universidade federal de sergipe prÓ-reitoria de pÓs …€¦ · obrigado, meus velhos, por sempre...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA E
CONSERVAÇÃO
FORMIGAS E O CÓDIGO FLORESTAL BRASILEIRO:
COMPARANDO ÁREAS DE PRESERVAÇÃO PERMANENTE (APP) E
RESERVA LEGAL (RL)
Rony Peterson Santos Almeida
Mestrado Acadêmico
São Cristóvão
Sergipe - Brasil
2015

ii
RONY PETERSON SANTOS ALMEIDA
FORMIGAS E O CÓDIGO FLORESTAL BRASILEIRO:
COMPARANDO ÁREAS DE PRESERVAÇÃO PERMANENTE (APP) E RESERVA
LEGAL (RL)
Dissertação apresentada ao Programa de Pós-Graduação em
Ecologia e Conservação da Universidade Federal de Sergipe,
como requisito parcial para obtenção do título de Mestre em
Ecologia.
Orientador: Prof. Dr. Leandro de Sousa Souto
Co-orientadora: Prof.ª Dr.ª Yana Teixeira dos Reis
São Cristóvão
Sergipe - Brasil
2015

iii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL
UNIVERSIDADE FEDERAL DE SERGIPE
Almeida, Rony Peterson Santos A447f Formigas e o Código Florestal Brasileiro : comparando Áreas
de Preservação Permanente (APP) e Reserva Legal (RL) / Rony Peterson Santos Almeida ; orientador Leandro de Sousa Souto. – São Cristóvão, 2015. 47 f. : il. Dissertação (mestrado em Ecologia e Conservação) –
Universidade Federal de Sergipe, 2015.
O 1. Mimecofauna. 2. Diversidade biológica. 3. Complexidade
ambiental. 4. Mata Atlântica. 5. Grupos funcionais. I. Souto, Leandro de Sousa, orient. II. Título
CDU: 595.796

iv

v
AGRADECIMENTOS
O homem é do tamanho do seu sonho. Logo, este é mais um que se realiza dentre
tantos que hei de realizar. Foram dois anos de dedicação e compromisso em uma pós-
graduação, tendo que me privar de muitas coisas e encarar desafios: tensão nas vésperas das
apresentações, campos conturbados, trabalho de bancada... isso tudo veio acompanhado de
boas surpresas em congressos, amigos, passeios, festas... vou sentir saudade de tudo isso!
Não posso terminar essa jornada sem agradecer àqueles que de forma direta ou
indireta formaram minha base. A priori, a indispensável proteção divina. Gratifico também o
exemplo e amor dos meus pais, Nivaldo e Lúcia, que sequer entendiam as “gaiolas de
passarinho” (mini-winkler) na garagem de casa, mas sempre priorizaram e me incentivaram
para que eu chegasse até aqui. Obrigado, meus velhos, por sempre estarem ao meu lado!
Brenda, valeu irmã, por aturar todos os dias que cheguei em casa cansado e “sem cabeça”.
Ademais, Gislaine, minha namorada e amiga, pela compreensão dos percalços que passei e
ainda assim sempre a apoiar a conclusão desse trabalho. Agradeço a minha família, aos meus
avós paternos Manoel e Helena e maternos Maria e Domigos (in memoriam). A todos os tios,
em especial a Nete, Neide, Vânia e Fina, por toda a ajuda e principalmente pelas palavras de
carinho e a tio Loro e Luciano pela presença e apoio constante nessa caminhada. Aos meus
padrinhos Wilson e Bernadete pela preocupação de sempre. E também aos primos.
Ao meu orientador, Leandro Sousa-Souto, por todo apoio e preocupação, alguém que
me espelho genuinamente. Um verdadeiro paizão em todos os momentos que precisei, sempre
com disponibilidade e paciência para dúvidas e explicações. Obrigado pelas sugestões,
broncas, direcionamentos, brincadeiras e por aturar esse “menino do sítio”! A minha co-
orientadora, Yana Reis, pela paciência nas explicações sobre a morfotipagem do material,
quanta trabalheira! Pelas conversas e principalmente por auxiliar na minha formação
profissional. Juro que hoje sou menos “cabeça dura”! Com vocês aprendi muito e agradeço
por toda orientação, pela amizade e por acreditarem no meu potencial, sem dúvida são
exemplos de profissionais que quero seguir. Obrigado, chefia!
A todos do Laboratório de Entomologia (LABENTO) que me receberam tão bem,
pessoas que convivi grande parte desses dois anos e com as quais aprendi muito, Bianca
Ambrogi, Ana Paula, Abel, Ariely, Jusci, Arivânia, Vânia, Rafaella, Mariana, Meggie e
Philippe. A Arleu pela ajuda na estatística, ao Anderson Leite e Sinara Moreira pelas
conversas e dúvidas sobre os insetos em geral e a Andrea pela força na triagem do material.
Obrigado a todos!
Ao Programa de Pós-graduação em Ecologia e Conservação (PPEC) pela oportunidade
e aos professores pelo aprendizado oferecido, em especial a Gustavo Hirose e Marcelo Brito
pelas críticas construtivas nas disciplinas e a professora Adriana Bocchiglieri por emprestar o

vi
densiômetro utilizado nesse trabalho e por ser sempre tão atenciosa. As sugestões da banca de
qualificação, em nome de Adenir Teodoro e a Ana Paula Araújo e Frederico Neves, pela
presença na banca de defesa. A secretária, Juliana. Aos colegas de mestrado Pedro, Pablo,
Thomaz, Jeanne e Danyelly por toda bagagem que juntos formamos e Abel, obrigado pela
amizade e por toda discussão nesses dois anos, científica ou não, no laboratório ou fora dele,
valeu lindão!
A toda galera do LABEV (Laboratório de Biologia e Ecologia dos Vertebrados) com
quem convivi durante as pesquisas e campos, no nome do professor Eduardo Dias, e pela
liberação desse espaço nas triagens do material dessa dissertação, valeu Edu! Aos mestres-
amigos da graduação Paulo Maroti, Eduardo Dias, Ricardo Carmo e Márcio Guimarães pelas
oportunidades, confiança e incentivo dado. Um muito obrigado à turma de Ecologia III/2014-
2 pelo aprendizado que tivemos juntos nesse período e a todos os amigos que fiz na UFS,
agradeço a lealdade, conselhos e atenção.
A liberação das áreas onde esse estudo foi feito em nome de Dantas, Abércio e a Ari
juntamente com Agroindustrial Campo Lindo. A FAPITEC/SE pela bolsa de mestrado (ed. n°
01/2013) e auxílio financeiro do projeto geral (FAPITEC/SE/FUNTEC nº. 13/2012-NAP’s) e
ao ICMBio/SISBIO pela licença de coleta concedida.
A todos que ajudaram nas coletas e triagem do material: Sidieres, Daniela (Dany),
Hugo, Aryeli, Iúri, Márcio, Arleu, Bruno, Juliana (Ju), Vânia, Brenda, Adriano e ao Grupo
Ambientalista Dorense (GAD) nos nomes de Luciano e Micael. Ótimas lembranças comendo
“sandubex com morte lenta”, tirando jaca, carro quebrado na volta do campo, triagens
varando tarde/noite adentrando até a madrugada, chuva desmarcando coleta (uma, duas, três...
vezes), jararaca dentro da parcela, ataque de abelhas, Iúri “dando o sangue” durante o trabalho
e a perda do celular na mata (4 meses depois o celular é encontrado no banco do golzinho,
hein Sid? rsrs). Pô galera, o que seria de mim sem vocês? Só tenho a agradecer por toda
colaboração, aprendizado e pela amizade. Meu muito OBRIGADO, vocês são 10!
Agradecer também aos amigos de longas datas: Vinícius, Rodrigo, Floriano, Thiago,
Cicinho, Guedes e Anderson, por todo apoio e compreensão ao dividir comigo as felicidades
em cada encontro, em especial a Marcelo Fontes, por sua saúde (Melhoras irmão! Espero que
esteja conosco logo.). “Us Boa Cepa” sempre marcando presença!
Peço desculpas às pessoas não citadas, as quais não são menos importantes. Cada uma
cumpriu um papel essencial na minha caminhada. Agradeço por terem me ajudado a encontrar
não só uma profissão, mas uma paixão. A todos vocês dedico essa dissertação como forma de
reconhecimento, gratidão, respeito e carinho. Obrigado!

vii
“Que os vossos esforços desafiem as
impossibilidades, lembrai-vos de que as grandes
coisas do homem foram conquistadas do que
parecia impossível.”
Charles Chaplin

viii
RESUMO
O Novo Código Florestal Brasileiro (NCFB) atualmente em vigor apresenta alguns pontos
polêmicos em seu texto, em especial aqueles relacionados à redução ou substituição de
Reservas Legais (RLs) e Áreas de Proteção Permanentes (APPs) por vegetação não nativa ou
a compensação de uma dessas áreas pela outra, numa mesma propriedade. O NCFB portanto,
considera essas duas áreas estruturalmente similares e passíveis de abrigar e conservar a fauna
e flora regional. O presente trabalho visa analisar se RLs e APPs são, de fato, similares do
ponto de vista ecológico, tendo como itens de analise a complexidade ambiental existente nos
fragmentos e o uso de formigas como bioindicadores. Para tanto, foram realizadas coletas em
seis áreas, sendo três consideradas como RL e três como APP. Em cada área foi feito um
transecto e esse subdividido em 30 parcelas de 5 x 5 m, espaçadas 6 m. Em cada uma das
parcelas foi medida a profundidade da serapilheira, contabilizada a densidade de árvores e
medida a cobertura do dossel. Além disso, na área central de cada parcela foi retirada uma
amostra de 1m² de serapilheira para analise da mirmecofauna (riqueza, composição e grupos
funcionais). A riqueza de formigas, bem como de grupos funcionais foram testadas em
resposta às variáveis ambientais por meio de modelos lineares generalizados (GLMs) e a
composição entre as áreas verificada através da análise de similaridade (ANOSIM) e
escalonamento multidimensional não-métrico (NMDS). Foram coletadas 116 morfoespécies
de formigas, distribuídas em nove subfamílias e 42 gêneros. Não foi encontrada diferença na
riqueza de espécies de formigas entre RLs e APPs e, dentre as variáveis ambientais
analisadas, apenas houve relação positiva entre a riqueza e a porcentagem do dossel. Além
disso, não houve diferença na composição de formigas nem na riqueza de grupos funcionais e
estes resultados reforçam o texto atual do NCFB de que RLs ou APPs atuam de forma similar
na manutenção da biodiversidade local. Desta forma, utilizar a APP no cômputo da RL (ou
vice-versa) não implica na redução da diversidade local de formigas e consequentemente,
prejuízo de suas funções ecológicas e de interações mediadas por espécies desse grupo já que
essas áreas apresentam similaridade na sua estrutura e na diversidade de espécies.
Palavras chave: diversidade; complexidade ambiental; grupos funcionais; Mata Atlântica;
mirmecofauna.

ix
ABSTRACT
The New Brazilian Forest Code (NCFB) presents some controversial points in its text,
especially those related to the reduction or replacement of legal reserves (LRs) and Permanent
Protection Areas (PPAs) for non-native vegetation or the compensation of these areas by
another, on the same property. The NCFB therefore considers these two areas structurally
similar and able to house and preserve the fauna and regional flora. This work aims to analyze
the existence of similarity between LRs and PPAs, with focus on environmental complexity
on the fragments and the use of ants as bioindicators. For this purpose, samplings were taken
in six areas, three of them considered LR and three as PPA. In each area, one transect was
subdivided into 30 plots of 5 x 5m and 6 m spaced. In each plot, we measured plant litter
depth, density of trees and the canopy cover (%). Furthermore, in the central area of each plot,
we toke a 1m² samples of plant litter to analyze the ant fauna. The relationship between ant
richness and environmental variables were tested using generalized linear models (GLMS),
and the species composition between areas was checked through non-metric multidimensional
scaling analysis (NMDS). As a result, 116 ants morphospecies were collected, distributed in
nine subfamilies and 42 genera. No difference was found in species richness of ants between
LRs and PPAs and only percentage of canopy cover was significantly different. We also did
not find differences of ant assemblages or functional groups between LR and PPA and this
fact reinforces that the NCFB is coherent indicating that LR and PPA are similar in
maintaining of local biodiversity. Thus, using the PPA in the calculation of LR (or vice versa)
does not imply the reduction of local diversity of ants and consequently loss of ecological
functions and interactions mediated by species of this group as these areas have similarity in
structure and species diversity.
Keywords: diversity; environmental complexity; functional groups; Atlantic Forest; ant
fauna.

x
SUMÁRIO
RESUMO ................................................................................................................................ viii
ABSTRACT ............................................................................................................................. ix
SUMÁRIO ................................................................................................................................. x
INTRODUÇÃO ...................................................................................................................... 11
MATERIAL & MÉTODOS .................................................................................................. 15
Área de estudo .................................................................................................................... 15
Descrição dos fragmentos .................................................................................................. 16
Desenho amostral ............................................................................................................... 17
Amostragem de formigas .................................................................................................. 18
Grupos funcionais .............................................................................................................. 20
Análises dos dados ............................................................................................................. 20
Variáveis ambientais e riqueza de formigas entre áreas .............................................. 20
Composição de formigas entre áreas ............................................................................. 21
Diferenças entre grupos funcionais em RL e APP ....................................................... 22
RESULTADOS ....................................................................................................................... 23
Variáveis ambientais e riqueza de formigas entre áreas ................................................ 23
Composição de formigas entre áreas ............................................................................... 24
Diferenças entre grupos funcionais em RL e APP .......................................................... 26
DISCUSSÃO ........................................................................................................................... 28
CONCLUSÃO ......................................................................................................................... 35
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 36
ANEXO .................................................................................................................................... 44

11
INTRODUÇÃO
Hábitats com distintas fitofisionomias, relevo e litologia podem ser consideradas
similares do ponto de vista da sua função ecológica? Perguntas como essa, a princípio
requerem respostas complexas, entretanto, são muitas vezes tratadas de forma generalizada
por gestores ou tomadores de decisão, principalmente pelos órgãos governamentais. Esta
questão foi de certa forma abordada, por exemplo, por ambientalistas durante a reformulação
do Código Florestal Brasileiro (CFB) (Metzger, 2010).
O CFB foi regulamentado inicialmente através da Lei 4.771 de 1965, e pode ser
considerado como o primeiro mecanismo legal para a manutenção de ecossistemas naturais,
tendo por objetivo “a preservação de remanescentes de hábitats nativos, nos diferentes biomas
brasileiros” (Brasil, 1965). Com o passar dos anos, tornou-se necessário esclarecer alguns
pontos e adequar o CFB à realidade econômica e ambiental brasileira, sendo em 2010 iniciado
intenso debate a respeito dessa reforma. Tais discussões envolveram diferentes setores da
sociedade civil, e tiveram como maior ponto de divergência a importância e utilidade das
Áreas de Proteção Permanente (APPs) e Reserva Legal (RLs), para a manutenção da
biodiversidade, em agroecossistemas (Metzger, 2010; Lewinsohn et al., 2011).
De acordo com o CFB, uma APP é definida como a parte “protegida, coberta ou não
por vegetação nativa, com a função ambiental de preservar os recursos hídricos, a paisagem, a
estabilidade geológica e a biodiversidade, facilitar o fluxo gênico de fauna e flora, proteger o
solo e assegurar o bem-estar das populações humanas” (Brasil, 2012). São consideradas Áreas
de Preservação Permanente as faixas marginais de qualquer curso d’água natural perene e
intermitente, áreas no entorno dos lagos e lagoas naturais, no entorno das nascentes e dos
olhos d’água perenes, encostas ou partes destas com declividade superior a 45°, restingas,
manguezais, bordas dos tabuleiros ou chapadas, topo de morros, montes, montanhas e serras,
áreas em altitude superior a 1.800 (mil e oitocentos) metros e em veredas (Freitas, 2010;
Brasil, 2012; Lima et al., 2014).
Por outro lado, as RLs são áreas localizadas no interior de uma propriedade ou posse
rural, excetuada a APP, “necessárias ao uso sustentável dos recursos naturais; à conservação e
reabilitação dos processos ecológicos; à conservação da biodiversidade e ao abrigo e proteção
de fauna e flora nativas” (Freitas, 2010; Brasil, 2012). São consideradas áreas de Reserva
Legais 80% (oitenta por cento) do imóvel situado em área de florestas (Amazônia Legal),
35% (trinta e cinco por cento) do imóvel situado em área de Cerrado, 20% (vinte por cento)

12
em área de campos gerais e nas demais regiões do país (Caatinga e Mata Atlântica) (Freitas,
2010; Brasil, 2012; Lima et al., 2014).
Após a reforma, o Novo Código Florestal Brasileiro (NCFB), agora regulado pela Lei
nº 12.651, de 25 de maio de 2012 e alterado em 17 de outubro do mesmo ano, pela Lei nº
12.727, não modificou as definições, mas trouxe novidades para o uso da RL e APP. Apesar
do NCFB ter mantido as mesmas regras em relação a área que deve ser preservada nas
propriedades em relação à sua área total (20 a 80%, dependendo da região em questão),
permite agora a recomposição da RL com espécies não nativas e o computo da APP na área
de RL. A área abrangida por uma APP, localizada nas margens de cursos d’água, está
atualmente relacionada à largura da calha regular deste corpo d’água, ou seja, o canal por
onde corre regularmente as águas durante o ano, e não mais a partir do nível alcançado por
ocasião da cheia (leito maior) (Brasil, 2012). Considerando o regime fluvial ao longo do ano,
essa modificação significou uma efetiva redução da faixa de APP, podendo acarretar
alterações nas entradas de material orgânico e inorgânico no corpo d’água, com
consequências para todo o sistema aquático (Lima et al., 2014). Outro ponto relevante do
NCFB é a descaracterização de proteção as nascentes intermitentes, considerando apenas
aquelas produtoras de água de forma perene.
No NCFB, alguns autores afirmam que essas novas mudanças podem afetar a
biodiversidade como um todo, desta forma tanto APPs quanto RLs são fundamentais para a
manutenção das populações de inúmeras espécies, pois exercem funções complementares,
criando na paisagem corredores que permitem a conectividade entre populações previamente
isoladas (Metzger, 2010; Toledo et al., 2010; Galetti et al., 2010). Segundo Metzger (2010), a
diferença está no fato de que, as APPs atuam primariamente na proteção da estrutura
hidrológica e geológica dos agroecossistemas (controle de erosão e lixiviação de nutrientes),
ao passo que as RLs atuam no banco local de sementes, no abrigo e manutenção de agentes
polinizadores e de inimigos naturais de pragas, contribuindo para manutenção da diversidade
local.
Pode parecer óbvio que APP e RL são de fato ambientes distintos, com características
próprias e que ambas devem ser priorizadas. Entretanto, existem poucos estudos que
acompanhem e/ou demonstrem aspectos sobre a contribuição de tais áreas para a manutenção
de grupos específicos de animais e plantas (Metzger, 2010; Lima et al., 2014). Dessa forma,
não se pode precisar sobre os prejuízos para a biodiversidade local e/ou global, caso seja

13
autorizada, pela atual legislação, a redução das APP ou RL, ao ponto de não existir consenso
sobre se tais áreas são, de fato, similares e desta forma, do ponto de vista ecológico, podem
ser consideradas uma coisa só (Chan et al., 2006; Silva et al., 2010; Freitas, 2010; Toledo et
al., 2010, Marques et al., 2010). Essas dúvidas tornam-se mais pertinentes, tendo em vista que
pequenas modificações na estrutura do ambiente podem promover alterações em assembléias.
Por exemplo, Gomes et al., 2014, observaram mudanças na composição de formigas epigéicas
encontradas em uma RL existente em um fragmento de Mata Atlântica, onde o fragmento
preservado apresentou um assembleia diferente das áreas com índices de recuperação iniciais.
Dentre os biomas brasileiros, a Mata Atlântica foi um dos mais prejudicados pela
reforma do NCFB. Desde os anos 80, o complexo da Mata Atlântica vem sendo considerado
um dos hotspots da biodiversidade global (Myers et al., 2000; Ribeiro et al., 2009; 2011), em
virtude do seu histórico de ocupação ao longo do processo de colonização do país. Já é de
conhecimento geral que grande parte de sua área foi substituída por pastagens, áreas
agricultáveis ou urbanizadas ao longo dos anos, se tornando atualmente um ambiente
fragmentado, com pouca conectividade entre fragmentos remanescentes (Ribeiro et al., 2009;
Metzger, 2010; Aragão et al., 2011). Diante destes fatos, as áreas de APP e RL em Mata
Atlântica já eram reduzidas e permitir a supressão ou substituição da sua flora por vegetação
não nativa, se tornará uma ameaça para as populações de animais e plantas,
consequentemente, provocando maior perda de variabilidade genética e/ou extinção de
populações de diversos grupos (Ribeiro et al., 2009; Galetti et al., 2010; Toledo et al., 2010:
Freitas, 2010; Marques et al., 2010).
Os efeitos das alterações no habitat sobre a perda de espécies ou redução das
populações de formigas é bem documentadas na literatura. Na região tropical, a riqueza desse
grupo tende a ser menor em ambientes com baixa heterogeneidade e/ou complexidade
estrutural da vegetação (Leal et al., 2012; Matos et al., 2013, Dáttilo et al., 2013). Além disso,
foram observadas diferenças significativas na composição das espécies de formigas e de
outros grupos em relação à variação ambiental (Neves et al., 2010, Teodoro et al., 2010,
Matos et al., 2013, Dáttilo et al., 2013). Diversos autores têm demonstrado correlação entre
características estruturais dos habitats e padrões de comunidades de formigas (Andersen &
Majer, 2004; Neves et al., 2010; 2013; Castro, 2012; Dáttilo et al., 2013; Gomes et al., 2014).
Mesmo com o isolamento e perda de habitat em muitas comunidades ecológicas, as formigas

14
permanecem sendo um dos grupos dominantes, ocupando todos os espaços disponíveis,
exercendo um grande número de funções (Silva & Brandão, 1999).
Neste sentido, Brown-Jr (1991), analisa as respostas de diferentes grupos de insetos
aos fatores ambientais e suas características ecológicas e biogeográficas. Ele aplicou valores a
aos diferentes táxons usando alguns atributos, tais como: diversificação taxonômica e
ecológica, identificação acessível, presença de indivíduos em todos os meses do ano,
importância funcional no ecossistema, resposta visível aos distúrbios e baixa mobilidade
(hábito sedentário). Como resultado verificou que as formigas obtiveram os maiores valores
entre os insetos analisados, recebendo 21 pontos dentro de uma escala de 0 a 24 (Brown-Jr,
1991).
A partir dessa justificativa, diversos trabalhos utilizam formigas como indicadores
ecológicos, em diferentes abordagens como: sucessão vegetal (Braga et al., 2010; Neves et al.,
2010; Gomes et al., 2014), fragmentação de habitats (Leal et al., 2012) e impactos das
atividades humanas no ambiente (Delabie et al., 2006) como a mineração (Ribas et al., 2012)
e a agricultura (Pacheco et al., 2013). Desta forma, a utilização de formigas pode ser uma
ferramenta promissora para analisar e/ou detectar as possíveis diferenças entre RLs e APPs,
situadas em uma mesma propriedade. De acordo com alguns autores, diferenças na estrutura
do hábitat nessas áreas, podem refletir na riqueza e composição de espécies ou nos grupos
funcionais de formigas (Silvestre & Silva, 2001; Gomes et al., 2014; Leal et al., 2012). Um
exemplo desses grupos são as formigas de correição (nômades), que têm sido utilizadas para
demonstrar qualidade do habitat, uma vez que necessitam de extensas áreas de forrageamento
(Silvestre & Silva, 2001).
Por fim, o presente trabalho teve como objetivo verificar se Reservas Legais e Áreas
de Proteção Permanentes diferem quanto à sua complexidade ambiental ou se essas podem ser
consideradas áreas similares para manutenção da biodiversidade. Desta forma, foram testadas
as seguintes hipóteses: (1) a complexidade ambiental (profundidade de serapilheira, abertura
do dossel e abundância de árvores) irá diferir entre RL e APP; (2) áreas com maior
complexidade ambiental abrigam maior riqueza de formigas; (3) a composição de formigas de
serapilheira irá diferir entre RLs e APPs e (4) haverá diferença quanto aos grupos funcionais
de formigas presentes em cada área.

15
MATERIAL & MÉTODOS
Área de estudo
O presente estudo foi realizado entre fevereiro e novembro de 2013 e conduzido em
três diferentes fragmentos, presentes no domínio de Mata Atlântica sergipano, a saber:
Instituto Federal de Sergipe (-10.9033, -37.1985) em São Cristóvão, Reserva Creuza Ramos (-
10.6555, -37.3275) localizada entre os municípios de Itabaiana e Malhador e a Mata da Boa
Vistam (-10.4592, -37.1271) localizada entre os municípios de Nossa Senhora das Dores e
Capela (Figura 1). Esses municípios foram selecionados em virtude de sua importância como
região agrícola e por sua paisagem já possuir feições que permitem a identificação de
fragmentos de mata isolados em meio a uma matriz de monocultura, geralmente pastagens ou
cultivo de cana-de-açúcar (Aragão et al., 2011; Fundação SOS Mata Atlântica e Instituto
Nacional de Pesquisas Espaciais, 2013). Para a execução do experimento, em cada fragmento
foi escolhido duas áreas de acordo com a definição do NCFB, consideradas uma como RL e
outra como APP, as quais consideramos no presente estudo apenas as matas ciliares.
Figura 1: Propriedades particulares sob domínio de Mata Atlântica em Sergipe onde o estudo foi conduzido
(Coordenadas em GD-SAD69).

16
Descrição dos fragmentos
O Instituto Federal de Sergipe (IFS) apresenta uma área de 337 ha que compõem a
“Matinha” ou Reserva Legal da propriedade. No limite oeste a propriedade é cortada pelo Rio
Poxim-Açú, com aproximadamente quatro m de calha regular e com faixa de mata ciliar
(APP) variável de 20 m até uma continuidade com o fragmento da “Matinha”. No entorno da
área do IFS encontramos pastagens e cultivo de mandioca.
A Reserva Creuza Ramos (RCR) apresenta 72 ha de tamanho total onde cinco ha de
sua propriedade é utilizada para o cultivo não intensivo de banana e gado. Está situada em um
prolongamento da serra que compõe o Parque Nacional Serra de Itabaiana (PARNASI), um
dos poucos fragmentos preservados do estado. A propriedade é cortada pela rodovia estadual
SE-245, onde do lado sul encontramos a Reserva Legal que compreende uma mata de porte
primário a secundário e o “Sitio Velho” composto por mangueiras e jaqueiras abandonadas,
totalizando 60 ha. Do lado norte, encontramos a cede da propriedade e o Riacho Cambotá,
esse com aproximadamente um m de calha regular, largura de APP média de 30 m e
apresentando uma mata de porte secundário. Os fragmentos de mata são margeados por
pastagem e nas fronteiras da propriedade encontramos pequenos sítios de cultivo familiar e
pastagens.
A Mata da Boa Vista (MBV) apesenta 54 ha de tamanho total e compõe a Reserva
Legal da propriedade da usina de cana Agroindustrial Campo Lindo. É cortada por uma trilha
com placas informativas para o uso em trabalhos de Educação Ambiental e sofre pressão
antrópica principalmente do seu lado oeste (Gois-Santos et al., 2013), apesar disso é um
fragmento com vegetação secundária. Ao lado da mata, o canavial é cortado pelo Rio da
Aldeia, riacho com um m de calha regular, que apresenta APP da margem direita composta
por mata ciliar preservada, enquanto que a margem esquerda serve como pastagem (Silva,
2013). Os fragmentos de mata e a propriedade têm no seu entorno a pastagem e o cultivo de
cana de açúcar.
Essas informações foram registradas por observação “in loco” e/ou informações
passadas pelo proprietário ou pessoas que conhecem a área e visam um melhor
reconhecimento dos locais utilizados no presente trabalho.

17
Desenho amostral
Em cada área de RL e APP foram estabelecidas 30 parcelas de 25 m², espaçadas 6 m,
ao longo de um transecto de 324 m (Figura 2, A). Respeitou-se uma distância mínima de 20
metros entre o início do transecto e a borda da mata, a fim de tentar minimizar o efeito de
borda (Fisher, 1999), além disso, as parcelas foram demarcadas no centro da RL e da APP,
essa última demarcada a 15 m da margem do curso d’água em sua calha regular.
Figura 2: Representação esquemática das três primeiras parcelas do transecto de 324 m (30 amostras) espaçadas
6 m (A) e detalhe de uma parcela com o ponto de coleta das formigas bem como dos dados ecológicos avaliados
nesse estudo (B).
No centro de cada uma das 30 parcelas foi estabelecido um quadrate de 1 m² de onde
foi mensurada à profundidade média do folhiço e retirada toda à serapilheira (ver
“Amostragem de formigas” abaixo) (Figura 2, B). Após a coleta da serapilheira, foram feitas
leituras da cobertura do dossel com um densiômetro esférico convexo de Lemmon (Lemmon,
1957) (Figura 3, B), a norte, sul, leste e oeste. As leituras foram tomadas a 1 m do solo, por
uma mesma pessoa. Segundo Suganuma et al., (2008) essa metodologia apresenta resultado

18
satisfatório e vantagens quando comparado aos sistemas fotográficos com lentes de 8 mm
(“olho de peixe”).
Em cada parcela foi avaliada a densidade de árvores com diâmetro a altura do peito
(DAP) superior a 15 cm (Figura 3, A) (Dantas et al., 2010).
Figura 3: Retirada do diâmetro da altura do peito (DAP) das árvores com mais de 15 cm de circunferência
localizadas dentro da parcela de 25 m² (A) e densiômetro esférico convexo de Lemmon (B). Fotos: Arleu
Barbosa.
Amostragem de formigas
Para a coleta de formigas, foi retirado no centro de cada parcela uma amostra de 1 m²
de serapilheira (Figura 2, B), respeitando assim uma distância de 10 m entre pontos de coleta
(Baccaro et al., 2011). O material vegetal foi transferido para um peneirador com malha de
0,5 cm² para retirada do excesso de matéria orgânica e desalojamento dos exemplares de
formigas (Figura 4, A). Todo processo foi feito pelo menos 24 horas após a última chuva no
local, pois a eficiência da coleta é afetada com o folhiço úmido (Fisher, 1998). O material
peneirado foi transportado em saco de tecido e posteriormente transferindo ao saco de malha
do extrator de mini-winkler por 48 horas (Figura 4, B) (Delabie et al., 1999; Fisher, 2000;
Bestelmeyer et al., 2000).
A B

19
Figura 4: Material vegetal sendo separado com peneirador de grade de 0,5 cm² para retirada do excesso de
matéria orgânica e desalojamento inicial dos exemplares de formigas (A) e extrator de mini-winkler durante o
período de 48 horas para desalojamento final dos formicídeos (B). Fotos: Arleu Barbosa.
Após o período de 48h, o material foi levado para laboratório, devidamente triado
utilizando lupa estereoscópica e pinça, e os formicídeos acondicionados em Etanol a 70%. A
identificação foi até o menor nível taxonômico possível, para isso, foram utilizadas chaves de
identificação (Bolton 1994; 2003; Baroni-Urbani & de-Andrade, 2007) e por meio de
comparação com coleções de referência do Laboratório de Entomologia do Departamento de
Biologia da Universidade Federal de Sergipe. Foi feito duplicata do material testemunho e
depositados no Laboratório de Entomologia do Departamento de Biologia da Universidade
Federal de Sergipe, São Cristovão-SE e no Laboratório de Mirmecologia do Centro de
Pesquisa do Cacau, Ilhéus-BA.
A licença para coleta foi deferida pelo Sistema de Autorização e Informação em
Biodiversidade (SISBIO) do Instituto Chico Mendes de Conservação da Biodiversidade
(ICMBio) com o número 42620-1.
A B

20
Grupos funcionais
Foi utilizado a classificação de grupos funcionais proposta por Leal et al., (2012),
devido à similaridade entre os biomas, proximidade geográfica entre as áreas e a alta
diversidade taxonômica e ecológica das formigas de Mata Atlântica em outras classificações,
por exemplo, formigas “Especialistas Tropicais” (Andersen, 2010), utilizado em uma
classificação global.
Os grupos aceitos foram: (I) Crípticas predadoras – formigas pequenas que nidificam
no solo, folhas e troncos podres, onde se especializam em artrópodes vivos ou seus ovos; (II)
Crípticas onívoras - formigas pequenas que nidificam no solo, folhas e troncos podres, onde
são predadoras generalistas e catadores; (III) Epigéicas predadoras – espécies de médio a
grande porte que se alimentam na superfície de serapilheira e são predadoras especializadas
de outros artrópodes, (IV) Epigéicas onívoras – formigas de médio a grande porte que se
alimentam na superfície da serapilheira e são predadoras generalistas e catadoras; (V)
Arborícolas dominantes – espécies altamente ativas e agressivas com grandes colônias e
nidificação em árvores; (VI) Arborícolas subordinadas - outras espécies que nidificam em
árvores; (VII) Oportunistas – espécies não especializadas de baixa competitividade ou
associada a espécies invasoras; (VIII) Formigas nômades - altamente agressivass com
recrutamento legionário; (IX) Attini cortadoras de folhas - altamente ativas e agressivas,
polimórficas e que formam grandes colônias que usam as folhas para cultivar o fungo
simbionte; e (X) Attini não cortadoras de folhas – formigas monomórficas, não-agressivas,
com colônias de tamanho reduzido e usam outros materiais orgânicos para cultivar o fungo
simbionte.
As espécies coletadas e não citadas na lista de Leal et al., (2012) foram alocadas em
grupos seguindo a classificação de Delabie (2000).
Análises dos dados
Variáveis ambientais e riqueza de formigas entre áreas
A riqueza de formigas (variável resposta) em resposta ao tratamento (RL ou APP), à
cobertura do dossel, à profundidade da serapilheira e à densidade de árvores (variáveis
explicativas) foi testada por meio de modelos lineares generalizados (GLMs) (Crawley 2007).

21
Tendo em vista que as parcelas estão aninhadas (pseudoréplicas) em um conjunto maior
(transectos), que por sua vez estão aninhados em pares dentro de cada propriedade, utilizou-se
para as análises as médias dos valores de cada área, exceto para os valores de riqueza, que
foram utilizados os dados brutos. Assim, o modelo completo foi composto por seis repetições
(3 transectos para cada tratamento). Para testar se a riqueza de formigas era influenciada pelas
variáveis ambientais adotou-se a distribuição Poisson, após constatação de que a variável
explicativa não obedecia a uma distribuição normal.
Diferenças nos valores das variáveis ambientais (variável resposta) entre APP e RL
(variável explicativa) também foram testadas por meio de modelos lineares generalizados
(GLMs) (Crawley, 2007). Os GLMs foram realizados no software estatístico R (R
Development Core Team, 2014) em nível de significância de 5% (Crawley, 2007).
Composição de formigas entre áreas
Para se testar diferenças na composição das assembléias de formigas de serapilheira
entre as áreas (RL e APP) foi utilizada análise similaridade (ANOSIM) (Clarke, 1993) e uma
análise de escalonamento multidimensional não-métrico (NMDS), adotando-se ordenação
pelo índice de Jaccard (que considera a presença e ausência de espécies) (Gotelli et al., 2011).
Além do NMDS, adotou-se a análise de agrupamento (CLUSTER) utilizando-se a ordenação
pelo índice de Bray-Curtis, para isso considerou-se a frequência das espécies nas amostras
(n/30) em cada sítio. A análise de Cluster permite a ordenação hierárquica, indicando
possíveis grupos/subgrupos entre áreas e locais de amostragem (RL e APP) ao passo que a
NMDS utilizada permitiu apenas a comparação entre locais.
Além disso, uma análise de similaridade (ANOSIM) (Clarke, 1993) foi realizada para
se testar diferenças entre a composição de espécies entre as 6 áreas. Ao final desta análise,
utilizando-se o software estatístico R (R Development Core Team, 2014) com nível de
significância de 5%, valores de P e R foram estabelecidos e os padrões de similaridade entre
as assembléias definidos por meio destes valores (Hammer et al., 2001).

22
Diferenças entre grupos funcionais em RL e APP
Diferenças na riqueza de grupos funcionais em relação às áreas (RL e APP), bem
como as variáveis ambientais foram testadas usando modelos lineares generalizados (GLMs),
para isso foi utilizado a soma das presenças de cada grupo funcional nas amostras (Clarke,
1993). No GLM, a riqueza de grupos funcionais foi a variável resposta e as variáveis
utilizadas para estimar a complexidade ambiental (profundidade de serapilheira, abertura do
dossel e abundância de árvores) foram as variáveis explicativas. Um modelo completo, com
todas as variáveis foi primeiramente testado, excluindo-se, posteriormente, as variáveis não
significativas por contraste a posteriori. Todas as análises foram realizadas utilizando-se o
software estatístico R (R Development Core Team, 2014), com nível de significância de 5%.
23
RESULTADOS
Foram coletadas 116 morfoespécies, distribuídas em 42 gêneros (Anexo). As
morfoespécies estão inseridas em nove subfamílias, entre as quais a Myrmicinae foi a mais
diversificada (com 74 morfoespécies), seguida por Ponerinae (n = 17), Formicinae (n = 10) e
Ectatomminae (n = 5). Com relação aos gêneros amostrados, Pheidole, Strumigenys e
Solenopsis foram os mais diversificados com 23, 10 e 8 morfoespécies, respectivamente
(Anexo). A RL contribuiu com 86 morfoespécies enquanto a APP registrou 96, havendo entre
elas 66 morfoespécies em comum. Apesar da maior riqueza de morfoespécies, a quantidade
de gêneros foi igual nas duas áreas (n = 36).
Variáveis ambientais e riqueza de formigas entre áreas
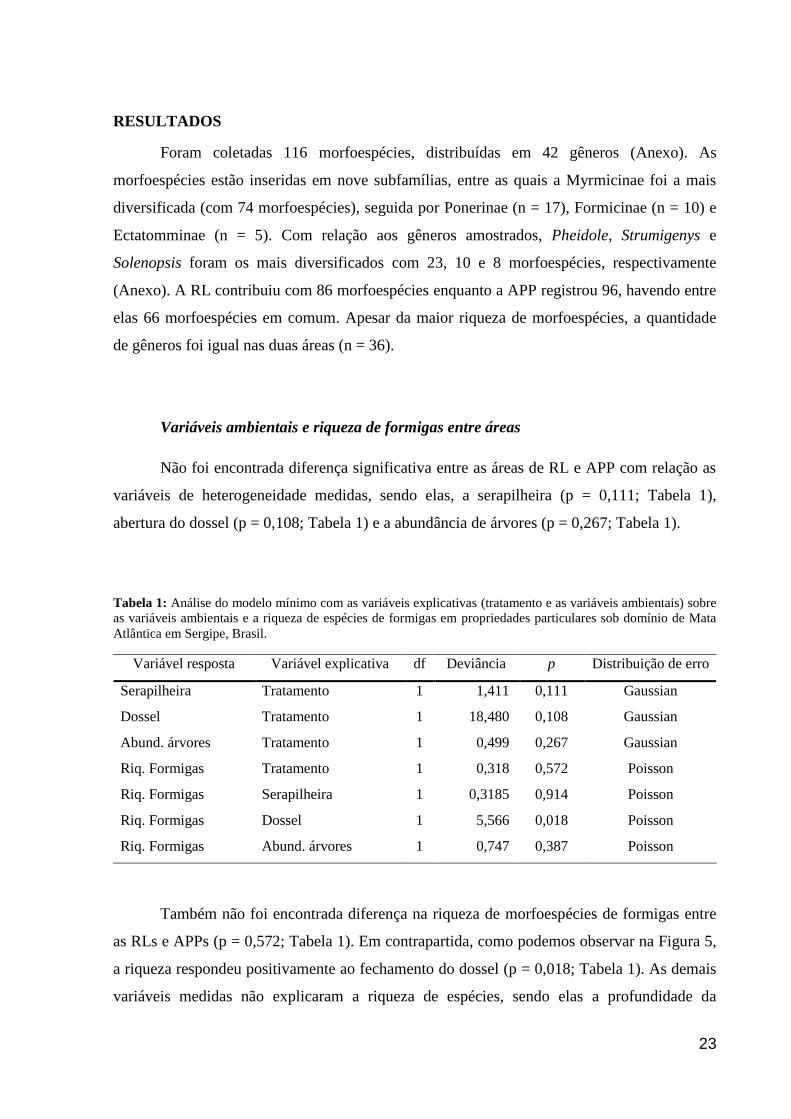
Não foi encontrada diferença significativa entre as áreas de RL e APP com relação as
variáveis de heterogeneidade medidas, sendo elas, a serapilheira (p = 0,111; Tabela 1),
abertura do dossel (p = 0,108; Tabela 1) e a abundância de árvores (p = 0,267; Tabela 1).
Tabela 1: Análise do modelo mínimo com as variáveis explicativas (tratamento e as variáveis ambientais) sobre
as variáveis ambientais e a riqueza de espécies de formigas em propriedades particulares sob domínio de Mata
Atlântica em Sergipe, Brasil.
Variável resposta Variável explicativa df Deviância p Distribuição de erro
Serapilheira Tratamento 1 1,411 0,111 Gaussian
Dossel Tratamento 1 18,480 0,108 Gaussian
Abund. árvores Tratamento 1 0,499 0,267 Gaussian
Riq. Formigas Tratamento 1 0,318 0,572 Poisson
Riq. Formigas Serapilheira 1 0,3185 0,914 Poisson
Riq. Formigas Dossel 1 5,566 0,018 Poisson
Riq. Formigas Abund. árvores 1 0,747 0,387 Poisson
Também não foi encontrada diferença na riqueza de morfoespécies de formigas entre
as RLs e APPs (p = 0,572; Tabela 1). Em contrapartida, como podemos observar na Figura 5,
a riqueza respondeu positivamente ao fechamento do dossel (p = 0,018; Tabela 1). As demais
variáveis medidas não explicaram a riqueza de espécies, sendo elas a profundidade da

24
serapilheira (p = 0,914; Tabela 1) e a cobertura do dossel (p = 0,387; Tabela 1) nos locais
amostrados.
Figura 5: Riqueza de formigas total em diferentes médias de aberturas do dossel nas APPs e RLs de
propriedades particulares sob domínio de Mata Atlântica em Sergipe, Brasil.
Composição de formigas entre áreas
Não houve diferença significativa entre as assembléias de formigas nas RLs em
comparação com as APPs (ANOSIM; R = 0,037; p = 0,6) (Figura 6), indicando que tais áreas
são similares não apenas na sua complexidade ambiental, mas também na diversidade local de
formigas.
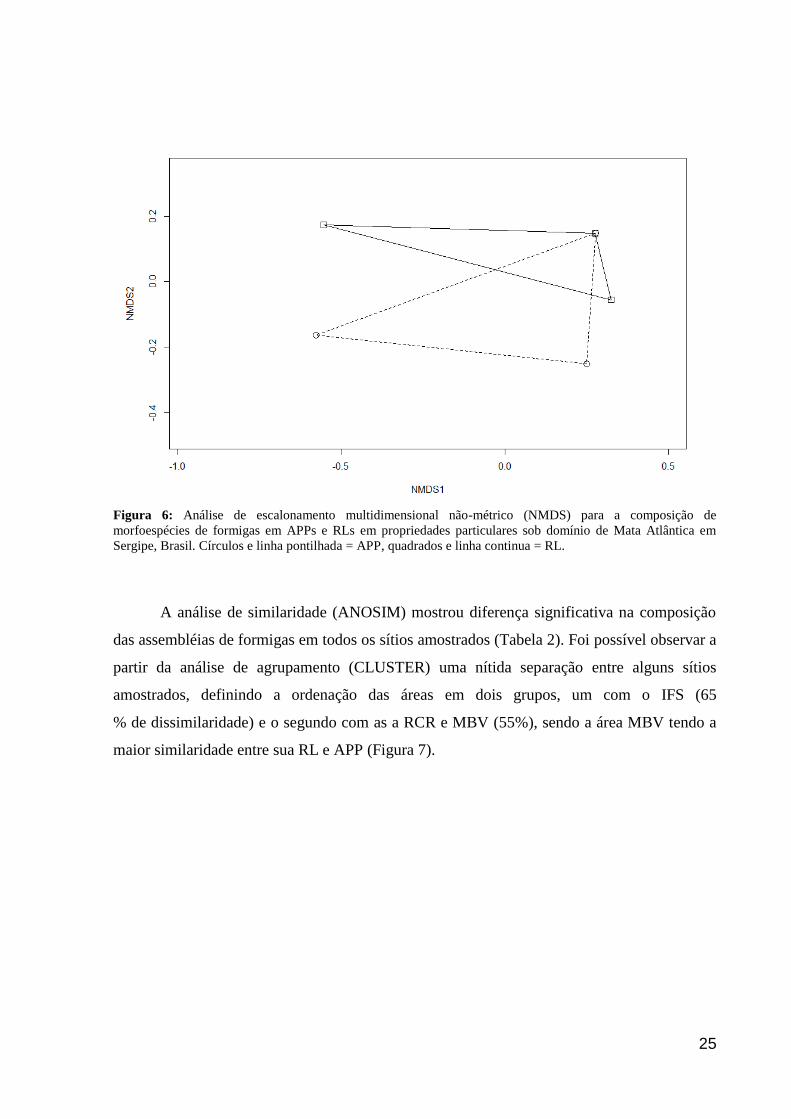
25
Figura 6: Análise de escalonamento multidimensional não-métrico (NMDS) para a composição de
morfoespécies de formigas em APPs e RLs em propriedades particulares sob domínio de Mata Atlântica em
Sergipe, Brasil. Círculos e linha pontilhada = APP, quadrados e linha continua = RL.
A análise de similaridade (ANOSIM) mostrou diferença significativa na composição
das assembléias de formigas em todos os sítios amostrados (Tabela 2). Foi possível observar a
partir da análise de agrupamento (CLUSTER) uma nítida separação entre alguns sítios
amostrados, definindo a ordenação das áreas em dois grupos, um com o IFS (65
% de dissimilaridade) e o segundo com as a RCR e MBV (55%), sendo a área MBV tendo a
maior similaridade entre sua RL e APP (Figura 7).

26
Tabela 2: Análise de similaridade (ANOSIM) para a composição de espécies de formigas em APPs e RLs e
propriedades particulares sob domínio de Mata Atlântica em Sergipe, Brasil.
Área 1 Área 2 Valor R Valor P
IFS-APP X IFS-RL 0.2650 < 0,01
IFS-APP X MBV-APP 0.5085 < 0,01
IFS-APP X MBV-RL 0.5198 < 0,01
IFS-APP X RCR-APP 0.3618 < 0,01
IFS-APP X RCR-RL 0.5552 < 0,01
IFS-RL X MBV-APP 0.3817 < 0,01
IFS-RL X MBV-RL 0.2916 < 0,01
IFS-RL X RCR-APP 0.2327 < 0,01
IFS-RL X RCR-RL 0.3945 < 0,01
MBV-APP X MBV-RL 0.1788 < 0,01
MBV-APP X RCR-APP 0.3611 < 0,01
MBV-APP X RCR-RL 0.2918 < 0,01
MBV-RL X RCR-APP 0.2719 < 0,01
MBV-RL X RCR-RL 0.2375 < 0,01
RCR-APP X RCR-RL 0.2531 < 0,01
Figura 7: Método de agrupamento (CLUSTER) utilizando a dissimilaridade de Bray-Curtis para a composição
de espécies de formigas em APPs e RLs e propriedades particulares sob domínio de Mata Atlântica em Sergipe,
Brasil.
Diferenças entre grupos funcionais em RL e APP
Foram encontrados 10 grupos funcionais nas três áreas amostradas (Figura 8) com
ausência de alguns grupos funcionais nas áreas de coleta como as Attini cortadoras de folhas
ausentes na RCR-RL e na MBV (RL e APP). Formigas nômades não foram coletadas nas
áreas do IFS–RL, RCR (RL e APP) e na MBV-APP. O IFS-RL também não apresentou as

27
Attini não cortadoras de folhas e a RCR-APP não apresentou o grupo das Arborícolas
dominantes. A área IFS-APP foi a única com registro de todos os grupos funcionais (Figura
8).
Figura 8: Frequência dos grupos funcionais de formigas em APPs e RLs e propriedades particulares sob
domínio de Mata Atlântica em Sergipe, Brasil.
Apesar da diferença nas frequências dos grupos funcionais entre as áreas e ausência de
alguns em determinados locais, não foi encontrada diferença entre os grupos funcionais das
RLs e APPs (F = 8,657; p = 0,208). Também não foi encontrada relação na riqueza de grupos
funcionais pelo volume de serapilheira (F = 86,012; p = 0,068), cobertura do dossel (F =
37,406; p = 0,103) e abundância de árvores (F = 48,733; p = 0,090).

28
DISCUSSÃO
Apesar da necessidade de se conservar e preservar fragmentos de vegetação, Reserva
Legal e Área de Proteção Permanente podem ser consideradas iguais, do ponto de vista
estrutural. Tais resultados trazem grandes implicações ecológicas para agroecossistemas e são
apoiados pela atual legislação vigente. Dentre as hipóteses estudadas, verificou-se que todas
foram refutadas. Alguns resultados podem ser atribuidos a fatores de escala pontual, que
determinam a riqueza e composição de formigas de serapilheira em diferentes ambientes. Por
exemplo, a riqueza de formigas respondeu a essas variáveis apenas em escala pontual (nas
parcelas). O fato de áreas de APP e RL apresentarem semelhanças estruturais (Tabela 1) abre
a possibilidade de se tratar as respectivas áreas de forma similar.
O número de morfoespécies de formigas no presente estudo foi maior do que em
outros trabalhos na Mata Atlântica no Brasil (62 espécies em Feitosa & Ribeiro, 2005; 110
espécies em Coelho & Ribeiro, 2006; 82 espécies em Gomes et al., 2014). Segundo Wilson
(1976), Pheidole e Solenopsis juntamente com Camponotus e Crematogaster, possuem a
maior diversidade de espécies e de hábitos, maior extensão de distribuição geográfica e de
abundância local, e, por isso, são considerados os gêneros mais prevalentes em escala global.
A alta diversidade de Stumigenys espécies (10 morfoespécies) encontrada no presente estudo
pode ser atribuído ao fato de adotarmos a sinonímia desse gênero com Pyramica (Baroni-
Urbani & de-Andersen, 2007). Essa sinonímia faz a representatividade do grupo aumentar
consideravelmente, por exemplo, no trabalho de Braga et al., (2010) o gênero Strumigenys (9
espécies coletadas) passaria de quarto para o segundo mais representado, atrás apenas de
Pheidole (19 espécies) e a frente de gêneros como Solenopsis (8 espécies) e Hypoponera (7
espécies).
A composição de formigas amostradas é composta pelas principais subfamílias e
gêneros encontrados para a Mata Atlântica em geral (Feitosa & Ribeiro, 2005; Coelho &
Ribeiro, 2006; Braga et al., 2010; Leal et al., 2012; Gomes et al., 2014) e outros ecossistemas
do Brasil como Caatinga (Leal, 2003; Neves, et al., 2013; Sousa-Souto, et al., 2014) e
Restinga (Gomes et al., 2010), ambos biomas amparados pelas mesmas regras do CF. Assim
como em trabalhos anteriores (Braga et al., 2010; Orsolon-Souza et al., 2011), a maior riqueza
da subfamília Myrmicinae neste estudo está relacionada à sua alta riqueza e abundância na
serapilheira da região neotropical (Silva & Brandão, 1999) bem como à sua adaptação a
diferentes ambientes (Fowler, 1991).

29
O fato da riqueza de formigas não ter diferido entre a RL e APP, pode ser explicado
por condições de microhabitats existentes em cada uma dessas áreas. Em outras palavras, cada
habitat independentemente possui um conjunto de variáveis que conferem disponibilidade de
recursos distintos (alimentação, espaço para nidificação, etc) (Queiroz et al., 2013) com
influência até do histórico da área (Kaspari, 2000). Neste sentido, a atenção deve-se voltar
para as especificidades de alguns grupos de formigas que acabam por limitar sua
sobrevivência em determinados habitats. Por exemplo, o gênero Prionopelta, foi registrado no
presente estudo apenas em APP e de acordo com a literatura está relacionado a ambientes
úmidos, como florestas ripárias (Brandão et al., 2009).
Analisando apenas a disponibilidade de recurso registrado nas APPs (maiores medias
de espessura de serapilheira e cobertura do dossel) o esperado seria que houvesse diferença da
riqueza entre as áreas (Leal et al., 2012). Entretanto, como mencionado anteriormente, neste
tipo de análise deve-se observar as especificidades das assembléias de formigas (Brandão et
al., 2009), uma vez que tanto as APPs como as RLs apresentam características similares de
pouca interferência antrópica. Dessa forma, os valores encontrados nas RLs, apesar de
menores, podem não representar uma “simplificação do hábitat” a ponto de interferir na
estrutura dessas assembléias (Neves et al., 2013; Queiroz et al., 2013). Assim, é possível que
as dinâmicas próprias existente em cada área permitem mudanças apenas na composição,
enquanto que os valores de riqueza permaneçam similares entre as áreas.
A cobertura do dossel foi usada como uma variável preditora da qualidade do habitat e
demonstrou ser um fator consistente para explicar a riqueza de formigas. Os resultados aqui
encontrados corroboram os existentes em estudos similares, que mostram o efeito positivo da
estrutura da vegetação sobre essas comunidades (Marinho et al., 2002; Coelho & Ribeiro,
2006; Braga et al., 2010; Leal et al., 2012; Queiroz et al., 2013; Neves et al., 2013). Efeitos
positivos da cobertura do dossel sobre a riqueza de formigas são esperados, pois o dossel
impede a entrada de raios solares e propicia maior umidade e uma vegetação
multiestratificada, ou seja, condições microclimáticas mais adequadas e características a Mata
Atlântica, quando comparado a áreas abertas. De acordo com Neves et al. (2013) a redução da
cobertura do dossel reduz a riqueza de formigas, pois altera as condições microclimáticas
locais bem como a qualidade e quantidade dos recursos ali disponíveis.
A quantidade e qualidade da matéria orgânica em decomposição no solo é um
importante preditor da abundância de espécies de artrópodes (Sayer et al., 2010), e por sua

30
vez serve como alimento para grupos de formigas epigéicas. Utilizando pitfalls de solo,
Queiroz et al. (2013) observaram que a heterogeneidade e peso da serapilheira foram as
variáveis que melhor explicaram a riqueza de espécies de formigas epigéicas, e associaram a
essas duas variáveis a disponibilidade de recursos, de espaço, alimentação e nidificação.
Apesar disso, no presente estudo, não foi encontrado relação entre a riqueza de formigas e o
volume da serapilheira, fato que pode está associado ao encontrado por Sobrinho &
Schoereder (2007), onde notaram uma maior profundidade da serapilheira em áreas de borda
de fragmentos com uma menor riqueza de espécies de formigas. É possível que as coletas,
principalmente na APP por essa sofrer efeito direto da radiação solar da borda, aumento da
temperatura e maior velocidade do vento, tenham mascarado uma possível relação entre
riqueza e serapilheira.
Já a abundância de árvores não foi um fator explicativo para a riqueza de formigas de
serapilheira, e diferiu de outros trabalhos (Leal et al., 2012; Neves et al., 2013). Uma
explicação para o presente resultado pode ser encontrada em Queiroz et al., (2013), onde foi
levantada a hipótese de que as formigas estão fortemente relacionadas às características do
habitat, como por exemplo, um maior fechamento do dossel para as epigéicas e riqueza e
densidade de árvores para formigas arborícolas. De certa forma, a densidade de árvores está
relacionada às duas outras medidas utilizadas no presente estudo (profundidade da
serapilheira e dossel). Entretanto, fatores não mensurados como riqueza, composição de
espécies arbóreas, porte e deciduidade poderiam elucidar e/ou explicar melhor os padrões
registrados para assembléias de formigas (Leal et al., 2012). Além disso, vale ressaltar, que
existem outros fatores que agem em diferentes escalas espaciais, e podem afetar a estrutura de
comunidades de formigas epigeicas, como a variação topográfica (Gunawardene et al., 2012),
tamanho do fragmento (Leal et al., 2012) e isolamento (Ribeiro et al., 2009; Freitas, 2010).
Tais fatores não foram abordados no presente trabalho e deve ser considerados em estudos
futuros, principalmente quando consideramos remanescentes florestais de RL e APP.
Através da análise de similaridade ficou evidente que as assembléias encontradas na
RL e APP são semelhantes, resultado que vai de encontro com a hipótese proposta, mas que
pode ser explicado pela proximidade dos locais amostrados ou pela conectividade entre os
fragmentos (Sobrinho & Schoereder, 2007). Esses resultados estão relacionados também ao
histórico de preservação na propriedade e por sua vez à especificidade de cada grupo ou a
espécie e seus recursos e/ou condições particulares do habitat encontrado nessas áreas (Silva

31
& Brandão, 1999; Brandão et al., 2009). De acordo com Leal et al. (2012), o isolamento em
fragmentos próximos influenciam a composição das espécies de formigas ali presente, por
vezes, motivados pela variação na estrutura do habitat, porém, o tempo de separação dessas
áreas de APP e RL podem não ter sido suficiente para uma modificação estrutural e das
assembléias desses ambientes amostrados.
No entanto, a partir do NMDS observa-se que parte da composição foi similar entre as
RLs e APPs. Estes resultados podem indicar a existência de conexão dentro da matriz
agrícola, possivelmente através de corredores de vegetação, ou o fato de no passado tais áreas
pertencerem a um fragmento contínuo. Dentro deste raciocínio, pode-se propor que áreas mais
próximas ou existentes em um único fragmento, independente se RL ou APP, apresentarão
composição semelhante, resultado observado para as áreas do IFS e da MBV na análise de
agrupamento. A semelhança entre as áreas mais próximas espacialmente pode reforçar a
importância da função da APP como corredor ecológico, para manutenção da biodiversidade
(Ribeiro et al., 2009; Galetti et al., 2010; Metzger, 2010; Toledo et al., 2010; Leal et al.,
2012).
Ainda analisando a análise de agrupamento, observa-se que no caso da RCR a resposta
da assembleia pareceu estar associada à complexidade do ambiente uma vez que a sua RL
apresenta valores da quantidade de serapilheira e cobertura do dossel próximo dos sítios
amostrados na MBV (dados não expostos aqui). Resultado similar foi encontrado também por
Pacheco et al. (2013) onde sugerem que as diferenças na estrutura do habitat entre RL e as
culturas adjacentes têm um efeito muito mais importante nas assembléias de formigas do que
a distância entre os locais de amostragem.
Os resultados obtidos a partir do uso dos grupos funcionais reforçam os resultados
encontrados para a composição de espécies e indicaram que não existe diferença entre APP e
RL, de forma geral Além disso, os grupos funcionais não responderam a nenhuma das
variáveis ambientais tomadas. Fato que vai de encontro ao encontrado na literatura, onde
alguns autores mostram essas variáveis influenciando os habitats de algumas guildas de
formigas, por vezes, causando a exclusão de grupos por não proporcionar condições
favoráveis (Silvestre & Silva, 2001; Leal et al., 2012). No presente estudo isso pode está
relacionado também à presença da maioria dos grupos funcionais nas áreas amostradas, com
uma variação de dois entre a menor e a que teve os 10 grupos de formigas amostrados.

32
Além da composição, os grupos funcionais passam a ser uma importante característica
de comparação entre duas comunidades, pois deixam de comparar apenas uma lista de
espécies e passam a observar um arranjo estrutural da assembleia de determinado local
(Silvestre & Silva, 2001). Dessa forma, os grupos encontrados em cada área apresentam suas
particularidades, essas podem está mantendo as funções ecológicas na propriedade em
equilíbrio uma vez que, assembléias de formigas na RL são, por vezes, complementadas na
APP (e vice versa). Esses dados reforçam a ideia de manter essas duas áreas como prioritárias
dentro da propriedade, visto que essas irão se complementar para manter as funções (Metzger,
2010). No entanto, esta tendência deve ser interpretada com cautela, pois não foi significativa
quando analisada em seu modelo geral. Sendo assim, as funções ecológicas dentro da
propriedade parecem estar relacionadas à “hipótese rebite” de Ehrlich e Ehrlich (1981), onde
essa pressupõe a estabilidade do ambiente baseada em um conjunto de espécies, essas,
divididas em subconjuntos menores desempenhando diferentes funções. A ausência de
algumas espécies pode não acarretar problemas para a manutenção das funções, mas um
conjunto delas pode desestabilizar o ambiente. Com isso, reforçamos a importância de se
manter essas duas áreas na propriedade rural, uma vez que os grupos funcionais são
aparentemente distintos em escala local, acarretando diferenças nas funções ecológicas
desempenhadas dentro da propriedade na RL e APP (Metzger, 2010).
Algumas assembléias podem ser tratadas isoladamente, por exemplo, houve uma
dominância dos grupos considerados generalistas e de acordo com os dados da literatura as
espécies oportunistas e generalistas por apresentarem maiores amplitudes nas dietas e
condições do ambiente podem ser mais facilmente registradas (Brandão et al., 2009; Galetti et
al., 2010; Toledo et al., 2010; Pacheco et al., 2013). Vale ressaltar que vários trabalhos
sugerem as formigas especializadas (por exemplo, Predadoras crípticas e Epigéicas
predadoras) como bioindicadoras da fauna edáfica e consequentemente da qualidade do
ambiente (Silvestre & Silva, 2001; Leal et al., 2012; Pacheco et al., 2013). Dentre esses
grupos, as Formigas nômades são importantes prediletores da qualidade do habitat, pois
algumas espécies necessitam de extensas áreas de forrageamento (Silvestre & Silva, 2001).
Apesar disso, sua ausência nas demais áreas pode ser atribuída à baixa densidade e/ou
limitação da metodologia utilizada na coleta desse grupo em específico (Orsolon-Souza et al.,
2011). Dessa forma, não foi utilizado como grupo resposta devido a sua baixa frequência
(Leal et al., 2012). A ausência do grupo das Attini não cortadoras de folhas na RL do IFS, foi

33
devido a um evento estocástico pontual, uma vez que se trata de um grupo abundante na
serapilheira e com representantes amostrados nessa localidade em outras coletas (piloto).
De maneira geral, APP e RL podem ser consideradas similares, situação condizente
com o NCFB. Os dados aqui encontrados demonstraram que APPs e RLs abrigam
semelhantes assembléias de formigas de serapilheira. Neste sentido, vale ressaltar que
artifícios legais, como a redução de uma delas para compensar a perda da outra,
aparentemente, não prejudicaria a biodiversidade de formigas. Pacheco et al. (2013) reforça a
manutenção da RL na propriedade em áreas de cerrado, por essas apresentarem entre 41-76%
de todas as espécies encontradas, quando analisada a área protegida e a utilizada para o
cultivo. No presente estudo, a exclusividade de cada área em comportar as espécies da
propriedade variou de 64 a 81% na RL e de 74 a 84% na APP, mostrando com isso que ambas
as áreas abrigam parcela significativa da diversidade de formigas local.
Em contrapartida, um argumento relevante e que reforça a manutenção de tais áreas, é
que todas elas estão inseridas no bioma de Mata Atlântica. De acordo com Ribeiro et al.,
(2009) aproximadamente 40% do que resta de Mata Atlântica encontra-se no formado de
fragmento, com menos de 100 ha, e tais fragmentos, como observado no presente estudo,
conseguem abrigar parcela significativa da biodiversidade local. Desta forma chama-se
atenção para a legislação vigente quanto aos meios e/ou ferramentas usadas para classificação,
definição e manutenção desses ambientes. Dados registrados na literatura indicam a
existências de diferenças cruciais nas interações e funções ecológicas desempenhadas por
outros grupos e espécies nesses fragmentos (Chan et al., 2006; Galetti et al., 2010; Silva et al.,
2010; Freitas, 2010; Toledo et al., 2010; Marques et al., 2010).
Outra justificativa é que o NCFB permite aos produtores, nas propriedades que abriga
mais de um tipo de vegetação nativa, escolher aquelas com menor heterogeneidade e
diversidade para servirem de RL, diminuindo desta forma a efetividade dessas na conservação
da biodiversidade. Por exemplo, extensas fazendas de cana-de-açúcar ou eucalipto no Brasil,
podem abrigar distintos ecossistemas ou áreas de transição, como floresta úmida, brejos,
cerrado (savanas), restingas e campos de altitude (Metzger, 2010). Ao se priorizar a
conservação de apenas um tipo de ambiente (geralmente o mais simples) como RL é possível
que muitas espécies especialistas dos demais ambientes sejam eliminadas (Galetti et al.,
2010). E destacamos ainda a importância da RL na mobilidade de animais nesses ambientes,

34
integrando fragmentos e reduzindo o isolamento de Unidades de Conservação através desses
“trampolins ecológicos” (Ribeiro et al., 2009; Galetti et al., 2010; Pacheco et al., 2013).
Apesar dos dados expostos encontrados na literatura, sugerimos, a partir dos nossos
dados, que o NCFB é condizente com a ideia de que APPs podem atuar como RLs e serem
efetivas ao compensar a perda de espécies, uma vez que trata-se de ambientes estruturalmente
similares para a conservação dessas áreas em propriedades localizadas no complexo de Mata
Atlântica em Sergipe.

35
CONCLUSÃO
O Novo Código Florestal Brasileiro foi o foco de polêmicas entre pesquisadores,
políticos e a sociedade civil, sendo criticado por possuir um texto que possivelmente
simplificava a questão da importância de Áreas de Proteção Permanente e Reservas Legais na
manutenção da biodiversidade em áreas dominadas pela paisagem agrícola. Entre os seus
artigos, está o que permite a substituição da RL pelo cômputo da APP como forma de
compensação e anistia por delitos ambientais ocorridos no passado. Apesar das críticas ao
NCFB, o presente estudo não encontrou diferenças marcantes na estrutura do ambiente para
essas áreas, indicando que o texto atual do NCFB é condizente no que se refere a uma
possível substituição da RL pelo cômputo da APP ou vice-versa.
Além disso, a fauna de formigas não diferiu entre as áreas (riqueza, composição e
quantidade de grupos funcionais), corroborando os dados encontrados para estrutura do
habitat. Os grupos funcionais apresentaram diferenças na sua frequência e ocorrência quando
analisados em escala de propriedades, e por apresentar apenas essa diferença, consideramos
que formigas de serapilheira respondem de forma similar aos dois ambientes amostrados. Esse
resultado tem um impacto importante sobre funções ecológicas e de interações mediadas por
formigas, uma vez que essas fazem parte da dieta de outros grupos e que talvez esses grupos
possam responder da mesma forma. Contudo, sugerimos priorizar áreas com um maior
fechamento do dossel, pois, RLs ou APPs com menor cobertura do dossel, pode significar
uma área com baixa riqueza de espécies.

36
REFERÊNCIAS BIBLIOGRÁFICAS
Andersen, A.N. & Majer, J.D. (2004). Ants show the way down under: Invertebrates as
bioindicators in land management. Front in Ecology and the Environ 2: 291-298.
Andersen, A.N. (2010). Functional groups in ant community ecology. In: Lach. L., Parr, C. &
Abbott, K. (Eds) Ant ecology (pp. 142–144). Oxford University Press, Oxford.
Aragão, R., Almeida, J.A.P., Figueiredo, E.E. & Srinivasan, V.S. (2011). Mapeamento do
potencial de erosão laminar na Bacia do Rio Japaratuba, SE, via SIG. Rev Bras de Eng Agríc
e Ambient, 15(7): 731-740.
Baccaro, F.B., Ketelhut, S.M. & Morais, J.W. (2011). Efeitos da distância entre iscas nas
estimativas de abundância e riqueza de formigas em uma floresta de terra-firme na Amazônia
Central. Acta Amazon, 41(1): 115-122.
Baroni-Urbani, C. & de-Andrade, M.L. (2007). The ant tribe Dacetini: limits and constituent
genera, with descriptions of new species (Hymenoptera, Formicidae). Annali del Museo
Civico di Storia Naturale “Giacomo Doria”, 99: 1-191.
Bestelmeyer, B.T., Agosti, D., Alonso, L.E., Brandão, C.R.F., Brown,W.L., Delabie, J.H.C. &
Silvestre, R. (2000). Field techniques for the study of ground-dwelling ants: An overview,
description, and evaluation. In Agosti, D.; Majer, J.D.; Alonso, L.E. & Schultz, T.R. (Eds.),
Ants: Standard methods for measuring and monitoring biodiversity (pp. 122-144).
Washington and London, Smithsonian Institution Press, 280 p.
Bolton, B. (1994). Identification guide to the ant genera of the world. Cambridge, Harvard
University Press, 222 p.
Bolton, B. (2003). Synopsis and classification of Formicidae. Mem Am Entomol Institute, 71:
1-370.
Braga, D.L., Louzada, J.N.C., Zanetti, R. & Delabie, J.H.C. (2010). Avaliação rápida da
diversidade de formigas em sistemas de uso do solo no sul da Bahia. Neotropical
Entomology, 39(4): 464-469.
Brandão, C.R.F., Silva, R.R. & Delabie, J.H.C. (2009). Formigas (Hymenoptera). In A.R.
Panizzi, J.R.P. Parra, (Eds.), Bioecologia e nutrição de insetos: base para o manejo integrado
de pragas (pp. 323-369). Embrapa Tecnológica, Brasília-DF, 1164 p.

37
Brasil. (1965). Presidência da República, Casa Civil, Subchefia para Assuntos Jurídicos. Lei
nº 4.771, de 15 de setembro de 1965. Diário Oficial [da] República Federativa do Brasil,
Brasília.
Brasil. (2012). Presidência da República, Casa Civil, Subchefia para Assuntos Jurídicos. Lei
nº 12.651, de 25 de maio de 2012. Diário Oficial [da] República Federativa do Brasil,
Brasília.
Brown-Jr, K.S. (1991). Conservation of neotropical environments: insects as indicators. In:
Collins N.M. & Thomas J.A. (Eds.). The conservation of insects and their habitats. Academic
Press, London, 450 p.
Castro, F.S., Gontijo, A.B., Castro, P.T.A. & Ribeiro, S.P. (2012). Annual and Seasonal
Changes in the Structure of Litter-Dwelling Ant Assemblages (Hymenoptera: Formicidae) in
Atlantic Semideciduous Forests. Psyche, 959715: 1-12.
Chan, K.M.A., Shaw, M.R., Cameron, D.R., Underwood, E.C. & Daily, G.C. (2006).
Conservation planning for ecosystem services. PLOS Biol, 4(11): 2138-2152.
Clarke, K.R. (1993). Non-parametric multivariate analysis of changes in community structure.
Austral Ecol, 18: 117-143.
Coelho, I.R. & Ribeiro, S.P. (2006). Environment Heterogeneity and Seasonal Effects in
Ground-Dwelling Ant (Hymenoptera: Formicidae) Assemblages in the Parque Estadual do
Rio Doce, MG, Brazil. Neotropical Entomology, 35(1): 19-29.
Crawley, M.J. (2007). Statistical computing – an introduction to data analysis using s-plus.
John Wiley & Sons, London, UK.
Dantas, T.V.P., Nascimento-Júnior, J.E., Ribeiro, A.S. & Prata, A.P.N. (2010). Florística e
estrutura da vegetação arbustivo-arbórea das Areias Brancas do Parque Nacional Serra de
Itabaiana/Sergipe, Brasil. Rev Bras Bot, 33(4): 575-588.
Dáttilo. W., Guimarães-Jr., P.R. & Izzo, T.J. (2013). Spatial structure of ant – plant
mutualistic networks. Oikos, 122: 1643–1648.
Delabie, J.H.C., Nascimento, I.C. & Mariano, C.S.F. (1999). Importance de l'agriculture
cacaoyère pour le maintien de la biodiversité: étude comparée de la myrmécofaune de
différents milieux du sud-est de Bahia, Brésil (Hymenoptera; Formicidae). In: Cocoa

38
Producer's Alliance, XII International Cocoa Research Conference, Cocoa producers'
Alliance, p. 23–30.
Delabie, J.H.C., Agosti, D. & Nascimento, I. (2000). Litter ant communities of the Brazilian
Atlantic rain forest region. In: Agosti, D., Majer, J., Alonso, L. & Schultz, T.R. (Eds)
Sampling ground-dwelling ants: case studies from the world’s rain forests. Curtin University
of Technology School of Environmental Biology Bulletin No. 18, Perth.
Delabie, J.H.C., Paim, V.R.L.M., Nascimento, I.C., Campiolo, S. & Mariano, C.S.F. (2006).
As Formigas como Indicadores Biológicos do Impacto Humano em Manguezais da Costa
Sudeste da Bahia. Neotropical Entomology, 35(5): 602-615.
Ehrlich, P.R. & Ehrlich, A.H. (1981). The rivet poppers. Not Man Apart, 2-15.
Feitosa, R.S.M. & Ribeiro, A.S. (2005). Mirmecofauna (Hymenoptera, Formicidae) de
serapilheira de uma área de Floresta Atlântica no Parque Estadual da Cantareira – São Paulo,
Brasil Biotemas, 18(2): 51-71.
Fisher, B.L. (1998). Ant diversity patterns along an elevational gradient in the Reserve
Specialed'Anjanaharibe-Sud and on the western Masoala Peninsula, Madagascar. Fieldiana
Zoology, 90: 39-67.
Fisher, B.L. (1999). Improving inventory efficiency: a case study of leaf-litter ant diversity in
Madagascar. Ecol Appl, 9(2): 714-731.
Fisher, B.L., Malsch, A.K.F., Gadagkar, R., Delabie, J.H.C., Vasconcelos H.L. & Majer, J.D.
(2000). Applying the ALL Protocol: Selected case studies. In Agosti, D.; Majer, J.D.; Alonso,
L.E. & Schultz, T.R. (Eds.), Ants: Standard methods for measuring and monitoring
biodiversity (pp. 207-214). Washington and London, Smithsonian Institution Press, 280 p.
Fowler, H.G., Forti, L.C., Brandão, C.R.F., Delabie, J.H.C. & Vasconcelos, H.L. (1991).
Ecologia nutricional de formigas. In Panizzi, A.R. & Parra, J.R.P, Ecologia nutricional de
insetos e suas implicações no manejo de pragas (pp. 131-223). São Paulo, Manole.
Freitas, A.V.L. (2010). Impactos potenciais das mudanças propostas no Código Florestal
Brasileiro sobre as borboletas. Biota Neotrop, 10(4): 53-58.
Fundação SOS Mata Atlântica e Instituto Nacional de Pesquisas Espaciais. (2013).
Divulgados novos dados do Atlas dos Remanescentes Florestais da Mata Atlântica. Publicado

39
terça-feira, 4 de Junho de 2013. http://www.inpe.br/noticias/noticia.php?Cod_Noticia=3299.
(Acessado dia: 22 de Janeiro, 2015).
Galetti, M., Pardini, R., Duarte, J.M.B., Silva, V.M.F., Rossi, A. & Peres, C.A. (2010)
Mudanças no Código Florestal e seu impacto na ecologia e diversidade dos mamíferos no
Brasil. Biota Neotrop, 10(4): 47-52.
Gois-Santos, L., Leite, J.L., Silva, G.S. & Santos, R.S. (2013). Levantamento faunístico de
térmitas (Insecta: Isoptera) existente em Remanescente de Mata Atlântica, Mata da Boa Vista,
Nossa Senhora das Dores, Sergipe. XVIII Encontro de Zoologia do Nordeste: Do Oceano ao
Sertão; Encontro de Zoologia do Nordeste, Maceió-AL, Livro de resumos e programa oficial,
2013, p 220.
Gomes, E.C.F., Ribeiro, G.T., Souza T.M.S. & Sousa-Souto, L. (2014). Ant assemblages
(Hymenoptera: Formicidae) in three different stages of forest regeneration in a fragment of
Atlantic Forest in Sergipe, Brazil. Sociobiology 61(3): 250-257.
Gomes, J.B.V., Barreto, A.C., Filho, M.M., Vidal, W.C.L., Costa, J.L.S., Oliveira-Filho, A.T.
& Curi, N. (2010). Relações entre atributos do Solo e Atividade de Formigas em Restingas.
Rev Bras Ciênc Solo, 34: 67-78.
Gotelli, N.J., Ellison, A.M., Dunn, R.R. & Sanders, N.J. (2011). Counting ants (Hymenoptera:
Formicidae): biodiversity sampling and statistical analysis for myrmecologists. Myrmecol
News, 15: 13-19.
Gunawardene, N.R., Majer, J.D. & Edirisinghe, J. P. (2012). Correlates of ant (Hymenoptera:
Formicidae) and tree species diversity in Sri Lanka. Myrmecol News, 17: 81-90.
Hammer, O., Harper, D.A.T. & Ryan, P.D. (2001). PAST: Palaeonthological Statistics
Software Package for education and data analysis. Palaeontologia Electronica, 4: 1-9.
Kaspari, M. (2000). A primer of ant ecology. In Agosti, D.; Majer, J.D.; Alonso, L.E. &
Schultz, T.R. (Eds.), Ants: Standard methods for measuring and monitoring biodiversity (pp.
207-214). Washington and London, Smithsonian Institution Press, 280 p.
Ketelhut, S.M. (2004). Ecologia de comunidades de formigas (Hymenoptera: Formicidae) na
várzea da Ilha de Marchantaria – município de Iranduba, Amazônia Central. Tese em Ciências

40
Biológicas - Programa de Pós-Graduação em Biologia Tropical e Recursos Naturais,
INPA/UA, Manaus, Amazônia. 218 p.
Leal, I.R. (2003). Diversidade de formigas em diferentes unidades da paisagem da Caatinga.
In I.R. Leal, M. Tabarelli & J.M. Silva (Eds.), Ecologia e conservação da Caatinga. (pp. 435-
460). Recife: Editora da Universidade Federal de Pernambuco.
Leal, I.R., Filgueiras, B.K.C., Gomes, J.P., Iannuzzi, L. & Andersen, A.N. (2012). Effects of
habitat fragmentation on ant richness and functional composition in Brazilian Atlantic forest.
Biodivers Conserv, 21: 1687–1701.
Lemmon, P.E. (1957). A new instrument for measuring forest overstory density. J Forest,
55(9): 667-668.
Lewinsohn, T.M., Metzger, J.P., Joly, C.A., Casatti, L., Rodrigues, R.R. & Martinelli, L.A.
(2011). Impactos potenciais das alterações propostas para o Código Florestal Brasileiro na
biodiversidade e nos serviços ecossistêmicos. http://www2.unesp.br/revista/wp-
content/uploads/2010/10/Biota-Fapesp-ABECO-Sintese-CFB-e-biodiversidade.pdf.
(Acessado dia: 10 de Dezembro, 2014).
Lima, A., Bensusan, N. & Russ, L. (2014). Código Florestal - Por um debate pautado em
ciência. Realização do Instituto de Pesquisa Ambiental da Amazônia. Brasília, dez. 2014, 76
p.
Marinho, C.G.S., Zanetti, R., Delabie, J.H.C., Schlindwein, M.S. & Ramos, L.S. (2002).
Diversidade de Formigas (Hymenoptera: Formicidae) da Serapilheira em Eucaliptais
(Myrtaceae) e Área de Cerrado de Minas Gerais. Neotrop Entomol, 31(2): 187-195.
Marques, O.A.V., Nogueira, C., Martins, M. & Sawaya, R.J. (2010). Impactos potenciais das
mudanças propostas no Código Florestal Brasileiro sobre os répteis brasileiros. Biota
Neotrop, 10(4): 39-42.
Martinelli L.A., Joly, C.A., Nobre, C.A. & Sparovek, G. (2010). A falsa dicotomia entre a
preservação da vegetação natural e a produção agropecuária. Biota Neotrop, 10(4): 323-330.
Matos, M.C.B., Sousa-Souto, L., Almeida, R.S. & Teodoro, A.V. (2013). Contrasting patterns
of species richness and composition of solitary wasps and bees (Insecta: Hymenoptera)
according to land-use. Biotropica, 45(1): 73-79.

41
Metzger, J.P. (2002). Bases biológicas para definição de Reservas Legais. Ciência Hoje, 31:
183-184.
Metzger, J.P. (2010). O Código Florestal Brasileiro tem base científica? Natureza &
Conservação, 8(1): 92-99.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., Fonseca, G.B.A. & Kent, J. (2000).
Biodiversity hotspots for conservation priorities. Nature, 403: 853-858.
Neves, F.S., Braga, R.F., Espírito-Santo, M.M., Delabie, J.H.C., Fernandes, G.W., Sánchez-
Azofeifa, G.A. (2010). Diversity of arboreal ants in a brazilian tropical dry forest: effects of
seasonality and successional stage. Sociobiology, 56(1): 177-194.
Neves, F.S., Queiroz-Dantas, K.S., Darocha, W.D. & Delabie, J.H.C. (2013). Ants of three
adjacent habitats of a transition region between the Cerrado and Caatinga biomes: The effects
of heterogeneity and variation in canopy cover. Neotrop Entomol, 42: 258–268.
Orsolon-Souza, G., Esbérard, C.E.L., Mayhé-Nunes, A.J., Vargas, A.B., Veiga-Ferreira, S. &
Folly-Ramos, E. (2011). Comparison between Winkler’s extractor and pitfall traps to estimate
leaf litter ants richness (Formicidae) at a rainforest site in southest Brazil. Braz. J. Biol.,
71(4): 873-880.
Pacheco, R. & Vasconcelos, H.L. (2012). Habitat diversity enhances ant diversity in a
naturally heterogeneous Brazilian landscape. Biodivers Conserv, 21:797–809.
Pacheco, R., Vasconcelos, H.L., Groc, S., Camacho, G.P. & Frizzo, T.L.M. (2013). The
importance of remnants of natural vegetation for maintaining ant diversity in Brazilian
agricultural landscapes. Biodivers Conserv, 22: 983–997.
Queiroz, A.C.M., Ribas, C.R. & França, F.M. (2013). Microhabitat characteristics that
regulate ant richness patterns: The importance of leaf litter for epigaeic ants. Sociobiology,
60(4): 367-373.
R Development Core Team (2014) R: a language and environment for statistical computing.
R foundation for statistical computing, Vienna, Austria. ISBN 3-900051-07-0. http://www.r-
roject.org.

42
Ribas, C.R., Solar, R.R.C., Campos, R.B.F., Schmidt, F.A., Valentim, C.L. & Schoereder,
J.H. (2012). Can ants be used as indicators of environmental impacts caused by arsenic? J
Insect Conserv, 16: 413–421.
Ribeiro, M.C., Martensen, A.C., Metzger, J.P., Tabarelli, M., Scarano, F. & Fortin, M.J.
(2011). The brazilian atlantic forest: a shrinking biodiversity Hotspot. In: Zachos, F.E &
Habel, J.C. (Eds.). Biodiversity Hotspots: Distribution and Protection of Conservation Priority
Areas. Springer-Verlag Berlin Heidelberg, 550 p.
Ribeiro, M.C., Metzger, J.P., Martensen, A.C., Ponzoni, F.L., Hirota, M.M. (2009). The
Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed?
Implications for conservation. Biol Conserv, 142: 1141–1153.
Sayer, E.J., Sutcliffe, L.M.E., Ross, R.I.C. & Tanner, E.V.J. (2010). Arthropod abundance
and diversity in a lowland tropical forest floor in Panama: the role of habitat space vs. nutrient
concentrations. Biotropica, 42(2): 194-200.
Silva, G.S. (2013). Guia de Campo Mata da Boa Vista. Projeto de conclusão do curso de
Ciências Biológicas – Lic. – da Universidade Federal de Sergipe sob orientação do Profº Dr.
Claudio Sérgio Lisi. São Cristóvão, Sergipe. 84 p.
Silva, R.R. & Brandão, C.R.F. (1999). Formigas (Hymenoptera: Formicidae) como
indicadoras da qualidade ambiental e da biodiversidade de outros invertebrados terrestres.
Biotemas, 12(2): 55-73.
Silva, S.T., Figueiredo, G.J.P., Leuzinger, M.D. & Neto, J.N. 2010. Código Florestal: Em
defesa das nossas florestas e do nosso futuro. In: Lavratti, P. & Prestes, V.B. (Orgs.). Direito e
Mudanças Climáticas – A Reforma do Código Florestal: Limites Jurídicos. São Paulo:
Instituto O Direito por um Planeta Verde (Direito e Mudanças Climáticas; 1) 189 p.
Sobrinho, T.G. & Schoereder, J.H. (2007). Edge and shape effects on ant (Hymenoptera:
Formicidae) species richness and composition in forest fragments. Biodivers Conserv, 16:
1459–1470.
Sousa-Souto, L., Santos, E.D.S., Figueiredo, P.M.F.G., Santos, A.J. & Neves, F.S. (2014). Is
there a bottom-up cascade on the assemblages of trees, arboreal insects and spiders in a
semiarid Caatinga? Arthropod-Plant Interactions, 8: 581–591.

43
Suganuma, M.S., Torezan, J.M.D., Cavalheiro, A.L., Vanzela, A.L.L. & Benato, T. (2008).
Comparando metodologias para avaliar a abertura do dossel e a luminosidade no sub-bosque
de um reflorestamento e uma floresta madura. Rev Árvore, 32(2): 377-385.
Teodoro, L.F.A., Eke, V.R. & Elphic, R.C. (2010). Spatial distribution of lunar polar
hydrogen deposits after KAGUYA (SELENE), Geophys. Res. Lett, 37, L12201.
Toledo, L.F., Carvalho-e-Silva, S.P., Sánchez, C., Almeida, M.A. & Haddad, C.F.B. (2010.)
A revisão do Código Florestal Brasileiro: impactos negativos para a conservação dos anfíbios.
Biota Neotrop, 10(4): 35-38.
Wilson, E.O. (1976). Which are the most prevalent ant genera? Stud Entomol, 19: 187-200.

44
ANEXO
Anexo: Lista, grupo funcional e número de ocorrências de cada uma das morfoespécies de formigas coletadas
em 30 amostras de mini-winkler nas APP e RL de propriedades sob domínio de Mata Atlântica em Sergipe.
Locais de coleta
IFS MBV RCR
Subfamílias ou morfoespécies/grupos
funcionais
A
P
P
R
L
A
P
P
R
L
A
P
P
R
L
AMBLYOPONINAE
Prionopelta sp.1 I - - 6 - - -
DOLICHODERINAE
Dolichoderus attelaboides (Fabricius,
1775) V - 2 - - - -
Dolichoderus cf. lutosus (Smith, 1858) V 2 - - - - -
Dolichoderus sp.1 V - 4 15 16 - 6
Dorymyrmex sp.1 VII 1 - - - - -
DORYLINAE
Neivamyrmex sp.1 VIII - - - 3 - -
Neivamyrmex sp.2 VIII 1 - - - - -
ECTATOMMINAE
Ectatomma edentatum Roger, 1863 VII 6 4 - - - -
Ectatomma tuberculatum (Olivier,
1792) VII - 1 4 2 5 3
Gnamptogenys gracili (Santschi, 1929) IV - - 5 4 3 4
Gnamptogenys horni (Santschi, 1929) IV - - 3 - 1 -
Gnamptogenys cf. sulcata (Smith,
1858) IV - - - - 1 -
FORMICINAE
Acropyga sp.1 II - - 1 1 5 -
Brachyrmymex sp.1 VII - - 3 13 7 7
Brachyrmymex sp.2 VII - - - - - 4
Camponotus nevagrande Mayr, 1870 IV - - - - 4 1
Camponotus pilosa (Olivier, 1792) IV 6 2 - - 6 -
Camponotus sp.3 IV 5 2 1 4 - 1
Camponotus sp.5 IV - 1 - - - -
Nylanderia sp.1 VII 2 11 25 25 14 25
Nylanderia sp.2 VII - - - - 1 3
Paratrechina sp.1 VII - 3 1 1 - -
MYRMICINAE
Acanthognathus cf. ocellatus Mayr,
1887 I - - - - 1 -
Apterostigma sp.1 X - - 2 - - -
Atta sexdens (Linnaeus, 1758) IX 2 1 - - 1 -
Carebara (grupo lignata) sp.1 II - 1 10 11 7 16
Carebara cf. brevipilosa Fernández,
2004 II - - 3 3 2 4
Cephalotes cf. maculatus (Smith, 1876) VI 1 - - - - -
Cephalotes cf. minutus (Fabricius,
1804) VI 2 1 1 - - -
Cephalotes cf. pusillus (Klug, 1824) VI - 2 - - - - Cephalotes cf. umbraculatus (Fabricius, 1804) VI - 1 - - - -

45
(Anexo, continuação...)
Locais de coleta
IFS MBV RCR
Subfamílias ou morfoespécies/grupos
funcionais
A
P
P
R
L
A
P
P
R
L
A
P
P
R
L
Crematogaster sp.1 VI 12 1 15 4 7 15
Crematogaster sp.2 VI 2 - - - - - Cyphomyrmex cf. rimosus (Spinola, 1851) X 13 - 7 2 - -
Cyphomyrmex sp.2 X - - 2 5 3 8
Hylomyrma cf. balzani (Emery, 1894) IV 2 - - - - -
Hylomyrma sp.1 IV - - 1 - - 1
Lachnomyrmex cf. victori Feitosa &
Brandão, 2008 IV - - - - - 1
Megalomyrmex sp.1 IV - - 4 3 - 2
Monomorium sp.1 VI - 1 - - - -
Mycocepurus cf. smithii (Forel, 1893) X - - - 4 - 4
Myrmicocrypta sp.1 X - - - - - 1
Nesomyrmex sp.1 IV 1 - - - - -
Nesomyrmex sp.2 IV - - - - - 1
Nesomyrmex sp.3 IV - - 1 - - -
Nesomyrmex sp.4 IV - 1 - - - -
Octostruma iheringi (Emery, 1888) I 4 - 4 3 8 3
Octostruma sp.2 I - - 2 1 - -
Pheidole sp.1 IV 7 3 22 21 14 24
Pheidole sp.2 IV 8 14 18 11 12 16
Pheidole sp.3 IV 22 3 6 6 7 12
Pheidole sp.4 IV 4 1 20 4 6 9
Pheidole sp.5 IV 16 3 5 1 - -
Pheidole sp.6 IV - - - - 20 5
Pheidole sp.7 IV - - 11 6 - 1
Pheidole sp.8 IV - - 2 17 - -
Pheidole sp.9 IV - - 1 - - 7
Pheidole sp.10 IV - - 1 - 4 1
Pheidole sp.11 IV - - 13 1 - -
Pheidole sp.12 IV - - - - 6 6
Pheidole sp.13 IV 6 - - - 1 -
Pheidole sp.14 IV 2 - - - 2 2
Pheidole sp.15 IV 3 - - - 1 -
Pheidole sp.16 IV 4 - - - - -
Pheidole sp.17 IV - - - - - 3
Pheidole sp.18 IV - - - - 1 1
Pheidole sp.19 IV 1 - - - - -
Pheidole sp.20 IV - - - - 1 -
Pheidole sp.21 IV - - - - 1 -
Pheidole sp.22 IV - - 1 - - -
Pheidole sp.23 IV - 2 - - - -
Rogeria cf. besucheti Kugler, 1994 IV 1 - - - - -
Rogeria cf. germaini Emery, 1894 IV - - - - 1 -
Sericomyrmex sp.1 X - - 6 7 6 4
Solenopsis sp.1 II 27 21 29 26 28 30
Solenopsis sp.2 II 13 6 16 18 20 25
Solenopsis sp.3 II 9 4 13 6 6 14
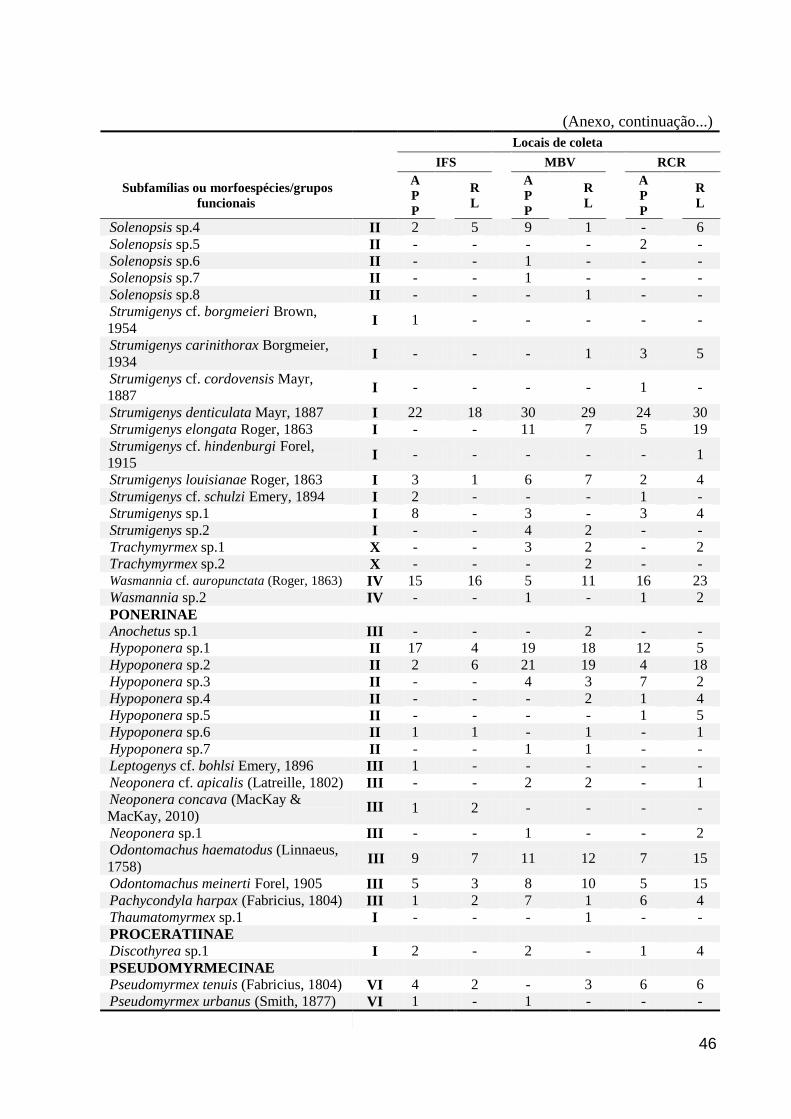
46
(Anexo, continuação...)
Locais de coleta
IFS MBV RCR
Subfamílias ou morfoespécies/grupos
funcionais
A
P
P
R
L
A
P
P
R
L
A
P
P
R
L
Solenopsis sp.4 II 2 5 9 1 - 6
Solenopsis sp.5 II - - - - 2 -
Solenopsis sp.6 II - - 1 - - -
Solenopsis sp.7 II - - 1 - - -
Solenopsis sp.8 II - - - 1 - -
Strumigenys cf. borgmeieri Brown,
1954 I 1 - - - - -
Strumigenys carinithorax Borgmeier,
1934 I - - - 1 3 5
Strumigenys cf. cordovensis Mayr,
1887 I - - - - 1 -
Strumigenys denticulata Mayr, 1887 I 22 18 30 29 24 30
Strumigenys elongata Roger, 1863 I - - 11 7 5 19
Strumigenys cf. hindenburgi Forel,
1915 I - - - - - 1
Strumigenys louisianae Roger, 1863 I 3 1 6 7 2 4
Strumigenys cf. schulzi Emery, 1894 I 2 - - - 1 -
Strumigenys sp.1 I 8 - 3 - 3 4
Strumigenys sp.2 I - - 4 2 - -
Trachymyrmex sp.1 X - - 3 2 - 2
Trachymyrmex sp.2 X - - - 2 - - Wasmannia cf. auropunctata (Roger, 1863) IV 15 16 5 11 16 23
Wasmannia sp.2 IV - - 1 - 1 2
PONERINAE
Anochetus sp.1 III - - - 2 - -
Hypoponera sp.1 II 17 4 19 18 12 5
Hypoponera sp.2 II 2 6 21 19 4 18
Hypoponera sp.3 II - - 4 3 7 2
Hypoponera sp.4 II - - - 2 1 4
Hypoponera sp.5 II - - - - 1 5
Hypoponera sp.6 II 1 1 - 1 - 1
Hypoponera sp.7 II - - 1 1 - -
Leptogenys cf. bohlsi Emery, 1896 III 1 - - - - -
Neoponera cf. apicalis (Latreille, 1802) III - - 2 2 - 1
Neoponera concava (MacKay &
MacKay, 2010) III 1 2 - - - -
Neoponera sp.1 III - - 1 - - 2
Odontomachus haematodus (Linnaeus,
1758) III 9 7 11 12 7 15
Odontomachus meinerti Forel, 1905 III 5 3 8 10 5 15
Pachycondyla harpax (Fabricius, 1804) III 1 2 7 1 6 4
Thaumatomyrmex sp.1 I - - - 1 - -
PROCERATIINAE
Discothyrea sp.1 I 2 - 2 - 1 4
PSEUDOMYRMECINAE
Pseudomyrmex tenuis (Fabricius, 1804) VI 4 2 - 3 6 6
Pseudomyrmex urbanus (Smith, 1877) VI 1 - 1 - - -
47
Grupos Funcionais: I - Crípticas predadoras; II - Crípticas onívoras; III - Epigéicas predadoras; IV - Epigéicas
onívoras; V - Arborícolas dominantes; VI - Arborícolas subordinadas; VII – Oportunistas; VIII - Formigas
nômades; IX - Attini cortadoras de folhas e; X - Attini não cortadoras de folhas.
(Anexo, continuação...)
Locais de coleta
Subfamílias ou morfoespécies/grupos
funcionais
IFS MBV RCR
A
P
P
R
L
A
P
P
R
L
A
P
P
R
L
Pseudomyrmex sp.2 VI - - - 1 - 2