frutose 1,6 bifosfato, um intermediÁrio da...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
PÓS-GRADUAÇÃO EM IMUNOLOGIA BÁSICA E APLICADA
FRUTOSE 1,6 BIFOSFATO, UM INTERMEDIÁRIO
DA GLICÓLISE, REGULA A PRODUÇÃO DE IL-
10 EM MACRÓFAGOS DE FORMA DEPENDENTE
DA VIA ADENOSINÉRGICA
Paula Ramos Viacava
Ribeirão Preto – SP
2015

PAULA RAMOS VIACAVA
FRUTOSE 1,6 BIFOSFATO, UM INTERMEDIÁRIO
DA GLICÓLISE, REGULA A PRODUÇÃO DE IL-
10 EM MACRÓFAGOS DE FORMA DEPENDENTE
DA VIA ADENOSINÉRGICA
Dissertação apresentada ao Programa de
Pós-graduação em Imunologia Básica e
Aplicada da Faculdade de Medicina de
Ribeirão Preto da Universidade São Paulo
como parte das exigências para obtenção
do título de Mestre em Ciência.
Área de concentração: Imunologia
Orientador: Prof. Dr. José Carlos Farias Alves Filho
Ribeirão Preto – SP
2015

AUTORIZO A REPRODUÇÃO E DIVULGAÇÃO TOTAL OU PARCIAL
DESTE TRABALHO, POR QUALQUER MEIO CONVENCIONAL OU
ELETRÔNICO, PARA FINS DE ESTUDO E PESQUISA, DESDE QUE
CITADA A FONTE.
FICHA CATALOGRÁFICA
Viacava, Paula Ramos
Frutose 1,6 bifosfato, um intermediário da glicólise, regula a
produção de IL-10 em macrófagos de forma dependente da via
adenosinérgica. Ribeirão Preto, 2015. 98 p; il.:30cm
Dissertação de mestrado, apresentada à Faculdade de Medicina de
Ribeirão Preto/USP. Área de concentração: Imunologia Básica e
Aplicada.
Orientador: Alves-Filho, José Carlos Farias.
1. Frutose 1,6- Bifosfato. 2. IL-10. 3. Adenosina. 4. ATP.
5. Via Glicolítica.

FOLHA DE APROVAÇÃO
PAULA RAMOS VIACAVA
FRUTOSE 1,6 BIFOSFATO, UM INTERMEDIÁRIO
DA GLICÓLISE, REGULA A PRODUÇÃO DE IL-
10 EM MACRÓFAGOS DE FORMA DEPENDENTE
DA VIA ADENOSINÉRGICA
Dissertação apresentada ao Programa de
Pós-graduação em Imunologia Básica e
Aplicada da Faculdade de Medicina de
Ribeirão Preto da Universidade São Paulo
como parte das exigências para obtenção
do título de Mestre em Ciência.
Área de concentração: Imunologia
Banca examinadora
________________________________________
Prof. Dr. José Carlos Farias Alves Filho
FMRP-USP
________________________________________
Prof. Dra. Vanessa Carregaro Pereira
FMRP-USP
________________________________________
Prof. Dr. Robson Coutinho Silva
UFRJ

Trabalho realizado no Laboratório de Inflamação e dor do Departamento de Farmacologia e
associado à área de Imunologia Básica e Aplicada da Faculdade de Medicina de Ribeirão Preto
– Universidade de São Paulo, com auxílio financeiro da FAPESP, CAPES e CNPq.

DEDICATÓRIA

Dedico este trabalho à minha Mãe,
Zoraide Conceição Maciel Ramos,
por ser não só a melhor mãe do
mundo, mas também o meu melhor
pai, minha melhor amiga, minha
conselheira, ou seja, meu alicerce.

AGRADECIMENTOS

AGRADECIMENTOS
Gostaria de agradecer a todos que contribuíram para execução deste trabalho.
Agradeço em especial:
Ao Prof. Dr. José Carlos Farias Alves Filho não só pela orientação, mas também pela
paciência, entusiasmo com o projeto e todos os ensinamentos que são muito valiosos para a
minha formação.
Aos Profs. Dr. Fernando de Queiróz Cunha, Dr. Thiago Cunha e Prof. Dr. Sérgio
Henrique Ferreira pelo auxilio, disponibilidade e contribuição para a realização deste trabalho.
À Profa. Dra. Vanessa Carregaro por aceitar ao meu convite de ser membro da banca
avaliadora, além de gentilmente ter cedido os camundongos A2AR KO.
Ao Porf. Robson Coutinho por disponibilizar parte do seu tempo para se dedicar como
membro avaliador da banca de mestrado.
À minha família que sempre acreditou e torceu por mim (TODOS). Especialmente Minha mãe,
Zoraide Ramos, meu pai, Vitor Viacava, irmãos Keililine, Vinicius e Fernando, pelo valioso
apoio e companheirismo, em todos os momentos da minha vida.
Ao Roberto Ferreira, por me mostrar que laços de sangue nem sempre são os que unem.
Ao Alexandre Ribeiro, por ser sempre muito compreensivo com a minha ausência e sempre
presente quando eu precisei.
À minha família “Ribeirão Pretana”, Maria Claudia e Angélica por fazerem do nosso humilde
apartamento um verdadeiro lar.
Aos colaboradores desse trabalho e também amigos João Paulo Mesquita, Flavio Protásio,
Daniele Nascimento, Raphael Ferreira, Priscila Tartari, Paulo Henrique Melo, Douglas

Prado, Annie Piñeros, Vanessa Borges, Tatiana Cecílio e Alexandre Kanashiro pela ajuda
na execução do projeto, correções na escrita dessa dissertação e momentos de amizade
inesquecíveis.
Aos amigos do Laboratório de Inflamação e Dor, com quem pude compartilhar momentos
únicos de aprendizado, alegrias, medos, insegurança, conquistas e incentivo no desenvolvimento
deste trabalho e pelas horas de lazer, não irei citar nomes para não cometer injustiças.
À Ana Kátia dos Santos, Diva Amabile Montanha de Sousa, Marcos, Sérgio Roberto Rosa,
Ieda Regina dos Santos Schivo pela amizade, convívio e auxilio técnico. Especialmente à
Giuliana Bertozi Francisco, por ter me passado valiosos ensinamentos de como trabalhar com
cultura de células, e por carinhosamente me adotar como filha.
Aos funcionários do biotério do Departamento de Genética, Júlio Siqueira, Dener e; e do
Departamento de Farmacologia Eliana Barros e Ronaldo pela atenção e cuidado com os animais,
que foi fundamental para a qualidade deste trabalho.
Aos amigos do programa de pós-graduação em Imunologia Básica e Aplicada, e do
Departamento de Farmacologia da FMRP-USP.
Aos professores do programa de pós-graduação em Imunologia Básica e Aplicada pelo empenho
constante para manter o nível de excelência de nosso curso.
À Ana Cristina S. Ferreira secretária da pós-graduação, pela amizade e ajuda incondicional.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional
de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do
Estado de São Paulo (FAPESP), Fundação de poio ao Ensino, Pesquisa e Assistência (FAEPA)
pela concessão dos recursos para a realização deste trabalho.

RESUMO

RESUMO
VIACAVA, P. R. Frutose 1,6 bifosfato, um intermediário da glicólise, regula a produção de
IL-10 em macrófagos de forma dependente da via adenosinérgica. Ribeirão Preto, 2015. 98
p. Dissertação (Mestrado) – Faculdade de Medicina de Ribeirão Preto, Universidade de São
Paulo.
Frutose 1,6-bifosfato (FBP) é um intermediário da via glicolítica, que quando administrada
exogenamente apresenta efeito protetor em modelos de injuria isquêmica, o qual foi associado a
uma sustentada glicólise e aumento da produção de ATP. Além disso, estudos prévios do nosso
grupo demonstraram que a FBP apresenta-se como um fármaco promissor na artrite
experimental, através da geração de adenosina extracelular e ativação subsequente do receptor
de adenosina A2A (A2AR). Neste trabalho, nós investigamos os mecanismos imunomoduladores
da FBP dependentes das vias adenosinérgica e glicolítica em macrófagos ativados. Os nossos
resultados mostram que a FBP apresenta redução dos níveis de IL-1β e IL-6, e um aumento
significativo (p<0,001) da produção de IL-10 em macrófagos derivados de medula óssea
(BMDMs) ativados com LPS. O aumento dos níveis de IL-10 é dependente da produção
adenosina (ADO), pois a presença de adenosina deaminase, enzima que metaboliza a ADO,
aboliu o efeito proporcionado pela FBP. A inibição farmacológica do A2AR bloqueou a
produção de IL-10 pela FBP, e este resultado não ocorreu com antagonistas dos receptores A1R,
A2BR, ou A3R. Corroborando esses dados, a FBP não apresentou efeitos em BMDMs
originados de camundongos deficientes para o A2AR. Além disso, a inibição de CD39 e CD73,
que promovem hidrólise de ATP em ADO, também atenuou a produção de IL-10 pela FBP. A
produção de ATP extracelular também encontra-se aumentada na presença de FBP, e esse
aumento é dependente da via glicolítica. Além disso, verificamos que a inibição da Pannexin1,
um canal de liberação de ATP, reduz a produção de IL-10 promovida pela FBP. Por fim, o
tratamento prévio com FBP foi capaz de aumentar os níveis da citocina IL-10 no soro, lavado
peritonial e pulmão, em modelo de endotoxemia peritonial induzida por LPS, de modo
dependente do receptor A2A. Em conjunto, estes dados sugerem o FBP é uma molécula-chave
no aumento da produção de IL-10, uma importante citocina anti-inflamatória, por mecanismo
dependente da produção, liberação e hidrólise do ATP em ADO via CD39 e CD73 em
macrófagos ativado, demonstrando ser um promissor fármaco no tratamento de doenças
inflamatórias.
Palavras-chave: Frutose 1,6-bifosfato, adenosina, IL-10.

ABSTRACT

ABSTRACT
VIACAVA, P. R. Fructose 1,6 bisphosphate, an intermediate of glycolysis, regulates IL-10
production in macrophage dependent by adenosinergic pathway. Ribeirão Preto, 2015. 98 p.
Master’ Degree – Faculdade de Medicina de Ribeirão Preto, Universidade de São Paulo.
Fructose 1,6-bisphosphate (FBP) is an intermediate in the glycolytic pathway, that when
administered exogenously introduced protective effect in models of ischemic injury, which was
associated with a sustained increase in glycolysis and ATP production. Besides our previous
studies show that FBP minimizes the effects of experimental arthritis by systemic generation of
extracellular adenosine and subsequent activation of adenosine receptor A2a (A2aR). The aim of
this study was to investigate the molecular mechanisms of the anti-inflammatory effect of FBP
in macrophages. To this end, macrophages derived from bone marrow (BMDM) or cell line
RAW 264.7 were treated with FBP 6h before LPS stimulation for 24 hours, the supernatant was
collected for quantification of cytokines by ELISA and ATP by luminescence, and cells was
collected for flow cytometry and RT-PCR. We found that the FBP decrease levels of IL-1β and
IL-6, whereas significant enhanced IL-10 production by LPS-activated BMDMs. The increased
IL-10 production was dependent on adenosine, since the adenosine deaminase presence
abolished the effect of increasing IL-10 provided by FBP or ATP. Pharmacological inhibition of
A2AR blocked the increased production of IL-10 for FBP, and failed to enhance IL-10
production in BMDMs originated from mice deficient for the A2AR. Moreover, the inhibition of
CD39 and CD73 also blocked the enhanced production of IL-10 by the FBP, even if their
presence does not alter the expression of these ectonucleotidases. While ATP production was
increased in the presence of FBP, and this increase is dependent on the glycolytic pathway. In
addition, we found that the inhibition of Pannexin1, an ATP-release channel, blocks the IL-10
induction by FBP. Finally, in a model of endotoxemia with LPS injection into the peritoneal
cavity, pre-treatment with FBP was able to increase the levels of cytokine IL-10 in serum,
peritoneal lavage and lung 2A receptor-dependent manner. Taken together, these data imply
FBP as a key molecule that increases the production, release and hydrolysis of ATP to
adenosine via CD39 / CD73 in activated macrophages, demonstrating an anti-inflammatory
promising.
Key words: Fructose 1,6-bisphosphate, adenosine, IL-10.

LISTA DE ABREVIAÇÕES
2-DG 2-Deoxy-D-glicose
A1R receptor de adenosina 1
A2AR receptor de adenosina 2A
A2BR receptor de adenosina 2B
A3R receptor de adenosina 3
ADA Adenosina deaminase
ADO Adenosina
ADP Adenosina difosfato
AHR receptor de hidrocarboneto de arilo
AMP Adenosina monofosfato
AMPc adenosina 3',5'-monofosfato cíclico
APC Células apresentadoras de antígenos
APCP adenosine 5'-(α,β-methylene)diphosphate inibidor de CD73
ARL ARL 67156 inibidor de cd39
ATP Adenosina trifosfato
BMDM Macrófagos derivados da medulla óssea
CBX Carbenoxolone
CD Cluster of differentiation
Con-A Concavalina A
CREB cAMP response element-binding protein
CSC 8-(3-Chlorostyryl)caffeine, antagonista de A2AR
CSF-1 colony stimulating factor 1
DAMP Damage-associated molecular pattern molecules
DHAP fosfato de dihidroxiacetona
DPCPX 8-Ciclopentil-1,3-dipropilxantina, antagonista de A1R
ERK Extracellular signal-regulated kinases
FIZZ1 Found in inflammatory zone protein

FBP Frutose 1,6 –bifosfato
IAA Iodoacetamide
IFN-γ Interferon gama
IL Interleucina
iNOS Óxido nítrico sintase induzível
IRF4 interferon-regulatory fator
G-CSF Fator Estimulador de Colônias de Granulócitos
GM-CSF Fator de estimulação de colónias de granulócitos-macrófagos
G3P D-gliceraldeído-3-fosfato
LPS Lipopolissacarídeo
MAP quinase Mitogen Activated Protein
MHC Complexo principal de histo-compatibilidade
MRC 1 Mannose receptor C type 1
MTX Metotrexato
MyD88 Myeloid differentiation primary response gene 88
NF-κB Fator nuclear kappa B
MRS MRS 1334, antagonista de A3R
MRS MRS 1754, antagonista de A2BR
NO Óxido nítrico
PBS Tampão salina-fosfato
PFK 1 Fosfofrutoquinase 1
PKA Proteína quinase A
PLC Fosfolipase C
PMNs leucócitos polimorfonucleares
PRRs Receptores de reconhecimento de padrões
SOCS 3 Suppressor of cytokine signaling 3
STAT Signal transducer and activator of transcription
Syk Spleen tyrosine kinase

TAM Macrófagos associados a tumores
TCR Receptores de células T
TGF-β Transforming Growth Factor beta
TLR Receptores de tipo Toll
TNF-α Fator de necrose tumoral alfa
TR1 células reguladoras do tipo 1
Treg Células T reguladoras

ÍNDICE DE FIGURAS
FIGURA 1: ESQUEMA DA VIA ADENOSINÉRGICA............................................................... 25
FIGURA 2: ESQUEMA DA VIA GLICOLÍTICA ....................................................................... 30
FIGURA 3: FBP REDUZ A PRODUÇÃO DE CITOCINAS PRÓ-INFLAMATÓRIAS EM
MACRÓFAGOS ESTIMULADOS POR LPS ........................................................................ 48
FIGURA 4: FBP AUMENTA A PRODUÇÃO DE IL-10 EM MACRÓFAGOS
ESTIMULADOS COM LPS .................................................................................................. 49
FIGURA 5: O AUMENTO DA PRODUÇÃO DE IL-10 INDUZIDO POR FBP É
DEPENDENTE DE ADENOSINA ........................................................................................ 51
FIGURA 6: O EFEITO DA FBP NA PRODUÇÃO DE IL-10 É DEPENDENTE DA
ATIVAÇÃO DOS RECEPTORES A2A PELA ADENOSINA ............................................... 53
FIGURA 7: MACRÓFAGOS DERIVADOS DE ANIMAIS DEFICIENTES DO RECEPTOR
A2A NÃO APRESENTAM O AUMENTO NA PRODUÇÃO DE IL-10 INDUZIDO PELA
FBP APÓS O ESTIMULO DE LPS ....................................................................................... 54
FIGURA 8: LPS AUMENTA AS EXPRESSÕES DOS RECEPTORES A2, MAS FBP
ALTERA SOMENTE A EXPRESSÃO DO RECEPTOR A2B. .............................................. 55
FIGURA 9: FBP NÃO ALTERA A EXPRESSÃO DE CD39 E CD73 .......................................... 57
FIGURA 10: EFEITO DA INIBIÇÃO DAS ECTONUCLEOTIDASES CD39 E CD73 NA
PRODUÇÃO DE IL-10 .......................................................................................................... 59
FIGURA 11: FBP EM MACRÓFAGOS RAW NÃO ALTERA A EXPRESSÃO DE CD39 E
CD73 APÓS ESTIMULO DE LPS ........................................................................................ 61
FIGURA 12: MACRÓFAGOS BMDM E RAW DIFEREM NO PADRÃO DE PRODUÇÃO
DE TNF-Α E IL-10 ................................................................................................................ 63
FIGURA 13: FBP INDUZ O AUMENTO DA GERAÇÃO DE ATP EM MACRÓFAGOS .......... 65
FIGURA 14: FBP AUMENTA A LIBERAÇÃO DE ATP EM MACRÓFAGOS
ESTIMULADOS COM LPS DE FORMA DEPENDENTE DA VIA GLICOLÍTICA ............ 68

FIGURA 15: FBP AUMENTA A LIBERAÇÃO DE ATP EM MACRÓFAGOS
ESTIMULADOS COM LPS DE FORMA DEPENDENTE DA VIA GLICOLÍTICA ............ 69
FIGURA 16: O ATP EXTRACELULAR AUMENTA A PRODUÇÃO DE IL-10 INDUZIDA
PELA FBP EM MACRÓFAGOS ESTIMULADOS COM LPS ............................................. 71
FIGURA 17: INIBIÇÃO DO CANAL PURINÉRGICO P2X7 NÃO ALTERA A PRODUÇÃO
DE IL-10. ............................................................................................................................... 73
FIGURA 18: FBP PROMOVE EFEITO ANTI-INFLAMATÓRIO, IN VIVO, EM MODELO
DE PERITONITE .................................................................................................................. 75
FIGURA 19: MECANISMO IMUNOMODULADOR DA FBP, ATRAVÉS DO AUMENTO
DA PRODUÇÃO DE IL-10, DE FORMA DEPENDENTE DA VIA GLICOLÍTICA E
ADENOSINÉRGICA. ............................................................................................................ 85

SUMÁRIO
1. INTRODUÇÃO..................................................................................................................................... 22
1.1. ADENOSINA ............................................................................................................................ 23
1.2. FRUTOSE 1,6- BIFOSFATO ..................................................................................................... 28
2. OBJETIVOS ..................................................................................................................................... 33
2.1 OBJETIVO GERAL ................................................................................................................... 34
2.2 OBJETIVOS ESPECÍFICOS ...................................................................................................... 34
3. MATERIAL E MÉTODOS .................................................................................................................. 35
3. MATERIAL E MÉTODOS ............................................................................................................ 36
3.1 ANIMAIS DE EXPERIMENTAÇÃO ........................................................................................ 36
3.2 PREPARO DAS SOLUÇÕES E REAGENTES .......................................................................... 36
3.3 DIFERENCIAÇÃO DE MACRÓFAGOS PROVENIENTES DA MEDULA ÓSSEA ................ 38
3.4 CULTURA DE MACRÓFAGOS DE LINHAGEM RAW 264.7 ................................................ 39
3.5 ENSAIOS COM FBP ................................................................................................................. 40
3.6 DETECÇÃO DE CITOCINAS PELO ENSAIO IMUNOENZIMÁTICO (ELISA) ..................... 41
3.7 PCR QUANTITATIVO EM TEMPO REAL .............................................................................. 42
3.8 CITOMETRIA DE FLUXO DE MACRÓFAGOS ...................................................................... 43
3.9 ENSAIO DE ATP ...................................................................................................................... 43
3.10 PROTOCOLO DE ENDOTOXEMIA ........................................................................................ 44
3.11 ANÁLISE ESTATÍSTICA ......................................................................................................... 45
4. RESULTADOS ..................................................................................................................................... 46
4.1 FBP ALTERA A PRODUÇÃO DE CITOCINAS POR MACRÓFAGOS ESTIMULADOS COM
LPS ....................................................................................................................................................... 47
4.2 O AUMENTO DA PRODUÇÃO DE IL-10 INDUZIDO POR FBP É DEPENDENTE DE
ADENOSINA E DO RECEPTOR A2A .................................................................................................. 50
4.3 ECTONUCLEOTIDASES CONTRIBUEM PARA O AUMENTO DA PRODUÇÃO DE IL-10
PROMOVIDO PELA FBP ..................................................................................................................... 56
4.4 FBP AUMENTA A PRODUÇÃO E LIBERAÇÃO DE ATP POR MACRÓFAGOS
ESTIMULADOS COM LPS DE FORMA DEPENDENTE DA VIA GLICOLÍTICA ............................ 64

4.5 ATP GERADO É FUNDAMENTAL PARA O AUMENTO DE IL-10, SEM ATIVAÇÃO DO
INFLAMASSOMA................................................................................................................................ 70
4.6 FBP PROMOVE EFEITO ANTI-INFLAMATÓRIO, IN VIVO, EM MODELO DE
ENDOTOXEMIA. ................................................................................................................................. 74
5. DISCUSSÃO ......................................................................................................................................... 76
6. CONCLUSÃO ....................................................................................................................................... 84
7. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................................. 86

1. INTRODUÇÃO

Introdução
23
1. INTRODUÇÃO
1.1. ADENOSINA
A adenosina (ADO) é um nucleosídeo de purina, formado pela ligação de uma adenina a
uma D-ribose, na forma fosfatada é conhecida como AMP, quando apresenta dois fosfatos é
chamada de ADP ou ATP quando possuir três fosfatos (Lehninger et al. 2007). Pode ser
encontrada em todos os tecidos, suas principais ações são descritas como regulação das funções
cardiovasculares e modulação do sistema nervoso central. Além disso, a ADO é gerada tanto
intracelular como extracelular, por fibroblastos, células epiteliais, endoteliais, plaquetas e células
musculares, embora também possa ser derivada do metabolismo extracelular de nucleosídeos
purínicos (Fredholm et al. 2000, Lappas et al. 2005, Sato et al. 2005).
A formação intracelular de adenosina ocorre através da hidrólise do AMP pela enzima
5’-nucleotidase, ou S-adenosilhomocisteína pela ação da S-adenosilhomocisteína hidrolase (Coe
et al. 1996). Transportadores bidirecionais são capazes de transportar ADO formada
intracelularmente para o ambiente extracelular ou vice-versa, esse nucleosídeo pode ser
transportado a favor do gradiente de concentração pelos transportadores ENTs (equilibrative
nucleoside transporter). Situações de hipóxia e inflamação levam ao consumo do ATP e
acúmulo de adenosina no interior da célula, o qual se difunde para o ambiente extracelular a
favor do gradiente de concentração por meio de ENTs. A ADO também pode ser transportada
através dos CNT (concentrative nucleoside transporter) que são transportadores dependentes da
concentração de Na+ agindo contra o gradiente de concentração e consequentemente
consumindo energia (Molina-Arcas et al. 2009).
O estresse celular pode levar a um aumento exacerbado de ATP ou ADP no interior da
célula, e esses nucleosídeos podem ser liberados para o ambiente extracelular através de

Introdução
24
apoptose ou necrose (Yegutkin 2008). Uma vez encontrados no meio extracelular, os
nucleosídeos medeiam seus efeitos através da sinalização de uma série de receptores
ionotrópicos P2X e receptores metabotrópicos P2Y, classificados pela sua afinidade em relação
ATP, ADP (Kronlage et al. 2010). Sabe-se que altos níveis de ATP no meio extracelular (1-
8mM) é caracterizado como um sinal de estresse celular, pois promove um perfil pró-
inflamatório via sinalização do receptor P2X7, ativando a plataforma do inflamassoma e
consequentemente a produção de IL-1β (Ferrari et al. 1997, Pelegrin & Surprenant 2006)
No entanto, altos níveis de ATP normalmente não são encontrados nos organismos
vivos, pois as concentrações de ATP extracelular são rigorosamente controladas pela ação de
ectoenzimas que hidrolisam o ATP para ADP e AMP, essa enzima é a NTPDase-1 (ecto-apirase)
também conhecida como CD39. Ao passo que, o AMP pode ser convertido em ADO através da
ação da ecto-5’-nucleotidase (CD73), essa via é uma das principais formas responsáveis pela
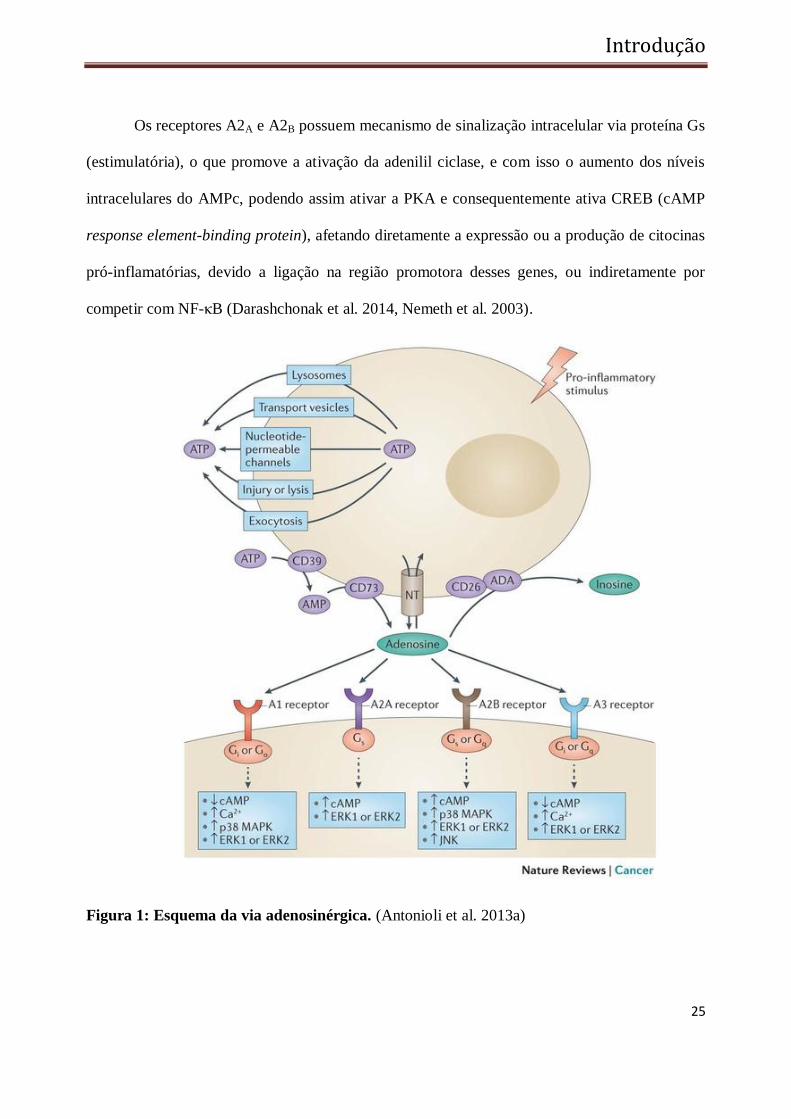
geração de ADO no meio extracelular. (Antonioli et al. 2013b, Eltzschig 2009) (Figura 1).
A ADO extracelular atua através de um conjunto de receptores (A1, A2A, A2B e A3),
esses receptores contêm sete domínios transmembrana acoplados a proteínas G (ligantes de GTP
intracelular) estimulatória (Gs) ou inibitória (Gi) da adenilato ciclase, enzima que cataliza a
formação ou diminuição dos níveis de cAMP (AMP cíclico) intracelular. Quando ocorre a
ativação de A1R ou A3R ocasiona na inibição da atividade da adenilil ciclase, pois esses
receptores estão acoplados a proteína Gi, e consequentemente, inibe a conversão do ATP em
AMPc e assim não ativando a via da proteína kinase A (PKA), além disso ocorre uma maior
liberação de Ca2+
o que aumenta a atividade da fosfolipase C (PLC) (Duro et al. 2014, Osada et
al. 1992, Pereira et al. 2010).
Introdução
25
Os receptores A2A e A2B possuem mecanismo de sinalização intracelular via proteína Gs
(estimulatória), o que promove a ativação da adenilil ciclase, e com isso o aumento dos níveis
intracelulares do AMPc, podendo assim ativar a PKA e consequentemente ativa CREB (cAMP
response element-binding protein), afetando diretamente a expressão ou a produção de citocinas
pró-inflamatórias, devido a ligação na região promotora desses genes, ou indiretamente por
competir com NF-κB (Darashchonak et al. 2014, Nemeth et al. 2003).
Figura 1: Esquema da via adenosinérgica. (Antonioli et al. 2013a)

Introdução
26
A primeira vez que relataram o A2AR como um importante modulador da resposta
inflamatória foi em modelo de lesão hepática, onde os pesquisadores demonstraram que a
administração de um agonista seletivo para A2AR pode prevenir o dano hepático, inibindo a
produção de TNF-α, IL-12 e IFN-γ, assim como a deleção do gene para A2AR demonstrou um
papel importante na redução da sobrevida dos animais submetidos a uma lesão hepática por
Concavalina-A (Con-A), pois a ausência desse receptor aumentou a produção de citocinas pró-
inflamatórias (Ohta & Sitkovsky 2001). Além disso, a administração de antagonistas ou a
deleção genética do receptor também promoveu a piora dos processos inflamatórios crônicos em
diferentes modelos experimentais, tais como: encefalomielite autoimune experimental (Mills et
al. 2012), miastenia gravis experimental autoimune (Li et al. 2012) e pleurisia induzida por
carragenina (Lapa et al. 2012).
Um dos genes em que a transcrição é induzida pelo AMPc é o da citocina
imunomoduladora IL-10, que possui um papel supressor do processo inflamatório (Montminy e
Bilezikjian, 1987; Zidek, 1999), Essa citocina apresenta uma estrutura homodimérica, sem
presença de carboidratos, constituída por uma cadeia polipleptídica estruturada em quatro α-
hélices (Stroud & Wells 2004). Pode ser produzida por linfócitos Th1, Th2, Th17, células T
reguladoras, células B, macrófagos, queratinócitos, células dendríticas, entre outras células
(Maynard & Weaver 2008, Moore et al. 2001, O'Garra & Vieira 2004, Roncarolo et al. 2006). A
IL-10 inibe a síntese de citocinas como TNF-α, IL-1β, IL-6, IL-8, IL-18 e IFN-γ, também é
quimioatrativa para linfócitos T CD8+ e participa da downregulação da secreção de IL-12, uma
citocina crítica para a diferenciação de Th1 (Fiorentino et al. 1991).
Recentemente, pesquisadores descreveram que outro fator de transcrição, o C/EBPβ
(CCAAT/enhancer-binding protein beta), também é responsável pela produção de IL-10 em
macrófagos expostos a Escherichia coli, esse efeito é decorrente da ativação de A2AR, que são

Introdução
27
mediados via ativação de MAPK (proteína quinase ativada por mitógeno) e PKC (proteína
quinase C) (Csoka et al. 2007).
Em macrófagos, a IL-10 consegue suprimir a produção de óxido nítrico e outros
mediadores inflamatórios como TNF-α, IL-1β, IL-6, IL-8 e GM-CSF (Saraiva & O'Garra 2010),
Essas células são conhecidas pela eficiência no killing de microrganismos através de fagocitose,
apresentação de antígenos ao linfócito T, e clearance do organismo por remoção das células
apoptóticas, reparo e remodelamento tecidual, além de apresentar importante papel resolutivo
nos processos inflamatórios. Originam-se a partir de um progenitor mielóide, compartilhado
também por granulócitos na medula óssea, circulam por vários dias no sangue periférico como
monócitos antes de alcançarem os tecidos e se diferenciarem em macrófagos (Gordon & Taylor
2005, Vanfurth 1985, Volkman 1985).
Quando macrófagos são ativados com LPS podem liberar ATP no meio extracelular, e
como sua membrana plasmática apresenta uma alta expressão das ectonucleotidases CD39 e
CD73, essas células são capazes de converter o ATP liberado à adenosina (Cohen et al. 2013).
Adenosina também bloqueia a ativação clássica de macrófagos, inibindo seus mecanismos
microbicidas (Csoka et al. 2012), alterando de um perfil pró-inflamatório para anti-inflamatório
(Hasko et al. 2008), inibindo a produção de espécies reativas do oxigênio (Cronstein et al. 1990)
e a fagocitose (Salmon & Cronstein 1990). Além disso, a ativação dos receptores A2 pela
adenosina pode diminuir a produção de TNF-α e aumentar a produção de IL-10 pelos
macrófagos, e também levando a diferenciação de macrófagos M2 (Csoka et al. 2012, Hasko et
al. 2000).
Uma das principais rotas para geração de ATP nas células e está presente em todos os
tipos de tecidos é através da via glicolítica. A glicólise é uma sequência metabólica composta

Introdução
28
por um conjunto de dez reações catalizadas por enzimas livres no citosol, na qual a glicose é
oxidada consumindo duas moléculas de ATP, para produzir duas moléculas de piruvato, quatro
moléculas de ATP e dois equivalentes reduzidos de NADH+, que serão introduzidos na cadeia
respiratória ou na fermentação (Lehninger et al. 2007).
Visto que a via glicolítica pode ser dividida didaticamente em duas fases, uma fase de
“investimento” em que ocorre o consumo de 2 mols de ATP para a formação de frutose1,6-
bifosfato (FBP), e a partir desse ponto da via uma fase de “produção” de 4 mols de ATP, com
um balanço final na produção de 2 mols de ATP (Markov 1986). Nesse sentido, torna-se
interessante investigar se a administração exógena de FBP pode apresentar um papel no aumento
da produção de ADO através da via glicolítica.
1.2. FRUTOSE 1,6- BIFOSFATO
A FBP é um carboidrato intermediário da via glicolítica, altamente energético. Trata-se
de uma molécula polar composta por um monossacarídeo frutose fosforilado em dois carbonos,
C-1 e C-6, produzida pela fosforilação da frutose-6-fosfato através da adição de um fosfato pela
enzima fosfofrutoquinase 1 (PFK1). A FBP é clivada pela enzima aldolase em D-gliceraldeído-
3-fosfato (G3P) e fosfato de dihidroxiacetona (DHAP), tendo uma participação fundamental na
rota metabólica da glicólise que culmina na produção de 4 mols de ATPs para cada molécula de
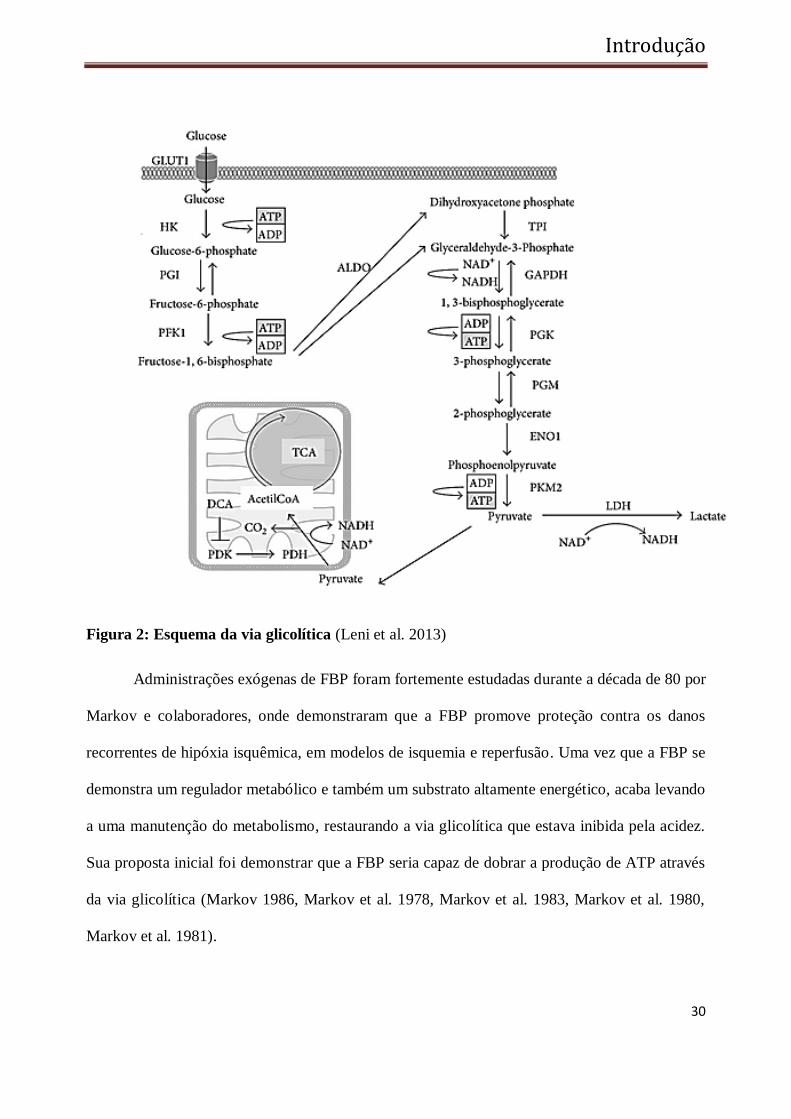
FBP. (Lehninger et al. 2007) (Figura 2).
Na via glicolítica, a FBP se encontra após as reações das enzimas hexoquinase e
fosfofrutoquinase, etapas essas que consomem ATP, e antes da piruvato quinase, sobre o qual a
FBP atua como um ativador alostérico. A produção de ATP é estimulada, pois a piruvato
quinase não será um oponente para sua metabolização, além de aumentar os níveis de ATP, a

Introdução
29
FBP também facilita na formação de ATP a partir do ADP (Bailey et al. 1968, Irving &
Williams 1973, Loguercio et al. 1996).
Esse metabólito é capaz de entrar na célula atingindo o citosol, e essa entrada de FBP no
meio intracelular faz com que aumente o fluxo glicolítico (Hassinen et al. 1991, Lazzarino et al.
1984, Nuutinen et al. 1991). Porém, ainda não foi identificado um transportador de membrana
próprio para o transporte de FBP do meio extracelular para o interior da célula. Sabe-se que a
FBP é uma molécula altamente polar, e que em pH fisiológico se encontra carregada. O
mecanismo de entrada da FBP no meio intracelular, provavelmente, seja a sua difusão pela
membrana de forma dose dependente (Ehringer et al. 2000), visto que ela tem a capacidade de
atravessar a membrana plasmática (Hardin & Roberts 1994).
Introdução
30
Figura 2: Esquema da via glicolítica (Leni et al. 2013)
Administrações exógenas de FBP foram fortemente estudadas durante a década de 80 por
Markov e colaboradores, onde demonstraram que a FBP promove proteção contra os danos
recorrentes de hipóxia isquêmica, em modelos de isquemia e reperfusão. Uma vez que a FBP se
demonstra um regulador metabólico e também um substrato altamente energético, acaba levando
a uma manutenção do metabolismo, restaurando a via glicolítica que estava inibida pela acidez.
Sua proposta inicial foi demonstrar que a FBP seria capaz de dobrar a produção de ATP através
da via glicolítica (Markov 1986, Markov et al. 1978, Markov et al. 1983, Markov et al. 1980,
Markov et al. 1981).

Introdução
31
Quando a FBP é administrada, apresenta baixos efeitos tóxicos, visto que em humanos
saudáveis não apresentaram efeitos adversos após receberem altas quantidades de FBP via
intravenosa (Markov et al. 2000). Além disso, a FBP apresenta propriedades anti-inflamatórias
como, em modelo de pleurisia induzida por carragenina, no qual, o tratamento com FBP
apresentou redução do edema de pata induzida por injeção de carragenina na região plantar
(Planas et al. 1993). Em adição, o tratamento com FBP reduziu a migração de leucócitos
polimorfonucleares (PMNs) para a cavidade pleural (Alves et al. 2004).
Em outros modelos como de infecção bacteriana e fúngica, a FBP também apresentou
efeitos anti-inflamatório. Ao passo que animais foram tratados com FBP apresentaram um
aumento na sobrevida em infecções por Escherichia coli, porém esse efeito da FBP foi anti-
inflamatório e não microbicida (Nunes et al. 2003). Efeitos semelhantes também foram
observados em infecções por Candida albicans. (Vianna Santos et al. 2012). Recentemente
nosso grupo demonstrou que o tratamento com FBP atenua os efeitos causados pela artrite
experimental, demonstrando ser uma nova estratégia terapêutica para essa doença (Veras et al.
2015).
Características anti-inflamatórias da FBP também foram observadas em estudos in vitro,
pois o tratamento com FBP inibiu a proliferação de células T (Nunes et al. 2004), além de
reduzir a produção de citocinas pró-inflamatórias (Lopes et al. 2006, Yin et al. 2008). Porém,
até o início desse século, muito pouco se sabia sobre seu mecanismo de ação, mesmo já sendo
descrito como um promissor agente terapêutico em vários modelos patológicos (Hardin et al.
2001).
As propriedades anti-inflamatórias da FBP podem estar fortemente relacionadas com o
aumento dos níveis de ADO extracelular. No primeiro trabalho que relacionou os efeitos

Introdução
32
produzidos pelas duas substâncias, não encontrou uma relação clara entre a produção de ADO e
FBP (Akimitsu et al. 1995). No entanto, em 2003 foi demonstrado que os efeitos anti-
inflamatórios observados com FBP são dependentes da ação da ADO. Em modelo de isquemia e
reperfusão, o tratamento com FBP apresentou um efeito protetor, porém com a administração de
adenosina deaminase (ADA), uma enzima que promove a desaminação da ADO para inosina, o
efeito protetor foi abolido (Sola et al. 2003). Veras e colaboradores demonstraram que o
tratamento com FBP em modelos de artrite experimental apresenta efeitos anti-inflamatórios
semelhantes aos descritos para metotrexato (MTX), demostrando uma forte dependência da
presença de ADO no tratamento com FBP (Veras et al. 2015).
Sabendo a importância da via glicolítica na produção de ATP e dos efeitos anti-
inflamatórios promovidos pela ADO. Nossa hipótese de trabalho foi que a FBP promoveria o
aumento de ATP extracelular em macrófagos ativados, que sofre ação das ectonucleotidases e
seria hidrolisado para adenosina, promovendo um efeito anti-inflamatório dependente das vias
adenosinérgica e glicolítica.

2. OBJETIVOS

Objetivos
34
2. OBJETIVOS
2.1 OBJETIVO GERAL
Investigar os mecanismos imunomoduladores da frutose 1,6 bifosfato (FBP) em
macrófagos ativados com LPS.
2.2 OBJETIVOS ESPECÍFICOS
2.2.1 Avaliar o efeito da FBP na produção de citocinas pro- e anti-inflamatórias;
2.2.2 Verificar participação da adenosina e dos seus receptores nos efeitos
imunomoduladores da FBP;
2.2.3 Verificar participação das ectonucleotidases, CD39 e CD73, nos efeitos
imunomoduladores da FBP;
2.2.4 Verificar participação da geração de ATP nos efeitos imunomoduladores da FBP;
2.2.5 Verificar a participação da via glicolítica nos efeitos imunomoduladores da FBP;
2.2.6 Avaliar a participação do receptor purinérgico nos efeitos imunomoduladores da
FBP;
2.2.7 Verificar os efeitos anti-inflamatórios da FBP in vivo em modelo de
endotoxemia.

3. MATERIAL E MÉTODOS

Material e Métodos
36
3. MATERIAL E MÉTODOS
3.1 ANIMAIS DE EXPERIMENTAÇÃO
No presente estudo, foram utilizados camundongos isogênicos da linhagem BALB/c e
C57BL/6, machos, com 6 semanas de idade. Esses camundongos foram adquiridos no Serviço
de Biotério da Prefeitura do Campus da USP-Ribeirão Preto. Também utilizamos camundongos
deficientes para o receptor de adenosina A2a (A2a-/-
) (background BALB/c), os quais foram
criados no Centro de Criação de Camundongos Especiais da FMRP USP. Mantemos todos os
animais em experimentação no biotério do Departamento de Farmacologia da FMRP-USP em
microisoladores de 30cmx20cmx12cm, forrados com maravalha, em temperatura de (23-25 °C)
e ciclo claro/escuro controlados, com livre acesso à ração e água. Mantemos no máximo 5
animais por microisolador. Os protocolos experimentais realizados neste trabalho estão de
acordo com os Princípios Éticos de Experimentação Animal, adotado pelo Colégio Brasileiro de
Experimentação Animal (lei 11794/ 2008).
3.2 PREPARO DAS SOLUÇÕES E REAGENTES
Solução de Ketamina e Xilazina:
Cetamina 10% (Agener União) .................................................................................... 1,25mg/mL
Xylazina 2% (Agener União) ......................................................................................... 0,5mg/mL
Salina 0.9 % q.s.p ................................................................................................................ 100mL
A solução estocada a 4 ºC.
Solução de Tampão Salina-fosfato (PBS) 10X (Solução Mãe):
Cloreto de Sódio (NaCl, Merck) ............................................................................................... 80g
Cloreto de Potássio (KCl, Merck)................................................................................................ 2g
Fosfato de Sódio dibásico (Na2HPO4, Merck)........................................................................ 11,5g

Material e Métodos
37
Fosfato de Potássio monobásico (KH2PO4, Merck)................................................................... 20g
Água deionizada Milli-Q q.s.p. ......................................................................................... 1000mL
A solução estocada a 4 ºC.
PBS 1X:
PBS10X ............................................................................................................................... 100mL
Água Milli-Q q.s.p. .............................................................................................................. 900mL
O pH ajustado para 7.2, filtrada e estocada à 4 °C.
PBS/EDTA (1mM):
PBS 1X q.s.p. ....................................................................................................................... 100mL
Etilenodiaminotetracético (EDTA, Merck) ........................................................................ 37,2mg
O pH ajustado para pH 7.2, filtrada e estocada a 4 °C.
Meio RPMI incompleto:
Hepes (C8H18N2O4S, Sigma) .................................................................................................. 2,38g
RPMI 1640 (Sigma) – 1 frasco ....................................................................................................
10,4g
Bicarbonato de sódio (NaHCO3, Vetec) ................................................................................ 2,20g
Água de Milli-Q q.s.p......................................................................................................... 1000mL
O pH ajustado para pH 7.2, filtrada e estocada a 4 °C.
Meio RPMI completo:
Soro Bovino Fetal inativado (SBF-I, Difco) ......................................................................... 10mL
Estreptomicina (10mg/mL)/penicilina (10 unidades) (antibiótico, Sigma) ............................. 1mL
Fungizona (antifúngico, Sigma) ........................................................................................... 120µL
L--Gultamina (200mM, Sigma) ............................................................................................ 1,3mL
Gentamicina (Sigma) ............................................................................................................ 100µL

Material e Métodos
38
RPMI incompleto q.s.p. ....................................................................................................... 100mL
A solução foi preparada no dia do uso.
Tampão de citometria de fluxo (Tampão de FACS):
Soro Bovino Fetal inativado (SBF-I, Difco) 2% ..................................................................... 2mL
PBS 1X q.s.p. ....................................................................................................................... 100mL
A solução foi preparada no dia do uso e utilizada gelada.
Tampão de bloqueio (ELISA)
Albumina derivada de soro bovino (BSA, Sigma) 1 % .............................................................. 1g
PBS 1X q.s.p. ....................................................................................................................... 100mL
A solução foi preparada no dia do uso.
Tampão de lavagem (ELISA) (PBS/T)
Teewn 20 (Sigma) 0,05 %.. ..................................................................................................... 1mL
PBS 1X q.s.p. ..................................................................................................................... 2000mL
A solução foi preparada no dia do uso.
Tampão de lise:
Cloreto de amônio (NH4Cl, Merck) ....................................................................................... 4,01g
EDTA (Labsynth) .................................................................................................................. 0,18g
Bicarbonato de sódio (NaHCO3, Vetec) ................................................................................ 0,42g
Água de Milli-Q q.s.p. ......................................................................................................... 500mL
O pH ajustado para pH 7.2, filtrada e estocada a 4 °C.
3.3 DIFERENCIAÇÃO DE MACRÓFAGOS PROVENIENTES DA MEDULA ÓSSEA
A diferenciação de macrófagos derivados da medula óssea (BMDM) foi realizada como
descrito previamente (Kurowska-Stolarska et al. 2009). Os animais foram sacrificados por
deslocamento cervical e após esse procedimento foi coletado o fêmur, as epífises foram cortadas

Material e Métodos
39
para exposição do canal medular. Para remoção da medula óssea, utilizamos uma agulha de 26-
gauge acoplada a uma seringa plástica de 3 mL que foi inserida na cavidade medular para
retirada da medula óssea através de flush utilizando meio de cultura RPMI-C. A suspensão de
medula óssea foi homogeneizada e centrifugada à 400g por 10 minutos à 4°C, o sobrenadante
foi desprezado, e as células ressuspendidas em 10mL de tampão de lise então novamente
centrifugada, o sobrenadante foi retirado cuidadosamente e o pelit ressuspendido em 10mL de
meio RPMI-C para retirada total do tampão de lise. As células foram centrifugadas, o
sobrenadante desprezado e então ressuspendidas em RPMI-C enriquecido com 10% de SFB-I e
20% do sobrenadante de cultura de L929, essas células foram colocadas em 4 placas de Petri
especiais (BD OPTILUX) contendo 10mL do meio para macrófagos e ficaram por 3 dias numa
estufa de CO2 à 37°C, adicionamos mais 10mL do meio de diferenciação para macrófagos e
retornaram para a estufa pro mais 4 dias.
Após uma semana na estufa as células foram destacadas com tampão fosfato salina
(PBS) gelado e com o auxílio de um suporte plástico (Cell Scraper, Corning NY) para a
raspagem das células aderentes, a solução contendo os macrófagos foi centrifugada a 400g por
10 minutos a 4°C, após o sobrenadante desprezado as células foram ressuspendida em RPMI-C
numa concentração de 200 mil células por mL para os ensaios, onde a diferenciação de
macrófagos será confirmada por citometria de fluxo.
3.4 CULTURA DE MACRÓFAGOS DE LINHAGEM RAW 264.7
Utilizamos a linhagem de macrófagos RAW 264.7 (RAW), obtida previamente por
doação do Prof. Dr. Dário Simões Zamboni. As células foram mantidas em nitrogênio líquido
(criopreservação) em solução de congelamento composta por RPMI-C com 10% de
Dimetilsulfóxido (DMSO) e 90% de soro fetal bovino. Para os ensaios de estimulação in vitro as

Material e Métodos
40
células serão descongeladas e expandidas em garrafas de cultura celular, em meio RPMI-C em
estufa incubadora umidificada à temperatura de 37°C.
Após a formação da monocamada de células e as mesmas atingirem confluência,
destacamos com PBS e auxílio de um suporte plástico (Cell Scraper, Corning NY) para
raspagem das células aderentes, o meio contendo as células foi centrifugado a 400g, durante 10
minutos a 4°C, descartamos o sobrenadante e ressuspendimos o pelit em RPMI-C.
3.5 ENSAIOS COM FBP
Nos ensaios, os macrófagos foram plaqueados em placas Costar (Sigma) de 96 poços
contendo (2 x 105 células/poço) e incubados por 1 hora na estufa de CO2 a 37°C para a aderência
das células na placa. Após adicionamos o tratamento de D-Fructose 1,6-bisphosphate trisodium
salt hydrate FBP (300µM, Sigma), Adenosine 5′-triphosphate disodium salt hydrate ATP
(100µM, Sigma), Adenosine 5′-diphosphate sodium salt ADP (100µM, Tocris) ou NECA
(10µM, Tocris) por 6 horas na estufa de CO2 a 37°C, juntamente com o tratamento, também
utilizamos Adenosina deaminase ADA (1U/mL, Sigma), 8-Ciclopentil-1,3-dipropilxantina
DPCPX (10µM, Sigma), 8-(3-Chlorostyryl)caffeine CSC (100µM, Sigma), MRS 1754 (1µM,
Tocris), MRS 1334 (10µM, Tocris), ARL 67156 trisodium salt ARL (200µM, Tocris),
adenosine 5'-(α,β-methylene)diphosphate APCP (100µM, Sigma), 2-Deoxy-D-glucose 2-DG
(1mg/mL, Sigma), STF-31 (10µM, Sigma), Iodoacetamide IAA (3 µM, Sigma), Carbenoxolone
disodium salt CBX (50µM, Sigma) e A-43807 (50µM, Sigma). Após o tratamento de 6 horas,
estimulamos as células com LPS (100ng/mL, Sigma) por 24 horas, então coletamos o
sobrenadante da cultura celular para dosagem de citocinas por ELISA.

Material e Métodos
41
3.6 DETECÇÃO DE CITOCINAS PELO ENSAIO IMUNOENZIMÁTICO (ELISA)
A determinação dos níveis de IL-10, IL-6, IL-1β e TNF-α murinos foram feitas pelo
método imunoenzimático (ELISA) utilizando kits DuoSet ELISA Development Systems (R&D
Systems) de acordo com as informações do fabricante. Placas de microtitulação (96 poços)
foram recobertas com 50 µl/ poço do anticorpo específico anti-IL-10, anti-IL6, anti-IL-1β e anti-
TNF-α, nas concentrações descritas pelo fabricante diluídos em PBS e incubados overnight a 4
ºC. As placas foram lavadas por quatro vezes com PBS/Tween-20 (0,05 % Sigma), as ligações
não-especificas foram bloqueadas com 100 µL de PBS contendo BSA 1% durante 1 hora a
temperatura ambiente. As placas foram lavadas 4 vezes com PBS/T e então adicionamos as
amostras e o padrão (curva-padrão) contendo as concentrações conhecidas de IL-10
(8000pg/mL), IL-6 (4000pg/mL), IL-1β (400 pg/mL) e TNF-α (4000pg/mL), foram colocadas
nas placas (50µL) e incubados overnight a 4 ºC. Após esse período, as placas foram lavadas com
PBS/T 4 vezes e adicionamos 50µL dos anticorpos biotinilados específicos para cada citocina
foram adicionados nas concentrações descritas pelo fabricante. Após duas horas, as placas foram
lavadas com PBS/T e o conjugado estreptavidina-peroxidase, na diluição de 1:40 foi adicionado
a cada poço e incubadas por 1 hora, novamente as placas foram lavadas com PBS/T e foi
adicionado 100µL do substrato TMB. A densidade ótica foi medida a 630 nm no
espectrofotômetro SpectraMAX 190 Microplate Reader (Molecular Devices) e os dados foram
analisados usando o software SoftMax Pro 5. A concentração de citocinas contidas nas amostras
foi calculada a partir de uma curva padrão com 12 pontos obtidos por diluição seriada. Os
resultados foram expressos em pg/mL.

Material e Métodos
42
3.7 PCR QUANTITATIVO EM TEMPO REAL
Para analisar a expressão gênica, o RNA total de macrófagos (1 x 106 células) derivados
da medula óssea de murinos foi extraído usando RNeasy Mini Kit 250 (Qiagen), de acordo com
as instruções do fabricante. O RNA total (1ug) foi utilizado para as reações de transcrição
reversa utilizando o kit High-CapacitycDNA Reverse Transcription (AppliedBiosystems) de
acordo com as instruções fornecidas pelo fabricante.
A expressão de RNAm para Il10 (NM_010548.2), A2A (adora 2a, NM_009630.3) e A2B
(adora2b, NM_007413.4) foi avaliada por RT-PCR em tempo real. O gene Gapdh
(NM_001289726.1) foi usado como controle endógeno. A reação quantitativa do PCR em tempo
real foi feita no Via7 Real-Time PCR (Life Technologies) usando o sistema de fluorescência
SYBR green (Applied Biosystems, Warrington, UK) ou TaqMan (AppliedBiosystems) para a
quantificação das amplificações.
Os resultados foram analisados através do método comparative de cycle threshold (CT).
Todos dados foram normalizados em relação aos valores de Gapdh, e a quantificação das
diferenças entre os grupos foi calculada de acordo com as instruções do fabricante. A expressão
gênica foi apresentada baseada na quantidade de vezes que aumentou em relação aos BMDMs
não-estimulados. Para realizar os qPCRs foram usados primers Taqman (AppliedBiosystems)
para A2A (adora 2a, Mm00802075_m1), A2B (adora2b, Mm00839292_m1), Gapdh
(Mm99999915_g1) e pares de primers SYBER Green (Life Technologies) mostrados na tabela
1.

Material e Métodos
43
Tabela – Primers utilizados nas reações de RT-PCR e suas respectivas sequências sense e anti-
sense:
3.8 CITOMETRIA DE FLUXO DE MACRÓFAGOS
Para caracterizar a expressão de CD39 e CD73 na população de BMDM ou RAW
realizamos as marcações com F4/80+ e CD39+ ou F4/80+ e CD73+.
Macrófagos pré-incubados 6 horas na presença ou ausência de 300 µM de FBP e
estimulados ou não com 100 ng/mL de LPS por 24 horas, coletamos os macrófagos 1x106
(células/tubo) incubadas com 1μL (1μg) de F4/80 FITC (eBioscience) e 1μL de CD39 ou CD73
ambos PE (eBioscience) (anticorpos monoclonais extracelulares), por 30 minutos, a 4 ºC. Após
a incubação, lavamos as amostras 2 vezes com 2 mL de PBS contendo 2 % de soro bovino fetal,
centrifugadas a 400g por 10 minutos e ressuspensas em 100 μL de solução de formaldeído a 1%
em PBS. Após esses procedimentos, adquirimos as amostras em FACSVerse (BD
Immunocytometry System, Franklin Lakes, NJ), e as análises realizadas no programa FCS
Express V5 (DE NOVO SOFTWARE).
3.9 ENSAIO DE ATP
Para avaliar a quantitativamente os níveis de ATP extracelular, dosamos no sobrenadante
das culturas de BMDMs ou RAW incubadas overnight com FBP e estimuladas por 3 horas com
100 ng/mL de LPS. A quantificação foi conduzida utilizando o ensaio luminescente ATPlite
Primers Sense Anti-sense
IL-10 5´-TGGACAACATACTGCT-3´ 5´-GGATCATTTCCGATAAGGCT-3´
GAPDH 5´-GGGTGTGAACCACGAGAAAT-3´ 5´-CCTTCCACAATGCCAAAGTT-3´

Material e Métodos
44
PerkinElmer conforme as recomendações do fabricante. Adicionamos numa placa preta de 96
poços 100 µL de amostra ou meio de cultura contendo uma diluição seriada do padrão (1x10-
5M), adicionamos 50 µL da solução de lise celular e aguardamos 5 minutos num shaker a 700
rpm. Após adicionamos 50 µL da solução substrato e novamente deixamos 5 minutos num
shaker a 700 rpm, 10 minutos após medimos a luminescência (Flexstation 3, Molecular
Devices), os valores expressos em nM.
3.10 PROTOCOLO DE ENDOTOXEMIA
Os animais submetidos à endotoxemia receberam duas injeções via intraperitoneal (i.p)
de FBP ou salina, a segunda injeção foi 23 horas após a primeira, na dose de 300mg/kg de
FBP. Após uma hora da última injeção de tratamento, os animais receberam uma injeção de
25mg/kg de LPS de bactéria Escherichia coli (Sigma-Aldrich). Transcorridas 6 horas após o
protocolo de endotoxemia, os animais foram anestesiados com Ketamina 1,25mg/mL e
Xylazina 0,5mg/mL com injeção subcutânea e então coletados o sangue pelo plexo retro-
orbrital, lavado peritoneal e pulmão.
Para a coleta do pulmão foi necessário o protocolo de perfusão cardíaca. Após a coleta
do lavado peritoneal foi realizada uma primeira incisão longitudinal da região púbica até a
incisura jugular para a retirada da pele e visualização dos músculos. Uma segunda incisão
longitudinal é realizada na região púbica até o processo xifóide para a retirada dos músculos
e visualização do diafragma. Em seguida, é realizada a abertura da caixa torácica pela
dissecação das partes laterais e ventral do diafragma e rebatimento da porção ventral do
gradil costal. Com a visualização do coração, injeta-se no ventrículo esquerdo uma agulha de
10mL com PBS e então coletamos o pulmão. As amostras foram congeladas e processadas
para as dosagens de citocinas por ELISA.

Material e Métodos
45
3.11 ANÁLISE ESTATÍSTICA
Os dados estão apresentados como média ± SEM de três experimentos independentes. A
significância estatística será estimada por teste t de Student para amostras não pareadas para
comparação entre dois grupos. Para comparação entre três ou mais grupos utilizamos ANOVA
de uma via, seguido pelo pós-teste de Bonferroni. Diferenças com p<0,05 serão consideradas
significativas. Para realizar as análises estatísticas utilizamos o GraphPadPrism 5.0 (GraphPad
Software).

4. RESULTADOS

Resultados
47
4. RESULTADOS
4.1 FBP ALTERA A PRODUÇÃO DE CITOCINAS POR MACRÓFAGOS ESTIMULADOS
COM LPS
No intuito de avaliar o potencial anti-inflamatório da FBP, macrófagos derivados da
medula óssea (BMDM), foram pré-tratados com FBP durante 6 horas e posteriormente
estimulados por LPS (100ng/mL) por 24 horas. Decorrido o período de estimulação, o padrão de
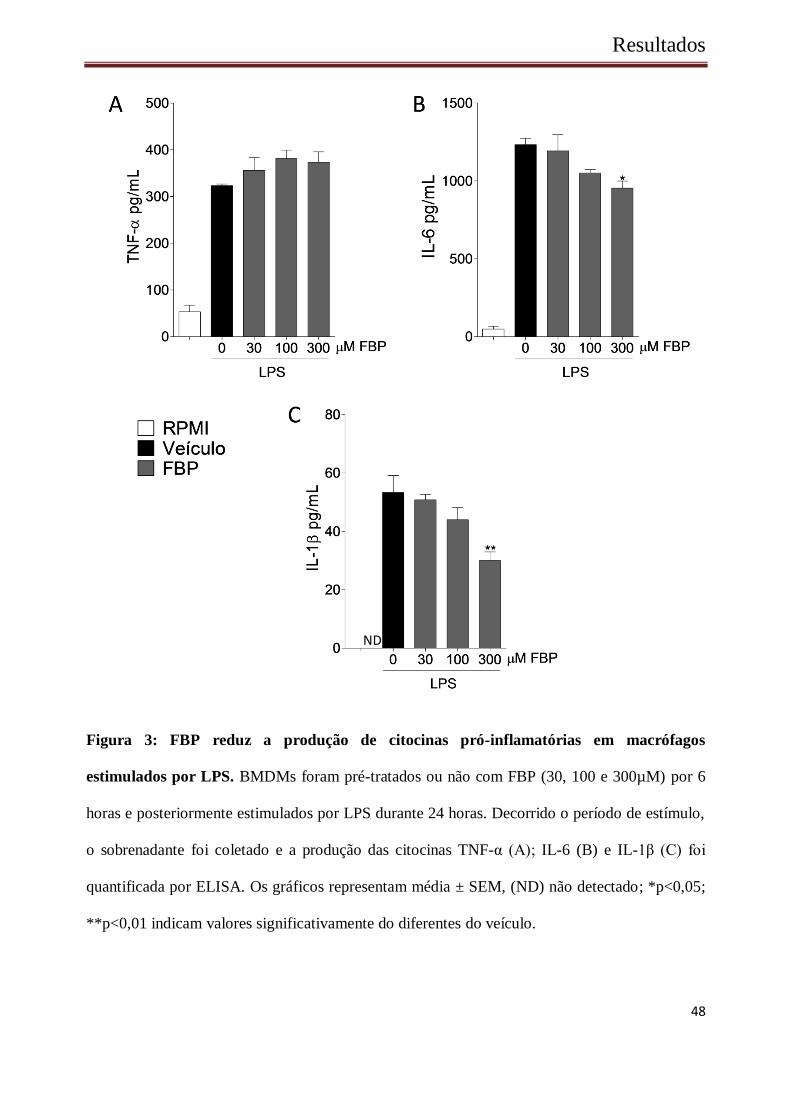
citocinas produzido por essas células foi quantificado por ELISA (Figura 3). Macrófagos
estimulados com LPS aumentaram a produção de citocinas, quando comparados com células não
estimuladas. O estímulo de LPS na presença de FBP, não alterou a produção de TNF-α entre os
grupos (Figura 3 A). No entanto, foi observado que o tratamento por FBP (300µM) ocasionou
uma redução significativa na produção de IL-6 (Figura 3 B) e IL-1β (Figura 3 C).
Além disso, observamos que macrófagos estimulados com LPS liberam IL-10, porém
apresentam um aumento acentuado da produção de IL-10 na presença de FBP (300µM) quando
comparados com o veículo (Figura 4 A). Com a finalidade de confirmar o aumento da expressão
de IL-10 induzido por FBP, a expressão de RNAm dessa citocina foi avaliada por RT-PCR.
Corroborando com os resultados de ELISA, a expressão do gene da IL-10 foi aumentada a partir
da terceira hora da incubação com o estímulo de LPS (6,9 - fold increase). Observamos um
aumento na expressão ainda maior no grupo que foi pré-tratado com FBP (8,8 - fold increase),
permanecendo aumentada no grupo tratado até 6 horas após a ativação celular. No entanto, após
12h de estímulo, os níveis de expressão do gene retornaram aos valores basais (Figura 4 B).
Tendo em vista o aumento significativo na produção de IL-10 proporcionado pela FBP o
próximo passo foi avaliar o papel de FBP na produção dessa citocina anti-inflamatória.
Resultados
48
Figura 3: FBP reduz a produção de citocinas pró-inflamatórias em macrófagos
estimulados por LPS. BMDMs foram pré-tratados ou não com FBP (30, 100 e 300µM) por 6
horas e posteriormente estimulados por LPS durante 24 horas. Decorrido o período de estímulo,
o sobrenadante foi coletado e a produção das citocinas TNF-α (A); IL-6 (B) e IL-1β (C) foi
quantificada por ELISA. Os gráficos representam média ± SEM, (ND) não detectado; *p<0,05;
**p<0,01 indicam valores significativamente do diferentes do veículo.
ND

Resultados
49
Figura 4: FBP aumenta a produção de IL-10 em macrófagos estimulados com LPS. (A)
BMDM foram incubados na presença de concentrações crescentes de FBP (0, 30, 100 e 300µM)
por 6h e então estimulados com LPS (100ng/mL) durante 24h, no sobrenadante celular foi
quantificada a concentração de IL-10 por ELISA. (B) BMDM foram incubados por 6 horas na
presença de RPMI ou FBP, então ativado com LPS por 3, 6 e 12 horas, onde foi quantificada a
expressão relativa do gene da IL-10 por PCR-RT normalizada por GADPH. Os gráficos
representam média ± SEM, *p<0,05; **p<0,01; *** p<0,001 indicam valores significativamente
do diferentes do veículo.

Resultados
50
4.2 O AUMENTO DA PRODUÇÃO DE IL-10 INDUZIDO POR FBP É DEPENDENTE DE
ADENOSINA E DO RECEPTOR A2A
Em 2003, Sola e colaboradores foram os primeiros a demonstrar que FBP apresenta
efeitos anti-inflamatórios dependentes da produção de adenosina. Em modelos de isquemia e
reperfusão, a FBP foi capaz de reduzir os níveis de atividade da enzima mieloperoxidase (MPO),
efeito que não foi observado na presença de adenosina deaminase (ADA) uma enzima que
converte a adenosina à inosina, que é um metabólito que não apresenta ações anti-inflamatórias,
demonstrando dependência da ação da adenosina (Sola et al. 2003). Recentemente, nosso
laboratório aprofundou os estudos iniciais de Sola e colaboradores, mostrando que FBP atenua
artrite experimental por um mecanismo dependente de adenosina e da sua ativação pelo receptor
A2A (Veras et al. 1015). Para avaliar a contribuição da adenosina sobre o efeito anti-inflamatório
da FBP, BMDM foram pré-incubados com FBP na presença de ADA.
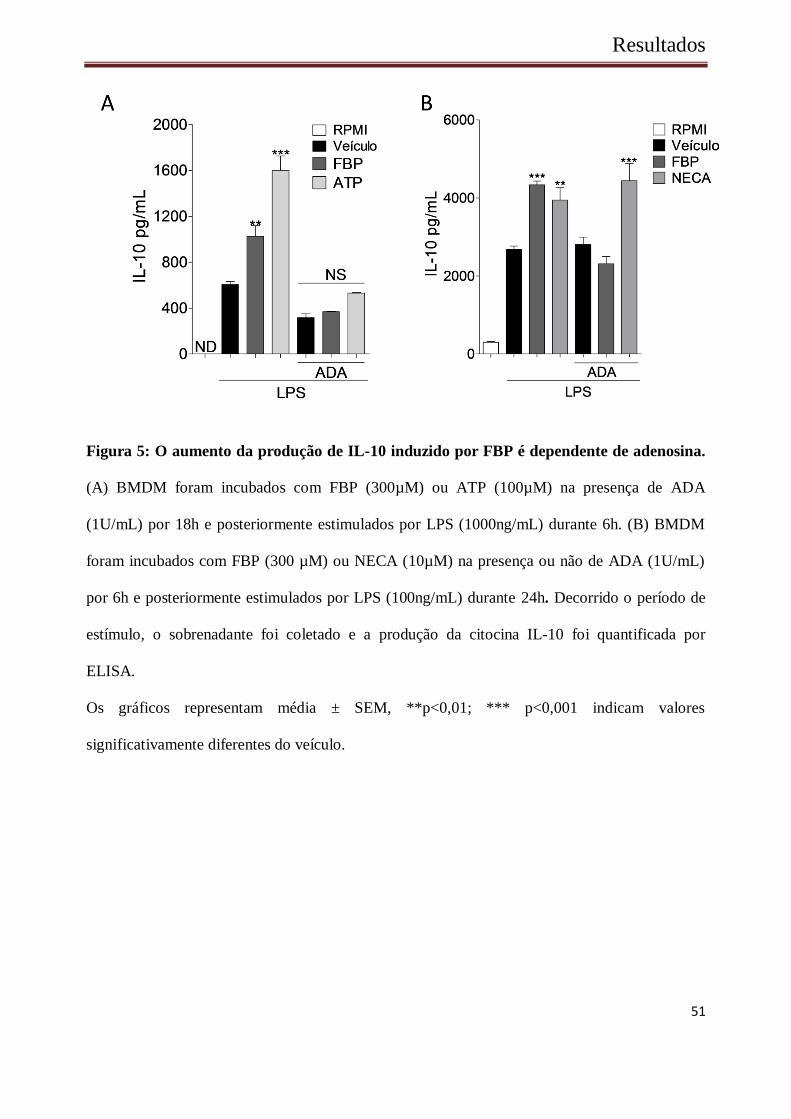
BMDM quando ativados com LPS produzem IL-10, A produção de IL-10 induzida por
LPS é aumentada na presença de FBP ou ATP. Quando a enzima ADA foi adicionada na cultura
de BMDMs juntamente com FBP ou ATP, o aumento na produção de IL-10 foi abolido (Figura
5 A). No entanto, na presença de um análogo de adenosina (NECA) o aumento na produção de
IL-10 por essas células é mantido, no entanto, o efeito inibitório de ADA não foi observado
nesse análogo (Figura 5 B). Juntos, esses resultados sugerem que o efeito da FBP na produção
de IL-10 pode ser dependente de adenosina.
Resultados
51
Figura 5: O aumento da produção de IL-10 induzido por FBP é dependente de adenosina.
(A) BMDM foram incubados com FBP (300µM) ou ATP (100µM) na presença de ADA
(1U/mL) por 18h e posteriormente estimulados por LPS (1000ng/mL) durante 6h. (B) BMDM
foram incubados com FBP (300 µM) ou NECA (10µM) na presença ou não de ADA (1U/mL)
por 6h e posteriormente estimulados por LPS (100ng/mL) durante 24h. Decorrido o período de
estímulo, o sobrenadante foi coletado e a produção da citocina IL-10 foi quantificada por
ELISA.
Os gráficos representam média ± SEM, **p<0,01; *** p<0,001 indicam valores
significativamente diferentes do veículo.

Resultados
52
Com a comprovação de que FBP induz a produção de IL-10 de maneira dependente de
adenosina, o próximo passado foi investigar qual o receptor de adenosina está envolvido nesse
fenômeno. Para isso, foram utilizados diversos antagonistas dos receptores de adenosina, tais
como, A1R (DPCPX), A2AR (CSC), A2BR (MRS1754) e A3R (MRS1334).BMDMs foram
tratados com os diferentes antagonistas juntamente com FBP ou NECA e posteriormente
estimulados por LPS. A2AR foi identificado como o único receptor capaz de alterar o padrão de
produção de IL-10 entre os grupos (Figura 6 B), sendo esta alteração ausente quando os outros
antagonistas foram utilizados, os quais não promovem alterações nas concentrações de IL-10
induzidas por FBP ou NECA (Figura 6 A, C e D). Para confirmar a participação do receptor A2A
na produção de IL-10 induzida por FBP, foram utilizados BMDM provenientes de animais
deficientes para esse receptor. Conforme representado na figura 9, à produção de IL-10 em
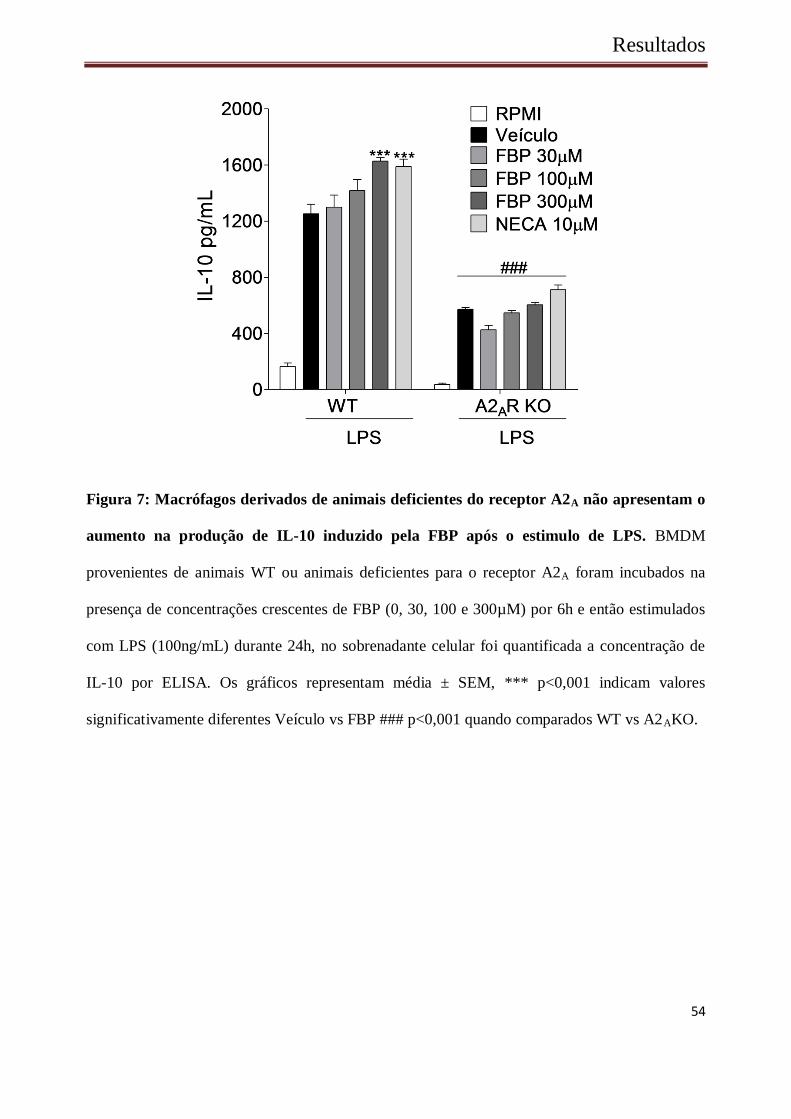
BMDMs de animais deficientes para o receptor A2A foi drasticamente reduzida quando
comparada a de macrófagos originados de animais WT (Figura 7).
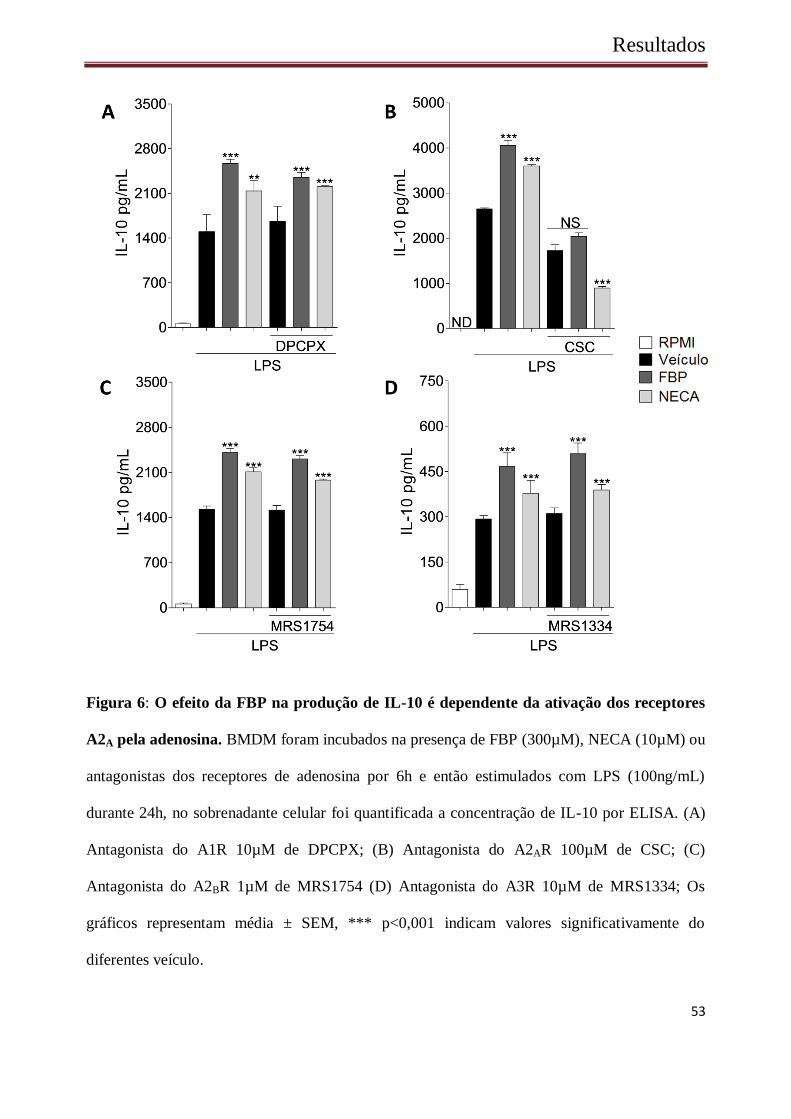
Resultados
53
Figura 6: O efeito da FBP na produção de IL-10 é dependente da ativação dos receptores
A2A pela adenosina. BMDM foram incubados na presença de FBP (300µM), NECA (10µM) ou
antagonistas dos receptores de adenosina por 6h e então estimulados com LPS (100ng/mL)
durante 24h, no sobrenadante celular foi quantificada a concentração de IL-10 por ELISA. (A)
Antagonista do A1R 10µM de DPCPX; (B) Antagonista do A2AR 100µM de CSC; (C)
Antagonista do A2BR 1µM de MRS1754 (D) Antagonista do A3R 10µM de MRS1334; Os
gráficos representam média ± SEM, *** p<0,001 indicam valores significativamente do
diferentes veículo.
Resultados
54
Figura 7: Macrófagos derivados de animais deficientes do receptor A2A não apresentam o
aumento na produção de IL-10 induzido pela FBP após o estimulo de LPS. BMDM
provenientes de animais WT ou animais deficientes para o receptor A2A foram incubados na
presença de concentrações crescentes de FBP (0, 30, 100 e 300µM) por 6h e então estimulados
com LPS (100ng/mL) durante 24h, no sobrenadante celular foi quantificada a concentração de
IL-10 por ELISA. Os gráficos representam média ± SEM, *** p<0,001 indicam valores
significativamente diferentes Veículo vs FBP ### p<0,001 quando comparados WT vs A2AKO.

Resultados
55
Sugerida à participação da adenosina e do receptor A2A na produção de IL-10 induzida
por FBP em macrófagos estimulados com LPS, o próximo objetivo foi avaliar se a FBP altera o
nível trascricional dos receptores A2A e A2B após o estímulo por LPS. A expressão dos
receptores foi avaliada por RT-PCR em BMDM tratados ou não com FBP (300µM) após 3, 6 e
12 hs do estímulo de LPS (100ng/mL). Conforme representado na figura 8 A e B, a expressão
do A2AR e A2BR é aumentada após o estimulo por LPS (2,7 e 3,5 - fold increase,
respectivamente), além disso, a expressão do receptor A2BR, mas não A2AR, encontrou-se
elevada nas células estimuladas com LPS quando na presença de FBP (3,9 - fold increase)
(Figura 8 B). Embora a expressão de A2BR tenha sido alterada na presença de FBP, o nível de
produção de IL-10 nessas células não foi alterado.
Figura 8: LPS aumenta as expressões dos receptores A2, mas FBP altera somente a
expressão do receptor A2B. BMDM foram incubados na presença de FBP (300µM) por 6h e
então estimulados com LPS (100ng/mL) durante (3, 6, 12h). A expressão de RNAm para
Adora2a, Adora2a foram analisadas por RT- PCR e normalizada em relação à expressão de
Gadph; *p<0,05; **p<0,01; ***p<0,001 quando comparados RPMI vs FBP.

Resultados
56
4.3 ECTONUCLEOTIDASES CONTRIBUEM PARA O AUMENTO DA PRODUÇÃO DE
IL-10 PROMOVIDO PELA FBP
Sabe-se que uma fonte comum para obtenção de adenosina no meio extracelular é a
hidrólise do ATP, com a ação das ectonucleotidases CD39 e CD73, que liberam os fosfatos do
ATP para a formação de ADP, AMP e o produto final dessa hidrólise é a formação da adenosina
(Regateiro et al. 2013). Macrófagos derivados da medula óssea expressam constitutivamente na
superfície celular CD39 e CD73 (Cohen et al. 2013, Zanin et al. 2012). Com base nesses relatos
anteriores, o próximo passo foi verificar se a FBP é capaz de alterar a expressão dessas
moléculas na superfície de macrófagos. Para isso, BMDM foram pré-incubados com FBP por 6
h e submetidos à análise da expressão das ectonucleotidases CD39 e CD73 por citometria de
fluxo. FBP não alterou a expressão de ambas as moléculas na superfície celular (Figura 9 A e
C), mesmo após o estímulo por LPS (Figura 9 B e D).

Resultados
57
Figura 9: FBP não altera a expressão de CD39 e CD73 após estimulo de LPS. BMDM
foram incubados na presença de FBP (300µM) por 6h e então estimulados com LPS (100ng/mL)
durante 24 horas. Histograma representativo da expressão de CD39 (A, B) e da expressão de
CD73 (C, D).
IsotipoRPMI + RPMIFBP + RPMIRPMI + LPSFBP + LPS
CD73
CD39
CD73
CD39
A
C
B
D

Resultados
58
Mesmo que a FBP não altere a expressão das ectonucleotidases CD39 ou CD73, a
participação dessas proteínas pode ser fundamental para a produção de adenosina e,
consequentemente, para o aumento da produção de IL-10 em macrófagos estimulados com LPS.
Assim, BMDMs foram pré-incubados por 6h com FBP (300µM) ou ADP (100µM) com ou sem
ARL (200µM) e APCP (100µM) inibidores de CD39 e CD73, respectivamente, em seguida as
células foram estimuladas por 24h com LPS e no sobrenadante quantificado as concentrações de
IL-10 por ELISA. Quando utilizamos inibidor de CD39 (ARL) juntamente com FBP perdemos o
efeito de aumento na produção de IL-10 (Figura 10 A), também encontramos um resultado
semelhante com o uso do inibidor de CD73 (APCP) perdendo novamente o aumento de IL-10
promovido pela FBP (Figura 10 B).

Resultados
59
Figura 10: Efeito da inibição das ectonucleotidases CD39 e CD73 na produção de IL-10.
BMDM foram incubados na presença de FBP (300µM) ou ADP (100µM) por 6h com ou sem
inibidores de CD39 ou CD73 e então estimulados com LPS (100ng/mL) durante 24 horas. No
sobrenadante celular foi quantificado a concentração de IL-10 por ELISA. (A) Inibidor de CD39
(ARL, 200µM); (B) Inibidor de CD73 (APCP, 100µM). No sobrenadante celular foi
quantificada a concentração de IL-10 por ELISA. Os gráficos representam média ± SEM; ND -
não detectado; *** p<0,001 indicam valores significativamente diferentes Veículo vs FBP.

Resultados
60
Conforme descrito na literatura, macrófagos da linhagem RAW 264.7 possuem baixa
expressão das ectonucleotidases CD39 e CD73 (Cohen et al. 2013, Murphy & Elliott 2014).
Para confirmar a ausência do papel de FBP na expressão dessas proteínas, observado em
BMDM, macrófagos da linhagem RAW 264.7 foram estimulados por FBP seguido de estímulo
por LPS e a expressão de CD39 e CD73 foi avaliada por citometria de fluxo. Corroborando com
os resultados anteriores, a expressão de CD39 e CD73 em células RAW 264.7 não foi alterada
pelo estímulo de FBP, independente da presença de LPS (Figura 11). No entanto, as células de
linhagem RAW 264.7 apresentaram uma expressão reduzida das ectonucleotidases CD39 e
CD73, quando comparados com a expressão encontrada nos macrófagos provenientes da medula
óssea.

Resultados
61
Figura 11: FBP em macrófagos RAW não altera a expressão de CD39 e CD73 após
estimulo de LPS. RAW foram incubados na presença de FBP (300µM) por 6h e então
estimulados com LPS (100ng/mL) durante 24 horas. Histograma representativo da expressão de
CD39 (A, B) e da expressão de CD73 (C, D).
CD73
IsotipoBMDM + RPMIRPMI + RPMIFBP + RPMIRPMI + LPSFBP + LPS
A
C
CD73
CD39
CD73
CD39
A
C
B
D

Resultados
62
Visto que, macrófagos RAW 264.7 possuem uma baixa expressão das ectonucleotidases
CD39 e CD73, provavelmente não demonstrariam o mesmo padrão na produção de citocinas
comparados às BMDMs. Para confirmar essa hipótese, a produção das citocinas TNF-α e IL-10
nos dois tipos celulares, BMDMs e RAW 264.7 foi avaliada. Para isso, os macrófagos foram
pré-incubados por 6h com FBP ou ADP e posteriormente estimulados por LPS durante 24h.
Conforme representado na figura 12, o estímulo por LPS levou a um aumento das concentrações
de TNF-α em BMDM, porém, o nível de produção desta citocina não foi alterado sob o estímulo
por FBP. No que concerne ao tratamento por ADP, observou-se que a produção de TNF-α foi
prejudicada nessas células (Figura 12 A). Por outro lado, macrófagos da linhagem RAW 264.7
apresentaram níveis de TNF-α mais elevados em comparação com BMDM, no entanto, esse
efeito foi observado em todos os grupos estimulados por LPS, de maneira independente aos
tratamentos com FBP ou ADP (Figura 12 B).
A quantificação de IL-10 reforça a hipótese de que FBP e ADP são capazes de regular
positivamente a produção dessa citocina em macrófagos originados da medula óssea (Figura 12
C). No entanto, não foram encontrados níveis detectáveis de IL-10 em macrófagos RAW 264.7
sob esses estímulos (Figura 12 D). Esses resultados sugerem que a produção de IL-10,
observada em BMDM estimulados por FBP pode ser dependente da expressão de
ectonucleotidases como a CD39 e a CD73.

Resultados
63
Figura 12: Macrófagos BMDM e RAW diferem no padrão de produção de TNF-α e IL-10.
BMDM e RAW foram incubados com FBP (300µM) ou ADP (100µM) por 6h e estimulados
com LPS (100ng/mL) durante 24h. No sobrenadante celular foi quantificada a concentração de
citocinas por ELISA. (A) BMDM, TNF-α; (B) RAW, TNF-α; (C) BMDM, IL-10; (D) RAW, IL-
10; Os gráficos representam média ± SEM, ND - não detectado; * p<0,05, *** p<0,001 indicam
valores significativamente diferentes do veículo.

Resultados
64
4.4 FBP AUMENTA A PRODUÇÃO E LIBERAÇÃO DE ATP POR MACRÓFAGOS
ESTIMULADOS COM LPS DE FORMA DEPENDENTE DA VIA GLICOLÍTICA
Tendo em vista que a produção de IL-10 induzida pela FBP em macrófagos de medula
óssea estimulados com LPS depende da degradação de ATP extracelular pelas
ectonucleotidases, o próximo passo foi avaliar se FBP promove aumento extracelular de ATP.
Conforme foi demonstrado nos experimentos anteriores, macrófagos da linhagem RAW 264.7
apresentaram uma baixa expressão de CD39 e CD73 (Figura 11), e também não produziam
níveis detectáveis de IL-10 (Figura 12 D). Sendo assim, estas células são uma boa ferramenta
para dosagem de ATP, pois, com a ausência das ectonucleotidases, não são capazes de hidrolisar
o ATP para ADP tornando a molécula passível de detecção sem necessidade de inibidores.
Macrófagos RAW 264.7 e BMDM foram incubados overnight na presença de FBP e
estimulados com LPS por 3 horas. Após o estimulo, os níveis de ATP foram determinados por
ensaio de bioluminescência. Em células RAW 264.7, a presença de FBP foi suficiente para
geração de ATP. O estímulo por LPS induziu aumento dos níveis desse nucleotídeo e, em
conjunto com FBP, esse aumento foi ainda mais pronunciado. (Figura 13 A). O uso de um
inibidor para CD39 (ARL) não afetou a produção de ATP por células Raw 264.7, o que era
esperado, devido ao fato de essas células não expressarem essa ectonucleotidase. Por outro lado,
em BMDM o estímulo por LPS só induziu a produção de níveis detectáveis de ATP quando na
presença de ARL. Além disso, maiores concentrações de ATP foram detectadas em células que
além do estimulo de LPS também receberam FBP (figura 13 B).

Resultados
65
Figura 13: FBP induz o aumento da geração de ATP em macrófagos. Macrófagos RAW (A)
ou BMDM (B) foram incubados com 300µM de FBP ou RPMI overnight e depois estimulados
com (100ng/mL) LPS por 3h. Adicionamos 1h antes do estimulo de LPS 200µM de ARL. No
sobrenadante celular foi quantificada a concentração de ATP. Os gráficos representam média ±
SEM *p<0,05; **p<0,01; ***p<0,001 indicam valores significativamente diferentes quando
comparados com o grupo RPMI; ou ##p<0,01 quando comparado com o grupo veículo.

Resultados
66
Uma vez demonstrado que o tratamento com FBP aumenta os níveis de IL-10 em
macrófagos, e essa produção aumentada é dependente de adenosina, agindo através do receptor
A2A. Também vimos que, a adenosina pode ser originada pela ação das ectonucleotidases CD39
e CD73 que retiram os fosfatos do ATP, mesmo a FBP não alterando as expressões dessas
moléculas, é capaz de aumentar a concentração extracelular de ATP, então o nosso próximo
passo foi avaliar se esse aumento de ATP é oriundo da via glicolítica.
A via glicolítica apresenta um saldo positivo na geração de ATP (Lehninger et al. 2007),
desse modo, o próximo passo foi verificar se o aumento na geração de ATP em macrófagos após
o estímulo por LPS seria intensificado na presença da FBP mesmo com a via glicolítica inibida.
Para esse fim, células RAW 264.7 foram pré-incubadas ou não com FBP overnight, e então
estimuladas por 3h por LPS (100ng/mL). Uma hora antes do estímulo, incubamos as células
com inibidores da via glicolítica, 2-deoxi-glicose (2-DG) um competidor da Hexoquinase
inibindo a formação de glicose-6-fosfato, ou STF-31 uma pequena molécula que bloqueia o
canal GLUT1, que permite a entrada de glicose no interior celular (Figura 14 A).
Após o estimulo com LPS, novamente observou-se um aumento produção de ATP,
intensificado pela presença de FBP. No entanto, quando as mesmas foram tratadas com
inibidores da via glicolítica, os níveis de ATP foram semelhantes aos valores basais (Figura 14
B, C), demonstrando um importante papel da via glicolítica na geração de ATP após a ativação
com LPS. No entanto, quando as células foram estimuladas por FBP a produção de ATP
permaneceu elevada, mesmo na presença de 2-DG e STF31, indicando que a entrada de FBP na
célula ocorre de maneira independente de GLUT1 e que essa proteína consegue agir
posteriormente a hexoquinase.
A fim de confirmar a dependência da via glicolítica na produção aumentada de ATP, a
glicólise foi inibida num ponto abaixo da FBP, utilizando Iodoacetamida (IAA), um conhecido

Resultados
67
inibidor da enzima gliceraldeído 3-fosfato desidrogenase (GAPDH) (Figura 15 A). Conforme
representado na figura 15 B, à presença do inibidor de GAPDH, foi capaz de abolir a produção
de ATP inclusive no grupo tratado com FBP. Por outro lado, observou-se que somente a FBP é
capaz de aumentar a produção de IL-10, visto que tanto a glicose quanto a frutose não alteraram
os níveis da produção de IL-10 em comparação com o grupo controle (Figura 15 C).
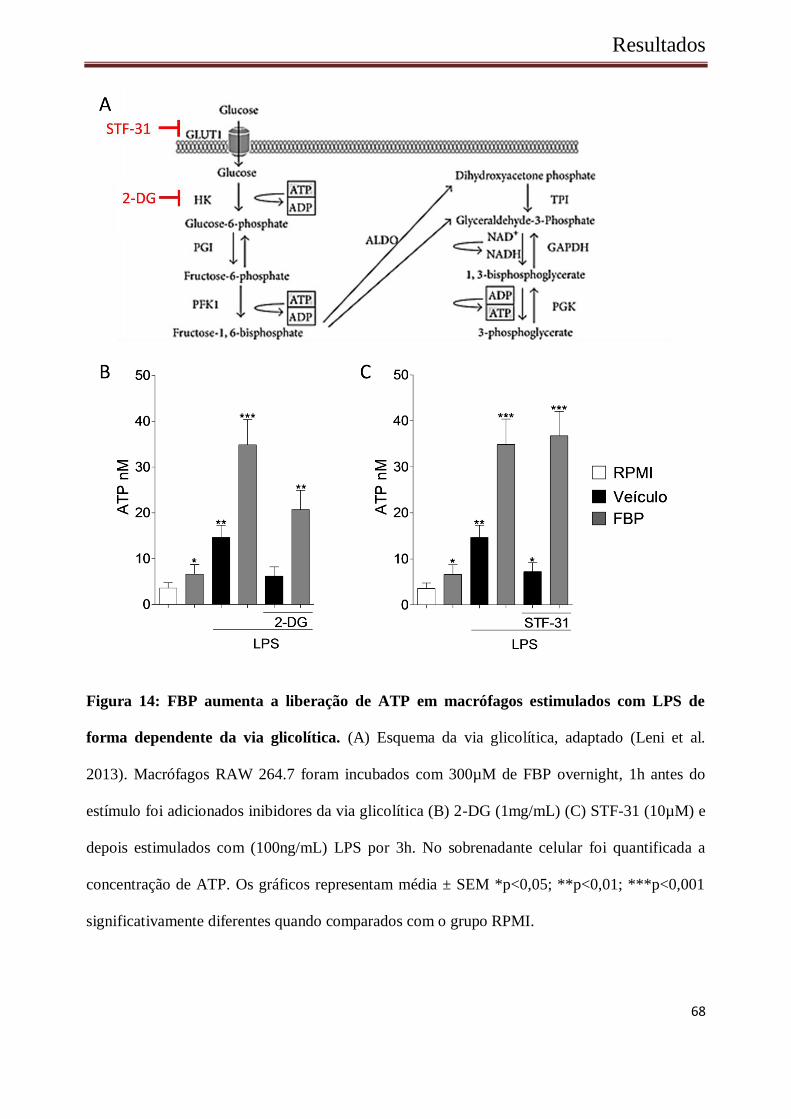
Resultados
68
Figura 14: FBP aumenta a liberação de ATP em macrófagos estimulados com LPS de
forma dependente da via glicolítica. (A) Esquema da via glicolítica, adaptado (Leni et al.
2013). Macrófagos RAW 264.7 foram incubados com 300µM de FBP overnight, 1h antes do
estímulo foi adicionados inibidores da via glicolítica (B) 2-DG (1mg/mL) (C) STF-31 (10µM) e
depois estimulados com (100ng/mL) LPS por 3h. No sobrenadante celular foi quantificada a
concentração de ATP. Os gráficos representam média ± SEM *p<0,05; **p<0,01; ***p<0,001
significativamente diferentes quando comparados com o grupo RPMI.

Resultados
69
Figura 15: FBP aumenta a liberação de ATP em macrófagos estimulados com LPS de
forma dependente da via glicolítica. (A) Esquema da via glicolítica, adaptado (Leni et al.
2013). (B) Macrófagos RAW foram incubados com 300µM de FBP overnight, 1h antes do
estímulo foi adicionado inibidores da via IAA (3 µM) e depois estimulados com (100ng/mL)
LPS por 3h. No sobrenadante celular foi quantificada a concentração de ATP. (C) BMDM foram
incubados com 300µM de FBP e então estimulados com LPS (100ng/mL) por 24 horas. No
sobrenadante quantificamos as concentrações de IL-10 por ELISA. Os gráficos representam
média ± SEM *p<0,05; **p<0,01; ***p<0,001 significativamente diferentes quando
comparados com o grupo (A) RPMI (B) Veículo.

Resultados
70
4.5 ATP GERADO É FUNDAMENTAL PARA O AUMENTO DE IL-10, SEM ATIVAÇÃO
DO INFLAMASSOMA.
O conjunto de resultados demonstrados no presente trabalho sugere que a produção de
IL-10 por macrófagos estimulados com LPS pode ser aumentada conforme a disponibilidade de
ATP e ADO, e que a FBP possibilita o aumento da expressão dessas moléculas. Também foi
demonstrado que esse efeito é dependente das ectonucleotidases CD39 e CD73, visto que a
molécula de ATP necessita da ação dessas enzimas para ser convertida em adenosina. Para
confirmar, se o efeito no aumento da produção de IL-10 proporcionado pela FBP é dependente
da formação de ATP intracelular, e se esse ATP necessita ser direcionado para o ambiente
extracelular.
Para responder essa questão, carbexolone (CBX), um bloqueador do canal de panexina-
1, responsável pela saída do ATP para o meio extracelular foi utilizado. Células RAW 264.7
foram tratadas ou não com CBX (50µM) 1h antes do estímulo, incubadas com FBP por 18h e,
posteriormente estimuladas com LPS (100ng/mL) por 3h. Um aumento nos níveis de ATP no
sobrenadante dessas células foi observado quando estas foram estimuladas por LPS. FBP
demonstrou-se capaz de intensificar esse aumento observado pelo estímulo de LPS, no entanto,
na presença do bloqueador CBX esse efeito foi abolido (Figura 16 A). Para avaliar os níveis de
IL-10, BMDM foram pré-incubados com FBP (300µM) ou ATP (100µM) e CBX (µM) por 6h e
posteriormente estimulados por LPS durante 24h. Corroborando com os dados anteriores,
observou-se que o tratamento com CBX foi capaz de reduzir a produção de IL-10 nas células
estimuladas na presença de FBP, enquanto que nenhuma alteração nos níveis dessa citocina foi
observada em células tratadas com ATP (Figura 16 B).

Resultados
71
Figura 16: O ATP extracelular aumenta a produção de IL-10 induzida pela FBP em
macrófagos estimulados com LPS. (A) Macrófagos RAW foram incubados com FBP (300µM)
overnight e depois estimulados com (100ng/mL) LPS por 3h. Adicionamos 1h antes do estimulo
CBX (50µM); (B) BMDM foram incubados com FBP (300µM) ou ATP (100µM) com CBX
(50µM) por 6h e estimulados com LPS (100ng/mL) durante 24h. No sobrenadante celular foi
quantificada a concentração de ATP por kit e IL-10 por ELISA. Os gráficos representam média
± SEM *p<0,05; **p<0,01; ***p<0,001 indicam valores significativamente diferentes quando
comparados com o grupo RPMI (A); ou com o grupo veículo (B).

Resultados
72
No ambiente extracelular, a molécula de ATP pode funcionar como um DAMP (Danger-
Associated Molecular Pattern) ao se ligar ao receptor purinérgico P2X7, ativando o
inflamassoma via NALP3 e caspase-1. Um dos sinais de ativação dessa via é a produção da
citocina IL-1β (Riteau et al. 2012). Com base nessas informações, o papel da via P2x7 na
produção aumenta de IL-10 na presença de FBP foi avaliado. Para isso, BMDM foram pré-
incubados por 6h com FBP ou ATP e com inibidor do canal purinérgico P2X7 (A-438079) e
posteriormente estimulados com LPS por 24h. Decorrido o período de incubação o sobrenadante
dessas células foi coletado e a produção de IL-10 e IL-1β foi quantificada por ELISA. Conforme
indicado na figura 17, a inibição do receptor purinérgico P2X7, BMDM estimulados na presença
de FBP mantiveram o aumento na produção de IL-10 mesmo na presença desse inibidor (Figura
17 A). O aumento da produção de IL-10 sob o estímulo de FBP é acompanhado por uma
acentuada redução dos níveis de IL-1β, que não foi alterada na presença do inibidor do canal
P2X7 (Figura 17 B).
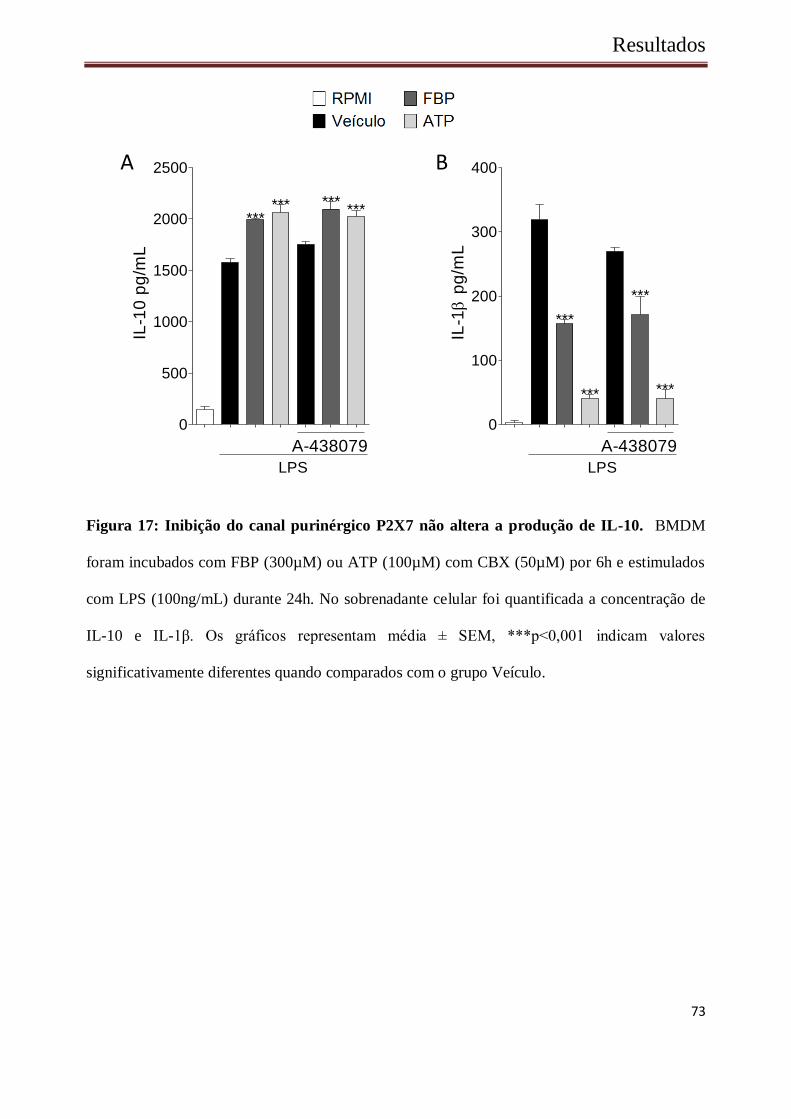
Resultados
73
Figura 17: Inibição do canal purinérgico P2X7 não altera a produção de IL-10. BMDM
foram incubados com FBP (300µM) ou ATP (100µM) com CBX (50µM) por 6h e estimulados
com LPS (100ng/mL) durante 24h. No sobrenadante celular foi quantificada a concentração de
IL-10 e IL-1β. Os gráficos representam média ± SEM, ***p<0,001 indicam valores
significativamente diferentes quando comparados com o grupo Veículo.
BA
0
100
200
300
400
LPS
***
***
A-438079
***
***
IL-1
pg
/mL
0
500
1000
1500
2000
2500
LPS
******
A-438079
******
IL-1
0 p
g/m
L

Resultados
74
4.6 FBP PROMOVE EFEITO ANTI-INFLAMATÓRIO, IN VIVO, EM MODELO DE
ENDOTOXEMIA.
Para confirmar o efetivo efeito anti-inflamatório da FBP, um modelo de endotoxemia foi
realizado. Para isso, camundongos C57/BL6, Balb/c e A2ARKO foram tratados com 300mg/kg
de FBP 24h e outra dose 1h antes da injeção de LPS, o desafio com LPS ocorreu com uma
injeção de (25mg/kg) na cavidade peritoneal. Soro, pulmão e lavado peritoneal forma coletados
6 horas após o estímulo com LPS, para dosagem das citocinas IL-10.
Camundongos que foram tratados previamente com FBP e submetidos à endotoxemia
apresentaram perfil anti-inflamatório, evidenciado pelo aumento das concentrações séricas de
IL-10 (Figura 18 A e D), bem como no lavado peritoneal (Figura 18 B e E) e no lavado
brônquio-alveolar (Figura 18 C e F). Por outro lado, quando camundongos nocautes para o
receptor A2A receberam esse tratamento, o aumento na produção de IL-10 não foi observado
(Figura 18 D, E e F).

Resultados
75
Figura 18: FBP promove efeito anti-inflamatório, in vivo, em modelo de peritonite. A
produção de citocinas foi avaliada 6 horas após injeção de 25mg/kg de LPS via i.p em
camundongos C57/BL6 (n=8) pré-tratados com FBP (300mg/kg, i.p.) 24 e 1 h antes do LPS.
Foram coletados soro (A,D), lavado peritoneal (B,E) e pulmão (C,F), processados para
mensuração dos níveis de IL-10. Camundongos C57/BL6 (A, B, C) Balb/c vs A2AR KO (D, E,
F).Os gráficos representam a média ± SEM. *P <0,05; **P<0,01 indica diferença significante
quando comparado salina vs FBP.

5. DISCUSSÃO

Discussão
77
4. DISCUSSÃO
FBP é um intermediário da via glicolítica que possui vários efeitos farmacológicos.
Neste sentido, alguns trabalhos científicos demonstraram a FBP como um promissor agente
terapêutico em doenças inflamatórias (Alves et al. 2004; Nunes et al. 2003, Planas et al. 1993;
Vianna Santos et al. 2012; Lopes et al. 2006, Yin et al. 2008), porém o mecanismo de ação não
está bem esclarecido. Por outro lado, nosso grupo de pesquisa demonstrou que a FBP possui
atividade anti-inflamatória em modelos experimentais de artrite (Veras 2014). O tratamento com
FBP foi capaz de reduzir a migração leucocitária, hipernocicepção articular e edema articular,
através do mecanismo dependente do aumento sérico de adenosina em camundongos (Veras et
al. 2015).
Podemos perceber com os nossos resultados, que a entrada de FBP na célula ocorre por
um transportador diferente que o da glicose, no caso GLUT-1, mas ainda não foi identificado um
transportador específico para a FBP, no entanto alguns estudos sugerem que a entrada dessa
molécula na célula provavelmente ocorra por difusão, e possa ser facilitado com concentrações
maiores do açúcar no meio extracelular (Ehringer et al. 2000, Hardin & Roberts 1994).
Sabemos que a FBP é capaz aumentar o fluxo glicolítico (Hassinen et al, 1991)
apresentando baixos efeitos tóxicos (Markov et al., 2000), além de possuir efeitos
farmacológicos com ação anti-inflamatória como na participação em diminuir a produção de
citocinas inflamatórias (Cuesta et al, 2006; Lopes et al, 2006; Yin et al, 2008). Também, já foi
demonstrado que o tratamento com FBP reduz a proliferação de linfócitos T (Nunes et al. 2004),
no entanto os mecanismos não estão bem esclarecidos. Sendo assim, no nosso trabalho,
encontramos alterações nos níveis das citocinas pró-inflamatórias como IL-6 e IL-1β, mas não
nas de TNF-α, durante a incubação com FBP em macrófagos. Porém, Yin e colaboradores
demonstraram que o pré-tratamento com FBP reduziu a produção de TNF-α, em macrófagos
alveolares estimulados com LPS (Yin et al. 2008).

Discussão
78
Vários fatores podem explicar diferenças encontradas em alguns estudos, como o tempo
de incubação, a linhagem celular e a concentração da droga. Por exemplo, a diferença entre os
resultados de Yin e colaboradores (2008) com os nossos foi provavelmente devido ao tratamento
com FBP em uma concentração aproximadamente 10 vezes maior que utilizada em nossos
experimentos. Além disso, em um trabalho com micróglia, um tipo de macrófago encontrado no
sistema nervoso, mostrou que a produção de citocinas foi dependente do tempo de incubação,
uma vez que, a redução dos níveis de TNF-α ocorreu a partir de 48 horas de estímulo, enquanto
de IL-10 modificaram com apenas 3 horas (Seo et al. 2004).
Demonstramos que a incubação com FBP aumenta os níveis de IL-10 em macrófagos.
Esta citocina apresenta características anti-inflamatórias inibindo a produção de diversos
mediadores pró-inflamatórios produzidos por macrófagos e células dendríticas ativados (Saraiva
& O'Garra 2010). Neste sentido, o efeito da IL-10 é dependente da ativação de STAT3, quando
ocorre a produção de IL-10 à quinase IKK é inibida, o que culmina na não ativação do NF-κB,
um importante fator de transcrição envolvido na produção de citocinas pró-inflamatórias
(Murray 2005).
Sola e colaboradores (2003) foram os primeiros a demonstrarem que os efeitos anti-
inflamatórios da FBP estão relacionados com alterações dos níveis endógenos de ADO (Sola et
al. 2003). Assim, em nosso trabalho demonstramos que além da FBP, o ATP, também pode
aumentar os níveis de IL-10 em macrófagos estimulados, de forma dependente de ADO. Uma
vez que o aumento da IL-10 foi abolido na presença de ADA, que faz a ADO virar inosina e não
apresentar efeitos anti-inflamatórios. Neste sentido, para comprovar que o aumento da produção
de IL-10 foi fortemente correlacionado com a atividade da ligação da adenosina em seu receptor,
pré tratamos os macrófagos com um análogo de adenosina chamado NECA. Com este
tratamento demonstramos que o análogo também foi capaz de aumentar a produção de IL-10. E

Discussão
79
como esperado, este efeito não foi revertido na presença da ADA, pois esta enzima é seletiva a
ADO e não ao seu análogo.
Em mamíferos a interação da ADO com os membros da família 1 de receptores
purinérgico (P1R) podem ter diferentes ações (Trevethick et al. 2008). Conhecidamente, a
interação da ADO pode ocorrer através de quatro diferentes receptores, que são o A1R, A2AR,
A2BR e A3R. Nos experimentos de Trevethick e colaboradores (2008), a resposta inflamatória
foi regulada com a administração endógena de ADO, uma vez que esta molécula reduziu o
infiltrado inflamatório, a liberação de citocinas pró-inflamatórias, como também o aumento da
produção de IL-10, por um mecanismo mediado pelo receptor A2A (Trevethick et al. 2008).
Desta forma, o próximo passo foi investigar qual o receptor de ADO estaria envolvido no
aumento da produção de IL-10. A utilização de antagonistas para os receptores de adenosina
(A1R, A2AR, A2BR e A3R), nos sugere que apenas A2AR foi capaz de alterar a produção de IL-
10, sendo esta alteração ausente quando os outros antagonistas foram utilizados. Em seguida,
confirmamos estes resultados com a utilização de macrófagos originados de camundongos
deficientes para esse receptor.
Deste modo, dados da literatura confirmam nossos achados, onde Vicenzi e
colaboradores (2013) demonstraram que agonistas para o receptor A2A podem promover o
aumento de IL-10 em pacientes com artrite. O mecanismo intracelular envolvido com o efeito
anti-inflamatório está relacionado com a ativação do A2AR, através da estimulação da adenilato
ciclase, a produção de AMPc, o aumento da PKA e fosforilação de CREB, que culminaram na
inibição da via do NF-kB. Consequentemente, observaram redução na produção de mediadores,
tais como TNF-α, IL-1β e IL-6 e aumento IL-10 (Varani et al. 2011). Recentemente nosso grupo
também demonstrou uma dependência dos efeitos da FBP através do A2AR, onde o bloqueio
desse receptor aboliu completamente os efeitos promovidos pelo tratamento com FBP (Veras et
al. 2015).

Discussão
80
Os macrófagos quando não ativados apresentam uma baixa expressão dos receptores A2
de adenosina, porém ligantes dos receptores tipo Toll, são ótimos indutores da expressão
receptores de ADO (Elson et al. 2013, Streitova et al. 2010). Apesar do estímulo com LPS
induzir a expressão dos receptores A2, o mesmo não ocorre com o tratamento com FBP que não
alterou a expressão do receptor A2A em nossos resultados, o qual é o receptor que apresenta a
maior afinidade para ADO em comparação com os outros receptores (Bruns et al. 1986). Dados
na literatura demonstram que as citocinas do tipo Th1 podem levar ao aumento da expressão dos
receptores A2B (Khoa et al. 2003). No nosso estudo, demonstramos que a expressão do A2B
também foi aumentada nos macrófagos estimulados com LPS. Além disso, o tratamento com
FBP aumentou a expressão de A2B nos macrófagos, mesmo que ele não esteja desempenhando
um papel relevante na produção de IL-10. Porém, essa alteração na expressão através da FBP
necessita de maiores investigações para ser esclarecida.
Os macrófagos derivados da medula óssea expressam constitutivamente na superfície
celular o CD39 e CD73 (Cohen et al. 2013, Zanin et al. 2012). Sabe-se que em condições
fisiológicas as concentrações de ADO extracelular são relativamente constantes (Chen & Chang
2013). No entanto, esses níveis fisiológicos podem ser alterados através do catabolismo do ATP,
no qual é uma das principais vias de produção de ADO no meio extracelular onde ocorre a
hidrólise do ATP pela ação das ectonucleotidases CD39 e CD73, que convertem o ATP
extracelular em ADO (Antonioli et al. 2013b). Neste sentindo, em nossos experimentos, a FBP
não alterou a expressão de CD39 e CD73 em BMDM ou RAW 264.7 estimulados com LPS,
sugerindo que os efeitos promovidos pela droga são dependentes do aumento dos níveis de ATP
extracelular. Em relação à expressão das ectonucleotidases, alguns fatores são capazes de
aumentar a expressão dessas moléculas na superfície celular, como por exemplo, o tratamento
com 8-Bromoadenosine 3',5'-cyclic monophosphate (8-Br-cAMP) para CD39 (Baek et al. 2013)
e IL-4 para CD39 e CD73 (Zanin et al. 2012).

Discussão
81
As ectonucleotidases exibiram uma participação bastante relevante na produção de IL-10
em macrófagos tratados com FBP, pois com a utilização de inibidores para essas moléculas, o
efeito proporcionado pela FBP foi abolido. Cohen e colaboradores (2013) também encontrou um
resultado semelhante, onde macrófagos tratados com ATP na presença do inibidor de CD39
reduz a produção de IL-10 (Cohen et al. 2013). Estudos com Tregs, também demonstraram a
importância do CD73 na produção de IL-10, pois células deficientes para essa ectonucleotidases
produziram menores níveis de IL-10 (Ehrentraut et al. 2013).
Em células RAW 264.7 não detectamos a produção de IL-10, na presença ou não de
FBP, pois estas apresentam baixa expressão das ectonucleotidases, no qual são importantes para
conversão do ATP para ADO (Cohen et al. 2013). Contudo, Nemeth e colaboradores (2005)
detectaram níveis de IL-10 em células RAW264.7 estimuladas com LPS (Nemeth et al. 2005),
provavelmente pela concentração do estimulo utilizado, uma vez que o grupo citado acima
utilizou uma concentração de LPS cerca de 100 vezes maior do que utilizado em nossos
experimentos. Além disso, vimos que essas células apresentaram uma alta produção de TNF-α
quando comparadas com os macrófagos originados da medula óssea.
Em nossos experimentos demonstramos que a FBP promove a produção de ATP
extracelular, porém Markov e colaboradores já haviam demonstrado que o tratamento exógeno
de FBP era capaz de alterar os níveis de ATP, indicando que a FBP teria a capacidade de
sustentar a glicólise e aumentar a produção de ATP em um ambiente de hipóxia (Markov 1986,
Markov et al. 1983, Markov et al. 1981). Contudo, esse dado citado nos levou a questionar se o
aumento na produção de ATP gerado pelo tratamento com FBP acontecia de forma dependente
da glicólise.
Nesse contexto, demonstramos que há uma dependência da via glicolítica no aumento da
produção de ATP promovida pela FBP. Quando inibimos a via glicolítica em reações acima da
FBP vimos que a célula ativada liberava uma quantidade de ATP, no entanto, a presença de FBP

Discussão
82
na cultura fez com que os níveis de ATP aumentassem mesmo com a via glicolítica inibida. Por
fim, quando inibimos a via glicolítica num ponto abaixo da FBP a produção de ATP foi
reduzida, o que nos leva a sugerir que a FBP entra na célula e que o aumento de ATP promovido
pela FBP é dependente de sua interação com a via glicolítica, um achado nunca demonstrado na
literatura.
Vários investigadores reportaram que após a administração com FBP houve aumento na
produção de ATP em diversos órgãos como rins, cérebro, fígado e coração (Aiub et al. 2003,
Antunes et al. 2006, Bickler & Buck 1996). Para que ocorra um efeito anti-inflamatório, o ATP
necessita ser transportado para o meio extracelular através de um canal específico, o canal
Panexina-1, e assim convertido em ADO pela ação das ectonucleotidases CD39 e CD73. Este é
um canal iônico transmembrana com uma grande condutância e um poro que permite a
passagem do ATP (Elliott et al. 2009). Nesse sentido, demonstramos que o ATP foi liberado
para o meio extracelular, através do canal de pannexin-1, sendo que a presença de ATP
extracelular foi imprescindível para que a FBP promovesse seus efeitos.
O ATP extracelular em excesso pode funcionar como um DAMP ao se ligar ao receptor
purinérgico P2X7, ativando o inflamassoma via NALP3 e caspase-1 (Ferrari et al. 1997). As
concentrações do ATP liberado por células ativadas ou lesadas para meio extracelular é um
ponto crítico na participação como componente imunomodulador ou durante a ativação do
inflamassoma (Ferrari et al. 1997). Os nossos dados demonstram que a FBP promove aumento
das concentrações de ATP, no entanto esse aumento não foi suficiente para que ocorresse a
ativação da plataforma do inflamassoma, uma vez que a inibição do receptor P2X7 não alterou
os efeitos anti-inflamatórios promovidos pela FBP. Com isso, sugerimos que o ATP foi
rapidamente convertido em ADO e assim não poderia se ligar ao receptor P2X7.
Adicionalmente, em nossas condições experimentais, a FBP reduziu os níveis de IL-1β, um
marcador clássico da ativação do inflamassoma (Davis et al. 2011)

Discussão
83
Nossos resultados obtidos de experimentos in vitro foram validados em um modelo
experimental de endotoxemia, onde camundongos C57/BL6 submetidos a um tratamento prévio
com FBP apresentam um aumento da produção de IL-10 no soro, lavado da cavidade peritoneal
e também no homogenato de pulmão. Corroborando os dados in vitro, vimos que o receptor A2A
contribui para o efeito anti-inflamatório promovido pela FBP, pois em animais deficientes para
este receptor o pré-tratamento com FBP não apresentou o aumento na produção de IL-10.
Em suma, demonstramos que a FBP possui mecanismo anti-inflamatório pelo aumento
de ATP extracelular dependente da via glicolítica, e consequentemente um aumento na produção
de ADO, via CD39 e CD73. Esse mecanismo de aumento de ADO é semelhante ao que acontece
com o MTX, que é a primeira droga de escolha para o tratamento de doenças reumáticas. No
entanto, a FBP apresenta algumas vantagens em relação ao MTX, principalmente por não
apresentar reações adversas no organismo (Kremer et al. 1994, Markov et al. 2000). Com isso, a
FBP poderá, futuramente, ser inserida como um potencial agente terapêutico para tratamento de
doenças inflamatórias.

6. CONCLUSÃO

Conclusão
85
5. CONCLUSÃO
Como conclusão do conjunto de resultados deste trabalho, a FBP apresentou um
aumento na produção de IL-10, uma citocina anti-inflamatória, através de um mecanismo
dependente da liberação de ATP pela via glicolítica. O ATP, produzido no interior da célula, é
transportado para o meio extracelular através do canal de Panexin-1. Esse ATP no meio
extracelular sofre ação das ectonucleotidases CD39 e CD73, que retiram os fosfatos do ATP,
formando ADP, AMP e por fim ADO. A adenosina formada se liga ao receptor A2A aumentando
a produção de IL-10 (Figura 19).
Figura 19: Mecanismo imunomodulador da FBP, através do aumento da produção de IL-
10, de forma dependente da via glicolítica e adenosinérgica.
2x

7. REFERÊNCIAS
BIBLIOGRÁFICAS

Referências
87
6. REFERÊNCIAS BIBLIOGRÁFICAS
Aiub CAF, Bortolini R, Azambuja AA, Alves JCF, Nunes FB, de Oliveira JR (2003):
Alterations in the indexes of apoptosis and necrosis induced by galactosamine in the liver of
Wistar rats treated with fructose-1,6-bisphosphate. Hepatology Research 25, 83-91
Akimitsu T, White JA, Carden DL, Gute DC, Korthuis RJ (1995): Fructose-1,6-
diphosphate or adenosine attenuate leukocyte adherence in postischemic skeletal-muscle.
American Journal of Physiology-Heart and Circulatory Physiology 269, H1743-H1751
Alves JCF, Santos RCV, Castaman TA, de Oliveira JR (2004): Anti-inflammatory
effects of fructose-1,6-bisphosphate on carrageenan-induced pleurisy in rat. Pharmacological
Research 49, 245-248
Antonioli L, Blandizzi C, Pacher P, Hasko G (2013a): Immunity, inflammation and
cancer: a leading role for adenosine. Nature Reviews Cancer 13, 842-857
Antonioli L, Pacher P, Vizi ES, Hasko G (2013b): CD39 and CD73 in immunity and
inflammation. Trends in Molecular Medicine 19, 355-367
Antunes N, Martinusso CA, Takiya CM, da Silva AJR, de Ornellas JFR, Elias PR, Leite
M, Cardoso LR (2006): Fructose-1,6 diphosphate as a protective agent for experimental
ischemic acute renal failure. Kidney International 69, 68-72
Baek AE, Kanthi Y, Sutton NR, Liao H, Pinsky DJ (2013): Regulation of ecto-apyrase
CD39 (ENTPD1) expression by phosphodiesterase III (PDE3). Faseb Journal 27, 4419-4428
Bailey E, Stirpe F, Taylor CB (1968): regulation of rat liver pyruvate kinase - effect of
preincubation ph copper ions fructose 1,6-diphosphate and dietary changes on enzyme activity.
Biochemical Journal 108, 427-&

Referências
88
Bickler PE, Buck LT (1996): Effects of fructose-1,6-bisphosphate on glutamate release
and ATP loss from rat brain slices during hypoxia. Journal of Neurochemistry 67, 1463-1468
Bruns RF, Lu GH, Pugsley TA (1986): Characterization of the A2 adenosine receptor
labeled by H-3 neca in rat striatal membranes. Molecular Pharmacology 29, 331-346
Chen C-J, Chang S-J (2013): Purinergic Signaling during Inflammation. New England
Journal of Medicine 368, 1260-1260
Coe IR, Dohrman DP, Constantinescu A, Diamond I, Gordon AS (1996): Activation of
cyclic AMP-Dependent protein kinase reverses tolerance of a nucleoside transporter to ethanol.
Journal of Pharmacology and Experimental Therapeutics 276, 365-369
Cohen HB, Briggs KT, Marino JP, Ravid K, Robson SC, Mosser DM (2013): TLR
stimulation initiates a CD39-based autoregulatory mechanism that limits macrophage
inflammatory responses. Blood 122, 1935-1945
Cronstein BN, Daguma L, Nichols D, Hutchison AJ, Williams M (1990): the adenosine
neutrophil paradox resolved - human neutrophils possess both A1 and A2 receptors that promote
chemotaxis and inhibit O-2- generation, respectively. Journal of Clinical Investigation 85, 1150-
1157
Csoka B, Nemeth ZH, Virag L, Gergely P, Leibovich SJ, Pacher P, Sun C-X, Blackburn
MR, Vizi ES, Deitch EA, Hasko G (2007): A(2A) adenosine receptors and C/EBP beta are
crucially required for IL-10 production by macrophages exposed to Escherichia coli. Blood 110,
2685-2695
Csoka B, Selmeczy Z, Koscso B, Nemeth ZH, Pacher P, Murray PJ, Kepka-Lenhart D,
Morris SM, Jr., Gause WC, Leibovich SJ, Hasko G (2012): Adenosine promotes alternative
macrophage activation via A(2A) and A(2B) receptors. Faseb Journal 26, 376-386

Referências
89
Darashchonak N, Koepsell B, Bogdanova N, von Versen-Hoeynck F (2014): Adenosine
A(2B) receptors induce proliferation, invasion and activation of cAMP response element
binding protein (CREB) in trophoblast cells. Bmc Pregnancy and Childbirth 14
Davis BK, Wen H, Ting JPY (2011): The Inflammasome NLRs in Immunity,
Inflammation, and Associated Diseases. Annual Review of Immunology, Vol 29 29, 707-735
Duro E, Pallai A, Koeroeskenyi K, Sarang Z, Szondy Z (2014): Adenosine A(3)
receptors negatively regulate the engulfment-dependent apoptotic cell suppression of
inflammation. Immunology Letters 162, 292-301
Ehrentraut H, Clambey ET, McNamee EN, Brodsky KS, Ehrentraut SF, Poth JM, Riegel
AK, Westrich JA, Colgan SP, Eltzschig HK (2013): CD73(+) regulatory T cells contribute to
adenosine-mediated resolution of acute lung injury. Faseb Journal 27, 2207-2219
Ehringer WD, Niu WY, Chiang B, Wang OL, Gordon L, Chien S (2000): Membrane
permeability of fructose-1,6-diphosphate in lipid vesicles and endothelial cells. Molecular and
Cellular Biochemistry 210, 35-45
Elliott MR, Chekeni FB, Trampont PC, Lazarowski ER, Kadl A, Walk SF, Park D,
Woodson RI, Ostankovich M, Sharma P, Lysiak JJ, Harden TK, Leitinger N, Ravichandran KS
(2009): Nucleotides released by apoptotic cells act as a find-me signal to promote phagocytic
clearance. Nature 461, 282-U165
Elson G, Eisenberg M, Garg C, Outram S, Ferrante CJ, Hasko G, Leibovich SJ (2013):
Induction of murine adenosine A(2A) receptor expression by LPS: analysis of the 5 ' upstream
promoter. Genes and Immunity 14, 147-153
Eltzschig HK (2009): Adenosine: An Old Drug Newly Discovered. Anesthesiology 111,
904-915

Referências
90
Ferrari D, Chiozzi P, Falzoni S, DalSusino M, Melchiorri L, Baricordi OR, DiVirgilio F
(1997): Extracellular ATP triggers IL-1 beta release by activating the purinergic P2Z receptor of
human macrophages. Journal of Immunology 159, 1451-1458
Fiorentino DF, Zlotnik A, Mosmann TR, Howard M, Ogarra A (1991): IL-10 inhibits
cytokine production by activated macrophages. Journal of Immunology 147, 3815-3822
Fredholm BB, Arslan G, Halldner L, Kull B, Schulte G, Wasserman W (2000): Structure
and function of adenosine receptors and their genes. Naunyn-Schmiedebergs Archives of
Pharmacology 362, 364-374
Gordon S, Taylor PR (2005): Monocyte and macrophage heterogeneity. Nature Reviews
Immunology 5, 953-964
Hardin CD, Roberts TM (1994): Metabolism of exogenously applied fructose-1,6-
bisphosphate in hypoxic vascular smooth-muscle. American Journal of Physiology-Heart and
Circulatory Physiology 267, H2325-H2332
Hardin CD, Lazzarino G, Tavazzi B, Di Pierro D, Roberts TM, Giardina B, Rovetto MJ
(2001): Myocardial metabolism of exogenous FDP is consistent with transport by a
dicarboxylate transporter. American Journal of Physiology-Heart and Circulatory Physiology
281, H2654-H2660
Hasko G, Kuhel DG, Chen JF, Schwarzschild MA, Deitch EA, Mabley JG, Marton A,
Szabo C (2000): Adenosine inhibits IL-12 and TNF-alpha production via adenosine A(2a)
receptor-dependent and independent mechanisms. Faseb Journal 14, 2065-2074
Hasko G, Linden J, Cronstein B, Pacher P (2008): Adenosine receptors: therapeutic
aspects for inflammatory and immune diseases. Nature Reviews Drug Discovery 7, 759-770

Referências
91
Hassinen IE, Nuutinen EM, Ito K, Nioka S, Lazzarino G, Giardina B, Chance B (1991):
Mechanism of the effect of exogenous fructose-1,6-bisphosphate on myocardial energy-
metabolism. Circulation 83, 584-593
Irving MG, Williams JF (1973): Kinetic studies on regulation of rabbit liver pyruvate-
kinase. Biochemical Journal 131, 287-301
Keller M, Rueegg A, Werner S, Beer H-D (2008): Active caspase-1 is a regulator of
unconventional protein secretion. Cell 132, 818-831
Khoa ND, Montesinos MC, Williams AJ, Kelly M, Cronstein BN (2003): Th1 cytokines
regulate adenosine receptors and their downstream signaling elements in human microvascular
endothelial cells. Journal of Immunology 171, 3991-3998
Kremer JM, Alarcon GS, Lightfoot RW, Willkens RF, Furst DE, Williams HJ, Dent PB,
Weinblatt ME (1994): Methotrexate for rheumatoid-arthritis - suggested guidelines for
monitoring liver toxicity. Arthritis and Rheumatism 37, 316-328
Kronlage M, Song J, Sorokin L, Isfort K, Schwerdtle T, Leipziger J, Robaye B, Conley
PB, Kim H-C, Sargin S, Schon P, Schwab A, Hanley PJ (2010): Autocrine Purinergic Receptor
Signaling Is Essential for Macrophage Chemotaxis. Science Signaling 3
Kurowska-Stolarska M, Stolarski B, Kewin P, Murphy G, Corrigan CJ, Ying S, Pitman
N, Mirchandani A, Rana B, van Rooijen N, Shepherd M, McSharry C, McInnes IB, Xu D, Liew
FY (2009): IL-33 Amplifies the Polarization of Alternatively Activated Macrophages That
Contribute to Airway Inflammation. Journal of Immunology 183, 6469-6477
Lapa FdR, da Silva MD, Cabrini DdA, Santos ARS (2012): Anti-inflammatory effects of
purine nucleosides, adenosine and inosine, in a mouse model of pleurisy: evidence for the role of
adenosine A(2) receptors. Purinergic Signalling 8, 693-704

Referências
92
Lappas CM, Rieger JM, Linden J (2005): A(2A) adenosine receptor induction inhibits
IFN-gamma production in murine CD4(+) T cells. Journal of Immunology 174, 1073-1080
Lazzarino G, Cattani L, Costrini R, Mulieri L, Candiani A, Galzigna L (1984): Increase
of intraerythrocytic fructose-1,6-diphosphate after incubation of whole human-blood with
fructose-1,6-diphosphate. Clinical Biochemistry 17, 42-45
Lehninger A, Nelson D, Cox M (2007): Princípios de Bioquimica, Único
Leni Z, Parakkal G, Arcaro A (2013): Emerging Metabolic Targets in the Therapy of
Hematological Malignancies. Biomed Research International
Li N, Mu L, Wang J, Zhang J, Xie X, Kong Q, Tang W, Yao X, Liu Y, Wang L, Wang
G, Wang D, Jin L, Sun B, Li H (2012): Activation of the adenosine A2A receptor attenuates
experimental autoimmune myasthenia gravis severity. European Journal of Immunology 42,
1140-1151
Loguercio C, Nardone G, Sicolo P, Cuomo R, Blanco CD, Budillon G (1996):
Intravenous load of fructose and fructose 1,6-diphosphate: Effects on uricemia in patients with
nonalcoholic liver disease. American Journal of Gastroenterology 91, 559-564
Lopes RP, Lunardelli A, Preissler T, Leite CE, Alves-Filho JCF, Nunes FB, de Oliveira
JR, Bauer ME (2006): The effects of fructose-1,6-bisphosphate and dexamethasone on acute
inflammation and T-cell proliferation. Inflammation Research 55, 354-358
Markov AK, Chilliard N, Lehan P, Hellems H (1978): Fructose 1-6 diphosphate - agent
for protection of acutely ischemic myocardium. Circulation 58, 98-98
Markov AK, Oglethorpe NC, Blake TM, Lehan PH, Hellems HK (1980): Hemodynamic,
electrocardiographic, and metabolic effects of fructose diphosphate on acute myocardial
ischemia. American Heart Journal 100, 639-646

Referências
93
Markov AK, Turner MD, Oglethorpe N, Neely WA, Hellems HK (1981): Fructose-1,6-
diphosphate - an agent for treatment of experimental endotoxin-shock. Surgery 90, 482-488
Markov AK, Oglethorpe N, Grillis M, Neely WA, Hellems HK (1983): Therapeutic
action of fructose-1,6-diphosphate in traumatic shock. World Journal of Surgery 7, 430-436
Markov AK (1986): Hemodynamics and metabolic effects of fructose 1-6 diphosphate in
ischemia and shock - experimental and clinical observations. Annals of Emergency Medicine
15, 1470-1477
Markov AK, Neely WA, Didlake RH, Terry J, Causey A, Lehan PH (2000): Metabolic
responses to fructose-1,6-diphosphate in healthy subjects. Metabolism-Clinical and
Experimental 49, 698-703
Maynard CL, Weaver CT (2008): Diversity in the contribution of interleukin-10 to T-
cell-mediated immune regulation. Immunological Reviews 226, 219-233
Mills JH, Kim D-G, Krenz A, Chen J-F, Bynoe MS (2012): A2A Adenosine Receptor
Signaling in Lymphocytes and the Central Nervous System Regulates Inflammation during
Experimental Autoimmune Encephalomyelitis. Journal of Immunology 188, 5713-5722
Molina-Arcas M, Javier Casado F, Pastor-Anglada M (2009): Nucleoside Transporter
Proteins. Current Vascular Pharmacology 7, 426-434
Moore KW, Malefyt RD, Coffman RL, O'Garra A (2001): Interleukin-10 and the
interleukin-10 receptor. Annual Review of Immunology 19, 683-765
Murphy P, Elliott M (2014): Apoptotic cells suppress TNF production by tissue resident
macrophages through a CD73-dependent mechanism (INM7P.433). The Journal of Immunology
192, 123.11-123.11

Referências
94
Murray PJ (2005): The primary mechanism of the IL-10-regulated anti inflammatory
response is to selectively inhibit transcription. Proceedings of the National Academy of Sciences
of the United States of America 102, 8686-8691
Nemeth ZH, Leibovich SJ, Deitch EA, Sperlagh B, Virag L, Vizi ES, Szabo C, Hasko G
(2003): Adenosine stimulates CREB activation in macrophages via a p38 MAPK-mediated
mechanism. Biochemical and Biophysical Research Communications 312, 883-888
Nunes FB, Graziottin CM, Alves JCF, Lunardelli A, Pires MGS, Wachter PH, De
Oliveira JR (2003): An assessment of fructose-1,6-bisphosphate as an antimicrobial and anti-
inflammatory agent in sepsis. Pharmacological Research 47, 35-41
Nunes FB, Alves JCF, Bastos CMA, Tessele PM, Caberlon E, Moreira KB, Ferreira TM,
de Oliveira JR (2004): Effect of the chlorpropamide and Fructose-1,6-bisphosphate on soluble
TNF receptor II levels. Pharmacological Research 49, 449-453
Nuutinen EM, Lazzarino G, Giardina B, Hassinen IE (1991): Effect of exogenous
fructose-1,6-bisphosphate on glycolysis in the isolated perfused rat-heart. American Heart
Journal 122, 523-527
O'Garra A, Vieira P (2004): Regulatory T cells and mechanisms of immune system
control. Nature Medicine 10, 801-805
Ohta A, Sitkovsky M (2001): Role of G-protein-coupled adenosine receptors in
downregulation of inflammation and protection from tissue damage. Nature 414, 916-920
Osada S, Mizuno K, Saido TC, Suzuki K, Kuroki T, Ohno S (1992): A new member of
the protein-kinase-c family, NPKC-theta, predominantly expressed in skeletal-muscle.
Molecular and Cellular Biology 12, 3930-3938

Referências
95
Pelegrin P, Surprenant A (2006): Pannexin-1 mediates large pore formation and
interleukin-1 beta release by the ATP-gated P2X(7) receptor. Embo Journal 25, 5071-5082
Pereira MR, Hang VR, Vardiero E, de Mello FG, Paes-de-Carvalho R (2010):
Modulation of A1 adenosine receptor expression by cell aggregation and long-term activation of
A2a receptors in cultures of avian retinal cells: involvement of the cyclic AMP/PKA pathway.
Journal of Neurochemistry 113, 661-673
Planas ME, Sanchez S, Gonzalez P, Deoliveira JR, Bartrons R (1993): Protective effect
of fructose-1,6-bisphosphate against carrageenan-induced inflammation. European Journal of
Pharmacology 237, 251-255
Regateiro FS, Cobbold SP, Waldmann H (2013): CD73 and adenosine generation in the
creation of regulatory microenvironments. Clinical and Experimental Immunology 171, 1-7
Riteau N, Baron L, Villeret B, Guillou N, Savigny F, Ryffel B, Rassendren F, Le Bert M,
Gombault A, Couillin I (2012): ATP release and purinergic signaling: a common pathway for
particle-mediated inflammasome activation. Cell Death & Disease 3
Roncarolo MG, Gregori S, Battaglia M, Bacchetta R, Fleischhauer K, Levings MK
(2006): Interleukin-10-secreting type 1 regulatory T cells in rodents and humans. Immunological
Reviews 212, 28-50
Salmon JE, Cronstein BN (1990): Fc-gamma-receptor-mediated functions in neutrophils
are modulated by adenosine receptor occupancy - A1 receptors are stimulatory and A2 receptors
are inhibitory. Journal of Immunology 145, 2235-2240
Saraiva M, O'Garra A (2010): The regulation of IL-10 production by immune cells.
Nature Reviews Immunology 10, 170-181

Referências
96
Sato A, Terata K, Miura H, Toyama K, Loberiza FR, Hatoum OA, Saito T, Sakuma I,
Gutterman DD (2005): Mechanism of vasodilation to adenosine in coronary arterioles from
patients with heart disease. American Journal of Physiology-Heart and Circulatory Physiology
288, H1633-H1640
Seo DR, Kim KY, Lee YB (2004): Interieukin-10 expression in lipopolysaccharide-
activated rnicroglia is mediated by extracellular ATP in an autocrine fashion. Neuroreport 15,
1157-1161
Sola A, Panés J, Xaus C, Hotter G (2003): Fructose-1,6-biphosphate and nucleoside pool
modifications prevent neutrophil accumulation in the reperfused intestine. Journal of Leukocyte
Biology 73, 74-81
Streitova D, Hofer M, Hola J, Vacek A, Pospisil M (2010): Adenosine A(1), A(2a),
A(2b), and A(3) Receptors in Hematopoiesis. 2. Expression of Receptor mRNA in Resting and
Lipopolysaccharide-Activated Mouse RAW 264.7 Macrophages. Physiological Research 59,
139-144
Stroud R, Wells J (2004): Mechanistic Diversity of Cytokine Receptor Signaling Across
Cell Membranes. scince signaling 2004, 17
Trevethick MA, Mantell SJ, Stuart EF, Barnard A, Wright KN, Yeadon M (2008):
Treating lung inflammation with agonists of the adenosine A(2A) receptor: promises, problems
and potential solutions. British Journal of Pharmacology 155, 463-474
Vanfurth R (1985): Monocyte production during inflammation. Comparative
Immunology Microbiology and Infectious Diseases 8, 205-211
Varani K, Padovan M, Vincenzi F, Targa M, Trotta F, Govoni M, Borea PA (2011):
A(2A) and A(3) adenosine receptor expression in rheumatoid arthritis: upregulation, inverse

Referências
97
correlation with disease activity score and suppression of inflammatory cytokine and
metalloproteinase release. Arthritis Research & Therapy 13
Veras FP 2014: Estudo dos mecanismos antiinflamatórios da frutose1,6-difosfato na
artrite experimental: papel do receptor de adenosina A2a e das ectonucleotidases CD39 E CD73,
Universidade de São Paulo, Faculdade de Medicina de Ribeirão Preto da Universidade de São
Paulo 94 pp
Veras, F. P. , Peres, R. S. , Saraiva, A. L. L. , Pinto, L. G. , Louzada-Junior, P. , Cunha,
T. M. , Paschoal, J. A. R. , Cunha, F. Q. & Alves-Filho, J. C. (2015)Fructose 1,6-bisphosphate, a
high-energy intermediate of glycolysis, attenuates experimental arthritis by activating anti-
inflammatory adenosinergic pathway. Sci. Rep. 5, 15171.
Vianna Santos RC, Moresco RN, Pena Rico MA, Garcia Susperregui AR, Luis Rosa J,
Bartrons R, Ventura F, Mario DN, Alves SH, Tatsch E, Kober H, de Mello RO, Scherer P, Dias
HB, de Oliveira JR (2012): Fructose-1,6-Bisphosphate Reduces the Mortality in Candida
albicans Bloodstream Infection and Prevents the Septic-Induced Platelet Decrease. Inflammation
35, 1256-1261
Volkman A (1985): Citation classic - the origin of macrophages from bone-marrow in
the rat. Current Contents/Life Sciences, 15-15
Yegutkin GG (2008): Nucleotide- and nucleoside-converting ectoenzymes: Important
modulators of purinergic signalling cascade. Biochimica Et Biophysica Acta-Molecular Cell
Research 1783, 673-694
Yin H, Jin X-b, Gong Q, Yang H, Hu L-y, Gong F-l, Zhu J-y (2008): Fructose-1,6-
diphosphate attenuates acute lung injury induced by lipopolysaccharide in mice. International
Immunopharmacology 8, 1842-1847

Referências
98
Zanin RF, Braganhol E, Bergamin LS, Ingrassia Campesato LF, Zanotto Filho A,
Fonseca Moreira JC, Morrone FB, Sevigny J, Chitolina Schetinger MR, de Souza Wyse AT,
Oliveira Battastini AM (2012): Differential Macrophage Activation Alters the Expression
Profile of NTPDase and Ecto-5 '-Nucleotidase. Plos One 7