associaÇÃo das biotÉcnicas: aspiraÇÃo folicular guiada por ... · tua alma: mãe, pai... ......
TRANSCRIPT

UNIVERSIDADE ESTADUAL PAULISTA“JÚLIO MESQUITA”
FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIASCÂMPUS DE JABOTICABAL
ASSOCIAÇÃO DAS BIOTÉCNICAS: ASPIRAÇÃOFOLICULAR GUIADA POR ULTRA-SONOGRAFIA E
SUPEROVULAÇÃO NA PRODUÇÃO IN VITRO E IN VIVODE EMBRIÕES BOVINOS
Andréa Renesto
Médica Veterinária
JABOTICABAL – SÃO PAULO – BRASIL
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

ii
Agosto de 2004UNIVERSIDADE ESTADUAL PAULISTA
“JÚLIO MESQUITA”FACULDADE DE CIÊNCIAS AGRÁRIAS E VETERINÁRIAS
CÂMPUS DE JABOTICABAL
ASSOCIAÇÃO DAS BIOTÉCNICAS: ASPIRAÇÃOFOLICULAR GUIADA POR ULTRA-SONOGRAFIA E
SUPEROVULAÇÃO NA PRODUÇÃO IN VITRO E IN VIVODE EMBRIÕES BOVINOS
Andréa Renesto
Orientador: Profa. Dra. Lia de Alencar Coelho
Dissertação de Mestradoapresentada à Faculdade deCiências Agrárias e Veterinárias– Unesp, campus de Jaboticabal,como parte das exigências paraa obtenção do título de Mestreem Medicina Veterinária(Reprodução Animal).
JABOTICABAL – SÃO PAULO – BRASIL
Agosto de 2004
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

iii
DADOS CURRICULARES DO AUTOR
ANDRÉA RENESTO – Nascida aos 10 dias de março de 1977 em Pereira
Barreto – SP. Graduou-se em Medicina Veterinária na Universidade do Oeste
Paulista – UNOESTE – no período de Janeiro de 1995 a Dezembro de 1999. No
período de Fevereiro de 2000 a Agosto de 2001 cursou a Especialização Lato
Sensu em Reprodução Animal, na Universidade do Oeste Paulista – UNOESTE.
Em fevereiro de 2001 ingressou na Especialização Lato Sensu em Julgamento
das Raças Zebuínas, da Faculdade de Agronomia e Zootecnia de Uberaba –
FAZU, obtendo o título de Especialista em Julgamento das Raças Zebuínas em
Dezembro de 2001. Graduou-se como Jurada efetiva das Raças Zebuínas através
do Colégio Técnico de Jurados das Raças Zebuínas da Associação Brasileira dos
Criadores de Zebu, no período de Junho de 1999 a Outubro de 2001. Ingressou
no Programa de Mestrado em Medicina Veterinária, Área de Concentração
Reprodução Animal, pela faculdade de Ciências Agrárias e Veterinárias – UNESP
- Jaboticabal-SP, em março de 2002.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

iv
Agradeço a Deus por essa existência, pela coragem,
vontade e principalmente pela força
nos momentos mais difíceis...
Dedico este trabalho aos meus pais!
É mais uma prova do sinônimo de amor e carinho que
vocês me proporcionaram durante toda minha existência.
.... Vocês que abriram mão de muitos sonhos em função
dos meus... Não há palavras que expressem minha
gratidão e meu amor!!!
“ Se um dia já homem feito e realizado, sentires que aterra cedeu a teus pés, e que tuas obras morreram, quenão há ninguém a tua volta para te estender a mão,esquece a tua maturidade, passa pela tua mocidade,volta a tua infância e balbucia, entre lágrimas eesperanças, as últimas palavras que sempre estarão natua alma: Mãe, Pai...
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

v
Rui Barbosa
Aos meus irmãos, que tanto amo, Silvana e Éden:
Obrigada pelo apoio incessante e por acreditarem emmim, mesmo nas horas em que eu cheguei a duvidar!!
“ À medida que o nosso amor progride, mais nos convencemosde que Deus existe e de que a alma é imortal”.
(Dostoievski)
Ao meu querido amigo Irineu Gonçalves Filho;
“ A imortalidade de que se reveste a natureza humanafaz o homem sempre presente.
Presente pela cultura que transmitiu.Presente pela amizade que conquistou.
Presente pelo exemplo que legou.Sempre presente porque o homem é educador.”
(Dostoievski)
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

vi
Ofereço:
A Profa Dra. Lia de Alencar Coelho
Ao Prof. Dr. Joaquim Mansano Garcia
A Profa Dra. Caliê Castilho
Obrigada ... pela orientação,valiosos ensinamentos,
pela confiança e inestimável apoio,pela amizade e convivência enriquecedora...
À vocês, minha eterna admiração!!Em especial a minha querida amiga Caliê... não tenho
palavras de agradecimento, admiração e respeito...
“ É muito melhor ousar fazer coisas grandiosas,triunfar gloriosamente, mesmo que com alguns fracassos nomeio do caminho, do que se igualar aquela pobres almas que
não aproveitam nem sofrem muito,pois vivem na penumbra cinza de quem não sabe o que é a
vitória nem a derrota”(Theodore Roosevelt)
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

vii
“ ...E encontrei liberdade e segurança
em minha loucura:
a liberdade da solidão e
a segurança de não ser compreendido,
pois aqueles que nos compreendem
nos escravizam de algum modo...”
(Gibran Khalil Gibran)
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

viii
AGRADECIMENTOS
A realização deste trabalho só foi possível graças à colaboração direta e indireta
de muitas pessoas. Manifesto minha eterna gratidão a todas elas!!
À Universidade Estadual Paulista, Faculdade de Ciência Agrárias e Veterinárias
de Jaboticabal, e ao departamento de Medicina Veterinária Preventiva e
Reprodução Animal – DRA pelas instalações, condições de trabalho e
oportunidade oferecida.
À Universidade do Oeste Paulista pelo incentivo a pesquisa, por ceder os animais
da segunda fase do trabalho e colocar a disposição às instalações da Fazenda
Experimental, em nome da amiga Profa Dra. Caliê Castilho, que me guiou no
caminho da pesquisa. Não tenho palavras de agradecimento... é só sentimento!!!
À Gilmar Garcia que gentilmente forneceu os animais e toda instalação da
Agropecuária J Garcia para os experimentos da primeira fase do trabalho.
“ A arte suprema do mestre consiste em despertar alegria, provocando curiosidade
pelo conhecimento criativo. A única finalidade da educação deve consistir em
preparar indivíduos que pensem e ajam como pessoas – independentes e livres”.
(Albert Einstein)
À todos os funcionários que me acompanharam no campo, ajudando
intensamente na realização desse trabalho. Fica aqui minha gratidão, vocês foram
fundamentais para o projeto...e, foram... incansáveis!!
Aos meus companheiros de trabalho e amigos, Dr. Irineu Gonçalves Filho, Dr.
Valdecir Marin Júnior e Dr. Marcelo Moura, inicialmente pela amizade e por
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

ix
acreditarem em mim e posteriormente pela confiança, compreensão e força...
Vocês fizeram parte da realização desse sonho e com vocês aprendo mais a cada
dia os principais valores da vida: ética, companheirismo, conhecimento, respeito,
seriedade e amizade! “... Amigos que se tornaram parceiros, parceiros que se
tornaram amigos. Sementes plantadas e regadas com respeito, admiração e muita
amizade!! “
Ao meu querido e doce JV. Obrigada pela sinceridade, pelos olhos cheio de
carinho pela beleza e grandeza dos seus sentimentos. Você foi fundamental em
um momento da minha vida em que cheguei a achar que as pessoas eram só
maldade e que a vida era repleta de injustiças e incompreensão.
“ Parece que tudo já foi dito entre nós. Agora só vale a pena dizer da saudade que
você deixou...” (Carlos Drummond de Andrade)
A todos meus familiares que mesmo distantes sempre me apoiaram e torceram
por mim em mais essa etapa da minha vida.
A todos os professores e funcionários do Departamento de Reprodução Animal
pela amizade, convívio harmonioso e colaboração. Em especial a Bel e a Roberta
pela paciência e atenção.
Ao Médico Veterinário e amigo José Otávio Folino pelo auxílio na colheita de
embriões, pela ajuda em um dos momentos mais difíceis e tão engrandecedor que
passei na minha vida e principalmente pela amizade! A você e sua esposa Milene,
nunca esquecerei o carinho, a amizade, o apoio, o incentivo, as palavras de fé, de
esperança... pela mão amiga estendida, pelo estímulo e conforto...
Ao Médico Veterinário Fábio Mustafá pelo auxílio no desenvolvimento dos
trabalhos na Fazenda Experimental. Obrigado companheiro!!
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

x
Aos funcionários da Avanti Consultoria Pecuária, Carol, Regina e Jú! A realização
desse trabalho contou diretamente com a colaboração de vocês... por me
apoiarem e principalmente pela compreensão da minha inconstância de humor. E,
também aos funcionários da Nelore, Adriana e Maurílio! Obrigada pela força!!
Aos pós-graduandos de Jaboticabal, em especial a Déia e Gabriel que foram
extremamente prestativos quando precisei de ajuda. Companheirismo é uma
qualidade de poucos... Obrigado companheiro!!! Ao Max, Cris, Vi, Letícia, Éricles...
A minha amiga irmã Ka (Karina Ueda Stringuetta). Você, mesmo distante, sentia
quando eu precisava de uma palavra amiga e de um conforto e mesmo quando eu
não queria falar você sempre tinha uma palavra de carinho. Obrigada por você
nunca desistir de mim!!!
A minha amiga irmã Lê (Alessandra Reis).... sempre presente em todos os
momentos em que precisei, sempre pronta para ouvir! Obrigada pelos sábios
conselhos e por me fazer ver, por muitas vezes, a força que existe em mim e que
Deus nos dá a todo instante. Obrigada pela amizade e pelos momentos de alegria
fraterna que sempre marcaram nossa convivência... Você que une a ação ao
sentimento e ao pensamento! Muito obrigada!!!
À minha amiga querida, Janete (Fernanda Prata Cunha)... Muito obrigada pela
força e por se preocupar tanto comigo. Você tem sido uma grande amiga...
sabemos que podemos contar uma com a outra!!! “Toghethersss... foreverssss....”
À minha eterna amiga Maria de Lamare... nem tenho palavras para agradecer toda
força que já me deu. Obrigada pelo querer bem e pela confiança! Saiba que você
é companheira ímpar... parceira cem por cento!
Àos meus amigos: Lu Kahale, Flor (Fabiana), Verinha, Baraldi, Talita Medeiros, Li,
Gabi, Dani Bolota, Dipilik (Bárbara), Dani Tenca, Lala, Dudu, Fer Rezek, Tiago,
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xi
Kel, Gui, SI, Déia Basso, Bidu, Elaine, Ricardo Porção, Mary, Dedé, Thá, Morcega
(Kelma), Paulinha, Flá, Ana Laura, D.Lenita, Mav... enfim a todos MUIIITTOOO
OBRIGADA PELO APOIO E PRINCIPALMENTE PELA AMIZADE!!
SUMÁRIO
I. INTRODUÇÃO ........................................................................................................... 15
II. REVISÃO DA LITERATURA..................................................................................... 17
III. MATERIAL E MÉTODOS......................................................................................... 25
Local do Experimento.............................................................................................. 25
Animais ..................................................................................................................... 25
Tratamentos.............................................................................................................. 25
Aspiração Folicular.................................................................................................. 28
Lavagem e seleção dos oócitos ............................................................................. 28
Transporte de oócitos ............................................................................................. 29
Maturação in vitro (MIV) .......................................................................................... 29
Fecundação in vitro (FIV) ........................................................................................ 29
Cultivo in vitro (CIV)................................................................................................. 30
Colheita de embriões............................................................................................... 30
Manipulação e Avaliação Morfológica dos Embriões TE ..................................... 30
Análise Estatística ................................................................................................... 30
IV. RESULTADOS......................................................................................................... 32
V. DISCUSSÃO ............................................................................................................. 38
VI. CONCLUSÃO .......................................................................................................... 41
VII. REFERÊNCIAS BIBLIOGRÁFICAS....................................................................... 42
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xii
ASSOCIAÇÃO DAS BIOTÉCNICAS: ASPIRAÇÃO FOLICULAR GUIADA POR
ULTRA-SONOGRAFIA E SUPEROVULAÇÃO NA PRODUÇÃO IN VITRO E IN
VIVO DE EMBRIÕES BOVINOS
RESUMO – Visando otimizar o uso alternado das biotecnologias TE e OPU-
FIV o presente trabalho teve como objetivo avaliar em vacas da raça Nelore o
efeito do início da superovulação ovariana (SOV) 48 a 60 horas após a aspiração
folicular em fase aleatória do ciclo estral (OPU 1) e avaliar a produção in vitro de
embriões oriundos de oócitos aspirados (OPU 2) em diferentes momentos da
primeira onda folicular induzida após a SOV. As fêmeas (n=23) foram submetidas
a OPU 1, em fase aleatória do ciclo estral, seguida pela inserção de implante de
progestágeno (Crestar®). Em torno de 48 a 60 horas após a aspiração iniciou-se a
SOV, utilizando-se FSH (Folltropin®) 180 mg por vaca dividida em 8 doses,
durante 4 dias consecutivos. Durante 6a e 7a aplicações de FSH foi administrada
500 µg de PGF2α (Preloban®) e o Crestar foi retirado durante a administração da
sétima dose de FSH. Após 12 horas do término da SOV, as fêmeas receberam
25mg de LH exógeno (Lutropin-V®) para causar ovulação e realizar a IA com
tempo fixo. Foram realizadas 2 inseminações com intervalo de 12 horas, sendo
que a 1ª inseminação ocorreu 12 horas após a administração de LH. Sete dias
após a 1ª IA as fêmeas foram coletadas. No dia da colheita dos embriões as
fêmeas foram divididas em 6 tratamentos, T(0-144) (n=4), T(48-120) (n=5), T(48-
96) (n=4), T(72-96) (n=3), T(96-72) (n=4), T(96-48) (n=3) que variaram de acordo
com o momento da administração de PGF2α em relação a colheita de embriões e
aspiração na primeira onda após a SOV. A aplicação de PGF2α foi realizada no
dia da colheita (dia 0), 48, 72 ou 96 horas após a colheita e a aspiração 144, 96,
72 ou 48 horas após a PGF2α. Não houve diferença significativa na % de oócitos
aspirados e embriões produzidos com a alternância das biotecnologias OPU e TE.
No entanto, foi observada diferença significativa (p>0,05) no número de oócitos
recuperados entre os tratamentos, sendo que a recuperação no T(96-48) foi
significativamente menor quando comparada ao T(48-96) e T(72-96),
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xiii
demonstrando que o momento da aspiração muito próxima da PGF2α (48 horas
após) prejudicou a recuperação de oócitos. Concluí-se que a superovulação não
afeta a recuperação de oócitos e a produção in vitro de blasctocistos quando a
OPU é realizada a partir de 48 horas após a PGF2α na onda subseqüente a
superovulação podendo ser utilizada para o incremento na produção de embriões
de vacas da raça Nelore.
Palavras chaves: aspiração folicular, competência oocitária, dinâmica folicular,
nelore, superovulação
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xiv
ASSOCIATION OF THE BIOTECHNIQUES: ULTRASOUND-GUIDED
TRANSVAGINAL FOLLICULAR ASPIRATION AND SUPEROVULATION IN
PRODUCTION IN VITRO AND IN VIVO OF BOVINE EMBRYOS
SUMMARY - Aiming at to optimize the alternating use of the biotechnologies
multiple ovulation and embryo transfer (MOET) and Ovum Pick Up and in vitro
fertilization (OPU-FIV) the present work had as objective to evaluate in Nelore
donors breed the effect of the beginning of the superovulation (SOV) at 48-60
hours after the follicular aspiration in randomly phase of the estrous cycle (OPU 1)
and to evaluate the in vitro embryo development of recovered oocytes (OPU 2) at
different moments of the first folicular wave after the SOV. The females (n=23)
were submitted to a OPU 1, in randomly phase of the estrous cycle, followed for
the insertion of progesterone (Crestar®). Around 48 - 60 hours after the aspiration
the superovulaction was induced with FSH (Folltropin®) 180 mg/ cow divided in 8
doses, during 4 consecutive days. Luteolytis was induced with two doses of PGF2á
(Preloban®, 500 µg), during the sixth and seventh infections of FSH and Crestar
was removed during the administration of the seventh dose of FSH. 12 hours after
the end of the SOV, the females received 25mg of LH (Lutropin-V®) to cause
ovulation. Two Artificial inseminations with fixed time were done with interval of 12
hours and the first occurred 12 hours after the LH administration. Seven days after
1ªIA the embryo were collected and all cows divided in 6 treatments, T(0-144)
(n=4), T(48-120) (n=5), T(48-96) (n=4), T(72-96) (n=3), T(96-72) (n=4), T(96-48)
(n=3) that accordance with the injection of PGF2 á , embryo recovery and
aspiration in the first wave after the SOV. The PGF2 á application was carried in
the embryo recovery (day 0), 48, 72 or 96 hours after and aspiration 144, 96, 72 or
48 hours after the PGF2 á. There was no significant difference in % of oocytes
recovery and embryos development with the alternation of the biotechnologies
OPU and SOV. However, significant difference (p>0,05) in the number of oocytes
recovery between the treatments was observed. The recovery in T(96-48) was
significantly lesser when compared with T(48-96) and T(72-96), demonstrating that
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xv
the moment of the aspiration very next to the PGF2á (48 hours after) harmed the
oocytes recovery. The results of the present study concluded that the
superovulation not affect the oocytes recovery and the embryo development when
the OPU is carried 48 hours after the injection of PGF2á in the subsequent wave
after the superovulation and may to be used for the increment of embryos yield of
Nelore donors.
key words: superovulation, follicular aspiration, oocyte competence, follicular
dynamic, Nelore breed.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xvi
I. INTRODUÇÃO
No atual contexto de evolução da produtividade na pecuária nacional,
associado às evoluções científicas e tecnológicas, várias biotecnologias ligadas a
reprodução animal vem sendo desenvolvidas e aprimoradas com o intuito de
aumentar a eficiência reprodutiva, maximizando a produção de animais
geneticamente superiores, visando o aproveitamento deste material genético para
obtenção do maior número de descendentes, em um curto período de tempo.
Seguindo a evolução das principais biotecnologias adotadas e trabalhadas
no Brasil, é importante ressaltar, inicialmente, o papel da Inseminação Artificial
(IA), sendo a primeira biotecnologia adotada nos sistemas de produção brasileiros
que visa a multiplicação genética de touros de alto valor. Com a introdução de
esquemas de ovulações múltiplas, recuperação e transferência de embriões, mais
conhecida como Multiple Ovulation and Embryo Transfer (MOET), junto com a
criopreservação de embriões na década de 80, a bovinocultura passou a ter em
mãos ferramentas para aumentar o número de gestações provenientes de fêmeas
de alto mérito genético (RODRIGUES, 2001). A produção embrionária através da
TE é uma biotécnica mundialmente difundida e vem apresentando um crescimento
acentuado onde mais de 500.000 embriões são colhidos e transferidos ou
congelados anualmente (THIBIER, 2000).
A Fertilização In Vitro (FIV), por sua vez, é considerada a terceira geração
de biotecnologia aplicada ao Melhoramento Genético, após a IA e a TE. No início
da década de 90, com a introdução da aspiração folicular guiada por ultra-
sonografia (Ovum Pick-up) seguida pela produção in vitro de embriões (OPU-PIV),
a expectativa no incremento da produtividade das fêmeas aumentou.
Na bovinocultura brasileira, essas tecnologias também vêm sendo
amplamente utilizadas e adaptadas ao nosso sistema de produção e, ao longo da
sua evolução, tem revelado bons índices reprodutivos, confirmando ser um
processo economicamente viável, bastante divulgado e empregado na pecuária
moderna.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xvii
No entanto, segundo BO et al., (2000) alguns inconvenientes podem ser
observados nos programas de superovulação como a necessidade de iniciar o
tratamento hormonal em momento determinado do ciclo estral e pouca
consistência na produção de embriões viáveis pelas doadoras, sobretudo quando
se considera que 20% a 30% das doadoras não produzem nenhum embrião
transferível. A variabilidade na produção de embrião pode ser influenciada por
fatores relacionados com o tratamento superovulatório ou em maior grau por
fatores individuais associados às características da dinâmica folicular ovariana
(BO et al., 1995; BO et al., 2000) ou condição ovariana no momento da
superovulação (MONNIAUX et al., 1983; DIAZ, 2001).Sem contar que repetidos
tratamentos superovulatórios em novilhas e vacas afetam a fertilidade podendo
causar cistos e dificuldade para estabelecimento de prenhez posterior (GALLI et
al., 2003).
A FIV pode contribuir como uma alternativa para a superovulação e maior
produção de embriões por unidade de tempo quando comparada a TE. No
entanto, mesmo com os progressos obtidos nos processos de maturação in vitro
(MIV), fecundação in vitro (FIV) e cultivo in vitro (CIV) para o desenvolvimento
embrionário, esta técnica ainda apresenta algumas desvantagens que precisam
ser melhoradas e solucionadas. As taxas de blastocistos bovinos oriundos de
oócitos maturados e fertilizados in vitro têm permanecido relativamente estáticas
na última década, onde somente em torno de um terço (LONERGAN, et al., 1994;
THOMPSON; DUGANIZICH, 1996; DAYAN, et al., 2000) dos oócitos selecionados
morfologicamente antes de serem submetidos a MIV resultam em embriões
viáveis.
Tendo em vista que as biotecnologias MOET e OPU-PIV são de extrema
importância para aumentar a velocidade de ganho genético do rebanho nacional, o
presente trabalho visando a otimização pelo uso alternado destas biotecnologias
testou:
- o efeito do início da superovulação (SOV) 48 a 60 horas após a aspiração
folicular em fase aleatória do ciclo estral – aspiração referência (OPU 1);
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xviii
- a produção in vitro de embriões oriundos de oócitos aspirados (OPU 2)
em diferentes momentos da primeira onda folicular após a SOV.
II. REVISÃO DA LITERATURA
O ciclo estral (CE) bovino é compreendido pelos eventos reprodutivos que
se apresentam entre dois períodos de receptividade sexual (cio). As fêmeas da
espécie bovina ao atingirem a puberdade exibem comportamento estral em média
a cada 21 dias, variando de 17 a 24 dias tanto em raças européias (ROBINSON;
SHELTON, 1991; WRIGHT; MALMO, 1992; BO; ADAMS; MAPLETOFT; 2000)
quanto zebuínas (GALINA; ARTUR, 1990; PINHEIRO, et al., 1998; FIGUEIREDO,
et al., 1997; GAMBINI, et al., 1999) até que a prenhez se estabeleça.
Durante o ciclo estral ocorrem importantes alterações no córtex ovariano
que incluem crescimento e atresia de vários folículos antrais até o aparecimento
do folículo ovulatório, bem como a formação e lise do corpo lúteo (CASTILHO,
1999). O crescimento folicular na espécie bovina exibe padrão contínuo de
crescimento e atresia dos folículos ovarianos (MATTON, et al., 1981; WOOLUMS;
PETTER, 1994) que se inicia na vida fetal, passa pela puberdade (EVANS, et al.,
1997) e continua na vida reprodutiva até a seniIidade (HAFEZ, 1993). Muitos
estudos com ultra-sonografias seriadas foram realizados e demonstraram que
durante o ciclo estral de novilhas ou vacas zebuínas e européias ocorrem duas
(PIERSON; SIROIS; SAVIO, 1988; GINTHER; KNOPF, 1989; FIGUEIREDO, et al.,
1997) ou três (SAVIO, et al., 1988; GAMBINI, et al., 1998; BARROS, et al., 1993;
FIGUEIREDO, et al., 1997; CASTILHO, et al., 2000) ondas de crescimento
folicular.
Este processo contínuo de crescimento e regressão dos folículos, que leva
ao crescimento do folículo pré-ovulatório, é conhecido como dinâmica folicular,
enquanto que o padrão de crescimento e atresia de um grupo de folículos
ovarianos é denominado onda de crescimento folicular (LUCY, et al., 1992). Cada
onda folicular é composta por 3 fases: recrutamento ou emergência, seleção e
dominância (SIROIS, 1990).
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xix
A emergência da primeira onda folicular é observada em torno de um dia e
meio após a ovulação (ADAMS, et al., 1992; GINTHER, et al., 1997) quando um
conjunto de folículos antrais dependentes de FSH começa a se desenvolver. Foi
observado um aumento na concentração plasmática de FSH que antecede 1 a 2
dias a emergência de cada onda folicular (ADAMS, et al., 1992; GIBONS;
GINTHER, 1999). Em torno de 3 a 4 dias após a emergência da onda o FSH reduz
para níveis basais e o futuro folículo dominante é selecionado, continua seu
crescimento, enquanto o restante dos folículos detém seu crescimento ou
regridem e são chamados de subordinados (BODENSTEINER, et al. 1996;
GINTHER, et al., 1997; GINTHER, et al., 2000).
A fase de seleção do folículo dominante durante a primeira onda folicular foiobservada por GINTHER et al. (1996), como a divergência na taxa de crescimento entre ofuturo folículo dominante (FD) e o maior folículo subordinado (FS) 2,8 dias após aemergência da onda quando o futuro folículo dominante apresentou 8,5 mm de diâmetro e omaior subordinado 7,2 mm. CASTILHO; RENESTO; GARCIA (2003), estudando aseleção folicular em novilhas da raça Nelore observaram que em média 84 horas após aovulação o maior folículo subordinado medindo 5,5 mm cessa seu crescimento, enquanto ofuturo dominante com 7,5 mm continua a se desenvolver. Nas novilhas da raça Nelore, 24horas antes da divergência no crescimento folicular foi observada a menor concentraçãoplasmática de FSH.
Existe uma relação funcional entre a concentração de estradiol e FSH em novilhas.O aumento ou decréscimo na concentração de estradiol no início da divergência resulta emdecréscimo ou aumento, respectivamente, na concentração de FSH (KULICK, et al., 1999;GINTHER, et al., 2000b).
Provavelmente o responsável pela queda do FSH antes da seleção é a produção deestradiol e inibina, sobretudo pelas células da granulosa do folículo dominante, que atuaminibindo a liberação de FSH pela hipófise anterior (FINDLAY; CLARKE, 1987). Asconcentrações de FSH são mantidas em níveis basais até o folículo dominante da primeiraonda perder sua dominância, resultando em aumento nos níveis de FSH e subseqüenteemergência da segunda onda folicular (BODENSTEINER, et. al., 1996).
Alguns trabalhos têm demonstrado que qualquer folículo saudável ou emcrescimento é capaz de se tornar dominante. O aumento da concentração sérica de FSH,decorrente da destruição do folículo dominante após a divergência, resulta na dominânciado maior folículo subordinado (KO, et al., 1991, GINTHER, et al., 2000). Da mesmaforma, um folículo aleatoriamente escolhido dentre um grupo de folículos de 5 mm, noinício da onda folicular, pode ser direcionado para a dominância após a aspiração de todosos outros da onda (GIBBONS; GINTHER, 1997).
Tratamentos com FSH no início do desenvolvimento folicular estimulam muitosfolículos a obterem diâmetro de dominância (ADAMS, et al., 1994), sendo à base dosprotocolos de superovulação com FSH para colheita de embriões.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xx
A superovulação é uma eficiente técnica para se obter várias progênies de
fêmeas geneticamente superiores e pode ser realizada mediante aplicação
seriada de FSH no início da 2a onda folicular, ou seja, entre os dias 9 a 12 do ciclo
estral (GOULDING, et al., 1990). A resposta ovariana a superovulação depende
do número de folículos presentes no ovário sensíveis a gonadotrofina e do estágio
da onda folicular no momento em que a aplicação de FSH é iniciada
(DRIANCOUT, 2001). No entanto, um dos principais inconvenientes observados
nos programas de superovulação é a necessidade de iniciar o tratamento
hormonal em momento determinado do ciclo estral (BO, et al., 2000).
Outro inconveniente da SOV é a baixa consistência na produção de
embriões viáveis pelas doadoras, um terço das doadoras tratadas não respondem
a SOV, por outro lado um terço produz em média 3 embriões e somente o terço
restante resulta em grande número de embriões transferidos (GALLI, et al., 2003).
Esta variação na resposta, têm-se transformado em fator limitante para o uso em
larga escala da tecnologia (ARMSTRONG, 1993; MAPLETOFT; PIERSON, 1993,
BO, et al., 1995). Dentre os fatores que podem ser responsabilizados por tais
variações, podemos citar a influencia do tipo, o grau de pureza, a dose e o
esquema de administração do hormônio (BECKER; PINHEIRO, 1986;
MAPLETOFT; PIERSON, 1994), estado nutricional da doadora (YAAKUB, et al.,
1999; SIDDIQUI, et al., 2002), histórico reprodutivo, idade, estação do ano, efeito
de repetidas superovulações (MAPLETOFT; PIERSON, 1994), raça, Bos indicus
quando comparada a raças européias (BARROS; NOGUEIRA, 2001), além da
condição folicular no início do tratamento superovulatório (MONNIAUX, et al.,
1983; GRASSO, et al., 1989; PIERSON; GINTHER, 1988; BUNGARTZ;
NIEMMAN, 1994, BO, et al., 1995) que pode estar diretamente relacionada a fase
ideal para se iniciar a estimulação hormonal exógena.
A estimulação hormonal com FSH pode ser iniciada antes da emergência
da onda folicular ou no dia da emergência (GOULDING, et al., 1990; NASSER, et
al., 1993; ADAMS; ROBERTS, et al., 1994; BO, et al., 1995; STOCK, et al., 1996),
ou seja, antes dos folículos subordinados iniciarem seu processo de atresia (BO,
et al., 1995).
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxi
Segundo PIERSON et al. (1988), GRASSO et al. (1989), GUIBAULT et al.
(1991) e ADAMS et al. (1993), o início de tratamentos com gonadotrofinas na
presença de um folículo dominante ou após a seleção do folículo dominante
resulta em uma redução da resposta superovulatória.
BERGFELT et al. (1994) e MERTON et al. (2002), iniciaram a
superovulação 48 e 38 a 46 horas, respectivamente, após a aspiração e obtiveram
melhores resultados.
Em vista dessas informações, uma alternativa para se obter uma melhor
resposta superovulatória é controlar a dinâmica folicular (BO, et al., 2000), ou a
emergência da nova onda folicular em programas de transferência de embriões. A
sincronização da emergência da onda folicular tem como objetivo remover o efeito
supressivo do folículo dominante e iniciar uma nova onda (BO, et al.,1995; 2000),
situação que ocorre normalmente em animais entre os dias 8º e 12° após a
detecção do estro.
Existem diferentes métodos pelos quais se pode controlar a dinâmica
folicular em bovinos. Estudos demonstraram que a eliminação do folículo
dominante, por métodos físicos ou hormonais, resultam em aumento de FSH e
desta forma emergência de uma nova onda folicular num período de tempo
conhecido (ADAMS, et al., 1992; BO; GHINTHER, et al., 1996; MIHM, et al.,
1997). A possibilidade de utilização de estradiol (BO, et al., 1993,1994;
CARRIERE, et al., 1995) ou estradiol associado a progestágenos (BO, et al., 1991;
1996) para sincronizar a onda folicular, vem sendo muito utilizado em protocolos
de superovulação em bovinos (BO, et al., 1995b, 1996; BROADBENT, et al., 1995;
ANDRADE, et al., 2002a,b,c; OLIVEIRA, et al., 2002). Quando a progesterona é
usada associada ao estrógeno, a emergência da nova onda folicular ocorre ao
redor de 4 ou 5 dias após o tratamento ( BO, et al., 1995, 1996; CACCIA; BO,
1998; MAPLETOFT, et al., 1999). BO et al. (2000), demonstraram que o início do
tratamento com gonadotrofinas 4 dias após o tratamento com progesterona e
estradiol resulta em resposta superovulatória semelhante a iniciada na emergência
da segunda onda folicular ou entre os dias 8º e 12º do ciclo estral.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxii
O mesmo foi observado em vacas da raça Nelore, onde trabalhos utilizando
métodos hormonais para sincronizar a onda folicular para início da SOV
demonstraram que não há comprometimento da morfologia do embrião ou taxa de
gestação, quando comparado ao protocolo padrão e elimina a necessidade de
observar o cio, permitindo a superovulação de grande número de doadoras em
curto período de tempo (AZEVEDO; COELHO,1991; ANDRADE; OLIVEIRA, et al.,
2002; ANDRADE, et al., 2003).
Além dos métodos hormonais empregados para promover a sincronização
da emergência da onda folicular em vacas, o método físico através da aspiração
folicular guiada por ultra-sonografia transvaginal vem sendo utilizado com
sucesso.
BERGFELT et al. (1994), realizaram aspiração folicular de todos os folículos
maiores ou iguais a 5 mm de diâmetro, em estágio aleatório do ciclo estral, e
constataram aumento no nível sérico de FSH em 24 horas e emergência da nova
onda folicular em torno de 48 horas após a aspiração folicular. O mesmo foi
constatado por trabalhos com aspiração somente do folículo dominante, tanto em
novilhas Bos taurus (AMARIDIS, et al., 1999; ADAMS, et al., 1994; BERGFELT,
1994) quanto Bos indicus (BURATINE, et al., 2000).
SHAW et al. (1999) e MERTON (2003), estudando procedimentos de
superovulação com aspiração prévia do folículo dominante observaram aumento
no índice de embriões viáveis de 3,9 a 5,9. GRADELA et al. (2000), avaliando em
vacas Nelore a resposta da SOV iniciada na presença ou ausência do FD e após a
aspiração do FD observaram que a resposta superovulatória das doadoras não
diferiu entre os grupos, mas a taxa de viabilidade embrionária foi maior nos grupos
sem FD (69,4%) e FD aspirado (68,99%) quando comparados ao grupo com FD
presente (48,54%).
No início da década de 90, com a introdução da Ovum Pick-up, seguida
pela produção in vitro de embriões (OPU-PIV), a expectativa no incremento da
produtividade das fêmeas aumentou. A aplicação da OPU-FIV é de grande
importância para a multiplicação rápida de animais, e exibe algumas vantagens
quando comparada com a MOET. NIBART et al. (1995), demonstraram que se
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxiii
pode obter até 18 gestações em três meses utilizando aspiração folicular,
enquanto com a TE clássica, no mesmo período, seria possível obter apenas 5
gestações. Desta forma pela OPU-FIV é possível produzir 2 a 3 vezes mais
embriões por unidade de tempo quando comparado à TE convencional. Outras
vantagens da OPU-PIV são a obtenção de oócitos em qualquer fase do ciclo estral
sem tratamento gonadotrófico (MERTON, et al., 2003), possibilitando realizar
repetida recuperação de oócitos de animais vivos sem trauma aparente do trato
reprodutivo, permitindo, desta forma, a produção de embriões oriundos de vacas
senis (BROGLIATTI; ADAMS, 1996), com patologias reprodutivas adquiridas
(LOONEY, et al., 1994; HASLER, et al., 1995; RODRIGUES; GARCIA, 2000),
prenhes, durante o primeiro trimestre de gestação (BUNGARTZ, et al., 1995;
MEINTJES, et al., 1995) ou em novilhas pré-púberes (ARMSTRONG, et al.;
ADAMS, 1994; BROGLIATTI, 1996). Outra vantagem particular da FIV em relação
a outras biotecnologias é a maximização do uso do sêmen, permitindo maior
produção de embriões com doses de alto valor comercial e inclusive sêmen
sexado (FABER, et al., 2003).
Entretanto, as taxas de blastocistos bovinos oriundos de oócitos maturados
e fertilizados in vitro têm permanecido relativamente estáticas na última década,
somente em torno de um terço (LONERGAN, et al., 1994, THOMPSON;
DUGANIZICH, 1995), 15 a 20% (HENDRIKSEN, et al., 2000) ou 35% (DAYAN,
2001) dos oócitos selecionados morfologicamente antes de serem submetidos a
MIV resultam em embriões viáveis. Segundo PEIXER et al. (1995), além dos
baixos índices de blastocistos, a produção in vitro de embriões ainda apresenta
algumas limitações: qualidade biológica dos embriões, dificuldades na
criopreservação dos embriões e dos oócitos, menor viabilidade dos oócitos obtidos
de bezerras em relação aos de vacas e novilhas, e o custo, que é superior ao
embrião produzido pela transferência de embriões (TE).
Buscando obter maior número de embriões em curto espaço de tempo, os
protocolos têm geralmente consistido de uma ou duas aspirações semanais
usando animais superovulados ou não (LOONEY, et al., 1994; BOLS;
BUNGARTZ, et al., 1995; HASLER, et al., 1995; STUBBINGS, 1995). Embora a
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxiv
aspiração realizada 2 vezes por semana aumente o número de folículos, oócitos
recuperados e embriões produzidos (GIBBONS, et al., 1994), a alta freqüência e o
curto intervalo entre as aspirações afeta negativamente o número de oócitos
(HASLER, et al., 1995; BOUSQUET, et al., 1999) a longo prazo.
Têm sido reportadas taxas de recuperação de oócitos em torno de 2 a 3 por
doadora, por sessão de aspiração (PIETERSE, et al., 1988), 4,7 (BOUSQUET, et
al., 1999), 9,7 (DAYAN, et al., 2000) e acima de 11 oócitos (BORDIGNON, et al.,
1997) .
Existe também uma ampla variação na produção de oócitos e embriões,
KRUIP et al. (1994), relataram variação entre as doadoras de 9 a 26% e HASLER
et al. (1995), variação entre 4% a 33%. DAYAN et al. (2000a), observaram
variação entre 14 e 54% na produção de embriões em 20 doadoras submetidas a
OPU-FIV.
Essa variação pode estar diretamente relacionada ao fato de que o oócito
para ser fertilizado in vivo é obtido através da ovulação de um folículo saudável,
durante uma fase específica do ciclo estral e os oócitos aspirados para a produção
in vitro são obtidos de folículos em diferentes estágios do desenvolvimento
(LONERGAN, et al., 1994; HENDRIKSEN, et al., 2000), em fases distintas do ciclo
estral (MACHATKOVÁ, et al., 1996), portanto expostos a diferentes concentrações
de FSH, podendo levar a uma diminuição na competência oocitária
(MACHATKOVA, et al., 2004).
Vários estudos têm demonstrado que a fase do ciclo estral e inclusive do
desenvolvimento folicular influenciam a competência oocitária, uma vez que o
diâmetro do folículo afeta diretamente o oócito (MACHATKOVA, et al., 2004). Em
alguns estudos, oócitos provenientes de folículos maiores se desenvolvem melhor
in vitro, onde se observou aumento na produção de embriões de oócitos aspirados
de folículos médios e grandes quando comparado a folículos pequenos (PAVLOK,
et al., 1992; LOONERGAN, et al., 1994; ARLOTTO, et al., 1996; HAGEMANN, et
al., 1999b). Provavelmente devido a fase de dominância do ciclo estral onde o
folículo dominante afeta negativamente a competência oocitária dos folículos
subordinados induzindo avanço no estádio de atresia (HENDRIKSEN, et al.,
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxv
2000). No entanto, CAROLAN et al. (1996), SENEDA et al. (2001), não
observaram influência do diâmetro folicular sobre a competência do oócito,
analisando oócitos aspirados de folículos pequenos e grandes.
MACHATKOVA et al. (1996), comparando a aspiração em diferentes
estádios do ciclo estral, demonstraram que oócitos colhidos nos dias 14 a 16 do
ciclo apresentavam melhores índices de competência para o desenvolvimento até
balstocisto quando comparados aos aspirados nos dias 7, 8 e 9. Em outro estudo,
oócitos colhidos de folículos subordinados nos dias 2, 3 e principalmente no dia 7
da onda, demonstram menor competência que aqueles recuperados no dia 5
(VASSENA, 2003). HAGEMAN (1999), MACHATKOVA et al. (2000), concluem
que o desenvolvimento até balstocisto é significativamente maior em oócitos
colhidos durante a fase de crescimento folicular comparado à fase de dominância
independente do diâmetro do folículo, porém a competência oocitária tendeu a
aumentar em oócitos oriundos de folículos maiores.
CASTILHO; GARCIA (2003), estudando a fase da 1ª onda e o diâmetro
folicular sobre a competência oocitária em Nelore observaram que não houve
efeito do diâmetro folicular sobre a produção de balstocistos, mas houve interação
do diâmetro com a fase folicular e concluíram que a competência é melhor nos
folículos pequenos no início da onda e melhora nos folículos maiores com o
avanço da onda. O mesmo foi observado por MACHATKOVA et al. (2004) em Bos
taurus.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxvi
III. MATERIAL E MÉTODOS
Local do Experimento
Os experimentos para colheita de material (oócitos e embriões) foram
desenvolvidos na Agropecuária J. Garcia localizada no município de Regente Feijó
– SP e Fazenda Experimental da Unoeste localizada no município de Presidente
Bernardes – SP. Posteriormente os oócitos foram enviados ao Laboratório de
Biotecnologia do Departamento de Medicina Veterinária Preventiva e Reprodução
Animal da Faculdade de Ciências Agrárias e Veterinária (FCAV) da Universidade
Estadual Paulista (UNESP), Câmpus de Jaboticabal – SP, para produção in vitro
(PIV) dos embriões.
Animais
Foram utilizadas 23 fêmeas adultas da raça Nelore (Bos taurus indicus), com idadeentre quatro a nove anos, pesando em média 500 Kg. As fêmeas, em perfeitas condiçõessanitárias e reprodutivas foram mantidas em pasto de Brachiaria decumbens, com acesso aágua e sal mineral Ad libitum.
Tratamentos
As fêmeas foram submetidas a uma aspiração folicular – aspiração folicularreferência (OPU 1) em fase aleatória do ciclo estral, seguida pela inserção de implante deliberação lenta de progestágeno (Crestar® Intervet International B.V-Boxmeer-Holanda).Em torno de 48 a 60 horas após a aspiração iniciou-se o tratamento de supererovulaçãoovariana (SOV) utilizando-se FSH (Folltropin® Vetrepharm Inc. Ontário, Canadá) na dosetotal de 180 mg por vaca, com duração de quatro dias consecutivos, em oito aplicações(quadro 1). Durante a sexta e sétima aplicação de FSH foi administrada por via IM 500 µgde PGF2α (Preloban® Intervet International B.V-Boxmeer-Holanda) e o implante deprogestágeno foi retirado durante a administração da sétima dose de FSH.
Quadro 1. Esquema de superovulação com FSH:
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxvii
Horário Dia 1(FSH)
Dia 2(FSH)
Dia 3
(FSH/ PGF2α)
Dias4
(FSH/ PGF2α)
7:00 h 40mg 20mg 20mg 10mg + 500 µg19:00 h 40mg 20mg 20mg + 500 µg 10mg
Aproximadamente 12 horas após o término da SOV, as fêmeas receberam 25mg deLH exógeno (Lutropin-V® Vetrepharm, Beleville, Ontário, Canadá) para causar ovulação erealizar a IA com tempo fixo. Foram realizadas 2 inseminações com intervalo de 12 horas,sendo que a 1ª inseminação ocorreu 12 horas após a administração de LH, utilizando sêmencongelado da raça nelore em palhetas de 0,5 mL, descongelado de acordo com o critérioindicado pelas centrais (36°C por 30’).
No dia da colheita dos embriões as fêmeas foram divididas em seis tratamentos quevariaram de acordo com o momento da aplicação de PGF2α após a colheita de embrião(Dia 0) e a aspiração folicular (OPU2) realizada após a SOV (figura 1). Os tratamentosrealizados foram:
T (0-144) – aplicação de PGF2á imediatamente após a colheita de embriões easpiração 144 horas após a administração de PGF2á;
T (48-120) – aplicação de PGF2á 48 horas após a colheita de embriões e aspiração120 horas após a administração de PGF2á;
T (48-96) – aplicação de PGF2á 48 horas após a colheita de embriões e aspiração96 horas após a administração de PGF2á;
T (72-96) – aplicação de PGF2á 72 horas após a colheita de embriões e aspiração96 horas após a administração de PGF2á;
T (96-72) – aplicação de PGF2á 96 horas após a colheita de embriões e aspiração72 horas após a administração de PGF2á;
T (96-48) – aplicação de PGF2á 96 horas após a colheita de embriões e aspiração48 horas após a administração de PGF2á
A PGF2α foi administrada em diferentes momentos para avaliar se haveria efeito dafase da primeira onda folicular na segunda aspiração folicular (OPU 2) e avaliar se estasegunda aspiração seria afetada pela prévia superovulação.
Tratamento – T(0-144) (n=4)
144 horasDia 0 OPU 2PGF2á
Tratamento – T(48-120) (n=5)
48 horas 120 horas
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxviii
Dia 0 PGF2á OPU 2
Tratamento – T(48-96) (n=4)
48 horas 96 horasDia 0 PGF2á OPU 2
Tratamento – T(72-96) (n=3)
72 horas 96 horasDia 0 PGF2á OPU 2
Tratamento – T(96-72) (n=4)
96 horas 72 horasDia 0 PGF2á OPU 2
Tratamento – T(96-48) (n=3)
96 horas 48 horasDia 0 PGF2á OPU 2
Figura 1. Representação esquemática dos tratamentos realizados após a colheita deembriões em 23 fêmeas nelore.D 0 – dia da colheita dos embriões/ n – número de animais
Aspiração Folicular
O procedimento de aspiração folicular foi realizado utilizando-se equipamento deultra-som Aloka SSD-500 com transdutor microconvexo de 5 mHz (UST 974-5) conectadoa guia de biópsia adaptado por Chuck Bolland, com agulhas 18 G, (Cook VBOAS 1855) elinha de aspiração (Cook VBOA 18L) em tubos de centrífuga de 50 mL. A pressão devácuo foi obtida com uma bomba Cook V-MAR 5000, ajustada entre 72 e 78 mmHg.
Para evitar movimentos peristálticos e desconforto ao animal foi feita anestesiaepidural com 5 mL de Lidocaína a 2% (Pearson®) e em seguida o transdutor foi inseridoaté o fundo vaginal e, com o auxílio da manipulação transretal, os ovários foramposicionados para obtenção de uma boa visualização na tela do ultra-som. Os folículos aserem aspirados foram posicionados no percurso da linha de punção indicada na tela do
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxix
ultra-som e quando a agulha se aproximou do folículo a ser aspirado, o pedal da bomba devácuo foi pressionado e o folículo aspirado (NIBART et al., 1995), o mesmo procedimentofoi repetido em todos os folículos visíveis de cada ovário. A lavagem da agulha e o meio derecebimento dos oócitos foi composto de DPBS (Dulbeco Modificado - Nutricell) acrescidode 5,0 UI/mL de heparina sódica (Liquemine ) e 50 mg/mL de Gentamicina (Gentocin ).
Lavagem, seleção dos oócitos
O material aspirado foi transferido para o filtro de colheita de embriões
(EmCom®) e lavado com a mesma solução utilizada na aspiração. O sedimento
restante no filtro foi observado em placas de Petri e efetuado a busca e contagem
dos oócitos e posterior classificação da qualidade. Os oócitos foram classificados
de acordo com sua morfologia (número de camadas de células do cumullus e
aspecto do citoplasma) em graus I, II e III (GI, GII e GIII), oócitos sem cumullus
(s/c), expandidos (exp), degenerados (deg) e atrésicos (atr), segundo LONERGAN
(1992). Os oócitos considerados viáveis foram classificados como GI, GII e GIII,
lavados em solução TCM 199 Hepes (Gibco) suplementado com 10% SFB
(Gibco), 50 µg de gentamicina, 2,2 µg de piruvato e transportados em criotubos
(Corning®) contendo meio de maturação em banho Maria a 35°C.
Transporte dos oócitos
Os oócitos foram transportados em meio de maturação composto por meio
TCM 199 Bicarbonato, suplementado com 10% SFB, 50UI de hCG/mL, 0,5 µg/mL
de FSH, 1µg/mL de estradiol, 2,2µg/mL de piruvato, 70 µg/mL de amicacina em
atmosfera de 5% de CO2 e 5% de O2. O tempo médio de transporte variou de 6 a
8 horas, não ultrapassando 8 horas do início da aspiração na primeira vaca até a
chegada dos oócitos ao laboratório.
Maturação in vitro (MIV)
Chegando no laboratório os oócitos foram lavados três vezes em meio de lavagemTCM-199 Hepes, suplementado com 10% de SFB e 70 µg de amicacina. Em seguida,foram transferidos para placas de Petri, em microgotas de 100 µL de meio de maturação,
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxx
que consistiu de meio TCM 199 Bicarbonato, suplementado com 10% SFB, 50UI dehCG/mL, 0,5 µg/mL de FSH, 1µg/mL de estradiol, 2,2µg/mL de piruvato, 70 µg/mL deamicacina. Os oócitos permaneceram incubados por 24 horas a 38,5°C em atmosfera de 5%de CO2 em ar.
Fecundação in vitro (FIV)
Os oócitos maturados foram lavados três vezes em meio de fecundação
TALP-FIV e transferidos para microgotas de 100µL de meio de fecundação Tyrode
modificado (TALP) suplementado com 10 µg/mL de heparina e 160 µL da solução
PHE. O sêmen congelado foi separado em Gradiente de Percoll 90 e 45%,
submetido a uma força de centrifugação de 900g durante 30 minutos. O sedimento
foi recuperado e avaliado quanto ao volume, concentração e motilidade
espermática. A concentração final foi ajustada para 25 x 106 espermatozóides
vivos por mL, de modo que, ao adicionar 4µL do sêmen em cada microgota de
100µL de meio TALP-FIV-Gotas, obteve-se a concentração final de 100x103
espermatozóides vivos por gota. Posteriormente, foram incubados em temperatura
de 39°C por 18 a 20 horas, com atmosfera de 5% de CO2 em ar, para a
fecundação.
Cultivo in vitro (CIV)
Após o tempo de fecundação, os zigotos foram lavados por três vezes em meio SOFe as estruturas transferidas para microgotas com 100 µL de meio de cultivo, recobertas poróleo mineral, permanecendo nestas por um período de 6 a 8 dias até os zigotos atingirem osestádios de mórula (Mo) e blastocisto (Bl). O meio de cultivo foi renovado em cadamicrogota no terceiro e quinto dia (feeding) e no 6º e 7º dia foi observado odesenvolvimento embrionário.
Colheita de embriões
Sete dias após a primeira IA, os embriões foram colhidos pelo método não cirúrgicocomo descrito por NEWCOMB et al. (1978), utilizando-se Solução Salina FosfatadaTamponada (DPBS – Dulbecco Modificado-Cutilab), contendo 1% de Soro Fetal Bovino
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxi
(SFB), ou seja, 10 mL de SFB em 1 litro de DPBS, ambos a 37°C e recuperados em filtropróprio (EmCom ).
Manipulação e Avaliação Morfológica dos Embriões TE
Os embriões foram rastreados e transferidos para Placa de Petri contendo DPBSenriquecido com 10% de SFB. Posteriormente foi avaliado o estádio de desenvolvimentodo embrião (mórula, blastocisto, blastocisto em expansão e balstocisto eclodido)(LINDNER & WRIGHT-JR, 1983). Quanto ao formato e integridade da zona pelúcida e amorfologia foram classificados em normais, degenerados, não fertilizados e sem zonapelúcida (BOLAND et al., 1978).
Análise Estatística
Os dados referentes à aspiração folicular realizada após a superovulação
(OPU 2) foram avaliados pelo teste do Qui-Quadrado. Para verificar se houve
diferença com relação aos resultados da OPU 2 quando comparamos com os
resultados da OPU 1 (referência), utilizou-se um delineamento inteiramente
casualizado
utilizando o procedimento GLM (General Linear Model) no software
Statystical Analysis System (SAS), seguido pelo teste de Duncan para estabelecer
a comparação entre médias. Neste caso, as variáveis analisadas (percentual de
oócitos viáveis e a taxa de blastocistos) foram obtidas a partir das diferenças com
relação aos resultados da OPU 2 quando comparados com os resultados da OPU
1 (referência).
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com
xxxii
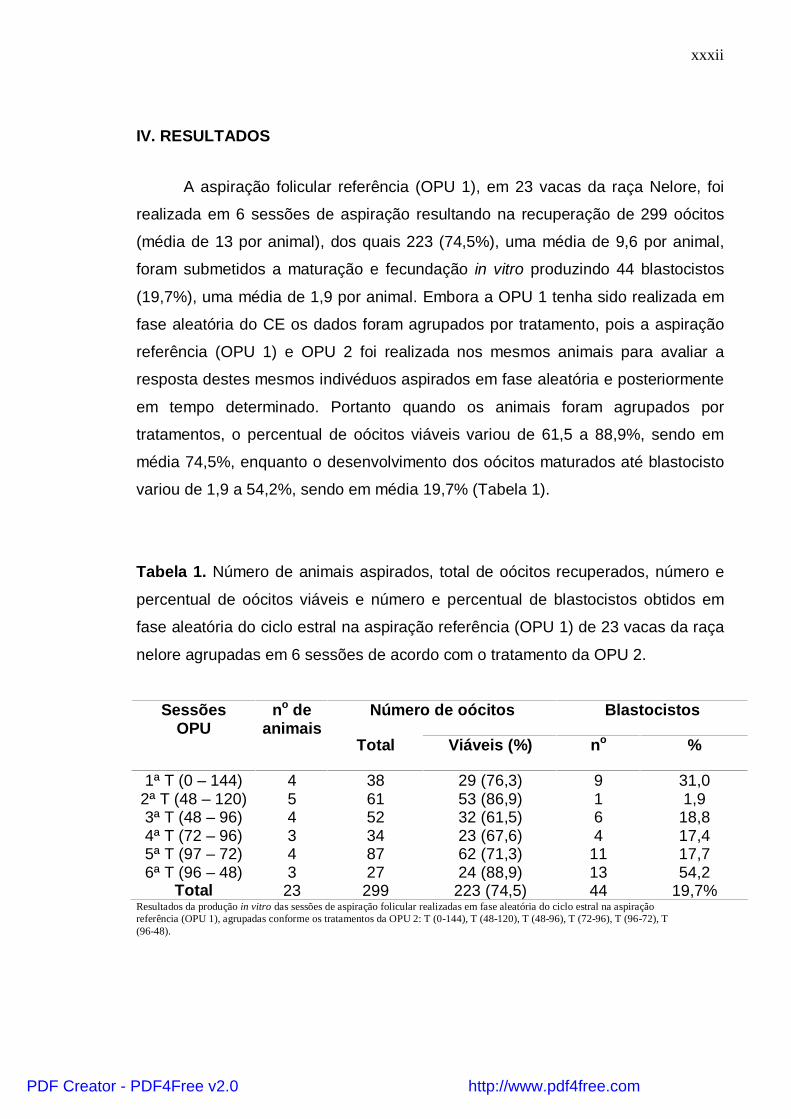
IV. RESULTADOS
A aspiração folicular referência (OPU 1), em 23 vacas da raça Nelore, foi
realizada em 6 sessões de aspiração resultando na recuperação de 299 oócitos
(média de 13 por animal), dos quais 223 (74,5%), uma média de 9,6 por animal,
foram submetidos a maturação e fecundação in vitro produzindo 44 blastocistos
(19,7%), uma média de 1,9 por animal. Embora a OPU 1 tenha sido realizada em
fase aleatória do CE os dados foram agrupados por tratamento, pois a aspiração
referência (OPU 1) e OPU 2 foi realizada nos mesmos animais para avaliar a
resposta destes mesmos indivéduos aspirados em fase aleatória e posteriormente
em tempo determinado. Portanto quando os animais foram agrupados por
tratamentos, o percentual de oócitos viáveis variou de 61,5 a 88,9%, sendo em
média 74,5%, enquanto o desenvolvimento dos oócitos maturados até blastocisto
variou de 1,9 a 54,2%, sendo em média 19,7% (Tabela 1).
Tabela 1. Número de animais aspirados, total de oócitos recuperados, número e
percentual de oócitos viáveis e número e percentual de blastocistos obtidos em
fase aleatória do ciclo estral na aspiração referência (OPU 1) de 23 vacas da raça
nelore agrupadas em 6 sessões de acordo com o tratamento da OPU 2.
Número de oócitos BlastocistosSessõesOPU
no deanimais
Total Viáveis (%) no %
1ª T (0 – 144) 4 38 29 (76,3) 9 31,02ª T (48 – 120) 5 61 53 (86,9) 1 1,93ª T (48 – 96) 4 52 32 (61,5) 6 18,84ª T (72 – 96) 3 34 23 (67,6) 4 17,45ª T (97 – 72) 4 87 62 (71,3) 11 17,76ª T (96 – 48) 3 27 24 (88,9) 13 54,2
Total 23 299 223 (74,5) 44 19,7%Resultados da produção in vitro das sessões de aspiração folicular realizadas em fase aleatória do ciclo estral na aspiraçãoreferência (OPU 1), agrupadas conforme os tratamentos da OPU 2: T (0-144), T (48-120), T (48-96), T (72-96), T (96-72), T(96-48).
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxiii
Após a aspiração folicular referência (OPU 1) as mesmas fêmeas foram
superovuladas para colheita in vivo de embriões e aspiradas novamente (OPU 2)
em diferentes momentos da 1º onda folicular após a superovulação (Tabela 2). Do
total de 252 oócitos recuperados (média de 10,9 por animal), 73,8% ou 186 (média
de 8,1 por animal) foram classificados morfologicamente como viáveis e
submetidos à maturação e fecundação in vitro, resultando em 17,2% ou 32
blastocistos (média de 1,4 por animal).
O número de oócitos recuperados e produção de blastocistos (total e
percentual) foram afetados (P<0,05) pelo momento da aplicação de PGF2α e a
segunda aspiração folicular (OPU 2) e podem ser observados na tabela 2.
Tabela 2. Número de animais, total de oócitos aspirados, número e percentual de
oócitos viáveis e número e percentual de blastocistos obtidos de vacas da raça
nelore aspiradas em diferentes momentos da 1ª onda folicular após a
superovulação (OPU 2) nos tratamentos T (0-144), T (48-120), T (48-96), T (72-
96), T (96-72) e T (96-48).
Número de oócitos BlastocistosTratamento no deanimais
Total Viáveis (%) no %
T (0 – 144) 4 41 29 (70,7) b 14 48,3 aT (48 – 120) 5 58 52 (89,7) a 6 11,5 bT (48 – 96) 4 61 41 (67,2) bc 6 14,6 bT (72 – 96) 3 21 17 (81,0) ab 2 11,8 bT (96 – 72) 3 56 42 (75,0) ab 3 7,1 bT (96 – 48) 4 15 05 (33,3) c 1 20,0 ab
Total 23 252 186 (73,8%) 32 17,2%Letras diferentes diferem entre si dentro da coluna (p<0,05). Tratamentos:
T (0-144) – PGF2á imediatamente após a colheita de embriões, aspiração 144 horas após a injeção de PGF2á
T (48-120) – PGF2á 48 horas após a colheita de embriões e aspiração 120 horas após a injeção de PGF2á
T (48-96) – PGF2á 48 horas após a colheita de embriões e aspiração 96 horas após a injeção de PGF2á
T (72-96) – PGF2á 72 horas após a colheita de embriões e aspiração 96 horas após a injeção de PGF2á
T (96-72) – PGF2á 96 horas após a colheita de embriões e aspiração 72 horas após a injeção de PGF2á
T (96-48) - PGF2á 96 horas após a colheita de embriões e aspiração 48 horas após a injeção de PGF2á
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxiv
O percentual de oócitos viáveis foi menor (P<0,05) no T (96-48) (33,3%),
quando a PGF2α foi aplicada 96 horas depois da colheita de embriões e 48 horas
após foi realizada a OPU 2, comparando aos demais tratamentos. No tratamento
T (48-120), a PGF2á foi administrada 48 horas após a colheita de embriões e 120
horas após foi realizada a OPU 2 e obteve-se o maior percentual de oócitos
viáveis entre os tratamentos (89,7%), diferindo dos tratamentos T (0-144), T (48-
96) e T (96-48) que obtiveram 70,7%, 67,2% e 33,3%, respectivamente. Os
tratamentos T (72-96) e T (96-72) não diferiram entre si (P<0,05) do tratamento T
(48-120) quanto ao número de oócitos viáveis.
Com relação à taxa de embriões produzidos in vitro, o melhor tratamento
observado foi o T (0-144) com 48,3% de blastocisto, que diferiu dos tratamentos T
(48-120), T (48-96), T (72-96) e T (96-72) e não diferiu (P<0,05) do T (96-48).
(Tabela 2)
Uma vez que os animais foram submetidos a duas aspirações foliculares
com um tratamento superovulatório intermediário foi estabelecida uma
comparação da produção in vitro entre a OPU 1 (referência) e OPU 2 como
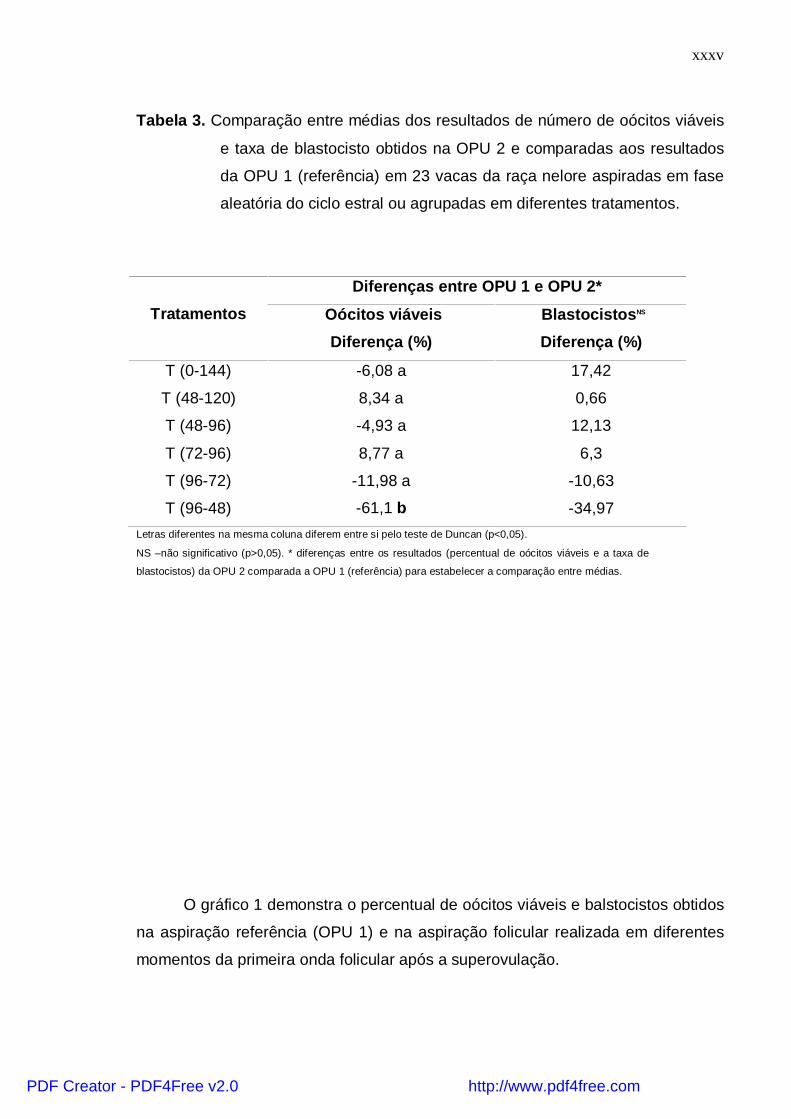
demonstra a tabela 3, onde observa-se que a produção de embriões no T (96-48)
foi significativamente menor (p<0,05) quando comparada aos demais tratamentos,
os quais não deferiram entre si (p>0,05).
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com
xxxv
Tabela 3. Comparação entre médias dos resultados de número de oócitos viáveis
e taxa de blastocisto obtidos na OPU 2 e comparadas aos resultados
da OPU 1 (referência) em 23 vacas da raça nelore aspiradas em fase
aleatória do ciclo estral ou agrupadas em diferentes tratamentos.
Diferenças entre OPU 1 e OPU 2*
Tratamentos Oócitos viáveis
Diferença (%)
BlastocistosNS
Diferença (%)
T (0-144) -6,08 a 17,42
T (48-120) 8,34 a 0,66
T (48-96) -4,93 a 12,13
T (72-96) 8,77 a 6,3
T (96-72) -11,98 a -10,63
T (96-48) -61,1 b -34,97Letras diferentes na mesma coluna diferem entre si pelo teste de Duncan (p<0,05).
NS –não significativo (p>0,05). * diferenças entre os resultados (percentual de oócitos viáveis e a taxa de
blastocistos) da OPU 2 comparada a OPU 1 (referência) para estabelecer a comparação entre médias.
O gráfico 1 demonstra o percentual de oócitos viáveis e balstocistos obtidos
na aspiração referência (OPU 1) e na aspiração folicular realizada em diferentes
momentos da primeira onda folicular após a superovulação.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxvi
OPU1 e OPU2 - % oócitos e Bl
76,3
86,9
61,567,6
88,9
71,370,7
89,7
67,2
81
33,3
75
31
1,9
18,8 17,4
54,2
17,7
48,3
11,5 14,6 11,8
20
7,1
0
10
20
30
40
50
60
70
80
90
100
0 - 144 48 - 120 48 - 96 72 - 96 96 - 48 96 - 72
Tratamentos
%oó
cito
svi
ávie
s/%
Bl
% oócitos viáveis - OPU 1 % oócitos viáveis - OPU 2 % Bl viáveis - OPU 1 % Bl viáveis - OPU 2
Gráfico1. Resultado da % média de oócitos viáveis recuperados e taxa de
blastocistos obtidos de oócitos aspirados em fase aleatória do ciclo
estral na aspiração referência (OPU 1) e em diferentes momentos
da primeira onda folicular após o tratamento superovulatório (OPU
2), realizado em 23 fêmeas da raça nelore, agrupadas nos
tratamentos T(0-144), T (48-120), T(48-96), T(72-96), T(96-72) e T
(96-48), de acordo com o tempo de administração de PGF2á após
a colheita de embriões e o momento da segunda aspiração após a
SOV (OPU 2).
A produção in vivo de embriões (TE) nas 23 vacas da raça Nelore resultou
em 72 embriões (3,1 por animal), dos quais 73,6% ou 53 (2,3 por animal) foram
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxvii
classificados morfologicamente como viáveis e 26,3% ou 19 (3,1 por animal) foram
classificados como não viáveis (Tabela 4).
Tabela 4. Número de animais, número e média total de embriões recuperados,
número de embriões não viáveis, número, média e percentual de
embriões viáveis produzidos in vivo.
Produção de embriões in vivoSessões n Total e
médiaNão
viáveisViáveise média
% Viáveis
1ª T (0 – 144) 4 17 (4,2) 1 16 (4,0) 94,12ª T ( 48 – 120) 5 7 (1,4) 6 1 (0,2) 14,33ª T (48 – 96) 4 12 (3,0) 1 11 (2,7) 91,74ª T (72 – 96) 3 1 (0,3) 1 0 (0,0) 0,05ª T 96 – 72) 4 19 (4,7) 7 12 (3,0) 63,26ª T (96 – 48) 3 16 (5,3) 3 13 (4,3) 81,3
Total 23 72 (3,1) 19 (26,3%) 53 (2,3) 73,6%Produção in vivo de embriões realizada após a aspiração referência (OPU 1) e que precedeu a OPU 2.
V. DISCUSSÃO
No presente estudo com vacas da raça nelore a aspiração referência (OPU
1) realizada em fase aleatória do CE resultou em uma média 9,6 oócitos viáveis e
na OPU 2 a média, independente do tratamento, foi de 8,1 oócitos. Quando os
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxviii
animais foram agrupados por tratamentos o percentual de oócitos viáveis variou
de 33,3% no T (96-48) a 89,7% no T (48-120).
Essas médias obtidas são semelhantes as taxas de recuperação de oócitos
observadas por CASTILHO, (2003), SENEDA, et al., (2001) e DAYAN et al.,
(2000), 9,0, 9,2 e 9,7, respectivamente.
Com relação à taxa de blastocistos, na OPU 2, embora não houve diferença
significativa, os tratamentos que resultaram em maior taxa de desenvolvimento
embrionária foram o T (0 – 144) com 48,3% e T (48-120) com 11,5 % dos oócitos
maturados e fecundados atingindo o estágio de blastocisto. O desenvolvimento
embrionário em todos os tratamentos da OPU 2 variou de 7,1% a 48,3% e na OPU
1, excluindo o valor de 1,9%, variou de 17,4% a 54,2% e ampla variação também
foi observada por KRUIP et al. (1994), 9 a 26% e HASLER et al. (1995), 4% a 33%
e DAYAN et al. (2000a), 14 e 54%.
Na produção in vivo de embriões, iniciada 48 a 60 horas após a OPU em
fase aleatória do CE, obtivemos uma média de 2,3 embriões por animal, que não
diferiu da média nacional nas doadoras zebuínas observada por BECKER et al. e
RODRIGUES et al., (1988), ALVIM et al., 2000; ANDRADE et al., 2000; FONSECA
et al., 2000; NOGUEIRA et al., 2000; PENNA et al., 2000; PINTO et al., 2000) que
variou de 2,4 a 9,9 embriões. GRADELA et al. (2000), também não observaram
aumento de estruturas quando realizaram a SOV na ausência do FD, embora
houve melhora na viabilidade embrionária.
Quando analisamos por tratamentos obtivemos as melhores médias de
embriões produzidos in vivo nos tratamentos T (0-144) e T (96-48), 4,0 e 4,3, as
taxas intermediárias nos tratamentos T (48-96) e T (96-72), 2,7 e 3,0 e ausência
de produção nos tratamentos T (48-120) e T (72-96) que de acordo com GALLI
(2003) confirmam a baixa consistência na produção in vivo de embriões, sendo
que um terço das doadoras tratadas não respondem a SOV, um terço produz em
média 3 embriões e somente o terço restante resulta em grande número de
embriões transferidos.
Quando comparamos os tratamentos realizados na OPU 2, observou-se
que a recuperação no T(96-48), 5 oócitos (33,3%), foi significativamente menor
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xxxix
que os demais tratamentos, demonstrando que o momento da aspiração folicular
muito próxima a administração de PGF2α apenas 48 horas prejudicou a
recuperação dos oócitos. È importante ressaltar que a resposta a superovulação
desses animais foi boa e a presença de muitos CLs nos ovários pode ter
dificultado a recuperação dos oócitos na OPU 2, reforçando o fato que a aspiração
realizada muito próxima a administração de PGF2α pode prejudicar a posterior
recuperação de oócitos. Por outro lado podemos observar que nos tratamentos
em que a aspiração folicular foi realizada a partir de 96 horas da aplicação de
PGF2á as taxas de oócitos não variaram, demonstrando que a recuperação e a
produção in vitro após a SOV é possível sem prejudicar os índices de produção.
Além da possibilidade da diminuição da recuperação dos oócitos em função
da presença de vários CLs nos ovários , outro fator que pode ter afetado
diretamente essa recuperação é a ausência da emergência da onda folicular até
48 horas após a colheita de embriões. Segundo ADAMS et. al (1992), FORTUNE
et al. (1993), GIBONS, GINTHER (1999), em torno de 24 a 36 horas após a
ovulação ocorre o crescimento de um grupo de folículos que é caracterizado como
a emergência da onda folicular no CE normal. No entanto, não existe na literatura
trabalhos que demonstrem o tempo de emergência da onda folicular após
superovulação, mas no presente trabalho podemos afirmar que até 48 horas após
a PGF2á não houve emergência da onda folicular e esta provavelmente ocorre
mais tarde em animais superovulados, pois acima de 72 horas observamos
aumento no percentual de oócitos recuperados e acima de 96 horas não houve
comprometimento da taxa de oócitos recuperados e produção de blastocistos.
É importante ressaltar que assim como no T (0-144) obteve-se a maior taxa
de blastocistos produzidos in vitro, também observou-se maior taxa de blastocistos
produzidos in vivo, demonstrando que a SOV além de não ter prejudicado a
recuperação na OPU 2, ainda gerou bom índice na produção de embriões in vivo.
Os resultados de recuperação de oócitos e taxa de balstocistos das
aspirações não variaram significativamente entre a OPU 1 e OPU 2, concluindo
que é possível realizar a alternância das biotecnologias sem prejudicar os índices
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xl
de produção, desde que a OPU ocorra após a lise dos CLs presentes nos ovários
após a superestimulação.
VI. CONCLUSÃO
- a aspiração folicular realizada muito próxima à colheita de embriões
diminui significativamente a recuperação de oócitos;
- a superovulação realizada a partir de 96 horas após a aplicação de
PGF2α em animais previamente superovulados para colheita de embriões in vivo
não afeta a recuperação de oócitos e a produção in vitro de blasctocistos;
- é possível realizar a alternância das biotecnologias sem prejudicar
significativamente os índices de produção.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xli
VII. REFERÊNCIAS BIBLIOGRÁFICAS
ADAMS, G.P.; MATTERI, R.L.; KASTELIC, J.P.; et al. Association Between
Surges of Follicle-stimulating Hormone and The Emergence of Folicular Waves in
Heifers. J. Reprod. Fert., v. 94, p. 177-188, 1992.
ADAMS, G.P.; MATTERI, R.L.; GINTHER, O.J. Effect of progesterone on ovarian
follicles, emergencies of follicular waves and circulating follicle-stimulating hormone in
heifers. J. Reprod. Fert., v. 96, p. 627-40, 1992.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xlii
ADAMS, G.P.; KOT, K.; SMITH, C.A.; GINTHER, O.J. Selection of a dominant follicleand suppression of follicular growth in heifers. Animal Reprod. Science, v. 30, p. 259-271, 1993.
ADAMS, G.P. Control of ovarian follicular wave dynamics in cattle: implications for
synchronization and superstimulation. Theriogenology, v. 41, p. 19-24, 1994.
ADAMS, G.P.; EVANS, A.C.O.; RAWLINGS, N.C. Folicular waves and circulating
gonadotrophins in 8-month-old prepubertal heifers. J. Reprod. Fertil., v. 100, p.
27-33, 1994.
ALVIM, M.T.T.; MATOS, S.P.M.; SALIBA, W.P.; BELISÁRIO, H.; BRANDÃO, F.Z.;
PALHARES, M.S.; SILVA FILHO, J.M. Effect of two different commercial products
(Pluset vs Folltropin) on the superovulatory response and embryo production in
Nelore breed cows. Arquivos da Faculdade de Veterinária UFRGS, v. 28, Suppl
1, p. 201, abst., 2000.
AMARIDIS, G.S.; ROBERTSON, L.; REID, S.; BOYD, J.S.; O’ SHAUGHNESSY,
P.J.; JEFFCOATE, I.A. Plasma estradiol, FSH and LH concentration after
dominant follicle aspiration in the cow. Theriogenology, v. 52, p. 995-1003, 1999.
ANDRADE, J.C.O.; OLIVEIRA, M.A.L.; SANTOS FILHO, A.S.; LIMA, P.F. Use of
Synchro-Mate B (SMB) and of Controlled internal drug releasing device (CIDR) to
synchronize the superovulation in Nelore donors. Arquivos da Faculdade de
Veterinária UFRGS, v. 28, Suppl 1, p. 205, abst., 2000.
ANDRADE, J.C.O.; OLIVEIRA, M.A.L.; LIMA, P.F.; SANTOS FILHO, A.S.; PINA,
V.M. Use of steroid hormone treatments prior to superovulation in Nelore donors.
Anim. Reprod. Sci., v. 69, p. 09-14, 2002.
ANDRADE, J.C.O.; OLIVEIRA, M.A.L.; LIMA, P.F.; GUIDO, S.I.; TENÓRIO FILHO,
F.; AZEVEDO, J.B.; SOUZA, B.P.A.A.; PINA, V.M.R.; BELLO, A.C.; OLIVEIRA,
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xliii
L.R.S. Superovulatory response of Nelore donors treated with FSH-o and FSH-p.
Rev. Bras. Reprod. Anim., v. 27, p. 104-106, 2002a.
ANDRADE, J.C.O.; OLIVEIRA, M.A.L.; LIMA, P.F.; SANTOS FILHO, A.S.; GUIDO,
S.I.; TENÓRIO FILHO, F.; CAVALCANTI NETO, C.C. Use of CIDR prior to
superovulation of Nelore donors in differents stages of the estrus cycle:
preliminaries results. Rev. Bras. Reprod. Anim., v. 26, p. 224-226, 2002b.
ANDRADE, J.C.O.; OLIVEIRA, M.A.L.; LIMA, P.F.; SANTOS FILHO, A.S.; PINA,
V.M.R. Use of steroid hormone treatments prior to superovulation in Nelore donors.
Anim. Reprod. Sci., v. 69, p. 9-14, 2002c.
ANDRADE, J.C.O.; OLIVEIRA, M.A.L.; LIMA, P.F.; GUIDO, S.I.; BARTOLOMEU,
C.C.; TENÓRIO FILHO, F.; PINA, V.M.R.; IUNES-SOUZA, T.C.; PAULA, N.R.O.;
FREITAS, J.C.C. The use of steroid hormones in superovulation of Nelore donors
at different stages of estrous cycle. Anim. Reprod. Sci., v. 77, p. 117-125, 2003.
ARLOTTO, T.; SCHWARTZ, J.L.; FIRST, N.L.; LIEBFRIED-RUTLEDGE, M.L.
Aspects of follicle and oocyte stage that affect in vitro maturation and development
of bovine oocyte. Theriogenology, v. 45, p. 943-956, 1996.
ARMSTRONG, D.T. Recent advances in superovulation of cattle.
Theriogenology, v. 55, p. 7-23, 1993.
ARMSTRONG, D.T.; IRVINE, B.J.; EARL, C.R.; McLEAN, D.; SEAMARK, R.F.Gonadotropins stimulations regiments for follicular aspiration and in vitro embryoproductions from calf oocytes. Theriogenology, v. 42, p. 1227-1236, 1994.
AZEVEDO, N.; COELHO, S.G. Particularidades na transferência de embriões
zebuínos. In: Congresso Brasileiro de Reprodução Animal. Anais. p. 33-45. Belo
Horizonte, MG, 1991.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xliv
BARROS, C.; NOGUEIRA, M. Embryo Transfer In Bos Indicus Cattle.
Theriogenology, v. 56, p. 1483-1496, 2001.
BARROS, C.M.; FIGUEIREDO R.A.; PAPA, F.O.; ROCHA, G. Follicular growth in
Nelore cows. (Bos indicus) after PGF2α administration. J. Anim. Sci., v. 71, supl,
1993.
BECKER, W.; PINHEIRO, L. Ovarian response to superovulation in Nelore cows
(Bos Taurus indicus). Theriogenology, v. 25, p. 785-793, 1986.
BECKER, W.A.P.; MORAES, G.V.; PINHEIRO, L.E.L.; RODRIGUES, C.F.M.;
CARVALHO, C. Quali-quantitative evaluation of bovine embryo transfer: II –
Hormonal effects. Revista do Centro de Ciências Rurais UFSM, 18 (Suppl), 43
abst., 1988.
BERGFELT, D.R.; LIGHTFOOT, K.C.; ADAMS, G.P. Ovarian synchronization
following ultrasound-guided transvaginal follicle ablation in heifers.
Theriogenology, v. 42, p. 895-907, 1994.
BERGFELT, D.R.; LIGHTFOOT, K.C.; ADAMS, G.P. Ovarian dynamics following
ultrasound-guided transvaginal follicle ablation in heifers. Theriogenology, v. 42,
p. 895-907, 1994a.
BO, G.A.; PIERSON, R.A.; MAPLETOFT, R.J. The effect of estradiol valerate on
follicular dynamics and superovulatory response in cows with syncro-mate-B
implants. Theriogenology, v. 36, p. 169-183, 1991.
BO, G.A.; ADAMS, G.P.; NASSER, L.F.; PIERSON, R.A.; MAPLETOFT, R.J.
Effects of estradiol valerate on ovarian follicles, emergence of follicular waves and
circulating gonadotropins in heifers. Theriogenology, v. 40, p. 225-239, 1993.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xlv
BO, G.A.; ADAMS, G.P.; PIERSON, R.A.; TRIBULO, H.E.; CACCIA, M.;
MAPLETOFT, R.J. Follicular waves dynamics after estradiol -17 â treatment of
heifers with or without a progestogen implant. Theriogenology, v. 41, p. 1555-
1569, 1994.
BO, G.A.; ADAMS, G.P.; CACCIA, M.; MARTINEZ, M.; PIERSON, R.A.;
MAPLETOFT, R.J. Ovariam follicular wave emergence after treatment with
progestogen and estradiol in cattle. Anim. Reprod. Sci., v. 39, p. 139-204, 1995.
BO, G.A.; ADAMS, G.P.; PIERSON, R.A.; MAPLETOFT, R.J. Exogenous control of
follicular wave emergence in cattle. Theriogenology, v. 43, p. 32-40, 1995b.
BO, G.A.; BERGFELT, D.R.; MAPLEOFT, R.J. Follicle wave dynamics and
superovulation en cattle: recent advances and practical experience. Arq. Fac. Vet.
UFRGS, v. 24, p. 31-52, 1996
BO, G.A.; BERGFELT, D.R.; MAPLETOFT, R.J. Follicle wave dynamics and
superovulation in cattle: recent advances and pratical experience. Arq. Fac. Vet.
UFRGS, v. 24, p. 31-52, 1996.
BÓ, G.A.; ADAMS, G.P.; MAPLETOFT, R.J. Dinámica Folicular Ovariana en el
Bovino. In: Simpósio sobre controle farmacológico do ciclo estral em ruminantes,
2000, São Paulo: Departamento de reprodução animal: Faculdade de Medicina
Veterinaria e Zootecnia, Anais, p. 12 –34, 2000.
BODENSTEINER, K.J.; KOT, K.; WILTBANK, M.C.; GINTHER, O.J. Synchronization
of emergence of follicular wave in cattle. Theriogenology, v. 45, p. 1115-28, 1996.
BOLAND, M.P.; GOULDING, D.; ROCHE, J.F. Alternatives gonadotrophins for
superovulation in cattle. Theriogenology, v. 35, p. 5-17, 1991.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xlvi
BOLS, P.E.J.; VANDENHEEDE, J.M.M.; VAN SOOM, A.; Transvaginal ovum-pich-
up (OPU) in the cow: a new disposable needle guindance system.
Theriogenology, v. 43, p. 677-687, 1995.
BORDIGNON, V.; MORIN, N.; DUROCHER, J.; BOUSQUET, D.; SMITH, L.C.
GnRH improves the recovery rate and the in vitro developmental competence of
oocytes obtained by transvaginal follicular aspiration from superstimulate heifers.
Theriogenology, v. 48, p. 291-298, 1997.
BOUSQUET, D.; TWAGIRAMUNGU, H.; MORIN, N.; BRISSON, C.; CRBONNEAU,
G.C.; DUROCHER, J. In Vitro Production in Cow: An Effective Alternative to the
Conventional Embryo Production Approach. Theriogenology, v. 51, p. 59-70,
1999.
BROADBENT, P.J.; GEBBIE, F.E.; DOLMAN, D.F.; WATT, R.G.; KING, M.E.;
HIGGINS, L.C. Superovulatory responses in cattle pre-treated with estradiol and
progestagen. Theriogenology, v. 43. p. 176, 1995.
BROGLIATTI, G.M.; ADAMS, G.P. Ultrasound-guined transvaginal oocyte
collection in prepurbetal calves. Theriogenology, v. 45, p. 1163-1176, 1996.
BUNGARTZ, L.; NIEMANN, H. Assessment of the presence of a dominant follicle
and selection of dairy cows suitable for superovulation by a single ultrasound
examination. J. Reprod. Fert., v. 101, p. 583-591, 1994.
BUNGARTZ, L.; LUCAS-HAHN, A.; RATH, D.; NIEMANN, H. Collection of oocytes
from cattle via follicular aspiration aided by ultrasound with or without gonadotropin
pretreatment and in different reproductive stages. Theriogenology, v. 43, n. 667-
675,
1995.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xlvii
BURATINI Jr, J.; PRICE, C.A.; VISINTIN, J.A.; BO, G.A. Effects of dominant follicle
aspiration and treatment with recombinant bovine somatotropin (BST) on ovarian
follicular development in Nelore (Bos indicus) heifers. Theriogenology, v. 54, p.
421-431, 2000.
CACCIA, M.; BO, G.A. Follicle emergence following treatment of CIDR-B implanted
beef cows with estradiol benzoate and progesterone. Theriogenology, v. 49, p.
341, 1998.
CAROLAN, C.; LONERGAN, P.; MONGET, P.; MONNIAUX, D.; MERMILLOD, P.
Effect of follicle size and quality on the ability of follicular fluid to support
cytoplasmic maturation of bovine oocytes. Mol. Reprod. Dev., v. 43, p. 477-483,
1996.
CARRIERE, P.D.; AMAYA, D.; LEE, B. Ultrasonography and endocrinology of
ovarian disfunctions induced in heifers with Estradiol Valerate. Theriogenology, v.
43, p. 1061-1076, 1995.
CASTILHO, C. Desenvolvimento folicular e sincronização da ovulação em
novilhas da raça Girolando. Dissertação (Mestrado), 94 p., Unesp, Botucatu,
1999.
CASTILHO C.; GAMBINI, A.L.G.; FERNANDES, P.; TRINCA, L.A.; TEIXEIRA,
A.B.; BARROS, C.M. Synchronization of ovulation in crossbred dairy heifers using
gonadotropin-releasing hormone agonist, prostaglandin F2 á and human chorionic
ganadotropin or estradiol benzoate. Brazilian Journal of Medical and Biological
Research, v. 33, p. 91-101, 2000.
CASTILHO, C. Divergência no crescimento folicular em novilhas da raça
nelore e sua influência sobre a competência oocitária para o
desenvolvimento embrionário In vitro. Tese (Doutorado) – Faculdade de
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xlviii
Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Jaboticabal,
2003.
CASTILHO, C.; GARCIA, J.M. Influência do crescimento folicular sobre a
competência oocitária para o desenvolvimento embrionário in vitro em novilhas da
raça nelore. ACTA Scientciae Veterinare, v. 31, (suplemento), Porto Alegre:
UFRGS, 2003.
CASTILHO, C.; RENESTO A. Divergência no crescimento folicular em novilhas da
raça nelore. ACTA Scientciae Veterinare, v. 31, (suplemento), Porto Alegre:
UFRGS, 2003.
CHUPIN, D.; COMBARNOUS, Y.; PROCUREUR, R. Antagonistic effect of LH on
FSH induced superovulation in cattle. Theriogenology, v. 21, p. 229, 1984
COELHO, S.G.; AZEVEDO, N.A. Particularidades na transferência de embriões
zebuínos. In Congresso Brasileiro de Reprodução Animal, 9, Belo Horizonte,
Anais, p. 33-45, 1991.
DAYAN, A.; WATANABE, M.R.; WATANABE, Y.F. Fatores que interferem na
produção comercial de embriões FIV. Arq. Faculd. Vet. UFRGS, v. 28, p. 181-
185, 2000.
DAYAN, A., WATANABE, M.R., WATANABE, Y.F. Fatores que interferem na
produção comercial de embriões FIV. Arq. Fac. Vet. UFRGS, v. 28,n.1, p. 181-
185, 2000a.
DAYAN, A. Fatores que interferem na produção de embriões bovinos
mediante aspiração folicular e fecundação in vitro. Dissertação (Mestrado),
55p., Unesp, Botucatu, 2001.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

xlix
DIAZ, T. Effects of the Persistent Dominant Follicle on the Ability of Follicle
Stimulating Hormone to Induce Follicle Development and Ovulatory Responses. J.
Dairy Science, v. 84, n. 1, p. 88-99, 2001.
DRIANCOURT, M. Regulation of Ovarian Follicular Dynamics in Farm Animal.
Implications for Manipulation of Reproduction. Theriogenology, v. 55, p. 1211-
1231, 2001.
FABER, D.C.; MOLINA, J.A.; OHLRICHS, C.L..; VANDER WAAG, D.F.; FERRÉ,
L.B. Commercialization of animal biotechnology. Theriogenology, v. 59, p. 125-
138, 2003.
FIGUEIREDO, R. A.; BARROS, C.M.; PINHEIRO, O.L.; SOLER, J.M.P. Ovarian
Follicular Dnamics in Nelore Breed (Bos indicus) Cattle. Theriogenology, v. 47, p.
1484-1505, 1997.
FINDLAY, J.K.; CLARKE, I.J. Regulation of the secretion of FSH In domestic
ruminants. J. Reprod. Fertil., v. 34, p. 27-37, 1987.
FONSECA, J.F.; SILVA FILHO, J.M.; PINTO NETO, A.; BELISÁRIO, H.; SALIBA,
W.P.; ALVIM, M.T.T. Different embryonic stages recovered from zebu donors.
Arquivos da Faculdade de Veterinária UFRGS, v. 28, Suppl. 1, p. 263, abst.,
2000.
FORTUNE, J.E.; HANSEL, W. Concentrations of steroids and gonadotropins in
follicular fluid from normal heifers primed for superovulation. Biol. Reprod., v. 32,
p. 1069-1079, 1985.
GALINA, C.; ARTHUR, G. Review on Cattle Reproduction in the Tropics. Part 4.
Oestrus Cycles. Animal Breed. Abst., v. 58, p. 697-707, 1990.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

l
GALLI, C.; DUCHI, R.; GROTTI, G.; TURINI, P.; PONDERATO COLLEONI, S.;
LAGUTINA, I.; LAZZARI, G. Bovine embryo technologies. Theriogenology, v. 59,
p. 599-616, 2003.
GAMBINI, A.L.G.; MOREIRA, M.B.P.; CASTILHO, C.; BARROS, C.M. Dinâmica
follicular e sincronização da ovulação em vacas da raça Gir. Rev. Bras. Reprod.
Anim., v. 22, p. 201-210, 1999.
GIBBONS, J.R.; BEAL, W.E.; KRISHER, R.L.; FABER, E.G.; PEARSON, R.E.;
GWAZDAUSKAS, F.C. Effects of once versus twice weekly transvaginal follicular
aspiration on bovine oocyte recovery and embryo development. Theriogenology,
v. 42, p. 405, 1994.
GIBBONS, J.R.; WILTBANK, M.C.; GINTHER, O.J. Functional interrelationships
between follicles greater than 4 mm and the follicle-stimulating hormone surge in
heifers. Biol. Reprod., v. 57, p. 1066-73, 1997.
GIBBONS, J.; WILBANK, M.; GINTHER, O.J. Relationship Between Follicular
Development and the Decline in the Follicle-Stimulation Hormone Surge in
HEIFERS. Biol. of Reprod., v. 60, p. 72-77, 1999.
GINTHER, O.J.; KNOPF, L.; KASTELIC, J.P. Temporal associations among ovarian
events in cattle during oestrous cycles with two and three follicular waves. J. Reprod.
Fertil., v. 887, p. 223-30, 1989.
GINTHER, O.J.; WILTBANK, M.C.; FRICKE, P.M.; GIBBONS, J.R.; KOT, K.
Selection of the dominant follicle in cattle. Biol. Reprod., v. 55, p. 1187-94, 1996.
GINTHER, O.J.; KOT, K.; KULICK, L.J.; WILTBANK, M.C. Emergence and
desviation of follicle during the development of follicular waves in cattle.
Theriogenology, v. 48, p. 75-87, 1997.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

li
GINTHER, O.J.; BERGFELD, D.R.; KULICK, L.J.; KOT, K. Selection of the
dominant follicle in cattle: establishment of follicle deviation in less than 8 hours
through depression of FSH concentration. Theriogenology. v. 52, p. 1079-1093,
1999.
GINTHER, O.J;, BERFELT, D.R.; KULICK, L.J.; KOT, K. Selection of the dominant
follicle in cattle: role of estradiol. Biol. Reprod., v. 63, p. 383-389, 2000.
GINTHER, O.J.; BERGFELD, D.R.; KULICK, L.J.; KOT, K. Selection of the
dominant follicle in cattle: role of estradiol. Biol Reprod., v. 63, p. 383-389, 2000b.
GINTHER, O.J.; BERFELT, D.R.; BEG, M.A.; KOT, K. Follicle selection in
monovular species. Biol. Reprod., v. 65, p. 638-647, 2001.
GOULDING, D.; WILLIANS, D.H.; DUFFY, P.; BOLAND, M.P.; ROCHE, J.F.
Superovulation in heifers given FSH initiated either at Day 2 or at mid luteal phase
of the estrous cycle. Theriogenology, v. 33, p. 239, 1990.
GRADELA, A.; ESPER, C.R.; MATOS, S.P.M.; LANZA, J.A.; DERAGON, L.A.G.;
MALHEIROS, R.M. Dominant follicle by ultrasound guided transvaginal aspiration
and superovulatory response in Nellore cows. Arq. Bras. Med. Vet. Zootec., v. 52,
n. 1, p. 53-58, 2000.
GRASSO, F.; GUILBAULT, L.A.; ROY, G.L. The influence of the presence of a
dominant follicle at the time of inititation of a superovulatory treatment on
superovulatory responses in cattle. Theriogenology, v. 31, p. 199, 1989.
GUILBAULT, L.A.; GRASSO, F.; LUSSIER, J.G.Decreased superovulatory
responses in heifers superovulated in the presence of a dominant follicle. J.
Reprod. Fert., v. 91, p. 81-89, 1991.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

lii
HAFEZ, E.S.E.; Reproduction in farm animal, 6ª edição, p. 573, 1995.
HAGEMANN, L.J.; BEAUMONT, S.E.; BERG, M.; DONNINSON, M.J.; LEDGARD,
A.;
PETERSON, A.J.; SCHURMANN, A.; TERVIT, H.R. Development during sIVP of
bovine
oocytes from dissected follicules: interative effects of cycle stage, follicle size and
atresia. Mol. Reprod. Dev., v. 53, p. 451-458, 1999.
HAGEMANN, L.J.; BEAUMONT, S.E.; BERG, M.; DONNINSON, M.J.; LEDGARD,
A.; PETERSON, A.J.; SCHURMANN, A.; TERVIT, H.R. Development during sIVP
of bovine oocytes from dissected follicles: interative effects of estrous cycle stage,
follicle size and atresia. Mol reprod Dev., v. 53, p. 451-458, 1999b.
HASLER, J.F.; HENDRERSON, W.B.; HURTGEN, P.J.; JIN, Z.Q.; McCAULEY,
A.D.; MOWER, S.A.; NEELY, B.; SHUEY, L.S.; STOKES, J.E.; TRIMMER, S.A.
Production, freezing and transfer of bovine IVF embryos and subsequent calving
results. Theriogenology, v.43, p.141-152, 1995.
HENDRIKSEN, P.J.M.; VOS P.L.A.M. Bovine follicular development and its effect
on the in vitro competence of oocytes. Theriogenology, v. 53, p. 11-20, 2000.
HYTTEL, P.; CALLESEN, H.; GREVE, T. Oocyte maturation and sperm transport
in superovulated cattle. Theriogenology, v. 35, n. 1, p. 91-108, 1991.
KNOPF, L.; KASTELIC, J.P.; SCHALLENBERGER, E. Ovarian follicular dynamics
in heifers: test of two wave hypotesis by ultrasonically monitoring individual
follicles. Domestic Animal Endocrinology, v. 6, p. 111-119, 1989.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

liii
KO, J.C.H.; KASTELIC, J.P.; DEL CARMO, M.R. Effects of a dominant follicle on
ovarian follicular dynamics during the estrus cycle in heifers. J. Reprod. Fert., v.
91, p. 511-51, 1991.
KRUIP, T.A.M.; BONI, R.; WURTH, Y.A.; ROELOFSEN, M.W.M.; PIETERSE, M.C.
Potential use of ovum pick-up for embryo production and breeding in cattle.
Theriogenology, v. 42, p. 675-684, 1994.
KULICK. A.C. Follicular and Hormonal Dynamics During the First Follicular Wave
in Heifers. Theriogenology, v. 52, p. 913-921,1999.
LONERGAN, P.; MONAGHAN, P.; RIZOS, D.; BOLAND, M.P.; GORDON,I. Efect
of follicle size on bovine oocyte quality and development competence following
maturation, fertilization and culture in vitro. Mol. Reprod. Dev., v. 37, p. 48-53,
1994.
LOONEY, C.R.; LINDSEY, B.R.; GONSETH, C.L.; JOHNSON, D.L. Commercial
aspects of oocyte retrieval and in vitro fertilization (IVF) for embryo production in
problem cows. Theriogenology, v. 41, p. 67-72, 1994.
LUCY, M.C.; SAVIO, J.D.; BADINGA, L.; DE LA SOTA, R.L.; THATCHER, W.W.
Factors that affect ovarian follicular dynamics in cattle. J. Anim. Sci., v. 70, p. 3615-
26, 1992.
MACHATKOVÁ, M.; JOKESOVÁ, E.; PETELÍKOVÁ, J.; DVORÁCEK, V.
Developmental competence of bovine embryo derived from oocytes collected at
various stages of the estrous cycle. Theriogenology, v. 45, p. 801-810, 1996.
MACHATKOVA, M.; JOKESOVA, E.; HORKY, F.; KREPELOVA, A. Utilization of
the growth phase of the first follicular wave for bovine oocyte collection improves
blastocyst production. Theriogenology, v. 54, p. 543-550, 2000.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

liv
MACHATKOVA, M.; KRAUSOVA, K.; JOKESOVA, E.; TOMANEK, M.
Developmental competence of bovine oocytes: effects of follicle size and the phase
of follicular wave on in vitro embryo production. Theriogenology, v. 61, p. 229-
335, 2004.
MAPLETOFT, R.J.; R.A. Featured article: Factors affecting superovulation in the
cow: pratical considerations. IETS Embryo Transfer Newsletter, v. 11, p. 15-24,
1993.
MAPLETOFT, R.J.; PIERSON, R.A. Recruitment of Follicles for Superovulation.
Comp. Cont. Ed., v. 16, p. 127-141, 1994.
MAPLETOFT, R.J.; MARTINEZ, M.F.; ADAMS, G.P.; KASTELIC, J.; BURNLEY,
C.A. The effect of estradiol on follicular wave emergence and superovulatory
response in norgestomet-impanted cattle. Theriogenology, v. 51, p. 411, 1999.
MATTON, P.; ADELAKOUN, V.; COUTURE, Y.; DUFOUR, J.J. Growth and
replacement of the bovine ovarian follicles during the estrous cycle. J. Anim. Sci., v.
52, p. 813-20, 1981.
MERTON J.S.; ROOS A.P.W.; MULLAART, E.; RUIGH L.; KAAL, L.; VOS,
P.L.A.M.; DIELEMANl. Factors affecting oocyte quality and quantity in commercial
application of embryo technologies in the cattle breeding industry.
Theriogenology, v. 59, n. 2, p. 651-674, 2003.
MEINTJES, M.; BELLOW, M.S.; BROUSSARD, J.R.; PAUL J.B.; GODKE, R.A.
Transvaginal aspiration of oocytes from hormone-treated pregnant beef for in vitro
fertilization. J. Anim. Sci., v. 73, p. 967-974, 1995.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

lv
MIHM, M.; GOOD, T.E.M.; IRELAND, J.L.H.; KNIGHT P.G.; ROCHE, J.F. Decline
in serum FSH concentration alters key intrafollicular growth factors involved in
selection of the dominant follicle in heifers. Biol. Reprod., v. 57, p. 1328-1337,
1997.
MONNIAUX, D.;, CHUPIN D.; SAUMANDE, J. Superovulatory responses of cattle.
Theriogenology. v. 19, p. 55-81, 1983.
MONNIAUX, D.; CHUPIN, D.; SAUMANDE, J. Superovulatory responses of cattle.
Theriogenology, v. 19, p. 55-81, 1983.
NASSER, L.; ADAMS, G.P.; BO, G.A.; MAPLEOFT, R.J. Ovarian superstimulatory
response relative to follicular wave emergence in heifers. Therigenology, v. 40, p.
713-724, 1993.
NIBART, M.; SILVA PEIXER, M.; THUARD, J.M.; DURANT, M.; GUYADER-JOLY,
C.; PONCHON, S.; MARQUANT-LE GUIENNE, B.; HUMBLOT, P. Embryo
production by OPU and IVF in dairy cattle. In: Réunion A.E.T.E., XI, Hannover,
Proceedings, p. 216, 1995.
NOGUEIRA, M.F.G.; BARROS, B.J.P.; ANDREUSSI, P.A.T.; D’OCCHIO, M.J.;
BARROS, C.M. Embryo recovery and pregnancy rates after delay of ovulation and
fixed-time insemination in superestimulated Nelore cows. Theriogenology, v. 53,
p. 503, abst., 2000.
NOGUEIRA, M.F.G. Superovulation and timed artificial insemination, in
Nelore cows. Tese (Mestrado), p. 68, Faculdade de Medicina Veterinária e
Zootecnia, Unesp, Botucatu, 2001.
OLIVEIRA, M.A.L.; ANDRADE, J.C.O.; LIMA, P.F.; SANTOS FILHO, A.S.;
GUIDO,S.I.; CAVALCANTI NETO, C.C.; TENÓRIO FILHO, F.; OLIVEIRA, L.R.S.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

lvi
Influência da idade sobre a resposta superovulatória de doadoras Nelore.
Resultados preliminares. Ver. Bras. Reprod. Anim., v. 26, p. 246-248, 2002.
PAVLOK, A.; LUCANS-HAHN; NIEMANN, H. Fertilization and developmental
competence of bovine oocytes derived from different categories of antral follicules.
Mol.
Reprod. Dev., v. 31, p. 63-67, 1992.
PEIXER M.A.S., SOUZA R.V., RUMPF R., DE BEM A.R., NETO M.A.P.
Nascimento dos primeiros produtos de FIV da raça Nelore no cenargem.
Zootecnia, Nova Odessa, v. 32, p. 49, 1995.
PENNA, V.M. The MOET nucleus of improvement of the Guzerá breed. Arquivos
da Faculdade de Medicina Veterinária UFRGS, v. 28, Suppl. 1, p. 167-174,
2000.
PIERSON, R.A. Ultrasonic imaging of the ovaries and utero in cattle.
Theriogenology, v. 29, p. 21-37, 1998.
PIERSON, R.A.; GINTHER, O.J. Follicular popilations during the estrous cycle in
heifers. III Time of selection of the ovulatory follicle. Anim. Reprod. Sci., v. 16, p.
81-95, 1988.
PIETERSE, M.C.; KAPPEN, K.A.; KRUIP, T.A.M.; TAVERNE, M.A.M. Aspiration of
bovine oocytes during transvaginal ultrasound scanning of the ovaries.
Theriogenology, v. 30, p. 751-761, 1988.
PINHEIRO, O.L.; BARROS, C.M.; FIGUEREDO, R.A.; VALLE,; E.R. DO,
ENCARNAÇÃO, R.O.; PADOVANI, C.R. Estrous Behavior and Estrous-to-
ovulation interval in Nelore Cattle (Bos indicus) With Natural Estrous or induced
With Prostaglandin F2 á or Norgestomet and Estradiol Valerato. Therigenology, v.
49, p. 667-681, 1998.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

lvii
PINTO NETO, A.; SILVA FILHO, J.M.; FONSECA, J.F.; MOTA, M.F.; BELISÁRIO,
H.; PARDINI, W.S.; ALVIM, M.T.T. Performance of donor cows from Nelore breed
in na embryo transfer program. Arquivos da Faculdade de Medicina Veterinária
UFRGS, p. 28, Suppl. 1, v. 311, abst., 2000.
ROBERTS, A.J.; GRIZZLE, J.M.; ECHTERNKAMP, S.E. Follicular development
and superovulation response in cows administered multiple FSH injections early in
the estrous cycle. Theriogenology, v. 42, p. 917-929, 1994.
ROBINSON, T.; SHELTON, J. Reproduction in Cattle, In: CUPPS, P.
Reproduction in Domestic Animals. 4ª Edicion. San Diego: Academic Press, p.
445-470, 1991.
RODRIGUES, C.F.M.; MORAES, G.V.; BECKER, W.A.P.; CARVALHO, C.;
PINHEIRO, L.E.L. Quali-quantitative evaluation of bovine embryo transfer: I –
General means in Zebu and European breeds. Revista do Centro de Ciências
Rurais UFSM, v. 18, Suppl. 1, p. 42, abst., 1988.
RODRIGUES, C.F.M.; GARCIA ,J.M. Fecundação in vitro em bovinos: aplicação
comercial. Arq. Fac. Vet. UFRGS, v. 28, p. 186-187, 2000.
RODRIGUES, J. L. Transferencia de Embriões Bovinos - Historico e Perspectivas
Atuais. Revista Brasileira De Reprodução Animal, v. 25, n. 2, p. 102-107, 2001.
SAUMANDE, J.; BATRA, S.K. Superovulation in the cow: comparison of
oestradiol-17â and progesterona patterns in plasma and milk of cows induced to
superovulote, relatioonships eith ovarian responses. J. of Endocrin., v. 107, n. 2,
p. 259-264, 1985
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

lviii
SENEDA, M.M.; ESPER, C.R.; GARCIA, J.M.; de OLIVEIRA, J.A.; VANTINI, R.
Relationship between follicle size and ultrasound-guided transvaginal oocyte
recovery. An. Reprod. Sci., v. 67, p. 37-43, 2001.
SHAW, D.W. Recovery rates and embryo quality following dominant follicle
ablation in superovulated cattle. Theriogenology, v. 53, p. 1521-1528, 1999.
SIDDIQUI, M.A.R.; SHAMSUDDIN, M.; BHUIYAN, M.M.U.; AKBAR, M.A.;
KAMARUDDIN, K.M. Effect of feeding and body condition score on multiple
ovulation and embryo production in zebu cows. Reprod. Dom. Anim., v. 37, p. 37-
41, 2002.
SIROIS, J.; FORTUNE, J.E. Ovarian follicular dynamics during the estrous cycle in
heifers monitored by real-time ultrasonography. Biol. Reprod., v. 39, p. 308-317,
1988.
SIROIS, J.; FORTUNE, J.E. Lengthening of the bovine estrous cycle with low levels
of exogenous progesterone: a model for studying ovarian follicular dominance.
Endocrinology, v. 127, p. 916-25, 1990.
STOCK, A.E.; ELLIGTON, J.E.; FORTUNE, J.E.A. A dominant follicle does not
affect follicular recruitment by superovulatory doses of FSH in cattle but can inhibit
ovulation. Theriogenology, v. 45, p. 1091-1102, 1996.
STUBBINGS, R.B.; WALTON, J.S. Effect of ultrasonically-guided follicle aspiration
on estrous cycle and follicular dynamics in Holstein cows. Theriogenology, v. 43,
p. 705-712, 1995.
THIBIER, M. The IETS statistics of embryo transfers in livestock in the world for the year1999: A new record for bovune in vivo-derived embryos transferred. Embryo TransferNewsletter, v. 18, p. 24-28, 2000.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com

lix
THOMPSON, J.G.; DUGANZICH, D. Analysis of culture systems for bovine in vitroembryo production reported in abstractw of the Proceedings of the International EmbryoTransfer Society (1991-1995). Theriogenology, 45:195, 1996.
VASSENA, R.; MAPLETOFT, R.J.; ALLODI, S.; SINGH, J.; ADAMS,
G.P.Morphology and developmental competence of bovine oocytes relative to
follicular status. Theriogenology, v. 60, p. 923-932, 2003.
WOOLUMS, A.R.; PETER, A.T. Cystic ovarian condition in cattle: Part I.
Folliculogenesis and ovulation. Compend. Cont. Educ. Article, v. 16, p. 935-43,
1994.
WRIGHT, P.; MALMO, J. Pharmacological Manipulation of Fertility. Appl. Food
Anim. Pract., v. 8, p. 57-89, 1992.
YAAKUB, H.; O’CALLAGHAN, D.; BOLAND, M.P. Effect of type and quantity of
concentrates on superovulation and embryo yield in beef heifers. Theriogenology,
v. 51, p. 1259-1266, 1999.
PDF Creator - PDF4Free v2.0 http://www.pdf4free.com