dinÂmica do banco de sementes do solo em …livros01.livrosgratis.com.br/cp000695.pdf · aos...
TRANSCRIPT

MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL DE PELOTAS FACULDADE DE AGRONOMIA ELISEU MACIEL
DINÂMICA DO BANCO DE SEMENTES DO SOLO
EM ECOSSISTEMA CAMPESTRE SOB UTILIZAÇÃO
AGROPECUÁRIA COM SOJA E AZEVÉM ANUAL
ENG. AGR. M.Sc. FERNANDA COSTA MAIA
Tese apresentada à Universidade Federal de Pelotas, sob orientação do Prof. Dr. Manoel de Souza Maia, como parte das exigências do Programa de Pós-Graduação em Ciência e Tecnologia de Sementes, para a obtenção do título de Doutor em Ciências.
PELOTAS Rio Grande do Sul – Brasil
Agosto de 2005

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
MINISTÉRIO DA EDUCAÇÃO
UNIVERSIDADE FEDERAL DE PELOTAS FACULDADE DE AGRONOMIA ELISEU MACIEL
DINÂMICA DO BANCO DE SEMENTES DO SOLO
EM ECOSSISTEMAS CAMPESTRES SOB UTILIZAÇÃO
AGROPECUÁRIA COM SOJA E AZEVÉM ANUAL
ENG. AGR. M.Sc. FERNANDA COSTA MAIA
Tese apresentada à Universidade Federal de Pelotas, sob orientação do Prof. Dr. Manoel de Souza Maia, como parte das exigências do Programa de Pós-Graduação em Ciência e Tecnologia de Sementes, para a obtenção do título de Doutor em Agronomia.
PELOTAS Rio Grande do Sul – Brasil
Agosto de 2005

iii
COMITÊ DE ORIENTAÇÃO
Orientador:
Manoel de Souza Maia Eng. Agr., Dr., Prof. Adjunto do Departamento de Fitotecnia
Faculdade de Agronomia Eliseu Maciel – FAEM
Universidade Federal de Pelotas – UFPel
Co-orientadores:
Orlando Antônio Lucca Filho Eng. Agr., Dr., Prof. Adjunto do Departamento de Fitotecnia
Faculdade de Agronomia Eliseu Maciel – FAEM
Universidade Federal de Pelotas – UFPel
Renato Borges de Medeiros Eng. Agr., Dr., Prof. Adjunto do Departamento de Plantas Forrageiras e Agrometeorologia
Faculdade de Agronomia
Universidade Federal do Rio Grande do Sul - UFRGS
COMISSÃO EXAMINADORA
__________________________________________________ Francisco Amaral Villela Eng. Agrícola, Dr., Prof. Adjunto do Departamento de Física – IFM – UFPel
__________________________________________________ Orlando Antônio Lucca Filho Eng. Agr., Dr., Prof. Adjunto do Departamento de Fitotecnia – FAEM – UFPel
__________________________________________________ Renato Borges de Medeiros Eng. Agr., Dr., Prof. Adjunto do Departamento de Plantas Forrageiras e Agrometeorologia – FA – UFRGS
__________________________________________________ José Pedro Pereira Trindade Eng. Agr., Dr., Pesquisador EMBRAPA Pecuária Sul

iv
Quem não compreende um olhar, tampouco compreenderá uma longa explicação...
MÁRIO QUINTANA

v
AGRADECIMENTOS
Ao Programa de Pós-Graduação em Ciência e Tecnologia de Sementes pela
oportunidade de realizar o curso de Doutorado.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
pelo suporte financeiro ao projeto de pesquisa que resultou nessa tese.
A Fundação Estadual de Pesquisa Agropecuária (FEPAGRO), que permitiu a
realização deste trabalho de pesquisa.
Ao Professor Manoel de Souza Maia pela orientação, conhecimento, amizade e
companheirismo, como orientador e como pai, que sempre foi um grande incentivador e
a quem admiro pessoal e profissionalmente.
Ao Prof. Dr. Orlando Antônio Lucca Filho pelas importantes contribuições,
incentivo e amizade.
Aos professores e amigos Dr. Renato Borges de Medeiros e Dr. Valério De Patta
Pillar, da Universidade Federal do Rio Grande do Sul, que prestaram importantes
colaborações ao trabalho.
As biólogas e amigas Renée Bekker e Irma Knevel, grandes orientadoras durante
o estágio proporcionado por este curso e pela Coordenação de Aperfeiçoamento de
Pessoal de Nível Superior (CAPES) e sempre amigas, que deram contribuições
fundamentais para os artigos desta tese.

vi
Aos estagiários Rogério Berton, Leandro Caetano e Silvia Simioni,
fundamentais para a concretização deste trabalho, e aos funcionários do Laboratório
Didático de Análise de Sementes da UFPel.
Aos meus familiares, que sempre forneceram todo o apoio material e espiritual
para que pudesse finalizar este curso. A minha mãe, também pelo esforço nas correções
de linguagem do texto.
Aos meus amigos e amigas que souberam entender a importância desse trabalho
para mim e que, inclusive, contribuíram na concretização do mesmo.
A todos que direta ou indiretamente contribuíram de alguma forma para a
realização desse trabalho.
Aos meus pais e minha irmã, a quem dedico este trabalho.

vii
ÍNDICE
LISTA DE TABELAS..................................................................................................... ix
LISTA DE FIGURAS....................................................................................................... x
RESUMO....................................................................................................................... xiii
ABSTRACT.................................................................................................................... xv
1. INTRODUÇÃO GERAL ........................................................................................... 1
2. DINÂMICA DO BANCO DE SEMENTES DO SOLO DE AZEVÉM ANUAL
EM SISTEMA DE SEMEADURA DIRETA COM SOJA
RESUMO.......................................................................................................................... 7
ABSTRACT...................................................................................................................... 8
INTRODUÇÃO................................................................................................................ 9
MATERIAL E MÉTODOS............................................................................................ 11
RESULTADOS .............................................................................................................. 19
DISCUSSÃO .................................................................................................................. 26
CONCLUSÕES .............................................................................................................. 31
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 32
3. LONGEVIDADE DE SEMENTES DE AZEVÉM ANUAL EM CONDIÇÕES
NATURAIS
RESUMO........................................................................................................................ 35
ABSTRACT.................................................................................................................... 36
INTRODUÇÃO.............................................................................................................. 37
MATERIAL E MÉTODOS............................................................................................ 39

viii
RESULTADOS .............................................................................................................. 41
DISCUSSÃO .................................................................................................................. 43
CONCLUSÕES .............................................................................................................. 47
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 48
4. ALTERAÇÕES NO BANCO DE SEMENTES DO SOLO DE UM
ECOSSISTEMA CAMPESTRE PROVOCADAS POR NOVE ANOS DE
MANEJO AGROPECUÁRIO INTENSIVO E POTENCIAL DE
RECUPERAÇÃO DA VEGETAÇÃO NATIVA
RESUMO........................................................................................................................ 51
ABSTRACT.................................................................................................................... 52
INTRODUÇÃO.............................................................................................................. 53
MATERIAL E MÉTODOS............................................................................................ 57
RESULTADOS .............................................................................................................. 61
DISCUSSÃO .................................................................................................................. 71
CONCLUSÕES .............................................................................................................. 79
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 80
ANEXO .......................................................................................................................... 85
5. CONSIDERAÇÕES FINAIS ................................................................................... 88
6. REFERÊNCIAS BIBLIOGRÁFICAS.................................................................... 94

ix
LISTA DE TABELAS
DINÂMICA DO BANCO DE SEMENTES DO SOLO DE AZEVÉM ANUAL EM
SISTEMA DE SEMEADURA DIRETA COM SOJA
1. Diferenças de tamanho do banco de sementes do solo de azevém anual (0-10 cm) nas áreas de 3, 6, 8 e 9 anos de manejo................................................................................. 20
2. Diferenças de conteúdos de argila, matéria orgânica, fósforo, potássio, alumínio e cálcio em cada área da série temporal (n = 4), representando tempos de implantação do sistema agropastoril com semeadura direta sobre campo natural................................... 20
3. Número de sementes na superfície do solo, por metro quadrado, nas séries temporais de 6, 8 e 9 anos de manejo .............................................................................................. 24
4. Modelos de regressão polinomial dos dados com transformação logarítmica do banco de sementes do solo para cada série temporal (P<0,001) ............................................... 27
ALTERAÇÕES NO BANCO DE SEMENTES DO SOLO DE UM
ECOSSISTEMA CAMPESTRE PROVOCADAS POR NOVE ANOS DE
MANEJO AGROPECUÁRIO INTENSIVO E POTENCIAL DE
RECUPERAÇÃO DA VEGETAÇÃO NATIVA
1. Espécies comuns e características dos bancos de sementes do solo de 0 a 5 cm, em cada série temporal, com base na freqüência (%). Letras diferentes significam diferenças significativas na composição do banco de sementes (P<0,05)...................... 63
2. Espécies comuns e características dos bancos de sementes do solo de 5 a 10 cm, em cada área da série temporal, com base na freqüência (%). Letras diferentes significam diferenças significativas na composição do banco de sementes (P<0,05)...................... 63

x
LISTA DE FIGURAS
DINÂMICA DO BANCO DE SEMENTES DO SOLO DE AZEVÉM ANUAL EM
SISTEMA DE SEMEADURA DIRETA COM SOJA
1. Pastagem de azevém anual durante o mês de junho, com altura de aproximadamente 20 cm............................................................................................................................... 12
2. Condição da pastagem de azevém anual durante o inverno no momento da retirada dos animais ..................................................................................................................... 13
3. Azevém anual dessecado por glifosato no início de dezembro .................................. 13
4. Condição da pastagem de azevém anual no final do ciclo da soja em março ............ 14
5. Área da pastagem de azevém anual após a colheita da soja no fim de abril .............. 14
6. Amostras de solo utilizadas na análise do banco de sementes do solo....................... 16
7. Germinação das sementes de azevém anual retiradas das amostras de solo em sala climatizada ...................................................................................................................... 16
8. Diagrama de ordenação do BSS azevém (0-10 cm) ao longo do ano nas parcelas (A, B, C e D) das séries temporais de 6, 8 e 9 anos de manejo (n = 12). As variáveis pH, matéria orgânica (MO), potássio (K), sódio (Na) e cálcio (Ca) no solo apresentam correlação acima de 0,52 com o eixo X. O eixo horizontal representa 53,8% e o vertical 33,1% da variação do BSS de azevém ao longo do ano ................................................. 21
9. Diferenças no tamanho do banco de sementes do solo de azevém anual nos primeiros 5 cm de solo, ao longo do ano, nas séries temporais com 6 (A), 8 (B) e 9 (C) anos de manejo (P<0,05).............................................................................................................. 23

xi
10. Dinâmica da vegetação de azevém anual nas séries temporais de 6 (A), 8 (B) e 9 (C) anos de manejo................................................................................................................ 25
11. Curvas obtidas por análise de regressão polinomial do banco de sementes do solo de azevém anual, coletado de setembro a agosto de 2003, nas diferentes séries temporais......................................................................................................................... 27
LONGEVIDADE DE SEMENTES DE AZEVÉM ANUAL EM CONDIÇÕES
NATURAIS
1. Alterações observadas nas sementes no experimento de enterrio e diferenças significativas (P<0,05) entre os diversos períodos de exumação das mesmas, considerando-se o conjunto das variáveis estudadas ...................................................... 42
ALTERAÇÕES NO BANCO DE SEMENTES DO SOLO DE UM
ECOSSISTEMA CAMPESTRE PROVOCADAS POR NOVE ANOS DE
MANEJO AGROPECUÁRIO INTENSIVO E POTENCIAL DE
RECUPERAÇÃO DA VEGETAÇÃO NATIVA
1. Tamanho do BSS das 30 espécies com maior quantidade de sementes nas duas profundidades de solo estudadas, nas quatro áreas da série temporal. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Juncus bufonius, Lolium multiflorum, Gamochaeta spp., Kyllinga brevifolia, Sisyrinchium micranthum, Digitaria ciliaris, Plantago spp., Cuphea carthagenensis, Centela asiatica, Paspalum nicorae, Trifolium repens, Eragrostis neesii, Juncus cf. effusus, Lythrum hyssopifolium, Cerastium rivulare, Cyperus eragrostis, Kyllinga odorata, Fimbristylis diphylla, Cyperaceae 1, Centunculus minimus, Conyza cf. floribunda, Digitaria adscendens ...................................................................................................................... 62
2. Espécies com maior BSS nas duas profundidades de solo estudadas, na área com 3 anos de manejo. As espécies estão identificadas segundo duas as primeiras letras dos epítetos genérico e específico: Juncus bufonius, Gamochaeta spp., Plantago spp., Digitaria ciliaris, Coniza cf. floribunda, Digitaria adscendens, Kyllinga brevifolia, Sisyrinchium micranthum, Cuphea carthagenensis, Lolium multiflorum, Eragrostis neesii, Cyperus reflexus, Paspalum nicorae, Cerastium glomeratum, Trifolium repens, Eclipta megapotamica, Fimbristylis autumnalis, Cyperaceae 1 .................................... 64
3. Espécies com maior BSS nas duas profundidades de solo estudadas, na área com 6 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Gamochaeta spp., Lolium multiflorum, Sisyrinchium micranthum, Centela asiática, Juncus bufonius, Kyllinga brevifolia, Eragrostis neesii, Centunculus minimus, Hydrocotyle exigua, Plantago spp., Soliva pterosperma, Hypoxis decumbens, Cerastium glomeratum, Oxalis cf. conorrhiza, Kyllinga vaginata, Lotus subbiflorus, Eleusine tristachya, Cyperus cf. cayennensis, Juncus capillaceus, Kyllinga odorata............................................................................................................................ 65

xii
4. Espécies com maior BSS nas duas profundidades de solo estudadas, na área com 8 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Lolium multiflorum, Gamochaeta spp., Kyllinga odorata, Juncus bufonius, Cyperus eragrostis, Juncus capillaceus, Kyllinga brevifolia, Fimbristylis dyphilla, Cyperaceae 1, Sisyrinchium micranthum, Juncus dichotomus, Oxalis cf. conorrhiza, Centela asiática, Amaranthus lividus ssp. Polygonoides, Glandularia selloi, Kyllinga vaginata, Centunculus minimus, Dichondra sericea, Hypoxis decumbens, Cyperaceae 2 ................................................................................ 66
5. Espécies com maior BSS nas duas profundidades de solo estudadas, na área com 9 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Juncus bufonius, Kyllinga brevifolia, Digitaria ciliaris, Lolium multiflorum, Sisyrinchium micranthum, Juncus cf. effusus, Cerastium rivulare, Plantago spp., Trifolium repens, Cuphea carthagenensis, Kyllinga odorata, Lythrum hyssopifolium, Juncus dichotomus, Eragrostis neesii, Fimbristylis autumnalis, Centunculus minimus, Cyperaceae 1, Poa annua, Gamochaeta spp. ............................ 67
6. Diagrama de ordenação da composição do BSS de 0 a 5 cm nas parcelas das quatro áreas da série temporal (3, 6, 8 e 9 anos de manejo) e linha de tendência. As espécies identificadas segundo as duas primeiras letras dos epítetos genérico e específico, em itálico: Amaranthus lividus ssp. polygonoides, Briza minor, Centela asiatica, Conyza bonariensis, C. cf. floribunda, Cuphea carthagenensis, Dichondra sericea, Digitaria ciliaris, Eclipta megapotamica, Fimbristylis autumnalis, Juncus bufonius, Lolium multiflorum, Panicum dichotomiflorum e Poa annua apresentam correlação acima de 0,50 com pelo menos um dos eixos de ordenação. O eixo horizontal representa 64,6% e o vertical 16,9% da variação da composição do BSS da série ....................................... 69

xiii
Dinâmica do banco de sementes do solo em ecossistema campestre sob utilização agropecuária com soja e azevém anual. Autora: Eng. Agr. M.Sc. Fernanda Costa Maia. Orientador: Prof. Dr. Manoel de Souza Maia (UFPEL). Projeto financiado pelo CNPq.
RESUMO. O presente trabalho de tese consta de três artigos. O primeiro e o segundo
focam, respectivamente, a dinâmica e a longevidade das sementes de azevém anual
(Lolium multiflorum Lam.) no solo, em rotação com soja em sistema de semeadura
direta na palha, atualmente muito difundido na região. O terceiro artigo procura
quantificar possíveis alterações na composição e tamanho do banco de sementes do solo
(BSS) ao longo de diferentes séries temporais, buscando inferir sobre o potencial de
recuperação da vegetação campestre através do seu banco de sementes. A soja é
tradicionalmente cultivada por dois anos e, após, a área permanece mais dois anos em
pousio. As quatro áreas estudadas correspondem a diferentes séries temporais,
representando diferentes tempos de implementação desta prática de manejo. Em duas
áreas a soja foi semeada; as outras duas permaneceram em pousio. Em cada uma das
quatro parcelas consecutivas (25 x 20 m) de cada área, foram retiradas 10 amostras de
solo de 5 cm de diâmetro, mensalmente, durante um ano e divididas em duas
profundidades (0-5 e 5-10 cm), para a análise do BSS de azevém. No meio de cada
parcela, a dinâmica da pastagem também foi avaliada, utilizando-se três quadros
contíguos de 0,5 x 0,5 m para contagem de plantas e inflorescências/infrutescências, e 3
amostras da superfície do solo com 10 cm de diâmetro, para contagem de plântulas.
Para o estudo da longevidade da espécie, foram enterrados, próximo a uma das parcelas,
a 5 cm de profundidade, sacos de náilon contendo solo esterilizado e 100 sementes. Em
intervalos de três ou quatro meses, quatro sacos foram sendo exumados; as sementes
foram contadas e submetidas a testes de germinação e de viabilidade. Com uma das
coletas realizadas durante o outono (maio), foi avaliado o BSS como um todo, através
da germinação das sementes das amostras de solo em casa de vegetação, e posterior
identificação das espécies de plantas. O estudo apontou para um BSS de azevém anual
de caráter transitório, já que a maior parte das sementes não persistiu no solo por mais
de um ano. Verifica-se, com base nesses dois estudos, que a maioria das sementes se
dispersa durante o fim da primavera e início do verão (dezembro), com alta dormência
primária, germinam no outono, quando as condições de temperatura e disponibilidade
de água tornam-se adequadas. A dinâmica do BSS mostra padrões estruturais no tempo,
com um período de “armazenamento” das sementes no solo durante o verão, devido a

xiv
uma alta dormência primária, um período de “exaustão”, durante o outono e um período
de “transição”, durante o inverno e a primavera. Até o fim do outono, o BSS é
praticamente exaurido. O efeito da área cultivada e da área em pousio apresentou maior
influência no BSS de azevém do que o número de anos em que ela se repete,
principalmente em virtude da sua transitoriedade, que tornam os distúrbios de cada ano
mais importantes do que suas conseqüências de longo prazo. O estabelecimento da
pastagem por ressemeadura natural é totalmente dependente do recrutamento anual das
sementes que estão no solo, em densidades que variam de 3.363 a 22.835 sementes.m-2.
Quanto às sementes das demais espécies espontâneas, observou-se que os distúrbios
decorrentes do fato de a área estar em pousio ou com o cultivo da soja também
apresentaram maior influência no BSS do que o tempo de implementação do manejo.
Houve um predomínio de sementes de espécies de monocotiledôneas anuais, em termos
quantitativos, e de perenes, em termos qualitativos. Aponta-se aqui para uma possível
influência do uso de herbicidas na redução da densidade e da riqueza de espécies do
banco de sementes. A cada ano, possivelmente o controle químico provoca um aparente
reinício da sucessão na comunidade vegetal, embora as sementes de algumas espécies
“escapem” e, lentamente, dêem lugar ao processo de regeneração. Observou-se maior
concentração de sementes nos primeiros 5 cm de solo. Conforme aumenta o tempo de
utilização da área, o tamanho e a riqueza do BSS diminuem, com tendência de
substituição de espécies perenes por espécies anuais, em termos quantitativos, e o
conseqüente desaparecimento de espécies nativas, de importante valor forrageiro, após 6
anos.
Palavras-chave: Lolium multiflorum, Glycine max, semeadura direta, sementes
enterradas, ecologia de sementes, campo natural.

xv
Soil seed bank dynamic in fields with soybean and annual ryegrass. Author: Agr. Eng. M.Sc. Fernanda Costa Maia. Supervisor: Prof. Dr. Manoel de Souza Maia (UFPEL). Project supported by CNPq.
ABSTRACT. This thesis is composed by three articles. The first and the second paper
are related to the dynamic and longevity of annual ryegrass seeds in the soil, in no
tillage systems in rotation with soybean, that have been growing in the South of Brazil.
The third one aims to quantify possible changes in the soil seed bank (SSB) of the area,
and to determine the regeneration potential of the natural grassland communities from
the seed bank. Soybean is commonly cultivated for two years and then is left in follow
for another two years. The sites correspond to four areas, representing different periods
of management, two of them with soybean and the other two resting. FFoouurr consecutive
plots (25 x 20 m) were designed in each area. Ten soil samples with 5 cm of diameter
were taken every month during one year and divided in two depths (0-5 cm and 5-10
cm). In the middle of each plot, the vegetation dynamic was also evaluated. The number
of plants and inflorescences were counted in three contiguous quadrats (0.5 x 0.5 m)
and the number of seedlings on three soil cores with 10 cm of diameter, per plot. For the
longevity experiment, bags of nylon screen containing sterilised soil with 100 seeds
each one were randomly buried in the field, 5 cm deep. Every three or four months, four
nylon bags were exhumed. Seeds were counted and tested by germination and viability.
With the soil cores collected in the autumn (May), the whole SSB was analysed using
the direct germination method in a greenhouse. The SSB of annual ryegrass was
characterized as being transitory, due to the majority of the seeds does not persist viable
in the soil for more than one year. The results of these two papers showed that the great
part of seed dispersion occurs at the end of spring and beginning of summer
(December), with high primary dormancy, and then they germinate in the autumn, when
temperature conditions and water requirements were suitable. SSB dynamic showed
structural patterns in time, with a “storage period” in summer, an “exhausting period”
during autumn and a “transition period” in winter and spring. At the end of the autumn,
SSB is practically exhausted. The effect of the soybean presence had more influence on
annual ryegrass SSB than the number of cropping years, mainly due to its transitory
character that make the disturbs of each year more important than their long-term
consequences. The establishment of the pasture by natural re-sowing is totally
dependent of the annual recruitment of seeds from the SSB, in densities that vary from
3,363 to 22,835 seeds.m-2. About the other spontaneous grassland species it was

xvi
observed that crop disturbance have higher influence in SSB than the accumulated
effect of cropping age in the fields. There were more seeds of annual monocot species in
quantitative terms and perennials in qualitative terms. The study pointed to a possible
influence of desiccant herbicides on SSB species richness and density reduction. Each
year, it seems that chemical control induces a sort of re-colonization, i.e., the succession
comes back to the beginning, but some species “escape” and, slowly, give place to the
regeneration process. Most of the collected seeds were at the first 5 cm of soil. As the
utilization years of the area proceeds, SSB size and species richness decrease, tending to
the substitution of perennial species by annuals in quantitative terms and the consequent
disappearance of important native species with forage value just after 6 years.
Key-words: Lolium multiflorum, Glycine max, no tillage, buried seeds, seed ecology,
grassland.

INTRODUÇÃO GERAL
No Rio Grande do Sul (RS), a vegetação campestre ocupa cerca de 10,5 milhões
de hectares, contribuindo com mais de 90% da alimentação dos cerca de 14 milhões de
bovinos e 5 milhões de ovinos criados no estado (UCPEL/ITEPA, 2001). Além disso,
constitui a base de sustentação de outros herbívoros domésticos e dos componentes da
fauna selvagem, que representam, sem dúvida, um condicionante da vegetação natural
hoje presente na área. São campos cuja produção de forragem é considerada excelente
para o período de primavera-verão, em função da diversidade de espécies e da qualidade
das mesmas, se comparadas com as pastagens naturais de qualquer outra região do
mundo. Segundo Boldrini (1997), essa vegetação é formada por, aproximadamente, 400
espécies de Poaceae e 150 de Fabaceae.
Mesmo assim, os ecossistemas de pastagem natural encontram-se em processo
de permanente degradação, em decorrência do superpastejo e/ou pastejo seletivo, da
utilização excessiva do fogo ou da simples substituição por lavouras, como acontece,
geralmente, em todo o mundo. Em 1942, cerca de 46% do estado do Rio Grande do Sul
era coberto por essa formação vegetal (Rambo, 1942), enquanto que atualmente ela
ocupa cerca de 37% do território (IBGE, 1997).
Embora se disponha de um número relativamente grande de informações sobre
identificação e classificação das plantas ocorrentes em pastagens nativas, pouco se sabe
sobre sua dinâmica e seus efeitos na restauração dessas áreas após cultivo intensivo,
principalmente com relação ao efeito de herbicidas aplicados nos sistemas de produção.

2
A distribuição da produção dos campos no RS ao longo do ano é sazonal, devido
à ocorrência de baixas temperaturas durante o inverno, com o predomínio de espécies
subtropicais (ciclo estival). Entre as práticas mais utilizadas pelos produtores para
contornar este problema está a introdução de espécies exóticas de inverno, com manejo
direcionado para a ressemeadura natural da espécie introduzida, no sentido de reduzir
custos da produção de forragem.
No caso particular do sistema de integração lavoura-pecuária, com azevém
(Lolium multiflorum Lam.) e soja (Glycine max L.) em semeadura direta na palha,
muitos produtores vêm obtendo bons resultados.
Entretanto, ainda não está bem esclarecido por quanto tempo essas áreas
permanecerão com elevados índices de produtividade e como ocorre a dinâmica das
sementes de azevém no solo que proporciona o estabelecimento de áreas de pastagem
baseadas exclusivamente na ressemeadura desta espécie.
Além das estratégias de dispersão, outro fator que faz parte dos mecanismos de
sobrevivência das espécies é a distribuição da germinação por determinado período de
tempo, prevenindo o desaparecimento ou substituição da espécie, devido às condições
ambientais adversas logo após a germinação. Portanto, nem todas as sementes
produzidas e dispersadas em determinado ano germinarão no ano seguinte. Muitas
destas sementes podem permanecer dormentes ou ter a germinação inibida após caírem
no solo (Mayer & Poljakoff-Mayber, 1989).
As sementes que caem no solo e permanecem dormentes após a disseminação
formam os chamados “banco de sementes do solo” (BSS). Segundo Milberg (1992), o
BSS é um reflexo da vegetação passada do local, sendo que as espécies não mais
presentes podem às vezes persistir como sementes no solo por décadas.
A persistência das espécies no ambiente está associada, entre outros fatores, às
suas estratégias de colonização, o que envolve o banco de sementes armazenadas no
solo (Miles, 1978), componente importante da dinâmica da vegetação. A existência de
uma numerosa e rica população de sementes no solo pode ter profundos efeitos na
dinâmica genética e populacional das plantas de uma determinada comunidade vegetal
(Cook, 1980), sendo um dos mecanismos de sobrevivência das espécies, ao longo do
tempo. Este conjunto de sementes, juntamente com estruturas vegetativas, assegura, no
caso dos ecossistemas agropastoris, a perenidade da maioria das espécies que
constituem sua diversidade florística.

3
Além da importância genética e produtiva, o BSS pode apresentar papel
fundamental na recuperação de áreas que sofreram processos de distúrbio drásticos,
sendo necessários entendimentos referentes ao manejo, conservação e influência dos
BSS na manutenção da diversidade florística e da sustentabilidade social e ecológica
desses ecossistemas.
Para plantas perenes, a semente é uma alternativa de perpetuação, mas não é tão
importante quanto para espécies anuais, onde a semente dormente é o elo crucial entre
as gerações (Harper, 1977). Sementes de espécies perenes geralmente não permanecem
viáveis no solo tanto quanto as anuais (Roberts, 1970; Pettit & Froend, 2001), que
podem ter ciclos de vida extremamente longos (Cook, 1980). O número de sementes
viáveis no solo geralmente diminui com a redução na freqüência de distúrbio e o
aumento do ciclo de vida das espécies que formam a vegetação (Thompson, 1978;
Symonides, 1986).
Várias evidências apontam que espécies do gênero Lolium tendem a não formar
BSS persistentes (Douglas, 1965; Thompson & Grime, 1979; Howe & Chancellor,
1983; López-Mariño et al., 2000), indicando que a ressemeadura observada em campos
melhorados decorre basicamente de sementes depositadas no solo na estação anterior.
Muitas espécies que acumulam grande número de sementes no solo apresentam-
se em estado de dormência ou, pelo menos, em populações parcialmente compostas por
sementes dormentes. O fato de essas espécies apresentarem um grande número de
sementes de vida longa, aliado ao polimorfismo com respeito à dormência ou ao
desenvolvimento de dormência secundária, apresenta fluxos de germinação por um
longo período de tempo. Como essas plantas produzem um grande número de sementes
pequenas, muitas, provavelmente, acabam penetrando no solo através de rachaduras ou
canais de origem biológica e distribuindo-se verticalmente a partir dos primeiros
centímetros superiores do perfil do solo (Bryant, 1989).
A composição do banco de sementes depende da produção e composição das
comunidades vegetais anterior e atual, bem como da longevidade das sementes de cada
espécie, sob as condições locais (López-Mariño et al., 2000). Em seu trabalho, Stocklin
& Fischer (1999) demonstraram que a longevidade das sementes está altamente
correlacionada com o índice de desaparecimento de determinadas espécies campestres.
A longevidade das sementes é, portanto, um importante tópico para práticas de manejo
que visem recuperação de áreas degradadas (Bekker et al., 1998a; Bekker et al., 1998b;
Bakker & Berendse, 1999), revelando a possível vulnerabilidade da vegetação à

4
extinção, inclusive à de seus bancos de sementes do solo, a necessidade de sua
conservação e possibilidades de regeneração (Bekker et al., 1998c). Entretanto, pouco
se sabe quanto aos efeitos das condições ambientais sobre a longevidade e
sobrevivência de sementes em condições naturais. Esta relação entre a composição do
banco de sementes e a vegetação é particularmente importante em áreas que sofrem
diferentes tipos de manejo (Fredrickson & Taylor, 1982, apud López-Mariño, 2000).
A importância do conhecimento da ecologia das sementes e dos bancos de
sementes para o entendimento da função e estrutura das comunidades tem sido
mencionada por muitos autores (Heydecker, 1973; Harper, 1977; Roberts, 1981; Fenner,
1985, 2000; Leck et al., 1989; Bekker, 1998; Medeiros, 2000). Em seu trabalho de tese,
Bekker (1998) menciona recentes revisões referentes a BSS (Vyvey, 1989a,b; Bernhardt
& Poschlod, 1993) e uma compilação sobre longevidade de sementes (Milberg, 1990).
Uma compilação de vários estudos sobre BSS é apresentada por Thompson et al.
(1997). Atualmente, quase não há área da ecologia vegetal moderna na qual os BSS não
estejam implicados, muitos deles com relevância direta para a ecologia de recuperação
de espécies e comunidades (Bakker et al., 1996).
Darwin, em 1859, foi provavelmente o primeiro a publicar informações
relacionadas a BSS quando, no livro A Origem das Espécies, descreve a presença de
sementes na lama do fundo de um lago. A partir de 1970, os pesquisadores se
preocuparam em estudar as relações dos BSS com a dinâmica vegetal e o seu papel na
manutenção da diversidade florística e na recuperação de ecossistemas degradados.
Estudos com a finalidade de avaliar o efeito do distúrbio (freqüência, intensidade e
amplitude) sobre a dinâmica vegetal e suas implicações com os BSS nos diferentes
ecossistemas, incluindo florestas naturais (Kellman, 1970), florestas plantadas (Hill &
Stevens, 1981), margens de lagos (Keddy & Reznicek, 1982), pântanos (Whigham et
al., 1979), dunas (Bekker et al., 1999), desertos (Reichman, 1975; Nelson & Chew,
1977; Henderson et al., 1978) e solos agrícolas (Roberts & Ricketts, 1979; Graham &
Hutchings, 1988b; Bakker & Berendse, 1999; López-Mariño et al., 2000; Vanasse &
Leroux, 2000; Favreto, 2004) têm sido realizados. Também tem se estudado o papel dos
BSS na revegetação de áreas de minas (Archbold, 1980) e florestas devastadas pelo
fogo (Archbold, 1979), demografia vegetal (Sarukhán, 1974), preservação de espécies
raras (Baskin & Baskin, 1978) e reconstrução da história vegetal (Van der Valk &
Davis, 1979). Entretanto, segundo Cook (1980), a principal carga de informação em

5
relação ao número de sementes dormentes veio de trabalhos de pesquisadores
interessados no controle de plantas concorrentes em solos cultivados.
Recentemente, devido a preocupações com a intensa degradação antrópica,
cresceu o interesse em estudar BSS com o objetivo de desenvolver modelos de predição
da sucessão vegetal mais realistas (Medeiros, 2000). Pesquisadores de vários países têm
demonstrado estreitas relações entre os BSS e a dinâmica populacional das espécies e
muitos são os trabalhos que abordam aspectos relacionados ao BSS em ecossistemas
campestres (Milberg, 1992; Boccanelli & Lewis, 1994; Bakker et al., 1997; Bekker et
al., 1997; Kalamees & Zobel, 1997; Morgan, 1998; Favreto et al., 2000; Lodge, 2001;
Maia et al., 2003; Maia et al., 2004). Apesar da reconhecida influência dos BSS na
manutenção da dinâmica populacional de ambientes de campo nativo, muito pouca
importância tem sido dada aos mesmos por parte das instituições de pesquisa do Brasil.
Os impactos do uso da terra e práticas de manejo são diferentes na vegetação e
no BSS. Os distúrbios causados pela intensificação da agricultura alteram a composição
da vegetação da área (Zanin et al., 1997), levando a um decréscimo na riqueza de
espécies, incluindo o desaparecimento de espécies características de campo.
Sendo assim, a composição de espécies do BSS no local cultivado é influenciada
pela exploração agrícola (Bakker et al., 1997; Medeiros & Steiner, 2002; Favreto,
2004). O preparo reduzido do solo, por exemplo, deixa as sementes próximas da
superfície. Segundo Yenish et al. (1992), em sistemas de semeadura direta na palha,
60% de todas as sementes presentes no perfil do solo encontram-se localizadas entre 0-1
cm de profundidade, com poucas sementes abaixo de 10 cm.
A utilização maciça de herbicidas em sistemas de semeadura direta na palha
pode estar comprometendo a heterogeneidade da florística de uma vasta região de
campos. Se as aplicações de herbicidas se sucederem a cada estação de crescimento e a
cada ano e ainda, se formulações combinadas de herbicidas forem utilizadas, é possível
pensar na hipótese de que os efeitos sobre as complexas cadeias biológicas existentes
nestes ecossistemas superem a sua resiliência, ou seja, sua habilidade para retornar à sua
composição e estrutura originais ou muito próximas destas (Holling, 1973; Goldsmith,
1978).
Pesquisas sobre a dinâmica populacional desses campos melhorados e suas
relações com os BSS a médio e longo prazo devem ser propostas. Para isto, trabalhos
abrangendo levantamentos do BSS em todas as estações do ano e observações da
dinâmica populacional da vegetação, aliados ao estudo da longevidade das sementes em

6
condições naturais, permitirão um melhor entendimento sobre a dinâmica da vegetação
campestre sob utilização agropecuária e inferências sobre o potencial de recuperação da
mesma. Tudo isto visando oferecer informações para a adoção de práticas de manejo
que otimizem a utilização econômica desses campos e assegurem a sua sustentabilidade.

7
Dinâmica do banco de sementes do solo de azevém anual em sistema de semeadura direta com soja. Autora: Eng. Agr. M.Sc. Fernanda Costa Maia. Orientador: Prof. Dr. Manoel de Souza Maia (UFPel). Projeto financiado pelo CNPq.
RESUMO. O estudo buscou caracterizar a dinâmica da população de sementes de
azevém anual (Lolium multiflorum Lam.) em sistema de semeadura direta com soja, em
diferentes áreas, representando séries temporais (3, 6, 8 e 9 anos de manejo). No sul do
Brasil, em áreas de integração de agricultura e pecuária, utiliza-se cada vez mais o
sistema de semeadura direta de cultivos de verão, como a soja. A cultura de cobertura
de inverno é pastejada e manejada para ressemeadura natural. As quatro áreas estudadas
correspondem as séries temporais, representando tempos de implementação desta
prática de manejo. Em duas áreas a soja foi semeada e as outras duas permaneceram em
pousio. Quatro parcelas consecutivas (25 x 20 m) foram desenhadas em cada área.
Foram retiradas 10 amostras de solo de 5 cm de diâmetro, em cada parcela,
mensalmente, durante um ano e divididas em duas profundidades (0-5 e 5-10 cm). No
meio de cada parcela, a dinâmica da pastagem também foi avaliada, utilizando-se três
quadros contíguos de 0,5 x 0,5 m, para contagem de plantas e inflorescências, e 3
amostras da superfície do solo, com 10 cm de diâmetro, para contagem de plântulas. A
dinâmica do banco de sementes do solo mostra padrões estruturais no tempo, com um
período de “armazenamento” das sementes no solo durante o verão, um período de
“exaustão” durante o outono e um período de “transição” durante o inverno e a
primavera. O estabelecimento da pastagem por ressemeadura natural é totalmente
dependente do recrutamento anual das sementes que estão no solo. A influência das
práticas de manejo utilizadas na área no banco de sementes do solo é mais importante
do que o tempo de implementação destas práticas. As áreas onde a soja foi semeada
apresentaram maiores quantidades de sementes no solo. A maior parte das sementes
supera a dormência e germina no fim do verão e início do outono, mostrando um banco
de sementes do solo tipicamente transitório, mas com uma pequena proporção de
sementes persistente.
Palavras-chave: Lolium multiflorum, sementes enterradas, sementes de espécies
forrageiras, plantio direto.

8
Soil seed bank dynamic of annual ryegrass in no tillage system with soybean. Author: Agr. Eng. M.Sc. Fernanda Costa Maia. Supervisor: Prof. Dr. Manoel de Souza Maia (UFPEL). Project supported by CNPq.
ABSTRACT. The study aimed to characterise annual ryegrass (Lolium multiflorum
Lam.) seed population dynamic in no tillage systems in rotation with soybean, in
different chronosequences (3, 6, 8 and 9 years-old). In the South of Brazil, integration
of soil cultivation and cattle grazing in the same area has been used, in no tillage
system, managing natural re-sowing of the grass. The soybean is traditionally cultivated
for two years and after that the area stays resting for two years. The four sites
correspond to the chronosequences, representing different periods of management, two
of them with soybean and the other two resting. FFoouurr consecutive plots (25 x 20 m)
were designed in each area. Ten soil samples with 5 cm of diameter were taken every
month during one year and divided in two depths (0-5 cm and 5-10 cm). In the middle
of each plot, the vegetation dynamic was also evaluated. The number of plants and
inflorescences were counted in three contiguous quadrats (0.5 x 0.5 m) and the number
of seedlings on three soil cores with 10 cm of diameter, per plot. Soil seed bank
dynamic shows structural patterns in time, with a “storage period” in summer, an
“exhausting period” during autumn and a “transition period” in winter and spring. The
establishment of the pasture by natural re-sowing is totally dependent of the annual
recruitment of seeds from the soil seed bank. The influence of the management
practices in the seed bank is more important than the number of years that these
practices have been implemented. Sites where soybean was sown showed the biggest
soil seed banks. Most part of the seeds overcome dormancy and germinate in the end of
summer and beginning of autumn, showing a typically transitory soil seed bank, but
with a small proportion of persistent seeds.
Key words: Lolium multiflorum, buried seeds, seeds of forage species, no tillage system.

INTRODUÇÃO
A distribuição da produção dos campos no sul do Brasil ao longo do ano é
tipicamente sazonal, devido à ocorrência de baixas temperaturas durante o inverno, que
prejudica o crescimento da vegetação campestre, composta predominantemente por
espécies subtropicais (Maia et al., 2003). No estado do Rio Grande do Sul, tal situação
provoca uma perda de aproximadamente 5% do total do rebanho (Maia & Primo, 1995).
O estado é um dos grandes responsáveis pela exportação brasileira de carne, sendo o
Brasil o maior exportador mundial de carne bovina desde 2003 (IBGE, 2005). Entre as
práticas mais utilizadas pelos produtores para contornar o problema da sazonalidade da
produção dos campos está a introdução de espécies exóticas de inverno, com manejo
direcionado para a ressemeadura natural da espécie introduzida, visando reduzir custos
na produção de forragem.
O azevém anual, devido à dormência de suas sementes e ao seu valor nutritivo
como alimento, é uma das gramíneas mais utilizadas no sul do Brasil em sistemas
rotativos com cultivos de verão, tanto como forrageira quanto como cobertura protetora
do solo; dispersando suas sementes antes da semeadura da soja, permite uma
ressemeadura natural. Entretanto, ainda não está claro por quanto tempo estes campos
permanecem com os altos índices de produtividade observados atualmente e como
acontece a dinâmica das sementes no solo, em áreas baseadas exclusivamente em
ressemeadura natural.

10
No caso particular da sucessão azevém e soja em semeadura direta, muitos
produtores vêm obtendo bons resultados com relação a esta prática, embora faltem
estudos que qualifiquem as decisões inerentes aos “custos de oportunidade” da
utilização do azevém para pastejo e manejo para a ressemeadura (Medeiros, 2000).
Ayala (2002) considerou importante o entendimento dos padrões de emergência e
viabilidade das sementes no solo, como uma forma de aumentar fortemente os índices
de ativação do banco de sementes de algumas espécies de importância forrageira.
O banco de sementes pode ter um importante papel nos sistemas produtivos,
tornando viável a utilização da pastagem (Silva, 2004). O sucesso econômico da
introdução de uma espécie forrageira em campo nativo depende de quanto tempo esta
espécie permanece produzindo. Em geral, a persistência está fortemente relacionada
com a adoção de adequadas práticas de manejo e com a capacidade de ressemeadura
natural através do banco de sementes do solo (Marañon, 1995; Medeiros, 2000).
Alguns autores apontam que muitas gramíneas cultivadas, incluindo Lolium spp.,
não formam bancos de sementes persistentes (Douglas, 1965; Thompson & Grime,
1979; Howe & Chancellor, 1983; López-Mariño, 2000), indicando que a ressemeadura
observada em campos melhorados decorre basicamente de sementes depositadas no solo
na estação anterior.
A grande maioria dos estudos descreve a composição, diversidade e densidade
do banco de sementes, mas poucos explicam como estes aspectos acontecem e quão
rapidamente eles se modificam (Fenner & Thompson, 2005). Pesquisas sobre dinâmica
da população desses campos melhorados e sua relação com o banco de sementes do solo
a médio e longo prazo devem ser propostas. Para isto, estudos que objetivem
caracterizar o banco de sementes em todas as estações do ano permitirão uma melhor
compreensão da dinâmica da vegetação campestre sob utilização agrícola e inferências
sobre o potencial de recuperação da mesma, oferecendo informações que possibilitem a
adoção de práticas de manejo agrícola, otimizando, assim, a utilização econômica
desses campos e assegurando sua sustentabilidade.
O presente estudo buscou caracterizar a dinâmica da população de sementes de
azevém anual em sistema de semeadura direta em rotação com soja, dentro de diferentes
séries temporais.

MATERIAL E MÉTODOS
1. Área de estudo
O trabalho foi desenvolvido no sul do Brasil, na Estância Santa Eulália, região
sudeste do estado do Rio Grande do Sul, município de Pelotas, situada a 31º 38’ 00”S e
52º 27’ 00”O.
O clima da região é considerado subtropical, com quatro estações do ano bem
definidas, precipitações pluviais totais anuais de 1.300 a 1.500 mm, bem distribuídas ao
longo do ano, temperaturas médias anuais de 17 ºC e possível ocorrência de geadas no
período de abril a setembro. A área apresenta relevo suavemente ondulado, solo com
textura argilosa, não bem drenado, com boas características químicas e físicas, profundo
e com excessiva compactação superficial; também apresenta forte variação nos níveis
de fósforo e potássio, de acordo com a profundidade; pH varia de 4,4 a 5,1.
A Estância Santa Eulália possui uma área física de 2500 ha, sendo 850 ha
destinados à semeadura direta de soja e azevém anual em sucessão, por ressemeadura
natural, para pastejo de gado bovino. Os quatro potreiros em estudo medem 100 ha,
aproximadamente, cada um representando um tempo de implantação do sistema de
manejo acima citado (série temporal), sendo estes 3, 6, 8 e 9 anos. Quatro parcelas
consecutivas, de 25 x 20 m cada, em faixa, foram demarcadas em cada série temporal.

12
O local foi escolhido de modo a estar situado em posições intermediárias do relevo e
com boa representação do sistema de manejo utilizado.
2. Sistema de semeadura direta na palha de azevém anual e soja
Na implantação do sistema, o azevém anual foi primeiramente semeado (30
kg.ha-1) em campo nativo dessecado. A semeadura não foi mais realizada nos outros
anos. Os potreiros são divididos com cerca elétrica e utilizados de forma rotativa para
pastejo de gado bovino, com capacidade média de 1,5 unidade animal.ha-1.ano-1,
alcançando uma produtividade comercial de aproximadamente 450 kg.ha-1.ano-1 de peso
vivo. O período de pastejo é usualmente de junho (fim do outono), conforme a FIGURA
1, a outubro (primavera), quando os animais são vendidos ou retirados para outra área
(FIGURA 2). Neste momento, são aplicados 50 kg.ha-1 de uréia, para favorecer a
recuperação das plantas, a produção de sementes e a formação de um adequado BSS,
capaz de garantir a ressemeadura natural com uma adequada população de plantas (600
plantas.m-2). No início de dezembro (fim da primavera), conforme a FIGURA 3, é feita
uma aplicação de glifosato (1,25 a 2,0 kg.ha-1 de produto comercial), permitindo a
semeadura da soja, 10 a 15 dias depois. A soja é colhida no fim de abril (FIGURA 4),
quando as plantas de azevém anual já se encontram vegetando na área, com alturas que
podem atingir 15 cm (FIGURA 5).
FIGURA 1. Pastagem de azevém anual durante o mês de junho, com altura de aproximadamente 20 cm.

13
FIGURA 2. Condição da pastagem de azevém anual durante o inverno, no momento da retirada dos animais.
FIGURA 3. Azevém anual dessecado por glifosato, no início de dezembro.

14
FIGURA 4. Condição da pastagem de azevém anual, no final do ciclo da soja, em março.
FIGURA 5. Área da pastagem de azevém anual após a colheita da soja, no fim de abril.
A soja é habitualmente cultivada por dois anos consecutivos e depois a área
permanece dois anos seguidos em pousio. No período em que a área está em pousio, o
campo natural modificado é pastejado e roçado durante o verão e o azevém anual é
pastejado durante o inverno e manejado visando continuar o sistema de ressemeadura
natural.

15
Durante o período de estudo, a soja estava sendo cultivada em duas áreas (6 e 8
anos de manejo em semeadura direta); as outras duas se encontravam em pousio (3 e 9
anos).
Antes da aplicação do glifosato para a semeadura da soja nas áreas com 6 e 8
anos, algumas sementes foram coletadas, para verificação do grau de maturidade através
do conteúdo de água.
Variáveis de fertilidade do solo de cada parcela foram analisadas.
3. Dinâmica do banco de sementes do solo
Dez amostras de solo (10 cm de profundidade x 5 cm de diâmetro) foram
retiradas em cada uma das quatro parcelas (FIGURA 6), totalizando quarenta amostras
em cada série temporal, aproximadamente a cada 30 dias, durante um ano (de dezembro
de 2002 a dezembro de 2003). Um total de 1.963 cm3 foi amostrado em cada parcela,
estando de acordo com estudos japoneses, que recomendam um volume mínimo de solo
de 400 a 600 cm3 para uma adequada estimativa do banco de sementes de campos e
pastagens (Numata et al., 1964; Hayashi & Numata, 1971).
Cada amostra foi dividida em duas profundidades (0 a 5 cm e 5 a 10 cm) e
passadas por peneiras de perfuração de 9,52 mm, para remoção de restos vegetais e
pedras, de 1,68 mm e de 0,84 mm. O resíduo concentrado, contendo as sementes, foi
examinado e as sementes de azevém anual contadas e removidas. As sementes foram
colocadas em bandejas de alumínio, contendo solo esterilizado da própria área mantido
úmido, em sala com temperatura controlada, para verificação da viabilidade (FIGURA
7). Aquelas que não germinaram foram submetidas ao teste de tetrazólio (Brasil, 1992).
As unidades amostrais foram compostas pelas médias dos resultados das dez
amostras de cada parcela, em cada profundidade.

16
FIGURA 6. Amostras de solo utilizadas na análise do banco de sementes do solo.
FIGURA 7. Germinação das sementes de azevém anual retiradas das amostras de solo, em sala climatizada.
A série de 3 anos de manejo foi amostrada somente de dezembro de 2002 a maio
de 2003, quando então não foi mais considerada no experimento, em virtude de nova
semeadura de azevém e correção da superfície do solo realizada pelo produtor, já que a
área onde a parcela se localizava apresentou problemas de alagamento e poucas
sementes sobreviveram.

17
4. Dinâmica da população de plantas
No interior de cada uma das três parcelas de cada série temporal, três quadros
contíguos, de 0,5 x 0,5 m (0,25 m2), foram utilizados para um levantamento mensal da
população de plantas de azevém, durante um ano, determinando-se o número de plantas
e de inflorescências/infrutescências, totalizando doze repetições em cada área. A média
dos valores obtidos nos três quadros em cada uma das três parcelas determinou cada
unidade amostral. De acordo com o número de inflorescências/infrutescências por metro
quadrado, observado em cada área, e o peso de mil sementes (Brasil, 1992), no fim de
novembro e início de dezembro (fim da primavera), foi determinado o número potencial
de sementes que poderiam fazer parte do banco de sementes do solo; este cálculo foi
feito através de uma estimativa do número de sementes produzidas por metro quadrado.
Os números de plântulas e de sementes na superfície do solo foram
determinados em três amostras de solo de 10 cm de diâmetro, por parcela, totalizando
doze repetições em cada série temporal, sendo cada unidade amostral formada pelos
valores médios obtidos em cada parcela.
5. Análise estatística
Análise de variância simples (ANOVA) foi realizada com os dados do banco de
sementes do solo de azevém, para a identificação de diferenças entre as séries
temporais, em cada período de coleta e entre os períodos, dentro de cada série, através
do aplicativo SPSS, utilizando-se o teste de Tukey, com 5% de probabilidade. Um teste
de homogeneidade de variâncias (Levene), também no aplicativo SPSS, foi realizado
antes da análise de variância (α de 5%). Análise de regressão também foi feita com os
mesmos dados e utilizando-se o mesmo aplicativo, buscando-se um modelo para melhor
visualização da dinâmica do BSS ao longo do ano.
Foi realizada análise multivariada dos dados do banco de sementes de azevém e
de fertilidade do solo. O teste de aleatorização dos dados de fertilidade do solo foi
realizado com o aplicativo MULTIV (Pillar, 1997), utilizando-se, como medida de
semelhança entre unidades amostrais, o Índice de Gower, já que as variáveis

18
apresentavam diferentes unidades de medida. Com este teste buscou-se identificar
diferenças significativas de fertilidade do solo das quatro séries avaliadas.
O teste de homogeneidade de variância e ANOVA também foram realizados
com estes dados, para verificar quais variáveis foram responsáveis pelas diferenças
observadas, utilizando-se o Teste de Tukey (α de 5%) do aplicativo SPSS.
Através do aplicativo SYNCA (Pillar & Orlóci, 1993), foi realizada análise de
ordenação dos dados do banco de sementes, plotando-se também, no diagrama, as
variáveis de solo com maiores índices de correlação com pelo menos um dos eixos da
ordenação. O diagrama de ordenação elaborado permitiu visualizar o posicionamento,
no espaço, de cada unidade amostral, considerando-se a quantidade de sementes de
azevém no solo, ao longo de todo o ano, e as variáveis de solo que podem influenciar
esta disposição.

RESULTADOS
No período com maior BSS, a série com 3 anos de manejo apresentou o menor
número de sementes viáveis no solo (3.363 sementes.m-2), seguida pela área com 9 anos
(6.623 sementes.m-2), ambas sem soja. As séries com 6 e 8 anos apresentaram os
maiores bancos de sementes, com 22.835 e 18.824 sementes.m-2, respectivamente,
conforme demonstrado na TABELA 1, considerando-se as duas profundidades
estudadas. Os períodos em que foram coletados os maiores números de sementes no
solo foram o início de dezembro (fim da primavera), para as séries com 6 e 9 anos, e o
fim de janeiro (verão), para as com 3 e 8 anos. Estas diferenças com relação aos
períodos devem-se, provavelmente, à retirada dos animais dos potreiros, que ocorre de
modo escalonado, pelo produtor, em função da comercialização. Se forem incorporados
a esses valores os números de sementes não viáveis, o tamanho do banco de sementes
pode variar entre 12.341 e 28.873 sementes.m-2.
Considerando-se as coletas nas quais foram verificados os maiores números de
sementes no solo e o número potencial de sementes que poderiam vir a fazer parte do
BSS, determinado através do número de inflorescências/infrutescências e o peso de mil,
a série com 3 anos de manejo apresentou um banco de sementes 56% menor; a série
com 6 anos, um banco de sementes 9% maior; com 8 anos, 4% maior e com 9 anos, um
banco 44% menor. Mais de 90% das sementes foram encontradas nos primeiros 5 cm de
solo.

20
TABELA 1. Diferenças de tamanho do banco de sementes do solo de azevém anual (0-10 cm) nas séries temporais de 3, 6, 8 e 9 anos de manejo.
Período de amostragem 3 anos 6 anos 8 anos 9 anos Setembro - 13 a 77 a 13 a Outubro - 89 a 89 a 38 a
Novembro - 22.835 c 6.203 b 2.458 a Dezembro - 13.831 b 13.398 b 6.623 a
Janeiro 3.363 a 13.194 b 18.824 b 4.941 a Fevereiro 1.605 b 3.248 b 7.909 c 319 a
Março 561 a 2.356 b 8.801 c 904 ab Abril 357 a 1.478 b 4.712 b 503 a Maio 64 a 599 a 624 a 102 a Junho - 318 a 1.541 a 217 a Julho - 166 a 128 a 38 a
Agosto - 13 a 77 a 13 a Letras diferentes na linha indicam diferenças significativas de médias a 5% de probabilidade (ANOVA seguida do Teste de Tukey) dos dados com transformação logarítmica.
Considerando-se separadamente os períodos com maior número de sementes no
solo, as áreas sem soja foram significativamente diferentes das áreas onde a soja foi
cultivada no verão.
As variáveis de composição química do solo em cada série foram
significativamente distintas. Os contrastes entre as áreas com e sem soja também
mostraram diferenças significativas de fertilidade. De acordo com o teste de
homogeneidade e análise de variância, os solos de todas as séries temporais
apresentaram diferenças significativas no conteúdo de argila, fósforo, potássio, alumínio
e cálcio, como pode ser observado na TABELA 2. O teor de matéria orgânica não
apresentou diferença significativa entre as áreas estudadas; as outras variáveis (pH,
sódio e magnésio) não foram consideradas, já que as variâncias não eram homogêneas,
de acordo com o teste de Levene.
TABELA 2. Diferenças de conteúdos de argila, matéria orgânica, fósforo, potássio, alumínio e cálcio em cada área da série temporal (n = 4), representando tempos de implantação do sistema agropastoril com semeadura direta sobre campo natural.
3 anos 6 anos 8 anos 9 anos Argila (%) 13ab 17c 15,75bc 11a
Matéria orgânica (%) 4,31a 3,38a 3,69a 4,23a Fósforo (mg/L) 10,43ab 7,33a 19,7b 11,28ab
Potássio (mg/L) 68a 135,3b 181,3b 53a
Alumínio (me/100 mL) 0,9b 0,3a 0,1a 0,08a
Cálcio (me/100 mL) 3,4a 4,05ab 5,1bc 5,48c
Letras diferentes na linha indicam diferenças significativas de médias (ANOVA seguida do Teste de Tukey).

21
O diagrama de ordenação da FIGURA 8 ilustra as diferenças do BSS de azevém
ao longo do ano, incorporando as doze coletas realizadas, e da composição química do
solo de três séries temporais (6, 8 e 9 anos). O solo da série com 3 anos de manejo
apresentou os maiores índices de alumínio e matéria orgânica; os das séries temporais
de 6 e 8 anos apresentaram os maiores conteúdos de argila; o da série de 8 anos, os
maiores índices de potássio, fósforo e magnésio, enquanto que o solo da série de 9 anos
apresentou os maiores índices de sódio e maior pH. Através da FIGURA 8 pode-se
observar possíveis influências do teor de potássio na constituição do banco de sementes
de azevém das parcelas das séries temporais de 6 e 8 anos de manejo, ambas com
cultura da soja presente no ano de avaliação, e do pH, teores de cálcio, sódio e matéria
orgânica do solo na constituição do banco das parcelas da série de 9 anos.
8D 8C8B
8A
K
CaMO
NapH 9D9C9B
9A
6D
6C
6A
6B
-20000
-15000
-10000
-5000
0
5000
10000
15000
20000
-30000 -20000 -10000 0 10000 20000 30000
FIGURA 8. Diagrama de ordenação do BSS azevém (0-10 cm) ao longo do ano nas parcelas (A, B, C e D) das séries temporais de 6, 8 e 9 anos de manejo (n = 12). As variáveis pH, matéria orgânica (MO), potássio (K), sódio (Na) e cálcio (Ca) no solo apresentam correlação acima de 0,52 com o eixo X. O eixo horizontal representa 53,8% e o vertical, 33,1% da variação do BSS de azevém ao longo do ano.
A observação da dinâmica da população de plantas de azevém anual possibilitou
um maior entendimento sobre o potencial anual de sementes que viriam constituir ou
não o BSS da área e sobre possíveis flutuações nesse fluxo de sementes anualmente
produzido.

22
No fim do verão (fim de fevereiro e março), a dormência da maior parte das
sementes que estavam no solo foi superada e, conseqüentemente, verificou-se uma
diminuição de 49% a 82% no tamanho do banco de sementes, o que coincidiu com o
início do aumento no número de plântulas. No primeiro mês do outono (abril), mais de
90% das sementes haviam germinado. Em maio, o banco de sementes tornou-se
insignificante e o número de plântulas atingiu o máximo, diminuindo posteriormente.
Um expressivo número de plantas foi observado durante este período, aumentando até o
início do inverno e diminuindo, gradativamente, até estabilizar-se logo após esse
período. No início da primavera (fim de setembro), as primeiras flores aparecem. No
fim da primavera (fim de novembro e dezembro), a deposição das sementes no solo
recomeça.
Houve diferenças significativas entre as séries temporais quanto ao tamanho do
banco de sementes de 0 a 10 cm de profundidade, do fim da primavera, quando a
dispersão das sementes se inicia, ao início do outono (pico de germinação). Não houve
diferenças do fim do outono até a próxima primavera, quando quase nenhuma semente
era encontrada no solo (TABELA 1). As sementes que se encontravam de 5 a 10 cm de
profundidade não apresentaram diferenças significativas entre as séries estudadas.
A FIGURA 9 mostra as diferenças graduais no tempo (entre as diversas coletas
ao longo do ano) do tamanho do banco de sementes nos primeiros 5 cm de solo, dentro
das séries temporais de 6, 8 e 9 anos, separadamente. As sementes que estavam de 5 a
10 cm de profundidade não foram consideradas, porque as variâncias das amostras não
foram homogêneas (Teste de Levene). Mesmo não sendo considerada em algumas
análises e nas FIGURAS 9 e 10, por não apresentar a totalidade das coletas ao longo do
ano, a série de 3 anos também apresenta diferenças significativas entre o tamanho do
banco de sementes no verão (de dezembro a março) e no outono (coleta de maio),
quando foi realizada a última coleta, neste caso.

23
0
5000
10000
15000
20000
25000
Set. Out. Nov. Dez. Jan. Fev. Mar. Maio Jun. Jul. Ag.
Período de amostragem
Sem
ente
s.m
-2
0
5000
10000
15000
20000
25000
Set. Out. Nov. Dez. Jan. Fev. Mar. Maio Jun. Jul. Ag.
Período de amostragem
Sem
ente
s.m-2
0
5000
10000
15000
20000
25000
Set. Out. Nov. Dez. Jan. Fev. Mar. Maio Jun. Jul. Ag.
Período de amostragem
Sem
ente
s.m
-2
FIGURA 9. Diferenças no tamanho do banco de sementes do solo de azevém anual nos primeiros 5 cm de solo, ao longo do ano, nas séries temporais com 6 (A), 8 (B) e 9 (C) anos de manejo (P<0,05).
Nas séries de 6 e 8 anos, houve diferenças significativas entre o período no qual
as plantas estavam florescendo, que se iniciou no fim do inverno e início da primavera
(setembro), onde quase nenhuma semente podia ser encontrada no solo, e o período no
qual iniciou-se a dispersão, no fim da primavera (fim de novembro). Durante o outono,
C
B
A
a ab
c
de de
cde cde bcd abc
a
de
de
cde cde
a a c
c c
c c bc bc ab a
abc a
a ab
e
bcd abc
abc

24
quando a maioria das sementes germina, algumas ainda se encontravam no solo (de
maio a julho). Depois de maio, quando o maior número de plântulas foi observado
(FIGURA 9), o banco de sementes começou a diminuir, tendo uma drástica redução em
junho e julho, chegando a zero em agosto.
Essas diferenças também foram significativas para a série temporal de 9 anos,
onde a soja não foi cultivada durante o período de estudo, mas a diminuição do banco
iniciou-se mais cedo (fim de março). Entretanto, o pico no número de plântulas também
ocorreu em maio (FIGURA 10).
Muitas sementes se encontravam na superfície do solo e sobre a cobertura
vegetal, conforme demonstra a TABELA 3, acompanhando a mesma dinâmica do banco
de sementes. Estas foram consideradas no total da profundidade de 0 a 5 cm.
TABELA 3. Número de sementes na superfície do solo, por metro quadrado, nas séries temporais de 6, 8 e 9 anos de manejo.
Período de amostragem 6 anos 8 anos 9 anos Setembro 0 0 0 Outubro 0 0 0
Novembro 14.621 9.263 7.915 Dezembro 104.692 173.924 117.541
Janeiro 97.594 130.029 69.582 Fevereiro 135.272 135.934 56.574
Março 172.950 141.839 43.566 Abril 159.450 221.684 254.075 Maio 145.950 270.292 21.608 Junho 0 48.650 0 Julho 0 0 0
Agosto 0 0 0

25
FIGURA 10. Dinâmica da vegetação de azevém anual nas séries temporais de 6 (A), 8 (B) e 9 (C) anos de manejo.
02000400060008000
10000120001400016000
1 2 3 4 5 6 7 8 9 10 11 12
Período de amostragemN
úmer
o.m
-2
Plantas Plântulas Inflorescências
0
20004000
60008000
1000012000
14000
16000
1 2 3 4 5 6 7 8 9 10 11 12
Período de amostragem
Núm
ero.
m-2
Plantas Plântulas Inflorescências
02000400060008000
10000120001400016000
1 2 3 4 5 6 7 8 9 10 11 12
Período de amostragem
Núm
ero.
m-2
Plantas Plântulas Inflorescências
A
B
C
Set. Out. Nov. Dez. Jan. Fev. Mar. Abr. Maio Jun. Jul. Ag.
Set. Out. Nov. Dez. Jan. Fev. Mar. Abr. Maio Jun. Jul. Ag.
Set. Out. Nov. Dez. Jan. Fev. Mar. Abr. Maio Jun. Jul. Ag.

DISCUSSÃO
As espécies anuais associadas a sistemas de semeadura direta normalmente
apresentam um banco de sementes transitório, germinando no outono e apresentando
uma dinâmica previsível (Silva, 2004), como o azevém anual nesse caso. Considerando
que a longevidade da espécie também é curta em condições naturais (Thompson et al.,
1997; ver também Artigo II), o padrão do banco de sementes do solo observado é
totalmente compreensível, onde quase todas as sementes iniciam a germinação até seis
meses após a dispersão.
O modelo cúbico mostrou ser o que melhor descreve a dinâmica do banco de
sementes do solo de azevém em todas as séries temporais e profundidades (FIGURA 11
e TABELA 4). É possível identificar três períodos distintos nesses modelos. O primeiro
período ocorre na primavera (setembro e outubro), quando as plantas iniciam o
florescimento e a formação das sementes, com praticamente nenhuma semente presente
no solo. O segundo período vai do fim da primavera até o verão (de novembro a março),
quando a dispersão e um pequeno período de dormência no solo ocorrem. Neste
período, assinalado na FIGURA 11, observa-se um alto número de sementes no solo e
também diferenças significativas no tamanho do banco de sementes entre as três séries
temporais. Com relação à dormência, Bazzigalupi (1982) relata ausência de germinação
de sementes de azevém anual imediatamente após a colheita, aumentando
gradativamente até alcançar o pico após três meses de armazenamento, o que coincide
com as condições no campo de início de germinação. O terceiro período inicia-se
quando começa a germinação, no outono (abril), mostrando uma forte redução no
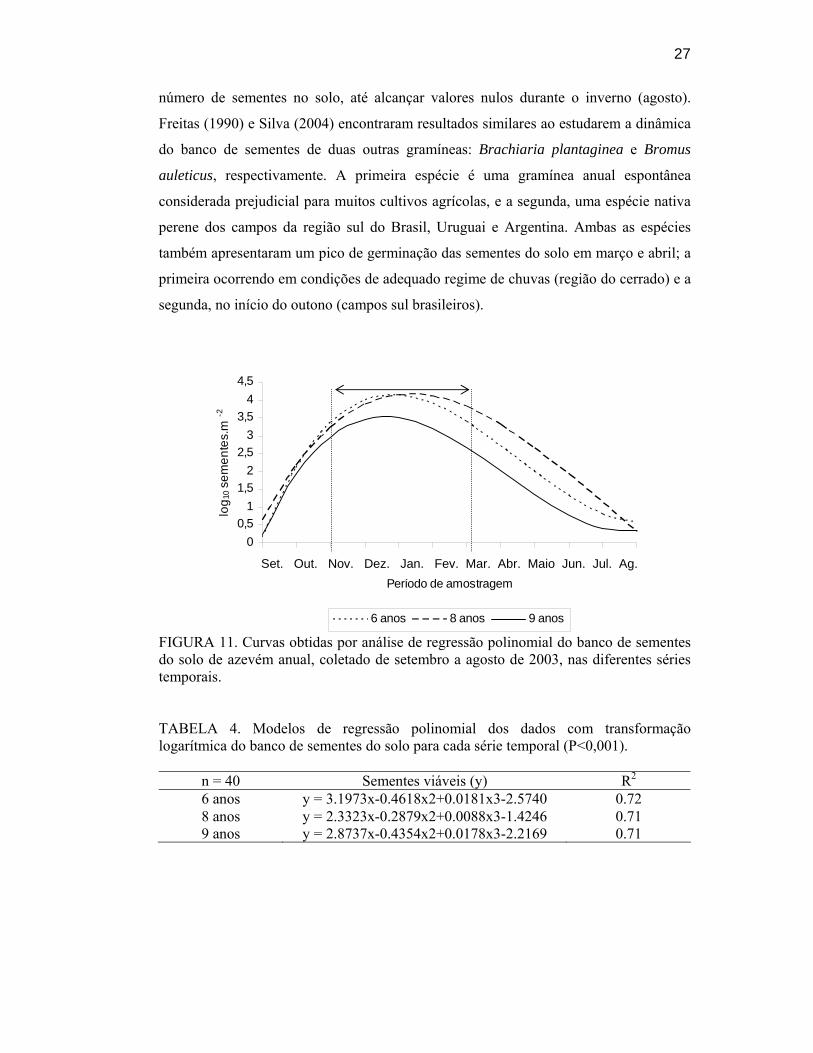
27
número de sementes no solo, até alcançar valores nulos durante o inverno (agosto).
Freitas (1990) e Silva (2004) encontraram resultados similares ao estudarem a dinâmica
do banco de sementes de duas outras gramíneas: Brachiaria plantaginea e Bromus
auleticus, respectivamente. A primeira espécie é uma gramínea anual espontânea
considerada prejudicial para muitos cultivos agrícolas, e a segunda, uma espécie nativa
perene dos campos da região sul do Brasil, Uruguai e Argentina. Ambas as espécies
também apresentaram um pico de germinação das sementes do solo em março e abril; a
primeira ocorrendo em condições de adequado regime de chuvas (região do cerrado) e a
segunda, no início do outono (campos sul brasileiros).
FIGURA 11. Curvas obtidas por análise de regressão polinomial do banco de sementes do solo de azevém anual, coletado de setembro a agosto de 2003, nas diferentes séries temporais.
TABELA 4. Modelos de regressão polinomial dos dados com transformação logarítmica do banco de sementes do solo para cada série temporal (P<0,001).
n = 40 Sementes viáveis (y) R2 6 anos y = 3.1973x-0.4618x2+0.0181x3-2.5740 0.72 8 anos y = 2.3323x-0.2879x2+0.0088x3-1.4246 0.71 9 anos y = 2.8737x-0.4354x2+0.0178x3-2.2169 0.71
00,5
11,5
22,5
33,5
44,5
1 2 3 4 5 6 7 8 9 10 11 12
Período de amostragem
log 1
0 sem
ente
s.m
-2
6 anos 8 anos 9 anos
Set. Out. Nov. Dez. Jan. Fev. Mar. Abr. Maio Jun. Jul. Ag.

28
Este padrão foi observado em ambas as profundidades (0 a 5 e 5 a 10 cm).
Mesmo sendo um sistema de semeadura direta, a dinâmica das sementes que se
encontravam de 5 a 10 cm de profundidade no solo não foi estável, mostrando o mesmo
decréscimo no fim do outono (por volta de maio), quando nenhuma semente foi
encontrada, e o mesmo aumento no fim da primavera (fim de novembro), quando as
sementes foram dispersas, mas, claro, em menor escala, devido à pequena quantidade de
sementes encontrada nesta profundidade. Isto provavelmente se deve à máquina
utilizada na semeadura da soja, que provoca distúrbios somente nas linhas de
semeaduras, com distâncias que variam de 40 a 60 cm. A soja é semeada a 2 cm de
profundidade e o fertilizante é colocado de 5 a 7 cm, permitindo às sementes que estão
na superfície e nos primeiros centímetros de solo penetrarem mais profundamente.
Pequenas diferenças nas séries temporais das áreas com e sem soja, puderam ser
observadas durante o período de amostragem. A dinâmica do banco de sementes foi a
mesma, mas o decréscimo verificado na série sem soja (9 anos) se iniciou mais cedo
(fim de março), já que a sombra da cultura não existia neste caso. Entretanto, o pico no
número de plântulas também foi em maio (FIGURA 10), como nas outras séries
temporais. Neste período do ano, a vegetação campestre nativa ainda estava cobrindo a
área, cobertura esta formada por espécies rasteiras e prostradas, decorrentes do intenso
pastejo a que a área foi submetida. Neste ambiente, as plântulas que emergiram durante
o verão estavam diretamente expostas à forte radiação solar e morreram no estágio
inicial de vida.
A sobrevivência da semente e o tamanho do banco de sementes do solo podem
depender das condições de enterrio e de hábitat, idade da semente, densidade e
predação, além de outros fatores como herbivoria, que afetam o fluxo de entrada de
sementes no solo (Alexander & Schrag, 2003). A hipótese inicial do presente trabalho
era de que o tempo de manejo realizado no local de estudo seria o principal fator a
afetar o tamanho do banco de sementes, com a série temporal de 9 anos apresentando o
maior número de sementes no solo. Entretanto, as práticas de manejo utilizadas em cada
área foram mais importantes que o fator tempo. As áreas com soja tiveram grande
influência no BSS de azevém. A cultura da soja cria um microclima, que protege as
plântulas da gramínea no início do seu desenvolvimento, como sombreamento durante o
período final de verão, reduzindo o índice de mortalidade das plântulas.

29
Além disso, a aplicação de fertilizantes na soja promove o melhoramento da
fertilidade do solo, o que se manifesta no desenvolvimento das plantas de azevém.
Conforme relatado anteriormente, algumas das variáveis de solo analisadas promoveram
diferenças significativas nos índices de fertilidade das diferentes séries, afetando
principalmente as sem soja, como a de 9 anos. O nitrato, por exemplo, é citado como
promotor da germinação de algumas sementes não dormentes (Fenner & Thompson,
2005), além de ser uma importante fonte de nutrientes e contribuir para o rápido
crescimento das plântulas.
A resposta do nitrato é interpretada como um removedor de dormência para
alguns autores (Pons, 1989), bem como um promotor da germinação em sementes não
dormentes para outros (Fenner & Thompson, 2005). O conteúdo de nitrato interno da
semente possui uma clara correlação com a germinabilidade (Bouwmeester et al.,
1994). Entretanto, o nitrato é rapidamente lixiviado da semente para o solo, fazendo
com que o conteúdo endógeno da mesma tenha pouca importância ecológica em
comparação com a sua sensibilidade aos íons de nitrato externos (Murdoch & Carmona,
1993 apud Fenner & Thompson, 2005; Bouwmeester et al., 1994).
A resposta ao nitrato pode ser usada pela semente como um mecanismo para a
detecção de espaços vazios na vegetação, através da redução deste nutriente, provocada
pelo consumo das plantas, do efeito do distúrbio e da estação do ano (Pons, 1989). Em
muitos casos, a resposta de germinação ao nitrato é influenciada por outros fatores como
a luz e a flutuação de temperatura (Probert et al., 1987). Como estes três fatores (nitrato,
luz e temperatura) se alteram, simultaneamente, quando um espaço é criado na
vegetação, uma resposta a todos eles possivelmente formaria um mecanismo de
detecção eficiente destes espaços.
Uma plântula recém emergida também está em grande desvantagem em relação
à população de plantas já estabelecida pela captura de recursos antes da formação de
suas raízes e expansão das folhas (Fenner & Thompson, 2005), como provavelmente
ocorre na área de estudo coberta por vegetação nativa. Schafer & Chilcote (1970, apud
Baskin & Baskin, 2000) relatam ocorrência de germinação de sementes enterradas de
azevém anual, seguida de morte das plântulas antes mesmo da emergência, o que
também foi observado neste trabalho. A disponibilidade de pequenos e poucos sítios
favoráveis ao estabelecimento também pode limitar o recrutamento.

30
Essas condições refletem a situação observada no campo, onde as áreas com soja
apresentaram maiores BSS do que as áreas sem a cultura. Aquelas tiveram melhores
condições nutricionais, de acordo com a análise de fertilidade do solo; principalmente,
maiores níveis de potássio e fósforo, e também mais espaços disponíveis para o
estabelecimento das plântulas de azevém, já que o controle de plantas concorrentes foi
realizado sistematicamente, com grande eficiência. O diagrama da FIGURA 10
demonstra exatamente a influência do teor de potássio do solo, por exemplo, no banco
de sementes das séries de 6 e 8 anos, as quais receberam adubação potássica para o
cultivo da soja.
Como se pode observar, a ressemeadura natural do azevém anual é totalmente
dependente do recrutamento anual de sementes; além disso, a influência das práticas de
manejo adotadas é muito maior do que o número de anos de adoção das mesmas.
Pequenos atrasos e sazonalidades na germinação estão freqüentemente
associados a alguma forma de dormência (Thompson, 2000). O presente estudo revelou
que a maior parte das sementes de azevém superou a dormência e germinou no fim do
verão e início do outono.
Muitos modelos vêm sendo publicados na tentativa de descrever e classificar os
diferentes tipos de bancos de sementes (Csontos & Tamas, 2003). Neste trabalho foi
utilizado o modelo proposto por Thompson et al. (1997), permitindo classificar o
azevém anual como uma espécie que apresenta um BSS que varia de transitório
(sementes viáveis no solo por menos de um ano) a pouco persistente (sementes viáveis
no solo por um a cinco anos), semelhantemente a resultados obtidos em outros países
(Douglas, 1965; Howe & Chancellor, 1983; López-Mariño et al., 2000). Conforme
Thompson & Grime (1979), este padrão majoritariamente transitório do banco de
sementes está associado aos distúrbios estacionais previsíveis.

CONCLUSÕES
A dinâmica do banco de sementes de azevém anual apresenta padrão estrutural
no tempo, com um período de “armazenamento” durante o verão, um período de
“exaustão” durante o outono e um período de “transição” durante o inverno e a
primavera.
O estabelecimento da pastagem através de ressemeadura natural depende do
recrutamento anual de sementes de azevém do banco de sementes do solo.
A influência das práticas de manejo no banco de sementes de azevém é muito
mais importante do que o período de tempo de aplicação destas práticas.
O tamanho do banco de sementes de azevém está positivamente associado ao
cultivo da soja, cuja integração possibilita uma maior sobrevivência de plântulas e
antecipação na produção de forragem.
A maior parte das sementes de azevém supera a dormência e germina no fim do
verão e início do outono, mostrando um banco de sementes que varia de transitório a
pouco persistente.

REFERÊNCIAS BIBLIOGRÁFICAS
ALEXANDER, H.M.; SCHRAG, A.M. Role of soil seed banks and newly dispersed seeds in population dynamics of the annual sunflower, Helianthus annuus. Journal of Ecology, v.91, p.987-998, 2003.
AYALA, W. Rol del banco de semillas en la persistencia de pasturas majoradas. In: REUNIÓN DE GRUPO TÉCNICO EN FORRAJERAS DEL CONO SUR – ZONA CAMPOS. SISTEMAS DE PRODUCCIÓN: CAMINOS PARA UNA INTEGRACIÓN SUSTENTABLE, 19., 2002, Mercedes, Argentina. Memórias... Mercedez: INTA, 2002. p.188.
BASKIN, C.C.; BASKIN, J.M. Seeds: ecology, biogeography, and evolution of dormancy and germination. 2 ed., San Diego: Academic Press, 2000. 666p.
BAZZIGALUPI, O. Efeito da época de colheita sobre o rendimento e a qualidade de sementes de azevém anual. 1982. 74f. Dissertação (Mestrado em Ciência e Tecnologia de Sementes) – Faculdade de Agronomia Eliseu Maciel, Universidade Federal de Pelotas, Pelotas, 1982.
BOLDRINI, I.I. Campos do Rio Grande do Sul: caracterização fisionômica e problemática ocupacional. Boletim do Instituto de Biociências/UFRGS, v. 56, p. 1-39, 1997.
BOUWMEESTER, H.J.; DERKS, L.; KEIZER, J.L.; KARSSEN, C.M. Effects of endogenous nitrate content of Sisymbrium officinale seeds on germination and dormancy. Acta Botanica Neerlandica, v.43, p.39-50, 1994.
BRASIL. Regras para Análise de Sementes. Brasília: Ministério da Agricultura e Reforma Agrária, 1992. 365 p.
CSONTOS, P.; TAMAS, J. Comparisons of soil seed bank classification systems. Seed Science Research, v.13, p.101-111, 2003.
DOUGLAS, G. The weed flora of chemically-renewed lowland swards. Journal of British Grassland Society, v.20, p.91-100, 1965.

33
FENNER, M.; THOMPSON, K. The ecology of seeds. Cambridge: Cambridge University Press, 2005. 250p.
FREITAS, R.R. Dinâmica do banco de sementes em uma comunidade de plantas daninhas com aspectos da germinação e dormência de sementes de capim-marmelada (Brachiaria plantagineai (Link) Hitc.). 1990. 98f. Dissertação (Mestrado em Agronomia) – Faculdade de Agronomia, Universidade Federal de Lavras, Lavras, 1990.
HAYASHI, I.; NUMATA, M. Viable buried-seed population in the Miscanthus- and Zoysia- type grasslands in Japan: Ecological studies on the buried-seed population in the soil related to plant succession. VI. Japonese Journal of Ecology, v.20, p.243-252, 1971.
HOWE, C.D.; CHANCELLOR, R.J. Factors affecting the viable seed content of soils beneath lowland pastures. Journal of Applied Ecology, v.20, p.915-922, 1983.
IBGE. /www.ibge.gov.br/ Recuperado em 2005.
LÓPEZ-MARIÑO, A.; LUIS-CALABUIG, E.; FILLAT, F.; BERMUDEZ, F.F. Floristic composition of established vegetation and the soil seed bank in pasture communities under different traditional management regimes. Agriculture Ecosystems & Environment, v.78, n.3, p.273-282, 2000.
MAIA, F.C.; MEDEIROS, R.B.; PILLAR, V.P.; FOCHT, T.; CHOLLET, D.M.S.; OLMEDO, M.O.M. Composição, riqueza e padrão de variação do banco de sementes do solo em função da vegetação de um ecossistema de pastagem natural. Iheringia (Sér. Bot.), v.58, n.1, p.61-80, 2003.
MAIA, M.S.; PRIMO, A.T. Cadeia forrageira para a região sul. In: FEDERACITE (org.). Cadeias forrageiras regionais. Porto Alegre: Caramuru, 1995. p.106-132.
MARAÑON, T. Ecologia de los bancos de semilla em el suelo: uma revisión de estúdios españoles. Pastos, v.25, n.1, p.3-25, 1995.
MEDEIROS, R.B. Bancos de sementes no solo e dinâmica vegetal. In: REUNIÃO DO GRUPO TÉCNICO REGIONAL DO CONE SUL (ZONA CAMPOS) EM MELHORAMENTO E UTILIZAÇÃO DE RECURSOS FORRAGEIROS DAS ÁREAS TROPICAL E SUBTROPICAL, 18., 2000, Guarapuava. Memórias... Guarapuava: UFPR, 2000. p.61-87.
NUMATA, M.; AOKI, K.; HAYASHI, I. Ecological studies on the buried-seed population as related to plant succession. II-Particularly on the pioneer stage dominated by Ambrosia elatior. Japonese Journal of Ecology, v.14, p.224-227, 1964.
PILLAR, V.P. Multivariate exploratory analysis and randomization testing with MULTIV. Coenoses, v.12, p.145-148, 1997.
PILLAR, V.P.; ORLÓCI, L. Character-based community analysis: the theory and an application program. The Hague: SPB, 1993. 270p. (Ecological Computation Series; v.5).
PONS, T.L. Breaking of seed dormancy by nitrate as a gap detection mechanism. Annals of Botany, v.63, p.139-143, 1989.

34
PROBERT, R.J.; GAJJAR, K.H.; HASLARN, I.K. The interactive effects of phytochrome, nitrate and thiourea on the germination response to alternating temperatures in seeds of Ranunculus sceleratus L.: a quantal approach. Journal of Experimental Botany, v.38, p.1012-1025, 1987.
THOMPSON, K. The functional ecology of seed banks. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI, 2000. p.215-235.
THOMPSON, K.; GRIME, J.P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. Journal of Ecology, v.67, p.893-921, 1979.
THOMPSON, K.; BAKKER, J.P.; BEKKER, R.M. The soil seed banks of North West Europe: methodology, density and longevity. Cambridge: Cambridge University Press, 1997.
UCPEL/ITEPA. Vinte e cinco anos de agropecuária na zona sul – RS: levantamento estatístico 1975/2000. Pelotas: Educat, 2001. 606p.

35
Longevidade de sementes de azevém anual em condições naturais. Autora: Eng. Agr. M.Sc. Fernanda Costa Maia. Orientador: Prof. Dr. Manoel de Souza Maia (UFPel). Projeto financiado pelo CNPq.
RESUMO. O interesse do trabalho foi estudar a longevidade das sementes de azevém
anual (Lolium multiflorum Lam.) em condições naturais como uma importante
informação ao estudo da sua dinâmica nos sistemas de semeadura direta na palha
utilizados no sul do Brasil. A espécie é manejada visando a ressemeadura natural,
permitindo a alimentação do gado durante a estação fria, em uma pastagem implantada
com custos reduzidos. O estudo da dinâmica do banco de sementes de azevém que vem
sendo realizado permitiu a classificação do banco de sementes da espécie como
transitório ou persistente a curto prazo. Entretanto, poucas informações estão
disponíveis sobre o efeito das condições ambientais na longevidade e sobrevivência das
sementes em condições naturais, especialmente quanto ao tempo que as não germinadas
podem permanecer viáveis no solo. Em fevereiro de 2003, vinte sacos de náilon
contendo solo esterilizado e 100 sementes foram aleatoriamente enterrados em área de
campo a 5 cm de profundidade. Em intervalos de três ou quatro meses, quatro sacos
foram sendo desenterrados. As sementes foram contadas e submetidas a testes de
germinação e tetrazólio. Inicialmente, as sementes apresentaram uma alta dormência
primária, a qual foi rapidamente superada. Após 108, 226, 326, 565 e 732 dias de
enterrio não houve diferenças significativas nas condições das sementes, já que a
dormência secundária das que não germinaram durante o outono foi baixa. A última
coleta foi a única que apresentou diferença significativa das demais devido à acentuada
queda na viabilidade das sementes. Em função da reduzida quantidade de sementes que
permaneceram viáveis após 732 dias de enterrio, o banco de sementes do solo foi
classificado como transitório, evidenciando que em pastagens anuais estes bancos
transitórios apresentam papel fundamental na regeneração da espécie.
Palavras-chave: Lolium multiflorum, viabilidade, dormência, sementes enterradas.

36
Longevity of annual ryegrass seeds in natural conditions. Author: Agr. Eng. M.Sc. Fernanda Costa Maia. Supervisor: Prof. Dr. Manoel de Souza Maia (UFPEL). Project supported by CNPq.
ABSTRACT. The interest was the seed longevity dynamic of annual ryegrass (Lolium
multiflorum Lam.) in natural conditions as an important tool to explain its dynamic in
no tillage systems used in the South of Brazil. The species is commonly managed for
natural resowing and, in this way, allows cattle grazing with reduced costs during the
winter time. We have already studied the soil seed bank dynamic of the pasture in the
first chapter, where we could classify the seed bank of the species as transient or short-
term persistent. However, there is little concern about the effects of the environmental
conditions on the longevity and survival of seeds in natural conditions to get better
information about how long these seeds that do not germinate can survive in the soil. In
February of 2003, twenty bags of nylon screen containing sterilised soil with 100 seeds
in each were randomly buried in the field, 5 cm deep. Around every three or four
months, four sacks were exhumed. Seeds were counted and tested by a germination and
tetrazolium tests. The seeds showed high primary dormancy, that was overcame very
fast. After 108, 226, 326, 565 and 732 days of burial there were no significant
differences as the secondary dormancy of the seeds that did not germinate in autumn
was not high. The last exhumation period was significantly different from the others due
to the strong decay on seed viability. As few seeds remained viable after 732 days, the
soil seed bank was classified as transient, being evident that in annual pastures the
transitory seed banks have a main role in the regeneration of the species.
Key words: Lolium multiflorum, viability, dormancy, buried seeds.

INTRODUÇÃO
As sementes constituem uma importante alternativa para a perpetuação das
plantas, notadamente para espécies anuais, cujas sementes dormentes são o elo crucial
entre as gerações (Harper, 1977). Sementes de espécies perenes normalmente não
permanecem viáveis no solo por longos períodos como as anuais (Pettit & Froend,
2001), que podem ter ciclos de vida extremamente longos (Cook, 1980). O número de
sementes viáveis no solo normalmente diminui com a redução da freqüência dos
distúrbios (impactos sobre a vegetação do ecossistema) e o aumento da longevidade da
vegetação (Symonides, 1986).
Muitas espécies que acumulam um grande número de sementes no solo possuem
sementes dormentes ou, pelo menos, populações parcialmente dormentes. O fato de
estas espécies apresentarem alto número de sementes de vida longa, aliado aos
polimorfismos com relação à dormência ou ao desenvolvimento de uma dormência
secundária, permite fluxos de germinação por longos períodos de tempo. Como essas
plantas, em geral, produzem um grande número de pequenas sementes, muitas delas
provavelmente penetram no solo e distribuem-se verticalmente nas primeiras camadas
(Bryant, 1989).
A composição do banco de sementes do solo depende da produção e da
composição das comunidades vegetais passada e presente, bem como da longevidade
das sementes de cada espécie, em condições locais (López-Mariño et al., 2000).
Stocklin & Fischer (1999) demonstraram que a longevidade das sementes está altamente

38
correlacionada com o índice de extinção de algumas espécies campestres. A
longevidade da semente é, portanto, um importante tópico na proposição de práticas de
manejo para recuperação de áreas degradadas (Bekker et al., 1998a; Bekker et al.,
1998b; Bekker & Berendse, 1999), revelando a possível vulnerabilidade da vegetação à
extinção, incluindo seu banco de semente do solo, a necessidade de sua conservação e
possibilidades de regeneração (Bekker et al., 1998c). Essa relação entre a composição
do banco de sementes do solo e a vegetação é muito importante em áreas submetidas a
diferentes práticas de manejo (Fredrickson & Taylor, 1982 apud López-Mariño, 2000).
No caso do azevém anual (Lolium multiflorum Lam.) no sul do Brasil, o
interesse foi sobre a longevidade de suas sementes em condições naturais, como um
importante tópico para a compreensão da sua dinâmica em sistemas de semeadura
direta. A espécie é manejada neste sistema visando-se à ressemeadura natural,
permitindo, desta forma, uma redução nos custos de implantação da pastagem de
inverno, para o gado utilizado neste sistema de integração lavoura e pecuária. A
dinâmica do banco de sementes do solo desta espécie já foi abordada no primeiro
capítulo, onde se pôde classificá-la como uma espécie que apresenta um banco de
sementes transitório a pouco persistente. Entretanto, existe necessidade de mais
informações sobre os efeitos das condições ambientais na longevidade e sobrevivência
das sementes em condições naturais, o que permitiria saber por quanto tempo estas
sementes que não germinam na próxima estação de crescimento podem sobreviver no
solo.
Este estudo teve por objetivo estudar a viabilidade das sementes de azevém
anual, em condições naturais.

MATERIAL E MÉTODOS
O estudo foi desenvolvido na Fazenda Santa Eulália, no sudeste do estado do
Rio Grande do Sul, município de Pelotas, na região sul do Brasil, 31º 38’ 00”S e 52º 27’
00”O, de fevereiro de 2003 a setembro de 2004.
O clima da região é considerado subtropical, com quatro estações do ano bem
definidas, chuvas bem distribuídas ao longo do ano (1.300 a 1.500 mm), temperaturas
médias anuais de 17 ºC e possível ocorrência de geadas no período de abril a setembro
(15 a 20 vezes). A área apresenta relevo suavemente ondulado e solos de textura
argilosa, não bem drenados, com boas características químicas e físicas, profundos e
com camada superficial compactada. O solo apresenta forte variação nos índices de
fósforo e potássio, de acordo com a profundidade. O pH varia de 4,4 a 5,1.
Em fevereiro de 2003, vinte sacos de náilon com solo esterilizado, contendo 100
sementes de azevém anual com 80% de poder germinativo, foram aleatoriamente
enterrados a cerca de 5 cm de profundidade.
Aproximadamente a cada três meses, quatro sacos foram desenterrados. As
sementes foram contadas e avaliadas pelo teste de germinação, em caixas gerbox, com
temperatura constante de 20 ºC, segundo as Regras para Análise de Sementes (Brasil,
1992). As que não germinaram dentro de 14 dias foram submetidas ao teste de tetrazólio
(Brasil, 1992), para verificação da viabilidade.

40
Foram realizadas análises multivariadas nos dados através do aplicativo
MULTIV 2.3.4. (Pillar, 1997; Pillar, 1994-2004). A medida de semelhança utilizada foi
a distância euclidiana e um teste de aleatorização foi realizado para identificar
diferenças significativas entre os tempos de enterrio, a 5% de probabilidade.

RESULTADOS
Como se pode ver na FIGURA 1, houve diferenças significativas entre as
condições iniciais da semente e após exumação, mas não entre estas, exceto após 226 e
565 dias de enterrio.
Em fevereiro, as sementes apresentaram um alto índice de dormência (72%);
somente 8% germinaram. De acordo com o teste de tetrazólio, cerca de 80% das
sementes enterradas estavam viáveis.
Após 108 dias de enterrio, 52% das sementes não foram mais detectadas,
provavelmente devido à germinação, mas também predação ou degradação, já que 28%
de plântulas foram detectadas nas amostras. Somente 16% das sementes ainda se
encontravam presentes e viáveis, apresentando alta qualidade, retratada pelo resultado
do teste de germinação, que evidenciou 80% de poder germinativo. Um baixo nível de
dormência foi verificado nesse período, com um pequeno aumento nas exumações
restantes (FIGURA 1).
Após 326 dias de enterrio, o índice de germinação em campo diminuiu
consideravelmente e o número de sementes não viáveis aumentou. O número de
sementes viáveis ficou praticamente o mesmo, mas a qualidade diminuiu (73% de poder
germinativo).
Após 565 dias, o número de sementes viáveis diminuiu ainda mais (11%), assim
como a qualidade (67% de sementes germinadas). O número de sementes não viáveis
diminuiu fortemente. Isto provavelmente deve-se ao fato de a identificação das

42
sementes danificadas nas amostras não ser mais possível e a maioria ter passado para a
categoria denominada “perda não identificada”.
Cerca de 732 dias após o enterrio, somente 12% do número inicial de sementes
foi encontrado, sendo 2% destas viáveis. O percentual de sementes dormentes se
manteve praticamente estável (5%). A qualidade das sementes encontradas, entretanto,
declinou bastante, ficando ao redor de 24% de germinação. Das sementes submetidas ao
teste de germinação, 50% delas não germinaram devido a algum tipo de dormência
secundária.
A FIGURA 1 retrata a situação inicial das sementes e as encontradas após cada
período de exumação, evidenciando as diferenças significativas entre estes períodos,
considerando todas as variáveis analisadas.
FIGURA 1. Alterações observadas nas sementes no experimento de enterrio e diferenças significativas (P<0,05) entre os diversos períodos de exumação das mesmas, considerando-se o conjunto das variáveis estudadas.
0%10%20%30%40%50%60%70%80%90%
100%
0 108 226 326 565 732
Dias de enterrio
Viáveis Dormentes Não viáveis Germinadas no campo Perda não identificada
a bc b bcd c d

DISCUSSÃO
As sementes no solo podem se encontrar nos mais variados estádios fisiológicos.
Embora sementes não dormentes apresentem uma leve tendência a serem menos
persistentes, praticamente todas as combinações de persistência e dormência existem
(Thompson et al., 2003).
A superação da dormência não deve ser confundida com germinação, pois são
processos completamente diferentes, que não acontecem na mesma escala de tempo e
dependem de diferentes condições ambientais (Benech-Arnold et al., 2000). As
sementes de azevém anual apresentaram alta dormência primária, que foi rapidamente
superada, provavelmente devido às altas temperaturas do verão. Posteriormente,
germinaram. A dormência secundária das sementes que não germinaram no outono não
foi muito alta, quando submetidas às condições requeridas para a germinação, durante a
condução do teste em laboratório.
A dormência das sementes de azevém anual pode ter causas ambientais ou
genéticas, podendo ser agravada por muitos fatores desconhecidos, que variam no
tempo e espaço (Jensen & Pierpoint, 1960).
A maior parte da diminuição observada no número de sementes encontradas nas
exumações após 108 dias foi, provavelmente, devido à germinação. As sementes de
azevém anual apresentam dormência até aproximadamente quatro meses após a
maturidade fisiológica, alcançando máximos valores de germinação somente após o
terceiro mês (Jensen & Pierpoint, 1961; Bazzigalupi, 1982), o que coincide com a

44
estação do ano do primeiro período de exumação. Neste período também foi possível a
observação do maior número de plântulas em área de pastagem no local (Artigo 1).
Entretanto, a predação e a degradação das sementes também podem ter contribuído com
este resultado. As conseqüências dos padrões de predação, que são muito variáveis, na
dinâmica da população de plantas são potencialmente altas (Crawley, 2000). Em seus
estudos, Hulme (1996) mostrou que sementes relativamente grandes como Festuca
pratensis, Lolium perenne e Trifolium pratense, que apresentam tamanho e forma
similares a L. multiflorum, foram as sementes mais predadas por pequenos mamíferos
após o enterrio, comparando-se com outras espécies.
Os resultados após 226 e 565 dias de enterrio foram significativamente
diferentes, provavelmente devido à alta percentagem de germinação em campo, no
primeiro caso, e alta perda não identificada, no segundo. A alta percentagem de
germinação em campo pode ser uma conseqüência do fato de alguns sacos não se
encontrarem mais na sua posição original no solo, mas muito perto da superfície,
provavelmente devido à ação de algum animal. Os outros períodos não apresentaram
diferenças significativas, tornando evidente que a superação da dormência primária e o
estímulo à germinação pelas altas temperaturas do verão, além do baixo grau de
dormência secundária, permitem a germinação da maior parte das sementes que estão
enterradas ainda no outono.
Roberts (1988) reconhece três processos fisiológicos distintos nas sementes
afetados pela temperatura: i) temperatura, junto com o grau de umidade, determina o
índice de deterioração em todas as sementes; ii) temperatura afeta o grau de remoção da
dormência, em sementes secas, e o padrão de modificações na dormência de sementes
úmidas; iii) em sementes não dormentes, a temperatura determina o grau de germinação.
Quando as sementes de espécies anuais de inverno, como o azevém, estão em
dormência primária, logo após a dispersão, a germinação está restrita, de forma
característica, a estreitas amplitudes de baixas temperaturas (Baskin & Baskin, 2000;
Probert, 2000). Durante o período de fim de primavera e início de verão, as
temperaturas do solo normalmente são maiores do que a temperatura máxima favorável
à germinação e, portanto, as sementes não germinam. Durante o período subseqüente,
mais quente, nas condições normalmente secas de verão, modificações pós-maturação
resultam em um aumento na temperatura máxima para a germinação. Durante o início
do outono, quedas na temperatura do solo coincidem com a amplitude de temperatura

45
requerida para a germinação, além de outras condições favoráveis, como adequada
disponibilidade de água no solo (Probert, 2000).
Segundo os dados da Estação Agroclimatológica de Pelotas, durante o mês de
fevereiro de 2003, a temperatura média chegou a 24,2 °C, com máxima de 34,2 °C e
mínima de 14,2 °C. Até o mês de maio, as temperaturas caíram para uma média de 16
°C e em junho, para 14,4 °C. Nos meses de março, abril e junho, a precipitação pluvial
foi maior que as normais para a região (143,8 e 151,8 mm, respectivamente). Estas
condições adequadas de temperatura e disponibilidade de água até o mês de maio
provavelmente favoreceram a germinação da maioria das sementes.
Estudando o efeito da germinação em espécies campestres no Japão, cuja
germinação era restrita ao período de primavera até o início do verão, Washitani &
Matsuda (1990) contataram que o tempo de emergência não estava correlacionado com
a presença de dormência ou de requerimentos para remoção da dormência, mostrando
que a mesma, normalmente, possui pouca influência na determinação do tempo de
germinação.
Tem sido comprovado que altas temperaturas durante o processo de maturação
reduzem a dormência das sementes de algumas espécies. Wiesner & Grabe (1972)
verificaram que cultivares de azevém anual apresentam diferentes potencialidades para
dormência, influenciadas pelas temperaturas registradas durante a maturação. Embora a
germinação no campo seja promovida, certamente, pelas temperaturas alternadas, na
maioria das espécies, experimentos laboratoriais sugerem que as sementes, na verdade,
respondem somente ao aumento da temperatura (Fenner & Thompson, 2005).
A interação da temperatura com o nitrato e a luz pode ser responsável pela
regulação dos ciclos de dormência de várias espécies (Hilhorst & Karssen, 2000). No
modelo proposto por Hilhorst (1993, apud Hilhorst & Karssen, 2000), a cadeia que leva
à germinação inicia em um receptor protéico na membrana, que fica exposto, pela ação
da temperatura na fluidez da membrana celular. Isto facilita a ligação do nitrato,
resultando em um aumento na afinidade da proteína com o fitocromo. Uma vez ligado
ao fitocromo, o complexo do receptor protéico é ativado e induz o sinal ao processo que
leva, ao final, à germinação da semente. Este modelo, embora seja somente uma
especulação a respeito do assunto, é importante por integrar os três principais fatores
ambientais (luz, temperatura e nitrato) no mecanismo da germinação. O local e o

46
período nos quais as sementes foram enterradas parecem ter refletido as condições
necessárias para o início desse processo.
Poucas sementes permaneceram viáveis por mais de um ano e meio no solo,
com um índice de germinação de 67%. McDonald et al. (1996) observaram um
comportamento similar para L. perenne, mencionando que a espécie apresenta tanto
bancos de sementes transitórios quanto persistentes a curto prazo (viáveis de 1 a 5
anos). A maioria das observações para azevém anual classificam-no como possuindo
um banco de sementes transitório, embora possam também ser encontradas
classificações que falem até em persistentes a curto e a longo prazo (Thompson et al.,
1997).
Desta forma, é possível perceber evidências de que tanto o solo como o clima
podem influenciar a longevidade das sementes (Bekker et al., 1998b; Blaney &
Kotanen, 2001; Cavieres & Arroyo, 2001). Entretanto, esta influência não é
determinante, pois, conforme Fenner & Thompson (2005) mencionam em seu livro, a
persistência é, sobretudo, uma característica da semente, que pode ser ou não
modificada pelas condições ambientais.
Como restaram poucas sementes ainda aptas a germinar após as exumações e
como os níveis de dormência foram baixos, o estudo concorda com os trabalhos de
Thompson & Grime (1979), na Inglaterra, e de D’Angela et al. (1988), na Argentina,
que classificaram o azevém anual como uma espécie com banco de sementes transitório,
em pastagens submetidas a distúrbios sazonais (seca e pastejo, por exemplo), fazendo
parte dos mecanismos regenerativos, onde mortalidades na população estabelecida de
plantas são anualmente recuperadas durante a estação do ano particularmente favorável.
Assim sendo, é evidente que, em pastagens anuais, os bancos de sementes
transitórios possuem papel fundamental na regeneração das espécies, juntamente com a
porção mais persistente de sementes (Levassor, 1990). Com esta informação, pode-se
dizer que os sistemas de semeadura direta da região, manejados visando à ressemeadura
natural da gramínea de inverno, são vulneráveis e dependentes da produção prévia de
sementes, já que o período de dormência da espécie não é tão longo quanto se previa, e
somente poucas sementes permanecem no solo ao longo do ano.

CONCLUSÕES
Uma fração das sementes de azevém anual, dispersa no fim da primavera,
mantém-se dormente no verão, sendo que as sobreviventes germinam no outono após a
remoção da dormência, provavelmente pela ação das altas temperaturas do verão e das
condições adequadas do outono.
A dormência secundária do azevém anual é muito baixa e poucas sementes
permanecem viáveis no solo ao longo do ano.
Como poucas sementes da espécie estavam aptas a germinar após mais de um
ano de enterrio, o banco de sementes foi classificado, neste estudo, como transitório.
O sistema de semeadura direta de soja sobre azevém anual, manejado para
ressemeadura natural e pastejo da gramínea, é totalmente dependente da produção anual
de sementes, já que estas não são aptas a permanecerem viáveis por longos períodos no
solo.

REFERÊNCIAS BIBLIOGRÁFICAS
BAKKER, J.P.; BERENDSE, F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends in Ecology and Evolution, v.14, n.2, p.63-68, 1999.
BASKIN, C.C.; BASKIN, J.M. Seeds: ecology, biogeography, and evolution of dormancy and germination. 2 ed., San Diego: Academic Press, 2001. 666p.
BAZZIGALUPI, O. Efeito da época de colheita sobre o rendimento e a qualidade de sementes de azevém anual. Pelotas, 1982. 74f. Dissertação (Mestrado em Ciência e Tecnologia de Sementes) – Programa de Pós-Graduação em Ciências Agrárias, Universidade Federal de Pelotas, 1982.
BEKKER, R.M.; KNEVEL, I.C.; TALLOWIN, J.B.R.; TROOST, E.M.L.; BAKKER, J.P. Soil nutrient input effects on seed longevity: a burial experiment with fen-meadow species. Functional Ecology, v.12, n.4, p.673-682, 1998a.
BEKKER, R.M.; OOMES, M.J.M.; BAKKER, J.P. The impact of groundwater level on soil seed bank survival. Seed Science Research, v.8, n.3, p.399-404, 1998b.
BEKKER, R.M.; SCHAMINEE, J.H.J.; BAKKER, J.P.; THOMPSON, K. Seed bank characteristics of Dutch plant communities. Acta Botanica Neerlandica, v.47, n.1, p.15-26, 1998c.
BENECH-ARNOLD, R.L.; SÁNCHEZA, R.A.; FORCELLA, F.; KRUKA, B.C., GUERSAA, C.M. Environmental control of dormancy in weed seed banks in soil. Field Crops Research, v.67, n.2, p.105-122, 2000.
BLANEY, C.S.; KOTANEN, P.M. Effects of fungal pathogens on seeds of native and exotic plants: a test using congeneric pairs. Journal of Applied Ecology, v.38, p.1104-1013, 2001.
BRASIL. Regras para análise de sementes. Brasília: Ministério da Agricultura e Reforma Agrária, 1992. 365p.
BRYANT, J.A. Fisiologia da Semente. São Paulo: EPU, 31 (Temas de Biologia), 1989. 86p.

49
CAVIERES, L.A.; ARROYO, M.T.K. Persistent soil seed banks in Phacelia secunda (Hydrophyllaceae): experimental detection of variation along an altitudinal gradient in the Andes of central Chile (33 degrees S). Journal of Ecology, v.89, p.31-39, 2001.
COOK, R. The biology of seeds in the soil. In: SOLBRIG, O.T. (ed.). Demography and evolution in plant populations. Botanical Monographs, v.15, 1980. p.107-129.
CRAWLEY, M.J. Seed predators and plant population dynamics. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2ed., Wallingford: CABI, 2000. p.157-191.
D’ANGELA, E.; FACELLI, J.M.; JACOBO, E. The role of permanent soil seed bank in early stages of a post-agricultural succession in the Inland Pampa, Argentina. Vegetatio, v.74, p.39-45, 1988.
FENNER, M.; THOMPSON, K. The ecology of seeds. Cambridge: Cambridge University Press, 2005. 250p.
HARPER, J.L. Population biology of plants. London: Academic Press, 1979. p.33-111.
HILHORST, H.W.M.; KARSSEN, C.M. Effect of chemical environment on seed germination. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI, 2000. p.293-309.
HULME, P.E. Herbivory, plant regeneration and species coexistence. Journal of Ecology, v.84, p.609-615, 1996.
JENSEN, L.; PIERPOINT, M. Dormancy in common ryegrass. Newsletter Association of Seed Analysis, v.35, n.4, p.31-33, 1960.
JENSEN, L.; PIERPOINT, M. Survey of post harvest dormancy in Oregon ryegrass samples. Proc. Association of Seed Analysis, n.51, p.178-183, 1961.
LEVASSOR, C.; ORTEGA, M.; PECO, B. Seed bank dynamics of Mediterranean pastures subjected to mechanical disturbance. Journal of Vegetation Science, v.1, p.339-344, 1990.
LÓPEZ-MARIÑO, A.; LUIS-CALABUIG, E.; FILLAT, F.; BERMUDEZ, F.F. Floristic composition of established vegetation and the soil seed bank in pasture communities under different traditional management regimes. Agriculture Ecosystems and Environment, v.78, n.3, 2000. p.273-282.
MC DONALD, A.W.; BAKKER, J.P.; VEGELIN, K. Seed bank classification and its importance for the restoration of species-rich flood-meadows. Journal of Vegetation Science, v.7, p.157-164, 1996.
PETTIT, N.E.; FROEND, R.H. Availability of seed for recruitment of riparian vegetation: a comparison of a tropical and a temperate river ecosystem in Australia. Australian Journal of Botany, v.49, n.4, p.515-528, 2001.
PILLAR, V.P. Multivariate exploratory analysis and randomization testing with MULTIV. Coenoses, v.12, p.145-148, 1997.
PILLAR, V.P. MULTIV: aplicativo para análise multivariada e testes de hipóteses – versões para Macintosh e Windows. Departamento de Ecologia da Universidade Federal do Rio Grande do Sul, Porto Alegre, 1994-2004.

50
PROBERT, R.J. The role of temperature in seed dormancy and germination. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI, 2000. p.261-292.
STOCKLIN, J.; FISCHER, M. Plants with longer-lived seeds have lower local extinction rates in grassland remnants 1950-1985. Oecologia, v.120, n.4, p.539-543, 1999.
SYMONIDES, E. Seed banks in old-field successional ecosystems. Ekologia Polska, v.34, n.3, p.29, 1986. (Polish Journal of Ecology)
THOMPSON, K.; GRIME, J.P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. Journal of Ecology, v.67, p.893-921, 1979.
THOMPSON, K.; BAKKER, J.P.; BEKKER, R.M. The soil seed banks of North West Europe: methodology, density and longevity. Cambridge: Cambridge University Press, 1997.
THOMPSON, K.; CERIANI, R.M.; BAKKER, J.P.; BEKKER, R.M. Are seed dormancy and persistence in soil related? Seed Science Research, v.13, p.97-100, 2003.
WASHITANI, I.; MATSUDA, M. A comparative study of the germination characteristics of seeds from a moist tall grassland community. Functional Ecology, v.4, p.543-557, 1990.

51
Alterações no banco de sementes do solo de um ecossistema campestre provocadas por nove anos de manejo agropecuário intensivo e potencial de recuperação da vegetação nativa. Autora: Eng. Agr. M.Sc. Fernanda Costa Maia. Orientador: Prof. Dr. Manoel de Souza Maia (UFPel). Projeto financiado pelo CNPq.
RESUMO. Os campos naturais na região sul do Brasil ocupam grande parte do
território e são a principal fonte de alimentos dos rebanhos bovino e ovino, além de
apresentar um alto valor ecológico, pela riqueza de espécies. Entretanto, estes campos
vêm sofrendo processo de degradação devido ao pastejo intenso, ao fogo e à
substituição por áreas de lavoura. Entre os cultivos utilizados, a soja é o que apresenta
um crescimento contínuo, ocupando áreas que antes eram formadas por ecossistemas
campestres. O objetivo deste estudo foi conhecer a dinâmica do banco de sementes do
solo (BSS) em um ecossistema de campo natural após 3, 6, 8 e 9 anos de cultivo de soja,
em sistemas de semeadura direta na palha, em rotação com azevém anual, submetido a
pastejo, buscando inferir sobre o potencial de recuperação da vegetação através do BSS.
As áreas são cultivadas com soja por dois anos e depois permanecem dois anos em
pousio. Quatro parcelas consecutivas (25 x 20 m) foram desenhadas em cada área.
Foram retiradas 10 amostras de solo de 5 cm de diâmetro, em cada parcela, divididas
em duas profundidades (0-5 e 5-10 cm). Cerca de 70% das sementes encontravam-se
nos primeiros 5 cm do solo. Os BSS das quatro séries apresentaram diferenças
significativas, entretanto, a trajetória da cenosere modelada não foi unidirecional. Os
distúrbios decorrentes do fato da área estar em pousio ou com soja apresentaram maior
influência no BSS do que o tempo de implantação do manejo. Predominaram sementes
de espécies de monocotiledôneas anuais, em termos quantitativos, e de perenes, em
termos qualitativos. O BSS da área com 9 anos de manejo apresentou a menor riqueza
de espécies, mas também a maior densidade de sementes no solo. Ao longo da cenosere,
há uma diminuição no tamanho e na riqueza do BSS, com uma substituição de espécies
perenes por anuais, em termos quantitativos, e o desaparecimento de importantes
espécies campestres nativas, já após os seis anos de manejo. Há uma tendência de que a
cada ano o controle químico provoca o reinício da sucessão na comunidade vegetal, mas
algumas espécies “escapam” e, lentamente, dão lugar ao processo sucessional.
Palavras-chave: sementes enterradas, semeadura direta, campo natural, soja, azevém anual.

52
Soil seed bank changes in a grassland ecosystem due to nine years of intensive agricultural and cattle management, and recovery potential of the vegetation. Author: Agr. Eng. M.Sc. Fernanda Costa Maia. Supervisor: Prof. Dr. Manoel de Souza Maia (UFPEL). Project supported by CNPq.
ABSTRACT. In the South of Brazil, natural grasslands occupies the major part of the
territory and are the main source of forage for ruminants, having also an important
ecological value represented by its high species richness. However, these vegetation
have been suffering great degradation process due to selective or intensive grazing, fire
and substitution by crop fields. Soybean is the main one that is replacing the natural
grassland. The study aimed to evaluate the soil seed bank (SSB) dynamic of a natural
grassland after 3, 6, 8 and 9 years of intensive disturbance by grazing and soybean
cultivation in no tillage system with annual ryegrass, trying to determine the potential of
recolonization from the seed bank. Soybean is traditionally cultivated for two years and
after that the area stays resting for two years. FFoouurr consecutive plots (25 x 20 m) were
designed in each area. Ten soil samples with 5 cm of diameter were taken every month
during one year and divided in two depths (0-5 cm and 5-10 cm).Almost 70% of the
seeds were in the first 5 cm of soil. The seed bank of all coenoseres showed significant
differences, however, the trajectory of the simulated coenosere was not unidirectional.
Disturbances due to the presence of soybean or follow in the warm season had more
influence in the seed bank, than the time factor. There were more seeds of annual
monocot species in quantitative terms and perennial species in qualitative terms. The
soil seed bank after 9 years of management had the smallest species richness, but the
highest density. Throughout the simulated coenosere, there was a decrease in the soil
seed bank size and species richness, with the replacement of perennial species by
annuals in quantitative terms, and the disappearance of important native grasses in 6
years. Each year, it seems that the chemical weed control restarts the succession, but
some species escape and slowly give place to a succession process.
Key-words: buried seeds, no tillage system, natural grassland, soybean, annual ryegrass.

INTRODUÇÃO
A paulatina degradação dos campos de pastagens naturais no mundo, em
decorrência do superpastejo e da simples substituição de áreas de pastagens nativas por
lavouras, tem provocado a erosão genética destes ecossistemas (Pott, 1989). No sul do
Brasil, particularmente no estado do Rio Grande do Sul (RS), este tipo de formação
vegetal também se encontra em processo de degradação. Em 1942, cerca de 46% da
área do estado era coberta pela vegetação campestre (Rambo, 1942); atualmente,
somente 37% da área é coberta por este tipo de vegetação (Boldrini, 1997), o que
corresponde a aproximadamente 10,5 milhões de hectares (IBGE, 1997).
Os campos naturais da região contribuem com mais de 90% da alimentação dos
cerca de 14 milhões de bovinos e 5 milhões de ovinos criados no estado
(UCPEL/ITEPA, 2001), além de constituir a base de sustentação de outros herbívoros
domésticos e dos componentes da fauna selvagem, que representam, sem dúvida, um
condicionante da vegetação hoje presente na área. São campos cuja produção de
forragem é considerada excelente para o período de primavera/verão, em função da
diversidade de espécies e da qualidade das mesmas, se comparadas com as pastagens
naturais de qualquer outra região do mundo, sendo formados por aproximadamente 400
espécies de Poaceae e 150 de Fabaceae (Boldrini, 1997).
A distribuição da produção dos campos no RS ao longo do ano é tipicamente
sazonal devido à ocorrência de baixas temperaturas durante o inverno, prejudicando o
crescimento da vegetação campestre, onde predominam espécies subtropicais de ciclo
estival. Entre as práticas mais utilizadas pelos produtores para contornar este problema

54
está a introdução de espécies exóticas de inverno, com manejo direcionado para a
ressemeadura natural da espécie introduzida, no sentido de reduzir os custos de
produção de forragem.
A adoção de sistemas agropastoris na região vem crescendo nos últimos anos,
possibilitando a utilização integral e intensiva da área durante o ano todo. Uma das
integrações mais utilizadas é a semeadura direta de soja (Glycine max L.) sobre os
restos culturais de azevém anual (Lolium multiflorum Lam.). Esta prática permite aos
produtores utilizarem a gramínea não só como cultura de cobertura do sistema, mas
também como pastagem de inverno para o gado bovino e ovino. A soja é uma das
culturas agrícolas mais cultivadas no país atualmente. O Brasil é o segundo maior
produtor mundial de soja e o RS é responsável por cerca de 25% da produção nacional
(IBGE, 2005).
Embora se disponha de um número relativamente grande de informações sobre
identificação e classificação das plantas ocorrentes em pastagens nativas, há
necessidade de maiores informações sobre sua dinâmica e os efeitos na restauração
destas áreas após cultivos intensivos, como os descritos anteriormente. Principalmente,
com relação ao efeito de herbicidas aplicados no sistema de produção.
O banco de sementes do solo (BSS) constitui um componente importante da
dinâmica vegetal, sendo um dos mecanismos de sobrevivência das espécies ao longo do
tempo. Este conjunto de sementes, juntamente com estruturas vegetativas, assegura, no
caso dos ecossistemas agropastoris, a perenidade da maioria das espécies que
constituem sua diversidade florística. A existência de uma grande população de
sementes no solo pode ter profundos efeitos na dinâmica genética e populacional das
plantas da superfície (Cook, 1980).
Além da importância produtiva, o BSS pode representar papel fundamental na
recuperação de áreas que sofreram processos de distúrbio drásticos. Para tal são
necessários entendimentos referentes ao manejo, conservação e influência dos BSS na
manutenção da diversidade florística e da sustentabilidade social e ecológica desses
ecossistemas.
A composição do banco de sementes depende da produção e composição das
comunidades vegetais anterior e atual, bem como da longevidade das sementes de cada
espécie, sob as condições locais (López-Mariño et al., 2000). Esta relação entre a

55
composição do banco de sementes e a vegetação é particularmente importante em áreas
que sofrem diferentes tipos de manejo (Fredrickson & Taylor, 1982, apud López-
Mariño, 2000).
Os impactos do uso da terra e das práticas de manejo são diferentes na vegetação
e no BSS. Os distúrbios causados pela intensificação da agricultura alteram a
composição da vegetação da área (Zanin et al., 1997), levando a um decréscimo na
riqueza de espécies, incluindo o desaparecimento de espécies características de campo.
A utilização maciça de herbicidas em sistemas de semeadura direta, pode estar
comprometendo a heterogeneidade da florística de uma vasta região de campos. Se as
aplicações de herbicidas se sucederem a cada estação de crescimento e a cada ano, e se
formulações combinadas de herbicidas forem utilizadas, é possível pensar na hipótese
de que os efeitos sobre as complexas cadeias biológicas existentes nestes ecossistemas
superem a sua resiliência, ou seja, sua habilidade para retornar à sua composição e
estrutura originais, ou muito próximas destas (Holling, 1973; Goldsmith, 1978).
Recentemente, devido a preocupações com a intensa degradação antrópica,
cresceu o interesse em estudar os BSS, com o objetivo de desenvolver modelos mais
realistas de predição da sucessão vegetal (Medeiros, 2000). Pesquisadores de vários
países têm demonstrado estreitas relações entre os BSS e a dinâmica populacional das
espécies. Muitos são os trabalhos que abordam aspectos relacionados ao BSS, em
ecossistemas campestres (Milberg, 1992; Boccanelli & Lewis, 1994; Bakker et al.,
1997; Bekker et al., 1997; Kalamees & Zobel, 1997; Morgan, 1998; Favreto et al.,
2000; Lodge, 2001; Maia et al., 2003 e 2004). Apesar da reconhecida influência dos
BSS na manutenção da dinâmica populacional de ambientes de campo nativo, pouca
importância tem sido dada aos mesmos, por parte das instituições de pesquisa do Brasil.
Pesquisas sobre a dinâmica populacional dos campos melhorados e suas relações
com os BSS a médio e longo prazo devem ser propostas. Para isto, trabalhos
abrangendo levantamentos do BSS em todas as estações do ano, e observações da
dinâmica populacional da vegetação, aliados a estudos de longevidade das sementes em
condições naturais, permitirão uma melhor compreensão sobre a dinâmica da vegetação
campestre, sob utilização agropecuária e inferências sobre o potencial de recuperação da
mesma. Deste modo, serão disponibilizadas informações que possibilitem a adoção de
práticas de manejo otimizadoras da utilização econômica desses campos, assegurando a
sua sustentabilidade.

56
Os objetivos deste estudo foram conhecer o tamanho e composição do BSS e
determinar alterações do mesmo ao longo do tempo, em ecossistema de campo natural
submetido a distúrbio por cultivo agrícola e pastejo, em sistemas de semeadura direta de
soja em azevém anual, em diferentes áreas, representando séries temporais, buscando
inferir sobre o potencial de recuperação da vegetação campestre através do seu banco de
sementes.

MATERIAL E MÉTODOS
1. Área de estudo
O trabalho foi desenvolvido no sul do Brasil, na Estância Santa Eulália, na
região sudeste do estado do Rio Grande do Sul, município de Pelotas, situado a 31º 38’
00”S e 52º 27’ 00”O.
O clima da região é considerado subtropical, com quatro estações do ano bem
definidas, precipitações pluviais totais anuais, de 1.300 a 1.500 mm, bem distribuídas ao
longo do ano, temperaturas médias anuais de 17 ºC e possível ocorrência de geadas no
período de abril a setembro. A área apresenta relevo suavemente ondulado e solo com
textura argilosa, não bem drenado, com boas características químicas e físicas, profundo
e com excessiva compactação superficial. O solo também apresenta forte variação nos
níveis de fósforo e potássio, de acordo com a profundidade. O pH varia de 4,4 a 5,1.
A Estância Santa Eulália possui uma área física de 2500 ha, sendo 850 ha destinados
à semeadura direta de soja e azevém anual em sucessão, por ressemeadura natural, para
pastejo de gado bovino. Os quatro potreiros em estudo medem 100 ha,
aproximadamente, cada um representando um tempo de implantação do sistema de
manejo acima citado (série temporal), sendo estes 3, 6, 8 e 9 anos. Quatro parcelas
consecutivas, de 25 x 20 m cada, em faixa, foram demarcadas em cada série temporal.
O local foi escolhido de modo a estar situado em posições intermediárias do relevo e
com boa representação do sistema de manejo utilizado.

58
Variáveis indicadoras da fertilidade do solo de cada área foram analisadas em
laboratório.
2. Sistema de semeadura direta na palha de azevém anual e soja
O azevém anual foi primeiramente semeado (30 kg.ha-1) em campo nativo
dessecado. A semeadura não foi mais realizada nos anos subseqüentes. Os potreiros são
divididos com cerca elétrica e utilizados de forma rotativa para pastejo de gado bovino,
com capacidade média de 1,5 unidade animal.ha-1.ano-1, alcançando uma produtividade
comercial de aproximadamente 450 kg. ha-1.ano-1 de peso vivo. O período de pastejo é
usualmente de junho (fim do outono) a outubro (primavera), quando os animais são
vendidos ou retirados para outra área, e o nitrogênio, na forma de uréia, é aplicada para
favorecer a recuperação das plantas para a ressemeadura natural (50 kg.ha-1). No início
de dezembro (fim da primavera), é feita uma aplicação de glifosato (1,25 a 2,0 kg.ha-1
de produto comercial), permitindo a semeadura da soja, 10 a 15 dias depois. Ela é
tradicionalmente cultivada por dois anos sucessivos, e depois a área permanece mais
dois anos em pousio, sendo mantido o pastejo. A cultura é colhida no fim de abril,
quando as plantas de azevém anual já se encontram vegetando na área, com alturas que
podem atingir 15 cm, conforme observado no campo.
3. Dinâmica do banco de sementes do solo
Dez amostras de solo (10 cm de profundidade x 5 cm de diâmetro) foram
retiradas em cada uma das quatro parcelas, totalizando quarenta amostras em cada série
temporal, no mês de maio de 2003. Cada amostra foi dividida em duas profundidades (0
a 5 cm e 5 a 10 cm). As amostras foram passadas por peneiras de perfuração de 9,52
mm, para remoção de restos vegetais e pedras. O resíduo concentrado, contendo todas
as sementes da amostra, foi distribuído em camadas sobre solo esterilizado da área, com
igual volume de vermiculita (Favreto et al., 2000), em bandejas de alumínio colocadas
em casa de vegetação. As plântulas foram então identificadas, contadas e removidas. As

59
plântulas não identificadas foram transplantadas para recipientes maiores e cultivadas,
até que estruturas reprodutivas permitissem sua identificação.
Foram realizados dois ciclos de germinação, com a finalidade de forçar a
exaustão do BSS. A fim de remover a dormência entre cada ciclo de germinação, as
bandejas permaneceram secas por um período de sete dias (Favreto et al., 2000; Maia et
al., 2003 e 2004).
4. Análise estatística
Na análise dos dados foram adotados procedimentos de análise multivariada, que
englobam síntese das informações obtidas, busca exploratória de padrões de variação do
BSS, correlações com fatores ambientais e estudo de efeitos específicos por testes de
aleatorização, através dos aplicativos MULTIV (Pillar, 1997) e SYNCSA (Pillar &
Orlóci, 1993).
Através do teste de aleatorização (Pillar & Orlóci, 1996), disponibilizado pelo
aplicativo MULTIV, utilizando-se como medida de semelhança a distância euclidiana
entre as unidades amostrais, buscou-se detectar diferenças significativas na composição
do BSS e de variáveis químicas do solo das quatro séries, nas duas profundidades
avaliadas, com base em um índice de significância de 5%. Os dados do BSS de 0 a 5 cm
foram transformados vetorialmente, através da normalização, dentro de variáveis, e os
de 5 a 10 cm foram transformados com raiz quadrada.
O aplicativo SYNCSA forneceu a freqüência e a dominância das espécies em
cada série, em cada uma das profundidades. Também foram realizadas análises de
ordenação das unidades amostrais, através de coordenadas principais, utilizando-se
como medida de semelhança a distância euclidiana, e correlação das variáveis de solo
com os eixos da ordenação. Os diagramas de ordenação elaborados permitiram uma
visualização do posicionamento de cada unidade amostral no espaço, considerando-se a
composição e tamanho do banco de sementes, em cada uma das profundidades
estudadas, e as variáveis de solo que podem influenciar esta disposição.

60
Com este mesmo aplicativo, utilizaram-se as séries temporais estudadas na
tentativa de elaboração de modelo baseado em cadeias de Markov (Orlóci et al., 1993;
Quadros & Pillar, 2001). Para isto, os bancos de sementes de diferentes idades, das
diferentes séries, representaram uma série temporal modelada, tratada como cenosere,
ou seja, o conjunto de estados do BSS no tempo.

RESULTADOS
1. Tamanho e composição do BSS
Foram encontradas 96 espécies nas quatro séries temporais (ANEXO), 30 delas
exclusivamente nos primeiros 5 cm e 11, de 5 a 10 cm de profundidade. Cerca de 70%
das sementes estavam na profundidade de 0 a 5 cm, representando um valor médio de
786.098 sementes.m-2. Aproximadamente 52% das espécies presentes no BSS eram
perenes, e 34% eram anuais. As outras espécies apresentaram variação relativamente a
duração de vida, conforme as condições ambientais nas quais estão vegetando, sendo
10% perenes ou anuais e 3% anuais ou bianuais1. As espécies perenes predominaram
em ambas as profundidades estudadas. Entretanto, a maior quantidade de sementes foi
de espécies anuais.
A maioria das espécies encontradas pertence às famílias Poaceae, Asteraceae,
Cyperaceae e Juncaceae. As 30 espécies com maior BSS em todas as séries temporais,
nas duas profundidades estudadas, estão identificadas na FIGURA 1. As famílias
Poaceae, Cyperaceae e Apiaceae predominaram na profundidade de 0 a 5 cm. Espécies
das famílias Amaranthaceae, Caryophillaceae, Hypoxidaceae, Iridaceae, Lamiaceae,
Lythraceae, Malvaceae, Plantaginaceae, Polygonaceae, Rubiaceae, Solanaceae,
Urticaceae e Verbenaceae foram encontradas exclusivamente na profundidade de 0 a 5
1 Espécies que podem apresentar duração de vida tanto anual como perene, dependendo das condições ambientais nas quais se encontrem vegetando.

62
cm. A família Juncaceae predominou na profundidade de 5 a 10 cm, e espécies das
famílias Primulaceae e Scrophulariaceae foram exclusivas desta profundidade.
A riqueza em cada área foi semelhante, ao redor de 60 espécies, excetuando-se a
área com 9 anos de manejo, que apresentou 49 espécies, ou seja, uma redução de 18,3%.
Os bancos de sementes das séries temporais apresentaram diferença
significativa, nas duas profundidades. Entre as duas profundidades estudadas, também
se verificaram diferenças significativas nos bancos das quatro séries. Além disso, foi
possível verificar uma interação altamente significativa entre o número de anos de
implantação do manejo e a profundidade do BSS.
As espécies comuns e as características dos BSS das séries temporais, nas
profundidades de 0 a 5 e 5 a 10 cm, estão apresentadas nas TABELAS 1 e 2,
respectivamente. Observa-se que a maioria das espécies comuns às quatro áreas é anual.
FIGURA 1. Tamanho do BSS das 30 espécies com maior quantidade de sementes, nas duas profundidades de solo estudadas, nas quatro séries temporais. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Juncus bufonius, Lolium multiflorum, Gamochaeta spp., Kyllinga brevifolia, Sisyrinchium micranthum, Digitaria ciliaris, Plantago spp., Cuphea carthagenensis, Centela asiatica, Paspalum nicorae, Trifolium repens, Eragrostis neesii, Juncus cf. effusus, Lythrum hyssopifolium, Cerastium rivulare, Cyperus eragrostis, Kyllinga odorata, Fimbristylis diphylla, Cyperaceae 1, Centunculus minimus, Conyza cf. floribunda, Digitaria adscendens.
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
Jubu
Lom
u
Gas
p
Kybr
Sim
i
Dic
i
Plsp
Cuc
a
Cea
s
Pan
i
Trre
Ern
e
Jucf
Lyhy Cer
i
Cye
r
Kyod FIdi
Cyp
Cem
i
Coc
f
Dia
d
Sem
ente
s.m-2
0-5 cm 5-10 cm

63
TABELA 1. Espécies comuns e características dos bancos de sementes do solo de 0 a 5 cm, em cada série temporal, com base na freqüência (%). Letras diferentes significam diferenças significativas na composição do banco de sementes (P<0,05).
3 anosa
(n = 4) 6 anosb
(n = 4) 8 anosc
(n = 4) 9 anosd
(n = 4) Espécies comuns: freqüência de 100% em mais de uma área.
Lolium multiflorum 100 100 100 100 Gamochaeta spp. 100 100 100 100
Kyllinga brevifolia 100 100 100 100 Sisyrinchium micranthum 100 100 100 100
Digitaria ciliaris 100 100 100 100 Juncus bufonius 100 100 100 100
Cerastium glomeratum 100 100 100 75 Plantago spp. 100 100 25 100
Centunculus minimus 100 75 75 100 Eragrostis neesii 100 50 50 100
Oxalis cf. conorhiza 75 100 100 75 Espécies características: freqüência de 100% em uma das áreas.
Fimbristylis autumnalis 100 75 25 75 Centela asiatica 50 100 25 Sida rhombifolia 50 100 25 Trifolium repens 75 75 50 100
Juncus dichotomus 75 50 50 100 Eclipta megapotamica 75 50 100 Lythrum hyssopifolium 75 100
TABELA 2. Espécies comuns e características dos bancos de sementes do solo de 5 a 10 cm, em cada série temporal, com base na freqüência (%). Letras diferentes significam diferenças significativas na composição do banco de sementes (P<0,05).
3 anosa
(n = 4) 6 anosb
(n = 4) 8 anosc
(n = 4) 9 anosd
(n = 4) Espécies comuns: freqüência de 100% em mais de uma área.
Sisyrinchium micranthum 100 100 100 100 Juncus bufonius 100 100 75 100 Digitaria ciliaris 100 75 100 50 Gamochaeta spp. 50 100 100 25
Espécies características: freqüência de 100% em uma das áreas. Centunculus minimus 100 50 75 75
Fimbristylis autumnalis 100 25 50 Juncus dichotomus 100 25 75 25 Kyllinga brevifolia 100 50 75 50
Oxalis cf. conorhiza 75 100 75 50 Hydrocotyle exígua 25 100 25 25

64
Na série de 3 anos de manejo foram encontradas 60 espécies, com 71% das
sementes nos primeiros 5 cm de profundidade. Cerca de 60% das 50 espécies
encontradas no BSS de 0 a 5 cm de profundidade, nesta área, pertencem às famílias
Poaceae (13), Asteraceae (8), Cyperaceae (5) e Juncaceae (4), com predominância de
perenes (49%). Cerca de 49% das espécies possuem duração de vida perene, 33% anual,
13% anual ou perene e 4% anual ou bianual, nesta profundidade. As espécies da família
Juncaceae foram as que apresentaram maior BSS, neste caso (235.355 sementes.m-2),
seguidas pelas de Poaceae (130.924 sementes.m-2), Asteraceae (110.292 sementes.m-2)
e Cyperaceae (80.405 sementes.m-2). Na profundidade de 5 a 10 cm foram encontradas
38 espécies (pequena predominância de anuais), nessa mesma área, sendo 54% delas
das famílias Cyperaceae (8 espécies e 40.501 sementes.m-2), Asteraceae (6 espécies,
56.547 sementes.m-2), Poaceae (6 espécies, 77.263 sementes.m-2), que também
apresentou o maior número de sementes no solo nesta profundidade, e Juncaceae (4
espécies, 40.160 sementes.m-2). Neste caso, 46% das espécies possuem duração de vida
anual. As 20 espécies com maior número de sementes no solo nas duas profundidades
estudadas, na série de 3 anos de manejo, estão identificadas na FIGURA 2.
FIGURA 2. Espécies com maior BSS, nas duas profundidades de solo estudadas, na série temporal de 3 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Juncus bufonius, Gamochaeta spp., Plantago spp., Digitaria ciliaris, Coniza cf. floribunda, Digitaria adscendens, Kyllinga brevifolia, Sisyrinchium micranthum, Cuphea carthagenensis, Lolium multiflorum, Eragrostis neesii, Cyperus reflexus, Paspalum nicorae, Cerastium glomeratum, Trifolium repens, Eclipta megapotamica, Fimbristylis autumnalis, Cyperaceae 1.
0
50000
100000
150000
200000
250000
300000
Jubu
Gas
p
Plsp Dic
i
Coc
f
Dia
d
Kyb
r
Sim
i
Cuc
a
Lom
u
Ern
e
Cyr
e
Pan
i
Ceg
l
Trre
Ecm
e
Fiau
Cy1
Cea
s
Cob
o
Sem
ente
s.m-2
0-5 cm 5-10 cm

65
Na série de 6 anos de manejo foram encontradas 58 espécies, com 75% das
sementes nos primeiros 5 cm de profundidade. Cerca de 58% das 49 espécies
encontradas no BSS de 0 a 5 cm de profundidade, nesta área, pertencem às famílias
Cyperaceae (10), Poaceae (9), Asteraceae (5) e Juncaceae (5). As espécies da família
Poaceae foram as que apresentaram maior BSS, neste caso (101.886 sementes.m-2),
seguidas pelas de Asteraceae (88.131 sementes.m-2), Cyperaceae (49.840 sementes.m-2)
e Iridaceae (41.518 sementes.m-2). Na profundidade de 5 a 10 cm foram encontradas 36
espécies, nesta mesma área, sendo 61% delas das famílias Cyperaceae (8 espécies e
18.001 sementes.m-2), Poaceae (8 espécies, 10.190 sementes.m-2) e Juncaceae (6
espécies, 19.359 sementes.m-2). As três espécies de Asteraceae encontradas nesta
profundidade apresentaram o maior BSS, com uma média de 37.188 sementes.m-2. As
espécies perenes predominaram em ambas as profundidades, principalmente na maior.
As vinte espécies com maior número de sementes no solo, nas duas profundidades
estudadas, nesta área, estão mostradas na FIGURA 3.
FIGURA 3. Espécies com maior BSS, nas duas profundidades de solo estudadas, na série de 6 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Gamochaeta spp., Lolium multiflorum, Sisyrinchium micranthum, Centela asiática, Juncus bufonius, Kyllinga brevifolia, Eragrostis neesii, Centunculus minimus, Hydrocotyle exigua, Plantago spp., Soliva pterosperma, Hypoxis decumbens, Cerastium glomeratum, Oxalis cf. conorrhiza, Kyllinga vaginata, Lotus subbiflorus, Eleusine tristachya, Cyperus cf. cayennensis, Juncus capillaceus, Kyllinga odorata.
Na série de 8 anos de manejo foram encontradas 60 espécies, com 78% das
sementes nos primeiros 5 cm de profundidade. Cerca de 59% das 45 espécies
encontradas no BSS de 0 a 5 cm de profundidade, nesta área, pertencem às famílias
0
20000
40000
60000
80000
100000
120000
Gas
p
Lom
u
Sim
i
Cea
s
Jubu
Kyb
r
Ern
e
Cem
i
Hye
x
Pls
p
Sop
t
Hyd
e
Ceg
l
Oxc
f
Kyv
a
Losu Eltr
Cyc
f
Juca
Kyo
d
Sem
ente
s.m-2
0-5 cm 5-10 cm

66
Cyperaceae (12), Poaceae (7) e Asteraceae (7). As espécies da família Poaceae foram
as que apresentaram maior BSS, neste caso (293.939 sementes.m-2), seguidas pelas de
Cyperaceae (69.453 sementes.m-2) e Asteraceae (65.972 sementes.m-2). Na
profundidade de 5 a 10 cm foram encontradas 36 espécies nesta mesma área, sendo 60%
delas das famílias Cyperaceae (7 espécies e 31.584 sementes.m-2), Poaceae (7 espécies,
18.170 sementes.m-2), Juncaceae (4 espécies, 21.736 sementes.m-2) e Apiaceae (4
espécies, 14.263 sementes.m-2). Houve predominância de espécies perenes em ambas as
profundidades, principalmente na de 5 a 10 cm. A FIGURA 4 mostra as 20 espécies
com maior número de sementes no solo, nas duas profundidades estudadas, nesta área.
0
50000
100000
150000
200000
250000
300000
350000
Lom
u
Gas
p
Kyod
Jubu
Cye
r
Juca
Kybr FIdi
Cy1
Sim
i
Judi
Oxc
f
Cea
s
Amli
Gls
e
Kyva
Cem
i
Dis
e
Hyd
e
Cy2
Sem
ente
s.m-2
0-5 cm 5-10 cm
FIGURA 4. Espécies com maior BSS, nas duas profundidades de solo estudadas, na série de 8 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Lolium multiflorum, Gamochaeta spp., Kyllinga odorata, Juncus bufonius, Cyperus eragrostis, Juncus capillaceus, Kyllinga brevifolia, Fimbristylis dyphilla, Cyperaceae 1, Sisyrinchium micranthum, Juncus dichotomus, Oxalis cf. conorrhiza, Centela asiática, Amaranthus lividus ssp. Polygonoides, Glandularia selloi, Kyllinga vaginata, Centunculus minimus, Dichondra sericea, Hypoxis decumbens, Cyperaceae 2.
Na área com 9 anos de manejo foram encontradas 49 espécies, com 70% das
sementes nos primeiros 5 cm de profundidade. Cerca de 55% das 42 espécies
encontradas no BSS de 0 a 5 cm de profundidade, nesta área, pertencem às famílias
Poaceae (7), Cyperaceae (6), Asteraceae (5) e Juncaceae (5). As espécies da família
Juncaceae foram as que apresentaram maior BSS, neste caso (203.686 sementes.m-2),
seguidas pelas de Poaceae (165.393 sementes.m-2) e Cyperaceae (114.451 sementes.m-
2). As 5 espécies de Asteraceae apresentaram um dos menores BSS (27.424 sementes.

67
m-2) em comparação com outras famílias como Lythraceae, Iridaceae, Caryophillaceae,
Plantaginaceae e Fabaceae, que apresentaram mais sementes no solo. Na profundidade
de 5 a 10 cm foram encontradas 26 espécies nesta mesma área, sendo 47% delas das
famílias Cyperaceae (9 espécies e 196.723 sementes.m-2) e Apiaceae (3 espécies, 9.680
sementes.m-2). As espécies de Cyperaceae foram as que apresentaram maior BSS nesta
profundidade, seguidas de 2 espécies de Juncaceae (63.848 sementes.m-2) e de
Sisyrinchium micranthum (18.085 sementes.m-2), da família Iridaceae. Sementes de
espécies com duração de vida anual apresentaram pequena predominância nos primeiros
5 cm de solo, enquanto que as perenes predominaram na maior profundidade (5 a 10
cm). As 20 espécies com maior número de sementes no solo nas duas profundidades
estudadas, nesta área, podem ser visualizadas na FIGURA 5.
0
50000
100000
150000
200000
250000
300000
Jubu
Kybr Dic
i
Lom
u
Sim
i
Jucf
Cer
i
Plsp
Trre
Cuc
a
Kyod
Lyhy Judi
Erne
Fiau
Cem
i
Cy1
Poan
Gas
p
NãI
d
Sem
ente
s.m-2
0-5 cm 5-10 cm
FIGURA 5. Espécies com maior BSS, nas duas profundidades de solo estudadas, na série de 9 anos de manejo. As espécies estão identificadas segundo as duas primeiras letras dos epítetos genérico e específico: Juncus bufonius, Kyllinga brevifolia, Digitaria ciliaris, Lolium multiflorum, Sisyrinchium micranthum, Juncus cf. effusus, Cerastium rivulare, Plantago spp., Trifolium repens, Cuphea carthagenensis, Kyllinga odorata, Lythrum hyssopifolium, Juncus dichotomus, Eragrostis neesii, Fimbristylis autumnalis, Centunculus minimus, Cyperaceae 1, Poa annua, Gamochaeta spp.

68
2. Dinâmica do BSS
As chamadas cadeias de Markov são utilizadas com o propósito de modelar tanto
o passado quanto o futuro. Uma cenosere é puramente markoviana quando a descrição
de seus estados podem ser derivados dos estados anteriores e da matriz transicional
conhecida, que expressa o nível pelo qual cada população cede lugar a outra, quando a
cenosere se move de um de seus estados para o próximo (anos). O modelo seria
utilizado para a predição da trajetória do BSS, extrapolando para um futuro de 20 anos
as tendências observadas, e abreviando-se o tempo necessário para observação dos
efeitos do manejo utilizado na área sobre o BSS e a recuperação do ecossistema natural.
Em virtude de o modelo markoviano exigir intervalos regulares de observação, foi
realizada a interpolação dos dados anteriormente à análise, obtendo-se uma série
modelada de 3 a 9 anos, em intervalos de um ano. Posteriormente, verifica-se se a
cenosere é estacionária, ou seja, se mantém seu alvo em estado constante, ou dinâmica
(em movimento), ou seja, se vai reajustando o alvo de tempos em tempos; neste último
caso, a cenosere é dita caótica, onde cada estado (ano) transita para outro através de
uma matriz de transição diferente, não podendo ser expressa pelo modelo markoviano.
O estado de caos pode ser simulado por permutações aleatórias das unidades amostrais
observadas, gerando valores de estresse (Orlóci et al., 1993). O estresse aumenta
conforme as transições se tornam caóticas. A hipótese nula testada é de que a cenosere é
indireta, não direcionada, caótica. Neste caso, o valor de estresse obtido foi alto, com
P>0,10. Desta forma, como a proporção dos valores de estresse simulados menor ou
igual ao observado foi maior que o α (0,10) assumido, a hipótese nula foi aceita e o
modelo não pôde ser utilizado para representar a trajetória caótica do BSS.
Através da análise de ordenação, pode-se visualizar a semelhança entre o BSS
das áreas com 3 e 9 anos de manejo e entre o das áreas com 6 e 8 anos, o que evidenciou
a trajetória dinâmica e instável do BSS da área referida anteriormente.
Os solos das quatro áreas apresentaram diferenças significativas de fertilidade. O
contraste entre os solos das áreas com soja e das em pousio apresentaram diferenças
significativas (P<0,05), principalmente devido à adubação utilizada na área cultivada.

69
O diagrama de ordenação da FIGURA 6 ilustra a composição do BSS das
parcelas estudadas, na profundidade de 0 a 5 cm, juntamente com as espécies de maior
correlação com os eixos da ordenação. O diagrama ilustra apenas o banco de sementes
dos primeiros 5 cm de profundidade de solo, devido à grande predominância das
sementes nesta camada. Verifica-se que os bancos das séries de 3 e 9 anos de
implantação, ambas com campo natural modificado no momento da coleta e onde não
foram feitas aplicações de herbicida, apresentaram alta correlação com as espécies Briza
minor, Conyza cf. floribunda, Cuphea carthagenensis, Eclipta megapotamica, Digitaria
ciliaris, Fimbristylis autumnalis, Juncus bufonius, Panicum dichotomiflorum e Poa
annua e com os teores de alumínio e sódio no solo. Os bancos de sementes das séries de
6 e 8 anos de manejo, ambas com cultura da soja no momento da coleta, caracterizaram-
se pela presença de sementes de Amaranthus lividus, Centela asiatica, Dichondra
sericea e Lolium multiflorum, e pelos teores de magnésio, fósforo e potássio no solo.
FIGURA 6. Diagrama de ordenação da composição do BSS de 0 a 5 cm, nas parcelas das quatro séries temporais (3, 6, 8 e 9 anos de manejo), e linha de tendência. As espécies identificadas, segundo as duas primeiras letras dos epítetos genérico e específico, em itálico: Amaranthus lividus ssp. polygonoides, Briza minor, Centela asiatica, Conyza bonariensis, C. cf. floribunda, Cuphea carthagenensis, Dichondra sericea, Digitaria ciliaris, Eclipta megapotamica, Fimbristylis autumnalis, Juncus bufonius, Lolium multiflorum, Panicum dichotomiflorum e Poa annua apresentam correlação acima de 0,50 com pelo menos um dos eixos de ordenação. O eixo horizontal representa 64,6%, e o vertical, 16,9% da variação da composição do BSS da série.
3D
9D9A
6A6B
6D3A
6C
8A8D
8B
8C
9C 9B3B3C
JubuPoanBrmiPadi Cocf
FiauCoboDici
EcmeCuca
Ceas
AmliDise
Lomu
-200000
-150000
-100000
-50000
0
50000
100000
150000
-300000 -200000 -100000 0 100000 200000 300000 400000

70
Sementes de Amaranthus lividus e Dichondra sericea foram encontradas
somente na série de 8 anos de manejo, enquanto que de Briza minor, Conyza cf.
floribunda e Panicum dichotomiflorum foram encontradas somente no início da
cenosere (3 anos). Sementes de Conyza bonariensis foram encontradas nas séries de 3 e
6 anos somente. Sementes das espécies Cuphea carthagenensis, Digitaria ciliaris e
Juncus bufonius estavam presentes no banco de sementes de todas as áreas, aumentando
no decorrer da cenosere. Sementes de Eclipta megapotamica e Fimbristylis autumnalis
também foram encontradas em todas as áreas, mas diminuíram conforme aumentava o
tempo de implantação do manejo. Sementes de Poa annua mantiveram-se em número
constante no solo no decorrer da cenosere, até os 9 anos. O banco de sementes de
Lolium multiflorum (azevém anual) foi maior nas séries de 6 e 8 anos (ver Artigo I).
A presença de algumas espécies como aveia-preta (Avena strigosa) e trevo
vermelho (Trifolium pratense), na série de 3 anos de manejo, deve-se a uma semeadura
realizada após o primeiro cultivo de soja. As sementes de aveia-preta foram encontradas
somente na profundidade de 5 a 10 cm e as de trevo vermelho, em ambas,
predominando nos primeiros 5 cm. O trevo branco (T. repens) também foi semeado
nessa mesma época, mas se perpetuou por mais tempo, estando presente em toda a
cenosere, nas duas profundidades.
Outras espécies como Borreria capitata, Cerastium rivulare, Crysanthemum
myconis, Gratiola peruviana, Hydrocotyle verticillata, Hypochaeris chillensis, Hypoxis
decumbens, Juncus dichotomus, J. dombeyanus, J. cf. effusus, Kyllinga brevifolia, K.
odorata, K. vaginata, Lythrum hyssopifolium, Oxalis sp., Panicum decipiens, P.
racemosum, Rhynchospora rugosa, Sisyrinchium micranthum, Solanum americanum,
Soliva pterosperma e Trifolium repens aumentaram seus bancos de sementes ao longo
da cenosere. Por outro lado, espécies como Baccharis trimera, Briza minor, Chaptalia
nutans, Conyza bonariensis, C. cf. floribunda, Cyperus spp., Digitaria adscendens,
Fimbristylis dichotoma, F. diphylla, Lotus subbiflorus, Luzula campestris, Panicum
aristellum, P. dichotomiflorum, Paspalum nicorae, P. urvillei, Pluchea sagittalis,
Polygonum hydropiperoides, Setaria sphacelata, Verbena litoralis e Wahlenbergia
linarioides tendem a desaparecer conforme os anos de manejo repetido.
As demais espécies tendem a diminuir seus bancos de sementes na cenosere.

DISCUSSÃO
Os principais distúrbios que influenciaram as diferenças dos BSS das quatro
áreas foram o uso agrícola, a intensa densidade de plântulas de azevém anual,
direcionada pelo manejo praticado no local e o pastejo, tanto do azevém, no inverno,
como do campo natural modificado, no verão. A influência do tempo de manejo parece
não ter sido tão marcante na composição e tamanho do banco de sementes quanto as
práticas agrícolas utilizadas a cada dois anos e o pastejo constante. Menalled et al.
(2001) fizeram observação semelhante com a composição de espécies na vegetação
infestante de cultivos agrícolas, que variou em função do manejo utilizado, mas sem
uma influência do tempo de avaliação.
Em virtude dessa trajetória caótica da cenosere do banco de sementes, não se
pôde utilizar o modelo markoviano inicialmente proposto. Este nível de caos observado
pode ser resultante principalmente da hipótese levantada no parágrafo anterior, além das
diferenças de relevo e de solo das áreas estudadas, que parecem ser preponderantes
sobre o tempo de implementação do manejo.
As menores diferenças observadas entre o BSS das áreas de 3 e 9 anos e o das
áreas de 6 e 8 anos de manejo também evidenciam essa impossibilidade de modelagem
unidirecional. As áreas com 3 e 9 anos de manejo se encontravam em situação de pousio
de segundo ano, no momento da coleta, onde o único distúrbio, além do pastejo do
azevém anual com baixa densidade e do campo natural, era a roçada mecânica;
enquanto isso, as áreas com 6 e 8 anos encontravam-se com a cultura da soja, quando
também foram realizadas diversas outras práticas de distúrbio, como adubação do solo e

72
uso de herbicidas, seguindo-se a emergência do BSS de azevém anual com altíssima
população, pastejado com alta carga animal, no período de inverno até meados de
novembro.
Em virtude do manejo intensivo utilizado, com agricultura e pecuária,
simultaneamente, houve um predomínio no BSS de espécies monocotiledôneas anuais,
em termos quantitativos, e de espécies de monocotiledôneas perenes, em termos
qualitativos. Sementes de espécies perenes geralmente não permanecem viáveis no solo
tanto quanto as anuais (Roberts, 1970; Pettit & Froend, 2001), que podem ter ciclos de
vida extremamente longos (Cook, 1980), entretanto, esta não é uma regra geral.
Segundo Fenner & Thompson (2005), alguns hábitats como os campos não são
necessariamente associados com uma persistência alta ou baixa das sementes,
principalmente porque tais comunidades vegetais são compatíveis com uma ampla gama
de regimes de distúrbios (pastejo, seca, fogo, etc.). Neste caso, os distúrbios promovidos
por este tipo de manejo agropecuário intensivo parecem estar favorecendo espécies com
bancos de sementes transitórios. Os BSS transitórios podem ter sua composição e
tamanho fortemente modificados de um ano para o outro. Em virtude disto, o fato de a
área se encontrar em um período de pousio ou de cultivo de soja pode apresentar uma
influência mais marcante no BSS do que o tempo de implementação deste manejo.
Esta influência também pode ser observada com relação ao tamanho do BSS. As
áreas onde a soja estava sendo cultivada no momento (6 e 8 anos) apresentaram as
menores quantidades de sementes no solo. Amplas variações observadas na densidade
do banco de sementes também foram relatadas por Roberts & Neilson (1981),
Schweizer & Zimdahl (1984), Cardina et al. (1991), Mayor & Dessaint (1998), Vanasse
& Leroux (2000), Menalled et al. (2001) e Medeiros & Steiner (2002), que trabalharam
em sistemas de cultivo com uso de herbicidas. Um resultado comum destes estudos foi
que o uso de herbicidas e outras práticas de manejo podem reduzir a densidade do banco
de sementes, assim como ocorreu nas áreas de 6 e 8 anos, onde herbicidas foram
utilizados intensa e recentemente no controle de plantas indesejáveis. A alta densidade
observada na área de 9 anos pode-se dever ao fato de há dois anos não estarem sendo
feitas aplicações de herbicidas ou/e a oscilações de curto prazo, que provocam
diferenças na dormência das sementes, devido a condições climáticas específicas do
período, conforme a hipótese relatada por Menalled et al. (2001). Mayor & Dessaint
(1998) também concordam com esta hipótese de forte influência ambiental

73
determinando a germinabilidade do banco de sementes. Os autores observaram que a
densidade do banco de sementes pode variar em mais de 100%, de um ano para outro.
O fenômeno conhecido como “dinâmica de espaços abertos na comunidade
vegetal” tem sido descrito por vários autores (Daubenmire, 1988; Glenn-Lewin & Van
der Maarel, 1992; Bullock, 2000). Os distúrbios causados pelo uso de herbicidas,
durante o cultivo da soja, e pelo pastejo promovem o aparecimento de solo descoberto e
uma maior heterogeneidade espacial e temporal (Denslow, 1987), formando microsítios
de recrutamento constante (Boldrini, 1993) e diminuindo o tamanho do BSS, conforme
evidenciado nas áreas de 6 e 8 anos, mas determinando um possível aumento da riqueza
florística da vegetação (Focht, 2001; Maia et al., 2003). Em ambientes com alta
freqüência de sítios regenerativos, o número de espécies, de início de sucessão aumenta,
determinando uma maior diversidade de espécies pela redução da presença de espécies
dominantes (Connell, 1980).
É a imprevisibilidade do distúrbio, ao invés da sua severidade, que seleciona as
sementes persistentes (Peco et al, 1998; Fenner & Thompson, 2005). Distúrbios
moderados podem promover a diversidade de espécies pela redução da dominância das
mais vigorosas, prevenindo a exclusão competitiva, assim como distúrbios mais
drásticos podem reduzir a diversidade, prevenindo a regeneração de espécies de vida
longa (Fenner & Thompson, 2005). A área com 9 anos demonstrou claramente esta
tendência, pois foi a que apresentou a menor riqueza de espécies. Além disto, neste
caso, houve um aumento na riqueza de espécies anuais em comparação com as outras
áreas, promovendo uma equiparação com o número de espécies perenes.
Isto evidencia o maior tempo de utilização agropecuária ao qual a área vem
sendo submetida, onde as espécies anuais na vegetação tendem a adquirir uma maior
importância, pela sua relação com a severidade do distúrbio em termos temporais
amplos (Zanin et al., 1997). Esta relação pode-se refletir no banco de sementes, que
também possui a tendência de aumentar o número de sementes de espécies anuais e a
correspondência com a vegetação, quanto maior o distúrbio (Lewis, 1973; Harper, 1977;
Levassor et al., 1990; Boccanely & Lewis, 1994; Fenner & Thompson, 2005).
Entretanto, esta hipótese deve ser vista com ressalva, neste caso, já que o efeito do
tempo em uma escala mais ampla é questionável, e o fato de predominarem espécies
anuais pode somente refletir uma diminuição dos espaços abertos na vegetação quando
cessa o cultivo da soja e a área entra em pousio.

74
A diminuição brusca na riqueza de espécies do banco de sementes, a partir de
nove anos, pode estar também relacionada a variáveis de fertilidade do solo. A partir da
composição de espécies encontrada no banco de sementes, das parcelas com 3 anos de
manejo (53 espécies), verificou-se uma queda já no próximo período, que seriam as
áreas com 6 anos (46 espécies). Posteriormente, parece que o efeito da fertilidade do
solo, devido às adubações sucessivas realizadas durante o cultivo da soja, altera
novamente a composição de espécies, com um novo aumento na riqueza (53).
O diagrama de ordenação da FIGURA 6 ilustra a relação da composição do
banco de sementes das parcelas das séries de 6 e 8 anos com os solos mais férteis,
evidenciada pelas altas correlações com as variáveis fósforo, potássio e magnésio. O
teor de fósforo é reportado como possuindo um maior efeito sobre a composição de
espécies do que sobre a densidade (Albrecht & Auerswald, 2003). Já na área de 9 anos,
que se encontrava no segundo ano sem adubação pelo fato de estar em pousio, foram
encontradas somente 43 espécies em um solo com maiores teores de alumínio e sódio,
apesar de apresentar a maior densidade de sementes.
Albrecht & Auerswald (2003) correlacionam altos números de sementes com
solos de baixa fertilidade. A presença de compostos orgânicos, devido à grande
concentração de raízes das plantas de azevém (4.000 a 10.000 plantas.m-2), também
pode influenciar na redução do tamanho do BSS, devido à germinação. O etileno, por
exemplo, é um composto constituinte da atmosfera do solo, que é produzido por muitos
tipos de bactérias. Fenner & Thompson (2005) mencionam o composto como sendo um
estimulador da germinação de algumas poucas espécies. O significado ecológico do
etileno do solo para a germinação das sementes no campo não é ainda conhecido. Um
sinergismo entre o etileno e o nitrato é visto em algumas espécies (Saini et al., 1986). A
habilidade de responder a ambos os compostos simultaneamente pode possibilitar à
semente mecanismos de detecção do fluxo de atividade microbiana, indicativo de solos
com recente perturbação (Fenner & Thompson, 2005).
Bakker et al (1997) enfatizam que a composição de espécies do BSS no local
cultivado é influenciada pela exploração agrícola. Sistemas de semeadura direta, em
comparação com sistemas de manejo de solo tradicionais, com lavração e gradagem,
apresentam uma certa sucessão ecológica em direção a comunidades mais maduras, com
mais espécies perenes presentes na vegetação, em termos qualitativos, e, em termos
quantitativos, a comunidades características de espécies pioneiras de sucessão

75
secundária (Zanin et al., 1997; Favreto, 2004), como observado neste estudo (maior
riqueza de espécies perenes e maior quantidade de sementes de espécies anuais no
BSS). Menalled et al. (2001) também relatam sobre a predominância de sementes de
espécies de monocotiledôneas anuais no solo, principalmente da família Poaceae, em
sistemas de semeadura direta.
O preparo reduzido do solo também deixa as sementes próximas da superfície.
Observou-se que 70 a 78% das sementes encontravam-se nos primeiros 5 cm de
profundidade, indicando que o estabelecimento da vegetação natural através de
sementes de camadas mais profundas é pouco provável. Segundo Yenish et al. (1992),
em sistemas de semeadura direta, 60% de todas as sementes presentes no perfil do solo
encontram-se localizadas entre 0-1 cm de profundidade, com poucas sementes abaixo de
10 cm. Em ecossistemas naturais de campo, vários estudos também mencionam uma
predominância das sementes nas camadas superiores, como em Bekker et al. (1999).
O manejo empregado no local visa favorecer a ressemeadura natural do azevém
anual. Quando não se considera esta espécie nas avaliações, o banco de sementes das
áreas com soja sofrem grandes alterações nas proporções de espécies anuais e perenes,
inclusive passando a um domínio de perenes na área com 8 anos de manejo, em ambas
as profundidades, e na profundidade de 5 a 10 cm, na área com 9 anos. Na profundidade
de 5 a 10 cm, mesmo considerando-se o banco de sementes de azevém anual, as
sementes de espécies perenes passaram a predominar sobre as de anuais, nas áreas de 8
e 9 anos. Em virtude da intensa relação conhecida de espécies anuais na vegetação em
ambientes com alto grau de distúrbio como os agrícolas, como a área com 8 anos (Zanin
et al, 1997; Fenner & Thompson, 2005), o recrutamento pode ter sido estimulado pelo
grande número de espaços abertos, causando uma redução na quantidade de sementes
de espécies anuais no solo. Bullock (2000) relata vários estudos que evidenciaram uma
maior emergência plântulas de espécies com sementes pequenas, em espaços abertos na
vegetação, do que de sementes grandes. Sabe-se que espécies anuais tendem a ter
sementes pequenas (Fenner & Thompson, 2005), o que é outra evidência para a hipótese
de um maior recrutamento destas.
Mesmo sendo um sistema de manejo de semeadura direta, a presença de uma
maior quantidade de sementes de espécies perenes em maiores profundidades, pode
indicar este grande recrutamento de sementes de espécies anuais mencionado, nessas
áreas que sofreram distúrbios por mais tempo, com um provável predomínio destas na

76
vegetação. Nos primeiros centímetros, na área com 9 anos, devido ao tempo de pousio
(segundo ano no momento da coleta) e à existência de poucos espaços abertos na
vegetação campestre natural, o recrutamento das espécies, entre estas as anuais, na
superfície diminuiu, sendo somente visualizado em maiores profundidades.
Outra possível interpretação é de que devido ao maior tempo sem ocorrer
revolvimento do solo, as espécies perenes estejam sendo favorecidas. Entretanto, esta
última hipótese é pouco provável, devido às práticas de controle de plantas indesejáveis,
com herbicida, que são realizadas durante o período em que as áreas estão sendo
cultivadas com soja.
Torna-se evidente que, conforme o distúrbio diminui, mesmo que por alguns
anos somente, parece haver um direcionamento para um banco de sementes de espécies
perenes, com menor densidade, possivelmente de caráter transitório. Isto pode refletir
uma estratégia de sobrevivência da vegetação, com rápida reocupação dos espaços
abertos de solo provocados pelo cultivo da soja ou pastejo intenso. A resiliência
observada, entretanto, pode ocasionar grande erosão genética, pois o pastejo intenso é
mantido e, logo em seguida (após dois anos), o cultivo da soja retorna, alterando
novamente a composição e tamanho do BSS. Alguns trabalhos apontam o banco de
sementes como sendo o principal responsável pela recolonização de áreas campestres
após distúrbio (Zimmergren, 1980; Pakeman et al., 1998).
O tamanho do banco de sementes geralmente diminui com a redução da
freqüência de distúrbio e o aumento da longevidade das espécies que formam a
vegetação (Thompson, 1978; Symonides, 1986). O fato de as duas áreas em pousio (3 e
9 anos de manejo) apresentarem os maiores BSS é outro indício para o que foi afirmado
anteriormente sobre o caráter transitório do mesmo, sendo determinado pelas práticas de
manejo.
Segundo o projeto RADAMBRASIL (Teixeira et al., 1986), a vegetação da
região da Serra do Sudeste, onde se localiza a área de estudo, corresponde à savana,
com arbustos e árvores isoladas no estrato superior. O estrato inferior é constituído
especialmente por gramíneas. Entre as várias espécies que ocorrem na região estão
Adesmia punctata, Andropogon lateralis, A. selloanus, A. ternatus, Aristida jubata, A.
filifolia, A. spegazzini, A. circinalis, A. vernustula, Axonopus affinis, Baccharis
coridifolia, B. trimera, Briza subaristata, Chloris polydactyla, Clitoria nana,
Desmodium incanum, Eriosema tacuaremboense, Eryngium horridum, Galactia neesii,

77
Hypochaeris spp., Hypoxis decumbens, Lathyrus pubescens, Luziola peruviana,
Paspalum compressifolium, P. notatum, P. nicorae, P. urvillei, Piptochaetium
montevidense, Schizachyrium tenerum, Rhynchosia diversifolia, Stipa filifolia e
Trifolium polymorphum (Boldrini, 1997).
Observou-se que a maioria das espécies encontradas no BSS não são espécies
campestres e sim de plantas consideradas invasoras (Brachiaria plantaginea, Solanum
sisymbrifolium, S. americanum, Sida spp., etc.). Isto pode-se dever ao longo período
pelo qual a área vem sendo submetida ao cultivo da soja. Algumas sementes de espécies
campestres podem ser inclusive, de plantas não mais presentes na vegetação (Milberg,
1992), mas, embora muitas possuam uma dispersão limitada (Verkaar et al., 1983;
Spence, 1990), a dispersão recente de áreas adjacentes não pode ser excluída.
É importante mencionar que a entrada do gado bovino não se deu de forma
constante e uniforme em todas as áreas, estando dependente da condição da pastagem, o
que pode influenciar a dispersão zoocórica e o depósito de sementes no solo. Sabe-se
que a maior parte das sementes são dispersas próximas à planta-mãe (Wilson &
Traveset, 2000), mas outra parte importante é ingerida pelos herbívoros, transportadas e
expulsas nos excrementos. Marañon (1995) relata vários estudos sobre o papel das
sementes com dispersão endozoocórica, em campos espanhóis. Estes estudos
contabilizaram mais de 20 sementes.cm-2 de 46 espécies campestres no esterco das
vacas, durante a primavera, o que presume um banco concentrado de 200.000
sementes.cm-2 no solo. Malo & Suárez (1995) concluíram que uma vaca pode dispersar
cerca de 300.000 sementes por dia. O potencial de recuperação da vegetação pelo BSS
pode ser drasticamente afetado com o tempo de utilização da área, principalmente pela
diminuição da riqueza das espécies que formam o banco de sementes, conforme
evidenciado na área de 9 anos de manejo, mais intensivamente utilizada em escala
temporal. Em sistemas de cultivo onde o uso de herbicidas é a ferramenta principal de
controle de plantas indesejáveis, a diversidade de espécies no banco de sementes pode
ser drasticamente afetada (Menalled et al., 2001; Medeiros & Steiner, 2002).
As diferenças significativas entre os bancos de sementes das quatro séries
confirmam certa alteração na composição de espécies e número de sementes ao longo
do tempo, conforme relatado anteriormente. Entretanto, da mesma forma como ocorreu
em Bekker et al. (1997), a maioria das variáveis de solo quantificadas não explica de
forma significativa a variação observada no banco de sementes das quatro áreas, exceto

78
fósforo, potássio, magnésio, alumínio e sódio, que apresentaram uma boa correlação
com a variação do BSS. Embora essas variáveis possam aparentemente parecer de
reduzida significância no processo, elas podem ser, ao contrário, de alta relevância e
determinantes na resposta da vegetação.
O aumento na abundância de sementes de espécies anuais em sistemas de
semeadura direta na palha, com aplicação constante de herbicida, também vem sendo
observado em vários trabalhos (Forcella & Lindstrom, 1988; Buhler & Oplinger, 1990;
McCloskey et al., 1996; Swanton et al., 1999; Menalled et al., 2001). Já no início da
avaliação, após 3 anos de implementação do manejo utilizado na fazenda, não se
verificou a presença de várias espécies citadas por Boldrini (1997), como características
dos campos da região no BSS. Algumas espécies das famílias Cyperaceae e Poaceae
aumentaram seus bancos de sementes, enquanto que famílias como Asteraceae e
Apiaceae apresentaram forte diminuição.
Conforme progrediu a cenosere, o tamanho e a riqueza do banco de sementes
diminuiu, com uma substituição de espécies perenes por espécies anuais, em termos
quantitativos, e o conseqüente desaparecimento de espécies nativas de importante valor
forrageiro, como as do gênero Paspalum, já após 6 anos de manejo.
De acordo com estudo relatado por Zanin et al. (1997) sobre a dinâmica da
vegetação de plantas indesejáveis sob diferentes sistemas de cultivo, a combinação da
ausência e redução dos distúrbios causados pelo controle químico com herbicidas
dessecantes e competição com as plantas de azevém anual, capazes de produzirem
sucessões secundárias anualmente, afetou o banco de sementes. Isto ocorreu,
provavelmente, como um reflexo da alteração ocorrida na comunidade vegetal, em duas
diferentes direções evolutivas: uma tipicamente sucessional e outra de recolonização
repetida. A cada ano, o controle químico provoca o reinício da sucessão na comunidade
vegetal, mas algumas espécies “escapam” e, lentamente, dão lugar ao processo
sucessional.

CONCLUSÕES
A condição atual de manejo da área (pousio ou cultivo da soja) apresenta maior
influência na composição e tamanho do BSS do que o tempo de implementação dos
manejos.
A redução do distúrbio pelo pousio, mesmo que somente por dois anos, induz a
um direcionamento para um banco de sementes de espécies perenes, com menor
densidade, possivelmente de caráter transitório, visando a uma rápida reocupação dos
espaços abertos no solo, provocados pelo cultivo agrícola ou pelo pastejo intenso.
Ao longo da cenosere há uma diminuição no tamanho e na riqueza do banco de
sementes, com uma substituição de espécies perenes por espécies anuais, em termos
quantitativos, e o desaparecimento de importantes espécies campestres nativas, já após
seis anos de manejo.
A maior quantidade de sementes encontra-se nos primeiros 5 cm de
profundidade do solo.
Embora o BSS das quatro séries temporais tenha apresentado diferenças
significativas, a trajetória da cenosere não é unidirecional.
Predominaram nos bancos de sementes, em termos quantitativos, espécies de
monocotiledôneas anuais e, em termos qualitativos, de monocotiledôneas perenes.
As áreas com solos mais férteis, com recente aplicação de fertilizantes,
apresentaram uma maior riqueza, enquanto as áreas menos férteis apresentaram maior
densidade de sementes no solo.

REFERÊNCIAS BIBLIOGRÁFICAS
ALBRECHT, H.; AUERSWALD, K. Arable weed seedbanks and their relation to soil properties. Aspects of Applied Biology, v.69, p.11-20, 2003. (Seedbanks: Determination, Dynamics & Management)
BAKKER, J.P.; BAKKER, E.S.; RÓSEN, E.; VERWEIJ, G.L. The soil seed bank of undisturbed and disturbed dry limestone grassland on Öland (Sweden). Zeitschrift für Ökologie und Naturschutz, v.6, p.9-18, 1997.
BEKKER, R.M.; VERWEIJ, G.L.; SMITH, R.E.N.; REINE, R.; BAKKER, J.P.; SCHNEIDER, S. Soil seed banks in European grasslands: does land use affect regeneration perspectives? Journal of Applied Ecology, v.34, p.1293-1310, 1997.
BEKKER, R.M.; LAMMERTS, E.J.; SCHUTTER, A.; GROOTJANS, A.P. Vegetation development in dune slacks: the role of persistent seed banks. Journal of Vegetation Science, v.10, p.745-754, 1999.
BOCCANELLI, S.I.; LEWIS, J.P. The seed bank of old pampean prairie and its relation with the standing vegetation. Pesquisa Agropecuária Brasileira, v.29, p.1833-1840, 1994.
BOLDRINI, I.I. Dinâmica de vegetação de uma pastagem natural sob diferentes níveis de oferta de forragem e tipos de solos, Depressão Central, RS. 1993. 262f. Tese (Doutorado em Zootecnia) – Faculdade de Agronomia, Universidade Federal do Rio Grande do Sul, Porto Alegre, 1993.
BOLDRINI, I.I. Campos do Rio Grande do Sul: caracterização fisionômica e problemática ocupacional. Boletim do Instituto de Biociências/UFRGS, v.56, p.1-39, 1997.
BUHLER, D.D.; OPLINGER, E.S. Influence of tillage systems on annual weed densities and control in solid-seeded soybean (Glycine max). Weed Science, v.38, p.158-165, 1990.
BULLOCK, J.M. Gaps and seedling colonization. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI Publishing, 2000. p.375-395.

81
CARDINA, J.; REGNIER, E.; HARRISON, K. Long-term tillage effects on seed banks in three Ohio soils. Weed Science, v.39, p.186-194, 1991.
COOK, R. The biology of seeds in the soil. In: SOLBRIG, O.T. (ed.). Demography and evolution in plant populations. Botanical Monographs, v.15, p.107-129, 1980.
CONNEL, J.H. Diversity and coevolution of competition, or the ghost of competition past. Oikos, v.35, p.131-138, 1980.
DAUBENMIRE, R.F. Ecology of fire in grasslands. Advances on Ecological Research, v.5, p.109-266, 1988.
DENSLOW, J.S. Tropical rainforest gaps and tree species diversity. Annual Review of Ecology and Systematics, v.18, p.431-451, 1987.
FAVRETO, R. Vegetação espontânea e banco de sementes do solo em área agrícola estabelecida sobre campo natural. 2004. 116 f. Dissertação (Mestrado em Ecologia) - Centro de Ecologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, 2004.
FAVRETO, R.; MEDEIROS, R.B.; PILLAR, V.D.D. Composição do banco de sementes do solo de um campo natural sujeitos a intensidades de pastejo e posições de relevo. In: REUNIÃO DO GRUPO TÉCNICO REGIONAL DO CONE SUL (ZONA CAMPOS) EM MELHORAMENTO E UTILIZAÇÃO DE RECURSOS FORRAGEIROS DAS ÁREAS TROPICAL E SUBTROPICAL, 18., 2000, Guarapuava, Anais... Guarapuava: UFPR, 2000. p.233-235.
FENNER, M., THOMPSON, K. The ecology of seeds. Cambridge: Cambridge University Press, 2005. 250p.
FOCHT, T. Padrões espaciais em comunidades vegetais de um campo pastejado e suas relações com fatores de ambiente. 2001. 157f. Dissertação (Mestrado em Ecologia) – Centro de Ecologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, 2001.
FORCELLA, F.; LINDSTROM, M.J. Weed seed populations in ridge and conventional tillage. Weed Science, v.36, p.500-503, 1988.
GLENN-LEWIN, D.C.; VAN DER MAAREL, E. Patterns and processes of vegetation dynamics. In: GLENN-LEWIN, D.C.; PEET, R.K.; VEBLEN, T.T. (eds.). Plant Succession: Theory and prediction. Londres: Chapman & Hall, 1992, p.11-59.
GOLDSMITH, E. Complexity and stability in the real world. The Ecologist Quarterly, Winter, n.4, p.305-316, 1978.
HOLLING, C.S. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics, v.4, p.1-23, 1973.
IBGE. \www.ibge.gov.br\ Recuperado em 1997.
IBGE. \www.ibge.gov.br\ Recuperado em 2005.
KALAMEES, R.; ZOBEL, M. The seed bank in an Estonian calcareous grassland: comparison of different successional stages. Folia Geobotanica, v.32, n.1, p.1-14, 1997.
LODGE, G.M. Studies of soil seedbanks in native and sown pastures in northern New South Wales. Rangeland Journal, v. 23, n. 2, p. 204-223, 2001.

82
LÓPEZ-MARIÑO, A., LUIS-CALABUIG, E., FILLAT, F., BERMUDEZ, F.F. Floristic composition of established vegetation and the soil seed bank in pasture communities under different traditional management regimes. Agriculture Ecosystems & Environment, v.78, n.3, p.273-282, 2000.
MAIA, F.C., MEDEIROS, R.B., PILLAR, V.P., FOCHT, T., CHOLLET, D.M.S., OLMEDO, M.O.M. Composição, riqueza e padrão de variação do banco de sementes do solo em função da vegetação de um ecossistema de pastagem natural. Iheringia (Sér. Bot.), v. 58, n. 1, p. 61-80, 2003.
MAIA, F.C.; MEDEIROS, R.B.; PILLAR, V.P.; FOCHT, T. Soil seed bank variation patterns according to environmental factors in a natural grassland. Revista Brasileira de Sementes, v.26, n.2, p.126-137, 2004.
MALO, J.E.; SUÁREZ, F. Herbivorous mammals as seed dispersers in a Mediterranean dehesa. Œcologia, v.104, p.246-255, 1995.
MARAÑON, T. Ecología de los bancos de semilla en el suelo: una revisión de estúdios españoles. Revista Pastos, v.25, n.1, p.3-25, 1995.
MAYOR, J.P.; DESSAINT, F. Influence of weed management strategies on soil seedbank diversity. Weed Research, v.38, p.95-105, 1998.
McCLOSKEY, M.; FIRBANK, L.G.; WATKINSON, A.R.; WEBB, D.J. The dynamics of experimental arable weed communities under different management practices. Journal of Vegetation Science, v.7, p.799-808, 1996.
MEDEIROS, R.B. Bancos de sementes no solo e dinâmica vegetal. IN: REUNIÃO DO GRUPO TÉCNICO REGIONAL DO CONE SUL (ZONA CAMPOS) EM MELHORAMENTO E UTILIZAÇÃO DE RECURSOS FORRAGEIROS DAS ÁREAS TROPICAL E SUBTROPICAL, 18., 2000, Guarapuava. Memórias... Guarapuava: UFPR, 2000. p.61-87.
MEDEIROS, R.B.; STEINER, J.J. Influence of temperate grass seed rotation systems on weed seed soil bank composition. Revista Brasileira de Sementes, v.24, n.1, 2002.
MENALLED, F.D.; GROSS, K.L.; HAMMOND, M. Weed aboveground and seedbank community responses to agricultural management systems. Ecological Applications, v.11, n.6, p.1586-1601, 2001.
MILBERG, P. Seed bank in a 35-year-old experiment with different treatments of a semi-natural grassland. Acta Œcologica, v.13, n.6, p.743-752, 1992.
MORGAN, J.W. Composition and seasonal flux of the soil seed bank of species-rich Themeda triandra grasslands in relation to burning history. Journal of Vegetation Science, v.9, n.2, p.145-156, 1998.
PAKEMAN, R.J.; ATTWOOD, J.P.; ENGELEN, J. Sources of plants colonizing experimentally disturbed patches in an acidic grassland, in eastern England. Journal of Ecology, v.86, p.1032-1041, 1998.
PECO, B.; ORTEGA, M.; LEVASSOR, C. Similarity between seed bank and vegetation in Mediterranean grassland: a predictive model. Journal of Vegetation Science, v.9, p.815-828, 1998.
PETTIT, N.E., FROEND, R.H. Availability of seed for recruitment of riparian vegetation: a comparison of a tropical and a temperate river ecosystem in Australia. Australian Journal of Botany, v.49, n.4, p.515-528, 2001.

83
PILLAR, V.P. Multivariate exploratory analysis and randomization testing with MULTIV. Coenoses, v. 12, p. 145-148, 1997.
PILLAR, V.P.; ORLÓCI, L. Character-based community analysis: the theory and an application program. The Hague: SPB, 1993. 270p. (Ecological Computation Series; v.5).
PILLAR, V.P.; ORLÓCI, L. On randomization testing in vegetation science: multifactor comparisons of relevé groups. Journal of Vegetation Science, v. 7, p. 585-592, 1996.
POTT, A. O papel da pastagem na modificação da vegetação climax. In: FAVORETO, V.; RODRIGUES, L.R.A. (eds.). SIMPÓSIO SOBRE ECOSSISTEMA DE PASTAGENS, 1989, Jaboticabal. Anais... Jaboticabal: FUNEP, 1989. p.43-68.
QUADROS, F.L.F.; PILLAR, V.P. Dinâmica vegetacional em pastagem natural submetida a tratamentos de queima e pastejo. Ciência Rural, v. 31, n. 5, p. 863-868, 2001.
RAMBO, B. A fisionomia do Rio Grande do Sul. Porto Alegre: Of. Gráf. da Imprensa Oficial, 1942. 394p.
ROBERTS, H.A. Viable weed seeds in cultivated soils. Report of the National Vegetable Research Station, 25-38p., 1970.
ROBERTS, H.A.; NEILSON, J.E. Changes in the soil seed bank of four long-term crop/herbicide experiments. Journal of Applied Ecology, v.18, p.661-668, 1981.
SAINI, H.S.; BASSI, P.K.; SPENCER, M.S. Use of ethylene and nitrate to break seed dormancy of common lambsquarters (Chenopodium album). Weed Science, v.34, p.502-506, 1986.
SCHWEIZER, E.E.; ZIMDAHL, R.L. Weed seed decline in irrigated soil after six years of continuous corn (Zea mays) and herbicides. Weed Science, v.32, p.76-83, 1984.
SPENCE, J.R. Seed rain in grassland, herfield, snowbank, and fellfield in the alpine zone, Craigieburn Range, South Island, New Zealand. New Zealand Journal of Botany, v. 28, p. 439-450, 1990.
SWANTON, C.J.; SHRESTHA, A.; ROY, R.C.; BALL-COELHO, B.R.; KNEZEVIC, S.Z. Effect of tillage systems, N, and cover crop on the composition of weed flora. Weed Science, v.47, p.454-461, 1999.
SYMONIDES, E. Seed banks in old-field successional ecosystems. Ekologia Polska, v.34, n.3, p.29, 1986. (Polish Journal of Ecology)
TEIXEIRA, M.B.; COURA NETO, A.B.; PASTORE, U.; RANGEL FILHO, L.R. Vegetação: as regiões fitoecológicas, sua natureza e seus recursos econômicos; estudo fitogeográfico. In: FUNDAÇÃO INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. IBGE. http//ibge.gov.br. 1999. Levantamento de recursos naturais. Rio de Janeiro: IBGE. v.33, p.541-620, 1986.
THOMPSON, K. The occurrence of buried viable seeds in reaction to environmental gradients. Journal of Biogeography, v.5, p.425-430, 1978.
UCPEL/ITEPA. Vinte e cinco anos de agropecuária na zona sul – RS: levantamento estatístico 1975/2000. Pelotas: Educat, 2001. 606p.

84
VANASSE, A.; LEROUX, G.D. Floristic diversity, size, and vertical distribution of the weed seedbank in ridge and conventional tillage systems. Weed Science, v.48, p.454-460, 2000.
VERKAAR, H.J.; SCHENKEVELD, A.D.; VAN DE KLASHORTS, M.P. The ecology of short-lived forbes in chalk grasslands: dispersal of seeds. New Phytology, v.95, p.335-344, 1983.
WILSON, M.F.; TRAVESET, A. The ecology of seed dispersal. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI Publishing, 2000. p. 85-110.
YENISH, J.P.; DOLL, J.D.; BUHLER, D.D. Effect of tillage on vertical distribution and viability of weed seed in soil. Weed Science, v.40, p.429-433, 1992.
ZANIN, G.; OTTO, S.; RIELLO, L.; BORIN, M. Ecological interpretation of weed flora dynamics under different tillage systems. Agriculture Ecosystems & Environment, v.66, n.3, p.177-188, 1997.
ZIMMERGREN, D. The dynamics of seed banks in an area of sandy soil in southern Sweden. Botaniska Notiser, v.133, p.633-641, 1980.

85
ANEXO
1. Número de sementes por metro quadrado das espécies encontradas no BSS ao longo da cenosere, de 3 a 9 anos, nas profundidades de 0-5 e de 5-10 cm.
0 - 5 cm 5 - 10 cm Espécie 3 6 8 9 3 6 8 9
Agrostis montevidensis 0 0 0 0 0 255 0 0 Amaranthus lividus ssp. polygonoides 0 0 4330 0 0 0 2802 0 Apium leptophyllum 0 0 0 0 0 0 255 0 Avena strigosa 0 0 0 0 255 0 0 0 Baccharis trimera 0 0 0 0 0 255 0 0 Borreria capitata 0 0 764 255 0 0 0 0 Brachiaria plantaginea 0 255 0 0 0 0 0 0 Briza minor 255 0 0 0 0 0 0 0 Bulbostylis capillaris 0 2038 0 255 2547 2038 0 1783 Centela asiatica 2038 29292 0 509 5094 7641 1528 764 Centunculus minimus 7641 11971 3057 10698 8915 4075 3311 5604 Cerastium glomeratum 16556 7132 2292 1528 764 4840 255 255 Cerastium rivulare 2038 4330 255 19103 2547 0 0 2802 Chaptalia nutans 255 0 255 0 509 0 0 0 Conyza bonariensis 9934 509 0 0 0 0 0 0 Conyza cf. floribunda 255 0 0 0 9424 0 0 0 Crysanthemum myconis 0 0 0 255 0 0 0 0 Cuphea carthagenensis 16047 255 0 33113 509 255 0 1274 Cyperus cf. cayennensis 0 2547 255 0 0 764 0 0 Cyperus eragrostis 0 0 5349 0 0 0 0 0 Cyperus laetus 0 0 0 0 0 0 255 0 Cyperus meyenianus 0 0 255 0 0 0 0 0 Cyperus reflexus 8915 1528 0 1274 0 0 0 0 Dichondra sericea 0 0 1274 0 0 509 2038 0 Digitaria adscendens 0 255 0 0 9424 0 0 0 Digitaria ciliaris 50433 3057 3821 64697 29547 1019 3057 509 Eclipta megapotamica 9934 0 764 7132 255 255 0 0 Eleocharis maculosa 0 255 0 0 0 0 0 0 Eleocharis minima 0 0 255 0 0 255 0 0
86
ANEXO 1 (cont.) 0 - 5 cm 5 - 10 cm Espécie
3 6 8 9 3 6 8 9 Eleusine tristachya 255 2802 1274 0 0 255 764 0 Eragrostis lugens 0 0 0 0 0 255 0 0 Eragrostis neesii 18849 8151 764 15792 0 509 1274 0 Fimbristylis autumnalis 12990 1019 255 6368 7896 509 0 7387 Fimbristylis dichotoma 509 1783 0 0 1528 1528 255 0 Fimbristylis diphylla 0 0 2547 0 0 1019 0 0 Gamochaeta spp. 61640 73867 59348 9424 6368 35150 9170 764 Glandularia selloi 0 0 1274 0 0 0 0 0 Gratiola peruviana 255 0 0 255 0 0 0 1528 Hydrocotyle exigua 764 5349 1274 255 255 7896 1528 1274 Hydrocotyle verticillata 0 0 0 0 0 0 0 764 Hypochaeris albiflora 0 255 255 0 0 0 255 0 Hypochaeris chillensis 255 255 0 1019 0 0 0 0 Hypoxis decumbens 764 4840 1274 0 0 509 2038 1019 Juncus bufonius 225166 29037 5858 167601 29801 10188 5094 100611 Juncus capillaceus 0 3821 1783 1019 1783 1019 5094 0 Juncus dichotomus 2038 1274 3057 1783 2038 255 2802 5349 Juncus dombeyanus 0 764 255 255 0 1019 0 0 Juncus cf. effusus 4840 509 0 23434 3566 764 255 31839 Kyllinga brevifolia 34641 18594 5094 89404 13245 1274 7641 20886 Kyllinga odorata 0 1274 5604 764 0 0 4585 20122 Kyllinga vaginata 0 4075 2038 0 255 255 1274 509 Lolium multiflorum 20377 69791 281966 64697 4075 2038 6368 2802 Lotus subbiflorus 0 2038 0 0 0 0 0 0 Luzula campestris 255 0 0 0 0 255 0 0 Lythrum hyssopifolium 0 2802 0 21905 0 0 1019 0 Mecardonia tenella 0 0 0 0 0 0 509 0 Ornithopus micranthus 0 0 0 0 0 0 255 0 Oxalis cf. conorrhiza 2292 8660 6113 4840 1783 6368 2292 1783 Oxalis corniculata 0 0 0 255 0 509 0 0 Panicum aristellum 255 0 0 0 0 0 0 0 Panicum decipiens 255 0 0 1019 0 0 0 0 Panicum dichotomiflorum 509 0 0 0 509 0 0 0 Panicum racemosum 0 0 0 3311 0 0 0 0 Paspalum nicorae 8406 0 0 0 0 0 0 0 Paspalum urvillei 764 0 0 0 0 0 0 0 Piptochaetium montevidense 0 0 0 0 0 0 255 0 Plantago spp. 52216 10443 764 34131 5858 255 0 0 Pluchea sagittalis 255 0 255 0 509 0 0 0 Poa annua 4840 255 255 4840 0 0 0 0 Polygonum hydropiperoides 764 0 0 0 0 0 0 0 Polygonum cf. punctatum 6113 255 0 2292 509 0 0 0 Relbunium richardianum 764 0 0 255 0 0 255 0 Rhynchospora globularis 0 0 0 0 1783 0 255 764 Rhynchospora microcarpa 0 0 255 0 255 0 0 0 Rhynchospora rugosa 0 0 0 0 0 0 0 255 Rhynchospora tenuis 0 0 255 0 0 0 0 0 Richardia brasiliensis 0 0 1019 0 0 0 0 0 Setaria sphacelata 509 0 0 0 0 0 0 0 Sida rhombifolia 509 0 2038 509 255 0 764 0 Sida sp. 0 255 0 0 0 0 0 0

87
ANEXO 1 (cont.) 0 - 5 cm 5 - 10 cm Espécie
3 6 8 9 3 6 8 9 Sisyrinchium micranthum 32094 41518 8151 44575 17066 10188 7641 18085 Solanum americanum 0 255 1274 255 0 0 0 0 Soliva pterosperma 0 5094 509 4330 255 0 0 0 Sporobolus indicus 0 764 509 0 0 255 255 0 Trifolium pratense 764 0 0 0 509 0 0 0 Trifolium repens 11462 3311 764 33367 7132 0 1019 764 Urtica dioica 0 255 0 0 0 0 0 0 Verbena litoralis 764 255 0 0 0 0 0 0 Wahlenbergia linarioides 255 255 0 0 0 0 0 0 Campanulaceae 255 255 0 764 1528 0 0 0 Cyperaceae 1 9679 255 2292 9934 5858 0 0 1528 Cyperaceae 2 0 0 0 0 0 0 1019 0 Lamiaceae 255 0 0 0 0 0 0 0 Poaceae 1783 255 255 1019 1274 255 255 0 Indeterminada 1 255 0 0 1019 0 0 0 0 Indeterminada 2 764 0 0 3566 0 0 0 0

CONSIDERAÇÕES FINAIS
Conforme mencionado por Bekker (1998), nos anos 90, os bancos de sementes
finalmente foram aceitos como parte da comunidade vegetal, como primeiramente
sugerido por Major & Pyott (1966).
Os BSS de comunidades campestres são pouco estudados no Brasil, onde
predominam estudos em ecossistemas florestais e em ambientes agrícolas; este último
com a abordagem da área de plantas concorrentes daninhas. Espera-se que esse trabalho
possa contribuir com algumas informações sobre a dinâmica e a funcionalidade dos
bancos de sementes de comunidades campestres alteradas pelo cultivo agrícola, como
um importante recurso a ser observado neste ecossistema.
O primeiro e o segundo artigo focaram a dinâmica e a longevidade das sementes
de azevém anual no solo, respectivamente, em sistemas de semeadura direta, em rotação
com soja, atualmente muito difundidos na região. Muitas controvérsias podem ser vistas
com relação à classificação do banco de sementes desta e de várias outras espécies, no
tocante à sua longevidade. Em Thompson et al. (1997), há uma compilação de dados
sobre os BSS de uma ampla gama de espécies do noroeste da Europa, entre elas, o
azevém anual, mostrando classificações, para esta última, que variam desde um banco
de sementes persistente, até um transitório. As informações obtidas nesse trabalho
concordam com a maioria dos estudos retratados pelo autor, apontando a espécie como
apresentando um banco de sementes de caráter transitório, já que a maior parte das
sementes não persistiu no solo por mais de um ano (somente 11% das sementes foram
encontradas após 565 dias de enterrio). Pouco mais de dois anos após o enterrio,

89
somente 2% das sementes foram encontradas. Verifica-se, com base nestes dois estudos,
que a maioria das sementes é dispersa, durante o fim da primavera e início do verão
(dezembro), com alta dormência primária, e germinam no outono, quando as condições
de temperatura e disponibilidade de água tornam-se adequadas. Até o fim do outono, o
BSS é praticamente exaurido.
A prática de manejo utilizada, com dois anos de cultivo de soja e dois anos de
pousio, apresentou maior influência no tamanho do BSS de azevém do que o número de
anos nos quais esta prática vem sendo utilizada, principalmente em virtude da sua
transitoriedade, que tornam os distúrbios de cada ano mais importantes do que suas
conseqüências de longo prazo.
Apesar do referido manejo ser bem sucedido em termos produtivos,
principalmente por proporcionar duas fontes de renda para o produtor – uma oriunda da
lavoura e outra da produção de carne –, diminuindo os custos de implantação da
pastagem e antecipando o pastejo, em virtude da intensividade e do manejo visando a
ressemeadura natural da gramínea, esse caráter transitório das sementes de azevém no
solo evidencia a necessidade da reposição anual do BSS para assegurar pastagens
subseqüentes. As sementes são depositadas na estação anterior, em densidades que
variam de 3.363 a 22.835 sementes.m-2. Isto representa uma quantidade de sementes
equivalente a 67 até 456 kg.ha-1, considerando-se um peso de 1000 sementes de 2
gramas, o que está bem acima das quantidades normais recomendadas para semeadura,
que variam de 25 a 30 kg.ha-1.
A área cultivada com soja vem crescendo a cada ano na região e no país como
um todo. Cada vez mais áreas de campo estão sendo ocupadas com essa cultura,
levando ao gradativo desaparecimento de espécies campestres.
O terceiro artigo procurou quantificar possíveis alterações na composição e
tamanho do BSS da área, buscando inferir sobre o potencial de recuperação da
vegetação campestre, através do seu banco de sementes.
Alguns estudos têm demonstrado que o BSS possui um papel significativo na
colonização após pequenos distúrbios, enquanto outros mostram que este papel é
irrelevante (Thompson, 2000).

90
Bakker (1989) e Van der Valk & Verhoeven (1988), por exemplo, apontam para
um possível potencial regenerativo do BSS, em casos onde as espécies tenham
desaparecido da vegetação estabelecida. A maioria das espécies campestres tende a
formar bancos de sementes que variam de transitórios a persistentes de curto prazo,
normalmente dominados por espécies pioneiras persistentes, por longos períodos, e
espécies ruderais (Bakker, 1989). Durante a sucessão, não ocorrendo distúrbios no solo,
as espécies ruderais podem permanecer presentes no banco de sementes por um longo
período de tempo, mas quando elas desaparecem da vegetação estabelecida e das áreas
adjacentes, a acumulação das sementes no solo cessa e, com o tempo, a dominância
dessas no banco de sementes termina (Chancellor, 1986). As espécies pioneiras e
ruderais, entretanto, podem influenciar negativamente as perspectivas de regeneração
das comunidades campestres, pela rápida ocupação de espaços abertos na vegetação,
aumentando seu poder competitivo sobre as espécies alvo (Graham & Hutchings,
1988b; Strykstra et al., 1998).
Utilizando uma comunidade alvo, Bekker (1998) mediu quantitativamente o
potencial de recuperação da vegetação pelo BSS e, assim como Kirkham & Kent
(1997), concluiu que o potencial de regeneração de comunidades campestres degradadas
pode diminuir conforme o passar do tempo, e que processos de drenagem e de
fertilização podem afetar a sobrevivência das sementes no solo.
Kalamees & Zobel (2002) argumentam que parte do problema está nos
diferentes métodos experimentais. Alguns pesquisadores têm criado espaços vazios por
remoção da vegetação, mas causaram pouco distúrbio no solo, observando assim uma
pequena importância do BSS (Bullock et al., 1994; Edwards & Crawley, 1999a). Um
experimento totalmente fatorial, em um campo de pedra calcária, com solo alterado a
uma profundidade de 10 cm, revelou que 36% dos colonizadores dos espaços de 10 x 10
cm originaram-se do BSS, 46%, da chuva de sementes (mas somente de 12% das
plantas a mais de 0,5 m do espaço vazio) e 18% invadiram vegetativamente (Kalamees
& Zobel, 2002). Pakeman et al. (1998) encontraram resultados similares em um campo
de solo ácido, no leste da Inglaterra. Edwards & Crawley (1999b) sugerem que, em
locais com distúrbio, o recrutamento, normalmente, seria dominado pela corrente chuva
de sementes. Entretanto, na ausência desta, o distúrbio aumentou o recrutamento
somente nas espécies com bancos de sementes persistentes.

91
Alguns experimentos vêm confirmando a pouca importância da chuva de
sementes na colonização de pequenos espaços abertos, na vegetação campestre (Marks
& Mohler, 1985; Milberg, 1993). Os trabalhos de Zimmergren (1980) e Pakeman et al.
(1998) apontam o banco de sementes como sendo o principal responsável pela
recolonização de áreas campestres, após distúrbio. Estes resultados sugerem que o
banco de sementes pode ter uma contribuição fundamental na regeneração, quando da
ausência de distúrbio ou se o distúrbio ocorreu em um período que houve chuva de
sementes pequena.
Desta forma, fica claro que o padrão e a diversidade do BSS pode depender dos
eventos após a dispersão, pelo menos tanto quanto da dispersão por si mesma. Tais
eventos podem ter efeitos altamente idiossincráticos, tanto nos padrões de composição,
quanto espacial, dos BSS.
O clima, nutrientes e herbivoria são os principais determinantes da composição
dos campos e comunidades relacionadas (Thompson, 2000). A maioria dos estudos tem
concluído que o crescimento vegetativo, a chuva de sementes e o banco de sementes
persistente estão todos envolvidos, em vários graus, na colonização de espaços abertos,
na vegetação campestre. Embora o banco de sementes persistente tenha freqüentemente
a menor importância numérica, como fonte de colonizadores, sua contribuição pode ser
dramaticamente alterada por uma sutil mudança na severidade do distúrbio e na locação
e tempo de criação do espaço aberto (Thompson et al., 1996).
Alguns hábitats, incluindo muitos, mas nem todos os campos, apresentam
oportunidades sazonais razoáveis para o estabelecimento de plântulas, normalmente, no
fim do período seco ou de baixas temperaturas. Tais oportunidades são melhores
aproveitadas por um BSS transitório, com germinação mais ou menos sincronizada com
o início do período favorável para o desenvolvimento vegetativo das espécies em
questão. Estes tipos de bancos de sementes estão associados à previsibilidade da
ocorrência de distúrbios.
Neste caso, os principais distúrbios são o cultivo da soja, a alta densidade de
plântulas de azevém (já que a área é manejada para propiciar a ressemeadura dessa
espécie, com grande densidade de sementes no solo) e o pastejo. O cultivo da soja
implica não só na presença da cultura, mas, também, em distúrbios causados pelo uso de
herbicidas dessecantes e fertilização.

92
A influência na composição e tamanho do BSS, das práticas de manejo
utilizadas a cada dois anos, com o cultivo da soja, e o pastejo constante, foi maior do
que o tempo de manejo, assim como em Menalled et al. (2001). Houve um predomínio
de espécies monocotiledôneas anuais ,quantitativamente, e de perenes, qualitativamente.
O BSS da área de estudo apresentou uma trajetória dinâmica, não sendo possível
sua representação através do “modelo markoviano”. Estas variações, possivelmente,
devem-se às diferentes estratégias de sobrevivência das espécies em virtude do manejo
alternado e da provável presença de um grande banco de sementes transitório, que pode
apresentar grandes variações de densidade de um ano para outro (Roberts & Neilson,
1981; Schweizer & Zimdahl, 1984; Cardina et al., 1991; Mayor & Dessaint, 1998;
Vanasse & Leroux, 2000; Menalled et al., 2001). Além disso, a entrada do gado bovino
nas áreas dependeu da condição da pastagem de cada local, ou seja, não se deu
uniformemente, o que pode ter influído na dispersão zoocórica das sementes e,
conseqüentemente, no depósito das mesmas no solo.
Aponta-se aqui para uma possível influência do uso de herbicidas, na redução da
densidade e da riqueza de espécies do banco de sementes, assim como nos estudos
acima citados. A maior presença de espaços abertos no solo, nas áreas com soja,
promove um recrutamento constante, que contribuiu para a diminuição do BSS. A cada
ano, o controle químico parece provocar o reinício da sucessão na comunidade vegetal,
com o “escape” de algumas espécies que, gradativamente, seguem o processo
sucessional.
Estudos recentes têm examinado a associação da vegetação e do BSS com
variáveis relacionadas à fertilidade do solo (Favreto, 2004). A influência da fertilidade
do solo também pode ser observada, com maiores bancos de sementes nas áreas com
solos menos férteis, assim como em Albrecht & Auerswald (2003).
Conforme o distúrbio diminuiu, houve uma tendência à formação de um banco
de sementes de espécies perenes, com menor densidade, possivelmente de caráter
transitório, provavelmente, visando a rápida reocupação de espaços abertos no solo,
provocados pelo cultivo da soja ou pastejo intenso.
A maior quantidade de sementes anuais, assim como a maior concentração de
sementes nos primeiros centímetros de solo, em sistema de semeadura direta, com uso
de herbicida, foram observadas também por vários autores (Forcella & Lindstrom,

93
1988; Buhler & Oplinger, 1990; Yenish et al,.1992; McCloskey et al., 1996; Bekker et
al., 1999; Swanton et al., 1999; Menalled et al., 2001; Favreto, 2004).
Apesar de não constarem as características do banco de sementes de uma área de
vegetação nativa, já após três anos de manejo, não se verifica a presença de muitas das
espécies campestres características da região.
Conforme aumenta o tempo de utilização da área, o tamanho e a riqueza do BSS
diminuem, com uma tendência de substituição de espécies perenes por espécies anuais,
em termos quantitativos, e o conseqüente desaparecimento de nativas de importante
valor forrageiro, como as do gênero Paspalum, após 6 anos.
Sendo assim, é importante que sejam feitos estudos por um maior período de
tempo em áreas agropastoris, enfocando a dinâmica da vegetação, do banco de sementes
e da chuva de sementes, abordando aspectos relativos à dispersão, para que possam ser
obtidos resultados mais completos, contribuindo na geração de indicações para o uso
sustentável e para a recuperação desse importante recurso natural que são os campos,
que ocupam grande parte da região.

REFERÊNCIAS BIBLIOGRÁFICAS
ALBRECHT, H.; AUERSWALD, K. Arable weed seedbanks and their relation to soil properties. Aspects of Applied Biology, v.69, p.11-20, 2003. (Seedbanks: Determination, Dynamics & Management)
ARCHBOLD, O.W. Buries viable propagules as a factor in postfire regeneration in northern Saskatchewan. Canadian Journal of Botany, v.57, p.54-58, 1979.
ARCHBOLD, O.W. Seed input as a factor in the regeneration of strip-mine wastes in Saskatchewan. Canadian Journal of Botany, v.58, p.1490-1495, 1980.
BAKKER, J.P. Nature management by grazing and cutting: on the ecological significance of grazing and cutting regimes applied to restore species-rich grassland communities in The Netherlands. Dordrecht: Kluwer Academic Publishers, 1989.
BAKKER, J.P.; POSCHLOD, P.; STRIJKSTRA, R.J.; BEKKER, R.M.; THOMPSON, K. Seed banks and seed dispersal: important topics in restoration ecology. Acta Botanica Neerlandica, v.45, p.461-490, 1996.
BAKKER, J.P.; BAKKER, E.S.; ROSÉN, E.; VERWEIJ, G.L. The soil seed bank of undisturbed and disturbed dry limestone grassland on Öland (Sweden). Zeitschrift für Ökologie and Naturschutz, v.6, p.9-18, 1997.
BAKKER, J.P.; BERENDSE, F. Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends in Ecology and Evolution, v.14, n.2, p.63-68, 1999.
BASKIN, J.M.; BASKIN, C.C. The seed bank in a population of an endemic plant species and its ecological significance. Biological Conservation, v.14, p.125-130, 1978.
BEKKER, R.M. The ecology of soil seed banks in grassland ecosystems. 1998. 192f. Tese (Doutorado em Ecologia Vegetal) – Universidade de Groningen, Haren, Holanda, 1998.
BEKKER, R.M.; VERWEIJ, G.L.; SMITH, R.E.N.; REINE, R.; BAKKER, J.P.; SCHNEIDER, S. Soil seed banks in European grasslands: does land use affect regeneration perspectives? Journal of Applied Ecology, v.34, p.1293-1310, 1997.

95
BEKKER, R.M.; KNEVEL I.C.; TALLOWIN, J.B.R.; TROOST, E.M.L.; BAKKER, J.P. Soil nutrient input effects on seed longevity: a burial experiment with fen-meadow species. Functional Ecology, v.12, n.4, p.673-682, 1998a.
BEKKER, R.M.; OOMES, M.J.M.; BAKKER, J.P. The impact of groundwater level on soil seed bank survival. Seed Science Research, v.8, n.3, p.399-404, 1998b.
BEKKER, R.M.; SCHAMINEE, J.H.J.; BAKKER, J.P.; THOMPSON, K. Seed bank characteristics of Dutch plant communities. Acta Botanica Neerlandica, v.47, n.1, p.15-26, 1998c.
BEKKER, R.M.; LAMMERTS, E.J.; SCHUTTER, A.; GROOTJANS, A.P. Vegetation development in dune slacks: the role of persistent seed banks. Journal of Vegetation Science, v.10, p.745-754, 1999.
BERNHARDT, K.G.; POSCHLOD, P. Diasporenbanken im Boden als Vegetationsbestandteil. Teil 1: Europa. Excerpta Botanica Section B, v.29, p.241-260, 1993.
BOCCANELLI, S.I.; LEWIS, J.P. The seed bank of old pampean prairie and its relation with the standing vegetation. Pesquisa Agropecuária Brasileira, v.29, p.1833-1840, 1994.
BOLDRINI, I.I. Campos do Rio Grande do Sul: caracterização fisionômica e problemática ocupacional. Boletim do Instituto de Biociências/UFRGS, v.56, p.1-39, 1997.
BRYANT, J.A. Fisiologia da Semente. São Paulo: EPU, 1989. 86p. (Temas de Biologia, v.31)
BUHLER, D.D.; OPLINGER, E.S. Influence of tillage systems on annual weed densities and control in solid-seeded soybean (Glycine max). Weed Science, v.38, p.158-165, 1990.
BULLOCK, J.M.; HILL, B.C.; DALE, M.P.; SILVERTOWN, J. An experimental study on the effects of sheep grazing on vegetation change in a species-poor grassland and the role of seedling recruitment into gaps. Journal of Applied Ecology, v.31, p.493-507, 1994.
CARDINA, J.; REGNIER, E.; HARRISON, K. Long-term tillage effects on seed banks in three Ohio soils. Weed Science, v.39, p.186-194, 1991.
CHANCELLOR, R.J. Decline of arable weed seeds during 20 years in soil under grass and the periodicity of seedling emergence after cultivation. Journal of Applied Ecology, v.23, p.631-637, 1986.
COOK, R. The biology of seeds in the soil. In: SOLBRIG, O.T. (ed.). Demography and evolution in plant populations. Botanical Monographs, v.15, p.107-129, 1980.
DOUGLAS, G. The weed flora of chemically-renewed lowland swards. Journal of British Grassland Society, v.20, p.91-100, 1965.
EDWARDS, G.R.; CRAWLEY, M.J. Herbivores, seed banks and seedling recruitment in mesic grasslands. Journal of Ecology, v.87, p.423-435, 1999a.
EDWARDS, G.R.; CRAWLEY, M.J. Effects of disturbance and rabbit grazing on seedling recruitment of six mesic grassland species. Seed Science Research, v.9, p.145-156, 1999b.

96
FAVRETO, R.; MEDEIROS, R.B.; PILLAR, V.D. Composição do banco de sementes do solo de um campo natural sujeitos a intensidades de pastejo e posições de relevo. In: REUNIÃO DO GRUPO TÉCNICO REGIONAL DO CONE SUL (ZONA CAMPOS) EM MELHORAMENTO E UTILIZAÇÃO DE RECURSOS FORRAGEIROS DAS ÁREAS TROPICAL E SUBTROPICAL, 18., 2000, Guarapuava. Anais... Guarapuava: UFPR, 2000. p.233-235.
FAVRETO, R. Vegetação espontânea e banco de sementes do solo em área agrícola estabelecida sobre campo natural. 2004. 116 f. Dissertação (Mestrado em Ecologia), Centro de Ecologia, Universidade Federal do Rio Grande do Sul, Porto Alegre, 2004.
FENNER, M. (ed.). Seed Ecology. Londres: Chapman & Hall, 1985.
FENNER, M. (org.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI, 2000. 410p.
FORCELLA, F.; LINDSTROM, M.J. Weed seed populations in ridge and conventional tillage. Weed Science, v.36, p.500-503, 1988.
GOLDSMITH, E. Complexity and stability in the real world. The Ecologist Quarterly, Winter, n.4, p.305-316, 1978.
GRAHAM, D.J.; HUTCHINGS, M.J. Estimation of the seed bank of a chalk grassland ley established on former arable land. Journal of Applied Ecology, v. 25, p. 241-252, 1988a.
GRAHAM, D.J.; HUTCHINGS, M.J. A field investigation of germination from the seed bank of a chalk grassland ley on former arable land. Journal of Applied Ecology, v.25, p.253-263, 1988b.
HARPER, J.L. Population biology of plants. Londres: Academic Press, 1977. p.33-111.
HENDERSON, C.B.; PETERSEN, K.E.; REDAK, A. Spatial and temporal patterns in the seed bank na vegetation of a desert grassland community. Journal of Ecology, v.76, p.717-728, 1978.
HEYDECKER, W. (ed.). Seed Ecology. Pensylvania: Pensylvania State University Press. 1973.
HILL, M.O.; STEVENS, P.A. The density of viable seed in soils of forest plantations in upland Britain. Journal of Ecology, v.69, p.693-709, 1981.
HOLLING, C.S. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics, v.4, p.1-23, 1973.
HOWE, C.D.; CHANCELLOR, R.J. Factors affecting the viable seed content of soils beneath lowland pastures. Journal of Applied Ecology, v.20, p.915-922, 1983.
IBGE. \www.ibge.gov.br\ Recuperado em 1997.
KALAMEES, R.; ZOBEL, M. The seed bank in an Estonian calcareous grassland: comparison of different successional stages. Folia Geobotanica, v.32, n.1, p.1-14, 1997.
KALAMEES, R.; ZOBEL, M. The role of the seed bank in gap regeneration in a calcareous grassland community. Ecology, v.83, p.1017-1025, 2002.
KEDDY, P.A.; REZNICEK, A.A. The role of seed banks in the persistence of Ontario´s coastal plain flora. American Journal of Botany, v.69, p.13-22, 1982.

97
KELLMAN, M.C. The viable seed content of some forest soil in coastal British Columbia. Canadian Journal of Botany, v.48, p.1383-1385, 1970.
KIRKHAM, F.W.; KENT, M. Soil seed bank composition in relation to the above-ground vegetation in fertilized and unfertilized hay meadows on a Somerset peat moor. Journal of Applied Ecology, v.34, p.889-902, 1997.
LECK, M.A.; PARKER, V.T.; SIMPSON, R.L. (eds.). Ecology of Soil Seed Banks. San Diego: Academic Press, 1989.
LODGE, G.M. Studies of soil seedbanks in native and sown pastures in northern New South Wales. Rangeland Journal, v.23, n.2, p.204-223, 2001.
LÓPEZ-MARIÑO, A.; LUIS-CALABUIG, E.; FILLAT, F.; BERMUDEZ, F.F. Floristic composition of established vegetation and the soil seed bank in pasture communities under different traditional management regimes. Agriculture Ecosystems and Environment, v.78, n.3, p.273-282, 2000.
MAIA, F.C.; MEDEIROS, R.B.; PILLAR, V.P.; FOCHT, T.; CHOLLET, D.M.S.; OLMEDO, M.O.M. Composição, riqueza e padrão de variação do banco de sementes do solo em função da vegetação de um ecossistema de pastagem natural. Iheringia (Sér. Bot.), v.58, n.1, p.61-80, 2003.
MAIA, F.C.; MEDEIROS, R.B.; PILLAR, V.P.; FOCHT, T. Soil seed bank variation patterns according to environmental factors in a natural grassland. Revista Brasileira de Sementes, v.26, n.2, p.126-137, 2004.
MAJOR, J.; PYOTT, W.T. Buried viable seeds in two Californian bunch grass sites and their bearing on the definition of a flora. Vegetatio, v.13, p.253-282, 1966.
MARKS, P.L.; MOHLER, C.L. Succession after elimination of buried seeds from a recently plowed field. Bulletin of the Torrey Botanical Club, v.112, p.376-382, 1985.
MAYER, A.M.; POLJAKOFF-MAYBER, A. The germination of seeds. 4 ed., Londres: Pergamon Press, 1989.
MAYOR, J.P.; DESSAINT, F. Influence of weed management strategies on soil seedbank diversity. Weed Research, v.38, p.95-105, 1998.
McCLOSKEY, M.; FIRBANK, L.G.; WATKINSON, A.R.; WEBB, D.J. The dynamics of experimental arable weed communities under different management practices. Journal of Vegetation Science, v.7, p.799-808, 1996.
MEDEIROS, R.B. Bancos de sementes no solo e dinâmica vegetal. In: REUNIÃO DO GRUPO TÉCNICO REGIONAL DO CONE SUL (ZONA CAMPOS) EM MELHORAMENTO E UTILIZAÇÃO DE RECURSOS FORRAGEIROS DAS ÁREAS TROPICAL E SUBTROPICAL, 18., 2000, Guarapuava. Memórias... Guarapuava: UFPR, 2000. p.61-87.
MEDEIROS, R.B.; STEINER, J.J. Influence of temperate grass seed rotation systems on weed seed soil bank composition. Revista Brasileira de Sementes, v.24, n.1, 2002.
MENALLED, F.D.; GROSS, K.L.; HAMMOND, M. Weed aboveground and seedbank community responses to agricultural management systems. Ecological Applications, v.11, n.6, p.1586-1601, 2001.
MILBERG, P. Hur länge kan ett frö leva? Svensk Botanisk Tidskrift, v. 84, p. 323-352, 1990.

98
MILBERG, P. Seed bank in a 35-year-old experiment with different treatments of a semi-natural grassland. Acta Oecologica - International Journal of Ecology, v.13, n.6, p.793-752, 1992.
MILBERG, P. Seed bank and seedlings emerging after soil disturbance in a wet semi-natural grassland in Sweden. Annales Botanici Fennici, v.30, p.9-13, 1993.
MILES, J. Vegetation Dynamics. Londres: Chapman and Hall, 1978. 80p.
MORGAN, J.W. Composition and seasonal flux of the soil seed bank of species-rich Themeda triandra grasslands in relation to burning history. Journal of Vegetation Science, v.9, n.2, p.145-156, 1998.
NELSON, J.F.; CHEW, R.M. Factors affecting seed reserves in the soil of a Mojave Desert ecosystem. American Midland Naturalist, v.97, p.300-320, 1977.
PAKEMAN, R.J.; ATTWOOD, J.P.; ENGELEN, J. Sources of plants colonizing experimentally disturbed patches in an acidic grassland, in eastern England. Journal of Ecology, v.86, p.1032-1041, 1998.
PETTIT, N.E.; FROEND, R.H. Availability of seed for recruitment of riparian vegetation: a comparison of a tropical and a temperate river ecosystem in Australia. Australian Journal of Botany, v.49, n.4, p.515-528, 2001.
RAMBO, B. A fisionomia do Rio Grande do Sul. Porto Alegre: Of. Gráf. da Imprensa Oficial, 1942. 394p.
REICHMAN, O.J. Relation of desert rodent diets to available resources. Journal of Mammalogy, v.56, p.731-751, 1975.
ROBERTS, H.A. Viable weed seeds in cultivated soils. Report of the National Vegetable Research Station, 1970. 25-38p.
ROBERTS, H.A. Seed banks in soils. In: COAKER, T.H. (ed.). Advances in Applied Biology, v.6, p.1-55, 1981.
ROBERTS, H.A.; RICKETTS, M.E. Quantitative relationships between the weed flora after cultivation and the seed population in the soil. Weed Research, v.19, p.269-275, 1979.
ROBERTS, H.A.; NEILSON, J.E. Changes in the soil seed bank of four long-term crop/herbicide experiments. Journal of Applied Ecology, v.18, p.661-668, 1981.
SARUKHÁN, J. Studies on plant demography: Ranunculus repens L., R. bulbosus L., R. acris L. 2. Reproductive strategies and seed population dynamics. Journal of Ecology, v.62, p.151-177, 1974.
SCHWEIZER, E.E.; ZIMDAHL, R.L. Weed seed decline in irrigated soil after six years of continuous corn (Zea mays) and herbicides. Weed Science, v.32, p.76-83, 1984.
STOCKLIN, J.; FISCHER, M. Plants with longer-lived seeds have lower local extinction rates in grassland remnants 1950-1985. Oecologia, v.120, n.4, p.539-543, 1999.
STRYKSTRA, R.J.; BEKKER, R.M.; BAKKER, J.P. Assessment of dispersule availability: its practical use in restoration management. Acta Botanica Neerlandica, v.47, p.57-70, 1998.

99
SWANTON, C.J.; SHRESTHA, A.; ROY, R.C.; BALL-COELHO, B.R.; KNEZEVIC, S.Z. Effect of tillage systems, N, and cover crop on the composition of weed flora. Weed Science, v.47, p.454-461, 1999.
SYMONIDES, E. Seed banks in old-field successional ecosystems. Ekologia Polska, Lomianki, v.34, p.3-29, 1986. (Polish Journal of Ecology)
THOMPSON, K. The occurrence of buried viable seeds in reaction to environmental gradients. Journal of Biogeography, v.5, p.425-430, 1978.
THOMPSON, K. The functional ecology of seed banks. In: FENNER, M. (ed.). Seeds: the ecology of regeneration in plant communities. 2 ed., Wallingford: CABI, 2000. p.215-235.
THOMPSON, K.; GRIME, J.P. Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats. Journal of Ecology, v.67, p.893-921, 1979.
THOMPSON, K.; HILLIER, S.H.; GRIME, J.P.; BOSSARD, C.C.; BAND, S.R. A functional analysis of a limstone grassland community. Journal of Vegetation Science, v.7, p.371-380, 1996.
THOMPSON, K.; BAKKER, J.P.; BEKKER, R.M. The soil seed banks of North West Europe: methodology, density and longevity. Cambridge: Cambridge University Press, 1997.
UCPEL/ITEPA. Vinte e cinco anos de agropecuária na zona sul – RS: levantamento estatístico 1975/2000. Pelotas: Educat, 2001. 606p.
VANASSE, A.; LEROUX, G.D. Floristic diversity, size, and vertical distribution of the weed seedbank in ridge and conventional tillage systems. Weed Science, v.48, p.454-460, 2000.
VAN DER VALK, A.G.; DAVIS, B. A reconstruction of the recent vegetational history of a prairie marsh. Eagle Lake. Iowa. its seed bank. Aquatic Botany, v.6, p.29-51, 1979.
VAN DER VALK, A.G.; VERHOEVEN, J.T.A. The restoration of quaking fens from floating forests: the role of the seed bank and understorey species. Vegetatio, v.76, p.3-13, 1988.
VYVEY, Q. Bibliographical review on buried viable seeds in the soil. Excerpta Botanica Section B, v.26, p.311-320, 1989a.
VYVEY, Q. Bibliographical review on buried viable seeds in the soil. Excerpta Botanica Section B, v.27, p.1-52, 1989b.
WHIGHAM, D.F.; SIMPSON, R.L.; LECK, M.A. The distribution of seeds, seedlings, and established plants of arrow arum (Peltandra virginica (L.) (Kunth)) in a freshwater tidal wetland. Bulletin of the Torrey Botanical Club, v.106, p.193-199, 1979.
YENISH, J.P.; DOLL, J.D.; BUHLER, D.D. Effect of tillage on vertical distribution and viability of weed seed in soil. Weed Science, v.40, p.429-433, 1992.
ZANIN, G.; OTTO, S.; RIELLO, L.; BORIN, M. Ecological interpretation of weed flora dynamics under different tillage systems. Agriculture Ecosystems & Environment, v.66, n.3, p.177-188, 1997.
ZIMMERGREN, D. The dynamics of seed banks in an area of sandy soil in southern Sweden. Botaniska Notiser, v.133, p.633-641, 1980.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo