
solução de Ficoll-PaqueTM PLUS, d=1,078 g/mL (GE Healthcare Bio-Sciences AB, Uppsala,
Sweden). Logo após, as amostras foram centrifugadas a 640 x g por 30 minutos a 20°C para
separação da camada de células mononucleares. As células presentes na interfase do gradiente

SOARES, L.S. Material e Métodos
30
de separação foram cuidadosamente coletadas, transferidas para um novo tubo e lavadas duas
vezes pela adição de 50 mL de PBS seguida de centrifugação a 400 x g por 10 min a 20°C.
Por fim, ressuspendeu-se o precipitado celular em 5 mL de meio RPMI-1640 incompleto e
procedeu-se à contagem das células em câmara de Neubauer, utilizando o corante azul de
Tripan para análise da viabilidade celular.
4.3. Ensaio imunoenzimático para quantificação das moléculas CD14 e CD163 solúveis
no plasma
As amostras de plasma separadas no dia da coleta e armazenadas a -20°C foram
descongeladas e centrifugadas a 1000 x g por 10 min a 10°C. Em seguida, utilizou-se 100 µL
da amostra previamente diluída para dosagem por ELISA das moléculas CD14 (diluição de
1000x) e CD163 (diluição de 100x) solúveis com a utilização de kit específico (DuoSet
ELISA Development System, R&D). As concentrações foram determinadas empregando-se
anticorpos de captura, citocinas-padrão e anticorpos associados à biotina e amplificados com
estrepto-avidina-peroxidase. Como substrato foi utilizado tetrametilbenzidina (TMB) e a
reação bloqueada adicionando-se ácido sulfúrico 1M aos poços. A leitura das amostras foi
realizada em espectrofotômetro a 450 nm.
4.4. Quantificação de citocinas e quimiocinas no plasma
Para determinação da quantidade de citocinas e quimiocinas presentes no plasma foi
empregada a plataforma multiplex customizada (MILLIPLEX MAP magnetic bead-based
multi-analyte panels, Cat #HCYTOMAG-60K, Merck Millipore®) e analisada em leitor de
placas próprio (Luminex Multiplexing Instrument - EMD Millipore, Luminex Corporation,
Austin, TX, USA). Segundo instruções do fabricante, foram detectadas as seguintes citocinas:
IFN-γ, IL-10, IL-1β, IL-6, TNF-α, IFN-α2, IL-12p70, IL-17A, IL-4, IL-5; e as quimiocinas:
MCP-1, RANTES, IP-10, MIP-1α, MIP-1β, e GM-CSF. As amostras foram adicionadas às
placas específicas para o ensaio e diluídas em tampão fornecido pelo kit e, após a incubação
com microesferas magnéticas fluorescentes recobertas por anticorpos de captura específicos
para as citocinas listadas, realizou-se a leitura das amostras no equipamento. Os dados
adquiridos foram analisados por meio do software Milliplex Analyst v3.5 (Millipore;
VigeneTech Inc., Boston, EUA) utilizando uma curva logística de três parâmetros.

SOARES, L.S. Material e Métodos
31
4.5. Análise fenotípica das células NK e suas subpopulações por citometria de fluxo
A partir de 5x105 células mononucleares do sangue periférico realizou-se a marcação de
moléculas de superfície para caracterização das células NK e suas subpopulações. Para tanto,
as células foram resssuspendidas em 100µL de PBS contendo 2% de SBF e a esse volume foi
adicionado 20µL da solução de anticorpos previamente diluída. Foram utilizados os seguintes
anticorpos: CD3 PerCPCy5.5 (clone SK7), CD56 APC (clone NCAM 16.2), CD16 PE-Cy7
(clone 3G8), NKp46 PE (clone 9E2), NKB1 FITC (clone DX9), NKAT2 PE (clone DX27) e
CD158a FITC (clone HP-3E4), adquiridos da BD Biosciences. Os anticorpos foram titulados
para utilização da melhor diluição no experimento, sendo os volumes padronizados para cada
um de: 5 µL (CD3), 3µL (CD56, NKp46, NKB1, NKAT2 e CD158a) e 2µL (CD16). A
marcação foi realizada por 25 min a 4°C, no escuro. Após o tempo de incubação, adicionou-
se 2 mL de PBS contendo 2% de Soro Bovino Fetal (SBF) em cada tubo. Os tubos foram
centrifugados por 10 min a 400 x g, 4°C e o sobrenadante foi descartado. Em seguida, as
células foram fixadas em 200 µL de PBS com 1% de formaldeído e mantidas a 4°C até o
momento da aquisição. Foram adquiridos 100.000 eventos de cada amostra no citômetro
FACS Canto II (BD Biosciences, San Diego, EUA) e a análise realizada no software FlowJo
versão 7.6.5.
4.6. Separação das células NK por esferas magnéticas
Após obtenção das células mononucleares do sangue periférico, a suspensão celular foi
utilizada como amostra para separação magnética das células NK. Para tanto, após contagem,
as células mononucleares foram ressuspendidas em solução tampão de PBS livre de Ca2+ e
Mg2+ contendo 2mM de EDTA e 0,5% de SBF na proporção de 40 µL para cada 107 células
totais. Em seguida, acrescentou-se 10 µL da solução de anticorpos humanos monoclonais
anti-CD56 acoplados a esferas magnéticas (MACS, Miltenyi Biotec Inc., Auburn CA, USA)
para cada 107 células totais e incubou-se a 4°C por 15 min. A seguir, as células foram lavadas
pela adição de 2 mL do tampão descrito acima e centrifugadas a 400 x g por 10min.
Descartou-se o sobrenadante, ressuspendeu-se o precipitado em 500 µL do tampão para cada
108 células totais e procedeu-se à separação magnética (seleção positiva), segundo instruções
do fabricante utilizando as colunas LS (Miltenyi Biotec Inc., Auburn CA, USA).
As células NK obtidas foram contadas em câmara de Neubauer para análise do
rendimento do método de separação e utilizadas nos ensaios subsequentes. Para verificar a

SOARES, L.S. Material e Métodos
32
pureza da separação, foi realizada citometria de fluxo empregando-se os anticorpos CD3
PerCPCy5.5 (clone SK7) e CD56 APC (clone NCAM 16.2). Na figura 2 abaixo está
demonstrada pureza obtida de um dos experimentos realizados. Observa-se que as células NK
obtidas após separação correspondiam a 74,3% (Q1) das células dentro da população de
linfócitos.
Figura 2. Análise da pureza das células NK obtidas após separação magnética.
4.7. Análise fenotípica das células NK por PCR em Tempo Real
Após obtenção das células NK, o precipitado celular foi armazenado a -80°C até
posterior extração do RNA total utilizando-se o reagente TRIzol (Ambion®) segundo
instruções do fabricante. O RNA extraído foi quantificado no fluorímetro Qubit 2.0 e
estocado a -80ºC.
O RNA obtido foi tratado com Turbo DNase (Ambion®) segundo o protocolo do
fabricante. O DNA complementar foi sintetizado utilizando-se o High Capacity cDNA
Reverse Transcription Kit (Applied Biosystems). A partir de 50 ng da amostra de RNA
convertido em cDNA, foi realizada a reação em cadeia da polimerase em tempo real
empregando-se a sonda TaqMan® Fast da Applied Biosystems. As moléculas quantificadas
por PCR em tempo real foram: CD69 (Hs00934033_m1), Lamp1 (Hs00174766_m1), NCR3
(Hs00394809_m1), KLRC1 (Hs00970274_m1). Para análise da expressão gênica, empregou-

SOARES, L.S. Material e Métodos
33
se o método comparativo ou 2-ΔΔ
Ct [80] e utilizou-se os seguintes genes constitutivos: GAPDH
e β-actina.
4.8. Avaliação funcional das células NK por ensaio de citotoxicidade
4.8.1. Cultivo das células alvo
Para realização do ensaio de citotoxicidade, utilizou-se como célula alvo a linhagem
celular K-562 (ATCC N° CCL-243™). Essa linhagem celular foi gentilmente cedida pela
Profa. Dra. Fabíola Attié de Castro da FCFRP/USP. As células foram cultivadas em meio
RPMI-1640 acrescido de 10% de SBF, em estufa úmida a 37°C e 5% de CO2. No dia anterior
ao ensaio, realizou-se o subcultivo destas células em placa de 24 poços contendo 5 x 105
células/mL por poço.
4.8.2. Ensaio de citotoxicidade
O ensaio de citotoxicidade foi realizado utilizando-se o “CytoTox 96 Non-Radioactive
Cytotoxicity Assay” (Promega), segundo instruções do fabricante. Para isso, células NK
purificadas (efetoras) foram cultivadas com as células alvo (células K-562) em placas de 96
poços de fundo U sob diferentes razões entre células efetoras e células alvo (20:1; 10:1 e 5:1).
O ensaio foi realizado em triplicata num volume total de 100 µL/poço. A porcentagem de
citotoxicidade foi calculada a partir da quantificação da enzima lactato desidrogenase (LDH)
liberada no meio de cultura após lise celular, segundo a fórmula:
%citotoxicidade = Experimental − Efetora espontânea − Alvo Espontânea
Alvo máxima − Alvo espontânea x 100
4.8.3. Quantificação de citocinas e quimiocinas em sobrenadante de cultura
O sobrenadante da cultura do ensaio de citotoxicidade descrito acima foi coletado para
quantificação das citocinas e quimiocinas. Para tanto, utilizou-se o “Human
Cytokine/Chemokine Magnetic Bead Panel Kit” (Cat #HCYTOMAG-60K, Millipore) e
realizou-se o ensaio segundo instruções do fabricante. Foram detectadas as seguintes
citocinas: IFN-γ, IL-10, IL-1β, IL-6, TNF-α, IFN-α2, IL-12p70, IL-17A, IL-4, IL-5; e as
quimiocinas: MCP-1, RANTES, IP-10, MIP-1α, MIP-1β, e GM-CSF. A fluorescência foi

SOARES, L.S. Material e Métodos
34
quantificada no equipamento Magpix (Luminex Corporation) segundo instruções do
fabricante e os resultados foram analisados no software Analyst.
4.8.4. Quantificação da produção de espécies reativas de oxigênio por
quimioluminescência
A produção de espécies reativas de oxigênio (ROS) foi avaliada por meio de
quimioluminescência amplificada por Luminol (Sigma). As células NK purificadas foram
divididas em dois tubos, sendo que cada um recebeu 1x105 células. Foram adicionados aos
tubos 10% do volume total de Luminol ([ ] final de 100 µM) para realização da leitura.
Utilizou-se como controle positivo da produção de ROS, a estimulação das células com PMA
a 10-7M. O controle de produção espontânea (negativo) foi realizado sem a adição de
estímulo. A leitura foi realizada em luminômetro (AutLumat LB 953 Multi-Tube
Luminometer from Berthold) no Laboratório de Imunologia e Citologia dos Fluidos
Biológicos da FCFRP/USP, sob responsabilidade da Profa. Dra. Cleni Mara Marzocchi
Machado.
4.9. Avaliação de mudanças epigenéticas relacionadas às células NK
4.9.1. Determinação do padrão de metilação global do DNA cromossômico
O DNA genômico foi extraído das células NK purificadas utilizando-se o “PureLink
Genomic DNA kit” (Invitrogen, Life Technologies), segundo instruções do fabricante. Em
seguida, o DNA foi quantificado no fluorímetro Qubit 2.0. Foram utilizadas 50 ng da amostra
de DNA genômico de cada indivíduo para quantificação do perfil de metilação global relativa
utilizando-se o “Imprint Methylated DNA Quantification kit” (Sigma-Aldrich), conforme
instruções do fabricante. Os cálculos de porcentagem relativa da metilação global foram
realizados considerando-se o controle positivo de DNA 100% metilado fornecido pelo kit.
4.9.2. Avaliação do padrão de metilação gene-específico
Após análise do fenótipo de células NK por citometria de fluxo e PCR em Tempo Real,
definiu-se a molécula KIR2DL2 como alvo para estudo da metilação gene-específico. A

SOARES, L.S. Material e Métodos
35
estratégica utilizada foi clonagem e posterior sequenciamento do DNA pelo método de
Sanger, conforme descrito por Gao et al. em 2009 [81], com modificações.
4.9.2.1.Análise da sequência genômica referente ao gene KIR2DL2
Inicialmente, realizou-se a amplificação da região analisada por Gao et al., antes da
modificação do DNA por bissulfito, pela técnica de PCR. A reação foi feita utilizando-se 1
µL da amostra de DNA genômico previamente extraído (50 ng/µL) e o seguintes reagentes:
0,1 µL de Taq DNA Polymerase (Invitrogen); 1,25 µL de tampão; 0,25 µL de MgCl2
(50mM); 0,25 µL de cada primer a 10µM (2DL2-gen F e 2DL2-gen R); 0,25 µL de dNTP
(10mM) e 9,65 µL de H2O Nuclease-Free. As amostras foram submetidas a reação de PCR no
termociclador no Veriti Thermal Cycler (ThermoFisher Scientific) com a seguinte
programação: 94°C por 5 minutos; 35 ciclos de 94°C por 30 segundos, 56°C por 30 segundos
e 72°C por 30 segundos; e extensão final a 72°C por 10 minutos. Após amplificação,
analisou-se o resultado em gel de agarose a 1,5% (Apêndice -Figura suplementar 01) corado
com SYBR Safe DNA Gel Stain (Thermo Fisher Scientific). As respectivas sequências de
cada primer estão demonstradas abaixo:
2DL2-gen F: 5’ – GTACTAGAAACTCAAGCAGG – 3’ 2DL2-gen R: 5’ – ACACGCCATGCTGACGACCA – 3’
Em seguida, extraiu-se os produtos de PCR a partir do gel de agarose utilizando-se o
“Illustra GFX PCR DNA and Gel Band Purification Kit” (Ge Heatlhcare), segundo instruções
do fabricante. O DNA purificado foi quantificado no fluorímetro Qubit 2.0 e analisado em gel
de agarose a 1,5% corado com SYBR Safe DNA Gel Stain (Thermo Fisher Scientific).
A partir de 2 µL da amostra de DNA purificado, realizou-se a reação de
sequenciamento do DNA pelo método de Sanger utilizando o “BigDye Terminator v3.1 Cycle
Sequencing Kit” (ThermoFisher Scientific). A reação foi feita no equipamento ABI Prism
3130 Genetic Analyzer do Núcleo de Serviços em Biotecnologia da Fundação Hemocentro de
Ribeirão Preto. Os cromatogramas e as sequências foram obtidas com auxílio do programa
FinchTV, comparadas com a sequência referência depositada no banco de dados GenBank
(sequência referência NM_014219.2) e alinhadas utilizando-se o programa MultAlin [82] e
Clustal.

SOARES, L.S. Material e Métodos
36
4.9.2.2. Quantificação da metilação do gene KIR2DL2 após modificação por bissulfito
Após avaliação in silico das sequências genômicas relativas à região analisada do gene
KIR2DL2 de cada amostra, realizou-se a reação de modificação por bissulfito utilizando-se o
EZ DNA Methylation Gold Kit (Zymo Research), segundo instruções do fabricante. Utilizou-
se 300 ng de DNA genômico total para cada reação e o DNA modificado foi eluído no
volume final de 20 µL.
Após a modificação por bissulfito, amplificou-se a região específica por meio de PCR
empregando-se os seguintes reagentes: 0,05 µL de Platinum Taq DNA Polymerase High
Fidelity (Invitrogen, 5U/µL); 1,25 µL de tampão; 0,5 µL de MgSO4 (50mM); 0,25 µL de
dNTP (10mM); 0,25 µL de cada primer (2DL2-bis Fa/Ra) e 8,95 µL de H2O Nuclease-Free.
Utilizou-se 0,5 µL da amostra de DNA modificado por bissulfito em cada reação. As
configurações do termociclador foram as seguintes: 94°C por 1 minuto e 30 segundos; 35
ciclos de 94°C por 45 segundos, 53°C por 45 segundos e 68°C por 45 segundos; e extensão
final a 68°C por 1 minuto e 30 segundos. Para a clonagem do fragmento amplificado acima,
realizou-se a PCR aumentando a quantidade proporcional de cada reagente para o volume
final de 100 µL. O tamanho de 452 pb do fragmento amplificado foi verificado em gel de
agarose a 1,5% (Apêndice - Figura suplementar 02) corado com SYBR Safe DNA Gel Stain
(Thermo Fisher Scientific). Os primers foram desenhados com auxílio do software on line
MethPrimer [83]. As sequências dos respectivos primers estão demonstradas abaixo:
2DL2-bis Fa: 5’ – GAAGAAGAGTTTGAATTTTAGA – 3’
2DL2-bis Ra: 5’ – CCTATATCTCCAACTCTAAACC – 3’
A reação de clonagem foi realizada utilizando-se o TOPO TA Cloning kit (pCR 2.1 –
TOPO vector, ThermoFisher Scientific). Para tanto, inicialmente adicionou-se a cauda poli-A
nas extremidades do fragmento amplificado por meio da incubação do produto de PCR
amplificado acima com a Taq DNA Polymerase (Invitrogen) a 72°C por 10 minutos. Em
seguida, os produtos da PCR foram purificados utilizando-se o kit “Wizard SV Gel and PCR
Clean-up System” (Promega). O DNA foi eluído em 20 µL e quantificado no fluorímetro
Qubit 2.0. Para reação de clonagem, incubou-se 1 µL do DNA (7 ng/µL) com 1 µL do vetor
TOPO (10 ng/µL), 1µL da solução de sal e 3 µL de H2O Nuclease-Free por 30 minutos a
temperatura ambiente.

SOARES, L.S. Material e Métodos
37
Todo o volume da ligação acima foi utilizado na transformação de células de E. coli
Top10 competentes, as quais foram plaqueadas em meio LB ágar suplementado com
canamicina, visto que o vetor TOPO pCR 2.1 apresenta o gene de resistência a esse
antibiótico. A seleção dos clones recombinantes foi feita a partir da PCR de colônias, na qual
o molde consistiu de uma solução de bactérias “palitadas” a partir da placa de cultura,
dissolvidas em 20 µL de água estéril Nuclease-free e fervidas por 5 minutos. Para a reação,
utilizou-se: 0,1 µL de Taq DNA Polymerase (Invitrogen); 1,25 µL de tampão; 0,25 µL de
MgCl2 (50mM); 0,25 µL de cada primer a 10µM (2DL2-bis Fa/Ra); 0,25 µL de dNTP
(10mM) e 9,65 µL de H2O Nuclease-Free.
O produto da PCR foi analisado em gel de agarose a 1,5% e a partir desse resultado,
extraiu-se os plasmídios das colônias que apresentaram banda de amplificação de 452 pb
(Apêndice - Figura suplementar 03), relativa ao tamanho do fragmento analisado por Gao et
al. O DNA obtido foi quantificado no fluorímetro Qubit 2.0, diluído a 105 ng/ µL e enviado
para sequenciamento no Centro de Pesquisas sobre o Genoma Humano e Células-Tronco da
USP/SP, utilizando nas reações o primer universal M13 Forward (Life Technologies).
As sequências foram obtidas com auxílio do programa FinchTV e a qualidade das bases
analisadas no programa computacional PHRED [84]. Os pontos de metilação foram alinhados
pelo programa Clustal Omega (EMBL-EBI). A região amplificada compreendeu o estudo do
percentual de metilação de 19 dinucleotídeos CpG como destacado na figura 3 abaixo. A
comparação entre as sequências tratadas com bissulfito e as de referência foram realizadas
com o auxílio do programa QUMA – Quantification tool for Methylation Analysis [85]. As
análises de bioinformática foram realizadas com o auxílio da Profa. Dra. Silvana Giuliatti do
Departamento de Genética da FMRP/USP, Dr. Murilo Racy Soares e Dra. Maria Gabriela
Fontanetti Rodrigues.
Figura 3. Sequência genômica analisada do KIR2DL2 quanto ao perfil de metilação. Estão destacados em amarelo os 19 dinucleotídeos CpG analisados, compreendidos entre as regiões -271 a +82. As regiões de pareamento dos primers utilizados estão destacadas em vermelho.

SOARES, L.S. Material e Métodos
38
4.9.3. Quantificação da expressão de enzimas que promovem as alterações epigenéticas
Extraiu-se o RNA total a partir das amostras de células NK purificadas dos diferentes
grupos, conforme descrito nas seções anteriores, utilizando-se o reagente TRIzol (Ambion®).
A partir de 30 ng da amostra de RNA convertido em cDNA foi realizada a reação em cadeia
da polimerase em tempo real empregando-se o RT2 Profiler PCR Array customizado pela
Qiagen para análise da expressão das enzimas envolvidas na ativação da transcrição gênica
(ATF2, HAT1, KAT2B, KAT5, CARM1, PRMT5, SMYD3, SUV39H1, ASH1L, KMT2C,
KMT2E, SETD1B, KDM5B, KDM6B) utilizando-se o RT2 SYBR Green PCR Master Mix,
segundo instruções do fabricante. Para cálculo da expressão gênica, empregou-se os seguintes
genes constitutivos: GAPDH e β-actina.
E para determinação da expressão das enzimas envolvidas na repressão da transcrição
gênica, utilizou-se o “TaqMan→ Array Human DNA Methylation & Transcriptional
Repression kit” (Applied Biosystems, Foster City, EUA). Foram analisadas as seguintes
enzimas: DNMT1, DNMT3A, DNMT3B, HDAC1, HDAC2, HDAC3, HDAC4, HDAC5,
HDAC6, HDAC7, HDAC8, HDAC9, HDAC10, HDAC11, MBD2, MBD3 e MECP2. Para
cálculo da expressão gênica, empregou-se os seguintes genes constitutivos: GAPDH, ACTB,
18S, HPRT1, GUSB, RPLPO, HMBS e B2M.
A partir dos resultados de Ct (cycle treshold) obtidos acima, calculou-se os níveis de
expressão relativa utilizando-se o método comparativo ou 2-ΔΔ
Ct [80].
4.9.4. Imunoprecipitação da cromatina
A partir de 1x106 células NK purificadas foi realizada a fixação da estrutura DNA-
proteína com 55 µL de formaldeído a 18,5% em 1 mL de PBS, durante 10 minutos a 37°C.
Em seguida, adicionou-se 100 µL por tubo da solução de glicina concentrada 10 vezes
([]FINAL= 125mM) e incubou-se a temperatura ambiente por 5 minutos. Centrifugou-se a
amostra a 3400 x g por 5 minutos a 4°C e descartou-se o sobrenadante. Realizou-se duas
lavagens seguidas do precipitado celular com 500 µL de PBS acrescido da solução de
inibidores de protease (Sigma) diluída 1:100 e centrifugou-se a 3400 x g por 5 minutos a 4°C.
Ao final, descartou-se o sobrenadante e armazenou-se o precipitado celular a -80°C.
A lise das células foi realizada pela adição de 50 µL de tampão de lise (1% SDS, 10
mM EDTA, 50 mM Tris-HCl pH 8,1) e 150 µL de tampão de diluição (0,01% SDS; 1,1%

SOARES, L.S. Material e Métodos
39
Triton X-100; 1,2 mM EDTA; 16,7 mM Tris-HCl pH8,1 e 167 mM NaCl) acrescidos dos
inibidores de protease (1:100). Homogeneizou-se a amostra com auxílio do vórtex e incubou-
se no gelo por 10 minutos. Cada amostra foi dividida em dois tubos contendo 100 µL cada
para ser realizada a etapa de sonicação utilizando-se o Bioruptor Pico Sonication System
(Diagenode). Conforme padronização prévia (Apêndice - Figura suplementar 04), realizou-se
11 ciclos de 15 segundos-on e 90 segundos-off a 4°C. Em seguida, centrifugou-se a amostra a
12000 x g por 20 minutos a 4°C e transferiu-se o sobrenadante para um novo tubo.
Após a centrifugação, o sobrenadante contendo a cromatina foi diluído com tampão de
diluição (200 µL para cada anticorpo), e uma alíquota (5% em volume) foi reservada para
indicar a quantidade de DNA de cada amostra (input). As frações de cromatina restantes
foram submetidas à imunoprecipitação e incubadas com os anticorpos específicos, overnight a
4°C, com suave rotação. Foram utilizados os seguintes anticorpos: normal mouse IgG (1µg,
Millipore), anti-histone H3K27me3 (4µg, Abcam) e anti-histone H3K4me3 (4µg, Abcam).
Após incubação com os anticorpos específicos, adicionou-se 50 µL da solução de
esferas de agarose com DNA de esperma de salmão por tubo e incubou-se por 1 hora a 4°C
sob rotação. Realizou-se as lavagens do precipitado com os diferentes tampões (low-salt,
high-salt, LiCl e TE) por 5 minutos a 4°C, sob agitação, seguida de centrifugação a 100 x g
por 1 minuto. Os complexos imunes foram eluídos pela adição de tampão de eluição e a
ligação DNA-proteína foi revertida após incubação a 62°C por 2 horas com proteinase K. O
DNA livre foi purificado por protocolo padrão de precipitação por fenol/clorofórmio/álcool
isoamílico e quantificado no fluorímetro Qubit 2.0.
O DNA obtido após imunoprecipitação da cromatina foi utilizado como amostra na
PCR em Tempo Real para quantificação de regiões específicas das moléculas de IFN-γ e
TNF-α, previamente descritas como alvo de modificações de histona [86, 87]. Foram
utilizados 2 µL da amostra de DNA imunoprecipitado em cada reação, empregando-se o
reagente SYBR Green Master Mix (Life Technologies), segundo instruções do fabricante. A
reação foi realizada no aparelho StepOnePlus Real-Time PCR System (Life Technologies)
conforme as seguintes configurações: 94°C por 10 minutos, 50 ciclos de 94°C por 20
segundos e 60°C por 1 minuto, seguida da curva de dissociação ao final. A análise foi
realizada segundo a fórmula: 100*2^(input ajustado – Ct(IP)), onde o input ajustado refere-se
ao valor de input corrigido pela diluição inicial e o IP, à amostra de cada anticorpo
empregado.

SOARES, L.S. Material e Métodos
40
4.10. Análise estatística
Os dados obtidos foram analisados utilizando-se o programa GraphPad Prism 6.0 (San
Diego, CA, EUA) e as diferenças foram consideradas estatisticamente significativas quando p
< 0,05. Considerando a natureza não paramétrica do conjunto de dados, verificado pelo teste
de Kolmogorov-Smirnov, empregou-se o teste de Mann-Whitney para comparações entre dois
grupos ou o teste de Kruskal-Wallis, para comparações múltiplas.
Com o objetivo de avaliar a associação entre duas variáveis distintas, realizou-se o teste
de correlação de Spearman. As correlações significativas foram compiladas no software
Cytoscape (versão 3.2.1; http://www.cytoscape.org). As redes foram construídas usando o
layout de círculo e as conexões entre as variáveis designadas como linhas, com espessura de
acordo com o índice de correlação (r). As correlações foram classificadas em negativa (r < 0),
fraca (r ≤ 0,35), moderada (0,36 ≥ r ≤ 0,67) e forte (r ≥ 0,68), de acordo com estudos prévios
[88].
Para análise do perfil de citocinas e quimiocinas presentes no plasma ou produzidas
pelas células NK, realizou-se o cálculo da mediana global de cada molécula, o qual foi
utilizada como ponto de corte na classificação dos indivíduos em altos ou baixos produtores
para cada citocina ou quimiocina. Esses dados foram utilizados para gerar diagramas, nos
quais os indivíduos classificados como altos produtores foram destacados e sua frequência
determinada para cada indicador. A partir desses resultados, construiu-se gráficos de radar
para demonstrar as respectivas frequências dos indivíduos com níveis elevados para cada
molécula analisada.

SOARES, L.S.
41
5. Resultados

SOARES, L.S. Resultados
42
5. Resultados
5.1. Características da população em estudo
Com o objetivo de caracterizar fenotipicamente e funcionalmente as células NK de
pacientes infectados com HIV-1 submetidos ou não ao tratamento antirretroviral, foram
obtidas 28 amostras de sangue periférico dos indivíduos HIV-1+ virgens de tratamento (grupo
HIV-1+), 29 amostras daqueles em uso regular dos antirretrovirais (grupo TARV) e 30
amostras do grupo controle, como descrito em Materiais e Métodos.
Na tabela 1 estão resumidos os principais dados de cada grupo considerando os
parâmetros de gênero, idade, contagem de linfócitos T CD4+, carga viral, tempo aproximado
de infecção, assim como o esquema terapêutico empregado nos pacientes do grupo TARV e o
respectivo tempo de tratamento. Os três grupos apresentam características de idade e
distribuição de gêneros semelhantes, com a observação de que o grupo TARV compreende
indivíduos de idade um pouco mais avançada, sendo a mediana da idade de 46 anos, o que
reflete as características dessa população em estudo.
Desde dezembro de 2013, o Ministério da Saúde recomenda o início imediato da terapia
antirretroviral em pessoas vivendo com HIV/AIDS, independente da contagem de linfócitos T
CD4+ e estágio clínico da doença, na perspectiva de diminuir a transmissão do HIV, dentre
outros fatores. Dessa forma, o grupo HIV-1+ compreendeu indivíduos recentemente
infectados, sendo a mediana do tempo de infecção de 5,3 meses quando comparado ao grupo
TARV com 156,4 meses. O tempo aproximado de infecção foi determinado a partir do
primeiro resultado positivo para o teste de ELISA. Além disso, os pacientes do grupo HIV-1+
apresentavam menor contagem de linfócitos T CD4+, sendo a mediana de 415,5 células/mm3,
em relação ao grupo TARV com mediana de 705 células/mm3.
Os indivíduos do grupo TARV incluídos no estudo possuíam carga viral indetectável
(<40 cópias/mL) e estavam sob determinado esquema terapêutico a no mínimo seis meses,
sendo a mediana do tempo de tratamento atual de 72,3 meses. Os pacientes desse grupo foram
subdivididos de acordo com a terapia a qual foram submetidos, sendo a primeira linha de
tratamento, preconizada pelo Ministério da Saúde, composta por dois Inibidores da
Transcriptase Reversa Análogos Nucleosídeos ou Nucleotídeos (ITRN/ITRNt) associados a
um Inibidor da Transcriptase Reversa Não Análogo de Nucleosídeo (ITRNN). Na segunda
linha de tratamento, há substituição do ITRNN por um Inibidor de Protease (IP) e esta terapia
é selecionada baseada em fatores clínicos inerentes ao paciente.

SOARES, L.S. Resultados
43
Tabela 1. Características da população em estudo. Controle HIV-1+ TARV n 30 28 29
Idade 32,0 33,5 46,0
(27,5 - 54) (23 - 43,3) (40,5 - 52,5) Gênero M = 21 M = 21 M = 14
F = 9 F = 7 F = 15
Esquema terapêutico - - ITRN + ITRNN (n = 17)
ITRN + IP/r (n = 12)
Carga viral (cópias/mL) - 21638 < 40
(4272 - 117414)
Células T CD4+ (céls/mm3) não determinado 415,5 705
(214,5 - 640,3) (597,5 - 932)
Tempo aproximado de infecção (meses) - 5,3 156,4
(2 - 29,5) (73,2 - 193,6)
Tempo de tratamento (meses) - - 72,3
(27 - 114,9) Os dados referentes a idade, carga viral, células T CD4+, tempo de infecção e de tratamento estão expressos como mediana (percentil 25-75).

SOARES, L.S. Resultados
44
5.2. Análise de moléculas relacionadas à progressão da doença
Inicialmente, com o intuito de caracterizar a progressão da infecção por HIV-1,
realizou-se a quantificação das citocinas e quimiocinas presentes no plasma, assim como das
moléculas CD14 e CD163 solúveis. Para tanto, após obtenção das amostras de sangue
periférico dos indivíduos dos grupos controle, HIV-1+ virgens de tratamento e TARV,
procedeu-se à separação do plasma, o qual foi armazenado à -20°C e posteriormente
descongelado para dosagem das citocinas e quimiocinas por plataforma Multiplex, e das
moléculas CD14 e CD163 por ELISA, como descrito na seção de materiais e métodos.
5.2.1 Quantificação das moléculas CD14 e CD163 solúveis no plasma
A quantidade de CD163 solúvel no plasma dos indivíduos infectados com HIV-1 e
virgens de tratamento foi significativamente maior quando comparada aos grupos controle e
TARV. Quanto à molécula sCD14, os grupos HIV-1+ e TARV apresentaram aumento
significativo em relação aos indivíduos controle, sendo a mediana do grupo TARV maior que
o HIV-1+, embora não estatisticamente significativa (Figura 4).

SOARES, L.S. Resultados
45
Figura 4. Pacientes HIV-1+ apresentam níveis plasmáticos elevados de sCD163 e sCD14 plasmáticos. Quantificação das moléculas CD14 e CD163 solúveis no plasma dos indivíduos controle, HIV-1+ virgens de tratamento e TARV. O plasma foi obtido do sangue periférico e a análise quantitativa foi realizada por meio de ensaio imunoenzimático (ELISA). As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05; ***p<0,001; ****p<0,0001.

SOARES, L.S. Resultados
46
5.2.2. Quantificação de citocinas e quimiocinas do plasma
Na figura 5, estão demonstrados os níveis de citocinas pró-inflamatórias, aquelas mais
relacionadas à resposta TH2 (IL-4 e IL-5) e reguladora (IL-10) quantificadas no plasma dos
indivíduos dos diferentes grupos.
No grupo HIV-1+, observou-se aumento significativo de TNF-α quando comparado aos
grupos controle e TARV. Os níveis plasmáticos da citocina IL-10 também estavam
aumentados neste grupo, sendo este aumento estatisticamente significativo em relação aos
pacientes HIV-1+ sob tratamento antirretroviral (grupo TARV). As demais citocinas
quantificadas não apresentaram diferenças significativas entre os grupos analisados (Figura
5).
Quanto aos níveis plasmáticos das diferentes quimiocinas analisadas, verificou-se maior
quantidade de IP-10 no grupo HIV-1+, em relação aos grupos controle e TARV. Os
indivíduos infectados e tratados também apresentaram aumento significativo desta quimiocina
quando comparado aos controles. Por outro lado, a molécula GM-CSF apresentou diminuição
significativa no grupo TARV. E por fim, a quimiocina CCL5, envolvida no recrutamento de
leucócitos para os sítios inflamatórios e que também atua como fator supressor do HIV,
estava diminuída no grupo HIV-1+, sendo essa diminuição estatisticamente significativa em
relação ao grupo TARV, o qual parece restaurar os níveis dessa quimiocina próximos aos do
controle (Figura 6).

SOARES, L.S. Resultados
47
Figura 5. Quantificação de citocinas no plasma dos indivíduos controle, HIV-1+ virgens de tratamento e TARV. O plasma foi obtido do sangue periférico e a análise quantitativa foi realizada utilizando-se microesferas magnéticas na plataforma Magpix®. O limite de detecção de cada citocina foi <5pg/mL. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo e cada ponto representa um individuo. *p<0,05; **p<0,01 e ****p<0,0001.
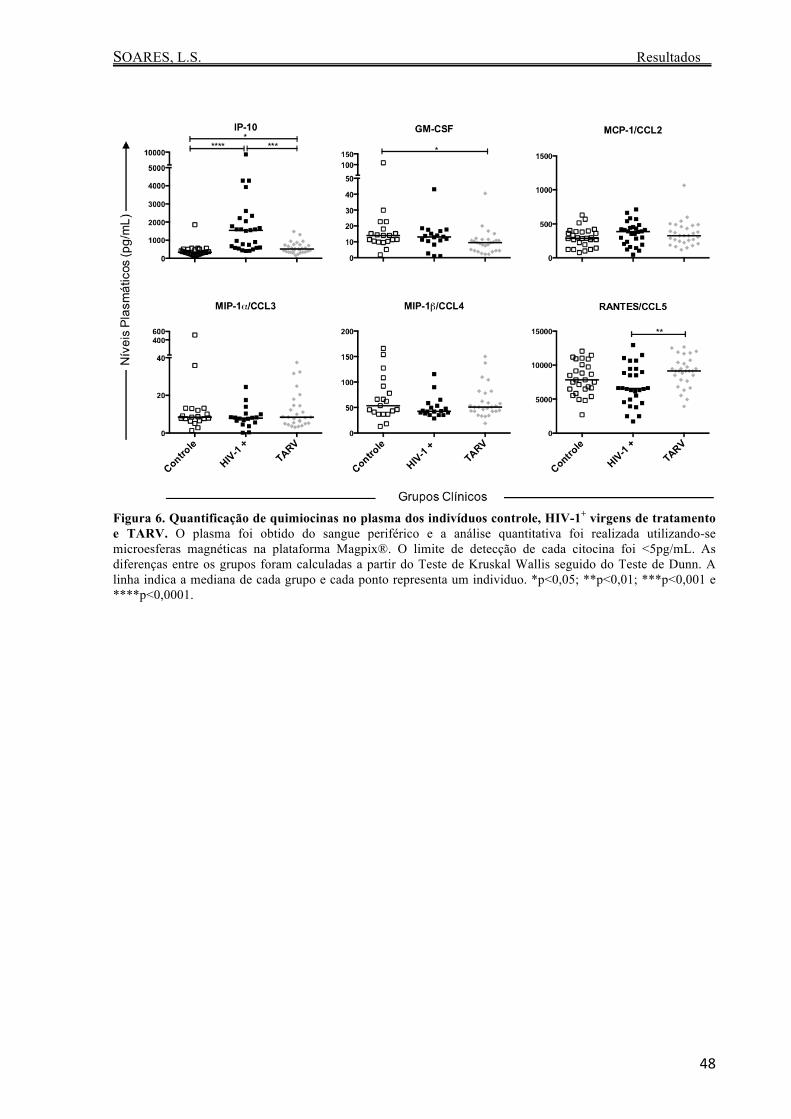
SOARES, L.S. Resultados
48
Figura 6. Quantificação de quimiocinas no plasma dos indivíduos controle, HIV-1+ virgens de tratamento e TARV. O plasma foi obtido do sangue periférico e a análise quantitativa foi realizada utilizando-se microesferas magnéticas na plataforma Magpix®. O limite de detecção de cada citocina foi <5pg/mL. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo e cada ponto representa um individuo. *p<0,05; **p<0,01; ***p<0,001 e ****p<0,0001.

SOARES, L.S. Resultados
49
5.2.3. Correlação entre os marcadores de ativação do plasma
Para compreender melhor a progressão da infecção pelo HIV-1+, além de analisarmos
os níveis plasmáticos das diferentes moléculas, faz-se necessário o estudo da correlação entre
essas variáveis e os parâmetros empregados na rotina clínica (contagem de linfócitos T CD4+
e carga viral). Dessa forma, a partir dos resultados obtidos por meio dessa análise no grupo
HIV-1+, nota-se correlação negativa entre a contagem de linfócitos T CD4+ e a viremia, ou
seja, à medida que ocorria aumento da carga viral no individuo infectado, o número de células
T CD4+ diminuía (Figura 7).
Quando correlacionamos a viremia com as citocinas IL-10 e TNF-α, e a quimiocina IP-
10, as quais estavam estatisticamente aumentadas no grupo HIV-1+, observou-se forte
correlação positiva, confirmando em nossos dados a relação dessas moléculas como
indicadores da progressão da doença.

SOARES, L.S. Resultados
50
Figura 7. Correlação entre os marcadores plasmáticos e o aumento da viremia em pacientes HIV-1+. Associação entre os principais marcadores plasmáticos da progressão da infecção por HIV-1+ foi feita a partir da determinação da concentração de cada variável no grupo HIV-1+ e então calculou-se o coeficiente de correlação de Spearman entre as variáveis. Nos gráficos estão representados os valores de r e p, respectivamente.

SOARES, L.S. Resultados
51
A partir dos resultados dos mediadores solúveis quantificados no plasma, caracterizou-
se o perfil imunológico de cada grupo. Para tanto, calculou-se a frequência de indivíduos
definidos como altos produtores a partir da mediana global de cada marcador, como
representado na figura 8. No diagrama, os quadrados preenchidos correspondem aos valores
superiores à mediana global do respectivo analito, e em branco, aqueles com valores
inferiores à mediana. Cada linha representa um individuo daquele grupo e as moléculas
quantificadas estão dispostas em colunas. Ao final de cada coluna, estão demonstradas as
frequências de altos produtores para aquele determinado analito.
No grupo controle, observou-se que a frequência de altos produtores para a maioria dos
marcadores plasmáticos analisados foi inferior ou próxima a 50%, exceto para IL-17 e GM-
CSF, em que 63% dos indivíduos controles apresentaram níveis elevados de ambos acima da
mediana global (5,13 e 11,4 pg/mL, respectivamente). Quanto às moléculas associadas à
progressão da infecção por HIV como TNF-α, IP-10 e IL-10, apenas 22, 15 e 48%,
respectivamente, dos indivíduos controles apresentaram quantidade maior que a mediana
global.
De forma semelhante ao grupo controle, o grupo TARV também apresentou frequências
de altos produtores inferiores ou próximas a 50%, exceto para sCD14, RANTES e IL-5 (77,
67 e 60%, respectivamente). Enquanto os níveis de altos produtores para TNF-α, IP-10 e IL-
10 foram de: 47, 47 e 33% , respectivamente. Em resumo, nota-se que a maioria dos pacientes
do grupo TARV demonstram diminuição do perfil inflamatório sistêmico, mas ainda exibem
um painel de marcadores plasmáticos diferente daquele observado para os indivíduos não
infectados (grupo controle).
Por sua vez, o grupo compreendendo os indivíduos infectados pelo HIV-1+ e virgens de
tratamento, apresentou frequência maior do que 50% de altos produtores para 11 dos 18
mediadores solúveis analisados. É interessante notar que os maiores valores foram observados
para os seguintes marcadores: sCD163, TNF-α, IP-10 e IL-10 (76%, 75%, 86% e 64%,
respectivamente). Dentre as moléculas analisadas no grupo HIV-1+, verificou-se que a menor
frequência de indivíduos considerados altos produtores foi observada para a quimiocina
CCL5, em que somente 32% dos pacientes deste grupo apresentaram níveis plasmáticos
maior que a mediana global de 8505 pg/mL.

SOARES, L.S. Resultados
52
Figu
ra 8
. Fre
quên
cia
de in
divi
duos
car
acte
riza
dos c
omo
alto
s pro
duto
res p
ara
os d
ifere
ntes
med
iado
res p
lasm
átic
os. O
s di
agra
mas
das
freq
uênc
ias
fora
m c
onst
ruíd
os
utili
zand
o-se
o v
alor
da
med
iana
glo
bal d
e ca
da m
edia
dor s
olúv
el c
omo
pont
o de
cor
te p
ara
mar
car c
ada
indi
vidu
o co
mo
“alto
s” o
u “b
aixo
s” p
rodu
tore
s par
a ca
da a
nalit
o. O
s qu
adra
ntes
mar
cado
s em
pre
to c
orre
spon
dem
aos
val
ores
sup
erio
res
à m
edia
na g
loba
l do
resp
ectiv
o an
alito
, e e
m b
ranc
o aq
uele
s co
m v
alor
es in
ferio
res
à m
edia
na. A
baix
o de
ca
da q
uadr
o es
tão
repr
esen
tado
s a
freq
uênc
ia (%
) de
indi
vidu
os q
ue a
pres
enta
m n
ívei
s el
evad
os d
os m
edia
dore
s e
os d
ados
rele
vant
es (≥
50%
) est
ão su
blin
hado
s.

SOARES, L.S. Resultados
53
5.3. Avaliação fenotípica das células NK e suas subpopulações
5.3.1. Determinação da frequência de células NK e a distribuição de suas
subpopulações
A quantidade de células NK e suas subpopulações no contexto da infecção por HIV-1
foi determinada por meio da técnica de citometria de fluxo a partir da utilização dos
anticorpos: anti-CD3, anti-CD56 e anti-CD16, previamente titulados. As células NK foram
definidas dentro da população de linfócitos determinada de acordo com tamanho e
granularidade e que possuíam a marcação CD3-CD56+.
Na figura 9 estão demonstrados os valores de porcentagem e número das células NK
obtidas a partir da população em estudo. A mediana relativa à frequência de células NK do
grupo controle foi de 9,6% comparada aos valores de 7,3 e 5,8% nos grupos HIV-1+ e
TARV, respectivamente. A diminuição observada nos pacientes infectados pelo HIV-1+,
independente do tratamento, foi estatisticamente significativa em relação aos indivíduos
controles. Quanto ao número de células NK, observou-se diminuição estatisticamente
significativa no grupo HIV-1+, comparado aos grupos controle e TARV. Não se observou
diferença significativa quanto aos esquemas terapêuticos empregados (dados não mostrados).
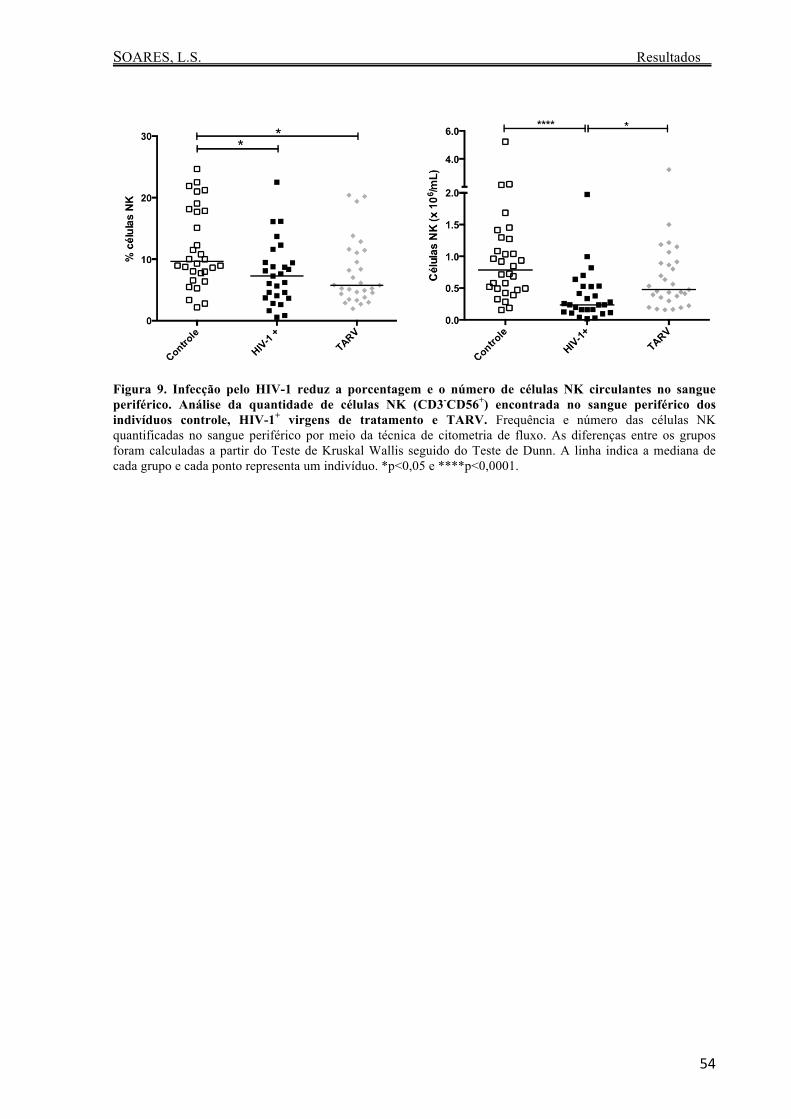
SOARES, L.S. Resultados
54
Figura 9. Infecção pelo HIV-1 reduz a porcentagem e o número de células NK circulantes no sangue periférico. Análise da quantidade de células NK (CD3-CD56+) encontrada no sangue periférico dos indivíduos controle, HIV-1+ virgens de tratamento e TARV. Frequência e número das células NK quantificadas no sangue periférico por meio da técnica de citometria de fluxo. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05 e ****p<0,0001.

SOARES, L.S. Resultados
55
Conforme dados da literatura, as subpopulações de células NK foram classificadas de
acordo com a intensidade de expressão das moléculas CD56 e CD16 [89]. Dessa forma, foram
determinadas três subpopulações de células NK: CD56brightCD16-, CD56dimCD16+ e CD56-
CD16+ (Figura 10A), sendo que cada uma é descrita por possuir funções específicas,
justificando sua análise durante a infecção pelo HIV-1.
A subpopulação de células NK de fenótipo CD56dimCD16+, caracterizada por sua
elevada capacidade citotóxica, estava diminuída significativamente no grupo HIV-1+ em
relação ao controle, evidenciando umas das disfunções encontradas nos pacientes infectados e
não tratados (Figura 10B, C). Essa subpopulação corresponde à maioria das células NK
encontradas no sangue periférico, sendo a mediana de sua frequência semelhante nos grupos
controle e TARV (87,6 e 81,7%, respectivamente).
Quanto às células NK CD56brightCD16-, descritas por possuírem função predominante
de produção de citocinas, notou-se diminuição no grupo HIV-1+, embora não significativa,
com valor de mediana relativo à frequência dessa subpopulação de 3,9%. Assim como
observado para as células NK CD56dimCD16+, os indivíduos controle e do grupo TARV
mantiveram quantidades próximas quanto a essa outra subpopulação, sendo a mediana de 5,9
e 5,5% do total de células NK encontradas no sangue periférico, respectivamente (Figuras
10B,C).
E, por fim, a frequência de células NK de fenótipo CD56-CD16+ apresentou aumento
significativo nos indivíduos HIV-1+, independente do tratamento antirretroviral (Figura
10B,C). Essa subpopulação é denominada não funcional por expressar altos níveis de
receptores inibitórios, secretar menor quantidade citocinas e possuir defeitos na lise de células
alvo infectadas. A porcentagem destas células NK não funcionais manteve-se elevada no
grupo TARV, embora ainda menor que o grupo HIV-1+.
Dessa forma, sugere-se redução da função de defesa desempenhada pelas células NK
durante a infecção por HIV-1+, em decorrência da alteração da distribuição de suas
subpopulações, principalmente nos indivíduos recentemente infectados e que não iniciaram a
terapia antirretroviral. O tratamento, por sua vez, parece reestabelecer parcialmente as
proporções de células NK, exceto para a subpopulação não funcional que permanece elevada.
Não foram observadas diferenças quanto aos esquemas terapêuticos do grupo TARV.

SOARES, L.S. Resultados
56
Figura 10. Alteração da distribuição das subpopulações de células NK nos pacientes HIV-1+. A) Estratégia para análise das subpopulações de células NK por citometria de fluxo. B) Gráficos representativos da distribuição das subpopulações de células NK dos grupos controle, HIV-1+ e TARV. C) Frequência das subpopulações de células NK. (O total de células NK foi definido como a soma das três subpopulações analisadas). As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. **p<0,01; ***p<0,001; ****p<0,0001.

SOARES, L.S. Resultados
57
5.3.2. Análise da expressão dos receptores de ativação e inibição das células NK
Além de alterar a proporção das subpopulações de células NK, a infecção por HIV-1
também está associada à modificação da expressão dos receptores nestas células. Dessa
forma, com o objetivo de complementar a caracterização fenotípica das células NK na
população em estudo, realizou-se a quantificação da expressão dos receptores de ativação e
inibição por citometria de fluxo e PCR em Tempo Real.
Inicialmente, verificou-se a expressão dos receptores inibitórios: CD158a (KIR2DL1),
NKAT2 (KIR2DL2/L3) e NKB1 (KIR3DL1). Não foi observada diferença significativa
quanto à porcentagem de células NK expressando esses receptores entre os três grupos
estudados (Figura 11A-C), com exceção do grupo TARV que demonstrou diminuição na
frequência de células NK positivas para CD158a em relação ao grupo controle (Figura 11A).
Essa redução também foi verificada quanto ao número de células NK CD158a+ nos pacientes
HIV-1+, independente do tratamento antirretroviral (Figura 11D).
Notou-se ainda diminuição estatisticamente significativa no número de células NK
positivas para NKAT2 e NKB1 no grupo HIV-1+ comparado ao controle (Figura 11E, F),
refletindo em parte a redução do número total de células NK nos pacientes desse grupo. Em
contrapartida, a mediana da intensidade de fluorescência (MIF) da molécula NKAT2 estava
aumentada no grupo HIV-1+ em relação aos grupos controle e TARV (Figura 11H). Dessa
forma, conclui-se que poucas células NK do grupo HIV-1+ expressavam o receptor NKAT2,
mas com alta densidade na superfície celular como demonstrado pela mediana da intensidade
de fluorescência desse receptor. Os outros receptores não apresentaram diferença significativa
quanto ao MIF (Figura 11 G-I), embora verifique-se relativo aumento da mediana da
molécula NKB1 no grupo TARV (Figura 11-I).
Uma vez identificadas alterações na porcentagem e número de células NK expressando
os receptores de inibição entre os grupos avaliados, investigou-se também se entre as
diferentes subpopulações de células NK havia variação na expressão desses receptores.
Observou-se que a expressão dos receptores inibitórios nas subpopulações de células NK foi
diferente somente para a molécula CD158a, como demonstrado na figura 12. A redução na
expressão desse receptor foi estatisticamente significativa na subpopulação citotóxica
(CD56dimCD16+) do grupo TARV. Por sua vez, o grupo de pacientes que receberam IP em
seu esquema terapêutico apresentaram aumento da expressão de CD158a na subpopulação
não funcional em níveis próximos ao grupo controle.

SOARES, L.S. Resultados
58
Figura 11. Diminuição da quantidade de células NK positivas para os receptores de inibição. A, B, C) Porcentagem de células NK que expressam as moléculas: CD158a, NKAT2 e NKB1, respectivamente. D, E, F) Número de células NK que expressam as moléculas: CD158a, NKAT2 e NKB1, respectivamente. G, H, I) Mediana da intensidade de fluorescência das moléculas: CD158a, NKAT2 e NKB1 na superfície de células NK, respectivamente. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05; **p<0,01 e ***p<0,001.

SOARES, L.S. Resultados
59
Figura 12. Modulação da expressão do receptor de inibição CD158a nas diferentes subpopulações de células NK de acordo com o tratamento do paciente. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn para comparações múltiplas, e Teste de Mann-Whitney para comparações entre dois grupos. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05.

SOARES, L.S. Resultados
60
Outro conjunto de moléculas envolvidas na modulação da resposta das células NK são
os receptores de ativação. Nesse sentido, quantificou-se a expressão da molécula NKp46
pertencente à família dos receptores de citotoxicidade natural (Figura 13 A-C). Verificou-se
diminuição significativa no número de células NK expressando a molécula NKp46 nos grupos
HIV-1+ e TARV (Figura 13B). Interessantemente, a mediana da intensidade de fluorescência
deste receptor nas células NK do grupo HIV-1+ foi estatisticamente maior em relação ao
grupo controle (Figura 13C). Esse resultado demonstra que apesar de poucas células NK
expressarem esse receptor no grupo HIV-1+, a expressão na superfície celular estava
aumentada, sugerindo maior estado de ativação dessas células.
Quanto à análise da expressão da molécula NKp46 nas diferentes subpopulações de
células NK, observou-se aumento desse receptor na superfície das células NK
CD56brightCD16- nos indivíduos infectados, independente do tratamento, sendo significativo
no grupo TARV (Figura 14). Por outro lado, notou-se diminuição dessa molécula,
principalmente nos pacientes tratados, quanto à subpopulação citotóxica (CD56dimCD16+),
embora não significativa. Interessantemente, as células NK descritas como não funcionais
apresentaram redução estatisticamente significativa da expressão de NKp46 nos grupos HIV-
1+ e TARV+, sendo essa diminuição ainda mais pronunciada no grupo TARV (Figura14).
Com o objetivo de complementar a análise fenotípica das células NK quanto à
expressão das moléculas de ativação, realizou-se a quantificação da expressão gênica das
moléculas: CD69, NCR3 (NKp30) e KLRC1 (NKG2a) pela técnica de PCR em Tempo Real.
Na figura 15, estão demonstrados os resultados da expressão relativa das amostras dos grupos
HIV-1+ e TARV, sendo o grupo controle utilizado como amostra de referência para o cálculo
da expressão relativa. Não foi observada diferença estatisticamente significativa quanto às
moléculas analisadas entre os grupos.
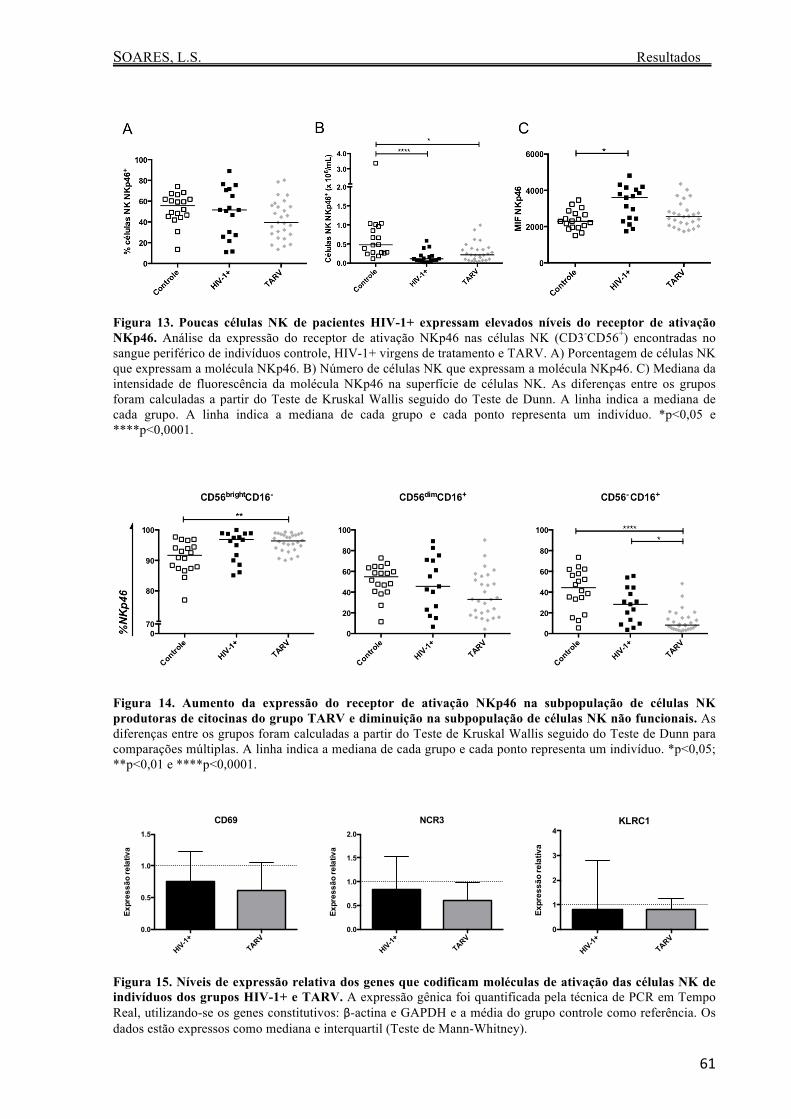
SOARES, L.S. Resultados
61
Figura 13. Poucas células NK de pacientes HIV-1+ expressam elevados níveis do receptor de ativação NKp46. Análise da expressão do receptor de ativação NKp46 nas células NK (CD3-CD56+) encontradas no sangue periférico de indivíduos controle, HIV-1+ virgens de tratamento e TARV. A) Porcentagem de células NK que expressam a molécula NKp46. B) Número de células NK que expressam a molécula NKp46. C) Mediana da intensidade de fluorescência da molécula NKp46 na superfície de células NK. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn. A linha indica a mediana de cada grupo. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05 e ****p<0,0001.
Figura 14. Aumento da expressão do receptor de ativação NKp46 na subpopulação de células NK produtoras de citocinas do grupo TARV e diminuição na subpopulação de células NK não funcionais. As diferenças entre os grupos foram calculadas a partir do Teste de Kruskal Wallis seguido do Teste de Dunn para comparações múltiplas. A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05; **p<0,01 e ****p<0,0001.
Figura 15. Níveis de expressão relativa dos genes que codificam moléculas de ativação das células NK de indivíduos dos grupos HIV-1+ e TARV. A expressão gênica foi quantificada pela técnica de PCR em Tempo Real, utilizando-se os genes constitutivos: β-actina e GAPDH e a média do grupo controle como referência. Os dados estão expressos como mediana e interquartil (Teste de Mann-Whitney).
HIV-1+TARV
0.0
0.5
1.0
1.5
Expr
essã
o re
lativ
a
CD69
HIV-1+TARV
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
NCR3
HIV-1+TARV
0
1
2
3
4
Expr
essã
o re
lativ
a
KLRC1

SOARES, L.S. Resultados
62
5.4. Avaliação funcional das células NK
5.4.1. Análise da citotoxicidade das células NK purificadas
Para avaliar a função das células NK obtidas após separação magnética, realizou-se o
ensaio de citotoxicidade, utilizando como célula alvo a linhagem celular K-562. A
porcentagem de citotoxicidade foi calculada a partir da quantificação da enzima lactato
desidrogenase liberada no meio de cultura após lise celular e segundo fórmula detalhada na
seção de materiais e métodos. No ensaio, foram utilizadas diferentes razões de células
efetoras em relação à célula alvo (razão E:A). Conforme demonstrado na figura 16, foi
possível diferenciar a porcentagem de citotoxicidade da célula NK entre as diferentes razões e
observou-se tendência de diminuição da função citotóxica nas células do grupo HIV-1+. Por
outro lado, os valores de citotoxicidade do grupo TARV estavam próximos aos do controle,
sugerindo recuperação parcial da função citotóxica nos indivíduos infectados após tratamento
antirretroviral.

SOARES, L.S. Resultados
63
Figura 16. Ensaio de citotoxicidade realizado a partir da quantificação da enzima lactato desidrogenase. Células NK purificadas (efetoras) dos grupos controle, HIV-1+ virgens de tratamento e TARV foram cultivadas com células alvo (K-562) em diferentes razões entre células efetoras e células alvo (20:1; 10:1; 5:1), considerando a concentração inicial de 5x103 células alvo. A porcentagem de citotoxicidade foi calculada a partir da quantificação da enzima lactato desidrogenase (LDH) liberada no meio de cultura após lise celular. Os dados estão expressos como média ± SEM (Teste de Kruskal Wallis seguido do Teste de Dunn).
0
20
40
60
80
100
NK:K-562
% C
itoto
xici
dade
ControleHIV-1 +
20:1
10:1 5:1
TARV

SOARES, L.S. Resultados
64
5.4.2. Quantificação das citocinas e quimiocinas no sobrenadante do ensaio de
citotoxicidade
Com o objetivo de complementar a caracterização funcional das células NK dos grupos
controle, HIV-1+ virgem de tratamento e TARV, realizou-se a quantificação de citocinas e
quimiocinas liberadas no sobrenadante de cultura durante o ensaio de citotoxicidade. Pela
plataforma Multiplex foram detectadas as seguintes citocinas: TNF-α, IL-6, IL-1β, IL-12p70,
IFN-γ, IFN-α2, IL-5, IL-4, IL-10 (Figuras 17 e 18); e as quimiocinas: RANTES (CCL5),
MCP-1 (CCL2), IP-10, MIP-1α (CCL3), MIP-1β (CCL4) e GM-CSF (Figura 19).
Os resultados obtidos a partir das amostras coletadas do grupo TARV demonstram
aumento significativo na produção da maioria das citocinas pró-inflamatórias (TNF-α, IL-6,
IFN-γ, IFN-α2 e IL-12p70) pelas células NK dos indivíduos desse grupo quando comparados
ao controle e/ou HIV-1+ virgens de tratamento (Figura 17). Essa maior produção de citocinas
inflamatórias pode ter favorecido o aumento na porcentagem de citotoxicidade observada no
grupo TARV no experimento descrito anteriormente.
Por outro lado, nota-se que as principais citocinas relacionadas à função das células NK
estão diminuídas no grupo HIV-1+, inclusive em relação ao grupo controle, embora essa
diferença não tenha sido estatisticamente significativa. A mediana da produção de TNF-α no
grupo HIV-1+ foi de 192,9 pg/mL comparado aos valores de 400,9 e 1031 pg/mL nos grupos
controle e TARV, respectivamente. Quanto à citocina IFN-γ, essa redução foi de 4,8 vezes em
relação ao grupo controle e de aproximadamente 10 vezes em relação ao grupo TARV
considerando os valores de mediana. A produção das citocinas IL-12p70 e IFN-α2 também
estava consideravelmente diminuída no grupo HIV-1+ com valores de mediana 2,9 e 1,5
vezes menores que os indivíduos controles, respectivamente.
Concomitantemente ao aumento na produção de citocinas pró-inflamatórias pelas
células NK do grupo TARV, verificou-se também elevada produção de IL-10, IL-4 e IL-5
(Figura 18). A mediana da quantidade de IL-10 foi de 8,2 pg/mL comparada aos valores de
2,2 pg/mL no grupo controle e 1,1 pg/mL no grupo HIV-1+. Embora os níveis de produção
das citocinas IL-4 e IL-5 tenham sido baixos, os valores correspondentes à mediana foram
respectivamente: 4,3 e 1,7 vezes maior no grupo TARV do que no grupo HIV-1+.
Quanto à análise da produção das diferentes quimiocinas, observou-se aumento
significativo de RANTES e MCP-1 no grupo TARV em relação ao grupo HIV-1+ e em
relação aos grupos HIV-1+ e TARV, respectivamente. As outras quimiocinas analisadas

SOARES, L.S. Resultados
65
também estavam aumentadas nos pacientes infectados e tratados, embora não
significativamente (Figura 19). Vale ressaltar a diminuição na produção de RANTES/CCL5
pelas células NK do grupo HIV-1+, sendo a mediana de 52,1 pg/mL. Enquanto nos grupos
controle e TARV, os valores observados foram de 167 e 461,2 pg/mL, evidenciando outra
possível disfunção das células NK nos indivíduos recentemente infectados e que ainda não
iniciaram a terapia antirretroviral.

SOARES, L.S. Resultados
66
Figura 17. Células NK de indivíduos do grupo TARV apresentam maior capacidade de produção de citocinas pró-inflamatórias. Células NK purificadas (efetoras) dos grupos controle, HIV-1+ virgem de tratamento e TARV foram cultivadas com células alvo (K-562) na proporção 20:1, e a liberação de citocinas no sobrenadante foi avaliada quantitativamente utilizando-se microesferas magnéticas na plataforma Magpix®. O limite de detecção de cada citocina foi <5pg/mL. Os dados estão expressos como mediana e interquartil (Teste de Kruskal Wallis seguido do Teste de Dunn). A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05; **p<0,01 e ***p<0,001.
Figura 18. Células NK de indivíduos do grupo TARV apresentam maior capacidade de produção de citocinas do perfil TH2 (IL5, IL-4) e regulador (IL-10). Células NK purificadas (efetoras) dos grupos controle, HIV-1+ virgem de tratamento e TARV foram cultivadas com células alvo (K-562) na proporção 20:1, e a liberação de citocinas no sobrenadante foi avaliada quantitativamente utilizando-se microesferas magnéticas na plataforma Magpix®. O limite de detecção de cada citocina foi <5pg/mL. Os dados estão expressos como mediana e interquartil (Teste de Kruskal Wallis seguido do Teste de Dunn). A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05 e **p<0,01.
TNF-α
0
500
1000
1500
2000
pg/m
L
**
IFN-γ
Controle
HIV-1 +
TARV0
5
10
15
20
pg/m
L ***
IL-12p70
0
1
2
3
4
pg/m
L
**
IL-6
Controle
HIV-1 +
TARV0
500
1000
1500
pg/m
L
*
IFN-α2
0
5
10
15
20
25
pg/m
L
*****
IL-1β
Controle
HIV-1 +
TARV0
100
200
300
400
500
pg/m
L
IL-10
Controle
HIV-1
+TARV
0
10
20
30
40
pg/m
L
***
IL-4
Controle
HIV-1
+TARV
0
2
4
6
8
10
pg/m
L
**
IL-5
Controle
HIV-1
+TARV
0.0
0.1
0.2
0.3
0.4
pg/m
L
*

SOARES, L.S. Resultados
67
Figura 19. Aumento da produção de quimiocinas pelas células NK dos indivíduos do grupo TARV. Células NK purificadas (efetoras) dos grupos controle, HIV-1+ virgem de tratamento e TARV foram cultivadas com células alvo (K-562) na proporção 20:1, e a liberação de citocinas no sobrenadante foi avaliada quantitativamente utilizando-se microesferas magnéticas na plataforma Magpix®. O limite de detecção de cada citocina foi <5pg/mL. Os dados estão expressos como mediana e intervalo interquartil (Teste de Kruskal Wallis seguido do Teste de Dunn). A linha indica a mediana de cada grupo e cada ponto representa um indivíduo. *p<0,05 e **p<0,01.
IP-10
0
10
20
30
40
50
200
300pg
/mL
MCP-1
Controle
HIV-1 +
TARV0
500
1000
1500
2000
pg/m
L
***
GM-CSF
0
10
20
30
40
pg/m
L
MIP-1α
Controle
HIV-1
+TARV
0
500
1000
1500
2000
2500
pg/m
L
RANTES
0
200
400
600
800
1000
pg/m
L
*
MIP-1β
Controle
HIV-1
+TARV
0
500
1000
1500
2000
pg/m
L

SOARES, L.S. Resultados
68
5.4.3. Correlações entre os parâmetros que caracterizam as disfunções das células NK
nos indivíduos HIV-1+
A partir dos dados obtidos da quantificação de citocinas e quimiocinas no sobrenadante
do ensaio de citotoxicidade, nota-se um perfil de maior ativação das células NK provenientes
dos indivíduos do grupo TARV em relação aos outros grupos. Para compreender melhor essa
observação, realizou-se a análise da frequência de indivíduos considerados altos produtores
para as diferentes moléculas estudadas considerando a mediana global de cada mediador.
Dessa forma, construiu-se os gráficos de radar demonstrados na figura 20, evidenciando o
estado de ativação do grupo TARV, no qual a frequência de altos produtores foi maior de
50% para a maioria dos analitos. Por outro lado, o grupo HIV-1+ sugere uma disfunção
generalizada nas células NK dos indivíduos desse grupo, visto que a frequência de pacientes
com altos níveis de produção das diferentes citocinas estava consideravelmente menor do que
o grupo controle.

SOARES, L.S. Resultados
69
Figu
ra 2
0. R
epre
sent
ação
do
perf
il de
pro
duçã
o de
cito
cina
s pe
las
célu
las
NK
dos
gru
pos
cont
role
, HIV
-1+
e TA
RV
. As
freq
uênc
ias
fora
m
calc
ulad
as u
tiliz
ando
-se
o va
lor
da m
edia
na g
loba
l de
cada
med
iado
r so
lúve
l co
mo
pont
o de
cor
te p
ara
defin
ição
dos
indi
vídu
os c
omo
alto
s ou
ba
ixos
pro
duto
res
em re
laçã
o a
dete
rmin
ado
anal
ito.

SOARES, L.S. Resultados
70
Além de investigarmos as variações individuais das moléculas quantificadas, faz-se
necessário o estudo da correlação entre as mesmas para melhor compreensão do cenário
doença versus hospedeiro. Percebe-se que muitas correlações positivas são formadas, sendo
que o maior número delas é encontrado no grupo controle (Figura 21). Enquanto no grupo
HIV-1+, ocorre perda de grande parte dessas correlações e formação de novas. Por exemplo, a
quimiocina GM-CSF apresentou correlação positiva de moderada a forte com 12 outras
citocinas/quimiocinas (IFN-α2, IFN-γ, IL-12p70, IL-1β, IL-6, TNF-α, IP-10, MCP-1, MIP-
1α, MIP-1β, IL-5 e IL-10) no grupo controle. Todas essas correlações foram perdidas no
grupo HIV-1+, e formaram-se novas conexões com as moléculas IL-17 e IL-1β.
Interessantemente, houve recuperação parcial no grupo TARV das correspondências
inicialmente observadas no grupo controle para essa quimiocina.
Para a citocina IFN-γ, importante na função desempenhada pelas células NK, à medida
que sua produção aumentava, ocorria também aumento das quimiocinas MIP-1α e MIP-1β,
quimioatraentes relevantes na atração de outros leucócitos durante o processo inflamatório.
Essa correlação observada no grupo controle foi perdida nos grupos HIV-1+ e TARV, sendo
que, interessantemente, quando os pacientes são tratados, o número de conexões formadas
considerando a citocina IFN-γ foi o menor observado entre os três grupos.
A quimiocina RANTES apresentou o menor número de correlações entre os outros
analitos nos três grupos. Nos indivíduos controles, verificou-se correlação positiva forte com
IP-10 e moderada com IL-17. Enquanto no grupo HIV-1+, essas correspondências foram
perdidas e observou-se somente associação fraca com IL-10. Nos pacientes sob tratamento,
esta quimiocina não teve correlação significativa com nenhuma outra citocina/quimiocina,
embora sua produção tenha sido significativamente maior neste grupo (Figura 19).
Outra molécula com pequeno número de correlações formadas foi a quimiocina IP-10.
No grupo HIV-1+, esse marcador perdeu todas as seis conexões positivas existentes com as
moléculas: IL-6, IL-17, GM-CSF, RANTES, MIP-1α e MIP-1β, as quais foram observadas
no grupo controle. Essas conexões não foram reestabelecidas nos pacientes tratados, nos quais
surgiram diferentes associações com IL-5 e IFN-γ.
Esses resultados evidenciam a importância de estudarmos as correlações entre as
diferentes moléculas e não somente sua expressão de forma individual. Isso porque mais
importante que a quantidade de determinada citocina e/ou quimiocina é a interação existente
entre elas no contexto da infecção, e o resultado de tal interação como sendo benéfico ou não
para o hospedeiro. Nota-se claramente os diferentes perfis de células NK entre os grupos

SOARES, L.S. Resultados
71
analisados, sendo que no grupo HIV-1+, esta célula aparenta estar não funcional, e mesmo
após o tratamento antirretroviral, a homeostase não é restaurada.
Outra associação relevante observada nos pacientes infectados pelo HIV-1+ foi entre as
citocinas/quimiocinas produzidas pelas células NK e o tempo aproximado de infecção nesses
indivíduos. Na figura 22, verifica-se correlação positiva significativa entre o tempo de
infecção e as principais moléculas produzida pelas células NK, independente do tratamento
antirretroviral. Ou seja, quanto maior o tempo em que o individuo encontra-se infectado pelo
HIV-1+, maiores são os níveis de produção das moléculas: IFN-α2, IFN-γ, TNF-α, IL-12, IL-
6, MCP-1, IL-10 e IL-5, sugerindo maior estado de ativação dessas células. Essa observação
corrobora com a definição atual da infecção por HIV-1 como doença inflamatória crônica
destacando aspectos que devem ser levados em consideração quanto a sua implicação,
principalmente naqueles pacientes que convivem com o vírus há vários anos.

SOARES, L.S. Resultados
72
Figu
ra 2
1. R
ede
de in
tera
ções
ent
re a
s ci
toci
nas
e qu
imio
cina
s pr
oduz
idas
pel
as c
élul
as N
K. A
aná
lise
da re
de m
ostra
cor
rela
ções
sig
nific
ativ
as
(p<0
,05)
em
tod
as a
s va
riáve
is,
calc
ulad
as p
or m
eio
da c
orre
laçã
o de
Spe
arm
an e
ntre
cad
a pa
r de
mol
écul
as e
são
rep
rese
ntad
as p
or l
inha
s no
s di
fere
ntes
gru
pos
clín
icos
est
udad
os. A
for
ça d
e co
rrel
ação
foi
det
erm
inad
a pe
lo v
alor
de
“r”.
Os
tipos
de
inte
raçõ
es f
oram
def
inid
os c
omo:
for
te
(pre
to),
mod
erad
a (v
erde
) e fr
aca
(ver
mel
ho p
ontil
hado
).
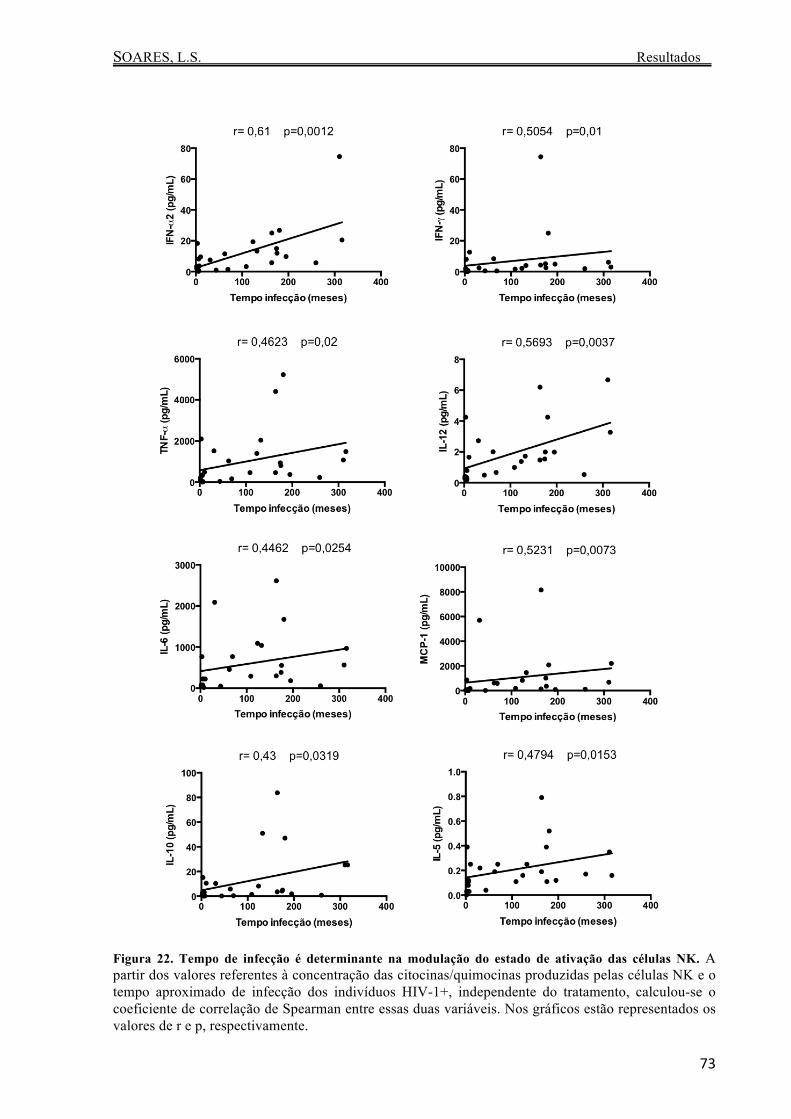
SOARES, L.S. Resultados
73
Figura 22. Tempo de infecção é determinante na modulação do estado de ativação das células NK. A partir dos valores referentes à concentração das citocinas/quimocinas produzidas pelas células NK e o tempo aproximado de infecção dos indivíduos HIV-1+, independente do tratamento, calculou-se o coeficiente de correlação de Spearman entre essas duas variáveis. Nos gráficos estão representados os valores de r e p, respectivamente.

SOARES, L.S. Resultados
74
5.4.4. Determinação da produção de granzima B pelas células NK
Dentre as principais funções descritas para as células NK está a função citotóxica. Para
executar tal função, após ativação da célula ocorre liberação de moléculas importantes como
granzimas e perforinas. As granzimas estão envolvidas na ativação de caspases no interior da
célula-alvo e consequente indução da apoptose. Nesse sentido, avaliou-se a produção de
granzima B pelas células NK dos diferentes grupos clínicos estudados mediante o estímulo
com IL-2. No entanto, não se observou diferença significativa entre os grupos como
demonstrado na figura 23, com valores de mediana similares entre os grupos controle e
TARV (188,7 e 264,5 pg/mL, respectivamente). No grupo HIV-1+, a mediana da produção de
granzima B foi de 624,2 pg/mL, embora seja uma quantidade significativa, ocorreu grande
variação neste grupo, como demonstrado pela dispersão dos valores na figura 23.

SOARES, L.S. Resultados
75
Figura 23. Determinação da produção de granzima B pelas células NK. As células NK obtidas após separação por microesferas magnéticas foram incubadas com IL-2 (200U/mL) por 72 horas. Após esse período, coletou-se o sobrenadante e quantificou-se a produção de granzima B por ELISA. Boxplot dos valores relativos à produção de granzima pelas células NK, em que os dados estão expressos como mediana e intervalo interquartil (Teste de Kruskal Wallis seguido do Teste de Dunn).
Controle
HIV-1+TARV
0
500
1000
1500
2000
2500
Gra
nzim
a B
(pg/
mL)

SOARES, L.S. Resultados
76
5.4.5. Quantificação da produção de espécies reativas de oxigênio pelas células NK
O estresse oxidativo celular pode ser indicativo do fenômeno de senescência celular e é
facilmente avaliado pela quantificação de espécies reativas de oxigênio (ROS – reactive
oxygen species) por quimioluminescência. Considerando que na senescência, a célula também
perde sua função, realizou-se a quantificação de ROS por quimioluminescência na população
em estudo, para verificarmos se as células dos indivíduos infectados pelo HIV-1 estavam não
funcionais devido a esse fenômeno.
Como demonstrado na figura 24, observou-se tendência no aumento da produção de
ROS pelas células NK do grupo TARV (quando estimulada com PMA), com valor de
mediana 1,7 vezes maior que a do grupo controle e aproximadamente 2 vezes maior que a do
grupo HIV-1+. Esse resultado sugere a ocorrência do fenômeno de imunosenescência nestas
células e que pode estar correlacionado devido ao maior estado de ativação verificado nesse
grupo pela produção de citocinas e quimiocinas durante o ensaio de citotoxicidade.

SOARES, L.S. Resultados
77
Figura 24. Quantificação de espécies reativas de oxigênios liberadas por células NK de indivíduos dos grupos controle, HIV-1+ e TARV espontaneamente ou após estímulo com PMA. As células NK foram purificadas e estimuladas com PMA a 10-7 M. Os resultados estão expressos em mediana e intervalo interquartil da área sob a curva de quimioluminescência do Luminol em células NK após 3500 segundos de estímulo.
0
2×106
4×106
6×106
PMA - + - + - +
Áre
a so
b a
curv
aControleHIV-1+TARV

SOARES, L.S. Resultados
78
5.5. Caracterização das modificações epigenéticas relacionadas às disfunções das células
NK
5.5.1. Determinação do padrão de metilação global do DNA das células NK
Considerando as alterações fenotípicas e funcionais observadas nas células NK de
indivíduos HIV-1+ submetidos ou não ao tratamento antirretroviral, procedeu-se à
investigação de mudanças epigenéticas que pudessem estar correlacionadas à resposta imune
das células NK nestes indivíduos. Para tanto, inicialmente analisou-se o perfil de metilação
global do DNA purificado de células NK dos grupos controle, HIV-1+ e TARV como
demonstrado na figura 25. Apesar da mediana da porcentagem de metilação relativa ter sido
semelhante entre os grupos, quando os pacientes HIV-1+ foram analisados conforme a
progressão da doença, nota-se aumento na frequência de metilação do DNA das células NK
dos indivíduos em estágio avançado da doença (carga viral > 10.000 cópias/mL e células T
CD4+ < 200/mm3), mais especificamente dos pacientes HIV15 e HIV18, como destacado na
figura.
Por outro lado, observa-se semelhança no perfil de metilação global do DNA das
células NK dos indivíduos do grupo TARV em relação ao controle. Considerando que as
células NK do grupo TARV encontram-se mais ativadas, essa observação parece refletir em
parte na menor porcentagem de metilação global do DNA em relação ao grupo HIV-1+,
sendo que essa diferença deve ser mais pronunciada quanto à metilação dos genes específicos
como TNF-α , IFN-γ, IFN-α2 e RANTES (não analisadas neste trabalho). Não foi vista
diferença no perfil de metilação quanto aos diferentes esquemas terapêuticos do grupo TARV.
Esses dados sugerem fortemente a relação de alterações epigenéticas em genes da
resposta imune nas células NK com as modificações fenotípicas e funcionais observadas
nestas células nos pacientes HIV-1+ e TARV, que podem ser resultado direto dos
mecanismos de escape induzidos pelo vírus no hospedeiro assim como do contexto
inflamatório em que essas células se encontram.
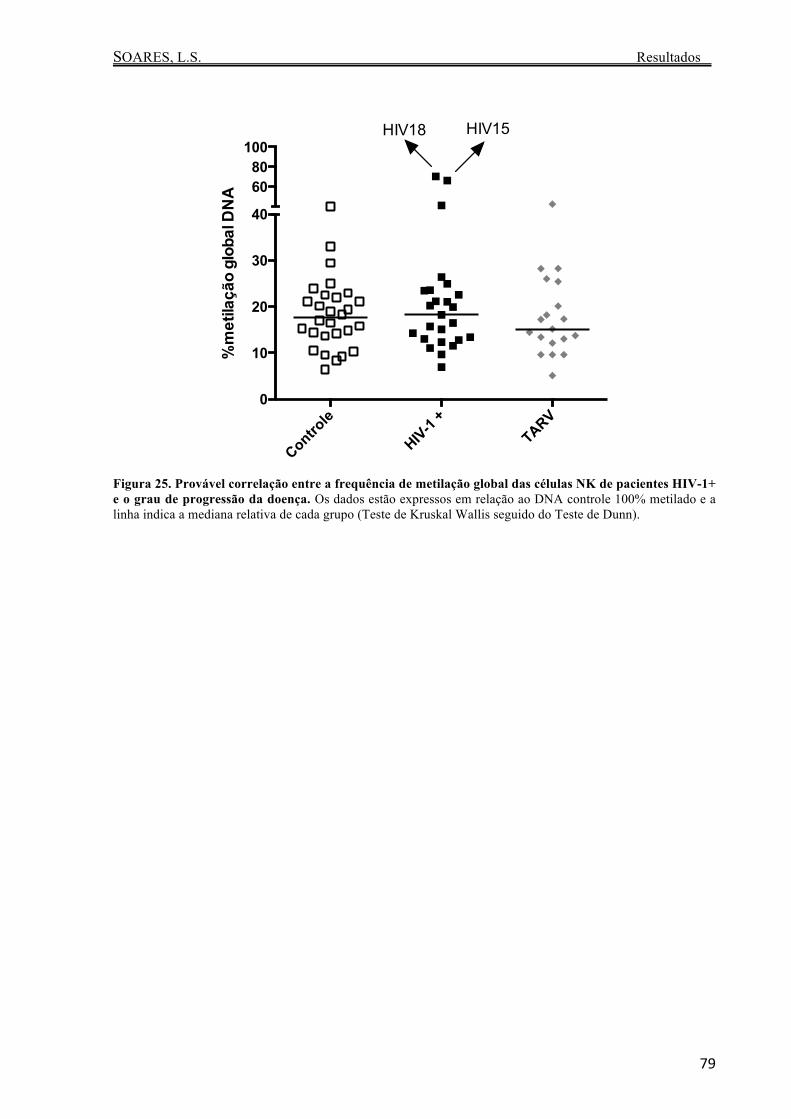
SOARES, L.S. Resultados
79
Figura 25. Provável correlação entre a frequência de metilação global das células NK de pacientes HIV-1+ e o grau de progressão da doença. Os dados estão expressos em relação ao DNA controle 100% metilado e a linha indica a mediana relativa de cada grupo (Teste de Kruskal Wallis seguido do Teste de Dunn).
Controle
HIV-1 +
TARV0
10
20
30
40
6080
100HIV15HIV18
%m
etila
ção
glob
al D
NA

SOARES, L.S. Resultados
80
5.5.2. Avaliação do padrão de metilação específico do gene KIR2DL2/L3
Considerando que os mecanismos epigenéticos, em especial a metilação, possuem
grande importância na regulação da expressão dos receptores do tipo KIR e que eles estavam
alterados nas células NK analisadas, prosseguiu-se à investigação da porcentagem de
metilação específica na região promotora do gene KIR2DL2. Interessantemente, as células
NK positivas para esse marcador estavam diminuídas em número no grupo HIV-1+, no
entanto os valores de MIF estavam aumentados em relação aos grupos controle e TARV.
Com o intuito de correlacionar esses resultados com possíveis modificações epigenéticas,
realizou-se a análise da metilação após modificação do DNA por bissulfito, com posterior
clonagem e sequenciamento do DNA pelo método de Sanger.
Inicialmente, verificou-se a sequência de DNA genômico da região analisada antes da
reação de modificação do DNA por bissulfito para confirmação da sequência e investigação
de possíveis polimorfismos, os quais devem ser levados em consideração para correto
alinhamento das sequências em análise posterior. Os resultados obtidos após sequenciamento
do DNA pelo método de Sanger e os respectivos alinhamentos com a sequência referência do
genoma estão demonstrados na figura suplementar 05 (Apêndice). As amostras foram
escolhidas para análise da metilação com base nos níveis de expressão da molécula NKAT2.
Dentre os indivíduos investigados, observou-se a ocorrência de dois polimorfismos na
sequência do paciente HIV11 do grupo HIV-1+ (Figura 26). O primeiro polimorfismo
(rs10414999) envolveu a troca da base Adenina por uma Guanina. Enquanto o segundo
polimorfismo identificado (rs10415421) referiu-se à troca de uma Citosina por uma Adenina.
Ambos polimorfismos encontram-se mapeados na região promotora do gene KIR2DL2
(sequencia referencia GenBank NM_014219.2).
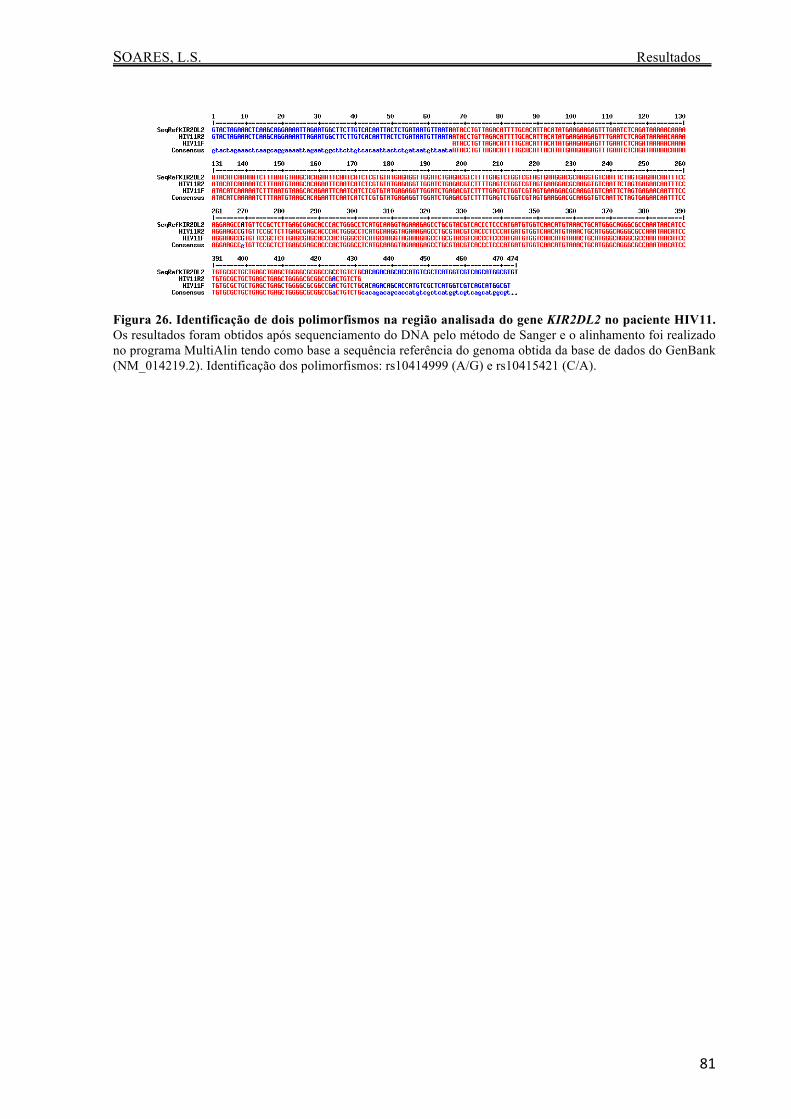
SOARES, L.S. Resultados
81
Figura 26. Identificação de dois polimorfismos na região analisada do gene KIR2DL2 no paciente HIV11. Os resultados foram obtidos após sequenciamento do DNA pelo método de Sanger e o alinhamento foi realizado no programa MultiAlin tendo como base a sequência referência do genoma obtida da base de dados do GenBank (NM_014219.2). Identificação dos polimorfismos: rs10414999 (A/G) e rs10415421 (C/A).

SOARES, L.S. Resultados
82
Após verificação das sequências de DNA antes da modificação por bissulfito, realizou-
se a clonagem do fragmento de interesse e posterior sequenciamento de no mínimo oito
clones positivos de cada amostra para quantificação da porcentagem de metilação. Foram
analisadas o total de 19 dinucleotídeos CpG localizados na posição física do cromossomo 19:
54738176-54738672 (ou -271 a +81) do gene KIR2DL2, tendo como referência a base de
dados do GenBank. Para quantificação da frequência de metilação, os pontos de metilação
foram alinhados com auxílio dos programas Clustal e MultiAlin como demonstrado na figura
suplementar 06 (Apêndice).
Na figura 27 estão demonstrados os resultados representativos de cada grupo quanto ao
perfil de metilação dos clones sequenciados. Nota-se que a maioria dos dinucleotídeos CpG
estão metilados como representado pelo círculo preenchido. A média da porcentagem de
metilação foi semelhante entre os grupos, sendo de 74,5; 76,5 e 75,3% nos grupos controle,
HIV-1+ e TARV, respectivamente (Figura 28).
Em resumo, não foi observada diferença significativa entre os níveis de metilação total
da região analisada entre os grupos (Figura 28). Quanto às frequências de metilação pontuais
também não foi verificada diferença significativa entre os grupos como demonstrado pelos
gráficos representativos na figura 29.

SOARES, L.S. Resultados
83
Figura 27. Gráficos representativos dos níveis de metilação na região analisada do gene KIR2DL2. O perfil de metilação para cada posição CpG está representada pelos círculos não preenchidos (não metilado) e preenchidos (metilado). Os números referem-se à posição física de cada CpG em relação ao sítio de inicio da transcrição.

SOARES, L.S. Resultados
84
Figura 28. Maior porcentagem de metilação total na região analisada do gene KIR2DL2. A avaliação da metilação foi feita após clonagem e sequenciamento do DNA pelo método de Sanger seguida do alinhamento com a sequência referência do gene em questão e análise no software QUMA.
Figura 29. Frequências de metilação pontuais dos dinucleotídeos CpG na região -271 a +81 do gene KIR2DL2. Os gráficos são representativos dos grupos controle, HIV-1+ e TARV, respectivamente.
Controle
HIV-1+TARV
0
20
40
60
80
100
%M
etila
ção
tota
l - K
IR2D
L2

SOARES, L.S. Resultados
85
5.5.3. Análise da frequência de enzimas que promovem as modificações epigenéticas
Em resumo, os mecanismos epigenéticos envolvidos na regulação gênica podem ser
classificados como ativador transcricional ou silenciador gênico. Na primeira análise do perfil
epigenético, quantificou-se a porcentagem de metilação global e específico do DNA das
células NK, sendo que essa modificação está associada à repressão gênica. Com o objetivo de
continuar essa caracterização, procedeu-se à quantificação das enzimas envolvidas nas
alterações que promovem a transcrição (Figura 30) e a repressão gênica (Figuras 31 e 32).
Para tanto, analisou-se por PCR em tempo real a expressão dessas enzimas nas células NK
dos grupos controle, HIV-1+ e TARV, sendo que os dados estão expressos considerando-se o
grupo controle como amostra de referência.
Dentre as enzimas que promovem as alterações relacionadas à transcrição gênica estão
as histonas acetil-transferases. Essas enzimas estão envolvidas na acetilação de histonas,
promovendo o remodelamento da cromatina e regulação dos processos nucleares. Não foram
constatadas diferenças significativas entre as histonas acetil-transferases analisadas (ATF2,
HAT1, KAT2B, KAT5) nas células NK dos pacientes infectados pelo HIV-1+, independente
do tratamento (Figura 30).
Outro grupo de enzimas quantificadas foram as histonas metil-transferases (CARM1,
PRMT5, SMYD3, SUV39H1, ASH1L, KMT2C, KMT2E e SETD1B). Enquanto a metilação
do DNA está sempre associada à repressão gênica, a metilação de histonas pode promover a
ativação da transcrição gênica. A expressão dessas enzimas também não se modificou nas
células NK dos diferentes indivíduos estudados como demonstrado na figura 30.
E, por fim, as histonas demetilases KDM5B e KDM6B também não modificaram sua
expressão nos grupos HIV-1+ e TARV. Embora seja importante ressaltar a maior expressão
de KDM6B nos pacientes infectados, independente do tratamento, com valores de mediana
3,6 e 2,2 vezes maior que os indivíduos controles nos grupos HIV-1+ e TARV,
respectivamente. Essa enzima está envolvida especificamente na demetilação da lisina 27 da
histona H3 e possui importante função na resposta inflamatória e diferenciação de
macrófagos.
Quanto às enzimas envolvidas na repressão gênica, realizou-se a quantificação de uma
classe de proteínas que atuam por meio da remoção de grupos acetilas das lisinas de histonas
denominadas histonas deacetilases (HDAC). Foram analisadas um total de 11 HDACs (Figura
31) nas células NK provenientes dos grupos controle, HIV-1+ e TARV, sem alteração
significativa da expressão nestes três grupos.

SOARES, L.S. Resultados
86
Outro conjunto de enzimas analisadas foram as DNA metil-transferases (DNMT1, 3A e
3B), MECP2 (methyl CpG binding protein 2), MBD2 e 3 (methyl-CpG binding domain
protein), RBBP4 e 7 (Retinoblastoma binding protein), CHD4 (Chromodomain helicase DNA
binding protein 4), SIN3A, TRDMT1 (TRNA aspartic acid methyltransferase 1), SAP18 e 30.
Não se verificou alteração estatisticamente significativa quanto à expressão dessas proteínas
nos diferentes grupos analisados, com a observação de tendência de aumento de CHD4 no
grupo TARV (Figura 32). Essa enzima representa um dos componentes principais do
complexo de deacetilases e atua no remodelamento do nucleossomo, sendo sua função
fundamental na repressão da transcrição gênica.
As enzimas analisadas estão envolvidas na formação de complexos protéicos com
funções biológicas importantes na regulação gênica. Os resultados obtidos quanto às
expressões de cada proteína indicam que, a princípio, as células NK circulantes nos pacientes
infectados pelo HIV-1+ não apresentam alterações epigenéticas no seu estado basal, quanto
ao nível de enzimas promotoras dessas modificações.

SOARES, L.S. Resultados
87
Figura 30. Níveis de expressão relativa dos genes que codificam as enzimas promotoras de modificações epigenéticas relacionadas à transcrição gênica. A expressão gênica foi quantificada pela técnica de PCR-Array em Tempo Real, utilizando-se os genes constitutivos: β-actina e GAPDH e a amostra controle como referência. Os dados estão expressos como mediana e intervalo interquartil.
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
ATF2
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
CARM1
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
ASH1L
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
KDM5B
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
HAT1
0.0
0.5
1.0
1.5
2.0Ex
pres
são
rela
tiva
PRMT5
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
KMT2C
012345678
Expr
essã
o re
lativ
a
KDM6B
HIV-1+TARV
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
KAT2B
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
SMYD3
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
KMT2E
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
KAT5
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
SUV39H1
0.0
0.5
1.0
1.5
2.0
Expr
essã
o re
lativ
a
SETD1B

SOARES, L.S. Resultados
88
Figura 31. Níveis de expressão relativa dos genes que codificam as enzimas promotoras de modificações epigenéticas relacionadas à repressão gênica: Histonas Deacetilases (HDAC). A expressão gênica foi quantificada pela técnica de PCR-Array em Tempo Real. Os dados estão expressos como mediana e intervalo interquartil.
0
1
2
3
4
Expr
essã
o re
lativ
aHDAC1
0.0
0.5
1.0
1.5
2.0
2.5
Expr
essã
o re
lativ
a
HDAC5
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC9
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC2
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC6
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC10
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC3
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC7
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC11
HIV-1 +TARV
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC4
0
1
2
3
4
Expr
essã
o re
lativ
a
HDAC8

SOARES, L.S. Resultados
89
Figura 32. Níveis de expressão relativa dos genes que codificam as enzimas promotoras de modificações epigenéticas relacionadas à repressão gênica. A expressão gênica foi quantificada pela técnica de PCR-Array em Tempo Real. Os dados estão expressos como mediana e intervalo interquartil.
0
1
2
3
4
Expr
essã
o re
lativ
aDNMT1
0
1
2
3
4
Expr
essã
o re
lativ
a
MBD2
0
1
2
3
4
Expr
essã
o re
lativ
a
SAP18
0
1
2
3
4
Expr
essã
o re
lativ
a
DNMT3A
0
1
2
3
4
Expr
essã
o re
lativ
a
MBD3
0
1
2
3
4
Expr
essã
o re
lativ
a
SAP30
HIV-1 +TARV
0
1
2
3
4
Expr
essã
o re
lativ
a
DNMT3B
0
1
2
3
4
Expr
essã
o re
lativ
a
RBBP4
0
1
2
3
4
Expr
essã
o re
lativ
a
SIN3A
0
1
2
3
4
Expr
essã
o re
lativ
a
RBBP7
0
1
2
3
4
Expr
essã
o re
lativ
a
TRDMT1
0
1
2
3
4
Expr
essã
o re
lativ
a
MECP2
0
1
2
3
Expr
essã
o re
lativ
a
CHD4

SOARES, L.S. Resultados
90
5.5.4. Investigação de modificações epigenéticas associadas às histonas em genes da
resposta imune de células NK
Os resultados descritos no tópico anterior demonstraram que a maioria das enzimas
envolvidas nas modificações de histonas não estão alteradas nas células NK durante a
infecção por HIV-1+. Dessa forma, investigou-se se essas alterações já teriam ocorrido nas
regiões promotoras dos genes relativos às citocinas que apresentaram diferença de expressão
entre os grupos estudados.
Dentre as modificações de histonas, avaliou-se a trimetilação da lisina 27 na histona H3
(H3K27me3) associada à repressão da transcrição gênica e a trimetilação da lisina 4 na
histona H3 (H3K4me3) relacionada à ativação. Para tanto, realizou-se o ensaio de
imunoprecipitação da cromatina utilizando anticorpos específicos para essas modificações e
posterior quantificação da região imunoprecipitada por PCR em Tempo Real. As sequências
analisadas eram relativas à região promotora das citocinas TNF-α e IFN-γ, previamente
descritas na literatura por influenciarem a transcrição gênica dessas proteínas.
Na figura 33 estão demonstrados os resultados obtidos quanto à citocina TNF-α por
alguns individuos de cada grupo. Se compararmos a porcentagem de DNA imunoprecipitado
relativo à quantidade de amostra inicial (% input), constata-se aumento da modificação
H3K27me3, relacionada à repressão em detrimento daquela associada à ativação (H3K4me3)
em todos os indivíduos analisados, exceto pelo CT19, que apresentou maior ativação do que
repressão. Devido à variação entre os indivíduos, não foi possível a observação de um padrão
entre os grupos analisados e a consequente correlação com os níveis de produção dessa
citocinas pelas células NK.
Quanto ao perfil de modificação de histonas observada na região promotora da citocina
IFN-γ, também não foram constatadas diferenças significativas, com aparente enriquecimento
de alterações associadas à repressão (H3K27me3) em detrimento daquelas relacionadas à
ativação (H3K4me3) em todos os grupos analisados (Figura 34).
Esses resultados não explicam o aumento na produção de citocinas pelas células NK no
grupo TARV como esperado. No entanto, justificam em parte a menor quantidade dessas
citocinas no grupo HIV-1+, embora os indivíduos controle também tenham apresentado perfil
de histonas mais relacionado à repressão. Além disso, analisou-se somente uma pequena
porção da região promotora, sendo que podem ocorrer modificações de histonas associadas a
regiões próximas às que foram amplificadas e que possuam maior influência na transcrição

SOARES, L.S. Resultados
91
das proteínas quantificadas. Nesse sentido, mais estudos devem ser realizados para
compreensão dos mecanismos epigenéticos diretamente relacionados à alteração de expressão
das diferentes moléculas nos indivíduos HIV-1+, submetidos ou não ao tratamento
antirretroviral, e a implicação de tais alterações a longo prazo.

SOARES, L.S. Resultados
92
Figura 33. Regulação epigenética via modificações de histonas associadas à região promotora da citocina TNF-α. Após separação por esferas magnéticas, 1x106 células NK foram empregadas no ensaio de imunoprecipitação da cromatina utilizando anticorpos específicos paras modificações de histonas associadas à repressão (H3K27me3) e transcrição (H3K4me3) gênicas. As regiões imunoprecipitadas foram quantificadas por PCR em Tempo Real utilizando-se primers específicos para a região promotora dessa proteína.
Figura 34. Regulação epigenética via modificações de histonas associadas à região promotora da citocina IFN-γ Após separação por esferas magnéticas, 1x106 células NK foram empregadas no ensaio de imunoprecipitação da cromatina utilizando anticorpos específicos paras modificações de histonas associadas à repressão (H3K27me3) e transcrição (H3K4me3) gênicas. As regiões imunoprecipitadas foram quantificadas por PCR em Tempo Real utilizando-se primers específicos para a região promotora dessa proteína.
CT16CT19
CT24CT29
HIV12HIV14
HIV17
TARV06
TARV15
TARV18
TARV210.0
0.5
1.0
1.5
2.0
51015
% In
put
H3K27me3 - TNF-α
CT16CT19
CT24CT29
HIV12HIV14
HIV17
TARV06
TARV15
TARV18
TARV210.0
0.5
1.0
1.5
2.0
51015
% In
put
H3K4me3 - TNF-α
CT16CT19
CT24CT29
HIV12HIV17
HIV19
TARV06
TARV15
TARV18
TARV210.0
0.5
1.0
1.5
2.0
345
% In
put
H3K27 - IFN-γ
CT16CT19
CT24CT29
HIV12HIV17
HIV19
TARV06
TARV15
TARV18
TARV210.0
0.5
1.0
1.5
2.0
345
% In
put
H3K4 IFN-γ

SOARES, L.S.
93
6. Discussão

SOARES, L.S. Discussão
94
6. Discussão
A infecção por HIV-1 tem sido caracterizada como doença inflamatória crônica devido
aos elevados níveis de citocinas pró-inflamatórias encontradas no plasma dos indivíduos
infectados [90], assim como pela ativação de células T, principalmente durante a infecção
aguda [91], e posterior evolução para estado de disfunção e exaustão dessas células com a
cronicidade da infecção [92]. Com os avanços da terapia antirretroviral, os indivíduos
infectados pelo HIV têm controlado mais eficientemente o desenvolvimento de infecções
oportunistas e a progressão da doença para o quadro de imunodeficiência. No entanto, em
detrimento do aumento da expectativa de vida, associada ao novo status imunológico após o
tratamento, esses indivíduos têm apresentado maior incidência de doenças não relacionadas à
AIDS, como doenças cardiovasculares, osteoporose, diabetes, e alguns tipos de câncer, as
quais geralmente são associadas ao processo de envelhecimento.
Apesar do tratamento antirretroviral diminuir significativamente a viremia, os
reservatórios virais não são completamente eliminados e a integridade de mucosa não é
inteiramente reestabelecida, permanecendo o estímulo ao sistema imunológico. Essa
observação levantou a hipótese de que a infecção por HIV-1 poderia estar relacionada à
aceleração do envelhecimento natural [32, 93]. Além disso, considerando que em pacientes
tratados, a progressão da doença tem sido mais relacionada às células da imunidade inata [94],
tem crescido o número de estudos objetivando maior compreensão do envolvimento dessas
células na patogênese do HIV. Nesse sentido, este trabalho descreveu as principais alterações
fenotípicas e funcionais de células natural killer na infecção por HIV-1 e a correlação com
modificações epigenéticas nessas células. Embora estas últimas tenham sido sutis, em
conjunto com as disfunções observadas, sugerimos possível grau de exaustão nas células NK
provenientes dos indivíduos HIV-1+ recentemente infectados, em paralelo ao estado de
imunosenescência celular naqueles sob tratamento antirretroviral regular há no mínimo seis
meses.
Os resultados obtidos a partir da análise plasmática da população em estudo
demonstram o perfil pró-inflamatório característico nos pacientes infectados pelo HIV-1+.
Esse perfil é evidente nos indivíduos recentemente infectados (grupo HIV-1+) como visto
pelos maiores níveis de sCD163, TNF-α, IP-10 e IL-10 (Figuras 4, 5 e 6). O aumento dos
níveis plasmáticos de CD163 solúvel está relacionado à clivagem dessa molécula a partir da
membrana de monócitos e/ou macrófagos, após ativação do receptor do tipo Toll, entre
outros, como forma de diminuir a ativação dos monócitos e a inflamação [95]. Em 2011,

SOARES, L.S. Discussão
95
Burdo e colaboradores [96] observaram pela primeira vez a associação desse marcador com a
replicação do HIV e sugeriram a utilização do sCD163 como um dos fatores preditivos de
progressão da doença.
As moléculas TNF-α, IP-10 e IL-10 estão entre o conjunto de citocinas descritas
previamente como aumentadas sistemicamente nos pacientes HIV-1+, especificamente
naqueles considerados progressores rápidos, os quais apresentam diminuição da contagem de
linfócitos T CD4+ abaixo de 350 células/mm3 em até doze meses após a infecção [31], assim
como naqueles que ainda não iniciaram a terapia antirretroviral [29]. Diversas células são
consideradas fontes dessas moléculas, sendo que em pacientes virêmicos a maior quantidade
de TNF-α é produzida por monócitos e sua produção excessiva compromete a integridade da
barreira epitelial favorecendo ainda mais a indução da inflamação sistêmica [97]. A produção
de IL-10 pelas células T reguladoras também está relacionada à progressão da doença, pois
seus altos níveis em decorrência do excesso de inflamação acabam por inibir o
desenvolvimento da resposta imune adaptativa protetora durante a infecção [98]. E por fim, a
quimiocina IP-10 foi destacada como fator preditivo, melhor indicador do prognóstico do
paciente do que a carga viral e a contagem de linfócitos T CD4+, logo ao inicio da infecção
[31]. As células T são consideradas a principal fonte de IP-10 em resposta ao estímulo de IFN
do tipo I, e a principal função dessa quimiocina é o recrutamento de células CXCR3+ para os
linfonodos, amplificando a inflamação e atraindo novos alvos para infecção pelo HIV [99],
justificando sua utilização na análise da progressão da doença.
Outra importante ferramenta para análise da progressão da infecção pelo HIV é a
quantificação da expressão das moléculas CD38 e HLA-DR, principalmente nas células T
CD8+ [100, 101]. Nesse sentido, um estudo recente também demonstrou a correlação entre a
expressão desses marcadores nas células NK e a progressão da doença [102]. Embora essas
moléculas não tenham sido analisadas em nosso estudo, realizamos a quantificação de outros
marcadores como o CD69 (Figura 15). Dados na literatura geralmente demonstram aumento
da expressão de CD69 nas células NK, caracterizando-as como ativadas fenotipicamente, mas
não funcionais [51]. Em nossos resultados, nos indivíduos HIV-1+ virgens de tratamento, a
expressão de CD69 estava ligeiramente aumentada, embora não significativa. Observou-se
discreto aumento dos receptores de ativação NKp30 e NKG2A em relação aos grupos
controle e TARV (Figura 15), no entanto os resultados obtidos não nos permitem tirar mais
conclusões nesse sentido, sendo necessária também a quantificação dessas proteínas, além de
sua expressão gênica.

SOARES, L.S. Discussão
96
Diversos fatores estão relacionados à inflamação sistêmica observada nos pacientes
HIV-1+, principalmente naqueles recentemente infectados e que ainda não iniciaram a terapia
antirretroviral como visto em nossos resultados. Dentre as causas estão a expansão de células
T HIV-específicas, ativação de células inatas em resposta aos ligantes de TLR, diminuição de
células T reguladoras, ocorrência de co-infecções, e principalmente a ruptura da integridade
de barreira epitelial da mucosa intestinal [103]. Esse último fenômeno favorece a translocação
de produtos microbianos como LPS, sendo que um de seus ligantes é a molécula CD14, além
do TLR4 [104]. Dessa forma, a quantificação de CD14 solúvel no plasma funciona como
medida indireta da translocação microbiana, e em nossos resultados, esse aumento foi
progressivo nos pacientes sob tratamento antirretroviral (Figura 4), assim como demonstrado
por Burdo et al. [96] e Sandler et al. [105]. Essas observações sugerem a não recuperação da
integridade da mucosa intestinal mesmo após o tratamento e evidenciam a importância do
início imediato da terapia antirretroviral, como recomendado atualmente.
Diante desse cenário inflamatório sistêmico encontrado nos pacientes HIV-1+, a
caracterização dos mecanismos envolvidos, em níveis celular e molecular, é essencial para
compreensão da patogênese do HIV e subsequente implementação de estratégias
translacionais visando a melhoria da qualidade de vida desses pacientes. Nesse sentido, os
linfócitos T, monócitos e macrófagos estão entre as células mais estudadas durante a infecção,
principalmente pela correlação positiva entre seu estado de ativação e a progressão da doença
[96, 100], e por serem os principais reservatórios virais [106, 107]. Em paralelo, as células
NK também são importante personagem neste cenário, sendo que seu perfil de ativação
também está correlacionado com a progressão da doença [102]. Desta forma, os resultados
aqui apresentados, especialmente aqueles relacionados a maior produção de citocinas
inflamatórias nos pacientes do grupo TARV, os quais possuíam maior tempo de infecção,
reforçam seu papel chave na resposta contra o HIV.
A maioria dos estudos com células NK durante a infecção por HIV-1 descrevem
alterações quanto à expressão dos receptores de ativação e inibição, assim como as variações
nas proporções das subpopulações destas células, verificadas por meio da técnica de
citometria de fluxo [50, 51, 108]. A partir da utilização dessa técnica, confirmamos neste
trabalho a diminuição da frequência e número de células NK nos pacientes infectados por
HIV-1+ e a recuperação parcial, embora não significativa, do número dessas células nos
pacientes tratados (Figura 9). Essa diminuição também já foi descrita por outros grupos de
pesquisa [51, 55, 109]. No entanto, existem resultados que demonstram aumento inicial das
células NK devido à expansão da subpopulação CD56dimCD16+ [108]. Esse aumento foi

SOARES, L.S. Discussão
97
relatado durante a fase aguda, em que os pacientes ainda não apresentavam resultado positivo
para o teste de ELISA, mas possuíam vírus detectável no organismo. Em nossos resultados,
não foi possível observar esse fenômeno, provavelmente devido aos diferentes tempos de
infecção em que os pacientes se encontravam, e considerando a mediana do tempo de
infecção de 5,3 e 156,4 meses nos grupos HIV-1+ e TARV, respectivamente, nota-se que
grande parte dos pacientes analisados não estavam mais durante a fase aguda da infecção.
Proporcionalmente, a queda no número de células NK da subpopulação citotóxica foi a
principal responsável pela diminuição significativa do total de células NK no grupo HIV-1+
(Figura 10). Em contrapartida, a subpopulação não funcional CD56-CD16+ permaneceu
elevada, inclusive nos pacientes HIV-1+ sob tratamento antirretroviral, sendo que nessa
subpopulação observou-se diminuição significativa do receptor de ativação NKp46,
característico das células NK (Figura 14). A expansão dessas células anérgicas foi observada
há cerca de 20 anos e está relacionada em grande parte à desregulação funcional apresentada
pelas células NK durante os diferentes estágios da infecção por HIV [54]. Com nossos
resultados, mostramos que essa desregulação foi mais evidente no grupo HIV-1+, como visto
pela menor porcentagem de citotoxicidade contra a linhagem tumoral K-562 e diminuição da
produção de citocinas pelas células NK desse grupo (Figuras 16-19). A redução da função
citotóxica em indivíduos com carga viral detectável também foi reportada por Mavilio et al.
[50], por meio da análise da expressão da molécula CD107a na superfície das células NK
como indicativo do fenômeno de desgranulação.
Interessantemente, as células NK do grupo HIV-1+ demonstraram certa responsividade
ao estímulo com IL-2 como observado pela produção de granzima B por essas células,
embora a diferença entre os grupos não tenha sido significativa (Figura 23). Vale ressaltar que
essa maior produção de granzima B foi observada naqueles pacientes com quantidade de
células NK CD56brightCD16- acima da mediana do grupo HIV-1+ (dados não mostrados). Um
fator bastante interessante e essencial ao entendimento deste resultado é que essa
subpopulação é a única a expressar constitutivamente o receptor heterotrimérico de alta
afinidade para a citocina IL-2, o que a torna responsiva mesmo a quantidades mínimas de IL-
2 [110]. Em nossos resultados, esse fato pode estar relacionado à maior produção de granzima
B pelas células NK especificamente nesses indivíduos.
Além da liberação de granzimas que estão relacionadas à indução de apoptose nas
células infectadas, as células NK também são importantes produtoras de citocinas que
auxiliam as demais células do sistema imune em suas funções essenciais. Enquanto os
indivíduos do grupo HIV-1+ exibiram retração da resposta funcional das células NK, os

SOARES, L.S. Discussão
98
pacientes do grupo TARV apresentaram recuperação da função citotóxica, acompanhada de
elevada produção de citocinas e quimiocinas por essas células, sendo que para alguns analitos,
como IFN-α2 e MCP-1, a produção foi significativamente maior até mesmo em relação ao
grupo controle (Figuras 17 e 19). Esse fenótipo mais ativado do grupo TARV sugeriu, a
principio, restauração da função das células NK em decorrência do tratamento antirretroviral.
No entanto, após analisarmos o perfil de produção de citocinas e quimiocinas por meio do
gráfico de radar e as correlações existentes entre essas moléculas (Figuras 20 e 21), nos
deparamos com um fenótipo totalmente diferente do grupo controle e, sendo assim, distante
do perfil inicial dessas células NK antes da infecção pelo HIV. Portanto, apesar do tratamento
antirretroviral reduzir a viremia e auxiliar na recuperação parcial dos níveis de linfócitos T
CD4+, estes pacientes apresentam inflamação residual comprometendo a completa restauração
da função das células NK.
Segundo a literatura, o fenômeno de exaustão ocorre após exposição excessiva e
contínua a determinado estímulo [111], fato que ocorre durante o início da infecção pelo HIV-
1 nos indivíduos virêmicos. Em paralelo, existem dois outros eventos denominados anergia e
imunosenescência, os quais também estão intimamente relacionados à disfunção da resposta
imune. Na anergia, o estímulo é subótimo; enquanto na senescência, a causa está associada à
permanência do estímulo de forma repetitiva mesmo após o desenvolvimento da resposta
imune primária. Este último compara-se ao cenário encontrado nos pacientes do grupo
TARV, que mesmo após tratamento antirretroviral, apresentam inflamação residual e
replicação viral persistente, principalmente naqueles locais denominados santuários
anatômicos, devido à baixa penetração das drogas antirretrovirais, como: mucosa intestinal,
linfonodos, trato genital e sistema nervoso central [112].
O perfil super ativado das células NK do grupo TARV corrobora com a ideia de
imunosenescência nas células desses pacientes. Embora não tenham sido analisados neste
estudo os marcadores característicos de senescência, como atividade da telomerase e tamanho
dos telômeros [113], os resultados obtidos quanto à produção de ROS, que estava elevada
(Figura 24), são outro forte indicativo da ocorrência desse fenômeno nas células NK do grupo
TARV. Isto porque o aumento do estresse oxidativo celular está entre os fatores
característicos da senescência celular [114]. Além disso, observou-se diminuição da expressão
do receptor de inibição CD158a na subpopulação de células NK citotóxicas do grupo TARV
(Figura 12) e maior expressão do receptor de ativação NKp46 na subpopulação
CD56brightCD16- deste grupo (Figura 14), o que pode ter favorecido o maior estado de

SOARES, L.S. Discussão
99
ativação dessas células e a subsequente maior capacidade citotóxica e de produção de
citocinas.
A primeira evidência clínica do envelhecimento celular durante a infecção por HIV
refere-se ao acúmulo de células T CD8+ que perderam a expressão do correceptor CD28 e
aumentaram a expressão da molécula CD57. Essas células possuem capacidade reduzida de
produção de IL-2 e proliferação, além de apresentarem encurtamento do comprimento dos
telômeros [115]. No entanto, os estudos ainda não são conclusivos quanto à associação desse
fenótipo celular à imunosenescência, assim como sua relação com a progressão da doença.
Além disso, é importante ressaltar que a molécula CD57 foi inicialmente descrita em células
NK e é considerada um marcador de maturação final dessas células e não necessariamente um
marcador de imunosenescência [116]. Entretanto, há relatos do acúmulo de células NK
CD57+ com o avanço da idade, e também em hepatites virais [117, 118]. Dessa forma, mais
estudos são necessários com o objetivo de compreender as implicações do perfil relacionado à
senescência na infecção por HIV, visto que os pacientes HIV+ tem aumentado sua
expectativa de vida ao mesmo tempo em que passaram a apresentar maior incidência de co-
morbidades, que são mais frequentes em indivíduos idosos.
Considerando que a imunosenescência caracteriza-se como um estado de inflamação
crônica sistêmica semelhante ao observado nos pacientes HIV-1+, existem diversos fatores
que podem estar relacionados à indução desse fenômeno. Entre eles estão: a própria infecção
viral, a terapia antirretroviral, assim como a ocorrência de co-infecções [93]. Dentre os
antirretrovirais utilizados no tratamento da infecção por HIV-1, os inibidores da transcriptase
reversa como o AZT (zidovudina) e Tenofovir inibem a atividade da telomerase in vitro e
estão relacionados ao encurtamento do telômero, evidenciando a influência também do
tratamento antirretroviral na senescência celular além da infecção per se [119, 120]. Além
disso, co-infecções também favorecem a ativação do sistema imune nos pacientes HIV-1+,
como a co-infecção com CMV, que mesmo em indivíduos assintomáticos estava associada ao
aumento da ativação de células T e da replicação viral nos pacientes HIV-1+, tratados ou não
[121]. Nos pacientes analisados em nosso estudo, a maioria possuía anticorpos IgG positivo
para o CMV (dados não mostrados), no entanto não temos como afirmar se esses indivíduos
apresentavam a infecção ativa no período em que foram realizadas as coletas e não fica clara a
sua relação com o estado de ativação das células NK nesses pacientes.
Enquanto nas células NK do grupo TARV sugerimos o estado de imunosenescência, as
células do grupo HIV-1+ apresentaram características mais condizentes com o perfil de
exaustão celular. Dentre as propriedades observadas nesse sentido estão a menor porcentagem

SOARES, L.S. Discussão
100
de citotoxicidade contra a linhagem tumoral K-562 (Figura 16) e reduzida produção de
citocinas (Figura 17-19). Outro aspecto analisado por nós e que também está relacionado à
exaustão celular foi a expressão de receptores de inibição nestas células (Figura 11). Embora
o número de células NK positivas para os três KIR analisados tenha diminuído no grupo HIV-
1+, observou-se aumento significativo da mediana da intensidade de fluorescência para o
receptor NKAT2. Em contrapartida, foi interessante notar que essas células também possuíam
maior expressão do receptor de ativação NKp46 (Figura 13), o que contradiz com o perfil de
exaustão celular, pois nesse estado, as células diminuem a expressão de marcadores
característicos das células NK como o NKp46. Portanto, para confirmação da hipótese aqui
sugerida, outras moléculas devem ser quantificadas como PD-1 e Tim-3, que apresentam forte
correlação com o fenômeno de exaustão celular [111]. Em resumo, verificamos neste trabalho
que o estado funcional das células NK nas diferentes fases da doença (infecção recente versus
crônica associada ao tratamento antirretroviral) encontra-se alterado e parece estar associado à
progressão da doença. Nos pacientes do grupo TARV que estão infectados há mais tempo, as
células NK modificam seus perfis de ativação, evoluindo de um padrão de exaustão como
observado na infecção recente para a imunosenescência.
Após analisarmos os efeitos que a infecção pelo HIV induziu nas células NK dos
pacientes investigados, nossa principal hipótese era de que essas alterações fenotípicas e
funcionais das células NK discutidas até aqui seriam resultado de modificações epigenéticas
nestas células. Isto porque sabemos que infecções podem alterar o estado funcional do sistema
imune e já foi mostrado que modificações epigenéticas estariam envolvidas nesta
programação celular. A primeira análise feita nesse sentido foi a quantificação da metilação
global do DNA em células NK obtidas do sangue periférico dos indivíduos infectados pelo
HIV-1, em tratamento antirretroviral ou não (Figura 25). Interessantemente, observou-se
aumento significativo da metilação global do DNA naqueles pacientes virgens de tratamento e
que estavam em estágio avançado da doença, com carga viral maior que 10 000 cópias/mL e
células T CD4+ menor que 200 por mm3. Esses resultados sugerem uma relação de causa e
efeito entre a maior porcentagem de metilação do DNA e o estado disfuncional das células
NK desses pacientes, em que a não responsividade dessas células poderia ser explicada pela
menor expressão de moléculas essenciais para execução de sua função durante a infecção
viral.
Ainda existem poucos estudos que investigam alterações epigenéticas em decorrência
da infecção por HIV-1, visto que a maioria está relacionada à análise de modificações
associadas à latência viral. Recentemente, foi publicado um trabalho com gêmeos

SOARES, L.S. Discussão
101
monozigóticos em que um deles estava infectado pelo HIV-1 e o outro não [122]. A partir da
quantificação da metilação global em células mononucleares do sangue periférico, notou-se
que o gêmeo infectado possuía maior percentual de metilação global em comparação ao
gêmeo saudável, corroborando com nossos dados. Outro trabalho também demonstrou
resultado semelhante utilizando modelo de infecção in vitro, com aumento da porcentagem de
metilação global após 12 horas de cultura das células mononucleares com o HIV-1 [123].
Considerando a provável influência do mecanismo de metilação do DNA no fenótipo e
função das células NK dos pacientes HIV-1+, realizou-se a análise do perfil de metilação
gene-específico. Para tanto, quantificou-se a porcentagem de metilação das ilhas CpG
presentes na região promotora do gene KIR2DL2. Esse receptor foi quantificado por
citometria de fluxo por meio da utilização do anticorpo anti-NKAT2, sendo que poucas
células NK do grupo HIV-1+ expressavam esse receptor em grande quantidade como
observado pela mediana da intensidade de fluorescência dessa molécula (Figura 11). Para
quantificação da metilação gene-específico, empregou-se a técnica de modificação por
bissulfito seguida da clonagem e sequenciamento do DNA de no mínimo oito clones de cada
amostra pelo método de Sanger. No entanto, não se observou diferença significativa quanto à
porcentagem de metilação da região analisada entre os diferentes grupos (Figuras 27-29).
Embora um estudo prévio tenha relacionado o maior percentual de metilação dessa região do
gene KIR2DL2 com a menor expressão da proteína [81], pode ser que, em nosso contexto,
outras regiões e/ou até mesmo outros mecanismos estejam envolvidos na regulação da
expressão desse receptor nas células NK dos indivíduos HIV-1+.
Interessantemente, a partir dos resultados obtidos do sequenciamento de DNA antes
da modificação por bissulfito, observou-se a ocorrência de dois polimorfismos na sequência
analisada do gene KIR2DL2 de um dos pacientes do grupo HIV-1+ (HIV11) (Figura 26).
Esses dois SNPs foram previamente descritos em estudos relacionados à história da evolução
e adaptação de populações [124]. No entanto, não foram encontrados trabalhos relacionando
esses polimorfismos com alterações da função de células NK. Dessa forma, não podemos
afirmar se essa troca de bases na região intrônica do KIR2DL2 teria alguma implicação na
função dessas células e consequentemente na resposta imune desse indivíduo contra o HIV.
Embora dados da literatura demonstrem que dentre os polimorfismos, as trocas de base única
(SNPs – single nucleotide polymorphims) são os mais frequentes e geralmente estão
localizados ao longo de todo o genoma, sendo relativamente estáveis e variando entre as
populações estudadas [125].

SOARES, L.S. Discussão
102
Outra abordagem realizada nesse estudo com objetivo de investigar as alterações
epigenéticas, foi a quantificação da expressão gênica, pelas células NK, de enzimas que
promovem essas modificações, ou seja, que estão relacionadas à ativação ou repressão da
transcrição gênica. Apesar de não ter sido observada diferença significativa para nenhuma das
enzimas avaliadas, algumas delas apresentaram sua expressão relativamente aumentada nos
indivíduos infectados em comparação aos controles (Figuras 30-32). Dentre essas enzimas
está a KDM6B, envolvida na demetilação da lisina 27 da histona 3, propiciando a transcrição
gênica e também está relacionada a respostas inflamatórias ao participar da diferenciação de
macrófagos [126]. Outras enzimas que estavam ligeiramente aumentadas foram as histonas
deacetilases 4, 5, 10 e 11. Estas últimas promovem a deacetilação de histonas e favorecem a
repressão da transcrição gênica [127].
Embora não tenhamos observado diferenças significativas quanto à expressão das
enzimas envolvidas na indução das modificações epigenéticas, podemos sugerir que essas
alterações não estejam associadas às disfunções observadas nas células NK dos pacientes
HIV-1+ ou ainda que, no momento analisado, essas células já teriam retornado ao nível de
expressão basal dessas enzimas. Além disso, não investigamos se as células NK dos
indivíduos estudados estavam infectadas com o HIV, o que poderia exercer efeito mais
significativo na modulação dessas enzimas como observado por alguns autores. Em modelos
de infecção in vitro foi relatado aumento da histona deacetilase 2 (HDAC2) assim como da
DNMT1, ambas associadas ao silenciamento gênico [128, 129], o que pode favorecer tanto a
indução da latência viral quanto a diminuição da função da célula infectada. Nos dois casos,
essas modificações estariam relacionadas a um mau prognóstico para o paciente HIV-1+.
Considerando a hipótese de que algumas modificações epigenéticas já teriam ocorrido
nas células NK e que por isso as enzimas não apresentaram mudanças significativas,
analisamos também o padrão de metilação das histonas associadas à sequência de DNA de
algumas das citocinas diferencialmente expressas nos grupos HIV-1+ e TARV(Figuras 33 e
34). Embora as regiões analisadas após imunoprecipitação da cromatina com anticorpos
específicos para as modificações de ativação (H3K4me3) e repressão (H3K27me3) tenham
sido previamente associadas com mudanças na expressão do IFN-γ e TNF-α [86, 87], em
nossos resultados não observamos diferenças significativas entre os indivíduos testados.
Apesar disso, não podemos afirmar que esse perfil de histonas não está relacionado à menor
expressão dessas citocinas no grupo HIV-1+ e maior no grupo TARV, visto que outras
regiões desses e de outros genes também precisariam ser quantificadas.

SOARES, L.S. Discussão
103
De maneira geral, as células NK circulantes do sangue periférico de pacientes HIV-1+
apresentaram mudanças epigenéticas sutis quando comparadas às notórias alterações
funcionais e fenotípicas encontradas nestas células. Grande parte da literatura existente sobre
células NK e modificações epigenéticas está associada à regulação da expressão dos
receptores característicos destas células durante seu desenvolvimento. Dessa forma, este é um
dos primeiros trabalhos a investigar a existência de modificações epigenéticas ex vivo, em
células NK provenientes diretamente de pacientes infectados pelo HIV-1. Mais estudos
devem ser realizados nesse sentido com o intuito de compreendermos detalhadamente a
relação desta população celular com a patogênese do HIV, e sobretudo com o fenômeno de
senescência evidenciado. Com este trabalho e o conjunto de dados aqui discutidos, temos o
intuito de promover avanços na área de ciência básica, os quais poderão, de forma
translacional, influenciar na terapia do paciente. Isto porque tão importante quanto a redução
da carga viral, está o controle da inflamação crônica em seus níveis celular e molecular, assim
como é de suma relevância o reestabelecimento das funções de células do sistema imune inato
que cooperam para manter o organismo vigilante, como as células natural killer.

SOARES, L.S.
104
7. Conclusões

SOARES, L.S. Conclusões
105
7. Conclusões
Neste trabalho demonstramos a associação entre o perfil inflamatório sistêmico de
pacientes infectados pelo HIV-1+ e as alterações de fenótipo e função de células NK. Nos
pacientes recentemente infectados e virgens de tratamento, as células NK apresentaram perfil
sugestivo de exaustão imune, com retração da produção de citocinas e função citotóxica.
Naqueles indivíduos HIV-1+ sob estágio avançado da doença, essas disfunções foram mais
pronunciadas e parecem estar correlacionadas ao maior grau de metilação do DNA nessas
células. Por outro lado, células NK de pacientes sob tratamento antirretroviral e com maior
tempo de infecção apresentaram um padrão de ativação indicativo do estágio de
imunosenescência, evidenciado pela elevada produção de citocinas e maior estresse oxidativo
nestas células. E por fim, a maioria das modificações epigenéticas observadas nas células NK
foram sutis, sugerindo o envolvimento de outros mecanismos na mudança fenotípica e
funcional dessas células durante a infecção pelo HIV-1.

SOARES, L.S.
106
8. Referências Bibliográficas

SOARES, L.S. Referências Bibliográficas
107
8. Referências Bibliográficas 1. (CDC), C.f.D.C., Update on acquired immune deficiency syndrome (AIDS)--United States. MMWR Morb Mortal Wkly Rep, 1982. 31(37): p. 507-8, 513-4. 2. Masur, H., et al., An outbreak of community-acquired Pneumocystis carinii pneumonia: initial manifestation of cellular immune dysfunction. N Engl J Med, 1981. 305(24): p. 1431-8. 3. Hymes, K.B., et al., Kaposi's sarcoma in homosexual men-a report of eight cases. Lancet, 1981. 2(8247): p. 598-600. 4. (CDC), C.f.D.C., A cluster of Kaposi's sarcoma and Pneumocystis carinii pneumonia among homosexual male residents of Los Angeles and Orange Counties, California. MMWR Morb Mortal Wkly Rep, 1982. 31(23): p. 305-7.
5. Gallo, R.C., et al., Isolation of human T-cell leukemia virus in acquired immune deficiency syndrome (AIDS). Science, 1983. 220(4599): p. 865-7.
6. Gelmann, E.P., et al., Proviral DNA of a retrovirus, human T-cell leukemia virus, in two patients with AIDS. Science, 1983. 220(4599): p. 862-5.
7. Barré-Sinoussi, F., et al., Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science, 1983. 220(4599): p. 868-71.
8. Rabson, A.B. and M.A. Martin, Molecular organization of the AIDS retrovirus. Cell, 1985. 40(3): p. 477-80.
9. Sanchez-Pescador, R., et al., Nucleotide sequence and expression of an AIDS-associated retrovirus (ARV-2). Science, 1985. 227(4686): p. 484-92.
10. Wain-Hobson, S., et al., Nucleotide sequence of the AIDS virus, LAV. Cell, 1985. 40(1): p. 9-17.
11. Pépin, J., The origins of AIDS: from patient zero to ground zero. J Epidemiol Community Health, 2013. 67(6): p. 473-5.
12. Gao, F., et al., Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature, 1999. 397(6718): p. 436-41.
13. Hirsch, V.M., et al., An African primate lentivirus (SIVsm) closely related to HIV-2. Nature, 1989. 339(6223): p. 389-92.
14. Wright, K., AIDS therapy. First tentative signs of therapeutic promise. Nature, 1986. 323(6086): p. 283.
15. Adamson, C.S. and E.O. Freed, Recent progress in antiretrovirals--lessons from resistance. Drug Discov Today, 2008. 13(9-10): p. 424-32.
16. Galvão, J., Brazil and access to HIV/AIDS drugs: a question of human rights and public health. Am J Public Health, 2005. 95(7): p. 1110-6.
17. Allers, K., et al., Evidence for the cure of HIV infection by CCR5Δ32/Δ32 stem cell transplantation. Blood, 2011. 117(10): p. 2791-9.
18. Lampton, L.M., Functional HIV cure achieved in Mississippi. J Miss State Med Assoc, 2013. 54(4): p. 94.
19. Liu, C., et al., HIV-1 functional cure: will the dream come true? BMC Med, 2015. 13(1): p. 284.

SOARES, L.S. Referências Bibliográficas
108
20. Krakower, D.S., S. Jain, and K.H. Mayer, Antiretrovirals for primary HIV prevention: the current status of pre- and post-exposure prophylaxis. Curr HIV/AIDS Rep, 2015. 12(1): p. 127-38.
21. UNAIDS, World AIDS Day - Fact sheet 2015. 2015. 22. Saúde, M.d., Boletim epidemiológico HIV-AIDS. 2015: Brasília.
23. Lackner, A.A., M.M. Lederman, and B. Rodriguez, HIV pathogenesis: the host. Cold Spring Harb Perspect Med, 2012. 2(9): p. a007005.
24. Brandenberg, O.F., et al., The HIV-1 Entry Process: A Stoichiometric View. Trends Microbiol, 2015. 23(12): p. 763-74.
25. Salazar-Gonzalez, J.F., et al., Genetic identity, biological phenotype, and evolutionary pathways of transmitted/founder viruses in acute and early HIV-1 infection. J Exp Med, 2009. 206(6): p. 1273-89. 26. Broder, C.C. and R.G. Collman, Chemokine receptors and HIV. J Leukoc Biol, 1997. 62(1): p. 20-9. 27. Haase, A.T., Early events in sexual transmission of HIV and SIV and opportunities for interventions. Annu Rev Med, 2011. 62: p. 127-39. 28. Klatt, N.R., et al., Immune activation and HIV persistence: implications for curative approaches to HIV infection. Immunol Rev, 2013. 254(1): p. 326-42. 29. Espindola, M.S., et al., Dysregulated Immune Activation in Second-Line HAART HIV+ Patients Is Similar to That of Untreated Patients. PLoS One, 2015. 10(12): p. e0145261. 30. Brenchley, J.M., et al., Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med, 2006. 12(12): p. 1365-71. 31. Liovat, A.S., et al., Acute plasma biomarkers of T cell activation set-point levels and of disease progression in HIV-1 infection. PLoS One, 2012. 7(10): p. e46143. 32. Deeks, S.G., R. Tracy, and D.C. Douek, Systemic effects of inflammation on health during chronic HIV infection. Immunity, 2013. 39(4): p. 633-45. 33. Streeck, H., D. van Bockel, and A. Kelleher, T-cell responses in primary HIV-1 infection. Curr Opin HIV AIDS, 2008. 3(1): p. 52-9. 34. Altfeld, M., et al., DCs and NK cells: critical effectors in the immune response to HIV-1. Nat Rev Immunol, 2011. 11(3): p. 176-86. 35. Berger, B.J., F. Hussain, and K. Roistacher, Bacterial infections in HIV-infected patients. Infect Dis Clin North Am, 1994. 8(2): p. 449-65. 36. Biron, C.A., More things in heaven and earth: defining innate and adaptive immunity. Nat Immunol, 2010. 11(12): p. 1080-2. 37. Paust, S., B. Senman, and U.H. von Andrian, Adaptive immune responses mediated by natural killer cells. Immunol Rev, 2010. 235(1): p. 286-96. 38. Kiessling, R., E. Klein, and H. Wigzell, "Natural" killer cells in the mouse. I. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur J Immunol, 1975. 5(2): p. 112-7.
39. Spits, H., et al., Innate lymphoid cells--a proposal for uniform nomenclature. Nat Rev Immunol, 2013. 13(2): p. 145-9.

SOARES, L.S. Referências Bibliográficas
109
40. Jost, S. and M. Altfeld, Control of human viral infections by natural killer cells. Annu Rev Immunol, 2013. 31: p. 163-94. 41. Robertson, M.J. and J. Ritz, Biology and clinical relevance of human natural killer cells. Blood, 1990. 76(12): p. 2421-38. 42. Lanier, L.L., Up on the tightrope: natural killer cell activation and inhibition. Nat Immunol, 2008. 9(5): p. 495-502. 43. Höglund, P. and P. Brodin, Current perspectives of natural killer cell education by MHC class I molecules. Nat Rev Immunol, 2010. 10(10): p. 724-34. 44. Vivier, E., et al., Functions of natural killer cells. Nat Immunol, 2008. 9(5): p. 503-10.
45. Brenner, B.G., et al., Natural killer cell function in patients with acquired immunodeficiency syndrome and related diseases. J Leukoc Biol, 1989. 46(1): p. 75-83.
46. Brenner, B.G., C. Gryllis, and M.A. Wainberg, Role of antibody-dependent cellular cytotoxicity and lymphokine-activated killer cells in AIDS and related diseases. J Leukoc Biol, 1991. 50(6): p. 628-40. 47. Ahmad, A. and J. Menezes, Defective killing activity against gp120/41-expressing human erythroleukaemic K562 cell line by monocytes and natural killer cells from HIV-infected individuals. AIDS, 1996. 10(2): p. 143-9.
48. Portales, P., et al., Interferon-alpha restores HIV-induced alteration of natural killer cell perforin expression in vivo. AIDS, 2003. 17(4): p. 495-504.
49. De Maria, A., et al., The impaired NK cell cytolytic function in viremic HIV-1 infection is associated with a reduced surface expression of natural cytotoxicity receptors (NKp46, NKp30 and NKp44). Eur J Immunol, 2003. 33(9): p. 2410-8. 50. Mavilio, D., et al., Natural killer cells in HIV-1 infection: dichotomous effects of viremia on inhibitory and activating receptors and their functional correlates. Proc Natl Acad Sci U S A, 2003. 100(25): p. 15011-6.
51. Vieillard, V., et al., Specific phenotypic and functional features of natural killer cells from HIV-infected long-term nonprogressors and HIV controllers. J Acquir Immune Defic Syndr, 2010. 53(5): p. 564-73. 52. Mavilio, D., et al., Characterization of CD56-/CD16+ natural killer (NK) cells: a highly dysfunctional NK subset expanded in HIV-infected viremic individuals. Proc Natl Acad Sci U S A, 2005. 102(8): p. 2886-91.
53. Oliva, A., et al., Natural killer cells from human immunodeficiency virus (HIV)-infected individuals are an important source of CC-chemokines and suppress HIV-1 entry and replication in vitro. J Clin Invest, 1998. 102(1): p. 223-31. 54. Hu, P.F., et al., Natural killer cell immunodeficiency in HIV disease is manifest by profoundly decreased numbers of CD16+CD56+ cells and expansion of a population of CD16dimCD56- cells with low lytic activity. J Acquir Immune Defic Syndr Hum Retrovirol, 1995. 10(3): p. 331-40. 55. Fauci, A.S., D. Mavilio, and S. Kottilil, NK cells in HIV infection: paradigm for protection or targets for ambush. Nat Rev Immunol, 2005. 5(11): p. 835-43. 56. Bernstein, H.B., et al., CD4+ NK cells can be productively infected with HIV, leading to downregulation of CD4 expression and changes in function. Virology, 2009. 387(1): p. 59-66.

SOARES, L.S. Referências Bibliográficas
110
57. Collins, K.L., et al., HIV-1 Nef protein protects infected primary cells against killing by cytotoxic T lymphocytes. Nature, 1998. 391(6665): p. 397-401. 58. Cohen, G.B., et al., The selective downregulation of class I major histocompatibility complex proteins by HIV-1 protects HIV-infected cells from NK cells. Immunity, 1999. 10(6): p. 661-71.
59. Martin, M.P., et al., Epistatic interaction between KIR3DS1 and HLA-B delays the progression to AIDS. Nat Genet, 2002. 31(4): p. 429-34.
60. Martin, M.P., et al., Innate partnership of HLA-B and KIR3DL1 subtypes against HIV-1. Nat Genet, 2007. 39(6): p. 733-40.
61. Alter, G., et al., HLA class I subtype-dependent expansion of KIR3DS1+ and KIR3DL1+ NK cells during acute human immunodeficiency virus type 1 infection. J Virol, 2009. 83(13): p. 6798-805. 62. Uhrberg, M., Shaping the human NK cell repertoire: an epigenetic glance at KIR gene regulation. Mol Immunol, 2005. 42(4): p. 471-5. 63. Scott-Algara, D., et al., Cutting edge: increased NK cell activity in HIV-1-exposed but uninfected Vietnamese intravascular drug users. J Immunol, 2003. 171(11): p. 5663-7. 64. Portela, A. and M. Esteller, Epigenetic modifications and human disease. Nat Biotechnol, 2010. 28(10): p. 1057-68. 65. Gómez-Díaz, E., et al., Epigenetics of host-pathogen interactions: the road ahead and the road behind. PLoS Pathog, 2012. 8(11): p. e1003007. 66. Schneider, R., et al., Histone H3 lysine 4 methylation patterns in higher eukaryotic genes. Nat Cell Biol, 2004. 6(1): p. 73-7. 67. Cao, R. and Y. Zhang, The functions of E(Z)/EZH2-mediated methylation of lysine 27 in histone H3. Curr Opin Genet Dev, 2004. 14(2): p. 155-64. 68. Biel, M., V. Wascholowski, and A. Giannis, Epigenetics--an epicenter of gene regulation: histones and histone-modifying enzymes. Angew Chem Int Ed Engl, 2005. 44(21): p. 3186-216.
69. Wen, H., et al., Epigenetic regulation of dendritic cell-derived interleukin-12 facilitates immunosuppression after a severe innate immune response. Blood, 2008. 111(4): p. 1797-804. 70. Wen, H., et al., Dendritic cells at the interface of innate and acquired immunity: the role for epigenetic changes. J Leukoc Biol, 2008. 83(3): p. 439-46. 71. Ishii, M., et al., Epigenetic regulation of the alternatively activated macrophage phenotype. Blood, 2009. 114(15): p. 3244-54. 72. Dixon, S.E., et al., A decade of epigenetic research in Toxoplasma gondii. Mol Biochem Parasitol, 2010. 173(1): p. 1-9. 73. Chookajorn, T., et al., Epigenetic memory at malaria virulence genes. Proc Natl Acad Sci U S A, 2007. 104(3): p. 899-902. 74. Marinus, M.G. and J. Casadesus, Roles of DNA adenine methylation in host-pathogen interactions: mismatch repair, transcriptional regulation, and more. FEMS Microbiol Rev, 2009. 33(3): p. 488-503.

SOARES, L.S. Referências Bibliográficas
111
75. Pennini, M.E., et al., Mycobacterium tuberculosis 19-kDa lipoprotein inhibits IFN-gamma-induced chromatin remodeling of MHC2TA by TLR2 and MAPK signaling. J Immunol, 2006. 176(7): p. 4323-30.
76. Marazzi, I., et al., Suppression of the antiviral response by an influenza histone mimic. Nature, 2012. 483(7390): p. 428-33.
77. Ay, E., et al., Epigenetics of HIV infection: promising research areas and implications for therapy. AIDS Rev, 2013. 15(3): p. 181-8.
78. Donahue, D.A. and M.A. Wainberg, Cellular and molecular mechanisms involved in the establishment of HIV-1 latency. Retrovirology, 2013. 10: p. 11.
79. Deeks, S.G., HIV: Shock and kill. Nature, 2012. 487(7408): p. 439-40. 80. Livak, K.J. and T.D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods, 2001. 25(4): p. 402-8. 81. Gao, X.N., et al., Demethylating treatment suppresses natural killer cell cytolytic activity. Mol Immunol, 2009. 46(10): p. 2064-70. 82. Corpet, F., Multiple sequence alignment with hierarchical clustering. Nucleic Acids Res, 1988. 16(22): p. 10881-90. 83. Li, L.C. and R. Dahiya, MethPrimer: designing primers for methylation PCRs. Bioinformatics, 2002. 18(11): p. 1427-31. 84. Ewing, B., et al., Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res, 1998. 8(3): p. 175-85. 85. Kumaki, Y., M. Oda, and M. Okano, QUMA: quantification tool for methylation analysis. Nucleic Acids Res, 2008. 36(Web Server issue): p. W170-5. 86. Morinobu, A., Y. Kanno, and J.J. O'Shea, Discrete roles for histone acetylation in human T helper 1 cell-specific gene expression. J Biol Chem, 2004. 279(39): p. 40640-6. 87. Lee, J.Y., et al., Histone acetylation and chromatin conformation are regulated separately at the TNF-alpha promoter in monocytes and macrophages. J Leukoc Biol, 2003. 73(6): p. 862-71.
88. Shannon, P., et al., Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res, 2003. 13(11): p. 2498-504.
89. Cooper, M.A., T.A. Fehniger, and M.A. Caligiuri, The biology of human natural killer-cell subsets. Trends Immunol, 2001. 22(11): p. 633-40.
90. Stacey, A.R., et al., Induction of a striking systemic cytokine cascade prior to peak viremia in acute human immunodeficiency virus type 1 infection, in contrast to more modest and delayed responses in acute hepatitis B and C virus infections. J Virol, 2009. 83(8): p. 3719-33.
91. Papagno, L., et al., Immune activation and CD8+ T-cell differentiation towards senescence in HIV-1 infection. PLoS Biol, 2004. 2(2): p. E20.
92. El-Far, M., et al., T-cell exhaustion in HIV infection. Curr HIV/AIDS Rep, 2008. 5(1): p. 13-9.
93. Appay, V. and A.D. Kelleher, Immune activation and immune aging in HIV infection. Curr Opin HIV AIDS, 2016. 11(2): p. 242-9.

SOARES, L.S. Referências Bibliográficas
112
94. Hunt, P.W., et al., Gut epithelial barrier dysfunction and innate immune activation predict mortality in treated HIV infection. J Infect Dis, 2014. 210(8): p. 1228-38. 95. Weaver, L.K., et al., Pivotal advance: activation of cell surface Toll-like receptors causes shedding of the hemoglobin scavenger receptor CD163. J Leukoc Biol, 2006. 80(1): p. 26-35. 96. Burdo, T.H., et al., Soluble CD163 made by monocyte/macrophages is a novel marker of HIV activity in early and chronic infection prior to and after anti-retroviral therapy. J Infect Dis, 2011. 204(1): p. 154-63.
97. Dutertre, C.A., et al., Pivotal role of M-DC8⁺ monocytes from viremic HIV-infected patients in TNFα overproduction in response to microbial products. Blood, 2012. 120(11): p. 2259-68.
98. Estes, J.D., et al., Premature induction of an immunosuppressive regulatory T cell response during acute simian immunodeficiency virus infection. J Infect Dis, 2006. 193(5): p. 703-12. 99. Foley, J.F., et al., Roles for CXC chemokine ligands 10 and 11 in recruiting CD4+ T cells to HIV-1-infected monocyte-derived macrophages, dendritic cells, and lymph nodes. J Immunol, 2005. 174(8): p. 4892-900.
100. Giorgi, J.V., et al., Shorter survival in advanced human immunodeficiency virus type 1 infection is more closely associated with T lymphocyte activation than with plasma virus burden or virus chemokine coreceptor usage. J Infect Dis, 1999. 179(4): p. 859-70. 101. Sodora, D.L. and G. Silvestri, Immune activation and AIDS pathogenesis. AIDS, 2008. 22(4): p. 439-46. 102. Kuri-Cervantes, L., et al., Activation of NK cells is associated with HIV-1 disease progression. J Leukoc Biol, 2014. 96(1): p. 7-16. 103. Paiardini, M. and M. Müller-Trutwin, HIV-associated chronic immune activation. Immunol Rev, 2013. 254(1): p. 78-101. 104. Gioannini, T.L. and J.P. Weiss, Regulation of interactions of Gram-negative bacterial endotoxins with mammalian cells. Immunol Res, 2007. 39(1-3): p. 249-60. 105. Sandler, N.G., et al., Plasma levels of soluble CD14 independently predict mortality in HIV infection. J Infect Dis, 2011. 203(6): p. 780-90. 106. Chomont, N., et al., HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat Med, 2009. 15(8): p. 893-900. 107. Campbell, J.H., et al., The importance of monocytes and macrophages in HIV pathogenesis, treatment, and cure. AIDS, 2014. 28(15): p. 2175-87. 108. Alter, G., et al., Sequential deregulation of NK cell subset distribution and function starting in acute HIV-1 infection. Blood, 2005. 106(10): p. 3366-9. 109. Tarazona, R., et al., Selective depletion of CD56(dim) NK cell subsets and maintenance of CD56(bright) NK cells in treatment-naive HIV-1-seropositive individuals. J Clin Immunol, 2002. 22(3): p. 176-83.
110. Caligiuri, M.A., et al., Functional consequences of interleukin 2 receptor expression on resting human lymphocytes. Identification of a novel natural killer cell subset with high affinity receptors. J Exp Med, 1990. 171(5): p. 1509-26. 111. Wherry, E.J. and M. Kurachi, Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol, 2015. 15(8): p. 486-99.

SOARES, L.S. Referências Bibliográficas
113
112. Massanella, M., R. Fromentin, and N. Chomont, Residual inflammation and viral reservoirs: alliance against an HIV cure. Curr Opin HIV AIDS, 2016. 11(2): p. 234-41. 113. Akbar, A.N. and S.M. Henson, Are senescence and exhaustion intertwined or unrelated processes that compromise immunity? Nat Rev Immunol, 2011. 11(4): p. 289-95. 114. Passos, J.F. and T. Von Zglinicki, Oxygen free radicals in cell senescence: are they signal transducers? Free Radic Res, 2006. 40(12): p. 1277-83. 115. Effros, R.B., et al., Shortened telomeres in the expanded CD28-CD8+ cell subset in HIV disease implicate replicative senescence in HIV pathogenesis. AIDS, 1996. 10(8): p. F17-22. 116. Nielsen, C.M., et al., Functional Significance of CD57 Expression on Human NK Cells and Relevance to Disease. Front Immunol, 2013. 4: p. 422. 117. Fagnoni, F.F., et al., Expansion of cytotoxic CD8+ CD28- T cells in healthy ageing people, including centenarians. Immunology, 1996. 88(4): p. 501-7. 118. Béziat, V., et al., CMV drives clonal expansion of NKG2C+ NK cells expressing self-specific KIRs in chronic hepatitis patients. Eur J Immunol, 2012. 42(2): p. 447-57. 119. Hukezalie, K.R., et al., In vitro and ex vivo inhibition of human telomerase by anti-HIV nucleoside reverse transcriptase inhibitors (NRTIs) but not by non-NRTIs. PLoS One, 2012. 7(11): p. e47505.
120. Leeansyah, E., et al., Inhibition of telomerase activity by human immunodeficiency virus (HIV) nucleos(t)ide reverse transcriptase inhibitors: a potential factor contributing to HIV-associated accelerated aging. J Infect Dis, 2013. 207(7): p. 1157-65. 121. Freeman, M.L., et al., CD8 T-Cell Expansion and Inflammation Linked to CMV Coinfection in ART-treated HIV Infection. Clin Infect Dis, 2016. 62(3): p. 392-6. 122. Zhang, Y., S.K. Li, and S.K. Tsui, Genome-wide analysis of DNA methylation associated with HIV infection based on a pair of monozygotic twins. Genom Data, 2015. 6: p. 12-5. 123. Maricato, J.T., et al., Epigenetic modulations in activated cells early after HIV-1 infection and their possible functional consequences. PLoS One, 2015. 10(4): p. e0119234. 124. Lachance, J., et al., Evolutionary history and adaptation from high-coverage whole-genome sequences of diverse African hunter-gatherers. Cell, 2012. 150(3): p. 457-69. 125. Shastry, B.S., SNP alleles in human disease and evolution. J Hum Genet, 2002. 47(11): p. 561-6. 126. Satoh, T., et al., The Jmjd3-Irf4 axis regulates M2 macrophage polarization and host responses against helminth infection. Nat Immunol, 2010. 11(10): p. 936-44. 127. Biel, M., V. Wascholowski, and A. Giannis, Epigenetics--an epicenter of gene regulation: histones and histone-modifying enzymes. Angew Chem Int Ed Engl, 2005. 44(21): p. 3186-216.
128. Blazkova, J., et al., CpG methylation controls reactivation of HIV from latency. PLoS Pathog, 2009. 5(8): p. e1000554.
129. Youngblood, B. and N.O. Reich, The early expressed HIV-1 genes regulate DNMT1 expression. Epigenetics, 2008. 3(3): p. 149-56.

SOARES, L.S.
114
APÊNDICE

SOARES, L.S.
115
APÊNDICE A – FIGURAS SUPLEMENTARES
Figura suplementar 01. Amplificação por PCR da região referente ao gene KIR2DL2 antes da modificação do DNA por bissulfito. O fragmento amplificado corresponde a 474 pb e foi analisado em gel de agarose a 1,5%. (C- grupo controle, H- grupo HIV-1+ e T- grupo TARV; MM- marcador molecular 100pb).
Figura suplementar 02. Amplificação por PCR da região referente ao gene KIR2DL2 após a modificação do DNA por bissulfito. O fragmento amplificado corresponde a 452 pb e foi analisado em gel de agarose a 1,5%. (MM- marcador molecular 1Kb Plus).

SOARES, L.S.
116
Figura suplementar 03. Amplificação por PCR da região referente ao gene KIR2DL2 a partir dos plasmídeos purificados dos clones positivos. O fragmento amplificado corresponde a 452 pb e foi analisado em gel de agarose a 1,5%. (MM- marcador molecular 1Kb Plus) A numeração acima de cada gel refere-se à identificação dos clones positivos para o fragmento analisado em cada amostra.
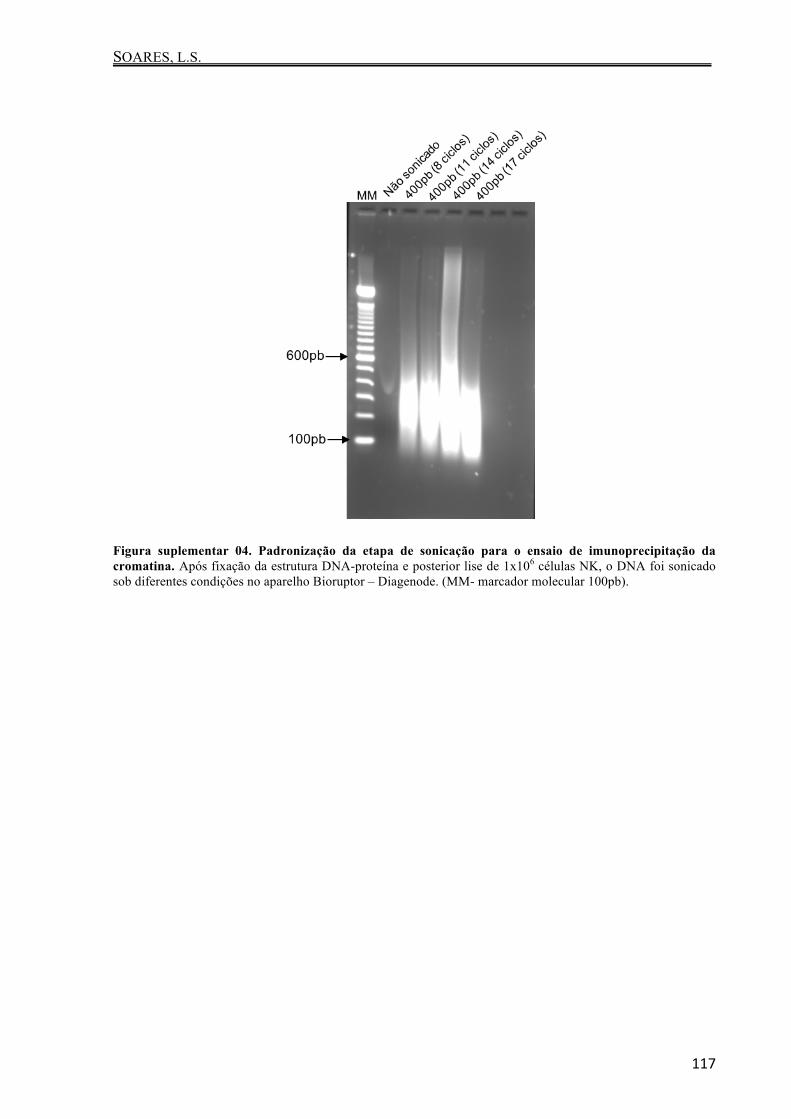
SOARES, L.S.
117
Figura suplementar 04. Padronização da etapa de sonicação para o ensaio de imunoprecipitação da cromatina. Após fixação da estrutura DNA-proteína e posterior lise de 1x106 células NK, o DNA foi sonicado sob diferentes condições no aparelho Bioruptor – Diagenode. (MM- marcador molecular 100pb).

SOARES, L.S.
118
Figura suplementar 05a. Sequenciamento da região referente ao gene KIR2DL2 antes da modificação por bissulfito no grupo controle. O alinhamento foi realizado com auxílio da ferramenta MultiAlin e tendo com base a sequência referência da base de dados GenBank (NM_014219.2).

SOARES, L.S.
119
Figura suplementar 05b. Sequenciamento da região referente ao gene KIR2DL2 antes da modificação por bissulfito no grupo HIV-1+. O alinhamento foi realizado com auxílio da ferramenta MultiAlin e tendo com base a sequência referência da base de dados GenBank (NM_014219.2).

SOARES, L.S.
120
Figura suplementar 05c. Sequenciamento da região referente ao gene KIR2DL2 antes da modificação por bissulfito no grupo TARV. O alinhamento foi realizado com auxílio da ferramenta MultiAlin e tendo com base a sequência referência da base de dados GenBank (NM_014219.2).

SOARES, L.S.
121
Figura suplementar 06a. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra CT12. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.
SOARES, L.S.
122
Figura suplementar 06b. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra CT16. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
123
Figura suplementar 06c. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra CT18. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
124
Figura suplementar 06d. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra HIV11. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
125
Figura suplementar 06e. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra HIV15. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
126
Figura suplementar 06f. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra HIV18. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
127
Figura suplementar 06g. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra TARV04. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.
SOARES, L.S.
128
Figura suplementar 06h. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra TARV05. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
129
Figura suplementar 06i. Sequenciamento dos clones positivos para o fragmento do gene KIR2DL2 da amostra TARV12. O alinhamento foi realizado com auxílio das ferramentas MultiAlin e Clustal, tendo com base a sequência predita in silico após conversão por bissulfito.

SOARES, L.S.
130
ANEXOS

SOARES, L.S.
131
ANEXO I – MANUSCRITO PARA PUBLICAÇÃO
NATURAL KILLER CELLS EXHIBIT FEATURES OF
IMMUNOSENESCENCE IN LONG-TERM HIV-1 TREATED PATIENTS
Luana Silva Soares1, Milena Sobral Espíndola1, Fabiana Albani Zambuzi1, Leonardo Judson
Galvão de Lima1, Valdes Roberto Bollela2, Fabiani Gai Frantz1,*.
1 – Laboratório de Imunologia e Epigenética, Faculdade de Ciências Farmacêuticas de
Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil; 2 – Faculdade de
Medicina de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil.
*Corresponding author: Fabiani Gai Frantz, DACTB, Faculdade de Ciências Farmacêuticas
de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil. ZIP Code: 14041-
903. Tel: +55 1633150241; e-mail: [email protected]
Key words: natural killer cells, HIV infection, activation state, immunosenescence.
ABSTRACT
HIV-1 infection is one of the main concerns regarding infectious disease worldwide.
Nowadays, it is recognized as an inflammatory chronic disease and affects many immune
cells such as the natural killer cells. Our aim in this work was to characterize the dysfunctions
of NK cells during HIV-1 infection as a result of cell senescence. In our study population,
there was an expansion of the non-functional NK cell subset, despite of antiretroviral
treatment, that is also seen in the elderly people. NK cells from HAART group had increased
cytotoxic function and overall cytokine production, even higher than the non-infected
individuals. This higher activated state associated to increased production of oxygen reactive
species by NK cells from this group confirm our hypothesis of immunosenescence in these
cells. Although other markers of senescence might be investigated to better understand this
phenomenon during HIV-1 infection creating perspectives of new interventions for restoration
of immune cell homeostasis.

SOARES, L.S.
132
Introduction
The Acquired Immune Deficiency Syndrome (AIDS) was first recognized as a disease
in the early 80s [1] and since then, many research groups have studied this pathology aiming
its effective cure and prevention. The etiologic agent is the Human Immunodeficiency Virus
(HIV) that have been originated from a virus that infects primates (Simian Immunodeficiency
Virus - SIV) [2]. Nowadays, HIV infection is recognized as an inflammatory chronic disease
in parallel with a dysfunctional immune system [3]. Despite the advance of antiretroviral
therapy and the subsequent increase in life expectancy, the HIV-1+ patients have shown
higher occurrence of non-AIDS related diseases such as cardiovascular and liver diseases,
osteoporosis, cancers and metabolic syndromes. As these pathologies are usually seen in the
elderly people, nowadays HIV infection has been associated to acceleration of natural aging
[4].
The pro-inflammatory state seen in HIV infected individuals is mainly due to the
disruption of intestinal mucosa and the translocation of microbial products from the gut to the
blood and tissues [5]. In this context, many cells of the immune system have shown higher
activation status, such as CD8 T cells, monocytes and macrophages, and that was correlated
to disease progression [6, 7]. A recent study also reported an activated phenotype of natural
killer cells in HIV+ patients even on ART by analyzing the expression of surface markers [8].
Considering that NK cells are one of the pivotal cells of innate immune system, the
understanding of its role and dysfunctions during HIV infection is of great importance.
Natural killer cells constitute nearly 10% of peripheral blood lymphocytes [9], and most
studies have reported changes in NK cell frequency and subset distribution during HIV
infection [10-12]. As the infection progresses, phase described as chronic, a non-functional
NK cell subset characterized as CD3-CD56-CD16+ accumulates, impairing the function of the
total NK cells [13]. Also, the increased expression of inhibitory receptors in parallel with a
decrease of activating receptors renders the NK cells unresponsive and not functional during
HIV infection [10]. It is not known whether the phenotypic and functional changes seen in the
NK cell population during HIV infection is due to the direct infection of these cells or if is a
consequence of the overall disease.
The distinct phenotype and function of NK cells during HIV-1 infection may occur as a
result of genetic, epigenetic and environmental factors. In this work, we are particularly
interested in analyzing the function of these cells and some markers of immunosenescence
that might be related to disease progression. Also, we investigated some epigenetic

SOARES, L.S.
133
modifications that could be involved in the alterations of these cell’s function during HIV-1
infection.
Our results clearly showed a higher activation status of NK cells from HIV-1+ treated
patients. This phenotype was very similar to that seen in elderly people, also reinforced by the
expansion of the non-functional NK subset and the higher oxidative stress in these cells. On
the other hand, these alterations might not be directly due to epigenetic modifications, as
enzymes involved in these process were not changed in our study.
Materials and Methods
Blood samples and study population
Samples from 28 adult subjects ART-naïve and 29 on ART were collected at the Hospital das
Clínicas de Ribeirão Preto – São Paulo, Brazil. Control samples (n = 30) were obtained from
Fundação Hemocentro de Ribeirão Preto – São Paulo, Brazil (Table 1). The individuals were
from both sexes and aged between 18 and 65 years. Patients on ART were included in the
study considering that viremia was undetectable for at least six months. All of the participants
gave written, informed consent, and the study was evaluated and accepted by the Ethics
Committee of the Hospital das Clínicas de Ribeirão Preto (Process HCRP # 9818/2012).
Flow Cytometry
After collection of blood samples (40 mL), peripheral blood mononuclear cells were obtained
by gradient density separation. The total PBMC was then counted in a Neubauer chamber,
and the cell viability was evaluated with Tripan blue 0.5% staining. Thereafter, 1 x 105 cells
were stained for surface markers at 4°C for 25 min using the following fluorochrome-labeled
mAb: anti-CD3 PerCPCy5.5 (clone SK7), anti-CD56 APC (clone NCAM 16.2) and anti-
CD16 PE-Cy7 (clone 3G8). Next, the cells were washed and fixed in 1% paraformaldehyde.
At least 100,000 events were acquired from each sample in a FACSCantoTM II flow cytometer
(BD Biosciences, San Diego, USA) using the FACSDiva 6.1.3 software for data acquisition.
Data were analyzed on lymphocyte population defined by size and granularity using FlowJo
software version 7.6.5 (Tree Star Inc., Oregon, USA)
Evaluation of NK cell cytotoxicity
NK cells were purified from PBMC by positive selection using CD56 MicroBeads (MACS,
Miltenyi Biotec Inc., Auburn CA, USA) according to manufacturer’s instructions. Then, NK
cells were co-cultured with K-562 (ATCC N° CCL-243™) as a target cell in different effector

SOARES, L.S.
134
to target ratios for four hours. NK cytotoxicity was measured using the CytoTox 96 Non-
Radioactive Cytotoxicity Assay (Promega) by quantifying the lactate dehydrogenase release
in the supernatant following manufacturer’s protocol.
Quantification of cytokines and chemokines produced by NK cells
The production of cytokines and chemokines by NK cells was quantified by a customized
multiplex kit (16-plex, EMD Millipore Corporation, Billerica, Massachusetts, USA) in the
supernatant of the cytotoxicity assay. The multiplex plate was read in a fluorescent bead-
based instrument (Luminex® MAGPIX® System; Luminex Corporation, Austin, Texas,
USA) and analyzed using the Milliplex Analyst software v3.5 ((Millipore; VigeneTech Inc.,
Boston, Massachusetts, USA) with a three-parameter logistic curve fit.
Assessment of oxidative stress of NK cells
After purification of NK cells by CD56 MicroBeads, oxidative stress was quantified by
monitoring the induction of NADPH oxidase activity in a luminol-dependent
chemiluminescence assay. For this, 1 x 105 NK cells were stimulated with 10-7M PMA or not
(spontaneous) and 100µM Luminol was used. The reaction was recorded with a luminometer
(EG & G Berthold, AutoLumat Plus LB 953, Bad Wildbad, Germany). The area under the
time-course curve of chemiluminescence (AUC) was calculated for each individual.
Measurement of enzymes that promote epigenetic modifications
Total RNA was extracted from NK cells with Trizol Reagent (Ambion®) and treated with
Turbo DNase (Ambion) according to manufacturer’s instructions. Total RNA was quantified
in a Qubit 2.0 Fluorometer (Thermo Fisher Scientific) and then reverse transcribed into cDNA
using the High Capacity cDNA Reverse Transcription kit (Applied Biosystems). 30ng of total
RNA after reverse transcription was applied in the RT2 Profiler PCR Array for quantification
of enzymes involved in the activation of gene transcription (ATF2, HAT1, KAT2B, KAT5,
CARM1, PRMT5, SMYD3, SUV39H1, ASH1L, KMT2C, KMT2E, SETD1B, KDM5B,
KDM6B). Relative expression of each enzyme was calculated by using the 2-ΔΔ
Ct method
considering the control group as reference. And GAPDH and beta-actin as housekeeping
genes.

SOARES, L.S.
135
ChIP Assay
The chromatin immunoprecipitation (ChIP) procedure was performed starting from 1 x 106
purified NK cells. DNA-protein structure was cross-linked by 1% paraformaldehyde for 10
minutes at 37°C. Then, cells were washed with PBS plus protease inhibitors at 400 x g for 10
min at 4°C and lysed in 200 µL SDS Lysis Buffer. The lysate was sonicated to obtain DNA
fragments around 400 bp using a Bioruptor Pico Sonication System (Diagenode) under the
following conditions: 11 times for periods of 15 seconds on and 90 seconds off. Next,
samples were centrifuged and the supernatant containing chromatin was diluted, and an
aliquot (5% volume) was separated as the input DNA in each sample. The remaining
chromatin fractions were precleared with salmon sperm DNA/protein A agarose beads
followed by immunoprecipitation with the following antibodies: anti-histone H3K27me3
(Abcam), anti-histone H3K4me3 (Abcam) and normal mouse IgG (Millipore) overnight at
4°C with gentle rotation. Cross-linking was reversed for 2 hours at 62°C associated with
proteinase K digestion. DNA was purified by standard phenol/chloroform and ethanol
precipitation and was subjected to real-time PCR. Primers for the promoter regions of IFN-γ
and TNF-α were described elsewhere [14, 15].
Results
Characteristics of the study population
The characteristics of the individuals included in this study were described in the Material and
Methods section and detailed in table 1. The HIV-1+ infected patients were separated in two
groups: HIV-1+ (naïve-treated) and HAART (HIV-1+ patients on two distinct antiretroviral
regimens). Patients on HAART had undetectable levels of plasma viral load and a median of
72,3 months of treatment. On the other side, HIV-1+ naïve-treated patients were recently
infected with a median of 5,3 months of infection compared to 156,4 months in the HAART
group.
Change in the distribution of NK cell subsets in HIV-1 infected individuals
Our first aim in this work was to characterize the distribution of NK cell subsets in our study
population. The figure 1a shows the gating strategy used to calculate the proportion of NK
cell subsets. NK cells were analyzed in the lymphocyte population defined by size and
granularity, and by the expression of CD56 and lack of expression of the CD3 molecule. The

SOARES, L.S.
136
proportion of NK cell subsets were determined considering the differential expression of the
markers CD56 and CD16. Considering this characterization, we observed that the cytotoxic
NK cell subset (CD3-CD56dimCD16+) was significant reduced only in the HIV-1+ group
compared to the control one (Figures 1b, c). Although, the NK cell subset described as more
related to cytokine production (CD3-CD56brightCD16-) showed no differences between the
groups. On the other hand, the non-functional NK cell subset (CD3-CD56-CD16+) was
increased in the HIV-1+ patients, despite of antiretroviral treatment. These results clearly
indicated that the antiretroviral treatment did not restore the proportions of NK cell subsets.
And that patients recently infected should have greater loss of effector response mediated by
NK cells against HIV-1.
Impaired function of NK cells from treatment-naïve patients
We next investigated if the change in the distribution of NK cell subsets in HIV-1+ patients
could have an affect on the function of these cells. So, we evaluated the cytotoxic function of
NK cells against a target cell line that down-regulates the expression of HLA-class I (K-562).
The percentage of NK cytotoxicity was reduced in the treatment-naïve patients compared to
control and treated groups, although it was not statistically significant (Figure 2).
Interestingly, the HAART group demonstrated a trend of higher cytotoxicity function even
than the control individuals, first suggesting a more activated phenotype of NK cells from
these patients.
NK cell activation correlates with time of infection
To better characterize the activation phenotype of NK cells beyond the expression of surface
molecules, we quantified the cytokine and chemokine production in the supernatant of the
cytotoxicity assay by a Multiplex Platform. While NK cells from the HIV-1+ group produced
small amounts of all the molecules analyzed, NK cells from the HAART group exhibited
increased levels of pro-inflammatory cytokines, chemokines and also of regulatory and anti-
inflammatory cytokines (Figure 3). For some molecules, such as IFN-α2, MCP-1 and IL-10,
the median production by NK cells from treated patients was even higher than the control
individuals. Although the expression of enzymes related to activation of transcription were
very similar between the groups, suggesting that other mechanisms might be related to the
modulation of these cytokines (Figuure S1).
The super-activated state of NK cells from the HAART group was better seen after the
calculation of frequency of higher producers for each molecule considering the overall

SOARES, L.S.
137
median of every analyte. The radar charts shown in Figure 4 clearly demonstrated that more
than 50% of all the patients in the HAART group were higher producers for all the molecules
quantified. On the other hand, treated-naïve patients showed a contraction of the NK cell
effector response. When we analyzed the presence of histone marks related to activation
(H3K4me3) or repression (K3K27me3) of transcription in the promoter region of the
cytokines IFN-γ and TNF-α, that were differentially expressed, we didn’t see any difference
between the groups (Figure S2). Although other regions must be investigated in order to
understand the mechanisms that are driving these modifications in NK cells during HIV-1
infection.
We next asked ourselves if this activated phenotype was a result of drug therapy or time of
infection. We didn’t see any correlation between the cytokine production by NK cells from
the HAART group and the time of treatment (data not shown). But we did see a positive
correlation between the levels of some cytokines and the time of infection, irrespective of
patient’s treatment as demonstrated in figure 5. The higher association was seen between the
levels of IFN-α2 and time of infection (r = 0.61, p = 0.0012). The molecules TNF-α and IL-
10, that are usually associated to disease progression, were also positive correlated to time of
infection. These results suggest that antiretroviral treatment does not restore effector response
of NK cells during HIV-1 infection and that as longer as the patient is infected, their NK cells
progresses to a more activated state. However, considering that this activation is even higher
than the control group, this phenotype may not be a good prognostic for the patient.
Increased NK cell activation is associated to oxidative stress and an immunosenescence
phenotype
Taken together the accumulation of the non-functional NK cell subset, the higher activated
phenotype of these cells as the infection goes by, and the evidence in the literature that HIV-1
infection accelerates natural aging, we speculated that NK cells were evolving to a
immunosenescence phenotype. To evaluate this phenomena, we quantified the production of
reactive oxygen species by NK cells either spontaneously or stimulated with PMA. NK cells
from the HAART group produced increased levels of ROS compared to the other groups
(Figure 6). Other markers of cellular senescence must be evaluated in order to better
characterize this phenotype, but our results strongly suggest that this might be the case of NK
cells during HIV-1 infection.

SOARES, L.S.
138
Discussion
HIV infection is an inflammatory chronic disease that has no cure, except for the Berlin
patient, and so patients have to be treated for all their lifetime [3]. Despite the increase in
survival, HIV-infected individuals have shown acceleration of natural aging and subsequently
higher occurrence of comorbidities related to senescence [4]. In this context, we
demonstrated, in our work, that natural killer cells from HIV-1 infected patients exhibited
characteristics of immunosenescence such as an increased activation state and oxidative
stress, mainly in the long-term treated patients.
Considering that immunosenescence is an inflammatory state [16] similar to what is seen in
HIV-1+ patients, there are many factors related to the induction of this phenomena. Among
them are: residual virus replication, antiretroviral therapy, as well as occurrence of co-
infections [4]. For example, the antiretrovirals AZT and Tenofovir are involved on the
inhibition of telomerase activity in vitro and are related to telomere shortening [17, 18]. Also,
co-infections like the co-infection with Cytomegalovirus can promote T cell activation and
virus replication during HIV-1 infection [19].
Although in our study the treated patients had no detectable viremia, it is well known that
antiretroviral drugs do not penetrate well in tissues called sanctuary anatomic such as
intestinal mucosa, lymph nodes, genital tract and central nervous system, thus favoring
residual replication of the virus [20]. Also, some studies [21], including ours [22], had
demonstrated that HIV-1+ patients on treatment had elevated levels of plasma inflammatory
markers. Considering that cellular senescence cause is associated to the presence of a constant
stimulus even after the development of a primary immune response [16, 23], we can conclude
that this is the case during HIV-1 infection after long-term treatment.
One of the first evidences of the immunosenescence phenotype in our study was the
expansion of the non-functional NK cell subset in HIV-1+ individuals, despite of
antiretroviral treatment, as usually seen in the elderly people [24]. This expansion is well
related in the literature, but until know no one had correlated to aging process in HIV-1
infection. Many studies usually investigate surface markers of differentiation in NK cells such
as CD57 [25], but this phenotype is not clearly related to senescence.
Another characteristic of immunosenescence is an increased effector function as shown in our
results by the higher production of overall cytokines by NK cells from the HAART group.
This enhanced function was even seen in the cytotoxicity assay and correlated to time of
infection, raising the question that this phenomenon might not only be a result of
antiretroviral drugs. On the other hand, patients that were recently infected demonstrated

SOARES, L.S.
139
signs of NK cells exhaustion, mainly because of reduced effector response, although other
markers might be investigated.
To confirm our hypothesis of NK cell senescence in the HAART group, we quantified the
oxidative stress in these cells. There was an increased production of reactive oxygen species
by NK cells from the HAART group even when these cells were not stimulated. At a glance,
our results clearly demonstrated the development of NK cell senescence during HIV-1
infection, mainly in the long-term treated patients. This finding contributes to the
understanding of HIV pathogenesis and also creates new strategies of intervention aiming the
restoration of immune cell homeostasis, specifically of natural killer cells.
Authorship
LSS, FGF, VRB: conceived and designed the study; LSS, MSE, FAZ, LJGL: conducted the
experiments; LSS, FGF: were involved in writing the manuscript.
Acknowledgments
This work was supported by São Paulo Research Foundation (FAPESP, grants
#2011/24026-3; #2011/12199-0).
The authors are grateful to Caroline Fontanari and Thaisa da Silva Araújo for their
technical support.
Disclosures The authors declare no conflict of interest.

SOARES, L.S.
140
References 1. (CDC), C.f.D.C., Update on acquired immune deficiency syndrome (AIDS)--United States. MMWR Morb Mortal Wkly Rep, 1982. 31(37): p. 507-8, 513-4. 2. Gao, F., et al., Origin of HIV-1 in the chimpanzee Pan troglodytes troglodytes. Nature, 1999. 397(6718): p. 436-41. 3. Klatt, N.R., et al., Immune activation and HIV persistence: implications for curative approaches to HIV infection. Immunol Rev, 2013. 254(1): p. 326-42. 4. Appay, V. and A.D. Kelleher, Immune activation and immune aging in HIV infection. Curr Opin HIV AIDS, 2016. 11(2): p. 242-9. 5. Brenchley, J.M., et al., Microbial translocation is a cause of systemic immune activation in chronic HIV infection. Nat Med, 2006. 12(12): p. 1365-71. 6. Papagno, L., et al., Immune activation and CD8+ T-cell differentiation towards senescence in HIV-1 infection. PLoS Biol, 2004. 2(2): p. E20. 7. Hunt, P.W., et al., Gut epithelial barrier dysfunction and innate immune activation predict mortality in treated HIV infection. J Infect Dis, 2014. 210(8): p. 1228-38. 8. Kuri-Cervantes, L., et al., Activation of NK cells is associated with HIV-1 disease progression. J Leukoc Biol, 2014. 96(1): p. 7-16. 9. Robertson, M.J. and J. Ritz, Biology and clinical relevance of human natural killer cells. Blood, 1990. 76(12): p. 2421-38. 10. Mavilio, D., et al., Natural killer cells in HIV-1 infection: dichotomous effects of viremia on inhibitory and activating receptors and their functional correlates. Proc Natl Acad Sci U S A, 2003. 100(25): p. 15011-6.
11. Vieillard, V., et al., Specific phenotypic and functional features of natural killer cells from HIV-infected long-term nonprogressors and HIV controllers. J Acquir Immune Defic Syndr, 2010. 53(5): p. 564-73. 12. Alter, G., et al., Sequential deregulation of NK cell subset distribution and function starting in acute HIV-1 infection. Blood, 2005. 106(10): p. 3366-9. 13. Hu, P.F., et al., Natural killer cell immunodeficiency in HIV disease is manifest by profoundly decreased numbers of CD16+CD56+ cells and expansion of a population of CD16dimCD56- cells with low lytic activity. J Acquir Immune Defic Syndr Hum Retrovirol, 1995. 10(3): p. 331-40. 14. Morinobu, A., Y. Kanno, and J.J. O'Shea, Discrete roles for histone acetylation in human T helper 1 cell-specific gene expression. J Biol Chem, 2004. 279(39): p. 40640-6. 15. Lee, J.Y., et al., Histone acetylation and chromatin conformation are regulated separately at the TNF-alpha promoter in monocytes and macrophages. J Leukoc Biol, 2003. 73(6): p. 862-71.
16. Wherry, E.J. and M. Kurachi, Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol, 2015. 15(8): p. 486-99.
17. Hukezalie, K.R., et al., In vitro and ex vivo inhibition of human telomerase by anti-HIV nucleoside reverse transcriptase inhibitors (NRTIs) but not by non-NRTIs. PLoS One, 2012. 7(11): p. e47505.

SOARES, L.S.
141
18. Leeansyah, E., et al., Inhibition of telomerase activity by human immunodeficiency virus (HIV) nucleos(t)ide reverse transcriptase inhibitors: a potential factor contributing to HIV-associated accelerated aging. J Infect Dis, 2013. 207(7): p. 1157-65.
19. Freeman, M.L., et al., CD8 T-Cell Expansion and Inflammation Linked to CMV Coinfection in ART-treated HIV Infection. Clin Infect Dis, 2016. 62(3): p. 392-6.
20. Massanella, M., R. Fromentin, and N. Chomont, Residual inflammation and viral reservoirs: alliance against an HIV cure. Curr Opin HIV AIDS, 2016. 11(2): p. 234-41.
21. Liovat, A.S., et al., Acute plasma biomarkers of T cell activation set-point levels and of disease progression in HIV-1 infection. PLoS One, 2012. 7(10): p. e46143.
22. Espíndola, M.S., et al., Dysregulated Immune Activation in Second-Line HAART HIV+ Patients Is Similar to That of Untreated Patients. PLoS One, 2015. 10(12): p. e0145261.
23. Akbar, A.N. and S.M. Henson, Are senescence and exhaustion intertwined or unrelated processes that compromise immunity? Nat Rev Immunol, 2011. 11(4): p. 289-95.
24. Solana, R., et al., Shaping of NK cell subsets by aging. Curr Opin Immunol, 2014. 29: p. 56-61.
25. Nielsen, C.M., et al., Functional Significance of CD57 Expression on Human NK Cells and Relevance to Disease. Front Immunol, 2013. 4: p. 422.

SOARES, L.S.
142
Tables and Figures
Table 1. Characteristics of the study population
Control HIV-1+ HAART
n 30 28 29 Age 32,0 33,5 46,0
(27,5 - 54) (23 - 43,3) (40,5 - 52,5) Gender M = 21 M = 21 M = 14
F = 9 F = 7 F = 15
Treatment - - NRTI + NNRTI
(n = 17)
NRTI + PI
(n = 12) Viral load (copies/mL) - 21638 < 40
(4272 - 117414)
CD4+ T cells (cells/mm3) Not determined
415,5 705
(214,5 - 640,3) (597,5 - 932)
Months of infection - 5,3 156,4
(2 - 29,5) (73,2 - 193,6)
Months on ART - - 72,3
(27 - 114,9) The data for age, viral load, CD4+ T cell count, time of infection and treatment were expressed as median (25-75
percentil).
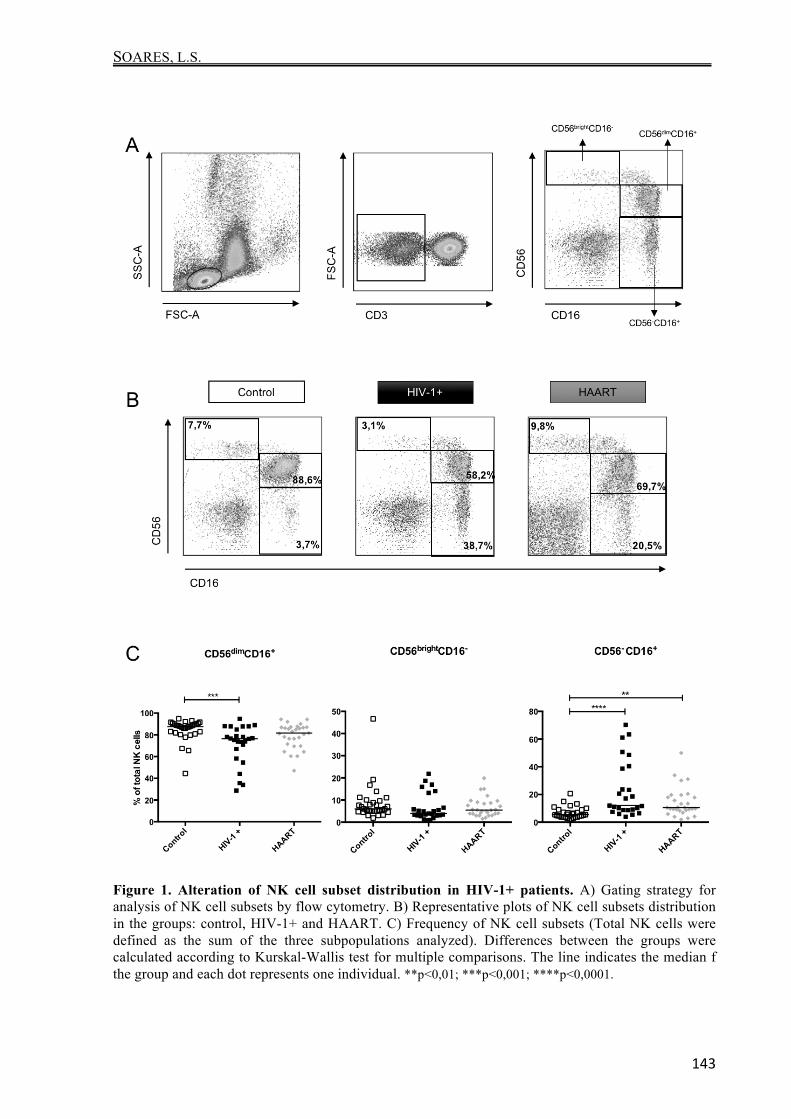
SOARES, L.S.
143
Figure 1. Alteration of NK cell subset distribution in HIV-1+ patients. A) Gating strategy for analysis of NK cell subsets by flow cytometry. B) Representative plots of NK cell subsets distribution in the groups: control, HIV-1+ and HAART. C) Frequency of NK cell subsets (Total NK cells were defined as the sum of the three subpopulations analyzed). Differences between the groups were calculated according to Kurskal-Wallis test for multiple comparisons. The line indicates the median f the group and each dot represents one individual. **p<0,01; ***p<0,001; ****p<0,0001.

SOARES, L.S.
144
Figure 2. Increased cytotoxic function of NK cells from HAART group. NK cells were purified from the blood and co-cultured with a target cell (K-562) in different effector to target ratios. The percentage of cytotoxicity was calculated from the quantification of lactate dehydrogenase enzyme released in the supernatant. Data are expressed as mean ± SEM. (Kruskal-Wallis Test for multiple comparisons).
0
20
40
60
80
100
NK:K-562
% C
itoto
xici
ty
ControlHIV-1 +
20:1
10:1 5:1
HAART
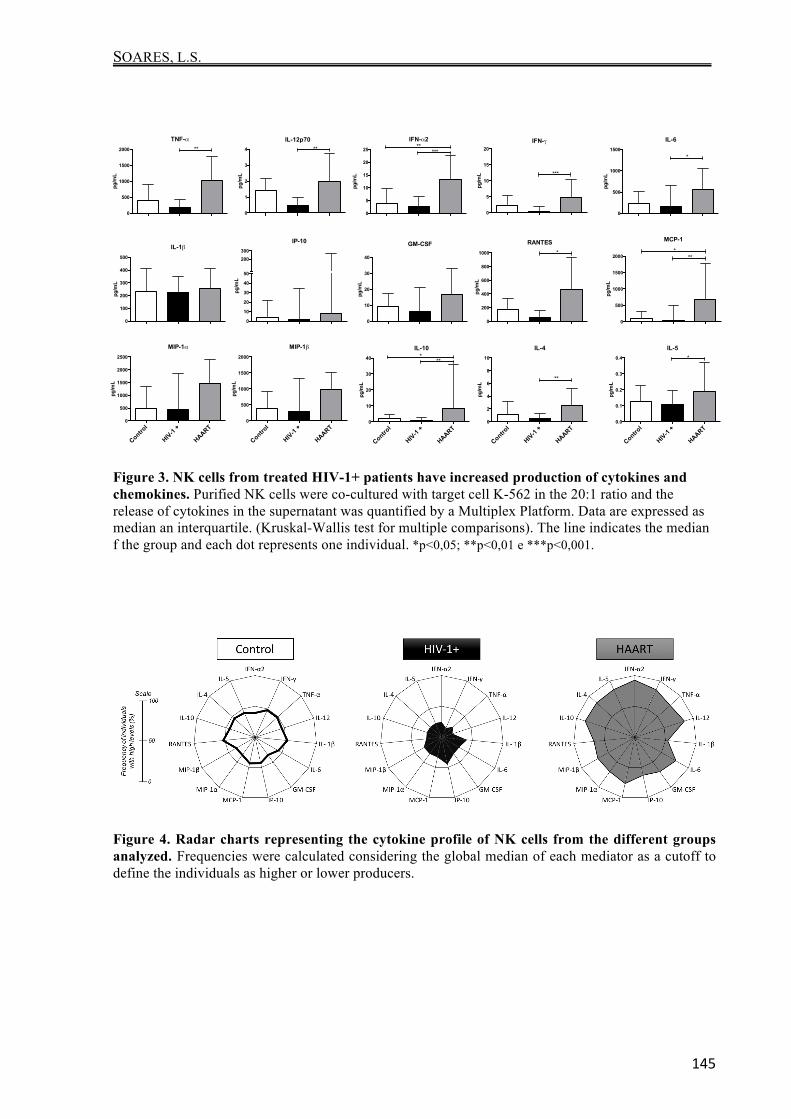
SOARES, L.S.
145
Figure 3. NK cells from treated HIV-1+ patients have increased production of cytokines and chemokines. Purified NK cells were co-cultured with target cell K-562 in the 20:1 ratio and the release of cytokines in the supernatant was quantified by a Multiplex Platform. Data are expressed as median an interquartile. (Kruskal-Wallis test for multiple comparisons). The line indicates the median f the group and each dot represents one individual. *p<0,05; **p<0,01 e ***p<0,001.
Figure 4. Radar charts representing the cytokine profile of NK cells from the different groups analyzed. Frequencies were calculated considering the global median of each mediator as a cutoff to define the individuals as higher or lower producers.
TNF-α
0
500
1000
1500
2000
pg/m
L
**IL-12p70
0
1
2
3
4
pg/m
L
**IFN-α2
0
5
10
15
20
25
pg/m
L
*****
IFN-γ
0
5
10
15
20
pg/m
L ***
IL-6
0
500
1000
1500
pg/m
L
*
IL-1β
0
100
200
300
400
500
pg/m
L
IP-10
0
10
20
30
40
50
200
300
pg/m
LGM-CSF
0
10
20
30
40
pg/m
L
RANTES
0
200
400
600
800
1000
pg/m
L
*
MCP-1
0
500
1000
1500
2000
pg/m
L
***
MIP-1α
Control
HIV-1
+
HAART0
500
1000
1500
2000
2500
pg/m
L
MIP-1β
Control
HIV-1
+
HAART0
500
1000
1500
2000
pg/m
L
IL-10
Control
HIV-1
+
HAART0
10
20
30
40pg
/mL
***
IL-4
Control
HIV-1
+
HAART0
2
4
6
8
10
pg/m
L
**
IL-5
Control
HIV-1
+
HAART0.0
0.1
0.2
0.3
0.4
pg/m
L
*

SOARES, L.S.
146
Figure 5. Time of infection is crucial in the modulation of the activation state of NK cells. Spearman correlation coefficient was calculated between the cytokine/chemokine levels and time of infection, independent of treatment.
0 100 200 300 4000
20
40
60
80
Months of infection
IFN
-α2
(pg/
mL)
r= 0,61 p=0,0012
0 100 200 300 4000
2000
4000
6000
Months of infection
TNF-α
(pg/
mL)
r= 0,4623 p=0,02
0 100 200 300 4000
1000
2000
3000
Months of infection
IL-6
(pg/
mL)
r= 0,4462 p=0,0254
0 100 200 300 4000
20
40
60
80
100
Months of infection
IL-1
0 (p
g/m
L)
r= 0,43 p=0,0319
0 100 200 300 4000
20
40
60
80
Months of infection
IFN
-γ (p
g/m
L)
r= 0,5054 p=0,01
0 100 200 300 4000
2
4
6
8
Months of infection
IL-1
2 (p
g/m
L)
r= 0,5693 p=0,0037
0 100 200 300 4000
2000
4000
6000
8000
10000
Months of infection
MC
P-1
(pg/
mL)
r= 0,5231 p=0,0073
0 100 200 300 4000.0
0.2
0.4
0.6
0.8
1.0
Months of infection
IL-5
(pg/
mL)
r= 0,4794 p=0,0153

SOARES, L.S.
147
Figure 6. Increased oxidative stress in NK cells from HIV-1+ treated patients. Purified NK cells were stimulated with PMA 10-7M or not and the reactive oxygen species production was quantified by chemoluminescence. Data are expressed as median and interquartile range of the area under the curve of the chemoluminescence of luminol after 3500 seconds of stimulus.
0
2×106
4×106
6×106
PMA - + - + - +
Are
a un
der c
urve
ControlHIV-1+HAART

SOARES, L.S.
148
Figure S1. Relative expression levels of enzymes that promote epigenetic modifications related to activation of transcription. Gene expression was quantified by PCR-Array, considering the housekeeping genes: actin-b and GAPDH and the control group as reference. Data are expressed as median and interquartile range.
ATF2
0.0
0.5
1.0
1.5
Fold
cha
nge
HAT1
0.0
0.5
1.0
1.5
Fold
cha
nge
KAT2B
0.0
0.5
1.0
1.5
Fold
cha
nge
KAT5
0.0
0.5
1.0
1.5
Fold
cha
nge
CARM1
0.0
0.5
1.0
1.5
Fold
cha
nge
PRMT5
0.0
0.5
1.0
1.5Fo
ld c
hang
eSMYD3
0.0
0.5
1.0
1.5
Fold
cha
nge
SUV39H1
0.0
0.5
1.0
1.5
Fold
cha
nge
ASH1L
0.0
0.5
1.0
1.5
Fold
cha
nge
KMT2C
0.0
0.5
1.0
1.5
Fold
cha
nge
KMT2E
0.0
0.5
1.0
1.5Fo
ld c
hang
e
SETD1B
0.0
0.5
1.0
1.5
Fold
cha
nge
KDM5B
0.0
0.5
1.0
1.5
Fold
cha
nge
KDM6B
0
1
2
3
4
5 HAARTHIV-1+
Fold
cha
nge
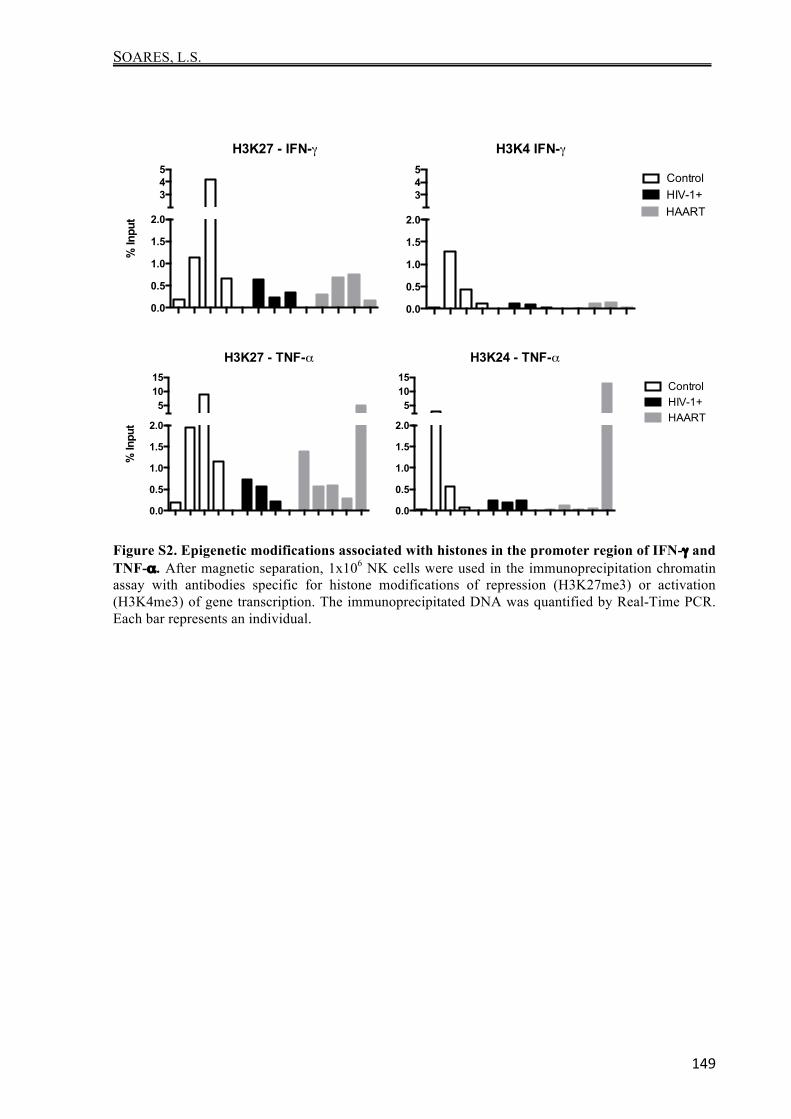
SOARES, L.S.
149
Figure S2. Epigenetic modifications associated with histones in the promoter region of IFN-γ and TNF-α. After magnetic separation, 1x106 NK cells were used in the immunoprecipitation chromatin assay with antibodies specific for histone modifications of repression (H3K27me3) or activation (H3K4me3) of gene transcription. The immunoprecipitated DNA was quantified by Real-Time PCR. Each bar represents an individual.
0.0
0.5
1.0
1.5
2.0
345
% In
put
H3K27 - IFN-γ
0.0
0.5
1.0
1.5
2.0
345
H3K4 IFN-γ
HAARTHIV-1+Control
0.0
0.5
1.0
1.5
2.0
51015
% In
put
H3K27 - TNF-α
0.0
0.5
1.0
1.5
2.0
51015
H3K24 - TNF-α
ControlHIV-1+HAART

SOARES, L.S.
150
ANEXO II – Aprovação do Comitê de Ética em Pesquisa para Seres Humanos

SOARES, L.S.
151
ANEXO III – Aprovação do Centro de Saúde Escola para coleta de amostras

SOARES, L.S.
152
ANEXO IV – Termo de Consentimento Livre e Esclarecido – Indivíduos Controles
UNIVERSIDADE DE SÃO PAULO - Campus Universitário Monte Alegre HOSPITAL DAS CLÍNICAS DA FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
NOME DA PESQUISA: “CARACTERIZAÇÃO DE CÉLULAS NATURAL KILLER DE PACIENTES HIV+ E ESTUDO DE ALTERAÇÕES EPIGENÉTICAS RELACIONADAS AS
SUAS FUNÇÕES E FENÓTIPO”. PESQUISADOR RESPONSÁVEL: MSc. Luana Silva Soares Profª. Drª. Fabiani Gai Frantz
Gostaríamos de convidá-lo a participar deste projeto de pesquisa. Este projeto faz parte de
uma linha de pesquisa que é desenvolvida pela Faculdade de Farmácia da USP de Ribeirão
Preto em conjunto com os pesquisadores do Laboratório de Biologia Molecular do
Hemocentro de Ribeirão Preto e da Unidade de Doenças Infecciosas do Hospital das
Clínicas da FMRP-USP, com o objetivo de avaliar aspectos imunológicos a partir da análise
do sangue de pacientes infectados com o vírus HIV.
A sua participação é voluntária. Para participar, é necessário ler atentamente este documento e ouvir nossas explicações em caso de dúvidas. Se estiver interessado em
participar, solicitaremos que assine este documento. Caso não queira participar, o seu
atendimento no Hemocentro de Ribeirão Preto continuará o mesmo, sem prejuízo nenhum.
O que é célula natural killer? As células natural killer são células de defesa do corpo importantes no combate a
doenças infecciosas e tumorais.
O que são alterações epigenéticas?
As alterações epigenéticas constituem modificações associadas ao DNA e que podem
influenciar na função das células no corpo humano.
Porque esta pesquisa está sendo feita?
Esta pesquisa trará benefícios para o entendimento de como as células de defesa de
um indivíduo infectado com HIV funcionam em comparação com as células de indivíduos
que não estejam infectados com o vírus (controle negativo). Além disso, os dados dessa

SOARES, L.S.
153
pesquisa poderão ser úteis para o desenvolvimento de um teste para melhorar o tratamento
de pacientes infectados com HIV.
Para a realização deste trabalho, precisamos da coleta de sangue de pessoas que NÃO estejam infectadas com o vírus HIV-1 e que também não estejam infectadas com fungos e bactérias oportunistas. Será importante para utilizar como controle negativo nesta pesquisa. A pessoa que participar desta pesquisa deverá doar 40 mL de sangue. Esta quantidade de
sangue retirada não será prejudicial à saúde. A coleta de sangue será realizada no braço
durante o processo de doação de sangue juntamente com as amostras usualmente coletadas,
com seringas e agulhas descartáveis por um profissional capacitado. Não há riscos
previsíveis decorrentes da pesquisa para o doador, a não ser o desconforto da picada da
agulha na coleta de sangue.
Não haverá nenhuma despesa e nenhum ganho em dinheiro. Afirmamos que os
dados obtidos serão guardados de maneira sigilosa e que os nomes dos participantes não
serão divulgados em nenhum momento, e os mesmos serão informados dos resultados
finais da pesquisa. Esta pesquisa não trará benefício direto ao sujeito envolvido, ficando a
critério do mesmo a desistência em participar da pesquisa a qualquer momento, sem
nenhum prejuízo ao seu seguimento hospitalar.
Qualquer dúvida sobre esta pesquisa, entrar em contato com Luana Silva Soares
pelo telefone (16) 3602-4189 ou (16) 99719-4270.
Eu,_______________________________________________________________________
RG n°: ____________________________, autorizo a retirada de 40 mL do meu sangue da
veia do braço para a realização de pesquisa. Fui informado (a) sobre o procedimento e que
o volume retirado não causará dano a minha saúde. Fui informado (a) também que a
finalidade dessa pesquisa é de entender como o vírus se comporta e promove o
aparecimento das infecções.
Ribeirão Preto, _____ de _________________ de 20__.
_______________________________ _________________________________
Assinatura do paciente – Registro MSc. Luana Silva Soares (Pesquisador Responsável)

SOARES, L.S.
154
ANEXO V – Termo de Consentimento Livre e Esclarecido – Pacientes HIV-1+
UNIVERSIDADE DE SÃO PAULO - Campus Universitário Monte Alegre HOSPITAL DAS CLÍNICAS DA FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
TERMO DE CONSENTIMENTO LIVRE E ESCLARECIDO
NOME DA PESQUISA: “CARACTERIZAÇÃO DE CÉLULAS NATURAL KILLER DE
PACIENTES HIV+ E ESTUDO DE ALTERAÇÕES EPIGENÉTICAS RELACIONADAS AS SUAS FUNÇÕES E FENÓTIPO”.
PESQUISADOR RESPONSÁVEL: MSc. Luana Silva Soares Profª. Drª. Fabiani Gai Frantz Gostaríamos de convidá-lo a participar deste projeto de pesquisa. Este projeto faz parte de
uma linha de pesquisa que é desenvolvida na Faculdade de Farmácia da USP de Ribeirão
Preto em conjunto com os pesquisadores do Laboratório de Biologia Molecular do
Hemocentro de Ribeirão Preto e da Unidade de Doenças Infecciosas do Hospital das
Clínicas da FMRP-USP, com o objetivo de avaliar aspectos imunológicos a partir da análise
do sangue de pacientes infectados com o vírus HIV.
A sua participação é voluntária. Para participar, é necessário ler atentamente este documento e ouvir nossas explicações em caso de dúvidas. Se estiver interessado em
participar, solicitamos que assine este documento. Caso não queira participar, o seu
atendimento no ambulatório continuará o mesmo, sem prejuízo nenhum para o seu
tratamento.
O que é célula natural killer? As células natural killer são células de defesa do corpo importantes no combate a doenças
infecciosas e tumorais.
O que são alterações epigenéticas? As alterações epigenéticas constituem modificações associadas ao DNA e que podem
influenciar na função das células no corpo humano.
Como você, paciente, vai participar da pesquisa? A pessoa que participar desta pesquisa deverá doar 40 mL de sangue. Esta quantidade de
sangue retirada não será prejudicial à saúde. A coleta de sangue será realizada no braço,
com seringas e agulhas descartáveis por um profissional capacitado. Não há riscos

SOARES, L.S.
155
previsíveis decorrentes da pesquisa para o doador, a não ser o desconforto da picada da
agulha na coleta de sangue.
Qual o objetivo desta pesquisa? Esta pesquisa trará benefícios para o entendimento de como as células de defesa de um
indivíduo infectado com HIV funcionam em comparação com as células de indivíduos que
não estejam infectados com o vírus (controle negativo). Além disso, os dados dessa
pesquisa poderão ser úteis para o desenvolvimento de um teste para melhorar o tratamento
de pacientes infectados com HIV.
Não haverá nenhuma despesa e nenhum ganho em dinheiro. Afirmamos que os dados
obtidos serão guardados de maneira sigilosa e que os nomes dos participantes não serão
divulgados em nenhum momento, e os mesmos serão informados dos resultados finais da
pesquisa. Esta pesquisa não trará benefício direto ao sujeito envolvido, ficando a critério do
mesmo a desistência em participar da pesquisa a qualquer momento, sem nenhum prejuízo
ao seu seguimento hospitalar.
Qualquer dúvida sobre esta pesquisa, entrar em contato com Luana Silva Soares
pelo telefone (16) 3602-4189 ou (16) 99719-4270.
Eu,_______________________________________________________________________
RG n°: ____________________________, autorizo a retirada de 40 mL do meu sangue da
veia do braço esquerdo para a realização de pesquisa. Fui informado (a) sobre o
procedimento e que o volume retirado não causará dano a minha saúde. Fui informado (a)
também que a finalidade dessa pesquisa é de entender como o vírus se comporta e
promove o aparecimento das infecções.
Ribeirão Preto, _____ de _________________ de 200__.
________________________________ ________________________________
Assinatura do paciente – Registro MSc. Luana Silva Soares (Pesquisador Responsável)