universidade federal de goiÁs escola de veterinÁria e ... · entre as espécies domésticas de...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
O PAPEL DAS GONADOTROFINAS HIPOFISÁRIAS NO CICLO
ESTRAL EQUINO
Kele Amaral Alves
Orientadora: Profª Dra. Maria Lúcia Gambarini
Goiânia
2011

ii
KELE AMARAL ALVES
O PAPEL DAS GONADOTROFINAS HIPOFISÁRIAS NO CICLO
ESTRAL EQUINO
Seminário apresentado junto à
Disciplina Seminários Aplicados do
Programa de Pós-Graduação em
Ciência Animal da Escola de Veterinária
da Universidade Federal de Goiás
Nível: Doutorado
Área de Concentração: Produção Animal
Linha de pesquisa: Biotecnologia e eficiência reprodutiva animal
Orientadora Profª. Dra. Maria Lúcia Gambarini – UFG Comitê de Orientação Prof. Dr. José Octavio Jacomini – UFU Prof. Dr. Benedito Dias de O. Filho – UFG
GOIÂNIA
2011

iii
SUMÁRIO
1 INTRODUÇÃO.............................................................................................. 01
2 REVISÃO DE LITERATURA......................................................................... 03
2.1 Composição e Mecanismos de Produção ................................................. 03
2.2 Mecanismos de ação................................................................................. 06
2.3 Mecanismos de Regulação e Controle...................................................... 10
2.4 Contribuição Biotecnológica....................................................................... 12
3 CONSIDERAÇÕES FINAIS…………………………………………………….. 14
REFERÊNCIAS ........................................................…………………………... 15

iv
LISTA DE FIGURAS
FIGURA 1 – Concentração plasmática de melatonina de acordo com o
período do dia, evidenciando o pico de produção no período noturno. .......... 03
FIGURA 2 – Ilustração anatômica dos vasos portais entre hipotálamo e
hipófise anterior e associação entre células neurossecretoras do
hipotálamo, células endócrinas da hipófise, hormônios liberadores e
inibidores do hipotálamo e hormônios da hipófise. ......................................... 04
FIGURA 3 - Foto hemisfério cerebral equino, comprovando a ligação entre
hipotálamo e hipófise.......................................................................................... 05
FIGURA 4 - Microfotografias das zonas de projeções citoplasmáticas em
folículo ovariano .............................................................................................. 07
FIGURA 5 - Ilustração dos aspectos das concentrações hormonais durante
o desvio usando um modelo de dois folículos................................................. 09
FIGURA 6 - Folículo pré-ovulatório equino. O colchete evidencia o ápice
folicular com a parede delgada. As setas brancas demonstram a parede
espessa fora do ápice...................................................................................... 10
FIGURA 7 - Imagens ultrassonográficas de folículos equinos dominantes
com Doppler colorido, demonstrando a vascularização de folículos
dominantes equinos......................................................................................... 12

v
LISTA DE TABELAS
TABELA 1 – Número de ovócitos equinos maturados in vitro por 24 horas com gonadotrofinas de origem suína, classificados de acordo com o estágio de maturação nuclear...................................................................................... 13

1. INTRODUÇÃO
A espécie equina acompanha o histórico evolutivo da humanidade
mesmo não tendo a produção de alimentos como característica principal de sua
criação. Entre as espécies domésticas de grande porte os cavalos são os únicos
que podem ser considerados animais de companhia na cultura ocidental.
Os aspectos relacionados a equideocultura revelam diversas
particularidades da espécie, desde nutricionais, cicatriciais, comportamentais e
reprodutivos sendo que este último tem desempenho ínfimo quando se trata de
produtividade, rendendo o título de menor fertilidade entre as espécies
domésticas para as éguas. Esta informação ressalta a necessidade de melhor
entendimento de todos os fatores envolvidos com a fisiologia reprodutiva desta
espécie.
Estas fêmeas são poliéstricas estacionais e exibem atividade ovariana
máxima durante a primavera e verão. Nos meses de inverno, essa atividade é
reduzida e conhecida como anestro sazonal (HANS, 1986). A causa determinante
deste comportamento é a duração do período luz/dia, mas outros fatores como
nutrição, temperatura e estado sanitário também podem alterar os padrões de
ciclicidade (ANDRADE, 1986). Em resposta ao aumento da luminosidade diária,
ocorre o início da estação ovulatória nas éguas, pois receptores presentes no
nervo óptico estão ligados ao hipotálamo que estimula a glândula hipófise e
executa modificações na secreção de gonadotrofinas hipofisárias (hormônio
folículo estimulante e hormônio luteinizante) as quais estimularão os ovários
(GINTHER, 1992).
PALMER & JOUSSET (1975), descreveram o ciclo estral da égua com
duração média 21 dias, sendo 14 dias de diestro (fase luteal) e sete dias de estro
(fase folicular). Porém, devido a duração da fase folicular (dois a 15 dias), existe
grande variação entre os ciclos, mais longos durante o período transicional de
primavera e mais curtos no final da estação ovulatória (GINTHER, 1974). O efeito
de altos níveis de estrógeno sistêmico associados com o desenvolvimento dos
folículos determina o comportamento característico do cio na fase folicular. Por
outro lado, o comportamento de diestro é atribuído aos altos níveis sistêmicos de

2
progesterona associados com o desenvolvimento do corpo lúteo, caracterizando a
fase luteal (BERGFELT, 2000).
O processo contínuo de crescimento e regressão folicular nos ovários
dos mamíferos é denominado dinâmica folicular, que ocorre em forma de ondas
foliculares sincronizadas, recrutando grupos de folículos sensíveis ao hormônio
folículo estimulante (FSH), seguido pela seleção do folículo dominante em
detrimento dos demais que entram no processo de atresia. O folículo dominante
normalmente continua crescendo e secreta elevada quantidade de estrógeno e
quando sensibilizado pelo padrão de liberação do hormônio luteinizante (LH)
responde ao estímulo ovulatório (DRIANCOURT, 2001).
Os fatores intrafoliculares, tais como fatores de crescimento, peptídeos,
receptores de gonadotrofinas, fatores angiogênicos e esteróides, determinam o
controle do desvio folicular. As gonadotrofinas produzidas na hipófise exercem
ações fundamentais na vida reprodutiva das éguas, seja estimulando o
crescimento folicular (FSH) ou a ovulação (LH) (KOHEK & LATRONICO, 2001).
O sucesso da manipulação do ciclo estral e a implementação de
biotecnologias reprodutivas dependem do esclarecimento dos mecanismos de
desenvolvimento folicular, assim como da seleção do folículo dominante, com o
intuito de propiciar a multiplicação de indivíduos zootecnicamente superiores.
Este seminário revisou os trabalhos sobre mecanismos de produção,
liberação, ação e utilização das gonadotrofinas hipofisárias relacionados
diretamente com o ciclo ovariano na espécie equina.

3
2 REVISÃO DE LITERATURA
2.1 Composição e Mecanismos de Produção
O ciclo anual previsto de alteração na duração dos dias (fotoperíodo) é
o principal sinal de adaptação estacional, e permite que os organismos
expressem respostas adaptativas em antecipação a mudanças ambientais
favoráveis. Nos mamíferos a informação fotoperiódica está relacionada com a
secreção do hormônio melatonina pela glândula pineal, o qual tem pico produtivo
no período noturno (Figura 1) e atua no sistema neuroendócrino, produzindo
mudanças na endocrinologia, anatomia e fisiologia, afetando o comportamento, o
balanço energético e o sistema reprodutivo (MORGAN & HAZLERIGG, 2008).
Figura1- Concentração plasmática de melatonina de acordo com o período do dia,
evidenciando o pico de produção no período noturno.
Fonte: ARENDT (1988).

4
O solstício de verão traz dias com incidência luminosa maior, a qual, na
espécie equina, incide pela rotina ocular, estimula seus receptores, conectando
com a glândula pineal e inibindo a síntese do hormônio melatonina, o que
funciona como estímulo positivo das atividades hipotalâmicas (HAFEZ et al.,
1995). O hipotálamo é o órgão responsável por converter os sinais neurológicos
originados em estímulos externos e internos em descargas hormonais, e um de
seus produtos é o GnRH (hormônio liberador de gonadotrofinas hipofisárias)
composto peptídico com 10 aminoácidos (GIGLI et al., 2006). O GnRH é
armazenado na eminência média até que ocorra uma despolarização neuronal e o
hormônio entre nos capilares fenestrados e chegue a hipófise via vasos do
sistema portal (Figura 2) (HART et al., 1984).
Figura 2- Ilustração anatômica dos vasos portais entre hipotálamo e hipófise
anterior e associação entre células neurossecretoras do hipotálamo, células
endócrinas da hipófise, hormônios liberadores e inibidores do hipotálamo e
hormônios da hipófise.
Fonte: http://wwwmomalu.blogspot.com/

5
A ligação entre hipotálamo hipófise, demonstrada na figura 3, tem
papel importante por permitir que quantidades mínimas de hormônios exerçam
ações diretamente nas células da hipófise anterior, antes de se diluírem na
circulação sistêmica (SENGER, 2003)
Figura 3- Foto hemisfério cerebral equino, comprovando a ligação entre
hipotálamo e hipófise.
Fonte:Departamento de Anatomia Animal, Faculdade de Zootecnia de
Uberaba
As células endócrinas da hipófise anterior recebem o estímulo dos
receptores de GnRH e iniciam a produção das gonadotrofinas hipofisárias FSH
(hormônio folículo estimulante) e LH (hormônio luteinizante), que são compostos
glicoprotéicos com mesmo peso molecular e com duas subunidades diferenciais,
designadas como α e β. A subunidade α é específica de cada espécie e é
essencialmente igual em todos os hormônios hipofisários. Em contraste, a
subunidade β confere a cada substância sua função biológica específica
(MCKINNON & VOSS, 1993).
IRVINE & ALEXANDER (1988) relatam que os pulsos de secreção
gonadotrófica não podem ser detectados em amostras sanguíneas periféricas,

6
devido a longa meia vida circulatória das gonadotrofinas manter um pool contínuo
das mesmas no sangue, mascarando os pulsos individuais. Os mesmos autores
em 1993, afirmam que a secreção de FSH na égua pode ser uni ou bimodal,
sendo que o padrão bimodal pode ser observado somente no período de
transição da primavera e o início da temporada reprodutiva com um primeiro pico
no final do estro e outro na metade do diestro. Já o padrão unimodal ocorre em
plena temporada reprodutiva e o pico único só ocorre na metade do diestro.
O LH tende a ter um padrão bimodal de secreção no início da estação
reprodutiva com um pulso a cada dois dias. A concentração sérica aumenta nos
intervalos pré-ovulatórios, atingindo os níveis necessários para desencadear a
ovulação (IRVINE et al., 2000).
2.2 Mecanismos de Ação
O ciclo funcional ovariano depende totalmente das gonadotrofinas
hipofisárias, uma vez que a remoção da hipófise causa depressão da fertilidade e
leva os ovários a um estágio juvenil (MCKINNON & VOSS, 1993).
Os mecanismos que controlam a ativação dos folículos permanecem
pouco elucidados, principalmente em animais de interesse zootécnico (BINELLI et
al., 2009).A função do folículo ovariano é proporcionar um ambiente ideal para a
manutenção da viabilidade, bem como, o crescimento e maturação do ovócito
(GONÇALVES et al., 2001).
De acordo com VAN DEN HURK & ZHAO (2005), a partir do momento
em que ocorre a formação do antro, os folículos passam a ser denominados
terciários ou antrais. Este fluido se origina nos folículos equinos de 0,2 a 0,4 mm
de diâmetro e pode servir como fonte de substâncias regulatórias derivadas do
sangue e de secreções das células foliculares.
O hormônio folículo estimulante é necessário para o recrutamento dos
folículos antrais. Sob a influência desta gonadotrofina as células da granulosa se
multiplicam por mitose o que aumenta o número de camadas ao redor do ovócito
e consequentemente a quantidade de fluido produzido pelas mesmas. As células
da teca interna dos folículos terciários respondem ao estímulo do LH produzindo
andrógenos e quando o FSH estimula a ativação da enzima aromatase nas

7
células da granulosa ocorre a transformação dos andrógenos em estradiol
(HAFEZ , 2004).
O FSH também está envolvido no aumento da vascularização do
folículo dominante, o que permite uma maior captação de nutrientes, além de
estimular a formação da cavidade antral e a expressão dos receptores para LH
nas células da granulosa do folículo pré-ovulatório (SIROIS & FORTUNE, 1990).
Segundo ALBERTINI et al. (2001), a proliferação e diferenciação das
células da granulosa influencia diretamente a maturação do ovócito com a
secreção de mediadores parácrinos, e através de extensões destas células na
zona pelúcida, denominadas processos citoplasmáticos transzonais (TZP) onde
junções do tipo “gap” permitem transporte bidirecional de moléculas reguladoras
as quais interpretam as mensagens geradas pelos receptores gonadotróficos
(Figura 4).
Figura 4- Microfotografias das zonas de projeções citoplasmáticas em folículo
ovariano.
Fonte: ALBERTINI et al. (2001)
A função do LH é reiniciar a meiose do ovócito no folículo pré-
ovulatório onde se une a receptores de membrana nas células da teca e da
granulosa para desencadear a ovulação e controlar o desenvolvimento e
manutenção do corpo lúteo. Este hormônio produz um aumento do AMPc por

8
meio da estimulação da enzima adenil ciclase, promovendo a conversão do
colesterol em pregnolona que participa ativamente da ovulação (GINTHER, 1992).
O hormônio luteinizante da égua tem uma maior concentração de carboidratos e
de ácido siálico o que o torna diferente das demais espécies (BOUSFIELD et al.,
1987). Nestas fêmeas, as elevadas concentrações de LH estão temporariamente
associadas com o desvio e representadas por uma parte distinta do grande pico
pré-ovulatório do hormônio luteinizante nestas espécies (GASTAL et al., 1997).
Assim, essa primeira elevação dos índices de LH é seguida pela expressividade
de seus receptores nas células da granulosa e representa um dos eventos iniciais
para a sequência que leva ao desvio durante o declínio do FSH (GINTHER et
al.,2001).
O termo onda folicular compreende um conjunto de fenômenos
foliculares que obedecem a sequência de recrutamento, seleção, dominância,
ovulação ou atresia. O estudo realizado por GINTHER et al. (2003) demonstrou
que o recrutamento folicular na égua ocorre em dois tipos de ondas: maiores
(desenvolvimento de folículos dominantes e subordinados) e menores (folículos
maiores não atingem diâmetro para dominância). As concentrações de LH são
maiores nas ondas maiores, suportando a idéia de que o folículo dominante
necessita de um estímulo adicional desta gonadotrofina
Após a emergência, os folículos de uma onda entram em fase de
comum de crescimento, envolvendo em média de 7-11 folículos. O surgimento de
cada onda é temporariamente associada com o surgimento do FSH e o seu pico
de concentração ocorre quando o folículo maior atinge, aproximadamente 13 mm
de diâmetro, para depois começar a declinar (GINTHER, 2005).
O crescimento paralelo dos folículos finaliza quando o maior folículo
alcança um diâmetro de 22 mm e o segundo maior alcança 19 mm, a partir de
então, o maior folículo geralmente torna-se dominante e continua a crescer
enquanto os demais se tornam subordinados e apresentam uma interrupção ou
redução do crescimento, caracterizando o fenômeno de desvio folicular (GASTAL
et al., 1999).
O mecanismo pelo qual ocorre a dominância folicular é denominado
seleção e exerce um efeito inibitório indireto sobre as estruturas subordinadas,

9
ocorrendo no fim da fase de crescimento quando o folículo dominante cresce
continuadamente e os demais regridem (BEG & GINTHER, 2006).
Figura 5- Ilustração dos aspectos das concentrações hormonais durante o desvio
usando um modelo de dois folículos.
Fonte: GINTHER et al. (2001)
A ovulação é definida como ponto auge de uma série de mecanismos
bioquímicos, fisiológicos e morfológicos complexos desencadeados pela elevação
do LH, e tem como resultado a liberação do ovócito. Atualmente os processos
ovulatórios aceitos são de contração muscular e o mecanismo bioquímico.
Segundo ESPEY (1994), a contração é produzida por fibras de
colágeno e musculares presentes na teca folicular externa, mas os eventos que
coordenam a ação das enzimas proteolíticas e as trocas vasculares são
evidentemente mais importantes. O aumento pré-ovulatório do LH aumenta a
expressão das enzimas proteolíticas plasmina e procolagenase. A plasmina
aumenta a ativação do plasminogênio e a procolagenase se converte em
10
colagenase, e ambas irão romper o colágeno da teca externa, diminuindo a
tensão da parede folicular. A medida que acontece a degradação do tecido
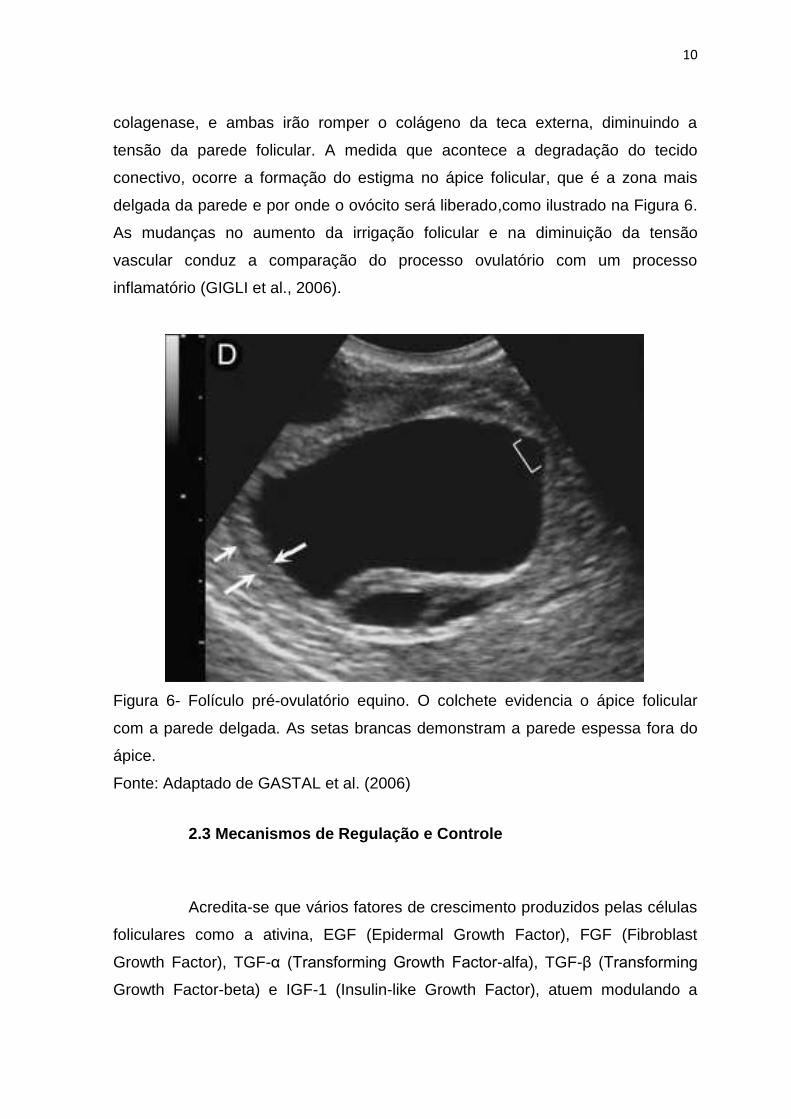
conectivo, ocorre a formação do estigma no ápice folicular, que é a zona mais
delgada da parede e por onde o ovócito será liberado,como ilustrado na Figura 6.
As mudanças no aumento da irrigação folicular e na diminuição da tensão
vascular conduz a comparação do processo ovulatório com um processo
inflamatório (GIGLI et al., 2006).
Figura 6- Folículo pré-ovulatório equino. O colchete evidencia o ápice folicular
com a parede delgada. As setas brancas demonstram a parede espessa fora do
ápice.
Fonte: Adaptado de GASTAL et al. (2006)
2.3 Mecanismos de Regulação e Controle
Acredita-se que vários fatores de crescimento produzidos pelas células
foliculares como a ativina, EGF (Epidermal Growth Factor), FGF (Fibroblast
Growth Factor), TGF-α (Transforming Growth Factor-alfa), TGF-β (Transforming
Growth Factor-beta) e IGF-1 (Insulin-like Growth Factor), atuem modulando a

11
ação das gonadotrofinas FSH e LH, controlando a foliculogênese,
esteroidogênese e atresia folicular (LEITÃO, 2009).
A inibina atua diminuindo a síntese e a liberação de FSH. Seu efeito na
secreção do hormônio folículo estimulante é lento, enquanto a redução na síntese
do mesmo sofre ação rápida (MCKINNON & VOSS, 1993). Estudos realizados por
DONADEU & GINTHER (2002) apontaram um aumento considerável na
concentração de inibina nos folículos maiores entre 16 e 19mm o que sugere a
participação no desvio folicular e no subsequente crescimento diferencial do
folículo dominante.
A principal função da ativina na regulação do hormônio folículo
estimulante circulante pode ser por aumentar a quantidade disponível deste
hormônio na hipófise anterior pronto para liberação. Esta glicoproteína induz a
formação de células da granulosa, aumentando a expressão dos receptores de
FSH. Outra glicoproteína isolada do fluido folicular eqüino com atividade tanto
inibitória da secreção de FSH como de neutralização da ativina, constituindo um
grande modulador da ação do FSH (BEG & GINTHER, 2006).
O estradiol aumenta a atividade da enzima aromatase nas células da
granulosa, promovendo a expressão de receptores de gonadotrofinas,
aumentando a sensibilidade do folículo ao LH (GURGEL et al. 2008). Os estudos
de GASTAL et al. (1999) relatam que a eliminação do folículo mais desenvolvido,
resulta em diminuição dos índices de estradiol e inibina e aumento nos níveis de
FSH, relacionando o controle desta gonadotrofina ao estradiol.
De acordo com ZULU et al. (2002), o fator semelhante a insulina IGF-1
interage com o FSH no estímulo da produção de estradiol pelas células da
granulosa, resultando em sinergismo com as gonadotrofinas hipofisárias para
estimular o desenvolvimento folicular. Da mesma forma, o fator de crescimento de
endotélio vascular promove a angiogênese nos folículos dominantes,
proporcionando suporte preferencial de nutrientes, gonadotrofinas e fatores de
crescimento aos mesmos (DONADEU & WATSON 2007) Utilizando a técnica de
Doppler colorido GASTAL (2006) evidenciou o aumento do fluxo sanguíneo nos
maiores folículos após a seleção (Figura 7).

12
Figura 7- Imagens ultrassonográficas de folículos equinos dominantes com
Doppler colorido, demonstrando a vascularização de folículos dominantes
equinos.
Fonte: GASTAL et al. (2006)
2.4 Contribuição biotecnológica
Considerando o extenso papel das gonadotrofinas hipofisárias no
processo de desenvolvimento folicular e maturação ovocitária in vivo, deduz-se
que sua participação nos processos de diversas biotecnologias reprodutivas, seja
imprescindível.
BORGES et al. (1998) utilizaram diferentes doses de FSH, LH ou
ambos combinados em meio de maturação in vitro de ovócitos equinos e conclui
que tanto a associação FSH/LH como somente o FSH, proporcionaram uma taxa
de maturação nuclear ovocitária significativamente maior que o grupo controle
(Tabela 1). Em concordância, DELL’ ÁQUILA et al. (2004) testaram o efeito da

13
presença de FSH na maturação nuclear de cumulus oóforos equinos e relatam
maior expansão das estruturas na presença da gonadotrofina.
Tabela 1- Número de ovócitos equinos maturados in vitro por 24 horas com gonadotrofinas de origem suína, classificados de acordo com o estágio de maturação nuclear.
Tratamentos n MII/TI MI/AI QVG VG SE/DEG
n % n % n % n % n %
Controle 98 21 22,8a
35 38,0 12 13,0 6 6,0 18 20,0
LH 108 35 32,4a
41 38,0 9 8,0 9 8,0 13 12,0
FSH 102 55 53,9b
23 23,0 8 8,0 4 4,0 12 12,0
LH/FSH 106 59 55,6b
23 22,0 5 5,0 6 6,0 13 12,0
Letras diferentes na mesma coluna (p<0,05) MII/TI – Metáfase II / Telófase I MI/AI – Metáfase I / Anáfase I QVG – Quebra da vesícula germinativa VG – Vesícula germinativa SEG/DEG – Sem estrutura / Degenerados Adaptado de BORGES et al. (1998).
Conforme ALMEIDA et al. (2010), a transferência de embriões tem
impacto no agronegócio do cavalo, com alteração de preços dos animais, no
entanto, a técnica ainda é relativamente ineficiente na espécie equina em relação
a bovina, devido a dificuldades de superovulação das éguas. Vários hormônios
tem sido testados a fim de melhorar os índices de coleta de embriões, dentre os
quais se destacam o FSH e o extrato de pituitária equina (EPE) com resultados
mais consistentes (LIRA et al., 2009).
O estudo de BONIN et al (2010) destacam o EPE como precursor de
gonadotrofinas hipofisárias e sua aplicação exógena aumenta a incidência de
múltiplas ovulações em éguas e incrementou a taxa de recuperação embrionária
por ciclo.
A janela de sincronização entre a ovulação da doadora e da receptora
equina é entre +1 e -3 dias e constitui uma técnica realizada de maneira
relativamente simples em éguas cíclicas. Em todos os protocolos, monitora-se o

14
crescimento folicular por ultrassonografia e utiliza-se precursores de FSH e LH
(GnRH ou EPE) para induzir a ovulação nas receptoras dentro de 48 horas da
inseminação da doadora (BERGFELT, 2000).
3. CONSIDERAÇÕES FINAIS
A fisiologia reprodutiva das éguas é composta de esquemas complexos
e integrados e que ainda possuem alguns pontos inexplicados ligados direta ou
indiretamente com as gonadotrofinas hipofisárias.
Os artigos científicos revisados esclarecem os aspectos de produção e
função destas gonadotrofinas, mas os aspectos relacionados aos receptores das
mesmas e suas características durantes as fases de desenvolvimento folicular e
maturação in vitro intrigam a curiosidade científica, levantando a hipótese de estar
neles a resposta para a baixa eficiência das fêmeas equinas em programas
reprodutivos.

15
REFERÊNCIAS 1.ALBERTINI, D. F., COMBELLES, C. M. H., BENECCHI, E., CARABATSOS, M.J. Cellular basis for paracrine regulation of ovarian follicle development. Reproduction, v. 121, p. 647-653, 2001. 2.ALMEIDA, F. Q., SILVA, V. P. Progresso científico em equideocultura na 1ª década do século XXI. Revista Brasileira De Zootecnia, v. 39, p. 119-129, 2010. 3.ANDRADE, L. S. Fisiologia e manejo da reprodução equina. Ed.Recife. 1986, 338p. 4.BEG, M. A., GINTHER, O, J. Follicle selection in cattle and horses: role of intrafollicular factors. Reproduction Review, v. 132, p. 365-377, 2006. 5.BERGFELT, D. R. Estrous synchronization. In: SAMPER, J.C. Equine breeding management and artificial insemination. Philadelphia: Saunders, Cap.12, p.169-170, 2000. 6.BINELLI, M., PORTELA, V. M., MURPHY, B. D. Dinâmica ovariana e eficiência reprodutiva: estado da arte. Congresso Brasileiro de Reprodução Animal, 18, 2009, Belo Horizonte, MG. Anais ... Belo Horizonte: CBRA, 2009. 7.BONIN, B. F., DELL’AQUA, J. A. J., FIORATTI, E. G., ALVARENGA, M. A. Efeito do tratamento com extrato de pituitária equina na resposta ovariana e eficiência reprodutiva de éguas idosas em programa de transferência de embriões. Revista Veterinária e Zootecnia, v. 17, p. 94-103, 2010. 8.BORGES, J. M. L., RUBIN, M. I. B., SILVA, C. A. M., GONÇALVES, P. B. D.,RIECK, A. C. Influência das gonadotrofinas na regulação da maturação nuclear de oócitos equinos. Ciência Rural, v.28, p.293-297, 1998. 9.BOUSFIELD, G. R., LIU, W. K., SUGINO, H., WARD, D. N. Structural studies on equine glycoprotein hormones. The Journal of Biological Chemistry, v. 262, p. 8610-8620, 1987. 10.DELL’ÁQUILA, M. E., CAILLAUD, M., MARIATO, F., MARTORIATI, A., GERARD, N., AWDI, G., MINOIA, P., GOUDET, G. Cumullus expansion, nuclear maturation on conexin 43 ciclooxigenase-2 and FSH receptor mRNA expression in equine cumullus-oocyte cultured in the presence of FSH and precursors for hialuronic acid synthesis. Reproductive Biology and Endocrinology, v. 2, p. 1-13, 2004. 11.DONADEU, F. X., GINTHER, O. J. Follicular waves and circulating concentration of gonadotrophins, inhibin and oestradiol during the anovulatory season in mares. Reproduction, v. 124, p. 875-885, 2002.

16
12.DONADEU, F. X., WATSON, E. D. Seasonal changes in ovarian activity: lessons learnt from the horse. Animal Reproduction Science, v. 100, p. 225-242, 2007. 13.DRIANCOURT, M. A. Regulation of ovarian follicular dynamics in farm animals: implications for manipulation of reproduction. Theriogenology, v. 55, p. 1211-1239, 2001. 14.ESPEY, L. L. Current status of the hipothesis that mammalian ovulation in comparable to an inflammatory reaction. Biology of Reproduction, v. 50, p. 233-238, 1994. 15.GASTAL, E. L., GASTAL, M. O., BERGFELT, D. R., GINTHER, O. J. Role of diameter differences among follicles in selection of a future dominant follicle in mares. Biology of Reproduction, v. 57, p. 1320-1327, 1997. 16.GASTAL, E. L., GASTAL, M. O., WILTBANK, M., GINTHER, O. J. Follicle deviation and intrafollicular and systemic estradiol concentration in mares. Biology of Reproduction, v. 61, p. 31-39, 1999. 17.GASTAL, E. L., GASTAL, M. O., GINTHER, O. J. Relationships of changes in B-mode echotexture and colour Doppler signal in the wall of the preovulatory follicle to changes in systemic oestradiol concentrations and the effects of human chorionic gonadotrophin in mares. Reproduction Research, v. 131, p. 699-709, 2006. 18.GIGLI, I., RUSSO, A., AGUERO, A. Consideraciones sobre La dinâmica ovárica em equino, bovino e camelydos sudamericanos. In Veterinary, v. 8, p. 183-204, 2006. 19.GINTHER, O. J. Occurrence of anoestrous, estrus, diestrus and ovulation over 12 monthperiod in mares. American Journal of Veterinary Research, v. 35, p. 1173-1179, 1974. 20.GINTHER, O. J. Reproductive biology of the mare: basic and applied aspects, 2ed, Wisconsin: Equiservices Publishing. 1992. 642p. 21.GINTHER, O. J., BEG, M. A., BERGFELT, D. R., DONADEU, F. X., KOT, K. Follicle selection in monovular species. Biology of Reproduction, v. 65, p. 638-647, 2001. 22.GINTHER, O. J., BEG, M. A., DONADEU, F. X., BERGFELT, D. R. Mechanism of follicle deviation in monovular farm species. Animal Reproduction Science, v. 78, p. 239-257, 2003. 23.GINTHER, O. J., GASTAL, E. L., GASTAL, M. O., BEG, M. A. Regulation of circulating gonadotropins by the negative effects ovarian hormones in mares. Biology of Reproduction, v. 73, p. 315-323, 2005.

17
24.GONÇALVES, P. B. D.; FIGUEIREDO, J. R.; FREITAS, V. J. F. Biotécnicas Aplicadas à Reprodução Animal. São Paulo: Editora Varela, 2001. 340p. 25.GURGEL, J. R. C., VIANA, C. H. C., PEREZ, E. G. A., NICHI, M. Dinâmica follicular em éguas: aspectos intrafoliculares. Revista Brasileira de Reprodução Animal, v. 32, p. 122-132, 2008. 26.HAFEZ, E. S. E. Reprodução Animal. 6.ed. São Paulo: Manole, 1995, 582p. 27.HAFEZ, E. S. E. Reprodução Animal. 7.ed. São Paulo: Manole, 2004, 513p. 28.HANNS, J. W. Equine diseases: a textbook for students and practitioners. Berlin: Paul Parey, 1986, 48p. 29.HART, P. J., SQUIRES, E. L., IMEL, K. J., NETT, T. M. Seasonal variation in hypothalamic content of gonadotropin release hormone (GnRH), pituitary receptors for GnRH and pituitary content of luteinizing hormone and follicle stimulating hormone in the mare. Biology of Reproduction, v. 30, p. 155-162, 1984. 30.IRVINE, C. H. G., ALEXANDER, S. L. Secretion rates on short term patterns of gonadotropin releasing hormone, FSH and LH in the normal stallion in the breeding season. Journal of Endocrinology, v. 117, p. 197-206, 1988. 31.IRVINE, C. H. G., ALEXANDER, S. L., MCKINNON, A. O. Reproductive hormone profiles in mares during the autumn transition as determined by collection of jugular blood at 6h intervals throughout ovulatory and anovulatory cycles. Journal of Reproduction and Fertility, v. 118, p. 101-109, 2000. 32.KOHEK, M. B. F., LATRONICO, A. C. O papel dos receptores das gonadotrofinas na reprodução feminina. Arquivo Brasileiro de Endocrinologia Metabólica, v. 45, p. 369-374, 2001. 33.LEITÃO, C. C. F., BRITO, I. R., FROTA, I. M. A., SILVA, J. R. V. Importância dos fatores de crescimento locais na regulação da foliculogênese ovariana em mamíferos. Acta Scientiae Veterinariae, v. 37, p. 215-224, 2009. 34.LIRA, R. A.; PEIXOTO, G. C. X., SILVA, A. R. Transferência de embrião em equinos: revisão. Acta Veterinaria Brasilica, v. 3, n. 4, p.132-140, 2009. 35.MCKINNON, A. O., VOSS, J. L. Equine Reproduction. 1ed. Philadelphia: Lea e Febiger, 1993, 1137p. 36.MORGAN, P. J., HAZLERIGG, D. G. Photoperiodic signaling through the melatonin receptor turns full circle. Journal of Neuroendocrinology, v. 20, p. 820-826, 2008.

18
37.MURPHY, B. A., MARTIN, A. M., FURNEY, P., ELLIOT, J. Absence of a serum melatonin rhythm under acutely extended darkness in the horse. Journal of Circadian Rhythms, v. 9, p. 1-8, 2011. 38.PALMER, E., JOUSSET, B. Urinary oestrogen and plasma progesterone levels in non pregnant mares. Journal of Reproduction and Fertility, v. 23, p. 213-221, 1975. 39.SENGER, P. L. Pathways to Pregnancy and Parturition. 2ed. Washington: Current Conceptions, 2003, 368p. 40.SIROIS, J., FORTUNE, J. E. Lengthening the bovine estrous cycle with low levels of exogenous progesterone: A model for studying ovarian follicular dominance. The Endocrine Society, v. 127, p. 159-166, 1990. 41.VAN DEN HURK, R., ZHAO, J. Formation of mammalian oocytes and their growth, differentiation and maturation within ovarian follicles. Theriogenology, v. 63, p. 1717–1751, 2005. 42.ZULU, V. C., NAKAO, T., SAWAMUKAI, Y. Insulin growth factor I as a possible hormonal mediator of nutritional regulation of reproduction in cattle. Journal of Veterinarian Medicine Science, v. 64, p. 657-665, 2002.