efeitos do canabidiol e escitalopram sobre as … · ansiedade e capaz de reduzir a latência para...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
DEPARTAMENTO DE FARMACOLOGIA
EFEITOS DO CANABIDIOL E ESCITALOPRAM SOBRE AS DIFERENTES
ETAPAS DO PROCESSO NEUROGÊNICO HIPOCAMPAL ADULTO EM
CAMUNDONGOS CRONICAMENTE ESTRESSADOS
VINÍCIUS DETONI LOPES
RIBEIRÃO PRETO
2018

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE MEDICINA DE RIBEIRÃO PRETO
DEPARTAMENTO DE FARMACOLOGIA
VINÍCIUS DETONI LOPES
Efeitos do canabidiol e escitalopram sobre as diferentes etapas do processo neurogênico
hipocampal adulto em camundongos cronicamente estressados
RIBEIRÃO PRETO
2018
Dissertação apresentada à
Faculdade de Medicina de Ribeirão
Preto da Universidade de São Paulo
para obtenção do grau de Mestre
em Ciências. Área de concentração:
Farmacologia.
Orientador: Profª Drª Alline
Cristina de Campos

Autorizo a reprodução e divulgação total ou parcial deste trabalho, por qualquer meio
convencional ou eletrônico, para fins de estudo e pesquisa, desde que citada a fonte.
Lopes, Vinícius Detoni
Efeitos do canabidiol e escitalopram sobre as diferentes etapas do
processo neurogênico hipocampal adulto em camundongos cronicamente
estressados. Ribeirão Preto, 2018.
Dissertação de Mestrado, apresentada à Faculdade de Medicina de
Ribeirão Preto/USP. Área de concentração: Farmacologia.
Orientador: Campos, Alline Cristina de
1.Neurogênese. 2. Canabidiol. 3. Estresse 4. Neuroplasticidade

FOLHA DE APROVAÇÃO
Nome: LOPES, Vinícius Detoni
Título: Efeitos do canabidiol e escitalopram sobre as diferentes etapas do processo
neurogênico hipocampal adulto em camundongos cronicamente estressados.
Aprovado em:___/___/2018
Banca Examinadora
Profª Drª Alline Cristina de Campos FMRP/USP
Julgamento: __________________ Assinatura: ___________________
Membro 1: ___________________ Instituição: ___________________
Julgamento: __________________ Assinatura: ___________________
Membro 2: ___________________ Instituição: ___________________
Julgamento: __________________ Assinatura: ___________________
Membro 3: ___________________ Instituição: ___________________
Julgamento: __________________ Assinatura: ___________________
Dissertação apresentada à
Faculdade de Medicina de Ribeirão
Preto da Universidade de São Paulo
para obtenção do grau de Mestre
em Ciências. Área de concentração:
Farmacologia.

Dedico este trabalho à minha avó
Otília, por ter tido um papel
fundamental na minha educação.

AGRADECIMENTOS
Agradeço primeiramente aos meus pais Carlos e Regina pelo apoio incondicional durante toda
a minha formação. Desconheço dois seres humanos com tamanha capacidade de amar e ser
acalento sempre quando preciso. Compreender a distância é algo sempre muito difícil e tem
seus preços. Obrigado por entenderem minha necessidade de buscar novos horizontes e por
apostarem na minha educação. Amo vocês!
À minha irmã Cecília pelos anos de discussão que sem dúvidas me fizeram enxergar as coisas
com mais clareza e maturidade. Mesmo que os caminhos tenham por hora, nos distanciado,
você sempre foi uma palavra amiga nos momentos mais difíceis.
À minha orientadora Alline Campos por ter me aceito nesta empreitada. Sou um profundo
admirador das suas qualidades, como pesquisadora, mas principalmente como ser humano.
Obrigado por sempre me estimular a ir além e ter participado dessa etapa tão engrandecedora
da minha vida.
Aos meus colegas de laboratório Davi, Paula, Jordan, Mari, Dri, Gabriel, Karla, Rafa, Edu,
Erika, Melissa, Fran e Cláudia pelos inesquecíveis anos de convivência e pela ajuda na
conclusão deste trabalho. Podem ter certeza que levo um pouco de cada um de vocês em
minha jornada.
À Fernanda e a Giu por terem sido equilíbrio em momentos fundamentais do nosso
crescimento coletivo de grupo e pela amizade sincera.
Á todo o laboratório do Professor Francisco Guimarães pela paciência em ter nos acolhido
durante todo o tempo e pela solicitude marcante de todos com que tive o prazer de conviver.
Ao laboratório da professora Elaine Del Bel por ter disponibilizado gentilmente o uso do
microscópio.
À Gabi por ter se tornado um refúgio na minha vida. Obrigado pelos inesquecíveis bons
momentos e por ter “segurado a barra” durante os momentos difíceis. Obrigado também por
participar deste momento, com sua ajuda científica valiosa.
Aos meus amigos de Ribeirão Preto, em especial ao Cassiano e Atlante por terem
proporcionado bons momentos fora do ambiente universitário. Vocês foram fundamentais
durante todo este processo. Seja para debater de forma muito saudável temas dos mais

variados possíveis bebendo uma cerveja, seja para oferecer uma palavra amiga nos momentos
difíceis.
Aos meus amigos de São João Nepomuceno, amigos de infância, por terem me proporcionado
momentos que me fizessem esquecer por um instante que seja da rotina.
Aos meus amigos da República Megazord de Diamantina pelos 5 anos de convivência.
Criamos laços e meu crescimento, sem dúvidas, passa por esse marcante momento da minha
vida, quando com 18 anos decidi seguir meu caminho e permanecer mesmo que longe da
família, em Diamantina.
Agradeço a todos os mestres verdadeiros que participaram da minha formação, em especial ao
professor Disney que me incentivou a encarar essa jornada desafiadora.
Aos meus amados tios e tias pelo carinho e apoio que recebi durante minha jornada.
A toda pós-graduação do departamento de Farmacologia da FMRP da Universidade de São
Paulo, na qual incluo os professores, técnicos, secretários e alunos pela seriedade,
competência e paixão em realizar suas funções. Fazer parte disso é motivo de muito orgulho.
A CAPES pelo fomento e ao instituto L’oreal e FAPESP pelo financiamento.

”
“A alegria não chega apenas no
encontro do achado, mas faz parte do
processo da busca. E ensinar e
aprender não pode dar-se fora da
procura, fora da boniteza e da alegria”.
Paulo Freire

RESUMO

RESUMO
DETONI, V.L. Efeitos do canabidiol e escitalopram sobre as diferentes etapas do
processo neurogênico hipocampal adulto em camundongos cronicamente estressados.
2018. Dissertação (Mestrado). Faculdade de Medicina de Ribeirão Preto- Universidade de São
Paulo, Ribeirão Preto, 2018.
O conhecimento atual sugere que a neurogênese adulta é um mecanismo de plasticidade
importante para a manutenção da homeostasia cerebral e conservado no cérebro de
mamíferos. A neurogênese adulta pode ser modulada negativamente por exposição
prolongada ao estresse e positivamente por fármacos antidepressivos. Nesse sentido,
utilizamos o modelo de estresse crônico imprevisível (CUS) para mimetizar em animais o
impacto da exposição crônica ao estresse sobre o comportamento e sobre a neurogênese
adulta. A administração crônica de antidepressivos pode atenuar o aparecimento de respostas
comportamentais em animais submetidos ao CUS, embora exista uma considerável latência
para o surgimento desses efeitos. Recentemente, o canabidiol (CBD) emergiu como um novo
fármaco com potencial terapêutico para tratamento de transtornos psiquiátricos, como a
ansiedade e capaz de reduzir a latência para início dos efeitos em relação aos antidepressivos
clássicos, sendo os mecanismos até então pouco compreendidos. Portanto, o objetivo deste
trabalho foi investigar se o tratamento com CBD ou escitalopram (ESC) durante 7 ou 14 dias
é capaz de atenuar respostas comportamentais do tipo ansiogênica em animais submetidos ao
CUS e observar se esse efeito poderia correlacionar-se a diferentes mecanismos relacionados
à neurogênese adulta. Para isso, camundongos C57BL/6 machos foram divididos em 6
grupos. Os animais de cada grupo receberam somente um tratamento: CBD (30mg/Kg), ESC
(20mg/Kg) ou veículo diariamente durante 7 (n=9-10=grupo) ou 14 dias (n=6-10/grupo) de
maneira independente e foram submetidos ou não ao CUS durante o mesmo período. Um dia
após o último tratamento, os animais foram submetidos ao teste do Novelty Supressed
Feeding (NSF). Além disso, foram realizadas análises de proliferação celular (Ki67), da
sobrevivência celular (BrdU) e de marcadores associados a distintos tipos celulares (Sox2 e
DCX). Os resultados obtidos no presente trabalho sugerem que o CBD é capaz de prevenir os
efeitos do estresse crônico em protocolos de 7 e 14 dias de tratamento, enquanto que o
antidepressivo escitalopram previne os efeitos do estresse apenas no protocolo de 14 dias.
Além disso, o estresse não alterou significativamente a taxa de sobrevivência celular em
ambos protocolos, mas promoveu redução da proliferação celular em 7 dias. Além disso,
observou-se uma redução do número de células DCX+ no grupo dos animais
estressados/veículo em relação ao grupo de animais controle/veículo no protocolo de 7 dias,
diferença esta não encontrada no protocolo de 14 dias. Com relação ao efeito dos tratamentos,
encontramos que o CBD ou ESC não previnem a redução da proliferação celular mediada
pelo estresse, mas previnem a redução de células DCX+ no protocolo de 7 dias. Os resultados
encontrados no presente trabalho sugerem que o CBD pode prevenir os efeitos do estresse em
um menor período de tempo quando comparado ao ESC. Porém, o presente trabalho não
demonstrou alteração significativa nos parâmetros relacionados à neurogênese, tampouco
modulação diferencial entre os fármacos.
Palavras-chave: neurogênese, canabidiol, antidepressivo, latência.

ABSTRACT

ABSTRACT
Effects of Cannabidiol and escitalopram in different stages of adult hippocampal
neurogenesis in n chronically stressed mice. 2018. Master Thesis. Faculdade de Medicina
de Ribeirão Preto- Universidade de São Paulo, Ribeirão Preto, 2018
Current knowledge suggests that adult neurogenesis is an important plasticity mechanism for
the maintenance of cerebral homeostasis and conserved in the mammalian brain. Adult
neurogenesis can be modulated negatively by prolonged exposure to stress and positively by
antidepressant drugs. In the present study, we expose mice to the unpredictable chronic stress
paradigm (CUS) and evaluated the impact of chronic exposure to stress on behavior. Chronic
administration of antidepressants attenuate CUS-induced behavioral abnormalities in animals,
although there is a considerable latency for the appearance of the effects of these drugs.
Recently, cannabidiol (CBD) has emerged as a putative new drug with for the treatment of
psychiatric disorders, such as anxiety. Therefore, the objective of this study was to investigate
whether the treatment with CBD or escitalopram (ESC) for 7 or 14 days is able to attenuate
CUS-induced anxiogenic response in mice and, if its effects are correlated with mechanism
involving adul type in animals submitted to the CUS and to observe if this effect could
correlate to different related mechanisms to adult neurogenesis. Male C57BL6 mice were
divided in 6 groups. Each animal of every experimental group received: CBD (30mg / kg),
ESC (20mg / kg) or vehicle daily for 7 (n = 9-10 = group) or 14 days (n = 6-10 / group) and
were submitted or not to the CUS during the same period. One day after the last treatment,
animals were submitted to the Novelty Supressed Feeding (NSF) test. In addition, analyzes of
cell proliferation (Ki67), cell survival (BrdU) and markers associated with different cell types
(Sox2 and DCX) were performed. The results obtained in the present work suggest that CBD
is able to prevent the effects of chronic stress in protocols of 7 and 14 days of treatment,
whereas the antidepressant escitalopram prevents the effects of stress only in the protocol of
14 days. In addition, stress did not significantly alter the rate of cell survival in both protocols,
but promoted reduction of cell proliferation in 7 days. In addition, a reduction in the number
of DCX + cells in the group of stressed animals / vehicle was observed in relation to the group
of control / vehicle animals in the 7-day protocol, a difference not found in the 14-day
protocol. Regarding the effect of treatments, we found that CBD or ESC did not prevent the
reduction of cell proliferation mediated by stress, but prevented the reduction of DCX + cells
in the 7-day protocol. The results found in the present study suggest that CBD can prevent the
effects of stress in a shorter window when compared to ESC. These effects however seem not
to be related to adult hippocampal neurogenesis.
Key- words: neurogenesis, canabidiol, escitalopram, anxiety.

SUMÁRIO

SUMÁRIO
Sumário 1. Introdução.......................................................................................................................... 16
1.1. Neurogênese adulta .................................................................................................... 16
1.2. Conceito e nichos neurogênicos................................................................................. 18
1.3. Neurogênese Adulta: Participação na manutenção de importantes funções cerebrais e
evidências de fatores que podem exercer influência positiva ou negativa. .......................... 21
1.4. Estresse, transtornos psiquiátricos, antidepressivos e neurogênese adulta ................ 23
1.5. Canabinoides- Breve histórico, atuação sobre o sistema endocanabinoide ............... 27
1.6. Canabidiol (CBD) ...................................................................................................... 30
1.6.1. Química do CBD ................................................................................................ 30
1.6.2. Efeitos clínicos do CBD ..................................................................................... 31
1.6.3. Farmacodinâmica e farmacocinética do CBD .................................................... 33
1.6.4. Efeitos pré-clínicos observados com o CBD........................................................... 34
2. Objetivos ........................................................................................................................... 37
2.1. Objetivo geral ............................................................................................................ 37
2.2. Objetivos específicos ................................................................................................. 37
3. Material e métodos ............................................................................................................ 39
3.1. Animais ...................................................................................................................... 39
3.2. Drogas ........................................................................................................................ 39
3.3. Procedimento de estresse ........................................................................................... 39
3.4. Teste comportamental ................................................................................................ 40
3.5. Perfusão e preservação da amostra ............................................................................ 40
3.6. Delineamento experimental ....................................................................................... 41
3.6.1. Delineamento experimental ................................................................................ 41
3.6.2. Protocolo experimental ....................................................................................... 42
3.7. Preparação dos tecidos para a técnica de imunohistoquímica ................................... 43
3.7.1. Determinação de células doblecortin (DCX) positivas por imunohistoquímica ..... 43
3.7.2. Determinação de células BrdU positivas por imunohistoquímica .......................... 44
3.7.3. Determinação de células Ki67 positivas para imunohistoquímica ......................... 45
3.8. Imunofluorescência para detecção de BrdU/SOX2 ................................................... 45
3.9. Análise estatística ...................................................................................................... 45
4. Resultados ............................................................................................................................. 47

4.1. Protocolo experimental de 7 dias .................................................................................. 47
4.1.1. Efeito do tratamento repetido com CBD ou escitalopram (7 dias) sobre a taxa de
sobrevivência celular, em animais controle e cronicamente estressados .............................. 49
4.1.2. Efeito do tratamento repetido com CBD ou escitalopram (7 dias) sobre a taxa de
proliferação celular, em animais controle e cronicamente estressados................................. 52
4.1.3. Efeito do tratamento repetido com CBD ou Escitalopram (7 dias) sobre a expressão
de células DCX+ .................................................................................................................... 55
4.2 Protocolo experimental de 14 dias .................................................................................. 59
4.2.1 Efeito do tratamento repetido (14 d) sobre a expressão de células BrdU+ .................. 61
4.2.2 Efeito do tratamento repetido (14 d) sobre a expressão de células DCX+ e sobre a
migração celular. ................................................................................................................... 64
4.2.3. Efeito do tratamento repetido (14 d) sobre a expressão de células SOX2+ ................ 68
5. Discussão .............................................................................................................................. 72
6. Conclusão ............................................................................................................................. 84
7. Referências ........................................................................................................................... 86

LISTA DE FIGURAS
Figura 1- Nichos neurogênicos no cérebro adulto de mamíferos em corte coronal.
Figura 2- Etapas da neurogênese hipocampal adulta com respectivos marcadores que
caracterizam determinado tipo celular e características de cada fase.
Figura 3- Estresse mecanismos de adaptação.
Figura 4- Esquema simplificado ilustrando a sinalização endocanabinoide.
Figura 5- Estrutura química do canabidiol.
Figura 6- Delineamento experimental. Protocolo de 7 dias.
Figura 7- Delineamento experimental. Protocolo de 14 dias.
Figura 8- Efeitos comportamentais do tratamento repetido com o CBD e escitalopram em
animais controle ou estressados e submetidos ao paradigma do Novelty Supressed Feeding.
Time point: 7 dias.
Figura 9- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a sobrevivência de células BrdU +/0,1mm2 de giro denteado do hipocampo.
Time point: 7 dias.
Figura 10- Análise correlacional entre o efeito comportamental obtido no NSF e o número de
células BrdU+/0,1mm2 de giro denteado. Time point: 7 dias.
Figura 11- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a proliferação celular no giro denteado do hipocampo. Células
Ki67+/0,1mm2. Time point: 7 dias.
Figura 12- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a quantidade de células DCX + /0,1mm2 no giro denteado do hipocampo.
Time point: 7 dias.

Figura 13- Análise correlacional entre o efeito comportamental obtido no NSF e o número de
células DCX+/0,1mm2 de giro denteado. Time point: 7 dias.
Figura 14- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a porcentagem de células DCX+ em estágio de migração em relação ao
número de células DCX+ totais no giro denteado do hipocampo. Time point: 7 dias.
Figura 15- Efeitos comportamentais do tratamento repetido com o CBD e escitalopram em
animais controle ou estressados e submetidos ao paradigma do Novelty Supressed Feeding.
Time point: 14 dias.
Figura 16- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a quantidade de células BrdU + /0,1mm2 de giro denteado do hipocampo.
Time point: 14 dias.
Figura 17- Análise correlacional entre o efeito comportamental obtido no NSF e o número de
células BrdU+/0,1mm2 de giro denteado. Time point: 14 dias.
Figura 18- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a quantidade de células DCX + /0,1mm2 de giro denteado do hipocampo.
Time point: 14 dias.
Figura 19- Análise correlacional entre o efeito comportamental obtido no NSF e o número de
células DCX+/0,1mm2 de giro denteado. Time point: 14 dias.
Figura 20- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a porcentagem de células DCX+ em estágio de migração em relação ao
número de células DCX+ totais no giro denteado do hipocampo. Time point: 14 dias.
Figura 21- Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou
estressados sobre a quantidade de células SOX2 + /mm2 de giro denteado do hipocampo.
Time point: 14 dias.

LISTA DE SIGLAS
SNC- Sistema Nervoso Central
NSE- neuron specific enolase
BrdU- 5-bromo-2’-deoxiuridina
SVZ- Zona subventricular
SGZ- Zona subgranular
GD- giro denteado
OB- bulbo olfatório
RMS- cadeia migratória rostral
Str- estriado
LV- ventrículo lateral
Cx- Córtex
Hip- Hipocampo
DCX- doublecortin
GFAP- glial fibrillary acidic protein
eixo HPA- eixo hipotálamo-hipofisário-adrenal
AD- antidepressivo
GC- glicocorticoides
CUS- chronic unpredictable stress
CBD- Canabidiol
Δ9-THC- 1- delta-9-trans-tetrahidrocanabinol
CB1- receptor canabinoide 1
CB2- receptor canabinoide 2
v.o- via oral
GPR55- receptor 55 acoplado à proteína G
TRPV1- receptor vaniloide tipo 1
PPARγ- receptor ativado por proliferador de peroxissoma
Tmax- Tempo no qual a concentração é máxima no plasma
t1/2- tempo de meia-vida
CYP450- citocromo P450
mTOR-mammalian target of rapamycin
PFA- Paraformaldeído
TBS- Tris-buffered saline
BSA- bovine sérum albumine
DAB- 3,3’ diaminobenzidina
HCl- ácido clorídrico
M- molar
ANOVA- Análise de variância
SEM- erro padrão da média
NS- Não estressado
ESC- Escitalopram

INTRODUÇÃO

16
1. Introdução
“Nos centros adultos, os caminhos nervosos são algo fixo, terminado, imutável. Tudo
pode morrer, nada pode ser regenerado. É para a ciência do futuro mudar, se possível, este
severo decreto.” (Ramón e Cajal, 1928). Esta frase do grande neurocientista Santiago Ramón
e Cajal contida em sua pioneira obra “Degeneração e regeneração do Sistema Nervoso
Central”, explicitou o entendimento à época acerca da capacidade mutável do Sistema
Nervoso Central. Embora o autor propusesse que seria necessária uma população fixa de
células neuronais no cérebro adulto para manter a estabilidade funcional da circuitaria
cerebral adulta, Cajal, concomitantemente, demonstrava sua inquietude em relação a este
paradigma vigente na época, de que o cérebro adulto seria algo imutável, incapaz de ser
alterado. Coube à ciência do futuro, como referido pelo visionário e célebre neurocientista,
alterar este severo decreto. Nas últimas décadas, evidências crescentes nos levaram a
“redescobrir” o cérebro e a admirar a sua grande capacidade dinâmica e plástica. Dentre os
processos plásticos que ocorrem no cérebro adulto, a neurogênese, evidenciada pelos
trabalhos de Joseph Altman, na década de 60 do século anterior, ofereceu uma nova
perspectiva ao demonstrar experimentalmente que novos neurônios poderiam ser
incorporados no cérebro adulto de ratos (Altman e Das, 1965). O pioneirismo de Altman
possibilitou o estudo deste fenômeno, de sua dinâmica e de suas funções no cérebro adulto,
alterando, de forma instigante os rumos da neurociência.
1.1.Neurogênese adulta
Historicamente, os primeiros relatos de que a neurogênese poderia ser um evento
conservado em cérebros adultos foram divulgados por Altman (1963). O pesquisador
demonstrou por meio da técnica de autorradiografia, a existência de células marcadas com
timidina H-3
(marcador que acopla-se durante a replicação do DNA de células em
proliferação) no neocórtex e em células granulares do hipocampo de ratos de 4 meses de idade
e em gatos de idade desconhecida. Porém, somente em 1965, o mesmo pesquisador publicou
seus resultados demonstrando evidências que sugeriam a existência de neurogênese pós-natal
no hipocampo de ratos de 1, 4, 6 e 8 meses de idade (Altman e Das, 1965). Kaplan e Hinds
em 1977, também reportaram que novos neurônios seriam formados no bulbo olfatório e giro
denteado de ratos adultos (4 meses de idade), através da análise da incorporação do mesmo
marcador utilizado por Altman. Porém, a utilização de micrografia eletrônica para análise

17
permitiu, segundo os autores, distinguir as células neurogliais de outras populações celulares.
As descobertas de Altman e de Kaplan e Hinds, embora revolucionárias, permaneceram como
fonte de debate por vários anos, em grande parte, em virtude da limitação de técnicas
disponíveis na época que comprovassem a existência de neurogênese adulta (Rakic, 1985).
Além disso, não era possível determinar o papel desses novos neurônios no cérebro adulto ou
se os mesmos integrariam-se funcionalmente no sistema nervoso central (SNC). Nesse
sentido, Paton e Notebohm (1984) foram pioneiros em demonstrar que estes novos neurônios
em aves canoras eram capazes de se integrar à rede sináptica pré-existente. Outro trabalho
relevante para os rumos do entendimento da neurogênese como um evento conservado no
cérebro adulto foi desenvolvido por Cameron e colaboradores (1993). Os autores
demonstraram que após duas semanas de administração de timidina H3 em ratos adultos,
muitas células haviam incorporado esse marcador e que muitas delas foram imunorreativas
para NSE (neuron specific enolase), que é um marcador específico da linhagem neuronal.
Além disso, os autores, a partir do estudo de time points determinados (1 h, 1, 2, 3 e 4
semanas), conseguiram determinar a posição destas células no giro denteado do hipocampo e
sugerir que muitos dos neurônios recém-formados migravam durante o seu desenvolvimento
para a camada granular do giro denteado.
O crescente avanço de técnicas moleculares como a microscopia confocal e de novos
marcadores celulares, tais como o análogo da timidina 5-bromo-2`-deoxiuridina (BrdU), que
se incorpora ao DNA das células que estão na fase S do ciclo celular, permitiram um grande
avanço no entendimento deste evento. Tais evoluções, possibilitaram sequencialmente a
descoberta da neurogênese adulta em primatas (Gould et al., 1997) e a existência da
neurogênese adulta humana, evidenciada pelo trabalho de Eriksson e colaboradores (1998). A
partir de análises post mortem no cérebro de pacientes que haviam recebido BrdU como um
marcador para diagnóstico de tumores, os autores observaram que, através da técnica de
imunofluorescência para marcação específica de populações neuronais (NeuN, calbindina ou
NSE) que haviam sido coexpressos com BrdU, novos neurônios eram gerados no giro
denteado do hipocampo de seres humanos adultos. Atualmente observa-se um grande
interesse pelo estudo da neurogênese adulta em seres humanos. Embora fonte de debates,
diversos autores consideram a existência deste processo em seres humanos adultos
(Kempermann et al., 2018; Ernst et al., 2014; Spalding et al., 2013; Bergmann et al., 2015).

18
1.2.Conceito e nichos neurogênicos
Segundo definições de Gage (2000) e Kempermann et al. (2015) a neurogênese adulta
não pode ser descrita como meramente um evento e sim um processo altamente dinâmico,
sujeito à regulação por vários estímulos fisiológicos, patológicos e farmacológicos que
culmina com o surgimento de novos neurônios no cérebro adulto, a partir de células tronco
progenitoras, localizados em nichos bem definidos (Gage, 2000; Kempermann et al., 2015). O
conhecimento atual reconhece a existência de 2 nichos neurogênicos sob condições
fisiológicas no cérebro de mamíferos, a zona subventricular dos ventrículos laterais (SVZ) e a
zona subgranular (SGZ) do giro denteado da formação hipocampal (Ming e Song, 2011)
(Figura 1).
Além da presença de células tronco progenitoras dotadas de capacidades de auto-
renovação e multipotência, fundamentalmente, os nichos possuem um microambiente
favorável à ocorrência do processo, como uma população de células gliais (astrócitos e
micróglia), contatos específicos entre diferentes tipos celulares, a presença de células
endoteliais e uma complexa matriz extracelular (Doetsch, 2003; Conover e Notti, 2008). A
neurogênese adulta, embora seja caracterizada por um processo complexo compreendido por
vários passos e que sofre a regulação de diversos fatores, pode ser dividida genericamente em
4 etapas principais: fase de proliferação, migração, diferenciação e maturação. Inicialmente,
Figura 1- Bátiz et al., 2016. Nichos neurogênicos no cérebro adulto de mamíferos em corte
coronal. A SVZ e SGZ são atualmente definidos como nichos neurogênicos no cérebro adulto de
mamíferos. Em destaque é representada a zona subgranular (SGZ) do giro denteado (DG). Legenda:
OB: bulbo olfatório; RMS: cadeia migratória rostral; Str: estriado; SVZ: zona subventricular; LV:
ventrículo lateral; Cx: córtex; Hip: Hipocampo. Copyright © 2016 Bátiz, Castro, Burgos, Velásquez,
Muñoz, Lafourcade, Troncoso-Escudero and Wyneken.

19
células precursoras com propriedades gliais (Tipo1-GD; ou tipo B1-SVZ) podem sofrer
divisões simétricas, gerando duas células filhas semelhantes ou sofrer divisões assimétricas,
originando uma célula filha, que pode ser um progenitor amplificado transitório (Tipo 2a-GD;
ou tipo C-SVZ) e uma célula semelhante a ela. Após esta fase proliferativa, as células podem
migrar longas distâncias, através da cadeia migratória rostral (SVZ) ou curtas distâncias
através da camada granular (GD), nos quais diferenciam-se em neuroblastos (Tipo 2b e Tipo 3
GD; Tipo A-SVZ) (Kempermann et al., 2015).
Importante enfatizar que essas populações celulares podem assumir determinada
heterogeneidade em seus respectivos nichos durante todo o processo de neurogênese,
possivelmente possuindo funções específicas (Alvarez-Buylla et al., 2008). Caso sobrevivam
a todo esse processo, podem gerar novos neurônios imaturos e integrar-se às sinapses pré-
existentes na camada granular do giro denteado ou no bulbo olfatório (em roedores), onde
assumem a forma de células granulares glutamatérgicas, no GD, ou interneurônios
GABAergicos, no bulbo olfatório. Nesta fase, ocorre a maturação de espinhos dendríticos e
aumento da plasticidade sináptica (Kempermann et al., 2015). Ressalta-se, no entanto, que
todos os condicionantes deste processo não estão ainda completamente elucidados, embora
possivelmente, a sinalização presente no nicho neurogênico mediada por fatores de
crescimento e neurotransmissores permitam uma complexa regulação, por exemplo, entre
quiescência/proliferação de células tronco progenitoras e diferenciação entre
neurônios/células gliais (Conover e Notti, 2008).
Uma estratégia utilizada para se ter um indício do estágio da neurogênese em que a
célula se encontra é o uso de marcadores neurais que são expressos em distintas populações
celulares. Por exemplo, células precursoras amplificadas nas regiões neurogênicas são
negativas para GFAP, portanto a dupla marcação de SOX2/GFAP pode ser utilizada para
diferenciação entre células progenitoras e precursoras amplificadas (Suh et al., 2007). Após a
fase proliferativa, as células se diferenciam e caso sobrevivam, expressam, dentre outros
marcadores, doublecortin (DCX), proteína associada à microtúbulos que é expressa somente
na linhagem neuronal e que está intimamente relacionada ao processo de migração e de
iniciação da diferenciação neuronal (células 2b e 3). No caso da neurogênese adulta que
ocorre no hipocampo, após sofrer integração sináptica na camada granular do giro denteado,
os neurônios expressam calbindina correspondendo à fase de maturação pós-mitótica tardia. A
expressão destes marcadores pode ser avaliada, portanto, por técnicas moleculares,
ferramentas de particular importância para o presente estudo. Sinteticamente, a expressão de
alguns marcadores celulares e de características que compreendem as distintas fases do

20
processo de neurogênese adulta são exibidos esquematicamente abaixo (Figura 2). Em
seguida, na tabela 1 são atribuídas funções para marcadores de fundamental interesse ao nosso
estudo e sua expressão celular.
Figura 2. Modificado de Kempermann et al., 2015. Etapas da neurogênese hipocampal adulta
com respectivos marcadores que caracterizam determinado tipo celular e características de cada
fase.

21
Marcador Função Expressão
celular
durante a
neurogênese
adulta no SGZ
Referências
GFAP (glial
fibrillary acidic
protein)
Suporte de força
mecânica em astrócitos.
Células progenitoras
positivas para GFAP
podem gerar tipos
celulares específicos
durante a neurogênese
adulta.
Células radiais
gliais ( Tipo 1)
Kriegstein e
Gotz (2003)
SOX 2 Fator de transcrição que
codifica um motivo de
ligação de DNA
altamente conservado,
possuindo papel vital
durante distintos
estágios do
desenvolvimento de
mamíferos. São capazes
de gerar células
diferenciadas (divisões
assimétricas e células
idênticas (divisões
simétricas).
Células tronco
progenitoras
(Tipo 1) e
células
progenitoras
amplificadas
(Tipo 2 a)
Suh et al.,
2007
DCX
(doublecortin)
Proteína que facilita a
polimerização de
microtúbulos
Células tipo 2b
e células do
tipo 3
(neuroblastos).
Brown et al.,
2003
1.3.Neurogênese Adulta: Participação na manutenção de importantes funções
cerebrais e evidências de fatores que podem exercer influência positiva ou
negativa.
A neurogênese adulta é um processo regulador da homeostasia cerebral em eventos
fundamentais, tais como memória e aprendizado e processos emocionais (Sahay et al., 2008).
Tabela 1- Biomarcadores moleculares na neurogênese adulta, suas funções e expressão celular
em tipos celulares durante a neurogênese adulta na SGZ do giro denteado.

22
É bem descrito que fatores como o enriquecimento ambiental, atividade física voluntária
(Sakalem et al., 2017), bem como algumas classes de fármacos, como os antidepressivos
(Mallberg et al., 2000) exercem efeitos pró-neurogênicos, enquanto que fatores como
envelhecimento e exposição ao estresse podem promover um prejuízo sobre a neurogênese
adulta (Duman et al; 2006). Além disso, sua disfunção é um dos possíveis fatores relacionado
com a neurobiologia dos transtornos psiquiátricos, tais como depressão maior e ansiedade
(Schoenfeld e Cameron, 2015).
Neste sentido, trabalhos pioneiros desenvolvidos por Gould e colaboradores (1992),
demonstraram que a administração de glicocorticoides promove uma redução de células
granulares precursoras no hipocampo de ratos adultos. Posteriormente, o mesmo grupo
demonstrou que a exposição de um primata não humano a um estresse social (animal intruso)
promoveu uma redução da proliferação de células granulares no hipocampo adulto destes
animais (Gould et al., 1998).
Outro importante estudo demonstrou que o aumento da neurogênese hipocampal
adulta per se, através da deleção do gene pró-apotótico Bax de células tronco progenitoras e
sua progênie no cérebro adulto, é suficiente para reduzir comportamentos relacionados à
ansiedade e depressão em animais tratados cronicamente com corticosterona, um modelo bem
delineado de estresse (Hill et al., 2015).
Algumas classes de fármacos, também possuem papel facilitador sobre a neurogênese
adulta. Neste sentido, Mallberg e colaboradores (2000) foram pioneiros em demonstrar que o
tratamento crônico, mas não agudo, com antidepressivos aumentou significativamente o
número de células marcadas com BrdU no giro denteado do hipocampo de ratos adultos. Este
aumento da proliferação evidenciado com BrdU, somado a evidências de colocalização com
marcadores específicos da linhagem neuronial, indicou um efeito pró- neurogênico. Outro
ponto importante destacado, refere-se à latência encontrada para o início dos efeitos sobre a
neurogenese adulta: somente o tratamento crônico com AD foi capaz de gerar os efeitos
positivos sobre a proliferação celular. Este achado era condizente com o tempo necessário
para a ação terapêutica clínica observada para os antidepressivos, evidenciando o possível
papel de processos plásticos, como a neurogênese adulta para o efeito desta classe de
fármacos (Mallberg et al., 2000).
A participação da neurogênese hipocampal para o efeito comportamental observado
em animais após a utilização crônica de fármacos antidepressivos foi sugerida
experimentalmente no estudo de Santarelli e colaboradores (2003). Foi observado, que após a
indução da morte de células progenitoras no hipocampo por meio da irradiação com raios-X,

23
o efeito comportamental observado após o tratamento de 28 dias com fluoxetina ou
imipramina era abolido (Santarelli et al., 2003). Porém, como o próprio autor enfatizou,
apesar do fortalecimento da hipótese neurogênica para a ação dos AD, não se poderia excluir
a hipótese que outras consequências da irradiação do hipocampo com raios X poderiam ter
contribuído para a perda de ação dos AD. Existem, portanto, fatores extrínsecos que
influenciam o processo neurogênico de forma negativa, como o estresse ou de maneira
positiva como, por exemplo, os fármacos antidepressivos (Encinas et al., 2006), os quais serão
enfatizados em nossa investigação.
1.4. Estresse, transtornos psiquiátricos, antidepressivos e neurogênese adulta
Evidências atuais indicam a importante participação do estresse sobre a neurogênese
hipocampal adulta. Porém, apesar da atual compreensão, em uma perspectiva histórica, o
estudo do estresse, de seus efeitos sobre o corpo e das respostas adaptativas geradas para
contraposição de seus efeitos são seculares e sofreram ao longo do tempo modificações que
permitiram o aprimoramento de sua compreensão. Portanto, é necessário destacar duas
importantes teorias que lançaram bases fundamentais do entendimento de como o organismo
responderia quando desafiado por um estressor (Jackson, 2014).
A primeira, desenvolvida por Walter Cannon indicou que quando o organismo sofria
determinada exposição a um insulto, mecanismos de retroalimentação seriam ativados
visando a manutenção da estabilidade do meio interno, o que definiu como homeostasia.
Como exemplo, quando exposto a uma situação potencialmente perigosa, o corpo responderia
através da ativação da glândula adrenal, que ao liberar adrenalina, geraria respostas
fisiológicas necessárias para a reação de luta ou fuga. Essa resposta, portanto, visaria ao
reestabelecimento da homeostasia, fundamental para a sobrevivência (de I Brito, 2017). Outra
teoria foi desenvolvida por Hans Selye, responsável pela popularização do conceito de
estresse. Segundo o mesmo, quando o corpo é exposto a um determinado estressor,
desenvolve-se uma resposta fisiológica não específica a este insulto, dentre elas a ativação do
eixo hipotalâmico-hipofisário-adrenal (eixo HPA) e a consequentemente liberação de cortisol.
Esta resposta por sua vez, seria estabelecida através de um padrão trifásico compreendido por
uma fase inicial de alarme, seguido por uma fase de resistência ou adaptação, podendo levar
eventualmente à exaustão ou morte (Szabo et al., 2017). O estresse, portanto, poderia ser
definido como uma resposta adaptativa necessária à sobrevivência, porém, a partir de uma
exposição prolongada, doenças físicas e transtornos mentais poderiam ser desenvolvidos (Mc

24
Ewen, 2000).
Teorias mais contemporâneas sobre o conceito de estresse consideram que a resposta
ao estresse pode ter determinado padrão de especificidade, sendo condicionada pelo tipo de
estressor, sua duração, à percepção do organismo ao insulto e a habilidade para enfretamento.
As alterações naturais dentro de determinados parâmetros, que culminam com a estabilidade
das respostas fisiológicas nesse contexto, são atualmente definidas como alostase.
Cronicamente, exposição a estressores pode prejudicar a coordenação entre respostas
neuroendócrinas e comportamentais, gerando uma sobrecarga alostática que pode levar ao
desenvolvimento de respostas mal adaptativas e, consequentemente, a transtornos
psiquiátricos (Mc Ewen, 2000).
O importante papel do hipocampo, além das já referidas estruturas hipotalâmicas sobre
a regulação deste processo, já são estudadas desde a década de 60. Neste período, estudos
utilizando perda de função indicaram que lesões hipocampais levavam a um aumento nos
níveis basais de glicocorticoides (GC) (Fendler et al., 1961; Knigge, 1961), indicando que
estas regiões eram importantes para a regulação basal da liberação de GC. Além deste tipo de
regulação, foi evidenciado que a lesão do subiculum ventral, principal fonte de eferências
hipocampais no núcleo paraventricular, prejudicava a regulação do eixo HPA, gerando
hipersecreção de GC e aumento da biossíntese do hormônio liberador de corticotropina
(Herman et al., 1998). Estes efeitos foram também acompanhados por alterações
comportamentais nos animais mediante a exposição ao estresse agudo, fortalecendo o papel
importante do hipocampo na modulação da resposta frente ao estresse (Herman et al., 1998).
O estresse crônico, somado às condições citadas acima podem, portanto, prejudicar o
feedback negativo exercido pelo hipocampo sobre o eixo HPA (Mizoguchi et al., 2003).
Embora o papel do hipocampo sobre a regulação do estresse tenha sido extensamente
estudado, o papel da neurogênese hipocampal adulta sobre a modulação desta resposta foi
elucidado apenas recentemente. Estudo de Snyder e colaboradores (2011) sugeriu que em
animais transgênicos com capacidade neurogênica reduzida, o estresse agudo promovia uma
resposta mais acentuada em relação a secreção de corticosterona quando comparado a animais
controle, indicando que mecanismos de feedback negativos sobre o eixo HPA poderiam estar
comprometidos. Evidenciando ainda o importante papel da neurogênese na atenuação dos
efeitos do estresse crônico, recente estudo demonstrou que o aumento da neurogênese
induzido por meio da ablação do gene pró-apoptótico ibax sobre um promotor de nestina, era
capaz de reverter o déficit induzido pelo estresse sobre a neurogênese adulta (Culig et al.,
2017).

25
.
Experimentalmente, em animais, os efeitos do estresse podem ser estudados sobre
diversas perspectivas, dentre elas, o efeito da exposição a eventos estressantes sobre
manifestações comportamentais, bem como sobre eventos fisiológicos, por exemplo,
relacionados à plasticidade cerebral. Neste sentido, a aplicação do modelo de estresse crônico
imprevisível, tradução do inglês de chronic unpredictable stress (CUS) é capaz de gerar uma
diversidade de efeitos comportamentais e alterações fisiológicas que são paralelos a sintomas
de depressão. Essa exposição mimetiza o que ocorre em seres humanos, suportando a boa
validade de face do modelo (Willner et al., 2017). Além disso, a utilização do CUS justifica-
se por sua boa validade preditiva, ou seja, os animais são responsivos ao tratamento com
fármacos como, por exemplo, ao tratamento crônico com fármacos antidepressivos (Santarelli
et al., 2003).
Os efeitos negativos do estresse sobre a neurogênese hipocampal adulta, podem em
Figura 3. Estresse mecanismos de adaptação. Eventos estressantes são acompanhados de respostas
adaptativas visando o reestabelecimento da homeostase. Porém, de acordo com o tempo e a intensidade
de exposição a estes eventos, podem ser geradas respostas mal adaptativas que podem culminar no
desenvolvimento de transtornos psiquiátricos. O hipocampo é uma estrutura importante na regulação
destes processos e pode ser afetado negativamente pela exposição repetida ao estresse.

26
parte ser prevenidos ou revertidos com a utilização de fármacos antidepressivos. Estudo de
Morais e colaboradores (2014) evidenciaram que a aplicação do estresse crônico imprevisível
por 7 semanas promoveu uma redução no número de células BrdU colocalizadas com o
marcador de neuroblastos PSA-NCAM. Este efeito foi acompanhado por um comportamento
do tipo pró-depressivo nestes animais. O tratamento com fluoxetina, por sua vez, administrada
durante as últimas 3 semanas do período de aplicação do estresse, foi capaz de atenuar este
efeito comportamental e reverter o déficit sobre a neurogênese adulta. Analogamente, outro
estudo demonstrou que a fluoxetina (10mg/Kg) administrada por 5 semanas é capaz de
promover uma atenuação dos efeitos do CUS em camundongos no splash test e Novelty
Supressed Feeding. Os efeitos comportamentais foram acompanhados por um aumento no
número de células BrdU+, o que neste trabalho indicou um aumento de proliferação celular
(Surget et al., 2011). Seguindo a mesma compreensão, dados indicaram que a administração
do antidepressivo escitalopram (5 ou 10mg/Kg) por 4 semanas em ratos foi capaz de reverter
a redução de proliferação celular no giro denteado destes animais e o sintoma de anedonia
após a aplicação do estresse crônico variado aplicado inicialmente por 2 semanas e,
posteriormente por 4 semanas em combinação com o tratamento. (Jayatissa et al., 2006).
Embora diversos estudos demonstrem o efeito pró-neurogênico do tratamento crônico
com AD, dados são ainda escassos em relação a influencia dos AD em determinadas etapas da
neurogênese adulta. Nesse sentido, dados indicam que o tratamento crônico com fluoxetina
estimula a maturação e plasticidade sináptica de células granulares hipocampais e aumenta a
arborização dendrítica de células DCX+ (Wang et al., 2008; Wang et al., 2013). A fim de
determinar o período crítico no qual a fluoxetina induziria a maturação, Amellem e
colaboradores (2017) encontraram que o tratamento crônico com fluoxetina (28 dias) acelera
a maturação de novos neurônios em camundongos da linhagem C57BL6 quando comparado
ao tratamento com veículo, promovendo a maturação de células DCX+ (neurônios jovens)
para células calbindina+ (neurônios maduros) em um tempo anterior. Embora pertencente à
mesma classe da fluoxetina (inibidores seletivos da receptação de serotonina), o escitalopram
possui algumas diferenças em termos farmacodinâmicos (Pastoor et al., 2014), o que pode
condicionar respostas comportamentais e moleculares diferentes ainda pouco elucidadas.
Em termos estruturais, o escitalopram é uma formulação que contém apenas o
enantiômero S- do citalopram. Em um estudo pioneiro, foi demonstrado que tanto o
escitalopram, quanto o citalopram eram efetivos em reverter o fenótipo pró-depressivo em
ratos, após a aplicação do estresse crônico imprevisível (CUS). Porém, aparentemente o
escitalopram possuía um início mais rápido de ação. Neste mesmo estudo, essa hipótese foi

27
confirmada clinicamente, tendo o escitalopram eficácia comprovada no tratamento da
depressão e um efeito mais rápido que o citalopram (Montgomery et al., 2001). Foi
demonstrado também que o escitalopram possui uma alta afinidade pelo transportador de
serotonina, o que também poderia contribuir para o seu melhor perfil na clínica (Owens et al.,
2001). Além disso, testes clínicos posteriores confirmaram seu potencial no tratamento de
manifestações de ansiedade (Spadone, 2002). Cipriani e colabadoradores (2009, 2018)
demonstraram, através de um estudo de revisão sistemática com metanálise que o
escitalopram possui melhores perfis de aceitabilidade e alta taxa de responsividade na prática
clínica.
Embora recentes avanços no manejo de transtornos de ansiedade e de transtornos de
depressão tenham sido feitos com a incorporação de novos antidepressivos com melhores
perfis de eficácia e aceitabilidade e com menores incidências de efeitos adversos (Cipriani et
al., 2018), clinicamente ainda são observadas algumas limitações inerentes a essa classe de
fármacos. Algumas das mais importantes referem-se à alta taxa de refratariedade ao
tratamento (Pierz e Thase, 2014) e à latência observada para o início dos efeitos clínicos
(Frazer e Bemansour, 2002; Cipriani et al., 2018). Esta latência, por sua vez, pode estar
relacionada à capacidade de modulação de eventos neuroplásticos, como a neurogênese adulta
em diferentes etapas e tempos.
Sustenta-se, portanto, a necessidade de se estudar alternativas viáveis para o
tratamento destes transtornos. Evidências indicam que a manipulação do sistema
endocanabinoide, através de fármacos sintéticos ou através de fitocanabinoides como o
canabidiol (CBD) em testes pré-clínicos, é igualmente capaz de modular efeitos
comportamentais relacionados à ansiedade e à depressão (Zanelatti et al., 2010, Campos et al.,
2013) e seu mecanismo parece estar relacionado à regulação da neurogênese hipocampal
adulta (Wolf et al., 2010; Campos et al., 2013). Porém, de maneira semelhante aos AD,
estudos de como os fármacos canabinoides atuam sobre etapas específicas da neurogênese
hipocampal adulta são escassos e, portanto, de interesse para o presente estudo.
1.5. Canabinoides- Breve histórico, atuação sobre o sistema endocanabinoide
Os primeiros registros da utilização da Cannabis datam aproximadamente do século II
a.c, em uma passagem da importante obra da Cultura Antiga Chinesa, intitulada Livro dos
Ritos. Nesta obra, é relatado o importante papel que a fibra do cânhamo, extraída da
Cannabis, possuía na fabricação de roupas e sapatos, reduzindo a dependência do homem na
utilização de peles de animais para o vestuário (Abel, 1980). A utilização das fibras do

28
cânhamo também impulsionou a descoberta do papel que de fato se configurou como um
objeto fundamental de desenvolvimento da sociedade moderna. Embora possivelmente
utilizada com finalidades médicas durante um período mais antigo, o primeiro relato concreto
do uso da Cannabis está contido no manual padrão chinês sobre drogas, conhecido como Pen
Ts’ao. As indicações eram para fadiga menstrual, gota, reumatismo, malária, béri-béri,
constipação e distração, doenças que segundo a medicina chinesa, estavam associados a uma
perda do yin corporal. O vasto conhecimento à época já acumulado sobre a maconha, fez com
que os chineses fossem os primeiros a conhecer as suas propriedades psicoativas, seguido de
outras culturas como os povos que habitavam a Índia. Relatos em sânscrito, atribuída aos
indianos, incluso, referem-se a preparações com Cannabis realizadas com proporções bem
definidas destinadas para o uso em guerras, a fim de fortalecer a mente e abolir o pânico
provocado pela batalha (Abel, 1980)
Embora as fibras do cânhamo já fossem utilizadas pelos povos europeus, africanos e
posteriormente pelos americanos levadas pelos conquistadores europeus, a introdução da
maconha na Medicina Ocidental somente aconteceu no SÉC XIX, sendo atribuída ao médico
irlandês William O’Shaughnessy, pioneiro nos estudos sobre as propriedades medicinais da
Cannabis. O marco ocorreu em 1843, quando o médico publicou um resumo de seus estudos
sobre a droga em diversas afecções como reumatismo e epilepsia, despertando a atenção de
médicos europeus interessados em adotar o tratamento em suas práticas médicas. Porém,
dada a variabilidade de efeitos promovidos pela maconha, muitos médicos à época receberam
com cautela esses estudos, levando grupos de químicos a tentar isolar e identificar os
componentes ativos da planta.
Em 1890, um grupo de químicos da Universidade de Cambridge conseguiu isolar um
extrato relativamente puro que denominaram canabinol. Impulso importante, no entanto, foi
dado somente aproximadamente 70 anos depois com os trabalhos dos israelistas Gaoni e
Mechoulam, que identificaram estruturalmente os dois principais componentes da Cannabis
sativa, o canabidiol (CBD), isolado inicialmente em 1940 por Adams et al., (1940)
(Mechoulam e Shvo, 1963) e o 1- delta-9 trans- tetrahidrocanabinol (Δ9-THC), principal
componente psicoativo presente na planta (Gaoni e Mechoulam, 1965; Gaoni e Mechoulam,
1967). Outras substâncias, além das mais conhecidas Δ9-THC e CBD foram também
extraídas, identificadas e denominadas conjuntamente de fitocanabinoides. A elucidação das
novas substâncias, somado ao conhecimento milenar do uso terapêutico da Cannabis
impulsionaram a investigação de mecanismos relacionados à sua ação, culminando na
descoberta do Sistema Endocanabinoide em meados da década de 80.

29
A descoberta do sistema endocanabinoide, embora relativamente recente, foi baseada
em um acúmulo de evidências que apontaram para a existência de um conjunto de receptores
(CB1 e CB2) (Matsuda et al., 1990; Munro et al., 1993) e de enzimas de síntese e degradação
de neuromoduladores que seriam produzidos endogenamente sob demanda. O impulso inicial
para a pesquisa deste sistema foi dado pela tentativa de se elucidar os mecanismos pelos quais
a Cannabis sativa elicitava seus efeitos no sistema nervoso central. A descoberta e elucidação
estrutural do THC, possibilitou que as pesquisas avançassem consideravelmente, com a
síntese de análogos sintéticos que possibilitaram o estudo do sistema até então desconhecido.
A busca pela elucidação da “neurotransmissão canabinoide” rompeu paradigmas ao
indicar que a neurotransmissão do novo sistema não se daria da maneira clássica, mas por
meio da modulação da liberação de outros neurotransmissores (Cadogan, et al. 1997; Gifford
e Ashby 1996).
Embora o sistema endocanabinoide atue sob demanda regulando a atividade de outros
sistemas através da síntese e liberação dos eCB mencionados, os receptores e enzimas do
sistema endocanabinoide também podem ser alvo para os canabinoides sintéticos e
fitocanabinoides extraídos da planta Cannabis sativa. Apesar de por muito tempo, grande
parte da atenção científica tenha sido dispensada ao ∆9-tetrahidrocanabinol, principal
componente psicotomimético da C. sativa, o canabidiol (CBD) emergiu ao longo dos últimos
anos, tendo seu estudo grande enfoque atualmente, como uma droga promissora para o
tratamento de transtornos psiquiátricos.

30
1.6. Canabidiol (CBD)
1.6.1. Química do CBD
O CBD é um dos mais abundantes compostos naturalmente produzidos pela planta do
gênero Cannabis. Quimicamente é classificado como um composto terpernofenólico formado
por descarboxilação do precursor ácido canabidiólico sob influência do calor. Naturalmente, é
produzido como enantiômero (-)CBD, sendo os seus níveis de produção influenciados por
condições ambientais como temperatura do solo, índice de precipitação e temperatura
ambiental (Sikora et al., 2011). Rotas sintéticas também foram desenvolvidas em laboratórios
de síntese orgânica (Mechoulam et al., 2002), gerando diferentes fontes de disponibilidade
para o CBD. Em condições laboratoriais, por exemplo, sob condições ácidas, o CBD pode ser
convertido a Δ9-THC (Mechoulam et al., 2002) ao passo que espontaneamente essa
conversão parece não ocorrer.
Figura 4. de Oliveira RW et al. (2018). Esquema simplificado ilustrando a sinalização endocanabinoide. O
sistema endocanabinoide é composto pelos endocanabinoides 2-AG e AEA, biossentetizados nesta
representação a partir de aumento de Ca 2+
intracelular; pelas enzimas de síntese dos eCB: DAGL
(diacilglicerol lipase), NAPE (N-araquidonoil PE); enzimas de degradação dos eCB: FAAH e MAGL e
receptores CB1 e CB2. Além disso, os astrócitos e a micróglia podem exercer essa comunicação biredicional
neste sistema.

31
1.6.2. Efeitos clínicos do CBD
Historicamente, a utilização da Cannabis com propósitos clínicos data de séculos. O
impulso dado com o isolamento e elucidação estrutural do CBD e dada a necessidade de se
encontrar tratamentos alternativos a diversas doenças e transtornos mentais, fizeram com que
o estudo pormenorizado do CBD ganhasse bastante enfoque durante a segunda metade do
século XX. Um dos primeiros estudos clínicos realizados com esse composto foi
desenvolvido pelo grupo brasileiro liderado pelo professor Elisaldo Carlini da Universidade
de São Paulo que administrou CBD (66-67% pureza) nas doses de 200-300mg (v.o) a
pacientes com epilepsia generalizada secundária por um período de 4-5 meses. Dos 8
pacientes que receberam CBD, 4 demonstraram melhora clínica significativa, efeito que não
ocorreu em pacientes que já estavam recebendo medicações antiepiléticas, mas que não
haviam recebido CBD (Cunha et al., 1980). Outros estudos com enfoque da ação do CBD sob
manifestações da epilepsia foram publicados demonstrando o potencial do CBD isolado na
redução da frequência mensal de convulsões motoras por 12 semanas em crianças e adultos
jovens com epilepsia severa (Devinsky et al., 2016), mantendo-se com taxas semelhantes ao
longo de 96 semanas de tratamento (Szaflarski et al., 2018). Estudos também foram
conduzidos através da administração de extratos de Cannabis com alta proporção de CBD,
obtendo resultados promissores em relação à redução de manifestações da epilepsia em
crianças e adolescentes (Gedde e Maa, 2013; Tzadok et al., 2016).
Estudos com o CBD como potencial droga ansiolítica também foram realizados
clinicamente. O primeiro deles, também foi realizado por um grupo de pesquisa brasileiro, no
qual foi demonstrado que o CBD diminuía alguns dos efeitos ansiogênicos gerados pela
administração de altas doses de Δ9-THC (Karniol et al., 1974). Outro estudo confirmou os
Figura 5. Estrutura química do canabidiol

32
resultados previamente obtidos e demonstrou que a administração concomitante de CBD
(1mg/Kg) era capaz de atenuar alguns dos efeitos relacionados à ingestão de altas doses de
Δ9-THC (0,5mg/Kg), dentre eles o efeito ansiogênico (Zuardi et al., 1982). Em 1993, o
mesmo grupo demonstrou que a administração de CBD (300mg) reduzia a ansiedade de
voluntários submetidos ao teste simulado de falar em público, sugerindo que o CBD possuía
propriedades ansiolíticas em indivíduos submetidos a situações de estresse (Zuardi et al.,
1993). A fim de determinar como os efeitos do CBD eram mediados em nível central, Crippa
e colaboradores (2004) valendo-se de técnica de neuroimagem demonstraram que o CBD
além de diminuir a ansiedade subjetiva promovida pela própria aplicação do teste, exercia
seus efeitos através da ação em áreas cerebrais límbicas e pré-límbicas. Posteriormente,
Bergamaschi e colaboradores (2011) demonstraram que o CBD na dose de 600mg era capaz
de reduzir manifestações relacionadas à ansiedade em pacientes com Transtorno de Ansiedade
Social Generalizada, que, todavia, nunca haviam recebido tratamento para a condição. As
pesquisas relatadas acima denotam o potencial do CBD para o tratamento de transtornos
relacionados à ansiedade.
Estudos clínicos com o CBD também foram conduzidos em transtornos psicóticos.
Embora os resultados encontrados sejam conflitantes (McGuire et al., 2018; Boggs et al.,
2018), alguns estudos evidenciaram que o canabinoide administrado durante 4-6 semanas
poderia ser efetivo para o tratamento da esquizofrenia, através da melhora dos sintomas
psicóticos, somado a uma baixa incidência de efeitos adversos (Leweke et al., 2012; McGuire
et al., 2018). No estudo de Leweke e colaboradores, (2012) este efeito correlacionou-se
clinicamente a um aumento dos níveis de anandamida (AEA) (Leweke et al., 2012).
Atualmente o CBD é comercializado em combinação com o THC (1:1) na forma de
spray de administração oral em uma formulação farmacêutica de spray (Sativex®) para o
tratamento da espasticidade refratária moderada a severa em pacientes com esclerose
múltipla, proporcionando melhora clinicamente relevante. (Markovà et al., 2018; Keating,
2017).
Além das aplicações mencionadas, o CBD vem sendo estudado para o tratamento da
síndrome de retirada a drogas de abuso como Cannabis (Crippa et al., 2013; Trigo et al.,
2018) e nicotina (Morgan et al., 2013). Todos estes estudos sinalizam a uma grande tendência
para o estudo dos canabinoides nas mais diversas condições clínicas. Dados da literatura
(Bergamaschi et al., 2011) endossam ainda que o CBD é seguro e bem tolerado nas doses
testadas, mesmo em voluntários saudáveis (Martin-Santos et al., 2012), não tendo sido
demonstrados efeitos adversos significativos em pacientes administrados com dose de até

33
1500mg/Kg por via oral (Zuardi et al., 1995). Porém, dada a multiplicidade de efeitos, faz-se
necessário estudar os mecanismos pelos quais o CBD e outros canabinoides exercem seus
efeitos.
1.6.3. Farmacodinâmica e farmacocinética do CBD
Atualmente, sabe-se que o CBD pode produzir ações farmacológicas múltiplas, dada a
sua capacidade de atuar em diversos receptores presentes em distintas estruturas do sistema
nervoso central e dado o seu efeito dose dependente (Campos et al., 2012) . Em relação ao
sistema endocanabinoide, dados experimentais obtidos a partir de estudos de binding
demonstram que o CBD possui baixa afinidade pelos receptores CB1 e CB2 (Showalter et al.,
1996; Bisogno et al., 2001).
Estudo recente indicou que o CBD pode atuar como um modulador alostérico negativo
sobre os receptores CB1 (Lapraire et al., 2015; Tham et al., 2018). De maneira intrigante, no
entanto, diversos resultados indicaram que os efeitos do CBD eram revertidos por agonistas
inversos do receptor CB1 e que eram ausentes em animais knockouts para o receptor CB1
(Casarotto et al., 2010; Campos et al., 2013). Dado esse fato, Bisogno e colaboradores (2001)
observaram que o CBD possuía a capacidade de inibir a enzima de degradação da
anandamida, a FAAH, aumentando, portanto, a sinalização mediada pelo endocanabinoide,
por exemplo, em CB1. Outros alvos para o CBD já foram elucidados, como o receptor 5-
HT1A, através de estudos de binding (Russo et al., 2005) e a partir de evidências obtidas de
experimentos comportamentais (Zanelati et al., 2010; Sonego et al., 2016; Campos et al.,
2013), o receptor órfão GPR55 (Ryberg et al., 2007), os receptores µ e ƍ opióides, nos quais
atua como modulador alostérico (Kathmann et al., 2006), receptor vanilóide TRPV1, o qual
parece ser estimulado e dessensibilizado pelo CBD (Bisogno et al., 2001; Ianotti et al., 2014;
De Petrocellis et al., 2011) e receptores PPARγ (Esposito et al., 2011).
Em relação aos parâmetros farmacocinéticos do CBD, sabe-se que possui baixa
solubilidade em água e absorção errática pelo trato gastrointestinal. Além disso, sofre
significativo metabolismo de primeira passagem no fígado, o que reduz sua
biodisponibilidade a partir da administração oral (Devinsky et al., 2014). Em camundongos,
após a administração i.p, o Tmax foi atingido após 1 hora no cérebro e 2 horas no plasma. O
tempo de meia vida t1/2 aparente no plasma e no cérebro não sofreram variações significativas
e foram de aproximadamente 4,5 a 5 horas (Deiana et al., 2012). Como outros canabinoides, o
CBD é metabolizado pelo fígado por enzimas do CYP450 (Ujváry e Hanus, 2016).

34
Importante mencionar que variações interindividuais na expressão e função de enzimas do
CYP450 podem afetar a farmacocinética do CBD e seus metabólitos (Ujváry e Hanus, 2016).
1.6.4. Efeitos pré-clínicos observados com o CBD
Estudos pré-clínicos também confirmam o potencial do CBD, tendo sido
demonstrados efeitos do tipo antipsicótico (Moreira e Guimarães, 2005; Long et al., 2006),
efeitos anticonvulsivantes (Vilela et al., 2017) e do tipo ansiolítico em modelos como
labirinto em cruz elevado (Onaivi et al., 1990; Guimarães et al., 1990), teste de conflito de
Vogel (Campos et al., 2008; Moreira et al., 2006) e novelty supressed feeding (Campos et al.,
2013).
Recentemente, foi demonstrado que a administração crônica de CBD por 14 dias
(3mg/Kg) reduziu o tempo de imobilidade no tail suspension test em relação ao grupo
controle, indicando um comportamento do tipo antidepressivo (Schiavon et al., 2016). Outro
estudo indicou que o efeito do tipo antidepressivo promovido pelo CBD em camundongos
Swiss machos (7-8 semanas de idade) no teste do nado forçado modula a via de sinalização
mTOR (Sartim et al., 2018) e dependente dos níveis cerebrais de serotonina (Sales et al.,
2018).
Outro aspecto importante diz respeito à possível capacidade do CBD em exercer
efeitos comportamentais rápidos. Com relação a este fator, estudo de Linge e colaboradores
(2016) demonstrou que o CBD administrado por 7 dias é capaz de induzir um efeito rápido e
sustentado do tipo antidepressivo via aumento da sinalização cortical de serotonina e
glutamato. No entanto, os mecanismos pelos quais o CBD pode promover seus efeitos e
possivelmente acelerar as manifestações comportamentais, permanecem como fonte de
investigação. Embora grande número de informações tenham sido geradas, devido o grande
potencial que pode advir deste composto, estudos adicionais são necessários para se investigar
outros mecanismos que podem estar associados aos efeitos pré-clínicos do CBD, dado o seu
comprovado efeito pleiotrópico. Pesquisas recentes demonstraram que o CBD, assim como os
AD também atua sobre processos neuroplásticos, dentre eles a neurogênese hipocampal
adulta. Os mecanismos pelos quais o CBD interfere sobre a neurogênese adulta, no entanto,
permanecem sob investigação. Um dos estudos realizados identificou que o CBD exerce
efeitos pró-neurogênicos através do envolvimento do receptor PPARγ (Esposito et al., 2011).
Além disso, promove o aumento da sobrevivência neuronial e de outros estágios da

35
neurogênese adulta (Esposito et al., 2011, Wolf et al., 2010), através de um mecanismo
dependente da sinalização via receptor CB1.
No estudo citado de Wolf et al., 2010 foi demonstrado que o CBD administrado a
camundongos por meio da dieta por 6 semanas, é capaz de promover um aumento de células
Brdu/NeuN positivas 7 dias após a injeção de BrdU, indicando que o CBD pode acelerar a
maturação neuronal. Estes efeitos neurogênicos mediados por CBD foram dependentes da
sinalização via receptor CB1. Na ausência deste receptor, no entanto, o CBD promovia um
aumento da proliferação celular, mas a capacidade de diferenciação neuronal era reduzida.
Em outro recente estudo, foi demonstrado que o tratamento repetido com CBD (14
dias) foi capaz de atenuar os efeitos da aplicação do estresse crônico imprevisível no teste do
Novelty Supressed Feeding e concomitantemente, reverter o déficit neurogênico após a
aplicação do mesmo. Os autores demonstraram que este efeito do tipo ansiolítico foi
dependente da sinalização mediada pelos receptores CB1 e CB2 e que o efeito pró-
neurogênico foi mediado através da sinalização via CB2. Além disso, foi observado que o
CBD é capaz de agir em outros processos neuroplásticos, como no aumento da expressão de
proteínas sinápticas em relação aos animais submetidos ao estresse e aumento do
remodelamento dendrítico (Fogaça et al., 2018). Campos e colaboradores (2013)
demonstraram que o tratamento crônico com CBD por 14 dias é capaz de prevenir o
comportamento do tipo ansiogênico em animais submetidos ao modelo de estresse crônico
imprevisível no modelo de labirinto em cruz elevado e no paradigma do Novelty suspressed
feeding (NSF) (Campos et al., 2013). Não obstante, os autores demonstraram ainda que os
efeitos ansiolíticos do CBD em camundongos cronicamente estressados são ausentes em
animais transgênicos com capacidade neurogênica adulta nula ou bastante reduzida (Campos
et al., 2013).
Embora os autores não tenham comparados os efeitos do CBD com os antidepressivos
clássicos, o efeito em 14 dias poderia indicar uma possível redução de latência em relação ao
tratamento com imipramina e fluoxetina (geralmente após 21 dias- Santarelli et al., 2003). O
presente trabalho consiste, portanto, em testar a hipótese de que o CBD é capaz de produzir
efeitos comportamentais em animais cronicamente estressados em uma latência menor do que
o antidepressivo escitalopram. Além disso, objetiva-se analisar se a redução da latência
observada possui relação com mecanismos da neurogênese hipocampal adulta, analisando-se
as etapas e marcadores celulares relacionados à neurogênese.

36
OBJETIVOS

37
2. Objetivos
2.1. Objetivo geral
Analisar o papel do CBD em comparação direta ao antidepressivo escitalopram sobre os
efeitos ansiolíticos do CBD em animais cronicamente estressados, bem como os mecanismos
pró-neurogênicos possivelmente associados.
2.2.Objetivos específicos
. Analisar o efeito ansiolítico do tratamento repetido (7 dias) com CBD ou escitalopram em
animais submetidos ou não ao protocolo de estresse crônico durante o mesmo período.
Analisar o efeito do estresse e dos tratamentos sobre a sobrevivência celular, testado pela
incorporação de BrdU no time point de 7 dias.
. Identificar por meio da análise de DCX (doblecortin) o efeito dos tratamentos em diferentes
condições sobre o número de células (Tipo 2b e Tipo 3) comprometidas com o fenótipo
neuronial no time points de 7 dias.
. Identificar por meio da análise de células Ki67+ o efeito do estresse e dos tratamentos sobre
a proliferação celular.
. Analisar o efeito ansiolítico do tratamento repetido (14 dias) com CBD ou escitalopram em
animais submetidos ou não ao protocolo de estresse crônico durante o mesmo período.
. Analisar a expressão de células BrdU+ no time point de 14 dias. Essa análise permite
identificar células que sobreviveram durante o time point (14 dias) testado pela incorporação
de BrdU.
. Identificar a expressão de células SOX2+ no time point de 14 dias. Essa análise permite
identificar células precursoras amplificadas (Tipo 2 a) neste time point.

38
. Identificar por meio da análise de DCX (doblecortin) o efeito dos tratamentos em diferentes
condições sobre o número de células (Tipo 2b e Tipo 3) comprometidas com o fenótipo
neuronial no time points de 14 dias.
MATERIAL E MÉTODOS

39
3. Material e métodos
3.1.Animais
Os experimentos foram conduzidos com camundongos machos selvagens C57BL/6
(n=120), originados do biotério central da Universidade de São Paulo (USP)/ Campus de
Ribeirão Preto/SP com aproximadamente 6 semanas e utilizados no início do protocolo
experimental com aproximadamente 9-12 semanas de idade. Após a chegada, os
camundongos foram alocados randomicamente em caixas de acrílico (30, 20, 13 cm) em 5
animais/grupo com acesso ad libitum à água e comida e viveram em um ciclo 12X12 horas
claro/escuro. A cama dos animais consistiu em aproximadamente 2 cm de maravalha, trocada
uma vez na semana. Além disso, foram mantidos em biotério com temperatura controlada
(aproximadamente 23ºC). Os protocolos experimentais foram previamente submetidos à
análise da Comissão de Ética no Uso de Animais (CEUA) da Universidade local e aprovados
(Protocolo nº 56/2017).
3.2.Drogas
Foram utilizados escitalopram (Prati Donaduzzi®) (20mg/Kg) dissolvido em salina
0,9% (Dong et al., 2016) e canabidiol (BSPG®) (30mg/Kg) dissolvido em 2% de Tween 80 e
salina 0,9% (Campos et al., 2013). As drogas foram preparadas imediatamente antes da
utilização em ambiente estéril e os animais foram tratados por via intraperitoneal (10ml/Kg)
de acordo com o grupo experimental uma vez ao dia durante todo o protocolo experimental,
preferencialmente, 2 horas após o estressor diário (Campos et al., 2013). O grupo controle
recebeu veículo (2% Tween 80 e salina 0,9%). Além dos tratamentos citados, os animais
receberam injeção de BrdU (200mg/Kg) em dose única no primeiro dia de protocolo
experimental.
3.3.Procedimento de estresse
Os animais de cada grupo experimental foram submetidos ao regime de estresse crônico
imprevisível, definição traduzida do inglês de Chronic Unpredictable Stress (CUS), modelo
modificado proposto por Katz e Hersch et al., 1981. Os animais foram expostos a diferentes
estressores de maneira randomizada, sendo que apenas 1 estressor diário foi utilizado. Os
estressores aplicados foram: 15 minutos de natação forçada a 25ºC, maravalha molhada (24

40
horas), inversão do ciclo claro/escuro, inclinação da caixa (overnight), intervalos inconstantes
de inversão de ciclo durante o dia (24 horas), estresse de restrição em tubo falcon por 2 horas
e privação de comida por 24 horas, sendo este o estressor utilizado em comum para os grupos
controle e estressado, de maneira a ser necessário para a aplicação do teste comportamental.
Durante o protocolo de estresse, foram registrados os pesos dos animais.
3.4.Teste comportamental
No 8º ou 15º dia de protocolo experimental, os animais foram submetidos ao
paradigma da Supressão da Alimentação pela Novidade, ou do inglês, Novelty Supressed
Feeding (NSF). O modelo em questão, desenvolvido por Bodnoff e colaboradores (1988)
baseia-se na privação de alimento do animal durante 24 horas. Decorrido este período, o
animal é exposto a uma caixa transparente de acrílico (50x50x20 cm), contendo serragem
cobrindo o fundo da caixa, localizada em uma sala aos quais nunca havia sido exposto. Nesta
caixa foi introduzido no seu centro um pellet de ração sobre um suporte de plástico, focados
por uma luz incidente e a caixa colocada em ambiente ausente de qualquer outra fonte de
luminosidade. Os animais previamente habituados por cerca de 2 horas em sala diferente da
de aplicação do procedimento, foram introduzidos individualmente em uma das extremidades
da caixa e foi medida a latência para que o animal iniciasse o consumo do alimento dentro de
um tempo limite de 10 minutos. Este modelo cria, portanto, um conflito entre uma tendência
natural para comer após a privação de comida e aversão etológica ao novo, à luz incidente e
ao ambiente desprotegido conferido pelo centro da caixa de teste (Campos et al., 2013). Logo
em seguida, os animais foram colocados individualmente em sua home cage e foi avaliada a
latência para que o camundongo comesse, bem como o consumo total de ração dentro de um
período de 5 minutos. Todos os testes comportamentais foram registrados por meio de
filmagem.
3.5.Perfusão e preservação da amostra
No dia seguinte ao teste comportamental, os animais previamente anestesiados com
um volume de 10ml/Kg de ketamina (150mg/Kg) e xilazina (8mg/Kg) foram perfundidos por
via transcardíaca com salina tamponada em tampão fosfato (30mL) seguidos pela infusão de
paraformaldeído (PFA) 4% previamente filtrado (30mL). Os animais foram decapitados e os
encéfalos removidos. Posteriormente, os encéfalos foram pós-fixados overnight em PFA 4% e

41
crioprotegidos em solução de sacarose 30% por aproximadamente 4 dias. Após estes
procedimentos, a sacarose foi removida e os encéfalos congelados em isopentana/vapor de
gelo seco (-30°C) por aproximadamente 30 segundos e armazenados em freezer -80°C até o
momento dos cortes em criostato (Leica, Alemanha). Os animais foram escolhidos
aleatoriamente e, posteriormente é realizada a análise correlacional obtido entre os dados
comportamentais e os resultados das técnicas moleculares.
3.6.Delineamento experimental
No início do protocolo experimental (dia 1), todos os animais receberam uma injeção
i.p única de BrdU 200mg/Kg e foram ou não submetidos ao protocolo de estresse crônico
imprevisível durante 7 ou 14 dias. Durante o mesmo período os animais receberam tratamento
i.p com veículo, CBD 30mg/Kg ou Escitalopram 20mg/Kg. No 8º ou 15º dia de protocolo, os
animais foram submetidos ao teste comportamental (NSF) e perfundidos no dia seguinte para
a remoção dos encéfalos para análises moleculares (Figura 5 e Figura 6).
3.6.1. Delineamento experimental
No experimento de 7 dias, os animais foram divididos randomicamente de forma
aleatória entre 6 grupos; animais não estressados (NS) tratados com veículo, CBD 30mg/Kg
ou Escitalopram 20m/Kg e animais submetidos ao protocolo de estresse crônico imprevisível
tratados com veículo, CBD 30mg/Kg ou Escitalopram 20mg/Kg. Inicialmente, todos os
grupos experimentais foram constituídos por um n=10 por grupo. Porém, reduções neste
número ocorreram durante o experimento e são explicitados nos resultados. Os experimentos
foram conduzidos de maneira independente
Tratamento
Condição
Veículo CBD 30mg/Kg Escitalopram
(20mg/Kg)
NS NS/Vei (n=9) NS/CBD (n=10) NS/ESC (n=10)
CUS CUS/Vei (n=10) CUS/CBD (n=9) CUS/ESC (n=10)
Tabela 2. Grupos experimentais. Protocolo de 7 dias. Todos os experimentos foram conduzidos de
forma independente com n=9/grupo NS/Vei e CUS/CBD e n=10 demais grupos em um total de 6
grupos experimentais.

42
No experimento de 14 dias, os animais foram divididos randomicamente de forma aleatória
entre 6 grupos; animais não estressados (NS) tratados com veículo, CBD 30mg/Kg ou
Escitalopram 20m/Kg e animais submetidos ao protocolo de estresse crônico imprevisível
tratados com veículo, CBD 30mg/Kg ou Escitalopram 20mg/Kg. Inicialmente, todos os
grupos experimentais foram constituídos por um n=10 por grupo. Porém, reduções neste
número ocorreram durante o experimento e são explicitados nos resultados. Os experimentos
foram conduzidos de maneira independente
Tratamento
Condição
Veículo CBD 30mg/Kg Escitalopram
(20mg/Kg)
NS NS/Vei (n=10) NS/CBD (n=10) NS/ESC (n=10)
CUS CUS/Vei (n=6) CUS/CBD (n=10) CUS/ESC (n=10)
3.6.2. Protocolo experimental
No início do protocolo experimental (dia 1), todos os animais receberam uma injeção i.p
única de BrdU 200mg/Kg e foram ou não submetidos ao protocolo de estresse crônico
imprevisível durante 7 ou 14 dias. Durante o mesmo período de aplicação do estresse, os
animais receberam tratamento diário único i.p com veículo, CBD 30mg/Kg ou Escitalopram
20mg/Kg. No 8º ou 15º dia de protocolo, os animais foram submetidos ao teste
comportamental (NSF) e perfundidos no dia seguinte para a remoção dos encéfalos para
análises moleculares (Figura 6 e Figura 7).
Tabela 3. Grupos experimentais. Protocolo de 14 dias. Todos os experimentos foram conduzidos de
forma independente com n=6/grupo CUS/Vei e n=10 demais grupos em um total de 6 grupos
experimentais.

43
3.7.Preparação dos tecidos para a técnica de imunohistoquímica
Previamente os cérebros foram cortados em criostato (-25°C) em seções de 30µm
(bregma 1,34mm- 2,7mm- Paxinos e Franklin, 2012) de maneira a se ter representatividade
ântero-posterior do hipocampo. Os cortes foram colocados em placas de 24 poços (8 poços
por animal contendo aproximadamente 8 cortes em cada um) contendo solução anticongelante
e armazenados em freezer -20°C até o momento do protocolo. Os cortes para
imunohistoquímica foram selecionados em placa de Petri de acordo com a estrutura desejada
(hipocampo) e colocados em poço de placa de 24 poços.
3.7.1. Determinação de células doblecortin (DCX) positivas por imunohistoquímica
Os cortes foram lavados por 3 vezes em solução de TBS 50mM por 15 minutos e em
seguida foi realizada a recuperação antigênica com tampão citrato (10mM, pH=6) em estufa a
70°C por 30 minutos. Foi realizada novamente a lavagem do tecido, porém utilizou-se TBS +
(TBS 50mM + 0.1% soro albumina bovina- BSA + Triton 100X 0.125%) como solução de
Figura 6. Delineamento experimental. Protocolo de 7 dias
Figura 7. Delineamento experimental. Protocolo de 14 dias

44
lavagem para permeabilização do tecido. O bloqueio foi realizado por 2 horas com solução de
bloqueio (TBS 50mM + 1% BSA + Triton 100X 0.25%) para em seguida, os cortes serem
incubados com o anticorpo primário anti-Doblecortin 1:200 (Goat- Santa Cruz®) diluído em
solução de TBS +. A incubação foi realizada por 16 horas em agitador orbital a uma
temperatura de 18°C. Posteriormente, os cortes foram lavados em solução de TBS+ e o
anticorpo secundário anti-Goat 1:1000 incubado por 1 hora (Vector Labs). Procedeu-se nova
lavagem e posterior incubação do complexo avidina-biotina por 1 hora (1:1000). Após nova
lavagem, os cortes foram incubados em solução de 3,3’ diaminobenzidina DAB por
aproximadamente 20 minutos. Os cortes foram, por fim, lavados com TBS para posterior
montagem das lâminas (mínimo de 6 cortes por lâmina) e diafanização. Ao término dos
procedimentos, os cortes foram montados em lâmina/lamínula com auxílio do meio de
montagem Permount e as análises foram realizadas por microscopia de luz convencional
(Olympus) (Campos et al. 2013). Foram quantificados 16 giros denteados em 8 cortes de
hipocampo, localizados entre as seguintes coordenadas: -1.22 a -2.26mm (baseadas do atlas
Paxinos e Franklin). As células DCX positivas foram quantificadas utilizando-se a objetiva de
40X. Além do número total de células, células que tivesse seu corpo celular localizado na
cama granular do giro denteado foram consideradas em estado de migração. O total de células
foi expresso através da quantidade total de células positivas/ mm2 de giro denteado (Fogaça et
al., 2018).
3.7.2. Determinação de células BrdU positivas por imunohistoquímica
O protocolo foi realizado como descrito anteriormente, excetuando-se o processo de
recuperação antigênica, no qual foi utilizado HCl (2M) em estufa a 37°C por 30 minutos para
procedimentos de desnaturação do DNA, resfriamento imediato a 10ºC por 10 minutos, com
posterior lavagem dos cortes com solução de ácido Bórico (1M- pH= 8.9). A incubação do
anticorpo primário, foi realizada com anti-BrdU 1:100 (Mouse- Hybridoma Bank, Iowa
University diluído em solução de TBS + e o anticorpo secundário utilizado foi o anti-Mouse
1:500 por 1 hora (Vector Labs). Durante a montagem das lâminas, além da diafanização, os
cortes foram tratados por 1 minuto com hematoxilina de Mayer. As células BrdU positivas
foram quantificadas utilizando-se a objetiva de 20X e o total de células foi expresso através da
quantidade total de células positivas/ mm2 de giro denteado (Fogaça et al., 2018).

45
3.7.3. Determinação de células Ki67 positivas para imunohistoquímica
O protocolo foi realizado de maneira semelhante ao protocolo para determinação de
células DCX+. No entanto, a incubação de anticorpo primário, foi realizada com anti-Ki67
1:100 (Rabbit) diluído em solução de TBS + e o anticorpo secundário utilizado foi anti-rabbit
(1:500) incubado por 1 hora (Vector Labs). Durante a montagem das lâminas, além da
diafanização, os cortes foram tratados por 1 minuto com hematoxilina de Mayer. As células
Ki67 positivas foram quantificadas utilizando-se a objetiva de 20X e o total de células foi
expresso através da quantidade total de células positivas/0,1 mm2 de giro denteado.
3.8.Imunofluorescência para detecção de BrdU/SOX2
O processo foi realizado de maneira semelhante ao protocolo para imunohistoquímica
para detecção de BrdU. Porém, os anticorpos primários utilizados foram anti-SOX2 1:500
(Rabbit-Millipore) e anti-BrdU (rat- Abcam) 1:100 preparados na mesma solução de
incubação de TBS+. Os anticorpos secundários utilizados foram Alexa 594 (anti-rat-
Invitrogen) e Alexa 647 (anti-rabbit- Invitrogen). Os cortes foram montados em lâminas no
mesmo dia em ambiente escuro em meio Fluormalte (Eletron Microscopy) e selados com
esmalte para posterior análise. As células BrdU+
foram contadas em objetiva de 20X e o total
de células expresso através do número total de células/0,1mm2. As células SOX2
+, por sua
vez foram determinadas através do programa ImageJ® (NCBI) em objetiva de 10X.
3.9.Análise estatística
Os dados foram testados quanto à normalidade (teste Kolmorov-Smirnov) e à
homogeneidade de variâncias (teste de Levene). No caso de cumprimento destes pré-
requisitos, a análise dos resultados do teste comportamental foi realizada por two-way
ANOVA (tratamento x condição) seguida de teste post hoc de Duncan. Foi adotado um valor
de α de 0,05 como nível de significância estatístico. Nos testes moleculares procedeu-se a
mesma análise estatística anteriormente descrita, a fim de identificar possível interação entre
os fatores condição (Controle x CUS) e tratamento (Veículo, CBD 30mg/Kg, Escitalopram
20mg/Kg). Foi realizado posteriormente análise de one-way ANOVA, seguido de post hoc de
Duncan para identificação de qual (is) dos grupos experimentais eram diferentes
estatisticamente entre si.

46
RESULTADOS

47
4. Resultados
4.1. Protocolo experimental de 7 dias
No protocolo comportamental de 7 dias apresentado, a análise de two-way ANOVA
revelou diferença significativa no fator tratamento (CBD 30mg/Kg/ ESC 20mg/Kg/ veículo, F
(2,58)=5,896, p<0,05) e embora bastante próxima da significância estatística, não foi
encontrado diferença significativa no fator condição (controle vs estresse F= (1,58) = 3,577,
p=0,06). Além disso, as análises indicaram que houve interação entre condição (animais
submetidos ao protocolo de estresse crônico imprevisível ou animais não estressados) e
tratamento (F (2,58) = 3,733 , p<0,05). Análises post hoc indicaram que os animais do grupo
controle tratados com escitalopram 20mg/Kg apresentaram significativamente uma menor
latência para comer no novo ambiente em relação aos animais controle tratados com veículo.
Além disso, no grupo estressado cronicamente, a administração repetida de canabidiol
(30mg/Kg/ 7 dias), mas não de escitalopram, reduziu a latência para comer no novo ambiente
quando comparado ao grupo veiculo. Nenhum efeito foi observado no consumo de alimento
na home cage, indicando que o estresse, tampouco nenhum dos tratamentos alterou a ingesta
alimentar dos animais. Neste protocolo, um animal do grupo NS/Veículo foi excluído da
análise por ter encontrado ração na maravalha durante o teste no novo ambiente,
prejudicando a geração de conflito no animal e, portanto, inviabilizando sua utilização.

48
Figura 8. Efeitos comportamentais do tratamento repetido com o CBD e escitalopram em animais controle ou estressados e submetidos ao paradigma do Novelty Supressed Feeding. Time point: 7 dias A) Latência em segundos (s) que os animais gastam para se alimentarem em um novo ambiente. Tempo de corte do teste: 10 minutos. Dados são expressos como média±SEM. n=9- grupo NS/Veículo ou n=10- demais grupos. p<0,05 em relação ao grupo Controle/Veículo; # p<0,05 em relação ao grupo CUS/Veículo. (ANOVA de duas vias; ANOVA de uma via seguido de post hoc de Duncan). B) Consumo na home cage após o tratamento repetido com o CBD ou escitalopram em animais controle ou estressados e submetidos ao paradigma do Novelty Supressed Feeding. Consumo de alimento em gramas (g) durante 5 minutos. Time point de 7 dias. Dados são expressos como média±SEM. n=9 - grupo NS/Veículo e grupo CUS/CBD; ou n=10 - demais grupos.
A
B

49
4.1.1. Efeito do tratamento repetido com CBD ou escitalopram (7 dias) sobre a taxa de
sobrevivência celular, em animais controle e cronicamente estressados
A taxa de sobrevivência celular foi determinada neste protocolo a partir da análise da
quantidade de células BrdU+ por 0,1mm
2 de área no giro denteado dos animais submetidos ao
tratamento repetido com CBD ou escitalopram, submetidos ou não ao protocolo de estresse
crônico imprevisível. A administração de BrdU, que incorpora-se ao DNA de células que
estão na fase S do ciclo celular, no primeiro dia de protocolo, permite a análise da
sobrevivência celular decorridos os 7 dias de protocolo, permitindo observar se algum dos
tratamentos ou a aplicação do estresse modulam a taxa de sobrevivência celular.
A análise estatística por two-way ANOVA não indicou efeito significativo do fator
tratamento ou condição. Além disso, não foi observada interação entre os fatores tratamento e
condição. Mediante os efeitos comportamentais encontrados neste time point explorados no
tópico anterior, realizamos análises de correlação de Pearson entre a latência dos animais para
se alimentarem no novo ambiente e o respectivo efeito sobre o número de células BrdU+.
Realizamos a análise entre todos os animais, sem distinção entre os grupos, visando obter a
informação se o comportamento do animal é diretamente relacionado a um efeito sobre a taxa
de sobrevivência celular. Também realizamos as mesmas análises correlacionais distinguindo
os grupos pelo fator condição (controle x submetidos ao estresse) e pelo fator tratamento
(véiculo, CBD, escitalopram). Não encontramos, todavia, nenhum efeito correlacional pela
análise global (Figura 10) ou pela distinção entre os grupos sugerida acima (ANEXO B)

50
Figura 9. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a sobrevivência de células BrdU
+/0,1mm
2 de giro denteado do hipocampo. Time point: 7 dias. Dados
são expressos como média±SEM. n=4 grupo CUS/Escitalopram ou n=5 demais grupos.
Figura 10. Análise correlacional entre o efeito comportamental obtido no NSF e o número de células BrdU
+/0,1mm
2 de giro denteado. Time point: 7 dias. Foi realizada análise por correlação de Pearson entre o
efeito observado no comportamento para cada animal, independentemente do grupo experimental e o respectivo efeito molecular. n=28 animais.

51
Imag
ens
rep
rese
nta
tivas
do
gir
o d
ente
ado
do
s gru
po
s ex
per
imen
tais
no
tim
e p
oin
t d
e 7
dia
s. F
oi
uti
liza
da
técnic
a d
e im
uno
his
toq
uím
ica
par
a a
mar
caçã
o d
as
célu
las
Brd
U+,
que
estã
o e
vid
enci
adas
po
r m
arca
ção
de
DA
B.
Alé
m d
isso
, nest
e p
roto
colo
, as
cél
ula
s fo
ram
co
ntr
aco
rad
as c
om
Hem
ato
xil
inad
e M
ayer
. A
s fo
tos
fora
m t
irad
as e
m a
um
ento
de
20
X e
co
m o
s m
esm
os
par
âm
etr
os
de
tem
po
de
exp
osi
ção
e b
alan
ço d
e b
ranco
.

52
4.1.2. Efeito do tratamento repetido com CBD ou escitalopram (7 dias) sobre a taxa de
proliferação celular, em animais controle e cronicamente estressados
A proliferação celular foi determinada neste protocolo a partir da análise da
quantidade de células Ki67+ por 0,1mm2 de área no giro denteado dos animais submetidos ao
tratamento repetido com CBD ou escitalopram, submetidos ou não ao protocolo de estresse
crônico imprevisível. O Ki67 é um antígeno nuclear expresso durante todos os estágios do
ciclo celular, exceto na fase G0, ou seja, é expresso durante todas as fases proliferativas do
ciclo celular (Fisher et al., 2002).
A análise por two way ANOVA indicou diferença estatisticamente significativa no
fator condição F (1,17)=30,43, p<0,05) e no fator tratamento F(2,17)= 3,69, p=0,05, mas não
indicou interação entre condição e tratamento. O teste post hoc de Duncan, aplicado após o
teste de one way ANOVA, demonstrou diferença significativa entre o grupo NS/Veículo e
CUS/Veículo, no qual o estresse reduziu a taxa de proliferação celular. Em contrapartida,
nenhum dos tratamentos empregados (CBD ou Escitalopram), foram capazes de reverter a
redução da proliferação celular promovida pelo estresse crônico. Além disso, não alteraram a
taxa de proliferação celular nos animais controle.

53
Figura 11. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a proliferação celular no giro denteado do hipocampo. Células Ki67
+/0,1mm
2. Time point: 7 dias. Dados
são expressos como média±SEM. n=4 grupo CUS/Escitalopram ou n=5 demais grupos.

54
Imag
ens
rep
rese
nta
tivas
do
gir
o d
ente
ado
do
s gru
po
s ex
per
imen
tais
no
tim
e p
oin
t d
e 7
dia
s. F
oi
uti
liza
da
técnic
a d
e im
uno
his
toq
uím
ica
par
a a
mar
caçã
o d
as
célu
las
Ki6
7+,
que
estã
o e
vid
enci
adas
po
r m
arca
ção
de
DA
B.
Nes
te p
roce
dim
ento
, as
cél
ula
s fo
ram
co
ntr
aco
rad
as c
om
hem
ato
xil
ina
de
Mayer
. A
s fo
tos
.fo
ram
tira
das
em
au
mento
de
20
X e
co
m o
s m
esm
os
par
âm
etr
os
de
tem
po
de
exp
osi
ção
e b
alan
ço d
e b
ranco
.

55
4.1.3. Efeito do tratamento repetido com CBD ou Escitalopram (7 dias) sobre a
expressão de células DCX+
Após os testes comportamentais, os animais foram anestesiados e perfundidos para a
remoção dos encéfalos para a posterior realização de técnicas de imunohistoquímica. A DCX
+ é um marcador expresso somente em tipos celulares já comprometidos com o fenótipo
neuronial (células 2b e tipo 3). A análise de células que expressam este marcador permite
inferir se o estresse ou algum dos tratamentos influenciam sobre a etapa da neurogênese
relacionada à diferenciação neuronial.
A análise por two-way ANOVA não indicou diferença estatisticamente significativa
entre nenhum dos fatores testados (condição: Controle, CUS) ou (tratamento: Veículo, CBD
30mg/Kg ou Escitalopram 20mg/Kg) sobre a quantidade de células DCX+ localizadas no giro
denteado do hipocampo. Ademais, não houve interação entre os fatores condição e
tratamento.
A partir de uma análise por teste t de amostras independentes, foi encontrado diferença
estatisticamente significativa entre o grupo de animais controle tratados com veículo e o
grupo de animais submetidos ao estresse crônico tratados com veículo (teste t de amostras
independentes; t(8)= 2,273, p=0,05). A análise gráfica indica, portanto, uma redução no
número de células DCX+ no grupo submetido ao estresse tratado com veículo em relação ao
grupo controle submetido ao mesmo tratamento (Figura 12).
Foi realizado teste de correlação de Pearson entre os dados comportamentais
individuais obtidos e os resultados obtidos nas análises moleculares correspondentes em
relação ao número total de células DCX+ expressas no giro denteado do hipocampo. Foi
realizada também análise correlacional, somente entre o grupo de animais submetidos ou não
ao estresse independentemente do tratamento e entre os animais submetidos aos diferentes
tratamentos independentemente da condição, a fim de observar se o comportamento
apresentado após a exposição ao estresse ou a algum dos tratamentos influencia diretamente
no número de células DCX+. Nenhuma das análises realizadas, no entanto, indicou correlação
entre os dados (Figura 13) (ANEXO).
A análise por two- way ANOVA não encontrou diferença estatisticamente significativa
em nenhum dos fatores (Condição, Tratamento) ou interação entre estes fatores, em relação à
porcentagem de células DCX+ em estágio de migração (Figura 14).

56
Figura 12. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a quantidade de células DCX
+ /0,1mm
2 no giro denteado do hipocampo. Time point: 7 dias. Dados são
expressos como média±SEM. n=4 grupo CUS/Escitalopram ou n=5 demais grupos. *p<0,05 em relação ao grupo Controle/Veículo (teste t de amostras independentes).
Figura 13. Análise correlacional entre o efeito comportamental obtido no NSF e o número de células DCX
+/0,1mm
2 de giro denteado. Time point: 7 dias. Foi realizada análise por correlação de Pearson entre o
efeito observado no comportamento para cada animal, independentemente do grupo experimental e o respectivo efeito molecular. n=28 animais.

57
Figura 14. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a porcentagem de células DCX
+ em estágio de migração em relação ao número de células DCX
+ totais
no giro denteado do hipocampo. Time point: 7 dias. Dados são expressos como média±SEM. n= 4/grupo CUS/ESC; n=5/demais grupos.

58
VE
ÍCU
LO
C
BD
E
SC
CONTROLE CUS
Imag
ens
rep
rese
nta
tivas
do
gir
o d
ente
ado
do
s gru
po
s ex
per
imen
tais
no
tim
e p
oin
t d
e 7
dia
s. F
oi
uti
liza
da
técnic
a d
e im
uno
his
toq
uím
ica
par
a a
mar
caçã
o d
as
célu
las
DC
X+,
que
estã
o e
vid
enci
adas
po
r m
arca
ção
de
DA
B.
As
foto
s fo
ram
tir
adas
em
au
men
to d
e 1
0X
e c
om
os
mes
mo
s p
arâm
etro
s d
e te
mp
o d
e e
xp
osi
ção
e
bal
anço
de
bra
nco
.
250
µm

59
4.2 Protocolo experimental de 14 dias
No protocolo comportamental de 14 dias, a análise por two way ANOVA indicou
haver interação entre os fatores condição x tratamento (F (2,55) = 4,778 , p<0,05). Neste
sentido, análises post hoc apontaram que os animais do grupo CUS tratados com veículo
tiveram uma maior latência para se alimentar no novo ambiente em relação aos animais não
estressados tratados com veículo, ou seja, o estresse crônico foi capaz de gerar uma resposta
do tipo ansiogênica neste teste.
A análise nos indica também que os animais submetidos ao estresse crônico, porém
tratados com CBD 30mg/Kg ou escitalopram 20mg/Kg, durante 14 dias, apresentaram uma
menor latência para se alimentar no novo ambiente em relação ao grupo estressado tratado
com veículo. Este resultado indica, portanto, que neste time point, ambos tratamentos foram
ansiolíticos (Figura 15A).
De maneira semelhante, nenhum efeito foi observado no consumo de alimento na
home cage, indicando que o estresse, tampouco nenhum dos tratamentos alterou a ingesta
alimentar dos animais, eliminando este viés das análises (Figura 15B). Durante a aplicação
deste protocolo de estresse crônico, 4 animais do grupo CUS/ veículo morreram após a
aplicação do estresse de maravalha molhada. Após este estressor, foi observada considerável
perda de peso nos animais o que pode ter justificado o ocorrido. Dessa maneira, este grupo é
apresentado com menor n experimental em relação aos demais grupos.
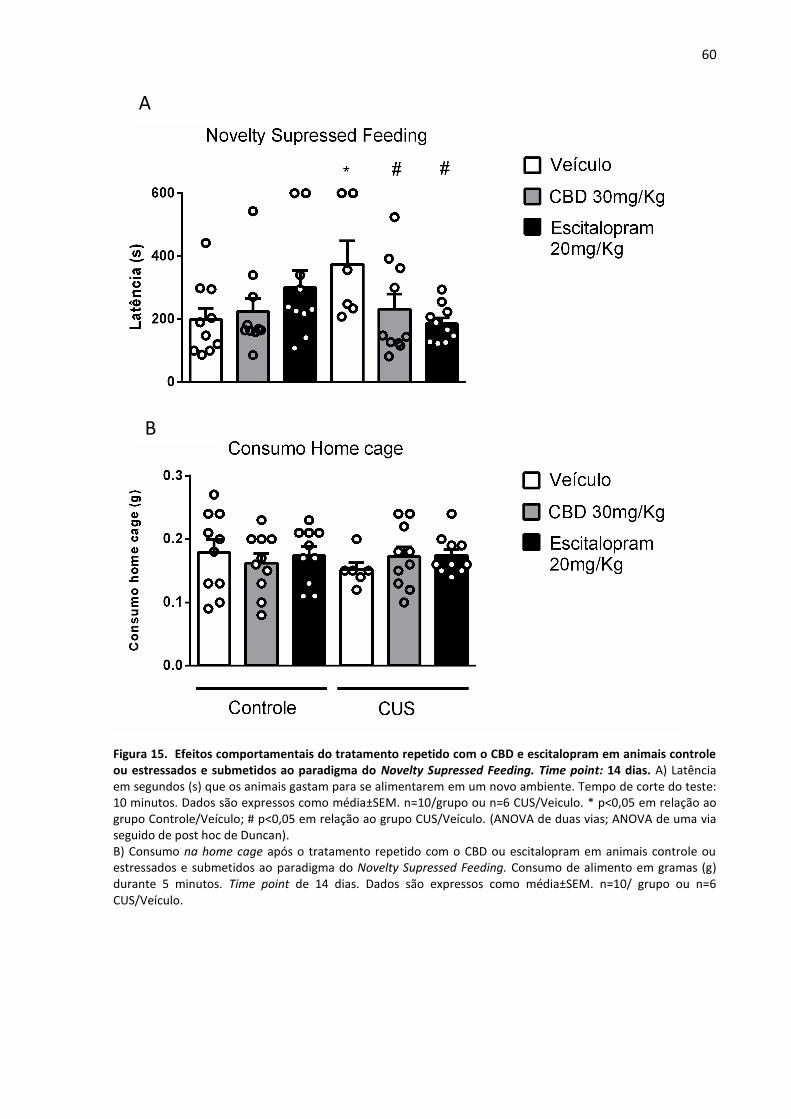
60
Figura 15. Efeitos comportamentais do tratamento repetido com o CBD e escitalopram em animais controle ou estressados e submetidos ao paradigma do Novelty Supressed Feeding. Time point: 14 dias. A) Latência em segundos (s) que os animais gastam para se alimentarem em um novo ambiente. Tempo de corte do teste: 10 minutos. Dados são expressos como média±SEM. n=10/grupo ou n=6 CUS/Veiculo. * p<0,05 em relação ao grupo Controle/Veículo; # p<0,05 em relação ao grupo CUS/Veículo. (ANOVA de duas vias; ANOVA de uma via seguido de post hoc de Duncan). B) Consumo na home cage após o tratamento repetido com o CBD ou escitalopram em animais controle ou estressados e submetidos ao paradigma do Novelty Supressed Feeding. Consumo de alimento em gramas (g) durante 5 minutos. Time point de 14 dias. Dados são expressos como média±SEM. n=10/ grupo ou n=6 CUS/Veículo.
A
B

61
4.2.1 Efeito do tratamento repetido (14 d) sobre a expressão de células BrdU+
A fim de determinar se o tratamento repetido com CBD ou Escitalopram nas
condições testadas (Controle ou Veículo) altera a sobrevivência celular, utilizamos a técnica
de incorporação por BrdU. Este composto foi administrado no primeiro dia de protocolo
experimental e acopla-se às células que se encontram na fase S do ciclo celular, permitindo a
análise da sobrevivência celular decorridos os 14 dias de protocolo.
Análise por two way ANOVA não indicou diferença estatisticamente significativa em
nenhum dos fatores testados (Condição= Controle ou CUS; Tratamento= Veículo, CBD ou
Escitalopram). Ademais, a mesma análise não indicou interação entre condição e tratamento.
Este resultado indica que neste experimento, a sobrevivência celular não foi alterada por
nenhuma das condições ou tratamentos testados (Figura 16).
A análise de correlação individual entre os valores comportamentais obtidos e os
dados moleculares do número de células BrdU+, não apontou nenhum tipo de correlação neste
protocolo (Figura 17). Foi realizada também análise correlacional, somente entre o grupo de
animais submetidos ou não ao estresse independentemente do tratamento e entre os animais
submetidos aos diferentes tratamentos independentemente da condição, a fim de observar se o
comportamento apresentado após a exposição ao estresse ou a algum dos tratamentos
influencia diretamente sobre a sobrevivência celular.
Nenhuma das análises, no entanto, indicaram correlação estatisticamente significativa
(ANEXO D).

62
Figura 16. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a quantidade de células BrdU
+ /0,1mm
2 de giro denteado do hipocampo. Time point: 14 dias. Dados
são expressos como média±SEM. n=4 grupo Controle/CBD ou n=5 demais grupos.
Figura 17. Análise correlacional entre o efeito comportamental obtido no NSF e o número de células BrdU
+/0,1mm
2 de giro denteado. Time point: 14 dias. Foi realizada análise por correlação de Pearson entre o
efeito observado no comportamento para cada animal, independentemente do grupo experimental e o respectivo efeito molecular. n=29 animais.

63
VE
ÍCU
LO
C
BD
E
SC
CONTROLE CUS
Imag
ens
rep
rese
nta
tivas
do
gir
o d
ente
ado
do
s gru
po
s ex
per
imen
tais
no
tim
e p
oin
t d
e 1
4 d
ias.
Fo
i uti
liza
da
técnic
a d
e im
uno
fluo
resc
ênci
a p
ara
a m
arca
ção
das
célu
las
Brd
U+.
Fo
i uti
liza
do
nes
te p
roto
colo
o a
nti
corp
o p
rim
ário
anti
-Brd
U (
rat-
Ab
cam
) e
o a
nti
corp
o s
ecund
ário
Ale
xa
® 5
94
(an
ti-r
at-
Invit
rogen).
As
foto
s
.fo
ram
tir
adas
em
au
men
to d
e 2
0X
e c
om
os
mesm
os
par
âm
etr
os
de
tem
po
de
exp
osi
ção
.
75 µ
m

64
4.2.2 Efeito do tratamento repetido (14 d) sobre a expressão de células DCX+ e sobre a
migração celular.
Com relação à expressão de células DCX/mm2 de giro denteado, 14 dias de protocolo
experimental, foi encontrada diferença estatisticamente significativa no fator condição (F
(1,29) = 5,591 , p<0,05). Este resultado indica que o estresse promoveu uma redução do
número de células DCX+
em relação aos animais controle, independente do fator tratamento.
Análise de ANOVA one way seguida de teste post hoc de Duncan, não indicou diferença
estatisticamente entre o grupo de animais estressados tratados com veículo e entre os animais
controle submetidos ao mesmo tratamento. A análise indicou ainda que o tratamento repetido
com CBD nos animais controle por 14 dias, aumenta significativamente o número de células
DCX+ no hipocampo de camundongos adultos. (Figura 18)
O tratamento com CBD 30mg/Kg ou Escitalopram 20mg/Kg, por sua vez, nos animais
estressados não altera a quantidade de células que expressam DCX+ (Figura 18)
A análise estatística das células DCX+ localizadas na camada granular do giro
denteado, o que sugere um estágio de célula em migração, não indicou diferença entre
nenhum dos fatores testados (condição ou tratamento) ou interação entre os mesmos (Figura
20).
Não foi observado neste experimento nenhuma correlação entre os dados
comportamentais obtidos no teste do NSF com os dados moleculares de expressão de DCX+
em nenhum dos grupos de tratamento ou em nenhuma das condições testadas (Figura 19)
(ANEXO C)

65
Figura 18. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a quantidade de células DCX
+ /0,1mm
2 de giro denteado do hipocampo. Time point: 14 dias. Dados são
expressos como média±SEM. n= 5/grupo. *p<0,05 em relação ao grupo Controle/Veículo (ANOVA one way seguido de post hoc de Duncan)
Figura 19. Análise correlacional entre o efeito comportamental obtido no NSF e o número de células DCX
+/0,1mm
2 de giro denteado. Time point: 14 dias. Foi realizada análise por correlação de Pearson entre o
efeito observado no comportamento para cada animal, independentemente do grupo experimental e o respectivo efeito molecular (expressão de células DCX
+). n=30 animais.

66
Figura 20. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a porcentagem de células DCX
+ em estágio de migração em relação ao número de células DCX
+ totais
no giro denteado do hipocampo. Time point: 14 dias. Dados são expressos como média±SEM. n= 5/grupo.

67
Imag
ens
rep
rese
nta
tivas
do
gir
o d
ente
ado
do
s gru
po
s ex
per
imen
tais
no
tim
e p
oin
t d
e 1
4 d
ias.
Fo
i uti
liza
da
técnic
a d
e im
uno
his
toq
uím
ica
par
a a
mar
caçã
o
das
cél
ula
s D
CX
+,
que
estã
o e
vid
enci
adas
po
r m
arca
ção
de
DA
B.
As
foto
s fo
ram
tir
adas
no
au
mento
de
10
X.
CONTROLE CUS V
EÍC
UL
O
CB
D
ES
C
250
µm

68
4.2.3. Efeito do tratamento repetido (14 d) sobre a expressão de células SOX2+
Analisamos a expressão de células SOX 2 +/mm 2
de giro denteado, a fim de se
analisar o efeito do estresse e dos tratamentos administrados sobre a população de célula
progenitora (Tipo 1 e Tipo 2a). Decorridos 14 dias de protocolo experimental, a análise por
two-way ANOVA não indicou diferenças estatisticamente significativas entre nenhum dos
fatores testados (condição ou tratamento). Além disso, a mesma análise não demonstrou
interação entre os fatores condição e tratamento. Este resultado demonstra que o estresse ou
os tratamentos empregados não promovem alteração no número de células SOX2+
em relação
aos animais controle (Figura 21).

69
Figura 21. Efeitos do tratamento repetido com o CBD e escitalopram em animais controle ou estressados sobre a quantidade de células SOX2
+ /mm
2 de giro denteado do hipocampo. Time point: 14 dias. Dados são
expressos como média±SEM. n= 4/grupo Controle/CBD e grupo Controle/Escitalopram.

70
Imag
ens
rep
rese
nta
tivas
do
gir
o d
ente
ado
do
s gru
po
s ex
per
imen
tais
no
tim
e p
oin
t d
e 1
4 d
ias.
Fo
i uti
liza
da
técnic
a d
e im
uno
fluo
resc
ênci
a p
ara
a m
arca
ção
das
célu
las
SO
X 2
+.
Fo
i uti
liza
do
nes
te p
roto
colo
o a
nti
corp
o p
rim
ário
anti
-SO
X2
(ra
bb
it-
Mil
lip
ore
) e
o a
nti
corp
o s
ecund
ário
Ale
xa
® 6
47
(an
ti-r
abb
it-
Invit
rogen).
As
foto
s .f
ora
m t
irad
as e
m a
um
ento
de
10
X e
co
m o
s m
esm
os
par
âmet
ros
de
tem
po
de
exp
osi
ção
.

71
DISCUSSÃO

72
5. Discussão
Os resultados obtidos no presente trabalho sugerem que o CBD é capaz de prevenir
os efeitos do estresse crônico em protocolos de 7 e 14 dias de tratamento, enquanto que o
antidepressivo escitalopram preveniu os efeitos do estresse apenas no protocolo de 14 dias.
Nossos resultados indicaram que o estresse não gerou alterações significativas na taxa de
sobrevivência celular, medida pela incorporação de BrdU decorridos 7 ou 14 dias de
protocolo experimental. No entanto, o estresse promoveu uma significativa redução do
número de células Ki67+ em 7 dias, o que se configura como uma redução da proliferação
celular. Além disso, observou-se uma redução do número de células DCX+ no grupo dos
animais estressados/veículo em relação ao grupo de animais controle/veículo no protocolo de
7 dias, diferença esta não encontrada no protocolo de 14 dias. Por fim, o estresse não
promoveu alteração significativa no número de células SOX 2+ no protocolo de 14 dias. Com
relação ao efeito dos tratamentos, encontramos que o CBD ou o Escitalopram não previnem a
redução da proliferação celular mediada pelo estresse, mas previnem a redução de células
DCX+ no protocolo de 7 dias.
Considerando os efeitos deletérios do estresse crônico sobre a neurogênese adulta,
como mencionado na introdução do presente estudo, utilizamos o modelo de estresse crônico
imprevisível. Na medida em que os animais são expostos a diversos estressores de maneira
imprevisível e com alta intensidade, podem ao longo do tempo, desenvolver alterações
comportamentais e fisiológicas associadas com estados emocionais, mimetizando o que
ocorre em seres humanos, sendo, portanto, um modelo de boa validade de face (Willner,
2017). Além disso, a utilização do CUS justifica-se por sua boa validade preditiva, ou seja, os
animais são responsivos ao tratamento crônico com fármacos, no caso em questão, fármacos
antidepressivos (Surget et al., 2011) e o CBD (Campos et al., 2013; Fogaça et al., 2018).
Originalmente, a resultante comportamental após a aplicação do CUS era determinada pelo
desenvolvimento de um sintoma conhecido como anedonia (Willner, 2017), no qual o animal
perdia o interesse por atividades outrora prazerosas como o consumo de soluções adocicadas.
Porém, ao longo do tempo, observou-se que este modelo promovia uma diversidade de
alterações comportamentais, além do sintoma clássico de anedonia, como o de provocar nos
animais comportamentos defensivos relacionados à “ansiedade”. (Papp et al., 2017; Surget et
al., 2008; Campos et al., 2013). Desta maneira, permitiu-se que novos testes fossem
associados ao CUS, visando à observação de outras alterações comportamentais que não
somente o comportamento do tipo depressivo.

73
Em nosso estudo, os animais experimentais foram expostos a estressores de diferentes
intensidades e natureza (físicos, psicológicos) de forma independente durante 7 ou 14 dias e
receberam ou não tratamento (CBD 30mg/Kg ou Escitalopram 20mg/Kg) durante toda a
aplicação do protocolo. Para as análises comportamentais, no presente trabalho, optou-se pela
aplicação do Novelty Supressed Feeding, em virtude, igualmente de sua boa validade de face
e por permitir determinar temporalmente o efeito comportamental gerado após a
administração de determinado fármaco (Campos et al., 2013). Experimentalmente, o NSF
evidencia respostas comportamentais somente após o tratamento crônico com antidepressivos,
sendo possível dissociar, portanto, de um efeito gerado após a administração aguda (Bodnoff
et al., 1988). Foi demonstrado igualmente por nosso grupo, que o CBD induz alterações
comportamentais no NSF somente após tratamento repetido, não induzindo efeitos agudos
(Campos, dados não publicados).
A partir da análise comportamental no nosso protocolo de 7 dias, é possível observar
que a aplicação do estresse durante este período não foi capaz de gerar efeito do tipo
ansiogênico no NSF, embora haja uma tendência clara para uma significância estatística do
fator condição (Controle x CUS). Observando o gráfico (Figura 8A) é possível constatar a alta
dispersão dos dados entre o grupo de animais controle tratados com veículo, atendo-se ao fato
de que aproximadamente 1/3 dos animais não comeram durante o tempo limite de teste (600
segundos). Esse fator pode ter comprometido a análise do efeito do estresse, na medida em
que o teste é afetado pelas condições basais dos animais, refletindo a imensa complexidade
existente em análises comportamentais. Neste caso, portanto, consideramos baseados nos
testes estatísticos, não ser possível afirmar que os animais não foram susceptíveis aos efeitos
da aplicação do estresse, mesmo com um período de aplicação relativamente curto (7 dias).
Clinicamente, o manejo de transtornos psiquiátricos como ansiedade e depressão,
impõe desafios complexos a profissionais de saúde. Fármacos antidepressivos, por exemplo,
possuem limitações clínicas clássicas como alta incidência de efeitos adversos, alta taxa de
refratariedade e uma alta latência para o início dos efeitos clínicos (Cipriani et al., 2018). No
nosso trabalho, utilizamos o escitalopram, pois segundo alguns autores, promove efeitos
clínicos em um tempo inferior a outros antidepressivos (Kasper et al., 2006), está associado a
maiores taxas de aceitabilidade e por ser em alguns transtornos, mais eficaz na remissão dos
sintomas (Cipriani et al., 2018). Além disso, devido a seu relativo curto tempo de
comercialização em relação a outros antidepressivos, como a fluoxetina, julgamos ser
interessante a geração de mais dados pré-clínicos, visando à obtenção de novas informações
quanto a seu mecanismo de ação.

74
Embora o escitalopram possa apresentar algumas vantagens em relação a outros
antidepressivos, o uso desta classe de fármacos ainda é associado a limitações como
refratariedade, incidência considerável de eventos adversos e considerável latência clínica
(Wade e Friis, 2006), prejudicando o tratamento de transtornos geradores de um alto grau de
inabilidade, como ansiedade e depressão. Portanto, inicialmente nosso trabalho prestou-se a
investigar se os efeitos comportamentais gerados após administração repetida do CBD,
surgiriam anteriormente aos antidepressivos. Ao retomar a análise comportamental de 7 dias
(Figura 8 a) , observamos que o CBD 30mg/Kg atenua significativamente os efeitos do
estresse, enquanto que no mesmo período, o antidepressivo escitalopram 20mg/Kg não é
capaz. Nossos achados sugerem, portanto, que o CBD pode prevenir os efeitos
comportamentais do estresse com uma latência ainda inferior ao testado por Campos et al.,
(2013) e Fogaça et al., (2018) (7 dias vs 14 dias). No caso do grupo de animais tratados com
escitalopram e submetidos ao estresse repetido (7 dias), nota-se a presença de duas
populações distintas de resposta ao tratamento. Embora possa ser aventada a hipótese da
limitação do modelo em gerar uma resposta a partir de um tratamento de 7 dias com
antidepressivos, pode ser observado que o escitalopram, na mesma dose, gerou um efeito do
tipo ansiolítico nos animais não estressados. Portanto, na nossa concepção, trata-se de uma
resposta dependente da condição (Controle X CUS), ou seja, o escitalopram demanda um
maior período para exercer seu efeito quando os animais são submetidos a um desafio, no
caso o estresse, o que foi observado, posteriormente no protocolo de 14 dias (Figura 15 A).
Resultados não publicados de nosso grupo indicam que o Escitalopram na dose de 10mg/kg
administrado nos últimos 7 dias de protocolo em camundongos C57BL6 submetidos ao
estresse crônico imprevisível ou no estresse crônico de derrota social (10 dias), também não
foi capaz de gerar um efeito do tipo ansiolítico no teste do Novelty Supressed Feeding
(Scarante, 2018), embora a mesma dose seja ansiolítica após 21 dias de tratamento (Aguiar et
al., dados não publicados). Assim como no presente trabalho, Scarante (2018) observou a
grande dispersão dos dados dos grupos controle, o que corrobora com nossos achados.
Diferenças interindividuais dos animais no padrão de resposta a antidepressivos também
podem explicar a ocorrência de animais que respondem ao AD e que não respondem, o que
também se correlaciona a achados clínicos (Einarson, 2004).
No protocolo comportamental de 14 dias, observamos que a aplicação do estresse
crônico foi capaz de gerar um comportamento do tipo ansiogênico nos animais tratados com
veículo, o que pode ser evidenciado pelo aumento da latência para comer no novo ambiente
quando comparado aos animais controle submetidos ao mesmo tratamento (Figura 15 A).

75
Essa resposta indica que o estresse foi aplicado durante um tempo necessário, com a
intensidade suficiente e com a imprevisibilidade necessária. Esse achado evidencia uma das
vantagens inerentes ao modelo do CUS sobre outros modelos de estresse, como por exemplo
os homotípicos, na medida em que reduzem a possibilidade de habituação ao estresse
(Willner, 2017). Neste mesmo experimento, observamos que tanto o CBD, quanto o
escitalopram, administrados repetidamente (14 dias) foram capazes de atenuar o efeito da
aplicação do estresse crônico, ou seja, promovem um efeito do tipo ansiolítico.
Diferentemente do observado durante o tratamento repetido por 7 dias, pode-se observar que a
administração do escitalopram estabeleceu nos animais um padrão de comportamento
homogêneo (Figura 15 A), o que corrobora com a ideia de que este AD necessita de um maior
tempo de administração para promover alterações suficientes para contrapor a resposta
induzida pelo estresse, assim como observado para outros antidepressivos inibidores da
recaptação de serotonina (Amellen et al., 2018). O CBD por sua vez, permaneceu sendo capaz
de atenuar a resposta ao estresse. Estes resultados comportamentais indicam, portanto, que
neste modelo, o CBD é capaz de gerar uma resposta comportamental do tipo ansiolítica em
um tempo inferior ao antidepressivo escitalopram e de maneira sustentada.
Realizamos, análises moleculares em ambos protocolos (7 e 14 dias) a fim de
observar o padrão de resposta molecular relacionado à sobrevivência celular e a diferenciação
neuronial frente a exposição ao estresse e aos tratamentos empregados. Além disso,
observamos se os efeitos comportamentais foram acompanhados de alterações moleculares,
através de análises correlacionais.
Estudos demonstram que somente cerca de 50% de novas células granulares
sobrevivem após as primeiras semanas no giro denteado de roedores (Dayer et al., 2003),
porém fatores ambientais como exposição ao estresse e tratamentos farmacológicos podem
modular diferencialmente este processo (Schoenfeld e Gould, 2013). No entanto, através de
análises moleculares dos time points de 7 e 14 dias, não observamos efeitos do estresse ou
algum dos tratamentos sobre alterações na sobrevivência celular, medida pela incorporação de
BrdU administrado no primeiro dia de protocolo (Figuras 9 e 16).
Grande parte dos estudos examina o efeito do estresse sobre a proliferação celular.
Neste sentido, diversos trabalhos demonstram que a aplicação do estresse crônico em
diferentes espécies, promovem um impacto negativo sobre a taxa de proliferação celular, seja
pelo modelo de derrota social (Czeh et al., 2001; Czeh et al., 2002; Simon et al., 2005; Mitra
et al., 2006; Ferragud et al., 2010), modelo de estresse de restrição (Veena et al., 2009; Pham
et al., 2003) ou estresse crônico imprevisível (Xu et al., 2009). No entanto, na literatura, os

76
efeitos do estresse sobre a sobrevivência celular são apresentados como passíveis de maior
variação e parecem ser dependentes de fatores como o tipo de estresse aplicado, o tempo de
exposição ao (s) estressor (es), tipo de animal utilizado, linhagem, entre outros (Tanti et al.,
2013).
Ilustrando estas observações, estudo de Snyder e colaboradores (2009) demonstraram
que o estresse de restrição aplicado por 14 dias em camundongos machos C57 adultos,
promove um aumento da sobrevivência celular em relação ao grupo controle. No entanto, o
mesmo efeito não foi observado após 21 dias de aplicação do mesmo estressor, não sendo
observada diferença significativa em relação ao controle (Snyder et al., 2009), efeito
encontrado também no estudo de Heine e colaboradores (2004) em ratos adultos. Em outro
protocolo, no entanto, foi demonstrado que a aplicação do CUS por 3 semanas em ratos
adultos, machos da linhagem Sprague-Dawley reduz a sobrevivência celular decorridos 14
dias de aplicação de BrdU (Koo e Duman, 2008). Resultados semelhantes indicaram que a
aplicação do mesmo protocolo anteriormente citado, por 4 semanas, promove uma redução da
sobrevivência celular em camundongos da linhagem Balb/cBYJ (Tanti et al., 2013), resultante
encontrada também por Fogaça e colaboradores (2018) em camundongos C57BL6 expostos
ao CUS por 14 dias. Visando investigar se diferentes linhagens de camundongos (C57BL6;
BALB/cJ e DBA/2J), responderiam de maneira diferente ao estresse em relação à taxa de
sobrevivência celular, um estudo encontrou que decorridas 2 semanas de aplicação do
protocolo do CUS, todas as linhagens estudas, tiveram uma redução da taxa de sobrevivência
celular (Mineur et al., 2007). Portanto, é interessante observar a grande variabilidade de
respostas relacionadas à modulação da sobrevivência celular, decorrentes da exposição ao
estresse crônico.
Até o momento, não há registros de estudos que realizaram a aplicação do CUS por 7
dias e estudaram seus efeitos sobre a sobrevivência celular em camundongos da linhagem
utilizada em nosso estudo. No entanto, estudo de Mitra e colaboradores (2006) encontrou que
a aplicação do estresse de derrota social por 7 dias em camundongos machos adultos da
linhagem C57BL6, promove uma redução da proliferação celular. É possível, portanto, que o
CUS (7 dias) não promova efeito sobre a sobrevivência celular, como observado no nosso
trabalho, podendo, no entanto influenciar sobre os processos de proliferação celular. Portanto,
diante desta hipótese analisamos a expressão do marcador endógeno Ki67, expresso em todas
as fases do ciclo, excetuando-se G 0. Neste sentido, encontramos que o CUS (7 dias) é capaz
de reduzir a proliferação celular (Figura 11). Esta redução foi encontrada por outros
pesquisadores utilizando este modelo de estresse, porém com um maior tempo de aplicação

77
(Xu et al., 2009). Assim, esse resultado é inédito em mostrar que o CUS aplicado por 7 dias é
capaz de promover alterações moleculares sobre a taxa de proliferação celular em
camundongos C57BL6. A observação do número de células progenitoras dotadas de alta
capacidade proliferativa, como as células do tipo 2a, positivas para SOX 2, neste time point,
permitirá também inferir sobre os efeitos do estresse sobre este tipo celular.
Os efeitos do estresse crônico imprevisível de 14 dias sobre a sobrevivência celular em
camundongos machos C57BL6, como anteriormente citado, são divergentes tendo um estudo
demonstrado um aumento deste fator (Snyder et al., 2009), enquanto outro demonstrou uma
redução da sobrevivência celular (Fogaça et al., 2018). No estudo de Snyder, no entanto, não
foi realizada análise de nenhuma resultante comportamental ou de algum parâmetro
fisiológico após a aplicação do CUS. Esse dado faz com que não seja possível afirmar que o
estresse crônico aplicado tenha sido suficiente para gerar o fenótipo “estressado”. Nesse
sentido, nosso estudo e o estudo de Campos e colaboradores (2013), foram capazes de
identificar que o estresse crônico aplicado por 14 dias gera manifestações comportamentais
relacionadas à ansiedade. Entretanto, diferentemente de Campos et al. e Fogaça et al. (2018),
não encontramos nenhum efeito do estresse sob o parâmetro de sobrevivência celular.
Realizamos, portanto, análises correlacionais para identificar se o comportamento nos animais
experimentais está relacionado diretamente ao efeito de sobrevivência celular. No entanto,
não foi observado nenhum tipo de correlação (Figura 17). Isso sugere que outros efeitos
relacionados à neurogênese adulta podem ter influenciado a resposta comportamental.
Nossos resultados indicam também que o efeito do tipo ansiolítico promovido pelo
CBD em ambos time points (7 e 14 dias) e pelo escitalopram no time point de 14 dias não
possuem relação com a sobrevivência celular. Embora a análise global da sobrevivência seja
fundamental, é importante mencionar que o estresse e o tratamento farmacológico podem
afetar diferencialmente a sobrevivência celular entre populações celulares distintas (Tanti et
al., 2013), condicionando o balanço entre morte e sobrevivência destas populações (Wolf et
al., 2010). Ademais, os tratamentos podem alterar a dinâmica da neurogênese adulta, por
exemplo, aumentando a sobrevivência de determinado tipo celular em relação a outro,
visando o reestabelecimento da homeostasia em animais submetidos ao estresse.
Para a investigação do exposto, utilizamos técnicas imunoistoquímicas, em ambos
time points. No time point de 7 dias realizamos análises de dupla marcação para GFAP/SOX2,
objetivando analisar a população de células tronco progenitoras (tipo 1), que podem expressar
GFAP e SOX2 e células precursoras amplificadas (tipo 2a) que não expressam GFAP, mas
expressam SOX2 (Suh et al., 2007). Analisamos também, a população de células expressando

78
o marcador DCX+ intimamente relacionado ao fenótipo neuronial imaturo (tipo 2b e tipo 3).
No protocolo de 14 dias, realizamos a análise da expressão de células SOX2 positivas,
visando um enfoque sobre a população de células precursoras e de células DCX positivas,
visando analisar o processo de diferenciação neuronial.
Em relação ao número de células DCX+, foi encontrado que a aplicação do estresse
durante 7 dias reduziu significativamente esta população em relação ao grupo controle tratado
com veículo (Figura 12) . Esse efeito pode estar relacionado à expressão diferencial de DCX +
em diferentes tipos celulares (célula 2b e célula tipo 3). Uma das possíveis hipóteses é a de
que o estresse possa ter impactado essa população celular, que pode ser mais sensível às
variações homeostáticas, uma vez que deixaram recentemente o nicho neurogênico.
Embora dados já estabeleçam o importante efeito do estresse sobre a taxa de
proliferação celular (Schoenfeld e Gould, 2003), estudos indicando quais populações são
afetadas ainda são escassos. Alguns trabalhos demonstram que o estresse crônico gera um
impacto negativo sobre a população de células DCX+. Van Bokhoven e colaboradores (2011)
evidenciaram que o estresse de derrota social aplicado por 3 meses em ratos adultos, promove
uma redução na quantidade destas células. Analogamente, Oomen et al. (2007) e Dagyte et al.
(2011) demonstraram que o CUS de 21 dias e de 5 semanas, respectivamente, também
promovem uma redução de células com este fenótipo celular em ratos. Outro estudo
evidenciou que o CUS (14 dias) em camundongos machos C57BL6, reduz significativamente
o número de células DCX+, alterando também a complexidade dendrítica destas células
(Fogaça et al., 2018).
A observação dos efeitos gerados pelo tratamento repetido com CBD em um curto
prazo (7 dias) em relação à neurogênese, também são inéditos. De maneira interessante, nosso
trabalho demonstrou que o tratamento com CBD poderia atenuar os efeitos da aplicação do
CUS em 7 dias, porém o mecanismo pelo qual o CBD é capaz de gerar este efeito rápido,
permanece como fonte de debates. No nosso estudo, demonstramos que o CBD não promoveu
um aumento significativo do número de células DCX+ em relação aos animais estressados
tratados com veículo, mas preveniu a redução promovida pela aplicação do estresse (Figura
12). Ou seja, o CBD pode estar favorecendo a estabilização das células neste fenótipo
específico, mediante o insulto sofrido, a fim de garantir a manutenção da homeostase.
Este efeito de manutenção desta população celular pode ter fundamental importância,
já que este tipo celular pode estar promovendo respostas importantes para “frear” a resposta
ao estresse. Os mecanismos pelos quais esse evento poderia ocorrer necessita de mais
investigações, mas já é demonstrado que os neurônios jovens podem condicionar respostas

79
eletrofisiológicas diferentes das células granulares maduras, possuindo um menor limiar de
disparo e gerando uma resposta de LTP de maior amplitude (Piatti et al., 2011; Schmidt-
Hieber et al., 2004). Além disso, os neurônios recebem distintos padrões de conectividade de
acordo com seu grau de desenvolvimento no giro denteado (gabaérgico x glutamatérgico)
(Esposito et al., 2005; Ge et al., 2006). Essas diferenças fazem com que os neurônios jovens
difiram em relação a diversas propriedades dos neurônios maduros já integrados à rede
sináptica (Esposito et al., 2005; Piatti et al., 2011). Nesse sentido, pode não ser necessário que
um neurônio complete todo o processo de maturação para influenciar em respostas
importantes no giro denteado, na medida em que quando todo o seu processo de maturação é
completado e o mesmo se integra às redes sinápticas, não há diferença no padrão de resposta
sináptica em relação aos demais neurônios maduros (Laplagne et al., 2006; Mongiat and
Schinder, 2011).
É importante destacar, no entanto, que a ausência de marcação por colocalização com
BrdU, não nos permite distinguir células formadas após o início do tratamento e células que já
existiam anteriormente. Os dados apresentados acerca do efeito da administração repetida de
CBD por 7 dias sobre a neurogênese adulta, mediante condições de estresse, ainda não são
explorados na literatura. Sua investigação posterior pode elucidar importantes mecanismos
relacionados à possível ação comportamental rápida observada.
No protocolo experimental de 14 dias, encontramos que o CBD promoveu um
aumento significativo no número de células DCX+
nos animais não estressados (Figura 18),
efeito semelhante ao encontrado por Campos e colaboradores (2013). Este efeito, como
observado, não foi acompanhado de um aumento na sobrevivência celular. É interessante
observar, portanto, que embora este processo de diferenciação neuronial tenha sido
aumentado, o CBD não promoveu efeito comportamental nos animais não submetidos ao
estresse. Portanto, os efeitos de aumento de uma população celular per se pode, de fato, estar
dissociado de um efeito comportamental.
Neste protocolo não foi observada diferença significativa quanto ao número de células
DCX+ do grupo de animais estressados tratados com veículo e de animais controle tratados
com veículo (Figura 18). No entanto, foi observada significância estatística do fator condição.
Ou seja, o grupo de animais submetidos ao CUS possui menor quantidade dessas células em
relação aos animais controle. Não foi observado, no entanto, efeito do tratamento com CBD
ou escitalopram nos animais estressados em relação a este parâmetro. É importante mencionar
que a exposição ao estresse e a administração de fármacos é capaz de gerar uma vasta gama
de respostas neuroplásticas, podendo influenciar na complexidade de espinhos dendríticos, na

80
sinaptogênese, dentre outros fatores (Fogaça et al., 2018; Morais et al., 2014). Portanto,
embora o CBD não tenha promovido um efeito pró-neurogênico neste tipo celular específico,
seu efeito do tipo ansiolítico, pode estar relacionado a uma multiplicidade de eventos que em
conjunto, foram hábeis para prevenir os efeitos deletérios do estresse .
Neste sentido, Fogaça e colaboradores (2018) demonstraram que o CBD na dose de
30mg/Kg em animais submetidos ao CUS, reverte a redução da densidade de espinhos
dendríticos no giro denteado do hipocampo e a redução no tamanho dos dendritos. Estes
efeitos sobre a complexidade sináptica podem ter contribuído, neste protocolo para o efeito
comportamental observado.
Outra análise realizada no trabalho de Fogaça e colaboradores (2018) e testada no
presente trabalho, refere-se à análise da migração celular. As células DCX+ são proteínas que
estão associadas à microtúbulos e que participam do processo de migração das células da
camada subgranular do giro denteado para a camada granular sobrejacente (Brown et al.,
2003). Portanto, analisamos a porcentagem de células DCX+
localizadas na camada granular
do GD em relação ao número total de células com este fenótipo (Figura 20). Nossos
resultados, no entanto, não indicaram nenhum efeito do estresse ou dos tratamentos
empregados sobre este parâmetro. Estes resultados são conflitantes ao encontrado por Fogaça
e colaboradores (2018), na medida em que os autores demonstraram que o CUS (14 dias),
promove uma redução do número de células em migração em relação ao controle e que o
CBD (30mg/Kg) é capaz de reverter este efeito. Essa abordagem, no entanto, ainda é pouco
utilizada e em termos funcionais, sem a devida análise morfológica, pode não ser possível
distinguir entre diferentes tipos celulares que expressam DCX+. Portanto, é necessária a
criação de mecanismos que permitam identificar diferentes tipos celulares neste estágio, na
medida em que são células que possivelmente estão em uma fase mais posterior do processo
de neurogênese hipocampal adulta e que, portanto, podem possuir diferentes padrões de
conectividade com outras regiões do GD (Kempermann et al., 2015).
Outra hipótese é a de que o tratamento com os fármacos CBD e escitalopram durante
14 dias possa estar modulando os níveis de proteínas sinápticas em regiões como o córtex pré-
frontal e o hipocampo. Scarante (2018) abordou esta questão e demonstrou que a combinação
de CBD e escitalopram em doses inefetivas quando administrados isoladamente, aumentam o
nível da proteína sinaptofisina. Essa proteína é expressa na membrana vesicular e sua
presença exclusiva em terminais pré-sinápticos, a configura como um importante marcador.
Visando ainda a observação de outras etapas da neurogênese adulta, realizamos a
análise da expressão do marcador SOX 2 expresso em células após 14 dias de protocolo

81
experimental. Este marcador está intimamente relacionado à células precursoras (Células do
tipo 1 e 2a). A ausência de efeitos do estresse crônico sobre este tipo celular, sugere que o
mecanismo pelo qual o estresse promova os efeitos comportamentais deletérios, possa estar
dissociado de um efeito direto sobre a população de células precursoras (Figura 21). Além
disso, nenhum dos tratamentos promoveu alteração significativa sobre este parâmetro, o que
nos faz inferir que a manutenção deste pool celular, possa ser importante para a homeostasia
do giro denteado.
O antidepressivo escitalopram, assim como o CBD, foi capaz de atenuar os efeitos
comportamentais do estresse no protocolo de 14 dias. A administração deste AD não gerou
efeito estatisticamente significativo no número de células DCX+ em relação ao grupo
estressado/veículo, semelhante ao observado para o CBD (Figura 18). O escitalopram pode ter
promovido um acúmulo de alterações neuroplásticas, cujo efeito, na presente investigação não
foi necessariamente associado a um aumento propriamente dito da neurogênese adulta. Estudo
de Wang et al., 2008 demonstra o exposto. Os autores demonstraram que o tratamento crônico
(28 dias) com fluoxetina, um AD pertencente à mesma classe do escitalopram, gera um efeito
comportamental do tipo ansiolítico no NSF. Este efeito foi acompanhado por um aumento da
arborização dendrítica de neurônios imaturos positivos para DCX. Em adição, o mesmo
tratamento reduziu o número de células DCX+ em relação ao controle, com um aumento
concomitante na expressão de marcadores de neurônios maduros. Portanto, o efeito sobre a
quantidade de células DCX+ não necessariamente está relacionado a um efeito sobre sua
complexidade estrutural. O mesmo autor discute que a maturação neuronial promovida pela
fluoxetina, somado a efeitos sobre a propriedade de neurônios jovens, podem ser necessários
para os seus efeitos ansiolíticos e contribuir para a latência para o início dos efeitos
comportamentais e clínicos (Wang et al., 2008), o que também pode ocorrer no caso do
escitalopram.
Outros autores demonstraram resultados semelhantes ao anteriormente citado. No
estudo de Amellen e colaboradores (2017) demonstrou-se que o tratamento crônico com
fluoxetina (28 dias) promovia uma aceleração da maturação neuronial em relação aos animais
controle. Será fundamental para a complementação do presente trabalho que, posteriormente
seja observada a expressão de marcadores mais relacionados à maturação neuronial, como,
por exemplo, NeuN e calbindina (Kempermann et al., 2015).
É possível, portanto, que os mecanismos pelos quais o CBD e o escitalopram exerçam
seus efeitos sobre a neurogênese adulta sejam diferentes. Porém, o uso de mais marcadores
celulares que permitam uma abordagem mais individualizada de cada etapa da neurogênese e

82
abordagens mais sofisticadas, como a utilização de animais transgênicos, por exemplo,
animais k.o para a proteína DCX+, permitirão uma melhor compreensão sobre esta possível
modulação diferencial. Será possível também, investigar se os efeitos destas drogas sobre o
comportamento estão diretamente relacionados a algum tipo específico de população celular.
É fundamental, no entanto, mencionar que os efeitos neuroplásticos não são
independentes e podem ocorrer simultaneamente durante condições de estresse ou durante o
tratamento farmacológico. Portanto, é possível que não exista uma dependência estrita de
mecanismos relacionados à neurogênese para o efeito comportamental de fármacos como os
antidepressivos ou o CBD (David et al., 2009; Bessa et al., 2009). No estudo de Bessa et al.,
(2009), por exemplo, a ação de antidepressivos foram independentes de um efeito direto sobre
a neurogênese adulta, porém, associados ao remodelamento dendrítico.
Os estudos conduzidos através da retirada de populações de células progenitoras,
como nos estudos de Santarelli et al. (2003) e Campos et al. (2013) evidenciam que a
neurogênese é importante para o efeito comportamental da fluoxetina e do CBD,
respectivamente. Porém, assumindo a existência de interdependência dos processos
neuroplásticos, essa abordagem pode ter alterado toda a complexidade dos processos
fisiológicos do cérebro adulto, tornando o animal mais susceptível ao efeito do estresse e
incapaz de revertê-lo pela administração farmacológica. Essa difícil análise correlacional
entre comportamento e alterações neuroplásticas é explorada em uma frase do cientista Gerd
Kempermann (2015).

83
CONCLUSÃO

84
6. Conclusão
Os resultados do presente estudo indicaram que em animais estressados, o canabidiol é
capaz de gerar efeito comportamental do tipo ansiolítico de maneira mais rápida que o
antidepressivo escitalopram (7 vs 14 dias). Essa resposta não foi acompanhada de uma
modulação sobre os parâmetros neurogênicos testados no presente estudo (proliferação,
sobrevivência celular, modulação sobre a célula progenitora SOX2+
ou migração celular). No
entanto, a prevenção da redução de células DCX+ promovida pelo estresse a partir do
tratamento por 7 dias com CBD e ESC podem fornecer indícios de que a manutenção da
homeostase, seja um processo importante para a contraposição ao efeito deletério do estresse.
Portanto, neste estudo não encontramos indícios de que o efeito comportamental mais
rápido promovido pelo CBD esteja relacionado a uma modulação diferencial, quando
comparado ao escitalopram, sobre os parâmetros neurogênicos testados. Outras análises, no
entanto, devem ser realizadas, a fim de determinar temporalmente os efeitos do estresse e do
tratamento com CBD e antidepressivos sobre a neurogênese hipocampal adulta. Além disso,
permanece por ser testada a influência do estresse e destes tratamentos sobre outras fases que
compreendem a neurogênese hipocampal adulta como o balanço entre quiescência e ativação
e o processo de maturação neuronial.

85
REFERÊNCIAS

86
7. Referências
Abel, EL. Marihuana. The First Twelve Thousand Years, 1980.
Adams, R.; Hunt, M.; Clark, JH. Structure of cannabidiol, a product isolated from the
marihuana extract of Minnesota wild hemp. Journal of the American Chemical Society, v. 62,
n.1, p.196–200, 1940.
Altman J. Autoradiographic investigation of cell proliferation in the brains of rats and
cats.The Anatomical record, v. 145, n. 4, p. 573-591, 1963.
Altman, J.; Das, GD. Autoradiographic and histological evidence of postnatal hippocampal
neurogenesis in rats. The Journal of comparative neurology, v.123, n. 3, p. 319-335, 1965.
Alvarez-Buylla, A. et al. The heterogeneity of adult neural stem cells and the emerging
complexity of their niche. Cold Spring Harbor symposia on quantitative biology, v. 73, p.
357-365, 2008.
Amellen, I. et al. A critical period for antidepressant-induced acceleration of neuronal
maturation in adult dentate gyrus. Translational Psychiatry, v. 7, n. 9, p. 1235, 2017.
Bátiz, LF. et al. Exosomes as Novel Regulators of Adult Neurogenic Niches. Frontiers in
cellular neuroscience, v. 9, 2016.
Bergamaschi, MM. et al. Cannabidiol Reduces the Anxiety Induced by Simulated Public
Speaking in Treatment-Naïve Social Phobia Patients. Neuropsychopharmacology, v. 36, n. 6,
p. 1219-1226, 2011.
Bergmann, O.; Spalding, KL.; Frisén, J. Adult Neurogenesis in Humans. Cold Spring Harbor
perspectives in biology, v. 7, n. 7, 2015.

87
Bessa, JM. et al. The mood-improving actions of antidepressants do not depend on
neurogenesis but are associated with neuronal remodeling. Molecular psychiatry, v. 14, n. 8,
p. 764-773,739, 2009.
Bisogno, T. et al. Molecular targets for cannabidiol and its synthetic analogues: effect on
vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide.
British Journal of Pharmacology, v. 134, n. 4, p. 845-852, 2001.
Bodnoff, SR. et al. The effects of chronic antidepressant treatment in an animal model of
anxiety. Pyschopharmacology (Berl.), v. 95, n. 3, p.298-302, 1988.
Boggs, DL. et al. The effects of cannabidiol (CBD) on cognition and symptoms in outpatients
with chronic schizophrenia a randomized placebo controlled trial. Psychopharmacology
(Berl), v. 235, n. 7, p. 1923-1932, 2018.
Brito, I.; Haddad, H. A formulação do conceito de homeostase por Walter Cannon. Filosofia e
História da Biologia, v. 12, n. 1, p. 99-113, 2017.
Brown, JP. et al. Transient expression of doublecortin during adult neurogenesis. The Journal
of comparative neurology, v. 467, n. 1, p. 1-10, 2003.
Cadogan, AK. et al. Influence of Cannabinoids on Electrically Evoked Dopamine Release and
Cyclic AMP Generation in the Rat Striatum, v. 69, n. 3, p. 1131-1137, 1997.
Cameron, HA. et al. Differentiation of newly born neurons and glia in the dentate gyrus of the
adult rat. Neuroscience, v. 56, n. 2, p. 337-344, 1993.
Campos, AC. et al. Multiple mechanisms involved in the large-spectrum therapeutic potential
of cannabidiol in psychiatric disorders. Philosophical transactions of the Royal Society of
London, v. 367, n. 1607, p. 3364-3378, 2012.

88
Campos, AC. et al. The anxiolytic effect of cannabidiol on chronically stressed mice depends
on hippocampal neurogenesis: involvement of the endocannabinoid system. The international
journal of neuropyschopharmacology, v. 16, n. 6, p. 1407-1419, 2013.
Campos, AC.; Guimarães, FS. Involvement of 5HT1A receptors in the anxiolytic-like effects
of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacology
(Berl), v. 199, n. 2, p. 223-230, 2008.
Casarotto, PC. et al. Cannabidiol inhibitory effect on marble-burying behaviour: involvement
of CB1 receptors. Behavioural pharmacology, v. 21, n. 4, p. 353-358, 2010.
Cipriani, A. et al. Comparative efficacy and acceptability of 12 new-generation
antidepressants: a multiple-treatments meta-analysis. Lancet, v. 373, n. 9665, p. 746-758,
2009.
Cipriani, A. et al. Comparative efficacy and acceptability of 21 antidepressant drugs for the
acute treatment of adults with major depressive disorder: a systematic review and network
meta-analysis. Lancet, v. 391, n. 10128, p. 1357-1366, 2018.
Conover, JC.; Notti, RQ. The neural stem cell niche. Cell and tissue research, v. 331, n. 1, p.
211-224, 2008.
Crippa, JA. et al. Cannabidiol for the treatment of cannabis withdrawal syndrome: a case
report. Journal of clinical pharmacy and therapeutics, v. 38, n. 2, p. 162-164, 2013.
Crippa, JA. et al. Effects of cannabidiol (CBD) on regional cerebral blood flow.
Neuropsychopharmacology, v. 29, n.2, p. 417-426, 2004.

89
Culig, L. et al. Increasing adult hippocampal neurogenesis in mice after exposure to
unpredictable chronic mild stress may counteract some of the effects of stress.
Neuropharmacology, v. 126, p. 179-189, 2017.
Cunha, JM. et al. Chronic administration of cannabidiol to healthy volunteers and epileptic
patients. Pharmacology, v. 21, n. 3, p. 175-185, 1980.
Czéh, B. et al. Chronic psychosocial stress and concomitant repetitive transcranial magnetic
stimulation: effects on stress hormone levels and adult hippocampal neurogenesis. Biological
psychiatry, v. 52, n. 11, p. 1057-1065.
Czéh, B. et al. Stress-induced changes in cerebral metabolites, hippocampal volume, and cell
proliferation are prevented by antidepressant treatment with tianeptine. Proceedings of the
National Academy of Sciences of the USA, v. 98, n. 22, p. 12796-12801, 2001.
Dagyte, G. et al. Agomelatine reverses the decrease in hippocampal cell survival induced by
chronic mild stress. Behavioural Brain research, v. 218, n. 1, p. 121-128, 2011.
David, DJ. et al. Neurogenesis-dependent and -independent effects of fluoxetine in an animal
model of anxiety/depression. Neuron, v. 62, n. 4, p. 479-493, 2009.
Dayer, AG. et al. Short-term and long-term survival of new neurons in the rat dentate gyrus.
The journal of comparative neurology, v. 460, n. 4, p. 563-572, 2003.
De Oliveira, RW. et al. Cannabinoid signalling in embryonic and adult neurogenesis: possible
implications for psychiatric and neurological disorders. Acta Psychiatrica, p. 1-16, 2018.
De Petrocellis, L. et al. Effects of cannabinoids and cannabinoid-enriched Cannabis extracts
on TRP channels and endocannabinoid metabolic enzymes. British Journal of Pharmacology,
v. 163, n. 7, p. 1479-1494, 2011.

90
Deiana, S. et al. Plasma and brain pharmacokinetic profile of cannabidiol (CBD),
cannabidivarine (CBDV), Δ⁹-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats
and mice following oral and intraperitoneal administration and CBD action on obsessive-
compulsive behaviour. Psychopharmacology (Berl), v. 219, n. 3, p. 859-873, 2012.
Devinsky, O. et al. Cannabidiol in patients with treatment-resistant epilepsy: an open-label
interventional trial. Lancet Neurology, v. 15, n. 3, p. 270-278, 2016.
Devinsky, O. et al. Cannabidiol: pharmacology and potential therapeutic role in epilepsy and
other neuropsychiatric disorders. Epilepsia, v. 55, n. 6, p. 791-802, 2014.
Doetsch, F. A niche for adult neural stem cells. Current opinion in genetics & development, v.
13, n. 5, p. 543-550, 2003.
Duman, RS.; Monteggia, LM. A neurotrophic model for stress-related mood disorders.
Biological psychiatry, v. 59, n. 12, p. 1116-1127, 2006.
Einarson, TR. Evidence based review of escitalopram in treating major depressive disorder in
primary care. International clinical psychopharmacology, v. 19, n.5, p. 305-310.
Encinas, JM.; Vaahtokari, A.; Enikolopov, G. Fluoxetine targets early progenitor cells in the
adult brain. Proceedings of National Academy of Sciences of the United States of America, v.
103, n. 21, p. 8233-8238, 2006.
Eriksson, PS. et al. Neurogenesis in the adult human hippocampus. Nature medicine, v. 4, n.
11, p. 1313-1317, 1998.
Ernst, A. et al. Neurogenesis in the striatum of the adult human brain. Cell, v. 156, n. 5, p.
1072-1083, 2014.

91
Esposito, G. et al. Cannabidiol reduces Aβ-induced neuroinflammation and promotes
hippocampal neurogenesis through PPARγ involvement. PLoS One, v. 6, n. 12, 2011.
Esposito, MS. et al. Neuronal differentiation in the adult hippocampus recapitulates
embryonic development. The journal of neuroscience, v. 25, n. 44, p. 10074-10086, 2005.
Fendler, K.; Karmos, G.; Telegdy, G. The effect of hippocampal lesion on pituitary-adrenal
function. Europe PMC, v. 20, p. 293-297, 1961.
Ferragud, A. et al. Enhanced habit-based learning and decreased neurogenesis in the adult
hippocampus in a murine model of chronic social stress. Behavioural brain research, v. 210, n.
1, p. 134-139, 2010.
Fogaça, MV. et al. The anxiolytic effects of cannabidiol in chronically stressed mice are
mediated by the endocannabinoid system: Role of neurogenesis and dendritic remodeling.
Neuropharmacology, v. 135, p. 22-33, 2018.
Frazer, A.; Benmansour, S. Delayed pharmacological effects of antidepressants. Molecular
psychiatry, v. 7, 2002.
Gage, FH. Mammalian neural stem cells. Science, v. 287, n. 5457, p. 1433-1438, 2000.
Gedde, M.; Maa, E. Whole Cannabis Extract of High Concentration Cannabidiol May Calm
Seizures in Highly Refractory Pediatric Epilepsies. American Epilepsy Society, 67th Annual
Meeting, 2013.
Gifford, AN.; Ashby, CR Jr. Electrically evoked acetylcholine release from hippocampal
slices is inhibited by the cannabinoid receptor agonist, WIN 55212-2, and is potentiated by
the cannabinoid antagonist, SR 141716A. The journal of pharmacology and experimental
therapeutics, v. 277, n. 3, p. 1431-1436, 1996.

92
Gould, E. et al. Adrenal hormones suppress cell division in the adult rat dentate gyrus. The
Journal of neuroscience, v. 12, n. 9, p. 3642-3650, 1992.
Gould, E. et al. Neurogenesis in the dentate gyrus of the adult tree shrew is regulated by
psychosocial stress and NMDA receptor activation. The journal of neuroscience, v. 17, n. 7, p.
2492-2498, 1997.
Gould, E. et al. Proliferation of granule cell precursors in the dentate gyrus of adult monkeys
is diminished by stress. Proceedings of National Academy of Sciences of the United States of
America, v. 95, n. 6, p. 3168-3171, 1998.
Guimarães, FS. et al. Antianxiety effect of cannabidiol in the elevated plus-maze.
Psychopharmacology (Berl), v. 100, n. 4, p. 558-559, 1990.
Heine, VM. et al. Suppressed proliferation and apoptotic changes in the rat dentate gyrus after
acute and chronic stress are reversible. The European Journal of Neuroscience, v. 19, n. 1, p.
131-144, 2004.
Herman, JP.; Dolgas, CM.; Carlson, SL. Ventral subiculum regulates hypothalamo-pituitary-
adrenocortical and behavioural responses to cognitive stressors. Neuroscience, v. 86, n. 2, p.
449-459, 1998.
Hill, AS.; Sahay, A.; Hen, R. Increasing Adult Hippocampal Neurogenesis is Sufficient to
Reduce Anxiety and Depression-Like Behaviors. Neuropsychopharmacology, v. 40, n. 10, p.
2368-2378, 2015.
Ianotti, FA. et al. Nonpsychotropic plant cannabinoids, cannabidivarin (CBDV) and
cannabidiol (CBD), activate and desensitize transient receptor potential vanilloid 1 (TRPV1)
channels in vitro: potential for the treatment of neuronal hyperexcitability. ACS chemical
neuroscience, v. 5, n. 11, p. 1131-1141, 2014.

93
Jackson, M. Stress, Shock, and Adaptation in the Twentieth Century. Chapter 1, Evaluating
the Role of Hans Selye in the Modern History of Stress, Rochester (NY), University of
Rochester Press, 2014.
Jayatissa, MN. et al. Hippocampal cytogenesis correlates to escitalopram-mediated recovery
in a chronic mild stress rat model of depression. Neuropyschopharmacology, v. 31, n. 11, p.
2395-2404, 2006.
Kaplan, MS.; Hinds, JW. Neurogenesis in the adult rat: electron microscopic analysis of light
radioautographs. Science, v. 197, n. 4308, p. 1092-1094, 1977.
Karniol, IG. et al. Cannabidiol interferes with the effects of delta 9 - tetrahydrocannabinol in
man. European Journal of pharmacology, v. 28, n. 1, p. 172-177, 1974.
Kasper, S. et al. Onset of action of escitalopram compared with other antidepressants: results
of a pooled analysis. International clinical psychopharmacology, v. 21, n. 2, p. 105-110, 2006.
Kathmann, M. et al. Cannabidiol is an allosteric modulator at mu- and delta-opioid receptors.
Naunyn-Schmiedebergs archives of pharmacology, v. 372, n. 5, p. 354-361, 2006.
Katz, RJ.; Hersh, S. Amitriptyline and scopolamine in an animal model of depression.
Neuroscience and biobehavioral reviews, v. 5, n. 2, p. 265-271, 1981.
Keating, GM. Delta-9-Tetrahydrocannabinol/Cannabidiol Oromucosal Spray (Sativex®): A
Review in Multiple Sclerosis-Related Spasticity. Drugs, v. 77, n. 5, p. 563-574, 2017.
Kempermann, G. et al. Human Adult Neurogenesis: Evidence and Remaining Questions. Cell
stem cell, v. 23, n. 1, p. 25-30, 2018.
Kempermann, G.; Song, H.; Gage, FH. Neurogenesis in the Adult Hippocampus. Cold Spring
Harbor perspectives in biology, v. 7, n. 9, 2015.

94
Knigge, KM. Adrenocortical Response to Stress in Rats with Lesions in Hippocampus and
Amygdala. Experimental Biology and Medicine, v. 108, n.1, p. 18-21, 1961.
Koo, JW.; Duman, RS. IL-1β is an essential mediator of the antineurogenic and anhedonic
effects of stress. Proceedings of the National Academy of Sciences of the USA, v. 105, n. 2,
p. 751-756, 2008.
Kriegstein, AR.; Gotz, M. Radial glia diversity: a matter of cell fate. Glia, v. 43, n. 1, p. 37-
43, 2003.
Laplagne, DA. et al. Functional convergence of neurons generated in the developing and adult
hippocampus. PLoS Biology, v. 4, n. 12, 2006.
Lapraire, RB. et al. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1
receptor. British Journal of Pharmacology, v. 172, n. 20, p. 4790-4805, 2015.
Leweke, FM. et al. Cannabidiol enhances anandamide signaling and alleviates psychotic
symptoms of schizophrenia. Translational Psychiatry, v. 2, n.3, 2012.
Linge, R. et al. Cannabidiol induces rapid-acting antidepressant-like effects and enhances
cortical 5-HT/glutamate neurotransmission: role of 5-HT1A receptors. Neuropharmacology,
v. 103, p. 16-26.
Long, LE.; Malone, DT.; Taylor, DA. Cannabidiol reverses MK-801-induced disruption of
prepulse inhibition in mice. Neuropsychopharmacology, v. 31, n. 4, p. 795-803, 2006.
Mallberg, JE. et al. Chronic antidepressant treatment increases neurogenesis in adult rat
hippocampus. The Journal of neuroscience, v. 20, n. 24, p. 9104-9110, 2000.

95
Markovà, J. et al. Sativex® as add-on therapy vs. further optimized first-line ANTispastics
(SAVANT) in resistant multiple sclerosis spasticity: a double-blind, placebo-controlled
randomised clinical trial. The International journal of neuroscience, v. 13, p. 1-10, 2018.
Martin-Santos, R. et al. Acute effects of a single, oral dose of d9-tetrahydrocannabinol (THC)
and cannabidiol (CBD) administration in healthy volunteers. Current pharmaceutical design,
v. 18, n. 32, p. 4966-4979, 2012.
McEwen, BS. Allostasis and allostatic load: implications for neuropsychopharmacology.
Neuropsychopharmacology, v. 22, n. 2, p. 108-124, 2000.
McGuire, P. et al. Cannabidiol (CBD) as an Adjunctive Therapy in Schizophrenia: A
Multicenter Randomized Controlled Trial. The American journal of psychiatry, v. 175, n. 3, p.
225-231, 2018.
Mechoulam, R.; Gaoni, Y. A Total Synthesis of dl-Δ1-Tetrahydrocannabinol, the Active
Constituent of Hashish. Journal of the American Chemical Society, v. 87, n. 14, p. 3273-3275,
1965.
Mechoulam, R.; Gaoni, Y. The absolute configuration of delta-1-tetrahydrocannabinol, the
major active constituent of hashish. Tetrahedron letters, v. 12, p. 1109-1111, 1967.
Mechoulam, R.; Hanus, L. Cannabidiol: an overview of some chemical and pharmacological
aspects. Part I: chemical aspects. Chemistry and Physics of Lipids, v. 121, p. 35-43, 2002.
Mechoulam, R.; Shvo, Y. Hashish. I. The structure of cannabidiol. Tetrahedron, v. 19, n. 12,
p. 2073-2078, 1963.
Mineur, YS.; Belzung, C.; Crusio, WE. Functional implications of decreases in neurogenesis
following chronic mild stress in mice. Neuroscience, v. 150, n. 2, p. 251-259, 2007.

96
Ming, GL.; Song, H. Adult neurogenesis in the mammalian brain: significant answers and
significant questions. Neuron, v. 70, n. 4, p. 687-702, 2011.
Mitra, R. et al. Social stress-related behavior affects hippocampal cell proliferation in mice.
Physiology & Behavior, v. 89, n. 2, p. 123-127, 2006.
Mizoguchi, K. et al. Chronic stress attenuates glucocorticoid negative feedback: involvement
of the prefrontal cortex and hippocampus. Neuroscience, v. 119, n. 3, p. 887-897, 2003.
Mongiat, LA.; Schinder, AF. Adult neurogenesis and the plasticity of the dentate gyrus
network. The European Journal of Neuroscience, v. 33, n. 6, p. 1055-1061, 2011.
Montgomery, SA. et al. Escitalopram (S-enantiomer of citalopram): clinical efficacy and
onset of action predicted from a rat model. Pharmacology & Toxicology, v. 88, n. 5, p. 282-
286, 2001.
Morais, M. et al. The effects of chronic stress on hippocampal adult neurogenesis and
dendritic plasticity are reversed by selective MAO-A inhibition. Journal of
Psychopharmacology, v. 28, n. 12, p. 1178-1183, 2014.
Moreira, FA.; Aguiar, DC.; Guimarães, FS. Anxiolytic-like effect of cannabidiol in the rat
Vogel conflict test. Progress in neuro-psychopharmacology & biological psychiatry, v. 30, n.
8, p. 1466-1471, 2006.
Moreira, FA.; Guimarães, FS. Cannabidiol inhibits the hyperlocomotion induced by
psychotomimetic drugs in mice. European journal of pharmacology, v. 512, n.2-3, p. 199-205,
2005.
Morgan, CJ. et al. Cannabidiol reduces cigarette consumption in tobacco smokers:
preliminary findings. Addictive behaviors, v. 38, n. 9, p. 2433-2436, 2013.

97
Onaivi, ES.; Green, MR.; Martin, BR. Pharmacological characterization of cannabinoids in
the elevated plus maze. The journal of pharmacology and experimental therapeutics, v. 253, n.
3, p. 1002-1009, 1990.
Oomen, CA. et al. Brief treatment with the glucocorticoid receptor antagonist mifepristone
normalizes the reduction in neurogenesis after chronic stress. The European Journal of
Neuroscience, v. 26, n. 12, p. 3395-3401, 2007.
Owens, MJ.; Knight, DL.; Nemeroff, CB. Second-generation SSRIs: human monoamine
transporter binding profile of escitalopram and R-fluoxetine. Biological psychiatry, v. 50, n.
5, p. 345-350, 2001.
Papp M. et al. Antidepressant, anxiolytic and pro-cognitive effects of subacute and chronic
ketamine in the chronic mild stress model of depression. Behavioural Pharmacology, v. 28, n.
1, p. 1-8, 2017.
Pastoor, D.; Gobburu, J. Clinical pharmacology review of escitalopram for the treatment of
depression. Expert opinion on drug metabolism & toxicology, v. 10, n. 1, p. 121-128, 2014.
Paton, JA.; Nottebohm, FN. Neurons generated in the adult brain are recruited into functional
circuits. Science, v. 225, n. 4666, p. 1046-1048, 1984.
Paxinos, G.; Franklin, K. Paxinos and Franklin's the Mouse Brain in Stereotaxic Coordinates.
4ª edição. Academic Press, 2012.
Pham, K. et al. Repeated restraint stress suppresses neurogenesis and induces biphasic PSA-
NCAM expression in the adult rat dentate gyrus. The European journal of neuroscience, v. 17,
n. 4, p. 879-886, 2003.

98
Piatti, VC. et al. The timing for neuronal maturation in the adult hippocampus is modulated
by local network activity. The journal of neuroscience, v. 31, n. 21, p. 7715-7728, 2011.
Pierz, KA.; Thase, ME. A review of vilazodone, serotonin, and major depressive disorder.
The primary care companion for CNS disorders, v. 16, n.1, 2014.
Rakic, P. Limits of neurogenesis in primates. Science, v. 227, n. 4690, p. 1054-1056, 1985.
Ramon y Cajal, S. Degeneration and regeneration of the nervous system. Oxford, England:
Clarendon Press,1928.
Russo, EB. et al. Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochemical
research, v. 30, n. 8, p. 1037-1043, 2005.
Ryberg, E. et al. The orphan receptor GPR55 is a novel cannabinoid receptor. British Journal
of Pharmacology, v. 152, n. 7, p. 1092-1101, 2007.
Sahay, A.; Hen, R. Hippocampal neurogenesis and depression. Novartis Foundation
symposium, v. 289, p. 152-160, 2008.
Sakalem, ME. et al. Environmental enrichment and physical exercise revert behavioral and
electrophysiological impairments caused by reduced adult neurogenesis. Hippocampus, v. 27,
n. 1, p. 36-51, 2017.
Sales, AJ. et al. Cannabidiol Induces Rapid and Sustained Antidepressant-Like Effects
Through Increased BDNF Signaling and Synaptogenesis in the Prefrontal Cortex. Molecular
neurobiology, p. 1143-1444, 2018.
Santarelli, L. et al. Requirement of hippocampal neurogenesis for the behavioral effects of
antidepressants. Science, v.301, n. 5634, p. 805-809, 2003.

99
Sartim, AG. et al. Hippocampal mammalian target of rapamycin is implicated in stress-coping
behavior induced by cannabidiol in the forced swim test. Journal of psychopharmacology, v.
32, n. 8, p. 922-931, 2018.
Scarante, FF. Efeitos comportamentais e neuroplásticos da combinação de escitalopram e
canabinoides. Dissertação (Mestrado em Farmacologia)- Faculdade de Medicina de Ribeirão
Preto, Universidade de São Paulo. Ribeirão Preto, p. 175, 2018.
Scheonfeld, TJ.; Cameron, HA. Adult neurogenesis and mental illness.
Neuropsychopharmacology, v. 40, n. 1, p. 113-128, 2015.
Schiavon, AP. et al. Influence of single and repeated cannabidiol administration on emotional
behavior and markers of cell proliferation and neurogenesis in non-stressed mice. Progress in
neuro-psychopharmacology & biological psychiatry, v. 64, p. 27-34, 2016.
Schimidt-Hieber, C.; Jonas, P.; Bischofberger, J. Enhanced synaptic plasticity in newly
generated granule cells of the adult hippocampus. Nature, v. 429, n. 6988, p. 184-187, 2004.
Schoenfeld, TJ.; Gould, E. Differential effects of stress and glucocorticoids on adult
neurogenesis. Current topics in behavioral neuroscience, v. 15, p. 139-164, 2013.
Showalter, VM. et al. Evaluation of binding in a transfected cell line expressing a peripheral
cannabinoid receptor (CB2): identification of cannabinoid receptor subtype selective ligands.
The journal of pharmacology and experimental therapeutics, v. 278, n. 3, p. 989-999, 1996.
Sikora, V.; Berenji, J.; Latkovic, D. Influence of agroclimatic conditions on content of main
cannabinoids in industrial hemp (Cannabis sativa L. ). Genetika, v. 43, n. 3, p. 449 -456,
2011.

100
Simon, M.; Czeh, B.; Fuchs, E. Age-dependent susceptibility of adult hippocampal cell
proliferation to chronic psychosocial stress. Brain research, v. 1049, n. 2, p. 244-248, 2005.
Snyder, JS. et al. Adult hippocampal neurogenesis buffers stress responses and depressive
behaviour. Nature, v. 476, n. 7361, p. 458-461, 2011.
Snyder, JS. et al. The effects of exercise and stress on the survival and maturation of adult-
generated granule cells. Hippocampus, v. 19, n. 10, p. 898-906, 2009.
Sonego, AB. et al. Cannabidiol attenuates haloperidol-induced catalepsy and c-Fos protein
expression in the dorsolateral striatum via 5-HT1A receptors in mice. Behavioural brain
research, v. 309, p. 22-28, 2016.
Spadone, C. [Effects of escitalopram on anxiety symptoms in depression]. L’Enchephale, v.
28, n. 5, p. 461-465, 2002.
Spalding, KL. et al. Dynamics of hippocampal neurogenesis in adult humans. Cell, v. 153, n.
6, p. 1219-1227, 2013.
Suh, H. et al. In vivo fate analysis reveals the multipotent and self-renewal capacities of
Sox2+ neural stem cells in the adult hippocampus. Cell Stem Cell, v. 1, n.5, p. 515-528, 2007.
Surget, A. et al. Antidepressants recruit new neurons to improve stress response regulation.
Moleclar psychiatry, v. 16, n. 12, p. 1177-1188, 2011.
Surget, A. et al. Drug-dependent requirement of hippocampal neurogenesis in a model of
depression and of antidepressant reversal. Biological Psychiatry, v. 64, n. 4, p. 293-301, 2008.
Szabo, S. et al. "Stress" is 80 Years Old: From Hans Selye Original Paper in 1936 to Recent
Advances in GI Ulceration. Current pharmaceutical design, v. 23, n. 27, p. 4029-4041, 2017.

101
Szaflarski, JP. et al. Long‐term safety and treatment effects of cannabidiol in children and
adults with treatment‐resistant epilepsies: Expanded access program results. Epilepsia, v. 59,
n. 8, p. 1540-1548, 2018.
Tanti, A. et al. Region-dependent and stage-specific effects of stress, environmental
enrichment, and antidepressant treatment on hippocampal neurogenesis. Hippocampus, v. 23,
n. 9, p. 797-811, 2013.
Tham, M. et al. Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol-
dimethylheptyl at the type 1 and type 2 cannabinoid receptors. British Journal of
Pharmacology, 2018.
Trigo, JM. et al. Nabiximols combined with motivational enhancement/cognitive behavioral
therapy for the treatment of cannabis dependence: A pilot randomized clinical trial. PLos One,
v. 13, n. 1, 2018.
Tzadok, M. et al. CBD-enriched medical cannabis for intractable pediatric epilepsy: The
current Israeli experience. Seizure, v. 35, p. 41-44, 2016.
Ujváry, I.; Hanus, L. Human Metabolites of Cannabidiol: A Review on Their Formation,
Biological Activity, and Relevance in Therapy. Cannabis and cannabinoid research, v. 1, n. 1,
p.90-101, 2016.
Van Bokhoven, P. et al. Reduction in hippocampal neurogenesis after social defeat is long-
lasting and responsive to late antidepressant treatment. The European Journal of
Neuroscience, v. 33, n. 10, p. 1833-1840, 2011.
Veena, J. et al. Exposure to enriched environment restores the survival and differentiation of -
new born cells in the hippocampus and ameliorates depressive symptoms in chronically
stressed rats. Neuroscience Letters, v. 455, n. 3, p. 178-182, 2009.

102
Vilela, LR. et al. Anticonvulsant effect of cannabidiol in the pentylenetetrazole model:
Pharmacological mechanisms, electroencephalographic profile, and brain cytokine levels.
Epilepsy and behavior, v. 75, p. 29-35, 2017.
Wade, A.; Friis Andersen, H. The onset of effect for escitalopram and its relevance for the
clinical management of depression. Current medical research and opinion, v. 22, n. 11, p.
2101-2110, 2006.
Wang, JW. et al. Chronic fluoxetine stimulates maturation and synaptic plasticity of adult-
born hippocampal granule cells. The Journal of neuroscience, v. 28, n. 6, p. 1374-1384, 2008.
Willner, P. The chronic mild stress (CMS) model of depression: History, evaluation and
usage. Neurobiology of Stress, v. 6, p. 78-93, 2017.
Wolf, SA. et al. Cannabinoid receptor CB1 mediates baseline and activity-induced survival of
new neurons in adult hippocampal neurogenesis. Cell communication and signaling, v. 8,
n.12, 2010.
Xu, Y. et al. Curcumin reverses impaired cognition and neuronal plasticity induced by chronic
stress. Neuropharmacology, v. 57, n. 4, p. 463-471, 2009.
Zanelatti, TV. et al. Antidepressant-like effects of cannabidiol in mice: possible involvement
of 5-HT1A receptors. British journal of pharmacology, v. 159, n. 1, p. 122-128, 2010.
Zuardi, AW. et al. Action of cannabidiol on the anxiety and other effects produced by delta 9-
THC in normal subjects. Psychopharmacology (Berl), v. 76, n. 3, p. 245-250, 1982.
Zuardi, AW. et al. Antipsychotic effect of cannabidiol. The journal of clinical psychiatry, v.
56, n. 10, p. 485-486, 1995.

103
Zuardi, AW. et al. Effects of ipsapirone and cannabidiol on human experimental anxiety.
Journal of psychopharmacology, v. 7, p. 82-88, 1993.

104
ANEXOS
ANEXO A) Análises correlacionais entre os valores de latência para comer no novo
ambiente no teste do NSF e o número de células DCX+/0,1mm
2 no time point de 7 dias.
Análises de correlação de Pearson entre os valores de latência observados no NSF e as
respectivas quantidades de células DCX+/0,1 mm
2 no giro denteado do hipocampo após 7 dias
de protocolo. Testamos neste experimento as seguintes correlações: A) somente entre os
animais não submetidos ao estresse; B) entre os animais submetidos ao estresse; C) Entre os
animais tratados com veículo independentemente da condição; D) entre os animais tratados
com CBD independentemente da condição e E) entre os animais tratados com escitalopram
independente da condição.
A
B
E
D
C

105
ANEXO B
Análises de correlação de Pearson entre os valores de latência observados no NSF e as
respectivas quantidades de células BrdU+/0,1 mm
2 no giro denteado do hipocampo após 7
dias de protocolo. Testamos neste experimento as seguintes correlações: A) somente entre os
animais não submetidos ao estresse; B) entre os animais submetidos ao estresse; C) Entre os
animais tratados com veículo independentemente da condição; D) entre os animais tratados
com CBD independentemente da condição e E) entre os animais tratados com escitalopram
independente da condição.
A
B
E
D
C

106
ANEXO C) Análises correlacionais entre os valores de latência para comer no novo
ambiente no teste do NSF e o número de células DCX+/0,1mm
2 no time point de 14 dias.
Análises de correlação de Pearson entre os valores de latência observados no NSF e as
respectivas quantidades de células DCX+/0,1 mm
2 no giro denteado do hipocampo após 14
dias de protocolo. Testamos neste experimento as seguintes correlações: A) somente entre os
animais não submetidos ao estresse; B) entre os animais submetidos ao estresse; C) Entre os
animais tratados com veículo independentemente da condição; D) entre os animais tratados
com CBD independentemente da condição e E) entre os animais tratados com escitalopram
independente da condição.
A
B
E
D
C

107
ANEXO D) Análises correlacionais entre os valores de latência para comer no novo
ambiente no teste do NSF e o número de células BrdU+/0,1mm
2 no time point de 14 dias.
Análises de correlação de Pearson entre os valores de latência observados no NSF e as
respectivas quantidades de células BrdU+/0,1 mm
2 no giro denteado do hipocampo após 14
dias de protocolo. Testamos neste experimento as seguintes correlações: A) somente entre os
animais não submetidos ao estresse; B) entre os animais submetidos ao estresse; C) Entre os
animais tratados com veículo independentemente da condição; D) entre os animais tratados
com CBD independentemente da condição e E) entre os animais tratados com escitalopram
independente da condição.
A
B
E
D
C