dissertação de mestrado aspectos ecológicos da ...‡Ão... · figura 2. a) vista panorâmica da...
TRANSCRIPT

Universidade Federal de Ouro Preto
Instituto de Ciências Exatas e Biológicas
Departamento biodiversidade, evolução e meio ambiente – DEBio
Programa de Pós-Graduação em Ecologia de Biomas Tropicais
Dissertação de Mestrado
Aspectos ecológicos da ocorrência de Aedes (Stegomyia) aegypti
(Linnaeus,1762) e Aedes (Stegomyia) albopictus (Skuse, 1984)
(DIPTERA:CULICIDAE) em áreas verdes urbanas e residenciais
MICHELLE CRISTINE PEDROSA
Ouro Preto, Minas Gerais
Março de 2013


Catalogação: [email protected]
P372a Pedrosa, Michelle Cristine.
Aspectos ecológicos da ocorrência de Aedes (Stegomyia) aegypti
(Linnaeus, 1762) e Aedes (Stegomyia) albopictus (Skuse, 1984)
(DIPTERA:CULICIDAE) em áreas verdes urbanas e residenciais
[manuscrito] / Michelle Cristine Pedrosa - 2013.
85f.: il., color; graf.; tab.; mapas.
Orientador: Prof. Dr. Sérvio Pontes Ribeiro.
Dissertação (Mestrado) - Universidade Federal de Ouro Preto.
Instituto de Ciências Exatas e Biológicas. Departamento de Biodiversidade,
Evolução e Meio Ambiente. Programa de Pós-Graduação em Ecologia de
Biomas Tropicais.
Área de concentração: Evolução e Funcionamento de Ecossistemas.
1. Aedes aegypti - Teses. 2. Aedes albopictus - Teses. 3. Vetores de
doença - Teses. 4. Entomologia - Teses. 5. Ecologia urbana (Biologia) -
Mariana (MG) - Teses. I. Ribeiro, Sérvio Pontes. II. Universidade Federal
de Ouro Preto. III. Título.
CDU: 574.4(815.1):616.928.8

Dedico este trabalho à meu marido e aos meus pais

“O conhecimento nos faz responsáveis.”
Che Guevara
“A tarefa não é tanto ver aquilo que ninguém viu,
mas pensar o que ninguém ainda pensou
sobre aquilo que todo mundo vê”.
Arthur Schopenhauer

AGRADECIMENTOS
Como o tempo passou rápido! Muitas pessoas fizeram parte dessa fase, contribuindo
direta ou indiretamente para a realização do trabalho.
Agradeço à algumas pessoas em especial: Alexandre, meus pais e ao Sérvio. À
Alexandre, meu companheiro, em todos os sentidos, os meus agradecimentos...você foi
essencial da primeira à última etapa. Foi biólogo em campo e em laboratório, motorista,
revisor, debatedor de ideias, psicólogo...foi tudo o que podia ser e tudo o que eu precisava que
fosse. E sem nunca esquecer de me dar os puxões de orelha e o carinho necessários...
Mãe e pai, como não agradecê-los? Apoiaram-me mesmo sem no início terem
compreendido o que eu estava fazendo...cederam o carro para quando não havia o transporte,
“financiaram” alguns materiais da pesquisa, carregaram minhas larvas pra casa só para que eu
pudesse triar e ficar pertinho de vocês algumas vezes...assim como em todas as outras etapas
da minha vida, vocês foram (e são!) muito importantes! Aos meus irmãos, cunhadas, sobrinho
(as), sogro, sogra o meu agradecimento pelo apoio.
Agradeço ao orientador, amigo e “cumpadre”, Sérvio. Se não fosse você ter acolhido
lá em 2008 “a menina que queria trabalhar com vetores” essa dissertação não estaria sendo
entregue hoje, ao menos não na UFOP. Obrigada Sérvio pelo grande ensinamento teórico-
prático da ciência e da vida...ser orientada (ou desorientada em alguns momentos rsrsr) por
você me amadureceu muito como profissional! Considero você um exemplo em nossa
profissão pela dedicação, eficiência e honestidade.
Ah o “lab.”! Quantas saudades já sinto e tenho certeza que sentirei...quanto coisa
vivenciada, quantas amizades construídas e aprendizado adquirido! Só não sentirei saudades
de ficar limpando o “dossel” durante horas e ganhar uma bela dor de coluna por dias! Luna,
Luana, Núbia, Babi, Cinthia, Raquel, Zé, Flavinho, Alice, Roiber, Caxelove, e Gi, jamais
esquecerei dos bons momentos que vivemos alí! Agradeço em especial às “meninas” sempre
parceiras: me lembrarei para sempre das paçocas, das belas risadas da Cinthia ou
simplesmente do “convite” de Lulu e Nubita para irmos lá pra fora papear!
Aos remanescentes do 05.2, Adriele, Fifi, Glênia, Guilherme, Hudson, Água Moli e
Amanda obrigada pela companhia, pelos cafés no fim da tarde...pizzas no O Passo...não posso
deixar de agradecer em especial ao Guilherme, por ter sempre me ajudado com boa vontade,
compromisso e simpatia! E à Adriele, bem é até difícil falar...Dri amiga, obrigada por tudo!
Você me ajudou em tudo o que pôde: me deu abrigo, foi pra campo várias vezes comigo, me

apoiou quando precisei. Já disse e repito, você tornou minha vida mais alegre enquanto estive
aqui!
À turma do mestrado pela companhia e risadas, em especial à Fê que se tornou uma
amiga desde o primeiro dia em que nos vimos...
Ao prof. Álvaro Eiras pela colaboração em todos estes anos! Seu apoio possibilitou a
realização deste trabalho! Obrigada também à todos do LABEq e ECOVEC!
À Raquel (Royal) e Maíra colegas mosquitológas, muito obrigada por toda
colaboração, troca de artigos e conselhos!
Agradeço ao Dr. Aldo Malavasi e ao MSc. Danilo Carvalho pelas correções,
compreensão e incentivo. E à todos da Moscamed, em especial Luiza, Miriam e Maylen pela
torcida!
Ao Talvani por permitir a “entrada” dos meus mosquitos malucos em seu
laboratório...e ao Dr. Sérgio Caldas pela colaboração na FUNED.
Ao Júlio Fontenelle (Julinho!) pela colaboração na parte inicial deste trabalho.
À todos estagiários que “passaram por este projeto”: Léo, Lucas, Laís, Luana, Ana e
Márcia, muito obrigada!
Ao meu sobrinho Álef pela boa vontade constante em ajudar e por tentar fazer com
que meu experimento desse certo!
À profa. Michelly da UNIVASF-PE que permitiu o acesso ao laboratório para que eu
pudesse identificar o restante das amostras...
Ao secretário Rubens, sempre simpático, pela ajuda constante!
Agradeço a cada morador de Mariana e Ouro Preto que confiaram à essa estranha os
quintais de suas casas...
Agradeço à UFOP e à Ouro Preto por esta etapa tão especial da minha vida.
À CAPES pelo apoio financeiro.

LISTA DE FIGURAS
Introdução
Figura 1. Classificação de Áreas de Vulnerabilidade para ocorrência de Dengue por
Unidades Federadas, Brasil, 2010/2011. Fonte: Ministério da Saúde
http://portal.saude.gov.br/portal/arquivos/pdf/ntaval_vulerpid_dengue_1011.pdf.
Modificado em 26/08/2012................................................................................
16
Figura 2. Estágios de desenvolvimento do mosquito Aedes. Fonte: UNICAMP
(2001)...........................................................................................................................
17
Capítulo I
Figura 1. Espaço verde urbano utilizado no estudo em Ouro Preto. Foto:
Modificada do Google Earth, Michelle Pedrosa, 2012.................................................
30
Figura 2. a) Vista panorâmica da área verde urbana utilizada para o estudo em
Ouro Preto. b, c e d) vista interna da área de estudo. Foto: Alexandre Cortez do
Amaral, 2009.................................................................................................................
30
Figura 3. Área verde utilizada no estudo na cidade de Mariana. Foto: Modificada
do Google Earth, Michelle Pedrosa, 2012....................................................................
31
Figura 4. a) Vista panorâmica da área verde urbana utilizada para o estudo em
Ouro Preto. b, c e d) vista interna da área de estudo. Foto: Alexandre C. do Amaral,
2009..............................................................................................................................
31
Figura 5.Esquema do desenho amostral em cada área verde urbana utilizada de
Mariana e Ouro reto, Minas Gerais................................................................................
32
Figura 6. Proporção de cada espécie coletada em ambas áreas verdes de Mariana e
Ouro Preto, Minas Gerais..............................................................................................
36
Figura 7. Distribuição da abundância de Aedes aegypti em cada coleta nas áreas
verdes de Ouro Preto e Mariana....................................................................................
36
Figura 8. Distribuição da abundância de Aedes albopictus em cada coleta na áreas
verdes de Ouro Preto e Mariana....................................................................................
37
Figura 9. Relação entre a abundância de Aedes aegypti e Ae. albopictus
considerando todas as amostras de diferentes cidades e datas na escala de
ovitrampa. Observar que as ovitrampas com maiores densidades de Ae. albopictus
são as com zero Ae. aegypti..........................................................................................
39
Figura 10.Gráfico de dispersão entre a abundância média por coleta de Aedes e a
pluviosidade (atrasada em 15 dias) em ambos anos de coleta em Mariana, Minas
Gerais...........................................................................................................................
40
Figura 11. Gráfico de dispersão entre a abundância média por coleta de Aedes e a
pluviosidade (atrasada em 15 dias) em ambos anos de coleta em Ouro Preto, Minas
Gerais............................................................................................................................
40
Figura 12. Gráfico de dispersão entre a abundância de Aedes e a pluviosidade
(atrasada em 15 dias) em ambos anos de coleta em Mariana e Ouro Preto, Minas
Gerais............................................................................................................................
41
Capítulo II
Figura 1. Esquema representando o delineamento amostral em cada cidade, com os
pontos amostrais devidamente distribuídos..................................................................
55
Figura 2. Visualização dos pontos amostrais utilizados no estudo em Ouro Preto.
Observar que pontos inseridos no círculo azul correspondem aos biótopos I e II.
Pontos inseridos no círculo laranja correspondem ao biótopo III. Imagem
modificada do Google Earth em 14/03/2013................................................................
56

Figura 3. Visualização dos pontos amostrais utilizados no estudo em Mariana.
Observar que pontos inseridos no círculo azul correspondem aos biótopos I e II.
Pontos inseridos no círculo laranja correspondem ao biótopo III. Imagem
modificada do Google Earth em 14/03/2013................................................................
56
Figura 4. Distribuição das espécies, na forma adulta, entre os biótopos durante a
estação chuvosa em Mariana, Minas Gerais.................................................................
61
Figura 5. Distribuição das espécies, na forma adulta, entre os biótopos durante a
estação chuvosa em Ouro Preto, Minas Gerais.............................................................
61
Figura 6. Distribuição das espécies, na forma imatura, entre os biótopos durante a
estação chuvosa em Mariana, Minas Gerais.................................................................
62
Figura 7. Distribuição das espécies, na forma imatura, entre os biótopos durante a
estação chuvosa em Ouro Preto, Minas Gerais.............................................................
63
APÊNDICE E ANEXO
Figura 1.a) Gráfico de amplificação gerado pelas amostras de Aedes em que
apenas os controles positivos (da extração e da PCR) demonstraram amplificação.
b) Curva de “melting” gerada após os 40 ciclos de amplificação, onde se observa
um Tm em torno de 76,8oC (apenas para os controles positivos) esperado para o
produto de DENV
amplificado........................................................................................................
83
Figura 2. Ovitrampas utilizadas nos estudos dos capítulos I e II.a) A armadilha é
vista lateralmente. b) A armadilha é vista por cima. Foto: Michelle C. Pedrosa,
2009..............................................................................................................................
85
Figura 3. MosquiTRAP®
utilizadas no estudo do capítulo II. a) Armadilha montada,
pronta para uso. Fonte: https://www.ufmg.br/online/arquivos/Dengue_armadilha-
thumb.jpg acesso em 17/03/2013. b) Armadilha desmontada, com seus componentes
expostos. Sendo: I, parte superior da armadilha; II, parte inferior (armazenamento
de água); III, cartao adesivo; IV AtrAedes® e V, tela. Fonte: Laila Heringer
Costa............................................................................................................................. .
85

LISTA DE TABELAS
Capítulo I
Tabela 1. Número de imaturos coletados em ovitrampas em Mariana e Ouro
Preto, cidades de Minas Gerais, entre Fevereiro de 2009 e Maio de 2009.................
35
Tabela 2. Número de imaturos coletados em ovitrampas em Mariana e Ouro
Preto, cidades de Minas Gerais, em Fevereiro de 2012..............................................
35
Tabela 3. Positividade das palhetas e coexistência de Ae. aegypti e Ae. albopictus
nas cidades de Mariana e Ouro Preto, Minas Ferais...................................................
38
Tabela 4. Análise da distribuição espacial dos ovos nas palhetas............................. 38
Capítulo II
Tabela 1. Tabela de contingência apresentando a abundância total das espécies em
cada biótopo e a porcentagem de diminuição da abundância das espécies entre a
estação chuvosa e seca nas cidades de Mariana, Minas Gerais..................................
60
Tabela 2. Tabela de contingência apresentando a abundância total das espécies em
cada biótopo e a porcentagem de diminuição da abundância das espécies entre a
estação chuvosa e seca na cidade de Ouro Preto, Minas Gerais.................................
60
Tabela 3. Valores absolutos de abundância das espécies em cada habitat, na
estação chuvosa, com as diferentes metodologias utilizadas......................................
63
Tabela 4. Frequência de ocorrência de Aedes nas armadilhas, por habitat, durante
as quatro semanas de coleta da estação chuvosa e porcentagem de ocorrência em
relação ao número total de armadilhas utilizadas.......................................................
64

SUMÁRIO
APRESENTAÇÃO DO TRABALHO................................................................... 11
INTRODUÇÃO GERAL........................................................................................... 12
CAPÍTULO I: Aspectos ecológicos dos vetores da dengue em
áreas verdes urbanas: um estudo da ocorrência de vetores nas
cidades de Mariana e Ouro Preto, Minas Gerais..........................
24
RESUMO ................................................................................................................... 25
ABSTRACT............................................................................................................... 26
1.INTRODUÇÃO....................................................................................................... 27
2.MATERIAL E MÉTODOS..................................................................................... 29
2.1.Área de estudo .................................................................................................... 29
2.2 Coleta de dados .................................................................................................... 32
2.3.Análises estatísticas.............................................................................................. 34
3.RESULTADOS...................................................................................................... 34
4.DISCUSSÃO ......................................................................................................... 41
5. CONSIDERAÇÕES EPIDEMIOLÓGICAS......................................................... 46
Capítulo II: Padrões de distribuição espacial e sazonal de Aedes
(Stegomyia) aegypti (Linnaeus, 1762) e Aedes (Stegomyia)
albopictus (Skuse, 1894) (DIPTERA: CULICIDAE) em
biótopos urbanos ..............................................................................................
48
RESUMO................................................................................................................... 49
ABSTRACT............................................................................................................... 50
1. INTRODUÇÃO...................................................................................................... 51
2. MATERIAL E MÉTODOS.................................................................................... 53
2.1. Área de estudo..................................................................................................... 53
2.2. Delineamento amostral........................................................................................ 54
2.2. Coleta de dados................................................................................................... 56
2.5. Dados abióticos.................................................................................................... 58
2.6. Análises estatísticas 59
3.RESULTADOS....................................................................................................... 59
4.DISCUSSÃO........................................................................................................... 65
REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 70
APÊNDICE- Inquérito Virológico ......................................................................... 81
ANEXO...................................................................................................................... 86

11
APRESENTAÇÃO DO TRABALHO
Prever e controlar a trajetória de um sistema biológico de evolução rápida, composto
por vários agentes, entre eles parasitas, hospedeiros e vetores é algo desafiador (Codeço
et al., 2010). O estudo da ecologia de vetores, considerado como o pilar central da
vigilância entomológica (Donalísio & Glasser, 2002), visa contribuir para que tal desafio
seja menor, pois busca conhecer os fatores ecológicos determinantes para que os insetos
vetores sejam capazes de atuarem como agentes de transmissão de parasitas ao ser
humano (Codeço et al., 2010). Estratégias eficientes para a prevenção de doenças
vetoriais podem ser construídas a partir de um maior número de informações sobre as
mudanças comportamentais dos agentes transmissores (Codeço et al., 2010). Assim,
diante das características dos vetores e de sua capacidade de adaptação às alterações
ambientais, é esperado que os quadros epidemiológicos das infecções por eles
transmitidas apresentem mudanças constantes (Forattini, 1998).
O dengue é hoje a arbovirose com maior importância no planeta, com cerca de 2,5
bilhões de pessoas sob risco de infecção, especialmente em países tropicais. Como o
vetor é o único elemento vulnerável da cadeia de transmissão da dengue (Tauil, 2002),
estudos relativos a diversos aspectos da ecologia dos vetores Ae. aegypti e Ae. albopictus,
tem sido realizados. Nestes estudos tem-se buscado desvendar os comportamentos e
hábitos preferenciais desses mosquitos na natureza e no espaço habitado pelo ser humano,
devido à grande capacidade de adaptação que apresentam diante as diversas
características sociais, urbanas e ambientais (Donalísio & Glasser, 2002).
Para o controle vetorial, a compreensão dos padrões de ocorrência em cidades com
altos níveis de infestação dos vetores é essencial. Todavia, para a prevenção, é possível
que seja igualmente importante o entendimento destes padrões em locais onde a
infestação das espécies transmissoras é baixa, pois é a partir da infestação mínima que as
estratégias devem ser colocadas em prática a fim de minimizar o risco de surtos
epidêmicos. Assim, neste trabalho, buscou-se compreender aspectos ecológicos da
ocorrência de Ae. aegypti e Ae. albopictus em situações não usuais, como em áreas verdes
inseridas em ambiente urbano e ainda em cidades com baixa infestação destes vetores
(Índice de Infestação Predial <1), sendo uma delas com elevada altitude. Pretendeu-se
verificar se ambas espécies podem ser encontradas em áreas verdes urbanas destas
cidades e ainda, se os padrões espaciais e sazonais de ocorrência dessas espécies em

12
cidades com baixa infestação são semelhantes aos padrões encontrados em cidades com
maiores índices de infestação.
As próximas subdivisões desta introdução são considerações teóricas sobre a dengue
e seus vetores, bem como sobre o biótopo estudado: a área verde urbana. Foram
desenvolvidos dois estudos entre os anos de 2009 e 2012. Nos capítulos que se seguem,
tem-se metodologia, resultados e discussão de cada trabalho desenvolvido, sendo o
primeiro capítulo um estudo desenvolvido por completo em áreas verdes urbanas de duas
cidades de Minas Gerais nos anos de 2009 e 2012. Já o capítulo II, desenvolvido no ano
de 2011, abrangeu tanto as áreas verdes urbanas quanto as áreas residencial e comercial
das mesmas cidades.
INTRODUÇÃO GERAL
1. Antecedentes históricos da dengue
A dengue é uma doença febril aguda, que possui um vírus como seu agente
etiológico, sendo conhecidos quatro sorotipos causadores da doença: DENV-1, DENV-2,
DENV-3 e DENV-4 (WHO, 2012). Atualmente é a doença vetorial mais importante no
continente americano e coloca em constante ameaça a saúde de milhões de pessoas que
residem em áreas urbanas, suburbanas e rurais. O vírus da dengue é um arbovírus, palavra
derivada da expressão inglesa “ARthropod BOrne VIRuses”. Tal expressão denomina
um grupo de vírus que é mantido no meio ambiente através da transmissão biológica
entre hospedeiros vertebrados e artrópodes hematófagos como mosquitos e carrapatos,
entre outros (CDC, 2005). A transmissão do vírus ocorre principalmente através da
picada da fêmea do mosquito Aedes aegypti (Linnaeus, 1762) (DIPTERA:CULICIDAE),
entretanto outra espécie do mesmo gênero, Aedes albopictus (Skuse 1894)
(DIPTERA:CULICIDAE), recentemente encontrado na América é reconhecida como
vetor secundário da doença no continente asiático (Tauil, 2001), além de ter sido
considerado o transmissor primário durante um recente surto no Havaí (MMWR, 2002;
Effler et al., 2005).
De acordo com Gubler (1997) a epidemia de dengue é uma doença muito antiga,
cuja história é caraterizada por epidemias periódicas, muitas vezes raras. As primeiras
notificações de epidemias de dengue ocorreram nos anos de 1779 e 1780 nos continentes

13
asiático, africano e norte-americano (CDC, 2000; Mahmood, 2006). No século seguinte,
ocorreram quatro grandes epidemias entre 1827 e 1897 no Caribe e sul dos Estados
Unidos (Ehrenkranz et al. apud Pinheiro e Nelson 1997). Nas décadas de 1940 e 1950 as
epidemias tiveram seu reinício no sudeste da Ásia durante e após a Segunda Guerra
Mundial (Maciel et al. 2008), se expandindo posteriormente, para o restante do globo,
incluindo o continente Americano (Pinheiro & Nelson, 1997; Maciel et al., 2008).
Durante muitas décadas, um programa iniciado em 1946 pela Organização Pan-
Americana da Saúde (OPAS), foi realizado no intuito de erradicar Aedes aegypti,
prevenindo epidemias de febre amarela urbana e dengue nos países da América
continental. Entretanto, no início dos anos 70, a não continuidade do programa em países
que já haviam conquistado a erradicação acarretou em uma lenta reinfestação pelo vetor,
sendo que em 1987, quase todos os países já apresentavam Ae. aegypti em seu território
(Vasconcelos et al., 1999). Ainda nos anos 70, o DENV-1 foi reintroduzido nas Américas
e houveram epidemias na América do Sul (por exemplo, na Venezuela) e do Norte, bem
como no Caribe (Maciel et al., 2008), ocorrendo a a re-emergência da doença (Guzman et
al., 2003) e ainda, houve a primeira epidemia documentada clínica e laboratorialmente,
no início da década de 80 no Brasil, no estado de Roraima pelo DENV-1 e DENV-4
(Osanai et al., 1983;Tauil, 2001).
Depois de cinco anos sem casos confirmados da doença, ocorreu uma epidemia
(DENV1) no Rio de Janeiro e em sequência, várias epidemias em cidades bastante
povoadas nas regiões sudeste e nordeste do país (Siqueira et al., 2005). A localização e o
intenso fluxo de pessoas na cidade do Rio de Janeiro contribuíram para a dispersão da
doença para o Nordeste, São Paulo, Minas, Mato Grosso do Sul e, posteriormente, outros
estados, incluindo Goiás, na década de 1990 (Maciel, 1999). Cem mil casos de dengue
foram registrados em 1987 (DENV1) e 1991 (DENV2), provavelmente pela introdução
dos diferentes sorotipos virais (Siqueira et al., 2005).
Siqueira et al. (2005), através da análise de dados da dengue, encontraram um padrão
epidemiológico no Brasil, durante os últimos 20 anos, que divide a incidência da doença
em dois períodos distintos:
1. Fase inicial, entre 1986 e 1993, representada por ondas epidêmicas em áreas
localizadas.
2. Padrão nacional epidêmico e endêmico de circulação viral, entre 1994 e 2002.

14
Entre 1994 e 2002, cerca de 2,826,948 casos de dengue foram notificados,
indicando uma tendência de aumento na incidência da doença. Grande parte dos casos de
dengue ocorreram em áreas metropolitanas, apesar de vários surtos terem ocorrido em
áreas urbanas menores em 25 dos 27 estados brasileiros. Neste segundo período, a
vigilância sobre o vetor Ae. aegypti foi estendida cobrindo 69,7% dos municípios do país
em 1996 e 89,6% em 2002. A partir deste sistema inferiu-se que a porcentagem de
municípios infestados variou de 44,5% em 1996 a 58,3% em 2002 (Siqueira et al., 2005).
1.1- Dengue: Situação atual
Na atualidade mais de 100 países com uma população combinada de mais de 2,5
bilhões de pessoas estão em risco de infecção pela doença (Gómez-Dantés & Willoquet,
2009). Houve um aumento exponencial do número de casos de dengue notificados à
Organização Mundial da Saúde - World Health Organization- (WHO, Maciel et al.; 2008)
e ainda, expansão geográfica das áreas de transmissão da doença (WHO 2010; Guzman et
al., 2010 ). Atualmente, todos os quatro sorotipos do vírus da dengue (DENV-1 a 4)
circulam nos continentes asiáticos, africano e americano, incluindo Brasil (M.S., 2010b).
Nas Américas a dengue tem apresentado uma propensão à ascensão, com picos
epidêmicos cada vez maiores que se repetem a cada 3–5 anos de forma regular. Mais de
30 países do continente americano registram atualmente casos de dengue em seu
território e o Brasil está entre os países que apresentam o maior número de casos (San
Martín & Brathwaite-Dick, 2007).
Desde o final da década de 90 as epidemias de dengue se constituem numa das
maiores prioridades de saúde pública no Brasil (S.V.S, 2004) que têm apresentado um
padrão sazonal de incidência da doença, com maiores índices nos meses mais quentes e
úmidos (janeiro a maio), acompanhando a estação chuvosa (M.S., 2006). Além disso, a
maioria dos casos de dengue que se concentravam nos grandes centros urbanos,
apresentou modificação entre os anos de 2007 e 2008, verificando-se o aumento da
incidência em municípios pequenos e médios (M.S., 2007 e 2008; Maciel et al., 2008.).
Em 2009, o Ministério da Saúde em estreita cooperação com outros órgãos e
diversas iniciativas, elaborou as Diretrizes Nacionais para a Prevenção e Controle de
Epidemias de Dengue, no intuito de orientar os gestores e técnicos na adequação dos
planos de contingência estaduais, regionais, metropolitanos ou locais (M.S., 2010b). Na
figura 1, por exemplo, pode ser encontrada uma matriz, elaborada pelo Ministério da

15
Saúde e Secretaria de Vigilância em Saúde/MS, que revela áreas vulneráveis para
ocorrência de dengue no Brasil entre 2010 e 2011. Tal matriz fornece subsídios para
auxiliar na delimitação das áreas que necessitam de maior intensificação das ações de
combate ao vetor (M.S., 2010b).
Na Figura 1 pode-se observar a pequena quantidade de estados com baixo risco de
transmissão e que Minas Gerais está classificada como uma região de alto risco de
transmissão da dengue.
Na dinâmica da dengue o vetor é fundamental sendo, até o momento, o único
elemento vulnerável a uma medida preventiva na cadeia de transmissão da doença (Tauil,
2002). Somando-se a isso o fato de ainda não existir uma vacina preventiva eficaz
disponível (Tauil, 2002), a luta antivetorial é a principal estratégia de controle do dengue
tanto no Brasil, quanto nos demais países onde a doença está presente (Costa, 2008).
Como toda doença vetorial, a ocorrência da dengue está diretamente relacionada
com a ecologia e distribuição do seu vetor, ecologia dos hospedeiros, variáveis
ambientais e a forma como se dá a ocupação do espaço natural pelo homem (Forattini,
1992). Fatores estruturais e conjunturais como concentração humana e aumento da
mobilidade dos grupos populacionais (proporcionada pelo maior acesso aos meios de
transporte) também podem potencializar a propagação da doença, favorecendo a
expansão e a manutenção da circulação do vírus e seus vetores (Maciel, 2008).

16
Figura 1. Classificação de Áreas de Vulnerabilidade para ocorrência de Dengue por
Unidades Federadas, Brasil, 2010/2011. Fonte: Ministério da Saúde
http://portal.saude.gov.br/portal/arquivos/pdf/nt_aval_vul_epid_dengue_verao_10_11.pdf
Modificado em 26/08/2012.
2. Considerações importantes sobre a ecologia dos vetores da dengue
Ochlerotatus, Stegomyia, Howardina e Protomacleaya são subgêneros de Aedes,
pertencentes à família Culicidae, existentes no Brasil (Kingh & Stone, 1978 apud
(Consoli & Lourenço-de-Oliveira, 1998). Entretanto, apenas os subgêneros Stegomyia e
Ochlerotatus reúnem espécies de Aedes com importância epidemiológica (Consoli &
Lourenço-de-Oliveira, 1998).
Aedes (Stegomyia) aegypti e Ae. (Stegomyia) albopictus são espécies exóticas na
América do Sul, homólogas e simpátricas, sendo que Ae. aegypti foi introduzido no
Brasil provavelmente na época do tráfego de escravos e Ae. albopictus foi notificado pela
primeira vez em 1986 no país (Forattini 1986; Consoli & Lourenço 1998).

17
Figura 2. Estágios de desenvolvimento do mosquito Aedes. Fonte: UNICAMP (2001)
O desenvolvimento do mosquito do gênero Aedes dura cerca de 8 a 10 dias (Fig.
2) e possui do mesmo modo que todos os culicídeos, uma fase aquática e uma fase
terrestre durante o ciclo de vida. A primeira fase possui três estádios de
desenvolvimento: ovo, larva e pupa, sendo que a fase terrestre equivale ao mosquito . Da
oviposição à fase adulta decorrem em média 10 dias em condições favoráveis de
temperatura e disponibilidade de alimentos. Os ovos são depositados individualmente
pelas fêmeas nas paredes internas de recipientes , próximos à superfície da água, embora
as posturas possam ser realizadas diretamente na água (Gomes et al., 2006; Madeira et
al., 2002) . Entretanto, aceita-se que as espécies prefiram ovipor em locais onde haja
água limpa, mas que possam utilizar também criadouros que disponibilizem água rica em
matéria orgânica. A eclosão do ovo ocorre após o desenvolvimento do embrião (2 a 3
dias em condições adequadas de umidade e temperatura) após o contato com a água. Se o
período necessário para o desenvolvimento embrionário ocorrer em ambiente úmido, os

18
ovos podem permanecer por mais de um ano em locais secos, sendo portanto, resistentes
à dessecação (Consoli & Lourenço-de-Oliveira, 1998).
Após a eclosão, emergem as larvas que apresentam quatro estádios larvais (L1,
L2, L3 e L4) e em seguida ao desenvolvimento larval tem início a fase de pupa. Neste
estágio, o imaturo não se alimenta e seu desenvolvimento é consideravelmente
influenciado pela temperatura. Estes mosquitos passam de forma rápida pela fase
imatura, já que todo o ciclo inteiro de vida deve ser acontecer antes que o líquido do
criadouro seque por completo. Em condições ideais (27 a 38oC) o mosquito emerge entre
um e três dias e estes, machos e fêmeas, se alimentam de néctar de fluidos açucarados de
variadas fontes, porém as fêmeas são fortemente hematófagas e atacam durante o dia
(Consoli & Lourenço-de-Oliveira, 1998).
Aedes aegypti e Ae. albopictus podem coexistir na mesma região e utilizar
criadouros análogos, colonizando com rapidez locais onde as condições são vantajosas
para sua proliferação (Forattini 2002). A dispersão destas espécies pode ser rápida
dependendo da pouca disponibilidade de locais para oviposição (Braks et al., 2003;
Forattini 2002; Fantinatti et al., 2007; Lounibos 2002).
2.1 Aedes aegypti
A fêmea do mosquito Ae. aegypti, principal responsável pela transmissão do
dengue, possui atividade hematofágica diurna, é adaptada ao ambiente urbano (Taveira et
al., 2001), sendo portanto, sinantrópica. É consenso que a partir da população silvestre,
devido à destruição dos hábitats naturais, uma linhagem da espécie teria sido selecionada
positivamente, originando mosquitos adaptados às áreas alteradas e que posteriormente
teriam encontrado nos aglomerados humanos ambiente adequado para se estabelecer
(Christophers, 1960, Natal, 2002). Um dos fatos que corroboram tal afirmação é o de que
no continente africano ainda podem ser encontrados Ae. aegypti utilizando criadouros
naturais como ocos de árvores e outras cavidades (Crovello & Hacker, 1972). Apesar de
em algumas áreas do Velho Mundo Ae. aegypti poder ser registrado distante dos
aglomerados humanos, no Brasil a espécie é encontrada nos locais de maior densidade
humana e raramente em ambientes semi-silvestres ou onde a população humana é
menos numerosa (Consoli & Lourenço-de-Oliveira, 1998).
A adaptação do mosquito às áreas urbanas e, portanto, domiciliares, teria sido
favorecida pela possibilidade de utilizar artefatos produzidos pelo homem (vasos de

19
planta, garrafas, caixas d’água, pneus, etc.) para a reprodução (Natal, 2002). A
preferência da espécie por criadouros artificiais faz com que a concentração populacional
originada pela urbanização e a grande utilização de recipientes artificiais na atualidade,
contribuem para a crescente proliferação do mosquito nos centros urbanos das regiões
tropicais e subtropicais do planeta (Gadelha, 1985, Tauil 2001).
No Brasil, o vetor foi considerado eliminado em 1957, reintroduzido em 1967,
novamente erradicado em 1973, mas acredita-se que em 1976 tenha ocorrido a
reintrodução da espécie (Catão, 2009). A crescente expansão da infestação de Ae.
aegypti no Brasil é um agravante no que diz respeito ao controle populacional e
consequentemente da incidência do dengue. Além disso, o controle vetorial tem sido
dificultado pelo fato do mosquito apresentar grande capacidade de adaptação a diferentes
condições ambientais consideradas não comuns para a espécie (Tauil, 2002). Pode-se
citar como exemplo a presença de larvas em recipientes com água poluída (Tauil, 2002)
e ainda o registro da espécie em locais de altitude elevada (fator limitante para o vetor)
onde ocorreram inclusive, algumas epidemias como no México em 1988, em Taxco, a
1.735m (primeira epidemia notificada em altitudes maiores que 1.700m) e em
Tlayacapan, 1.630m (Herrera, 1992).
A relação entre a densidade populacional da espécie e fatores abióticos tem se
mostrado diretamente proporcional, principalmente no que se refere à pluviosidade e
temperatura. Apesar de Ae. aegypti conseguir manter uma população considerável
durante as estações menos chuvosas, se reproduzindo em criadouros semipermanentes e
independentes das chuvas, como caixas d'água, cisternas e tonéis, é na estação chuvosa
que a abundância aumenta significativamente (Consoli & Lourenço-de-Oliveira, 1998).
Além disso, a relação direta entre abundância de Aedes e temperatura é comumente
descrita na literatura: para Ae. aegypti, a taxa de metabolismo aumenta nos meses
quentes e seu ciclo reprodutivo se torna mais rápido, durando até oito dias. O aumento da
temperatura torna o período extrínseco de incubação do vírus no inseto mais rápido
(Halstead, 2008).
As fêmeas de Ae. aegypti realizam hematofagia nos horários diurnos,
principalmente ao amanhecer e pouco antes do crepúsculo, entretanto pode picar o
homem e animais domésticos, a qualquer hora do dia e mais raramente, à noite, caso o
hospedeiro se aproxime de seu abrigo. O hábito diurno também é apresentado pelos
machos, que seguem as fêmeas em seus abrigos na tentativa de efetuar a cópula e

20
também de obter alimento como substâncias açucaradas (Consoli & Lourenço-de-
Oliveira, 1998). Aedes aegypti é habilidosa a escapar de ser morta por sua vítima durante
o repasto sanguíneo: qualquer movimento da vítima faz o mosquitos abandoná-la
rapidamente e após, pode atacá-la novamente ou procurar nova fonte de alimentação. Tal
fato é importante do ponto de vista epidemiológico, já que apenas uma fêmea infectada
pode alimentar-se rapidamente por diversas vezes, em diferentes hospedeiros,
disseminando o vírus para um maior número de pessoas (Consoli & Lourenço-de-
Oliveira, 1998).
2.2.Aedes albopictus
Aedes albopictus é uma espécie proveniente das florestas do Sudeste Asiático
descrita originalmente na Índia (Skuse, 1894 apud Gratz, 2004). Ocorre naturalmente em
locais de clima temperado e tropical e atualmente pode ser encontrado na Ásia temperada
e tropical, Europa, Norte e América do Sul, Austrália, Nova Guiné, nas Ilhas Marianas,
Havaí, Benin, em Madagascar (Reunião e Mauricus) e no oeste do Irã (Consoli &
Lourenço-de-Oliveira, 1998; Paupy et al. 2009).
A introdução nas Américas ocorreu em 1985, ocupando áreas ao sul dos Estados
Unidos, sendo que em 1986 ocorreram seus primeiros registros no Brasil nos estados do
Rio de Janeiro e Minas Gerais (Forattini, 1986; Consoli & Lourenço-de-Oliveira, 1998).
Acredita-se que a entrada do mosquito no país ocorreu através do comércio marítimo de
minério de ferro (pelos portos no Espírito Santo, se adentrando pelo país via estrada de
ferro, no vale do rio Doce) (Consoli & Lourenço-de-Oliveira, 1998). Desde então, Ae.
albopictus têm se expandido rapidamente pelo território brasileiro, estando presente em
quase todos estados do país (Santos, 2003). A população dessa espécie introduzida no
Brasil parece ser originária do Japão e apresenta distinções biológicas se comparada à
cepa invasora da América do Norte (Consoli & Lourenço-de-Oliveira, 1998).
A espécie tem mostrado capacidade de dispersão e adaptação considerável ao
ambiente humano, colonizando praticamente todos os continentes através da dispersão
ativa e passiva (Borges, 2001). Sendo menos antropofílica quando comparada a Ae.
aegypti (Tauil, 2002), a espécie é a responsável pela transmissão da dengue no sudeste da
Ásia (Borges, 2001) e também na última década no Havaí (MMWR, 2002; Effler et al.,
2005).

21
Aedes albopictus apresenta grandes semelhanças com Ae. aegypti como por
exemplo, ter hábitos diurnos e ter sua densidade fortemente influenciada pela
pluviosidade. Entretanto exibe algumas diferenças consideráveis: a primeira espécie, se
dispersa com facilidade no ambiente rural, semisilvestre e silvestre, sendo habitualmente
encontrado em áreas com baixa densidade humana, onde a segunda é inexistente ou rara.
Em locais povoados, apesar de estar presente no domicílio humano, é mais frequente no
peridomicílio. Além da divergência sobre habitats preferenciais, há discrepância também
quanto ao hospedeiro: Ae. albopictus é mais eclético em relação à Ae. aegypti, sendo o
ser humano e aves os hospedeiros mais frequentes. Além disso, diferentemente de Ae.
aegypti, Ae. albopictus é capaz de se adaptar à temperaturas frias
tornando-se dormente durante o inverno em regiões temperadas (Paupy et al., 2009),
enquanto a primeira evita temperaturas mais baixas (Consoli & Lourenço-de-Oliveira,
1998).
É um mosquito generalista que tem demonstrado capacidade de utilizar variados
tipos de criadouros, naturais (internódios de bambu, ocos em árvores e cascas de frutas) e
artificiais (pneus, vidros, pratos sob vasos de plantas, recipientes plásticos, vasos de
cemitério etc.) desde que encontre condições favoráveis (Consoli & Lourenço-de-
Oliveira, 1998).
Aedes albopictus é considerado como um vetor secundário nas Américas (Tauil,
2002), mas devido a algumas características da espécie, principalmente capacidade
adaptativa e de dispersão, muitos estudos foram desenvolvidos com intuito de avaliar a
potencialidade da espécie como vetor de doença nas Américas. A partir desses estudos, o
mosquito foi considerado como um vetor experimental de várias arboviroses, dentre elas,
as doenças causadas por flavivírus como febre amarela, encefalite japonesa, febre do
Nilo ocidental e dengue. Sob condições laboratoriais a população de Ae. albopictus
existente no Brasil demonstrou capacidade de transmitir o vírus do dengue, da febre
amarela e da encefalite eqüina venezuelana (Alencar et al., 2008). Porém, no país
existem até o momento, dois registros de indivíduos naturalmente infectados com o vírus
dengue: em Minas Gerais (Serufo et al., 1993) e Fortaleza ( Martins et al. 2012).
Aedes albopictus e Ae. aegypt podem co-ocorrer em várias áreas, compartilando,
muitas vezes, os mesmos criadouros. Porém, a coexitencia pode ser uma situação
transitória que resultrá no deslocamento de uma das espécies. A diminuição ou o

22
deslocamento populacional de espécies residentes tem sido observada em vários países
após a introdução de Ae. albopictus (Paupy et al. 2009).
3.Áreas verdes urbanas.
Alterações imprevisíveis provocadas pelo homem no ambiente natural formam
um ecossistema antrópico com suas características próprias. Como é um ambiente
complexo, o número de nichos e habitats que se oferecem a adaptabilidade das
populações é variável neste sistema (Forattini, 1996). Como resultado da formação de um
ecossistema antrópico, é possível perceber o aparecimento de áreas verdes urbanas. Essas
áreas podem ser consideradas elementos essenciais à qualidade de vida da população na
medida em que oferecem benefícios diretos e indiretos tanto para os habitantes das
cidades contemporâneas quanto para o ambiente de modo geral (Amaral, 2009).
Os espaços verdes podem ter diversas finalidades, entre elas a de contribuir para a
qualidade de vida, o lazer, a preservação ambiental e à própria sociabilidade. Além de tais
benefícios, esses fatores contribuem para atenuar a paisagem urbana (Loboda & Angelis,
2005) e atuam na absorção de ruídos, diminuem a sensação do calor do sol, atenuam a
sensação de opressão do ser humano com relação às grandes construções; filtra partículas
sólidas em suspensão no ar e colabora para a formação e o aprimoramento do senso
estético, entre tantos outros benefícios (Loboda & Angelis, 2005).
Fragmentos de mata em área urbana podem desempenhar variadas funções
ecológicas, mas apesar de sua incontestável relevância na formação de ambientes de lazer
e proteção ambiental (Navarro-Silva, 2004), é possível que tais espaços quando inseridos
em áreas alteradas possam agrupar condições favoráveis aos processos adaptativos dos
insetos em relação a suas interações com humanos (Taipe-Lagos & Natal, 2003).
Nesse contexto, além dessas áreas poderem atuar como abrigo para mosquitos s
provenientes de outras áreas (Silva, & Lozovei, 1996), sua proximidade com
aglomerados humanos torna-os mais vulneráveis às interferências antrópicas negativas
como despejo de lixo e entulhos de construção civil, e esses artefatos, desde que
retenham água, podem funcionar como criadouros artificiais para espécies de dípteros da
família Culicidae. É importante considerar ainda, a existência de criadouros naturais
nesses espaços que também podem ser úteis para a procriação de Culicidae (Zequi, et al.,
2005).

23
Diante de tais fatos, percebe-se que há oferta de diferentes microambientes nesses
espaços que podem ser úteis ao desenvolvimento de variadas espécies de mosquitos.
Portanto, a investigação de culicídeos nesses locais pode revelar a presença de espécies
com importância epidemiológica (Tissot & Navarro-Silva, 2004) além de avaliar o
impacto da atividade humana na composição das espécies (Frankie, 1978). Porém, de
acordo com Petschow (2008) poucos estudos sobre a diversidade da fauna de Culicidae
têm sido realizados em parques ou outro tipo de áreas de preservação que contenham
fragmentos de mata e que estejam inseridos no perímetro urbano ou periurbano.

24
Capítulo I
Aspectos ecológicos dos vetores da dengue em áreas verdes
urbanas: um estudo da ocorrência de vetores nas cidades de
Mariana e Ouro Preto, Minas Gerais

25
RESUMO
Aedes aegypti e Ae. Albopictus sofrem efeitos consideráveis, mas contrários, da
urbanização e cobertura vegetal. Este estudo verificou se estas espécies estão presentes
em áreas verdes inseridas e avaliou os aspectos dessa ocorrência em cidades com
diferenças altitudinais e climáticas de uma mesma região. O estudo foi desenvolvido em
duas áreas verdes de Ouro Preto e Mariana, Minas Gerais, com um total de 60 ovitrampas
instaladas durante 15 semanas de 2009 e 3 semanas em 2012. A presença de Ae. aegypti e
Ae. albopictus foi detectada, sendo coletados 4.793 imaturos no total. Aedes albopictus
foi a espécie mais abundante (80,5 %) e Mariana apresentou a maior abundância dos
culicídeos. O Índice de Positividade de Ovitrampa (IPO) máximo foi de 86% para ambas
cidades e a distribuição dos ovos nas palhetas revelou disposição agregada. Em geral,
locais com elevada abundância de Ae. albopictus também tiveram grande número de
indivíduos de Ae. aegypti. Embora seja clara a relação de coexistência, foi observada uma
separação espacial de habitats dentro destes locais favoráveis, sugerindo competição
interespecífica por sítios de oviposição. A proporção das espécies nas áreas verdes foi
similar em ambas cidades. Pluviosidade explicou o aumento de abundância apenas de
Ae. albopictus. Os vetores ocorreram em áreas verdes urbanas mesmo em cidades sem
casos de dengue e com altitude elevada. A elevada abundância de Ae. albopictus e a
possível relação competitiva deste com Ae. aegypti leva a crer que áreas verdes possam
ter um papel na desaceleração de infestações urbanas. A compreensão dos padrões de
ocorrência dessas espécies em situações pouco estudadas e em locais com recente
estabelecimento pode ser importante para a prevenção da dengue. O monitoramento de
áreas verdes,é necessário para auxiliar na prevenção e controle da doença.
Palavras chave: Aedes aegypti; Aedes albopictus; entomologia urbana; diferenças
climáticas; Mariana, Ouro Preto.

26
ABSTRACT
Aedes aegypti and Ae. albopictus suffer considerable effects, but contrary,
urbanization and vegetation cover. This study examined whether these species are present
in green areas entered and evaluated aspects of this occurrence in cities with altitudinal
and climatic differences within the same region. The study was conducted in two areas of
green Ouro Preto and Mariana, Minas Gerais, with a total of 60 ovitraps installed for 15
weeks in 2009 and 3 weeks in 2012. The presence of Ae. aegypti and Ae. albopictus was
detected and collected 4793 total immature. Aedes albopictus was the most abundant
species (80.5%) and Mariana had the highest abundance of Culicidae. The Positivity
Index of ovitrap (IPO) maximum was 86% for both cities and the distribution of eggs in
the reeds revealed aggregate provision. In general, sites with high abundance of Ae.
albopictus were also large numbers of individuals of Ae. aegypti. While it is clear the
relationship of coexistence, we observed a spatial separation of habitats within these
favorable locations, suggesting interspecific competition for oviposition sites. The
proportion of species in the green areas was similar in both cities. Rain explained the
increased abundance of only Ae. albopictus. The vectors occurred in urban green areas
even in towns without dengue cases and high altitude. The high abundance of Ae.
albopictus and the possible relationship of this with competitive Ae. aegypti suggests that
green spaces can play a role in slowing urban infestations. Understanding the patterns of
occurrence of these situations poorly studied species and locations with recent
establishment may be important for the prevention of dengue. The monitoring of green
areas, is needed to help prevent and control the disease.
Keywords: Aedes aegypti, Aedes albopictus, urban entomology; climatic differences;
Mariana, Ouro Preto.

27
1.INTRODUÇÃO
O mosquito Aedes aegypti é o responsável pela transmissão da dengue no Brasil
e a partir de sua re-infestação no país na década de 70, se disseminou por todos os estados
brasileiros (Tauil, 2002). Além disso, desde os anos 80 encontra-se no território brasileiro
a espécie Ae. albopictus, que apesar de não ser considerada um vetor natural da doença
no país, é o responsável pela transmissão do vírus dengue no Sudeste da Ásia (Borges,
2001). A espécie foi também a transmissora do vírus durante surto ocorrido em 2001 no
Havaí (Effler et al, 2005).
Aedes aegypti possui característica antropofílica, com hábitos essencialmente
domésticos e utiliza, em sua maioria, criadouros artificiais para se reproduzir (Pontes
1994). O encontro de seus imaturos em residências de ambientes urbanos e suburbanos é
frequente, especialmente onde há alta concentração populacional humana (Braks et al.
2003; Consoli & Lourenço-de-Oliveira 1994; Honório et al. 2009a; Lourenço-de-Oliveira
2004; Pontes 1994). Aedes albopictus utiliza os mesmos criadouros artificiais somados à
criadouros naturais, como ocos de árvores e buracos em rochas (Consoli & Lourenço-de-
Oliveira 1994; Pontes 1994). A espécie é capaz de sobreviver e reproduzir em uma maior
gama de ambientes, não sendo sua distribuição limitada a apenas ao domicílio e
peridomicílio, como ocorre comumente com Aedes aegypti. A espécie é encontrada ainda
em ambientes onde prevalecem áreas abertas com vegetação, nas zonas urbanas,
suburbanas, bem como na zona rural (Braks, 2003; Consoli & Lourenço-de-Oliveira,
1994; Honório et al., 2009b; Lourenço-de-Oliveira, 2004; Pontes, 1994). Diante de tais
características é possível inferir que apesar dessas espécies apresentarem nichos
ecológicos análogos (Serpa, 2008) a urbanização e o grau de cobertura vegetal podem
exercer efeitos contrários na frequência dessas espécies (Braks et al. 2003; Carbajo et al.
2006; Honório et al. 2009b).
Áreas verdes urbanas surgem por planejamento, ou por mera consequência do
uso e ocupação irregular do solo pelo homem (Forattini, 1996). Áreas verdes inseridas em
ambiente urbano são elementos essenciais à qualidade de vida da população e sua
relevância é incontestável (Amaral, 2009; Navarro-Silva, 2004). Sob o ponto de vista
biológico, acredita-se que espaços verdes quando inseridos em áreas alteradas possam
reunir condições oportunas aos processos adaptativos dos insetos em relação a suas
interações com humanos (Taipe-Lagos & Natal, 2003). Assim, a detecção de vetores ou
de potenciais vetores de doenças em espaços verdes inseridos em ambiente urbano pode

28
ser importante para gerar informações que contribuirão para futuras implicações no
controle de doenças (Guimarães, 2000). Todavia, quando se considera os estudos de
vetores, existe um número pouco expressivo de trabalhos sobre ocorrência de vetores em
áreas verdes urbanas, podendo-se citar trabalhos realizados em cidades como Rio de
Janeiro-RJ (Lourenço-de-Oliveira et al., 2003; Maciel-de-Freitas et al., 2006; Honório et
al. 2009), Fortaleza-CE (Alencar, 2009) e Governador Valadares-MG (Morais, 2011).
Apesar das já conhecidas preferências de Ae. aegypti e Ae. albopictus em relação à
cobertura vegetal, se torna importante compreender o papel desses espaços naturais para a
ocorrência das espécies, na medida em que eles também compõe o ambiente urbano.
Além dos padrões de uso do solo e da vegetação, outras variáveis ambientais
também podem estar diretamente relacionadas com a ocorrência dos Aedes (Forattini,
1992). A temperatura influencia nas atividades de repasto sanguíneo das fêmeas dos
mosquitos, na longevidade e no período de incubação extrínseco do vírus (Donalísio &
Glasser, 2002), além de interferir na distribuição geográfica de populações vetoras de
doenças, permitindo estabelecer limites para ocorrência de espécies (Glasser & Gomes,
2002). Outra variável ambiental importante é a precipitação pluviométrica, pois o
acúmulo de água aumenta o número de potenciais criadouros, levando a um aumento na
densidade de criadouros disponíveis no extradomicílio e consequentemente, elevando a
abundância dos vetores (Gubler, 1998; Glasser & Gomes 2003). A altitude também é
indicada como um fator limitante na reprodução do vetor, porém algumas epidemias já
foram registradas a 1.735m de altitude no México, a 1.735m e a 1.630m, ambas no
México (Herrera et al., 1992; Donalísio & Glasser, 2002). Diante do atual cenário de
mudanças climáticas é esperado um aumento na incidência de doenças vetoriais em
montanhas tropicais, devido à facilitação da imigração mediante a uma maior frequência
e constância de climas mais amenos (Barcellos et al. 2009; Cortez, 2004; Mendonça,
2003).
Neste contexto, Ouro Preto e Mariana, duas cidades do interior de Minas Gerais,
despertam interesse para o estudo dessas espécies. Apesar de próximas (12 km entre si),
elas possuem diferenças climáticas marcantes, devido à maior altitude de Ouro Preto em
relação à Mariana. Além disso, Ouro Preto não apresentou casos autóctones de dengue
até o ano de 2010 e a eventual ocorrência de Aedes na cidade deve então representar
eventos recentes de colonização. Compreender aspectos da ocorrência dessas espécies em
ambientes naturais inseridos na zona urbana, em cidades com diferenças altitudinais e

29
climáticas pode elucidar aspectos importantes da ocorrência destes vetores em diferentes
contextos ecológicos regionais. A identificação de biótopos preferenciais pode contribuir
para o maior conhecimento sobre o padrão de distribuição das populações de mosquitos
(Glasser & Gomes, 2002).
Este trabalho investigou a hipótese de que as duas espécies de Aedes serão
encontradas e que suas abundâncias irão responder de forma diferente ao habitat e às
condições climáticas. A predição é que Ae. albopictus seja a espécie predominante em
áreas verdes e que Mariana apresente maior abundância de Aedes como um todo devido
às condições climáticas. Outra hipótese investigada foi a de que há uma relação
competitiva entre Ae. albopictus e Ae. aegypti nestes ambientes. Para tanto, a predição é
que as espécies coexistam nos locais de estudo, mas que haja competição nos criadouros.
2. MATERIAIS E MÉTODOS
2.1. Área de estudo
Mariana (20° 22′ 40″ de latitude sul e 43° 24′ 57″ longitude) e Ouro Preto
(Latitude 20º23'08" S e Longitude 43º30'29" O) são municípios do estado de Minas
Gerais distantes aproximadamente 14 km entre si e cerca de 110 km e 96 km da capital
Belo Horizonte, respectivamente. O clima na região é do tipo Cwb pela classificação de
Köppen, com precipitações abundantes no verão entre 1000 mm e 2000 mm (Gomes,
1998). Ouro Preto, cidade situada à 1100 m , possui temperaturas médias anuais entre
17,4°C e 19,8°C (Gomes, 1998) e Mariana à 750 m de altitude , possui temperaturas
médias estão entre 18°C e 22° C (Ozório, 2000).
Duas áreas verdes localizadas nestas cidades foram utilizadas no estudo (figuras
1 a 4). A área situada no centro histórico de Ouro Preto tem aproximadamente 1,4ha e
apresenta uma vegetação composta por - palmiteiros, bambuzais e espécies nativas e
exóticas de árvores frutíferas, plantas ornamentais, além de árvores de grande porte como
Chorisia sp., além de uma abundante população de “lírios-do-brejo” (Amaral, 2009). A
área verde em Mariana, com cerca de 2,0ha está localizada no bairro Rosário e sua
vegetação é composta predominantemente por eucaliptos, capins, bananeiras, dentre
outras árvores frutíferas. Ambas as áreas são margeadas por ruas e residências, sendo que
em Mariana há uma maior quantidade de edificações circundando o espaço.

30
Figura 1. Espaço verde urbano utilizado no estudo em Ouro Preto. Foto:
Modificada do Google Earth, Michelle C. Pedrosa, 2012.
Figura 2. a) Vista panorâmica da área verde urbana utilizada para o estudo em Ouro
Preto. b, c e d) vista interna da área de estudo. Foto: Alexandre Cortez do Amaral,
2009.

31
Figura 3. Área verde utilizada no estudo na cidade de Mariana. Foto: Modificada
do Google Earth, Michelle C. Pedrosa, 2012.
Figura 4. a) Vista panorâmica da área verde urbana utilizada para o estudo em Mariana. b, c e d)
vista interna da área de estudo. Foto: Alexandre Cortez do Amaral, 2009.
a b
c d

32
2.2. Coleta de dados
A coleta de imaturos foi realizada utilizando 60 ovitrampas: armadilhas de
oviposição caracterizadas como recipiente plástico, preto-fosco com capacidade de
700ml, onde prende-se verticalmente, por meio de um clips, uma palheta de madeira
(Bonat, 2008) (ver anexo ). A armadilha de oviposição é um método mais sensível do que
a coleta ativa de larvas (utilizada para vigilância entomológica no Brasil), mas não é
válido para quantificar as populações do vetor (Focks et al., 2003, Gama et al., 2007).
Entretanto, tal metodologia pode fornecer estimativas relativas das populações de Aedes
nos locais estudados.
Cada ovitrampa foi amarrada em árvores, arbustos e outros locais que
possibilitassem sua suspensão a aproximadamente 1,5m do solo e em local sombreado.
Nas bordas de cada área verde (não foram instaladas armadilhas no interior da
mata) foram utilizados três pontos (B1, B2 e B3), distantes aproximadamente 30m da
calçada mais próxima. Cada bloco distava cerca de 50 metros um do outro (exceto os
Blocos 1 e 2 da área verde em Ouro Preto, distantes 20m). Em cada ponto foram
instaladas 10 ovitrampas, com distância estimada de 1m entre elas, totalizando 30
ovitrampas por área verde. Para melhor compreensão do desenho amostral utilizado veja
a figura 10. Vale ressaltar que o formato de cada área verde foram respeitadas no desenho
abaixo (figura 5), bem como a distribuição de cada bloco de ovitrampas.
Figura 5.Esquema do delineamento amostral em cada área verde urbana utilizada de
Mariana e Ouro Preto, Minas Gerais.

33
Ocorreram 15 coletas semanais entre 18 de fevereiro e 28 de maio de 2009 e em
uma segunda campanha, três coletas foram realizadas entre 15 e 29 fevereiro do ano de
2012. Após cada coleta, as palhetas de madeira e água contida nas ovitrampas eram
armazenadas a fim de eclodir os ovos presentes. Depois de no mínimo três dias, as
amostras eram triadas e as larvas, após o 2° instar larval e pupas, eram mortos para
posterior identificação taxonômica, com auxílio dos seguintes de manuais de
identificação.
Dados de pluviosidade diária, referentes à cidade de Mariana, foram cedidos
pela Gerência Geral de Desenvolvimento e Planejamento do Sudeste - GEPDS/DIFS -
Complexo Mariana da empresa VALE, sendo que a estação meteorológica está situada na
mina de Timbopeba, localizada aproximadamente 15km da cidade. Os dados relativos à
cidade de Ouro Preto foram cedidos pelo Setor de Defesa Civil da prefeitura da cidade,
que tem sua estação meteorológica localizada no Bairro São Cristovão.
Para avaliar a eficiência das ovitrampas na captura dos Aedes foi estimado o
Índice de Positividade de Ovitrampa (IPO) que é uma relação entre o número de
armadilhas positivas e o número de armadilhas instaladas, onde a abundância é
desconsiderada (GOMES 1998 e 2002).
IPO = Armadilhas positivas *100
Armadilhas instaladas
A distribuição espacial dos ovos também foi avaliada a fim de se verificar se há
aleatoriedade ou agregação, de acordo com a fórmula:
I = (Variância)
Média
Abaixo segue-se a descrição do significado dos valores obtidos na análise de
variância (Pereira et al. 2004, Fantinatti et al. 2007; Rabinovich 1980):
I=1: disposição espacial aleatória
I<1: disposição espacial regular ou uniforme
I>1: disposição agregada

34
2.3. Análises estatísticas
Para análises de correlação entre a abundância e precipitação, um atraso de duas
semanas neste dado foi utilizado para que houvesse concordância com o início das chuvas
e o tempo de desenvolvimento do mosquito (Honório 2009).
A análise de variância de abundância de espécies entre habitats e cidades foram
concentradas nos dados de 2009. Os dados de 2012 foram utilizados para verificar a
permanência das espécies três anos após a primeira coleta e para a análise de correlação
entre abundância de Aedes e precipitação. Dados faltantes foram compensados pela
aleatorização de amostras e geração de médias entre locais ou datas, preservando o
balanço do delineamento.
Uma vez que os dados brutos de contagem seguem a distribuição de Poisson,
gerou-se a média dos blocos logaritmizadas, escala na qual, em concordância com o
Teorema Central do Limite, a distribuição de frequência dos dados foi normal. Utilizou-
se teste t de Student para comparar a variação das espécies entre as cidades e um qui-
quadrado (X2) para verificar se a proporção das espécies foi semelhante em cada cidade.
Foi realizada uma ANOVA Bifatorial de medidas repetidas com nível de significância de
5% para verificar a influência das datas de coleta e das áreas verdes sobre a abundância
das espécies e ainda, uma ANCOVA, para analisar se houve relação positiva entre
abundância e precipitação. A Correlação de Spearman foi usada a fim de averiguar o que
ocorre com ambas as espécies na escala de ovitrampas.
3.RESULTADOS
No ano de 2009 foram coletados 4.250 imaturos (ovos, larvas e pupas) das quais
19,5% foram de Ae. aegypti e 80,5% de Ae. albopictus (figura 6 e tabela I). Entretanto,
foram capturados outros 247 imaturos de culicídeos, Culex sp. (N= 206) e Aedes sp.
(N=41) em 2009 e 36 Culex sp. e 2 Aedes sp. em 2012 que não serão avaliados neste
trabalho.
Em 2012 um total de 543 imaturos foram coletados, sendo 145 indivíduos de
Ae. aegypti e 398 de Ae. Albopictus (Tabela 2). Em 2009, para Mariana, as coletas com
maior sucesso de captura de Aedes (N= 1334), ocorreram no mês de março seguido do
mês de abril (N=1233). Em Ouro Preto, o inverso pôde ser observado, visto que as
35
coletas mais abundantes aconteceram no mês de abril (N=777), seguido do mês de março
(N=329).
A abundância total de Aedes variou entre as datas de coleta (ANOVA, F 1;174
=4,76; p <0,001; figuras 7 e 8) e entre as cidades, sendo encontrados mais indivíduos em
Mariana (ANOVA, F 1:174 = 18,54; p <0,01; figura 7).
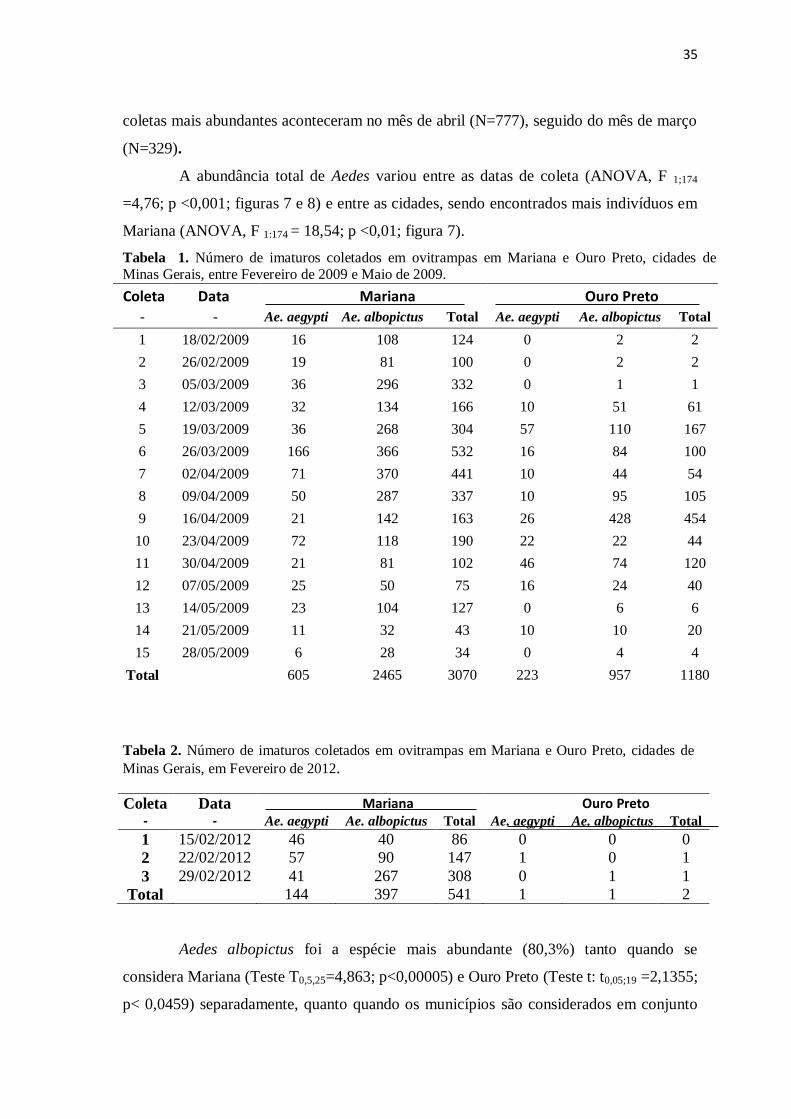
Tabela 1. Número de imaturos coletados em ovitrampas em Mariana e Ouro Preto, cidades de Minas Gerais, entre Fevereiro de 2009 e Maio de 2009.
Tabela 2. Número de imaturos coletados em ovitrampas em Mariana e Ouro Preto, cidades de
Minas Gerais, em Fevereiro de 2012.
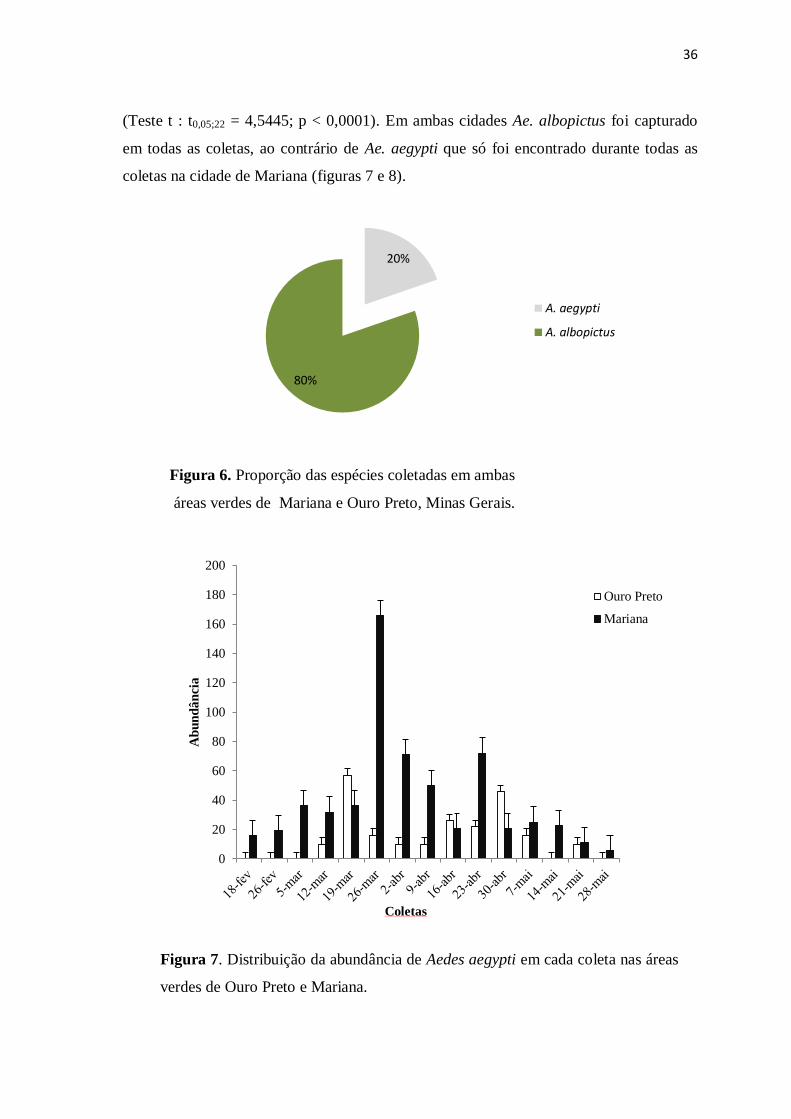
Aedes albopictus foi a espécie mais abundante (80,3%) tanto quando se
considera Mariana (Teste T0,5,25=4,863; p<0,00005) e Ouro Preto (Teste t: t0,05;19 =2,1355;
p< 0,0459) separadamente, quanto quando os municípios são considerados em conjunto
Coleta Data Mariana Ouro Preto
- - Ae. aegypti Ae. albopictus Total Ae. aegypti Ae. albopictus Total
1 18/02/2009 16 108 124 0 2 2
2 26/02/2009 19 81 100 0 2 2
3 05/03/2009 36 296 332 0 1 1
4 12/03/2009 32 134 166 10 51 61
5 19/03/2009 36 268 304 57 110 167
6 26/03/2009 166 366 532 16 84 100
7 02/04/2009 71 370 441 10 44 54
8 09/04/2009 50 287 337 10 95 105
9 16/04/2009 21 142 163 26 428 454
10 23/04/2009 72 118 190 22 22 44
11 30/04/2009 21 81 102 46 74 120
12 07/05/2009 25 50 75 16 24 40
13 14/05/2009 23 104 127 0 6 6
14 21/05/2009 11 32 43 10 10 20
15 28/05/2009 6 28 34 0 4 4
Total 605 2465 3070 223 957 1180
Coleta Data Mariana Ouro Preto - - Ae. aegypti Ae. albopictus Total Ae. aegypti Ae. albopictus Total
1 15/02/2012 46 40 86 0 0 0
2 22/02/2012 57 90 147 1 0 1
3 29/02/2012 41 267 308 0 1 1
Total 144 397 541 1 1 2
36
(Teste t : t0,05;22 = 4,5445; p < 0,0001). Em ambas cidades Ae. albopictus foi capturado
em todas as coletas, ao contrário de Ae. aegypti que só foi encontrado durante todas as
coletas na cidade de Mariana (figuras 7 e 8).
Figura 6. Proporção das espécies coletadas em ambas
áreas verdes de Mariana e Ouro Preto, Minas Gerais.
Figura 7. Distribuição da abundância de Aedes aegypti em cada coleta nas áreas
verdes de Ouro Preto e Mariana.
20%
80%
A. aegypti
A. albopictus
0
20
40
60
80
100
120
140
160
180
200
Ab
un
dâ
ncia
Coletas
Ouro Preto
Mariana

37
Figura 8. Distribuição da abundância de Aedes albopictus em cada coleta na áreas verdes
de Ouro Preto e Mariana.
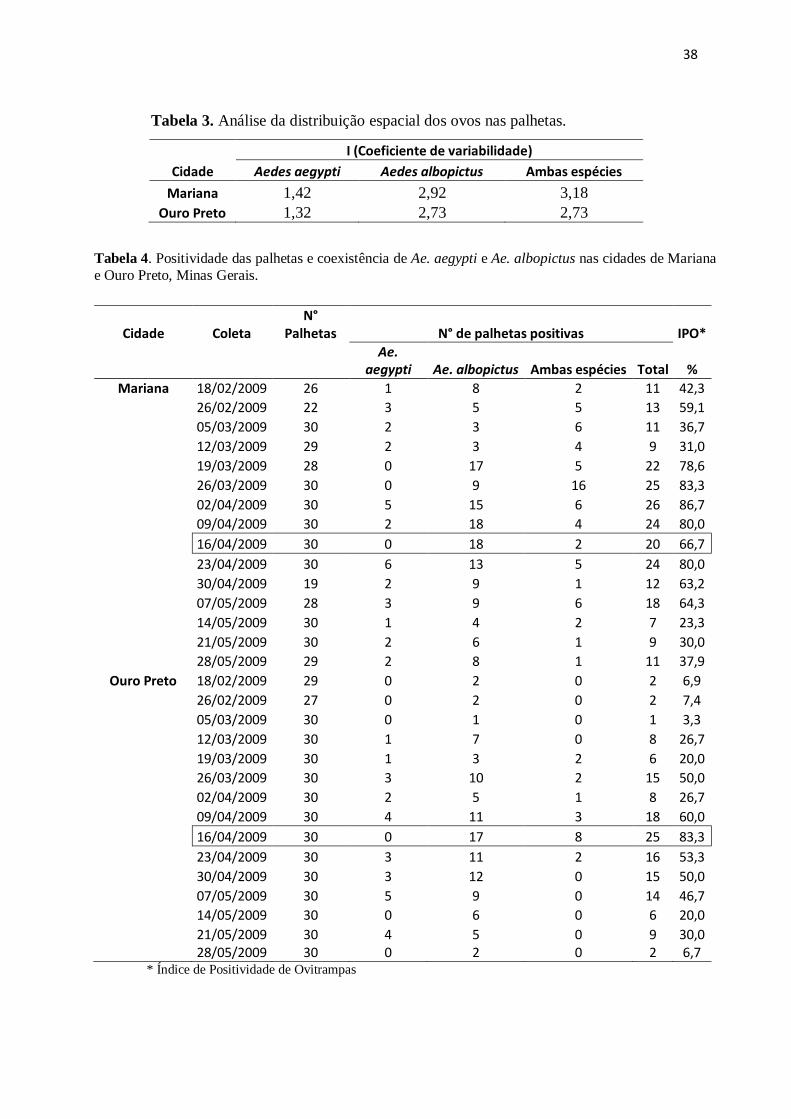
Em relação à distribuição espacial dos ovos nas armadilhas, foi observada uma
distribuição agregada tanto para Ae. aegypti quanto para Ae. albopictus quando
analisados separadamente, bem como quando as espécies foram analisadas em conjunto
(Tabela 4). A proporção encontrada de cada espécie nas áreas verdes não diferiu entre as
cidades (qui-quadrado= 0,35; gl=1; p>0,05) já que as proporções tanto de Ae. aegypti
(20,1% e 18,6 %) quanto de Ae. albopictus (79,9% e 81,4%) foram semelhantes em
Mariana e Ouro Preto, respectivamente.
A maior positividade das armadilhas (IPO) foi alcançada em Mariana, com
exceção de uma amostra (Tabela 4). A positividade máxima atingida foi cerca de 86%
para ambas as cidades, entretanto em datas diferentes. O menor IPO ocorreu em Ouro
Preto, quando apenas 3,3% das ovitrampas foram positivas. Valor consideravelmente
abaixo do menor IPO para Mariana que foi 23,3%. A co-ocorrência das espécies
aconteceu em 84 das 389 palhetas positivas.
0
20
40
60
80
100
120
140
160
180
200
Ab
un
dân
cia
Coletas
Ouro Preto
Mariana
38
Tabela 3. Análise da distribuição espacial dos ovos nas palhetas.
Cidade
I (Coeficiente de variabilidade)
Aedes aegypti Aedes albopictus Ambas espécies
Mariana 1,42 2,92 3,18
Ouro Preto 1,32 2,73 2,73
Tabela 4. Positividade das palhetas e coexistência de Ae. aegypti e Ae. albopictus nas cidades de Mariana
e Ouro Preto, Minas Gerais.
* Índice de Positividade de Ovitrampas
Cidade Coleta N°
Palhetas N° de palhetas positivas IPO*
Ae.
aegypti Ae. albopictus Ambas espécies Total %
Mariana 18/02/2009 26 1 8 2 11 42,3
26/02/2009 22 3 5 5 13 59,1
05/03/2009 30 2 3 6 11 36,7
12/03/2009 29 2 3 4 9 31,0
19/03/2009 28 0 17 5 22 78,6
26/03/2009 30 0 9 16 25 83,3
02/04/2009 30 5 15 6 26 86,7
09/04/2009 30 2 18 4 24 80,0
16/04/2009 30 0 18 2 20 66,7
23/04/2009 30 6 13 5 24 80,0
30/04/2009 19 2 9 1 12 63,2
07/05/2009 28 3 9 6 18 64,3
14/05/2009 30 1 4 2 7 23,3
21/05/2009 30 2 6 1 9 30,0
28/05/2009 29 2 8 1 11 37,9
Ouro Preto 18/02/2009 29 0 2 0 2 6,9
26/02/2009 27 0 2 0 2 7,4
05/03/2009 30 0 1 0 1 3,3
12/03/2009 30 1 7 0 8 26,7
19/03/2009 30 1 3 2 6 20,0
26/03/2009 30 3 10 2 15 50,0
02/04/2009 30 2 5 1 8 26,7
09/04/2009 30 4 11 3 18 60,0
16/04/2009 30 0 17 8 25 83,3
23/04/2009 30 3 11 2 16 53,3
30/04/2009 30 3 12 0 15 50,0
07/05/2009 30 5 9 0 14 46,7
14/05/2009 30 0 6 0 6 20,0
21/05/2009 30 4 5 0 9 30,0
28/05/2009 30 0 2 0 2 6,7

39
Houve uma relação positiva entre as abundâncias de Ae. aegypti e Ae. albopictus
quando se considera cada local de oviposição (R2=0,8) não havendo separação temporal
de habitats (Figura 9). Assim, locais e datas favoráveis para uma espécie também foram
favoráveis para a outra, compartilhando inclusive, ovitrampas para a postura de ovos.
Porém, os locais com maior oviposição de Ae. albopictus foram em armadilhas onde não
se verificou ovos de Ae. aegypti. Além disso, Ae. albopictus parece ter tido preferência
por ovipositar em criadouros onde não se verificava a presença de Ae.egypti (figura 9).
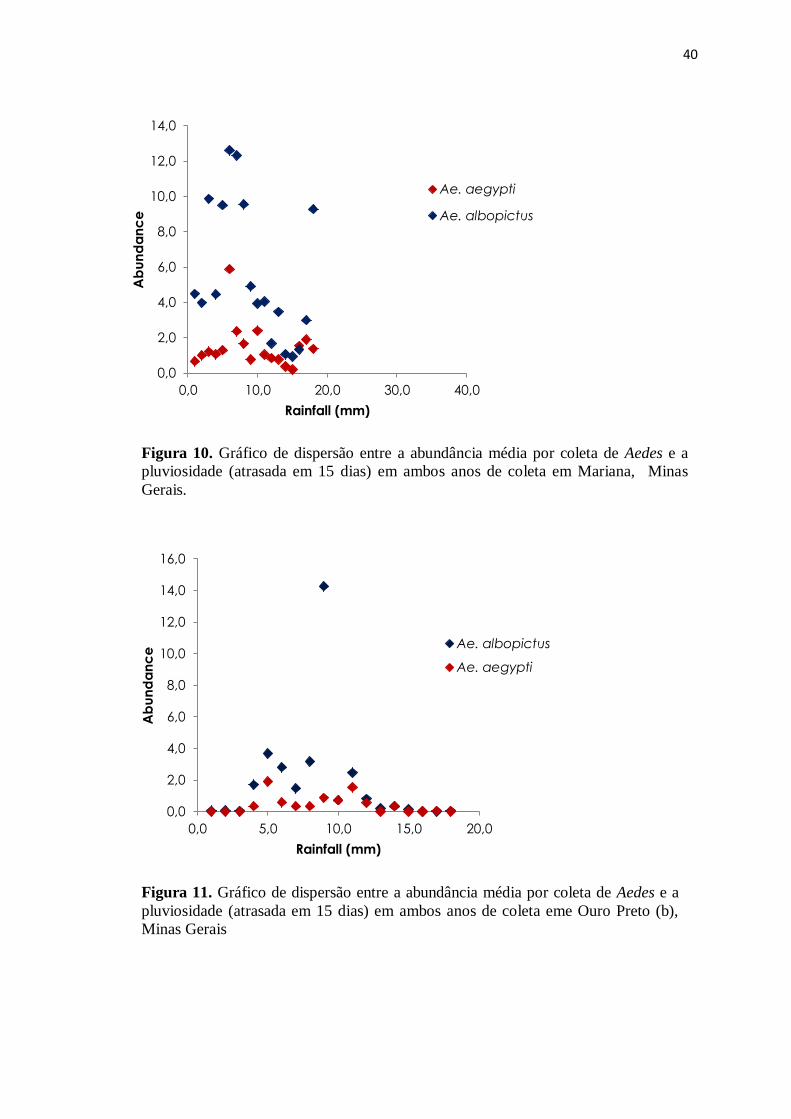
Considerando as cidades juntas e em ambos os anos de coleta, apenas Ae.
albopictus demonstrou associação positiva com a pluviosidade (ANCOVA F1;103= 8,58;
p<0,01). Foi também notável que esta relação foi mais evidente em Mariana (figura 10,11
e 12), sendo que o efeito das cidades também apareceu tanto para Ae. albopictus
(ANCOVA F1;103= 13,07; p<0,01) quanto para Ae. aegypti (ANCOVA F1;103= 12,98;
p<0,01).
Figura 9. Relação entre a abundância de Aedes aegypti e Ae. albopictus
considerando todas as amostras de diferentes cidades e datas na escala de
ovitrampa (observar que as ovitrampas com maiores densidades de Ae.
albopictus são as com zero Ae. aegypti).
05
101520253035404550556065707580859095
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75
Ab
un
dân
cia
Ae.
alb
op
ictu
s
Abundância de Ae. aegypti
40
Figura 10. Gráfico de dispersão entre a abundância média por coleta de Aedes e a
pluviosidade (atrasada em 15 dias) em ambos anos de coleta em Mariana, Minas
Gerais.
Figura 11. Gráfico de dispersão entre a abundância média por coleta de Aedes e a
pluviosidade (atrasada em 15 dias) em ambos anos de coleta eme Ouro Preto (b),
Minas Gerais
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
0,0 10,0 20,0 30,0 40,0
Ab
un
da
nc
e
Rainfall (mm)
Ae. aegypti
Ae. albopictus
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
16,0
0,0 5,0 10,0 15,0 20,0
Ab
un
da
nc
e
Rainfall (mm)
Ae. albopictus
Ae. aegypti
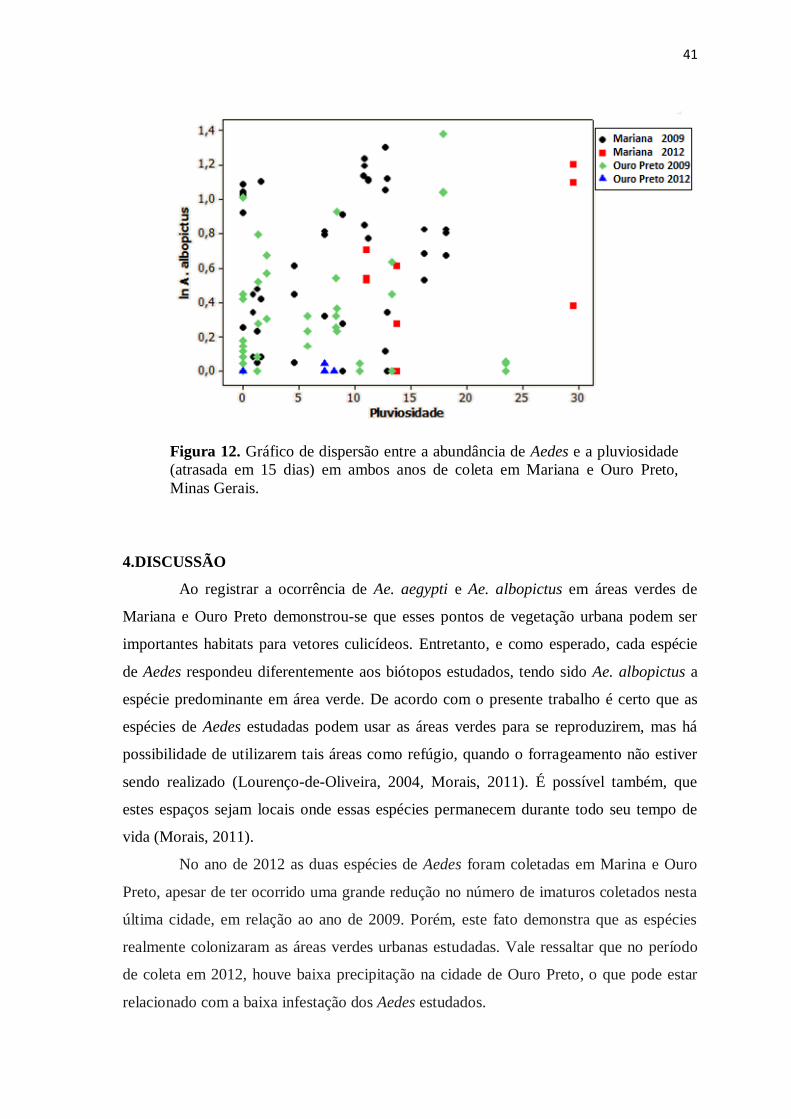
41
Figura 12. Gráfico de dispersão entre a abundância de Aedes e a pluviosidade
(atrasada em 15 dias) em ambos anos de coleta em Mariana e Ouro Preto,
Minas Gerais.
4.DISCUSSÃO
Ao registrar a ocorrência de Ae. aegypti e Ae. albopictus em áreas verdes de
Mariana e Ouro Preto demonstrou-se que esses pontos de vegetação urbana podem ser
importantes habitats para vetores culicídeos. Entretanto, e como esperado, cada espécie
de Aedes respondeu diferentemente aos biótopos estudados, tendo sido Ae. albopictus a
espécie predominante em área verde. De acordo com o presente trabalho é certo que as
espécies de Aedes estudadas podem usar as áreas verdes para se reproduzirem, mas há
possibilidade de utilizarem tais áreas como refúgio, quando o forrageamento não estiver
sendo realizado (Lourenço-de-Oliveira, 2004, Morais, 2011). É possível também, que
estes espaços sejam locais onde essas espécies permanecem durante todo seu tempo de
vida (Morais, 2011).
No ano de 2012 as duas espécies de Aedes foram coletadas em Marina e Ouro
Preto, apesar de ter ocorrido uma grande redução no número de imaturos coletados nesta
última cidade, em relação ao ano de 2009. Porém, este fato demonstra que as espécies
realmente colonizaram as áreas verdes urbanas estudadas. Vale ressaltar que no período
de coleta em 2012, houve baixa precipitação na cidade de Ouro Preto, o que pode estar
relacionado com a baixa infestação dos Aedes estudados.

42
As espécies responderam ainda às cidades com climas diferentes, apesar de não
se poder afirmar que a maior abundância encontrada em Mariana seja resultado apenas da
temperatura. Porém, a incidência relativamente pequena de ambas as espécies em Ouro
Preto, associado ao fato de que registros de casos autóctones de dengue somente
ocorreram posteriormente à realização deste estudo (Setor de Zoonoses Municipal) são
fortes indícios da colonização recente do gênero nesta cidade montana de mais de 1000
metros de altitude.
Dentre as espécies encontradas, Ae. albopictus foi a espécie mais abundante nas
duas áreas verdes estudadas. A maior abundância dessa espécie pode ser explicada pelo
fato desta apresentar hábitos tipicamente mais silvestres em relação à Ae. aegypti, além
de ser registrado em áreas de altitudes mais elevadas, desenvolvendo-se bem em
temperaturas diversas (de 15°C até 30°C) (Eiras, 2005). Tal faixa de temperatura abrange
tanto a cidade de Mariana quanto a de Ouro preto, apesar da segunda apresentar
temperaturas médias menores em relação à primeira. A presença de Ae. aegypti nas áreas
estudadas também é de extrema relevância, já que é entendido como um mosquito com
hábitos essencialmente domésticos (Gadelha, 1985; Pontes, 1994; Eiras 2005).
Entretanto, embora seja uma espécie atualmente sinantrópica, existem populações no
continente africano com alguma afinidade por ambientes silvestres (Pontes, 1994; Eiras
2005), local de origem da espécie, onde foram encontrados Ae. aegypti utilizando
criadouros naturais como ocos de árvores, buracos em rochas e outras cavidades
(Crovello & Hacker, 1972).
Mariana foi o município em que as espécies foram mais abundantes, o que pode
estar relacionado à temperatura do local, já que a cidade está localizada em uma região de
menor altitude em relação à Ouro Preto, apresentando temperaturas médias mais
elevadas. É comum encontrar na literatura descrições sobre a relação positiva entre
abundância de Aedes e a temperatura, sendo que para Ae. aegypti por exemplo, a taxa de
metabolismo aumenta nos meses quentes e seu ciclo reprodutivo se torna mais rápido (em
até oito dias) (Consoli & Lourenço-de-Oliveira 1994). Aedes albopictus também
apresenta diminuição do período de desenvolvimento de seus estágios imaturos como
resultado do aumento da temperatura (Calado et al., 2002). Contudo, apesar de Mariana
apresentar maior abundância total de Aedes e tendo as áreas verdes características
fisionômicas distintas, as proporções de Ae. aegypti e Ae. albopictus coletados nos dois
municípios foram semelhantes.

43
A distribuição dos ovos nas palhetas durante todo o estudo apresentou padrão
agregado, tanto para Ae. aegypti quanto para Ae. albopictus, em ambas cidades, o que
corrobora com estudo de Fantinatti et al. (2007) em que foi detectado um alto padrão de
agregação para as mesmas espécies analisadas em conjunto em áreas urbanas no Paraná.
Sabe-se que, em laboratório, fêmeas de Ae. aegypti obrigadas a reter seus ovos (por
exemplo, devido à indisponibilidade de criadouros), ao receberem o substrato de
oviposição realizam grande parte da postura em um mesmo local (Chadee, 2010). Assim,
os resultados sugerem que as fêmeas destes mosquitos possam estar com a demanda de
criadouros limitada nos locais onde forrageiam (Fantinatti et al. 2007) e portanto, tiveram
na ovitrampa uma possibilidade de realizar postura. Neste contexto, não é possível
afirmar que haja uma baixa demanda de criadouros (naturais) nos locais estudados,
porém, o fato de que tal padrão foi encontrado em armadilhas (que foram oferecidas em
número maior do que o necessário), leva a crer que o mesmo aconteça com os criadouros
naturais, pois apesar das fêmeas poderem distribuir seus ovos em todas as ovitrampas do
bloco, elas não o fizeram.
É sabido que chuvas auxiliam no desenvolvimento dos mosquitos pelo aumento
do número de criadouros disponíveis (Gomes, 2002). No presente trabalho, a correlação
entre a abundância de Ae. albopictus e pluviosidade foi evidenciada, embora o mesmo
não tenha acontecido para Ae. aegypti. Fato semelhante ocorreu em trabalho realizado por
Serpa e et al. (2006) em Potim, SP. Vale ressaltar que a pluviosidade demonstrou ser
mais influente na abundância de Ae. albopictus em Mariana do que Ouro Preto. Fato que
talvez possa ser explicado tanto pelas características dos espaços verdes quanto pela
notável diferença de temperatura entre as cidades. A área verde de Ouro Preto possui
vegetação mais densa, o que somado à maior umidade da cidade como um todo, torna-o
notavelmente mais úmido do que o espaço verde em Mariana. Assim, os criadouros
podem se manter viáveis por mais tempo (disponibilidade de água) e a chuva não seria
um fator essencial e sim, complementar em Ouro Preto. Honório et al. (2009a) analisaram
a distribuição temporal de Ae. aegypti em três bairros da cidade do Rio de Janeiro, sendo
que o efeito da pluviosidade sobre a abundância de mosquitos foi positivo apenas em uma
das áreas com um com um longo atraso de três a cinco semanas na precipitação. Assim, a
pluviosidade demonstra ser importante em áreas onde os sítios de oviposição são
principalmente produzidos por precipitação. Os efeitos positivos da precipitação com um
atraso nas semanas podem ser atribuídos principalmente ao impacto da chuva sobre a

44
dinâmica da população, pois produz novos sítios de oviposição e estimula a eclosão de
ovos já existentes no ambiente (Honório et al. 2009a).
A maior abundância de Ae. albopictus do que de Ae. aegypti corrobora com
Urbinatti (2004), que descreveu padrão semelhante no Parque Ecológico do Tietê, com
área de 14.000 ha dentro da cidade de São Paulo. Nesse estudo, as coletas foram
realizadas utilizando nove criadouros naturais e 20 artificiais (larvitrampas e ovitrampas)
e Ae. albopictus foi a espécie mais abundante em ambos. Nas 10 ovitrampas utilizadas,
foram coletados no período de um ano 1.571 imaturos de Ae. albopictus. Em estudo
realizado por Lourenço-de-Oliveira et al (2004) em parte da também urbana Floresta da
Tijuca, cidade do Rio de Janeiro, foram capturados 25.783 imaturos, sendo que a ampla
maioria também correspondia a Ae. albopictus (83.7%) do total de culicídeos. Honório et
al. (2009b), realizou estudo em áreas com diferentes graus de cobertura vegetal no
campus da FIOCRUZ RJ e encontrou predominância de Ae. albopictus nas 80 ovitrampas
instaladas principalmente nas áreas com alta e média cobertura vegetal.
Embora ainda não esteja claro que a presença de Ae. albopictus interfira na
dinâmica de transmissão do dengue nas Américas, a sua interação com Ae. aegypti requer
atenção pois ambas espécies se desenvolvem nos mesmos criadouros artificiais (Honório
& Lourenço-de-Oliveira, 2001; Urbinatti, 2004). Aedes albopictus têm sido considerado
um melhor competidor, tanto em laboratório quanto em campo, na fase aquática em
relação à Ae. aegypti ( Juliano, 1998; Juliano et al., 2004). Na Flórida, EUA, por
exemplo, a relação de competição entre essas espécies tem sido consideravelmente
estudada, pois Ae. albopictus parece deslocar Ae. aegypti em áreas suburbanas e rurais,
apesar de Ae. aegypti permanecer dominante em alguns ambientes urbanos onde Ae.
albopictus está presente ( Juliano,1998; Hornby et al. 1994). Contudo, outros trabalhos
mais antigos mostram que Ae. aegypti pode ser um competidor superior em condições
laboratoriais (Black et al., 1989, Ho et al., 1989).
Em suporte à hipótese competitiva no presente trabalho, Ae. aegypti e Ae.
albopictus tiveram respostas populacionais semelhantes às condicionantes ambientais em
ambas as cidades, já que o aumento da abundância de uma espécie foi correlacionado
com o da outra. Todavia, a separação espacial de habitats de postura apareceu diversas
vezes, principalmente para grandes posturas de Ae. albopictus realizadas em locais onde
Ae. aegypti não o fez. Em trabalho realizado por Martins et al. (2010) em Fortaleza-CE,
resultado semelhante foi obtido nas 1391 amostras no ambiente urbano em que, Ae.

45
albopictus, mesmo com um expressivo sucesso na infestação, depositou seus ovos nos
criadouros em que Ae. aegypti estava ausente. Já as fêmeas grávidas de Ae. aegypti
demonstraram atração por água contendo Ae. albopictus (Sucharit et al., 1978),
provavelmente devido aos menores efeitos que a competição interespecífica causa em
relação à competição intra-específica de alta densidade (Serpa, et al. 2008). Espécies em
competição apresentam distribuições diferentes, quando consideradas em uma escala de
resolução mais fina, mesmo que estejam coexistindo em escala espacial (Townsend et al.,
2006). Tal fato é demonstrado quando a separação espacial de habitats em diversos
momentos foi evidenciada, mesmo que a separação temporal de habitats não tenha sido
aparente. A separação espacial de habitats pode acarretar na redução da força de
competição entre esses culicídeos, porém, é possível que a própria separação seja
resultado da competição gerada pela coexistência das espécies (ausência de separação
temporal de habitats). A possível relação competitiva de Ae. albopictus com Ae. aegypti
nas áreas verdes urbanas torna plausível cogitar que estes locais possam contribuir para a
desaceleração de infestações urbanas.
Estudo realizado em cidades de Minas Gerais demonstrou não existir relação
entre a captura das espécies de Aedes estudadas com as distâncias dos fragmentos e áreas
com arborização urbanas situadas ao redor das armadilhas utilizadas (Morais 2011).
Porém, neste trabalho a presença destes vetores nas áreas verdes urbanas foi registrada,
mas suas preferêcias, a distribuição, a sazonalidade da ocorrência e as consequências da
oviposição nestes locais ainda não foram totalmente compreendidas. Para que as áreas
verdes não sejam consideradas obstáculos para o controle vetorial nas cidades, acredita-se
que o monitoramento e a manutenção das áreas verdes urbanas seja o mais indicado, bem
como ações de conscientização da população, na tentativa de eliminar o descarte de lixo
nesses locais. Tais práticas são sugeridas já que o combate ao vetor nestes ambientes
naturais seria algo complexo e talvez irrealizável, devido às diversas interações bióticas
que ali ocorrem e que poderiam ser afetadas.
O presente trabalho mostrou que espécies vetoras podem ocorrer em áreas
verdes urbanas, mesmo em cidades sem ocorrência de dengue. Mesmo que a
interpretação deste fato possa não ser alarmante, é importante lembrar que as espécies de
Aedes encontradas são exóticas e estão em constante processo de invasão Brasil adentro.
Assim, a elucidação da conjuntura local em relação à infestação dos vetores pode
contribuir para a criação de diretrizes primárias de planos para prevenção da dengue. De

46
forma complementar, a compreensão dos padrões de ocorrência dessas espécies em
situações pouco estudadas e em locais em que a população do vetor ainda é,
possivelmente, incipiente, também podem ser importantes para a prevenção da doença,
principalmente se considerada a tendência de invasão dos ecossistemas montanos prevista
(Cortez, 2004; Mendonça, 2003) com as mudanças climáticas globais .
5. CONSIDERAÇÕES EPIDEMIOLÓGICAS
A presença de Ae. aegypti nas áreas estudadas é relevante já que é considerado em
nosso país como vetor primário do dengue e no passado, como vetor da febre amarela
urbana (Urbinatti, 2004). Além disso, é preciso atentar-se que há primatas não humanos
(gênero Callithrix) nas duas áreas estudadas, que podem atuar como reservatórios do
vírus da febre amarela. Tais primatas fazem parte do ciclo silvestre da doença juntamente
com mosquitos dos gêneros Haemagogus e Sabethes. Caso estes mosquitos estejam
presentes no local e o ciclo silvestre seja rompido pela entrada do homem na área verde,
poderá ter início o ciclo da febre amarela urbana, na qual Ae. aegypti é o principal vetor.
É importante citar a possibilidade de existir um grande contingente de pessoas não
vacinadas contra a doença, o que levaria ao ressurgimento da febre amarela urbana em
locais infestados pela dengue, como detectado por Massad et al. (2003) no estado de São
Paulo.
Do ponto de vista epidemiológico a infestação de Ae. albopictus nas duas áreas
verdes estudadas pode representar um problema para a saúde pública, se considerado o
potencial da espécie de atuar como vetor de várias arboviroses, dentre elas a dengue
(Hawley, 1988; Borges 2001; Martins et al 2012). Recentemente, foi registrada a primeira
evidência natural da transmissão vertical do dengue em populações de Ae. albopictus
coletados em Fortaleza (CE) (Martins et al. 2012), ou seja, os indivíduos estudados
estavam infectados no momento em que foram coletados. Neste contexto, a possibilidade
de transmissão do vírus dengue pela espécie não deve ser desconsiderada, mesmo que
ainda não existam casos confirmados no Brasil (Martins et al. 2012).
Marcondes & Tauil (2011) ressaltam a possibilidade de ocorrência de ciclo
silvestre de dengue, mencionando algumas ocorrências pelo mundo. Dentre elas, a
documentação de ciclo silvestre da doença envolvendo macacos no oeste da África e na
Península da Malásia (Gubler, 2004) e a infecção por vírus dengue em alguns tatus,
veados, porcos-espinhos, roedores e marsupiais na Guiana Francesa (Thoisy et al., 2004).

47
Segundo os autores, mesmo que surtos de dengue com transmissão silvestre não tenham
sido registrados, esta possibilidade deve ser considerada, sendo indicado estudos com
mosquitos e mamíferos em áreas silvestres com diversos graus de preservação.
AGRADECIMENTOS
Aos moradores de Mariana e Ouro Preto que permitiram o acesso às suas casas. À
Alexandre Cortez, Leonardo Rodrigues e Lucas Ramos, pela colaboração em campo. Ao
Setor de zoonoses de ambas cidades. À Defesa Civil de Ouro Preto e à Vale/Mariana. Ao
LEEIDS da UFOP e LABEq da UFMG. À CAPES pelo apoio financeiro. SPR é bolsista
pesquisador do CNPq.

48
Capítulo II
Padrões de distribuição espacial e sazonal de Aedes (Stegomyia)
aegypti (Linnaeus, 1762) e Aedes (Stegomyia) albopictus (Skuse,
1894) (DIPTERA: CULICIDAE) em biótopos urbanos

49
RESUMO
Aedes aegypti e Ae. albopictus são vetores do dengue que apresentam nichos ecológicos
análogos. A segregação espacial destes culicídeos tem sido utilizada como uma possível
explicação para a coexistência dessas espécies em várias regiões. Este estudo teve como
objetivo a análise da distribuição espacial e sazonal de Ae. aegypti e Ae. albopictus, e
ainda, realizar inquérito virológico (DENV) nos indivíduos coletados. O estudo foi
realizado em dois municípios de Minas Gerais, Mariana e Ouro Preto, sendo que cada
um deles, foram utilizadas uma área verde urbana e dezesseis edificações, em diferentes
bairros. Os locais foram selecionados de acordo com o grau de cobertura vegetal e de
adensamento de casas, abrangendo assim, três biótopos distintos. A área verde foi
denominada biótopo I, os pontos situados em locais próximos à área verde e com menor
adensamento de edificações foram agrupados no biótopo II. O biótopo III reuniu pontos
localizados onde o adensamento de casas era maior e a cobertura vegetal, menor.As
coletas com duração de um mês cada, ocorreram nas estações chuvosa e seca. As
amostras de Aedes foram coletadas a partir da instalação de ovitrampas e
MosquiTRAPs®. Temperatura foi mensurada com termohigrômetro de campo e dados
pluviométricos cedidos pela VALE Mariana e Defesa Civil de Ouro Preto. As amostras
foram encaminhadas à FUNED realização da análise virológica. No total, foram
coletados 111 mosquitos e 618 imauturos de Ae. aegypti e 44 s e 484 imaturos de Ae.
albopictus em ambas cidades e estações. Devido à baixa abundancia na estação seca, as
análises foram realizadas apenas com os dados da estação chuvosa. A coocorrência das
espécies aconteceu em 6,9% e 20,6% das armadilhas em Mariana e em 2,9% e 17,8%
em Ouro Preto, em MosquiTRAPs®
e ovitrampas respectivamente. Na forma adulta,
apenas Ae. aegypti variou em abundância entre os biótopos, demonstrando
predominância no biótopo mais urbanizado. Os imaturos apresentaram diferentes
abundâncias entre os biótopos, sendo Ae. aegypti prevalente no biótopo mais urbanizado
e Ae. albopictus nas áreas verdes e residenciais com menor grau de adensamento
residencial. A abundância de Ae. aegypti não foi correlacionada com os fatores abióticos
em nenhuma de suas formas, porém Ae. albopictus apresentou correlação positiva com a
temperatura em todas seus estágios de vida. Não houve uma espécie mais abundante
quando se considera os s, porém, Ae.aegypti foi a espécie que apresentou maior
abundância na forma imatura. O Índice de Positividade de Ovitrampa (IPO) se mostrou
maior em Mariana nos biótopos I e II (60% e 57,2%, respectivamente) e em Ouro Preto
no biótopo III (59,4%). O vírus dengue não foi detectado em nenhuma amostra. Aedes
aegypti demonstrou preferência pelo ambiente mais urbanizado e Ae. albopictus pela
área de mata ou com maior arborização, mas ambos foram encontrados em todos os
biótopos, coocorrendo inclusive, em muitas armadilhas. Apenas Ae. albopictus, se
mostrou sensível à temperatura e ambas espécies, principalmente Ae. albopictus,
tiveram uma expressiva queda em sua abundância na estação seca, fato semelhante a
outros estudos. A compreensão destes padrões de ocorrência das espécies pode ser útil
na elaboração das práticas de controle e prevenção da dengue. Além disso, a elucidação
destes padrões em locais com baixa infestação dos Aedes e provável invasão recente,
também pode contribuir para a prevenção da doença, pois ajudam a esclarecer a
ocorrência das espécies durante o seu processo inicial de estabelecimento em um bairro,
cidade ou região.
Palavras chave: coocorrência, segregação espacial, temperatura, pluviosidade, DENV,
Mariana, Ouro Preto.

50
ABSTRACT
Aedes aegypti and Ae. albopictus are vectors of dengue that have similar
ecological niches. Spatial segregation of Culicidae has been used as a possible
explanation for the coexistence of these species in several regions. This study aimed to
analyze the spatial and seasonal distribution of Ae. aegypti and Ae. albopictus, and also
perform virological investigation (DENV) in sampled individuals. The study was
conducted in two municipalities of Minas Gerais, Ouro Preto and Mariana, and each one
of them we used a green urban and 16 buildings in different neighborhoods. The sites
were selected according to the degree of plant cover and density of houses, thus covering
three different biotopes. The green area was called biotope I, the points at locations close
to green areas with lower density buildings were grouped in biotope II. The biotope III
gathered points located where the density of houses and vegetation cover was bigger,
smaller. The collections last a month each in the rainy and dry seasons. Aedes samples
were collected using ovitraps and MosquiTRAPs®. Temperature was measured with a
hygrometer field and rainfall data obtained from ceded by VALE Mariana and civil
defense of Ouro Preto.. The samples were sent to FUNED achievement of virological
analysis. A total of 111 mosquitoes were collected and 618 imauturos of the Ae. aegypti
and 44 adults and immatures of 484 Ae. albopictus in both towns and stations. Due to the
low abundance in the dry season, the analyzes were performed with the data of the rainy
season. The co-occurrence of species was due 6.9% and 20.6% of the traps in Mariana
and 2.9% and 17.8% in Ouro Preto, for MosquiTRAPs ® and ovitraps, respectively. For
the adult form, only Ae. aegypti varied in abundance among habitats, demonstrating the
predominance in the biotope more urbanized. The immature showed different abundances
between biotopes, and Ae. aegypti prevailed in more urban biotope and Ae. albopictus in
residential and green areas with less houses density. Aedes aegypti abundance was not
correlated with abiotic factors in any of its forms, but Ae. albopictus correlated positively
with temperature in all life stages. There was a species more abundant when considering
adults, however, A.aegypti was the species with the highest abundance in immature form.
The Positivity Index of Ovitrap (IPO) was larger in the biotopes Mariana I and II (60%
and 57.2%, respectively) and Ouro Preto in the biotope III (59.4%). Dengue virus was not
detected in any sample. The results showed a significant variation in patterns of spatial
and seasonal distribution of Ae. aegypti and Ae. albopictus, regardless of town. Aedes
aegypti demonstrated preference for more urban environment and Ae. albopictus by green
areas, but both were found in all habitats, as well as co-occurred in some tramps.. In
relation to abiotic factors, only Ae. albopictus, was sensitive to both temperature had a
significant drop in their abundance during the dry season, a fact similar to other studies.
The understanding of these patterns of species occurrence can be useful in the
development of practical control and prevention of dengue. Furthermore, the elucidation
of these standards in areas of low infestation of Aedes recent and probable invasion, may
also contribute to the prevention of disease, because they help to clarify the occurrence of
the species during their initial establishment in a neighborhood, city or region.
Keywords: co-occurrence; spatial segregation; temperature; rainfall; DENV; Mariana;
Ouro Preto.

51
1. INTRODUÇÃO
O dengue é hoje a arbovirose mais importante do planeta, cuja etiologia e
mecanismos de transmissão são bem conhecidos (Tauil, 2002). A doença é causada por
um vírus que pertence ao gênero Flavivirus (Gubler, 1998) e é transmitida geralmente,
pelos mosquitos Aedes aegypti e Ae. albopictus, sendo o primeiro seu principal vetor
(Tauil, 2002).
Aedes aegypti e Ae. albopictus são espécies exóticas que invadiram a região
neotropical e que apresentam nichos ecológicos análogos ( Forattini, 1999; Serpa &
Voltolini, 2008). Os s de Ae. aegypti são em sua maioria domiciliados, sendo a habitação
humana o local onde as fêmeas exercem a hematofagia, se abrigam e encontram
criadouros (Forattini, 2002). As fêmeas demonstram baixa tendência a se afastarem do
ambiente antrópico em que são encontradas e utilizam a maior parte do tempo disponível
procurando possíveis criadouros (Forattini, 2002). A distância de dispersão que a fêmea
adulta alcança dependerá da acessibilidade dos recursos, principalmente fonte de
alimentação sanguínea e locais para reprodução, e será curta caso tais recursos estejam
acessíveis nas proximidades (Forattini, 2002; Natal, 2002; Russel et al., 2005). Estudos
de dispersão com marcação-soltura-recaptura têm mostrado diferentes distâncias médias
de dispersão que variam entre 56m e 78m, ou até mesmo entre 28m e 199m (Muir & Kay
1998; Harrington et al., 2005; Russel et al, 2005). Todavia, a dispersão máxima pode
estar entre 594 m até aproximadamente a 800m na busca por sítios de oviposição
(Honório et al., 2003), ou ainda por uma área de pelo menos 840 metros de diâmetro
(Reiter et al., 1995).
Aedes albopictus possui características ainda silvestres e portanto, possui menor
grau de domiciliação em relação à Ae. aegypti (Forattini, 1986). Apesar da espécie
também utilizar criadouros artificiais, ela é perfeitamente capaz de sobreviver sem
artefatos humanos (Moore, 1999), utilizando criadouros naturais, alimentando-se de
néctar e de sangue de animais silvestres (Borges, 2001). Tais possibilidades
proporcionam à espécie maiores chances de dispersão e colonização, levando a ocupação
de diversificados ambientes (Borges, 2001). Como a preferência de Ae. albopictus é por
locais onde haja alto grau de cobertura vegetal (Braks et al. 2003; Honório et al. 2009;
Lima-Camara, 2010), as características ambientais naturais se tornam fundamentais para
determinação das áreas onde a espécie se estabelecerá (Moore, 1999). Neste contexto,
diversos estudos têm demonstrado que a infestação urbana de Ae. albopictus depende da

52
vegetação (de porte arbóreo) presente em locais habitados, parques e bosques (Urbinatti,
2004).
Ambas espécies podem utilizar os mesmos criadouros artificiais, sendo possível a
co-ocorrência nos sítios de oviposição (Forattin, 2002; Braks et al., 2003), o que
pressupõe a existência de competição intra e interespecífica (Braks et al., 2003; Serpa et
al, 2008). Devido a tal fato e dependendo de fatores regionais, tem-se observado a
possibilidade de exclusão competitiva entre as mesmas (O’Meara et al., 1995; Forattini,
2002,). Aedes albopictus têm sido deslocado por Ae. aegypti de centros urbanos do
sudeste da Ásia, onde a primeira é nativa (Gilotra e col 1967 apud Forattini, 2002;
Hawley 1988). No EUA a possibilidade de exclusão competitiva têm sido relatada, já que
na Flórida e sudeste do país a abundância de Ae. aegypti tem declinado
concomitantemente à expansão de Ae. albopictus (O’Meara et al, 1993 e 1995; Harrison
et al. 1998). Contudo, há alguns locais no sudeste do EUA em que as espécies ainda
coexistem (O’Meara et al., 1995; Juliano et al., 2002). A persistência de Ae. aegypti em
determinadas áreas e habitats leva a crer que fatores abióticos como temperatura e
pluviosidade, bem como a estrutura macroscópica do habitat e composição química da
água nos criadouros podem influenciar o processo competitivo (Juliano et al. 2002 e
2004).
As invasões biológicas desafiam o entendimento dos processos bióticos e abióticos
que orientam a distribuição e abundância das espécies (Juliano et al., 2004). Somado a
isso, o fato de ambas espécies poderem ter a preferência relacionada ao tipo de cobertura
de solo (Braks et al, 2003; Carbajo et al. 2006) ou a fatores socioeconômicos da
população (Santos 1999; Tauil, 2001; Ferreira & Neto, 2007), torna complexa a
compreensão dos padrões de ocorrência desses vetores.
Dentro de uma mesma cidade, podem existir diferentes tipos de habitats, sendo
importante portanto compreender como ocorre a distribuição dos Aedes nesses ambientes.
Tal entendimento seria relevante para as ações de controle, caso fosse possível determinar
se os padrões de distribuição e de co-ocorrência observados em determinadas áreas,
pudessem ser generalizados para outros locais em que as espécies estão (Braks et al.,
2003). Além disso, a elucidação dos padrões de ocorrência de Ae. aegypti e Ae albopictus
em locais com baixa infestação e provável invasão recente, também pode contribuir para
maior eficácia nas medidas de prevenção e controle da dengue, na medida em que ajudam
a esclarecer o processo inicial de estabelecimento em um bairro, cidade ou região.

53
Segundo Braks et al. (2003), no Brasil existem poucos registros sobre as relações
espaciais e de segregação de habitats entre Ae. albopictus e Ae. aegypti. Assim, o
presente estudo tem como objetivo a análise da distribuição espacial e sazonal de Ae.
aegypti e Ae. albopictus, verificando a prevalência e coexistência das espécies em
diferentes biótopos: espaço verde urbano, área plenamente urbanizada e área
intermediária entre os dois habitats citados, em cidades com baixo índice de infestação
(Índice de Infestação Predial<1 Comunicação pessoal Setor Zoonoses das cidades
estudadas), localizadas no interior de Minas Gerais.
Este estudo investiga as hipóteses de que o padrão da distribuição é diferente entre
os biótopos, mas semelhante entre as cidades. A predição é que o grau de cobertura
vegetal influenciará na distribuição das espécies, independente da cidade estudada. Além
disso, teve-se como hipótese que ambas espécies são correlacionadas com temperatura e
pluviosidade, predizendo a maior abundância total na estação chuvosa e um declínio na
estação seca. Por fim, investiga-se a hipótese de que na estação seca a ocorrência das
espécies no espaço verde reduzirá drasticamente, devido à previsão de diminuição de
criadouros disponíveis.
2. MATERIAL E MÉTODOS
2.1.Área de estudo
Este estudo foi realizado nos municípios de Mariana (20° 22′ 40″ de latitude sul
e 43° 24′ 57″ longitude) e Ouro Preto (Latitude 20º23'08" S e Longitude 43º30'29" O),
distantes aproximadamente 14 km entre si e localizados aproximadamente a 100km de
Belo Horizonte. Tais municípios estão inseridos em uma área de Minas Gerais onde o
clima na região é do tipo Cwb de acordo com a classificação de Koppen, com estações
seca e chuvosa bem definidas, verões suaves concentrando cerca de 90% da
precipitação anual (Gomes, 1998). Ouro Preto, cidade situada à 1100 m de altitude,
possui temperaturas médias anuais entre 17,4°C e 19,8°C (Gomes, 1998) e Mariana à
750 m, possui temperaturas médias entre 18°C e 22°C (Ozório, 2000).
Para o estudo foram utilizadas uma área verde urbanas em cada cidade, além de
residências situadas em diferentes bairros na sede dos municípios (figuras 1,2 e 3). A área
verde em Ouro Preto é uma área particular, situada no centro histórico da cidade, com
aproximadamente 1,4 hectares. Algumas edificações utilizadas no estudo pertencem ao

54
bairro Centro, mesmo bairro onde está situada a área verde e outras estão situadas no
bairro Antônio Dias (figura 2). Em Mariana, a área verde utilizada possui
aproximadamente 2,0 ha e está localizada no bairro Rosário. A área pertence à prefeitura
da cidade sendo completamente contornada por residências, tornando o acesso possível
apenas através das mesmas. Residências localizadas neste bairro foram utilizadas, bem
como algumas edificações no bairro Centro (figura 3). Devido às dificuldades de acesso
ao interior do espaço verde em Mariana, decidiu-se por não utilizar a região central de
ambos espaços verdes estudados.
2.2. Delineamento amostral
No ano de 2011 ocorreram duas campanhas, a primeira na estação chuvosa (março
e abril) e a segunda na estação seca (setembro e outubro), com duração de cinco semanas
cada. Para tal, foi utilizado o desenho amostral apresentado na figura 1. Pontos amostrais
foram distribuídos aleatoriamente tanto nas áreas verdes quanto em residências. Tal
seleção ocorreu da seguinte forma: a partir do ponto de entrada da área verde foram
selecionados outros três pontos na tentativa de formar uma cruz (com 4 pontos) nas
bordas do fragmento. Estando cada ponto a aproximadamente 30m da rua ou calçada na
extremidade da área (ver figura 2, bloco arborizado). A partir de cada ponto selecionado,
a primeira edificação localizada, dentro do possível, em linha reta logo após o fim da área
verde era escolhida, ou seja, a residência localizada limítrofe do espaço verde. Partindo
do último ponto escolhido, a próxima residência distante 100m era selecionada. Os locais
foram selecionados com a utilização do programa Google Earth, considerando sempre a
ideia inicial de que os pontos amostrais deveriam formar uma cruz. Vale ressaltar que a
utilização de cada edificação deveria ser consentida pelo proprietário e portanto, em casos
em que o imóvel melhor localizado não pôde ser utilizado, a construção vizinha àquele
ponto foi a escolhida.
Para o estudo foi necessário que as coletas abrangessem diferentes paisagens e
contextos dentro de uma mesma localidade. Assim, denominou-se por biótopo cada área
com características semelhantes que podem ter importância na ecologia dos vetores
estudados. Os biótopos definidos foram caracterizados da seguinte forma:
Biótopo I: Área verde propriamente dita, com quatro pontos amostrais por cidade;
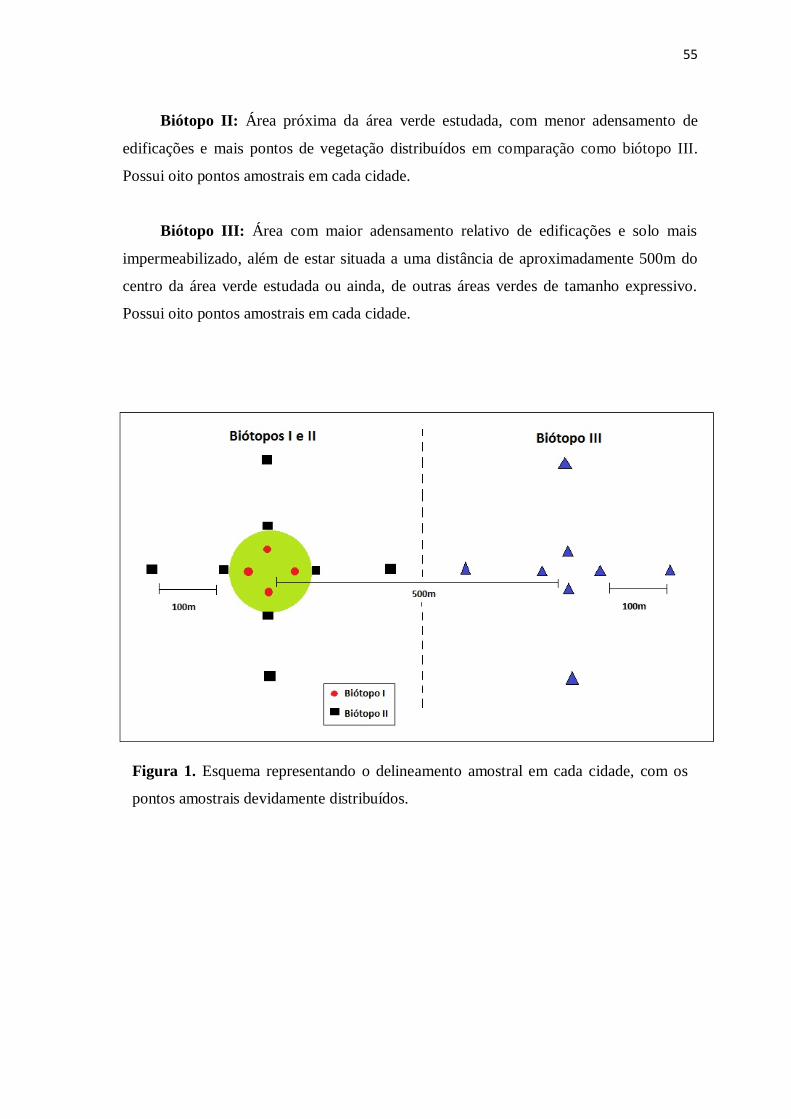
55
Biótopo II: Área próxima da área verde estudada, com menor adensamento de
edificações e mais pontos de vegetação distribuídos em comparação como biótopo III.
Possui oito pontos amostrais em cada cidade.
Biótopo III: Área com maior adensamento relativo de edificações e solo mais
impermeabilizado, além de estar situada a uma distância de aproximadamente 500m do
centro da área verde estudada ou ainda, de outras áreas verdes de tamanho expressivo.
Possui oito pontos amostrais em cada cidade.
Figura 1. Esquema representando o delineamento amostral em cada cidade, com os
pontos amostrais devidamente distribuídos.

56
Figura 2. Visualização dos pontos amostrais utilizados no estudo em Ouro Preto. Observar que
pontos inseridos no círculo azul correspondem aos biótopos I e II. Pontos inseridos no círculo
laranja correspondem ao biótopo III. Imagem modificada do Google Earth em 14/03/2013.
Figura 3. Visualização dos pontos amostrais utilizados no estudo em Mariana. Observar que pontos
inseridos no círculo azul correspondem aos biótopos I e II. Pontos inseridos no círculo laranja
correspondem ao biótopo III. Imagem modificada do Google Earth em 14/03/2013.
2.3. Coleta de dados
Durante o estudo, dois modelos de armadilhas, ovitrampa e MosquiTRAP®
(ver
anexo), foram empregados. A primeira, também conhecida como armadilha de

57
oviposição, é utilizada para coleta de imaturos especialmente ovos de mosquitos (ver
anexo I). A armadilha usada consistiu em um recipiente plástico preto-fosco com
capacidade de 700ml, ao qual se prendeu verticalmente uma palheta de madeira Eucatex
com 10cm de comprimento, por 3cm de largura e 0,5cm de espessura. Além da palheta, a
ovitrampa continha cerca de 300ml de água de abastecimento. Importante ressaltar que o
objetivo era amostrar passivamente, assim refletindo a densidade real na área, portanto,
optou-se por não utilizar a infusão de gramíneas. Cada armadilha foi acomodada de modo
que permanecesse suspensa entre 1m e 1,5m do solo e em local sombreado, quando
possível.
A MosquiTRAP® é uma armadilha para captura de mosquitos s, principalmente
fêmeas grávidas e raramente paridas e nulíparas, já que utiliza um atraente de oviposição,
o AtrAedes® (Ecovec Ltda., Belo Horizonte, Brasil). A armadilha consiste em um frasco
preto-fosco, dividida em duas partes, cuja parte inferior possui capacidade para 300ml de
água. Possui ainda uma tela para evitar a postura e acima do nível da água coloca-se um
cartão (onde os mosquitos capturados ficam presos pela patas) com o atraente sintético de
oviposição (Resende et al., 2010). Cada armadilha permaneceu com o mesmo cartão
adesivo e atraente durante toda a coleta, salvo algumas exceções em que houve a
necessidade de substituí-los. Estudos de campo mostraram que o MosquiTRAP® é mais
sensível que a pesquisa larvária, mas possui menor sensibilidade que a ovitrampa.
Entretanto, os dados obtidos semanalmente pela MosquiTRAP®
mostraram
comportamento semelhante ao da ovitrampa, possibilitando a verificação da flutuação
populacional dos vetores na área (Resende & Eiras, 2009). As armadilhas foram
deixadas no solo e procurou-se sempre abrigá-las da chuva, apesar da impossibilidade
disto ocorrer nas áreas verdes. Além disso, como a água contida em cada MosquiTRAP®
não era constantemente substituída, foi adicionado à agua o larvicida Temephos cedido
pelo setor de zoonoses da cidade de Ouro Preto.
Em cada ponto amostral nas áreas verdes foram instaladas uma ovitrampa e uma
MosquiTRAP®. Já nas residências, foram utilizadas uma ovitrampa e duas
MosquiTRAP®, na tentativa de abranger diferentes ambientes mesmo dentro de uma só
residência (exemplo: varanda e quintal), quando possível. Assim, no estudo foram
empregadas 40 ovitrampas e 72 MosquiTRAPs® por campanha, considerando ambos
municípios.

58
Entre 14/03/2011 e 13/04/2011 ocorreram quatro coletas semanais em cada cidade,
sempre obedecendo um intervalo de sete dias para cada nova coleta. Para ovitrampas, a
cada semana, água e palhetas eram coletadas e imediatamente colocadas em pote de
500ml devidamente identificados. A cada coleta, 300ml de água era adicionado a cada
armadilha, bem como nova palheta de Eucatex. Todos os potes foram levados ao
Laboratório de Ecologia Evolutiva de Dossel e Sucessão Natural. Adicionava-se água de
abastecimento nos potes até que a palheta ficasse embebida (para a eclosão dos ovos).
Após a triagem do material, as larvas eram acondicionadas em potes J10 e quando
atingissem maior estágio de maturação (de L2 a L4), eram repassadas para Placa de Petri
e então, resfriadas em freezer -20° C por cerca de 4 minutos para que se tornasse possível
a identificação taxonômica. As larvas já identificadas, e ainda vivas, foram
acondicionadas em eppendorf (procedimento realizado em ambiente estéril), enumeradas
e armazenadas no freezer -80°C do Laboratório de Doença de Chagas-UFOP (ver
apêndice)
As MosquiTRAPs®
sempre foram vistoriadas nos mesmos dias em que as
ovitrampas. Durante a vistoria a armadilha era aberta e caso algum díptero estivesse
grudado no cartão adesivo, este era identificado com auxílio de lupa de mão e em seguida
colocado em potes J10. A água da armadilha era verificada e caso houvesse larvas ou
pupas essas eram retiradas e levadas ao laboratório para posterior identificação.
Entretanto tal coleta de imaturos não consta como captura da MosquiTRAP®.
Em laboratório os mosquitos das espécies Aedes aegypti e Ae. albopictus eram
temporariamente (durante algumas horas) armazenados em freezer -20°C e
posteriormente submetidos ao mesmo procedimento realizado com as larvas (descrito
acima) para serem acondicionados no freezer 80°C (ver apêndice).
2.5. Dados abióticos
A temperatura foi mensurada durante cada dia de coleta, em todas as campanhas,
com a utilização de termohigrômetro digital de campo (Kestrel0®). Os dados referentes à
pluviosidade diária da cidade de Mariana foram cedidos pela Gerência Geral de
Desenvolvimento e Planejamento Sudeste - GEPDS/DIFS - Complexo Mariana da
empresa VALE, sendo que a estação meteorológica está situada na mina de Timbopeba,
localizada aproximadamente 15km da cidade, porém na mesma altitude. Os dados
relativos à cidade de Ouro Preto foram cedidos pelo Setor de Defesa Civil da prefeitura

59
da cidade, sendo que sua estação é localizada no bairro São Cristovão. Para as análises,
os dados de pluviosidade foram atrasados em duas semanas em relação à semana de
coleta e os dados de temperatura em apenas uma semana.
2.6.Análises estatísticas
Utilizou-se tabela de contingência para relacionar a abundância de Aedes obtida
na estação chuvosa, com a abundância na estação seca, e testada com o uso de qui
quadrado (X2). Valores médios por amostra foram utilizados para testar os efeitos de
“espécie”, co-variáveis abióticas e “habitats”, o que foi realizado com modelos de
ANOVA e regressões lineares simples, respectivamente, além de teste t de Student para
comparar variação entre cidades. Dados que escapavam aos pressupostos de
homoscedasticidade e normalidade, foram logaritimizados. Dados cuja distribuição de
frequência distava à distribuição normal em qualquer escala de interesse para este estudo,
foram analisados com uso de modelos não paramétricos de Kruskal-Wallis.
3. RESULTADOS
Na estação chuvosa, foram coletados 66 mosquitos s e 388 imaturos de Ae.
aegypti e 28 mosquitos e 321 de Ae. albopictus em Mariana. Em Ouro Preto, 36 s e 170
de Ae. aegypti foram coletados, e ainda 16 mosquitos e 144 imaturos de Ae. albopictus.
Além disso, foram coletados 80 imaturos de Culex sp., que não foram considerados nas
análises dos dados.
Na estação seca, apenas Ae. aegypti foi encontrado na forma adulta, sendo
coletados oito indivíduos em Mariana e apenas um em Ouro Preto. Quanto a forma
imatura, 59 Ae. aegypti e 19 Ae. albopictus foram coletados em Mariana e apenas um
indivíduo da primeira espécie em Ouro Preto. Além disso, foram capturados 23 s e 68
imaturos de Culex sp nas duas cidades e estações, porém estes dados não foram
utilizados neste estudo (Tabelas 1 e 2).
Uma drástica redução na abundância de indivíduos entre as diferentes estações foi
detectada principalmente nas áreas verdes de ambas as cidades (Tabelas 1 e 2). Aedes
albopictus não foi encontrado no período de seca no município de Ouro Preto em nenhum
estágio de vida, e em Mariana, só foi encontrado na forma imatura em quantidade
consideravelmente menor, quando comparado à estação chuvosa. A abundância de

60
imaturos observada na estação seca foi menor do que na chuvosa, tanto em Mariana
(X2=146,1; gl=2; p<0,05), quanto em Ouro Preto (X
2=8,23; gl=2; p<0,05). O mesmo
padrão foi observado para os mosquitos s de Mariana (X2 =6,10; gl=2; ; p<0,05) e de
Ouro Preto (X2 =6,02; gl=2; p<0,05).
Tabela 1. Tabela de contingência apresentando a abundância total das espécies em cada biótopo e a
porcentagem de diminuição da abundância das espécies entre a estação chuvosa e seca nas cidades de
Mariana, Minas Gerais, no ano de 2011.
Tabela 2. Tabela de contingência apresentando a abundância total das espécies em cada biótopo e a
porcentagem de diminuição da abundância das espécies entre a estação chuvosa e seca na cidade de Ouro
Preto, Minas Gerais.
Mariana foi a cidade com maior abundância total dos Aedes (ANOVA: F1,72=5.59;
p<0,05), mas nenhuma espécie foi mais abundante que a outra na forma adulta, tanto
quando as cidades são consideradas em conjunto (Teste t; gl=12; p<0,05) ou
separadamentente (Teste t; gl= 24; p<0,05. Na forma imatura, houve predominância
apenas de Ae. aegypti quando ambas cidades foram consideradas (Teste t; gl=12;p
<0,05).
Foram detectadas diferenças nos padrões de distribuição de ambas formas de vida
entre os biótopos.
Espécie Forma
de vida
I II III Total I II III Total I II III
Ae. aegypti Imaturo 22 88 278 388 0 8 51 59 100 91,7 84,5
Ae. albopictus Imaturo 58 179 84 321 0 2 17 19 100 98,9 83,2
Total 80 267 362 709 0 10 68 78 _ _ _
Ae. aegypti Adulto 0,5 11,5 19,5 31,5 0 1 3 4 100 92,0 86,7
Ae. albopictus Adulto 1,5 11 4,5 17 0 0 0 0 100 100 100
Total 2 22,5 24 48,5 0 1 3 4 _ _ _
Abundância na estação chuvosa Abundâncida na estação seca
Biótopos Biótopos Biótopos
% de diminuição da abundância
Espécie Forma
de vida
I II III Total I II III Total I II III
Ae. aegypti Imaturo 2 68 100 170 0 0 1 1 100 100 99
Ae. albopictus Imaturo 7 72 65 144 0 0 0 0 100 100 100
Total 9 140 165 314 0 0 1 1 _ _ _
Ae. aegypti Adulto 0 4,5 13,5 18 0 0 0,5 0,5 0 100 96,4
Ae. albopictus Adulto 0 6,25 2 8,25 0 0 0 0 0 100 100
Total 0 10,75 15,5 26,25 0 0 0,5 0,5 _ _ _
Abundância na estação chuvosa Abundância na estação seca % de diminuição da abundância
Biótopos Biótopos Biótopos
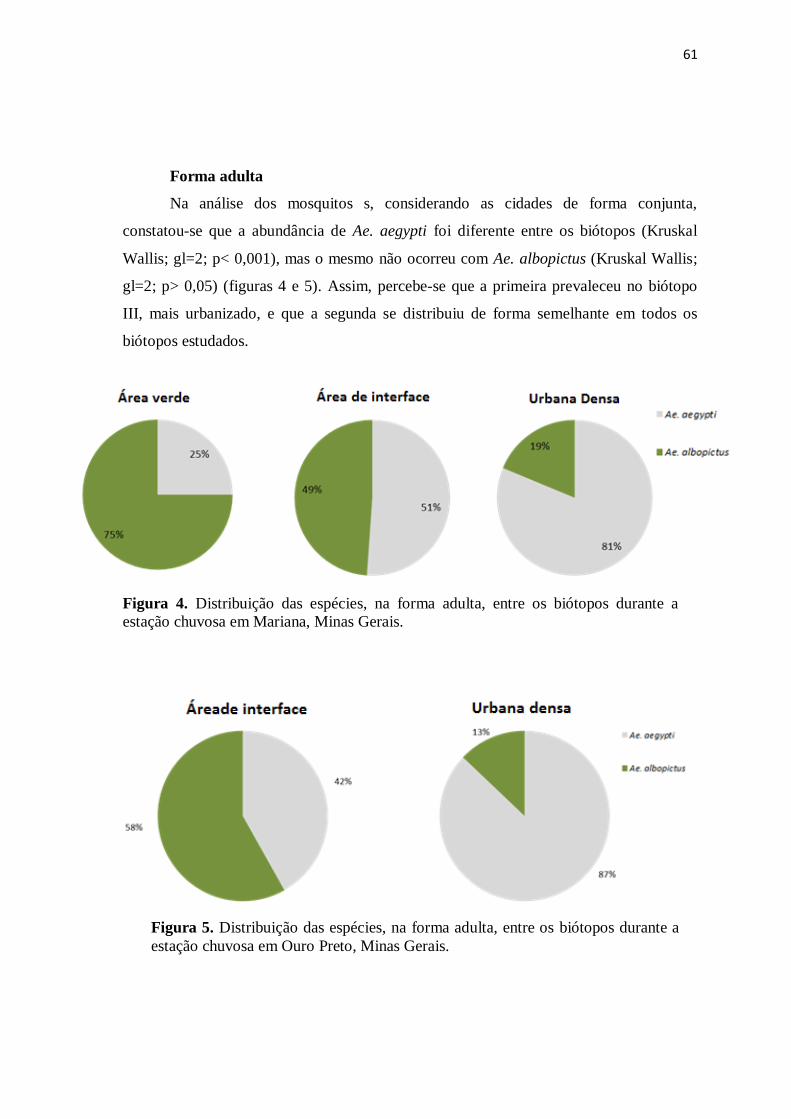
61
Forma adulta
Na análise dos mosquitos s, considerando as cidades de forma conjunta,
constatou-se que a abundância de Ae. aegypti foi diferente entre os biótopos (Kruskal
Wallis; gl=2; p< 0,001), mas o mesmo não ocorreu com Ae. albopictus (Kruskal Wallis;
gl=2; p> 0,05) (figuras 4 e 5). Assim, percebe-se que a primeira prevaleceu no biótopo
III, mais urbanizado, e que a segunda se distribuiu de forma semelhante em todos os
biótopos estudados.
Figura 4. Distribuição das espécies, na forma adulta, entre os biótopos durante a
estação chuvosa em Mariana, Minas Gerais.
Figura 5. Distribuição das espécies, na forma adulta, entre os biótopos durante a
estação chuvosa em Ouro Preto, Minas Gerais.
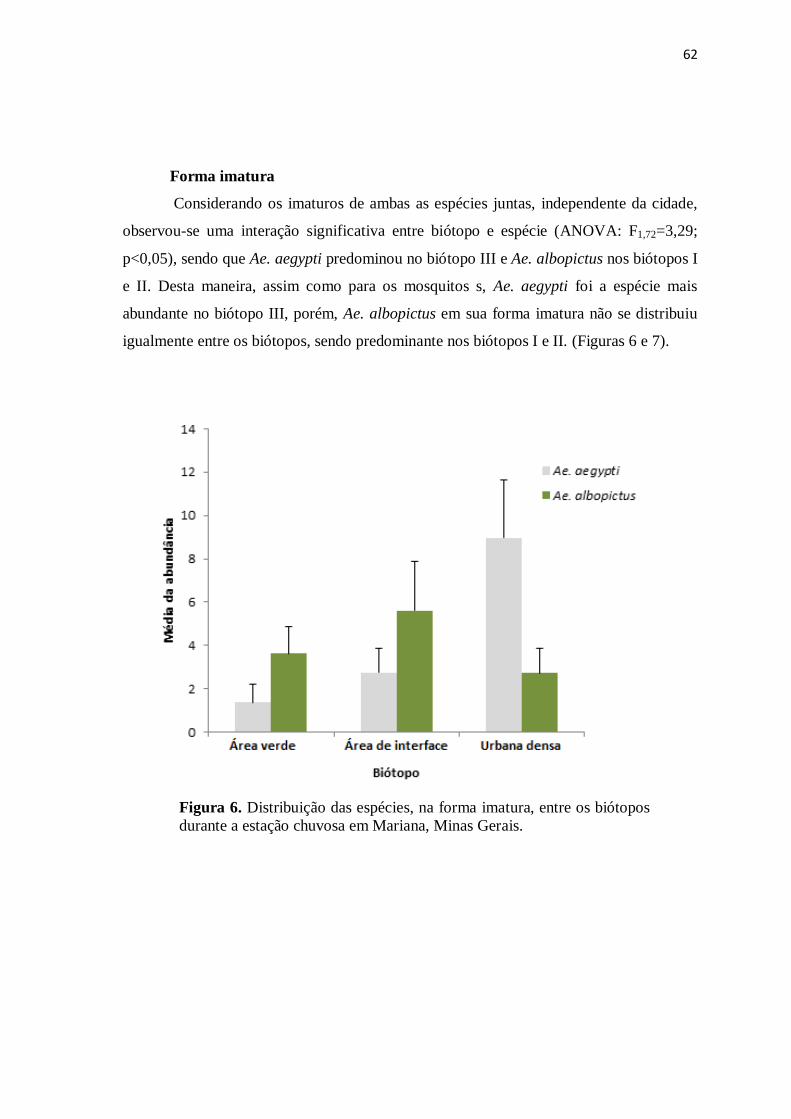
62
Forma imatura
Considerando os imaturos de ambas as espécies juntas, independente da cidade,
observou-se uma interação significativa entre biótopo e espécie (ANOVA: F1,72=3,29;
p<0,05), sendo que Ae. aegypti predominou no biótopo III e Ae. albopictus nos biótopos I
e II. Desta maneira, assim como para os mosquitos s, Ae. aegypti foi a espécie mais
abundante no biótopo III, porém, Ae. albopictus em sua forma imatura não se distribuiu
igualmente entre os biótopos, sendo predominante nos biótopos I e II. (Figuras 6 e 7).
Figura 6. Distribuição das espécies, na forma imatura, entre os biótopos
durante a estação chuvosa em Mariana, Minas Gerais.

63
Figura 7. Distribuição das espécies, na forma imatura, entre os biótopos
durante a estação chuvosa em Ouro Preto, Minas Gerais.
Tabela 3. Valores absolutos de abundância das espécies em cada habitat, a
estação chuvosa, com as diferentes metodologias utilizadas.
*Espécie mais abundante capturada pela ovitrampa em cada habitat.
**Espécie mais abundante capturada pela Mosquitrap em cada habitat.
Quando se compara os valores absolutos de abundância das espécies (Tabela 3),
percebe-se que os resultados encontrados com as diferentes armadilhas são análogos, já
que a espécie predominante em cada biótopo na forma imatura, também prevaleceu na
Cidade Habitat
Ae. aegypti Ae. albopitcus Ae. aegypti Ae. albopitcus
I 2 7 * 0 0
Ouro Preto II 68 73 * 4,5 6,25 **
III 100 * 65 13,5 ** 2
I 22 58* 0,5 1,5**
Mariana II 67 155* 10 11**
III 145* 70 15** 4
Ovitrampa Mosquitrap

64
forma adulta. Ou seja, independente da armadilha utilizada, Ae. albopictus foi sempre
predominante nos biótopos I e II e Ae. aegypti no biótopo III.
Tabela 4. Frequência de ocorrência de Aedes nas armadilhas, por habitat, durante as quatro semanas de coleta
da estação chuvosa e porcentagem de ocorrência em relação ao número total de armadilhas utilizadas.
A co-ocorrência de Ae. aegypti e Ae. albopictus (Tabela 4) em Mariana foi
semelhante nos habitats II (sete ovitrampas) e III (seis ovitrampas), o que corresponde à
coexistência das espécies em 43,7% e 37,5% das ovitrampas positivas, respectivamente.
Já em Ouro Preto, houve mais casos de co-ocorrência (47,7% das ovitrampas positivas)
no habitat III do que no habitat II (33,3% ovitrampas positivas). Ainda nesta cidade, no
habitat I, todos Aedes capturados co-ocorreram em uma única armadilha.
Mas o mesmo não pôde ser observado para os imaturos, pois em ambas as cidades
a co-ocorrência foi detectada em mais casos do que a ocorrência solitária das espécies.
O Índice de Positividade de Ovitrampa (IPO) calculado para cada biótopo,
somando-se todas as coletas da estação chuvosa, mostrou-se maior em Mariana nos
biótopos I e II, sendo igual a 60% e 57,2% respectivamente, em relação à Ouro Preto:
6,25% e 38,8%. Contudo, o mesmo não ocorre no habitat III, já que o IPO foi maior em
Ouro Preto (59,4%) do que em Mariana (53,3%).
Habitat Ae. aegypti a
Ae. albopictusb
Co-ocorrênciac
Armadilhas positivasd
Ae. aegypti a
Ae. albopictusb
Co-ocorrênciac
Armadilhas positivasd
1 1 1 0 3 0 0 0 0
2 9 2 4 19 3 3 2 10
3 11 1 6 20 9 2 2 15
Porcentagem 14,6 2,8 6,9 29,2 8,5 3,6 2,9 17,6
Habitat Ae. aegypti a
Ae. albopictusb
Co-ocorrênciac
Armadilhas positivasd
Ae. aegypti a
Ae. albopictusb
Co-ocorrênciac
Armadilhas positivasd
1 1 6 2 9 0 0 1 1
2 1 6 7 16 1 5 4 12
3 6 3 6 16 6 4 9 19
Porcentagem 11,0 18,8 20,6 56,2 8,9 11,4 17,8 40,5
MosquiTRAP®
Ovitrampa
Mariana Ouro Preto
Mariana Ouro Preto
Ae. aegypti a -número de armadilhas em que foram encontrados somente Ae. aegypti.
Ae. albopictusb -número de armadilhas em que foram encontrados somente Ae. albopictus.
Co-ocorrênciac -número de armadilhas em que ambas espécies foram encontradas.
Armadilhas positivasd
– Número total de armadilhas positivas por habitat, incluindo positivas com
Culex sp. apenas ou ainda com Aedes e Culex sp.

65
Fatores abióticos
Aedes aegypti, tanto na forma alada (F1;23= 0,157 p>0,05; F1;23= 0,158 p>0,05)
quanto na imatura (F1;23= 0,059 p>0,05; F1;23= 0,059 p>0,05), não apresentou correlação
com a pluviosidade ou com a temperatura, respectivamente. Todavia, Ae. albopictus,
apesar de também não ter sido correlacionado com a pluviosidade em nenhuma de suas
formas, mostrou que sua abundância possui relação com a temperatura quando
(y=0,0101x-0,2187, R2=0,218, F1;20=5,30; p< 0,05) e quando imaturo (y=0,0041x-0,0888,
R2=0,2237; F1;20=5,48; p<0,05).
4.DISCUSSÃO
Os resultados mostram que Aedes aegypti e Ae. albopictus responderam ao grau
de cobertura vegetal e de adensamento urbano. A maior abundância de Ae. aegypti em
áreas com uma urbanização mais intensa e de Ae. albopictus em locais com maior
cobertura vegetal, mostram a existência de um padrão de distribuição que também é
descrito em outros estudos (Braks et al. 2003; Carabajo et al., 2006; Lourenço-de-
Oliveira et al., 2004; Maciel-de-Freitas et al., 2006; Serpa et al, 2006; Honório et al.,
2009b). Apesar da segregação de habitats revelada, o padrão sazonal das espécies foi
parecido, uma vez que ambas se mostraram sensíveis à transição da estação chuvosa
para a seca. Contudo, Ae. albopictus apresentou maior sensibilidade às condições
ambientais resultantes do período seco. Assim, enquanto Ae. aegypti demonstra uma
maior estabilidade de resposta à variação climática, (Honório et al., 2009b), Ae.
albopictus revela uma maior redução de sua população perante a ausência de chuvas. O
que provavelmente, deve-se à menor resistência de seus ovos à dessecação (Juliano et
al., 2002) e à diminuição de sítios de oviposição disponíveis, já que os criadouros
naturais utilizados podem não conter água em consequência da ausência de chuvas.
Durante a estação chuvosa, os imaturos de ambas espécies foram comuns em
todos habitats e cidades, mas o mesmo não foi observado com as formas adultas pois no
biótopo arborizado em Ouro Preto elas não foram encontradas. Na estação seca, além da
redução na abundância dos imaturos ocorrida nos dois municípios, o de Ae. albopictus
não foi encontrado em nenhuma área verde. Variações similares de abundância em
resposta positiva à estação chuvosa foram detectadas em outros estudos (Mogi et al.,

66
1988; Serpa et al., 2006; Honório et al., 2009b), incluindo trabalhos realizados em zonas
de transição (mata-área urbana) no Brasil (Honório et al., 2009).
Os estágios de vida de Aedes responderam diferentemente aos biótopos urbanos.
Aedes aegypti foi o único a se correlacionar com os biótopos, apresentando baixa
abundância no habitat arborizado, enquanto Ae. albopictus , se distribuiu de forma
uniforme entre os locais. A distribuição dos imaturos foi heterogênea entre os biótopos
estudados, independentemente da cidade, corroborando a principal hipótese de que os
diferentes biótopos afetam a abundância das espécies. Aedes aegypti foi claramente
predominante nas áreas com maior adensamento de edificações. Já Ae. albopictus foi
mais abundante no biótopo II, apesar de ser esperada sua prevalência nas áreas verdes
urbanas, diante de suas características ecológicas e principalmente se considerada sua
abundância nas mesmas áreas verdes no ano de 2009 (Pedrosa et al., dados ainda não
publicados) e ainda, em estudo semelhante na Paraíba (Leandro, 2012).
O grau de cobertura vegetal tem demonstrado influência sobre a distribuição de
ambas as espécies, mas de maneiras opostas (Braks et al., 2003; Honório et al., 2009;
Leandro, 2012; Lourenco-De-Oliveira et al.,2004). Assim, ao ser mostrado que as
abundâncias de Ae. albopictus e Ae. aegypti foram relacionadas com o tipo de biótopo, é
admissível acreditar que os fatores utilizados na definição dos biótopos estudados, como
grau de cobertura vegetal e de adensamento residencial, influenciem a distribuição das
espécies. O padrão espacial encontrado foi semelhante ao verificado em outras cidades
brasileiras como Rio de Janeiro e Nova Iguaçú-RJ, que possuem índices de infestação
mais altos em relação às cidades de Mariana e Ouro Preto (Braks et al., 2003; Lima-
Câmara et al., 2006; Honório et al., 2009). Tal distribuição espacial também foi
detectada em João Pessoa-PB (Leandro, 2012) e na Tailândia (Tsuda et al., 2006).
O padrão de ocorrência sugere que Ae. albopictus viva principalmente nas áreas
verdes e residenciais com maior arborização, mas que visite áreas com maior densidade
de casas. Já Ae. aegypti parece viver em uma malha urbana mais densa, bem como em
locais residenciais com menor cobertura vegetal, mas visitando as áreas verdes urbanas.
Admite-se que Ae. albopictus foi originalmente uma espécie que forrageava e se
reproduzia nas bordas das florestas, mas que passou a adaptar-se ao ambiente
doméstico nos vários locais em que ocorre pelo mundo (Lima-Camara et al., 2006).
Presume-se que Ae. albopictus evite áreas urbanizadas, ou que essas áreas acarretem em

67
baixo crescimento populacional e ainda que Ae. aegypti evite áreas rurais, ou que essas
áreas gerem pequeno crescimento de sua população (Braks et al., 2003).
A segregação espacial tem sido usada para tentar explicar a coexistência de Ae.
aegypti e Ae. albopictus em vários regiões (Hawley 1988; Barrera, 1996; Honório &
Lourenço de Oliveira, 2001; Braks et al., 2003). A possibilidade de coexistência destas
espécies ocorre quando o ambiente favorece a fase adult de uma espécie e a fase
aquática da outra (Braks et al., 2003). Além disso, períodos com clima secos e quentes
atenuam a competição interespecífica na fase aquática, devido à maior mortalidade dos
ovos de Ae. albopictus pela dessecação, o que facilita a coexisência local de ambas
espécies, já que favorece Ae. aegypti neste período (Juliano et al., 2002).
Neste trabalho foi observado que Ae. albopictus quando adulto, parece forragear
em todos os habitats, porém com sua reprodução (cujo resultado é a forma imatura) não
ocorreu o mesmo. Sua reprodução no ambiente mais urbanizado pode não ser oportuna,
pois a espécie tem demonstrado preferência por ovipositar em locais onde Ae. aegypti
não o fez (Pedrosa et al dados ainda não publicados). Tal fato sugerem que um ambiente
mais propício para a reprodução de Ae. aegypti, pode não ser conveniente para Ae.
albopictus realizar a oviposição, porém pode ser favorável seu forrageamento.
Apesar de ter sido detectada uma variação na distribuição espacial de Ae.
albopictus e Ae. aegypti, as espécies co-ocorreram nos sítios de oviposição em todos os
biótopos, mas em quantidades diferentes de armadilhas. A co-ocorrência destas espécies
no mesmo criadouro foi descrita em diversos estudos (Schultz,1989; Honório &
Lourenço-de- Oliveira, 2001; Juliano et al., 2004). O mesmo foi detectado ainda por
Braks et al. (2003) sendo relatado um alto e inesperado, nível de co-ocorrência em
habitats urbanos, suburbanos e rurais de cidades brasileiras e do EUA. No trabalho de
Braks et al. (2003) a co-ocorrência diferiu entre os países estudados, sendo habitat-
dependente no estado do Rio de Janeiro, Brasil, e cidade-dependente na Flórida, Estados
Unidos. Considerando as cidades do Rio de Janeiro e Nova Iguaçú onde o trabalho foi
realizado, as duas espécies co-ocorreram em 61% das ovitrampas. No presente estudo,
Ae. albopictus e Ae. aegypti co-ocorreram nas armadilhas de s e principalmente nas
armadilhas de imaturos. A coocorrência foi evidenciada em 20,6% e 17,8% das
ovitrampas em Mariana e Ouro Preto, respectivamente. Estes valores foram inferiores à
outras observações, mas poucos estudos tem relatado altas co-ocorrências destas duas
espécies no mesmo criadouro (Schultz, 1989; Braks et al.,2003). É plausível considerar

68
que a baixa infestação das cidades estudadas possa acarretar em um maior número de
criadouros disponíveis e portanto, na diminuição da co-ocorrência das espécies.
Contudo, a menor co-ocorrência nos criadouros enfatiza a existência de uma variação da
distribuição espacial destas espécies no estudo.
Estudos em campo e laboratório mostraram que na competição larval (em um
mesmo criadouro) entre Ae. aegypti e Ae. albopictus, a segunda espécie exibiu maior
capacidade competitiva (Barrera, 1996; Juliano, 1998; Braks et al., 2004). No Brasil, a
mesma espécie também revelou vantagem competitiva em experimento realizado em
campo no Rio de Janeiro (Braks et al, 2003). Mas Serpa et al. (2008), mostraram que
Ae. aegypti apresentou maior capacidade de competição quando em densidade larvária
intermediária, apesar de em alta densidade larvária a maior sobrevivência ter sido de Ae.
albopictus. Aedes aegypti tem demonstrado ser mais afetado pela competição
intraespecífica do que pela competição interespecífica com Ae. albopictus (Sucharit e et
al.,1980; Serpa et al., 2008).
A incidência do dengue está fortemente relacionada com as estações chuvosas,
altas temperaturas, vento e altitude (Donalísio & Glasser, 2002). Além disso, a
pluviosidade pode influenciar no aumento populacional dos mosquitos estudados já que
a precipitação possibilita a eclosão dos ovos e a formação de novos criadouros. Neste
trabalho, a abundância de Ae. aegypti não apresentou relação com nenhum dos fatores
abióticos analisados, diferente do que foi encontrado por Honório et al. (2009), que
verificou correlação tanto com a precipitação quanto com a temperatura (atrasada em
uma semana) e ainda, por Serpa et al. (2006), em que a espécie revelou associação
positiva com a última variável. A temperatura, quando abaixo do limiar (22°C- 24°C)
parece contribuir para o declínio da população de mosquitos Ae. aegypti (Honório et al.
2009).
A abundância de Ae. albopictus também não se correlacionou com a pluviosidade,
ao contrário do encontrado por Serpa et al. (2006) e em trabalho realizado nas mesmas
cidades em anos anteriores (Pedrosa et al. dados ainda não publicados). Entretanto, neste
último, as coletas ocorreram exclusivamente em áreas verdes, o que pode explicar a
dependência da chuva para preencher os criadouros. Esta espécie apresentou relação de
sua abundância com a temperatura, corroborando com estudo realizado em campo (Serpa
et al., 2006) e laboratório (Calado & Navarro-Silva, 2002). Calado & Navarro-Silva
(2002) mostraram que a temperatura influenciou as atividades de hematofagia e

69
oviposição de Ae. albopictus em laboratório, bem como a velocidade de desenvolvimento
das formas imaturas que se revelou inversamente proporcional à temperatura. Os mesmos
autores verificaram que as baixas temperaturas parecem atuar como fator limitante ao
crescimento populacional desta espécie sob condições de laboratório. Deste modo, a
ocorrência frequente de temperaturas abaixo deste limiar, pode ser um fator limitante à
distribuição espacial e temporal das populações de Ae. albopictus (Calado & Navarro-
Silva 2002). É possível que os fatores abióticos contribuam para a segregação de habitats
(Julliano et al., 2002; Lounibos et al. 2002; Braks et al., 2004) já que as áreas urbanas
podem ter temperatura mais elevada que as áreas periféricas (McIntyre, 2000; Braks et al.
2004).
As medidas de abundância das duas espécies são importantes para a eclucidação de
mecanismos relacionados com a distribuição espacial, que por sua vez, são úteis para as
práticas de controle dos vetores (Braks et al., 2003). De forma complementar, a
compreensão dos padrões de ocorrência das espécies em locais com baixa infestação dos
Aedes e com provável invasão recente, contribui para o esclarecimento do cenário do
estabelecimento do vetor em um novo local, colaborando com a elaboração de estratégias
que contenham a abundância e a dispersão dos vetores do dengue.
AGRADECIMENTOS
Aos moradores de Mariana e Ouro Preto que permitiram o acesso às suas residências. Às
estagiárias Laís Sato, Luna Ferreira, Márcia Regina F. Luzia e Ana Cláudia A. Carneiro e
à MSc Adriele Magalhães pela apoio em campo. À Guilherme de P. Costa pela ajuda em
laboratório. Ao Setor de Zoonoses de ambas cidades. À Secretaria de Turismo e Defesa
Civil de Ouro e à Vale/Mariana. Ao LEEIDS e Lab. de Doenças de Chagas da UFOP,
LABEq da UFMG. À ECOVEC. À Dra. Alzira Cecílio e Dr. Sérgio Caldas da FUNED.
À CAPES pelo apoio financeiro. SPR é bolsista pesquisador do CNPq.

70
REFERÊNCIAS BIBLIOGRÁFICAS
Alencar CHM 2008. Infestação pelo Aedes albopictus (Skuse), em criadouros naturais e
artificiais encontrados em áreas verdes na cidade de Fortaleza-CE5. Dissertação de
mestrado. Universidade Federal do Ceará.
Amaral AC 2009. Ecologia histórico - botânica de um quintal e aspectos histórico-sócio-
ambientais de espaços verdes no centro histórico de Ouro Preto – MG. Monografia,
Departamento de Biodiversidade, Evolução e Meio Ambiente, ICEB, Universidade
Federal de Ouro Preto.
Barcelos C,Monteiro AMV, Corvalán C, Gurgel HC, Carvalho MS, Artaxo P, Hacon S,
Ragoni V 2009. Mudanças climáticas e ambientais e as doenças infecciosas: cenários e
incertezas para o Brasil* Epidemiol. Serv. Saúde, Brasília 18(3):285-304.
Barrera R 1996. Competition and resistance to starvation in larvae of container-
inhabiting Aedes mosquitoes. Ecol Entomol 21:117-27
Black WC, Rai KS, Turco BJ, Arroyo DC 1989. Laboratory study of competition
between United States strains of Aedes albopictus and Aedesaegypti (Diptera: Culicidae).
J. Med. Entomol. 26: 260-271.
Borges SMAA 2001. Importância epidemiológica do Aedes albopictus nas américas.
Dissertação de Mestrado, Departamento de Epidemiologia da Faculdade de Saúde
Pública, Universidade de São Paulo.
Braks MAH, Honório NA, Oliveira LR, Juliano SA, Lounibos LP 2003. Convergent
habitat segregation of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in
southeastern Brazil and Florida. J Med Entomol 40:785-94.
Braks MAH, Honório NA, Lounibos LP, Lourenço-de-Oliveira R, Juliano AS 2004.
Interespecific competition between two invasive species of container mosquitoes, Aedes
aegypti and Aedes albopictus (Diptera: Cullicidae), in Brasil. Annuals of the
Entomological Society of America 97:130-139.
Calado DC, Navarro-Silva MA. 2002. Exigências térmicas de Aedes (Stegomyia)
albopictus Skuse, 1894 (Diptera, Culicidae) em condições de laboratório. Revista
Brasileira de Entomologia. 46: 547-551
Câmara FP, Gomes AF, Santos GT, Câmara DCP 2009. Clima e epidemias de dengue no
estado do Rio de Janeiro. Revista da Sociedade Brasileira de Medicina Tropical. 42:137-
140.
Carbajo AE, Curto SI, Schweigmann NJ 2006. Spatial distribution pattern of oviposition
in the mosquito Aedes aegypti in relation to urbanization in Buenos Aires: Southern
fringe bionomics of an introduced vector. Medical and Veterinary Entomology 20, 209–
218.
Catão RF, Guimarães RB 2009. Espaço e Dengue: Uma análise miltoniana em geografia
da saúde. 12O Encuentro de Geógrafos de América Latina, Montevideo,
Uruguay.Availablefrom:http://egal2009.easyplanners.info/area07/7095_Castro_Catao_Ra
fael.pdf

71
Central Disease Control (CDC) 2000. http://www.cdc.gov/ncidod/dvbid/arbor/arbdet.htm
Acessado em 23/11/2012.
Central Disease Control (CDC) 2005. http://www.cdc.gov/ncidod/dvbid/arbor/arbdet.htm
Acessado em 05/05/2013.
Chadee DD 2010. The diel oviposition periodicity of Aedes aegypti (L.) (Diptera:
Culicidae) in Trinidade, West Indies: effects of forced egg retention. Bulletin of
Entomological Research 100: 599-603.
Christophers R 1960. Aedes aegypti (L.). The yellow fever mosquito. Cambridge Univ.
Press. Cambridge.
Codeço C, Ribeiro K, Figueiró R, Honório N 2010. Interações ecológicas e doenças
transmissíveis. Oecolologia. Australis, 14(3): 588-590.
Consoli RAGB, Lourenço-de-Oliveira R 1994. Principais mosquitos de importância
sanitária no Brasil. Rio de Janeiro: Editora Fiocruz. 225p.
Consoli RAGB, Oliveira LR 1998. Principais mosquitos de importância sanitária no
Brasil. Rio de Janeiro: Fiocruz, 228p.
Correa PRL, França E, Bagutchi TF 2005. Infestação pelo Aedes aegypti e ocorrência da
dengue em Belo Horizonte, Minas Gerais. Revista de Saúde Pública, São Paulo vol.39.
Cortez H 2004 . Série: Consciência e Meio Ambiente/Tema: Aquecimento Global e
Água.
Costa AC da, Santos IGC dos, Barbosa MG da 2009. Detecção e tipagem de vírus
dengue em Aedes aegyptiv(Diptera: Culicidae) na Cidade de Manaus, Estado do
Amazonas. Revista da Sociedade Brasileira de Medicina Tropical, 42(6):677-681.
Costa FS, Silva JJ, Souza CM, Mendes J 2008. Dinâmica populacional de Aedes aegypti
(L) em área urbana de alta incidência de dengue. Revista da Sociedade Brasileira de
Medicina Tropical 41:309-312.
Crovello TJH Hacker CS 1972. Evolutionary strategies in life table characteristics among
feral and urban strains of Aedes aegypti (L.). Evolution V.26, P.185-96
Donalísio MR, Glasser CM 2002. Vigilância entomológica e controle de vetores do
dengue. Rev. Brasileira de Epidemiologia 5:259-272.
Effler PV, Pang L, Kitsutani P,Vorndam V, Nakata M, Ayers T, Elm J,Tom T,Reiter P,
Rigau-Perez JG, Hayes JM, Mills K, Napier M, Clark CG, Gubler DJ 2005. Dengue
Fever, Hawaii, 2001–2002. Emerging Infectious Diseases 11(5) 742-749.
Eiras AE 2005. Culicidae. In Parasitologia Humana, Neves DP, Melo AL de, Linardi PM
& et al, 1nd ed., Editora Atheneu.pp. 355-36
Epstein PR, Diaz HF, Elias S, Grabherr G, Graham NE, Martens EJM, Thompson EM,
Susskind J 1998. Biological and physical signs of climate change: Focus on mosquito-
born diseases. Bull. Am. Meteorol. Soc. 79(3), 409–417.

72
Fantinatti ECS, Duqye JEL, Silva AM, Navarro-Silva M 2007. Abundância e agregação
de ovos de Aedes aegypti (Linnaeus) e Aedes albopictus (Skuse) (Diptera: Culicidae) no
norte e noroeste do Paraná. Neotropical Entomology 36(6).
Ferreira AC, Neto FC 2007. Infestação de área urbana por Aedes aegypti e relação com
níveis socioeconômicos. Rev Saúde Pública 2007;41(6):915-22.
Ferreira CP, Yang, H.M., 2003. Estudo Dinâmico da População de Mosquito Aedes
Forattini OP 1992. Ecologia, epidemiologia e sociedade. São Paulo: Ed. Artes Médicas,
EDUSP.
Focks DA 2003. A review of entomological sampling methods and indicators for
dengue vectors. WHO, Genebra, 40p.
Forattini OP 1986. Identification of Aedes (Stegomyia) albopictus (Skuse) in Brazil.
Revista de Saúde Pública, São Paulo, 20:244-245.
Forattini OP 1992. Ecologia, Epidemiologia e Sociedade. São Paulo: Ed. Artes Médicas,
EDUSP.
Forattini OP 1996. Culicidologia Médica, Vol 1: Princípios gerais, morfologia,
glossário taxonômico. São Paulo: Editora da Universidade De São Paulo. 546p.
Forattini OP, Marques GRAM, Kakitani I, Brito M de, Sallum MAM 1998. Significado
epidemiológico dos criadouros de Aedes albopictus em bromélias. Rev. Saúde Pública
32(2):186-8.
Forattini OP 2002. Culicidologia Médica. Identificação, Biologia e Epidemiologia. São
Paulo: Editora da Universidade de São Paulo. Vol 2. 860p.
Frankie GW, Ehler LE 1978. Ecology of insects in urban environments. Ann Revista
Entomológica; 23:367-87.
Gadelha DP, Toda AT 1985. Biologia e comportamento do Aedes aegypti. Revista
Brasileira de Malariologia e Doenças Tropicais. 37: 29-36.
Glasser CM & Gomes AC 2002. Clima e sobreposição da distribuição de Aedes aegypti
e Aedes albopictus na infestação do estado de São Paulo. Rev Saúde Pública 36:166-
72.
Gomes AC 1998. Medidas dos níveis de infestação urbana para Aedes (Stegomyia)
aegypti e Aedes (Stegomyia) albopictus. Informe Epidemiológico do SUS 7(3):49-57.
Gomes AC 2002. Informe Epidemiológico do SUS 2002 -Vigilância Entomológica
11(2):79 – 90.
Gomes A dos S, Sciavico CJ de S, Eiras AE 2006. Periodicidade de oviposição de fêmeas
de Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae) em laboratório e campo. Revista
da Sociedade Brasileira de Medicina Tropical, vol. 39 (4): 327-332.
Gómez-Dantés H, Willoquet JR 2009. Dengue in the Americas: challenges for prevention
and control Cad. Saúde Pública, Rio de Janeiro, 25 Sup 1:S19-S31.

73
Gratz NG 2004. Critical review of the vector status of Aedes albopictus. Medical and
Veterinary Entomology ,18:3, 215–227.
Gubler 2004. The changing epidemiology of yellow fever and dengue, 1900 to 2003: full
circle?
Gubler DJ, 2004. Comp. Immun. Microbiol. Infect. Dis. 27 (2004) 319–330
Gubler DJ 1998. Dengue and Dengue Hemorrhagic Fever: Its history and resurgence as a
global public health problem. In: Gubler D.J. & Kuno G. Dengue and dengue
hemorrhagic fever. New York: Cab International. Clinical Microbiology Reviews 11
(3):480-496.
Guimaraes AE, Gentile C, Lopes CM, Sant’anna A, Jovita AM 2000. Ecologia de
mosquito00s (Diptera: Culicidae) em áreas do Parque Nacional da Serra da Bocaina,
Brasil. I - Distribuição por habitat. Revista de Saúde Pública, São Paulo 34 (3): 243-50.
Guzman MG, Kouri G 2003. Dengue and dengue hemorrhagic fever in the Americas:
Lessons and Challenges. Journal of Clinical Virology 27: 1-13.
Guzman A, Isturiz RE 2010. Update on the global spread of dengue. Int. J. Antimicrob.
Agents, v. 36, n. l, p. 40-42.
Halstead SB 2008. Dengue Virus–Mosquito Interactions. Annu. Rev. Entomol,
53:273–91.
Harrington LC, Scott TW, Lerdthusnee K, Coleman RC, Costero A, Clark GG, Jones JJ,
Kitthawee S, Kittayapong P, Sithiprasasna R, Edman JD 2005. Dispersal of the dengue
vector Aedes aegypti within and between rural communities. The American Society of
Tropical Medicine and Hygiene Trop. Am. J. Med. Hyg., 72(2) pp. 209–220.
Harrison BA, White PB, Powell EE, Hickman JR 1998. North Carolina mosquito records.
Uncommon Aedes and Anopheles (Diptera: Culicidae) Journal of the American Mosquito
Control Association, I4(2):165-172, 1998.
Hawley WA 1988. The biology of Aedes albopictus. Journal of the American Mosquito
Control Association. 4 (Suplemento 1):2-39.
Herrera BE, Prevots DR, Sarate ML, Silva I, Sepúlveda AJ 1992. First reported of
classical dengue fever at 1700 meters above the sea in Guerrero State México. The
American Journal Of Tropical Medicine And Hygiene, 46(6): 649-53.
Ho BC, Ewert A, Chow LM 1989. Interspecific competition among Aedes aegypti, Ae.
albopictus, and Ae. triseriatus (Diptera: Culicidae): larval development in mixed cultures.
J. Med. Entomol26: 615-623
Honório NA, Lourenço-de-Oliveira R 2001. Frequência de larvas e pupas de Aedes
aegypti e Aedes albopictus em armadilhas, Brasil. Rev. Saúde Pública. 34: 385- 391.
Honório NA, Silva WC, Leite PJ, Gonçalves JM, Lounibos LP, Lourenço-de-Oliveira R
2003. Dispersal of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in an urban

74
endemic dengue area in the state of Rio de Janeiro, Brazil. Mem Inst Oswaldo Cruz, Rio
de Janeiro, Vol. 98(2): 191-198.
Honório NA, Codeço CT, Alves FC, Magalhães MAFM, Lourenço-de-Oliveira R 2009a.
Temporal distribution of Aedes aegypti in diferente districts of Rio de Janeiro, Brazil,
measured by two types of traps. J Med Entomol (in press).
Honório NA, Castro MG, Barros FSM, Magalhães MAFM, Sabroza PC 2009b. The
spatial distribution of Aedes aegypti and Aedes albopictus in a transition zone, Rio de
Janeiro, Brazil. Cad. Saúde Púlica, Rio de Janeiro, 25(6):1203-1214.
Honório NA, Lourenço-de-Oliveira R 2001. Frequência de larvas e pupas de Aedes
aegypti e Aedes albopictus em armadilhas, Brasil. Rev. Saúde Pública. 34: 385- 391.
Juliano SA 1998. Species introduction and replacement among mosquitoes:
interspecific resource competition or apparent competition. Ecology 79:255–268
Juliano SA, O’Meara GF, Morrill JR, Cutwa MM 2002. Desiccation and thermal
tolerance of eggs and the coexistence of competing mosquitoes. Oecologia (Heidelbg)
130: 458-469.
Juliano SA, Lounibos LP, O’Meara GF 2004. A field test for competitive effects of
Aedes albopictus on Aedes aegypti in south Florida: Differences between sites of
coexistence and exclusion? Oecologia 139:583-593.
Juliano SA. Species introduction and replacement among mosquitoes: interspecific
resource competition or apparent competition? Ecology 1998; 79:255-68.
Leandro, RS 2012. Competição e dispersão de Aedes (Stegomyia) aegypti (Linnaeus,
1762) e Aedes (Stegomyia) albopictus (Skuse, 1894) (DIPTERA: CULICIDAE) em áreas
de ocorrência no município de João Pessoa- PB. Dissertação de Mestrado, Universidade
Estadual da Paraíba.
Lima-Camara TN, Honório NA, Lourenço-de-Oliveira R 2006. Freqüência e distribuição
espacial de Aedes aegypti e Aedes albopictus (Diptera: Culicidae) no Rio de Janeiro,
Brasil. Cadernos de saúde pública, 22: 2079-2084.
Lima-Camara TN 2010 Activity patterns of Aedes aegypti and Aedes albopictus
(DIPTERA: CULICIDAE) under natural and artificial conditions. Oecologia Australis
14(3): 737-744.
Loboda CR, Aneglis, BLD de 2005. Áreas Verdes Públicas Urbanas: Conceitos, Usos E
Funções.
Lounibos LP 2002. Invasions by insect vectors of human disease. Annu. Rev. Entomol.
47: 233 - 266.
Lourenço-de-Oliveira R, Castro MG, Braks MA, Lounibos LP 2004. The invasion of
urban forest by dengueNvectors in Rio de Janeiro. J. Vector Ecol. 29: 94-100.

75
Maciel I 1999. Avaliação epidemiológica do dengue no município de Goiânia no Período
de 1994 a 1997. Mestrado. Instituto de Patologia Tropical e Saúde Pública. Universidade
Federal de Goiás: Goiânia.119 pp.
Maciel IJ, Júnior JBS, Martelli CM 2008. Epidemiologia e desafios no controle do
dengue. Revista de Patologia Tropical. 37: 111-118
Maciel-de-Freitas R, Brocki Neto R, Gonçalves JM, Codeço CT, Lourenço-de-Oliveira R
2006. Movement of Dengue Vectors Between the Human Modified Environment
and an Urban Forest in Rio de Janeiro. J. Med. Entomol. 43(6), 1112-1120.
Maciel-de-Freitas R, Eiras AE, Lourenço-de-Oliveira R 2006. Field evaluation of
effectiveness of the BG-Sentinel, a new trap for capturing adult Aedes aegypti
(Diptera:Culicidae). Mem Inst Oswaldo Cruz, p. 321-25.
Madeira NG, Macharelli CA, Carvalho LR 2002. Variation of the Oviposition
Preferences of Aedes aegypti in Function of Substratum and Humidity. Mem. Inst.
Oswaldo Cruz Vol. 97(3): 415-420.
Mahmood S 2006. Dengue: an Epidemic is larged a failure in public health
administration! The role of 23. Dhaka City Corporation, DCC of Bangladesh. World
Health & Population. http://www.longwoods.com/product.php?productid=17900
acessado em 12/08/2012.
Marcondes CB, Tauil PL 2011. Dengue silvestre: devemos nos preocupar? Revista da
Sociedade Brasileira de Medicina Tropical 44(2):263-264.
Marques GRAM, Serpa LLN, Brito M 2009. Chave de identificação de gêneros de
culicídeos. Laboratório de Culicídes- SUCEN-Taubaté-SP.
Marques GRAM, Santos RC, Forattini OP 2001. Aedes albopictus em bromélias de
ambiente antrópico no estado de São Paulo, Brasil. Revista de Saúde Pública, São Paulo,
35: 243-248.
Martins VEP , Alencar CHM, Facó PEG, Dutra RF, Alves CR , Pontes RJS, Guedes MIF
2010. Distribuição espacial e características dos criadouros de Aedes albopictus e Aedes
aegypti em Fortaleza, estado do Ceará. Revista da Sociedade Brasileira de Medicina
Tropical 43(1):73-77.
Martins VEP, Alencar CH, Kamimura MT, Araújo FMC de, Simone SG de, Fireman R,
Izabel DM, Guedes F. 2012. Occurrence of Natural Vertical Transmission of Dengue-2
and Dengue-3 Viruses in Aedes aegypti and Aedes albopictus in Fortaleza, Ceará, Brazil.
Plos One. 7: 1-9.
Massad E, Brattini MN, Coutinho FAB, Lopez LF 2003.Dengue e risco da reintrodução
da febre amarela urbana no Estado de São Paulo. Rev Saúde Pública 37(4):477-84
McIntyre, N E 2000. Ecology of urban arthropods: a reviewand a call to action.
Ann.Entomol. Soc. Am. 93: 825-835.

76
McMichael AJ, Woodruff RE, Hales S. 2006 Climate Change And Human Health:
Present And Future Risks. The Lancet; 367:859-869.
Mendonça F 2003. Aquecimento global e saúde: uma perspectiva geográfica– notas
introdutórias Francisco Mendonça. Terra Livre São Paulo. Ano19 - vol. I - n. 20205-221
Ministério da Saúde (MS) 2006. Abordagens Espaciais na Saúde Pública, Brasília.
Fundação Oswaldo Cruz, volume 1.
Ministério da Saúde (MS) 2007. Introdução à Estatística Espacial para a Saúde
Pública,mBrasília. Fundação Oswaldo Cruz, volume 3.
Ministério da Saúde (M.S.) 2010a. Isolamento do sorotipo DENV 4 em Roraima /
Brasil. Nota técnica CGPNCD/DEVEP/SVS. Brasília. Available from
:http://www.saude.gov.br 20/12/ 2012.
Ministério da Saúde (M.S.) 2010b. Identificação de Áreas de Maior Vulnerabilidade para
Ocorrência de Dengue no período de Transmissão 2010/2011 com vistas a subsidiar a
intensificação das ações de controle. Nota técnica N.º 118 / 2010
CGPNCD/DEVEP/SVS/MS Fonte: Ministério da Saúde
http://portal.saude.gov.br/portal/arquivos/pdf/nt_aval_vul_epid_dengue_verao_10_11.pdf
Modificado em 26/08/2012.
Mortality and Morbidity Weekly Reports (MMWR) 2002.Imported Dengue United
States, 1999 and 2000. 51, 281–283.
Mogi M, Choochote W, Khamboonruang C, Suwanpanit P 1990. Applicability of
presence-absence and sequential sampling for ovitrap surveillance of Aedes (Diptera:
Culicidae) in Chiang Mai, northern Thailand. J Med Entomol 27:509-514.
Moore CG 1999. Aedes albopictus in the United States: current status and prospects for
further spread. J Am Mosq Control Assoc 15:221-227.
Morais, MM 2011. A proporção de Mata Atlântica remanescente no entorno de áreas
urbanas relaciona-se à incidência de dengue? Dissertação de Mestrado, Universidade
Federal de Minas Gerais.
Muir E, Kay BH 1998. Aedes aegypti Survival and dispersal estimated by mark-
release-recapture in northern Australia. Am. J. Trop. Med. Hyg. 58, 277-282.
Natal D 2002. Bioecologia do Aedes aegypti. Biológico, São Paulo, 64(2):205-207.
Navarro-Silva MA, Barbosa AA, Calado D 2004. Atividade de Mansônia Spp.
(Mansoniini, Culicidae) em fragmento florestal na área urbana de Curitiba, Paraná,
Brasil. Revista Brasileira de Zoologia 21(2): 24-247.
O’Meara GF, Gettman AD, Evans Jr., Curtis GA 1993. The spread of Aedes albopictus in
Flórida. reprinted from american entomologist, 39:3.

77
O’Meara, GF, Evans LF, Gettman AD, Cuda JP 1995. Spread of Aedes albopictus and
decline of Aedes aegypti (Diptera: Culicidae) in Florida. J. Med. Entomol. 32: 554-562.
Osanai CH, Travassos-da-Rosa APA, Amaral S, Passos ACD, Tauil PL 1983. Surto de
dengue em Boa Vista, Roraima. Revista do Instituto de Medicina Tropical de São Paulo,
1:53- 54.
Ozório TF 2000. Potencial de uso da serapileira na recueração de áreas degradas por
mineraçaõ de ferro, Mariana-MG. 62f. (Mestrado em Ciência Florestal) Universidade
Federal de Viçosa - MG.
Paupy C, Delatte H, Bagny L, Corbel V, Fontenille D 2009. Aedes albopictus, an
arbovirus vector: From the darkness to the light. Microbes and Infection, 11 1177- 1185.
Pereira MFA, Boiça Jr. AL, Barbosa J.C 2004. Distribuição espacial de Bemisia tabaci
(Genn.) Biótipo B (Hemiptera: Aleyrodidae) em feijoeiro (Phaseolus vulgaris L.).
Neotrop. Entomol. 33: 493-498.
Petschow LA, Zequi JAC (2008). Programa de vigilância de insetos vetores:
levantamento de imaturos de mosquito (Diptera: Culicidae) em fragmento florestal da
área periurbana, Cornélio Procópio, Paraná. II Encontro de Extensão da UniFil,
Filadélfia, Paraná.
Pinheiro F, Nelson M 1997. Re-emergence of dengue and emergence of dengue
haemorrhagic fever in the Americas. Dengue Bulletin 21: 16-24
Pontes RJS, Ruffino-Netto A 1994. Dengue em localidade urbana da região sudeste do
Brasil: Aspectos Epidemiológicos. Revista de Saúde Pública, São Paulo 28(3).
Rabinovich, J.E. 1980. Introducción a la ecología de poblaciones animales. México,
Continental, 313p.
Rai, KS 1986. Genetics Of Aedes Albopictus. J Am Mosq Control Assoc, 2(4): 429-36
Reiter P, Amador MA, Anderson RA, Clark GG 1995. Short report: Dispersal of Aedes
aegypti in a urban area after blood feeding as demonstrated by rubidium-marked eggs.
Am J Trop Med Hyg 52: 177-179.
Resende MC, Eiras AE 2009. Estudo multicêntrico do uso da armadilha MosquiTRAP
para captura de Aedes aegypti e geração de índices de vigilância entomológica.
Dissertação de Mestrado, Universidade Federal de Minas Gerais.
Resende MC, Silva IM, Eiras AE 2010. Avaliação da operacionalidade da armadilha
mosquitrap no monitoramento de Aedes aegypti. Epidemiol. Serv. Saúde, Brasília,
19(4):329-338.
Russell RC, Webb CE, Williams CR, Ritchie SA 2005. Medical and veterinary
entomology mark–release–recapture study to measure dispersal of the mosquito Aedes
aegypti in Cairns, Queensland, Australia. Volume 19, Issue 4, pages 451–457.
San Martín JBD. La estrategia de gestión integrada para la prevención y el control del
dengue en la región de las Américas. Rev Panam Salud Publica 21: 55-63, 2007.

78
Santos M 1999. A Natureza do Espaço: técnica e tempo: razão e emoção. São Paulo:
Hucitec.
Santos RC 2003. Atualização da distribuição de Aedes albopictus no Brasil (1997-2002).
Revista de Saúde Pública, São Paulo, 37(5):671-673.
Schultz GW 1989. Cemetery vase breeding of dengue vectors in Manila, Republic of the
Philippines. J. Am. Mosq. Control Assoc. 5: 508-513.
Secretaria de Vigilância em Saúde 2004. Situação da prevenção e controle das doenças
transmissíveis no Brasil: Uma análise da situação de saúde, SAÚDE BRASIL. Available
from: http://portal.saude.gov.br/portal/arquivos/pdf/capitulo6_sb.pdf 17/10/2012
Serpa LLN, Costa KVRM, Kakitani I, Voltolini JC 2006.Variação Sazonal de Aedes
aegypti e Aedes albopictus no município de Potim, São Paulo. Revista de Saúde Pública
Revista de Saúde Pública 40:1101-1105.
Serpa LLM, Kakitani I, Voltolini JC 2008. Competição entre larvas de Aedes aegypti e
Aedes albopictus em laboratório. Revista da Sociedade Brasileira de Medicina Tropical
41(5):479-484.
Serufo JC, Oca HM, Tavares VA, Souza AM, Rosa RV, Jamal MC 1993. Isolation of
dengue type 1 from larvae of Aedes albopictus in the Campos Altos city, state of Minas
Gerais, Brazil. Memórias Do Instituto Oswaldo Cruz; 88: 503-504.
Sucharit S, Watanasak T, Vutikes S, Viraboonchai S 1978. Interactions Between Larvae of
Aedes aegypti and Aedes albopictus in mixed experimental populations. 9(1) 93-97
Silva MAN, Lozovei AL 1998. Mosquitos (Diptera, Culicidae) capturados com isca
humana em área preservada de Curitiba, Paraná. Revista Brasileira de Zoologia, Curitiba,
15: 965-976.
Siqueira Júnior JB, Martelli CM, Coelho GE, Simplicio AC, Hatch DL 2005. Dengue and
dengue hemorrhagic fever, Brazil, 1981-2002. Emerg Infect Dis 11: 48-53.
Taipe-Lagos CB, Natal D 2003. Abundância de Culicídeos em área metropolitana
preservada e suas implicações epidemiológicas. Revista de Saúde Pública. São Paulo 37(3)
275-279.
Tauil PL 2001. Urbanização e ecologia do dengue. Caderno de Saúde Pública
17(Suplemento): 99-102.
Tsuda Y, Suwonkerd W, Chawprom S, Prajakwong S, Takagi M 2006. Different spatial
distribution of Aedes aegypti and Aedes albopictus along an urban–rural gradient and the
relating environmental factors examined in three villages in northern thailand. Journal of
the American Mosquito Control Association 22(2):222-228.
Tauil PL 2002. Aspectos críticos do controle do dengue no Brasil. Caderno de Saúde
Pública, Rio de Janeiro 18(3):867-871.

79
Taveira LA, Fontes LR, Natal D 2001. Manual de diretrizes e procedimentos no controle
do Aedes aegypti. Ribeirão Preto: Prefeitura Municipal de Ribeirão Preto. Availablefrom:
http://www.ribeiraopreto.sp.gov.br/ssaude/vigilancia/vigep/dengue/vetor-1.pdf acesso em
19/10/2012.
Teixeira MG, Costa MCN, Barreto ML, Barreto FR 2001. Epidemiologia do dengue em
Salvador-Bahia, 1995-1999. Revista da Sociedade Brasileira de Medicina Tropical 34(3):
269-274.
Thoisy B, Dussart P, Kazanji M 2004.Wild terrestrial rainforest mammals as potential
reservoirs for flaviviruses (yellow fever, dengue 2 and St Louis encephalitis viruses) in
French Guiana. Trans R Soc Trop Med Hyg 98:409-412.
Tissot AC, Navarro-Silva MA 2004. Preferência por hospedeiro e estratificação de
Culicidae (Díptera) em área de remanescente florestal do Parque Ecológico do Iguaçu,
Curitiba Paraná, Brasil. Revista Brasileira de Zoologia 21 (4): 877–886.
Townsend CR, Begon M, Harper JL 2006. Fundamentos em Ecologia.2nd ed, Artmed, pp
239- 247
Urbinatti PR 2004. Observações ecológicas de Aedes albopictus (Diptera: Culicidae) em
áreas de proteção ambiental e urbana da periferia na grande São Paulo. Tese de Doutorado,
Departamento de Epidemiologia, Unidade Faculdade de Saúde Pública (FSP),
Universidade de São Paulo.
Vasconcelos PR, Rosa APAT, Pinheiro FP, Rodrigues SG, Rosa EST, Cruz ACR, Rosa
JFST 1999. Aedes aegypti, Dengue and re-urbanization of yellow fever in Brazil and
other South American countries - past and present situation and future perspectives.
Dengue Bulletin 23: 55-66.
Werneck MS, Pedralli G, Koenig R, Gieseke LF 2000. Florística e estrutura de três trechos
de uma floresta semidecídua na Estação Ecológica do Tripuí, Ouro Preto, MG. Revista
Brasileira de Botânica 23:97-106.
World Health Organization (WHO) 2010. Health topics – Dengue. Available from:
http://www.who.int/topics/dengue/en/ 17/10/2012
World Health Organization 2012. Dengue y dengue hemorrágico. Nota descriptiva N° 117
ttp://www.who.int/mediacentre/factsheets/fs117/es/ acessado em 28/04/2013.
Zequi JAC, Lopes J, Medri IM 2005. Imaturos de Culicidae encontrados em recipientes
instalados em mata residual no município de Londrina, Paraná, Brasil. Revista Brasileira
de Zoologia 22(3): 656–661.

80
APÊNDICE
No território brasileiro existem apenas dois registros de infeccção natural com
vírus dengue em Aedes albopictus (Serufo et al. 1993; Martins et al. 2012). Além disso,
ainda não há um consenso sobre a densidade mínima de Ae. aegypti que permita a
transmissão do vírus (Halstead, 2008), o que torna importante o conhecimento antecipado
dos locais com circulação de mosquitos infectados. Tal informação pode contribuir para
estratégias mais adequadas de controle e prevenção da doença (Costa et al., 2009). Neste
contexto, buscou-se complementar o estudo, verificando se há presença do vírus nos
vetores de Aedes coletados que foram coletados.
Inquérito Virológico (PCR em Tempo Real)
Todos os mosquitos e larvas coletados no ano de 2011, na primeira fase (estação
chuvosa) e na segunda fase (estação seca) deste estudo, foram armazenados em freezer a -
80 °C e posteriormente encaminhados à Fundação Ezequiel Dias (FUNED) para a
triagem molecular das amostras positivas para o Dengue virus (DENV). Ao todo, foram
coletados 182 mosquitos e 46 larvas, os quais foram agrupados em pools de acordo com a
localidade de captura. Cada pool continha entre 1 e 3 mosquito e entre 5 e 10 larvas.
As amostras foram transportadas, em gelo seco, para o Serviço de Biotecnologia e
Saúde (SBS) da FUNED, onde foram, inicialmente, submetidas à extração de RNA pelo
método do Trizol de acordo com as recomendações do fabricante. O Trizol LS Reagent
(Invitrogen) utilizado é composto por uma solução monofásica de fenol e isotiocianato de
guanidina. Durante a homogeneização ou lise da amostra, a integridade do RNA foi
mantida enquanto as células foram rompidas e os componentes celulares dissolvidos. A
adição de clorofórmio, seguida por centrifugação, permitiu a separação da solução em
uma fase orgânica e uma fase aquosa contendo o RNA, o qual foi recuperado por
precipitação induzida pelo isopropanol. Durante a extração de RNA, foram utilizados
controle negativo (um Ae. aegypti sabidamente não infectado pelo DENV) e controle
positivo (um Ae. aegypti acrescido de 5 µL de 1x104 PFU/mL do DENV-2).
Para a síntese de cDNA, foi utilizado o High-Capacity cDNA Reverse
Transcription Kit (Applied Biosystems). As reações continham 2,0 µL de 10X RT
Buffer, 0,8 µL de 25X dNTP Mix (100 mM), 2,0 µL de 10X RT Random Primers, 1,0 µL
de MultiScribe™Reverse Transcriptase, 1,0 µL de RNase Inhibitor, 3,2 µL de Nuclease-

81
free H2O e 10 µL de RNA. As reações foram levadas ao termociclador (Veriti 96 Well
Thermal Cycler, Applied Biosystems), e submetidas às temperaturas de: 25 °C por 10
minutos, 37 °C por 120 minutos e 85 °C por 5 minutos, seguido de resfriamento a 4 oC e
acondicionamento a -20 oC até serem requeridas para a PCR em tempo real.
Após a realização da Transcrição Reversa, foi realizado o PCR em tempo real
para triagem das amostras positivas para DENV. A reação de PCR continha 2 μL de
cDNA, 0,50 μM de oligonucleotídeos iniciadores específicos para a região 5’UTR de
DENV (5’UTR-S: 5’-AGTTGTTAGTCTACGTGGACCGA-3’ e 5’UTR-C: 5’-
CGCGTTTCAGCATATTGAAAG-3’ (InvitrogenTM) (Aquino e cols., 2006), o qual
amplifica um produto de 120 pb com temperatura de melting (Tm) em torno de 77oC (+
0,8 oC), 5 μL de Sybr®Green PCR Mastermix (Applied Biosystems), e água ultra pura
para um volume final de 10 μL. As reações foram distribuídas em tubos de 0,1 mL
(MicroAmp™Fast Reaction Tube With Cap), centrifugadas por 2 minutos a 200 x g e
levadas ao termociclador StepOnePlus (Apllied Biosystems). O programa de
termociclagem consistiu de aquecimento a 95 oC por 10 minutos, 40 ciclos de 95
oC por
15 segundos e 60 oC por 1 minuto, com aquisição da fluorescência a 60
oC. A
amplificação foi imediatamente seguida por um programa de melting com desnaturação
inicial a 95 oC por 15 segundos, resfriamento a 60
oC por 1 minuto e aumento gradual de
temperatura de 0,3 oC/s até 95
oC. Como controle negativo da PCR, foi utilizado água no
lugar de cDNA, e como controle positivo foi utilizado o cDNA proveniente de larvas
naturalmente infectadas, do banco de amostras da Intituição. No caso de existirem
amostras positivas, seria realizada posteriormente a tipagem do vírus (DENV-1, DENV2,
DENV3, DENV4) também +-através de PCR em tempo real, utilizando-se primers
específicos para cada sorotipo viral.
Resultados
Todas as amostras de larvas e mosquitos analisadas apresentaram resultados da
PCR em tempo real negativos para o DENV. A figura 1 apresenta dois gráficos típicos de
PCR em tempo real gerados neste estudo. A figura 8a apresenta o gráfico de amplificação
onde se observa a detecção de material genético apenas nos controles positivos (da
extração e da PCR) e ausência de amplificação para as demais amostras (linha basal). A
figura 8b apresenta a curva de dissociação (Melt Curve) do produto gerado após os 40
ciclos de amplificação, onde se observa um pico em torno de 76,8 oC correspondendo ao
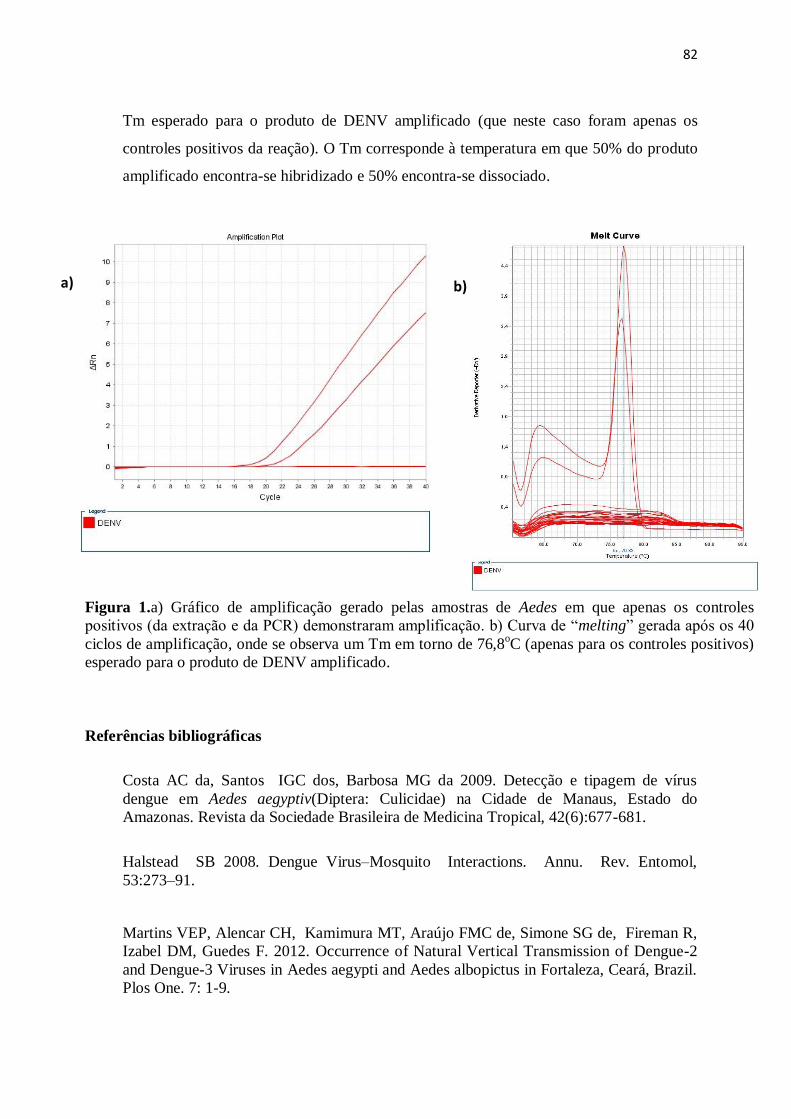
82
Tm esperado para o produto de DENV amplificado (que neste caso foram apenas os
controles positivos da reação). O Tm corresponde à temperatura em que 50% do produto
amplificado encontra-se hibridizado e 50% encontra-se dissociado.
Figura 1.a) Gráfico de amplificação gerado pelas amostras de Aedes em que apenas os controles
positivos (da extração e da PCR) demonstraram amplificação. b) Curva de “melting” gerada após os 40
ciclos de amplificação, onde se observa um Tm em torno de 76,8oC (apenas para os controles positivos)
esperado para o produto de DENV amplificado.
Referências bibliográficas
Costa AC da, Santos IGC dos, Barbosa MG da 2009. Detecção e tipagem de vírus
dengue em Aedes aegyptiv(Diptera: Culicidae) na Cidade de Manaus, Estado do
Amazonas. Revista da Sociedade Brasileira de Medicina Tropical, 42(6):677-681.
Halstead SB 2008. Dengue Virus–Mosquito Interactions. Annu. Rev. Entomol,
53:273–91.
Martins VEP, Alencar CH, Kamimura MT, Araújo FMC de, Simone SG de, Fireman R,
Izabel DM, Guedes F. 2012. Occurrence of Natural Vertical Transmission of Dengue-2
and Dengue-3 Viruses in Aedes aegypti and Aedes albopictus in Fortaleza, Ceará, Brazil.
Plos One. 7: 1-9.
a) b)

83
Serufo JC, Oca HM, Tavares VA, Souza AM, Rosa RV, Jamal MC 1993. Isolation of
dengue type 1 from larvae of Aedes albopictus in the Campos Altos city, state of Minas
Gerais, Brazil. Memórias Do Instituto Oswaldo Cruz; 88: 503-504.
AGRADECIMENTOS
Aos moradores de Mariana e Ouro Preto que permitiram o acesso às suas residências e à
Secretaria de Turismo de Ouro Preto. Às estagiárias Laís Sato, Luna Ferreira, Márcia
Regina F. Luzia e Ana Cláudia A. Carneiro e à MSc Adriele Magalhães pela apoio em
campo. À Guilherme de P. Costa pela ajuda em laboratório. Ao LEEIDS e Lab. de
Doenças de Chagas da UFOP, LABEq da UFMG. À ECOVEC. À Dra. Alzira Cecílio e
Dr. Sérgio Caldas da FUNED. À CAPES pelo apoio financeiro. SPR é bolsista
pesquisador do CNPq.

84
ANEXO
Figura 2. Ovitrampas utilizadas nos estudos dos capítulos I e II.a) A armadilha é vista
lateralmente. b) A armadilha é vista por cima. Foto: Michelle C. Pedrosa, 2009.
Figura 3. MosquiTRAP®
utilizadas no estudo do capítulo II. a) Armadilha montada, pronta
para uso. Fonte: https://www.ufmg.br/online/arquivos/Dengue_armadilha-thumb.jpg
acessado em 17/03/2013. b) Armadilha desmontada, com seus componentes expostos. Sendo:
I, parte superior da armadilha; II, parte inferior (armazenamento de água); III, cartao
adesivo; IV AtrAedes® e V, tela. Fonte:Laila Heringer Costa.
a b
a) )