avaliação da reação em cadeia da polimerase (pcr) em...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO INSTITUTO DE MICROBIOLOGIA PROFESOR PAULO DE GÓES
LAURA MARIA ABREU OLIVEIRA
Avaliação da Reação em Cadeia da Polimerase (PCR) em Amostras de Fezes para o Diagnóstico
da Infecção pelo Schistosoma mansoni
Rio de Janeiro Março/2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

LAURA MARIA ABREU OLIVEIRA
Avaliação da Reação em Cadeia da Polimerase (PCR) em Amostras de Fezes para o Diagnóstico da Infecção pelo Schistosoma mansoni
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Microbiologia do Instituto de Microbiologia Professor Paulo de Góes, Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Mestre em Ciências Biológicas (Microbiologia).
Orientadores: Dr. José Mauro Peralta Dra. Margareth Maria Lessa Gonçalves
Rio de Janeiro 2007

Avaliação da Reação em Cadeia da Polimerase (PCR) em Amostras de Fezes
para o Diagnóstico da Infecção pelo Schistosoma mansoni
Laura Maria Abreu Oliveira
Orientadores: José Mauro Peralta e Margareth Maria Lessa Gonçalves
Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências Biológicas (Microbiologia), Instituto de Microbiologia Professor Paulo de Góes, da Universidade Federal do Rio de Janeiro – UFRJ, como parte dos requisitos necessários à obtenção do título de Mestre em Ciências (Microbiologia)
Aprovada por:
___________________________________ Prof a Dr a Maria Helena da Silva - Presidente
Universidade Federal do Rio de Janeiro
___________________________________ Prof. Dr. Walter Martin R. Oelemann - Revisor
Universidade Federal do Rio de Janeiro
___________________________________ Prof a Dr a Norma Suely O. Santos
Universidade Federal do Rio de Janeiro
___________________________________ Prof. Dr. Arnaldo Maldonado Junior
Fundação Oswaldo Cruz
Rio de Janeiro Março/2007

Oliveira, Laura Maria Abreu. Avaliação da Reação em cadeia da Polimerase (PCR) em amostras de fezes para o diagnóstico da infecção pelo Schistosoma mansoni . Laura Maria Abreu Oliveira. – Rio de Janeiro: UFRJ/ IMPPG, 2007. p.92. Orientadores: José Mauro Peralta e Margareth Maria Lessa Gonçalves Dissertação (Mestrado) – UFRJ/ Instituto de Microbiologia Professor Paulo de Góes-IMPPG/ Programa de Pós-graduação em Ciências Biológicas (Microbiologia), 2007. Referências bibliográficas: f. 79 - 89 1- Índices da infecção pelo Schistosoma mansoni, 2- Avaliação dos métodos de diagnóstico da infecção pelo Schistosoma mansoni, 3- Avaliação clínica dos pacientes com esquistossomose mansônica. I- Peralta, José Mauro. II- Universidade Federal do Rio de Janeiro. Instituto de Microbiologia (IMPPG), Programa de Pós-Graduação em Ciências Biológicas (Microbiologia). III- Título.

Dedico este trabalho aos meus Pais
(Celeste e Manuel), pelo amor, apoio e
dedicação incondicional em todos os momentos
da minha vida.
À minha estimada irmã Fernanda
pelo amor, ensinamentos eternos, força,
vitalidade, incentivo e amizade.

Agradecimentos
Ao Dr. José Mauro Peralta, Pesquisador Chefe do Laboratório de Sorologia do Departamento de Imunologia do Instituto de Microbiologia Professor Paulo de Góes-IMPPG/UFRJ, pela oportunidade de trabalhar em seu grupo, pela excelente orientação, constante apoio, amizade e paciência na realização desta dissertação.
A Dra. Margareth Maria Lessa Gonçalves, pelo apoio, presteza constante no decorrer
deste trabalho, palavras amigas, paciência e confiança, que tornaram este período muito mais construtivo e agradável.
Ao IMPPG e à Fiocruz pela estrutura e condições necessárias ao desenvolvimento deste
trabalho. Ao CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), pelo
financiamento deste trabalho. À Helena Carneiro dos Santos, pela amizade e incentivo, que não mediu esforços nas
horas difíceis para realizar este trabalho. Ao Professor Walter Oelemann, pelo incentivo, além da valiosa contribuição durante o
desenvolvimento deste trabalho. Ao Dr. Franklin. D. Rumjanek e a técnica Marta Freire Finotte, do Laboratório de
Bioquímica e Biologia Molecular do Schistosoma mansoni, pela doação de ovos do parasito para a realização da padronização da PCR.
A Dra. Lúcia Martins Teixeira, do Laboratório de Biotecnologia do Departamento de
Microbiologia Médica do IMPPG, pela amizade, pelo incentivo, além da valiosa contribuição na tradução de meu texto para o inglês.
Ao Carlos Ausberto Bonnetti de Souza pela amizade e pelo apoio técnico. À turma do Laboratório 046, turma da amizade, turma da alegria, turma do almoço,
turma do apoio, vocês são demais!!! (Regina Peralta, Giovani Veríssimo, Mauro Jorge Castro, Nathalia Motta, Diva Monteiro, Jorge Luiz Gonçalves, Adriana Neves, Tiana Gonçalves, Adriana Sudré, Maria José Ponce, Aline Caseca, Eduardo Volotão e Ronald Ferreira da Costa Neto).
Aos colegas Magali Barreto e Marisa Soares do Departamento de Biologia do Instituto
Oswaldo Cruz-Fiocruz, pela amizade e pelo incentivo durante todo o desenvolvimento desta dissertação.
Aos professores, coordenadores e secretárias do Curso de Pós-graduação do IMPPG. A todos os pacientes que colaboraram com este trabalho, espero que possam ser
beneficiados no futuro, com estas investigações.

A todos que ajudaram, de alguma forma, mas não foram citados aqui por falha minha. E por fim, a Deus, meu grande amigo que em todos os momentos me guia pelos
melhores caminhos, restabelece minhas esperanças e sonhos, dá vida à minha vida e está sempre presente nos momentos em que tive meus problemas, nos meus desafios e nas minhas alegrias.
Não teria sido possível desenvolver este trabalho sem a amizade e o auxílio de muitas
pessoas e instituições. Em muitas ocasiões, pequenos gestos valeram o mundo. Muito obrigada.

Resumo Avaliação da Reação em Cadeia da Polimerase (PCR) em Amostras de Fezes para o Diagnóstico da Infecção pelo Schistosoma mansoni.
Laura Maria Abreu Oliveira
Orientadores: José Mauro Peralta e Margareth Maria Lessa Gonçalves.
Resumo da Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências Biológicas (Microbiologia), do Instituto de Microbiologia Professor Paulo de Góes, da Universidade Federal do Rio de Janeiro, 2007. Este estudo teve como objetivo avaliar a PCR como alternativa ao diagnóstico da
esquistossomose mansônica em indivíduos portadores de baixa carga parasitária, de áreas de
baixa endemicidade ou expostos ocasionalmente ao Schistosoma mansoni. Cinco grupos de
pacientes foram estudados: 16 com esquistossomose que tinham o exame coprológico positivo
com eliminação de menos de 10 ovos por grama de fezes (OPG); 18 com eliminação maior
que 10 OPG; 22 com sorologia reativa através do ELISA-IgG contra antígeno solúvel de
membrana de vermes S. mansoni adultos e sem eliminação de ovos; 25 portadores de outras
parasitoses intestinais e 21 indivíduos com resultado parasitológico negativo. Foram avaliados
dois procedimentos de extração do DNA fecal e dois pares de oligonucleotídeos iniciadores,
um originado de seqüência altamente repetitiva ao longo do genoma do S. mansoni e outro de
uma seqüência da região conservada do RNA ribossomal do parasito. Os resultados
observados entre os indivíduos esquistossomóticos mostraram uma alta sensibilidade da PCR,
sendo detectado DNA do parasito em 91% das amostras analisadas (31/34). Não houve
amplificação em três amostras de fezes que apresentavam carga parasitária abaixo de 10 OPG.
A especificidade do teste para ambos os iniciadores foi de 100%. No grupo dos soropositivos,
observamos que 59% (13/22) das amostras fecais analisadas apresentaram DNA do S.
mansoni, corroborando os resultados da sorologia. Em vista dos resultados obtidos, foi
padronizada a PCR-Multiplex utilizando os dois pares de iniciadores. A técnica apresentou
uma concordância de 100% com as PCR convencionais. Um novo grupo de pacientes
esquistossomóticos (grupo VI) que compreendia indivíduos que forneceram até três amostras
fecais colhidas em dias diferentes, foi avaliado pela PCR-Multiplex. Nesse grupo, havia
pacientes com apenas uma amostra fecal contendo ovos do parasito e pacientes com mais de
uma amostra positiva na coproscopia. Com o emprego da PCR-Multiplex, o DNA do parasito
pôde ser detectado na maioria das amostras com resultados negativos pela coproscopia.
Entretanto, houve um caso em que uma das amostras com coproscopia positiva de um mesmo

indivíduo, foi negativa pela PCR-Multiplex. Foi avaliado também um reagente de extração
comercial para obtenção de DNA de amostra fecal. Os resultados obtidos reproduziram
aqueles observados anteriormente com os protocolos padronizados em nosso laboratório. Os
resultados alcançados demonstraram que a PCR-Multiplex pode ser uma importante
ferramenta para detecção da infecção pelo S.mansoni em indivíduos que eliminem pouca
quantidade de ovos nas fezes. Além disso, com a determinação da infecção através da
detecção de DNA do S. mansoni naquelas amostras de fezes dos indivíduos apenas
soropositivos para a esquistossomose, surge uma nova possibilidade para a confirmação dos
resultados obtidos pelo ELISA-IgG. Portanto, estudos nesta direção devem ser incentivados e
ampliados.
Schistosoma mansoni, esquistossomose, baixa carga parasitária,diagnóstico,PCR

Abstract .Evaluation of the Polymerase Chain Reaction in Fecal Samples for the Diagnosis of Infection by Schistosoma mansoni.
Laura Maria Abreu Oliveira
Advisors: José Mauro Peralta e Margareth Maria Lessa Gonçalves Abstract da Dissertação de Mestrado submetida ao Programa de Pós-Graduação em Ciências
Biológicas (Microbiologia), do Instituto de Microbiologia Professor Paulo de Góes, da
Universidade Federal do Rio de Janeiro, 2007
The aim of the present study was to evaluate the PCR as an alternative tool for the diagnosis of schistosomiasis mansoni in individuals with low parasitic load from areas of low endemicity or under occasional risk of infection by Schistosoma mansoni. Five groups of patients were studied: 16 individuals with schistosomiasis that had a positive stool exam with elimination of less than 10 eggs per gram of feces (EPG); 18 individuals with egg elimination high than 10 EPG; 22 individuals with reactive ELISA-IgG against S. mansoni soluble membrane antigen and negative coproscopy; 25 individuals with other intestinal parasites and 21 individuals with negative parasitological exam. Two procedures for DNA extraction from fecal samples and two different pairs of primers were evaluated. One of the primer pairs was designed to amplify a highly repeated 121-basepair sequence of S. mansoni and the other was designed to amplify Schistosoma 28S rDNA. The results obtained with stool samples from individuals with schistosomiasis showed a high sensibility for the PCR, since S. mansoni DNA was detected in 91% (31/34) of the samples analyzed. No amplification was observed in three stool samples from individuals excreting below 10 EPG. The specificity of the test for both pairs of primers was 100%. In the group of seropositive individuals, S. mansoni DNA was detected in 59% (13/22) of fecal samples, corroborating the serological results. In view of the results obtained, a Multiplex-PCR using the two pairs of primers was standardized. This technique presented an agreement of 100% with the conventional PCR. A new group (group VI), composed by individuals with schistosomiasis that collected three fecal samples in different days, was evaluated by the PCR-Multiplex. In this group, there were patients with one stool sample containing S. mansoni eggs and patients with more than a positive sample in the coproscopy. DNA of the parasite was detected by the PCR-Multiplex in the majority of the fecal samples that were negative by coproscopy in this group. However, one of the samples with positive coproscopy from the same individual, was negative by the PCR-Multiplex. A commercial kit for extraction of fecal DNA was also evaluated. The results reproduced those previously observed with the protocols standardized in our laboratory. Overall, the results indicated that PCR-Multiplex can be an important tool for detection of the S. mansoni infection in individuals excreting few eggs in feces. Moreover, the determination of the infection through the detection of S. mansoni DNA in stool samples from seropositive individuals represented a new possibility to confirm the results of ELISA-IgG for schistosomiasis. Therefore, studies in this direction should be encouraged and extended.
Schistosoma mansoni, schistosomiasis, low parasitic load, diagnosis, PCR.

Lista de Abreviaturas, Siglas e Unidades
BSA ..........
CAA .........
CCA .........
DNA .........
dATP ........
dCTP ........
dGTP ........
dTTP .........
DMSO ......
ELISA ......
fg ..............
FNS ..........
g ................
IBGE ........
IgA ...........
IgG ...........
IgM ...........
kDa ...........
Mb ............
mL ............
mM ….......
Bovine serum albumin
Circulating Anodic Antigen
Circulating Cathodic Antigen
Deoxyribonucleic Acid
Deoxiadenosine triphosphate
Deoxicytidine triphosphate
Deoxiguanosine triphosphate
Deoxithymidine triphosphate
Dimetilsulfóxido
Enzyme-linked immunosorbent assay
fentograma
Fundação Nacional de Saúde
grama
Instituto Brasileiro de Geografia Estatística
Imunoglobulina A
Imunoglobulina G
Imunoglobulina M
Quilodalton
Megabases (106 bases)
Mililitro
Milimolar

MRE .........
N ...............
OMS .........
OPG ..........
pb ..............
PBS ...........
PCR ..........
pg ..............
pH .............
PM ……...
PAGE rpm SMMA .....
TE ….........
TMB …….
U …….…..
v/v ………
μg ………
μL ……….
μM ………
Síndrome Mielorradicular
Normalidade
Organização Mundial de Saúde
Ovos por grama de fezes
Pares de base
Tampão fosfato-salina “Phosphate-buffered saline”
Reação em cadeia da polimerase “Polymerase chain reaction”
Picograma
Potencial hidrogeniônico
Peso Molecular
Eletroforese em gel de poliacrilamida – Polyacrylamide gel electrophoresis Rotação por minuto
Antígeno solúvel de membrana de verme adulto
Tris (hidroximetil) aminometano, EDTA (ácido etilenodiaminotetracético)
3, 3’,5, 5’ tetramethylbenzidine
Unidade
volume/volume
Micrograma
Microlitro
Micromolar

Sumário
INTRODUÇÃO
I- A esquistossomose no mundo e no Brasil ...........................................................
II- Ciclo de vida do Schistosoma mansoni ..............................................................
III- A patologia da esquistossomose mansônica ....................................................
IV- Diagnóstico laboratorial da esquistossomose mansônica ..............................
V- A técnica da reação em cadeia da polimerase aplicada à Esquistossomose ..
OBJETIVOS ................................................................................................................
MATERIAIS & MÉTODOS ......................................................................................
I- Área de estudo ......................................................................................................
II- Amostragem ........................................................................................................
II.1. Grupo de estudo ............................................................................................
II.2. Amostras controles .......................................................................................
III- Técnica coprológica ...........................................................................................
IV- Técnica sorológica .............................................................................................
IV.1. Preparação do antígeno SMMA utilizado no teste de ELISA-IgG .........
IV.2. Ensaio imunoenzimático (ELISA-IgG) ......................................................
V- Diagnóstico molecular ........................................................................................
V.1. Extração do DNA de Schistosoma mansoni ................................................
V.1.1. Protocolo de extração segundo Pitcher, Saunders & Owen (1989) .
V.1.2. Protocolo de extração de Pitcher, Saunders & Owen (1989), com modificações propostas por Silva et al. (1999) ..............................................
V.1.3. Protocolo de extração através de um método comercial ..................
V.2. Reação em cadeia da polimerase .................................................................
12
13
15
17
23
28
29
29
31
31
33
33
34
35
36
37
37
37
39
41
42

V.2.1. Protocolo de amplificação de uma região repetitiva ao longo do genoma do Schistosoma mansoni ...................................................................
V.2.2- Protocolo de amplificação do gene que codifica o DNA ribossomal do Schistosoma mansoni ..................................................................................
V.2.3- Protocolo de amplificação do Multiplex- PCR ..................................
V.3. Teste para a presença de inibidores da PCR ..............................................
V.4. Detecção do produto amplificado ................................................................
RESULTADOS ............................................................................................................
DISCUSSÃO ................................................................................................................
CONCLUSÕES ...........................................................................................................
REFERÊNCIAS BIBLIOGRÁFICAS ......................................................................
ANEXOS ......................................................................................................................
42
43
44
45
45
47
67
77
79
90

Introdução
I- A esquistossomose no mundo e no Brasil
A esquistossomose é uma parasitose causada por helmintos do gênero Schistosoma
presente entre a humanidade desde a antiguidade. Há registros que comprovam a presença de
ovos de Schistosoma em múmias egípcias datando de 3500 a.C. No entanto, a infecção foi
descrita pela primeira vez em 1852, por Bilharz, que observou a presença de vermes adultos
em múmias egípcias, nomeando-os de Distoma haematobia. Posteriormente, em 1858, o
gênero foi modificado para Schistosoma por Weinland e a espécie passou a ser denominada
de Schistosoma haematobium. Em 1902, Patrick Manson descreveu uma outra espécie que
eliminava ovos com espículas laterais. Porém, somente em 1907, Sambon, designou esta
espécie de Schistosoma mansoni em homenagem à descrição feita por Manson. No Brasil, a
espécie S. mansoni foi descrita pela primeira vez por Pirajá da Silva, em um estudo realizado
no Estado da Bahia em 1908 (apud Pessôa & Martins, 1982).
As espécies do gênero Schistosoma de maior interesse à saúde pública são: S.
japonicum (endêmico em países do Sudeste Asiático e do Pacífico Ocidental), S. mansoni
(endêmico em países da África, da América do Sul e nas Antilhas); S. hematobium (endêmico
na África e na região oriental do Mediterrâneo); S. intercalatum (endêmico em países da
África Central e Ocidental); S. mekongi (endêmico na região da Bacia do rio Mekong entre
Laos e Camboja). As esquistossomoses afetam mais de 200 milhões de pessoas em todo o
mundo (Rey, 2001).
No Brasil, ocorre apenas a espécie S. mansoni, capaz de infectar seres humanos e
alguns roedores silvestres como o Nectomys squamipes. Estima-se que aproximadamente 2,5
milhões de brasileiros estejam infectados e outros 25 milhões estejam expostos ao risco de

infecção pelo parasito (Katz & Peixoto, 2000).
A definição do grau de endemicidade nas áreas onde existe a doença baseia-se na
intensidade de infecção que, por sua vez, é estimada pela média geométrica do número de
ovos de S. mansoni freqüentemente eliminado pelos indivíduos parasitados. Assim, segundo a
Organização Mundial de Saúde (OMS, 1993), áreas de baixa endemicidade estão na faixa de
1-100 ovos eliminados por grama de fezes (OPG), áreas de média endemicidade estão na
faixa de 101-400 OPG e as áreas de alta endemicidade encontram-se na faixa acima de 400
OPG.
Nos municípios dos estados de Pernambuco, Alagoas, Sergipe e Bahia observam-se
as maiores prevalências de infecção pelo S. mansoni. No estado de Minas Gerais são
encontradas áreas de alta e média endemicidades da doença, enquanto que áreas de baixa
endemicidade são citadas nos estados do Pará, Maranhão, Ceará, Espírito Santo, Rio de
Janeiro, São Paulo, Paraná, Santa Catarina e, mais recentemente, no Rio Grande do Sul (Rey,
2001). O controle da doença nestas áreas tem ficado sob responsabilidade da Fundação
Nacional de Saúde-FNS, que vem realizando inquéritos parasitológicos anuais e tratamento
dos indivíduos parasitados.
O Estado do Rio de Janeiro apresenta áreas de baixa endemicidade ou mesmo focos
isolados de esquistossomose, com prevalências de infecção geralmente abaixo dos 4% e
cargas parasitárias que não ultrapassam 100 OPG entre os indivíduos infectados (Doumenge
et al., 1987; Rey, 2001).
II- Ciclo de vida do Schistosoma mansoni
O S. mansoni apresenta diferentes formas evolutivas: ovo, miracídio, cercária,
esquistossômulo e verme adulto. Os vermes adultos apresentam dimorfismo sexual e,

habitualmente, são encontrados nas vênulas do sistema porta. Os ovos medem cerca de 150
μm e possuem uma característica marcante que é a presença de espícula lateral (Rey, 2001).
A infecção pelo S. mansoni ocorre pela penetração de cercárias na pele do indivíduo.
Durante a penetração, as cercárias perdem a cauda transformando-se em esquistossômulos
que inicialmente migram na derme. Estes passam para a corrente sanguínea atingindo o
pulmão e, posteriormente, o sistema porta intra-hepático, onde amadurecem até o estágio de
vermes adultos (machos e fêmeas) e acasalam-se. Os casais de vermes migram para as
vênulas da mucosa e submucosa intestinais, onde as fêmeas liberam seus ovos que passam
para o lúmen do intestino e são excretados nas fezes. Os ovos podem ser detectados nas fezes
cerca de 40 dias após a infecção. Depois de eliminados nas fezes, os ovos eclodem após o
contato com a água-doce e liberam os miracídios que penetram no hospedeiro intermediário,
um molusco do gênero Biomphalaria. No molusco ocorrem transformações morfológicas e
processos de multiplicação assexuada que dão origem aos esporocistos primários. Estes
sofrem novas transformações originando os esporocistos secundários, que finalmente, dão
origem as cercárias. Estas então, saem do molusco e nadam até o encontro do hospedeiro
definitivo susceptível (Figura 1).
No Brasil, apenas três espécies de moluscos do gênero Biomphalaria são
susceptíveis à infecção pelo S. mansoni: B. glabrata, B. straminea e B. tenagophila (Rey,
2001).

Figura 1: Ciclo evolutivo do Schistosoma mansoni. (CDC, http://www.dpd.cdc.gov/html/Imagelibrary/Schistosomiasis)
III- A patologia da esquistossomose mansônica
Em relação ao aspecto clínico da doença, a infecção pelo S. mansoni na maioria dos
casos é assintomática. A susceptibilidade do hospedeiro humano à infecção ou à doença
depende de uma série de fatores, como: o número de cercárias que penetram pela pele (carga
infectante), características do hospedeiro (idade e estado nutricional), a freqüência de
ocorrência de reinfecções e o grau de imunidade desenvolvido ao longo do tempo (Rey,
2001). Os sintomas da doença, sob o ponto de vista clínico da esquistossomose, podem ser
divididos em fase aguda e fase crônica.

Na fase aguda da infecção, pode ocorrer a dermatite cercariana que é uma inflamação
da pele no local da penetração da cercária. Essa reação dura cerca de dois dias e depois
regride. Esse mesmo processo de inflamação pode ocorrer no pulmão, durante a etapa de
migração dos esquistossômulos. Nessa fase podem ocorrer ainda sintomas como: febre alta,
tosse, cefaléia, esplenomegalia, linfadenopatia, diarréia e quadros de alergia.
Na fase crônica da doença, a lesão típica é o granuloma formado ao redor dos ovos
retidos nos tecidos do indivíduo infectado. Pode haver progressão para fibrose, culminando na
hipertensão do sistema porta. Variações clínicas podem ocorrer nessa fase, gerando três
principais formas da doença: intestinal (predominante), hepatointestinal e hepatoesplênica.
A forma intestinal ocorre após a regressão dos sintomas da fase aguda e apresenta
sintomas geralmente inespecíficos: perda de apetite, desconforto abdominal, pequenos
quadros diarréicos ou prisão de ventre e, em alguns casos, ocorrência de eliminação de fezes
sanguinolentas. Essa forma coincide geralmente com a detecção de ovos do parasito nas fezes
do hospedeiro.
Na forma hepatointestinal, os sintomas são, em alguns casos, mais acentuados que os
da forma intestinal. A principal diferença entre essas duas formas da doença está no
surgimento de lesões hepáticas.
A forma hepatoesplênica caracteriza-se pelo comprometimento progressivo do
fígado e do baço. Os principais sintomas relatados são: indisposição, má digestão, sensação de
plenitude gástrica, inapetência e emagrecimento. Nessa fase predominam as manifestações
decorrentes da hipertensão do sistema porta, como a presença de varizes esofágicas, cuja
ruptura pode causar hemorragias fatais. Ocorre geralmente hematêmese que vai estar
associada ao grau de hipertensão porta. Os quadros hemorrágicos podem ser acompanhados
de febre, anemia, edema e ascite. A superfície do fígado apresenta cicatrizes de fibrose além
de veias dilatadas e colaterais que são visíveis na parte interna da parede abdominal. Essa

forma clínica, embora seja um processo raro, é a causa mais comum de morbidade da infecção
pelo S. mansoni (Warren, 1984; Prata, 1987; El-Ridi et al., 1997; Rey, 2001).
É possível ainda que os ovos de S. mansoni sejam encontrados em outras
localizações não habituais como bexiga, sistema nervoso central e ovário (Rey, 2001).
O sistema nervoso central pode ser acometido pela deposição assintomática de ovos
de S. mansoni. Na forma sintomática, o indivíduo parasitado apresenta um quadro de
síndrome mielorradicular (MRE). A neuroesquistossomose constitui importante causa de
mielopatia não traumática em regiões endêmicas. (Nascimento-Carvalho & Moreno-Carvalho,
2005). Entretanto, sua prevalência não é conhecida, provavelmente devido ao
desconhecimento desta patologia (Santos et al., 2001).
Oxaminiquine e praziquantel são os quimioterápicos utilizados no tratamento da
esquistossomose. O oxaminiquine é um derivado das tetraidroquileinas produzido por síntese
biológica que tem efeito anticolinérgico, causando aumento na motilidade do parasito e
inibindo a síntese de ácidos nucléicos. Praziquantel é um derivado de pirazino-isoquinolina
que afeta principalmente as fêmeas. Causa alterações no tegumento e redução da concentração
de glutationa do verme (FNS, 1998).
IV- Diagnóstico laboratorial da esquistossomose mansônica
O diagnóstico laboratorial tem sido realizado, sobretudo, por métodos parasitológicos
e anátomo-parasitológicos, através da demonstração de ovos de S. mansoni nas fezes ou
tecidos.
Os métodos parasitológicos utilizados no diagnóstico dessa doença podem ser de
dois tipos: qualitativos (apenas detectam a presença de ovos dos parasitos nas fezes) ou

quantitativos (detectam a presença dos ovos e também permitem a contagem do número de
ovos por grama de fezes).
A técnica de sedimentação espontânea descrita por Lutz (1919) e modificada por
Hoffman, Pons & Janer (1934), é o método qualitativo mais comumente empregado, que
permite a identificação e a diferenciação dos ovos do helminto em viáveis ou não viáveis.
Dentre os métodos quantitativos existem as técnicas de Bell, Stoll, Simões Barbosa
(Rey 2001) e Kato, posteriormente modificado por Katz, Chaves & Pelegrino (1972). Essas
técnicas permitem a visualização, contagem e estimativa do número de ovos do helminto em
determinada quantidade de fezes, servindo assim para avaliar também a intensidade de
infecção do indivíduo positivo (FNS, 1998).
O método Kato-Katz tem sido o exame de escolha para o diagnóstico da
esquistossomose mansônica, sendo recomendado pela OMS por ser barato, simples e
específico. Porém, estudos têm demonstrado que em áreas de baixa endemicidade, onde a
eliminação de ovos está abaixo dos 100 OPG, o método perde na sensibilidade de detecção da
infecção. Deste modo, há a necessidade de implementação de ferramentas alternativas de
diagnóstico para evitar possíveis resultados falso-negativos (Doumenge et al., 1987; FNS,
1994; Rey, 2001).
Outro método parasitológico é a técnica de eclosão de miracídios, descrita por
Fulleborn em 1921. Este método consiste em colocar a amostra fecal em recipiente contendo
água à determinada temperatura e expô-la a uma fonte luminosa para se observar, depois de
algum tempo, a olho nu ou com auxílio de lupa, a presença de miracídios liberados dos ovos
que porventura existam nessa amostra (FNS, 1998).
No diagnóstico anátomo-parasitológico da doença é realizada biopsia, que consiste
na retirada de fragmentos da mucosa retal (mais utilizado) ou de fígado e seu exame para
detecção de ovos ou de granulomas. A contagem de ovos na biopsia recebe o nome de

oograma. O resultado do oograma é expresso em ovos por grama de tecido retal biopsiado e,
assim, permite a avaliação da intensidade de infecção. Essa técnica costuma ser mais
empregada quando repetidos exames de fezes resultam negativos ou na avaliação de
eficiência terapêutica, através de oogramas seriados, por se tratar de um método invasivo e
bastante incômodo, necessitando, inclusive, de instrumental mais sofisticado em ambiente
hospitalar e pessoal especializado. A biopsia hepática é usada quando a doença se apresenta
clinicamente grave e não se consegue confirmar a esquistossomose ou diferenciá-la de outras
hepatopatias (FNS, 1998).
Como a identificação do parasito é crucial para a eficiência dos programas de
controle da esquistossomose e diante da subestimação da prevalência da doença pelo
diagnóstico coprológico, alguns países como a Venezuela, Suriname e algumas ilhas do
Caribe, que apresentam áreas de baixa endemicidade com indivíduos apresentando menor
intensidade de infecção, têm incorporado métodos imunológicos aos seus programas de
controle (Bergquist, 1992; Noya et al., 1998).
As técnicas de imunodiagnóstico podem ser utilizadas tanto na detecção de antígenos
circulantes do Schistosoma como na detecção de anticorpos produzidos contra o parasito.
O método da intradermorreação foi amplamente empregado no diagnóstico da
doença durante algum tempo. Esse método baseia-se na resposta celular do hospedeiro à
presença do parasito. A técnica consistia na inoculação, por via intradérmica, de pequena
quantidade de solução composta por água e extrato de vermes adultos ou cercárias e a
observação de pápula formada no local. Seu uso, porém, foi limitado pela constatação de que
era pouco sensível em crianças e tinha pouca utilidade na determinação da prevalência da
esquistossomose, além da permanência do resultado positivo depois da cura da doença e a
reatividade cruzada com outros helmintos (FNS, 1998).
A aplicação do imunodiagnóstico baseado em ensaios para detecção de anticorpos

produzidos pelo hospedeiro no curso da esquistossomose, teve início em 1909, quando
Fujinami & Nakamura descreveram a reação de fixação do complemento para o diagnóstico
da doença (apud Rabello, 1997). Prosseguiu com Fairley em 1919, que usou um preparado de
hepatopâncreas de caramujos infectados como antígeno na reação de fixação do
complemento. Depois disto, em 1927, Fairley & Williams desenvolveram um teste cutâneo
empregando o mesmo preparado antigênico. Desde então, uma série de testes imunológicos
tem sido estudada para uso no diagnóstico da esquistossomose (Brandt & Finch, 1946; Vogel
& Minning, 1949; Kagan & Oliver-Gonzales, 1950; Oliver-Gonzales, 1954; Sadun, Williams
& Anderson, 1960; Coudert et al., 1968; Hoshino, Camargo & Corrêa, 1970; Wilson, Sulzer
& Walls, 1974; Huldt et al., 1975; Mclaren et al., 1978; Nash, 1978; Nash, Lunde & Cheever,
1981; Deelder et al., 1980, 1989; Taylor & Wells, 1984; Maddison et al., 1985; Kanamura,
Hoshino-Shimizu & Silva, 1992).
Muitas preparações antigênicas foram produzidas a partir dos diferentes estágios
evolutivos do helminto. Freqüentemente são mais usados antígenos provenientes de vermes
adultos e ovos, podendo ser proteínas, glicoproteínas, polissacarídeos, além de alguns
constituintes preparados com atividade enzimática. Até o momento as técnicas de
imunofluorescência indireta, “Western blot” e principalmente o Enzyme Linked Immuno
Sorbent Assay (ELISA) têm sido mais aplicadas ao diagnóstico da doença (Tsang, Peralta &
Simons, 1983; Ruppel, Diesfield & Pother,1985; Ruppel et al.,1985; Idris & Ruppel, 1988;
Rossi, Tsang & Picher, 1991; Lima et al., 1993, 1998; Rabello et al., 1993; Silva et al., 1993;
Tsang & Wilkins, 1991, 1997; Valli et al., 1997; Camargo-Neves et al., 1998; Gomes et al.,
1998; Tarp, Black & Petersen, 2000; Van Gool et al., 2002; Burlandy-Soares et al., 2003;
Makarova et al., 2003, 2005; Sulahian et al., 2005; Gonçalves et al., 2006).
Através da técnica de imunofluorescência indireta em cortes de verme adulto, foi
observado que os anticorpos IgA aparecem na fase aguda da doença, enquanto IgM e IgG

aparecem tanto na fase aguda como crônica. Anticorpos IgA, IgG, IgM e IgE apresentam uma
fluorescência interna localizada na estrutura compatível com o intestino do verme adulto,
demonstrando um padrão focal de fluorescência. Esta reação caracteriza um diagnóstico de
infecção aguda. Por outro lado, o padrão de fluorescência de IgG é difuso, em todo o parasito.
Na infecção crônica, os anticorpos IgM sofrem uma redução em seus títulos de quase dez
vezes, mas permanece positiva (Kanamura et al., 1979, 1991). Uma hipótese razoável para
essa persistência de IgM, seria o contínuo estímulo por antígenos de natureza polissacarídica,
oriundos do tubo digestivo do S. mansoni, regurgitados na corrente sangüínea do hospedeiro
(Nash, 1978; Attallah et al. 1999).
Tsang, Peralta & Simons (1983), descreveram pela primeira vez a técnica de
“Western blot” aplicada ao diagnóstico da esquistossomose usando diferentes preparações
antigênicas do S. mansoni e amostras de soros de pacientes procedentes de áreas endêmicas
distintas, mostrando que o diagnóstico imunológico da doença apresentava grande potencial e
abrindo caminho para as pesquisas sobre os componentes antigênicos que compunham os
extratos preparados a partir do verme.
Dentre as provas sorológicas usadas na detecção da infecção pelo S. mansoni, o
ELISA é o mais amplamente empregado nos países onde a doença é endêmica. Em 1979,
Lunde, Ottesen & Cheever, realizaram um dos primeiros ELISA aplicados ao diagnóstico da
esquistossomose. Os autores analisaram amostras de soro de pacientes com esquistossomose
aguda e crônica, e amostras de soro obtidas de animais (macacos) experimentalmente
infectados pelo S. mansoni. Foram preparados antígenos a partir de ovos, cercárias e vermes
adultos de S. mansoni. Foi observada reatividade tanto nos soros de seres humanos como de
animais nos ensaios com as três preparações antigênicas. Os soros dos pacientes com infecção
aguda e os soros dos animais referentes ao período de dois meses de infecção, reagiram
fortemente com o antígeno de cercária. Por outro lado, os soros de pacientes com doença

crônica e dos animais, colhidos após quatro meses da infecção, reagiram mais fortemente com
o antígeno de verme adulto. Os pesquisadores concluíram que a diferença observada nas
respostas aos antígenos preparados apresentava potencial diagnóstico para distinção entre
infecção aguda ou crônica pelo S. mansoni.
Posteriormente, o padrão de detecção dos anticorpos IgA, IgM e IgG no decurso da
esquistossomose, revelado pela imunofluorescência indireta, pode ser confirmado também nos
ensaios imunoenzimáticos. Assim, a IgA figura como marcador da fase aguda da doença (três
meses) e a IgM e/ou IgG como marcadores de fase crônica.
Contudo, a permanência de anticorpos depois do tratamento da doença tem sido a
principal desvantagem desses ensaios.
Okabe e Tanaka, em 1961, foram os primeiros a observar que antígenos secretados
ou excretados pelo parasito poderiam ter potencial para o diagnóstico da infecção. Esses
antígenos se originam no tubo digestivo do parasito sendo continuamente regurgitados na
corrente sanguínea do hospedeiro (Nash, 1978; Attalah et al., 1999). Estudos posteriores
demonstraram diferentes antígenos derivados de Schistosoma, mas as pesquisas têm sido
focadas particularmente em duas proteoglicanas, que já foram parcialmente caracterizadas por
suas estruturas químicas: CAA (antígeno anódico circulante de 30kDa) e CCA (antígeno
catódico circulante de 70kDa), denominadas de acordo com seu potencial de comportamento
migratório em imunoeletroforese (Deelder et al., 1976; Bergwerff et al., 1994; Hamilton,
Klinker & Doenhoff, 1998).
As abordagens para detecção dos antígenos circulantes CAA e CCA geralmente
envolvem o ELISA de captura empregando anticorpos monoclonais. Apesar de serem capazes
de detectar infecção ativa, as técnicas que utilizam esses antígenos geralmente não são
espécie-específicas e não têm apresentado boa sensibilidade em casos de infecções leves.
Aparentemente existe correlação positiva entre os níveis de antígenos circulantes e as taxas

de excreção de ovos. Além disso, é necessária uma etapa adicional de pré-tratamento da
amostra de soro ou de urina antes da realização do teste (De Jonge et al., 1988; Van Lieshout
et al., 1995; Doenhoff, Chiodini & Hamilton, 2004).
V- A técnica da reação em cadeia da polimerase aplicada à esquistossomose
Diante da baixa sensibilidade do exame parasitológico de fezes e dos ensaios baseados na detecção de antígenos nas áreas de baixa endemicidade, e pela incapacidade dos testes baseados na detecção de anticorpos determinarem infecção ativa, a técnica molecular baseada na reação em cadeia da polimerase (PCR) vem sendo cogitada como mais uma alternativa de diagnóstico da esquistossomose mansônica.
A PCR tem sido utilizada como uma ferramenta alternativa no diagnóstico de outras parasitoses intestinais (Entamoeba histolytica, Cryptosporidium parvum, Enterocytozoon bieneusi, Cyclospora cayetanensis, Giardia lamblia), demonstrando ser sensível e específica (Gibbons-Matthews & Prescott, 2003; Gonin & Trudel, 2003; Verweij et al., 2003, 2004; Subrungruang et al., 2004).
O método de PCR permite a amplificação “in vitro” de uma seqüência específica do
DNA-alvo que pode ser um simples gene ou uma parte dele. A PCR tem mostrado ser
específica e sensível para o diagnóstico de doenças infecto-parasitárias (Saiki et al., 1988,
Singh, 1997). A técnica fundamenta-se em ciclos repetitivos de três reações que ocorrem em
diferentes temperaturas de incubação. Em um mesmo microtubo com reagentes termoestáveis
e seqüências específicas de DNA a serem amplificadas, consiste na etapa de desnaturação das
fitas do DNA-alvo, seguida pela etapa em que ocorrerá a hibridização das seqüências de
iniciadores e terminando com a etapa na qual ocorre a extensão da região flanqueada pelos
iniciadores. Em condições ótimas, essas reações ocorrem na presença de iniciadores
(pequenas seqüências de DNA sintetizadas para serem complementares às seqüências
conhecidas do DNA-alvo) e de grande quantidade dos quatro desoxirribonucleotídeos
trifosfatados (dATP, dCTP, dGTP e dTTP). Utiliza-se ainda uma enzima termoestável (Taq
DNA-polimerase) e solução tampão contendo magnésio.
Com o advento do projeto genoma do S. mansoni, desenvolvido pela OMS, novas

perspectivas na identificação de alvos para quimioterápicos e vacinas e para o
desenvolvimento do diagnóstico têm sido implementadas. Esse projeto vem realizando o
seqüênciamento do genoma do parasito. O S. mansoni tem genoma haplóide de 270 mega-
bases (Mb) com 34% de guanina + citosina, constando de oito pares de cromossomas. O
genoma é constituído de 15.000 a 25.000 genes, sendo que 40% destes genes apresentam
seqüências altamente repetitivas; o restante representa uma cópia simples ou pequeno gene
familiar (LoVerde et al., 2004). Estas seqüências repetitivas têm sido utilizadas na PCR para o
diagnóstico da infecção pelo S. mansoni em amostras fecais.
Inicialmente em 1991, Hamburger et al., descreveram seqüências altamente
repetitivas ao longo do genoma do S. mansoni. Mais tarde, em 1998, os autores utilizaram
essas sequências na padronização da PCR para detectar DNA de cercárias em coleções
hídricas. Contudo, em 1997, foram publicados os primeiros trabalhos sobre a detecção de
DNA específico de Schistosoma para determinação da infecção em moluscos hospedeiros
intermediários do parasito (Hanelt et al., 1997; Janotti-Passos et al., 1997).
Pontes, Dias-Neto & Rabello (2002), padronizaram um ensaio de PCR utilizando
amostras de soro e de fezes humanas, sendo possível detectar até um fentograma de DNA do
parasito. Neste ensaio as seqüências-alvo empregadas foram as mesmas originalmente
descritas por Hamburger et al em 1991. A PCR para detecção da infecção pelo S. mansoni em
amostra fecal foi capaz de diagnosticar DNA do parasito a partir de baixas cargas como 2,4
OPG, demonstrando uma sensibilidade dez vezes maior que a da técnica de Kato-Katz.
Posteriormente, os autores empregaram a PCR para diagnóstico num maior número de
indivíduos, residentes numa área endêmica para esquistossomose (Comercinho, Minas
Gerais). A prevalência da infecção mostrada pela coproscopia de três amostras fecais
diferentes foi de 30,9%, enquanto que pela análise de uma amostra fecal através da PCR foi
de 38,1%. Exames de fezes complementares nas doze pessoas diagnosticadas apenas pela

PCR revelaram que cinco delas eliminavam ovos do S.mansoni. Tomando por base o exame
parasitológico das fezes como padrão-ouro, a PCR apresentou 96,7% de sensibilidade e 88%
de especificidade (Pontes et al., 2003).
A detecção de fragmentos de DNA do S. mansoni através da PCR tem se mostrado
satisfatoriamente sensível e específica, tanto quando testada com amostras provenientes de
animais (modelos experimentais) ou de seres humanos (Hamburger et al., 2001; Sandoval et
al., 2006a; Suzuki et al., 2006). Mais recentemente, mostrou-se capaz de determinar a espécie
de Schistosoma (S. mansoni, S. haematobium, S. japonicum, S. intercalatum ou S. bovis)
responsável pela infecção esquistossomótica além de diagnosticar a doença (Sandoval et al.,
2006b).
Entretanto, essa técnica não foi empregada em áreas de baixa endemicidade nem em
amostras de fezes de pacientes que apresentavam apenas reatividade sorológica para a
esquistossomose sem a confirmação parasitológica.
A PCR vem demonstrando ser um método sensível e específico no diagnóstico de
parasitoses intestinais, no entanto, apresenta algumas dificuldades quanto ao processo de
extração do DNA em fezes, devido às peculiaridades do material fecal. Nas fezes existe
grande quantidade de substâncias capazes de inibir a ação da enzima (Taq polimerase) que é
necessária para a realização do processo. As principais substâncias que interferem na reação
de PCR são: sais biliares, bilirrubina, etanol, complexos de polissacarídeos, etc (Deuter et al.,
1995; Monteiro et al., 1997; Wilson, 1997; Vandenberg & Oorschot, 2002).
Para a PCR ser realizada, é necessário um método de extração de DNA que seja simples e rápido e produza DNA altamente puro. A literatura relata que a extração de DNA a partir de material fecal é um processo complexo, em decorrência da co-extração de substâncias inibidoras e da própria presença de resíduos contaminantes oriundos do processo de extração. A remoção total das substâncias inibidoras nem sempre é fácil (Deuter et al., 1995; Wilson, 1997; Vandenberg & Oorschot, 2002). Porém, esses mesmos autores argumentam que é possível identificar a presença desses inibidores na amostra, introduzindo na reação controles de amplificação ou mesmo realizando a reação de amplificação em duplicata. Outra sugestão dos pesquisadores como estratégia para detectar a presença de inibidores na amostra amplificada, é a contaminação proposital (“ Spiking”) da mesma com

DNA extraído de ovos obtidos de fígado de camundongos infectados por S. mansoni, após realização do processo de perfusão.
Variações da PCR clássica têm sido alvo de várias pesquisas, com o objetivo de
desenvolver metodologias mais sensíveis para o diagnóstico da esquistossomose, utilizando
vários iniciadores dirigidos a seqüência de DNA do Schistosoma, sobretudo, S. mansoni
(Gobert et al., 2005; Sandoval et al., 2006a, 2006b; Suzuki et al., 2006).
A PCR-Multiplex emprega vários pares de iniciadores que hibridizam com várias
seqüências genômicas presentes na amostra. Deste modo, pode ser utilizada como estratégia
da co-amplificação de seqüências repetitivas localizadas na região de minisatelite do DNA
mitocondrial de ovo de Schistosoma e de uma seqüência do genoma do verme adulto.
Suzuki et al. (2006), desenvolveram um ensaio para detecção da infecção pelo
S.mansoni através do “Touchdown-PCR”. Neste ensaio, após a etapa de desnaturação, inicia-
se a etapa de hibridização em uma determinada temperatura que vai sendo reduzida
gradualmente a cada período de três ciclos. Ao atingir a temperatura mais baixa de
hibridização, dentro das condições ótimas para a reação, são realizados 20 ciclos nos quais
ocorre a amplificação. A padronização foi realizada com amostras de soro de camundongos
infectados pelo S. mansoni, sendo possível detectar DNA do parasito já na segunda semana
após a infecção desses animais. Além disso, pôde ser empregada a amostra de soro
diretamente no ensaio, descartando a etapa de extração de DNA. Os autores comentam que o
teste mostrou-se útil ao diagnóstico precoce da infecção esquistossomótica.
Nosso objetivo foi avaliar a PCR como método alternativo para complementação do
diagnóstico da esquistossomose mansônica em indivíduos provenientes de áreas de baixa
endemicidade ou naqueles expostos ocasionalmente ao helminto, adquirindo baixa carga
parasitária. A escolha por esta metodologia se deu pela demonstração da viabilidade de se
amplificar o DNA de S. mansoni em amostras de fezes com alta sensibilidade e especificidade
através de técnicas simples para extração do DNA e de um ensaio de amplificação rápido (em

torno de duas horas) (Pontes, Dias-Neto & Rabello, 2002; Pontes et al., 2003).

Objetivos
Geral
Avaliar a técnica de reação em cadeia da polimerase (PCR) como alternativa de
diagnóstico da esquistossomose mansônica em indivíduos provenientes de áreas de baixa
endemicidade da doença ou naqueles expostos ocasionalmente ao Schistosoma mansoni,
adquirindo baixa carga parasitária.
Específicos
1. Empregar a técnica de PCR para detecção de fragmentos de DNA de S. mansoni em
amostras de fezes de indivíduos com baixa carga parasitária;
2. Comparar os resultados da PCR com métodos de diagnóstico coprológico e sorológico;
3. Padronizar a técnica PCR-Multiplex para detecção de fragmentos de DNA de S. mansoni
em amostras de fezes, com o intuito de aumentar a sensibilidade do diagnóstico molecular,
a partir da amplificação simultânea de múltiplas regiões-alvo;
4. Avaliar um protocolo comercial de extração de DNA de amostra fecal.

Materiais & Métodos
I- Área de estudo
Este estudo representa como parte do projeto “Avaliação de testes rápidos para o
diagnóstico da esquistossomose”, desenvolvido pelo Departamento de Biologia do Instituto
Oswaldo Cruz, tendo sido aprovado pelo Comitê de Ética em Pesquisa da Fiocruz (CAAE –
0035.1.011.009-05) (Anexo 1).
Sumidouro localiza-se na região Serrana, há aproximadamente 180 quilômetros da
cidade do Rio de Janeiro. Segundo o IBGE (http:// www.ibge.gov.br/cidadesat), abriga uma
população de 14.927 habitantes e abrange desde áreas tipicamente urbanas até áreas rurais.
Desde o início do século XIX, sua economia baseia-se na agropecuária.
Este Município tem sérios problemas no tratamento e na distribuição de água tratada,
assim como nos sistemas de coleta de lixo e de esgoto. Esta região possui como principal
fonte de abastecimento de água o rio Paquequer que possui um percurso de 29 Km dentro do
município e é um afluente do rio Paraíba do Sul. Os escoamentos de esgoto da cidade
desembocam ao longo dos córregos, efluentes e afluentes do Rio Paquequer.
As localidades nas quais o projeto se desenvolveu foram Soledade (figura 2) e
Encanto (figura 3), comunidades rurais onde predomina o cultivo de lavouras. Existem vários
córregos nestas localidades e a única espécie de hospedeiro intermediário do S.mansoni
observada, foi o molusco Biomphalaria glabrata (Coelho da Silva, Soares & Barreto., 1997).

Figura 2: Localidade rural de Soledade, uma das áreas estudadas do Município de Sumidouro (RJ), endêmico para a esquistossomose mansônica.
Figura 3: Localidade rural de Encanto, uma das áreas estudadas do Município de Sumidouro (RJ), endêmico para a esquistossomose mansônica.

II– Amostragem
II.1- Grupo de estudo
Foram selecionadas 102 amostras de fezes e de sangue de moradores que
concordaram em participar deste estudo através da assinatura de informe concensual (Anexo
2). No caso de menores de 21 anos, o informe foi assinado pelo responsável. Cada
participante recebeu um frasco sem conservante para colheita das amostras fecais. Foram
colhidos de cada participante entre 5 a 10 mL de sangue venoso em tubos para coleta sem
anticoagulante (Vacutainer-Becton Dickinson, Ind. Cirúrgicas Ltda., Juiz de Fora, Minas
Gerais, Brasil).
As amostras de fezes foram processadas na Fiocruz, sendo submetida à técnica
coprológica Kato-Katz.
Após o processamento das amostras fecais, uma alíquota foi enviada para nosso
laboratório (UFRJ) juntamente com as amostras de soro e conservadas a –20°C até o
momento do uso. A triagem sorológica foi realizada através do ensaio ELISA para detecção
de anticorpos IgG, empregando antígeno solúvel de membrana de S. mansoni adulto.
Após a realização da coprologia e da sorologia, conforme os resultados obtidos,
foram selecionados cinco grupos nos quais as alíquotas de fezes enviadas ao nosso laboratório
e congeladas puderam ser distribuídas:
[Grupo I]
Dezoito amostras de fezes de indivíduos com presença de ovos do S. mansoni e
sorologia reativa, sendo incluídos neste grupo apenas indivíduos eliminando mais que 10
OPG (ovos por grama de fezes).

[Grupo II]
Dezesseis amostras de fezes de indivíduos com presença de ovos do parasito e
sorologia reativa, sendo incluídos neste grupo apenas indivíduos eliminando menos que 10
OPG.
[Grupo III]
Vinte e duas amostras de fezes de indivíduos com resultado positivo na sorologia
para a esquistossomose sem confirmação parasitológica da infecção.
[Grupo IV]
Vinte e cinco amostras de fezes de indivíduos com resultados negativos na
coproscopia e na sorologia para a esquistossomose, mas com coproscopia positiva para outras
parasitoses intestinais.
[Grupo V]
Vinte e uma amostras de fezes de indivíduos com resultados negativos na
coproscopia e na sorologia para a esquistossomose.
Estas amostras de fezes foram colhidas na localidade de Soledade e compuseram o
painel de teste para a análise do desempenho da PCR na detecção da infecção pelo S.
mansoni. Posteriormente, amostras dos grupos I e II foram também analisadas através da
Multiplex-PCR.
Foi selecionado um sexto grupo composto por indivíduos esquistossomóticos
provenientes da localidade de Encanto, que concordaram em fornecer mais duas ou três
amostras de fezes. Estas amostras foram analisadas apenas através do PCR-Multiplex.
[Grupo VI]

Vinte amostras de fezes referentes à colheita de dois ou três exames de dias
diferentes provenientes de oito indivíduos esquistossomóticos.
II.2- Amostras controles
Na etapa inicial de padronização da PCR foram utilizadas 14 amostras de fezes
provenientes de indivíduos de área endêmica que apresentavam ovos de S. mansoni (carga
parasitária entre 1 e 112 OPG) e outras duas amostras de fezes de indivíduos de área
endêmica com reatividade sorológica para a infecção pelo S. mansoni e exame parasitológico
negativo.
Foram testadas também amostras de fezes contaminadas artificialmente com ovos de
S.mansoni. A partir de uma amostra de fezes de indivíduo não residente em área endêmica
para esquistossomose e comprovadamente negativo pelo exame parasitológico para S.mansoni
e para outras helmintíases, foram utilizadas alíquotas de cerca de 0,2 g contaminadas com 1,
3, 5, 10 ou 25 ovos de S. mansoni.
Foi realizada ainda a extração de DNA de ovos de S. mansoni imersos em tampão
fosfato-salina PBS pH 7,2 (Na2HPO4 0,0075 M; NaH2 PO4 0,0025 M; NaCl 0,15 M), sendo
preparadas alíquotas contendo 1, 3, 5, 10 e 25 ovos do parasito por mililitro de PBS.
Foram usados 100 μL de cada material para a realização da extração de DNA.
Os ovos de S. mansoni foram cedidos pelo Laboratório de Bioquímica e Biologia
Molecular do Instituto de Bioquímica Médica da UFRJ, obtidos através do processamento de
fígados retirados de camundongos infectados após a realização da perfusão destes animais.

III- Técnica Coprológica
Toda a etapa da coprologia foi realizada por laboratoristas do Departamento de
Biologia do Instituto Oswaldo Cruz, Fiocruz.
As amostras de fezes foram analisadas pelo método parasitológico Kato-Katz (Katz,
Chaves & Pelegrino,1972). Este método permite a visualização e contagem dos ovos por
grama de fezes (OPG), fornecendo uma avaliação da carga parasitária e da intensidade da
infecção do indivíduo parasitado.
Uma porção de fezes (cerca de 43mg) foi aplicada sobre lâmina de vidro para
coproscopia através de uma tela e uma placa de plástico contendo um orifício central de
diâmetro (6mm) e espessura (3cm x 4cm x 1,37mm) padronizados. As lamínulas usadas
foram embebidas em solução de verde-malaquita com glicerina. Foram preparadas duas
lâminas por cada amostra processada.
O OPG referente a cada indivíduo foi calculado como a média aritmética do número
de ovos encontrados nas lâminas examinadas, conforme a seguinte fórmula:
no de ovos lâmina1 + no de ovos lâmina2 + ... = resultado × 24 = OPG
no total de lâminas
Exemplo: durante a realização da coproscopia de um paciente, o laboratorista preparou e
leu quatro lâminas. Ele contou, respectivamente, dois ovos, nenhum, oito ovos e dois ovos
em cada lâmina examinada. Ao aplicar a fórmula para estimativa da carga parasitária do
indivíduo, chegou-se ao resultado de 72 OPG.
2 + 0 + 8 + 2 = 3 × 24 = 72 OPG
4
IV- Técnica Sorológica

A triagem sorológica foi feita através do ensaio imunoenzimático (“Enzyme Linked
Immunosorbent Assay-ELISA”) para detecção de anticorpos IgG, de acordo com o protocolo
descrito por Gonçalves et al. (2006). O antígeno empregado no ELISA corresponde ao
antígeno solúvel de membrana de verme adulto (SMMA), que foi preparado seguindo o
método descrito por Silva et al. (1992).
IV.1- Preparação do Antígeno SMMA utilizado no teste de ELISA- IgG
A preparação do SMMA foi realizada com vermes S. mansoni adultos, obtidos
através de perfusão do sistema porta de camundongos após 50 dias de infecção. Estes animais
foram gentilmente cedidos pela pesquisadora Dra. Lygia dos Reis Correia do Departamento
de Malacologia do Instituto Oswaldo Cruz/Fiocruz.
Depois de recuperados, os vermes foram lavados em solução salina fisiológica e
homogeneizados no Potter (PYREX Laboratory Glassware, Corning, EUA), em banho de
gelo. Para facilitar a homogeneização dos vermes foram acrescentados até 10mL de tampão
Tris-HCl pH 8,0 (0,05M de Tris).
O material foi submetido a centrifugações sucessivas com velocidades diferenciadas
em temperatura de 4oC, conforme procedimento descrito por Silva (1992). O último
sobrenadante constituiu o antígeno solúvel de membrana de S. mansoni/SMMA.
Antes de usar o SMMA no ELISA, foi feita a determinação de sua concentração
protéica através do reagente comercial “BCA Protein assay reagent” (Pierce, Rockford,
Illinois, EUA), sendo seguida a orientação do fabricante. A curva-padrão foi estabelecida em
cada ensaio, utilizando como proteína padrão uma solução de albumina bovina na
concentração de 1mg/mL (BSA, Sigma Chem. Co, EUA).

IV.2- Ensaio imunoenzimático (ELISA-IgG)
O SMMA foi utilizado no ELISA na concentração de 5μg/mL. Foram utilizadas
placas de poliestireno de microtitulação (Maxisorp, Nunc-Immuno modules, Nalge Nunc
International, Denmark). Para a adsorção do antígeno na placa, houve um período de
incubação de uma hora a 37oC e mais 18 horas a 4oC. Foram realizadas três lavagens com
tampão fosfato-salina, PBS pH 7,2 contendo 0,3% de Tween 20 para retirada do excesso de
antígeno não adsorvido. Em cada cavidade da placa foram adicionados 100μL de solução
bloqueadora (PBS-Tween e 5% de leite desnatado - Molico, Nestlé, Araçatuba, SP, Brasil). A
placa foi incubada por 30 minutos a 37oC. Seguiu-se uma série de três lavagens com PBS-
Tween.
As amostras de soro foram diluídas 1:100 em solução bloqueadora e 100μL das
diluições adicionadas a cada cavidade. Após incubação de 30 minutos a 37oC, a placa foi
lavada oito vezes com PBS-Tween. Foram adicionados 100μL da diluição 1:6.000 em solução
bloqueadora do conjugado (IgG de cabra anti-IgG humana ligada à peroxidase, Biolab
Diagnóstica AS, RJ, Brasil). Seguiu-se uma nova incubação de 30 minutos a 37oC e,
posteriormente, nova série de oito lavagens.
A reação foi revelada com solução de 100μL TMB (3,3’,5,5’ tetrametilbenzidina,
Sigma) preparado na concentração de 10mg/mL de DMSO (dimetilsulfóxido, Sigma), 2μL de
peróxido de hidrogênio H2O2 a 30% adicionados a 10mL de tampão citrato-fosfato pH 5,0
(citrato de sódio 0,1M; fosfato de sódio 0,2M). Cada cavidade recebeu 100μL da solução
reveladora e a placa foi mantida no escuro por 20 minutos à temperatura ambiente. A reação
foi interrompida com a adição, em cada cavidade, de 100μL de ácido sulfúrico a 2N.
A leitura das densidades ópticas das reações foi efetuada em espectofotômetro para
placas de microtitulação (BioRad, Model 3550 UV), na faixa de 450nm.
Para determinação do limiar de reatividade (cut-off) do ELISA, foram incluídos nos

ensaios, soros controles positivos (duplicatas) e negativos (triplicatas). A média do controle
negativo foi subtraída da média do controle positivo, sendo o resultado dividido por três que
representou o fator de ajuste do teste.
Os resultados do ELISA foram expressos em unidade imunoenzimática (unidade de
ELISA), que reflete a porcentagem da reatividade da amostra de soro testada em relação ao
cut-off. As densidades ópticas obtidas para cada uma das amostras de soro testadas foram
divididas pelo valor do cut-off do referido ensaio.
Foram aceitos resultados com valores de cut-off no intervalo de 0,35-0,45.
V- Diagnóstico Molecular
V.1- Extração do DNA de Schistosoma mansoni
Nesta etapa foram utilizados três métodos diferentes de extração de DNA em
amostra fecal, descritos na literatura.
V.1.1- Protocolo de extração segundo Pitcher, Saunders & Owen (1989)
Este protocolo foi utilizado com ligeiras modificações na etapa de preparação da
amostra fecal (Figura 4).

Figura 4: Extração de DNA conforme protocolo descrito por Pitcher, Saunders & Owen (1989).
A amostra fecal foi diluída utilizando 0,2g de fezes em 1mL de água destilada.
Inicialmente, foram adicionados 500μL da solução de tiocianato de guanidina 5M a 100μL da
suspensão fecal diluída num microtubo (Microtubes MCT-060-C, Axygen Scientific Inc.,
EUA). A mistura foi agitada manualmente e incubada à temperatura ambiente por 10 minutos,
sendo o lisado resfriado no gelo. Foram acrescentados 250μL de acetato de amônio 7,5M.
O material foi, então, homogeneizado várias vezes por inversão e incubado no gelo
Clorofórmio/ Álcool isoamílico
Tris-EDTA
Isopropanol
Etanol 70 %
100µL fezes
Sol. Tiocianato de
Guanidina 5M Resfriar lisado no
gelo
Centrifugar 10 min.
Transferir fase aquosa
Centrifugar 2 min.
Lavar Sedimento 3X
Acetato de amônia
Agitar/ Resfriar no gelo
Secar o DNA 37°C/ 24h
DNA extraído

por mais 10 minutos. Foram adicionados 500μL de clorofórmio-álcool isoamílico (24:1 v/v),
seguido de nova agitação manual e centrifugação por 16.000 xg (Centrífuga 5415c,
Eppendorf, Alemanha) durante 10 minutos.
A fase aquosa resultante foi transferida para um novo microtubo, onde foram
acrescentados 340μL de isopropanol para precipitar o DNA. O microtubo foi agitado por
inversão e centrifugado a 3.448 xg por dois minutos. O sobrenadante foi descartado e o
sedimento lavado com etanol a 70% (v/v) por cinco vezes. O microtubo permaneceu em
estufa a 37oC por 24 horas. Após a secagem, o sedimento ou DNA foi dissolvido em 50μL de
tampão Tris-EDTA pH 8,0 e o microtubo incubado a 37oC por uma hora.
O DNA extraído foi armazenado a –20oC até o momento da amplificação. Para cada
reação de extração foi incluída uma amostra fecal positiva pelo método parasitológico Kato-
Katz com carga parasitária de 15 OPG.
V.1.2- Protocolo de extração de Pitcher, Saunders & Owen (1989), com modificações propostas por Silva et al. (1999)
Foi elaborado um protocolo (Figura 5), baseado em alguns dos procedimentos
descritos tanto por Pitcher, Saunders & Owen em (1989), como por Silva et al. (1999).
Resumidamente, em nosso protocolo, utilizou-se um grama de fezes não preservada e
estocada a –20ºC, dissolvida em água deionizada. Alíquotas de 300µL foram preparadas e
depois centrifugadas duas vezes a 16.000 xg durante cinco minutos. Esta suspensão fecal
(300μL) foi submetida à etapa de lise mecânica e química, adicionando 0,1 g de pérolas de
vidro (106 microns). Na etapa de precipitação de DNA foram acrescentados 340μL de
isopropanol e 1µg/µL de glicogênio. O glicogênio foi empregado na etapa de precipitação
porque tem a finalidade de melhorar a recuperação de DNA em amostras que apresentem
provavelmente pouca quantidade.

Acetato de amônia
Clorofórmio / Álcool isoamílico
Figura 5: Protocolo de extração de DNA modificado (Pitcher, Saunders & Owen, 1989; Silva et al., 1999).
Tris-EDTA
Isopropanol +
Glicogênio
Etanol 70 %
0,1 g de pérolas de vidro +
Sol. Tiocianato de Guanidina 5M
300µL fezes em PBS 13400g X 5’
Resfriar lisado no gelo
Centrifugar/10 min.
Transferir a fase aquosa
Centrifugar /2 min.
Lavar Sedimento 3X
Secar o DNA a 37°C/ 24h
DNA extraído
Agitar/ Resfriar no gelo

V.1.3- Protocolo de extração através de um método comercial
Foi utilizado um método comercial específico para extração de DNA de fezes
(QIAamp DNA stool Mini Kit, Qiagen, Alemanha) seguindo-se os procedimentos descritos
pelo fabricante.
A extração de DNA foi realizada colocando 200µL de fezes em microtubo mantido
no gelo. Adicionou-se 1,4 mL do tampão ASL e o microtubo foi agitado no agitador de tubos
(Super-Mixer, Lab-Line Instruments Inc., EUA) por um minuto até ficar completamente
homogeneizada. A suspensão foi aquecida por cinco minutos a 95ºC, novamente agitada no
agitador de tubos por 15 segundos e centrifugada a 16.000 xg durante um minuto.
Da fase aquosa resultante, foi transferido 1,2 mL para um novo microtubo e o
sedimento foi descartado. Foi acrescentado um comprimido do InhibitEX em cada amostra e
imediatamente agitada durante um minuto ou até o tablete ser completamente dissolvido. A
suspensão foi incubada por um minuto em temperatura ambiente para permitir que os
inibidores fossem absorvidos pela matriz InhibitEX.
Após a incubação, a amostra foi centrifugada a 16.000 xg durante três minutos. O
sobrenadante foi novamente transferido para outro microtubo e centrifugado a 16.000 xg por
mais três minutos. Em um novo microtubo foi, então, colocado 15µL de proteinase K, 200µL
da amostra e 200µL do tampão AL, sendo agitado durante 15 segundos. Após essa etapa, a
amostra foi incubada a 70ºC por 10 minutos.
Na etapa seguinte, adicionou-se 200µL de etanol (96%-100%) ao lisado e novamente
misturou-se no agitador de tubos (vortex) por um minuto. Cuidadosamente o sobrenadante foi
transferido para a coluna QIAamp e centrifugado a 16.000 xg durante um minuto. A coluna
foi colocada em um novo microtubo descartando-se o filtrado. Foram adicionados 500µL do
tampão AW1 e, em seguida, nova centrifugação a 16.000 xg durante um minuto. Este
procedimento foi repetido utilizando-se 500µL do tampão AW2. A amostra foi centrifugada a

16.000 xg durante mais três minutos. O tubo contendo o filtrado foi novamente descartado. A
coluna QIamp foi transferida para um outro microtubo no qual foi pipetado 200µL do tampão
AE. A coluna foi incubada por um minuto na temperatura ambiente e depois centrifugada a
16.000 xg por mais um minuto.
O eluato extraído (DNA) foi armazenado a –20oC até o momento da amplificação.
Para cada reação de extração foi incluída uma amostra fecal positiva pelo método
parasitológico Kato-Katz com carga parasitária de 15 OPG.
V.2- Reação em cadeia da polimerase
Foram desenvolvidas e otimizadas duas PCR clássicas, seguindo os protocolos
descritos por Nuñez et al. (2001) e Sandoval et al. (2006a). Posteriormente, foi padronizado o
Multiplex-PCR.
Para cada protocolo de PCR foram incluídos: controle positivo/DNA (DNA de
S.mansoni, cedido pelo Laboratório de Bioquímica e Biologia Molecular do Instituto de
Bioquímica Médica da UFRJ, que foi extraído de ovos do parasito obtidos de fígados de
camundongos infectados); controle positivo/fezes (amostra fecal positiva pelo método
parasitológico Kato-Katz com carga parasitária de 15 OPG); controle negativo (extração de
amostra fecal de um indivíduo comprovadamente negativo para S. mansoni pelo exame
parasitológico de fezes); Branco da reação (todos os reagentes usados na mistura, exceto o
DNA que foi substituído por água ultra-pura).

V.2.1- Protocolo de amplificação de uma região repetitiva ao longo do genoma do Schistosoma mansoni
Foram utilizados pares de oligonucleotídeos iniciadores (“primers”) descritos por
Hambuger et al. (1991), SMEGG1- 5’GATCTAACCGACCAACC3’ e SMEEG2-
5’ATATTAACGCCCACGCTCTC3’, a partir de seqüências de uma região altamente
repetitiva ao longo do genoma do S. mansoni. O fragmento amplificado é composto de 110
pares de bases (pb).
A reação foi realizada em 30 ciclos, onde cada ciclo apresentou uma fase de
desnaturação a 94°C por 30 segundos, uma fase de hibridização a 55oC por 30 segundos e
uma fase de extensão a 72oC por 30 segundos. Antes do primeiro ciclo ocorreu uma fase de
pré-desnaturação a 94°C por três minutos e após o último ciclo, uma fase de extensão final a
72°C por sete minutos. Na etapa de amplificação do DNA foi utilizado o termociclador
geneAmp PCR System 2400 (Applied - Biosistems, CA, EUA).
O volume final da mistura de reação foi de 50 μl, contendo: tampão 20mM Tris-HCl
pH 8,4; 50 mM KCl; 1,5 mM MgCl2; 250 μM de cada nucleotídeo (dNTPs); 1,25 unidades
(U) de Taq DNA-polimerase (Invitrogen Life technologies, USA); 0,1% albumina de soro
bovino (BSA– Sigma Chem. Co., EUA); 25 pmoles de cada “iniciador”e 2μL do DNA
extraído.
V.2.2- Protocolo de amplificação do gene que codifica o DNA ribossomal do Schistosoma mansoni
Um fragmento de 350pb oriundo da subunidade menor do DNA ribossomal do
S.mansoni foi amplificado através da utilização dos iniciadores SmF-
5’GAGATCAAGTGTGACAGTTTTGC3’ e SmR- 5’ACAGTGCGCGCGTCGTAA GC3’,
descritos por Sandoval et al. (2006a). A mistura de reação teve um volume final de 50 μL,
contendo: tampão 20mM Tris-HCl pH 8,4; 50 mM KCl; 3mM MgCl2; 2,5 U Taq polimerase;

20μM de cada “iniciador”; 0,5mM de cada nucleotídeo (dNTPs) (Invitrogen Life
technologies, USA) e 2μL de DNA extraído.
A reação foi realizada em 35 ciclos, onde cada ciclo apresentou uma fase de
desnaturação a 94°C por 20 segundos, uma fase de hibridização a 65oC por 20 segundos e
uma fase de extensão a 72oC por 30 segundos. Antes do primeiro ciclo ocorreu uma fase de
pré-desnaturação a 94°C por três minutos e após o último ciclo, uma fase de extensão final a
72°C por sete minutos. Para a amplificação do DNA foi usado o mesmo termociclador.
V.2.3- Protocolo de amplificação do PCR-Multiplex
A padronização da reação de amplificação foi realizada seguindo os procedimentos
descritos por Henegariu et al. (1997), com algumas modificações. As seqüências alvo foram
amplificadas utilizando os iniciadores (SMEGG1 5’GATCTAACCGACCAACC3’, SMEEG2
5’ATATTAACGCCCACGCTCTC3’; SmF 5’GAGATCAAGTGTGACAGTTTTGC3’ e
SmR 5’ACAGTGCGCGCGTCGTAA GC-3’), específicos para S. mansoni, descritos acima
nos itens V.2.1 e V.2.2.
O volume final da mistura de reação foi de 50 μl, contendo: tampão 20mM Tris-HCl
pH 8,4; 50mM KCl; 3mM MgCl2; 10 pmoles de cada “iniciador”; 250µM de cada nucleotídeo
(dNTPs) e 1,25 U de Taq platinum DNA-polimerase(Invitrogen); 0,1% de albumina de soro
bovino (BSA – Sigma) e 2μl de DNA extraído.
A etapa de amplificação foi realizada em 30 ciclos, onde cada ciclo apresentou uma
fase de desnaturação a 94°C por 30 segundos, uma fase de hibridização a 60oC por 30
segundos e uma fase de extensão a 72oC por 30 segundos. Antes do primeiro ciclo ocorreu
uma fase de pré-desnaturação a 94°C por quatro minutos e após o último ciclo, ocorreu a fase
de extensão final a 72°C por sete minutos. Foi utilizado o mesmo modelo de termociclador.

V.3- Teste para a presença de inibidores da PCR
A presença de inibidores da PCR foi avaliada através da contaminação de amostras
de fezes com DNA de S. mansoni (“spiking”) (Silva et al., 1997). Esse método foi utilizado
com as amostras que persistiram negativas depois de terem sido analisadas com os diferentes
protocolos de extração e pares de iniciadores. Como resultado, é esperado que o DNA de S.
mansoni empregado como contaminante das amostras, seja amplificado mostrando a ausência
de substâncias inibidoras ou que não ocorra amplificação de nenhum produto, sinalizando a
interferência dos inibidores. Foi utilizado nessa reação de amplificação, 1µL (5fg) de DNA de
S. mansoni juntamente com 1µL do DNA extraído das amostras negativas.
V.4- Detecção do produto amplificado
Inicialmente foi empregada a eletroforese em gel de agarose 2% (Invitrogen) para
visualização do produto amplificado. Devido a maior sensibilidade de detecção do produto
amplificado em gel de poliacrilamida (PAGE), foi decidido posteriormente, realizar a
eletroforese utilizando um sistema contínuo contendo 12% de acrilamida e dodecil sulfato de
sódio no gel de separação e a corrida foi feita em um sistema vertical (Mini Protean II Dual
Slab cell. Bio-Rad), como descrito por Laemmli (1970) para proteínas.
O produto amplificado foi aplicado juntamente com o corante azul de bromofenol, na
proporção de 9μL do produto para 1μL do corante. Foram aplicados cerca de 5μL do
marcador de tamanho molecular (100 pb DNA ladder, Invitrogen). O gel foi, então, submetido
à tensão de 120 volts por aproximadamente duas horas. Após a corrida eletroforética, o gel foi
corado pela prata.
A coloração do gel pelo nitrato de prata seguiu o protocolo descrito por Herring et al.

(1982). O gel foi colocado em 100 mL da solução fixadora (etanol absoluto 10%, ácido
acético glacial 0,5%) e mantido na geladeira por 12 horas. Após esse período, a solução
fixadora foi desprezada e o gel lavado rapidamente em água destilada. Foi acrescentada a
solução de nitrato de prata 0,16% (peso/volume) e o gel foi mantido no escuro sob agitação
por 30 minutos. Em seguida, o gel foi lavado em água destilada. Foi adicionada a solução
reveladora (hidróxido de sódio 0,75M, formaldeído 0,75%) e mantido sob agitação até o
aparecimento das bandas, quando a solução reveladora pôde ser desprezada. O gel foi
novamente lavado em água destilada e acrescentou-se a solução neutralizante (ácido acético
glacial 5%). Após 10 minutos sob agitação na solução neutralizante, o gel foi lavado em água
destilada.
Terminada a coloração do gel, iniciou-se sua secagem. Folhas de papel celofane e o
gel foram mergulhados numa solução de metanol a 50%, acrescida de glicerol a 0,5%. Após
20 minutos, o gel foi embrulhado no papel celofane, tomando-se o cuidado de evitar a
formação de bolhas de ar e permaneceu pelo menos 24 horas em temperatura ambiente para a
sua secagem.

RESULTADOS
Na fase inicial da padronização da PCR foi utilizado o protocolo de extração de
DNA proposto por Pitcher, Saunders & Owen (1989) com algumas modificações, e o par de
iniciadores originalmente descrito por Hamburger et al., (1991). Estes iniciadores hibridizam
com uma região altamente repetitiva ao longo do genoma do parasito (SMEGG1/SMEEG2).
O produto amplificado (“amplicon”) foi de 110 pb. Observa-se também a presença de outros
fragmentos de DNA com tamanhos superiores a 110 pb, que são, na verdade, bandas
amplificadas em decorrência da grande quantidade de oligonucleotídeos livres,
complementares de uma seqüência repetitiva de DNA. Das 14 amostras de fezes positivas
para ovos de S.mansoni, a PCR detectou a presença de DNA do parasito em todas elas. Foi
detectada também a presença de DNA do S.mansoni nas duas amostras de fezes negativas
pelo método de Kato-Katz. Estes resultados estão sumarizados na tabela 1. O produto gerado
de 110 pb pela amplificação do par de iniciadores SMEGG1/SMEEG2 pode ser observado em
gel de agarose 2% (Figura 6).
A seguir, a sensibilidade da PCR foi determinada empregando amostras fecais
negativas que foram contaminadas com diferentes quantidades de ovos de S. mansoni,
variando de um a 25 ovos em 100 µL de fezes. Foi possível detectar o produto amplificado de
110 pb em amostras fecais contaminadas com até um ovo (Figura 7). A partir dessa etapa, os
produtos amplificados foram detectados através do gel de poliacrilamida a 12% corados pela
prata, por apresentar uma maior sensibilidade quando comparado ao gel de agarose corado
pelo brometo de etídio.

Tabela 1: Resultados dos métodos Kato-Katz e PCR das 16 amostras de fezes controles provenientes de indivíduos residentes no Município de Sumidouro (Rio de Janeiro).
Indivíduos Kato-Katz
OPG a
PCR
A1 Positivo 112 Positivo
A2 Positivo 15 Positivo
A3 Positivo 13 Positivo
A4 Positivo 5 Positivo
A5 Positivo 5 Positivo
A6 Positivo 5 Positivo
A7 Positivo 4 Positivo
A8 Positivo 4 Positivo
A9 Positivo 4 Positivo
A10 Positivo 4 Positivo
A11 Positivo 4 Positivo
A12 Positivo 3 Positivo
A13 Positivo 1 Positivo
A14 Positivo 1 Positivo
A15 Negativo - Positivo
A16 Negativo - Positivo
a OPG: ovos por grama de fezes. 1 2 3 4 5 6 7 8 9 10 11
110 bp

Figura 6: Produto amplificado da PCR utilizando o par de iniciadores SMEGG1/ SMEEG2. 1- padrão de tamanho molecular ( 2- controle negativo 3- controle positivo, 4- branco da reação (sem DNA), 5-11 amostras clínicas.Gel corado pelo corante brometo de etidium
1 2 3 4 5 6 7 8 9
110 pb
Figura 7: Produtos amplificados utilizando o par de iniciadores SMEGG-1/SMEEG-2. Avaliação da sensibilidade da PCR utilizando amostras de fezes contaminadas com diferentes números de ovos de S. mansoni. Eletroforese em gel de acrilamida a 12% corado pela prata. 1- amostra de fezes negativa contaminada com 25 ovos de S. mansoni, 2- amostra de fezes negativa contaminada com 10 ovos de S. mansoni, 3- amostra de fezes negativa contaminada com cinco ovos de S.mansoni, 4- amostra de fezes negativa contaminada com três ovos de S. mansoni, 5- amostra de fezes negativa contaminada com um ovo de S. mansoni, 6- controle negativo, 7- branco da reação (sem DNA), 8- controle positivo, 9- Padrão de tamanho molecular.
Outra estratégia utilizada para avaliar a sensibilidade da PCR foi o teste com
amostras experimentais em que os ovos de S. mansoni foram acrescentados em PBS nas
concentrações de um, três, cinco, dez e 25 ovos para cada 100 µL de PBS. Nestas amostras foi

possível detectar a fração amplificada de 110 pb até a concentração de um ovo/100µL (Figura
8). Não foi observada diferença em relação à intensidade das bandas destes fragmentos
amplificados em comparação com aqueles obtidos nas amostras fecais contaminadas com
ovos de S.mansoni.
1 2 3 4 5 6 7 8 9
110 pb
Figura 8: Produtos amplificados através da PCR utilizando o par de iniciadores SMEGG-1/SMEEG-2. Avaliação da sensibilidade da PCR utilizando amostras de PBS contaminadas com diferentes números de ovos de S.mansoni. Eletroforese em gel de acrilamida a 12% corado pela prata. 1– padrão de tamanho molecular, 2- controle positivo, 3- controle negativo, 4- Branco da reação (sem DNA), 5- um ovo de S. mansoni/100μL PBS 6- três ovos de S.mansoni/100μL PBS, 7- cinco ovos de S. mansoni/100μL PBS, 8- 10 ovos de S. mansoni/100μL PBS, 9- 25 ovos de S.mansoni/100μL PBS.

A validação dessa técnica foi realizada empregando 102 amostras de fezes obtidas de
indivíduos residentes em Sumidouro e utilizando o protocolo de extração de DNA proposto
por Pitcher, Saunders & Owen (1989) com algumas modificações, e o par de iniciadores
originalmente descrito por Hamburger et al., (1991). Para o estudo, as amostras analisadas
foram divididas em cinco grupos, como descrito no item II.1 do material e métodos.
O grupo I foi composto de 18 amostras de fezes de indivíduos com carga parasitária
acima de 10 OPG e reatividade no ELISA-IgG. A PCR detectou a presença do fragmento de
DNA de S. mansoni em 44% (8) das amostras. Os resultados estão sumarizados na tabela 2.
No grupo II, constituído de 16 amostras de fezes de indivíduos com carga parasitária
menor que 10 OPG e reatividade ELISA-IgG, a PCR detectou a presença de DNA de
S.mansoni em 25% (4) das amostras. Os resultados encontram-se na tabela 3.
No grupo III, composto de 22 amostras de fezes de indivíduos com exame
parasitológico negativo e sorologia positiva para a infecção pelo S. mansoni, a PCR detectou a
presença de DNA de S. mansoni em apenas uma das amostras.
No grupo IV, constituído de 21 amostras de fezes de indivíduos com exames
parasitológico e sorológico negativos para a infecção pelo S. mansoni, e no grupo V,
constituído de 25 amostras de fezes de indivíduos portadores de outras parasitoses e com
sorologia negativa para S. mansoni, a PCR foi negativa em todas as amostras. Esses
resultados indicam uma especificidade de 100% para a PCR.

Tabela 2: Resultados da PCR nas amostras de fezes de indivíduos com carga parasitária acima de 10 OPG e sorologia positiva.
Indivíduos OPG a ELISA -IgG PCR
SMEGG1/SMEEG2
B1 2451 Positivo Positivo
B2 270 Positivo Positivo
B3 174 Positivo Positivo
B4 141 Positivo Positivo
B5 132 Positivo Negativo
B6 90 Positivo Positivo
B7 81 Positivo Negativo
B8 66 Positivo Positivo
B9 52 Positivo Negativo
B10 30 Positivo Positivo
B11 30 Positivo Positivo
B12 18 Positivo Negativo
B13 18 Positivo Negativo
B14 18 Positivo Negativo
B15 16 Positivo Negativo
B16 15 Positivo Negativo
B17 12 Positivo Negativo
B18 12 Positivo Negativo
a OPG: ovos por grama de fezes.
Tabela 3: Resultados da PCR nas amostras de fezes de indivíduos com carga parasitária abaixo de 10 OPG e sorologia positiva.

Indivíduos OPG a ELISA-IgG PCR SMEGG1/SMEEG2
C1 3 Positivo Positivo
C2 3 Positivo Negativo
C3 9 Positivo Negativo
C4 3 Positivo Negativo
C5 6 Positivo Positivo
C6 3 Positivo Positivo
C7 3 Positivo Negativo
C8 3 Positivo Negativo
C9 9 Positivo Positivo
C10 3 Positivo Negativo
C11 6 Positivo Negativo
C12 3 Positivo Negativo
C13 6 Positivo Negativo
C14 6 Positivo Negativo
C15 6 Positivo Negativo
C16 9 Positivo Negativo
a OPG: ovos por grama de fezes.
Baseado nos resultados obtidos para os dois grupos de indivíduos esquistossomóticos
(grupos I e II) e para o grupo com somente sorologia reativa para a infecção pelo S. mansoni
(grupo III), apenas 12 amostras e uma amostra, respectivamente, apresentaram a PCR positiva.
Portanto, a metodologia empregada não apresentou uma boa sensibilidade para o diagnóstico
da esquistossomose. Assim, decidimos fazer algumas modificações nas etapas de extração e
usar um novo par de iniciadores, recentemente descrito (Sandoval et al., 2006a). Na etapa de
extração, aumentou-se o volume de fezes de 100µL para 300µL, adicionou-se uma etapa de
homogeneização das fezes empregando pérolas de vidro e na etapa de precipitação do DNA
acrescentou-se glicogênio (Figura 9).

Figura 9: Extração de DNA com as modificações realizadas seguindo os protocolos de Pitcher, Saunders & Owen (1989) e Silva et al. (1999).
A determinação da sensibilidade desse protocolo, empregando os dois pares de
iniciadores, foi realizada utilizando as amostras de fezes humanas contaminadas artificialmente
com um a 25 ovos do parasito. A sensibilidade obtida foi a detecção de apenas um ovo, em
ambas as amplificações (Figuras 10a-b). Posteriormente, este protocolo foi avaliado
Tris-EDTA
Acetato de amônia
Solução de Tiocianato de Guanidina M5
Clorofórmio/ Álcool isoamílico
Isopropanol
Etanol 70 %
300 µl de fezes/ PBS 13400g X 5’
O,1g pérolas de vidroLise mecânica Guanidina 5M
1μL de glicog
Modificações no protocolo de extração:
100 µL de fezes
Resfria o lisado no gelo
Agitar e Resfriar no gelo
Centrifugar por 10 min.
Transferir a fase aquosa
ênio
Centrifugar por 2 min
Lavar o Sedimento 3X
Secar DNA a 37°C/ 24h
DNA extraído

empregando o mesmo painel de 102 amostras de fezes, divididas em cinco grupos, conforme
descrito em material e métodos.
A
1 2 3 4 5 6 7 8 9 10
110 pb
B
1 2 3 4 5 6 7 8 9
350 pb
Figura 10: Eletroforese em gel de acrilamida a 12% corado pela prata. Avaliação da sensibilidade da PCR com o método de extração modificado, utilizando amostras de fezes contaminadas com diferentes números de ovos de S. mansoni. A- Produto amplificado utilizando o par de iniciadores SMEEG-1/SMEGG-2. 1- padrão de tamanho molecular, 2- controle positivo 3-controle negativo, 4- Branco da reação (sem DNA), 5- amostra de fezes negativa contaminada com um ovo de S. mansoni, 6- amostra de fezes negativa contaminada com três ovos de S. mansoni, 7- amostra de fezes negativa contaminada com cinco ovos de S.mansoni, 8- controle negativo, 9- amostra de fezes negativa contaminada com 25 ovos de S. mansoni 10- Branco da reação (sem DNA). B- Produto amplificado utilizando o par de iniciadores SmF/SmR. 1- padrão de tamanho molecular, 2- controle negativo, 3- controle positivo, 4- Branco da reação (sem DNA), 5- amostra de fezes negativa contaminada com um ovo de S. mansoni, 6- amostra de fezes negativa contaminada com três ovos de S.mansoni, 7- amostra de fezes negativa contaminada com cinco ovos de S.mansoni, 8- Branco da reação (sem DNA), 9- amostra de fezes negativa contaminada com 25 ovos de S. mansoni.
No grupo I, todas as 18 amostras de fezes de indivíduos que apresentavam carga
parasitária acima de 10 OPG e sorologia positiva para S. mansoni, testadas pela PCR
apresentaram as bandas de 110 pb e/ou 350 pb. Dessas, 14 amostras apresentaram
concordância nos resultados da PCR, sendo positivas quando usado tanto o par de iniciadores
SMEGG1/SMEEG2 como SmF/SmR. Duas amostras foram positivas para

SMEGG1/SMEEG2 e negativas quando usado SmF/SmR. Outras duas amostras foram
positivas apenas na PCR empregando SmF/SmR (Tabelas 4 e 5).
O grupo II, constituído de 16 indivíduos com carga parasitária abaixo de 10 OPG e
sorologia positiva para S. mansoni, apresentou 81% (13) das amostras de fezes positivas na
PCR com dois ou pelo menos um dos pares de iniciadores. Em oito amostras de fezes,
utilizando tanto SMEGG1/SMEEG2 como SmF/SmR, a PCR foi positiva. Uma amostra desse
grupo foi positiva apenas na PCR com o par de iniciadores SMEGG1/SMEEG2. Quatro
amostras foram positivas somente quando empregado o par de iniciadores SmF/SmR. Três
amostras foram negativas em ambas as PCRs convencionais (Tabelas 6 e 7).

Tabela 4: Resultados das duas PCR convencionais em amostras de fezes de indivíduos com carga parasitária acima de 10 OPG e sorologia positiva.
Indivíduos OPG a ELISA -IgG PCR 1 SMEGG1/SMEEG2
PCR 2 SmF/SmR
B1 2451 Positivo Positivo Positivo
B2 270 Positivo Positivo Positivo
B3 174 Positivo Positivo Positivo
B4 141 Positivo Positivo Positivo
B5 132 Positivo Negativo Positivo
B6 90 Positivo Positivo Positivo
B7 81 Positivo Positivo Positivo
B8 66 Positivo Positivo Positivo
B9 52 Positivo Positivo Positivo
B10 30 Positivo Positivo Positivo
B11 30 Positivo Positivo Negativo
B12 18 Positivo Positivo Positivo
B13 18 Positivo Positivo Positivo
B14 18 Positivo Negativo Positivo
B15 16 Positivo Positivo Positivo
B16 15 Positivo Positivo Positivo
B17 12 Positivo Positivo Negativo
B18 12 Positivo Positivo Positivo a OPG: ovos por grama de fezes.
Tabela 5: Comparação dos resultados obtidos nas duas PCR convencionais com os diferentes pares de iniciadores em amostras de fezes de indivíduos

com carga parasitária acima de 10 OPG e sorologia positiva.
SMEGG1/SMEEG2 PCR
SmF/SmR Positivo Negativo Total
Positivo 14 2 16
Negativo 2 0 2
Total 16 2 18
Tabela 6: Resultados das duas PCR convencionais em amostras de fezes de indivíduos com carga parasitária abaixo de 10 OPG e sorologia positiva.
Indivíduos
OPG a
ELISA-IgG PCR
SMEGG1/SMEEG2PCR
SmF/SmR C1 3 Positivo Positivo Positivo
C2 3 Positivo Negativo Positivo
C3 9 Positivo Positivo Positivo
C4 3 Positivo Positivo Positivo
C5 6 Positivo Negativo Positivo
C6 3 Positivo Positivo Negativo
C7 3 Positivo Negativo Negativo
C8 3 Positivo Positivo Positivo
C9 9 Positivo Positivo Positivo
C10 3 Positivo Negativo Positivo
C11 6 Positivo Positivo Positivo
C12 3 Positivo Negativo Negativo
C13 6 Positivo Positivo Positivo
C14 6 Positivo Positivo Positivo
C15 6 Positivo Negativo Negativo
C16 9 Positivo Negativo Positivo
a OPG: ovos por grama de fezes.
Tabela 7: Comparação dos resultados obtidos nas duas PCR convencionais com os diferentes pares de iniciadores em amostras de fezes de indivíduos com carga parasitária abaixo de 10 OPG e sorologia positiva.

SMEGG1/SMEEG2 PCR
SmF/SmR Positivo Negativo Total
Positivo 8 4 12
Negativo 1 3 4
Total 9 7 16
No intuito de verificar a presença de substâncias inibidoras da Taq polimerase nas
reações de amplificação, três amostras de fezes de indivíduos que eliminavam ovos do
S.mansoni e negativas na PCR, foram contaminadas com 5fg de DNA de S. mansoni
(“spiking”). Todas as amostras testadas tiveram o DNA de S. mansoni amplificado pela PCR,
servindo, portanto, como um indicador da ausência de interferência de inibidores.
No grupo III, constituído de 22 amostras de fezes de indivíduos com sorologia
positiva sem a confirmação parasitológica da infecção pelo S. mansoni, ambas as reações de
PCR detectaram a presença de DNA do parasito em 54,5% (12) das amostras. Seis amostras
de fezes foram positivas em ambos os testes: PCR com SMEGG1/SMEEG2 e PCR com
SmF/SmR. Uma amostra de fezes foi positiva apenas na PCR com o par de iniciadores
SMEGG1/SMEEG2. Seis amostras de fezes foram positivas somente quando o par de
iniciadores SmF/SmR foi usado. Nove amostras de fezes mantiveram-se negativas nos dois
testes de PCR (Tabelas 8 e 9).
Tabela 8: Resultados nas duas PCR convencionais em amostras de fezes de indivíduos com exame parasitológico negativo e sorologia positiva.

Indivíduos
ELISA–IgG
PCR 1
SMEGG1/SMEEG2
PCR 2
SmF/SmR
D1 Positivo Positivo Positivo
D2 Positivo Positivo Positivo
D3 Positivo Positivo Positivo
D4 Positivo Positivo Positivo
D5 Positivo Positivo Positivo
D6 Positivo Positivo Positivo
D7 Positivo Positivo Negativo
D8 Positivo Negativo Positivo
D9 Positivo Negativo Positivo
D10 Positivo Negativo Positivo
D11 Positivo Negativo Positivo
D12 Positivo Negativo Positivo
D13 Positivo Negativo Positivo
D14 Positivo Negativo Negativo
D15 Positivo Negativo Negativo
D16 Positivo Negativo Negativo
D17 Positivo Negativo Negativo
D18 Positivo Negativo Negativo
D19 Positivo Negativo Negativo
D20 Positivo Negativo Negativo
D21 Positivo Negativo Negativo
D22 Positivo Negativo Negativo
Tabela 9: Comparação dos resultados obtidos nas duas PCR convencionais com os diferentes pares de iniciadores em amostras de fezes de indivíduos com exame parasitológico negativo e sorologia positiva.
PCR SMEGG1/SMEEG2

SmF/SmR Positivo Negativo Total
Positivo 6 6 12
Negativo 1 9 10
Total 7 15 22
Em vista dos resultados obtidos, decidimos padronizar uma PCR-Multiplex
utilizando os dois pares de iniciadores em um único tubo de reação. Inicialmente, para a
padronização da PCR-Multiplex foram utilizadas as amostras fecais negativas contaminadas
com um a 25 ovos de S.mansoni. Conforme demonstrado na figura 11, ambos os produtos de
110 pb e de 350 pb correspondentes aos pares de iniciadores SMEGG1/SMEEG2 e
SmF/SmR respectivamente, foram detectados.
1 2 3 4 5 6 7 8 9 10
350 pb 110 pb
Figura 11: Avaliação da sensibilidade da PCR-Multiplex com amostras de fezes contaminadas com diferentes números de ovos de S. mansoni. Eletroforese em gel de acrilamida a 12% corado pela prata. 1– padrão de peso molecular, 2- controle positivo, 3- controle negativo, 4- Branco da reação (sem DNA), 5– amostra de fezes negativa contaminada com um ovo de S.mansoni, 6- amostra de fezes negativa contaminada com três ovos de S.mansoni, 7- amostra de fezes negativa contaminada com cinco ovos de S.mansoni, 8- controle negativo, 9- amostra de fezes negativa contaminada com 25 ovos de S. mansoni, 10- Branco da reação (sem DNA).
Para avaliar a PCR-Multiplex, foram selecionadas oito amostras de fezes de
indivíduos esquistossomóticos pertencentes aos grupos I e II, que apresentavam diferentes
combinações de resultados nas PCR convencionais e na sorologia. Na tabela 10, pode-se

observar que houve uma concordância de 100% entre os resultados obtidos pela PCR-
Multiplex e PCR convencionais.
Tabela 10: Comparação dos resultados das PCR convencionais com os da PCR-Multiplex em amostras de fezes de oito indivíduos esquistossomóticos com diferentes cargas parasitárias.
PCR-Multiplex
Indivíduos
OPG a
PCR
SMEGG1/ SMEEG2
PCR
SmF/SmR SMEGG1/
SMEEG2 SmF/SmR
B5 132 Negativo Positivo Negativo Positivo
B7 81 Positivo Positivo Positivo Positivo
B10 30 Positivo Positivo Positivo Positivo
B17 12 Positivo Negativo Positivo Negativo
C16 9 Negativo Positivo Negativo Positivo
C7 3 Negativo Negativo Negativo Negativo
C12 3 Negativo Negativo Negativo Negativo
C15 6 Negativo Negativo Negativo Negativo
a OPG: ovos por grama de fezes
Em nosso estudo nos deparamos com a ausência das bandas diagnósticas na PCR de
três amostras de fezes que haviam sido positivas para a presença de ovos do S. mansoni.
Tratava-se de amostras de fezes do grupo de indivíduos com carga parasitária abaixo de 10
OPG. Resolvemos avaliar, não só a influência da baixa carga parasitária no resultado do

diagnóstico, mas também a questão da eliminação intermitente de ovos do parasito pelos
hospedeiros. Para isto, foram analisadas as amostras de fezes selecionadas no grupo VI
através da PCR-Multiplex.
Analisando os dados apresentados na tabela 11, referentes ao grupo VI, observamos
como realmente os indivíduos esquistossomóticos apresentaram uma variabilidade de carga
parasitária e, conseqüentemente, de resultados nos testes de diagnóstico. Considerando-se os
pacientes 6.1 e 6.5, apenas uma das amostras de fezes analisadas pelo método Kato-Katz
apresentou ovos de S. mansoni, com cargas de 24 OPG e 12 OPG respectivamente. Todas as
amostras de fezes de cada um destes pacientes, analisadas pela PCR-Multiplex foram
positivas. Isso demonstrou que provavelmente havia uma quantidade muito baixa de material
do parasito nestas amostras fecais para detecção pela coproscopia, mas suficiente para o
diagnóstico pela PCR. Já em relação aos pacientes 6.7 e 6.8, pudemos observar que
novamente houve concordância entre a coproscopia e a PCR em determinadas amostras de
fezes e que também a PCR continuou revelando amostras falso-negativas do exame de fezes
pela técnica de Kato-Katz. Todavia, ocorreu que em uma das amostras de fezes de cada um
desses pacientes, tanto a coproscopia como a PCR foram negativas. Nos pacientes 6.3, 6.4 e
6.6 sempre houve concordância entre os resultados da coproscopia com a PCR-Multiplex.
Foram observadas nestes indivíduos cargas parasitárias mais altas. Todavia, uma amostra foi
positiva no exame pelo método Kato-Katz e negativa pela PCR-Multiplex (paciente 6.2).
Tabela 11: Resultados do exame coprológico e da PCR-Multiplex em amostras de fezes obtidas de diferentes dias de oito indivíduos esquistossomóticos residentes na localidade de Encanto (Sumidouro, RJ).
PCR-Multiplex
Indivíduos
Amostras em
diferentes dias
OPG aSmF/SmR SMEGG1/
SMEEG2
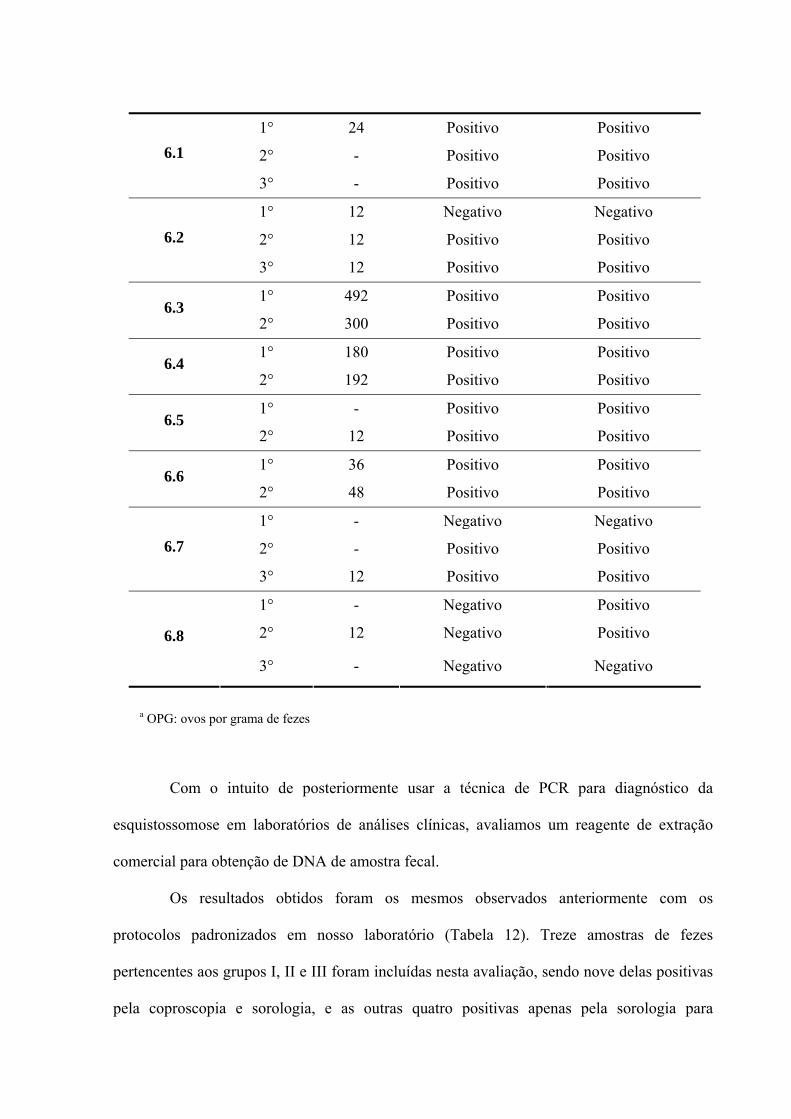
1° 24 Positivo Positivo
2° - Positivo Positivo 6.1
3° - Positivo Positivo
1° 12 Negativo Negativo
2° 12 Positivo Positivo 6.2
3° 12 Positivo Positivo
1° 492 Positivo Positivo 6.3
2° 300 Positivo Positivo
1° 180 Positivo Positivo 6.4
2° 192 Positivo Positivo
1° - Positivo Positivo 6.5
2° 12 Positivo Positivo
1° 36 Positivo Positivo 6.6
2° 48 Positivo Positivo
1° - Negativo Negativo
2° - Positivo Positivo 6.7
3° 12 Positivo Positivo
1° - Negativo Positivo
2° 12 Negativo Positivo 6.8
3° - Negativo Negativo
a OPG: ovos por grama de fezes
Com o intuito de posteriormente usar a técnica de PCR para diagnóstico da
esquistossomose em laboratórios de análises clínicas, avaliamos um reagente de extração
comercial para obtenção de DNA de amostra fecal.
Os resultados obtidos foram os mesmos observados anteriormente com os
protocolos padronizados em nosso laboratório (Tabela 12). Treze amostras de fezes
pertencentes aos grupos I, II e III foram incluídas nesta avaliação, sendo nove delas positivas
pela coproscopia e sorologia, e as outras quatro positivas apenas pela sorologia para

S.mansoni. As onze amostras de fezes, que haviam sido positivas com os pares de iniciadores
SMEGG1/SMEEG2 e/ou SmF/SmR, mostraram positividade na PCR-Multiplex com o DNA
extraído conforme o protocolo comercial. Duas amostras de fezes de indivíduos apenas
soropositivos foram positivas tanto com o nosso protocolo de extração de DNA
desenvolvido em laboratório como com o protocolo comercial. As outras duas amostras de
fezes de indivíduos soropositivos permaneceram sempre negativas.

Tabela 12: Comparação dos resultados da PCR-Multiplex após a extração de 13 amostras de fezes através de reagente comercial com os da PCR obtidos nos diferentes protocolos para detecção da infecção pelo Schistosoma mansoni.
Indivíduos OPG a
PCR SMEGG1/ SMEEG2
PCR SmF/SmR
PCR-Multiplex
PCR-Multiplex extração comercial
B1 2451 Positivo Positivo NR b Positivo
B17 12 Positivo Negativo Positivo Positivo
C11 06 Positivo Positivo NR Positivo
B10 30 Positivo Positivo Positivo Positivo
C16 09 Negativo Positivo Positivo Positivo
C15 06 Negativo Negativo NR Negativo
B3 174 Positivo Positivo NR Positivo
C2 09 Negativo Positivo NR Positivo
B5 132 Negativo Positivo Positivo Positivo
D3 0 Positivo Positivo NR Positivo
D7 0 Positivo Negativo NR Positivo
D8 0 Negativo Positivo NR Positivo
D14 0 Negativo Negativo NR Negativo
a OPG: ovos por grama de fezes. b NR – não realizado

Discussão
Nos últimos anos, houve uma mudança no perfil da esquistossomose em algumas
regiões do Brasil devido à implementação de intervenções de controle desde 1976. Estas
medidas de controle tiveram um impacto incontestável sobre a prevalência, a intensidade de
infecção e morbidade. Todavia, no Estado do Rio de Janeiro ainda ocorre a persistência da
transmissão e quadros de infecções leves. A infecção por S. mansoni é considerada leve
quando o hospedeiro elimina menos que 100 ovos do parasito por grama de fezes. Em virtude
desses baixos índices de intensidade de infecção tem sido necessário empregar-se métodos de
diagnóstico de alta sensibilidade.
A reação de PCR vem demonstrando resultados satisfatórios no diagnóstico de
muitas parasitoses intestinais (Gibbons-Matthews & Prescott, 2003; Gonin & Trudel, 2003;
Verweij et al., 2003; Subrungruang et al., 2004), inclusive, mais recentemente, para a
esquistossomose (Pontes et al., 2003; Sandoval et al., 2006b). Todavia, a PCR foi empregada
para detecção de DNA de S. mansoni em áreas de média e de alta endemicidades (Pontes et
al., 2003).
Nenhum estudo com amostra de fezes obtidas de pacientes de áreas de baixa
endemicidade foi realizado. Nosso objetivo foi justamente avaliar a PCR para a detecção de
DNA de S. mansoni em amostras fecais de indivíduos que apresentem carga parasitária
reduzida residentes em áreas de baixa endemicidade.
Na padronização da PCR, nossos resultados alcançaram 100% de sensibilidade. De
fato, pôde ser detectada a banda diagnóstica de 110 pb em todas as amostras de fezes que
continham ovos de S. mansoni. Além disso, a PCR detectou a presença do DNA do parasito
em amostras de fezes negativas pelo método Kato-Katz. Foram observados, ainda, alguns

fragmentos de DNA com tamanhos moleculares acima de 110 pb. Sabemos que na reação de
PCR existe uma grande quantidade de oligonucleotídeos livres, que são complementares a
uma seqüência de DNA repetitiva agrupadas em arranjo consecutivo no genoma de S.
mansoni. Assim, a PCR amplifica fragmentos de DNA com peso molecular variado.
Esses resultados preliminares nos estimularam a passar para a etapa de aplicação da
técnica em amostras de fezes de indivíduos moradores de uma área de baixa endemicidade.
Em nosso estudo, inicialmente usamos um método de extração descrito por Pitcher,
Saunders & Owen (1989) e o par de iniciadores SMEGG1/SMEEG2 (Hamburger et al.,
1991). A opção por esse método de extração partiu dos resultados obtidos em nosso
laboratório por Santos et al. (aceito para publicação) que avaliou a PCR-Multiplex e um
conjunto comercial de reagentes–ELISA para detecção e diferenciação simultânea Entamoeba
histolytica e Entamoeba dispar. A PCR-Multiplex padronizada no laboratório foi capaz de
identificar quase a totalidade dos indivíduos infectados revelando um limiar de detecção de
cinco trofozoítos por 100 microlitros de suspensão fecal. O teste de PCR foi mais eficiente do
que o exame parasitológico realizado com uma amostra de fezes e seu resultado foi
concordante com o ensaio de detecção de antígeno de E. histolytica pelo ELISA.
Entretanto, quando esse protocolo de extração foi empregado com as amostras de
fezes provenientes de área endêmica para a esquistossomose, os resultados não foram tão
bons como aqueles demonstrados por Santos et al. (aceito para publicação). No grupo dos
indivíduos com ovos de S. mansoni nas fezes acima e abaixo de 10 OPG, a PCR detectou
DNA do parasito em menos da metade das amostras fecais analisadas, enquanto no grupo dos
indivíduos apenas soropositivo para esquistossomose, somente uma amostra de fezes
apresentou o produto amplificado de 110 pb. Quanto à especificidade do teste, nenhuma das
amostras de fezes dos indivíduos sem a presença da infecção pelo S. mansoni apresentou a
banda diagnóstica.

A baixa sensibilidade dos resultados pode ter sido devida às dificuldades na obtenção
de DNA em amostras de fezes, uma vez que substâncias inibidoras da Taq polimerase
poderiam estar presentes nas fezes e inibir a ação da enzima (Deuter et al., 1995; Holland et
al., 2000; Vandenberg & Oorschot et al., 2002). Até o momento, não existe um consenso
quanto ao método de extração ideal e universal, visto que vários fatores presentes nas fezes
podem interferir ou atuar como substâncias inibidoras como ácidos biliares, hemoglobina,
bilirrubina e carboidratos (Monteiro et al., 1997; Wilson., 1997; Silva et al., 1999). Estas
interferências são extremamente variáveis entre populações diferentes, talvez em decorrência
dos hábitos alimentares que possam estar influenciando no tipo e na quantidade de substâncias
inibidoras presentes no material fecal e até mesmo pela não uniformidade na composição das
próprias fezes, conforme a dieta, estado de saúde e flora intestinal de cada indivíduo (Silva, et
al., 1999; Holland et al., 2000; Li, Hullar & Lampe, 2007).
Não podemos descartar também a possibilidade de que no final do processo de
extração seja obtido um baixo rendimento de DNA em decorrência da baixa quantidade de
ovos de S.mansoni na amostra. Além disso, os ovos de S. mansoni possuem uma membrana
altamente robusta e compacta, dificultando, assim, a liberação de DNA nas fezes.
Na tentativa de melhorar a obtenção de DNA, decidimos alterar algumas etapas de
extração de DNA. Nesse novo protocolo, utilizamos 300 µL de fezes diluídas em água, ao
invés de 100 µL. Nossa intenção era de aumentar as chances de ter ovos do parasito naquela
alíquota processada. Todavia, estaríamos aumentando, também, o percentual de substâncias
inibidoras. Assim, para diminuir a ação dos inibidores, as amostras de fezes diluídas foram
lavadas três vezes. Além disso, pérolas de vidro foram acrescentadas ao sedimento fecal e
ressuspensas em solução de tiocianato de guanidina, aumentando a possibilidade lise dos ovos
do parasito. A etapa de lise mecânica favorece a liberação do DNA e, conseqüentemente
maior quantidade da molécula, devido à avidez do DNA pela superfície de vidro (Marko,

Chipperfield & Birnboim, 1982; Boom et al., 1990). Como última modificação, foi
adicionado glicogênio à fase aquosa contendo o DNA, para auxiliar na precipitação do mesmo
(Teixeira, 2000).
Depois de realizadas alterações no protocolo de extração, foi feita, também, uma
modificação no processo de amplificação da PCR, acrescentando a BSA na mistura da reação.
Segundo Al-Sound & Radstrom (2000), algumas substâncias, tal como a BSA, quando usadas
simultaneamente na reação de amplificação, comportam-se como facilitadores da reação, pois
captam substâncias inibidoras das amostras fecais e, conseqüentemente, produzindo um
aumento na qualidade da amplificação. A BSA como proteína “capta” os inibidores.
A PCR foi novamente realizada com o par de iniciadores SMEGG1/SMEEG2. No
momento em que estávamos padronizando este novo protocolo de extração, surgiu um
trabalho aplicando um par de iniciadores direcionados para uma região da subunidade 28S do
DNA ribossomal do S. mansoni (Sandoval et al., 2006a). Os autores enfocaram o estudo na
aplicação da PCR em amostras de urina de camundongos. Os resultados obtidos na PCR com
as amostras de urina foram comparados com aqueles obtidos na PCR e no exame
parasitológico pela técnica de Kato-Katz com amostras de fezes e com o ELISA-IgG em
amostras de soro desses mesmos animais. Empregando esta metodologia, os autores
demonstraram que a PCR em urina seria uma ferramenta útil no diagnóstico da
esquistossomose aguda. Em vista desses dados, decidimos avaliar esse mesmo par de
iniciadores, SmF/SmR, empregando as amostras de DNA extraídas pelo novo protocolo de
extração e amplificação. A PCR foi realizada com o mesmo painel de amostras clínicas que
havíamos selecionado para o estudo. Os resultados observados entre os indivíduos
esquistossomóticos (Tabelas 4 e 6) mostraram que as PCRs quando analisadas em conjunto,
apresentaram uma alta sensibilidade para o diagnóstico da esquistossomose. O fragmento de
DNA de 110 pb ou 350 pb foi detectado em 91% das amostras analisadas (31/34), não houve

amplificação em três amostras de fezes que apresentavam carga parasitaria abaixo de 10 OPG.
Durante a execução desse estudo, as amostras fecais foram primeiramente analisadas
pelo método Kato-Katz e uma parte da amostra foi guardada para a realização da reação de
PCR. Em se tratando de infecções leves, pode ter ocorrido que no preparo de lâminas para
coproscopia tenham sido retirados os poucos ovos presentes na amostra.
Sabemos da possibilidade de resultados falso-negativos da PCR devido a presença de
substâncias inibidoras nas fezes. Entretanto, esse problema pode ter sido contornado quando
modificamos o protocolo de extração. Porém, para nos certificarmos disto, realizamos o
procedimento de contaminação das três amostras de fezes negativas com 5fg de DNA de
S.mansoni (“spiking”) como teste controle de inibidores (Silva et al., 1997, 1999). O resultado
demonstrou não haver a interferência de inibidores nas amostras.
É importante salientar que a qualidade das amostras é um fator crucial para um bom
desempenho da PCR. Por mais cuidados que tivéssemos com o transporte das amostras fecais
da área endêmica para o laboratório, persiste a possibilidade de degradação do DNA (Pontes
et al., 2003). Outro aspecto importante a ser considerado, principalmente diante da escassez
de ovos do parasito, é a quantidade de fezes utilizadas pelos métodos de diagnóstico. Foram
preparadas duas lâminas de cada amostra, totalizando cerca de 86mg de fezes examinadas
pela técnica de Kato-Katz. Em princípio, parece que para a extração do DNA, uma quantidade
maior de fezes estaria sendo usada (1g), mas vale lembrar que essa alíquota foi diluída em
água e, finalmente, empregado um volume de 300µL, que corresponderia a uma porção bem
menor que aquela empregada pela técnica parasitológica.
Quando analisamos isoladamente a eficiência de cada iniciador nas reações das PCR
(Tabelas 5-7), observamos que houve resultados discordantes entre nove, das quais seis foram
negativas, quando usados o par de iniciadores SMEGG1/SMEEG2, e positivas para o par
SmF/SmR. Com as outras três amostras fecais foi constatado o inverso, visualizando o

produto amplificado de 350pb referente ao par de iniciadores SmF/SmR e não a banda
diagnóstica de 110pb referente ao par de SMEGG1/SMEEG2.
Estes resultados demonstraram, à primeira vista, que o iniciador SmF/SmR foi mais
eficiente quando comparado com o SMEGG1/SMEEG2. Sabemos que é muito difícil
determinar qual o fator responsável pela discordância entre os resultados das reações de PCR,
porém, não podemos deixar de mencionar que o protocolo de amplificação empregando
SmF/SmR foi padronizado em condições de alta estringência quando comparado com o
protocolo para o par de iniciadores do SMEGG1/SMEEG2.
O ciclo térmico da amplificação tem um efeito substancial na sensibilidade e
especificidade da PCR. Partindo-se do fato de que a etapa de desnaturação tenha ocorrido de
forma eficiente, o sucesso completo da reação de amplificação, dependerá da hibridização do
par de iniciadores com o seu alvo, da atividade ótima da enzima, da concentração de dNTPs e
da constituição do DNA a ser amplificado (Elnifro et al., 2000).
Em nosso estudo, a temperatura de hibridização e a concentração de MgCl2 usadas
no protocolo de amplificação quando o par SMEGG1/SMEEG2 foi empregado, podem ter
afetado o rendimento da reação, apesar de se tratar de uma seqüência altamente repetitiva ao
longo do genoma do S. mansoni. Suzuki et al. (2006), observaram melhores resultados de
sensibilidade e de especificidade com o mesmo par de iniciadores, SMEGG1/SMEEG2,
quando empregaram temperatura diferente. Os autores usaram a técnica Touchdown-PCR, na
qual é mantida uma temperatura de anelamento mais alta em comparação a que seria ideal
(maior estringência) nos ciclos iniciais da reação, evitando, assim, a formação de produtos
inespecíficos. Isto permite que o DNA-alvo seja predominante durante o restante dos ciclos da
amplificação. Os autores realizaram a etapa de hibridização, utilizando a temperatura de 60ºC,
que foi gradualmente diminuída a cada três ciclos para 50ºC. Quando a temperatura de
anelamento chegou aos 50ºC, foram realizados os vinte ciclos restantes.

Por outro lado, existe ainda a questão do limiar de detecção apresentado por cada par
de iniciadores. Pontes, Dias-Neto & Rabello (2002) demonstraram que o par
SMEGG1/SMEEG2 foi capaz de amplificar o DNA do S. mansoni até a concentração de um
fentograma presente na amostra testada. Os autores que descreveram o par SmF/SmR
(Sandoval et al., 2006a) mencionaram o limiar de 1.9 picogramas de DNA presente na
amostra, para que ocorra a amplificação do alvo. As amostras analisadas continham pouca
quantidade de ovos que acarretaria na menor quantidade de DNA disponível. Possivelmente,
nas três amostras de fezes negativas pela PCR com o par de iniciadores SmF/SmF houvesse
menor concentração de DNA do que o limiar de detecção exigido pelo teste.
Em nosso estudo avaliamos a PCR como um método de diagnóstico mais sensível de
identificação direta do S. mansoni. Dentro do painel de amostras clínicas analisadas havia um
grupo de indivíduos com resultado positivo para a esquistossomose apenas pela sorologia.
Ainda existem muitas controvérsias e indefinições a respeito da especificidade da sorologia
(Rabello, 1997; Hamilton, Klinkert & Doenhoff ., 1998; Doenhoff, Chiodini & Hamilton.,
2004).
Entretanto, ao avaliarmos os resultados da PCR entre os indivíduos com sorologia
positiva, observamos que 59% das amostras fecais (13/22) apresentaram DNA do S.mansoni
(Tabela 8). Como a PCR detectou fragmentos de DNA do S. mansoni nestas amostras,
garantindo a confirmação parasitológica da infecção, os resultados da sorologia foram
corroborados.
Dentre as amostras de fezes dos indivíduos soropositivos, ocorreu que entre aquelas
positivas pela PCR, algumas apresentaram ambos produtos de 110pb e 350pb, referentes aos
dois pares de iniciadores usados, enquanto, outras seis mostraram apenas uma das duas
bandas diagnósticas.

As possíveis causas para explicar a presença de apenas uma das bandas diagnósticas
nos testes de PCR, já foram discutidas anteriormente, e quanto às amostras de fezes restantes,
nas quais não foram observados os produtos amplificados de 100pb e/ou 350pb, existe ainda a
questão da oviposição intermitente do parasito.
Em relação à especificidade da PCR, com o novo protocolo de extração e o emprego
dos dois pares de iniciadores, novamente nenhuma das amostras de fezes de indivíduos sem a
infecção pelo S. mansoni apresentou alguma banda diagnóstica.
Em vista dos resultados obtidos, decidimos padronizar uma reação de PCR-
Multiplex, pois estaríamos empregando os dois pares de iniciadores, SMEGG1/SMEEG2 e
SmF/SmR, em uma única reação. Nesse caso, a etapa de hibridização foi realizada
empregando condições de alta estringência.
Do painel de amostras de fezes usado para avaliação das duas PCR convencionais,
oito foram escolhidas para a primeira avaliação da PCR-Multiplex, incluindo aquelas três
amostras das quais não se amplificou nenhuma das bandas diagnósticas. Os resultados da
PCR-Multiplex reproduziram àqueles encontrados nas PCR convencionais. Novamente as três
amostras de fezes que haviam sido anteriormente negativas, mantiveram o resultado (Tabela
10).
Devido aos resultados discordantes entre as reações de PCR e a coproscopia no
grupo de pacientes com eliminação menor que 10 OPG, decidimos estudar um novo grupo de
pacientes (grupo VI). Este grupo compreendia indivíduos que forneceram até três amostras
fecais colhidas em dias diferentes e cada uma das amostras de um mesmo indivíduo foi
avaliada separadamente.
Dessa forma, tínhamos pacientes com somente uma amostra positiva e outros com
mais de uma amostra positiva no exame parasitológico de fezes, através do método de Kato-
Katz. Esse novo grupo de amostras fecais foi, então, avaliado pela PCR-Multiplex (Tabela

11). Podemos observar que várias dessas amostras, que tiveram resultado negativo na
coproscopia, foram PCR positivas, apresentando os dois fragmentos de DNA amplificados.
Em uma amostra com coproscopia positiva, a PCR foi negativa (paciente 6.2). Entretanto,
esse fato só ocorreu com uma amostra desse indivíduo, sendo as outras duas positivas em
ambas as análises.
Possivelmente, neste caso, tenha acontecido que, depois da realização da
coproscopia, através da técnica de Kato-Katz, os poucos ovos de S. mansoni presentes na
amostra tenham sido retirados e a alíquota analisada pela PCR-Multiplex fosse realmente
negativa. Em contrapartida, a PCR-Multiplex foi bem mais sensível que a coproscopia, visto
que se manteve positiva naqueles casos em que o exame pelo Kato-Katz mostrava um
resultado falso negativo, em indivíduos esquistossomóticos.
Uma vez que pretendemos utilizar esta metodologia em laboratórios de análises
clínicas, decidimos avaliar um método de extração comercial para a etapa de obtenção do
DNA da amostra fecal. Para este propósito, foi escolhido o reagente QIAamp (Qiagen)
recomendado para extração de DNA de amostra fecal. Em uma análise preliminar,
processamos treze amostras de fezes pertencentes aos grupos I, II e III, sendo que, nove eram
positivas no exame parasitológico, pelo método de Kato-Katz, e quatro eram positivas apenas
pela sorologia para S. mansoni.
Com o método de extração comercial 85% das amostras de fezes testadas (11/13)
foram positivas na PCR-Multiplex. Essas mesmas onze amostras já haviam sido positivas
anteriormente, quando os pares de iniciadores foram testados separadamente. Duas amostras
de indivíduos, com apenas sorologia positiva para infecção pelo S. mansoni, mantiveram
negativas em todos os testes de PCR, mesmo com a PCR-Multiplex empregando a extração
comercial (Tabela 12).

Levando-se em consideração que a PCR reproduziu os resultados encontrados
quando foram feitas as PCR convencionais, com os dois pares de iniciadores separadamente,
nota-se que, nosso protocolo de extração apresentou eficiência similar ao da extração
comercial.
Os resultados obtidos neste estudo demonstraram que a PCR-Multiplex pode ser uma
importante ferramenta para detecção da infecção pelo S. mansoni em indivíduos que eliminem
pouca quantidade de ovos nas fezes. Além disso, com a determinação da infecção através da
detecção de DNA do S. mansoni naquelas amostras de fezes dos indivíduos apenas
soropositivos para a esquistossomose, surge uma nova possibilidade para a confirmação dos
resultados obtidos pelo ELISA-IgG. Em vista desses resultados, os indivíduos da área
endêmica que tiveram a parasitose confirmada através da PCR foram tratados. Estes
indivíduos serão acompanhados através de novos exames de fezes e de sangue para a
verificação de cura. Isso nos dará a chance, inclusive, de avaliar a PCR não só como mais um
critério para elucidar casos de suspeita da esquistossomose nos quais a coproscopia deixe
dúvidas, mas também como uma nova ferramenta para a investigação da eficiência da
terapêutica.

Conclusões
Como conclusões deste estudo, podemos destacar que:
1. A extração de DNA de amostras de fezes dos indivíduos com infecção pelo
S.mansoni, empregando as técnicas de Pitcher, Saunders & Owen (1989) e Silva et al.
(1999), modificada, foi superior à descrita originalmente;
2. O emprego de um dos pares de iniciadores, com esta técnica de extração, mostrou
uma alta sensibilidade para a detecção da infecção pelo S. mansoni. Entretanto, o
somatório dos resultados das duas reações de PCR apresentou ainda uma maior
sensibilidade;
3. A técnica de PCR para a detecção de fragmentos de DNA de S. mansoni nas fezes
apresentou 100% de sensibilidade quando empregada em amostras com mais de dez
ovos por grama de fezes e 81% de sensibilidade em amostras com menos de dez ovos
por grama de fezes;
4. A técnica de PCR para a detecção de fragmentos de DNA de S. mansoni nas fezes foi
positiva em 54,5% das amostras de indivíduos que apresentavam somente a sorologia
reativa;

5. A técnica de PCR para a detecção de fragmentos de DNA de S. mansoni nas fezes
apresentou 100% de especificidade;
6. A PCR-Multiplex padronizada com os dois pares de iniciadores apresentou resultados
similares àqueles obtidos com o emprego dos iniciadores individualmente;
7. O emprego de uma técnica comercial para extração de DNA de fezes, apresentou a
mesma eficiência da técnica de Pitcher, Saunders & Owen, (1989) e Silva et al. (1999)
modificada;
8. As técnicas de PCR convencional e multiplex têm grande potencial para se tornarem
métodos de diagnóstico referencial na confirmação da infecção pelo S.mansoni, através da
detecção do DNA do parasito, adicionalmente a coproscopia, sobretudo, considerando-se
sua utilização para indivíduos portadores de infecções leves.

Referências Bibliográficas
AL-SOUD, W.A. & RADSTROM, P. Effects of amplification facilators on diagnostic PCR in the presence of blood, feces, and meat. J. Clin. Microbiol. 38(12):4463-4470, 2000
ATTALLAH, A.M.; ISMAIL, H.; EL MASRY, S.A.; RIZK, H.; HANDOUSA, A.; EL BENDARY, M.; TABIL, A.; EZZAT, F. Rapid detection of a Schistosoma mansoni circulating antigen excreted in urine of infected individuals by using a monoclonal antibody. J. Clin. Microbiol., 37(2): 354-357, 1999.
BERGQUIST, N.R. Present aspects of immunodiagnosis of schistosomiasis. Mem. Inst. Oswaldo Cruz, 87(4): 29-38, 1992.
BERGWERFF, A.A.; VAN DAM, G.J.; ROTMANS, J.P.; DEELDER, A.M.; KAMERLING, J.P.; VLIEGENTHART, J.F. The immunologically reactive part of immunopurified circulating anodic antigen from Schistosoma mansoni is a threonine-linked polysaccharide consisting of --> 6)-(beta-D-GlcpA-(1->3)-beta-D-GalpNAc-(1->) repeating units. J. Biol. Chem., 269(50): 31510-7, 1994.
BOOM, R.; SOL, C.J.; SALIMANS, M.M.; JANSEN, C.L.; WERTHEIM-VAN DILLEN, P.M.; VAN DER NOORDAA, J. Rapid and simple method for purification of nucleic acids. J. Clin.Microbiol, 28 (3): 495-503, 1990.
BRANDT, J.L. & FINCH, E.P. A simple flocculation slide test for diagnosis of schistosomiasis. Am. J. Clin. Pathol., 10: 141-152, 1946.
BURLANDY-SOARES, L.C.; DIAS, L.C.S.; KANAMURA, H.Y.; OLIVEIRA, E.J.; CIARAVOLO, R.M. Schistosomiasis mansoni: follow-up of control program based on parasitologic and serologic methods in a Brazilian community of low endemicity. Mem. Inst. Oswaldo Cruz, 98(6): 853-859, 2003.
CDC, http://www.dpd.cdc.gov/html/Imagelibrary/Schistosomiasis CAMARGO-NEVES, V.L.F.; KANAMURA, H.Y.; VELLOSA, S.A.G.; GAGIONI, C.; DIAS, L.C.S. IgM antibodies to Schistosoma mansoni gut-associated antigens for the study of schistosomiasis in Ribeirão Pires, São Paulo. Mem. Inst. Oswaldo Cruz, 93(1): 273-278, 1998
COELHO da SILVA, C.L.P.A.; SOARES M.S.; BARRETO,. G.M. Occurrence of Biomphalaria tenagophila and disappearance of Biomphalaria straminea in Paracambi, RJ, Brazil. Mem. Inst. Oswaldo Cruz, 92 (1): 37-38, 1997. (research note)

COUDERT, J.; GARIN, J.P.; AMBROISE-THOMAS, P.; POTHIER, M.A. Diagnostic serologic par immunofluorescences sur coupes à la congelation d’infectious à Schistosoma mansoni, S. haematobium et S. intercalatum. Acta Trop., 25: 109-132, 1968.
DE JONGE, N.; GRYSEELS, B.; HILBERATH, G.W.; POLDERMAN, A.M.; DEELDER, A.M. Detection of circulating anodic antigen by ELISA for seroepidemiology of schistosomiasis mansoni. Trans. R. Soc. Trop. Med. Hyg., 82: 591-594, 1988.
DEELDER, A.M.; KLAPPE, H.T.; VAN DEN AARDWEG, G.J.; VAN MEERBEKE, E.H. Schistosoma mansoni: demonstration of two circulating antigens in infected hamsters. Exp Parasitol., 40 (2):189-97, 1976.
DEELDER, A.M.; KORNELIS, D.; MAKBIN, H.N.; NOORDPOOL, H.N.; CODFRIED, R.M.; ROTMANS, J.P.; DOSTBURG, B.F.J. Applicability of different antigens preparations in the enzyme-linked immunosorbent assay for schistosomiasis mansoni. Am. J. Trop. Med. Hyg., 29: 401-410, 1980.
DEELDER, A.M.; ZEYLVAR, R.J.M.; FILLIE, Y.E.; ROTMANS, J.P.; DUCHENNE, W. Recognation of gut-associated antigens by immunoglobulin M in the direct florescent antibody test for schistosomiasis mansoni. Trans. R. Soc. Trop. Med. Hyg., 83: 364-367, 1989.
DEUTER, R.; PIETSCH, S.; HERTER, S.; MÜLLER, O. A method for preparation of fecal DNA suitable for PCR. Nucl. Acids Res., 23(18): 3800-3801, 1995.
DOENHOFF, M.J.; CHIODINI, P.L.; HAMILTON, J.V. Specific and sensitive diagnosis of schistosome infection: can it be done with antibodies ? Trends Parasitol., 20(1): 35-39, 2004.
DOUMENGE, J.P.; MOTT, K.E.; CHEUNG, C.; VILLENAVE, D.; CHAPUIS, O.; PERRIN, M.F.; REAUD-THOMAS, G. Atlas of the global distribution of schistosomiasis. Geneve, OMS/WHO, 1987. 400p.
EL-RIDI, R.; OZAKI, T.; INABA, T.; ITO, M.; KAMIYA, H. Schistosoma mansoni oviposition in vitro reflects worm fecundity in vivo: individual-, parasite age- and host-dependent variations. Inter. J. Parasitol., 27(4):381-7, 1997.
ELNIFRO, E.M.; ASHSHI, A.M.; COOPER, R.; KLAPPER, P.E. Multiplex PCR: optimization and application in diagnostic virology. Clin Microbiol Rev.,13(4):559-70, 2000.
FAIRLEY, N.H. The discovery of a specific complement fixation test for bilharziasis and its practical application to clinical medicine. J. R. Army Med. Corps., 32: .449-460, 1919.
FAIRLEY, N.H. & WILLIAMS, F.F. A preliminary report on an intradermal reaction in schistosomiasis. Med. J. Austral., 2: 811-818, 1927.

FNS. FUNDAÇÃO NACIONAL DE SAÚDE. Controle da esquistossomose: diretrizes técnicas. 1. ed. Brasília, Ministério da Saúde, 1994. 85p.
FNS. FUNDAÇÃO NACIONAL DE SAÚDE. Controle da esquistossomose: diretrizes técnicas. 2. ed. Brasília, Ministério da Saúde, 1998. 70p.
GIBBONS-MATTHEWS, C. & PRESCOTT, A.M. Intra-isolate variation of Cryptosporidium parvum small subunit ribosomal RNA genes from human hosts in England. Parasito.l Res., 90(6):439-44, 2003.
GOBERT, N.G.; CHAI, M.; DUKE, M.; McMANUS, P.D. Copro-PCR based detection of Schistosoma eggs using mitochondrial DNA markers. Mol. Cell. Probes, 19: 250-254, 2005.
GOMES, Y.M.; GOMES, F.C.;FURTADO, V.C.; TEIXEIRA, K.M.; SOUZA, W.V.; ABATH, F.G.C. IgE and IgG4 antibodies in subjects reinfected with Schistososma mansoni in an endemic area of northeast Brazil. Mem. Inst. Oswaldo Cruz, 93(1): 183-184, 1998.
GONÇALVES, M.M.L.; BARRETO, M.G.M.; PERALTA, R.H.S.; GARGIONI, C.; GONÇALVES, T.; IGREJA, R.P.; SOARES, M.S.; PERALTA, J.M. Immunoassays as an auxiliary tool for the serodiagnosis of Schistosoma mansoni infection in individuals with low intensity of egg elimination. Acta tropica, 100(1): 24-30, 2006.
GONIN, P. & TRUDEL, L. Detection and Differentiation of Entamoeba histolytica and Entamoeba dispar isolates in clinical samples by PCR and Enzyme-Linked. Immunosorbent Assay. J. Clin. Microbiol., 41(1): 237-241, 2003.
HAMBURGER, J.; TURETSKI, T.; KAPELLER, I.; DERESIEWIC, R. Highly repeated hort DNA sequences in the genome of Schistosoma mansoni recognized by a species- pecific probe. Mol. Bioch. Parasitol., 44: 73-80, 1991.
HAMBURGER, J.; YU-XIN, X.; RAMZY, M.R.; JOURDANE, J.; RUPPEL, A. Development and laboratory evaluation of a polymerase chain reaction for monitoring Schistosoma mansoni infestation of water. Am. J. Trop. Med Hyg., 59(3): 468-473, 1998.
HAMBURGER, J.; HE-NA.; ABBASI, I.; RAMZY, R.M.; JOURDANE, J.; RUPPEL. A.polymerase chain reaction assay based on a highly repeated sequence of Schistosoma haematobium: a potential tool for monitoring schistosome-infested water. Am. J. Trop. Med Hyg, 65(6): 907-11, 2001.
HAMILTON, J.V.; KLINKERT, M.; DOENHOFF, M.J. Diagnosis of schistosomiasis: antibody detection, with notes on parasitological and antigen detection methods. Parasitol., 117: 41-57, 1998.
HANELT, B; ADEMA, C.M; MANSOUR, M.H; LOKER, E.S. Detection of Schistosoma mansoni in Biomphalaria using nested PCRJ. Parasitol; 83(3):387-94. 1997

HENEGARIU, O.; HEEREMA, N.A.; DLOUHY, S.R.; VANCE, G. H.; VOGT, P.H. Multiplex PCR: Critical parameters and step-by-step protocol. BioTech., 23(3): 504-511, 1997.
HERRING, A.J.; INGLIS, N.F.; OJHE, C.K.; SNODGRASS, D.P. & MENZIES, D. Rapid diagnosis of rotavirus infection by direct detection of a viral nucleic acid in silver-stained polyacrylamide gel. Clin. Microbiol., 16: 473-477, 1982.
HOFFMAN, W.A.; PONS, J.A.; JANER, J.L. Sedimentacion concentracion method in schistosomiasis mansoni. J. Publ. Health Trop. Med., 9: 283-298, 1934.
HOLLAND, J.L.; LOQUIE,L.; SIMOR, A.E.; LOQUIE, M.. PCR detection of Escherichia coli O157:H7 directly from stools: Evaluation of comercila extraction methods for purifying fecal DNA. J. Clin. Microbiol, 38(11):4108-4113, 2000.
HOSHINO, S.; CAMARGO, M.E.; CORRÊA, N.S. A hemaggluttination test for Manson’s schistosomiasis using chromium chloride, formalin treated human erythocytes, sensitized with worm extracts. Rev. Inst. Med. Trop. São Paulo, 12: 192-194, 1970.
HULDT, G.; LAGERGUIST, B.; PHILLIPS, T.; DRAPER, C.C.; VOLLER, A. Detection of antobodies in schistosomiasis by enzyme-linked immunosorbent assay (ELISA). Ann. Trop. Med. Parasitol., 69: 483-488, 1975.
IDRIS, M.A.; RUPPEL, A. Diagnostic Mr 31/3200 Schistosoma mansoni proteins (Sm 31/32): Reaction with sera from Sudanese patients infected with S. mansoni or S. haematobium. J. Helminthology, 62: 92-101, 1988.
JANNOTTI-PASSOS, L.K; VIDIGAL, T.H; DIAS-NETO E; PENA, S.D; SIMPSON, A.J; DUTRA. W.O; SOUZA, C.P; CARVALHO-PARRA, J.F. PCR amplification of the mitochondrial DNA minisatellite region to detect Schistosoma mansoni infection in Biomphalaria glabrata snails. J. Parasitol., 83(3):395-9, 1997.
KAGAN, I.G. & OLIVER-GONZALEZ, J. Preliminary studies on hemaggluttination in schistosomiasis. J. Parasitol., 41: 26, 1950.
KANAMURA, H.Y.; HOSHINO-SHIMIZU, S.; CAMARGO, M.E.; SILVA, L.C. Class- specific antibodies and fluorescent staining paterns in acute and chronic forms of schistosomiasis mansoni. Am. J. Trop. Med. Hyg., 28: 242-248, 1979.
KANAMURA, H.Y.; SILVA, R.M.; RABELLO, A.L.T.; ROCHA, R.S.; KATZ, N. Anticorpos séricos IgA no diagnóstico da fase aguda da esquistossomose mansoni humana. Rev. Inst. Adolfo Lutz, 51(1/2): 101-104, 1991.
KANAMURA, H.Y.; HOSHINO-SHIMIZU, S.; SILVA, L.C. Schistosoma mansoni cercarie and schistosomulum antigens in serodiagnosis of schistosomiasis. Bull. Pan. Amer. Health. Org., 26: 220-229, 1992.

KATZ, N.; CHAVES, A.; PELLEGRINO, J. A simple device for quantitative stool thick-smear technique in schistosomiasis mansoni. Rev. Inst. Med. trop. São Paulo, 14: 397-400, 1972.
KATZ, N. & PEIXOTO, S.V. Análise crítica da estimativa do número de portadores de esquistossomose no Brasil. Rev. Soc. Bras. Med. Trop., 33(3): 303-308, 2000.
LAEMMLI, U.K. Clevage of structural proteins during the assembly of head of bacteriophage T4. Nature, 227: 680-685, 1970.
LI, F; HULLAR, M.A; LAMPE, J.W. Optimization of terminal restriction fragment lenght polymorphism (TRFLP) analysis of human gut microbiota.. J. Microbiol Methods, 68(2): 303-11, 2007.
LIMA, D.M.C.; ABRANTES-LEMOS, C.P.; HOSHINO-SHIMIZU, S.; KANAMURA, H.Y.; da SILVA, L.C.; VELLOSA, S.A.G. Schistosoma mansoni low infection. Evaluation of serologic assays. In: International Symposion of Schistosomiasis. Rio de Janeiro, 1993.
LIMA, V.L.C.; GUERCIO, V.M.S.; RANGEL, O.; KANAMURA, H.Y.; DIAS, L.C.S. Immunofluorescence test on Schistosoma mansoni worm paraffin sections (IgM-IFT) for the study schistosomiasis transmission in Campinas, São Paulo, Brazil. Mem. Inst. Oswaldo Cruz, 93(1): 283-288, 1998.
LOVERDE, P.T.; HIRAI, H.; MERRICK, J.M.; LEE, N.H.; EL-SAYED, N. Schistosoma mansoni genome project: an update. Parasitol. Int., 53: 183-192, 2004.
LUNDE, M.N.; OTTESEN, E.A.; CHEEVER, A.W. Serologice differences between acute and chronic schistosomiasis mansoni detected by enzyme-linked immunosorbent assay (ELISA). Am. J. Trop. Med. Hyg., 28(1): 87-91, 1979.
LUTZ, A. O Schistosomum e a schistossomose segundo observações feitas no Brasil. Mem. Inst. Oswaldo Cruz, 11: 121-150, 1919.
MADDISON, S.A.; SLEMENDA, S.B.; TSANG, V.C.W.; POLLARD, R.A. Serodiagnosis of Schistososma mansoni with microsomal adult worm antigen in an enzyme-linked immunosorbent assay using a standard curve developed with a reference serum pool. Am. J. Trop. Med. Hyg., 34: 484-494, 1985.
MAKAROVA, E.; GOES, T.S.; MARCATTO, A.L.M.; LEITE, M.F.; GOES, A.M. Serological differentiation of acute and chronic schistosomiasis using Schistosoma mansoni recombinant protein RP26. Parasitol. Inter., 52: 269-279, 2003.
MAKAROVA, E.; GOES, T.S.; LEITE, M.F.; GOES, A.M. Detection of IgG binding to Schistosoma mansoni recombinant protein RP26 is a sensitive and specific method for acute schistosomiasis diagnosis. Parasitol. Inter., 54: 69-74, 2005.

McLAREN, M.L.; DRAPER, C.C.; ROBERTS, J.M.; MINTER-GOEDBLOED, E.; LIGTHART, T.; TEESDALE, C.H.; AMIN, M.A.; OMER, A.H.S.; BARTLETT, A.; VOLLER, A. Studies on the enzyme linked immunosorbent assay (ELISA) for Schistosoma mansoni infections. Ann. Trop. Med. Parasitol., 72: 243-253, 1978.
MARKO, M.A..; CHIPPERFIELD, R & BIRNBOIM, H.C. A procedure for the large scale isolation of highly purified plasmid DNA using alkaline extraction and binding to glass powder. Anal Biochem.,. 121: 382-387,1982.
MONTEIRO, L.; BONNEMAISON, D.; VEKRIS, A.; PETRY, K.G.; BONNET, J.; VIDAL, R.; CABRITA, J.; MEGRAUD, F. Complex Polysaccharides as PCR Inhibitors in Feces Helicobacter pylory Model. J. Clin. Microbiol., 35(4): 995-998, 1997.
NASCIMENTO-CARVALHO, M. & MORENO-CARVALHO,O.A. Neuroschistosomiasis due to Schistosoma mansoni: a review of pathogenesis, clinical syndromes and diagnostic approaches. Rev Inst Med Trop São Paulo., 47(4):179-84, 2005.
NASH, T.E. Antibody response to polysaccharide antigen present in the schistosome gut. Am. J. Trop. Med. Hyg., 27: 938-943, 1978.
NASH, T.E.; LUNDE, M.N.; CHEEVER, A.W. Analysis and antigenic activity of a carbohydrate fraction derived from adult Schistosoma mansoni. J. Immunol., 126: 805-810, 1981.
NOYA, O.; NOYA, B.A.; BALLEN, D.; ZERPA, N.; COLMENARES, C.; LOSADA, S.; BERMUDEZ, H. Use of synthetic peptides derived from adult worm proteins of Schistosoma mansoni, in the diagnosis of schistosomiasis. Mem. Inst. Oswaldo Cruz, 93(1):157-158, 1998.
NUÑEZ, Y. O.; FERNÁNDEZ, M.A.; TORRES-NÚNEZ, D.; SILVA, J.A.; MONTANO, I.; MAESTRE, J.L.; FONTE, L.. Multiplex polymerase chain reaction amplification and differention of Entamoeba histolytica and Entamoeba dispar DNA from stool samples. Am. J. Trop. Med. Hyg, 64(5/6): 293-297, 2001
OKABE, K. & TANAKA, T. Urine precipitation reaction for schistosomiasis japonica. Kurume Med. J., 8: 24-29, 1961.
OLIVER-GONZALEZ, J. Anti-egg precipitins in the serum of humans infected with Schistosoma mansoni. J. Infect. Dis., 95: 86-91, 1954.
OMS. ORGANIZAÇÃO MUNDIAL DA SAÚDE (World Health Organization). The control of schistosomiasis: second report of the WHO Expert Committee. Technical Report Series, 830, Geneva, OMS/WHO, 1993. 97p.
PESSÔA, S.B. & MARTINS, A.V. Trematódeos parasitas do sistema sangüíneo – Schistosoma mansoni. In: Pessôa Parasitologia Médica. 11. ed. Rio de Janeiro, Guanabara Koogan, 1982. Cap. 35, p. 361-381.

PITCHER, D.G.; SAUNDERS, N.A.; OWEN, R.J. Rapid extraction of bacterial genomic DNA with guanidine thiocyanate. Lett. Appl. Microbiol., 4: 1513-1521, 1989.
PONTES, L.A.; DIAS-NETO, E.; RABELLO, A. Detection by polymerase chain reaction of Schistosoma mansoni DNA in human serum and feces. Am. J. Trop. Med. Hyg., 66(2): 157-162, 2002.
PONTES, L.A.; OLIVEIRA, M.C.; KATZ, N.; DIAS-NETO, E.; RABELLO, A. Comparison of a polymerase chain reaction and the Kato-Katz technique for diagnosing infection with Schistosoma mansoni. Am. J. Trop. Med. Hyg., 68(6): 652-656, 2003.
PRATA, A. Schistosomiasis mansoni in Brazil. Baill. Clin. Trop. Med. Comm. Dis., 2: 349-369, 1987.
RABELLO, A.T.L.; GARCIA, M.M.A.; GOMES, P.F.; DIAS NETO, E.; ROCHA, R.S.; KATZ, N. Dot-dye-immunoassay and dot-ELISA for the serological differentiation of acute and chronic schistosomiasis mansoni using keyhole limpet haemocyanin as antigen. Trans. R. Trop. Med. Hyg., 87: 279-281, 1993.
RABELLO, A.L.T. Diagnosing schistosomiasis. Mem Inst. Oswaldo Cruz, 92(5): 669-676, 1997.
REY, L. Schistosoma e esquistossomose: epidemiologia e controle. In: Parasitologia. 3. ed. Rio de Janeiro, Guanabara Koogan, 2001. Cap. 35, p. 455-479.
ROSSI, C.L.; TSANG, V.C.W.; PICHER, J.B. Rapid low-technology field and laboratory applicable enzyme-linked immunosorbent assays for immunodiagnosis of Schistosoma mansoni. J. Clin. Microbiol., 29: 1836-1841, 1991.
RUPPEL, A.; DIESFELD, H.J.; ROTHER, U. Western blot analysis of Schistosoma mansoni antigens with sera of schistosomiasis patients: diagnostic potential of an adult schistosome polypeptide. Clin. Exp. Immunol., 62:499-506, 1985a.
RUPPEL, A.; ROTHER, U; VONGERICHTEN, H.; LUCIUS, R.; DIESFELD, H.J. Schistosoma mansoni: Western blot analysis of adult worm proteins. Exp. Parasitol., 60: 195-206, 1985b.
SADUN, E.H.; WILLIAMS, J.S.; ANDERSON, R.I. Fluorescent antibody technic for sero-diagnosis of schistosomiasis in humans. Proc. Soc. Exp. Biol. Med., 105: 289-291, 1960.
SAIKI, R.K.; GELFAND, D.; STOFFEL, S.; SCHARF, S.; HIGUICHI, R.; HORN, G.T.; MULLIS, K.B.; ERLICH, H.A. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science, 239: 487-491, 1988.
SANDOVAL, N.; SILES-LUCAS, M.; ABAN, J.L.; PEREZ–ADELLANO, J.L.; GARATE, T.; MURO, A. Schistosoma mansoni: A diagnostic approach to detect acute schistosomiasis infection in a murine model by PCR. Exp. Parasitol., 114(2):84-8, 2006a.

SANDOVAL, N.; SILES-LUCAS, M.; PEREZ-ARELLANO, J.L.; CARRANZA, C.; PUENTE, S.; LOPEZ-ABAN, J.; MURO, A. A new PCR-based approach for the specific amplification of DNA from different Schistosoma species applicable to human urine samples. Parasitology, 133(Pt 5): 581-7, 2006b.
SANTOS, E.C; CAMPOS, G. B; DINIZ, A.C; LEAL, J.C; ROCHA,M.O.C. Perfil clínico e critérios diagnósticos da mielorradiculopatia esquistossomótica. Arq Neuropsiquiatr, 59(3-B): 772-777, 2001.
SANTOS, H.L.C; PERALTA, R.H.S; MACEDO, H.W; BARRETO, M.G.M; PERALTA, M.P. COMPARISON OF MULTIPLEX-PCR AND ANTIGEN DETECTION FOR DIFFERENTIAL DIAGNOSIS ENTAMOEBA HISTOLYTICA. BRAZILIAN J INFECT DIS, ACEITO PARA PUBLICAÇÃO. SILVA, A.J. Desenvolvimento e Avaliação de ensaios imunoenzimáticos para o diagnóstico sorológico da esquistossomose mansônica. Rio de Janeiro, 1992. 109p. Dissertação (Mestrado), Instituto de Microbiologia, Universidade Federal do Rio de Janeiro-UFRJ.
SILVA, A.J; PIUVEZAM, M.R.; MOURA, H.; MADDISON, S.; PERALTA, J.M. Rapid competitive enzyme linked immunosorbent assay using monoclonal antibody reacting with a 15 kilodalton tegumental antigen of Schistosoma mansoni for serodiagnosis of schistosomiasis. J. Clin. Microbiol., 31: 2315-2319, 1993.
SILVA, A.J.; BORNAY-LLINARES, F.J.; DEL AGUILA DE LA PUENTE, C.D.E.L. A.; MOURA, H; PERALTA, J.M.; SOBOTTKA, I.; SCHWARTZ, D.A.; VISVESVARA, G.S.; SLEMEND, S.B.; PIENIAZEK, N.J. Diagnosis of Enterocytozoon bieneusi (microsporidia) infections by polymerase chain reaction in stool samples using primers based on the region coding for small-subunit ribosomal RNA. Arch Pathol Lab Med., 121(8):874-9, 1997.
SILVA, A.J.; BORNAY-LLINARES, F.J.; MOURA, I.N.; SLEMENDA, S.B.; TUTTLE, J.L.; PIENIAZEK, N.J. .Fast and reliable extraction of protozoan parasite DNA from fecal specimens. Mol. Diagn.,4(1):57-64, 1999.
SINGH, B. Molecular Methods for Diagnosis and Epidemiological Studies of Parasitic Infections. Int. J. Parasitol., 27(10): 1135-1145, 1997.
SUBRUNGRUANG, I.; MUNGTHIN, M.; CHAVALITSHEWINKOON-PETMITR, P.; RANGSIN, R.; NAAGLOR, T.; LEELAYOOVA, S. Evaluation of DNA extraction and PCR methods for detection of Enterocytozoon bienusi in stool specimens. J. Clin. Microbiol.,3490-3494, 2004.
SULAHIAN, A.; GARIN, Y.J.F.; IZRI, A.; VERRET, C.; DELAUNAY, P.; VAN GOOL, T.; DEROUIN, F. Development and evaluation of a Western blot kit for diagnosis of schistosomiasis. Clin. Diagn. Lab. Immunol., 12(4): 548-551, 2005.

SUZUKI, T; OSADA, Y; KUMAGAI, T; HAMADA, A; OKUZAWA, E; KANAZAWA, T. Early detection of Schistosoma mansoni infection by touchdown PCR in a mouse model. Parasitol Int. 55(3):213-8. Epub 2006
TARP, B., BLACK, F.T.; PETERSEN, E. The immunofluorescence antibody test (IFAT) for the diagnosis of schistosomiasis used in a non-endemic area. Trop. Med. Inter. Health, 5(3): 185-191, 2000.
TAYLOR, D.W. & WELLS, P.Z. Isolation and antigen analysis of surface tegument membranes from schistosomula of Schistosoma mansoni. Parasitol., 89: 495-510, 1984.
TEIXEIRA, M.G.M. Diagnóstico da infecção chagásica: aplicação e avaliação de métodos sorológicos e da reação em cadeia da polimerase em amostras clínicas de pacientes chagásicos crônicos. Rio de janeiro, 2000. Dissertação (Doutorado), Instituto Oswaldo Cruz, Fundação Oswaldo Cruz-FIOCRUZ.
TSANG, V.C.W.; PERALTA, J.M.; SIMONS, R.A. The enzyme-linked immuno-electrotranfer blot techniques (EITB) for studying the specificities of antigens and antibodies separated by gel electrophoresis. Methods Enzymol., 92: 377-391, 1983.
TSANG, V.C.W. & WILKINS, P.P. Immunodiagnosis of schistosomiasis. Clinics Lab. Med., 11(4): 1029-1039, 1991.
TSANG, V.C.W. & WILKINS P.P. Immunodiagnosis of schistosomiasis. Immunol. Investigations, 26(1,2): 175-188, 1997.
VALLI, L.C.P.; KANAMURA, H.Y.; SILVA, R.M.; SILVA, M.I.P.G.; VELLOSA, S.A.G.; GARCIA, E.T. Efficacy of an enzyme-linked immunosorbent assay in the diagnosis of and serologic distinction between acute and chronic Schistosoma mansoni infection. Am. J. Trop. Med. Hyg., 57(3): 258-362, 1997.
VAN GOOL, T.; VETTER, H.; VERVOOT, T.; DOENHOFF, M.J.; WETSTEYN, J.; OVERBOSCH, D. Improved serodiagnosis of imported schistosomiasis using a combination of a commercial indirect haemagglutination test (IHA) with Schistosoma mansoni adult worm antigens and an enzyme-linked immunosorbent assay (ELISA) with S. mansoni egg antigens. J. Clin. Microbiol., 40: 3432-3437, 2002.
VAN LIESHOUT L.; PANDAY U.G.; DE JONGEN N.; KRIJGER, F.W.; OOSTBURG, B.F.J.; POLDERMAN, A.M.; DEELDER, A.M. Immunodiagnosis of schistosomiasis mansoni in a low endemic area in Surinam by determination of the circulating antigens CAA and CCA. Acta Trop., 59: 19-29, 1995.
VANDENBERG, N. & OORSCHOT, V.R.A.H. Extraction of Human Nuclear DNA from Feces Samples Using the QIAamp DNA Stool Mini Kit. J. Forensic. Sci., 47(5): 993-995, 2002.

VERWEIJ, J.J.; LAEIJENDECKER, D.; BRIENEN, E.A.; VAN,L.L.; POLDERMAN, A.M. Detection of Cyclospora cayetanensis in travelers returning from the tropics and subtropics using microscopy and real-time PCR. Int. J. Med. Microbiol., 293(2-3): 199-202, 2003.
VERWEIJ, J.J.; BLAGÉ, R.A.; TEMPLETON, K.; SCHINKEL, J.; BRIENEN, E. A.T.; ROOYEN, M.A.A.; LIESHOUT, L.V.; POLDERMAN, A.M. Simultaneous detection of Entamoeba histolytica, Giardia lamblia and Crytosporidium parvus in Fecal Samples by Using Multiplex Real –Time PCR. J. Clin. Microbiol., 42(3): 1220-1223, 2004.
VOGEL, H. & MINNING, W. Hüllenbildung bei Bilharzia-Cercarien im Serum bilharzia-infizierter Tiere und Menschen. Zentrbl. Bakt. Parasitk, 153: 91-105, 1949.
WARREN, K.S. The kinetics of hepatosplenic schistosomiasis. Semin. Liver Dis., 4:293-300, 1984.
WILSON, M.; SULZER, A.J.;WALLS, K.W. Modified antigens in the indirect immunofluorescence test for schistosomiasis. Am. J. Trop. Med. Hyg., 23: 1072-1076, 1974.
WILSON, I.G. Inhibition and Facilitation of Nucleic Acid Amplification. Appl. Envirol.
Microbiol., 63(10): 3741-3751, 1997.

Anexos
Anexo 1: Documento de aprovação pelo Comitê de Ética em Pesquisa da Fiocruz.
Andamento do projeto - CAAE - 000029/0011-02
Título do Projeto de Pesquisa IDENTIFICAçãO DE DEMANDAS DE SANEAMENTO, DE INFORMAçãO E DE SENSIBILIZAçãO POPULAR, ATRAVéS DE INDICADORES BIOLóGICOS E SOCIAIS, PARA ENFRENTAMENTO DA ESQUISTOSSOMíASE E OUTRAS DOENçAS DE ORIGEM SóCIO-AMBIENTAL: O MUNICíPIO DE SUMIDOURO COMO áREA MODELO.
Situação Data Inicial no CEP Data Final no CEP Data Inicial na CONEP Data Final na CONEP Aprovado no CEP 05/11/2002 00:00:00 05/11/2002 00:00:00 05/11/2002 00:00:00 05/11/2002 00:00:00
Descrição Data Documento Nº do Doc Origem
1 - Envio da Folha de Rosto pela Internet 14/05/2002 00:00:00 Folha de Rosto FR-251 Pesquisador
2 - Recebimento de Protocolo pelo CEP (Check-List) 24/05/2002 14:44:32 Folha de Rosto 000029/0011-02 CEP
3 - Protocolo Pendente no CEP 27/06/2002 12:47:31 Folha de Rosto 182/02 CEP
4 - Protocolo Aprovado no CEP 05/11/2002 10:12:32 Folha de Rosto 182/02 CEP

Anexo 2: Informe concensual para participação nos inquéritos coprológico e sororlógico da
esquistossomose mansônica.
TERMO DE CONSENTIMENTO
Eu, ...................................................................................................concordo
em participar do trabalho que a equipe da FIOCRUZ, está realizando na minha
comunidade para prevenir e combater as verminoses. Sei que os perquisadores da
Fiocruz vão precisar me fazer perguntas para descobrir se eu ou alguém da minha
família tem vermes, onde "pegou os verme" e se algum de nós tem sintomas de
doenças. Vão precisar saber, também, que doenças cada pessoa da minha casa já
teve, como são as condições de vida da minha comunidade e o que eu penso dos
problemas de saúde e de meio-ambiente do bairro e do município. Para saber tudo
isso, eles vão me pedir para fazer exames de fezes vão conversar comigo, fazer
debates e perguntas.
Eu sei que fazendo apenas 1 exame de fezes a chance de o exame acusar
que a pessoa tem certos vermes é bem pequena. Por isso vai ser necessário que
cada um de nós faça 3 exames de fezes, em dias diferentes. Sei, também, que
posso pesquisar através do sangue, se tenho esquistossomose ou não. Sei que o
exame feito com o sangue não é aquele exame comum, que informa se a pessoa
tem anemia, colesterol alto ou diabetes; é ume exame apenas para saber se a
pessoa tem ou já teve a esquistossomose.
Estou ciente de que ninguém é obrigado a participar dos exames de sangue e
de fezes e nem precisa responder aos questionários, e de que temos o direito de
desistir de participar a qualquer momento.
Data / /
Assinatura ____________________________________________________

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo