mestrado 2012 - ri.ufs.br · direções, ocorrendo, então, uma grande competição pelo mesmo...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SERGIPE
PALEOFAUNA E PALEOAMBIENTES DO PLEISTOCENO
SUPERIOR NO MUNICÍPIO DE JAGUARARI, NORTE DA BAHIA
MÁRCIA CRISTINA TELES XAVIER
MESTRADO 2012

ii
UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
NÚCLEO DE PÓS-GRADUAÇÃO EM ECOLOGIA E CONSERVAÇÃO
MESTRADO EM ECOLOGIA E CONSERVAÇÃO
PALEOFAUNA E PALEOAMBIENTES DO PLEISTOCENO
SUPERIOR NO MUNICÍPIO DE JAGUARARI, NORTE DA BAHIA
AUTORA: MÁRCIA CRISTINA TELES XAVIER
ORIENTADOR: PROF. DR. ADAUTO DE SOUZA RIBEIRO
CO-ORIENTADOR: PROF. DR. MARIO ALBERTO COZZUOL
SÃO CRISTOVÃO, SERGIPE
FEVEREIRO, 2012

iii
FICHA CATALOGRÁFICA ELABORADA PELA BIBLIOTECA CENTRAL
UNIVERSIDADE FEDERAL DE SERGIPE
Xavier, Márcia Cristina Teles
X3p Paleofauna e paleoambientes do pleistoceno superior no
município de Jaguarari, norte da Bahia / Márcia Cristina
Teles Xavier ; orientador Adauto de Souza Ribeiro. – São
Cristóvão, 2012.
110 f. ; Il.
Dissertação (mestrado em Ecologia e Conservação)–
Universidade Federal de Sergipe, 2012.
O
1. Reconstrução paleoambiental. 2. Megafauna. 3.
Matéria orgânica do solo. 4. Carvão - Fragmentos. 5.
Educação patrimonial I. Ribeiro, Adauto de Souza, orient.
II. Título.
CDU: 551.583.7

iv

v
Agradecimentos
Hoje as palavras fogem e é com grande emoção que escrevo os agradecimentos,
última parte do meu trabalho. Gostaria de agradecer a todos que estiveram presentes neste
processo, especialmente:
A meu bom Deus, por ter me permitido chegar até aqui e ter proporcionado todo
equilíbrio para concluir mais esta etapa da minha vida;
A minha família, que soube suportar a saudade, distância e por transmitir doses
diárias de coragem e carinho;
Ao meu querido fêssor Adauto, por ter aceitado orientar o meu trabalho científico e
durante todo o curso, teve um carinho e atenção muito maior que professor/orientador. Foi
em todas as caronas, conversas, conselhos e até nas broncas que encontrei um amigo, um
parceiro e até o acolhimento de um pai. Fêssor meu muito obrigada, como sempre disse, o
senhor é meu orgulho!
Ao Mário Cozzuol que me recebeu tão bem em seu laboratório e partilha os sonhos
futuros;
Ao fêssor Cláudio Lisi por toda dedicação e acreditar tanto em mim. Por ter
abraçado meu projeto e trabalhado com tanto afinco, por me apresentar o fascinante mundo
da dendroecologia e muitas vezes, por ter sido meu orientador/coorientador... Eu prometo
que darei uma réplica da manchadinha, rsrs;
Aos professores revisores que contribuíram com as correções/sugestões para uma
melhor qualidade do trabalho: Kleberson Porpino, Edison Oliveira, Maria Helena Zucon,
Renato Faria, Cláudio Lisi, Susy Gouveia, Edilma Andrade, Maria Angélica;
A minhas queridas amigas Fran e Chris, por ter sido a minha família fora de casa e
ter compartilhado comigo as suas famílias. Sem elas tudo seria mais difícil. Se nada desse
certo, só o fato de ter conhecido vocês já teria valido a pena;
Ao Bruno, colega de orientação, companheiro nas mesmas angustias e
preocupações, por todas as risadas e momentos de “lavar a roupa suja”, rsrs;
A Érica pela amizade construída, por partilhar muitos momentos bons, por todas as
conversas (aquelas de caráter crítico) e por ser esta baiana arretada. Só uma baiana pra
entender outra baiana rsrs;

vi
Aos amigos de turma, especialmente Anny, Fábio, Higor Vieira, Daniel por todas
as “alterações”;
Aos amigos de laboratório, Patrício, Taiguã, Raone, Herlânia, Mônica, Jefferson,
Carolzinha, Douglas, Renato, Mariana, Francisco, Leonardo, Nayane, Ana Paula, Rodrigo,
Lucas, Priscila, Marcelo, Daniel, por todos os cafés, almoços no Resun;
As minhas estagiárias Mônica e Cleice por ter dedicado tanto tempo no meu
trabalho, pela vontade em ajudar e a torcida que tudo desse certo;
Aos amigos que mesmo de longe, estiveram bem perto com todo carinho: Mário
Dantas, Cris ruiva, Dan, Rubens, Juan, Sarinha, Adriana, Sandra, Naiara, Emília, D.
Alzenir, Harlem, Taty, Taci, Jô, Silvia;
A todos da Secretaria do Meio Ambiente em Jaguarari, especialmente ao secretário
Antônio Marcos pela confiança depositada e disposição, à Glécia, Robson, Clécio, sem o
apoio de vocês, não teria conseguido;
Ao senhor Celso, por ter permitido que a pesquisa fosse feita em sua propriedade;
A todos os moradores de Lajedo II em especial ao Devan, Neguinho, Sr. Geovane,
Renato e D. Quinininha;
A Juliana por ter dividido comigo o seu espaço e durante mais de um ano ter me
aguentado, rsrs. Sei que não sou tão fácil assim, e você só reclamou dos banhos
demorados, rsrs, você é um doce! Mimimi pra você!
A Juliana Cordeiro, pela amizade, pelas conversas e companhia na lanchonete
durante o almoço totalmente fora de horário;
A Cristiana por ter contribuído mais uma vez na minha pesquisa, por toda a
colaboração do Laboratório de Arqueologia e Paleontologia da Universidade do Estado da
Bahia;
Ao CNPQ pela bolsa concedida e ao PROCAD-NF: 23032-042965/2008-11 pelo
apoio concedido;
Ao programa de Pós-graduação em Ecologia e Conservação da Universidade
Federal de Sergipe;
E a todos que contribuíram para que esta etapa fosse concluída com sucesso!

vii
“... Refugie-se no tempo, antes que as horas passem...”.
Márcia Xavier

viii
À minha tia Dina (in memoriam),
que tanta falta faz!

ix
SUMÁRIO
RESUMO ................................................................................. Erro! Indicador não definido.
ABSTRACT ............................................................................ Erro! Indicador não definido.
Introdução Geral ............................................................................................................... 12
1.1. Considerações Gerais e Objetivos ............................................................................ 13
1.2. Quaternário: Período de Mudanças. ......................................................................... 17
1.3. Mastofauna Pleistocênica da Bahia: Histórico das pesquisas. ................................. 21
1.4. Paleoecologia e suas implicações na dinâmica ambiental da Caatinga. ................... 26
Capítulo 1. Megafauna Neopleistocênica da Caatinga ................................................... 35
Capítulo 2. Dinâmica de Vegetação e Clima no Quaternário recente .......................... 55
Capítulo 3. Considerações Finais ..................................................................................... 91
Apêndice ............................................................................................................................. 93
Anexo ................................................................................................................................ 108

x
RESUMO
Resumo: Estudos de reconstrução paleoambiental (vegetação e clima) são mais
abundantes nas regiões Sul, Sudeste e Central do Brasil utilizando pólen depositado em
sedimentos lacustres e turfeiras. Por outro lado, na região Nordeste, trabalhos análogos
são mais escassos. A partir deste tipo de estudo é possível inferir sobre os processos
ecológicos associados às comunidades vegetais e animais. Neste trabalho buscou-se
entender a dinâmica paleoambiental ocorrida em um afloramento fossilífero do tipo tanque
localizado no município de Jaguarari, o qual faz parte da Microrregião de Senhor do
Bonfim – Centro-Oeste da Bahia, em domínio de Caatinga. Para tanto, coletou-se fósseis
de mamíferos neopleistocênicos; solo e plantas para análises isotópicas (13
C). A fauna
identificada é composta por 6 taxa: Eremotherium laurillardi; Notiomastodon platensis;
Toxodontinae; Felidae; Panochthus greslebini; Equus (Amerhippus) neogeus.
Interpretações paleoambientais das características ecológicas dos taxa sugerem que este
conjunto faunístico estava associado há um ambiente com predomínio de áreas abertas em
associação com fisionomias mais fechadas. Os resultados isotópicos apontam para
estabilidade vegetacional na região até o presente, sem troca de vegetação C3 para C4 nos
locais estudados. A presença de fragmentos de carvão soterrados em várias profundidades
do solo sugere a ocorrência de paleoincêndios durante todo o Holoceno, com a possível
influência de um clima mais seco para a região. Estes incêndios podem ser de natureza
antrópica, já que foram encontrados alguns vestígios de ocupação de populações antigas na
região. A dinâmica da vegetação observada para Jaguarari, até a profundidade de solo
estudada, sugere tendência a uma vegetação arbórea dominante, desde aproximadamente
15.000 anos AP. com retorno a um clima mais úmido e provavelmente similar ao atual. A
região estudada, possivelmente, não sofreu modificação acentuada na vegetação no médio
Holoceno, quando do estabelecimento de períodos mais secos, permanecendo com áreas de
vegetação de floresta. Durante os trabalhos de escavação dos fósseis, sentiu-se a
necessidade de desenvolver um trabalho de Educação Patrimonial junto à população do
povoado de Lajedo II, para subsidiar ações de conservação dos sítios paleontológicos. Os
resultados mostraram que esta ferramenta é um instrumento capaz de fornecer informações
sobre patrimônio, noções de conservação, leis que normatizam estas práticas, as sanções
legais previstas, como também, proporcionar a construção de novos conceitos e uma
aprendizagem significativa na comunidade alvo.
Palavras Chave: Reconstrução paleoambiental; Megafauna; Matéria orgânica do solo;
Fragmentos de carvão; Educação patrimonial.

xi
ABSTRACT
Abstract: Numerous studies of paleo-environmental reconstruction (vegetation and
climate) have been carried out in southern, southeastern, and central Brazil, based on the
pollen deposited in lacustrine and peatbog sediments. Such studies are scarce in the
Brazilian Northeast, however. Studies of this type permit the development of inferences on
the ecological processes affecting local plant and animal communities. The present study
aimed to understand the paleo-environmental dynamics of a tank-type fossil outcrop in the
municipality of Jaguarari, part of the Senhor do Bonfim micro-region in central-western
Bahia state, located within the Caatinga biome. Fossils of neopleistocene mammals were
collected, together with samples of soil and plants for isotopic analyses (13
C). Six
mammalian taxa were identified: Eremotherium laurillardi, Notiomastodon platensis,
Toxodontinae, Felidae, Panochthus greslebini, and Equus (Amerhippus) neogeus. The
paleo-environmental interpretation of the ecological characteristics of these taxa indicated
that this assemblage was found with a predominantly open environment, associated with
areas of denser habitat. The isotopic analysis indicated that the vegetation of the area has
been stable up to the present day, with no exchange of C3 to C4 at the study sites. The
presence of charcoal fragments at different depths in the soil indicates the occurrence of
wildfires throughout the Holocene, possibly as a consequence of a relatively much drier
climate. The wildfires may have been anthropogenic, given that the evidence was found in
association with vestiges of ancient human populations. The dynamics of the vegetation
observed at the site to the depth excavated indicated a predominance of arboreal vegetation
since approximately 15,000 BP, finally returning to a more humid climate similar to that of
the present day. The vegetation of the study region may not have suffered major
modifications of the vegetation during the drier periods of the mid-Holocene, with forest
formations predominating. During excavations, the need for a patrimonial education
program was perceived, in order to promote measures for the conservation of
paleontological sites, together with the local population of Lajedo II village. The results of
this initiative indicated that this approach is a useful tool for the generation of information
on natural patrimony, basic concepts of conservation, the legal basis for this process and
the possible sanctions that enforce it, as well as contributing to the development of new
knowledge and perspectives in the local community.
Key words: Paleoenvironmental reconstruction; Megafauna; Organic matter on soil;
Charcoal fragments; Patrimonial education.

xii
Introdução Geral

13
1.1. Considerações Gerais e Objetivos
Há sessenta milhões de anos a América do Sul, estava isolada geograficamente, assim
como acontece atualmente com a Austrália, e os registros fósseis de mamíferos que teriam
surgido há 150 milhões de anos antes dessa data, mostram que estes se tornavam cada vez
mais variados e numerosos. Estavam isolados e teriam se originado aqui ou para cá teriam
emigrado procedentes da América do Norte, antes de nosso continente se tornar uma ilha
(CARTELLE, 1994).
Na América do Sul mudanças geográficas ocorreram com elevação dos Andes há 35
milhões de anos, e com a formação de algumas ilhas, até que por volta de 1,8 milhão de
anos houve o estabelecimento de uma ponte intercontinental, e a então ilha da América do
Sul passou a estar ligada a América do Norte por meio da América Central (WEBB, 1985;
CARTELLE, 1994; WOODBURNE, 2010). Estas pontes-de-terra (land bridges)
facilitaram a passagem da biota, permitindo migração entre as duas Américas em ambas as
direções, ocorrendo, então, uma grande competição pelo mesmo alimento e espaço
(território) (SALGADO-LABOURIAU, 1994).
Este “grande intercâmbio faunístico” possibilitado por um corredor contínuo após a
formação do istmo do Panamá facilitou as idas e vindas de faunas diferentes entre as
Américas do Norte e do Sul. Tatus e preguiças conseguiram avançar com relativo sucesso
para o norte, sendo bem sucedidos com o intercâmbio os mamíferos setentrionais: cavalos,
camelídeos, antas, carnívoros (ursos, quatis, felinos, cães) e proboscídeos levando à
extinção boa parte de seus similares sulinos (LOPES, 2004).
Contudo, por volta de dez, doze mil anos ocorreu na América do Sul e outras partes do
mundo, uma grande extinção, onde numerosas espécies aparentemente bem adaptadas, em
um tempo geológicamente curto desapareceram do continente, coincidindo com a
instabilidade climática e diminuição da temperatura. Estes fatores teriam provocado
variações ambientais como: aumento de áreas de floresta ao longo dos rios, desertificações
nas planícies, diminuição de pastagens nos cerrados (pelo menos temporariamente) e em
conseqüência a isso, redução gradual de indivíduos e nascimentos em algumas espécies
animais que apresentavam um grau elevado de especialização, sobretudo quanto ao regime
alimentar (CARTELLE, 1999)
Outros autores mencionam também a possibilidade de doenças terem ocasionado
grandes mortandades entre estes grandes animais (e.g. FERIGOLO, 1999). Nesse caso o

14
homem poderia ter contribuído, pois, pode ter introduzido na América animais domésticos
que contaminaram animais selvagens com suas doenças. Um terceiro fator pode ter sido a
caça predatória que teria levado à extinção estes animais (FUNARI & NOELLI, 2002;
CIONE et al. 2003).
Logo, é provável que os três fatores tenham atuado de maneira conjunta, pois houve,
efetivamente, mudança climática com a diminuição da área dos campos e cerrados –
habitats originais desses grandes animais – concomitante à expansão da ocupação humana,
que pode ter espalhado doenças bem como extinguido o número desses animais por meio
das caças.
Os estudos da paleofauna na America do Sul desde os achados de Charles Darwin no
Uruguai e Argentina atraíram não só a atenção da Europa para a fauna extinta no
continente americano, como também o estabelecimento da Paleontologia como ciência na
metade do século XVIII a partir dos trabalhos de Georges Cuvier (CARTELLE, 1994).
Desde então numerosos trabalhos têm sido feitos buscando identificar e descrever esta
fauna, assim como as razões que levaram a sua extinção. No Brasil tem-se encontrado
muitos fósseis de mamíferos pleistocênicos em vários estados, a exemplo os estudos
realizados por Cástor Cartelle na Bahia e Minas Gerais; Carlos de Paula Couto em quase
todo o Brasil, e Peter Lund em Minas Gerais.
No Nordeste há registros da ocorrência desta fauna em todos os estados, a exemplo de
alguns trabalhos realizados em: Alagoas (e.g. ALVIM, 1939; SILVA, 2001); Bahia (e.g.
CARTELLE, 1987; BELTRÃO & LOCKS, 1989; LESSA, 2005, XAVIER et al., 2008);
Ceará (e.g. MORAES, 1924; PAULA-COUTO, 1953; XIMENES, 1996; XIMENES &
MACHADO, 2004; XIMENES, 2006;); Maranhão (e.g. ALVIM, 1939; SIMPSON &
PAULA-COUTO, 1957); Paraíba (e.g. BERQVIST, 1993; COSTA et al., 1993;
BERQVIST et al.,1997; BARRETO et al., 2003); Pernambuco (e.g. ALVIM, 1939;
ROLIM, 1974; SILVA, 2001; OLIVEIRA et al., 2009; OLIVEIRA et al., 2010); Piauí
(e.g. GUÉRIN et al., 1993; FAURE et al., 1999); Rio Grande do Norte (e.g. OLIVEIRA et
al., 1984; PORPINO & SANTOS, 1997; PORPINO, 2000; SANTOS et al., 2001;
PORPINO & SANTOS, 2003; PORPINO et al., 2004); Sergipe (e.g. GOES et al., 2002;
DANTAS et al., 2003; DANTAS & ZUCON, 2007).
Algumas localidades onde foram encontrados fósseis da megafauna pleistocênica no
Nordeste do Brasil foram plotadas em um mapa por Viana et al. (2007), onde pode ser
vista a distribuição destas espécies por toda a região (Figura 1) sendo destacadas na figura
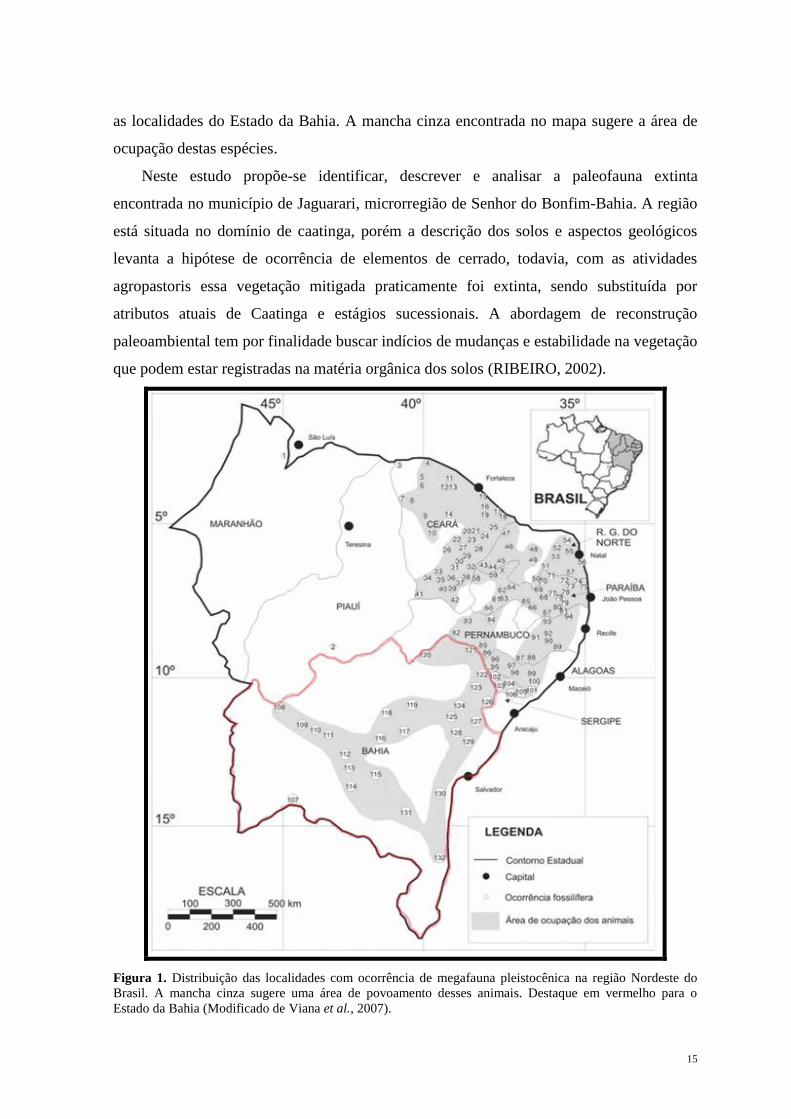
15
as localidades do Estado da Bahia. A mancha cinza encontrada no mapa sugere a área de
ocupação destas espécies.
Neste estudo propõe-se identificar, descrever e analisar a paleofauna extinta
encontrada no município de Jaguarari, microrregião de Senhor do Bonfim-Bahia. A região
está situada no domínio de caatinga, porém a descrição dos solos e aspectos geológicos
levanta a hipótese de ocorrência de elementos de cerrado, todavia, com as atividades
agropastoris essa vegetação mitigada praticamente foi extinta, sendo substituída por
atributos atuais de Caatinga e estágios sucessionais. A abordagem de reconstrução
paleoambiental tem por finalidade buscar indícios de mudanças e estabilidade na vegetação
que podem estar registradas na matéria orgânica dos solos (RIBEIRO, 2002).
Figura 1. Distribuição das localidades com ocorrência de megafauna pleistocênica na região Nordeste do
Brasil. A mancha cinza sugere uma área de povoamento desses animais. Destaque em vermelho para o
Estado da Bahia (Modificado de Viana et al., 2007).

16
Segundo Da Silva (2001) a microrregião de Senhor do Bonfim apresenta-se como uma
das áreas de maior potencial paleontológico do Estado da Bahia, sendo nos ambientes de
carste (cavernas) e tanques onde se depositaram grande parte das jazidas fossilíferas
pleistocênicas do Estado.
No entanto, a maior parte dos estudos está voltada para o ambiente de caverna, por
exemplo, a Toca Da Barriguda e a Toca da Boa vista no município de Campo Formoso,
onde se conhece um número maior de espécies para este tipo de ambiente.
Para os tanques localizados na região, a realidade já é diferente. Existem poucos
estudos acadêmicos, poucas coletas com o critério científico adequado, embora seja do
conhecimento da maioria que o material fóssil está sendo perdido. Esta perda é ocasionada,
tanto por agentes naturais, que tendem a degradar naturalmente os restos fósseis, como
também pela ação antrópica, onde em suas atividades agrícolas de forma não intencional
(desmatamento, preparo da terra para plantio, escavação de reservatórios de água contendo
fósseis, etc), danificam o material; ou de maneira mais perniciosa, ao realizar a coleta
indiscriminada das ossadas sem nenhum critério ou registro científico, descontextualizando
os achados.
Esta retirada do material fóssil do contexto tafonômico e estatigráfico, muitas vezes é
realizada pelos moradores que vivem no entorno do sítio, onde curiosos e desejosos de
maiores informações, removem-nos de forma indiscriminada.
Em estudos anteriores Xavier et al. (2008) constataram que a megafauna pleistocênica
composta de oito táxons de 07 herbívoros e 01 carnívoro identificados: Eremotherium
laurillardi (Lund, 1842) Cartelle & Bohórquez, 1982; Glyptodon clavipes Owen, 1839;
Panochthus greslebini Castellanos, 1941; Panochthus jaguaribensis Moreira, 1965;
Palaeolama major Liais, 1872; Stegomastodon waringi Holand, 1920; Toxodon platensis
Owen, 1840; Família Felidae Gray, 1821, foram encontrados em tanques da microrregião
de Senhor do Bonfim abrangendo os municípios: Andorinha, Antonio Gonçalves,
Caldeirão Grande, Campo Formoso, Filadélfia, Jaguarari, Pindobaçu, Ponto Novo, Senhor
do Bonfim.
Sendo assim, considerando que estudos adicionais para áreas de tanque se fazem
necessários, este trabalho busca: ampliar os registros tanto para a microrregião como para o
Estado; caracterizar a megafauna quanto aos seus hábitos alimentares e o ambiente onde
vivia para estabelecer sua paleoecologia, paleocronologia e conseqüentemente conhecer os

17
ambientes pretéritos e inferir se a diminuição da biodiversidade local levou a uma
diminuição da estabilidade ambiental.
A partir dessas novas informações, verificar se há indícios de mudança na paisagem no
local estudado, e em caso positivo, sugerir os possíveis agentes de mudanças, as hipóteses
que justificam estes eventos e sugerir para onde estas mudanças apontam atualmente.
Com objetivo secundário de desenvolver na comunidade local uma ação de
conservação do material estudado, propôs-se um experimento de educação patrimonial no
povoado de Lajedo II buscando transmitir conceitos básicos sobre paleontologia, fósseis da
megafauna e medidas de conservação.
1.2. Quaternário: Período de Mudanças.
Desnoyers (1829) foi o primeiro a propor o termo Quaternário para diferenciar os
estratos sobrejacentes aos sedimentos do Terciário da Bacia de Paris, sendo redefinido
posteriormente por Reboull (1833) visando incluir todos os estratos caracterizados por
vestígios de flora e fauna, cujos similares poderiam ainda hoje ser encontrados em vida
(MOURA, 2009).
Embora não existindo concordância geral entre os pesquisadores, considera-se que
este período tenha iniciado há 2,6 milhões de anos, sendo dividido em Pleistoceno e
Holoceno (este compreendendo os últimos 10 mil anos) (Tabela 1). O Quaternário
apresenta-se como o período que contém a maior quantidade de evidências paleoecológicas
e a história da civilização humana com as suas intervenções sobre os outros ecossistemas.
Não é possível afirmar que todas as paisagens atuais são datadas apenas do Quaternário,
pois algumas delas conservam traços de outros períodos como o Paleógeno e Neógeno
podendo chegar até o Cretáceo, no entanto, raras são as regiões que estiveram sujeitas a
condições estáveis, sob a ação de um único sistema morfoclimático durante o Cenozóico
nos último milhão de anos (AB’ SABER, 1969).
Sabe-se que por todo o planeta, a intensidade das variações climáticas ocorridas
durante o Quaternário produziu efeitos nas taxas de intemperismo e pedogênese, nos
regimes fluviais e nível dos oceanos, e na distribuição ecológica dos seres vivos, forçados a
migrações e adaptações a condições mutáveis. Dessas contínuas modificações nas
condições ambientais resultaram transformações mundiais na paisagem. Sendo assim, a

18
análise geomorfológica dos ambientes atuais constitui a base para a compreensão da
seqüência evolutiva da paisagem no passado geológico recente (MOURA, 2009).
Tabela 1. Escala do Tempo Geológico - 2009, baseada na International Commission on Stratigraphy (ICS)
(modificado de Silva-Filho, 2011).
Éon Era Período Época Milhões de Anos
Fanerozóico Cenozóico
Quaternário Holoceno 0,01 à atualidade
2,6 a 0,01 Pleistoceno
Neógeno Plioceno
Mioceno
5,3 a 2,6
23,0 á 5,3
Paleógeno
Oligoceno
Eoceno
Paleoceno
33,9 a 23,0
55,8 a 33,9
65,5 a 55,8
Estas mudanças ambientais, tanto espaciais quanto temporais, estão refletidas em um
complexo mosaico de paisagens, seqüências sedimentares, vestígios de floras, faunas e
artefatos humanos (LOWE & WALKER, 1984).
A distribuição da área florestal brasileira sofreu modificações cíclicas no tempo e no
espaço, sendo periodicamente substituída por formações vegetais abertas (Cerrado,
Caatinga). Nas épocas climáticas favoráveis sua expansão foi máxima, desempenhando
papel importante na intemperização química das rochas (BIGARELLA et al., 1994).
Durante o quaternário, as flutuações paleoclimáticas promoveram deslocamentos
espaciais das formações vegetais, embora, no tocante ao patrimônio florístico no
Pleistoceno, os grandes tipos de cobertura vegetal teriam sido os mesmo de hoje. O mesmo
não aconteceu em relação à fauna, onde as alterações foram maiores ocasionando a
extinção de numerosas espécies, entre elas, os grandes mamíferos terrestres característicos
deste período (BIGARELLA et al., 1994).
Estas mudanças foram possíveis porque durante o Pleistoceno ocorreram grandes
pulsações climáticas com períodos de temperatura muito baixas (glaciações) e períodos
mais quentes (interglaciações). Uma glaciação, por sua vez, pode apresentar episódios
menores, referidos como estádios e interestádios. Os estádios correspondem aos avanços
das geleiras e os interestádios, às fases de recuo dessas geleiras, causados pela elevação da
temperatura (MENDES, 1984; COX & MOORE, 1994; SALGADO-LABOURIAU, 1994).
No início do último episódio glacial pleistocênico (Würm – Wisconsin Superior), o
paleoespaço geográfico era igual ou similar ao atual, mas com o avanço da glaciação

19
desintegrou-se o caráter tropical dos climas até então predominantes na fase pré-Würm. A
implantação desta última grande época glacial foi marcada pela regressão das florestas e
ampliação das formações vegetais abertas intertropicais e subtropicais, em conseqüência da
expansão compartimentada dos climas secos sazonários, determinados pelo avanço das
correntes frias, até latitudes bem mais baixas do que as de suas extremidades atuais.
(BIGARELLA et al., 1994).
Estes intervalos glaciais e interglaciais constituem unidades geoclimáticas que
recebem nomes diferentes de acordo com as regiões em que foram descritas, sendo que, no
Brasil utiliza-se a descrição usada na América do Norte. Considera-se que em todas as
regiões do mundo, esse interglacial ou Holoceno iniciou-se há 12.000 anos chegando aos
dias atuais, mesmo em regiões que não tiveram influência direta das glaciações as quais,
ainda sim, sofreram alterações com períodos áridos ou de baixas temperaturas (Salgado-
Labouriau, 1994).
Sendo assim, profundas mudanças climáticas ou as pequenas flutuações do clima
desempenharam um papel importante no desenvolvimento da paisagem atual. Nos últimos
anos tem-se percebido que nas épocas frias do Quaternário, em grande parte das regiões
subtropicais e tropicais, teria ocorrido uma diminuição sensível da pluviosidade ou
modificação da distribuição das chuvas, generalizando-se condições de semi-aridez e
mesmo aridez, enquanto as épocas úmidas corresponderiam aos interglaciais
(BIGARELLA et al., 1994).
Durante o clima úmido desenvolveu-se um manto de decomposição formado,
principalmente, por alteração química, sobre o qual vicejou formações florestais, onde no
sul do Brasil, este manto de intemperismo atinge uma espessura usual de 5 a 15m, ou
mesmo mais, chegando ao norte do Paraná, nas imediações de Londrina a 60m. Observa-se
na transição do clima úmido para um mais seco a regressão da floresta, sendo substituída
por uma cobertura vegetal menos densa (aberta), do tipo cerrado ou Caatinga. Sob a
cobertura florestal, os solos eram protegidos contra a erosão acelerada o que não vai
acontecer sob o novo tipo de vegetação. Neste novo regime, onde as chuvas são
concentradas, os rios tendem a ser intermitentes, com grandes variações na sua descarga
havendo um progressivo aumento na carga sólida dos rios, resultante da erosão acelerada
das vertentes desprotegidas (BIGARELLA et al., 1994).
Durante os episódios de semi-aridez, as florestas ficaram restritas a refúgios enquanto
as condições climáticas permitiram sua sobrevivência. Contudo, a retração do revestimento

20
vegetal com a exposição do solo a um regime diferente de chuvas fortes e concentradas,
fez com que o manto de intemperismo formado sob condições climáticas úmidas, fosse
removido pela erosão mecânica das fases semi-áridas, ao fim do qual, a região sofreu
aplainamentos laterais (pedimentação) (KRAMER, 2002).
Para Ab’ Saber (1973), o modelo de transformação da paisagem brasileira durante o
Quaternário baseia-se no fato de que algumas rampas de pedimento estiveram
condicionadas às bruscas mudanças climáticas ocorridas no passado; e após o fim da era
glacial e o aumento da temperatura e das chuvas, as florestas foram se restabelecendo
lentamente entre 9,5 e 8,5 Ka AP, de acordo com as condições ambientais de cada região.
Crowley (1984) reconhece que apenas uma pequena quantidade dos eventos
deposicionais que caracterizam os ambientes atuais são preservadas. O registro sedimentar
é dominado por eventos raros de alta magnitude, devido à ocorrência de hiatos (fases de
não deposição e/ou de remoção pela erosão), e por isso, muitas feições identificadas em
ambientes atuais não se preservam como registro sedimentar.
Segundo Schumm (1977) a natureza incompleta de tal registro é inerente à natureza
episódica dos processos de erosão/sedimentação (conceito intimamente ligado à questão de
equilíbrio/desequilíbrio e tempo). Desta forma, a associação entre a natureza do registro
sedimentar e a dinâmica de evolução da paisagem seria a base para a reconstrução dos
períodos de estabilidade e instabilidade ambiental, fundamentação básica da
Geomorfologia do Quaternário.
Os mais importantes avanços no conhecimento do curto espaço de tempo envolvido
pelo Quaternário puderam ser obtidos a partir do desenvolvimento do método de datação
pelo Carbono 14 e estudos do fundo oceânico, onde por meio deste último, foi possível
identificar um registro sedimentar praticamente não perturbado, bastante extenso e quase
contínuo de todo o período (MOURA, 2009).
A freqüência e a amplitude das oscilações climáticas registradas dentro de um episódio
de tempo bastante curto fazem com que o Quaternário contraste bastante com os outros
períodos geológicos. A quantidade de eventos climáticos registrados nos sedimentos
oceânicos, mais do que o dobro dos glaciais e interglaciais distinguidos nos depósitos
continentais, torna este período mais que a simples ocorrência de fases quentes e frias
distribuídas por todo o registro geológico (FISCHER, 1982).
A teoria amplamente aceita para justificar a regularidade e a freqüência das oscilações
climáticas quaternárias foi elaborada por Milankovitch (1941) e baseia-se na admissão de

21
que as variações de temperatura da superfície terrestre estão relacionadas a modificações
periódicas de magnitudes variadas na órbita e no eixo terrestre. Estes mecanismos
combinados afetariam a quantidade de radiação solar recebida e seriam responsáveis pelos
efeitos climáticos.
1.3. Mastofauna Pleistocênica da Bahia: Histórico das pesquisas.
Na Bahia os estudos com material fóssil para afloramentos fossilíferos do tipo tanque
são descritos na literatura desde Paula-Couto (1953) e Simpson & Paula-Couto (1957).
Para Souza-Cunha (1966), tanques são fendas naturais, em rochas cristalinas, de
paredes íngremes e de contorno elíptico, cuja formação está relacionada à ação de águas
turbulentas no trajeto dos rios, contendo fragmentos de rochas que desgastam a superfície
cristalina. Paula-Couto (1953) sugere que este atrito constante nas fendas aumenta
progressivamente de tamanho influenciado pelo intemperismo químico do próprio rio.
Trabalhos mais recentes (BATISTA, 2002; DANTAS & TASSO, 2007; DANTAS &
ZUCON, 2007; XAVIER et al., 2008) apontam outras localidades, como também
identificam e descrevem uma fauna mais diversa do que se conhecia.
Também fazem parte dos estudos paleontológicos do Estado, trabalhos realizados em
cavernas sendo a Toca da Boa Vista a mais extensa caverna conhecida no Hemisfério Sul
(AULER, 1999) e 16ª dentre as mais extensas do mundo, possuindo 84 km de galerias
mapeadas. A Toca da Barriguda (segunda maior caverna do Brasil - com 19,5 km), a Toca
do Calor de Cima, Toca do Pitu e Toca do Morrinho, são as que mais se destacam e
constituem um sítio de grande valor científico para o Estado (AULER & SMART, 1999).
Dentre os trabalhos já realizados na Bahia podemos destacar as pesquisas realizadas
em Mairi, Central e Xique-Xique. Nestas localidades foram encontrados ossos e dentes
fósseis de mamíferos coletados pelas equipes do Museu de Arqueologia e Etnologia
(UFBA), Museu Geológico do Estado da Bahia (COM/SME), Museu Nacional (UFRJ) e
da seção de Paleontologia (DGM/DNPM). Do material coletado dois conjuntos faunísticos
das idades distintas dentro do Pleistoceno foram individualizados, sendo um mais novo que
o outro. A fauna mais recente é composta por representantes da fauna atual (cavalos,
porcos-do-mato, etc.) e outra, mais antiga, com formas já extintas (Eremotérios,
Toxodontes, etc) (LAGE, 1985).

22
Na Gruta das Onças em Caatinga do Moura, Jacobina (BA), em pesquisa realizada
pela equipe de Paleontologia da PUC-MG, foi encontrada grande quantidade de ossos de
Eremotherium laurillardi (Lund) (maior número de peças), Pampatherium paulacoutoi
Cartelle e Bohórquez, 1984, Nothrotherium maquinense Lydekker, 1889, Hoplophorus
euphractus (Lund, 1839), Smilodon populator (Lund, 1842) e restos, por identificar, de
Milodontídeo (CARTELLE & BELTRÃO, 1985).
Em Janeiro de 1993 foram coletados um molar superior e diversas peças esqueletais
na Toca da Boa Vista em associação com restos de Tayassu peccari (Fisher), Mazana sp.
e Smilodon populator Lund. Após comparação com esqueletos de Lama guanicoe (Muller)
e Lama glama (Linneus) e de Palaeolama major (Liais), as ossadas foram identificadas
como provavelmente pertencentes a, Lama guanicoe (CARTELLE, 1993).
Cartelle & Ferreira (1993) identificaram Brachyteles aracnoides (Géoffroy) em Morro
do Chapéu a partir da região rostral do crânio; em Campo Formoso identificaram
Brachyteles brasiliensis (Lund) baseado em numerosos ossos do esqueleto pós-craniano e
dois crânios completos com a maior parte da dentição. Os crânios pertencem a espécimes
macho e fêmea e a espécie extinta é 25% maior que o atual muriqui e apresenta-se como o
maior tamanho entre os primatas brasileiros.
Cartelle & Abuhid (1993) ainda a partir de material procedente de grutas da Bahia,
identificam espécies de Quirópteros fósseis: Mormoops aff. M. megalophylla*, Pteronotus
aff. P. gymnonotus*, Tonatia bidens*, Phylostomus hastatus, Chrotopterus auritus*,
Loncophylla mordax, Artibeus jamaicensis*, Desmodus rotundus, Desmodus draculae*,
Eptesicus brasiliensis, Eumops sp., Tadarida brasiliensis e Molossus molossus. Para os
autores as espécies assinaladas com asterisco (*) foram encontradas associadas à fauna
extinta de mamíferos enumerada por Lund em Lagoa Santa. As demais são identificadas
como primeiro registro fóssil para o Brasil.
Em 1989 no sítio arqueológico Toca do Aragão situado na Fazenda Aragolândia,
Município de Central foram retirados 715 kg de ossos, contendo centenas de osteodermos.
Em 1991 foram obtidos 1.500kg de ossos em uma área de 4m2, e a partir dos osteodermos
coletados foram identificadas as seguintes espécies: Pampatherium humboldti, Holmesina
sp. (Dasypodoidea); Panochthus sp., Hoplophorus sp., Glyptodon sp. (Glyptodontoidea),
distribuídas por todo o sítio e em todos os níveis estratigráficos onde foi encontrado o
material ósseo (LOCKS et al., 1993).

23
No ano de 1997, vários trabalhos foram feitos na Bahia. Abuhid (1997) estudou o
material procedente de uma gruta calcárea localizada no nordeste da Bahia, identificando
estes restos como sendo de Glossotherium aff. G. lettsomi. Vucetich (1997); Cartelle
(1997) e Monteiro (1997) registraram roedores caviomorfos para o Pleistoceno tardio da
Bahia; Locks et al. (1997) encontraram na região arqueológica de Central, uma associação
de mamíferos fósseis sub-recentes; Cartelle (1997) identificou a espécie Arctotherium
brasiliense, baseado em fósseis encontrados na Toca da Barriguda, Campo Formoso,
Bahia; Ferreira (1997) estudou restos de Stegomastodon waringi encontrados em
Nordestina durante escavações realizadas para aprofundamento de uma cacimba
pertencentes e o exemplar pertence ao IGEO-UFBA.
Lessa et al. (1997) analisaram restos fósseis de mamíferos carnívoros procedentes de
três grutas calcárias Toca dos Ossos, Brejões e Boa Vista (todas na Bahia), onde foram
reportados 15 crânios, diversos fragmentos mandibulares e ossos pós-cranianos e vários
dentes isolados, sendo possível identificar 12 (doze) espécies, sendo a maioria delas
pertencentes à Família Felidae (Smilodon populator, Felis tigrina, Felis pardalis, Felis
yagouaroundi, Felis concolor e Panthera onca), e as demais pertencentes à Família
Canidae (Protocyon troglodytes e Cerdocyon thous, à Família Mustelidae, Conepatus
semistriatus e Eira barbara), à Família Ursidae (Arctotherium brasiliensis) e à Família
Procyonidae (Procyon cancrivorus). Destas espécies S. populator, P. troglodytes e A. (P)
brasiliensis são espécies pleistocênicas extintas e as restantes são consideradas recentes,
porém ocorrentes também no Pleistoceno. Juntamente com estes carnívoros o autor
também coletou restos de Eremotherium laurillardi, Nothrotherium maquinense,
Palaeolama major e Xenorhinotherium bahiense.
Perônico et al. (2000) estudaram cropólitos de mamíferos Quaternários encontrados
nas cavernas da Toca dos Ossos e Brejões, ambos na Bahia, e Alberdi et al. (2002)
descreveram o crânio de Notiomastodon platensis (=Stegomastodon waringi) encontrado
no município de Pains, Minas Gerais, e ossos encontrados na Tocados Ossos, Bahia.
Viana et al. (2007) realizam um inventário da distribuição geográfica da megafauna
pleistocênica no Nordeste Brasileiro e apresentam um mapa de localidades (Figura 1). Os
dados referentes a nome dos municípios, numeração destes no mapa, localidades,
referencias usadas pelos autores e citadas neste trabalho contam na Tabela 2. Para o Estado
da Bahia existem outras localidades onde já foram coletados materiais os quais não
aparecem plotados no mapa, sendo listados neste trabalho.
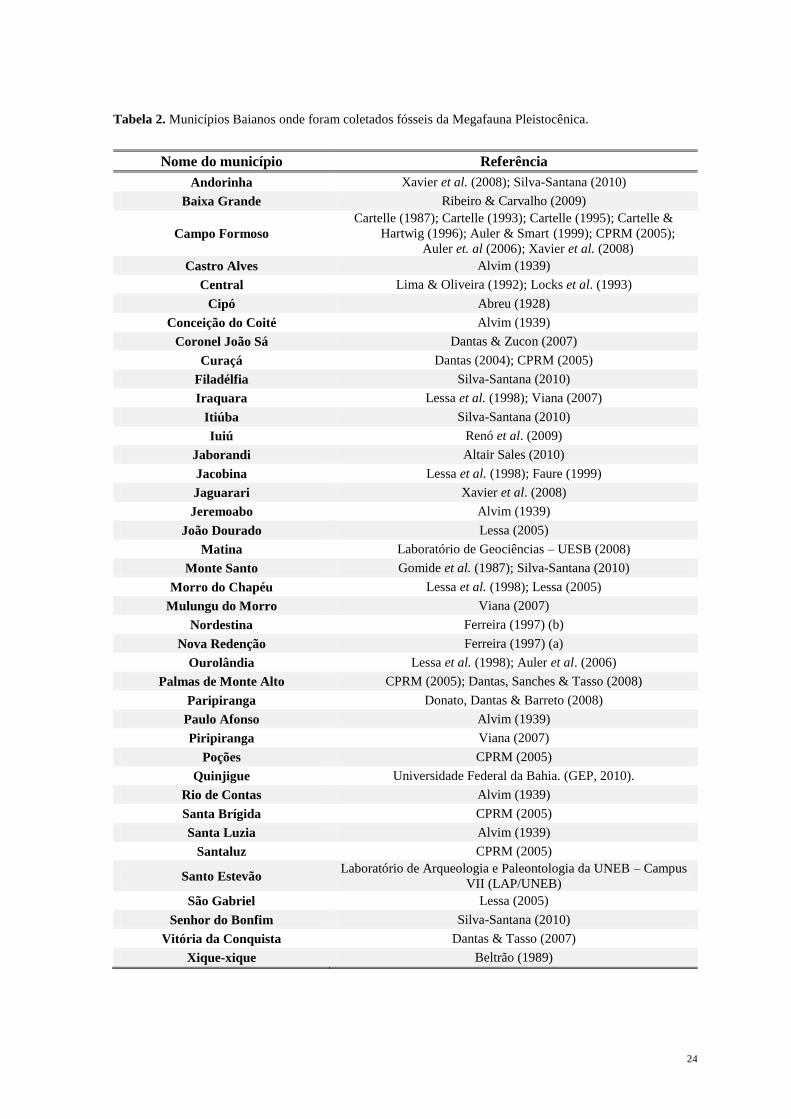
24
Tabela 2. Municípios Baianos onde foram coletados fósseis da Megafauna Pleistocênica.
Nome do município Referência
Andorinha Xavier et al. (2008); Silva-Santana (2010)
Baixa Grande Ribeiro & Carvalho (2009)
Campo Formoso
Cartelle (1987); Cartelle (1993); Cartelle (1995); Cartelle &
Hartwig (1996); Auler & Smart (1999); CPRM (2005);
Auler et. al (2006); Xavier et al. (2008) Castro Alves Alvim (1939)
Central Lima & Oliveira (1992); Locks et al. (1993)
Cipó Abreu (1928)
Conceição do Coité Alvim (1939)
Coronel João Sá Dantas & Zucon (2007)
Curaçá Dantas (2004); CPRM (2005)
Filadélfia Silva-Santana (2010)
Iraquara Lessa et al. (1998); Viana (2007)
Itiúba Silva-Santana (2010)
Iuiú Renó et al. (2009)
Jaborandi Altair Sales (2010)
Jacobina Lessa et al. (1998); Faure (1999)
Jaguarari Xavier et al. (2008)
Jeremoabo Alvim (1939)
João Dourado Lessa (2005)
Matina Laboratório de Geociências – UESB (2008)
Monte Santo Gomide et al. (1987); Silva-Santana (2010)
Morro do Chapéu Lessa et al. (1998); Lessa (2005)
Mulungu do Morro Viana (2007)
Nordestina Ferreira (1997) (b)
Nova Redenção Ferreira (1997) (a)
Ourolândia Lessa et al. (1998); Auler et al. (2006)
Palmas de Monte Alto CPRM (2005); Dantas, Sanches & Tasso (2008)
Paripiranga Donato, Dantas & Barreto (2008)
Paulo Afonso Alvim (1939)
Piripiranga Viana (2007)
Poções CPRM (2005)
Quinjigue Universidade Federal da Bahia. (GEP, 2010).
Rio de Contas Alvim (1939)
Santa Brígida CPRM (2005)
Santa Luzia Alvim (1939)
Santaluz CPRM (2005)
Santo Estevão Laboratório de Arqueologia e Paleontologia da UNEB – Campus
VII (LAP/UNEB)
São Gabriel Lessa (2005)
Senhor do Bonfim Silva-Santana (2010)
Vitória da Conquista Dantas & Tasso (2007)
Xique-xique Beltrão (1989)

25
Para a Microrregião de Senhor do Bonfim (antes chamada de Vila da Rainha), Spix
e Martius em 1817-1820 em sua expedição pelo Brasil relatam achados da megafauna
pleistocênica (“animais antidiluvianos”):
Aqui e acolá, nas vargens rasas, existe camada de argila avermelhada de três a
quatro pés de altura, que os sertanejos costumam cavar até à profundidade de
alguns pés, formando caldeirões e tanques, para conservarem água da chuva; e
neles aparecem numerosas ossadas de animais antidiluvianos, porém geralmente
tão fragmentados e dispersas, que dificilmente se pode contar com o achado de
um esqueleto completo. Os ossos que aqui tivemos ocasião de colecionar em
estado de poderem ser reconhecidos foram um maxilar inferior, uma vértebra
dorsal e parte de uma omoplata de mastodonte. Outras partes, especialmente
costelas e fêmures, são aqui freqüentemente desenterradas pelos sertanejos; nós,
no entanto, não tivemos essa sorte, pois todos os pedaços que nos aparecem
estavam muito fragmentados. Também perto da |vizinha Fazenda Barriga Mole,
segundo contam os moradores, foram desenterrados dos tanques uns ossos
semelhantes, e, conforme dizem, aparece às vezes em grande quantidade. Ativas
investigações neste sertão tão pouco conhecido enriqueceriam provavelmente os
conhecimentos sobre animais antediluvianos e ainda de muitos outros fatos
importantes (SPIX & MARTIUS, 1976, p.192).
Levantamentos recentes para esta microrregião foram realizados equipe do LAP-
UNEB. Esses trabalhos de registros de sítios e coletas ainda não foram publicados estando
disponíveis sob a forma de relatórios em Da Silva (2001; 2002). Uma monografia foi
elaborada por Batista (2002) a qual incluiu a localização dos sítios visitados à época e
informações sobre as coletas efetuadas até então pelo Laboratório.
Xavier et al. (2008) organiza a coleção da paleofauna do Laboratório, coletando
quatro espécimes novos, reunindo com os existentes, sendo um total de oito, os quais
foram posteriormente identificados, descritos, catalogados e acondicionados, compondo a
coleção didática do LAP-UNEB, Tabela 3.
Silva-Santana et al. (2010) realizou um estudo visando levantar informações sobre a
Megafauna Pleistocênica para o Estado da Bahia, listando as localidades onde esta fauna já
foi encontrada.

26
Tabela 3. Municípios que compõe a Microrregião de Senhor do Bonfim, com as espécies da Megafauna
Pleistocênicas encontrada em cada localidade. Destaque em verde para os municípios onde ainda não foi
coletado material fóssil.
Taxons
An
do
rin
ha
An
tôn
io
Go
nça
lves
*
Ca
mp
o
Fo
rmo
so
Fil
ad
élfi
a
Itiú
ba
Ja
gu
ara
ri
Pin
do
ba
çu*
Sen
ho
r d
o
Bo
nfi
m
Um
bu
ran
as*
Eremotherium laurillardi x x x x x
Gliptodon clavipes x
Panochthus greslebini x
Panochthus jaguaribensis x
Palaeolama major x x
Smilodon populator x
Stegomastodon waringi x x x x
Toxodon platensis x
* Municípios não apresentam registro de espécies coletadas para a Megafauna Pleistocênica.
1.4. Paleoecologia e suas implicações na dinâmica ambiental da Caatinga.
O período Quaternário caracteriza-se por apresentar fauna e flora composta por formas
predominantemente viventes, além de ser também a idade de surgimento do homem na
Terra (SUGUIO, 1999).
Dois importantes registros palinológicos destacam as mudanças vegetacionais e
climáticas da região da Caatinga nordestina: o registro continental da Caatinga do campo
de dunas fósseis do médio rio São Francisco/BA (DE OLIVEIRA et al., 1999), com
sedimentos datados em 10.990 anos A.P. e o registro de sedimentos marinhos, obtidos a
cerca de 90 km ao leste da cidade de Fortaleza/CE que indica que a vegetação semi-árida
de Caatinga predominou entre 42.000 e 8.500 anos A.P (BEHLING et al., 2000).
A partir de datação com potássio-argônio e paleomagnetismo, verificou-se que toda
flora moderna já existia desde o início do período, sendo os megafósseis de plantas, os
grãos de pólen, os esporos de pteridófitas, os foraminíferos e as diatomáceas achadas em
sedimentos quaternários, os mesmos dos atuais podendo ser relacionados com os gêneros
modernos, sendo possível desta forma, reconstruir os ecossistemas, estudar a sucessão da

27
vegetação de uma região e observar o seu comportamento frente às mudanças e oscilações
climáticas (SALGADO-LABOURIAU, 1994).
Com os animais vertebrados o processo não se deu da mesma forma, já que a maioria
dos grandes mamíferos terrestres extinguiu-se. Algumas espécies de tigre dente-de-sabre,
de mamute, de mastodonte, de preguiça e outros, ainda existiam há uns dez mil anos atrás.
O número de extinções é considerável e tem sido objeto de muita especulação
(SALGADO-LABOURIAU, 1994).
Visando entender os motivos que levaram a estas extinções e transformações,
atualmente tem-se lançado mão de várias técnicas de investigação, sendo uma delas os
estudos envolvendo isótopos estáveis. Estes estudos são possíveis já que determinadas
fontes específicas de alimentos tem relações com os isótopos estáveis de carbono (13
C =
razão entre 13
C/12
C permil, PDBellemita) e nitrogênio (15
N), e a composição isotópica da
dieta é incorporada a frações de tecidos do corpo, tais como colágeno e preserva-se por
milhões de anos. Devido a isso, é possível estudar a relação dos isótopos estáveis de
carbono e nitrogênio nos tecidos ósseos na reconstrução de paleodietas (SCHOENINGER,
1995).
Os isótopos de carbono provenientes da Matéria Orgânica do Solo (MOS) também
auxiliam no entendimento destes estudos, uma vez que, além de fornecerem registros das
mudanças de vegetação no passado, os valores isotópicos não são modificados com o
tempo (CERLING et al., 1989).
De posse destas informações é possível determinar a origem da MOS de plantas C3,
C4, a mistura de plantas (C3 e C4) ou CAM com base nas razões isotópicas do carbono
13C, enquanto que a datação por
14C nos permite estabelecer a cronologia.
Outra técnica que permite elucidar estas questões é a Datação por Ressonância por
SPIN Eletrônico, a qual baseia-se nos efeitos da radiação sobre o material, assim como na
determinação da concentração de elétrons desemparelhados (KINOSHITA & BAFFA,
2005).
Rastreando estes marcadores, é possível identificar a dieta destes animais e
conseqüentemente, o tipo vegetacional existente no período, e juntamente com
informações sobre morfologia dos dentes, proporção dos ossos e biomecânica do aparato
mastigatório inferir sobre a paleoecologia das espécies (BARGO et al., 2000; 2006), uma
vez que as dietas dos animais influenciam a morfologia, comportamento, interações

28
ecológicas e a história evolucionária dos mesmos (GANNES et al., 1998) e podem
responder ao status nutricional, taxa de turnover e rota biosintética (KOCK, et al. 1994).
Referências Bibliográficas
Ab’ Saber, A.N. 1969. Gênese das vertentes pendentes em inselbergs no nordeste
brasileiro. Inst. Geogr., Univ. S. Paulo, Notas prévias, Geomorfologia, São Paulo,
14:6-8.
Ab’ Saber, A.N. 1973. A organização natural das paisagens inter e subtropicais brasileiras.
Geomorfologia, 41: 1-39.
Abreu, S. F. 1928. Um capítulo da Geographia do Ceará. In: REUNIÃO DA
SOCIEDADE BRASILEIRA DE GEÓGRAFOS. Recursos e Minerais, Rio de Janeiro,
vol. 33: p.145-180.
Alberdi, M.T.; Prado, J.L.; Cartelle, C. 2002. El registro de Stegomastodon (Mammalia,
Gomphotheriidae) en el Pleistoceno Superior de Brasil. Revista Española de
Paleontologia 17(2): 217-235.
Alvim, G.F. 1939. Jazigos Brasileiros de Mamíferos Fósseis. Notas Preliminares e Estudos
18: 8-16.
Auler, A.S. 1999. Karst Evolution and Palaeoclimate of Eastern Brazil. PhD Thesis,
University of Bristol.
Auler, A.S.; Smart, P.L. 1999. Toca da Boa Vista, Bahia - A maior caverna do Hemisfério
Sul. In: Schobbenhaus,C.; Campos,D.A.; Queiroz,E.T.; Winge,M.; Berbert-Born,M.
(Edit.) Sítios Geológicos e Paleontológicos do Brasil.
Auler, A. S., Piló, L. B.; Esperto, P. L.; Wang, X.; Hoffmann, D.; Richards, D. A.;
Edwards, R. L.; Neves, W. A.; & Cheng, H. 2006. U-series dating and taphonomy of
Quaternary vertebrates from Brazilian caves. Palaeogeography, palaeoclimatology,
palaeoecology. Revista do d’aquesta dos sumaris de Altres, 240(3-4): 508-522.
Bargo, M.S.; de Iuliis, G.; Vizcaíno, S.F. 2006. Hypsodonty in Pleistocene ground sloths.
Acta Palaeontol. Pol. 51(1): 53-61.
Bargo, M.S.; Vizcaíno, S.F.; Archuby, F.M.; Blanco, R.E. 2000. Limb bone proportions,
strength and digging in some Lujanian (late Pleistocene – early Holocene) Mylodontid
ground sloths (Mammalia, Xenarthra). Journal of Vertebrate Paleontology 20(3): 601-
610.
Barreto, A.M.F.; Almeida, J.A.C.; Silva, F.M.; Kinoshita, A.; Baffa, O. & Macário, J.
2003. Datação por EPR em dentes de Haplomastodon e Xenorhinotherium de Lagoa
de Dentro, Puxinanã, Paraíba. In: CONGRESSO BRASILEIRO DE
PALEONTOLOGIA, 18, Brasília, 2003. Boletim de Resumos, Brasília: Universidade
de Brasília, p.60.
Batista, I.C.M. 2002. Levantamentos e registros paleontológicos de megamamíferos
pleistocênicos na microregião de Senhor do Bonfim – Bahia. 24p. Monografia – curso
de Licenciatura em Ciências Biológicas, Universidade do Estado da Bahia, Senhor do
Bonfim.
Behling, H.; Arz, H.W.; Patzold, J.; Wefer, G. 2000. Late Quaternary vegetational and
climate dynamics in northeastern Brazil, inferences from marine core GeoB 3104-1.
Quaternary Science Reviews 19: 981-994.

29
Beltrão, M.C.M.C. & Locks, M. 1989. Pleistocene fauna from the “Toca da Esperança”
site, (Archaeollogical-Region of Central, Bahia, Brazil): Mammals – nº 1. In:
CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 11, Curitiba, 1989. Anais do
Congresso, vol. 1: 685- 697.
Berqvist, L. P.; Gomide, M.; Cartelle, C. & Capilla, R. 1997. Faunas locais de mamíferos
pleistocênicos de Itapipoca/Ceará, Taperoá/Paraíba e Campina Grande/Paraíba. Estudo
comparativo, bioestratinômico e paleoambiental. Revista Universidade de Guarulhos –
Geociências 2(6): 23-32.
Berqvist, L.P. 1993. Jazimentos pleistocênicos do Estado da Paraíba e seus fósseis. Revista
Nordestina de Biologia 8(2): 143-158.
Bigarella, J.J. 1994. Estrutura e origem das paisagens tropicais e subtropicais / João José
Bigarella, Rosemari Dora Becker, Gilberto Friedenreich dos Santos, Everton Passos,
Kenitiro Suguio. Florianópolis: Ed. UFSC, 3v.
Cartelle, C. & Abuhid, V.S. 1993. Chiroptera do Pleistoceno Final-Holoceno da Bahia.
In:13º Congresso Brasileiro de Paleontologia, São Leopoldo, 19 a 26 de Setembro de
1993, 1º Simpósio Paleontológico do Cone Sul: Companhia de Pesquisa de Recursos
Minerais - Companhia de Pesquisa de Recursos Minerais - CPRM. p. 145.
Cartelle, C. & Beltrão, M.C.M.C. 1985. Notícia Prévia Sobre o Achado do “Homem de
Lagoa Santa” na Bahia. In: IX Congresso Brasileiro de Paleontologia, Fortaleza, 1 a 7
de Setembro de 1985. Fortaleza: Sociedade Brasileira de Paleontologia. p. 148.
Cartelle, C. & Ferreira, M. A. C. 1993. Nota Prévia Sobre Primatas Pleistocênicos
Brasileiro. In: 13º Congresso Brasileiro de Paleontologia, São Leopoldo, 19 a 26 de
Setembro de 1993, 1º Simpósio Paleontológico do Cone Sul. São Leopoldo:
Companhia de Pesquisa de Recursos Minerais - Companhia de Pesquisa de Recursos
Minerais CPRM. p. 140.
Cartelle, C. & Hartwigt, W. C. A new extinct primate among the Pleistocene megafauna of
Bahia, Brazil Proc. Natl. Acad. Sci. USA Vol. 93, pp. 6405-6409, June 1996
Evolution.
Cartelle, C. 1987. Um pequeno urso-de-face-curta Arctotherium brasiliense, da Bahia. In:
CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 15, São Pedro, 1997. Boletim
de Resumos, São Pedro: UNESP, p.130.
Cartelle, C. 1993. Presença de Lama (Artiocactyla, Camelidae) no Pleistoceno Final-
Holoceno da Bahia. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 13, e
SIMPÓSIO PALEONTOLÓGICO DO CONE SUL, 1, São Leopoldo, 1993. Boletim
de Resumos, Rio Grande do Sul, p.139.
Cartelle, C. 1994. Tempo Passado: Mamíferos do Pleistoceno em Minas Gerais. Belo
Horizonte: Acesita.
Cartelle, C. 1995. A fauna local de mamíferos pleistocênicos da Toca da Boa Vista
(Campo Formoso, BA). Tese para professor titular, Universidade Federal de Minas
Gerais.
Cartelle, C.1997. Um pequeno urso-de-face-curta, Arctotherium brasiliense, da Bahia. In:
CONGRESO BRASILEIRO DE PALEONTOLOGIA, 15., São Pedro, SP. Boletim de
resumos. p. 130.
Cartelle, C. 1999. Pleistocene mammals of the Cerrado and Caatinga of Brazil. In:
EISENBERG, J.F. & REDFORD, K.H. (eds.). Mammals of the Neotropics. The
University of Chicago Press, p. 27-46.

30
Cerling, T.E.; Quade, J.; Wang, Y.; Bowman, J.R. 1989. Carbon Isotopes In Soils And
Paleosols As Ecology And Paleoecology Indicators. Nature 341:138-139.
Cione et al., 2003. The borken Zig-Zag: Late Cenozoic large mammal and tortoise
extinction in South America. Revista del Museo Argentino de Ciencias Naturales 5(1):
1-19.
Congresso Brasileiro de Paleontologia, São Pedro, Agosto, 1997. São Pedro: UNESP.
p.122, 1997.
Costa, A.C.D.; Apoluceno, D.M.; Machado, M.F.; Santos, M.F.C.F. & Trindade, R.I.F.
1993. Registro de um Megaterídeo na Fazenda Algodão de Jandaíra, Município de
Pocinhos (PB). In: SIMPÓSIO DE GEOLOGIA DO NORDESTE, 15, Natal, 1993.
Boletim de Resumos, Natal, p.19.
Cox, C.B. & Moore, P.D. 1994. Biogeography. 5. Ed. Oxford: Blackwel Scientific
Publications, 326p.
CPRM – Serviço Geológico do Brasil: Projeto Cadastro de Fontes de Abastecimento por
Água Subterrânea, Diagnóstico do Município de Jaguarari Estado da Bahia /
Organizado [por] Ângelo T. Vieira, Felicíssimo Melo, Hermínio Brasil V. Lopes,
Hermínio Brasil V. Lopes, José C. Viégas Campos, José T Guimarães, Juliana M. da
Costa, Luís Fernando C. Bomfim, Pedro Antonio de A. Couto, Sara Maria Pinotti
Benvenuti . Salvador: CPRM/PRODEEM, 2005. 14p + anexos
Crowley, K.D. 1984. Filtering of depositional events and the completeness of sedimentary
sequences. J. Sediment. 54(1): 127-136.
Da Silva, C.C. 2001. Relatório de Pesquisas: Arqueologia e Paleontologia – semestres
2000.2, 2001.1 e 2001.2. Laboratório de Geociências. Departamento de Educação-
Campus VII/UNEB, Senhor do Bonfim, 31p.
Da Silva, C.C. 2002. Relatório de Pesquisas em Arqueologia e Paleontologia - semestres
2002.1 e 2002.2. Laboratório de Arqueologia e Paleontologia. Departamento de
Educação-Campus VII/UNEB, Senhor do Bonfim, 8p.
Dantas, M.A.T. & Tasso, M.A. de L. 2007. Megafauna do Pleistoceno final de Vitória da
Conquista, Bahia: taxonomia e aspectos tafonômicos. Scientia Plena 3(3):30-36.
Dantas, M. A. T.; Sanches, A. L. & Tasso, M. A. L. de. 2008. Nota sobre a ocorrência de
fósseis da megafauna do Pleistoceno final – Holoceno em Palmas de Monte Alto,
Bahia, Brasil Revista de Geologia, Vol. 21, nº 1, 109-114.
Dantas, M.A.T. & Zucon, M.H. 2007. Occurrence of Catonyx cuvieri (Lund, 1839)
(Tardigrada, Scelidotheriinae) in Late Pleistocene – Holocene of Brazil. Revista
Brasileira de Paleontologia 10(2):129-132.
Dantas, M.A.T.; Andrade, L.; Zucon, M.H. & Teodósio, C. 2003. Novo achado de
megafauna do Pleistoceno fi nal de Sergipe – Fazenda Elefante, Gararu. In:
CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 18, Brasília, 2003. Boletim de
Resumos, Brasília: UnB, p.115.
Dantas, M.A.T., 2004, Os fósseis da megafauna pleistocênica do Instituto Histórico e
Geográfico de Sergipe. Canindé, 4: 383-393.
De Oliveira, P.E. de; Barreto, A.M.F.; Suguio, K. 1999. Late Pleistocene / Holocene
climatic and vegetational history of the brazilian Caatinga: the fossil dunes of the
middle São Francisco river. Palaeogeography, Palaeoclimatology, Palaeoecology
152: 319-337.

31
Desnoyer, J. 1829. Observations sur um ensemble de dépôts Marins plus recents que lês
terrains tertiaires du bassin de La Seine et constituant une formation geologique
distincte; precedes d’ un appercu de La non-simultaneité dês bassins tertiaires. Ann.
Sci. Nat. 16: 171-214, 402-91.
Donato, C. R. ; Dantas, M. A. T. ; Barreto, E. A. S. . Ocorrência de Morcegos Fósseis
(Myotis sp.) na Caverna do Bom Pastor, Paripiranga, Bahia, Brasil.. In: VI Simpósio
Brasileiro de Paleontologia de Vertebrados, 2008, Ribeirão Preto - SP. Paleontologia
em Destaque: Boletim Informativo da Sociedade Brasileira de Paleontologia, 2008. p.
85-86.
Faure, M.; Guérin, C. & Parenti, F. 1999. Découverte d’une mégafaune holocène à la Toca
do Serrote do Artur (aire archéologique de São Raimundo Nonato, Piauí, Brésil).
Comptes rendus de l’Académie des sciences 329(6): 443-448.
Ferigolo, J. 1999. Late Pleistocene South-American land mammal extinctions: the
infection hypotheses. Quaternary of South America and Antarctic Peninsula.
Ferreira, M. T. G. M. 1997. Haplomastodon waringi (Holland,1920) Procedente de
Nordestina,BA. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 15, São
Pedro, 1997. Boletim de Resumos, São Pedro: UNESP, p.133. (b)
Ferreira, M. T. G. M. 1997. Nota sobre a Presença de Megalonichidae sp. No Poço Azul,
BA. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 15, São Pedro, 1997.
Boletim de Resumos, São Pedro: UNESP, p.121. (a)
Fischer, A.G. 1982. Long-term climatic oscillations recorded in stratigraphy. In: Studies
geophysics: Climate in Earth history. National Acad. Press. PP. 97 – 104.
Funari, P.P. & Noelli, 2002. Pré-História do Brasil. 1 ed. São Paulo: Contexto, 110p.
Gannes, L.Z.; Del-Rio, C.M.; Koch, P. 1998. Natural abundance variations in stable
isotopes and their potential uses in animal physiological ecology. Comp. Biochem.
Physiol 119 A (3), 725-737.
Goes, F.A.S.; Vieira, F.S.; Zucon, M.H.; Cartelle, C. & Teodósio, C. 2002. Ocorrência de
mamíferos pleistocênicos em Sergipe, Brasil. Arquivos do Museu Nacional, 60(3):199-
206.
Gomide, M.; Berqvist, L. P. & Rego, D. D. 1987. O “Tigre dente-de-sabre” (Smilodon
poulator) de Itapipoca, Ceará. In: CONGRESSO BRASILEIRO DE
PALEONTOLOGIA, 10, Rio de Janeiro, Anais, Rio de Janeiro, vol.1. p.197-204.
Guérin, C.; Curvello, M. A.; Faure, M.; Hugueney, M. & Mourer-Chauviré. C. 1993. La
Faune Pleistocene du Piaui (Nordeste du Bresil): implications paleoecologiques et
biochronologiques. Quaternaria Nova 3: 303-341.
Kinoshita, A. & Baffa, O. 2005. Datação por Ressonância do Spin Eletrônico. Canindé 6.
Kock, P.L.; Fogel, M.L.; Tuross, N. 1994. Tracing the diets of fossil animals using stable
isotopes. In: Lajtha, K.; Michener, R.H. (Ed). Stable Isotopes in Ecology and
Enviromental Science. Oxford: Blackwel Scientific Publications. cap4. p.63-92.
Kramer, V.M.S. 2002. Mudanças ambientais climáticas durante o quaternário na região do
alto curso do rio Paraná, Akrópolis, Umuarama, v10, n°1 e 2, jan./jun., 2002
Lage, A.C.T. 1985 Observações sobre a Paleontologia do Plesitoceno da Região
centro_ociedental do estado da Bahia, In: IX CONGRESSO BRASILERIO DE
PALEONTOLOGIA, Fortaleza, 1 a 7 de setembro de 1985. Fortaleza: Sociedade
Brasileira de Paleontologia. P. 143.

32
Lessa, G. 2005. Novas ocorrências fossilíferas na APA Gruta dos Brejões/Vereda do
Romão Gramacho, Bahia, Brasil. In: CONGRESSO LATINO-AMERICANO DE
PALEONTOLOGIA DE VERTEBRADOS, 2, Rio de Janeiro, 2005. Boletim de
Resumos, Rio de Janeiro: Museu Nacional, p151.
Lessa, G.; Cartelle, C.; Faria, H. & Goncálves, P. R. 1997. Novos Achados de Mamíferos
Carnívoros do Pleistoceno Final-Holoceno em Grutas Calcárias do Estado da Bahia.
In: 15º Congresso Brasileiro de Paleontologia, São Pedro, Agosto de 1997. São Pedro:
p. 129.
Lessa, G.; Cartelle, C.; Faria, H. D. & Gonçalves, P. R., 1998. Novos achados de
mamíferos carnívoros do pleistoceno final – Holoceno em grutas calcárias do Estado
da Bahia, Acta geológica leopoldensia, XXI (46/47): 157-169.
Lima, L. C.; Oliveira, M. P. de. 1992. Pleistocene Gastropods from Toca da Esperança,
Municipality of Central, State of Bahia, Brasil. Mem. Inst. Oswaldo Cruz, Rio de
Janeiro, Vol. 87 (3): 423-427, jul./sep.
Locks, M.; Beltrão, M.C.M.C. & Cherfan, A.G. 1993. Região arqueológica de Central,
Bahia, nº 6 – Dasypodoidea e Glyptodontoidea (Mammalia-Edentata). In:
CONGRESSO DE PALEONTOLOGIA, 13, e SIMPÓSIO PALEONTOLÓGICO DO
CONE SUL, 1, São Leopoldo, 1993. Boletim de Resumos, Rio Grande do Sul, p.147.
Lopes, R.J. 2004. Terra de Gigantes. Scientific American. 5: 82-89.
Lowe, J.J. & Walker, M.J. 1984. Reconstructing Quaternary environments. New York,
Longman. 389p.
Mendes, J.C. 1984. Aspectos da Estratigrafia do Quaternário. In: Queiroz, T.A. (Ed.).
Elementos de Estratigrafia. São Paulo, EDUSP, pp. 468-496.
Milankovitch, M. 1941. Kanon der Erdbestrahlung und Seine Anwendung anf das
Eiszeitenproblem. Acad. Roy. Seibe, Belgrado , v.13, Sec. Sci. Math. Nat, 331, 1941.
Moraes, L.J. 1924. Serras e Montanhas do Nordeste. Rio de Janeiro, Inspectoria Federal de
Obras Contra as Secas, 1924. Coleção Mossoroense, 35. Vol. 1. (série I. D.58). 1077p.
Monteiro, C. A. 1997. Novas ocorrências de mamíferos pleistocênicos no norte de Minas
Gerais, Brasil. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 15., São
Pedro, SP. Boletim de resumos. P. 104.
Moura, J.R. da S. de. 2009. Geomorfologia do Quaternário. In Guerra, A.J.T. & Cunha,
S.B. (orgs.). Geomorfologia: uma atualização de bases e conceitos. 9ª Ed. – Rio de
Janeiro: Bertrand Brasil, 474p.
Nikiforova, K.V. & Krasnov, I.I. 1976. Stratigraphic scheme of upper Pliocene and
Quaternary deposits in the european parto f the U.R.S.S. in IGPC – INQUA.
Quaternary glaciations in the Northern Hemisphere. pp. 339-347.
Oliveira, L.D.D.; Santos, C.L.A. & Santos, M.F.C.F. 1984. Nova ocorrência de Panochtus
greslebini Castellanos, 1914, no Estado do Rio Grande do Norte. In: SIMPÓSIO DE
GEOLOGIA DO NORDESTE, 11, Natal, 1984. Atas, Natal, SBG, p.346-351.
Oliveira, E.V., Barreto, A.L.M.F., Silva, A.R., 2009. Aspectos sistemáticos,
paleobiogeográficos e paleoclimáticos dos mamíferos quaternários de Fazenda Nova,
PE, nordeste do Brasil. GAEA 5 (2), 75–85.
Oliveira, E.V., Porpino, K., de, O., Barreto, A.M.F., 2010. On the presence of
Glyptotherium in the Late Pleistocene of Northeastern Brazil, and the status of
“Glyptodon” and “Chlamydotherium”. Paleobiogeographic implications. Neues
Jahrbuch fur Geologie und Palaontologie-Abhandlungen 258/3, 353–363.

33
Paula Couto, C. de. 1953. Paleontologia Brasileira: Mamíferos. Rio de Janeiro: Instituto
Nacional do Livro.
Perônico, C.; Pilar, A.; Souza, R. De C. M.; Becho, D P. 2000. Estudos de cropólitos de
mamíferos Quatrenários preservados nas cavernas Toca dos Ossos e Brejões – BA.
Revista Brasileira de Paleontolgia, nº 2, p.114.
Porpino, K. de O. 2000. Sobre a ocorrência de Glyptodon clavipes Owen, 1939 e
Glyptodon reticulatus Owen, 1845 no estado do Rio Grande do Norte. In: SIMPÓSIO
BRASILEIRO DE PALEONTOLOGIA DE VERTEBRADOS, 2., Rio de Janeiro, RJ.
Boletim de resumos. P. 48.
Porpino, K. de O.; Santos, M.F. 1997. Mamíferos pleistocênicos de Lagoa do Santo, Rio
Grande do Norte – Brasil. In: CONGRESSO BRASILEIRO DE PALEONTOLOGIA,
16.; São Pedro, SP. Boletim de resumos. p. 116.
Porpino, K. de O.; Santos, M.F.. 2003. Novos registros de artiodactyla e perissodactyla
para o Lajedo da escada, Baraúna/RN. In: CONGRESSO BRASILEIRO DE
PALEONTOLOGIA, 18.; Brasília, DF. Boletim de resumos. p. 226.
Porpino, K. de O.; Santos, M.F.C.F. dos; Bergqvist, L.P. 2004. Registros de Mamíferos
Fósseis no Lajedo De Soledade, Apodi, Rio Grande do Norte, Brasil. Revista
Brasileira de Paleontologia 7(3):349-358.
Reboull, H. 1833. Geólogie de La période Quaternaire et introduction a l’histoire
ancienne. Paris, Levrault. 222p.
Renó, R.; Duque, T. R. F.; Lucon, T. N.; Levy, M. de O. Pinto; Silva, C. M. T. da; Simões,
P. R. 2009. Prospecção e caracterização Espeleológica da serra de Iuiu, Iuiu-BA.
ANAIS do XXX Congresso Brasileiro de Espeleologia Montes Claros MG.
Ribeiro, A.S., 2002. Dinâmica paleoambiental da vegetação e clima durante o Quaternário
tardio em domínios da mata Atlântica, brejo do semi-árido e cerrado nordestinos,
utilizando isótopos do carbono da matéria orgânica do solo e das plantas. Ph.D.
dissertation. University of São Paulo, Piracicaba (Brazil). 192 pp.
Ribeiro, R. da C. & Carvalho, 2009 I. de S. Megafauna do Quaternário tardio de Baixa
Grande, Bahia, Brasil, V.32 p. 42-50.
Rolim, J.L. 1974. Paleontologia e estratigrafia do Pleistoceno continental do Nordeste
Brasileiro – “Formação Cacimbas”. Programa de Pós-graduação em Geologia,
Universidade Federal do Rio Grande do Sul, Dissertação de Mestrado em Geociências.
Salgado-Labouriau, M.L. 1994. História ecológica da Terra. São Paulo: Edgard Blucher.
Santos, M.F.C.F.; Bergqvist, L.P. & Filho, F.P.L. 2001. Fósseis Pleistocênicos da Ravina
do Leon, Lajedo de Soledade, Apodi/RN. In: SIMPÓSIO DE GEOLOGIA DO
NORDESTE, 19, Natal, 2001. Atas, Natal, p.79.
Schoeninger M.J. 1995. Stable isotope studies in human evolution. Evol. Anthropology 4:
83-98.
Schumm, S.A. 1977. The Fluvial System. New York, John Wilwey & Sons, 338p.
Silva, J.L.L. 2001. Tafonomia em Mamíferos Pleistocênicos: caso da planície colúvio-
aluvionar de Maravilha-AL. Programa de Pós-graduação em Geologia, Universidade
Federal de Pernambuco, Dissertação de Mestrado em Geociências, 96p.
Silva-Filho, R.A. da. 2011. História Geológica da Bahia. Salvador: CBPM.
Silva-Santana, C. De C.; Santana, H. A. Silva, V. de C. 2010. Levantamentos
Paleontológicos, Arqueológicos e Programa de Patrimônio Histórico e Cultural em

34
Itapura, Miguel Calmon, Bahia. Relatório – Diagnóstico. Contratante: Química Geral
do Nordeste. Miguel Calmon, Maio de 2010. 113 p.
Simpson, G.G.; Paula-Couto, C. de. 1957. The Mastodonts of Brazil. Bulletin of American
Museum of Natural History 112(2): 1-65.
Souza-Cunha, F.L. de. 1966. Explorações paleontológicas no Pleistoceno do Rio Grande
do Norte. Arquivos do Instituto de Antropologia “Câmara Cascudo” 2(1-2): 1-46.
Spix & Martius. 1976. Viagem pelo Brasil: 1817-1820. São Paulo: Melhoramentos.
Suguio, K. 1999. Geologia do Quaternário e mudanças ambientais, passado + presente =
futuro?. São Paulo: Paulo’s comunicação e artes gráficas.
Viana, M.S.S.; Ximenes, C.L.; Rocha, L.A. de S.; Chaves, A.P. de P.; Oliveira, P.V. de.
2007. Distribuição Geográfica da Megafauna Pleistocênica no Nordeste Brasileiro. In
Carvalho, I. de S. (Ed.). Paleontologia: Cenários de Vida. Editora Interciência.
Vucetich, M, G. 1997. Los roedores caviomorfos del Pleistoceno Final del Estado de Bahia
(Brasil). CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 15., São Pedro, SP.
Boletim de resumos. P. 127.
Webb, S.D. 1985. Late Cenozoic mammal dispersals between the Americas. In: F.G.
Stehli, & S.D.Webb (eds.) The Great American Biotic Interchange, Plenum Press, p.
357-386.
Woodburne, M.O., 2010. The Great American Biotic Interchange: Dispersals,Tectonics,
Climate, Sea Level and Holding Pens. Journal of Mammalian Evolution 17(4), 245-
264.
Woldstedt, P. 1958. Das einzeitalter: grundlinien einer geologie es Quartärs – Band II.
Stutgart. Ferdinand Enke Verlag, 438p.
Xavier, M.C.T.; Dantas, M.A.T.; Santana, C.C. da S. 2008. A coleção de fósseis da
megafauna pleistocênica do Laboratório de Arqueologia e Paleontologia da
Universidade do Estado da Bahia, Senhor do Bonfim, Bahia. Paleontologia em
Destaque 23(61): 13-14.
Ximenes , C.L. 2006. Novas Ocorrências de Fósseis de Megafauna no Neo-Quaternário do
Estado do Ceará, Brasil. In: REUNIÃO ANUAL REGIONAL DA SOCIEDADE
BRASILEIRA DE PALEONTOLOGIA, Sobral, 2006. Boletim de Resumos, Sobral-
Ce, Fundação Paleontológica Phoenix, p.25.
Ximenes, C. L. 1996. Geologia de uma área localizada na porção Norte dos Municípios de
Tururu e Itapipoca, CE, e a caracterização de seus jazigos fossilíferos. Universidade
Federal do Ceará, Relatório de Graduação em Geologia.
Ximenes, C.L. & Machado, D.A.N. 2004. Diagnóstico paleontológico da Província
Espeleológica de Ubajara, Estado do Ceará. In: ENCONTRO BRASILEIRO DE
ESTUDOS DO CARSTE, 1, Belo Horizonte, 2004. Resumos do encontro, Belo
Horizonte, p.40.

35
Capítulo 1. Megafauna Neopleistocênica da Caatinga

36
Interpretações Paleoecológicas da Megafauna Neopleistocênica da Microrregião de
Senhor do Bonfim – BA em uma área de Caatinga
Interpretations paleoecological and occurrence of Megafauna Neopleistocênica in
Bioma Caatinga of the Microregion of Senhor do Bonfim – Bahia, Brazil.
Abstract In the Brazilian state of Bahia, studies of fossils from tank-like deposits date
back to the 1950s, with the published work of Carlos de Paula Couto. The Senhor do
Bonfim micro-region is one of the richest fossil-producing areas of Bahia, with large
deposits of Pleistocene fossils in karst formations and tanks. Six taxa of neopleistocene
mammals were recorded in a tank in Lajedo II village in the municipality of Jaguarari –
Eremotherium laurillardi, Notiomastodon platensis, Toxodontinae indet., Felidae indet.,
Panochthus greslebini, and Equus (Amerhippus) neogeus. The paleo-environmental
interpretation of the ecological characteristics of these taxa indicated that this assemblage
was found with predominantly open habitats, associated with areas of denser vegetation.
Key words: Neopleistocenic Megafauna; Tank; Bahia.
Resumo Na Bahia os estudos com material fóssil para afloramento fossilífero do tipo
tanque são descritos na literatura desde os trabalhos de Carlos de Paula Couto na década de
1950. A microrregião de Senhor do Bonfim se apresenta como uma das áreas de maior
potencial paleontológico do Estado da Bahia, sendo nos ambientes de carste e tanques onde
se depositaram grande parte das jazidas fossilíferas pleistocênicas. Para o município de
Jaguarari foram registrados fósseis de mamíferos neopleistocênicos, localizados em um
tanque no povoado de Lajedo II com a ocorrência de seis taxa: Eremotherium laurillardi;
Notiomastodon platensis; Toxodontinae indet.; Felidae indet.; Panochthus greslebini;
Equus (Amerhippus) neogeus. Interpretações paleoambientais das características
ecológicas dos taxa sugerem que este conjunto faunístico estava associado há um ambiente
com predomínio de áreas abertas em associação com fisionomias mais fechadas.
Palavras Chave: Megafauna Neopleistocênica; Tanque; Bahia.

37
1. Introdução
Na Bahia, os estudos com material fóssil em afloramentos fossilíferos do tipo
tanque são descritos na literatura desde a década de 1950 (PAULA-COUTO, 1953;
SIMPSON & PAULA-COUTO, 1957).
Na expedição Viagem pelo Brasil, Spix e Martius (1817-1820) relatam achados
fósseis em tanques naturais na microrregião de Senhor do Bonfim, antes chamada de Vila
da Rainha.
A microrregião de Senhor do Bonfim está situada na porção Centro - Oeste da
Bahia, pertencendo ao Território Piemonte Norte do Itapicuru e é composta por nove
municípios: Andorinha, Antonio Gonçalves, Caldeirão Grande, Campo Formoso,
Filadélfia, Jaguarari, Pindobaçu, Ponto Novo, Senhor do Bonfim (Figura 01).
Segundo Silva (2001) essa microrregião se apresenta como uma das áreas de maior
potencial paleontológico do Estado da Bahia, sendo nos ambientes de carste e tanques onde
se depositaram grande parte das jazidas fossilíferas pleistocênicas do Estado.
Deste modo, o presente trabalho teve como objetivos: registrar a ocorrência de
fósseis de seis táxons da megafauna pleistocênica para um tanque no município de
Jaguarari, Bahia e discutir a paleoecologia destes táxons.
2. Materiais e Métodos
Em Janeiro de 2011 foi realizada a coleta de material fóssil no afloramento
fossilífero do tipo tanque (Tanque Lajedo II –10°21’17”S, 40°16’19”W), localizado no
povoado Lajedo II, município de Jaguarari – BA. Os fósseis coletados compõem a
Coleção Didática do Laboratório de Arqueologia e Paleontologia da Universidade do
Estado da Bahia – UNEB (LAP-UNEB).
Os fósseis foram comparados com exemplares da coleção de Paleontologia do
Museu de Ciências Naturais da Pontifícia Universidade Católica de Minas Gerais
(PUC/MG) e do Laboratório de Paleontologia da Universidade Federal de Sergipe
(LPUFS). Constam de elementos mandibulares e algumas peças bastante fragmentadas do
esqueleto pós-craniano que apresentam diferentes tonalidades de coloração. Essa diferença
sugere variações nos padrões mineralógicos, impregnação sofrida pelo material e tempo de
exposição na superfície do tanque.

38
A sistemática dos Tardigrada segue Cartelle & De Iuliis (1995); dos
Proboscidea segue Mothé et al. (2011); dos Notoungulata segue Miño-Boilini et al. (2006);
dos Carnivora segue Paula-Couto (1979); dos Cingulata segue Porpino et al. (2004); dos
Perissodactyla segue Prado & Alberdi (2008). A descrição das peças segue, sempre que
possível Cartelle (1992).
3. Sistemática
TARDIGRADA Latham & Davies in Forster, 1795
MEGATHERIOIDEA Gray, 1821
MEGATHERIIDAE Gray 1821
EREMOTHERIUM Spillmann 1948
Eremotherium laurillardi (Lund, 1842) Cartelle & Bohórquez, 1982
Figura 02 (A-L), Figura 03 (E e G)
Material Referido: LAP 040 Porção proximal da costela; Fragmentos de ramo mandibular
LAP 104 e 105; Corpos de vértebra LAP 108, 109, 118, 126, 127, 128, 129, 130;
Cuneiforme LAP 111; Falange proximal/medial do dedo III LAP 113; Lunar LAP 114;
Navicular LAP 122; Fragmentos de molar LAP 059, 154, 155, 156, 157, 158, 159, 160 e
161.
Descrição: No fragmento do ramo mandíbular LAP 104 (Figura 02 - A) percebe-se uma
porção do alvéolo dentário, com formato quadrangular. Está ausente toda a sínfise.
O fragmento de molar LAP 155 possui formato quadrangular em corte transversal
como característico dos Megatheriinae
A falange proximal medial do dedo III direito LAP 113 (Figura 02 – F, I, L) está
desgastada, principalmente em vista dorsal. Na face ventral, a faceta articular distal
encontra-se parcialmente conservada, apresentando fratura na porção ungueal. Devido ao
desgaste ser maior na porção proximal da falange não observa-se claramente a concavidade
intermédia articular para a quilha.
Observa-se na coossificação entre a falange proximal e média, bem como
medialmente, uma faixa articular e a superfície articular distal em formato de carretel,
como diagnóstico em Eremotherium.

39
O Navicular LAP 122 (Figura 03 – E) apresenta em vista proximal a borda medial
convexa e a lateral côncava. A metade dorsal em formato de domo semi-esférico está
desgastada, expandido látero-dorsalmente. Percebe-se uma pequena porção da metade
plantar.
Comentários: Cartelle & Bohorquez (1982) estabelecem Eremotherium laurillardi como
uma espécie Panamericana. Segundo revisões taxonômicas, comparada com outras
preguiças, esta espécie apresenta uma ampla distribuição geográfica, que vai desde Nova
Jersey na América do Norte, em direção ao sul, passando pela Flórida e ao longo da costa
do Golfo do México, América Central, América do Sul desde o Norte do Peru na Costa do
Pacífico (PUJOS & SALAS, 2004) até o Rio Grande do Sul na Costa Atlântica
(CARTELLE & DE IULIIS, 1995).
Eremotherium laurillardi possivelmente tinha preferencias climáticas por
ambientes tropicais e subtropicais, o que pode ser inferido pelo seu registro limitado nos
Estados Unidos, indicando provavelmente, o limite norte de sua distribuição
(MCDONALD & LUNDELIUS, 2009).
Segundo Spillmann (1948 apud Paula Couto, 1979), Eremotherium, assim como as
demais preguiças terrícolas, seriam animais que viveriam em estepes secas, alimentando-se
de plantas xerófilas, com hábitos particularmente xilófagos.
Além de E. Laurillardi, cujos fósseis já foram encontrados em quase todos os
estados brasileiros (CARTELLE, 1994; OLIVEIRA et al., 2002), outra espécie pretencente
a subfamília Megatheriinae é encontradas no Brasil, Megatherium americanum Cuvier,
1796, com registros no Rio Grande do Sul.
Como características atribuídas à espécie, podemos citar molariformes com formato
prismático, retangulares, hipsodontes, sem esmalte e na superfície de mastigação,
acentuada aresta transversal. O material aqui descrito (e.g. dente com raiz aberta -
hipsodonte) confirma o registro realizado anteriormente.

40
PROBOSCIDEA Illiger, 1811
GOMPHOTHERIIDAE Hay, 1922
Notiomastodon platensis (Ameghino, 1888)
Figura 03 (A-D)
Material Referido: Corpos de vértebras LAP 115, 116, 121, 131; Fragmentos de molar
LAP 163, 167, 168; Fragmento de incisivo LAP 058, 169, 170 e 171.
Descrição do material: Nos fragmentos de molar LAP 163 Figura (A e D) e 167 (Figura
03 – A) observam-se cúspides pouco desgastadas, cônicas, compostas de colinas
transversais, com a presença do conulido do interlófido (?) em LAP 167.
Comentários: Em revisão recente dos Gonfotérios Sul Americanos Mothé et al. (2011)
sugerem para este táxon uma nova combinação, sugerindo o binômio Notiomastodon
platensis (Ameghino, 1888).
Desta forma, há uma redução do número de espécies dos gonfotérios na América do
Sul, passando para dois gêneros e duas espécies, apresentando distribuição geográfica ao
longo do Equador, Venezuela, Colômbia, Peru, Paraguai, Chile, Argentina, e Uruguai
(PRADO et al., 2001; GADENS-MARCON, 2008).
Como características atribuídas à espécie pode-se citar dentes com um padrão
quadrangular, com superfície oclusal apresentando duas séries longitudinais de cúspides
cônicas, compostas por colinas transversais, que permitem quando do desgaste destas,
formarem duas fileiras de figuras trifoliadas. O material aqui descrito (e.g. molar com
cúspides cônicas, com colinas transversais) apresenta a morfologia para a espécie.

41
NOTOUNGULATA Roth, 1901
TOXODONTIA Owen, 1853
TOXONDONTIDAE Owen, 1845
TOXODONTINAE (Owen 1845) Trouessart, 1898
Figura 03 (H-J)
Material Referido: Fragmento de molar LAP 165;
Descrição: O molar LAP 165 (Figura 03 – H, I, J) está fragmentado. Em vista transversal,
observa-se forma triangular, onde na face interna localiza-se o sulco anterior que divide o
molar em dois lóbulos, um anterior e outro posterior.
Na face externa (ectolofo), observa-se uma elevação próxima ao ângulo anterior do
dente, sendo percebida apenas nos M1 e M2. O dente é bilobado lingualmente sendo o
sulco interlobular recoberto por uma faixa de esmalte, característico para os toxodontídeos.
Comentários: Toxodontes eram um grupo de grandes mamíferos herbívoros endêmicos na
América do Sul e Neotrópico durante o Cenozoíco. Desde o início das descobertas fósseis
na metade do século 19 (por exemplo, OWEN, 1840), o clado está registrado para o
Pleistoceno como o mega herbívoro Toxodon difundido para a América do Sul
(MACFADDEN, 2005).
Esta ordem é a mais diversificada dentro dos ungulados nativos (BOND, 1986,
1999; BOND et al., 1995).
Ecologicamente estavam relacionados com ambientes abertos, cercados de corpos
d’água e hábitos pastadores (OLIVEIRA, 1999; CZERWONOGORA et al., 2003),
possuindo uma dieta com preferência por gramíneas abrasivas (MACFADDEN, 2005),
com crescimento contínuo dos dentes (CARTELLE, 1994).
A presença deste gênero já foi constatada em depósitos pleistocênicos da Argentina,
Uruguai, Paraguai, Bolívia e Brasil, pelo menos (PAULA COUTO, 1979).
Alguns autores, consideram para o Brasil a ocorrencia de três espécies
pleistocênicas de Toxodontíneos, sendo: Toxodon platensis Owen, 1840; Trigodonops
lopesi (Roxo, 1921) e Mixotoxodon larensis van Frank, 1957 (PAULA-COUTO, 1982;
CARTELLE, 1992, 1994).

42
O fóssil é aqui atribuído a um Toxodontíneo, baseado nas características do dente
bilobado lingualmente com sulco interlobular recoberto por uma faixa de esmalte.
CARNIVORA Bowdich, 1821
FELIDAE Fischer, 1817
Material Referido: Falange LAP 020.
Descrição: A falange proximal (LAP 020) apresenta um bom estado de conservação, com
suas extremidades distal e proximal presentes.
Comentários: Para a América reconhecem-se três espécies pertencentes ao gênero
Smilodon (Smilodon gracilis Cope, 1888; Smilodon fatalis Leidy, 1868; Smilodon
populator Lund, 1842) e uma pertencente ao gênero Homotherium (H. venezuelensis
Rincón, Prevosti & Parra, 2011) (Rincón et al., 2011).
Os primeiros registros de mamíferos carnívoros pleistocênicos no Brasil foram
feitos por Lund (1843) e revisados por Winge (1895-96), baseados em material coletado
em cavernas calcárias do Estado de Minas Gerais (LESSA et al., 1998).
Cartelle & Abuhid (1989) desconsideram a subdivisão da espécie S. populator em
subespécies, tendo como base somente a diferenciação de tamanho. Estes concluem que a
forma norte-americana e a sul americana, na verdade, seriam coespecíficas.
Alguns autores (e.g., KURTÉN & ANDERSON, 1980), no entanto aceitam a
diferenciação especificas entre as espécies, utilizando o nome de S. fatalis Leidy, 1869,
para o Norte da América, dada a sua prioridade sobre S. californicus Bovard, 1907.
Recentemente, S. gracilis Cope, 1880 foi reconhecido com um novo taxon, de posição
filogenética mais basal em relação às outras espécies do gênero, restrita ao final do
Plioceno - Pleistoceno Médio da América do Norte (KÜRTEN & WERDELIN, 1990).
Na América do Sul, foram encontrados fósseis de S. Populator no Equador, Peru,
Bolívia, Brasil, Argentina e Venezuela (BERTA, 1985; ZURITA et al., 2004; RINCÓN,
2006).

43
Para a Bahia foram registrados fósseis de Smilodon nos municípios de Campo
Formoso (CARTELLE, 1992); Ourolândia (CARTELLE, 1992); Caatinga do Moura –
Jacobina (CARTELLE & BOHÓRQUEZ, 1982, 1985; CARTELLE & ABUHID, 1989;
CARTELLE, 1992).
O fóssil é aqui atribuído a um Felidae em virtude de suas dimensões avantajadas,
que contrasta, visivelmente, com peças homólogas de outros Carnivora do Pleistoceno
final-Holoceno.
CINGULATA Illiger, 1811
GLYPTODONTOIDEA Simpson, 1931
GLYPTODONTIDAE Burmeister, 1879
HOPLOPHORINAE Weber, 1925
PANOCHTINI Simpson, 1945
Panochthus greslebini Castellanos, 1941
Figura 03 (F)
Material Referido: Osteodermo de carapaça LAP 162
Descrição: O osteodermo de carapaça LAP 162 (Figura 03 - F) encontra-se desgastado na
sua face externa, na qual se pode ver, parcialmente, uma ornamentação homogênea
consistindo de pequenas figuras irregulares, separadas por sulcos estreitos e rasos,
distribuídas desordenadamente, rugosas e sem a presença de uma figura central maior.
O diâmetro das figuras varia de 5 a 7 mm e a largura dos sulcos não ultrapassa 2
mm, a espessura do osteodermo varia de 10 mm a 25 mm transversalmente. Esta variação
de 15 mm encontrada na espessura do osteodermo deve-se ao fato deste estar danificado na
porção interna. Percebe-se dorsalmente nos sulcos, e ventralmente em todo osteodermo,
forames de tamanhos diminutos.
Comentários: O gênero Panochthus, o mais conhecido entre os animais da tribo
Panochtini Simpson, 1945, se distribui cronologicamente por todo o Pleistoceno
compreendendo espécies de grande tamanho (PAULA-COUTO, 1979).

44
Além do Nordeste brasileiro e Rio Grande do Sul, também possui registros no Quaternário
tardio da Argentina, Bolívia, Paraguai, Peru e Uruguai (PAULA-COUTO, 1979; PORPINO, 1999).
Habitava as savanas e alimentava-se de gramíneas (RANZI, 2000).
Para a Bahia foram registrados fósseis de Panochthus em Ourolândia (PAULA
COUTO, 1947), Itiuba (PAULA COUTO, 1957) e Jacobina (CARTELLE, 1992).
O fóssil é aqui atribuído a Panochthus cf. greslebini baseando-se na ornamentação
composta por um aglomerado de figurinhas de pequeno tamanho, de número variável,
distribuídas sem orientação preferencial, sem figura central maior e separadas entre si por
sulcos estreitos com pouca profundidade.
Ordem PERISSODACTYLA Owen, 1848
Família EQUIDAE Gray, 1821
Equus (Amerhippus) neogeus Lund, 1840
Figura (K)
Material Referido: m2 direito LAP 166.
Descrição: No fragmento m2 direito LAP 166 (Figura 03 - K) observa-se desgaste na
superfície oclusal no metacônido e metastílido e ausência de raiz.
O duplo laço é composto por metacônido e metastílido unidos, onde o primeiro é
arredondado e o segundo anguloso, concordando com o referido para a espécie. Possui
dimensões de 28 mm ântero-posterior e 17 mm línguo-vestibular.
Comentários: O registro de eqüídeos na América do Sul está bem documentado
(ALBERDI & PRADO, 1992; 1995) pela presença dos gêneros Hippidion Owen, 1869 e
Equus Hoffstetter, 1950. O primeiro está regitrado desde o Plioceno tardio até o Holoceno,
o segundo a partir do Pleistoceno médio. Conhecem-se cinco espécies de Equus
(Amerhippus): neogeus, insulatus, lasallei, andinum e santaelenae (PRADO & ALBERDI,
1994), que estão distribuídas na Colômbia, Equador, Bolívia e Brasil (BRIZUELA, 2005).
Equus (A.) neogaeus é a espécie de maior tamanho dentro do subgênero e encontra-
se registrada nas zonas de planícies da província de Buenos Aires e no Brasil, associadas à
ambientes abertos de pastagens xerófilas e solos compactos. Esta característica a diferencia

45
das outras espécies, as quais apresentam extremidades comparativamente mais robustas
(BRIZUELA, 2005).
No Brasil, Equus, tem sido registrado para os estados de Piauí, Ceará, Paraíba,
Pernambuco, Bahia, Minas gerais, Mato Grosso do Sul e Rio Grande do Sul (KERBER &
OLIVEIRA, 2008).
Para a Bahia foram registrados fósseis de Equídeos para Pedra Vermelha
(HOLLAND, 1920), Ourolândia (PAULA COUTO, 1947), Xique-Xique (PAULA
COUTO, 1953), Morro do Chapéu (CAMPOS & CAMPOS, 1976), Central (BELTRÃO &
LOCKS, 1989) e Santana (CARTELLE et al., 1989).
O fóssil é aqui atribuído a Equus (Amerhippus) neogeus com base nas
características encontradas no dente acima descrito, que apresenta caracteres diagnosticos
para a espécie.
Interpretações Paleoecológicas
É possível que a mudança na estrutura e composição do ecossistema cerrado-
caatinga, tenha ocorrido através da diminuição do ciclo pluvial e do aumento do ciclo seco,
durante a mudança climática no final do Pleistoceno, provocando variações nos
ecossistemas (CARTELLE, 1992).
Os táxons aqui identificados viviam em condições ambientais distintas das
existentes atualmente no sertão nordestino, onde predomina o clima semi-árido, altas
temperaturas, e uma cobertura vegetal escassa, composta predominantemente por plantas
xerófilas (Ribeiro & Carvalho, 2009).
A diminuição na área de Caatinga teria sido influenciada pelo aumento da
precipitação entre 15.500 a 11.800 anos, favorecendo o estabelecimento de um período
mais úmido e frio que o encontrado atualmente, contribuindo assim, para a expansão das
Florestas Amazônica e Atlântica, unidas através de corredores (DE OLIVEIRA et al.,
1999; BEHLING et al., 2000).
Do ponto de vista paleoambiental, E. laurillardi, Equus (Amerhippus) neogaeus e
Toxodontinae são associados a um habitat do tipo savana (com áreas recobertas por
gramíneas) em associação com fisionomias mais fechadas (borda de floresta) e
temperaturas mais baixas. Este fato implica em um clima mais úmido do que o atual em
certos momentos do Pleistoceno final (ARAÚJO JÚNIOR & PORPINO, 2007; RIBEIRO

46
& CARVALHO, 2009). Estas condições ambientais estariam de acordo com o modelo para
o Pleistoceno do Brasil Intertropical Extra-Amazônico proposto por Cartelle (1999).
Eremotherium laurillardi apresentava hábito pastador/ramoneador (BERGQVIST,
1993; DE IULLIS et al., 2000); enquanto P. greslebini seria pastador (Vizcaíno, 2000).
Notiomastodon platensis e Toxodontinae seriam animais de grande tamanho e
hábitos pastadores ou pastadores/ramoneadores (MACFADDEN, 2005; PRADO et al.,
2005) e ocorreriam em regiões semelhantes. Baseado em sua morfologia dental, N.
platensis apresentaria uma alimentação mista, consistindo em gramíneas, folhas e galhos
de arbustos (C3 e C4) (CARTELLE, 1992; SÁNCHEZ et al., 2003); enquanto que os
Toxodontinae alimentavam-se de gramíneas abrasivas (MACFADDEN, 2005).
A extinção dos grandes mamíferos, em função da hipótese de mudança ambiental
em todo o Brasil intertropical durante o Pleistoceno final-Holoceno (CARTELLE, 1992),
juntamente com desaparecimento de espécies de médio porte, são os possíveis fatores
envolvidos na extinção de predadores como os Felidae (PORPINO et al., 2004).
Sánches et al., (2006) sugerem a partir de análises isotópicas de carbono e oxigênio
amostradas de esmalte dentário dos táxons Hippidion Owen, 1869 e Equus (Amerhippus)
Hoffstetter, 1950, que os eqüídeos do Pleistoceno Médio apresentavam estratégias
oportunistas de forrageio. Estas podem ter contribuído para que os tornassem aptos a
viverem em diversos hábitats, e ao final do Pleistoceno teriam convergido para uma dieta
mais seletiva, restringindo seus hábitats.
Equus (amerhippus) neogaeus contém praticamente as mesmas características
morfológicas que os cavalos pastadores atuais (ALMEIDA, 2009), sendo considerado
pastador adaptado à corrida em áreas abertas (ALBERDI et al., 2003), alimentando-se de
plantas C4 (KOHN et al., 2005).
Em virtude a presença de alguns animais, tais como E. laurillardi e P. greslebini,
estes podem ser localizados no Neopleistoceno-Holoceno (PORPINO et al., 2004), ou
equivalente ao Lujanense, já que E. (A.) neogeus é considerado como um fóssil-guia do
Lujanense - Biozona Equus (Amerhippus) neogeus, no começo do Neopleistoceno (CIONE
& TONNI, 1999).

47
Conclusões
Registra-se pela primeira vez, a ocorrência de Eremotherium laurillardi (Lund,
1842) Cartelle & Bohórquez, 1982; Notiomastodon platensis (Ameghino, 1888); Felidae;
Toxodontinae; Panochthus greslebini e Equus (Amerhippus) neogeus para o municipio de
Jaguarari - BA.
A partir deste conjunto fossilífero analizado, sugere-se condições ambientais
diversas da encontrada atualmente para a região, provavelmente semelhantes àquelas
inferidas por Cartelle (1999) para o Pleistoceno do Brasil Intertropical Extra-Amazônico,
onde, durante o Pleistoceno final - Holoceno, a região compreenderia áreas abertas de
savana associadas, possivelmente, com fisionomias mais fechadas, tipo florestas.
Referencias Bibliográficas
Alberdi, M. T. y Prado, J. L. 1992. El registro de Hippidion Owen, 1869 y Equus
(Amerhippus) Hoffstetter, 1950 (Mammalia, Perissodactyla) en América del Sur.
Ameghiniana 29(3): 265 - 284.
Alberdi, M. T. y Prado, J. L. 1995. Los Équidos de América del Sur. Em Alberdi, M. T.,
Leone G. y Ton, E. P. (eds.), Evolución biológica y climática de la Región Pampeana
durante los últimos cinco millones de años, p. 295 - 308, Madrid.
Alberdi, M. T., Cartelle, C. & Prado, J. L. 2003. El registro Pleistoceno de Equus
(Amerhippus) e Hippidion (Mammalia, Perissodactyla) de Brasil. Consideraciones
paleoecológicas y biogeográficas. Ameghiniana, 40(2): 173-196.
Almeida, C.B.; Sicuro, F.L; Avilla, L.S. 2009 Paleoecologia dos Equídeos Plesitocênicos
Brasileiros (Mammalia, perissodactyla, Equidae): Uma análise
ecomorfológicacomparativa de suas regiões rostrais. Anais do IX Congresso de
Ecologia do Brasil, 13 a 17 de Setembro de, São Lourenço – MG
Araújo Junior H.I., Porpino K.O. 2007. Mamíferos fósseis da Fazenda Lágea Formosa, São
Rafael, Rio Grande do Norte, Brasil: Interpretações paleoecológicas. In: VIII
Congresso de Ecologia do Brasil. Boletim de resumos. Anais, p. 1-2.
Behling, H.; Arz, H.W.; Tzold, J.R.P.; Wefer, G. 2000. Late Quaternary vegetational and
climate dynamics in northeastern Brazil, inferences from marine core GeoB 3104-1.
Quaternary Science Reviews, 19:981-994.
Beltrão, M. C. M. C. & Locks, M. 1989. Pleistocene fauna from the “Toca da Esperança”
site, (Archaeollogical-Region of Central, Bahia, Brazil): Mammals – nº 1. In:
CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 11, Curitiba, 1989. Anais do
Congresso, vol. 1: 685- 697.
Berqvist, L. P. 1993. Jazimentos pleistocênicos do Estado da Paraíba e seus fósseis.
Revista Nordestina de Biologia, 8(2): 143-158.

48
Berta, A. 1985. The status of Smilodon in North and SouthAmerica. Contribution in
Science of the Natural History Museum of Los Angeles County 370:219–227.
Bond, M., Cerdeño, E. P. & López, G. 1995. Los ungulados nativos de América de Sur. In:
Evolución climática y biológica de la región Pampeana durante los últimos cinco
millones de años. Un ensayo de correlación con el Mediterráneo occidental (Eds. M.
T. Alberdi, G. Leone & E. P. Tonni). Monografías del Museo de Ciencias Naturales,
Consejo Superior de Investigaciones Científi cas, Madrid, 12, 259-275.
Bond, M. 1986. Los ungulados fósiles de Argentina: evolución y paleoambientes. Actas IV
Congreso Argentino Paleontología, Bioestratigrafía, Mendoza, 2, 173-185.
Bond, M. 1999. Quaternary native ungulates of Southern South America. A synthesis. In:
Quaternary of South America and Antarctic Peninsula (Eds. J. Rabassa & M.
Salemme). Centro Austral de Investigaciones Científicas and Universidad Nacional de
la Patagonia, Ushuaia, Tierra de Fuego, 12, 177-205.
Brizuela, Rafael Rodríguez, 2005 Presencia de Equus (Amerhippus) neogeus Lund em el
yacimiento paleontológico de Pehuen Có (Pleistoceno tardío de la provincia de
Buenos Aires) y su significado bioestratigráfico Revista de la Asociación Geológica
Argentina, 60 (3): 613-615
Campos, D. de a., Campos, D. R. B., 1976 Localidades fossilíferas da Folha de Brasília –
SD 23. In: Bruni, M. A. L., Carta geológica do Brasil ao milionésimo: folha Brasília.
Brasília. DNPM, p. 141-144.
Cartelle, C.& Abuhid, V.S., 1989. Novos espécimes de Smilodon populator (Lund, 1842),
Carnívora, Machairodontinae: morfologia e conclusões taxonômicas. In:
CONGRESSO BRASILEIRO DE PALEONTOLOGIA, 11, 1989. Anais, Curitiba,
UFPR, p. 607-620.
Cartelle, C. & Bohórquez, G.A., 1982, Eremotherium laurillardi (Lund, 1842). Parte I.
Determinação específica e dimorfismo sexual. Iheringia, Série Geológica 7: 45-63.
Cartelle, C. & Bohórquez, G.A. 1985. Pampatherium paulacoutoi, uma nova espécie de
tatu gigante da Bahia, Brasil (Edentata, Dasypodidae). Revista Brasileira de Biologia,
2(4):229-254.
Cartelle, C. & De Iuliis, G. 1995. Eremotherium laurillardi: the Panamerican late
Pleistocene Megatheriid sloth. Journal of Vertebrate Paleontology, 15(4):830-841.
Cartelle, C. 1999. Pleistocene mammals of the Cerrado and Caatinga of Brazil. In:
EISENBERG, J.F. & REDFORD, K.H. (eds.). Mammals of the Neotropics. The
University of Chicago Press, p. 27-46.
Cartelle, c. brant. W & piló, b. 1989 A Gruta do Túnel de Santana (BA): morfogênese e
paleontologia. In: XI Congresso Brasileiro de Paleontologia. Curitiba. Anais, 1:593-
606.
Cartelle, C. 1992. Edentata e megamamíferos herbívoros extintos da toca dos ossos
(Ourolândia, BA). Programa de Pós-graduação em Morfologia, Universidade Federal
de Minas Gerais, Tese de Doutorado, 516pp.
Cartelle, C. 1994. Tempo Passado: Mamíferos do Pleistoceno em Minas Gerais. Belo
Horizonte: Palco, 131 p.
Cione, A.L. & Tonni, E.P. 1999. Biostratigraphy and chronological scale of upper-most
Cenozoic in the Pampean Area, Argentina. In: J. Rabassa & M. Salemme (eds.)
Quaternary of South America and Antarctic Peninsula, A.A. Balkema Publishers, p.
23-52.

49
Czerwonogora, A., De Esteban-Trivigno, S. & Fariña, R. A. 2003. Hábitos alimentícios de
ungulados pleistocenos sudamericanos: un enfoque ecomorfológico. Ameghiniana
(Resúmenes), 40, 83R.
De Iullis, G., Bargo, M. S. & Vizcaíno, S. F. 2000. Variation in skull morphology and
mastication in the fossil giant armadillos Pampatherium spp. And allied genera
(Mammalia: Xenarthra: Pampatheriidae), with coments on their systematics and
distribution. Journal of Paleontology Vertebrate. 20(4): 743- 754.
De Oliveira, P.E. de; Barreto, A.M.F.; Suguio, K. 1999. Late Pleistocene / Holocene
climatic and vegetational history of the brazilian Caatinga: the fossil dunes of the
middle São Francisco river. Palaeogeography, Palaeoclimatology, Palaeoecology
152: 319-337.
Gadens Marcon, G.T. 2008. Contribuição ao estudo dos Proboscidea (Mammalia,
Gomphotheriidae) do Quaternário do Estado do Rio Grande do Sul, Brasil. Revista
UnG – Geociências, 7(1):93-109.
Holland, w. j., 1920 Fossil mammals collected at Pedra Vermelha. Bahia, Brasil, by
Gerald A. Waring. Annals of the Carnegie Museum, 13 (1-2):224-232.
Kerber, L. & Oliveira, E.V. 2008. Presença de Equus (Perissodactyla, Equidae) e
Neuryurus (Cingulata, Glyptodontidae), no Quaternário do município de Quaraí, oeste
do Rio Grande do Sul. Caderno de Pesquisa, Sér. Biol., 20(3):18-25.
Kohn, M.J., McKay, M.P., and Knight, J.L. 2005. Dining in the Pleistocene: who’s on the
menu? Geology 33:649–652.
Kurtén, B., and Anderson, E., 1980, Pleistocene mammals of North America: New York,
Columbia University Press, 442 p.
Kurtén, B., and L. Werdelin. 1990. Relationships between North and South American
Smilodon. Journal of Vertebrate Paleontology 10:158–169.
Lessa, G.; Cartelle, C.; Faria, H. D. & Gonçalves, P. R., 1998. Novos achados de
mamíferos carnívoros do pleistoceno final – Holoceno em grutas calcárias do Estado
da Bahia, Acta geológica leopoldensia, XXI (46/47): 157-169,
MacFadden, B.L. 2005. Diet and habitat of toxodont megaherbivores (Mammalia,
Notoungulata) from the late Quaternary of South and Central America. Quaternary
Research, 64: 113-124.
Mcdonald, H. Gregory & Lundelius, JR Ernest L. 2009 The giant ground sloth
Eremotherium laurillardi (Xenarthra, Megatheriidae) in Texas. Albright, L. B. III, ed.,.
Papers on Geology, Vertebrate Paleontology, and Biostratigraphy in Honor of Michael
O. Woodburne. Museum of Northern Arizona Bulletin 65, Flagstaff, Arizona.
Miño Boilini, A.R., Cerdeño, E. & Bond, M. 2006. Revisión del género Toxodon Owen,
1837 (Notoungulata: Toxodontidae) en el Pleistoceno de las provincias de Corrientes,
Chaco y Santa Fe, Argentina. Revista Española de Paleontología, 21(2):93-103.
Mothé, D.; Avilla, L.S.; Cozzuol, M.A. & Winck, G.R. no prelo. Revision of the
Quaternary South America lowland gomphotheres (Mammalia: Proboscidea:
Gomphotheriidae). Quaternary International. doi:10.1016/j.quaint.2011.05.018
Oliveira, E. 1999. Quaternary vertebrate and climates from southern Brazil. In: Quaternary
of South America and Antarctic Peninsula (Eds. J. Rabassa & M. Salemme). Centro
Austral de Investigaciones Científi cas y Universidad Nacional de La Patagonia,
Ushuaia, Tierra de Fuego, 5, 61-75.

50
Oliveira, E.V.; Dutra, T. & Zeltzer, F. 2002. Megaterídeos (Mammalia, Xenarthra) do
Quaternário de Caçapava do Sul, Rio Grande do Sul, com considerações sobre a flora
associada. Geologia Colombiana, 27:77-86.
Paula Couto, C. de, 1947. Contribuição para o estudo de Hoplophorus euphractus Lund,
1839. Summ Brasiliensis Geologiae, 1 (4):33-53.
Paula Couto, C de, 1957, Sobre um Gliptodonte do Brasil. Divisão de Geologia e
Mineralogia. DNPM. Boletim, 165:1-37.
PAULA COUTO, C. de. 1953 Paleontologia Brasileira: mamíferos. Rio de Janeiro:
Biblioteca Cientifica Brasileira, 516p..
Paula-Couto, C. de 1979. Tratado de Paleomastozoologia.Rio de Janeiro, Academia
Brasileira de Ciências, 590 p.
Paula Couto, C. de. 1982. Fossil mammals from the Cenozoic of Acre, Brazil. V.
Notoungulata Nesodontinae (II) Haplodontheriinae and Liptoterna, Pyrotheria and
Astrapotheria (II). Iheringia, Série Geológica, 7:5-43.
Porpino, K.O. & Bergqvist, L.P. 2002. Novos achados de Panochthus (Mammalia,
Cingulata, Glyptodontoidea) no nordeste do Brasil. Revista Brasileira de
Paleontologia 4: 51-62.
Porpino, K. O., Santos, M. F. C. F. & Bergqvist, L. P. 2004. Registro de mamíferos fósseis
no Lajedo de Soledade, Apodi, Rio Grande do Norte, Brasil. Revista Brasileira de
Paleontologia, 7(3): 349-358.
Porpino, K.O. 1999. Estudo dos Cingulata (Xenarthra, Mammalia) fósseis depositados no
Museu Câmara Cascudo, Natal-RN. Programa de Pós-Graduação em Zoologia,
Universidade Federal do Rio de Janeiro, Dissertação de Mestrado, 138 p.
Prado, J. L. & Alberdi, M. T. 2008. Restos de Hippidion y Equus (Amerhippus)
procedentes de las Barrancas de San Lorenzo, Pleistoceno tardío (Provincia de Santa
Fé, Argentina). [Hippidion and Equus (Amerhippus) from the San Lorenzo cliff, late
Pleistocene (Santa Fe province, Argentina).] Revista Española de Paleontología, 23
(2), 225-236. ISSN 0213-6937.
Prado, J. L. y Alberdi, M. T. 1994. A quantitative review of the horses Equus from South
America. Paleontology, 37: 459-481.
Prado, J.L. et al., 2001. The Pleistocene Gomphotheres (Proboscidea) from South America:
diversity, habitats and feeding ecology. In: THE WORLD OF ELEPHANTS –
INTERNACIONAL CONGRESS, Rome. Annals…p. 337-340.
Prado, J. L., Alberdi, M. T., Azanca, B., Sánchez, B. & Frassinetti, D. 2005. The
Pleistocene Gomphotheriidae (Proboscidea) from South America. Quaternary
International, 126- 128: 21-30.
Pujos, F. & Salas, R. 2004. A new species of Megatherium (Mammalia: Xenarthra:
Megatheriidae) from the Pleistocene of Sacaco and Tres Ventanas, Peru. Paleontology
47(3): 579-604.
Ranzy, A. 2000. Paleoecologia da Amazônia – Megafauna do Pleistoceno. Florianópolis,
Editora da UFSC, 101 p.
Ribeiro, Ricardo da Costa & Carvalho, Ismar de Souza 2009 Megafauna do Quaternário
tardio de Baixa Grande, Bahia, Brasil Anuário do Instituto de Geociências – UFRJ -
Vol. 32 - 2 / p. 42-50

51
Rincón, A.D.; Prevostri, F.J. & Parra, G.E. 2011. New saber-toothed cat records (Felidae:
Machairodontinae) for the Pleistocene of Venezuela, and the great American biotic
interchange. Journal of Vertebrate Paleontology, 31(2):468-478.
Rincón A.D. 2006. A first record of the Pleistocene saber-toothed cat Smilodon populator
Lund, 1842 (Carnivora: Felidae: Machairodontinae) from Venezuela. Ameghiniana,
43(2):499-501.
Sanchez, B., Prado, J.L., Alberdi, M.T. 2006. Ancient feeding, ecology and extinction of
Pleistocenic horses from the Pampean Region, Argentina. Ameghiniana, 43(2): 427 -
436,
Sánchez, Begoña; Prado, José Luis; Alberdi, María Teresa 2003 Paleodiet, ecology, and
extinction of Pleistocene gomphotheres (Proboscidea) from the Pampean Region
(Argentina), Coloquios de Paleontología, Vol. Ext. 1. 617-625.
Silva, C. C. 2001. Relatório de Pesquisas: Arqueologia e Paleontologia – semestres 2000.2,
2001.1 e 2001.2. Laboratório de Geociências. Departamento de Educação-Campus
VII/UNEB, Senhor do Bonfim, 31p.
Simpson, G.G.; Paula-Couto, C. de. 1957. The Mastodonts of Brazil. Bulletin of American
Museum of Natural History 112(2): 1-65.
Spix & Martius. 1976. Viagem pelo Brasil: 1817-1820. São Paulo: Melhoramentos.
Vizcaíno, S.F. 2000. Vegetation Partitioning among Lujanian (Late- Pleistocene/Early
Holocene) armored herbivores in the Pampean Region. Current Research in the
Pleistocene, 17:135-137.
Zurita, A.E.; Carlini, A.A.; Scillato-Yané, G.J.; Tonni, E.P. 2004. Mamíferos extintos del
Cuaternário de la provincia del Chaco (Argentina) y su relación com aquéllos del este
de la región pampeana y de Chile. Revista Geológica de Chile, 31(1):65-87.

52
Figura 01. Mapa de localização da Microrregião de Senhor do Bonfim, com visualização dos nove
municípios. (Modificado de SEI, 2007).
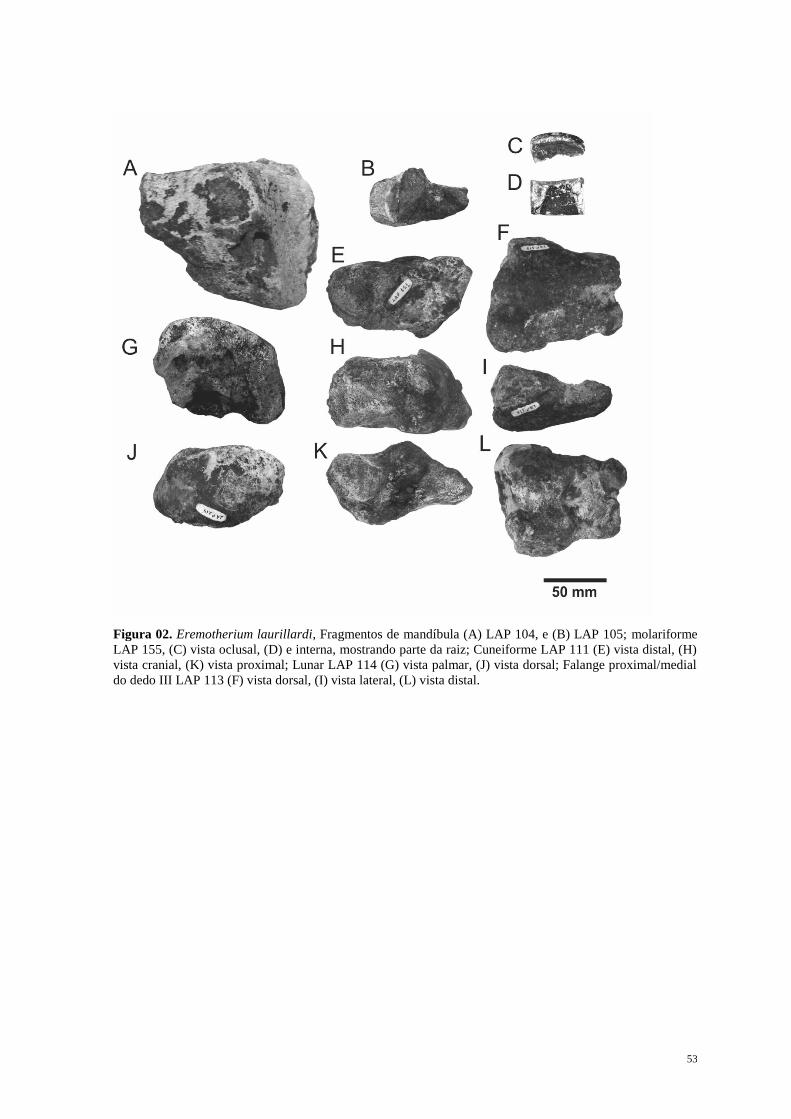
53
Figura 02. Eremotherium laurillardi, Fragmentos de mandíbula (A) LAP 104, e (B) LAP 105; molariforme
LAP 155, (C) vista oclusal, (D) e interna, mostrando parte da raiz; Cuneiforme LAP 111 (E) vista distal, (H)
vista cranial, (K) vista proximal; Lunar LAP 114 (G) vista palmar, (J) vista dorsal; Falange proximal/medial
do dedo III LAP 113 (F) vista dorsal, (I) vista lateral, (L) vista distal.

54
Figura 03. Notiomastodon platensis, Fragmento de molar LAP 163 (A), vista oclusal, (D) vista labial;
fragmento de incisivo LAP 169 (B) vista transversal, (C) vista rostral; Eremotherium laurillardi LAP 122 (E)
vista proximal, (G) vista lateral; Panochthus greslebini LAP 162 (F) osteodermo isolado; Toxodontinae
Fragmento de molar LAP 165 (H) vista rostral, (I) vista oclusal, (J) vista labial; Equus (Amerhippus) neogeus
LAP 166 (K) vista oclusal.

55
Capítulo 2. Dinâmica de Vegetação e Clima no
Quaternário recente

56
DINÂMICA DE VEGETAÇÕES E CLIMA NO QUATERNÁRIO RECENTE EM
JAGUARARI – BA INFERIDA A PARTIR DOS ISÓTOPOS DE CARBONO DA
MATÉRIA ORGÂNICA DO SOLO
DYNAMICS OF VEGETATIONS AND CLIMATE IN THE RECENT QUATERNARY IN
JAGUARARI - BA INFERRED FROM CARBON ISOTOPES OF THE SOIL ORGANIC
MATTER
Abstract: Numerous studies of paleo-environmental reconstruction (vegetation and
climate) have been carried out in southern, southeastern, and central Brazil, based on the
pollen deposited in lacustrine and peatbog sediments. Such studies are scarce in the
Brazilian Northeast, however, which stimulated the development of the present study,
using carbon isotopes (13
C) from organic material found in the soil (MOS) and plants, to
provide systematic inferences on climatic variation. Soil and plant samples were collected
for isotopic analysis from three areas in the municipality of Jaguarari, located in the
Caatinga biome. The isotopic data (13
C) indicate that the region’s vegetation has been
stable in the past, and up to the present day, with no exchange of C3 to C4 at any of the
study sites. The presence of charcoal fragments at different depths in the soil indicates the
occurrence of wildfires throughout the Holocene, possibly as a consequence of a relatively
much drier climate. These wildfires may have been anthropogenic, given that the charcoal
was found in association with vestiges of the region’s ancient human populations. The
dynamics of the vegetation observed in Jaguarari to the depth excavated correspond to that
found in other studies in the Brazilian Northeast, which indicate a predominance of
arboreal vegetation since approximately 15,000 BP, eventually returning to a more humid
climate similar to that of the present day. The vegetation of the study region may not have
suffered major modifications of the vegetation during the drier periods of the mid-
Holocene, with forest formations predominating.
Key words: Caatinga; Stable isotopes; Organic matter on soil..
Resumo: Estudos de reconstrução paleoambiental (vegetação e clima) no Brasil vêm sendo
desenvolvidos em várias regiões, principalmente Sul, Sudeste e Central, utilizando pólen
depositado em sedimentos lacustres e turfeiras. Na região Nordeste, os trabalhos são mais
escassos, o que motivou o desenvolvimento deste estudo utilizando os isótopos do carbono
(13
C) da matéria orgânica do solo (MOS) e das plantas, com inferências às variações
climáticas. Foram amostrados solos e plantas para análises isotópicas em três áreas no
município de Jaguarari, em um domínio de Caatinga. Os dados isotópicos (13
C) indicam
estabilidade vegetacional na região até o presente, sem troca de vegetação C3 para C4 nos
locais estudados. A presença de fragmentos de carvão soterrados em várias profundidades
do solo sugere a ocorrência de paleoincêndios durante todo o Holoceno, com a possível
influência de um clima mais seco para a região. Estes incêndios podem ser de natureza
antrópica, já que foram encontrados alguns vestígios de ocupação de populações antigas na
região. A dinâmica da vegetação observada para Jaguarari, até a profundidade de solo
estudada, se em concordância com outros estudos no Nordeste, sugere uma vegetação
arbórea dominante, desde aproximadamente 15.000 anos AP. com um retorno a um clima
mais úmido e provavelmente similar ao atual.
Palavras Chave: Caatinga; Isótopos estáveis; Matéria orgânica do solo.

57
1. Introdução
Estudos de reconstrução paleoambiental (vegetação e clima) no Brasil,
principalmente nas regiões Sul, Sudeste e Central, têm sido mais intensamente
desenvolvidos desde o início da década de 1990. Nestes trabalhos têm-se utilizado pólens
depositados em sedimentos lacustres e turfeiras (FERRAZ-VICENTINI, 1993; LEDRU,
1993; NEVES & LORSCHEITTER, 1995; LEDRU et al., 1996, 1998; BEHLING,
1995a,1997a; FERRAZ-VICENTINI & SALGADO-LABOURIAU, 1996; BARBERI et
al, 2000), isótopos de carbono da matéria orgânica do solo (MOS) (DESJARDINS et al.,
1996; PESSENDA et al., 1996a, 1996b, 1998a, 1998b, 1998c) e dos fragmentos de carvão
encontrados nos solos (DESJARDINS et al., 1996; PESSENDA et al., 1996a; GOUVEIA
et al., 1999, 2002; SANTOS et al., 2000).
Embora os estudos paleoecológicos do Quaternário apresentem um considerável
avanço, as paleovegetações e paleoclimas presentes nos Nordeste brasileiro durante o
Pleistoceno tardio e Holoceno permanecem praticamente desconhecidos. A dificuldade em
encontrar áreas permanentemente alagadas ou sedimentos de turfa não oxidados em
domínios fitogeográficos de floresta de Caatinga em regiões Semi-Áridas representa um
dos principais obstáculos ao desenvolvimento de estudos paleoecológicos baseados na
palinologia nesta região (DE OLIVEIRA et al., 1999).
Conti e Furlan (2003) afirmam que o mosaico geobotânico brasileiro é originário do
resultado da expansão e retração das florestas, cerrados e caatingas, provocados pela
alternância de climas úmidos e secos nas regiões tropicais, durante os períodos glaciais do
Quaternário.
Esta dinâmica é ocasionada pelas diferentes capacidades de adaptação ao ambiente,
onde plantas C4 são mais resistentes a um regime de baixa disponibilidade hídrica e são
típicas de vegetações mais abertas (maior intensidade luminosa) (EHLERINGER, 1978;
NORDT et al., 1994) que plantas C3 de florestas tropicais (EHLERINGER, 1991).
O mecanismo de fixação do carbono nas plantas C3 foi descrito por Calvin &
Benson (1948). No ciclo de Calvin como foi denominado, uma molécula de CO2 é
incorporada a uma pentose com dois átomos de fósforo, a ribulose-1-5-difosfato (RUDP).
A enzima RUDP-carboxilase é fundamental na ligação da pentose ao gás CO2, formando
duas moléculas de 3-fosfoglicerato (3PGA) com 3 átomos de carbono, razão pela qual esta
via é chamada de ciclo C3.

58
As plantas de ciclo C4 foram descritas por Hatch & Slack (1966) ao descobrirem
que existia outra via de pela qual o CO2 pode ser incorporado. Neste outro mecanismo, o
ácido axalacético com quatro átomos de carbono, na presença de CO2 reage com o
fosfopiruvato (PEP), que possui três átomos de carbono. Essa reação é mediada por uma
enzima chamada de PEP-carboxilase. A capacidade dos tecidos das plantas C4 de
concentrarem CO2 atmosferico nos sítios de produção de carboidratos (céluas da bainha
vascular) é um dos parâmetros mais importantes na diferenciação das plantas C3 e C4.
Uma terceira via de fixação de CO2 é conhecida como Metabolismo Ácido
Crassuláceo (CAM), e evoluiu em muitas plantas suculentas de clima árido. Nessas
plantas, os ácidos málico e isocítrico acumulam-se nas folhas durante à noite e são
novamente convertidos em CO2 na presença de luz. As plantas CAM abrem os estômatos à
noite, quando a temperatura e a evapotranspiração diminuem, favorecendo o aumento da
umidade relativa da camada de ar que envolve a planta. O CO2 é fixado pela via C4 e é
acumulado em forma de malato. Durante o dia os estômatos permanecem fechados e o
carbono acumulado é utilizado na fotossíntese, aumentando a eficiência na utilização da
água (RAVEN et al., 1978).
As plantas C3 apresentam valores de 13
C mais negativos do que as plantas C4
(SMITH & EPSTEIN, 1971; SMITH & BROWN, 1973) e os mecanismos dos ciclos
fotossintéticos foram estudados através da composição isotópica 13
C/12
C, independente dos
estudos da fisiologia ou da anatomia da planta. Estabeleceu-se que a variação nos valores
da relação 13
C/12
C era tanto interespecífica quanto intraespecífica, podendo assim, ser
utilizada para entender o histórico dos metabolismos e do ambiente de vida das plantas
(WINTER et al., 1976; EHLERINGER, 1978).
Os valores de 13
C das plantas C3 variam de -32‰ a -22‰, com uma média de -
27‰, estando presente em aproximadamente 85% das espécies vegetais terrestres, desde as
florestas boreias até os trópicos. Nas plantas C4, os valores de 13
C variam de -9‰ a -17‰,
com uma média de -13‰, compreendendo 5% de todas as espécies vegetais, sendo que
ocorrem em 18 famílias e metade são espécies de gramíneas tropical e subtropical que
cobrem extensas áreas de savanas, campos e pântanos salgados. As espécies de plantas
CAM, são geralmente suculentas, estão distribuídas em 30 famílias, tais como
Crassulaceae, Euphorbiaceae, Orchidaceae, Bromeliaceae, Cactaceae. As espécies CAM
facultativas apresentam valores de 13
C de -10‰ a -28‰, enquanto que as espécies CAM
obrigatórias os valores isotópicos são comparáveis aos das plantas C4 (BOUTTON, 1996).

59
A utilização dos isótopos do carbono da matéria orgânica do solo (MOS) é uma
fundamental ferramenta na caracterização de vegetações atuais e passadas (BOUTTON,
1991), uma vez que, os métodos convencionais para estudo da dinâmica da vegetação atual
utilizam os parâmetros estruturais e fitossociológicos para medir os estágios sucessionais
das comunidades vegetacionais (GLENN-LEWIS et al., 1992).
Utilizando-se análises isotópicas durante a dissolução e precipitação da calcita em
solos sob condições de semi-aridez, inferiu-se que o clima no Nordeste foi mais úmido por
volta de 26.000 e 20.000 anos Antes do Presente (AP) (DEVER et al., 1987).
Czaplewski & Cartelle (1998) em cinco cavernas na Caatinga no Nordeste da
Bahia, encontraram 27 taxa de Quirópteras fósseis. Datação por 14
C do húmero de
Mormops megalophyla revelou uma idade de 20.060 anos AP e, em outra caverna, os
coprólitos e o crânio de Desmodus rotundus revelaram uma idade de aproximadamente
12.000 anos AP para os fósseis estudados. A distribuição e abundância destes fósseis,
exceto uma espécie extinta Desmodus vampire, estão relacionadas com florestas e cavernas
com alta umidade, sugerindo que as cavernas estudadas foram mais úmidas durante o
Pleistoceno tardio do que no presente.
Para um campo de dunas fósseis no Vale do Icatu, localizado entre Barra e Pilão
Arcado (BA) ou médio São Francisco, constatou-se a ocorrência de sedimentos de origem
eólica que foram datados por termoluminescência. Os resultados indicaram estabilidade
nos períodos de geração das dunas depositadas entre 10.000 e 1.000 anos AP (TATUMI et
al., 1998).
Em estudos palinológicos também no Vale do Rio Icatu, região semi-árida da
Bahia, De Oliveira et al., (1999) sugeriram mudanças na vegetação e clima durante os
últimos 11.000 anos AP para a região. Para os autores, os taxa de espécies de florestas da
Amazônia, Atlântica e de montanha, indicam um período muito úmido e de temperaturas
mais reduzidas no final do Pleistoceno (10.990 – 10.540 anos AP).
De 10.540 a 8.910 anos AP a vegetação dominada por Mauritia vinifera sugere um
progressivo aumento de temperatura e alta umidade. De 8.910 a 6.790 anos AP houve um
progressivo declínio de taxa de floresta e aumento de taxa de Caatinga e Cerrado. Até
4.535 anos AP houve um retorno a um mosaico de vegetações composta de floresta de
galeria, caatinga e cerrado, indicando um clima mais úmido. Desde então, houve um
aumento nos taxa de caatinga e de cerrado, caracterizando uma diminuição de umidade e o

60
declínio da floresta de galeria, aspecto encontrado atualmente (DE OLIVEIRA et al.,
1999).
Na região de Lage dos Negros e Abreus, norte da Bahia, Auler e Smart (2001)
estudaram depósitos de travertino fóssil e espeleotemas subaquáticos em cavernas secas da
caatinga. Esta região atualmente apresenta um dos menores índices de precipitação do
Nordeste, em torno de 490 mm anuais. Utilizando técnicas isotópicas (datação 230
Th/234
U),
os autores verificaram que durante o Quaternário ocorreram elevações no nível da água
subterrânea. O travertino foi depositado em duas fases distintas, uma fase datada em
40.000 anos AP e outra entre 21.400 anos AP até o início do Holoceno. Em duas cavernas
distintas foi datada a formação de depósitos de calcita em espeleotemas. A idade de
145.000 anos AP foi associada ao penúltimo período glacial e as de 21.000 e 17.300 anos
AP ao Último Máximo Glacial (UMG).
Estes resultados evidenciam períodos úmidos durante o UMG, contrastando com a
aridez proposta por estudos realizados na Amazônia e sudeste do Brasil. Pelo menos nas
duas últimas glaciações, o semi-árido nordestino estaria sob condições úmidas (AULER &
SMART, 2001).
A hipótese de dinâmica da paleovegetação na região Nordeste sob influência do
semi-árido e das florestas Atlântica e Amazônica necessita de uma melhor compreensão,
uma vez que a região central do Nordeste, dominada pela vegetação de caatinga,
caracteriza-se por apresentar uma grande diversidade de espécies adaptadas à semi-aridez e
ao relevo local. O domínio do semi-árido, aproximadamente 1.000.000 km2, forma um
polígono de aridez que separa duas florestas tropicais, a mata Atlântica da Amazônia. Na
transição entre a caatinga e região Amazônia ocorrem várias subformações vegetais
indefinidas de cerrado e caatinga, provavelmente associadas às mudanças climáticas
passadas (RIBEIRO, 2002).
Neste estudo, iniciou-se o estudo da dinâmica da vegetação atual e passada durante
o Pleistoceno tardio e Holoceno, utilizando os isótopos de carbono da matéria orgânica de
solos coletados em áreas com vegetação de floresta e capoeira, no domínio de Caatinga –
região Centro-Oeste da Bahia.
Nas duas áreas de estudo efetuou-se a caracterização isotópica das espécies vegetais
predominantes e procurou-se associá-las à dinâmica da vegetação atual.

61
2. Materiais e Métodos
2.1 Características geomorfológicas e climáticas da área de estudo
As amostras de solo e plantas foram coletadas em um Latossolo Amarelo (LA) no
Sítio Poço Velho, em três pontos de amostragem com vegetações distintas. Dois destes
pontos encontrava-se em uma floresta hipoxerófila de Caatinga arbórea, e o terceiro em
uma vegetação de capoeira, localizados numa propriedade particular que dista 8 km da
sede do município de Jaguarari (10° 15’ 00” S e 40° 11’ 00” W).
O município de Jaguarari possui clima semi-árido e está incluído no “Polígono das
Secas” (CPRM, 2005). Possui tipos variados de solos, sendo possível identificar:
planossolos, argissolos e neossolos litólicos eutróficos, neossolos regolíticos e nitossolos
distróficos. Apresenta vegetação variegada composta por contato cerrado-floresta
estacional e as Caatingas arbóreas abertas ou densas, ambas com ou sem palmeiras e,
ainda, o contato Caatinga-floresta estacional. O relevo local é do tipo serrano, passando a
baixadas e pediplano sertanejo, cortados pelos rios Jaguarari e do Brejo, que servem às
bacias Itapicuru e São Francisco (CPRM, 2005).
A sede do município tem altitude de 650 metros e coordenadas geográficas
10°15’00”de latitude sul e 40°11’00” de longitude oeste, e geologicamente caracteriza-se
pela ocorrência de rochas cristalinas do Complexo Itapicuru, Mairi, Tanque Novo-Ipirá e
Caraíba, além de também serem encontrados corpos graníticos e granitóides. É possível
encontrar no município três tipos de coberturas quaternárias: coberturas detrito-lateríticas,
formadas por areia com níveis de cascalho e argila e crosta laterítica, ocorrendo em
segmentos isolados nas porções central e sul do município; brecha calcífera e calcrete na
porção noroeste; e, em menor proporção, depósitos aluvionares recentes, relacionados ao
riacho da Barrinha (CPRM, 2005).
2.2 Análises do Solo
2.2.1 Amostragem e preparo do solo
Em um dos pontos de amostragem, foi feita uma trincheira de aproximadamente
150 cm x 150 cm x 300 cm de profundidade, de onde foram coletadas amostras de solo a
cada 10 cm de profundidade, sendo aproximadamente 10 kg por camada, no sentido das

62
camadas inferiores para as superiores (Figura 01). As amostras foram acondicionadas em
sacos plásticos transparentes e devidamente identificadas.
Para amostras mais profundas na trincheira (até 420 cm) utilizou-se perfuração a
trado. Estas amostras foram acondicionadas em potes plásticos devidamente identificados e
possuíam capacidade de até 250 g de solo. Foram coletadas no sentido das camadas mais
superiores para as mais inferiores. Estas amostras são aqui denominas “solo da trincheira”.
Em um segundo ponto de amostragem, foi feita apenas a perfuração a trado até 200
cm de profundidade. Estas amostras (denominadas neste trabalho “solo da capoeira”)
foram acondicionadas em potes plásticos devidamente identificados e possuíam capacidade
de até 250 g de solo. Foram coletadas no sentido das camadas mais superiores para as mais
inferiores (Figura 01).
O terceiro ponto de coleta foi em uma área de floresta, no local onde foi encontrado
um artefato lítico (machadinha semilunar). Embora o encontro tenha sido acidental, e
inicialmente fora das áreas de amostragem, optou-se por fazer uma coleta de solo para
comparação com os outros locais estudados. O perfil do solo estava exposto, e percebia-se
uma grande quantidade de fragmentos de carvão presente nas camadas (Figura 01). Fez-se
uma coleta de aproximadamente 10 kg de solo por camada, no sentido das camadas
inferiores para as superiores. Essas últimas amostras (denominadas neste trabalho “solo da
machadinha”) foram acondicionadas em sacos plásticos transparentes e devidamente
identificadas.
Não foi necessário um período prolongado de secagem das amostras, já que estas
estavam praticamente secas, mesmo nas camadas mais profundas, apresentando apenas 1%
de umidade, medida com auxilio do aparelho HigroFarm – Falker 2010.
No laboratório as amostras foram passadas em peneiras de malha grossa para o
destorroamento e obtenção da terra fina seca ao ar (TFSA). Durante o processo retirou-se,
com o auxílio de uma pinça metálica, restos de raízes, vegetação, animais presentes e
fragmentos de carvão. O peneiramento foi realizado em peneiras com malha de 2 mm para
análises granulométricas e a 0,2 mm para análises isotópicas e de carbono orgânico total.
Ao término de cada peneiramento por camada, os utensílios utilizados (peneiras,
bacias, pinças) eram lavados em água corrente com detergente neutro, água destilada, e
colocados para secar em estufa a 60° C por 1h, na tentativa de evitar a mistura de carbono
das diferentes camadas. Só depois desta etapa, seguia-se com o peneiramento da camada
seguinte. Para tanto, utilizou-se três conjuntos metálicos.

63
Figura 01. Pontos de amostragem de solos no Município de Jaguarari. (A) Vista superior da trincheira na
área de vegetação de floresta; (B) Delimitação das camadas no interior da trincheira; (C) Camadas já
demarcadas e com a presença de carvões, destacados em amarelo; (D) Vista superior do perfil da trincheira
evidenciando as camadas de solo que já foram retiradas; (E) Perfil do solo sob vegetação de floresta, onde foi
encontrada a machadinha, destaque em amarelo para os carvões encontrados no solo e que se desprenderam
das camadas, quando da limpeza do perfil; (F) Coleta de solo na área de vegetação de campo utilizando
perfuração a trado.

64
2.2.2 Análises químicas e físicas do solo
As análises físico-químicas e granulométricas das amostras de solo foram
realizadas no Instituto Tecnológico e de Pesquisas do Estado de Sergipe – ITPS. Enviaram-
se aproximadamente 250 g de solo das amostras coletadas até 300 cm na trincheira e 420
cm por perfuração a trado na trincheira, tendo assim todo o perfil de 0-420 cm.
2.2.3 Determinação do Carbono Orgânico Total do solo
Partindo-se de 8 a 11 g de TFSA peneirada a 0,2 mm, foi determinado o carbono
orgânico total, que será reportado como Carbono Total (C-total), no Laboratório de
Isótopos Estáveis – Centro de Energia Nuclear na Agricultura da Universidade de São
Paulo. Os resultados apresentados referem-se à medida de três determinações, com
precisão de 1 a 2%, sendo expressos em % de C.
2.2.4 Determinação da razão 13
C/12
C (13
C) das amostras de solo
Foram enviados aproximadamente, 11g de TFSA, peneirada a 0,2 mm, ao
Laboratório de Isótopos Estáveis – Centro de Energia Nuclear na Agricultura da
Universidade de São Paulo, para a determinação das razões isotópicas 13
C/12
C em um
espectrômetro de massas ANCA-SL, Europa Scientific. Os resultados estão expressos pela
unidade relativa “ ”, determinada em relação ao padrão internacional PDB:
(1) 1000R
RRC
PDB
PDBamostra13
Onde a razão isotópica expressa a abundância relativa entre 13
C/12
C. Os resultados
apresentados referem-se à média de três determinações, com precisão de 0,2‰.

65
2.3 Análises dos Fragmentos de Carvão
2.3.1 Amostragem e preparo dos carvões
Com o auxílio de uma pinça de aço, os fragmentos de carvão foram coletados
durante o peneiramento das amostras de solo por camada, e colocados dentro de potes
etiquetados.
2.3.2 Análise do 14
C dos carvões
Para a determinação da idade 14
C dos fragmentos de carvão, foram enviadas ao
Laboratório de Carbono 14 do Centro de Energia Nuclear na Agricultura da Universidade
de São Paulo amostras que apresentaram, por camada, um peso superior a 5g, perfazendo
um total de 6 amostras.
Amostras de carvão com peso inferior a 5g por camada serão enviadas para os EUA
para serem analisadas por AMS e complementarem os dados.
2.4 Análises da Vegetação
2.4.1 Amostragem e preparo da vegetação
A amostragem de plantas foi realizada na área de floresta e na capoeira. Coletaram-
se amostras para análise isotópica (13
C) e para identificação botânica.
As amostras destinadas para análise isotópica foram lavadas com água destilada,
secas em estufa a 60° C, moídas, peneiradas a 0,2 mm e armazenadas em potes plásticos
com a devida identificação da espécie.
Utilizou-se para as análises isotópicas apenas as folhas das plantas, coletadas a uma
altura de 1,60 cm para as espécies lenhosas, e as mais externas, para as cactáceas, que eram
rasteiras.
2.4.2 Determinação da razão 13
C/12
C (13
C) das amostras de plantas
Enviaram-se aproximadamente 1 g de amostras de plantas, secas, moídas e
peneiradas ao Laboratório de Isótopos Estáveis – Centro de Energia Nuclear na Agricultura
da Universidade de São Paulo, para a determinação da razão isotópica 13
C/12
C em um

66
espectrômetro de massas ANCA-SL, Europa Scientific. Os resultados estão expressos pela
unidade relativa “ ”, determinada em relação ao padrão internacional PDB.
3. Resultados e Discussão
3.1 Análises químicas do solo.
As características químicas do solo da trincheira encontram-se na Tabela 01 e nas
Figuras 02 e 03.
Os valores de pH em água para o solo da trincheira foram classificados como de
acidez elevada, ao longo de todo o perfil (KIEHL, 1979; COCHRANE, 1984). Os valores
ficaram compreendidos entre 4,32 a 4,78, e apresentam uma distribuição normal (Figura
02).
Valores de pH compreendidos entre 3,5 a 5,5 foram encontrados e estudados por
Gomes (1996) para três Latossolos e um Podzólico Vermelho-Amarelo localizados no
estado de Rondônia.
A acidez do solo pode estar diretamente relacionada à quantidade de matéria
orgânica presente. Os valores ficaram compreendidos na faixa de 2,45 a 11,5 g/dm3.
Observou-se um decréscimo da matéria orgânica em relação à profundidade do
solo, contudo, devido ao aporte orgânico no solo, o pH ácido se manteve constante em todo
o perfil.
Figura 02. Variação do pH do solo em água (H20) em função da profundidade (0 – 4,20 cm) em Floresta
Hipoxerófila de Caatinga Arbórea, sitio Tanque Velho, Jaguarari – Bahia.
4.2 4.3 4.4 4.5 4.6 4.7 4.8 4.9
pH
0
1
2
3
4
5
6
7
8
9
Fre
qu
en
cia
Ob
se
rva
da
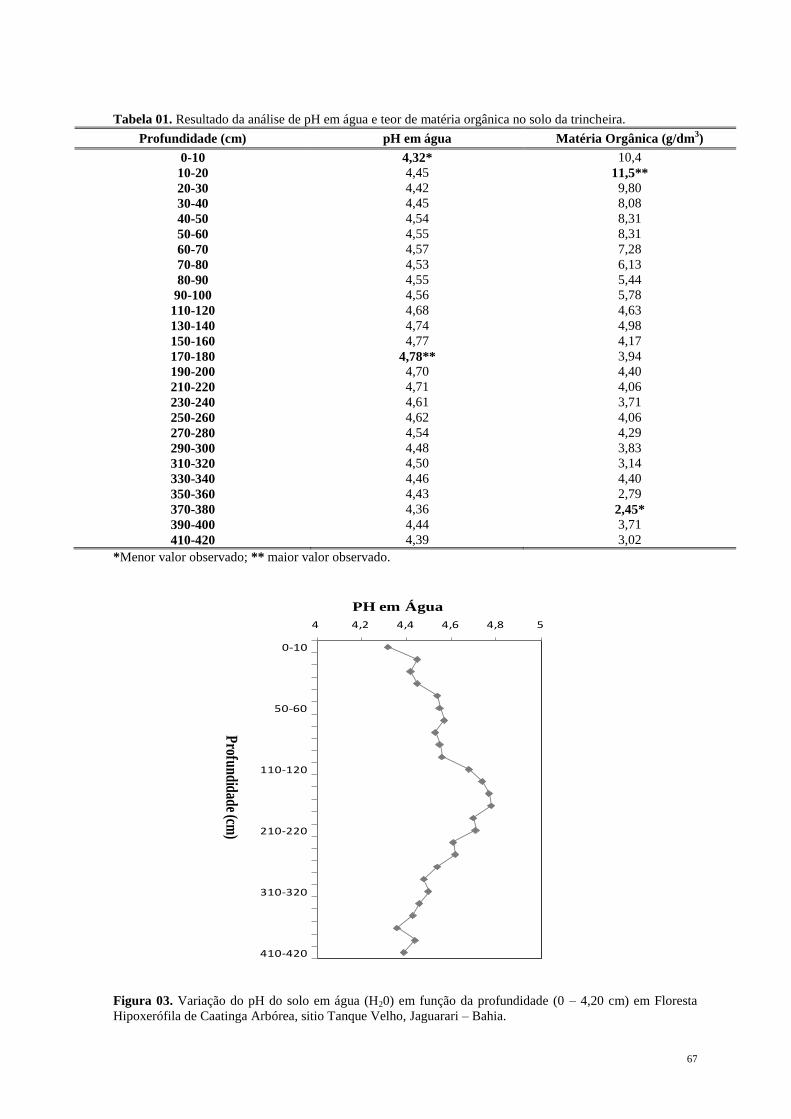
67
Tabela 01. Resultado da análise de pH em água e teor de matéria orgânica no solo da trincheira.
Profundidade (cm) pH em água Matéria Orgânica (g/dm3)
0-10 4,32* 10,4
10-20 4,45 11,5**
20-30 4,42 9,80
30-40 4,45 8,08
40-50 4,54 8,31
50-60 4,55 8,31
60-70 4,57 7,28
70-80 4,53 6,13
80-90 4,55 5,44
90-100 4,56 5,78
110-120 4,68 4,63
130-140 4,74 4,98
150-160 4,77 4,17
170-180 4,78** 3,94
190-200 4,70 4,40
210-220 4,71 4,06
230-240 4,61 3,71
250-260 4,62 4,06
270-280 4,54 4,29
290-300 4,48 3,83
310-320 4,50 3,14
330-340 4,46 4,40
350-360 4,43 2,79
370-380 4,36 2,45*
390-400 4,44 3,71
410-420 4,39 3,02
*Menor valor observado; ** maior valor observado.
Figura 03. Variação do pH do solo em água (H20) em função da profundidade (0 – 4,20 cm) em Floresta
Hipoxerófila de Caatinga Arbórea, sitio Tanque Velho, Jaguarari – Bahia.
4 4,2 4,4 4,6 4,8 5
0-10
50-60
110-120
210-220
310-320
410-420
PH em Água
Profundidade (cm
)
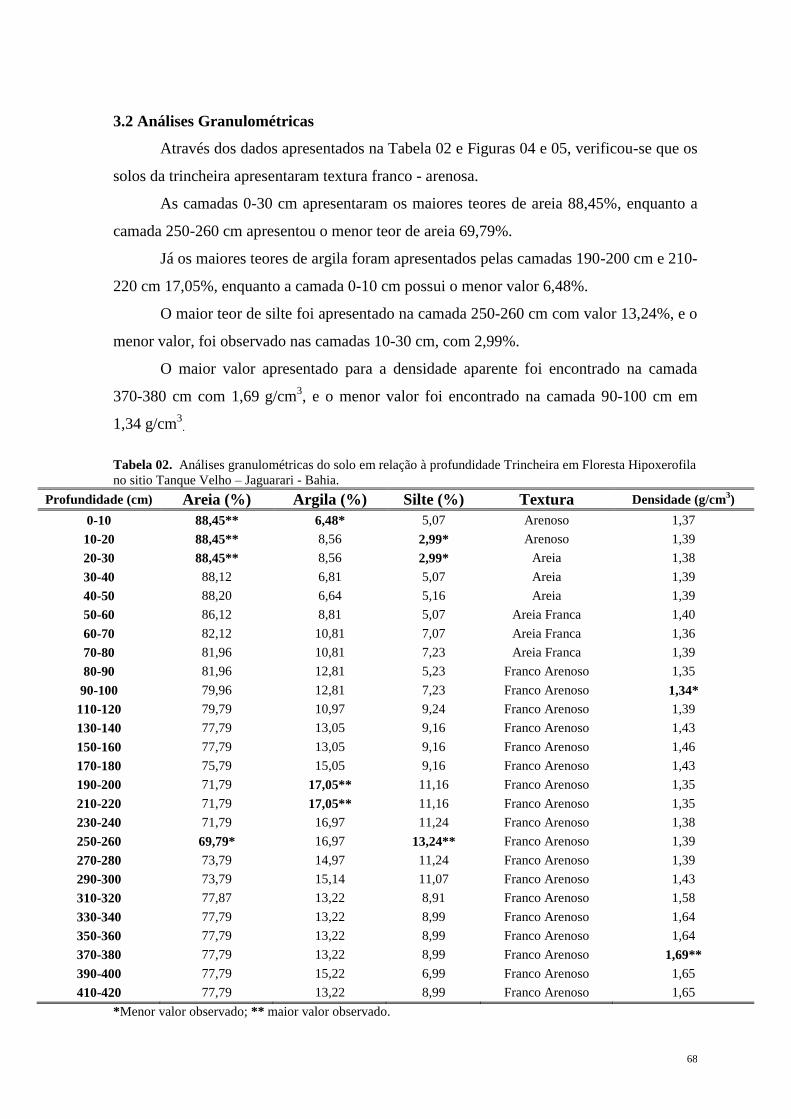
68
3.2 Análises Granulométricas
Através dos dados apresentados na Tabela 02 e Figuras 04 e 05, verificou-se que os
solos da trincheira apresentaram textura franco - arenosa.
As camadas 0-30 cm apresentaram os maiores teores de areia 88,45%, enquanto a
camada 250-260 cm apresentou o menor teor de areia 69,79%.
Já os maiores teores de argila foram apresentados pelas camadas 190-200 cm e 210-
220 cm 17,05%, enquanto a camada 0-10 cm possui o menor valor 6,48%.
O maior teor de silte foi apresentado na camada 250-260 cm com valor 13,24%, e o
menor valor, foi observado nas camadas 10-30 cm, com 2,99%.
O maior valor apresentado para a densidade aparente foi encontrado na camada
370-380 cm com 1,69 g/cm3, e o menor valor foi encontrado na camada 90-100 cm em
1,34 g/cm3
.
Tabela 02. Análises granulométricas do solo em relação à profundidade Trincheira em Floresta Hipoxerofila
no sitio Tanque Velho – Jaguarari - Bahia.
Profundidade (cm) Areia (%) Argila (%) Silte (%) Textura Densidade (g/cm3)
0-10 88,45** 6,48* 5,07 Arenoso 1,37
10-20 88,45** 8,56 2,99* Arenoso 1,39
20-30 88,45** 8,56 2,99* Areia 1,38
30-40 88,12 6,81 5,07 Areia 1,39
40-50 88,20 6,64 5,16 Areia 1,39
50-60 86,12 8,81 5,07 Areia Franca 1,40
60-70 82,12 10,81 7,07 Areia Franca 1,36
70-80 81,96 10,81 7,23 Areia Franca 1,39
80-90 81,96 12,81 5,23 Franco Arenoso 1,35
90-100 79,96 12,81 7,23 Franco Arenoso 1,34*
110-120 79,79 10,97 9,24 Franco Arenoso 1,39
130-140 77,79 13,05 9,16 Franco Arenoso 1,43
150-160 77,79 13,05 9,16 Franco Arenoso 1,46
170-180 75,79 15,05 9,16 Franco Arenoso 1,43
190-200 71,79 17,05** 11,16 Franco Arenoso 1,35
210-220 71,79 17,05** 11,16 Franco Arenoso 1,35
230-240 71,79 16,97 11,24 Franco Arenoso 1,38
250-260 69,79* 16,97 13,24** Franco Arenoso 1,39
270-280 73,79 14,97 11,24 Franco Arenoso 1,39
290-300 73,79 15,14 11,07 Franco Arenoso 1,43
310-320 77,87 13,22 8,91 Franco Arenoso 1,58
330-340 77,79 13,22 8,99 Franco Arenoso 1,64
350-360 77,79 13,22 8,99 Franco Arenoso 1,64
370-380 77,79 13,22 8,99 Franco Arenoso 1,69**
390-400 77,79 15,22 6,99 Franco Arenoso 1,65
410-420 77,79 13,22 8,99 Franco Arenoso 1,65
*Menor valor observado; ** maior valor observado.
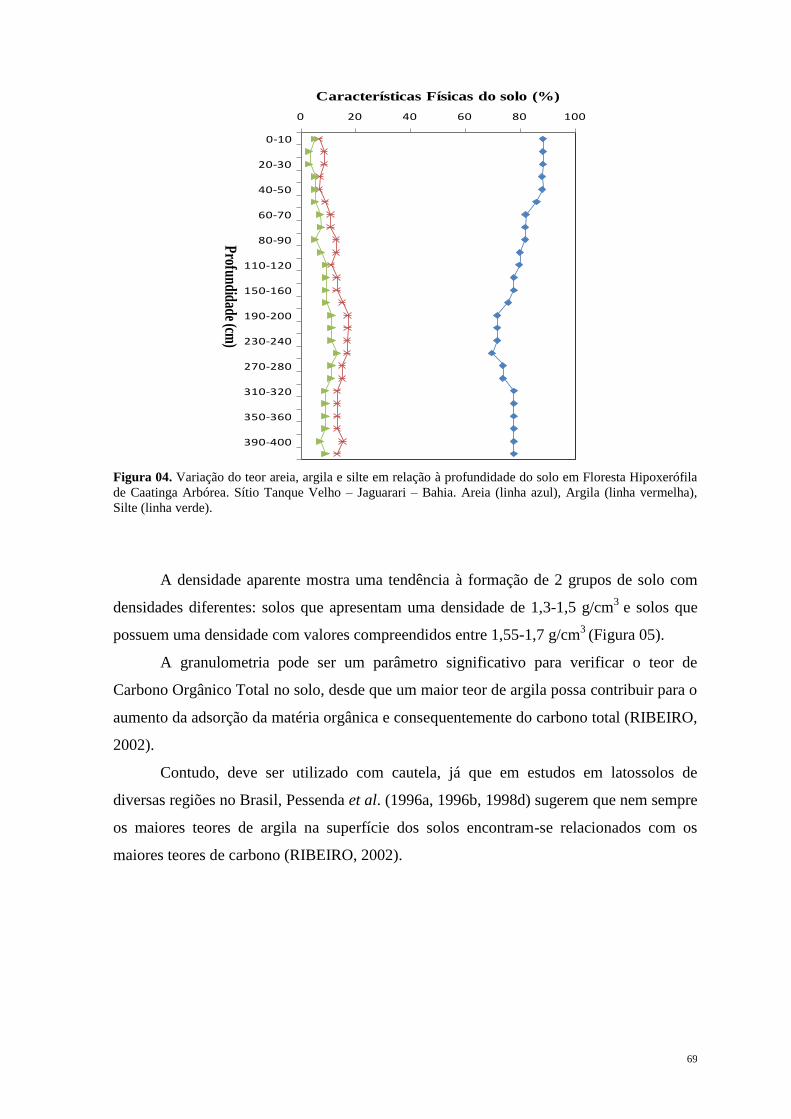
69
Figura 04. Variação do teor areia, argila e silte em relação à profundidade do solo em Floresta Hipoxerófila
de Caatinga Arbórea. Sítio Tanque Velho – Jaguarari – Bahia. Areia (linha azul), Argila (linha vermelha),
Silte (linha verde).
A densidade aparente mostra uma tendência à formação de 2 grupos de solo com
densidades diferentes: solos que apresentam uma densidade de 1,3-1,5 g/cm3
e solos que
possuem uma densidade com valores compreendidos entre 1,55-1,7 g/cm3
(Figura 05).
A granulometria pode ser um parâmetro significativo para verificar o teor de
Carbono Orgânico Total no solo, desde que um maior teor de argila possa contribuir para o
aumento da adsorção da matéria orgânica e consequentemente do carbono total (RIBEIRO,
2002).
Contudo, deve ser utilizado com cautela, já que em estudos em latossolos de
diversas regiões no Brasil, Pessenda et al. (1996a, 1996b, 1998d) sugerem que nem sempre
os maiores teores de argila na superfície dos solos encontram-se relacionados com os
maiores teores de carbono (RIBEIRO, 2002).
0 20 40 60 80 100
0-10
20-30
40-50
60-70
80-90
110-120
150-160
190-200
230-240
270-280
310-320
350-360
390-400
Características Físicas do solo (%)
Profundidade (cm
)
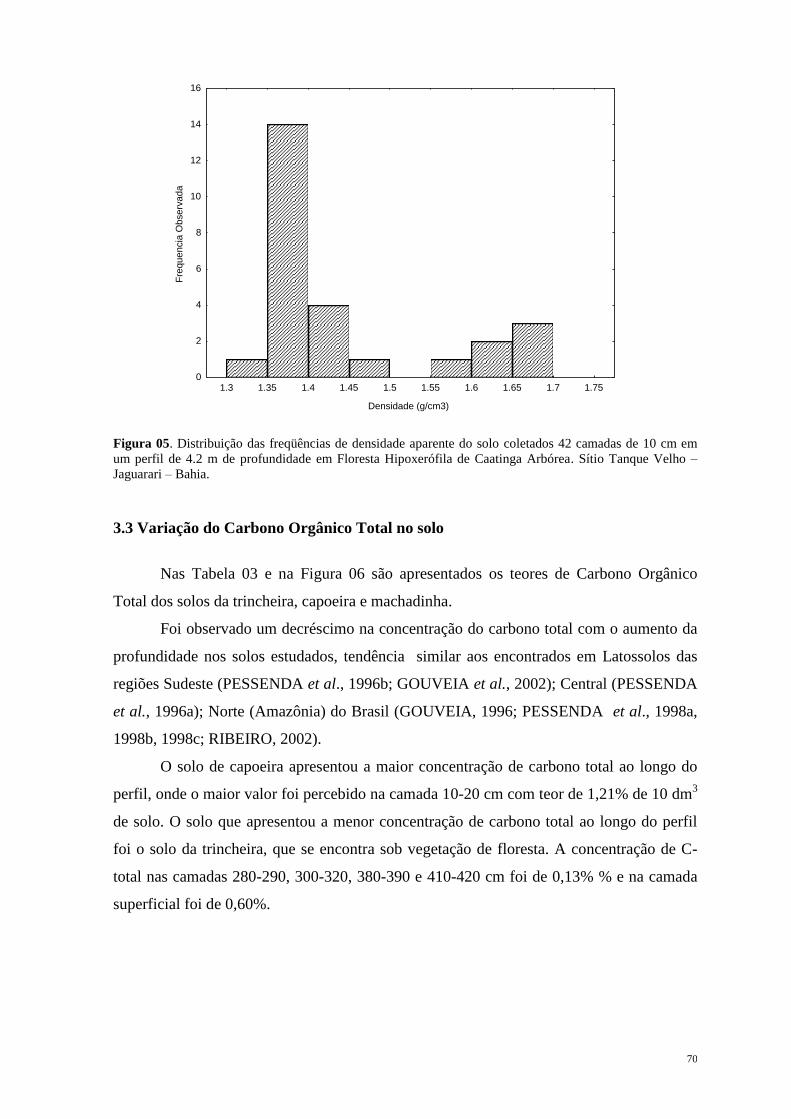
70
Figura 05. Distribuição das freqüências de densidade aparente do solo coletados 42 camadas de 10 cm em
um perfil de 4.2 m de profundidade em Floresta Hipoxerófila de Caatinga Arbórea. Sítio Tanque Velho –
Jaguarari – Bahia.
3.3 Variação do Carbono Orgânico Total no solo
Nas Tabela 03 e na Figura 06 são apresentados os teores de Carbono Orgânico
Total dos solos da trincheira, capoeira e machadinha.
Foi observado um decréscimo na concentração do carbono total com o aumento da
profundidade nos solos estudados, tendência similar aos encontrados em Latossolos das
regiões Sudeste (PESSENDA et al., 1996b; GOUVEIA et al., 2002); Central (PESSENDA
et al., 1996a); Norte (Amazônia) do Brasil (GOUVEIA, 1996; PESSENDA et al., 1998a,
1998b, 1998c; RIBEIRO, 2002).
O solo de capoeira apresentou a maior concentração de carbono total ao longo do
perfil, onde o maior valor foi percebido na camada 10-20 cm com teor de 1,21% de 10 dm3
de solo. O solo que apresentou a menor concentração de carbono total ao longo do perfil
foi o solo da trincheira, que se encontra sob vegetação de floresta. A concentração de C-
total nas camadas 280-290, 300-320, 380-390 e 410-420 cm foi de 0,13% % e na camada
superficial foi de 0,60%.
1.3 1.35 1.4 1.45 1.5 1.55 1.6 1.65 1.7 1.75
Densidade (g/cm3)
0
2
4
6
8
10
12
14
16
Fre
quencia
Observ
ada
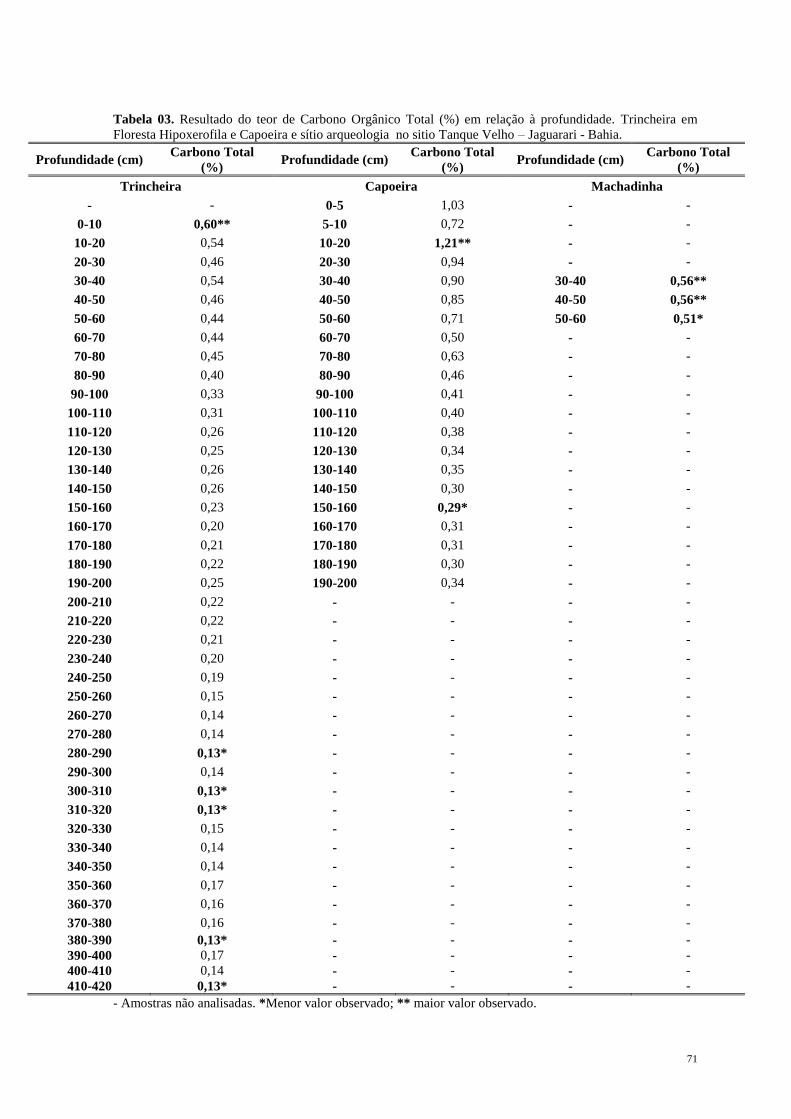
71
Tabela 03. Resultado do teor de Carbono Orgânico Total (%) em relação à profundidade. Trincheira em
Floresta Hipoxerofila e Capoeira e sítio arqueologia no sitio Tanque Velho – Jaguarari - Bahia.
Profundidade (cm) Carbono Total
(%) Profundidade (cm)
Carbono Total
(%) Profundidade (cm)
Carbono Total
(%)
Trincheira Capoeira Machadinha
- - 0-5 1,03 - -
0-10 0,60** 5-10 0,72 - -
10-20 0,54 10-20 1,21** - -
20-30 0,46 20-30 0,94 - -
30-40 0,54 30-40 0,90 30-40 0,56**
40-50 0,46 40-50 0,85 40-50 0,56**
50-60 0,44 50-60 0,71 50-60 0,51*
60-70 0,44 60-70 0,50 - -
70-80 0,45 70-80 0,63 - -
80-90 0,40 80-90 0,46 - -
90-100 0,33 90-100 0,41 - -
100-110 0,31 100-110 0,40 - -
110-120 0,26 110-120 0,38 - -
120-130 0,25 120-130 0,34 - -
130-140 0,26 130-140 0,35 - -
140-150 0,26 140-150 0,30 - -
150-160 0,23 150-160 0,29* - -
160-170 0,20 160-170 0,31 - -
170-180 0,21 170-180 0,31 - -
180-190 0,22 180-190 0,30 - -
190-200 0,25 190-200 0,34 - -
200-210 0,22 - - - -
210-220 0,22 - - - -
220-230 0,21 - - - -
230-240 0,20 - - - -
240-250 0,19 - - - -
250-260 0,15 - - - -
260-270 0,14 - - - -
270-280 0,14 - - - -
280-290 0,13* - - - -
290-300 0,14 - - - -
300-310 0,13* - - - -
310-320 0,13* - - - -
320-330 0,15 - - - -
330-340 0,14 - - - -
340-350 0,14 - - - -
350-360 0,17 - - - -
360-370 0,16 - - - -
370-380 0,16 - - - -
380-390 0,13* - - - -
390-400 0,17 - - - -
400-410 0,14 - - - -
410-420 0,13* - - - -
- Amostras não analisadas. *Menor valor observado; ** maior valor observado.
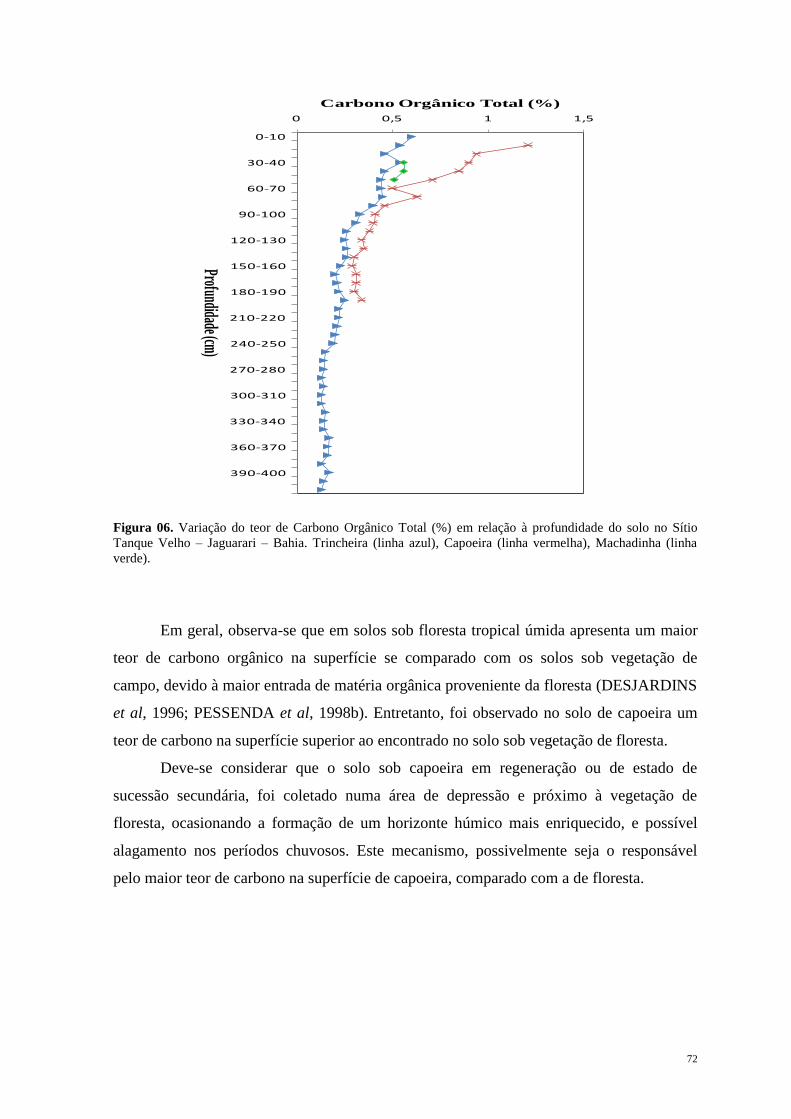
72
Figura 06. Variação do teor de Carbono Orgânico Total (%) em relação à profundidade do solo no Sítio
Tanque Velho – Jaguarari – Bahia. Trincheira (linha azul), Capoeira (linha vermelha), Machadinha (linha
verde).
Em geral, observa-se que em solos sob floresta tropical úmida apresenta um maior
teor de carbono orgânico na superfície se comparado com os solos sob vegetação de
campo, devido à maior entrada de matéria orgânica proveniente da floresta (DESJARDINS
et al, 1996; PESSENDA et al, 1998b). Entretanto, foi observado no solo de capoeira um
teor de carbono na superfície superior ao encontrado no solo sob vegetação de floresta.
Deve-se considerar que o solo sob capoeira em regeneração ou de estado de
sucessão secundária, foi coletado numa área de depressão e próximo à vegetação de
floresta, ocasionando a formação de um horizonte húmico mais enriquecido, e possível
alagamento nos períodos chuvosos. Este mecanismo, possivelmente seja o responsável
pelo maior teor de carbono na superfície de capoeira, comparado com a de floresta.
0 0,5 1 1,5
0-10
30-40
60-70
90-100
120-130
150-160
180-190
210-220
240-250
270-280
300-310
330-340
360-370
390-400
Carbono Orgânico Total (%)
Profundidade (cm)

73
3.4 Inferencia paleoincendios com ocorrência de fragmentos de carvão nos solo
Na Tabela 04, e nas Figuras 07, 08 e 09 são apresentados os resultados das massas
de carvão em função da profundidade dos solos da trincheira, capoeira e machadinha.
A presença de fragmentos de carvão foi observada em todos os pontos coletados
nas áreas da trincheira, capoeira e da machadinha, desde a superfície até as camadas mais
profundas, evidenciando a ocorrência de incêndios durante provavelmente todo o
Holoceno. Ressalta-se que o solo da área de estudo era arenoso, o que eventualmente
poderia ter facilitado o recobrimento dos fragmentos pela fauna local do solo.
Nas camadas superficiais foram encontradas quantidades significativas de
fragmentos de carvão, principalmente na área onde foi encontrada a machadinha.
No solo da trincheira observa-se que massas de até 22,92 g de carvão por 10 kg de
solo foram encontradas até aproximadamente 90 cm. Massas de pouco mais de 1g/10 kg
foram encontradas até 160 cm, e de 170 cm até a profundidade máxima, a massa de carvão
foi inferior a 1g/10 kg de solo.
Na área de capoeira todos os valores da massa de carvão encontrados no perfil
foram inferiores a 1 g/10 kg de solo. Mesmo efetuada a coleta das amostras por meio de
trado manual, um dos fatores da quantidade reduzida de carvão amostrada, os fragmentos
apareceram em todo o perfil amostrado.
A maior quantidade de massa de carvão foi amostrada no local onde a machadinha
foi encontrada, um total de 127,94 g de carvão para as três camadas analisadas.
Na área de capoeira foi encontrada uma porção de uma panela de cerâmica com
vários fragmentos de carvão. Os carvões foram coletados e serão feitas análises isotópicas
de 13
C e 14
C para verificar o tipo de vegetação que estava sendo utilizada pela população,
a idade do material, e a localização temporal da população para a região.
A partir da determinação das taxas de recobrimento destes solos após a datação por
14C dos fragmentos de carvão, será possível inferir sobre os paleoincêndios na região.
Saber se a área onde foi encontrada a machadinha era um local de queima de alimentos
(coivara), ou se estes fragmentos de carvão poderão ser associados a incêndios ocorridos
no passado, há pelo menos algumas centenas de anos, coincidindo com períodos mais
secos para o Holoceno.
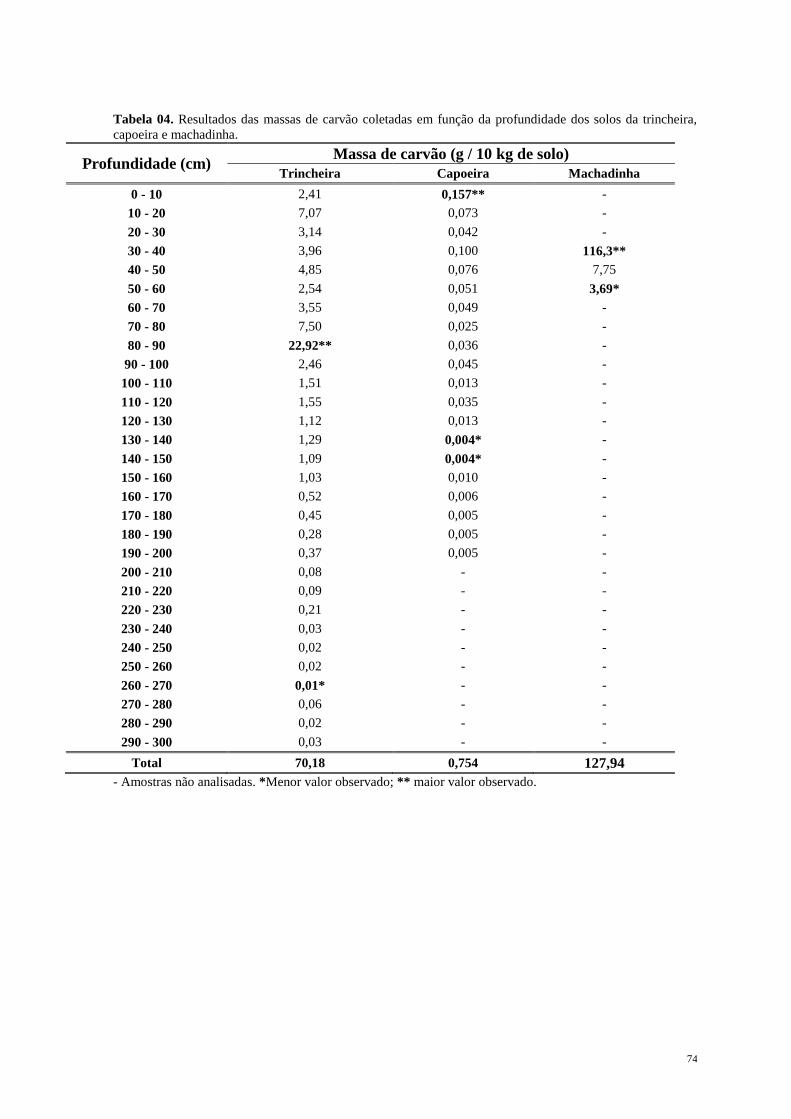
74
Tabela 04. Resultados das massas de carvão coletadas em função da profundidade dos solos da trincheira,
capoeira e machadinha.
- Amostras não analisadas. *Menor valor observado; ** maior valor observado.
Profundidade (cm) Massa de carvão (g / 10 kg de solo)
Trincheira Capoeira Machadinha
0 - 10 2,41 0,157** -
10 - 20 7,07 0,073 -
20 - 30 3,14 0,042 -
30 - 40 3,96 0,100 116,3**
40 - 50 4,85 0,076 7,75
50 - 60 2,54 0,051 3,69*
60 - 70 3,55 0,049 -
70 - 80 7,50 0,025 -
80 - 90 22,92** 0,036 -
90 - 100 2,46 0,045 -
100 - 110 1,51 0,013 -
110 - 120 1,55 0,035 -
120 - 130 1,12 0,013 -
130 - 140 1,29 0,004* -
140 - 150 1,09 0,004* -
150 - 160 1,03 0,010 -
160 - 170 0,52 0,006 -
170 - 180 0,45 0,005 -
180 - 190 0,28 0,005 -
190 - 200 0,37 0,005 -
200 - 210 0,08 - -
210 - 220 0,09 - -
220 - 230 0,21 - -
230 - 240 0,03 - -
240 - 250 0,02 - -
250 - 260 0,02 - -
260 - 270 0,01* - -
270 - 280 0,06 - -
280 - 290 0,02 - -
290 - 300 0,03 - -
Total 70,18 0,754 127,94

75
Figura 07. Distribuição das massas de carvão em relação à profundidade no perfil da Trincheira.
Figura 08. Distribuição das massas de carvão em relação à profundidade no perfil da Capoeira.
0 5 10 15 20 25
20 - 30
50 - 60
80 - 90
110 - 120
140 - 150
170 - 180
200 - 210
230 - 240
260 - 270
290 - 300
Massa de carvão
(g / 10 kg de solo)
Profundidade (cm)
0 0,05 0,1 0,15 0,2
10-20 cm
30 - 40
50 - 60
70 - 80
90 - 100
110 - 120
130 - 140
150 - 160
170 - 180
190 - 200
Massa de carvão
(g/10 kg de solo)
Pro
fun
did
ad
e (cm)
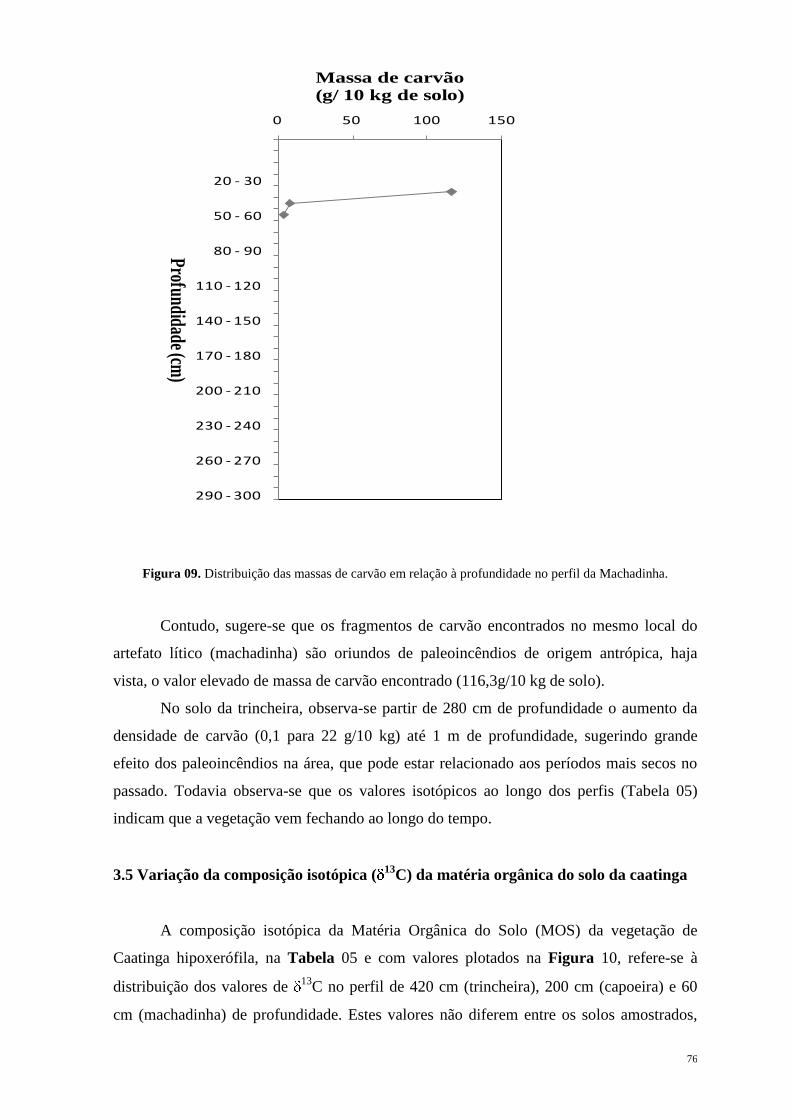
76
0 50 100 150
20 - 30
50 - 60
80 - 90
110 - 120
140 - 150
170 - 180
200 - 210
230 - 240
260 - 270
290 - 300
Massa de carvão
(g/ 10 kg de solo)
Profundidade (cm
)
Figura 09. Distribuição das massas de carvão em relação à profundidade no perfil da Machadinha.
Contudo, sugere-se que os fragmentos de carvão encontrados no mesmo local do
artefato lítico (machadinha) são oriundos de paleoincêndios de origem antrópica, haja
vista, o valor elevado de massa de carvão encontrado (116,3g/10 kg de solo).
No solo da trincheira, observa-se partir de 280 cm de profundidade o aumento da
densidade de carvão (0,1 para 22 g/10 kg) até 1 m de profundidade, sugerindo grande
efeito dos paleoincêndios na área, que pode estar relacionado aos períodos mais secos no
passado. Todavia observa-se que os valores isotópicos ao longo dos perfis (Tabela 05)
indicam que a vegetação vem fechando ao longo do tempo.
3.5 Variação da composição isotópica (13
C) da matéria orgânica do solo da caatinga
A composição isotópica da Matéria Orgânica do Solo (MOS) da vegetação de
Caatinga hipoxerófila, na Tabela 05 e com valores plotados na Figura 10, refere-se à
distribuição dos valores de 13
C no perfil de 420 cm (trincheira), 200 cm (capoeira) e 60
cm (machadinha) de profundidade. Estes valores não diferem entre os solos amostrados,

77
seguindo a mesma tendência de enriquecimento isotópico em função da profundidade. A
tendência de enriquecimento de 13
C com a profundidade foi de -27‰ com sinal isotópico
da vegetação atual entre 0 - 0.5 cm, representado por uma Caatinga hipoxerófila densa. A
vegetação de Caatinga hiperxerófita apresentou valor isotópico de -24‰ ou mais aberta até
a uma profundidade de 300 cm. Boutton (1979), Cerri et al. (1994), Pessenda et al. (1996a,
1998b) atribuem esse fracionamento isotópico devido a decomposição da matéria orgânica
da liteira, seguido dos processos de mineralização da MOS e de processos associados com
a formação do húmus nos solos o qual esse fracionamento pode variar entre 0 a 2‰ .
Estudos realizados em florestas úmidas e cerrados no Brasil, em caso de mudança
ou troca de vegetação, como vistos na Amazônia de Rondônia, Brasil Central e Maranhão,
a variação é bem acentuada chegando a uma amplitude de 4‰.
O sinal isotópico da vegetação estudada variou em -28,03‰ e um desvio padrão de
6.04 ‰ com uma amplitude de -12‰ a -34‰, todavia esse valor médio é input de
Matéria Orgânica atual, o que não difere significativamente do sinal da liteira -27‰. Os
valores de 13
C obtidos para as camadas ao longo do perfil dos solos variaram entre -24 a -
27‰, são característicos de vegetação com predomínio de plantas de ciclo fotossintético
C3 tendendo a formação florestal mais fechada.
.
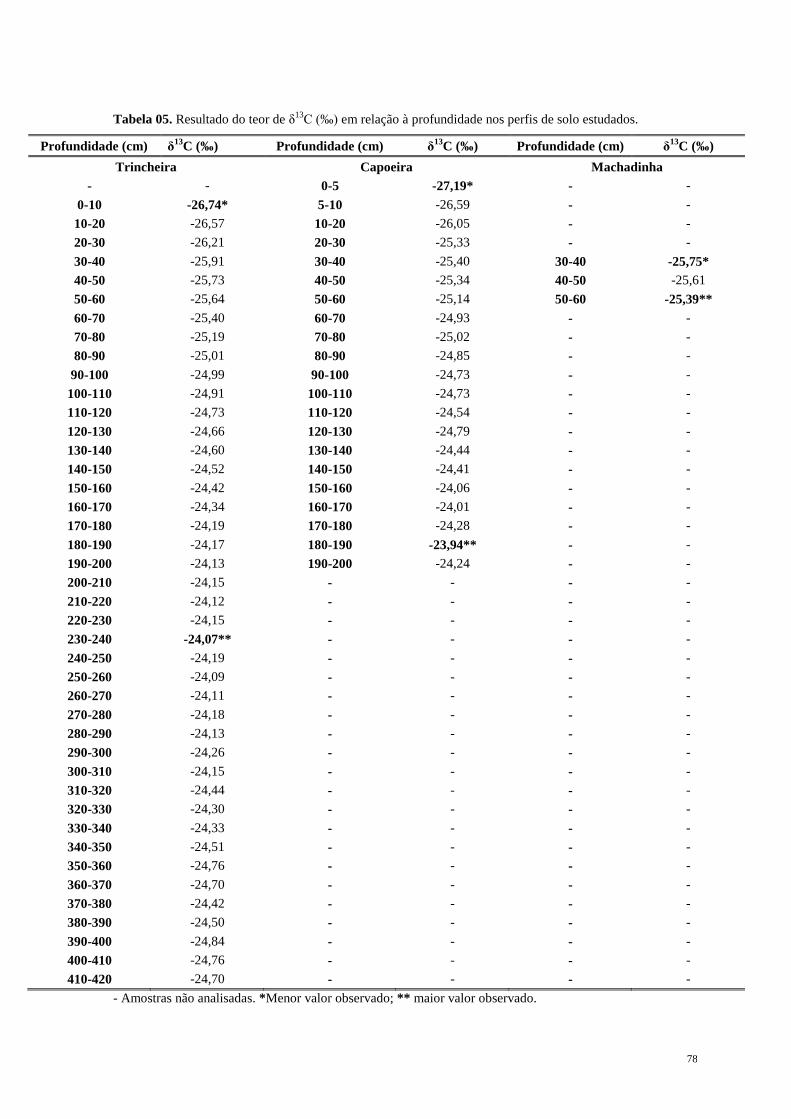
78
Tabela 05. Resultado do teor de δ13
C (‰) em relação à profundidade nos perfis de solo estudados.
Profundidade (cm) δ13
C (‰) Profundidade (cm) δ13
C (‰) Profundidade (cm) δ13
C (‰)
Trincheira Capoeira Machadinha
- - 0-5 -27,19* - -
0-10 -26,74* 5-10 -26,59 - -
10-20 -26,57 10-20 -26,05 - -
20-30 -26,21 20-30 -25,33 - -
30-40 -25,91 30-40 -25,40 30-40 -25,75*
40-50 -25,73 40-50 -25,34 40-50 -25,61
50-60 -25,64 50-60 -25,14 50-60 -25,39**
60-70 -25,40 60-70 -24,93 - -
70-80 -25,19 70-80 -25,02 - -
80-90 -25,01 80-90 -24,85 - -
90-100 -24,99 90-100 -24,73 - -
100-110 -24,91 100-110 -24,73 - -
110-120 -24,73 110-120 -24,54 - -
120-130 -24,66 120-130 -24,79 - -
130-140 -24,60 130-140 -24,44 - -
140-150 -24,52 140-150 -24,41 - -
150-160 -24,42 150-160 -24,06 - -
160-170 -24,34 160-170 -24,01 - -
170-180 -24,19 170-180 -24,28 - -
180-190 -24,17 180-190 -23,94** - -
190-200 -24,13 190-200 -24,24 - -
200-210 -24,15 - - - -
210-220 -24,12 - - - -
220-230 -24,15 - - - -
230-240 -24,07** - - - -
240-250 -24,19 - - - -
250-260 -24,09 - - - -
260-270 -24,11 - - - -
270-280 -24,18 - - - -
280-290 -24,13 - - - -
290-300 -24,26 - - - -
300-310 -24,15 - - - -
310-320 -24,44 - - - -
320-330 -24,30 - - - -
330-340 -24,33 - - - -
340-350 -24,51 - - - -
350-360 -24,76 - - - -
360-370 -24,70 - - - -
370-380 -24,42 - - - -
380-390 -24,50 - - - -
390-400 -24,84 - - - -
400-410 -24,76 - - - -
410-420 -24,70 - - - -
- Amostras não analisadas. *Menor valor observado; ** maior valor observado.
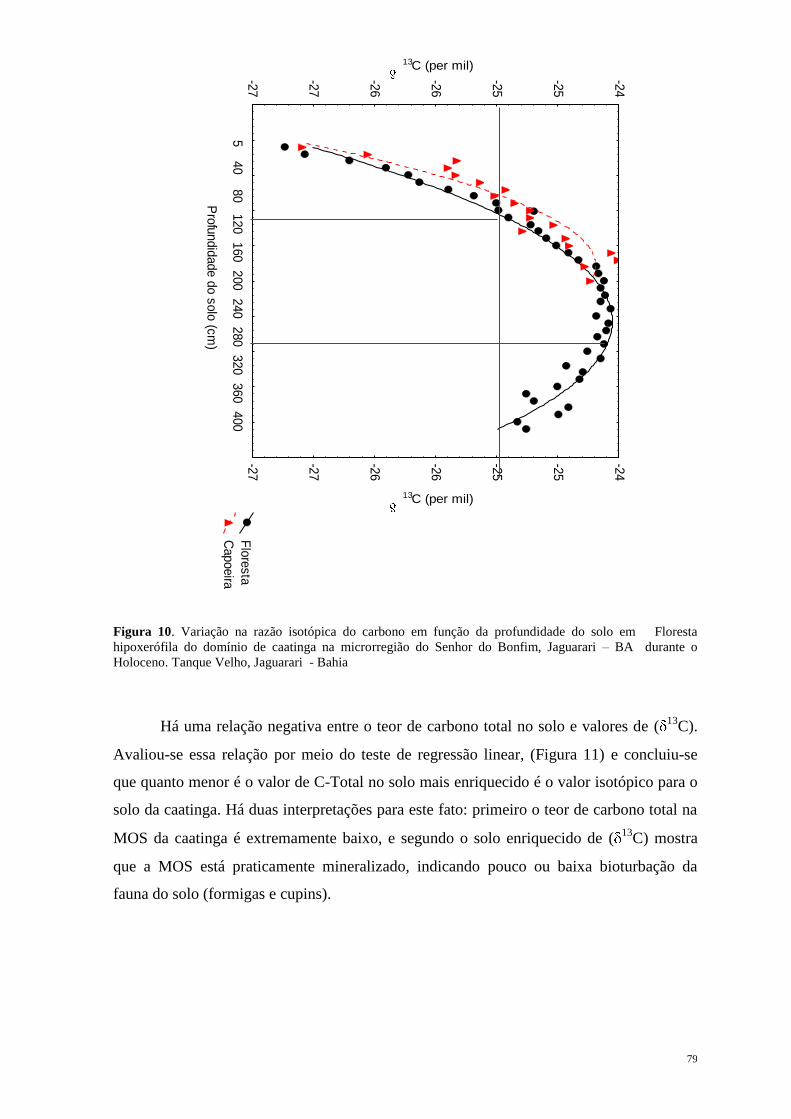
79
Figura 10. Variação na razão isotópica do carbono em função da profundidade do solo em Floresta
hipoxerófila do domínio de caatinga na microrregião do Senhor do Bonfim, Jaguarari – BA durante o
Holoceno. Tanque Velho, Jaguarari - Bahia
Há uma relação negativa entre o teor de carbono total no solo e valores de (13
C).
Avaliou-se essa relação por meio do teste de regressão linear, (Figura 11) e concluiu-se
que quanto menor é o valor de C-Total no solo mais enriquecido é o valor isotópico para o
solo da caatinga. Há duas interpretações para este fato: primeiro o teor de carbono total na
MOS da caatinga é extremamente baixo, e segundo o solo enriquecido de (13
C) mostra
que a MOS está praticamente mineralizado, indicando pouco ou baixa bioturbação da
fauna do solo (formigas e cupins).
Flo
resta
Capoeira
540
80
120
160
200
240
280
320
360
400
Pro
fundid
ade d
o s
olo
(cm
)
-27
-27
-26
-26
-25
-25
-24
13C (per mil)
-27
-27
-26
-26
-25
-25
-24
13C (per mil)
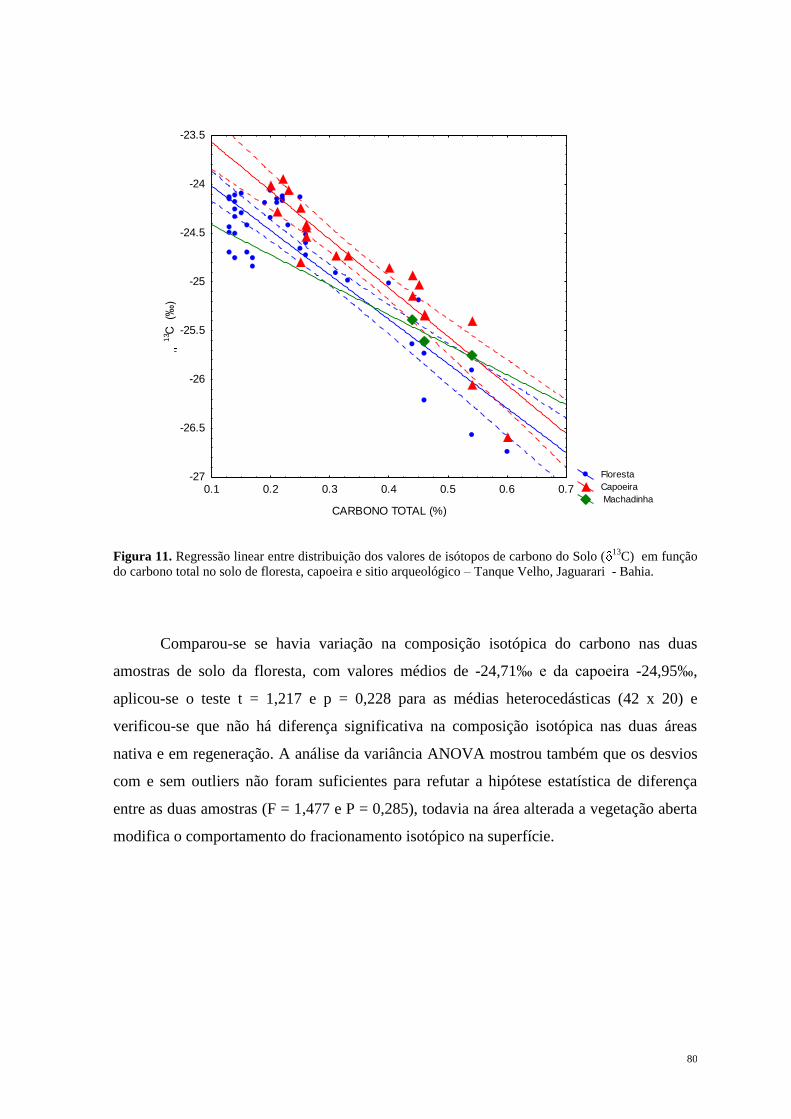
80
Figura 11. Regressão linear entre distribuição dos valores de isótopos de carbono do Solo (13
C) em função
do carbono total no solo de floresta, capoeira e sitio arqueológico – Tanque Velho, Jaguarari - Bahia.
Comparou-se se havia variação na composição isotópica do carbono nas duas
amostras de solo da floresta, com valores médios de -24,71‰ e da capoeira -24,95‰,
aplicou-se o teste t = 1,217 e p = 0,228 para as médias heterocedásticas (42 x 20) e
verificou-se que não há diferença significativa na composição isotópica nas duas áreas
nativa e em regeneração. A análise da variância ANOVA mostrou também que os desvios
com e sem outliers não foram suficientes para refutar a hipótese estatística de diferença
entre as duas amostras (F = 1,477 e P = 0,285), todavia na área alterada a vegetação aberta
modifica o comportamento do fracionamento isotópico na superfície.
13 C
(‰
)
Floresta
Capoeira
Machadinha0.1 0.2 0.3 0.4 0.5 0.6 0.7
CARBONO TOTAL (%)
-27
-26.5
-26
-25.5
-25
-24.5
-24
-23.5
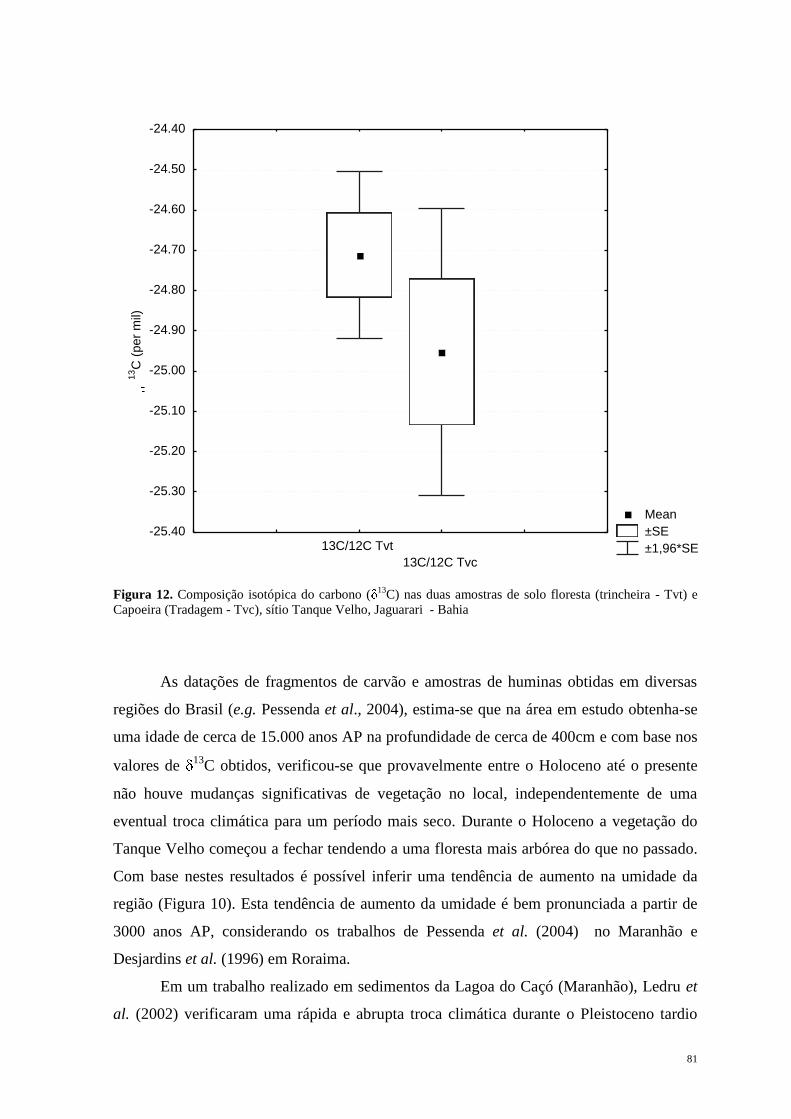
81
Figura 12. Composição isotópica do carbono (13
C) nas duas amostras de solo floresta (trincheira - Tvt) e
Capoeira (Tradagem - Tvc), sítio Tanque Velho, Jaguarari - Bahia
As datações de fragmentos de carvão e amostras de huminas obtidas em diversas
regiões do Brasil (e.g. Pessenda et al., 2004), estima-se que na área em estudo obtenha-se
uma idade de cerca de 15.000 anos AP na profundidade de cerca de 400cm e com base nos
valores de 13
C obtidos, verificou-se que provavelmente entre o Holoceno até o presente
não houve mudanças significativas de vegetação no local, independentemente de uma
eventual troca climática para um período mais seco. Durante o Holoceno a vegetação do
Tanque Velho começou a fechar tendendo a uma floresta mais arbórea do que no passado.
Com base nestes resultados é possível inferir uma tendência de aumento na umidade da
região (Figura 10). Esta tendência de aumento da umidade é bem pronunciada a partir de
3000 anos AP, considerando os trabalhos de Pessenda et al. (2004) no Maranhão e
Desjardins et al. (1996) em Roraima.
Em um trabalho realizado em sedimentos da Lagoa do Caçó (Maranhão), Ledru et
al. (2002) verificaram uma rápida e abrupta troca climática durante o Pleistoceno tardio
13C
(p
er
mil)
Mean
±SE
±1,96*SE 13C/12C Tvt
13C/12C Tvc
-25.40
-25.30
-25.20
-25.10
-25.00
-24.90
-24.80
-24.70
-24.60
-24.50
-24.40

82
(11.600 até aproximadamente 10.000 anos AP), relacionado com o evento “Younger
Dryas” ocorrido no Hemisfério Norte. Os autores consideram que a temperatura pode ter
diminuído cerca de 2°C neste intervalo, assim como observaram a presença significativa de
fragmentos de carvão em todo o Holoceno, indicando a ocorrência de paleoincêndios que
foram associados ao domínio de uma vegetação arbórea mais aberta.
Embora não se possa fazer inferências mais precisas pela ausência dos resultados
das datações por 14
C, sugere-se que as interpretações paleoclimáticas do presente trabalho
são comparáveis às publicadas por Ledru et al. (2002), onde verifica-se na profundidade
máxima (400cm) uma vegetação mais fechada e a possível existência de uma vegetação
mais aberta em 350-170 cm, com o retorno da umidade em 170-0 cm (Figura 10).
Pode-se relacionar também as análises paleoclimáticas do presente trabalho com o
estudo de reconstrução paleoambiental realizado por De Oliveira et al., (1999) em uma
turfeira no norte da Bahia, no Vale do Rio Icatu. Segundo os autores, no final do
Pleistoceno (10.990-10.540 anos AP), espécies características de floresta da Amazônia,
Atlântica e de montanha indicam um período muito úmido e temperaturas mais reduzidas.
De 10.540 a 8.910 anos AP, a vegetação dominada por Mauritia vinifera sugeriu um
progressivo aumento de temperatura e com alta umidade. De 8.910 a 6.790 AP houve um
progressivo declínio de taxa de floresta e aumento de taxa de caatinga e cerrado. Até 4.535
anos AP houve um retorno a um mosaico de vegetação composta de floresta de galeria,
caatinga e cerrado, indicando um clima mais úmido. De 4.535 até o presente houve um
aumento no taxa de caatinga e de cerrado, caracterizando uma diminuição de umidade e o
declínio da floresta de galeria, aspecto encontrado no ambiente atual.
Verifica-se que as interpretações ambientais do presente trabalho são similares com
as obtidas no trabalho de De Oliveira et al. (1999), onde um provável período seco
predominou na região norte da Bahia desde o início do Holoceno (cerca de 9.000 anos
AP.) até o Holoceno médio (4.000-3000 anos AP), vindo a seguir um período mais úmido
e provavelmente similar ao atual.
Vale ressaltar que no final do Pleistoceno/início do Holoceno foram inferidas
situações distintas a nível paleoclimático nas regiões sul/sudeste do Brasil. Dados
isotópicos de solos de Salitre (MG) (PESSENDA et al., 1996a, 1998d), Londrina (PR) e
Piracicaba (SP) (PESSENDA et al., 1996b, 1998d) e Jaguariúna (SP) (GOUVEIA et al.,
1999, 2002) indicaram a presença significativa de gramíneas desde cerca de 11.000 anos
AP até cerca de 4.000 anos AP, associado a presença de um clima mais seco. Dados

83
palinológicos de sedimentos lacustres também indicaram um clima seco em período
similar em localidades ao sul do Brasil (LEDRU et al., 1996; BEHLING, 1997, 1998).
Desta forma, enquanto na região sul do Brasil por volta de 11.000 anos AP até
8.000 anos AP havia o predomínio de gramíneas e clima seco em vários locais, em regiões
do nordeste e no sul da Amazônia as florestas dominavam associadas a um provável clima
úmido. A partir de aproximadamente 8.000-7.000 anos AP, um clima seco também se
instalou nas regiões nordeste e no sul e centro da Amazônia, modificando a partir de
aproximadamente 4.000-3.000 anos AP para um clima mais úmido em conjunto com
outros locais da região sul.
Esta hipótese poderá ser corroborada com as datações dos carvões encontrados nas
camadas no perfil do solo, mas, os valores dos isótopos de 13
C da MOS sugerem esta
tendência para o local de estudo.
3.6 Resultado das análises isotópicas (13
C) foliares e Carbono total (%)
Os resultados referentes à composição isotópica da vegetação nas áreas da
trincheira, capoeira e machadinha são representados na Tabela 06 e Figuras 13 e 14.
Foram coletadas 26 amostras de plantas, sendo identificadas 7 espécies, 12 gêneros
e as demais não foram identificadas.
22 plantas foram identificadas como de ciclo fotossintético C3, correspondendo a
85% das amostras analisadas, com os valores 13
C variando entre -32 a -24‰, enquanto as
plantas de ciclo fotossintético CAM corresponderam a 15% (n=4) das amostras analisadas,
com valores isotópicos variarando de -12 a -16‰ indicando mistura de plantas CAM
obrigatórias e facultativas (Tabela 6, Figura 14).
O enriquecimento isotópico da MOS em relação a profundidade pode ser explicado
pela alteração ao longo do tempo, da composição isotópica da vegetação como
consequência das variações recentes no conteúdo de 13
C no CO2 atmosférico; uma possível
decomposição diferencial dos componentes bioquímicos das plantas ou ainda, um
fracionamento isotópico durante a mineralização da MOS (DESJARDINS et al., 1996).
Os valores de C-Total apresentaram uma distribuição normal (Figura 13).
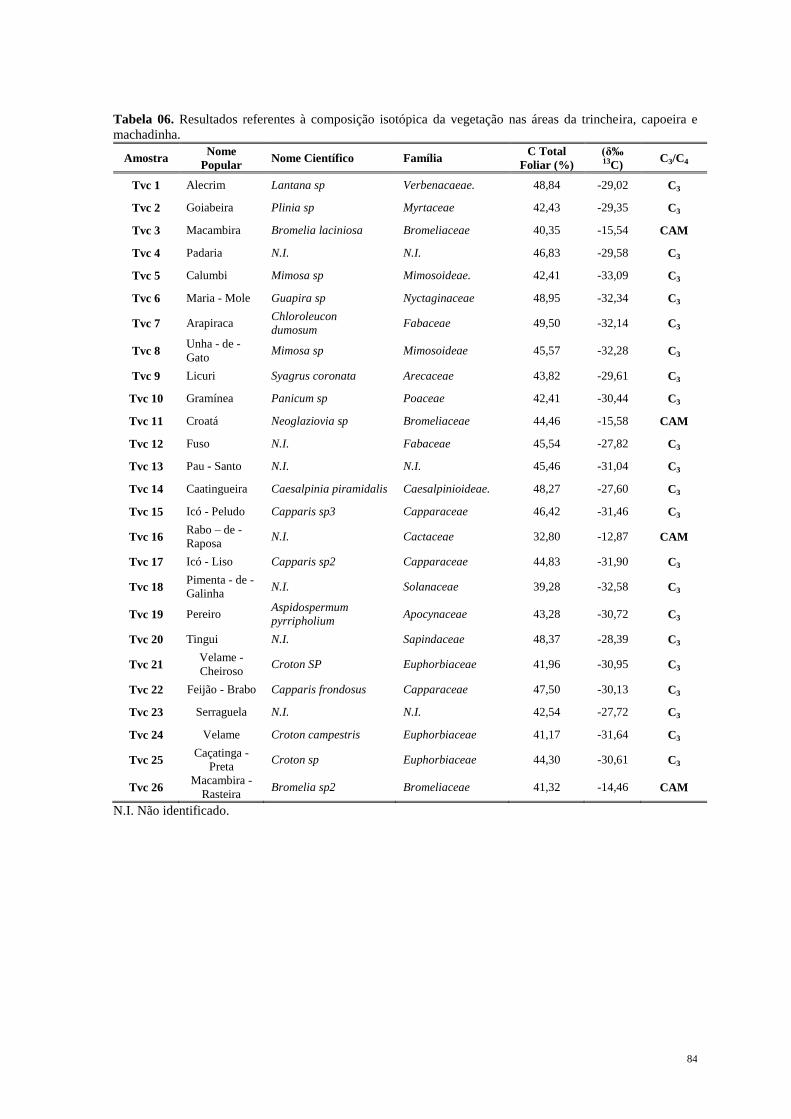
84
Tabela 06. Resultados referentes à composição isotópica da vegetação nas áreas da trincheira, capoeira e
machadinha.
Amostra Nome
Popular Nome Científico Família
C Total
Foliar (%)
(δ‰ 13C)
C3/C4
Tvc 1 Alecrim Lantana sp Verbenacaeae. 48,84 -29,02 C3
Tvc 2 Goiabeira Plinia sp Myrtaceae 42,43 -29,35 C3
Tvc 3 Macambira Bromelia laciniosa Bromeliaceae 40,35 -15,54 CAM
Tvc 4 Padaria N.I. N.I. 46,83 -29,58 C3
Tvc 5 Calumbi Mimosa sp Mimosoideae. 42,41 -33,09 C3
Tvc 6 Maria - Mole Guapira sp Nyctaginaceae 48,95 -32,34 C3
Tvc 7 Arapiraca Chloroleucon
dumosum Fabaceae 49,50 -32,14 C3
Tvc 8 Unha - de -
Gato Mimosa sp Mimosoideae 45,57 -32,28 C3
Tvc 9 Licuri Syagrus coronata Arecaceae 43,82 -29,61 C3
Tvc 10 Gramínea Panicum sp Poaceae 42,41 -30,44 C3
Tvc 11 Croatá Neoglaziovia sp Bromeliaceae 44,46 -15,58 CAM
Tvc 12 Fuso N.I. Fabaceae 45,54 -27,82 C3
Tvc 13 Pau - Santo N.I. N.I. 45,46 -31,04 C3
Tvc 14 Caatingueira Caesalpinia piramidalis Caesalpinioideae. 48,27 -27,60 C3
Tvc 15 Icó - Peludo Capparis sp3 Capparaceae 46,42 -31,46 C3
Tvc 16 Rabo – de -
Raposa N.I. Cactaceae 32,80 -12,87 CAM
Tvc 17 Icó - Liso Capparis sp2 Capparaceae 44,83 -31,90 C3
Tvc 18 Pimenta - de -
Galinha N.I. Solanaceae 39,28 -32,58 C3
Tvc 19 Pereiro Aspidospermum
pyrripholium Apocynaceae 43,28 -30,72 C3
Tvc 20 Tingui N.I. Sapindaceae 48,37 -28,39 C3
Tvc 21 Velame -
Cheiroso Croton SP Euphorbiaceae 41,96 -30,95 C3
Tvc 22 Feijão - Brabo Capparis frondosus Capparaceae 47,50 -30,13 C3
Tvc 23 Serraguela N.I. N.I. 42,54 -27,72 C3
Tvc 24 Velame Croton campestris Euphorbiaceae 41,17 -31,64 C3
Tvc 25 Caçatinga -
Preta Croton sp Euphorbiaceae 44,30 -30,61 C3
Tvc 26 Macambira -
Rasteira Bromelia sp2 Bromeliaceae 41,32 -14,46 CAM
N.I. Não identificado.
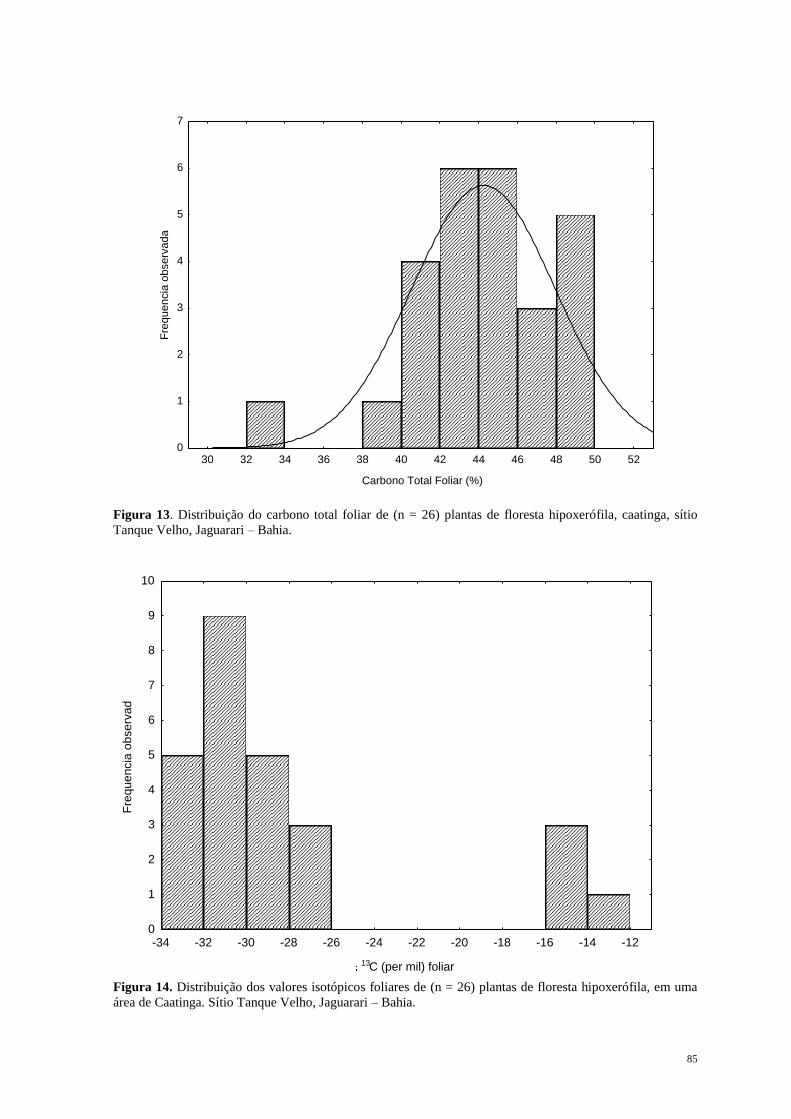
85
30 32 34 36 38 40 42 44 46 48 50 52
Carbono Total Foliar (%)
0
1
2
3
4
5
6
7
Fre
quen
cia
observ
ada
Figura 13. Distribuição do carbono total foliar de (n = 26) plantas de floresta hipoxerófila, caatinga, sítio
Tanque Velho, Jaguarari – Bahia.
-34 -32 -30 -28 -26 -24 -22 -20 -18 -16 -14 -12
13C (per mil) foliar
0
1
2
3
4
5
6
7
8
9
10
Fre
qu
en
cia
ob
se
rva
d
Figura 14. Distribuição dos valores isotópicos foliares de (n = 26) plantas de floresta hipoxerófila, em uma
área de Caatinga. Sítio Tanque Velho, Jaguarari – Bahia.

86
4. Considerações Finais
Os solos analisados apresentaram textura principalmente franca-arenosa e arenosa.
No perfil do solo da trincheira foi observado valores de pH na faixa de 4,32 a 4,78,
sendo considerado solo ácido, e valores de matéria orgânica que permitem considerá-lo
como horizonte orgânico.
Observou-se um decréscimo da concentração de carbono orgânico total com o
aumento da profundidade nos solos estudados. O solo que apresentou o maior teor de
carbono nas camadas superficiais estava localizado sob capoeira e em uma área mais baixa.
A vegetação amostrada para a área estudada foi composta de 85% de plantas do
ciclo fotossintético C3 e 15% de plantas do ciclo C4.
O solo que apresentou uma maior variação nos valores de 13
C da MOS foi o
encontrado sob vegetação de capoeira, com -27,19‰ na superfície e -24,24‰ na camada
mais profunda. Uma variação de 2,95 unidades de .
O solo que apresentou a segunda maior variação no teor de 13
C da MOS foi o
encontrado sob vegetação de floresta, com -26,74‰ na superfície e -24,70‰ na camada
mais inferior, representando uma variação de 2,04 unidades de 13
C.
Os resultados isotópicos mostram estabilidade vegetacional no local de estudo sem
troca de vegetação. É possível que tenha ocorrido no local, em períodos durante o
Holoceno e presente, a mudança de uma vegetação arbórea menos densa ou mistura de
plantas C3 e CAM com predomínio da primeira, sugerindo que provavelmente durante todo
o Holoceno um clima semelhante ao atual ocorreu na região de Jaguarari.
Possivelmente não houve na região estudada, modificação acentuada na vegetação
no Holoceno. Mesmo durante os períodos mais secos no continente, quando os
ecossistemas de cerrado e o sul da Amazônia abriram (Pessenda et al., 1998a, Gouveia
2000), na região de Jaguarari a cobertura vegetal permaneceu estável.
A razão isotópica 13
C mostra-se eficiente na caracterização de ocorrências de
mudanças de vegetações e, associada à datação radiocarbônica, mais eficaz nos estudos
paleoambientais.

87
Referências Bibliográficas
Auler AS, Smart PL. 2001. Late Quaternary paleoclimate in semi-arid northeastern Brazil
from U-series dating of travertine and water table speleothems. Quaternary Research
55: 159–167.
Barberi, M., Salgado-Laboariau, M.L., Suguio, K., 2000. Paleovegetation and
palaeoclimate of Veredas de Águas Emendadas, Q central Brazil. Journal of South
American Earth Sciences 13, 241– 254.
Behling, H., 1995A. Investigations into the Late Pleistocene and Holocene history of
vegetation and climate in Santa Catarina (S Brazil). Vegetation History and
Archaeobotany 4 (3), 127–152.
Behling, H., 1997. Late Quaternary vegetation, climate and fire history from the tropical
mountain region of Morro de Itapeva. SE Brazil. Palaeogeography, Palaeoclimatology,
Palaeoecology 129, 407 - 422.
Behling, H. Late Quaternary vegetacional and climatic changes in Brazil. Review of
Paleobotany and Palynology, v. 99, p. 143-156,1998.
Boutton, T.W., 1991. Stable carbon isotope ratios ofnatural materials. II. Atmospheric,
terrestrial, marine and freshwater environments. In: Coleman, D.C., Fry, B. (Eds.),
Carbon Isotope Techniques. Academic Press, New York, pp. 173–185.
Boutton, T.W. 1996. Stable carbon isotope ratios of soil organic matter and their use as
indicators of vegetation and climate change. In: BOUTTON, T.W.; YAMASAKI, S.I.
(Ed.) Mass spectrometry of soils. New York: Marcel Dekker, p.47-82.
Calvin, M.; Benson, A. A. 1948 The path of carbon in photosynthesis. Science, v. 107, p.
476-480.
Cocharane, t. t. 1984 Amazônia: computerized overview of the climate, landscape and soils
resources. Interciência, v.9, n.5, p.,.
Conti, J. B.; Furlan, S. A. (2003). Geoecologia: O Clima, os Solos e a Biota. In: ROSS, J.
S. (Org.). Geografia do Brasil. 4. ed. 1 reimp. São Paulo: Editora da Universidade de
São Paulo.
CPRM – 2005. Serviço Geológico do Brasil: Projeto Cadastro de Fontes de Abastecimento
por Água Subterrânea, Diagnóstico do Município de Jaguarari Estado da Bahia /
Organizado [por] Ângelo T. Vieira, Felicíssimo Melo, Hermínio Brasil V. Lopes,
Hermínio Brasil V. Lopes, José C. Viégas Campos, José T Guimarães, Juliana M. da
Costa, Luís Fernando C. Bomfim, Pedro Antonio de A. Couto, Sara Maria Pinotti
Benvenuti . Salvador: CPRM/PRODEEM. 14p + anexos
Czaplewksi, N. J.; Cartelle,C. 1998. Pleistocene bats from cave deposits in Bahia, Brazil.
Journal of Mammalogy , 79:784-803.
De Oliveira, P.E. de; Barreto, A.M.F.; Suguio, K. 1999. Late Pleistocene / Holocene
climatic and vegetational history of the brazilian Caatinga: the fossil dunes of the
middle São Francisco river. Palaeogeography, Palaeoclimatology, Palaeoecology
152: 319-337.
Desjardins, t.; volkoff b.; andreux F.; cerri c.c. 1991 Distribution Du carbone total de
l’isotope 13
C dans de Sols Ferrallitiques di Brésil. Cahiers. Orstom. Série. Pédologie.
Vol 26, n. 4, p.343-348.

88
Desjardins, T., Filho, A.C., Mariotti, A., Chauvel, A., Girardin, C., 1996. Changes of the
forest–savanna boundary in Brazilian Amazonia during the Holocene as revealed by
soil organic carbon isotope ratios. Oecologia 108, 749– 756.
Dever, L., Fonte. C.; Riché. G. 1987 Isotopic approach to calcite dissolution and
recipitacion is soils inder semi-arid conditions. Chemical Geology, v.66, p.307-314.
Ehleringer, J. R. 1978 Implications of quantum yield differences on distributions of C3 and
C4 grasses.Oecologia, v.31, p. 255-267.
Ehleringer, J.R. 13
C/12
C fractionation and its utility in terrestrial plant studies. In Colleman,
D.C.; Fry, b. (Eds). Carbon Isotope Techniques, San Diego: Academic Press, p. 187 –
200, 1991.
Ferraz-Vicentini, K.R. 1993 Análise palinológica de uma veredaem Cromínia, GO.
Brasília,. 87p. Dissertação (Mestrado) – Departamento de Ecologia, Universidade de
Brasília.
Ferraz-Vincentini, K.R., Salgado-Labouriau, M.L., 1996. Palynological analysis of a palm
swamp in Central Brazil. Journal of South American Earth Sciences 9, 207 - 219.
Glenn-Lewis, P.; Peet, R.K.; Veblen, T.T. 1992. Plant sucession: theory and prediction.
Chapman and Hall, London.
Gomes, B. M.; 1996. Estudo paleoambiental no estado de Rondônia utilizando datação por
14C e razão 13C/12C da matéria orgânica do solo. Piacicaba,. 106p. Dissertação
(mestrado) – Centro de Energia Nuclear na Agricultura, Universidade de São Paulo.
Gouveia, S. E. M. 1996 Estudos das alterações de paleovegetações na Amazônia Central,
utilizando a datação radiocarbônica e razão 13
C/12
C da matéria orgânica do solo.
Piracicaba,. 75p. Dissertação (Mestrado) – Centro de Energia Nuclear na Agricultura,
Universidade de São Paulo.
Gouveia, S.E.M., Pessenda, L.C.R., Boulet, R., Aravena, R., Scheel- Ybert, R., 1999.
Isótopos do carbono dos carvões e da matéria orgânica do solo em estudos de mudança
de vegetação e clima no Quaternário e da taxa de formação de solos do estado de São
Paulo. Anais da Academia Brasileira de Cieˆncias 71 (4), 969–980.
Gouveia, S.E.M., Pessenda, L.C.R., 2000. Datation par le 14C de charbons inclus dans le
sol pour l’etude du role de la remonte´e biologique de matie´re et Du colluvionnement
dans la formation de latosols de l’etat de Sa˜o Paulo, Bre´ sil. Comptes Rendus de
l’Acade´mie des Sciences de Paris, Serie IIA Earth and Planetary Science 330 (2),
133–138.
Gouveia, S.E.M., Pessenda, L.C.R., Aravena, R., Boulet, R., Scheel-Ybert, R., Bendassoli,
J.A., Ribeiro, A.S., Freitas, H.A., 2002. Carbon isotopes in charcoal and soils in
studies of paleovegetation and climate changes during the late Pleistocene and the
Holocene in the southeast and centerwest regions of Brazil. Global and Planetary
Change 33 (1–2), 95– 106.
Hatch, M. D.; Slach, C. R. 1966. Photosynthesis by sugarcane leaves: A new carboxylation
reaction and the pathway of sugar formation. Biochemestry Journal, v.101, p.103-111.
Kiehl, E.J., 1979. Manual de edafologia; relações solo/planta. Ceres, São Paulo.
Ledru, M. P. 1993 Late Quaternary and climatic changes in Central Brazil. Quaternary
Research, v.39, p.90-98.
Ledru, M.-P., Braga, P.I.S., Soubie`s, F., Fournier, M., Martin, L., Suguio, K., Turcq, B.,
1996. The last 50,000 years in the Neotropics (Southern Brazil): evolution of

89
vegetation and climate. Palaeogeography, Palaeoclimatology, Palaeoecology 123,
239–257.
Ledru, M.-P., Bertaux, J., Sifeddine, A., Suguio, K., 1998. Absence of last glacial
maximum records in lowland tropical forest. Quaternary Research 49, 233– 237.
Ledru, M.-P., Mourguiart, P., Ceccantini, G., Turcq, B., Sifeddine, A., 2002. Tropical
climates in the game of two hemispheres revealed by abrupt climatic change. Geology
30 (3), 275– 278.
Neves, P.C.P.; Lorscheitter, M.L. 1995 Upper Quaternary paleoenvironments in the
northern coastal plain of Rio Grande do Sul, Brazil. Quaternary of South America and
Antartic Peninsula, v.9,p.39-67.
Nordt, l.c.; boutton, t.w.; hallmark, c.t.; Waters, mr. 1994 Late Quaternary vegetation
changes in Central Texas based on the isotopic composition of organic carbon.
Quaternary Research, v.41, p. 109-120,
Pessenda, L.C.R.; Aravena, R.; Melfi, A.J.; Telles, E. C. C.; Boulet, R.; Valencia, E.P.E.,
Tomazello, M. 1996a. The use of carbon isotopes (C-13, C-14) in soil to evaluate
vegetation changes during the Holocene in central Brazil. Radiocarbon v.38, n.2,
p.191–201.
Pessenda, L.C.R.; Valencia, E.P.E.; Camargo, P. B.; Telles, E. C. C.; Martinelli, L.A.;
Cerri, C.C.; Avarena, R.; Rozanski, K. 1996b. Natural radiocarbon measurements in
brazilian soils developed on basic rocks. Radiocarbon, v.38, n.2, p.203-208.
Pessenda, L.C.R., Gouveia, S.E.M., Aravena, R., Gomes, B.M., Boulet, R., Ribeiro, A.S.,
1998a. 14C dating and stable carbon isotopes of soil organic matter in Forest savanna
boundary areas in the southern Brazilian Amazon region. Radiocarbon 40 (2), 1013–
1022.
Pessenda, L.C.R., Gomes, B.M., Aravena, R., Ribeiro, A.S., Boulet, R., Gouveia, S.E.M.,
1998b. The carbon isotope record in soils along a forest–cerrado ecosystem transect:
implications for vegetation changes in the Rondonia state, southwestern Brazilian
Amazon region. The Holocene 8 (5), 631–635.
Pessenda, L.C.R.; Lisi, C.S.; Gouveia, S.E.M 1998c Datação por 14C. In Pessenda, L.C.R.
Laboratório de 14C. Técnicas e aplicação paleoambientais. Piracicaba: CENA/US,
cap.1, p.3-5.
Pessenda, L.C.R., Valencia, E.P.E., Aravena, R., Telles, E.C.C., Boulet, R., 1998d.
Paleoclimate studies in Brazil using carbon isotopes in soils. In: Wasserman, J.C.,
Silva-Filho, E.V., Villas- Boas, R. (Eds.), Environmental Geochemistry in the Tropics,
Lecture Notes in Earth Sciences., Vol. 72. Springer, Berlin/New York, pp. 7–16.
Pessenda, L.C.R., Ribeiro, A.S., Gouveia, S.E.M., Aravena, R., Boulet, R., Bendassolli,
J.A., 2004. Vegetation dynamics during the Late Pleistocene in the Barreirinhas
region, Maranhaão State, Northeastern Brazil, based on carbon isotopes in soil organic
matter. Quaternary Research 62, 183–193.
Raven, P.H.; Evert, R. V.; Curtis, H. 1978. Biologia Vegetal. Rio de Janeiro: Guanabara
Koogan.
Ribeiro, A.S., 2002. Dinâmica paleoambiental da vegetação e clima durante o Quaternário
tardio em domínios da mata Atlântica, brejo do semi-árido e cerrado nordestinos,
utilizando isótopos do carbono da matéria orgânica do solo e das plantas. Ph.D.
dissertation. University of São Paulo, Piracicaba (Brazil). 192 pp.

90
Santos, G.M.; Gomes, P.R.S.; Anjos, R.M.; Cordeiro, R.C.; Turcq, B.J.; Sifeddine, A.; Di
Tada, M.L.; Cresswell, R.G.; Fifield, L.K. 2000. 14C AMS dating of fires in the
central Amazon rain forest. Nuclear Instruments and Methods in Physics Research,
172: 761-766.
Smit, B. N.; Brown, W.V. The Kranz Syndrome in the gramineae as indicated by carbon
isotope ratios. American Journal of Botany, v.60, n.6, p.505-513, 1973.
Smith, B. N.; Epstein, S. Two categories of 13
C/12
C ratios for higer plants. Plant
Physiology, v.47, p.380-384, 1971.
Tatumi, S.H.; Nagatomo, T.; Dengupta, D.; Watanabe, s.; barrteo, a.m.f.; suguio, K. 1998
Termoluminescence dating of eolian sediments from São Francisco River. State of
Bahia, Brazil. Radiation Effects and Defects in Solids, v.146, n.1-4, p. 285-295.
Winter, K.; Troughton, J. H.; Card, K.A. 1976. 13
C values of grass species collected in the
Northern Sahara Desert. Oecologia, v.25, p.115-123.

91
Capítulo 3. Considerações Finais

92
1. Foram descritas pela primeira vez, para o municipio de Jaguarari – BA, a ocorrência de
seis espécies de mamíferos pleistocênicos: Eremotherium laurillardi; Notiomastodon
platensis; Panochthus greslebini; Equus (Amerhippus) neogeus; Toxodontinae; Felidae.
2. A partir do conjunto fossilífero estudado, sugere-se condições ambientais semelhantes
àquelas inferidas por Cartelle (1999) para o Pleistoceno do Brasil Intertropical Extra-
Amazônico.
3. Durante o Pleistoceno final - Holoceno, a região de Jaguarari compreenderia áreas
abertas de savana associadas, possivelmente, com fisionomias mais fechadas, tipo
florestas.
4. A vegetação amostrada para a área estudada foi composta de 85% de plantas do ciclo
fotossintético C3 (vegetação de floresta) e 15% de plantas do ciclo C4 (vegetação de
campo).
5. A matéria orgânica do solo encontrada nas áreas estudadas sugere estabilidade
vegetacional, sem troca de vegetação, podendo ter ocorrido no local, em períodos
passados, uma vegetação arbórea menos densa ou mistura de plantas C3 e C4, com
predomínio da primeira.
6. Provavelmente no Holoceno, um clima semelhante ao atual ocorreu na região de
Jaguarari.
7. Na região estudada, possivelmente, não houve modificação acentuada na vegetação no
médio Holoceno, quando do estabelecimento de períodos mais secos, permanecendo com
áreas de vegetação de floresta.
8. A Educação Patrimonial, no presente trabalho, mostrou-se um instrumento capaz de
fornecer informações sobre patrimônio, noções de conservação, leis que normatizam estas
práticas, sanções legais previstas, como também, proporcionar a construção de novos
conceitos e uma aprendizagem significativa.
9. Recomenda-se, a aplicação Trabalhos de Educação Patrimonial em sítios de ocorrência
de fósseis, para subsidiar ações de conservação nos locais.

93
Apêndice

94
Educação Patrimonial como instrumento de conservação de um afloramento
fossilífero numa área de Caatinga1
Resumo
Os fósseis são importantes registros de restos de animais ou de vegetais pretéritos e de suas
atividades que se conservam nas rochas. Contudo, nem sempre recebem o tratamento
adequado, sendo manipulados de forma indevida ocasionando a perda de informações que
poderiam auxiliar nas interpretações ambientais. O presente trabalho teve como objetivo
discutir com os moradores do povoado Lajedo II, no município de Jaguarari, microrregião
de Senhor do Bonfim-BA a noção de patrimônio, a importância de se preservar o material
fossilífero, bem como as medidas que estes podem vir a tomar, quando da descoberta de
novos achados. Os resultados obtidos apontam a mudança de conceitos dos moradores e a
adoção de práticas que visam à conservação dos fósseis encontrados no afloramento
fossilífero tipo tanque numa área de Caatinga.
Resumen
Los fósiles son importantes registros de restos animales o vegetales pretéritos, y de sus
actividades que se conservan en rocas. Sin embargo, no siempre reciben el tratamiento
adecuado, siendo manipulados de forma indebida ocasionando la perdida de información
que podrían auxiliar en las interpretaciones ambientales. El presente trabajo tuvo como
objetivo discutir con los residentes del poblado Lajeado II, en el municipio de Jaguarari,
microrregión de Senhor do Bomfim-BA la noción de patrimonio, la importancia de
preservar el material fosilífero, también los cuidados a tomar en caso nuevos
descubrimientos. Los resultados obtenidos apuntan a un cambio de concepto de los mismos
y la adopción de prácticas que visan la conservación de los fósiles encontrados en los
afloramientos fosilíferos del tipo tanque en la región de Caatinga.
Palavras-chave: Conservação, Educação Patrimonial, Caatinga.
1 Artigo publicado nos Anais do V Colóquio Internacional “Educação e Contemporaneidade”, setembro de
2011.

95
1. Introdução
Este trabalho é resultado de algumas atividades desenvolvidas durante o processo de
escavação de um afloramento fossilífero tipo tanque numa área de Caatinga, para coleta de
fósseis e fará parte da dissertação de mestrado da autora, compondo um dos capítulos
voltados para a Educação.
O desenvolvimento de atividades voltadas para o ensino de Educação Patrimonial
surgiu da necessidade de fornecer subsídios para a implementação de ações de conservação
dos fósseis encontrados no tanque Lajedo II, povoado Lajedo II, localizado no município
de Jaguarari, que compõe a microrregião de Senhor do Bonfim – BA.
Nesta área, fósseis de mamíferos neopleistocênicos foram removidos do sítio
paleontológico ao longo de alguns anos sem nenhum critério científico, e transportados
sem registro há outras localidades, dificultando hoje trabalhos que visam identificar esta
fauna, bem como fazer a reconstrução paleoambiental da área.
Essa prática é prejudicial ao desenvolvimento de uma pesquisa mais sistemática, pois
impossibilita a datação do material, um estudo tafonômico mais elaborado e até mesmo,
identificação da flora existente na época, que mesmo não estando presente, pode ser
estudada pela presença dos pólens encontrados junto ao material sedimentar.
A promoção da descontextualizando dos achados se deu a partir de descoberta
acidental, quando moradores do povoado na tentativa de aumentar a capacidade de
armazenamento de água do tanque, ao fazer a limpeza do material sedimentar que este
continha, localizou os fósseis nas camadas mais profundas.
Após a remoção dos fragmentos fossilíferos do tanque, moradores locais, bem como
curiosos recém - chegados, atraídos pela “novidade” se apropriaram de partes de fosseis
retendo-os para si e dispersando de maneira irreversível, o material que deveria ser objeto
de estudo, preservação e educação das gerações futuras.
Por desconhecimento da importância histórica e cultural do material estes foram
manipulados indevidamente e importantes informações paleontológicas foram perdidas.
Desta forma, o presente trabalho teve como objetivo discutir com a população a noção
de patrimônio; discutir a importância de se preservar o material fossilífero, bem como as
medidas que eles podem vir a tomar, quando da descoberta de novos achados.

96
Para alcançar tais objetivos, foi ministrada uma palestra, exposição de um
documentário mostrando os animais inseridos nos seus habitas e a realização de uma
oficina com a confecção, em massa de modelar, de miniaturas das espécies estudadas.
1.1. Um pouco de paleontologia
A paleontologia (gr. Palaios = antigo + ontos = ser + logos = estudo) pode ser definida
como a ciência que estuda os fósseis, isto é, restos ou vestígios de animais ou de vegetais
que viveram antes dos tempos históricos e que se conservam nas rochas (MENDES, 1977).
A palavra fóssil deriva do termo “fossilis” referido pela primeira vez por Plínio (séc. I
a.C), a sua raiz “fossus”, particípio passado de “foldere” (cavar), significa literalmente “o
que se extrai cavando” (CACHÃO, 2005).
Esta terminologia ainda hoje é utilizada e compreende os restos de organismos como
ossos e dentes, tecidos moles, estruturas biogênicas formados à partir das atividades dos
seres quando vivos, como por exemplo pegadas, perfurações, escavações, coprólitos, ovos
e gastrólitos (TIMM; KISCHLAT, 2005).
A paleontologia é considerada como ciência só a partir do século XIX, mas estudos e
observações paleontológicas não são recentes, podendo ser observadas interpretações
desde a antiguidade clássica, onde os filósofos utilizavam os fósseis para obter as
informações que precisavam (MENDES, 1977).
Hoje é sabido que o clima é um fator que atua diretamente no processo de fossilização
e este é sem dúvidas, um dos condicionantes que mais influenciam o ambiente. O clima
pode permanecer mais estável ou variar com o tempo, e exerce influência na distribuição
geográfica e diversificação dos organismos (MENDES, 1988).
Com a instalação de climas mais secos ou, mesmo, áridos, houve, ainda no Pleistoceno
ou talvez já no sub-Recente, profunda modificação na geografia regional, com o
desaparecimento de tais lagoas e entulhamentos das respectivas bacias (AB’SABER,
1956).
De acordo com Souza-Cunha (1966) o termo cacimba não é propriamente um
acidente natural e sim, resultado do trabalho humano, geralmente de escavações no fundo
seco de antigas lagoas que ocupavam depressões topográficas naturais do terreno; e tanque,
seria o resultado de estruturas resultantes de fendas naturais, em rochas cristalinas, de
paredes íngremes e de contorno elíptico.

97
Neste trabalho, o afloramento fossilífero encontrado no povoado Lajedo II trata-se
de um tanque (Figura 01).
Sendo que os restos esqueletais cranianos e pós-cranianos encontrados no local de
estudo são atribuídos a megafauna, que segundo Bergqvist (2002) a definição mais
apropriada para mamíferos fósseis seria: todos aqueles Sinapsidas cuja articulação entre o
crânio e a mandíbula é feita pelos ossos escamosal e dentário.
Embora os ossos craniais sejam de difícil conservação, pois são muito frágeis e se
quebram com facilidade, com algumas exceções encontra-se um crânio inteiro, resta
classificá-los baseados em outras características.
Também é possível Identificar os mamíferos por meio de seus outros ossos, tais como
os ossos longos, que são bem característicos ao grupo, as vértebras, como também a sua
dentição. A identificação por meio destes outros materiais é bastante eficiente, pois ossos
longos, vértebras e dentes abundam nos registros fossilíferos, haja vista, são formados por
partes duras onde a sua fossilização é possível e fica-se preservado por mais tempo
(PAULA-COUTO, 1979).
1.2. Patrimônio e Educação Patrimonial
Conceito de patrimônio normalmente está relacionado com a herança paterna, ou seja,
aos bens materiais transmitidos de pai para filho. A extensão do seu uso tutela e protege
antiguidades nacionais às quais era atribuído significado para a história da nação
(TEIXEIRA et al., 2004, p. 02).
Embora o uso deste termo tenha sido ampliado, geralmente está relacionado aos bens
culturais de um povo, associado às produções humanas, não apenas os bens imóveis, mas
também os bens móveis, tais como acervos de museus e documentos textuais (TEIXEIRA
et al., 2004, p. 02).
Extrapolando um pouco este conceito, os achados paleontológicos são considerados
como patrimônio nacional, uma vez que para regulamentar as questões do patrimônio
brasileiro no que se refere aos fósseis, foram criados uma série de leis, decretos-leis e
portarias para assegurar a proteção destes (BRASIL, 1988).
Para tanto, na Constituição da Republica Federativa do Brasil quando da sua
promulgação em 1988, foi criado um capitulo próprio no que se refere ao Meio Ambiente e
em 12.02.1998, dez anos depois, foi aprovada a Lei 9605 – Lei de Crimes Ambientais, que

98
dispõe sobre as sanções penais e administrativas aplicáveis às condutas e atividades lesivas
ao meio ambiente.
Esta lei é influente por:
representar significativo avanço na tutela do ambiente, pro inaugurar uma
sistematização das sanções administrativas e por tipificar organicamente os
crimes ecológicos” (BOCCASIUS-SIQUEIRA, 2002).
Os artigos 20º, 23º e 24º da Constituição de 1988 indicam que os fósseis são bens da
União e que o estado é responsável na defesa do patrimônio natural do país. Os jazidos
fossilíferos são enquadrados como bens da União disposto nos incisos I, IX e X do artigo
20º.
Como conceito de registro de vida do passado, preservado graças a um processo de
mineralização, pode-se enquadrar os fósseis no inciso IX e /ou X deste mesmo artigo:
Artigo 20º. São bens da União:
I - os que atualmente lhe pertencem e os que lhe vieram a ser atribuídos;IX - os
recursos minerais, inclusive os do subsolo; X - as cavidades naturais
subterrâneas e os sítios arqueológicos e pré-históricos.
Artigo 23º. É competência comum da União, dos Estados, do Distrito Federal e
dos Municípios: III - proteger os documentos, as obras e outros bens de valor
histórico, artístico e cultural, os monumentos, as paisagens notáveis e os sítios
arqueológicos; IV - impedir a evasão, a destruição e descaracterização de obras
de arte e de outros bens de valor histórico, artístico e cultural;
Artigo 24º. Compete à União, aos Estados e ao Distrito Federal legislar
corretamente sobre: VII - proteção ao patrimônio histórico, cultural, turístico e
paisagístico; VIII - responsabilidade por dano ao meio ambiente, ao consumidor,
a bens e direitos de valor artístico, estético, histórico, turístico e paisagístico.
Artigo 216º. Constitui patrimônio cultural brasileiro os bens de natureza material
e imaterial, tomados individualmente ou em conjunto, portadores de referência à
identidade, à ação, à memória dos diferentes grupos formadores da sociedade
brasileira, nos quais se incluem: V - os conjuntos urbanos e sítios de valor
histórico, paisagístico, artístico, arqueológico, paleontológico, ecológico e
científico (BRASIL, 1988).
Diante do exposto, é notório que os estudos e as atividades que envolvam os fósseis
devem ser realizados por um paleontólogo, que são profissionais preparados a
desenvolverem a atividade específica de intervenção. Estas ações podem ser de caráter
científico, educacional e cultural, sendo reguladas nas conformidades e requisitos
estabelecidos por leis próprias e ligada a instituições sérias de pesquisa.
No tocante à Educação Patrimonial, estes profissionais devem promover um ensino
(formal e não formal) que possibilite a construção de conceitos e práticas que visem à
preservação e conservação do material fóssil.

99
O que nos faz pensar que “A prática educativa exige comunicação em múltiplas
linguagens e, também, exige a competência de conhecer a multiplicidade de significados
de significações dessas linguagens (FREIRE, 2001)”.
Podendo assim, evitar uma exploração geradora de impactos e em desacordo com a
legislação vigente ou com as técnicas que são indispensáveis para um melhor
aproveitamento das informações.
Em se tratando de fósseis é considerado impacto toda extração ilegal, feita de forma
não criteriosa realizada por pessoas não capacitadas, tratamento inapropriado das peças e
acondicionamento irregular do material.
Onde a Lei nº 9.605 de 12 de fevereiro de 1998 prevê sanções para crimes ambientais,
protegendo assim, o patrimônio natural:
Seção III. Da Poluição e outros Crimes Ambientais.
Artigo 55º. Executar pesquisa, lavra ou extração de recursos minerais sem a
competente autorização, permissão, concessão ou licença, ou em desacordo com
a obtida. Pena: detenção, de seis meses a um ano, e multa (BRASIL, 1998).
2. Metodologia
A escavação paleontológica no sítio Lajedo II ocorreu nos dias 17 a 21 de Janeiro de
2011. Esta tinha como objetivo o resgate de ossadas de mamíferos neopleistocênicos
encontrados no interior do tanque, como também, fragmentos localizados no entorno deste.
Os moradores do povoado haviam sido informados uma semana antes, que durante os
próximos dias, trabalhos paleontológicos de cunho científico estariam acontecendo.
Na data acima citada, antes de dar início aos trabalhos de escavação, a população do
povoado Lajedo II foi reunida em frente à casa de uma moradora e em seguida,
apresentado o plano de trabalho que seria desenvolvido ao longo da semana.
No primeiro e segundo dia, ao longo dos trabalhos de escavação, sem a presença de
questionários, algumas perguntas foram feitas aos moradores buscando perceber o nível de
entendimento que eles tinham sobre o que estudava a paleontologia, o que eram os fósseis
ali encontrados, que espécies animais eles achavam que pertenciam às ossadas, e quais os
cuidados que eles deveriam ter para salvaguardar estes achados.

100
Embora não tenha sido aplicado questionário, as perguntas dirigidas aos morados
haviam sido elaboradas com antecedência, buscando integrar os novos conhecimentos de
Ensino de Ciências e Paleontologia; Educação Patrimonial; Aprendizagem Significativa e
Novas Tecnologias.
Para tanto, foram utilizadas como referencias livros, artigos científicos nos temas de
interesse, revistas, leituras e informações sobre a história do município de Jaguarari – BA e
sobre o povoado Lajedo II.
O método utilizado na obtenção dos dados foi a pesquisa qualitativa, pois esta tem
como objetivo verificar a eficiência dos questionamentos como forma de compreensão
particular do objeto de estudo (MARTINS; BICUDO, 1989). Optou-se por não aplicar
questionário na tentativa de não coibir e influenciar as respostas dos moradores, como
também, por acreditar que perguntas feitas de forma informal, poderiam facilitar as
respostas e aproximá-los da pesquisadora.
Ao término do segundo dia, novamente os moradores foram reunidos. Foram
mostradas algumas imagens dos animais que existiram na região no período pleistocênico e
algumas informações fornecidas, tais como a ecologia destes.
Estas informações foram antecipadas, pois, estes estavam bastante curiosos em relação
ao conteúdo da palestra do dia seguinte, como também, bastante surpresos com tudo que já
tinha sido dito sobre os achados que estavam acontecendo durante as escavações.
À medida que os fragmentos fósseis eram resgatados, informações sobre as peças
eram passadas, tais como, a que possível animal pertencia, o tamanho apresentado, o
habito alimentar que possuíam. Sempre ressaltando que, dados conclusivos só seriam
obtidos quando estas informações fossem confrontadas com as existentes na literatura
especializada.
No terceiro dia de escavação, os trabalhos foram encerrados mais cedo e a população
do povoado foi reunida em frente casa de uma moradora, a fim de assistirem a uma
palestra sobre a Megafauna Pleistocênica e Educação Patrimonial. Com o auxilio de um
DataShow a palestra desenvolvida utilizando-se o software da Microsoft – PowerPoint,
versão 2007 foi projetada na parede da casa. A palestra continha hipertextos apresentando
figuras estáticas e animadas, como também um pequeno documentário mostrando uma
reconstrução destes animais em seus habitats.

101
A palestra contou com a presença dos representantes oficiais da Secretaria do Meio
Ambiente do município de Jaguarari - BA, sendo o Secretário e Diretores do Meio
Ambiente, como também, com a presença dos Líderes Comunitários do povoado Lajedo II.
No quarto dia, foi ministrada uma oficina para que eles confeccionassem utilizando
massa de modelar, os animais que mais tinham chamado a atenção. Eles poderiam utilizar
como referencia tanto as imagens projetadas durante a palestra, quanto os animais que
apareceram no vídeo. Embora esta prática fosse destinada as crianças, houve a participação
da maioria dos adultos presentes no local.
À medida que estes iam confeccionando seu animal, eles estavam sendo entrevistados
sobre as impressões da palestra do dia anterior e de todas as atividades que estava
acontecendo ao longo dos dias e questionados sobre a escolha do animal. Todas as
perguntas e respostas foram filmadas com o auxilio de uma câmera filmadora digital.
Quando todos os moradores participantes na oficina terminaram de confeccionar seu
animal, houve uma votação para escolher quais as três melhores produções. Em seguida
foi realizada uma premiação simbólica do primeiro, segundo e terceiro colocado (Figura
02).
Ao término das escavações no quinto dia, cada morador envolvido nos trabalhos de
escavação e nas atividades desenvolvidas ao longo da semana, foi entrevistado de forma
individual e perguntado sobre as impressões que tiveram e se tinham sugestões para
trabalhos futuros.
Durante os cinco dias de atividades desenvolvidas no povoado, tanto atividades mais
técnicas (escavações realizadas dentro do tanque e na sua área de entorno e palestra),
quanto à atividade lúdica (oficina), foram filmadas para composição de um documentário e
fotografadas para uma exposição.
O documentário bem como a exposição das fotos das atividades realizadas, ainda será
produzido.
3. Resultados e Discussão
Os resultados das atividades de Educação Patrimonial foram satisfatórios, pois ao
término dos trabalhos pôde-se perceber que os moradores do povoado Lajedo II haviam
modificado e adquirido novos conceitos sobre os temas discutidos.

102
O ato pedagógico pode, então, ser definido como uma atividade sistemática de
interação entre seres sociais, tanto no nível do intrapessoal como no nível da
influencia do meio, interação essa que se configura numa ação exercida sobre
sujeitos visando provocar neles mudanças tão eficazes que os tornem elementos
alvos desta própria ação exercida (ARANHA, 1996).
Esta aquisição de novos conceitos foi perceptível quando novamente perguntados
sobre os temas, os moradores já apresentavam resposta com os conteúdos abordados nas
atividades desenvolvidas.
Foram repetidas as mesmas perguntas, tais como: “O que são fósseis?”, “Qual o
processo de formação dos fósseis?”, “Que medidas podem ser tomadas para que haja
conservação dos fósseis como do sítio paleontológico que eles estão inseridos?”,
“Sabendo que eles são considerados patrimônio, podem ser comercializados”, “Quando
novas descobertas na região fossem realizadas, que tipo de atitude eles deveriam tomar?”.
Ficou evidenciada também, uma aprendizagem significativa, quando os participantes
deram abertamente suas opiniões sobre o objeto de estudo e a partir de suas respostas,
pode-se perceber que estas validavam a metodologia proposta.
Esta mudança de conceitos e atitude vista nos moradores é resultado da total
integração nas atividades. Os participantes se mostraram interessados em todo o momento,
tanto em atuar ativamente na escavação, na coleta dos fósseis, quanto nas atividades
voltadas para Educação Patrimonial.
Durante a palestra se mantiveram atentos o tempo inteiro, tecendo entre si comentários
sobre tudo que foi visto. Mostraram-se surpresos ao ver as imagens dos animais, como
também em saber do tamanho que eles apresentavam.
E ficaram curiosos ao perceber que na região existiram animais tão diferentes do que
existe hoje, principalmente descobrir a presença de alguns que hoje só existem em regiões
mais frias.
Assim, ficaram evidentes as mudanças que a região sofreu, porque foi possível a
formação dos fósseis naquela área, porque estes podem ser encontrados nos dias de hoje, e
a necessidade de conservação destes achados.
Estes puderam concluir que dificilmente as condições que originaram os fósseis da
megafauna neopleistocênica naquele sítio serão repetidas em outro lugar, e que os animais
que ali são encontrados, guardam informações particulares que precisam ser estudadas,
para possibilitar o entendimento das mudanças ocorridas na região.

103
Lamentaram profundamente o desconhecimento dos fósseis enquanto patrimônio e da
sua importância para os estudos de paleontologia, e afirmaram que se fosse sabido na
época dos achados o que eles sabiam hoje, não só não teriam deixados que curiosos
levassem o material, como também, teriam dado um tratamento diferente as peças.
Sabendo destas singularidades, tinham consciência da necessidade de proteção do
sítio, bem como as medidas que poderiam tomar, quando da descoberta de novos sítios,
haja vista, a região ser rica em afloramentos fossilíferos.
Uma das atitudes informadas foi procurar a Secretária do Meio Ambiente no
município de Jaguarari - BA, onde esta entraria em contato com os profissionais
competentes em estudá-los; como também, estes se dirigissem a universidade mais
próxima, sendo a Universidade do Estado da Bahia – UNEB Campus VII, procurando a
responsável pelo Laboratório de Arqueologia e Paleontologia – LAPUNEB, até mesmo,
informarem a pesquisadora autora do presente trabalho.
Estas medidas deverão ser adotas sempre que novos achados forem realizados, e que
jamais estes comercializassem as peças, pois irão incorrer em um crime ambiental e serão
penalizados por esta prática.
Um dos resultados observações que mais chamou atenção foi em relação valorização
do povoado. Os moradores sentiram-se privilegiados em relação à pesquisa e mostraram
profundo interesse em conservar o local, pois perceberam mais que a importância do
material histórico ali contido, a construção de uma identidade local.
Durante os trabalhos houve uma integração muito forte entre a pesquisadora e os
moradores, mas, mais ainda entre os próprios moradores, que incentivados com as
descobertas, traçavam metas e formas de organização para manter os ensinamentos
adquiridos presente no seu cotidiano.
Foi percebido o interesse em algumas crianças de aprofundar os estudos em
paleontologia quando chegassem à universidade.
Também foram desconstruídos alguns conceitos errôneos que haviam sido passados
aos moradores. Para estes, as ossadas ali encontradas eram pertencentes aos dinossauros.
Durante a palestra e conversas informais foi explicada a diferença entre os dinossauros e os
mamíferos ali encontrados, como também a diferenças entre os períodos geológicos que
cada espécie estava situada.

104
Outro conceito equivocado que estes possuíam, era que os fósseis possuíam
propriedades medicinais e que desta forma, podiam se utilizados na produção de
medicamentos naturais ou tivessem qualquer propriedade curativa.
Durante a oficina, os moradores mostraram-se mais falantes, até porque, já se sentiam
mais seguros quanto aos assuntos abordados, e através desta, puderam expressar suas
observações.
Chamaram a atenção algumas miniaturas confeccionadas por eles. Em uma delas foi
representado um carnívoro predando um herbívoro mostrando a compreensão da interação
presa/predador. Em outras, ficou evidenciado a estrutura corporal, e até o comportamento,
onde foi representado como o animal se mantinha em pé como forma de afugentar
predadores.
4. Considerações Finais
Com base no que foi exposto, fica evidente que trabalhos de Educação Patrimonial
precisam ser realizados em sítios de ocorrência de fósseis, para subsidiar ações de
conservação nos locais.
Estas ações serão possíveis e mais eficazes, quando as informações disponibilizadas
aos moradores de cada região, forem postas em práticas, uma vez, serem eles, geralmente,
as primeiras pessoas a ter contato com os fósseis.
Desta forma, é necessário que o conhecimento seja passado de forma clara e objetiva,
buscando a compreensão de todos, aproximando-os do meio científico e fazendo parcerias
com estes.
Assim, a Educação Patrimonial é um instrumento capaz de fornecer informações sobre
patrimônio, noções de conservação, leis que normatizam estas práticas, as sanções legais
previstas, como também, proporcionar a construção de novos conceitos e uma
aprendizagem significativa.
Os fósseis coletados da megafauna neopleistocênica no povoado Lajedo II, município
de Jaguarari, na microrregião de Senhor do Bonfim – BA, serão depositados e integrarão a
Coleção Didática do Laboratório de Arqueologia e Paleontologia da Universidade do
Estado da Bahia – Campus VII.

105
5. Agradecimentos
Sou especialmente agradecida aos moradores do povoado de Lajedo II pelo carinho e
total participação nas atividades, onde sem eles este trabalho não seria possível; ao
Secretário do Meio Ambiente do município de Jaguarari- BA, Antônio Marcos Gomes do
Nascimento pelo apoio logístico; a Glécia Gama pelo acolhimento; a Robson Marques pelo
auxílio nas atividades em campo, incentivo e apoio durante todo o processo; a Juan Pedro
pelo apoio irrestrito; ao CNPq pela bolsa de fomento à pesquisa.
Referencias Bibliográficas
Ab’saber, A.N. 1956. Depressões periféricas e depressões semi-áridas no Nordeste do
Brasil. Bol. São Paulo, Paulista Geogr. 22: 3-18.
Aranha, M.L. de A. 1996. Filosofia da Educação. 2ª ed. Rev. e ampl. São Paulo: Moderna.
Bergqvist, L.P. 2002. Os fósseis da bacia Sergipe-Alagoas: A megafauna pleistocênica.
Phoenix 4(43).
Boccasius-Siqueira, A. 2002. O Direito Ambiental na Legislação Brasileira – um
contributo para o resgate da história. Revista eletrônica do Mestrado em Educação
Ambiental 9.
BRASIL. 2004. Constituição da República Federativa do Brasil. Barueri, São Paulo:
Manole, 2004.
BRASIL. 1998. Lei nº 9.605 de 12 de Fevereiro de 1998. Protege o patrimônio Cultural.
Brasília, DF.
Cachão, M. 2005. Paleontologia, Curso técnico. Departamento de Geologia, Faculdade de
Ciências, Universidade de Lisboa.
Martins, J. & Bicudo, M.A.V. 1989. A pesquisa qualitativa em psicologia: fundamentos e
recursos básicos. São Paulo: Moraes.
Mendes, J.C. 1988. Paleontologia Básica. 1 ed., São Paulo: T. A. Queiroz, editora da
Universidade de São Paulo. p. 347.
Mendes, J.C. 1977. Paleontologia Geral. Rio de janeiro, Livros Técnicos e Científicos;
São Paulo, Ed. Da Universidade de São Paulo.
Paula Couto, C. de. 1979. Tratado de Paleomastozoologia. Rio de Janeiro: Academia
Brasileira de Ciências, p. 590.

106
Paulo Freire. 2001. Vida e obra / Organizado por Ana Inês Souza... [et al].- São Paulo:
Expressão Popular.
Souza-Cunha, F.L. de. 1966. Explorações paleontológicas no Pleistoceno do Rio Grande
do Norte. Arquivos do Instituto de Antropologia “Câmara Cascudo” 2(1-2): 1-46.
Teixeira, S.; Moraes, A.; et alli. Educación Patrimonial: aprendiendo a conocer el
Patrimônio Cultural. In: (Anais) “Primeras Jornadas Del Mercosur y Segundas
Bonarenses sobre Patrimonio Cultural y Vida Cotidiana”. La Plata - Argentina (Texto
publicado integralmente em CD-Rom, ISBN 987-21148-6-2). Novembro/2004.
Timm, L. de L.; Kischlat, E.E. 2005. Considerações sobre áreas de estudo em
paleontologia, métodos de coleta e preparação de vertebrados fósseis. Caderno La
Salle Xi 2(1): 39-48.
Figura 01. Tanque do Sítio Lajedo II, no povoado de Lajedo II, município de Jaguarari -
BA. (Foto: Márcia Xavier, 2011).

107
Figura 02. Miniaturas feitas em massa de modelar, confeccionadas pelos moradores do
povoado Lajedo II durante a oficina ministrada. A - miniaturas sendo entregues para
votação. B - votação das três melhores miniaturas. (Fotos: Márcia Xavier, 2011).

108
Anexo

109

110