prospecção e caracterização de linhagens fúngicas ... · de enzimas lignocelulolíticas para a...
TRANSCRIPT

Universidade Estadual de Montes Claros
Programa de Pós-graduação Stricto Sensu em Ciências Biológicas
PROSPECÇÃO E CARACTERIZAÇÃO DE LINHAGENS
FÚNGICAS PRODUTORAS DE COMPLEXOS ENZIMÁTICOS
LIGNOCELULOLÍTICOS PROVENIENTES DE RESÍDUOS DE
AMBIENTE CANAVIEIRO
Maria Lúcia Gomes e Souza
Montes Claros, Minas Gerais
2011

ii
Maria Lúcia Gomes e Souza
PROSPECÇÃO E CARACTERIZAÇÃO DE LINHAGENS
FÚNGICAS PRODUTORAS DE COMPLEXOS ENZIMÁTICOS
LIGNOCELULOLÍTICOS PROVENIENTES DE RESÍDUOS DE
AMBIENTE CANAVIEIRO
Prof. Dr. Dario Alves de Oliveira – Orientador
Prof. Dr. Henrique Maia Valério – Co-orientador
Dissertação apresentada ao Programa de
Pós-Graduação Stricto Sensu em Ciências
Biológicas da Universidade Estadual de
Montes Claros como requisito necessário para
conclusão do curso de Mestrado em Ciências
Biológicas.
Montes Claros, Minas Gerais
2011

iii
SUMÁRIO
Pág
RESUMO....................................................................................................... v
ABSTRACT.................................................................................................... vi
LISTA DE FIGURAS...................................................................................... vii
LISTA DE TABELAS...................................................................................... ix
1 INTRODUÇÃO............................................................................................ 1
2 REVISÃO BIBLIOGRÁFICA........................................................................ 4
2.1 Fontes de energia – resíduos lignocelulósicos......................................... 4
2.1.1 Celulose................................................................................................. 6
2.1.2 Hemicelulose......................................................................................... 9
2.1.3 Lignina................................................................................................... 10
2.2 Hidrólise da lignocelulose......................................................................... 10
2.3 Sistemas enzimáticos............................................................................... 12
2.3.1 Celulases............................................................................................... 12
2.3.1.1 Produção de celulases....................................................................... 14
2.3.2 Hemicelulases....................................................................................... 17
2.3.3 Ligninases............................................................................................. 18
2.4 Microrganismos celulolíticos..................................................................... 18
2.4.1 Aspectos ecológicos das comunidades que degradam a celulose....... 21
3 MATERIAL E MÉTODOS............................................................................ 23
3.1 Local de coleta......................................................................................... 23
3.2 Isolamento dos microrganismos............................................................... 24
3.3 Manutenção das linhagens....................................................................... 25
3.4 Pré seleção dos microrganismos lignocelulolíticos.................................. 25
3.5 Fermentação dos microrganismos isolados ativos................................... 27

iv
3.5.1 Quantificação dos microrganismos durante as fermentações............... 30
3.6 Extração das enzimas.............................................................................. 30
3.7 Determinação das atividades e concentrações enzimáticas do
complexo celulolítico e hemicelulolítico..........................................................
31
3.7.1 Atividade Carboximetilcelulase – CMCase........................................... 31
3.7.2 Atividade FPase.................................................................................... 31
3.7.3 Atividade glicosidase......................................................................... 32
3.7.4 Atividade hemicelulase (xilanase)......................................................... 32
3.7.5 Cálculo da atividade enzimática........................................................... 32
3.8 Identificação taxonômica......................................................................... 34
3.9 Análise estatística..................................................................................... 34
4 RESULTADOS E DISCUSSÃO................................................................... 35
4.1 Isolamento dos fungos filamentosos........................................................ 35
4.2 Manutenção e viabilidade dos isolados.................................................... 38
4.3 Seleção dos fungos com atividade lignocelulolítica................................. 39
4.4 Análise da atividade celulolítica e hemicelulolítica................................... 43
4.5 Identificação taxonômica.......................................................................... 57
5 CONCLUSÕES E PERSPECTIVAS........................................................... 58
6 REFERÊNCIAS BIBLIOGRÁFICAS............................................................ 60
APÊNDICES .................................................................................................. 68
ANEXOS......................................................................................................... 78

v
RESUMO
O potencial bioenergético do Brasil está representado em grande parte
pelo setor sucroalcooleiro, pois a cana-de-açúcar constitui uma fonte de
energia renovável e abundante. Além do aproveitamento do caldo para a
produção de etanol e do emprego do bagaço para fins energéticos, os
polissacarídeos (principalmente a celulose) podem ser liberados por hidrólises
enzimáticas para serem fermentados a etanol e outros produtos de maior valor
agregado. O elevado custo das enzimas celulolíticas representa o maior
desafio para a viabilidade do processo. Atualmente os principais fornecedores
de enzimas lignocelulolíticas para a indústria são os fungos. O conhecimento
da diversidade e potencialidade dos fungos do ambiente é ainda pouco
explorado. Portanto, neste trabalho, fungos filamentosos foram isolados e
selecionados do solo de ambiente canavieiro da região Norte do Estado de
Minas Gerais, com o objetivo de obter linhagens com potencial para produção
de enzimas lignocelulolíticas. Foram coletadas amostras de solo e resíduos de
cana-de-açúcar nos períodos pós-estiagem e pós-chuvas. As coletas foram
realizadas em pontos estrategicamente pré-selecionados, perfazendo um total
de 16 amostras. Foram isolados 230 morfotipos dos quais 124 identificados
como celulolíticos, através de testes qualitativos, destes 5 também
hemicelulolíticos e 7 lignolíticos. Os fungos Trichoderma reesei RUT C-30
(ATCC 56765) e Aspergillus awamori B.361 U2/1 foram empregados como
referência de linhagens produtoras de celulases e hemicelulases. Em relação à
análise enzimática quantitativa, foram testados em fermentação submersa em
frascos sob agitação mecânica e em fermentação semi-sólida em frascos sob
cultivo estacionário, 19 morfotipos celulolíticos. Destes, 5 morfotipos foram
identificados taxonomicamente, 2 a nível de gênero e 3 espécies, sendo eles:
Coriolopsis sp. (basidiomiceto); Aspergillus cf. phoenicis; Penicillium raistrickii;
Penicillium angulare e Penicillium variabile. Em cultivo submerso com lactose, o
Coriolopsis sp. apresentou maior atividade endoglucanase (CMCase 7,8946
UI/mL); e os Penicillium angulare e Penicillium raistrickii, maior atividade
FPase, 0,7103 e 0,6631UI/mL respectivamente. O Penicillium variabile se
destacou por apresentar maior atividade -Glicosidase 1,3356 UI/mL, maior
que o controle Aspergillus awamori nas mesmas condições de cultivo. A
atividade hemicelulósica (xilanase) foi melhor apresentada pelo Penicillium
raistrickii, 5,2161UI/mL. O bagaço de cana-de-açúcar se apresentou como bom
substrato e indutor em cultivo submerso para atividade CMCase produzida
pelos Penicillium raistrickii e Penicillium angulare, bem como para todas as
enzimas analisadas produzidas pelo Coriolopsis sp., comparando-se com o
cultivo com lactose ou farelo de trigo.
Palavras-chave: celulase, isolamento, resíduos, fungos.

vi
ABSTRACT
The bioenergetic potential of Brazil is represented largely by the alcohol and
sugar sector, because the sugar-cane is a source of renewable and abundant
energy. Besides the use of the sugar-cane juice for ethanol production and use
of bagasse for energy purposes, the polysaccharides (mainly cellulose) can be
released by enzymatic hydrolysis to be fermented into ethanol and other
products with higher added value. The high cost of cellulolytic enzymes
represents the biggest challenge to the viability of the process. Currently, the
fungi are the main producers of industrial enzymes lignocellulolytic. The
Knowledge of the diversity and potential of the fungi from the environment is still
a little explored. Therefore, in this study, filamentous fungi were isolated and
selected from sugarcane soil environment of the northern region of Minas
Gerais state, with the aim of obtaining strains with potential for production of
enzymes lignocellulolytic. The samples were collected from soil and waste of
sugar cane in post-drought and post-rains periods. The Samples were collected
at pre-selected points strategically for a total of 16 samples. 230 morphotypes
were isolated of which 124 identified as cellulolytic, through qualitative tests,
and of these 5 also hemicellulolytic and 7 lignolytics. The fungus Trichoderma
reesei RUT C-30 ATCC 56765 and Aspergillus awamori B.361 U2/1 were used
as reference of producer strains of cellulases and hemicellulases. In relation to
the quantitative enzymatic analysis, were tested in submerged fermentation in
mechanical shaken flasks and in solid state fermentation in stationary culture
flasks, 19 morphotypes cellulolytic. Of these, five morphotypes were identified
taxonomically, 2 genus and 3 species, namely: Coriolopsis sp. (Basidiomycete),
Aspergillus cf. phoenicis, Penicillium raistrickii; Penicillium angulare and
Penicillium variabile. In submerged culture with lactose, the Coriolopsis
sp. presented the highest endoglucanase activity (CMCase 7.8946 IU/ mL), and
the Penicillium angulare and Penicillium raistrickii, increased activity FPase,
0.7103 and 0.6631 IU/ mL respectively. The Penicillium variabile stood out due
to its higher activity -glucosidase 1.3356 IU/ mL, greater than the Aspergillus
awamori control in the same cultivation conditions Hemicellulosic activity
(xylanase) was better presented by Penicillium raistrickii, 5.2161 IU/
mL. Sugarcane bagasse is presented as a good substrate and inducer in
culture submerged for CMCase activity produced by Penicillium angulare and
Penicillium raistrickii as well as for all analysed enzymes produced by
Coriolopsis sp., compared with cultivation with lactose or wheat bran.
Keywords: Cellulase, isolation, waste, fungi

vii
LISTA DE FIGURAS
Figura 1 – Composição da lignocelulose ......................................... Pág. 6
Figura 2 – Estrutura da celulose (A) ligações -glicosídicas. (B)
Estrutura esquemática de uma fibrila ...............................................
Pág. 7
Figura 3 – Estrutura das fibras de celulose ...................................... Pág. 8
Figura 4 – Sinergismo entre endoglucanases, celobiohidrolases e
-glicosidases ...................................................................................
Pág. 13
Figura 5 – pH de cada ponto de coleta nos 2 períodos sazonais ... Pág. 37
Figura 6 – Correlação da diversidade fúngica com pH nos 2
períodos sazonais ............................................................................
Figura 7 – Fotos de halos de degradação em meio sintético com
CMC ou xilana .................................................................................
Pág. 37
Pág. 40
Figura 8 – Quadro representativo dos IEs dos fungos filamentosos
isolados ............................................................................................
Pág. 42
Figura 9 - Concentração enzimática de endoglucanase (CMCase)
e proteína total, obtidas do cultivo dos isolados em meio Mandels
& Weber com lactose.......................................................................
Pág. 44
Figura 10 - Concentração enzimática de exoglucanase (FPase) e
proteína total, obtidas do cultivo dos isolados em meio Mandels &
Weber com lactose..........................................................................
Pág. 45
Figura 11 - Concentração enzimática de -glicosidase e proteína
total, obtidas do cultivo dos isolados em meio Mandels & Weber
com lactose......................................................................................
Pág. 46
Figura 12 - Concentração enzimática de xilanase e proteína total,
obtidas do cultivo dos isolados em meio Mandels & Weber com
lactose...............................................................................................
Pág. 47
Figura 13 - Comparativo meio Mandels & Weber (1969) com
lactose e com bagaço. Maiores concentrações CMCase (UI/mL)
obtidas em cultivo submerso por até 120 h......................................
Figura 14 - Comparativo meio Mandels & Weber (1969) com
Pág. 48

viii
lactose e com bagaço. Maiores concentrações FPase (UI/mL)
obtidas em cultivo submerso por até 120 h.....................................
Figura 15 - Comparativo meio Mandels & Weber (1969) com
lactose e com bagaço. Maiores concentrações de -glicosidase
(UI/mL) obtidas em cultivo submerso por até 120 h.........................
Figura 16 - Comparativo meio Mandels & Weber (1969) com
lactose e com bagaço. Maiores concentrações de xilanase (UI/mL)
obtidas em cultivo submerso por até 120 h.....................................
Pág. 48
Pág. 49
Pág. 50
Figura 17 - Cultivo submerso meio Breccia (1995) modificado com
farelo de trigo ou bagaço 1,5 % e 3,0 % do isolado 67. Controle
sem farelo ou bagaço. Concentração enzimática máxima
obtida...............................................................................................
Pág. 51
Figura 18 - Cultivo submerso meio Breccia (1995) modificado com
farelo de trigo ou bagaço 1,5 % e 3,0 % do isolado 87. Controle
sem farelo ou bagaço. Concentração enzimática máxima
obtida.................................................................................................
Figura 19 - Concentração CMCase em cultivo semi sólido por 33
dias, comparativo substrato farelo de trigo e bagaço.......................
Pág. 52
Pág. 53
Figura 20 - Concentração FPase em cultivo semi sólido por 33
dias, comparativo substrato farelo de trigo e bagaço.......................
Pág. 53
Figura 21 - Concentração -glicosidase em cultivo semi sólido por
33 dias, comparativo substrato farelo de trigo e bagaço..................
Figura 22 - Concentração xilanase em cultivo semi sólido por 33
dias, comparativo substrato farelo de trigo e bagaço.......................
Figura 23 – Concentração média máxima de enzimas produzidas
pelo isolado S87 em cultivo semi-sólido nos diferentes substratos
(bagaço 3,0%; farelo de Trigo 3,% e mistura bagaço 1,5% e farelo
de trigo 1,5%)....................................................................................
Figura 24 – Concentração média máxima de enzimas produzidas
pela referência A. awamori em cultivo semi-sólido nos diferentes
substratos (bagaço 3,0%; farelo de Trigo 3,% e mistura bagaço
1,5% e farelo de trigo 1,5%)..............................................................
Pág. 54
Pág. 54
Pág. 55
Pág. 55

ix
LISTA DE TABELAS
Tabela 1 – Composição do meio sintético CMC 1% ........................ Pág. 25
Tabela 2 – Composição do meio para detecção de lacase.............. Pág. 26
Tabela 3 – Composição do meio Mandels & Weber (1969)
modificado (pH 5,0)...........................................................................
Pág. 28
Tabela 4 – Composição do meio Breccia et al. (1995) modificado .. Pág. 28
Tabela 5 – Composição da solução nutritiva (pH 5,0) para meio
semi-sólido .......................................................................................
Pág. 29
Tabela 6 – Número de fungos filamentosos (x104 UFC/mL) ........... Pág. 35
Tabela 7 – Diversidade de morfotipos encontrados por ponto de
coleta ................................................................................................
Pág. 36
Tabela 8 – Número de fungos filamentosos com atividade
enzimática ........................................................................................
Pág. 41
Tabela 9 – Seleção de fungos celulolíticos com IE ≥ 3,5 ................. Pág. 42
Tabela 10 – Identificação taxonômica de 5 fungos celulolíticos ...... Pág. 56

AGRADECIMENTOS
À Universidade Estadual de Montes Claros, UNIMONTES pela oportunidade de
participar do Programa de Pós Graduação em Ciências Biológicas.
À BIOMM SA, em especial ao diretor científico Luciano Vilela, pela permissão e
apoio em participar do programa de pós-graduação enquanto funcionária da
empresa.
À FINEP pelo financiamento e apoio ao projeto maior “Desenvolvimento de
processo para a obtenção industrial de misturas enzimáticas celulolíticas
destinadas à produção de biocombustíveis a partir de biomassa” no qual este
trabalho está inserido.
Ao Dr. Dario Alves de Oliveira, pela orientação, confiança e acolhimento, meus
sinceros agradecimentos.
Ao co-orientador Dr. Henrique Maia Valério, pela amizade, orientação,
incentivo, apoio e valorosas discussões e correções ao longo deste trabalho.
Ao Dr. Torquato Gonçalves da Fonseca pelo apoio, interesse, disponibilidade
do seu precioso tempo e concordância na coleta de material de sua
propriedade.
Ao IBAMA pela autorização de acesso e de remessa de amostra de
componente do patrimônio genético.
À Profa. Dra. Elba Bon pelo envio e permissão do uso das cepas referência
utilizadas neste trabalho.
À Escola técnica de Montes Claros (FEMC) por ceder o laboratório de
Bioquímica e Microbiologia à BIOMM para desenvolvimento do projeto FINEP,
no qual este trabalho foi realizado.
Às colegas de pós-graduação Leide e Patrícia, pelo companheirismo, por tornar
o período de execução dos créditos obrigatórios do programa menos
estressante.
Aos queridos amigos e alunos de iniciação científica da FACIT (Clara, Patrícia,
Bárbara(s), Mariana, Helbert e Laiane) os quais contribuíram significativamente

para a realização deste trabalho por alguns períodos ou durante todo o projeto
e pelos momentos de descontração, meus sinceros agradecimentos.
Ao José Carlos pela valiosa colaboração no estoque dos fungos isolados.
Aos meus colegas da BIOMM, pelo apoio e amizade e em especial à Heloísa
Tunes pelo constante incentivo e contribuição com a estatística.
Aos meus queridos amigos, sempre presentes mesmo que de longe, Andréa,
Rosi, Mônica, Simone, Élder, Denise, Lúcia, Luiz, Janete, Henrique, Fernanda,
Gustavo, Mauro, Alessandra e os mais recentes Dayane e José Carlos, muito
obrigada pela torcida, carinho, alegrias e momentos de descontração.
Aos responsáveis pela minha iniciação na pesquisa científica, Dr. Marcos
Mares Guia (in memoriam) e Dr. Joseph Ernest Thiemann, meu eterno
reconhecimento.
À minha família, queridas irmãs que sempre me incentivaram, obrigada pelo
amor incondicional e carinho.
Aos meus pais, que já estão em outro plano, mas que sempre sinto por perto,
por todo o amor e orientação para a vida.
Ao meu querido Arthur, meu orgulho e minha alegria.
E meus sinceros agradecimentos a todas as pessoas e instituições que
tornaram de alguma forma possível a realização deste trabalho.

1
1. INTRODUÇÃO
O desenvolvimento de tecnologia para produção de energia alternativa é
criticamente importante devido à crescente produção dos gases de efeito
estufa causado pelo aumento do uso de combustíveis fósseis, fator
preponderante no esgotamento das reservas e aumento dos preços do
petróleo. A bioconversão da biomassa lignocelulósica tem significativas
vantagens ambientais e econômicas sobre outras estratégias de energia
alternativa, pois sendo a lignocelulose o principal componente estrutural de
todas as plantas, corresponde ao material orgânico renovável mais abundante
do planeta (DASHTBAN et al., 2009; PANDEY et al., 2000; GOMEZ et al.,
2008; LIN & TANAKA, 2006).
A lignocelulose consiste de três principais componentes: celulose,
hemicelulose e lignina, os quais variam em proporção de espécie para espécie.
Em média os resíduos agrícolas contêm de 20 a 60% de celulose, 20 a 30% de
hemicelulose e 15 a 30% de lignina. O bagaço da cana-de-açúcar contém
cerca de 25 a 40% de celulose e o restante de hemicelulose (20 a 35%) e
lignina (15 a 35%) (COWLING & KIRK, 1976).
Desses três polímeros a celulose é a principal fonte de açúcares
fermentos-alvos em etanol, pois é constituída de polímeros de glicose, que uma
vez hidrolisada é facilmente fermentada em etanol pela levedura
Saccharomyces cerevisiae em processos industriais aprovados e com bom
desempenho (WARZWODA et al., 2006).
O processo de produção de etanol no Brasil ocorre pela via de
fermentação alcoólica do caldo de cana-de-açúcar e gera como resíduos a
vinhaça, a palha e o bagaço, os quais são parcialmente utilizados através da
queima para geração de energia (vapor e elétrica) e para adubação das áreas
de plantio. Parte considerável deste subproduto não é utilizada, e causa
transtornos e problemas no que se refere a sua destinação e processamento
(SILVA et al., 2009).
A área ocupada pela cana-de-açúcar no Brasil e destinada ao setor
sucroalcooleiro alcança 8,1 milhões de hectares na safra 2010/11, ou 9,2% a
mais do que no ciclo anterior. A previsão de produção total de cana-de-açúcar

2
a ser moída pela indústria sucroalcooleira no país na safra 2010/11 é de 664,33
milhões de toneladas. Deste total de cana-de-açúcar a ser esmagada, cerca de
54,6% (362,8 milhões de toneladas) se destina à produção de 28,5 bilhões de
litros de álcool. Já a outra parte, cerca de 45,4% (301,6 milhões de toneladas),
se destina à produção de 38,7 milhões de toneladas de açúcar. Baseado nesta
previsão, cerca de 150 milhões de toneladas de bagaço e 120 milhões de
toneladas de palha serão gerados (CONAB, 2010).
A produção de etanol de segunda geração é obtida através da
transformação dos materiais lignocelulósicos, que se destacam como fontes
energéticas, como bagaço e palha de cana-de-açúcar, em etanol. Uma
tonelada de cana contém a energia equivalente a 1,2 barris de petróleo, sendo
que cerca de 1/3 dessa energia está armazenada quimicamente no caldo
(açúcares) e o restante na biomassa de cana: metade no bagaço e metade na
palha, aproximadamente. A produção de etanol através da utilização de
resíduos lignocelulósicos pode representar um adicional significativo de 15% na
produção (GOES, 2008).
A bioconversão dos substratos lignocelulósicos pode ser feita por muitos
microrganismos, dentre bactérias, actinomicetos, protozoários e fungos,
através da produção de celulases. Aqueles microrganismos os quais produzem
níveis apropriados do complexo enzimático em ação conjunta e sinérgica,
destacando-se: exoglucanases (celobiohidrolases), endoglucanases,
celobiases (ß-glicosidase) e fenoloxidases, são capazes de transformar os
resíduos lignocelulósicos em compostos de fácil assimilação para o
metabolismo (VALASKOVÁ & BALDRIAN, 2006, LYND et al., 2002, KUMAR et
al., 2008).
As celulases foram investigadas inicialmente há várias décadas atrás
para bioconversão de biomassa, abrindo caminho para pesquisas de
aplicações industriais das enzimas em ração animal, alimentos, têxteis,
detergentes e na indústria de papel. Com a diminuição dos combustíveis
fósseis e o aumento da necessidade de encontrar fontes alternativas para
combustíveis e energia renovável, existe uma renovação do interesse na
bioconversão da biomassa lignocelulósica usando celulases e outras enzimas
(SUKUMARAN et al., 2005).

3
As pesquisas voltadas para os mecanismos enzimáticos da celulose e
os problemas envolvidos na conversão direta da biomassa lignocelulósica em
produtos úteis por meio de enzimas isoladas ou de microorganismos
lignocelulolíticos tem aumentado nas últimas décadas (SCHWARZ, 2001).
Ressalta-se que o custo atual das enzimas existentes no mercado ainda
é a maior limitação para o emprego potencial destas enzimas na hidrólise de
resíduos lignocelulósicos e implantação desta tecnologia (KUMAR et al., 2008,
SUKUMARAN et al., 2005, WYMAN, 1994).
Os microrganismos utilizados nas fermentações industriais devem
preencher uma série de requisitos a fim de tornar operacional o processo do
qual participam (HESSELTINE & HAYNES, 1973).
Este trabalho teve como objetivo principal buscar na natureza cepas
potencialmente interessantes do ponto de vista industrial para possibilitar o
desenvolvimento de processos tecnológicos mais competitivos.

4
2. REVISÃO BIBLIOGRÁFICA
2.1 Fontes de energia - resíduos lignocelulósicos
O carbono atmosférico foi capturado por plantas há milhões de anos
atrás, pelo processo da fotossíntese, e passado o tempo foi manifestado em
petróleo bruto e carvão. No entanto, desde a revolução industrial, nós temos
usado muito destas fontes de energia, causando excessiva liberação de
carbono de volta para a atmosfera. Assim, nos últimos 150 anos os níveis de
CO2 na atmosfera aumentaram de 280 para 380 ppm. Em troca, isto tem sido
potencialmente a causa da elevação das temperaturas ao redor do mundo
levando às mudanças climaticas globais (DASHTBAN et al., 2009, GOMEZ et
al. 2008).
A produção global de petróleo bruto é prevista para diminuir 5 vezes até
o ano de 2050. Baseado nos cálculos do World Energy Council (WEC), o
consumo mundial de energia primária é aproximadamente equivalente a 12
bilhões de toneladas de carvão/ano. Os cálculos das Nações Unidas têm
mostrado que a população mundial irá aumentar para cerca de 10 bilhões de
pessoas em 2050 o que aumentará a demanda de energia para equivalente de
no mínimo 24 bilhões de toneladas de carvão/ano (2x o consumo de hoje)
dependendo econômica, social e politicamente do desenvolvimento
(DASHTBAN et al., 2009).
O aumento do consumo de energia, o esgotamento dos combustíveis
fósseis e aumento das preocupações com o meio ambiente se tornaram fortes
justificativas para geração de energia aplicada ao uso de biocombustíveis. Os
resíduos lignocelulósicos são produzidos em larga escala por diferentes
indústrias incluindo florestal, polpa e papel, agricultura e de alimentos, além de
diversos resíduos do lixo sólido municipal e resíduos animais. Significantes
esforços, muitos dos quais têm tido sucesso, tem sido realizados para
converter estes resíduos lignocelulósicos em produtos de valor como
biocombustíveis, químicos e alimentação animal. Estes materiais de potencial
valor foram tratados no passado em muitos países como lixo, e ainda hoje em

5
alguns países desenvolvidos, o que levanta muitas preocupações ambientais
(RAGAUSKAS et al., 2006, PRASAD et al., 2007).
O etanol como combustível é usado em mistura (10 a 22%) com a
gasolina ou puro (95% etanol e 5% água). Por ser um combustível oxigenado
que contém 35% de oxigênio, reduz a emissão de partículas e NOx da
combustão. Com a escassez das reservas de petróleo, aumento da poluição do
ar e acúmulo de CO2 na atmosfera, o etanol terá um papel significativo na
matriz energética do planeta (LIN & TANAKA, 2006, PRASAD et al., 2007).
Um dos cultivos comerciais de maior importância em todo o mundo, a
cana-de-açúcar, ocupa mais de 20 milhões de hectares, com destaque para o
Brasil, que, com uma área plantada de mais de 7 milhões de hectares,
corresponde a cerca de 42% do total produzido. Grande quantidade de
resíduos é gerada no processamento da cana-de-açúcar para a produção de
álcool. Para cada tonelada de cana, aproximadamente 250 a 280 Kg de bagaço
é produzido.
Grandes investimentos estão sendo efetuados para viabilizar a produção
de combustível a partir de celulose, estimando-se que, em 2020, apenas nos
EUA, cerca de 30 bilhões de litros de álcool poderiam ser obtidos dessa fonte.
Será necessário investir recursos elevados em PD&I, especialmente na
hidrólise da celulose (EMBRAPA, 2006).
A lignocelulose é um material orgânico e principal componente estrutural
das plantas, além de ser o produto principal da fotossíntese em ambientes
terrestres, e a mais abundante fonte de energia renovável produzida na
biosfera - cerca de 100 bilhões de toneladas seca/ano (ZHANG, 2006).
A composição da lignocelulose consiste de 3 principais componentes:
celulose, hemicelulose e lignina (Figura 1). Em adição, pequenas quantidades
de outros materiais como proteínas e pectina tem sido encontrados em
resíduos lignocelulósicos, em diferentes proporções, dependendo da fonte. A
quebra da biomassa lignocelulósica envolve a formação de longas cadeias de
polissacarídeos, principalmente celulose e hemicelulose, e subseqüente
hidrólise destes polissacarídeos em cadeias de açúcares compostas por 5 e 6
carbonos. Na produção de biocombustível, estes açúcares podem ser

6
convertidos em bioetanol através do processo de fermentação (DASHTBAN et
al., 2009).
Figura 1 – Desenho esquemático da estrutura da lignocelulose e seus componentes
De acordo com BON et al. (2008), dentre os resíduos agroindustriais de
composição lignocelulósica de maior importância destacam-se: bagaço e palha
de cana-de-açúcar; sabugo e palha de milho; palhas de trigo e arroz, restos de
madeira processada e lixo baseados em papel.
O desenvolvimento de tecnologias para efetivamente converter resíduos
florestais e de agricultura de baixo custo para açúcares fermentáveis oferece
um potencial para gerar benefícios de interesse nacional através de: melhorar a
estratégica de segurança; diminuir os déficits comerciais; salutar a economia
rural; melhorar a qualidade do ambiente; exportar tecnologia; e fornecer fonte
de energia sustentável (ZHANG, 2006).
2.1.1 - Celulose
A celulose é o principal constituinte de todo o material das plantas,
dominante na agricultura, é a molécula orgânica mais abundante na Terra,
representante de cerca de 1,5x1012 ton de biomassa total anual produzida
através da fotossíntese especialmente nos trópicos, e é considerada uma fonte
quase inesgotável de matéria prima para diferentes produtos (SUKUMARAN et
al., 2005, DASHTBAN et al., 2009).
Fonte: Sticklen (2008)
Pectina
Microfibrila
de celulose
Hemicelulose
Proteínas
solúveis
Lignina

7
Na natureza podemos encontrar a celulose quase que exclusivamente
na parede celular das plantas, apesar de ser produzida por alguns animais (ex.
tunicatos) e poucas bactérias. A despeito de grandes diferenças na
composição e na estrutura anatômica da parede celular de várias plantas, um
alto conteúdo celulósico – cerca de 35 a 50% do peso seco da planta – é uma
característica geral (LYND et al., 2002).
Sua estrutura molecular consiste em um biopolímero linear de moléculas
de anidroglucopiranose, conectados por ligações -1,4-glicosídicas (Figura 2A).
Figura 2 – Estrutura da celulose. (A) Ligações -glucosídicas. (B) Estrutura esquemática de
uma fibrila.
O polímero é insolúvel em água e de massa molecular que varia entre
50 mil e 2,5 milhões de Daltons, dependendo da origem da amostra. A celulose
é composta por unidades monoméricas de celobiose, a qual é formada pela
junção de duas moléculas de glicose seguida da eliminação da água através
das hidroxilas ligadas ao carbono 1 e 4 (FILHO, 2008).
Independentemente da sua orientação, as cadeias são rígidas por
pontes de hidrogênio intra e inter cadeias. Segmentos adjacentes se
sobrepõem entre si e são unidos por fracas ligações de Van der Waals.
Aproximadamente 30 moléculas individuais de celulose são montadas
em grandes unidades conhecidas como fibrilas elementares (protofibrilas), as
Fonte: Béguin & Aubert (1994)
Celobiose Glicose
Região cristalina Região cristalina
Região amorfa

8
quais são empacotadas em grandes unidades chamadas microfibrilas, e estas
por sua vez são montadas nas conhecidas fibras de celulose (Figura 3).
Figura 3 – Estrutura das fibras de celulose
A natureza cristalina da celulose implica em uma organização estrutural
na qual todos os átomos são fixados em posições distintas em relação ao
outro. Uma importante característica do arranjo cristalino é que os
componentes moleculares das microfibrilas individuais são ―empacotados
hermeticamente‖ para prevenir a penetração não só de enzimas, mas também
de pequenas moléculas como a água (BÉGUIN & AUBERT, 1994).
Apesar de a celulose formar estruturas cristalinas distintas, as fibras de
celulose naturais não são puramente cristalinas. O grau de cristalinidade é
variável, deixando regiões amorfas com variados graus de organização das
fibrilas (Figura 2B). Além das regiões amorfas e cristalinas as fibras de celulose
contêm vários tipos de irregularidades, tais como dobras ou torções das
Fonte: Wyman,C.E. 2009
Cultura de bioenergia
Celulose
Células
Hemicelulose
Lignina
microfibrila
Moléculas
de açúcar Glicose

9
microfibrilas, ou vazios tais como microporos da superfície, buracos grandes e
capilares. Alguns microporos e capilares são suficientemente grandes para
permitir a penetração, em alguns casos, de enzimas celulolíticas (LYND et al.,
2002, DASHTBAN et al., 2009).
Na natureza, em alguns casos, como no algodão, a celulose está
presente em um estado quase puro, enquanto na maioria dos casos, as fibras
de celulose são embebidas na matriz de outros polímeros estruturais,
principalmente hemiceluloses e lignina, os quais compreendem,
respectivamente, 20 a 35 e 5 a 30% do peso seco das plantas. Apesar dessa
proporção ter variações de acordo com o tipo e a maturidade da planta, estes
polímeros são a estrutura dominante e limitante da velocidade e do grau de
utilização da biomassa não tratada (SUKUMARAN et al., 2005).
2.1.2 - Hemiceluloses
As hemiceluloses, o segundo componente mais abundante da biomassa
lignocelulósica, são polímeros heterogêneos de pentoses (incluindo xilose e
arabinose), hexoses (principalmente manose, menos glucose e galactose),
ácidos urônicos e grupos acetila. Sua estrutura apresenta ramificações e
cadeias laterais que interagem facilmente com a celulose, dando flexibilidade
ao agregado (DASHTBAN et al., 2009).
A composição das hemiceluloses é muito variável na natureza e
depende da fonte da planta, as madeiras em geral possuem de 20 a 30% de
hemicelulose na composição geral, enquanto que nas gramíneas esse valor é
de 20 a 40% (BORTOLAZZO, 2011).
A porção de hemiceluloses nas plantas contribuem significativamente
como potencial para a produção de combustíveis a partir da lignocelulose. No
entanto, espécies de leveduras e bactérias comumente usadas na produção de
etanol não são muito eficientes na metabolização de açúcares pentose tais
como xilose e arabinose, e do contrário, estes açúcares podem inibir a
atividade desses organismos. Uma das abordagens para superar este
problema em particular é a identificação na natureza ou engenheiramento de
microrganismos que possam usar eficientemente as pentoses (GOMEZ, 2008).

10
2.1.3 - Lignina
A lignina, o terceiro principal polímero heterogêneo dos resíduos da
lignocelulose, geralmente contém 3 álcoois aromáticos incluindo álcool
cumárico, coniferílico e sinapílico.
Segundo BON et al. (2008), a lignina representa um dos maiores
estoques de carbono/energia da natureza e é ainda, o maior depósito natural
de estruturas químicas aromáticas, constituindo-se em uma fonte potencial de
valiosos insumos para a indústria química.
A lignina, junto com a hemicelulose e a pectina, preenche os espaços
entre as fibrilas de celulose e além de atuar como material ligante entre os
componentes da parede celular, age como barreira e previne a penetração de
enzimas lignocelulolíticas no interior da estrutura lignocelulósica.
Mesmo presente em quantidades menores em relação à celulose, a
lignina confere limitação suficiente para retardar, ou mesmo impedir
completamente, a atuação microbiana sobre o material. Não
surprendentemente, a lignina é o principal componente recalcitrante do material
lignocelulósico a degradar (DASHTBAN et al., 2009).
2.2 - Hidrólise da Lignocelulose
A hidrólise enzimática da celulose na natureza é comumente um
processo incompleto e lento. Embora em intervalos de 48 horas, observam-se
exemplos de consumação satisfatória, como a da flora microbiana do rúmen
bovino, hidrolisando de 60-65% da celulose disponível e também de cupins
capazes de assimilar até 90% de celulose da madeira (FILHO, 2008). Enquanto
isso, em sistemas mais complexos, como o apodrecimento de uma árvore no
solo, essa degradação pode levar meses para ser completada (SCHWARZ,
2001).
A conversão inicial da biomassa em açúcares é o gargalo chave no
processo de produção de biocombustíveis e novas soluções biotecnológicas
são necessárias para melhorar sua eficiência, o que pode reduzir o custo global
da produção de bioetanol (BON et al., 2008).

11
Uma estratégia promissora para eficiente utilização de resíduos
lignocelulolíticos é a hidrólise microbiana e fermentação dos açúcares
resultantes para produção dos metabólitos desejados ou produção de
biocombustível.
A utilização da biomassa lignocelulósica é mais complexa do que a
celulose pura, não somente devido à complexa composição com a presença de
hemicelulose e lignina, o que impede às celulases de acessarem o substrato
eficientemente (ZHANG et al., 2006), mas também por causa da diversa
arquitetura das células das plantas. O tecido das plantas difere tremendamente
com respeito ao tamanho e organização. Assim, em adição às restrições
impostas pela estrutura da celulose, limitações adicionais são impostas pela
difusão e transporte do agente celulolítico ao local de ataque. Estas restrições
podem limitar fortemente a utilização em alguns habitats (LYND et al, 2002).
A molécula de celulose é muito estável, com meia vida de 5-8 milhões
de anos para clivar as ligações -glicosídicas a 25ºC, enquanto o processo de
degradação da celulose dirigido pelas enzimas é muito mais rápido e vital para
retornar o carbono dos sedimentos para a atmosfera (ZHANG et al., 2006).
Dessa maneira, para utilização industrial, a conversão efetiva da
lignocelulose recalcitrante a açúcares fermentáveis requer um pré-tratamento o
qual deve ao mesmo tempo produzir uma polpa celulósica com elevada
acessibilidade e reatividade da fibra aos agentes hidrolíticos ácidos ou
enzimáticos (digestibilidade), garantir adequada recuperação das pentoses,
além de limitar a geração de compostos inibidores aos microrganismos usados
na fermentação e às enzimas. Adicionalmente, aspectos associados ao uso de
catalisadores de baixo custo, reciclagem de insumos e geração de subprodutos
de alto valor agregado a partir da lignina caracterizam sistemas de pré-
tratamento eco-eficientes (BONOMI, 2007).
ZHANG et al. (2006) e BON et al. (2008) comentam os diferentes
métodos aplicáveis ao pré-tratamento da lignocelulose. Alta temperatura e
ácido foram usados inicialmente para degradar quimicamente a celulose e
ainda são usados no pré-tratamento de resíduos lignocelulósicos em escala
industrial. Entretanto esta abordagem é cara, lenta e ineficiente. Além do mais,
o rendimento global do processo de fermentação será diminuído por causa

12
deste pré-tratamento pela liberação de inibidores tais como ácidos fracos,
compostos furano e fenólicos. Outras tecnologias propostas de pré-tratamento
da lignocelulose propostos incluem, moagem, irradiação, explosão da fibra por
amônia, explosão a vapor, reciclagem de percolação com amônia e cal, ação
de solventes orgânicos e tratamento com fungos da podridão branca.
Não existe um tratamento único aplicável a todas as biomassas e um
mesmo pré-tratamento pode ser aplicado com diferentes graus de severidade.
De acordo com o pré-tratamento utilizado pode ocorrer: a remoção da
hemicelulose ou da lignina, em proporções variáveis; a redução da
cristalinidade da celulose; e o aumento da porosidade (BON et al., 2008).
A degradação microbiana dos resíduos celulósicos e dos produtos
derivados resulta da ação corrigida de várias enzimas principalmente das
celulases as quais são produzidas por muitos microrganismos e incluem
diferentes classificações enzimáticas. O principal desafio na conversão da
biomassa a bioetanol é alcançar rendimentos que tornam seu custo competitivo
aos combustíveis fósseis (SUKUMARAN et al., 2005).
2.3 - Sistemas enzimáticos
2.3.1 - Celulases
Na natureza, a degradação da biomassa celulósica é realizada por
misturas de enzimas hidrolíticas conhecidas como celulases. As celulases
hidrolisam a celulose (ligações -1,4-D-Glucan) e produzem como principal
produto glicose, celobiose e celo-oligossacarides. Existem 3 principais tipos de
enzimas celulolíticas: as exoglucanases formadas pela Celodextrinases (1,4-b-
D-glicana-glicano-hidrolase, EC 3.2.1.74) e pela Celobiohidrolase (CBH ou 1,4-
-D-glicana-celobio-hidrolase, EC 3.2.1.91); as endoglucanases (EG ou endo-
1,4, -D-glican 4-glicano-hidrolase, EC 3.2.1.4); e as Celobiases ou -
glicosidase (BG- EC 3.2.1.21) (BÉGUIN & AUBERT,1994).
O mecanismo mais aceito para hidrólise enzimática da celulose envolve
ação sinergística pela endoglucanase, exoglucanase ou celobiohidrolase, e -
glicosidase. Endoglucanases hidrolizam randomicamente em locais internos
amorfos as ligações -1,4-glucosídicas intramoleculares acessíveis das

13
cadeias de celulose para produzir oligossacarídeos de vários comprimentos e
consequentemente novos finais de cadeia. Exoglucanases clivam as cadeias
de celulose nos finais redutores ou não redutores para liberar celobiose solúvel
ou glicose. Exoglucanases podem também agir na celulose microcristalina,
presumivelmente descascando as cadeias de celulose da estrutura
microcristalina. As -glucosidases hidrolizam celodextrinas solúveis e celobiose
a glicose a fim de eliminar a inibição por celobiose. Estes 3 processos de
hidrólise ocorrem simultaneamente (Figura 4).
Figura 4 – Sinergismo entre endoglucanases, celobiohidrolases e -glicosidases
A ação combinada de endoglucanases e exoglucanases modifica as
características da superfície da celulose ao longo do tempo, resultando em
Fonte: Béguin & Aubert, 1994
Endoglucanases
Região
cristalina
Região amorfa Adsorção de celulases
Celobiohidrolases
-glicosidases

14
rápidas mudanças na taxa de hidrólise (BÉGUIN & AUBERT,1994, LYND et al,
2002, ZHANG et al., 2006).
Para a caracterização de preparações brutas de celulases com relação à
sua atividade de endoglucanases ou exoglucanases são utilizados substratos
diferentes, ressaltando-se, entretanto, que a sinergia existente entre dois tipos
de enzima impede uma quantificação precisa. Para a atividade de
endoglucanase utiliza-se, como substrato, derivado de celulose substituída
como a carboximetilcelulose (CMC), que é solúvel. A enzima ataca o polímero
em uma forma randômica, produzindo uma mudança rápida no grau de
polimerização. Após a reação enzimática é determinada a formação de
açúcares redutores; sendo esta atividade conhecida como CMCase. A medida
da atividade enzimática utilizando como substrato o papel de filtro Whatman
nº1 (FPase) é comumente utilizada como referência para determinação da
atividade celulósica global tanto em trabalhos acadêmicos quanto para
preparações enzimáticas comerciais (GHOSE, 1987).
A atividade endoglucanase também pode ser detectada facilmente por
exame de ―halos‖ em placas de ágar sólido usando CMC como substrato,
seguido por coloração por Vermelho Congo e lavagem. Altas taxas de hidrólise
geralmente resultam em grandes halos. Esse método é semi-quantitativo, e
está bem adaptado ao monitoramento de um grande número de amostras. Não
é surpresa que tantos relatos de exemplos de endoglucanases têm sido
selecionados pelo uso do método Vermelho Congo/CMC (KIM et al., 2000,
WANG et al., 2005; ZHANG et al., 2006; BORTOLLAZO, 2011).
2.3.1.1. Produção de celulases
As celulases são enzimas relativamente caras e têm uso comercial
significativo no mercado mundial: na indústria textil para amolecimento do
algodão e acabamento do brim; no mercado de detergentes para o cuidado da
cor, limpeza e anti-resíduos; na industria alimentícia para trituração; e nas
industrias de polpa e papel, no descoloramento, melhoria de drenagem e
modificação da fibra (BHAT & BHAT, 1997).

15
O mercado de celulase tem expectativa de expandir muito quando as
celulases forem usadas para hidrolisar material celulósico pré-tratado à
açúcares, o qual pode ser fermentado a produtos como bioetanol e produtos
biológicos produzidos em larga escala. Por exemplo, o mercado de celulase
potencial tem sido estimado ser mais alto que US$400 milhões por ano se as
celulases forem usadas para hidrolisar palha de milho no meio oeste dos
Estados Unidos (ZHANG et al., 2006). Este cenário do mercado representa um
aumento de cerca de 33% no total do mercado de enzimas industriais dos
Estados Unidos. O grande potencial de mercado e o importante papel das
celulases na bioenergia emergente e bioprodutos industriais representam uma
motivação maior para desenvolver preparações melhores de celulase para
hidrólise da celulose da parede celular das plantas (ZHANG et al., 2006,
HIMMEL et al., 2007).
As celulases são enzimas induzíveis e alguns desafios na sua produção
industrial envolvem o desenvolvimento de bioprocessos adequados e meio de
fermentação, ao lado de identificação de substratos e indutores baratos
(SUKUMARAN et al., 2005, RECZEY et al., 1996).
Aproximadamente 90% de todas as enzimas industriais são produzidas
através da fermentação submersa, frequentemente usando microrganismos
geneticamente otimizados. Por outro lado, quase todas as enzimas podem ser
também produzidas através de fermentação semi-sólida usando
microrganismos selvagens. A fermentação semi-sólida pode ser biologicamente
vantajosa para alguns microrganismos, uma vez que as condições de cultivo se
assemelham mais ao seu habitat natural. Tais vantagens incluem alta
produtividade da fermentação, menor repressão catabólica, baixa demanda de
água e, portanto, baixa demanda de esterilidade devido à baixa atividade da
água, cultivo de microrganismos que requerem um suporte sólido e cultivo
misto de vários fungos (HOLKER et al., 2004).
Sob o ponto de vista econômico e ambiental, a seleção de resíduos
agroindustriais que atuem como substratos e possam ser utilizados em cultivo
semi-sólido de microrganismos celulolíticos, se faz interessante (SINGHANIA et
al., 2007).

16
A grande disponibilidade de biomassa lignocelulósica de baixo custo,
como a do bagaço de cana-de-açúcar, no Brasil, estimula a busca por
microrganismos mais eficientes no uso desse resíduo (BASSO, et al., 2010).
De acordo com ZHANG et al. (2006), estratégias baseadas em celulases
que farão a biorefinaria processar com maior economia incluem: aumento da
produtividade em volume da enzima comercial, produção de enzimas usando
substratos baratos, produzindo preparações enzimáticas com maior
estabilidade para processos específicos, e produção de celulases com mais
alta atividade específica nos substratos sólidos.
Recentemente as empresas de biotecnologia Genencor International e
Novozymes Biotech tem relatado o desenvolvimento da tecnologia que tem
reduzido o custo do processo de celulose a etanol de US$5,40 por galão de
etanol para aproximadamente 20 centavos por galão de etanol. Para tal foram
necessárias 2 principais estratégias: melhoria econômica na produção de
celulase para reduzir o custo por grama de enzima como, por exemplo, através
de meio de cultivo mais barato e sistema indutor alternativo, e melhoria na
performance da enzima celulase para reduzir a quantidade de enzima
necessária para alcançar equivalente hidrólise por coquetéis e aperfeiçoamento
do componente. A afirmação de expressiva redução no custo não tem sido
largamente aceita porque a mistura de celulase foi testada somente para um
substrato lignocelulósico pré-tratado específico e não pode ser aplicado a
outras lignoceluloses pré-tratadas (ZHANG, et al., 2006).
Sistemas celulolíticos completos são produzidos por diferentes gêneros
e espécies de fungos e bactérias. Atualmente a maioria das celulases
comerciais são produzidas por espécies de Trichoderma e Aspergillus. O
sistema enzimático dos fungos Trichoderma tem sido estudado extensivamente
e mostra insuficiente atividade -glicosidase para a eficiente sacarificação da
celulose. Altos níveis de -glicosidase são importantes para a completa
conversão da celulose devido à inibição por celobiose, das atividades exo e
endoglucanases. Desta forma a alta produção de -glicosidase por algumas
espécies de Aspergillus complementam o sistema (GARCIA-KIRCHNER et al.,
2005).

17
A modificação genética dos produtores de celulases para melhorar a
atividade da celulase tem sido um longo caminho para proporcionar melhores
produtores com altos títulos de enzimas, mas ainda assim a economia de
produção de celulases tem de ser melhorada para a produção comercial de
etanol a partir da biomassa (SUKUMARAN et al., 2005).
2.3.2 Hemicelulases
As endoxilanases (EC 3.2.1.8) são as hemicelulases mais estudadas.
Para a completa hidrólise das hemiceluloses, ainda é necessária a ação das
enzimas desramificadoras dos grupos laterais ligados à cadeia principal de
xilana: -1,4-xilosidase, -glicuronosidase, -arabinosidase e acetil xilana
estearase (KROGH et al., 2004).
De acordo com BON et al. (2008), o desenvolvimento comercial de
hemicelulases para a hidrólise de materiais lignocelulósicos não está tão
avançado quanto o de celulases, pois as preparações celulásicas comerciais
correntes têm sido principalmente desenvolvidas para a hidrólise de biomassa
pré-tratada com ácido diluído, em que a hemicelulose é removida antes da
sacarificação de celulose. No entanto, com o desenvolvimento de pré-
tratamentos totalmente enzimáticos, as hemicelulases serão compulsoriamente
requeridas.
Além da atuação das enzimas celulolíticas, da -glicosidase e das
xilanases, estão recebendo atenção crescente as enzimas acessórias que
possuem papel facilitador para a degradação da celulose. Assim é que
pectinases, tanases e feruloil esterase atuam na desconstrução da biomassa
através da degradação de diferentes ligações covalentes intra e inter-
poliméricas. Existe muito interesse atualmente na enzima feruloil esterase que
rompe ligações entre a lignina e a hemicelulose, facilitando o acesso das
celulases à fibra de celuloses. Assim sendo, misturas enzimáticas eficientes na
desestruturação da biomassa precisam apresentar um conjunto de atividades
enzimáticas acessórias além da atividade das enzimas típicas do complexo
celulolítico e xilanolítico (BON at al.,2008; KANG et al., 2004).

18
2.3.3 Ligninases
Apesar da necessidade de investigação de vários de seus aspectos, a
degradação da lignina pode ser entendida como um processo multienzimático
por reação não específica, resultante da ação coordenada de uma série de
enzimas ligninolíticas intra e extracelulares: peroxidases – lignina peroxidase
(Lip) e manganês peroxidase (Mnp); lacases – fenol oxidase e outras oxidases
produtoras de peróxido de hidrogênio e de metabólitos intermediários de baixa
massa molecular as quais, desestabilizam as ligações da macromolécula,
causando assim seu colapso (FILHO, 2008).
As aplicações mais estudadas dessas enzimas, especialmente as
lacases, incluem a deslignificação de polpas na indústria de papel e celulose,
branqueamento de corantes têxteis, degradação de efluentes, modificação de
biopolímeros e o uso dessas enzimas como um componente na fabricação de
detergentes (DASHTBAN et al., 2009).
2.4 Microrganismos celulolíticos
Em 1880 já era observada a presença de fungos contaminando o
algodão em fábricas de tecidos (ABOU-ZEID & EL- DIWANY, 1978).
A partir da década de 60 descobriu-se que preparados de enzimas
extracelulares eram responsáveis pela ação hidrolítica, despertando o interesse
pelos fungos. A idéia de aproveitar essas enzimas na conversão de resíduos
celulósicos em produtos de interesse alimentar e energético surgiu em 1973 e
em 1979 a equipe do Laboratório de Pesquisas em Natick, Massachussets,
anunciou o isolamento de cepas mutantes de Trichoderma reesei com poder
hidrolítico aproximadamente vinte vezes superior ao da cepa nativa
(BORTOLLAZO, 2011).
Os microrganismos celulolíticos são encontrados entre os grupos
taxonômicos mais variáveis. A maioria entre eubactéria e fungos, mas
microrganismos anaeróbicos também têm sido identificados no rúmen bovino.
Podem ser encontrados em todas as biotas onde resíduos celulósicos são
acumulados. Eles geralmente ocorrem em populações mistas compreendendo

19
espécies celulolíticas e não celulolíticas, as quais interagem sinergisticamente
(BÉGUIN & AUBERT, 1994).
Os fungos são agentes conhecidamente decompositores de matéria
orgânica em geral e de substratos celulósicos em particular e ocorrem no solo,
colonizando vegetais, suas raízes e resíduos, com importante função de
reciclagem de nutrientes. Na natureza, existe uma grande variedade de
microrganismos que produzem celulases, entretanto apenas alguns são
conhecidos como verdadeiros celulolíticos, isto é, são capazes de degradar a
celulose natural (LYND et al.,2002)
Entre os organismos que degradam a celulose e outras fibras da parede
celular das plantas através da produção de altos níveis de múltiplas enzimas,
os mais estudados incluem: espécies fúngicas – Trichoderma, Humicola,
Penicillium, Aspergillus; bactérias - Bacillus, Pseudomonas, Cellulomonas
Cellovibrio e Cytophaga; e actinomicetos – Streptomyces e Actinomucor.
Entretanto, os microrganismos comercialmente explorados para preparações
de celulases são principalmente limitados ao T. reesei, H. insolens, A. niger,
Thermomonospora fusca, Bacillus sp., e poucos outros organismos
(SUKUMARAN et al., 2005).
O Trichoderma reesei foi descoberto por ser a causa do desgaste de
roupas e tendas durante a segunda guerra mundial (ABOU-ZEID & EL-
DIWANY, 1978). O T. reesei selvagem e seu melhor mutante produtor de
celulase extracelular (RUT C-30) produzem pequenas quantidades de -
glicosidase, o que inibe a hidrólise da celulose devido ao acúmulo de produto
final (celobiose) (DASHTBAN et al., 2009).
Espécies de Aspergillus são conhecidas como bons produtores de -
glicosidase (GARCIA-KIRCHNER et al., 2005).
Os microrganismos utilizam de diferentes ações para hidrolisar
efetivamente a celulose, o que ocorre em partículas insolúveis ou embebidas
dentro de polímeros de hemicelulose e lignina. Os fungos filamentosos
celulolíticos (e bactérias actinomicetos) têm a habilidade de penetrar nos
substratos celulósicos através das extensões das hifas (SUKUMARAN et al.,
2005).

20
Apesar desta discussão focar principalmente a ação de hidrólise do
sistema enzimático na celulose, essa ação deve se realizar também na
hemicelulose e as enzimas hemicelulases são comumente co-produzidas por
microrganismos celulolíticos. Adicionalmente os fungos produtores de enzimas
lignolíticas são muitos e incluem espécies além do ascomiceto T. reesei,
basidiomicetos incluindo fungo da podridão branca (P. chrysosporium), da
podridão marrom (Fomitopsis palustris) e finalmente poucas espécies
anaeróbias (Orpinomyces sp.) os quais degradam a celulose no trato intestinal
de ruminantes (DASHTBAN et al., 2009).
Muitas são as espécies de fungos da podridão branca, sendo a maioria
destas Basidiomycotina, seguidas por alguns Ascomycotina. A degradação da
lignina por estes fungos da podridão branca é mais rápida do que a causada
por outros microrganismos. O crescimento destes fungos diminui em condições
limitadas de nitrogênio e carbono, e a atividade de enzimas lignolíticas aparece
como forma de metabolismo secundário (BORTOLLAZO, 2011).
Os fungos da podridão marrom degradam a celulose e hemicelulose
sem alterar a lignina. No entanto, basidiomicetos de podridão marrom são
conhecidos por serem responsáveis por grave deterioração das paredes das
células lenhosas. Portanto, os fungos chamados de podridão marrom
efetivamente digerem componentes celulósicos sem a remoção da lignina. O
que retarda ataques microbianos em hidratos de carbono lenhosos
(BORTOLLAZO, 2011).
Os fungos da podridão branda, geralmente pertencentes às divisões
Ascomicotina e Deuteromicotina, são fungos que podem causar a degradação
da madeira de forma suave, de aparência úmida. Estes fungos colonizam e
degradam madeiras duras de alta umidade, mas também degradam madeiras
moles, porém com uma taxa de hidrólise mais lenta quando comparada com
fungos de podridão branca e marrom, e são mais eficientes na
despolimerização de compostos sintéticos de lignina. Em contraste com outros
tipos de apodrecimento, o índice de deterioração decorrente da podridão mole
é diretamente proporcional à quantidade de nitrogênio e por isso ela é mais
ativa na madeira que está em contato com o solo ou com a água que
contenham este nutriente em solução (FILHO, 2008).

21
A aplicação de microrganismos como os fungos, por exemplo, espécies
de fungos termofílicos tais como Sporotrichum thermophile, Thermoascus
aurantiacus e Thielavia terrestris têm sido propostos como bons candidatos
para bioconversão de resíduos lignocelulósicos para açúcares e oferecem um
grande potencial a ser explorado em escala industrial. A aplicação de fungos
termofílicos em escala industrial também permite a economia de energia
porque o custo da refrigeração após o pré-tratamento por vapor é evitado e as
taxas de sacarificação são melhoradas. Estes fungos têm sido mostrados como
produtores de celulases e degradam a celulose natural; entretanto a atividade
enzimática de organismos termofílicos é geralmente baixa comparada a fungos
mesofílicos como T. reesei (DASHTBAN et al., 2009).
2.4.1 Aspectos ecológicos das comunidades que degradam a
celulose
Como um processo microbiano, a utilização da celulose está sujeita a
condições químicas e físicas do ambiente. Os efeitos da temperatura são
particularmente dramáticos. Uma comparação das espécies celulolíticas
através da máxima taxa de crescimento revela uma forte dependência da
temperatura de crescimento. Dois outros parâmetros ambientais, pH e
potencial redox, afetam a taxa e extensão de utilização da celulose (GAUTAM
et al., 2010).
Diferentes habitats em que a celulose é amplamente disponível, pelas
suas diferentes características (disponibilidade de água, de oxigênio, potencial
redox, variabilidade da temperatura e nível de nutrientes), têm promovido o
desenvolvimento de estratégias de utilização da celulose, que diferem na
arquitetura e apresentação da enzima, velocidade e extensão da celulólise,
atividades hidrolíticas acessórias, destino dos produtos de hidrólise e
interações entre microorganismos celulolíticos e não celulolíticos (BELL et al.,
2009).
A maioria, ou talvez todos os microrganismos que provavelmente
desempenham um papel importante na hidrólise da celulose na natureza
evoluíram estratégias que aproximam a celulose à superfície da célula e

22
possibilitam ao organismo celulolítico o ―primeiro acesso‖ aos produtos de
hidrólise (BELL et al., 2009; LYND et al., 2002).
Nos solos, a celulose está disponível primeiramente na forma de
serrapilheira (material vegetal morto) que é relativamente recalcitrante, devido
ao alto conteúdo de lignina das plantas terrestres. As necessidades nutricionais
para crescimento das espécies celulolíticas incluem a disponibilidade de
nitrogênio, fósforo e enxofre, além de macro e micro nutrientes padrão e várias
vitaminas. Assim, uma falta de nitrogênio fixado e outros nutrientes podem
secundariamente limitar o crescimento microbiano (LYND et al., 2002).
A estratégia dos fungos para a celulólise envolve celulases
extracelulares que trabalham ao lado do sistema de enzimas lignolíticas do
qual requer eficiência de produção contínua de espécies reativas de oxigênio
(por ex. pelas peroxidases associadas). De acordo com VARDAVAKIS (1989),
a atividade celulolítica diminui com a profundidade do solo, sugerindo que a
utilização da celulose é predominantemente um processo aeróbico, e a bactéria
celulolítica primária isolada foi da espécie Cytophaga, apesar de linhagens de
Bacillus e Cellulomonas também terem sido isoladas. Em compostagem de
estrume de gado, bactérias filamentosas do gênero Micromonospora,
Cytophaga, e Sporocytophaga foram numericamente as isoladas mais
abundantes e fungos foram muito menos abundantes (GODDEN &
PENNINCKX, 1984).
De acordo com RUEGGER & TAUK-TORNISIELO (2004) fungos
produtores de celulases, na natureza, não ocupam o mesmo nicho ecológico
em cultura pura, onde não existe competição, ao contrário, eles interagem com
outros organismos celulolíticos ou não e com aqueles que degradam vários
polímeros, microrganismos os quais nem sempre são revelados pelas técnicas
de isolamento em laboratório. Essa associação é responsável pela completa
mineralização dos substratos celulósicos nos ecossistemas onde estes
organismos estão inseridos.

23
3. MATERIAL E MÉTODOS
3.1 Local de coleta
O local escolhido para a coleta de amostras pertence à propriedade
particular rural (Fazenda São Lucas), com área de plantio de cana-de-açúcar
com produção estimada entre 400 a 500 toneladas/ano. A área faz parte da
comunidade de Inhaúma, parte da antiga Fazenda Lagoa Grande, Distrito de
São Roberto de Minas, Município de São João da Lagoa, região norte de MG,
S 16º 44′ 16,2", W 44º 12′ 36,5", coordenadas X 584.193 Y 8.149.280 (Mapa no
ANEXO 1).
O local de coleta foi estabelecido seguindo as determinações do
Conselho de Gestão do Patrimônio Genético (CGEN) sob autorização de
acesso e remessa de amostra de componente do patrimônio genético nº
014/2009 concedida pelo IBAMA (ANEXO 1).
As amostras foram coletadas ao final do período de estiagem (mês de
Setembro/09) e após o período de chuvas (mês de Abril/10). Foram coletados
resíduos de bagaço e palha de cana-de-açúcar, bem como porções do solo
(até 15 cm de profundidade), provenientes de oito ambientes pré-selecionados
na propriedade.
Os pontos de coleta foram concentrados em duas regiões da fazenda
(destacadas no mapa – Anexo 1) e foram definidos para ambos os períodos e
assim descritos e caracterizados:
Ponto 01 - Piquete 2 – Canavial existente a 10 anos . Resíduo de cana no solo
até 15 cm de profundidade.
Ponto 02 - Piquete 2 – Raiz da cana e solo até 15 cm de profundidade na base
do colmo das plantas.
Ponto 03 – Canavial da Tiririca - existente a 4 anos. Solo (superfície) de palha
de cana.
Ponto 04 – Canavial da Tiririca até 15 cm de profundidade do solo do canavial

24
Ponto 05 – Área de confinamento do gado – Bagaço da cana-de-açúcar,
superfície do solo abaixo do cocho de alimentação do gado.
Ponto 06 – Casa de ração animal, área coberta. Monte de cana-de-açúcar
picada.
Ponto 07 – Engenho – área de moagem de cana a céu aberto. Parte interior do
monte de bagaço de cana-de-açúcar.
Ponto 08 – Engenho – área de moagem de cana a céu aberto. Parte superior
do monte de cana – bagaço de cana-de-açúcar ―verde‖.
Cerca de 100 a 200 g de cada amostra foram coletadas com auxílio de
uma pá de jardim, armazenadas em sacos plásticos devidamente identificados
e acondicionados em caixa de isopor até completar o transporte ao laboratório.
3.2 Isolamento dos microrganismos
Os fungos filamentosos foram isolados a partir do processamento das
amostras em meio de cultura BDA (2% Ágar, 20% Batata, 2% Dextrose),
estrategicamente preparado com pH acidificado para 5,0 acrescido de
antibiótico tetraciclina 100 g/mL.
O processamento das amostras consistiu em diluição seriada (1/10) a
partir de 10g do material de cada ponto de coleta, em solução salina estéril
(NaCl 0,9%) e após homogeneização através de agitação por 1 hora. Em
seguida foram realizadas medida e registro do pH de cada amostra e
plaqueamento de alíquotas de 100 l das diluições 10-3, 10-4 e 10-5, incubadas
a 28ºC ±2 durante 7-9 dias em BDA.
Após o crescimento dos fungos as colônias individualizadas foram
contabilizadas, plaqueadas e diferenciadas, com base nos caracteres
macroscópicos das mesmas, em meio BDA, e submetidas novamente à
incubação durante 7 dias à 28ºC ± 2.
A técnica de microcultivo em lâmina foi utilizada para observação
microscópica das estruturas morfológicas reprodutivas típicas dos fungos
filamentosos (RIDELL, 1950).

25
3.3 Manutenção das linhagens
O estoque para manutenção das linhagens foi preparado em tubos de
ensaio com ágar BDA inclinado, mantidas a 4ºC e renovado a cada 6 meses e
também de acordo com Castellani (1967) do seguinte modo: após o
crescimento da colônia em meio BDA, discos de aproximadamente 0,7 cm de
diâmetro contendo micélio foram transferidos para tubos de ensaio com tampa
contendo água estéril e armazenados a 4ºC.
3.4 Pré-seleção dos microrganismos lignocelulolíticos
Os morfotipos isolados foram pré-selecionados baseando-se na
morfologia macroscópica e microscópica e por testes em meio sólido para
avaliação de atividade lignocelulolítica.
Os fungos filamentosos foram selecionados como produtores de
enzimas celulolíticas, especificamente endoglucanase, pela análise em placa
de Petri com meio sintético acrescido de 1% (p/v) CMC (Carboximetilcelulose)
como única fonte de carbono e substrato indutor de celulases (Tabela 1).
Componentes Concentração (g.L-1
)
NaNO3 3,0
K2HPO4 1,0
MgSO4
KCl
FeSO4 . 7H2O
CMC
Ágar
0,5
0,5
0,01
10,0
20,0
Tabela 1 – Composição do meio sintético CMC 1% (RUEGGER & TAUK-TORNISIELO, 2004)
Os inóculos foram feitos com utilização de palitos estéreis em 3 pontos
equidistantes no centro das placas de Petri. Estas foram incubadas por 2-4 dias

26
a 28ºC ± 2 e em seguida submetidas a choque térmico por 16 h a 50ºC. Após
esse período, foram adicionados 10 mL de solução corante Vermelho Congo
(0,5 g.L-1) em tampão Tris HCl 0,1 M, pH 8,0. Após 30 minutos a solução foi
descartada e as placas lavadas com 5 mL de solução de Cloreto de sódio
(NaCl) 0,5 M neste mesmo tampão. Os diâmetros das colônias e dos halos
produzidos foram medidos com paquímetro (MULLINGS, 1985; RUEGGER &
TAUK-TORNISIELO, 2004; NOGUEIRA & CAVALCANTI, 1996).
O choque térmico a 50ºC por 16 horas antes da revelação com o corante
permite rápida hidrólise da celulose e a seleção de espécies que produzem
enzimas celulolíticas termoestáveis (NOGUEIRA & CAVALCANTI, 1996).
Através do cálculo do Índice Enzimático, de acordo com Nogueira e
Cavalcanti, 1996, (IE = diâmetro da colônia + diâmetro do halo formado/
diâmetro da colônia), foram selecionados aqueles morfotipos com maiores
valores de IE, para na sequência, serem avaliados quantitativamente pela
produção de enzimas (celulases e hemicelulases) em frascos agitados.
A indicação de morfotipos produtores de hemicelulase (xilanase)
também foi realizada em placa de Petri com a utilização do mesmo meio
sintético usado para revelar endoglucanases, sendo o substrato CMC
substituído pela Xilana solúvel (1% p/v). Da mesma forma o halo de
degradação foi revelado com o Vermelho Congo.
A análise para detecção da produção de enzimas ligninolíticas dos
isolados em meio sólido com bagaço de cana peneirado a 1,19 mm, foi
realizada com Guaiacol como descrito na Tabela 2.
Componentes Concentração
Bagaço de cana de açúcar 2,0 g.L-1
Guaiacol 0,1 mL.L-1
Ágar 16,0 g.L-1
Tabela 2 – Composição do meio para detecção de lacase (FILHO, 2008)
Da mesma maneira que o teste em placa com CMC, o inóculo foi feito
com a utilização de palitos estéreis para fazer o repique. As placas foram
incubadas a 28ºC± 2 por até 7 dias para visualização do halo avermelhado ao

27
redor da colônia formado pela oxidação do guaiacol causado pela ação das
enzimas fenoloxidases, mais especificamente lacases (OKINO et al., 2000;
FILHO, 2008).
3.5 Fermentação dos microrganismos isolados ativos
Uma vez pré-selecionados os fungos isolados os quais apresentaram
maiores índices enzimáticos, estes foram submetidos à fermentação submersa
em condição aeróbica em frascos (Erlenmeyers) agitados e aqueles isolados
os quais apresentaram maiores valores de atividade enzimática foram
escolhidos para novos testes em fermentação submersa e fermentação semi-
sólida em cultivo estacionário.
Cepas conhecidas como boas produtoras de celulases, Trichoderma
reesei RUT C-30 (ATCC 56765) e Aspergillus awamori B.361 U2/1, gentilmente
cedidas pela Dra. Elba Pinto da Silva Bon do Laboratório de Tecnologia
Enzimática do Instituto de Química da UFRJ, foram utilizadas como referência.
KHALID et al., 2006 apontam que vários meios de cultura contendo
todos os nutrientes essenciais, são úteis para os estudos de fungos, embora
nenhum deles representem o substrato ideal. Suas deficiências são evidentes,
pois certos fungos não são cultiváveis em meios artificiais.
Sabendo-se da importância das fontes de carbono, nitrogênio e fósforo,
assim como a taxa C/N oferecida e elementos traço, os meios MANDELS &
WEBER (1969) e BRECCIA et al. (1995) modificados foram utilizados neste
trabalho (Tabelas 3 e 4).
O uso de Tween 80 teve como objetivo aumentar a solubilidade do meio
e, portanto o acesso ao substrato. A adição de Tween 80 no meio de cultura
dobrou os rendimentos de -glicosidase em culturas submersas de Aspergillus
niger C-6 (GARCIA-KIRCHNER et al., 2005).

28
Componentes Concentração
Uréia - CO(NH2)2 0,3 g/L
Sulfato de amônio - (NH4)2SO4 1,4 g/L
Fosfato de Potássio monobásico - KH2PO4 0,2 g/L
Cloreto de Cálcio - CaCl2 0,3 g/L
Sulfato de Magnésio heptahidratado - MgSO4.7H2O 0,3 g/L
Sulfato Ferroso heptahidratado - FeSO4.7H2O 0,005 mg/L
Cloreto de Cobalto hexahidratado - CoCl2.6H2O
Sulfato de Manganês tetrahidratado - MnSO4.4H2O
0,02 mg/L
0,0016mg/L
Sulfato de Zinco heptahidratado - ZnSO4.7H2O 0,0014 mg/L
Extrato de Levedura 0,6% (p/v)
Tween 80 0,5 mL/L
Fonte de Carbono*1 3,0% (p/v)
Tabela 3 – Composição do meio Mandels & Weber (1969) modificado (pH 5,0) para cultivo
submerso.
Componentes Concentração
Nitrito de sódio - NaNO3 1,2 g/L
Fosfato de Potássio monobásico - KH2PO4 6,0 g/L
Fosfato de Potássio dibásico – K2HPO4 3,0 g/L
Cloreto de Cálcio - CaCl2 . 2H2O 0,05 g/L
Sulfato de Magnésio heptahidratado - MgSO4.7H2O 0,2 g/L
Sulfato Ferroso heptahidratado - FeSO4.7H2O 0,005 mg/L
Cloreto de Cobalto hexahidratado - CoCl2.6H2O Sulfato de Manganês tetrahidratado - MnSO4.4H2O
0,02 mg/L 0,0016mg/L
Sulfato de Zinco heptahidratado - ZnSO4.7H2O 0,0014 mg/L
Extrato de Levedura 1,2% (p/v)
Tween 80 0,5 mL/L
Fonte de Carbono*1 3,0% (p/v)
Tabela 4 – Composição do meio BRECCIA et al. (1995) modificado (pH 5,0).
Nota (*1): A fonte de carbono usada nestes meios pode ser lactose ou bagaço de cana-de-açúcar ou
farelo de trigo.

29
O micélio fúngico foi utilizado como inóculo na forma de discos de 7 mm
de diâmetro em meio Mandels & Weber (1969) modificado. O bagaço de cana-
de-açúcar moída ou lactose comercial foram avaliados como fonte de
carbono*1 e substrato indutor da expressão enzimática.
O bagaço de cana-de-açúcar utilizado nos meios, seja submerso ou
semi-sólido, não foi submetido a nenhum pré-tratamento específico, a não ser
pelo processo de esterilização por 20 minutos a 121ºC.
O meio Breccia et al. (1995) modificado (Tabela 4) com farelo de trigo
3%, foi utilizado para os testes de fermentação submersa, por vezes
substituindo o farelo de trigo por bagaço de cana-de-açúcar, como fonte de
carbono*1 e indutor da produção de enzimas.
Os frascos foram esterilizados a 121ºC por 15 minutos, e posteriormente
os inóculos foram incubados a 28ºC ± 2 durante 5 a 10 dias sob agitação 150
rpm.
O cultivo em meio semi-sólido foi utilizado como alternativo e comparativo ao
cultivo submerso e seguiu os mesmos procedimentos para inóculo tal como
descrito acima, no entanto o meio foi composto por 20 mL de solução nutritiva
(Tabela 5), e o bagaço de cana ou o farelo de trigo como substrato indutor do
mesmo modo que para fermentação submersa, porém em maior proporção,
30g por frasco Erlenmeyer de 500 mL de capacidade, acrescidos de 10 g de
casca de arroz para possibilitar maior aeração do meio.
Componentes Concentração
Fosfato de amônio dibásico - (NH4)2HPO4 2,5 g/L
Sulfato de magnésio heptahidratado - MgSO4.7H2O 0,5 g/L
Cloreto de potássio - KCl 0,5 g/L
Tween 80 0,5 mL/L
Tabela 5 – Composição da solução nutritiva (pH 5,0) para meio semi-sólido.
Os frascos que continham a solução nutritiva e o substrato indutor foram
esterilizados a 121ºC por 15 minutos e após os inóculos feitos foram incubados
a 28ºC ± 2 durante cerca de 30 ou mais dias em cultivo estacionário.

30
3.5.1 Quantificação dos microrganismos durante fermentações
Para os cultivos sob substratos solúveis, a concentração celular é
comumente medida diretamente via filtração e subsequentemente pela
determinação de peso de matéria seca ou indiretamente via densidade ótica ou
contagem de células. Devido à presença de substratos insolúveis a aplicação
dessas técnicas neste estudo não foi possível.
Os fungos foram observados e determinados de acordo com o padrão
adaptado da ASTM (American Society for Testing Materials), Standard Methods
G21-96 (2002), que apresentaram as seguintes faixas de crescimento:
(-) ausência de crescimento;
(+) pouco crescimento com turvação de pequenos fragmentos de micélio
lançados no meio;
(++) moderado crescimento com surgimento de ―pellet‖ fino na superfície do
meio;
(+++) ótimo crescimento de massa micelial da metade a todo o frasco.
3.6 Extração das enzimas
Em seguida aos procedimentos de incubação, todas as amostras e
controles foram processas, tal como se segue:
Passo 1 - Foram recuperadas amostras (10-20 mL) em duplicata, de cada
frasco Erlenmeyer para testes enzimáticos subseqüentes, a cada 5 dias para
cultivo semi-sólido e a cada 24 hs para cultivo submerso.
Passo 2 - As alíquotas retiradas foram submetidas à centrifugação refrigerada
por 10 min. a 18.000 rpm. O sobrenadante foi utilizado para avaliar a atividade
das enzimas lignocelulolíticas além das medidas de pH e dosagem de proteína
extracelular total (de acordo com BRADFORD, 1976).
As amostras do cultivo em meio semi-sólido tiveram 5,0 g de material
coletado o qual teve 50 mL de água destilada estéril adicionada, agitada por 30
minutos e então centrifugada e recuperada como descrito no passo 1.

31
3.7 Determinação das atividades e concentrações enzimáticas do
complexo celulolítico e hemicelulolítico.
3.7.1 Atividade Carboximetilcelulase – CMCase
Foi determinada de acordo com a metodologia padrão descrita pela
IUPAC (GHOSE, 1987). O método se baseia na dosagem da concentração de
açúcares redutores liberados durante a degradação do substrato
Carboximetilcelulose (CMC).
O meio reacional foi formado por 0,250 mL de uma solução de CMC 4%
p/v em tampão citrato de sódio 50 mM pH 4,8 e 0,250 mL do sobrenadante das
culturas (extrato enzimático). A mistura reacional foi incubada a 50ºC, durante
10 minutos, sob agitação. A reação enzimática foi interrompida pela adição
imediata de 0,5 mL de DNS (ácido 3,5-dinitro salicílico). O reagente DNS, além
de interromper a reação enzimática, possibilita a realização da dosagem da
concentração de açúcares redutores produzidos pela ação enzimática, obtida
após fervura por 5 minutos a 96ºC e após resfriamento dos tubos, feita leitura
em espectrofotômetro em absorbância 540nm (MILLER,1959) (Esquema no
Apêndice A).
3.7.2 Atividade FPase (exoglucanase)
Foi determinada de acordo com a metodologia padrão descrita pela
IUPAC (GHOSE, 1987). O método se baseia na dosagem da concentração de
açúcares redutores liberados durante a degradação de uma fita de papel de
filtro.
O meio reacional foi formado por 0,5 mL do sobrenadante das culturas,
1,0 mL de tampão citrato de sódio 50 mM pH 4,8 e uma tira de papel filtro
Whatmann nº1 medindo 1,0 x 6,0 cm (aproximadamente 50 mg). A mistura
reacional foi incubada a 50ºC, durante 60 minutos, sob agitação. A reação foi
interrompida pela adição de 0,5 mL de DNS a um tubo contendo 0,5 mL da
mistura reacional. A determinação da concentração de açúcares redutores foi

32
realizada em seguida, como já descrita no item 3.7.1 (Esquema no Apêndice
B).
3.7.3 Atividade -glicosidase
Foi determinada de acordo com a metodologia padrão descrita pela
IUPAC (GHOSE, 1987). O método se baseia na determinação da concentração
de glicose liberada quando a enzima hidrolisa a celobiose.
O meio reacional foi formado por 0,5 mL do sobrenadante das culturas e
0,5 mL de solução de substrato (celobiose 15 mM em tampão citrato de sódio
pH 4,8, 50mM). A mistura reacional foi incubada a 50ºC por 30 minutos, sob
agitação. A reação foi terminada por imersão dos tubos em água fervente por 5
minutos. A determinação da glicose foi realizada utilizando-se o kit para análise
de Glicose GOD-POD® da Biosystems (Esquema no Apêndice C).
3.7.4 Atividade hemicelulase (Xilanase)
A atividade da enzima xilanase foi determinada segundo MILLER (1959).
A reação consiste na mistura contendo 0,1 mL de sobrenadante das culturas,
0,2 mL de solução de 1% de xilana em 0,05 M de tampão acetato pH 5,0. A
mistura reacional foi incubada a 50º C por 30 minutos, e o sistema enzima-
substrato foi agitado periodicamente para manter a xilana em suspensão. A
reação foi interrompida pela adição de 0,3 mL da solução de ácido 3,5-
dinitrosalicílico (DNS). A determinação da concentração de açúcares redutores,
tendo como padrão a xilana, foi realizada em seguida, como já descrita no item
3.7.1. (Esquema no Apêndice D).
3.7.5 Cálculo da atividade enzimática
A atividade enzimática é expressa em UI (μmol de produto açúcar
redutor liberado / minuto), calculada conforme equação:

33
UI = diluição x Abs. X Fator ( mol/mL) x Vol (mL)
Tempo (min)
Onde:
Diluição = Fator de diluição, apenas se for necessário diluir o sobrenadante
(solução de enzima);
∆Abs = Diferença da absorbância medida ao final da reação e no início da
reação enzimática;
Fator = fator de correção obtido da curva padrão de glicose ou xilana (para
dosagens de Endoglucanases e Exoglucanases);
Vol = Volume total da mistura reacional em mL;
Tempo = Tempo da reação em minutos.
Determinação da concentração enzimática
A concentração da enzima é expressa em UI/mL, calculado conforme equação:
[Enz]= diluição x Abs. X Fator ( mol/mL) x Vol. 1 (mL)
Tempo (min) x Vol. 2 (mL)
Onde:
Diluição = Fator de diluição, apenas se for necessário diluir o sobrenadante
(solução de enzima);
∆Abs = Diferença da absorbância medida ao final da reação e no início da
reação enzimática;
Fator = fator de correção obtido da curva padrão de glicose ou xilana (para
dosagens de Endoglucanases e Exoglucanases);
Vol. 1 = Volume total da mistura reacional em mL;
Tempo = Tempo da reação em minutos;
Vol. 2 = Volume da amostra em mL.
Para o cálculo da -glicosidase o fator de correção foi obtido pela
amostra padrão nas dosagens de β-glicosidase utilizando o kit GOD-POD®

34
(glucose oxidase/peroxidase) da Biosystems, onde a concentração do padrão é
igual a 5,55mM/L de glicose e calculado pela fórmula abaixo.
Fator = ([glicose padrão]) / (Abs. Padrão) = 5,55 / (Abs. Padrão)
3.8 Identificação taxonômica
A identificação taxonômica de 5 linhagens fúngicas celulolíticas, pré-
selecionadas por apresentarem altos valores de IE, foram realizadas pelo
Centro Pluridisciplinar de Pesquisas Químicas, Biológicas e Agrícolas (CPQBA)
da Unicamp. As metodologias usadas foram a extração do DNA genômico,
amplificação da região D1/D2 (DNAr 28S) ou da região ITS (―Internal
Transcribed Spacer‖ – região espaçadora de transcrição interna) através de
PCR, seqüenciamento e análise filogenética (Anexo 2).
3.9 Análise estatística
Os testes estatísticos foram realizados com o auxílio do programa
Microsoft Office Excel e utilização do programa Minitab para análise de
variância (ANOVA) e teste de Tukey, com nível de significância 5%, p<0,05.

35
4. RESULTADOS E DISCUSSÃO
4.1 Isolamento dos fungos filamentosos
Após o devido processamento das amostras e período de incubação de
7-9 dias em meio BDA, os resultados revelaram elevado número de morfotipos
filamentosos, contabilizados em UFC (unidades formadoras de colônias)
(Tabela 6), bem como alta diversidade de morfotipos por ponto de coleta
(Tabela 7).
A adição de antibiótico e a correção do pH do meio para 5,0 foram
importantes para inibir o crescimento de bactérias e leveduras. As diluições
10-3 e 10-4 foram satisfatórias para obtenção de até 100 UFC/placa. A análise
microscópica através de lâminas preparadas por microcultivo (RIDELL, 1950)
corroborou para a identificação das estruturas morfológicas específicas dos
fungos. Algumas fotos estão registradas no apêndice E.
Ponto de coleta 1ª coleta (seca) 2ª coleta (chuva)
P01 230 1
P02 30 40
P03 30 30
P04 150 30
P05 30 100
P06 50 30
P07 10 300
P08 850 300
Total 1380 831
Tabela 6 – Número de fungos filamentosos (x104 UFC /mL) por ponto de coleta e período
correspondente.

36
Ponto de coleta 1ª coleta (seca) 2ª coleta (chuva)
P01 23 10
P02 45 4
P03 12 6
P04 15 9
P05 10 15
P06 11 7
P07 8 3
P08 22 31
Total 145 85
Tabela 7 – Diversidade de morfotipos encontrados por ponto e período de coleta.
Os resultados sugerem que a sazonalidade pode ter influenciado o
número de UFC’s de fungos filamentosos encontrados. Na 1ª coleta (período
pós-estiagem) a riqueza de morfotipos foi cerca de 40% maior. Esta ocorrência
pode ser devido à maior deposição de folhas e ramos no solo no período,
favorecendo a presença de maior número de microrganismos.
O menor número de morfotipos encontrados na estação chuvosa pode
ser explicado pela lixiviação dos esporos dos fungos para uma área mais
profunda do solo e também pelo aumento de substâncias alelopáticas. Estes
resultados corroboram os apresentados por CARNEIRO (2000) que estudou
solo de Cerrado e de áreas cultivadas no Distrito Federal, que confirmam a
ocorrência de um maior número de UFC's no início da estação chuvosa
seguido pelo seu declínio quando o índice pluviométrico atinge valores
elevados.
A diversidade de microrganismos do solo depende da química, da
textura e da capacidade do solo de manter a umidade. O sucesso de um fungo
alcançar e colonizar uma área do solo se deve principalmente a sua
capacidade saprobiótica, expressa por crescimento rápido do micélio, produção
de esporos, presença de um eficiente e extenso sistema de poder enzimático e

37
tolerância a antibióticos, salinidade, e metais pesados, compostos
químicos/fungicidas e temperatura (KHALID et al, 2006).
O total de morfotipos isolados nas duas coletas foi de 230 e a maior
diversidade encontrada no período de estiagem, cerca de 40% maior que no
período de chuvas, também encontra suporte na maior riqueza de substratos
para utilização dos fungos, específicamente no ponto de coleta 2, o qual se
refere à amostra de raiz e colmo da cana. Sugere-se que outros fatores podem
corroborar para a maior diversidade (BELL et al., 2009), como a maior faixa de
pH nos pontos de coleta na 1ª coleta (Figura 5) e o pH mais ácido como
favorável à maior diversidade fúngica (Figura 6).
Figura 5 – pH de cada ponto de coleta nos 2 períodos sazonais
Figura 6 – Correlação da diversidade fúngica com pH, na 1ª (A) e 2ª (B) coleta.

38
De acordo com o pH medido nas amostras de solo em cada ponto de
coleta, sugere-se que os maiores valores nos pontos 5 e 6 da 1ª coleta,
correspondentes à área do cocho dos animais e casa de ração,
respectivamente, foram conseqüência da mistura do bagaço de cana com uréia
e sulfato de amônio, utilizados como aditivos na alimentação animal da
propriedade (informação do proprietário/comunicação pessoal).
Os valores de pH mais baixos nos pontos 7 e 8, referentes ao monte de
bagaço de cana-de-açúcar, parte interna e superior, respectivamente,
provavelmente resultam do processo de fermentação e degradação do bagaço.
Moderada tendência de correlação entre a diversidade e o pH foi
observada e registrada nos gráficos A e B (Figura 6). Houve maior diversidade
de morfotipos encontrados em pontos de coleta de menores valores de pH.
Preliminarmente pode-se inferir que a diferença nos pHs entre os 2
períodos de coleta assim como as modificações provocadas no meio, sejam
naturais (mudanças do clima, sazonalidade, levando a variação do substrato)
ou impostas por ação humana tais como práticas culturais (adubação, irrigação
gradeio, etc), de uma forma ou de outra poderão influenciar positiva ou
negativamente na riqueza e abundância dos microrganismos encontrados,
assim como na expressão enzimática dos mesmos em condições
edafoclimáticas diferenciadas.
KHALID et al. (2006), verificaram em estudo de fungos celulolíticos de
solo, que além dos fatores abióticos da região, as características/habilidades
dos fungos (esporulação, taxa de crescimento, sistema enzimático,
preferências alimentares e tolerância à condições de estresse) são
determinantes na frequência e distribuição fúngica.
4.2 Manutenção e viabilidade dos isolados
O procedimento para estoque dos morfotipos em tubos com BDA
inclinado a 4ºC e renovação a cada seis meses se mostrou eficiente para
manter as culturas estáveis e livres de contaminação, assim como as culturas
mantidas pelo método Castellani (1969). No entanto, essas só foram avaliadas
em período de 40 dias de estocagem (Apêndice F).

39
4.3 Seleção dos fungos com atividade celulolítica
Nenhum método de isolamento e seleção de fungos do solo é 100%
eficiente. CAVALCANTI & MAIA (1994) usaram discos de papel de filtro, os
quais cobriam as amostras de solo coletadas e dessa forma foram colonizados
pelos fungos celulolíticos do solo da região sob estudo, posteriormente
transferidos para placas de Petri e mantidos em câmara úmida.
Subsequentemente as colônias fúngicas desenvolvidas foram transferidas para
meio Sabouraud com antibiótico e em segundo repique para meio BDA. Dessa
forma obtiveram o isolamento e seleção de espécies celulolíticas.
THEATER & WOOD (1982) mostraram uma forte interação entre o
Vermelho Congo e os polissacarídeos (1-4) ligados em unidades D-
glucopiranosil fornecendo a base para um ensaio simples e sensível para a
detecção de colônias de bactérias produtoras de celulase.
Para ZHANG et al. (2006), atividades endoglucanase podem ser
facilmente detectadas em placas com CMC (carboximetilcelulose) e corantes
que são adsorvidos somente por polissacrídeos de cadeia longa. Os autores
consideram este método semi-quantitativo e bem adequado para monitorar
grande número de amostras.
No presente trabalho, a seleção dos fungos celulolíticos foi realizada
mediante crescimento dos isolados em placa de Petri com meio acrescido de
CMC e coloração com Vermelho Congo. De acordo com este método foi
possível visualizar a formação de halos de degradação da carboximetilcelulose
em fragmentos menores que celohexose, à qual o Vermelho Congo não se liga,
devido à ação de endoglucanases (Figura 7).
O número de fungos filamentosos com atividade celulolítica revelados
em placa com CMC e Vermelho Congo, nos dois períodos sazonais avaliados
neste trabalho, seguiu a mesma composição de morfotipos totais, ou seja, na
estação seca a presença de fungos celulolíticos foi cerca de duas vezes maior
que na estação chuvosa (Tabela 8).

40
Figura 7 – Fotos de halos de degradação (região clara) indicadores de ação endoglucanase
nos isolados S88, S123, S74 e S87 em meio sintético com CMC 1% e ação de xilanase da
amostra 87 em meio sintético com xilana 1%. Índice enzimático - valor IE entre parênteses.
No início da estação seca a proporção de matéria orgânica é maior,
favorecendo a colonização dos fungos celulolíticos (VANCE & CHAPIN, 2001).
No total foram 124 morfotipos celulolíticos para 230 morfotipos inicialmente
isolados, ou seja, 54%. JAHANGER et al. (2005), obtiveram 67,83% de fungos
produtores de celulase dos fungos isolados do ambiente.
CAVALCANTI & MAIA (1994), no isolamento de fungos celulolíticos de
solo aluvial de zona semi-árida do nordeste do Brasil, observaram pequeno
número de espécies e correlacionaram este fato a pouca matéria orgânica
presente no solo da região.
Isolado S88 (IE = 5,4) S123 (IE=5,8) S74 (IE = 4,0)
S87 (IE = 4,10) S87 (substrato xilana)
xilana CMC
CMC CMC CMC CMC

41
Ponto de coleta 1ª coleta 2ª coleta
P01 12 1
P02 20 2
P03 8 4
P04 10 6
P05 9 13
P06 10 6
P07 5 3
P08 9 6
Total 83 41
Tabela 8 – Número de fungos filamentosos com atividade celulolítica por ponto de coleta e
período correspondente.
Diante do alto número de isolados (124 morfotipos) que apresentaram
halos de degradação do CMC, utilizou-se o critério do tamanho do halo (maior
ou igual a 3,5) para selecionar alguns morfotipos para cultivo em frascos e
posterior quantificação da atividade enzimática, tendo como referência os
valores de ―Índice Enzimático‖ dos fungos controle.
Obteve-se o ―Índice Enzimático‖ (IE) através do cálculo da soma dos
valores dos diâmetros da colônia e do halo de degradação, dividido pelo valor
do diâmetro da colônia, de acordo com NOGUEIRA & CAVALCANTI (1996)
(Apêndice G). Os fungos controle Trichoderma reesei e Aspergillus awamori,
apresentaram IE de 2,86 e 3,2, respectivamente.
A figura 8 e a tabela 9 representam os IEs dos isolados nos pontos e nos
dois períodos de coleta, e a indicação daqueles que apresentaram IE ≥ 3,5,
perfazendo um total de 27 isolados. Destes, 19 isolados foram escolhidos
aleatoriamente (amostras sublinhadas no apêndice G), submetidos ao
crescimento em frascos e tiveram as enzimas CMCase, FPase, -glicosidase e
xilanase quantificadas.

42
P8P7P6P5P4P3P2P1
6
5
4
3
2
P8P7P6P5P4P3P2P1
2009
PC
IE
2010
3,5
Indice Enzimático
2009 - 1ª coleta - período seco 2010 - 2ª coleta - período chuvoso
PC = Ponto de coleta
Figura 8 - Quadro representativo dos IEs dos fungos filamentosos isolados nos pontos de
coleta (PC = P1 a P8) na 1ª e 2ª coletas, mostrando o ponto de corte IE ≥ 3,5.
Ponto de coleta 1ª coleta 2ª coleta Total
P01 2 0 2
P02 7 0 7
P03 1 3 4
P04 3 2 5
P05 1 1 2
P06 2 0 2
P07 2 0 2
P08 1 2 3
Total 19 8 27
Tabela 9 – Número de fungos celulolíticos selecionados com IE ≥ 3,5 por ponto de coleta e
período correspondente.

43
O teste em placa para atividade lignolítica revelou halo de oxidação do
guaiacol nos isolados S09, S29, S42, S71, S72, S73 e S87. Alguns registros
dos halos formados podem ser observados no apêndice H. Não foi feita análise
quantitativa de lignase. No entanto, todos os 124 isolados cresceram no meio
contendo ágar, bagaço e guaiacol (FILHO, 2008), indicando a potencialidade
do bagaço como substrato e a seleção dos fungos para estudos mais
aprofundados.
O teste para atividade hemicelulolítica (xilanase) em placa, com a
utilização da xilana como substrato e Vermelho Congo, revelou halos de
degradação somente nos isolados S42, S48, S72, S73 e S87 (Figura 7).
4.4 Análise da atividade celulolítica e hemicelulolítica
Os 19 morfotipos isolados selecionados com IE ≥ 3,5, foram submetidos
ao cultivo em frascos agitados, em meio MANDELS & WEBER (1969) com
lactose por 144 horas a 30ºC. As concentrações enzimáticas máximas (média
de duplicata) de CMCase, FPase, -glicosidase e xilanase, bem como a
dosagem de proteína extracelular total estão representadas nas figuras 9, 10,
11 e 12, respectivamente.
Em relação à dosagem de proteína extracelular total, observa-se que
alguns perfis de concentração enzimática de uma dada enzima se mostram
correlacionados à concentração de proteína total, o que sugere que
provavelmente aquela enzima específica é o componente protéico dominante
no extrato. No entanto, muitos extratos analisados não mostram essa relação,
pois se tratam de extratos brutos. ZHANG et al. (2007), em estudos de
fermentação submersa de Trichoderma reesei RUT C-30 verificaram essas
distorções em se tratando de coquetéis enzimáticos não purificados. Portanto,
as demais análises comparativas neste trabalho foram realizadas
considerando-se especialmente as concentrações enzimáticas e não as
atividades específicas obtidas sob as diferentes condições de cultivo dos
isolados.

44
Figura 9 - Concentração enzimática de endoglucanase (CMCase) e dosagem de proteína total
(linha contínua), obtidas do cultivo dos isolados em meio Mandels & Weber com lactose,
máxima alcançada em até 144 h. Barra indicativa desvio padrão. Médias com a mesma letra
não diferem entre si, segundo teste de Tukey (p<0,05).
Observa-se que o isolado S87 apresentou concentração enzimática
CMCase destacada em relação aos demais (7,89 UI/mL), mas
significativamente menor que o produzido pela linhagem referência T. reesei
(9,10 UI/mL).
O teste de degradação de CMC em placa revelou halo de degradação
para todos os isolados, sendo halos maiores que o revelado pelo T. reesei (IE
2,86). Os isolados S21, S48, S67, por exemplo, revelaram halos que levaram
ao IE de 4,0, no entanto esses morfotipos não mostraram altas concentrações
de CMCase, sugerindo que esta técnica não é válida para análise quantitativa
de enzima endoglucanase, não se revelando um bom preditor. Pelos resultados
há indicações de que não existe correspondência entre tamanho de halo e
proporção da concentração enzimática entre os isolados avaliados neste
estudo.
RUEGGER & TAUK-TORNISIELO (2004), predizem que o índice
enzimático pode ser utilizado, somente como uma medida útil para selecionar
linhagens dentro de uma mesma espécie ou como um parâmetro simples e
rápido para selecionar mutantes.
F
E E
E E E E E E E
B
A
C
D D D D
F F F F F

45
Figura 10 - Concentração enzimática de exoglucanase (FPase) e dosagem de proteína total
(linha contínua), obtidas do cultivo dos isolados em meio Mandels & Weber com lactose,
máxima alcançada em até 144 h. Barra indicativa desvio-padrão. Médias com a mesma letra
não diferem entre si, segundo teste de Tukey (p<0,05).
Os isolados S48 e S93 mostraram maiores concentrações da enzima
FPase, 0,7103 e 0,6631 UI/mL, respectivamente, no entanto os valores foram
significativamente menores que o apresentado pela referência T. reesei.
Observa-se que os demais isolados apresentaram atividades FPase
estatisticamente semelhantes ao A. awamori ou maiores, nestas condições de
cultivo.
Segundo HAHN-HÄGERDAL et al., (1991) a produção de FPase por
T.reesei RUT C-30 em cultivo submerso relatada na literatura está entre 0,25 a
6,4 UI/mL.
RUEGGER & TAUK-TORNISIELO (2004) fazem algumas considerações
ao final do estudo de fungos filamentosos celulolíticos isolados do solo da
Estação Ecológica de Juréia-Itatins, São Paulo, das quais se podem
compartilhar algumas neste trabalho. De acordo com estes autores, a produção
de celulases depende do tipo de substrato utilizado e o uso de distintos
métodos para avaliar a atividade enzimática torna difícil a comparação com os
resultados obtidos por diferentes autores.
G
A
B B C C D
F F F F F E E
H
G
G G G
G G
G

46
Figura 11 - Concentração enzimática de -glicosidase e dosagem de proteína total (linha
contínua), obtidas do cultivo dos isolados em meio Mandels & Weber com lactose, máxima
alcançada em até 144 h. Barra indicativa desvio-padrão. Médias com a mesma letra não
diferem entre si, segundo teste de Tukey (p<0,05).
Altos níveis de -glicosidase são importantes para a completa conversão
da celulose, devido à inibição pela celobiose, das atividades celobiohidrolase e
endoglucanase. Espécies de Aspergillus são conhecidas como bons produtores
de -glicosidase (GARCIA-KIRCHNER et al., 2005).
O isolado S67 se destacou pela expressão da -glicosidase (1,3356
UI/mL), ultrapassando inclusive (cerca de 75% maior) a concentração desta
enzima produzida pelo fungo referência A. awamori, nestas condições de
cultivo.
Ainda, nota-se que vários isolados não expressaram a -glicosidase.
A
B
C C
C C
D D D D D D
D D D D D D
D D D

47
Figura 12 - Concentração enzimática de xilanase e dosagem de proteína total (linha contínua),
obtidas do cultivo dos isolados em meio Mandels & Weber com lactose, máxima alcançada em
até 144 h. Barra indicativa desvio-padrão. Médias com a mesma letra não diferem entre si,
segundo teste de Tukey (p<0,05).
Vários isolados expressaram a enzima xilanase nestas condições de
cultivo, mas o isolado S93 se destacou com 5,2161 UI/mL, atividade
estatisticamente maior que o T. reesei. No entanto, dos isolados selecionados
a priori de acordo com teste em placa com xilana e Vermelho Congo, somente
os isolados S87 e S93 revelaram halo de degradação, sendo que os demais
isolados não mostraram nenhum halo, sugerindo que a referida técnica para
seleção prévia de fungos xilanolíticos, pode não ser adequada.
Aqueles isolados os quais se destacaram no teste preliminar de cultivo
submerso em meio MANDELS & WEBER (1969) com lactose (S21, S48, S67,
S87 e S93), foram submetidos a avaliações de expressão das enzimas
utilizando outros substratos indutores e também cultivo semi-sólido.
Com a utilização do bagaço em cultivo submerso no meio MANDELS &
WEBER (1969) por até 120 horas e comparando-se com a lactose, observa-se
que a expressão de enzimas não segue um padrão, dependendo do fungo e da
enzima em questão (Figuras 13, 14, 15 e 16).
A
B
C C C D D D
E E E E E E E
E
E
E E E E

48
O bagaço de cana-de-açúcar se mostrou melhor indutor que a lactose
para síntese de CMCase, para os isolados S48 (6,06 UI/mL), S93 (4,03 UI/mL),
A. awamori (6,22 UI/mL) e T. reesei (11,48 UI/mL) (Figura 13).
Figura 13 - Comparativo produção de CMCase (UI/mL) pelos isolados S21, S48, S67, S87,
S93 e controles A. awamori e T.reesei em cultivo submerso em meio Mandels & Weber (1969)
com lactose e com bagaço por 120 h. Barra indicativa desvio-padrão.
Para síntese de FPase, o bagaço de cana não foi capaz de induzir
maiores concentrações em relação à lactose a nenhum isolado testado (Figura
14).
Figura 14 - Comparativo produção de FPase (UI/mL) pelos isolados S21, S48, S67, S87, S93 e
controles A.awamori e T.reesei em cultivo submerso em meio Mandels & Weber (1969) com
lactose e com bagaço por 120 h. Barra indicativa desvio-padrão.

49
Em relação à atividade -glicosidase, apenas o isolado S93 expressou
melhor (0,35 UI/mL) na presença de bagaço, sendo que para o isolado S67 o
uso do bagaço reduziu muito (de 1,34 pra 0,54 UI/mL) a expressão da enzima.
Para as demais amostras a expressão da enzima -glicosidase produzida em
meio MANDELS & WEBER (1969) com lactose ou bagaço, não foi expressiva
(Figura 15).
Figura 15 - Comparativo produção de -glicosidase (UI/mL) pelos isolados S21, S48, S67,
S87, S93 e controles A.awamori e T.reesei em cultivo submerso em meio Mandels & Weber
(1969) com lactose e com bagaço por 120 h. Barra indicativa desvio-padrão.
O bagaço foi melhor indutor de xilanase para o isolado S48 e para os
controles A. awamori e T. reesei. O isolado S21 teve pior expressão de
xilanase quando em meio com bagaço e os demais isolados não apresentaram
valores estatisticamente diferentes entre a expressão em meio com bagaço ou
lactose. (Figura 16).

50
Figura 16 - Comparativo produção de xilanase (UI/mL) pelos isolados S21, S48, S67, S87, S93
e controles A.awamori e T.reesei em cultivo submerso em meio Mandels & Weber (1969) com
lactose e com bagaço por 120 h. Barra indicativa desvio-padrão.
O bagaço de cana-de-açúcar não foi bom indutor enzimático em meio
MANDELS & WEBER modificado (1969) para a maioria dos isolados testados.
Considerando a relação C/N, o meio BRECCIA et al. (1995) modificado fornece
mais carbono pela maior concentração de extrato de levedura e também pelo
farelo de trigo que representa ótima fonte de carbono. Portanto, os isolados
S67 e S87, os quais para nenhuma enzima apresentaram melhor expressão
com bagaço, foram avaliados em meio BRECCIA et al. (1995) modificado, com
utilização de farelo de trigo ou bagaço como substrato e indutor (em diferentes
concentrações).
Para o isolado S67, o farelo de trigo se mostrou melhor indutor para
todas as enzimas avaliadas, em relação ao bagaço, sendo que a -glicosidase
e xilanase foram expressas diante de menor concentração de farelo de trigo
(1,5%) e ainda assim melhor do que o bagaço (Figura 17).

51
Figura 17 - Concentração enzimática máxima obtida em cultivo submerso em meio Breccia
(1995) modificado com farelo de trigo ou bagaço 1,5 % e 3,0 % do isolado 67. Barra indicativa
desvio-padrão. Controle sem farelo ou bagaço.
No entanto, somente a xilanase produzida na presença de farelo de trigo
1,5% foi um pouco maior que em meio MANDELS & WEBER (1969) com
lactose. Todas as demais enzimas foram produzidas em maiores
concentrações na presença de lactose.
Com estes resultados pode-se sugerir preliminarmente que os
substratos insolúveis de maneira geral não foram melhores indutores que a
lactose solúvel. Há de se levar em consideração ainda a diferença da
proporção C/N entre os meios testados.
De forma oposta, o isolado S87 apresentou maiores concentrações de
todas as enzimas avaliadas quando em presença de bagaço (3,0%) em meio
BRECCIA et al. (1995) modificado. E ainda, observa-se que o farelo de trigo
em meio Breccia não foi melhor indutor que a lactose em meio MANDELS &
WEBER (1969) (Figura 18).

52
Figura 18 - Concentração enzimática máxima obtida em cultivo submerso em meio Breccia
(1995) modificado com farelo de trigo ou bagaço 1,5 % e 3,0 % do isolado S87. Barra indicativa
desvio-padrão. Controle sem farelo ou bagaço.
Estes resultados são uma pequena amostra de que não há um único
substrato indutor de celulases e hemicelulases que possa ser padronizado a
todos os fungos celulolíticos. O estudo deve ser aprofundado levando em conta
as necessidades nutricionais do fungo produtor em questão.
Apesar de 90% das enzimas serem produzidas industrialmente através
de fermentação submersa, a fermentação semi-sólida tem sido utilizada com
diferentes resíduos agroindustriais como palha de arroz (KHAN et al., 2007),
farelo de arroz (LATIFIAN et al., 2007), farelo de trigo (CAMASSOLA &
DILLON, 2007), bagaço de cana-de-açúcar (MASSADEH et al., 2001;
MUTHUVELAYUDHAM & VIRUTHAGIRI, 2006). Esse bioprocesso tem sido
apontado como alternativa para mitigar problemas de poluição ambiental
(SOCCOL & VANDENBERGHE, 2003).
Existe uma grande variedade de fungos e substratos utilizados para
produção de celulases. KANG et al. (2004) estudaram a produção de celulases
e hemicelulases por Aspergillus niger KK2 em palha de arroz e farelo de trigo.
AGUIAR & MENEZES (2000) avaliaram a atividade enzimática de A. niger IZ-9
em bagaço de cana e CAMASSOLA & DILLON (2007) utilizaram Penicillium
echinulatum em bagaço de cana-de-açúcar e farelo de trigo.

53
Em nosso estudo sob cultivo semi-sólido, comparando-se o uso de
bagaço e farelo de trigo como substrato, avaliado por 33 dias, observa-se nas
Figuras 19, 20, 21 e 22 que o bagaço não induziu melhor a expressão das
enzimas para nenhum isolado avaliado em relação ao farelo de trigo.
Figura 19 – Concentração máxima de CMCase obtida em cultivo semi-sólido por 33 dias,
comparativo substrato farelo de trigo e bagaço. Barra indicativa desvio-padrão.
Figura 20 – Concentração máxima de FPase obtida em cultivo semi-sólido por 33 dias,
comparativo substrato farelo de trigo e bagaço. Barra indicativa desvio-padrão.

54
Figura 21 – Concentração máxima de -glicosidase obtida em cultivo semi-sólido por 33 dias,
comparativo substrato farelo de trigo e bagaço. Barra indicativa desvio-padrão.
Figura 22 – Concentração máxima de xilanase obtida em cultivo semi-sólido por 33 dias,
comparativo substrato farelo de trigo e bagaço. Barra indicativa desvio-padrão.
O isolado S87 quando avaliado em cultivo submerso apresentou maiores
atividades enzimáticas na presença de bagaço, no entanto quando em cultivo
semi-sólido o comportamento foi contrário.
Na avaliação deste isolado S87 frente à composição do meio semi-sólido
com mistura de bagaço e farelo de trigo, observa-se que o farelo de trigo puro

55
confirmou ser melhor substrato e indutor de enzimas sob as condições de
cultivo semi-sólido (Figura 23).
Figura 23 – Concentração média máxima de enzimas produzidas pelo isolado S87 em cultivo
semi-sólido nos diferentes substratos (bagaço 3,0%; farelo de trigo 3,0% e mistura bagaço
1,5% e farelo de trigo 1,5%). Barra indicativa desvio-padrão.
O mesmo foi observado para o Aspergillus awamori (Figura 24).
Figura 24 - Concentração média máxima de enzimas produzidas pela referência A. awamori
em cultivo semi-sólido nos diferentes substratos (bagaço 3,0%; farelo de trigo 3,0% e mistura
bagaço 1,5% e farelo de trigo 1,5%). Barra indicativa desvio-padrão.

56
Nota-se que tanto o isolado S87 como o controle A. awamori produziram
baixas concentrações de FPase (0,2 a 1,6 UI/g de substrato), assim como em
outros experimentos, também para outros isolados, observou-se reduzidas
concentrações de FPase. KROGH et al. (2004) em screening do gênero
Penicillium para produção de enzimas celulolíticas e xilanolíticas, comparando-
os com T. reesei RUT C-30 encontraram concentrações de FPase máximas na
faixa de 0,68 UI/mL. O autor pondera que fungos com baixa atividade FPase
devem conter enzimas específicas com propriedades interessantes, tais como
atividades específicas mais altas para enzimas únicas ou baixa inibição pelo
produto (repressão catabólica).
De acordo com HOLKER et al. (2004), as principais desvantagens de um
cultivo semi-sólido são principalmente devido a formação de gradientes de
temperatura, pH, concentração de substrato ou oxigênio dissolvido durante o
cultivo, as quais dificultam o controle sob quantidade limitada de água.
Melhorias relativas ao pré-tratamento do bagaço, à composição da
solução nutriente e o volume (nível de umidade) desta no cultivo semi-sólido
são pertinentes para continuidade dos estudos.

57
4.5 Identificação taxonômica
Os 5 fungos celulolíticos isolados, os quais, em experimento em frasco,
apresentaram concentrações enzimáticas expressivas, foram identificados
taxonomicamente, sendo 3 isolados a nível de espécie pertencentes ao gênero
Penicillium e 2 isolados a nível de gênero, espécie não confirmada.
Isolados Identificação
S21 Aspergillus sp. cf. phoenicis
S48
S67
S87
Penicillium angulare
Penicillium variabile
Coriolopsis sp. (basidiomiceto)
S93
Penicillium raistrickii
Tabela 10 - Identificação taxonômica de cinco fungos celulolíticos
O fungo basidiomiceto Coriolopsis sp. apresentou 99% de similaridade
de sequência com o fungo Coriolopsis polyzona.
O relatório da identificação taxonômica realizada pelo CPQBA da
Unicamp encontra-se no anexo 2.

58
5. CONCLUSÕES E PERSPECTIVAS
- Os resultados evidenciam o grande potencial da biodiversidade fúngica
existente em ambientes canavieiros.
- O método de seleção de morfotipos produtores de endoglucanases através da
revelação do halo de degradação com Vermelho Congo em meio sintético
acrescido de CMC (carboximetilcelulose), se mostrou eficiente sob o ponto de
vista qualitativo, mas não confere relação do tamanho do halo com
concentração enzimática.
- A variação do mesmo método com utilização de xilana para seleção de
morfotipos produtores de xilanase não foi consistente, diferindo da análise dos
isolados quando submetidos ao cultivo submerso e quantificação da enzima.
- O método de seleção de morfotipos lignolíticos em placa em meio com
guaiacol revelou resultados positivos, devendo ser confirmado por
quantificação das lignases em cultivo submerso em futuros trabalhos.
- O bagaço de cana-de-açúcar se mostrou um substrato adequado para o
cultivo de alguns fungos isolados e consequentemente um bom indutor de
celulases, em condições específicas de meio de cultura e forma de cultivo,
como por exemplo, o isolado S87 (Coriolopsis sp.) em cultivo submerso e meio
Breccia et al. modif (1995) com bagaço 3,0%, expressou melhor todas as
enzimas analisadas, fato econômica e ecologicamente importante para
utilização de resíduos da indústria sucroalcooleira.
- O meio semi-sólido com bagaço não se mostrou eficiente para indução das
enzimas dos isolados, nem mesmo do A. awamori.
- O isolado S67 (Penicillium variabile) produziu melhor concentração enzimática
de -glicosidase que o A. awamori, conhecidamente bom produtor desta
enzima, em diversas condições de cultivo e substratos (lactose, bagaço ou

59
farelo de trigo). Como em se tratando de uma cepa selvagem, pode-se inferir
que existem reais chances de melhoramento da produção desta enzima, em
trabalhos futuros.
- O isolado S93 (Penicillium raistrickii) também deve ser submetido a estudos
complementares e aprofundados, pois indicou boa expressão de xilanase,
melhor do que A. awamori e T. reesei em fermentação submersa com lactose.
- O estoque de fungos filamentosos em ágar (BDA) inclinado a 4ºC com
repiques a cada 6 meses, comprovou ser uma técnica eficiente para manter a
viabilidade e pureza das cepas.
- Um banco de 124 morfotipos celulolíticos foi obtido com este trabalho para
futuros estudos para obtenção de maior conhecimento sobre a biodiversidade e
as funções exercidas por estes microrganismos em ambientes terrestres e de
suas interações com fatores bióticos e abióticos dentro de um ecossistema, ou
ainda, estudos de hidrólise enzimática de resíduos lignocelulósicos para etanol
de 2ª geração ou outros usos das enzimas expressas por estes fungos na
biotecnologia.

60
6. REFERÊNCIAS BIBLIOGRÁFICAS
ABOU-ZEID, A. A. & EL- DIWANY, A. I. Cellulose-decomposing Fungi. Zbl.
Bakt. II. Abt., Bd. 133, S.647-656. 1978.
AGUIAR, C. L. & MENEZES, T. J. B. Produção de celulases e xilanase por
Aspergillus niger IZ, usando fermentação submersa sobre bagaço de cana de
açúcar. Boletim Centro de Pesquisa de Processamento de Alimentos, v.
18, p.57-70, 2000.
ASTM G21 – 96. Standard Practice for Determining Resistance of Synthetic
Polymeric Materials to Fungi. 2002.
BASSO, T. P., GALLO, C. R., BASSO, L. C. Atividade celulolítica de fungos
isolados de bagaço de cana de açúcar e madeira em decomposição. Pesq.
Agropec. Bras., Brasília, Vol. 45, n.11, p.1282-1289. 2010.
BÉGUIN, P. & AUBERT, J-P. The biological degradation of cellulose. FEMS
Microbiology Reviews Vol. 13, p. 25-58. 1994.
BELL, C. W., ACOSTA-MARTINEZ, V., McINTYRE, N. E,, COX, S., TISSUE, D.
T., ZAK, J. C. Linking microbial community structure and function to seasonal
differences in soil moisture and temperature in a Chihuahuan Desert Grassland.
Microb. Ecol. Vol. 58, p.827-842. 2009.
BHAT, M. K. & BHAT, S. Cellulose degrading enzymes and their potential
industrial applications. Biotechnology Advances. Vol. 15, Nº 3/4, p. 583-620.
1997.
BON, E. P. S., FERRARA, M. A., CORVO, M. L. Enzimas em Biotecnologia:
produção, aplicação e mercado. Ed. Interciência – Rio de Janeiro, RJ. 2008.
BONOMI, A. Workshop hidrólise de material lignocelulósico. 2007. Disponível em
www.apta.sp.gov.br/cana/.../Relatorio Acesso em 29 maio, 2010.
BORTOLAZZO, N. G. – Isolamento e seleção de fungos celulolíticos para
hidrólise enzimática do bagaço de cana-de-açúcar . Dissertação de mestrado.

61
Escola Superior de Agricultura ―Luiz de Queiroz‖ – Universidade de São Paulo,
Piracicaba, 2011.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding.
Anal. Biochem. Vol. 72, p. 248.1976.
BRECCIA, J. D., CASTRO, G. R., BAIGARI, M. D., SIÑERIZ, F. Screening of
xylanolitic bacteria using a colour plate method. J. Appl. Bacteriol. V. 78, p.
469-472. 1995.
CAMASSOLA, M. & DILLON, A. J. P. Production of cellulases and
hemicellulases by Penicillium echinulatum grown on pretreated sugar cane
bagasse and wheat bran in solid-state fermentation. Journal of Applied
Microbiology. Vol. 103, p.2196-2204. 2007.
CASTELLANI, A. Maintenance and cultivation of the common pathogenic fungi
of man in sterile distilled water. Further researches. J. Trop. Med. Hyg. Vol. 70,
p.181-184. 1967.
CARNEIRO, M. A. C. Características bioquímicas do solo em duas
cronossequências de reabilitação em áreas de mineração de bauxita. p.166.
Tese (Doutorado) - Departamento Ciência do Solo, Universidade Federal de
Lavras. 2000.
CAVALCANTI, M. A. & MAIA, L. C. Cellulolytic fungi isolated from an alluvial
soil in a semi-arid area of the northeast of Brazil. Rev. Microbiol. São Paulo,
Vol 25 (4), p. 251-4. 1994.
COMPANHIA NACIONAL DE ABASTECIMENTO – CONAB. Disponível em:
www.conab.gov.br. Acesso em 29 mai. 2010.
COWLING, E. B. & KIRK, T. K. Properties of cellulose and lignocellulose
materials and substrats for enzimatic conversion processes. Biotechnology
and Bioengineering Symposium. Vol.6, p. 95-123. 1976.

62
DASHTBAN, M., SCHRAFT, H., QIN, W. – Fungal bioconversion of
lignocellulosic residuos; opportunities & perspectives. International Journal of
Biological Sciences. Vol. 5 (6), p. 578-595. 2009.
EMBRAPA Informação Tecnológica. Plano Nacional de Agroenergia 2006-
2011. Ministério da Agricultura, Pecuária e Abastecimento, Secretaria de
Produção e Agroenergia. 2. ed. rev. - Brasília, DF:, 110p. 2006.
FILHO, J. M. M. A. Análise enzimática de fungos lignocelulolíticos cultivados
em vinhaça e bagaço de cana-de-açúcar. Dissertação de Mestrado. Escola
Superior de Agricultura Luiz de Queiroz – Universidade de São Paulo,
Piracicaba, 2008.
GARCIA-KIRCHNER, O., SEGURA-GRANADOS, M., RODRÍGUEZ-PASCUAL,
P. Effect of media composition and growth conditions on production of -
glucosidase by Aspergillus niger C-6. Applied Biochemistry and
Biotechnology. Vol. 121-124, p. 347-360. 2005.
GAUTAM, S. P., BUNDELA, P. S., PANDEY, A. K., KHAN, J., AWASTHI, M. K.,
SARSAIYA, S. Optimization for the production of cellulose enzyme from
municipal solid waste residue by two novel cellulolytic fungi. Biotechnology
Research International. Vol. 2011, 8 pages. 2010.
GHOSE, T. K. Measurement of cellulose activities. Pure & Appl. Chem. Vol.
59(2), p. 257-268. 1987.
GODDEN, B. & PENNINCKX, M. J. Identification and evolution of the cellulolytic
microflora present during composting of cattle manure: on the role of
actinomycetes sp. Ann. Microbiol. Vol. 135B, p. 69-78. 1984.
GOES, T. A energia que vem da cana-de-açúcar. Artigo disponível em
www.embrapa.com.br Acessado em 29 mai. 2010.
GOMEZ, L. D., STEELE-KING, C. G., MCQUEEN-MASON, S. J. – Sustainable
liquid biofuels from biomass: the writing’s on the walls. New phytologist. Vol.
178, p. 473-485. 2008.

63
HAHN-HÄGERDAL, B., LINDÉN, T., SENAC, T., SKOOG, K. Ethanolic
fermentation of pentoses in lignocellulose hydrolysates. Appl Biochem
Biotechnol 28/29:131–141. 1991.
HESSELTINE, C. W. & HAYNES, W. C. Sources and management of micro-
organisms for the development of a fermentation industry. Prog. Ind.
Microbiol. Vol. 12 (0), p. 1-46. 1973.
HIMMEL, M. E., DING, S. –Y., JOHNSON, D. K., ADNEY, W. S., NIMLOS, M.
R., BRADY, J. W., FOUST, T.D. Biomass recalcitrance: Engineering plants and
enzymes for biofuels production. Science. Vol. 315. 2007.
HOLKER, U., HOFER, M., LENZ, J. Biotechnological advantages of laboratory-
scale solid-state fermentation with fungi. Appl. Microbiol Biotechnol. Vol 64,
p.175-186. 2004.
JAHANGEER, S., KHAN, N., JAHANGER, S., SOHAIL, M., SHAHZAD, S.,
AHMED, A., KHAN, S.A. Screening and characterization of fungal cellulases
isolated the native environmental source. Pak. J. Bot. Vol. 37 (3), p. 739-748.
2005.
KANG, S. W., PARK, Y. S., LEE, J. S., HONG, S. I., KIM, S. W. Production of
cellulases and hemicellulases by Aspergillus niger KK2 from lignocellulosic
biomass. Bioresource Technology. Vol. 91, p.153-156. 2004.
KHALID, M., YANG, W., KISHWAR, N., RAJPUT, Z. I., ARIJO, A. G. Study of
cellulolytic soil fungi and two nova species and new medium. Journal of
Zhejiang University Science B. Vol. 7(6), p.459-466. 2006.
KHAN, M. H., ALI, S., FAKHRU’L-RAZI, A., ALAM, Z. Use of fungi for the
bioconversion of rice straw into cellulase enzyme. Journal of Environmental
Science and Health, Part B: pesticides, Food Contaminants, and
Agricultural Wastes. Vol. 42, p. 381-386. 2007.
KIM, Y. S., JUNG, H. C., PAN, J. G. Bacterial cell surface display of an enzyme
library for selective screening of improved cellulose variants. Applied
Environmental Microbiology. Vol. 66, p. 788-93. 2000.

64
KROGH, K. B. R., MØRKEBER, A., JØRGENSEN, H., FRISVAD, J. C.,
OLSSON, L. Screening genus Penicillium for products of cellulolytic and
xylanolytic enzymes. Applied Biochemistry and Biotechnology. Vol. 113-
116, p. 389. 2004.
KUMAR, R., SINGH, S., SINGH, O. V. – Bioconversion of lignocellulosic
biomass: biochemical and molecular perspectives. J. Ind. Microbiol.
Biotechnol. Vol. 35, p. 377-391. 2008.
LATIFIAN, M., HAMIDI-ESFAHANI, Z., BARZEGAR, M. Evaluation of culture
conditions for cellulose production by two Trichoderma reesei mutants under
solid-state fermentation conditions. Bioresource Technology. Vol. 98, p. 3634-
3637. 2007.
LIN, Y. & TANAKA, S. Ethanol fermentation from biomass resources: current
state and prospects. Appl Microbiol Biotechnol. Vol. 69, p. 627-642. 2006.
LYND, L. R., WEIMER, P. J., ZYL, W. H., PRETORIUS, I. S. Microbial cellulose
utilization: Fundamentals and Biotechnology. Microb. Molec. Biol. Rewies.
Vol. 66 (3), p. 506-577. 2002.
MANDELS, M. & WEBER, J. Production of cellulases. Adv. Chem. Ser. Vol.
95, p. 391-414. 1969.
MASSADEH, M. I., YUSOFF, W. M. W., OMAR, O., KADER, J. Synergism of
cellulose enzymes in mixed culture solid substrate fermentation.
Biotechnology Letters. Vol. 23, p.1771-1774. 2001.
MILLER, L. Use of a dinitrosalicylic acid reagent for determination of reducing
sugar. Anal. Chem. Vol. 31, p. 426-428. 1959.
MULLINGS, R. Measurement of saccharification by cellulases. Enzyme and
Microbial Technology. Vol. 7, p. 586-591. 1985.
MUTHUVELAYUDHAM, R. & VIRUTHAGIRI, T. Fermentative production and
kinetics of cellulose protein on Trichoderma reesei using sugarcane bagasse
and rice straw. African Journal of Biotechnology. Vol. 5, p. 1873-1881. 2006.

65
NOGUEIRA, E. B. S. & CAVALCANTI, M. A. Q. Cellulolytic fungi isolated from
processed oats. Revista de Microbiologia. Vol. 27, p.7-9. 1996.
OKINO, L. K., MACHADO, K. M. G., FABRIC, C., BONOMI, V. L. R. Ligninolytic
activity of tropical rainforest basidiomycetes. World J. of Microbiol. Biotech.
vol. 16, p. 889-893. 2000.
PANDEY, A., SOCCOL, C. R., NIGAM, P., SOCCOL, V. T., VANDENBERGHE,
L. P. S., MOHAN, R. Biotechnological potential of agro-industrial residues. II:
cassava bagasse. Bioresource Technology, Amsterdam, vol. 74, p. 81-87.
2000.
PRASAD, S., SINGH, A., JOSHI, H. C. Ethanol as an alternative fuel from
agricultural, industrial and urban residues. Resources, Conservation and
recycling. Vol. 50, p.1-39. 2007.
RAGAUSKAS, A. J., WILLIAMS, C. K., DAVISON, B. H., BRITOVSEK, G.,
CAIRNEY, J., ECKERT, C. A., FREDERICK Jr., W. J., HALLETT, J. P., LEAK,
D. J., LIOTTA, C. L., MIELENZ, J. R., MURPHY, R., TEMPLER, R.,
TSCHAPLINSKI, T. The Path Forward for Biofuels and Biomaterials - Science
27 January Vol 311. 2006.
RECZEY, K., SZENGYEL, Z. S., EKLUND, R., ZACCHI, G. Cellulase
production by T. reesei. Biores Technol. Vol. 57, p. 25-30. 1996.
RIDELL, R. W. Permanent stained mycological preparation obtained by slide
culture. Mycologia. Vol. 42, p. 265-270. 1950.
RUEGGER, M. J. S., TAUK-TORNISIELO, S. M. Atividade da celulase de
fungos isolados do solo da Estação Ecológica de Juréia-Itatins, São Paulo,
Brasil. Revista Brasileira de Botânica. Vol. 27, nº2, p.205-211. 2004.
SCHWARZ, W. The cellulosome and cellulose degradation by anaerobic
bactéria. Applied Microbiology and Biotechnology. Vol. 56 (5-6), p.634-649.
2001.
SILVA, A. S., WEBB, C., FILHO, E. X. F., BON, E. P. S., ELEUTHERIO, E. C.
A., GOTTSCHALK, L. M. F., MOREIRA, L. R. S., PEREIRA, M. D., FERRARA,

66
M. A., TEIXEIRA, R. S. S. Composição de enzimas, uso da composição na
hidrólise enzimática de material lignocelulósico, processo de produção de
enzimas que degradam a fração de polissacarídeos da biomassa, processo de
produção de álcool utilizando a composição de enzimas. Patente publicada em
2009 sob o nº PI0705744-OA2.
SINGHANIA, R. R., SUKUMARAN, R. K., PANDEY, A. Improved cellulose
production by Trichoderma reesei RUT C-30 under SSF through process
optimization. Applied Biochemistry and Biotechnology. V. 142, p.60-70.
2007.
SOCCOL, C. R. & VANDENBERGHE, L. P. S. Overview of applied solid-stae
fermentation in Brazil. Biochemical Engineering Journal. Vol. 13, p.205-218.
2003.
STICKLEN, M. B. – Plant genetic engineering for biofuel production: towards
affordable cellulosic ethanol. Nature Reviews. Vol. 9, p. 433 – 443. 2008.
SUKUMARAN, R. K., SINGHANIA, R. R., PANDEY, A. Microbial cellulases –
Production, applications and challenges. Journal of Scientific & Industrial
Research. Vol. 64, p. 832-844. 2005.
TEATHER, R. M. & WOOD, P. J. Use of Congo red-polysaccharide interactions
in enumeration and characterization of cellulolytic bacteria from the bovine
rumen. Applied and Environmental Microbiology. Vol. 43, p.777-780. 1982.
VALASKOVÁ, V. & BALDRIAN, P. Estimation of bound and free fractions of
lignocellulose-degrading enzymes of wood-rotting fungi Pleurotus ostreatus,
Trametes versicolor and Pictoporus betulinus. Research in Microbiology. Vol.
157(2), p.119-124. 2006.
VANCE, E. D. & CHAPIN III, F. S. Substract limitations to microbial activity in
taiga forest floors. Soil Biology & Biochemistry. Vol 33, p.173-188. 2001.
VARDAVAKIS, E. Seasonal fluctuations of aerobic cellulolytic bacteria, and
cellulose and respiratory activities in a soil profile under a forest. Plant soil. Vol.
115, p.145-150. 1989.

67
WANG, T., LIU, X., YU, Q., ZHANG, X., QU, Y., GAO, P. Direct evolution for
engineering pH profile of endoglucanase III from Trichoderma reesei. Biomol
Eng. Vol. 22, p. 89-94. 2005.
WARZWODA, M., BALLERINI, D., MONOT, F. Processo de produção de
enzimas celulolíticas e hemicelulolíticas, utilizando os resíduos de destilação de
fermentação etanólica de hidrolisados enzimáticos de materiais (ligno)-
celulósicos. Patente depositada em 2006 sob o nº PI0600409-1ª.
WYMAN, C. E. Ethanol from lignocellulosic biomass — technology, economics,
and opportunities. Biores Technol. Vol. 50(1), p.16. 1994.
WYMAN, C. E. Cellulosic biomass could help meet California's transportation
fuel needs. California Agriculture. Vol. 63(4), p.185-190. 2009.
ZHANG, Y. H. P., HIMMEL, M. E., MIELENZ, J. R. Outlook for cellulose
improvement: Screening and selection strategies. Biotechnology Advances.
Vol. 24, p.452-481. 2006.

68
APÊNDICE A - Resumo esquemático do ensaio da atividade CMCase
TUBO BRANCO 0,250 mL tampão citrato + 0,250 mL substrato CMC
TUBOS TEMPO ZERO (T0) 0,250 mL tampão citrato + 0,250 mL amostra enzima
TUBOS AMOSTRA (T10) 0,250 mL substrato CMC + 0,250 mL amostra enzima REAÇÃO ENZIMÁTICA 50ºC / 10 MINUTOS
TODOS OS TUBOS + 0,500 mL reagente DNS
REAÇÃO DNS 100ºC / 5 MINUTOS
BANHO DE GELO
TODOS OS TUBOS + 6,5 mL água destilada
ESPECTROFOTÔMETRO A540nm

69
APÊNDICE B - Resumo esquemático do ensaio da atividade FPase
TUBO BRANCO 0,500 mL tampão citrato
TUBOS TEMPO ZERO (T0) 1,0 mL tampão citrato + 0,500 mL amostra enzima
TUBOS AMOSTRA (T60) PAPEL FILTRO + 1,0 mL tampão citrato +
0,500 mL amostra enzima REAÇÃO ENZIMÁTICA 50ºC / 60 MINUTOS
Transferir dos TUBOS AMOSTRA (T60) para outros tubos de ensaio
0,500 mL
TODOS OS TUBOS + 0,500 mL reagente DNS
REAÇÃO DNS 100ºC / 5 MINUTOS
BANHO DE GELO
TODOS OS TUBOS + 6,5 mL água destilada
ESPECTROFOTÔMETRO A540nm

70
APÊNDICE C - Resumo esquemático do ensaio da atividade β- glicosidase
TUBOS TEMPO ZERO (T0) 0,500 mL tampão citrato + Bioanalisador ou Kit GOD-POD 0,500 mL amostra enzima
TUBOS AMOSTRA (T30) 0,500 mL substrato celobiose
+ 0,500 mL amostra enzima REAÇÃO ENZIMÁTICA 50ºC / 30 MINUTOS FINALIZAR A REAÇÃO 100ºC / 5 MINUTOS TODOS OS TUBOS GELO DETERMINAÇÃO DE GLICOSE PELO BIOANALISADOR DIRETA
OU DETERMINAÇÃO DE GLICOSE PELO KIT GOD-POD
TUBO BRANCO 1,0 mL reagente GOD-POD
TUBOS PADRÃO 1,0 mL do reagente GOD-POD +
10 µl do padrão GOD-POD
TUBOS TEMPO ZERO (T0) 1,0 ml do reagente GOD-POD +
10 µl da mistura reacional T0
TUBOS AMOSTRA (T30) 1,0 ml do reagente GOD-POD +
10 µl da mistura reacional T30 REAÇÃO GOD-POD TEMP. AMBIENTE / 10 MINUTOS
ESPECTROFOTÔMETRO A500nm

71
APÊNDICE D - Resumo esquemático do ensaio da atividade Xilanase
TUBO BRANCO 0,150 mL tampão acetato
TUBOS BRANCO Substrato 0,050 mL tampão acetato + 0,100 mL substrato Xilana
TUBOS TEMPO ZERO (T0) 0,100 mL tampão acetato + 0,050 mL amostra enzima
TUBOS AMOSTRA (T30) 0,100 mL substrato Xilana + 0,050 mL amostra enzima REAÇÃO ENZIMÁTICA 50ºC / 30 MINUTOS
TODOS OS TUBOS + 0,300 mL reagente DNS
REAÇÃO DNS 100ºC / 10 MINUTOS
BANHO DE GELO
TODOS OS TUBOS + 1,5 mL água destilada
ESPECTROFOTÔMETRO A540nm

72
APÊNDICE E – Visualização da morfologia de alguns isolados ao microscópio
óptico Olympus modelo BX41 (aumento 1000x). Câmera fotográfica Canon
modelo EOS 5D digital Lentes EF 24-70mm e/ou câmera Sony DSC-W310.
Coloração com azul de metileno 0,02 g/L.
S123
S67 P. variabile
S15 S93 P. raistrickii
A.awamori A.awamori
S21 Aspergillus sp.
S48 P. angulare

73
APÊNDICE F – Estoque dos fungos
BDA inclinado (4ºC) Castellani (1969) - Submersos em água (4ºC)
APÊNDICE H – Indicação de presença de enzimas lignolíticas (lacase) por oxidação
do Guaiacol.
S71 S42
S09 S29
S87

74
APÊNDICE G - Atividade celulolítica (IE) dos fungos filamentosos isolados, cultivados
em meio sintético com CMC 1% durante 3 a 4 dias. Øh = diâmetro do halo (mm); Øc =
diâmetro da colônia (mm); IE = Índice enzimático.
Isolados Ponto (coleta) Øh Øc IE
S01 P01 (1ª) 21 10 3,10 S02 P01 (1ª) 8 5 2,60 S03 P01 (1ª) 13 7 2,86 S04 P01 (1ª) 45 25 2,80 S05 P01 (1ª) 12 3 5,00 S06 P01 (1ª) 10 6 2,67 S07 P01 (1ª) 10 8 2,25 S08 P01 (1ª) 10 8 2,25 S09 P01 (1ª) 7 3 3,33 S10 P01 (1ª) 10 8 2,25 S11 P01 (1ª) 9 6 2,50 S12 P01 (1ª) 5 2 3,50 S13 P02 (1ª) 17 9 2,89 S14 P02 (1ª) 18 9 3,00 S15 P02 (1ª) 20 7 3,86 S16 P02 (1ª) 24 7 4,43 S17 P02 (1ª) 18 9 3,00 S18 P02 (1ª) 18 9 3,00 S19 P02 (1ª) 20 7 3,86 S20 P02 (1ª) 14 5 3,80 S21 P02 (1ª) 15 5 4,00 S22 P02 (1ª) 7 5 2,40 S23 P02 (1ª) 13 6 3,17 S24 P02 (1ª) 20 14 2,43 S25 P02 (1ª) 12 3 5,00 S26 P02 (1ª) 14 4 4,50 S27 P02 (1ª) 8 6 2,33 S28 P02 (1ª) 11 5 3,20 S29 P02 (1ª) 17 8 3,13 S30 P02 (1ª) 10 5 3,00 S31 P02 (1ª) 10 5 3,00 S32 P02 (1ª) 9 5 2,80 S33 P03 (1ª) 7 4 2,75 S34 P03 (1ª) 10 6 2,67 S35 P03 (1ª) 16 10 2,60 S36 P03 (1ª) 9 4 3,25 S37 P03 (1ª) 9 5 2,80 S38 P03 (1ª) 7 4 2,75 S39 P03 (1ª) 12 8 2,50 S40 P03 (1ª) 15 5 4,00 S41 P04 (1ª) 13 4 4,25 S42 P04 (1ª) 15 4 4,75 S43 P04 (1ª) 17 9 2,89 S44 P04 (1ª) 10 7 2,43 S45 P04 (1ª) 13 11 2,18
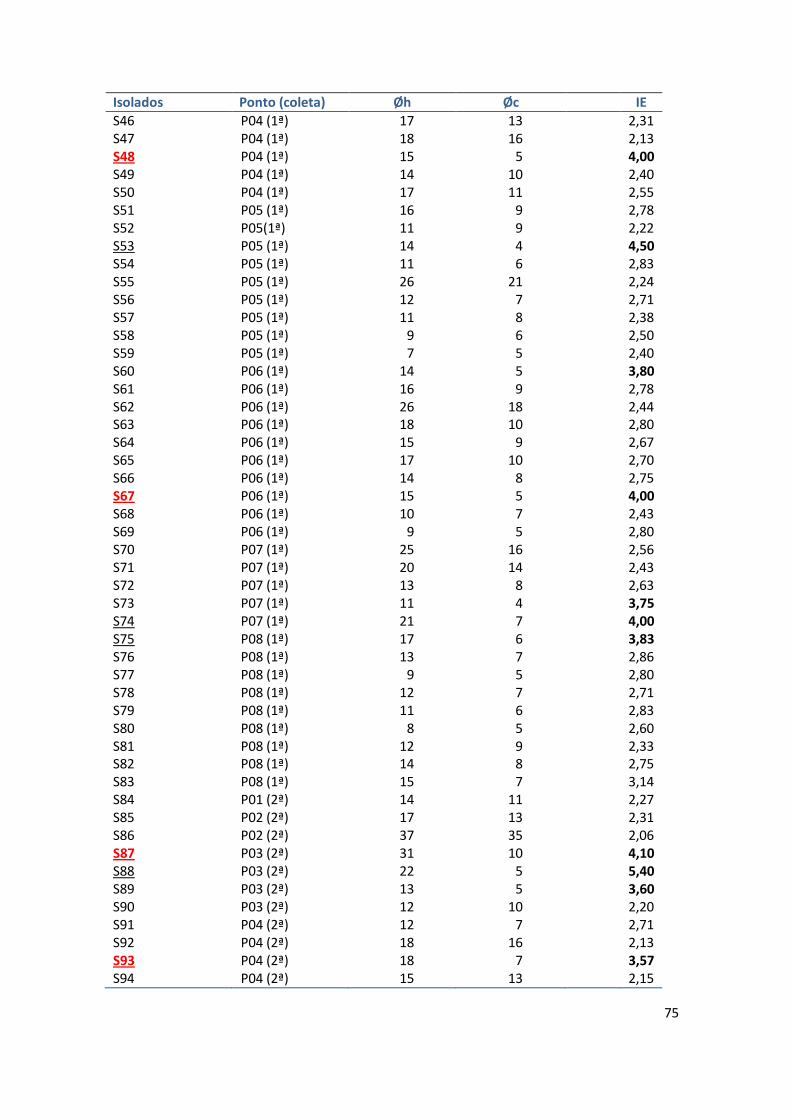
75
Isolados Ponto (coleta) Øh Øc IE
S46 P04 (1ª) 17 13 2,31 S47 P04 (1ª) 18 16 2,13 S48 P04 (1ª) 15 5 4,00 S49 P04 (1ª) 14 10 2,40 S50 P04 (1ª) 17 11 2,55 S51 P05 (1ª) 16 9 2,78 S52 P05(1ª) 11 9 2,22 S53 P05 (1ª) 14 4 4,50 S54 P05 (1ª) 11 6 2,83 S55 P05 (1ª) 26 21 2,24 S56 P05 (1ª) 12 7 2,71 S57 P05 (1ª) 11 8 2,38 S58 P05 (1ª) 9 6 2,50 S59 P05 (1ª) 7 5 2,40 S60 P06 (1ª) 14 5 3,80 S61 P06 (1ª) 16 9 2,78 S62 P06 (1ª) 26 18 2,44 S63 P06 (1ª) 18 10 2,80 S64 P06 (1ª) 15 9 2,67 S65 P06 (1ª) 17 10 2,70 S66 P06 (1ª) 14 8 2,75 S67 P06 (1ª) 15 5 4,00 S68 P06 (1ª) 10 7 2,43 S69 P06 (1ª) 9 5 2,80 S70 P07 (1ª) 25 16 2,56 S71 P07 (1ª) 20 14 2,43 S72 P07 (1ª) 13 8 2,63 S73 P07 (1ª) 11 4 3,75 S74 P07 (1ª) 21 7 4,00 S75 P08 (1ª) 17 6 3,83 S76 P08 (1ª) 13 7 2,86 S77 P08 (1ª) 9 5 2,80 S78 P08 (1ª) 12 7 2,71 S79 P08 (1ª) 11 6 2,83 S80 P08 (1ª) 8 5 2,60 S81 P08 (1ª) 12 9 2,33 S82 P08 (1ª) 14 8 2,75 S83 P08 (1ª) 15 7 3,14 S84 P01 (2ª) 14 11 2,27 S85 P02 (2ª) 17 13 2,31 S86 P02 (2ª) 37 35 2,06 S87 P03 (2ª) 31 10 4,10 S88 P03 (2ª) 22 5 5,40 S89 P03 (2ª) 13 5 3,60 S90 P03 (2ª) 12 10 2,20 S91 P04 (2ª) 12 7 2,71 S92 P04 (2ª) 18 16 2,13 S93 P04 (2ª) 18 7 3,57 S94 P04 (2ª) 15 13 2,15

76
Isolados Ponto (coleta) Øh Øc IE
S95 P04 (2ª) 10 5 3,00 S96 P04 (2ª) 20 5 5,00 S97 P05 (2ª) 11 7 2,57 S98 P05 (2ª) 12 6 3,00 S99 P05 (2ª) 7 5 2,40 S100 P05 (2ª) 10 5 3,00 S101 P05 (2ª) 12 5 3,40 S102 P05 (2ª) 8 5 2,60 S103 P05 (2ª) 14 6 3,33 S104 P05 (2ª) 44 39 2,13 S105 P05 (2ª) 12 5 3,40 S106 P05 (2ª) 20 5 5,00 S107 P05 (2ª) 15 8 2,88 S108 P05 (2ª) 6 4 2,50 S109 P05 (2ª) 5 4 2,25 S110 P06 (2ª) 11 7 2,57 S111 P06 (2ª) 10 5 3,00 S112 P06 (2ª) 11 7 2,57 S113 P06 (2ª) 8 6 2,33 S114 P06 (2ª) 13 7 2,86 S115 P06 (2ª) 6 5 2,20 S116 P07 (2ª) 15 13 2,15 S117 P07 (2ª) 20 17 2,18 S118 P07 (2ª) 15 9 2,67 S119 P08 (2ª) 16 9 2,78 S120 P08 (2ª) 35 33 2,06 S121 P08 (2ª) 13 6 3,17 S122 P08 (2ª) 9 7 2,29 S123 P08 (2ª) 24 5 5,80 S124 T. reesei A. awamori
P08 (2ª) Referência Referência
10 13 16
3 7 7
4,33 2,86 3,2

77
APÊNDICE J - Dados para estatística
CMCase Fpase -glicosidase xilanase
Amostras (UI/mL) PT (mg/mL) (UI/mL) PT (mg/mL) (UI/mL) PT (mg/mL) (UI/mL) PT (mg/mL)
S05 1,986 0,219 0,371 0,205 0,0150 0,219 2,519 0,219
S05 1,886 0,219 0,289 0,205 0,0104 0,219 2,574 0,219
S15 0,704 0,053 0,163 0,064 0,0127 0,064 0,500 0,064
S15 0,754 0,053 0,157 0,064 0,0116 0,064 0,472 0,064
S16 1,509 0,05 0,126 0,082 0,0185 0,082 0,219 0,082
S16 1,710 0,05 0,195 0,082 0,0150 0,082 0,205 0,082
S19 2,816 0,064 0,069 0,054 0,0173 0,064 0,329 0,054
S19 2,841 0,064 0,119 0,054 0,0104 0,064 0,246 0,054
S21 2,967 0,083 0,509 0,076 0,1573 0,083 2,690 0,594
S21 2,866 0,083 0,522 0,076 0,1583 0,083 2,785 0,594
S25 1,056 0,06 0,038 0,06 0,0104 0,06 0,465 0,056
S25 1,257 0,06 0,063 0,06 0,0127 0,06 0,329 0,056
S26 1,358 0,053 0,465 0,214 0,0173 0,214 0,657 0,214
S26 1,760 0,053 0,427 0,214 0,0139 0,214 0,698 0,214
S40 1,659 0,075 0,088 0,075 0,0112 0,075 0,596 0,076
S40 1,710 0,075 0,107 0,075 0,0134 0,075 0,637 0,076
S48 2,313 0,077 0,698 0,077 0,1450 0,079 1,506 0,077
S48 2,464 0,077 0,723 0,077 0,1490 0,079 2,471 0,077
S53 0,955 0,046 0,157 0,084 0,0145 0,046 0,151 0,046
S53 0,805 0,046 0,189 0,084 0,0156 0,046 0,192 0,046
S67 1,609 0,077 0,515 0,071 1,4543 0,072 0,671 0,071
S67 1,609 0,077 0,515 0,071 1,2168 0,072 0,726 0,071
S74 2,263 0,052 0,239 0,058 0,0078 0,052 0,370 0,058
S74 2,263 0,052 0,264 0,058 0,0123 0,052 0,383 0,058
S75 0,955 0,059 0,013 0,059 0,0173 0,059 0,534 0,059
S75 0,981 0,059 0,050 0,059 0,0162 0,059 0,589 0,059
S87 7,794 0,077 0,289 0,079 0,1250 0,131 2,677 0,134
S87 7,995 0,077 0,314 0,079 0,1390 0,131 2,656 0,134
S88 3,822 0,051 0,157 0,063 0,0243 0,063 0,507 0,063
S88 3,721 0,051 0,145 0,063 0,0220 0,063 0,534 0,063
S93 2,665 0,057 0,660 0,062 0,1330 0,043 5,237 0,062
S93 2,715 0,057 0,666 0,062 0,1350 0,043 5,196 0,062
S96 1,861 0,045 0,182 0,058 0,0173 0,058 2,341 0,058
S96 2,011 0,045 0,214 0,058 0,0173 0,058 2,273 0,058
S106 2,263 0,059 0,145 0,059 0,0162 0,065 0,698 0,065
S106 2,112 0,059 0,195 0,059 0,0150 0,065 0,739 0,065
S123 1,609 0,061 0,132 0,08 0,0123 0,061 0,753 0,08
S123 1,861 0,061 0,126 0,08 0,0123 0,061 0,835 0,08
A.awamori 0,955 0,054 0,233 0,036 0,3130 0,090 1,711 0,067
A.awamori 1,106 0,054 0,201 0,036 0,3300 0,090 2,368 0,067
T. reesei 8,6732 0,3890 1,8516 0,3890 0,0164 0,5490 4,4936 0,5490
T. reesei 9,5178 0,3890 1,8678 0,3890 0,0154 0,5490 4,6992 0,5490

78
ANEXO 1 – Autorização IBAMA CGEN e mapa do local de coleta

79

80
Visualização Google Earth – Fazenda São Lucas, Distrito de São Roberto de
Minas, Município de São João da Lagoa, MG.
S 16º44’16,2” W 44º12’36,5”
ANEXO 2 – Relatório CPQBA Identificação taxonômica
Fazenda São Lucas

81
Mapa da Fazenda São Lucas
Regiões de seleção de locais de coleta circuladas em vermelho

82
ANEXO 2 – Relatório CPQBA Identificação taxonômica

83
NOTA: A amostra identificada como Aspergillus tubingensis não faz parte
deste trabalho de dissertação.

84

85

86

87

88

89

90

91

92

93

94

95

96