impacto da administraÇÃo intranasal de ocitocina no
TRANSCRIPT

UNIVERSIDADE PRESBITERIANA MACKENZIE
JULIA HORTA TABOSA DO EGITO
IMPACTO DA ADMINISTRAÇÃO INTRANASAL DE OCITOCINA NO
COMPORTAMENTO SOCIAL DE EMPATIA À DOR E PERCEPÇÃO DE
AMEAÇA
São Paulo 2018

JULIA HORTA TABOSA DO EGITO
IMPACTO DA ADMINISTRAÇÃO INTRANASAL DE OCITOCINA NO
COMPORTAMENTO SOCIAL DE EMPATIA À DOR E PERCEPÇÃO DE
AMEAÇA
Tese apresentada ao Programa de Pós-Graduação em Distúrbios do Desenvolvimento da Universidade Presbiteriana Mackenzie como requisito para à obtenção do título de Doutora em Distúrbios do Desenvolvimento.
Orientadora: Prof.a Dr.a Ana Alexandra Caldas Osório Co-orientador: Prof. Dr. Paulo Sérgio Boggio
São Paulo 2018



AGRADECIMENTOS
Aos meus pais Ana Lúcia de Moraes Horta Tabosa do Egito e Eryberto Steves
Tabosa do Egito, minha avó Maria de Lourdes Moraes Horta, à minha segunda mãe
Ana Maria da Cruz de Oliveira e ao meu querido namorado Adalto Romualdo pelo
constante apoio e carinho que torna nossa união familiar mais íntima e respeitosa a
cada dia e por me incentivarem a realizar meus sonhos.
À minha orientadora, Prof.a Dra. Ana Alexandra Caldas Osório, agradeço a
confiança, diversas oportunidades e o suporte constante que me permitiram
amadurecer pessoal e profissionalmente. Ao co-orientador deste trabalho, Prof. Dr.
Paulo Sérgio Boggio, que me orientou desde o início de minha trajetória e me ensinou
a ter um rigor científico. A Prof.a Dra. Simone Shamay-Tsoory agradeço pela
experiência de ter frequentado seu laboratório e aprendido tanto com suas orientações
e equipes.
A toda equipe do Laboratório de Neurociência Cognitiva e Social que por anos
trabalhamos juntos e de onde guardo lembranças especiais e significativas amizades.
Em especial, aos alunos Gabrielle Ferreira, Manoela Gonçalves, Ana Cláudia Freire,
Merikmar Lomar, Fernando da Silva Reis, Gabriela Pezzatti, Marianne Cremonesi,
Tiago Cortés, Isabella Quilici, Luiza Mendes e Mayara Barros que me ofereceram
todo o suporte no processo de coleta de dados e que tornaram o dia-a-dia mais leve
diante de tantos desafios e aprendizados. À Samara Dina, que produziu e editou os
estímulos apresentados na tarefa de avaliação explícita de empatia à dor física. À
Ruth Lyra pelo auxílio em diversos momentos críticos desse estudo.
À secretária do programa de Pós-Graduação Dani Gomes que pacientemente
me instruiu e apoiou.
Às Doutoras Olivia Morgan Lapenta e Cláudia Valasek por toda a
disponibilidade e auxílio referente à técnica de Estimulação Magnética Transcraniana.
Ao Prof. Dr. Eder de Carvalho Pincinato, por disponibilizar um espaço no
Laboratório de Análises Clínicas da Universidade Presbiteriana Mackenzie que
permitiu manter em condições adequadas os materiais das coletas de dados. Ao
Técnico Bruno Silva do Laboratório de Bioquímica, pelo suporte em preparar as
monodoses utilizadas desse estudo. Ao Técnico Murilo Corrêa pelo auxílio logístico
no Laboratório de Análises Clínicas.

Aos técnicos em programação, Rani Amit Bar-On, e em estatística, Michael
Nevat, membros do Social and Affective Neuroscience Laboratory na Universidade
de Haifa (Israel) que me auxiliaram incansavelmente a organizar os dados referentes
aos Potenciais Evocados Motores e a analisar os dados desse projeto.
À minha prima Lorraine Egito, pelas trocas constantes durante essa fase e por
ter me ajudado a tornar mais clara a minha apresentação.
À Prof.a Dra. Miriam Oliveira Ribeiro e ao Prof. Dr. Óscar Gonçalves, pelas
importantes contribuições na etapa de qualificação. Agradeço ainda a todos os
membros da banca pela generosidade da leitura do trabalho final e pelas relevantes
considerações.
Por fim, agradeço a todos os colaboradores de pesquisa que contribuíram para
o desenvolvimento desse trabalho.

“Ninguém nasce odiando outra pessoa pela cor de sua pele, por sua origem ou ainda
por sua religião. Para odiar, as pessoas precisam aprender; e se podem aprender a
odiar podem ser ensinadas a amar”.
Nelson Mandela

Apoio:
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES
Fundação de Amparo à Pesquisa do Estado de São Paulo - FAPESP
- Número do processo do presente estudo: 2016/17242-5 - Estudo vinculado ao processo: 2014/06777-0 (Programa
Jovens Pesquisadores em Centros Emergentes)

RESUMO
O papel da ocitocina (OT) enquanto promotora indiscriminada de cognição social e
prosocialidade tem sido questionado por resultados que indicam alguns efeitos
antissociais, como inveja e etnocentrismo. Assim, o objetivo do presente trabalho foi
avaliar na mesma amostra o impacto da administração intranasal de OT em
comportamentos pró-sociais como, empatia à dor no outro, e anti-sociais como,
percepção de ameaça. Para incluir a dimensão de pertencimento ao grupo, avaliamos
as respostas de participantes autodeclarados Brancos diante de alvos membros do
endogrupo (Branco) e do exogrupo (Negro). Tratou-se de estudo duplo-cego, placebo-
controlado, randomizado e inter-sujeitos, no qual participaram 107 estudantes
universitários, do sexo masculino, Brancos e destros. No primeiro dia de coleta, foram
verificados os critérios de exclusão e administrados questionários para a avaliação de
empatia e viés racial. No segundo dia de coleta, os participantes auto-administraram
uma das soluções distribuídas de maneira randomizada (OT ou placebo [PL]). Após
uma espera de 40 minutos, foram administradas tarefas computadorizadas de empatia
explícita à dor (avaliação da intensidade de dor percebida durante observação de mãos
Brancas, Negras e Roxas – controle de cor – em situações neutras ou potencialmente
dolorosas) e percepção de ameaça (identificação de homens Brancos e Negros
armados em contextos sociais). No terceiro dia de coleta, os participantes inalaram a
mesma substância e foram submetidos a avaliação de empatia implícita à dor (durante
observação de vídeos de mãos Brancas, Negras e Roxas sendo tocadas por um
cotonete ou penetradas por uma seringa). Durante essa última fase, foram obtidos
registros de potencial evocado motor na mão direita, enquanto a bobina da
Estimulação Magnética Transcraniana era posicionada sobre o córtex motor primário
no hemisfério esquerdo do participante. Quanto à tarefa de percepção de ameaça,
resultados da ANOVA mista revelaram uma interação significativa para Etnia x
Grupo, F(1,105)=5.44, p=.022, η2=.049, indicando que o grupo PL apresentou maior
taxa de Rejeição Correta (abstendo-se de atirar em alvos desarmados) para alvos
Brancos em comparação aos Negros, enquanto as taxas de rejeição correta para alvos
Brancos e Negros foram equivalentes no grupo OT. Assim, a ocitocina reduziu os
falsos alarmes para os alvos Negros via uma melhora da discriminação entre alvos
armados ou não, reduzindo a percepção de ameaça diante do exogrupo. Em relação à

empatia explícita à dor, resultados da ANOVA mista revelaram uma atribuição de dor
de maior intensidade em contextos dolorosos com alvos Negros em comparação aos
alvos Brancos. Implicitamente, os níveis de contágio sensóriomotor diante de alvos
Brancos e Negros foram equivalentes. Em síntese, resultados da nossa amostra
Brasileira apontam para um efeito prosocial da OT durante avaliação de percepção de
ameaça especificamente para o exogrupo, mas nenhum efeito dessa substância nas
avaliações de empatia explícita e implícita modulada por pistas raciais. Em linhas
gerais, as evidências desse estudo estão de acordo com a abordagem interacionista e
ampliam o conhecimento sobre a interação entre pertença ao endo- e exogrupo (em
relação ao grupo étnico) e a presença ou ausência de sinais contextuais de ameaça nos
efeitos da OT na cognição e comportamento social. Estudos futuros devem examinar
se estes resultados são aplicáveis no gênero feminino e em outras sociedades.
Palavras-chave: Ocitocina, Administração Intranasal, Empatia, Percepção de
Ameaça, Viés Racial

ABSTRACT
The role of oxytocin (OT) as an indiscriminate promoter of social cognition and pro-
sociality has come under question due to results indicating antisocial effects, such as
envy or ethnocentrism. Thus, the current study aimed to evaluate in the same sample
the impact of intranasal administration of OT in pro-social behaviors, such as
empathy for pain, and anti-social behaviors, as threat perception. In order to include
the dimension of belonging to the group, we evaluated the responses of self-reported
White participants to members of the ingroup (White) and outgroup (Black). It was a
double-blind placebo-controlled, randomized, and inter-subject study, in which 107
White, right-handed, undergraduate Brazilian males participated. On the first day of
collection, the exclusion criteria were verified and questionnaires were administered
to assess empathy and racial bias. On the second day, participants self-administered
the randomly assigned solution (Groups: OT or placebo [PL]). After a 40-minute
waiting period, computerized tasks of explicit empathy for pain (assessment of pain
intensity perceived during observation of White, Black, Purple – color control – hands
in neutral or potentially painful situations) and threat perception (identification of
White and Black armed men in social contexts) were administered. On the third day
of collection, participants inhaled the same substance and underwent the evaluation of
implicit empathy for pain (assessment of pain intensity perceived during observation
of videos depicting White, Black, Purple hands being touched by a Q-tip or penetrated
by a syringe). During this last phase, records of motor evoked potential in the right
hand were obtained while the Transcranial Magnetic Stimulation coil was positioned
over the primary motor cortex in the participant's left hemisphere. Regarding threat
perception task, results of a Mixed ANOVA revealed a significant Race*Group
interaction, F(1,105)=5.44, p=.022, η2=.049, indicating that the PL group presented
higher Correct Rejection rates (i.e., refraining from shooting unarmed targets) for
White than Black targets, whereas no difference between these targets’ rates was
found in the OT group. Thus, oxytocin reduced false alarms facing Black targets
Via improvement of the discrimination between armed and non-armed targets,
reducing the perceived threat. Regarding explicit empathy for pain, results from a
mixed ANOVA revealed that higher pain intensity was attributed in painful contexts
to Black targets compared to White targets. On the implicit assessment, the levels of

sensory-motor contagion between White and Black targets were the same. In sum,
results from our Brazilian sample point to a prosocial effect of OT on threat
perception specifically for outgroup, but no effect on explicit or implicit empathy
modulated by racial clues. Overall, this study supports the interactionist approach and
offers insights on the interaction between ingroup/outgroup membership (specifically
race) and the presence/absence of contextual threat to expand understanding of the
effects of OT on social cognition and behavior. Future studies could examine whether
these findings apply to female and to other societies as well.
Keywords: Oxytocin, Intranasal administration, Empathy, Threat Perception, Racial Bias.

12
LISTA DE ILUSTRAÇÕES
Figura 1 - Modelo interacionista do efeito social da ocitocina em humanos. (Figura retirada da revisão de BARTZ, ZAKI, BOLGER, OCHSNER, 2011; traduzida pela autora da presente pesquisa).
26
Figura 2 - Ilustração dos estímulos utilizados na Tarefa de avaliação das respostas empáticas explícitas durante observação de situações dolorosas ou neutras com mãos Brancas, Negras e Roxas
42
Figura 3 - (A) Desenho experimental com duas condições de sequências possíveis (homem-alvo Negro ou Branco) apresentados durante a Tarefa do Atirador em Primeira Pessoa (CORRELL et al. (2002); (B) Condições experimentais que foram randomicamente apresentadas: alvo Negro com objeto inócuo / com objeto ameaçador ou alvo Branco com objeto inócuo / com objeto ameaçador
44
Figura 4 - Ilustração dos estímulos apresentados em cada bloco experimental do IAT racial
46
Figura 5 - (A) Desenho experimental da Tarefa de avaliação do contágio sensório-motor ao observar dor no outro e (B) estímulos originais de Avenanti Stimuli, Sirigu & Aglioti (2010)
47
Figura 6 - Da esquerda para a direita: eletrodos utilizados; eletrodos posicionados para registro de FDI; configuração completa de montagem dos eletrodos de EMG, ponto preto corresponde ao “pólo +” (protuberância do músculo-musculaturas alvos) e ponto cinza corresponde ao “pólo –” (articulação - eletrodo de referência); posicionamento da bobina de EMT sobre M1.
49
Figura 7 - (A) Distribuições normais hipotéticas dos resultados com a presença (armas) ou ausência (objeto inócuo) de sinais. Tais distribuições são construídas separadamente entre os alvos Negros e Brancos para a Tarefa do Atirador em Primeira Pessoa. A linha vertical representa o limiar do critério de decisão do colaborador. Valores acima do limiar indicam resposta de “atirar” no alvo, enquanto que valores a baixo ou iguais ao critério indicam respostas de “não atirar” no alvo; (B) Condições prováveis de respostas em Paradigmas de Detecção de Sinal (figura baseada nas ilustrações do material do curso de Psicologia da Percepção da Universidade do Colorado, ministrada pelo Prof. Lewis O. Harvey)
54
Figura 8 - (a) Taxa de Erro por cada tipo de estímulo no TAPP 58 Figura 9 - Latência de Resposta por cada tipo de estímulo no TAPP 59 Figura 10 - Taxa de Rejeição Correta por cada Grupo no TAPP.
59
Figura 11 - (a) Comparação dos resultados de cBranco, cNegro e d’Branco, d’Negro entre amostras Norte-Americana e Brasileira
60
Figura 12 - (A) Sensibilidade e (B) Critério de Decisão por cada tipo de 61

13
estímulo Figura 13 - Avaliação de explícita de empatia à dor imaginada na própria perspectiva
63
Figura 14 - Avaliação de explícita de empatia à dor percebida na perspectiva do outro
63
Figura 15 - Média da diferença de amplitude entre potencial evocado motor (MEP) das situações de [dor] – [neutra], registro no músculo alvo FDI. (A) Amostra Brasileira (B) Amostra Italiana (resultados extraídos do estudo de AVENANTI et al., 2010). Não foram observados efeitos significativos de inibição sensório-motor para nenhum dos alvos (teste-t para uma amostra contrastando com 0, não indicou contágio sensório-motor nem diante de alvos Negro (p= .109) e nem de Branco (p= .835)
65

14
LISTA DE TABELAS
Tabela 1 - Fluxograma do processo de coleta de dados e critérios de exclusão 37
Tabela 2 - Caracterização dos colaboradores por Grupo e comparação por Teste-t para amostras independentes
56
Tabela 3 – Dados brutos da amplitude média dos MEPs (desvio-padrão) em mV, comparando as amostras Brasileira e Italiana
66

15
LISTA DE ABREVIAÇÕES
MAM – Músculo Abdutor do Mínimo (ADM, do inglês Abductor Digiti Minimum)
ATV – Área Tegmentar Ventral
IDB – Inventário de Depressão de Beck (BDI, do inglês Beck Depression Inventory)
DA – Dopamina
EMG – Eletromiógrafo
EMRI – Escala Multidimensional de Reatividade Interpessoal
EMT – Estimulação Magnética Transcraniana
PER – Potencial Evocado Relacionado à Evento (ERP, do inglês Evoked Related Potential)
ETCC – Estimulação Transcraniana por Corrente Contínua
PID – Primeiro Interósseo Dorsal (FDI, do inglês First Dorsal Interosseous)
IRMF – Imagem de Ressonância Magnética Funcional (fMRI, do inglês Functional Magnetic
Resonance Imaging)
TAI racial – Tarefa de Associação Implícita racial (IAT, do inglês Implicit Association Task)
LM – Limiar Motor
M1 – Córtex Motor Primário
PEM(s) – Potencial Evocado Motor (MEP, do inglês Motor Evoked Potential)
OT – Ocitocina
OTR – Receptor de ocitocina
PL – Placebo
POE – Posição Ótima no Escalpe
SI – Área somatosensorial Primária
SII – Área somatosensorial Secundária
TAPP – Tarefa do Atirador na Primeira Pessoa (FPST, First Person Shooter Task)
UPM – Universidade Presbiteriana Mackenzie
VP – Vasopressina

16
SUMÁRIO
1.INTRODUÇÃO ....................................................................................................................17 2.HIPÓTESES..........................................................................................................................37 3.OBJETIVOS ........................................................................................................................39 4.MÉTODO .............................................................................................................................40 4.1.PARTICIPANTES .............................................................................................................40 4.2.DESCRIÇÃO DOS INSTRUMENTOS UTILIZADOS....................................................42
4.2.1. Escalas e Questionários ..............................................................................................42 4.2.1.1. Questionário Sócio-Demográfico e Rastreio dos critérios de exclusão para a administração de ocitocina ................................................................................................42 4.2.1.2. Inventário de Depressão de Beck .........................................................................42 4.2.1.3. Rastreio dos critérios de Exclusão para a aplicação da Estimulação Magnética Transcraniana ....................................................................................................................42 4.2.1.4. Inventário de Lateralidade de Edinburgh .............................................................43 4.2.1.5. Escala Multidimensional de Reatividade Interpessoal ........................................43 4.2.1.6. Escala de Preconceito Flagrante e Sutil ...............................................................43 4.2.1.7. Questionário de Agressividade de Buss-Perry .....................................................44 4.2.1.8. Questionário de experiências com diferentes grupos étnicos ..............................44
4.2.2. Tarefas Computadorizadas (programadas no E-Prime, versão 2.10).........................44 4.2.2.1. Tarefa de avaliação das respostas empáticas explícitas durante observação de situações dolorosas ou neutras...........................................................................................45 4.2.2.2. Tarefa do Atirador na Primeira Pessoa / Dilema do Policial................................46 4.2.2.3. Tarefa de Associação Implícita racial...................................................................48 4.2.2.4. Tarefa de avaliação das respostas implícitas de contágio sensório-motor durante observação de situações dolorosas ou neutras.................................................................. 50
4.2.3. Técnica.........................................................................................................................53 4.2.3.1. Estimulação Magnética Transcraniana.................................................................53
4.2.4. Amostras para inalação...............................................................................................54 4.2.4.1. Solução com Ocitocina sintética (OT) e solução Placebo (PL)...........................54
4.3.PROCEDIMENTO.............................................................................................................54 4.4.ANÁLISE DOS DADOS....................................................................................................56 4.5.ASPECTOS ÉTICOS..........................................................................................................60 5.RESULTADOS......................................................................................................................61 6.DISCUSSÃO.........................................................................................................................73 7.REFERÊNCIAS ....................................................................................................................91

17
1. INTRODUÇÃO
1.1. Origem da ocitocina, formas de medição e administração
A ocitocina (OT) é um nonapeptídeo sintetizado pelo hipotálamo, especificamente nos
núcleos paraventricular e supraóptico, que projetam axônios para partes posteriores da
hipófise (neurohipófise) bem como para outras áreas cerebrais. A partir daí, a OT endógena é
liberada na circulação e atua de duas formas: a) como hormônio, liberado na corrente
sanguínea para afetar a atividade de células em alvos corporais periféricos; ou b) como
neurotransmissor, ou seja, mensageiro químico que modula sinais entre neurônios de distintas
regiões do cérebro. A OT é amplamente conhecida pelo seu papel hormonal quando ela é
liberada na circulação sistêmica em resposta à sucção, durante a lactação, e favorecendo a
estimulação rítmica da musculatura lisa do útero para auxiliar a expulsão do feto, durante o
trabalho de parto (INSEL, YOUNG e WANG, 1997). Ainda, os dendritos dos núcleos
hipotalâmicos liberam o peptídeo diretamente no cérebro, em estruturas onde a OT tem um
papel neuromodulador capaz de regular processos como a cognição social (ASPÉ-SÁNCHEZ
et al., 2016), que envolve o estreitamento de vínculos na relação mãe-bebê (TAYLOR et al.,
1994). Dentre as regiões no cérebro com maior densidade de receptores de OT destaca-se a
amígdala (LOUP et al., 1991; BAUMGARTNER et al., 2008; DELGADO, 2008), estrutura
subcortical altamente envolvida no processamento sócio-emocional, na aprendizagem de
medo e em mecanismos de recompensa (PHELPS e LEDOUX, 2005; KOSCIK e TRANEL,
2011).
Os experimentos sobre os efeitos comportamentais da OT são possíveis a partir de
duas abordagens principais – a avaliação de marcadores endógenos e a administração
exógena. Relativamente à primeira alternativa, a maior parte dos estudos recorreram à análise
da OT plasmática (LANCASTER et al., 2015). Efetivamente, existem evidências que
apontam para a liberação sincronizada do peptídeo de forma central e periférica, em roedores
(WOTJAK et al., 1998; ROSS & YOUNG, 2009), e evidência de correlação positiva entre os
níveis de OT do líquido cefalorraquidiano e do plasma, em humanos (CARSON et al., 2015).
Além disso, os níveis de OT plasmática variam entre os indivíduos e isso parece impactar nos
seus perfis comportamentais, como mostram estudos com amostras clínicas e normativas.
Referente às amostras clínicas, existem evidências preliminares que descrevem níveis
inferiores de OT plasmática em crianças e adultos com autismo, sugerindo um papel causal
das alterações ocitocinérgicas nos problemas apresentados (MODAHL et al., 1998); e maiores

18
concentrações de OT plasmática em pessoas com Síndrome de Williams (SW), sendo que a
medida basal da concentração desse peptídeo nos pacientes com SW foi correlacionada
positivamente com medidas de aproximação social excessiva e negativamente com
comportamentos de adaptação social (DAI et al., 2012). Ainda, observam-se diferentes níveis
plasmáticos de OT em relação a distintos perfis de pacientes com Esquizofrenia, inclusive,
pesquisadores verificaram que níveis menores de OT nesses pacientes estavam associados a
um pior desempenho em tarefas de reconhecimento de emoções faciais (GOLDMAN et al.,
2008). Em relação às amostras com colaboradores saudáveis, Zak, Kurzban e Matzner (2005)
analisaram o plasma de adultos de ambos os sexos e constataram que quando os
colaboradores recebem dinheiro em um jogo econômico, há maior liberação de OT endógena
diante de situações em que eles consideram-se dignos de confiança por parte de outro ser
humano, em comparação ao ganho monetário aleatório e programado por um computador.
Por outro lado, a OT exógena artificialmente sintetizada, pode ser administrado pela
via intravenosa (PIETROWSKY et al., 1996) ou intranasal (SCHULZE et al., 2011).
Atualmente, a maioria das pesquisas têm realizado a administração intranasal de OT em
humanos, uma vez que existem evidências da sua ação neuromodulatória e porque seu manejo
é mais fácil em comparação às demais alternativas (BORN et al., 2002; PETROVIC et al.,
2008; ECKSTEIN et al., 2014). Atualmente, limitações metodológicas impedem os
pesquisadores de aferir ou manipular as concentrações de OT diretamente no cérebro humano
(MCCALL e SINGER, 2012). Porém, há evidências de que a administração intranasal deste
neuropeptídeo (em contraste com a via intravenosa) é capaz de induzir alterações replicáveis
ao nível do comportamento (NABER et al., 2010), e da percepção (THEODORIDOU et al.,
2009), sugerindo a penetração da barreira hematoencefálica. Todavia, destacam-se alguns
efeitos de interferência e viés que podem ocorrer nos experimentos com essa forma de
administração. Para um melhor controle desses vieses nos dados, deve-se ter especial atenção
à necessidade de desenhos experimentais duplo-cegos, à distribuição de gênero dos
colaboradores e tipo de solução placebo selecionada (salina ou solução com os ingredientes
do spray de OT exceto pelo nonapeptídeo) (EISENEGGER et al., 2010). Adiante
apresentaremos em detalhe os principais estudos que recorreram à administração intranasal de
OT para modulação do comportamento social em humanos. Em seguida, passamos a
apresentar dados comparativos que ampararam a área de estudo dos efeitos centrais da OT.
1.2. Estudos com ocitocina em roedores

19
Desde os anos 90 do século XX, diversos estudos com roedores sugerem que a OT
pode gerar efeitos pró-sociais. Inicialmente, observou-se que a OT tem um papel central nos
comportamentos maternos de cuidado da prole (INSEL & YOUNG, 2001; RILLING e
YOUNG, 2014), sendo que roedores com prejuízos na liberação de OT apresentam redução
no comportamento maternal (JIN et al., 2007). Adicionalmente a isso, outros estudos
verificaram uma redução na expressão dos receptores de OT no cérebro de roedores
poligâmicos quando comparados aos monogâmicas (INSEL e SHAPIRO, 1992). Evidências
indicam que em ratos monogâmicos, a OT interage com o sistema dopaminérgico
favorecendo a formação de laços entre os parceiros (NUMAN e YOUNG, 2016). A dopamina
(DA) e OT são neuromoduladores cujos neurônios estão presentes na área tegumentar ventral
(ATV)/substância negra e hipotálamo, respectivamente. Os efeitos sinérgicos dessas
substâncias são de grande interesse para os estudos da área, pois as projeções dos neurônios
de OT e DA direcionam-se para regiões como córtex pré-frontal, núcleo accumbens e estriado
com a finalidade de regular comportamentos sociais e afiliativos (NEUMANN, 2009). Xiao et
al (2017) descreveram os mecanismos básicos subjacentes à interação destes
neurotransmissores, onde a OT parece aumentar diretamente a atividade dos neurônios
dopaminérgicos na ATV e inibir indiretamente, pelo recrutamento de interneurônios
gabaérgicos, os neurônios dopaminérgicos na substância negra. A partir destas regiões, as
projeções dopaminérgicas da ATV seguem para o hipocampo e núcleo accumbens (LOVE,
2014), além do córtex pré-frontal e putâmen, favorecendo comportamentos de interesse social
(GUNAYDIN et al., 2014). Em paralelo, as projeções dopaminérgicas da substância negra
seguem para o córtex pré-frontal e putâmen reduzindo os comportamentos exploratórios
irrelevantes, pelo controle da atividade motora (PATEL et al., 2012).
Outro sistema que também regula os comportamentos sociais com a OT é o sistema da
vasopressina (AVP), substância produzida pelos núcleos supraóptico e paraventricular do
hipotálamo. No sistema nervoso central, AVP e OT parecem ter funções antagônicas,
principalmente em relação à regulação amigdalar durante o controle da ansiedade. Enquanto
AVP potencializa a resposta de medo, a OT exerce um efeito ansiolítico. Porém, ainda não é
claro como ocorre o equilíbrio entre a AVP e OT na regulação do sistema dopaminérgico.
Quando a OT liga-se ao receptor de OT (OTR) esta ligação ativa a proteína G a qual
determina modificações em toda a célula. Os receptores da OT e da AVP seguem o caminho
pela via da fosfolipase C (BURBACH et al., 1995), sendo que sua expressão e disponibilidade
geralmente depende do grau de desenvolvimento cerebral do indivíduo. A dessensibilização
ou redução da atividade do segundo mensageiro podem ainda ocasionar uma down-regulation

20
do receptor de OT (SHAW, 1996). Além disso, para explorar a relação causal entre a OT e os
padrões comportamentos, pesquisadores produziram animais com nocaute para a OT (OTKO)
ou para o receptor para OT (OTRKO), sendo que os resultados dessas pesquisas confirmaram
que a OT é fundamental para o comportamento materno, e para os comportamentos sociais
(PEDERSEN et al., 2006).
Quando a professora Peggy Mason estudou com sua equipe os comportamentos de
empatia entre ratos, observaram-se resultados bastante interessantes, os quais em conjunto
revelaram que o comportamento de oferecer ajuda ao outro requer familiaridade com esse
indivíduo (BEN-AMI BARTAL et al., 2014). Essa evidência foi testada com um experimento
onde um rato livre estava na mesma gaiola de um outro rato preso em um pequeno
compartimento. O rato livre, ao observar um outro rato preso, parece ficar ansioso e
estressado porque ele se identifica com as reações advindas dessa situação e costuma
esforçar-se para liberar o rato preso. O diferencial dessas pesquisas envolveu o estudo do
impacto do grau de familiaridade entre os ratos para a promoção da expressão de
comportamentos de ajuda. Assim, os resultados indicam que os ratos livres ajudam ratos
desconhecidos que sejam de sua mesma espécie (provavelmente devido a um viés social que
direciona os ratos a ajudar apenas aqueles de seu próprio grupo) e não ajudam aqueles de uma
espécie diferente. Porém, se o rato livre tiver uma breve convivência significativa ou for
criado com indivíduos de uma espécie diferente da sua de origem, ele ajudará inclusive esses
indivíduos, mesmo que eles não sejam similares. Caso o rato livre nunca tenha convivido com
membros de sua espécie, ele não os ajudará futuramente. Em outras palavras, a familiaridade
entre os ratos parece ser um elemento fundamental para guiar os comportamentos de ajuda
mas, um ambiente social diverso parece permitir que os indivíduos auxiliem uns aos outros,
mesmo que eles sejam diferentes. Curiosamente, ratos livres que receberam ansiolíticos
tiveram uma motivação reduzida para auxiliar qualquer outro rato, provavelmente porque há
um bloqueio do efeito de estresse e de emoções negativas (BEN-AMI BARTAL et al., 2014).
Em suma, os dados disponíveis sugerem que os sistemas ocitocinérgico e
dopaminérgico tendem a controlar-se mutuamente devido à disponibilidade de seus receptores
porém, a maneira como isso acontece não é completamente clara (CHARLET e
GRINEVICH, 2017). Estudos atuais ainda buscam compreender em detalhes a relação entre
as doses e as respostas decorrentes da administração de OT. Além disso, estudos com
indivíduos nocaute para o receptor de OT (OTRKO) apresentam consequências piores em
comparação aos animais com nocaute para a OT, sugerindo que outros ligantes, além da OT,
devem ativar esse receptor. Em linhas gerais, fatores ambientais modulam as via

21
ocitocinérgica. Então OT pode sim contribuir na expressão de comportamentos protetivos ao
endogrupo porém, parece que a exposição a diferentes contextos ambientais devem modular
o sistema ocitocinérgico de modo que respostas positivas socialmente podem ser eliciadas
diante de membros do exogrupo. Até o momento, diversos estudos clínicos de OT intranasal
em pacientes humanos com prejuízos sociais já foram desenvolvidos e parecem promissores.
1.3. Estudos com ocitocina em humanos
A reputação de hormônio do amor que a OT adquiriu em decorrência dos estudos com
humanos foi por muito tempo fortalecida entre o público leigo e científico. É comum em
revistas não científicas uma apresentação simples e entusiasmada sobre o tema. Uma
ilustração é a matéria da Revista Veja Hormônio da felicidade: saiba como estimular seus
efeitos (abril, 2017), que introduz os neurotransmissores que compõem o “quarteto da
felicidade”, são eles: endorfina, serotonina, dopamina e, em destaque, ocitocina. Em linhas
gerais, a matéria apresenta endorfina como a substância que regula as respostas à dor e os
sentimentos de unidade grupal; a serotonina relacionada com os sentimentos de solidão e
sintomatologia depressiva; a dopamina como substância moduladora do prazer; e a ocitocina
relacionada aos comportamentos que favorecem o vínculo social. termina com a
recomendação da exposição a estímulos externos específicos para favorecer reações químicas
que contribuam para a produção e liberação destes neurotransmissores.
No âmbito da pesquisa científica, os efeitos da OT são analisados mais
cuidadosamente e em maior detalhe. Assim, evidências de pesquisas em humanos revelam
que a OT tem um efeito inibidor no núcleo dorsal da rafe, principal região de síntese de
serotonina, e em regiões como a amígdala, ínsula e córtex orbitofrontal, regulando assim a
interação social (MOTTOLESE et al., 2014). Além disso, verifica-se que a OT parece
promover comportamentos e competências sócio-emocionais positivas, como a capacidade de
inferir estados emocionais a partir de faces (DOMES, et al., 2007; DI SIMPLICIO et al.,
2009), a confiança interpessoal (KOSFELD et al., 2005; BAUMGARTNER et al., 2008), a
empatia (HURLEMANN et al., 2010), a fidelidade nas relações amorosas (SCHEELE et al.,
2012), a generosidade e o altruísmo (ZAK, STANTON e AHMADI, 2007; DE DREU et al.,
2010). Estes elementos em conjunto são necessários para o desenvolvimento de
relacionamentos emocionais saudáveis, o que é crucial para a sobrevivência das espécies.
As pesquisas com humanos são relativamente recentes, portanto, ainda são necessários
novos estudos para esclarecemos os mecanismos subjacentes à influência de diferentes

22
contextos e os efeitos de distintos parâmetros de administração da OT nos comportamentos
sociais, como variações no tempo de espera entre a administração de OT e a coleta, dosagem
de OT e o efeito moderador dos níveis de OT endógena em relação à administração de OT
exógena. A maioria dos estudos revelam que a inalação de OT parece estar associada a efeitos
positivos e de melhora de habilidades de cognição social e da prosocialidade (KOSFELD et
al., 2005; DOMES et al., 2007; MARSH et al., 2010; HURLEMANN et al., 2010;
DECLERCK et al., 2010). Porém, a revisão de Bartz, Zaki, Bolger & Ochsner (2011) revelou
que esses efeitos da OT são de magnitude fraca a moderada, e não forte como até então se
acreditava, onde aproximadamente metade dos estudos de prosocialidade não apresentam o
efeito esperado após administração de OT e cerca de 60% dos dados encontrados podem ser
explicados por diferenças individuais e situacionais (ver inclusive revisão de SHAMAY-
TSOORY e ABU-AKE, 2015).
As descobertas controversas acerca dos efeitos da OT desafiam os pesquisadores a
explorar o impacto de diferentes contextos na prosocialidade, considerada anteriormente
como uma resposta indiscriminada. Estudos recentes têm constatado que o tipo de relação que
se estabelece com o outro (considerando o distanciamento/proximidade social) é capaz de
determinar o impacto da OT (STALLEN et al., 2012). É evidente que questões religiosas,
econômicas, étnicas, entre outras, interferem nas relações interpessoais assim como na
identidade grupal, definindo demarcações sociais de quem compõe o nós (endogrupo) e os
outros (exogrupo). Considerando tais diferenças grupais, um estudo conduzido com homens
holandeses (Brancos) verificou que a administração de OT propiciou sentimentos de
preferência a Etnia de origem (etnocentrismo), em detrimento de grupos étnicos distintos (DE
DREU et al., 2011). Por outro lado, um estudo que analisou a medida de potencial evocado
relacionado a evento (ERP, sigla em inglês) junto de uma pequena amostra Asiática, verificou
uma resposta neuronal aumentada do potencial P2 após a visualização de faces com
expressões de dor quando estas pertenciam a indivíduos Asiáticos, em comparação com
Caucasianos (SHENG et al., 2013). Os autores verificaram ainda que a administração
intranasal de OT acentuava este padrão, promovendo um aumento da resposta neuronal
apenas diante de estímulos da mesma Etnia (endogrupo). Além disso, uma interessante meta-
análise de Van IJzendoorn e Bakermans-Kranenburg (2012) observou que a administração
intranasal de OT aumenta a habilidade de reconhecer expressões faciais e eleva a confiança
entre pessoas do endogrupo porém, não do exogrupo.
As controvérsias sustentam-se diante de evidências que reportam inclusive
modulações de emoções negativas advindas da administração desse peptídeo. Ilustrações

23
disso incluem os efeitos anti-sociais reportados como o aumento de comportamentos de inveja
(SHAMAY-TSOORY et al., 2009) e de agressividade (NE’EMAN et al., 2016). Na pesquisa
de Shamay-Tsoory e colaboradores (2009) foi aplicada uma tarefa onde os colaboradores
escolhiam uma entre três possíveis portas que surgiam na tela do computador. Assim, em cada
rodada ou eles ganhavam mais pontos do que o jogador oponente, ou o seu oponentes
ganhava mais pontos do que eles. Após administração de OT, na primeira situação descrita os
colaboradores sinalizaram, por uma escala Likert, sentir maior exaltação/arrogância de terem
recebido mais pontos em comparação ao oponente; enquanto que quando os colaboradores
recebiam menos pontos, eles sinalizavam sentir mais inveja, em comparação ao grupo que
havia recebido para inalação uma substância placebo. No estudo de Ne’eman et al. (2016), os
colaboradores realizavam um jogo monetário em que eles poderiam apresentar um dentre três
tipos de respostas após provocação (e.g. um jogador fictício que retirava pontos do
colaborador): (1) retirar pontos do jogador fictício – resposta agressiva, (2) aumentar os seus
próprio pontos – resposta individualista – e (3) dividir seus pontos com o jogador fictício –
resposta colaborativa. Os resultados revelaram que quem recebeu OT, aumentou a taxa de
respostas agressivas. Ambos autores justificam a ocorrência de modulação de emoções
relacionadas a sentimentos negativos pela OT uma vez que essa substância promove um
aumento atencional do colaborador à saliência dos agentes sociais, resultando em um aumento
de comportamentos anti-sociais, como a inveja e a agressividade em contextos competitivos.
Isso refuta a ideia de que a OT sempre eleva a frequência de comportamentos pró-sociais em
todos os contextos.
Além disso, outros estudos revelam que elevadas concentrações de OT endógena
correlacionam-se com dificuldade de relacionamento (TABAK et al., 2011). Esses resultados
de modulações de emoções negativas foram observados tanto em estudos com ratos, como
com humanos. Por exemplo, Ferris e colaboradores (1992) verificaram que a administração de
OT em ratos aumentou a frequência de comportamentos agressivos relacionados à
preservação de território e da prole. Apesar das evidências descritas acima, ainda são escassos
os estudos sobre os efeitos da OT em relação a diferentes grupos étnicos. Além disso, a
maioria dos estudos citados foram conduzidos nos Estados Unidos e na Europa, o que indica a
necessidade de nova pesquisa em diferentes culturas.
Diante desse panorama, tem crescido o interesse da neurociência social, em
compreender os mecanismos neurobiológicos de funcionamento da OT. Isso porque as
evidências de contribuições da OT em processos de cognição social com modelos animais
(DONALDSON e YOUNG, 2008) e com humanos (HEINRICHS, VON DAWANS,

24
DOMES, 2009), motivam pesquisadores a considerar o potencial terapêutico da OT em
diferentes grupos clínicos com alterações psiquiátricas, como a esquizofrenia (ABU-AKEL et
al, 2014a), e com distúrbios do desenvolvimento, como o autismo (DADDS et al., 2014). Tal
compreensão poderia favorecer consideravelmente intervenções psicoterapêuticas com
pessoas com déficits de empatia, pois em conjunto com essas ações, a OT poderá catalisar
processos de sensibilidade ao outro nas relações interpessoais favorecendo, por exemplo,
componentes envolvidos no compartilhamento de emoções, na flexibilidade mental ou na
auto-regulação. Todavia, o crescente número de pesquisas na área que têm revelado alguns
resultados controversos às descobertas primordiais, nos lembra da necessidade de cautela
quanto a conclusões sobre os efeitos exclusivamente benignos da OT e quanto à possível
implementação individualizada do uso desse neuropeptídeo como agente terapêutico. Quando
Churchland e Winkielman (2012) discutem sobre a improvável possibilidade de que a OT
tenha uma ação tão ampla nos processos de cognição social, eles ainda nos lembram que as
pesquisas aplicadas necessitam de precisão quanto ao seu uso clínico e a descrição dos efeitos
da OT. Pode ser que a OT venha a ser tão versátil quanto outras drogas, tal como a aspirina,
que apenas após décadas de estudo, pode ser aplicada em diferentes doses e frequências para a
promoção de efeitos distintos, que incluem um alívio de uma cefaleia, um potente efeito anti-
inflamatório ou até mesmo, forte inibição na agregação plaquetária. Desta forma, a aspirina
passou a fazer parte do arsenal terapêutico na neuroclínica, reumatologia e na cardiologia,
respectivamente.
1.4. Efeito biológicos da ocitocina
Após administrada a substância, a OT é suficientemente absorvida pela mucosa nasal,
sendo que seu efeito modulatório é estabelecido gradativamente alcançando seu estado de
equilíbrio geralmente após 40 minutos. Além disso, a ação da OT tem um período de latência
estimado de 30-60 minutos. Segundo a bula da OT sintética, dados pré-clínicos não revelam
riscos especiais para humanos baseados em estudos convencionais de dose única de
toxicidade. Considerando as possíveis reações adversas descritas, destacam-se como sintomas
de ocorrência rara: cefaleia, taquicardia e náusea (SYNTOCINON® (ocitocina), Novartis
Biociências SA).
Com o decorrer das pesquisas, diferentes modelos teóricos buscaram explicar os
mecanismos de funcionamento neurobiológico da OT. A maioria dos estudos relatam efeitos
pró-sociais da OT, como a aproximação social, gerando o favorecimento de vínculo

25
interpessoal. Dois mecanismos explicativos que defendem um efeito indiscriminado da OT,
consideram que esta deve promover (a) a redução da ansiedade social (CAMPEBELL et al.,
2010) ou (b) o aumento da motivação afiliadora (MCDONALD E MCDONALD, 2010). A
hipótese da redução da ansiedade como justificativa do efeito social da OT baseia-se, por
exemplo, na evidência de Kirsch et al. (2005) a qual demonstra que a OT é capaz de regular a
reatividade em humanos, por modular a atividade amigdalar diante de situações que geram
medo; e de reduzir a ansiedade social, permitindo que as pessoas tomem riscos que de outra
forma evitariam. Em relação à hipótese da motivação afiliadora, esta embasa-se nas
evidências de que a OT promove comportamentos de confiança e cooperação entre as pessoas
(KOSFELD et al., 2005), devido a um viés positivo generalizado em relação ao estímulo
social. No estudo de Gamer et al. (2010), com ressonância magnética funcional, foi
constatado que a OT diminui a atividade de sub-regiões da amígdala diante de estímulos
negativos e aumenta a atividade diante de estímulos positivos, o que permitiria justificar
ambas as hipóteses acima. Entretanto, tais hipóteses apresentam limitações para a
compreensão de muitos dos dados inconsistentes observados na área.
Outros pesquisadores passaram a questionar essas hipóteses sobre o efeito social
indiscriminado da OT e consideram se após sua inalação, a interação com indivíduos de
distintos grupos gera (c) ativação de recursos atencionais distintos (em função do membro
observado), com o aumento da saliência social de estímulos alvos no contexto (SHAMAY-
TSOORY et al., 2009). Esse modelo propõe que a OT aumenta diversas emoções, não apenas
as emoções positivas. Acredita-se que dessa forma existiria um aumento de reações agressivas
em relação ao exogrupo em contextos ameaçadores (NE’EMAN et al., 2016), além de uma
pré-disposição seletiva a emoções positivas em relação ao endogrupo em contextos
cooperativos. Essa terceira hipótese de alteração do processamento / saliência de um estímulo
social visual foi proposta a partir das evidências de que a OT aumenta a funcionalidade da
amígdala posterior e dos colículos superiores (GAMER et al., 2010) produzindo reflexos no
processamento de estímulos sociais de natureza positiva bem como no direcionamento
atencional.
Esse modelo da Hipótese de Saliência Social baseia-se na interação entre os sistemas
ocitocinérgico e dopaminérgico (DA) diante de pistas sociais externas (SHAMAY-TSOORY
e ABU-AKEL, 2016). Assim, as características pessoais e funções biológicas homeostáticas
determinam a disponibilidade de dopamina tônica, a qual modula o sistema ocitocinérgico
que regula a responsividade do sistema dopaminérgico. A saliência de um estímulo é
inicialmente processada pela amígdala e núcleo accumbens, que em seguida transmitem

26
informação para a ATV, a qual regula a modulação de dopamina. Assim, a amígdala medeia o
processamento de estímulos sociais positivos e negativos, promovendo a integração dos
sistemas em questão e comunicando-se, por projeções aferentes e eferentes, a regiões corticais
e sub-corticais. Toda essa interação descrita favorece a percepção de estímulos sociais e o
redirecionamento atencional (ROSENFELD, LIEBERMAN e JARSKOG, 2011). Os efeitos
comportamentais da OT parecem depender da relevância do estímulo social porém, mais
pesquisas com paradigmas atencionais são necessárias para melhor se entender o papel da OT
no aumento da saliência social.
Por fim, (d) a abordagem interacionista (BARTZ, ZAKI, BOLGER, OCHSNER,
2011) considera que os efeitos da OT são moderados por fatores contextuais e características
individuais e que todos os mecanismos descritos acima de funcionamento da OT podem
acontecer independentemente ou não (Figura 1). Essa última proposta auxilia a compreensão
sobre os efeitos sociais da OT, pois entende o que os estudos apresentam como
inconsistências, como verdadeiras “pistas” de quando a OT favorece a cognição social e o
comportamento pró-social nos indivíduos.
Figura 1 - Modelo interacionista do efeito social da ocitocina em humanos. De acordo com o modelo, a administração exógena (e.g. intranasal) de ocitocina influencia uma série de resultados relacionados à cognição social (e.g. reconhecimento de emoção e empatia) e pró-socialidade (e.g. percepção e comportamentos de confiança) mas esses efeitos da ocitocina são geralmente moderados por características da situação social na qual a ocitocina é administrada e/ou o indivíduo o qual a ocitocina é administrada. Os moderadores situacionais podem incluir dificuldades com as características da tarefa, a valência do estímulo, bem como a familiaridade do oponente, confiabilidade e pertencimento ao endo- em comparação ao exogrupo. Os moderadores da diferença individual podem incluir características comportamentais como proficiência em cognição social, traços de diferenças individuais como apego ansioso; essas diferenças individuais devem refletir uma variação natural no sistema de ocitocina endógeno (e.g. nível de ocitocina plasmático ou polimorfismo do gene receptor da

27
ocitocina). Adicionalmente as variáveis moderadoras, o efeito da ocitocina na cognição social e na prosocialidade provavelmente são mediados por um ou mais de um processo de mecanismo básico; especificamente por: redução da ansiedade social, aumento da motivação social / afiliação e/ou aumento da saliência de pistas sociais. Como indicado pelas linhas pontilhadas que conectam os hipotéticos mecanismos, esses mecanismos podem trabalhar independentemente ou em conjunto para mediar o efeito da ocitocina na cognição social e na prosocialidade. Por exemplo, a ocitocina pode aumentar a saliência de uma pista social reduzindo a ansiedade social e permitindo que pessoas com ansiedade social se atentem às pistas sociais que antes elas provavelmente evitariam. A ocitocina pode inclusive aumentar a saliência de uma pista social pela ativação de motivos afiliativos que devem aumentar a atenção da pessoa a informações sociais no ambiente, trabalhando para promover ou impedir tal objetivo. Finalmente, a ocitocina pode aumentar o direcionamento afiliativo / aproximação social pela diminuição da ansiedade social. (Figura retirada da revisão de BARTZ, ZAKI, BOLGER, OCHSNER, 2011, p. 305; traduzida pela autora da presente pesquisa).
De forma a testar adequadamente os modelos teóricos de funcionamento da OT,
defendemos que será necessário avaliar nos mesmos indivíduos dimensões comportamentais
(a) pró- e (b) anti-social. Assim, no âmbito da presente pesquisa focaremos no estudo do
papel da OT na regulação de comportamentos empáticos (pró-social), bem como seu efeito
em relação a reações emocionais negativas (anti-social), como a detecção de ameaça. Além
disso, no sentido de incluir a dimensão de pertencimento ao grupo, o presente estudo testará
as respostas em relação quer a membros do endogrupo (mesma etnia do participante), como a
membros do exogrupo (etnia distinta). Estamos cientes de que é possível que no Brasil a
divisão proposta entre os grupos étnicos citados (colaboradores Brancos avaliando situações
com alvos Brancos e Negros) não apresente resultados tão distintos entre si, pois a história da
constituição da população Brasileira é marcada pela miscigenação interétnica. Todavia, uma
vez que ainda se observam situações cotidianas de discriminação entre esses grupo étnicos,
nota-se a complexidade dessa relação e faz-se necessário o seu estudo. Então, optamos por
oferecer OT (versus uma solução placebo) aos colaboradores e em seguida replicar tarefas
clássicas na área da neurociência que avaliam esses comportamentos.
Investir na replicabilidade de estudos em neurociência é um desafio porém, uma
prioridade para promover maior confiabilidade dos resultados produzidos no campo científico
e assim, melhor embasar as conclusões da área (BUTTON et al., 2013). A competitividade e a
exigência por elevado nível de publicação dificultam a divulgação de resultados negativos ou
menos inovadores. A contaminação por falsos-positivos na área da neurociência pode ser alta,
como ocorre nas pesquisas biomédicas em que há apenas cerca de 25% de taxa de
replicabilidade (PRINZ, SCHLANGE, ASADULLAH, 2011; BEGLEY e ELLIS, 2012). A
importância da replicação pode ser considerada particularmente crítica na área de estudo do
viés racial, dadas as importantes diferenças que podem existir entre sociedades/regiões (com
distintas composições étnicas), não se podendo assumir que os resultados obtidos em um
contexto serão universais. Além disso, algumas regras sociais tem sido modificadas com o

28
passar dos anos, sendo recentemente promovida uma maior convivência entre indivíduos de
diferentes crenças e grupos étnicos, o que tende a aumentar a representatividade social e o
respeito aos direitos humanos. Esse processo de convivência e respeito favorece a
modificação das associações estereotípicas em relação aos distintos grupos (MIZAEL et al.,
2016) porém, muitas vezes seleciona comportamentos mais sutis de discriminação, os quais
costumam ser mais aceitos socialmente (PETTIGREW e MEERTENS, 1995). Essas
mudanças comportamentais desafiam os estudos a aprimorarem formas de registro das
atitudes que se tem em relação ao outro. Para além do uso de métodos clássicos de avaliação
(como exclusiva aplicação de tarefas comportamentais e questionários), o recurso a métodos
de investigação em neurociência, bastante sensíveis, como ressonância, medidas oculares,
eletroencefalograma, estimulação cerebral, entre outros, permitem avaliar com maior precisão
o processamento de respostas automáticas e controladas, além de demonstrar o envolvimento
de circuitarias neurais específicas dos mais diversos processos cognitivos durante ativação e
controle de um comportamento enviesado.
1.5. O papel da ocitocina no processo de empatia à dor
Apesar do termo empatia ser amplamente usado, ele costuma ser erroneamente
empregado pois é muitas vezes confundido com conceitos como compaixão, simpatia, teoria
da mente, entre outros (SINGER & DECETY, 2011). De fato, é considerado um “termo
guarda-chuva” que desperta grande interesse em pesquisadores da área da neurociência social,
sendo que mesmo sem uma definição única, denota em linhas gerais a capacidade de um
indivíduo entender o estado mental do outro. Já na década de 80, a habilidade empática era
compreendida como uma experiência vicária e um construto multidimensional que combina
componentes cognitivos e afetivos (DAVIS, 1983) e pesquisas sobre as bases neuronais da
empatia têm contribuído consideravelmente para sua definição (KENNEDY & ADOLPHS,
2012). Simone Shamay-Tsoory, uma importante pesquisadora da área, propôs um modelo que
divide a empatia entre uma dimensão emocional e outra dimensão cognitiva (SHAMY-
TSOORY et al., 2009). A primeira envolve o compartilhamento de emoções entre as pessoas
(experiência do contágio emocional) e a outra dimensão permite o entendimento dos
pensamentos e intenções dos outros a partir do movimento de adotar o ponto de vista alheio
que possibilitar inferências precisas sobre o que se passa com o outro (DECETY &
JACKSON, 2004; SHAMY-TSOORY et al., 2009; ABU-AKEL et al., 2014b). Essa
habilidade tem um papel crucial no processo de interação social humana, permitindo a

29
conexão entre os indivíduos, uma vez que favorece a compreensão da intenção da ação do
outro, de suas emoções, de sentimentos e de pensamentos (SINGER & DECETY, 2011;
DECETY & SVETLOVA, 2012). É possível avaliar a empatia explícita e implicitamente.
Todavia, inúmeras evidências revelam que as respostas explícitas, em relação às medidas
implícitas, são fortemente influenciadas por fatores como desejabilidade social (NETO, 2015)
e que por conta disso, é possível que se verifiquem resultados contrastantes quanto a essas
formas de avaliação.
No modelo neurocientífico de compreensão da empatia de Preston e de Waal (2002),
foi proposto que quando imaginamos ou quando observamos o estado mental de uma outra
pessoa, nosso sistema nervoso automaticamente reage promovendo ativação de redes neurais
semelhantes a circuitaria ativa no observador. Isso foi constatado inclusive em estudos
subsequentes conduzidos com ressonância magnética funcional, em que se verificou uma
ativação compartilhada de redes neuronais específicas durante experiências infligidas em si e
no outro. Na mesma linha, Keysers e colaboradores (2004), por exemplo, observaram uma
ativação do córtex somatossensorial quando os colaboradores eram tocados e quando
observavam o toque feito no outro.
Desde 2004, diversas pesquisas sobre o domínio afetivo da empatia envolvendo o
estudo da dor foram desenvolvidas visando a compreensão de como respondemos de maneira
empática aos outros. A empatia à dor física, por exemplo, é um dos mecanismos mais básicos
e refere-se ao mal-estar subjetivo resultante da exposição à dor do outro (SHAMAY-
TSOORY et al., 2013). Segundo a Associação Internacional para o Estudo da Dor (2018), dor
é definida como uma experiência sensitiva e emocional desagradável associada a dano
tecidual real ou potencial, que tem uma elevada importância evolutiva para alertar os
indivíduos a interromper ou afastar-se do estímulo causador, promovendo assim sua proteção.
Quanto ao processamento da dor, sabe-se que ele é composto pelas dimensões sensitivo-
discriminativa (percepção e detecção do estímulo nocivo em relação a intensidade,
localização e duração) e afetivo-motivacional (integração da informação dolorosa com a
experiência afetiva que regula as respostas comportamentais e o humor). Esta é uma resposta
complexa em que informações dos neurônios aferentes primários nociceptivos seguem até a
medula espinal onde neurônios de segunda ordem conduzem essas informações para regiões
supra-espinais. Então, as projeções do trato espinotalâmico vão para estruturas do troco
encefálico (tálamo e córtex) para iniciar o processamento das dimensões do estímulo
doloroso. As informações de dor dos núcleos talâmicos seguem para o córtex alcançando
principalmente áreas somatosensoriais primárias (SI) e secundárias (SII). Estas áreas enviam

30
projeções para o córtex parietal posterior que integra elementos do estímulo doloroso com o
contexto em que ele ocorre. Além disso, outras projeções talâmicas seguem para o córtex
insular (regula o processamento de informação do estado interno do corpo) e para o córtex
cingulado anterior (integra respostas afetivas, cognitivas e sociais). Por fim, o córtex pré-
frontal é ativado para promover uma avaliação e planejamento de consequências futuras da
situação vivenciada (OLIVEIRA e SILVA, 2009).
Vários estudos de neuroimagem mostraram que as áreas ativadas pela observação de
dor física alheia se sobrepõem às áreas ativadas durante a própria experiência de dor física
(SINGER et al., 2004; DECETY e LAMM, 2006; DECETY, ECHOLS e CORRELL, 2010;
LAMM, DECETY, & SINGER, 2011). Uma das pesquisas iniciais com uma interessante
proposta metodológica foi conduzida por Singer e colaboradores (2004) onde casais iam ao
laboratório e qualquer um dos dois poderia receber um choque como estímulo doloroso
enquanto que apenas um deles estava num equipamento de neuroimagem para avaliação das
mudanças na ativação de regiões cerebrais. Assim, os autores avaliaram a experiência de dor
em si mesmo e em seu parceiro. Resultados dessa pesquisa revelaram que para ambas as
condições de dor, a codificação desse estímulo gerou uma ativação da mesma circuitaria
cerebral, que envolve essencialmente o córtex insular e cingulado. Estudos de diversas
pesquisas conseguiram observar que regiões posteriores da ínsula estavam envolvidas na
experiência pessoal de dor - representações primárias, e que regiões anteriores eram ativadas
diante da empatia a dor no outro - representações secundárias relacionadas com o afeto
negativo antecipatório em si e no outro (DECETY & LAMM, 2009; SINGER & DECETY,
2011).
Ainda é importante entender sob quais circunstâncias reagimos de maneira empática
com o outro. A resposta empática à dor a nível cerebral é modulada por diversos fatores como
o direcionamento atencional para situação dolorosa (GU & HAHN, 2007), a intensidade do
estímulo doloroso (AVENANTI et al., 2006), a relação afetiva entre as pessoas (SINGER et
al., 2004) e a experiência que se tem com situações dolorosas, como o caso dos médicos em
comparação a não médicos (CHENG et al., 2007). Destacam-se outros estudos que revelam
que apesar da natureza eminentemente automática dos processos empáticos, a empatia à dor
física no outro é inclusive modulada por fatores contextuais, como pistas relativas ao estatuto
de pertença ao grupo social (TARRANT, DAZELEY e COTTOM, 2009). Algumas
ilustrações disso envolvem evidências de que participantes Brancos e Negros apresentam
maior contágio sensório motor ao observar situações dolorosas infligidas em membros do
endogrupo em comparação a membros do exogrupo (AVENANTI, SIRIGU e AGLIOTI,

31
2010) e outros achados que revelam que os participantes foram mais empático à dor em
pessoas que contraíram AIDS por transfusão de sangue em comparação a quem contraiu a
doença por uso de drogas (DECETY, ECHOLS e CORRELL, 2010).
Em relação as pesquisas nessa linha com OT, considera-se que o peptídeo seriam um
facilitador de respostas empáticas afetivas e cognitivas. Assim, é possível que após inalação
de OT os colaboradores fiquem mais sensíveis aos estímulos dolorosos no outro. No estudo
de Shamay-Tsoory et al. (2013), por exemplo, evidências revelaram que a administração de
OT é capaz de promover um aumento na avaliações subjetivas da dor física no outro, mesmo
sendo este um membro do exogrupo. Porém, Bos e colaboradores (2015) verificaram em um
estudo com ressonância, que a OT reduziu a atividade neuronal da circuitaria da dor (ínsula e
regiões sensório-motoras) durante observação de dor física no outro, independentemente da
etnia do alvo observado. Porém, os autores defendem que novas pesquisas ainda precisam ser
conduzidas pois nesse estudo não foram coletadas medidas subjetivas de empatia à dor após a
inalação de OT e a reduzida amostra pode não ter sido suficiente para captar efeitos étnicos.
Ainda não é claro o efeito da OT quando contrastamos os resultados da percepção
dolorosa em si e no outro, pois diferentes pesquisas sócio-endocrinológica demonstram
evidências contrárias. Exemplos disso, são os estudos de Jackson et al. (2006) e Abu-Akel et
al. (2014) em que os autores avaliaram o efeito da OT na percepção empática da dor em si
mesmo vs. na perspectiva do outro com a mesma tarefa experimental. Por um lado, no estudo
Norte-Americano de Jackson et al. (2006) eles convidaram colaboradores Brancos a avaliar a
intensidade de dor sentida em alvos Brancos desconhecidos. Enquanto que, no estudo
conduzido em Israel por Abu-Akel et al. (2014) os autores aplicaram a tarefa de empatia à dor
apresentando antes de cada situação o nome do colaborador (judeu israelita) ou um outro
nome tipicamente judeu israelita. Ao considerarmos diferenciações étnicas, pode-se dizer que
nos dois estudos os colaboradores interagiam com alvos do endogrupo. Porém, no primeiro
estudo ao observar as situações dolorosas os colaboradores sentiam-se mais desconfortáveis
(Jackson et al., 2006) e no segundo estudo, os colaboradores sentiam que o desconforto
gerado no alvo do estímulo observado foi maior do que o seu próprio (Abu-Akel et al., 2014).
Dessa forma, evidencia-se que a OT medeia a definição das barreiras entre si e o outro, mas
novos estudos precisam ser desenvolvidos para esclarecer esse fenômeno.
O efeito das pistas étnicas na observação de um alvo sofrendo estimulação dolorosa,
pode ser implicitamente avaliada com o uso da Estimulação Magnética Transcraniana (EMT).
Essa técnica é um método não invasivo utilizada em diversos tópicos de pesquisa para duas
finalidades (a) a neuromodulação de áreas cerebrais específicas ou (b) a avaliação da

32
fisiologia cortical e vias motoras descendentes. No âmbito do presente trabalho utilizamos
esta técnica para a segunda finalidade descrita, onde acessamos o parâmetro de pulso único de
EMT mais comum, o potenciais evocados motores (MEPs, sigla em inglês). Os MEPs nesse
estudo ocorrem durante a observação de situações dolorosas e neutras infligida em alvos de
diferentes grupos étnicos. Isso foi possível, pois a bobina de EMT produz um campo
magnético ao seu redor que em contato com o crânio induz um potencial de ação no tecido
cerebral subjacente. Essa ativação do córtex motor gera uma resposta de contração observável
no músculo correspondente (BARKER et al., 1985). Como ilustração, podemos destacar o
estudo de Avenanti, Sirigu e Aglioti (2010) que se propõem medir a intensidade das respostas
empáticas durante observação de situação de dor física. Esses autores verificaram um efeito
de supressão motora quando os colaboradores observaram estimulação dolorosa na mão de
membros do seu grupo étnico (endogrupo), e excitação motora quando a mão pertencia a
membros do grupo étnico distinto (exogrupo). O estudo de Azevedo e colaboradores (2012)
expande os achados de Avenanti, Sirigu e Aglioti (2010), pois com a mesma tarefa eles
avaliam os dados de ressonância magnética funcional e de respostas autonômicas diante
observação de dor no outro. Em linhas gerais, verificou-se reatividade empática enviesada
para a própria Etnia, uma vez que foi observada maior dilatação pupilar e ativação da ínsula
anterior quando o estímulo doloroso era infligido a membro do endogrupo em comparação ao
exogrupo.
A proposta do presente estudo envolve a replicação da tarefa desenvolvida por
Avenanti, Sirigu e Aglioti (2010), adicionando a administração de OT, o que possibilitará
importantes contribuições para a testagem dos efeitos comportamentais empáticos seletivos
desse neuropeptídeo em relação a diferentes grupos étnicos. Adicionalmente, o presente
estudo será o primeiro a avaliar o impacto da OT na empatia à dor física na mesma etnia vs.
etnia distinta em uma amostra ocidental.
1.6. O papel da ocitocina no processo de detecção de ameaça
A detecção de ameaça é uma habilidade necessária para a proteção dos indivíduos
pois, os prepara para lutar ou fugir de potenciais perigos. Assim, evidências revelam que
durante as interações sociais existe uma tendência de favorecimento dos membros do
endogrupo em detrimento aos membros do exogrupo (TAJFEL et al, 1971). Provavelmente,
isso ocorre devido a necessidade de segurança que os membros do endogrupo costumam
compartilhar, em contraste com o que ocorre com membros do exogrupo. Pesquisas nas áreas

33
da psicologia social e das neurociências têm permitido identificar como variáveis pessoais
(internas) e sociais (externas) influenciam esse viés perceptivo nas respostas
comportamentais. Assim, estas evidências explicam situações cotidianas, como a formação de
grupos entre os adolescentes, comportamentos protetivos de mães com seus bebês em relação
a potenciais ameaças, a rotina de policiais que devem ser precisos em agir diante de alvos
ameaçadores, entre outras situações. O fato é que, se tomarmos as ações policiais como
exemplo, nota-se que o controle cognitivo necessário em seu dia-a-dia durante sua tomada de
decisão por vezes falha, ocasionando certas injustiças sociais. Sendo que esta falha perceptiva
sustenta-se em tendências pessoais de comportamentos impulsivos ou nos estereótipos étnicos
constituídos sócio, cultural e biologicamente que regulam nossa percepção.
Para tanto, foram desenvolvidas diversas tarefas computadorizadas que permitem
estudar essa temática em contextos laboratoriais. Uma dessas relevantes tarefas é conhecida
como a Tarefa do Atirador em Primeira Pessoa (TAPP; First-person-shooter-task, FPST na
sigla em inglês) onde os colaboradores observam diferentes cenários, como praça, campo de
futebol e biblioteca, até que em um determinado momento surge na tela um alvo humano
(homem Branco ou Negro, em diferentes posições corporais) segurando um objeto que pode
ser uma arma ou um objeto inócuo (e.g. carteira, celular ou latinha de refrigerante)
(CORRELL et al., 2002). Os colaboradores são instruídos a identificar da maneira mais rápida
possível os homens que seguram armas, e que assim representam uma ameaça social. Os
resultados dessa pesquisa conduzida com frequência nos Estados Unidos, confirmaram o
panorama observado fora dos laboratórios. Assim, os colaboradores – universitários em sua
grande maioria Brancos – foram mais rápidos e precisos em atirar em alvos armados quando
estes eram homens Negros. Ainda, eles foram mais rápidos e precisos em não atirar quando os
alvos eram homens Brancos. Dessa forma, as pesquisas no contexto Norte-Americano,
revelaram uma tendência de uma associação (viés) negativa atribuída aos Negros, como sendo
mais violentos e ameaçadores que os Brancos (CORRELL et al., 2002), sendo observado na
vida real, onde os Negros costumam ser mais abordados e atingidos injustamente por
policiais.
Nessa mesma linha de pesquisa, Payne (2001) desenvolveu a Tarefa de Identificação
de Armas, os resultados desse estudo revelaram que os colaboradores tendem a identificar
erradamente ferramentas como sendo armas com maior frequência quando estas eram
precedidas por faces de homens Negros em comparação às faces de Brancos. O autor ainda
destaca a relevâncias desse dado no cotidiano quando pensamos que os estereótipos negativos
comumente atribuídos aos Negros podem gerar graves injustiças sociais, como testemunhos

34
enviesados, em que a acusação de pessoas Negras como criminosas tende a ser frequente. No
estudo I de Correll, Park, Judd, Wittenbrink (2007) foi testado o impacto do desempenho da
TAPP pela manipulação da associação entre grupo étnico e estereótipo de ameaça via
apresentação de uma matéria de jornal com uma descrição detalhada de um crime. Metade da
amostra desse estudo leu uma matéria onde o criminoso era Negro e a outra metade da
amostra leu a mesma matéria alterando apenas a identidade étnica do criminoso, que dessa
vez era Branco. Os resultados revelaram que a exposição a matéria com um criminoso Negro
aumentou o viés dos colaboradores em atirar em alvos Negros durante a TAPP. Além disso,
os autores verificaram que a prática melhora o desempenho dos universitários na TAPP.
Entretanto, esse efeito positivo não foi observado durante a replicação dessa tarefa, após 48
horas. Tais evidências em conjunto nos fazem pensar nos vieses compartilhados socialmente e
em como podemos modulá-los a longo prazo.
Uma análise minuciosa da identificação de alvos ameaçadores pela Teoria de
Detecção de Sinal (HARVEY, 1992), considera que os processos decisórios de atirar ou não
em um alvo envolvem (a) ser sensível à discriminação de alvos armados ou não
(independentemente da Etnia do alvo) e (b) um limiar dos critérios pessoais que regulam a
predisposição do colaborador em atirar em alguém (CORRELL et al., 2002). Curiosamente,
verifica-se que os colaboradores parecem apresentam menos critérios cognitivos no processo
de decisão de atirar diante de um alvo Negro em comparação a um alvo Branco
(GREENWALD, OAKES, E HOFFMAN, 2003), provavelmente devido aos estereótipos dos
Negros que, de forma automática, costumam ser associadas (no contexto Norte-Americano)
ao conceito de perigo. Assim, tendem a ser mais imediatas as respostas aos estímulos que
confirmam os estereótipos culturalmente vigentes.
Evidentemente, o treinamento de habilidades específicas pode modular o controle das
respostas apresentadas. Desse modo, Correll et al. (2007) compararam o desempenho na
tarefa do TAPP entre (a) amostras de policiais, de um departamento de polícia local e outro
nacional, (b) uma amostra de civis da comunidade em que esses policiais servem e (c)
universitários. Os resultados revelaram que os policiais apresentaram melhor desempenho em
comparação com os civis e universitários, julgando de forma mais rápida e precisa as
respostas de atirar e de não atirar. Porém, todos os grupos amostrais apresentam maior
latência para avaliar alvos que não são comumente associados (ex. Negros não armados e
Brancos armados) em comparação a avaliação de alvos comumente associados (ex. Negros
armados e Brancos não armados) na cultura Norte-Americana. Maiores latências de respostas
sinalizam que nos Estados Unidos a associação implícita entre Branco e Arma é mais fraca

35
em comparação a associação entre Negro e Arma. Diversas pesquisas foram então
desenvolvidas para melhor compreender as variáveis que orientam os colaboradores nos
processos decisórios nesses contextos (CORRELL et al., 2002; PLANT e PERUCHE, 2005).
Assim, nota-se a complexidade desse fenômeno, pois eventuais falhas do controle cognitivo
em regular atitudes preconceituosas ocorrem, apesar do exigente treinamento que os policiais
recebem para discriminar alvos ameaçadores.
Como já referido, o processamento neurobiológico da percepção de ameaça é
inicialmente dependente da amígdala, sendo que vários estudos evidenciaram uma maior
ativação dessa estrutura em resposta à visualização de faces do exogrupo étnico em
comparação a faces de membros do endogrupo (HART et al., 2000; PHELPS et al., 2000).
Dessa forma, essa estrutura cerebral tem um relevante papel em decodificar importantes
informações sociais. A OT é capaz de modular a ativação da amígdala, reduzindo a percepção
de estímulos ameaçadores tais como cenas não-sociais de ameaça (KIRSCH et al., 2005), ou
faces zangadas e com medo (DOMES, HEINRICHS, GLASCHER et al., 2007). Além disso,
Labuschagne et al. (2010) observaram que a OT foi capaz de reduzir a hiperativação
amigdalar em resposta à visualização de faces de medo em pacientes com Transtorno de
Ansiedade Social. Porém, até o momento nenhum estudo foi conduzido com o intuito de
analisar o efeito da OT em uma tarefa que reúna a identificação de estímulos ameaçadores em
contextos com pistas étnicas em uma amostra marcada pela miscigenação interétnica.
Pretendemos, com o presente estudo, contribuir para a clarificação sobre a possível
especificidade dos efeitos pró-endogrupo e/ou anti-exogrupo da OT, analisando o impacto da
sua administração durante o processo de identificação de armas comparando situações com
homens Brancos e Negros.
A escolha por avaliar os efeitos das pistas étnicas nas interações sociais é de grande
relevância para o contexto Brasileiro devido às consequências até hoje vivenciadas do
processo de socialização entre Negros e Brancos. O Brasil foi a última nação americana a
abolir a escravidão que era vigente há quase quatro séculos. Ainda hoje, identificam-se atos
preconceituoso no país porém, as pessoas não costumam assumir tal viés étnico (dados da
pesquisa conduzida pela Abril/MindMinders em 2017). Essa lacuna em relação à
conscientização da discriminação baseada na cor da pele, evidencia a necessidade de falarmos
sobre racismo e, especificamente, sobre como estes gestos apresentam-se diariamente de
forma velada e indireta. Segundo o censo do Instituto Brasileiro de Geografia e Estatística
(IBGE, 2014), 54% dos respondentes da pesquisa censitária nacional declararam-se pretos ou
pardos. Esse panorama faz do Brasil o segundo país com maior concentração de pessoas

36
Negras no mundo (apenas atrás da Nigéria). Atualmente, a renda média de pretos e pardos
continua aproximadamente metade da renda de Brancos e, além disso, os Negros sofrem com
outras consequências, como a baixa acessibilidade a educação de qualidade e ao ensino
superior, dificuldades no acesso a saneamento básico e maior disponibilidade à trabalhos de
baixa qualificação (segundo IBGE, 2014).
Em 1989 entrou em vigor a Lei do Racismo, a qual prescreve que o racismo é um
crime inafiançável e imprescritível. Todavia, na prática as ações intolerantes para com os
Negros costumam apenas serem enquadradas como crime de injúria racial, sendo que muitas
pessoas pretas e pardas revelam que não denunciam tais atitudes preconceituosas (GAMA,
2017). Esse cenário pode ser justificado pelo fato de que não temos no Brasil distinções
culturais tão evidentes entre Brancos e Negros, nem demarcações territoriais, como existiram
em outros países (por exemplo durante Apartheid na África do Sul). Talvez por isso,
minimizamos os impactos de certas atitudes discriminativas e não priorizamos o cumprimento
de ações afirmativas em relação aos Negros. A elaboração deste trabalho permitirá inclusive
discutir os efeitos atuais decorrentes desse processo histórico de relação entre esses grupos
étnicos, em um panorama atual de incentivo à convivência com as diferenças.

37
2. HIPÓTESES
De acordo com o escopo da literatura, a maioria dos resultados recentes tendem a
confirmar os modelos da Saliência Social dos estímulos e da Abordagem Interacionista como
os mecanismos neurobiológicos mais completos para se compreender os efeitos da OT em
humanos. Considerando isto, a seguir são descritas as hipóteses do presente estudo relativas
aos possíveis efeitos da OT na modulação de (a) respostas empáticas e de (b) percepção de
ameaça, diante dos estímulos étnicos do endogrupo e exogrupo. Vale ressaltar que esse estudo
se trata de um único experimento pois, avaliamos conjuntamente os efeitos pró- e anti-sociais
da OT nos comportamentos sociais em uma mesma amostra. Não é esperado que os grupos
experimentais, OT e PL, diferenciem-se em relação às avaliações explícitas de auto-relato das
competências empáticas, nível de agressividade e viés racial (resultados dos questionários).
Também não é esperado que a inalação de OT module a excitabilidade cortical em relação à
medida de linha de base (registro com EMT). Todavia, espera-se que o desempenho
comportamental dos colaboradores nas tarefas computadorizadas propostas seja:
(a) Quanto aos comportamentos de percepção de ameaça, espera-se diante do endogrupo
um forte perfil pró-social (efeito teto) que não esta sujeito a modulação pela OT.
Ainda, diante do exogrupo a inalação de OT pode alterar mecanismos neurais de
atenção aos estímulos (evidências anteriores que revelam modulação da atividade da
amígdala e colículos superiores) enquanto deve promover uma redução da ansiedade
social, gerando redução do viés racial negativo para com esses alvos.
Ha: aumento da acurácia na identificação de armas diante de alvos do exogrupo após
inalação de OT.
(b) Quanto aos comportamentos empáticos explícitos, a tendência em apresentar
comportamentos politicamente corretos costuma ser forte em pesquisas que envolvem
contextos de estudo de preconceito racial e esse fator deve sobrepor-se aos modestos
efeito comportamentais modulados pela OT.
Hb1: maior empatia explícita diante de membros do exogrupo.

38
Hb2: ausência de modulação dos comportamentos empáticos explícitos pela OT.
(c) Quanto aos comportamentos empáticos implícitos, evidências anteriores
demonstraram um aumento da empatia afetiva em relação ao endogrupo quando
comparado com o exogrupo e a OT costuma fortalecer esse efeito basal.
Hc: aumento da intensidade do contágio sensório-motor durante a observação de
situações de dor física infligida em membros do endogrupo após inalação de OT.

39
3. OBJETIVOS
3.1. OBJETIVO GERAL
O objetivo do presente trabalho consiste em avaliar a ocorrência de viés racial positivo
(efeito pró-social) e negativo (efeito anti-social) em universitários durante interações sociais
com o endo- e exogrupo e verificar se a administração intranasal de ocitocina é capaz de
modular os efeitos basais.
3.2. OBJETIVOS ESPECÍFICOS
- Explorar a universalidade de fenômenos encontrados a partir da comparação dos
resultados do grupo placebo obtidos nessa pesquisa e das evidências verificadas nas
pesquisas originais de percepção de ameaça (Correll et al., 2002), empatia explícita
(Jackson, Meltzoff, Decety, 2005) e empatia implícita (Avenanti, Sirigu, Aglioti,
2010).
- Analisar os efeitos pró- e anti-sociais modulados pela ocitocina em uma tarefa de
identificação de ameaças entre membros do endogrupo e do exogrupo.
- Analisar modulação das respostas empáticas explícitas pela ocitocina em uma tarefa
para avaliar a intensidade de dor física infligida em membros do endogrupo e do
exogrupo.
- Analisar modulação das respostas empáticas implícitas de contágio sensório-motor
pela ocitocina em uma tarefa de observação de dor física infligida em membros do
endogrupo e do exogrupo.
- Discutir se o padrão de respostas empáticas explícitas e implícitas tem mesma direção
ou direções opostas.
- Discutir de forma integrada os resultados pró- e anti-sociais observados pela
modulação da OT.

40
4. MÉTODO
4.1. PARTICIPANTES
Cento e setenta e cinco estudantes universitários participaram da coleta de dados do
presente estudo. Na Tabela 1 verifica-se o fluxograma do processo de aquisição de dados,
assim como detalhes em relação à exclusão de colaboradores. A amostra final incluída para a
análise reuniu os dados de 128 colaboradores (dimensão previamente determinada com base
numa análise de poder em que d = .05, α = .05; β = .80). Realizamos uma entrevista inicial
guiada pelo Questionário Sócio-Demográfico que desenvolvemos para explorar os critérios de
inclusão estabelecidos como necessários: ausência de doenças psiquiátricas (além de
aplicação do BDI para verificar especificamente sintomatologia depressiva) ou médicas
(incluindo alterações do funcionamento endócrino – avaliação por auto-relato do
colaborador), ausência do uso de medicações que interferissem no sistema nervoso central ou
que interagissem com a ocitocina sintética (verificação das possíveis interações
medicamentosas no site http://www.webmd.com/drugs/2/drug-32789/oxytocin-
bulk/details/list-interaction-medication), não ter consumo de tabaco superior a cinco cigarros
por dia ou fazer uso abusivo de álcool ou drogas.
A homogeneidade da presente amostra envolveu colaboradores homens, destros e
autodeclarados brancos (pela pigmentação de sua pele e pela representatividade dos membros
da família de origem), não tendo sido possível selecionar uma amostra com homogeneidade
étnica visto que os colaboradores não foram avaliados quanto a sua ascendência (e.g. latina,
europeia, americana, entre outras). A amostra foi composta exclusivamente por homens, uma
vez que os níveis endógenos de OT plasmática das mulheres variam ao longo do ciclo
menstrual (ALTEMUS et al., 2001), dificultando o controle de uma das variáveis centrais
deste estudo. Adicionalmente, uma vez que a maioria dos estudos mencionados na seção de
referencial teórico têm amostras totalmente masculinas, nossa escolha por esse grupo facilita
a comparação e discussão dos resultados. No sentido de testar as questões relativas à empatia
em relação ao endogrupo versus exogrupo e por questões de poder estatístico e de
conveniência, foram recrutados apenas indivíduos Branco, desta forma, selecionamos um
grupo étnico homogêneo a partir do qual nos foi permitido fazer inferências mais fiáveis.
Finalmente, recrutamos apenas colaboradores destros uma vez que diferenças de lateralidade
poderiam gerar vieses nos desempenhos das tarefas computadorizadas realizadas e, além
disso, porque na coleta do DIA 3 avaliou-se o contágio sensório-motor nos colaboradores
enquanto eles observavam mãos direitas sendo tocadas por diferentes objetos. Assim, caso

41
incluíssemos colaboradores com diferentes perfis de lateralidade poderíamos ter um fator de
confusão devido à possibilidade de modulação da sua auto-percepção em relação à mão
direita.
Os colaboradores incluídos na análise tinham em média 21.20 anos de idade (DP =
2.65, mín.-máx.: 18-29 anos; Coef. de variação: 12.51 %) e 93.08% cursavam o Ensino
Superior. Sessenta e quatro sujeitos foram alocados (aleatoriamente) ao Grupo que fez a
inalação de ocitocina sintética (OT), e outros sessenta e quatro colaboradores receberam para
inalação uma solução placebo contendo soro fisiológico. Os anúncios de convocação para as
coletas de dados ocorreram via redes sociais, sendo que os colaboradores receberam créditos
escolares pela participação.
Tabela 1 - Fluxograma do processo de coleta de dados e critérios de exclusão
Período de coletas de dados: Dezembro/2016 - Outubro/2017 (~1000 horas de coleta) Total de coletas realizadas: 175 Exclusões: 47 colaboradores (100%) 11 / 23.40% - Passaram mais do que 4 meses entre a coleta dos dias 1 e 2 para a coleta do dia 3 11 / 23.40% - Não quiseram mais participar do estudo 9 / 19.15% - Pontuaram mais do que 18 pontos no BDI 4 / 8.51% - Tomam medicações que interferem no sistema nervoso central 3 / 6.38% - Tiveram muita dificuldade em compreender as tarefas propostas 2 / 4.25% - Possuíam metais no corpo, ex. pino, marca-passo, etc. 2 / 4.25% - Apresentaram grande grau de ansiedade visivelmente (e fumavam mais do que 5 cigarros por dia) 1 / 2.13% - Era canhoto 1 / 2.13% - Usou uma amostra que foi preparada com uma concentração abaixo do necessário 1 / 2.13% - Fez cateterismo 1 / 2.13% - Teve traumatismo craniano há menos de 4 meses
Incluídos para as coletas dos DIAS 1, 2 e 3: 128 colaboradores (64 em cada Grupo) Dados analisados das coletas no DIA 2: • Tarefa de empatia explícita - 110 colaboradores (incluído apenas quem apresentou ±2 DP em relação à média de intensidade dolorosa percebida: OT = 53; PL = 57) • Tarefa do Atirador em Primeira Pessoa - 107 colaboradores (incluído apenas quem apresentou taxa de omissão < 20%: OT = 50; PL = 57) Coletas de dados do DIA 3 que foram interrompidas: 11 colaboradores (100%) 7 / 63.63% - MEPs não encontrados 4 / 36.36% - Desconforto durante a coleta de dados que impediu continuação Número de colaboradores, com pelo menos 44% dos registros de MEPs adequados para análise (8 MEPs entre os 18 pulsos liberados por categoria de análise): 73 colaboradores (distribuição nos grupos experimentais: OT = 31; PL = 42)

42
4.2. DESCRIÇÃO DOS INSTRUMENTOS UTILIZADOS
4.2.1. Escalas e Questionários
4.2.1.1. Questionário Sócio-Demográfico e Rastreio dos critérios de exclusão para a
administração de ocitocina (OT): questionário que possibilitou a caracterização dos
colaboradores, pois nele se questionou a idade, escolaridade, curso universitário realizado,
histórico de doenças e se o sujeito fazia uso de alguma medicação na época da coleta. As
questões do instrumento de rastreio dos critérios de exclusão para a inalação foram
organizadas para o presente trabalho com base nas informações destacadas por outras
pesquisas na área de administração hormonal via nasal, especificamente a ocitocina. Quanto
aos critérios de exclusão para a inalação da OT, buscamos verificar com 15 questões se o
colaborador tinha alergia à ocitocina sintética ou aos excipientes do syntocinon, se cumpriu as
duas horas de jejum necessárias, evitando a ingestão de café e o consumo de cigarros no dia
da coleta, além de evitar o uso de qualquer droga lícita ou ilícita pelos menos 24 horas antes
do momento da coleta de dados. Ainda, perguntamos se o sujeito estava com congestão nasal
por gripe ou alergia e se recentemente havia feito uso de corticóides ou realizou cirurgia de
correção de desvio de septo nos últimos 40 dias.
4.2.1.2. Inventário de Depressão de Beck (BDI, sigla em inglês; BECK et al., (1961); validada
para o português por GORENSTEIN e ANDRADE, 1998): Escala do tipo Likert de quatro
pontos, que contém 21 questões que avaliam a presença e intensidade de sintomatologia
depressiva nos colaboradores. A escala tem uma gradação de 0-63 pontos, sendo que a nota
de corte para definição de sintomas depressivos de nível moderado a severo varia entre os
perfis amostrais e diferentes culturas. Para a amostra Brasileira em geral, pontuações acima de
18 são consideradas clínicas (ver manual em português produzido por Cunha, p.12, 1996).
4.2.1.3. Rastreio dos critérios de Exclusão para a aplicação da Estimulação Magnética
Transcraniana (EMT): Instrumento aplicado pelo pesquisador, com dez perguntas que
rastrearam a existência de critérios de exclusão para a aplicação da EMT, especificamente,
verificou-se a presença de implantes metálicos no corpo, ocorrência de traumatismos
cranianos e histórico de epilepsia ou convulsões. Ainda perguntávamos se os colaboradores já
haviam participado de pesquisas que utilizaram estimulação cerebral, como Estimulação
Transcraniana por Corrente Contínua (tDCS) ou Estimulação Magnética Transcraniana

43
(EMT). Respostas afirmativas a uma ou mais questões não necessariamente indicavam
contraindicação absoluta aos procedimentos. Apenas quatro colaboradores foram de fato
excluídos devido a presença de implantes metálicos e traumatismos craniano. Porém, os riscos
e benefícios foram cuidadosamente analisados e discutidos caso a caso. Duração estimada de
aplicação de dois minutos.
4.2.1.4. Inventário de Lateralidade de Edinburgh (OLDFIELD, 1971): Instrumento que avalia
a dominância dos colaboradores em atividades específicas. Assim, o colaborador indicou com
qual mão (esquerda ou direita) ou pé ele comumente realiza dez atividades motoras cotidianas
específicas, como escrever, arremessar, escovar os dentes, entre outras. O uso desse
instrumento teve como objetivo controlar possíveis vieses de tempo de reação para
apresentação das respostas durante as tarefas computadorizadas e justificou-se ainda pelo fato
da tarefa de empatia à dor física apresentar estímulos dolorosos sendo aplicados à mão direita.
Desta forma, espera-se que o contágio sensório-motor pela observação desses estímulos seja
mais eficaz em pessoas destras, na medida em que estas observaram estimulação visualmente
congruente com sua mão dominante. A aplicação durou em torno de três minutos.
4.2.1.5. Escala Multidimensional de Reatividade Interpessoal (EMRI; DAVIS (1983):
Instrumento que avalia o perfil empático dos respondentes. No presente projeto utilizou-se a
versão traduzida e adaptada para o Brasil (SAMPAIO et al., 2011), que contém 26 itens
divididos nas seguintes subescalas: Angústia Pessoal – itens 27, 24, 23, 18, 20 e 15 (ex. “Fico
apreensivo em situações emergenciais”); Consideração Empática – itens 3, 5, 6, 10, 12, 13 e
17 (ex. “Costumo me emocionar com as coisas que vejo acontecer aos outros”); Tomada de
Perspectiva – itens 4, 7, 8, 9, 21, 22 (ex. “Tento compreender o argumento dos outros”); e
Fantasia – itens 1, 2, 14, 16, 19, 25, 26 (ex. “Sinto emoções de um personagem de filme como
se fossem minhas próprias emoções”). Os colaboradores indicaram seu grau de concordância
com cada afirmação por meio de uma escala Likert de cinco pontos, a qual variava de 1 =
"não me descreve bem" a 5 = "descreve-me muito bem". Apenas o item 19 apresentou uma
cotação invertida. A aplicação durou aproximadamente cinco minutos.
4.2.1.6. Escala de Preconceito Flagrante e Sutil (PETTIGREW e MEERTENS, 1995; versão
traduzida para o português por GIRÃO, DINA, & OSÓRIO, 2015): A versão em português da
Escala de Preconceito Flagrante e Sutil foi traduzida e adaptada pela nossa equipe de
laboratório porém, esta versão está em processo de validação. A escala avalia duas formas de

44
expressão de preconceito racial, o flagrante (direto) e o sutil (indireto). O instrumento é
composto por 20 enunciados, sendo 10 itens relacionados ao preconceito flagrante (ex. “Os
Negros conseguem empregos que deveriam ser dos Brancos”) e 10 itens ao preconceito sutil
(ex. “Algumas pessoas não se esforçam o suficiente. Se os Negros se esforçassem mais, eles
seriam tão bem sucedidos quanto os Brancos”), sendo que para as respostas utilizou-se a
escala Likert de quatro pontos, a qual variava de 1 = "discorda fortemente" a 5 = "concorda
fortemente". Quanto maior o escore, maior o perfil preconceituoso do colaborador. A
aplicação durou em torno de cinco minutos.
4.2.1.7. Questionário de Agressividade de Buss-Perry (BUSS e PERRY (1992): A versão em
português desse questionário (GOUVEIA et al., 2008) é composta por 26 itens que avaliam
quatro dimensões da agressividade: física – itens 1, 2, 5, 6, 14, 15, 21 e 22 (ex: “Se alguém
me bater, eu bato nele de volta”); verbal – itens 10, 16, 20 e 26 (ex: “Não consigo ficar calado
(a) quando as pessoas discordam de mim”); raiva – itens 3, 8, 11, 12, 13 e 17 (ex: “Alguns
amigos dizem que sou cabeça quente”); e hostilidade – itens 4, 7, 9, 18, 19, 23, 24 e 25 (ex:
“Quando as pessoas são muito gentis, duvido de suas intenções”). Os colaboradores
respondiam utilizando uma escala Likert de cinco pontos que variava de 1 = “Discordo
totalmente” a 5 = “Concordo totalmente” para indicar seu grau de concordância com cada
item. Quanto maior o escore, maior o perfil agressivo do colaborador. O preenchimento dessa
escala durou cerca de cinco minutos.
4.2.1.8. Questionário de experiências com diferentes grupos étnicos: Questionário construído
para o presente estudo com o intuito de avaliar: o grau de contato e intimidade dos
colaboradores com pessoas de diferentes grupos étnicos; os principais estereótipos
relacionados às pessoas Brancas e Negras (de acordo com a opinião do próprio colaborador e
o que ele acredita que a cultura Brasileira tem de referência desses grupos); e avaliação da
existência de preferências explícitas dos colaboradores em relação a esses grupos étnicos.
Instrumento composto por 29 itens, com questões para serem respondidas ora por escalas
Likert, ora por questões abertas. Instrumento em processo de validação sem que tenha sido
estabelecido a priori critérios para nota de corte. Seu preenchimento durou em média 35
minutos.
4.2.2. Tarefas Computadorizadas (programadas no E-Prime, versão 2.10)

45
4.2.2.1. Tarefa de avaliação das respostas empáticas explícitas durante observação de
situações dolorosas ou neutras (Adaptada de JACKSON, MELTZOFF, DECETY, 2005):
Essa tarefa foi baseada na proposta do estudo de Jackson, Meltzoff, Decety (2005) onde são
apresentadas diferentes situações potencialmente dolorosas ou situações neutras sendo que o
colaborador deve avaliar o quanto de dor ele percebe que a pessoa da imagem sentiu em cada
situação. Os estímulos originais não puderam ser usados, pois o presente estudo se propõe
avaliar diferenças nas respostas empáticas em contextos com pistas raciais. Assim, foi
necessária a criação de novos estímulos apenas com mãos. Estas mãos eram de coloração
Branca e Negra. Tirámos fotos de algumas situações equivalentes ao estudo original e criámos
ainda outras situações. Uma mesma situação era apresentada ora com modelos de mãos
Brancas, ora com modelos de mãos Negras, sendo que estas surgiram em ordem randomizada
durante a aplicação da tarefa. Ainda, considerando a proposta do estudo de Avenanti, Sirigu,
Aglioti (2010), que incluiu um controle de cor de mão das situações apresentadas, inserimos
estímulos das mesmas situações, com mãos Roxas. As mãos foram editadas para produzir a
coloração púrpura no programa Adobe Photoshop CC 2016, sendo todas baseadas nas mãos
Negras, assim teríamos uma mão característica de um exogrupo pela sua forma e pela cor.
Foram apresentadas no total 54 imagens (ver exemplos na Figura 2) coloridas digitais (18 de
cada condição Branca, Negra e Roxa, metade delas em situações potencialmente dolorosas e
outra metade nas situações neutras correspondentes), com resolução de 1280x800 píxeis. As
imagens foram tiradas em um ângulo que promovem uma perspectiva em primeira pessoa e
simulam situações do cotidiano (ex. passar roupa, cortar legumes, abrir um armário, etc.).
Foram representados vários tipos de dor (mecânica, térmica e de pressão) e as mãos exibidas
foram de modelos masculinos. Nessa tarefa avalia-se tendência dos colaboradores de
perceberem uma intensidade dolorosa no outro e em si. Para isso, os estímulos permaneciam
na tela por cinco segundos e, em seguida, surgiam duas perguntas, por um segundo e meio
cada, que avaliavam (a) a intensidade da dor que os colaboradores acreditavam que a pessoa
da imagem estava sentindo (foco no outro) e (b) o quanto de desconforto eles mesmos
sentiram ao ver a imagem (foco em si). As respostas a esses estímulos foram dadas
considerando uma escala Likert que variou entre 1 (“sem dor”) a 9 (“pior dor possível”).
Instruções dessa tarefa encontram-se a baixo. A duração média dessa tarefa foi de 15 minutos.
Observe as imagens a seguir com muita atenção. Essas imagens podem mostrar mãos Brancas, Negras e Roxas em situações potencialmente dolorosas ou situações inofensivas. Você deve classificar cada uma das imagens usando a escala a seguir: 1 = sem dor a 9 = pior dor possível. É muito importante que

46
para avaliar essas imagens você se coloque no lugar da pessoa da imagem. Após analisar a imagem por alguns segundos, você deverá responder as seguintes questões sempre nessa ordem:
- Em sua opinião, quanto de dor a pessoa da imagem está sentindo? (1 = sem dor a 9 = pior dor possível)
- Em sua opinião, quanto de sensação dolorosa você sentiu ao ver essa imagem? (1 = sem dor a 9 = pior dor possível) Para essas questões não existem respostas certas ou erradas, mas sim respostas que melhor descrevem o que você sentiu ao ver cada imagem. Seja o mais sincero possível.
Figura 2 - Ilustração dos estímulos utilizados na Tarefa de avaliação das respostas empáticas explícitas durante observação de situações dolorosas ou neutras com mãos Brancas, Negras e Roxas
4.2.2.2. Tarefa do Atirador em Primeira Pessoa / Dilema do Policial (TAPP; CORRELL et al.,
2002): Nessa tarefa são apresentados 20 estímulos de diferente cenários (ex., praças, frente de
prédios, universidade, etc.) e 80 estímulos com os mesmos cenários onde se encontra um
homem-alvo, que pode ser da Etnia Branco (endogrupo) ou Negro (exogrupo). Cada alvo
apareceu na tela em uma de cinco possíveis posições corporais (ex. em pé de perfil com a mão
ao longo do corpo ou com a mão próxima peito, em pé posicionado lateralmente, agachado
com a mão próxima ao ombro ou com a mão próxima ao joelho) e segurava uma arma ou um
objeto inofensivo (ex. carteira, celular, lata de refrigerante). Em linhas gerais, os
colaboradores simularam estar no papel social de um policial e, então, julgavam se os
homens-alvo que surgiam nos cenários indicavam ameaça social (alvos armados) ou não
(alvos desarmados). Ver detalhes da instrução dessa tarefa abaixo.
De acordo com o desempenho na tarefa eram oferecidas recompensas ou penalidades
em pontos acumulados durante essa atividade. Estas pontuações eram baseadas nas
consequências sociais reais desse tipo de julgamento: indicação correta de não atirar em um

47
alvo desarmado gerava ganho de cinco pontos (rejeição correta), mas indicando atirar perdia-
se 20 pontos (alarme falso); indicação correta de atirar em resposta a um alvo armado
ganhava-se 10 pontos (acerto), mas indicando não atirar perdia-se 40 pontos (erro). Caso os
colaboradores não respondessem, isso resultaria em uma penalidade de 10 pontos. As
respostas poderiam ocorrer até 630 ms a partir da apresentação desses estímulos de cenário
com um homem-alvo. Essa janela de tempo justifica-se uma vez que o foco do estudo é
exatamente reduzir a variabilidade do tempo de reação e aumentar a ocorrência de erros para
então analisar o viés racial dos colaboradores (janela de tempo de resposta definida de acordo
com CORRELL et al., 2002). Ao término do julgamento de cada estímulo, o total de pontos
acumulados a cada rodada apareciam na tela. Para que os colaboradores não se habituassem
ao momento em que surgia o homem-alvo, foi apresentado um número variado de cenários
sem pessoas, antes da tela onde julgava-se o alvo. A sequência completa de telas era
composta pela seguinte ordem: de 1 a 4 cenários sem pessoas (com 500-1000 ms de
apresentação de cada cenário), cenário com o homem-alvo (500-1000 ms); tela com os pontos
acumulados (1000 ms); e ponto de fixação (500 ms) (ver Figura 3).
O número variado de cenários no início das sequências inclusive foi idealizado nessa
tarefa visando aumentar o engajamento dos colaboradores no jogo, pois assim eles não
saberiam a hora exata que apareceria o homem-alvo e aumentavam sua atenção durante todo o
experimento. Na perspectiva do participante, o homem-alvo aparece de repente na tela. A
tarefa programada em E-Prime, assim como os estímulos originais, foram obtidos no site
oficial do Dr. Correll (http://psych.colorado.edu/~jclab/FPST.html). O tempo estimado para
realização dessa tarefa foi de aproximadamente 15 minutos.
Para jogar esse jogo você irá imaginar que é um policial e portanto deverá decidir o mais rápido possível se os homens que surgem no monitor seguram uma Arma ou um Objeto Inofensivo. - Caso o homem da imagem segure uma Arma isso representa uma ameaça social, e você, como policial, precisa atirar nele. Para isso pressione a tecla “D” para ATIRAR. - Caso o homem da imagem segure um Objeto Inofensivo isso não representa uma ameaça social, e você, como policial, NÃO precisa atirar nele. Para isso pressione a tecla “L” para NÃO ATIRAR. O participante dessa pesquisa que tiver o melhor desempenho, ou seja, que julgar de forma mais exata e rápida possível, e acumular o máximo de pontos possível, ganhará um vale FNAC!!! Considerando as penalidades reais e sociais desses tipos de escolha, todas as suas respostas serão pontuadas seguindo os critérios abaixo: a) Atirar em um alvo ameaçador: +10 pontos

48
b) Não atirar em um alvo ameaçador: -40 pontos c) Atirar em um alvo não ameaçador: -20 pontos d) Rejeitar corretamente atirar em um alvo não ameaçador = +5 pontos e) Ser muito lento em apresentar uma resposta diante de um alvo = -10 pontos Você deverá memorizar essas pontuações, pois ao final da tarefa iremos lhe perguntar sobre esses valores de penalidades. É muito importante que você preste atenção à face dos homens que surgirão no monitor, pois ao final dessa tarefa iremos testar também a sua habilidade de reconhecer os alvos. Os colaboradores que responderem mais de 10 vezes de forma muito lenta serão excluídos da análise final.
Figura 3 - (A) Desenho experimental com duas condições de sequências possíveis (homem-alvo Negro ou Branco) apresentados durante a Tarefa do Atirador em Primeira Pessoa (CORRELL et al. (2002); (B) Condições experimentais que foram randomicamente apresentadas: alvo Negro com objeto inócuo / com objeto ameaçador ou alvo Branco com objeto inócuo / com objeto ameaçador
Ao final desse teste, os colaboradores realizaram uma Tarefa de Reconhecimento de faces dos homens apresentadas na atividade anterior. Sendo-lhe apresentada a seguinte instrução:
Indique quais das faces Negras e Brancas a seguir foram apresentadas no jogo anterior o Dilema do Policial. Ainda você deverá informar o grau de certeza de cada uma de suas respostas de acordo com a escala a seguir: 1 = pouca certeza a 5 = muita certeza.
4.2.2.3. Tarefa de Associação Implícita racial (IAT racial, sigla em inglês; GREENWALD et
al., 1998): Essa tarefa avalia as atitudes implícitas de viés racial, pela medida da força de
associação entre Conceitos e Atributos. Os Conceitos – Negro e Branco – são representados
por faces de pessoas dos grupos étnicos Negro e Branco. Os Atributos – Positivo e Negativo –
são representados por palavras de valência positiva e negativa, como felicidade, prazer,
horrível e agonia (Figura 4). Para avaliar a força da associação, calcula-se o d-Score baseado

49
nas respostas e tempo de reação dos colaboradores (GREENWALD, NOSEK, BANAJI,
2003). O d-Score é um valor que varia entre -2 e +2, sendo que valores negativos indicam
uma associação mais forte entre Branco com Atributos Positivos e Negros com atributos
Negativos. A tarefa é composta por cinco blocos; o primeiro bloco serve para avaliar a
capacidade de discriminação dos Conceitos e o segundo bloco a discriminação dos Atributos.
No terceiro bloco, momento de avaliação de associações automáticas comuns, os
colaboradores observavam faces e palavras no centro da tela e era instruídos a classificá-las
de acordo com os Conceitos e Atributos que apareciam agrupados nos cantos superiores da
tela, como Branco - Positivos (à esquerda) e Negros – Negativos (à direita). No quarto bloco a
posição dos Conceitos na tela aparecia de forma invertida em comparação ao bloco 1.
Finalmente, no quinto bloco era o momento de avaliação de associações automáticas
incomuns (similar aos terceiro bloco, exceto pelo fato de que a posição dos Conceitos foram
os mesmos do bloco quatro). Metade dos colaboradores realizaram a tarefa na ordem descrita
acima e o restante da amostra realizou a tarefa com os blocos três e cinco na ordem inversa. A
tarefa durou em torno de 15 minutos. Abaixo seguem as instruções apresentadas nessa tarefa.
Você será apresentado a um conjunto de palavras ou imagens para serem classificados em grupos usando os botões “B” e “N”. Classifique os itens o mais rápido que puder, cometendo o menor número de erros possível. Realizando o teste de forma lenta ou cometendo muito erros, resultará em um valor não interpretável. Bloco 1: Aperte “B” se a foto na tela corresponder a uma face da categoria “Branco” e “N” se a foto corresponder a uma face da categoria “Negro”. Bloco 2: Aperte “B” se a palavra na tela corresponder a uma valência da categoria “Positiva” e “N” se a palavra corresponder a uma valência da categoria “Negativa”. Bloco 3: Aperte “B” se a palavra ou a face na tela corresponder as categorias “Branco” ou “Positiva” e “N” se corresponder as categorias “Negro” ou “Negativa”. Bloco 4: Aperte “B” se a foto na tela corresponder a uma face da categoria “Negro” e “N” se a foto corresponder a uma face da categoria “Branco”.

50
Bloco 5: Aperte “B” se a palavra ou a face na tela corresponder as categorias “Negro” ou “Positiva” e “N” se corresponder as categorias “Branco” ou “Negativa”.
Figura 4 - Ilustração dos estímulos apresentados em cada bloco experimental do IAT racial
4.2.2.4. Tarefa de avaliação das respostas implícitas de contágio sensório-motor durante
observação de situações dolorosas ou neutras (AVENANTI, SIRIGU, AGLIOTI, 2010):
Replicamos essa tarefa que consistiu na apresentação inicial de uma cruz de fixação que foi
apresentada por 1 seg., seguida de um entre seis diferentes tipos de vídeos com 1.8 segundos
cada (estímulos originais foram disponibilizados pelo Dr. Alessio Avenanti) e finalizamos
com a apresentação de uma tela cinza por 7.2 seg. Esses vídeos mostravam situações
dolorosas ou neutras, respectivamente: (a) seringas (internamente com líquidos de diferentes
cores: vermelho, amarelo ou marrom) penetrando o primeiro interósseo dorsal (FDI, sigla em
inglês) da mão direita de homens ou (b) cotonete (com cabos de diferentes cores: amarelo,
azul ou branco) tocando esse mesmo músculo da mão direita de homens. Essas mãos
poderiam ser de coloração Branca (endogrupo), Negra (exogrupo) ou Roxa (controle de cor)
(ver Figura 5). As mãos Roxas resultaram da edição em programa gráfico (Photoshop) das
mãos Negras, conforme recomendação do estudo original. Nessa tarefa foram apresentados
seis blocos ordenados de maneira pseudo-randomizada, com 18 vídeos em cada bloco, sendo
que dentro do mesmo bloco mostrava-se apenas um tipo de vídeo (mão Branca-seringa / mão

51
Branca-cotonete / mão Negra-seringa / mão Negra-cotonete / mão Roxa-seringa / mão Roxa-
cotonete). As instruções da tarefa eram:
Mantenha seus braços relaxados durante o experimento e busque movimentar-se o mínimo possível. Assista, com muita atenção, a todos os vídeos a seguir. Ao final da apresentação de todos os vídeos, lhe faremos perguntas sobre o que você observou.
Durante a apresentação dos vídeos, programamos para ocorrer a liberação de um pulso
magnético pelo aparelho de Estimulação Magnética Transcraniana (EMT) que estava com sua
bobina posicionada sobre a área do córtex motor primário (M1) no hemisfério esquerdo, sobre
a região que corresponde ao controle da mão direita do colaborador. Utilizamos essa técnica
para medir alterações na excitabilidade cortical de representações córtico-espinais corporais
dos colaboradores durante a observação desses vídeos. O pulso poderia surgir a qualquer
momento entre os 200-600 ms antes do fim da apresentação do vídeo para evitar quaisquer
efeitos de priming que pudessem afetar o contágio sensório-motor medido pelo potencial
evocado motor (MEP) dos colaboradores. Ainda, programamos a tarefa de forma que a
distância entre um pulso magnético e outro fosse de 10 seg, uma vez que nosso objetivo não
era produzir uma neuromodulação. Os potenciais motores evocados foram registrados pelo
posicionamento de dois pares de eletrodos de um eletromiógrafo (EMG). O par de eletrodos
que coletava os registros do músculo alvo, nomeadamente o FDI, estava sobre a mesma
musculatura da mão do participante que era estimulada nos vídeos dessa tarefa. Além disso,
para avaliarmos a especificidade da resposta de contágio-sensório motor sobre FDI foi
necessário contrastar esses registros com os dados advindos do posicionamento de outro par
de eletrodos controle em uma musculatura neutra (que não sofria estimulação durante os
vídeos apresentados). Conforme montagem proposta por Avenanti, Sirigu & Aglioti (2010), o
músculo controle selecionado foi o Músculo Abdutor do Mínimo (ADM).

52
Figura 5 - (A) Desenho experimental da Tarefa de avaliação do contágio sensório-motor ao observar dor no outro e (B) estímulos originais de Avenanti Stimuli, Sirigu & Aglioti (2010)
Após a conclusão da Tarefa de avaliação do contágio sensório-motor ao observar dor
no outro solicitamos que os colaboradores respondessem três tipos de perguntas: primeiro
para avaliar, de forma explícita, a intensidade de dor que eles acreditavam que as pessoas dos
vídeos e eles mesmo sentiram durante cada uma das situações apresentas (usando uma escala
que variou de 1 = “nenhuma dor sentida” a 9 = “máximo de dor sentida”); segundo, perguntas
que avaliaram o grau de familiaridade visual e similaridade física entre a mão do colaborador
em relação as mãos dos vídeos (ver detalhes das instruções a baixo); e terceiro, perguntas
sobre as cores dos objetos apresentados nos vídeos, assim como perguntas sobre
características das mãos dos vídeos (definição do gênero e idade estimada) para avaliar o grau
de atenção dos colaboradores na tarefa proposta. A tarefa durou aproximadamente 80
minutos.
Antes de finalizar a tarefa hoje, você deverá observar uma vez mais cada tipo de vídeo da tarefa anterior para responder as seguintes questões: - Em sua opinião, quanto de dor a pessoa do vídeo sentiu em cada situação? (1
= sem nenhum efeito doloroso a 9 = máximo de efeito doloroso imaginado) - Em sua opinião, quanto de dor você sentiu ao ver cada situação? (1 = sem
nenhum efeito doloroso em mim a 9 = máximo de efeito doloroso imaginado) - Em sua opinião, quanto cada estímulo observado é visualmente familiar na
sua própria mão? (1 = inteiramente desconhecida a 9 = inteiramente familiar)
- Em sua opinião, quanto cada estímulo observado é fisicamente similar à sua própria mão? (1 = inteiramente diferente a 9 = inteiramente similar)

53
4.2.3. Técnica
4.2.3.1. Estimulação Magnética Transcraniana (EMT): No presente estudo, essa técnica foi
utilizada para a mensuração de excitabilidade cortical. O aparelho utilizado é da marca
Magstim Bistim, sendo que o pulso magnético foi realizado utilizando uma bobina em
formato de 8 com 70 mm de diametro. Desenvolvemos um material informativo para o uso
dessa técnica visando a padronização da aplicação em relação aos materiais necessários para
esse tipo de coleta, parâmetros de como encontramos a região cortical de interesse e
referências para o procedimento de extração de dados. Os dados eletromiográficos foram
coletados por pares de eletrodos posicionados no músculo alvo (FDI) e músculo controle
(ADM), conectados ao amplificador de 5 canais Myoquick 1400ME e registrados com o
software SystemPlus.
Realizámos o posicionamento dos dois pares de eletrodos em cada um dos músculos
por uma montagem do tipo protuberância do músculo-articulação (Figura 6), onde o “pólo +”
(protuberância do músculo) e o “pólo –” (articulação) distanciaram-se entre 2-4 cm
(BASHIR, PEREZ, HORVATH, & PASCUAL-LEONE, 2013). Os cabos desses eletrodos
permaneceram entrelaçados entre si durante todas as coletas para que anulassem seus campos
magnéticos, uma vez que eles não eram blindados. Os parâmetros no Sistem Plus incluíram
EMG coletado com uma amostragem de 10kHZ e filtro band-pass de 20Hz-2.5kHz
(AVENANTI, SIRIGU, AGLIOTI, 2010). O Myoquick, outro aparelho também utilizado
nesse sistema, possibilitou a amplificação e registro de medidas neurofisiológicas dos
potenciais evocados por EMT. A delimitação do hotspot, região cortical alvo, baseou-se em
Bashir, Perez, Horvath & Pascual-Leone (2013) que demonstraram que a distância
correspondente à FDI e à ADM é mínima, portanto passível de sobreposição. Assim, a
Posição Ótima no Escalpe (POE) no hemisfério esquerdo foi encontrada com a bobina
orientada com uma rotação de ~ 45 graus, no sentido anti-horário, a partir do sulco central.
Buscamos primeiramente determinar o Limiar Motor (LM) em ADM, musculatura que
necessitava de uma potência na EMT ligeiramente maior em comparação ao FDI. O LM foi
obtido considerando a menor intensidade da máquina de EMT na qual pelo menos 5 em 10
pulsos sobre a POE resultavam em um potencial evocado motor (MEP) de 50µV ou maior.
Seguindo os parâmetro do estudo que buscamos replicar, a intensidade do pulso magnético
aplicado durante a Tarefa de avaliação do contágio sensório-motor ao observar dor no outro
foi de 130% do limiar motor de repouso encontrado, para assim obtermos um sinal estável em
ambas musculaturas.

54
Figura 6 - Da esquerda para a direita: eletrodos utilizados; eletrodos posicionados para registro de FDI; configuração completa de montagem dos eletrodos de EMG, ponto preto corresponde ao “pólo +” (protuberância do músculo - musculaturas alvos) e ponto cinza corresponde ao “pólo –” (articulação - eletrodo de referência); posicionamento da bobina de EMT sobre M1.
4.2.4. Amostras para inalação
4.2.4.1. Solução com Ocitocina sintética (OT) e solução Placebo (PL): Os colaboradores
fizeram a autoadministração intranasal de uma dose de 24 UI de OT (três nebulizações, de 4
UI, em cada narina – concentração de OT baseada nos estudos da área referenciados nesse
projeto) ou de uma solução placebo (soro fisiológico), 40 minutos antes do início das tarefas
computadorizadas. Em ambos os grupos os colaboradores receberam a mesma embalagem
porém, no grupo placebo, esta possuía uma solução neutra em que a OT não estava presente.
As soluções foram preparadas em monodose no Laboratório de Bioquímica da Universidade
Presbiteriana Mackenzie (UPM), sendo que o farmacêutico responsável pelo preparo das
amostras foi quem determinou códigos genéricos para caracterizar cada tipo de embalagem
visando preservar o sigilo das amostras utilizadas. Apenas no momento de tabulação e análise
do dados as identificações das amostras foram reveladas.
4.4. PROCEDIMENTO
Assim que os colaboradores chegavam ao Laboratório de Neurociência Social e
Cognitiva da UPM eram recebidos pela equipe de coleta a qual lhes apresentavam uma
descrição completa das etapas do presente estudo, sendo-lhes então solicitada a assinatura do
Termo de Consentimento Livre e Esclarecido. Esse estudo consiste em um único estudo uma
vez que nosso objetivo foi avaliar efeitos pró- e anti-sociais da OT nos comportamentos
sociais na mesma amostra de colaboradores. A coleta ocorreu em três dias distintos com
duração média de 2h cada encontro, perfazendo cerca de 6h de participação de cada

55
colaborador no estudo (total de aproximadamente 770h de coleta de dados completa, apenas
considerando os participantes incluídos na amostra final). Para isso, no DIA 1 de coleta os
colaboradores preencheram as seguintes escalas e questionários nesta ordem: Questionário
Sócio-Demográfico e Rastreio dos critérios de exclusão para a administração de ocitocina;
Inventário Beck de Depressão; Rastreio dos critérios de Exclusão para a aplicação da
Estimulação Magnética Transcraniana; Inventário de Lateralidade de Edinburgh; Escala
Multidimensional de Reatividade Interpessoal; Escala de Preconceito Flagrante e Sutil;
Questionário de Agressividade de Buss-Perry; e Questionário de experiências com diferentes
grupos étnicos. Os colaboradores permaneciam sozinhos na sala de coleta e o pesquisador
sempre avisava que não avaliaríamos as respostas de uma forma individual, mas sim coletiva,
e que resguardaríamos a identidade do colaborador. Por fim, os colaboradores realizaram,
acompanhados pelo pesquisador, um treino das tarefas computadorizadas, pois no próximo
dia de coleta eles estariam sozinhos em sala e precisariam estar familiarizados com as tarefas
aplicadas.
As coletas de dados dos DIAS 2 e 3 ocorreram com uma distância máxima de 15
semanas (devido à logística de uso do espaço), entre as 12h e 16h, para controle de oscilações
hormonais normais que ocorrem ao longo do dia. Ainda, solicitou-se que os colaboradores
chegassem para a coleta com jejum de duas horas (exceto água), que não fumassem nem
tomassem café no dia da coleta, nem consumissem drogas lícitas ou ilícitas pelo menos 24
horas antes da coleta. Após conferência desses quesitos fundamentais para a devida aplicação
dos procedimentos previstos, os colaboradores fizeram a auto-administração de uma das duas
substâncias selecionadas, ocitocina ou solução placebo. O estudo foi duplo-cego, pois nem os
colaboradores, nem os pesquisadores tinham conhecimento sobre qual substância foi inalada.
Em seguida, eles permaneceram na sala de coleta sozinhos por 40 minutos sem realizar
qualquer atividade (incluindo consultar o celular) para que a OT alcançasse gradativamente
seu estado de equilíbrio, então buscávamos aplicar as tarefas de cada encontro em até 50
minutos, tempo em que ainda se observa seu efeito modulatório no cérebro (BORN et al.
2002). No DIA 2, quando o tempo de espera encerrava-se, um dos pesquisadores retornava à
sala para iniciar as tarefas computadorizadas previstas e saia da sala na sequência. Em uma
ordem randomizada pelo programa, foram apresentadas as seguintes atividades: Tarefa de
percepção explícita de dor no outro e Tarefa do Atirador em Primeira Pessoa / Dilema do
Policial. No final dessa coleta verificávamos se os colaboradores perceberam alguma
mudança corporal relacionada à inalação da substância recebida e qual foi o seu grau de
ansiedade durante o momento de espera (escala Likert entre 1 = “nada ansioso” a 5 = “muito

56
ansioso”). Antes dos colaboradores irem embora, era oferecido um lanche considerando o
tempo prolongado de jejum e agendávamos o dia da última coleta.
No DIA 3, a coleta era iniciada com a conferência dos quesitos fundamentais para a
nova auto-administração do mesmo tipo de substância inalada no DIA 2 de coleta. Os
colaboradores responderam à Tarefa de Associação Implícita racial e então, o pesquisador
posicionava os eletrodos do eletromiógrafo sobre as musculaturas de interesse da mão direita
dos colaboradores (FDI, alvo e ADM, controle). Além disso, foi realizada a etapa de
neuronavegação para a conferência da representação cortical do primeiro interósseo dorsal
(FDI), considerada a Posição Ótima no Escalpe (POE). A POE foi identificada com o uso da
EMT por meio de pulsos simples e registros da contração muscular por análise
eletromiográfica, sendo que a tarefa seria iniciada quando era possível induzir MEPs no
músculo controle (ADM) com amplitude de pico a pico > 50 µV em ao menos cinco entre dez
aplicações.
A partir disso, conduziu-se a medida de linha de base dos potenciais evocados motores
para registrar a excitabilidade cortical anterior à inalação e realização da tarefa
computadorizada. Em seguida, os colaboradores fizeram a inalação da substância selecionada
e aguardaram 40 minutos na sala de coleta sem realizar qualquer atividade. Após esse tempo,
conduzimos mais uma vez uma medida para avaliar a excitabilidade cortical pós-inalação,
para verificar possíveis impactos da inalação na intensidade de contração muscular sem a
apresentação de nenhum estímulo. Somente após essas etapas, foi aplicada a Tarefa de
avaliação do contágio sensório-motor ao observar dor no outro enquanto mediamos os MEPs
dos colaboradores durante a observação dos vídeos dessa tarefa. Por último, fizemos algumas
perguntas sobre essa tarefa para verificar o grau de atenção do colaborador durante o
experimento; avaliamos possíveis efeitos colaterais relacionados à inalação e verificamos o
grau de ansiedade dos colaboradores durante a espera. Finalmente, apresentamos uma breve
devolutiva (que abarcou uma explicação geral do estudo) e os colaboradores partilharam suas
impressões acerca da experiência conosco. Nesse momento avisamos que posteriormente
enviaríamos por e-mail um resumo dos principais achados do trabalho. Novamente
oferecemos um lanche e entregamos o comprovante de participação na pesquisa para a
validação de créditos escolares.
4.5. ANÁLISE DOS DADOS

57
A análise dos dados foi conduzida com o uso do pacote IBM SPSS Statistics 21.0,
respeitando-se a distribuição dos dados e estabelecendo para todos os testes estatísticos um
erro de α=5%. Quanto aos dados sócio-demográficos, estatística descritiva com valores de
média e desvio-padrão foram calculados para as escalas e questionários preenchidas, além de
terem sido conduzidos Testes-t para amostras independentes para comparar o perfil dos dois
grupos amostrais. Para explorar quais os principais estereótipos atribuídos à pessoas Brancas
e Negras, utilizamos um programa próprio para análise de dados textuais chamado ALCEST.
Assim, essa análise possibilitou um mapeamento dos três estereótipos comumente associados
a cada um desses grupos étnicos pelos colaboradores desse estudo. Teste de Qui-quadrado foi
conduzido para avaliar a efetividade da coleta duplo-cego e portanto averiguar se os
colaboradores identificaram a substância inalada.
Tarefa do Atirador em Primeira Pessoa. Os valores da latência das respostas corretas
sofreram uma transformação logarítmica para a condução das análises estatísticas porém, nos
gráficos apresentados na seção de resultados, seus valores foram apresentados em
milissegundos para facilitar a compreensão dos dados. Conduziu-se uma ANOVA mista 2
(Etnia: Negro, Branco) x 2 (Objeto: Arma, Não-Arma), com as variáveis dependentes taxa de
erro e latência de resposta corretas, fator entre-sujeito Grupos Experimentais (OT, PL), com
comparação entre médias do tipo LSD. Em todos os testes de diferenças conduzidos nessa e
nas demais tarefas, reportamos as magnitudes dos efeitos – ANOVA mista, ƞ: pequeno a
partir de .02; médio .13 e grande .26 (ESPÍRITO-SANTO & DANIEL, 2018).
Ainda, os dados dessa tarefa foram reorganizados de maneira a nos permitir conduzir
uma análise segundo a Teoria de Detecção de Sinal (HARVEY, 1992). Essa teoria propõem
uma análise em separado dos processos sensoriais e decisórios durante a execução dessa
tarefa. As variáveis analisadas destes processos são conhecidas como sensibilidade (d’) e
critério de decisão (c), as quais são decorrentes de cálculos com as medidas das taxas de
Atirar Corretamente e as taxas de Falsos Alarmes. Harvey (1992) assumiu que a sensibilidade
de discriminar alvos tem distribuições normais (ver Figura 7 para compreensão esquemática
da Teoria de Detecção de Sinais). Dessa forma, o índice de sensibilidade (d’) indica a
capacidade de discriminação de um sinal (no contexto do presente estudo, seriam os estímulos
de indivíduos segurando armas) em relação a ruídos (estímulos de indivíduos segurando
outros objetos que não armas). Assim, quanto melhor a sensibilidade do colaborador, mais
distantes serão as curvas de distribuições. Vale destacar que valores baixos de sensibilidade
produzem elevadas taxas de Falsos Alarmes (atirar em alvos desarmados) e elevadas taxas de

58
Perdas (não atirar em alvos armados). O ponto de encontro entre essas curvas caracteriza uma
limitação perceptiva do colaborador que, nesse momento, não consegue discriminar entre
alvos armados e desarmados. Esse viés perceptivo tem um limiar que varia de acordo com as
características pessoais de cada colaborador. Aqueles que apresentam uma maior
predisposição em atirar em um alvo, apesar dele estar desarmado, terão o critério de decisão
(c) com um menor limiar. Isso produz elevadas taxas de Falsos Alarmes e baixas taxas de
Perdas. Por outro lado, colaboradores com menor predisposição a atirar em alvos terão o
critério de decisão com um maior limiar. Testes-t para amostras independentes foram
conduzidas para comparar as medidas de sensibilidade e critério de decisão entre (a) as
amostras Norte-americanas (estudo original) e Brasileira; e em nosso estudo (b) comparando
as amostras OT e PL. Em todos os testes de diferenças conduzidos nessa e nas demais tarefas,
reportamos as magnitudes dos efeitos – Teste-t, d de Cohen: pequeno, a partir de 0.20, médio
0.50, e grande 0.80 (ESPÍRITO-SANTO & DANIEL, 2018).
Figura 7 – (A) Distribuições normais hipotéticas dos resultados com a presença (armas) ou ausência (objeto
inócuo) de sinais. Tais distribuições são construídas separadamente entre os alvos Negros e Brancos para a
Tarefa do Atirador em Primeira Pessoa. A linha vertical representa o limiar do critério de decisão do
SENSIBILIDADE
Distribuição de Ruído (Não-Arma)
Probabilidade
Percepção de Ameaça
Distribuição de Sinal (Arma)
CRITÉRIO DE DECISÃO
Não Atirar Atirar
Taxa Atirar Corretamente Taxa Não Atirar
Taxa Falso Alarme Taxa Rejeição Correta
Sinal presente
Sinal ausente
Não Atirar Atirar
A
B

59
colaborador. Valores acima do limiar indicam resposta de “atirar” no alvo, enquanto que valores a baixo ou
iguais ao critério indicam respostas de “não atirar” no alvo; (B) Condições prováveis de respostas em
Paradigmas de Detecção de Sinal (figura baseada nas ilustrações do material do curso de Psicologia da
Percepção da Universidade do Colorado, ministrada pelo Prof. Lewis O. Harvey)
Tarefa de empatia explícita à dor física no outro. ANOVA mista 2 (Perspectiva da dor
percebida: pelo Outro, em Si) x 3 (Cor da mão: Negro, Branco, Roxa) x 2 (Situação:
Dolorosa, Neutra), com a variável dependente intensidade de dor física percebida e fator
inter-sujeitos Grupo (OT, PL).
Tarefa de empatia implícita à dor física no outro. Para esta análise das medidas
implícitas de empatia, primeiramente foi necessário o pré-processamento dos dados de
potenciais motores evocados (MEPs) indicativos da intensidade de contágio sensório-motor
dos colaboradores ao observarem os vídeos de situações dolorosas ou neutras. As etapas
compostas nesta fase para cada colaborador envolviam: marcação no Sistem Plus dos pontos
de interesse do MEP, como momento de início (onset) e fim, além dos pontos mais altos e
baixos para determinação da amplitude (2h); extração dos valores referentes a estas
marcações para o Word (1h); e reorganização de dados no Excel (1h). No total, a atividade de
pré-processamento dos dados dos 128 colaboradores que participaram da coleta do dia 3
decorreu em aproximadamente em 512 horas, sendo que dez integrantes da equipe
contribuíram para essa etapa. A partir disso, um técnico, especialista em Tecnologia de
Informação, de uma equipe parceira do Social and Affective Neuroscience Laboratory,
University of Haifa (Israel) e a aluna responsável por esse trabalho desenvolveram um código
de programação no software Python para organizar os dados de MEPs visando as variáveis de
interesse para as análises estatísticas subsequentes. A criação desta linha de programação
demorou 32 dias, permitindo que posteriormente o processamento completo dos dados dos
128 participantes ocorresse em menos do que 1 hora. Dez por cento dos casos foram revistos
após a execução desse processamento para confirmar a inexistência de erros.
As medidas de MEP analisadas foram: (a) percentagem de MEPs salvos por categoria de
estímulos apresentados (assim definimos critérios para a exclusão de participantes nessa
tarefa); (b) latência do MEP (em ms); (c) o momento de início (onset, em ms); e (d) subtração
das amplitudes dos MEPs de situações dolorosas menos as situações neutras (análise do
contágio sensório-motor). Essas informações foram analisadas por ANOVAs mistas para
comparação das medidas de MEPs entre Momentos (com 2 níveis: pré- e pós-inalação) e em

60
seguida, uma análise 3 (Cor da mão: Negro, Branco, Roxa) x 2 (Situação: Dolorosa, Neutra) x
2 (Músculo: FDI, ADM) com Grupo como fator inter-sujeitos (OT, PL). Para avaliarmos
efeitos modulatórios da OT sobre o nível de contágio sensório-motor em FDI conduzimos
primeiramente três Testes-t para uma amostra contrastando os valores médios de cada
categoria de cor de mão com 0 e em seguida uma ANOVA mista com amplitudes dos MEPs
de situações de [dor] – [neutra] diante da observação de mãos Brancas e Negras como
variável dependente e Grupo (OT, PL) como fator inter-sujeitos.
4.6. ASPECTOS ÉTICOS
O estudo foi conduzido de acordo com os requerimentos do Comitê de Ética da
Universidade Presbiteriana Mackenzie e também baseado nas recomendações estabelecidas
na Declaração de Helsinki (1964), conforme emenda em Tóquio (1975), Veneza (1983) e
Hong- Kong (1989). A todos os colaboradores foram apresentados os objetivos e métodos
propostos nesse experimento, sendo que as coletas apenas iniciaram-se após eles terem
assinado o Termo de Consentimento Livre e Esclarecido. Os colaboradores foram
devidamente avisados que poderiam deixar o estudo a qualquer momento que desejassem sem
necessidade de justificativa e sem lhes causar qualquer prejuízo. Além disso, todas as
informações fornecidas foram estritamente sigilosas, e cada participante foi referenciado por
uma codificação de cinco dígitos (construída a partir da idade do colaborador no DIA 1 de
coleta e a posição deste considerando a ordem de coleta realizada). A coleta de dados do
presente estudo foi completada anteriormente à etapa de Qualificação do Doutorado devido a
extensa dedicação necessária para essa fase e considerando o curto período remanescente de
vigência da bolsa FAPESP. Todavia, nosso respaldo sustentou-se na aprovação pela FAPESP
da proposta metodológica presente e, evidentemente, pela aprovação pelo Comitê de Ética da
Universidade Presbiteriana Mackenzie e pela Plataforma Brasil (CAAE:
25874414.0.0000.0084).

61
5. RESULTADOS
5.1.Caracterização da amostra
Teste-t para amostras independentes não revelou diferenças estatisticamente
significativas entre os grupos em relação as variáveis de caracterização (ver detalhes na
Tabela 2). Os grupos PL e OT apresentaram um padrão de resultados semelhante, em que
observa-se: ausência de sintomas depressivos clinicamente relevantes; maior nível de contato
com pessoas Brancas em comparação a pessoas Negras; automatização da associação entre a
Etnia Branco com atributos positivos e Negro com atributos negativos; durante os 40 minutos
de espera após inalação da substância distribuída, o nível de ansiedade dos colaboradores era
de intensidade média; na avaliação de agressividade, os colaboradores revelaram que diante
de situações ameaçadoras eles têm mais comportamentos hostis e menos comportamentos de
agressividade verbal; na avaliação de empatia os colaboradores pontuaram maior nível de
consideração empática e menor de angústia pessoal; um perfil preconceito mais sutil do que
explicitamente evidente; e estereótipos favorecendo pessoas Brancas em detrimento de
pessoas Negras.
Teste de Qui-quadrado revelou que os colaboradores não diferenciaram o tipo de
substância inalada, χ2(1) = .32, p = .571. Nos dois grupos, a maioria dos colaboradores
indicou ter auto-administrado a substância placebo (69% do grupo PL e 63% do grupo OT).
Ainda, não se observaram diferenças estatisticamente significativas para a ordem de
apresentação dos instrumentos aplicados neste projeto.
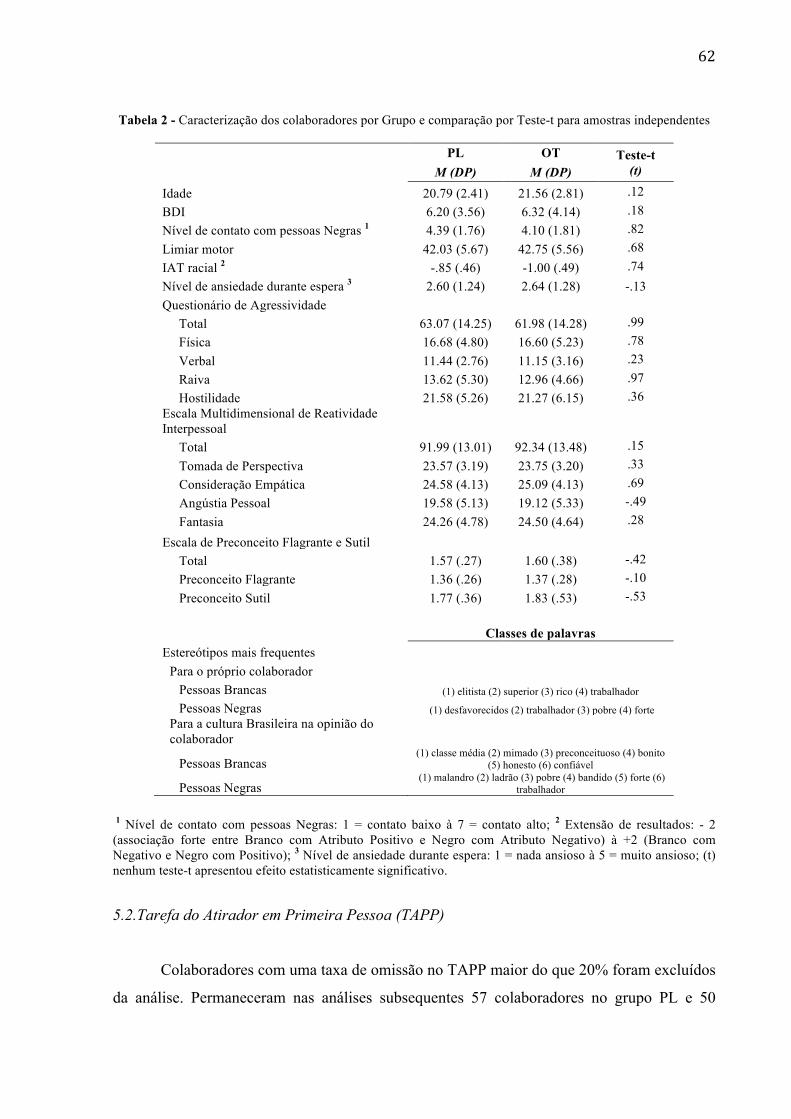
62
Tabela 2 - Caracterização dos colaboradores por Grupo e comparação por Teste-t para amostras independentes
PL OT Teste-t
(t)
M (DP) M (DP) Idade 20.79 (2.41) 21.56 (2.81) .12 BDI 6.20 (3.56) 6.32 (4.14) .18 Nível de contato com pessoas Negras 1 4.39 (1.76) 4.10 (1.81) .82 Limiar motor 42.03 (5.67) 42.75 (5.56) .68 IAT racial 2 -.85 (.46) -1.00 (.49) .74 Nível de ansiedade durante espera 3 2.60 (1.24) 2.64 (1.28) -.13 Questionário de Agressividade Total 63.07 (14.25) 61.98 (14.28) .99 Física 16.68 (4.80) 16.60 (5.23) .78 Verbal 11.44 (2.76) 11.15 (3.16) .23 Raiva 13.62 (5.30) 12.96 (4.66) .97 Hostilidade 21.58 (5.26) 21.27 (6.15) .36 Escala Multidimensional de Reatividade Interpessoal
Total 91.99 (13.01) 92.34 (13.48) .15 Tomada de Perspectiva 23.57 (3.19) 23.75 (3.20) .33 Consideração Empática 24.58 (4.13) 25.09 (4.13) .69 Angústia Pessoal 19.58 (5.13) 19.12 (5.33) -.49 Fantasia 24.26 (4.78) 24.50 (4.64) .28
Escala de Preconceito Flagrante e Sutil Total 1.57 (.27) 1.60 (.38) -.42 Preconceito Flagrante 1.36 (.26) 1.37 (.28) -.10 Preconceito Sutil 1.77 (.36) 1.83 (.53) -.53
Classes de palavras Estereótipos mais frequentes
Para o próprio colaborador Pessoas Brancas (1) elitista (2) superior (3) rico (4) trabalhador Pessoas Negras (1) desfavorecidos (2) trabalhador (3) pobre (4) forte
Para a cultura Brasileira na opinião do colaborador
Pessoas Brancas (1) classe média (2) mimado (3) preconceituoso (4) bonito
(5) honesto (6) confiável
Pessoas Negras (1) malandro (2) ladrão (3) pobre (4) bandido (5) forte (6)
trabalhador
1 Nível de contato com pessoas Negras: 1 = contato baixo à 7 = contato alto; 2 Extensão de resultados: - 2 (associação forte entre Branco com Atributo Positivo e Negro com Atributo Negativo) à +2 (Branco com Negativo e Negro com Positivo); 3 Nível de ansiedade durante espera: 1 = nada ansioso à 5 = muito ansioso; (t) nenhum teste-t apresentou efeito estatisticamente significativo.
5.2.Tarefa do Atirador em Primeira Pessoa (TAPP)
Colaboradores com uma taxa de omissão no TAPP maior do que 20% foram excluídos
da análise. Permaneceram nas análises subsequentes 57 colaboradores no grupo PL e 50
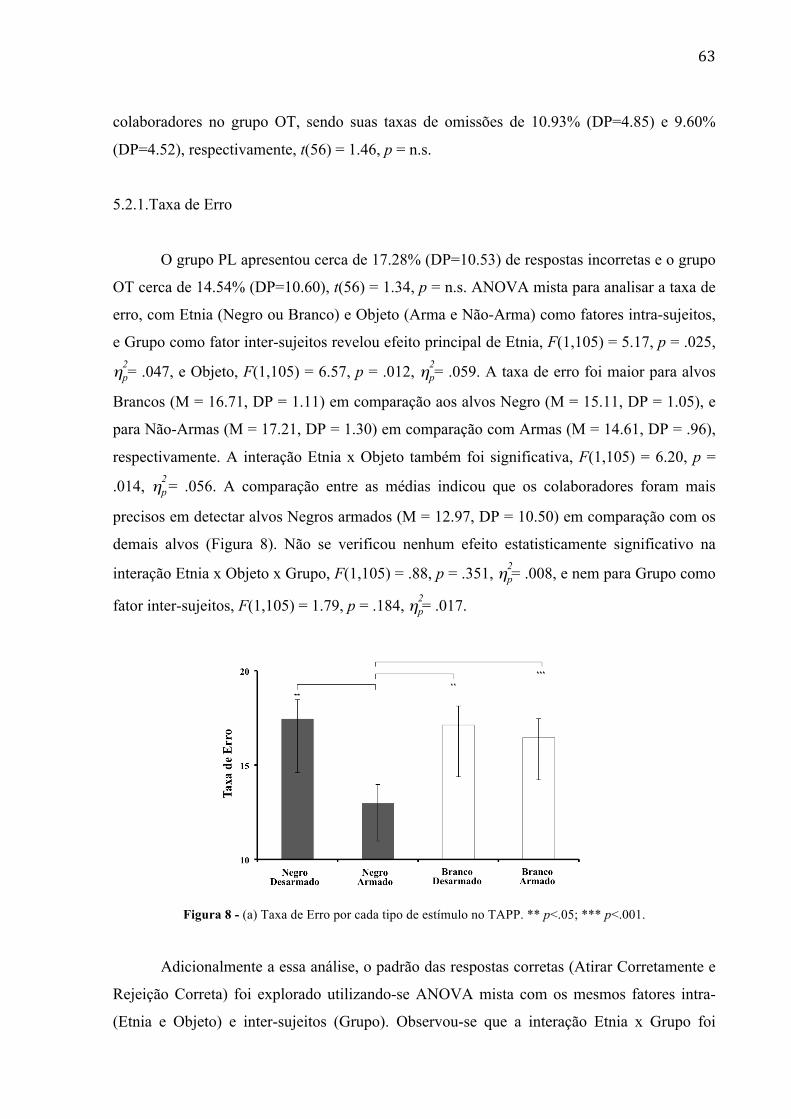
63
colaboradores no grupo OT, sendo suas taxas de omissões de 10.93% (DP=4.85) e 9.60%
(DP=4.52), respectivamente, t(56) = 1.46, p = n.s.
5.2.1.Taxa de Erro
O grupo PL apresentou cerca de 17.28% (DP=10.53) de respostas incorretas e o grupo
OT cerca de 14.54% (DP=10.60), t(56) = 1.34, p = n.s. ANOVA mista para analisar a taxa de
erro, com Etnia (Negro ou Branco) e Objeto (Arma e Não-Arma) como fatores intra-sujeitos,
e Grupo como fator inter-sujeitos revelou efeito principal de Etnia, F(1,105) = 5.17, p = .025,
ηp2 = .047, e Objeto, F(1,105) = 6.57, p = .012, ηp
2 = .059. A taxa de erro foi maior para alvos
Brancos (M = 16.71, DP = 1.11) em comparação aos alvos Negro (M = 15.11, DP = 1.05), e
para Não-Armas (M = 17.21, DP = 1.30) em comparação com Armas (M = 14.61, DP = .96),
respectivamente. A interação Etnia x Objeto também foi significativa, F(1,105) = 6.20, p =
.014, ηp2 = .056. A comparação entre as médias indicou que os colaboradores foram mais
precisos em detectar alvos Negros armados (M = 12.97, DP = 10.50) em comparação com os
demais alvos (Figura 8). Não se verificou nenhum efeito estatisticamente significativo na
interação Etnia x Objeto x Grupo, F(1,105) = .88, p = .351, ηp2 = .008, e nem para Grupo como
fator inter-sujeitos, F(1,105) = 1.79, p = .184, ηp2 = .017.
Figura 8 - (a) Taxa de Erro por cada tipo de estímulo no TAPP. ** p<.05; *** p<.001.
Adicionalmente a essa análise, o padrão das respostas corretas (Atirar Corretamente e
Rejeição Correta) foi explorado utilizando-se ANOVA mista com os mesmos fatores intra-
(Etnia e Objeto) e inter-sujeitos (Grupo). Observou-se que a interação Etnia x Grupo foi
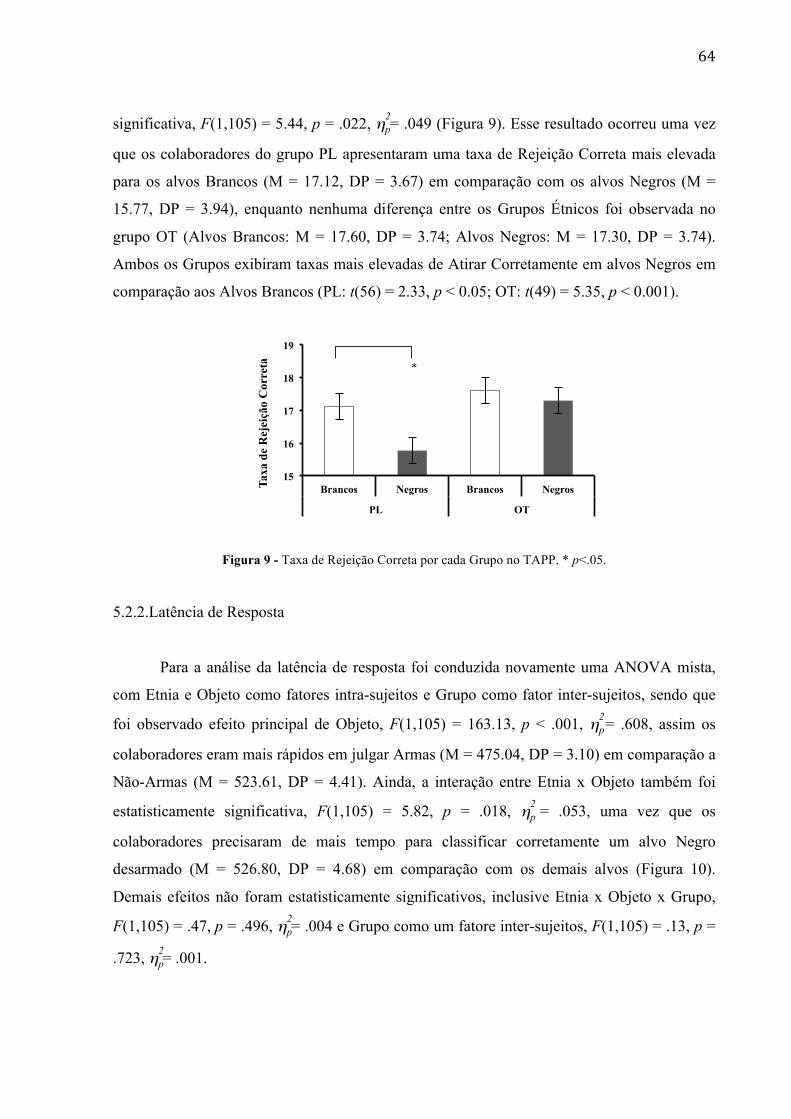
64
significativa, F(1,105) = 5.44, p = .022, ηp2 = .049 (Figura 9). Esse resultado ocorreu uma vez
que os colaboradores do grupo PL apresentaram uma taxa de Rejeição Correta mais elevada
para os alvos Brancos (M = 17.12, DP = 3.67) em comparação com os alvos Negros (M =
15.77, DP = 3.94), enquanto nenhuma diferença entre os Grupos Étnicos foi observada no
grupo OT (Alvos Brancos: M = 17.60, DP = 3.74; Alvos Negros: M = 17.30, DP = 3.74).
Ambos os Grupos exibiram taxas mais elevadas de Atirar Corretamente em alvos Negros em
comparação aos Alvos Brancos (PL: t(56) = 2.33, p < 0.05; OT: t(49) = 5.35, p < 0.001).
Figura 9 - Taxa de Rejeição Correta por cada Grupo no TAPP. * p<.05.
5.2.2.Latência de Resposta
Para a análise da latência de resposta foi conduzida novamente uma ANOVA mista,
com Etnia e Objeto como fatores intra-sujeitos e Grupo como fator inter-sujeitos, sendo que
foi observado efeito principal de Objeto, F(1,105) = 163.13, p < .001, ηp2 = .608, assim os
colaboradores eram mais rápidos em julgar Armas (M = 475.04, DP = 3.10) em comparação a
Não-Armas (M = 523.61, DP = 4.41). Ainda, a interação entre Etnia x Objeto também foi
estatisticamente significativa, F(1,105) = 5.82, p = .018, ηp2 = .053, uma vez que os
colaboradores precisaram de mais tempo para classificar corretamente um alvo Negro
desarmado (M = 526.80, DP = 4.68) em comparação com os demais alvos (Figura 10).
Demais efeitos não foram estatisticamente significativos, inclusive Etnia x Objeto x Grupo,
F(1,105) = .47, p = .496, ηp2 = .004 e Grupo como um fatore inter-sujeitos, F(1,105) = .13, p =
.723, ηp2 = .001.
15
16
17
18
19
Brancos Negros Brancos Negros
PL OT
Taxa
de
Rej
eiçã
o C
orre
ta
*
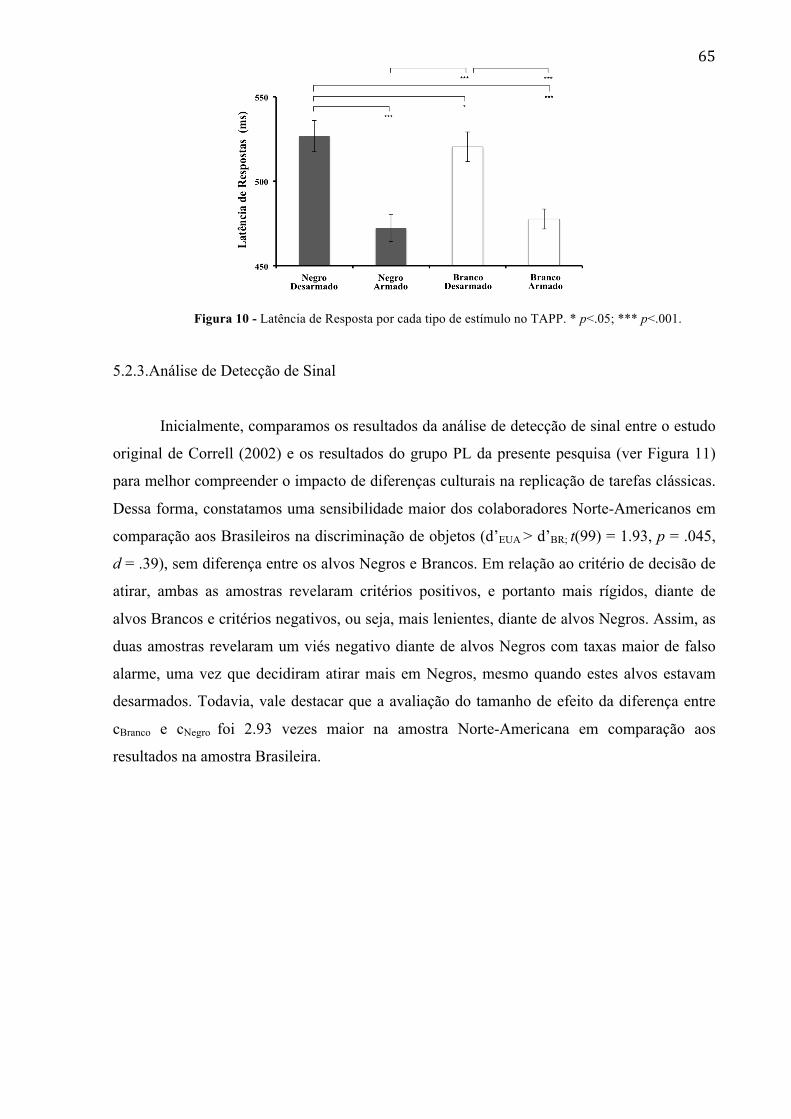
65
Figura 10 - Latência de Resposta por cada tipo de estímulo no TAPP. * p<.05; *** p<.001.
5.2.3.Análise de Detecção de Sinal
Inicialmente, comparamos os resultados da análise de detecção de sinal entre o estudo
original de Correll (2002) e os resultados do grupo PL da presente pesquisa (ver Figura 11)
para melhor compreender o impacto de diferenças culturais na replicação de tarefas clássicas.
Dessa forma, constatamos uma sensibilidade maior dos colaboradores Norte-Americanos em
comparação aos Brasileiros na discriminação de objetos (d’EUA > d’BR; t(99) = 1.93, p = .045,
d = .39), sem diferença entre os alvos Negros e Brancos. Em relação ao critério de decisão de
atirar, ambas as amostras revelaram critérios positivos, e portanto mais rígidos, diante de
alvos Brancos e critérios negativos, ou seja, mais lenientes, diante de alvos Negros. Assim, as
duas amostras revelaram um viés negativo diante de alvos Negros com taxas maior de falso
alarme, uma vez que decidiram atirar mais em Negros, mesmo quando estes alvos estavam
desarmados. Todavia, vale destacar que a avaliação do tamanho de efeito da diferença entre
cBranco e cNegro foi 2.93 vezes maior na amostra Norte-Americana em comparação aos
resultados na amostra Brasileira.

66
Figura 11 - (a) Comparação dos resultados de cBranco, cNegro e d’Branco, d’Negro entre amostras Norte-Americana e Brasileira. <.10; * p<.05.
Em seguida, os índices de sensibilidade d'Negro e d'Branco foram comparados, por Teste-t
para amostras em pares, para cada um dos Grupos Experimentais de nossa amostra
separadamente (Figura 12A). Essa comparação não revelou diferenças estatisticamente
significativas entre os d’ do grupo PL, t(56) = .27, p = .789, r = .04 (d'Negro: M = 2.08, DP =
1.17; d'Branco: M = 2.04, DP = 1.27), nem entre os d’ do grupo OT, t(49) = 1.66, p = .104, r =
.23 (d'Negro: M = 2.57, DP = 1.44; d'Branco: M = 2.30, DP = 1.25). Entretanto, a comparação,
por Teste-t para amostras independentes, entre os d'Negro dos grupos PL vs. OT revelou uma
diferença marginalmente significativa, t(105) = 1.95, p = .053, sendo que a OT tende a
melhorar a sensibilidade dos voluntários diante de alvos Negros (Figura 10A). Nenhuma
diferença foi encontrada na comparação entre os d'Branco, t(105) = 1.09, n.s. Assim como para
o critério de decisão, os resultados do Teste-t para amostras em pares revelou que cNegro (M = -
.12, SD = .37) foi significativamente menor do que cBranco (M = .02, SD = .36) no grupo PL,
t(56) = -2.32, p = 0.024, r = 0.30, o que replica os resultados originais de Correll et al (2002).
Além disso, essa diferença observada foi reduzida no grupo OT, t(49) = -1.63, p = .109, r =
.23 (cNegro: M = -.10, DP = .32; cBranco : M = .02, DP = .44), como pode ser observado na
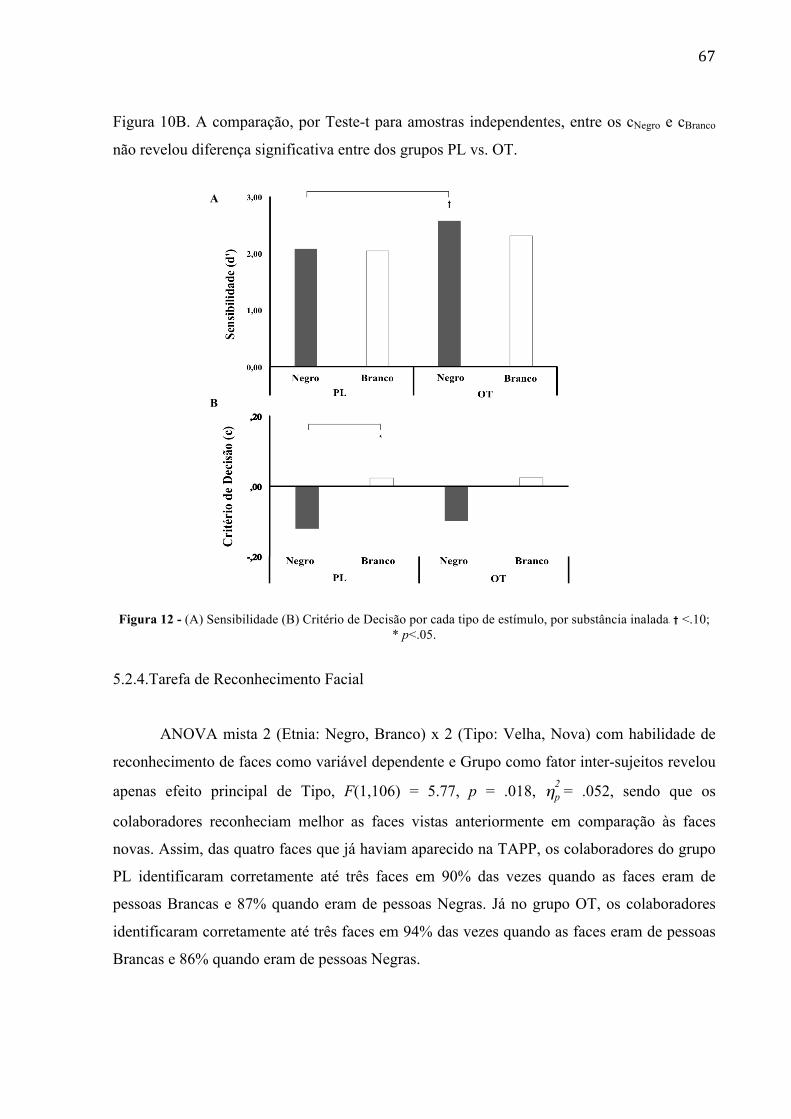
67
Figura 10B. A comparação, por Teste-t para amostras independentes, entre os cNegro e cBranco
não revelou diferença significativa entre dos grupos PL vs. OT.
Figura 12 - (A) Sensibilidade (B) Critério de Decisão por cada tipo de estímulo, por substância inalada. <.10; * p<.05.
5.2.4.Tarefa de Reconhecimento Facial
ANOVA mista 2 (Etnia: Negro, Branco) x 2 (Tipo: Velha, Nova) com habilidade de
reconhecimento de faces como variável dependente e Grupo como fator inter-sujeitos revelou
apenas efeito principal de Tipo, F(1,106) = 5.77, p = .018, ηp2 = .052, sendo que os
colaboradores reconheciam melhor as faces vistas anteriormente em comparação às faces
novas. Assim, das quatro faces que já haviam aparecido na TAPP, os colaboradores do grupo
PL identificaram corretamente até três faces em 90% das vezes quando as faces eram de
pessoas Brancas e 87% quando eram de pessoas Negras. Já no grupo OT, os colaboradores
identificaram corretamente até três faces em 94% das vezes quando as faces eram de pessoas
Brancas e 86% quando eram de pessoas Negras.
A
B

68
5.3.Tarefa de avaliação explícita de empatia à dor no outro
Inicialmente conduzimos uma análise exploratória e verificamos que alguns
colaboradores eram outliers apresentando médias de intensidade dolorosa percebida por
estímulo na tarefa adaptada de Jackson (2004) ±2 desvios-padrão. Considerando isso, esses
dados foram excluídos e permaneceram 57 colaboradores no grupo PL e 53 no grupo OT.
ANOVA mista 3 (Cor: Negro, Branco, Roxo) x 2 (Situação: Dor, Neutro) x 2
(Perspectiva: dor percebida no Outro, Em si) foi conduzida com intensidade de dor percebida
como variável dependente e Grupo (OT, PL) como fator inter-sujeitos. Esta análise revelou
efeito principal estatisticamente significativo para Etnia, F(2,216) = 3.53, p = .031, ηp2 = .032,
Situação, F(1,108) = 974.21, p < .001, ηp2 = .900, e Perspectiva, F(1,108) = 22.13, p < .001, ηp
2
= .170, sendo que os colaboradores imaginaram maior intensidade de dor nos contextos com
(a) alvos Negros em comparação aos alvos Roxos (p = .010), (b) em contextos dolorosos em
comparação aos neutros (p < .001) e (c) quando avaliavam a dor percebida no Outro versus a
dor percebida em Si (p < .001). Ainda foram observados efeitos de interação de Etnia x
Situação, F(2,216) = 4.02, p = .019, ηp2 = .036, em que as avaliações da intensidade de dor
sentida em situações dolorosas foram maiores para alvos Negros em comparação a alvos
Roxos (p = .003) e a alvos Brancos (p = .089). Além disso, ainda verificamos efeito
significativo para a interação de Situação x Perspectiva, F(1,108) = 22.20, p < .001, ηp2 = .170,
onde percebia-se maior intensidade de dor sentida em avaliações de situações dolorosas na
perspectiva do outro em comparação a si (p < .001). Importante ainda mencionar que não
observamos nenhum efeito de interação de Grupo, nem como fator inter-sujeito, F(1,108) =
.63, p = .430, ηp2 = .006, o que indica que a OT não modulou a avaliação da intensidade de dor
explícita (ver Figuras 13 e 14).

69
Figura 13 - Avaliação de explícita de empatia à dor percebida na própria perspectiva. Adicionou-se a linha pontilhada para facilitar a inspeção visual dos dados na comparação entre Perspectiva Em si (Figura 13) e no Outro (Figura 14). <.10; * p<.05; *** p<.001.
Figura 14 - Avaliação de explícita de empatia à dor percebida na perspectiva do outro. Adicionou-se a linha pontilhada para facilitar a inspeção visual dos dados na comparação entre Perspectiva Em si (Figura 13) e no Outro (Figura 14). τ <.10; * p<.05; *** p<.001.
5.4.Tarefa de avaliação implícita de empatia à dor no outro
Nessa seção de análise, verificamos grande variabilidade individual nas medidas de
MEPs porém, os testes de homogeneidade foram cumpridos, uma vez que os dados analisados
apresentaram medidas de Assimetria e Curtose dentro do intervalo de ±2. Apenas foram
incluídos os dados dos colaboradores que possuíam pelo menos oito MEPs válidos em todas
as categorias de análise, ou seja, diante de alvos Negros e Brancos nas situações dolorosas e
neutras. Este critério resultou na exclusão dos dados de 43% da amostra (55 colaboradores),
sendo que provavelmente isso possa ter ocorrido devido ao extenso processo de coleta do dia
3 e pelas movimentações corporais dos colaboradores o que impactava nos registros das

70
informações eletromiográficas. A partir disso, foram incluídos nas análises subsequentes
dados de 73 colaboradores (42 do grupo PL e 31 do grupo OT). Então, analisamos se os
Grupos apresentaram diferenças entre si em relação às características básicas dos MEPs
decorrentes dos registros em repouso comparando os momentos Pré- e Pós-inalação da
substância selecionada. Para isso, conduzimos, separadamente por músculos (FDI, ADM),
ANOVAs mistas comparando as variáveis dependentes: momento de início (onset), latência e
amplitude dos MEPs entre esses momentos, sendo Grupo o fator inter-sujeitos destas análises.
Nenhuma destas análises revelou efeito de interação estatisticamente significativa entre
Momento x Grupo, o que revelou ausência de modulação de OT nas medidas de linha de base.
Em seguida, iniciamos as análises relacionadas à tarefa de empatia implícita à dor
propriamente dita, seguindo a estrutura de análise do estudo original de Avenanti et al.
(2010). Uma análise geral desses dados, revelou diferenças significativas do padrão de
respostas entre FDI (músculo alvo) e ADM (músculo controle) durante os blocos críticos
(vídeos de situações dolorosas), F(1,28) = 37.11, p < .001, ηp2 = .570, onde as contrações no
músculo alvo FDI (M = 1.84, DP = .15) foram maiores em comparação às contrações no
músculo controle ADM (M = 1.19, DP = .13). Após esse resultado, foram explorados no
músculo alvo, efeitos relacionados aos vídeos de diferentes categorias que foram
apresentados, assim como o impacto das substâncias administradas nas respostas implícitas de
contração muscular. Dessa forma, ANOVA mista 2 (Cor da mão: Negro, Branco) foi
conduzida para as respostas em FDI da subtração das amplitudes dos MEPs de situações de
[dor] – [neutra] como variável dependente e Grupo (OT, PL) como fator inter-sujeitos. Esta
análise não revelou efeito principal para Cor da mão, F(1,71) = 1.80, p = .184, ηp2 = .025, nem
para Grupo, F(1,71) = .31, p = .581, ηp2 = .004 (ver estatística descritiva na Tabela 3 e ver
comparação entre as respostas das amostras Brasileira e Italiana, quanto aos dados de
contágio sensório-motor na Figura 15).
Para a análise do contágio sensório-motor do colaborador em relação a cada modelo
de mão apresentada na tarefa, três testes-t para uma amostra foram conduzidos separadamente
contrastando os valores médios de cada categoria de cor de mão com 0. Essa análise revelou
que nenhuma das médias das amplitudes de MEP foram estatisticamente significativa de 0,
mão Brancas: t(72)=-.21 , p= .835; Negras: t(72)= 1.62 , p= .109; Roxas: t(72)=-1.19 , p=
.239).
Analisou-se ainda a confiabilidade da mão controle de coloração Roxa com uma
ANOVA mista para comparar se há diferença entre o nível de contágio sensório-motor entre
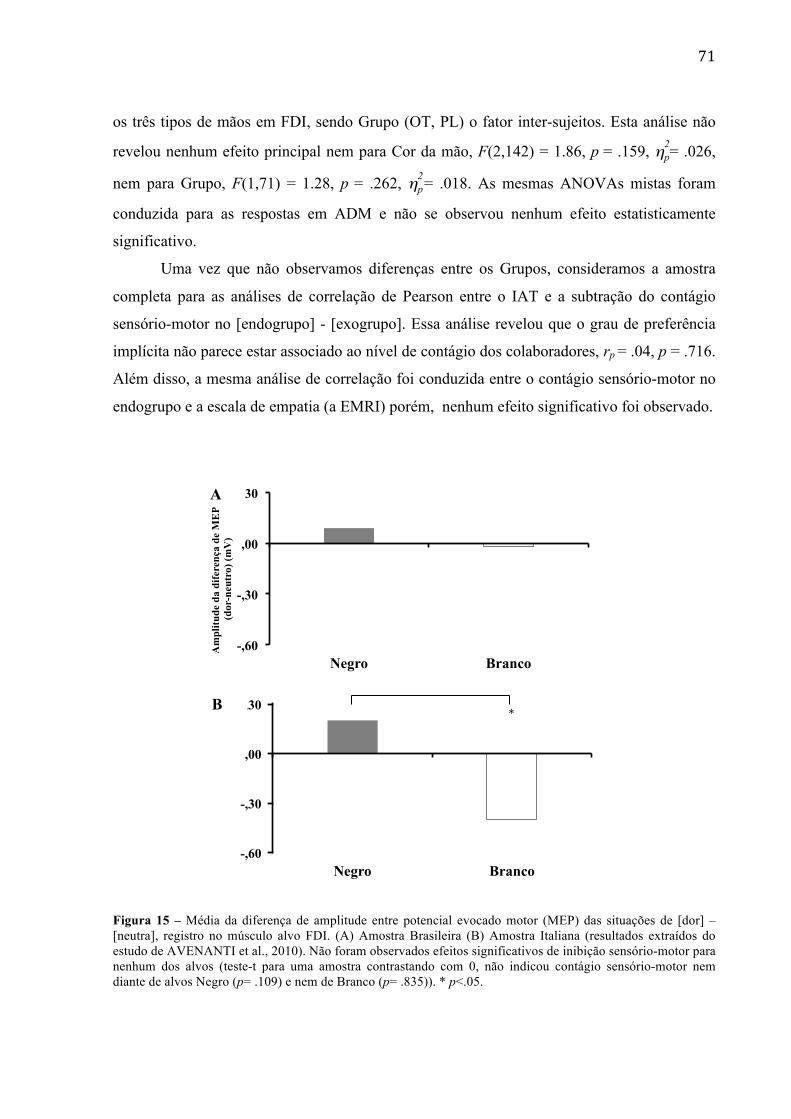
71
os três tipos de mãos em FDI, sendo Grupo (OT, PL) o fator inter-sujeitos. Esta análise não
revelou nenhum efeito principal nem para Cor da mão, F(2,142) = 1.86, p = .159, ηp2 = .026,
nem para Grupo, F(1,71) = 1.28, p = .262, ηp2 = .018. As mesmas ANOVAs mistas foram
conduzida para as respostas em ADM e não se observou nenhum efeito estatisticamente
significativo.
Uma vez que não observamos diferenças entre os Grupos, consideramos a amostra
completa para as análises de correlação de Pearson entre o IAT e a subtração do contágio
sensório-motor no [endogrupo] - [exogrupo]. Essa análise revelou que o grau de preferência
implícita não parece estar associado ao nível de contágio dos colaboradores, rp = .04, p = .716.
Além disso, a mesma análise de correlação foi conduzida entre o contágio sensório-motor no
endogrupo e a escala de empatia (a EMRI) porém, nenhum efeito significativo foi observado.
Figura 15 – Média da diferença de amplitude entre potencial evocado motor (MEP) das situações de [dor] – [neutra], registro no músculo alvo FDI. (A) Amostra Brasileira (B) Amostra Italiana (resultados extraídos do estudo de AVENANTI et al., 2010). Não foram observados efeitos significativos de inibição sensório-motor para nenhum dos alvos (teste-t para uma amostra contrastando com 0, não indicou contágio sensório-motor nem diante de alvos Negro (p= .109) e nem de Branco (p= .835)). * p<.05.
-,60
-,30
,00
,30
Negro Branco
*
A
B
-,60
-,30
,00
,30
Negro Branco
Am
plitu
de d
a di
fere
nça
de M
EP
(dor
-neu
tro)
(mV
)
A

72
Tabela 3 - Dados brutos da amplitude média dos MEPs (desvio-padrão) em mV, comparando as amostras
Brasileira e Italiana
Amostras
Brasileira M(DP)
Italiana M(DP)
FDI
Endogrupo
Dor 1.91 (1.34) 2.22 (1.31)
Neutro 1.93 (1.47) 2.68 (1.83)
Exogrupo
Dor 2.00 (1.28) 2.71 (1.79)
Neutro 1.91 (1.28) 2.55 (1.88)
ADM
Endogrupo
Dor 1.21 (1.06) 1.05 (0.74)
Neutro 0.98 (0.67) 1.21 (1.00)
Exogrupo
Dor 1.03 (0.82) 1.32 (1.10) Neutro 0.99 (0.75) 1.28 (0.98)

73
6. DISCUSSÃO
O presente estudo teve como principal objetivo avaliar efeitos comportamentais pró- e
anti-sociais pela administração intranasal de ocitocina (versus placebo) durante avaliação da
percepção de empatia à dor no outro e de detecção de ameaça, respectivamente, em contextos
com pistas étnicas, junto a jovens universitários Brancos do sexo masculino. Tanto quanto nos
foi possível apurar, tratou-se do primeiro estudo, realizado com amostra brasileira, que
simultaneamente avaliou eventuais efeitos pró- e anti-sociais da OT. Ainda, esse estudo
permitiu expandir resultados de pesquisas prévias, conduzidas em amostras Norte-Americanas
e Europeias, analisando um contexto de maior interação entre grupos étnicos.
De uma forma global, os resultados obtidos na nossa amostra, indicam que a
administração intranasal de OT não promoveu modulação das respostas empáticas implícitas
e explícitas à dor porém, surpreendentemente a OT reduziu a percepção de ameaça diante de
alvos do exogrupo, via aumento da rejeição correta em atirar diante de Negros desarmados na
TAPP. Esse conjunto de resultados foi observado a partir da administração intranasal de OT
seguida da replicação de tarefas amplamente conhecidas e validadas pela comunidade
científica desenvolvidas por JACKSON, 2004; AVENANTI, SIRIGU, AGLIOTI, 2010; e
CORRELL et al., 2002, o que nos permitiu explorar o impacto da OT durante processos de
sensibilização à dor do outro e identificação de estímulos ameaçadores entre membros de
grupos étnicos distintos. Discute-se a seguir a ausência de efeito observável em relação ao
endogrupo e o efeito pró-social específico com o exogrupo promovido pela OT considerando
ainda uma provável mediação de fatores como variabilidade individual e contexto social,
conforme propõe a Abordagem Interacionista.
Em síntese, evidências prévias em animais e em humanos demonstraram que a OT é
igualmente capaz de favorecer, uma ligação emocional e um distanciamento social. Ratas
recém paridas (elevada concentração de OT endógena) cuidam de sua prole, enquanto estão
mais sensíveis a potenciais ameaças externas para seus filhotes (FERRIS et al., 1992; INSEL
& YOUNG, 2001). A administração de OT em humanos, pode promover comportamentos
pró-sociais (LEE et al., 2009) porém, em situações específicas (ex. diante de uma
provocação), esse neuropeptídio pode aumentar a saliência do agente social e favorecer
comportamentos anti-sociais como a agressividade (NE’EMAN et al., 2016). Esses achados
embasaram nossas hipóteses. Todavia, os resultados observados na presente pesquisa
reforçam a complexidade da modulação dos comportamentos pró- e anti-sociais pela OT.
Paralelamente, as principais evidências dos estudos originais replicados nessa pesquisa

74
incluem uma forte associação entre Negros e ameaça (CORRELL et al., 2002 – estudo
Americano) e um menor contágio emocional de Brancos com Negros e vice-versa
(AVENANTI, SIRIGU, AGLIOTI, 2010 – estudo Italiano). Isso pode inclusive ter sido
observado porque nestes países há um baixo grau de miscigenação e uma convivência
limitada entre esses grupos étnicos. Considerando dados referente às distribuições
populacionais, por exemplo, vale destacar que o percentual de Negros na população dos
Estados Unidos (EUA) é 13.4% (Census Bureau, 2017), na Itália 15.2% (ISTAT, 2008) e no
Brasil 7.6% (IBGE de 2010). Porém, no Brasil 43.1% da população declara-se pardo. Nos
EUA e Itália não se utiliza a classificação “pardo”. Baseado em tais diferenças contextuais e
culturais, nossa expectativa era de que os resultados basais (grupo PL) das respostas pró- e
anti-sociais sejam diferenciados pela população Brasileira ser de um contexto com maior
interação entre grupos étnicos. Por isso, o impacto da OT poderia ser ainda mais
surpreendente. De fato, a modulação comportamental pela OT não promoveu um
favorecimento específico e indiscriminado para o endogrupo, mas favoreceu uma redução
(moderada) do viés perceptivo de ameaça associado ao exogrupo. Esses resultados são
relevantes para a comunidade científica porque nos permitem compreender melhor o impacto
de diferenças contextuais nos efeitos da OT. Além disso, tais contribuições podem
potencialmente orientar as intervenções terapêuticas com grupos clínicos. A seguir,
discutimos detalhadamente em diferentes sub-sessões os resultados obtidos nas avaliações da
(1) caracterização da amostra, (2) empatia (explícita e implícita) e (3) percepção de ameaça.
6.1. Avaliação da Caracterização da Amostra
Ao avaliar objetivamente atitudes e estereótipos em relação aos grupos étnicos por
escalas de auto-relato, notamos que os participantes revelam um predomínio de
comportamentos preconceituosos sutis e indiretos. Assim, mesmo em contextos
miscigenados, confirma-se que formas indiretas de preconceito (ex. brincadeiras ou uso
depreciativo das palavras “negro” e “preto”) tem sustentado a estratificação entre os grupos
étnicos com tentativas de atribuir desvantagens ao exogrupo, ao invés de comportamentos
discriminatórios explícitos (ex. acesso restrito a espaços sociais, como lugares específicos no
ônibus). Essas evidências são reforçadas quando analisamos os estereótipos atribuídos aos
Brancos e Negros pelos participantes. Nota-se que os participantes consideram, na própria
perspectiva, os Brancos como uma referência social. Enquanto, associam os Negros a uma
condição de desfavorecimento econômico e profissional. Quando os colaboradores indicam os

75
estereótipos reforçados pela cultura Brasileira, nota-se maior pluralidade de associações aos
Brancos, com atribuições de adjetivos positivos (ex. “bonito”, “honesto”, “confiável”) e
negativos (ex. “mimado” e “preconceituoso”). Todavia, nota-se ainda uma restrição
perceptiva em relação aos Negros, os quais são mais fortemente associados com adjetivos
negativos (ex. “malandro”, “ladrão”, “pobre”, “bandido”). É possível que esses padrões
estejam sendo sustentados por mensagens midiáticas e uma convivência ainda limitada entre
esses grupos mesmo no Brasil, em que ainda ocorre uma primazia do favorecimento do grupo
dos Brancos em detrimento de outros grupos étnicos, principalmente dos Negros. Dessa
forma, o processo histórico de maior relacionamento entre os grupos étnicos de Brancos e
Negros no Brasil fortalece, gradualmente, ações afirmativas e ampliação da representatividade
porém, não extinguiu estereótipos negativos relativos a esses grupos.
6.2. Avaliação de Empatia
Era de interesse do escopo desse trabalho analisar se existia uma congruência de
direção e intensidade de resultados entre as medidas empáticas implícitas e explícitas em uma
amostra que é marcada pelo processo de miscigenação. Além disso, estudos sobre o uso da
OT com finalidade terapêutica, geram a necessidade de investigações que comparem os
efeitos da OT nos processos empáticos explícitos e implícitos. Assim, o presente estudo
amplia a discussão sobre respostas empáticas à membros do endo- e exogrupo pois
observamos que independente da substância inalada pelo colaborador, implicitamente, não foi
verificado diferenças de contágio sensório-motor na responsividade aos alvos desses grupos, e
explicitamente, foi observado uma tendência dos colaboradores declararam perceber mais dor
em situações dolorosas quando estas ocorriam em alvos Negros em comparação aos Brancos.
Esse resultado de empatia explícita contradiz evidências atuais de neuroimagem que revelam
aumento da ativação da circuitaria de dor do colaborador apenas quando observamos
situações dolorosas em membros do endogrupo e redução da atividade do cíngulo anterior
diante de membros do exogrupo (XU et al., 2009).
Assim, nossos dados parecem evidenciar uma preocupação dos colaboradores em
explicitar uma postura favorável ao exogrupo. Esse tipo de observação já foi constatado em
antigas pesquisas sobre mudanças de atitudes raciais ao longo do tempo nos Estados Unidos.
Por exemplo, na década de 50, menos da metade dos entrevistados Brancos declararam que
Negros deveriam ter acesso a escolaridade, assim como os Brancos, e na década de 90 essa
porcentagem elevou-se para mais de 90% (SCHUMAN et al., 2001). Apesar dessa postura

76
mais favorável com o exogrupo na década de 90, ainda hoje não se observam grandes
mudanças sociais reais no contexto Norte-Americano. De forma equivalente, este mesmo
cenário é revelado no Brasil quando contrastamos uma aparente progressão social dos Negros
e Pardos com poucas mudanças sociais significativas (GOMES & MARLI, 2018 em Relatos e
Revista do IBGE, pp.15-19). Ilustrações disso estão disponíveis em dados censitários que
revelam uma taxa de analfabetismo em 2016, mais de duas vezes maior para Negros e Pardos
em comparação aos Brancos (PNAD Contínua 2016) e ainda uma maior dificuldade da
população Negra em ascender economicamente no Brasil (PNAD Contínua 2017).
Aparentemente, o padrão de dados que verificamos em nossa pesquisa sugere uma influência
de pressões sociais de que sejam apresentadas respostas “politicamente corretas” (não
discriminatórias) em detrimento das reais crenças. Além disso, durante a aplicação das tarefas
em laboratório fica logo evidente ao colaborador o fenômeno a ser estudado, o que pode
regular seus comportamentos explícitos. Fica em aberto a questão do quanto nossos dados de
empatia explícita podem sinalizar um real declínio do preconceito contra o Negro, pela
observada aproximação das respostas empáticas entre o endo- e exogrupo ou se de fato,
formas abertas de discriminação são cognitivamente controladas por não serem socialmente
aceitas.
Em relação ao efeito da OT sobre medidas explícitas de cooperação entre diferentes
grupos, Ma et al. (2015) publicaram um trabalho o qual evidenciou que a OT gerou um efeito
de aumento da preferência pelo endogrupo em um jogo econômico apenas com participantes
que apresentam um estilo cognitivo intuitivo (que preferem respostas rápidas e gerais) e
redução da preferência pelo endogrupo naqueles com estilo reflexivo (que preferem respostas
lentas e deliberadas). Em contraste a esses achados, em nossa pesquisa não observamos
modulação pela OT da preferência pelo endogrupo, apesar de também termos aplicado uma
tarefa que favorecia a apresentação de respostas resultantes de um controle cognitivo do estilo
reflexivo. Os resultados destoantes entre o estudo de Ma e colaboradores e o nosso podem
estar relacionados as diferenças nos procedimentos aplicados, uma vez que no primeiro
estudo ocorreu uma interação real (tarefa mais ecológica) onde explicitamente 10
participantes reais foram divididos em subgrupos definindo assim quem pertencia ao
endogrupo e ao exogrupo. Já em nosso estudo não estabelecemos subgrupos a priori e a
interação proposta não era real, mas sim por tarefas computadorizadas (previamente
programada pelo pesquisador). Na presente pesquisa, verificamos uma tendência de efeito de
maior sensibilidade dos participantes Brancos ao perceber dor em alvos Negros em
comparação aos alvos Brancos, independente da substância inalada. Assim, esses resultados

77
em conjunto confirmam nossas hipóteses Hb1 de que fatores como desejabilidade social
devem ter conduzidos os colaboradores a revelar maior empatia explícita diante de alvos do
exogrupo e nossa Hb2 a qual previa que a OT não seria capaz de modular as respostas de
empatia explícita porque estas são decorrentes de processamentos deliberados que favorecem
a apresentação de comportamentos socialmente aceitos, ocorrendo uma sobreposição desse
padrão de resposta aos modestos efeitos comportamentais promovidos pela OT.
Quanto à avaliação das respostas de empatia implícita, é importante destacar que esta
foi devidamente conduzida conforme recomendações de pesquisas anteriores (AVENANTI,
SIRIGU, AGLIOTI, 2010). Assim, a liberação dos pulsos de EMT foi programada para
ocorrer entre 200-600 ms antes do fim da apresentação dos estímulos, o que permitiu o
registro de modulações na excitabilidade cortical associada às respostas de contágio sensório-
motor. Isso foi necessário, pois a liberação dos pulsos no início dos estímulos geraria registros
de excitabilidade cortical associados com a ativação do sistema de neurônios espelho devido a
visualização de seringas ou cotonetes em movimento. Ainda, a comparação entre a magnitude
das medidas basais dos MEPs (ex. momento de início, latência e amplitude) registradas em
nosso estudo, em comparação as medidas do estudo original, foram equivalentes e com baixa
variabilidade, o que sinaliza uma utilização adequada da técnica de EMT.
Todavia, foi surpreendente a ausência de um padrão empático diferenciado para os
grupos étnicos observados, bem como a ausência de modulação das respostas empáticas
implícitas pela OT. Vários estudos em neurociência social indicam que a resposta neural de
empatia a dor no outro é modulada por fatores como grupo, sendo que os colaboradores
costumam apresentar modulação dos marcadores biológicos de forma mais favorável em
relação aos membros do endogrupo em comparação aos do exogrupo (XU et al., 2009;
AVENANTI, SIRIGU, AGLIOTI, 2010; SHENG et al., 2013). Adicionalmente, de acordo
com evidências que denotam que a OT tem um efeito especialmente positivo na
prosocialidade e cognição social diante de membros do endogrupo, era expectável que a OT
amplificasse o efeito basal descrito de empatia implícita a dor diante do endogrupo, assim
como ocorreu em evidências anteriores de SHENG et al., 2013. Essa resposta empática mais
evidente com o endogrupo diante de estimulação dolorosa foi constatada em um estudo
Chinês de neuroimagem que revelou maior ativação da circuitaria de dor em colaboradores
Asiáticos observando uma perfuração facial por uma seringa em membros de seu próprio
grupo étnico em comparação ao exogrupo (e.g. alvos Caucasianos) (XU et al., 2009). Outro
exemplo seria o estudo Italiano de Avenanti, Sirigu & Aglioti (2010), em que foi verificado
que os participantes Brancos e os Negros, apenas apresentaram contágio sensório-motor

78
durante observação de mãos de modelos do endogrupo sendo perfuradas por seringas e que
essa resposta empática não ocorria diante de membros do exogrupo na mesma situação.
Ainda, efeitos na mesma direção foram posteriormente descritos por Sheng et al. (2013) em
um estudo com EEG que evidenciou modulação das respostas empáticas pela OT com um
fortalecimento do efeito basal de maior amplitude do potencial relacionado a evento P2 ao
observar faces com expressão de dor, exclusivamente para os estímulos do endogrupo. Então,
avançamos com algumas hipóteses explicativas para a ausência de modulação pelo grupo
étnico e pela OT das respostas implícitas observadas em nosso estudo.
Primeiramente, é importante destacar que quando um colaborador observa dor no
outro, é provável que ocorra ativação da mesma circuitaria neuronal de quando a situação
dolorosa é vivenciada pelo colaborador (AVENANTI e AGLIOTI, 2006) e isso promoverá o
compartilhamento dessa experiência com o outro (SINGER & FRITH, 2005), favorecendo a
cognição social humana. Pesquisas descritas a seguir têm utilizado técnicas de neuroimagem
para dissociar os componentes sensórios e afetivos desse processamento de dor no outro. Os
estudos de Morrison et al. (2004), Singer et al. (2004) e Jackson et al. (2005), conduzidos com
fMRI, apontam para uma forte associação entre a observação de situações dolorosas e a
ativação da circuitaria de dor afetiva (que inclui córtex cingulado anterior e ínsula anterior).
Outras evidências ainda apontam para um efeito de menor magnitude quanto ao envolvimento
do córtex somatosensorial primário no processamento de dor (PEYRON et al., 2000).
Bushnell et al. (1999) sugeriu que o significado funcional da reatividade dessa região é uma
sinalização de que existe um aspecto sensório, além do afetivo, no processamento da dor.
Anos depois, Avenanti et al. (2005) e Fecteau et al. (2008) demonstraram que um pulso
simples de EMT sobre M1 é capaz de registrar uma supressão do potencial evocado motor
(MEP) pela observação de uma situação dolorosa no outro (e.g. agulha penetrando uma região
específica da mão). Esse resultado em conjunto com o estudo de Bufalari et al. (2007)
demonstraram o envolvimento de componentes sensório-motores durante o processamento de
empatia à dor. Sabe-se que o processamento de dor é uma resposta subjetiva e complexa
porém, ainda não é claro a universalidade desse efeito de supressão na região somatosensorial
durante observação de dor no outro. Em nosso estudo, por exemplo, obtivermos um
inesperado resultado de ausência do contágio sensório-motor, independente do grupo étnico
que estava sendo observado. Em contraste, Avenanti, Sirigu & Aglioti (2010) verificaram que
além do maior contágio para o endogrupo, registros de condutância da pele indicaram uma
reatividade emocional mais imediata diante do endogrupo em comparação ao exogrupo. Da
mesma forma, a resposta de taxa cardíaca foi reduzida diante de membros do endogrupo em

79
contraste ao exogrupo, indicando maior resposta de orientação em janelas temporais iniciais
(0-4 seg) para os membros do próprio grupo étnico do colaborador. Os registros periféricos
supracitados complementam os achados das medidas centrais de excitabilidade cortical por
adicionarem informações de respostas autonômicas durante a interação com o endo- e
exogrupo. Na presente pesquisa não tivemos a oportunidade de realizar tais registros
periféricos, sendo que provavelmente eles poderiam revelar diferenças entre endo- e exogrupo
que não foram captadas em nossas medidas de excitabilidade cortical. Todavia, vale destacar
que no estudo de Zuo & Han (2013) se verificou que durante o processamento de empatia à
dor, o grupo étnico de pertença não é um determinante intrínseco que promove viés racial,
pois dados de fMRI com Chineses criados em países ocidentais (e.g. Estado Unidos, Londres
e Canadá) e que se declaram Asiáticos, revelaram que eles apresentam reação empáticas
elevadas (níveis semelhantes de ativação neural) diante da observação de situações dolorosas
com alvos Caucasianos e Asiáticos. Então, diferentes estudos (SHENG & HAN, 2012; ZUO
& HAN, 2013), assim como o nosso demonstram que experiências interétnica podem atenuar
o viés racial de empatia à dor.
Alguns estudos verificam uma reatividade empática automática quando observamos
pessoas em situação de dor, mesmo que elas nos sejam desconhecidas (PRESTON e de
WAAL, 2002; DECETY e JACKSON, 2004). Entretanto, pesquisas adicionais apontam que a
intensidade das respostas de incorporação da dor do outro é modulada por variáveis como
similaridade e familiaridade entre o participante e o alvo observado (EBERHARDT, 2005;
XU et al., 2009). Dessa forma, estima-se intuitivamente um padrão de resposta crescente de
contágio sensório-motor, quanto maior for a familiaridade com o membro observado. Assim,
a subtração da amplitude do MEP resultante da observação de situação dolorosa menos da
situação neutra, seria capaz de gerar valores negativos (marcador biológico de contágio
sensório-motor) porém, o valor diante de alvos do exogrupo seria de menor magnitude
(indicação de menor contágio) e o valor diante de alvos do endogrupo de maior magnitude
(indicação de maior contágio). Todavia, Avenanti, Sirigu & Aglioti (2010) curiosamente
demonstraram que ao observar dor no outro há um evidente contágio sensório-motor apenas
se o alvo pertencer ao endogrupo (valor negativo da subtração acima), enquanto há uma
ausência de contágio diante de membros do exogrupo (valor positivo da subtração acima). O
padrão das respostas obtido em nosso estudo, contrapôs-se ao da pesquisa original Italiana
(AVENANTI, SIRIGU, AGLIOTI, 2010), pois em nossa pesquisa os valores da subtração dos
MEPs para o endogrupo e exogrupo não foram estatisticamente diferentes de 0 (zero),
sugerindo ausência de contágio sensório-motor para ambos os grupos. Vale destacar que

80
apesar das médias de subtração dos MEPs para o endogrupo e exogrupo não serem
estatisticamente diferentes entre si (magnitude reduzida em relação ao estudo original),
observamos que estas apresentam a mesma direção do estudo Italiano. Isso refutou nossa Hc
que previa respostas diferenciadas de direção e magnitude entre esses grupos étnicos. Assim,
a marcante diferença na magnitude desses dados em relação aos dados Italianos pode
objetivamente evidenciar o impacto de diferenças culturais resultantes da maior miscigenação
e convivência entre Brancos e Negros no Brasil que aproxima as respostas observadas entre
os membros do endo- e exogrupo.
Hipóteses alternativas podem ser levantadas para compreender esses resultados de
nosso estudo. Uma delas envolve o fato dos participantes poderem ter percebido as situações
dolorosas apresentadas como estímulos de fraca intensidade e com isso observamos um efeito
global de ausência da modulação das respostas implícitas diante dos grupos étnicos. Outra
hipótese é que a evidente redução da reatividade corticoespinal às situações dolorosas com o
endogrupo, em comparação ao estudo original Italiano, ocorreu porque com o decorrer da
tarefa os participantes podem ter antecipado reações de desconforto causado por essas
situações e modificavam seu direcionamento atencional, gerando uma menor reação de
supressão a esses estímulos. Adicionar medidas de registro ocular durante essa tarefa teria nos
auxiliado na confirmação dessas hipóteses, pois isso nos permitiria avaliar as regiões de
fixação do olhar no estímulo apresentado e o nível de engajamento cognitivo relacionado ao
processamento dessa informação pelas medidas de diâmetro pupilar. Paralelamente a essas
considerações, há a possibilidade de que maior supressão diante do endogrupo seja um sinal
de comportamento etnocêntrico porém, parece existir uma atenuação desse perfil na amostra
Brasileira em comparação a Italiana, pela primeira ser mais miscigenada. Isso pode ser
ilustrado pela análise da diferença dos dados brutos da amplitude média dos MEPs de [dor] -
[neutro] para o endogrupo, onde verifica-se um valor para a amostra Italiana 23 vezes maior
do que para a amostra Brasileira.
Além disso, evidências complementares revelam que a inibição corticoespinal da mão
correspondente que sofre uma estimulação dolorosa é acompanhada pela facilitação
corticoespinal da mão oposta como uma estratégia de favorecer comportamentos reativos de
defesa à observação de uma situação de sofrimento, como buscar escapar da situação em
questão (AVENANTI et al., 2009). Ainda, o sistema de ressonância de dor parece ser bastante
sensível às informações sensoriais básicas observadas, como localização e intensidade, pois
não há uma modulação da amplitude dos MEPs registrados no músculo da mão quando o
observador visualiza estimulações no pé ou em um tomate sendo perfurado por uma seringa

81
(AVENANTI et al., 2005). Da mesma forma, no presente estudo verificamos tal modulação
dos MEPs na musculatura alvo, FDI e não na musculatura controle, ADM.
No estudo Italiano, a ausência de contágio apenas ocorreu para membros do exogrupo
e esta foi justificada por uma provável interferência de processos cognitivos de
desumanização diante desse grupo (HASLAM, 2006; FISKE, 2009). Tal processo de
desumanização inicia-se com as dimensões de categorização social que são embasadas na
percepção de traços de proximidade e competência do outro (ex. moradores de rua são
considerados distantes e pouco competentes; enquanto cônjuges são considerados próximos e
muito competentes). Essa categorização pode, por um lado, distanciar as pessoas da
possibilidade de apreciação da mente do outro, ou por outro lado, favorecer o processamento
das experiências internas do outro, como intenções, pensamentos e sentimentos, porque
acreditamos que o outro tem uma mente como nós – mentalização (EPLEY, 2009). Estudos
de ressonância revelam que grupos estigmatizados costumam não modular a ativação de
circuitarias cerebrais específicas como ocorre com membros do endogrupo (AMODIO e
FRITH, 2006; LIEBERMAN, 2007). Assim, quanto mais hostil for o observador em relação a
um grupo social, menor sua capacidade de mentalização sobre o outro (PHELPS et al., 2000;
CIKARA, SAID, EBERHARDT, FISKE, 2009). Esse mesmo processo de desumanização
pode contextualizar o porquê verificamos em nossa amostra que a subtração da amplitude do
MEP resultante da observação de situação dolorosa menos da situação neutra foi positiva
(indicação de ausência de contágio) para o exogrupo. Caso os participantes Brancos
desumanizem os alvos Negros, eles não sentirão empatia ao observar alvos Negras em
situação de dor física, assim, não se observa uma supressão corticoespinal da representação
cerebral da musculatura correspondente nos participantes Brancos. A maior convivência entre
Brancos e Negros no Brasil pode ter sido o fator que reduziu a magnitude desse efeito em
comparação ao estudo Italiano. Portanto, os resultados obtidos no presente estudo sustentam a
ideia de que a ativação de circuitarias neurais envolvidas na cognição social é influenciada
pelo contexto social.
No seu estudo, Avenanti e colaboradores (2010) destacam ainda que novas pesquisa
precisam explorar se a ausência de contágio diante o exogrupo ocorreu porque os alvos não
eram familiares ou porque eles eram evidentemente diferentes do endogrupo. Outra
importante observação presente no estudo Italiano original, envolve o fato de ter sido
verificada uma reatividade empática maior para a mão Roxa em comparação a Negra. Em
nosso estudo verificamos novamente a mesma direção de dados, mas uma menor magnitude
desses resultados que não gerou diferenças estatisticamente significativas entre essas

82
condições. Caso as diferenças tivessem alcançado significância, tal revelaria que é possível
existir um contágio sensório-motor com quem não nos é familiar, contanto que a ele não
sejam ativados automaticamente estereótipos negativos (efetivamente, nenhuma categoria
social deve ter sido associada à mão Roxa). Porém, o padrão de resultados que encontramos
foi de equivalência estatística para mãos Brancas, Negras e Roxas, sugerindo que a coloração
da pele não parece ser uma característica com forte impacto nos processos de contágio
sensório motor, na nossa amostra.
Outra contribuição dos estudos de Avenanti, Sirigu & Aglioti (2010) e Sheng et al.
(2013) é a constatação de que a resposta cerebral de empatia à dor é positivamente associada
ao grau de preferência implícita ao endogrupo. Entretanto, no presente estudo não
constatamos correlação significativa entre a medida de contágio sensório-motor com a
preferência étnica implícita (medida pela TAI), nem com a capacidade empática (medida pela
EMRI). Isso aconteceu apesar de verificarmos na Tarefa de Associação Implícita (TAI) uma
maior automatização da associação da categoria “Branco” com atributos positivos e “Negro”
com atributos negativos. Ainda, vale destacar que na avaliação de empatia os participantes
apresentaram maior nível de característica empática cognitiva (elevados escores nas
subescalas de fantasia e tomada de perspectiva) e na avaliação da empatia afetiva, eles apenas
apresentam nível elevado de preocupação empática, mas não de angústia pessoal. Dessa
forma, os participantes parecem estar aptos em colocar-se no lugar do outro, seja para
entender seus pensamentos e sentimentos (tomada de perspectiva), como para identificar-se
imaginativamente com personagens fictícios (fantasia), ainda, motivam-se para ajudar o outro
(preocupação empática) porém, a amostra da presente pesquisa revelou menor nível de
incômodo ou ansiedade pessoal quando estão diante de situações de emergência com o outro
(angústia pessoal). Este último resultado pode complementar a explicação do porque os
colaboradores não apresentaram um evidente contágio sensório-motor, uma vez que relataram
menor desconforto pessoal diante da observação à dor do outro, o que pode gerar
consequências de uma menor tendência a motivação pessoal em ajudar o outro.
Em suma, observamos uma ausência de modulação das medidas empáticas explícita e
implícita entre os grupos étnicos pela OT. Quanto as medidas explícitas, discutimos a
tendência dos participantes Brancos a uma compensação favorável ao exogrupo (no caso,
alvos Negros) que parece estar relacionada as pressões contextuais de apresentação de
respostas socialmente aceitas. Algumas hipóteses ainda foram levantadas sobre a ausência de
modulação das medidas explícitas e implícitas, envolvendo (a) mudança do direcionamento
atencional por uma antecipação do estímulo doloroso ou (b) impacto da elevada

83
miscigenação no Brasil que: pode ter reduzido tendências etnocêntricas a favor do endogrupo
enquanto, pode ter reduzido o efeito de desumanização em relação aos membros do exogrupo,
como descrito em outros estudos. O presente estudo não foi capaz de esclarecer o que mais
contribuiu no processamento de dor no outro. Ainda esses dados sinalizam que estudos que
descrevem o papel da OT como um promotor de efeitos pró-sociais indiscriminados e
universais devem ser ponderados, uma vez que diferentes variáveis inter e intrapessoais nos
modelos de análise parecem esclarecer os efeitos da OT na cognição e comportamento social.
6.2. Avaliação da Percepção de Ameaça
Percepção de ameaça é uma resposta vicária necessária para sobrevivência e
preservação que pode promover comportamentos agressivos com a finalidade de autodefesa.
Na vida cotidiana, esse tipo de percepção embasa diversos conflitos entre grupos sociais
distintos, evidenciados por ações discriminativas, conflitos étnicos, entre outros (BOWLES,
2009). Ameaças reais ou imaginadas podem gerar comportamentos agressivos (reativos ou
proativos) em relação a um exogrupo, o que não deixa de ser uma forma indireta de defesa do
endogrupo (ARROW, 2007). Diversas pesquisas exploram os efeitos modulatórios da OT
sobre a percepção de ameaça e comportamentos agressivos, sendo que até o momento não há
grande consistência em relação aos efeitos biológicos que sustentam os resultados observados.
Assim, o objetivo do presente trabalho foi contribuir para o esclarecimento sobre o impacto
da OT nas respostas de percepção de ameaça em contexto de interação entre grupos étnicos
distintos.
Enfatizaremos nessa discussão diferenças culturais entre os dados das amostras Norte-
Americana (estudo original) e Brasileira durante a Tarefa do Atirador em Primeira Pessoa
(TAAP; CORRELL et al., 2002), para compreender os impactos da OT nos comportamentos
observados. Os principais resultados analisados são: (a) taxa de acurácia e tempo de reação,
(b) taxa de rejeição correta (não atirar em alvos desarmados) e (c) medidas de sensibilidade e
critério de decisão, variáveis da Teoria de Detecção de Sinal. Em linhas gerais, uma análise
do desempenho global na tarefa revelou que os participantes Brancos na amostra Brasileira
apresentam maior precisão na identificação de alvos Negros armados e maior latência de
resposta para classificar alvos Negros desarmados, assim como as amostras Norte-
Americanas (CORRELL, PARK, & WITTERNBRINK et al., 2002; CORRELL, URLAND,
& ITO, 2002; GREENWALD, OAKES, & HOFFMAM, 2003). Esses resultados sinalizam
que os participantes de ambos os contextos parecem ter maior expectativa em deparar-se com

84
um alvo Negro armado do que com um alvo Branco nessa condição. Em outras palavras, há
um viés negativo na decisão de atirar diante de membros do exogrupo dos Negros. Isso
reafirma que o estereótipo racial influencia a percepção de ameaça facilitando as respostas
diante de alvos do tipo estereótipos-congruentes (Brancos não armados e Negros armados),
mas inibindo as respostas para alvos estereótipos-incongruentes (Brancos armados e Negros
não armados). Na Tarefa de Identificação de Armas, que apresenta uma estrutura semelhante
a TAAP, Payne (2006) verificou que com 200 ms a mais para a classificação de objetos (ex.
armas ou objetos inócuos), os participantes já são capazes de apresentar maior controle
cognitivo, regulando consideravelmente a predisposição a identificar armas após a observação
de faces Negras. Outras manipulações experimentais devem favorecer esse tipo de controle
durante a TAAP. Na presente pesquisa, por exemplo, verificamos que após a inalação da OT
ocorreu um aumento do limiar do critério de decisão de atirar diante de alvos Negros via
aumento da taxa de Rejeições Corretas (evitar atirar em alvos desarmados) seletivamente para
o exogrupo. Essa evidência indica que nossa Ha foi confirmada, pois a OT foi capaz de
interferir nos processos decisórios, gerando redução de falsos alarmes diante do exogrupo.
Distintos efeitos biológicos promovidos pela OT podem explicar a redução de falsos
alarmes seletivamente para o exogrupo, como a redução da ansiedade social (CAMPBELL et
al., 2010) ou o aumento da saliência do agente social (SHAMAY-TSOORY et al., 2009). Por
um lado, pesquisas em animais (EBITZ et al., 2013) e humanos (LABUSCHAGNE et al.,
2010) demonstraram que a OT pode ter um efeito ansiolítico e que isso deve ser evidenciado
durante a interação social pela redução da percepção de ameaça social. Respectivamente a
esses estudos, constata-se que a OT ameniza o comportamento de vigilância social em
macacos e reduz a atividade amigdalar diante de faces de raiva em pessoas com ansiedade
social generalizada. Considerando o que observamos na presente pesquisa, esse efeito
ansiolítico poderá ter favorecido comportamentos pró-sociais como redução da percepção de
que Negros desarmados sinalizam ameaça social. A seletividade do efeito da OT para esses
alvos pode estar relacionada ao fato de que eram esses estímulos que mais provocavam
ansiedade social nos participantes existindo, portanto, margem para modulação. Por outro
lado, a Hipótese de Saliência Social (SHAMAY-TSOORY et al., 2009) também pode explicar
esses resultados ao considerar a provável modulação da amígdala e dos colículos superiores
pela OT, que promovem aumento do direcionamento atencional aos agentes sociais. Assim, o
efeito pró-social da OT em relação ao exogrupo poderá assentar em uma maior acurácia na
identificação de ameaça proveniente de indivíduos Negros, por via da redução da taxa de

85
falso alarme, da tendência de melhoria na discriminação de alvos armados e desarmados e da
diminuição da tendência de atirar contra alvos Negro.
Todavia, considerando o exposto, podemos dizer que a Abordagem Interacionista
(BARTZ et al., 2011) é a explicação que melhor descreve os efeitos biológicos produzidos
pela OT em nossa pesquisa pois os possíveis efeitos acima podem ter ocorrido de forma
independente ou em conjunto. Além disso, essa abordagem sinaliza que também se deve
considerar o impacto de elementos moderadores como o contexto (ex. dificuldade com
características da tarefa, uma vez que na TAAP é solicitado um rápido processo decisório
incomum aos participantes) e variabilidades individuais (ex. características comportamentais
como nível de agressividade dos participantes) nos nossos resultados. Assim, na condição
placebo, os participantes podem ter apresentado maior ansiedade social diante de alvos
Negros pelo fato destes serem culturalmente mais associados à ameaça em comparação aos
Brancos e porque os participantes estão cientes das pressões sociais de que não se deve
apresentar comportamentos discriminatórios. Em contraste, para aqueles que inalaram OT,
essa substância pode ter favorecido o aumento da saliência social diante de alvos Negros ao
reduzir a ansiedade social durante a decisão de atirar. Isso deve ter promovido um maior
direcionamento atencional às pistas sociais relevantes, no caso os membros do exogrupo,
gerando provavelmente uma redução da taxa de falsos alarmes.
Então, de forma geral, em nossa amostra verificou-se que a OT modulou
seletivamente – ao invés de indiscriminadamente – uma tendência pró-social de redução da
percepção de ameaça associada ao exogrupo. Outro estudo que apresentou evidências na
mesma direção foi o de Shamay-Tsoory et al. (2013). Nessa pesquisa, os autores observaram
que a OT atenuou o viés empático à dor física de israelitas judeus em relação ao endogrupo,
via um aumento do relato de empatia à dor física em palestinos (exogrupo adversário).
Algumas diferenças entre o estudo de Shamay-Tsoory e o nosso precisam ser sinalizadas em
relação a amostra, estímulos selecionados e design experimental para entendermos em que
medida estes estudos se complementam. Shamay-Tsoory e colaboradores realizaram a
pesquisa com homens (n=37) e mulheres (n=18) em um design intra-sujeitos, enquanto que
nosso estudo envolveu coleta exclusiva com homens (n=107) em um design inter-sujeitos.
Apesar de nossa amostra ter sido maior, não avaliamos efeitos de interação de gênero e
ocitocina em relação a percepção de ameaça, o que se recomenda ser explorado em estudos
futuros. Adicionalmente, esses estudos selecionaram diferentes estímulos de exogrupo, o que
contribui com as discussões sobre diferenciação entre si-outro e distanciamento social na
modulação de nossos vieses perceptivos. Assim, Shamay-Tsoory analisou respostas de

86
empatia à dor em membros de um exogrupo neutro (alvos Europeus) versus um exogrupo
adversário (alvos Palestinos), enquanto que em nosso estudo analisamos respostas de
percepção de ameaça em relação a membros de um exogrupo muito miscigenado com o
endogrupo do participante (alvos Negros).
A modulação comportamental pela OT é um processo complexo, pois observamos um
efeito pró-social seletivo para exogrupo, enquanto outros estudos revelam resultados
diferentes. Certos comportamentos agressivos com o exogrupo diante de uma ameaça real ou
imaginada, podem sugerir uma indireta defesa do endogrupo (ARROW, 2007) e nesse sentido
a OT pode amplificar essa atitude. Por exemplo, no estudo de Shamay-Tsoory et al. (2009)
verificou-se que a OT promoveu inveja quando se perde um jogo e promove um efeito sutil de
exaltação quando se ganha. Já De Dreu et al. (2010) observaram efeitos da OT mais
marcantes com reações divergentes para grupos de pertença distintos. Os autores exploraram
os efeitos da OT em contextos com pistas étnicas (alvos Holandeses vs. Árabes), revelando
que essa substância foi capaz de promover - em um jogo econômico - a coesão do endogrupo,
pelo aumento da confiança e cooperação (maior alocação de recursos financeiros), enquanto
modulou uma agressividade reativa / defensiva diante da ameaça representada pelo exogrupo.
Assim, a OT pareceu promover, em uma amostra de holandeses, o etnocentrismo diante do
endogrupo (DE DREU et al., 2011), favorecendo a unidade do endogrupo com
comportamentos de cooperação, enquanto elevou as tendências anti-sociais em relação aos
membros do exogrupo (DE DREU & KRET 2016). Para além do papel da experiência e
contato com membros de outras raças (acima citado), deve-se ainda considerar o possível
impacto da variabilidade individual (ex. perfil ansioso) para compreendermos os efeitos
promovidos pela OT. Ilustrações disso envolvem pesquisas que indicam que a OT gera
aumento dos comportamentos agressivos em pessoas com baixo nível de ansiedade
(PFUNDMAIR et al., 2018) e promove redução modesta da agressividade em pessoas com
elevado nível de ansiedade (CAMPBELL & HAUSMANN, 2013). Assim, a modulação do
comportamento favorável ao endogrupo pela OT pode não acompanhar uma depreciação do
exogrupo (CHEN et al., 2011). Inclusive se observam estudos recentes que revelam a
promoção de um efeito pró-social de redução do medo social após inalação desse
nonapeptídeo (NEUMANN & SLATTERY, 2016).
Em suma, o efeito pró-social da OT ao endogrupo não parece ser indiscriminado em
todos os contextos (SINGER et al., 2008). A partir disso, as revisões de Jong e Neumann
(2017) e Neumann e Slattery (2016) discutem as expectativas no campo científico de que a
administração da OT sintética possa vir a tornar-se uma intervenção complementar ao

87
tratamento de quadros psiquiátricos caracterizados por comportamentos anti-sociais e de
ansiedade. Considerando a inconsistência dos resultados publicados, são necessários mais
estudos no sentido de esclarecer as oportunidades e limites do uso terapêutico da OT. Uma
contribuição do presente estudo foi demonstrar que a OT é capaz de modular a percepção de
ameaça e os comportamentos agressivos, mas a magnitude e direção desse efeito variam
conforme as pistas raciais.
Finalmente, a análise das variáveis destacadas na Teoria de Detecção de Sinal
proporciona relevantes conclusões. A primeira delas é que os participantes Norte-Americanos
são mais hábeis em discriminar armas em comparação aos Brasileiros. Isso foi demonstrado
na comparação dos dados de sensibilidade entre o estudo original de Correll (2002) e os
resultados do grupo PL na presente pesquisa (d’EUA > d’BR). Essa maior sensibilidade dos
Norte-Americanos poderá estar associada à maior presença de armas de fogo na população
em geral, o que amplia sua experiência com esse tipo de objeto. Porém, devemos também
considerar que a legalidade na aquisição de armas de fogo nos EUA promove uma
normalização do seu uso, o que por sua vez pode favorecer a ocorrência de comportamentos
agressivos no cotidiano (RODRIGUES, JABLONSKI E ASSMAR, 2000), vide a ocorrência
sistemática de tragédias em escolas e espaços públicos. Dessa forma, maior contato com arma
de fogo pode favorecer uma capacidade mais apurada de identificação desse objeto, assim
como revelado em estudos anteriores que policiais apresentam melhor desempenho na TAAP
(com menor viés racial na média) em comparação a universitários e membros da comunidade
em geral (CORRELL et al., 2007; MA & CORRELL, 2011).
A segunda conclusão desse estudo é que participantes Brancos Norte-Americanos e
Brasileiros controlam mais sua decisão de atirar quando estão diante de membros do
endogrupo em relação aos exogrupo. Verificamos isso pois ambas as amostras revelaram
critério de decisão de atirar positivo, ou seja, mais rígido e deliberado, diante de alvos
Brancos e critério de decisão negativo, mais leniente e automático, diante de alvos Negros
(cBranco > cNegro). Os resultados obtidos na nossa amostra indicam que, no Brasil, apesar da
maior convivência entre os grupos étnicos em questão, ainda há certo viés negativo diante de
alvos Negros uma vez que se atira neles mesmo quando estes estão desarmados (falso
alarme). Ainda assim, ressaltamos que a magnitude desse efeito para os Norte-Americanos é
2.93 vezes maior em comparação aos Brasileiros. Alguns elementos precisam ser
considerados para compreendermos tais diferenças culturais em relação ao processo de
decisão de atirar. Interessantes evidências com participantes Brancos Norte-Americanos
revelam que eles são mais sensíveis as manipulações experimentais, como apresentação de

88
estímulos priming de Brancos criminosos, pois isso eleva a tendência deles de atirar em alvos
Brancos na TAAP, o que não acontece com os alvos Negros porque eles comumente são
considerados ameaçadores independente dos estímulos priming apresentados (CORRELL et
al., 2007). Além disso, foi demonstrado anteriormente que quanto menos prototípicos foram
os alvos Brancos (ex. pessoas de cor de pele clara e características faciais típicas de Brancos),
maior é a probabilidade de decisões de atirar nos alvos (MA & CORRELL, 2011).
Transpondo esses achados para o contexto social Brasileiro verificamos que nosso estudo
pode adicionar relevantes contribuições para compreendermos esses complexos fenômenos
em uma amostra com maior miscigenação e convivências entre os grupos étnicos em questão.
Assim, no Brasil tem-se mais contato com pessoas desses grupos étnicos e com pessoas de
famílias interraciais, o que amplia nossa capacidade de sensibilização ao outro e transforma a
valência e intensidade de alguns estereótipos étnicos. Provavelmente por isso, na nossa
amostra de indivíduos Brasileiros a tendência do critério de decisão de atirar em alvos Negros
foi mais próxima do limiar de alvos Brancos e essa tendência foi quase três vezes menor que a
registrada por Correll nos Estados Unidos.
6.3. Limitações e Estratégias Futuras
Apesar dos cuidados metodológicos empregados na condução da presente pesquisa,
alguns aspectos poderão eventualmente limitar as interpretações dos resultados obtidos.
Relativamente ao controle por placebo, vale ressaltar que o grupo PL inalou uma solução
salina e o grupo experimental inalou solução de OT que incluía em sua composição solução
salina, além de outros excipientes (ácido cítrico, clorobutanol, cloreto de sódio, fosfato de
sódio dibásico, glicerol, metilparabeno, propilparabeno, sorbitol e água purificada, conforme
bula ANVISA / Novartis). Esse estudo poderia ter maior validade interna e externa caso a
única diferença entre as amostras inaladas fosse a presença ou ausência da molécula de OT,
conforme realizado em algumas pesquisas (HURLEMANN et al. 2010; COHEN &
SHAMAY-TSOORY, 2017). Entretanto, a solução salina (soro fisiológico) também é
habitualmente utilizada em pesquisas recentes (ECKSTEIN et al., 2014; NE’EMAN et al.
2016) sendo econômica e de fácil acesso. Porém, por ser familiar à maioria das pessoas, não
podemos excluir a possibilidade de os participantes identificarem de qual grupo eles fizeram
parte. Todavia, verificamos que isso não ocasionou um viés em nossa amostra uma vez que os
participantes não foram capazes de discriminar a substância inalada. Ainda, é necessário
destacar que a via de administração intranasal limita o controle da dosagem disponibilizada e

89
a capacidade de absorção da substância. Porém, apesar do provável impacto desses fatores na
modulação pela OT, essa é uma via amplamente empregada e que evidencia efeitos
comportamentais (NABER et al., 2010; THEODORIDOU et al., 2009).
Outra limitação, diz respeito à composição homogênea da nossa amostra – seja em
termos de gênero e grupo étnico (totalidade de homens Brancos), faixa etária (maioria jovens
adultos, entre 18-29 anos), nível socioeconômico (a pesquisa foi conduzida em uma
universidade particular, onde a maioria dos participantes deve ser de nível sócio-econômico
médio e médio-alto) e região de proveniência (totalidade de São Paulo). Assim, os resultados
obtidos não permitem gerar conclusões definitivas sobre os efeitos da OT sobre os
comportamentos de empatia e percepção de ameaça para a população Brasileira. São
necessários estudos adicionais com amostras mais diversificadas. Quanto à avaliação de
empatia implícita, a perda de 43% da amostra pode sinalizar limitações procedimentais desse
estudo durante essa coleta de dados específica. As pesquisas da área não costumam descrever
claramente a taxa de perda amostral dos experimentos que envolvem o uso de EMT e
registros de MEPs. Por isso, se faz necessário estudos futuro que explore limites aceitáveis de
perda amostral com essa técnica para aprimorarmos esse tipo de coleta. Por fim, de acordo
com os objetivos da presente pesquisa optamos por uma testagem com dosagem única,
entretanto, para fim de generalizações dos dados visando intervenções terapêuticas, avaliações
continuadas são fundamentais para oferecer resultados quanto à dependência da modulação
das respostas relativamente à dose administrada.
Na replicação de tarefas clássicas, verificamos que a magnitude dos resultados foi
consideravelmente menor em nossa amostra Brasileira em relação às amostras Norte-
Americana e Italiana. Assim, estudos futuros devem considerar adaptação das tarefas
selecionadas, especialmente quando aplicadas em populações com maior miscigenação, para
investigar efeitos de menor magnitude. Possibilidades de estudos sobre os comportamentos
pró- e anti-sociais devem utilizar estímulos que enfatizem a percepção sensorial da dor (ex.
com o uso de realidade virtual ou com a aplicação de ilusões perceptivas, como a Ilusão da
Mão de Borracha (LIRA et al., 2017) ou desenvolver tarefas mais ecológicas, que podem
gerar resultados mais evidentes oriundos, por exemplo, de interações interpessoais reais.
6.4. Considerações Finais
As habilidades de empatia e percepção de ameaça social são cruciais para favorecer o
vínculo e a proteção. Nesse trabalho, os efeitos da OT nessas habilidades foram avaliados em

90
contextos com pistas raciais, junto a uma amostra proveniente de um contexto com alto
percentual de miscigenação (comparativamente a estudos anteriores). Verificamos que nas
avaliações implícita e explícita, a OT não modulou a empatia à dor apresentada pelos
participantes, independentemente da etnia dos alvos. Por outro lado, os participantes que
inalaram a substância apresentaram redução do viés perceptivo de ameaça social, apenas
diante de membros do exogrupo. Isso indica que, dependendo do contexto analisado, a
inalação da OT sintética pode promover um efeito pró-social específico (ainda que modesto)
na diminuição de ameaça associada a indivíduos do exogrupo. Adicionalmente, foi
interessante verificar que os dados de empatia implícita e percepção de ameaça na amostra
Brasileira apresentaram direção semelhante às amostras Italiana e Norte-Americana porém,
com magnitude menor. Os resultados destacados nessa pesquisa são encorajadores quanto ao
papel de alguns fatores para a redução de comportamentos agressivos diante de membros do
exogrupo – nomeadamente a administração intranasal de OT e a maior convivência entre
membros de diferentes grupos étnicos. Em suma, o presente estudo oferece contribuições para
o crescente número de pesquisas que têm questionado os supostos efeitos universais da OT,
sublinhando a necessidade de contemplar as complexas interações entre variáveis individuais
e sociais na compreensão dos efeitos da substância na cognição e comportamento social
humano.

91
7. REFERÊNCIAS
ABU-AKEL, A.; FISCHER-SHOFTY, M.; LEVKOVITZ, Y.; DECETY, J.; SHAMAY-TSOORY, S. The role of oxytocin in empathy to the pain of conflictual out-group members among patients with schizophrenia. Psychol Med, v. 44, p. 3523–3532, 2014a. ABU-AKEL, A.; PALGI, S.; KLEIN, E.; DECETY, J.; SHAMAY-TSOORY, S. Oxytocin increases empathy to pain when adopting the other- but not the self-perspective. Social Neuroscience, p.1-9, 2014b. ALTEMUS, M.; REDWINE, L. S.; LEONG, Y. M.; FRYE, C. A.; PORGES, S. W.; e CARTER, C. S. Responses to laboratory psychosocial stress in postpartum women. Psychosomatic Medicine, v. 63, p. 814 – 821, 2001. AMODIO, D. M., & FRITH, C. D. Meeting of minds: The medial frontal cortex and social cognition. Nat. Rev. Neurosci., v. 7, p. 268–277, 2006. ANDARI, E., et al. Promoting social behavior with oxytocin in high-functioning autism spectrum disorders. Proceedings of the National Academy of Sciences, v. 107, n. 9, p. 4389-4394, 2010. ARROW, H. The Sharp End of Altruism. Science, v. 318, p. 581–582, 2007. ASPÉ-SÁNCHEZ, M.; MORENO, M.; RIVERA, M. I.; ROSSI, A.; & EWER, J. Oxytocin and Vasopressin Receptor Gene Polymorphisms: Role in Social and Psychiatric Traits. Front. Neurosci., 2016. AVENANTI, A., BUETI, D., GALATI, G., AGLIOTI, S. Transcranial magnetic stimulation highlights the sensorimotor side of empathy for pain Nat. Neurosci., v. 8, p. 955-960, 2005. AVENANTI, A.; PALUELLO, I. M.; BUFALARI, I.; & AGLIOTI, S. M. Stimulus-driven modulation of motor-evoked potentials during observation of others' pain. NeuroImage, v. 32, p. 316-324, 2006. AVENANTI, A., MINIO-PALUELLO, I., BUFALARI, I., AGLIOTI, S. M. The pain of a model in the personality of an onlooker: Influence of state-reactivity and personality traits on embodied empathy for pain. Neuroimage, v. 44, p. 275–283, 2009. AVENANTI, A.; SIRIGU, A.; AGLIOTI, S. M. Racial bias reduces empathic sensorimotor resonance with other-race pain. Current Biology, v. 20, n. 11, p. 1018-1022, 2010. AZEVEDO, R.T., MACALUSO, E., AVENANTI, A., SANTANGELO, V., CAZZATO, V., AGLIOTI, S.M. Their pain is not our pain: brain and autonomic correlates of empathic resonance with the pain of same and different race individuals. Hum Brain Mapp, v. 34, n.12, p. 3168-81, 2013. BAKERMANS-KRANENBURG, M. J., & VAN IJZENDOORN, M. H. Oxytocin receptor (OXTR) and serotonin transports (5-HTT) gene associated with observed parenting, Soc. Cogn. Affect. Neurosci., v. 3, p. 128-134, 2008.

92
BARTZ, J.A.; ZAKI, J.; BOLGER, N.; OCHSNER, K.N. Social effects of oxytocin in humans: context and person matter. Trends Cogn Sci., v. 15, n. 7, p. 301-309, 2011. BARKER, A.T., JALINOUS, R., FREESTON, I.L. Non-invasive magnetic stimulation of human motor cortex. Lancet 1, p. 1106–1107, 1985. BASHIR, S.; PEREZ, J.; HORVATH, J. C.; PASCUAL-LEONE, A. Differentiation of motor cortical representation of hand muscles by navigated mapping of optimal EMT current directions in healthy subjects. Journal Clinic Neurophysiology, v. 30, n. 4, p. 390–395, 2013. BAUMGARTNER, T. et al. Oxytocin shapes the neural circuitry of trust and trust adaptation in humans. Neuron, v. 58, n. 4, p. 639-650, 2008. BECK, A.T.; WARD, C.H.; MENDELSON, M.; MOCK, J.; ERBAUGH, J. An inventory for measuring depression. Arch Gen Psychiatry, v. 4, p. 561-71, 1961. BEN-AMI BARTAL I., RODGERS D.A., BERNARDEZ SARRIA M.S., DECETY J., MASON P. Pro-social behavior in rats is modulated by social experience. eLife, v. 3, 2014. BEGLEY, C. G., ELLIS, L. M. Drug development: raise standards for preclinical cancer research. Nature, v. 483, p. 531–533, 2012. BORN, J.; LANGE, T.; KERN, W.; McGREGOR, G. P.; BICKEL, U.; FEHM, H. L. Sniffing neuropeptides: a transnasal approach to the human brain. Nature Neuroscience, v. 5, n. 6, p. 514-516, 2002. BOS, P. A., MONTOYA, E. R., HERMAN, E. J., KEYSERS, C., HONKABG, J. Oxytocin reduces neural activity in the pain circuitry when seeing pain in others. NeuroImage, v. 113, p. 217-224, 2015. BUFALARI, I., APRILE, T., AVENANTI, A., DI RUSSO, F., AGLIOTI, S.M. Empathy for pain and touch in the human somatosensory cortex. Cereb. Cortex, v. 17, p. 2553–2561, 2007. BURBACH, J.P.H., ADAN, R.A.H., LOLAIT, S.J., VAN LEEUWEN, F.W., MEZEY, E., PALKOVITS, M., BARBERIS, C. Molecular neurobiology and pharmacology oh the vasopressina, oxytocin receptor Family. Cell. Mol. Neurobiol., v. 15, p.573-595, 1995. BUSHNELL, M. C., DUNCAN, G. H., HOFBAUER, R. K., HA, B., CHEN, J. I., CARRIER, B. Pain perception: Is there a role for primary somatosensory cortex? Proc. Natl. Acad. Sci., v. 96, p. 7705–7709, 1999. BUSS, A. H.; PERRY, M. The aggression questionnaire. Journal of Personality and Social Psychology, v. 63, p. 452-459, 1992. BUTTON, K. S., IOANNIDIS, J. P. A., MOKRYSZ, C., NOSEK, B. A., FLINT, J., ROBINSON, E. J. S., MUNAF, M. S. Power failure: why small sample size undermines the reliability of neuroscience. Nature Reviews, v. 14, p. 365-376, 2013.

93
BOWLES, S. Did Warfare Among Ancestral Hunter-Gatherers Affect the Evolution of Human Social Behaviors? Science, v. 324, n. 5932, pp. 1293-1298, 2009. CAMPBELL, A. Oxytocin and Human Social Behavior. Personality and Social Psychology Review, v. 14, n. 3, p. 281-295, 2010. CAMPBELL, A., HAUSMANN, M. Effects of oxytocin on women's aggression depend on state anxiety. Aggressive behavior., v. 39, n. 4, p. 316-322, 2013. CARSON, D. S., BERQUIST, S. W., TRUJILLO, T. H., GARNER, J.P., HANNAH, S. L., HYDE, S. A., SUMIYOSHI, R. D., JACKSON, L. P., MOSS, J. K., STREHLOW, M. C., CHESHIER, S. H., PARTAP, S., HARDAN A. Y., PARKER, K. J. Cerebrospinal fluid and plasma oxytocin concentrations are positively correlated and negatively predict anxiety in children. Molecular Psychiatry, v. 20, p. 1085–1090, 2015. CHARLET, A., GRINEVICH, V. Oxytocin Mobilizes Midbrain Dopamine toward Sociality. Neuron, v. 95, n. 2, p. 235-237, 2017. CHENG, Y.; LIN, C.; LIU, H. L.; HSU, Y.; LIM, K.; HUNG, D.; et al. Experience modulates the perception of pain in others. Current Biology, v. 17, p. 1708-1713, 2007. CHENA, F. S., KUMSTAA, R., DAWANSA, B., MONAKHOVB, M., EBSTEINB, R. P., HEINRICHS. M. Common oxytocin receptor gene (OXTR) polymorphism and social support interact to reduce stress in humans. PNAS, p.1-6, 2011. CHURCHLAND, P. S., WINKIELMAN, P. Modulating Social Behavior with Oxytocin: How does it work? What does it mean? Horm Behav., v. 61, n. 3, p. 392–399, 2012. COHEN, D., SHAMAY-TSOORY, S. G. Oxytocin regulates social approach. Soc Neurosci., v. 17, p. 1-8, 2017. CIKARA, M., C.P. SAID, J.L. EBERHARDT & S.T. FISKE. From subjects to objects: Sexist attitudes and instrumental processing of sexualized women. Paper presented to Society for Personality and Social Psychology, Tampa, 2009. CORRELL, J.; PARK, B.; JUDD, C. M.; & WITTENBRINK, B. The police officer's dilemma: using ethnicity to disambiguate potentially threatening individuals. Journal of personality and social psychology, v. 83, n. 6, p. 1314, 2002. CORRELL, J., URLAND, G. R., ITO, T. A. Event-related potentials and the decision to shoot: The role of threat perception and cognitive control. Journal of Experimental Social Psychology, v. 42, 120–128, 2006. CORRELL, J.; PARK, B.; JUDD, C. M.; & WITTENBRINK, B. The Influence of Stereotypes on Decisions to Shoot. European Journal of Social Psychology, v. 37, p. 1102-1117, 2007. CORRELL, J., WITTENBRINK, B., PARK, B., JUDD, C.M., SADLER, M. Across the Thin Blue Line: Police Officers and Racial Bias in the Decision to Shoot. Journal of Personality and Social Psychology. 92, 1006–1023, 2007.

94
CUNHA, J. A., PRIEB, R. G. G., GOULART, P. M., LEMES, R. B. O uso do inventario de Beck para avaliar depressão em universitários. Psico, v. 27, n. 1, p. 107-115, 1996. DADDS, M. R.; MACDONALD, E.; CAUCHI, A.; WILLIAMS, K.; LEVY, F.; BRENNAN, J. Nasal oxytocin for social deficits in childhood autism: A randomized controlled trial. J Autism Dev Disord, v. 44, p. 521–531, 2014. DAI, L.; CARTER, C. S.; YING, J.; BELLUGI, U.; POURNAJAFI-NAZARLOO, H.; KORENBERG, J. R. Oxytocin and Vasopressin Are Dysregulated in Williams Syndrome, a Genetic Disorder Affecting Social Behavior. PLoS ONE, v. 7, n. 6, 2012. DAVIS, M.H. Measuring individual differences in empathy: evidence for a multidimensional approach. Journal of personality and social psychology, v. 44, n. 1, p. 113-136, 1983. DECLERCK, C.H. et al. Oxytocin and cooperation under conditions of uncertainty: the modulating role of incentives and social information. Horm. Behav., v. 57, p. 368–374, 2010. DELGADO, M. R. Fool me once, shame on you; fool me twice, shame on oxytocin. Neuron, v. 58, n. 4, p. 470-471, 2008. DE DREU, C. K.; GREER, L. L.; VAN KLEEF, G. A.; SHALVI, S.; HANDGRAAF, M. J. Oxytocin promotes human ethnocentrism. Proc Natl Acad Sci U S A, 108, p. 1262–1266, 2010. DE DREU, C. K., et al. The Neuropeptide Oxytocin Regulates Parochial Altruism in Intergroup Conflict Among Humans. Science, v. 328, n. 5984, p. 1408-1411, 2010. DE DREU, C. K., KRETAC, M. Oxytocin Conditions Intergroup Relations Through Upregulated In-Group Empathy, Cooperation, Conformity, and Defense. Biological Psychiatry, v. 79, n. 3, p.165-173, 2016. DE DREU, C. K., GREER, L. L., VAN KLEEF, G. A., SHALVI, S., HANDGRAAF, M. J. Oxytocin promotes human ethnocentrism. Proc. Natl. Acad. Sci. U.S.A. v. 108, p. 1262–1266, 2011. DECETY, J.; & ECHOLS, S. C.; & CORRELL, J. The blame game: The effects of responsibility and social stigma on empathy for pain. Journal of Cognitive Neuroscience, v. 22, p. 985-997, 2010. DECETY, J.; JACKSON, P. L. The functional architecture of human empathy. Behavioral and Cognitive Neuroscience Reviews, v. 3, n. 2, p. 71-100, 2004. DECETY, J.; LAMM, C. Empathy versus personal distress: Recent evidence from social neuroscience. In J. DECETY & W. ICKES (Eds.), The social neuroscience of empathy (p. 199-213). Cambridge, MA: MIT Press, 2006. DECETY, J.; SVETLOVA, M. Putting together phylogenetic and ontogenetic perspectives on empathy. DeveloMEPntal Cognitive Neuroscience, v. 2, n. 1, p. 1-24, 2012

95
DI SIMPLICIO, M., MASSEY-CHASE, R., COWEN, P. J., HARMER, C. J. Oxytocin enhances processing of positive versus negative emotional information in healthy male volunteers. Journal of Psychopharmacology, v. 23, n. 3, p. 241-248, 2009. DONALDSON, Z. R.; YOUNG, L. J. Oxytocin, vasopressin, and the neurogenetics of sociality. Science, v. 322, p. 900–904, 2008. DOMES, G., HEINRICHSD, M., GLÄSCHER, J., BÜCHEL, C., BRAUS, D. F., HERPERTZA, S. C. Oxytocin Attenuates Amygdala Responses to Emotional Faces Regardless of Valence. Biological Psychiatry, v. 62, n. 10, p. 1187-1190, 2007. DOMES, G. et al. Oxytocin improves “mind-reading” in humans. Biological Psychiatry, v. 61, n. 6, p. 731-733, 2007. ECKSTEIN, M., SCHEELE, D., WEBER, K., STOFFEL-WAGNER, B., MAIER, W., AND HURLEMANN, R. Oxytocin Facilitates the Sensation of Social Stress. Human Brain Mapping, p. 1-10, 2014. EISENEGGER, C.; NAEF, M.; SNOZZI, R.; HEINRICHS, M.; FEHR, E. Prejudice and truth about the effect of testosterone on human bargaining behaviour. Nature, v. 463, p. 356-360. 2010. EBERHARDT, J.L. Imaging race. Am. Psychol., v. 60, p. 181–190, 2005. EPLEY, N. In press. Mind perception. In Handbook of Social Psychology, 5th edn. S.T. Fiske, D.T. Gilbert & G. Lindzey, Eds. John Wiley. New York, NY. EBITZ, R. B., WATSON, K. K., PLATT, M. L. Oxytocin blunts social vigilance in the rhesus macaque. Proc. Natl. Acad. Sci. U.S.A. v. 110, p. 11630–11635, 2013. ESPÍRITO-SANTO H, DANIEL, F. Calcular e apresentar tamanhos do efeito em trabalhos científicos (3): Guia para reportar os tamanhos do efeito para análises de regressão e ANOVAs. Revista Portuguesa de Investigação Comportamental e Social v. 4, p. 43-60, 2018. FECTEAU, S., PASCUAL-LEONE, A., THEORET, H. Psychopathy and the mirror neuron system: Preliminary findings from a non-psychiatric sample. Psychiatry Res. v. 160, p. 137–144, 2008. FERRIS, C. F.; FOOTE, K. B.; MELTSER, H. M.; PLENBY, M. G.; SMITH, K. L.; INSEL, T. R. Oxytocin in the amygdala facilitates maternal aggression. Ann N Y Acad Sci, v. 652, p. 456–457, 1992. FISKE, S. T. From dehumanization and objectification to rehuma- nization: Neuroimaging studies on the building blocks of empathy. Ann. N. Y. Acad. Sci., v. 1167, p. 31–34, 2009. GAMA, R. Uma verdade inconveniente. Revista Veja, São Paulo, p.79, 16 novembro de 2017.

96
GAMER, M.; et al. Different amygdala subregions mediate valence-related and attentional effects of oxytocin in humans. Proc. Natl. Acad. Sci. U.S.A., v. 107, p. 9400-9405, 2010. GORENSTEIN, C.; ANDRADE, L. H. S. G. Inventário de depressão de Beck: propriedades psicométricas da versão em português. Rev Psiq Clin, v. 25, n. 5, p. 245-50, 1998. GOUVEIA, V. V.; CHAVEZ, C. M. C.; PEREGRINO, R. R.; CASTELLO BRANCO, A. O.; & GONÇALVES, M. P. Medindo a agressão: o Questionário de Buss-Perry. Arquivos Brasileiros de Psicologia, v. 60, n. 3, 2008. GOLDMAN, M., et al. Diminished plasma oxytocin in schizophrenic patients with neuroendocrine dysfunction and emotional deficits. Schizophrenia research, v. 98, n. 1, p. 247-255, 2008. GREENWALD, A. G., MCGHEE, D.E., SCHWARTZ, J. L. K. Measuring individual differences in implicit cognition: The implicit association test. Journal of Personality and Social Psychology, v. 74, n. 6, p. 1464-1480, 1998. GREENWALD, A. G.; NOSEK, B. A.; BANAJI, M. R. Understanding and using the implicit association test: I. An improved scoring algorithm. Journal of personality and social psychology, v. 85, n. 2, p. 197, 2003. GREENWALD, A. G.; OAKES, M. A.; HOFFMAN, H. G. Targets of discrimination: Effects of race on responses to weapons holders. Journal of Experimental Social Psychology, n. 39, v.4, p.399-405, 2003. GU, X.; & HAN, S. Attention and reality constraints on the neural processes of empathy for pain. NeuroImage, v. 36, p. 256-267, 2007. GUNAYDIN, L.A., GROSENICK, L., FINKELSTEIN, J.C., KAUVAR, I.V., FENNO, L.E., ADHIKARI, A., LAMMEL, S., MIRZABEKOV, J.J., AIRAN, R.D., ZALOCUSKY, K.A., et al. Natural neural projection dynamics underlying social behavior. Cell, v. 157, p. 1535-1551, 2014. HART, A. J., WHALEN, P. J., SHIN, L. M., MCINERNEY, S. C., FISCHER, H., RAUCH, S. L. Differential response in the human amygdala to racial outgroup vs ingroup face stimuli. NeuroRepor, v.11, n. 11, p. 2351–2354, 2000. HARVEY, L. O., Jr. (1992). The critical operating characteristic and the evaluation of expert judgment. Organizational Behavior & Human Decision Processes, v. 53, n. 2, p. 229. HASLAM, N. Dehumanization: An integrative review. Pers. Soc. Psychol. Rev., v. 10, p. 252–264, 2006. HEINRICHS, M.; VON DAWANS, B.; DOMES, G. Oxytocin, vasopressin, and human social behavior. Front Neuroendocrinol, v. 30, p. 548–557, 2009. HURLEMANN, R.; PATIN, A.; ONUR, O. A.; COHEN, M. X.; BAUMGARTNER, T.; METZLER, S.; KENDRICK, K. M. Oxytocin enhances amygdala-dependent, socially

97
reinforced learning and emotional empathy in humans. Journal of Neuroscience, v. 30, n. 14, p. 4999-5007, 2010. INSEL, T.R.; YOUNG, L.; WANG, Z. Central oxytocin and reproductive behaviours. Rev Reprod, v. 2, p. 28–37, 1997. INSEL, T. R., AND YOUNG, L. J. The neurobiology of attachment. Nature Reviews, v. 2, p. 129-136, 2001. INSEL, T.R., SHAPIRO, L.E. Oxytocin receptor distribution reflects social organization in monogamous and polygamous voles. Proc. Natl. Acad. Sci., v. 89, p. 5981-5985, 1992. JACKSON, P. L.; BRUNET, E.; MELTZOFF, A. N.; DECETY, J. Empathy examined through the neural machanisms involved in imagining how I feel versus how you feel pain. Neuropsychologia, v. 44, n. 5, p. 752-761, 2006. JACKSON, P. L.; MELTZOFF, Andrew N.; DECETY, Jean. How do we perceive the pain of others? A window into the neural processes involved in empathy. Neuroimage, v. 24, n. 3, p. 771-779, 2005. JIN, D, et al. CD38 is critical for social behaviour by regulating oxytocin secretion. Nature, v. 446, p. 41–45, 2007. JONG, T. R., NEUMANN, I. D. Oxytocin and Aggression. Curr Topics Behav Neurosci, p. 1-18, 2017. KENNEDY, D.P., ADOLPHS, R. The social brain in psychiatric and neurological disorders. Trends Cogn Sci., v. 16, p. 559-72, 2012. KEYSERS, C.; WICKER, B.; GAZZOLA, V.; ANTON, J. L.; FOGASSI, L.; GALLESE, V. A touching sight: SH/PV activation during the observation and experience of touch. Neuro, v. 42, p. 335-346, 2004. KIRSCH, P. et al. Oxytocin modulates neural circuitry for social cognition and fear in humans. J. Neurosci, v. 25, p. 11489-11493, 2005. KOSFELD, M.; et al. Oxytocin increases trust in humans. Nature, v. 435, p. 673-676, 2005. KOSCIK, T. R.; TRANEL, D. The human amygdala is necessary for developing and expressing normal interpersonal trust. Neuropsychologia, v. 49, n. 4, p. 602-611, 2011. LAMM, C., DECETY, J., SINGER, T. Meta-analytic evidence for common and distinct neural networks associated with directly experienced pain and empathy for pain. NeuroImage, v. 54, n. 3, , p. 2492-2502, 2011. LABUSCHAGNE, I., PHAN, K. L., WOOD, A., ANGSTADT, M., CHUA, P., HEINRICHS, M., NATHAN, P. J. Oxytocin attenuates amygdala reactivity to fear in generalized social anxiety disorder. Neuropsychopharmacology, v. 35, n. 12, p. 2403, 2010.

98
LANCASTER, K.; CARTER, C. S.; POURNAJAFI-NAZARLOO, H.; KARAOLI, T.; LILLARD, T. S.; JACK, A.; DAVIS, J. M.; MORRIS, J. P.; CONNELLY, J. J. Plasma oxytocin explains individual differences in neural substrates of social perception. Front. Hum. Neurosci. v. 9, p. 1-7, 2015. LEE, H. J., MACBETH, A. H., PAGANI, J. H., YOUNG, W. S.Oxytocin: the great facilitator of life. Prog Neurobiol. v. 88, n. 2, p.127-51, 2009. LIEBERMAN, M. D. Social cognitive neuroscience: A review of core processes. Ann. Rev. Psychol., v. 58, p. 259–289, 2007. LIU, W., PAPPAS, G.D., CARTER, C.S. Oxytocin receptors in brais cortical regions are reduced in haploinsufficient (+/-) reler mice. Neurol. Res., v. 27, p. 339-345, 2005. LOUP, F. et al. Localization of high-affinity binding sites for oxytocin and vasopressin in the human brain. An autoradiographic study. Brain research, v. 555, n. 2, p. 220-232, 1991. LOVE, T.M. Oxytocin, motivation and the role of dopamine. Pharmacol Biochem Behav, v. 119, p. 49-60, 2014. MA, D. S.; CORRELL, J. Target prototypicality moderates racial bias in the decision to shoot. Journal of Experimental Social Psychology, n. 47, v.2, p. 391-396, 2011. MA, Y., LIU, Y., RAND, D. G., HEATHERTON, T. F., HAN, S. Opposing Oxytocin Effects on Intergroup Cooperative Behavior in Intuitive and Reflective Minds. Neuropsychopharmacology. v. 40, n. 10, p. 2379-87, 2015. MA, D., CORRELL, J. Target prototypicality moderates racial bias in the decision to shoot. Journal of Experimental Social Psychology, v. 47, n. 2, p. 391-396, 2011. MACDONALD, K.; MACDONALD, T. M. The peptide that binds: a systematic review of oxytocin and its prosocial effects in humans. Harvard review of psychiatry, v. 18, n. 1, p. 1-21, 2010. MARSH, A.A. et al. Oxytocin improves specific recognition of positive facial expressions. Psychopharmacology (Berl), v. 209, p. 225–232, 2010. MCCALL, C.; SINGER, T. The animal and human neuroendocrinology of social cognition, motivation and behavior. Nature neuroscience, v. 15, n. 5, p. 681-688, 2012. MIZAEL, M. T., ALMEIDA, J. H., SILVEIRA, C. C., DE ROSE, J. C. Changing Racial Bias by Transfer of Functions in Equivalence Classes. Psychol Rec, v. 66, p. 451–462 , 2016. MODAHL et al. Plasma oxytocin levels in autistic children. Biol. Psychiatry, v. 43, p. 270-277, 1998. MORRISON, I., LLOYD, D., DI PELLEGRINO, G., ROBERTS, N. Vicarious responses to pain in anterior cingulate cortex: is empathy a multisensory issue? Cogn. Affect. Behav. Neurosci., v. 4, p. 270-278, 2004.

99
MOTTOLESE, R., REDOUTÉ, J., COSTES, N., BARS, D. L., SIRIGU, A. Switching brain serotonin with oxytocin. PNAS, p. 1-6, 2014. NABER, F. et al. Intranasal oxytocin increases fathers’ observed responsiveness during play with their children: a double-blind within-subject experiment. Psychoneuroendocrinology, v. 35, n. 10, p. 1583-1586, 2010. NE'EMAN, R. et al. Intranasal administration of oxytocin increases human aggressive behavior. Hormones and behavior, v. 80, p. 125-131, 2016. NETO, F. O paradigma do teste de associação Implícita. Revista E-Psi, v. 5, n. 2, 1-15, 2015. NEUMANN, I.D. The advantange of social living: brain neuropeptides mediate the benefical consequences of sex and motherhood. Front. Neuroendocrinol, v. 30, p. 483-496, 2009. NEUMANN, I. D., SLATTERY, D. A. Oxytocin in General Anxiety and Social Fear: A Translational Approach. Biological Psychiatry, v. 79, n. 3, p. 213-221, 2016. NUMAN, M., YOUNG, L. J. Neural mechanisms of mother–infant bonding and pair bonding: Similarities, differences, and broader implications. Hormones and Behavior, v. 77, p. 98-112, 2016. NISHIMORI, K., YOUNG, L.J., GUN, Q.X., WANG, Z.X., INSEL, T.R. Oxytocin is required for nursing, but is not essentioal for parturition or reproductive behavior. Proc. Natl. Acad. Sci., v. 93, p. 11699-11704, 1996. OLDFIELD, R. C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia, v. 9, n. 1, p. 97-113, 1971. OLIVEIRA, L. M. e SILVA, L. F. S. Mecanismos neurais e modulação da dor. Dor: princípios e prática. p. 235-242, 2009. PAYNE, B. K. Prejudice and perception: The role of automatic and controlled processes in misperceiving a weapon. Journal of Personality and Social Psychology, v. 81, n. 2, p. 181-192, 2001. PAYNE, B. K. Weapon Bias: Split-Second Decisions and Unintended Stereotyping. Current Directions in Psychological Science, v. 15, p. 287, 2006. PATEL, J.C., ROSSIGNOL, E., RICE, M.E. e MACHOLD, R.P. Opposing regulation of dopaminergic activity and exploratory motor behavior by forebrain and brainstem colonergic circuits. Nat. Commun, v. 3, p. 1172 , 2012. PEDERSEN, C. A., VADLAMUDI, S. V., BOCCIA, M. L., AMICO, J. A. Maternal behavior deficits in nulliparous oxytocin knockout mice. Genes, Brain and Behavior, v. 5, p. 274–281, 2006. PETTIGREW, T. F.; MEERTENS, R. W. Subtle and blatant prejudice in Western Europe. European journal of social psychology, v. 25, n. 1, p. 57-75, 1995.

100
PETROVIC, P., KALISCH, R., SINGER, T., AND DOLAN, R. J. Oxytocin Attenuates Affective Evaluations of Conditioned Faces and Amygdala Activity. The Journal of Neuroscience, v. 28, p. 6607– 6615, 2008. PEYRON, R., LAURENT, B., GARCIA-LARREA, L. Functional imaging of brain responses to pain. A review and meta-analysis.) Neurophysiol. Clin., v. 30, p. 263-288, 2000. PFUNDMAIRA, M., REINELTA, A. C., DEWALL, N., FELDMANN, L. Oxytocin strengthens the link between provocation and aggression among low anxiety people. Psychoneuroendocrinology, v. 93, p. 124-132, 2018. PIETROWSKY, R.; STRTIBEN, C.; MOLLER, M.; FEHM, H. L.; & BORN, J. Brain Intravenous for a Direct in Humans Potential Changes after Intranasal vs. Administration of Vasopressin: Evidence Nose-Brain Pathway for Peptide Effects. Biol Psychiatry, v. 39, p. 332-340, 1996. PLANT, E. A.; PERUCHE, B. M. The consequences of race for police officers' responses to criminal suspects. Psychological Science, n. 16,v.3, p.180-183, 2005. PHELPS, E. A., O'CONNOR, K. J., CUNNINGHAM, W. A., FUNAYAMA, E. S., GATENBY, J. C., GORE, J. C., BANAJI, M. R. Performance on Indirect Measures of Race Evaluation Predicts Amygdala Activation. Journal of Cognitive Neuroscience, v. 12, n. 5, p.729-738, 2000. PHELPS, E. A.; LEDOUX, J. E. Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron, v. 48, n. 2, p. 175-187, 2005. PRESTON, S. D.; & de WAAL, F. B. M. Empathy: its ultimate and proximate bases. Behavioral and Brain Science, v. 25, p. 1-27, 2002. PRINZ, F., SCHLANGE, T., ASADULLAH, K. Believe it or not: how much can we rely on published data on potential drug targets? Nature Rev. Drug Discov., v. 10, p. 712, 2011. RAGNAUTH, A.K., GOODWILLIE, A., BREWER, C., MUGLIA, L.J., PFAFF, D.W., KOW, L.M. Vasopressin stimulates ventromedial hypothalamic neurons via oxytocin receptors in oxytocin gene knockout male and female mice. Neuroendocrinology, v. 80, p.92-99, 2004. RILLING, J. K., YOUNG, L. J. The biology of mammalian parenting and its effect on offspring social development. Science, v. 345, n. 6198, p. 771-776, 2014. RUBIN, L. H. et al. Peripheral oxytocin is associated with reduced symptom severity in schizophrenia. Schizophr. Res., v. 124, p. 13-21, 2010. RODRIGUES, A., ASSMAR, E. M. L., JABLONKI, B.. Comportamento anti-social: a agressão. In RODRIGUES, Aroldo; ASSMAR, Eveline Maria Leal; JABLONKI, Bernado. Psicologia Social. 20. ed. Petrópolis: Vozes, 2001.

101
ROSENFELD, LIEBERMAN e JARSKOG. Oxytocin, dopamine, and the amygdala: A neurofunctional modelo f social cognitive déficits in schizophrenia. Schizophr Bull, v. 37, p. 1077-1087, 2011. ROSS, H. E.; YOUNG, L. J. Oxytocin and the neural mechanisms regulating social cognition and affiliative behavior. Front Neuroendocrinol, v. 30, p. 534–547, 2009. SAMPAIO, L. R.; GUIMARÃES, P. R. B.; DOS SANTOS CAMINO, C. P.; FORMIGA, N. S.; & MENEZES, I. G. Estudos sobre a dimensionalidade da empatia: tradução e adaptação do Interpersonal Reactivity Index (IRI). Psico, v. 42, n. 1, 2011. SHAMAY-TSOORY, S. G.; FISCHER, M.; DVASH, J.; HARARI, H.; PERACH-BLOOM, N.; LEVKOVITZ, Y. Intranasal administration of oxytocin increases envy and schadenfreude (gloating). Biol Psychiatry, v. 66, p. 864–870, 2009. SHAMAY-TSOORY e ABU-AKEL. The Social salience Hypothesis of Oxytocin. Biological Psychiatry, v. 79, p. 194-202, 2016. SHAMAY-TSOORY, S. G., ABU-AKEL, A., PALGIA, S., SULIEMAN, R., FISCHER-SHOFTY, M., LEVKOVITZ, Y., DECETY, J. Giving peace a chance: Oxytocin increases empathy to pain in the context of the Israeli–Palestinian conflict. Psychoneuroendocrinology, v. 38, n. 12, p. 3139-3144, 2013. SHAW, C. A. Age-dependent expression of receptor properties and function in CNS development. In: SHAW, C.A. (Ed.), Receptor dynamics in neural development. CRC Press, New York, pp-3-17, 1996. SHENG, F.; LIU, Y.; ZHOU, B.; ZHOU, W.; HAN, S. Oxytocin modulates the racial bias in neural responses to others' suffering. Biol Psychol, v. 92, p. 380–386, 2013. SCHULZE et al. Oxytocin increases recognition of masked emotional faces. J. Psyneuen, 2011. SCHUMAN, H. et al. Racial attitudes in America: trends and interpretations. 1ª edição 1997. Cambridge, MA, Harvard University Press, 2001. SCHEELE, D., STRIEPENS, N., GÜNTÜRKÜN, O., DEUTSCHLÄNDER, S., MAIER, W., KENDRICK, K. M., HURLEMANN, R. Oxytocin Modulates Social Distance between Males and Females. Journal of Neuroscience, v. 32, n. 46, p. 16074-16079, 2012. SHENG, F., HAN, S. Manipulations of cognitive strategies and intergroup relationships reduce the racial bias in empathic neural responses. NeuroImage, v. 61, p. 786–797, 2012. SINGER, T.; DECETY, J. Social Neuroscience of Empathy (2011). In DECETY, J.; CACIOPPO, J. T. (Eds.), The Oxford handbook of social neuroscience (pp. 551-564). Oxford, New Your: Oxford University Press. SINGER, T.; SEYMOUR, B.; O'DOHERTY, J.; KAUBE, H.; DOLAN, R. J.; &FRITH, C. D. Empathy for pain involves the affective but not sensory components of pain. Science, v. 303, p. 1157-1162, 2004.

102
SINGER, T., FRITH, C. The painful side of empathy. Nature Neuroscience, v. 8, p. 845-846, 2005. SINGER, T.; SNOZZI, R.; BIRD, G.; PETROVIC, P.; SILANI, G.; HEINRICHS, M.; DOLAN, R. J. Effects of oxytocin and prosocial behavior on brain responses to direct and vicariously experienced pain. Emotion, v. 8, n. 6, p. 781-79, 2008. STALLEN, M.; DE DREU, C. K.; SHALVI, S.; SMIDTS, A.; SANFEY, A. G. The herding hormone: Oxytocin stimulates in-group conformity. Psychol Sci, v. 23, p. 1288–1292, 2012. TABAK, B. A.; MCCULLOUGH, M. E.; SZETO, A.; MENDEZ, A. J.; MCCABE, P. M. Oxytocin indexes relational distress following interpersonal harms in women. Psychoneuroendocrinology, v. 36, p. 115–122, 2011. TAJFEL, H., TURNER, J.C. An integrative theory of intergrup conflict. In: AUSTIN, W.G., WORCHEL, S., editors. The social psychology of intergroup relations. Monterey, CA: Brooks-Cole, 1979. TAYLOR, A.; LITTLEWOOD, J.; ADAMS, D.; DORÉ, C.; & GLOVER, V. Serum cortisol levels are related to moods of elation and dysphoria in new mothers. Psychiatry Research, v. 54, p. 241-247, 1994. TARRANT, M., DAZELEY S., COTTOM, T. Social categorization and empathy for outgroup members. Social Psychology, v. 48, n. 3, p. 427-446, 2009. TEIXEIRA, C.M., MARTIN, E.D., SAHUN, I., MASACHS, N., PUJUDAS, L., CORVELO, A., BOSCH, C., ROSSI, D., MARTINEZ, A., MALDONADO, R., DIERSSEN, M., SORIANO, E. Overexpression of reelen prevents the manisfestation of behavioral phenotypes related to schizophrenia and bipolar disorder. Neuroendocrinilogy, v. 36, p. 2395-2405, 2005. THEODORIDOU, A. et al. Oxytocin and social perception: oxytocin increases perceived facial trustworthiness and attractiveness. Hormones and behavior, v. 56, n. 1, p. 128-132, 2009. VAN IJZENDOORN, M. H.; BAKERMANS-KRANENBURG, M. J. A sniff of trust: Meta-analysis of the effects of intranasal oxytocin administration on face recognition, trust to in-group, and trust to out-group. Psychoneuroendocrinologu, v. 37, p. 438-443, 2012. XIAO, L., PRIEST, M.F., NASENBENY, J., LU, T., KOZOROVITSKIY, Y. Biased Oxytocinergic Modulation of Midbrain Dopamine Systems. Neuron, v. 95, p.368-384, 2017. XU, X., ZUO, X., WANG, X., HAN, S. Do you feel my pain? Racial group membership modulates empathic neural responses. J. Neurosci. v. 29, p. 8525–8529, 2009. WOTJAK, C. T.; GANSTER, J.; KOHL, G.; HOLSBOER. F.; LANDGRAF, R.; ENGELMANN, M. Dissociated central and peripheral release of vasopressin, but not oxytocin, in response to repeated swim stress: new insights into the secretory capacities of peptidergic neurons. Neuroscience, v. 85, p. 1209–1222, 1998.

103
ZAK, P. J.; KURZBAN, R.; MATZNER, W. T. Oxytocin is associated with human trustworthiness. Hormones and behavior, v. 48, n. 5, p. 522-527, 2005. ZAK, J. P., STANTON, A. A., AHMADI, S. Oxytocin Increases Generosity in Humans. Plos One, v. 2, n. 11, 2007. ZHONG, S. et al. U-shaped relation between plasma oxytocin levels and behavior in the trust game. PLoS ONE, v. 7, n. 12, 2012.
ZUO, X., HAN, S. Cultural experiences reduce racial bias in neural responses to others’
suffering. Cult. Brain, v. 1, n. 1, p. 34–46, 2013.