estudo do envolvimento dos receptores da …‡Ão... · receptores da adenosina e espironolactona...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE OURO PRETO
NÚCLEO DE PESQUISA EM CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
ESTUDO DO ENVOLVIMENTO DOS RECEPTORES DA
ADENOSINA SOBRE A AÇÃO CARDIOPROTETORA DA
ESPIRONOLACTONA E EPLERENONA EM
CARDIOMIÓCITOS DE RATO
Ouro Preto – MG
2016

GABRIELA DE OLIVEIRA FARIA
ESTUDO DO ENVOLVIMENTO DOS RECEPTORES DA ADENOSINA
SOBRE A AÇÃO CARDIOPROTETORA DA ESPIRONOLACTONA E
EPLERENONA EM CARDIOMIÓCITOS DE RATO
Orientador: Prof. Dr. Mauro César Isoldi
Ouro Preto – MG
2016
Dissertação apresentada ao Programa de Pós-Graduação em
Ciências Biológicas do Núcleo de Pesquisas em Ciências
Biológicas da Universidade Federal de Ouro Preto, como parte
integrante dos requisitos para obtenção do Título de Mestre em
Ciências Biológicas, área de concentração Bioquímica
Metabólica e Fisiológica.



Dedico este trabalho aos meus pais Agenor e Iolanda,
à minha irmã Aline e ao Gustavo pelo amor e apoio
durante essa caminhada. Dedico também à Milla pela
ajuda e solidariedade durante todo o tempo.

Agradecimentos
Agradeço primeiramente a Deus pelo dom da vida e pelas bênçãos concedidas.
À minha família, em especial ao meu pai Agenor por me proporcionar incondicional
apoio, orientação e incentivo para realizar meus sonhos, sempre com sabedoria e pés no chão. À
minha mãe Iolanda, minha irmã Aline pelo apoio nas horas difíceis e por compartilhar as alegrias
nas vitórias.
Agradeço de forma muito especial ao Gustavo que, mesmo de longe, desde o início dessa
caminhada me apoiou em cada situação adversa, me dando seus sábios conselhos, ombro amigo
sempre disponível, carinho e amor que foram imprescindíveis para conseguir continuar a
caminhar. Sua paciência e apoio foram fundamentais para chegar ao fim dessa trajetória. Muito
obrigada amor!
Agradeço a meu orientador Prof. Dr. Mauro César Isoldi pela oportunidade de iniciar no
trabalho científico. Com sua filosofia de trabalho aprendi e cresci muito nesses anos de
laboratório. Meu imenso obrigada à Milla, minha “mãe” na ciência. Milla, foi um prazer
trabalhar com você! Passamos muitos apertos juntas, mas com certeza aprendemos e
amadurecemos muito! Serei sempre grata por sua disponibilidade para ajudar, ensinar, fazer
companhia que seja. Você é um exemplo de dedicação e amizade e sentirei sua falta. Obrigada
por tudo!
Obrigada a toda equipe do Laboratório de Hipertensão pelos ensinamentos, ajuda sempre
disponíveis e pela boa companhia. Durante meus 6 anos de LH, o laboratório foi uma segunda
casa para mim e vocês, definitivamente colaboraram para isso tornando meus dias muito mais
agradáveis.
Muito obrigada ao prof. Dr. Milton Hércules Guerra de Andrade pelos ensinamentos e
colaboração, e a todo pessoal do Laboratório de Enzimologia e Proteômica pela receptividade
e boa vontade durante os períodos em que lá estive preparando amostras para os experimentos
de HPLC e Espectometria de Massas. Milton, sem sua ajuda não sei se conseguiria chegar aos
resultados. Agradeço também à Ananda Lima Sanson, técnica de nível superior do Laboratório
de Caracterização Molecular e Espectrometria de Massas da UFOP, responsável pela leitura
das minhas amostras no equipamento. Muito obrigada!

Agradeço ao professor Dr. Luís Carlos Crocco Afonso e ao Laboratório de
Imunoparasitologia pela disponibilidade de empréstimo de equipamentos. Agradeço de forma
muito especial à professora Dra. Silvana de Queiroz Silva, demais professores e equipe do
Laboratório de Biologia e Tecnologia de Microrganismos (LBTM) da UFOP pelo apoio e
empréstimos realizados. Me sinto praticamente parte dessa equipe! Muito obrigada!
Agradeço aos professores do NUPEB que de alguma forma contribuíram para a realização
deste trabalho tirando dúvidas, dando conselhos ou até mesmo emprestando algum material
necessário para a execução dos experimentos. Agradeço ainda ao Centro de Ciência Animal da
UFOP, em especial à Lorena, pelo cuidado correto para com os animais, proporcionando um
bom trabalho.
À FAPEMIG e ao CNPq pelo apoio financeiro, e à Fundação Gorceix, em especial à
Aline que tanto ajudou durante o processo de aquisição de materiais e controle dos gastos do
projeto, muito obrigada.
Agradeço também à Ana Cláudia, Karininha, Jéssica, Renatinha, Laila e Brab que,
nesses últimos 2 anos, em momentos diferentes, foram mais do que colegas de república e sim
amigas e companheiras para todas as horas. Com vocês dividi minhas lutas, vitórias, alegrias e
tristezas. Vocês são muito importantes pra mim!
Meu sincero obrigada ao “PIVICantina” pelos momentos descontraídos, sempre com
discussões de alto nível, apesar de bem mais raros que na época da graduação. Ao nosso professor
orientador Dr. Fábio, à Ana Maria Sampaio Rocha, eterna Miss Mariana, Brab, Léozinho,
Felipe, Nalaura, Silêncio e Bruninha à distância. Ter vocês como amigos é uma honra.
Passamos muitos momentos sensacionais juntos e, com certeza vocês serão o maior motivo de
saudade que já sinto em não estar mais em Ouro Preto. Amocês demais da conta!!!
Brab, te agradeço de forma especial, pois, além da parceria (sempre certa e eficaz) nos
trabalhos desde os tempos da graduação, enfrentamos o desafio do mestrado juntas. Você se
tornou uma grande amiga e, convivendo com você no dia-a-dia aprendi que a vida é muito mais
do que fazemos dentro da universidade. Obrigada por me ouvir nos momentos difíceis, por sua
sinceridade nas opiniões e sincera disposição a ajudar sempre, enfim, pelo companheirismo que
foi mais importante para mim do que possa imaginar. Muito obrigada!
A todos que, direta ou indiretamente, contribuíram para a realização desse trabalho e para
minha formação, meu muito obrigada.

Resumo
Adenosina, e os fármacos espironolactona e eplerenona são conhecidos cardioprotetores.
Fármacos com esse perfil costumam mimetizar a ação da adenosina, que pode interagir com
cardiomiócitos através de quatro receptores (A1R, A2AR, A2BR e A3R). Nosso objetivo foi testar
a ação de dois conhecidos cardioprotetores, os antagonistas do receptor MR espironolactona e
eplerenona, assim como da adenosina e seus antagonistas como controle. Culturas primárias de
cardiomiócitos de ratos neonatos da linhagem Wistar foram utilizadas em ensaios onde foram
simuladas as condições de pré- e pós-condicionamentos isquêmicos. Nossos resultados
demonstraram que, por meio de análise de viabilidade celular por MTT, os danos sofridos pelos
cardiomiócitos, impostos pela isquemia, foram igualmente revertidos por espironolactona,
eplerenona e pela adenosina. Entretanto, ao associa-los a antagonistas dos receptores da
adenosina A1R, e A3R no pré-condicionamento e A2AR e A2BR no pós-condicionamento, não
houve diferença significativa na proteção das células em condições isquêmicas. Ao dosar a
adenosina por espectrometria de massas nas amostras de cardiomiócitos submetidos a isquemia,
observamos adenosina detectada e mostramos, pela primeira vez, que houve aumento na
concentração de adenosina nas células tratadas com espironolactona no pré-condicionamento e
no tratamento com eplerenona no pós-condicionamento farmacológico. As amostras do grupo
normóxia, no entanto, não apresentaram adenosina detectada. A análise conjunta dos nossos
resultados comprova o efeito protetor da espironolactona, eplerenona e adenosina em condições
de isquemia, além disso, na presença desses antagonistas do MR houve uma maior
disponibilidade de adenosina, agente associado a todo um quadro direto e indireto de proteção
cardíaca em estados de isquemia. Esses resultados nos levam a crer que há mediação entre a ação
da espironolactona e eplerenona com a adenosina, contudo parece ocorrer por outro mecanismo
que não via ligação aos receptores da adenosina. Adicionalmente, nossos resultados são
consequentes de uma análise aguda, feita 18 horas após os tratamentos, podendo não ter sido
tempo suficiente para observar os resultados consequentes dessa possível mediação de ação entre
receptores da adenosina e espironolactona e eplerenona.
Se por um lado a reperfusão é necessária para a sobrevivência do tecido, por outro a
reperfusão em si pode também causar danos teciduais, fato denominado lesão de reperfusão.

Abstract
Adenosine as well as eplerenone and spironolactone display cardioprotective effects. Drugs
with this profile tend to mimic the actions of adenosine, which interact with cardiomyocytes via
four receptors (A1R, A2ARs, A2BR and A3R). Primary neonatal rat cardiomyocyte cultures from
Wistar rat were used in trials where the conditions of pre- and postischemic conditioning was
simulated. Our aim was test the cardioprotective effect of two known MR antagonists,
spironolactone and eplerenone, as well as adenosine and its antagonist as a control. Per cell
viability analysis by MTT, our results demonstrated that the damage sustained by
cardiomyocytes, imposed by ischemia, were equally reversed by spironolactone, eplerenone and
by adenosine. However, by associating them to A1R and A3R antagonists at preconditioning and
A2ARs and A2BR at postconditioning, there was no significant difference in protecting cells in
ischemic conditions. Adenosine was dosed by mass spectrometry in the samples of
cardiomyocytes underwent ischemia, where adenosine was detected and, for the first time, we
show there was an increase of the concentration of adenosine in cells treated with spironolactone
at preconditioning and posttreatment with eplerenone as pharmacological postconditioning.
However, samples of normoxia group showed no detected adenosine. The analysis of our results
demonstrates the protective effect of spironolactone, eplerenone and adenosine in ischemic
conditions. In addition, the presence of these MR antagonists there was a greater availability of
adenosine, agent associated directly and indirectly with protection to the heart ischemic. These
results lead us to believe that there is a mediation between the action of spironolactone and
eplerenone with adenosine however it seems to occur by other mechanism than via binding to
adenosine receptors. In addition, our results are consequent of an acute analysis made 18 hours
after treatment and may not have been enough time to observe the consequent results of possible
mediation action of adenosine receptors and spironolactone and eplerenone.

Lista de Ilustrações
Figura 1: Fotos de dois corações de porcos submetidos ao infarto evidenciando a área infartada
em cor mais clara (coloração trifenil tetrazólio). ........................................................................ 20
Figura 2: Esquema ilustrativo dos ciclos de isquemia-reperfusão do pré-condicionamento
isquêmico. ................................................................................................................................... 21
Figura 3: Esquema da ativação da via RISK induzida por receptor opióide. ............................ 22
Figura 4: Esquema ilustrativo dos ciclos de isquemia-reperfusão do pós-condicionamento
isquêmico. ................................................................................................................................... 23
Figura 5: Vias de sinalização desencadeadas no pré- e pós-condicionamento isquêmico. ....... 24
Figura 6: Esquema resumindo as vias de sinalização envolvidas no condicionamento isquêmico
..................................................................................................................................................... 25
Figura 7: Estrutura química da adenosina. ................................................................................ 26
Figura 8: Esquema ilustrativo das vias de produção e degradação da adenosina. ..................... 27
Figura 9: Vias de sinalização envolvidas na ativação dos receptores de adenosina. ................. 28
Figura 10: Fórmulas estruturais dos antagonistas do receptor mineralocorticóide da aldosterona.
..................................................................................................................................................... 30
Figura 11: Cromatograma e espectro de massas obtidos da amostra “espironolactona” do pré-
condicionamento. ........................................................................................................................ 47
Figura 12: Cromatograma e espectro de massas obtidos da amostra “controle” do pré-
condicionamento. ........................................................................................................................ 48
Figura 13: Cromatograma e espectro de massas obtidos da amostra “controle” do pós-
condicionamento. ........................................................................................................................ 48
Figura 14: Cromatograma e espectro de massas obtidos da amostra “controle” normóxia. ..... 49

Lista de Gráficos
Gráfico 1: Análise do dano isquêmico mediante exposição dos cardiomiócitos a condições de
isquemia ..................................................................................................................................... 46
Gráfico 2: Análise da ação da espironolactona, eplerenona e adenosina no pré-condicionamento
e no pós-condicionamento farmacológico .................................................................................. 50
Gráfico 3: Análise da ação dos antagonistas dos receptores A1R e A3R da adenosina no pré-
condicionamento farmacológico ................................................................................................ 51
Gráfico 4: Análise da ação dos antagonistas dos receptores A2AR e A2BR da adenosina no pós-
condicionamento farmacológico ................................................................................................ 51
Gráfico 5: Análise da ação dos antagonistas dos receptores A1R e A3R da adenosina associados
à espironolactona no pré-condicionamento farmacológico ........................................................ 52
Gráfico 6: Análise da ação dos antagonistas dos receptores A2AR e A2BR da adenosina associados
à espironolactona no pós-condicionamento farmacológico ........................................................ 53
Gráfico 7: Análise da ação dos antagonistas dos receptores A1R e A3R da adenosina associados
à eplerenona no pré-condicionamento farmacológico ................................................................ 53
Gráfico 8: Análise da ação dos antagonistas dos receptores A2AR e A2BR da adenosina associados
à eplerenona no pós-condicionamento farmacológico ................................................................ 54

Lista de Tabelas
Tabela 1: Composição da solução de BRDU [10-2M] (5-Bromo-2'-deoxiuridina) .................... 38
Tabela 2: Composição do meio de cultura normal ..................................................................... 38
Tabela 3: Composição da solução tampão de digestão .............................................................. 38
Tabela 4: Composição do meio de cultura para isquemia .......................................................... 38
Tabela 5: Composição da solução de parada para evitar degradação da adenosina .................. 38
Tabela 6: Concentração de adenosina detectada nos cromatogramas para amostras dos grupos
normóxia, pré-condicionamento e pós-condicionamento ........................................................... 49
Tabela 7: Informações gerais dos antagonistas do MR, espironolactona, eplerenona, canrenona
e finerenona ................................................................................................................................ 74

Lista de Abreviaturas
A1R Receptor da adenosina subtipo A1R
A2AR Receptor da adenosina subtipo A2AR
A2BR Receptor da adenosina subtipo A2BR
A3R Receptor da adenosina subtipo A3R
AC Adenilato Ciclase
ADA Adenosinas deaminase
ADO Adenosina
Akt Serina/Treonina Quinase (ou Proteína Quinase B – PKB)
AMP Adenosina monofosfato
AMPc Adenosina monofosfato cíclico
ANOVA Análise de variância
AP1 Ativador de proteína 1
ATP Adenosina trifosfato
AVE Acidente Vascular Encefálico
BAY60-6583 Agonista do receptor A2A da adenosina
BAY94-8862 Antagonista do MR Finerenona
BRDU 5-Bromo-2'-deoxyuridine
Ca2+ Íon cálcio
CCA Centro de Ciência Animal
CCPA Agonista do receptor A1 da adenosina
CEUA Comitê de Ética de Uso Animal
CI-IB-MECA Agonista do receptor A3R da adenosina
CO2 Dióxido de Carbono
COX-2 Ciclooxigenase (também conhecida como prostaglandina G/H sintetase)
DPCPX Antagonista do receptor A1R da adenosina
DAG Diacilglicerol
DMEN Dulbeco´s Modified Eagle Medium
DMSO Dimetilsulfóxido
ECA Enzima conversora de angiotensina
eNOS Óxido nítrico sintase endotelial

EPHESUS Eplerenone Heart failure na Survival study – Principal estudo clínico sobre
eplerenona
ERK 1/2 Quinase regulada por sinais extracelulares tipo 1/2
FMUSP Faculdade de Medicina da Universidade de São Paulo
Gi Proteína G inibitória
GMPc Guanosina 3’,5’-monofosfato cíclico
Gq Proteína G tipo q
GR Receptor glicocorticoide
Gs Proteína G estimulatória
H+ Íon hidrogênio
HC Hospital das Clínicas
HPLC-MS High performance liquid chromatography- mass spectrometry – Cromatografia
Líquida de Alta Performance – Espectometria de Massas
HSPs Heat shock protein – Proteínas de choque térmico
IAM Infarto Agudo do Miocárdio
IC Insuficiência cardíaca
InCor Instituto do Coração
IP3 Inositol trifosfato
I-R Isquemia-Reperfusão
KATP Canais de potássio sensíveis a ATP
LDH Enzima Lactato Desidrogenase
MAPK Proteína quinase ativada por mitógeno
MEK 1/2 Proteína quinase ativada por mitógeno tipo 1/2
MR Receptor mineralocorticoide
MRS 1523 Antagonista do receptor A3R da adenosina
MRS 1706 Antagonista do receptor A2BR da adenosina
MTT Brometo de 3-4,5-dimetil-tiazol-2-il-2,5-difeniltetrazólio
m/z Massa/carga
N2 Gás nitrogênio
n.d. Não detectado
NFκB Fator de transcrição NF-kappaB
NO Óxido nítrico
NrF2 Fator nuclear eritróide 2 relacionado ao fator 2
O2 Gás oxigênio

p38 Subfamília de proteína quinase MAPK
p70S6K Proteína S6 quinase 1
PCI Pré-condicionamento isquêmico
PCIR Pré-condicionamento remoto
PI3K Enzima fosfatidilinositol-3-quinase
PIP2 Fosfatidilinositol 4,5-bifosfato
PKA Proteína quinase A
PKC Proteína quinase C
PLC Fosfolipase C
PTK Proteína tirosina quinase
PTPM Poros de transição de permeabilidade mitocondrial
q.s.p Quantidade suficiente para
RAF Proteína quinase específica para Ser/Thr
RALES Randomized Aldosterone Evaluation Study – Principal estudo clínico sobre
espironolactona
RISK Reperfusion Injury Salvage Kinase – Quinases de salvamento da lesão de
reperfusão
ROS Espécie reativa de oxigênio
rpm Rotações por minuto
SAH S-adenosil-homocisteína
SCH58261 Antagonista do receptor A2A da adenosina
SD Linhagem de rato – Spreague Dawley
SEM Erro padrão da média
SRAA Sistema Renina-Angiotensina-Aldosterona
STAT Transdutor de sinal e ativador de transcrição
T1/2 Tempo de meia-vida
tr Tempo de retenção
UFOP Universidade Federal de Ouro Preto
ZM 241385 Antagonista do receptor A2AR da adenosina

Sumário
1- INTRODUÇÃO .................................................................................................................. 18
1.1- Epidemiologia ...................................................................................................................... 19
1.1. Condicionamento Isquêmico ..................................................................................... 20
1.2. Adenosina .................................................................................................................... 26
1.3. Receptores da adenosina ............................................................................................ 28
1.4. Antagonistas do receptor mineralocorticoide da aldosterona (MR) ..................... 29
1.5. Antagonistas do receptor mineralocorticóide (MR), receptores de adenosina e o
condicionamento isquêmico .................................................................................................. 31
2- OBJETIVOS ....................................................................................................................... 34
OBJETIVO GERAL ................................................................................................................... 35
3- METODOLOGIA ............................................................................................................... 36
3.1. Animais ........................................................................................................................ 37
3.2. Cultura Primária de Cardiomiócitos........................................................................ 37
3.3. Soluções Utilizadas nos Ensaios ................................................................................ 37
3.4. Desenho experimental ................................................................................................ 39
3.5. Indução da isquemia in vitro ..................................................................................... 40
3.6. Validação do método de isquemia in vitro ................................................................ 41
3.7. Determinação da adenosina em cardiomiócitos ventriculares de ratos neonatos sob
condições de isquemia ........................................................................................................... 42
3.8. Avaliação da possível ação da espironolactona e eplerenona mediada pelos receptores
da adenosina (A1R, A2AR, A2BR e A3R) em condições de isquemia .................................. 43
3.9. Análise estatística ........................................................................................................... 44
4- RESULTADOS .................................................................................................................. 45
4.1. Validação do método de Isquemia in vitro .......................................................................... 46
4.2. Determinação da adenosina em cardiomiócitos ventriculares de ratos neonatos sob
condições de isquemia ........................................................................................................... 46

4.3. Espironolactona e eplerenona x Antagonistas da adenosina em condições de isquemia
................................................................................................................................................. 50
5- DISCUSSÃO ...................................................................................................................... 55
6- CONCLUSÃO .................................................................................................................... 60
7- REFERÊNCIAS ................................................................................................................. 62
8- ANEXOS ............................................................................................................................ 73

18
1- INTRODUÇÃO

Introdução
19
1.1- Epidemiologia Doenças cardiovasculares estão entre as doenças não contagiosas que mais levam a óbito
segundo a Organização Mundial da Saúde. Estimativa feita no ano de 2012 mostrou que das 56
milhões de mortes no mundo, cerca de 36 milhões foram consequentes de doenças não
contagiosas como diabetes (4%), doenças respiratórias crônicas (10,7%), câncer (21,7%) e
doenças cardiovasculares (46,2% ou 17,6 milhões de pessoas). Uma projeção estima que o
número de mortes por doenças cardiovasculares passará de 17,6 milhões ocorridas em 2012 para
22,2 milhões no ano de 2030 (WHO – World Health Organization, 2014).
Dados do Ministério da Saúde mostram que as doenças cardiovasculares foram
responsáveis por 29,4% das mortes no Brasil, isto é, 308 mil pessoas morreram principalmente
por infarto e acidente vascular encefálico (AVE) no ano de 2007 (Portal da Saúde). Segundo o
DATASUS/MS, o número de óbitos causados por doenças cardiovasculares entre 2006 a 2013
foi de 3.153.175, equivalente a 40 mortes por segundo (Portal da Sociedade Brasileira de
Cardiologia – Cardiômetro).
Diante destes números, é evidente que doenças cardiovasculares são um problema de saúde
pública importante e, estudos voltados à prevenção e tratamento são essenciais para reduzir a
morbidade e mortalidade dos pacientes.
Segundo Nicolau e colaboradores (1999), a hipertensão é um dos fatores de risco mais
importantes para o desenvolvimento de infarto agudo do miocárdio (IAM) (Nicolau et al., 1999).
O coração infartado tem seu tecido lesionado podendo levar à arritmia e insuficiência cardíaca.
No caso específico do IAM, clinicamente, a meta é restabelecer o fluxo sanguíneo para a área
isquêmica o mais rapidamente possível para evitar maiores danos celulares que poderiam ser
provocados pela isquemia. Se por um lado a reperfusão é necessária para a sobrevivência do
tecido, por outro a reperfusão em si pode também causar danos teciduais, fato denominado lesão
de reperfusão. Os danos provocados pelo processo de isquemia-reperfusão atingem as
mitocôndrias de forma significativa, nas quais ocorre redução na síntese de ATP, aumento no
acúmulo de cálcio e acentuada geração de espécies reativas de oxigênio, acarretando em lesões
e morte celular (Silva Jr. et al., 2002). Dessa forma, também já foi demonstrado que células
viáveis, mesmo após a isquemia, no ato da reperfusão sofrem necrose, o que pode contribuir para
até 50% da área infartada final, mitigando os benefícios da reperfusão (Bulluck e Hausenloy,
2015).

Introdução
20
Encontrar uma forma de tornar as células cardíacas resistentes à morte por isquemia-
reperfusão (I-R) é um alvo terapêutico pertinente para promover cardioproteção. Dentre as
formas de minimizar esses efeitos estão o uso de antagonistas do receptor mineralocorticoide da
aldosterona (MR) e o condicionamento isquêmico.
1.1. Condicionamento Isquêmico
Condicionamento isquêmico é o termo usado, atualmente, que engloba estratégias
cardioprotetoras endógenas, baseadas em ciclos breves de isquemia-reperfusão, sejam elas
aplicadas diretamente no coração como no pré-condicionamento isquêmico e no pós-
condicionamento isquêmico, ou indiretamente como é o caso do pré-condicionamento remoto.
Além da completa reperfusão, o condicionamento isquêmico é conhecido como a intervenção
mais poderosa na redução da área infartada (Figura 1) (Hausenloy e Yellon, 2016).
Figura 1: Fotos de dois corações de porcos submetidos ao infarto evidenciando a área infartada em cor mais clara
(coloração trifenil tetrazólio). A primeira imagem mostra um coração onde não houve proteção e a segunda de um
coração protegido. Ao lado, um gráfico mostra a área infartada em relação ao fluxo sanguíneo residual. Fonte:
(Heusch, 2015).
1.1.1. Pré-condicionamento isquêmico (PCI)
Descrito pela primeira vez em 1986 por Murry e colaboradores, o pré-condicionamento
isquêmico (PCI) é caracterizado por breves episódios de isquemia seguida de reperfusão,
realizados antes de um episódio isquêmico longo. Estudando cães, Murry e colaboradores
demonstraram que era possível proteger o coração dos danos causados pelo infarto ao executar
4 ciclos de 5 minutos de oclusão da artéria coronária esquerda, intercalados por 5 minutos de
reperfusão realizados antes da oclusão prolongada (Fig. 2). Este procedimento foi capaz de
reduzir 75% do dano promovido pela isquemia em comparação ao grupo controle, não submetido
aos ciclos de isquemia-reperfusão (Murry et al., 1986).

Introdução
21
Figura 2: Esquema ilustrativo dos ciclos de isquemia-reperfusão do pré-condicionamento isquêmico.
Após sua descoberta, a função protetora do PCI foi demonstrada em todas as espécies
testadas, como camundongos, ratos, coelhos, gatos, cães, ovelhas, macacos e corações humanos
(Hausenloy e Yellon, 2016).
Nos primeiros anos após a descoberta do pré-condicionamento isquêmico, a hipótese mais
aceita era a de que o PCI protege por melhorar o equilíbrio metabólico durante o período
isquêmico. Vários estudos examinaram os efeitos metabólicos e iônicos do PCI em corações e
observaram que o miocárdio apresenta menor concentração do nucleotídeo adenina, um aumento
substancial de fosfato de creatina, excesso de glicose intracelular e déficit contrátil (Murry et al.,
1991; Jennings et al., 2001). Adicionalmente, durante o período isquêmico prolongado,
cardiomiócitos pré-condicionados exibem uma taxa mais lenta de depleção de ATP, assim como
de lactato e acúmulo de H+. Por meio desses resultados, os autores concluíram que a redução da
procura de energia é um componente essencial para o mecanismo de proteção cardíaca no pré-
condicionamento (Jennings et al., 2001).
Tem sido descrito que o pré-condicionamento atua por duas janelas de cardioproteção
distintas, caracterizadas como precoce e tardia devido ao tempo que perdura a proteção
promovida em cada uma delas. O pré-condicionamento precoce ocorre imediatamente após os
estímulos de isquemia-reperfusão e dura de 2 a 3 horas. O pré-condicionamento tardio, por sua
vez, tem início entre 12 e 24 horas mais tarde e dura entre 48 e 72 horas (Hausenloy e Yellon,
2016).
Atualmente, sabe-se que durante os breves ciclos de isquemia-reperfusão promovidos
durante o pré-condicionamento, é iniciada a produção de uma série de autacóides como a
adenosina, bradicinina, endotelina e opióides, nos cardiomiócitos. A ligação desses autacóides a
seus respectivos receptores presentes na membrana plasmática das células cardíacas, transmite
sinais para a mitocôndria que promoverão a cardioproteção. Espécies reativas de oxigênio (ROS)
são geradas na mitocôndria e ativam proteínas quinases tais como a Akt (serina/treonina quinase),

Introdução
22
ERK1/2 (proteína quinase regulada por sinal extracelular), PKC (proteína quinase C) e tirosina
quinase (PTK). Esse processo permite que o efeito cardioprotetor possa durar de 2 a 3 horas,
caracterizando assim o pré-condicionamento precoce (Hausenloy e Yellon, 2016).
Estímulos químicos liberados endogenamente por pequenas moléculas, tais como íons
cálcio, espécies reativas de oxigênio (ROS), espécies reativas de nitrogênio e sulfeto de
nitrogênio também eliciam a cardioproteção (Heusch, 2015). Íons de cálcio extracelulares podem
atuar por meio da adenosina e pela ativação da PKC, no entanto, aparentam possuir um papel
menor no pré-condicionamento (Heusch, 2015), uma vez que, segundo Wallbridge e
colaboradores, o uso de antagonistas de cálcio não interfere no PCI (Wallbridge et al., 1996;
Heusch, 2015).
No pré-condicionamento tardio, proteínas quinase ativam fatores de transcrição como a
proteína ativadora-1 (AP-1), o fator de transcrição nuclear NF-κB, o fator NrF2 (fator nuclear
eritróide 2 relacionado ao fator 2) e o transdutor de sinal e ativador de transcrição (STAT) 1/3.
Estes fatores de transcrição levam à síntese de mediadores como a prostaglandina (G/H) sintetase
(também conhecida como COX-2), proteínas heat shock (Hsp72) e óxido nítrico sintase induzível
que induzem o efeito cardioprotetor entre 12-24h após o estímulo do PCI (Heusch, 2015).
Vias de sinalização também têm sido descritas como atuantes na prevenção da lesão de
reperfusão, dentre elas pode-se citar a via RISK (Reperfusion Injury Salvage Kinase). A via
quinase de salvamento da lesão de reperfusão (RISK) é um grupo de quinases pró-sobrevivência
que incluem as proteínas Akt e ERK 1/2 (figura 3). Quando ativada no início da reperfusão, esta
via é capaz de promover cardioproteção (Hausenloy e Yellon, 2010).
Figura 3: Esquema da ativação da via RISK induzida por receptor opióide. Fonte: (Luna-Ortiz et al., 2011).

Introdução
23
Estudos in vitro também têm sido realizados simulando isquemia-reperfusão ao expor
células cardíacas mantidas em meio pobre em nutrientes a uma atmosfera de N2 ou mistura gasosa
composta por 95% de N2 e 5% de CO2. Dessa forma, é possível mimetizar a isquemia e buscar
compreender diretamente nas células, os mecanismos envolvidos no condicionamento
isquêmico. Safran e colaboradores, por meio de ensaios in vitro, avaliaram o papel dos receptores
A1R e A3R da adenosina em cardiomiócitos de ratos e concluíram que ambos receptores,
ativados, são capazes de atenuar a lesão de miócitos durante a hipóxia (Safran et al., 2001).
1.1.2. Pós-condicionamento isquêmico
Apesar de sua eficácia em proteger o coração, o pré-condicionamento isquêmico possui
uma grande desvantagem: a intervenção deve ser feita antes da isquemia prolongada, tornando-
o inviável em casos de infarto agudo do miocárdio. Felizmente, estudos subsequentes ao PCI
demonstraram também que a aplicação de pequenos ciclos de isquemia-reperfusão no início do
período de reperfusão também é capaz de reduzir a área do infarto e da apoptose de modo similar
ao pré-condicionamento (Zhao et al., 2003); (Crisostomo et al., 2006). A esse fenômeno deu-se
o nome de pós-condicionamento isquêmico – Fig. 4.
Figura 4: Esquema ilustrativo dos ciclos de isquemia-reperfusão do pós-condicionamento isquêmico.
Zhao e colaboradores demonstraram que o pós-condicionamento confere uma miríade de
efeitos cardioprotetores, como a redução dos níveis de edema do miocárdio, diminuição do
estresse oxidativo, acúmulo de neutrófilos polimorfonucleares, e função endotelial preservada
(Zhao et al., 2003; Hausenloy e Yellon, 2016).
O pós-condicionamento compartilha algumas vias de sinalização do PCI (tais como as vias
RISK), no entanto, possui mecanismos de ação particulares. Por exemplo, a reperfusão gradual
promovida pelo pós-condicionamento atrasa o restabelecimento do pH intracelular, o que pode
contribuir para a inibição da abertura dos poros de transição de permeabilidade mitocondrial
(PTPM) em corações pós-condicionados (Hausenloy e Yellon, 2016). A inibição de PTPM
mantém a integridade da mitocôndria, evitando assim a liberação de proteínas pró-apoptóticas.

Introdução
24
A cascata de sinalização promovida pela ligação da adenosina ao receptor, resulta na
ativação de fosfolipase C (PLC). Paralelamente, a hidrólise do fosfatidilinositol 4,5-bifosfato
(PIP2) produz inositol trifosfato (IP3) e diacilglicerol (DAG). A PLC ativa o DAG formado e
então, a proteína quinase C (PKC) que é ativada diretamente pelo DAG e indiretamente por Ca2+.
PKC pode promover proteção pela ativação dos canais KATP mitocondriais ou ativação da
proteína tirosina quinase (PTK), que ativa proteínas quinases ativadas por mitógeno (MAPK),
família de proteínas conhecida pela promoção da cardioproteção – Fig. 5 (Crisostomo et al.,
2006).
Figura 5: Vias de sinalização desencadeadas no pré- e pós-condicionamento isquêmico. Fonte: (Crisostomo et al.,
2006)
1.1.3. Pré-condicionamento remoto (PCIR)
O pré-condicionamento remoto recebe atenção especial na prática clínica, pois, em
oposição ao pré- e ao pós-condicionamento que exigem uma intervenção direta no coração, no
pré-condicionamento remoto o estímulo é realizado em um órgão ou tecido “distante”
(Hausenloy e Yellon, 2016).
Semelhante ao pré-condicionamento clássico, Przyklenk e Whittaker propuseram em 1994
o pré-condicionamento remoto (PCIR) ou “pré-condicionamento à distância”. Com base em um
modelo matemático e em dados experimentais, os pesquisadores levantaram a hipótese de que
breves episódios de isquemia-reperfusão aplicados em tecidos ou órgãos distantes, tais como

Introdução
25
músculo esquelético, rins e mesentério, são capazes de provocar um fenótipo de proteção no
coração, tornando o músculo cardíaco resistente a um episódio longo de isquemia subsequente
(Przyklenk e Whittaker, 2011).
Evidências têm indicado que o efeito cardioprotetor subjacente promovido pelo pré-
condicionamento remoto, ocorre por uma via neuro-humoral central. A reperfusão do órgão
remoto promove a proteção cardíaca por meio de uma “lavagem” que transporta a substância ou
o fator humoral gerado pelo PCIR até o coração – Fig. 6 (Hausenloy e Yellon, 2016).
Figura 6: Esquema resumindo as vias de sinalização envolvidas no condicionamento isquêmico. Fonte:
(Hausenloy e Yellon, 2016) .
1.1.4. Condicionamento farmacológico
Apesar dos benefícios notórios promovidos pelos três tipos de condicionamento isquêmico,
a saber, PCI, PCIR e pós-condicionamento, na maior parte das vezes necessitam de intervenção
cirúrgica para serem executados, o que não os isenta de riscos. Como alternativa, já foi
demonstrado que a perfusão de algumas substâncias antes da isquemia prolongada ou durante a
reperfusão, podem desencadear alterações celulares capazes de proteger o coração. A esse
fenômeno chamou-se condicionamento farmacológico, podendo também ser dividido em pré- e
pós-condicionamento, conforme o momento em que é realizado. Tanto o pré- quanto o pós-

Introdução
26
condicionamento farmacológico são capazes de reduzir a necrose e apoptose no miocárdio
durante isquemia-reperfusão (Li et al., 2015).
Assim como o PCI e o pós-condicionamento isquêmico compartilham de mecanismos de
ação semelhantes, no pré- e pós-condicionamento farmacológico também ocorrem semelhanças
funcionais. Por exemplo, tem sido relatado que o pré-condicionamento por meio de sevoflurano
melhora a função ventricular e recupera o miocárdio, e seu uso no pós-condicionamento reduz
arritmias da reperfusão (Tian et al., 2013).
A ativação de mediadores farmacológicos durante isquemia-reperfusão pode mimetizar ou
mesmo intensificar os efeitos do condicionamento isquêmico. Desvendar os mecanismos de
atuação desses agentes de condicionamento farmacológico pode abrir caminhos importantes para
a prática clínica.
Dentre as substâncias envolvidas no condicionamento farmacológico, pode-se citar a
adenosina, bradicinina, eritropoietina, óxido nítrico (NO) (Gulati e Singh, 2014), opióides e
antagonistas do receptor mineralocorticoide (MR) da aldosterona como espironolactona e
eplerenona (Chai, Garrelds, Arulmani, et al., 2005; Chai, Garrelds, De Vries, et al., 2005).
1.2. Adenosina
A adenosina (ADO) é um autacóide que, assim como a bradicinina, é liberado a partir do
endotélio e células intersticiais durante os ciclos de isquemia-reperfusão no condicionamento
isquêmico (Heusch, 2015). Constituído por uma adenina ligada a uma ribose (Fig. 7), esse
nucleosídeo endógeno é um dos moduladores plausíveis para a proteção do miocárdio isquêmico
(Safran et al., 2001). Possui papel fundamental no controle da homeostase (Cunha, 2001), é um
vasodilatador potente (Berne, 1964; Sollevi, 1986) e neuromodulador importante na regulação
da transmissão sináptica e excitabilidade neuronal do sistema nervoso central (Cunha, 2001;
Rahman, 2009). Em 1985, foi descoberto que adenosina exerce efeitos cardioprotetores diretos
sobre os miócitos (Ely et al., 1985).
Figura 7: Estrutura química da adenosina. Fonte: http://www.universitario.com.br/noticias/n.php?i=16780

Introdução
27
A adenosina é formada a partir de AMP pela família de 5'-nucleotidases – Fig. 8 (Chiu e
Freund, 2014). No miocárdio, pode ser produzida pela desfosforilação do AMP promovida
intracelularmente pela ação da endo-5’-nucleotidase, localizada no citosol (Sommerschild e
Kirkebøen, 2000). A ecto-5’-nucleotidase, por sua vez, é responsável por desfosforilar o AMP
no espaço intersticial, gerando a maior parte da adenosina extracelular. Outra maneira de síntese
intracelular da adenosina é pela clivagem da S-adenosil-homocisteína (SAH), no entanto é menos
comum (Chiu e Freund, 2014). A formação de adenosina a partir de SAH é catalisada pela S-
Adenosil-homocisteína hidrolase, no entanto, a taxa de hidrólise de SAH durante a isquemia não
aumenta muito, sugerindo que esta via não seja fonte significativa de produção de adenosina no
tecido cardíaco nessas condições (Sommerschild e Kirkebøen, 2000).
Além da síntese de adenosina, os processos de degradação podem estar envolvidos no
aumento de sua disponibilidade. Sua degradação é feita pelas enzimas adenosina quinase e
adenosina deaminase (ADA), que convertem a adenosina em inosina por uma desaminação
irreversível – Fig. 8 (Chiu e Freund, 2014).
Figura 8: Esquema ilustrativo das vias de produção e degradação da adenosina. Fonte: (Latini e Pedata, 2001).
A adenosina atua via receptores transmembrana acoplados a proteínas G, divididos em 4
subtipos: A1R, A2AR, A2BR e A3R. Estudos têm demonstrado que todos os receptores da

Introdução
28
adenosina são expressos no coração e estão envolvidos na cardioproteção. Além disso, há
evidências de que todos podem ser expressos em miócitos ventriculares (Zhan et al., 2011), no
entanto pouco se sabe quanto à suas ações.
1.3. Receptores da adenosina
A ligação da adenosina a seus respectivos receptores exerce efeitos por meio da regulação
de AMPc (Chiu e Freund, 2014). No entanto, esses efeitos ocorrem de maneira diferente entre os
receptores. Enquanto os receptores A1R e A3R, acoplados respectivamente a proteínas Gi e Gq,
inibem adenilato ciclase (AC), receptores A2AR e A2BR, acoplados a proteína Gs, ativam AC,
levando à formação de AMPc e subsequente ativação de PKA (proteína quinase A dependente
de AMPc) – Fig. 9. A adenosina também ativa PKC diretamente, que por sua vez, ativa a via
RISK protegendo contra lesão de reperfusão (Heusch, 2015).
Figura 9: Vias de sinalização envolvidas na ativação dos receptores de adenosina. Fonte:
https://sites.google.com/a/unicam.it/diego-dal-ben/activities-and-collaborations
Sabe-se que os receptores A1R e A3R são ativados antes da isquemia, portanto, exercem
seus efeitos cardioprotetores no PCI. Em oposição, os efeitos protetores de A2AR e A2BR
aparecem durante a reperfusão, atuando assim, no pós-condicionamento isquêmico (Zhan et al.,
2011).
Estudos in vitro simularam a ação dos receptores de adenosina no PCI e pós-
condicionamento. Conforme descrito por Leshem-Lev e colaboradores, a ativação dos receptores
A1R e A3R por seus respectivos agonistas CCPA e CI-IB-MECA, protege os cardiomiócitos de
rato contra hipóxia por meio de fosforilação da MAPK p38, o que ocorre downstream à abertura
dos canais KATP da mitocôndria (Leshem-Lev et al., 2010). A sinalização promovida durante a

Introdução
29
ativação do receptor A1R ocorre por meio de sua associação a proteína Gi e efetores como a
MAPK (proteína quinase ativada por mitógeno), Akt e ERK 1/2 (St Hilaire et al., 2009).
O agonista altamente seletivo do receptor A2AR, BAY 60-6583, é capaz de reduzir a área
infartada de corações de coelho quando aplicado durante a reperfusão. O agonista dos receptores
A1R/A2R da adenosina, 5’-(N-ethylcarboxamido) aplicado durante a reperfusão limitou a área de
infarto, enquanto os antagonistas SCH58261 e MRS1706 dos receptores A2AR e A2BR,
respectivamente foram capazes de bloquear esse efeito (Xi et al., 2009).
Muitos aspectos da ação dos receptores de adenosina na I-R ainda são pouco
compreendidos. É consenso que receptores A1R e A3R são cardioprotetores quando ativados
antes da isquemia e receptores A2AR e A2BR, quando ativados durante a reperfusão. Muitas
questões ainda não estão claras, incluindo os mecanismos de sinalização ativados e se esses são
todos mediados via ação direta sobre os cardiomiócitos.
1.4. Antagonistas do receptor mineralocorticoide da aldosterona (MR)
Pacientes que sofrem infarto do miocárdio apresentam elevados níveis de aldosterona e
têm associado a isso, um aumento de fibrose, hipertrofia, morte celular, inflamação e estresse
oxidativo (Qin et al., 2003; Hayashi et al., 2008).
Aldosterona é um hormônio mineralocorticoide sintetizado na zona glomerulosa da supra-
renal sendo o efetor terminal da cascata de sinalização do Sistema Renina-Angiotensina-
Aldosterona (SRAA). Entretanto, elevados níveis plasmáticos de aldosterona resultam em
complicações vasculares, renal, cardíaca e metabólica independente da pressão sanguínea.
A grande quantidade de efeitos deletérios promovidos pela aldosterona sobre o sistema
cardiovascular tem motivado estudos sobre inibidores do receptor mineralocorticóide (MR).
Dois grandes estudos clínicos já realizados, o RALES (Randomized ALdosterone
Evaluation Study) e o EPHESUS (EPlerenone HEart failure and SUrvival Study), mostraram
que os antagonistas do MR reduzem a mortalidade em pacientes com insuficiência cardíaca e
disfunção ventricular esquerda após infarto. Esses estudos foram realizados com os dois
principais antagonistas do MR disponíveis comercialmente: espironolactona e eplerenona, e têm
apresentado excelentes resultados no tratamento de pacientes com disfunção ventricular esquerda
e evidência clínica de insuficiência cardíaca (IC) após infarto (Pitt et al., 1999; Pitt et al., 2003).

Introdução
30
Existem, atualmente, quatro principais antagonistas do receptor mineralocorticoide:
Espironolactona (Aldactone®), Eplerenona (Inspra®), Canrenona e o recém-sintetizado
Finerenona (BAY 94-8862) (Figura 10). Cada antagonista possui particularidades quanto às suas
características farmacológicas e ação (Tabela 7).
Figura 10: Fórmulas estruturais dos antagonistas do receptor mineralocorticóide da aldosterona. Fonte: Wikipedia
Espironolactona, o primeiro antagonista do MR produzido, é um anti-hipertensivo
poupador de potássio sintetizado na década de 1960. Em seu efeito antagonista a espironolactona
atua como um inibidor competitivo da aldosterona, contudo possui capacidade para se ligar a
receptores progesterônicos e androgênicos, promovendo efeitos adversos tais como
ginecomastia, alterações no ciclo menstrual e diminuição da libido (Delyani, 2000). Além de seu
papel na redução da pressão sanguínea, estudos clínicos têm mostrado efeitos anti-inflamatórios
(Zhang et al., 2014) e antifibróticos da espironolactona (Hermidorff et al., 2015; Zhao et al.,
2015).
Sintetizada a partir da espironolactona em 2002, eplerenona é o princípio ativo do fármaco
Inspra® (Núñez et al., 2006) que atua como um inibidor específico para o receptor de aldosterona
(Specific Aldosterone Receptor Antagonist – SARA). Em oposição à espironolactona, a
eplerenona apresenta maior seletividade pelo receptor mineralocorticóide e baixa afinidade pelos
receptores de progesterona (GR < 1%) e de andrógenos (0,1%). Essa maior seletividade pelo MR
e menor afinidade para os receptores esteroidais reduz os efeitos sexuais secundários observados
no tratamento a longo prazo com espironolactona, como ginecomastia, alteração no ciclo
menstrual, impotência, mastodinia e sangramento vaginal (Funder, 2013), tornando a eplerenona
mais segura para tratamento crônico. Adicionalmente, a eplerenona possui meia-vida (T1/2 = 4 –
6 h) maior que a espironolactona (T1/2 = 1,3 – 1,4 h) (Mcmahon, 2001), porém tem se mostrado
menos potente (Zannad et al., 2011).

Introdução
31
Estudos têm mostrado que a eplerenona atenua a síntese de colágeno induzida pela
corticosterona, exercendo, assim como a espironolactona, uma função de prevenir a fibrose
cardíaca (Omori et al., 2014; Hermidorff et al., 2015).
Canrenona é um metabólito ativo da espironolactona usado como diurético que possui
efeitos antiandrogênicos, demonstrados em pacientes com cirrose hepática (Andriulli et al.,
1989). É um fármaco de preço acessível e não tem apresentado limitações em sua prescrição nos
países em que está disponível. Estudos têm mostrado que, além de sua ação bloqueadora da
aldosterona, canrenona também reduz a progressão da resistência à insulina e de
microalbuminúria na diabetes tipo 2 e outras doenças relacionadas (Armanini et al., 2014).
Derosa e colaboradores demonstraram que canrenona é capaz de reduzir a pressão sanguínea em
pacientes com síndrome metabólica. Maiores estudos, porém, são necessários a respeito desse
metabólito da espironolactona (Derosa et al., 2013).
O antagonista Finerenona (BAY 94-8862) foi sintetizado em 2012 como um antagonista
não-esteroidal que possui maior seletividade ao MR que a espironolactona e maior afinidade
comparado à eplerenona. Um grande estudo clínico (ARTS – MinerAlocorticoid Receptor
Antagonist Tolerability Stud: NCT01345656) tem sido realizado para demonstrar a eficácia da
finerenona. Pitt e colaboradores demonstraram que doses de finerenona 5-10 mg/dia eram pelo
menos tão eficientes quanto a espironolactona 25 ou 50 mg/dia em diminuir biomarcadores do
estresse hemodinâmico em pacientes com insuficiência cardíaca, fração de ejeção ventricular
esquerda reduzida e doença renal crônica moderada. Entretanto, esse resultado foi associado a
menor incidência de hipercalemia e falha renal (Pitt et al., 2013). Por ser um composto recém-
sintetizado, mais estudos são necessários para comprovar a eficácia desse antagonista.
O presente trabalho é focado nos antagonistas do receptor mineralocorticoide
espironolactona e eplerenona por serem os medicamentos de uso mais comum na prática clínica.
1.5. Antagonistas do receptor mineralocorticóide (MR), receptores de
adenosina e o condicionamento isquêmico
Elevados níveis plasmáticos de aldosterona resultam em complicações vasculares, renal,
cardíaca e metabólica independente da pressão sanguínea. O tratamento de tais pacientes é feito
por meio do uso de antagonistas do receptor mineralocorticoide (MR) da aldosterona. Estudos
clínicos têm demonstrado que baixas doses desses antagonistas reduzem a morbidade e
mortalidade em pacientes com insuficiência cardíaca e infarto do miocárdio, apesar dos níveis
normais de aldosterona no plasma (Mccurley e Jaffe, 2012).

Introdução
32
A espironolactona foi originalmente desenvolvida para neutralizar os efeitos do hormônio
aldosterona, em especial os efeitos relacionados à excreção de potássio. Contudo, durante
décadas no mercado sendo usada para esse fim, começou-se a observar um papel secundário para
a espironolactona em relação aos diuréticos mais potentes (Nappi e Sieg, 2011).
Ao ligar-se ao receptor MR, a espironolactona promove efeitos benéficos sobre a
morfologia e função do ventrículo esquerdo em ratos submetidos a infarto do miocárdio
(Nagatomo et al., 2014). O estudo RALES demonstrou que a espironolactona associada ao
inibidor da enzima conversora de angiotensina (ECA), reduz significativamente a morbidade e
mortalidade em pacientes com insuficiência cardíaca congestiva grave (Ma et al., 2014).
Estudo realizado por Sohn e colaboradores demonstrou que em corações de filhotes de rato
da linhagem Spreague Dawley (SD) a expressão da proteína p38 (subfamília de proteína quinase
ativada por mitógeno responsável por regular vários processos celulares como diferenciação
celular, apoptose, respostas à inflamação e crescimento celular) foi diminuída significativamente
após administração de 200mg/kg de espironolactona, além de reduzir a apoptose de
cardiomiócitos nesses ratos (Sohn et al., 2010).
Trabalhos experimentais e clínicos demonstraram que a eplerenona promove acentuada
proteção cardíaca, cerebral, vascular e renovascular na isquemia cardíaca após infarto do
miocárdio (Rudolph et al., 2004). Dentre estes podemos citar os trabalhos nos quais o tratamento
por longo prazo com eplerenona promoveu uma melhoria da função ventricular e vascular em
ratos cronicamente infartados (Fraccarollo et al., 2003; Schäfer et al., 2003).
Estudo prévio em nosso laboratório, avaliou a ação da espironolactona e eplerenona de
maneira comparativa, na presença e na ausência da aldosterona. Os resultados mostraram que
ambos promoveram proliferação de cardiomiócitos e reduziram a proliferação de fibroblastos,
mesmo na ausência do referido hormônio, sugerindo uma via de sinalização independente do
antagonismo sobre aldosterona (Hermidorff et al., 2015).
Possíveis vias de sinalização evocadas por esses fármacos também foram analisadas e
constatou-se que, tanto espironolactona quanto eplerenona foram capazes de aumentar os níveis
de Ca2+, GMPc e a atividade de ERK 1/2. Também reverteram a ação da aldosterona sobre a
atividade de PKC e ERK 1/2. Curiosamente, apenas espironolactona aumentou os níveis de
AMPc. Nossos dados apoiam o fato de que, para além da aldosterona, tanto espironolactona
quanto eplerenona possuem respostas rápidas (não genômicas) (Hermidorff et al., 2015).

Introdução
33
Existem evidências consistentes de que antagonistas do MR agem sobre as células
cardíacas, independentemente do bloqueio da ação da aldosterona. No entanto, o mecanismo pelo
qual eles promovem cardioproteção ainda é pouco compreendido.
Chai e colaboradores em estudos com corações isolados de ratos submetidos a isquemia e
reperfusão, relataram que antagonistas do MR administrados antes da isquemia reduziram a área
isquêmica em relação ao grupo controle, sugerindo que, de alguma forma, esses antagonistas
promovem pré-condicionamento farmacológico (Chai, Garrelds, Arulmani, et al., 2005; Chai,
Garrelds, De Vries, et al., 2005).
Em estudo subsequente, também realizado em coração isolado de ratos submetidos a I-R,
Schmidt e colaboradores demonstraram que antagonistas do MR também foram capazes de
reduzir a área infartada quando administrados antes da reperfusão (pós-condicionamento), e esses
efeitos foram bloqueados por um antagonista não-seletivo de receptores da adenosina. Neste
mesmo estudo, o efeito cardioprotetor do canrenoato de potássio (metabólito ativo da
espironolactona) foi bloqueado em camundongos knock-outs pela enzima ecto-5’-nucleotidase
(enzima envolvida na síntese de adenosina) (Schmidt et al., 2010).
Estas evidências somadas ao que se conhece sobre os efeitos cardioprotetores da adenosina
no coração, nos permite sugerir que a ação direta de antagonistas de MR sobre o coração pode
estar sendo mediada por uma interação com a adenosina, seja via seus receptores de membrana
ou atuando sobre a disponibilidade de adenosina.

34
2- OBJETIVOS

Objetivos
35
OBJETIVO GERAL Avaliar o efeito cardioprotetor da espironolactona e eplerenona sobre cardiomiócitos de
ratos submetidos à isquemia-reperfusão in vitro, na presença e na ausência de antagonistas dos
receptores A1R, A2AR, A2BR e A3R da adenosina.
OBJETIVOS ESPECÍFICOS
1. Indução de isquemia in vitro (isquemia-reoxigenação).
2. Caracterização do dano isquêmico:
2.1. Avaliação da citotoxicidade por meio de ensaio por kit de LDH
3. Determinação da adenosina em cardiomiócitos ventriculares de ratos neonatos
3.1. Identificação do nucleosídeo por Espectrometria de Massas
4. Avaliação da possível ação da espironolactona e eplerenona mediada pelos receptores da
adenosina (A1R, A2AR, A2BR e A3R) em condições de isquemia nos miócitos cardíacos.
4.1. Análise da viabilidade celular por meio de ensaio por MTT

36
3- METODOLOGIA

Metodologia
37
3.1. Animais
Para obtenção de cardiomiócitos, foram utilizados ratos neonatos da linhagem Wistar,
fêmeas, pesando em média 5g. Estes permaneceram juntamente com a mãe até o dia do
experimento.
Os animas foram provenientes do Centro de Ciência Animal (CCA) da Universidade
Federal de Ouro Preto, com os protocolos CEUA-UFOP de número 2012/22 e 2016/07. As
progenitoras foram acomodadas uma por gaiola, com seus respectivos filhotes, mantidas em
ambiente silencioso, com temperatura controlada, em ciclos de 12 horas de período diurno (das
07:00 às 19:00 horas) e 12 horas de período noturno (das 19:00 às 07:00 horas) mantidos
artificialmente. Receberam água e ração Purina “ad libitum”. Todas as recomendações do Comitê
de Ética em Pesquisa Animal (CEUA-UFOP) foram seguidas, conforme protocolo em anexo.
3.2. Cultura Primária de Cardiomiócitos
O protocolo de extração de células cardíacas foi gentilmente cedido pelo Prof. Dr. Eduardo
M. Krieger (InCor – HC – FMUSP). Resumidamente, foram utilizados 10 ratos neonatos em
cada cultura, os quais foram higienizados com álcool 70% e decapitados. Os corações foram
removidos e lavados em solução DMEM enriquecida com glicose 5mg/mL. Em seguida
selecionou-se os ventrículos que foram fragmentados e, posteriormente digeridos
enzimaticamente em solução tampão de digestão (Tabela 3), agitados em banho maria a 37oC em
4 ciclos de 20 min. A suspensão de células resultante desta etapa foi centrifugada com soro fetal
bovino (LGC Biotecnologia) a 1200rpm/10min. O pellet foi ressuspendido com meio de cultura
DMEM enriquecido com 5% de soro fetal bovino, 10% de soro de cavalo, 1% de
penicilina/estreptomicina e 0,1% de inibidor de fibroblasto BRDU (5-Bromo-2’-deoxiuridina –
Sigma-Aldrich) (Tabela 2). As células foram então distribuídas em placa de 24 poços e incubadas
em estufa de CO2 (5%) a 37ºC por 60min para adesão preferencial dos fibroblastos e aumentar
assim, a pureza da cultura. O sobrenadante desta etapa (cardiomiócitos) foi retirado e distribuído
em uma nova placa de 24 poços. A placa foi acondicionada em estufa de CO2 (5%) a temperatura
constante de 37ºC por 48 horas para adesão completa das células (Barreto-Chaves et al., 2000).
3.3. Soluções Utilizadas nos Ensaios

Metodologia
38
Tabela 1: Composição da solução de BRDU [10-2M] (5-Bromo-2'-deoxiuridina)
Componentes Quantidade
Água Milli-Q 165 mL
BRDU 0,5 g
Tabela 2: Composição do meio de cultura normal
Componentes Quantidade (q.s.p)
DMEN (Sigma-Aldrich) 42,5 mL
Soro de cavalo (LGC) 5,0 mL
Soro fetal bovino (LGC) 2,5 mL
Penicilina-estreptomicina (LGC) 0,5 mL
Solução de BRDU [10-2M] 50 µL
Tabela 3: Composição da solução tampão de digestão
Componentes Quantidade (q.s.p)
Solução de DMEM + Glicose 5mg/mL 50 mL
Colagenase tipo II (Sigma-Aldrich) 10 mg
Pancreatina (Sigma-Aldrich) 30 mg
Tabela 4: Composição do meio de cultura para isquemia
Componentes Quantidade (q.s.p)
DMEN (Sigma-Aldrich) 42,5 mL
Glicose 250 mg
Penicilina-estreptomicina (LGC) 0,5 mL
Solução de BRDU [10-2M] 50 µL
Tabela 5: Composição da solução de parada para evitar degradação da adenosina
Componentes Quantidade (q.s.p)
Dipyridamole 75 µM
EHNA 15 µM
EDTA 3 mM
Solução salina isotônica (pH 6) 1 L
Todas as soluções utilizadas para cultivo e manutenção das culturas de cardiomiócitos
foram esterilizadas em membrana Millipore 0,22 µm.

Metodologia
39
3.4. Desenho experimental
As culturas foram divididas nos grupos Normóxia, Pré-condicionamento e Pós-
condicionamento farmacológico e os tratamentos foram igualmente administrados em ambos
grupos com as concentrações demonstradas na lista a seguir. Para facilitar a compreensão, os
antagonistas seletivos usados de cada receptor da adenosina foram identificados nessa lista (itens
de 6 a 9), contudo ao longo do trabalho serão referidos apenas como “antagonista do receptor
A1R”, ou simplesmente “ant. A1R”, por exemplo.
1. Controle normóxia: somente as células mantidas em condições normais
2. Controle isquemia: somente as células mantidas em condições de isquemia
3. Células tratadas com Espironolactona [10-9 M]
4. Células tratadas com Eplerenona [10-8 M]
5. Células tratadas com Adenosina [10-8 M]
6. Células tratadas com antagonista do receptor A1R (DPCPX [10-6 M])
7. Células tratadas com antagonista do receptor A2AR (ZM 241385 [10-9 M])
8. Células tratadas com antagonista do receptor A2BR (MRS 1754 [10-6 M])
9. Células tratadas com antagonista do receptor A3R (MRS 1523 [10-6 M])
10. Células tratadas com ant. A1R [10-6 M] + ant. A3R [10-6 M]
11. Células tratadas com ant. A2AR + ant. A2BR [10-6 M]
12. Células tratadas com Adenosina [10-8 M] + ant. A1R [10-6 M]
13. Células tratadas com Adenosina [10-8 M] + ant. A3R [10-6 M]
14. Células tratadas com Adenosina [10-8 M] + ant. A1R [10-6 M] + ant. A3R [10-6 M]
15. Células tratadas com Adenosina [10-8 M] + ant. A2AR [10-9 M]
16. Células tratadas com Adenosina [10-8 M] + ant. A2BR [10-6 M]
17. Células tratadas com Adenosina [10-8 M] + ant. A2AR [10-9 M] + ant. A2BR [10-6 M]
18. Células tratadas com Espironolactona [10-9 M] + ant. A1R [10-6 M]
19. Células tratadas com Espironolactona [10-9 M] + ant. A3R [10-6 M]
20. Células tratadas com Espironolactona [10-9 M] + ant. A1R [10-6 M] + ant. A3R [10-6 M]
21. Células tratadas com Espironolactona [10-9 M] + ant. A2AR [10-9 M]
22. Células tratadas com Espironolactona [10-9 M] + ant. A2BR [10-6 M]
23. Células tratadas com Espironolactona [10-9 M] + ant. A2AR [10-9 M] + ant. A2BR [10-6 M]
24. Células tratadas com Eplerenona [10-8 M] + ant. A1R [10-6 M]
25. Células tratadas com Eplerenona [10-8 M] + ant. A3R [10-6 M]

Metodologia
40
26. Células tratadas com Eplerenona [10-8 M] + ant. A1R [10-6 M] + ant. A3R [10-6 M]
27. Células tratadas com Eplerenona [10-8 M] + ant. A2AR [10-9 M]
28. Células tratadas com Eplerenona [10-8 M] + ant. A2BR [10-6 M]
29. Células tratadas com Eplerenona [10-8 M] + ant. A2AR [10-9 M] + ant. A2BR [10-6 M]
3.5. Indução da isquemia in vitro
Para avaliar a ação da espironolactona e eplerenona, como possíveis agentes de pré- e pós-
condicionamento farmacológico, foi necessário mimetizar isquemia in vitro. Isquemia é
caracterizada pela falta de sangue local que leva à deficiência na distribuição de O2 e nutrientes
na área afetada. Sendo assim, ao induzir isquemia in vitro, as células foram expostas por 2 horas
a uma mistura gasosa composta por 95% de N2 – responsável por substituir o O2 do meio – e 5%
de CO2. Associado a isso, as células também foram tratadas com meio de cultura pobre em
nutrientes, o qual é livre de soro, mas acrescido de 5mg/mL de glicose apenas para manutenção
dos cardiomiócitos (Safran et al., 2001).
Visando controlar a eficácia da indução das células à isquemia, em todos os ensaios foram
preparadas duas placas, uma submetida a condições ideais de nutrição e oxigênio para controle,
e uma para o ensaio de isquemia. Dessa forma, para melhor compreensão, demos os nomes a
essas placas de grupo normóxia e grupo isquemia. É importante salientar que, as células do grupo
normóxia se mantiveram em meio de cultura enriquecido normalmente, uma vez que o meio
pobre em nutrientes se faz necessário apenas para as condições de isquemia.
Em relação aos tratamentos farmacológicos, foram feitos dois grupos: o pré-
condicionamento, para as células tratadas antes da exposição ao nitrogênio, e o grupo pós-
condicionamento, para as células tratadas no ato da “reperfusão”. Dessa forma, foi preparada
uma placa para cada grupo exposto a condições de isquemia e as células do grupo normóxia,
também receberam os tratamentos respeitando os grupos de tratamento pré- e pós-
condicionamento para controle.
3.5.1. Preparo das placas
Após 48 horas de incubação (conforme sessão 3.2), os cardiomiócitos foram soltos das
placas utilizando-se TrypLE™ Express (tríplex - tripsina sintética). Para tanto, todo o meio de
cultura foi retirado e em seguida foi acrescentado 350μL de tríplex em cada poço para que as
células perdessem a adesão com a placa. Esse tríplex contendo células foi, então, retirado e
centrifugado a 1020rpm/10min, o sobrenadante foi descartado e as células do grupo normóxia

Metodologia
41
foram ressuspendidas em meio de cultura normal enquanto as do grupo isquemia foram
ressuspendidas em meio de cultura próprio para isquemia.
As células foram contadas em câmara de Neubauer e distribuído o equivalente a 105 células
em cada poço em duas placas de 96 poços. As placas foram acondicionadas em estufa de CO2
(5%) a temperatura constante de 37ºC por 24 horas para adesão completa das células.
3.5.2. Tratamentos
As células do grupo pré-condicionamento (tanto das placas isquemia quanto normóxia)
receberam o tratamento com espironolactona 10-9 M e eplerenona 10-8 M, cujas concentrações
foram definidas em ensaio de curva de concentração previamente realizado, e foram incubadas
por 2h para ação dos fármacos.
Após esse período, as placas isquemia foram colocadas em câmara de hipóxia por 2h em
estufa contendo uma mistura de N2 (95%) e CO2 (5%) a 37ºC. As placas normóxia foram
mantidas em estufa a 37ºC e 5% de CO2.
Em seguida, para a reperfusão as células foram expostas a condições normais de oxigênio.
Imediatamente, o grupo pós-condicionamento foi tratado com os fármacos espironolactona 10-9
M e eplerenona 10-8 M. As células do grupo pré-condicionamento tiveram o meio de cultura
substituído por meio de cultura normal (DMEM enriquecido de 10% de soro de cavalo e 5%
SFB). As placas foram incubadas em estufa a 37ºC e 5% de CO2 por 2 horas para ação dos
fármacos.
Para que ambos os grupos fossem expostos pelo mesmo tempo ao tratamento, passadas as
duas horas de tratamento, o meio de cultura contendo os fármacos do grupo Pós-
condicionamento foi removido e substituído por meio normal. Essa troca de meio ocorreu para
todos os grupos (normóxia e isquemia) para evitar diferença devido à renovação do meio. As
células foram incubadas em estufa a 37ºC e 5% de CO2 por 16 horas e posteriormente prosseguiu-
se com os experimentos.
3.6. Validação do método de isquemia in vitro
Para comprovar se a metodologia de indução das células a isquemia é eficiente, realizou-
se a caracterização do dano isquêmico feita por meio de kit de Detecção de Citotoxicidade (LDH)
(Roche – Catálogo: 11.644.793.001). Trata-se de um ensaio colorimétrico para quantificação de
lise e morte celular, baseado na medida da atividade da enzima lactato desidrogenase (LDH)

Metodologia
42
liberada do citosol no sobrenadante pelas células danificadas. A lactato desidrogenase é uma
enzima que atua em condições de anaerobiose, quando catalisa a redução do piruvato em lactato,
utilizando o NADH produzido na fase oxidativa, regenerando o NAD+ necessário para a glicólise
na ausência de oxigênio (Lehninger et al., 2005). Dessa forma, quanto maior o dano celular
promovido pelo ensaio de isquemia in vitro, maior a quantidade de LDH liberado.
Após o tratamento das células conforme citado na sessão 3.5.2, procedeu-se o ensaio de
detecção de citotoxicidade por LDH conforme recomendações do fabricante do kit.
3.7. Determinação da adenosina em cardiomiócitos ventriculares de ratos
neonatos sob condições de isquemia
3.7.1. Preparo das amostras
Para determinar se há presença de adenosina nos cardiomiócitos ventriculares, procedeu-
se normalmente o ensaio de indução à isquemia como descrito na seção 3.5. Imediatamente após
as duas horas de exposição das células ao gás nitrogênio, coletou-se o meio sobrenadante das
células do grupo pré-condicionamento. As amostras do pós-condicionamento foram tratadas
conforme descrição na seção 3.5 e sua coleta foi realizada imediatamente após as 2h de incubação
da espironolactona e eplerenona.
Um total de 300 µL de amostra foi coletado para cada tratamento e adicionadas em tubos
de vidro contendo 150 µL de solução de parada (descrita na tabela 5) para evitar a degradação
da adenosina. Posteriormente, adicionou-se 900 µL de acetona P.A. para precipitação de
proteínas, agitou-se em vórtex e, em seguida, centrifugou-se por 10’ a 5.000 rpm. O sobrenadante
foi retirado e transferido para um novo tubo de vidro. A acetona presente nas amostras foi retirada
por evaporação com auxílio de fluxo de gás nitrogênio.
As amostras foram então concentradas em liofilizador (BenchTop ™ “K” Series – VirTis).
O precipitado obtido foi ressuspendido na solução da fase móvel, composta por água e metanol
com ácido fórmico 0,1%. Posteriormente, procedeu-se a leitura.
3.7.2. Análise por espectrometria de massa
O ensaio foi realizado no equipamento LC-MS Shimadzu 8040, coluna C18 (150 x 2 mm)
3µ, Phenomenex. A fase móvel utilizada foi composta por água e metanol com ácido fórmico
0,1% e a eluição por gradiente de 5% a 70% de metanol.

Metodologia
43
Observação: Foi separado um tubo apenas com solução de parada para ser lido juntamente com
as amostras para avaliar sua interferência na detecção e subtraí-la.
3.8. Avaliação da possível ação da espironolactona e eplerenona mediada pelos
receptores da adenosina (A1R, A2AR, A2BR e A3R) em condições de isquemia
Para avaliar se a ação protetora da espironolactona e eplerenona é mediada por receptores
da adenosina em condições de isquemia, realizou-se a determinação da viabilidade celular pós-
exposição com MTT (Sigma-Aldrich – Catálogo: 298-3-1).
O MTT (brometo de 3-4,5-dimetil-tiazol-2-il-2,5-difeniltetrazólio) é um sal tetrazólio de
cor amarela que, quando metabolizado forma cristais de formazan de cor azul. Esse processo
ocorre na cadeia respiratória mitocondrial de células metabolicamente ativas, dessa forma, a
quantidade de formazan formado é proporcional à quantidade de células viáveis. A análise é feita
por espectrofotometria.
Para o ensaio de viabilidade, as células devem estar aderidas em microplaca, sendo assim,
a indução da isquemia foi feita diretamente nas placas de ensaio. Após o preparo das células
conforme citado na sessão 3.2, os tratamentos foram realizados (sessão 3.5.2), com o diferencial
da presença dos antagonistas dos receptores da adenosina. As células do grupo pré-
condicionamento foram tratadas com os antagonistas dos receptores A1R e A3R (Zhan et al.,
2011), foram incubadas por 20 min para ligação dos antagonistas a seus receptores, em seguida
adicionou-se adenosina, espironolactona ou eplerenona conforme desenho experimental pré-
estabelecido (sessão 3.4). O tratamento do grupo pós-condicionamento seguiu o mesmo padrão
realizado no grupo pré-condicionamento, contudo, foram utilizados antagonistas para os
receptores A2AR e A2BR da adenosina (Zhan et al., 2011).
Após o tratamento, o meio de cultura foi retirado e adicionou-se 100µL de solução MTT
aos poços e incubou-se por 2h conforme protocolo. Após esse período, o MTT foi removido e
adicionou-se 100µL de DMSO aos poços. As placas foram agitadas em agitador orbital por 5min,
e mantidas sem agitação por mais 5min para estabilização da cor. Em seguida, as placas foram
lidas em leitor de placa (Victor™ X3 - Multimode Plate Reader – PerkinElmer) no comprimento
de onda de 530nm.

Metodologia
44
3.9. Análise estatística
A análise estatística dos resultados foi realizada por meio do software GraphPad Prism®
(versão 6.01). Utilizou-se ANOVA para testar as diferenças entre os grupos e entre os
tratamentos, seguido de pós-teste de Tukey para determinação das diferenças significativas entre
os grupos. Adotou-se o nível de significância p < 0,05 para todas as análises.

45
4- RESULTADOS

Resultados
46
4.1. Validação do método de Isquemia in vitro Nos ensaios de validação do método de indução dos cardiomiócitos a isquemia,
observamos que a técnica utilizada foi eficaz uma vez que a liberação de LDH foi elevada de
maneira significativa no grupo isquemia em relação ao controle normóxia (Gráfico 1).
G r u p o s
Pe
rc
en
tua
l d
e L
DH
lib
era
do
0
5 0
1 0 0
1 5 0
2 0 0
C o n tro le N o rm ó x ia
C o n tro le Is q u e m ia
- S ig n if ic a tiv a m e n te d ife re n te d o C o n tro le Is q u e m ia
Gráfico 1: Análise do dano isquêmico mediante exposição dos cardiomiócitos a condições de isquemia. Percentual
de LDH liberado em cardiomiócitos expostos a isquemia comparados a células em condições ideais (controle
normóxia). Resultado de três experimentos independentes realizados em triplicatas, n = 3 e expressos em média ±
SEM, p<0,05.
4.2. Determinação da adenosina em cardiomiócitos ventriculares de ratos
neonatos sob condições de isquemia
Para investigar a presença da adenosina em cardiomiócitos ventriculares submetidos a
isquemia, amostras dos seguintes tratamentos foram analisadas: controle, espironolactona,
eplerenona e adenosina, dos grupos normóxia, pré-condicionamento e pós-condicionamento. A
fim de simplificação, estão representados os cromatogramas apenas dos controles de cada grupo
e uma tabela geral (Tabela 6) demonstrando a área dos picos obtidos para todos os tratamentos.
Na figura 10 são apresentados um cromatograma e um espectro obtidos para confirmar a
presença da adenosina ou de algum outro composto de massa molecular semelhante ao da
adenosina que pudesse gerar falso positivo na análise da amostra. Para tanto, foi selecionada uma
das amostras aleatoriamente (espironolactona – pré-condicionamento) e a ela adicionou-se 10µL
de solução padrão de adenosina [1 mg/L] à amostra para separar os picos. O resultado está
demonstrado abaixo e foi usado para comparar com os espectros das demais amostras.

Resultados
47
4,0
4,5
5,0
5,5
6,0
6,5
7,0
7,5
0
10000
20000
30000
40000
50000
60000
70000
80000
268,1
0>
119,0
5(+
)268,1
0>
136,0
5(+
)
Adenosina/5,757
Esp
ectro co
ntam
inan
te (tr: 5,5
01
min
) E
spectro
da ad
enosin
a (tr: 5,7
57 m
in)
60,0
65,0
70,0
75,0
80,0
85,0
90,0
95,0
100,0
105,0
110,0
115,0
120,0
125,0
130,0
m/z
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
8,0
Inte
n.(x1
0,0
00)
119,1
136,1
57,1
60,0
65,0
70,0
75,0
80,0
85,0
90,0
95,0
100,0
105,0
110,0
115,0
120,0
125,0
130,0
m/z
0,0
1,0
2,0
3,0
4,0
5,0
6,0
7,0
Inte
n.(x1
0,0
00)
136,1
119,1
57,1
Fig
ura
11
: Cro
mato
gra
ma e e
spectro
de m
assas o
btid
os d
a am
ostra “esp
irono
lactona” d
o p
ré-cond
icionam
ento
. Os p
icos d
estacado
s no
espectro
à esquerd
a ap
resentam
a relação m
assa/carga
(m/z) refere
nte ao
conta
min
ante n
a am
ostra, e
nq
uanto
no
espectro
à direita o
s pico
s em
destaq
ue rep
resenta
m a
aden
osin
a com
os frag
mento
s iônico
s esp
erado
s. Entre p
arênteses, é d
em
on
strada
a carga p
ositiv
a adq
uirid
a duran
te a ionização
.

Resultados
48
Na figura 11 estão representados o cromatograma e o espectro obtidos da amostra controle
do grupo pré-condicionamento. Ao comparar aos espectros demonstrados na figura 10 é possível
comprovar a presença de adenosina pelo padrão dos picos semelhantes aos demonstrados no
espectro de detecção do nucleosídeo de interesse.
Figura 12: Cromatograma e espectro de massas obtidos da amostra “controle” do pré-condicionamento. Os picos
destacados apresentam a relação massa/carga (m/z) referente à adenosina com os fragmentos iônicos esperados Entre
parênteses, é demonstrada a carga positiva adquirida durante a ionização.
De forma semelhante ao resultado encontrado no controle do grupo pré-condicionamento,
no controle do pós-condicionamento também foi detectada presença de adenosina (Figura 12).
Figura 13: Cromatograma e espectro de massas obtidos da amostra “controle” do pós-condicionamento. Os picos
destacados apresentam a relação massa/carga (m/z) referente à adenosina com a massa molecular esperada. Entre
parênteses, é demonstrada a carga positiva adquirida durante a ionização.
60,0 65,0 70,0 75,0 80,0 85,0 90,0 95,0 100,0 105,0 110,0 115,0 120,0 125,0 130,0 m/z
0,00
0,25
0,50
0,75
1,00
1,25
1,50
1,75
2,00
Inten. (x10,000)
136,1
119,1
57,1
60,0 65,0 70,0 75,0 80,0 85,0 90,0 95,0 100,0 105,0 110,0 115,0 120,0 125,0 130,0 m/z
0,00
0,25
0,50
0,75
1,00
Inten. (x10,000)
136,1
119,1
57,1

Resultados
49
Em oposição aos resultados encontrados nos grupos submetidos a isquemia, o controle
normóxia não apresentou presença de adenosina (Figura 13).
Figura 14: Cromatograma e espectro de massas obtidos da amostra “controle” normóxia. Os picos destacados
apresentam a relação massa/carga (m/z) referente ao contaminante com a massa molecular semelhante à esperada
para a adenosina. Entre parênteses, é demonstrada a carga positiva adquirida durante a ionização.
A tabela 6 mostra as concentrações de adenosina detectadas em cada amostra. É possível
observar diferenças significativas nos valores. A amostra tratada com espironolactona no pré-
condicionamento apresentou maior concentração de adenosina em relação ao tratamento com
eplerenona e ao controle. Em oposição, no pós-condicionamento a maior concentração ocorreu
no tratamento com eplerenona em relação à espironolactona e ao controle.
Tabela 8: Concentração de adenosina detectada nos cromatogramas para amostras dos grupos normóxia, pré-
condicionamento e pós-condicionamento.
Amostra Concentração de Adenosina (mg/L)
Pré-condicionamento Pós-condicionamento Normóxia
Controle 0,00332 0,00180 n.d.
Adenosina 0,00744 0,00129 n.d.
Espironolactona 0,00478 0,00096 n.d.
Eplerenona 0,00317 0,00223 n.d.
Legenda: n.d. = não detectado
60,0 65,0 70,0 75,0 80,0 85,0 90,0 95,0 100,0 105,0 110,0 115,0 120,0 125,0 130,0 m/z
0,0
2,5
5,0
7,5
Inten. (x10,000)
119,1
136,1
57,1

Resultados
50
4.3. Espironolactona e eplerenona x Antagonistas da adenosina em condições de
isquemia
Para testar nossa hipótese de interação dos antagonistas do receptor mineralocorticoide
com receptores da adenosina no condicionamento isquêmico, tratamos os cardiomiócitos
ventriculares com espironolactona, eplerenona, adenosina e fizemos associações dos mesmos aos
antagonistas dos quatro receptores, A1R, A2AR, A2BR e A3R, da adenosina. Os tratamentos foram
realizados para os grupos pré- e pós-condicionamento.
Conforme demonstrado no gráfico 2, os tratamentos com espironolactona, eplerenona e
adenosina foram capazes de proteger as células submetidas a isquemia, mantendo a viabilidade
estatisticamente igual ao controle não tratado sob normóxia. Esse resultado foi mantido tanto no
pré-condicionamento quanto no pós-condicionamento.
G r u p o s
Via
bil
ida
de
ce
lula
r (
%)
P ré -co n d ic
ion a m
e n to
P ó s -co n d ic
ion a m
e n to
0
5 0
1 0 0
1 5 0
C o n tro le N o rm ó x ia
C o n tro le Is q u e m ia
E s p iro
E p le
A D O
- S ig n ific a tiv a m e n te d ife re n te d o C o n tro le N o rm ó x ia
- S ig n if ic a tiv a m e n te d ife re n te d o C o n tro le Is q u e m ia
Gráfico 2: Ação da espironolactona, eplerenona e adenosina no pré- e no pós-condicionamento farmacológico.
Percentual de cardiomiócitos viáveis após isquemia, tratados com espironolactona [10-9 M], eplerenona [10-8 M] e
adenosina [10-8 M] como agentes de pré- e pós-condicionamento farmacológico comparados a cardiomiócitos não
tratados expostos a isquemia (controle isquemia) e a células em condições ideais (controle normóxia). Resultado de
três experimentos independentes realizados em triplicatas, n = 3 e expressos em média ± SEM, p<0,05.
4.3.1. Adenosina
Pré-condicionamento – Adenosina
Os receptores A1R e A3R da adenosina são conhecidos por atuar predominantemente no
pré-condicionamento. Ao tratar as células com os antagonistas dos receptores A1R e A3R,
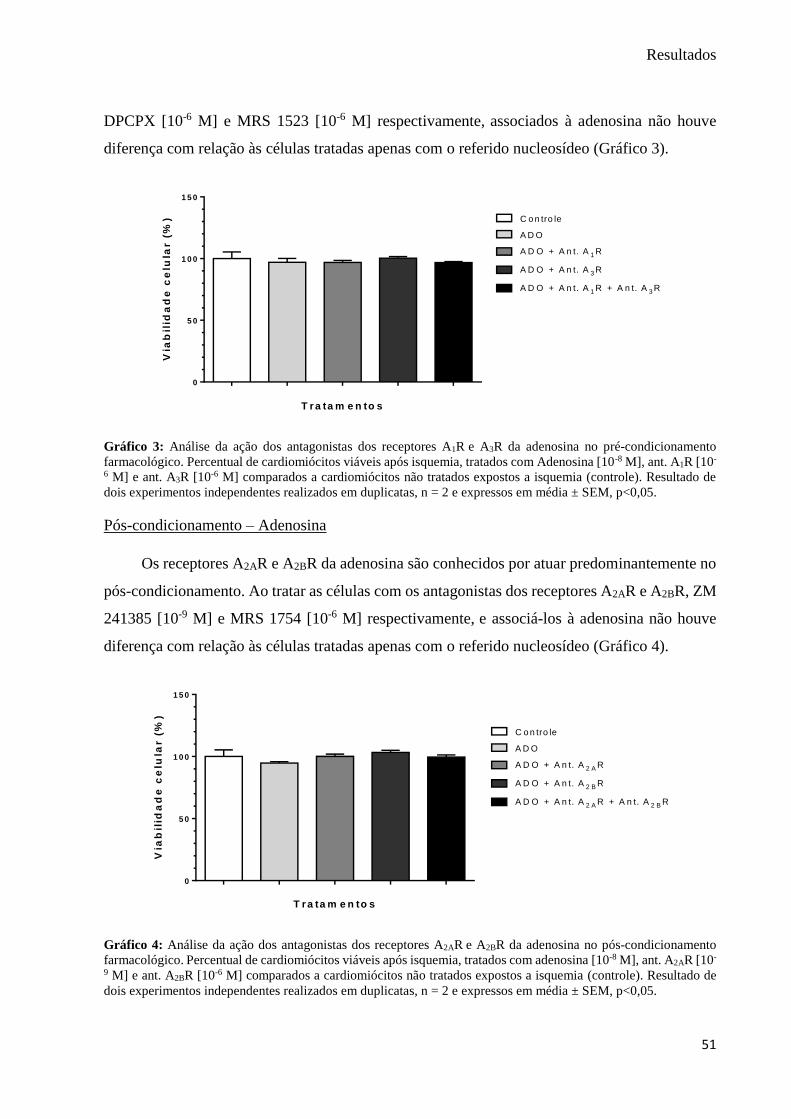
Resultados
51
DPCPX [10-6 M] e MRS 1523 [10-6 M] respectivamente, associados à adenosina não houve
diferença com relação às células tratadas apenas com o referido nucleosídeo (Gráfico 3).
T r a ta m e n to s
Via
bil
ida
de
ce
lula
r (
%)
0
5 0
1 0 0
1 5 0
C o n tro le
A D O
A D O + A n t. A 1 R
A D O + A n t. A 3 R
A D O + A n t. A 1 R + A n t. A 3 R
Gráfico 3: Análise da ação dos antagonistas dos receptores A1R e A3R da adenosina no pré-condicionamento
farmacológico. Percentual de cardiomiócitos viáveis após isquemia, tratados com Adenosina [10-8 M], ant. A1R [10-
6 M] e ant. A3R [10-6 M] comparados a cardiomiócitos não tratados expostos a isquemia (controle). Resultado de
dois experimentos independentes realizados em duplicatas, n = 2 e expressos em média ± SEM, p<0,05.
Pós-condicionamento – Adenosina
Os receptores A2AR e A2BR da adenosina são conhecidos por atuar predominantemente no
pós-condicionamento. Ao tratar as células com os antagonistas dos receptores A2AR e A2BR, ZM
241385 [10-9 M] e MRS 1754 [10-6 M] respectivamente, e associá-los à adenosina não houve
diferença com relação às células tratadas apenas com o referido nucleosídeo (Gráfico 4).
T r a ta m e n to s
Via
bil
ida
de
ce
lula
r (
%)
0
5 0
1 0 0
1 5 0
C o n tro le
A D O
A D O + A n t. A 2 A R
A D O + A n t. A 2 B R
A D O + A n t. A 2 A R + A n t. A 2 B R
Gráfico 4: Análise da ação dos antagonistas dos receptores A2AR e A2BR da adenosina no pós-condicionamento
farmacológico. Percentual de cardiomiócitos viáveis após isquemia, tratados com adenosina [10-8 M], ant. A2AR [10-
9 M] e ant. A2BR [10-6 M] comparados a cardiomiócitos não tratados expostos a isquemia (controle). Resultado de
dois experimentos independentes realizados em duplicatas, n = 2 e expressos em média ± SEM, p<0,05.

Resultados
52
4.3.2. Espironolactona
Pré-condicionamento – Espironolactona
Ao tratarmos os cardiomiócitos com antagonista dos receptores A1R e A3R associados à
espironolactona não houve diferença com relação às células tratadas apenas com o referido
antagonista do MR ou espironolactona + Adenosina (Gráfico 5).
T r a ta m e n to s
Via
bil
ida
de
ce
lula
r (
%)
0
5 0
1 0 0
1 5 0
C o n tro le
E s p iro
E s p iro + A D O
E s p iro + A n t. A 1 R
E s p iro + A n t. A 3 R
E s p iro + A n t. A 1 R + A n t. A 3 R
Gráfico 5: Análise da ação dos antagonistas dos receptores A1R e A3R da adenosina associados à espironolactona
no pré-condicionamento farmacológico. Percentual de cardiomiócitos viáveis após isquemia, tratados com
espironolactona [10-9 M], adenosina [10-8 M], ant. A1R [10-6 M] e ant. A3R [10-6 M] comparados a cardiomiócitos
não tratados expostos a isquemia (controle). Resultado de dois experimentos independentes realizados em
duplicatas, n = 2 e expressos em média ± SEM, p<0,05.
Pós-condicionamento – Espironolactona
Os tratamentos dos cardiomiócitos com os antagonistas dos receptores A2AR e A2BR
associados à espironolactona não mostraram diferença significativa na viabilidade celular no pós-
condicionamento farmacológico, conforme representado no gráfico 6.

Resultados
53
T r a ta m e n to s
Via
bil
ida
de
ce
lula
r (
%)
0
5 0
1 0 0
1 5 0
C o n tro le
E s p iro
E s p iro + A D O
E s p iro + A n t. A 2 A R
E s p iro + A n t. A 2 B R
E s p iro + A n t. A 2 A R + A n t. A 2 B R
Gráfico 6: Análise da ação dos antagonistas dos receptores A2AR e A2BR da adenosina associados à espironolactona
no pós-condicionamento farmacológico. Percentual de cardiomiócitos viáveis após isquemia, tratados com
espironolactona [10-9 M], adenosina [10-8 M], ant. A2AR [10-9 M] e ant. A2BR [10-6 M] comparados a cardiomiócitos
não tratados expostos a isquemia (controle). Resultado de dois experimentos independentes realizados em
duplicatas, n = 2 e expressos em média ± SEM, p<0,05.
4.3.3. Eplerenona
Pré-condicionamento – Eplerenona
Ao tratarmos os cardiomiócitos com antagonistas dos receptores A1R e A3R associados à
eplerenona não houve diferença com relação às células tratadas apenas com o referido fármaco
ou eplerenona + adenosina (Gráfico 7). Observa-se pelo gráfico que houve uma tendência a maior
viabilidade das células tratadas com eplerenona e com eplerenona + adenosina, ainda que não
tenha apresentado diferença estatística dos demais tratamentos.
T r a ta m e n to s
Via
bil
ida
de
ce
lula
r (
%)
0
5 0
1 0 0
1 5 0
C o n tro le
E p le
E p le + A D O
E p le + A n t. A 1 R
E p le + A n t. A 3 R
E p le + A n t. A 1 R + A n t. A 3 R
Gráfico 7: Análise da ação dos antagonistas dos receptores A1R e A3R da adenosina associados à eplerenona no
pré-condicionamento farmacológico. Percentual de cardiomiócitos viáveis após isquemia, tratados com eplerenona
[10-8 M], adenosina [10-8 M], ant. A1R [10-6 M] e ant. A3R [10-6 M] comparados a cardiomiócitos não tratados
expostos a isquemia (controle). Resultado de dois experimentos independentes realizados em duplicatas, n = 2 e
expressos em média ± SEM, p<0,05.

Resultados
54
Pós-condicionamento – Eplerenona
Eplerenona associada aos antagonistas dos receptores A2AR e A2BR não apresentou
diferença na viabilidade com relação às células tratadas apenas com o referido antagonista do
MR ou com eplerenona + adenosina (Gráfico 8) no pós-condicionamento farmacológico. Assim
como observado no pré-condicionamento, é possível notar uma tendência no gráfico a maior
viabilidade das células tratadas com eplerenona + adenosina comparado aos demais tratamentos,
embora não tenha havido diferença estatística para comprovar se há alguma relação.
T r a ta m e n to s
Via
bil
ida
de
ce
lula
r (
%)
0
5 0
1 0 0
1 5 0
C o n tro le
E p le
E p le + A D O
E p le + A n t. A 2 A R
E p le + A n t. A 2 B R
E p le + A n t. A 2 A R + A n t. A 2 B R
Gráfico 8: Análise da ação dos antagonistas dos receptores A2AR e A2BR da adenosina associados à eplerenona no
pós-condicionamento farmacológico. Percentual de cardiomiócitos viáveis após isquemia, tratados com eplerenona
[10-8 M], adenosina [10-8 M], ant. A2AR [10-9 M] e ant. A2BR [10-6 M] comparados a cardiomiócitos não tratados
expostos a isquemia (controle). Resultado de dois experimentos independentes realizados em duplicatas, n = 2 e
expressos em média ± SEM, p<0,05.

55
5- DISCUSSÃO

Discussão
56
No ano de comemoração de 30 anos da descoberta do pré-condicionamento isquêmico por
Murry e colaboradores (1986), ainda há muito a ser explorado para compreender esse poderoso
mecanismo de cardioproteção. Sabe-se que a adenosina é liberada em condições de isquemia e
atua no pré-condicionamento e no pós-condicionamento via receptores denominados A1R, A2AR,
A2BR e A3R. Adicionalmente, estudos têm demonstrado que todos os receptores da adenosina
são expressos no coração e estão envolvidos na cardioproteção (Zhan et al., 2011). Antagonistas
do MR, como espironolactona e eplerenona, também atuam como agentes de proteção durante
isquemia-reperfusão, sendo descritos como agentes de pré- e pós-condicionamento
farmacológico (Chai, Garrelds, Arulmani, et al., 2005; Chai, Garrelds, De Vries, et al., 2005).
Estas evidências nos motivaram a testar, comparativamente, a ação cardioprotetora da
espironolactona e eplerenona, bem como da adenosina perante seus antagonistas como controle.
Realizamos ensaios onde, inicialmente, foi estabelecida a metodologia de indução da
isquemia in vitro. Para mimetizar a isquemia, o método adotado associou subnutrição e
deslocamento do oxigênio do ambiente das células (Safran et al., 2001), técnica que se mostrou
eficaz, uma vez que, ao testar a citotoxicidade por meio de kit de lactato desidrogenase,
observamos que a atividade da enzima LDH aumentou significativamente nas células do grupo
isquemia em comparação às mantidas em condições normais de nutrição e oxigênio (gráfico 1).
Esse resultado pode ser explicado pelo fato de a lactato desidrogenase ser uma enzima que atua
em condições de anaerobiose, quando catalisa a redução do piruvato em lactato, utilizando o
NADH produzido na fase oxidativa, regenerando o NAD+ necessário para a glicólise na ausência
de oxigênio (Lehninger et al., 2005).
Com a metodologia de isquemia in vitro estabelecida, prosseguimos para a determinação
da presença da adenosina por espectometria de massas nos cardiomiócitos submetidos a isquemia
e tratados com espironolactona e eplerenona. Conforme exposto nas figuras 10, 11, 12 e 13, e na
tabela 6, demonstramos que nosso nucleosídeo de interesse foi detectado em todas as amostras
submetidas a isquemia, e, como esperado, nenhuma amostra do grupo normóxia apresentou
adenosina detectada. Esses resultados corroboram com estudos prévios em que determinaram a
presença da adenosina em cardiomiócitos, endotélio e células intersticiais durante os ciclos de
isquemia-reperfusão no condicionamento isquêmico (Heusch, 2015).
Entretanto, nosso trabalho mostra, pela primeira vez, que o tratamento com espironolactona
como agente de pré-condicionamento farmacológico levou a maior concentração de ADO em
relação ao tratamento com eplerenona e ao controle. Por outro lado, no pós-condicionamento a

Discussão
57
concentração da adenosina foi maior no tratamento com eplerenona quando comparada à
espironolactona e ao controle. Esses dados têm sido confirmados por um estudo do grupo, que
vêm sendo realizado em paralelo e têm demonstrado resultados semelhantes (dados não
mostrados). Adicionalmente, nossos resultados somam-se aos estudos de Schmidt e
colaboradores que, utilizando um modelo de infarto do miocárdio em rato, demonstraram que o
efeito cardioprotetor do canrenoato de potássio (metabólito ativo da espironolactona) foi
completamente abolido quando na ausência da enzima ecto-5’-nucleotidase, responsável pela
catalisação da conversão de AMP em adenosina. O mesmo ocorreu com camundongos knock-
outs para o receptor A2BR. O efeito da eplerenona também foi completamente abolido na
presença do antagonista não específico do receptor de adenosina, 8-p-sulphophenyltheophylline,
em corações perfundidos isolados de ratos (Schmidt et al., 2010). A análise, em conjunto, desses
dados demonstra que tanto espironolactona quanto eplerenona parecem estar relacionadas com o
aumento da disponibilidade de adenosina nos cardiomiócitos tanto no pré- quanto no pós-
condicionamento.
Esses resultados nos motivaram a prosseguir com nossa hipótese, e testar se a ação de
antagonistas do MR sobre o coração pode estar sendo mediada por uma interação com a
adenosina via seus receptores de membrana. Avaliamos então, o efeito cardioprotetor da
espironolactona e eplerenona a curto prazo sobre cardiomiócitos de ratos submetidos à isquemia-
reperfusão in vitro, na presença e na ausência de antagonistas dos receptores A1R, A2AR, A2BR e
A3R da adenosina.
Tanto a espironolactona, como a eplerenona e a adenosina foram capazes de prevenir (pré-
condicionamento) como reverter (pós-condicionamento) os danos impostos pela isquemia aos
cardiomiócitos, visto que a viabilidade nas células tratadas por elas foi semelhante ao grupo
controle normóxia (sem tratamento e em condições normais de oxigenação) e maior que o grupo
controle isquemia (sem tratamento, mas em condições isquêmicas), como mostrado pelo ensaio
de viabilidade celular (MTT) no gráfico 4. Esses dados confirmam a efetividade desses fármacos
e da adenosina em proteger as células sob condições isquêmicas conforme já descrito por Chai e
colaboradores (2005) e Schmidt et al., 2010.
Em contraposição, ao associar espironolactona e eplerenona a antagonistas dos receptores
da adenosina A1R, e A3R no pré-condicionamento e A2AR e A2BR no pós-condicionamento, dada
a ação preferencial desses receptores em cada um dos tipos de condicionamento isquêmico (Zhan
et al., 2011), não obtivemos resultados significativos no bloqueio da proteção das células

Discussão
58
(gráficos 5, 6, 7, 8, 9 e 10). A localização dos antagonistas da adenosina no coração ainda é
controversa. Há relatos da presença dos receptores em ventrículos, A1R (Auchampach e Bolli,
1999), A2AR (Norton et al., 1999), A2BR (Liang e Haltiwanger, 1995) e A3R (Strickler et al.,
1996). Entretanto, estudando os receptores a nível molecular, Robin e colaboradores, mostraram
por PCR em tempo real que a expressão dos receptores A1R, A2AR e A2BR foram maiores no
átrio em relação ao ventrículo, enquanto A3R foi igualmente expresso em ambos (Robin et al.,
2011). Koscsó e colaboradores, por sua vez, mostraram que a expressão do receptor A3R no
miocárdio é baixa, apesar de agonistas do A3R terem efeitos cardioprotetores em modelos de
isquemia-reperfusão (Koscsó et al., 2011). Esses estudos nos levam a crer que, provavelmente,
não houve uma resposta mais acentuada em relação ao bloqueio dos receptores em nossos
tratamentos, devido ao fato de usarmos cardiomiócitos ventriculares.
Por outro lado, contrapondo nossos resultados no pós-condicionamento, Grube e
colaboradores comprovaram, também por PCR em tempo real, a presença do receptor A2BR em
cardiomiócitos ventriculares isolados de rato, e demonstraram que a ativação deste receptor
protege fortemente os corações perfundidos da lesão de isquemia-reperfusão e o uso do
antagonista MRS 1754 bloqueou essa proteção (Grube et al., 2011).
Adicionalmente, alguns estudos vêm mostrando grande interação entre os receptores,
apesar da predominância de A1R e A3R no pré-condicionamento e de A2AR e A2BR no pós-
condicionamento. Analisando corações isolados e perfundidos de camundongos selvagens e
knock-outs para A2AR e A2BR, Zhan e colaboradores mostraram que ambos receptores são
necessários na cardioproteção promovida pelo receptor A1R, indicando uma interação entre os
subtipos de receptores da adenosina (Zhan et al., 2011). Essa interação também foi demonstrada
durante isquemia em células H9c2(2-1) (Urmaliya et al., 2009). Múltiplos resultados
consistentes, com diferentes espécies e modelos experimentais indicam interação entre os
receptores A1R e A3R antes da isquemia na proteção e para A2AR e A2BR durante a reperfusão
(Mcintosh e Lasley, 2012). Em nossos experimentos, a associação dos receptores não mostrou
diferença significativa comparada aos demais tratamentos.
Neste ponto, acreditamos que, apesar da comprovada participação da espironolactona e
eplerenona em proteger os cardiomiócitos sob isquemia, e sua evidente influência sobre a
disponibilidade de adenosina, nossos resultados não nos permite afirmar se a cardioproteção por
eles promovida é mediada via receptores da adenosina. É possível que o efeito cardioprotetor da
espironolactona e eplerenona no condicionamento isquêmico venha numa segunda fase, no

Discussão
59
entanto, diante de nossos resultados, admitimos que há possibilidade de o protocolo experimental
escolhido em nosso trabalho não ter sido o ideal para comprovar nossa hipótese. Essas
considerações nos levam a duas possíveis explicações:
1. Pré- e pós-condicionamento compartilham de vias de sinalização (com algumas
particularidades entre elas), que incluem PKC, receptor da aldosterona (MR), PI3K/AKT e ERK
(Schmidt et al., 2010). Sabidamente, espironolactona e eplerenona também atuam por essas vias,
sendo ainda capazes de aumentar a atividade da ERK 1/2 e elevar os níveis de AMPc (Hermidorff
et al., 2015). Adicionalmente, sabendo da ação dos antagonistas de MR sobre a atividade da
enzima ecto-5’-nucleotidase (Schmidt et al., 2010), sugerimos que espironolactona e eplerenona
possam estar atuando na promoção de substrato para formação de adenosina durante a isquemia.
Isto é, ao tratar as células com espironolactona e eplerenona, estimula-se o aumento dos segundos
mensageiros importantes na proteção, entre eles o AMPc. Com maior disponibilidade de AMPc,
a enzima ecto-5’-nucleotidase tem mais substrato para clivar o AMP em adenosina durante a
isquemia (Eckle et al., 2007). A adenosina, por sua vez, pode então atuar via seus receptores para
promover a proteção contra os danos causados por isquemia-reperfusão.
2. Espironolactona e eplerenona possuem ação proliferativa de cardiomiócitos e
antifibrótica, como demonstrado em trabalhos prévios do nosso laboratório. Tanto
espironolactona quanto eplerenona se mostraram capazes de promover proliferação de
cardiomiócitos após 24h e reverter a proliferação de fibroblastos promovida pela aldosterona
após 48h do tratamento (Hermidorff et al., 2015). Por nosso modelo de estudo ser agudo (ensaios
realizados 18h após tratamentos), é possível que nosso método não tenha sido suficiente para
observar os efeitos consequentes de uma possível ação dos antagonistas do MR associados aos
receptores da adenosina.

60
6- CONCLUSÃO

Conclusão
61
A cardioproteção promovida por antagonistas do receptor mineralocorticoide da
aldosterona é bem fundamentada e, estudos vêm demonstrando que espironolactona e eplerenona
atuam também como agentes de condicionamento farmacológico, protegendo o coração contra
lesão de reperfusão. A proteção promovida pela adenosina também é bem estabelecida durante
eventos de isquemia-reperfusão em várias espécies e modelos experimentais. Nosso trabalho
mostrou pela primeira vez, de forma comparativa, a ação da espironolactona e eplerenona
associadas à adenosina e seus receptores.
Com base em nossos resultados concluímos que:
1. Cardiomiócitos ventriculares de ratos são igualmente protegidos contra danos
promovidos por isquemia tanto por espironolactona e eplerenona, quanto por adenosina.
2. Espironolactona e eplerenona aumentam a concentração de adenosina presente em
cardiomiócitos ventriculares de ratos após isquemia, no pré-condicionamento e no pós-
condicionamento farmacológico.
3. Apesar da evidente proteção promovida por espironolactona e eplerenona durante a
isquemia, nossos dados não nos permite afirmar se esta ocorre via ligação aos receptores
da adenosina, uma vez que o tratamento das células com antagonistas dos receptores
A1R, A2AR, A2BR e A3R associados a espironolactona e eplerenona não alterou a
viabilidade celular quando comparado ao tratamento apenas com os fármacos. Outros
experimentos serão necessários para esclarecer essa questão.
A análise conjunta dos nossos resultados nos levam a crer que há relação entre a ação da
espironolactona e da eplerenona com a adenosina, contudo se faz necessário aprofundar ainda
mais no conhecimento desse mecanismo para compreendê-lo. É possível que a cardioproteção
promovida pela espironolactona e eplerenona durante a isquemia ocorra mediada pelos
receptores da adenosina, no entanto, nossos resultados são consequentes de uma análise aguda,
feita 18 horas após os tratamentos, podendo não ter sido tempo suficiente para observar os efeitos
consequentes dessa possível relação. Além disso, não se pode descartar a possibilidade de que a
ação desses cardioprotetores possa ocorrer por outro mecanismo, talvez por propiciar a ação da
enzima ecto-5’-nucleotidase.

62
7- REFERÊNCIAS

Referências
63
ANDRIULLI, A. et al. Canrenone and androgen receptor-active materials in plasma of cirrhotic
patients during long-term K-canrenoate or spironolactone therapy. Digestion, v. 44, n. 3, p. 155-
62, 1989. ISSN 0012-2823. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/2697627 >.
ARMANINI, D. et al. Aldosterone receptor blockers spironolactone and canrenone: two
multivalent drugs. Expert Opin Pharmacother, v. 15, n. 7, p. 909-12, May 2014. ISSN 1744-
7666. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/24617854 >.
AUCHAMPACH, J. A.; BOLLI, R. Adenosine receptor subtypes in the heart: therapeutic
opportunities and challenges. Am J Physiol, v. 276, n. 3 Pt 2, p. H1113-6, Mar 1999. ISSN 0002-
9513. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/10070100 >.
BARRETO-CHAVES, M. L.; HEIMANN, A.; KRIEGER, J. E. Stimulatory effect of
dexamethasone on angiotensin-converting enzyme in neonatal rat cardiac myocytes. Braz J Med
Biol Res, v. 33, n. 6, p. 661-4, Jun 2000. ISSN 0100-879X. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/10829094 >.
BERNE, R. M. METABOLIC REGULATION OF BLOOD FLOW. Circ Res, v. 15, p.
SUPPL:261-8, Aug 1964. ISSN 0009-7330. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/14206313 >.
BULLUCK, H.; HAUSENLOY, D. J. Ischaemic conditioning: are we there yet? Heart, v. 101,
n. 13, p. 1067-77, Jul 2015. ISSN 1468-201X. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/25887783 >.
CHAI, W. et al. Genomic and nongenomic effects of aldosterone in the rat heart: why is
spironolactone cardioprotective? Br J Pharmacol, v. 145, n. 5, p. 664-71, Jul 2005. ISSN 0007-
1188. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/15834444 >.

Referências
64
______. Nongenomic effects of aldosterone in the human heart: interaction with angiotensin II.
Hypertension, v. 46, n. 4, p. 701-6, Oct 2005. ISSN 1524-4563. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/16144984 >.
CHIU, G. S.; FREUND, G. G. Modulation of neuroimmunity by adenosine and its receptors:
metabolism to mental illness. Metabolism, v. 63, n. 12, p. 1491-8, Dec 2014. ISSN 1532-8600.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/25308443 >.
CRISOSTOMO, P. R. et al. Preconditioning versus postconditioning: mechanisms and
therapeutic potentials. J Am Coll Surg, v. 202, n. 5, p. 797-812, May 2006. ISSN 1072-7515.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16648020 >.
CUNHA, R. A. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous
system: different roles, different sources and different receptors. Neurochem Int, v. 38, n. 2, p.
107-25, Feb 2001. ISSN 0197-0186. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/11137880 >.
DELYANI, J. A. Mineralocorticoid receptor antagonists: the evolution of utility and
pharmacology. Kidney Int, v. 57, n. 4, p. 1408-11, Apr 2000. ISSN 0085-2538. Disponível em:
< http://www.ncbi.nlm.nih.gov/pubmed/10760075 >.
DEROSA, G. et al. Effects of canrenone in patients with metabolic syndrome. Expert Opin
Pharmacother, v. 14, n. 16, p. 2161-9, Nov 2013. ISSN 1744-7666. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/23984806 >.
ECKLE, T. et al. Cardioprotection by ecto-5'-nucleotidase (CD73) and A2B adenosine receptors.
Circulation, v. 115, n. 12, p. 1581-90, Mar 2007. ISSN 1524-4539. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/17353435 >.

Referências
65
ELY, S. W. et al. Functional and metabolic evidence of enhanced myocardial tolerance to
ischemia and reperfusion with adenosine. J Thorac Cardiovasc Surg, v. 90, n. 4, p. 549-56, Oct
1985. ISSN 0022-5223. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/4046621 >.
FRACCAROLLO, D. et al. Additive improvement of left ventricular remodeling and
neurohormonal activation by aldosterone receptor blockade with eplerenone and ACE inhibition
in rats with myocardial infarction. J Am Coll Cardiol, v. 42, n. 9, p. 1666-73, Nov 2003. ISSN
0735-1097. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/14607457 >.
FUNDER, J. W. Mineralocorticoid receptor antagonists: emerging roles in cardiovascular
medicine. Integr Blood Press Control, v. 6, p. 129-38, 2013. ISSN 1178-7104. Disponível em:
< http://www.ncbi.nlm.nih.gov/pubmed/24133375 >.
GRUBE, K. et al. Evidence for an intracellular localization of the adenosine A2B receptor in rat
cardiomyocytes. Basic Res Cardiol, v. 106, n. 3, p. 385-96, May 2011. ISSN 1435-1803.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21246204 >.
GULATI, P.; SINGH, N. Pharmacological evidence for connection of nitric oxide-mediated
pathways in neuroprotective mechanism of ischemic postconditioning in mice. J Pharm
Bioallied Sci, v. 6, n. 4, p. 233-40, Oct 2014. ISSN 0976-4879. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/25400405 >.
HAUSENLOY, D. J.; YELLON, D. M. The second window of preconditioning (SWOP) where
are we now? Cardiovasc Drugs Ther, v. 24, n. 3, p. 235-54, Jun 2010. ISSN 0920-3206.
______. Ischaemic conditioning and reperfusion injury. Nat Rev Cardiol, Feb 2016. ISSN 1759-
5010. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/26843289 >.
HAYASHI, H. et al. Aldosterone nongenomically produces NADPH oxidase-dependent reactive
oxygen species and induces myocyte apoptosis. Hypertens Res, v. 31, n. 2, p. 363-75, Feb 2008.
ISSN 0916-9636. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18360057 >.

Referências
66
HERMIDORFF, M. M. et al. Non-genomic effects of spironolactone and eplerenone in
cardiomyocytes of neonatal Wistar rats: do they evoke cardioprotective pathways? Biochem Cell
Biol, v. 93, n. 1, p. 83-93, Feb 2015. ISSN 1208-6002. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/25488178 >.
HEUSCH, G. Molecular basis of cardioprotection: signal transduction in ischemic pre-, post-,
and remote conditioning. Circ Res, v. 116, n. 4, p. 674-99, Feb 2015. ISSN 1524-4571.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/25677517 >.
JENNINGS, R. B. et al. Metabolism of preconditioned myocardium: effect of loss and
reinstatement of cardioprotection. J Mol Cell Cardiol, v. 33, n. 9, p. 1571-88, Sep 2001. ISSN
0022-2828. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11549338 >.
LATINI, S.; PEDATA, F. Adenosine in the central nervous system: release mechanisms and
extracellular concentrations. J Neurochem, v. 79, n. 3, p. 463-84, Nov 2001. ISSN 0022-3042.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11701750 >.
LEHNINGER, A. L.; NELSON, D. L.; COX, M. M. Lehninger principles of biochemistry. 4th
ed. New York, N.Y. ; Basingstoke: W.H. Freeman, 2005. ISBN 9780716743392 : ¹39.99
0716743396 : ¹39.99.
LESHEM-LEV, D. et al. Adenosine A(1) and A (3) receptor agonists reduce hypoxic injury
through the involvement of P38 MAPK. Mol Cell Biochem, v. 345, n. 1-2, p. 153-60, Dec 2010.
ISSN 1573-4919. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20730620 >.
LI, W. et al. Pharmacological preconditioning and postconditioning with nicorandil
attenuates ischemia/reperfusion-induced myocardial necrosis and apoptosis in
hypercholesterolemic rats.: Experimental and Therapeutic Medicine. 10: 2197–2205 p. 2015.

Referências
67
LIANG, B. T.; HALTIWANGER, B. Adenosine A2a and A2b receptors in cultured fetal chick
heart cells. High- and low-affinity coupling to stimulation of myocyte contractility and cAMP
accumulation. Circ Res, v. 76, n. 2, p. 242-51, Feb 1995. ISSN 0009-7330. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/7834835 >.
LUNA-ORTIZ, P. et al. Myocardial postconditioning: anaesthetic considerations. Arch Cardiol
Mex, v. 81, n. 1, p. 33-46, 2011 Jan-Mar 2011. ISSN 1405-9940. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/21592890 >.
MA, J. et al. Effects of low-dose spironolactone combined with irbesartan on cardiac
hypertrophy induced by pressure overload in rats. Am J Transl Res, v. 6, n. 6, p. 809-19, 2014.
ISSN 1943-8141. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/25628791 >.
MCCURLEY, A.; JAFFE, I. Z. Mineralocorticoid receptors in vascular function and disease.
Mol Cell Endocrinol, v. 350, n. 2, p. 256-65, Mar 2012. ISSN 1872-8057. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/21723914 >.
MCINTOSH, V. J.; LASLEY, R. D. Adenosine receptor-mediated cardioprotection: are all 4
subtypes required or redundant? J Cardiovasc Pharmacol Ther, v. 17, n. 1, p. 21-33, Mar 2012.
ISSN 1940-4034. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21335481 >.
MCMAHON, E. G. Recent studies with eplerenone, a novel selective aldosterone receptor
antagonist. Curr Opin Pharmacol, v. 1, n. 2, p. 190-6, Apr 2001. ISSN 1471-4892. Disponível
em: < http://www.ncbi.nlm.nih.gov/pubmed/11714095 >.
MURRY, C. E.; JENNINGS, R. B.; REIMER, K. A. Preconditioning with ischemia: a delay of
lethal cell injury in ischemic myocardium. Circulation, v. 74, n. 5, p. 1124-36, Nov 1986. ISSN
0009-7322. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/3769170 >.

Referências
68
______. New insights into potential mechanisms of ischemic preconditioning. Circulation, v.
84, n. 1, p. 442-5, Jul 1991. ISSN 0009-7322. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/2060119 >.
NAGATOMO, Y. et al. Significance of AT1 receptor independent activation of
mineralocorticoid receptor in murine diabetic cardiomyopathy. PLoS One, v. 9, n. 3, p. e93145,
2014. ISSN 1932-6203. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/24664319 >.
NAPPI, J. M.; SIEG, A. Aldosterone and aldosterone receptor antagonists in patients with
chronic heart failure. Vasc Health Risk Manag, v. 7, p. 353-63, 2011. ISSN 1178-2048.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/21731887 >.
NICOLAU, J. C. et al. Infarto do miocárdio em hipertensos.: HiperAtivo. 6: 38-41 p. 1999.
NORTON, G. R. et al. Adenosine A1 receptor-mediated antiadrenergic effects are modulated
by A2a receptor activation in rat heart. Am J Physiol, v. 276, n. 2 Pt 2, p. H341-9, Feb 1999.
ISSN 0002-9513. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/9950832 >.
NÚÑEZ, L. C. et al. Mecanismo de acción de la eplerenona. AL., E. Revista Española de
Cardiologia. 6: 31-47 p. 2006.
OMORI, Y. et al. Glucocorticoids Induce Cardiac Fibrosis via Mineralocorticoid Receptor in
Oxidative Stress: Contribution of Elongation Factor Eleven-Nineteen Lysine-Rich Leukemia
(ELL). Yonago Acta Med, v. 57, n. 3, p. 109-16, Sep 2014. ISSN 0513-5710. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/25349466 >.
PITT, B. et al. Safety and tolerability of the novel non-steroidal mineralocorticoid receptor
antagonist BAY 94-8862 in patients with chronic heart failure and mild or moderate chronic
kidney disease: a randomized, double-blind trial. Eur Heart J, v. 34, n. 31, p. 2453-63, Aug
2013. ISSN 1522-9645. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/23713082 >.

Referências
69
______. Eplerenone, a selective aldosterone blocker, in patients with left ventricular dysfunction
after myocardial infarction. N Engl J Med, v. 348, n. 14, p. 1309-21, Apr 3 2003. ISSN 1533-
4406 (Electronic)
0028-4793 (Linking). Disponível em: <
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt=Citation&li
st_uids=12668699 >.
______. The effect of spironolactone on morbidity and mortality in patients with severe heart
failure. Randomized Aldactone Evaluation Study Investigators. N Engl J Med, v. 341, n. 10, p.
709-17, Sep 1999. ISSN 0028-4793. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/10471456 >.
PRZYKLENK, K.; WHITTAKER, P. Remote ischemic preconditioning: current knowledge,
unresolved questions, and future priorities. J Cardiovasc Pharmacol Ther, v. 16, n. 3-4, p. 255-
9, 2011 Sep-Dec 2011. ISSN 1940-4034. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/21821525 >.
QIN, W. et al. Transgenic model of aldosterone-driven cardiac hypertrophy and heart failure.
Circ Res, v. 93, n. 1, p. 69-76, Jul 2003. ISSN 1524-4571. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/12791709 >.
RAHMAN, A. The role of adenosine in Alzheimer's disease. Curr Neuropharmacol, v. 7, n. 3,
p. 207-16, Sep 2009. ISSN 1875-6190. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/20190962 >.
ROBIN, E. et al. Adenosine A1 receptor activation is arrhythmogenic in the developing heart
through NADPH oxidase/ERK- and PLC/PKC-dependent mechanisms. J Mol Cell Cardiol, v.
51, n. 6, p. 945-54, Dec 2011. ISSN 1095-8584. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/21907719 >.

Referências
70
RUDOLPH, A. E.; ROCHA, R.; MCMAHON, E. G. Aldosterone target organ protection by
eplerenone. Mol Cell Endocrinol, v. 217, n. 1-2, p. 229-38, Mar 2004. ISSN 0303-7207.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/15134822 >.
SAFRAN, N. et al. Cardioprotective effects of adenosine A1 and A3 receptor activation during
hypoxia in isolated rat cardiac myocytes. Mol Cell Biochem, v. 217, n. 1-2, p. 143-52, Jan 2001.
ISSN 0300-8177. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11269659 >.
SCHMIDT, K. et al. Cardioprotective effects of mineralocorticoid receptor antagonists at
reperfusion. Eur Heart J, v. 31, n. 13, p. 1655-62, Jul 2010. ISSN 1522-9645. Disponível em:
< http://www.ncbi.nlm.nih.gov/pubmed/20028693 >.
SCHÄFER, A. et al. Addition of the selective aldosterone receptor antagonist eplerenone to
ACE inhibition in heart failure: effect on endothelial dysfunction. Cardiovasc Res, v. 58, n. 3,
p. 655-62, Jun 2003. ISSN 0008-6363. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/12798439 >.
SILVA JR., O. D. C. E. et al. Aspectos básicos da lesão de isquemia e reperfusão e do pré-
condicionamento isquêmico. Acta Cirúrgica Brasileira. 17: 96-100 p. 2002.
SOHN, H. J. et al. Aldosterone modulates cell proliferation and apoptosis in the neonatal rat
heart. J Korean Med Sci, v. 25, n. 9, p. 1296-304, Sep 2010. ISSN 1598-6357. Disponível em:
< http://www.ncbi.nlm.nih.gov/pubmed/20808672 >.
SOLLEVI, A. Cardiovascular effects of adenosine in man; possible clinical implications. Prog
Neurobiol, v. 27, n. 4, p. 319-49, 1986. ISSN 0301-0082. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/3538187 >.
SOMMERSCHILD, H. T.; KIRKEBØEN, K. A. Adenosine and cardioprotection during
ischaemia and reperfusion--an overview. Acta Anaesthesiol Scand, v. 44, n. 9, p. 1038-55, Oct
2000. ISSN 0001-5172. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11028722 >.

Referências
71
ST HILAIRE, C. et al. Mechanisms of induction of adenosine receptor genes and its functional
significance. J Cell Physiol, v. 218, n. 1, p. 35-44, Jan 2009. ISSN 1097-4652. Disponível em:
< http://www.ncbi.nlm.nih.gov/pubmed/18767039 >.
STRICKLER, J.; JACOBSON, K. A.; LIANG, B. T. Direct preconditioning of cultured chick
ventricular myocytes. Novel functions of cardiac adenosine A2a and A3 receptors. J Clin Invest,
v. 98, n. 8, p. 1773-9, Oct 1996. ISSN 0021-9738. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/8878427 >.
TIAN, Y. S. et al. Pharmacological postconditioning with diazoxide attenuates
ischemia/reperfusion-induced injury in rat liver. Exp Ther Med, v. 5, n. 4, p. 1169-1173, Apr
2013. ISSN 1792-0981. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/23596486 >.
URMALIYA, V. B. et al. Cardioprotection induced by adenosine A1 receptor agonists in a
cardiac cell ischemia model involves cooperative activation of adenosine A2A and A2B
receptors by endogenous adenosine. J Cardiovasc Pharmacol, v. 53, n. 5, p. 424-33, May 2009.
ISSN 1533-4023. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/19333129 >.
WALLBRIDGE, D. R. et al. No attenuation of ischaemic preconditioning by the calcium
antagonist nisoldipine. J Mol Cell Cardiol, v. 28, n. 8, p. 1801-10, Aug 1996. ISSN 0022-2828.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/8877789 >.
XI, J. et al. Adenosine A2A and A2B receptors work in concert to induce a strong protection
against reperfusion injury in rat hearts. J Mol Cell Cardiol, v. 47, n. 5, p. 684-90, Nov 2009.
ISSN 1095-8584. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19695259 >.
ZANNAD, F. et al. Eplerenone in patients with systolic heart failure and mild symptoms. N
Engl J Med, v. 364, n. 1, p. 11-21, Jan 2011. ISSN 1533-4406. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/21073363 >.

Referências
72
ZHAN, E.; MCINTOSH, V. J.; LASLEY, R. D. Adenosine A₂A and A₂B receptors are both
required for adenosine A₁ receptor-mediated cardioprotection. Am J Physiol Heart Circ
Physiol, v. 301, n. 3, p. H1183-9, Sep 2011. ISSN 1522-1539. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/21743001 >.
ZHANG, L. et al. The aldosterone receptor antagonist spironolactone prevents peritoneal
inflammation and fibrosis. Lab Invest, v. 94, n. 8, p. 839-50, Aug 2014. ISSN 1530-0307.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/24862968 >.
ZHAO, H. et al. Inhibitory effects of spironolactone on myocardial fibrosis in spontaneously
hypertensive rats. Genet Mol Res, v. 14, n. 3, p. 10315-21, 2015. ISSN 1676-5680. Disponível
em: < http://www.ncbi.nlm.nih.gov/pubmed/26345970 >.
ZHAO, Z. Q. et al. Inhibition of myocardial injury by ischemic postconditioning during
reperfusion: comparison with ischemic preconditioning. Am J Physiol Heart Circ Physiol, v.
285, n. 2, p. H579-88, Aug 2003. ISSN 0363-6135. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/12860564 >.
WHO – World Health Organization. Global Status Report on Noncommunicable Diseases 2014.
p. 9-10. 2014. Disponível em: http://www.who.int/mediacentre/factsheets/fs355/en/
Portal Brasil. Doenças cardiovasculares causam quase 30% das mortes no País. Disponível em:
www.brasil.gov.br/saude/2011/09/doencas-cardiovasculares-causam-quase-30-das-mortes-no-
pais Acesso em 20/04/15 às 18:00 horas.
Portal da Sociedade Brasileira de Cardiologia – Cardiômetro. Disponível em:
www.cardiometro.com.br Acesso em 09/03/16 às 19:30 horas.

73
8- ANEXOS

Anexos
74
Ta
bel
a 9
: In
form
açõ
es g
erai
s d
os
anta
go
nis
tas
do
MR
, es
pir
ono
lact
ona,
ep
lere
no
na,
can
reno
na
e fi
ner
eno
na.

Anexos
75

Anexos
76