caracterizaÇÃo limnolÓgica e produtividade secundÁria...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE MINAS GERAIS
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PÓS-GRADUAÇÃO EM ECOLOGIA, CONSERVAÇÃO E MANEJO DE VIDA SILVESTRE
CARACTERIZAÇÃO LIMNOLÓGICA E PRODUTIVIDADE
SECUNDÁRIA DAS PRINCIPAIS ESPÉCIES DE
MICROCRUSTÁCEOS EM DOIS BRAÇOS DOS RESERVATÓRIOS
DE TRÊS MARIAS E FURNAS, MINAS GERAIS, BRASIL.
SOFIA LUIZA BRITO
Belo Horizonte, Minas Gerais
2010

SOFIA LUIZA BRITO
CARACTERIZAÇÃO LIMNOLÓGICA E PRODUTIVIDADE
SECUNDÁRIA DAS PRINCIPAIS ESPÉCIES DE
MICROCRUSTÁCEOS EM DOIS BRAÇOS DOS RESERVATÓRIOS
DE TRÊS MARIAS E FURNAS, MINAS GERAIS, BRASIL.
Orientadora: Profa. Dra. Paulina Maria Maia-Barbosa
Co-Orientador: Prof. Dr. Ricardo Motta Pinto-Coelho
Belo Horizonte, Minas Gerais
2010
Tese apresentada ao Curso de Ecologia, Conservação e Manejo da Vida Silvestre da Universidade Federal de Minas Gerais, como um dos requisitos à obtenção do título de Doutor em Ecologia.

Ao meu amado Cláudio:
...você sabe.

AGRADECIMENTOS
À todas as Deusas, especialmente a das Águas que me protegeu ao longo de todas as coletas e a das
Ciências que sempre me inspirou ao longo de minha carreira de bióloga.
Ao curso de Pós-Graduação em Ecologia, Conservação e Manejo de Vida Silvestre – ECMVS, que
juntamente com a USFISH proporcionaram esta oportunidade.
À secretaria do curso, em especial Joyce, Fred, Cristiane e Mary, por serem os facilitadores de todo
este processo.
Ao Projeto Parques Aqüícolas (SEAP/SECTES/FUNDEP-UFMG), pelo apoio financeiro e logístico.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq, pela concessão da
bolsa de estudos e pela taxa de bancada, indispensável para realização das coletas.
À banca avaliadora, pela colaboração para o crescimento deste trabalho.
À Profa. Dra. Paulina Maria Maia-Barbosa, por me ajudar a alcançar mais este degrau na minha
carreira de bióloga e por ensinar a ser cada vez mais profissional.
Ao Prof. Dr. Ricardo Motta Pinto-Coelho, pelo convite para participar do projeto Parques Aqüícolas
e ampliar meus horizontes através dos grandes reservatórios.
A todos os professores do curso PG-ECMVS, especialmente a Profa. Dra. Arnola Rietzler e Profa.
Dra. Cláudia Jacobi, permitindo meu amadurecimento ao longo do curso, bem como serem uma fonte
inspiradora para meu trabalho.
A todos os membros dos laboratórios de Ecologia do Zooplâncton, de Gestão Ambiental de
Reservatórios e de Limnologia, especialmente: Atenágoras, Filipe e Alessandra, companheiros de coleta
insubstituíveis, Simone e Cid, por auxiliarem tanto em laboratório quanto em campo, Rosinha, Cristiane,
Akemi, Meire, Vanessa e Ericson, pela enorme ajuda, por ouvirem meus desabafos e acima de tudo por me
aturarem.
Aos barqueiros, seu Pico em Três Marias e seu Alberico em Furnas, por nos guiarem pelos “mares”
de Minas Gerais.
A duas famílias incríveis que me adotaram enquanto estive hospedada em suas pousadas, durante o
período de coleta: César, Edilamar e Ana Júlia, da pousada Recanto das Buganvílias em Três Marias e
Simonides e Luci, da pousada Bella Minas, em Furnas. Obrigada pelo enorme carinho e tratamento em
regime de engorda!
À minha mãe, Lenita; meu irmão e minha cunhada, Élcio e Cláudia e ao meu sobrinho, Herbert, que
nasceu quando eu pleiteava uma vaga no curso de doutorado, foi crescendo junto com este trabalho e sempre
foi motivo de alegria e descontração, me fazendo esquecer as intempéries do caminho.
E especialmente ao meu amado Cláudio, grande companheiro da jornada da vida, meu maior
entusiasta, um dos mais sacrificados ao longo das coletas, que acreditou em mim quando eu já não
acreditava mais. Muito obrigada pela companhia, críticas e compreensão e me perdoe pelas faltas.

SUMÁRIO
RESUMO 1
ABSTRACT 2
INTRODUÇÃO GERAL 3
Referências Bibliográficas 8
ARTIGO 1 - Zooplâncton como Indicador de Condições Tróficas em Dois Braços dos Reservatórios de Três Marias e Furnas, Minas Gerais, Brasil. 12
Resumo/Abstract 12
Introdução 13
Material e Métodos 14
Resultados 17
Discussão 23
Referências Bibliográficas 27
ARTIGO 2 – Regressões Peso-Comprimento e Flutuação da Biomassa das Principais Espécies de Microcrustáceos em Dois Braços dos Reservatórios de Três Marias e Furnas, Minas Gerais, Brasil. 33
Resumo/Abstract 33
Introdução 34
Material e Métodos 35
Resultados 39
Discussão 43
Referências Bibliográficas 48
ARTIGO 3 - Produtividade Secundária das Principais Espécies de Microcrustáceos em Dois Braços dos Reservatórios de Três Marias e Furnas, Minas Gerais, Brasil. 52
Resumo/Abstract 52
Introdução 53
Material e Métodos 54
Resultados 58
Discussão 62
Referências Bibliográficas 66
DISCUSSÃO GERAL 69
CONCLUSÕES 72
Referências Bibliográficas 73
ANEXOS 74

1
RESUMO
Os inúmeros reservatórios construídos no Brasil, inicialmente com o objetivo principal de geração de energia elétrica, foram aos poucos diversificando seus usos para atender as necessidades das atividades desenvolvidas na bacia hidrográfica. Atualmente, na maioria deles, a água também é utilizada para irrigação, abastecimento, turismo e pesca. O atual governo, visando incrementar a produção aqüícola e otimizar o uso dos reservatórios no país propôs, através Instrução Normativa Interministerial nº. 6, de 28 de maio de 2004, repassar aos produtores rurais de baixa renda autorizações para uso da água pública para a produção de pescados em tanques-rede. Neste contexto, o “Estudo Técnico-Científico Visando a Delimitação de Parques Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG” (Convênio 8713, SEAP/SECTES/FUNDEP-UFMG) se propôs a selecionar e indicar áreas com águas oligotróficas dentro destes reservatórios para a instalação de tanques-redes para cultivo de tilápias, e estimar a capacidade suporte destas áreas, com o intuito de minimizar os impactos deste tipo de atividade sobre a qualidade das águas. Considerando a importância do zooplâncton na transferência de energia entre os níveis tróficos, além de seu papel como indicadores de qualidade de água, este trabalho teve como objetivos, descrever esta comunidade em termos de composição, biomassa e produtividade secundária, em braços dos reservatórios de Três Marias e Furnas, como suporte para a avaliação da sustentabilidade do potencial pesqueiro nestes ambientes. Apesar de considerados oligotróficos, estes reservatórios apresentaram diferenças marcantes na comunidade zooplanctônica. No reservatório de Três Marias predominaram ciclopóides e cladóceros de pequeno tamanho corporal, e em densidades elevadas (valores médios do zooplâncton total de 23.721 org.m-³ e 90.872 org.m-³ nos períodos de seca e chuva, respectivamente) caracterizando este ambiente como mesotrófico. Em Furnas (valores médios do zooplâncton total de 9.022 org.m-³ e 40.434 org.m-³ nos períodos de seca e chuva, respectivamente) calanóides e cladóceros de maior tamanho corporal contribuíram de forma mais significativa para o zooplâncton, sugerindo águas de melhor qualidade neste reservatório. Apesar de copépodes contribuirem com a maior parte da biomassa de microcrustáceos (entre 68 e 80%), as espécies de cladóceros apresentaram maiores taxas de produtividade e razão P:B em ambos os reservatórios. Cladóceros são considerados presas preferenciais de peixes, por sua maior digestibilidade e menor capacidade de fuga, se comparados aos copépodes. A comunidade zooplanctônica natural terá uma grande importância como fonte de alimento adicional para as formas juvenis de tilápias, podendo diminuir sua mortalidade no início de cada ciclo de criação. Além disso, uma vez conhecidas a composição em espécies e a estrutura desta comunidade, bem como a magnitude da biomassa e da produtividade secundária, alterações nestes parâmetros, em decorrência da atividade proposta, permitirão adequações e um manejo mais adequado, visando à conservação da qualidade das águas destes reservatórios ao longo de todo o empreendimento. PALAVRAS-CHAVE: comunidade zooplanctônica, microcrustáceos, biomassa, produtividade secundária, reservatórios tropicais, parques aqüícolas

2
ABSTRACT
Limnologic Characterization and Secondary Productivity of the Main Microcrustacean Species in Two Arms of Três Marias and Furnas Reservoirs, Minas Gerais, Brazil. The large number of reservoirs built in Brazil with the initial aim of electric energy generation, were slowly diversified in order to supply other needs of the activities developed in the water basin. Currently, the water is mainly used for irrigation, water supply, tourism and fishing. In order to increase the aquaculture production and optimize the use of the reservoirs in the country, the current government proposed the Inter-ministerial Instruction number 6, May, 28, 2004, to the transference of authorizations of public water use for fish production in cage farming to the low-income rural producers. Thus, the “Technical-Scientific Study Aiming the Delimitation of Aquaculture Areas in the Lakes of Furnas and Três Marias Reservoirs – MG, Brazil” was proposed to select areas with oligotrophic waters within these reservoirs to install the cage farming for the cultivation of tilapia and estimate these areas’ support capacity, with the objective of minimizing the impact of these kind of activities on water quality. Considering the importance of zooplankton on energy transference between the trophic levels, as well as it’s roll as water quality indicator, this study had as objectives describe this community in terms of composition, biomass and secondary productivity, in the reservoirs of Furnas and Três Marias, as a support for the evaluation of the sustainability of the fishing potential in these environments. Although these reservoirs may be considered oligotrophic, they show important differences in the zooplankton community. In the Três Marias reservoir, there’s a predominance of small body-sized cyclopoids and cladocerans in high densities (mean values of total zooplankton of 23,721 org.m-³ and 90,872 org.m-³ in dry and rainy periods, respectively), characterizing this environment as mesotrophic. In the Furnas reservoir (mean values of total zooplankton of 9,022 org.m-³ and 40.434 org.m-³ in dry and rainy periods, respectively), larger bodied calanoids and cladocerans contribute more significantly to the zooplankton, suggesting better quality waters. Although the copepods contribute with most of the microcrustacean biomass (between 68 and 80%), the species of cladocerans showed the higher productivity and P:B ratios in both reservoirs. Cladocera are considered the favourite prey by the fish due to its higher digestability and lower ability of escaping, when compared to the copepods. The natural zooplankton community will be very important for the young tilapias as an additional food source, being able to decrease their mortality at the beginning of each creation cycle. Furthermore, once the composition in species and the structure of this community, as well as the biomass magnitude and secondary productivity are well-known, changes in these parameters, due to the proposed activity, will allow a more appropriate management, aiming at the conservation of the quality of water at these reservoirs throughout all the joint enterprise. KEY-WORDS: zooplankton community, microcrustacean, biomass, secondary productivity, tropical reservoirs, aquaculture areas

3
INTRODUÇÃO GERAL
Devido às extensas bacias localizadas em rios de planalto, a maior parte da energia elétrica
produzida no Brasil é de origem hídrica. Desta forma, cerca de 73% da energia elétrica brasileira
(107.796 MW) é gerada por 530 usinas hidrelétricas (UHEs) e pequenas centrais hidrelétricas
(PCHs) (ANEEL, 2010). Com a criação da Eletrobrás (Centrais Elétricas Brasileiras S/A) na década
de 1960, a expansão do setor foi caracterizada pela implantação de grandes projetos hidrelétricos
como Itaipu (1966), Sobradinho (1974), Tucuruí (1976) e Ilha Solteira (1978). O Brasil aproveita
atualmente 25% de seu potencial hidroelétrico inventariado (metade do potencial não explorado se
encontra na bacia Amazônica), enquanto outros países já esgotaram os seus próprios potenciais.
A maior parte das usinas hidroelétricas do Brasil se encontra nos Estados de Minas Gerais,
São Paulo e Paraná, utilizando recursos da bacia do rio Paraná, quase 60% do potencial instalado no
país. A bacia do rio São Francisco corresponde a 15% do potencial instalado (ANEEL, 2010).
Localizadas nas regiões Sudeste e Sul, estas bacias estão entre as mais densamente povoadas e
intensamente exploradas pela agropecuária. Além da geração hidrelétrica, outros usos se agregaram
com o tempo: irrigação, abastecimento, navegação, turismo e pesca (Straškraba & Tundisi, 2000).
O atual governo, numa tentativa de incrementar a produção pesqueira e aumentar a geração
de empregos e renda, lançou as normas para autorizações de uso dos espaços físicos em corpos
d’água de domínio da União através da Instrução Normativa Interministerial nº. 6, de 28 de maio de
2004 (Secretaria Especial de Aqüicultura e Pesca – SEAP, na época ainda vinculada ao Ministério
da Agricultura e Pecuária). A partir dela, o atual Minístério da Pesca e Aqüicultura (antiga SEAP),
repassará aos produtores interessados as autorizações para uso da água pública para a produção de
pescados, e organizará os parques aqüícolas nas diversas regiões do país. Com relação aos
reservatórios, o Projeto de Produção de Peixes em Grandes Lagos selecionou sete para esta primeira
etapa do projeto: Furnas e Três Marias (MG), Sobradinho (BA), Tucuruí (PA); Serra da Mesa (GO),
Ilha Solteira (SP) e Itaipu (PR) (Diretoria de Desenvolvimento da Aqüicultura do Ministério de
Pesca e Aqüicultura - MPA) (MPA, 2009).
O projeto Parques Aqüícolas (Estudo Técnico-Científico Visando a Delimitação de Parques
Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG, Convênio 8713,
SEAP/SECTES/FUNDEP-UFMG) foi criado com o objetivo de atender a esta política de
reestruturação do setor de pesca e aqüicultura, buscando otimizar e ampliar o uso dos grandes
reservatórios no Estado de Minas Gerais. O projeto visou, numa fase inicial, o levantamento de
informações sobre a qualidade das águas destes reservatórios através de parâmetros físicos,
químicos e biológicos. Numa segunda etapa a capacidade suporte (em tanques rede por área) foi
estimada para áreas selecionadas nos dois reservatórios (Parques Aqüícolas, 2007a e b) onde “lotes”

4
de água de 1.500 m² foram demarcados e serão concedidos pelo período de 20 anos a produtores
rurais com renda até cinco salários mínimos (MPA, 2009). A preferência por áreas ou braços
oligotróficos, bem como das regiões mais profundas dos reservatórios, garantirá a longa vida do
projeto, evitando alterações na composição em espécies e aumento da biomassa da comunidade
fitoplanctônica, além do comprometimento das trocas gasosas e da penetração de luz causada pela
colonização excessiva de macrófitas (Parques Aqüícolas, 2006a e b).
Há um grande volume de trabalhos realizados em reservatórios, de modo que generalizações
sobre sua estrutura física, dinâmica de nutrientes e padrões das comunidades bióticas são
encontrados na literatura: Thornton et al. (1990); Pinto-Coelho et al. (1994); Tundisi et al. (1995);
Henry (1999); Tundisi & Straškraba (1999) e Straškraba & Tundisi (2000); Rodrigues et al. (2003);
Rodrigues et al. (2005). Da mesma forma, padrões da comunidade zooplanctônica em reservatórios
são conhecidos através de vários trabalhos como os de Marzolf (1990); Matsumura-Tundisi (1999);
Rocha et al. (1995); Rocha et al. (1997); Rocha et al. (1999); Lansac-Tôha et al. (2003); Lansac-
Tôha et al. (2005).
A comunidade zooplanctônica possui uma grande importância ecológica nos ambientes
aquáticos atuando como elo entre o nível dos produtores (fitoplâncton) e os níveis tróficos
superiores. Tem grande importância na estruturação e funcionamento do ecossistema aquático,
desempenhando um papel fundamental na transferência de energia e decomposição de matéria
orgânica, ou através de interações como herbivoria e predação, (Bozelli & Huszar, 2003).
Alguns autores têm procurado relacionar a composição e densidade das espécies com o grau
de trofia do ambiente (Gannon & Stemberger, 1978; Sendacz, 1984; Matsumura-Tundisi et al.,
2002 e Ferdous & Muktadir, 2009). Sob condições de enriquecimento de nutrientes, espécies de
menor tamanho, com ciclo de vida mais curto, dietas mais amplas e taxas de reprodução mais
rápidas (bactérias, protozoários e rotíferos) geralmente predominam na comunidade zooplanctônica.
Com relação aos Copepoda, espécies da ordem Calanoida têm sido associadas a ambientes
oligotróficos, enquanto que Cyclopoida têm predominado em ambientes meso-eutróficos (Tundisi
et al., 1988; Rocha et al., 1995).
A biomassa é considerada o peso de matéria viva existente num dado momento podendo ser
expressa por unidade de área ou volume (Begon et al., 2006). É um importante parâmetro das
comunidades que permite inferências sobre a cadeia trófica, ciclagem de matéria e fluxo energia
além de fornecer a real contribuição de cada espécie nos processos ecológicos (McCauley, 1984).
Dependendo da abordagem a ser adotada, a densidade descreve bem a estrutura do zooplâncton,
mas se o objetivo é comparar ambientes e quantificar as interações tróficas, a biomassa é a melhor
variável para descrever esta comunidade (Bozelli & Huzsar, 2003).

5
Desta forma, apesar de geralmente Rotifera ser um grupo numericamente dominante,
microcrustáceos contribuem com maior proporção na biomassa, comprovando que estudos de
abundância não refletem a real estrutura da comunidade zooplanctônica (Pace, 1986; Matsumura-
Tundisi et al., 1989; Rocha et al., 1995). Zimmer et al. (2001) comparando a comunidade
zooplanctônica de áreas úmidas no centro-oeste de Minnesota, não observou diferença significativa
entre a estrutura de tamanho de corpo destas comunidades, mas sim entre a composição
taxonômica, biomassa e produção.
No entanto, a biomassa de um organismo, num determinado momento, pode não refletir as
taxas de produção de novo material ou taxas nas quais a energia é processada no ecossistema, uma
vez que porções consideráveis podem ser eliminadas do ambiente, por exemplo, por predação
(Lampert & Sommer, 2007). Portanto, a produtividade é uma medida realística da contribuição de
cada componente no fluxo de energia da comunidade (Melão & Rocha, 2000).
A produtividade secundária do zooplâncton faz parte de um amplo esquema de
movimentação de matéria e energia através do ecossistema aquático, baseado nas atividades de
indivíduos e populações animais (Edmondson, 1974). Trata-se da “biomassa acumulada pelas
populações heterotróficas por unidade de tempo. A produtividade secundária do zooplâncton é a
soma do crescimento (incluindo mudas e os produtos sexuais) de todos os indivíduos da população”
(Sipaúba-Tavares & Rocha, 2001).
Estudos de produtividade secundária são importantes, pois permitem o entendimento do
ciclo de matéria e fluxo de energia na comunidade, detectando os efeitos de impactos ambientais,
(como eutrofização, introdução de espécies exóticas) (Rocha et al., 1997), sobre suas populações,
auxiliando no manejo racional dos recursos hídricos e inferindo sobre o sucesso de uma espécie
numa comunidade ou ecossistema (Downing & Rigler, 1984). Através da comparação com outros
estudos de biomassa e produtividade secundária pode-se ainda avaliar a regularidade nas
comunidades ecológicas, fornecendo subsídios para o manejo de ecossistemas aquáticos (Rietzler et
al., 2004).
Apesar da importância das informações obtidas através dos dados de biomassa e
produtividade secundária para a compreensão da dinâmica da matéria e energia dos ecossistemas
aquáticos, estudos desta natureza são ainda escassos em regiões tropicais. A tabela 1 sumariza os
principais estudos realizados no país envolvendo biomassa e/ou produtividade. Analisando a tabela,
fica evidente o predomínio de estudos de produtividade secundária nos corpos d’água do Estado de
São Paulo (tipicamente subtropicais). A maioria destes estudos procurou comparar a produtividade
secundária de espécies zooplanctônicas entre períodos de chuva e seca dentro de um mesmo
ambiente.

6
Tabela 1 – Estudos sobre biomassa e produtividade secundária do zooplâncton realizados no Brasil.
*=Trabalhos que envolveram ambos os parâmetros. Demais trabalhos forneceram biomassa, peso
seco ou regressões peso-comprimento.
Ambiente Grupo/Espécies Autores
Reservatório do Broa (SP) Argyrodiaptomus furcatus Rocha & Matsumura-Tundisi (1984)*
Lagos Dom Helvécio, Carioca e Amarela (MG)
Copepoda, Cladocera Matsumura-Tundisi & Tundisi (1986)
Reservatório do Broa (SP) Copepoda, Cladocera, Rotifera Matsumura-Tundisi et al. (1989)
Reservatório de Monjolinho (SP) Copepoda, Cladocera, Rotifera Okano (1994)*
Lago Batata (PA) Cladocera Maia-Barbosa (2000)*
Lagoa Dourada (SP) Copepoda, Cladocera, Rotifera Melão & Rocha (2000, 2004, 2006)*
Represa Lobo-Broa (SP) Rotifera Peláez-Rodríguez, & Matsumura-Tundisi (2002)*
Reservatório Barra-Bonita (SP) Copepoda, Cladocera, Rotifera Santos-Wisniewski et al. (2002)*
Santos-Wisniewski & Rocha (2007)*
Lagoa do Nado (MG) Chaoborus brasiliensis Bezerra-Neto & Pinto Coelho (2002)*
Lagoas Coqueiral e Camargo (SP) Cladocera Panarelli (2004)*
Reservatório de Salto Grande (SP) Copepoda, Cladocera, Rotifera Rietzler et al. (2004)*
Res. Ponte Nova e Guarapiranga (SP) Copepoda, Cladocera, Rotifera Sendacz et al. (2006)
Laboratório Ceriodaphnia silvestrii Santos et al. (2006)*
Rio Caeté (PA) Pseudodiaptomus spp Magalhães et al. (2007)
Lago Monte Alegre (SP) Copepoda, Cladocera, Chaoborus brasiliensis
Castilho-Nol & Arcifa (2007)
Res. Segredo, Mourão e Iraí (PR) Copepoda, Cladocera, Rotifera Blettler & Bonecker (2007),
Bonecker et al. (2007)
Reservatório Bariri (SP) Copepoda, Cladocera, Rotifera González et al. (2008)
Em Minas Gerais, onde a maioria dos corpos d’água é de clima tropical, apesar da enorme
malha hídrica e grande número de usinas hidrelétricas (35 grandes e médios reservatórios, 17.270
MW representando 18,6% da energia gerada no país), apenas um estudo foi registrado envolvendo
Chaoborus brasiliensis de uma lagoa periurbana, a Lagoa do Nado (Bezerra-Neto & Pinto Coelho,
2002).
Por se tratar de um processo avaliado através de medidas discretas, e por esta comunidade
apresentar grandes variações ao longo do tempo, a metodologia a ser empregada deve minimizar os
erros na estimativa da produtividade secundária (Edmondson, 1974). Intervalos curtos de
amostragem, que contemplem o período de reposição das espécies, são recomendados. Portanto,
amostragem em dias consecutivos ou alternados parece ser mais apropriada para um estudo acurado
da produção secundária (Melão & Rocha, 2000).
Segundo Winberg et al. (1965) o método do incremento da biomassa é o mais adequado para
a estimativa da produtividade secundária. Este método baseia-se na soma dos incrementos diários
em peso, para cada estágio de desenvolvimento, idade ou classe de tamanho e requer o
conhecimento destas variações durante todo o ciclo de vida do indivíduo, desde a eclosão até a
morte.

7
Quando estes dados não estão disponíveis, ou quando sua obtenção não é possível,
estimativa da produção pode ser obtida através do produto da biomassa (B) da população e da taxa
de crescimento, ou seja, da taxa finita de natalidade (β) (Hart, 1987). Este enfoque, aplicado
anteriormente para populações em equilíbrio com uma distribuição de idade estável, tem sido usado
para populações que violam o pressuposto do estado de equilíbrio, oferecendo uma estimativa
aproximada da produção.
A partir da estimativa de biomassa e produtividade do zooplâncton é possível avaliar a
importância desta comunidade como recurso alimentar para os níveis tróficos superiores, uma vez
que trata-se de um alimento rico em ácidos graxos e vitaminas, mais completo em termos de
nutrientes do que rações, e essencial para a sobrevivência de peixes nos estágios iniciais de
desenvolvimento (Sipaúba-Tavares & Rocha 2001; Roche & Rocha, 2005).
Se considerarmos que no mundo milhões de pessoas vivem direta ou indiretamente de
produtos animais de água doce ou salgada (cerca de 97 milhões de toneladas de pescado por ano –
FAO, 2004), esta comunidade possui um grande valor econômico, pois, a produção de peixes e
invertebrados depende da qualidade da alimentação nos estágios larvais cuja mortalidade pode
chegar a 90% nesta fase (Sá Júnior, 1994).
OBJETIVOS
Com base nas considerações feitas acima, este trabalho compara a densidade, biomassa e
produtividade secundária das espécies de microcrustáceos mais abundantes dos reservatórios de
Três Marias e Furnas (MG); e assim pretende determinar a importância da comunidade
zooplanctônica para a sustentabilidade do potencial pesqueiro nestes reservatórios. Neste sentido,
esta tese foi dividida em três capítulos: o primeiro descreve a comunidade zooplanctônica dos
reservatórios de Três Marias e Furnas e propõe seu uso na caracterização do estado trófico; o
segundo e terceiro capítulos, apresentam as estimativas da biomassa e da produtividade secundária
das principais espécies de microcrustáceos, e os possíveis fatores responsáveis pelas variações
registradas. Na discussão geral, os dados obtidos nesta tese, bem como outros estudos realizados no
contexto do projeto Parques Aqüicolas, são apresentados a fim de se avaliar o impacto da criação de
peixes em tanques-rede nas comunidades aquáticas e a importância do monitoramento destas
comunidades em longo prazo.

8
REFERÊNCIAS BIBLIOGRÁFICAS
ANEEL – Agência Nacional de Energia Elétrica. 2010. BIG - Banco de Informações de Geração.
http://www.aneel.gov.br/aplicacoes/capacidadebrasil/capacidadebrasil.asp Acesso: 13 abr. 2010. BEGON, M., TOWNSEND, C.R. & HARPER., J.L. 2006. Ecology: from individuals to
ecosystems. Oxford: Blackwell Publishing. 4ª ed. 759p. BEZERRA-NETO, J. F. & PINTO-COELHO, R.M. 2002. Population dynamics and secondary
production of Chaoborus brasiliensis (Diptera: Chaoboridae) in a small tropical reservoir: Lagoa do Nado, Minas Gerais - Brazil. Acta Limnol. Bras., 14(3): 61-72.
BLETTLER, M.C.M. & BONECKER, C.C. 2007. Longitudinal distribution of microcrustacean biomass in three tropical reservoirs (Paraná State, Brazil). Acta Scientiarum, 29(3):297-304.
BONECKER, C.C.; NAGAE, M.Y.; BLETTLER, M.C.M.; VELHO, L.F.M. & LANSAC-TÔHA, F.A. 2007. Zooplankton biomass in tropical reservoirs in southern Brazil. Hydrobiologia, 579:115-123.
BOZELLI, R. L. & HUSZAR, V. L. M. 2003. Comunidades Fito e Zooplanctônicas Continentais em Tempo de Avaliação. LIMNOtemas. Sociedade Brasileira de Limnologia, 3:1-32.
CASTILHO-NOLL, M.S.M. & ARCIFA, M.S. 2007. Length-weight relationships for zooplanktonic species of a tropical Brazilian lake: Lake Monte Alegre. Acta Limnol. Bras., 19(1): 93-100.
DOWNING, J.A. & RIGLER, F.H. (eds.) 1984. A Manual on Methods for the Assessment of Secondary Productivity in Fresh Waters. IBP Hand Book 17, Blackwell Scientific Publishing. 2ª ed. 501p.
EDMONDSON, W. T. 1974. Secondary Production. Verh. Internat. Verein. Limnol., 20:229-272. FAO - Food and Agriculture Organization of the United Nations. 2004. The State of Food
Insecurity in the World. 6a ed. Rome: United Nations. 40p. FERDOUS, Z. & MUKTADIR, A.K.M. 2009. A Review: Potentiality of Zooplankton as
Bioindicator. American Journal of Applied Sciences, 6(10): 1815-1819. GANNON, J. E. & STEMBERGER, R. 1978. Zooplankton (especially crustaceans and rotifers) as
indicators of water quality. Trans. Am. Microsc. Soc., 97:16-35. GONZÁLEZ, E.J., MATSUMURA-TUNDISI, T. & TUNDISI, J.G. 2008. Size and dry weight of
main zooplankton species in Bariri reservoir (SP, Brazil). Braz. J. Biol., 68(1):69-75. HART, R. C. 1987. Population dynamics and production of five crustacean zooplankters in a
subtropical reservoir during years of contrasting turbidity. Freshwater Biol., 18:287-318. HENRY, R. (ed.) 1999. Ecologia dos Reservatórios: Estrutura, Função e Aspectos Sociais.
Botucatu: FUNDIBIO, FAPESP. 799p. LAMPERT, W. & SOMMER,U. 2007. Limnoecology. The Ecology of Lakes and Streams. 2nd ed.
Oxford: Oxford University Press. 324p. LANSAC-TÔHA, F. A., VELHO, L. F. M., TAKAHASHI, E. M., NAGAE, M. Y., PEDROSO, M.
M., GARCIA & D. L. PEREIRA, D. G. 2003. Riqueza e Abundância da Comunidade Zooplanctônica em Reservatórios do Estado do Paraná. In: Rodrigues, L.; Agostinho, A. A.; Gomes, L. C. & Thomaz, S. M. (eds). Anais do Workshop Produtividade em Reservatórios e Bioindicadores. Maringá: Nupélia. 281p.
LANSAC-TÔHA, F. A.; BONECKER, C.C. & VELHO, L. F. M. 2005. Estrutura da Comunidade Zooplanctônica em Reservatórios. In: Rodrigues, L.; Thomaz, S.M.; Agostinho, A.A. & Gomes, L.C. (eds.) Biocenoses em Reservatórios: padrões espaciais e temporais. Rima: São Carlos. 333p.
MAGALHÃES, A., COSTA, R. M., LIANG, T. H., PEREIRA, L. C. C. & RIBEIRO, M. J. S. 2006. Spatial and Temporal Distribution in Density and Biomass of Two Pseudodiaptomus Species (Copepoda: Calanoida) in the Caeté River Estuary (Amazon Region - North of Brazil). Braz. J. Biol., 66(2A): 421-430.

9
MAIA-BARBOSA, P. M. 2000. Ecologia de Cinco Espécies de Cladóceros de um Lago Amazônico Impactado por Rejeito de Bauxita (Lago Batata, Pará-Brasil). Universidade Federal do Rio de Janeiro. 218p. Tese.
MARZOLF, G.R. 1990. Reservoir as Environments for Zooplankton. In: Thornton, K. W.; Kimmel, B.L. & Payne, F.E. (eds.) Reservoir Limnology: Ecological Perspectives. New York: John Wiley & Sons. 246p.
MATSUMURA-TUNDISI, T. & TUNDISI, J.G. 1986. Biomass and Zooplankton Community Structure of Three Lakes of River Doce Valley (Minas Gerais - Brazil). The Fifth Japan-Brazil Symposium on Science and Tecnology. Tokyo, Japan.
MATSUMURA-TUNDISI, T., RIETZLER, A.C. & TUNDISI, J.G. 1989. Biomass (dry weight and carbon content) of plankton crustacea from Broa reservoir (São Carlos, SP, Brazil) and its fluctuations across on year. Hydrobiologia, 179:229-236.
MATSUMURA-TUNDISI, T.; TUNDISI, J. G. & ROCHA, O. 2002. Zooplankton diversity in eutrophic systems and its relation to the occurrence of cyanophycean blooms. Verh. Internat. Verein. Limnol. 28: 671-674.
MCCAULEY, E. 1984. The estimation of the abundance and biomass of zooplankton in samples. In: Downing J.A. & F.H. Rigler (eds.) A Manual on Methods for Assessment of Secondary Productivity in Freshwaters. IBP Hand Book 17, Blackwell Scientific Publishing. 2ª ed. 501p.
MELÃO, M. G. & ROCHA, O. 2000. Productivy of zooplankton in a tropical oligotrophic reservoir over short periods of time. Verh. Internat. Verein. Limnol., 27:2879-2887.
MELÃO, M. G. & ROCHA, O. 2004. Life history, biomass and production of two planktonic cyclopoid copepods in a shallow subtropical reservoir. J. Plankton Res.,, 26(8):909-923.
MELÃO, M. G. & ROCHA, O. 2006. Life history, populations dynamics, standing biomass and production of Bosminopsis deitersi (Cladocera) in a shallow tropical reservoir. Acta Limnol. Bras., 18(4):433-450.
MPA - Ministério da Pesca e Aqüicultura. 2009. Aqüicultura no Brasil. http://www.presidencia.gov.br/estrutura_presidencia/seap/. Acesso em: 29 jun. 2009.
OKANO, Y. W. 1994. Análise da estrutura e dinâmica populacional da comunidade zooplanctônica de um reservatório artificial (reservatório Monjolinho, São Carlos, SP). Universidade Federal de São Carlos. 128p. Tese.
PACE, M. L. 1986. An empirical analyses of zooplankton community size structure across lake trophic gradients. Limnology and Oceanography, 31(1):45-55.
PANARELLI, E. A. 2004. Flutuações mensais da comunidade zooplanctônica e dinâmica das populações de Cladocera em lagoas marginais, na região de transição Rio Paranapanema – Represa Jurumirim (SP). Universidade Estadual Paulista. 247p. Tese.
PARQUES AQÜÍCOLAS - Estudo Técnico-Científico Visando a Delimitação de Parques Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG. 2006a. Ranqueamento de Áreas Alvo e Delimitação de Polígonos Reservatório de Três Marias (Fase 1). BeloHorizonte: UFMG. 13p.
PARQUES AQÜÍCOLAS - Estudo Técnico-Científico Visando a Delimitação de Parques Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG. 2006b. Ranqueamento de Áreas Alvo e Delimitação de Polígonos Reservatório de Furnas (Fase 1). BeloHorizonte: UFMG. 17p.
PARQUES AQÜÍCOLAS - Estudo Técnico-Científico Visando a Delimitação de Parques Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG. 2007a. Relatório de Identificação de Áreas Técnicamente Adequadas para a Instalação de Parques Aqüícolas. Reservatório de Três Marias. Belo Horizonte: UFMG. 311p.
PARQUES AQÜÍCOLAS - Estudo Técnico-Científico Visando a Delimitação de Parques Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG. 2007b. Relatório de Identificação de Áreas Tecnicamente Adequadas para a Instalação de Parques Aqüícolas. Reservatório de Furnas. Belo Horizonte: UFMG. 348p.

10
PELÁEZ-RODRÍGUEZ, M. & MATSUMURA-TUNDISI, T., 2002. Rotifer Production in a Shallow Artificial Lake (Lobo-Broa Reservoir, SP, Brazil). Braz. J. Biol., 62(3): 509-516.
PINTO-COELHO, R.M.; GIANI, A. & VON SPERLING, E. (Eds.) 1994. Ecology and Human impact on lakes and reservoirs in Minas Gerais with special reference to future development and management strategies. Belo Horizonte: SEGRAC. 193p.
RIETZLER, A.C.; ROCHA, O. & ESPINDOLA, E. L. G. 2004. Produção Secundária de Espécies do Zooplâncton do Reservatório de Salto Grande, Estado de São Paulo. In: Espindola, E. L.G., Leite, M.A., Dornfeld, C.B. (eds.) Reservatório de Salto Grande (Americana-SP): caracterização, impactos e propostas de manejo. São Carlos: Rima.
ROCHA, O. & MATSUMURA-TUNDISI, T., 1984. Biomass and production of Argyrodiaptomus furcatus a tropical calanoid copepod in Broa reservoir, southern Brazil. Hydrobiologia, 113: 307-311.
ROCHA, O.; MATSUMURA-TUNDISI, T. & SAMPAIO, E. V. 1997. Phytoplankton and zooplankton community structure and production as related to trophic state in some Brazilian lakes and reservoirs. Verh. Internat. Verein Limnol., 26:599-604.
ROCHA, O.; MATSUMURA-TUNDISI, T.; ESPÍNDOLA, E.; ROCHE, K. & RIETZLER, A. 1999. Ecological Theory Applied to Reservoir Zooplankton. In: Tundisi, J.G. & Straškraba, M. (eds.) Theoretical Reservoir Ecology and its Applications. São Carlos: IIE. 585p.
ROCHA, O.; SENDACZ, S. & MATSUMURA-TUNDISI, T. 1995. Composition, Biomass and Productivity of Zooplankton in Natural Lakes and Reservoirs of Brazil. In: Tundisi, J. G.; Bicudo, C. E. M. & Matsumura-Tundisi, T. (eds) Limnology in Brazil. Rio de Janeiro: Brazilian Academy of Sciences, Brazilian Limnological Society. 376p.
ROCHE, F. K. & ROCHA, O. 2005. Aspectos de predação por peixes em lagos e represas, com enfoque na planctivoria. In: Roche, F. K. & Rocha, O. (eds.) Ecologia Trófica de Peixes com ênfase na planctivoria em ambientes lênticos de água doce no Brasil. São Carlos: Rima. 146p.
RODRIGUES, L.; AGOSTINHO, A. A.; GOMES, L. C. & THOMAZ, S. M. (eds). 2003. Anais do Workshop Produtividade em Reservatórios e Bioindicadores. Maringá: Nupélia. 281p.
RODRIGUES, L.; THOMAZ, S.M.; AGOSTINHO, A.A. & GOMES, L.C. (eds.) 2005. Biocenoses em Reservatórios: padrões espaciais e temporais. São Carlos: Rima. 333p.
SÁ JUNIOR, W. P. 1994. Production of Planktonic Biomass for Feed of Alevins at the Furnas Hydrobiology and Hatchery Station. In: Pinto-Coelho, R.M.; Giani, A. & von Sperling, E. (eds) Ecology and Human impact on lakes and reservoirs in Minas Gerais with special reference to future development and management strategies. Belo Horizonte: SEGRAC. 193p.
SANTOS, M.A.P.F., MELÃO, M.G.G. & LOMBARDI, A.T. 2006. Life history characteristics and production of Ceriodaphnia silvestrii Daday (Crustacea, Cladocera) under different experimental conditions. Acta Limnol. Bras., 18(2):199-212.
SANTOS-WISNIEWSKI, M.J. & ROCHA, O. 2007. Spatial distribution and secondary production of Copepoda in a tropical reservoir: Barra Bonita, SP, Brazil. Braz. J. Biol., 67(2): 223-233.
SANTOS-WISNIEWSKI, M.J., MATSUMURA-TUNDISI, T. & ROCHA, O. 2002. Spatial and Temporal Patterns of Zooplankton Biomass and Secondary Production in Barra Bonita Reservoir, Brazil. 4th International Conference on Reservoir Limnology and Water Quality, August 12-16, 2002, extended Abstrats. p: 303-306.
SENDACZ, S., CALEFFI, S. & SANTOS-SOARES, J. 2006. Zooplankton Biomass of Reservoirs in Different Trophic Conditions in the State of São Paulo, Brazil. Braz. J. Biol., 66(1B): 337-350.
SENDACZ, S. 1984. A study of the zooplankton community of Billings Reservoir – São Paulo In: Dumont, H. & Tundisi, J.G. (eds.) Developments in Hydrobiology – Tropical Zooplankton. W.Junk Publishers, p. 121-128.
SIPAÚBA-TAVARES, L. H. & ROCHA, O. 2001. Produção de Plâncton (Fitoplâncton e Zooplâncton) para alimentação de organismos aquáticos. São Carlos: Rima. 106p.
STRAŠKRABA, M. & TUNDISI, J.G. (eds.) 2000. Diretrizes para o Gerenciamento de Lagos. Volume 9: Gerenciamento da Qualidade da Água de Represas. São Carlos: ILEC, IIE. 300p.

11
THORNTON, K. W.; KIMMEL, B.L. & PAYNE, F.E. 1990. Reservoir Limnology: Ecological Perspectives. New York: John Wiley & Sons. 246p.
TUNDISI, J. G.; BICUDO, C. E. M. & MATSUMURA-TUNDISI, T. (eds.) 1995. Limnology in Brazil. Rio de Janeiro: Brazilian Academy of Sciences, Brazilian Limnological Society. 376p.
TUNDISI, J. G.; T. MATSUMURA-TUNDISI; R. HENRY; O. ROCHA & K. HINO. 1988. Comparação do estado trófico de 23 reservatórios do Estado de São Paulo: Eutrofização e manejo. In: Tundisi, J.G. (ed.) Limnologia e manejo de represas. vol. 1. Tomo 1. São Paulo: Série Monografias em Limnologia/USP.
TUNDISI, J.G. & STRAŠKRABA, M. (eds.) 1999. Theoretical Reservoir Ecology and its Applications. São Carlos: IIE. 585p.
WINBERG, G. G.; PECHEN, G. A. & SHUSHKINA, E. A. 1965. Prodution of Planktonic crustaceans in three lakes of different type. Zoologicheskii Zhurnal, 44: 676-687.
ZIMMER, K. D.; HANSON, M. A.; BUTLER, M. G. & DUFFY, W. G. 2001. Size distribution of aquatic invertebrates in two prairie wetlands, with and without fish, with implications for community production. Freshwater Biol., 46:1373-1386.

12
ARTIGO 1 – ZOOPLÂNCTON COMO INDICADOR DE CONDIÇÕES TRÓFICAS EM DOIS BRAÇOS DOS RESERVATÓRIOS DE TRÊS MARIAS E FURNAS, MINAS GERAIS, BRASIL. RESUMO As variáveis físicas e químicas da água, bem como a composição e a estrutura da comunidade zooplanctônica dos reservatórios de Três Marias e Furnas (MG) foram comparados com o objetivo de caracterizar estes ambientes quanto ao seu estado trófico. Maiores valores de condutividade elétrica, concentrações de clorofila-a, sólidos totais e orgânicos em suspensão e nitrogênio total foram registrados no reservatório de Três Marias, enquanto maior transparência da água e concentrações de nitrito e nitrato foram observadas em Furnas (p<0,000). Maiores densidades do zooplâncton sempre foram obtidas no reservatório de Três Marias e no período de chuvas (p<0,000) com valores médios na seca e chuva em Três Marias de 23.721 org.m-³ e 90.872 org.m-³ e em Furnas de 9.022 org.m-³ e 40.434 org.m-³, respectivamente, sendo Copepoda o grupo dominante em ambos os reservatórios, principalmente devido aos estágios juvenis (náuplios e copepoditos). Dentre os cladóceros, bosminídeos ocorreram com maior abundância em Três Marias, e dafinídeos e sidídeos no reservatório de Furnas. Em valores absolutos e relativos, a contribuição de rotíferos foi maior no reservatório de Três Marias em relação ao de Furnas. Apesar do Índice de Estado Trófico, baseado na transparência da água, concentrações de clorofila-a e fósforo total, apontar os dois ambientes como oligotróficos, no reservatório de Três Marias, as maiores densidades da comunidade zooplanctônica bem como o predomínio de ciclopóides e cladóceros de menor tamanho corporal como bosminídeos caracterizam este ambiente como mesotrófico. No reservatório de Furnas, espécies de maior porte como calanoídes, dafinídeos e sidídeos caracterizam as águas deste ambiente como de melhor qualidade. PALAVRAS-CHAVE: zooplâncton, indicador de qualidade de água, estado trófico, reservatórios tropicais ABSTRACT Zooplankton as an Indicator of Trophic Conditions in Two Arms of Três Marias and Furnas Reservoirs, MG, Brazil. The physical and chemical variables of water, as well as the composition and structure of the zooplankton community of Três Marias and Furnas reservoirs (MG, Brazil) were compared in order to characterize these environments in relation to their trophic state. Higher values of electric conductivity, chlorophyll-a concentrations, total solids and suspended organic matter and total nitrogen were registered in Três Marias reservoir, while higher water transparence and nitrite and nitrate concentrations were observed in Furnas (p<0.000). Higher zooplankton densities were always obtained at Três Marias reservoir and during the rain period (p<0.000) with average values in dry and rain periods in Três Marias of 23,721 org.m-3 and 90,872 org.m-3 and in Furnas of 9,022 org.m-3 and 40,434 org.m-3, respectively. Copepoda was the dominant group in both reservoirs, mainly due to the younger stages (nauplii and copepodids). Within the cladoceran, bosminids were more abundant in Furnas reservoir. According to absolute and relative values, the contribution of rotifers was higher in Três Marias reservoir, in relation to Furnas. Although the Trophic State Index, based in water transparency, chlorophyll-a concentrations and total phosphorous, shows both environments as oligotrophic, in Três Marias reservoir, the higher densities of the zooplankton community as well as the predominance of cyclopoids and smaller body-sized species such as bosminids characterize this environment as mesotrophic. In Furnas reservoir, larger species such as calanoids, daphnids and sidids characterize the better water quality of this environment. KEY-WORDS: zooplankton, water quality indicator, trophic state, tropical reservoirs

13
INTRODUÇÃO
Os reservatórios constituem uma complexa rede interativa entre os organismos (espécies,
populações, comunidades) e o seu ambiente físico e químico. Esta rede é dinâmica e resulta de um
processo permanente de resposta às funções de força climatológicas e aos efeitos produzidos pela
manipulação do sistema na barragem (Henry, 1999).
A eutrofização é um problema crescente nos reservatórios brasileiros uma vez que a
urbanização e industrialização (fontes pontuais) e o intenso uso do entorno em atividades agro-
pastoris (fontes difusas) têm comprometido a qualidade de suas águas. Em longo prazo, levam à
perda da diversidade biológica, diminuição da vida útil dos reservatórios e de estoques pesqueiros
(Straškraba & Tundisi, 2000). Neste contexto, estudos sobre variáveis físicas, químicas e biológicas
de suas águas são fundamentais para avaliar os impactos destas atividades e auxiliar na proposição
de medidas preventivas e mitigatórias.
A estrutura e abundância da comunidade zooplanctônica, bem como sua distribuição
espacial, são influenciadas por fatores abióticos e pelas interações entre as espécies. Geralmente em
ecossistemas aquáticos tropicais se observa maior abundância de rotíferos e cladóceros (Esteves,
1998); contudo modificações nas condições físicas e químicas da água podem alterar a composição
da comunidade fitoplanctônica, ou seja, do alimento disponível, alterando também a estrutura da
comunidade zooplanctônica (Harper, 1992, Kozlowsky-Suzuki & Bozelli, 2002).
Apesar do zooplâncton não depender diretamente dos nutrientes para sua sobrevivência, já
que são afetados pela qualidade e quantidade de algas, bactérias e detritos, o estado trófico de um
ambiente pode influenciar a riqueza, estrutura, tamanho de corpo e produtividade da comunidade
zooplanctônica (Sládeček, 1958, McCauley & Kalff, 1981, Harper, 1992, Lathrop & Carpenter,
1992).
Segundo Gannon & Stemberger (1978) uma vez que muitos ambientes não exibem
claramente atributos de oligotrofia ou eutrofia, o zooplâncton pode ser usado como indicador destas
características intermediárias. Contudo, devido à ampla distribuição de algumas espécies, estes
autores sugerem o uso da abundância relativa dentro da comunidade como um indicador mais
sensível. Desta forma, padrões como aqueles observados por Matsumura-Tundisi et al. (2000) no
sistema em cascata do médio e baixo Tietê (SP), onde a dominância de Rotifera sobre Copepoda em
reservatórios eutróficos e o inverso em reservatórios mesotróficos ou oligotróficos, têm sido
descritos para a comunidade zooplanctônica.
A ictiofauna também pode influenciar na estrutura da comunidade zooplanctônica uma vez
que peixes planctívoros consomem indivíduos de maior tamanho corporal promovendo a

14
dominância de pequenos copépodes e cladóceros (Carpenter et al., 1985) mesmo que a biomassa de
microcrustáceos não seja alterada (Quirós & Boveri, 1999).
Além disso, visando ampliar o uso e aumentar o potencial pesqueiro dos reservatórios de
Três Marias e Furnas, o projeto Parques Aqüícolas (SEAP/SECTES/FUNDEP-UFMG), no qual
este trabalho está inserido, visou à delimitação de áreas para produção pesqueira através de tanques-
rede. Neste projeto, variáveis abióticas e bióticas foram medidas para estimar a capacidade suporte
de áreas selecionadas. Os conhecimentos levantados sobre o zooplâncton auxiliarão, em fase
posterior, não só pelo uso destes organismos como importante fonte alimentar, bem como no
monitoramento da qualidade de água, sendo utilizados para tanto como possíveis bioindicadores,
mitigando os impactos da criação de peixes sobre toda a comunidade aquática.
Neste contexto, o objetivo deste trabalho foi avaliar a composição e a densidade da
comunidade zooplanctônica em alguns braços dos reservatórios de Três Marias e Furnas (MG), e a
possibilidade do uso desta comunidade como indicadora de qualidade de água destes reservatórios,
através da relação de sua estrutura com as variáveis ambientais medidas.
MATERIAL E MÉTODOS
ÁREA DE ESTUDOS
Os reservatórios das Usinas Hidrelétricas (UHEs) de Três Marias (18°12’S e 45°15’W) e
Furnas (46°19’W e 20°40’S) foram criados há mais de 40 anos para geração de energia elétrica,
sendo também utilizados para recreação, pesca amadora e profissinal, irrigação e abastecimento. O
reservatório da UHE de Três Marias localiza-se no alto rio São Francisco, Centro-Oeste do Estado
de Minas Gerais. Concluído em 1960, atualmente o assoreamento é o principal impacto que
compromete a capacidade geradora de energia e a qualidade da água, seguido pela substituição da
vegetação ripária por monoculturas de Eucaliptus spp e pecuária extensiva (Sampaio & López,
2003). O reservatório de Furnas está localizado na bacia do rio Grande, Sul do Estado de Minas
Gerais, sendo o braço norte formado pelo rio Grande e o braço sul pelo rio Sapucaí. O reservatório
foi concluído em 1962 tendo, atualmente, como principais impactos atividades como a
agropecuária, monoculturas, o despejo de efluentes domésticos e o lançamento de resíduos sólidos e
agrotóxicos (Nogueira et al., 2008). Algumas características morfométricas dos dois reservatórios
são apresentadas na tabela 1.
Em cada reservatório foram escolhidos como pontos amostrais dois braços com
características diferentes de entorno (figura 1). Desta forma, no reservatório de Três Marias foram
selecionados os braços Barrão (Zmáx=21,8 metros, com vegetação original do tipo Cerrado) e

15
Extrema (Zmáx=20,9 metros, com monocultura de Eucaliptus spp), localizados no município de
Morada Nova de Minas. No reservatório de Furnas foram selecionados os braços Varjão
(Zmáx=17,5 metros, com nascente localizada no Parque Municipal do Paredão e entorno com
monocultura de café) e Mendonça (Zmáx=25,5 metros, com pecuária em pastagem nativa),
localizados no município de Guapé. Estes braços foram selecionados a fim de caracterizar as
diferentes feições do entorno destes reservatórios.
Tabela 1 – Características morfométricas dos reservatórios das UHEs de Três Marias e Furnas
(MG). Fonte: CEMIG, 2009 e FURNAS, 2009.
UHE Três Marias UHE Furnas Área Inundada 1.100 km2 1.440 Km² Volume Útil 15,27 x 109 m³ 17,21 x 109 m³ Vazão Média 700 m³.s-1 800 m³.s-1
Tempo Médio de Residência 120 dias 160 dias Profundidade Máxima 75 m 90 m Profundidade Média 12 m 13 m Capacidade Geradora 396 MW 1126 MW
Figura 1 – Reservatórios de Três Marias (esquerda, 1:10.000) e Furnas (direita, 1:20.000) (MG)
com as setas apontando os braços utilizados para as coletas. Modificado de Miranda & Coutinho,
2004.
PROCEDIMENTOS

16
As amostras foram coletadas a cada dois dias, durante quatro semanas, em dois períodos de
seca (julho/agosto de 2006 e julho/agosto de 2007) e dois de chuvas (janeiro/março de 2007 e
janeiro/março de 2008), considerados os mais contrastantes em termos de variações de temperatura
e precipitação. As coletas foram realizadas no ponto mais profundo da região limnética de cada
braço.
Em todas as coletas foi medida a transparência da água (disco de Secchi) e realizados perfis
de temperatura (°C), oxigênio dissolvido (mg.L-1) e condutividade (�S.cm-1) com auxílio das
sondas YSI30 e YSI55. Semanalmente foi também coletada água, da sub-superfície para
determinação das concentrações de clorofila-a, fósforo total, nitrogênio total, nitrito, nitrato,
amônio, sólidos totais em suspensão e sua fração orgânica. As amostras foram analisadas em
laboratório, seguindo recomendações de Lorenzen (1967), Golterman et al. (1978) e Mackereth et
al. (1978). Para nitrogênio total as concentrações foram determinadas apenas para o segundo ano de
coletas seguindo as recomendações de Wetzel & Likens, (1991) e Strickland & Parsons, (1972). O
Índice Estado Trófico (IET) foi calculado segundo Carlson (1977), modificado por Toledo et al.
(1983).
Para as coletas quali-quantitativas da comunidade zooplanctônica, foram realizados arrastos
com rede de plâncton de 68�m de abertura de malha. Devido à presença de “paliteiros” (vegetação
original não removida) nos pontos de coleta, os arrastos foram realizados apenas na zona eufótica,
determinada a partir da leitura do disco de Secchi. Os organismos coletados foram anestesiados com
água mineral gaseificada, corados com corante vital Rosa de Bengala e fixados com solução
neutralizada de formol 4%. Sub-amostras de 1,0 mL foram contadas sob microscópio ótico
Olympus (modelo CBA), em câmaras de Sedgewick-Rafter. Os dados apresentados correspondem
ao número de organismos por m3. Foram considerados os três grupos principais da comunidade
zooplanctônica: Rotifera, Cladocera e Copepoda. A metodologia para contagem das amostras
seguiu as recomendações de Bottrell et al., (1976).
A fim de relacionar a estrutura da comunidade zooplanctônica e o estado trófico dos
reservatórios foram calculados o Quociente Brachionus:Trichocerca (QB/T), segundo Sládeček
(1983), e a razão Calanoida/Cyclopoida, segundo Tundisi et al. (1988). A diversidade foi avaliada
através do índice de Shannon-Wiener (Magurran, 1988). Também foram realizadas correlações de
Pearson entre as variáveis ambientais e a densidade da comunidade zooplanctônica (Sampaio,
2002).
Para testar as diferenças das variáveis ambientais e da densidade da comunidade
zooplanctônica entre braços, reservatórios e períodos de coleta (seca e chuva) foi empregada uma
Análise de Variância bi-fatorial, seguida do teste de Tukey para detectar as diferenças. Quando os
pressupostos de normalidade e homogeneidade de variâncias não foram atendidos, foi realizado

17
teste não paramétrico (Mann-Whitney) (Sampaio, op. cit.). Para análise das variáveis ambientais foi
empregada uma Análise de Componentes Principais, onde as variáveis foram transformadas (log10)
a fim de atender aos pressupostos de linearidade da ordenação (Valentin, 2000). Todos os testes
foram realizados com auxílio do programa Statistica 7.0 (StatSoft).
RESULTADOS
VARIÁVEIS AMBIENTAIS
A transparência da coluna d’água sempre foi maior no período de seca em ambos os
reservatórios (F=636,4; p<0,000) (Tabela 2), com valores maiores sempre registrados no
reservatório de Furnas (F=247,3; p<0,000) (Anexo 1). A coluna d’água apresentou-se
desestratificada nos períodos secos e estraficada nos chuvosos (Anexo 2), com diferenças de
temperatura significativas entre os períodos (F=203,8; p<0,000). Durante o período de
desestratificação, as concentrações de oxigênio dissolvido e a condutividade elétrica não
apresentaram diferenças ao longo da coluna d’água (Anexos 3 e 4). Durante a estratificação, foi
observada hipóxia e menores valores de condutividade elétrica nas maiores profundidades. Valores
de oxigênio dissolvido apresentaram diferenças significativas apenas entre os períodos seca e chuva
(F=12,3; p=0,003) e a condutividade elétrica apenas entre os reservatórios (F=153,9; p<0,000)
sendo os maiores valores registrados em Três Marias (Tabela 2). Para a transparência, temperatura,
oxigênio dissolvido e condutividade elétrica as diferenças não foram significativas entre braços de
um mesmo reservatório (F=1,594; p=0,2094). No segundo período de chuva (janeiro/março de
2008), devido à menor precipitação neste ano (Tabela 2; SIMGE/IGAM, 2010), houve uma menor
redução na transparência no reservatório de Furnas e maior concentração de íons na coluna d’água
no reservatório de Três Marias. Os valores de pH, medidos entre 2006 e 2007, durante o projeto
Parques Aqüícolas por Pinto-Coelho (2006), oscilaram entre 8,0 e 8,5 no reservatório Três Marias e
7,3 e 8,3 no reservatório de Furnas, não apresentando diferenças significativas (F = 1,746;
p=0,2109).
Clorofila-a apresentou maiores concentrações no reservatório de Três Marias (F= 103,1;
p<0,000) e durante o período chuvoso (F=52,5; p<0,000) (Tabela 2) (Anexo 5). Sólidos totais e
orgânicos em suspensão sempre apresentaram maiores valores no reservatório de Três Marias
(F=10,3; p<0,000 e F=16,7; p<0,000 respectivamente), à exceção do período de chuvas de 2008
quando a menor precipitação aumentou a concentração destas variáveis no reservatório de Furnas
(F=8,17; p=0,007 e F=9,00; p<0,000, respectivamente). As diferenças não foram significativas
entre braços de um mesmo reservatório (F=0,1333; p=0,7163).
18
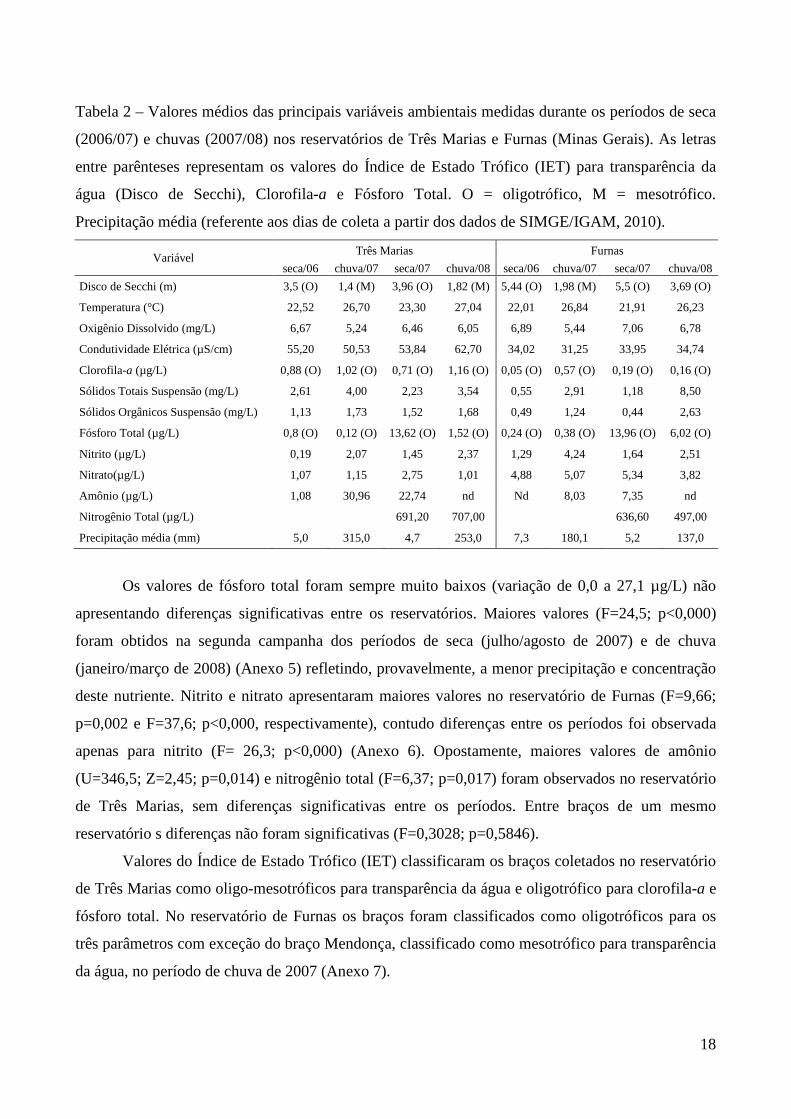
Tabela 2 – Valores médios das principais variáveis ambientais medidas durante os períodos de seca
(2006/07) e chuvas (2007/08) nos reservatórios de Três Marias e Furnas (Minas Gerais). As letras
entre parênteses representam os valores do Índice de Estado Trófico (IET) para transparência da
água (Disco de Secchi), Clorofila-a e Fósforo Total. O = oligotrófico, M = mesotrófico.
Precipitação média (referente aos dias de coleta a partir dos dados de SIMGE/IGAM, 2010).
Variável Três Marias Furnas
seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08
Disco de Secchi (m) 3,5 (O) 1,4 (M) 3,96 (O) 1,82 (M) 5,44 (O) 1,98 (M) 5,5 (O) 3,69 (O)
Temperatura (°C) 22,52 26,70 23,30 27,04 22,01 26,84 21,91 26,23
Oxigênio Dissolvido (mg/L) 6,67 5,24 6,46 6,05 6,89 5,44 7,06 6,78
Condutividade Elétrica (µS/cm) 55,20 50,53 53,84 62,70 34,02 31,25 33,95 34,74
Clorofila-a (µg/L) 0,88 (O) 1,02 (O) 0,71 (O) 1,16 (O) 0,05 (O) 0,57 (O) 0,19 (O) 0,16 (O)
Sólidos Totais Suspensão (mg/L) 2,61 4,00 2,23 3,54 0,55 2,91 1,18 8,50
Sólidos Orgânicos Suspensão (mg/L) 1,13 1,73 1,52 1,68 0,49 1,24 0,44 2,63
Fósforo Total (µg/L) 0,8 (O) 0,12 (O) 13,62 (O) 1,52 (O) 0,24 (O) 0,38 (O) 13,96 (O) 6,02 (O)
Nitrito (µg/L) 0,19 2,07 1,45 2,37 1,29 4,24 1,64 2,51
Nitrato(µg/L) 1,07 1,15 2,75 1,01 4,88 5,07 5,34 3,82
Amônio (µg/L) 1,08 30,96 22,74 nd Nd 8,03 7,35 nd
Nitrogênio Total (µg/L) 691,20 707,00 636,60 497,00
Precipitação média (mm) 5,0 315,0 4,7 253,0 7,3 180,1 5,2 137,0
Os valores de fósforo total foram sempre muito baixos (variação de 0,0 a 27,1 µg/L) não
apresentando diferenças significativas entre os reservatórios. Maiores valores (F=24,5; p<0,000)
foram obtidos na segunda campanha dos períodos de seca (julho/agosto de 2007) e de chuva
(janeiro/março de 2008) (Anexo 5) refletindo, provavelmente, a menor precipitação e concentração
deste nutriente. Nitrito e nitrato apresentaram maiores valores no reservatório de Furnas (F=9,66;
p=0,002 e F=37,6; p<0,000, respectivamente), contudo diferenças entre os períodos foi observada
apenas para nitrito (F= 26,3; p<0,000) (Anexo 6). Opostamente, maiores valores de amônio
(U=346,5; Z=2,45; p=0,014) e nitrogênio total (F=6,37; p=0,017) foram observados no reservatório
de Três Marias, sem diferenças significativas entre os períodos. Entre braços de um mesmo
reservatório s diferenças não foram significativas (F=0,3028; p=0,5846).
Valores do Índice de Estado Trófico (IET) classificaram os braços coletados no reservatório
de Três Marias como oligo-mesotróficos para transparência da água e oligotrófico para clorofila-a e
fósforo total. No reservatório de Furnas os braços foram classificados como oligotróficos para os
três parâmetros com exceção do braço Mendonça, classificado como mesotrófico para transparência
da água, no período de chuva de 2007 (Anexo 7).
19
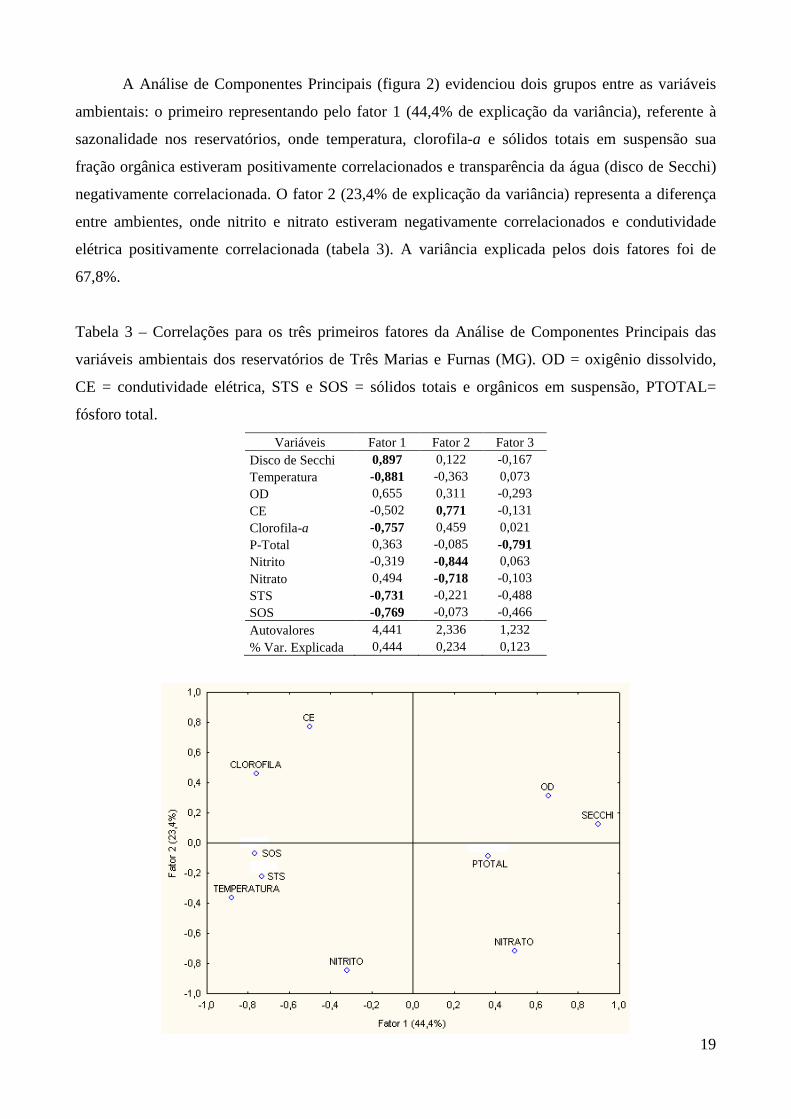
A Análise de Componentes Principais (figura 2) evidenciou dois grupos entre as variáveis
ambientais: o primeiro representando pelo fator 1 (44,4% de explicação da variância), referente à
sazonalidade nos reservatórios, onde temperatura, clorofila-a e sólidos totais em suspensão sua
fração orgânica estiveram positivamente correlacionados e transparência da água (disco de Secchi)
negativamente correlacionada. O fator 2 (23,4% de explicação da variância) representa a diferença
entre ambientes, onde nitrito e nitrato estiveram negativamente correlacionados e condutividade
elétrica positivamente correlacionada (tabela 3). A variância explicada pelos dois fatores foi de
67,8%.
Tabela 3 – Correlações para os três primeiros fatores da Análise de Componentes Principais das
variáveis ambientais dos reservatórios de Três Marias e Furnas (MG). OD = oxigênio dissolvido,
CE = condutividade elétrica, STS e SOS = sólidos totais e orgânicos em suspensão, PTOTAL=
fósforo total.
Variáveis Fator 1 Fator 2 Fator 3 Disco de Secchi 0,897 0,122 -0,167 Temperatura -0,881 -0,363 0,073 OD 0,655 0,311 -0,293 CE -0,502 0,771 -0,131 Clorofila-a -0,757 0,459 0,021 P-Total 0,363 -0,085 -0,791 Nitrito -0,319 -0,844 0,063 Nitrato 0,494 -0,718 -0,103 STS -0,731 -0,221 -0,488 SOS -0,769 -0,073 -0,466 Autovalores 4,441 2,336 1,232 % Var. Explicada 0,444 0,234 0,123
20
Figura 2 – Análise de Componentes Principais para as variáveis ambientais dos reservatórios de
Três Marias e Furnas (MG). OD = oxigênio dissolvido, CE = condutividade elétrica, STS e SOS =
sólidos totais e orgânicos em suspensão, PTOTAL= fósforo total.
COMUNIDADE ZOOPLANCTÔNICA
Foram identificadas no total, 62 espécies no reservatório de Três Marias (5 pertencentes a
Copepoda, 11 a Cladocera e 46 a Rotifera) e 65 espécies em Furnas (7 de Copepoda, 15 de
Cladocera e 43 de Rotifera).
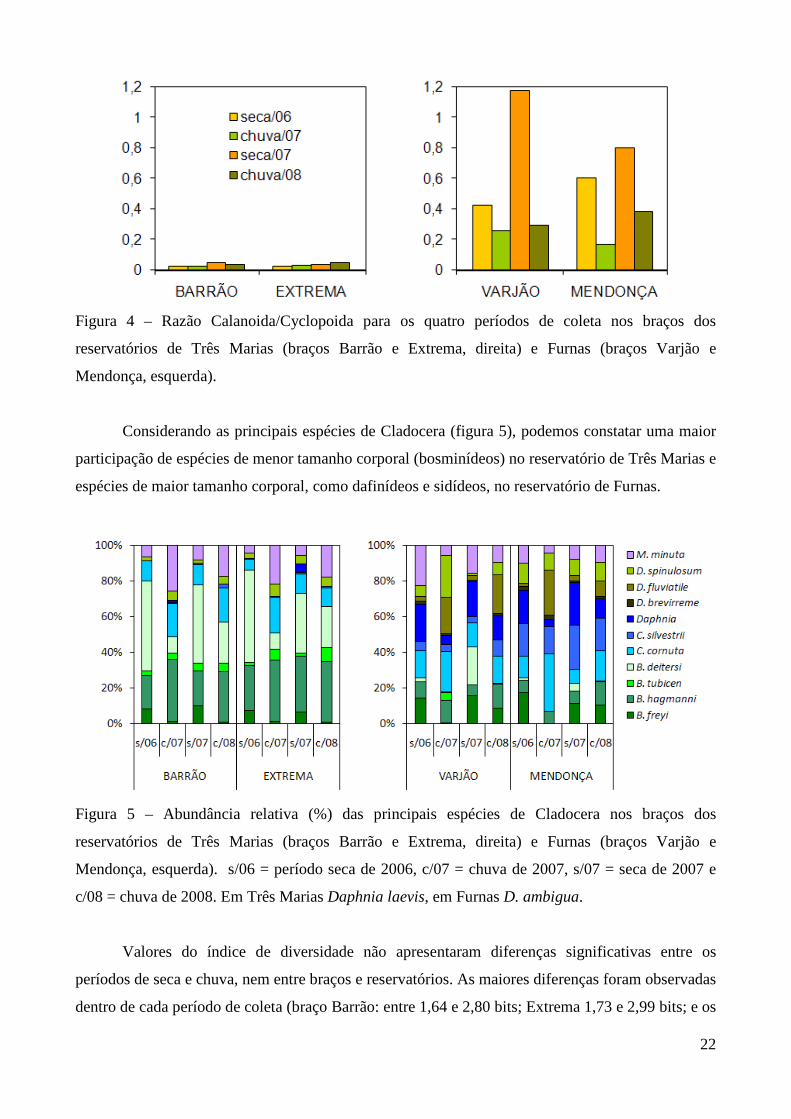
Maiores densidades sempre foram observadas no reservatório de Três Marias (F=18,6;
p<0,0001) e no período chuvoso (F=132,5; p<0,000) em ambos os reservatórios (valores médios na
seca e chuva em Três Marias de 23.721 org.m-³ e 90.872 org.m-³ e em Furnas de 9.022 org.m-³ e
40.434 org.m-³, respectivamente) (Figura 3 e Anexo 8). Entre os braços, maiores densidades foram
observadas no braço Barrão durante o período de seca, e no braço Extrema durante o período
chuvoso para o reservatório de Três Marias, contudo as diferenças não foram significativas
(t=0,0584; p=0,953). Em Furnas, o braço Mendonça apresentou maiores densidades em ambos os
períodos (t= 2,062; p=0,0416).
Figura 3 – Densidade média (org.m-3.103) e abundância relativa (%) dos principais grupos
zooplanctônicos e náuplios nos braços dos reservatórios de Três Marias (braços Barrão e Extrema,

21
acima) e Furnas (braços Varjão e Mendonça, abaixo). s/06 = período seca de 2006, c/07 = chuva de
2007, s/07 = seca de 2007 e c/08 = chuva de 2008.
No reservatório de Três Marias, o grupo dominante foi Copepoda, representando em média
50,6% (31.022 org.m-³) da comunidade zooplanctônica, sendo 34% (20.171 org.m-³) correspondente
às formas naupliares. Em seguida observa-se Rotifera, 40,1% e Cladocera 9,3% (densidades médias
de 20.569 e 5.561 org.m-³, respectivamente) (Figura 3 e Anexo 9). A espécie mais abundante foi
Thermocyclops minutus representando em média 15,5% (10.505 org.m-³, copepoditos e adultos) da
comunidade zooplanctônica. Dentre os Rotifera, destacam-se Bdelloidea, Conochilus coenobasis,
Collotheca sp, Hexarthra intermedia, Keratella americana, K. cochlearis, Polyarthra sp, Ptygura
sp e Sinantherina sp cada espécie participando com pelo menos 5% (2.787 org.m-³) em um dos
períodos (Anexos 12 a 19). Bosmina hagmanni, Bosminopis deitersi, Ceriodaphnia cornuta e
Moina minuta foram as espécies mais abundantes de Cladocera, representando entre 2 e 5% (1.683
org.m-³ em média) da abundância em pelo menos um dos períodos.
No reservatório de Furnas, Copepoda também foi o grupo dominante da comunidade
zooplanctônica, representando em média 62,5% (16.373 org.m-³) sendo 38,4% (10.329 org.m-³)
correspondente aos náuplios, seguido por Rotifera 20,6% e Cladocera 16,9% (densidades médias de
4.617 e 3.738 org.m-³, respectivamente) (Anexo 9 e Figura 3). Thermocyclops minutus e
Notodiaptomus henseni foram as espécies mais abundantes neste reservatório, representando em
média 16,5% (7.598 org.m-³, copepoditos e adultos) e 6,6% (2.041 org.m-³, copepoditos e adultos)
do zooplâncton, respectivamente. Dentre os Rotifera, Brachionus mirus, Conochilus coenobasis,
Collotheca spp e Lecane proiecta foram as espécies mais abundantes (1.130 org.m-³ em média)
(Anexos 20 a 27) e dentre os Cladocera predominaram Bosmina freyi, B. hagmanni, Bosminopsis
deitersi, Ceriodaphnia cornuta, C. silvestrii, Daphnia ambigua, Diaphanosoma fluviatile, D.
spinulosum e Moina minuta perfazendo entre 2,0 e 5,2% (862 org.m-³ em média) durante o período
de amostragem.
O Quociente Brachionus:Trichocerca foi de 1,25 para o reservatório de Três Marias,
classficando-o como mesotrófico (1 < QB/T < 2) e 0,80 para o reservatório de Furnas, classificando
como oligotrófico (QB/T < 1).
A razão Calanoida/Cyclopoida (figura 4) evidenciou o predomínio de indivíduos da ordem
Cyclopoida (valores menores que 1), especialmente no reservatório de Três Marias, indicando uma
condição meso-eutrófica. Mesmo que nem sempre a razão tenha alcançado valores maiores que 1
no reservatório de Furnas, ela sempre foi maior neste ambiente especialmente no período seca,
indicando uma condição mais oligotrófica.
22
Figura 4 – Razão Calanoida/Cyclopoida para os quatro períodos de coleta nos braços dos
reservatórios de Três Marias (braços Barrão e Extrema, direita) e Furnas (braços Varjão e
Mendonça, esquerda).
Considerando as principais espécies de Cladocera (figura 5), podemos constatar uma maior
participação de espécies de menor tamanho corporal (bosminídeos) no reservatório de Três Marias e
espécies de maior tamanho corporal, como dafinídeos e sidídeos, no reservatório de Furnas.
Figura 5 – Abundância relativa (%) das principais espécies de Cladocera nos braços dos
reservatórios de Três Marias (braços Barrão e Extrema, direita) e Furnas (braços Varjão e
Mendonça, esquerda). s/06 = período seca de 2006, c/07 = chuva de 2007, s/07 = seca de 2007 e
c/08 = chuva de 2008. Em Três Marias Daphnia laevis, em Furnas D. ambigua.
Valores do índice de diversidade não apresentaram diferenças significativas entre os
períodos de seca e chuva, nem entre braços e reservatórios. As maiores diferenças foram observadas
dentro de cada período de coleta (braço Barrão: entre 1,64 e 2,80 bits; Extrema 1,73 e 2,99 bits; e os

23
braços Varjão e Mendonça entre 1,81 e 2,85 bits). Quando calculados por grupo (Anexo 28),
diferenças significativas entre os reservatórios foram observadas para Cladocera (F=86,558;
p<0,0001), Rotifera (F=15,004; p<0,0001) e Copepoda (F= 258,41; p<0,0001). Para Cladocera e
Copepoda maiores valores do índice de diversidade ocorreram no reservatório Furnas (1,89 e 0,63
bits respectivamente), enquanto Rotifera apresentou maiores valores em Três Marias (2,28 bits).
DISCUSSÃO
O efeito da sazonalidade foi evidente nos dois reservatórios, com maiores valores de
transparência e concentração de oxigênio dissolvido no período de seca, e de temperatura, clorofila-
a, e sólidos totais e orgânicos em suspensão no período chuvoso. Este mesmo padrão foi relatado
para outros reservatórios como São Simão (Pinto-Coelho et al., 2005) e Salto Grande (Zanata &
Espíndola, 2002), onde valores de fósforo total e turbidez também foram maiores no período de
chuvas. Além disso, diferenças entre os ambientes ficaram nítidas com maiores valores de
condutividade elétrica, clorofila-a, íon amônio, nitrogênio total e sólidos totais e orgânicos em
suspensão no reservatório de Três Marias. Em Furnas apenas a transparência da água e as
concentrações de nitrito e nitrato foram maiores. Esta diferença é um reflexo do processo de
assoreamento que se desenvolve na bacia do rio São Francisco como um todo (Tundisi et al., 1993;
Sampaio & López, 2003). Apesar de possuírem a mesma idade, as dimensões e morfologia do
reservatório de Furnas provavelmente contribuiram para a diluição de nutrientes como fósforo e
nitrogênio, amenizando os efeitos que estes poderiam ter sobre a comunidade planctônica, e
permitiram que o reservatório se mantivesse em estado oligotrófico (Delgado, 1999; Assireu et al.,
2007).
Um estudo realizado no reservatório de Furnas (também no município de Guapé) sobre a
comunidade fitoplanctônica mostrou que as cianobactérias apesar de representarem, em
porcentagem de células, mais de 50% da comunidade fitoplanctônica, em termos de biovolume, não
ultrapassaram 20%, sendo Chlorophyceae, Bacillariophyceae, e Crysophyceae os grupos mais
representativos (Campos, 2008). Espécies de cianobactérias são importantes componentes do
fitoplâncton de lagos eutróficos, enquanto ambientes menos eutrofizados são caracterizados por
comunidades mais diversas representadas principalmente por espécies das classes Chlorophyta,
Crysophyta e Pyrrophyta (Esteves, 1998).
Apesar de classificado como oligotrófico para clorofila-a pelo IET, no reservatório de Três
Marias (em coletas também realizadas nos mesmos braços deste trabalho) foi registrada a
dominância de cianobactérias, que representaram até 86,7% do fitoplâncton (14.803 ind.mL-1),
seguidas de Cryptophyceae, 7,88% (1.347 ind.mL-1), e Bacillariophyceae 1,97% (184 ind.mL-1).

24
Crysophyecae, Chlorophyceae e Zygnemaphyceae não atingiram 1% (Pinto-Coelho & Greco,
2007a). Além disso, durante o presente trabalho, florações foram constantemente observadas em
todas as coletas do período de chuva no reservatório de Três Marias. No geral, as densidades de
cianobactérias em Furnas foram cerca de seis vezes menores: 2.251 ind.mL-1 do que em Três
Marias (Pinto-Coelho & Greco, 2007b).
A riqueza de espécies zooplanctônicas registrada nos reservatórios de Três Marias e Furnas
é condizente com o observado para outros reservatórios do Brasil, especialmente aqueles com idade
e área semelhantes, como por exemplo, Jurumirim e Barra Bonita (ambos em São Paulo), com 60 e
57 espécies, respectivamente (Rocha et al., 1999). A maior riqueza de Rotifera também é um padrão
recorrente em ambientes aquáticos tropicais (Matsumura-TundisI, 1999; Starling, 2001; Lansac-
Tôha et al., 2003) sendo no caso dos reservatórios, atribuída à hidrodinâmica que remove
indivíduos das regiões litorânea e profunda para o ambiente limnético, notadamente gêneros como
Lecane, Platyas, Lepadella, Colurella e Cephalodella (Velho et al., 2005).
Por se tratar de um índice quantitativo, não levando em conta a composição em espécies
(Buckland et al., 2005), a diversidade foi calculada por grupo, a fim de evidenciar as diferenças
entre os reservatórios. Neste caso, por apresentar maior número de espécies, com proporções mais
bem distribuídas, o reservatório de Furnas apresentou maiores valores de diversidade para
Cladocera e Copepoda, enquanto em Três Marias os valores foram maiores para Rotifera. Neste
ambiente, a dominância de poucas espécies, B. deitersi e B. hagmanni entre os cladóceros (figura 5)
e T. minutus entre copépodes, explicam os menores valores obtidos para este índice.
Maiores densidades de zooplâncton no período de chuvas geralmente são atribuídas às
maiores temperaturas neste período (que reduziriam o tempo de geração das espécies), bem como
maior disponibilidade de alimento (algas e sólidos em suspensão) (Sampaio et al., 2002; Melão et
al., 2005). De fato, correlações entre as densidades do zooplâncton e temperatura da água (r=0,806),
concentrações de clorofila-a (r=0,612), sólidos totais (r=0,576) e orgânicos (r=0,589) em suspensão
foram positivas e significativas (p<0,0001) em ambos os reservatórios. Para os nutrientes, as
correlações não foram significativas e apenas para a transparência da água foi negativa (r=-0,747)
uma vez que a maior transparência é o reflexo de menos material em suspensão nos meses de seca,
e provavelmente menos alimento disponível.
Com relação à abundância relativa, diferente do observado por vários autores (Arcifa, 1984;
Rocha et al., 1995; Nogueira, 2001; Sampaio et al., op.cit, Almeida et al., 2009) que relatam
Rotifera como o grupo numericamente dominante em reservatórios, em Três Marias e Furnas
copépodes foram mais abundantes, especialmente devido aos estágios naupliares. Este predomínio
de Copepoda (especialmente por náuplios e copepoditos) também foi observado por Espíndola et al.
(2000) no reservatório de Tucuruí (TO), por Ramos et al. (2008) em Emborcação (MG), por

25
Takahashi et al. (2009) no reservatório de Corumbá (GO) e por Lansac-Tôha et al. (2005) em
grande parte de 30 reservatórios do Paraná. Segundo estes últimos autores e Panarelli et al. (2003),
o maior tempo de residência da água pode ter favorecido este grupo devido seu ciclo de vida mais
longo. De acordo com Velho et al. (2005) a dominância de rotíferos em reservatórios não deve ser
considerada regra geral. Além disso, devido às grandes dimensões dos reservatórios aqui estudados,
e pelo fato dos pontos coletados não se localizarem no eixo central, mas dentro de braços, o
predomínio de copépodes pode ser explicado pela menor interferência da dinâmica de manejo do
reservatório (alterações na vazão, por exemplo) sobre estas espécies, que precisam de habitats mais
“constantes” para completarem seu ciclo mais longo, e são menos oportunistas do que os rotíferos
(Velho et al., 2001; Takahashi et al., op. cit.).
Contudo, Rotífera foi o segundo grupo mais abundante sendo sua dominância em
reservatórios geralmente atribuída à instabilidade deste tipo de ambiente bem como sua estratégia
de vida (r-estrategistas) (Matsumura-Tundisi, 1999), e amplo espectro alimentar (Pourriot, 1977).
López & Sampaio (2003), em trabalho realizado no reservatório de Três Marias,
encontraram a mesma estrutura da comunidade zooplanctônica, com predomínio de T. minutus (de
30 a 40%), H. intermedia, B. patulus, Ptygura libera, K. americana e C. coenobasis, além de B.
deitersi, M. minuta, C. cornuta e B. hagmanni como as espécies de Rotifera e Cladocera mais
abundantes.
Copépodes da ordem Cyclopoida são considerados onívoros, com habilidade de manipular
partículas maiores que 20µm (microfitoplâncton), sendo T. minutus capaz de se alimentar de
colônias de cianobactérias como Microcystis, Botryococcus e Aphanocapsa (Matsumura-Tundisi et
al., 1997), sendo esta uma das possíveis razões para sua dominância em Três Marias.
Já no reservatório de Furnas, o fitoplâncton de melhor qualidade, associado à maior
transparência e baixas concentrações de nutrientes totais, permitem que espécies de maior tamanho
corporal ocorram em maiores densidades. Pinto-Coelho et al., (2003) demonstraram que o
fitoplâncton de menor qualidade nutricional, associado à toxicidade de florações de Microcystis,
comuns também no reservatório de Três Marias, foram responsáveis pelo colapso de Daphnia
laevis em um reservatório urbano eutrófico (reservatório da Pampulha, MG). Alguns autores
(Tundisi et al., 2008; Almeida et al.; 2009) observaram que espécies de Calanoida apresentaram
maior abundância em ambientes com baixa condutividade elétrica (em Três Marias os valores desta
variável são quase o dobro de Furnas). De fato Rull de Aguila (2001) também encontrou maior
abundância de calanóides e cladóceros de grande porte (Daphnia, Diaphanosoma e Moina) próximo
à barragem do reservatório de Furnas, onde condições oligotróficas predominam.
Considerando o efeito da ictiofauna sobre o zooplâncton, em Três Marias, Anchoviella
vaillanti, uma espécie zooplanctívora (Peret, 2004), é o principal item alimentar de Acestrorhynchus

26
britskii e A. lacustris (peixes-cachorro), Pachyurus squamipennis (corvina), Cichla ocellaris
(tucunaré), piscívoros que controlam as populações de peixes forrageios. Desta forma A. vaillanti é
uma espécie-chave nas interações tróficas, envolvendo peixes e os demais organismos aquáticos
(Gomes & Verani, 2003; Peret, op. cit.). Já em Furnas, Santos & Formagio (2007) afirmam que
apenas quatro das 53 espécies de peixes registradas ingerem principalmente invertebrados aquáticos
(tecamebas, rotíferos, nematóides, microcrustáceos, moluscos e pequenas larvas de insetos), sendo
três delas consideradas bentófagas e uma onívora. Desta forma a presença de uma espécie
verdadeiramente zooplanctívora em Três Marias pode ser outro fator caracterizando espécies
zooplanctônicas de menor tamanho corporal neste reservatório.
Zooplâncton como Indicador de Qualidade de Água
Considerando que nos dois reservatórios as amostras foram realizadas fora do eixo central,
em áreas de menor influência do manejo do reservatório, o aumento na abundância relativa de
rotíferos em Três Marias, onde este grupo representou 40,1% da comunidade zooplanctônica
quando comparado com a contribuição deste mesmo grupo em Furnas (média 20,6%) (em termos
absolutos a densidade de rotíferos é quase cinco vezes maior no primeiro), parece ser um melhor
indicador de enriquecimento do que a presença de espécies em particular, como já salientado por
Gannon & Stemberger, (1978) e Matsumura-Tundisi et al., (2000). Soma-se a este fato, o resultado
obtido para o quociente Brachionus:Trichocerca, que também apontou mesotrofia para Três Marias
e oligotrofia para Furnas. Além disso, maiores densidades de B. angularis, B. falcatus, B.
calyciflorus, H. intermedia, K. cochlearis, C. coenobasis, Polyarthra sp., e indivíduos da ordem
Bdellloidea, apontadas como típicas de ambientes meso-eutróficos (Attayde & Bozelli, 1998,
Duggan et al., 2001; Kutikova, 2003) foram registradas no reservatório de Três Marias.
A diferença na abundância de Copepoda Calanoida entre os dois reservatórios, também pode
ser explicada pelo estado trófico. De acordo com vários autores (Gannon & Stemberger, 1978;
Pace, 1986; Tundisi et al., 1988 e Rocha et al., 1995) espécies da ordem Calanoida parecem mais
adaptadas a condições oligotróficas, enquanto Cyclopoida são mais abundantes em águas eutróficas.
Mesmo entre Cyclopoida, T. minutus é considerado típico de ambientes oligotróficos-mesotróficos
enquanto seu congênere T. decipiens (cujas densidades foram baixas em ambos os reservatórios)
domina em ambientes mais eutrofizados (Reid, 1989; Landa et al. 2007).
Por fim a maior abundância de pequenos cladóceros, notadamente bosminídeos, no
reservatório de Três Marias, reflete o favorecimento destas espécies em ambientes enriquecidos
(Esteves & Sendacz, 1988). A presença de algas coloniais e filamentosas que podem emaranhar no
aparato filtrador de grandes cladóceros, como Daphnia e Diaphanosoma (Lampert, 1987),

27
explicaria suas menores densidades em Três Marias enquanto as mesmas poderiam ser favorecidas
pelo fitoplâncton do reservatório de Furnas, onde predominam grupos como clorofíceas e
crisofíceas, de melhor qualidade nutricional.
Neste sentido, em ambientes oligotróficos, com maior transparência da água e menores
valores de condutividade elétrica e nutrientes, o nanofitoplâncton é a fração dominante, permitindo
que o zooplâncton herbívoro, representado por filtradores como calanóides e grandes cladóceros
(dafinídeoes e sidídeos), seja mais abundante (Tundisi et al., 1988; Rietzler et al., 2002; Sampaio et
al., 2002). Por outro lado, ambientes com concentrações mais elevadas de detritos e nutrientes
(meso-eutróficos), favorecem o crescimento de bactérias e protozoários, importante fonte de
alimento para pequenos filtradores como náuplios, rotíferos e pequenos cladóceros (bosminídeos).
Além disso, o predomínio de algas coloniais e filamentosas (cianobactérias) passa a favorecer
espécies de ciclopóides, que por seus hábitos raptoriais são capazes de quebrar colônias e
filamentos e aproveitar o microzooplâncton como recurso alimentar (Esteves & Sendacz, 1988;
Rocha et al., 1995; Matsumura-Tundisi et al., 2000 e 2002; Coelho-Botelho, 2003).
A partir dos dados levantados neste estudo, pode-se concluir que apesar de considerados
oligotróficos (Sampaio & López, 2003; Figueredo, 2000) e possuírem idade semelhante, estes dois
reservatórios apresentam diferenças marcantes na comunidade zooplanctônica. O reservatório de
Três Marias exibe uma comunidade zooplanctônica característica de ambientes mesotróficos, com
predomínio de ciclopóides e pequenos cladóceros dentre os microcrustáceos e maiores densidades
de rotíferos. Por outro lado, a maior participação de espécies de maior porte (calanóides, dafinídeos
e sidídeos) em Furnas, poderia ser um indicador do controle do fitoplâncton pela herbivoria,
caracterizando um ambiente com águas de melhor qualidade.
AGRADECIMENTOS
À toda a equipe dos laboratórios de Ecologia do Zooplâncton, de Gestão Ambiental de
Reservatórios e de Limnologia, que direta ou indiretamente ajudaram na realização deste trabalho.
Ao Projeto Parques Aqüícolas (Convênio 8713 SEAP/SECTES/FUNDEP-UFMG) e ao CNPq
(Processo n° 142079/2006-5) pelo financiamento do projeto.
REFERÊNCIAS BIBLIOGRÁFICAS
ALMEIDA, VLS., DANTAS, E.W., MELO-JÚNIOR, M., BITTENCOURT-OLIVEIRA, MC. &
MOURA, AN. 2009. Zooplanktonic community of six reservoirs in northeast Brazil. Braz. J. Biol., 69(1): 57-65.

28
ARCIFA, M.S. 1984. Zooplankton composition of ten reservoirs in southern Brazil. Hydrobiologia, 113:137-145.
ASSIREU, A.; ROLAND, F.; NOVO, E.; BARROS, N.O.; STECH, J.L. & PACHECO, F.S. 2007. Existe relação entre a complexidade geométrica do entorno dos reservatórios e a variabilidade espacial dos parâmetros limnológicos? Anais do XIII Simpósio Brasileiro de Sensoriamento Remoto, INPE, Florianópolis, p. 3263-3269.
ATTAYDE, J. L. & BOZELLI, R. 1998. Assessing the indicator properties of zooplankton assemblages to disturbance gradients by canonical correspondence analysis. Can. J. Fish. Aquatic. Sci. 55: 1789-1797.
BOTTRELL, H.H.; DUNCAN, A.; GLIWICZ, Z.M.; GRYGIEREK, E.; HERZIG, A.; HILLBRICHT-ILKOWSKA, A.; KURASAUA, H.; LARSSON, P. & WEGLENSKA, T. 1976. A review of some problems in zooplankton production studies. Norw. J. Zoo., 24: 419-456.
BUCKLAND, S.T., MAGURRAN,A.E., GREEN,R.E. & FEWSTER, R.M. 2005. Monitoring change in biodiversity through composite indices. Phil. Trans. R. Soc. B. 360:243-254.
CAMPOS, C.A. 2008. Estudo da diversidade de Cyanobacteria do reservatório de Furnas (MG, Brasil) por meio de caracterização morfológica e molecular. Universidade Federal de Minas Gerais. 81p. Dissertação.
CARLSON, R.E. A trophic state index for lakes. Limnology and Oceanography, 22(2): 361-369, 1977.
CARPENTER, S. R.; KITCHELL, J.F. & HODGSON, J. R. 1985. Cascading Trophic Interactions and Lake Productivity. Bioscience, 35(10):634-639.
CEMIG - Compania Energética de Minas Gerais. 2009. Usinas da CEMIG. http://www.cemig.com.br/institucional/usinas/11g.asp. Acesso em: 16 set. 2009.
COELHO-BOTELHO, M. J. 2003. Dinâmica da comunidade zooplanctônica e sua relação com o grau de trofia em reservatórios. In: IV Simpósio e IV Reunião de Avaliação do Programa Biota/FAPESP, Águas de Lindóia. Resumo expandido de Mini-Curso.
DELGADO, P. C. S. 1999. Variação sazonal e espacial da comunidade fitoplanctônica do Reservatório de Furnas, município de Guapé (MG). Universidade Federal de Minas Gerais. 106p. Dissertação.
DUGGAN, I.C., GREEN, J.D. & THOMASSON, K. 2001. Do rotifers have potential as bio-indicators of lake trophic state? Verh. Internat. Verein Limnol., 27: 3497-3502.
ESPÍNDOLA, E.L.G., MATSUMUTA-TUNDISI, T., RIETZLER, A.C. & TUNDISI, J.G. 2000 Spatial Heterogeneity of the Tucuruí Reservoir (State of Pará, Amazonia, Brazil) and the Distribuition of Zooplanktonic Species. Braz. J. Biol., 60 (2) p. 179-194.
ESTEVES, F. A. 1998. Fundamentos de Limnologia. 2ª ed. Rio de Janeiro: Interciência. 602p. ESTEVES, K. E. & SENDACZ, S. 1988. Relações entre a biomassa do zooplâncton e o estado
trófico de reservatórios do Estado de São Paulo. Acta Limnol. Bras., 11: 587-604. FIGUEREDO, C. C. 2000. Efeitos da tilápia (Oreochromis niloticus) nas características físicas e
químicas e estrutura da comunidade fitoplanctônica do Reservatório da Usina Hidrelétrica de Furnas (MG). Universidade Federal de Minas Gerais. 150p. Dissertação.
FURNAS – Furnas Centrais Elétricas S.A. 2009. Sistema Furnas. http://www.furnas.com.br/hotsites/sistemafurnas. Acesso em: 16 set. 2009.
GANNON, J. E. & STEMBERGER, R. 1978. Zooplankton (especially crustaceans and rotifers) as indicators of water quality. Trans. Am. Microsc. Soc., 97:16-35.
GOLTERMAN, H.L., CLYMO, R.S. & OHNSTAD, M.A.M. 1978. Methods for physical and chemical analysis of freshwaters. Oxford: Blackwell Scientific Publications. 166p.
GOMES, J.H.C. & VERANI, J.R. 2003. Alimentação de espécies de peixes do reservatório de Três Marias. In: Godinho, H. P. & Godinho, A. L. (org.) Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Hrizonte: PUCMinas. 468p.
HARPER, D. 1992. Eutrophication of Freshwaters. Principles, problems and restoration. London: Chapman & Hall. 327p.

29
HENRY, R. 1999. Ecologia dos Reservatórios: Estrutura, Função e Aspectos Sociais. Botucatu: FUNDIBI, FAPESP. 799p.
KOZLOWSKY-SUZUKI, B. & BOZELLI, R. L. 2002. Experimental Evidence of the Effect of Nutrient Enrichment on the Zooplankton in a Brazilian Coastal Lagoon. Braz. J. Biol., 62(4B): 835-846.
KUTIKOVA, L.A. 2003. Bdelloid Rotifers (Rotifera, Bdelloidea) as a Component of Soil and Land Biocenoses. Biology Bulletin, 30(3):271–274.
LAMPERT, W., 1987, Laboratory studies on zooplankton cyanobacteria interactions. N.Z. J. Mar. Freshwater Research, 21: 483-490.
LANDA, G.G.; BARBOSA, F.A.R.; RIETZLER, A.C. & MAIA-BARBOSA, P.M. 2007. Thermocyclops decipiens (Kiefer, 1929) (Copepoda, Cyclopoida) as Indicator of Water Quality in the State of Minas Gerais, Brazil. Brazilian Archives of Biology and Technology, 50(4):695-705.
LANSAC-TÔHA, F. A., VELHO, L. F. M., TAKAHASHI, E. M., NAGAE, M. Y., PEDROSO, M. M., GARCIA, D. L. & PEREIRA, D. G. 2003. Riqueza e Abundância da Comunidade Zooplanctônica em Reservatórios do Estado do Paraná. In: Rodrigues, L.; Agostinho, A. A.; Gomes, L. C. & Thomaz, S. M. (eds). Anais do Workshop Produtividade em Reservatórios e Bioindicadores. Maringá: Nupélia. 281p.
LANSAC-TÔHA, F. A.; BONECKER, C.C. & VELHO, L. F. M. 2005. Estrutura da Comunidade Zooplanctônica em Reservatórios. In: Rodrigues, L.; Thomaz, S.M.; Agostinho, A.A. & Gomes, L.C. (eds.) Biocenoses em Reservatórios: padrões espaciais e temporais. São Carlos: Rima. 333p.
LATHROP, R. C. & CARPENTER, S. R. 1992. Zooplankton and their relationship to phytoplankton. In: J.F. Kitchell (ed.). Food web management: a case study of Lake Mendota. New York: Springer-Verlag. 553p.
LÓPEZ, C. M. & SAMPAIO, E.V. 2003. A comunidade zooplanctônica no reservatório de Três Marias e no trecho do São Francisco a jusante. In: Godinho, H. P. & Godinho, A. L. (org.) Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Hrizonte: PUCMinas. 468p.
LORENZEN, C. J. 1967. Determination of chlorophyll and pheO-pigments: spectrophotometric equations. Limnology and Oceanography, 12:343-346.
MACKERETH, F.J.H., HERON, J. & TALLING, J.F. 1978. Water analysis and some revised methods for limnologists. New York: Freshwater Biological Association, 36. 120p.
MAGURRAN, A. E. 1988. Ecological Diversity and Its Measurement. New Jersey: Princeton University Press. 192p.
MATSUMURA-TUNDISI, T.; ROCHA, O. & TUNDISI, J. G. 1997. Carbon Uptake by Scolodiaptomus corderoi and Thermocyclops minutus Feeding on Different Size Fractions of Phytoplankton from Lake Dom Helvécio. In: Tundisi, J.G. & & Saijo, Y. (eds) Limnological Studies on the Rio Doce Valley Lakes, Brazil. Rio de Janeiro: Brazilian Academy of Sciences, University of São Paulo, School of Engineering at São Carlos, Center for Water Resources and Applied Ecology. 513p.
MATSUMURA-TUNDISI, T., 1999, Diversidade de zooplâncton em represas do Brasil. In: R. Henry (ed.) Ecologia de reservatórios: estrutura, função e aspectos sociais. Botucatu: FUNDIBIO, FAPESP. 799p.
MATSUMURA-TUNDISI, T.; LUZIA, A.P. & TUNDISI, J.G. 2000. Estado Trófico dos Reservatórios em Cascata do Médio e Baixo Tietê (SP) e Manejo para o Controle da Eutrofização. In: STRAŠKRABA, M. & TUNDISI, J.G. (eds.) Diretrizes para o Gerenciamento de Lagos. Volume 9: Gerenciamento da Qualidade da Água de Represas. São Carlos: ILEC, IIE. 300p.
MATSUMURA-TUNDISI, T.; TUNDISI, J. G. & ROCHA, O. 2002. Zooplankton diversity in eutrophic systems and its relation to the occurrence of cyanophycean blooms. Verh. Internat. Verein. Limnol. 28: 671-674.

30
MCCAULEY, E. & KALFF, J. 1981. Empirical Relationships between Phytoplankton and Zooplankton Biomass in lakes. Canadian Journal of Fisheries and Aquatic Science, 38:458-463.
MELÃO, M.G., ROCHA, O., & ROCHE, F. K. 2005. Produtividade, Biomassa e Flutuações Populacionais e interações biológicas da Comunidade Planctônica e Suas Implicações na Transferência de Energia na Cadeia Alimentar de um Reservatório Raso e oligotrófico. In: Roche, F. K. & Rocha, O. (eds.) Ecologia Trófica de Peixes com ênfase na planctivoria em ambientes lênticos de água doce no Brasil. São Carlos: Rima. 146p.
MIRANDA, E. E. & COUTINHO, A. C. (Coord.) 2004. Brasil Visto do Espaço. Campinas: Embrapa Monitoramento por Satélite. http://www.cdbrasil.cnpm.embrapa.br Acesso em: 16 set. 2009.
NOGUEIRA, M. G. 2001. Zooplankton composition, dominance and abundance as indicators of environmental compartmentalization in Jurumirim Reservoir (Paranapanema River), São Paulo, Brazil. Hydrobiologia, 455:1-18.
NOGUEIRA, D.J.; CASTRO, S.C. & RIGOLIN DE SÁ, O. 2008. Avaliação da qualidade da água no reservatório UHE Furnas - MG, utilizando as brânquias de Pimelodus maculatus (LACÈPÉDE, 1803) como biomarcador de poluição ambiental. Ciência et Praxis, 1(1): 15-20.
PACE, M. L. 1986. An empirical analyses of zooplankton community size structure across lake trophic gradients. Limnology and Oceanography, 31(1):45-55.
PANARELLI, E.; CASANOVA, S.M.C.; NOGUEIRA, M.G.; MITSUKA, P.M. & HENRY, R. 2003. A Comunidade Zooplanctônica ao Longo de Gradientes Longitudinais no Rio Paranapanema/Represa de Jurumirim (São Paulo, Brasil). In: Henry, R. (ed.) Ecótonos nas Interfaces dos Ecossistemas Aquáticos. São Carlos: Rima, p. 129-160.
PANARELLI, E. A. 2004. Flutuações mensais da comunidade zooplanctônica e dinâmica das populações de Cladocera em lagoas marginais, na região de transição Rio Paranapanema – Represa Jurumirim (SP). Universidade Estadual Paulista. 247p. Tese.
PERET, A.M. 2004. Dinâmica da alimentação de peixes piscívoros da represa de Três Marias (MG). Universidade Federal de São Carlos. 61p. Dissertação.
PINTO-COELHO, R.M. 2006. Ranqueamento de Áreas Alvo e Delimitação de Polígonos (Fase I). Projeto Parques Aqüícolas, UFMG/SECTES/SEAP. Belo Horizonte. 54p.
PINTO-COELHO, R.M., BEZERRA-NETO, J.F., GIANI, A., MACEDO, C.F., FIGUEIREDO, C.C. & CARVALHO, E.A. 2003. The collpase of a Daphnia laevis (Birge, 1878) population in Pampulha reservoir, Brazil. Acta Limnol. Bras., 15(3):53-70.
PINTO-COELHO, R.; AZEVEDO, L.; RIZZI, P.; BEZERRA-NETO, J. & ROLLA, M. 2005. Origens e Efeitos do Aporte Externo de Nutrientes em um Reservatório Tropical de Grande Porte: Reservatório de São Simão (MG/GO). In: Rodrigues, L.; Thomaz, S.M.; Agostinho, A.A. & Gomes, L.C. (eds.) Biocenoses em Reservatórios: padrões espaciais e temporais. São Carlos: Rima. 333p.
PINTO-COELHO, R.M. & GRECO, M.K.B. 2007a. Relatório de Estudos Ambientais e Regularização do Parque Aqüícola São Francisco-1, Reservatório de Três Marias. Projeto Parques Aqüícolas, UFMG/SECTES/SEAP. Belo Horizonte.183p.
PINTO-COELHO, R.M. & GRECO, M.K.B. 2007b. Relatório de estudos ambientais e Regularização do Parque Aqüícola Guapé-1, Reservatório de Furnas. Projeto Parques Aqüícolas, UFMG/SECTES/SEAP. Belo Horizonte. 254p.
POURRIOT, R. 1977. Food and feeding habits of Rotifera. Arch. Hydrobiol. Beih. Ergebn. Limnol., 8: 243-260.
QUIRÓS, R. & BOVERI, M.B. 1999. Fish Effects on Reservoir Trophic Relationships. In: TUNDISI, J.G. & STRAŠKRABA, M. (eds.) 1999. Theoretical Reservoir Ecology and its Applications. São Carlos: IIE. 585p.
RAMOS, J.D.; MELLO, H.O.O. & LIMA, L. 2008. Análise da composição, abundância e distribuição vertical das populações de Rotifera, Cladocera e Copepoda, no reservatório de Emborcação (Araguari - MG). Em Extensão, 7(2):80-94.

31
REID, J. W. 1989. The distribution of species of the genus Thermocyclops (Copepoda, Cyclopoida) in the western hemisphere, with description of T. parvus, new species. Hydrobiologia, 175: 149-174.
RIETZLER, A. C., MATSUMURA-TUNDISI, T. & TUNDISI, J. G. 2002. Life Cycle, Feeding and Adaptive Strategy Implications on the Co-occurrence of Argyrodiaptomus furcatus and Notodiaptomus iheringi in Lobo-Broa Reservoir (SP, Brazil). Braz. J. Biol., 62(1): 93-105.
ROCHA, O.; SENDACZ, S. & MATSUMURA-TUNDISI, T. 1995. Composition, Biomass and Productivity of Zooplankton in Natural Lakes and Reservoirs of Brazil. In: Tundisi, J. G.; Bicudo, C. E. M. & Matsumura-Tundisi, T. (eds.) Limnology in Brazil. Rio de Janeiro: Brazilian Academy of Sciences, Brazilian Limnological Society. 376p.
ROCHA, O.; MATSUMURA-TUNDISI, T.; ESPÍNDOLA, E.L.G.; ROCHE, K.F. & RIETZLER, A.C. 1999. Ecological Theory Applied to Reservoir Zooplankton. In: Tundisi, J.G. & Straskraba, M. (eds.) Theoretical Reservoir Ecology and its Applications. São Carlos: IIE. 585p.
RULL DE AGUILA, L. M. 2001. Gradiente trófico no Rio Sapucaí (reservatório de Furnas- MG): relação com a distribuição do zooplâncton e os usos do solo. Universidade Federal de Minas Gerais. 75p. Dissertação.
SAMPAIO, I.B.M. 2002. Estatística Aplicada à Experimentação Animal. Belo Horizonte: Fundação de Ensino e Pesquisa em Medicina Veterinária e Zootecnia. 2ª ed. 256p.
SAMPAIO, E. V., ROCHA, O., MATSUMURA-TUNDISI, T. & TUNDISI, J. G. 2002. Composition and Abundance of Zooplankton in the Limnetic Zone of Seven Reservoirs of the Paranapanema River, Brazil. Braz. J. Biol., 62(3): 525-545.
SAMPAIO, E.V. & LÓPEZ, C. M. 2003. Limnologias física, química e biológica da represa de Três Marias e do São Francisco. In: Godinho, H. P. & Godinho, A. L. (org.) Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Hrizonte: PUCMinas. 468p.
SIMGE/IGAM – Sistema Meteorológicos e de Recursos Hídricos de Minas Gerais/Instituto Mineiro de Gestão das Águas. 2010. Base da Dados Meteorológicos Diários. http://www.simge.mg.gov.br/. Acesso em: 25 jan. 2010.
SLÁDEČEK, V. 1958. A Note on the Phytoplankton-Zooplankton Relationship. Ecology, 39(3):547-549. Sládeček, 1958,
SLÁDEČEK, V. 1983. Rotifers as indicators of water quality. Hydrobiologia, 100:169-201. STARLING, F.L.R.M. 2001. Comparative Study of the Zooplankton Composition of Six
Lacustrine Ecosystems in Central Brazil During the Dry Seasonal. Braz. J. Biol., 60(1): 101-111. STRAŠKRABA, M. & TUNDISI, J.G. (eds.) 2000. Diretrizes para o Gerenciamento de Lagos.
Volume 9: Gerenciamento da Qualidade da Água de Represas. São Carlos: ILEC, IIE. 300p. STRICKLAND, J.D.H. & PARSONS, T.R. 1972. A Pratical Handbook of Seawater Analysis.
Fisheries Research Board, Canada. 310p. TAKAHASHI, E.M., LANSAC-TÔHA, F.A., DIAS, D. J., BONECKER, C.C. & VELHO, L.F.M. 2009. Spatial variations in the zooplankton community from the Corumbá Reservoir, Goiás State, in distinct hydrological periods. Aca Sienctiarum. Biological Sciences, 31(3):227-234. TOLEDO, A., TALARICO, M., CHINEZ, S. J., & AGUDO, E. G. 1983. A aplicação de modelos
simplificados para a avaliação do processo de eutrofização em lagos e reservatórios tropicais. Anais do Congresso Brasileiro de Engenharia Sanitária e Ambiental, 12, Camboriú. 1-34p.
TUNDISI, J. G.; T. MATSUMURA-TUNDISI; R. HENRY; O. ROCHA & K. HINO. 1988. Comparação do estado trófico de 23 reservatórios do Estado de São Paulo: Eutrofização e manejo. In: Tundisi, J.G. (ed.) Limnologia e manejo de represas. vol. 1. Tomo 1. São Paulo: Série Monografias em Limnologia/USP.
TUNDISI, J.G., MATSUMURA-TUNDISI, T. & CALIJURI, M.C. 1993. Limnology and management of reservoirs in Brazil. In: Straškraba, M.; Tundisi, J.G. & Duncan, A. Comparative Reservoir Limnology and Water Quality Management. Dordrecht: Kluwer Academic Publishers. 291p.

32
TUNDISI, J.G., MATSUMURA-TUNDISI, T. & ABE, D.S. 2008. The ecological dynamics of Barra Bonita (Tietê River, SP, Brazil) reservoir: implications for its biodiversity. Braz. J. Biol., 68(4, Suppl.): 1079-1098
VALENTIN, J.L. 2000. Ecologia Numérica. Uma introdução à Análise Multivariada de Dados Ecológicos. Rio de Janeiro: Interciência. 117p.
VELHO, L. F. M.; LANSAC-TOHA, F. A.; BONECKER, C. C.; BINI, L.M. & RSSA, D.C. 2001 The longitudinal distribution of copepods in Corumbá Reservoir, State of Goiás, Brazil. Hydrobiologia, 453/454: 385-391.
VELHO, L. F. M.; LANSAC-TOHA, F. A. & BONECKER, C. C. 2005. Distribuição Longitudinal da Comunidade Zooplanctônica em Reservatórios. In: Rodrigues, L.; Thomaz, S.M.; Agostinho, A.A. & Gomes, L.C. (eds.) Biocenoses em Reservatórios: padrões espaciais e temporais. São Carlos: Rima. 333p.
WETZEL, R. G. & LIKENS, G. E., 1991, Limnological Analyses. 2th ed. New York: Springer-Verlag.
ZANATA, L. H. & ESPÍNDOLA, E. L. G. 2002. Longitudinal Processes in Salto Grande Reservoir (Americana, SP, Brazil) and its Influence in the Formation of Compartment System. Braz. J. Biol., 62(2): 347-361.

33
ARTIGO 2 – REGRESSÕES PESO-COMPRIMENTO E FLUTUAÇÃO DA BIOMASSA DAS PRINCIPAIS ESPÉCIES DE MICROCRUSTÁCEOS EM DOIS BRAÇOS DOS
RESERVATÓRIOS DE TRÊS MARIAS E FURNAS, MINAS GERAIS, BRASIL. RESUMO Este trabalho teve por objetivo determinar as equações peso-comprimento para as principais espécies de microcrustáceos dos reservatórios de Três Marias e Furnas (MG), bem como avaliar a flutuação de sua biomassa. Para este estudo, foram utilizadas as seguintes espécies Thermocyclops minutus, Bominopsis deitersi, Bosmina hagmanni, Ceriodaphnia cornuta e Moina minuta no reservatório de Três Marias, e no reservatório de Furnas, além destas, foram também consideradas Notodiaptomus henseni, Daphnia ambigua, Ceriodaphnia silvestrii, Diaphanosoma spinulosum, D. fluviatile e Bosmina freyi. O peso seco foi obtido em microbalança, para cada classe de tamanho (no caso de Cladocera) ou estágios de desenvolvimento (Copepoda), sendo as regressões peso-comprimento consideradas adequadas, uma vez que diferenças entre o peso seco observado e o estimado não foram significativas (p>0,928). Diferente da densidade, maiores valores de biomassa de microcrustáceos foram observados no reservatório de Furnas (p=0,04) (médias de 5,76 mgPS.m-3 e 20,36 mgPS.m-3 e em Três Marias: 3,75 mgPS.m-3 e 18,14 mgPS.m-3, nos períodos de seca e chuva, respectivamente) e durante a estação chuvosa em ambos os reservatórios (p<0,000). Em termos de biomassa, T. minutus foi a espécie mais importante no reservatório de Três Marias, enquanto no reservatório de Furnas, N. henseni contribuiu na mesma proporção. Neste último reservatório, a maior biomassa de cladóceros foi explicada pela contribuição de espécies de maior tamanho corporal como D. ambigua, C. silvestrii, D. spinulosum e D. fluviatile. Mesmo considerados oligotróficos, foram observadas difererenças marcantes na biomassa de microcrustáceos estimada, que reforçaram a importância deste parâmetro como descritor da real contribuição de cada espécie na comunidade. PALAVRAS-CHAVE: regressões peso-comprimento, biomassa, microcrustáceos, reservatórios tropicais ABSTRACT Lenght-Weight Relationships and Biomass Fluctuation of the Main Microcrustacean Species in Two Arms of Três Marias and Furnas Reservoirs, MG, Brazil. This paper had as an objective to determine the weight-length equations for the main microcrustacean species of Três Marias and Furnas reservoirs (MG, Brazil), as well as evaluate the fluctuation of their biomass. For this study, the following species were used: Thermocyclops minutus, Bominopsis deitersi, Bosmina hagmanni, Ceriodaphnia cornuta and Moin minuta in Três Marias reservoir and, in Furnas reservoirs, together with these, we also considered: Notodiaptomus henseni, Daphnia ambigua, Ceriodaphnia silvestrii, Diaphanosoma spinulosum, D. fluviatile and Bosmina freyi. The dry weight was obtained in a microbalance for each size class (for Cladocera) or the development stages (Copepoda), with the appropriate length-weight relationships, since the differences between the observed and the estimated dry weight weren’t significant (p>0.928). Higher microcrustacean biomass values were observed in Furnas reservoir (p=0.04) (averages of 5,76 mgDW.m-3 and 20,36 mgDW.m-3, and in Três Marias: 3.75 mgDW.m-3 and 18.14 mgDW.m-3, during the dry and rainy periods, respectively) and during the rainy periods in both reservoirs (p<0.000). As for biomass, T. minutus was the most important species in Três Marias reservoir, whereas in Furnas reservoir, N. henseni contributed in the same proportion. In the later reservoir, the higher cladoceran biomass was explained by the contribution of larger body-sized species, such as D. ambigua, C. silvestrii, D. spinulosum and D. fluviatile. Even though they were considered oligotrophic, there were significant differences in the estimated microcrustacean biomass, which reinforces the importance of this parameter in the description of the real contribution of each species in the community. KEY-WORDS: lenght-weight relationships, biomass, microcrustacean, tropical reservoirs

34
INTRODUÇÃO
A maioria dos estudos sobre a comunidade zooplanctônica nos ecossistemas aquáticos
brasileiros envolve apenas a densidade numérica destes organismos. Entretanto, por apresentar
grande variação em tamanho (de µm a cm), esta comunidade é melhor descrita pela biomassa
(McCauley, 1984; Rocha et al., 1995; Maia-Barbosa, 2000; Bozelli & Huszar, 2003). Neste sentido,
grupos que contribuem significativamente em termos numéricos, como rotíferos e protozoários têm
pouca importância em termos de biomassa, sendo copépodes e cladóceros mais representativos
neste parâmetro (Pace, 1986; Matsumura-Tundisi et al., 1989; Melão et al., 2005; Bonecker et al.,
2007). Além disso, devido ao seu maior tamanho corporal em relação aos rotíferos, microcrustáceos
são presas preferenciais de peixes, predadores essencialmente visuais (Sá Júnior, 1994).
Contudo, a necessidade de equipamentos de alta precisão é um dos principais motivos para o
reduzido número de estudos de biomassa zooplanctônica, e explica porque muitos trabalhos
utilizam regressões peso-comprimento descritas na literatura (Esteves & Sendacz, 1988; Bezerra-
Neto & Pinto-Coelho, 2002; Panarelli, 2004; Pinto-Coelho et al., 2005, Corgosinho & Pinto-
Coelho, 2006). Na maioria das vezes, estas relações são generalizações para grandes grupos e
baseadas em espécies de clima temperado. Entretanto, efeitos espécie-específicos (duração do ciclo
de vida, capacidade de assimilação, estado nutricional), variações ambientais (temperatura,
poluição, fatores geográficos e climáticos, nutrientes, quantidade e qualidade do alimento,
predação) e particularidades metodológicas (preservação ou não dos organismos, tempo de
preservação, critérios para medição) podem afetar a estimativa de regressão peso-comprimento de
cada espécie (Bottrell et al., 1976; McCauley, 1984; Blettler & Bonecker, 2006).
Recentemente, com o desenvolvimento da pesquisa no país e aquisição de microbalanças,
estudos sobre relações peso-comprimento (Maia-Barbosa & Bozelli, 2005; Santos et al., 2006;
Castilho-Noll & Arcifa, 2007) e peso seco e biomassa (Melão & Rocha, 2004 e 2006; Sendacz et
al., 2006; Magalhães et al., 2006; Blettler & Bonecker, 2007; Bonecker et al., 2007; González et al.,
2008; Rosa, 2008) têm aumentado. Desta forma, pode-se dizer que esta última década se
caracterizou por um incremento nas estimativas da biomassa de microcrustáceos em ambientes
tropicais, especialmente em ecossistemas aquáticos brasileiros.
Trabalhos que considerem valores de biomassa são importantes porque permitem comparar
ambientes diferentes entre si, uma vez que fornecem uma unidade comum para avaliar os grupos
zooplanctônicos. Geralmente, ambientes mais enriquecidos apresentam maiores valores de
biomassa em oposição aos menos eutrofizados (Pace, 1986; Esteves & Sendacz, 1988). Soma-se a
este fato a importância de estudos de biomassa para estimativa da produtividade secundária do
zooplâncton, outro parâmetro com poucos estudos no país.

35
Além disso, visando ampliar o uso e aumentar o potencial pesqueiro dos reservatórios de
Três Marias e Furnas, o projeto Parques Aqüícolas (SEAP/SECTES/FUNDEP-UFMG), no qual
este trabalho está inserido, visou à delimitação de áreas para produção pesqueira através de tanques-
rede. Neste projeto, variáveis abióticas e bióticas foram medidas para estimar a capacidade suporte
de áreas selecionadas. Os conhecimentos levantados sobre o zooplâncton auxiliarão, em fase
posterior, não só pelo uso destes organismos como importante fonte alimentar, bem como no
monitoramento da qualidade de água, mitigando os impactos da criação de peixes sobre toda a
comunidade aquática.
Neste contexto, este trabalho teve por objetivo estabelecer as equações peso-comprimento
para as principais espécies de microcrustáceos de dois reservatórios tropicais, Três Marias e Furnas
(MG) bem como avaliar a flutuação da biomassa destas espécies em alguns braços nestes dois
ambientes, nos períodos de seca e chuva.
MATERIAL E MÉTODOS
ÁREA DE ESTUDOS
Os reservatórios das Usinas Hidrelétricas (UHEs) de Três Marias (18°12’S e 45°15’W) e
Furnas (46°19’W e 20°40’S) foram criados há mais de 40 anos para geração de energia elétrica,
sendo também utilizados para recreação, pesca amadora e profissinal, irrigação e abastecimento. O
reservatório da UHE de Três Marias localiza-se no alto rio São Francisco, Centro-Oeste do Estado
de Minas Gerais. Concluído em 1960, atualmente o assoreamento é o principal impacto que
compromete a capacidade geradora de energia e a qualidade da água (Sampaio & López, 2003),
seguido pela substituição da vegetação ripária por monoculturas de Eucaliptus spp e pecuária
extensiva. O reservatório de Furnas está localizado na bacia do rio Grande, Sul do Estado de Minas
Gerais, sendo o braço norte formado pelo rio Grande e o braço sul pelo rio Sapucaí. O reservatório
foi concluído em 1962 tendo, atualmente, como principais impactos atividades como a
agropecuária, monoculturas, o despejo de efluentes domésticos e o lançamento de resíduos sólidos
(Nogueira et al., 2008). Algumas características morfométricas dos dois reservatórios são
apresentadas na tabela 1.
Em cada reservatório foram escolhidos como pontos amostrais dois braços com
características diferentes de entorno (figura 1). Desta forma, no reservatório de Três Marias foram
selecionados os braços Barrão (Zmáx=21,8 metros, com vegetação original do tipo Cerrado) e
Extrema (Zmáx=20,9 metros, com monocultura de Eucaliptus spp), localizados no município de
Morada Nova de Minas. No reservatório de Furnas foram selecionados os braços Varjão

36
(Zmáx=17,5 metros, com nascente localizada no Parque Municipal do Paredão e entorno com
monocultura de café) e Mendonça (Zmáx=25,5 metros, com pecuária em pastagem nativa),
localizados no município de Guapé. Estes braços foram selecionados a fim de caracterizar as
diferentes feições do entorno destes reservatórios.
Tabela 1 – Características morfométricas dos reservatórios das UHEs de Três Marias e Furnas
(MG). Fonte: CEMIG, 2009 e FURNAS, 2009.
UHE Três Marias UHE Furnas Área Inundada 1.100 km2 1.440 Km² Volume Útil 15,27 x 109 m³ 17,21 x 109 m³ Vazão Média 700 m³.s-1 800 m³.s-1
Tempo Médio de Residência 120 dias 160 dias Profundidade Máxima 75 m 90 m Profundidade Média 12 m 13 m Capacidade Geradora 396 MW 1126 MW
Figura 1 – Reservatórios de Três Marias (esquerda, 1:10.000) e Furnas (direita, 1:20.000) (MG)
com as setas apontando os braços utilizados para as coletas. Modificado de Miranda & Coutinho,
2004.
PROCEDIMENTOS
As amostras foram coletadas a cada dois dias, durante quatro semanas, em dois períodos de
seca (julho/agosto de 2006 e julho/agosto de 2007) e dois de chuvas (janeiro/março de 2007 e

37
janeiro/março de 2008), considerados os mais contrastantes em termos de variações de temperatura
e precipitação. As coletas foram realizadas no ponto mais profundo da região limnética de cada
braço.
Para as coletas quali-quantitativas da comunidade zooplanctônica, foram realizados arrastos
com rede de plâncton de 68�m de abertura de malha. Devido à presença de “paliteiros” (vegetação
original não removida) nos pontos de coleta, os arrastos foram realizados apenas na zona eufótica,
determinada a partir da leitura do disco de Secchi. Os organismos coletados foram anestesiados com
água mineral gaseificada, corados com corante vital Rosa de Bengala e fixados com solução
neutralizada de formol 4%. Sub-amostras de 1,0 mL foram contadas sob microscópio ótico
Olympus (modelo CBA), em câmaras de Sedgewick-Rafter. Os dados apresentados correspondem
ao número de organismos por m3. Nesta fase, foram quantificados separadamente, os estágios de
náuplios, copepoditos e adultos das espécies de Copepoda.
Para este estudo foram consideradas apenas as espécies de microcrustáceos mais abundantes
(aquelas que contribuíram com pelos menos 5% da comunidade durante os períodos amostrados):
Thermocyclops minutus, Bominopsis deitersi, Bosmina hagmanni, Ceriodaphnia cornuta e Moina
minuta no reservatório de Três Marias, e no reservatório de Furnas além destas, foram também
consideradas Notodiaptomus henseni, Daphnia ambigua, Ceriodaphnia silvestrii, Diaphanosoma
spinulosum, D. fluviatile e Bosmina freyi (Artigo 1).
Para o estabelecimento das regressões peso-comprimento foram utilizados espécimes do
período de seca de 2006, medindo-se pelo menos os 30 primeiros indivíduos de cada estágio de
desenvolvimento (Bird & Praire, 1985), através de ocular micrometrada. Para Copepoda, as
medidas foram realizadas do cefalotórax até o abdômen, excetuando-se a furca; no caso de
Cladocera, as medidas foram tomadas da cabeça até a base da carapaça, excetuando-se o espinho da
mesma. No caso de Daphnia ambigua, a medida compreendeu da base do elmo até a base da
carapaça. Desta forma, foi possível determinar as classes de tamanho e a contribuição percentual de
neonatas, juvenis e adultos das espécies de Cladocera. Os espécimes foram lavados por três vezes
em água destilada e transferidos para estufa a 600C por 24 horas em cadinhos de alumínio
previamente pesados. Após a secagem, os cadinhos foram transferidos para dessecador com sílica-
gel para resfriamento, e logo após pesados (Bottrell et al., 1976; McCauley, 1984). As pesagens
foram realizadas em microbalança Sartorius (modelo SE2, precisão de 0,1µg) sendo os resultados
expressos em peso seco (µg PS). Apesar da literatura advertir sobre a perda de cerca de 10 a 40%
do peso para organismos fixados (Dumont et al., 1975; McCauley, op cit; Blettler & Bonecker,
2006) nenhuma correção foi adotada neste estudo.

38
Os valores do peso médio para cada classe de tamanho (Cladocera), ou estágio de
desenvolvimento (Copepoda), foram utilizados para calcular a regressão entre peso-comprimento
após transformação logarítmica do peso (W em µgPS) e comprimento (L em mm), de acordo com a
fórmula de Bottrell et al. (1976):
LnW = Lna + b LnL
Onde:
a = estimativa do intercepto
b = estimativa da inclinação.
Utilizando-se as regressões peso-comprimento e os comprimentos obtidos para as demais
coletas, foi possível estimar a flutuação da biomassa pelo produto da densidade (org.m-3) e do peso
seco médio (µgPS) (Winberg & Duncan, 1971) para os reservatórios de Três Marias e Furnas. Os
valores de biomassa foram expressos em mgPS. m-3.
As diferenças nos valores de comprimento e peso seco foram verificadas através do teste T
de Student, enquanto as diferenças nos valores de biomassa entre braços, reservatórios e períodos de
coleta (seca e chuva), através de uma Análise de Variância bi-fatorial, seguida do teste de Tukey
(Sampaio, 2002). Para estes testes e as regressões peso-comprimento foi utilizado o programa
Statistica 7.0 (StatSoft).
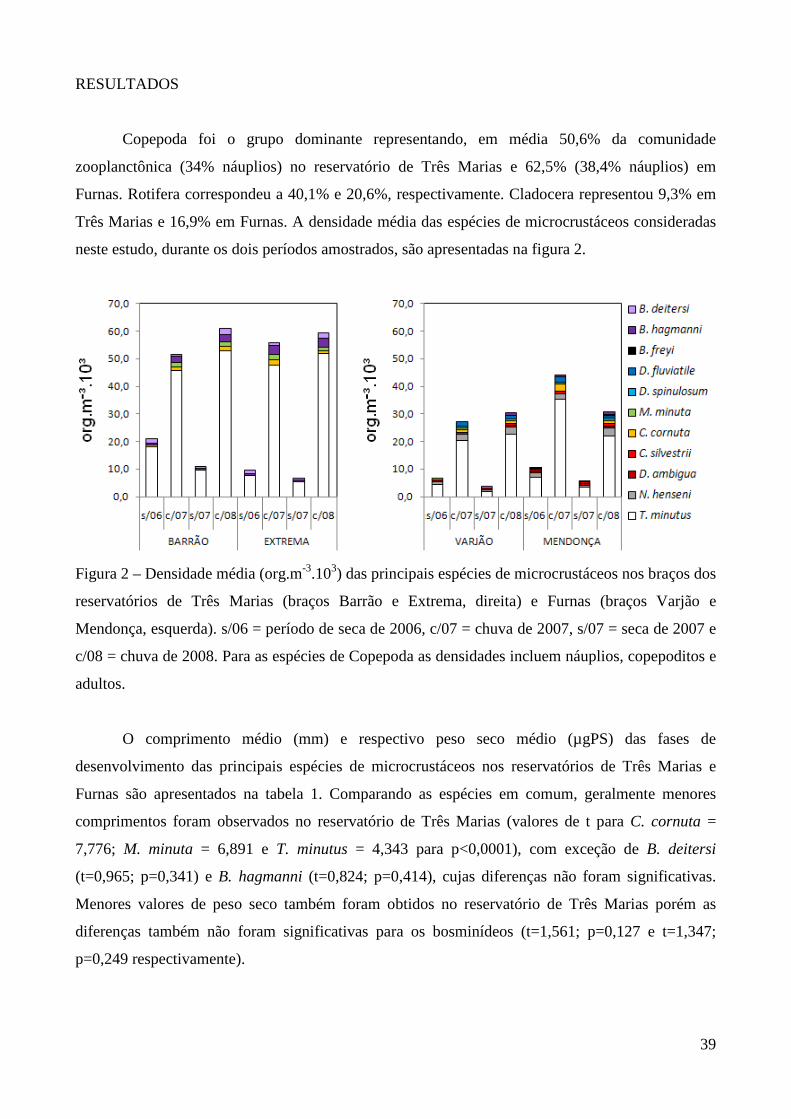
39
RESULTADOS
Copepoda foi o grupo dominante representando, em média 50,6% da comunidade
zooplanctônica (34% náuplios) no reservatório de Três Marias e 62,5% (38,4% náuplios) em
Furnas. Rotifera correspondeu a 40,1% e 20,6%, respectivamente. Cladocera representou 9,3% em
Três Marias e 16,9% em Furnas. A densidade média das espécies de microcrustáceos consideradas
neste estudo, durante os dois períodos amostrados, são apresentadas na figura 2.
Figura 2 – Densidade média (org.m-3.103) das principais espécies de microcrustáceos nos braços dos
reservatórios de Três Marias (braços Barrão e Extrema, direita) e Furnas (braços Varjão e
Mendonça, esquerda). s/06 = período de seca de 2006, c/07 = chuva de 2007, s/07 = seca de 2007 e
c/08 = chuva de 2008. Para as espécies de Copepoda as densidades incluem náuplios, copepoditos e
adultos.
O comprimento médio (mm) e respectivo peso seco médio (µgPS) das fases de
desenvolvimento das principais espécies de microcrustáceos nos reservatórios de Três Marias e
Furnas são apresentados na tabela 1. Comparando as espécies em comum, geralmente menores
comprimentos foram observados no reservatório de Três Marias (valores de t para C. cornuta =
7,776; M. minuta = 6,891 e T. minutus = 4,343 para p<0,0001), com exceção de B. deitersi
(t=0,965; p=0,341) e B. hagmanni (t=0,824; p=0,414), cujas diferenças não foram significativas.
Menores valores de peso seco também foram obtidos no reservatório de Três Marias porém as
diferenças também não foram significativas para os bosminídeos (t=1,561; p=0,127 e t=1,347;
p=0,249 respectivamente).
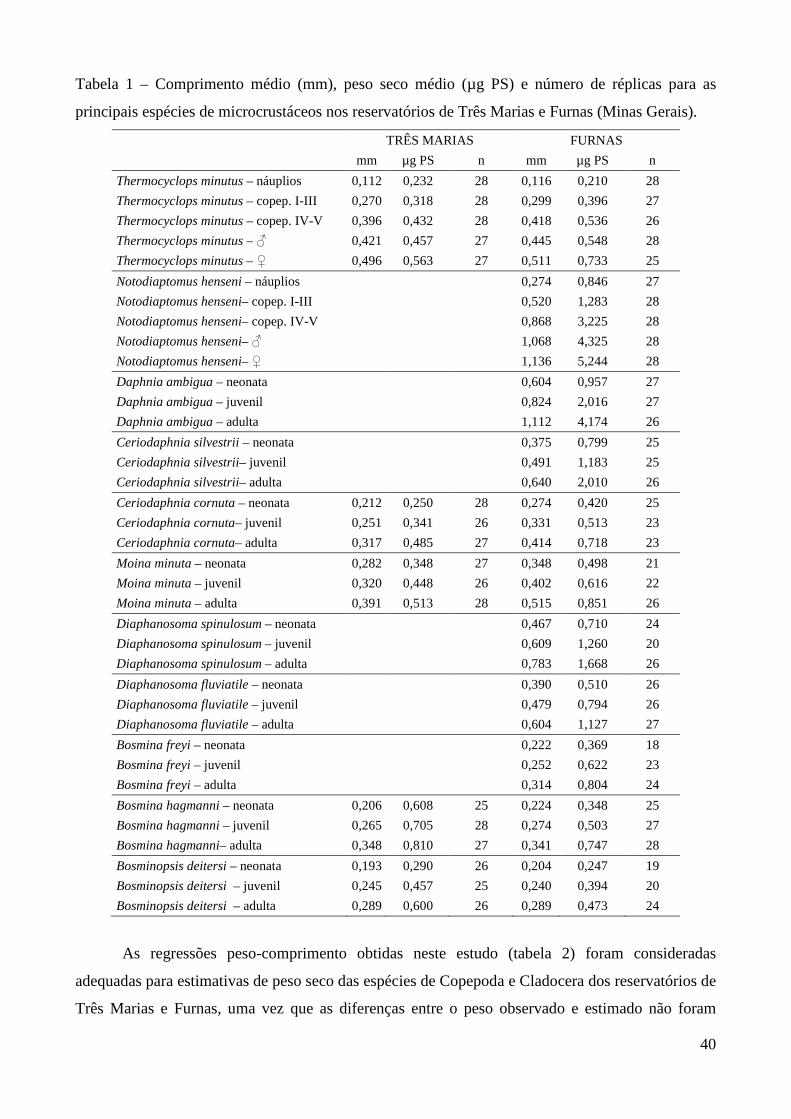
40
Tabela 1 – Comprimento médio (mm), peso seco médio (µg PS) e número de réplicas para as
principais espécies de microcrustáceos nos reservatórios de Três Marias e Furnas (Minas Gerais).
TRÊS MARIAS FURNAS
mm µg PS n mm µg PS n
Thermocyclops minutus – náuplios 0,112 0,232 28 0,116 0,210 28
Thermocyclops minutus – copep. I-III 0,270 0,318 28 0,299 0,396 27
Thermocyclops minutus – copep. IV-V 0,396 0,432 28 0,418 0,536 26
Thermocyclops minutus – ♂ 0,421 0,457 27 0,445 0,548 28
Thermocyclops minutus – ♀ 0,496 0,563 27 0,511 0,733 25
Notodiaptomus henseni – náuplios 0,274 0,846 27
Notodiaptomus henseni– copep. I-III 0,520 1,283 28
Notodiaptomus henseni– copep. IV-V 0,868 3,225 28
Notodiaptomus henseni– ♂ 1,068 4,325 28
Notodiaptomus henseni– ♀ 1,136 5,244 28
Daphnia ambigua – neonata 0,604 0,957 27
Daphnia ambigua – juvenil 0,824 2,016 27
Daphnia ambigua – adulta 1,112 4,174 26
Ceriodaphnia silvestrii – neonata 0,375 0,799 25
Ceriodaphnia silvestrii– juvenil 0,491 1,183 25
Ceriodaphnia silvestrii– adulta 0,640 2,010 26
Ceriodaphnia cornuta – neonata 0,212 0,250 28 0,274 0,420 25
Ceriodaphnia cornuta– juvenil 0,251 0,341 26 0,331 0,513 23
Ceriodaphnia cornuta– adulta 0,317 0,485 27 0,414 0,718 23
Moina minuta – neonata 0,282 0,348 27 0,348 0,498 21
Moina minuta – juvenil 0,320 0,448 26 0,402 0,616 22
Moina minuta – adulta 0,391 0,513 28 0,515 0,851 26
Diaphanosoma spinulosum – neonata 0,467 0,710 24
Diaphanosoma spinulosum – juvenil 0,609 1,260 20
Diaphanosoma spinulosum – adulta 0,783 1,668 26
Diaphanosoma fluviatile – neonata 0,390 0,510 26
Diaphanosoma fluviatile – juvenil 0,479 0,794 26
Diaphanosoma fluviatile – adulta 0,604 1,127 27
Bosmina freyi – neonata 0,222 0,369 18
Bosmina freyi – juvenil 0,252 0,622 23
Bosmina freyi – adulta 0,314 0,804 24
Bosmina hagmanni – neonata 0,206 0,608 25 0,224 0,348 25
Bosmina hagmanni – juvenil 0,265 0,705 28 0,274 0,503 27
Bosmina hagmanni– adulta 0,348 0,810 27 0,341 0,747 28
Bosminopsis deitersi – neonata 0,193 0,290 26 0,204 0,247 19
Bosminopsis deitersi – juvenil 0,245 0,457 25 0,240 0,394 20
Bosminopsis deitersi – adulta 0,289 0,600 26 0,289 0,473 24
As regressões peso-comprimento obtidas neste estudo (tabela 2) foram consideradas
adequadas para estimativas de peso seco das espécies de Copepoda e Cladocera dos reservatórios de
Três Marias e Furnas, uma vez que as diferenças entre o peso observado e estimado não foram
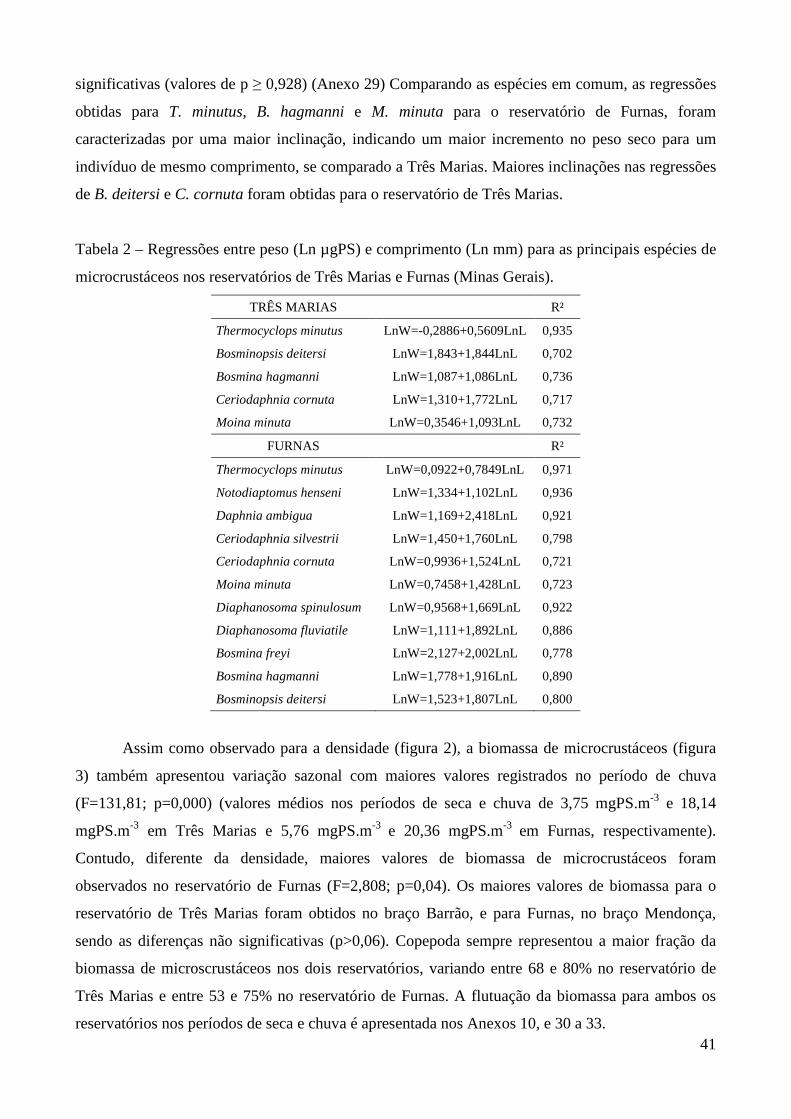
41
significativas (valores de p ≥ 0,928) (Anexo 29) Comparando as espécies em comum, as regressões
obtidas para T. minutus, B. hagmanni e M. minuta para o reservatório de Furnas, foram
caracterizadas por uma maior inclinação, indicando um maior incremento no peso seco para um
indivíduo de mesmo comprimento, se comparado a Três Marias. Maiores inclinações nas regressões
de B. deitersi e C. cornuta foram obtidas para o reservatório de Três Marias.
Tabela 2 – Regressões entre peso (Ln µgPS) e comprimento (Ln mm) para as principais espécies de
microcrustáceos nos reservatórios de Três Marias e Furnas (Minas Gerais).
TRÊS MARIAS R²
Thermocyclops minutus LnW=-0,2886+0,5609LnL 0,935
Bosminopsis deitersi LnW=1,843+1,844LnL 0,702
Bosmina hagmanni LnW=1,087+1,086LnL 0,736
Ceriodaphnia cornuta LnW=1,310+1,772LnL 0,717
Moina minuta LnW=0,3546+1,093LnL 0,732
FURNAS R²
Thermocyclops minutus LnW=0,0922+0,7849LnL 0,971
Notodiaptomus henseni LnW=1,334+1,102LnL 0,936
Daphnia ambigua LnW=1,169+2,418LnL 0,921
Ceriodaphnia silvestrii LnW=1,450+1,760LnL 0,798
Ceriodaphnia cornuta LnW=0,9936+1,524LnL 0,721
Moina minuta LnW=0,7458+1,428LnL 0,723
Diaphanosoma spinulosum LnW=0,9568+1,669LnL 0,922
Diaphanosoma fluviatile LnW=1,111+1,892LnL 0,886
Bosmina freyi LnW=2,127+2,002LnL 0,778
Bosmina hagmanni LnW=1,778+1,916LnL 0,890
Bosminopsis deitersi LnW=1,523+1,807LnL 0,800
Assim como observado para a densidade (figura 2), a biomassa de microcrustáceos (figura
3) também apresentou variação sazonal com maiores valores registrados no período de chuva
(F=131,81; p=0,000) (valores médios nos períodos de seca e chuva de 3,75 mgPS.m-3 e 18,14
mgPS.m-3 em Três Marias e 5,76 mgPS.m-3 e 20,36 mgPS.m-3 em Furnas, respectivamente).
Contudo, diferente da densidade, maiores valores de biomassa de microcrustáceos foram
observados no reservatório de Furnas (F=2,808; p=0,04). Os maiores valores de biomassa para o
reservatório de Três Marias foram obtidos no braço Barrão, e para Furnas, no braço Mendonça,
sendo as diferenças não significativas (p>0,06). Copepoda sempre representou a maior fração da
biomassa de microscrustáceos nos dois reservatórios, variando entre 68 e 80% no reservatório de
Três Marias e entre 53 e 75% no reservatório de Furnas. A flutuação da biomassa para ambos os
reservatórios nos períodos de seca e chuva é apresentada nos Anexos 10, e 30 a 33.
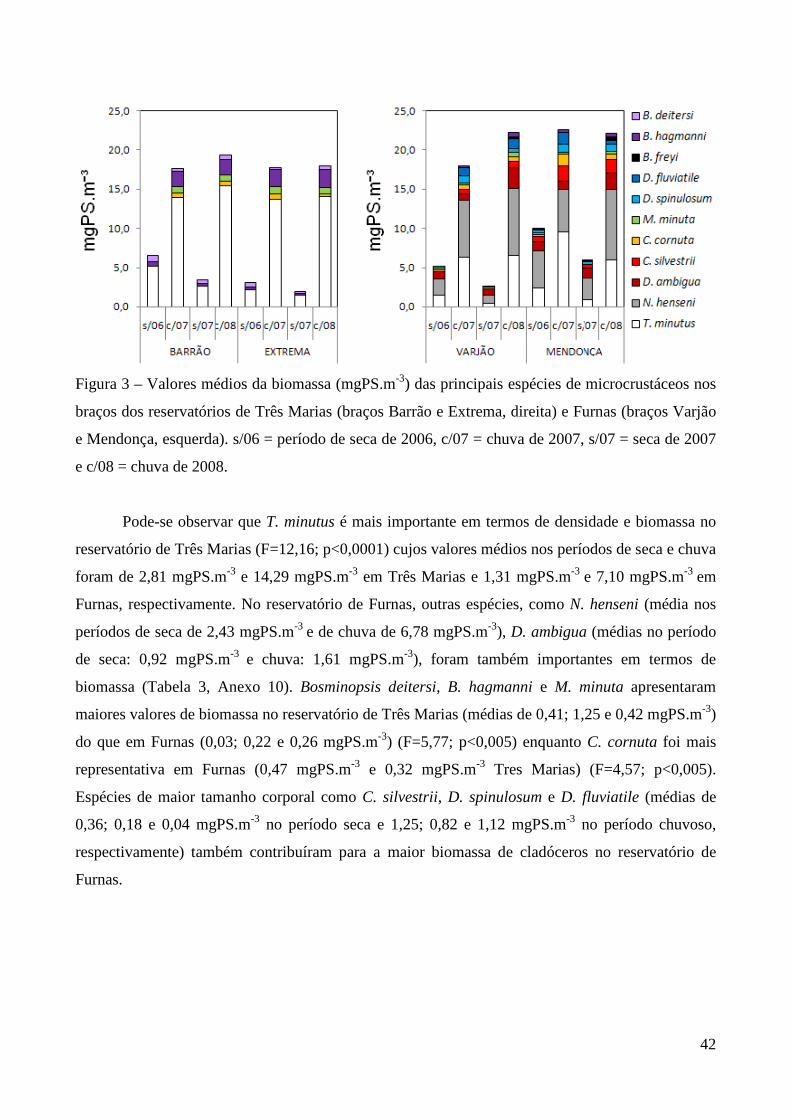
42
Figura 3 – Valores médios da biomassa (mgPS.m-3) das principais espécies de microcrustáceos nos
braços dos reservatórios de Três Marias (braços Barrão e Extrema, direita) e Furnas (braços Varjão
e Mendonça, esquerda). s/06 = período de seca de 2006, c/07 = chuva de 2007, s/07 = seca de 2007
e c/08 = chuva de 2008.
Pode-se observar que T. minutus é mais importante em termos de densidade e biomassa no
reservatório de Três Marias (F=12,16; p<0,0001) cujos valores médios nos períodos de seca e chuva
foram de 2,81 mgPS.m-3 e 14,29 mgPS.m-3 em Três Marias e 1,31 mgPS.m-3 e 7,10 mgPS.m-3 em
Furnas, respectivamente. No reservatório de Furnas, outras espécies, como N. henseni (média nos
períodos de seca de 2,43 mgPS.m-3 e de chuva de 6,78 mgPS.m-3), D. ambigua (médias no período
de seca: 0,92 mgPS.m-3 e chuva: 1,61 mgPS.m-3), foram também importantes em termos de
biomassa (Tabela 3, Anexo 10). Bosminopsis deitersi, B. hagmanni e M. minuta apresentaram
maiores valores de biomassa no reservatório de Três Marias (médias de 0,41; 1,25 e 0,42 mgPS.m-3)
do que em Furnas (0,03; 0,22 e 0,26 mgPS.m-3) (F=5,77; p<0,005) enquanto C. cornuta foi mais
representativa em Furnas (0,47 mgPS.m-3 e 0,32 mgPS.m-3 Tres Marias) (F=4,57; p<0,005).
Espécies de maior tamanho corporal como C. silvestrii, D. spinulosum e D. fluviatile (médias de
0,36; 0,18 e 0,04 mgPS.m-3 no período seca e 1,25; 0,82 e 1,12 mgPS.m-3 no período chuvoso,
respectivamente) também contribuíram para a maior biomassa de cladóceros no reservatório de
Furnas.
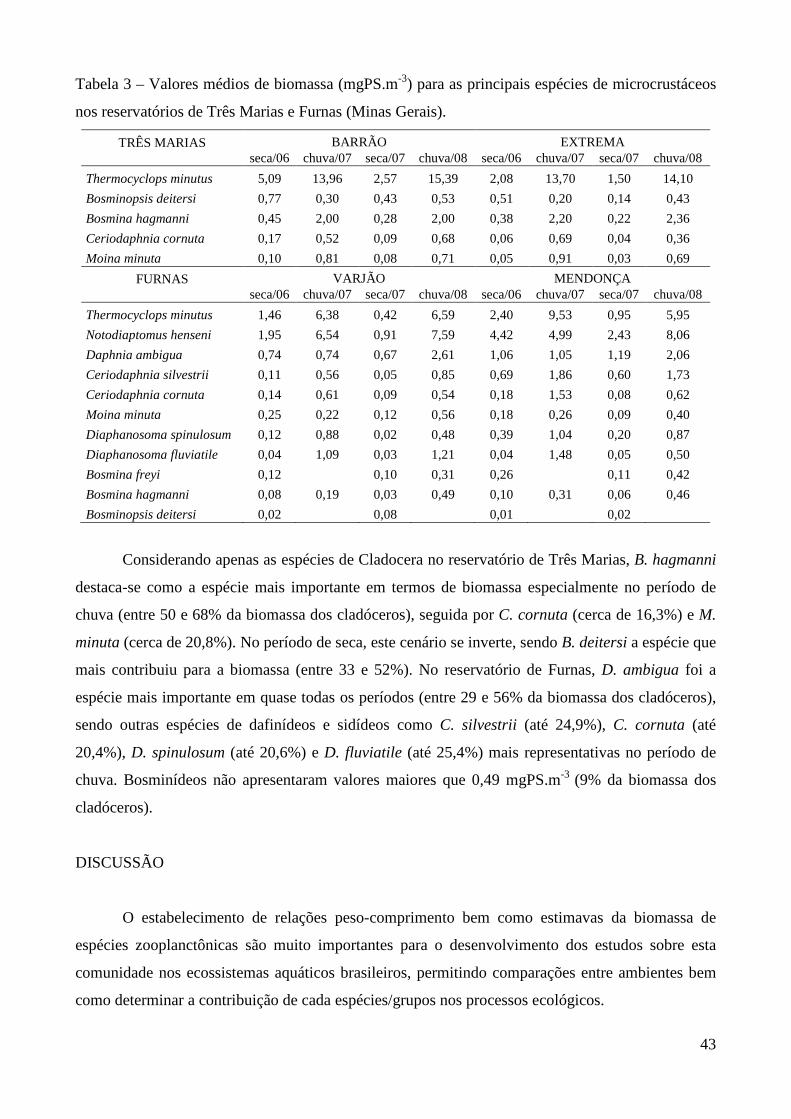
43
Tabela 3 – Valores médios de biomassa (mgPS.m-3) para as principais espécies de microcrustáceos
nos reservatórios de Três Marias e Furnas (Minas Gerais).
TRÊS MARIAS
BARRÃO EXTREMA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08
Thermocyclops minutus 5,09 13,96 2,57 15,39 2,08 13,70 1,50 14,10
Bosminopsis deitersi 0,77 0,30 0,43 0,53 0,51 0,20 0,14 0,43
Bosmina hagmanni 0,45 2,00 0,28 2,00 0,38 2,20 0,22 2,36
Ceriodaphnia cornuta 0,17 0,52 0,09 0,68 0,06 0,69 0,04 0,36
Moina minuta 0,10 0,81 0,08 0,71 0,05 0,91 0,03 0,69
FURNAS
VARJÃO MENDONÇA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08
Thermocyclops minutus 1,46 6,38 0,42 6,59 2,40 9,53 0,95 5,95
Notodiaptomus henseni 1,95 6,54 0,91 7,59 4,42 4,99 2,43 8,06
Daphnia ambigua 0,74 0,74 0,67 2,61 1,06 1,05 1,19 2,06
Ceriodaphnia silvestrii 0,11 0,56 0,05 0,85 0,69 1,86 0,60 1,73
Ceriodaphnia cornuta 0,14 0,61 0,09 0,54 0,18 1,53 0,08 0,62
Moina minuta 0,25 0,22 0,12 0,56 0,18 0,26 0,09 0,40
Diaphanosoma spinulosum 0,12 0,88 0,02 0,48 0,39 1,04 0,20 0,87
Diaphanosoma fluviatile 0,04 1,09 0,03 1,21 0,04 1,48 0,05 0,50
Bosmina freyi 0,12 0,10 0,31 0,26 0,11 0,42
Bosmina hagmanni 0,08 0,19 0,03 0,49 0,10 0,31 0,06 0,46
Bosminopsis deitersi 0,02 0,08 0,01 0,02
Considerando apenas as espécies de Cladocera no reservatório de Três Marias, B. hagmanni
destaca-se como a espécie mais importante em termos de biomassa especialmente no período de
chuva (entre 50 e 68% da biomassa dos cladóceros), seguida por C. cornuta (cerca de 16,3%) e M.
minuta (cerca de 20,8%). No período de seca, este cenário se inverte, sendo B. deitersi a espécie que
mais contribuiu para a biomassa (entre 33 e 52%). No reservatório de Furnas, D. ambigua foi a
espécie mais importante em quase todas os períodos (entre 29 e 56% da biomassa dos cladóceros),
sendo outras espécies de dafinídeos e sidídeos como C. silvestrii (até 24,9%), C. cornuta (até
20,4%), D. spinulosum (até 20,6%) e D. fluviatile (até 25,4%) mais representativas no período de
chuva. Bosminídeos não apresentaram valores maiores que 0,49 mgPS.m-3 (9% da biomassa dos
cladóceros).
DISCUSSÃO
O estabelecimento de relações peso-comprimento bem como estimavas da biomassa de
espécies zooplanctônicas são muito importantes para o desenvolvimento dos estudos sobre esta
comunidade nos ecossistemas aquáticos brasileiros, permitindo comparações entre ambientes bem
como determinar a contribuição de cada espécies/grupos nos processos ecológicos.

44
Os valores de tamanho de corpo e peso seco obtidos para as espécies de microcrustáceos dos
reservatórios de Três Marias e Furnas estiveram na mesma faixa de variação daqueles obtidos por
diversos autores em outros ecossistemas aquáticos brasileiros. Comparados ao peso seco médio
obtido para B. hagmanni, B. deitersi, C. cornuta e M. minuta no Lago Batata (PA) por Maia-
Barbosa & Bozelli (2005), verifica-se que as espécies de Três Marias e Furnas apresentaram pesos
geralmente maiores do que os estimados para cladóceros da estação natural daquele lago, contudo
menores do que os valores da estação impactada com rejeito de bauxita. Segundo os autores, a
ingestão de partículas de rejeito e a retenção deste material na parte interna das valvas (impossível
de ser removido durante a preparação para pesagem) seriam as razões para o maior peso dos
indivíduos da estação impactada.
Daphnia ambigua apresentou maiores valores de tamanho de corpo, porém menor peso seco
médio que o observado por Castilho-Noll & Arcifa (2007) no lago Monte Alegre (SP) (peso seco
médio de indivíduos pequenos, médios e grandes: 1,45; 4,64 e 8,27µg, respectivamente). O lago
Monte Alegre é considerado um ambiente eutrófico, portanto, indivíduos desta espécie devem
apresentar melhor estado nutricional e conseqüentemente maior peso seco. Contudo, valores de
peso seco similares aos registrados neste estudo foram relatados por Sendacz et al. (2006) nos
reservatórios de Ponte Nova (oligotrófico) e Guarapiranga (eutrófico) (SP).
Bosminopsis deitersi apresentou menores valores de tamanho de corpo e peso seco médio
comparado aos adultos dos reservatórios do Broa (0,61µg) (Matsumura-Tundisi et al., 1989) e da
Lagoa Dourada (Melão & Rocha, 2006) (peso seco de neonatas, juvenis e adultas: 0,23; 0,55 e
0,70µg, respectivamente). Contudo comparações com estes ambientes devem ser feitas com cautela,
uma vez que foram utilizados indivíduos não preservados em formol.
Moina minuta, Ceriodaphnia cornuta e C. silvestrii dos reservatórios estudados
apresentaram menores valores de peso seco médio em relação aos valores de Ponte Nova e
Guarapiranga (Sendacz et al., op. cit.), contudo esta última espécie apresentou valores maiores do
que os obtidos por González et al. (2008) para o reservatório de Bariri (SP) (peso seco médio de
adultos: 1,44µg). O comprimento e peso seco de C. silvestrii registrados por Santos et al. (2006)
(neonatas, juvenis e adultos de 1,337; 2,113 e 5,335 µg, respectivamente) foram maiores que os
obtidos para os reservatórios de Três Marias e Furnas. Contudo os indivíduos daquele estudo foram
criados em laboratório sendo o peso estimado a partir de indivíduos não preservados em formol. No
reservatório do Bariri, Diaphanosoma spinulosum e Bosmina hagmanni apresentaram maiores
valores do peso seco médio de adultos (1,897 e 2,233 µg, respectivamente) (González et al., op.
cit.). No reservatório do Broa (Matsumura-Tundisi et al., op. cit.), copepoditos (I-V) (420 µm e
0,430 µg) e machos e fêmeas (600µm e 0,780 µg) de Thermocyclops minutus apresentaram maiores

45
valores de tamanho de corpo e peso seco médio do que o observado nos reservatórios de Três
Marias e Furnas. Para esta espécie também foram pesados indivíduos não preservados em formol.
Estas diferenças no tamanho de corpo e peso seco, relatadas para os microcrustáceos,
refletem as respostas das espécies às variações das condições dos ecossistemas aquáticos (Dumont
et al., 1975; Boersma & Vijverberg, 1996; Blettler & Bonecker, 2006).
A relação entre peso e comprimento para espécies zooplanctônicas foi confirmada neste
estudo. Em alguns trabalhos, não foi observada linearidade nesta relação (Magalhães et al., 2006;
González et al., 2008), contudo os mesmos não empregaram transformações logarítmicas,
procedimento que torna as regressões peso-comprimento mais robustas devido à linearização dos
dados (Blettler & Bonecker, op.cit.).
Considerando a flutuação da biomassa nos reservatórios de Três Marias e Furnas, maiores
valores foram observados no período de chuva, sendo este padrão também observado por Rosa
(2008) em cinco reservatórios do sistema Furnas S.A. e por Melão et al. (2005) no reservatório
Lagoa Dourada. Em alguns estudos, maiores valores foram obtidos no período de seca (Rocha &
Matsumura-Tundisi, 1984; Sendacz et al., 2006, Santos-Wisniewski & Rocha, 2007) e em outros,
este padrão muitas vezes não é evidente devido à diferente contribuição dos grupos zooplanctônicos
entre os períodos de chuva e seca (Rietzler et al., 2004; Corgosinho & Pinto-Coelho, 2006).
Contudo em todos os trabalhos, os maiores valores de biomassa sempre foram associados a locais
ou períodos com maior disponibilidade de alimento (nutrientes, clorofila-a, sólidos em suspensão).
De fato, nos reservatórios de Três Marias e Furnas, maiores valores de clorofila-a e sólidos em
suspensão foram observados no período de chuva (Artigo 1), contudo correlações significativas
com a biomassa de microcrustáceos foram obtidas apenas para sólidos totais e orgânicos (r=0,560 e
0,547; p<0,0001; respectivamente). Nos reservatórios estudados esta pode ser uma importante fonte
de alimentar, não algal, para as espécies analisadas, pois segundo Marzolf (1990), apenas o
fitoplâncton não satisfaz todos os requerimentos energéticos do zooplâncton. Rull de Aguila (2001)
e Pinto-Coelho et al. (2005) afirmam que quando as correlações entre clorofila-a e biomassa do
zooplâncton são fracas, detritos são mais importantes do que a produção primária.
A dominância de T. minutus em termos de densidade e biomassa no reservatório de Três
Marias pode ser explicada pelo tipo de alimento predominante neste ambiente, as cianobactérias. T.
minutus é capaz de se alimentar de colônias de cianobactérias como Microcystis, Botryococcus e
Aphanocapsa (Matsumura-Tundisi et al., 1997). Contudo, pelo fato de constituírem uma fonte
alimentar de baixo valor nutricional (Rietzler & Espíndola, 1998) o maior consumo deste grupo de
algas por T. minutus em Três Marias pode ser uma das causas do menor tamanho e peso corporal
dos organismos em relação à mesma espécie em Furnas.

46
Os maiores valores de biomassa no reservatório de Furnas podem ser atribuídos às espécies
de maior tamanho corporal como calanóides (N. henseni), dafinídeos (D. ambigua e C. silvestrii) e
sidídeos (D. spinulosum e D. fluviatile) cujos pesos secos são bem maiores que o de pequenos
cladóceros como os bosminídeos, predominantes em Três Marias. Além disso, Castilho-Noll &
Arcifa (2007) observaram que apesar de menor em tamanho de corpo, D. ambigua apresentou maior
peso corporal que D. gessneri provavelmente por apresentar corpo mais arredondado, refletindo em
maior biomassa. O mesmo fato pode ser atribuído a B. hagmanni em Três Marias, cujo maior peso
seco torna esta espécie de cladócero a que mais contribuiu em termos de biomassa em oposição a B.
deitersi, que dominou em termos numéricos. Desta forma podemos constatar que mesmo entre os
microcrustáceos, a biomassa representa melhor a estrutura da comunidade do que a densidade
numérica.
Os valores de biomassa de microcrustáceos observados os reservatórios de Três Marias e
Furnas são próximos aos observados em outros reservatórios oligotróficos como os de Segredo e
Mourão (PR) (médias de 6,8 mgPS.m-3) com valores oscilando entre 0,03 e 20,26 mgPS.m-3 (Bletter
& Bonecker, 2007). Em Ponte Nova (SP), os valores oscilaram entre 9,264 e 9,794 mgPS.m-3 para
Cladocera e 0,357 e 1,657 mgPS.m-3 para Copepoda (Sendacz et al., 2006). Na Lagoa Dourada
(SP), Melão et al. (2005) apresentam valores médios de biomassa entre 3,496 e 10,037 mgPS.m-3
para Cladocera (sendo B. deitersi a principal espécie) e 0,852 e 5,956 mgPS.m-3 para Copepoda
(representados por Tropocyclops prasinus e Mesocyclops longisetus).
No reservatório do Broa (SP), médias anuais de biomassa de microcrustáceos estiveram
compreendidas entre 10 e 15 mgPS.m-3 com máximos de 60 a 80 mgPS.m-3 registrados nos meses
mais quentes do ano (Matsumura-Tundisi et al., 1989). Na época em que o estudo foi realizado, o
reservatório era considerado mesotrófico e os microcrustáceos representados por A. furcatus,
Bosminidae e Moinidae.
As médias anuais de biomassa estimadas para as principais espécies de Cladocera e
Copepoda nos lagos naturais Dom Helvécio e Carioca (MG) foram de 26,57 e 10,88 mgPS.m-3,
respectivamente (Matsumura-Tundisi & Tundisi, 1986 apud Rocha et al., 1995), quando no
primeiro lago dominavam duas espécies de Calanoida (Argyrodiaptomus furcatus e Scolodiaptomus
corderoi) e no segundo Thermocyclops minutus.
No lago Batata (PA) impactado por rejeito de bauxita, menores valores de biomassa de
cladóceros ocorreram na estação natural, entre 2,69 e 11,04 mgPS.m-3, enquanto na estação
impactada, maiores valores foram registrados: de 24,61 a 38,9 mgPS.m-3 (Maia-Barbosa, 2000).
Ambientes mais eutrofizados geralmente apresentam maiores valores de biomassa: no
reservatório de Iraí (PR) os valores de biomassa de microcrustáceos oscilaram entre 7,14 e 261,52
mgPS.m-3 (média de 101,5 mgPS.m-3) (Bletter & Bonecker, 2007). Em Salto Grande (SP) para

47
Cladocera os valores estiveram compreendidos entre 21,4 e 122,5 mgPS.m-3 e para Copepoda entre
117,0 e 319,0 mgPS.m-3 (Rietzler et al., 2004). No reservatório de Guarapiranga (SP), valores entre
37,72 e 69,59 mgPS.m-3 foram registrados para Cladocera e entre 6,48 e 236,18 mgPS.m-3 para
Copepoda (Sendacz et al., 2006). Na lagoa da Pampulha (MG), reservatório eutrófico urbano, para
Cladocera os valores médios estiveram compreendidos entre 40,42 e 572,16 mgPS.m-3 e para
Copepoda entre 11,34 e 379,51 mgPS.m-3 (Pinto-Coelho et al., 2005). Em Barra Bonita, Santos-
Wisniewski et al. (2002) observaram valores de biomassa de Cladocera e Copepoda em torno de 50
mgPS.m-3 (período chuvoso) e 100 mgPS.m-3 (período de seca) sendo este último grupo a maior
fração da biomassa. Em Monjolinho, considerado meso-eutrófico, Okano (1994 apud Rocha et al.,
1995) observou valores médios de biomassa de 4,008 gPS.m-3 para Cladocera e 2,560 gPS.m-3 para
Copepoda.
Mesmo em ambientes naturalmente eutróficos como a lagoa Amarela (Zmáx = 2 metros,
densamente colonizada por macrófitas e em estágio final de sucessão) maiores valores de biomassa
foram também obtidos, com média anual de 173,85 mgPS.m-3 para microcrustáceos (Matsumura-
Tundisi & Tundisi, 1986 apud Rocha et al., 1995).
Diversos autores têm apontado uma relação positiva entre biomassa zooplanctônica e trofia
dos ecossistemas aquáticos tanto tropicais (Esteves & Sendacz, 1988; Rocha et al., 1995; Rocha et
al., 1997) quanto temperados (Pederson et al., 1976; Pace, 1986). Estes autores consideram as
diferenças na quantidade e qualidade do fitoplâncton como fonte de alimento; na composição e
densidade da comunidade zooplanctônica entre ambientes, bem como no estado nutricional dos
indivíduos como outros fatores que afetam a biomassa.
Mesmo considerados oligotróficos, os reservatórios de Três Marias e Furnas apresentaram
diferenças marcantes na biomassa de microcrustáceos. Em Três Marias Thermocyclops minutus,
considerada como uma espécie comum em ecossistemas aquáticos brasileiros (Reid, 1989) foi a
espécie mais importante em termos de densidade e representando até 80% da biomassa.
Bosminopsis deitersi, B. hagmanni, C. cornuta e M. minuta foram pouco representativas neste
ambiente, ao contrário do observado para estas espécies em outros lagos e reservatórios brasileiros
(Maia-Barbosa, 2000; Melão & Rocha, 2006; Sendacz et al., 2006). Em Furnas, Copepoda também
foi o grupo dominante em densidade e biomassa, contudo N. henseni contribuiu na mesma
proporção que T. minutus. A maior biomassa de Cladocera neste reservatório deve-se
principalmente às espécies de maior tamanho corporal como D. ambigua, C. silvestrii, D.
spinulosum e D. fluviatile, especialmente durante o período de chuva. Desta forma, devido à sua
maior oferta em termos de biomassa e maior qualidade nutricional (Santeiro & Pinto-Coelho, 2000;
Pinto-Coelho et al., 2005) os microcrustáceos de maior porte podem ser considerados uma fonte de

48
alimento mais adequada para alevinos bem como adultos de espécies zooplanctívoras neste
reservatório.
AGRADECIMENTOS
À toda a equipe dos laboratórios de Ecologia do Zooplâncton, de Gestão Ambiental de
Reservatórios e de Limnologia, que direta ou indiretamente ajudaram na realização deste trabalho.
Ao Projeto Parques Aqüícolas (Convênio 8713 SEAP/SECTES/FUNDEP-UFMG) e ao CNPq
(Processo n° 142079/2006-5) pelo financiamento do projeto.
REFERÊNCIAS BIBLIOGRÁFICAS
BEZERRA-NETO, J. F. & PINTO-COELHO, R.M. 2002. Population dynamics and secondary
production of Chaoborus brasiliensis (Diptera: Chaoboridae) in a small tropical reservoir: Lagoa do Nado, Minas Gerais - Brazil. Acta Limnol. Bras., 14(3): 61-72.
BIRD D.F. & PRAIRE Y.T. 1985. Practical guidelines for the use of zooplankton length-weight regression equations. J. Plankton Res., 7:955-960.
BLETTLER, M.C.M. & BONECKER, C.C. 2006. Avaliação da Biomassa de Microcrustáceos em Ambientes Aquáticos Continentais. Interciência, 31(8):591-597.
BLETTLER, M.C.M. & BONECKER, C.C. 2007. Longitudinal distribution of microcrustacean biomass in three tropical reservoirs (Paraná State, Brazil). Acta Scientiarum, 29(3):297-304.
BONECKER, C.C.; NAGAE, M.Y.; BLETTLER, M.C.M.; VELHO, L.F.M. & LANSAC-TÔHA, F.A. 2007. Zooplankton biomass in tropical reservoirs in southern Brazil. Hydrobiologia, 579:115-123.
BOERSMA, M. & VIJVERBERG, J. 1996. Food effects on life history traits and seasonal dynamics of Ceriodaphnia pulchella. Freshwater Biol., 35:25-34.
BOTTRELL, H.H.; DUNCAN, A.; GLIWICZ, Z.M.; GRYGIEREK, E.; HERZIG, A.; HILLBRICHT-ILKOWSKA, A.; KURASAUA, H.; LARSSON, P. & WEGLENSKA, T. 1976. A review of some problems in zooplankton production studies. Norw. J. Zoo., 24: 419-456.
BOZELLI, R. L. & HUSZAR, V. L. M. 2003. Comunidades Fito e Zooplanctônicas Continentais em Tempo de Avaliação. LIMNOtemas. Sociedade Brasileira de Limnologia, 3:1-32.
CASTILHO-NOLL, M.S.M. & ARCIFA, M.S. 2007. Length-weight relationships for zooplanktonic species of a tropical Brazilian lake: Lake Monte Alegre. Acta Limnol. Bras., 19(1):93-100.
CEMIG - Compania Energética de Minas Gerais. 2009. Usinas da CEMIG. http://www.cemig.com.br/institucional/usinas/11g.asp. Acesso em: 16 set. 2009.
CORGOSINHO, P.H.C. & PINTO-COELHO, R.M. 2006. Zooplankton biomass, abundance and allometric patterns along an eutrophic gradient at Furnas Reservoir (Minas Gerais, Brazil). Acta Limnol. Bras., 182):213-224.
DUMONT,H.J.; VAN DE VELDE, I. & DUMONT, S. 1975. The Dry Weight Estimate of Biomass in a Selection of Cladocera, Copepoda and Rotifera from the Plankton Peryphyton and benthos of Continental Waters. Oecologia, 19:75-97.
ESTEVES, K. E. & SENDACZ, S. 1988. Relações entre a biomassa do zooplâncton e o estado trófico de reservatórios do Estado de São Paulo. Acta Limnol. Bras., 11: 587-604.

49
FURNAS – Furnas Centrais Elétricas SA. 2009. Sistema Furnas. http://www.furnas.com.br/hotsites/sistemafurnas. Acesso em: 16 set. 2009.
GONZÁLEZ, E.J., MATSUMURA-TUNDISI, T. & TUNDISI, J.G. 2008. Size and dry weight of main zooplankton species in Bariri reservoir (SP, Brazil). Braz. J. Biol., 68(1):69-75.
MAGALHÃES, A., COSTA, R. M., LIANG, T. H.,PEREIRA, L. C. C. & RIBEIRO, M. J. S. 2006. Spatial and Temporal Distribution in Density and Biomass of Two Pseudodiaptomus Species (Copepoda: Calanoida) in the Caeté River Estuary (Amazon Region - North of Brazil). Braz. J. Biol., 66(2A): 421-430.
MAIA-BARBOSA, P. M. 2000. Ecologia de Cinco Espécies de Cladóceros de um Lago Amazônico Impactado por Rejeito de Bauxita (Lago Batata, Pará-Brasil). Universidade Federal do Rio de Janeiro. 218p. Tese.
MAIA-BARBOSA, P.M. & BOZELLI, R. 2005. Lenght-Weight Relatioships for Five Cladoceran Species in an Amazonian Lake. Brazilian Archives of Biology and Technology, 48(2):303-308.
MARZOLF, G.R. 1990. Reservoir as Environments for Zooplankton. In: Thornton, K. W.; Kimmel, B.L. & Payne, F.E. (eds.) Reservoir Limnology: Ecological Perspectives. New York: John Wiley & Sons. 246p.
MATSUMURA-TUNDISI, T. & TUNDISI, J.G. 1986. Biomass and Zooplankton Community structure of Three Lakes of River Doce Valley (Minas Gerais - Brazil). The Fifth Japan-Brazil Symposium on Science and Tecnology. Tokyo, Japan.
MATSUMURA-TUNDISI, T., RIETZLER, A.C. & TUNDISI, J.G. 1989. Biomass (dry weight and carbon content) of plankton crustacea from Broa reservoir (São Carlos, SP, Brazil) and its fluctuations across on year. Hydrobiologia, 179:229-236.
MATSUMURA-TUNDISI, T.; ROCHA, O. & TUNDISI, J. G. 1997. Carbon Uptake by Scolodiaptomus corderoi and Thermocyclops minutus Feeding on Different Size Fractions of Phytoplankton from Lake Dom Helvécio. In: Tundisi, J.G. & & Saijo, Y. (eds) Limnological Studies on the Rio Doce Valley Lakes, Brazil. Rio de Janeiro: Brazilian Academy of Sciences, University of São Paulo, School of Engineering at São Carlos, Center for Water Resources and Applied Ecology. 513p.
MCCAULEY, E. 1984. The estimation of the abundance and biomass of zooplankton in samples. In: Downing J.A. & F.H. Rigler (eds.) A Manual on Methods for Assessment of Secondary Productivity in Freshwaters. IBP Hand Book 17, Blackwell Scientific Publishing, 2ª ed. 501 p.
MELÃO, M. G. & ROCHA, O. 2004. Life history, biomass and production of two planktonic cyclopoid copepods in a shallow subtropical reservoir. J. Plankton Res., 26(8):909-923.
MELÃO, M. G. & ROCHA, O. 2006. Life history, populations dynamics, standing biomass and production of Bosminopsis deitersi (Cladocera) in a shallow tropical reservoir. Acta Limnol. Bras., 18(4):433-450.
MELÃO, M.G., ROCHA, O., & ROCHE, F. K. 2005. Produtividade, Biomassa e Flutuações Populacionais e interações biológicas da Comunidade Planctônica e Suas Implicações na Transferência de Energia na Cadeia Alimentar de um Reservatório Raso e oligotrófico. In: Roche, F. K. & Rocha, O. (eds.) Ecologia Trófica de Peixes com ênfase na planctivoria em ambientes lênticos de água doce no Brasil. São Carlos: Rima. 146p.
MIRANDA, E. E. & COUTINHO, A. C. (Coord.) 2004. Brasil Visto do Espaço. Campinas: Embrapa Monitoramento por Satélite. http://www.cdbrasil.cnpm.embrapa.br Acesso em: 16 set. 2009.
NOGUEIRA, D.J.; CASTRO, S.C. & RIGOLIN DE SÁ, O. 2008. Avaliação da qualidade da água no reservatório UHE Furnas - MG, utilizando as brânquias de Pimelodus maculatus (LACÈPÉDE, 1803) como biomarcador de poluição ambiental. Ciência et Praxis, 1(1): 15-20.
OKANO, Y. W., 1994. Análise da estrutura e dinâmica populacional da comunidade zooplanctônica de um reservatório artificial (reservatório Monjolinho, São Carlos, SP). Universidade Federal de São Carlos. 128p. Tese.
PACE, M. L. 1986. An empirical analyses of zooplankton community size structure across lake trophic gradients. Limnology and Oceanography, 31(1):45-55.

50
PANARELLI, E. A. 2004. Flutuações mensais da comunidade zooplanctônica e dinâmica das populações de Cladocera em lagoas marginais, na região de transição Rio Paranapanema – Represa Jurumirim (SP). Universidade Estadual Paulista. 247p. Tese.
PEDERSON, G. L., WELCH, E. B. & LITT, A. H. 1976. Plankton secondary productivity and biomass: their relation to lake trophic state. Hydrobiologia, 50(2):129 - 144.
PINTO-COELHO, R. M., BEZERRA-NETO, J. F. & MORAIS-JR., C. A.. 2005. Effects of Eutrophication on Size and Biomass of Crustacean Zooplankton in a Tropical Reservoir. Braz. J. Biol., 65(2): 325-338.
RIETZLER, A.C. & ESPÍNDOLA, E. L. G. 1998. Microcystis as a food source for copepods in a subtropical eutrophic reservoir. Verh. Internat. Verein. Limnol., 26:2001-2005.
RIETZLER, A.C.; ROCHA, O. & ESPINDOLA, E. L. G. 2004. Produção Secundária de Espécies do Zooplâncton do Reservatório de Salto Grande, Estado de São Paulo. In: Espindola, E. L.G., Leite, M.A., Dornfeld, C.B. (eds.) Reservatório de Salto Grande (Americana-SP): caracterização, impactos e propostas de manejo. São Carlos: Rima.
ROCHA, O. & MATSUMURA-TUNDISI, T., 1984. Biomass and production of Argyrodiaptomus furcatus a tropical calanoid copepod in Broa reservoir, southern Brazil. Hydrobiologia, 113: 307-311.
ROCHA, O.; SENDACZ, S. & MATSUMURA-TUNDISI, T. 1995. Composition, Biomass and Productivity of Zooplankton in Natural Lakes and Reservoirs of Brazil. In: Tundisi, J. G.; Bicudo, C. E. M. & Matsumura-Tundisi, T. (eds) Limnology in Brazil. Rio de Janeiro: Brazilian Academy of Sciences, Brazilian Limnological Society. 376p.
ROCHA, O.; MATSUMURA-TUNDISI, T. & SAMPAIO, E. V. 1997. Phytoplankton and zooplankton community structure and production as related to trophic state in some Brazilian lakes and reservoirs. Verh. Internat. Verein Limnol., 26:599-604.
ROSA, P.G. 2008. Comunidade Zooplanctônica de Cinco Reservatórios Tropicais do Sistema Furnas S.A (Brasil): Abundância e Biomassa de Carbono. Dissertação. UFJF. 113p.
RULL DE AGUILA, L. M. 2001. Gradiente trófico no Rio Sapucaí (reservatório de Furnas- MG): relação com a distribuição do zooplâncton e os usos do solo. Universidade Federal de Minas Gerais. 75p. Dissertação.
SÁ JUNIOR, W. P. 1994. Production of Planktonic Biomass for Feed of Alevins at the Furnas Hydrobiology and Hatchery Station. In: Pinto-Coelho, R.M.; Giani, A. & von Sperling, E. (eds) Ecology and Human impact on lakes and reservoirs in Minas Gerais with special reference to future development and management strategies. Belo Horizonte: SEGRAC. 193p.
SAMPAIO, I.B.M. 2002. Estatística Aplicada à Experimentação Animal. Belo Horizonte: Fundação de Ensino e Pesquisa em Medicina Veterinária e Zootecnia, 2ª ed. 256p.
SAMPAIO, E.V. & LÓPEZ, C. M. 2003. Limnologias física, química e biológica da represa de Três Marias e do São Francisco. In: Godinho, H. P. & Godinho, A. L. (org.) Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Hrizonte: PUCMinas. 468p.
SANTEIRO, R.M. & PINTO-COELHO, R.M. 2000. Efeitos de fertilização na biomassa e qualidade nutricional do zooplâncton utilizado para alimentação de alevinos na estação de Hidrobiologia de Piscicultura de Furnas, MG. Acta Sientiarum, 22(3):707-716.
SANTOS, M.A.P.F., MELÃO, M.G.G. & LOMBARDI, A.T. 2006. Life history characteristics and production of Ceriodaphnia silvestrii Daday (Crustacea, Cladocera) under different experimental conditions. Acta Limnol. Bras., 18(2):199-212.
SANTOS-WISNIEWSKI, M.J., MATSUMURA-TUNDISI, T. & ROCHA, O. 2002. Spatial and Temporal Patterns of Zooplankton Biomass and Secondary Production in Barra Bonita Reservoir, Brazil. 4th International Conference on Reservoir Limnology and Water Quality, August 12-16, 2002, extended Abstrats. p: 303-306.
SANTOS-WISNIEWSKI, M.J. & ROCHA, O. 2007. Spatial distribution and secondary production of Copepoda in a tropical reservoir: Barra Bonita, SP, Brazil. Braz. J. Biol., 67(2): 223-233.

51
SENDACZ, S., CALEFFI, S. & SANTOS-SOARES, J. 2006. Zooplankton Biomass of Reservoirs in Different Trophic Conditions in the State of São Paulo, Brazil. Braz. J. Biol., 66(1B): 337-350.
WINBERG G.G. & DUNCAN, A. 1971. Methods for the estimation of production of aquatic animals. New York: Academic Press. 175 pp.

52
ARTIGO 3 - PRODUTIVIDADE SECUNDÁRIA DAS PRINCIPAIS ESPÉCIES DE MICROCRUSTÁCEOS EM DOIS BRAÇOS DOS RESERVATÓRIOS DE TRÊS MARIAS E
FURNAS, MINAS GERAIS, BRASIL. RESUMO Considerando a importância da comunidade zooplanctônica na transferência de energia entre níveis tróficos, este trabalho teve como objetivo estimar as taxas de produtividade secundária das principais espécies de microcrustáceos em dois grandes reservatórios, Três Marias e Furnas (MG). Para este estudo considerou-se Thermocyclops minutus, Bominopsis deitersi, Bosmina hagmanni, Ceriodaphnia cornuta e Moina minuta no reservatório de Três Marias, e no reservatório de Furnas além destas espécies, foram também consideradas Notodiaptomus henseni, Daphnia ambigua, Ceriodaphnia silvestrii, Diaphanosoma spinulosum, D. fluviatile e Bosmina freyi. Com relação à produtividade total, maiores taxas foram obtidas para o período de chuva em ambos os reservatórios (p<0,000) com valores médios nos períodos de seca e chuva de 0,44 e 1,80 mgPS.m-3.d-1 para o reservatório de Três Marias e 1,05 e 3,10 mgPS.m-3.d-1 para o reservatório de Furnas, respectivamente. T. minutus foi a espécie mais importante em termos de densidade e biomassa no reservatório Três Marias, e M. minuta a que apresentou as maiores taxas de produtividade secundária, especialmente no período de chuva. Em Furnas, N. henseni e D. ambigua apresentaram as maiores taxas de produtividade em ambos os períodos, enquanto C. silvestrii, C. cornuta, D. spinulosum e D. fluviatile também se destacaram no período chuvoso. Valores da razão produtividade:biomassa (P:B) foram geralmente menores para os copépodes, sendo M. minuta a espécie que apresentou maiores valores em ambos os reservatórios. As maiores taxas de produtividade secundária de microcrustáceos no reservatório de Furnas sejam provavelmente o resultado de uma maior eficiência na transferência de energia entre níveis tróficos, devido à presença de espécies de fitoplâncton de maior qualidade nutricional neste ambiente. PALAVRAS-CHAVE: produtividade secundária, microcrustáceos, reservatórios tropicais ABSTRACT Secondary Productivity of the Main Microcrustacean Species in Two Arms of Três Marias and Furnas Reservoirs, MG, Brazil. Considering the importance of the zooplankton community in the energy transference between trophic levels, this paper had as objective to estimate the secondary productivity rates of the main microcrustacean species in two large tropical reservoirs, Três Marias and Furnas (MG, Brazil). For this study, we considered Thermocyclops minutus, Bominopsis deitersi, Bosmina hagmanni, Ceriodaphnia cornuta and Moina minuta in Três Marias reservoir and, in Furnas reservoir, together with these, we also considered: Notodiaptomus henseni, Daphnia ambigua, Ceriodaphnia silvestrii, Diaphanosoma spinulosum, D. fluviatile and Bosmina freyi. As for the total productivity, higher rates were obtained in the rainy period in both reservoirs (p<0.000), with average values during the dry and rainy periods of 0.44 and 1.80 mgPS.m-3.d-1 for Três Marias Reservoir and 1.50 and 3.10 mgPS.m-3.d-1 for Furnas Reservoir, respectively. T.minutus was the most important species in terms of density and biomass in Três Marias reservoir and M. minuta was the one which presented the highest secondary productivity rates, especially during the rainy period. In Furnas, N. henseni and D. ambigua presented the highest productivity rates in both periods, whereas C. silvestrii, C. cornuta, D. spinulosum and D. fluviatille were also important during the rainy period. Values of productivity:biomass ratio (P:B) were usually lower for the copepods; M. minuta was the species that presented the highest values in both reservoirs. The higher microcrustacean secondary productivity rates in Furnas Reservoir are probably the result of the higher efficiency in energy transference between trophic levels, due to the presence of better nutritional quality phytoplankton species in this environment. KEY-WORDS: secondary productivity, microcrustacean, tropical reservoirs

53
INTRODUÇÃO
A produtividade secundária é uma importante ferramenta para a compreensão das
comunidades biológicas, seja através de parâmetros reprodutivos e populacionais, ou da ciclagem
de matéria e do fluxo de energia, uma vez que fornece a quantidade de material novo formada,
permitindo estimar a porção da biomassa que é renovada por unidade de tempo (Lampert &
Sommer, 2007). Para o zooplâncton, a produtividade ganha uma importância especial, uma vez que
esta comunidade é caracterizada por espécies de ciclo de vida curto e de altas taxas reprodutivas
onde parâmetros como densidade e biomassa sozinhos, não refletem todas as respostas desta
comunidade às variáveis ambientais e às interações bióticas (Edmondson, 1974). Além disso, a
produtividade secundária auxilia em estudos de cadeias tróficas dada a importância do zooplâncton
na transferência de energia entre os produtores (fitoplâncton) e níveis tróficos superiores (peixes).
Diversos métodos são descritos na literatura para estimativa da produtividade secundária,
contudo em populações naturais é difícil precisar dietas de organismos filtradores e onívoros, bem
como a porção de biomassa que é perdida através de processos fisiológicos e da predação (Allan &
Castilho, 2007). Neste contexto, métodos populacionais indiretos como das coortes (Allen, 1951), e
da soma dos incrementos (Edmondson & Winberg, 1971) são comumente empregados. Outros
métodos envolvem o produto entre taxas de crescimento espécies-específicos e da biomassa (Hart,
1987).
Dentre os fatores que influenciam a produtividade estão características intrínsecas de cada
espécie, como biomassa, fecundidade, taxas de desenvolvimento e crescimento, resistência à fome,
vulnerabilidade a predação e competitividade. Além destes, fatores ambientais também influenciam
a produtividade secundária e devem ser considerados para o entendimento de suas variações ao
longo do tempo: regime climático, variáveis hidrológicas, características morfométricas do
ambiente (Peláez-Rodríguez, & Matsumura-Tundisi, 2002; Rietzler et al., 2004), além das
interações entre as espécies, particularmente entre o fitoplâncton (alimento disponível), e as
pressões de predação (Melão & Rocha, 2004).
Muitos dos valores de produtividade secundária do zooplâncton em ecossistemas tropicais
foram estimados para os grandes lagos africanos (Burgis, 1974; Hart & Allanson, 1975; Mavuti,
1994), mas apesar da escassez destes estudos em ecossistemas aquáticos brasileiros nas duas
últimas décadas (Rocha & Matsumura-Tundisi, 1984; Okano, 1994), o número de trabalhos tem
aumentado (Maia-Barbosa, 2000; Peláez-Rodríguez, & Matsumura-Tundisi, 2002; Melão &
Rocha, 2004 e 2006; Rietzler et al., 2004; Santos-Wisniewski & Rocha, 2007). Entretanto, a
maioria destes estudos se concentra em ambientes aquáticos do Estado de São Paulo. O presente

54
trabalho fornece dados inéditos para Minas Gerais, uma vez que estudos sobre produtividade
secundária para espécies de microcrustáceos não foram registrados para corpos d’água deste Estado.
Além disso, visando ampliar o uso e aumentar o potencial pesqueiro dos reservatórios de
Três Marias e Furnas, o projeto Parques Aqüícolas (SEAP/SECTES/FUNDEP-UFMG), no qual
este trabalho está inserido, visou à delimitação de áreas para produção pesqueira através de tanques-
rede. Neste projeto, variáveis abióticas e bióticas foram medidas para estimar a capacidade suporte
de áreas selecionadas. Os conhecimentos levantados sobre o zooplâncton auxiliarão, em fase
posterior, não só pelo uso destes organismos como importante fonte alimentar bem como no
monitoramento da qualidade de água, mitigando os impactos da criação de peixes sobre toda a
comunidade aquática.
Neste sentido, este trabalho teve como objetivo estimar as taxas de produtividade secundária
das principais espécies de microcrustáceos em alguns braços dos reservatórios de Três Marias e
Furnas (MG) além de avaliar os possíveis fatores responsáveis pela sua variação.
MATERIAL E MÉTODOS
ÁREA DE ESTUDOS
Os reservatórios das Usinas Hidrelétricas (UHEs) de Três Marias (18°12’S e 45°15’W) e
Furnas (46°19’W e 20°40’S) foram criados há mais de 40 anos para geração de energia elétrica,
sendo também utilizados para recreação, pesca amadora e profissinal, irrigação e abastecimento. O
reservatório da UHE de Três Marias localiza-se no alto rio São Francisco, Centro-Oeste do Estado
de Minas Gerais. Concluído em 1960, atualmente o assoreamento é o principal impacto que
compromete a capacidade geradora de energia e a qualidade da água (Sampaio & López, 2003),
seguido pela substituição da vegetação ripária por monoculturas de Eucaliptus spp e pecuária
extensiva. O reservatório de Furnas está localizado na bacia do rio Grande, Sul do Estado de Minas
Gerais, sendo o braço norte formado pelo rio Grande e o braço sul pelo rio Sapucaí. O reservatório
foi concluído em 1962 tendo, atualmente, como principais impactos atividades como a
agropecuária, monoculturas, o despejo de efluentes domésticos e o lançamento de resíduos sólidos
(Nogueira et al., 2008). Algumas características morfométricas dos dois reservatórios são
apresentadas na tabela 1.
Em cada reservatório foram escolhidos como pontos amostrais dois braços com
características diferentes de entorno (figura 1). Desta forma, no reservatório de Três Marias foram
selecionados os braços Barrão (Zmáx=21,8 metros, com vegetação original do tipo Cerrado) e
Extrema (Zmáx=20,9 metros, com monocultura de Eucaliptus spp), localizados no município de

55
Morada Nova de Minas. No reservatório de Furnas foram selecionados os braços Varjão
(Zmáx=17,5 metros, com nascente localizada no Parque Municipal do Paredão e entorno com
monocultura de café) e Mendonça (Zmáx=25,5 metros, com pecuária em pastagem nativa),
localizados no município de Guapé. Estes braços foram selecionados a fim de caracterizar as
diferentes feições do entorno destes reservatórios.
Tabela 1 – Características morfométricas dos reservatórios das UHEs de Três Marias e Furnas
(MG). Fonte: CEMIG, 2009 e FURNAS, 2009.
UHE Três Marias UHE Furnas Área Inundada 1.100 km2 1.440 Km² Volume Útil 15,27 x 109 m³ 17,21 x 109 m³ Vazão Média 700 m³.s-1 800 m³.s-1
Tempo Médio de Residência 120 dias 160 dias Profundidade Máxima 75 m 90 m Profundidade Média 12 m 13 m Capacidade Geradora 396 MW 1126 MW
Figura 1 – Reservatórios de Três Marias (esquerda, 1:10.000) e Furnas (direita, 1:20.000) (MG)
com as setas apontando os braços utilizados para as coletas. Modificado de Miranda & Coutinho,
2004.
PROCEDIMENTOS

56
As amostras foram coletadas a cada dois dias, durante quatro semanas, em dois períodos de
seca (julho/agosto de 2006 e julho/agosto de 2007) e dois de chuvas (janeiro/março de 2007 e
janeiro/março de 2008), considerados os mais contrastantes em termos de variações de temperatura
e precipitação. As coletas foram realizadas no ponto mais profundo da região limnética de cada
braço.
Para as coletas quali-quantitativas da comunidade zooplanctônica, foram realizados arrastos
com rede de plâncton de 68µm de abertura de malha. Devido à presença de “paliteiros” (vegetação
original não removida) nos pontos de coleta, os arrastos foram realizados apenas na zona eufótica,
determinada a partir da leitura do disco de Secchi. Os organismos coletados foram anestesiados com
água mineral gaseificada, corados com corante vital Rosa de Bengala e fixados com solução
neutralizada de formol 4%. Sub-amostras de 1,0 mL foram contadas sob microscópio ótico
Olympus (modelo CBA), em câmaras de Sedgewick-Rafter. Os dados apresentados correspondem
ao número de organismos por m3. Nesta fase, foram quantificados separadamente, os estágios de
náuplios, copepoditos e adultos das espécies de Copepoda. Em cada sub-amostra analisada foram
quantificadas as fêmeas ovadas, bem como o número de ovos/embriões por fêmeas.
Para este estudo foram consideradas apenas as espécies de microcrustáceos mais abundantes
(aquelas que contribuíram com pelos menos 5% da comunidade durante os períodos amostrados):
Thermocyclops minutus, Bominopsis deitersi, Bosmina hagmanni, Ceriodaphnia cornuta e Moina
minuta no reservatório de Três Marias, e no reservatório de Furnas além destas, foram também
consideradas Notodiaptomus henseni, Daphnia ambigua, Ceriodaphnia silvestrii, Diaphanosoma
spinulosum, D. fluviatile e Bosmina freyi (Artigo 1).
Neste trabalho a produtividade secundária foi estimada segundo Hart (1987), utilizando-se
da taxa finita de natalidade per capita, β, estimada pela fórmula:
β = E/N.De
Onde:
E = densidade de ovos
N = densidade da população
De = tempo de desenvolvimento embrionário
Para estimativa do tempo de desenvolvimento embrionário, os espécimes foram cultivados
em laboratório em placas de acrílico multiescavadas, contendo apenas água do reservatório filtrada
em rede de 20 µm. As culturas foram mantidas em câmara incubadora (Eletrolab modelo EL202) a
22 e 26°C ±1°C (temperaturas próximas às médias dos períodos de seca e chuva nos reservatórios)
e fotoperíodo de 12 horas. A água das culturas foi substituída diariamente e as observações

57
ocorreram a cada 12 horas, e as fêmeas foram acompanhadas desde a produção dos ovos até a sua
eclosão.
Para T. minutus, N. henseni, B. freyi, B. hagmanni e B. deitersi, cujo tempo de
desenvolvimento embrionário não pode ser estimado, foi utilizada a expressão matemática proposta
por Bottrell et al., (1976), para Cyclopoida, Calanoida e outros Cadocera, respectivamente, e que
relaciona o tempo de desenvolvimento embrionário com a temperatura.
Desta forma, a produtividade secundária (P) foi estimada pelo produto da taxa finita de
natalidade per capita (β) e da biomassa (B) (Artigo 2) (Hart, 1987):
P= β . B
Para comparação, a produtividade secundária das espécies de Copepoda também foi
calculada pelo método de Winberg et al. (1965):
P= Ne.∆ωe + Nn.∆ωn + Nc.∆ωc
Te Tn Tc
Onde:
P = produção em uma unidade de tempo
N = número de indivíduos
∆ω = incremento em peso em cada estágio de desenvolvimento
T = duração de cada estágio de desenvolvimento
e = ovos
n = náuplios
c = copepoditos
Nesta fórmula, assume-se o peso final de um ovo como o peso inicial de um náuplio (pois o
peso inicial de um ovo é considerado zero) e o peso médio de um adulto como o final do
crescimento de copepoditos. Os tempos de desenvolvimento pós embrionário dos copépodes foram
extraídos de Rietzler (1995) e Corrêa et al. (2009), respectivamente.
As diferenças nos valores de produtividade entre braços, reservatórios e períodos de coleta
(seca e chuva) foram testadas através de uma Análise de Variância seguida do teste de Tukey
(Sampaio, 2002), sendo utilizado o programa Statistica 7.0 (StatSoft).

58
RESULTADOS
O tempo de desenvolvimento embrionário seguiu o mesmo padrão para espécies
zooplanctônicas em geral, com maior duração em temperaturas mais baixas (22°C), e para espécies
com maior tamanho corporal (D. ambigua e D. spinulosum) (tabela 2).
Tabela 2 – Tempo de desenvolvimento embrionário (dias) determinado para as espécies de
Cladocera nos reservatórios de Três Marias e Furnas (MG).
Espécies TRÊS MARIAS FURNAS
22 ±1°C 26±1°C 22±1°C 26±1°C
Daphnia ambigua 2,50 2,00
Ceriodaphnia silvestrii 2,00 1,63
Ceriodaphnia cornuta 2,07 1,50 2,00 1,67
Moina minuta 1,94 1,55 2,00 1,64
Diaphanosoma spinulosum 2,33 1,55
Diaphanosoma fluviatile 1,67 1,50
Valores médios de produtividade das principais espécies de microcrustáceos dos
reservatórios de Três Marias e Furnas são apresentandos na tabela 3 e figura 2. A flutuação das
taxas de produtividade é apresentada nos Anexos 11 e 34 a 37. Em termos de densidade e biomassa
T. minutus foi a espécie mais importante no reservatório de Três Marias. Em Furnas, esta espécie
também dominou em termos numéricos, contudo N. henseni, dafinídeos e sidídeos, contribuiram de
forma mais significativa para a biomassa de microcrustáceos (Artigos 1 e 2).
Considerando a produtividade total, maiores taxas foram obtidas para o período de chuva em
ambos os reservatórios (F=28,19; p<0,000) com valores médios nos períodos de seca e chuva de
0,44 e 1,80 mgPS.m-3.d-1 para o reservatório de Três Marias e 1,05 e 3,10 mgPS.m-3.d-1 para o
reservatório de Furnas, respectivamente. Assim como a biomassa (Artigo 2), maiores taxas de
produtividade secundária foram obtidas no reservatório de Furnas (F=9,30; p<0,0001), com maiores
valores no braço Mendonça. Em Três Marias, os maiores valores foram encontrados no braço
Barrão, contudo, diferenças entre braços não foram significativas (p>0,981). Cladocera foi o grupo
que mais contribuiu para a produtividade, variando entre 42 e 81% no reservatório de Três Marias e
47 e 65% no reservatório de Furnas.

59
Figura 2 – Valores médios de densidade (org.m-3.103) (Artigo 1), biomassa (mgPS.m-3) (Artigo 2) e
produtividade (mgPS.m-3.d-1) das principais espécies de microcrustáceos nos braços dos
reservatórios de Três Marias (braços Barrão e Extrema, direita) e Furnas (braços Varjão e
Mendonça, esquerda). s/06 = período de seca de 2006, c/07 = chuva de 2007, s/07 = seca de 2007 e
c/08 = chuva de 2008.
Comparando as espécies em comum entre os reservatórios, à exceção de C. cornuta
(F=2,63; p>0,05), todas apresentaram maiores taxas no reservatório de Três Marias, sendo as
diferenças significativas. Para T. minutus (F=4,89; p<0,003) os valores médios no período de seca e
chuva foram de 0,18 e 0,62 mgPS.m-3.d-1 em Três Marias e 0,13 e 0,39 mgPS.m-3.d-1 em Furnas,
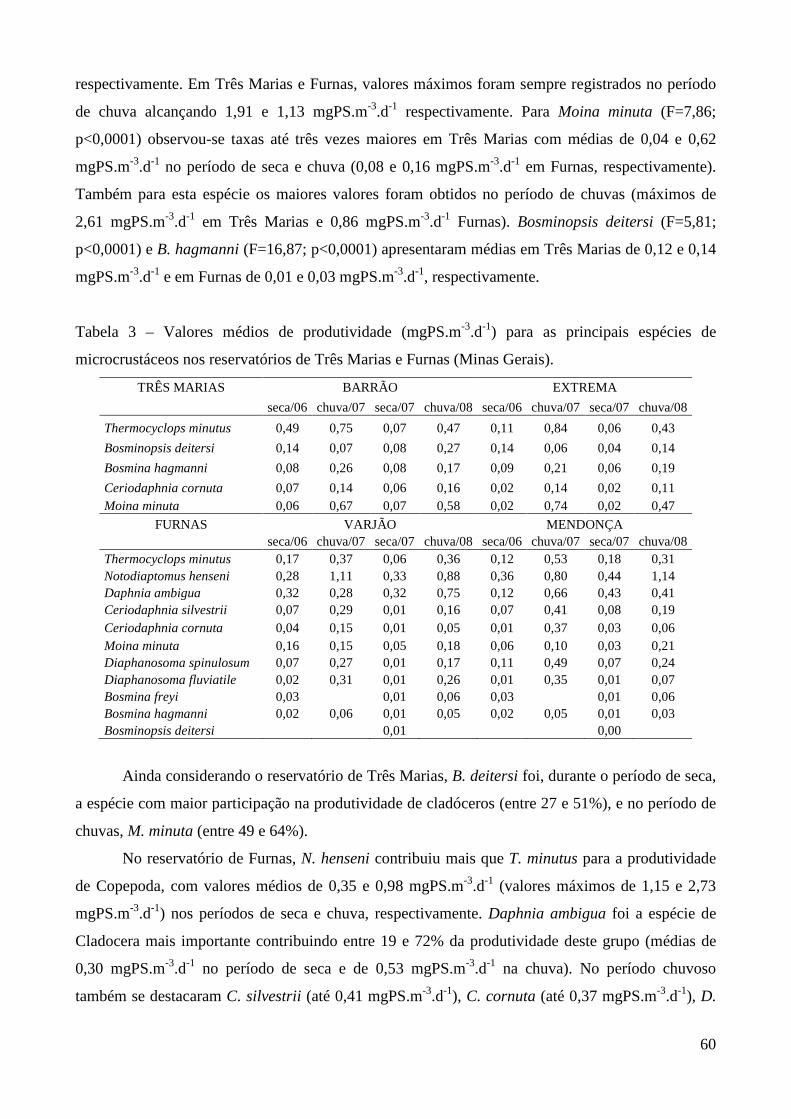
60
respectivamente. Em Três Marias e Furnas, valores máximos foram sempre registrados no período
de chuva alcançando 1,91 e 1,13 mgPS.m-3.d-1 respectivamente. Para Moina minuta (F=7,86;
p<0,0001) observou-se taxas até três vezes maiores em Três Marias com médias de 0,04 e 0,62
mgPS.m-3.d-1 no período de seca e chuva (0,08 e 0,16 mgPS.m-3.d-1 em Furnas, respectivamente).
Também para esta espécie os maiores valores foram obtidos no período de chuvas (máximos de
2,61 mgPS.m-3.d-1 em Três Marias e 0,86 mgPS.m-3.d-1 Furnas). Bosminopsis deitersi (F=5,81;
p<0,0001) e B. hagmanni (F=16,87; p<0,0001) apresentaram médias em Três Marias de 0,12 e 0,14
mgPS.m-3.d-1 e em Furnas de 0,01 e 0,03 mgPS.m-3.d-1, respectivamente.
Tabela 3 – Valores médios de produtividade (mgPS.m-3.d-1) para as principais espécies de
microcrustáceos nos reservatórios de Três Marias e Furnas (Minas Gerais).
TRÊS MARIAS BARRÃO EXTREMA
seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08
Thermocyclops minutus 0,49 0,75 0,07 0,47 0,11 0,84 0,06 0,43
Bosminopsis deitersi 0,14 0,07 0,08 0,27 0,14 0,06 0,04 0,14
Bosmina hagmanni 0,08 0,26 0,08 0,17 0,09 0,21 0,06 0,19
Ceriodaphnia cornuta 0,07 0,14 0,06 0,16 0,02 0,14 0,02 0,11 Moina minuta 0,06 0,67 0,07 0,58 0,02 0,74 0,02 0,47
FURNAS VARJÃO MENDONÇA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08 Thermocyclops minutus 0,17 0,37 0,06 0,36 0,12 0,53 0,18 0,31 Notodiaptomus henseni 0,28 1,11 0,33 0,88 0,36 0,80 0,44 1,14 Daphnia ambigua 0,32 0,28 0,32 0,75 0,12 0,66 0,43 0,41 Ceriodaphnia silvestrii 0,07 0,29 0,01 0,16 0,07 0,41 0,08 0,19 Ceriodaphnia cornuta 0,04 0,15 0,01 0,05 0,01 0,37 0,03 0,06 Moina minuta 0,16 0,15 0,05 0,18 0,06 0,10 0,03 0,21 Diaphanosoma spinulosum 0,07 0,27 0,01 0,17 0,11 0,49 0,07 0,24 Diaphanosoma fluviatile 0,02 0,31 0,01 0,26 0,01 0,35 0,01 0,07 Bosmina freyi 0,03 0,01 0,06 0,03 0,01 0,06 Bosmina hagmanni 0,02 0,06 0,01 0,05 0,02 0,05 0,01 0,03 Bosminopsis deitersi 0,01 0,00
Ainda considerando o reservatório de Três Marias, B. deitersi foi, durante o período de seca,
a espécie com maior participação na produtividade de cladóceros (entre 27 e 51%), e no período de
chuvas, M. minuta (entre 49 e 64%).
No reservatório de Furnas, N. henseni contribuiu mais que T. minutus para a produtividade
de Copepoda, com valores médios de 0,35 e 0,98 mgPS.m-3.d-1 (valores máximos de 1,15 e 2,73
mgPS.m-3.d-1) nos períodos de seca e chuva, respectivamente. Daphnia ambigua foi a espécie de
Cladocera mais importante contribuindo entre 19 e 72% da produtividade deste grupo (médias de
0,30 mgPS.m-3.d-1 no período de seca e de 0,53 mgPS.m-3.d-1 na chuva). No período chuvoso
também se destacaram C. silvestrii (até 0,41 mgPS.m-3.d-1), C. cornuta (até 0,37 mgPS.m-3.d-1), D.
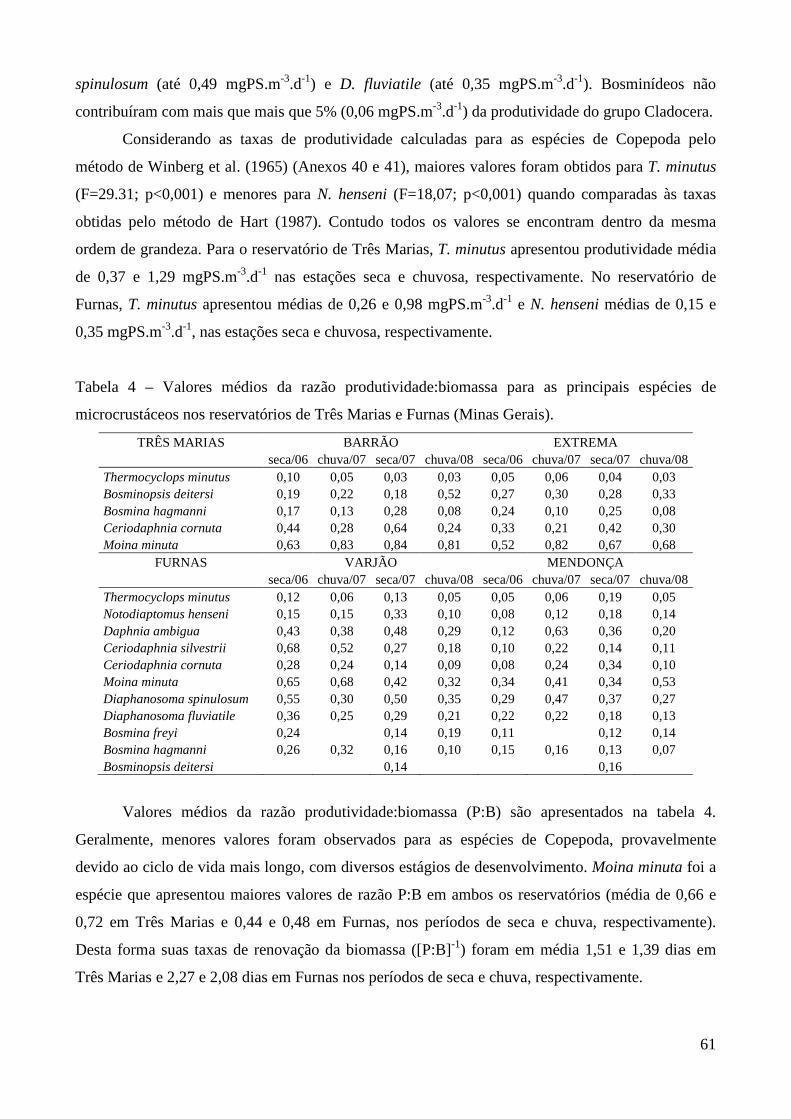
61
spinulosum (até 0,49 mgPS.m-3.d-1) e D. fluviatile (até 0,35 mgPS.m-3.d-1). Bosminídeos não
contribuíram com mais que mais que 5% (0,06 mgPS.m-3.d-1) da produtividade do grupo Cladocera.
Considerando as taxas de produtividade calculadas para as espécies de Copepoda pelo
método de Winberg et al. (1965) (Anexos 40 e 41), maiores valores foram obtidos para T. minutus
(F=29.31; p<0,001) e menores para N. henseni (F=18,07; p<0,001) quando comparadas às taxas
obtidas pelo método de Hart (1987). Contudo todos os valores se encontram dentro da mesma
ordem de grandeza. Para o reservatório de Três Marias, T. minutus apresentou produtividade média
de 0,37 e 1,29 mgPS.m-3.d-1 nas estações seca e chuvosa, respectivamente. No reservatório de
Furnas, T. minutus apresentou médias de 0,26 e 0,98 mgPS.m-3.d-1 e N. henseni médias de 0,15 e
0,35 mgPS.m-3.d-1, nas estações seca e chuvosa, respectivamente.
Tabela 4 – Valores médios da razão produtividade:biomassa para as principais espécies de
microcrustáceos nos reservatórios de Três Marias e Furnas (Minas Gerais).
TRÊS MARIAS BARRÃO EXTREMA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08 Thermocyclops minutus 0,10 0,05 0,03 0,03 0,05 0,06 0,04 0,03 Bosminopsis deitersi 0,19 0,22 0,18 0,52 0,27 0,30 0,28 0,33 Bosmina hagmanni 0,17 0,13 0,28 0,08 0,24 0,10 0,25 0,08 Ceriodaphnia cornuta 0,44 0,28 0,64 0,24 0,33 0,21 0,42 0,30 Moina minuta 0,63 0,83 0,84 0,81 0,52 0,82 0,67 0,68
FURNAS VARJÃO MENDONÇA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08
Thermocyclops minutus 0,12 0,06 0,13 0,05 0,05 0,06 0,19 0,05 Notodiaptomus henseni 0,15 0,15 0,33 0,10 0,08 0,12 0,18 0,14 Daphnia ambigua 0,43 0,38 0,48 0,29 0,12 0,63 0,36 0,20 Ceriodaphnia silvestrii 0,68 0,52 0,27 0,18 0,10 0,22 0,14 0,11 Ceriodaphnia cornuta 0,28 0,24 0,14 0,09 0,08 0,24 0,34 0,10 Moina minuta 0,65 0,68 0,42 0,32 0,34 0,41 0,34 0,53 Diaphanosoma spinulosum 0,55 0,30 0,50 0,35 0,29 0,47 0,37 0,27 Diaphanosoma fluviatile 0,36 0,25 0,29 0,21 0,22 0,22 0,18 0,13 Bosmina freyi 0,24 0,14 0,19 0,11 0,12 0,14 Bosmina hagmanni 0,26 0,32 0,16 0,10 0,15 0,16 0,13 0,07 Bosminopsis deitersi 0,14 0,16
Valores médios da razão produtividade:biomassa (P:B) são apresentados na tabela 4.
Geralmente, menores valores foram observados para as espécies de Copepoda, provavelmente
devido ao ciclo de vida mais longo, com diversos estágios de desenvolvimento. Moina minuta foi a
espécie que apresentou maiores valores de razão P:B em ambos os reservatórios (média de 0,66 e
0,72 em Três Marias e 0,44 e 0,48 em Furnas, nos períodos de seca e chuva, respectivamente).
Desta forma suas taxas de renovação da biomassa ([P:B]-1) foram em média 1,51 e 1,39 dias em
Três Marias e 2,27 e 2,08 dias em Furnas nos períodos de seca e chuva, respectivamente.

62
Ceriodaphnia cornuta e Bosminopsis deitersi também se destacaram no reservatório de Três
Marias com razão P:B de até 0,64 e 0,52 e taxas de renovação de 1,56 e 1,92 dias, respectivamente.
No reservatório de Furnas Daphnia ambigua (0,63), Ceriodaphnia silvestrii (0,68) e Diaphanosoma
spinulosum (0,55) também apresentaram maiores valores de razão P:B, sendo suas taxas de
renovação de 1,58; 1,47 e 1,81 dias, respectivamente.
DISCUSSÃO
Estudos envolvendo a produtividade secundária de espécies zooplanctônicas nos
ecossistemas aquáticos tropicais permitem elucidar a dinâmica destes ambientes uma vez que este
grupo é um elo importante entre organismos produtores e níveis superiores.
Os valores de tempo de desenvolvimento embrionário obtidos neste estudo foram muito
próximos ao de outros trabalhos realizados no Brasil. Para D. ambigua, Rocha & Matsumura-
Tundisi (1990 apud Melão 1999b) também obtiveram tempo de desenvolvimento de 2,0 dias, para
uma temperatura de 25°C.
Valores próximos também foram obtidos por Santos et al., (2006) para C. silvestrii, a 25°C,
variando entre 1,61 e 1,73 dias em tratamentos com diferentes meios e fontes de alimento. Para esta
mesma espécie, Rietzler (1998 apud Melão, 1999b) obteve valor maior para 20°C (2,32 dias) e
menor para 25°C (1,33 dias). Para C. cornuta, Melão (1997) observou tempos de desenvolvimento
de 3,24 e 1,66 dias (a 20 e 25°C, respectivamente); enquanto Maia-Barbosa (2000) observou 1,57
dias e para M. minuta 1,84 dias (a 25°C). Ambas as autoras encontraram valores muito próximos
para o tempo de desenvolvimento de B. deitersi a 25°C (1,21 e 1,10 dias, para Melão, 2006 e Maia-
Barbosa, op. cit., respectivamente).
As fórmulas para o tempo de desenvolvimento embrionário propostas por Bottrell et al.
(1976), que são generalizações para grandes grupos, não são consideradas adequadas para
cladóceros e copépodes de águas continentais brasileiras, típicamente tropicais. Os trabalhos
realizados no Brasil, apesar das diferentes condições nas quais foram desenvolvidos, utilizando
diferentes meios de cultura; água natural ou água reconstituída e com ou sem adição algas,
especialmente clorofíceas, geraram respostas próximas ntre si, considerando-se as mesmas espécies,
permitindo que padrões sejam delineados para as mesmas.
Vários autores (Margalef, 1983; Melão, 1999a) afirmam que temperatura e alimento
disponível são os principais fatores que afetam a produtividade do zooplâncton. Nos reservatórios
de Três Marias e Furnas, maiores valores de densidade, biomassa e produtividade foram observados
no período de chuvas. Temperaturas mais altas reduzem o tempo de geração e o tamanho de corpo
fazendo que os indivíduos alcancem mais rapidamente a idade reprodutiva e aumentando suas

63
densidades em poucas semanas (Gillooly, 2000; Gillooly et al., 2002). Além disso, a maior
disponibilidade de alimento (medida através da clorofila-a e sólidos totais e orgânicos em
suspensão) bem como um fitoplâncton de mais fácil digestibilidade (Artigo 1) permitiram um
melhor estado nutricional dos organismos, aumentando a proporção de fêmeas ovadas (Klein-
Breteler et al., 1990; Melão, 1999b; Santos et al., 2006), especialmente para as espécies de
Cladocera (Anexos 38 e 39).
Apesar das maiores densidades terem sido registradas no reservatório de Três Marias, a
maior produtividade das espécies de microcrustáceos foi observada no reservatório de Furnas. A
produtividade primária (estimada nos mesmos braços e nos mesmos períodos que o presente estudo)
apresentou valores muito mais elevados também no reservatório de Três Marias (entre 4,68 e
4.831,78 mgC.m-2.dia-1 e entre 4,27 e 100,89 mgC.m-2.dia-1 no reservatório de Furnas segundo
dados de Carvalhais-Júnior et al., 2007). Segundo Hillbricht-Ilkowska (1972, apud Pederson et al.,
1976) a eficiência na transferência de energia entre a produtividade primária e a secundária tende a
diminuir com o aumento do estado trófico uma vez que ambientes mais eutrofizados apresentam
altas densidades de cianobactérias e algas de maior tamanho. Apesar de considerado oligotrófico
(Sampaio & López, 2003), no reservatório de Três Marias, cianobactérias representaram mais de
80% do fitoplâncton (Artigo 1), sendo este grupo de algas de baixa digestibilidadde para o
zooplâncton (Lampert, 1987). No reservatório de Furnas, devido à maior contribuição de
clorofíceas, bacilariofíceas e crisofíceas (Campos, 2008), o zooplâncton poderia assimilar mais
eficientemente a energa produzida pelo fitoplâncton e apresentar maiores valores de produtividade
neste ambiente.
Dentre as espécies de cladóceros do reservatório de Três Marias, Moina minuta apresentou
os maiores valores de produtividade. A temperatura teve um papel importante, uma vez que a
proporção de fêmeas ovadas (média de 27%; máximo de 37%) e o número médio de ovos por
fêmea (média de 1,38 ovos, máximo de 2,0 ovos) foram maiores no período de chuva. Além disso,
a maior concentração de sólidos orgânicos dissolvidos no reservatório de Três Marias pode indicar
uma fonte adicional de alimento para esta espécie, como o observado para M. micrura por Ferrão-
Filho et al. (2005). Estes autores constataram que apenas esta espécie apresentou taxas de
crescimento similares ao tratamento controle, quando fontes de alimento diferentes foram ofertadas:
somente clorofíceas, somente séston ou séston + clorofíceas. Além disso, Sipaúba-Tavares & Rocha
(2001) apontam as espécies de Moina como as mais adequadas para aqüicultura devido à sua
facilidade de produção.
Bosminopsis deitersi participou com 43% da produtividade da estação impactada do lago
Batata (PA) (Maia-Barbosa, 2000), sendo a principal espécie no reservatório Lagoa Dourada (SP)
onde participou com 92,41 e 97,36% da produtividade (Melão & Rocha, 2006). Em Três Marias,

64
representou até 51% da produtividade no período de seca, estando sempre presente em todas as
amostras coletadas, ao contrário de Furnas, onde foi pouco abundante ou mesmo ausente. Melão &
Rocha (op. cit.) citam porcentagens entre 17 e 28% de fêmeas ovadas para esta espécie, sendo que
em Três Marias os valores variaram entre 8 e 30%.
Enquanto Maia-Barbosa (op.cit.) observou uma maior contribuição de B. hagmanni (0,88
mgPS.m-3.d-1) e D. birgei (0,77 mgPS.m-3.d-1) na produtividade secundária da estação natural do
lago Batata (PA), neste estudo (com exceção de B. deitersi no reservatório de Três Marias)
bosminídeos e C. cornuta foram as espécies que menos contribuíram para a produtividade
secundária.
Apesar da maior densidade e biomassa, T. minutus apresentou menores valores de
produtividade em ambos os reservatórios, provavelmente devido à menor prole em relação a outras
espécies de Copepoda. Segundo Reid e Pinto Coelho (1994), apesar de se tratar de uma espécie de
menor tamanho corporal, seu potencial reprodutivo é reduzido, com número médio de 5,7 ovos por
fêmea ovada. De fato, médias menores que a citada por aqueles autores foram registradas nos
reservatórios de Três Marias e Furnas (5,3 e 4,2 ovos por fêmea, respectivamente) (Anexo 39). No
reservatório Lagoa Dourada Tropocyclops prasinus, outra espécie de Cyclopoida com tamanho de
corpo semelhante, apresentou valores de produtividade maiores (até 2,8 mgPS.m-3.d-1 durante o
verão), e também maior número de ovos por fêmea, entre 10,7 e 12,7 (Melão & Rocha, 2004).
Notodiaptomus henseni, apesar de apresentar menor número de ovos (média de 2,4) sempre
apresentou maior proporção de fêmeas ovadas que T. minutus em Furnas (0,27 e 0,19,
respectivamente) contribuindo para maiores valores da taxa finita de natalidade per capita (β) e
conseqüentemente maiores taxas de produtividade, especialmente no período de chuvas.
As espécies de Cladocera de maior tamanho corporal que foram dominantes em biomassa e
produtividade no reservatório de Furnas (D. ambigua, C. silvestri, D. spinulosum e D. fluviatile)
geralmente investem menos em crescimento corporal, produzindo ninhadas maiores (Melão,
1999b), apresentando desta forma uma vantagem competitiva sobre as demais espécies de
cladóceros. Estas espécies também apresentaram as maiores razões produtividade:biomassa em
relação às demais espécies de microcrustáceos deste reservatório.
No geral, os valores de produtividade secundária, registrados para os reservatórios de Três
Marias e Furnas, estiveram dentro da mesma ordem de grandeza que os observados para outros
ambientes oligotróficos como a estação natural do lago Batata (PA), onde Maia-Barbosa (2000)
observou média anual de 2,72 mgPS.m-3.d-1 (média anual da estação impactada: 7,48 mgPS.m-3.d-1).
No reservatório Lagoa Dourada (SP), Melão et al. (2005) apresentaram para Cladocera, valores de
0,455 e 1,970 mgPS.m-3.d-1 e para Copepoda de 0,152 e 1,428 mgPS.m-3.d-1 (no inverno e verão,
respectivamente). Opostamente, Rietzler et al. (2004) encontraram valores no verão e inverno de

65
61,15 e 22,97 mgPS.m-3.d-1 para Cladocera e 46,22 e 53,55 mgPS.m-3.d-1 para Copepoda no
reservatório eutrófico de Salto Grande (SP). Esta relação entre produtividade do zooplâncton e
estado trófico é recorrente em outros ecossistemas aquáticos brasileiros, uma vez que ambientes
eutrofizados apresentam concentrações mais elevadas de nutrientes, detritos, bacterioplâncton,
biomassa algal e maiores taxas de produtividade primária, podendo sustentar uma maior produção
do zooplâncton (Rocha et al., 1995 e Rocha et al., 1997).
As diferenças nas taxas de produtividade obtidas pelos métodos de Winberg et al. (1965) e
Hart (1987) mostram que mais comparações devem ser realizadas antes de uma conclusão
definitiva. No método de Hart (op. cit.) toda a biomassa da população é utilizada no cálculo
enquanto no método de Winberg et al. (op. cit.) são utilizados os incrementos de peso em cada
estágio de desenvolvimento. Além disso, os estágios de desenvolvimento pós-embrionários
utilizados no cálculo foram obtidos em temperaturas diferentes das médias dos períodos de seca e
chuva observadas nos reservatórios de Três Marias e Furnas. Rietzler (1995) cultivou os copépodes
a 18, 23 e 28°C fornecendo como alimento protozoário e fitoplâncton, Corrêa et al. (2009) não
citaram no resumo as condições de temperatura e alimento utilizadas no cultivo, e neste trabalho
foram usadas 22 e 26°C, com água dos reservatórios filtrada em rede de 20 µm. Segundo Bottrell et
al. (1976) a temperatura representa 80 a 90% da variação no desenvolvimento embrionário, contudo
para o desenvolvimento pós-embrionário o alimento fornecido é tão importante quanto a
temperatura.
Considerando os valores de biomassa e produtividade avaliados neste estudo, constata-se
que Cladocera poderia ser uma melhor fonte de alimento para a ictiofauna, especialmente no
reservatório de Furnas, onde estes parâmetros apresentaram valores maiores. Além disso, este grupo
de microcrustáceos é considerado presa preferencial de peixes uma vez que Copepoda é conhecido
por sua capacidade de fuga através de movimentos rápidos e erráticos (Roche & Rocha, 2005).
Moina minuta em Três Marias, e dafinídeos e sidídeos em Furnas, são consideradas as melhores
espécies para aqüicultura não só por sua facilidade de cultivo (Sipaúba-Tavares & Rocha, 2001)
como também pelas maiores taxas de produtividade observadas neste estudo.
AGRADECIMENTOS
À toda a equipe dos laboratórios de Ecologia do Zooplâncton, de Gestão Ambiental de
Reservatórios e de Limnologia, que direta ou indiretamente ajudaram na realização deste trabalho.
Ao Projeto Parques Aqüícolas (Convênio 8713 SEAP/SECTES/FUNDEP-UFMG) e ao CNPq
(Processo n° 142079/2006-5) pelo financiamento do projeto.

66
REFERÊNCIAS BIBLIOGRÁFICAS
ALLAN, J.D. & CASTILHO, M.M. 2007. Stream Ecology. Structure and function of running waters. Dordrecht: Springer. 436p.
ALLEN, K.R. 1951. The Horowiki Stream. N. Z. Mar. Dept., Fish. Bull., 10. 238p. BOTTRELL, H.H.; DUNCAN, A.; GLIWICZ, Z.M.; GRYGIEREK, E.; HERZIG, A.;
HILLBRICHT-ILKOWSKA, A.; KURASAUA, H.; LARSSON, P. & WEGLENSKA, T. 1976. A review of some problems in zooplankton production studies. Norw. J. Zoo., 24: 419-456.
BURGIS, M. J. 1974. Revised estimates for the biomass and production of zooplankton in Lake George, Uganda. Freshwater Biol., 4, 535–541.
CAMPOS, C.A. 2008. Estudo da diversidade de Cyanobacteria do reservatório de Furnas (MG, Brasil) por meio de caracterização morfológica e molecular. Universidade Federal de Minas Gerais. 81p. Dissertação.
CARVALHAIS-JÚNIOR, A. C., MARTINS, F. G. & BARBOSA, F.A.R. 2007. Estimativa da Produtividade Primária Fitoplanctônica dos Reservatórios da UHE Três Marias e UHE Furnas. Projeto Parques Aqüícolas, UFMG/SECTES/SEAP. Belo Horizonte. 24p.
CEMIG - Compania Energética de Minas Gerais. 2009. Usinas da CEMIG. http://www.cemig.com.br/institucional/usinas/11g.asp. Acesso em: 16 set. 2009.
CORRÊA, A.C.G., MENDONÇA-GALVÃO, L., MAGALHÃES, D.M., NASCIMENTO, T.R., DALL’ASTTA, R.C. 2009. Ciclo de vida de Notodiaptomus cearensis Wright, 1936 (Copepoda, Calanoida). XII Congresso Brasileiro de Limnologia. Gramado, RS. p.316.
EDMONDSON W.T. & WINBERG, G.G. 1971. A manual on methods for the assessment of secondary productivity in fresh waters. Oxford: IBP Handbook Nº 17. Blackwell. 358 pp.
EDMONDSON, W. T. 1974. Secondary Production. Verh. Internat. Verein. Limnol., 20:229-272. FERRÃO-FILHO, A. S., ARCIFA, M. S. & FILETO, C. 2005. Influence of seston quantity and
quality on growth of tropical cladocerans. Braz. J. Biol., 65(1): 77-89. FURNAS – Furnas Centrais Elétricas SA. 2009. Sistema Furnas.
http://www.furnas.com.br/hotsites/sistemafurnas. Acesso em: 16 set. 2009. GILLOOLY, J.F. 2000. Effect of body size and temperature on generation time in zooplankton. J.
Plankton Res., 22, 241-251. GILLOOLY, J.; CHARNOV, E.; WETS, G.; SAVAGE, V. & BROWN, J. 2002. Effects of size and
temperature on developmental time. Nature, 417:70-73. HART, R.C. & ALLANSON, B.R. 1975. Preliminary estimates of production by a calanoid
copepod in subtropical Lake Sibaya. Verh. Internat. Verein. Limnol. 19:1434-1441. HART, R. C. 1987. Population dynamics and production of five crustacean zooplankters in a
subtropical reservoir during years of contrasting turbidity. Freshwater Biol., 18:287-318. KLEIN BRETELER, W. C. M.; SCHOGT, N. & GONZALES, S. R. 1990. On the role of food
quality in grazing and development of life stages, and genetic change of body size during cultivation of pelagic copepods. Journal of Experimental Marine Biology and Ecology, 135:177-189.
LAMPERT, W., 1987, Laboratory studies on zooplankton cyanobacteria interactions. N.Z. J. Mar. Freshwater Res., 21: 483-490.
LAMPERT, W. & SOMMER,U. 2007. Limnoecology. The Ecology of Lakes and Streams. 2nd ed. Oxford: Oxford University Press. 324p.
MAIA-BARBOSA, P. M. 2000. Ecologia de Cinco Espécies de Cladóceros de um Lago Amazônico Impactado por Rejeito de Bauxita (Lago Batata, Pará-Brasil). Universidade Federal do Rio de Janeiro. 218p. Tese.
MARGALEF, R. 1983. Limnologia. Barcelona: Omega. 1010pp. MAVUTI, K. M. 1994. Durations of development and production estimates by two crustacean
zooplankton species, Thermocyclops oblongatus Sars (Copepoda) and Diaphanosoma excisum

67
Sars (Cladocera), in Lake Naivasha, Kenya. In: Dumont, H. J., Green, J. and Masudire, H. (eds), Studies on the Ecology of Tropical Zooplankton. Hydrobiologia, 272, 27–38.
MELÃO, M. G. 1997. A comunidade planctônica (Fitoplâncton e Zooplâncton) e produtividade secundária do zooplâncton de um reservatório oligotrófico. Universidade Federal de São Carlos. Tese.
MELÃO, M. G. & ROCHA, O. 2004. Life history, biomass and production of two planktonic cyclopoid copepods in a shallow subtropical reservoir. J. Plankton Res., 26(8):909-923.
MELÃO, M. G. & ROCHA, O. 2006. Life history, populations dynamics, standing biomass and production of Bosminopsis deitersi (Cladocera) in a shallow tropical reservoir. Acta Limnol. Bras., 18(4):433-450.
MELÃO, M.G., ROCHA, O., & ROCHE, F. K. 2005. Produtividade, Biomassa e Flutuações Populacionais e interações biológicas da Comunidade Planctônica e Suas Implicações na Transferência de Energia na Cadeia Alimentar de um Reservatório Raso e oligotrófico. In: Roche, F. K. & Rocha, O. (eds.) Ecologia Trófica de Peixes com ênfase na planctivoria em ambientes lênticos de água doce no Brasil. São Carlos: Rima. 146p.
MELÃO, M.G.G. 1999a. A Produtividade Secundária do Zooplâncton: Métodos, Implicações e um estado na Lagoa Dourada. In: Henry, R. (ed) Ecologia dos Reservatórios: Estrutura, Função e Aspectos Sociais. Botucatu: FUNDIBIO, FAPESP. 799p.
MELÃO, M.G.G. 1999b. Desenvolvimento e aspectos reprodutivos de cladóceros e copépodes de águas continentais brasileiras. In: Pompêo, M.L.M. (ed) Perspectivas da Limnologia no Brasil. São Luís: União. 189p.
MIRANDA, E. E. & COUTINHO, A. C. (Coord.) 2004. Brasil Visto do Espaço. Campinas: Embrapa Monitoramento por Satélite. http://www.cdbrasil.cnpm.embrapa.br Acesso em: 16 set. 2009.
NOGUEIRA, D.J.; CASTRO, S.C. & RIGOLIN DE SÁ, O. 2008. Avaliação da qualidade da água no reservatório UHE Furnas - MG, utilizando as brânquias de Pimelodus maculatus (LACÈPÉDE, 1803) como biomarcador de poluição ambiental. Ciência et Praxis, 1(1): 15-20.
OKANO, Y. W., 1994. Análise da estrutura e dinâmica populacional da comunidade zooplanctônica de um reservatório artificial (reservatório Monjolinho, São Carlos, SP). Universidade Federal de São Carlos. 128p. Tese.
PELÁEZ-RODRÍGUEZ, M. & MATSUMURA-TUNDISI, T., 2002. Rotifer Production in a Shallow Artificial Lake (Lobo-Broa Reservoir, SP, Brazil). Braz. J. Biol., 62(3): 509-516.
PEDERSON, G. L., WELCH, E. B. & LITT, A. H. 1976. Plankton secondary productivity and biomass: their relation to lake trophic state. Hydrobiologia, 50(2):129 - 144.
REID, J. W. & PINTO-COELHO, R. M. 1994. Planktonic Copepoda of Furnas Reservoir: Initial Survey of Species (1993) and Review of Literature. In: Pinto-Coelho, R.M.; Giani, A. & von Sperling, E. (eds) Ecology and Human impact on lakes and reservoirs in Minas Gerais with special reference to future development and management strategies. Belo Horizonte: SEGRAC. 193p.
RIETZLER, A.C. 1995. Alimentação, ciclo de vida e análise da coexistência de espécies de Cyclopoida na Represa de Barra Bonita. Universidade Federal de São Carlos. 385p. Tese.
RIETZLER, A.C.; ROCHA, O. & ESPINDOLA, E. L. G. 2004. Produção Secundária de Espécies do Zooplâncton do Reservatório de Salto Grande, Estado de São Paulo. In: Espindola, E. L.G., Leite, M.A., Dornfeld, C.B. (eds.) Reservatório de Salto Grande (Americana-SP): caracterização, impactos e propostas de manejo. São Carlos: Rima.
ROCHA, O. & MATSUMURA-TUNDISI, T., 1984. Biomass and production of Argyrodiaptomus furcatus a tropical calanoid copepod in Broa reservoir, southern Brazil. Hydrobiologia, 113: 307-311.
ROCHA, O.; SENDACZ, S. & MATSUMURA-TUNDISI, T. 1995. Composition, Biomass and Productivity of Zooplankton in Natural Lakes and Reservoirs of Brazil. In: Tundisi, J. G.; Bicudo, C. E. M. & Matsumura-Tundisi, T. (eds) Limnology in Brazil. Rio de Janeiro: Brazilian Academy of Sciences, Brazilian Limnological Society. 376p.

68
ROCHA, O.; MATSUMURA-TUNDISI, T. & SAMPAIO, E. V. 1997. Phytoplankton and zooplankton community structure and production as related to trophic state in some Brazilian lakes and reservoirs. Verh. Internat. Verein Limnol., 26:599-604.
ROCHA, O.; MATSUMURA-TUNDISI, T.; ESPÍNDOLA, E.L.G.; ROCHE, K.F. & RIETZLER, A.C. 1999. Ecological Theory Applied to Reservoir Zooplankton. In: Tundisi, J.G. & Straskraba, M. (eds.) Theoretical Reservoir Ecology and its Applications. São Carlos: IIE. 585p.
ROCHE, F. K. & ROCHA, O. 2005. Aspectos de predação por peixes em lagos e represas, com enfoque na planctivoria. In: Roche, F. K. & Rocha, O. (eds.) Ecologia Trófica de Peixes com ênfase na planctivoria em ambientes lênticos de água doce no Brasil. São Carlos: Rima. 146p.
SAMPAIO, I.B.M. 2002. Estatística Aplicada à Experimentação Animal. Belo Horizonte: Fundação de Ensino e Pesquisa em Medicina Veterinária e Zootecnia. 2ª ed. 256p.
SAMPAIO, E.V. & LÓPEZ, C. M. 2003. Limnologias física, química e biológica da represa de Três Marias e do São Francisco. In: Godinho, H. P. & Godinho, A. L. (org.) Águas, peixes e pescadores do São Francisco das Minas Gerais. Belo Hrizonte: PUCMinas. 468p.
SANTOS, M.A.P.F., MELÃO, M.G.G. & LOMBARDI, A.T. 2006. Life history characteristics and production of Ceriodaphnia silvestrii Daday (Crustacea, Cladocera) under different experimental conditions. Acta Limnol. Bras., 18(2):199-212.
SANTOS-WISNIEWSKI, M.J. & ROCHA, O. 2007. Spatial distribution and secondary production of Copepoda in a tropical reservoir: Barra Bonita, SP, Brazil. Braz. J. Biol., 67(2): 223-233.
SIPAÚBA-TAVARES, L. H. & ROCHA, O. 2001. Produção de Plâncton (Fitoplâncton e Zooplâncton) para alimentação de organismos aquáticos. São Carlos: Rima. 106p.
WINBERG, G. C., PECHEN, G. A. AND SHUSHKINA, E. A. 1965. Production of planktonic crustaceans in three lakes of different type. Zool. Zh., 44, 676–687.

69
DISCUSSÃO GERAL
O projeto Parques Aqüícolas, visando incrementar a produção pesqueira em todo o país de
maneira rápida e a baixo custo para os produtores rurais, incentiva a criação de peixes em tanques-
rede, uma vez que neste tipo de produção, os custos para instalação e manutenção são muito
menores, se comparados à criação de peixes em viveiros em terra (MPA, 2009). A espécie de peixe
mais utilizada para cultivo em tanques-rede tem sido a tilápia do Nilo (Oreochromis niloticus)
devido a sua rusticidade, precocidade e hábito alimentar onívoro, boa aceitação de sua carne,
elevado valor comercial e excelente conversão alimentar (Sampaio & Braga, 2005; Marengoni,
2006). A criação em tanques-rede, por se tratar de um cultivo intensivo com elevada dependência
do fornecimento de ração, e realizada dentro de reservatórios, é considerada uma atividade
impactante podendo interferir rapidamente sobre a qualidade de água uma vez que os resíduos
(fezes, urina e sobras de ração) são lançados diretamente no ambiente aquático, sem possibilidade
de tratamento (Araripe et al., 2006).
Desta forma a estimativa da capacidade suporte e avaliação dos efeitos da criação em
tanques-rede nos reservatórios são fundamentais para a manutenção da qualidade de suas águas e da
sustentabilidade do empreendimento. Estudos sobre o efeito da criação de tilápias em tanques-rede
sobre a biota aquática ainda são escassos. Contudo merece destaque o trabalho realizado no
reservatório de Rosana (PR), onde as comunidades aquáticas foram monitoradas durante um ciclo
de criação de tilápias do Nilo, entre abril e agosto de 2006, nos braços do Corvo e Guairacá. Para o
fitoplâncton, Borges (2009) constatou o aumento na densidade e biomassa que foram atribuídos à
incorporação de nutrientes liberados pelo cultivo de tilápias. Segundo a autora, o estado trófico
(considerando o biovolume fitoplanctônico) passou de oligotrófico para hipereutrófico com
aumento na proporção de cianobactérias no decorrer do experimento.
Para o perifíton, Fonseca et al. (2009) observaram redução no número de espécies e na
abundância tanto nos pontos à montante quanto à jusante do experimento. No braço Guairacá,
diatomáceas e desmídias (favorecidas em ambientes com baixas concentrações de fósforo) tiveram
sua abundância reduzida e cianobactérias aumentaram sua contribuição em todos os pontos à
jusante dos tanques-rede. A comunidade zooplanctônica, analisada apenas no braço do Guairacá
(Dias, 2008), mostrou uma redução da riqueza logo após o início do experimento, com tendência ao
aumento no final. Rotíferos (cuja abundância foi relacionada à mesotrofia no final do experimento)
foram significativamente favorecidos pela instalação dos tanques-rede, enquanto cladóceros e
copépodes foram negativamente influenciados.
Para o reservatório de Furnas, os efeitos da criação de tilápias do Nilo em tanques-rede
sobre a qualidade da água foram avaliados por Rocha (2001) e sobre a comunidade fitoplanctônica

70
por Figueredo & Giani (2005). Considerando a qualidade da água não foram observadas diferenças
significativas para os diversos parâmetros amostrados, contudo foram constatados aumentos nos
valores de condutividade elétrica e concentrações de amônia nos pontos próximos aos tanques-rede.
Em mesocosmos de polietileno transparente onde as tilápias foram arraçoadas por duas semanas,
Rocha (op.cit.) observou aumento nas concentrações clorofila-a, amônio e fósforo total e
diminuição na transparência da água.
Com relação à comunidade fitoplanctônica, Figueredo & Giani (op.cit.) mantiveram juvenis
de tilápia em mesocosmos de polietileno transparente durante 28 dias sem adição de alimento. Em
todos os mesocosmos com tilápias, cianobactérias foram excluídas ou drasticamente reduzidas
enquanto Chlorophyta se tornou o grupo dominante. Segundo os autores, o efeito “top-down”
(herbivoria sobre algas filamenosas e coloniais) foi mais pronunciado que o “bottom-up” (aumento
na biomassa fitoplanctônica como consequência da excreção das tilápias).
Os dados obtidos nos reservatórios de Rosana (PR) e de Furnas (MG) mostram os cuidados
que se deve ter com a implantação empreendimento deste tipo, já que os reflexos sobre a qualidade
da água são rápidos e difíceis de serem revertidos. O monitoramento de longo prazo dos cultivos foi
recomendado para avaliar os reais efeitos desta atividade sobre a biota aquática e para que ajustes
(no arraçoamento, na densidade de peixes em cada gaiola ou número de tanques rede em cada lote
de água) possam ser feitos sempre que necessário.
Com relação à capacidade suporte, o Estudo Técnico-Científico Visando a Delimitação de
Parques Aqüícolas nos Lagos das Usinas Hidroelétricas de Furnas e Três Marias – MG, estimou a
capacidade suporte de 35.875 ton de peixes para o reservatório de Três Marias e 79.318 ton para o
reservatório de Furnas (Starling et al., 2007a e b).
A comunidade zooplanctônica tem um papel fundamental no início de cada ciclo de criação,
uma vez que juvenis de tilápia são adicionados aos tanques-rede com tamanhos entre 13 e 15 cm,
fase em que mesmo aceitando a ração, o plâncton ainda é considerado um alimento importante
(Sipaúba-Tavares et al., 2006, Negreiros et al., 2009). Uma vez que a ração não supre todas as
necessidades nutricionais, o zooplâncton continua sendo uma fonte rica em ácidos graxos e
vitaminas essenciais para o desenvolvimento das larvas de peixe (Sipaúba-Tavares & Rocha, 2001).
Em cultivos comerciais, a mortalidade de alevinos e peixes jovens pode chegar a 90%, sendo este
percentual reduzido quando além da ração é ofertado zooplâncton (Sá-Júnior, 1994). Nos
reservatórios de Três Marias e Furnas, os maiores valores de biomassa e produtividade de
cladóceros indicam uma fonte importante de alimento, tanto para as formas jovens de tilápias
quanto para toda a ictiofauna, devido à maior digestibilidade e qualidade nutricional deste grupo de
microcrustáceos (Roche & Rocha, 2005).

71
Ao longo de todo o empreendimento, o zooplâncton (somado a outros fatores abióticos e
bióticos) poderá auxiliar no monitoramento dos cultivos. Uma vez que este estudo levantou a
densidade de toda a comunidade, além da magnitude da biomassa e produtividade secundária de
microcrustáceos antes da implantação dos tanques-rede, comparações posteriores destes parâmetros
permitirão detectar impactos da criação de peixes (eutrofização, alterações na estrutura da
comunidade, diminuição na eficiência da transferência de energia) sobre a qualidade das águas dos
reservatórios.
O reservatório de Furnas, por apresentar águas de melhor qualidade, com maior
transparência, menores concentrações de nutrientes além de um zooplâncton de maior porte (sendo
melhor fonte alimentar), permitirá uma maior sustentatibilidade da criação de peixes em tanques-
rede do que o reservatório de Três Marias. Este fato é amparado pelo maior número de lotes de água
elencados pelo projeto e oferecidos pelo governo neste reservatório (1.527 lotes) em comparação ao
reservatório de Três Marias (543 lotes) (MPA, 2009).

72
CONCLUSÕES
- As diferenças nas variáveis ambientais, na densidade do zooplâncton, na biomassa e
produtividade dos microcrustáceos foram maiores entre reservatórios e sazonalmente (efeito da
temperatura e precipitação) do que entre braços dentro de um mesmo reservatório.
- A comunidade zooplanctônica mostrou-se como um importante indicador de qualidade de
água, uma vez que no reservatório de Três Marias o predomíno de ciclopóides e espécies de menor
tamanho corporal bosminídeos e altas densidades de rotíferos, caracterizou este ambiente como
mesotrófico. O reservatório de Furnas, por outro lado, apresentou maiores densidades de espécies
de maior porte como calanóides, dafinídeos e sidídeos, típicos de ambientes oligotróficos.
- Diferenças marcantes na biomassa de microcrustáceos foram observadas entre os dois
reservatórios. Em Três Marias Thermocyclops minutus, foi a espécie mais importante em termos de
densidade, representando até 80% da biomassa. Em Furnas, Copepoda também foi o grupo
dominante em densidade e biomassa, contudo N. henseni contribuiu na mesma proporção que T.
minutus. A maior biomassa de Cladocera neste reservatório deve-se principalmente às espécies de
maior tamanho corporal como D. ambigua, C. silvestrii, D. spinulosum e D. fluviatile,
especialmente durante o período de chuva.
- Apesar das maiores densidades terem sido registradas no reservatório de Três Marias,
maiores valores de biomassa e produtividade das espécies de microcrustáceos foram observados no
reservatório de Furnas. No reservatório de Três Marias, M. minuta apresentou as maiores taxas de
produtividade e razão P:B principalmente no período de chuvas. No reservatório de Furnas, N.
henseni apresentou maiores taxas de produtividade que T. minutus, enquanto M. minuta, D.
ambigua, C. silvestrii e D. spinulosum apresentaram as maiores razões P:B.
- Uma vez que a estrutura da comunidade zooplanctônica, bem como a biomassa e
produtividade de microcrustácos foram estimadas antes da implantação dos tanques-rede,
comparações com dados futuros permitirão avaliar o impacto desta atividade sobre a qualidade da
água nos braços dos reservatórios de Três Marias e Furnas.
- As regressões peso-comprimento obtidas para as espécies de microcrustáceos permitirão
que a biomassa e produtvidade sejam estimadas para outros ecossistemas aquáticos tropicais
brasileiros aumentando o conhecimento sobre a comunidade zooplanctônica neste tipo de ambiente.

73
REFERÊNCIAS BIBLIOGRÁFICAS
ARARIPE, M.N.B.A., SEGUNDO, L.F.F., LOPES, J.B. & ARARIPE, H.G.A. 2006. Efeito do
Cultivo de Peixes em Tanques Rede sobre o Aporte de Fósforo para o Ambiente. Rev. Cient. Prod. Anim., 8(2):56-65.
BORGES. P.A.F. 2009. Influência do cultivo experimental de tilápia (Oreochromis niloticus - Pisces, Cichlidae) em tanques-rede na qualidade da água de dois braços do reservatório da UHE Rosana: ênfase na comunidade fitoplanctônica. Programa de Pós-Graduação em Ecologia de Ambientes Aquáticos Continentais. Tese.
DIAS, J.D. 2008. Impacto da piscicultura em tanques-rede sobre a estrutura da comunidade zooplanctônica em um reservatório subtropical, Brasil. Programa de Pós-Graduação em Ecologia de Ambientes Aquáticos Continentais. Dissertação.
FIGUEREDO, C. C. & GIANI, A. 2005. Ecological interactions between Nile tilapia (Oreochromis niloticus, L.) and the phytoplanktonic community of the Furnas Reservoir (Brazil). Freshwater Biol., 50:1391-1403.
FONSECA, I.A., SIQUEIRA, N.S. & RODRIGUES, L. 2009. Algas perifíticas a montante e a jusante do local de instalação de tanques-rede em tributários do reservatório de Rosana, Estado do Paraná, Brasil. Acta Scientiarum, 31(2):135-141.
MARENGONI, N.G. 2006. Produção de tilápia do Nilo Oreochromis niloticus (linhagem chitralada), cultivada em tanques-rede, sob diferentes densidades de estocagem. Arch. Zootec. 55(210): 127-138.
MPA - Ministério da Pesca e Aqüicultura. 2009. Aqüicultura no Brasil. http://www.presidencia.gov.br/estrutura_presidencia/seap/. Acesso em: 29 jun. 2009.
NEGREIROS, N.F., ROJAS, N.E., ROCHA, O. & SANTOS WISNIEWSKI, M.J. 2009. Composition, diversity and short-term temporal fluctuations of zooplankton communities in fish culture ponds (Pindamonhangaba), SP. Braz. J. Biol.,69(3):785-794.
ROCHA, S.F. 2001. Influência do cultivo intensivo de tilápia (Oreochromis niloticus) em tanques-rede na qualidade da água do reservatório da Usina Hidrelétrica de Furnas – MG. Programa de Pós-Graduação em Ecologia, Conservação e Manejo de Vida Silvestre. Belo Horizonte. Dissertação. 150p. Dissertação.
ROCHE, F. K. & ROCHA, O. 2005. Aspectos de predação por peixes em lagos e represas, com enfoque na planctivoria. In: Roche, F. K. & Rocha, O. (eds.) Ecologia Trófica de Peixes com ênfase na planctivoria em ambientes lênticos de água doce no Brasil. São Carlos: Rima. 146p.
SÁ JUNIOR, W. P. 1994. Production of Planktonic Biomass for Feed of Alevins at the Furnas Hydrobiology and Hatchery Station. In: Pinto-Coelho, R.M.; Giani, A. & von Sperling, E. (eds) Ecology and Human impact on lakes and reservoirs in Minas Gerais with special reference to future development and management strategies. Belo Horizonte: SEGRAC. 193p.
SAMPAIO, J.M.C. & BRAGA, L.G.T. 2005. Cultivo de tilápia em tanques-rede na barragem do Ribeirão de Saloméa – Floresta Azul – Bahia. Rev. Bras. Saúde Prod. An., 6(2):42-52.
SIPAÚBA-TAVARES, L. H. & ROCHA, O. 2001. Produção de Plâncton (Fitoplâncton e Zooplâncton) para alimentação de organismos aquáticos. São Carlos: Rima. 106p.
SIPAÚBA-TAVARES, L.H., BACCARIN, A.E. & BRAGA, F.M. S. 2006. Limnological parameters and plankton community responses in Nile tilapia ponds under chicken dung and NPK (4-14-8) fertilizers. Acta Limnol. Bras.,18(3):335-346.
STARLING, F., PEREIRA., C.E. & ANGELINI, R. 2007a. Definição da capacidade suporte do reservatório de Três Marias para cultivo intensivo de peixes em tanques-redes. Projeto Parques Aqüícolas, UFMG/SECTES/SEAP. Belo Horizonte. 63p.
STARLING, F., PEREIRA., C.E. & ANGELINI, R. 2007b. Definição da capacidade suporte do reservatório de Furnas para cultivo intensivo de peixes em tanques-redes. Projeto Parques Aqüícolas, UFMG/SECTES/SEAP. Belo Horizonte. 62p.

74
ANEXOS
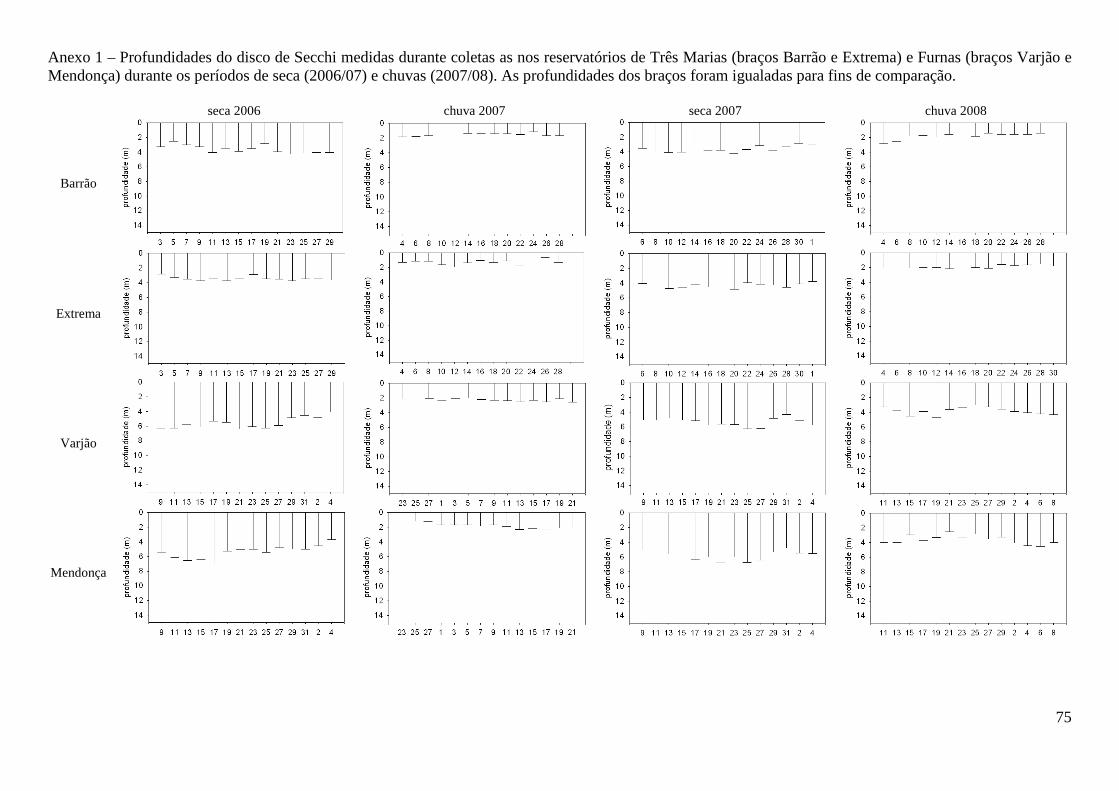
75
Anexo 1 – Profundidades do disco de Secchi medidas durante coletas as nos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuvas (2007/08). As profundidades dos braços foram igualadas para fins de comparação.
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
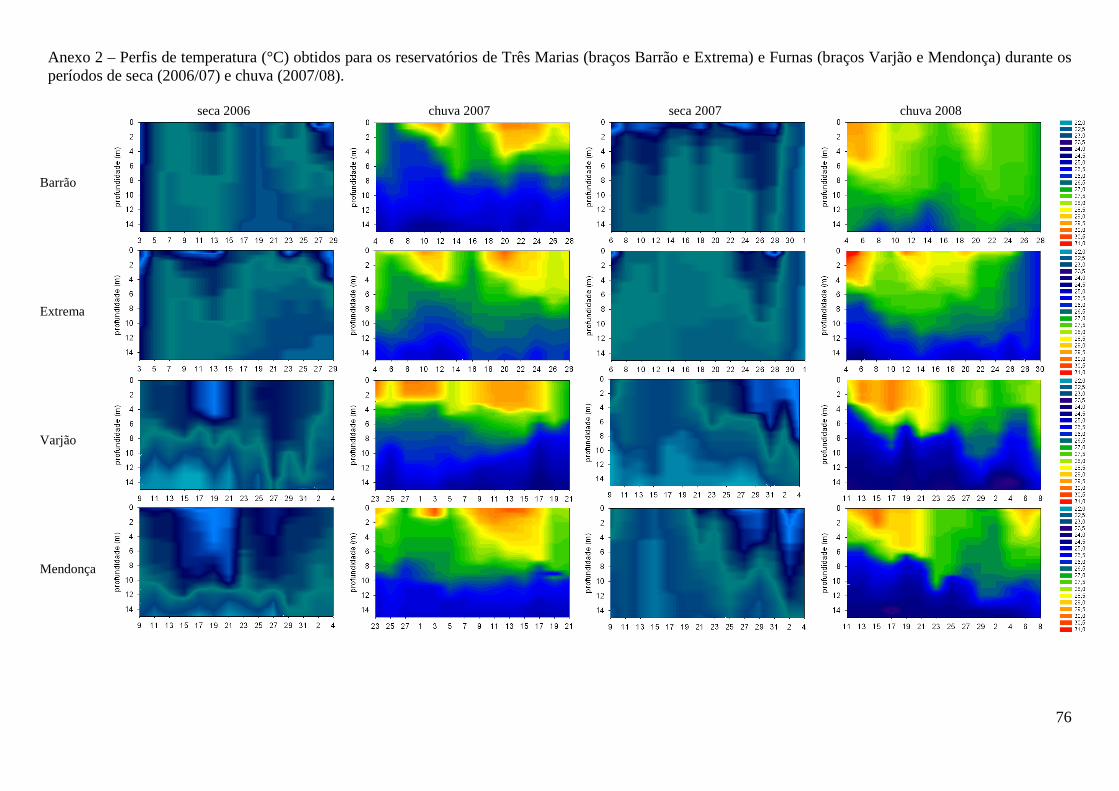
76
Anexo 2 – Perfis de temperatura (°C) obtidos para os reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
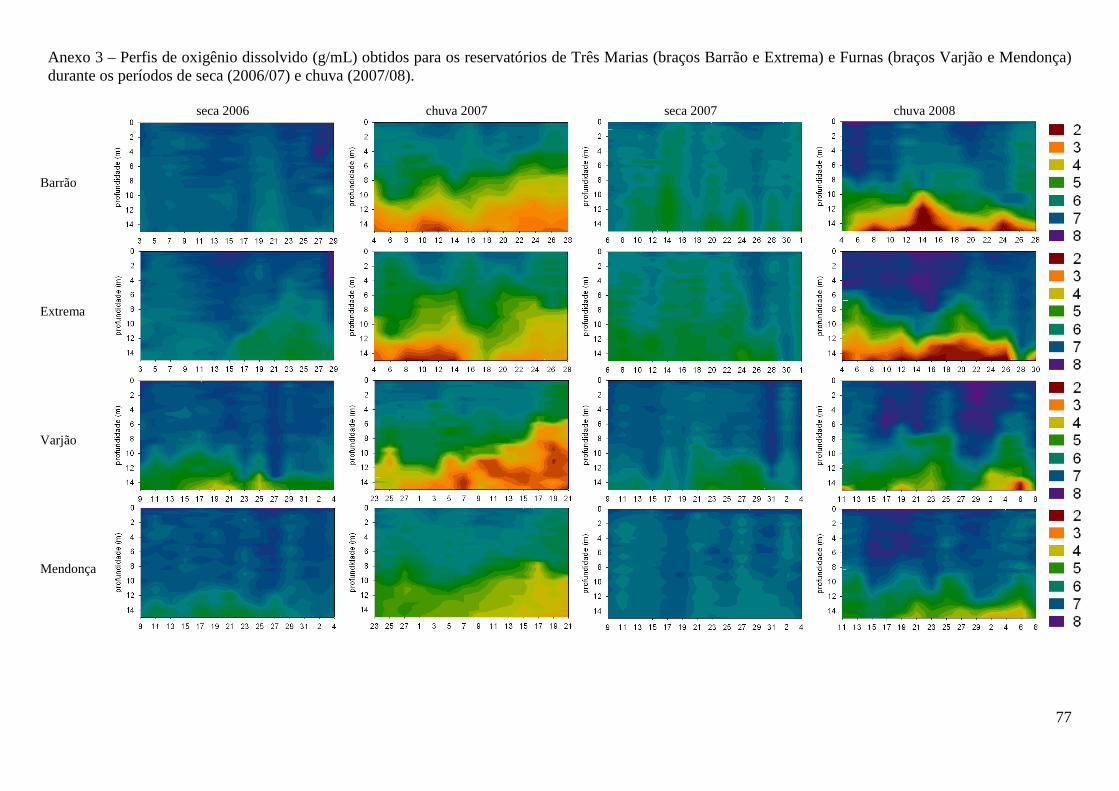
77
Anexo 3 – Perfis de oxigênio dissolvido (g/mL) obtidos para os reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça

78
Anexo 4 – Perfis de condutividade elétrica (µS/cm) realizados durante as coletas nos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08). Observação: notar valores de escala diferentes entre os reservatórios de Três Marias e Furnas.
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
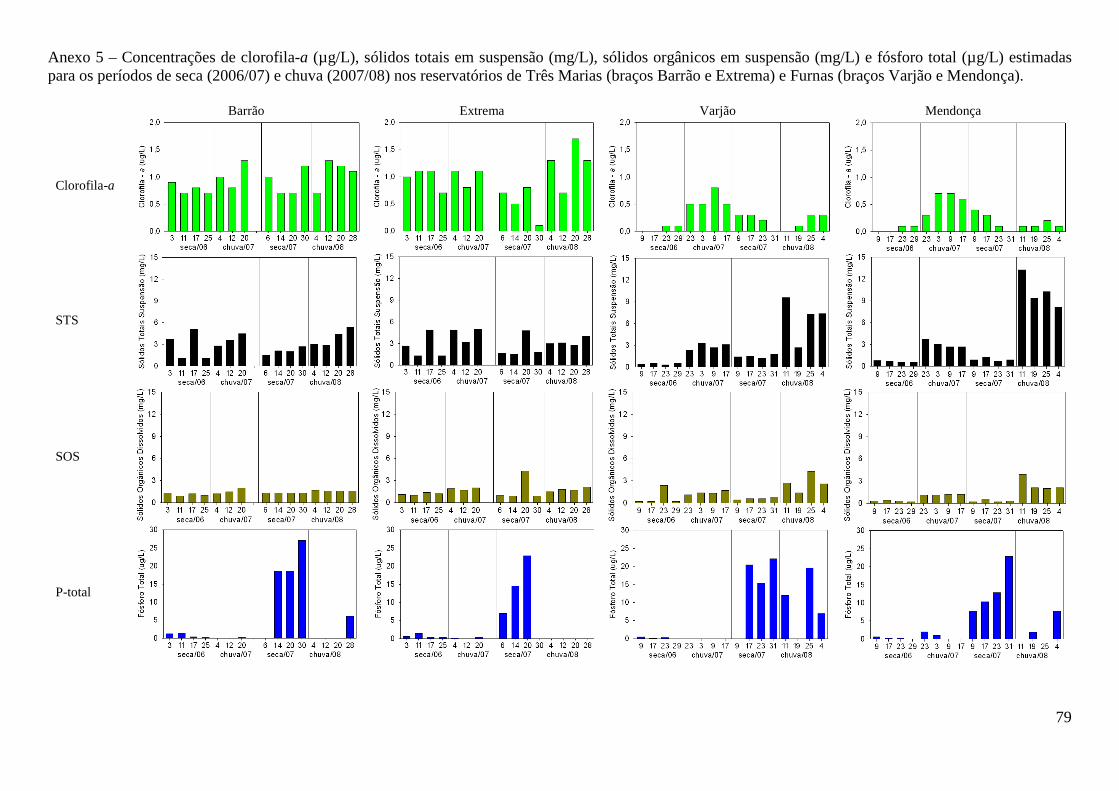
79
Anexo 5 – Concentrações de clorofila-a (µg/L), sólidos totais em suspensão (mg/L), sólidos orgânicos em suspensão (mg/L) e fósforo total (µg/L) estimadas para os períodos de seca (2006/07) e chuva (2007/08) nos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça).
Barrão Extrema Varjão Mendonça
Clorofila-a
STS
SOS
P-total
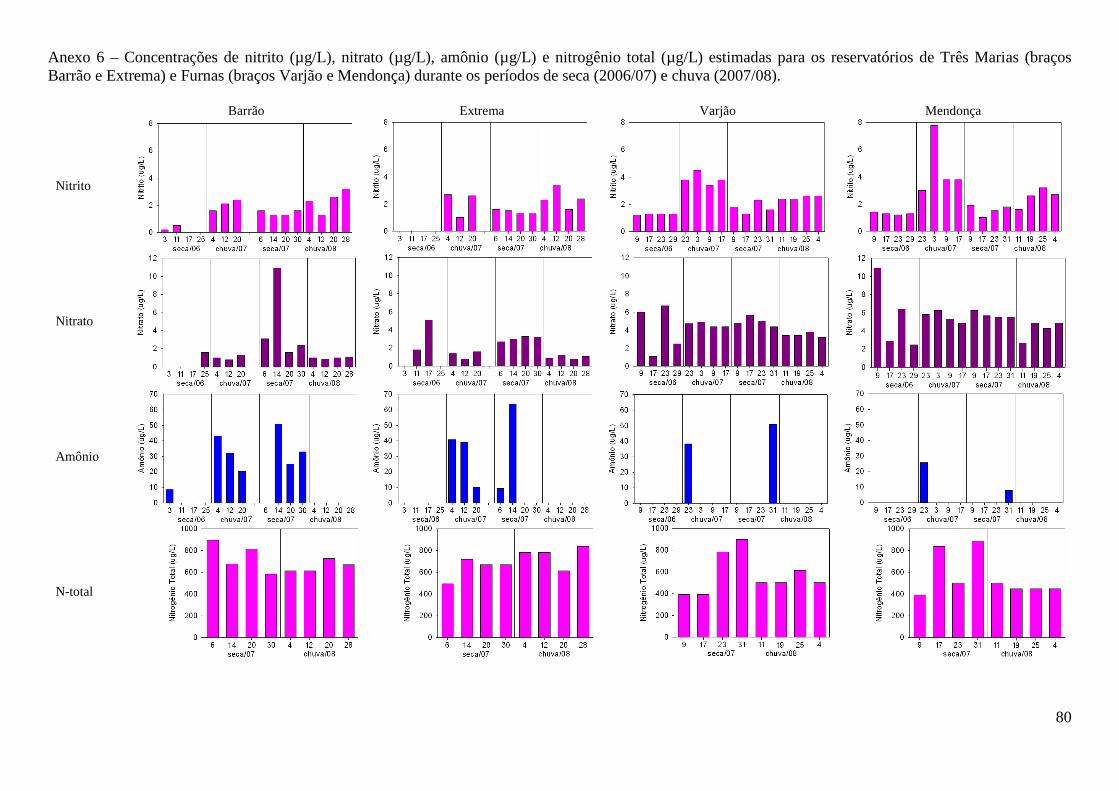
80
Anexo 6 – Concentrações de nitrito (µg/L), nitrato (µg/L), amônio (µg/L) e nitrogênio total (µg/L) estimadas para os reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
Barrão Extrema Varjão Mendonça
Nitrito
Nitrato
Amônio
N-total
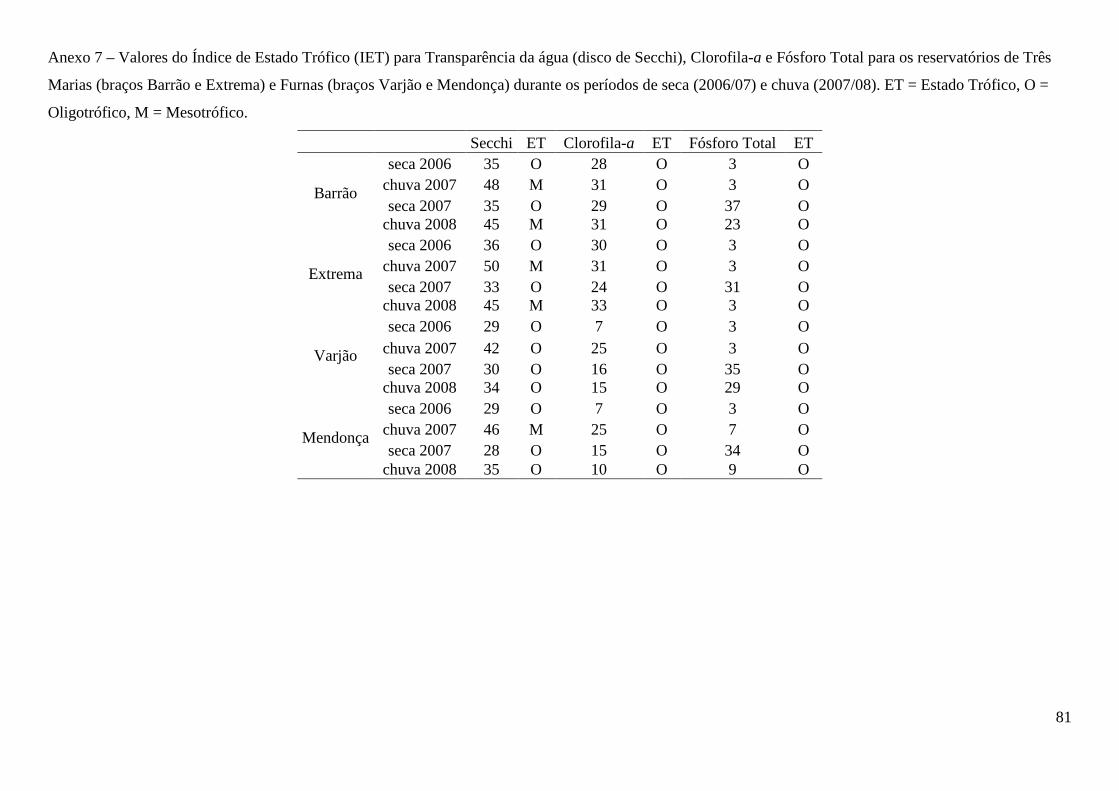
81
Anexo 7 – Valores do Índice de Estado Trófico (IET) para Transparência da água (disco de Secchi), Clorofila-a e Fósforo Total para os reservatórios de Três
Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08). ET = Estado Trófico, O =
Oligotrófico, M = Mesotrófico.
Secchi ET Clorofila-a ET Fósforo Total ET
Barrão
seca 2006 35 O 28 O 3 O chuva 2007 48 M 31 O 3 O seca 2007 35 O 29 O 37 O
chuva 2008 45 M 31 O 23 O
Extrema
seca 2006 36 O 30 O 3 O chuva 2007 50 M 31 O 3 O seca 2007 33 O 24 O 31 O
chuva 2008 45 M 33 O 3 O
Varjão
seca 2006 29 O 7 O 3 O chuva 2007 42 O 25 O 3 O seca 2007 30 O 16 O 35 O
chuva 2008 34 O 15 O 29 O
Mendonça
seca 2006 29 O 7 O 3 O chuva 2007 46 M 25 O 7 O seca 2007 28 O 15 O 34 O
chuva 2008 35 O 10 O 9 O
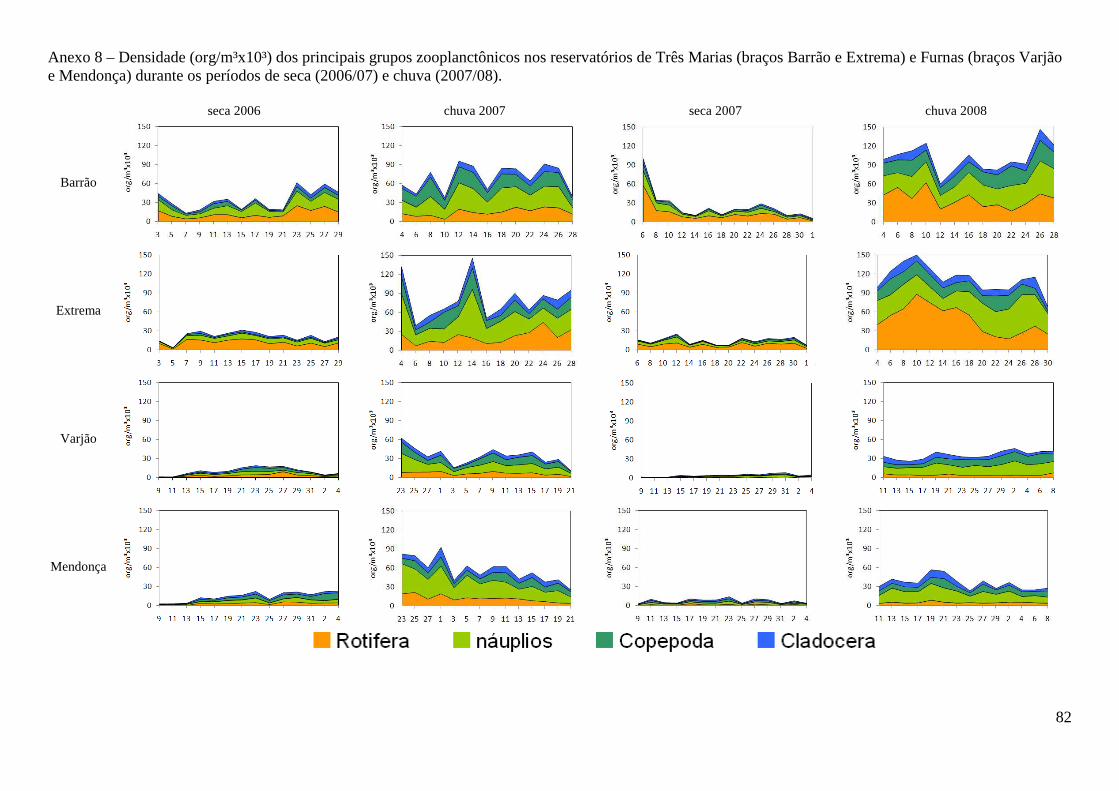
82
Anexo 8 – Densidade (org/m³x10³) dos principais grupos zooplanctônicos nos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
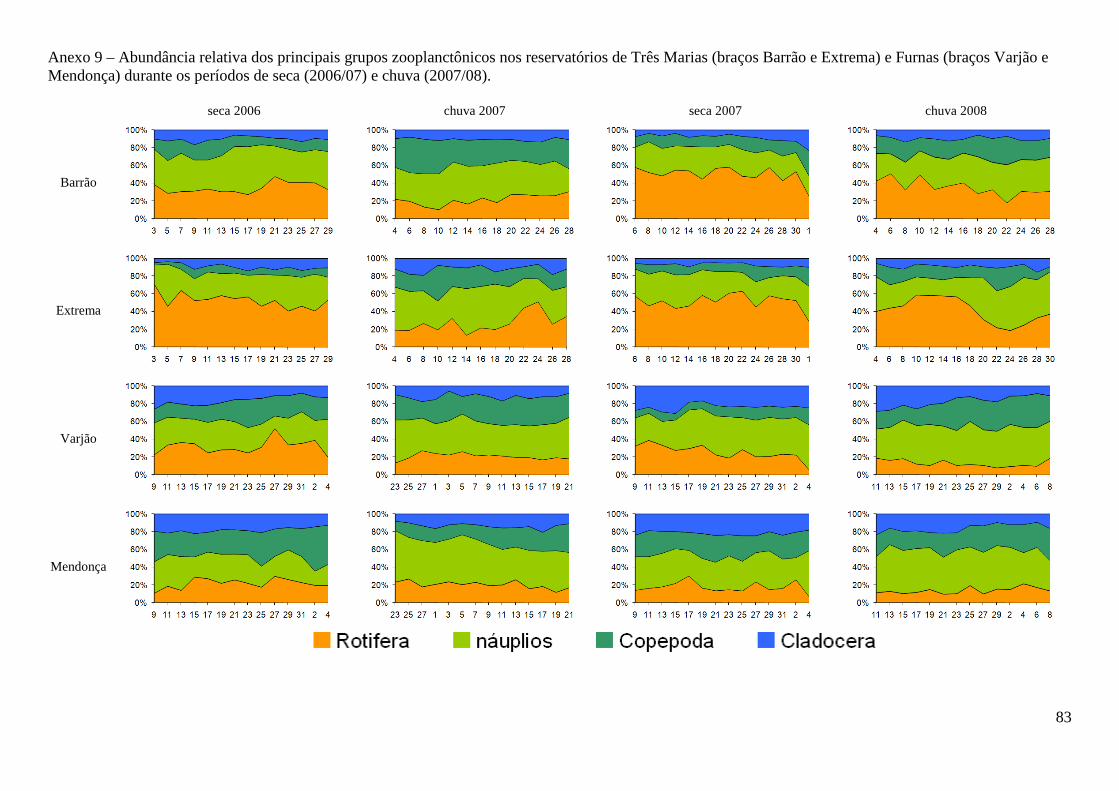
83
Anexo 9 – Abundância relativa dos principais grupos zooplanctônicos nos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
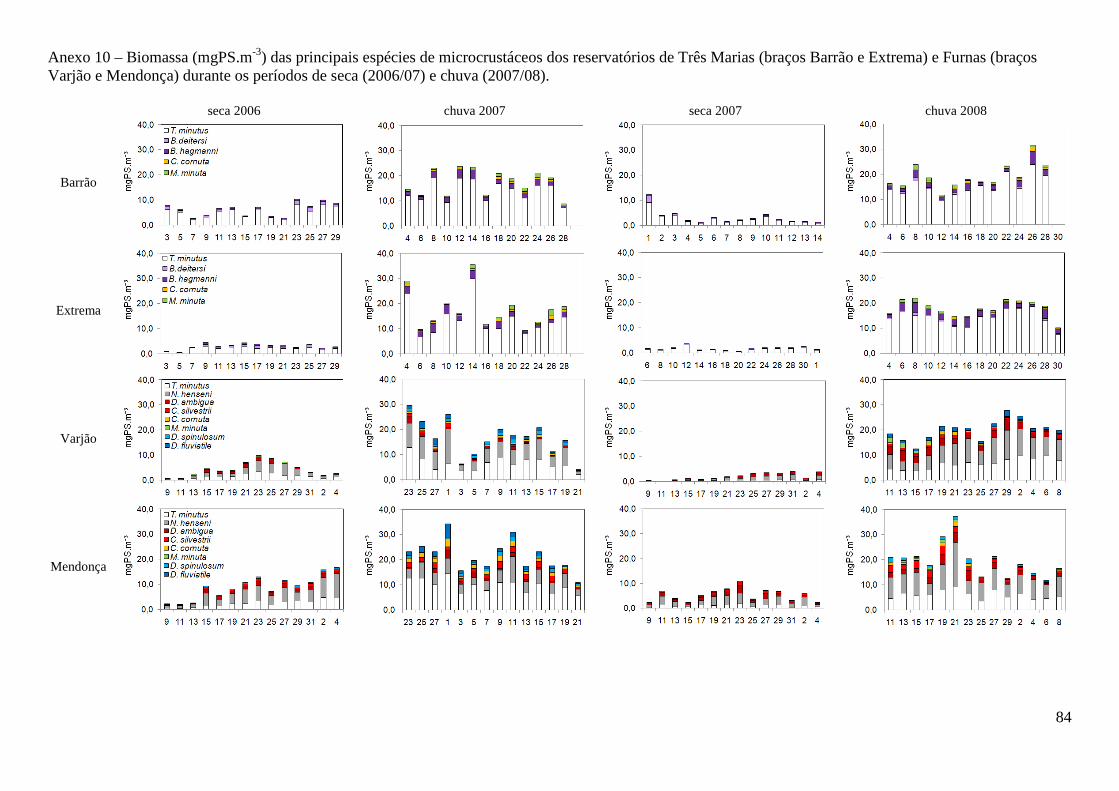
84
Anexo 10 – Biomassa (mgPS.m-3) das principais espécies de microcrustáceos dos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
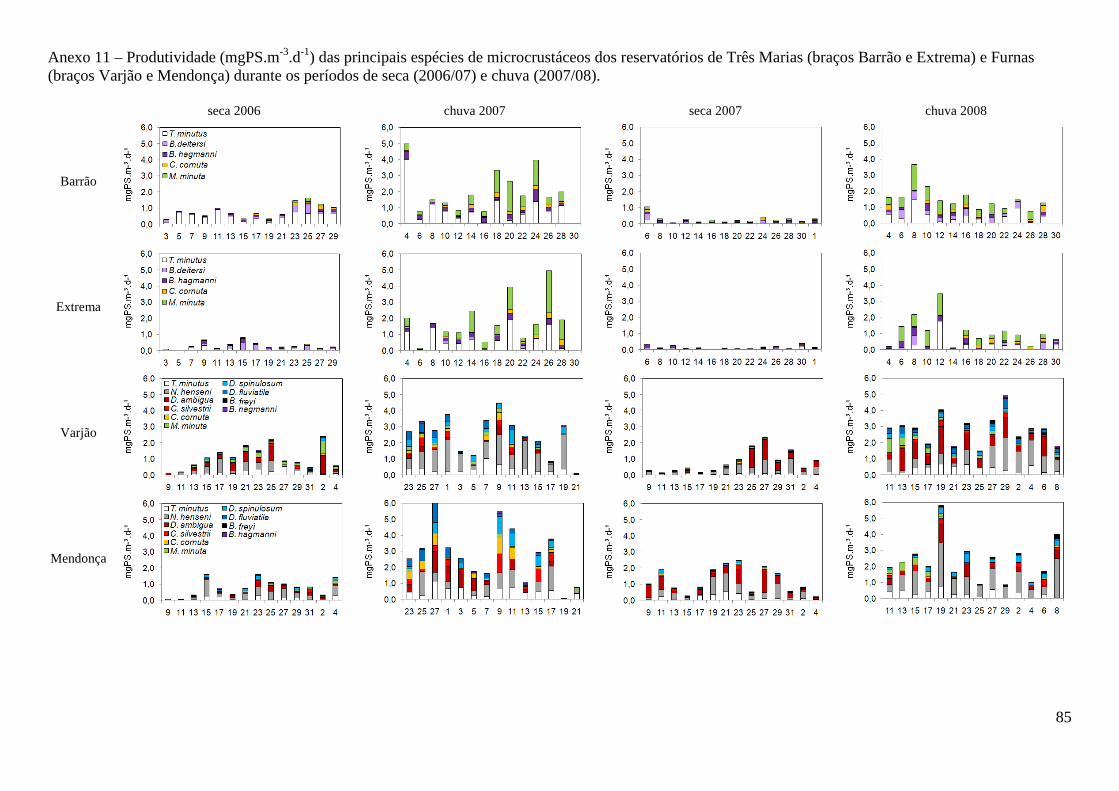
85
Anexo 11 – Produtividade (mgPS.m-3.d-1) das principais espécies de microcrustáceos dos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça) durante os períodos de seca (2006/07) e chuva (2007/08).
seca 2006 chuva 2007 seca 2007 chuva 2008
Barrão
Extrema
Varjão
Mendonça
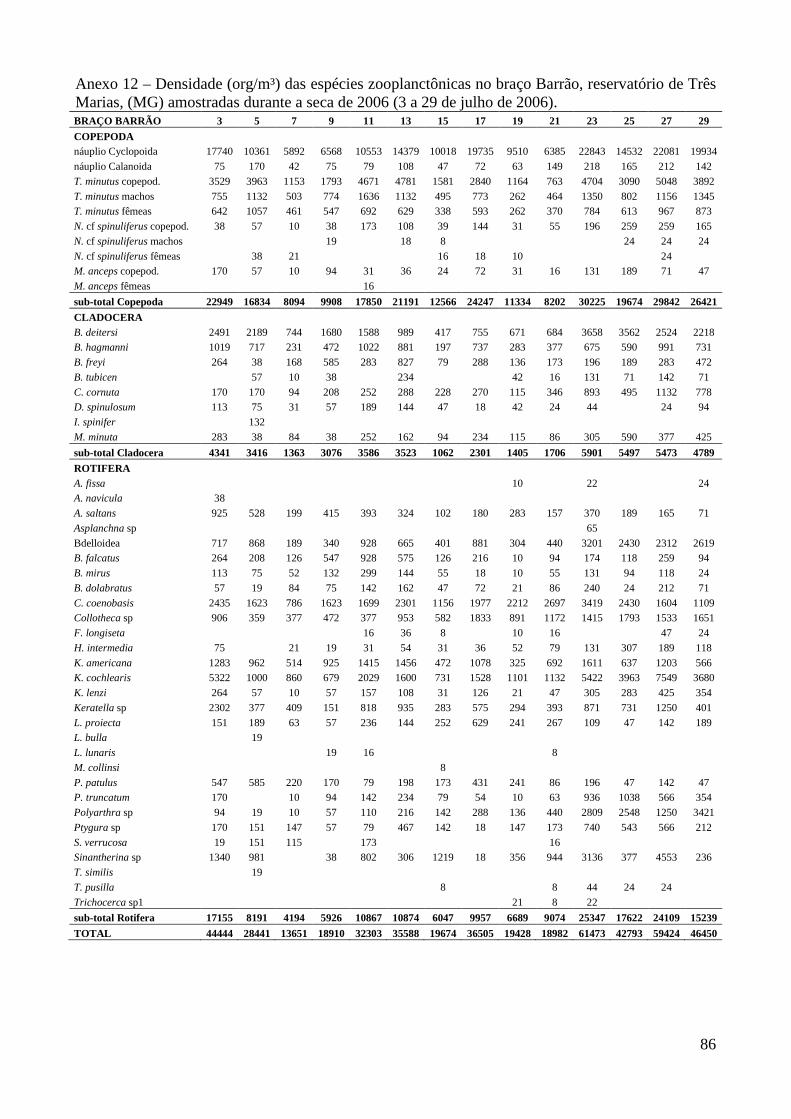
86
Anexo 12 – Densidade (org/m³) das espécies zooplanctônicas no braço Barrão, reservatório de Três Marias, (MG) amostradas durante a seca de 2006 (3 a 29 de julho de 2006). BRAÇO BARRÃO 3 5 7 9 11 13 15 17 19 21 23 25 27 29
COPEPODA náuplio Cyclopoida 17740 10361 5892 6568 10553 14379 10018 19735 9510 6385 22843 14532 22081 19934
náuplio Calanoida 75 170 42 75 79 108 47 72 63 149 218 165 212 142
T. minutus copepod. 3529 3963 1153 1793 4671 4781 1581 2840 1164 763 4704 3090 5048 3892
T. minutus machos 755 1132 503 774 1636 1132 495 773 262 464 1350 802 1156 1345 T. minutus fêmeas 642 1057 461 547 692 629 338 593 262 370 784 613 967 873
N. cf spinuliferus copepod. 38 57 10 38 173 108 39 144 31 55 196 259 259 165
N. cf spinuliferus machos 19 18 8 24 24 24
N. cf spinuliferus fêmeas 38 21 16 18 10 24 M. anceps copepod. 170 57 10 94 31 36 24 72 31 16 131 189 71 47
M. anceps fêmeas 16
sub-total Copepoda 22949 16834 8094 9908 17850 21191 12566 24247 11334 8202 30225 19674 29842 26421
CLADOCERA
B. deitersi 2491 2189 744 1680 1588 989 417 755 671 684 3658 3562 2524 2218 B. hagmanni 1019 717 231 472 1022 881 197 737 283 377 675 590 991 731
B. freyi 264 38 168 585 283 827 79 288 136 173 196 189 283 472
B. tubicen 57 10 38 234 42 16 131 71 142 71
C. cornuta 170 170 94 208 252 288 228 270 115 346 893 495 1132 778 D. spinulosum 113 75 31 57 189 144 47 18 42 24 44 24 94
I. spinifer 132
M. minuta 283 38 84 38 252 162 94 234 115 86 305 590 377 425
sub-total Cladocera 4341 3416 1363 3076 3586 3523 1062 2301 1405 1706 5901 5497 5473 4789
ROTIFERA
A. fissa 10 22 24 A. navicula 38
A. saltans 925 528 199 415 393 324 102 180 283 157 370 189 165 71
Asplanchna sp 65
Bdelloidea 717 868 189 340 928 665 401 881 304 440 3201 2430 2312 2619 B. falcatus 264 208 126 547 928 575 126 216 10 94 174 118 259 94
B. mirus 113 75 52 132 299 144 55 18 10 55 131 94 118 24
B. dolabratus 57 19 84 75 142 162 47 72 21 86 240 24 212 71
C. coenobasis 2435 1623 786 1623 1699 2301 1156 1977 2212 2697 3419 2430 1604 1109 Collotheca sp 906 359 377 472 377 953 582 1833 891 1172 1415 1793 1533 1651
F. longiseta 16 36 8 10 16 47 24
H. intermedia 75 21 19 31 54 31 36 52 79 131 307 189 118
K. americana 1283 962 514 925 1415 1456 472 1078 325 692 1611 637 1203 566 K. cochlearis 5322 1000 860 679 2029 1600 731 1528 1101 1132 5422 3963 7549 3680
K. lenzi 264 57 10 57 157 108 31 126 21 47 305 283 425 354
Keratella sp 2302 377 409 151 818 935 283 575 294 393 871 731 1250 401
L. proiecta 151 189 63 57 236 144 252 629 241 267 109 47 142 189 L. bulla 19
L. lunaris 19 16 8
M. collinsi 8
P. patulus 547 585 220 170 79 198 173 431 241 86 196 47 142 47 P. truncatum 170 10 94 142 234 79 54 10 63 936 1038 566 354
Polyarthra sp 94 19 10 57 110 216 142 288 136 440 2809 2548 1250 3421
Ptygura sp 170 151 147 57 79 467 142 18 147 173 740 543 566 212
S. verrucosa 19 151 115 173 16 Sinantherina sp 1340 981 38 802 306 1219 18 356 944 3136 377 4553 236
T. similis 19
T. pusilla 8 8 44 24 24
Trichocerca sp1 21 8 22
sub-total Rotifera 17155 8191 4194 5926 10867 10874 6047 9957 6689 9074 25347 17622 24109 15239
TOTAL 44444 28441 13651 18910 32303 35588 19674 36505 19428 18982 61473 42793 59424 46450
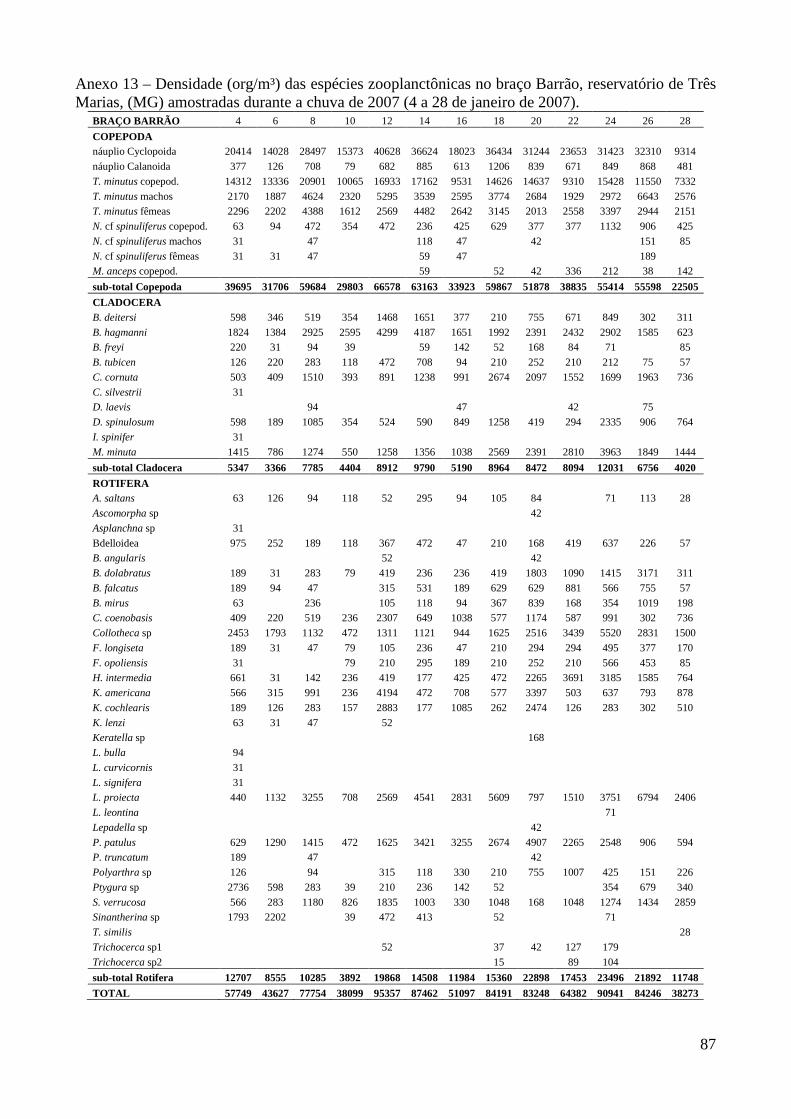
87
Anexo 13 – Densidade (org/m³) das espécies zooplanctônicas no braço Barrão, reservatório de Três Marias, (MG) amostradas durante a chuva de 2007 (4 a 28 de janeiro de 2007).
BRAÇO BARRÃO 4 6 8 10 12 14 16 18 20 22 24 26 28
COPEPODA náuplio Cyclopoida 20414 14028 28497 15373 40628 36624 18023 36434 31244 23653 31423 32310 9314
náuplio Calanoida 377 126 708 79 682 885 613 1206 839 671 849 868 481
T. minutus copepod. 14312 13336 20901 10065 16933 17162 9531 14626 14637 9310 15428 11550 7332
T. minutus machos 2170 1887 4624 2320 5295 3539 2595 3774 2684 1929 2972 6643 2576 T. minutus fêmeas 2296 2202 4388 1612 2569 4482 2642 3145 2013 2558 3397 2944 2151
N. cf spinuliferus copepod. 63 94 472 354 472 236 425 629 377 377 1132 906 425
N. cf spinuliferus machos 31 47 118 47 42 151 85
N. cf spinuliferus fêmeas 31 31 47 59 47 189 M. anceps copepod. 59 52 42 336 212 38 142
sub-total Copepoda 39695 31706 59684 29803 66578 63163 33923 59867 51878 38835 55414 55598 22505
CLADOCERA
B. deitersi 598 346 519 354 1468 1651 377 210 755 671 849 302 311
B. hagmanni 1824 1384 2925 2595 4299 4187 1651 1992 2391 2432 2902 1585 623 B. freyi 220 31 94 39 59 142 52 168 84 71 85
B. tubicen 126 220 283 118 472 708 94 210 252 210 212 75 57
C. cornuta 503 409 1510 393 891 1238 991 2674 2097 1552 1699 1963 736
C. silvestrii 31 D. laevis 94 47 42 75
D. spinulosum 598 189 1085 354 524 590 849 1258 419 294 2335 906 764
I. spinifer 31
M. minuta 1415 786 1274 550 1258 1356 1038 2569 2391 2810 3963 1849 1444
sub-total Cladocera 5347 3366 7785 4404 8912 9790 5190 8964 8472 8094 12031 6756 4020
ROTIFERA A. saltans 63 126 94 118 52 295 94 105 84 71 113 28
Ascomorpha sp 42
Asplanchna sp 31
Bdelloidea 975 252 189 118 367 472 47 210 168 419 637 226 57 B. angularis 52 42
B. dolabratus 189 31 283 79 419 236 236 419 1803 1090 1415 3171 311
B. falcatus 189 94 47 315 531 189 629 629 881 566 755 57
B. mirus 63 236 105 118 94 367 839 168 354 1019 198 C. coenobasis 409 220 519 236 2307 649 1038 577 1174 587 991 302 736
Collotheca sp 2453 1793 1132 472 1311 1121 944 1625 2516 3439 5520 2831 1500
F. longiseta 189 31 47 79 105 236 47 210 294 294 495 377 170
F. opoliensis 31 79 210 295 189 210 252 210 566 453 85 H. intermedia 661 31 142 236 419 177 425 472 2265 3691 3185 1585 764
K. americana 566 315 991 236 4194 472 708 577 3397 503 637 793 878
K. cochlearis 189 126 283 157 2883 177 1085 262 2474 126 283 302 510
K. lenzi 63 31 47 52 Keratella sp 168
L. bulla 94
L. curvicornis 31
L. signifera 31 L. proiecta 440 1132 3255 708 2569 4541 2831 5609 797 1510 3751 6794 2406
L. leontina 71
Lepadella sp 42
P. patulus 629 1290 1415 472 1625 3421 3255 2674 4907 2265 2548 906 594 P. truncatum 189 47 42
Polyarthra sp 126 94 315 118 330 210 755 1007 425 151 226
Ptygura sp 2736 598 283 39 210 236 142 52 354 679 340
S. verrucosa 566 283 1180 826 1835 1003 330 1048 168 1048 1274 1434 2859 Sinantherina sp 1793 2202 39 472 413 52 71
T. similis 28
Trichocerca sp1 52 37 42 127 179
Trichocerca sp2 15 89 104
sub-total Rotifera 12707 8555 10285 3892 19868 14508 11984 15360 22898 17453 23496 21892 11748
TOTAL 57749 43627 77754 38099 95357 87462 51097 84191 83248 64382 90941 84246 38273
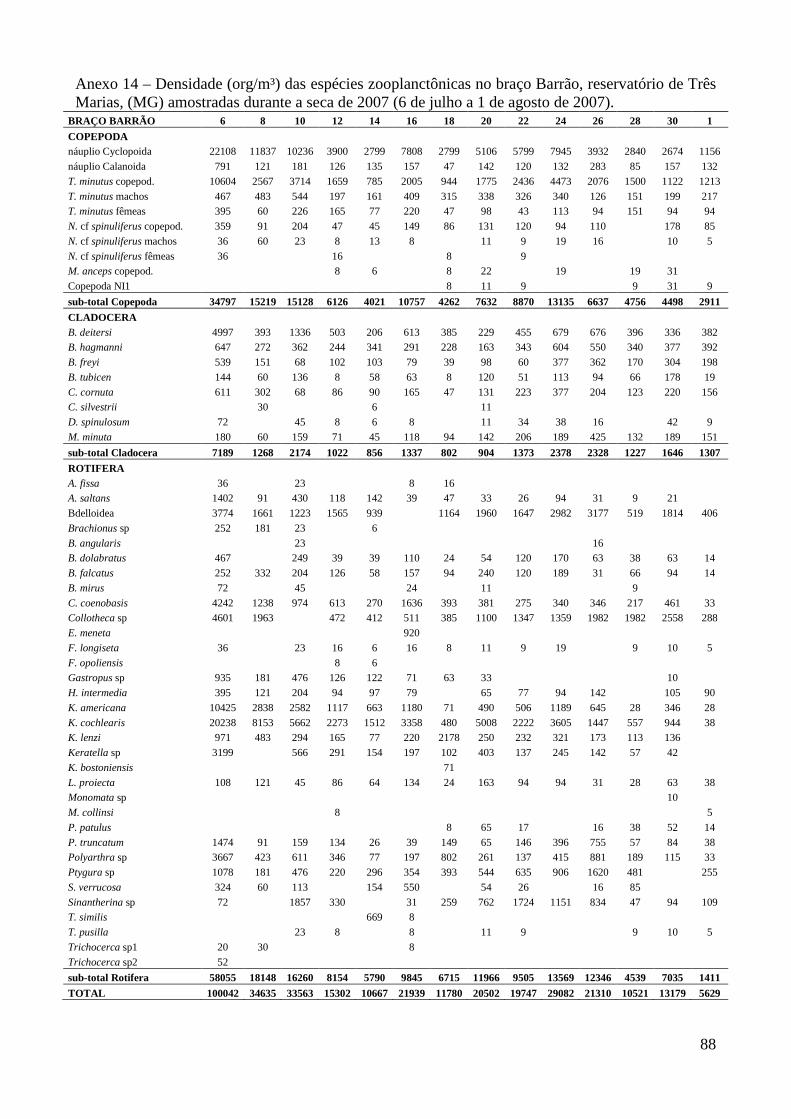
88
Anexo 14 – Densidade (org/m³) das espécies zooplanctônicas no braço Barrão, reservatório de Três Marias, (MG) amostradas durante a seca de 2007 (6 de julho a 1 de agosto de 2007).
BRAÇO BARRÃO 6 8 10 12 14 16 18 20 22 24 26 28 30 1
COPEPODA náuplio Cyclopoida 22108 11837 10236 3900 2799 7808 2799 5106 5799 7945 3932 2840 2674 1156
náuplio Calanoida 791 121 181 126 135 157 47 142 120 132 283 85 157 132
T. minutus copepod. 10604 2567 3714 1659 785 2005 944 1775 2436 4473 2076 1500 1122 1213
T. minutus machos 467 483 544 197 161 409 315 338 326 340 126 151 199 217 T. minutus fêmeas 395 60 226 165 77 220 47 98 43 113 94 151 94 94
N. cf spinuliferus copepod. 359 91 204 47 45 149 86 131 120 94 110 178 85
N. cf spinuliferus machos 36 60 23 8 13 8 11 9 19 16 10 5
N. cf spinuliferus fêmeas 36 16 8 9 M. anceps copepod. 8 6 8 22 19 19 31
Copepoda NI1 8 11 9 9 31 9
sub-total Copepoda 34797 15219 15128 6126 4021 10757 4262 7632 8870 13135 6637 4756 4498 2911
CLADOCERA
B. deitersi 4997 393 1336 503 206 613 385 229 455 679 676 396 336 382 B. hagmanni 647 272 362 244 341 291 228 163 343 604 550 340 377 392
B. freyi 539 151 68 102 103 79 39 98 60 377 362 170 304 198
B. tubicen 144 60 136 8 58 63 8 120 51 113 94 66 178 19
C. cornuta 611 302 68 86 90 165 47 131 223 377 204 123 220 156 C. silvestrii 30 6 11
D. spinulosum 72 45 8 6 8 11 34 38 16 42 9
M. minuta 180 60 159 71 45 118 94 142 206 189 425 132 189 151
sub-total Cladocera 7189 1268 2174 1022 856 1337 802 904 1373 2378 2328 1227 1646 1307
ROTIFERA
A. fissa 36 23 8 16 A. saltans 1402 91 430 118 142 39 47 33 26 94 31 9 21
Bdelloidea 3774 1661 1223 1565 939 1164 1960 1647 2982 3177 519 1814 406
Brachionus sp 252 181 23 6
B. angularis 23 16 B. dolabratus 467 249 39 39 110 24 54 120 170 63 38 63 14
B. falcatus 252 332 204 126 58 157 94 240 120 189 31 66 94 14
B. mirus 72 45 24 11 9
C. coenobasis 4242 1238 974 613 270 1636 393 381 275 340 346 217 461 33 Collotheca sp 4601 1963 472 412 511 385 1100 1347 1359 1982 1982 2558 288
E. meneta 920
F. longiseta 36 23 16 6 16 8 11 9 19 9 10 5
F. opoliensis 8 6 Gastropus sp 935 181 476 126 122 71 63 33 10
H. intermedia 395 121 204 94 97 79 65 77 94 142 105 90
K. americana 10425 2838 2582 1117 663 1180 71 490 506 1189 645 28 346 28
K. cochlearis 20238 8153 5662 2273 1512 3358 480 5008 2222 3605 1447 557 944 38 K. lenzi 971 483 294 165 77 220 2178 250 232 321 173 113 136
Keratella sp 3199 566 291 154 197 102 403 137 245 142 57 42
K. bostoniensis 71
L. proiecta 108 121 45 86 64 134 24 163 94 94 31 28 63 38 Monomata sp 10
M. collinsi 8 5
P. patulus 8 65 17 16 38 52 14
P. truncatum 1474 91 159 134 26 39 149 65 146 396 755 57 84 38 Polyarthra sp 3667 423 611 346 77 197 802 261 137 415 881 189 115 33
Ptygura sp 1078 181 476 220 296 354 393 544 635 906 1620 481 255
S. verrucosa 324 60 113 154 550 54 26 16 85
Sinantherina sp 72 1857 330 31 259 762 1724 1151 834 47 94 109 T. similis 669 8
T. pusilla 23 8 8 11 9 9 10 5
Trichocerca sp1 20 30 8
Trichocerca sp2 52
sub-total Rotifera 58055 18148 16260 8154 5790 9845 6715 11966 9505 13569 12346 4539 7035 1411
TOTAL 100042 34635 33563 15302 10667 21939 11780 20502 19747 29082 21310 10521 13179 5629
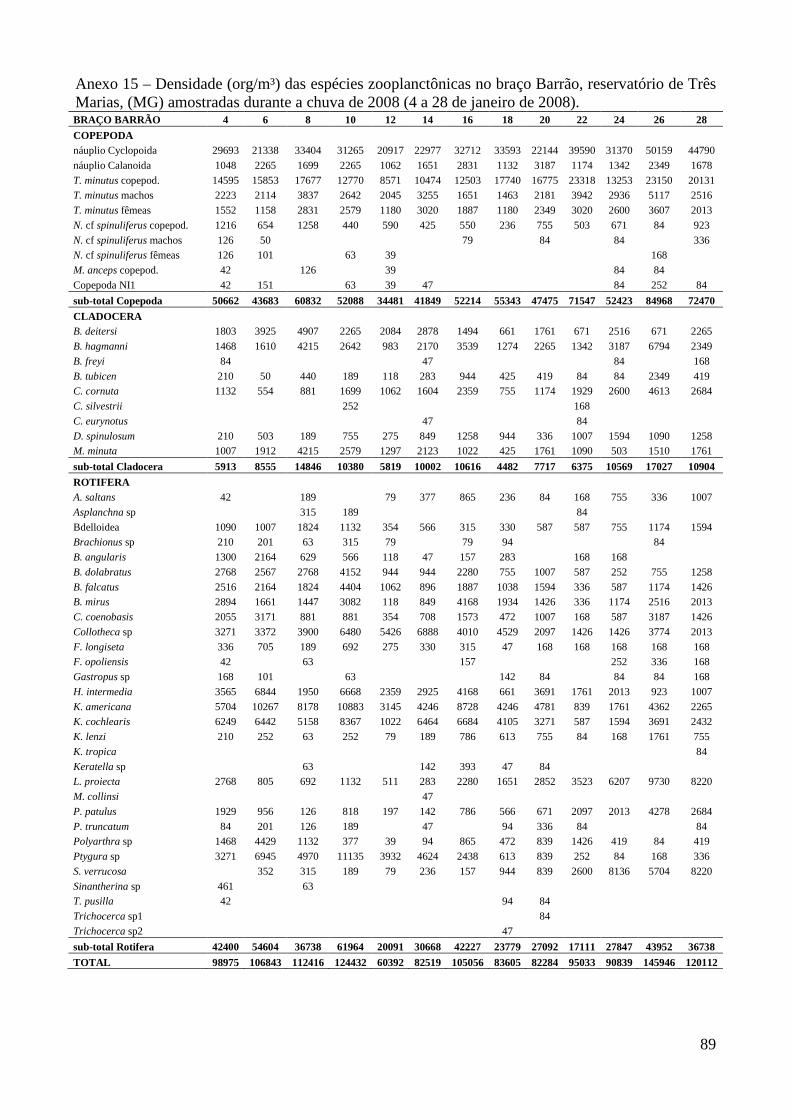
89
Anexo 15 – Densidade (org/m³) das espécies zooplanctônicas no braço Barrão, reservatório de Três Marias, (MG) amostradas durante a chuva de 2008 (4 a 28 de janeiro de 2008). BRAÇO BARRÃO 4 6 8 10 12 14 16 18 20 22 24 26 28
COPEPODA
náuplio Cyclopoida 29693 21338 33404 31265 20917 22977 32712 33593 22144 39590 31370 50159 44790
náuplio Calanoida 1048 2265 1699 2265 1062 1651 2831 1132 3187 1174 1342 2349 1678
T. minutus copepod. 14595 15853 17677 12770 8571 10474 12503 17740 16775 23318 13253 23150 20131 T. minutus machos 2223 2114 3837 2642 2045 3255 1651 1463 2181 3942 2936 5117 2516
T. minutus fêmeas 1552 1158 2831 2579 1180 3020 1887 1180 2349 3020 2600 3607 2013
N. cf spinuliferus copepod. 1216 654 1258 440 590 425 550 236 755 503 671 84 923
N. cf spinuliferus machos 126 50 79 84 84 336 N. cf spinuliferus fêmeas 126 101 63 39 168
M. anceps copepod. 42 126 39 84 84
Copepoda NI1 42 151 63 39 47 84 252 84
sub-total Copepoda 50662 43683 60832 52088 34481 41849 52214 55343 47475 71547 52423 84968 72470
CLADOCERA B. deitersi 1803 3925 4907 2265 2084 2878 1494 661 1761 671 2516 671 2265
B. hagmanni 1468 1610 4215 2642 983 2170 3539 1274 2265 1342 3187 6794 2349
B. freyi 84 47 84 168
B. tubicen 210 50 440 189 118 283 944 425 419 84 84 2349 419 C. cornuta 1132 554 881 1699 1062 1604 2359 755 1174 1929 2600 4613 2684
C. silvestrii 252 168
C. eurynotus 47 84
D. spinulosum 210 503 189 755 275 849 1258 944 336 1007 1594 1090 1258 M. minuta 1007 1912 4215 2579 1297 2123 1022 425 1761 1090 503 1510 1761
sub-total Cladocera 5913 8555 14846 10380 5819 10002 10616 4482 7717 6375 10569 17027 10904
ROTIFERA
A. saltans 42 189 79 377 865 236 84 168 755 336 1007
Asplanchna sp 315 189 84
Bdelloidea 1090 1007 1824 1132 354 566 315 330 587 587 755 1174 1594 Brachionus sp 210 201 63 315 79 79 94 84
B. angularis 1300 2164 629 566 118 47 157 283 168 168
B. dolabratus 2768 2567 2768 4152 944 944 2280 755 1007 587 252 755 1258
B. falcatus 2516 2164 1824 4404 1062 896 1887 1038 1594 336 587 1174 1426 B. mirus 2894 1661 1447 3082 118 849 4168 1934 1426 336 1174 2516 2013
C. coenobasis 2055 3171 881 881 354 708 1573 472 1007 168 587 3187 1426
Collotheca sp 3271 3372 3900 6480 5426 6888 4010 4529 2097 1426 1426 3774 2013
F. longiseta 336 705 189 692 275 330 315 47 168 168 168 168 168 F. opoliensis 42 63 157 252 336 168
Gastropus sp 168 101 63 142 84 84 84 168
H. intermedia 3565 6844 1950 6668 2359 2925 4168 661 3691 1761 2013 923 1007
K. americana 5704 10267 8178 10883 3145 4246 8728 4246 4781 839 1761 4362 2265 K. cochlearis 6249 6442 5158 8367 1022 6464 6684 4105 3271 587 1594 3691 2432
K. lenzi 210 252 63 252 79 189 786 613 755 84 168 1761 755
K. tropica 84
Keratella sp 63 142 393 47 84 L. proiecta 2768 805 692 1132 511 283 2280 1651 2852 3523 6207 9730 8220
M. collinsi 47
P. patulus 1929 956 126 818 197 142 786 566 671 2097 2013 4278 2684
P. truncatum 84 201 126 189 47 94 336 84 84 Polyarthra sp 1468 4429 1132 377 39 94 865 472 839 1426 419 84 419
Ptygura sp 3271 6945 4970 11135 3932 4624 2438 613 839 252 84 168 336
S. verrucosa 352 315 189 79 236 157 944 839 2600 8136 5704 8220
Sinantherina sp 461 63 T. pusilla 42 94 84
Trichocerca sp1 84
Trichocerca sp2 47
sub-total Rotifera 42400 54604 36738 61964 20091 30668 42227 23779 27092 17111 27847 43952 36738
TOTAL 98975 106843 112416 124432 60392 82519 105056 83605 82284 95033 90839 145946 120112
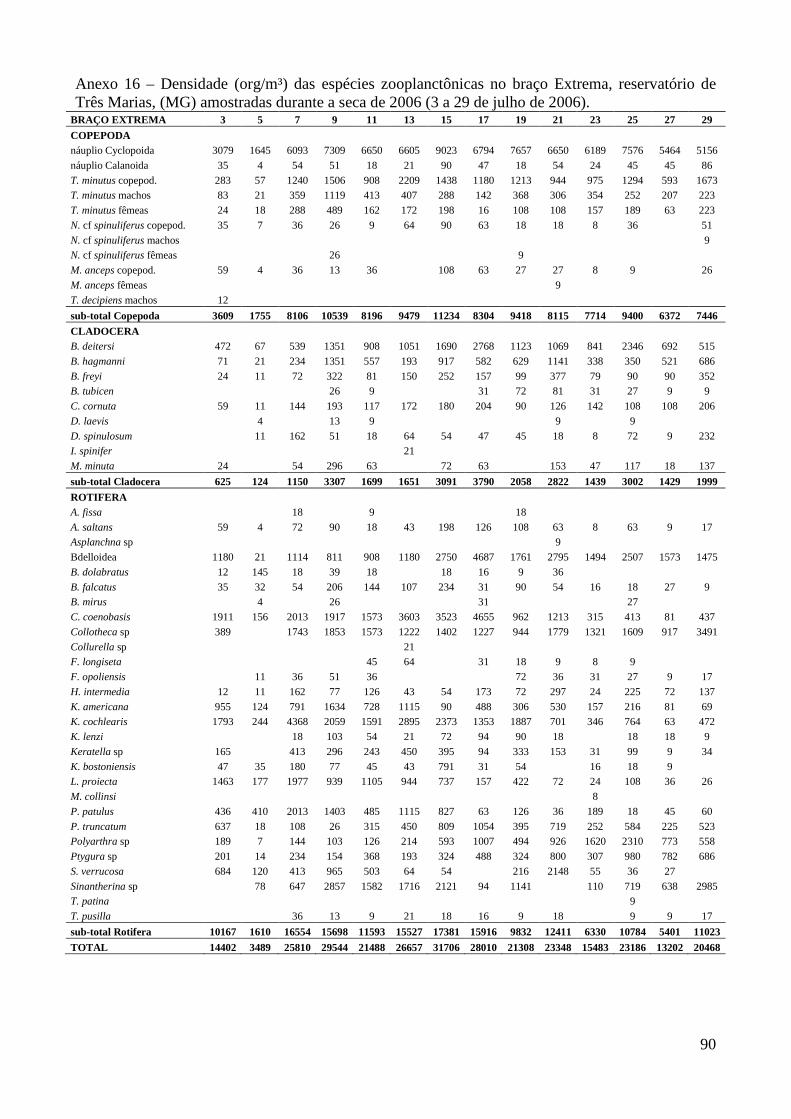
90
Anexo 16 – Densidade (org/m³) das espécies zooplanctônicas no braço Extrema, reservatório de Três Marias, (MG) amostradas durante a seca de 2006 (3 a 29 de julho de 2006).
BRAÇO EXTREMA 3 5 7 9 11 13 15 17 19 21 23 25 27 29
COPEPODA
náuplio Cyclopoida 3079 1645 6093 7309 6650 6605 9023 6794 7657 6650 6189 7576 5464 5156
náuplio Calanoida 35 4 54 51 18 21 90 47 18 54 24 45 45 86
T. minutus copepod. 283 57 1240 1506 908 2209 1438 1180 1213 944 975 1294 593 1673 T. minutus machos 83 21 359 1119 413 407 288 142 368 306 354 252 207 223
T. minutus fêmeas 24 18 288 489 162 172 198 16 108 108 157 189 63 223
N. cf spinuliferus copepod. 35 7 36 26 9 64 90 63 18 18 8 36 51
N. cf spinuliferus machos 9 N. cf spinuliferus fêmeas 26 9
M. anceps copepod. 59 4 36 13 36 108 63 27 27 8 9 26
M. anceps fêmeas 9
T. decipiens machos 12
sub-total Copepoda 3609 1755 8106 10539 8196 9479 11234 8304 9418 8115 7714 9400 6372 7446
CLADOCERA
B. deitersi 472 67 539 1351 908 1051 1690 2768 1123 1069 841 2346 692 515
B. hagmanni 71 21 234 1351 557 193 917 582 629 1141 338 350 521 686
B. freyi 24 11 72 322 81 150 252 157 99 377 79 90 90 352 B. tubicen 26 9 31 72 81 31 27 9 9
C. cornuta 59 11 144 193 117 172 180 204 90 126 142 108 108 206
D. laevis 4 13 9 9 9
D. spinulosum 11 162 51 18 64 54 47 45 18 8 72 9 232 I. spinifer 21
M. minuta 24 54 296 63 72 63 153 47 117 18 137
sub-total Cladocera 625 124 1150 3307 1699 1651 3091 3790 2058 2822 1439 3002 1429 1999
ROTIFERA
A. fissa 18 9 18
A. saltans 59 4 72 90 18 43 198 126 108 63 8 63 9 17 Asplanchna sp 9
Bdelloidea 1180 21 1114 811 908 1180 2750 4687 1761 2795 1494 2507 1573 1475
B. dolabratus 12 145 18 39 18 18 16 9 36
B. falcatus 35 32 54 206 144 107 234 31 90 54 16 18 27 9 B. mirus 4 26 31 27
C. coenobasis 1911 156 2013 1917 1573 3603 3523 4655 962 1213 315 413 81 437
Collotheca sp 389 1743 1853 1573 1222 1402 1227 944 1779 1321 1609 917 3491
Collurella sp 21 F. longiseta 45 64 31 18 9 8 9
F. opoliensis 11 36 51 36 72 36 31 27 9 17
H. intermedia 12 11 162 77 126 43 54 173 72 297 24 225 72 137
K. americana 955 124 791 1634 728 1115 90 488 306 530 157 216 81 69 K. cochlearis 1793 244 4368 2059 1591 2895 2373 1353 1887 701 346 764 63 472
K. lenzi 18 103 54 21 72 94 90 18 18 18 9
Keratella sp 165 413 296 243 450 395 94 333 153 31 99 9 34
K. bostoniensis 47 35 180 77 45 43 791 31 54 16 18 9 L. proiecta 1463 177 1977 939 1105 944 737 157 422 72 24 108 36 26
M. collinsi 8
P. patulus 436 410 2013 1403 485 1115 827 63 126 36 189 18 45 60
P. truncatum 637 18 108 26 315 450 809 1054 395 719 252 584 225 523 Polyarthra sp 189 7 144 103 126 214 593 1007 494 926 1620 2310 773 558
Ptygura sp 201 14 234 154 368 193 324 488 324 800 307 980 782 686
S. verrucosa 684 120 413 965 503 64 54 216 2148 55 36 27
Sinantherina sp 78 647 2857 1582 1716 2121 94 1141 110 719 638 2985 T. patina 9
T. pusilla 36 13 9 21 18 16 9 18 9 9 17
sub-total Rotifera 10167 1610 16554 15698 11593 15527 17381 15916 9832 12411 6330 10784 5401 11023
TOTAL 14402 3489 25810 29544 21488 26657 31706 28010 21308 23348 15483 23186 13202 20468
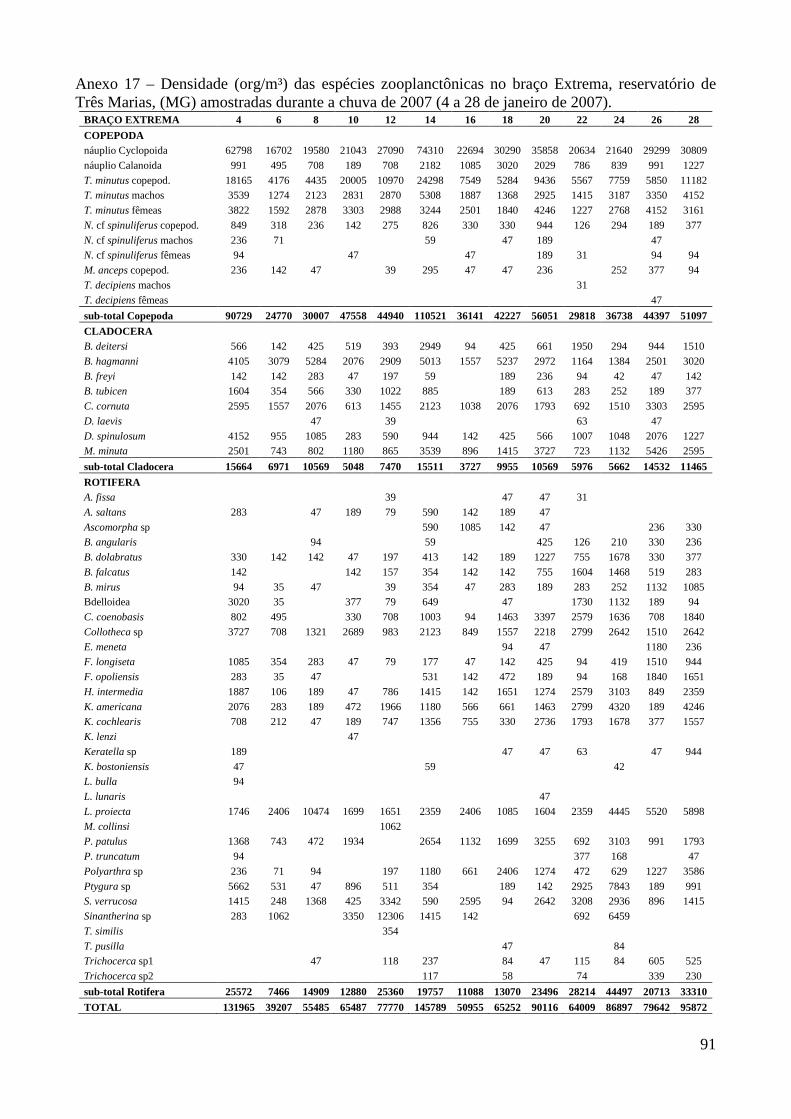
91
Anexo 17 – Densidade (org/m³) das espécies zooplanctônicas no braço Extrema, reservatório de Três Marias, (MG) amostradas durante a chuva de 2007 (4 a 28 de janeiro de 2007).
BRAÇO EXTREMA 4 6 8 10 12 14 16 18 20 22 24 26 28
COPEPODA
náuplio Cyclopoida 62798 16702 19580 21043 27090 74310 22694 30290 35858 20634 21640 29299 30809
náuplio Calanoida 991 495 708 189 708 2182 1085 3020 2029 786 839 991 1227
T. minutus copepod. 18165 4176 4435 20005 10970 24298 7549 5284 9436 5567 7759 5850 11182 T. minutus machos 3539 1274 2123 2831 2870 5308 1887 1368 2925 1415 3187 3350 4152
T. minutus fêmeas 3822 1592 2878 3303 2988 3244 2501 1840 4246 1227 2768 4152 3161
N. cf spinuliferus copepod. 849 318 236 142 275 826 330 330 944 126 294 189 377
N. cf spinuliferus machos 236 71 59 47 189 47 N. cf spinuliferus fêmeas 94 47 47 189 31 94 94
M. anceps copepod. 236 142 47 39 295 47 47 236 252 377 94
T. decipiens machos 31
T. decipiens fêmeas 47
sub-total Copepoda 90729 24770 30007 47558 44940 110521 36141 42227 56051 29818 36738 44397 51097
CLADOCERA
B. deitersi 566 142 425 519 393 2949 94 425 661 1950 294 944 1510
B. hagmanni 4105 3079 5284 2076 2909 5013 1557 5237 2972 1164 1384 2501 3020
B. freyi 142 142 283 47 197 59 189 236 94 42 47 142 B. tubicen 1604 354 566 330 1022 885 189 613 283 252 189 377
C. cornuta 2595 1557 2076 613 1455 2123 1038 2076 1793 692 1510 3303 2595
D. laevis 47 39 63 47
D. spinulosum 4152 955 1085 283 590 944 142 425 566 1007 1048 2076 1227 M. minuta 2501 743 802 1180 865 3539 896 1415 3727 723 1132 5426 2595
sub-total Cladocera 15664 6971 10569 5048 7470 15511 3727 9955 10569 5976 5662 14532 11465
ROTIFERA
A. fissa 39 47 47 31
A. saltans 283 47 189 79 590 142 189 47
Ascomorpha sp 590 1085 142 47 236 330 B. angularis 94 59 425 126 210 330 236
B. dolabratus 330 142 142 47 197 413 142 189 1227 755 1678 330 377
B. falcatus 142 142 157 354 142 142 755 1604 1468 519 283
B. mirus 94 35 47 39 354 47 283 189 283 252 1132 1085 Bdelloidea 3020 35 377 79 649 47 1730 1132 189 94
C. coenobasis 802 495 330 708 1003 94 1463 3397 2579 1636 708 1840
Collotheca sp 3727 708 1321 2689 983 2123 849 1557 2218 2799 2642 1510 2642
E. meneta 94 47 1180 236 F. longiseta 1085 354 283 47 79 177 47 142 425 94 419 1510 944
F. opoliensis 283 35 47 531 142 472 189 94 168 1840 1651
H. intermedia 1887 106 189 47 786 1415 142 1651 1274 2579 3103 849 2359
K. americana 2076 283 189 472 1966 1180 566 661 1463 2799 4320 189 4246 K. cochlearis 708 212 47 189 747 1356 755 330 2736 1793 1678 377 1557
K. lenzi 47
Keratella sp 189 47 47 63 47 944
K. bostoniensis 47 59 42 L. bulla 94
L. lunaris 47
L. proiecta 1746 2406 10474 1699 1651 2359 2406 1085 1604 2359 4445 5520 5898
M. collinsi 1062 P. patulus 1368 743 472 1934 2654 1132 1699 3255 692 3103 991 1793
P. truncatum 94 377 168 47
Polyarthra sp 236 71 94 197 1180 661 2406 1274 472 629 1227 3586
Ptygura sp 5662 531 47 896 511 354 189 142 2925 7843 189 991 S. verrucosa 1415 248 1368 425 3342 590 2595 94 2642 3208 2936 896 1415
Sinantherina sp 283 1062 3350 12306 1415 142 692 6459
T. similis 354
T. pusilla 47 84 Trichocerca sp1 47 118 237 84 47 115 84 605 525
Trichocerca sp2 117 58 74 339 230
sub-total Rotifera 25572 7466 14909 12880 25360 19757 11088 13070 23496 28214 44497 20713 33310
TOTAL 131965 39207 55485 65487 77770 145789 50955 65252 90116 64009 86897 79642 95872
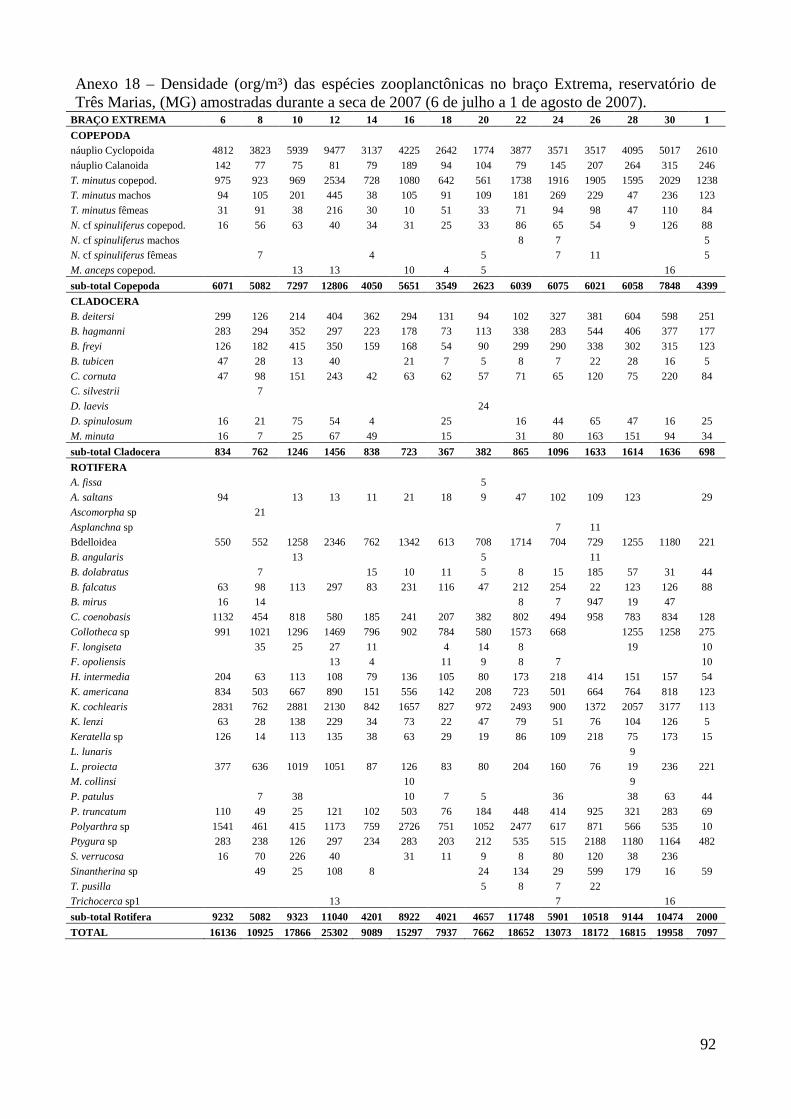
92
Anexo 18 – Densidade (org/m³) das espécies zooplanctônicas no braço Extrema, reservatório de Três Marias, (MG) amostradas durante a seca de 2007 (6 de julho a 1 de agosto de 2007).
BRAÇO EXTREMA 6 8 10 12 14 16 18 20 22 24 26 28 30 1
COPEPODA
náuplio Cyclopoida 4812 3823 5939 9477 3137 4225 2642 1774 3877 3571 3517 4095 5017 2610
náuplio Calanoida 142 77 75 81 79 189 94 104 79 145 207 264 315 246
T. minutus copepod. 975 923 969 2534 728 1080 642 561 1738 1916 1905 1595 2029 1238 T. minutus machos 94 105 201 445 38 105 91 109 181 269 229 47 236 123
T. minutus fêmeas 31 91 38 216 30 10 51 33 71 94 98 47 110 84
N. cf spinuliferus copepod. 16 56 63 40 34 31 25 33 86 65 54 9 126 88
N. cf spinuliferus machos 8 7 5 N. cf spinuliferus fêmeas 7 4 5 7 11 5
M. anceps copepod. 13 13 10 4 5 16
sub-total Copepoda 6071 5082 7297 12806 4050 5651 3549 2623 6039 6075 6021 6058 7848 4399
CLADOCERA
B. deitersi 299 126 214 404 362 294 131 94 102 327 381 604 598 251 B. hagmanni 283 294 352 297 223 178 73 113 338 283 544 406 377 177
B. freyi 126 182 415 350 159 168 54 90 299 290 338 302 315 123
B. tubicen 47 28 13 40 21 7 5 8 7 22 28 16 5
C. cornuta 47 98 151 243 42 63 62 57 71 65 120 75 220 84 C. silvestrii 7
D. laevis 24
D. spinulosum 16 21 75 54 4 25 16 44 65 47 16 25
M. minuta 16 7 25 67 49 15 31 80 163 151 94 34
sub-total Cladocera 834 762 1246 1456 838 723 367 382 865 1096 1633 1614 1636 698
ROTIFERA A. fissa 5
A. saltans 94 13 13 11 21 18 9 47 102 109 123 29
Ascomorpha sp 21
Asplanchna sp 7 11 Bdelloidea 550 552 1258 2346 762 1342 613 708 1714 704 729 1255 1180 221
B. angularis 13 5 11
B. dolabratus 7 15 10 11 5 8 15 185 57 31 44
B. falcatus 63 98 113 297 83 231 116 47 212 254 22 123 126 88 B. mirus 16 14 8 7 947 19 47
C. coenobasis 1132 454 818 580 185 241 207 382 802 494 958 783 834 128
Collotheca sp 991 1021 1296 1469 796 902 784 580 1573 668 1255 1258 275
F. longiseta 35 25 27 11 4 14 8 19 10 F. opoliensis 13 4 11 9 8 7 10
H. intermedia 204 63 113 108 79 136 105 80 173 218 414 151 157 54
K. americana 834 503 667 890 151 556 142 208 723 501 664 764 818 123
K. cochlearis 2831 762 2881 2130 842 1657 827 972 2493 900 1372 2057 3177 113 K. lenzi 63 28 138 229 34 73 22 47 79 51 76 104 126 5
Keratella sp 126 14 113 135 38 63 29 19 86 109 218 75 173 15
L. lunaris 9
L. proiecta 377 636 1019 1051 87 126 83 80 204 160 76 19 236 221 M. collinsi 10 9
P. patulus 7 38 10 7 5 36 38 63 44
P. truncatum 110 49 25 121 102 503 76 184 448 414 925 321 283 69
Polyarthra sp 1541 461 415 1173 759 2726 751 1052 2477 617 871 566 535 10 Ptygura sp 283 238 126 297 234 283 203 212 535 515 2188 1180 1164 482
S. verrucosa 16 70 226 40 31 11 9 8 80 120 38 236
Sinantherina sp 49 25 108 8 24 134 29 599 179 16 59
T. pusilla 5 8 7 22 Trichocerca sp1 13 7 16
sub-total Rotifera 9232 5082 9323 11040 4201 8922 4021 4657 11748 5901 10518 9144 10474 2000
TOTAL 16136 10925 17866 25302 9089 15297 7937 7662 18652 13073 18172 16815 19958 7097
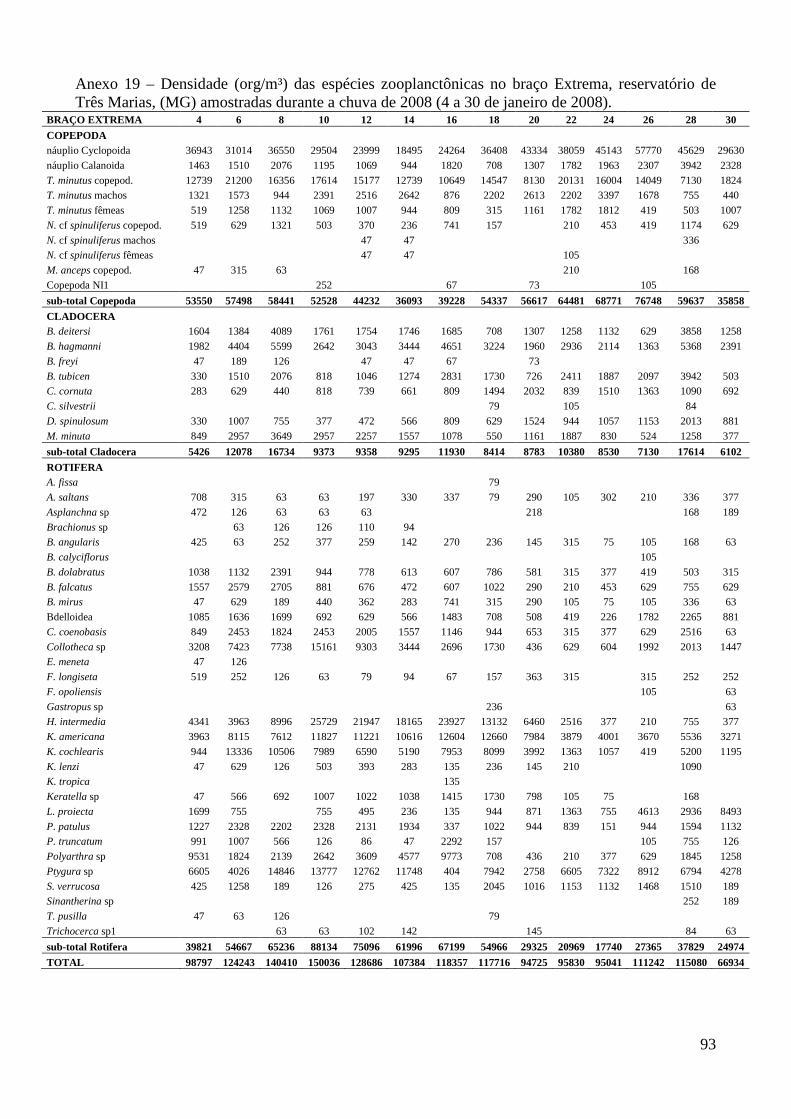
93
Anexo 19 – Densidade (org/m³) das espécies zooplanctônicas no braço Extrema, reservatório de Três Marias, (MG) amostradas durante a chuva de 2008 (4 a 30 de janeiro de 2008).
BRAÇO EXTREMA 4 6 8 10 12 14 16 18 20 22 24 26 28 30
COPEPODA
náuplio Cyclopoida 36943 31014 36550 29504 23999 18495 24264 36408 43334 38059 45143 57770 45629 29630
náuplio Calanoida 1463 1510 2076 1195 1069 944 1820 708 1307 1782 1963 2307 3942 2328
T. minutus copepod. 12739 21200 16356 17614 15177 12739 10649 14547 8130 20131 16004 14049 7130 1824 T. minutus machos 1321 1573 944 2391 2516 2642 876 2202 2613 2202 3397 1678 755 440
T. minutus fêmeas 519 1258 1132 1069 1007 944 809 315 1161 1782 1812 419 503 1007
N. cf spinuliferus copepod. 519 629 1321 503 370 236 741 157 210 453 419 1174 629
N. cf spinuliferus machos 47 47 336 N. cf spinuliferus fêmeas 47 47 105
M. anceps copepod. 47 315 63 210 168
Copepoda NI1 252 67 73 105
sub-total Copepoda 53550 57498 58441 52528 44232 36093 39228 54337 56617 64481 68771 76748 59637 35858
CLADOCERA B. deitersi 1604 1384 4089 1761 1754 1746 1685 708 1307 1258 1132 629 3858 1258
B. hagmanni 1982 4404 5599 2642 3043 3444 4651 3224 1960 2936 2114 1363 5368 2391
B. freyi 47 189 126 47 47 67 73
B. tubicen 330 1510 2076 818 1046 1274 2831 1730 726 2411 1887 2097 3942 503 C. cornuta 283 629 440 818 739 661 809 1494 2032 839 1510 1363 1090 692
C. silvestrii 79 105 84
D. spinulosum 330 1007 755 377 472 566 809 629 1524 944 1057 1153 2013 881
M. minuta 849 2957 3649 2957 2257 1557 1078 550 1161 1887 830 524 1258 377
sub-total Cladocera 5426 12078 16734 9373 9358 9295 11930 8414 8783 10380 8530 7130 17614 6102
ROTIFERA A. fissa 79
A. saltans 708 315 63 63 197 330 337 79 290 105 302 210 336 377
Asplanchna sp 472 126 63 63 63 218 168 189
Brachionus sp 63 126 126 110 94 B. angularis 425 63 252 377 259 142 270 236 145 315 75 105 168 63
B. calyciflorus 105
B. dolabratus 1038 1132 2391 944 778 613 607 786 581 315 377 419 503 315
B. falcatus 1557 2579 2705 881 676 472 607 1022 290 210 453 629 755 629 B. mirus 47 629 189 440 362 283 741 315 290 105 75 105 336 63
Bdelloidea 1085 1636 1699 692 629 566 1483 708 508 419 226 1782 2265 881
C. coenobasis 849 2453 1824 2453 2005 1557 1146 944 653 315 377 629 2516 63
Collotheca sp 3208 7423 7738 15161 9303 3444 2696 1730 436 629 604 1992 2013 1447 E. meneta 47 126
F. longiseta 519 252 126 63 79 94 67 157 363 315 315 252 252
F. opoliensis 105 63
Gastropus sp 236 63 H. intermedia 4341 3963 8996 25729 21947 18165 23927 13132 6460 2516 377 210 755 377
K. americana 3963 8115 7612 11827 11221 10616 12604 12660 7984 3879 4001 3670 5536 3271
K. cochlearis 944 13336 10506 7989 6590 5190 7953 8099 3992 1363 1057 419 5200 1195
K. lenzi 47 629 126 503 393 283 135 236 145 210 1090 K. tropica 135
Keratella sp 47 566 692 1007 1022 1038 1415 1730 798 105 75 168
L. proiecta 1699 755 755 495 236 135 944 871 1363 755 4613 2936 8493
P. patulus 1227 2328 2202 2328 2131 1934 337 1022 944 839 151 944 1594 1132 P. truncatum 991 1007 566 126 86 47 2292 157 105 755 126
Polyarthra sp 9531 1824 2139 2642 3609 4577 9773 708 436 210 377 629 1845 1258
Ptygura sp 6605 4026 14846 13777 12762 11748 404 7942 2758 6605 7322 8912 6794 4278
S. verrucosa 425 1258 189 126 275 425 135 2045 1016 1153 1132 1468 1510 189 Sinantherina sp 252 189
T. pusilla 47 63 126 79
Trichocerca sp1 63 63 102 142 145 84 63
sub-total Rotifera 39821 54667 65236 88134 75096 61996 67199 54966 29325 20969 17740 27365 37829 24974
TOTAL 98797 124243 140410 150036 128686 107384 118357 117716 94725 95830 95041 111242 115080 66934
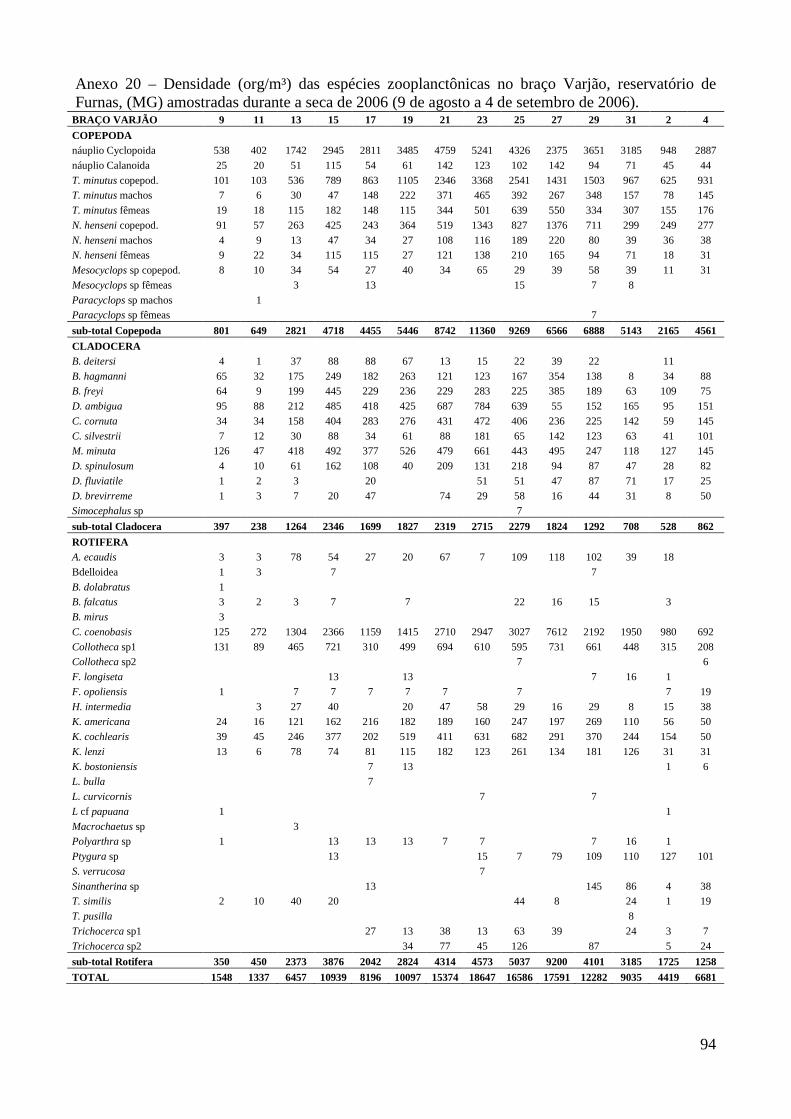
94
Anexo 20 – Densidade (org/m³) das espécies zooplanctônicas no braço Varjão, reservatório de Furnas, (MG) amostradas durante a seca de 2006 (9 de agosto a 4 de setembro de 2006).
BRAÇO VARJÃO 9 11 13 15 17 19 21 23 25 27 29 31 2 4
COPEPODA
náuplio Cyclopoida 538 402 1742 2945 2811 3485 4759 5241 4326 2375 3651 3185 948 2887
náuplio Calanoida 25 20 51 115 54 61 142 123 102 142 94 71 45 44
T. minutus copepod. 101 103 536 789 863 1105 2346 3368 2541 1431 1503 967 625 931 T. minutus machos 7 6 30 47 148 222 371 465 392 267 348 157 78 145
T. minutus fêmeas 19 18 115 182 148 115 344 501 639 550 334 307 155 176
N. henseni copepod. 91 57 263 425 243 364 519 1343 827 1376 711 299 249 277
N. henseni machos 4 9 13 47 34 27 108 116 189 220 80 39 36 38 N. henseni fêmeas 9 22 34 115 115 27 121 138 210 165 94 71 18 31
Mesocyclops sp copepod. 8 10 34 54 27 40 34 65 29 39 58 39 11 31
Mesocyclops sp fêmeas 3 13 15 7 8
Paracyclops sp machos 1 Paracyclops sp fêmeas 7
sub-total Copepoda 801 649 2821 4718 4455 5446 8742 11360 9269 6566 6888 5143 2165 4561
CLADOCERA
B. deitersi 4 1 37 88 88 67 13 15 22 39 22 11
B. hagmanni 65 32 175 249 182 263 121 123 167 354 138 8 34 88 B. freyi 64 9 199 445 229 236 229 283 225 385 189 63 109 75
D. ambigua 95 88 212 485 418 425 687 784 639 55 152 165 95 151
C. cornuta 34 34 158 404 283 276 431 472 406 236 225 142 59 145
C. silvestrii 7 12 30 88 34 61 88 181 65 142 123 63 41 101 M. minuta 126 47 418 492 377 526 479 661 443 495 247 118 127 145
D. spinulosum 4 10 61 162 108 40 209 131 218 94 87 47 28 82
D. fluviatile 1 2 3 20 51 51 47 87 71 17 25
D. brevirreme 1 3 7 20 47 74 29 58 16 44 31 8 50 Simocephalus sp 7
sub-total Cladocera 397 238 1264 2346 1699 1827 2319 2715 2279 1824 1292 708 528 862
ROTIFERA
A. ecaudis 3 3 78 54 27 20 67 7 109 118 102 39 18
Bdelloidea 1 3 7 7
B. dolabratus 1 B. falcatus 3 2 3 7 7 22 16 15 3
B. mirus 3
C. coenobasis 125 272 1304 2366 1159 1415 2710 2947 3027 7612 2192 1950 980 692
Collotheca sp1 131 89 465 721 310 499 694 610 595 731 661 448 315 208 Collotheca sp2 7 6
F. longiseta 13 13 7 16 1
F. opoliensis 1 7 7 7 7 7 7 7 19
H. intermedia 3 27 40 20 47 58 29 16 29 8 15 38 K. americana 24 16 121 162 216 182 189 160 247 197 269 110 56 50
K. cochlearis 39 45 246 377 202 519 411 631 682 291 370 244 154 50
K. lenzi 13 6 78 74 81 115 182 123 261 134 181 126 31 31
K. bostoniensis 7 13 1 6 L. bulla 7
L. curvicornis 7 7
L cf papuana 1 1
Macrochaetus sp 3 Polyarthra sp 1 13 13 13 7 7 7 16 1
Ptygura sp 13 15 7 79 109 110 127 101
S. verrucosa 7
Sinantherina sp 13 145 86 4 38 T. similis 2 10 40 20 44 8 24 1 19
T. pusilla 8
Trichocerca sp1 27 13 38 13 63 39 24 3 7
Trichocerca sp2 34 77 45 126 87 5 24
sub-total Rotifera 350 450 2373 3876 2042 2824 4314 4573 5037 9200 4101 3185 1725 1258
TOTAL 1548 1337 6457 10939 8196 10097 15374 18647 16586 17591 12282 9035 4419 6681
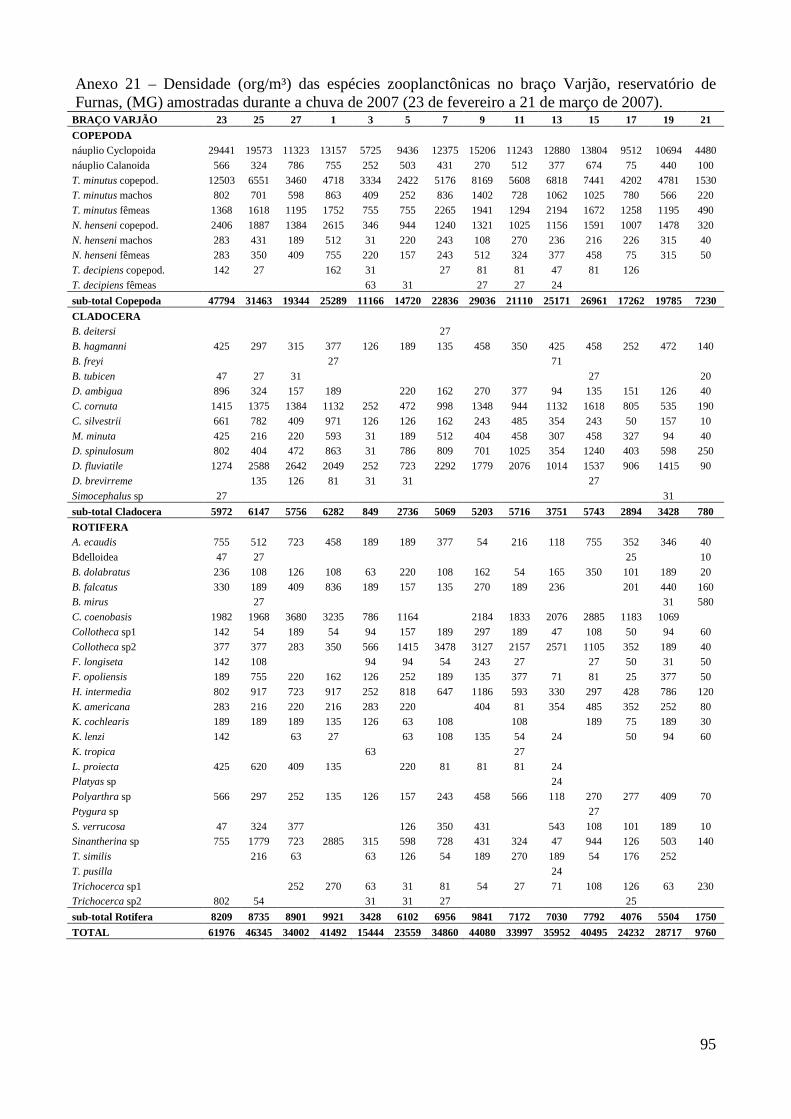
95
Anexo 21 – Densidade (org/m³) das espécies zooplanctônicas no braço Varjão, reservatório de Furnas, (MG) amostradas durante a chuva de 2007 (23 de fevereiro a 21 de março de 2007).
BRAÇO VARJÃO 23 25 27 1 3 5 7 9 11 13 15 17 19 21
COPEPODA
náuplio Cyclopoida 29441 19573 11323 13157 5725 9436 12375 15206 11243 12880 13804 9512 10694 4480
náuplio Calanoida 566 324 786 755 252 503 431 270 512 377 674 75 440 100
T. minutus copepod. 12503 6551 3460 4718 3334 2422 5176 8169 5608 6818 7441 4202 4781 1530 T. minutus machos 802 701 598 863 409 252 836 1402 728 1062 1025 780 566 220
T. minutus fêmeas 1368 1618 1195 1752 755 755 2265 1941 1294 2194 1672 1258 1195 490
N. henseni copepod. 2406 1887 1384 2615 346 944 1240 1321 1025 1156 1591 1007 1478 320
N. henseni machos 283 431 189 512 31 220 243 108 270 236 216 226 315 40 N. henseni fêmeas 283 350 409 755 220 157 243 512 324 377 458 75 315 50
T. decipiens copepod. 142 27 162 31 27 81 81 47 81 126
T. decipiens fêmeas 63 31 27 27 24
sub-total Copepoda 47794 31463 19344 25289 11166 14720 22836 29036 21110 25171 26961 17262 19785 7230
CLADOCERA B. deitersi 27
B. hagmanni 425 297 315 377 126 189 135 458 350 425 458 252 472 140
B. freyi 27 71
B. tubicen 47 27 31 27 20 D. ambigua 896 324 157 189 220 162 270 377 94 135 151 126 40
C. cornuta 1415 1375 1384 1132 252 472 998 1348 944 1132 1618 805 535 190
C. silvestrii 661 782 409 971 126 126 162 243 485 354 243 50 157 10
M. minuta 425 216 220 593 31 189 512 404 458 307 458 327 94 40 D. spinulosum 802 404 472 863 31 786 809 701 1025 354 1240 403 598 250
D. fluviatile 1274 2588 2642 2049 252 723 2292 1779 2076 1014 1537 906 1415 90
D. brevirreme 135 126 81 31 31 27
Simocephalus sp 27 31
sub-total Cladocera 5972 6147 5756 6282 849 2736 5069 5203 5716 3751 5743 2894 3428 780
ROTIFERA A. ecaudis 755 512 723 458 189 189 377 54 216 118 755 352 346 40
Bdelloidea 47 27 25 10
B. dolabratus 236 108 126 108 63 220 108 162 54 165 350 101 189 20
B. falcatus 330 189 409 836 189 157 135 270 189 236 201 440 160 B. mirus 27 31 580
C. coenobasis 1982 1968 3680 3235 786 1164 2184 1833 2076 2885 1183 1069
Collotheca sp1 142 54 189 54 94 157 189 297 189 47 108 50 94 60
Collotheca sp2 377 377 283 350 566 1415 3478 3127 2157 2571 1105 352 189 40 F. longiseta 142 108 94 94 54 243 27 27 50 31 50
F. opoliensis 189 755 220 162 126 252 189 135 377 71 81 25 377 50
H. intermedia 802 917 723 917 252 818 647 1186 593 330 297 428 786 120
K. americana 283 216 220 216 283 220 404 81 354 485 352 252 80 K. cochlearis 189 189 189 135 126 63 108 108 189 75 189 30
K. lenzi 142 63 27 63 108 135 54 24 50 94 60
K. tropica 63 27
L. proiecta 425 620 409 135 220 81 81 81 24 Platyas sp 24
Polyarthra sp 566 297 252 135 126 157 243 458 566 118 270 277 409 70
Ptygura sp 27
S. verrucosa 47 324 377 126 350 431 543 108 101 189 10 Sinantherina sp 755 1779 723 2885 315 598 728 431 324 47 944 126 503 140
T. similis 216 63 63 126 54 189 270 189 54 176 252
T. pusilla 24
Trichocerca sp1 252 270 63 31 81 54 27 71 108 126 63 230 Trichocerca sp2 802 54 31 31 27 25
sub-total Rotifera 8209 8735 8901 9921 3428 6102 6956 9841 7172 7030 7792 4076 5504 1750
TOTAL 61976 46345 34002 41492 15444 23559 34860 44080 33997 35952 40495 24232 28717 9760

96
Anexo 22 – Densidade (org/m³) das espécies zooplanctônicas no braço Varjão, reservatório de Furnas, (MG) amostradas durante a seca de 2007 (9 de agosto a 4 de setembro de 2007).
BRAÇO VARJÃO 9 11 13 15 17 19 21 23 25 27 29 31 2 4
COPEPODA
náuplio Cyclopoida 530 278 360 1420 1261 1569 1916 1966 2095 2089 3306 3262 1307 2402
náuplio Calanoida 33 11 14 64 30 59 66 60 82 63 101 101 42 118
T. minutus copepod. 31 8 24 85 88 71 142 110 248 126 381 283 129 217 T. minutus machos 8 6 6 19 25 33 54 53 72 44 60 72 52 38
T. minutus fêmeas 16 15 14 54 14 38 50 94 94 91 104 148 50 132
N. henseni copepod. 69 25 85 120 105 139 219 179 261 384 349 481 113 425
N. henseni machos 3 4 5 12 13 33 21 22 44 28 38 53 14 38 N. henseni fêmeas 20 6 16 17 9 24 35 31 50 69 41 75 30 104
M. anceps copepod. 3 2 19 9 21 12 9 3 25 25 19 16 9
M. anceps machos 3
M. anceps fêmeas 3 3 Copepoda NI 2
sub-total Copepoda 714 356 524 1812 1555 1986 2515 2526 2950 2919 4404 4498 1760 3482
CLADOCERA
B. deitersi 8 13 172 61 73 73 154 293 321 481 692 267 184
B. hagmanni 36 22 28 104 20 21 40 47 72 79 113 167 42 71 B. freyi 248 60 131 302 131 118 198 82 120 79 230 258 68 184
D. ambigua 38 23 75 90 49 64 142 267 355 384 337 355 154 429
C. cornuta 33 17 22 144 42 90 219 248 198 123 286 308 85 146
C. silvestrii 11 30 33 99 28 52 40 13 31 9 19 35 25 57 M. minuta 86 42 74 366 159 205 241 164 233 176 154 154 58 75
D. spinulosum 5 5 3 19 13 14 17 9 31 13 25 22 6 19
D. fluviatile 16 12 22 33 30 12 19 31 38 44 79 9 11 42
D. brevirreme 1 17 7 2 7 9 13 6 3 5 9
sub-total Cladocera 437 191 377 1059 453 561 875 815 1016 840 1135 1145 412 962
ROTIFERA A. ecaudis 5 5 38 17 9 21 25 35 25 31 16 17
Asplanchna sp 3 2
Bdelloidea 8 1 11 9 5 6 3 3 2
B. falcatus 2 C. coenobasis 335 206 263 550 396 774 550 472 1142 535 887 1205 310 38
Collotheca sp1 132 128 148 569 357 444 347 252 374 315 393 541 258 146
Collotheca sp2 2 5
Cephalodella sp 2 E. meneta
F. longiseta 6 1 2 5 6 19 9 6 16 5
F. opoliensis 2 2
Gastropus sp 2 H. intermedia 13 1 6 12 11 14 3 3 16 9 6
K. americana 6 4 2 5 5 14 7 9 9 22 6 9 6 5
K. cochlearis 35 12 9 17 35 33 41 44 94 91 28
K. lenzi 6 2 2 3 6 6 3 5 K. bostoniensis 2 2 6 2
L. bulla 3
L. leontina 2 3
L. curvicornis 3 Macrochaetus sp 2 3 3
Platyas sp 44
P. truncatum 2 3 5 6 2 21 6 16 57 35 13 9
Polyarthra sp 1 6 13 9 19 Ptygura sp 2 2
Trichocerca sp1 14 7 27 7 34 8 7 6 58 38 78 53 64 38
Trichocerca sp2 6 1 4 2 5 1 5 9 12 13 9 6 5
Trichocerca sp3 2
sub-total Rotifera 570 368 465 1193 876 1323 1017 821 1718 1063 1594 1969 709 264
TOTAL 1722 915 1366 4064 2884 3871 4407 4161 5685 4822 7134 7612 2881 4709
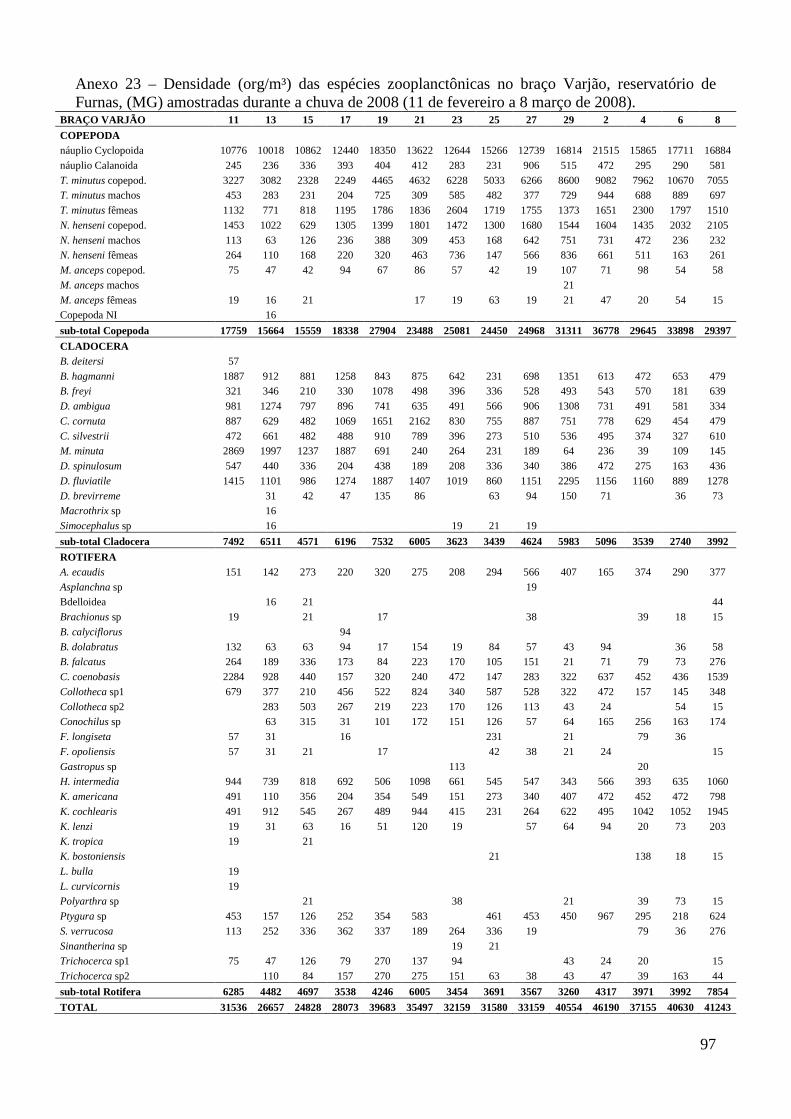
97
Anexo 23 – Densidade (org/m³) das espécies zooplanctônicas no braço Varjão, reservatório de Furnas, (MG) amostradas durante a chuva de 2008 (11 de fevereiro a 8 março de 2008).
BRAÇO VARJÃO 11 13 15 17 19 21 23 25 27 29 2 4 6 8
COPEPODA
náuplio Cyclopoida 10776 10018 10862 12440 18350 13622 12644 15266 12739 16814 21515 15865 17711 16884
náuplio Calanoida 245 236 336 393 404 412 283 231 906 515 472 295 290 581
T. minutus copepod. 3227 3082 2328 2249 4465 4632 6228 5033 6266 8600 9082 7962 10670 7055 T. minutus machos 453 283 231 204 725 309 585 482 377 729 944 688 889 697
T. minutus fêmeas 1132 771 818 1195 1786 1836 2604 1719 1755 1373 1651 2300 1797 1510
N. henseni copepod. 1453 1022 629 1305 1399 1801 1472 1300 1680 1544 1604 1435 2032 2105
N. henseni machos 113 63 126 236 388 309 453 168 642 751 731 472 236 232 N. henseni fêmeas 264 110 168 220 320 463 736 147 566 836 661 511 163 261
M. anceps copepod. 75 47 42 94 67 86 57 42 19 107 71 98 54 58
M. anceps machos 21
M. anceps fêmeas 19 16 21 17 19 63 19 21 47 20 54 15 Copepoda NI 16
sub-total Copepoda 17759 15664 15559 18338 27904 23488 25081 24450 24968 31311 36778 29645 33898 29397
CLADOCERA
B. deitersi 57
B. hagmanni 1887 912 881 1258 843 875 642 231 698 1351 613 472 653 479 B. freyi 321 346 210 330 1078 498 396 336 528 493 543 570 181 639
D. ambigua 981 1274 797 896 741 635 491 566 906 1308 731 491 581 334
C. cornuta 887 629 482 1069 1651 2162 830 755 887 751 778 629 454 479
C. silvestrii 472 661 482 488 910 789 396 273 510 536 495 374 327 610 M. minuta 2869 1997 1237 1887 691 240 264 231 189 64 236 39 109 145
D. spinulosum 547 440 336 204 438 189 208 336 340 386 472 275 163 436
D. fluviatile 1415 1101 986 1274 1887 1407 1019 860 1151 2295 1156 1160 889 1278
D. brevirreme 31 42 47 135 86 63 94 150 71 36 73 Macrothrix sp 16
Simocephalus sp 16 19 21 19
sub-total Cladocera 7492 6511 4571 6196 7532 6005 3623 3439 4624 5983 5096 3539 2740 3992
ROTIFERA
A. ecaudis 151 142 273 220 320 275 208 294 566 407 165 374 290 377
Asplanchna sp 19 Bdelloidea 16 21 44
Brachionus sp 19 21 17 38 39 18 15
B. calyciflorus 94
B. dolabratus 132 63 63 94 17 154 19 84 57 43 94 36 58 B. falcatus 264 189 336 173 84 223 170 105 151 21 71 79 73 276
C. coenobasis 2284 928 440 157 320 240 472 147 283 322 637 452 436 1539
Collotheca sp1 679 377 210 456 522 824 340 587 528 322 472 157 145 348
Collotheca sp2 283 503 267 219 223 170 126 113 43 24 54 15 Conochilus sp 63 315 31 101 172 151 126 57 64 165 256 163 174
F. longiseta 57 31 16 231 21 79 36
F. opoliensis 57 31 21 17 42 38 21 24 15
Gastropus sp 113 20 H. intermedia 944 739 818 692 506 1098 661 545 547 343 566 393 635 1060
K. americana 491 110 356 204 354 549 151 273 340 407 472 452 472 798
K. cochlearis 491 912 545 267 489 944 415 231 264 622 495 1042 1052 1945
K. lenzi 19 31 63 16 51 120 19 57 64 94 20 73 203 K. tropica 19 21
K. bostoniensis 21 138 18 15
L. bulla 19
L. curvicornis 19 Polyarthra sp 21 38 21 39 73 15
Ptygura sp 453 157 126 252 354 583 461 453 450 967 295 218 624
S. verrucosa 113 252 336 362 337 189 264 336 19 79 36 276
Sinantherina sp 19 21 Trichocerca sp1 75 47 126 79 270 137 94 43 24 20 15
Trichocerca sp2 110 84 157 270 275 151 63 38 43 47 39 163 44
sub-total Rotifera 6285 4482 4697 3538 4246 6005 3454 3691 3567 3260 4317 3971 3992 7854
TOTAL 31536 26657 24828 28073 39683 35497 32159 31580 33159 40554 46190 37155 40630 41243
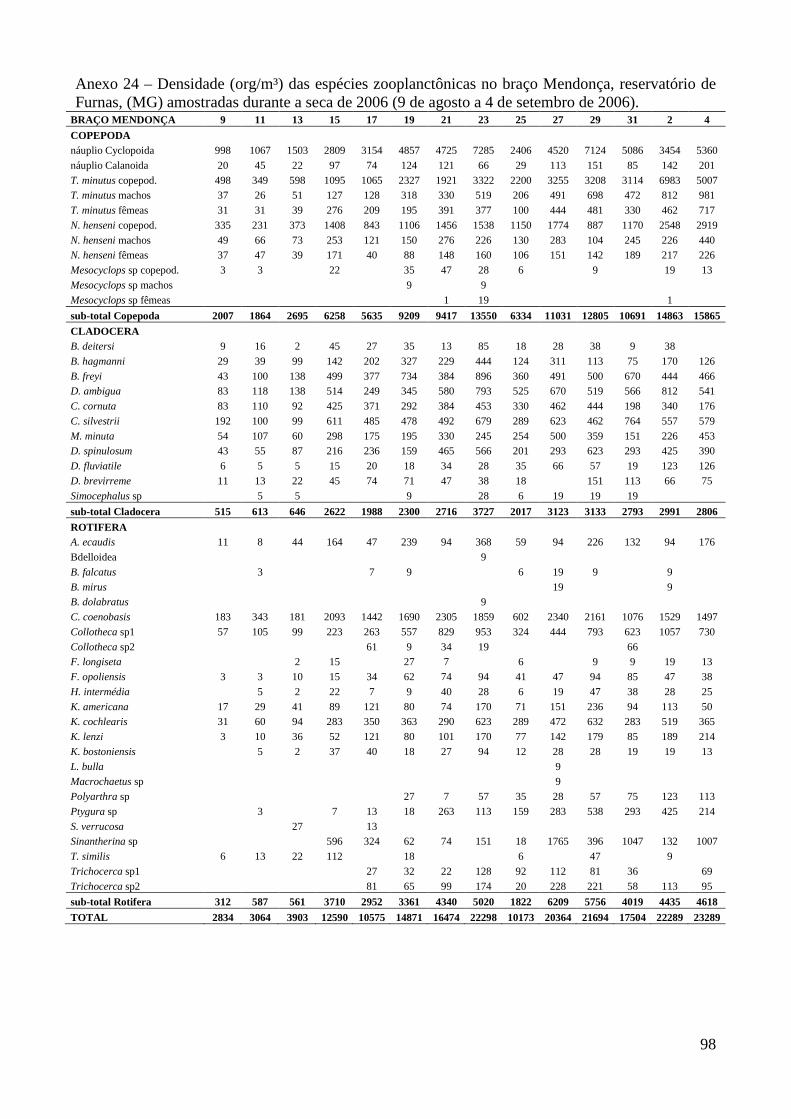
98
Anexo 24 – Densidade (org/m³) das espécies zooplanctônicas no braço Mendonça, reservatório de Furnas, (MG) amostradas durante a seca de 2006 (9 de agosto a 4 de setembro de 2006).
BRAÇO MENDONÇA 9 11 13 15 17 19 21 23 25 27 29 31 2 4
COPEPODA
náuplio Cyclopoida 998 1067 1503 2809 3154 4857 4725 7285 2406 4520 7124 5086 3454 5360
náuplio Calanoida 20 45 22 97 74 124 121 66 29 113 151 85 142 201
T. minutus copepod. 498 349 598 1095 1065 2327 1921 3322 2200 3255 3208 3114 6983 5007 T. minutus machos 37 26 51 127 128 318 330 519 206 491 698 472 812 981
T. minutus fêmeas 31 31 39 276 209 195 391 377 100 444 481 330 462 717
N. henseni copepod. 335 231 373 1408 843 1106 1456 1538 1150 1774 887 1170 2548 2919
N. henseni machos 49 66 73 253 121 150 276 226 130 283 104 245 226 440 N. henseni fêmeas 37 47 39 171 40 88 148 160 106 151 142 189 217 226
Mesocyclops sp copepod. 3 3 22 35 47 28 6 9 19 13
Mesocyclops sp machos 9 9
Mesocyclops sp fêmeas 1 19 1
sub-total Copepoda 2007 1864 2695 6258 5635 9209 9417 13550 6334 11031 12805 10691 14863 15865
CLADOCERA
B. deitersi 9 16 2 45 27 35 13 85 18 28 38 9 38
B. hagmanni 29 39 99 142 202 327 229 444 124 311 113 75 170 126
B. freyi 43 100 138 499 377 734 384 896 360 491 500 670 444 466 D. ambigua 83 118 138 514 249 345 580 793 525 670 519 566 812 541
C. cornuta 83 110 92 425 371 292 384 453 330 462 444 198 340 176
C. silvestrii 192 100 99 611 485 478 492 679 289 623 462 764 557 579
M. minuta 54 107 60 298 175 195 330 245 254 500 359 151 226 453 D. spinulosum 43 55 87 216 236 159 465 566 201 293 623 293 425 390
D. fluviatile 6 5 5 15 20 18 34 28 35 66 57 19 123 126
D. brevirreme 11 13 22 45 74 71 47 38 18 151 113 66 75
Simocephalus sp 5 5 9 28 6 19 19 19
sub-total Cladocera 515 613 646 2622 1988 2300 2716 3727 2017 3123 3133 2793 2991 2806
ROTIFERA A. ecaudis 11 8 44 164 47 239 94 368 59 94 226 132 94 176
Bdelloidea 9
B. falcatus 3 7 9 6 19 9 9
B. mirus 19 9 B. dolabratus 9
C. coenobasis 183 343 181 2093 1442 1690 2305 1859 602 2340 2161 1076 1529 1497
Collotheca sp1 57 105 99 223 263 557 829 953 324 444 793 623 1057 730
Collotheca sp2 61 9 34 19 66 F. longiseta 2 15 27 7 6 9 9 19 13
F. opoliensis 3 3 10 15 34 62 74 94 41 47 94 85 47 38
H. intermédia 5 2 22 7 9 40 28 6 19 47 38 28 25
K. americana 17 29 41 89 121 80 74 170 71 151 236 94 113 50 K. cochlearis 31 60 94 283 350 363 290 623 289 472 632 283 519 365
K. lenzi 3 10 36 52 121 80 101 170 77 142 179 85 189 214
K. bostoniensis 5 2 37 40 18 27 94 12 28 28 19 19 13
L. bulla 9 Macrochaetus sp 9
Polyarthra sp 27 7 57 35 28 57 75 123 113
Ptygura sp 3 7 13 18 263 113 159 283 538 293 425 214
S. verrucosa 27 13 Sinantherina sp 596 324 62 74 151 18 1765 396 1047 132 1007
T. similis 6 13 22 112 18 6 47 9
Trichocerca sp1 27 32 22 128 92 112 81 36 69
Trichocerca sp2 81 65 99 174 20 228 221 58 113 95
sub-total Rotifera 312 587 561 3710 2952 3361 4340 5020 1822 6209 5756 4019 4435 4618
TOTAL 2834 3064 3903 12590 10575 14871 16474 22298 10173 20364 21694 17504 22289 23289
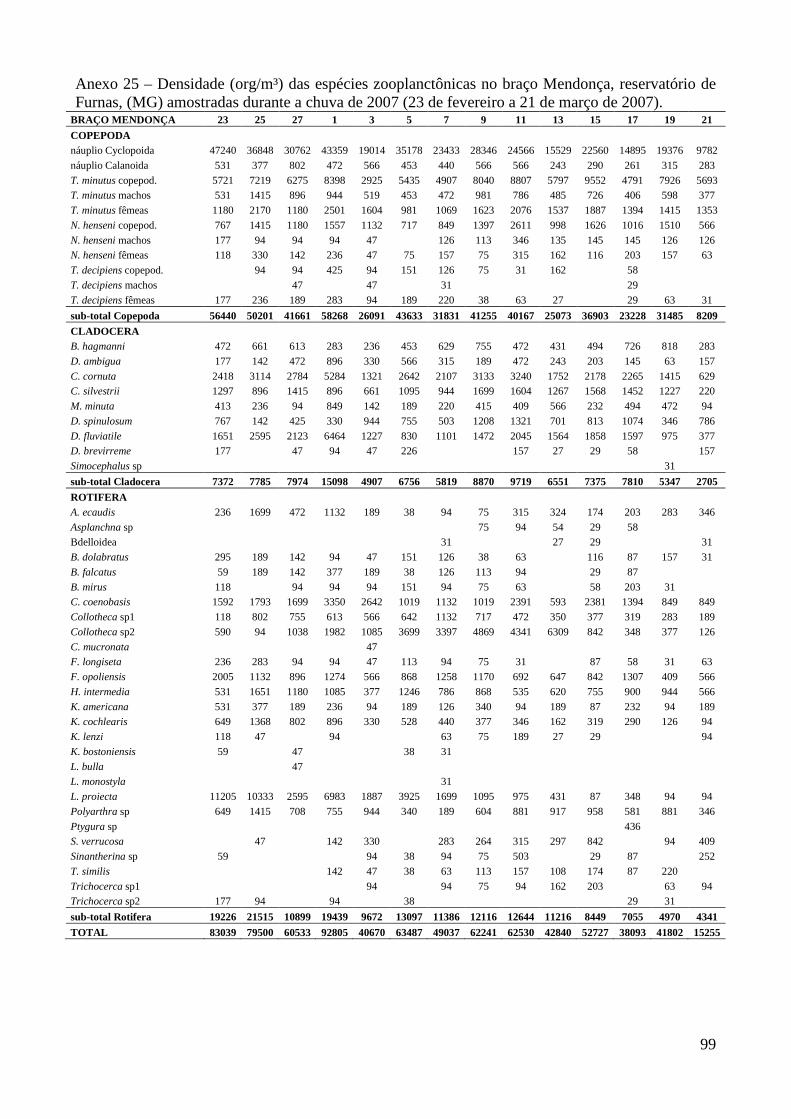
99
Anexo 25 – Densidade (org/m³) das espécies zooplanctônicas no braço Mendonça, reservatório de Furnas, (MG) amostradas durante a chuva de 2007 (23 de fevereiro a 21 de março de 2007).
BRAÇO MENDONÇA 23 25 27 1 3 5 7 9 11 13 15 17 19 21
COPEPODA
náuplio Cyclopoida 47240 36848 30762 43359 19014 35178 23433 28346 24566 15529 22560 14895 19376 9782
náuplio Calanoida 531 377 802 472 566 453 440 566 566 243 290 261 315 283
T. minutus copepod. 5721 7219 6275 8398 2925 5435 4907 8040 8807 5797 9552 4791 7926 5693 T. minutus machos 531 1415 896 944 519 453 472 981 786 485 726 406 598 377
T. minutus fêmeas 1180 2170 1180 2501 1604 981 1069 1623 2076 1537 1887 1394 1415 1353
N. henseni copepod. 767 1415 1180 1557 1132 717 849 1397 2611 998 1626 1016 1510 566
N. henseni machos 177 94 94 94 47 126 113 346 135 145 145 126 126 N. henseni fêmeas 118 330 142 236 47 75 157 75 315 162 116 203 157 63
T. decipiens copepod. 94 94 425 94 151 126 75 31 162 58
T. decipiens machos 47 47 31 29
T. decipiens fêmeas 177 236 189 283 94 189 220 38 63 27 29 63 31
sub-total Copepoda 56440 50201 41661 58268 26091 43633 31831 41255 40167 25073 36903 23228 31485 8209
CLADOCERA
B. hagmanni 472 661 613 283 236 453 629 755 472 431 494 726 818 283
D. ambigua 177 142 472 896 330 566 315 189 472 243 203 145 63 157
C. cornuta 2418 3114 2784 5284 1321 2642 2107 3133 3240 1752 2178 2265 1415 629 C. silvestrii 1297 896 1415 896 661 1095 944 1699 1604 1267 1568 1452 1227 220
M. minuta 413 236 94 849 142 189 220 415 409 566 232 494 472 94
D. spinulosum 767 142 425 330 944 755 503 1208 1321 701 813 1074 346 786
D. fluviatile 1651 2595 2123 6464 1227 830 1101 1472 2045 1564 1858 1597 975 377 D. brevirreme 177 47 94 47 226 157 27 29 58 157
Simocephalus sp 31
sub-total Cladocera 7372 7785 7974 15098 4907 6756 5819 8870 9719 6551 7375 7810 5347 2705
ROTIFERA
A. ecaudis 236 1699 472 1132 189 38 94 75 315 324 174 203 283 346
Asplanchna sp 75 94 54 29 58 Bdelloidea 31 27 29 31
B. dolabratus 295 189 142 94 47 151 126 38 63 116 87 157 31
B. falcatus 59 189 142 377 189 38 126 113 94 29 87
B. mirus 118 94 94 94 151 94 75 63 58 203 31 C. coenobasis 1592 1793 1699 3350 2642 1019 1132 1019 2391 593 2381 1394 849 849
Collotheca sp1 118 802 755 613 566 642 1132 717 472 350 377 319 283 189
Collotheca sp2 590 94 1038 1982 1085 3699 3397 4869 4341 6309 842 348 377 126
C. mucronata 47 F. longiseta 236 283 94 94 47 113 94 75 31 87 58 31 63
F. opoliensis 2005 1132 896 1274 566 868 1258 1170 692 647 842 1307 409 566
H. intermedia 531 1651 1180 1085 377 1246 786 868 535 620 755 900 944 566
K. americana 531 377 189 236 94 189 126 340 94 189 87 232 94 189 K. cochlearis 649 1368 802 896 330 528 440 377 346 162 319 290 126 94
K. lenzi 118 47 94 63 75 189 27 29 94
K. bostoniensis 59 47 38 31
L. bulla 47 L. monostyla 31
L. proiecta 11205 10333 2595 6983 1887 3925 1699 1095 975 431 87 348 94 94
Polyarthra sp 649 1415 708 755 944 340 189 604 881 917 958 581 881 346
Ptygura sp 436 S. verrucosa 47 142 330 283 264 315 297 842 94 409
Sinantherina sp 59 94 38 94 75 503 29 87 252
T. similis 142 47 38 63 113 157 108 174 87 220
Trichocerca sp1 94 94 75 94 162 203 63 94 Trichocerca sp2 177 94 94 38 29 31
sub-total Rotifera 19226 21515 10899 19439 9672 13097 11386 12116 12644 11216 8449 7055 4970 4341
TOTAL 83039 79500 60533 92805 40670 63487 49037 62241 62530 42840 52727 38093 41802 15255
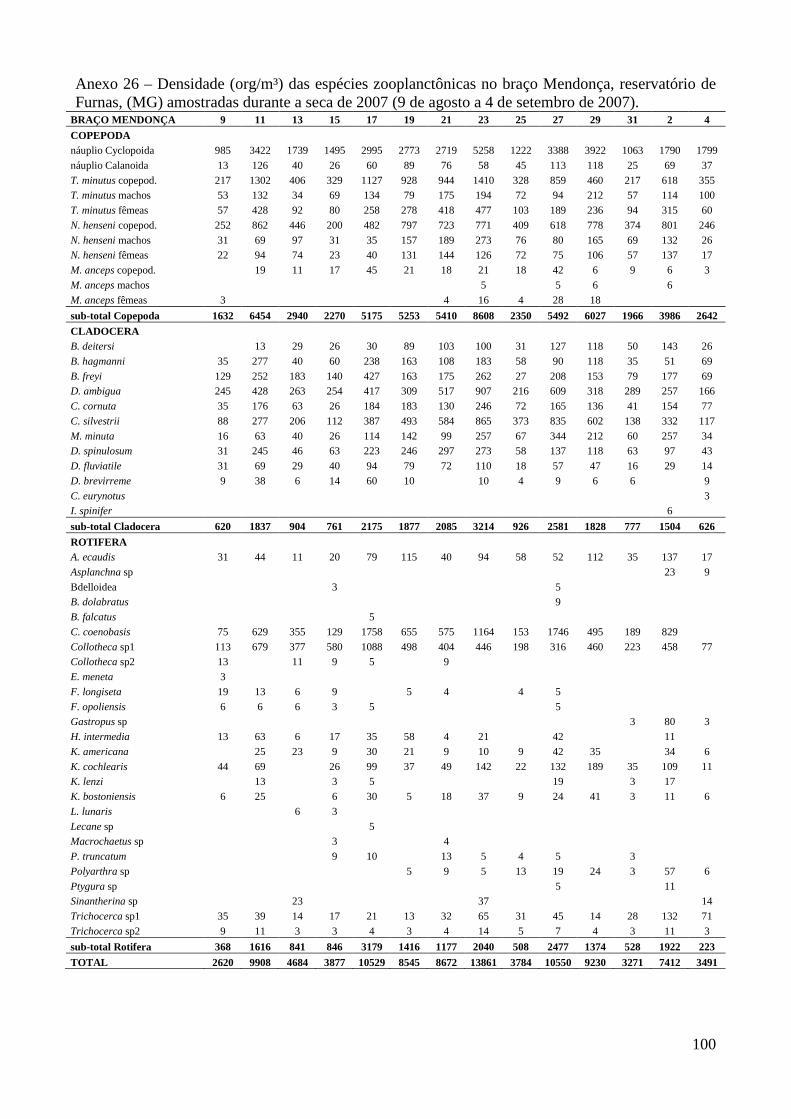
100
Anexo 26 – Densidade (org/m³) das espécies zooplanctônicas no braço Mendonça, reservatório de Furnas, (MG) amostradas durante a seca de 2007 (9 de agosto a 4 de setembro de 2007).
BRAÇO MENDONÇA 9 11 13 15 17 19 21 23 25 27 29 31 2 4
COPEPODA
náuplio Cyclopoida 985 3422 1739 1495 2995 2773 2719 5258 1222 3388 3922 1063 1790 1799
náuplio Calanoida 13 126 40 26 60 89 76 58 45 113 118 25 69 37
T. minutus copepod. 217 1302 406 329 1127 928 944 1410 328 859 460 217 618 355 T. minutus machos 53 132 34 69 134 79 175 194 72 94 212 57 114 100
T. minutus fêmeas 57 428 92 80 258 278 418 477 103 189 236 94 315 60
N. henseni copepod. 252 862 446 200 482 797 723 771 409 618 778 374 801 246
N. henseni machos 31 69 97 31 35 157 189 273 76 80 165 69 132 26 N. henseni fêmeas 22 94 74 23 40 131 144 126 72 75 106 57 137 17
M. anceps copepod. 19 11 17 45 21 18 21 18 42 6 9 6 3
M. anceps machos 5 5 6 6
M. anceps fêmeas 3 4 16 4 28 18
sub-total Copepoda 1632 6454 2940 2270 5175 5253 5410 8608 2350 5492 6027 1966 3986 2642
CLADOCERA
B. deitersi 13 29 26 30 89 103 100 31 127 118 50 143 26
B. hagmanni 35 277 40 60 238 163 108 183 58 90 118 35 51 69
B. freyi 129 252 183 140 427 163 175 262 27 208 153 79 177 69 D. ambigua 245 428 263 254 417 309 517 907 216 609 318 289 257 166
C. cornuta 35 176 63 26 184 183 130 246 72 165 136 41 154 77
C. silvestrii 88 277 206 112 387 493 584 865 373 835 602 138 332 117
M. minuta 16 63 40 26 114 142 99 257 67 344 212 60 257 34 D. spinulosum 31 245 46 63 223 246 297 273 58 137 118 63 97 43
D. fluviatile 31 69 29 40 94 79 72 110 18 57 47 16 29 14
D. brevirreme 9 38 6 14 60 10 10 4 9 6 6 9
C. eurynotus 3 I. spinifer 6
sub-total Cladocera 620 1837 904 761 2175 1877 2085 3214 926 2581 1828 777 1504 626
ROTIFERA
A. ecaudis 31 44 11 20 79 115 40 94 58 52 112 35 137 17
Asplanchna sp 23 9
Bdelloidea 3 5 B. dolabratus 9
B. falcatus 5
C. coenobasis 75 629 355 129 1758 655 575 1164 153 1746 495 189 829
Collotheca sp1 113 679 377 580 1088 498 404 446 198 316 460 223 458 77 Collotheca sp2 13 11 9 5 9
E. meneta 3
F. longiseta 19 13 6 9 5 4 4 5
F. opoliensis 6 6 6 3 5 5 Gastropus sp 3 80 3
H. intermedia 13 63 6 17 35 58 4 21 42 11
K. americana 25 23 9 30 21 9 10 9 42 35 34 6
K. cochlearis 44 69 26 99 37 49 142 22 132 189 35 109 11 K. lenzi 13 3 5 19 3 17
K. bostoniensis 6 25 6 30 5 18 37 9 24 41 3 11 6
L. lunaris 6 3
Lecane sp 5 Macrochaetus sp 3 4
P. truncatum 9 10 13 5 4 5 3
Polyarthra sp 5 9 5 13 19 24 3 57 6
Ptygura sp 5 11 Sinantherina sp 23 37 14
Trichocerca sp1 35 39 14 17 21 13 32 65 31 45 14 28 132 71
Trichocerca sp2 9 11 3 3 4 3 4 14 5 7 4 3 11 3
sub-total Rotifera 368 1616 841 846 3179 1416 1177 2040 508 2477 1374 528 1922 223
TOTAL 2620 9908 4684 3877 10529 8545 8672 13861 3784 10550 9230 3271 7412 3491
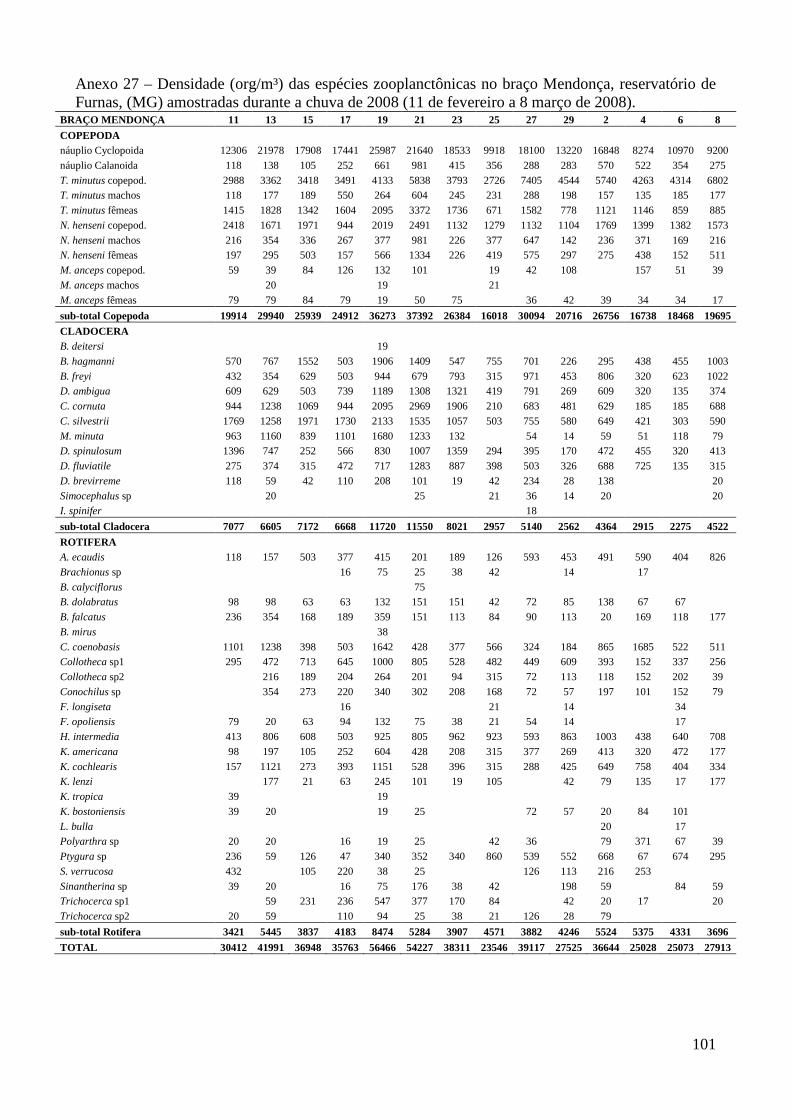
101
Anexo 27 – Densidade (org/m³) das espécies zooplanctônicas no braço Mendonça, reservatório de Furnas, (MG) amostradas durante a chuva de 2008 (11 de fevereiro a 8 março de 2008).
BRAÇO MENDONÇA 11 13 15 17 19 21 23 25 27 29 2 4 6 8
COPEPODA
náuplio Cyclopoida 12306 21978 17908 17441 25987 21640 18533 9918 18100 13220 16848 8274 10970 9200
náuplio Calanoida 118 138 105 252 661 981 415 356 288 283 570 522 354 275
T. minutus copepod. 2988 3362 3418 3491 4133 5838 3793 2726 7405 4544 5740 4263 4314 6802 T. minutus machos 118 177 189 550 264 604 245 231 288 198 157 135 185 177
T. minutus fêmeas 1415 1828 1342 1604 2095 3372 1736 671 1582 778 1121 1146 859 885
N. henseni copepod. 2418 1671 1971 944 2019 2491 1132 1279 1132 1104 1769 1399 1382 1573
N. henseni machos 216 354 336 267 377 981 226 377 647 142 236 371 169 216 N. henseni fêmeas 197 295 503 157 566 1334 226 419 575 297 275 438 152 511
M. anceps copepod. 59 39 84 126 132 101 19 42 108 157 51 39
M. anceps machos 20 19 21
M. anceps fêmeas 79 79 84 79 19 50 75 36 42 39 34 34 17
sub-total Copepoda 19914 29940 25939 24912 36273 37392 26384 16018 30094 20716 26756 16738 18468 19695
CLADOCERA
B. deitersi 19
B. hagmanni 570 767 1552 503 1906 1409 547 755 701 226 295 438 455 1003
B. freyi 432 354 629 503 944 679 793 315 971 453 806 320 623 1022 D. ambigua 609 629 503 739 1189 1308 1321 419 791 269 609 320 135 374
C. cornuta 944 1238 1069 944 2095 2969 1906 210 683 481 629 185 185 688
C. silvestrii 1769 1258 1971 1730 2133 1535 1057 503 755 580 649 421 303 590
M. minuta 963 1160 839 1101 1680 1233 132 54 14 59 51 118 79 D. spinulosum 1396 747 252 566 830 1007 1359 294 395 170 472 455 320 413
D. fluviatile 275 374 315 472 717 1283 887 398 503 326 688 725 135 315
D. brevirreme 118 59 42 110 208 101 19 42 234 28 138 20
Simocephalus sp 20 25 21 36 14 20 20 I. spinifer 18
sub-total Cladocera 7077 6605 7172 6668 11720 11550 8021 2957 5140 2562 4364 2915 2275 4522
ROTIFERA
A. ecaudis 118 157 503 377 415 201 189 126 593 453 491 590 404 826
Brachionus sp 16 75 25 38 42 14 17
B. calyciflorus 75 B. dolabratus 98 98 63 63 132 151 151 42 72 85 138 67 67
B. falcatus 236 354 168 189 359 151 113 84 90 113 20 169 118 177
B. mirus 38
C. coenobasis 1101 1238 398 503 1642 428 377 566 324 184 865 1685 522 511 Collotheca sp1 295 472 713 645 1000 805 528 482 449 609 393 152 337 256
Collotheca sp2 216 189 204 264 201 94 315 72 113 118 152 202 39
Conochilus sp 354 273 220 340 302 208 168 72 57 197 101 152 79
F. longiseta 16 21 14 34 F. opoliensis 79 20 63 94 132 75 38 21 54 14 17
H. intermedia 413 806 608 503 925 805 962 923 593 863 1003 438 640 708
K. americana 98 197 105 252 604 428 208 315 377 269 413 320 472 177
K. cochlearis 157 1121 273 393 1151 528 396 315 288 425 649 758 404 334 K. lenzi 177 21 63 245 101 19 105 42 79 135 17 177
K. tropica 39 19
K. bostoniensis 39 20 19 25 72 57 20 84 101
L. bulla 20 17 Polyarthra sp 20 20 16 19 25 42 36 79 371 67 39
Ptygura sp 236 59 126 47 340 352 340 860 539 552 668 67 674 295
S. verrucosa 432 105 220 38 25 126 113 216 253
Sinantherina sp 39 20 16 75 176 38 42 198 59 84 59 Trichocerca sp1 59 231 236 547 377 170 84 42 20 17 20
Trichocerca sp2 20 59 110 94 25 38 21 126 28 79
sub-total Rotifera 3421 5445 3837 4183 8474 5284 3907 4571 3882 4246 5524 5375 4331 3696
TOTAL 30412 41991 36948 35763 56466 54227 38311 23546 39117 27525 36644 25028 25073 27913
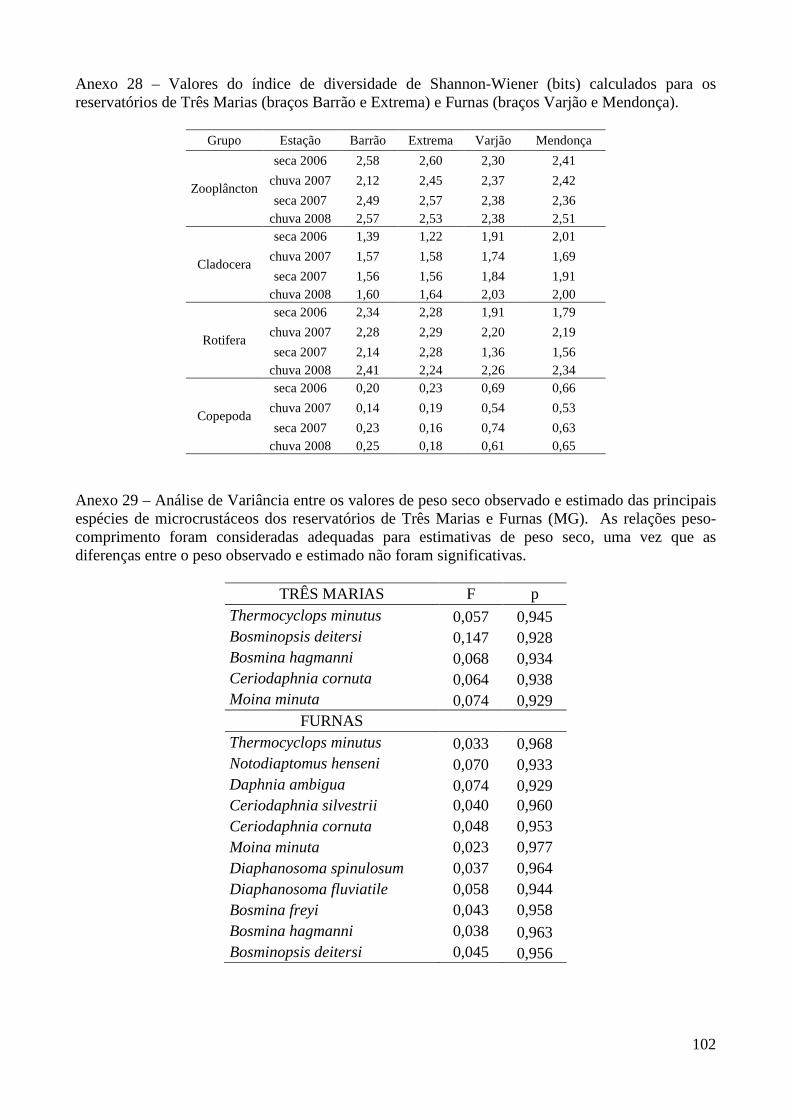
102
Anexo 28 – Valores do índice de diversidade de Shannon-Wiener (bits) calculados para os reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça).
Grupo Estação Barrão Extrema Varjão Mendonça
Zooplâncton
seca 2006 2,58 2,60 2,30 2,41
chuva 2007 2,12 2,45 2,37 2,42
seca 2007 2,49 2,57 2,38 2,36 chuva 2008 2,57 2,53 2,38 2,51
Cladocera
seca 2006 1,39 1,22 1,91 2,01
chuva 2007 1,57 1,58 1,74 1,69
seca 2007 1,56 1,56 1,84 1,91 chuva 2008 1,60 1,64 2,03 2,00
Rotifera
seca 2006 2,34 2,28 1,91 1,79
chuva 2007 2,28 2,29 2,20 2,19
seca 2007 2,14 2,28 1,36 1,56 chuva 2008 2,41 2,24 2,26 2,34
Copepoda
seca 2006 0,20 0,23 0,69 0,66
chuva 2007 0,14 0,19 0,54 0,53
seca 2007 0,23 0,16 0,74 0,63 chuva 2008 0,25 0,18 0,61 0,65
Anexo 29 – Análise de Variância entre os valores de peso seco observado e estimado das principais espécies de microcrustáceos dos reservatórios de Três Marias e Furnas (MG). As relações peso-comprimento foram consideradas adequadas para estimativas de peso seco, uma vez que as diferenças entre o peso observado e estimado não foram significativas.
TRÊS MARIAS F p Thermocyclops minutus 0,057 0,945 Bosminopsis deitersi 0,147 0,928 Bosmina hagmanni 0,068 0,934 Ceriodaphnia cornuta 0,064 0,938 Moina minuta 0,074 0,929
FURNAS Thermocyclops minutus 0,033 0,968 Notodiaptomus henseni 0,070 0,933 Daphnia ambigua 0,074 0,929 Ceriodaphnia silvestrii 0,040 0,960 Ceriodaphnia cornuta 0,048 0,953 Moina minuta 0,023 0,977 Diaphanosoma spinulosum 0,037 0,964 Diaphanosoma fluviatile 0,058 0,944 Bosmina freyi 0,043 0,958 Bosmina hagmanni 0,038 0,963 Bosminopsis deitersi 0,045 0,956
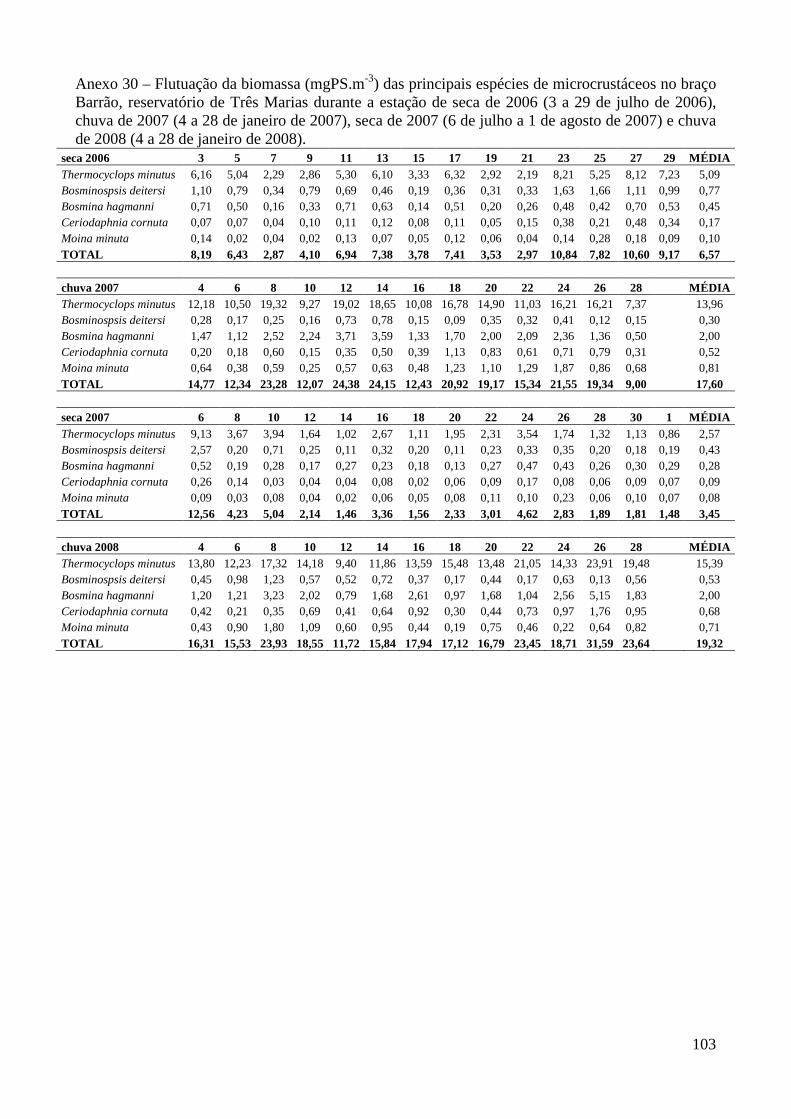
103
Anexo 30 – Flutuação da biomassa (mgPS.m-3) das principais espécies de microcrustáceos no braço Barrão, reservatório de Três Marias durante a estação de seca de 2006 (3 a 29 de julho de 2006), chuva de 2007 (4 a 28 de janeiro de 2007), seca de 2007 (6 de julho a 1 de agosto de 2007) e chuva de 2008 (4 a 28 de janeiro de 2008).
seca 2006 3 5 7 9 11 13 15 17 19 21 23 25 27 29 MÉDIA Thermocyclops minutus 6,16 5,04 2,29 2,86 5,30 6,10 3,33 6,32 2,92 2,19 8,21 5,25 8,12 7,23 5,09 Bosminospsis deitersi 1,10 0,79 0,34 0,79 0,69 0,46 0,19 0,36 0,31 0,33 1,63 1,66 1,11 0,99 0,77 Bosmina hagmanni 0,71 0,50 0,16 0,33 0,71 0,63 0,14 0,51 0,20 0,26 0,48 0,42 0,70 0,53 0,45 Ceriodaphnia cornuta 0,07 0,07 0,04 0,10 0,11 0,12 0,08 0,11 0,05 0,15 0,38 0,21 0,48 0,34 0,17 Moina minuta 0,14 0,02 0,04 0,02 0,13 0,07 0,05 0,12 0,06 0,04 0,14 0,28 0,18 0,09 0,10 TOTAL 8,19 6,43 2,87 4,10 6,94 7,38 3,78 7,41 3,53 2,97 10,84 7,82 10,60 9,17 6,57
chuva 2007 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA Thermocyclops minutus 12,18 10,50 19,32 9,27 19,02 18,65 10,08 16,78 14,90 11,03 16,21 16,21 7,37 13,96 Bosminospsis deitersi 0,28 0,17 0,25 0,16 0,73 0,78 0,15 0,09 0,35 0,32 0,41 0,12 0,15 0,30 Bosmina hagmanni 1,47 1,12 2,52 2,24 3,71 3,59 1,33 1,70 2,00 2,09 2,36 1,36 0,50 2,00 Ceriodaphnia cornuta 0,20 0,18 0,60 0,15 0,35 0,50 0,39 1,13 0,83 0,61 0,71 0,79 0,31 0,52 Moina minuta 0,64 0,38 0,59 0,25 0,57 0,63 0,48 1,23 1,10 1,29 1,87 0,86 0,68 0,81 TOTAL 14,77 12,34 23,28 12,07 24,38 24,15 12,43 20,92 19,17 15,34 21,55 19,34 9,00 17,60
seca 2007 6 8 10 12 14 16 18 20 22 24 26 28 30 1 MÉDIA Thermocyclops minutus 9,13 3,67 3,94 1,64 1,02 2,67 1,11 1,95 2,31 3,54 1,74 1,32 1,13 0,86 2,57 Bosminospsis deitersi 2,57 0,20 0,71 0,25 0,11 0,32 0,20 0,11 0,23 0,33 0,35 0,20 0,18 0,19 0,43 Bosmina hagmanni 0,52 0,19 0,28 0,17 0,27 0,23 0,18 0,13 0,27 0,47 0,43 0,26 0,30 0,29 0,28 Ceriodaphnia cornuta 0,26 0,14 0,03 0,04 0,04 0,08 0,02 0,06 0,09 0,17 0,08 0,06 0,09 0,07 0,09 Moina minuta 0,09 0,03 0,08 0,04 0,02 0,06 0,05 0,08 0,11 0,10 0,23 0,06 0,10 0,07 0,08 TOTAL 12,56 4,23 5,04 2,14 1,46 3,36 1,56 2,33 3,01 4,62 2,83 1,89 1,81 1,48 3,45
chuva 2008 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA Thermocyclops minutus 13,80 12,23 17,32 14,18 9,40 11,86 13,59 15,48 13,48 21,05 14,33 23,91 19,48 15,39 Bosminospsis deitersi 0,45 0,98 1,23 0,57 0,52 0,72 0,37 0,17 0,44 0,17 0,63 0,13 0,56 0,53 Bosmina hagmanni 1,20 1,21 3,23 2,02 0,79 1,68 2,61 0,97 1,68 1,04 2,56 5,15 1,83 2,00 Ceriodaphnia cornuta 0,42 0,21 0,35 0,69 0,41 0,64 0,92 0,30 0,44 0,73 0,97 1,76 0,95 0,68 Moina minuta 0,43 0,90 1,80 1,09 0,60 0,95 0,44 0,19 0,75 0,46 0,22 0,64 0,82 0,71 TOTAL 16,31 15,53 23,93 18,55 11,72 15,84 17,94 17,12 16,79 23,45 18,71 31,59 23,64 19,32
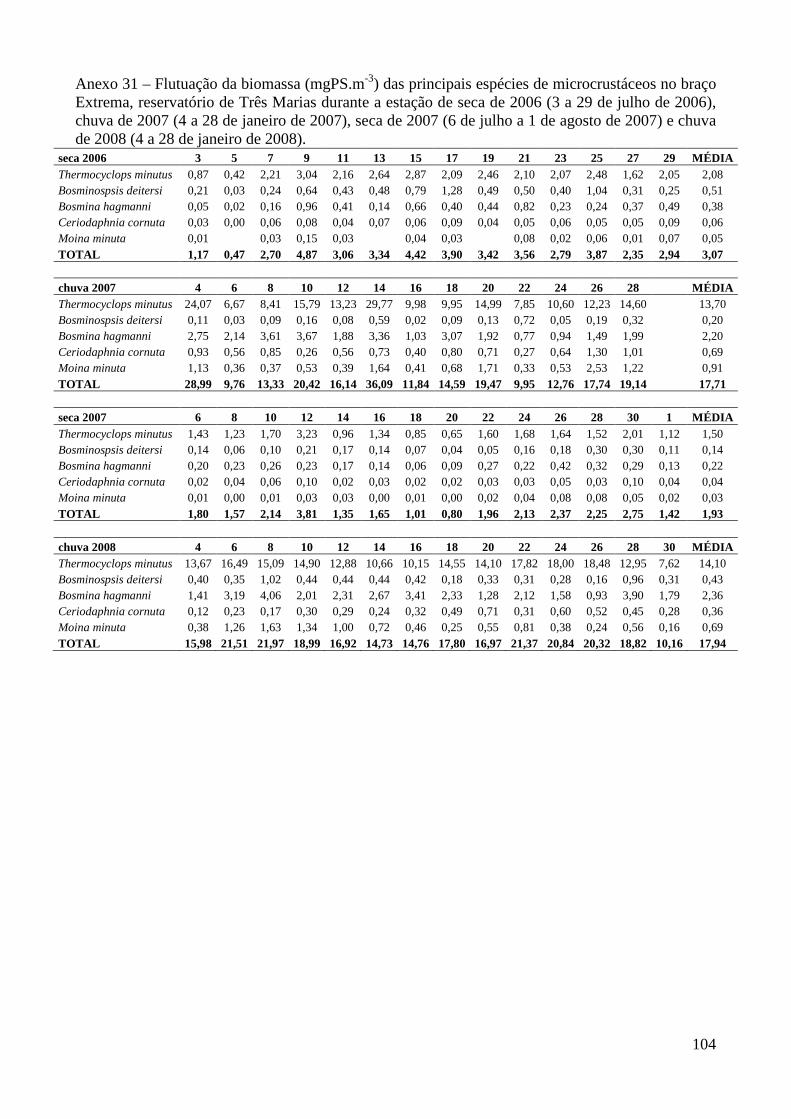
104
Anexo 31 – Flutuação da biomassa (mgPS.m-3) das principais espécies de microcrustáceos no braço Extrema, reservatório de Três Marias durante a estação de seca de 2006 (3 a 29 de julho de 2006), chuva de 2007 (4 a 28 de janeiro de 2007), seca de 2007 (6 de julho a 1 de agosto de 2007) e chuva de 2008 (4 a 28 de janeiro de 2008).
seca 2006 3 5 7 9 11 13 15 17 19 21 23 25 27 29 MÉDIA Thermocyclops minutus 0,87 0,42 2,21 3,04 2,16 2,64 2,87 2,09 2,46 2,10 2,07 2,48 1,62 2,05 2,08 Bosminospsis deitersi 0,21 0,03 0,24 0,64 0,43 0,48 0,79 1,28 0,49 0,50 0,40 1,04 0,31 0,25 0,51 Bosmina hagmanni 0,05 0,02 0,16 0,96 0,41 0,14 0,66 0,40 0,44 0,82 0,23 0,24 0,37 0,49 0,38 Ceriodaphnia cornuta 0,03 0,00 0,06 0,08 0,04 0,07 0,06 0,09 0,04 0,05 0,06 0,05 0,05 0,09 0,06 Moina minuta 0,01 0,03 0,15 0,03 0,04 0,03 0,08 0,02 0,06 0,01 0,07 0,05 TOTAL 1,17 0,47 2,70 4,87 3,06 3,34 4,42 3,90 3,42 3,56 2,79 3,87 2,35 2,94 3,07
chuva 2007 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA Thermocyclops minutus 24,07 6,67 8,41 15,79 13,23 29,77 9,98 9,95 14,99 7,85 10,60 12,23 14,60 13,70 Bosminospsis deitersi 0,11 0,03 0,09 0,16 0,08 0,59 0,02 0,09 0,13 0,72 0,05 0,19 0,32 0,20 Bosmina hagmanni 2,75 2,14 3,61 3,67 1,88 3,36 1,03 3,07 1,92 0,77 0,94 1,49 1,99 2,20 Ceriodaphnia cornuta 0,93 0,56 0,85 0,26 0,56 0,73 0,40 0,80 0,71 0,27 0,64 1,30 1,01 0,69 Moina minuta 1,13 0,36 0,37 0,53 0,39 1,64 0,41 0,68 1,71 0,33 0,53 2,53 1,22 0,91 TOTAL 28,99 9,76 13,33 20,42 16,14 36,09 11,84 14,59 19,47 9,95 12,76 17,74 19,14 17,71
seca 2007 6 8 10 12 14 16 18 20 22 24 26 28 30 1 MÉDIA Thermocyclops minutus 1,43 1,23 1,70 3,23 0,96 1,34 0,85 0,65 1,60 1,68 1,64 1,52 2,01 1,12 1,50 Bosminospsis deitersi 0,14 0,06 0,10 0,21 0,17 0,14 0,07 0,04 0,05 0,16 0,18 0,30 0,30 0,11 0,14 Bosmina hagmanni 0,20 0,23 0,26 0,23 0,17 0,14 0,06 0,09 0,27 0,22 0,42 0,32 0,29 0,13 0,22 Ceriodaphnia cornuta 0,02 0,04 0,06 0,10 0,02 0,03 0,02 0,02 0,03 0,03 0,05 0,03 0,10 0,04 0,04 Moina minuta 0,01 0,00 0,01 0,03 0,03 0,00 0,01 0,00 0,02 0,04 0,08 0,08 0,05 0,02 0,03 TOTAL 1,80 1,57 2,14 3,81 1,35 1,65 1,01 0,80 1,96 2,13 2,37 2,25 2,75 1,42 1,93
chuva 2008 4 6 8 10 12 14 16 18 20 22 24 26 28 30 MÉDIA Thermocyclops minutus 13,67 16,49 15,09 14,90 12,88 10,66 10,15 14,55 14,10 17,82 18,00 18,48 12,95 7,62 14,10 Bosminospsis deitersi 0,40 0,35 1,02 0,44 0,44 0,44 0,42 0,18 0,33 0,31 0,28 0,16 0,96 0,31 0,43 Bosmina hagmanni 1,41 3,19 4,06 2,01 2,31 2,67 3,41 2,33 1,28 2,12 1,58 0,93 3,90 1,79 2,36 Ceriodaphnia cornuta 0,12 0,23 0,17 0,30 0,29 0,24 0,32 0,49 0,71 0,31 0,60 0,52 0,45 0,28 0,36 Moina minuta 0,38 1,26 1,63 1,34 1,00 0,72 0,46 0,25 0,55 0,81 0,38 0,24 0,56 0,16 0,69 TOTAL 15,98 21,51 21,97 18,99 16,92 14,73 14,76 17,80 16,97 21,37 20,84 20,32 18,82 10,16 17,94
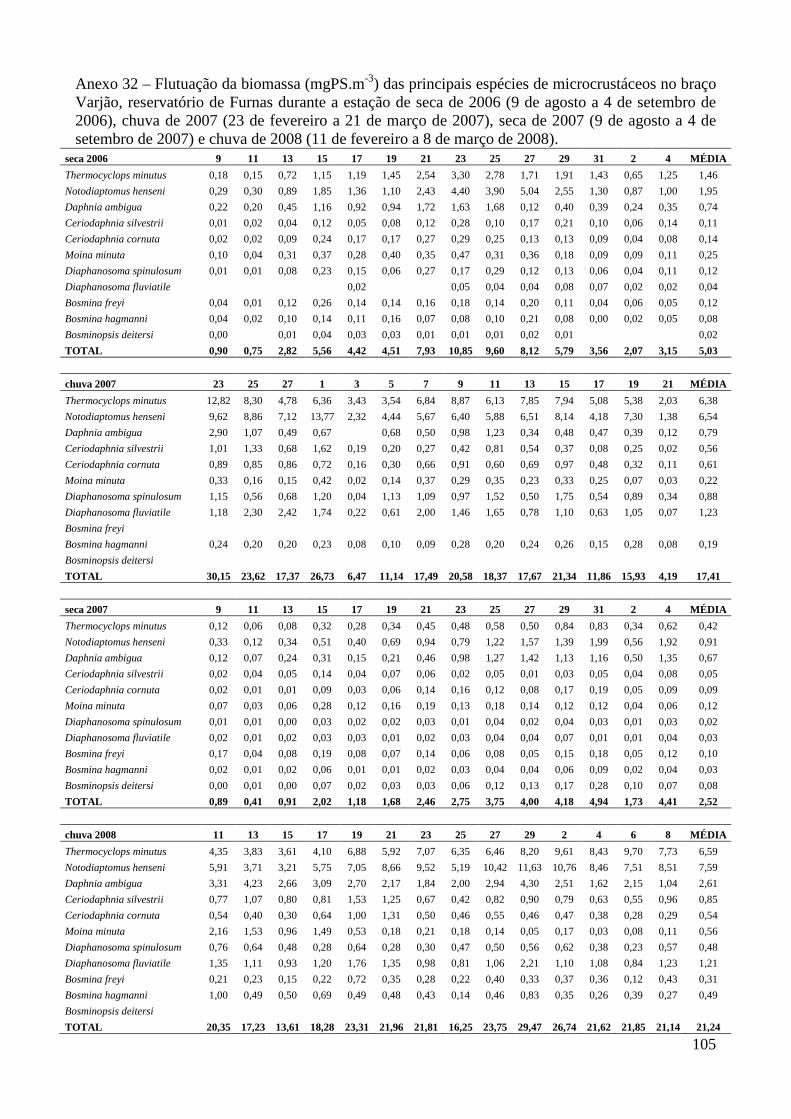
105
Anexo 32 – Flutuação da biomassa (mgPS.m-3) das principais espécies de microcrustáceos no braço Varjão, reservatório de Furnas durante a estação de seca de 2006 (9 de agosto a 4 de setembro de 2006), chuva de 2007 (23 de fevereiro a 21 de março de 2007), seca de 2007 (9 de agosto a 4 de setembro de 2007) e chuva de 2008 (11 de fevereiro a 8 de março de 2008).
seca 2006 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,18 0,15 0,72 1,15 1,19 1,45 2,54 3,30 2,78 1,71 1,91 1,43 0,65 1,25 1,46
Notodiaptomus henseni 0,29 0,30 0,89 1,85 1,36 1,10 2,43 4,40 3,90 5,04 2,55 1,30 0,87 1,00 1,95
Daphnia ambigua 0,22 0,20 0,45 1,16 0,92 0,94 1,72 1,63 1,68 0,12 0,40 0,39 0,24 0,35 0,74
Ceriodaphnia silvestrii 0,01 0,02 0,04 0,12 0,05 0,08 0,12 0,28 0,10 0,17 0,21 0,10 0,06 0,14 0,11
Ceriodaphnia cornuta 0,02 0,02 0,09 0,24 0,17 0,17 0,27 0,29 0,25 0,13 0,13 0,09 0,04 0,08 0,14
Moina minuta 0,10 0,04 0,31 0,37 0,28 0,40 0,35 0,47 0,31 0,36 0,18 0,09 0,09 0,11 0,25
Diaphanosoma spinulosum 0,01 0,01 0,08 0,23 0,15 0,06 0,27 0,17 0,29 0,12 0,13 0,06 0,04 0,11 0,12
Diaphanosoma fluviatile 0,02 0,05 0,04 0,04 0,08 0,07 0,02 0,02 0,04
Bosmina freyi 0,04 0,01 0,12 0,26 0,14 0,14 0,16 0,18 0,14 0,20 0,11 0,04 0,06 0,05 0,12
Bosmina hagmanni 0,04 0,02 0,10 0,14 0,11 0,16 0,07 0,08 0,10 0,21 0,08 0,00 0,02 0,05 0,08
Bosminopsis deitersi 0,00 0,01 0,04 0,03 0,03 0,01 0,01 0,01 0,02 0,01 0,02
TOTAL 0,90 0,75 2,82 5,56 4,42 4,51 7,93 10,85 9,60 8,12 5,79 3,56 2,07 3,15 5,03
chuva 2007 23 25 27 1 3 5 7 9 11 13 15 17 19 21 MÉDIA
Thermocyclops minutus 12,82 8,30 4,78 6,36 3,43 3,54 6,84 8,87 6,13 7,85 7,94 5,08 5,38 2,03 6,38
Notodiaptomus henseni 9,62 8,86 7,12 13,77 2,32 4,44 5,67 6,40 5,88 6,51 8,14 4,18 7,30 1,38 6,54
Daphnia ambigua 2,90 1,07 0,49 0,67 0,68 0,50 0,98 1,23 0,34 0,48 0,47 0,39 0,12 0,79
Ceriodaphnia silvestrii 1,01 1,33 0,68 1,62 0,19 0,20 0,27 0,42 0,81 0,54 0,37 0,08 0,25 0,02 0,56
Ceriodaphnia cornuta 0,89 0,85 0,86 0,72 0,16 0,30 0,66 0,91 0,60 0,69 0,97 0,48 0,32 0,11 0,61
Moina minuta 0,33 0,16 0,15 0,42 0,02 0,14 0,37 0,29 0,35 0,23 0,33 0,25 0,07 0,03 0,22
Diaphanosoma spinulosum 1,15 0,56 0,68 1,20 0,04 1,13 1,09 0,97 1,52 0,50 1,75 0,54 0,89 0,34 0,88
Diaphanosoma fluviatile 1,18 2,30 2,42 1,74 0,22 0,61 2,00 1,46 1,65 0,78 1,10 0,63 1,05 0,07 1,23
Bosmina freyi
Bosmina hagmanni 0,24 0,20 0,20 0,23 0,08 0,10 0,09 0,28 0,20 0,24 0,26 0,15 0,28 0,08 0,19
Bosminopsis deitersi
TOTAL 30,15 23,62 17,37 26,73 6,47 11,14 17,49 20,58 18,37 17,67 21,34 11,86 15,93 4,19 17,41
seca 2007 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,12 0,06 0,08 0,32 0,28 0,34 0,45 0,48 0,58 0,50 0,84 0,83 0,34 0,62 0,42
Notodiaptomus henseni 0,33 0,12 0,34 0,51 0,40 0,69 0,94 0,79 1,22 1,57 1,39 1,99 0,56 1,92 0,91
Daphnia ambigua 0,12 0,07 0,24 0,31 0,15 0,21 0,46 0,98 1,27 1,42 1,13 1,16 0,50 1,35 0,67
Ceriodaphnia silvestrii 0,02 0,04 0,05 0,14 0,04 0,07 0,06 0,02 0,05 0,01 0,03 0,05 0,04 0,08 0,05
Ceriodaphnia cornuta 0,02 0,01 0,01 0,09 0,03 0,06 0,14 0,16 0,12 0,08 0,17 0,19 0,05 0,09 0,09
Moina minuta 0,07 0,03 0,06 0,28 0,12 0,16 0,19 0,13 0,18 0,14 0,12 0,12 0,04 0,06 0,12
Diaphanosoma spinulosum 0,01 0,01 0,00 0,03 0,02 0,02 0,03 0,01 0,04 0,02 0,04 0,03 0,01 0,03 0,02
Diaphanosoma fluviatile 0,02 0,01 0,02 0,03 0,03 0,01 0,02 0,03 0,04 0,04 0,07 0,01 0,01 0,04 0,03
Bosmina freyi 0,17 0,04 0,08 0,19 0,08 0,07 0,14 0,06 0,08 0,05 0,15 0,18 0,05 0,12 0,10
Bosmina hagmanni 0,02 0,01 0,02 0,06 0,01 0,01 0,02 0,03 0,04 0,04 0,06 0,09 0,02 0,04 0,03
Bosminopsis deitersi 0,00 0,01 0,00 0,07 0,02 0,03 0,03 0,06 0,12 0,13 0,17 0,28 0,10 0,07 0,08
TOTAL 0,89 0,41 0,91 2,02 1,18 1,68 2,46 2,75 3,75 4,00 4,18 4,94 1,73 4,41 2,52
chuva 2008 11 13 15 17 19 21 23 25 27 29 2 4 6 8 MÉDIA
Thermocyclops minutus 4,35 3,83 3,61 4,10 6,88 5,92 7,07 6,35 6,46 8,20 9,61 8,43 9,70 7,73 6,59
Notodiaptomus henseni 5,91 3,71 3,21 5,75 7,05 8,66 9,52 5,19 10,42 11,63 10,76 8,46 7,51 8,51 7,59
Daphnia ambigua 3,31 4,23 2,66 3,09 2,70 2,17 1,84 2,00 2,94 4,30 2,51 1,62 2,15 1,04 2,61
Ceriodaphnia silvestrii 0,77 1,07 0,80 0,81 1,53 1,25 0,67 0,42 0,82 0,90 0,79 0,63 0,55 0,96 0,85
Ceriodaphnia cornuta 0,54 0,40 0,30 0,64 1,00 1,31 0,50 0,46 0,55 0,46 0,47 0,38 0,28 0,29 0,54
Moina minuta 2,16 1,53 0,96 1,49 0,53 0,18 0,21 0,18 0,14 0,05 0,17 0,03 0,08 0,11 0,56
Diaphanosoma spinulosum 0,76 0,64 0,48 0,28 0,64 0,28 0,30 0,47 0,50 0,56 0,62 0,38 0,23 0,57 0,48
Diaphanosoma fluviatile 1,35 1,11 0,93 1,20 1,76 1,35 0,98 0,81 1,06 2,21 1,10 1,08 0,84 1,23 1,21
Bosmina freyi 0,21 0,23 0,15 0,22 0,72 0,35 0,28 0,22 0,40 0,33 0,37 0,36 0,12 0,43 0,31
Bosmina hagmanni 1,00 0,49 0,50 0,69 0,49 0,48 0,43 0,14 0,46 0,83 0,35 0,26 0,39 0,27 0,49
Bosminopsis deitersi
TOTAL 20,35 17,23 13,61 18,28 23,31 21,96 21,81 16,25 23,75 29,47 26,74 21,62 21,85 21,14 21,24
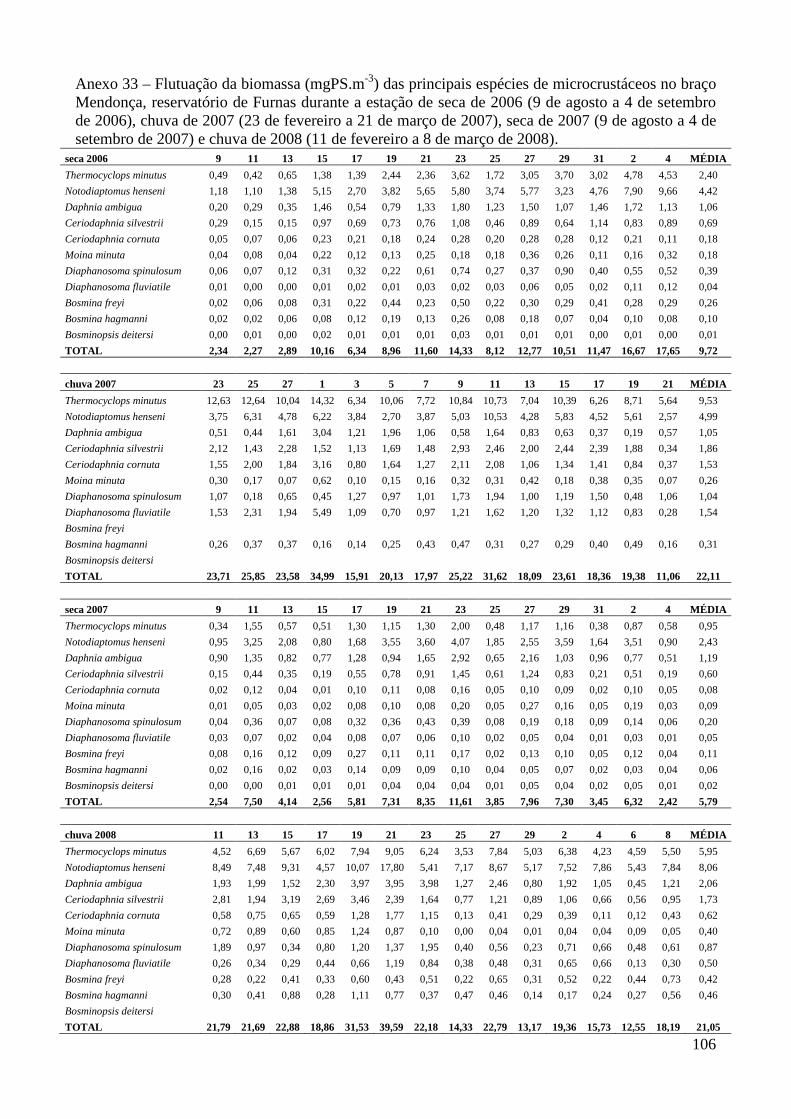
106
Anexo 33 – Flutuação da biomassa (mgPS.m-3) das principais espécies de microcrustáceos no braço Mendonça, reservatório de Furnas durante a estação de seca de 2006 (9 de agosto a 4 de setembro de 2006), chuva de 2007 (23 de fevereiro a 21 de março de 2007), seca de 2007 (9 de agosto a 4 de setembro de 2007) e chuva de 2008 (11 de fevereiro a 8 de março de 2008).
seca 2006 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,49 0,42 0,65 1,38 1,39 2,44 2,36 3,62 1,72 3,05 3,70 3,02 4,78 4,53 2,40
Notodiaptomus henseni 1,18 1,10 1,38 5,15 2,70 3,82 5,65 5,80 3,74 5,77 3,23 4,76 7,90 9,66 4,42
Daphnia ambigua 0,20 0,29 0,35 1,46 0,54 0,79 1,33 1,80 1,23 1,50 1,07 1,46 1,72 1,13 1,06
Ceriodaphnia silvestrii 0,29 0,15 0,15 0,97 0,69 0,73 0,76 1,08 0,46 0,89 0,64 1,14 0,83 0,89 0,69
Ceriodaphnia cornuta 0,05 0,07 0,06 0,23 0,21 0,18 0,24 0,28 0,20 0,28 0,28 0,12 0,21 0,11 0,18
Moina minuta 0,04 0,08 0,04 0,22 0,12 0,13 0,25 0,18 0,18 0,36 0,26 0,11 0,16 0,32 0,18
Diaphanosoma spinulosum 0,06 0,07 0,12 0,31 0,32 0,22 0,61 0,74 0,27 0,37 0,90 0,40 0,55 0,52 0,39
Diaphanosoma fluviatile 0,01 0,00 0,00 0,01 0,02 0,01 0,03 0,02 0,03 0,06 0,05 0,02 0,11 0,12 0,04
Bosmina freyi 0,02 0,06 0,08 0,31 0,22 0,44 0,23 0,50 0,22 0,30 0,29 0,41 0,28 0,29 0,26
Bosmina hagmanni 0,02 0,02 0,06 0,08 0,12 0,19 0,13 0,26 0,08 0,18 0,07 0,04 0,10 0,08 0,10
Bosminopsis deitersi 0,00 0,01 0,00 0,02 0,01 0,01 0,01 0,03 0,01 0,01 0,01 0,00 0,01 0,00 0,01
TOTAL 2,34 2,27 2,89 10,16 6,34 8,96 11,60 14,33 8,12 12,77 10,51 11,47 16,67 17,65 9,72
chuva 2007 23 25 27 1 3 5 7 9 11 13 15 17 19 21 MÉDIA
Thermocyclops minutus 12,63 12,64 10,04 14,32 6,34 10,06 7,72 10,84 10,73 7,04 10,39 6,26 8,71 5,64 9,53
Notodiaptomus henseni 3,75 6,31 4,78 6,22 3,84 2,70 3,87 5,03 10,53 4,28 5,83 4,52 5,61 2,57 4,99
Daphnia ambigua 0,51 0,44 1,61 3,04 1,21 1,96 1,06 0,58 1,64 0,83 0,63 0,37 0,19 0,57 1,05
Ceriodaphnia silvestrii 2,12 1,43 2,28 1,52 1,13 1,69 1,48 2,93 2,46 2,00 2,44 2,39 1,88 0,34 1,86
Ceriodaphnia cornuta 1,55 2,00 1,84 3,16 0,80 1,64 1,27 2,11 2,08 1,06 1,34 1,41 0,84 0,37 1,53
Moina minuta 0,30 0,17 0,07 0,62 0,10 0,15 0,16 0,32 0,31 0,42 0,18 0,38 0,35 0,07 0,26
Diaphanosoma spinulosum 1,07 0,18 0,65 0,45 1,27 0,97 1,01 1,73 1,94 1,00 1,19 1,50 0,48 1,06 1,04
Diaphanosoma fluviatile 1,53 2,31 1,94 5,49 1,09 0,70 0,97 1,21 1,62 1,20 1,32 1,12 0,83 0,28 1,54
Bosmina freyi
Bosmina hagmanni 0,26 0,37 0,37 0,16 0,14 0,25 0,43 0,47 0,31 0,27 0,29 0,40 0,49 0,16 0,31
Bosminopsis deitersi
TOTAL 23,71 25,85 23,58 34,99 15,91 20,13 17,97 25,22 31,62 18,09 23,61 18,36 19,38 11,06 22,11
seca 2007 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,34 1,55 0,57 0,51 1,30 1,15 1,30 2,00 0,48 1,17 1,16 0,38 0,87 0,58 0,95
Notodiaptomus henseni 0,95 3,25 2,08 0,80 1,68 3,55 3,60 4,07 1,85 2,55 3,59 1,64 3,51 0,90 2,43
Daphnia ambigua 0,90 1,35 0,82 0,77 1,28 0,94 1,65 2,92 0,65 2,16 1,03 0,96 0,77 0,51 1,19
Ceriodaphnia silvestrii 0,15 0,44 0,35 0,19 0,55 0,78 0,91 1,45 0,61 1,24 0,83 0,21 0,51 0,19 0,60
Ceriodaphnia cornuta 0,02 0,12 0,04 0,01 0,10 0,11 0,08 0,16 0,05 0,10 0,09 0,02 0,10 0,05 0,08
Moina minuta 0,01 0,05 0,03 0,02 0,08 0,10 0,08 0,20 0,05 0,27 0,16 0,05 0,19 0,03 0,09
Diaphanosoma spinulosum 0,04 0,36 0,07 0,08 0,32 0,36 0,43 0,39 0,08 0,19 0,18 0,09 0,14 0,06 0,20
Diaphanosoma fluviatile 0,03 0,07 0,02 0,04 0,08 0,07 0,06 0,10 0,02 0,05 0,04 0,01 0,03 0,01 0,05
Bosmina freyi 0,08 0,16 0,12 0,09 0,27 0,11 0,11 0,17 0,02 0,13 0,10 0,05 0,12 0,04 0,11
Bosmina hagmanni 0,02 0,16 0,02 0,03 0,14 0,09 0,09 0,10 0,04 0,05 0,07 0,02 0,03 0,04 0,06
Bosminopsis deitersi 0,00 0,00 0,01 0,01 0,01 0,04 0,04 0,04 0,01 0,05 0,04 0,02 0,05 0,01 0,02
TOTAL 2,54 7,50 4,14 2,56 5,81 7,31 8,35 11,61 3,85 7,96 7,30 3,45 6,32 2,42 5,79
chuva 2008 11 13 15 17 19 21 23 25 27 29 2 4 6 8 MÉDIA
Thermocyclops minutus 4,52 6,69 5,67 6,02 7,94 9,05 6,24 3,53 7,84 5,03 6,38 4,23 4,59 5,50 5,95
Notodiaptomus henseni 8,49 7,48 9,31 4,57 10,07 17,80 5,41 7,17 8,67 5,17 7,52 7,86 5,43 7,84 8,06
Daphnia ambigua 1,93 1,99 1,52 2,30 3,97 3,95 3,98 1,27 2,46 0,80 1,92 1,05 0,45 1,21 2,06
Ceriodaphnia silvestrii 2,81 1,94 3,19 2,69 3,46 2,39 1,64 0,77 1,21 0,89 1,06 0,66 0,56 0,95 1,73
Ceriodaphnia cornuta 0,58 0,75 0,65 0,59 1,28 1,77 1,15 0,13 0,41 0,29 0,39 0,11 0,12 0,43 0,62
Moina minuta 0,72 0,89 0,60 0,85 1,24 0,87 0,10 0,00 0,04 0,01 0,04 0,04 0,09 0,05 0,40
Diaphanosoma spinulosum 1,89 0,97 0,34 0,80 1,20 1,37 1,95 0,40 0,56 0,23 0,71 0,66 0,48 0,61 0,87
Diaphanosoma fluviatile 0,26 0,34 0,29 0,44 0,66 1,19 0,84 0,38 0,48 0,31 0,65 0,66 0,13 0,30 0,50
Bosmina freyi 0,28 0,22 0,41 0,33 0,60 0,43 0,51 0,22 0,65 0,31 0,52 0,22 0,44 0,73 0,42
Bosmina hagmanni 0,30 0,41 0,88 0,28 1,11 0,77 0,37 0,47 0,46 0,14 0,17 0,24 0,27 0,56 0,46
Bosminopsis deitersi
TOTAL 21,79 21,69 22,88 18,86 31,53 39,59 22,18 14,33 22,79 13,17 19,36 15,73 12,55 18,19 21,05

107
Anexo 34 – Flutuação da produtividade (mgPS.m-3.d-1) das principais espécies de microcrustáceos no braço Barrão, reservatório de Três Marias durante a estação de seca de 2006 (3 a 29 de julho de 2006), chuva de 2007 (4 a 28 de janeiro de 2007), seca de 2007 (6 de julho a 1 de agosto de 2007) e chuva de 2008 (4 a 28 de janeiro de 2008). seca 2006 3 5 7 9 11 13 15 17 19 21 23 25 27 29 MÉDIA Thermocyclops minutus 0,11 0,68 0,62 0,40 0,91 0,46 0,17 0,33 0,12 0,39 0,75 0,67 0,65 0,65 0,49 Bosminospsis deitersi 0,15 0,06 0,03 0,06 0,02 0,09 0,03 0,11 0,11 0,12 0,45 0,58 0,14 0,09 0,14 Bosmina hagmanni 0,03 0,03 0,06 0,07 0,06 0,05 0,11 0,05 0,06 0,07 0,15 0,19 0,08 Ceriodaphnia cornuta 0,02 0,01 0,06 0,05 0,08 0,02 0,04 0,14 0,08 0,21 0,11 0,07 Moina minuta 0,04 0,01 0,01 0,03 0,03 0,04 0,01 0,13 0,24 0,11 0,02 0,06 TOTAL 0,32 0,79 0,67 0,52 1,00 0,69 0,33 0,67 0,33 0,61 1,47 1,64 1,25 1,05 0,85
chuva 2007 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA Thermocyclops minutus 0,29 0,24 1,22 0,77 0,32 0,67 1,44 0,18 0,59 1,37 0,73 1,15 0,75 Bosminospsis deitersi 0,09 0,05 0,04 0,08 0,06 0,06 0,09 0,09 0,09 0,02 0,07 Bosmina hagmanni 0,42 0,30 0,12 0,20 0,07 0,23 0,34 0,13 0,31 0,16 0,77 0,19 0,14 0,26 Ceriodaphnia cornuta 0,07 0,02 0,09 0,06 0,21 0,06 0,33 0,17 0,20 0,22 0,20 0,07 0,14 Moina minuta 0,42 0,21 0,13 0,19 0,33 0,59 0,30 1,36 1,87 0,68 1,60 0,43 0,60 0,67 TOTAL 1,28 0,77 1,48 1,31 0,82 1,78 0,75 3,32 2,63 1,72 3,96 1,65 1,97 1,89
seca 2007 6 8 10 12 14 16 18 20 22 24 26 28 30 1 MÉDIA Thermocyclops minutus 0,28 0,03 0,05 0,07 0,02 0,05 0,04 0,04 0,02 0,07 Bosminospsis deitersi 0,33 0,07 0,03 0,09 0,01 0,03 0,03 0,05 0,05 0,16 0,05 0,08 0,03 0,08 Bosmina hagmanni 0,13 0,14 0,05 0,06 0,01 0,04 0,06 0,08 0,09 0,04 0,16 0,08 Ceriodaphnia cornuta 0,16 0,11 0,04 0,03 0,01 0,07 0,01 0,05 0,01 0,13 0,04 0,04 0,03 0,06 Moina minuta 0,17 0,08 0,01 0,04 0,03 0,06 0,06 0,08 0,10 0,09 0,02 0,07 0,07 TOTAL 1,06 0,32 0,07 0,29 0,13 0,22 0,13 0,21 0,16 0,43 0,23 0,30 0,13 0,32 0,35
chuva 2008 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA Thermocyclops minutus 0,55 0,27 1,48 0,53 0,09 0,23 0,50 0,24 0,41 0,94 0,38 0,51 Bosminospsis deitersi 0,14 0,60 0,49 0,28 0,25 0,21 0,41 0,07 0,37 0,19 0,41 0,03 0,12 0,27 Bosmina hagmanni 0,15 0,11 0,43 0,14 0,22 0,09 0,05 0,19 0,09 0,19 0,17 Ceriodaphnia cornuta 0,33 0,08 0,19 0,07 0,17 0,18 0,09 0,05 0,10 0,40 0,16 Moina minuta 0,44 0,66 1,62 0,87 0,87 0,39 0,58 0,40 0,66 0,27 0,12 0,49 0,18 0,58 TOTAL 1,61 1,64 3,66 2,30 1,42 1,22 1,76 0,85 1,22 0,92 1,46 0,71 1,28 1,70
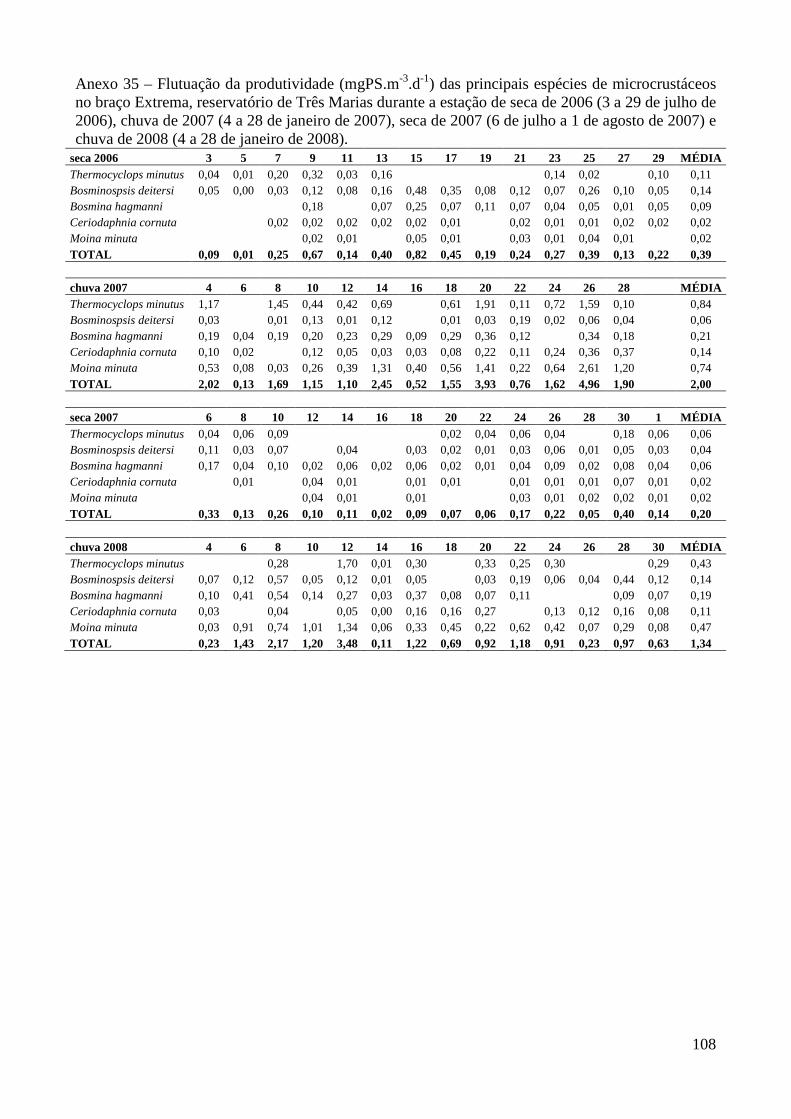
108
Anexo 35 – Flutuação da produtividade (mgPS.m-3.d-1) das principais espécies de microcrustáceos no braço Extrema, reservatório de Três Marias durante a estação de seca de 2006 (3 a 29 de julho de 2006), chuva de 2007 (4 a 28 de janeiro de 2007), seca de 2007 (6 de julho a 1 de agosto de 2007) e chuva de 2008 (4 a 28 de janeiro de 2008).
seca 2006 3 5 7 9 11 13 15 17 19 21 23 25 27 29 MÉDIA Thermocyclops minutus 0,04 0,01 0,20 0,32 0,03 0,16 0,14 0,02 0,10 0,11 Bosminospsis deitersi 0,05 0,00 0,03 0,12 0,08 0,16 0,48 0,35 0,08 0,12 0,07 0,26 0,10 0,05 0,14 Bosmina hagmanni 0,18 0,07 0,25 0,07 0,11 0,07 0,04 0,05 0,01 0,05 0,09 Ceriodaphnia cornuta 0,02 0,02 0,02 0,02 0,02 0,01 0,02 0,01 0,01 0,02 0,02 0,02 Moina minuta 0,02 0,01 0,05 0,01 0,03 0,01 0,04 0,01 0,02 TOTAL 0,09 0,01 0,25 0,67 0,14 0,40 0,82 0,45 0,19 0,24 0,27 0,39 0,13 0,22 0,39
chuva 2007 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA Thermocyclops minutus 1,17 1,45 0,44 0,42 0,69 0,61 1,91 0,11 0,72 1,59 0,10 0,84 Bosminospsis deitersi 0,03 0,01 0,13 0,01 0,12 0,01 0,03 0,19 0,02 0,06 0,04 0,06 Bosmina hagmanni 0,19 0,04 0,19 0,20 0,23 0,29 0,09 0,29 0,36 0,12 0,34 0,18 0,21 Ceriodaphnia cornuta 0,10 0,02 0,12 0,05 0,03 0,03 0,08 0,22 0,11 0,24 0,36 0,37 0,14 Moina minuta 0,53 0,08 0,03 0,26 0,39 1,31 0,40 0,56 1,41 0,22 0,64 2,61 1,20 0,74 TOTAL 2,02 0,13 1,69 1,15 1,10 2,45 0,52 1,55 3,93 0,76 1,62 4,96 1,90 2,00
seca 2007 6 8 10 12 14 16 18 20 22 24 26 28 30 1 MÉDIA Thermocyclops minutus 0,04 0,06 0,09 0,02 0,04 0,06 0,04 0,18 0,06 0,06 Bosminospsis deitersi 0,11 0,03 0,07 0,04 0,03 0,02 0,01 0,03 0,06 0,01 0,05 0,03 0,04 Bosmina hagmanni 0,17 0,04 0,10 0,02 0,06 0,02 0,06 0,02 0,01 0,04 0,09 0,02 0,08 0,04 0,06 Ceriodaphnia cornuta 0,01 0,04 0,01 0,01 0,01 0,01 0,01 0,01 0,07 0,01 0,02 Moina minuta 0,04 0,01 0,01 0,03 0,01 0,02 0,02 0,01 0,02 TOTAL 0,33 0,13 0,26 0,10 0,11 0,02 0,09 0,07 0,06 0,17 0,22 0,05 0,40 0,14 0,20
chuva 2008 4 6 8 10 12 14 16 18 20 22 24 26 28 30 MÉDIA Thermocyclops minutus 0,28 1,70 0,01 0,30 0,33 0,25 0,30 0,29 0,43 Bosminospsis deitersi 0,07 0,12 0,57 0,05 0,12 0,01 0,05 0,03 0,19 0,06 0,04 0,44 0,12 0,14 Bosmina hagmanni 0,10 0,41 0,54 0,14 0,27 0,03 0,37 0,08 0,07 0,11 0,09 0,07 0,19 Ceriodaphnia cornuta 0,03 0,04 0,05 0,00 0,16 0,16 0,27 0,13 0,12 0,16 0,08 0,11 Moina minuta 0,03 0,91 0,74 1,01 1,34 0,06 0,33 0,45 0,22 0,62 0,42 0,07 0,29 0,08 0,47 TOTAL 0,23 1,43 2,17 1,20 3,48 0,11 1,22 0,69 0,92 1,18 0,91 0,23 0,97 0,63 1,34
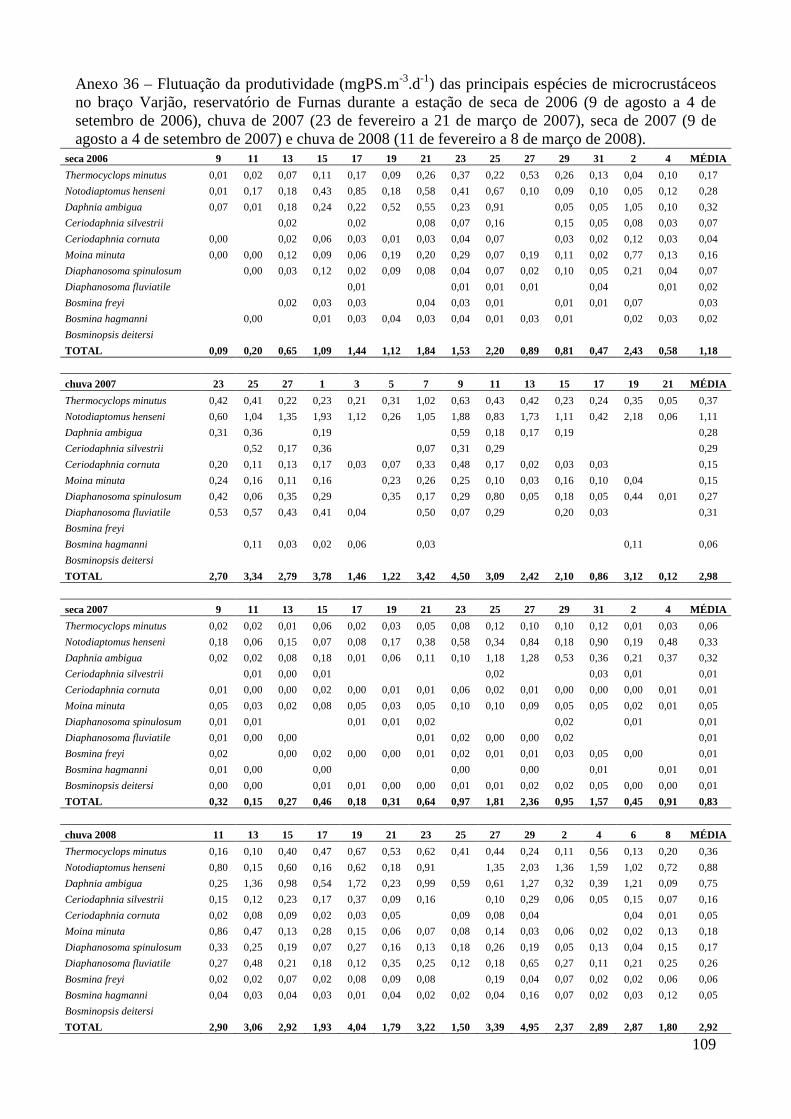
109
Anexo 36 – Flutuação da produtividade (mgPS.m-3.d-1) das principais espécies de microcrustáceos no braço Varjão, reservatório de Furnas durante a estação de seca de 2006 (9 de agosto a 4 de setembro de 2006), chuva de 2007 (23 de fevereiro a 21 de março de 2007), seca de 2007 (9 de agosto a 4 de setembro de 2007) e chuva de 2008 (11 de fevereiro a 8 de março de 2008).
seca 2006 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,01 0,02 0,07 0,11 0,17 0,09 0,26 0,37 0,22 0,53 0,26 0,13 0,04 0,10 0,17
Notodiaptomus henseni 0,01 0,17 0,18 0,43 0,85 0,18 0,58 0,41 0,67 0,10 0,09 0,10 0,05 0,12 0,28
Daphnia ambigua 0,07 0,01 0,18 0,24 0,22 0,52 0,55 0,23 0,91 0,05 0,05 1,05 0,10 0,32
Ceriodaphnia silvestrii 0,02 0,02 0,08 0,07 0,16 0,15 0,05 0,08 0,03 0,07
Ceriodaphnia cornuta 0,00 0,02 0,06 0,03 0,01 0,03 0,04 0,07 0,03 0,02 0,12 0,03 0,04
Moina minuta 0,00 0,00 0,12 0,09 0,06 0,19 0,20 0,29 0,07 0,19 0,11 0,02 0,77 0,13 0,16
Diaphanosoma spinulosum 0,00 0,03 0,12 0,02 0,09 0,08 0,04 0,07 0,02 0,10 0,05 0,21 0,04 0,07
Diaphanosoma fluviatile 0,01 0,01 0,01 0,01 0,04 0,01 0,02
Bosmina freyi 0,02 0,03 0,03 0,04 0,03 0,01 0,01 0,01 0,07 0,03
Bosmina hagmanni 0,00 0,01 0,03 0,04 0,03 0,04 0,01 0,03 0,01 0,02 0,03 0,02
Bosminopsis deitersi
TOTAL 0,09 0,20 0,65 1,09 1,44 1,12 1,84 1,53 2,20 0,89 0,81 0,47 2,43 0,58 1,18
chuva 2007 23 25 27 1 3 5 7 9 11 13 15 17 19 21 MÉDIA
Thermocyclops minutus 0,42 0,41 0,22 0,23 0,21 0,31 1,02 0,63 0,43 0,42 0,23 0,24 0,35 0,05 0,37
Notodiaptomus henseni 0,60 1,04 1,35 1,93 1,12 0,26 1,05 1,88 0,83 1,73 1,11 0,42 2,18 0,06 1,11
Daphnia ambigua 0,31 0,36 0,19 0,59 0,18 0,17 0,19 0,28
Ceriodaphnia silvestrii 0,52 0,17 0,36 0,07 0,31 0,29 0,29
Ceriodaphnia cornuta 0,20 0,11 0,13 0,17 0,03 0,07 0,33 0,48 0,17 0,02 0,03 0,03 0,15
Moina minuta 0,24 0,16 0,11 0,16 0,23 0,26 0,25 0,10 0,03 0,16 0,10 0,04 0,15
Diaphanosoma spinulosum 0,42 0,06 0,35 0,29 0,35 0,17 0,29 0,80 0,05 0,18 0,05 0,44 0,01 0,27
Diaphanosoma fluviatile 0,53 0,57 0,43 0,41 0,04 0,50 0,07 0,29 0,20 0,03 0,31
Bosmina freyi
Bosmina hagmanni 0,11 0,03 0,02 0,06 0,03 0,11 0,06
Bosminopsis deitersi
TOTAL 2,70 3,34 2,79 3,78 1,46 1,22 3,42 4,50 3,09 2,42 2,10 0,86 3,12 0,12 2,98
seca 2007 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,02 0,02 0,01 0,06 0,02 0,03 0,05 0,08 0,12 0,10 0,10 0,12 0,01 0,03 0,06
Notodiaptomus henseni 0,18 0,06 0,15 0,07 0,08 0,17 0,38 0,58 0,34 0,84 0,18 0,90 0,19 0,48 0,33
Daphnia ambigua 0,02 0,02 0,08 0,18 0,01 0,06 0,11 0,10 1,18 1,28 0,53 0,36 0,21 0,37 0,32
Ceriodaphnia silvestrii 0,01 0,00 0,01 0,02 0,03 0,01 0,01
Ceriodaphnia cornuta 0,01 0,00 0,00 0,02 0,00 0,01 0,01 0,06 0,02 0,01 0,00 0,00 0,00 0,01 0,01
Moina minuta 0,05 0,03 0,02 0,08 0,05 0,03 0,05 0,10 0,10 0,09 0,05 0,05 0,02 0,01 0,05
Diaphanosoma spinulosum 0,01 0,01 0,01 0,01 0,02 0,02 0,01 0,01
Diaphanosoma fluviatile 0,01 0,00 0,00 0,01 0,02 0,00 0,00 0,02 0,01
Bosmina freyi 0,02 0,00 0,02 0,00 0,00 0,01 0,02 0,01 0,01 0,03 0,05 0,00 0,01
Bosmina hagmanni 0,01 0,00 0,00 0,00 0,00 0,01 0,01 0,01
Bosminopsis deitersi 0,00 0,00 0,01 0,01 0,00 0,00 0,01 0,01 0,02 0,02 0,05 0,00 0,00 0,01
TOTAL 0,32 0,15 0,27 0,46 0,18 0,31 0,64 0,97 1,81 2,36 0,95 1,57 0,45 0,91 0,83
chuva 2008 11 13 15 17 19 21 23 25 27 29 2 4 6 8 MÉDIA
Thermocyclops minutus 0,16 0,10 0,40 0,47 0,67 0,53 0,62 0,41 0,44 0,24 0,11 0,56 0,13 0,20 0,36
Notodiaptomus henseni 0,80 0,15 0,60 0,16 0,62 0,18 0,91 1,35 2,03 1,36 1,59 1,02 0,72 0,88
Daphnia ambigua 0,25 1,36 0,98 0,54 1,72 0,23 0,99 0,59 0,61 1,27 0,32 0,39 1,21 0,09 0,75
Ceriodaphnia silvestrii 0,15 0,12 0,23 0,17 0,37 0,09 0,16 0,10 0,29 0,06 0,05 0,15 0,07 0,16
Ceriodaphnia cornuta 0,02 0,08 0,09 0,02 0,03 0,05 0,09 0,08 0,04 0,04 0,01 0,05
Moina minuta 0,86 0,47 0,13 0,28 0,15 0,06 0,07 0,08 0,14 0,03 0,06 0,02 0,02 0,13 0,18
Diaphanosoma spinulosum 0,33 0,25 0,19 0,07 0,27 0,16 0,13 0,18 0,26 0,19 0,05 0,13 0,04 0,15 0,17
Diaphanosoma fluviatile 0,27 0,48 0,21 0,18 0,12 0,35 0,25 0,12 0,18 0,65 0,27 0,11 0,21 0,25 0,26
Bosmina freyi 0,02 0,02 0,07 0,02 0,08 0,09 0,08 0,19 0,04 0,07 0,02 0,02 0,06 0,06
Bosmina hagmanni 0,04 0,03 0,04 0,03 0,01 0,04 0,02 0,02 0,04 0,16 0,07 0,02 0,03 0,12 0,05
Bosminopsis deitersi
TOTAL 2,90 3,06 2,92 1,93 4,04 1,79 3,22 1,50 3,39 4,95 2,37 2,89 2,87 1,80 2,92
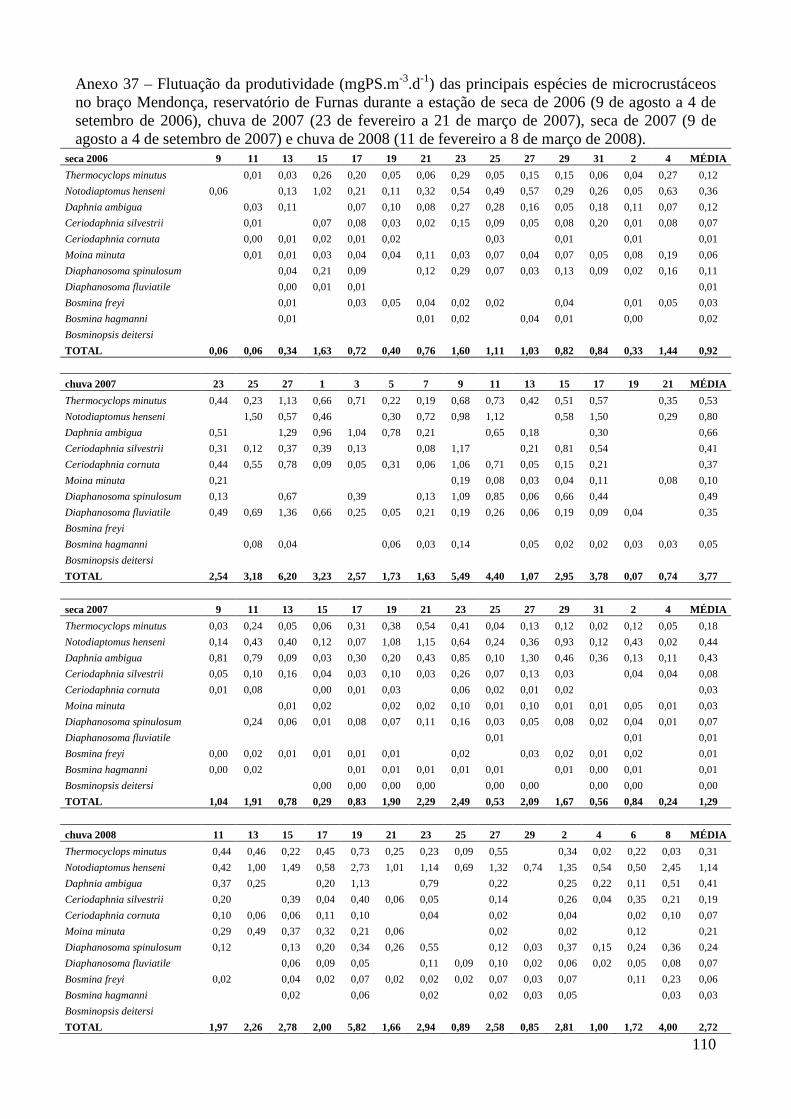
110
Anexo 37 – Flutuação da produtividade (mgPS.m-3.d-1) das principais espécies de microcrustáceos no braço Mendonça, reservatório de Furnas durante a estação de seca de 2006 (9 de agosto a 4 de setembro de 2006), chuva de 2007 (23 de fevereiro a 21 de março de 2007), seca de 2007 (9 de agosto a 4 de setembro de 2007) e chuva de 2008 (11 de fevereiro a 8 de março de 2008).
seca 2006 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,01 0,03 0,26 0,20 0,05 0,06 0,29 0,05 0,15 0,15 0,06 0,04 0,27 0,12
Notodiaptomus henseni 0,06 0,13 1,02 0,21 0,11 0,32 0,54 0,49 0,57 0,29 0,26 0,05 0,63 0,36
Daphnia ambigua 0,03 0,11 0,07 0,10 0,08 0,27 0,28 0,16 0,05 0,18 0,11 0,07 0,12
Ceriodaphnia silvestrii 0,01 0,07 0,08 0,03 0,02 0,15 0,09 0,05 0,08 0,20 0,01 0,08 0,07
Ceriodaphnia cornuta 0,00 0,01 0,02 0,01 0,02 0,03 0,01 0,01 0,01
Moina minuta 0,01 0,01 0,03 0,04 0,04 0,11 0,03 0,07 0,04 0,07 0,05 0,08 0,19 0,06
Diaphanosoma spinulosum 0,04 0,21 0,09 0,12 0,29 0,07 0,03 0,13 0,09 0,02 0,16 0,11
Diaphanosoma fluviatile 0,00 0,01 0,01 0,01
Bosmina freyi 0,01 0,03 0,05 0,04 0,02 0,02 0,04 0,01 0,05 0,03
Bosmina hagmanni 0,01 0,01 0,02 0,04 0,01 0,00 0,02
Bosminopsis deitersi
TOTAL 0,06 0,06 0,34 1,63 0,72 0,40 0,76 1,60 1,11 1,03 0,82 0,84 0,33 1,44 0,92
chuva 2007 23 25 27 1 3 5 7 9 11 13 15 17 19 21 MÉDIA
Thermocyclops minutus 0,44 0,23 1,13 0,66 0,71 0,22 0,19 0,68 0,73 0,42 0,51 0,57 0,35 0,53
Notodiaptomus henseni 1,50 0,57 0,46 0,30 0,72 0,98 1,12 0,58 1,50 0,29 0,80
Daphnia ambigua 0,51 1,29 0,96 1,04 0,78 0,21 0,65 0,18 0,30 0,66
Ceriodaphnia silvestrii 0,31 0,12 0,37 0,39 0,13 0,08 1,17 0,21 0,81 0,54 0,41
Ceriodaphnia cornuta 0,44 0,55 0,78 0,09 0,05 0,31 0,06 1,06 0,71 0,05 0,15 0,21 0,37
Moina minuta 0,21 0,19 0,08 0,03 0,04 0,11 0,08 0,10
Diaphanosoma spinulosum 0,13 0,67 0,39 0,13 1,09 0,85 0,06 0,66 0,44 0,49
Diaphanosoma fluviatile 0,49 0,69 1,36 0,66 0,25 0,05 0,21 0,19 0,26 0,06 0,19 0,09 0,04 0,35
Bosmina freyi
Bosmina hagmanni 0,08 0,04 0,06 0,03 0,14 0,05 0,02 0,02 0,03 0,03 0,05
Bosminopsis deitersi
TOTAL 2,54 3,18 6,20 3,23 2,57 1,73 1,63 5,49 4,40 1,07 2,95 3,78 0,07 0,74 3,77
seca 2007 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Thermocyclops minutus 0,03 0,24 0,05 0,06 0,31 0,38 0,54 0,41 0,04 0,13 0,12 0,02 0,12 0,05 0,18
Notodiaptomus henseni 0,14 0,43 0,40 0,12 0,07 1,08 1,15 0,64 0,24 0,36 0,93 0,12 0,43 0,02 0,44
Daphnia ambigua 0,81 0,79 0,09 0,03 0,30 0,20 0,43 0,85 0,10 1,30 0,46 0,36 0,13 0,11 0,43
Ceriodaphnia silvestrii 0,05 0,10 0,16 0,04 0,03 0,10 0,03 0,26 0,07 0,13 0,03 0,04 0,04 0,08
Ceriodaphnia cornuta 0,01 0,08 0,00 0,01 0,03 0,06 0,02 0,01 0,02 0,03
Moina minuta 0,01 0,02 0,02 0,02 0,10 0,01 0,10 0,01 0,01 0,05 0,01 0,03
Diaphanosoma spinulosum 0,24 0,06 0,01 0,08 0,07 0,11 0,16 0,03 0,05 0,08 0,02 0,04 0,01 0,07
Diaphanosoma fluviatile 0,01 0,01 0,01
Bosmina freyi 0,00 0,02 0,01 0,01 0,01 0,01 0,02 0,03 0,02 0,01 0,02 0,01
Bosmina hagmanni 0,00 0,02 0,01 0,01 0,01 0,01 0,01 0,01 0,00 0,01 0,01
Bosminopsis deitersi 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00
TOTAL 1,04 1,91 0,78 0,29 0,83 1,90 2,29 2,49 0,53 2,09 1,67 0,56 0,84 0,24 1,29
chuva 2008 11 13 15 17 19 21 23 25 27 29 2 4 6 8 MÉDIA
Thermocyclops minutus 0,44 0,46 0,22 0,45 0,73 0,25 0,23 0,09 0,55 0,34 0,02 0,22 0,03 0,31
Notodiaptomus henseni 0,42 1,00 1,49 0,58 2,73 1,01 1,14 0,69 1,32 0,74 1,35 0,54 0,50 2,45 1,14
Daphnia ambigua 0,37 0,25 0,20 1,13 0,79 0,22 0,25 0,22 0,11 0,51 0,41
Ceriodaphnia silvestrii 0,20 0,39 0,04 0,40 0,06 0,05 0,14 0,26 0,04 0,35 0,21 0,19
Ceriodaphnia cornuta 0,10 0,06 0,06 0,11 0,10 0,04 0,02 0,04 0,02 0,10 0,07
Moina minuta 0,29 0,49 0,37 0,32 0,21 0,06 0,02 0,02 0,12 0,21
Diaphanosoma spinulosum 0,12 0,13 0,20 0,34 0,26 0,55 0,12 0,03 0,37 0,15 0,24 0,36 0,24
Diaphanosoma fluviatile 0,06 0,09 0,05 0,11 0,09 0,10 0,02 0,06 0,02 0,05 0,08 0,07
Bosmina freyi 0,02 0,04 0,02 0,07 0,02 0,02 0,02 0,07 0,03 0,07 0,11 0,23 0,06
Bosmina hagmanni 0,02 0,06 0,02 0,02 0,03 0,05 0,03 0,03
Bosminopsis deitersi
TOTAL 1,97 2,26 2,78 2,00 5,82 1,66 2,94 0,89 2,58 0,85 2,81 1,00 1,72 4,00 2,72
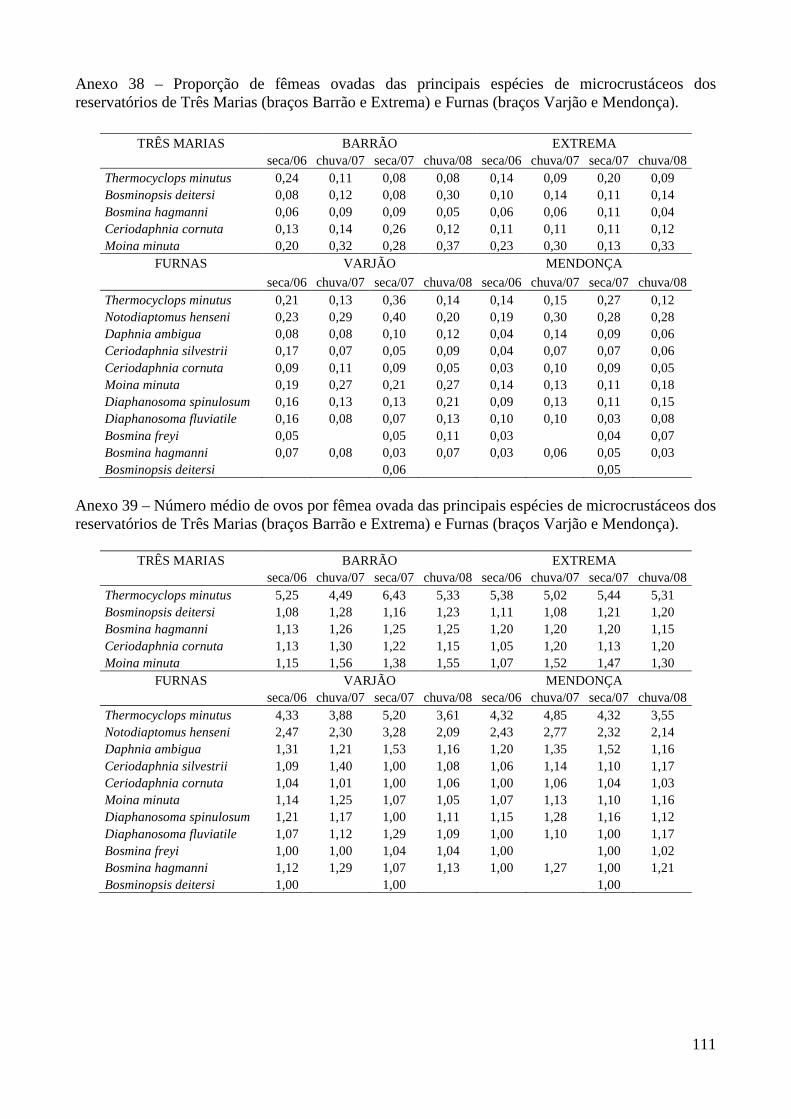
111
Anexo 38 – Proporção de fêmeas ovadas das principais espécies de microcrustáceos dos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça).
Anexo 39 – Número médio de ovos por fêmea ovada das principais espécies de microcrustáceos dos reservatórios de Três Marias (braços Barrão e Extrema) e Furnas (braços Varjão e Mendonça).
TRÊS MARIAS BARRÃO EXTREMA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08 Thermocyclops minutus 0,24 0,11 0,08 0,08 0,14 0,09 0,20 0,09 Bosminopsis deitersi 0,08 0,12 0,08 0,30 0,10 0,14 0,11 0,14 Bosmina hagmanni 0,06 0,09 0,09 0,05 0,06 0,06 0,11 0,04 Ceriodaphnia cornuta 0,13 0,14 0,26 0,12 0,11 0,11 0,11 0,12 Moina minuta 0,20 0,32 0,28 0,37 0,23 0,30 0,13 0,33
FURNAS VARJÃO MENDONÇA
seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08 Thermocyclops minutus 0,21 0,13 0,36 0,14 0,14 0,15 0,27 0,12 Notodiaptomus henseni 0,23 0,29 0,40 0,20 0,19 0,30 0,28 0,28 Daphnia ambigua 0,08 0,08 0,10 0,12 0,04 0,14 0,09 0,06 Ceriodaphnia silvestrii 0,17 0,07 0,05 0,09 0,04 0,07 0,07 0,06 Ceriodaphnia cornuta 0,09 0,11 0,09 0,05 0,03 0,10 0,09 0,05 Moina minuta 0,19 0,27 0,21 0,27 0,14 0,13 0,11 0,18 Diaphanosoma spinulosum 0,16 0,13 0,13 0,21 0,09 0,13 0,11 0,15 Diaphanosoma fluviatile 0,16 0,08 0,07 0,13 0,10 0,10 0,03 0,08 Bosmina freyi 0,05 0,05 0,11 0,03 0,04 0,07 Bosmina hagmanni 0,07 0,08 0,03 0,07 0,03 0,06 0,05 0,03 Bosminopsis deitersi 0,06 0,05
TRÊS MARIAS BARRÃO EXTREMA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08 Thermocyclops minutus 5,25 4,49 6,43 5,33 5,38 5,02 5,44 5,31 Bosminopsis deitersi 1,08 1,28 1,16 1,23 1,11 1,08 1,21 1,20 Bosmina hagmanni 1,13 1,26 1,25 1,25 1,20 1,20 1,20 1,15 Ceriodaphnia cornuta 1,13 1,30 1,22 1,15 1,05 1,20 1,13 1,20 Moina minuta 1,15 1,56 1,38 1,55 1,07 1,52 1,47 1,30
FURNAS VARJÃO MENDONÇA seca/06 chuva/07 seca/07 chuva/08 seca/06 chuva/07 seca/07 chuva/08 Thermocyclops minutus 4,33 3,88 5,20 3,61 4,32 4,85 4,32 3,55 Notodiaptomus henseni 2,47 2,30 3,28 2,09 2,43 2,77 2,32 2,14 Daphnia ambigua 1,31 1,21 1,53 1,16 1,20 1,35 1,52 1,16 Ceriodaphnia silvestrii 1,09 1,40 1,00 1,08 1,06 1,14 1,10 1,17 Ceriodaphnia cornuta 1,04 1,01 1,00 1,06 1,00 1,06 1,04 1,03 Moina minuta 1,14 1,25 1,07 1,05 1,07 1,13 1,10 1,16 Diaphanosoma spinulosum 1,21 1,17 1,00 1,11 1,15 1,28 1,16 1,12 Diaphanosoma fluviatile 1,07 1,12 1,29 1,09 1,00 1,10 1,00 1,17 Bosmina freyi 1,00 1,00 1,04 1,04 1,00 1,00 1,02 Bosmina hagmanni 1,12 1,29 1,07 1,13 1,00 1,27 1,00 1,21 Bosminopsis deitersi 1,00 1,00 1,00
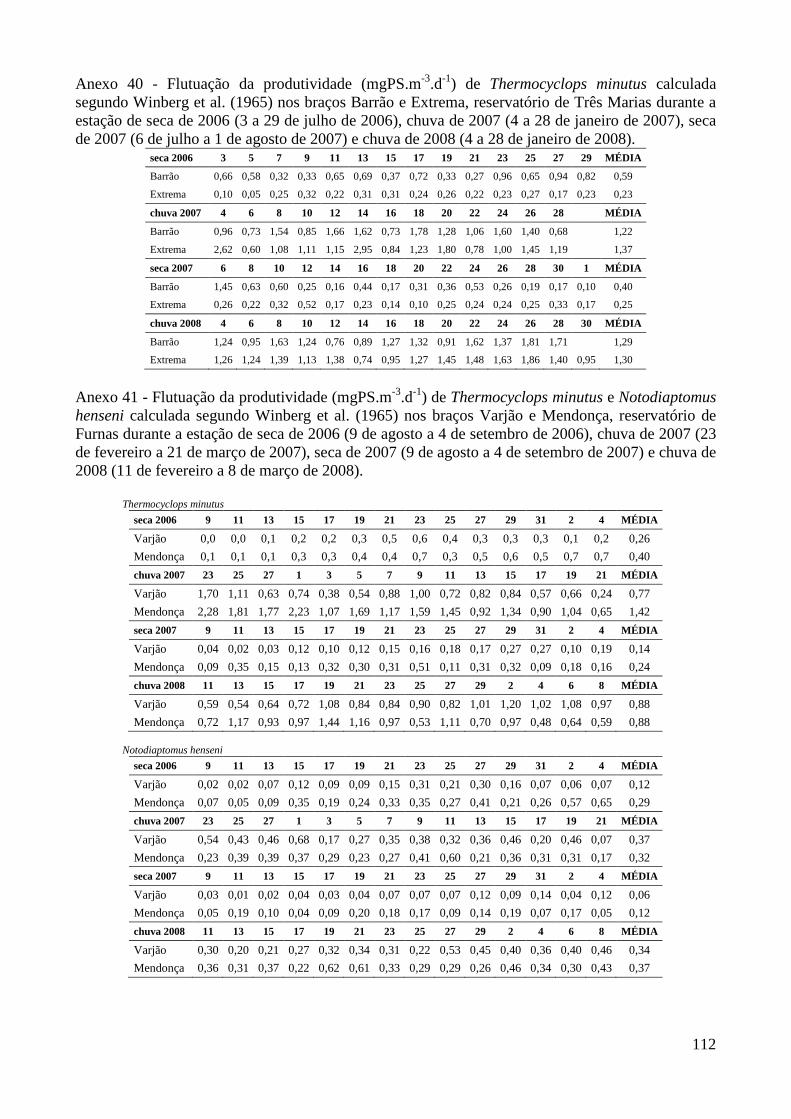
112
Anexo 40 - Flutuação da produtividade (mgPS.m-3.d-1) de Thermocyclops minutus calculada segundo Winberg et al. (1965) nos braços Barrão e Extrema, reservatório de Três Marias durante a estação de seca de 2006 (3 a 29 de julho de 2006), chuva de 2007 (4 a 28 de janeiro de 2007), seca de 2007 (6 de julho a 1 de agosto de 2007) e chuva de 2008 (4 a 28 de janeiro de 2008).
seca 2006 3 5 7 9 11 13 15 17 19 21 23 25 27 29 MÉDIA
Barrão 0,66 0,58 0,32 0,33 0,65 0,69 0,37 0,72 0,33 0,27 0,96 0,65 0,94 0,82 0,59
Extrema 0,10 0,05 0,25 0,32 0,22 0,31 0,31 0,24 0,26 0,22 0,23 0,27 0,17 0,23 0,23
chuva 2007 4 6 8 10 12 14 16 18 20 22 24 26 28 MÉDIA
Barrão 0,96 0,73 1,54 0,85 1,66 1,62 0,73 1,78 1,28 1,06 1,60 1,40 0,68
1,22
Extrema 2,62 0,60 1,08 1,11 1,15 2,95 0,84 1,23 1,80 0,78 1,00 1,45 1,19
1,37
seca 2007 6 8 10 12 14 16 18 20 22 24 26 28 30 1 MÉDIA
Barrão 1,45 0,63 0,60 0,25 0,16 0,44 0,17 0,31 0,36 0,53 0,26 0,19 0,17 0,10 0,40
Extrema 0,26 0,22 0,32 0,52 0,17 0,23 0,14 0,10 0,25 0,24 0,24 0,25 0,33 0,17 0,25
chuva 2008 4 6 8 10 12 14 16 18 20 22 24 26 28 30 MÉDIA
Barrão 1,24 0,95 1,63 1,24 0,76 0,89 1,27 1,32 0,91 1,62 1,37 1,81 1,71
1,29
Extrema 1,26 1,24 1,39 1,13 1,38 0,74 0,95 1,27 1,45 1,48 1,63 1,86 1,40 0,95 1,30
Anexo 41 - Flutuação da produtividade (mgPS.m-3.d-1) de Thermocyclops minutus e Notodiaptomus henseni calculada segundo Winberg et al. (1965) nos braços Varjão e Mendonça, reservatório de Furnas durante a estação de seca de 2006 (9 de agosto a 4 de setembro de 2006), chuva de 2007 (23 de fevereiro a 21 de março de 2007), seca de 2007 (9 de agosto a 4 de setembro de 2007) e chuva de 2008 (11 de fevereiro a 8 de março de 2008).
Thermocyclops minutus
seca 2006 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Varjão 0,0 0,0 0,1 0,2 0,2 0,3 0,5 0,6 0,4 0,3 0,3 0,3 0,1 0,2 0,26
Mendonça 0,1 0,1 0,1 0,3 0,3 0,4 0,4 0,7 0,3 0,5 0,6 0,5 0,7 0,7 0,40
chuva 2007 23 25 27 1 3 5 7 9 11 13 15 17 19 21 MÉDIA
Varjão 1,70 1,11 0,63 0,74 0,38 0,54 0,88 1,00 0,72 0,82 0,84 0,57 0,66 0,24 0,77
Mendonça 2,28 1,81 1,77 2,23 1,07 1,69 1,17 1,59 1,45 0,92 1,34 0,90 1,04 0,65 1,42
seca 2007 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Varjão 0,04 0,02 0,03 0,12 0,10 0,12 0,15 0,16 0,18 0,17 0,27 0,27 0,10 0,19 0,14
Mendonça 0,09 0,35 0,15 0,13 0,32 0,30 0,31 0,51 0,11 0,31 0,32 0,09 0,18 0,16 0,24
chuva 2008 11 13 15 17 19 21 23 25 27 29 2 4 6 8 MÉDIA
Varjão 0,59 0,54 0,64 0,72 1,08 0,84 0,84 0,90 0,82 1,01 1,20 1,02 1,08 0,97 0,88
Mendonça 0,72 1,17 0,93 0,97 1,44 1,16 0,97 0,53 1,11 0,70 0,97 0,48 0,64 0,59 0,88 Notodiaptomus henseni
seca 2006 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Varjão 0,02 0,02 0,07 0,12 0,09 0,09 0,15 0,31 0,21 0,30 0,16 0,07 0,06 0,07 0,12
Mendonça 0,07 0,05 0,09 0,35 0,19 0,24 0,33 0,35 0,27 0,41 0,21 0,26 0,57 0,65 0,29
chuva 2007 23 25 27 1 3 5 7 9 11 13 15 17 19 21 MÉDIA
Varjão 0,54 0,43 0,46 0,68 0,17 0,27 0,35 0,38 0,32 0,36 0,46 0,20 0,46 0,07 0,37
Mendonça 0,23 0,39 0,39 0,37 0,29 0,23 0,27 0,41 0,60 0,21 0,36 0,31 0,31 0,17 0,32
seca 2007 9 11 13 15 17 19 21 23 25 27 29 31 2 4 MÉDIA
Varjão 0,03 0,01 0,02 0,04 0,03 0,04 0,07 0,07 0,07 0,12 0,09 0,14 0,04 0,12 0,06
Mendonça 0,05 0,19 0,10 0,04 0,09 0,20 0,18 0,17 0,09 0,14 0,19 0,07 0,17 0,05 0,12
chuva 2008 11 13 15 17 19 21 23 25 27 29 2 4 6 8 MÉDIA
Varjão 0,30 0,20 0,21 0,27 0,32 0,34 0,31 0,22 0,53 0,45 0,40 0,36 0,40 0,46 0,34
Mendonça 0,36 0,31 0,37 0,22 0,62 0,61 0,33 0,29 0,29 0,26 0,46 0,34 0,30 0,43 0,37