homeostase e ambiente interno
TRANSCRIPT

1
Homeostase e Ambiente Interno
O conceito de Homeostase tem muito valor para o
estudo da fisiologia porque permite perceber o “porquê” e o
“como”, dos diversos mecanismos reguladores. O conceito
de Homeostase também pode ser uma boa forma de
diagnóstico médico. Em particular medidas do ambiente
interno, como as quantidades de substâncias dissolvidas no
sangue, onde desvios significantes em relação aos valores
normais, mostram que o Homeostase não foi mantida e que
essa pessoa se encontra doente. A quantificação dessas
medidas, combinadas com as observações clínicas,
permitem identificar defeitos particulares do mecanismo.
Para manter o meio interno constante, o corpo possui
diversos sensores para detectar alterações do “set point”. O
“set point” funciona como um termóstato para a temperatura
das nossas casas. Do mesmo modo, há um “set point” para
temperatura, concentração de glicose no sangue, tensão dos
tendões, etc. Quando o sensor detecta alterações nos valores
padrão, envia informação para o Sistema integrado, que
recebe informação dos diferentes sensores. O Sistema
integrado é uma região particular do cérebro ou da espinal
medula, mas em alguns casos pode ser um grupo
de células em glândulas endócrinas. As forças
relativas dos diferentes sensores são pesadas no
sistema integrado e ele responde através do
aumento ou diminuição da actividade de
efectores particulares, geralmente músculos ou
glândulas.
Se a temperatura do corpo exceder os 37ºC,
os sensores contidos no encéfalo vão detectar
isso e vão activar o sistema integrado,
estimulando a actividade dos efectores para
baixar a temperatura. Os agentes efectores
“defendem” os pontos de regulação dos desvios.
Quando a actividade dos efectores é influenciada
pelos efeitos que eles produzem, essa regulação dá-se no sentido negativo, ou reverso –
Feedback Negativo.
A natureza do desvio feedback negativo pode ser entendido através da analogia
com o termóstato e o ar condicionado. Quando o ar condicionado está ligado durante
algum tempo num quarto, a temperatura baixa consideravelmente. Quando isso
acontece, o ar condicionado vai desligar-se. O efector (ar condicionado) vai ligar
quando a temperatura estiver alta, o que vai fazer a temperatura baixar (produz
alterações negativas) que vão levar à desactivação do efector (ar condicionado desliga-
se). Assim é mantida uma certa constância na temperatura.
Fig. 1 – Ambiente interno constante
– Homeostase.
Fig. 2 – Detecção pelos sensores,
processamento da informação e resposta. –
Feedback negativo.

2
É importante que esse feedback negativo
seja um processo contínuo. Então, uma fibra
nervosa que faz parte dum mecanismo efector
pode sempre mostrar alguma actividade, e uma
hormona particular, que faz parte de outro
mecanismo efector, pode estar sempre presente
no sangue. A actividade do nervo e a
concentração da hormona podem diminuir em
resposta a alterações no ambiente interno numa
direcção (fig. 2), ou podem aumentar em
resposta a alterações em direcção oposta (fig.
3). Alteração ao normal para qualquer direcção
é compensada por alterações reversas na
actividade efectora.
A curva de feedback negativo
responde após variações ao “set
point” terem estimulado os
sensores, o ambiente interno nunca
é absolutamente constante. A
Homeostase concebeu o estado de
Constância Dinâmica, essas
condições são estabilizadas acima ou abaixo do “set point”. Essas condições podem ser
medidas quantitativamente: em graus Célsius para temperatura corporal; ou em
miligramas por decilitro para glicose no sangue. O “set point” pode ser tomado como
sendo o ponto médio das medições (fig. 4).
A concentração sanguínea de glicose, cálcio e outras substâncias é regulada por
um feedback negativo, envolvendo hormonas que promovem os efeitos opostos. Com a
insulina, por exemplo, quando os níveis de glicose sanguíneos estão baixos, outras
hormonas vão aumentar a glicemia. Do mesmo modo, o batimento cardíaco é
controlado por nervos fibrilares que produzem os efeitos contrários: a estimulação de
um grupo de nervos fibrilares aumenta o ritmo cardíaco; a estimulação de outro grupo
diminui o ritmo.
Feedback positivo
A manutenção do ambiente interno é mantida pelas acções dos agentes efectores
para compensar as alterações que servem como estímulo de activação, através do
feedback negativo. O oposto ocorre durante o feedback positivo, neste caso, a acção
dos efectores amplifica a alteração que os estimulou. Por exemplo, um termóstato que
trabalhe com um feedback positivo diminui ainda mais a temperatura, em resposta a
uma diminuição da temperatura.
Obviamente, a homeostase deve ser mantida preferencialmente por controlo
negativo e não por feedback positivo. No entanto, a eficácia de alguns mecanismos de
feedback negativo está ligada a mecanismos de feedback positivo para amplificar a
acção das suas respostas. A coagulação sanguínea, por exemplo, ocorre como resultado
de uma activação sequencial de factores de coagulação; a activação de um factor de
activação leva à activação de muitos, numa cascata de feedback positivo. Nesta via, uma
simples alteração é amplificada para formar um coágulo sanguíneo. No entanto,
formação do coágulo previne a perda de sangue, assim representa um complemento do
feedback negativo para restaurar a homeostase.
Fig. 3 – Detecção pelos efectores, diminuição do
estímulo.
Fig. 4 – Curva de feedback negativo mantêm o estado de
constância dinâmica, no ambiente interno.

3
Regulação Neurológica e Endócrina
A homeostase é mantida através de dois mecanismos reguladores: intrínsecos,
para a regulação dos órgãos; e extrínsecos, onde os órgãos são regulados por nervos e
pelo sistema endócrino. O sistema endócrino combina-se com o sistema nervoso na
regulação e integração das funções do corpo e manutenção da homeostase. O sistema
nervoso controla a secreção de muitas glândulas endócrinas, e algumas hormonas vão
afectar a função do sistema nervoso. Juntos, o sistema nervoso e o sistema endócrino,
regulam a actividade de muitos outros sistemas do corpo.
A regulação pelo sistema endócrino é activada pela secreção de reguladores
químicos, chamados hormonas. As hormonas são secretadas no sangue e percorrem
todo o corpo, passando por todos os órgãos. No entanto, apenas órgãos específicos vão
responder a uma hormona particular, são os chamados órgãos alvo da hormona.
As fibras nervosas que regulam os órgãos estão nos respectivos órgãos. Quando
estimuladas, essas fibras produzem impulsos nervosos electroquímicos que são
conduzidos desde a origem da fibra até ao seu fim, no órgão alvo enervado pela fibra.
Esses órgãos alvo podem ser músculos ou glândulas que têm funções na manutenção da
homeostase.
Controlo Feedback da Secreção de
Hormonas
As hormonas são secretadas em resposta a
um estímulo químico específico. O aumento na
concentração de glicose no plasma, por exemplo,
estimula o pâncreas a libertar insulina. As
hormonas são libertadas, apenas, em resposta a
uma estimulação nervosa e a uma estimulação por
outras hormonas.
A secreção de hormonas pode ser inibida
pelos seus efeitos, numa forma de feedback
negativo. A insulina libertada em resposta aos
níveis altos de glicose no sangue vai provocar uma
diminuição de glicose. Nesta situação há uma
inibição da secreção. Este sistema de controlo
fechado é chamado inibição por feedback
negativo.
As hormonas que regulam a manutenção da homeostase podem dividir-se em
grupos químicos:
Derivados de aminoácidos: por exemplo a
tiroxina, é produzida pela tiróide e deriva da
tirosina. Tem 3 ou 4 iodos incorporados, dai
se poder designar por T3 e T4,
respectivamente.
Proteínas: a insulina, sintetizada no pâncreas, deriva
da pró-insulina e é constituída por duas cadeias péptidicas.
Fig. 5 – Controlo por feedback negativo da
secreção de insulina e concentração de
glicose no sangue.

4
Esteróides, que são derivadas do colesterol.
São exemplos de homonas esteróides: a
progesterona e o estradiol (hormonas sexuais
femininas); cortisol (hormona animal); e a
testosterona (hormona sexual masculina).
Mecanismo de acção hormonal
As hormonas esteróides e tiroxinas permeiam a membrana plasmática e têm
receptores membranares, capazes de produzir mensageiros secundários intracelulares.
Por exemplo: a activação do receptor leva à produção cAMP, Ca2+
ou IP3; o sistema de
tirosina-cinase não envolve propriamente segundos mensageiros.
O efeito final da acção de algumas hormonas é
a activação da transcrição de genes, levando à síntese
de novas proteínas que não estavam a ser sintetizadas
nas células, anteriormente.
Algumas hormonas esteróides são moléculas
apolares que são transportadas no sangue com a
ajuda de proteínas transportadoras. Quando chega à
célula alvo, desliga da proteína cargo e passa pela
membrana até chegar ao seu receptor intracelular,
ligando-se a este. O complexo hormona-receptor
migra para o
núcleo, onde se
liga a um
determinado
local do DNA,
promovendo a
transcrição desse gene.
Nas hormonas sintetizadas na tiróide, o
mecanismo é o mesmo, mas os receptores
encontram-se no núcleo, e não livres no citoplasma.
Fig. 6 – Mecanismo de acção das
hormonas esteróides nas células alvo.
Fig. 7 – Mecanismo de acção das
hormonas tiróides nas células alvo.

5
Mas há hormonas que actuam através da acção de mensageiros secundários, por
exemplo:
A adrenalina, produzida nas
células do fígado, vai ligar-se
a um receptor que se encontra
na membrana plasmática.
Quando se liga ao receptor,
este vai activar um 2º
mensageiro, o cAMP. Este
mecanismo engloba a
activação de uma proteína G
com 3 subunidades (α, β e γ),
onde a subunidade α se
dissocia das restantes e vai
activar a enzima adenilato
ciclase, que catalisa a
produção de cAMP. Este vai
activar uma proteína cinase
(ligando-se à subunidade inibidora), que por sua vez, vai activar proteínas que
estavam inactivas na célula, fosforilando as enzimas que as vão activar ou
inibir.
Nestes casos diz-se que a hormona é um mensageiro primário.
Fosfolipase C: a hormona liga-se ao
receptor membranar, que através da
activação de uma proteína G, vai
activar a fosfolipase C (PLC), que por
sua vez vai activar o IP3. Este vai
estimular a libertação de Ca2+
do
retículo sarcoplasmático, através da
abertura de canais iónicos, aumentando
a concentração de Ca2+
, o que vai
causar a respostas na célula.
Os mensageiros secundários
intracelulares são o Ca2+
e o IP3.
A insulina liga-se a dois
receptores e promove a
dimerização deles. O dímero
forma-se no lado
citoplasmático e funciona
como cinase de resíduos de
tirosina. Ocorre então uma
auto-fosforilação e aumentam
os níveis de resíduos que
podem funcionar como sinal e
provocar uma cascata de
reacções.
Fig. 8 – Mecanismo de acção de uma hormona activadora de
cAMP.
Fig. 9 – Mecanismo de acção da fosfolipase C.
Fig. 10 – Receptor da insulina.

6
Hipófise: é uma pequena
glândula endócrina inserida no
hipotálamo, na base do encéfalo. É
geralmente dividida em duas partes: a
anterior e a posterior. O lóbulo
anterior secreta ACTH, TSH, FSH
LH, hormonas de crescimento e
prolactina. O posterior liberta
oxitocina e uma hormona anti-
diurética (ADH), que são produzidas
pelo hipotálamo.
Lóbulo posterior da hipófise:
neurónios com corpos celulares no
hipotálamo com axónios muito
compridos para o lóbulo da hipófise.
Aqui vai libertar duas hormonas:
Oxitocina – hormona
libertada no parto;
ADM – hormona anti-
diurética.
Lóbulo anterior da hipófise:
Neurónios com axónios mais curtos e
com terminais nervosos no pedúnculo
da glândula endócrina. Há substâncias
libertadas pelos neurónios do
hipotálamo que vão estimular a
produção de hormonas no lóbulo
anterior.
Fig. 11 - Hipófise
Fig. 12 – Lóbulo posterior da hipófise
Fig. 13 – Lóbulo anterior da hipófise

7
Fig. 14 – Organização do Sistema Nervoso.
O Sistema Nervoso (SN)
No sistema nervoso há detecção de estímulos que irão originar uma resposta, que é
processada pelo SNC e desencadeada através de agentes efectores – os músculos.
O SNC é constituído pelo encéfalo e pela espinal-medula, enquanto que o SNP é
constituído pelos nervos e respectivos gânglios.
O SNP pode dividir-se em duas vias:
Aferente (Sensorial) – que conduz a
informação desde os receptores para o
SNC.
Eferente – transporta a resposta para os
órgãos efectores: músculos ou
glândulas. É composto pelo:
o SN Somático – controla os músculos
esqueléticos.
o SN Autónomo – que controla
músculos lisos, cardíaco e glândulas.
Ainda se pode dividir em SN
Simpático e SN parassimpático, que
normalmente têm efeitos
antagónicos (contrários)
Por vezes considera-se também a
existência do SN Entérico, que existe no
tubo digestivo e que pode funcionar de
maneira autónoma. No entanto não vai ser
mencionado.
O SN é composto por Neurónios e por
Células da Glia. O bom funcionamento depende de
ambas as unidades.
Os Neurónios:
Apresentam tamanhos muito variados;
Podem ou não ter axónio;
Dendrites grandes ou pequenas.
A informação é recebida pelas dendrites, passa pelo
corpo celular e segue pelo axónio antes de ser
transferida para a célula seguinte.
Há muitos tipos de células da Glia:
Fig. 15 – Neurónios.
Fig. 16 – Diversidade de Células da Glia.

8
As Células de Schwann e os Oligodentrócitos formam a banda de mealina em
volta do axónio dos neurónios.
Os Astrócitos:
Dão suporte ao SNC;
Ajudam a formar a barreira protectora;
Liberta factores neurotróficos (estímulos
necessários para a sobrevivência das células
nervosas);
Captam neurotransmissores libertados pelos
neurónios (sabe-se hoje que o inverso também
acontece).
A Microglia tem função de defesa: metaboliza
substâncias agressivas para o SNC.
As Células Epidérmicas revestem espaços
internos do SNC.
As células do SN funcionam por impulsos
eléctricos que as percorrem.
Se colocarmos um eléctrodo dentro do axónio e outro de fora, verifica-se que há
diferença de potencial (geralmente, -70 mV) ou seja, no interior há mais cargas
negativas.
A membrana do axónio permite que se gere corrente eléctrica, uma vez que é
impermeável a iões.
Numa experiência com duas concentrações de
KCl, separadas por uma membrana permeável
apenas a cargas positivas. No princípio verifica-se
que o sistema não tem potencial, mas depois, pelo
gradiente químico, as moléculas de K+ passam pela
membrana. Essa transferência é compensada pelo
gradiente eléctrico, que vai gerar diferença de
potencial.
Potencial em equilíbrio de um ião (Equação
de Nernst):
O potencial de membrana é formado pelo transporte de iões através da membrana
plasmática. Isso pode acontecer por difusão simples ou por bombas de electrões.
Fig. 18 – Experiência com KCl.
Formação de potencial
Fig. 19 – Formação de potencial de membrana nos axónios.
Fig. 17 – Função das Células da Glia no
SNC.

9
Os iões mais importantes para a determinação de potenciais são: K+, Na
+ e Cl
-.
A Equação de Goldman mede a diferença de potencial:
P – permeabilidade da membrana ao ião
Tem em conta os três iões principais. No entanto, como a permeabilidade
da membrana para o Cl- é muito baixa, pode-se excluir a ultima parte da
equação (PCl[Cl-]int / PCl[Cl
-]ext).
As variações na permeabilidade da membrana para o Na+
são devidas à
abertura de canais iónicos de Na
+.
Geralmente, PK = 40 PNa.
A isto é chamado Potencial de Repouso, o que significa que a membrana não está
a ser estimulada (≈ -70 mV).
Por exemplo, se fizermos variar o PK ou o PNa, o potencial de membrana varia.
Assim como com a temperatura e com a concentração. A alteração do potencial de
membrana por alteração da permeabilidade é causada por canais eléctricos.
Quando há recepção de um estímulo que
excita as células, só o sentimos quando ele
chega ao nosso SNC.
As células do SN são excitáveis, ou seja,
quando há uma excitação causada por um
estímulo, há uma despolarização causada por
diminuição das cargas negativas do potencial de
membrana. No final há uma repolarização,
voltando aos valores normais de potencial de
membrana. A hiperpolarização acontece quando
há um aumento de cargas negativas.
Se tivermos 3 estímulos de diferentes
intensidades: quanto maior a intensidade desse
estímulo, maior é a despolarização. Se chegar ao
potencial limiar de despolarização, entra em
Potencial de Acção.
Fig. 21 – Alteração do potencial de
membrana, com estímulos diferentes.
Fig. 20 – Alterações do potencial de membrana.

10
Há vários tipos de receptores de estímulos:
Mecanoreceptores, quando, depois da
recepção de um estímulo, se abrem os canais
iónicos, por distensão da membrana,
permitindo o transporte de Na+, o que gera
despolarização do potencial de membrana.
Os paraplégicos também captam estímulos, mas estes não chegam ao SNC,
logo não se apercebem deles.
Quimioreceptores são específicos para
determinadas moléculas. Essa molécula é captada pelo
receptor e vai abrir o canal iónico, dando-se a
despolarização.
Fotoreceptores, funcionam ao contrário dos
outros receptores, abrem quando não há
estímulo. Ou seja, quando não há luz o canal
iónico está aberto, quando há luz o canal
iónico fecha. O estímulo gera uma
repolarização.
Para a Transdução Sensorial é necessário que haja estímulos e receptores.
Quando os receptores são activados há uma alteração da permeabilidade, que vai fazer
alterar o potencial de membrana, que pode gerar potencial de acção quando atinge o
potencial limiar.
Os Potenciais Graduais dependem da
intensidade do estímulo e diminuem a sua intensidade
à medida que se afasta do local onde foi captado.
Enquanto que os Potenciais de
Acção têm sempre a mesma
intensidade e não dependem da
intensidade do estímulo.
A despolarização, embora seja um processo rápido, demora algum tempo a
percorrer o neurónio.
Os potenciais de acção são gerados na zona de emparelhamento do axónio (zona
de iniciação ou zona de iniciação do axónio).
Fig. 22 – Potencias Graduais
Fig. 23 – Potenciais graduais vs. Potenciais de acção

11
Nos potenciais graduais podem gerar potencial de acção, que vai passar todo o
axónio. Isso depende apenas da intensidade do estímulo, tem de atingir o potencial
limiar.
O potencial de acção só se dá no axónio
porque aí os canais são sensíveis à voltagem,
enquanto que nas dendrites e no corpo celular
os canais são sensíveis a estímulos (químicos e
mecânicos).
Nos canais sensíveis à voltagem há uma
cancela de activação que controla a passagem
de substâncias. A abertura da cancela é
activada por um sensor de voltagem.
Enquanto que o canal de potássio tem uma cancela, o de sódio tem duas:
activação e inactivação:
No início, é necessário um
estímulo eléctrico que vai fazer com
que a membrana saia do repouso. 1Esse estímulo vai ser reconhecido
pelo sensor de voltagem que vai
influenciar a cancela de activação que
por sua vez vai activar a cancela que
inactivação (a cancela de inactivação
depende indirectamente da voltagem,
ao contrário da cancela de activação
que depende directamente) – fase de
rastreio. 2Posteriormente, a cancela de
activação também é activada,
deixando que o sódio entre na célula,
gerando despolarização. 3Quando a
cancela de inactivação inactiva, o sódio não pode entrar na célula e, como o potássio
está a sair, há uma repolarização. 4A cancela de activação inactiva (fecha). No entanto,
como ainda há saída de potássio para o espaço extracelular, porque a cancela do
potássio é mais lenta a inactivar, o que vai aumentar o número de cargas negativas na
célula, criando uma hiperpolarização.
Numa célula há vários canais de sódio
e de potássio e nem todos fecham ao mesmo
tempo. Isto leva a que a permeabilidade aos
iões aumente progressivamente até ser
máxima (é nessa altura que se atinge o maior
valor de potencial de acção). Depois, a
inactivação dos canais de sódio também é
gradual.
A partir de um certo momento, as
cancelas de inactivação inactivam,
terminando o ciclo.
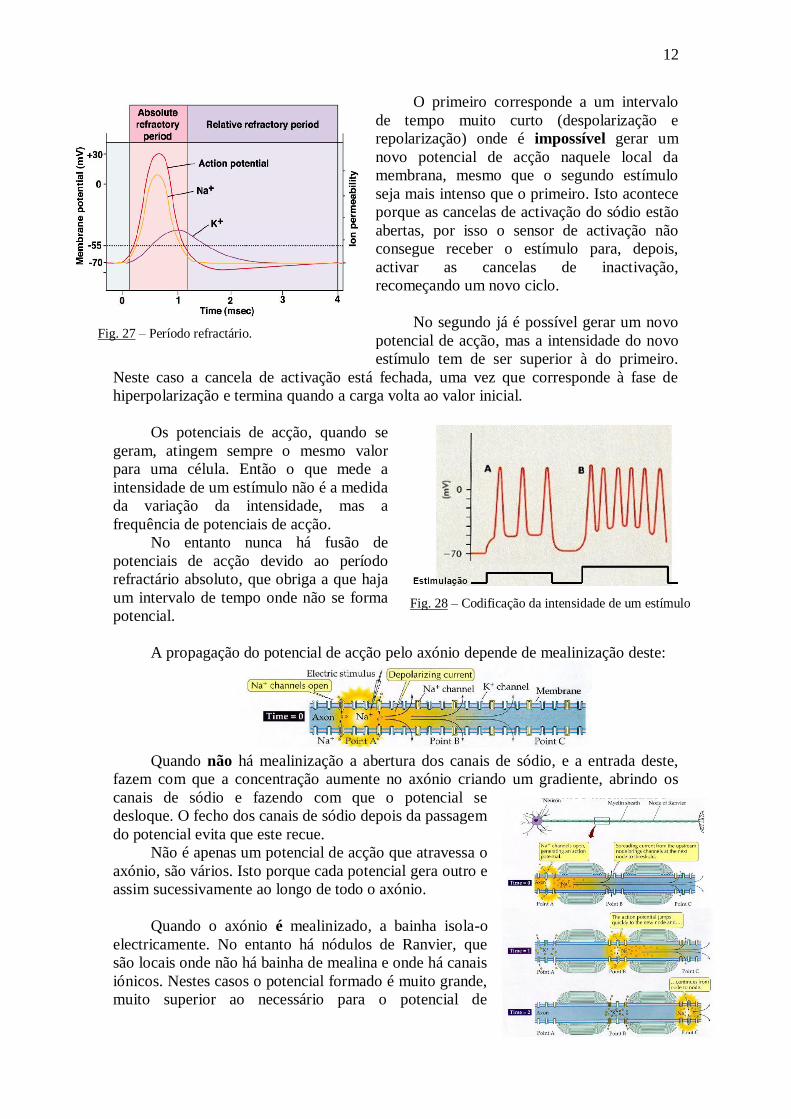
Há dois tipos de período refractário: o
absoluto e o relativo.
Fig. 24 – Canal iónico sensível à voltagem
Fig. 25 – Regulação das cancelas dos canais iónicos.
Fig. 26 – Movimento dos iões durante o potencial
de acção.
12
O primeiro corresponde a um intervalo
de tempo muito curto (despolarização e
repolarização) onde é impossível gerar um
novo potencial de acção naquele local da
membrana, mesmo que o segundo estímulo
seja mais intenso que o primeiro. Isto acontece
porque as cancelas de activação do sódio estão
abertas, por isso o sensor de activação não
consegue receber o estímulo para, depois,
activar as cancelas de inactivação,
recomeçando um novo ciclo.
No segundo já é possível gerar um novo
potencial de acção, mas a intensidade do novo
estímulo tem de ser superior à do primeiro.
Neste caso a cancela de activação está fechada, uma vez que corresponde à fase de
hiperpolarização e termina quando a carga volta ao valor inicial.
Os potenciais de acção, quando se
geram, atingem sempre o mesmo valor
para uma célula. Então o que mede a
intensidade de um estímulo não é a medida
da variação da intensidade, mas a
frequência de potenciais de acção.
No entanto nunca há fusão de
potenciais de acção devido ao período
refractário absoluto, que obriga a que haja
um intervalo de tempo onde não se forma
potencial.
A propagação do potencial de acção pelo axónio depende de mealinização deste:
Quando não há mealinização a abertura dos canais de sódio, e a entrada deste,
fazem com que a concentração aumente no axónio criando um gradiente, abrindo os
canais de sódio e fazendo com que o potencial se
desloque. O fecho dos canais de sódio depois da passagem
do potencial evita que este recue.
Não é apenas um potencial de acção que atravessa o
axónio, são vários. Isto porque cada potencial gera outro e
assim sucessivamente ao longo de todo o axónio.
Quando o axónio é mealinizado, a bainha isola-o
electricamente. No entanto há nódulos de Ranvier, que
são locais onde não há bainha de mealina e onde há canais
iónicos. Nestes casos o potencial formado é muito grande,
muito superior ao necessário para o potencial de
Fig. 27 – Período refractário.
Fig. 28 – Codificação da intensidade de um estímulo

13
despolarização atingir o nódulo seguinte. O potencial de acção não percorre todo o
axónio, a condução é saltatória.
A velocidade de passagem de um estímulo depende do diâmetro do axónio e da
existência de bainha de mealina.
Há situações onde a bainha de mealina é destruída, por exemplo na esclerose
múltipla. Nestes casos a corrente de despolarização pode perder-se na região de entre-
nódulos, fazendo diminuir a velocidade do potencial e, em casos extremos, pode
acontecer que a corrente que chega ao nódulo não seja necessária para gerar um novo
potencial, fazendo com que o estímulo se perca.
A informação dos estímulos tem de passar por várias células para poder chegar
SNC: pode haver uma rede neuronal, em que o neurónio recebe informação de vários
neurónios e/ou passa a informação para vários neurónios; ou então, pode haver um canal
único de neurónios, em que cada neurónio contacta apenas com outro neurónio e assim
sucessivamente.
Até ao princípio do século XX, pensava-se que a informação passava de neurónio
em neurónio apenas por sinapses eléctricas, onde as cargas eléctricas, sob a forma de
potenciais de acção, podem passar directamente de uma célula para outra através de
gap-junctions.
Mas em 1921, Loewi, fez uma experiência com dois
corações colocados em tinas diferentes, mas com contacto de
liquido banhante. Ambos os corações batiam a um determinado
ritmo cardíaco. Loewi estimulou o nervo vargus do coração 1,
que provocou uma diminuição do ritmo cardíaco deste.
No entanto, verificou-se que o ritmo do segundo coração
também diminuiu aquando da estimulação do nervo vargus do
coração 1. Como os corações apenas estavam a contactar
através do líquido banhante, concluiu-se que a informação do
estímulo passou através de mensageiros químicos para o
coração 2 e fez com
que este também
diminuísse o ritmo
cardíaco.
A este tipo de passagem de informação
chama-se sinapse química: quando o estímulo
chega à extremidade do axónio de um neurónio,
fundem-se com a membrana e são
libertados os neurotransmissores para a
fenda sináptica. Aí os neurotransmissores
vão ligar aos canais de sódio que estão
nas dendrites do neurónio seguinte,
activando-os. Com a entrada de sódio na
célula, gera-se uma despolarização
formando um novo potencial.
Fig. 29 – Experiência de Loewi.
Fig. 30 – Sinapse química vs Sinapse eléctrica
Tabela 1: Características das sinapses: eléctrica e química.
14
Nos terminais nervosos há canais de Ca2+
sensíveis à voltagem. Quando um
potencial eléctrico chega ao terminal vai activar os canais, fazendo aumentar a
concentração de cálcio. Este vai ligar-se a proteínas exocitóticas sensíveis ao Ca2+
,
activando-as, o que vai induzir a exocitose.
Após a abertura dos canais iónicos da célula seguinte, é necessário baixar os
níveis de neurotransmissores, sendo inactivados: degradados ou reabsorvidos pelo
sangue, células da Glia ou novamente pelo terminal nervoso.
Há muitos tipos
diferentes de
neurotransmissores, e estes
podem ter várias naturezas
químicas: podem ser
aminoácidos, aminas,
proteínas, etc.
A desregulação dos
neurotransmissores pode
originar doenças, como por
exemplo: Parkinson.
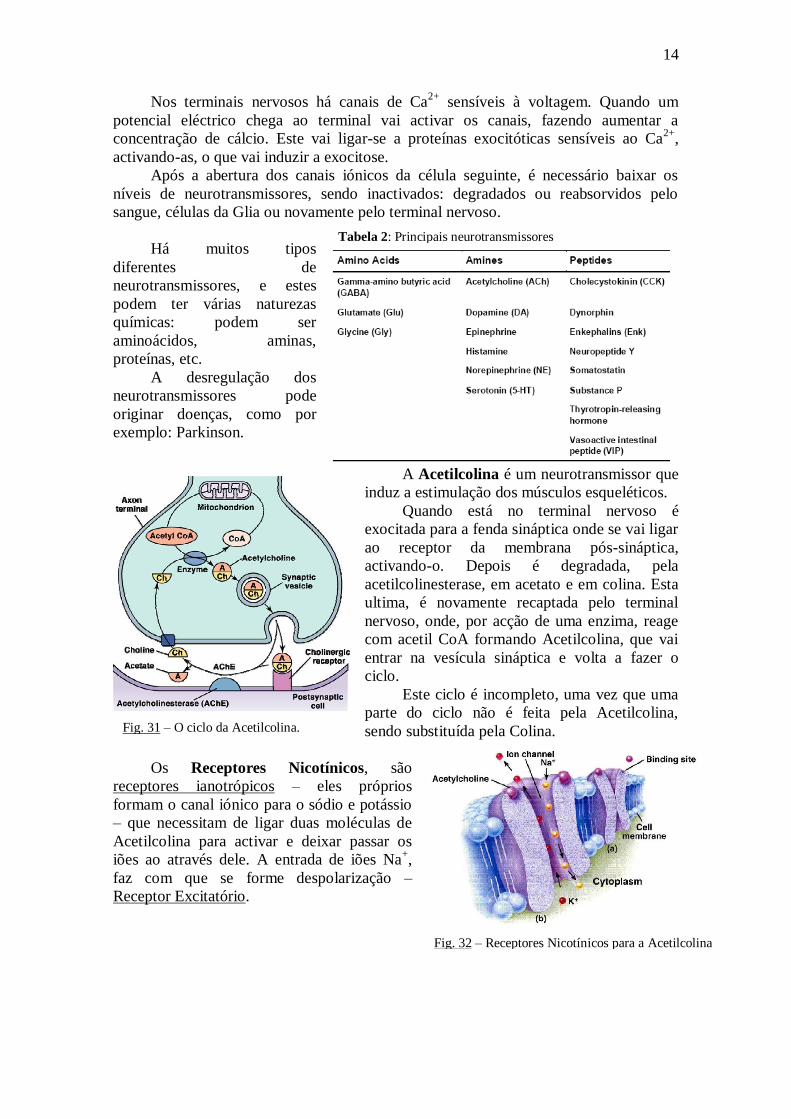
A Acetilcolina é um neurotransmissor que
induz a estimulação dos músculos esqueléticos.
Quando está no terminal nervoso é
exocitada para a fenda sináptica onde se vai ligar
ao receptor da membrana pós-sináptica,
activando-o. Depois é degradada, pela
acetilcolinesterase, em acetato e em colina. Esta
ultima, é novamente recaptada pelo terminal
nervoso, onde, por acção de uma enzima, reage
com acetil CoA formando Acetilcolina, que vai
entrar na vesícula sináptica e volta a fazer o
ciclo.
Este ciclo é incompleto, uma vez que uma
parte do ciclo não é feita pela Acetilcolina,
sendo substituída pela Colina.
Os Receptores Nicotínicos, são
receptores ianotrópicos – eles próprios
formam o canal iónico para o sódio e potássio
– que necessitam de ligar duas moléculas de
Acetilcolina para activar e deixar passar os
iões ao através dele. A entrada de iões Na+,
faz com que se forme despolarização –
Receptor Excitatório.
Tabela 2: Principais neurotransmissores
Fig. 31 – O ciclo da Acetilcolina.
Fig. 32 – Receptores Nicotínicos para a Acetilcolina

15
Os Receptores Muscarínicos ligam
apenas uma molécula de Acetilcolina. São
Receptores Metabotrópicos, uma vez que
quando a Acetilcolina se liga ao receptor, este
vai interagir com uma proteína G que por sua
vez vai activar os canais de potássio,
causando hiperpolarização – Receptor
Inibitório.
O facto de um receptor ser excitatório ou inibitório, não quer dizer que cause
apenas despolarização ou hiperpolarização, respectivamente.
Há substâncias, como a nicotina ou a muscarina, que quando se ligam a um
receptor conseguem desencadear a mesma resposta que a Acetilcolina. A estas
substâncias chama-se substâncias agónicas. Pelo contrário, as substâncias
antagónicas (como o curare e a atropina) ligam-se ao receptor e bloqueiam a sua acção.
É muito fácil afectar a transmissão nervosa e quer nós humanos, como os restantes
animais usam substâncias para regular o sistema nervoso (quer seja o seu ou o de outro
animal, predador ou presa). Por exemplo:
A toxina do botulismo (botax) é usada para reduzir as rugas de
expressão. Esta toxina bloqueia a exocitose da acetilcolina, por isso não
permite a contracção dos músculos que causam as rugas.
O curare é uma substância retirada da raiz das árvores e muito usada
pelos Índios para imobilizar as suas presas. Esta toxina bloqueia os
receptores para a acetilcolina, impedindo a contracção muscular, logo as
presas não se mexem.
α-bungarotoxina é uma neurotóxica existente no veneno das cobras, que
vai bloquear os receptores para a acetilcolina das suas presas, causando-
lhes imobilização.
A tetradotoxina existe no fugu (peixe muito comido no Japão), que vai
inibir os canais de sódio dependentes da voltagem nos indivíduos que o
ingerirem. Se os canais de sódio são bloqueados, os estímulos não podem
ser transportados, logo esse indivíduo fica paralisado e morre.
O gás dos nervos foi muito utilizado durante a 2ª Guerra Mundial. Esse
gás inibe a acetilcolinesterase não se permitindo a degradação da
acetilcolina, o que causa uma hiperestimulação, ou seja, os músculos não
consegue relaxar.
Nos neurotransmissores pode haver uma despolarização ou uma hiperpolarização
pós-sináptica, causada pelo aumento da permeabilidade ao Na+ e ao K
+ ou ao Cl
-,
respectivamente.
Fig. 33 – Receptor Muscarínico para a Acetilcolina.

16
Quando há uma rede neuronal com diferentes informações, temos de somar os
estímulos que são próximos temporalmente e só depois é que podemos se há ou não
formação de potencial de acção.
Fig. 34 – Simulação de potenciais pós-sinápticos.

17
Encéfalo Humano
Há vários mecanismos que protegem o
encéfalo de agressões externas: crânio e meninges.
Há três meninges: dura mater, aracnóide e pia
mater;
O líquido cefalorraquidiano é produzido
nos ventrículos – quatro cavidades existentes no
interior do encéfalo: duas laterais e duas centrais –
e vai envolver todo o encéfalo. Tem mais ou
menos a mesma função que o sangue, uma vez que
liberta para o encéfalo: glicose, oxigénio e
vitaminas e absorve produtos de excreção.
Funciona como almofada para os choques, uma
vez que absorve a energia deste.
É um líquido límpido, tanto que só a sua
observação é, desde logo, um diagnostico para
doenças, como meningite.
Fig. 35 – Constituição do encéfalo humano
Fig. 36 – Mecanismos de protecção:
A – Encéfalo; B- Medula Espinal.
A
B

18
Está sempre a ser produzido e absorvido, por isso se houver perda de líquido, por
exemplo numa anestesia epidoral, ele é rapidamente recuperado.
O facto de o encéfalo estar envolto em líquido retira-lhe algum do seu peso, caso
contrário poderia ser uma estrutura muito pesada em cima da coluna, o que poderia
facilmente causar lesões.
No encéfalo, os capilares estão revestidos por asterócitos e têm uma barreira
hematoencefálica que dificulta a troca de
substâncias. O que pode ser uma vantagem, por
exemplo quando há substâncias tóxicas em
circulação, uma vez que as células nervosas não
toleram grandes alterações no ambiente.
Mas há zonas em que estas barreiras
representam uma desvantagem, e em que ela não
está presente, como é o caso do lóbulo posterior da
hipófise e o centro de vómito, porque precisam de
saber se há determinadas substâncias em circulação
no organismo para reagirem.
Os doentes de Parkinson têm défice de
dopamina, mas esta não pode ser utilizada nos
tratamentos uma vez que não consegue atravessar a
barreira hematoencefálica. Por isso usa-se um precursor da dopamina (L-DOPA) que
consegue atravessar a barreira e, uma vez no encéfalo, vai formar dopamina.
O encéfalo está dividido em 2 hemisférios
separados por uma fenda. Cada hemisfério está
dividido em lóbulos. A comunicação entre os dois
hemisférios dá-se através do corpo caloso, onde
passam fibras
O facto de ser meio enrugado serve para
aumentar a superfície do córtex cerebral.
Há matéria de duas cores no
encéfalo: cinzenta e branca. A diferença
deve-se à existência ou não de mealina a
envolver os axónios.
Na parte mais periférica existe o córtex
cerebral, que é onde existe: as partes sensoriais
específicas (como: visual, gustativo e auditivo);
partes motoras primárias, que enviam informação
para os músculos para induzir o movimento; há
outra parte motora que é responsável pelos
movimentos mais complexos; as áreas de
associação existem no lóbulo frontal.
Fig. 37 – Barreira hematoencefálica
Fig. 38 – Corte transversal do encéfalo
Fig. 39 – Divisões do córtex cerebral.
Fig. 40 – Transmissão e processamento da
informação sensorial no sistema nervoso.

19
Fig. 41 – Divisões do córtex sensorial e motor.
Fig. 42 – Lateralização celular
Um estímulo só é sentido, e transformado em sensação, quando chega á zona
somatossensorial. Antes disso se houver reacção esta é espontânea.
O córtex sensorial está dividido por partes
do corpo. Uma zona é tanto maior quanto mais
terminais nervosos essa zona tiver.
Com o córtex motor passa-se a mesma
coisa, mas o tamanho da zona depende do número
de músculos que a correspondente parte do corpo
tem.
Sistema Nervoso Periférico Estímulos diferentes, mesmo que sejam captados na mesma zona, tomam vias
aferentes diferentes e vão ser processados em diferentes zonas do encéfalo. Nas vias
aferentes, a informação segue diferentes vias, consoante o órgão de origem.
O SN somático e o SN autónomo apresentam várias diferenças para a contracção
do músculo esquelético, a via necessita de passar por um neurónio que liberta
acetilcolina.
O SN autónomo necessita que haja dois
neurónios a libertar neurotransmissores: o neurónio
pré-ganglico e o neurónio pós-ganglico, separados
por um gânglio autónomo. Neste caso há
estimulação de um neurónio e inibição de outro.
Fig. 43 – SN somático e SN autónomo.

20
Os músculos estão constantemente a ser estimulados. A diferença está na
frequência de potenciais de acção gerados no neurónio. Se houver um aumento de
frequência de potenciais de acção dá-se a contracção muscular; quando a frequência de
potenciais diminui causa relaxamento muscular.
O SN Simpático tem uma função de
estimulação dos músculos, enquanto que o SN
Parassimpático inibe os músculos.
No SN Simpático há um neurónio pré-
ganglico curto e um pós-ganglico longo, ao
contrário do que acontece no SN Parassimpático.
Além disso saem de zonas diferentes da espinal-
medula.
Também há diferenças nos
neurotransmissores usados: a via Parassimpática é
feita por acetilcolina, enquanto que a via
Simpática é feita por acetilcolina no gânglio e por
epinefrina na célula alvo.
Há situações em que estes dois SN têm
funções semelhantes, por exemplo na produção de
saliva: ambos estimulam a produção de saliva,
mas com diferentes viscosidades.
Podem ainda ter funções de cooperativas.
Por exemplo no pâncreas o parassimpático
estimula a libertação de enzimas e o simpático
activa a contracção do intestino.
Há centros de controlo situados ao nível do SNC, onde partem as informações, de
locais diferentes consoante o estímulo. Por exemplo a estimulação e inibição do ritmo
cardíaco partem de locais diferentes, próximos, mas diferentes.
O Sistema Límbico é constituído por várias zonas do SNC, todas muito primárias,
e controla muitas das nossas emoções e comportamentos. O facto de haver poucas
ligações entre o sistema límbico e o córtex faz com
que haja comportamentos que não controlamos, como
comer, beber e sexo.
Estes comportamentos dão-nos prazer e
queremos repeti-los, porque eles vão activar o sistema
de recompensa, ou sistema mesolímbico
dopaminérgico que faz parte do sistema límbico. É
criada um estímulo na zona VTA que projecta o
estímulo dopaminérgico para o nucleus acumbens e
que depois migra para o córtex.
Este é o sistema que utiliza a dopamina como
neurotransmissor e funciona com a libertação deste.
Numa experiência com ratinho com a estimulação eléctrica de zona VTA ou
nucleus acumbens, ele vai repetir esse comportamento, por ele lhe dá prazer. Deixa
mesmo de comer e beber para estar constantemente a premir a alavanca que fornece o
estímulo.
Fig. 44 – SN Simpático e SN Parassimpático
Fig. 45 – Circuito de recompensa

21
Toxicodependência A cocaína vai actuar na zona de activação do circuito
de recompensa, inibindo a reabsorção de dopamina, o que
vai activar o circuito de recompensa.
O problema é que, um indivíduo sob acção desta
droga vai sofrer a uma redução da actividade do encéfalo.
Nos indivíduos viciados há uma redução, bastante drástica
das zonas activas, se ele estiver 10 dias sem consumir a
droga. Passados 100 dias sem consumo, há uma ligeira
recuperação, mas pode
nunca ficar com os
níveis normais.
Os Opiáceos (como a heroína, a morfina e a codeína) são substâncias derivadas
da papoila do ópio que actuam ao nível dos receptores de opiáceos. O nosso corpo
produz endomorfinas – opiáceos endogéneos.
A libertação de dopamina está refreada pelos GABA-receptores, que, na presença
de GABA libertado por outras células, vão ser activados.
Na presença de opiáceos, estes vão ser activados por morfina e vão ligar-se aos
seus receptores que, por sua vez irão inibir a
libertação de GABA. Por isso não vai haver
controlo na libertação de dopamina, havendo
uma libertação excessiva.
O consumo de opiáceos origina sensações
diferentes das reais, uma vez que actua, também, no
córtex sensorial, o que vai fazer com que as
sensações recebidas sejam deturpadas.
Os opiáceos podem ainda influenciar ou alterar
alguns factores de transcrição, o que vai alterar a
expressão de genes e síntese proteica.
Fig. 46 – Locais de acção da cocaína.
Fig. 47 – Mecanismos de acção da cocaína.
Fig. 48 – Efeitos da cocaína.
Fig. 49 – Acção dos Opiáceos.
Fig. 50 – Locais de acção dos Opiáceos.

22
Há diferenças entre os termos habituação ou tolerância, dependência e
viciação.
No primeiro o organismo deixa de responder à droga, necessitando de uma maior
quantidade para obter o mesmo efeito. A tolerância pode ocorrer por uma “down-
regulation” dos receptores para a substância, ou por um aumento das enzimas que
degradam essas substâncias. No segundo o organismo funciona bem na presença da
droga, caso contrário entra em distúrbios de abstinência: dores, comportamentos
violentos, etc. No terceiro há uma procura compulsiva da droga para causar boas
sensações. Normalmente a viciação surge antes da dependência.
O Ecstasy interfere com a seretonina, bloqueando os seus receptores, impedindo
que seja recaptada.
Actua ao nível do hipotálamo (redução
de apetite), redução de sensações, etc. Mas tem
muitos factores prejudiciais, como a
hipertermia, que causa desidratação exagerada,
o que facilita a ocorrência de arritmias e
insuficiências renais.
No dia seguinte há toma do
ecstasy, há pouca seretonina disponível
para ser libertada pelos terminais
nervosos, uma vez que não foi recaptada
por causa da acção da droga
anteriormente.
A longo prazo afecta
a existência de seretonina,
e de dopamina, nos
neurónios, causando danos
irremediáveis e até pode
causar Parkinson.
Fig. 51 – Mecanismos de acção do ecstasy.
Fig. 53 – Efeitos a curto prazo do Ecstasy.
Fig. 52 – Efeitos do ecstasy.
Fig. 54 – Níveis de transportadores
de seretonina.
Fig. 55 – Níveis de dopamina nos
neurónios depois do consumo de ecstasy.

23
Sentidos
Há sentidos especiais e sentidos somáticos:
Nos primeiros, há órgãos de captação específicos,
enquanto que, os segundos podem ser captados em todo
o corpo. Também é possível captar estímulos internos
do nosso corpo.
Para gerar uma sensação é necessário que seja
captado um estímulo que
forme potenciais de acção,
que depois são enviados
para uma determinada zona
do córtex e só aí tomamos consciência desse estímulo e
desencadeamos uma resposta. Se o estímulo não tiver
intensidade suficiente para formar potenciais de acção, a
sua intensidade vai perder-se progressivamente e não chega
ao córtex, logo não tomamos consciência dele.
Há casos em que podem ocorrer erros durante o
processamento ao nível do córtex.
Pode ainda acontecer que a sensação seja gerada por várias zonas do córtex.
Tacto Em toda a nossa pele há receptores sensoriais
(uns mais à superfície que outros). Os receptores mais
à superfície são activados por impulsos mais pequenos
que os receptores mais profundos, onde é necessário
que o impulso seja maior.
Por vezes basta o mexer de um pêlo para
haver uma sensação, isto porque este tem
muitos terminais nervosos na raiz. Este
mecanismo serve para detectar qualquer
movimento nas proximidades.
Os receptores cutâneos captam
diferenças de temperatura e não a temperatura
em si.
Os estímulos são processados no lado
oposto onde foram captados. Esta troca dá-se
já a nível superior.
Pode haver duas vias de transporte de um estímulo para o córtex: pela espinal-
medula ou pela via antero-lateral.
Fig. 57 – Mecanismo das sensações.
Fig. 58 – Receptores sensoriais cutâneos.
Fig. 59 – Transmissão e processamento da
informação sensorial no sistema nervoso
Fig. 56 - Sentidos

24
Há mecanismos que permitem determinar o local exacto onde um estímulo foi
recebido:
Por Inibição Lateral: Um estímulo pode
ser recebido por vários neurónios que
libertam neurotransmissores de segunda
ordem, mas o neurónio que recebe a maior
parte do estímulo liberta mais
neurotransmissores. Por sua vez o
neurónio de segunda ordem liberta
neurotransmissores inibidores que vão
inibir os outros neurónios. O efeito
inibitório dos outros neurónios também
existe, mas como são menos estimulados,
esse efeito inibitório é muito menor do que o do neurónio que foi mais
estimulado, não se fazendo sentir.
Por Campos Receptores: Quanto mais terminações
nervosas existirem numa zona, a sensação gerada é maior, do que
quando há apenas um único neurónio que ocupa uma área maior.
O que acontece é que dois estimulos recebidos em locais muito
próximos, podem ser sentidos como apenas um (fig. 61 B).
Para zonas diferentes da nossa pele, há
diferentes percepções dos estimulos. Por
exemplo nos lábios dois estímulos são
sentidos independentes se estiverem
afastados 0,2 cm, enquanto que nas costas
têm de estar afastados por 4 cm.
Pode acontecer também que os campos
receptores estejam ligados apenas a um
neurónio de segunda ordem, sentindo-se apenas um
estímulo nessa zona; ou podem estar ligados, cada um, a
um neurónio secundário.
O primeiro caso aumenta a sensibilidade aos impulsos,
uma vez que permite que haja somação destes.
Visão Nós, humanos, temos a visão limitada
a uma gama entre os 400 e os 700 nm – luz
visível.
Um estímulo mecânico não pode
gerar uma sensação visual. Mas, se a via de
transdução for activada, esse estímulo pode
ser sentido. Por exemplo: os boxers quando
levam um murro nos olhos têm visões,
apesar de terem os olhos fechados. Isto
acontece porque a energia mecânica
aplicada pelo murro vai activar o nervo
óptico.
Fig. 60 – Inibição lateral
Fig. 61 – Campos receptores
Fig. 62 – Discriminação de
dois estímulos.
Fig. 63 – Anatomia do glóbulo ocular.

25
No glóbulo ocular, a retina é onde se gera o
sinal, com a chegada de luz, e é enviado para o
córtex.
A íris é composta por duas camadas
musculares coradas: circular e radial. Em situações
de baixa luminosidade, a pupila dilata e está sob o
comando do SN simpático. O processo inverso é
comandado pela SN parassimpático.
A luz entra pela pupila e chega, invertida, à
retina.
A lente ou cristalino é o que nos permite
ver um objecto que está longe ou mais próximo:
quando está mais longe, os raios luminosos que
recebe são paralelos e o cristalino faz com eles
convirjam na retina; no caso de os objectos
estarem mais próximos, a luz não chega de forma
paralela, obrigando o cristalino a aumentar a sua
espessura para aumentar a convergência para a
retina. A esta capacidade de, o cristalino, alterar a
sua espessura chama-se acomodação.
Quando queremos ver um objecto que está perto, as fibras musculares relaxam,
permitindo ao cristalino ficar mais arredondado.
Quando o objecto está longe, as fibras contraem-se,
esticando e “empurrando” o cristalino, fazendo-o ficar
mais plano.
A capacidade
de acomodação do
cristalino diminui
com a idade.
As cataratas
afectam o cristalino,
tornando-o opaco.
A Retina é constituída por vários tipos de neurónios, além de células da glia.
Sendo que todas as células são importantes para o bom funcionamento do olho.
A luz passa por várias camadas de neurónios antes de chegar aos fotorreceptores,
excepto na Fóvea Central, onde as outras células foram como que “empurradas”. Esta é
a zona mais utilizada pela retina.
No ponto cego não há fotorreceptores, por isso não há percepção de uma imagem
que seja focada nessa zona. É nessa zona que chega o nervo óptico e vasos sanguíneos.
Fig. 64 – Imagem invertida na retina.
Fig. 65 – Acomodação.
Fig. 66 – Acomodação: alteração da
espessura do cristalino por contracção ou relaxamento das fibras musculares.
Fig. 67 – Gráfico da diminuição da
acomodação, com o aumento da idade.

26
São as Células Ganglionares que formam o
nervo óptico e levam a informação para fora do olho.
As Células Horizontais formam sinapses com
os fotorreceptores e com as células bipolares,
regulando a passagem de informação. Depois essa
informação passa para as Células Ganglionares e
sai do olho para o encéfalo, sendo que os seus
axónios vão formar o nervo óptico.
Há dois tipos de fotorreceptores:
Os Cones, que permitem ver com melhor
resolução e com cores. Existem em baixo
número: 6 milhões, no caso dos humanos.
Os bastonetes são muito mais numerosos (120
milhões) e fornecem imagens em condições de
baixa luminosidade, mas a resolução é baixa e
não detectam a cor.
Na zona da fóvea, existe quase só cones. Nas
zonas periféricas o número de cones diminui e
aumenta o número de bastonetes.
Durante a noite só usamos os bastonetes,
logo não usamos a fóvea. No entanto, quando há
boa luminosidade os bastonetes deixam de
responder, porque são muito sensíveis à luz.
Nós possuímos 120 M de bastonetes e 6 M
de cones, mas apenas temos 1,2 M de neurónios
ganglionares. Logo tem de haver convergência de neurónios, mas essa convergência não
é igual: nos cones a razão é quase de 1:1 enquanto que nos bastonetes é muito maior.
Fig. 70 – Camadas da Retina.
Fig 71 – Fotorreceptores: cones e bastonetes.
Fig. 68 – Organização laminar da retina. Fig. 69 – Fóvea Central.
Fig. 72 – Distribuição dos fotorreceptores na retina.
27
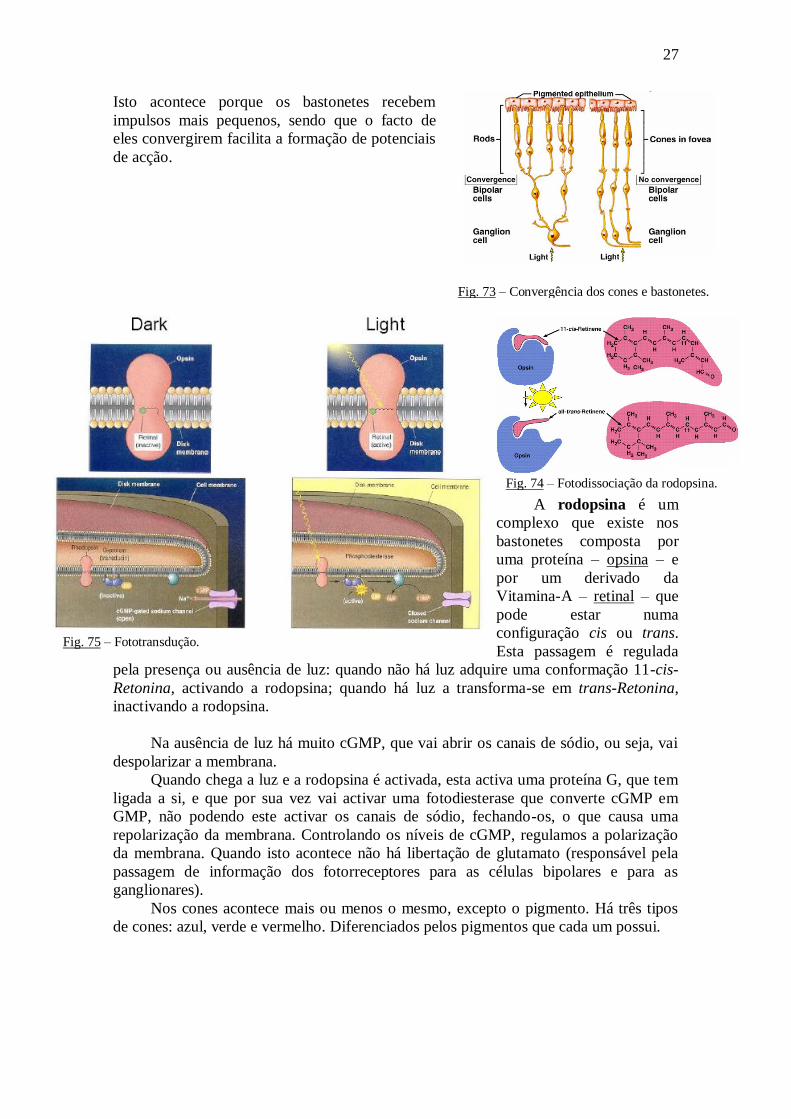
Isto acontece porque os bastonetes recebem
impulsos mais pequenos, sendo que o facto de
eles convergirem facilita a formação de potenciais
de acção.
A rodopsina é um
complexo que existe nos
bastonetes composta por
uma proteína – opsina – e
por um derivado da
Vitamina-A – retinal – que
pode estar numa
configuração cis ou trans.
Esta passagem é regulada
pela presença ou ausência de luz: quando não há luz adquire uma conformação 11-cis-
Retonina, activando a rodopsina; quando há luz a transforma-se em trans-Retonina,
inactivando a rodopsina.
Na ausência de luz há muito cGMP, que vai abrir os canais de sódio, ou seja, vai
despolarizar a membrana.
Quando chega a luz e a rodopsina é activada, esta activa uma proteína G, que tem
ligada a si, e que por sua vez vai activar uma fotodiesterase que converte cGMP em
GMP, não podendo este activar os canais de sódio, fechando-os, o que causa uma
repolarização da membrana. Controlando os níveis de cGMP, regulamos a polarização
da membrana. Quando isto acontece não há libertação de glutamato (responsável pela
passagem de informação dos fotorreceptores para as células bipolares e para as
ganglionares).
Nos cones acontece mais ou menos o mesmo, excepto o pigmento. Há três tipos
de cones: azul, verde e vermelho. Diferenciados pelos pigmentos que cada um possui.
Fig. 73 – Convergência dos cones e bastonetes.
Fig. 74 – Fotodissociação da rodopsina.
Fig. 75 – Fototransdução.

28
Nós vimos diferentes cores, quando há
activação diferente dos cones. Quando fixamos
uma imagem branca com um círculo vermelho e
passados alguns segundos o círculo desaparece,
nós vimos uma sombra azul-turquesa. Isto porque
a luz vermelha saturou os receptores de vermelho,
que não vão conseguir receber a cor vermelha,
nessa zona, quando a imagem fica branca.
Na cegueira das cores – daltonismo – os genes para os pigmentos vermelho e
verde estão no cromossoma X. No caso do homem, como só tem um cromossoma X, é
mais frequente. O gene para o azul existe noutro cromossoma, por isso esta mutação não
acontece.
Os gatos vêm melhor que os homens no
escuro porque: além de terem uma maior
flexibilidade da pupila, possuem um tapete
reflector depois dos fotorreceptores, que permite
que a luz passe novamente por estes. Enquanto
que em nós há uma camada preta que recebe toda
a luz depois de esta passar pelos fotorreceptores.
Em algumas células o glutamato dos bastonetes pode ir inactivar as células
bipolares na ausência de luz. Isto deve-se ao
efeito inibitório do glutamato e tem como
causa a existência de diferentes receptores
para o glutamato nas células bipolares: se os
receptores forem ianotrópicos, ao ligar
glutamato abrem canais de sódio causando
despolarização da membrana, ficando
inactivas. Quando a luz está presente, não há
glutamato para se ligar aos receptores,
inactivando os canais de sódio causando
repolarização da membrana e fazendo com
que as células bipolares libertem glutamato.
Fig. 76 – Sensibilidade dos diferentes cones.
Fig. 77 – A visão dos gatos.
Fig. 78 - Efeito inibitório do glutamato.

29
A luz influencia as sinapses entre os
fotorreceptores e as células bipolares: na sua ausência,
os canais de sódio estão abertos, a rodopsina está
inactiva e o glutamato é libertado do segmento interno
do bastonete para as células bipolares. Estas vão ser
inibidas ou estimuladas de acordo com os receptores de
glutamato. Na presença de luz, os canais de sódio estão
fechados, a rodopsina está activa, não ocorre libertação
de glutamato para as células bipolares. Estas tanto
podem libertar glutamato por serem inibidas ou por
serem suprimidas, dependendo dos receptores para o
glutamato.
Os humanos possuem visão binocular uma vez
que necessitam dos dois olhos para ter uma visão
tridimensional, com noção de profundidade. Isto
deve-se à sobreposição dos campos visuais. Os
predadores têm geralmente visão binocular, enquanto
que os herbívoros têm visão periférica (monocular).
A informação é transmitida a
partir do nervo óptico para os núcleos
geniculares laterais, em seguida saem do olho,
pelas fitas ópticas, chegam ao tálamo e só
depois ao córtex visual.
Ambos os olhos enviam a informação
para os
hemisférios,
tanto direito
como esquerdo.
As imagens
obtidas pela
metade direita e pela metade esquerda de cada olho são
enviadas para o hemisfério direito e esquerdo
respectivamente. A imagem que é projectada na retina
está invertida, permitindo-nos percepcionar a imagem
correctamente. A informação de todas as imagens que
não são projectadas na retina, tem de ser cruzada para o
hemisfério correcto. A informação cruza-se no quiasma
óptico e segue para o núcleo genicular lateral.
Nem toda a informação é enviada para o córtex
visual, uma pequena parte (20%) das fibras nervosas
vai para os colículas superiores, que permitem a
coordenação motora dos olhos, por exemplo, para
seguir objectos em movimento.
Fig. 79 – Efeito da luz nas sinapses
entre os fotorreceptores e as células
bipolares.
Fig. 80 – Condução do sinal visual.
Fig. 81 – Condução do sinal visual:
imagem monocular e binocular.
Fig. 82 – Córtex estriado (17) e áreas de
associação visual (18 e 19).

30
Paladar Os receptores para o paladar são
receptores químicos constituintes do botão
gustativo, que existe em zonas especializadas
da língua chamadas papilas, além do palato,
epiglote e da faringe, onde existem em menor
quantidade. São células epiteliares modificadas
e não neurónios.
Em cada botão gustativo há cerca de 50
células gustativas (receptores) e na extremidade
há umas microvilosidades, chamadas pêlos
gustativos, que se estendem para uma abertura
no epitélio chamada poro gustativo.
Os receptores dividem-se por diferentes zonas da língua:
Nos mecanismos de transdução do
salgado há entrada de sódio nas células,
causando a despolarização da membrana, e
que vai permitir a abertura dos canais de
Cálcio sensíveis à voltagem. Assim pode
haver libertação de neurotransmissores que
vão ser reconhecidos pelos neurónios
aferentes e vão levar a informação até ao
córtex cerebral.
No caso do mecanismo de
transdução do azedo, há entrada de H+, em
vez de sódio. Estas células podem, também,
atingir a despolarização membranar através
da inibição dos canais de potássio (se o
potássio não pode sair, gera-se
despolarização). Depois essa despolarização
vai activar os canais de Cálcio sensíveis à
voltagem, permitindo a libertação de
neurotransmissores (glutamato).
Fig. 83 – Botão gustativo.
Fig. 84 – Distribuição dos botões gustativos.
Fig. 85 – Mecanismo de transdução do salgado e do azedo.

31
Para a transdução do doce há receptores
metabotrópicos, que são activados pelo doce, e que vão activar
uma proteína G que, por sua vez, vai activar a enzima adenilato
ciclase para esta ir transformar o ATP em cAMP, que vai activar
uma proteína cinase A (PKA) para esta ir fosforilar os canais de
potássio, causando a despolarização da membrana por
diminuição da entrada de K+. Podendo, esta despolarização
activar os canais de cálcio, essenciais para a libertação de
neurotransmissores.
Para o
amargo há mais
do que uma via de
transdução
1. A substância
amarga vai ligar-
se aos receptores
ianotrópicos
existentes nos
canais de
potássio,
inactivando-os, o
que vai originar
uma
despolarização da
membrana. Essa
despolarização vai
activar os canais de
cálcio sensíveis à
voltagem,
permitindo a
entrada de cálcio
que vai ajudar a
formar as vesículas
sinápticas, para libertar os neurotransmissores.
2. A substância amarga liga-se a receptores metabotrópicos,
activando uma proteína G. Esta vai activar uma fosfolipase
C, que vai converter o PIP2 em IP3 que, por sua vez vai actuar
sobre as reservas de cálcio, fazendo com que este se liberte e
vá formar as vesículas sinápticas para libertar os
neurotransmissores.
Há quem considere um quinto sabor, o “umami” que em
japonês significa delicioso. O “umami” é um intensificador de
sabor, um aditivo de glutamato monossódico. O “Umami” vai
ligar-se a receptores ianotrópicos nos canais de sódio,
activando-os. A entrada de sódio na célula leva a uma
despolarização da membrana, indo activar os canais de cálcio
sensíveis à voltagem, fazendo com que este entre e permita
libertar os neurotransmissores.
Fig. 86 – Mecanismo de
transdução para o doce.
Fig. 87 – Mecanismos de transdução para o amargo.
Fig. 88 – Mecanismo de transdução para o Umami
(delicioso em Japonês).

32
O síndrome do restaurante chinês é causado por uma hiper estimulação, havendo
grandes quantidades de glutamato, que vão causar uma hiperactivação dos canais de
sódio. As pessoas sensíveis ao “umami” podem ficar mal dispostas.
No entanto foi realizada uma experiência
em que se viu que não há um mapa de sabores
assim tão linear, uma vez que o que os
receptores reconhecem alguns elementos, que
podem estar em substâncias de diferente
natureza. Consoante a natureza da substância
testada, o resultado não foi a que se esperava.
Os receptores que desencadearam resposta eram
diferentes dos que inicialmente se esperava,
como acontece com o HCl, nos gráficos, onde
há uma elevada resposta nos neurónios sensíveis
aos sais, por causa do ião Cl-.
Os estímulos recebidos nos receptores
gustativos passam pelo tálamo e vão para o
córtex gustativo.
Olfacto O epitélio olfactivo existe na parede
superior da cavidade nasal e é constituído por
células de suporte e por neurónios olfactivos
que possuem cílios que estão embebidos em
muco. É o único local do corpo humano em que
os neurónios estão em contacto directo com o
exterior.
O epitélio olfactivo está constantemente a
ser renovado.
As moléculas odoríferas vão
activar as proteínas receptoras de
odores – receptores metabotrópicos
– situadas na membrana plasmática,
que vão activar uma Proteína G, que
por sua vez vai activar a Adenilato
ciclase, indo esta formar cAMP a
parir de ATP para este ir activar os
canais de Cálcio e Sódio. Depois o
cálcio vai activar os canais de
Cloreto, que vão provocar a saída
deste ião, causando uma
despolarização da membrana,
permitindo a passagem do estímulo.
Fig. 89 – Preferências dos neurónios
receptores do paladar.
Fig. 90 – Epitélio olfactivo
Fig. 91 – Mecanismo de transdução olfactiva em vertebrados.

33
Esse estímulo vai ser enviado pelos axónios dos neurónios olfactivos, que entram
no bolbo olfactivo, onde passam a informação para as células mitrais ou para as células
tufadas. Estas, por sua vez, enviam a informação para o cérebro, através das vias
auditivas, e sinapsam com neurónios de associação no bolbo olfactivo. Os neurónios
de associação recebem também informação vinda do encéfalo, podendo modificar a
informação olfactiva antes de esta abandonar o bolbo olfactivo.
O olfacto é a única sensação que vai directamente para o córtex cerebral sem ter
de passar pelo tálamo. O córtex olfactivo localiza-se junto ao sulco lateral do cérebro e
divide-se em três partes:
Área olfactiva externa, que está envolvida na percepção consciente do
cheiro;
Área olfactiva interna, que é responsável pelas reacções emocionais e
viscerais aos odores. Tem ligação com o sistema límbico, através do qual
se liga ao hipotálamo.
Área olfactiva intermédia, possui neurónios que se estendem ao longo
das vias olfactivas até ao bolbo olfactivo, onde sinapsam com neurónios de
associação. É um mecanismo importante pelo qual a informação é
modulada no bolbo olfactivo.
Nós, humanos, conseguimos distinguir cerca de 10 000 odores diferentes. Mas
isto não significa que tenhamos 10 000 receptores diferentes, temos menos (embora não
se saiba quantos receptores temos ao certo).
Um determinado odor é originado pela activação simultânea de um dado número
de receptores.
Os ratinhos têm cerca de 1 000 receptores; enquanto que os cães têm muito mais
receptores, além de possuírem um epitélio maior, o que lhes confere uma capacidade
olfactiva maior.
Audição O som gera-se com a passagem de ondas
de grande amplitude, seguidas por ondas de
pequena amplitude.
A amplitude e a frequência dessas ondas
influenciam o som:
A frequência da vibração (ciclos/s)
mede o tom, em Hz. Quanto maior for
a frequência, mais alto é o tom do
som. Os sons mais frequentes ocorrem
entre 1000 e 4000 Hz, mas os
humanos conseguem ouvir frequências
entre os 20 e os 20000Hz. Mas há
animais com maior capacidade auditiva, por exemplo os cães conseguem ouvir
frequências até 40000 Hz.
A amplitude mede a intensidade (em decibéis
[dB]). O nosso limiar de audição é 0 dB. Uma conversa
normal corresponde a cerca de 50-60 dB e a música alta
corresponde a mais de 130 dB. É importante ter em
conta que uma elevação de 10 dB traduz um aumento de
10 vezes na intensidade, ou seja, um som com 60 dB é
106 mais intenso que outro com apenas 10dB).
Fig.92 – Formação do som.
Fig. 93 – Escala de decibéis.

34
O aparelho auditivo divide-se em 3
partes: ouvido externo, ouvido médio e
ouvido interno.
O ouvido externo é constituído pelo
pavilhão auditivo que liga ao tímpano pelo
canal auditivo. As ondas sonoras vão fazer
vibrar o tímpano, que por sua vez vai fazer
com que 3 ossículos, situados no ouvido
médio (martelo, bigorna e estribe) vibrem
também. O vibrar do martelo vai fazer
vibrar o bigorna, que por sua vez vai
transmitir a vibração ao estribe e que vai
fazer vibrar a janela oval, passando o
estímulo para o ouvido interno.
O ouvido médio tem um canal
que liga à garganta e que permite que o
ar no seu interior, esteja à mesma
pressão que o do exterior. O facto de
quando estamos no avião a subir
ficarmos com a sensação de ter os
ouvidos tapados é devido a uma
alteração de pressão no ouvido médio e
necessitamos de engolir em seco para o
ar voltar a entrar.
Os ossículos estão ligados ao
crânio por tendões, de modo a que
possam libertar energia, quando os
sons têm muita intensidade, para evitar
lesões.
A janela oval comunica com o tubo
vestibular que por sua vez comunica com
o tubo timpânico e são compostos pelo
mesmo líquido – perilinfa. No final do
tubo timpânico há uma membrana que
absorve os sons que chegam até lá –
janela redonda. Entre estes dois canais há
o canal coclear, que é constituído por
endolinfa. As ondas têm tendência a
encurtar caminho, passando através do canal
coclear para chegar ao canal timpânico,
fazendo movimentar a membrana tectorial.
Há medida que a frequência dos sons
aumenta, as ondas tendem a encurtar o seu
curso, atravessando o canal coclear mais
próximo da janela oval.
Fig. 94 – Aparelho auditivo.
Fig. 95 – Ouvido médio.
Fig. 96 – Condução do sinal sonoro.
Fig. 97 – Efeito de som com várias frequências na
membrana basilar.

35
Embebidos na membrana tectorial há cílios que se
movimentam com o movimento da membrana basilar. É
isto que permite activar a recepção da audição.
Se o movimento se der do mais pequeno para o
maior, há abertura dos canais de potássio, havendo
entrada de potássio, causando uma despolarização da
membrana.
No caso de o
movimento se dar do maior
para o mais pequeno há
inactivação dos canais de
potássio, não havendo
despolarização da
membrana.
A endolinfa é muito
diferente dos restantes
líquidos que banham as
nossas células. Neste caso
há uma elevada concentração de H+ e baixa concentração de Na
+, por isso a
despolarização é causada pela entrada de K+.
O reconhecimento dos diferentes sons depende do local onde foi detectado. Os
estímulos causados pelos sons são enviados através do nervo auditivo, passando pelo
tálamo, e vão para zonas distintas do córtex, consoante a sua frequência.
Se o som for muito forte pode provocar lesões nas células ciliadas e, como estas
células não têm capacidade de regeneração, essas lesões nunca são curadas.
Equilíbrio
Localizam-se nos nossos ouvidos
alguns órgãos responsáveis pelo nosso
equilíbrio, funcionando de uma forma
semelhante ao mecanismo de detecção de
sons.
Os Órgãos Otolíticos – Utrículo e
Sáculo – são capazes de reconhecer
diferentes acelerações.
Fig. 98 – Composição da Cóclea
Fig. 99 – Despolarização das células ciliares.
Fig. 100 – Cóclea e Aparelho Vestibular.

36
Os cílios dos órgãos otolíticos estão numa membrana otolítica que tem uma
grande inércia, por isso é muito sensível a acelerações: quando a aceleração aumenta a
membrana desliza para trás, permitindo-nos detectar a aceleração, devido à inibição dos
canais de potássio, uma vez que os cílios se deslocam na direcção do mais pequeno. No
caso de haver desaceleração acontece o oposto, a membrana desloca-se para a frente,
fazendo com que os cílios de desloquem para trás, activando os canais de potássio.
Os movimentos de rotação são
reconhecidos através de um mecanismo
semelhante, onde os cílios se deslocam
com o movimento da água em que
estão embebidos.
Um mecanismo semelhante a este
é também usado por peixes para
detectar deslocações da água.
Fig. 101 – Células Ciliares. Fig. 102 – Órgãos Otolíticos.
Fig. 103 – Ampola para detecção de movimentos de
rotação

37
Quando nós estamos a rodar e paramos de repente ficamos com dificuldade em
manter o equilíbrio. Isso acontece
porque o aparelho vestibular
fornece informação de movimento
e os olhos informam que estamos
parados, isso são informações
contraditórias, que fazem com que
o SNC não saiba como responder.
O contrário acontece
durante uma viagem, em que os
nossos olhos fornecem
informação de movimento rápido,
enquanto o aparelho vestibular diz
que estamos parados. Neste caso a
reacção pode dar enjoos.
Há animais que têm diferentes capacidades sensoriais que os humanos, por
exemplo:
Algumas espécies conseguem ver luz UV e IF;
Os elefantes conseguem comunicar através de ultrasons – sons de baixa
frequência, capazes de percorrer longas distâncias;
Algumas espécies utilizam a ecolocalização como modo de orientação e
procura de presas. Como é o caso de, por exemplo, golfinho e morcegos;
Alguns peixes conseguem detectar campos eléctricos.
Fig. 104 – Vias neuronais envolvidas na manutenção do
equilíbrio.