universidade federal do rio de janeiro centro de …livros01.livrosgratis.com.br/cp116909.pdf ·...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO CENTRO DE CIÊNCIAS DA SAÚDE
INSTITUTO DE BIOQUÍMICA MÉDICA Pós- Graduação em Química Biológica
CLARA RODRIGUES FERREIRA
ALTERAÇÕES EM PARÂMETROS BIOENERGÉTICOS
MITOCONDRIAIS PELO 3-BROMOPIRUVATO
Rio de Janeiro
2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

Rodrigues Ferreira, Clara
Alterações em parâmetros bioenergéticos mitocondriais pelo 3-bromopiruvato/ Clara Rodrigues Ferreira, Rio de Janeiro: UFRJ/IBqM, 2009.
xiv, 83 f.
Orientador: Antonio Galina Filho
Dissertação: Mestre em Ciências ( Química Biológica)
1. Cadeia Transportadora de Elétrons. 2. 3-Bromopiruvato. 3. Succinato
Desidrogenase. I. Galina, Antonio. II. Universidade Federal do Rio de
Janeiro, Instituto de Bioquímica Médica, Programa de Pós- graduação em
Química Biológica. III. Alterações em parâmetros bioenergéticos
mitocondriais pelo 3-bromopiruvato
FICHA CATALOGRÁFICA

ii
CLARA RODRIGUES FERREIRA
ALTERAÇÕES EM PARÂMETROS BIOENERGÉTICOS
MITOCONDRIAIS PELO 3-BROMOPIRUVATO
Dissertação apresentada ao Curso de
Pós-Graduação em Química Biológica da Universidade Federal do Rio de Janeiro, como requisito para obtenção do Grau de Mestre Modalidade Química Biológica.
Orientador: Prof. Dr. Antonio Galina Filho
Rio de Janeiro
2009

iii
CLARA RODRIGUES FERREIRA
ALTERAÇÕES EM PARÂMETROS BIOENERGÉTICOS
MITOCONDRIAIS PELO 3-BROMOPIRUVATO
Dissertação submetida ao corpo docente do Instituto de Bioquímica Médica da Universidade Federal do Rio de Janeiro – UFRJ, como parte dos requisitos necessários à obtenção do grau de Mestre em Ciências Biológicas modalidade Química Biológica. Aprovada em setembro de 2009 por: __________________________________________________________ DR. ANTONIO GALINA FILHO – ORIENTADOR PROFº ADJUNTO INSTITUTO DE BIOQUÍMICA MÉDICA/ UFRJ __________________________________________________________ DR. JOÃO BATISTA TEIXEIRA DA ROCHA PROFº ASSOCIADO DO DEPARTAMENTO DE QUÍMICA- CCNE/ UFSM __________________________________________________________ DR. JOSÉ ROBERTO MEYER FERNANDES PROFº ADJUNTO INSTITUTO DE BIOQUÍMICA MÉDICA/ UFRJ __________________________________________________________ DR. ROBSON DE QUEIROZ MONTEIRO PROFº ADJUNTO INSTITUTO DE BIOQUÍMICA MÉDICA/ UFRJ __________________________________________________________ DR. WAGNER SEIXAS DA SILVA PROFº ADJUNTO INSTITUTO DE BIOQUÍMICA MÉDICA/ UFRJ __________________________________________________________ DR. MARCELO EINICKER LAMAS PROFº ADJUNTO DO INSTITUTO DE BIOFÍSICA CARLOS CHAGAS FILHO/ UFRJ
Rio de Janeiro/2009

iv
Dedico este trabalho primeiramente à Deus por tudo. Aos meus pais e minha avó Maria (in memoriam) por sempre estarem ao
meu lado em todos os momentos.

v
AGRADECIMENTOS
A Deus em primeiro lugar, pois sem ele nada disso seria possível.
Ao meu pai, minha mãe e minha avó (que não está mais presente) pelo amor,
dedicação, companherismo, amizade e principalmente pela paciência.
A toda minha família, tios, tias, primos, primas que são essenciais na minha vida.
Aos meus irmãos do coração Rafaela, Marianna, Andréia, Fernanda, Dimitri, Elisa,
Laura, André e Vanessa por sempre estarem presentes na minha vida em todos os
momentos.
A todos os meus amigos que fiz no Instituto de Bioquímica Médica, sem dúvida
pessoas que somaram muito na minha trajetória.
Ao meu querido orientador Antonio Galina por tudo que me ensinou.
A toda equipe do laboratório antiga e atual Marne, Nattascha, Laudiene, Bruno,
André, Juliana, Ana Paula Pereira, Ana Paula Santiago, Vagner, Andreza, Douglas,
Murilo, Luiza e Daniele pelo carinho, amizade e ajuda.
A todas as pessoas importantes que já passaram pela minha vida e aos meus novos
amigos.

vi
RESUMO
Carcinoma hepatocelular (HCC) é um dos tumores malignos mais letais, apesar dos avanços científicos para detecção de HCC em estágio avançado. A maioria das lesões se desenvolve em pacientes com cirrose ou hepatite viral. Experimentos iniciais mostraram que o agente alquilante 3-Bromopiruvato (3-BrPA) tem atividade anti-tumoral baseada nas propriedades anti- proliferativas em células do hepatoma. Tem sido proposto que 3-BrPA causa distúrbios na glicólise e no consumo de oxigênio levando a uma diminuição na taxa de síntese de ATP. Apesar desta observação, o mecanismo bioquímico detalhado de ação do 3-BrPA na respiração mitocondrial em células tumorais e normais é desconhecido. Trabalhos recentes descreveram que 3-BrPA inibe a atividade da succinato desidrogenase (SDH) reagindo com grupamentos – SH, mas a influência no estado respiratório mitocondrial é desconhecido. O objetivo deste estudo foi investigar o efeito do 3-BrPA nos aparatos da fosforilação oxidativa da mitocôndria de fígado de camundongo; ex. na cadeia transportadora de elétrons (CTE) e a atividade da FoF1-ATPsintase. A atividade ATPase da FoF1-sintase foi somente parcialmente inibida (25%) por 1mM 3-BrPA. Nenhuma inibição na atividade ATPase foi detectada em concentrações mais baixas de 3-BrPA. Porém, a atividade da succinato desidrogenase (SDH) (complexo II) é inibida 50% em 100µM de 3-BrPA na presença ou ausência de ADP. A atividade da NADH: ubiquinona oxidoredutase (complexo I) não foi inibida por 500µM de 3-BrPA. Todavia, a pré- incubação da mitocôndria por 30 minutos com 3-BrPA causou uma grande inibição em 75 µM de 3-BrPA. As inibições observadas pelo 3-BrPA nas atividades dos complexos da CTE foram observadas na respiração e no potencial de membrana (ΔΨm). A inibição mais evidente da respiração e ΔΨm foi observada usando succinato como substrato. Em adição, quando a mitocôndria estava em um estado fosforilando ADP (estado 3), o efeito induzido pelo 3-BrPA foi maior. O agente desacoplador (FCCP), causando o mesmo nível de despolarização do estado 3, não foi capaz de aumentar inibição pelo 3-BrPA da respiração estimulada pelo succinato. Esses efeitos inibitórios do 3-BrPA foram revertidos ou atenuados pela glutationa reduzida (GSH) na mitocôndria de fígado de camundongo. Quando a respiração semelhante ao estado 3 foi induzida pela reação da HK de levedura, 3-BrPA inibiu mais rapidamente do que na ausência da reação catalisada pela HK. Os mesmos resultados foram observados usando mitocôndria de cérebro de camundongo que apresenta HK ligada a membranas externas. Esses resultados foram confirmados quando o carreador ANT foi parcialmente ou totalmente bloqueado por carboxiatractilosídeo. Esses resultados sugerem que HK-II ligada mitocondrialmente encontrada em tumores facilita a inibição pelo 3-BrPA em mitocôndrias de tumores através da ciclagem de ADP. A proteção causada pelo GSH sugere que importantes grupos - SH presentes no sítio catalítico da succinato desidrogenase são potencialmente alvos de alquilação promovida pelo 3-BrPA.

vii
ABSTRACT
Hepatocellular carcinoma (HCC) is one of the most lethal tumors despite of the scientific improvements for detection of HCC in advance it. The most injuries occur in patients with cirrhosis or viral-induced hepatitis. Early experiments showed that the alkylating agent 3-Bromopyruvate (3-BrPA) has been used as an anti-tumoral drug based on its anti-proliferative property in hepatomas cells. This effect has been proposed to occur by disturbance of glycolysis and respiration leading to a decreased rate of ATP synthesis. Despite of this observation, the detailed mechanism of action of 3-BrPA in mitochondrial respiration in normal and tumor cells is unknown. Previous works described that 3BrPA inhibits succinate dehydrogenase (SDH) activity reacting with SH groups, but the influence of mitochondrial respiration states is unknown. The aim of this study was investigate the effect of 3-BrPA on mice liver oxidative phosphorylation apparatus of mitochondria; i.e. electron transport chain (ETC) and FoF1-ATPsynthase activity. The FoF1-ATPase activity was only partially inhibited (25%) by 1mM 3-BrPA. No inhibition in ATPase activity was detected at lower 3-BrPA concentration. However, the succinate dehydrogenase (SDH) activity (complex II) is inhibited in 50 % at 100 µM 3-BrPA either in the presence or in the absence of ADP. The NADH: oxidoreductase activity (complex I) was not inhibited by 500 µM 3-BrPA. However, pre-incubation of mitochondria by 30 minutes with 3-BrPA caused a pronounced inhibition at 75 µM 3-BrPA. The inhibitions observed by 3-BrPA in the ETC complexes activities were coupled to respiration and membrane potential (ΔΨm). The most pronounced inhibition of respiration and ΔΨm was observed using succinate as substrate. In addition,when the mitochondria was in a phosphorylating ADP state (state 3), the effect induced by 3-BrPA was even more marked. The proton uncoupler (FCCP), set to cause the same level of ΔΨm depolarization of state 3, was not able to improve the 3-BrPA inhibition of respiration sustained by succinate. These inhibitory effects of 3-BrPA were reverted or attenuated by reduced glutathione (GSH) on mice liver mitochondria. When the respiration was induced by yeast HK reaction (state 3 like), 3-BrPA inhibition was faster than in the absence of HK catalyzed reaction. The same results were observed using mice brain mitochondria that present HK bound to the outer membranes. These results were confirmed when the ANT carrier was partially or totally blocked by carboxyatractiloside. These results suggest that mitochondrially bound HK type II found in tumors facilitates the 3-BrPA inhibition in tumors mitochondria through ADP re-cycling. The protection caused by GSH suggest that important –SH groups present at the catalytic site of succinate dehydrogenase are potentially target of alkylation promoted by 3-BrPA.

viii
LISTA DE ABREVIATURAS
ADP: adenosina 5'- difosfato ANT: adenine nucleotide translocator ATP: adenosina 5'- trifosfato
BSA: do inglês : bovine serum albumin COX: citocromo c oxidase CTE: cadeia transportadora de elétrons
DCIP: 2,6- diclorofenolindofenol EDTA: do inglês : Ethylenediamine tetraacetic acid
EGTA: do inglês :
ethylene glycol tetraacetic acid
FAD+
: do inglês : flavine adenine dinucleotide
FADH2 : do inglês : flavine adenine dinucleotide hydrogenase
FCCP: do inglês : Carbonylcyanide-4-(trifluoromethoxy)-phenylhydrazone
FDG: fluor-deoxiglicose FMN: flavina mononucleotídeo GSH: do inglês : Glutathione G-6-P: glicose 6- fosfato
HCC: hepatocarcinoma
HK: hexocinase HK- II: hexocinase tipo II NADH: do inglês: nicotinamide adenine dinucleotide hydrogenase NEM: N- etilmaleimida
PET: do inglês: positron emission tumography Pi: fosfato inorgânico
PMS: fenazina metilsulfato

ix
SDH: succinato desidrogenase TACE: do inglês: Transarterial chemoembolization Triton X-100: (t- Octilfenoxipolietoxietanol) VDAC: do inglês: Voltage-dependent anion
‘ channel
3-BrPA: 3-bromopiruvato

x
LISTA DE FIGURAS
Figura 1: Variações regionais na taxa de mortalidade causada pelo HCC 5
Figura 2: Método usado na administração de agentes quimioterapêuticos via
injeção intraarterial direta em tumor de fígado.
7
Figura 3: Estrutura e reatividade do 3-BrPA 15
Figura 4: Identificação das estruturas mitocondriais 19
Figura 5: Esquema representando a cadeia transportadora 22
Figura 6: Estados respiratórios 30
Figura 7: Atividade da NADH:Ubiquinona Oxidoredutase em presença
de 3BrPA 41
Figura 8: 3-BrPA não afeta o consumo de oxigênio quando
utilizados substratos do complexo IV
42
Figura 9: Efeito do 3-BrPA na hidrólise de ATP pela FoF1 44 -ATPase
Figura 10: Efeito do 3BrPA na atividade da succinato desidrogenase
na presença e na ausência de ADP
46
Figura 11: 3-BrPA diminui a formação do potencial de membrana
mitocondrial (ΔΨm) pela cadeia transportadora de elétrons(ETC)
49
Figura 12: Inibição do consumo de oxigênio é dependente do
estado respiratório mitocondrial
51
Figura 13: Efeito do 3-BrPA na despolarização do potencial de
membrana mitocondrial (ΔΨm) usando succinato
(substrato do complexo II) nos estados respiratórios 3 e 4
53
Figura 14: Diferenças na despolarização da formação potencial
de membrana mitocondrial (ΔΨm) usando substratos do
complexo I e II da cadeia transportadora de elétron (ETC)
na presença de 3-BrPA no estado 3
54
Figura 15: Glutationa reduzida (GSH) protege a dissipação do potencial
de membrana (ΔΨm) promovida pelo 3BrPA
56
Figura 16: Ciclagem ADP/ATP promovida pelo
complexo ANT.F0F1ATPsintase.HK aumenta a
sensibilidade da succinato desidrogenase ao 3-BrPA
58

xi
Figura 17: Despolarização parcial do potencial de membrana
Mitocondrial (ΔΨm) pelo ionóforo de próton FCCP não aumenta
a sensibilidade da succinato desidrogenase a inibição pelo
3-BrPA comparada a reciclagem de ADP pela atividade da HK
60
Figura 18: Importância da ciclagem de ADP no efeito do 3-BrPA 61
Figura 19: Correlação entre a taxa da respiração induzida pela atividade
da HK e a reatividade ao 3-BrPA
63

xii
SUMÁRIO
Resumo Vi Abstract Vii Lista de Abreviaturas Viii Lista de Figuras X 1- INTRODUÇÃO 2 1.1- Câncer 2 1.2- Epidemiologia do câncer de fígado 3 1.3- Hepatocarcinoma 4 1.4- Tratamento para o hepatocarcinoma 6 1.5- Aspectos gerais do metabolismo energético em células tumorais 8 1.6- Hexocinase 10 1.7- Hexocinase tipo II e células tumorais 12 1.8- 3-Bromopiruvato como um agente antitumoral promissor 13 1.9- Aspectos gerais da mitocôndria e fosforilação oxidativa 17 1.9.1- NADH: ubiquinona Oxidoredutase ou complexo I 23 1.9.2- Succinato- ubiquinona oxidoredutase ou complexo II 24 1.9.3- Ubiquinol: Citocromo c oxidoredutase, complexo citocromo
bc1
25
ou complexo III 1.9.4- Citocromo c oxidase ou complexo IV 26 1.9.5- ATP sintase ou complexo V 27 1.10- Estados respiratórios mitocônddriais 29 2- OBJETIVOS 32 2.1- Objetivo Geral 32 2.2- Objetivos Específicos 32 3- MATERIAIS E MÉTODOS 34 3.1- Animais 34 3.2- Isolamento de mitocôndria 34 3.3- Preparo da solução de 3-BrPA 35 3.4- Hidrólise de ATP 35 3.5- Consumo de oxigênio em mitocôndrias isoladas 35 3.6- Medidas do potencial de membrana mitocondrial (∆ψm 36 ) 3.7- Determinação da atividade da NADH: ubiquinona oxidoredutase 36 3.8- Ensaio da atividade da succinato desidrogenase 37 3.9- Quantificação de proteínas 38 3.10- Análise Estatística 38 4-RESULTADOS 40
4.1- Efeito do 3-BrPA nas atividades dos complexos I e IV da cadeia
transportadora de elétrons
40
4.2 - 3-BrPA não afeta a atividade da FoF1 43 - ATPsintase 4.3- 3-BrPA inibe a atividade da SDH 45 4.4- Inibição da formação do potencial de membrana (ΔΨm) e
consumo de oxigênio pelo 3-BrPA confirmam os efeitos na CTE
47
4.5- 3-BrPA reage diferentemente com os complexos da CTE e
depende dos estados respiratórios
50
4.6- Efeito protetor da glutationa reduzida (GSH) a inibição pelo 55

xiii
3BrPA 4.7- O efeito inibitório do 3-BrPA é modulado pelos estados
respiratórios da mitocôndria
57
5- DISCUSSÃO 65 6- CONCLUSÕES 71 7- REFERÊNCIAS 73

1- INTRODUÇÃO _______________________________________

Introdução |2
1- INTRODUÇÃO:
1.1- Câncer.
Apesar da enorme quantidade de estudos sobre o câncer durante a metade
do século passado por agências consolidadas através do mundo, a guerra contra
esta doença frequentemente fatal não se mostra iminente. Alguns estudos relatam
que atualmente um em cada dois homens e uma em cada três mulheres morre de
câncer. Embora durante o século passado um número de doenças tenham sido
seriamente encurtadas, o câncer está se tornando a doença número um no mundo
(Pedersen, 2007).
Câncer é caracterizado pelo crescimento descontrolado e propagação de
células anormais conhecido como neoplasias, que podem ser benignas ou malignas,
invadindo tecidos e órgãos podendo espalhar-se (metástase) para outras regiões do
corpo resultando em morte se essa propagação não for controlada.
Existem diferentes tipos de câncer correspondentes aos vários tipos de
células do corpo. Se o câncer tem início em tecidos epiteliais como pele ou
mucosas, é denominado carcinoma. Em tecidos conjuntivos como osso, músculo ou
cartilagem é chamado de sarcoma, linfa (linfoma), células neuroendrócrinas
(carcinóide), células geminativas (teratoma e seminoma), vasos sanguíneos
(hemangiossarcomas), vasos biliares (colangiossarcomas) e vasos linfáticos
(linfangiossarcomas)
Em 2005, de um total de 58 milhões de mortes ocorridas no mundo, o câncer
foi responsável por 7,6 milhões, o que representou 13%. Os principais tipos de
câncer com maior mortalidade foram: pulmão (1,3 milhão), estômago (cerca de 1
milhão), fígado (662 mil), cólon (655 mil) e mama (502 mil).
. O câncer é causado por ambos, fatores externos (tabaco,
organismos infecciosos, químicas e radiação) e fatores internos (mutações
herdadas, hormônios, condições imunes e mutações que ocorrem do metabolismo)
sendo que esses fatores podem agir juntos ou de maneira sequencial no
desenvolvimento da carcinogênese (American Cancer Society, 2009).

Introdução |3
Do total de óbitos por câncer ocorridos em 2005, mais de 70% ocorreram em
países de média ou baixa renda (World Health Organization, 2006). Estima-se que
em 2020 os números de casos novos anuais sejam da ordem de 15 milhões, tendo
cerca de 60% desses caos ocorrendo em países em desenvolvimento. Acredita-se
que um terço dos casos novos de câncer diagnosticados anualmente no mundo
poderia ser prevenido. Parkin et al. (2001) estimaram para o ano de 2000 que o
número de casos novos de câncer em todo o mundo seria maior que 10 milhões. No
Brasil, as estimativas para o ano de 2008 e válidas também para o ano de 2009
apontam que ocorrerão 466.730 novos diagnósticos (INCA, 2008).
1.2- Epidemiologia do câncer de fígado.
Dentre os tipos de câncer que estão apresentando um aumento na sua
incidência está o câncer de fígado. Este é dividido em duas categorias: o primário e
o secundário ou metastático (originado em outro órgão e que atinge também o
fígado). O termo "primário" é usado nos tumores originados no fígado, como o
hepatocarcinoma ou carcinoma hepatocelular (tumor maligno primário mais
frequente que ocorre em mais de 80% dos casos), o colangiocarcinoma (que
acomete os ductos biliares dentro do fígado), angiossarcoma (tumor do vaso
sangüíneo) e o hepatoblastoma (que acomete as crianças).
O câncer de fígado é o sexto tipo mais comum no mundo, apresentando
626.241 casos novos por ano. Sua incidência é 2,5 vezes maior em homens do que
em mulheres. Entre 1973 e 1997 a taxa de incidência aumentou em todas as partes
do mundo sendo que sua taxa de incidência para ambos, homens e mulheres, são
mais altas em regiões menos desenvolvidas (Kamangar et al., 2006).

Introdução |4
1.3- Hepatocarcinoma.
Dos quatro tipos de tumores primários de fígado citados acima, o carcinoma
hepatocelular (HCC) é o mais predominante. É um neoplasma comumente originado
de um fígado doente sendo um dos poucos cânceres com fatores de riscos bem
definidos (Bosch et al.,1999). A primeira descrição científ ica foi feita por
Eggel em 1901, mostrando o resultado de mais de 200 autópsias. É a
forma mais comum de câncer de fígado primário e um dos mais letais no mundo
(Saha et al., 2001), apresentando-se entre 85-90% dos cânceres primários de
fígado. É o quinto tipo mais comum no mundo e a terceira maior causa de
mortalidade (Parkin, 2001). Dentre as numerosas vítimas, não estão somente
aqueles com tumores primários que se desenvolvem diretamente no fígado, mas
aqueles com tumores secundários que frequentemente aumentam neste órgão
metabólico crítico como resultado de metástases de outros tecidos (Saha et al.,
2001). Em 80% dos casos, o HCC se desenvolve em fígados com cirrose que é o
fator que mais predispõe o desenvolvimento de tumores (Colombo, 2003).
Outros fatores de riscos para o desenvolvimento de HCC são doenças
crônicas causadas por infecção viral, hepatites B e C (Poynard et at.,1991). O HCC
apresenta diversas características epidemiológicas que incluem variações entre
regiões geográficas, raciais, grupos étnicos, sexo e aspectos ambientais (El-Serag e
Rudolph, 2007). Sua distribuição não é uniforme, e a maioria dos casos tem sido
observada na África subsaariana e Sudeste da Ásia (> 80%) onde a infecção pelo
vírus da hepatite B é comum juntamente com a ingestão de aflatoxina B
(micotoxinas) através de alimentos contaminados. Na América do Norte, América do
Sul, Oceania e Norte da Europa as taxas de incidências são menores apresentando
menos que 5 casos a cada 100.000 pessoas (Parkin, 2002, Liaw et al.,1986 e Sun et
al., 1999) (Figura 1).
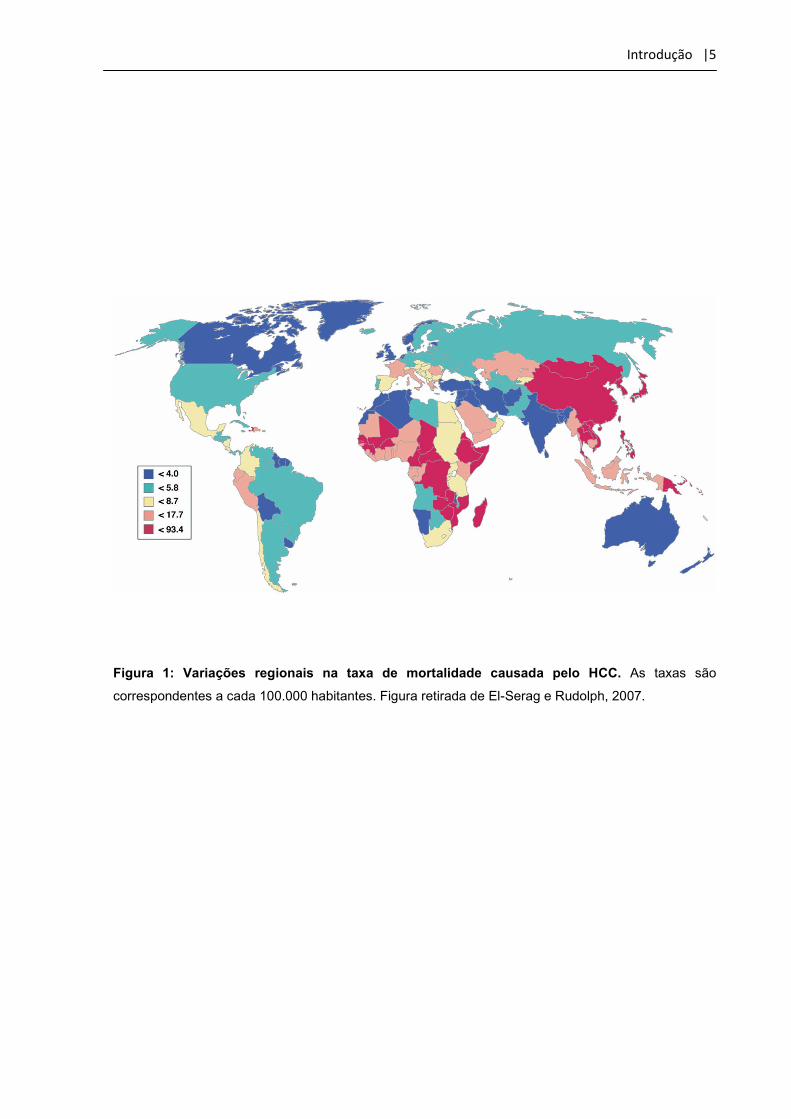
Introdução |5
Figura 1: Variações regionais na taxa de mortalidade causada pelo HCC. As taxas são
correspondentes a cada 100.000 habitantes. Figura retirada de El-Serag e Rudolph, 2007.

Introdução |6
1.4- Tratamento para o hepatocarcinoma.
Embora o HCC ocasione mais do que um milhão de mortes por ano, poucas
opções de tratamento estão disponíveis aos pacientes. O prognóstico para esses
pacientes é geralmente ineficiente, apresentando taxas de respostas baixas,
toxicidades severas e altas taxas de recorrência (Geschwind et al., 2002). Somente
uma minoria desses pacientes são candidatos a tratamentos de cura, tal como
excisão cirúrgica. Pouco menos de 10% dos pacientes sobrevivem cinco anos
depois do diagnóstico, e o período médio de sobrevivência é de quatro a seis meses
para pacientes com tumores não removíveis cirurgicamente (Okuda et al., 1985;
Rustgi, 1987; Di Bisceglie et al., 1988). Uma alternativa atualmente seria a
embolização e a quimioembolização.
Estas terapias levam vantagem pelo fato de que, tumores primários de fígado,
recebem a maior parte do suprimento sanguíneo da artéria hepática, ao contrário do
tecido do fígado normal que recebe predominantemente da veia porta (Breedis e Ko,
1954; Geschwind et al., 2000). Portanto, a embolização envolve o bloqueio da artéria
hepática alimentando o tumor com um material tipo resina misturado com uma base
de óleo (álcool polivinil em itiodol), deprivando o tumor de nutriente e oxigênio. A
quimioembolização arterial transcateter (TACE) refere-se ao mesmo procedimento,
mas com a inclusão de um ou mais agentes antitumorais (Ramsey et al., 2002).
Todavia, a artéria hepática seria um possível alvo para injeção de agentes
quimioterapêuticos em tumores de fígado embolizando-os (figura 2A e 2B). Estas
terapias têm sido amplamente investigadas e utilizadas em tratamentos de tumores
hepáticos. Os benefícios de TACE têm sido provados através de triagens clínicas
ocasionais, porém, a eficácia terapêutica é ainda limitada e a deterioração da função
do fígado é inevitável (Llovet et al., 2002). Todavia, uma nova estratégia para o
tratamento do HCC faz-se necessária.

Introdução |7
Figura 2: A e B método usado na administração de agentes quimioterapêuticos via injeção intraarterial direta em tumor de fígado. C, locais de ação do 3-BrPA na produção de ATP. Uma vez
dentro da célula tumoral, o 3-BrPA inibe a produção de ATP pela glicólise e fosforilação oxidativa.
Retirado de Geschwind et al., 2002.

Introdução |8
1.5- Aspectos gerais do metabolismo energético em células tumorais.
Em estudos bioquímicos e fisiológicos, células tumorais são geralmente
classificadas de acordo com a taxa de crescimento: lento, intermediário ou rápido
(Pedersen, 1978). Para tumores em animais experimentais, a taxa de crescimento é
determinada pelo tamanho e volume, taxa mitótica, grau de diferenciação e
incorporação de timidina (Weber, 2001). Exemplos de tumores de crescimento
rápido em camundongo incluem dois cânceres experimentais, tais como AS-30D e
carcinoma hepatocelular CH-252. Em tumores humanos, a classificação é baseada
nas características histológicas e estágio de desenvolvimento clínico. Todavia, pelo
estágio de desenvolvimento avançado e propriedades metastáticas, alguns tumores
considerados por ser de crescimento rápido são carcinoma ovariano, carcinoma da
tireóide e carcinoma uterino (Pedersen, 1978; Zu e Guppy, 2004).
Células tumorais exibem profundas diferenças histológicas, bioquímicas e
genéticas quando comparadas a tipos celulares originais e não transformadas. A
grande maioria dos tipos celulares de tumores de crescimento rápido sofre uma
grande modificação no metabolismo energético em comparação ao tecido de
origem. O mesmo se aplica a tumores experimentalmente desenvolvidos em
roedores.
A mais notória alteração do metabolismo energético em células tumorais é a
aumentada capacidade glicolítica mesmo na presença de uma alta concentração de
oxigênio (Warburg, 1930 e Pedersen, 1978). Foi observada em alguns tumores a
ocorrência de uma alta expressão de uma forma de hexocinase associada à
mitocôndria, que posteriormente foi identificada como sendo do tipo II (Nakashima et
al., 1988; Shinohara et al., 1991; Thelen e Wilson, 1991; Rempel e Mayer, 1994).
Esta enzima catalisa o primeiro passo na via glicolítica no qual o fosfato gama do
ATP é transferido para o carbono 6 da glicose. Propõe-se que a enzima teria um
papel significante em células altamente malignas na promoção do crescimento e
sobrevivência. Por esta razão, esta enzima representaria um dos possíveis alvos
para intervenção terapêutica.

Introdução |9
Outra justificativa para a glicólise aumentada em células tumorais com alta
taxa de proliferação, geralmente envolve as enzimas mitocondriais, na qual é
suposta por ser defeituosa ou menos efetiva do que enzimas glicolíticas na
competição por intermediários comuns (Warburg, 1930; Gosálvez et al., 1974).
Parlo e Coleman (1984), propuseram que a alta atividade glicolítica em
algumas células tumorais é causada pela disfunção mitocondrial a nível do ciclo de
Krebs, no qual resultaria numa baixa disponibilidade de equivalentes redutores para
a cadeia respiratória e portanto uma baixa fosforilação oxidativa, causando assim,
uma deficiência na produção de ATP que é muito importante para o crescimento do
tumor. Em tumores avançados "altamente glicolítico", é encontrado que a fonte de
energia (ATP) é derivada tanto da fosforilação oxidativa como da glicólise (Aisenberg
1961). Em um dos casos mais extremos estudados, a produção de ATP mitocondrial
foi estimada em aproximadamente 40% e a produção de ATP glicolítico em
aproximadamente 60% (Nakashima et al., 1984). Assim, a glicólise aumentada
contribuiria mais significativamente para a demanda energética por ATP. Isso é
contrastante com a maioria das células normais onde a mitocôndria é responsável
pela produção de 60% a 90% do ATP necessário a célula.
Todavia, estudos também determinaram que as taxas de oxidação de
piruvato, malato, citrato, acetoacetato e descarboxilação do acetato em células AS-
30D seriam similares a células não-tumorigênicas. De fato, as atividades de todas as
enzimas do ciclo de Krebs estavam 1-30 vezes mais alta em mitocôndria do
hepatoma AS-30D do que em mitocôndria de fígado normal (Dietzen e Davis, 1993).
Outras hipóteses para explicar a baixa eficiência do metabolismo oxidativo em
tumores altamente malignos seriam o baixo conteúdo mitocondrial por célula, cadeia
respiratória deficiente, sensibilidade aumentada do DNA mitocondrial ao estresse
oxidativo. Porém, estudos feitos em diversos tipos de hepatoma não mostraram
nenhuma diferença significativa quando comparados a células normais (Moreno-
Sánchez et al., 2007). De fato, existem linhagens de tumores nas quais certamente
exibem uma diminuída função mitocondrial, mas isso não se aplica a todos os tipos.

Introdução |10
Apesar de todas essas controvérsias, Muller et al., (1986) originalmente
propuseram que uma estratégia bioquímica para suprimir a proliferação acentuada
do tumor era o bloqueio simultâneo de ambos os modelos geradores de ATP
(glicólise e fosforilação oxidativa). Diversas drogas foram estudadas e uma delas
tem apresentado resultados satisfatórios no tratamento, o agente alquilante 3-
bromopiruvato (3-BrPA), que tem mostrado ser um inibidor em potencial da
hexocinase do tipo II (HK-II). Alguns estudos tem sido realizados sobre a ação do 3-
BrPA nesta enzima, mas há relatos da sua ação também na diminuição do consumo
de oxigênio em células do hepatoma que será explorado mais adiante (Ko et al.,
2001).
1.6- Hexocinase.
Tumores altamente malignos exibem a capacidade de metabolizar glicose a
lactato em taxas muito maiores do que células normais. Esta alta taxa é dependente
de níveis elevados de hexocinases. Em hepatomas com alta taxa de crescimento, a
hexocinase do tipo II (HK-II), uma forma capaz de se associar a membrana
mitocondrial externa, é a isoenzima predominantemente com níveis elevados
(Nakashima et al., 1988; Mathupala et al., 1995). Uma proposta de terapia
anticâncer seria o bloqueio do metabolismo energético através da inibição da HK-II.
O sufixo "cinase" (do inglês "Kinase") foi introduzido por Otto Meyerhof em
1927, quando este pesquisador verificou um grande aumento na taxa de ácido lático
ao adicionar glicose e extrato de levedura em homogenados de músculo. Meyerhof
decidiu buscar no extrato de levedura o fator protéico que promovia tal fenômeno.
Isolou uma fração protéica que em extratos de músculo era capaz de acelerar a
formação de lactato. Resolveu batizar a enzima como hexocinase (do inglês
hexokinase), ou enzima capaz de acelerar o uso da glicose no músculo (Kinase-
enzima que favorece a dinâmica (o metabolismo); hexose = açúcar de seis
carbonos. No inglês: hexokinase, no português hexocinase).

Introdução |11
Posteriormente Euler e Adler em 1935 verificaram a formação de hexose-
monofosfato quando misturavam hexose, ATP e íons de magnésio na fração
protéica purificada por Meyerhof. A hexocinase (EC 2.7.1.1) é uma enzima presente
em fungos, bactérias e todos os tecidos animais de vertebrados e invertebrados e
vegetais.
Hexocinase (ATP: D-hexose-6-fosfotransferase) catalisa o primeiro passo no
metabolismo da glicose:
Glicose + ATP → glicose-6- fosfato + ADP
A base para esta reação é o armazenamento da glicose-6-fosfato (G-6-P)
dentro da célula para sua utilização na via glicolítica, primeiramente para a geração
de energia (ATP) via glicólise e fosforilação oxidativa, metabolismo do glicogênio ou
a utilização desse metabólito para a via das pentoses sendo importante
principalmente para reações biossintéticas (Pedersen, 2007).
Em mamíferos há quatro isoformas diferentes de hexocinase (HK-I, -II, -III e –
IV ou glicocinase), nas quais diferem em propriedades cinéticas, expressão tecidual
e localização subcelular. Tipos I, II e III mostram alta afinidade pela glicose
apresentando o KM 250 vezes menor para glicose (KM = ~0.02 mM), são inibidas
pelo produto glicose-6-fostato (G-6-P). Todavia, a isoenzima do tipo IV tem afinidade
muito baixa pela glicose (KM
A isoforma tipo I é predominantemente encontrada no tecido cerebral
associada à membrana externa da mitocôndria através da porina. Também é
encontrada no seio, rim, retina e está presente também em alguns tumores
altamente glicolíticos (Kurokama et al., 1982; Parry et al., 1983; Arora et al., 1990;
Rempel et al., 1994), mas em níveis muito baixos relativo a Hexocinase do tipo II
(HK-II).
= 5 mM) sendo insensível a inibição por G-6-P
(Pedersen et al., 2002 e Pedersen, 2007). Baseado em análises de sequências
primária de aminoácidos, postula-se que o gene HK-IV, precursor das HKs sofreu
duplicação e fusão. Assim, a enzima HK-IV tem uma massa molecular de
aproximadamente 50 kDa enquanto as outras hexocinases tem uma massa
molecular de 100 kDa (Pedersen, 2007).

Introdução |12
A isoforma tipo II está presente no músculo esquelético, adipócitos e é
também a isoforma predominante em células tumorais. Esta isoforma também
apresenta a propriedade de se associar a mitocôndria. A isoforma III possui uma
localização perinuclear na região citoplasmática cerebelar do neurônio de Purkinje e
a isoforma do tipo IV é expressada em células do fígado (hepatócitos) e em células
pancreáticas recebendo o nome de glicocinase tendo uma localização nuclear ou
citosólica dependendo da disponibilidade de glicose no fígado, não se apresentando
associada à mitocôndria.
1.7- Hexocinase tipo II e células tumorais.
Em células tumorais de crescimento rápido, com exceção dos tumores
cerebrais nos quais possuem a HK-I como isoforma predominante, a HK-II é a
isoforma predominante e apresenta uma atividade de 20 a 306 vezes maior em
hepatomas de Novikoff, H19 e AS-30D do que em hepatócitos (Parry e Pedersen,
1983; Nakashima et al., 1988; Marin-Hernandez et al., 2006). Isto não é
característico de todos os tumores, mas sim de tumores com alta taxa de
proliferação pobremente diferenciados e que são altamente malignos.
A HK-II se liga a canais transmembranares formados pela proteína
denominada "porina" ou "VDAC" (canais de ânion voltagem dependente) inserida na
membrana mitocondrial externa (Nakashima et al., 1986). Esta interação reduz
marcadamente a sensibilidade da enzima à inibição pela G-6-P, provê acesso ao
ATP produzido pela mitocôndria e protege contra a degradação proteolítica (Rose e
Warms, 1982; Arora e Pedersen, 1988). Como já foi descrito pelo grupo do Dr.
Pedersen, essas propriedades combinadas juntamente com uma alta quantidade
desta enzima em tumores malignos resultam em uma rápida produção de G-6-P.
Este intermediário metabólico chave serve não somente como uma fonte de carbono
para biossíntese, que é essencial para o crescimento e rápida proliferação dos
tumores, mas também como substrato inicial para glicólise que gera ATP na
conversão à lactato.

Introdução |13
Células cancerígenas altamente malignas utilizam HK-II não somente para
protegê-las durante mudanças bruscas no estado metabólico e fisiológico, mas
também contra a morte celular. Tem sido demonstrado que a HK-II inibi a ligação de
BAX, uma proteína pró-apoptótica, a membrana mitocondrial externa inibindo a
liberação do citocromo c e, portanto, a apoptose (Pastorino et al., 2003). Como uma
forma de detecção, testes diagnósticos e PET (Tumografia de Emissão de Positron)
são importantes na identificação de tumores altamente glicolíticos com altos níveis
de HK-II. PET é uma técnica que provê imagens de processos metabólicos
utilizando FDG (18F-2-fluoro-2-deoxi-D-glicose), um análogo radiofarmacêutico da
glicose. Este é captado e fosforilado de maneira similar a glicose e a taxa de
captação pelas células tumorais é proporcional a atividade metabólica das mesmas.
(Rohren et al., 2004; Kapoor et al., 2004).
Considerando os vários papéis que a HK-II apresenta em células tumorais,
esta se tornou um alvo atrativo para intervenções terapêuticas. Geschwind em 2000,
propôs que HKII seria alvo de alquilação pelo 3-BrPA. Neste estudo, os autores
através de injeção intra-arterial deste composto, identificaram a HKII como alvo
principal até o momento, embora trabalhos já tenham sugerido sua ação na
fosforilação oxidativa (Ko et al., 2001, Pereira da Silva et al., 2009).
1.8- 3-Bromopiruvato como um agente antitumoral promissor.
Uma alternativa para interromper o crescimento de tumores caracterizados
como "altamente glicolíticos" e que demanda de quantidades significantes de ATP
seria a descoberta de um agente que fosse capaz de bloquear a formação de ATP
nas vias metabólicas das células tumorais.
Dessa forma, Ko e colaboradores em 2001 fizeram um exaustivo estudo de
vários compostos buscando por inibidores mais potentes e específicos de ambas as
vias, glicólise e fosforilação oxidativa. Neste estudo, foi usado o tumor VX2, que é
um tumor epidermóide de coelho induzido pelo vírus papiloma.

Introdução |14
Esse tumor tem sido descrito como um bom modelo para o estudo das
propriedades de crescimento e do sistema de vascularização encontrado em
tumores de fígado humano (Pauser et al., 1996; Geschwind et al., 2000).
Semelhante aos tumores de fígado, o tumor VX2 é alimentado através da artéria
hepática, enquanto o fígado é alimentado através da veia porta.
Dentre compostos testados, 3-Bromopiruvato (3-BrPA) mostrou-se um
potente inibidor das vias de síntese de ATP (Ko et al., 2001). Esse agente alquilante
agiria como uma potente droga antitumoral capaz de inibir a proliferação de células
do hepatoma (figura 3). Este composto é um análogo do lactato e baseado na
hipótese da similaridade estrutural ao lactato, é sugerido que entre nas células
cancerígenas pelo mesmo transportador que o lactado e induzir a depleção de ATP
(Ko et al., 2001; Geschwind et al., 2002) (figura 2C).

Introdução |15
d
Figura 3: Estrutura e reatividade do 3-BrPA segundo Ko et al., 2004.
Ácido lático Ácido Pirúvico 3-BrPA
Piruvilação da proteína (HK) (3-BrPA) HK HBr

Introdução |16
Com a descoberta do 3-BrPA como um inibidor da proliferação em modelos
de fatias de tumores VX2 em cultura, estudos foram realizados focando na
administração direta desse agente via injeção intra-arterial em tumores VX2
implantados no fígado de coelho para observar a capacidade de inibição do
crescimento (Geschwind et al., 2002). Os resultados obtidos comprovaram o grande
potencial desta droga em inibir a viabilidade das células tumorais com uma única
dose, pois comparado ao grupo controle (tumores não tratados com 3-BrPA), que
apresentou 100% das células viáveis, a droga foi capaz de reduzir a viabilidade das
células tumorais em 84% de células não viáveis sem causar qualquer dano no tecido
sadio.
Outro estudo também feito pelo grupo do Dr. Pedersen utilizou o hepatoma
AS-30D como modelo. Esse tumor é extensivamente estudado por ser altamente
glicolítico e apresentar altos níveis de HK-II associada à mitocôndria. Nesse estudo,
foi observado o efeito inibitório do 3-BrPA em tumor crescendo na cavidade
abdominal de ratos em diferentes estágios. A administração única da droga foi capaz
de inibir tumores moderadamente avançados até tumores sólidos avançados. Os
resultados obtidos mostraram a regressão e o completo desaparecimento dos
tumores entre 1-4 semanas sem qualquer recorrência meses após o tratamento e
sem necessidade de terapia adicional (Ko et al., 2004).
Análises histológicas do tecido hepático localizado perifericamente ao local da
administração do 3-BrPA seja por administração direta no tumor ou por injeção intra-
arterial, não apresentou nenhum dano. Esse resultado difere das terapias
convencionais, embolização ou quimioembolização descritos anteriormente, nos
quais se observa danos significantes no tecido próximo ao tecido maligno. Também
não foram observados danos aparentes em outros tecidos dos animais com tumor
de fígado tratados com 3-BrPA .
Até recentemente acreditava-se que o principal alvo da inibição pelo 3-BrPA
seria a HKII (Nakashima et al., 1988; Marin-Hernandez et al., 2006). Alguns estudos
demonstraram que 5 mM de 3-BrPA é capaz de inibir a reação da HK (Ko et al.,
2001). No entanto, 3-BrPA foi também descrito por ser um inibidor irreversível da
piruvato desidrogenase (Maldonado et al.,1972) e da succinato desidrogenase
(Sanborn et al., 1971; Kenney , 1975), da glutamato desidrogenase (Baker e Rabin,

Introdução |17
1969) e enzima málica (Chang e Hsu, 1973; Satterlee e Hsu, 1991) agindo então na
respiração mitocondrial. Assim como também de enzimas da via glicolítica como a
gliceraldeído 3-fosfato desidrogenase e a fosfoglicerato cinase (Jones et al., 1995).
Apesar destes estudos, o mecanismo de ação desta droga na respiração
mitocondrial de células normais e tumorais em concentrações mais baixas ainda não
foi investigado. O estudo a nível mitocondrial foi iniciado pelo nosso grupo utilizando
mitocôndrias isoladas de fígado sadio e células tumorais. Em células tumorais, foi
utilizado como modelo a HepG2 que apresenta um fenótipo glicolítico semelhante a
de outras células tumorais. Neste estudo, a HK mitocondrial não parece ser o alvo
preferencial do 3-BrPA quando doses baixas foram utilizadas. Outras enzimas da via
glicolítica parecem ser mais afetadas em concentrações mais baixas desta droga.
Porém, para estas concentrações, especulou-se que a succinato desidrogenase
possa ser um alvo promissor (Pereira da Silva et al., 2009).
Para uma melhor compreensão de como esta droga poderia estar agindo na
fisiologia mitocondrial, será apresentada uma breve revisão sobre a fosforilação
oxidativa e uma descrição dos complexos transportadores de elétrons localizados na
membrana interna mitocondrial que são o foco deste estudo.
1.9- Aspectos gerais da mitocôndria e fosforilação oxidativa.
As primeiras observações realizadas acerca das organelas que conhecemos
como mitocôndrias datam de meados do século XIX e foram feitas por diversos
citologistas que as descreveram como elementos granulares e inclusões presentes
no citoplasma de diferentes tipos celulares (Lehninger, 1965). Hoje, a mitocôndria é
considerada a principal organela transdutora de energia em células eucarióticas. É
nesta organela que ocorre a fase aeróbica do catabolismo das hexoses, ácidos
graxos e alguns aminoácidos que são oxidados a moléculas de acetil-coenzima A
(acetil-CoA) e alimentam as reações do ciclo dos ácidos tricarboxílicos (ciclo de
Krebs), gerando o potencial redutor a ser utilizado na fosforilação oxidativa. A
principal função mitocondrial é a conversão de energia armazenada em NADH e
FADH2 de processos metabólicos que ocorrem na matriz mitocondrial para a
molécula de ATP.

Introdução |18
São compostas de 2 compartimentos (espaço intermembrana e a matriz),
cada um com funções metabólicas específicas, demarcadas pelas membranas
mitocondriais internas e externas. A membrana externa envolve a organela,
enquanto a membrana interna envolve o espaço da matriz. O espaço intermediário é
localizado entre as duas membranas (figura 4). A membrana externa é impermeável
a moléculas maiores do que 1500 Da. O espaço intermembrana contém grupos
distintos de proteínas, incluindo o carreador de elétron móvel, citocromo c. A
membrana interna apresenta uma superfície de membrana maior com grande
quantidade de invaginações formando as cristas, que contêm a cadeia
transportadora de elétrons (CTE), aparatos da fosforilação e transportadores de
membrana, sendo porções da membrana interna que se aproximam da membrana
externa formando áreas de contato (Nicolay et al., 1990; Brdiczka e Wallimann,
1994). Essas áreas de contato participam da importação de proteínas, nucleotídeos
de adenina e ácidos graxos para a mitocôndria. Proteínas translocases,
translocadores de nucleotídeo de adenina (ANT), canais de ânion voltagem
dependente (VDAC), bem como enzimas de ativação e transporte de ácidos graxos,
são componentes dessas áreas de contato (Kerner e Hoppel, 2000).
Na CTE, os doadores de elétrons NADH e FADH2
Além do NADH e FADH
, são equivalentes redutores
formados na glicólise, oxidação de ácidos graxos e no ciclo do ácido cítrico. Essas
moléculas são ricas em energia, cada uma contendo um par de elétrons e possuindo
um grande potencial de transferência.
2
, três outros tipos de moléculas transportadoras de
elétrons funcionam na cadeia respiratória: uma quinona hidrofóbica (ubiquinona) e
dois tipos diferentes de proteínas que contêm ferro (citocromos e proteína ferro-
enxofre). Nas proteínas ferro-enxofre, o ferro está presente não no grupamento
heme, mas associados a átomos de enxofre inorgânico ou a átomos de enxofre de
resíduos de cisteína na proteína, ou a ambos.

Introdução |19
Figura 4: Identificação das estruturas mitocondriais. Retirada de
library.thinkquest.org/C004535/mitochondria.htlm.

Introdução |20
A transferência de elétrons de NADH ou FADH2 para oxigênio é acoplada ao
transporte ativo de prótons da matriz para o lado citosólico da membrana interna
formando um gradiente como descrito pela teoria quimiosmótica de Mitchell (Mitchell,
1976). Este gradiente, também conhecido como força próton motriz, é gerado pelos
três complexos respiratórios (I, III e IV) nos quais utilizam a energia livre liberada
durante o transporte de elétron para translocar prótons da matriz mitocondrial para o
espaço intermembrana. A força próton motriz tem dois componentes: o potencial de
membrana, no qual surge do movimento de cargas positivas através da membrana
interna (gradiente elétrico) e o gradiente de pH (gradiente químico). A membrana
interna da mitocôndria é impermeável aos prótons e eles voltam para matriz
somente através de canais específicos para prótons (Fo
O sistema de enzimas mitocondriais e moléculas carreadoras nas quais
transportam equivalentes redutores dos substratos ao oxigênio são coletivamente
conhecidos como sistema de transporte de elétron ou cadeia respiratória. A CTE é
composta de cinco complexos multienzimáticos nos quais suas características serão
brevemente exploradas: NADH: ubiquinona oxidoredutase, succinato- ubiquinona
oxidoredutase, ubiquinol: Citocromo c oxidoredutase, citocromo c oxidase e ATP
sintase.
). A força próton motriz que
direciona os prótons de volta para a matriz propicia a energia para a síntese de ATP
que é catalisada pela FoF1-ATPsintase (complexo V). Tanto a fosforilação oxidativa
como a teoria quimiosmótica estão representadas na figura 5.

Introdução |21
Fumarato
Figura 5: Esquema representando a cadeia transportadora de elétrons na membrana interna mitocondrial. A oxidação do piruvato gera NADH que é oxidado pelo complexo I. Este doa
elétrons para a coenzima Q que vai ser oxidada pelo complexo III. Assim, o complexo III reduz o
citocromo c. Citocromo c oxidase (complexo IV) transfere elétrons do citocromo c ao oxigênio
molecular. Succinato é oxidado pelo complexo II, reduzindo Q. Os elétrons do NADH e outros
substratos oxidáveis passam através de uma cadeia de carregadores arranjados na membrana
interna. O fluxo de elétrons é acompanhado por uma transferência de prótons através da membrana
produzindo um gradiente químico (∆pH) e um gradiente elétrico (ΔΨ), conhecido como força próton
motriz , sendo o intermediário para a síntese de ATP no modelo quimiosmótico estabelecido por
Mitchell, 1976. (adaptado do livro the mitochondrion, Lehninger,1965).
Matriz
Espaço Intermembrana
Citocromo c
Succinato
Complexo I Complexo II Complexo III Complexo IV Complexo V

Introdução |22
1.9.1- NADH: ubiquinona Oxidoredutase ou complexo I.
O complexo I é um dos pontos de entrada para os
elétrons na cadeia respiratória. Este se encontra localizado na
membrana interna mitocondrial e catalisa a transferência de
elétrons de NADH para o pool de ubiquinona. Os componentes
redox intrínsecos envolvem uma flavina mononucleotídeo ligada
não covalentemente (FMN), seis grupamentos ferro-enxofre (Fe-
S) detectáveis por ressonância paramagnética de elétrons (RPE)
e duas espécies de quinona distintas ligadas a proteína (Grigorieff, 1999).
NADH: ubiquinona oxidoretudase é o maior complexo e têm sido bem
estudado em células de mamíferos, plantas, fungos e bactéria. A composição das
subunidades de Escherichia coli, N.crassa e enzimas bovinas indica homologia entre
as subunidades apresentando uma massa molecular variando de 550KDa para a
enzima bacteriana a ~1000KDa na mitocôndria de coração bovina (Grigorieff, 1999).
Este complexo tem um centro comum de 14 subunidades onde são encontradas em
bactérias, enquanto em eucariotos apresentam subunidades adicionais com um total
de 45 em mitocôndrias de coração bovina (Vonck e Schäfer, 2009). Nesse
complexo, a etapa inicial é a transferência de elétrons do NADH para a flavina
mononucleotídeo (FMN) originando FMNH2
Em todos os organismos, o complexo I tem a forma de L e consiste de um
braço na membrana e outro braço periférico ou na matriz (Vonck e Schäfer, 2009).
Os elétrons são então transferidos do FMNH
(forma reduzida).
2 para uma série de centros ferro-
enxofre (Fe-S) que é o segundo grupo prostético da NADH desidrogenase e
exercem um papel crítico nas reações de redução em sistemas biológicos. O
complexo I tem diversos grupos tióis críticos e que são vulneráveis a modificações
oxidativas resultando em perda da atividade. A inibição do complexo I pode provocar
o escape de elétrons resultando na geração de estresse oxidativo, que leva a
inibição da enzima (Balijepalli et al., 1999).

Introdução |23
Os elétrons nos centros Fe-S são então encaminhados à coenzima Q
(ubiquinona) que depois de receber dois elétrons origina o ubiquinol (QH2
). A
transferência de elétrons está acoplada ao bombeamento de prótons da matriz para
o espaço intermembranar que contribui para a formação do gradiente de prótons
pelo complexo I e pelos complexos subsequentes na cadeia respiratória (citocromo
c: ubiquinona oxidoredutase ou complexo III e citocromo c oxidase ou complexo IV).
1.9.2- Succinato- ubiquinona oxidoredutase ou complexo II.
O complexo II (succinato-ubiquinona oxidoredutase (SQR);
EC 1.3.5.1) é uma enzima do ciclo do ácido cítrico integral da
membrana interna mitocondrial. Consiste de quatro subunidades
codificadas pelo DNA nuclear onde a maioria das estruturas e genes
foi conservada ao longo da evolução. Duas dessas subunidades
formam a succinato desidrogenase (SDH): a subunidade maior, uma
flavoproteína de 70 KDa (Fp; SDH A) contendo um sítio ativo
hipotético de FAD ligado covalentemente a parte da enzima e quatro
átomos de ferro não heme, e a proteína ferro-enxofre (Ip; SDH B,
30KDa), subunidade carregando 3 grupos ferro distintos. A SDH
está ancorada a membrana por duas subunidades adicionais, C e D (15 e 12.5 KDa
respectivamente) ligando um citocromo tipo- b, nas quais contém um único grupo
heme e os sítios de ligação para a ubiquinona. O complexo II faz parte tanto do ciclo
do ácido tricarboxílico quanto da cadeia respiratória mitocondrial catalisando a
transferência de seus elétrons a centros Fe-S e em seguida para a coenzima Q, a
fim de dar continuação a cadeia transportadora de elétrons. (Pierre e Agnes, 2002,
Pierre et al., 2002).

Introdução |24
Estudos prévios sugerem que o alvo de inibição da succinato desidrogenase
por 3-BrPA e N- etilmaleimida (NEM) seriam os grupamentos sulfidrilas em altas
concentrações (Sanborn et al., 1971 ). A participação de grupos sulfridilas na
oxidação de succinato em tecidos animais já foi demonstrada em trabalhos clássicos
(Hopkins et al., 1938). Desde então a sensibilidade da succinato desidrogenase aos
reagentes sulfidrilas foi registrada. Os grupos sulfidrilas estão envolvidos em uma
ligação estreita do inibidor oxalacetato a molécula da proteína e a uma espontânea
inativação da enzima. Estes grupos sulfidrilas estariam localizados na subunidade
contendo FAD+
da enzima. Foi demonstrado por Felberg, N. T. e Hollocher, T. C. em
1972 a incorporação de ['4C] NEM nesta mesma subunidade. Estes resultados
levam a conclusão que a inibição da enzima pelos reagentes alquilantes não é
devido a reação com o sítio ativo diretamente. Esses dados sugerem que os grupos
sulfidrilas estão envolvidos na primeira etapa da oxidação de succinato. Todavia,
muito pouco é conhecido sobre as propriedades químicas desses grupos na
molécula da enzima (Andrei et al., 1976).
1.9.3- Ubiquinol: Citocromo c oxidoredutase, complexo citocromo bc1
O complexo III é um complexo homodimérico,
apresentando 11 subunidades diferentes por monômero com
domínio substancial na matriz. O centro funcional é formado por
três subunidades: o citocromo b com seus dois hemes, a
proteína ferro-enxofre Riesk (na qual o átomo de Fe está
coordenado a dois resíduos de histidina em vez de dois
resíduos de cisteína) com seu centro 2Fe-2S e o citocromo c
ou complexo III.
1
com o seu heme. Esse complexo acopla a transferência de
elétrons do ubiquinol (QH2) para o citocromo c via carreadores
de elétrons (citocromo b562, b566, c1 e c) resultando no
transporte de prótons da matriz para o espaço intermembrana. O citocromo c é uma
proteína solúvel do espaço intermembranar. Após o seu único heme aceitar um
elétron do complexo III, o citocromo c se difunde ao complexo IV para doar o elétron
para um centro de cobre binuclear nesta enzima.

Introdução |25
1.9.4- Citocromo c oxidase ou complexo IV.
A citocromo c oxidase (COX) ou complexo IV
(ferrocitocromo c: oxigênio oxiredutase, EC 1.9.3.1) é a enzima
terminal da cadeia respiratória em eucariotos e alguns
procariotos. O complexo está presente na membrana agindo
como um dímero, tendo uma das suas partes voltada para o
espaço intermembrana, onde se apresenta mais inserida e a outra
voltada para a matriz. Apresenta um papel chave transportando
quatro elétrons de quatro moléculas de citocromo c para o
oxigênio molecular reduzindo-o a duas moléculas de H2
É uma proteína grande possuindo quatro subunidades em bactérias e em
mamíferos. Além das quatro subunidades contidas em bactérias, possui mais nove
totalizando treze subunidades (Vonck e Schäfer, 2008). Dentre as treze
subunidades, três são críticas para a função do transporte (Cox1p, Cox2p e Cox3p)
nas quais são codificadas pelo DNA mitocondrial e formam o centro ativo da enzima
representando a maior parte do complexo. As outras subunidades são codificadas
pelo genoma nuclear, traduzidas nos ribossomos citosólicos e transportadas para a
mitocôndria. Procariotos possuem três subunidades que apresentam homologia
significativa com as subunidades sintetizadas na mitocôndria (Khalimonchuk e
Rödel, 2005). Os centros catalíticos de todas as oxidases heme-cobre, incluindo a
COX, são altamente conservados a nível estrutural. Sobre os centros catalíticos
somente o centro Cu
O e
bombeando prótons (Khalimonchuk e Rödel, 2005). COXs são
membros de uma superfamília de heme e cobre contendo
oxidases terminais.
A será comentado por apresentar os grupamentos cisteínas
importantes na discussão do trabalho. O centro CuA está localizado no domínio
Cox1pA no lado externo da membrana mitocondrial interna e é o ponto de entrada
dos elétrons do citocromo c. A função do centro CuA é enviar elétrons para outros
centros redox localizados no Cox1pA (Khalimonchuk e Rödel, 2005).

Introdução |26
Contém dois íons cobre complexados aos grupos–SH de dois resíduos de
cisteína em um centro binuclear, além de conter dois resíduos de histidina, um de
metionina e um oxigênio carbonil. A subunidade a e a3 contém dois grupos heme e a
CuB outro íon. O heme a3 e o CuB formam um segundo centro binuclear que aceita
elétrons do heme a e então os transfere para o O2 ligado ao heme a3. Para cada
quatro elétrons que passam através desse complexo, a enzima consome quatro H+
da matriz convertendo O2 em H2
O. Ela também usa a energia dessa reação redox
para bombear um próton para o espaço intermembrana para cada elétron
transportado, aumentando o potencial eletroquímico.
1.9.5- ATP sintase ou complexo V.
A ATP sintase mitocondrial é uma ATPase do tipo F,
similar na estrutura e no mecanismo às ATP sintases de
cloroplastos e bactérias. Esse grande complexo enzimático da
membrana mitocondrial interna catalisa a formação de ATP
através de ADP e Pi acompanhado do retorno de prótons do
espaço intermembranas para a matriz. A ATP sintase apresenta
dois componentes distintos: F1, uma proteína periférica de
membrana, e Fo, uma proteína integral de membrana. A porção
é composta de 12 peptídeos estendidos através da membrana e
uma subunidade a. Está presente nas mitocôndrias,
cloroplastos, bactérias aeróbicas e fotossintéticas. A ATP
sintase translocadora de H+ é conhecida como FoF1
F
-ATPase (Nicholls e Fergunson,
2001).
1-ATPase é uma porção globular não membranar da enzima FoF1-
ATPsintase possuindo a mesma composição em todos organismos aeróbicos na
qual catalisa a síntese de ATP e H2O a partir de ADP e H2PO4 fazendo uso da força
próton motriz. Este também é capaz de bombear prótons através das membranas
usando a energia química de hidrólise de ATP.

Introdução |27
A “cabeça” da porção F1 é formada por três subunidades α e três subunidades
β entre as quais ficam três centros ativos. A “haste” entre Fo e F1 é formada por uma
subunidade γ e uma ε. Dois outros polipeptídeos, β e δ formam um “estabilizador”,
fixando as subunidades α e β em relação à porção Fo. A Fo contém uma via
condutora de prótons que opera junto a F1
O ciclo catalítico é dividido em três fases, as quais ocorrem em cada um dos
três centros ativos. Primeiramente, ADP e P
sintetizando de ATP e consumindo ΔΨm
ou hidrolizando ATP gerando ΔΨm dependendo das condições fisiológicas ou
experimentais.
i, se ligam formando uma ligação
anidrida e, em seguida o produto é separado. Cada vez que um próton flui através
do canal Fo
Já é bem estabelecido que os grupamentos sulfidrilas tenham um papel
importante na manutenção da integridade das membranas biológicas. E em
particular da membrana interna mitocondrial. Modificações dos grupamentos tióis da
membrana mitocondrial podem levar ao mau funcionamento de proteínas integrais
de membrana, como exemplo, os carreadores de fosfato e de nucleotídeos de
adenina que são consideravelmente diminuídos quando a mitocôndria é tratada com
agentes alquilantes (Lippe et al, 1988).
para a matriz, todos os três centros ativos passam de seus estados
momentâneos para o próximo. A energia do transporte de prótons primeiramente é
convertida em uma rotação da subunidade γ, que por outro lado, modifica
ciclicamente a conformação das subunidades α e β, em pos ição relativa com a
porção Fo, levando assim, à formação de ATP (Rohn S e Kroh LW, 2005).
Quanto a ATP sintase mitocondrial, o complexo enzimático na qual catalisa a
síntese e hidrólise de ATP, tem sido mostrado que grupos tióis pertecentes a Fo
estariam envolvidos no mecanismo de acoplamento entre Fo, a parte integrada a
membrana e F1,
a parte hidrofílica da enzima contendo os sítios catalíticos e estes
seriam modulados pela presença de ADP em proteínas de 25-27 kDa (subunidade b
ou Fo1) tornando-os livres através de mudanças conformacionais e susceptíveis a
reação com agentes redutores ou alquilantes interferindo na atividade da enzima
(Lippe et al, 1988, Dünschede et al., 2003).

Introdução |28
1.10- Estados respiratórios mitocondriais.
A produção de energia nas células envolve várias vias metabólicas ligadas
por intermediários comuns. A necessidade de ATP e a produção de precursores
para a construção de outras moléculas tornam essencial o controle destas vias.
Os fatores que controlam a respiração dependem de uma série de
características. A taxa de respiração é regulada pela concentração de ADP, Pi e o
produto ATP da fosforilação oxidativa acoplada ocorrendo na cadeia respiratória. O
requerimento de ADP para taxas máximas de respiração em mitocôndrias
fosforilando intactas foi primeiro estudado sistematicamente por Lardy e Wellman em
1952, que fizeram uma importante sugestão sobre a concentração de ADP que
poderia controlar a taxa de respiração dentro da célula.
A partir dos estudos feitos por Lardy e Wellman, Chance e Williams em 1956
sistematizaram as condições gerais que afetam a taxa respiratória de mitocôndrias
intactas. Estado 1 é a condição na qual ambos ADP e substrato respiratório estão
presentes. Estado 2 é a condição na qual somente o substrato respiratório está
ausente. O estado 3 refere-se a um aumento no conteúdo de ADP (aumento da
hidrólise de ATP devido o aumento da demanda energética) no qual aumenta a taxa
de fosforilação pelo complexo V, atenuando o gradiente de próton e estimulando o
transporte de elétron e o consumo de oxigênio. Estado 4 é quando uma baixa
concentração de ADP (baixa demanda energética) reduz a taxa de fosforilação
levando a um aumento do gradiente de próton através da membrana interna e
reduzindo a taxa de transporte de elétron. E finalmente o estado 5 é quando o
oxigênio está ausente (figura 6).

Introdução |29
Figura 6: Estados respiratórios. Estado 1, condição onde ambos ADP e substrato estão ausentes;
estado 2, somente o substrato respiratório está presente; estado 3, adição de ADP.ativa a fosforilação
oxidativa havendo consumo de oxigênio e fosforilação do ADP; estado 4, o ADP exógeno é
consumido e o consumo de oxigênio diminui e estado 5, onde a disponibilidade de oxigênio é zero.
Figura retirada de Chance e Williams, 1956.

2- OBJETIVOS ___________________________________

Objetivos |32
2- OBJETIVOS:
2.1- Objetivo Geral.
Apesar de diversos estudos terem investigado a ação do 3-BrPA em
hepatocarcinomas, todos eles identificaram efeitos somente sobre a glicólise, mais
especificamente na hexocinase pelo grupo do Dr. Peter Pedersen. Sua ação a nível
mitocondrial foi mostrada somente por Ko et al, 2001. Recentemente, nosso grupo
demonstrou que as doses de 3-BrPA que induzem a morte em tumores não são
capazes de causar qualquer inibição na atividade da HK mitocondrial (Pereira da
Silva et al., 2008) e que estas doses parecem ter efeito mais específico na atividade
da succinato desidrogenase. Apesar disso, estudos mais aprofundados eram
necessários para esclarecer a ação do 3-BrPA a nível mitocondrial analisando o seu
efeito na atividade das enzimas dos diferentes complexos da cadeia respiratória.
2.2- Objetivos Específicos.
Avaliar o efeito do 3-BrPA sobre a atividade catalítica dos Complexo I,
II, III, IV e Fo F1
Avaliar o efeito do 3-BrPA sobre os parâmetros bioenergéticos
mitocondriais em diferentes estados respiratórios (fosforilativo e não-fosforilativo)
usando diferentes substratos oxidáveis (substratos ligados a síntese de NADH e
substrato ligado a síntese de FADH
ATPase.
2
- Medidas de potencial de membrana (∆Ψ
.
m
- Consumo de oxigênio.
).
Estabelecer a cinética de inativação da respiração pelo 3-BrPA em
mitocôndrias isoladas de fígado e cérebro de camundongo.
Avaliar a relação causal entre a atividade de mitocôndrias exibindo
atividade hexocinase associadas ou não-associadas com a inibição da respiração
mitocondrial.

3- MATERIAIS E MÉTODOS _____________________________________________________________

Materiais e Métodos |34
3- MATERIAIS E MÉTODOS:
3.1- Animais.
Os animais utilizados nesta pesquisa foram mantidos e sacrificados segundo
as normas de biossegurança determinadas pelo comitê de ética da UFRJ. Foram
utilizados camundongos suíços disponibilizados pelos biotérios localizados no
Instituto de Bioquímica Médica e Instituto de Microbiologia da Universidade Federal
do Rio de Janeiro. Os animais foram mantidos em gaiolas plásticas forradas com
maravalha autoclavada em ambiente à 23ºC e ciclo de luz (12h/12h), recebendo
suplemento vitamínico semanal, ração apropriada e água filtrada.
3.2- Isolamento da mitocôndria.
Mitocôndrias de fígado e cérebro de camundongo suíço de aproximadamente
2 meses de idade foram isoladas por centrifugação diferencial como descrito por
Rosenthal et. al., 1987. Os camundongos foram sacrificados por deslocamento
cervical, o fígado foi rapidamente removido e colocado em 10 ml de tampão de
isolamento 1:10(v/v) gelado contendo: 225 mM manitol; 75 mM sacarose; 10 mM K-
Hepes; 1 mM EGTA; 0,1% BSA (livre de ácido graxo); pH 7,4). O tecido foi cortado
em pedaços pequenos e lavado pelo menos três vezes com o tampão de
isolamento. Um Potter-Elvehjem (Wheaton Sci, USA) de vidro de 30 ml foi utilizado
para a homogeneização manual em 11 ciclos do tecido. O homogenado foi então
centrifugado por 5 min a 600 g à 4ºC (Hitachi Himac SCR 20B,Tokyo, Japão)
utilizando o rotor RPR20-2. Depois da centrifugação o sobrenadante foi
recentrifugado por 10 min a 12000g à 4ºC. O precipitado foi ressuspendido em 10 ml
de tampão de isolamento e novamente centrifugado por 10 min à 12000g à 4ºC. O
sobrenadante foi descartado e o precipitado final ressuspendido em 300 µl de
tampão de isolamento. Para o isolamento de mitocôndrias de cérebro foram
utilizados os mesmos tampões e o mesmo procedimento tendo como diferencial
somente a primeira centrifugação, 3 min a 2000g à 4ºC.

Materiais e Métodos |35
3.3- Preparo da solução de 3-BrPA.
A solução de 3-BrPA (Sigma) foi preparada para uma concentração final de
25 mM em tampão Tris-HCl , pH 7.4. As soluções foram feitas minutos antes do
início de cada experimento.
3.4- Hidrólise de ATP.
Atividade da F1Fo-ATPase mitocondrial foi medida em 3 tempos 0, 30 e 60
minutos de incubação em um meio de reação contendo 20 mM Tris-HCl pH 7.5; 2
mM ATP e 10 mM MgCl2 a 37ºC na presença ou ausência de 1 mM 3-BrPA. A
reação foi iniciada pela adição da proteína (0,2 mg/ml concentração final). Como
controle foi feito o mesmo procedimento na presença de NaN3 que é um inibidor da
FoF1-ATPase mitocondrial e na ausência da droga. A medida da inibição da
hidrólise pelo 3-BrPA foi feita através da quantificação do fosfato inorgânico liberado
(Pi) pelo método de Fiske & Subbarow em 1925 através da atividade NaN3
sensível.
A reação de quantificação de Pi foi determinada no espectrofotômetro a 660nm.
3.5- Consumo de oxigênio em mitocôndrias isoladas.
O consumo de oxigênio foi monitorado através de respirometria de alta
resolução (OROBOROS® Oxygraph-2K, Instruments, Innsbruck, Austria) utilizando
mitocôndrias (0,2 mg/ml) de fígado e de cérebro de camundongo. Os ensaios foram
feitos em uma câmara contendo 2 ml de tampão de respiração (10 mM Tris-HCl pH
7.4; 320 mM manitol; 8 mM Pi-Tris; 4 mM MgCl2
; 0.08 mM EDTA e 1 mg/mL BSA
FFA; pH7,4 à 37ºC).

Materiais e Métodos |36
3.6- Medidas do potencial de membrana mitocondrial (∆ψm
O potencial de membrana mitocondrial foi avaliado em função de sinais
fluorescentes de um corante catiônico, safranina O. Este tem sua fluorescência
suprimida dentro de uma mitocôndria energizada, ou seja, que esteja mais positiva
no espaço intermembranas e menos na matriz (Akerman e Jarvisalo, 1980).
).
Foram incubadas 0.2 mg/ml de proteína mitocondrial no meio de respiração
padrão (vide seção 3.5) suplementado com 6,7 µM de safranina O. Utilizando-se
substratos, a medida da formação do potencial de membrana foi feita para os quatro
complexos: I, II, III e IV da cadeia transportadora de elétrons. Inicialmente utilizamos
em um fluorímetro Hitachi modelo F-3010 (Tokyo, Japan). Estudos posteriores foram
feitos somente para os complexos I e II em um fluorímetro (multi-mode microplate
reader spectrofluoremeter Spectra Max®
M5 (Molecular Devices) em uma placa de
96 poços. A fluorescência foi detectada no comprimento de onda de 495 nm de
excitação e 586 nm de emissão.O ensaio foi conduzido à 37ºC imediatamente após
o isolamento mitocondrial. As adições de substratos e reagentes são devidamente
indicadas na legenda da figura 13. Os ensaios foram feitos em diferentes
concentrações de 3-BrPA (0 – 1000µM).
3.7- Determinação da atividade da NADH: ubiquinona oxidoredutase.
A atividade da NADH: ubiquiquinona oxidoredutase foi determinada usando o
aceptor de elétrons ubiquinona-1 (CoQ1
NADH + H+ + ubiquinone-1 (CoQ1) → NAD+ + dihydroubiquinone-1 (CoQH2)
). A reação catalizada pelo complexo I segue
abaixo:
O tampão fosfato foi utilizado para o ensaio juntamente com 15 mM NADH, 1
mM KCN e 2 mM NaN3. Para a medida da atividade, a suspensão mitocondrial
estocada a -80ºC foi sonicada em dois ciclos de 20s no gelo (Bandelin Sonopuls HD
200 ultrasonicator (MS 73 probe tip; Berlin, Germany)).

Materiais e Métodos |37
A concentração final da proteína no ensaio foi de 0,1 mg/ml. A reação foi
iniciada com 0.1 mM coenzima Q1 após pré-incubações por 1 min com diferentes
concentrações de 3-BrPA (75, 150 e 500 µM) em um volume final de 1 ml. O tempo
de leitura foi de 3 min sendo a absorbância registrada a cada 15 segundos. Após
90s de reação, foi adicionado 5 µM rotenona para determinar a atividade insensível
a rotenona. A atividade real foi feita pela diferença da fração sensível pela
insensível. A leitura foi feita no espectrofotômetro a 340 nm e a diminuição linear da
absorbância devido à redução da coenzima Q1
foi então monitorada.
3.8- Ensaio da atividade da succinato desidrogenase.
A atividade da succinato desidrogenase foi determinada à temperatura
ambiente pelo ensaio DCIP-fenazina metilsulfato utilizando o tampão de respiração
contendo 10 mM Tris-HCl pH 7.4; 320 mM manitol; 8 mM Pi-Tris pH 7.4; 4 mM
MgCl2; 0.08 mM EDTA e 1 mg/mL BSA FFA; 4 mM NaNO3
, 10 mM DCIP; 0,2 mM
PMS e diferentes concentrações de 3-BrPA (0-1000 µM) em um volume final de 1ml.
A concentração final da proteína no ensaio foi de 0,1 mg/ml. Todas as amostras de
proteína foram tratadas com triton X-100 (0,1% concentração final). A reação foi
iniciada pela adição de diversas concentrações succinato após 1 min de pré-
incubação com as diferentes concentrações de 3-BrPA. O tempo de leitura foi de 3
min sendo a absorbância registrada a cada 15 segundos. A leitura foi feita no
espectrofotômetro a 600 nm e a diminuição linear da absorbância devido a oxidação
do DCIP foi então registrada. DCIP e PMS são aceptores de elétrons. Esses dois
compostos quando misturados desenvolvem uma coloração esverdeada que é
proporcional ao estado de oxidação do DCIP. Foram feitos brancos na ausência de
succinato.

Materiais e Métodos |38
3.9- Quantificação de proteínas.
A quantificação de proteínas utilizadas nos experimentos foi realizada pelo
método de Lowry et al. (1951).
3.10- Análise Estatística.
A análise estatística e a construção dos gráficos foram realizadas pelo
programa Origin® 8.0 (Originlab Corporation,USA).

4- RESULTADOS _______________________________________

Resultados |40
4- RESULTADOS:
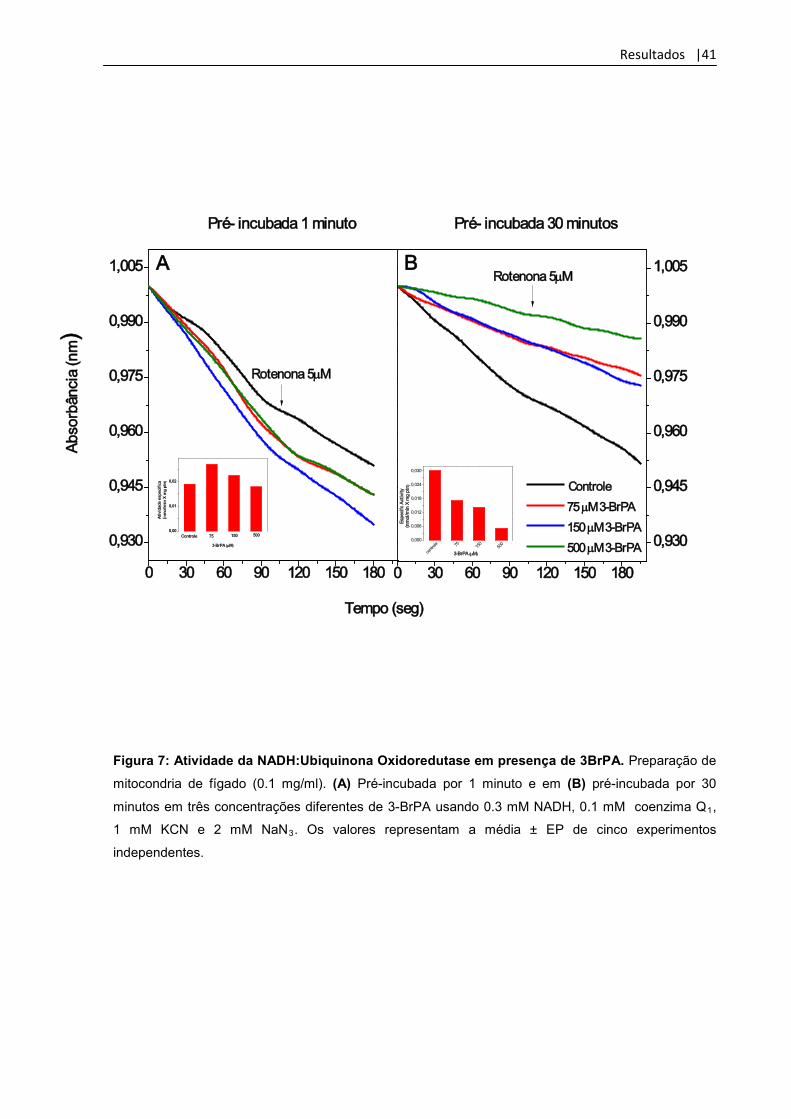
4.1- Efeito do 3-BrPA nas atividades dos complexos I e IV da cadeia transportadora de elétrons.
Os experimentos foram feitos em concentrações que variam de 0-1000 µM, já
que trabalhos anteriores sempre utilizaram concentrações acima de 1 mM. A
atividade da NADH: ubiquinona oxidoredutase ou complexo I foi medida em 3
concentrações diferentes da droga 0, 75, 150 e 500 µM com a fração mitocondrial de
fígado sonicada e pré-incubada por 1 e 30 min tendo a leitura da absorbância
medida a cada 15 s por 3 min. Não foi observado inibição da atividade em qualquer
concentração em 1 min. de incubação da partícula mitocondrial, embora apresente
um efeito inibitório principalmente quando usado 500 µM da droga mostrando ser
dependente de tempo quando medida em 30 min. de pré- incubação da partícula
mitocondrial com 3-BrPA. (figura 7). Todavia, quando medimos a atividade do
complexo IV através da taxa de consumo de oxigênio utilizando TMPD/ ascorbato
como substratos respiratórios na ausência ou presença de 100 µM 3-BrPA, não
observamos inibição na taxa do consumo de oxigênio neste complexo tanto no
estado 2 como no estado 3 (figura 8).
Resultados |41
0,00
0,01
0,02
500150
Ativ
idad
e es
pecí
fica
(nm
ol/m
in X
mg
ptn)
3-BrPA (µM)
Controle 75
0 30 60 90 120 150 180
0,930
0,945
0,960
0,975
0,990
1,005
Pré- incubada 30 minutos
B
Abso
rbân
cia
(nm)
Tempo (seg)
Controle 75 µM 3-BrPA 150 µM 3-BrPA 500 µM 3-BrPA
A
Pré- incubada 1 minuto
0 30 60 90 120 150 180
0,930
0,945
0,960
0,975
0,990
1,005
Rotenona 5µM
Rotenona 5µM
0,000
0,006
0,012
0,018
0,024
0,030
Espe
cifc
Act
ivity
(nm
ol/m
in X
mg
ptn )
50015
075
contr
ole
3-BrPA (µM)
Figura 7: Atividade da NADH:Ubiquinona Oxidoredutase em presença de 3BrPA. Preparação de
mitocondria de fígado (0.1 mg/ml). (A) Pré-incubada por 1 minuto e em (B) pré-incubada por 30
minutos em três concentrações diferentes de 3-BrPA usando 0.3 mM NADH, 0.1 mM coenzima Q1,
1 mM KCN e 2 mM NaN3
. Os valores representam a média ± EP de cinco experimentos
independentes.

Resultados |42
0
300
600
900
1200
1500
1800
Cons
umo
de O
2
(pm
ol O
2/ s
ec. m
g pt
n)
Basal TMPD/ Asc ADP 3-BrPA
Figura 8: 3-BrPA não afeta o consumo de oxigênio quando utilizados substratos do complexo IV. Mitocôndria de fígado de camundongo (0.2 mg/ml) foi usada na medida da taxa de consumo de
oxigênio quando 100 µM de 3-BrPA foi utilizado na presença de substratos do complexo IV ( 100 µM
TMPD/ 250 µM Ascorbato) e 1 mM de ADP. Os valores representam a média ± EP de três
experimentos independentes.

Resultados |43
4.2- 3-BrPA não afeta a atividade da FoF1
Em trabalhos prévios (Ko et al., 2001; Geschwind et al., 2002) demonstraram
uma grande baixa nos níveis de ATP em células tratadas com 150 µM de 3-BrPA.
Com o objetivo de avaliar se os efeitos da droga sob a quantidade de ATP celular
poderiam ser devido à inibição da síntese de ATP mitocondrial, resolvemos medir a
atividade da hidrólise de ATP de mitocôndria de fígado de camundongo. Avaliamos
o efeito de 1 mM 3-BrPA na hidrólise de ATP catalisada pelo complexo da FoF1-
ATPase sem pré- incubar ou pré-incubadando por 30 e 60 minutos na presença e
ausência de azida (inibidor do complexo V). O resultado obtido mostra uma pequena
inibição da atividade da FoF1-ATPase de aproximadamente 25%, sugerindo que o
3-BrPA não tem ação nesse sítio na concentração utilizada (figura 9).
- ATPsintase.

Resultados |44
0 30 600,00
0,05
0,10
0,15
µm
ol P
i/ m
g. 1
0min
. Controle 1 mM 3-BrPA
Tempo de pré-incubaçao (minutos)
Figura 9: Efeito do 3-BrPA na hidrólise de ATP pela FoF1-ATPase. Mitocôndria de fígado de
camundongo foi isolada por centrifugação diferencial. A atividade ATPase mitocondrial (FoF1-
ATPase) foi medida em um meio de reação descrito na seção 3.4 “metodologia” na presença ou
ausência de 5 mM NaN3 e 1 mM 3-BrPA a 37o
C. A reação foi iniciada adicionando proteína
mitocondrial (0,1 mg/ml) e a atividade medida em três tempos (0, 30 e 60 minutos) de pré- incubação.
A quantificação de Pi liberado foi determinada pelo método colorimétrico Fiske & Subbarow, 1925. Os
dados são referentes a média ± EP de quatro experimentos independentes.

Resultados |45
4.3- 3-BrPA inibe a atividade da SDH.
Estudos anteriores mostraram o efeito de 5 mM 3-BrPA na inibição da
succinato desidrogenase (Sanborn et al., 1971) porém essa concentração é
considerada elevada. A partir desses resultados prévios resolvemos testar o efeito
do 3-BrPA diretamente na atividade da enzima succinato desidrogenase (SDH)
através do ensaio fenazina metilsulfato (PMS) utilizando concentrações menores de
3-BrPA (ver materiais e métodos). Esse experimento foi realizado utilizando
diferentes concentrações de 3-BrPA na presença ou ausência de ADP. O resultado
obtido na figura 10 mostra que, SDH é fortemente inibida por 3-BrPA. O ADP não
influenciou na modulação da atividade de SDH pelo 3-BrPA, apesar de trabalhos
anteriores mostrarem que a succinato desidrogenase é regulada por nucleotídeos
(Affourtit et al., 2001).

Resultados |46
0 200 400 600 800 10000,0
0,2
0,4
0,6
0,8
1,0
Ativ
idad
e SD
H, t
axa
rela
tiva
(Vi/
Vo)
3 BrPA, uM
Sem ADP 1 mM ADP
Figura 10: Efeito do 3BrPA na atividade da succinato desidrogenase na presença e na ausência de ADP. A preparação de mitocôndria de fígado de camundongo (0.1 mg/ml) foi usada
para determinação da atividade da succinato desidrogenase (SDH) usando 0.05 mM DCIP, 0.2 mM
PMS, 0.1% (v/v) Triton X-100, 4 mM NaN3
e concentrações diferentes de 3-BrPA como indicado na
figura. Círculos abertos, representam o ensaio na presença de 1mM de ADP e círculos fechados sem
adição de ADP. A preparação foi pré-incubada por 1 minuto com 3-BrPA. Os valores são médias ±
EP de cinco experimentos independentes.

Resultados |47
Os resultados apresentados até o momento mostraram o efeito do 3-BrPA
diretamente sobre a atividade dos complexos da cadeia respiratória. Todavia, uma
avaliação mais realística dos efeitos desta droga requer medidas dos complexos
através da avaliação de parâmetros da fisiologia mitocondrial.
4.4 - Inibição da formação do potencial de membrana (ΔΨm) e consumo de oxigênio pelo 3-BrPA confirmam os efeitos na CTE.
Na tentativa de descobrir o mecanismo de ação do composto 3-BrPA na
fosforilação oxidativa, primeiramente testamos o quanto este agia nos complexos
respiratórios. Assim, investigamos seu efeito na formação do potencial de membrana
utilizando a sonda safranina O (ver materiais e métodos) para analisar cada
complexo separadamente com os seus respectivos substratos oxidáveis e inibidores
(legenda figura 11). Esse experimento foi realizado utilizando-se diferentes
concentrações de 3-BrPA variando de 0 a 1000 µM. Nosso objetivo também foi
achar uma concentração mínima de ação da droga já que em estudos prévios
(Sanborn et al., 1971), foi observado um efeito do 3-BrPA no complexo II porém em
uma dose maior (1 mM) que a provável em condições terapêuticas. A figura 11
mostra o efeito do 3-BrPA sobre os diferentes substratos respiratórios em
mitocôndria de fígado. Nela, é apresentada a atividade dos complexos I, II e IV
através da formação do ΔΨm (figura 11A).
Os ensaios foram feitos pré-incubando a fração mitocondrial (0,2 mg/ml) por 1
minuto com a droga antes da adição dos substratos. Na figura 11B observa-se
claramente o efeito de 250 µM 3-BrPA nos complexos, sendo o complexo II
majoritariamente afetado em relação ao complexo I e IV. O resultado mostrado na
figura 11B fica mais evidente na figura 11C, onde são mostrados os valores
máximos da inibição da formação do potencial de membrana para os complexos I, II
e IV em uma maneira dose dependente de 3-BrPA. Todavia, mostrando de uma
forma mais clara a inibição preferencial do complexo II que apresentou o IC50 de
170 µM, enquanto nessa concentração inibiu apenas 20% dos complexos I e IV.

Resultados |48
Para mostrar que o efeito da inibição do ΔΨm era realmente causado pelo 3-
BrPA, um ânion que poderia afetar a polaridade da membrana, foi feita uma medida
da formação do potencial de membrana utilizando o substrato do complexo II para
avaliar o efeito do 3-BrPA comparado com piruvato, seu análogo. Diante da
mitocôndria pré-energizada com 10 mM de succinato, foi feita uma titulação tanto
com 3-BrPA quanto com seu análogo piruvato (figura 11D). Os resultados obtidos
revelam que o efeito da inibição do ΔΨm pelo bombeamento de H+
provocado pelo
3-BrPA poderia ser pela inibição da atividade da SDH (figura 11C e 11D).

Resultados |49
FCCPSucc
Rot
2 min.
TMPD/Asc
Ant A
250 µM 3-BrPA
Mito+ Piruvato/Mal
Controle BA
FCCP
TMPD/Asc
Ant A
Succ
Rot
Mito+ Piruvato/Mal
10 AUF
Pir, µM4 min250
100150
50
250 200 150
100
3BrPir, µM
50
Succ
SuccSucc
Piruvato + Succ
Succ
40 AUF
D
200
3-BrPA + Succ
0 200 400 600 800 10000
20
40
60
80
100
% do
valor
máx
imo d
e ∆Ψ m
3- Bromopiruvato , µM
Piruvato/ Malato Succinato TMPD
C
Figura 11: 3-BrPA diminui a formação do potencial de membrana mitocondrial (ΔΨm) pela cadeia transportadora de elétrons(ETC). ΔΨm foi determinado pelo método de supressão de
fluorescência da safranina O usando o mesmo tampão usado na medida do consumo de oxigênio e
6,7μM safranina O. Perfil representativo da formação do ΔΨm pelos complexos I, II e IV (A). Painel
(B) gráfico representativo da inibição da formação do ΔΨm por 250µM 3-BrPA. No painel (C) Inibição
da formação do ΔΨm em uma maneira dose dependente depois de pré- incubar a mitocôndria com 3-
BrPA por 1 minuto e iniciar a reação usando os substratos oxidáveis para os diferentes complexos da
CTE. Para o complexo I (5 mM piruvato/malato), complexo II (10 mM succinato + rotenona 5 µM) e
complexo IV (0.3 mM TMPD/ 0.2 mM ascorbato + 5 µM rotenona + 1 µg/ml antimicina A). Estes
experimentos foram feitos com mitocôndrias isoladas de fígado (0.2 mg/ml). Os resultados
representam o efeito do 3-BrPA na mudança máxima no ΔΨm (B). No painel (D) dose resposta para
concentrações crescentes de 3-BrPA em uma mitocôndria pré-energizada com 10 mM succinato.
Como controle, foi adicionado a mesma quantidade de piruvato no lugar de 3 Br-PA. Os valores são
médias de quatro experimentos independentes.

Resultados |50
4.5- 3-BrPA reage diferentemente com os complexos da CTE e depende dos estados respiratórios.
Os efeitos observados do 3-BrPA sobre a formação do ΔΨm sugerem uma
ação inibitória sobre a atividade do complexo II. Entretanto, estas medidas envolvem
o estado de permeabilidade da membrana interna mitocondrial. Com o objetivo de
esclarecer se os efeitos do 3-BrPA afetam a atividade da SDH ou a permeabiblidade
de membrana, foram feitas medidas de consumo de oxigênio utilizando os
substratos para o complexo I e II . O consumo de O2
Estudos prévios (Sanborn et al., 1971; Maldonado et al., 1972), descreveram
o efeito desta droga na piruvato desidrogenase. Com base nessas informações,
avaliamos se o efeito do 3-BrPA que estávamos observando era em desidrogenases
ou era especificamente na piruvato desidrogenase. Para isso adicionamos
substratos para o complexo I com e sem piruvato (figuras 12C, 12D, 12E e 12F).
Verificamos que quando piruvato é adicionado junto aos outros subtratos o efeito
inibitório é maior do que na sua ausência (figuras 12E e 12F). Assim, o 3-BrPA age
na piruvato desidrogenase e favorece a inibição da respiração. Porém, também tem
efeito menor em outras enzimas como a NADH: ubiquinona oxidoredutase, malato
desidrogenase e glutamato desidrogenase em concentrações maiores. (figuras 12C
e 12D).
induzido por succinato
mostrou-se mais sensível a inibição pelo 3-BrPA (figuras 12A e 12B), onde em
concentrações baixas de 3-BrPA apresentam maior efeito comparadas com os
resultados utilizando substratos do complexo I, onde são necessárias concentrações
maiores para a inibição.

Resultados |51
0,0
0,3
0,6
0,9
1,2
estado 2 - succinato
0,0
0,3
0,6
0,9
1,2B
estado 3 - succinato 1min 10min 20min
0,0
0,3
0,6
0,9
1,2 C
0,0
0,3
0,6
0,9
1,2D
estado 3 - PMG
0 10 100 10000,0
0,3
0,6
0,9
1,2 E
estado 2 - MG
estado 2 - PMG
3BrPA (µM)
A
0 10 100 10000,0
0,3
0,6
0,9
1,2F
estado 3 - MG
Vi/V
o, c
onsu
mo
de o
xigê
nio
rela
tivo
Figura 12: Inibição do consumo de oxigênio é dependente do estado respiratório mitocondrial.
Mitocôndria de fígado de camundongo (0.2 mg/ml) foi usada na medida da taxa de consumo de
oxigênio. No painel (A) Efeito de diferentes concentrações do 3-BrPA na taxa respiratória na
presença de succinato sem ADP. No painel (B) Succinato com 1 mM ADP.Em (C), piruvato, malato e
glutamato sem ADP e (D) piruvato,malato e glutamato com 1 mM ADP. Nos painéis (E) e (F)
glutamato e malato sem e com 1 mM ADP respectivamente. Os valores representam quatro
experimentos independentes.

Resultados |52
Como mostrado na figura 12, no estado 3, as baixas concentrações de 3-
BrPA já são suficientes para prover a inibição. Isto sugere que o estado redox da
CTE poderia estar influenciando nessa potencialização através do potencial de
membrana (figura 13). Utilizando-se succinato como substrato do complexo II,
observamos que o 3-BrPA foi capaz de promover a despolarização (diminuição do
ΔΨm). Este efeito foi maior quando as mitocôndrias estavam no estado 3 (figura
13A) do que quando estavam no estado 4.
Porém, ao utilizarmos substratos para o complexo I, além de observarmos
uma inibição significativamente menor quando comparado ao complexo II, não
houve grande diferença frente aos diferentes estados respiratórios (estado 3 e
estado 4). Todavia, ao ser adicionado o succinato juntamente com os substratos do
complexo I observamos um aumento substancial na inibição da formação do ΔΨm
pelo 3-BrPA, sugerindo um efeito sinérgico ou potencializador (figura 14A e 14B).

Resultados |53
FCCP Oligomicina succinato 25uM 3-BrPA 50uM 3-BrPA 100uM 3-BrPA 1mM 3-BrPA
0 200 400 600 800 1000 12000
200
400
600
800B Succinato (estado 4)
Tempo (seg)
Succinato (estado 3)
Unid
ade
de F
luor
escê
ncia
da
Safra
nina
O
A
0 200 400 600 800 1000 12000
200
400
600
800
Figura 13: Efeito do 3-BrPA no potencial de membrana mitocondrial (ΔΨm) usando succinato (substrato do complexo II) nos estados respiratórios 3 e 4. O ΔΨm foi determinado pelo método
de fluorescência safranina O usando o mesmo tampão padrão usado para medidas de consumo de
oxigênio e 6,7 μM safranina O. Os painéis (A) e (B) são gráficos representativos mostrando o efeito
de diferentes concentrações de 3-BrPA na despolarização do potencial de membrana mitocondrial na
presença de succinato e ADP. No painel (A) presença de succinato com ADP (estado 3). No painel
(B) presença de succinato sem ADP (estado 4). Os valores são representativos de quatro
experimentos independentes.
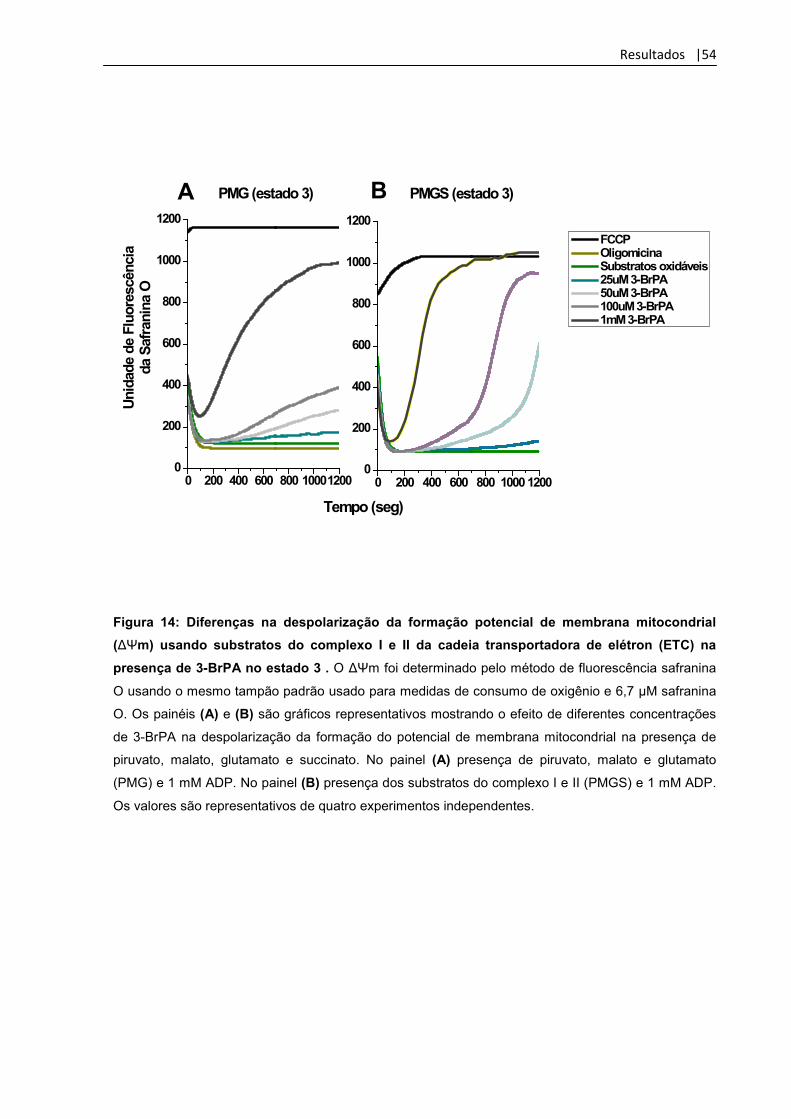
Resultados |54
FCCP Oligomicina Substratos oxidáveis 25uM 3-BrPA 50uM 3-BrPA 100uM 3-BrPA 1mM 3-BrPA
0 200 400 600 800 100012000
200
400
600
800
1000
1200
A PMGS (estado 3)PMG (estado 3)
Unid
ade
de F
luor
escê
ncia
da S
afra
nina
O
Tempo (seg)
B
0 200 400 600 800 1000 12000
200
400
600
800
1000
1200
Figura 14: Diferenças na despolarização da formação potencial de membrana mitocondrial (ΔΨm) usando substratos do complexo I e II da cadeia transportadora de elétron (ETC) na presença de 3-BrPA no estado 3 . O ΔΨm foi determinado pelo método de fluorescência safranina
O usando o mesmo tampão padrão usado para medidas de consumo de oxigênio e 6,7 μM safranina
O. Os painéis (A) e (B) são gráficos representativos mostrando o efeito de diferentes concentrações
de 3-BrPA na despolarização da formação do potencial de membrana mitocondrial na presença de
piruvato, malato, glutamato e succinato. No painel (A) presença de piruvato, malato e glutamato
(PMG) e 1 mM ADP. No painel (B) presença dos substratos do complexo I e II (PMGS) e 1 mM ADP.
Os valores são representativos de quatro experimentos independentes.

Resultados |55
4.6- Efeito protetor da glutationa reduzida (GSH) a inibição pelo 3BrPA.
Estudos prévios mostraram que o principal local da ação de agentes
alquilantes como 3-BrPA no complexo II, são em grupamentos tióis localizados no
sítio ativo da succinato desidrogenase, sugerindo que os resíduos sulfidrilas são
essenciais para a atividade desta enzima ( Sanborn et al., 1971; Maldonado et al.,
1972). A partir desses estudos testamos se o efeito observado do 3-BrPA poderia
ser revertido ou bloqueado pela adição de um agente redutor de grupamentos tióis
na succinato desidrogenase e na NADH: oxidoredutase. Para tal finalidade nós
usamos 2 mM GSH (agente redutor usado para estabilizar enzimas e proteínas que
possuem grupos sulfidrilas).
A figura 15A mostra o efeito do GSH na manutenção do potencial de
membrana em uma mitocôndria pré-energizada com 10 mM de succinato ou 5 mM
piruvato/malato na presença de diferentes concentrações de 3-BrPA e 1 mM de
ADP. Como controle foi utilizado o ionóforo de prótons FCCP. A figura 15B é um
gráfico representativo mostrando a proteção desse agente redutor. Estes resultados
sugerem que os grupos–SH presentes no sítio catalítico da succinato desidrogenase
e na estrutura da NADH: oxidoredutase seriam realmente os alvos potenciais da
alquilação promovida pelo 3-BrPA.

Resultados |56
0,0
0,1
0,2
0,3
3,57,0
10,5
3-BrPA, µM +GSH
100050
025
020
015010
07550
Taxa
de
desp
olar
izaçã
o do
∆Ψ
m(R
FU/s
ec x
0. 2
mg
ptn)
Succinato estado 3 Pir/Mal/Glut estado 3
FCCP
1000
sem
GSH
25
A
0 200 400 600 800 1000 12000
200
400
600
800
1000
FCCP Oligomicina 25mM 3-BrPA 50mM 3-BrPA 75mM 3-BrPA 100mM 3-BrPA 250µM 3-BrPA 500µM 3-BrPA 1mM 3-BrPA
Unid
ade
de F
luor
escê
ncia
d
a sa
frani
na O
Tempo (seg)
Figura 15: Glutationa reduzida (GSH) protege a dissipação do potencial de membrana (ΔΨm) promovida pelo 3BrPA. No painel (A) o ΔΨm foi determinado pelo método de fluorescência
safranina O usando o mesmo tampão para medir o consumo de oxigênio e 6,7 μM safranina O. GSH
2 mM foi adicionado previamente no meio de reação contendo a fração mitocondrial (0.2 mg/ ml) na
presença de 1 mM de ADP (estado 3) e diferentes concentrações de 3-BrPA utilizando os substratos
oxidáveis do complexo I (5 mM piruvato/malato e 2 mM glutamato) e complexo II (10 mM succinato +
rotenona 5 µM). 1 µM de FCCP e 1 µg/ml de oligomicina foram adicionados como controle. O painel
(B) é um gráfico representativo do efeito de GSH sobre a respiração usando succinato como
substrato. Os valores são representativos de três experimentos independentes.

Resultados |57
4.7- O efeito inibitório do 3-BrPA é modulado pelos estados respiratórios da mitocôndria.
Trabalhos prévios do nosso grupo demonstraram que a ciclagem de ADP
promovida pelo sistema ANT, FoF1ATPsintase, HK pode representar um importante
mecanismo de defesa antioxidante (da-Silva, 2004), inibição da apoptose e é
característico de várias linhagens tumorais. Nós decidimos investigar se a maior
sensibilidade da SDH ao 3-BrPA apresentada em estado 3 (1mM ADP) estaria
sendo mantida pela atividade do sistema ANT, FoF1ATPsintase, HK. Neste sentido,
facilitando a reação com 3-BrPA, já que em células de hepatocarcinomas há um
aumento da expressão de HKII associada a membrana externa da mitocôndria e
trabalhos anteriores descrevem um efeito majoritário nestes tipos celulares quando
comparados a hepatócitos na qual não apresentam HK associada a membrana
externa mitocondrial.
Neste ensaio, utilizamos além de mitocôndrias de hepatócitos, mitocôndrias
de cérebro de camundongo, uma vez que este tecido apresenta HK associada à
mitocôndria assim como as células tumorais. Nas figuras 16A e 16B estimulamos as
mitocôndrias de fígado e cérebro, respectivamente, com 10 mM de succinato e 200
µM de ADP. Após ter sido obtido o controle respiratório (estado 4), adicionamos 100
µM 3-BrPA e observamos a cinética de inibição do consumo de oxigênio total
provocada pela droga em ambas as preparações mitocondriais. Nas figuras 16C e
16D as mitocôndrias de fígado e cérebro respectivamente foram estimuladas de
forma semelhante como descrito acima.
Foi adicionado em ambas as preparações, 20 mM 2-DOG, que é um substrato
da HK análogo da glicose onde o produto dessa reação, 2-DOG-6P, não inibe a
enzima mantendo-a sempre ativa. Isso não acontece quando glicose é utilizada,
uma vez que há acúmulo de glicose-6-fosfato que inibe a reação da HK. Na figura
16C, a resposta da inibição quando 100 µM 3-BrPA foi adicionado foi semelhante a
figura 16A, pois mitocôndrias de hepatócitos não apresentam HK ligada a sua
membrana externa. Por outro lado, na figura 16D, quando 20 mM 2-DOG foi
adicionado ativando a HK e em seguida foi incluído 100 µM 3-BrPA, a resposta de
inibição do consumo de oxigênio foi imediata comparada a figura 16B onde a enzima
está inativa.
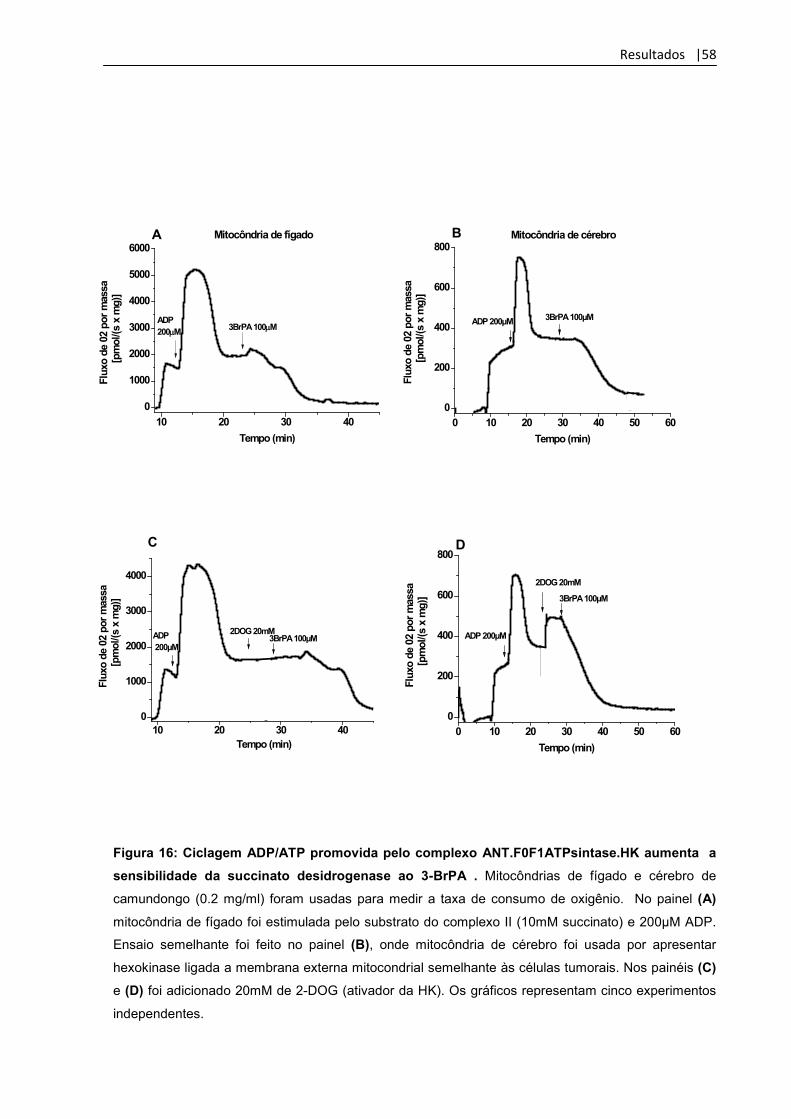
Resultados |58
10 20 30 400
1000
2000
3000
4000
5000
6000Mitocôndria de fígado
3BrPA 100µMADP 200µM
Flux
o de
02
por m
assa
[p
mol
/(s x
mg)
]
Tempo (min)
A
10 20 30 400
1000
2000
3000
4000
3BrPA 100µM2DOG 20mMADP
200µM
Tempo (min)
Flux
o de
02
por m
assa
[p
mol
/(s x
mg)
]
C
0 10 20 30 40 50 600
200
400
600
800
3BrPA 100µM
2DOG 20mM
ADP 200µM
Flux
o de
02
por m
assa
[p
mol
/(s x
mg)
]
Tempo (min)
D
0 10 20 30 40 50 600
200
400
600
800
3BrPA 100µMADP 200µM
Flux
o de
02
por m
assa
[p
mol
/(s x
mg)
]
Tempo (min)
Mitocôndria de cérebroB
Figura 16: Ciclagem ADP/ATP promovida pelo complexo ANT.F0F1ATPsintase.HK aumenta a sensibilidade da succinato desidrogenase ao 3-BrPA . Mitocôndrias de fígado e cérebro de
camundongo (0.2 mg/ml) foram usadas para medir a taxa de consumo de oxigênio. No painel (A)
mitocôndria de fígado foi estimulada pelo substrato do complexo II (10mM succinato) e 200µM ADP.
Ensaio semelhante foi feito no painel (B), onde mitocôndria de cérebro foi usada por apresentar
hexokinase ligada a membrana externa mitocondrial semelhante às células tumorais. Nos painéis (C) e (D) foi adicionado 20mM de 2-DOG (ativador da HK). Os gráficos representam cinco experimentos
independentes.

Resultados |59
Para confirmar que a maior sensibilidade observada no estado 3 seria
regulada pelo potencial de membrana como sugerido por resultados anteriores ou
pela presença do nucleotídeo, nós dissipamos parcialmente a formação do potencial
de membrana utilizando o ionóforo de prótons FCCP. O nível de dissipação por
FCCP foi ajustado de modo a ser o mesmo nível do induzido por 200 µM de ADP,
mimetizando a respiração em estado 3 porém, desacoplada (figura 17A). Na figura
17B, após a dissipação da formação do potencial de membrana, 50 µM de 3-BrPA
foi adicionado e uma leve inibição dependente de tempo foi observada.
Estes resultados são semelhantes aos resultados observados no estado 4. A
figura 17C sugere a partir dos resultados da figura 17A e 17B que a ciclagem de
ADP é fundamental para uma maior sensibilidade ao 3-BrPA e não o nível do ΔΨm e
nem a presença de nucleotídeos (ADP ou ATP). A ciclagem de ADP então
promoveria essa maior reatividade do 3-BrPA a SDH e demais complexos. Também
é sugerido nesta figura, que a HK ligada a mitocôndria de tumores coloca estas
mitocôndrias em um estado respiratório semelhante ao estado 3 permanente. Essa
característica, pelos resultados obtidos nesta dissertação, seria uma propriedade
facilitadora da reação inibitória do 3-BrPA as desidrogenases mitocondriais.
A importância do papel da ciclagem de ADP na potencialização do efeito do 3-
BrPA é melhor representada na figura 18. Neste ensaio, inibimos o ANT com
diferentes concentrações de carboxiatractilosídeo (figura 18).
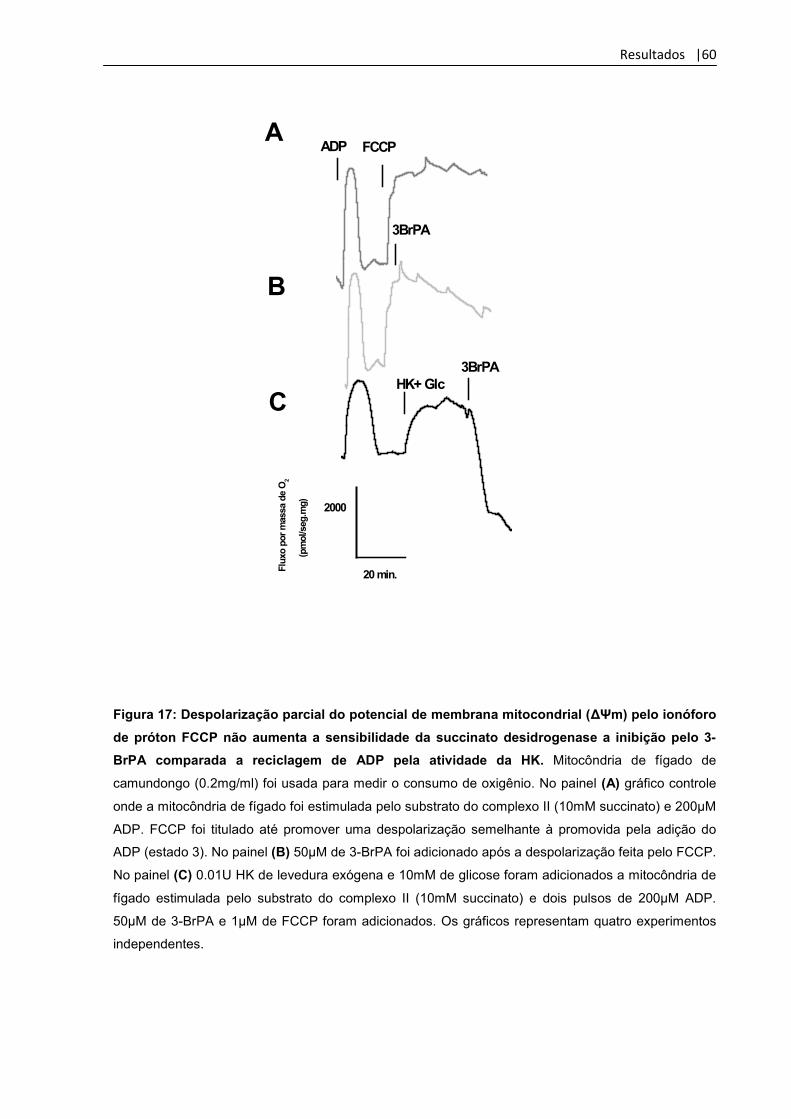
Resultados |60
3BrPA
FCCPADPA
B
C3BrPA
20 min.Flux
o po
r mas
sa d
e O
2
(pm
ol/s
eg.m
g) 2000
HK+ Glc
Figura 17: Despolarização parcial do potencial de membrana mitocondrial (ΔΨm) pelo ionóforo de próton FCCP não aumenta a sensibilidade da succinato desidrogenase a inibição pelo 3-BrPA comparada a reciclagem de ADP pela atividade da HK. Mitocôndria de fígado de
camundongo (0.2mg/ml) foi usada para medir o consumo de oxigênio. No painel (A) gráfico controle
onde a mitocôndria de fígado foi estimulada pelo substrato do complexo II (10mM succinato) e 200µM
ADP. FCCP foi titulado até promover uma despolarização semelhante à promovida pela adição do
ADP (estado 3). No painel (B) 50µM de 3-BrPA foi adicionado após a despolarização feita pelo FCCP.
No painel (C) 0.01U HK de levedura exógena e 10mM de glicose foram adicionados a mitocôndria de
fígado estimulada pelo substrato do complexo II (10mM succinato) e dois pulsos de 200µM ADP.
50µM de 3-BrPA e 1µM de FCCP foram adicionados. Os gráficos representam quatro experimentos
independentes.

Resultados |61
0
8000
16000
24000
0.5 µM
0.005 µM
0.1 µM
0.01 µM
A
3-BrPAGlic +HK Carboxi
ADP
0
8000
16000
24000
ADPCarboxi
Glic +HK
3-BrPA
E
D
C
B
0
8000
16000
24000
ADP
Carboxi
Glic +HK 3-BrPA
C
onsu
mo
de O
2
(pmol
O2/ s
ec. m
g pt
n)
0
8000
16000
24000
CarboxiGlic +HK
ADP
3-BrPA
10 20 30 400
8000
16000
240003-BrPA
ADP
Glic +HK
Controle
Tempo (min)
Figura 18: Importância da ciclagem de ADP no efeito do 3-BrPA. Mitocôndria de fígado de
camundongo (0.2mg/ml) foi usada para medir o consumo de oxigênio em diferentes concentrações de
carboxiatractlosídeo. O ensaio foi feito na presença de 10 mM succinato, 10 mM de glicose, 1 mM HK
de levedura e 100 µM 3-BrPA. No painel (A) mitocôndria de fígado na presença de 0,5µM de
carboxiatractilosídeo. No painel (B) 0,1 µM de carboxiatractilosídeo. Painel (C) 0,01 µM de
carboxiatractilosídeo. Painel (D) 0,005 µM de carboxiatractilosídeo e no painel (E) controle. Os
gráficos representam quatro experimentos independentes.

Resultados |62
Os resultados apresentados mostram a importância da ANT no efeito do 3-
BrPA corroborando com os resultados anteriores (figura 16 e 17). Em doses maiores
(0,5 µM de carboxiatractilosídeo) (figura 18A), a inibição pelo 3-BrPA foi mais lenta,
levando aproximadamente 13 minutos para inativar 50% da respiração induzida pela
hexocinase quando comparada ao controle na qual levou aproximadamente 3
minutos (figura 19). Além da importância da ciclagem de ADP, outro efeito curioso
que notamos foi a existência de 2 fases de inibição ( figura 18C e 18D), uma mais
lenta e outra mais rápida, sugerindo que a droga poderia está atuando em, pelo
menos 2 sítios, um de efeito imediato, que é apresentado quando o ANT está menos
inibido e outro em tempo maior, quando o trocador ANT esta mais inibido.

Resultados |63
0,000 0,01 0,1 1
2
4
6
8
10
12
14
T1/2
par
a a
inat
ivaç
ão d
e 50
%
da re
spira
ção
pelo
3-B
rPA
(min
.)
Carboxiatractilosídeo, µM
500
1000
1500
2000
2500
Tempo requerido para inativação de 50% da respiração pelo 3BrPA Respiração induzida pela hexocinase
Resp
iraçã
o in
duzi
da p
ela
hexo
cina
se
(
pmol
O2.
sec-1
. mg-1
)
Figura 19: Correlação entre a taxa da respiração induzida pela atividade da HK e a reatividade ao 3-BrPA. Mitocôndria de fígado de camundongo (0.2mg/ml) foi usada para medir o consumo de
oxigênio em diferentes concentrações de carboxiatractlosídeo. O ensaio foi feito na presença de 10
mM succinato, 10 mM de glicose, 1 mM HK de levedura e 100 µM 3-BrPA. Os valores são médias ±
EP de quatro experimentos independentes.

5- DISCUSSÃO ______________________________________

Discussão |65
5- DISCUSSÃO:
Diversos estudos propõem que o 3-BrPA poderia atuar como um potente
inibidor em certos tumores, particularmente em hepatocarcinomas. Entretanto, o
mecanismo bioquímico de ação desta droga não está completamente esclarecido.
Pedersen e colaboradores (Pedersen, 2007) têm enfatizado que o alvo celular
preferencial desta droga estaria na reação de fosforilação da glicose catalisada pela
hexocinase do tipo II presente em altos níveis nos carcinomas hepáticos. Os
tumores apresentam uma alta taxa glicolítica mesmo em presença de oxigênio,
efeito este observado por Otto Warburg em 1930. Vários mecanismos bioquímicos
têm sido propostos para explicar o fato de que a alta demanda energética requerida
pelo tumor é resultado de uma alta percentagem de síntese de ATP pela glicólise
(cerca de 60%,) e 40 % pela fosforilação oxidativa realizada pelas mitocôndrias
(Nakashima et al., 1984). Neste cenário a inibição do fluxo glicolítico em uma etapa
de grande controle sobre o fluxo desta via poderia explicar o potente efeito tóxico
desta droga sobre os tumores.
Todavia, estudos prévios já descreviam efeitos do 3-BrPA sobre a atividade
de preparações de enzimas mitocondriais tais como, piruvato desidrogenase e
succinato desidrogenase (Sanborn et al., 1971; Maldonado et al., 1972, Korotchkina
et al., 1999 e Acan e Ozer, 2001). Assim como também na glutamato desidrogenase
(Tunnicliff, 1978). Entretanto, nestes estudos preliminares as concentrações
experimentais utilizadas eram sempre bastante elevadas (entre 1 a 20 mM), sendo
de difícil significado terapêutico para justificar uma investigação detalhada sobre o
mecanismo de ação desta droga sobre a fosforilação oxidativa em um contexto
terapêutico. Porém, observações do grupo do Dr. Pedersen também confirmaram os
efeitos desta droga sobre o consumo de oxigênio (Ko et al., 2001) sugerindo um
outro potencial alvo de ação do 3-BrPA. De acordo com esses dados prévios, nosso
grupo iniciou um estudo detalhado analisando os possíveis alvos de ação do 3-
BrPA, tanto na glicólise como na respiração mitocondrial, em células de
hepatocarcinoma HepG2 (Pereira da Silva et al., 2009). Foi sugerido por Pereira da
Silva (2009) que um alvo importante do 3-BrPA seria a succinato desidrogenase,
independente da fonte de carbono (glicose ou glutamina) usado para o cultivo
celular. Interessantemente, a hexocinase não teve a sua atividade afetada pelo

Discussão |66
tratamento das células HepG2 com 3-BrPA, como sugerido por Pedersen e
colaboradores (Pereira da Silva et al., 2009).
Com base nos dados prévios da literatura (Sanborn et al., 1971; Maldonado et
al., 1972, Korotchkina et al., 1999 e Acan e Ozer, 2001 e Tunnicliff, 1978),
inicialmente foram feitas medidas diretas das atividades das enzimas do complexo II
(SDH), complexo I (NADH: oxidoredutase) em concentrações menores de 3-BrPA
que as estudadas previamente. Observamos que o 3-BrPA inibiu completamente a
atividade da SDH apresentando um IC50
Estes resultados foram confirmados quando analisamos mais profundamente
a ação do 3-BrPA em diferentes parâmetros mitocondriais ( medidas de potencial de
membrana e consumo de oxigênio). Na figura 11C, está mostrado que a formação
do potenxial de membrana (ΔΨm) é inibida em 50 % com 170 µM de 3-BrPA quando
o substrato é o succinato e em 20 % quando o substrato é ao nível de complexo I
(piruvato/malato) ou ao nível complexo IV (TMPD/ascorbado). Entretanto, a menor
inibição observada para os complexos I e IV para doses relativamente baixas do
agente alquilante não descarta um efeito inibitório sinérgico sobre a respiração ainda
maior em condições fisiológicas, ou seja, na presença tanto de piruvato como de
succinato como mostrado na figura 13, onde, quando o succinato foi adicionado
juntamente com os substratos do complexo I (piruvato, malato e glutamato) o efeito
sobre a despolarização da formação do ΔΨm foi potencializado. Este efeito sinérgico
sobre a respiração poderia depender da quantidade relativa de cada complexo na
CTE que determina a velocidade do fluxo total de consumo de oxigênio – limiar
inibitório de cada complexo sobre a respiração (Rossignol et al., 2003).
de aproximadamente 100 µM (figura 10).
Todavia nenhuma inibição foi observada na atividade da NADH: ubiquinona
oxidoredutase quando incubada por 1 minuto. Porém, quando as mitocôndrias de
fígado de camundongo são pré-incubadas por 30 minutos com a droga, observou-se
uma significativa inibição da atividade da NADH: ubiquinona oxidoredutase
mostrando ser dependente do tempo (figura 7). Nenhuma inibição da atividade do
complexo IV foi observada (figura 8), concluindo ser a succinato desidrogenase a
mais sensível a ação da droga.
Em trabalhos prévios foi mostrado uma queda dos níveis de ATP nas células
tumorais sob a ação do 3-BrPA (Ko et al., 2001; Geschwind et al., 2002). Este efeito

Discussão |67
não pode ser relacionado a uma inibição da atividade de síntese de ATP já que não
foi observado inibição da atividade ATPásica azida sensível da FoF1 ATPase (figura
9). Este dado sugere a queda nos níveis de ATP das células tumorais pode
depender de outros fatores. Uma possibilidade interessante é que a inibição da SDH
pelo 3-BrPA uma vez promovendo a queda do ΔΨm pode levar a FoF1 ATP sintase
a passar a funcionar como uma FoF1 ATPase na tentativa de restabelecer o ΔΨm
(notar que esta atividade não é inibida pelo 3-BrPA). O que facilitaria ainda mais o
decréscimo nos níveis intracelulares de ATP.
Os estudos de respirometria de alta resolução em mitocôndrias isoladas de
fígado de camundongo revelaram com maior clareza o caráter diferencial da
reatividade do 3BrPA frente aos diferentes estados respiratórios da mitocôndria
(figura 12). Os valores para inibição do consumo de oxigênio por 3-BrPA em
preparação de mitocôndrias isoladas foi maior no estado 3 quando comparado ao
estado 2 e 4 frente aos substratos do complexo I e complexo II. Estes dados
sugerem que talvez o fluxo de elétrons na CTE e a magnitude do ΔΨm possam estar
contribuindo para que a SDH se estabilize em uma conformação mais reativa ao 3-
BrPA. Em estados respiratórios não fosforilantes (estados 2 e 4), as unidades de
SDH não mobilizadas na transferência de elétrons poderiam existir em microestados
conformacionais de baixa reatividade ao 3-BrPA, como os revelados pelo maior
tempo de inibição (figuras 12, 16-19). Assim como na respiração, o efeito inibitório
do 3-BrPA sobre o ΔΨm foi mais pronunciado quando o ADP foi incluído ao meio
induzindo ao estado 3 da respiração (figura 13). Este efeito é bem claro quando
succinato é usado como substrato respiratório. Uma possível explicação para esta
maior inibição da SDH em estado 3 seria que a presença de nucleotídeos, tais
como, ADP e ATP, poderiam modular a atividade da SDH alterando a reatividade do
3-BrPA. De fato, já foi demonstrado que a SDH de planta tem a sua atividade
regulada por ADP e ATP e oxalacetato (Affourtit et al., 2001; Gutman, 1978). Na
figura 17, foi mostrado que a maior reatividade do 3-BrPA no estado 3 foi
independente do nível do ΔΨm, pois quando FCCP (ionóforo de prótons) foi utilizado
para despolarizar o potencial de membrana ao nível da despolarização equivalente a
obtida pelo estado 3 (ADP), não se observou uma reatividade tão rápida como a
observada pela ciclagem de ADP/ATP.

Discussão |68
Assim, a inibição por 3-BrPA da SDH poderia ser dependente do tipo de
nucleotídeo (ATP ou ADP) a da sua localização (dentro e/ou fora da matriz
mitocondrial) ou mesmo do fluxo de troca de ATP/ADP catalisada pelo ANT.
Entretanto, nós não fomos capazes de responder a estas questões no presente
estudo e outros experimentos serão conduzidos para se tentar esclarecer o papel
dos ATP/ADP na inibição da SDH causada pelo 3-BrPA.
Outro mecanismo possível para a inibição da SDH seria a reação do 3-BrPA
com os prováveis sítios de alquilação, resíduos de cisteína (-SH) e/ou treonina (-OH)
presentes na molécula da SDH e NADH-Oxidoredutase. De fato, Sanborn et al.,
1971 mostrou que a succinato desidrogenase de rato possui resíduos de cisteína no
sítio catalítico da enzima tornando assim a SDH mais reativa ao 3-BrPA. Estudos
demonstraram que a alquilação da succinato desidrogenase solúvel por NEM direto
no sítio ativo provoca a inibição da enzima (Kenney, 1975 e Vonogradov et al.,
1976). Observação importante foi o fato de que agentes redutores tal como GSH
protegem do efeito inibitório do 3-BrPA tanto sobre a respiração como para a
formação do ΔΨm usando succinato e piruvato/malato como substratos respiratórios
(figura 15) sugerindo que o estado redox da mitocôndria pode desempenhar também
um papel importante sobre a potência dos efeitos da droga. Sendo assim, uma
possibilidade seria que agentes redutores fisiológicos como a glutationa e NADPH2
Uma grande fração de tumores, inclusive hepatocarcinomas humanos,
apresenta uma grande quantidade de hexocinase associada à membrana
mitocondrial externa (Nakashima et al., 1988). Esta associação da HK a mitocôndria
acopla a glicólise extramitocondrial com a ativação da fosforilação oxidativa
intramitocondrial por um mecanismo de ciclagem de ADP (da-Silva, 2004). Nesta
situação, a glicose que entra no citoplasma recebe imediatamente um grupo fosforil
a partir do ATP gerado pela mitocôndria produzindo glicose 6-fosfato e enviando de
volta para a matriz mitocondrial o ADP que promove a ativação permanente da
respiração para um estado similar ao de estado 3. Outro fato curioso foi a diferença
de reatividade dos sítios de inibição, um lento e outro rápido sugerindo que o 3-BrPA
possam afetar o estado redox da respiração sobre certas condições metabólicas
influenciando a reatividade do 3-BrPA sobre a SDH.

Discussão |69
poderia estar agindo com 2 tipos de grupamentos sulfidrilas que estariam envolvidos
no sítio ativo da succinato desidrogenase, como já sugerido por LeQuôc et al., 1981.
A importância da ciclagem de ADP foi mostrada nas figuras 18 e 19, onde se
evidencia o papel crucial do ANT no tempo de inibição da respiração pelo 3-BrPA.
Assim sendo, mostramos que a presença favorável da HK nas preparações
mitocondriais de tumores levam a respiração mitocondrial deste tipo celular a
estados de ciclagem de nucleotídeos que se mostraram mais reativos ao 3-BrPA
culminando na inibição da respiração e colapso do ΔΨm. O colapso do ΔΨm
representa uma importante etapa na ativação das vias de morte celular. Desta
forma, se por um lado a localização preferencial da HK no VDAC funciona para os
tumores como um potente mecanismo de prevenção a morte, por outro, a atividade
da HK associada ao VDAC em tumores em que o efeito Warburg é proeminente, os
coloca em uma situação de vulnerabilidade ao agente alquilante 3-BrPA. Estes
achados podem representar um dos primeiros mecanismos descritos da fragilidade
do metabolismo energético das células cancerosas.

6- CONCLUSÃO ___________________________________

Conclusões |71
6- CONCLUSÕES.
Em mitocôndrias de fígado de camundongo 3-BrPA inibe tanto a taxa
de consumo de oxigênio e a formação do potencial de membrana (ΔΨm
)
principalmente a nível da succinato desidrogenase (complexo II).
O agente redutor GSH protege tanto o consumo de oxigênio o ΔΨm
da
inibição pelo 3-BrPA.
Na presença de outros substratos respiratórios, tais como glutamato,
malato e piruvato; a presença de succinato aumenta o efeito inibitório do 3-BrPA.
3-BrPA foi mais efetivo em dissipar o ΔΨm
e inibir o consumo de
oxigênio quando o ADP foi incluído no meio de reação.
A cinética de inativação da respiração pelo 3-BrPA é complexa sendo
mais rápida quando a mitochondria está sintetizando ATP e reciclando ADP pela
reação da hexoquinase.
A inativação da respiração sustentada por succinato pelo 3-BrPA não
é dependente do ΔΨm já que a sua dissipação parcial (ao mesmo nível do ADP)
induzida por ionóforo não foi capaz de causar uma reatividade equivalente. A
ciclagem de ADP/ATP pelo sistema do ANT, F0F1ATPsintase e HK afeta a
sensibilidade da succinato desidrogenase ao 3-BrPA .

7- REFERÊNCIAS __________________________________________

Referências |73
7- REFERÊNCIAS .
Acan NL; Ozer N. Modification of human erythrocyte pyruvate kinase by an active site-directed reagent: bromopyruvate. J. Enzyme Inhib, 16, 457-464, 2001.
Affourtit C; Krab K; Leach GR; Whitehouse DG; Moore AL. New insights into the regulation of plant succinate dehydrogenase. On the role of the protonmotive force. J Biol Chem
.,35, 32567- 32574, 2001.
Aisenberg AC. Studies on normal and neoplastic mitochondria. I. Respiration. Cancer Res., 21, 295- 303, 1961.
Akerman KE; Järvisalo JO. Effects of ionophores and metabolic inhibitors on the mitochondrial membrane potencial within isolated hepatocytes as measured with the safranine method. Biochem J, 192, 183-190, 1980.
American Cancer Society. Cancer facts and figures 2009. Atlanta: American Cancer Society; 2009.
Andrei DV; Eleonora VG; Victor VZ. Reactivity of the Sulfhydryl Groups of Soluble Succinate Dehydrogenase. Eur. J. Biochem. 63, 365-371 1976.
Arora KK; Pedersen PL. Functional significance of mitochondrial bound hexokinase in tumor cell metabolism: Evidence for preferential phosphorylation of glucose by intra-mitochondrially generated ATP. J. Biol. Chem. 263, 14422-14428, 1988.
Arora KK; Fanciulli M; Pedersen PL. Glucose phosphorylation in tumor cells. Cloning, sequencing and overexpression in active form of a full-length cDNA enconding a mitochondrial bindable form of hexokinase. J. Biol. Chem. 265, 6481-6488, 1990.

Referências |74
Baker JP; Rabin BR. Effects of bromopyruvate on the control and catalytic properties of glutamate dehydrogenase. Eur. J. Biochem., 11, 154- 159, 1969.
Balijepalli S; Annepu J; Boyd MR; Ravindranath, V. Effect of thiol modification on brain mitochondrial complex I activity. Neuroscience Lett., 203- 206, 1999.
Bosch X,;Ribes J, Borràs J. Epidemiology of primary liver cancer. Semin Liver Dis; 19: 271–85, 1999.
Brdiczka D; Wallimann T. The importance of the outer mitochondrial compartment in regulation of energy metabolism. Mol. Cell Biochem. 133-134, 69-83, 1994.
Breedis C; Young G. Blood supply in neoplasms. Am. J. Pathol. 30, 969-985, 1954.
Chance B; Williams GR. The respiratory chain and oxidative phosphorylation. Adv. Enzymol. 17, 65-134, 1956.
Chang GG; Hsu RY. The substrate analog bromopyruvate as a substrate, an inhibitor and a alkylating agent of malic enzyme of pigeon liver. Biochem. Biophys. Res. Commun., 55, 580- 587, 1973.
Colombo M. Risk groups and preventive strategies. In: Berr F,Bruix J, Hauss J, Wands J, Wittekind Ch, eds. Malignant livertumors: basic concepts and clinical management. Dordrecht: Kluwer Academic Publishers BV and Falk Foundation, 67–74,2003.
da-Silva WS, Gómez-Puyou A, de Gómez-Puyou MT, Moreno-Sanchez R, De Felice FG, de Meis L, Oliveira MF, Galina A. Mitochondrial bound hexokinase activity as a preventive antioxidant defense: steady-state ADP formation as a regulatory mechanism of membrane potential and reactive oxygen species generation in mitochondria. J Biol Chem., 38, 39846- 39855, 2004.

Referências |75
Nicholls DG; Ferguson SJ. Bioenergetics 3, 4º ed.London: Academic press, 2001. 297 p. cap. 7, p. 195-216.
Di Bisceglie AM; Rustgi VK; Hoofnagle JH; Dusheiko GM; Lotze MT. NIH conference. Hepatocellular carcinoma. Ann. Intern. Med. 108, 390-401, 1988.
Dietzen DJ; Davis EJ. Oxidation of pyruvate, malate, citrate, and cytosolic reducing equivalents by AS-30D hepatoma mitochondria. Arch. Biochem. Biophys. 305, 91-102, 1993.
Dünschede F, Zwicker K, Ackermann H, Zimmer G. ADP- and oligomycin-sensitive redox behavior of F0 b thiol in ATPsynthase depends on neighbored primary structure: Investigations using 14-C-labeled alpha lipoic acid. Biofactors, 19, 19-32, 2003.
El-Serag, HB; Mason, AC. Rising incidence of hepatocellular carcinoma in the United States. N. Engl. J. Med. 340, 745-750, 1999.
El- Serag, HB; Rudolph, KL. Hepatocellular carcinoma: epidemiology and molecular carcinogenegis. Gastroenterology, 132, 2557- 2576, 2007.
Euler HV; Adler EZ. Über die komponenten der dehydrasesysteme. 6. Dehyrierung von hexosen unter mitwirkung von adenosintriphosphorsäure.Physiol. Chem 235:122, 1935.
Felberg NT, Hollocher TC. Inactivation of succinate dehydrogenase by N-ethylmaleimide. Stoichiometry and chemistry. J Biol Chem
., 14, 4539-42, 1972.
Fink MP; Evans TW. Mechanisms of organ dysfunction in critical illness: report from a Round Table Conference held in Brussels. Intensive Care Med. 28, 369-375,2002.

Referências |76
Fiske, CF; Subbarow, Y. The colorimetric determination of phosphorus. J. Biol. Chem. 56,2, 375-400, 1925.
Geschwind JF; Artemov D; Abraham S; Omdal D; Huncharek C; McGee C; Arepally, A; Lambert, D; Venbrux, AC; Lund, GB. Chemoembolization of liver tumor in a rabbit model: Assessment of tumor cell death with diffusion-weighted MR imaging and histologic analysis. J. Vasc. Interv. Radiol. 11,1245-1255, 2000.
Geschwind JF; Ko YH; Toberson MS; Magee C; Pedersen PL. Novel therapy for liver cancer: direct intraarterial injection of a potent inhibitor of ATP production. Cancer Res. 3909-3913, 2002.
Gosálvez M; Perez-Garcia J; Weinhouse S. Competition for ADP between pyruvate kinase and mitochondrial oxidative phosphorylation as a control mechanism in glycolysis. Eur. J. Biochem. 46, 133-140, 1974.
Grigorieff, N. Structure of the respiratory NADH:ubiquinone oxidoreductase (complex I). Current Opinion in Structural Biology, 9, 476- 483, 1999.
Gutman M. Modulation of mitochondrial succinate dehydrogenase activity, mechanism and function. Mol Cell Biochem
., 1, 41- 60, 1978.
Instituto Nacional de Câncer; Ministério da Saúde. Estimativas 2008: Incidência de Câncer no Brasil, Rio de Janeiro (Brasil): INCA; 2007. Disponível em: http://www.inca.gov.br/estimativa/2008
Jones AR; Gillan L; Milmlow D. The anti- glycolitic activity of 3-bromopyruvate on mature boar spermatozoa in vitro. Contraception, 52, 317- 320, 1995.
Kamangar F, Dores GM, Anderson WF. Patterns of cancer incidence, mortality, and prevalence across five continents: defining priorities to reduce cancer disparities in different geographic regions of the world. J. Clin. Oncol. 24, 2137-2150, 2006.

Referências |77
Kapoor, V; McCook, BM; Torok, FS. An introduction to PET-CT imaging. Radiographics 24,523-543, 2004.
Kenney WC. The reaction of N-ethylmaleimide at the active site of succinate dehydrogenase. J Biol Chem
., 8, 3089- 3094, 1975.
Kenney WC. The reaction of N-ethylmaleimide at the active site of succinate dehydrogenase. J Biol Chem.
, 25, 3089- 3094, 1995.
Kenney WC. The reaction of N-ethylmaleimide at the active site of succinate dehydrogenase. J Biol Chem
., 8, 3089- 3094, 1975.
Kerner, J and Hoppel, C. Fatty acid import into mitochondria. Biochim. Biophys. Acta 1486, 1-17, 2000.
Khalimonchuk, O and Rödel, G. Biogenesis of cytochrome c oxidase. Mitochondrion, 5, 363- 388, 2005.
Ko, YH; Pedersen, PL; Geschwind, JF. Glucose catabolism in the rabbit VX2 tumor model for liver cancer: characterization and targeting hexokinase. Cancer Let.t 173, 1, 83-91,2001.
Ko, YH; Smith, BL; Wang, Y; Pomper, MG; Rini, DA; Torbenson, MS; Hullihen, J; Pedersen, PL. Advanced cancers: eradication in all cases using 3-bromopyruvate therapy to deplete ATP. Biochem. and Biophys. Res. Commun. 324, 269-275, 2004.
Korotchkina LG, Showkat AM E Patel MS. Involvement of alpha-cysteine-62 and beta-tryptophan-135 in human pyruvate dehydrogenase catalysis. Arch. Biochem. Biophys. 369, 277-287, 1999.

Referências |78
Kurokawa, M; Oda, S; Tsubotani, E; Fujiwara, H; Yokoyama, Y; Ishibashi, S. Characterization of hexokinase isoenzyme types I and II in ascites tumor mitochondrial membrane. Mol. Cell. Biochem. 45, 151-157, 1982.
Lardy HA; Wellman, H. Oxidative phosphorylations; rôle of inorganic phosphate and acceptor systems in control of metabolic rates. J Biol Chem
., 1, 215- 224, 1952.
Lê-Quôc, K.; Lê-Quôc, D.; Gaudemer, Y. Evidence for the existence of two classes of sulfhydryl groups essential for membrane-bound succinate dehydrogenase activity. Biochemistry, 20, 7, 1705-1710, 1981.
Lehninger AL. Molecular basis of structure and function. W. A. Benjamin, Inc. New York, 1, 1-14, 1965.
Lesnefsky, EJ and Hoppel, CL. Oxidative phosphorylation and aging. Ageing Res. Reviews 5, 402-433, 2006.
Liaw YF, Tai DI, Chu CM, et al. Early detection of hepatocellular carcinoma in patients with chronic type B hepatitis: a prospective study. Gastroenterology, 90, 263–67, 1986.
Lippe G, Sala FD, Sorgato MC. ATP Synthase Complex from Beef Heart Mitochondria : Role of the thiol group of the 25-kDa subunit of Fo in the coupling mechanism between Fo and F1. J. Biol. Chem. 263, 35, 18627-18634, 1988.
Llovet J; Real ML; Montana X. Arterial embolisation or chemoembolisation versus symptomatic treatment in patients with unresectable hepatocellular carcinoma: A randomized controlled trial. Lancet 359, 1734-1739, 2002.
Lowry OH; Rosebrough JN; Farr AL; Randall RJ. Protein measurements with the folin phenol reagent. J Biol. Chem 193, 265-275, 1951.

Referências |79
Maldonado ME; Oh K; Frey PA. J. Studies on Escherichia coli pyruvate dehydrogenase complex. I. Effect of bromopyruvate on the catalytic activities of the complex. Biol. Chem. 247, 2711-2716, 1972.
Marín-Hernandez A; Rodríguez-Henríquez S; Vital-Gonzalez PA; Flores-Rodríguez FL; Macias-Silva M; Sosa-Garrocho M; Moreno-Sánchez R. Determining and understanding the control of glycolysis in fast-growth tumor cells. Flux control by an over-expressed but strongly product-inhibited hexokinase. FESB J. 273, 1975-1988, 2006.
Mathupala SP; Rempel A; Pedersen PL. Glucose catabolism in cancer cells: Isolation, sequence, and activity of the promoter for type II hexokinase. J. Biol. Chem. 270, 16918-16925, 1995.
Meyerhof O. Die chemischen vorgange im muskel. Biochem. Z 183, 176-215, 1927.
Mitchell P. Possible molecular mechanisms of the protonmotive function of cytochrome systems. J. Theor. Biol. 62, 327-367, 1976.
Moreno-Sánchez R; Rodríguez-Enríquez S; Marín-Hernández A; Saavedra E. Energy metabolism in tumor cells. FESB J. 274, 1393-1418, 2007.
Muller M; Siems W; Buttgereit F; Dumdey R; Raport SM. Quantification of ATP- producing and consuming processes of Ehrlich ascites tumour cells. Eur. J Biochem 161, 701-705, 1986.
Nakashima RA; Mangan PS; Colombini M; Pedersen PL. Hexokinase receptor complex in hepatoma mitochondria: evidence from N,N'-dicyclohexylcarbodiimide-labeling studies for the involvement of the pore-forming protein VDAC. Biochemistry 25, 1015-1021, 1986.

Referências |80
Nakashima RA; Paggi MG; Pedersen PL. Contributions of glycolysis and oxidative phosphorylation to adenosine 5´- triphosphate production in AS-30D hepatomas cells. Cancer Res. 44, 5702-5706, Cancer Res. 44, 5702-5706, 1984.
Nakashima RA; Paggi MG; Scott LJ; Pedersen PL. Purification and characterization of a bindable form of mitochondrial bound hexokinase from the highly glycolytic AS-30D rat hepatoma cell line. Cancer Res. 48, 913-919, 1988.
Nelson, Davi L.; Cox, Michael M. Fosforilação Oxidativa e Fotofosforilação.In: .Lehninger Princípios de Bioquímica. 3º ed.São Paulo: Sarvier, 2002. 975 p. cap. 19, p. 515-526.
Nicolay K; Rojo M; Wallimann T; Demel R; Hovius R. The role of contact sites between inner and outer mitochondrial membrane in energy transfer. Biochim. Biophys. Acta 1018, 229-233, 1990.
Okuda K; Ohtsuki T; Obata H; Tomimatsu M; Okazaki N; Hasegawa H; et al. Natural history of hepatocellular carcinoma and prognosis in relation to treatment. Study of 850 patients. Cancer 56, 918-928, 1985.
Parkin DM. Global cancer statistics in the year 2000. Lancet Oncol 2:533–543, 2001.
Parkin DM. Cancer Incidence in five continents. IARC scientific publications volume VIII[No. 155]. 2002. Lyon: IARCPress.
Parry DM; Pedersen PL. Intracellular localization and properties of particulate hexokinase in the Novikoff ascites tumor. Evidence for an outer mitochondrial membrane location. J. Biol. Chem. 258, 10904-10912, 1983.
Parlo RA; Colemam PS. Enhanced rate of citrate export from cholesterol-rich hepatoma mitochondria. The truncated Krebs cycle and other metabolic

Referências |81
ramifications of mitochondrial mebrane cholesterol. J. Biol. Chem. 259, 9997-10003, 1984.
Pastorino JG; Hoek JB. Hexokinase II the integration of energy metabolism and control of apoptosis. Curr. Med. Chem. 10, 1535-1551, 2003.
Pauser S; Wagner S; Lippmann M; Rohlen U; Reszka R; Wolf KJ; Berger G. Evaluation of efficient chemoembolization mixtures by MR imaging therapy monitoring: An experimental atudy on the VX2 tumor in the rabbit liver. Cancer Res. 56, 1863-1867, 1996.
Pedersen PL; Mathupala S; Rempel A; Geschwind JF; Ko YH. Mitochondrial bound type II hexokinase: a key player in the growth and survival of many cancers and an ideal prospect for therapeutic intervention. Biochim. et Biophys. Acta 1555, 14-20, 2002.
Pedersen PL. Tumor mitochondria and the bioenergetics of cancer cells. Prog. Exp. Tumor Res. 22, 190-274, 1978.
Pedersen PL. The cancer cells "power plant" as promising therapeutic targets: An overview. J. Bioenerg. Biomembr., 2007.
Pereira da Silva AP; El-Bacha T; Kyaw N; dos Santos RS; da-Silva WS; Da Poian, AT; Galina A. Differential inhibition of energy- produncing pathways of HepG2 cells by 3- Bromopyruvate. Biochem J
., 3, 717- 726, 2009.
Pierre R; Agnes R. Inborn errors of complex II- Unusual human mitochondrial diseases. Biochimica et Biophysica Acta 1553,117-122, 2002.
Pierre R, Arnold M, Agnès R. Succinate dehydrogenase and human diseases: new insights into a well-known enzyme. European Journal of Human Genetics 10, 289-291, 2002.

Referências |82
Poynard T, Aubert A, Lazizi Y, Bedossa P, Hmelin B, Terris B, et al. Independent risk factors for hepatocellular carcinoma in French drinkers. Hepatology,13:896-901,1991.
Ramsey DE; Kernagis LY; Soulen MC; Geschwind JF. Chemoembolization of hepatocellular carcinoma. J. Vasc. Interv. Radiol. 13, S211-S221, 2002.
Rempel A; Bannasch P; Mayer D. Differents in expression and intracellular distribution of hexokinase isoenzymes in rat liver cells of differents transformation stages. Biochem. Biophys. Acta 1219, 660-668, 1994.
Rohn S, Kroh LW. Electron spin resonance--a spectroscopic method for determining the antioxidative activity. Mol Nutr Food Res
., 10, 898- 907, 2005.
Rohren EM; Turkington TG; Coleman RE. Clinical applications of PET in oncology. Radiology 231, 305-332, 2004.
Rose IA; Warms JVB. Stability of hexokinase II in vitro and in ascites tumor cells. Arch. Biochem. Biophys. 213, 625-634, 1982.
Rossignol R; Faustin B; Rocher C; Malgat M; Mazat JP; Letellier T. Mitochondrial threshold effects. Biochem. J. 370, 751-762, 2003.
Rous P; Kidd JG; Smith WE. Experiments on the cause of the rabbit carcinomas derived from virus-induced papillomas. II. Loss by the VX2 carcinomas of the power to immunize hosts against the papilloma virus. J. Exp. Med. 96, 159-174, 1952.
Rustgi VK. Epidemiology of hepatocellular carcinoma. Gastroenterol Clin. North Am 16, 545-551, 1987.
Saha S; Bardelli A; Guckhaults P; Velculescu VE; Rago C; St. Croix G; Romans KE; Choti MA; Lengauer C; Kinzler KW; Vogelstein BA. Phosphatase associated with metastasis of colorectal cancer. Science 294, 1343-1346, 2001.

Referências |83
Sanborn BM; Feldeberg NT; Hollocher TC. The inactivation of succinate dehydrogenase by bromopyruvate. Biochim. Biophys. Acta 227, 219-231, 1971.
Satterlee J; Hsu RY. Duck liver malic enzyme: sequence of a tryptic peptide containing the cysteine residue labeled by the substrate analog bromopyruvate. Biochim. Biophys. Acta, 1079, 247- 252, 1991.
Shinohara Y; Ichihara J; Terada H. Remarkably enhaceded expression of the type II hexokinase in rat hepatoma cell line AH130. FEBS Lett. 291, 55-57, 1991.
Syroeshkin AV, Vasilyeva EA, Vinogradov AD. ATP synthesis catalyzed by the mitochondrial F -F0 ATP synthase is not a reversal of its ATPase activity. FEBS Lett. 366, 29-32, 1995.
Sun Z, Lu P, Gail MH. Increased risk of hepatocellular carcinoma in male hepatitis B surface antigen carriers with chronic hepatitis whohave detectable aflatoxin metabolite M1. Hepatology,30: 379–83,1999.
Thele AP; Wilson, JE. Complete amino acid sequence of the type II isozyme of rat hexokinase, deduced from the cloned cDNA: comparison with a hexokinase from novikoff ascites tumor. Arch. Biochem. Biophys. 286, 645-651, 1991.
Tunnicliff G, Ngo TT. Mechanism of inactivation of brain glutamic decarboxylase by 3-bromopyruvate. Int. J. Biochem. 9, 249-252, 1978.
Vonck J; Schäfer E. Supramolecular organization of protein complexes in the mitochondrial inner membrane. Biochim. Biophys. Acta, 117- 124, 2009.
Vinogradov AD, Gavrikova EV, Goloveshkina VG. Kinetic and structural characteristics of succinate dehydrogenase components reacting with natural and artificial electron acceptors. Biokhimiia., 7, 155- 1168, 1976.

Referências |84
Weber, G. Ordered biochemical program of gene expression in cancer cells. Biochemistry (Moscou) 66, 1164-1173,2001.
Warburg, O. The Metabolism of Tumors. London Constable Co. Ltd, 1930.
Williams, GR. The respiratory chain and oxidative phosphorylation. Advan. Enzymol, 17, 65, 1956.
World Health Organization. Cancer. Fact sheet N°297. February 2006. [homepage on the Internet]. Geneva: World Health Organization; c2007 [cited 2007 oct. 10]. Available from: http://www.who.int/mediacentre/factsheets/fs297/en/print.html.
Zu XL; Guppy M. Cancer metabolism: facts, fantasy, and fiction. Biochem. Biophys. Res. Commun. 313, 459-465, 2004

Nome: Clara Rodrigues Ferreira Nascimento: 15/05/1983 Naturalidade: Rio de Janeiro Formação Acadêmica . Faculdade de Biologia – Modalidade Biologia do Desenvolvimento pela Universidade Fluminense, outubro 2003 a outubro 2007. . Mestrado em Química Biológica no Departamento de Bioquímica Médica – Instituto de Ciências Biomédicas – Universidade Federal do Rio de Janeiro. Orientação de Estudante
1. Andreza da Silva Lima – iniciação científica de janeiro/2008 a novembro/2009.
Comunicação em Congresso . 4 comunicações em congressos nacionais. Publicações 1. Cosentino-Gomes, Daniela, Russo-Abrahão, Thais, Fonseca-de-Souza, André Luiz, Rodrigues-Ferreira, C., Galina, Antonio, Meyer-Fernandes, José Roberto. (2009) Modulation of Trypanosoma rangeli ecto-phosphatase activity by hydrogen peroxide. Free Radical Biology & Medicine. , p.11 – 48.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo