i universidade federal do parÁ campus ... - ufpa… · maria cristina dos santos costa (ufpa -...
TRANSCRIPT

i
UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA INSTITUTO DE ESTUDOS COSTEIROS
CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL MESTRADO EM RECURSOS BIOLÓGICOS DA ZONA COSTEIRA AMAZÔNICA
A HERPETOFAUNA ASSOCIADA À ZONA COSTEIRA DA AMAZÔNIA BRASILEIRA E O PADRÃO DE DISTRIBUIÇÃO ESPAÇO TEMPORAL DE
ANUROS NA PENÍNSULA DE AJURUTEUA, BRAGANÇA-PA
ROSIVAN PEREIRA DA SILVA
Prof. Dr. MARCUS E. B. FERNANDES ORIENTADOR
BRAGANÇA-PA 2010

ii
UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA INSTITUTO DE ESTUDOS COSTEIROS
CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL MESTRADO EM RECURSOS BIOLÓGICOS DA ZONA COSTEIRA AMAZÔNICA
A HERPETOFAUNA ASSOCIADA À ZONA COSTEIRA DA AMAZÔNIA BRASILEIRA E O PADRÃO DE DISTRIBUIÇÃO ESPAÇO TEMPORAL DE
ANUROS NA PENÍNSULA DE AJURUTEUA, BRAGANÇA-PA
ROSIVAN PEREIRA DA SILVA Dissertação de Mestrado apresentada ao Curso de Mestrado em Biologia Ambiental (Área de Concentração: Ecologia de Ecossistemas Costeiros e Estuarinos), da Universidade Federal do Pará, Campus de Bragança, como parte dos requisitos necessários à obtenção do grau de Mestre em Biologia Ambiental.
Profº Dr. MARCUS E. B. FERNANDES ORIENTADOR
BRAGANÇA-PA 2010

iii

iv
UNIVERSIDADE FEDERAL DO PARÁ
CAMPUS UNIVERSITÁRIO DE BRAGANÇA INSTITUTO DE ESTUDOS COSTEIROS
CURSO DE PÓS-GRADUAÇÃO EM BIOLOGIA AMBIENTAL MESTRADO EM RECURSOS BIOLÓGICOS DA ZONA COSTEIRA AMAZÔNICA
A HERPETOFAUNA ASSOCIADA À ZONA COSTEIRA DA AMAZÔNIA BRASILEIRA E O PADRÃO DE DISTRIBUIÇÃO ESPAÇO TEMPORAL DE
ANUROS NA PENÍNSULA DE AJURUTEUA, BRAGANÇA-PA
Banca Examinadora
Dra. Maria Cristina dos Santos Costa (UFPA - ICB) (Titular)
Dr. José Antonio Renan Bernardi (Instituto Federal do Pará – IFPA) (Titular) ___________________________________________________________________ Dra. Gilda Vasconcellos de Andrade (Univ. Federal do Maranhão – UFMA) (Titular) Dr. Marcus Emanuel Barroncas Fernades (UFPA – IECOS) (Orientador)
BRAGANÇA-PA 2010

v
AGRADECIMENTOS
Esta dissertação é o resultado do trabalho em equipe, reunida pela amizade,
profissionalismo, interesse científico e curiosidade. Assim, só foi possível a partir da
ajuda mútua e colaboração de diversas pessoas. Certamente não conseguirei me
lembrar de todos. Para aqueles aqui não citados, minhas sinceras desculpas.
Agradeço a CAPES pela bolsa fornecida durante o curso.
Ao programa de Mestrado em Biologia Ambiental, IECOS, Campus de Bragança.
Ao Dr. Renan Bernardi por ter me apresentado a herpetofauna e ter dado
credibilidade em meus trabalhos e dividido comigo um pouco do seu conhecimento e
experiência acerca de répteis e anfíbios amazônicos.
Ao Dr. Marcus Fernades, que se dispôs a me orientar e acreditar na viabilidade do
trabalho, bem como está aberto a trabalhar com um grupo diferenciado de sua linha
de pesquisa.
Aos amigos Kelle, Folha, André Palmeira, Arthur, Tayana, Tarcio, Rafael, Lucia,
Andre, Marcio e Joely agradeço pela ajuda durante esta jornada, tendo enfrentado as
adversidades que com sem ajuda dos mesmos não teriam sido superadas.
Agradeço também aos motoristas e amigos Sr. Porisso, Antonio e José (Beraba) que
estiveram sempre a postos na execução de minhas tarefas.
Aos doutores, Ulf e Colin e ao Msc. Darlan que me ajudaram em momentos crucias
durante este trabalho.
Enfim, a todos que ajudaram nessa jornada.

vi
SUMÁRIO
AGRADECIMENTOS .......................................................................................... v
SUMÁRIO ............................................................................................................ vii
LISTA DE TABELAS .......................................................................................... viii
LISTAS DE FIGURAS.......................................................................................... ix
RESUMO 1
CAPÍTULO 1. INTRODUÇÃO GERAL ............................................................. 2
Origem e Características Gerais dos Anfíbios e Répteis ............................. 3
Riqueza e Distribuição da herpetofauna ...................................................... 6
A zona costeira da Amazônia brasileira ...................................................... 8
RÊFERENCIAS BIBLIOGRÁFICAS ............................................................... 9
CAPÍTULO 2. RÉPTEIS E ANFÍBIOS ASSOCIADOS À ZONA COSTEIRA
DO ESTADO DO PARÁ, AMAZÔNIA BRASILEIRA .....................................
16
RESUMO .............................................................................................................. 17
INTRODUÇÃO ................................................................................................... 18
MATERIAL E MÉTODOS .................................................................................. 19
Área de estudo ................................................................................................ 19
Coleta de dados .............................................................................................. 20
RESULTADOS E DISCUSSÃO .......................................................................... 21
Répteis ............................................................................................................ 21
Anuros ........................................................................................................... 26
REFERÊNCIAS BIBLIOGRÁFICAS .................................................................. 29
ANEXO 1. TAXALIST – TEXTO EM TXT COM INFORMAÇÕES
TAXONÔMICAS .................................................................................................
36
ANEXO 2. REGRAS PARA SUBMISSÃO DO CAPÍTULO 2 .......................... 37
CAPÍTULO 3. COMPOSIÇÃO E DISTRIBUIÇÃO ESPAÇO-TEMPORAL
DA ANUROFAUNA NO MOSAICO DE AMBIENTES DA PENÍNSULA DE
AJURUTEUA, BRAGANÇA – PARÁ, BRASIL ................................................
43
RESUMO .............................................................................................................. 44
INTRODUÇÃO .................................................................................................... 45
MATERIAL E MÉTODOS ................................................................................. 46
Área de estudo ............................................................................................... 46
Procedimento de campo e análise de dados .................................................. 50

vii
RESULTADOS .................................................................................................... 51
DISCUSSÃO ........................................................................................................ 58
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................. 61
ANEXO 3. REGRAS PARA SUBMISSÃO DO CAPÍTULO 3......................... 65
CAPÍTULO 4 – DISCUSSÃO GERAL .............................................................. 68
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................ 71

viii
LISTA DE TABELAS
Capítulo 2
Tabela 1. Lista das espécies de répteis e sua frequência de ocorrência (%) nos três
sítios de trabalho ao longo da península de Ajuruteua, Bragança – PA, segundo o
método da PLT.
Tabela 2. Lista das espécies de anfíbios e sua frequência de ocorrência (%) nos três
sítios de trabalho ao longo da península de Ajuruteua, Bragança – PA. (* com registros
no manguezal).
Capítulo 3
Tabela 1 - Distribuição de ocorrência das espécies registradas nos cinco pontos
amostrais ao longo da península de Ajuruteua, Bragança – PA. (+) presença e (-)
ausência. Ponto Amostral 1 – campos alagáveis com açudes no Taici, Ponto Amostral 2
– campos alagáveis na fazenda Salinas, Ponto Amostral 3 – lagoa na borda do
fragmento, Ponto Amostral 4 – poça temporária no interior do fragmento e Ponto
Amostral 5 – poças temporárias na borda da restinga. PA= Pontos Amostrais.
Tabela 2 – Valores estimados e observados de riqueza nos cinco pontos amostrais na
península de Ajuruteua, Bragança - PA. PA 1 – campos alagáveis com açudes no Taici,
PA 2 – campos alagáveis na fazenda Salinas, PA 3 – lagoa na borda do fragmento, PA 4
– poça temporária no interior do fragmento e PA 5 – poças temporárias na borda da
restinga.
Tabela 3 – Registro de atividade reprodutiva dos anuros nos pontos amostrais ao longo
de uma temporada reprodutiva na península de Ajuruteua, Bragança – PA. a= amplexo,
g= girinos, i= imagos, v= vocalização e (-) sem atividade reprodutiva.
Tabela 4 - Relação entre as variáveis abióticas e a riqueza de espécies nos pontos
amostrais P.A. 1 – campos alagáveis com açudes no Taici, P.A. 4 – poça temporária no
interior do fragmento e P.A. 5 – poças temporárias na borda da restinga, na península de
Ajuruteua, Bragança – PA. rs= Correlação de Spearman, A= vocalização, B= riqueza, t
= valor do teste t, p = nível de significância e n.s.= não significativo.

ix
LISTA DE FIGURAS
Capítulo 2
Figura 1. Mapa da área de estudo mostrando a localização dos sítios de trabalho: 1 =
Taici, 2 = Fazenda Salinas e 3 = Praia de Ajuruteua, ao longo da península de
Ajuruteua, Bragança-PA.
Figura 2. Diagrama climático apresentando a variação mensal da precipitação
pluviométrica e temperatura do ar para a península de Ajuruteua, Bragança-PA. A área
hachurada com linhas diagonais representa o período menos chuvoso (aqui denominado
de “estação seca”) e as áreas não hachuradas representam o período mais chuvoso (aqui
denominado “estação chuvosa”).
Figura 3. Espécies de escamados registrados na península de Ajuruteua, Bragança-PA:
Lagartos: Família Polychrotidae A, Anolis ortonii; Família Iguanidae B, Iguana iguana;
Família Sphaerodactylidae C, Gonatodes humeralis; Gekkonidae D, Hemidactylus
mabouia; Família Phyllodactylidae E, Thecadactylus rapicauda; Família Teiidae, F,
Cnemidophorus cryptus. Serpentes: Família Boidae: G, Corallus hortulanus; H, Boa
constrictor; Família Dipsadidae: I, Imantodes cenchoa; J, Liophis cobellus taeniogaster;
Família Colubridae: K, Leptophis ahaetulla.
Figura 4. Espécie de quelônio registrada na península de Ajuruteua, Bragança-PA.
Família Kinosternidae: Kinosternon scorpioides. A, vista dorsal e B, vista ventral.
Figura 5. Espécies de anuros registradas na península de Ajuruteua, Bragança-PA.
Família Hylidae: A, Dendropsophus nanus; B, Hypsiboas raniceps; C, Phyllomedusa
hypochondrialis; D, Trachycephalus venulosus; E, Scinax fuscomarginatus; F, Scinax
ruber e G, Scinax boesemani. Família Leptodactylidae: H, Leptodactylus fuscus; I,
Leptodactylus macrosternum; J, Leptodactylus labyrinthicus; K, Leptodactylus
mystaceus. Família Leiuperidae: L, Physalaemus ephipiffer; M, Pseudopaludicola sp. e
Família Bufonidae: N, Rhinella marina.

x
Capítulo 3
Figura 1. Mapa da área de estudo mostrando a localização dos sítios de trabalho: 1 =
Taici, 2 = Fazenda Salinas e 3 = Praia de Ajuruteua, ao longo da península de
Ajuruteua, Bragança-PA.
Figura 2 – Campos alagáveis com açudes abandonados no Ponto amostral 1 próximo
aos manguezais e áreas de terra-firme no Taici (Sítio 1), península de Ajuruteua,
Bragança – PA. A) campos alagáveis, B) e C) açudes abandonados.
Figura 3 – Pontos amostrais localizados na Fazenda Salinas (Sítio 2), no centro da
península de Ajuruteua, Bragança – PA. A) Ponto Amostral 2 - campos alagáveis, B)
Ponto Amostral 3 – lagoa e C) Ponto Amostral 4 - poça temporária.
Figura 4 - Poças temporárias em áreas abertas do Ponto amostral 5, localizadas nas
bordas da restinga na Praia de Ajuruteua (Sítio 3), península de Ajuruteua, Bragança –
PA.
Figura 5 - Dendrograma mostrando a similaridade entre os cinco pontos amostrais, com
base na presença/ausência de indivíduos das espécies registradas em pelo menos uma
atividade reprodutiva. P.A. 1 – campos alagáveis com açudes no Taici, P.A. 2 – campos
alagáveis na fazenda Salinas, P.A. 3 – lagoa na borda do fragmento, P.A. 4 – poça
temporária no interior do fragmento e P.A. 5 – poças temporárias na borda da restinga.
Figura 6 - Análise de correspondência entre os diferentes tipos de substratos de
vocalização e as espécies de anuros registradas na península de Ajuruteua, Bragança-
PA. Tipo 1: Solo; Tipo 2: Serrapilheira; Tipo 3: Gramíneas; Tipo 4: Vegetação
arbustiva; Tipo 5: Vegetação arbórea; Tipo 6: Nível da água; Tipo 7: Vegetação na
Água.
Figura 7 - Distribuição vertical dos hilídeos (cm) no Campo alagado com açudes no
Taici (Ponto Amostral 1) na península de Ajuruteua, Bragança-PA. 1- S.
fuscomarginatus, 2- D. nanus, 3- H. raniceps, 4- S. ruber e 5- P. hypochondrialis.

1
v
Resumo
Os répteis e anfíbios possuem registros nos ambientes costeiros em diversos países,
incluindo o Brasil. No entanto, nenhum estudo foi desenvolvido na região costeira da
Amazônia. A Península de Ajuruteua apresenta diferentes ambientes: manguezal,
restingas, campos salinos e fragmentos de floresta de terra-firme em áreas acima do
nível das inundações da maré. O presente estudo tem os objetivos de apresentar um
inventário da herpetofauna típica da zona costeira do estado do Pará, a composição e
distribuição espacial e temporal na temporada de vocalização da anurofauna ao longo
de uma estação reprodutiva na península. As amostragens ocorreram entre
julho/2008 e junho/2009, com intervalos quinzenais, em três sítios de trabalho,
utilizando-se a técnica de Procura Limitada por Tempo (PLT) e Encontros
Ocasionais, a PLT totalizou 1296 horas de campo. Foram coletados dados de
temperatura do ar, do solo e umidade do ar. A riqueza foi estimada por métodos não-
paramétricos, a similaridade quanto à presença de espécies em atividade reprodutiva
foi testada por uma análise de cluster. As variáveis abióticas foram correlacionadas
com a riqueza mensal e número de espécies vocalizando pelo coeficiente de
Spearman. Os substratos utilizados para vocalização foram categorizados quanto ao
tipo e altura de empoleiramento e associados com as espécies por uma análise de
Correspondência. A sobreposição na utilização vertical dos substratos foi testada por
uma análise de Kruskal-wallis. A herpetefauna na península é composta por 26
espécies, sendo 14 anuros e 12 espécies de répteis. Todas as espécies de anuros
estimadas foram registradas. S. ruber e L. macrosternum ocorreram em todos os
sítios, 11 espécies apresentaram atividade reprodutiva durante o período chuvoso,
quatro não foram registradas vocalizando. R. marina e P. hypochondrialis iniciaram
suas vocalizações no final do período seco, S. fuscomarginatus iniciou suas
vocalizações mais tardia entre todas as espécies. A umidade do ar apresentou
correlação com o número de espécies em atividade de vocalização. Diferenças
temporais na reprodução foram observadas nas espécies terrestres. Os substratos com
estratificação vertical foram exclusivamente utilizados por hilídeos e as famílias
Leptodactylidae, Leiuperidae e Bufonidae foram associadas aos substratos terrestres
e na água. Houve sobreposição na utilização vertical dos substratos.
Palavras-chave: Litoral, Herpetofauna, Ambientes Costeiros, Amazônia.

2
CAPÍTULO 1 INTRODUÇÃO GERAL

3
Origem e Características Gerais dos Anfíbios e Répteis
O termo “herpetofauna” é comumente utilizado para definir o grupo formado
pelos anfíbios e répteis, que apesar de formarem dois clados distintos de vertebrados,
possuem vários aspectos de sua biologia complementar, permitindo o seu estudo em
conjunto e através das mesmas técnicas ou similares (Zug et al., 2001).
Os anfíbios e os amniotas (répteis, aves e mamíferos) são originários dos
vertebrados de quatro patas com dígitos, os estegocéfalos, que apareceram cerca de 370
milhões de anos atrás no Devoniano. Posteriormente, em torno de 250 milhões de anos
atrás durante o Triássico, os lissanfíbios (cecílias, salamandras, anuros e seus aliados
fósseis) surgiram e exploram com sucesso uma grande variedade de hábitats, sempre
relacionados à água ou hábitats úmidos, sendo que é possível encontrar fósseis das
ordens atuais a partir do Jurássico Inferior (204 milhões de anos atrás) (Hutchins et al.,
2003). No Carbonífero (360 milhões de anos atrás), outro grupo divergente surgiu, os
antracossauros, os quais desenvolveram uma pele eficaz para reduzir a perda d’água
rápida e excessiva, permitindo sua existência em hábitats mais áridos e, dessa forma,
conquistando definitivamente o ambiente terrestre. A partir do Jurássico Superior (159
milhões de anos atrás) a ordem Squamata (escamados), que incluem lagartos e
serpentes, apareceu e tornou-se abundante em espécies desde então (Zug et al., 2001).
Os anfíbios e os répteis são animais ectotérmicos. Para os anfíbios, as taxas de
perda de água podem ser extremamente altas, pois a maioria das espécies absorve
grandes quantidades de água e produzem urina diluída. Em geral, possuem taxas
metabólicas baixas, enquanto a maioria dos répteis perde menos quantidade de água. Os
répteis, por sua vez, são capazes de reter mais água do que absorvê-la, pois produzem
urina relativamente concentrada, cujo conteúdo inclui ácido úrico, além de possuírem
taxas metabólicas mais elevadas do que as dos anfíbios (Zug et al., 2001; Wells, 2007).
Morfologicamente os anfíbios e répteis são muito diferentes. Os anuros são
especializados para o salto, com grandes patas traseiras, corpos encurtados, sem cauda e
com cabeça e olhos grandes. As salamandras e os tritões (ordem Urodela), por exemplo,
são alongados, com pernas anteriores e posteriores de igual tamanho e cauda longa. As
cecílias (ordem Gymnophiona) são anfíbios especializados para a vida subterrânea, com
o corpo alongado, sem pernas e olhos reduzidos (Hutchins et al., 2003; Wells, 2007).
Entre os répteis, a maioria possui dois pares de patas adaptadas para correr, rastejar ou
trepar, além da presença da cauda. Diversas espécies apresentam corpos robustos e

4
alguns lagartos possuem patas reduzidas, enquanto em outros as pernas estão ausentes,
como, por exemplo, em todas as serpentes e anfisbenas (Zug et al., 2001).
Os répteis e anfíbios, na sua maioria, possuem reprodução sexuada, mas há
reprodução assexuada em algumas espécies de anfíbios e lagartos; o desenvolvimento
pode ser direto ou indireto e a fecundação pode ser externa (na maioria dos anfíbios), ou
interna, o que é comum entre os répteis; há espécies que apresentam elevado esforço
reprodutivo no início da maturidade sexual e tempo de vida curto e as que apresentam
baixo esforço reprodutivo, maturidade sexual tardia e vida longa; o tamanho da prole e
o número de descendentes também são muito variáveis e, algumas espécies, parecem
estar relacionadas à disponibilidade de recursos, restrições morfológicas, ou até mesmo,
à possibilidade de mais de um fator (Zug et al., 2001).
Ambos os grupos apresentam oviparidade, ovoviparidade e viviparidade
(Stebbins e Cohen, 1997; Zug et al., 2001; Wells, 2007). Na maioria dos anuros ocorre
a fecundação externa, mas algumas espécies de anfíbios desenvolveram maneiras de
fertilizar os ovos internamente, resultando no nascimento de jovens semelhantes aos
adultos, não havendo estágios larvais aquáticos (Wells, 2007). Todos os crocodilianos,
tartarugas, tuataras e a maioria das cobras e lagartos ovipõem. Por outro lado, a
viviparidade evoluiu de forma independente em pelo menos 45 linhagens de lagartos e
35 linhagens de serpentes, onde a nutrição deriva inteiramente da mãe através de uma
placenta; nas ovovivíparas o desenvolvimento dos embriões é sustentado inteiramente
pela gema do ovo (lecitotrofia), assim como em espécies ovíparas (Zug et al., 2001).
Sabe-se que grande parte das espécies da herpetofauna tem reprodução sexuada, mas em
alguns poucos táxons a reprodução ocorre sem a contribuição genética do sexo
masculino, onde populações constituídas apenas de fêmeas apresentam reprodução
assexuada através da partenogênese, que é o desenvolvimento do óvulo na ausência de
um espermatozóide, sendo que toda a população é composta por fêmeas (Zug et al.,
2001).
O sucesso reprodutivo dos anfíbios e répteis é influenciado por diversos fatores,
tais como: modo de reprodução, cuidado parental, interações comportamentais,
localização da incubação e do sítio de desenvolvimento, cujos efeitos podem recair
sobre a sobrevivência e o desenvolvimento de ovos e larvas, bem como sobre a
morfologia (Zug et al., 2001). Os mesmos autores ressaltam que em alguns casos, estes
fatores podem recair até mesmo sobre a determinação do sexo da prole, como acontece
com os répteis, com machos nascem de ovos incubados em certas temperaturas e fêmeas

5
em outras (Zug et al., 2001). Em anfíbios e répteis de clima tempareado a reprodução
tendea ser sazonal. Já espécies que ocorrem em regiões tropicais apresentam ciclo
reprodutivo não sazonal, principalmente nos répteis. Fatores ambientais como seca e
chuva influenciam a reprodução de anfíbios e répteis (Crump, 1974; Hödl, 1990;
Duellman e Trueb, 1994; Ávila-Pires, 1995; Zimmerman e Simberloff, 1996; Pombal-
Jr, 1997; Stebbins e Cohen, 1997; Zug et al., 2001; Gottsberger e Gruber, 2004; Wells,
2007).
Os anfíbios desenvolveram as mais diversificadas estratégias reprodutivas, com
uma ampla variação na localização de sítios de oviposição, na estrutura de massas de
ovos e no desenvolvimento embrionário e larval (Crump, 1974; Hödl, 1990; Duellman e
Trueb, 1994; Zug et al., 2001; Hutchins et al., 2003; Wells, 2007). Os anuros possuem
cerca de 40 diferentes modos reprodutivos já descritos, sendo que o maior número de
espécies especializadas e os mais derivados modos de reprodução ocorrem nos trópicos
úmidos (Crump, 1974; Hödl, 1990; Zug et al., 2001; Wells, 2007). Na Amazônia, Hödl
(1990) identificou 18 modos reprodutivos para anuros, os quais foram divididos em
quatro categorias: a 1ª categoria agrupa as espécies que apresentam desenvolvimento
aquático, a 2ª apresenta o desenvolvimento semiterrestre, a 3ª engloba as espécies que
apresentam os modelos de reprodução terrestre e a 4ª as espécies cujo desenvolvimento
ocorre no dorso da fêmea.
Quanto à alimentação, os anfíbios e répteis apresentam diversas estratégias para
detectar, capturar e subjugar suas presas, desenvolvendo inúmeras adaptações
morfológicas e comportamentais. Na identificação das presas a visão é um importante
fator, geralmente considerando presas em movimento (Zug et al., 2001). No entanto,
algumas espécies são capazes de distinguir cores, enquanto outras utilizam pistas
químicas ou olfativas, podendo também ocorrer a detecção auditiva ou térmica, além
daquelas espécies, como os pipídeos, que podem perceber movimentos de presas
potenciais pela linha lateral (Stebbins e Cohen, 1997; Zug et al., 2001; Wells, 2007).
Os anfíbios e répteis adultos não comem necessariamente as mesmas presas
quando estão em fases larvais ou são juvenis. Os anfíbios que possuem larvas aquáticas
e adultos terrestres mudam, na grande maioria, de detritívoros (larvas) para insetívoros
(adultos). Por outro lado, entre as espécies de anfíbios e répteis, nas quais os juvenis
têm a mesma morfologia que os adultos, um grande componente de mudanças na dieta
está associado ao tamanho do indivíduo (Zug et al., 2001; Wells, 2007).

6
A dieta dos anuros consiste em uma grande variedade de insetos e outros
pequenos invertebrados, sendo a variedade de presas determinada pelo tamanho do
animal e pelo tamanho correspondente da presa (Hutchins et al., 2003). Alguns anuros
de grande porte alimentam-se de pequenos vertebrados, inclusive outros sapos, lagartos,
roedores e pássaros (Stebbins e Cohen, 1997; Zug et al., 2001; Hutchins et al., 2003;
Galatti et al., 2007; Wells, 2007). Os lagartos comem, principalmente, insetos e outros
invertebrados, mas alguns comem vertebrados, inclusive outros lagartos, enquanto
poucos são herbívoros. Já as serpentes, na sua maioria, possuem dieta baseada em
vertebrados e, com espécies especialistas em diversos grupos como os anuros, outras
serpentes, aves e invertebrados (Zug et al., 2001).
Outros fatores importantes na biologia de anfíbios e répteis são as interações
comportamentais, que estão relacionadas à reprodução, predação, alimentação, respostas
fisiológicas às doenças e etc. Seja qual for a sua frequência, tais interações requerem
alguma forma de comunicação, que geralmente ocorre através de uma variedade de
sentidos: visual, químico (nasal e vomeronasal), acústico e tátil (Hödl, 1977; Stebbins e
Cohen, 1997; Hödl e Amézquita, 2001; Zug et al., 2001; Hutchins et al., 2003; Wells,
2007). Os anuros são os únicos entre os anfíbios que elaboraram sinais acústicos.
Alguns possuem um repertório de vários tipos de chamadas que são usados para a
atração de fêmeas, defesa de territórios ao redor de sítios de vocalização ou de
territórios que contêm recursos atraentes às fêmeas (Hödl, 1977; Stebbins e Cohen,
1997; Hutchins et al., 2003; Wells, 2007).
Riqueza e distribuição da herpetofauna
As adaptações morfológicas e fisiológicas e as estratégias para a reprodução e
alimentação em anfíbios e répteis são o resultado de milhões de anos de evolução,
permitindo que esses vertebrados explorassem com sucesso uma grande variedade de
hábitats em quase todos os ambientes da terra, com exceção das regiões polares e do
mar aberto (Stebbins e Cohen, 1997; Zug et al., 2001; Hutchins et al., 2003; Lima et al.,
2006; Vitt et al., 2008).
O Brasil apresenta a maior riqueza de anfíbios e a segunda maior de répteis do
mundo com 875 espécies de anfíbios, das quais 847 são da ordem Anura, 27 da ordem
Gymnophiona e uma espécie representante da ordem Caudata (SBH, 2010). Os répteis,
apresentam cerca de 721 espécies, sendo 36 quelônios, seis jacarés, 241 lagartos, 67
anfisbênias e 371 serpentes, estando todas essas espécies distribuídas nos mais variados

7
biomas (Bérnils, 2010). Na Amazônia, os répteis da ordem Squamata pertencem ao
grupo mais numeroso com 325 espécies aproximadamente, sendo 180 espécies de
serpentes, 130 de lagartos e 15 anfisbênias. Já a Amazônia brasileira está representada
por cerca de 80% dessas espécies, sendo 149 espécies de serpentes, 94 de lagartos e 10
de anfisbênias. Quanto aos anfíbios, considerando os diversos gêneros que apresentam
dúvidas taxonômicas, estão presentes na Amazônia duas espécies de salamandras, 28 da
ordem Gymnophiona e 305 anfíbios anuros, ao passo que na Amazônia brasileira
ocorrem 221 espécies de anuros, nove da ordem Gymnophiona e duas espécies de
salamandras (Ávila-Pires et al., 2007).
Embora a riqueza de espécies seja alta na região amazônica, essas espécies não
ocorrem homogeneamente, existindo diferenças na distribuição da fauna de lagartos e
anfíbios entre as porções da Amazônia ocidental e oriental e, parcialmente, entre as
serpentes (Galatti et al., 2007; Ávila-Pires et al., 2009). Azevedo-Ramos e Galatti
(2002) analisaram a similaridade entre 28 localidades inventariadas na Amazônia
brasileira e demonstraram que a média da similaridade foi de 40%, indicando a presença
de diferentes assembléias de anfíbios por toda sua extensão.
Contudo, é importante ressaltar que a grande maioria dos trabalhos existentes
sobre herpetofauna foi realizada em ambientes no interior do continente (Crump, 1974;
Hödl, 1990; Ávila-Pires, 1995; Galatti et al., 2007; Ávila-Pires et al., 2009; Mas, por
outro lado, nas últimas cinco décadas, diversos trabalhos sobre a herpetofauna também
foram realizados em ambientes costeiros, ao redor do mundo. Esses estudos enfocaram
as diversas adaptações fisiológicas e osmorregulação em anuros e lagartos (Gordon et
al., 1961; Gordon e Tucker, 1968; Dicker e Elliott, 1970; Katz, 1973; Hatano et al.,
2001; Haramura, 2007), assim como a ocorrência, distribuição e biodiversidade da
herpetofauna nos diferentes ecossistemas costeiros (Burghardt e Rand, 1983; Dunson e
Mazzotti, 1989; Leitão, 1995; Conde, 1996; Platt et al., 1999; Fernandes, 2000; Boos,
2001; Lim et al., 2001; Bambaradeniya et al., 2002; Karns et al., 2002; Luiselli e Akani,
2002; Pauwels et al., 2004; Mcdiarmid e Savage, 2005; Greenberg et al., 2006;
Kacoliris et al., 2006; Kelly, 2006; Hansknecht, 2008; Kathiresan e Bingham, 2008;
Rojas-Runjaic et al., 2008; Luther e Greenberg, 2009).
Os estudos sobre a herpetofauna em ecossistemas costeiros no Brasil estão
centralizados nas regiões nordeste, sudeste e sul do país. Em geral, esses trabalhos
apresentam informações sobre herpetofauna em restinga (Freire, 1996; Seeiger et al.,
1998; Schineider e Teixeira, 2001; Teixeira, 2001; Teixeira e Schineider, 2002; Cascon

8
e Borges-Nojosa, 2003; Cunha-Barros et al., 2003; Carvalho e Araújo, 2004; Sluys et
al., 2004; Ferreira et al., 2005; Loebmann, 2005; Loebmann e Vieira, 2005; Carvalho et
al., 2007; Miranda, 2007; Rocha e Sluys, 2007; Loebmann e Mai, 2008; Rocha et al.,
2008a; Rocha et al., 2008b; Silva et al., 2008; Fernandes et al., 2009; Narvaes et al.,
2009; Quintela e Loebmann, 2009; Ferreira e Tonini, 2010). Alguns trabalhos, como o
de Rocha et al. (2005), por exemplo, demonstram que nas restingas do sudeste
brasileiro não só existe uma herpetofauna característica, mas também o registro de cinco
espécies de anfíbios e seis de répteis, todas endêmicas dessa região. Contudo, o
conhecimento da herpetofauna associado aos diversos ecossistemas litorâneos ainda é
muito inexpressivo, particularmente no que concerne à zona costeira da Amazônia
brasileira.
A zona costeira da Amazônia brasileira
O litoral da Amazônia brasileira abrange os estados do Amapá, Pará e a metade
ocidental do Maranhão, ocupando uma faixa de 2250 km entre o rio Oiapoque no
Amapá (5°N, 51°W) e a baía de São Marcos no Maranhão (2°S, 44°W), local onde
estão inseridos diversos ambientes, como praias, planícies de marés, pântanos salinos e
doces, estuários, manguezais, várzeas estuarinas, florestas de terra firme, lagoas,
lagunas, ilhas, rias, deltas e dunas (Souza-Filho, 2005; Pereira et al., 2009; Souza-Filho
et al., 2009).
Nessa linha de costa, somente o litoral nordeste do Pará perfaz 600 km de
extensão, desde a foz do rio Pará até a foz do rio Gurupi (Mendes, 2005; Souza-Filho,
2005). A formação vegetacional predominante dessa linha de costa é o manguezal,
caracterizado pela frequente inundação com águas salobras. Nos manguezais vivem
plantas e animais adaptados às condições mais adversas, resultantes de diferentes teores
de sal e oxigênio do solo, bem como dos elevados índices de temperatura e inundações
periódicas (Tomlinson, 1986; Hogarth, 1999). A planície costeira bragantina, onde o
presente estudo foi realizado, está inserida nessa faixa litorânea e estende-se da Ponta do
Maiaú até a foz do rio Caeté, perfazendo 40 km de linha de costa (Souza-Filho e El-
Robrini, 1996).
Considerando-se a ausência de informações sobre a fauna de anuros e répteis da
região costeira da Amazônia brasileira, o presente trabalho vem preencher parte dessa
lacuna apresentando informações inéditas sobre a distribuição desses dois grupos de
vertebrados, bem como sobre sua relação com o mosaico de ambientes típicos dessa

9
região. Com esse intuito, a presente dissertação foi concebida na forma de quatro
capítulos. O Capítulo 1 intitulado “INTRODUÇÃO GERAL”, versa sobre o tema
herpetofauna, apresentando uma síntese sobre a origem e as características
bioecológicas desses animais, bem como aspectos da sua distribuição e diversidade,
ressaltando sua estreita associação com os ambientes costeiros, inclusive aqueles
característicos da Amazônia brasileira.
O Capítulo 2 “RÉPTEIS E ANFÍBIOS ASSOCIADOS À ZONA COSTEIRA
DO ESTADO DO PARÁ, AMAZÔNIA BRASILEIRA” descreve a composição e a
distribuição da herpetofauna nos ambientes litorâneos do Estado do Pará. Através da
elaboração de uma lista de espécies associadas aos diversos ambientes da península de
Ajuruteua, município de Bragança, apresenta a mais recente revisão da herpetofauna
típica ocorrente na região costeira da Amazônia brasileira.
O Capítulo 3 “COMPOSIÇÃO E DISTRIBUIÇÃO ESPAÇO-TEMPORAL DA
ANUROFAUNA NO MOSAICO DE AMBIENTES DA PENÍNSULA DE
AJURUTEUA, BRAGANÇA – PARÁ, BRASIL” apresenta a riqueza e a similaridade
de espécies de anuros nos diferentes ambientes costeiros. Também apresenta sua
distribuição espaço-temporal no uso dos sítios e substratos de vocalização ao longo de
um ciclo anual.
O Capítulo 4 “DISCUSSÃO GERAL”, através de uma abordagem mais ampla,
elabora uma discussão sobre a herpetofauna ocorrente na costa amazônica brasileira.
Além do mais, enfatiza sua distribuição e resiliência dessa fauna às condições únicas
dos ecossistemas costeiros, enfocando as espécies de anuros, ressaltando o potencial
desses ambientes para a conservação e preservação desses animais.
Referências Bibliográficas
ÁVILA-PIRES, T. C. S. Lizards of Brazilian Amazônia (Reptilia: Squamata).
Zoologische Verhandelingen v. 1-706, p. 1-706, 1995.
ÁVILA-PIRES, T. C. S. et al. Herpetofauna da Amazônia. In: NASCIMENTO, L. B.;
OLIVEIRA, M. E. (Ed.). Herpetologia no Brasil II. Belo Horizonte: Sociedade
Brasileira de Herpetologia, 2007. p. 354.
______. Squamata (Reptilia) from four sites in southern Amazonia, with a
biogeographic analysis of Amazonian lizards. Boletim do Musel Paraense Emílio
Goeldi, Ciências Naturais v. 4, n. 2, p. 99-118, 2009.

10
AZEVEDO-RAMOS, C.; GALATTI, U. Patterns of amphibian diversity in Brazilian
Amazonia: conservation implications. Biological Conservation v. 103, p. 103-111,
2002.
BAMBARADENIYA, C. N. B. et al. An Assessment of the Status o f Biodiversity in
the Maduganga Mangrove Estuary. Occasional Papers of IUCN Sri Lanka v. 1, p. 1-49,
2002.
BÉRNILS, R. S. Brazilian reptiles-List of species Sociedade Brasileira de Herpetologia,
2010.
BOOS, H. E. A. The Snakes of Trinidad e Tobago. Texas: Texas A & M University
Press, College Station, 2001.
BURGHARDT, G. M.; RAND, A. S. Iguanas of the world: Their Behavior, Ecology, e
Conservation. New Jersey: Noyes Publications, 1983.
CARVALHO, A. L. G. D.; ARAÚJO, A. F. B. D. Ecologia dos Lagartos da Ilha da
Marambaia, RJ. Revista Universidade Rural: Série Ciências da Vida, Seropédica, RJ v.
24, n. 2, p. 159-165, 2004.
CARVALHO, A. L. G. D. et al. Lagartos da Marambaia, um remanescente insular de
Restinga e Floresta Atlântica no Estado do Rio de Janeiro, Brasil. Biota Neotropica v. 7,
n. 2, p. 221-226, 2007.
CASCON, P.; BORGES-NOJOSA, D. M. Anfíbios. In: CAMPOS, A. et al (Ed.). A
Zona Costeira do Ceará: Diagnóstico para a Gestão Integrada. Fortaleza: Gráfica e
Editora Pouchain Ramos, 2003. p. 125.
CONDE, J. E. A profile of laguna de Tacarigua, Venezuela: A tropical estuarine coastal
lagoon. . 21, n. 5, p. 282-292, 1996.
CRUMP, M. L. Reproductive Strategies in a Tropical Anuran Community. Kansas:
University of Kansas Printing Service, 1974. (Museum of Natural History, 61).
CUNHA-BARROS, M. et al. Patterns of infestation by chigger mites in four diurnal
lizard species from a Restinga habitat (Jurubatiba) of Southeastern Brazil. Brazilian
Journal of Biology v. 63, n. 3, p. 602-612, 2003.
DICKER, S. E.; ELLIOTT, A. B. Water uptake by the crab-eating frog Rana
cancrivora, as affected by osmotic gradients e by neurohypophysial hormones. Journal
Physiological v. 287, p. 119-132, 1970.
DUELLMAN, W. E.; TRUEB, L. Biology of Amphibians. Baltimore: The Johns
Hopkins University Press, 1994.

11
DUNSON, W. A.; MAZZOTTI, F. J. Salinity as a Limiting Factor in the Distribution of
Reptiles in Florida Bay: A Theory for the Estuarine Origin of Marine Snakes e Turtles.
Bulletin of Marine Science v. 44, n. 1, p. 229-244, 1989.
FERNANDES, M. E. B. Association of mammals with mangrove forests: a world wide
review. Boletim do Laboratório de Hidrobiologia v. 13, p. 83-108, 2000.
FERNANDES, M. E. B. et al. Gonatodes humeralis (NCH) Habitat Occurrence; Escape
Behavior. Herpetological Review v. 40, n. 2, p. 221-222, 2009.
FERREIRA, A. P. et al. Composição e estruturação da comunidade de serpentes da
restinga da praia de Panaquatira, Maranhão. Cd-rom do VII Congresso de Ecologia do
Brasil - Resumos expandidos, 2005.
FERREIRA, R. B.; TONINI, J. F. R. Living holed: Leptodactylus latrans occupying
crabs’ burrows. Herpetology Notes v. 3, n. 237-238, 2010.
FREIRE, E. M. X. Estudo ecológico e zoogeográfico sobre a fauna de lagartos (sauria)
das dunas de Natal, Rio Grande do Norte e da restinga de Ponta de Campina, Cabedelo,
Paraíba, Brasil. Revista Brasileira de Zoologia v. 13, n. 4, p. 903-921, 1996.
FROST, D. R. Amphibians Species of the World: an Online Reference. Version 5.4.
American Museum of Natural History, 2010.
GALATTI, U. et al. Anfíbios da Área de Pesquisa Ecológica do Guamá (APEG) e
Região de Belém. In: GOMES, J. I. et al (Ed.). Mocambo: Diversidade e Dinâmica
Biológica da Área de Pesquisa Ecológica do Guamá (APEG). Belém: MCT/Museu
Paraense Emílio Goeldi 2007. p. 456.
GORDON, M. S. et al. Osmotic regulation in the crab-eating frog (Rana cancrivora).
Journal Experimental Biology v. 38, p. 659-678, 1961.
GORDON, M. S.; TUCKER, V. A. Further Observations on the physiology of salinity
adaptation in the Crab-eating frog (Rana Cancrivora). Journal Experimental Biology v.
49, p. 185-193,1968.
GOTTSBERGER, B.; GRUBER, E. Temporal partitioning of reproductive activity in a
neotropical anuran community. Journal of Tropical Ecology v. 20, n. 1, p. 271-280,
2004.
GREENBERG, R. et al. Tidal Marshes: A Global Perspective on the Evolution e
Conservation of Their Terrestrial Vertebrates. BioScience v. 56, n. 8, p. 675–685, 2006.
HANSKNECHT, K. A. Lingual Luring by Mangrove Salt marsh Snakes (Nerodia
clarkii compressicauda). Journal of Herpetology v. 42, n. 1, p. 9-15, 2008.

12
HARAMURA, T. Salinity Tolerance of Eggs of Buergeria japonica (Amphibia, Anura)
Inhabiting Coastal Areas. Zoological Science v. 24, p. 820-823, 2007.
HATANO, F. H. et al. Thermal ecology e activity patterns of the lizard community of
the Restinga of Jurubatiba, Macaé, RJ. Revista Brasileira de Biologia v. 61, n. 2, p. 287-
294, 2001.
HÖDL, W. Call differences e calling site segregation in anuran species from Central
Amazonian floating meadows. Oecologia v. 28, p. 351-363, 1977.
______. Reproductive diversity in Amazonian lowland frogs. Fortschritte der Zoologie
v. 38, p. 41-60, 1990.
HÖDL, W.; AMÉZQUITA, A. Visual Signaling in Anuran Amphibians. In: RYAN, M.
J. (Ed.). Anuran Communication. Washington: Smithsonian Institution Press, 2001.
Cap.10. p. 252.
HOGARTH, P. The Biology of Mangroves. New York: Oxford University Press, 1999.
HUTCHINS, M. et al. Grzimek’s Animal Life Encyclopedia, Amphibians. Canadá: Galé
Group, 2003.
KACOLIRIS, F. et al. Herpetofauna, Coastal Dunes, Buenos Aires Province, Argentina
Check List v. 2, n. 3, 2006.
KARNS, D. R. et al. Ecology of oriental-australian rear-fanged water snakes
(Colubridae: Homalopsinae) in the pasir ris park mangrove forest, Singapore. The
Raffles Bulletin of Zoology v. 50, n. 2, p. 487-498, 2002.
KATHIRESAN, K.; BINGHAM, B. L. Biology of mangroves e mangrove ecosystems.
Advance in Marine Biology v. 40, p. 81-251, 2008.
KATZ, U. Studies on the adaptation of the toad Bufo viridis to high salinities: oxygen
consumption, plasma concentration e water content of the tissues. Journal Experimental
Biology V. 58, p. 785-796, 1973.
KELLY, L. Evolution's greatest survivior Crocodile. Sydney: Allen & Unwin, 2006.
LARA, R. J.; COHEN, M. C. L. Sensoriamento Remoto. In: FERNANDES, M. E. B.
(Ed.). Os Manguezais da Costa Norte Brasileira. Maranhão: Fundação Rio Bacanga,
2003. Cap.1. p. 11-28.
LEITÃO, S. N. A fauna do Manguezal. In: NOVELLI, Y. S. (Ed.). Manguezal,
Ecossistema entre a terra e o mar. São Paulo: Caribbean Ecological Research, 1995.
Cap.5. p. 64.

13
LIM, K. K. P. et al. A Guide to Mangroves of Singapore. In: Raffles Museum of
Biodiversity Research, The National University of Singapore & The Singapore Science
Centre, 2001.
LIMA, A. P. et al. Guia de sapos da Reserva Adolpho Ducke, Amazônia Central =
Guide to the frogs of Reserva Adolpho Ducke, Central Amazonia. Manaus: Áttema
Design Editorial, 2006.
LOEBMANN, D. Guia Ilustrado: Os anfíbios da região costeira do extremo sul do
Brasil. Pelotas: União Sul-Americana de Estudos da Biodiversidade, 2005.
LOEBMANN, D.; MAI, A. C. G. Amphibia, Anura, Coastal Zone, state of Piauí,
Northeastern Brazil. Check List v. 4, n. 2, p. 161-170, 2008.
LOEBMANN, D.; VIEIRA, J. P. Relação dos anfíbios do Parque Nacional da Lagoa do
Peixe, RS, Brasil. Revista Brasileira de Zoologia v. 22, n. 2, p. 339-341, 2005.
LUISELLI, L.; AKANI, G. C. An investigation into the composition, complexity e
functioning of snake communities in the mangroves of south-eastern Nigeria. African
Journal of Ecology v. 40, n. 3, p. 220-227, 2002.
LUTHER, D. A.; GREENBERG, R. Mangroves: A Global perspective on the evolution
e conservation of their terrestrial vertebrates. Bioscience v. 59, n. 7, p. 602-612, 2009.
MCDIARMID, R. W.; SAVAGE, J. M. The Herpetofauna of the Rincon Area,
Peninsula de Osa, Costa Rica, a Central American Lowland Evergreen Forest Site. In:
DONNELLY, M. A. et al (Ed.). Ecology e evolution in the tropics: a herpetological
perspective. Chicago: University of Chicago Press, 2005. Cap.16. p. 366-427.
MENDES, A. C. Geomorfologia e sedimentologia. In: FERNANDES, M. E. B. (Ed.).
Os manguezais da costa norte brasileira. Belém: Petrobrás/Fundação Rio Bacanga,
2005. p. 165.
MIRANDA, J. P. Ecologia e conservação de hepertofauna do Parque Nacional dos
Lençóis Maranhenses, Maranhão, Brasil. (2007). 140 f. Tese de Doutorado - Instituto
de Biologia Universidade Estadual de Campinas, Campinas, 2007.
NARVAES, P. et al. Composição, uso de hábitat e estações reprodutivas das espécies
de anuros da floresta de restinga da Estação Ecológica Juréia-Itatins, sudeste do Brasil.
Biota Neotropica, v. 9, n. 2, 2009. Disponível
em:<http://www.biotaneotropica.org.br/v9n2/en/abstract?article+bn02009022009>.
PAUWELS, O. S. G. et al. Reptiles of Loango National Park, Ogooué-Mariteme
Province, South-Western Gabon. Hamadryad v. 29, n. 1, p. 115-127, 2004.

14
PEREIRA, L. C. C. et al. The Brazilian Amazon Coastal Zone. Journal of Integrated
Coastal Zone Management v. 9, n. 2, p. 3-7, 2009.
PLATT, S. G. et al. Diversity, Observations, e Conservation of the Herpetofauna of
Turneffe, Leghthouse, e Glovers Atolls, Belize British Herpetological Society Bulletin
v. 66, p. 1-13, 1999.
POMBAL-JR, J. P. Distribuição espacial e temporal de anuros (Amphibia) em uma
poça permanente na Serra de Paranapiacaba, sudeste do Brasil. Revista Brasileira de
Biologia v. 57, n. 4, p. 583-594, 1997.
QUINTELA, F. M.; LOEBMANN, D. Os Répteis da região costeira do extremo sul do
Brasil. Pelotas: União Sul-Americana de Estudos da Biodiversidade, 2009.
ROCHA, C. F. D. et al. Effects of a fire on a population of treefrogs (Scinax cf. alter,
Lutz) in a restinga habitat in southern Brazil. Brazilian Journal of Biology v. 68, n. 3, p.
539-543, 2008a.
______. Frog species richness, composition e β-diversity in coastal Brazilian restinga
hábitats. Brazilian Journal of Biology v. 68, n. 1, p. 101-107, 2008b.
ROCHA, C. F. D.; SLUYS, M. V. Herpetofauna de Restingas. In: NASCIMENTO, L.
B.; OLIVEIRA, M. E. (Ed.). Herpetologia no Brasil II. Belo Horizonte: Sociedade
Brasileira de Herpetologia, 2007. p. 354.
ROCHA, C. F. D. et al. Endemic e threatened tetrapods in the Restingas of the
biodiversity corridors of Serra do Mar end of the central in Mata Atlântica in eastern
Brazil. Brazilian Journal of Biology v. 65, n. 1, p. 159-168, 2005.
ROJAS-RUNJAIC, F. J. M. et al. Amphibia, Anura, Hylidae, Scarthyla vigilans: Range
extensions e new state records from Delta Amacuro e Miranda states, Venezuela. .
Check List v. 4, n. 3, p. 301-303, 2008.
SBH. Brazilian amphibians–List of species. Sociedade Brasileira de Herpetologia, 2010.
SCHINEIDER, J. A. P.; TEIXEIRA, R. L. Relacionamento entre anfíbios anuros e
bromélias da Restinga de Regência, Linhares, Espírito Santo, Brasil. Iheringia, Série
Zoologia v. 91, p. 41-48, 2001.
SEEIGER, U. et al. Os Ecossistemas costeiros e marinhos do extremo sul do Brasil. Rio
Grande: Ecosciencia 1998.
SILVA, H. R. D. et al. Frogs of Marambaia: a naturally isolated Restinga e Atlantic
Forest remnant of southeastern Brazil. Biota Neotropica, v. 8, n. 4, p.167-174, 2008.
Disponível
em:<http://www.biotaneotropica.org.br/v8n4/en/abstract?inventory+bn01808042008>.

15
SLUYS, M. V. et al. Anfíbios da Restinga de Jurubatiba: Composição e História
Natural. In: ROCHA, C. F. D. et al (Ed.). Pesquisas de Longa Duração na Restinga de
Jurubatiba-Ecologia, História Natural e Conservação. São Carlos: Rima, 2004. Cap.9.
p. 376.
SOUZA-FILHO, P. W. M. et al. Environmental sensitivity index (ESI) mapping of oil
spill in the amazon coastal zone: the piatam mar project. Revista brasileira de Geofísica
v. 27, n. 1, p. 7-22, 2009.
SOUZA-FILHO, P. W. M. Costa de Manguezais de Macromaré da Amazônia: Cenários
Morfológicos, Mapeamento e Quantificação de Áreas usando dados de sensores
remotos. Revista brasileira de Geofísica v. 23, n. 4, p. 427-435, 2005.
SOUZA-FILHO, P. W. M.; EL-ROBRINI, M. Morfologia, processos de sedimentação e
litofácies dos ambientes morfo-sedimentares da planície costeira bragantina, nordeste do
Pará, Brasil. Geonomos v. 4, n. 2, p. 1-16, 1996.
STEBBINS, R. C.; COHEN, N. W. A Natural History of Amphibians. New Jersey:
Princeton University Press, 1997.
TEIXEIRA, R. L. Comunidade de lagartos da restinga de Guriri, São Mateus – ES,
Sudeste do Brasil. Atlântica v. 23, p. 77-84, 2001.
TEIXEIRA, R. L.; SCHINEIDER, J. A. P. The Occurrence of Amphibians in
Bromeliads from a Southeastern Brazilian Restinga habitat, with Special Reference to
Aparasphenodon brunoi (Anura, Hylidae). Brazilian Journal of Biology v. 62, n. 2, p.
263-268, 2002.
TOMLINSON, P. B. The Botany of Mangroves. Cambridge: Cambridge University
Press, 1986.
VITT, L. J. et al. Guia de Lagartos da Reserva Adolpho Ducke, Amazônia Central =
Guide to the Lizards of Reserva Adolpho Ducke, Central Amazônia. . Manaus: Áttema
Design Editorial, 2008.
WELLS, K. D. The Ecology e Behavior of Amphibians. Chicago: University of Chicago
Press, 2007.
ZIMMERMAN, B. L.; SIMBERLOFF, D. An historical interpretation of habitat use by
frogs in a Central Amazonian Forest. Journal of Biogeography v. 23, p. 27-46, 1996.
ZUG, G. R. et al. Herpetology an Intoductory biology of Amphibians e Reptiles.
Florida: Academic Press, 2001.

16
CAPÍTULO 2 RÉPTEIS E ANFÍBIOS ASSOCIADOS À ZONA COSTEIRA DO ESTADO DO
PARÁ, AMAZÔNIA BRASILEIRA

17
Resumo
A costa amazônica brasileira é extensa e possui os mais diferentes ecossistemas, onde
não há estudos com herpetofauna desenvolvido na região. Assim, o presente trabalho
tem o objetivo de apresentar um inventário da herpetofauna da zona costeira do Pará. A
partir das ocorrências determinou-se a freqüência das espécies em três sítios de trabalho.
As amostragens ocorreram entre julho/2008 e junho/2009. Utilizou-se a técnica de
Procura Limitada por Tempo (PLT), e Encontros Ocasionais. Registrou-se 26 espécies
na península, sendo 14 anuros e 12 répteis (6 lagartos, 5 serpentes e um quelônio). O
sítio 1 apresentou a maior riqueza e o sítio 3 menor. Os anuros L. macrosternum, L.
fuscus e S. ruber foram comum em todos os sítios. As serpentes foram consideradas
raras, já os lagartos apresentaram espécies raras em um determinado sítio e comum em
outro. Somente S. ruber entre os hilídeos ocorreu em todos os sítios, sendo os
leptodadctylídeos com maior numero de espécies ocorrendo em todos os sítios.
Palavras-chave: Herpetofauna, zona costeira, Amazônia

18
INTRODUÇÃO
Os répteis, incluindo crocodilos, jacarés, lagartos, serpentes e tartarugas, assim
como os anfíbios, já foram registrados em zonas costeiras em diversos países (Leitão
1995; Platt et al. 1999; Fernandes 2000; Boos 2001; Karns et al. 2002; Luiselli e Akani
2002; Pauwels et al. 2004; Kelly 2006; Kathiresan e Bingham 2008; Rojas-Runjaic et
al. 2008; Luther e Greenberg 2009). No Brasil, embora existam estudos que relatem a
composição da fauna de anfíbios e répteis em diferentes ambientes ao longo da linha de
costa brasileira, a maioria está concentrada nas regiões nordeste, sudeste e sul (Freire
1996; Hatano et al. 2001; Schineider e Teixeira 2001; Teixeira 2001; Teixeira e
Schineider 2002; Teixeira et al. 2002; Cunha-Barros et al. 2003; Carvalho e Araújo
2004; Mesquita et al. 2004; Rocha et al. 2004; Loebmann 2005; Rocha et al. 2005;
Carvalho et al. 2007; Rocha e Sluys 2007; Loebmann e Mai 2008; Rocha et al. 2008a;
Rocha et al. 2008b; Quintela e Loebmann 2009; Ferreira e Tonini 2010).
Contudo, o conhecimento da herpetofauna associado aos diversos ecossistemas
costeiros ainda é muito inexpressivo, particularmente na região amazônica, existindo
apenas dois trabalhos incluindo um réptil (Gonatodes humeralis – Fernandes et al.
2009) e um anfíbio (Leptodactylus fuscus – Brito et al. in press) em áreas de manguezal
no Estado do Pará.
É importante ressaltar que a costa amazônica brasileira é extensa e possui os
mais diferentes ecossistemas, os quais podem abrigar grande parte da fauna
herpetológica. A costa da Amazônia brasileira abrange os estados do Amapá, Pará e a
metade ocidental do Maranhão, compreendendo 2250 km, aproximadamente 27% do
litoral brasileiro (Awosika e Marone 2000; Mendes 2005; Souza-Filho et al. 2009).
Nessa faixa litorânea estão inseridos diversos ambientes, como: praias, planícies de
marés, pântanos salinos e doces, estuários, manguezais, floresta de várzea, florestas de
terra firme, lagoas, lagunas, ilhas, rias, deltas, dunas e restingas (Amaral et al. 2008;
Menezes et al. 2008; Pereira et al. 2009; Souza-Filho et al. 2009). O Pará possui cerca
de 600 km de costa, onde está inserida a zona costeira bragantina, que por sua vez, é
representada por diferentes unidades vegetacionais como: terra-firme, campo salino,
restinga e manguezal, sendo este último o principal tipo vegetacional (Souza-Filho e El-
Robrini 1996; Mendes 2005).
Com base nas dimensões regionais e na carência de informações sobre a
herpetofauna ocorrente ao longo da costa amazônica brasileira, o presente trabalho vem

19
contribuir para o conhecimento da herpetofauna costeira do Estado do Pará.
Adicionalmente, também são apresentados: i) os diferentes ambientes e suas respectivas
espécies ocorrentes e ii) a mais recente revisão da herpetofauna da zona costeira da
Amazônia brasileira.
MATERIAL E MÉTODOS
Área de estudo As amostragens foram realizadas no período de julho/2008 a junho/2009 com
intervalos quinzenais, em três sítios de trabalho localizados nos limites da Reserva
Extrativista Marinha Caeté-Taperaçu, península de Ajuruteua, Bragança - Pará. Os
sítios de trabalho estão inseridos ou localizados muito próximo dos ecossistemas que
formam a paisagem ao longo da península (Figura 1).
Sítio 1: Taici (01° 02’59,2”S e 046° 45’42,6”W) - paisagem composta por i)
manguezal misto [Rhizophora mangle L. e Avicennia germinans (L.) Stearn] inundada
periodicamente pelas marés, ii) campo inundado somente no período chuvoso e iii)
açudes abandonados;
Sítio 2: Fazenda Salinas (0°55’35,4"S e 46°40’13"W) - paisagem composta por
i) fragmentos de floresta de terra-firme (vegetação-relíquia), não estando sujeitos à
inundação pelas marés, mas apresentam formação de poças com água da chuva e ii)
campo inundado somente no período chuvoso;
Sítio 3: Praia de Ajuruteua (00°50’03,6”S e 46°36’05,9”W) – paisagem
composta por i) restinga com vegetação herbácea e arbustiva e ii) dunas, com presença
de vegetação herbácea e arbustiva.
De acordo com os dados meteorológicos para o ano amostrado, os níveis de
umidade relativa do ar ultrapassam 90%, com taxa de precipitação pluviométrica anual
de 3.613 mm e temperatura do ar média anual de 26°C. No presente trabalho, a estação
seca foi definida pelos meses que apresentaram valores de precipitação pluviométrica
inferiores a 100 mm, período de agosto a dezembro/2008, ao passo que a estação
chuvosa foi definida pelos meses com valores acima de 100 mm, período representado
pelos meses de julho/2008 e de janeiro a junho/2009 (Figura 2). Dados obtidos da
estação meteorológica automática (A226), localizada no município de Bragança-PA.

Figura 1. Mapa da área de estudo mostrando a localização dos sítios de trabalho: 1 = Taici, 2 = Fazenda Salinas e 3 =península de Ajuruteua, Bragança
Coleta de Dados As amostragens foram realizadas utilizando
Tempo (PLT) (Heyer et al.
1999), cujo registro dos espécimes é proveniente de outras atividades que não sejam as
de PLT. A procura ativa foi de seis horas/observador, totalizando 1296 horas de
atividade de campo. Os espécimes de anuros s
as serpentes de acordo com os trabalhos de
lagartos segundo Ávila-Pires (1995). Espécimes
Coleção de Zoologia do Campus de Bragança (CZB), localiz
PA.
A partir das ocorrências das espécies de an
um valor de frequência de encontro de cada espécie por sítio,
presença da espécie durante cada visita
utilizado para determinar: i)
das visitas) e ii) as espécies raras
iii) espécies intermediárias
2001; Neckel-Oliveira e Gordo 2004)
igura 1. Mapa da área de estudo mostrando a localização dos sítios de Taici, 2 = Fazenda Salinas e 3 = Praia de Ajuruteua, ao longo da
juruteua, Bragança-PA.
As amostragens foram realizadas utilizando-se a técnica de Procura Limitada por
et al. 1994) e através de encontros ocasionais (Martins
1999), cujo registro dos espécimes é proveniente de outras atividades que não sejam as
. A procura ativa foi de seis horas/observador, totalizando 1296 horas de
atividade de campo. Os espécimes de anuros seguiram a nomenclatura de Frost (2010),
de acordo com os trabalhos de Cunha e Nascimento (1978; 1993) e os
Pires (1995). Espécimes-testemunhos foram incorporados à
o de Zoologia do Campus de Bragança (CZB), localizada na cidade de Bragança
A partir das ocorrências das espécies de anuros, lagartos e serpentes foi criado
de frequência de encontro de cada espécie por sítio, baseado na
presença da espécie durante cada visita e o número total de visitas
: i) as espécies comuns - aquelas encontradas em mais de 50%
espécies raras - aquelas encontradas em menos de 20
iii) espécies intermediárias – encontradas entre 20 % > 50 % das visitas
Gordo 2004).
20
igura 1. Mapa da área de estudo mostrando a localização dos sítios de , ao longo da
rocura Limitada por
de encontros ocasionais (Martins e Oliveira
1999), cujo registro dos espécimes é proveniente de outras atividades que não sejam as
. A procura ativa foi de seis horas/observador, totalizando 1296 horas de
eguiram a nomenclatura de Frost (2010),
ento (1978; 1993) e os
testemunhos foram incorporados à
ada na cidade de Bragança-
, lagartos e serpentes foi criado
baseado na razão entre a
de visitas. Este valor foi
aquelas encontradas em mais de 50%
aquelas encontradas em menos de 20 % das visitas e
> 50 % das visitas (Hero et al.

21
Figura 2. Diagrama climático apresentando a variação mensal da precipitação
pluviométrica e temperatura do ar para a península de Ajuruteua, Bragança-
PA. A área hachurada com linhas diagonais representa o período menos
chuvoso (aqui denominado de “estação seca”) e as áreas não hachuradas
representam o período mais chuvoso (aqui denominado “estação chuvosa”).
RESULTADOS E DISCUSSÃO
Répteis Na península de Ajuruteua foram registradas 12 espécies de répteis, distribuídas
nas seguintes famílias: i) Iguanidae (n=1), Gekkonidae (n=1), Phyllodactylidae (n=1),
Polychrotidae (n=1), Sphaerodactylidae (n=1), Teiidae (n=1) (Figura 3); ii) Boidae
(n=2), Colubridae (n=1), Dipsadidae (n=2) (Figura 3) e iii) uma família de quelônio,
Kinostenidae (n=1) (Figura 4).

Figura 3. Espécies de escamados registrados na península de Ajuruteua, BragançaLagartos: A, Anolis ortoniiGonatodes humeralis (SphaerodactylidaeThecadactylus rapicaudaSerpentes: G, Corallus hortulanusJ, Liophis cobellus taeniogaster
Figura 4. Espécie de quelônio registrada na península de Ajuruteua, BragançaKinosternidae: dorsal e B, vista ventral.
Segundo Ávila-Pires
94 espécies de lagartos. Em se tratando de
região de 50.000 km2, incluindo a região bragantina, 87 espécies de serpentes já foram
registradas (Cunha e Nascimento 1978; 1993). Considerando apenas as serpentes, cinco
Figura 3. Espécies de escamados registrados na península de Ajuruteua, BragançaAnolis ortonii (Polychrotidae); B, Iguana iguana (Iguanidae
Sphaerodactylidae); D, Hemidactylus mabouia (GekkonidaeThecadactylus rapicauda (Phyllodactylidae); F, Cnemidophorus cryptus
Corallus hortulanus; H, Boa constrictor (Boidae); I, Imantodes cenchoataeniogaster (Dipsadidae); K, Leptophis ahaetulla (Colubridae
. Espécie de quelônio registrada na península de Ajuruteua, Bragança-PA. Família Kinosternidae: Kinosternon scorpioides. A, vista dorsal e B, vista ventral.
Pires et al. (2007), na Amazônia brasileira ocorrem, pelo
. Em se tratando de serpentes, somente no leste do Pará,
, incluindo a região bragantina, 87 espécies de serpentes já foram
imento 1978; 1993). Considerando apenas as serpentes, cinco
22
Figura 3. Espécies de escamados registrados na península de Ajuruteua, Bragança-PA: Iguanidae); C,
Gekkonidae); E, (Teiidae).
Imantodes cenchoa; Colubridae).
(2007), na Amazônia brasileira ocorrem, pelo menos,
somente no leste do Pará, uma
, incluindo a região bragantina, 87 espécies de serpentes já foram
imento 1978; 1993). Considerando apenas as serpentes, cinco

23
espécies foram registradas nos ambientes costeiros da península de Ajuruteua,
equivalente a 5,75% das espécies de serpentes que ocorrem na região leste do Pará
(Cunha e Nascimento 1978; 1993). No entanto, tais resultados tendem a ser alterados
com o aumento da abrangência das áreas inventariadas. Os trabalhos realizados nos
manguezais da Nigéria, por exemplo, ao longo de quatro anos de amostragens,
registraram 18 das 43 espécies de serpentes dos ambientes vizinhos, o que representa
41% do total das espécies, demonstrando o potencial dos ambientes costeiros para a
ocorrência de espécies desse grupo taxonômico (Luiselli e Akani 2002).
Boa contrictor (Linnaeus 1758) foi a única espécie não registrada pelo método
PLT, mas, no entanto, vários espécimes foram registrados atropelados na PA-458, que
liga Bragança à praia de Ajuruteua. Esta espécie de serpente também já foi registrada
em outras ocasiões na restinga e no manguezal, o que é um relato comum dos catadores
de caranguejo da península de Ajuruteua.
Segundo Cunha e Nascimento (1978), encontrar serpentes depende quase
sempre de sorte, pois elas movem-se continuamente de dia e de noite. Os mesmos
autores consideravam as vegetações de manguezal, campo e restinga como sendo de
quase nenhuma importância na ecologia desses animais. No entanto, Rocha et al. (2005)
relataram que Bothrops leucurus (Wagler 1824) é uma espécie endêmica de restingas no
Espírito Santo e Alagoas. Da mesma forma, diversos autores já registraram várias
espécies de serpentes em diferentes sistemas costeiros (Conde 1996; Seeiger et al. 1998;
Boss et al. 2001; Zug et al. 2001; Karns et al. 2002; Luiselli e Akani 2002; Ferreira et
al. 2005; Mcdiarmid e Savage 2005; Kacoliris et al. 2006; Rocha e Sluys 2007;
Hansknecht 2008; Kathiresan e Bingham 2008; Quintela e Loebmann 2009).
Quatro das cinco espécies de serpentes registradas no presente estudo foram
consideradas raras (Tabela 1), sendo poucos indivíduos registrados por espécies. No
entanto devemos considerar que todas as serpentes encontradas no presente estudo são
espécies frequentes em diversos ambientes da Amazônia (Cunha e Nascimento 1978;
1993; Neckel-Oliveira e Gordo 2004). Porém devido seus hábitos crípticos dificilmente
serão avistados e podem parecer que apresentam baixa densidade populacional.
Liophis cobellus taeniogaster foi considerada rara na península, tendo um
registro de juvenil no manguezal no início da estação chuvosa e um exemplar adulto
atropelado na altura do Km-21 da PA-458. A espécie Liophis cobellus cobellus
(Linnaeus 1758) é comum no manguezal de Trinidad e Tobago, onde é conhecida como
cobra-do-mangue (Boos 2001). Corallus hortulanus, foi a única espécie registrada nos
24
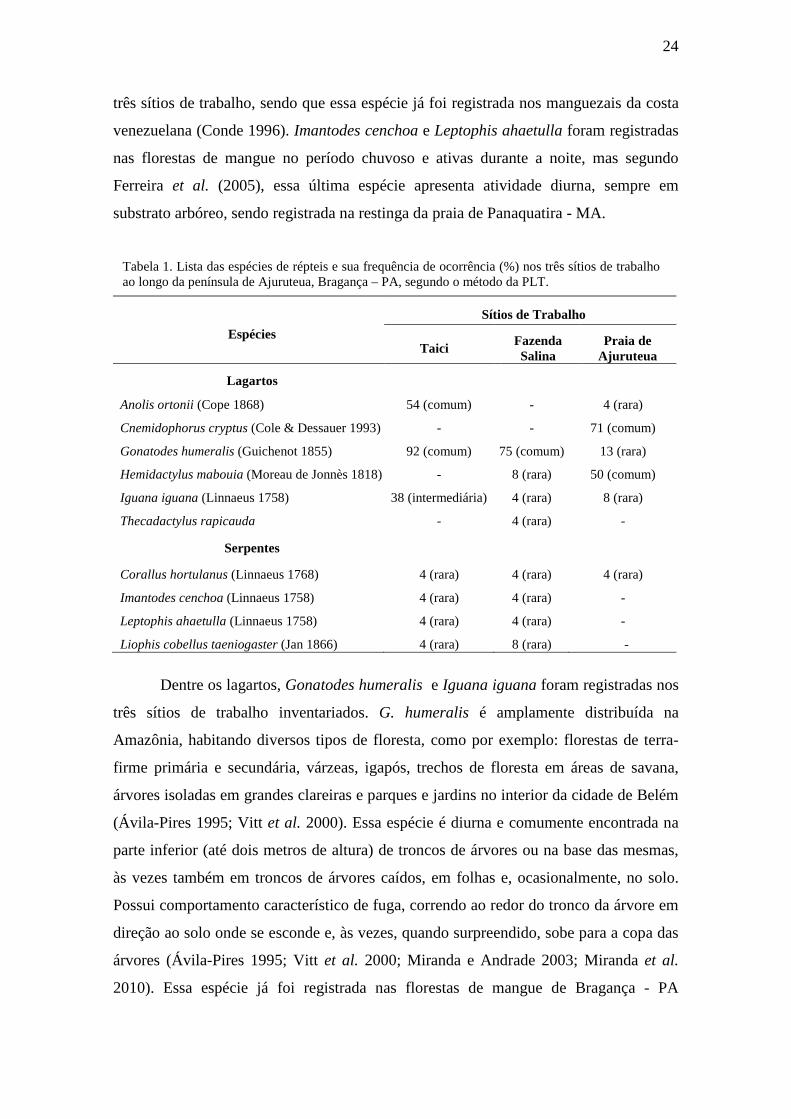
três sítios de trabalho, sendo que essa espécie já foi registrada nos manguezais da costa
venezuelana (Conde 1996). Imantodes cenchoa e Leptophis ahaetulla foram registradas
nas florestas de mangue no período chuvoso e ativas durante a noite, mas segundo
Ferreira et al. (2005), essa última espécie apresenta atividade diurna, sempre em
substrato arbóreo, sendo registrada na restinga da praia de Panaquatira - MA.
Tabela 1. Lista das espécies de répteis e sua frequência de ocorrência (%) nos três sítios de trabalho ao longo da península de Ajuruteua, Bragança – PA, segundo o método da PLT.
Dentre os lagartos, Gonatodes humeralis e Iguana iguana foram registradas nos
três sítios de trabalho inventariados. G. humeralis é amplamente distribuída na
Amazônia, habitando diversos tipos de floresta, como por exemplo: florestas de terra-
firme primária e secundária, várzeas, igapós, trechos de floresta em áreas de savana,
árvores isoladas em grandes clareiras e parques e jardins no interior da cidade de Belém
(Ávila-Pires 1995; Vitt et al. 2000). Essa espécie é diurna e comumente encontrada na
parte inferior (até dois metros de altura) de troncos de árvores ou na base das mesmas,
às vezes também em troncos de árvores caídos, em folhas e, ocasionalmente, no solo.
Possui comportamento característico de fuga, correndo ao redor do tronco da árvore em
direção ao solo onde se esconde e, às vezes, quando surpreendido, sobe para a copa das
árvores (Ávila-Pires 1995; Vitt et al. 2000; Miranda e Andrade 2003; Miranda et al.
2010). Essa espécie já foi registrada nas florestas de mangue de Bragança - PA
Espécies
Sítios de Trabalho
Taici Fazenda Salina
Praia de Ajuruteua
Lagartos
Anolis ortonii (Cope 1868) 54 (comum) - 4 (rara)
Cnemidophorus cryptus (Cole & Dessauer 1993) - - 71 (comum)
Gonatodes humeralis (Guichenot 1855) 92 (comum) 75 (comum) 13 (rara)
Hemidactylus mabouia (Moreau de Jonnès 1818) - 8 (rara) 50 (comum)
Iguana iguana (Linnaeus 1758) 38 (intermediária) 4 (rara) 8 (rara)
Thecadactylus rapicauda - 4 (rara) -
Serpentes
Corallus hortulanus (Linnaeus 1768) 4 (rara) 4 (rara) 4 (rara)
Imantodes cenchoa (Linnaeus 1758) 4 (rara) 4 (rara) -
Leptophis ahaetulla (Linnaeus 1758) 4 (rara) 4 (rara) -
Liophis cobellus taeniogaster (Jan 1866) 4 (rara) 8 (rara) -

25
(Fernandes et al. 2009). Na restinga esta espécie foi considerada rara, ao passo que na
Fazenda Salinas e no Taici a mesma foi considerada comum.
As espécies I. iguana e Anolis ortonii também já foram registradas no
manguezal. A primeira foi considerada rara na Fazenda Salinas e na Praia de Ajuruteua,
enquanto no Taici ocorreram em 38% das amostragens, com frequência intermediária.
Esta espécie já foi registrada em área de manguezal (Burghardt e Rand 1983). Por outro
lado, A. ortonii foi considerada rara na restinga e comum no manguezal. No México,
segundo Platt et al. (1999), esse gênero possui uma espécie, Anolis sagrei, que é
abundante nas florestas de mangue.
Hemidactylus mabouia foi registrada em duas áreas, sendo considerada rara na
Fazenda Salinas e comum na restinga da Praia de Ajuruteua, onde foi encontrada em
vegetação arbórea. A frequência de ocorrência na restinga está diretamente relacionada
ao fato de esta ser uma área antropizada, fator que tem favorecido a ampla distribuição e
ocorrência dessa espécie nos diferentes ambientes na Amazônia (Ávila-Pires 1995).
Cnemidophorus cryptus só ocorreu na restinga e foi considerada comum nesse
ecossistema. Essa espécie é comum em áreas urbanas e ambientes costeiros da
Amazônia (Ávila-Pires 1995).
O espécime de muçuã, Kinosternon scorpioides (Linnaeus-1766), foi registrado
posteriormente às coletas, enterrado em substrato lamoso entre a restinga e o
manguezal. Essa espécie é comumente encontrada em lagoas, lagos e rios lentos e
apresenta distribuição ampla ao longo de toda a Amazônia e América do Sul
(Duellman 1979; Zug et al. 2001).
Anuros
Na península de Ajuruteua ocorrem 14 espécies de anuros, distribuídos em
quatro famílias: Bufonidae (n=1), Hylidae (n=6), Leptodactylidae (n=5) e Leiuperidae
(n=2) (Figura 5) (Tabela 2). Todas são típicas de áreas abertas e apresentam ampla
distribuição na Amazônia (Crump 1971; Galatti et al. 2007; Frost 2010). De uma
maneira geral, o número de espécies de anuros registrado no presente trabalho não foi
elevado quando comparado a outros pontos da Amazônia brasileira (ex. 41 espécies em
Caxiuanã - Ávila-Pires e Hoogmoed 1997; Bernardi et al. 1999; 52 espécies em Belém -
Galatti et al. 2007; 31 espécies para as savanas na Amazônia - Neckel-Oliveira et al.
2000). Portanto, o inventário do presente trabalho sobre as espécies de anuros ainda
deve ser considerado preliminar, já que representa, em média, 35% das espécies

26
ocorrentes nos diversos pontos da Amazônia brasileira. No entanto, levando-se em
consideração as 26 espécies que ocorrem na região do município de Bragança, esse
valor percentual tende a aumentar, correspondendo a 54% do total de espécies
registrado. Assim, é de se esperar que um aumento no esforço amostral, utilizando os
diferentes ambientes ainda não inventariados ao longo do litoral amazônico, também
eleve o valor da riqueza das espécies associadas aos ambientes costeiros da Amazônia
brasileira.
Vários fatores podem contribuir para a baixa riqueza registrada no presente
estudo. Dentre eles pode-se citar a disponibilidade de sítios reprodutivos, que é um
fator-chave na estrutura da comunidade de anuros (Zimmerman e Simberloff 1996,
Neckel-Oliveira et al. 2000), assim como a baixa heterogeneidade de ecossistemas,
fornecendo poucos microambientes para a reprodução e alimentação das espécies.
Outro fator importante são as características dos ambientes costeiros, como é o caso do
manguezal, que encontra-se sob condições adversas e resultantes de diferentes teores de
sal e de oxigênio do solo, bem como dos elevados índices de temperatura e inundações
periódicas (Tomlinson 1986; Hogarth 1999; Souza-Filho e El-Robrini 1996). E, geral,
deve-se considerar que os ambientes salinos são umas das principais barreiras na
dispersão de grande parte das espécies de anfíbios, onde são submetidos à perda
excessiva de água e altas taxas de absorção de sódio e cloreto através da sua pele
permeável (Wells 2007).
Ainda que o manguezal não seja um ambiente favorável à maioria dos anfíbios,
ao menos três espécies (Fejervarya cancrivora (Gravenhorst, 1829), Buergeria japonica
(Hallowell, 1861) e Pseudepidalea viridis (Laurenti, 1768)) são tolerantes à alta
salinidade e apresentam adaptações fisiológicas a essas concentrações de sal em
diferentes continentes (Gordon et al. 1961; Gordon e Tucker 1968; Dicker e Elliott
1970; Zug et al. 2001; Haramura 2007; Wells 2007). Outras 22 espécies de anuros são
capazes de fazer uso de ambientes com salinidade moderada e também são encontradas
no ambientes salinos ao redor do mundo (Leitão 1995; Fernandes 2000; Zug et al. 2001;
Bambaradeniya et al. 2002; Mcdiarmid e Savage 2005; Venkataraman e Wafar L2005;
Wells 2007; Kathiresan e Bingham 2008). Dentre estas, três são consideradas comuns
nestes ambientes (Luther e Greenberg 2009; Kathiresan e Bingham 2008).

Figura 5. Espécies de anuros registradas na península de Ajuruteua, Bragança-PA. Família Hylidae: A, Hypsiboas ranicepsTrachycephalus venulosusruber e G, Leptodactylus fuscusLeptodactylus labyrinthicusLeiuperidae: L, Família Bufonidae: N,
No Brasil, seis espécies de anuros
(Loebmann e Mai 2008; Ferreira
atualizar essa lista, acresc
(Tabela 2). As espécies Dendropsophus nanus
fuscus, Leptodactylus macrosternum
Trachycephalus venulosus
manguezal, com maior frequência no período chuvoso
nanus e T. venulosus não foram consideradas comuns, sendo a última rara. Um
indivíduo adulto de Phyllomedusa hypochondrialis
Figura 5. Espécies de anuros registradas na península de Ajuruteua, PA. Família Hylidae: A, Dendropsophus nanus; B,
Hypsiboas raniceps; C, Phyllomedusa hypochondrialis; D, Trachycephalus venulosus; E, Scinax fuscomarginatus; F, Scinax
e G, Scinax boesemani. Família Leptodactylidae: H, Leptodactylus fuscus; I, Leptodactylus macrosternum; J, Leptodactylus labyrinthicus; K, Leptodactylus mystaceus. Família Leiuperidae: L, Physalaemus ephipiffer; M, Pseudopaludicola sp. e Família Bufonidae: N, Rhinella marina.
espécies de anuros já foram registradas em áreas de
Mai 2008; Ferreira e Tonini 2010), sendo quemas o presente trabalho
a lista, acrescentando mais nove espécies associadas a este ecossistema
Dendropsophus nanus, Hypsiboas raniceps
Leptodactylus macrosternum, Pseudopaludicola sp., Rhinella marina
foram registradas ativas principalmente durante a noite no
frequência no período chuvoso. Dentre essas espécies, apenas
não foram consideradas comuns, sendo a última rara. Um
Phyllomedusa hypochondrialis foi registrado dormindo durante o
27
Figura 5. Espécies de anuros registradas na península de Ajuruteua,
; B, ; D, Scinax
. Família Leptodactylidae: H, ; J,
ília sp. e
áreas de manguezal
sente trabalho vem
entando mais nove espécies associadas a este ecossistema
Hypsiboas raniceps, Leptodactylus
Rhinella marina e
foram registradas ativas principalmente durante a noite no
. Dentre essas espécies, apenas D.
não foram consideradas comuns, sendo a última rara. Um
dormindo durante o
28
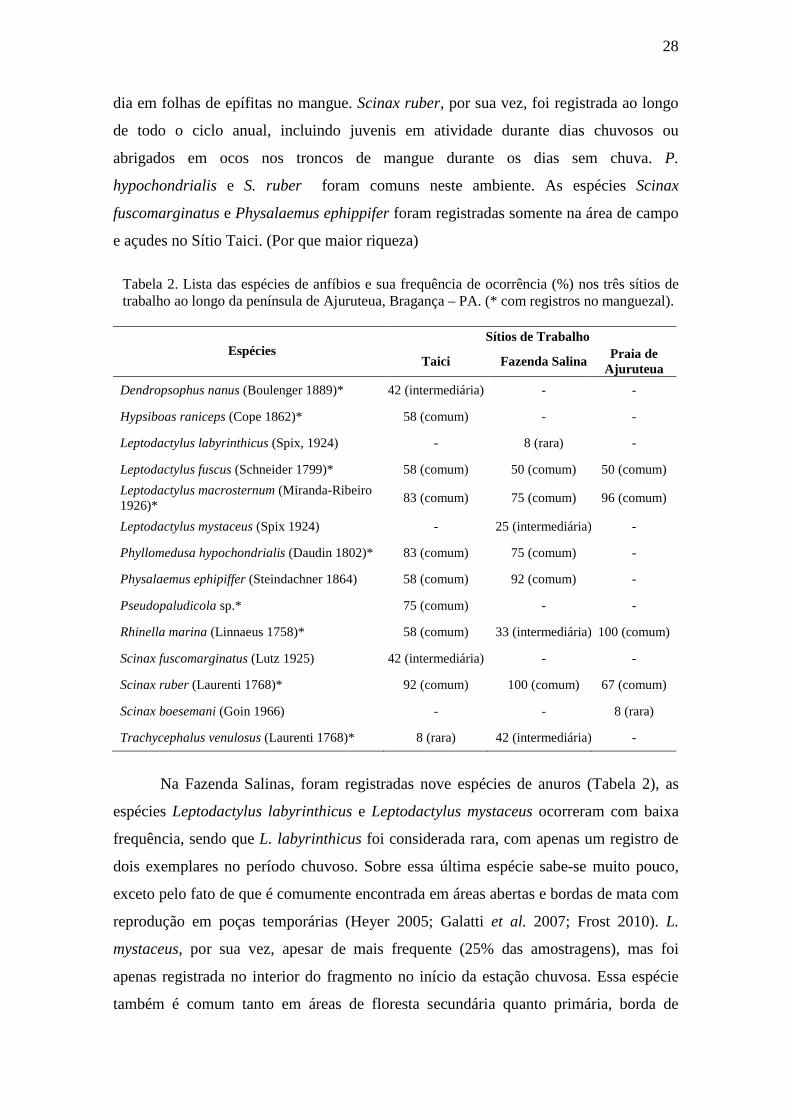
dia em folhas de epífitas no mangue. Scinax ruber, por sua vez, foi registrada ao longo
de todo o ciclo anual, incluindo juvenis em atividade durante dias chuvosos ou
abrigados em ocos nos troncos de mangue durante os dias sem chuva. P.
hypochondrialis e S. ruber foram comuns neste ambiente. As espécies Scinax
fuscomarginatus e Physalaemus ephippifer foram registradas somente na área de campo
e açudes no Sítio Taici. (Por que maior riqueza)
Tabela 2. Lista das espécies de anfíbios e sua frequência de ocorrência (%) nos três sítios de trabalho ao longo da península de Ajuruteua, Bragança – PA. (* com registros no manguezal).
Na Fazenda Salinas, foram registradas nove espécies de anuros (Tabela 2), as
espécies Leptodactylus labyrinthicus e Leptodactylus mystaceus ocorreram com baixa
frequência, sendo que L. labyrinthicus foi considerada rara, com apenas um registro de
dois exemplares no período chuvoso. Sobre essa última espécie sabe-se muito pouco,
exceto pelo fato de que é comumente encontrada em áreas abertas e bordas de mata com
reprodução em poças temporárias (Heyer 2005; Galatti et al. 2007; Frost 2010). L.
mystaceus, por sua vez, apesar de mais frequente (25% das amostragens), mas foi
apenas registrada no interior do fragmento no início da estação chuvosa. Essa espécie
também é comum tanto em áreas de floresta secundária quanto primária, borda de
Espécies Sítios de Trabalho
Taici Fazenda Salina Praia de
Ajuruteua
Dendropsophus nanus (Boulenger 1889)* 42 (intermediária) - -
Hypsiboas raniceps (Cope 1862)* 58 (comum) - -
Leptodactylus labyrinthicus (Spix, 1924) - 8 (rara) -
Leptodactylus fuscus (Schneider 1799)* 58 (comum) 50 (comum) 50 (comum)
Leptodactylus macrosternum (Miranda-Ribeiro 1926)*
83 (comum) 75 (comum) 96 (comum)
Leptodactylus mystaceus (Spix 1924) - 25 (intermediária) -
Phyllomedusa hypochondrialis (Daudin 1802)* 83 (comum) 75 (comum) -
Physalaemus ephipiffer (Steindachner 1864) 58 (comum) 92 (comum) -
Pseudopaludicola sp.* 75 (comum) - -
Rhinella marina (Linnaeus 1758)* 58 (comum) 33 (intermediária)100 (comum)
Scinax fuscomarginatus (Lutz 1925) 42 (intermediária) - -
Scinax ruber (Laurenti 1768)* 92 (comum) 100 (comum) 67 (comum)
Scinax boesemani (Goin 1966) - - 8 (rara)
Trachycephalus venulosus (Laurenti 1768)* 8 (rara) 42 (intermediária) -

29
floresta, floresta de várzea e se reproduzem em ninhos de espumas na serrapilheira
(Hödl 1990; Heyer e Rodrigues 2004; Galatti et al. 2007). As espécies T. venulosus e R.
marina foram registradas principalmente na borda do fragmento, ambas com dois
registros. As espécies L. fuscus, L. macrosternum, P. hypochondrialis e S. ruber foram
consideradas comuns, haja vista terem sido registradas em mais de 50% das
observações (Tabela 2).
No presente estudo, as dunas e restingas na praia de Ajuruteua apresentaram a
menor riqueza (n=5), sendo a espécie Scinax boesemani considerada rara. A literatura
aponta esses ambientes como aqueles que apresentam a maior riqueza de anuros na
costa brasileira, considerando o registro de 52 espécies entre os estados do Rio Grande
do Sul e Bahia e de cinco espécies endêmicas nas restingas da região sudeste (Sluys et
al. 2004; Loebmann 2005; Rocha et al. 2005; Rocha e Sluys 2007; Loebmann e Mai
2008; Rocha et al. 2008b; Silva et al. 2008). Segundo Rocha et al. (2008a) a riqueza e a
composição da assembléia de anuros possuem ampla variação ao longo das restingas da
costa leste brasileira e a taxa de mudança na composição de espécies é afetada, em
parte, pela distância geográfica entre as áreas.
Dentre as espécies consideradas comuns, isto é, presente em mais de 50% das
amostragens, L. fuscus, L. macrosternum, R. marina e S. ruber foram as que ocorreram
em todos os sítios amostrados (Tabela 1). R. marina e S. ruber foram registradas em
100 % das coletas no Sítio 3 e Sítio 2, respectivamente. Apesar dos hilídeos serem a
maioria das espécies ao longo da península, esperava-se uma distribuição mais
equivalente desta família nos ambientes representados. No entanto, apenas a espécie S.
ruber esteve em todos os sítios amostrados, ao passo que, os leptodadctylídeos
apresentaram duas espécies presentes em todos os sítios amostrados.
REFERÊNCIAS BIBLIOGRÁFICAS
Amaral, D.D., M.T. Prost, M.N.C. Bastos, S.V.C. Neto e J.U.M. Santos. 2008.
Restingas do litoral amazônico, estados do Pará e Amapá, Brasil. Boletim do Musel
Paraense Emílio Goeldi, Ciências Naturais 3(1): 35-67.
Ávila-Pires, T.C.S. 1995. Lizards of Brazilian Amazonian (Reptilia: Squamata).
Zoologische Verhandelingen 1-706(1-706.
Ávila-Pires, T.C.S. e M.S. Hoogmoed. 1997. The Herpetofauna; p. 389-401 In P.L.B.
Lisboa (ed.). Caxiuanã. Belém: Museu Paraense Emílio Goeldi.

30
Ávila-Pires, T.C.S., M.S. Hoogmoed e L.J. Vitt. 2007. Herpetofauna da Amazônia; p.
354 In L.B. Nascimento e M.E. Oliveira (eds.). Herpetologia no Brasil II. Belo
Horizonte: Sociedade Brasileira de Herpetologia.
Awosika, L. e E. Marone. 2000. Scientific needs to assess the health of the oceans in
coastal areas: a perspective of developing countries. Ocean & Coastal Management
43(781-791.
Bambaradeniya, C.N.B., S.P. Ekanayake, L.D.C.B. Kekulandala, R.H.S.S. Fernando,
V.A.P. Samarawickrama e T.G.M. Priyadharshana. 2002. An Assessment of the Status
o f Biodiversity in the Maduganga Mangrove Estuary. Occasional Papers of IUCN Sri
Lanka 1(1-49.
Bernardi, J.A.R., R.A. Estupiñán e U. Galatti. 1999. New Anuran Records from the
Floresta Nacional de Caxiuanã, Eastern Amazon, Brazil. Herpetological Review 30(3):
176-177.
Boss, H.E.A. 2001. The Snakes of Trinidad e Tobago. Texas: Texas A & M University
Press, College Station. 286 p.
Burghardt, G.M. e A.S. Rand. 1983. Iguanas of the world: Their Behavior, Ecology, e
Conservation. New Jersey: Noyes Publications. 491 p.
Carvalho, A.L.G.D. e A.F.B. Araújo. 2004. Ecologia dos Lagartos da Ilha da
Marambaia, RJ. Revista Universidade Rural: Série Ciências da Vida, Seropédica, RJ
24(2): 159-165.
Carvalho, A.L.G.D., A.F.B. Araújo e H.R. Silva. 2007. Lagartos da Marambaia, um
remanescente insular de Restinga e Floresta Atlântica no Estado do Rio de Janeiro,
Brasil. Biota Neotropica 7(2): 221-226.
Conde, J.E. 1996. A profile of laguna de Tacarigua, Venezuela: A tropical estuarine
coastal lagoon. . 21(5): 282-292.
Crump, M.L. 1971. Quantitative Analysis of Ecological Distribuition of tropical
herpetofauna. Occasional Papers Museum of Natural the University of Kansas 3(1-62.
Cunha-Barros, M., M. Van Sluys, D. Vrcibradic, C.A.B. Galdino, F.H. Hatano e C.F.D.
Rocha. 2003. Patterns of infestation by chigger mites in four diurnal lizard species from
a Restinga habitat (Jurubatiba) of Southeastern Brazil. Brazilian Journal of Biology
63(3): 602-612.
Cunha, O.R. e F.P. Nascimento. 1978. Ofídios da Amazônia X – As cobras da região
Leste do Pará. Belém: Museul Paraense Emílio Goeldi. 218 p.

31
Cunha, O.R. e F.P. Nascimento. 1993. Ofídios da Amazônia. As cobras da Região Leste
do Pará. Belém: Boletim do Museu Paraense Emílio Goeldi. 192 p.
Dicker, S.E. e A.B. Elliott. 1970. Water uptake by the crab-eating frog Rana
cancrivora, as affected by osmotic gradients e by neurohypophysial hormones. Journal
Physiological 287(119-132.
Duellman, W.E. 1979. The South American Herpetofauna: Its Origin, Evolution, e
Dispersal. Kansas: Museu of Natural History, The University of Kansas. 485 p.
Fernandes, M.E.B. 2000. Association of mammals with mangrove forests: a world wide
review. Boletim do Laboratório de Hidrobiologia 13(83-108.
Fernandes, M.E.B., A.O. Marciel, F.S. Santos, I.L.A.H.V. Linke e A.L. Ravetta. 2009.
Gonatodes humeralis (NCH) Habitat Occurrence; Escape Behavior. Herpetological
Review 40(2): 221-222.
Ferreira, A.P., E.M.S. Fialho and G.V. Andrade. 2005. Composição e estruturação da
comunidade de serpentes da restinga da praia de Panaquatira, Maranhão. VII Congresso
de Ecologia do Brasil - Resumos expandido.
Ferreira, R.B. e J.F.R. Tonini. 2010. Living holed: Leptodactylus latrans occupying
crabs’ burrows. Herpetology Notes 3:237-238.
Freire, E.M.X. 1996. Estudo ecológico e zoogeográfico sobre a fauna de lagartos
(sauria) das dunas de Natal, Rio Grande do Norte e da restinga de Ponta de Campina,
Cabedelo, Paraíba, Brasil. Revista Brasileira de Zoologia 13(4): 903-921.
Frost, D.R. 2010. Amphibians Species of the Wolrd: an Online Reference. Version 5.4.
Electronic Database accessible at
http://research.amnh.org/herpetology/amphibia/index.php. American Museum of
Natural History, New York, USA. Captured on 20 Abril 2010.
Galatti, U., R.A. Estupiñán, A.C.L. Dias e A.E.M. Travassos. 2007. Anfíbios da Área de
Pesquisa Ecológica do Guamá (APEG) e Região de Belém; p. 456 In J.I. G.M. Martins-
da-Silva, R.C. Viana e S.S. Almeida (eds.). Mocambo: Diversidade e Dinâmica
Biológica da Área de Pesquisa Ecológica do Guamá (APEG). Belém: MCT/Museu
Paraense Emílio Goeldi
Gordon, M.S., K. Schmidt-Neilson e H.M. Kelly. 1961. Osmotic regulation in the crab-
eating frog (Rana cancrivora). Journal Experimental Biology 38(659-678.
Gordon, M.S. e V.A. Tucker. 1968. Further Observations on the physiology of salinity
adaptation in the Crab-eating frog (Rana Cancrivora). Journal Experimental Biology
49(185-193.

32
Hansknecht, K.A. 2008. Lingual Luring by Mangrove Salt marsh Snakes (Nerodia
clarkii compressicauda). Journal of Herpetology 42(1): 9-15.
Haramura, T. 2007. Salinity Tolerance of Eggs of Buergeria japonica (Amphibia,
Anura) Inhabiting Coastal Areas. Zoological Science 24(820-823.
Hatano, F.H., V. D, C.A.B. Galdino, M. Cunha-Barros, R.C.F. D e M. Van Sluys. 2001.
Thermal ecology e activity patterns of the lizard community of the Restinga of
Jurubatiba, Macao, RJ. Revista Brasileira de Biologia 61(2): 287-294.
Hero, J.M., W.E. Magnusson, C.F.D. Rocha e C.P. Lateral. 2001. Ant predator
Defenses Inflhence the Distribution of Amphibian Prey Species in the Central Amazon
Rain Forest. Biotropica 33(1): 131-141.
Heyer, R.W., M.A. Donnelly, R.W. McDiarmid, L.C. Hayek e M.S. Foster. 1994.
Measuring e monitoring biological diversity: Standard methods for amphibians.
Washington: Smithsonian Institution Press. 364 p.
Hödl, W. 1990. Reproductive diversity in Amazonian lowland frogs. Fortschritte der
Zoologie 38(41-60.
Hogarth, P. 1999. The Biology of Mangroves. New York: Oxford University Press. 228
p.
Kacoliris, F., N. Horlent e J. Williams. 2006. Herpetofauna, Coastal Dunes, Buenos
Aires Province, Argentina Check List 2(3):
Karns, D.R., H.k. Voris e T.G. Goodwin. 2002. Ecology of oriental-australian rear-
fanged water snakes (Colubridae: Homalopsinae) in the pasir ris park mangrove
forest, Singapore. The Raffles Bulletin of Zoology 50(2): 487-498.
Kathiresan, K. e B.L. Bingham. 2008. Biology of mangroves e mangrove ecosystems.
Advance in Marine Biology 40(81-251.
Kelly, L. 2006. Evolution's greatest survivior Crocodile. Sydney: Allen & Unwin. 271
p.
Leitão, S.N. 1995. A fauna do Manguezal; p. 64 In Y.S. Novelli (eds). Manguezal,
Ecossistema entre a terra e o mar. São Paulo: Caribbean Ecoloical Research.
Loebmann, D. 2005. Guia Ilustrado: Os anfíbios da região costeira do extremo sul do
Brasil. Pelotas: União Sul-Americana de Estudos da Biodiversidade. 76 p.
Loebmann, D. e A.C.G. Mai. 2008. Amphibia, Anura, Coastal Zone, state of Piauí,
Northeastern Brazil. Check List 4(2): 161-170.

33
Luiselli, L. e G.C. Akani. 2002. An investigation into the composition, complexity e
functioning of snake communities in the mangroves of south-eastern Nigeria. African
Journal of Ecology 40(3): 220-227.
Luther, D.A. e R. Greenberg. 2009. Mangroves: A Global perspective on the evolution e
conservation of their terrestrial vertebrates. Bioscience 59(7): 602-612.
McDiarmid, R.W. e J.M. Savage. 2005. The Herpetofauna of the Rincon Area,
Peninsula de Osa, Costa Rica, a Central American Lowland Evergreen Forest Site; p.
366-427 In M.A. Donnelly B.I. Crother C. Guyer M.H. Wake e M.E. White (eds.).
Ecology e evolution in the tropics: a herpetological perspective. Chicago: University of
Chicago Press.
Mendes, A.C. 2005. Geomorfologia e sedimentologia; p. 13-31 In M.E.B. Fernandes
(ed.). Os manguezais da costa norte brasileira. Belém: Petrobrás/Fundação Rio
Bacanga.
Menezes, M.P.M., U. Berger e U. Mehlig. 2008. Mangrove vegetation in Amazonia: a
review of studies from the coast of Pará e Maranhão States, north Brazil. Acta
Amazonica 38(3): 403-420.
Mesquita, D.O., G.C. Costa e M.G. Zatz. 2004. Ecological aspects of the casque-headed
frog Aparasphenodon brunoi (Anura, Hylidae) in a Restinga habitat in southeastern
Brazil. Phyllomedusa 3(1): 51-59.
Miranda, J.P. e G.V. Andrade. 2003. Seasonality in Diet, Perch Use, e Reproduction of
the Gecko Gonatodes humeralis from Eastern Brazilian Amazon. Journal of
Herpetology 37(2): 433-438.
Miranda, J.P., A. Ricci-Lobão and C.F.D. Rocha. 2010. Influence of structural habitat
use on the thermal ecology of Gonatodes humeralis (Squamata: Gekkonidae) from a
transitional forest in Maranhão, Brazil. Zoologia 27(1): 35-39.
Neckel-Oliveira, S., W.E. Magnusson, A.P. Lima and A.L.K. Albernaz. 2000. Diversity
and distribution of frogs in an Amazonian savanna in Brazil. Amphibia-Reptilia 21(3):
317-326.
Neckel-Oliveira, S. e M. Gordo. 2004. Anfíbios, Lagartos e Serpentes do Parque
Nacional do Jaú; p. 173-185 In S.H. Borges S. Iwanaga C.C. Durigan e M.R. Pinheiro
(eds.). Janelas Para a Biodiversidade no Parque Nacional do Jaú: Uma Estratégia
Para o Estudo da Biodiversidade na Amazônia. Manaus: Ipiranga.
Pauwels, O.S.G., W.R. Branch e M. Burger. 2004. Reptiles of Loango National Park,
Ogooué-Mariteme Province, South-Western Gabon. Hamadryad 29(1): 115-127.

34
Pereira, L.C.C., J.A. Dias, J.A. Carmo e M. Polette. 2009. The Brazilian Amazon
Coastal Zone. Journal of Integrated Coastal Zone Management 9(2): 3-7.
Platt, S.G., J.C. Meerman e T.R. Rainwater. 1999. Diversity, Observations, e
Conservation of the Herpetofauna of Turneffe, Leghthouse, e Glovers Atolls, Belize
British Herpetological Society Bulletin 66(1-13.
Quintela, F.M. e D. Loebmann. 2009. Os Répteis da região costeira do extremo sul do
Brasil. Pelotas: União Sul-Americana de Estudos da Biodiversidade. 84 p.
Rocha, C.F.D., C.V. Ariani, V.A. Menezes e D. Vribradic. 2008a. Effects of a fire on a
population of treefrogs (Scinax cf. alter, Lutz) in a restinga habitat in southern Brazil.
Brazilian Journal of Biology 68(3): 539-543.
Rocha, C.F.D., H.G. Bergallo, J.P. Pombal-Jr, L. Geise, M.V. Sluys, R. Fernandes e U.
Caramaschi. 2004. Fauna de Anfíbios, Répteis e Mamíferos do estado do Rio de
Janeiro, Sudeste do Brasil. Publicações Avulsas do Museu Nacional 104(3-23.
Rocha, C.F.D., F.H. Hatano, D. Vrcibradic e M.V. Sluys. 2008b. Frog species richness,
composition e β-diversity in coastal Brazilian restinga hábitats. Brazilian Journal of
Biology 68(1): 101-107.
Rocha, C.F.D. e M.V. Sluys. 2007. Herpetofauna de Restingas; p. 354 In L.B.
Nascimento e M.E. Oliveira (eds.). Herpetologia no Brasil II. Belo Horizonte:
Sociedade Brasileira de Herpetologia.
Rocha, C.F.D., M. Van Sluys, H.G. Bergallo e M.A.S. Alves. 2005. Endemic e
threatened tetrapods in the Restingas of the biodiversity corridors of Serra do Mar end
of the central in Mata Atlântica in eastern Brazil. Brazilian Journal of Biology 65(1):
159-168.
Rojas-Runjaic, F.J.M., C.L. Barrio-Amorós, C.M. R., J.C. Señaris e I.C. Fedón. 2008.
Amphibia, Anura, Hylidae, Scarthyla vigilans: Range extensions e new state records
from Delta Amacuro e Miranda states, Venezuela. . Check List 4(3): 301-303.
Schineider, J.A.P. e R.L. Teixeira. 2001. Relacionamento entre anfíbios anuros e
bromélias da Restinga de Regência, Linhares, Espírito Santo, Brasil. Iheringia, Série
Zoologia 91(41-48.
Seeiger, U., C. Odebrechet e J.P. Castello. 1998. Os Ecossistemas costeiros e marinhos
do extremo sul do Brasil. Rio Grande: Ecosciencia 341 p.
Sluys, M.V., C.F.D. Rocha, F.H. Hatano, L. Boquimpani-Freitas e R.V. Marra. 2004.
Anfíbios da Restinga de Jurubatiba: Composição e História Natural; p. 376 In C.F.D.

35
Rocha, F.A. Esteves e F.R. Scarano (eds.). Pesquisas de Longa Duração na Restinga de
Jurubatiba-Ecologia, História Natural e Conservação. São Carlos: Rima.
Souza-Filho, P.W.M., M.T.R.C. Prost, F.P. Miranda, M.E.C. Sales, H.V. Borges, F.R.
Costa, E.F. Almeida e W.R.N. Junior. 2009. Environmental sensitivity index (ESI)
mapping of oil spill in the Amazon coastal zone: the Piatam mar project. Revista
Brasileira de Geofísica 27(1): 7-22.
Souza-Filho, P.W.M. e M. El-Robrini. 1996. Morfologia, processos de sedimentação e
litofácies dos ambientes morfo-sedimentares da planície costeira bragantina, nordeste do
Pará, Brasil. Geonomos 4(2): 1-16.
Teixeira, R.L. 2001. Comunidade de lagartos da restinga de Guriri, São Mateus-ES,
Sudeste do Brasil. Atlântica 23(77-84.
Teixeira, R.L. e J.A.P. Schineider. 2002. The Occurrence of Amphibians in Bromeliads
from a Southeastern Brazilian Restinga habitat, with special reference to
Aparasphenodon brunoi (Anura, Hylidae). Brazilian Journal of Biology 62(2): 263-268.
Teixeira, R.L., J.A.P. Schineider e G.I. Almeida. 2002. The occurrence of amphibians in
bromeliads from a southeastern Brazilian Restinga habitat, with special reference to
Aparasphenodon brunoi (Anura, Hylidae). Brazilian Journal of Biology 62(2): 263-268.
Tomlinson, P.B. 1986. The Botany of Mangroves. Cambridge: Cambridge University
Press. 419 p.
Venkataraman, k. e M. Wafar. 2005. Coastal e marine biodiversity of India. India
Journal of Marine Sciences 34(1): 57-75.
Vitt, L.J., R.A. Souza, S.S. Sartorius, T.C.S. Ávila-Pires, M.C. Espósito e W.L.
Montgomery. 2000. Comparative Ecology of Sympatric Gonatodes (Squamata:
Gekkonidae) in the Western Amazon of Brazil. Copeia 2000(1): 83-95.
Wells, K.D. 2007. The Ecology e Behavior of Amphibians. Chicago: University of
Chicago Press. 1148 p.
Zimmerman, B.L. and D. Simberloff. 1996. An historical interpretation of habitat use
by frogs in a Central Amazonian Forest. Journal of Biogeography 23(27-46.
Zug, G.R., L.J. Vitt e J.P. Caldwell. 2001. Herpetology an Intoductory biology of
Amphibians e Reptiles. Florida: Academic Press. 645 p.

36
ANEXO 1. TAXALIST – TEXTO COM INFORMAÇÕES TAXÔNOMICAS.
<taxaList> <taxon class="Amphibia" family="Bufonidae" genus="Rhinella" species="marina"/> <taxon class="Amphibia" family="Hylidae" genus="Dendropsophus" species="nanus"/> <taxon class="Amphibia" family="Hylidae" genus="Hypsiboas" species="raniceps"/> <taxon class="Amphibia" family="Hylidae" genus="Phyllomedusa" species="hypochondrialis"/> <taxon class="Amphibia" family="Hylidae" genus="Trachycephalus" species="venulosus"/> <taxon class="Amphibia" family="Hylidae" genus="Scinax" species="boesemani"/> <taxon class="Amphibia" family="Hylidae" genus="Scinax" species="fuscomarginatus"/> <taxon class="Amphibia" family="Hylidae" genus="Scinax" species="ruber"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="fuscus"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="macrosternum"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="mystaceus"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="labyrinthicus"/> <taxon class="Amphibia" family="Leiuperidae" genus="Physalaemus" species="ephipiffer"/> <taxon class="Amphibia" family="Leiuperidae" genus="Pseudopaludicola"/> <taxon class="Reptilia" family="Iguanidae" genus="Iguana" species="iguana"/> <taxon class="Reptilia" family="Gekkonidae" genus="Hemidactylus" species="mabouia"/> <taxon class="Reptilia" family="Phyllodactylidae" genus="Thecadactylus" species="rapicauda"/> <taxon class="Reptilia" family="Polychrotidae" genus="Anolis" species="ortonii"/> <taxon class="Reptilia" family="Sphaeroactylidae" genus="Gonatodes" species="humeralis"/> <taxon class="Reptilia" family="Teiidae" genus="Cnemidophorus" species="cryptus"/> <taxon class="Reptilia" family="Boidae" genus="Boa" species="Constrictor"/> <taxon class="Reptilia" family="Boidae" genus="Corallus" species="hortulanus"/> <taxon class="Reptilia" family="Colubridae" genus="Leptophis" species="ahaetulla"/> <taxon class="Reptilia" family="Dipsadidae" genus="Imantodes" species="cenchoa"/> <taxon class="Reptilia" family="Dipsadidae" genus="Liophis" species="cobellus"/> <taxon class="Reptilia" family="Kinosternidae" genus="kinosternon" species= "scorpioides/> </taxaList>

37
ANEXO 2. REGRAS PARA SUBMISSÃO DO CAPÍTULO.
GENERAL INFORMATION
Categories of papers:
There are two types of papers: Notes on Geographic Distribution (NGD) e Lists of
Species (LS). NGDs should provide reports of occurrence of one or a few species. SLs
generally report a species inventory from a given locality.
MANUSCRIPT STYLE e FORMAT
The manuscript must be submitted as Word document (.doc) or Rich Text Format (.rtf),
double-spaced throughout (including tables e references), 2.0 cm margins on 21.0 x
29.7 cm (A4) paper. Please use Times New Roman 12 e do not format table lines. All
pages should be numbered consecutively (except the cover page). Manuscripts must be
submitted in the order specified below for each manuscript category (NGD or SL).
Please indicate the category in the first line of the document.
INSTRUCTIONS FOR LISTS OF SPECIES (LS)
This section is dedicated to complete species inventory from a given locality. Maps,
color pictures e tables of species lists are recommended. SL should be organized as
follows:
Category of paper e short title (Cover page)
Indicate in the first line of the manuscript its category (NGD or LS). The second line
must contain a short title (up to 90 characters with spaces).
Title (Cover page)
Taxonomic group, locality (optional), State, Country.
e.g.: Anurans in bromeliads, Parque Estadual da Serra do Brigadeiro, state of Minas
Gerais, southeastern Brazil.
Author(s) (Cover page)
Name(s) with respective institution e author for correspondence (provide e-mail):
First Name e Surnames 1*, First Name e Surnames 2 e First Name e Surnames 1 1 University, Institute, Departament. Complete Address, Zip code (or CEP), City, State,
Country. 2 University, Institute, Departament. Complete Address, Zip code (or CEP), City, State,
Country. * Corresponding author. E-mail: author@company-or-university
Abstract (Second page forward)
Up to 150 words.

38
Introduction (Second page forward)
Brief.
Materials e Methods
Study site; Data Collection (any sort of permits/authorizations should be mentioned in
this section); Data Analysis (optional).
Results e Discussion
As concise e objective as possible.
Acknowledgements (optional)
Permits/authorizations should be mentioned in Materials e Methods, not in the
Acknowledgements.
Literature Cited
Journal titles should be in full, not abbreviated. Volume should be followed by issue
number in parenthesis in every journal reference. Cited publications should be included
in alphabetical order in the following formats (attention to the usage of upper e
lowercases, commas, semi-colon, brackets, spaces, italics e English words, ever):
Citing Check List articles:
e.g.: Arroyo-Rodriguez, V., J.C. Dunn, J. Benitez-Malvido e S. Mandujano. 2009.
Angiosperms, Los Tuxtlas Biosphere Reserve, Veracruz, Mexico. Check List 5(4): 787-
799.
Journal articles with usual volume e issue number:
e.g.: França, M.V.N. 1999. La extinción de los zorros (Foxidae: Foxtrotus spp.) de
España. Perro Negro 20(3): 251-265.
Two authors in a journal series:
e.g.: Westerman, A.C.B. e E.M. Wistuba. 2007. Born to be wild: Behaviour studies on
Steppe wolves. Canadian Canids 51(1): 25-27.
More than two authors in a journal series:
e.g.: Ferraz, M.J.O., P. Pinheiro, M.C. Wachowicz e L.M. Kozak. 2006. A new Wild
dog (Canidae: Archaeocerberus) from Madagascar with description of its disgusting
habits. South African Mastozoological News 33(6): 339-349.
Chapter in an edited volume:
e.g.: Wal, D. 2008. Die Anwesenheit von Hunden auf europächer Zirkus; p. 11-69 In L.
Paretto e C. Drummond (eds.). Der Zirkus als ein Ethik Ort. Hamburg: Fuchs Marke
Verlag.

39
Books:
e.g.: Felde, G. e M. Staveski. 2001. O lobo-guará nos campos do sul do Brasil.
Curitiba: Artes de Antes. 210 p.
e.g.: Beux, M.R. 1997. Red Tibetan fox (Vulpoides religiosa). Electronic Database
accessible at http://www.allcanids.org/species/ Vulpoides_religiosa.html. MCMK/Asian
Canid Group, Foxford University, UK. Captured on 14 February 2008.
Important remark: gray literature will not be accepted.
» "Gray Literature" is scientific or technical literature that is not available through the
usual bibliographic sources such as databases or indexes, i.e. that can not be found
easily through conventional channels such as regular scientific journals or the internet
(scientific open journals).
» Technical reports, pre-prints, committee reports, proceedings (conference, congress e
symposia), as well as unpublished works (Monographs, Dissertations e Thesis), will be
considered gray literature, even if available in the internet. If strictly necessary, the use
of gray literature information must be cited as "unpublished data" or "personal
communication". However, the acceptance of this kind of reference will depends on the
Area Editor's decision.
» Articles "in press/accepted" should be allowed only if the author has already the
formal/final acceptance from the editor. A journal document proving the acceptance can
be directed to Check List.
GENERAL INSTRUCTIONS
In-text reference to localities:
All names of localities should be in capitals, no matter the language, e not italicized;
e.g. Parque Nacional de Itatiaia, Espinhaço Range, Reunión Island.
In-text citations:
Author citations in the text must follow the pattern: "Lutz (1973)" or "(Sazima 1975)",
"Secor e Diamond (1998)", "(Abe e Fernandes 1977)", "Johansen et al. (1987)" or
"(Andrade et al. 2004)". Several publications must be cited in chronological order; e.g.
"(Lutz 1973; Sazima 1975; Secor e Diamond 1998)". Two or more publications by the
same author must be cited as "(Sazima 1975; 1976)" or "(Sazima 1974a, b; 1975)".
In-Text reference to tables e figures:
Tables e Figures should be numbered consecutively in Arabic numerals. In the text,
tables should be referred as Table 1, Tables 2 e 3, Tables 2-5; figures must be referred

40
as Figure 1, Figures 2 e 3, Figures 2-5, Figure 4A (not Fig. 1, figure 1, fig. 1, or Figure
4a).
Tables:
Tables must be formatted with horizontal, but not vertical rules. Do not format table
lines. Place all tables after Literature Cited, one table per page with its corresponding
heading.
Figures:
Each figure must be sent as a separate file. Please, try not to exceed 1 Mb per file (if
necessary use data compressors e send as .rar or .zip files). Use sans-serif font (such as
Arial) with enough size to allow good readiness. Please name your figure files with the
convention: "CorrespondingAuthorSurname_fig1". The use of professional software is
strongly recommended in order to get better results.
Figures sizes:
Two kinds of figures are allowed: one or two columns; exactly 87 mm (one column) or
180 mm (two columns) wide. Figures can not exceed 260 mm height. If possible, it is
preferable white color to the background.
Figures format:
Photographs e drawings must be sent as 150 or 300dpi (dots per inch, i.e.: pixel/inch) in
RGB, CYMK or Grayscale Color Mode e JPEG (.jpg) or TIFF (.tiff) formats.
Line vectors, as maps, must be sent as RGB, CYMK or Grayscale Color Mode e EPS
(.eps) files.
Figures must be merged into one single layer.
Nomenclature:
Authors are entirely responsible for correct species identifications. When first
mentioning a species name, provide its complete binomial name including the authority
e date of the species name. Scientific names must be in italics. Remember to italicize
the abbreviations "e.g.", "i.e.", e "et al." as well.
Time:
02:22 h; 14:55 h.
Distances e areas:
15 km²; 60 m; 20,760 ha.
Geographic coordinates:
05°44'00" S, 44°23'04" W.

41
Temperatures:
20 °C (note spacing)
Percentages:
15 % (note spacing).
SI Units:
Use SI units (quick downloads: International System of Units(SI), SI guide, SI rules e
style).
Proofs:
Check List will undertake proofreading of the original manuscript e send galley proofs
to corresponding author for final verification.
INSTRUCTIONS FOR THE .TXT FILE
After accepted for publication, the author(s) will have to send a file containing the
taxonomic information of the species treated in their articles - named "taxalist". It is
necessary to create an online database, which should be created in text editor software,
preferably in Word Pad. This file should be as complete as possible to facilitate retrieval
of articles after publication. All taxonomic information provided will be included in a
database, from where it will be able to search for articles. If your article deals with
Anurans e you do not include the word "Amphibia" in this file or in the title of the
manuscript, someone searching with the word "Amphibia" will not find the article. This
must be send as Text File (.txt) format.
<taxaList>
<taxon class="Actinopterygii" family="Acestrorhynchinae" genus="Acestrorhynchus"
species="falcatus"/>
<taxon class="Actinopterygii" family="Acestrorhynchinae" genus="Acestrorhynchus"
species="microlepis"/>
<taxon class="Actinopterygii" family="Acestrorhynchinae" genus="Acnodon"
species="normani"/>
<taxon class="Actinopterygii" family="Acestrorhynchinae" genus="Knodus"/>
</taxaList>
Important remarks: » The file must be saved in .txt format; » The symbols < = " " /> from the beginning to the end of each line can not be missing, or the information will not be retrievable from the database; » The first e last line of the file must be: <taxaList> e </taxaList>, respectively;

42
» Be careful with upper e lower cases - this file will be case sensitive. » When the Area Editor communicates you about the acceptance of the manuscript, please start the preparation of the taxalist, as it will be asked when you submited correct proofs. Taxalist preparation is time consuming, particularly for long Species Lists.
<taxaList> <taxon class="Amphibia" family="Bufonidae" genus="Rhinella" species="marina"/> <taxon class="Amphibia" family="Hylidae" genus="Dendropsophus" species="nanus"/> <taxon class="Amphibia" family="Hylidae" genus="Hypsiboas" species="raniceps"/> <taxon class="Amphibia" family="Hylidae" genus="Phyllomedusa" species="hypochondrialis"/> <taxon class="Amphibia" family="Hylidae" genus="Trachycephalus" species="venulosus"/> <taxon class="Amphibia" family="Hylidae" genus="Scinax" species="boesemani"/> <taxon class="Amphibia" family="Hylidae" genus="Scinax" species="fuscomarginatus"/> <taxon class="Amphibia" family="Hylidae" genus="Scinax" species="ruber"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="fuscus"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="macrosternum"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="mystaceus"/> <taxon class="Amphibia" family="Leptodactylidae" genus="Leptodactylus" species="labyrinthicus"/> <taxon class="Amphibia" family="Leiuperidae" genus="Physalaemus" species="ephipiffer"/> <taxon class="Amphibia" family="Leiuperidae" genus="Pseudopaludicola"/> <taxon class="Reptilia" family="Iguanidae" genus="Iguana" species="iguana"/> <taxon class="Reptilia" family="Gekkonidae" genus="Hemidactylus" species="mabouia"/> <taxon class="Reptilia" family="Phyllodactylidae" genus="Thecadactylus" species="rapicauda"/> <taxon class="Reptilia" family="Polychrotidae" genus="Anolis" species="ortonii"/> <taxon class="Reptilia" family="Sphaeroactylidae" genus="Gonatodes" species="humeralis"/> <taxon class="Reptilia" family="Teiidae" genus="Cnemidophorus" species="cryptus"/> <taxon class="Reptilia" family="Boidae" genus="Boa" species="Constrictor"/> <taxon class="Reptilia" family="Boidae" genus="Corallus" species="hortulanus"/> <taxon class="Reptilia" family="Colubridae" genus="Leptophis" species="ahaetulla"/> <taxon class="Reptilia" family="Dipsadidae" genus="Imantodes" species="cenchoa"/> <taxon class="Reptilia" family="Dipsadidae" genus="Liophis" species="cobellus"/> <taxon class="Reptilia" family="Kinosternidae" genus="kinosternon" species= "scorpioides/> </taxaList>

43
CAPÍTULO 3
COMPOSIÇÃO E DISTRIBUIÇÃO ESPAÇO-TEMPORAL DA ANUROFAUNA NO
MOSAICO DE AMBIENTES DA PENÍNSULA DE AJURUTEUA, BRAGANÇA –
PARÁ, BRASIL

44
Resumo
O presente trabalho tem o objetivo de inventariar a anurofauna em diferentes formações
vegetacionais, dando-se ênfase à riqueza e distribuição espaço-temporal desses animais
nos diversos microhábitats na península de Ajuruteua, Bragança-Pará. Cinco pontos
amostrais foram usados em três sítios de trabalho, através de busca ativa limitada por
tempo entre julho/2008 e junho/2009. Foram registradas 14 espécies de anuros, os sítios
tiveram todas as espécies esperadas registradas. S. ruber e L. macrosternum ocorreram
em todos os sítios, quatro espécies foram restritas ao sítio 1, duas no sítio 4 e uma no
sítio 5. 11 Espécies apresentaram atividade reprodutiva durante o período chuvoso,
quatro não foram registradas vocalizando, R. marina e P. hypochondrialis iniciaram
suas vocalizações no final do período seco, S. fuscomarginatus iniciou sua vocalização
mais tardia entre todas as espécies. A umidade do ar apresentou correlação com o
número de espécies em atividade de vocalização. Diferenças temporais na reprodução
foram observadas nas espécies terrestres. Os substratos com estratificação vertical
foram exclusivamente utilizados por hilídeos e as famílias Leptodactylidae, Leiuperidae
e Bufonidae foram associados aos substratos terrestres e na água. Houve sobreposição
na utilização vertical dos substratos. Os estimadores de riqueza mostraram que todos os
sítios de trabalho foram suficientemente amostrados, possibilitando a identificação de
todas as espécies estimadas. A maior riqueza no Ponto Amostral 1 parece ser decorrente
da sua proximidade aos ambientes de terra firme. Em geral, os pontos amostrais ao
longo da península parecem disponibilizar recursos suficientes para as atividades
reprodutivas de 11 das 14 espécies registradas no presente estudo. A umidade relativa
do ar demonstrou ser a variável mais correlacionada com as atividades reprodutivas das
espécies nos diferentes ambientes da península, certamente, por ter a capacidade de
variar de acordo com os microambientes presentes em cada sítio de trabalho ao longo da
península. As espécies da família Hylidae usam os substratos verticais em diferentes
alturas. No entanto, a análise estatística não mostrou diferença significativa na
estratificação vertical, evidenciando a sobreposição das espécies na partilha da
verticalização dos substratos.
Palavras-chave: Anura, Manguezal, Amazônia

45
INTRODUÇÃO
As características reprodutivas diferenciadas das espécies de anuros são um
fator-chave para a distribuição desses animais entre os mais diversos ambientes
amazônicos. Estudos realizados em diferentes localidades da Amazônia enfatizam que a
disponibilidade de hábitats reprodutivos temporários e permanentes é um dos principais
fatores que influênciam a distribuição dos anfíbios, tornando-se determinantes na
estrutura da comunidade desse grupo (Crump 1971, Hödl 1977, Zimmerman e
Bierregaard 1986, Zimmerman e Simberloff 1996, Neckel-Oliveira, et al. 2000).
Nas regiões tropicais com clima sazonal, a maioria das espécies de anuros
reproduz durante a estação chuvosa (Crump 1974, Hödl 1990, Zimmerman e Simberloff
1996, Zug, et al. 2001, Gottsberger e Gruber 2004, Menin, et al. 2008). Entretanto,
durante o período chuvoso, os períodos de atividades reprodutivas e a ocorrência das
espécies são bastante variáveis (Gottsberger e Gruber 2004, Duellman 2005).
As comunidades de anuros apresentam variações espaciais e temporais em
resposta a diversos fatores, podendo ser estes bióticos e ou abióticos (Crump 1971, Hödl
1977, Duellman 1978, Hödl 1990, Pombal-Jr 1997, Bernarde 2007, Giaretta, et al.
2008). Temporalmente, a partilha na utilização de ambientes reprodutivos nos anuros
está diretamente relacionada às condições físicas da região. Por exemplo, espécies que
se reproduzem em ambientes aquáticos temporários têm sua atividade reprodutiva
fortemente influenciada pelo aumento das águas das chuvas, que, por sua vez, propicia a
formação desses ambientes aquáticos (Crump 1974, Hödl 1977, Duellman 1978, Hödl
1990, Bernarde e Anjos 1999, Giaretta, et al. 2008).
Por outro lado, a partilha de ambientes reprodutivos reduz as interações
interespecíficas (Pombal-Jr 1997). Esse mesmo autor ressalta que, em se tratando de
espécies neotropicais, a sobreposição temporal no uso de microhábitat para vocalização
na estação chuvosa é muito comum. Gottsberger e Gruber (2004) demonstraram que as
espécies com reprodução terrestre podem apresentar maior atividade de vocalização no
início da estação chuvosa, enquanto aquelas com reprodução em ambientes aquáticos
podem não apresentar diferenças na atividade de vocalização ao longo da estação
chuvosa. Para Hödl (1977), a maioria das espécies encontradas no mesmo tipo de
vegetação apresenta vocalizações bem diferenciadas no que diz respeito à duração da
pulsação, ao intervalo entre cantos e ao padrão de notas.
Espacialmente, os anuros estão distribuídos em uma grande diversidade de
microambientes, explorando uma grande variedade deles como sítios reprodutivos

46
(Crump 1971, Duellman 1978, Gascon 1991, Zimmerman e Simberloff 1996, Stebbins e
Cohen 1997, Wells 2007). Segundo Heyer et al. (1990), espécies relacionadas
filogenéticamente tendem a apresentar similaridade ecológica em partes discretamente
diferentes do ambiente. Rossa-Feres e Jim (2001), por sua vez, apontaram para o fato de
que pode haver segregação espacial entre espécies taxonomicamente relacionadas que
vocalizam nos estratos verticais da vegetação, mas não entre aquelas espécies que
vocalizam na altura do chão.
De fato, o conhecimento dos fatores que determinam a distribuição de espécies
em escala local é fundamental para o entendimento dos processos de interação com o
meio, portanto, necessitam de uma investigação das características do ambiente onde
essas espécies ocorrem (Gascon 1991).
Na zona costeira paraense, o manguezal é a sua formação vegetacional
predominante (Mendes 2005). As florestas de mangue são constituídas por poucas
espécies arbóreas altamente adaptadas ao regime de marés e às condições abióticas
locais (Tomlinson 1986). Além do manguezal, outros tipos de formações vegetacionais
também compõem a paisagem nessa faixa costeira, como as restingas, os campos
salinos (marismas) e os fragmentos de floresta de terra firme que estão localizados
acima do nível de inundação das marés (Souza-Filho and El-Robrini 1996). De fato, o
conhecimento sobre esses ambientes é muito incipiente no que se refere à composição e
distribuição das espécies de anuros. O presente trabalho objetiva determinar a influência
de fatores climáticos (temperatura e umidade) na riqueza e abundância mensal das
espécies em atividade de vocalização, dando ênfase à riqueza e a distribuição espaço-
temporal desses animais nos diversos microhábitats utilizados ao longo de um ciclo
anual, mas principalmente durante a temporada reprodutiva, ao longo da península de
Ajuruteua, Bragança-Pará.
MATERIAL E MÉTODOS
Área de estudo
As amostragens foram realizadas no período de julho/2008 a junho/2009 com
intervalos quinzenais, em três sítios de trabalho localizados nos limites da Reserva
Extrativista Marinha Caeté-Taperaçu, península de Ajuruteua, Bragança - Pará. Os
sítios de trabalho estão inseridos ou localizados muito próximo dos ecossistemas que
formam a paisagem ao longo da península de Ajuruteua (Figura 1).

Figura 1. Mapa da área de estudo mostrando a localização dos sítios de trabalho: 1 = Taici, 2 = Fazenda Salinas e 3 = Praia de Ajuruteua, ao longo da península de Ajuruteua, Bragança
Uma vez que, na península não se encontram ambientes
doce, no presente estudo, foram selecionados cinco pontos amostrais em três sítios de
trabalho.
SÍTIO 1: TAICI (01°02’59,2”S e 046°45’42,6”W)
amostral foi selecionado.
Ponto Amostral 1 –
chuva e quatro açudes abandonados, medindo o maior 80x55x1,5 m e o menor
40x30x1,2 m (Figura 2). O nível mais alto da água nos açudes alcança cerca de 50 cm,
cujo abastecimento é feito somente pela chuva. Os açudes a
herbácea e arbustiva em suas bordas e, quando cheios, crescem em seu interior touceiras
de junco, Eleocharias sp. Esse ponto está localizado nas áreas de manguezal distante
cerca de 500 m da terra-firme, no início da península.
SÍTIO 2: FAZENDA SALINAS (0°55’35,4"S e 46°40’13"W)
foram selecionados três pontos amostrais.
Figura 1. Mapa da área de estudo mostrando a localização dos sítios de trabalho: 1 = Taici, 2 = Fazenda Salinas e 3 = Praia de Ajuruteua, ao longo da península de Ajuruteua, Bragança-PA.
Uma vez que, na península não se encontram ambientes permanentes com água
doce, no presente estudo, foram selecionados cinco pontos amostrais em três sítios de
SÍTIO 1: TAICI (01°02’59,2”S e 046°45’42,6”W) – nesse sítio apenas um ponto
– é caracterizado por campos alagáveis apenas por águas da
chuva e quatro açudes abandonados, medindo o maior 80x55x1,5 m e o menor
40x30x1,2 m (Figura 2). O nível mais alto da água nos açudes alcança cerca de 50 cm,
cujo abastecimento é feito somente pela chuva. Os açudes apresentam vegetação
herbácea e arbustiva em suas bordas e, quando cheios, crescem em seu interior touceiras
sp. Esse ponto está localizado nas áreas de manguezal distante
firme, no início da península.
FAZENDA SALINAS (0°55’35,4"S e 46°40’13"W)
foram selecionados três pontos amostrais.
47
Figura 1. Mapa da área de estudo mostrando a localização dos sítios de trabalho: 1 = Taici, 2 = Fazenda Salinas e 3 = Praia de Ajuruteua, ao longo
permanentes com água
doce, no presente estudo, foram selecionados cinco pontos amostrais em três sítios de
nesse sítio apenas um ponto
r campos alagáveis apenas por águas da
chuva e quatro açudes abandonados, medindo o maior 80x55x1,5 m e o menor
40x30x1,2 m (Figura 2). O nível mais alto da água nos açudes alcança cerca de 50 cm,
presentam vegetação
herbácea e arbustiva em suas bordas e, quando cheios, crescem em seu interior touceiras
sp. Esse ponto está localizado nas áreas de manguezal distante
FAZENDA SALINAS (0°55’35,4"S e 46°40’13"W) – nesse sítio

Ponto Amostral 2 –
sob influência das águas das marés e da chuva. São colonizados predominantem
juncos (Eleocharias sp.) e gramíneas (Figura 3A).
Figura 2 – Campos alagáveis com açudes abandonados no Ponto amostral 1 próximo aos manguezais e áreas de terraAjuruteua, Bragança – Ponto Amostral 3 –
acúmulo de água da chuva. Está localizada na borda de um fragmento de mata de terra
firme e possui, em seu entorno, vegetação arbustiva e arbórea. A margem da lagoa é
composta por uma camada não superior a 20 cm de serrapilheira, enquanto que a porção
em contato com os campos alagáveis é margeada por juncos do gênero
(Figura 3B).
Ponto Amostral 4 –
chuva. O fundo dessas poças é composto por serrapilheira e, em seu entorno, apresenta
vegetação arbustiva e arbórea. As poças estão localizadas no interior do maior
fragmento de terra-firme (cerca de 30 ha), situado entre uma área de manguezal e
manchas de campos alagáveis (F
SÍTIO 3: PRAIA DE AJURUTEUA (00°50’03,6”S e 46°36’05,9”W)
apenas um ponto amostral foi selecionado
Ponto Amostral 5
bordas da restinga próximo à praia de Ajuruteua (Figura
é caracterizado por uma vasta extensão de campos alagáveis,
sob influência das águas das marés e da chuva. São colonizados predominantem
sp.) e gramíneas (Figura 3A).
Campos alagáveis com açudes abandonados no Ponto amostral 1 próximo aos manguezais e áreas de terra-firme no Taici (Sítio 1), península de
– PA. A) campos alagáveis, B) e C) açudes abandonados.
– é uma pequena lagoa medindo 44x38x1,4 m e formada pelo
acúmulo de água da chuva. Está localizada na borda de um fragmento de mata de terra
firme e possui, em seu entorno, vegetação arbustiva e arbórea. A margem da lagoa é
a camada não superior a 20 cm de serrapilheira, enquanto que a porção
em contato com os campos alagáveis é margeada por juncos do gênero
– são poças temporárias formadas pelo acúmulo da água da
s poças é composto por serrapilheira e, em seu entorno, apresenta
vegetação arbustiva e arbórea. As poças estão localizadas no interior do maior
firme (cerca de 30 ha), situado entre uma área de manguezal e
manchas de campos alagáveis (Figura 3C).
SÍTIO 3: PRAIA DE AJURUTEUA (00°50’03,6”S e 46°36’05,9”W)
apenas um ponto amostral foi selecionado.
Ponto Amostral 5 – é caracterizado por poças temporárias a céu aberto nas
bordas da restinga próximo à praia de Ajuruteua (Figura 4).
48
é caracterizado por uma vasta extensão de campos alagáveis,
sob influência das águas das marés e da chuva. São colonizados predominantemente por
Campos alagáveis com açudes abandonados no Ponto amostral 1 firme no Taici (Sítio 1), península de
alagáveis, B) e C) açudes abandonados.
é uma pequena lagoa medindo 44x38x1,4 m e formada pelo
acúmulo de água da chuva. Está localizada na borda de um fragmento de mata de terra-
firme e possui, em seu entorno, vegetação arbustiva e arbórea. A margem da lagoa é
a camada não superior a 20 cm de serrapilheira, enquanto que a porção
em contato com os campos alagáveis é margeada por juncos do gênero Eleocharias
são poças temporárias formadas pelo acúmulo da água da
s poças é composto por serrapilheira e, em seu entorno, apresenta
vegetação arbustiva e arbórea. As poças estão localizadas no interior do maior
firme (cerca de 30 ha), situado entre uma área de manguezal e
SÍTIO 3: PRAIA DE AJURUTEUA (00°50’03,6”S e 46°36’05,9”W) – nesse sítio
é caracterizado por poças temporárias a céu aberto nas

As poças são formadas pelo acúmulo das águas da chuva e apresentam em suas
margens serrapilheira e vegetação herbácea dominada por
interior juncos (Eleocharis
Figura 3 – Pontos amostrais localizados na Fazenda da península de Ajuruteua, Bragança alagáveis, B) Ponto Amostral 3 temporária.
Figura 4 - Poças temporárias em áreas abertas do Ponto amostral 5, localinas bordas da restinga na Praia de Ajuruteua (Sítio 3), península de Ajuruteua, Bragança – PA.
As poças são formadas pelo acúmulo das águas da chuva e apresentam em suas
margens serrapilheira e vegetação herbácea dominada por Sporobolus
Eleocharis sp.) e aguapé.
Pontos amostrais localizados na Fazenda Salinas (Sítio 2), no centro da península de Ajuruteua, Bragança – PA. A) Ponto Amostral 2 alagáveis, B) Ponto Amostral 3 – lagoa e C) Ponto Amostral 4
Poças temporárias em áreas abertas do Ponto amostral 5, localinas bordas da restinga na Praia de Ajuruteua (Sítio 3), península de Ajuruteua,
49
As poças são formadas pelo acúmulo das águas da chuva e apresentam em suas
Sporobolus sp., e no seu
Salinas (Sítio 2), no centro - campos
lagoa e C) Ponto Amostral 4 - poça
Poças temporárias em áreas abertas do Ponto amostral 5, localizadas nas bordas da restinga na Praia de Ajuruteua (Sítio 3), península de Ajuruteua,

50
Procedimento de campo e análise de dados.
As amostragens foram realizadas utilizando-se a técnica de Procura Limitada por
Tempo (PLT) (Heyer et al. 1994, Martins e Oliveira 1999) e através da busca auditiva e
visual (Crump and Scott-Jr 1994). A procura ativa foi de seis horas/observador,
totalizando 1296 horas de atividade de campo. Os espécimes de anuros seguiram a
nomenclatura de Frost (2010). Espécimes-testemunhos foram incorporados à Coleção
de Zoologia do Campus de Bragança (CZB), localizada na cidade de Bragança-PA.
A riqueza de espécies foi estimada através dos métodos não-paramétricos:
Chao1, Chao2, Jackknife1 e Jackknife2, a partir de uma matriz elaborada com valores
de abundância mensal. O valor de abundância de uma dada espécie foi representado
pelo maior número de indivíduos registrados em qualquer uma das duas amostragens
quinzenais realizada por sítio de amostragen a cada mês. A escolha do maior registro
evita a contagem repetida dos indivíduos em cada ponto amostral (adaptado de Condrati
2009). Os resultados foram obtidos com o uso do programa BioDiversity Professional
2.0 (1997).
A similaridade na composição faunística entre os sítios foi obtida através da
análise de Cluster. Essa análise foi gerada a partir de uma matriz de similaridade com
uso do coeficiente de Jaccard. Foram utilizados registros de presença/ausência das
espécies com pelo menos um indício de reprodução nos sítios, como por exemplo,
vocalização, casais em amplexo, desovas, girinos e imagos. Essa análise foi realizada no
programa BioDiversity Professional 2.0 (1997).
Em cada sítio de trabalho um ponto amostral foi selecionado (Sítio 1 - ponto
amostral 1; Sítio 2 – ponto amostral 4 e Sítio 3 – ponto amostral 5), onde foram
coletados os seguintes dados: umidade relativa do ar (Termo-Higrômetro); temperatura
do ar, da água e do solo (Termômetro tipo vareta: TD-100). A relação destas variáveis
com a riqueza e abundância das espécies em atividade de vocalização foi testada através
da Correlação de Spearman, utilizando-se o programa BioEstat 5.0 (Ayres, et al. 2007).
Para caracterizar a distribuição espacial nos sítios de vocalização, foram
analisadas as seguintes variáveis:
a) tipo de substrato:
Tipo 1: Solo – animal registrado no chão úmido sem cobertura vegetal;
Tipo 2: Serrapilheira – animal abrigado ou sobre a serrapilheira;
Tipo 3: Gramíneas – animal registrado abrigado ou sobre touceiras de gramíneas;

51
Tipo 4: Vegetação arbustiva – animal registrado no extrato vertical até 150 cm do
nível do solo;
Tipo 5: Vegetação arbórea – animal registrado no extrato vertical acima de 150 cm
do nível do solo.
Tipo 6: Nível da água – animal parcialmente submerso ou flutuando agarrados à
vegetação aquática;
Tipo 7: Vegetação na Água – animal registrado ocupando estrato vertical acima do
nível da água em vegetação emergente e/ou vegetação marginal.
b) Verticalização no substrato: A distribuição vertical no substrato foi quantificada
pela determinação da altura (cm) em que os machos da família Hylidae vocalizam
no extrato vertical da vegetação, em relação ao nível do solo ou da água.
A relação entre as espécies e o uso vertical do substrato foi estudada apenas no
Campo algado e açudes no Taici. Isto se deve pelo fato desse ponto amostral apresentar
o maior número de espécies. A relação espécie/substrato de vocalização foi evidenciada
por meio de uma análise de correspondência (AC), baseada na frequência com que as
espécies foram registradas nos diferentes tipos de substrato. Esta análise foi realizada no
programa MVSP 3.1. Para verificar a existência de diferenças significativas na
distribuição do uso vertical do substrato foi realizado o teste não-paramétrico de
Kruskal-Wallis, através do pacote estatístico BioEstat 5.0 (Ayres et al. 2007).
RESULTADOS
A península de Ajuruteua abriga 14 espécies de anfíbios anuros, as quais estão
distribuídas em nove gêneros e quatro famílias: Bufonidae (n=1), Hylidae (n=6),
Leptodactylidae (n=5) e Leiuperidae (n=2) (Tabela 1). Os cinco pontos amostrais
estudados apresentaram variação na composição das espécies, sendo a maior riqueza
observada no Campo algado e açudes no Taici (ponto amostral 1), (n=11) e a menor no
Campo alagado na fazenda salinas (ponto amostral 2), (n=4). Os valores dos
estimadores de riqueza foram similares entre os pontos estudados, com exceção dos
estimadores Chao1 para o Campo alagado na fazenda salinas (ponto amostral 2) e
Jacknife2 para as Poças na borda da restinga (ponto amostral 5), onde a riqueza
observada foi de 80 e 83% das espécies esperadas, respectivamente (Tabela 2).
As espécies Leptodactylus macrosternum e Scinax ruber foram registradas em
todos os ambientes amostrados. Outras ocorreram de forma restrita em um único
ambiente. No Campo algado e açudes no Taici, por exemplo, ocorreram quatro espécies
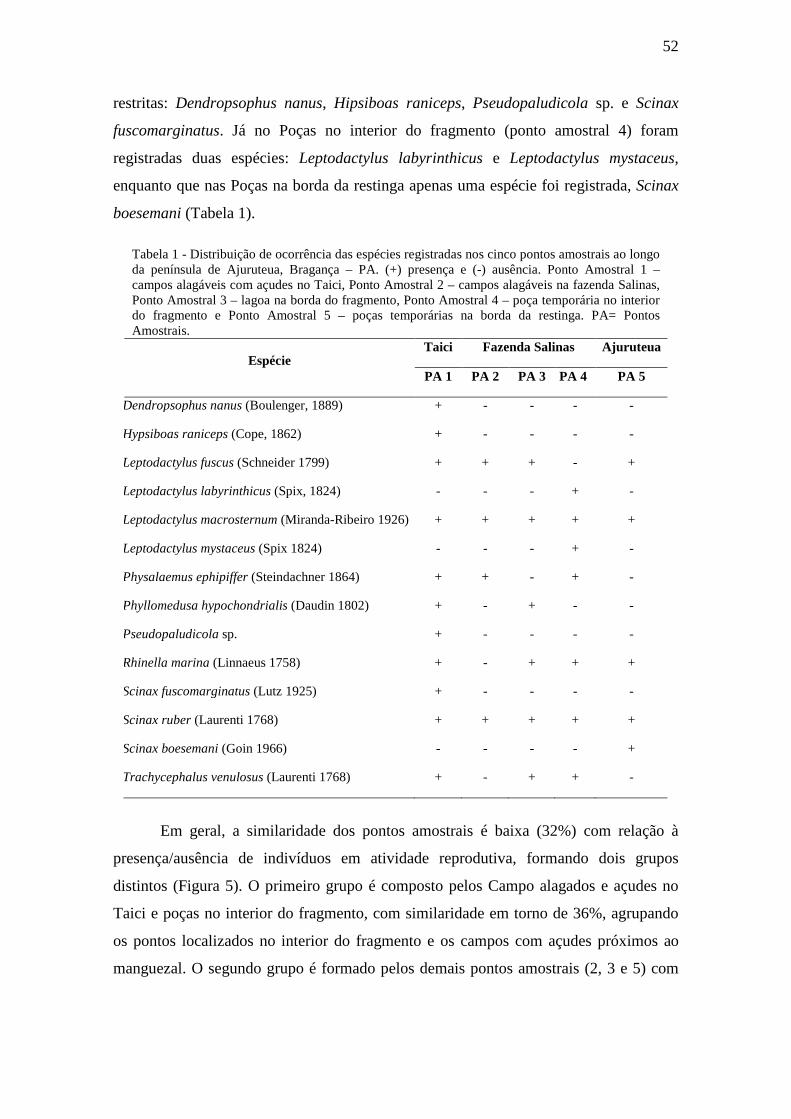
52
restritas: Dendropsophus nanus, Hipsiboas raniceps, Pseudopaludicola sp. e Scinax
fuscomarginatus. Já no Poças no interior do fragmento (ponto amostral 4) foram
registradas duas espécies: Leptodactylus labyrinthicus e Leptodactylus mystaceus,
enquanto que nas Poças na borda da restinga apenas uma espécie foi registrada, Scinax
boesemani (Tabela 1).
Tabela 1 - Distribuição de ocorrência das espécies registradas nos cinco pontos amostrais ao longo da península de Ajuruteua, Bragança – PA. (+) presença e (-) ausência. Ponto Amostral 1 – campos alagáveis com açudes no Taici, Ponto Amostral 2 – campos alagáveis na fazenda Salinas, Ponto Amostral 3 – lagoa na borda do fragmento, Ponto Amostral 4 – poça temporária no interior do fragmento e Ponto Amostral 5 – poças temporárias na borda da restinga. PA= Pontos Amostrais.
Espécie Taici Fazenda Salinas Ajuruteua
PA 1 PA 2 PA 3 PA 4 PA 5
Dendropsophus nanus (Boulenger, 1889) + - - - -
Hypsiboas raniceps (Cope, 1862) + - - - -
Leptodactylus fuscus (Schneider 1799) + + + - +
Leptodactylus labyrinthicus (Spix, 1824) - - - + -
Leptodactylus macrosternum (Miranda-Ribeiro 1926) + + + + +
Leptodactylus mystaceus (Spix 1824) - - - + -
Physalaemus ephipiffer (Steindachner 1864) + + - + -
Phyllomedusa hypochondrialis (Daudin 1802) + - + - -
Pseudopaludicola sp. + - - - -
Rhinella marina (Linnaeus 1758) + - + + +
Scinax fuscomarginatus (Lutz 1925) + - - - -
Scinax ruber (Laurenti 1768) + + + + +
Scinax boesemani (Goin 1966) - - - - +
Trachycephalus venulosus (Laurenti 1768) + - + + -
Em geral, a similaridade dos pontos amostrais é baixa (32%) com relação à
presença/ausência de indivíduos em atividade reprodutiva, formando dois grupos
distintos (Figura 5). O primeiro grupo é composto pelos Campo alagados e açudes no
Taici e poças no interior do fragmento, com similaridade em torno de 36%, agrupando
os pontos localizados no interior do fragmento e os campos com açudes próximos ao
manguezal. O segundo grupo é formado pelos demais pontos amostrais (2, 3 e 5) com

53
similaridade de 50%, agrupando as áreas com características de ambientes abertos,
localizados no interior da península.
Dentre as 14 espécies de anuros registradas no presente estudo, quatro não foram
observadas em atividade de vocalização (L. layirinthicus, L. macrosternum, S.
boesemani e T. venulosus) (Tabela 3). A maioria das espécies vocalizou no período
chuvoso, entre janeiro e junho/2009, apresentando variação o inicio das atividades ao
dos meses de atividade.
Tabela 2 – Valores estimados e observados de riqueza nos cinco pontos amostrais na península de Ajuruteua, Bragança - PA. PA 1 – campos alagáveis com açudes no Taici, PA 2 – campos alagáveis na fazenda Salinas, PA 3 – lagoa na borda do fragmento, PA 4 – poça temporária no interior do fragmento e PA 5 – poças temporárias na borda da restinga.
Estimado
Observado Chao1 Chao2 Jackknife1 Jackknife2
PA 1 11 9 10 10 11
PA 2 4 4 4 5 4
PA 3 7 6 6 5 7
PA 4 9 8 8 8 9
PA 5 6 4 5 5 5
As espécies terrestres apresentaram variação mais acentuada no período de
vocalização. Rhinella marina vocalizou por maior período dentre todas as espécies, com
registro de atividade ainda no período seco, a partir de dezembro e durante todo período
chuvoso. Leptodactylus mystaceus vocalizou por menor período, ficando restrito aos
dois meses no início do período chuvoso, sendo a única espécie ativa durante as chuvas
torrenciais. Physalaemus ephipiffer iniciou suas atividades reprodutivas logo após as
primeiras chuvas no campo alagado e na lagoa na borda da mata e, posteriormente,
ocupou todas as poças no interior do fragmento, substituindo totalmente a espécie L
mystaceus, no período chuvoso a partir da primeira quinzena do mês de março.

54
Figura 5 - Dendrograma mostrando a similaridade entre os cinco pontos
amostrais, com base na presença/ausência de indivíduos das espécies registradas
em pelo menos uma atividade reprodutiva. P.A. 1 – campos alagáveis com
açudes no Taici, P.A. 2 – campos alagáveis na fazenda Salinas, P.A. 3 – lagoa na
borda do fragmento, P.A. 4 – poça temporária no interior do fragmento e P.A. 5
– poças temporárias na borda da restinga.
As espécies P. ephipiffer e Leptodactylus fuscus também apresentaram variação
na sua atividade de vocalização em dias chuvosos ou nublados, iniciando sua
vocalização no final da tarde (das 16:00 às 17:00 hs). As demais espécies iniciaram suas
vocalizações sempre durante a noite ou no crepúsculo. Dentre os hilídeos, P.
hypochondrialis vocalizou por maior período, com registros a partir do fim do período
seco (dezembro) até o fim das amostragens (junho). A espécie S. fuscomarginatus
iniciou sua vocalização mais tardiamente do que as outras espécies, ocorrendo a partir
de março/2009 e mantendo-se em atividade até o fim das amostragens (Tabela 3).
A espécie L. macrosternum, apesar de presente em todos os pontos amostrados,
só apresentou atividade reprodutiva no Campo alagado e açudes no Taici, onde foram
encontrados casais em amplexo no açude no mês de janeiro, início da estação chuvosa,
sendo este evento registrado uma única vez. A atividade reprodutiva para
Trachycephalus venulosus ocorreu apenas pelo registro de dois juvenis. Por outro lado,
S. ruber apresentou atividade reprodutiva em todos os pontos amostrais. A espécie R.
marina não teve registro de atividade reprodutiva somente nas poças no interior do
fragmento. Está espécie teve seus girinos registrados em fevereiro co Campo alagado e
açudes no Taici e juvenis em abril e maio no referido local e na poças temporéris na

55
borda da restinga. Phyllomedusa hypochondrialis se reproduziu em todos ambientes em
que ocorreu (ponto amostrais 1, 3 e 4), com registro de massa com ovos em março nas
poças temporérias no interior do fragmento e imagos nos meses de março e abril na
referida área e no Campo algado e açude no Taici e na lagoa na borda do fragmento.
Tabela 3 – Registro de atividade reprodutiva dos anuros nos pontos amostrais ao longo de uma temporada reprodutiva na península de Ajuruteua, Bragança – PA. a= amplexo, g= girinos, i= imagos, v= vocalização e (-) sem atividade reprodutiva.
Espécies Dez Jan Fev Mar Abr Mai Jun
Dendropsophus nanus - v v v, a v v, i v
Hypsiboas raniceps - v v v v, i v, i v
Leptodactylus fuscus - v v v v v -
Leptodactylus macrosternum - a g - - - -
Leptodactylus mystaceus - v v i - - -
Physalaemus ephipiffer - v v v, a v, a v, i v, i
Phyllomedusa hypochondrialis v v v v, a v, i v, i v
Pseudopaludicola sp. - v v v v, i v v
Rhinella marina v v v, g v v, i v, i v
Scinax fuscomarginatus - - - v v v v
Scinax ruber - v v v v v v
Das três variáveis abióticas (umidade do ar, temperatura do ar e temperatura do
solo), apenas as duas últimas apresentaram correlação significativa com a riqueza de
espécies no Ponto amostral 5, sendo essas correlações negativas (rs= -0,92; t= -7,34;
p<0,01 e rs= -0,86; t= -0,53; p<0,01, respectivamente) (Tabela 4). A umidade do ar foi a
única variável significativamente correlacionada com o número de espécies em
atividade de vocalização nos três Pontos Amostrais, sendo essa correlação negativa no
Ponto Amostral 1 (rs= -0.66, t=-2.76, p<0,05; rs= 0.70, t=3.07, p< 0,01 e rs=0.86, t=5.39,
p< 0,01, nos pontos amostrais 1, 4 e 5, respectivamente). Da mesma forma, a umidade
relativa do ar também apresentou correlação com a riqueza mensal no Ponto Amostral 5
(rs=0,76, t=3,66 e p<0,001) (Tabela 4).

56
Tabela 4 - Relação entre as variáveis abióticas e a riqueza de espécies nos pontos amostrais P.A. 1 – campos alagáveis com açudes no Taici, P.A. 4 – poça temporária no interior do fragmento e P.A. 5 – poças temporárias na borda da restinga, na península de Ajuruteua, Bragança – PA. rs= Correlação de Spearman, A= vocalização, B= riqueza, t = valor do teste t, p = nível de significância e n.s.= não significativo.
Sítios Temperatura do Ar (°C) Temperatura do solo (°C) Umidade do ar (%)
A rs t p rs t p rs t p
PA 1 0,47 1,70 n.s. 0,54 2,01 n.s. -0,66 -2,76 0,05
PA 4 -0,30 -0,98 n.s. -0,40 -1,38 n.s. 0,70 3,07 <0,01
PA 5 -0,53 -1,98 n.s. -0,45 -1,61 n.s. 0,86 5,39 <0,01
B
PA 1 0,32 1,07 n.s. 0,56 2,14 n.s. -0,55 -2,08 n.s.
PA 4 0,09 0,28 n.s. 0,04 0,13 n.s. -0,01 -0,01 n.s.
PA 5 -0,92 -7,34 <0,01 -0,86 -5,23 <0,01 0,76 3,66 <0,01
Na utilização dos substratos para vocalização, a análise de Correspondência
(CA) evidenciou dois grupos distintos de espécies associados aos diferentes substratos
identificados (Figura 6). No eixo 1, com 42,2% da variação dos dados, agruparam-se os
substratos que apresentam estratificação vertical, com a família Hylidae,
exclusivamente, a eles associados. S. fuscomarginatus, D. nanus e H. raniceps
formaram associação, principalmente, na Vegetação na água (Tipo 7). S. ruber e P.
hypochondrialis agruparam Vegetação arbustiva e Vegetação arbórea (Tipo 4 e 5,
respectivamente). O Eixo 2 explicou 20,7% da variação dos dados, agrupando as
espécies comuns aos substratos terrestres (Tipo 1, 2 e 3) e ao nível da água (Tipo 6)
(Figura 5).
As espécies da família Leptodactylidae associaram-se, principalmente, aos
substratos terrestres (Tipo 1 e 2). A espécie L. mystaceus ficou restrita ao substrato
Serrapilheira (Tipo 2). As famílias Leiuperidae (P. ephippifer e Pseudopaludicola sp.) e
Bufonidae (R. marina), além de utilizarem os substratos terrestres, também utilizaram o
substrato no Nível da água (Tipo 6). A morfoespécie Pseudopaludicola sp. foi
comumente encontrada vocalizando em grupos de mais de 20 indivíduos abrigados sob
touceiras de gramíneas (Tipo 3) ou sobre a vegetação típica às margens das poças,
sempre parcialmente submersos (Tipo 6).
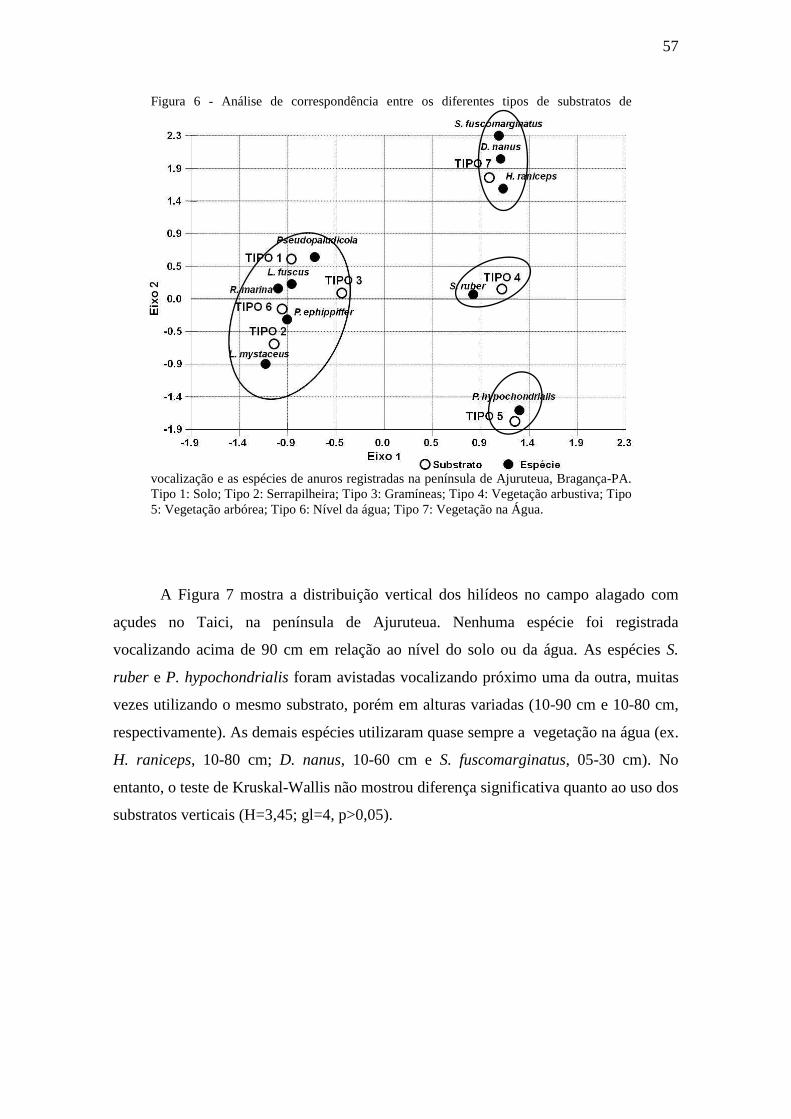
57
Figura 6 - Análise de correspondência entre os diferentes tipos de substratos de
vocalização e as espécies de anuros registradas na península de Ajuruteua, Bragança-PA. Tipo 1: Solo; Tipo 2: Serrapilheira; Tipo 3: Gramíneas; Tipo 4: Vegetação arbustiva; Tipo 5: Vegetação arbórea; Tipo 6: Nível da água; Tipo 7: Vegetação na Água.
A Figura 7 mostra a distribuição vertical dos hilídeos no campo alagado com
açudes no Taici, na península de Ajuruteua. Nenhuma espécie foi registrada
vocalizando acima de 90 cm em relação ao nível do solo ou da água. As espécies S.
ruber e P. hypochondrialis foram avistadas vocalizando próximo uma da outra, muitas
vezes utilizando o mesmo substrato, porém em alturas variadas (10-90 cm e 10-80 cm,
respectivamente). As demais espécies utilizaram quase sempre a vegetação na água (ex.
H. raniceps, 10-80 cm; D. nanus, 10-60 cm e S. fuscomarginatus, 05-30 cm). No
entanto, o teste de Kruskal-Wallis não mostrou diferença significativa quanto ao uso dos
substratos verticais (H=3,45; gl=4, p>0,05).

58
Figura 7 - Distribuição vertical dos hilídeos (cm) no Campo alagado com açudes no Taici (Ponto Amostral 1) na península de Ajuruteua, Bragança-PA. 1- S. fuscomarginatus, 2- D. nanus, 3- H. raniceps, 4- S. ruber e 5- P. hypochondrialis.
.
DISCUSSÃO
Os estimadores de riqueza mostraram que todos os sítios de trabalho foram
suficientemente amostrados, possibilitando a identificação de todas as espécies
estimadas. No entanto, as espécies não se distribuíram homogeneamente entre os pontos
amostrais. A maior riqueza no Campo alagado com açudes no Taici parece ser
decorrente da sua proximidade aos ambientes de terra firme. Tal proximidade possibilita
a entrada de um número maior de espécies devido à menor exposição às condições
salinas dos ambientes tipicamente costeiros, como o manguezal. Como os sítios 2 e 3
são mais distantes da massa continental, o deslocamento dos animais exige no mínimo
10 km ao longo do manguezal. A menor riqueza foi registrada no Ponto Amostral 2
(campo alagado), certamente em função dessa área ressecar e ficar sem cobertura
vegetal no período seco, disponibilizando poucos recursos para a alimentação e proteção
contra a dissecação dos anuros. Além do mais, essa área é constantemente alvo de
queimada nessa época do ano. Os outros pontos amostrais não têm disponibilidade de
água doce durante todo o ciclo anual e não são alagados pelas marés. Em compensação,
possuem cobertura vegetal, disponibilizando diversos microhábitats. Assim, o efeito dos

59
raios solares direto no solo é amenizado, funcionando como refúgio para as espécies que
habitam a parte central da península. De acordo com alguns autores (Heyer 1978,
Grandinetti e Jacobi 2005, Galatti, et al. 2007), a presença de ambientes abertos ou
antropizados é um fator importante a ser considerado para a ocorrência de espécies de
anuros. Na restinga da praia de Ajuruteua, por exemplo, além do ponto amostral
apresentar cobertura vegetal, de maneira adicional, a ação antrópica pode estar
favorecendo as espécies presentes, haja vista algumas espécies lá encontradas serem
comumente encontradas nesses tipos de ambientes.
Em geral, os pontos amostrais ao longo da península parecem disponibilizar
recursos suficientes para as atividades reprodutivas de 11 das 14 espécies registradas no
presente estudo, com exceção de L. labyrinthicus e S. boesemani que apresentaram
baixa densidade e não foram identificadas em atividade reprodutiva.
Hödl (1990), Heyer e Rodrigues (2004) e Galatti et al. (2007) apontam para o
fato de que pode haver exclusividade de espécies em ambientes de reprodução. Tal
comportamento territorial pode explicar a baixa similaridade entre os pontos amostrais
quanto à presença/ausência de espécies em atividade reprodutiva no presente estudo. L.
mystaceus, por exemplo, apresentou exclusividade na utilização de poças temporárias
no interior do fragmento de terra firme, que disponibilizaram serrapilheira em torno das
lagoas temporárias onde os machos constroem bacias para a deposição dos ovos em
ninhos de espuma. No presente estudo, indivíduos de L. mystaceus foram registrados
vocalizando apenas no início da estação chuvosa, durante um curto período e ao longo
de chuvas torrenciais ou após as mesmas. Este comportamento corrobora com a
informação de que essa espécie apresenta pico de vocalização no início da estação
chuvosa (Gascon 1991, Gottsberger e Gruber 2004, Borges e Juliano 2007), mas, por
outro lado, diverge do padrão de reprodução contínua na estação chuvosa, observado na
Amazonia Central, Manaus, por Lima et al. (2006).
Outras espécies também apresentaram exclusividade nos ambientes de
reprodução, como por exemplo: D. nanus, H. raniceps, S. fuscomargintus e
Pseudopaludicola sp. A espécie L. macrosternum, que mesmo estando presente em
todos os ambientes amostrados, só foi observada em atividades reprodutivas no Campo
alagado com açudes no Taici, com padrão reprodutivo explosivo no início do período
chuvoso, confirmando o observado por Dixon e Staton (1976) de que essa espécie
reproduz-se no início da estação chuvosa.

60
As espécies L. fuscus e P. ephipiffer apresentaram período de vocalização
semelhante ao registrado na literatura (Heyer 1978, Martins 1998, Grandinetti e Jacobi
2005, Galatti, et al. 2007, De-Carvalho, et al. 2008). No entanto, no presente estudo
houve variação do inicio da vocalização ao longo de dias chuvosos, ocorrendo ainda
pela parte da tarde. Entre os hilídeos, S. fuscomarginatus apresentou variação no início
das vocalizações divergindo no padrão de reprodução descrita para a espécie (Rossa-
Feres e Jim 2001, Toledo e Haddad 2005). É importante ressaltar que a distribuição e a
reprodução dos anuros tropicais são determinadas pelas taxas de pluviosidade (Crump
1971, 1974, Duellman 1979, Hödl 1990, Zimmerman e Simberloff 1996, Duellman
1999, Hogarth 1999, Gottsberger e Gruber 2004, Ávila-Pires, et al. 2007, Galatti, et al.
2007). No presente estudo a umidade relativa do ar demonstrou ser a variável mais
correlacionada com as atividades reprodutivas (vocalizações) apresentadas pelas
espécies nos diferentes ambientes da península, certamente, por ter a capacidade de
variar de acordo com os microambientes presentes em cada sítio de trabalho ao longo da
península.
Hödl (1977) e Zimmerman e Simberloff (1996) já demonstraram que a as
famílias são relacionadas a determinados tipos de substratos ou a altura que estas são
capazes de ocupar, estando fortemente associada com sua posição filogenética. No
presente estudo, a formação de dois grupos principais na utilização dos substratos
corrobora essas informações. Apenas L. mystaceus apresentou exclusividade na
utilização de substratos de vocalização entre as espécies terrestres, vocalizando apenas
na serrapilheira na margem das poças temporárias no fragmento de terra firme. A
completa substituição na utilização do substrato Serrapilheira, pela espécie P. ephipiffer
pode está relacionada à partição dos recursos disponíveis no ambiente, mas não há
informações suficientes para discutir nem suportar tal hipótese.
A espécie S. ruber esteve associada principalmente aos substratos arbustivos
devido não haver partilha com nenhuma espécie nestes substratos na restinga de
Ajuruteua, onde. Já P. hypochondrialis, que foi associada a Vegetaçõ arbórea (Tipo 5)
devido a frequência em que foram observados na Lagoa na borda da mata e nas Poças
temporárias no interior do fragmento (Pontos amostrais 3 e 4), onde esses substratos
apresentaram maior abundância.
As espécies da família Hylidae usam os substratos verticais em diferentes
alturas. No entanto, a análise estatística não mostrou diferença significativa na
estratificação vertical, evidenciando a sobreposição das espécies na partilha da

61
verticalização dos substratos, confirmando as observações feitas por vários autores que
já demonstraram a sobreposição de espécies no uso de substratos verticais para
atividades de vocalização (Hödl 1977, Heyer, et al. 1990, Gottsberger e Gruber 2004).
Por fim, é relevante ressaltar que as espécies de anuros aqui registradas demonstram ser
capazes de utilizar os recursos disponíveis na região costeira, não só para alimentação,
mas também para a reprodução. Considerando seus diferentes ambientes e suas
condições locais, a disponibilidade de hábitats como a Fazenda Salina pode estar
funcionando como refúgio para as espécies ao longo da península. Sendo que esses
hábitats são de extrema importância na distribuição e na manutenção das taxocenoses de
anuros.
REFERÊNCIAS BIBLIOGRÁFICAS
Ávila-Pires, T. C. S., M. S. Hoogmoed and L. J. Vitt. 2007. Herpetofauna da amazônia.
Pp. 12-43 in L. B. Nascimento and Oliveira, M. E. (eds.), Herpetologia no brasil
II . Belo Horizonte. Sociedade Brasileira de Herpetologia.
Ayres, M., A. J. M, D. L. Ayres and A. S. Santos (eds.). 2007. Bioestat. Versão 5.0.
Belém. Instituto de Desenvolvimento Sustentável Mamirauá, IDSM-MCT-
CNPq. 364 pp.
Bernarde, P. S. 2007. Ambientes e temporada de vocalização da anurofauna no
município de Espigão do Oeste, Rondônia, Sudoeste da Amazônia - Brasil
(Amphibia: Anura). Biota Neotropica 7: 87-92.
Bernarde, P. S. e L. D. Anjos. 1999. Distribuição espacial e temporal da anurofauna no
Parque Estadual Mata dos Godoy, Londrina, Paraná, Brasil (Amphibia: Anura).
Comunicações do Museu de Ciências e Tecnologia, Série Zoologia 12: 127-140.
Borges, F. J. A. e R. D. F. Juliano. 2007. Temporal e spatial distribution in an anuran
community in municipality of Morrinhos, Goiás, Brazil (Amphibia: Anura).
Neotropical Biology e Conservation 2: 21-27.
Condrati, L. H. 2009. Padrões de distribuição e abundância de anuros em áreas ripárias
e não ripárias de floresta de terra firme na reserva biológica do Uatumã
Amazônia central. Instituto Nacional de Pesquisas da Amazônia, Manaus.
Crump, M. L. 1971. Quantitative analysis of ecological distribution of tropical
herpetofauna. Occasional Papers Museum of Natural the University of Kansas
3: 1-62.

62
Crump, M. L. (eds.). 1974. Reproductive strategies in a tropical anuran community.
Kansas. University of Kansas Printing Service. 68 pp.
Crump, M. L. and N. J. Scott-Jr. 1994. Visual encounter surveys. Pp. 84-92 in W. R.
Heyer Donnelly, M. A. Mcdiarmid, R. W. Hayek, L. C. and Foster, M. S. (eds.),
Measuring and monitoring biological diversity: Standard methods for
amphibians. Washington. Smithsonian Institution Press.
De-Carvalho, C. B., E. B. D. Freitas e R. G. Faria1. 2008. História natural de
Leptodactylus mystacinus e Leptodactylus fuscus (Anura: Leptodactylidae) no
Cerrado do Brasil Central. Biota Neotropica 8: 105-115.
Duellman, W. E. (ed.). 1978. The biology of Equatorial Herpetofauna in Amazonian.
Laurence. Miscellaneous Publications Museum of Natural History. 352 pp.
Duellman, W. E. (ed.). 1979. The South American Herpetofauna: Its Origin, Evolution,
e Dispersal. Kansas. Museum of Natural History, The University of Kansas.
485 pp.
Duellman, W. E. 1999. Distribution patterns of amphibians in South America. Pp. 660
in W. E. Duellman (ed.), Patterns of distribution of amphibians. Baltimore. The
Johns Hopkins University Press.
Duellman, W. E. (ed.). 2005. Cusco Amazónico, the lives of amphibian e reptiles in an
Amazonian Rainforest. Ithaca. Cornell University Press. 472 pp.
Frost, D. R. (ed.). 2010. Amphibians species of the world: An online reference. Version
5.4. Url: http://research.amnh.org/herpetology/amphibia/index.php. Captured on
20 April 2010.
Galatti, U. R. A. Estupiñán, A. C. L. Dias e A. E. M. Travassos. 2007. Anfíbios da Área
de Pesquisa Ecológica do Guamá (APEG) e região de Belém. Pp. 456 in J. I.
Gomes Martins-Da-Silva, M. Viana, R. C. e Almeida, S. S. (eds.), Mocambo:
Diversidade e dinâmica biológica da Área de Pesquisa Ecológica do Guamá
(APEG). Belém. MCT/Museu Paraense Emílio Goeldi
Gascon, C. 1991. Population e community level analyses of species occurrences of
Central Amazonian rainforest tadpoles. Ecology 72: 1731-1746.
Giaretta, A. A., M. Menin, K. G. Facure, M. N. D. C. Kokubum e J. C. D. O. Filho.
2008. Species richness, relative abundance, e habitat of reproduction of
terrestrial frogs in the Triângulo Mineiro region, Cerrado biome,
Southeastern Brazil. Iheringia, Série Zoologia 98: 181-188.

63
Gottsberger, B. e E. Gruber. 2004. Temporal partitioning of reproductive activity in a
neotropical anuran community. Journal of Tropical Ecology 20: 271-280.
Grandinetti, L. e C. M. Jacobi. 2005. Distribuição estacional e espacial de uma
taxocenose de anuros (Amphibia) em uma área antropizada em Rio Acima -
MG. Lundiana 6: 21-28.
Heyer, R. W., M. A. Donnelly, R. W. Mcdiarmid, L. C. Hayek e M. S. Foster (eds.).
1994. Measuring e Monitoring Biological Diversity: Standard Methods for
Amphibians. Washington. Smithsonian Institution Press. 364 pp.
Heyer, W. R. 1978. Systematics of the group fuscus of the frog genus Leptodactylus
(Amphibia, Leptodactylidae). Natural History Museum of Los Angeles County,
Science Bulletin 29: 1-85.
Heyer, R. e M. T. Rodrigues (eds.). 2004. Leptodactylus mystaceus. Url:
www.iucnredlist.org. Captured on 21 Julho 2010.
Heyer, W. R., A. S. Rand, C. A. G. Cruz, O. L. Peixoto e C. E. Nelson. 1990. Frogs of
Boracéia. Arquivos de Zoologia, Museu de Zoologia da Universidade de São
Paulo 31: 231-410.
Hödl, W. 1977. Call differences e calling site segregation in anuran species from
Central Amazonian floating meadows. Oecologia 28: 351-363.
Hödl, W. 1990. Reproductive diversity in Amazonian lowland frogs. Fortschritte der
Zoologie 38: 41-60.
Hogarth, P. (ed.). 1999. The Biology of Mangroves. New York. Oxford University
Press. 228 pp.
Lima, A. P., W. E. Magnusson, M. Menin, L. K. Erdtmann, D. J. Rodrigues, C. Keller e
W. Hödl (eds.). 2006. Guia de sapos da reserva Adolpho Rucke, Amazônia
Central = Guide to the frogs of reserva Adolpho Ducke, Central Amazonia.
Manaus. Áttema Design Editorial. 168 pp.
Martins, M. 1998. Biologia reprodutiva de Leptodactylus fuscus em Boa Vista, Roraima
(Amphibia: Anura). Revista Brasileira de Biologia 48: 969-977.
Mendes, A. C. 2005. Geomorfologia e sedimentologia. Pp. 165 in M. E. B. Fernandes
(eds.), Os manguezais da costa norte brasileira. Belém. Petrobrás/Fundação Rio
Bacanga.
Menin, M., F. Waldez e A. P. Lima. 2008. Temporal variation in the abundance e
number of species of frogs in 10,000 ha of a forest in Central Amazônia, Brazil.
South American Journal of Herpetology 3: 68-81.

64
Neckel-Oliveira, S., W. E. Magnusson, A. P. Lima e A. L. K. Albernaz. 2000. Diversity
e distribution of frogs in an Amazonian savanna in Brazil. Amphibia-Reptilia 21:
317-326.
Pombal-Jr, J. P. 1997. Distribuição espacial e temporal de anuros (Amphibia) em uma
poça permanente na Serra de Paranapiacaba, sudeste do Brasil. Revista
Brasileira de Biologia 57: 583-594.
Rossa-Feres, D. C. e J. Jim. 2001. Similaridade do sítio de vocalização em uma
comunidade de anfíbios anuros na região noroeste do estado de São Paulo,
Brasil. Revista Brasileira de Zoologia 18: 439-454.
Souza-Filho, P. W. M. e M. El-Robrini. 1996. Morfologia, processos de sedimentação e
litofácies dos ambientes morfo-sedimentares da planície costeira bragantina,
nordeste do Pará, Brasil. Geonomos 4: 1-16.
Stebbins, R. C. e N. W. Cohen (eds.). 1997. A Natural History of Amphibians. New
Jersey. Princeton University Press. 316 pp.
Toledo, L. F. e C. F. B. Haddad. 2005. Acoustic repertoire e calling site of Scinax
fuscomarginatus (Anura, Hylidae). Journal of Herpetology 39: 455-464.
Tomlinson, P. B. (ed.). 1986. The botany of mangroves. Cambridge. Cambridge
University Press. 419 pp.
Wells, K. D. (ed.). 2007. The Ecology e Behavior of Amphibians. Chicago. University
of Chicago Press. 1148 pp.
Zimmerman, B. L. and R. O. Bierregaard. 1986. Relevance of the equilibrium theory of
island biogeography and species-area ralations to conservatios with a case from
amazonia. Journal Biogeography 13: 133-143.
Zimmerman, B. L. e D. Simberloff. 1996. An historical interpretation of habitat use by
frogs in a Central Amazonian forest. Journal of Biogeography 23: 27-46.
Zug, G. R., L. J. Vitt e J. P. Caldwell (eds.). 2001. Herpetology an Introductory Biology
of Amphibians e Reptiles. Florida. Academic Press. 645 pp.

65
ANEXO 3. REGRAS PARA SUBMISSÃO DO CAPÍTULO 3.
Phyllomedusa - Journal of Neotropical… Instructions to Authors MANUSCRIPT STYLE e FORMAT
Please put sentences in the active voice ("I did it"; "They did it") instead of the passive
voice ("It was done") to make it easy for readers to see who did what. Use the first
person ("I" or "we") to describe what you yourself did. Please double-space the entire
manuscript, including the references, with wide margins. Please use Times New Roman
12. PDF files are not acceptable. The pages of the manuscript should be numbered
consecutively. Manuscripts must include in the following order:
1) Title.
2) The author(s) name(s) with respective institution, e.g.
José Wellington Alves dos Santos1,2 , Roberta Pacheco Damasceno1,2 , Pedro
Luís Bernardo da Rocha2,3
1 Departamento de Zoologia, Instituto de Biociências, Universidade de São Paulo,
05508-900, São Paulo, SP, Brazil. E-mails: [email protected], [email protected].
2 Departamento de Zoologia, Instituto de Biologia, Universidade Federal da Bahia,
40170-210, Salvador, BA, Brazil. E-mail: [email protected].
3 Author for correspondence.
3) Abstract in English (up to 350 words), only for Articles. An additional abstract in
Portuguese (Resumo), Spanish (Resumen), French (Résumé), Italian (Riassunto), or
Deutsch (Zusammenfassung) could be included.
4) Keywords in English e (optional) in one of the languages above cited.
5) Introduction.
6) Materials e Methods (the editor encourages the authors to describe the study area e
include a location map).
7) Results.
8) Discussion.
9) Acknowledgements.
10) References.
11) Tables with their legends.
12) Figures with their legends.
Subtitles (items 5 to 8) should not be used in Short Communications. Please refer to a
recent issue for format details.

66
TABLES
Tables should be numbered consecutively in Arabic numerals e placed after the
References. Very large tables should be placed at the end of the article as Appendixes.
Tables should be formatted with horizontal, but not vertical, rules. In the text, tables
should be referred as Table 1, Tables 2 e 3, Tables 2-5.
FIGURES (INCLUDING COLOR PHOTOGRAPHS)
Figures should be numbered consecutively e placed after the Tables. Please embed them
in the text file. Figures should be referred to in the text as Figure 1, Figures 2 e 3,
Figures 2-5, Figure 4A (not Fig. 1, figure 1, fig. 1, or Figure 4a). After acceptance all
figures must be sent as separate files (not embedded in a text file) e their names should
follow the model: author(s) name(s) Figure X (e.g. Pritchard et al. Figure 1A). High
quality color or black e white photographs, or computer-generated figures (e.g. maps)
are preferable.
NOMENCLATURE
For the first mention of a species, give its complete binomial name; for taxonomic
papers give the authority e date. Scientific names must be shown in italics. Also italicize
words not adopted into the original language of the manuscript. Use SI units, e.g.
Time: "08:16 h", "17:52 h".
Distances e areas: "12 km 2 ", "55 m", "17,840 ha".
Geographic coordinates: "04 o 43'23'' S, 45 o 58'04'' W". Temperatures: 24°C.
CITATIONS
Author citations in the text must follow the pattern: "Vitt (2000)" or "(Wassersug
1975)". For two-author publications, both authors must be cited separated by "and"; e.g.
"Bastos e Haddad (1996)" or "(Zimmerman e Simberloff 1996)". For more than two
authors, only the first one must be cited followed by et al. (in italics); e.g. "Heyer et al.
(1990)" or "(Inger et al. 1986)". Several publications must be cited in chronological
order; e.g. "(Crump 1974, Duellman 1978, 1988, Duellman e Trueb 1986)". Two or
more publications by the same author must be cited as "(Vanzolini 1991, 1992)" or
"(Cadle 1984a, b, c, 1985)". Author names should be separated by "and", not by &.
REFERENCES
In the reference list, the cited publications must be included in alphabetical order in the
following format ("gray literature" must be avoided):

67
Journal articles with usual volume e issue number
Vanzolini, P. E. 1993. A new species of turtle, genus Trachemys, from the state of
Maranhão, Brazil (Testudines, Emydidae). Revista Brasileira de Biologia 55: 111-
125.
Two authors in a journal series
Zamudio, K. R. e H. W. Greene. 1997. Phylogeography of the bushmaster (Lachesis
muta: Viperidae): implications for Neotropical biogeography, systematics, e
conservation. Biological Journal of the Linnean Society 62: 421-442.
More than two authors in a journal series
Hero, J.-M., W. E. Magnusson, C. F. D. Rocha e C. P. Catterall. 2001. Antipredator
defenses influence the distribution of amphibian prey species in the Central Amazon
rain forest. Biotropica 33: 131-141.
Chapter in an edited volume
Hedges, S. B. 1999. Distribution patterns of amphibians in the West Indies. Pp. 211-254
in W. E. Duellman (ed.), Patterns of Distribution of Amphibians - a global
perspective. Baltimore e London. The Johns Hopkins University Press.
Unpublished M.S.c. Dissertation or Ph.D Thesis
Verdade, V. K. 2001. Revisão das espécies de Colostethus Cope, 1866 da Mata
Atlântica (Anura, Dendrobatidae). Unpublished M.Sc. Dissertation. Universidade de
São Paulo, Brazil.
Book
McDiarmid, R. W. e R. Altig (eds.). 1999. Tadpoles - the biology of anuran larvae.
Chicago e London. The University of Chicago Press. 633 pp.
Material from the World Wide Web
Frost, D. R. (ed.). 2000. Amphibian Species of the World - an online reference. URL:
http://reaserch.amnh.org/herpetology/amphibia/index.html. Captured on 23
December 2002.

68
CAPÍTULO 4
DISCUSSÃO GERAL

69
A riqueza da herpetofauna nos ambientes costeiros da Amazônia registrada no
presente trabalho representa densidade baixa em relação a áreas florestadas na
Amazônia. O manguezal é o ambiente florestado mais abundante na costa amazônica
brasileira e tem sua relevância para várias espécies de répteis e anuros, demonstrado que
seu papel para essa fauna associada é bastante subestimado.
Os diversos ambientes costeiros não só disponibilizam microhábitats para
alimentação da herpetofauna, mas também são capazes de gerar condições para a
reprodução de diversas espécies de anuros e répteis. A disponibilidade destes
microhábitats é um dos principais fatores que influência a distribuição e a manutenção
das taxocenoses de anuros e de répteis locais, tornando-se determinante na estrutura das
comunidades desses grupos (Hödl, 1977; Zimmerman e Simberloff, 1996; Neckel-
Oliveira et al., 2000; Grandinetti e Jacobi, 2005).
A grande maioria da herpetofauna associada ao manguezal na península de
Ajuruteua é são possívelmente visitantes. Contudo, ao menos uma espécie de lagarto
completa todo seu ciclo de vida nesse sistema, o lagarto Gonatodes humeralis. Desta
espécie já foi registrada a presença de fêmeas grávida, ovos inteiros e eclodidos em
cascas e troncos podres de árvores de mangue, além da presença de juvenis, indicando
não só a reprodução dessa espécie no manguezal, mas seu desenvolvimento. Segundo
Ávila-Pires (1995), essa espécie reproduz-se durante o ano inteiro. Miranda e Andrade
(2003) complementam essas informações com as observações de que fêmeas grávidas
ocorrem em maior número no período chuvoso no leste da Amazônia.
A utilização dos substratos para forrageio e o comportamento de fuga da espécie
no manguezal é similar aos descritos na literatura (Ávila-Pires, 1995; Vitt et al., 2000;
Miranda e Andrade, 2003; Miranda et al., 2010), sendo que Fernandes et al. (2009) já
observou esta espécie em comportamento de fuga mergulhando na água salobra do
manguezal e submergindo 10-15 cm a frente, um comportamento que ilustra uma
característica comportamental adaptativa a esse ambiente. No entanto, estudos mais
específicos com G. humeralis em florestas de mangue são necessários para esclarecer
suas adaptações a esse ambiente, até então inóspito para a maioria dos répteis.
Outro exemplo notável que fornece indícios da adaptabilidade da herpetofauna
na utilização de recursos costeiros é o anuro Leptodactylus macrosternum. Foi
identificado na dieta dessa espécie um exemplar de Pachygrapsus gracilis (Saussure,
1858), caranguejo típico dos manguezais bragantinos e três exemplares de pequenos
caranguejos do gênero Uca, além de vários exemplares de ácaros, coleópteros e

70
formigas como principais itens na sua dieta. No manguezal do Espírito Santo, Ferreira e
Tonini (2010) relataram que a espécie Leptodactylus latrans (Linnaeus 1758) alimenta-
se do caranguejo Uca, bem como faz uso das galerias de caranguejo abandonadas. Este
comportamento alimentar já foi relatado em manguezais da Ásia, onde a espécie
Fejervarya cancrivora (Gravenhorst 1829) tem sua dieta baseada em caranguejo (Wells,
2007). A utilização de uma ampla variedade de hábitats e recursos alimentares por L.
macrosternum trata-se de uma estratégia vantajosa, pois diminui a competição por
recursos com outras espécies.
Estudos focados em anuros típicos de ambientes salinos [ex. Fejervarya
cancrivora (Gravenhorst, 1829), Buergeria japonica (Hallowell, 1861) e Pseudepidalea
viridis (Laurenti, 1768), demonstram suas adaptações fisiológicas e osmorregulação,
sua elevada concentração plasmática de sódio, cloreto e uréia, além de uma diminuição
na taxa de produção de urina. Em F. cancrivora, a uréia corresponde a
aproximadamente 60% da elevação de concentração plasmática. Esta espécie
movimenta-se entre os ambientes salinos e doces, exigindo uma reposta relativamente
rápida a essas mudanças de concentração. Tais mudanças são controladas pela atuação
de hormônios, os quais regulam a produção de uréia e, de maneira complementar a
baixa produção de urina, sendo esses mecanismos similares na maioria das espécies
(Gordon et al., 1961; Gordon e Tucker, 1968; Dicker e Elliott, 1970; Haramura, 2007,
Wells 2007). Por outro lado, não existem informações acerca dos mecanismos
fisiológicos das espécies de anuros da Amazônia brasiliera que estão associados aos
ambientes salinos. Assim, faz-se necessário desenvolver estudos com enfoque
fisiológico para subsidiar discussões e conclusões sobre a capacidade de tolerância à
salinidade dessas espécies de anuros.
Os répteis, por sua vez, possuem diversas espécies que já desenvolveram
adaptações fisiológicas para esses ambientes, com espécies exclusivamente marinhas
(ex. serpentes, crocodilos e tartarugas), as quais desenvolveram capacidade de
eliminação de sal através de glândulas (Zug et al., 2001; Bambaradeniya et al., 2002;
Karns et al., 2002; Hansknecht, 2008; Kathiresan e Bingham, 2008). Espécies como
Amblyrhynchus cristatus (Bell, 1825) e Brachylophus fasciatus (Brongniart, 1800)
comumente expelem líquidos com sais pelas narinas, que são secretados
presumivelmente pelas glândulas nasais (Burghardt e Rand, 1983). Esses mesmos

71
autores relatam um comportamento similar para espécies do gênero Conolophus e para
três espécies de iguanídeos do gênero Sauromalus.
Na área de estudo do presente trabalho, a principal ameaça às comunidades de
herpetofauna é a perda de hábitat ou a descaracterização dos mesmos pelas atividades
antrópicas, como, por exemplo, as queimadas que ocorrem todos os anos nos campos
alagados, ameaçando também os fragmentos de terra firme no centro da península. Tais
fragmentos são de fundamental importância para a conservação das espécies de répteis e
anfíbios da península, especialmente os anuros para os quais esses fragmentos de mata
funcionam como refúgios. Na praia de Ajuruteua, as construções oriundas dos
assentamentos desordenados são a maior ameaça para a herpetofauna, visto que essas
construções têm, ao longo das décadas, avançado sobre a restinga e o manguezal.
Os resultados obtidos no presente estudo ressaltam alguns aspectos importantes
da história natural da herpetofauna, mas principalmente revela a necessidade de
preencher lacunas e a importância de se compreender melhor a composição e a
dinâmica populacional das assembléias de répteis e anfíbios nos ambientes costeiros da
Amazônia brasileira. É evidente que esses sistemas litorâneos têm sido pouco
estudados. Tal condição mostra a necessidade de se revelar os mecanismos responsáveis
pela sobrevivência das espécies dessa fauna sob condições adversas, em uma região
onde as forçantes ambientais aceleram a dinâmica costeira e as populações humanas
tradicionais vivenciam a corrida para o desenvolvimento urbano, reduzindo as áreas de
ocorrência desses animais.
REFERÊNCIAS BIBLIOGRÁFICAS
ÁVILA-PIRES, T. C. S. Lizards of Brazilian Amazônia (Reptilia: Squamata).
Zoologische Verhandelingen v. 1-706, p. 1-706, 1995.
BAMBARADENIYA, C. N. B. et al. An Assessment of the Status o f Biodiversity in
the Maduganga Mangrove Estuary. Occasional Papers of IUCN Sri Lanka v. 1, p. 1-49,
2002.
BURGHARDT, G. M.; RAND, A. S. Iguanas of the world: Their Behavior, Ecology,
and Conservation. New Jersey: Noyes Publications, 1983.

72
DICKER, S. E.; ELLIOTT, A. B. Water uptake by the crab-eating frog rana cancrivora,
as affected by osmotic gradients and by neurohypophysial hormones. Journal
Physiological v. 287, p. 119-132, 1970.
FERNANDES, M. E. B. et al. Gonatodes Humeralis (NCH) Habitat Occurrence;
Escape Behavior. Herpetological Review v. 40, n. 2, p. 221-222, 2009.
FERREIRA, R. B.; TONINI, J. F. R. Living holed: Leptodactylus latrans occupying
crabs’ burrows. Herpetology Notes v. 3, n. 237-238, 2010.
GORDON, M. S. et al. Osmotic regulation in the crab-eating frog (Rana cancrivora).
Journal Experimental Biology v. 38, p. 659-678, 1961.
GORDON, M. S.; TUCKER, V. A. Further Observations on the physiology of salinity
adaptation in the Crab-eating frog (Rana Cancrivora). Journal Experimental Biology v.
49, p. 185-193, 1968.
GRANDINETTI, L.; JACOBI, C. M. Distribuição estacional e espacial de uma
taxocenose de anuros (Amphibia) em uma área antropizada em Rio Acima - MG.
Lundiana v. 6, n. 1, p. 21-28, 2005.
HANSKNECHT, K. A. Lingual Luring by Mangrove Saltmarsh Snakes (Nerodia
clarkii compressicauda). Journal of Herpetology v. 42, n. 1, p. 9-15, 2008.
HARAMURA, T. Salinity Tolerance of Eggs of Buergeria japonica (Amphibia, Anura)
Inhabiting Coastal Areas. Zoological Science v. 24, p. 820-823, 2007.
HÖDL, W. Call differences and calling site segregation in anuran species from Central
Amazonian floating meadows. Oecologia v. 28, p. 351-363, 1977.
KARNS, D. R. et al. Ecology of oriental-australian rear-fanged water snakes
(colubridae: homalopsinae) in the pasir ris park mangrove forest, Singapore. The
Raffles Bulletin of Zoology v. 50, n. 2, p. 487-498, 2002.
KATHIRESAN, K.; BINGHAM, B. L. Biology of mangroves and mangrove
ecosystems. Advance in Marine Biology v. 40, p. 81-251, 2008.
LUTHER, D. A.; GREENBERG, R. Mangroves: A Global perspective on the evolution
and conservation of their terrestrial vertebrates. Bioscience v. 59, n. 7, p. 602-612,
2009.
MIRANDA, J. P.; ANDRADE, G. V. Seasonality in Diet, Perch Use, and Reproduction
of the Gecko Gonatodes humeralis from Eastern Brazilian Amazon. Journal of
Herpetology v. 37, n. 2, p. 433-438, 2003.

73
MIRANDA, J. P. et al. Influence of structural habitat use on the thermal ecoly of
Gonatodes humeralis (Squamata: Gekkonidae) from a transitional forst in Maranhão,
Brazil. Zoologia v. 27, n. 1, p. 35-39, 2010.
NECKEL-OLIVEIRA, S. et al. Diversity and distribution of frogs in an Amazonian
savanna in Brazil. Amphibia-Reptilia v. 21, n. 3, p. 317-326, 2000.
ROCHA, C. F. D. et al. Endemic and threatened tetrapods in the Restingas of the
biodiversity corridors of Serra do Mar end of the central in Mata Atlântica in eastern
Brazil. Brazilian Journal of Biology v. 65, n. 1, p. 159-168, 2005.
VITT, L. J. et al. Comparative Ecology of Sympatric Gonatodes (Squamata:
Gekkonidae) in the Western Amazon of Brazil. Copeia v. 2000, n. 1, p. 83-95, 2000.
WELLS, K. D. The Ecology and Behavior of Amphibians. Chicago: University of
Chicago Press, 2007.
ZIMMERMAN, B. L.; SIMBERLOFF, D. An historical interpretation of habitat use by
frogs in a Central Amazonian Forest. Journal of Biogeography v. 23, p. 27-46, 1996.
ZUG, G. R. et al. Herpetology an Intoductory biology of Amphibians and Reptiles.
Florida: Academic Press, 2001.