anÁlise da regulaÇÃo transcricional de genes de … · coisas tão belas? ... abstract vii...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS BIOLÓGICAS
DOUTORADO EM CIÊNCIAS BIOLÓGICAS
ANÁLISE DA REGULAÇÃO
TRANSCRICIONAL DE GENES DE Yersinia
EM Escherichia coli
Patrícia Rodrigues de Oliveira Haver
Recife/PE-BRASIL
2005

PATRÍCIA RODRIGUES DE OLIVEIRA HAVER
ANÁLISE DA REGULAÇÃO
TRANSCRICIONAL DE GENES DE Yersinia
EM Escherichia coli
Tese apresentada ao Curso de
Doutorado do Centro de Ciências
Biológicas da Universidade Federal
de Pernambuco, Área de
concentração em Genética.
Orientadora: Dra. Alzira Maria Paiva de Almeida
Co-Orientador: Dr. Osvaldo Pompílio de Melo Neto
Recife/PE-BRASIL
2005

Haver, Patrícia Rodrigues de Oliveira Análise da regulação transcricional de genes de Yersinia em Escherichia coli. / Patrícia Rodrigues de Oliveira Haver. – Recife: O Autor, 2005. 110 folhas : il., fig., tab.
Dissertação (doutorado) – Universidade Federal de Pernambuco. CCB. Ciências Biológicas. Genética de Microorganismos, 2005.
Inclui bibliografia e anexo.
1.YopH 2. SycH 3.Yersinia 4. H-NS 5. F1 6 Diagnóstico. Título. 579 CDU (2.ed.) UFPE 579.34 CDD (22.ed.) CCB – 2006 - 016


Trabalho desenvolvido no
Departamento de Microbiologia do
Centro de Pesquisas Aggeu
Magalhães (FIOCRUZ/MS), com o
suporte financeiro da FACEPE e da
CAPES/CNPq.

Triste? Eu? Não Eu não estou triste. Estou apenas pensando... Pensando em todas as maravilhas que vejo toda vez que olho o céu ou até mesmo que contemplo na terra. Então, por que pensar que estou triste se penso em coisas tão belas? Será porque vejo o homem se destruir em guerras e a tudo mais pela ganância? Será porque vejo um futuro tão cinza quanto o cinza de nossas próprias cinzas? É, eu poderia estar triste. Mas, não estou. Penso apenas que estou viva e isto é tudo. Lutarei por aquilo que acredito. Mas, o que me consola, é saber que desde que eu nasci, Tu me acordas com o raiar do dia e me ninas com o silêncio da noite. E, quando o dia chegar e eu me vir em Teus braços, será a minha vez de perguntar: E Você? Está triste?
Patrícia Haver

SUMÁRIO AGRADECIMENTOS I
LISTA DE FIGURAS E TABELAS II
LISTA DE ABREVIATURAS IV
RESUMO VI
ABSTRACT VII
INTRODUÇÃO 1
Revisão da literatura 3
1. Gênero Yersinia 3
1.1 Aspectos Moleculares de Yersiniae 6
1.2 Diagnóstico da peste 8
2. Escherichia coli enteropatogênica 9
2.1 Aspectos Moleculares 9
3. Sistema de Secreção Tipo III e seus elementos 12
4. Regulação Transcricional em Procariotas 20
4.1 Os elementos da transcrição 20
4.2 Modelos Clássicos de regulação transcricional 25
4.3 Fatores que influenciam a regulação transcricional 26
REFERÊNCIAS BIBLIOGRÁFICAS 29
JUSTIFICATIVA 42
OBJETIVOS 44
ARTIGOS 46
ARTIGO 1: Positioning of the Yersinia enterocolitica yopH promoter after the
Escherichia coli lac promoter induces a strong and regulated
expression of YopH in E. coli.
47
ARTIGO 2: Co-expressão das proteínas SycH e YopH de Yersinia
enterocolitica.
69
ARTIGO 3: Avaliação da YopH recombinante no diagnóstico sorológico da
peste
82
CONCLUSÕES 96
ANEXOS 97

AGRADECIMENTOS
• A Deus, que nunca me deixou desistir.
• A minha orientadora Dra. Alzira Almeida, por todo incentivo, apoio e
dedicação.
• Ao meu co-orientador Dr. Osvaldo Pompílio de Melo Neto, pela oportunidade e
orientação.
• As meninas de ouro: Marina Petribú e Mariana Chagas, que eu tive a honra de
orientar. Obrigada por toda a amizade e carinho.
• A amiga Maria Betânia Melo por todo o incentivo e amizade.
• Aos meus amigos do laboratório: Rafael Dhalia, Christian Reis, Éden Freire,
Cheila Bedor, Tamara De´Carli, Danielle Moura, Pollyanna Rocha, Rodrigo
Menezes, Rodolfo Katz, Francisco Cariri, Gerlane Souza, Soraya Silva, Franklin
Santos, Josiane Gomes, Wagner Luís.
• A todos os amigos que fazem parte do Departamento de Microbiologia: Dra.
Nilma Leal, Dra. Tereza Cristina Leal-Balbino, Dra. Marise Sobreira, Isaac
Martins, Yara Nakasawa, Silvana Santos, Fabiana Almeida, Lucas Rocha e
Fernanda Pimentel.
• A todos que fazem parte do Centro de Pesquisas Aggeu Magalhães.
• Ao CPqAM/FIOCRUZ, UFPE, CAPES e FACEPE, pelo suporte
técnico/científico/financeiro.
• Aos amigos da minha turma de doutorado (turma 2001), em especial a Fátima
Andrade, Leila Cabral e Vilma Loreto. Nós conseguimos!
• Ao Doutorado em Ciências Biológicas, em especial a Adenilda, por toda ajuda e
amizade.
• A todos os amigos do Departamento de Genética, por toda a amizade e
incentivo.
• A toda a minha família no Brasil e na Holanda, pelo apoio e suporte.
• A minha melhor amiga Eliane Bezerra, por ser essa pessoa maravilhosa e estar
sempre do meu lado.
• Ao meu marido Nicolaas Haver, pelo apoio, paciência e dedicação.
I

LISTA DE FIGURAS E TABELAS
REVISÃO BIBLIOGRÁFICA
Figura 1. Rotas de infecção das três espécies de Yersinia patogênicas. 5
Figura 2. Mapa do plasmídio pYV, de Yersinia enterocolitica. 7
Figura 3. Interações da EPEC com células epiteliais. 11
Figura 4. Injectisomo Ysc. 14
Figura 5. Diagrama dos sinais de secreção tipo III e funções das chaperonas. 17
Figura 6. Secreção das Yops pelo injectisomo Ysc e translocação através da
membrana da célula alvo.
19
Figura 7. Resumo das interações entre o promotor e a RNA polimerase. 24
ARTIGO 1
Figure 1. Scheme of the construct used in this study. 58
Figure 2. Expression of plasmid pTZ/YopH in E. coli. 59
Figure 3. Comparison of the expression levels of the yoph gene under the
control of different promoters.
60
Figure 4. SDS-PAGE and Western-blots showing the different levels of
expression of the YopH and fusion proteins at different
temperatures and O.D.s.
61
Figure 5. Expression of YopH protein in E. coli strains lacking the histone-
like proteins Hha, H-NS or both.
62
Figure 6. yopH promoter sequence used in this work as well as the
predicted curvature profile obtained using the "Wedge" software.
63
Table 1. Quantitative analysis of strains YK4122 (wild type) and YK4124
(hns-), in the beginning of the growth (DO = 0,2) and lag phase
(OV), shown at cels/ml.
64
II

ARTIGO 2
Figura 1. SDS/PAGE 17,5% mostrando a síntese da proteína SycH (seta),
em culturas de E. coli TG2 transformadas com os plasmídios
pBAD18/SycH (A) e pBAD33/SycH (B).
74
Figura 2. Western-blot mostrando a síntese da proteína YopH, em culturas
bacterianas de E. coli EPEC, transformadas com o plasmídeo
pTZ/YopH (seta).
75
Figura 3. Co-expressão das proteínas YopH (seta) e SycH (asterico). A-
SDS/PAGE, B- Western-blot.
76
Figura 4. Expressão da proteína YopH, em EPEC, na presença de glicose
2%, arabinose 2% e IPTG 0,1mM.
77
ARTIGO 3
Figura 1. Western-blot com soro de coelho anti-A1122 e a proteína F1
(linha 1) e proteínas totais da cepa P. CE 882 (linha 2); soro de
coelho anti-Y. pseudotuberculosis IP 32950 e a proteína F1 (linha
3).
87
Figura 2. Western-blot com proteínas totais da cepa P. PB 881 e soros
humanos HA+.
88
Figura 3. Proteínas totais de E. coli contendo o plasmídio pTZ/YopH
separadas por SDS/PAGE e coradas pelo Azul de Coomassie (A);
Western-blot com soro de coelho anti-YopH (B).
89
Figura 4. Western-blot com proteínas totais de E. coli contendo o plasmídio
pTZ/YopH e soros de coelho.
90
Figura 5. Western-blot das proteínas totais de E. coli contendo o plasmídio
pTZ/YopH e soros de coelho anti-YopH e soros humanos HA-.
91
III

LISTA DE ABREVIATURAS A/E: attaching/effacing
BFP: Fímbria de aderência
cAMP: Adenina monofosfato cíclica
CAP: Proteína ativadora de catabólito
DAG: Diacilglicerol
Dps: Proteína de ligação ao DNA em células da fase estacionária EPEC: Escherichia coli enteropatogênica
Esps: Proteínas secretadas de EPEC
F1: Fração 1
Fis: Fator para estimulação inversa
HA: Hemaglutinação
HI: Inibição da Hemaglutinação
H-NS: Proteína de estruturação do nucleóide similar as histonas
IHF: Fator de integração ao hospedeiro
IL-8: Interleucina 8
IP3: Inositol trifosfato
IPTG: Isopropil-tio-β-D-galactopiranosídeo
LA: Aderência localizada
LEE: Locus para enterócitos effacement”
M: “Microfold”
MLK: Miosina quinase de cadeia leve
NF-κB: Fator nuclear κB
OMS: Organização Mundial de Saúde
ORF: Quadro de leitura aberto
Pb: Pares de base
PIP2: Fosfatidilinositol-4,5-bifosfato
PKC: Proteína quinase C
PMNs: Leucócitos Polimorfonucleares
PP: Placas de Peyer
PTPase: Proteína tirosina fosfatase
pYV: Plasmídio de virulência de Yersinia
IV

SPI: Ilha de patogenicidade de Salmonella
StpA: Supressor do fenótipo A do mutante td
Sycs: Chaperona Yop específica
TNF-α: Fator de Necrose tumoral alfa
TTS: Secreção Tipo III
TTSS: Sistema de Secreção Tipo III
UP: anterior
Yops: Proteínas externa de Yersinia
Ysc: Secreção Yop
V

RESUMO
Bactérias do gênero Yersinia possuem um sistema de secreção tipo III, responsável pela
translocação de fatores protéicos conhecidos como Yops, para o interior de células
eucarióticas do hospedeiro. A proteína YopH é uma tirosina fosfatase que atua
desfosforilando moléculas sinalizadoras e impedindo a fagocitose em macrófagos. Para
uma secreção eficiente, ela necessita da presença da chaperona SycH que permite o seu
reconhecimento pela maquinaria de secreção. Com a finalidade de analisar a expressão
da proteína YopH em E. coli, foi construído um promotor híbrido formado pelo
promotor lac de Escherichia coli seguido do promotor yopH de Yersinia enterocolitica.
Este promotor foi capaz de direcionar uma forte expressão da YopH, em E. coli, e uma
síntese ótima de YopH foi observada com o aumento da temperatura (37oC) e em
culturas em fase de crescimento. A expressão da YopH foi investigada em linhagens
defectivas de E. coli para as proteínas homólogas às histonas, Hha e H-NS. A expressão
foi significantemente maior a 24oC que a 37oC, na ausência de H-NS, mas não na
ausência de Hha. Estes resultados são compatíveis com o papel da H-NS na regulação
da atividade do promotor yoph, possivelmente através da presença de curvas em sua
estrutura. Para determinar se a presença da chaperona SycH influenciaria na expressão
da YopH, em E. coli, a sequência codificadora para esta proteína foi amplificada e
clonada no plasmídio pBAD33. Foi observada a expressão da SycH em culturas de E.
coli, apenas na presença de arabinose. A co-expressão das proteínas YopH e SycH, em
EPEC, revelou uma redução na expressão da YopH, na presença de arabinose. O mesmo
foi observado em culturas de EPEC contendo apenas o plasmídio pTZ/YopH, onde
concentrações variadas de arabinose e glicose estariam influenciando a expressão da
YopH. Para verificar o potencial da proteína YopH no diagnóstico da peste, diversos
soros de pacientes suspeitos de peste e coelhos imunizados com Y. pestis foram testados
por Western-blot. Não houve reconhecimento da YopH nos soros testados.
Palavras chave: YopH, SycH, Yersinia, H-NS, F1, Diagnóstico
VI

ABSTRACT
Yersiniae possess a type III secretion system, responsible for the translocation of
proteins known as Yops, to the interior of the eukaryotic cells of the host. The YopH
protein is a tyrosine phosphatase that desphosphorylates signalling molecules and
impairs phagocytosis in macrophages. For its efficient secretion it needs the presence of
the chaperone SycH that allows its recognition by the secretion machinery. With the
purpose to analyse the expression of the YopH protein, a hybrid promoter was
constructed with the lac promoter of Escherichia coli followed by the yopH promoter of
Yersinia enterocolitica. This promoter was capable to direct a strong expression of the
YopH in E. coli, and an optimal synthesis of YopH was observed with increase in
temperature (37ºC) and in cultures in growth phase. The expression of the YopH was
investigated in E. coli strains defective in proteins homologous to the histones, Hha and
H-NS. The expression was significantly higher at 24ºC than to 37ºC, in the absence of
H-NS, but not Hha. These results are compatible with the role of the H-NS in the
regulation of the activity of the promoter yoph, possibly through the presence of curves
in its structure. To determine if the presence of the chaperone SycH would influence the
expression of the YopH, in E. coli, the sequence coding for this protein was amplified
and cloned in the plasmid pBAD33. The expression of SycH was observed in cultures
of E. coli, only in presence of arabinose. The co-expression of the proteins YopH e
SycH, in EPEC, resulted in reduction of the expression of the YopH, in presence of
arabinose. The same was observed in EPEC cultures with only the plasmid pTZ/YopH,
and concentration of arabinose and glucose would be acting in the expression of the
YopH. To evaluate the potential of the YopH protein in the diagnosis of plague, several
sera from plague suspect patients and rabbits immunized with Y. pestis strains had been
tested by Western-blot. The tested sera did not recognize YopH.
Keywords: YopH, SycH, Yersinia, H-NS, F1, Diagnostic
VII

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
INTRODUÇÃO
O sistema de secreção tipo III (TTSS) é um mecanismo utilizado por bactérias
Gram-negativas, para translocar proteínas diretamente no citosol de células eucarióticas.
Bactérias do gênero Yersinia e a Escherichia coli enteropatogênica (EPEC), utilizam
este aparato secretório para injetar toxinas no interior de macrófagos e leucócitos
polimorfonucleares (PMNs), como mecanismo de defesa contra a resposta imune
(HUECK, 1998).
O gênero Yersinia possui 3 espécies patogênicas para humanos e outros animais:
Y. pestis, o agente da peste; e a Y. enterocolitica e Y. pseudotuberculosis,
enteropatógenos responsáveis pela yersiniose. Para sobreviver e se multiplicar nos
tecidos linfóides, estas bactérias necessitam de um plasmídio de virulência chamado
pYV (Plasmid Yersinia Virulon), que codifica as proteínas responsáveis pelo TTSS, as
proteínas Yops (Yersinia Outer Proteins) e suas chaperonas Sycs (Specific Yop
Chaperone). Entre as principais Yops, encontra-se a YopH, uma tirosina fosfatase que
participa do bloqueio da fagocitose e indução da apoptose. Para a sua secreção, é
necessária uma chaperona específica, chamada SycH, no citoplasma bacteriano, que
previne a degradação das Yops, mantendo-as em conformação apropriada para a
secreção (CORNELIS, 2002a, 2002b; CORNELIS et al., 1998; PERRY;
FETHERSTON, 1997).
A patogenicidade de E. coli enteropatogênica é conferida pela presença de uma
ilha de patogenicidade, formada pela região LEE (locus for enterocyte effacement).
LEE é responsável pela codificação dos componentes do TTSS, as proteínas Esps
(EPEC-secreted proteins) e suas chaperonas. As Esps são fatores similares as Yops, que
promovem rearranjos no citoesqueleto da célula do hospedeiro e induzem a apoptose
(GROISMAN, 2001; WOLFF et al., 1998).
A regulação da transcrição é um dos principais mecanismos de controle da
expressão gênica em procariotas e alterações na estrutura do DNA podem modular a
ativação ou repressão de genes. A proteína H-NS (Histone-like nucleoid-structuring
protein) apresenta uma grande homologia com as histonas eucarióticas, sendo capaz de
interagir com regiões curvas do DNA. Em EPEC, essa proteína interage com sequências
regulatórias e influencia a expressão dos genes situados na região LEE. A expressão das
Yops, em Yersiniae, é ativada por temperatura (37oC) e controlada pela proteína virF,
1

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
também codificada no pYV. A síntese de virF é regulada pela proteína cromossomal
YmoA, que apresenta uma grande homologia com a proteína Hha e está envolvida na
modulação da expressão da toxina α-hemolisina, em E. coli (MACLEOD; JOHNSON,
2001; CORNELIS et al., 1998; MOURIÑO et al., 1996; PUENTE et al., 1996;
LAMBERT DE ROUVROIT et al., 1992; OWEN-HUGHES et al., 1992).
Neste trabalho, foi analisada a expressão da proteína YopH direcionada por um
promotor híbrido consistindo do promotor lac de E. coli seguido do promotor yopH de
Y. enterocolitica. Foram identificados múltiplos fatores que regulam a transcrição da
YopH, em diversas cepas de E. coli e em linhagens defectivas para as proteínas Hha e
H-NS. A expressão da proteína SycH foi realizada em E. coli enteropatogênica, e a co-
expressão da YopH e SycH também foi avaliada. A proteína YopH também foi
analisada quanto ao seu potencial para o diagnóstico da peste.
2

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Revisão da Literatura
1. Gênero Yersinia
Pertencente à família Enterobacteriaceae, o gênero Yersinia possui 11 espécies
das quais apenas 3 são patogênicas para animais e humanos: Y. pestis, Y.
pseudotuberculosis e Y. enterocolitica. Embora estas bactérias utilizem diferentes vias
de infecção e causem doenças distintas, elas possuem um organotropismo pelo tecido
linfático, onde proliferam como patógenos extracelulares, apesar da presença de células
imunes (FOULTIER et al., 2003; PERRY; FETHERSTON, 1997).
A Y. pestis é o agente etiológico da peste, doença primária de roedores que é
geralmente transmitida pela picada de uma pulga contaminada, podendo ocasionalmente
infectar outros mamíferos, inclusive o homem. A doença pode se apresentar em
diferentes formas clínicas, dependendo da via de transmissão e do curso da infecção
(Figura 1). Quando a infecção atinge os linfonodos regionais, a reação inflamatória
constitui um sinal clássico da forma bubônica da peste (bubão). A infecção pode evoluir
para o sangue periférico e causar septicemia, podendo atingir o baço, fígado e pulmões.
A forma mais perigosa é a peste pneumônica, uma vez que a disseminação ocorre via
aerossóis, além de ser mais letal por se espalhar rapidamente (GROISMAN, 2001;
PERRY; FETHERSTON, 1997).
Ao contrário da Y. pestis, que é um parasita obrigatório, Y. enterocolitica e Y.
pseudotuberculosis podem sobreviver fora dos seus hospedeiros como patógenos
alimentares. Elas são adquiridas por via oral, através da ingestão de alimentos e água
contaminadas, e provocam uma variedade de síndromes gastrointestinais. Y.
pseudotuberculosis é um patógeno que raramente causa doença em humanos e sua
manifestação mais comum é a linfadenite mesentérica, caracterizada por dor abdominal
e febre, geralmente benigna. A forma mais grave é a septicêmica, que é muito rara e
ocorre em pacientes com doenças subjacentes, tais como diabetes e cirrose hepática. Já
a Y. enterocolitica, que é a mais prevalente em humanos, causa uma grande variedade
de quadros clínicos que são determinadas em parte pela idade do hospedeiro, pelo
sorogrupo da cepa de Y. enterocolitica invasiva e pela condição imunológica do
hospedeiro (BERGMANN et al., 2001; CORNELIS et al., 1998; BOTTONE, 1997;
LJUNGBERG et al., 1995; CORNELIS et al., 1987).
3

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Para provocarem a infecção, Y. enterocolitica e Y. pseudotuberculosis precisam
cruzar a barreira intestinal (Figura 1), através de células epiteliais especializadas
chamadas células M. Estas células estão associadas com a camada epitelial linfóide e
com agregados de folículos linfóides chamados placas de Peyer (PP). Esta invasão leva
ao recrutamento de leucócitos polimorfonucleares, formação de micro-abscessos,
aparecimento de células apoptóticas, culminando na completa destruição da
citoarquitetura das Placas de Peyer. Os macrófagos restringem a multiplicação da
Yersinia nos tecidos do hospedeiro, permitindo o desenvolvimento de uma resposta
imune protetora. Entretanto, a Yersinia possui mecanismos que impedem a fagocitose,
inibindo o macrófago, ativando a apoptose e suprimindo a liberação de TNF-α (Fator de
necrose tumoral alfa) e outras citocinas (CORNELIS et al., 1998; AUTENRIETH;
FIRSCHING, 1996).
4

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 1 – Rotas de infecção das três espécies de Yersinia patogênicas. Y. pestis entra
no organismo do hospedeiro pela picada de uma pulga e espalha-se para os nódulos
linfáticos, onde forma bubões. Do sistema linfático, ela pode alcançar o sangue
periférico e eventualmente os pulmões, causando a peste pneumônica, que pode ser
transmitida via aerossóis. Y. enterocolitica e Y. pseudotuberculosis são patógenos
adquiridos por via oral que entram no hospedeiro via trato intestinal e passam através
das células M para as placas de Peyer (PP). Y. pseudotuberculosis pode também atingir
o sangue periférico. Fonte: Groisman (2001).
5

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
1.1 Aspectos Moleculares de Yersiniae
Grande parte da patogenicidade das três espécies de yersínias patogênicas
depende da presença do plasmídio pYV, com peso molecular de 70 kb (Figura 2). Este
plasmídio é responsável pela codificação da adesina YadA (inativa em Y. pestis); uma
lipoproteína YlpA; as proteínas antifagocíticas Yops; o aparato secretório Ysc (Yop
secretion); as chaperonas Sycs; e um sistema regulatório Lcr, que compreende os genes
vir (CORNELIS, 2002a, 2000; CORNELIS et al., 1998).
Cepas típicas de Y. pestis possuem, além do pYV, outros dois plasmídios de
virulência, pFra e pPst, responsáveis pelo aumento da virulência e transmissão pelo
vetor. O plasmídio pFra possui aproximadamente 100 kb e codifica uma proteína
capsular imunogênica F1 (Fração 1), que aumenta a resistência de Y. pestis contra
macrófagos e neutrófilos; e a toxina murina, envolvida na transmissão pelas pulgas. O
pPst possui cerca de 9,5 kb e codifica uma bacteriocina denominada pesticina, ativa
contra algumas cepas de Yersinia e E. coli; uma proteína que confere imunidade a
pesticina; um ativador de plasminogênio e uma coagulase, essenciais para a transmissão
da peste pela pulga (PUJOL; BLISKA, 2005; ANISIMOV et al., 2004; PERRY;
FETHERSTON, 1997).
No cromossomo das yersínias altamentes patogênicas, Y. pestis, Y. enterocolitica
biotipo 1B e Y. pseudotuberculosis sorogrupos I e III, foram identificadas proteínas
necessárias para a captação de ferro, mediada por um sideróforo denominado
Yersiniabactina. Essa região é conhecida como ilha de patogenicidade e apresenta um
conteúdo GC diferente do resto do cromossomo do hospedeiro, sugerindo uma origem
por transferência horizontal entre diferentes gêneros bacterianos (CARNIEL, 2001;
HACKER et al., 1997; FETHERSTON et al., 1992).
6

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 2 – Mapa do plasmídio pYV, de Yersinia enterocolitica. Estão
descritos os genes codificados pelo plasmídio e as funções de seus
produtos protéicos. Fonte: Groisman (2001).
7

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
1.2 Diagnóstico da peste
A peste está incluída entre as doenças reemergentes, em vista do aumento de
casos mundiais e da ocorrência de epidemias em vários países como Índia, Zaire,
Tanzânia, Madagascar, Equador e Peru. Devido à alta taxa de letalidade da Y. pestis e
seu potencial para o terrorismo biológico, a peste é classificada como doença de
notificação Classe I, pelo Regulamento Sanitário Internacional vigente. Em 2003, foram
notificados pela Organização Mundial de Saúde (OMS), 2.118 casos de peste com 182
óbitos em 9 países. Estes dados podem estar subestimados, devido a uma vigilância
inadequada e as dificuldades de diagnóstico laboratorial, sendo a prevalência
provavelmente maior nos países em desenvolvimento (WHO, 2004; INGLESBY et al.,
2000).
O diagnóstico laboratorial confirmatório de peste baseia-se no isolamento da Y.
pestis de material clínico ou pela demonstração de diferenças nos títulos de anticorpos
em amostras pareadas de soro. Para cultivo e isolamento do bacilo, amostras de sangue,
esputo ou aspirado de linfonodo devem ser coletadas antes do início do tratamento com
antibióticos. Algumas vezes, o crescimento da Y. pestis é mascarado por outras bactérias
presentes na amostra, tornando-se necessário recorrer à inoculação em camundongos
susceptíveis. Este procedimento requer laboratórios especializados e um acréscimo de 3
a 5 dias para conclusão do diagnóstico. Os sistemas de provas bioquímicas
automatizadas, usualmente empregados em laboratórios de diagnóstico, não têm se
mostrado confiáveis para a identificação de Y. pestis (PERRY; FETHERSTON, 1997).
A prova de hemaglutinação (HA) utiliza hemácias de carneiro sensibilizadas
com a proteína do envoltório capsular F1, para a detecção de anticorpos anti-F1 e vem
sendo usada há várias décadas para o diagnóstico e vigilância da peste humana e de
animais, em todos os focos do mundo. Esta técnica apresenta alguns inconvenientes
como: complexidade, emprego de reagentes perecíveis, baixa sensibilidade, entre
outros. Os resultados de HA positivos devem também ser confirmados pela prova de
inibição da hemaglutinação (HI). Amostras de soro pareadas (fase aguda e
convalescença) devem mostrar uma diferença de quatro títulos ou mais. Na maioria dos
pacientes, o aparecimento de anticorpos detectáveis pela HA ocorre dentro de 1 a 2
semanas do início dos sintomas, em alguns isto acontece mais cedo (5 dias) e outros não
produzem anticorpos hemaglutinantes (ARAGÃO et al., 2002; CHU, 2000; ALMEIDA
et al., 1989).
8

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
2. Escherichia coli enteropatogênica
E. coli enteropatogênica é a causa principal de diarréia infantil no mundo,
predominantemente em crianças menores de seis meses nos países desenvolvidos. Essa
doença é caracterizada por uma prolongada diarréia aquosa de severidade variada, com
vômito e febre baixa, freqüentemente acompanhada por perda de fluídos (BALDWIN,
1998; VALANCE; FINLAY, 2000).
Os mecanismos moleculares que provocam a diarréia ainda não estão bem
definidos, embora seja conhecido que a EPEC coloniza a superfície intestinal e exerce
seus efeitos patogênicos por ligação íntima com a superfície de enterócitos. Isto provoca
alterações histopatológicas específicas nos sítios de fixação conhecidas como A/E
(attaching/effacing), com dissolução da superfície intestinal e perda das
microvilosidades epiteliais. Nesses sítios, a sustentação da EPEC ocorre sobre projeções
em forma de pedestais, formadas por rearranjos no citoesqueleto de actina das células
do hospedeiro. Essa associação leva a uma perda da superfície absortiva, o que tem sido
associado à diarréia (CELLI et al., 2000; KAPER, 1998; DONNENBERG et al., 1997).
2.1 Aspectos Moleculares
A EPEC possui uma ilha de patogenicidade de 35 kb, formada pela região LEE
(locus for enterocyte effacement), que codifica cerca de 40 ORFs e apresenta um baixo
conteúdo G+C (38% em comparação aos 51% de E. coli). LEE pode ser dividida em
três regiões funcionais que codificam os componentes do sistema de secreção tipo III,
proteínas necessárias para uma íntima ligação com a célula hospedeira (Tir, CesT e
Intimina) e as proteínas Esps, que são secretadas pelo TTSS, com suas respectivas
chaperonas. As Esps são fatores específicos, similares as Yops das yersínias, que ativam
rearranjos no citoesqueleto da célula do hospedeiro e induzem apoptose (GROISMAN,
2001; WOLFF et al., 1998).
O desenvolvimento das lesões A/E pode ser dividido em três estágios (Figura 3).
O estágio inicial é caracterizado pela fixação da EPEC na superfície de células
epiteliais, caracterizada por uma aderência localizada (LA). A expressão de fatores de
aderência direciona a síntese de uma fímbria do tipo IV (BFP), associada com
interações interbacterianas. A formação de BFP, em cultura, ocorre durante a fase
exponencial de crescimento e pode ser modulada por concentrações de amônio e
9

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
temperatura (37oC). No segundo estágio, ocorre a secreção e translocação das proteínas
Esps, através do TTSS, responsáveis pela ativação dos caminhos de transdução de sinal.
A proteína Tir também é secretada e promove rearranjos no citoesqueleto da célula que
culminam na formação de um pedestal sob a EPEC. No terceiro estágio, a proteína
Intimina, que possui homologia com a invasina de Yersinia, promove uma fixação mais
íntima entre a EPEC e a célula do hospedeiro, através da interação com a proteína
translocada Tir, que é inserida na membrana eucariótica e serve como receptor para o
reconhecimento da Intimina (GROISMAN, 2001; KAPER, 1998; RAMER et al., 1996;
GIRON et al., 1991).
Os genes que codificam o TTS estão organizados em cinco unidades
policistrônicas (LEE1 a LEE5) que são reguladas pelo gene ler, excetuando LEE1. A
proteína Ler apresenta similaridade com a proteína H-NS, uma proteína histona-like que
regula negativamente vários fatores de virulência. Ler compete com H-NS pelas
mesmas sequências regulatórias localizadas antes da região –35 e regula positivamente
a expressão destes genes (BUSTAMANTE et al., 1999; ELLIOTT et al., 1998;
PUENTE et al., 1996).
10

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 3 – Interações da EPEC com células epiteliais. A fixação inicial das
microcolônias de EPEC na superfície da célula epitelial é mediada pela fímbria do tipo
IV, conhecida como BFP. Após a interação inicial, ocorre a secreção e translocação das
proteínas EspA, EspB, EspD e Tir, para o interior do citoplasma da célula do
hospedeirto, que é mediada pelo sistema de secreção tipo III. Proteínas EspA se
arranjam em uma grande organela filamentosa que é essencial para a translocação de
EspB. Após a translocação, Tir é fosforilada e inserida dentro da membrana plasmática,
onde liga a proteína de superfície externa Intimina. Neste estágio, Tir interage com
componentes do citoesqueleto incluindo actina, α-actina, talina e ezrin; e provoca
rearranjos no citoesqueleto que formam um pedestal rico em actina, localizado abaixo
da EPEC aderida. Proteínas translocadas disparam a ativação dos caminhos de
transdução de sinal que levam a uma resposta celular complexa. Aumento da atividade
da proteína associada a membrana Quinase C (PKC), estimula a secreção de Cl-.
Fosfolipase Cγ tirosina fosforilada converte fosfatidilinositol-4,5-bifosfato (PIP2) em
inositol trifosfato (IP3) e diacilglicerol (DAG), os quais estimulam a liberação de cálcio
intracelular (Ca2+) de estoques IP3 sensitivos. Ativação do fator transcricional NF-κB
inicia a transcrição de interleucina 8 (IL-8), a qual estimula a transmigração de células
polimorfonucleares. A ativação de miosina quinase de cadeia leve (MLK) leva a
fosforilação de junções compactas e aumento da permeabilidade intestinal. Fonte:
Groisman (2001).
11

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
3. Sistema de Secreção Tipo III e seus elementos
Para garantir a sobrevivência extracelular, as bactérias do gênero Yersinia e as
EPEC utilizam o sistema de secreção tipo III para secretar e translocar determinantes de
virulência no interior de células eucarióticas. Estas toxinas interferem com diversas
reações bioquímicas, bloqueando a fagocitose, suprimindo TNF-α e outras citocinas e
ativando a apoptose (CORNELIS, 2002b; CORNELIS et al., 1998; HUECK, 1998;
AUTENRIETH; FIRSCHING, 1996; ROSQVIST et al., 1994).
Mais de 20 sistemas TTS já foram descritos em patógenos de animais, plantas e
insetos; e as proteínas secretadas podem desempenhar papéis totalmente diferentes. A
bactéria Salmonella typhimurium, por exemplo, apresenta dois tipos de TTS (SPI para
“Salmonella pathogenicity island”), que atuam de maneira distinta na patogênese:
enquanto SPI-1 é necessária para a penetração inicial na mucosa intestinal, SPI-2 atua
nos estágios subseqüentes da infecção. Análises filogenéticas recentes demonstraram
que o TTS parece ter sido adquirido por transferência horizontal, uma vez que estes
sistemas são sempre encontrados em plasmídios, no caso das yersínias, ou em discretas
ilhas de patogenicidade cromossomais, como em EPEC (CORNELIS, 2002a; HUECK,
1998).
Antes do contato com células eucarióticas, bactérias incubadas na temperatura
do hospedeiro (37oC) produzem muitas unidades do TTS, presentes na superfície
bacteriana. Este aparato secretório é uma organela conhecida como injectisomo (Figura
4) que, em yersínia, é formada por 27 proteínas Ysc, e consiste de dois pares de anéis
ancorados na membrana interna e externa do envelope bacteriano e unidos por uma
haste central, formando o corpo basal. Esta estrutura suporta uma agulha oca com 10 nm
de espessura e 60 nm de largura, com um poro central de cerca de 50 Å, que se projeta
para o exterior da bactéria. Várias das proteínas Ysc que formam o corpo basal do
aparato secretório, compartilham significante homologia de aminoácidos com proteínas
do corpo basal do flagelo, sugerindo uma origem evolucionária comum (JOURNET et
al., 2003; ALDRIDGE; HUGHES, 2002; CORNELIS, 2002a; CORNELIS et al., 1998).
A parte mais interna do corpo basal (Figura 4) inclui a proteína YscN, uma
ATPase que se polimeriza para formar a parte mais baixa do cilindro; e as proteínas que
estão em contato com a membrana citoplasmática YscV, YscU, YscR, YscT, YscS e
YscQ. A parte distal é uma estrutura em forma de anel, formada pela polimerização da
proteína YscC, associada com a lipoproteína YscW. A parte externa do injectissomo é
12

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
composta das proteínas YscO, YscP, YscX e YscF, sendo este o principal constituinte
da agulha (CORNELIS, 2002a).
O comprimento do injectisomo é controlado geneticamente pela proteína YscP,
que controla o crescimento da agulha durante a formação do injectisomo. Linhagens de
Y. enterocolitica contendo grandes deleções no gene yscp, apresentaram estruturas com
comprimento indefinido. A complementação utilizando o gene yscp de Y. pestis, o qual
possui 90% de identidade, resultou na restauração do controle do comprimento e
formação de agulhas curtas. Dessa forma, foi observado que deleções afetando as
regiões N-terminal e C-terminal da proteína YscP, levam a perda de controle do
comprimento, provavelmente por servirem como âncoras. Além disso, YscP também
pode atuar como um sinal interno, determinando o comprimento ideal da agulha e
interrompendo a exportação de susbstratos como YscF e outros (THOMAS; FINLAY,
2004; JOURNET et al., 2003).
13

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 4 – Injectisomo Ysc. a. Representação esquemática do injectisomo Ysc
atravessando a membrana externa (OM), a camada peptídeoglicana (PG) e a membrana
citoplasmática (CM) da bactéria. O anel situado na OM é composto da secretina YscC,
associada pela lipoproteína YscW. YscJ é outra lipoproteína. YscF, YscO, YscP e YscX
são partes externas do injectisomo. YscF é o principal constituinte da agulha. YscV,
YscU, YscR, YscT e YscS como proteínas do corpo basal estão em contato com CM.
YscN é uma ATPase. YscQ está provavelmente localizada no cilindro interno. b.
Microscopia eletrônica mostrando as agulhas do injectisomo projetadas para o exterior
da Yersinia enterocolitica E40. Fonte: Cornelis (2002a).
14

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
O injectisomo Ysc é responsável pela secreção e translocação de proteínas
antifagocíticas (Yops) para o interior da célula do hospedeiro. A expressão dessas
proteínas ocorre pela ação da temperatura (37oC), mas a translocação só ocorre por
contato com células eucarióticas, diminuição de íons Ca2+ ou mutações em genes que
controlam a liberação das Yops (yopN, tyeA e lcrG). A expressão das Yops é controlada
por proteínas codificadas por vários loci chamados vir, que compõem uma região com
cerca de 20 Kb no pYV (IRIARTE et al., 1998; SARKER et al., 1998; SKRZYPEK;
STRALEY, 1993; FORSBERG et al., 1991; CORNELIS et al., 1986).
O virF é um regulador transcricional, da família AraC, que ativa os genes yop e
outros loci vir, quando a bactéria é colocada a 37ºC. Este ativador reconhece e se liga a
regiões específicas ricas em AT, localizadas antes do sítio de ligação da RNA
polimerase, que podem estar isoladas ou repetidas em orientação oposta. O VirF, por
sua vez, tem a sua síntese regulada tanto por temperatura como por uma proteína
chamada YmoA, que é codificada por um gene cromossomal e apresenta características
semelhantes às proteínas histonas de eucariotos, estando envolvida na compactação da
estrutura cromossomal do DNA (CORNELIS et al., 1998; CORNELIS et al., 1989;
WATTIAU, CORNELIS, 1994; LAMBERT DE ROUVROIT et al., 1992).
O reconhecimento específico das Yops pelo injectisomo é mediado por um sinal
de secreção (Figura 5), que foi inicialmente descrito nos primeiros 15 aminoácidos de
muitas Yops. Entretanto, substituições de aminoácidos em várias posições não afetaram
a eficácia do sinal de secreção e nenhuma homologia foi encontrada entre os resíduos de
várias Yops analisadas. Uma vez que as modificações alteravam apenas a proteína, foi
levantada a hipótese de que o sinal poderia estar na sequência do mRNA. Entretanto, as
dificuldades em distinguir similaridades de sequência ou estruturas secundárias entre
mRNAs de diferentes Yops, sugeriram a existência de um terceiro sinal de secreção
mediado pelas proteínas chaperonas específicas das Yops (HE et al., 2004;
RAMAMURTH; SCHNEEWIND, 2003).
As proteínas Sycs são moléculas acessórias e sua ausência resulta numa secreção
deficiente do substrato. Foram identificadas seis chaperonas: SycE, SycT, SycN, SycD,
SycH e SycO/ORF155; que atuam por reconhecimento e ligação de sítios localizados na
sequência de aminoácidos N-terminais, embora essa região não apresente muita
homologia entre as Yops. As Sycs tem a função de prevenir a degradação das Yops,
mantê-las em conformação apropriada e guiá-las do local de síntese até a maquinaria de
secreção; colaborando para uma secreção mais eficiente (FELDMAN; CORNELIS,
15

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
2003; CORNELIS, 2002b; ALDRIDGE; HUGHES, 2001; IRIARTE; CORNELIS,
1999; CORNELIS et al., 1998; IRIARTE; CORNELIS, 1998; WATTIAU et al., 1994;
WATTIAU; CORNELIS, 1993).
16

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 5 – Diagrama dos sinais de secreção tipo III e funções das chaperonas. Três
tipos de sinais de secreção foram descritos: 1, região 5’do RNA mensageiro (estrutura
secundária em forma de alça, indicada em azul); 2, chaperona ligando o domínio de
ligação da proteína efetora (chaperona homodimérica indicada por duas ferraduras e o
domínio de ligação indicado por uma curta linha preta); 3, região N-terminal do
peptídeo (indicado por uma curta linha vermelha). Ligação das chaperonas nas efetoras
cognatas permite a manutenção de uma conformação apropriada para a secreção,
prevenindo agregação e/ou formação de complexos no citoplasma bacteriano. Algumas
chaperonas podem ter função na regulação gênica. A associação das efetoras com as
chaperonas pode favorecer a secreção, sendo estas proteínas secretadas primeiro. A
efetora assume sua conformação final dentro da célula do hospedeiro e as chaperonas
permanecem no citoplasma bacteriano. Fonte: He et al. (2004).
17

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
As proteínas Yops incluem YopE, YopH, YopM, YopO (YpkA), YopP (YopJ) e
YopT, os quais são efetores intracelulares, YopB e YopD, que funcionam como
proteínas translocadoras e a YopK/YopQ, que funciona como fator modulador. As
proteínas YopB e YopD agem formando um poro na membrana da célula eucariótica,
através do qual outras Yops são translocadas para dentro do citosol da célula alvo. O
papel da YopK/YopQ, nesse processo, é o de controlar a quantidade de Yops secretadas
por influência no tamanho do poro induzido (CORNELIS, 2002b; ANDERSON;
SCHNEEWIND, 1999; HUECK, 1998).
O processo de “injeção” requer um contato muito íntimo entre as células, o que é
mediado por adesinas bacterianas e integrinas da célula do hospedeiro (Figura 6). As
proteínas YopB e YopD atuam promovendo uma desestabilização da membrana do
hospedeiro, permitindo que ocorra a perfuração da membrana e a passagem das Yops
efetoras. A função dessas proteínas efetoras é paralisar todos os mecanismos de defesa
celulares, contribuindo para o enfraquecimento da fagocitose por perturbar a dinâmica
do citoesqueleto (YopH, YopE, YopT, YopO); agir como GTPases monoméricas da
família Rho (YopE, YopT, YopO); promover a desfosforilação de adesões focais,
moléculas sinalizadoras e inibição da proliferação linfocítica (YopH); inibir resposta
inflamatória de macrófagos, células endoteliais e epiteliais e ativar a apoptose (YopP);
ou atuar diretamente no núcleo, como no caso da YopM (NAVARRO et al., 2005;
CORNELIS, 2002b; JURIS et al., 2002).
Entre as principais Yops, encontra-se a YopH, uma das mais poderosas proteínas
tirosina fosfatase conhecidas (PTPases), possuindo um peso molecular de 51 kDa, o que
lhe conferiu a sua primeira denominação, Yop51. O seu domínio C-terminal (resíduos
206-408) assemelha-se a fosfatases eucarióticas e contém uma alça de ligação ao fosfato
que inclui um importante resíduo de cisteína (Cys403), cuja mutação torna a enzima
inativa. O domínio N-terminal é bastante similar ao de outras Yops e contém um sinal
de secreção (resíduos 1-17) e o sítio de ligação a chaperona SycH. Sendo a fosforilação
um dos mecanismos básicos de modulação da atividade protéica, a atividade da YopH
representa um importante papel na resistência bacteriana ao sistema imune do
hospedeiro (NAVARRO et al., 2005; CORNELIS, 2002b; CORNELIS et al., 1998;
SORY et al., 1995; GUAN; DIXON, 1990).
18
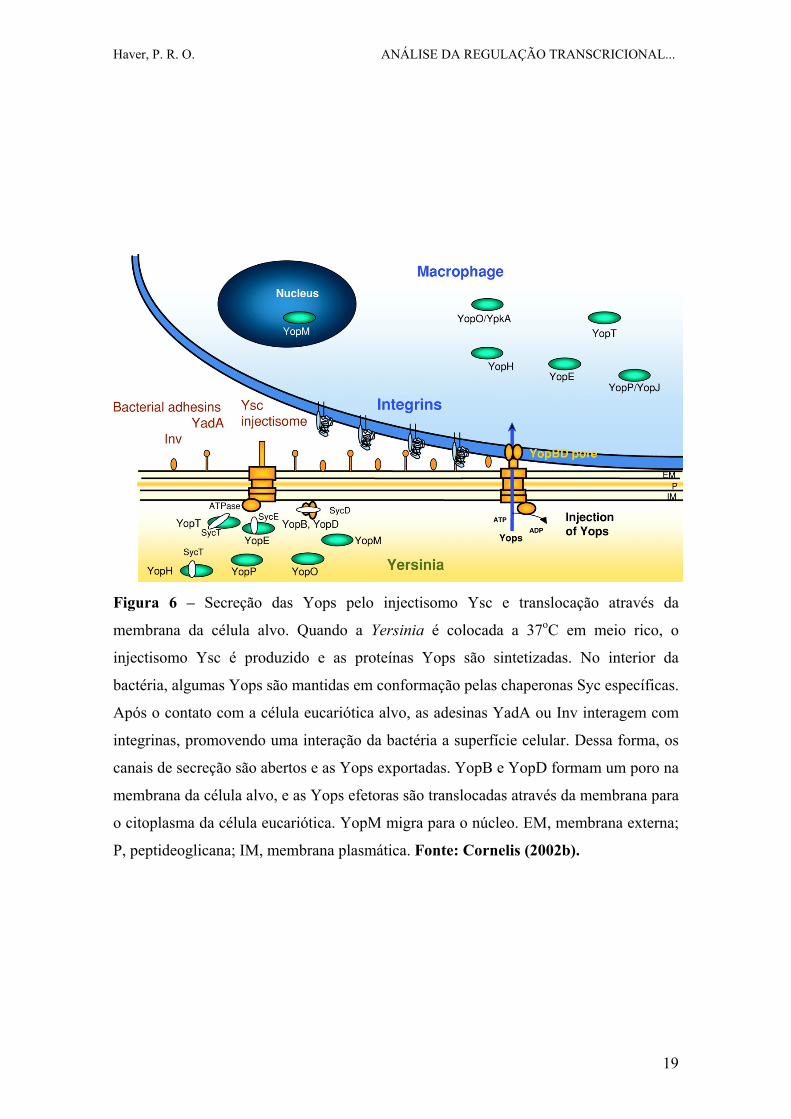
Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 6 – Secreção das Yops pelo injectisomo Ysc e translocação através da
membrana da célula alvo. Quando a Yersinia é colocada a 37oC em meio rico, o
injectisomo Ysc é produzido e as proteínas Yops são sintetizadas. No interior da
bactéria, algumas Yops são mantidas em conformação pelas chaperonas Syc específicas.
Após o contato com a célula eucariótica alvo, as adesinas YadA ou Inv interagem com
integrinas, promovendo uma interação da bactéria a superfície celular. Dessa forma, os
canais de secreção são abertos e as Yops exportadas. YopB e YopD formam um poro na
membrana da célula alvo, e as Yops efetoras são translocadas através da membrana para
o citoplasma da célula eucariótica. YopM migra para o núcleo. EM, membrana externa;
P, peptideoglicana; IM, membrana plasmática. Fonte: Cornelis (2002b).
19

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
4. Regulação Transcricional em Procariotas
Entre todos os mecanismos que controlam a expressão gênica, a regulação da
transcrição tem um papel predominante nas bactérias. Uma das razões é a existência de
um único tipo de RNA polimerase para transcrever todas as diferentes espécies de RNA,
sendo necessário regular sua especificidade. A outra, e talvez mais importante, é que em
bactérias, não há compartimentos como núcleo e citoplasma, e as reações acontecem de
forma simultânea. Conseqüentemente, enquanto uma parte do genoma está sendo
replicada, inicia-se a transcrição e a tradução do RNA nascente. Dessa forma, uma
regulação a nível transcricional é muito mais eficaz, impedindo que RNA’s
desnecessários sejam sintetizados (WAGNER, 2000).
4.1 Os elementos da transcrição
O processo de transcrição é caracterizado pela ligação da enzima RNA
polimerase a uma molécula de DNA dupla hélice, sendo uma das fitas de DNA utilizada
como a sequência molde para a produção de um RNA mensageiro o qual,
posteriormente, será traduzido em proteína (GRIFFITHS et al., 2002).
4.1.1 Região promotora
Para que ocorra a transcrição (Figura 7), é necessário que haja o reconhecimento
de seqüências específicas de DNA, as quais são chamadas de região promotora. A
estrutura geral dos promotores constitutivos é formada por quatro elementos principais:
dois hexâmeros nas posições –35 e –10 anterior ao sítio de início da transcrição, o DNA
espaçador de 17 pb situado entre eles, e elementos UP situados entre –60 e – 40. A
eficiência da transcrição é dependente do nível de correspondência com os elementos
–35 e –10, cujas seqüências são lidas na fita não codificante e correspondem a
TTGACA e TATAAT, respectivamente. Desvios do consenso, provocados por
substituições de um único nucleotídeo, levam a uma redução da atividade promotora
(WAGNER, 2000; deHASETH et al., 1998; HARLEY; REYNOLDS, 1987).
Um grupo especial de promotores funciona sem a presença de uma região –35
ou o envolvimento de proteínas ativadoras. Tais promotores são conhecidos como
promotores –10 estendidos, e parecem ser comuns em genes de virulência. Apresentam
20

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
a sequência TGNTATAAT, caracterizada por um motivo TG localizado antes da região
–10. Em alguns casos, a região –35 pode estar substituída por curvas de DNA;
nucleotídeos CCGACA em vez de TTGACA; e a presença da sequência CTGCAA
substituindo a região –35 (BALLESTEROS et al., 1998; DEHASETH et al., 1998;
DOMBROSKI, 1997; ESPINOSA-URGEL et al., 1996; WISE et al., 1996; KUMAR et
al., 1993; BOHANNON et al., 1991; VICENTE et al., 1991).
Seqüências flanqueadoras ricas em pares de base AT, situadas nas posições –40
e –60 foram reconhecidas em alguns promotores como um importante determinante
para a atividade promotora. Estas regiões foram chamadas de elementos UP e são
responsáveis por um aumento na eficiência da transcrição em aproximadamente 30
vezes. Isso parece ocorrer pela interação desses elementos com a subunidade α da RNA
polimerase, permitindo uma interação mais eficiente (deHASETH et al., 1998; ROSS et
al., 1993).
4.1.2 RNA polimerase
Descrita em 1959, a RNA polimerase é um complexo enzimático constituído por
quatro diferentes subunidades denominadas α, β, β´ e σ. A atividade catalítica reside no
core enzimático formado por duas subunidades α, uma β e uma β´ (α2ββ´), capazes de
promover uma correta elongação do RNA. Já a subunidade σ, é responsável pela
especificidade enzimática, ou seja, o reconhecimento de diferentes tipos de promotores,
não tendo participação na síntese do RNA (GRIFFITHS et al., 2002; ISHIHAMA,
2000; WAGNER, 2000; deHASETH et al., 1998).
A subunidade α apresenta um importante papel na ligação com sequências
anteriores a região promotora presentes em alguns tipos de promotores e na interação
com fatores transcricionais. Já as subunidades β e β´são os dois maiores componentes
da RNA polimerase, constituindo o cerne catalítico da RNA polimerase. A subunidade
β possui pelo menos três sítios de interação próximos ao início do RNA nascente e a
subunidade β´, apresenta um motivo semelhante a dedos de zinco (Zn finger-like) que
permite a ligação ao DNA. Na formação da RNA polimerase, uma molécula da
subunidade α interage com a subunidade β e a outra, com β´; não havendo interação
entre α e σ (WAGNER, 2000, ZHANG; DARST, 1998; HEYDUK et al., 1996).
21

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Existem sete tipos de subunidades sigma (σ), em E. coli, responsáveis por
direcionar a RNA polimerase para grupos de diferentes promotores, e com isso, mudar o
padrão de expressão gênica. O fator específico para o início da transcrição dos genes
constitutivos (housekeeping genes), durante o crescimento exponencial da bactéria, é o
produto do gene rpoD, o fator sigma 70 (σ70). Diferentes homólogos já foram
encontrados e todos compartilham quatro regiões com alto nível de conservação de
seqüência, que estão associadas a diferentes funções: ligação ao núcleo da RNA
polimerase, ligação com o DNA (reconhecimento dos elementos promotores da região
de consenso –10 e –35), desnaturação do DNA e interação com ativadores
transcricionais classe II (DOVE et al., 2003; GAAL et al., 2001; ISHIHAMA, 2000;
GROSS et al., 1998).
Como a especificidade da RNA polimerase depende da presença de fatores σ
específicos, a presença de fatores alternativos permite o reconhecimento de seqüências
promotoras especiais ou fora de consenso. Eles foram primeiro descritos em Bacillus
subtilis e em seus fagos SP01 e SP82, e foram relacionados com alterações do
crescimento do fago e mudanças na programação da transcrição, durante processos de
desenvolvimento como esporulação. Fatores σ específicos são requeridos, por exemplo,
para a transcrição de genes relacionados com a resposta de choque térmico (σH,σE), para
genes importantes da fase estacionária de crescimento (σS), para a expressão de genes
de fixação de nitrogênio (σN), expressão de proteína flagelar (σF), ou a formação de
esporos de bactérias Gram-positivas, como no caso do σF de B. subtilis
(HALDENWANG, 1995; ISHIHAMA, 1993).
Dentre os fatores alternativos existentes, o σS age como o regulator principal de
uma rede que controla a expressão de numerosos genes induzidos na fase estacionária,
em E. coli. A subunidade σS começa a ser acumulada na célula no início da fase
estacionária e a adição de fatores transcricionais específicos associados a uma alta
osmolaridade intracelular, parecem favorecer a transcrição de certos promotores. O fator
σS está envolvido na resposta ao stress oxidativo e osmótico e também é requerido para
genes de virulência, como nas bactérias patogênicas E. coli EHEC e Pseudomonas
aeruginosa (GAAL et al., 2001; BELTRAMETTI et al., 1999; SUH et al., 1999;
HENGGE-ARONIS, 1996; BARTH et al., 1995; HENGGE-ARONIS, 1993; LANGE;
HENGGE-ARONIS, 1991).
22

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Embora não exista uma seqüência consenso definida em promotores
reconhecidos pela subunidade σS, foi sugerido que eles reconheçam uma seqüência
similar à região –10 reconhecida pela σ70, chamada de região –10 estendida. Entretanto,
foi determinado que existe uma sobreposição nas seqüências promotoras reconhecidas
por σ70 e σS (in vitro), sugerindo um papel importante da região –35, pelo menos em
alguns promotores (GAAL et al., 2001; ESPINOSA-URGEL et al., 1996).
23

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 7 – Resumo das interações entre o promotor e a RNA polimerase. A figura
mostra um promotor com sequências consenso –10 e –35, que é reconhecido pelo fator
sigma70,. Em geral, quanto mais próximo um promotor está do consenso, mais forte
será sua atividade promotora. Também está demonstrado a sequência TGN localizada
antes da regiào –10; juntas, elas formam uma região –10 estendida no qual a atividade
promotora ocorre na sequência –35. O modelo da RNA polimerase mostra o contato
entre suas subunidades e o DNA promotor e não necessariamente reflete uma precisa
orientação das subunidades. Esta região da RNA polimerase liga sítios catalíticos de
DNA posteriores à região promotora. Contato entre RNA polimerase e DNA promotor
são demonstrados pelas linhas. Nem todos os contatos ocorrem pela interação promotor-
RNA polimerase, mas em alguns casos (incluindo a ativação de promotores por
proteínas regulatórias), no mínimo, um contato entre a região –10 é requerido. Fonte:
deHaseth et al. (1998).
24

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
4.2 Modelos Clássicos de regulação transcricional
O conceito de operon foi proposto por Jacob e Monod em 1961, e consiste de
uma região do DNA que inclui genes transcritos no mesmo mRNA; sequências
adjacentes necessárias para a transcrição, incluindo regiões promotoras e operadoras;
genes codificadores de fatores protéicos; e outras sequências envolvidas (SNYDER;
CHAMPNESS, 1997).
4.2.1 Operon Lac
O operon Lac é responsável pela síntese dos genes que degradam a lactose e sua
transcrição é iniciada pelo promotor lac, o qual é controlado por uma sequência
chamada operadora, palindrômica, localizada imediatamente após a região –10. Esta
região serve como sítio alvo para a ligação de uma proteína ao DNA, conhecida como
repressor lac (LacI ou LacR), produto do gene regulador lacI, localizado em um sítio
separado mas não muito distante dos genes lac (GRIFFITHS et al., 2002; VILAR et al.,
2003; WAGNER, 2000; SNYDER, CHAMPNESS, 1997).
O controle do operon Lac reside na ligação do repressor LacI na sequência
operadora, com uma afinidade muito maior do que a ligação da RNA polimerase,
bloqueando a transcrição. Entretanto, este mecanismo é reversível, uma vez que a
ligação de uma molécula indutora (alolactose e IPTG) diminui a afinidade do repressor
pela sequência operadora, através de modificações conformacionais alostéricas. O
aumento da transcrição também sofre influência da proteína ativadora de catabólito
chamada CAP que, na ausência de glicose, forma um complexo com cAMP, ligando
sequências específicas no DNA e aumentando a afinidade da RNA polimerase pela
região promotora (VILAR et al., 2003; SNYDER; CHAMPNESS, 2001; WAGNER,
2000; REZNIKOFF, 1992).
4.2.2 Operon araBAD
Um outro exemplo de regulação transcricional pode ser dado pelo operon
araBAD, que codifica um grupo de enzimas responsáveis pela conversão de L-arabinose
para D-xilulose-5-fosfato. A proteína repressora AraC apresenta homologia com a
proteína VirF, de Y. enterocolitica, e outros fatores transcricionais; dando origem a
25

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
família de reguladores semelhantes ao AraC. Esta proteína contém dois sítios de ligação
ao DNA, localizados na região C-terminal, e um sítio de dimerização, de localização N-
terminal. O gene responsável pela sua codificação é o araC, controlado pelo promotor
PC, que se encontra em orientação oposta ao promotor PBAD (GRIFFITHS et al., 2002;
WAGNER, 2000; BROWN; HOGG, 1972).
Para ativar a transcrição dos genes araBAD, a proteína AraC precisa estar ligada
ao indutor L-arabinose, o que ocorre em um domínio próximo a região N-terminal. Esta
ligação permite o reconhecimento de dois sítios formados por 17 pb orientados na
mesma direção, araI1 e araI2. Entretanto, na ausência do indutor, a proteína AraC ainda
é biologicamente ativa, diminuindo a afinidade por araI2 e passando a ligar o sítio
araO2, distante 210 pb do promotor, e que está localizado dentro da região
transcricional de AraC. Essa ligação induz a formação de uma curva de DNA, que
bloqueia a transcrição dos genes araBAD (WAGNER, 2000; HAMILTON; LEE, 1988).
4.3 Fatores que influenciam a regulação transcricional
A característica informacional do DNA não reside apenas na seqüência de
nucleotídeos, mas sua estrutura tri-dimensional também funciona como um sinal
determinante para a transcrição. Alterações na estrutura do DNA permitem que
proteínas reguladoras modulem a acessibilidade a um dado promotor, ativando ou
reprimindo a ligação da RNA polimerase, de acordo com estímulos ambientais. A
habilidade que essas proteínas têm para interagir com o promotor é um resultado direto
da sua afinidade por seqüências de DNA específicas na região promotora. Altas
temperaturas também podem induzir mudanças conformacionais, diminuindo a
afinidade destes fatores pelos sítios de ligação ou ainda modificando a estrutura da
cromatina, alterando curvas de DNA, o que facilita o acesso a regiões promotoras, e
estimula a transcrição (PÉREZ-MARTIN; LORENZO, 1997; RIPPE et al., 1995;
WATTIAU; CORNELIS, 1994; YAMADA et al., 1990).
4.3.1 Curvaturas e dobramentos de DNA
A presença de curvaturas e dobramentos antes dos promotores bacterianos e sua
influência na eficiência da transcrição foram descritas pela primeira vez no início de
1980 e hoje, são reconhecidas como um importante parâmetro na regulação gênica.
26

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Uma vez que a RNA polimerase envolve a molécula de DNA para promover uma
interação, tornam-se necessárias alterações na sua estrutura para formar o complexo
transcricional. Mudanças provocadas por forças externas ou ligação de proteínas são
geralmente chamadas de dobramentos (bends), e são opostas a curvaturas intrínsecas
(curves) que dependem apenas da seqüência de nucleotídeos. Alterações na temperatura
também podem estimular a formação de curvas no DNA, levando a modificações no
comportamento de proteínas regulatórias. Regiões contendo dobramentos podem estar
presentes tanto no núcleo do promotor como em regiões anteriores ao mesmo, de –40 a
–150, e são ricas em AT (DAME et al., 2001; WAGNER, 2000; PÉREZ-MARTIN;
LORENZO, 1997; HAGERMAN, 1990; YAMADA et al., 1990).
4.3.2 Fatores Transcricionais semelhantes às Histonas
Organismos procariotas, diferente de eucariotas, não possuem histonas para
compactar o DNA cromossômico, entretanto apresentam o seu material genético 1000
vezes condensado. Isso ocorre pela presença de proteínas envolvidas na compactação do
DNA bacteriano, chamadas proteínas “histone-like” ou proteínas nucleóides. Elas são
responsáveis pela modulação da estrutura dos cromossomos bacterianos, diferindo das
histonas eucarióticas por formarem complexos instáveis com o DNA. Além disso, são
responsáveis pela expressão de uma variedade de genes que são importantes para a
viabilidade celular, como síntese protéica e metabolismo; além de regular genes cuja
expressão varia em resposta a mudanças ambientais, como temperatura, osmolaridade e
genes que afetam a virulência (MACLEOD; JOHNSON, 2001; WAGNER, 2000).
As proteínas nucleóides mais importantes em E. coli são HU, IHF, H-NS, StpA,
Fis e Dps; e algumas compartilham homologia estrutural e sobreposição de funções. Por
exemplo, HU e IHF apresentam 45% das suas seqüências de aminoácidos idênticas e
possuem estruturas 3-D similares; já StpA compartilha 58% de identidade com H-NS,
sendo capaz de substituí-la em várias reações. Os níveis dessas proteínas variam
consideravelmente em E. coli onde, na fase exponencial, foram encontradas em maior
abundância Fis, HU, IHF, StpA e H-NS; e na fase estacionária, Dps, IHF e HU.
Experimentos de imunofluorescência mostram que as proteínas HU, IHF, H-NS, StpA e
Dps estão distribuídas de maneira uniforme dentro do nucleóide bacteriano
(MACLEOD; JOHNSON, 2001; AZAM et al., 2000; AZAM; ISHIHAMA, 1999;
DORMAN et al., 1999; ATLUNG; INGMER, 1997; WILLIAMS; RIMSKY, 1997).
27

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Entre as várias proteínas associadas com a organização do DNA, a H-NS
constitui o maior componente do nucleóide bacteriano. Ela é uma proteína neutra de
15,6 kDa; e apresenta uma estrutura similar às histonas de eucariotos. Possui dois
domínios funcionais distintos: um domínio responsável pela oligomerização entre
moléculas de H-NS, localizado na região N-terminal e com um baixo nível de
conservação; e um domínio C-terminal, responsável pela ligação com o DNA e
altamente conservado. A proteína H-NS regula a expressão gênica por interação com
estruturas curvas íntrinsecas do DNA, repressão de outras proteínas que regulam a
transcrição ou por indução de dobramentos de DNA (BERTIN et al., 2001; SMYTH et
al., 2000; FALCONI et al., 1998; OWEN-HUGHES et al., 1992; YAMADA et al.,
1990).
4.3.3 Hha
A proteína Hha pertence à mesma família da proteína YmoA, de Y.
enterocolitica, com a qual compartilha 82% de identidade na sequência de aminoácidos.
Em E. coli, esta proteína pode formar complexos com proteínas como a H-NS,
modulando a expressão da toxina α-hemolisina. Mutantes hha, quando submetidos a
baixas temperaturas ou alta osmolaridade, aumentam a expressão da hemolisina. Em Y.
enterocolitica, mutantes ymoA apresentam uma alteração na expressão das Yops e da
adesina YadA, quando submetidos a temperaturas baixas. Uma vez que a proteína H-NS
de E. coli apresenta domínios N-terminais idênticos a H-NS de Y. enterocolitica, a
formação de um complexo YmoA/H-NS pode modular também a expressão de genes de
virulência em yersínia. Dessa forma, tanto a proteína Hha como a proteína YmoA,
funcionam como repressores da expressão de genes bacterianos envolvidos em
processos patogênicos (MADRID et al., 2002; NIETO et al., 2002; MOURIÑO et al.,
1996; MIKULSKIS; CORNELIS, 1994; CARMONA et al., 1993; CORNELIS et al.,
1991; NIETO et al., 1991).
28

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
REFERÊNCIAS BIBLIOGRÁFICAS ALDRIDGE, P., HUGHES, K.T. Regulation of flagellar assembly. Current Opinion
Microbiology, v. 5, n. 2, p. 160-165, 2002.
ALDRIDGE, P.; HUGHES, K.T. How and when are substrates selected for type III
secretion. Trends in Microbiology, v. 9, n. 5, p. 209-214, 2001.
ALMEIDA, A. M. P.; BRASIL, D. P.; LEAL, N. C.; MELO, M. E. B.; REGO, R. V.
B.; ALMEIDA, C. R. Estudos bacteriológicos e sorológicos de um surto de peste no
Estado da Paraíba, Brasil. Memórias do Instituto Oswaldo Cruz RJ, v. 84, p. 249-256,
1989.
ANDERSON, D. M.; SCHNEEWIND, O. Yersinia enterocolitica type III secretion: an
mRNA signal that couples translation and secretion of YopQ. Molecular
Microbiology, v. 31, n. 4, p. 1139-1148, 1999.
ANISIMOV, P. A., LINDLER, L. E., PIER, G. B. Intraspecific diversity of Yersinia
pestis. Clinical Microbiology Reviews, v. 17, n. 2, p. 434-464, 2004.
ARAGÃO, A. I.; SEOANE, A. C.; LEAL, N. C.; ALMEIDA, A. M. P. Vigilância da
peste no Estado do Ceará: 1990-1999. Revista da Sociedade Brasileira de Medicina
Tropical, v. 35, p. 143-148, 2002.
ATLUNG, T.; INGMER, H. H-NS: a modulator of environmentally regulated gene
expression. Molecular Microbiology, v. 24, p. 7-17, 1997.
AUTENRIETH, I. B.; FIRSCHING, R. Penetration of M cells and destruction of
Peyer's patches by Yersinia enterocolitica: an ultrastructural and histological study.
Journal of Medical Microbiology, v. 44, p. 285-294, 1996.
29

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
AZAM, T.A.; ISHIRAMA, A. Twelve species of the nucleoid-associated protein from
Escherichia coli. Sequence recognition specificity and DNA binding affinity. Journal
of Biology Chemistry, v. 274, n. 46, p. 33105-33113, 1999.
AZAM, T. A.; IWATA, A.; NISHIMURA, A.; UEDA, S.; ISHIRAMA, A. Growth
phase-dependent variation in protein composition of the Escherichia coli nucleoid.
Journal of Bacteriology, v. 181, p. 6361-6370, 2000.
BALDWIN, T.J. The 18th C. L. Oakley Lecture: Pathogenicity of enteropathogenic
Escherichia coli. Journal Medical Microbiology, v. 47, n. 4, p. 283-293, 1998.
BALLESTEROS, M.; KUSANO, S.; ISHIRAMA, A.; VICENTE, M. The ftsQ1p
gearbox promoter of Escherichia coli is a major sigma S-dependent promoter in the
ddlB-ftsA region. Molecular Microbiology, v. 30, n. 2, p. 419-430, 1998.
BARTH, M.; MARSHALL, C.; MUFFLER, A.; FISCHER, D.; HENGGER-ARONIS,
R. Role of the histone-like protein H-NS in growth phase-dependent and osmotic
regulation of σS and many σS-dependent genes in Escherichia coli. Journal of
Bacteriology, v. 177, p. 3455-3464, 1995.
BELTRAMETTI, F.; KRESSE, A. U.; GUZMAN, C. A. Transcriptional regulation of
the esp genes of enterohemorrhagic Escherichia coli. Journal of Bacteriology, v. 181,
n. 11, p. 3409-3418, 1999.
BERGMANN, T. K.; VINDING, K.; HEY, H. Multiple hepatic abscesses due to
Yersinia enterocolitica infection secondary to primary haemochromatosis.
Scandinavian Journal of Gastroenterology, v. 36, n. 8, p. 891-5, 2001.
BERTIN, P.; HOMMAIS, F.; KRIN, E.; SOUTOURINA, O.; TENDENG, C.;
DERZELLE, S.; DANCHIN, A. H-NS and H-NS-like proteins in Gram-negative
bacteria and their multiple role in the regulation of bacterial metabolism. Biochimiestry
v. 83, n. 2, p. 235-241, 2001.
30

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
BOHANNON, D. E.; CONELL, N.; KEENER, J.; TORMO, A.; ESPINOSA-URGEL,
M.; ZAMBRANO, M. M.; KOLTER, R. Stationary-phase-inducible "gearbox"
promoters: differential effects of katF mutations and role of sigma 70. Journal of
Bacteriology, v. 173, n. 14, p. 4482-4492, 1991.
BOTTONE, E.J. Yersinia enterocolitica: the charisma continues. Clinical
Microbiology Reviews v. 10, p. 257-276, 1997.
BROWN, C. E.; HOGG, R. W. A second transport system for L-arabinose in
Escherichia coli B-r controlled by the araC gene. Journal of Bacteriology, v. 111, p.
606-613, 1972.
BUSTAMANTE, C.; GUTHOLD, M.; ZHU, X.; YANG, G. Facilitated target location
on DNA by individual Escherichia coli RNA polymerase molecules observed with the
scanning force microscope operating in liquid. Journal of Biology Chemistry, v. 274,
n. 24, p. 16665-16668, 1999.
CARMONA, M.; BALSALOBRE, C.; MUNOA, F.; MOURIÑO, M.; JUBETE, Y.; DE
LA CRUZ, F.; JUAREZ, A. Escherichia coli hha mutants, DNA supercoiling and
expression of the haemolysin genes from the recombinant plasmid pANN202-312.
Molecular Microbiology, v. 9, p. 1011-1018, 1993.
CARNIEL, E. The Yersinia high-pathogenicity island: an iron-uptake island. Microbes
Infection, v. 3, n. 7, p. 561-569, 2001.
CELLI, J.; DENG, W.; FINLAY, B. B. Enteropathogenic Escherichia coli (EPEC)
attachment to epithelial cells:exploiting the host cell cytoskeleton from the outside. Cell
Microbiology, v. 2, n. 1, p. 1-9, 2000.
CHU, M. Laboratory Manual of Plague Diagnostic Tests. CDC/WHO, 2000.
CORNELIS, G. R. The Yersinia Ysc-Yop type III' weaponry. Nature Review
Molecular Cell Biology, v. 3, n. 10, p. 742-752, 2002a.
31

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
CORNELIS, G. R. Yersinia type III secretion: send in the effectors. The Journal of
Cell Biology, v. 158, n. 3, p. 401–408, 2002b.
CORNELIS, G. R. Molecular and cell biology aspects of plague. Proceedings of the
National Academy of Science USA, v. 97, n. 16, p. 8778-8783, 2000.
CORNELIS GR; BOLAND A; BOYD AP; GEUIJEN C; IRIARTE M; NEYT C;
SORY MP; STAINIER I. The virulence plasmid of Yersinia, an antihost genome.
Microbiology and Molecular Biology Review, v. 62, p. 1315-1352, 1998.
CORNELIS, G. R.; SLUITERS, C.; DELOR, I.; GEIB, D.; KANIGA, K.; LAMBERT
DE ROUVROIT, C.; SORY, M. P.; VANOOTEGHEM, J. C.; MICHIELS, T. ymoA, a
Yersinia enterocolitica chromosomal gene modulating the expression of virulence
functions. Molecular Microbiology, v. 5, p. 1023-1034, 1991.
CORNELIS, G. R.; SLUITERS, C.; LAMBERT DE ROUVROIT, C.; MICHIELS, T.
Homology between VirF, the transcriptional activator of the Yersinia virulence regulon,
and AraC, the Escherichia coli arabinose operon regulator. Journal of Bacteriology v.
171, p. 254-262, 1989.
CORNELIS, G.; VANOOTEGEM, J. C.; SLUITERS, C. Transcription of the yop
regulon from Y. enterocolitica requires trans acting pYV and chromosomal genes.
Microbial Pathogenesis, v. 2, p. 367-379, 1987.
CORNELIS, G.; SORY, M. P.; LAROCHE, Y.; DERCLAYE, I. Genetic analysis of the
plasmid region controlling virulence in Yersinia enterocolitica O:9 by Mini-Mu
insertions and lac gene fusions. Microbial Pathogenesis, v. 1, p. 349-359, 1986.
DAME, R. T.; WYMAN, C; GOOSEN, N. Structural basis for preferential binding of
H-NS to curved DNA. Biochimiestry, v. 83, n. 2, p. 231-234, 2001.
DeHASETH, P. L.; ZUPANCIC, M. L.; RECORD, M. T. JR. RNA polymerase-
promoter interactions: the comings and goings of RNA polymerase. Journal of
Bacteriology, v. 180, n.12, p. 3019-3025, 1998.
32

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
DOMBROSKI, A. J. Recognition of the -10 promoter sequence by a partial polypeptide
of sigma70 in vitro. Journal of Biology Chemistry, v. 272, n. 6, p. 3487-3494, 1997.
DONNENBERG, M. S.; KAPER, J. B.; FINLAY, B. B. Interactions between
enteropathogenic Escherichia coli and host epithelial cells. Trends Microbiology, v. 5,
n. 3, p. 109-114, 1997.
DORMAN, C. J.; HINTON, J. C; FREE, A. Domain organization and oligomerization
among H-NS-like nucleoid-associated proteins in bacteria. Trends in Microbiology, v.
7, n. 3, p. 124-128, 1999.
DOVE, S. L.; DARST, S. A.; HOCHSCHILD, A. Region 4 of sigma as a target for
transcription regulation. Molecular Microbiology, v. 48, n. 4, p. 863-874, 2003.
ELLIOTT, S. J.; WAINWRIGHT, L. A.; MCDANIEL, T. K.; JARVIS, K. G.; DENG,
Y. K.; LAI, L. C.; MCNAMARA, B. P.; DONNENBERG, M. S.; KAPER, J. B. The
complete sequence of the locus of enterocyte effacement (LEE) from enteropathogenic
Escherichia coli E2348/69. Molecular Microbiology, v. 28, n. 1, p. 1-4, 1998.
ESPINOSA-URGEL, M.; CHAMIZO, C.; TORMO, A. A consensus structure for sigma
S-dependent promoters. Molecular Microbiology, v. 21, n. 3, p. 657-659, 1996.
FALCONI, M.; COLONNA, B.; PROSSEDA, G.; MICHELI, G.; GUALERZI, C. O.
Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-
dependent structural transition of DNA modulates accessibility of virF promoter to
transcriptional repressor H-NS. The EMBO Journal, v. 17, p. 7033-7043, 1998.
FELDMAN, M. F.; CORNELIS, G. R. The multitalented type III chaperones: all you
can do with 15 kDa. FEMS Microbiology Letters, v. 219, n. 2, p. 151-158, 2003.
FETHERSTON, J. D.; SCHUETZE, P.; PERRY, R. D. Loss of the pigmentation
phenotype in Yersinia pestis is due to the spontaneous deletion of 102 kb of
chromosomal DNA which is flanked by a repetitive element. Molecular Microbiology,
v. 6, n. 18, p. 2693-2704, 1992.
33

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
FORSBERG, A.; VIITANEN, A. M.; SKURNIK, M.; WOLF-WATZ, H. The surface-
located YopN protein is involved in calcium signal transduction in Yersinia
pseudoturberculosis. Molecular Microbiology, v. 5, p. 977-986, 1991.
FOULTIER, B.; TROISFONTAINES, P; VERTOMMEN, D.; MARENNE, M. N.;
RIDER, M.; PARSOT, C.; CORNELIS, G. R. Identification of substrates and
chaperone from the Yersinia enterocolitica 1B Ysa type III secretion system. Infection
Immunity, v. 71, n. 1, p. 242-253, 2003.
GAAL, T.; ROSS, W.; ESTREM, S. T.; NGUYEN, L. H.; BURGESS, R. R.;
GOURSE, R. L. Promoter recognition and discrimination by EsigmaS RNA
polymerase. Molecular Microbiology, v. 42, n. 4, p. 939-954, 2001.
GIRON, J. A.; HO, A. S.; SCHOOLNIK, G. K. An inducible bundle-forming pilus of
enteropathogenic Escherichia coli. Science, v. 254, n. 5032, p. 710-713, 1991.
GRIFFITHS, A. J. F.; MILLER, J. H.; SUZUKI, D. T.; LEWONTIN, R. C.;
GELBART, W. M. Introdução à genética. Guanabara Koogan, 7a ed., 2002.
GROISMAN, E. A. Principles of bacterial pathogenesis. Academic Press, 2001.
GROSS, C. A.; CHAN, C.; DOMBROSKI, A.; GRUBER, T.; SHARP, M.; TUPY, J.;
YOUNG, B. The functional and regulatory roles of sigma factors in transcription. Cold
Spring Harbor symposia on quantitative biology, v. 63, p. 141-155, 1998.
GUAN, K. L.; DIXON, J. E. Protein tyrosine phosphatase activity of an essential
virulence determinant in Yersinia. Science, v. 249, p. 553-556, 1990.
HACKER, J.; BLUM-OEHLER, G.; MUHLDORFER, I.; TSCHAPE, H. Pathogenicity
islands of virulent bacteria: structure, function and impact on microbial evolution.
Molecular Microbiology, v. 23, n. 6, p. 1089-1097, 1997.
HAGERMAN PJ. Sequence-directed curvature of DNA. Annual Review
Biochemistry, v. 59, p. 755-781, 1990.
34

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
HALDENWANG, W. G. The sigma factors of Bacillus subtilis. Microbiology Review,
v. 59, n. 1, p. 1-30, 1995.
HAMILTON, E.; LEE, N. Three binding sites for AraC protein required for
autoregulation of araC in Escherichia coli. Proceedings of the National Academy of
Science USA, v. 85, p. 1749-1753, 1988.
HARLEY, C. B.; REYNOLDS, R. P. Analysis of E. coli promoter sequences. Nucleic.
Acids Research, v. 15, n. 5, p. 2343-2361, 1987.
HE, S. Y.; NOMURAA, K.; WHITTAM, T. S. Type III protein secretion mechanism in
mammalian and plant pathogens. Biochimica et Biophysica Acta, v. 1694, p. 181–206,
2004.
HENGGE-ARONIS, R. Back to log phase: sigma S as a global regulator in the osmotic
control of gene expression in Escherichia coli. Molecular Microbiology, v. 21, n. 5, p.
887-893, 1996.
HENGGE-ARONIS, R. Survival of hunger and stress: the role of rpoS in early
stationary phase gene regulation in E. coli. The Cell, v. 72, n. 2, p. 165-168, 1993.
HEYDUK, T.; HEYDUK, E.; SEVERINOV, K.; TANG, H.; EBRIGHT, R. H.
Determinants of RNA polymerase alpha subunit for interaction with beta, beta', and
sigma subunits: hydroxyl-radical protein footprinting. Proceedings of the National
Academy of Science USA, v. 93, n. 19, p. 10162-10166, 1996.
HUECK, C. J. Type III protein secretion systems in bacterial pathogens of animals and
plants. Microbiology Molecular Biology Review, v. 62, p. 379-433, 1998.
INGLESBY, T. V.; DENNIS, D. T.; HENDERSON, D. A. Plague as a Biological
Weapon. Med Publ Health Man Jama, v. 283, p. 2281-2290, 2000.
35

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
IRIARTE, M.; CORNELIS, G. R. Identification of SycN, YscX, and YscY, three new
elements of the Yersinia Yop virulon. Journal of Bacteriology, v. 181, p. 675-680,
1999.
IRIARTE, M.; CORNELIS, G. R. YopT, a new Yersinia Yop effector protein, affects
the cytoskeleton of host cells. Molecular Microbiology, v. 29, n. 3, p. 915-929, 1998.
IRIARTE, M.; SORY, M. P.; BOLAND, A.; BOYD, A. P.; MILLS, S. D.;
LAMBERMONT, I.; CORNELIS, G. R. TyeA, a protein involved in control of Yop
release and in translocation of Yersinia Yop effectors. The EMBO Journal, v. 17, n. 7,
p. 1907-1918, 1998.
ISHIHAMA, A. Functional modulation of Escherichia coli RNA polymerase. Annual
Review Microbiology, v. 54, p. 499-518, 2000.
ISHIHAMA, A. Protein-protein communication within the transcription apparatus.
Journal of Bacteriology, v. 175, n. 9, p. 2483-2489, 1993.
JACOB, F.; MONOD, J. Genetic regulatory mechanisms in the synthesis of proteins.
Journal of Molecular Biology, v. 3, p. 318-356, 1961.
JOURNET, L.; AGRAIN, C.; BROZ, P.; CORNELIS, G. R. The needle length of
bacterial injectisomes is determined by a molecular ruler. Science, v. 302, n. 5, p. 1757-
1760, 2003.
JURIS, S. J.; SHAO, F.; DIXON, J. E. Yersinia effectors target mammalian signalling
pathways. Cell Microbiology, v. 4, n. 4, p. 201-211, 2002.
KAPER, J. B. EPEC delivers the goods. Trends in Microbiology, v. 6, p. 169-172,
1998.
KUMAR, A.; MALLOCH, R. A.; FUJITA, N.; SMILLIE, D. A.; ISHIHAMA, A.;
HAYWARD, R. S. The minus 35-recognition region of Escherichia coli sigma 70 is
36

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
inessential for initiation of transcription at an "extended minus 10" promoter. Journal
of Molecular Biology, v. 232, n. 2, p. 406-418, 1993.
LAMBERT DE ROUVROIT, C.; SLUITERS, C.; CORNELIS, G. R. Role of the
transcriptional activator VirF and temperature in the expression of the pYV plasmid
genes of Yersinia enterocolitica. Molecular Microbiology, v. 6, p. 379-388, 1992.
LANGE, R.; HENGGE-ARONIS, R. Identification of a central regulator of stationary-
phase gene expression in Escherichia coli. Molecular Microbiology, v. 5, n.1, p. 49-59,
1991.
LJUNGBERG, P.; VALTONEN, M.; HARJOLA, V. P.; KAUKORANTA-
TOLVANEN, S. S.; VAARA, M. Report of four cases of Yersinia pseudotuberculosis
septicemia and a literature review. European Journal Clinical Microbiology
Infection Disease, v. 14, p. 804-810, 1995.
MACLEOD, S. M.; JOHNSON, R. C. Control of transcription by nucleoid proteins.
Currents Opinions Microbiology, v. 4, p. 152-159, 2001.
MADRID, C.; NIETO, J. M.; PAYTUBI, S.; FALCONI, M.; GUALERZI, C. O.;
JUAREZ, A. Temperature- and H-NS-dependent regulation of a plasmid-encoded
virulence operon expressing Escherichia coli hemolysin. Journal Bacteriology, v. 184,
n. 18, p. 5058-5066, 2002.
MIKULSKIS, A. V.; CORNELIS, G. R. A new class of proteins regulating gene
expression in enterobacteria. Molecular Microbiology, v. 11, p. 77-86, 1994.
MOURIÑO, M.; MADRID, C.; BALSALOBRE, C.; PRENAFETA, A.; MUNOA, F.;
BLANCO, J.; BLANCO, M.; BLANCO, J. E.; JUAREZ, A. The Hha protein as a
modulator of expression of virulence factors in Escherichia coli. Infection
Immunology, v. 64, p. 2881-2884, 1996.
37

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
NAVARRO, L.; ALTO, N. M.; DIXON, J. E. Functions of the Yersinia effector
proteins in inhibiting host immune responses. Current Opinion in Microbiology, v. 8,
p. 21–27, 2005.
NIETO, J. M.; MADRID, C.; MIQUELAY, E.; PARRA, J. L.; RODRÍGUEZ, S.;
JUÁREZ, A. Evidence for direct protein-protein interaction between members of the
enterobacterial Hha/YmoA and H-NS families of proteins. Journal Bacteriology, v.
184, n. 3, p. 629-35, 2002.
NIETO, J. M.; CARMONA, M.; BOLLAND, S.; JUBETE, Y.; DE LA CRUZ, F.;
JUÁREZ, A. The hha gene modulates haemolysin expression in Escherichia coli.
Molecular Microbiology, v. 5, p. 1285-1293, 1991.
OWEN-HUGHES, T. A.; PAVITT, G. D.; SANTOS, D. S; SIDEBOTHAM, J. M.;
HULTON, C. S.; HINTON, J. C.; HIGGINS, C. F. The chromatin-associated protein H-
NS interacts with curved DNA to influence DNA topology and gene expression. The
Cell, v. 71, p. 255-265, 1992.
PEREZ-MARTIN, J.; DE LORENZO, V. Clues and consequences of DNA bending in
transcription. Annual Review Microbiology, v. 51, p. 593-628, 1997.
PERRY, R. D.; FETHERSTON, J. D. Yersinia pestis-etiologic agent of plague. Clinical
Microbiology Review, v. 10, p. 35-66, 1997.
PUENTE, J. L.; BIEBER, D.; RAMER, S. W.; MURRAY, W.; SCHOOLNIK, G. K.
The bundle-forming pili of enteropathogenic Escherichia coli: transcriptional regulation
by environmental signals. Molecular Microbiology, v. 20, n. 1, p. 87-100, 1996.
PUJOL, C.; BLISKA, J. B. Turning Yersinia pathogenesis outside in: subversion of
macrophage function by intracellular Yersiniae. Clinical Immunology, v. 114, p. 216–
226, 2005.
38

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
RAMAMURTH, K. S.; SCHNEEWIND, O. Substrate recognition by the Yersinia type
III protein secretion machinery. Molecular Microbiology, v. 50, n. 4, p. 1095-1102,
2003.
RAMER, S. W.; BIEBER, D.; SCHOOLNIK, G. K. BFPB, an outer membrane
lipoprotein required for the biogenesis of bundle-forming pili in enteropathogenic
Escherichia coli. Journal of Bacteriology, v. 178, n. 22, p. 6555-6563, 1996.
REZNIKOFF, W. S. Catabolite gene activator protein activation of lac transcription.
Journal of Bacteriology, v. 174, n. 3, p. 655-658, 1992.
RIPPE, K.; VON HIPPEL, P. H.; LANGOWSKI, J. Action at a distance: DNA-looping
and initiation of transcription. Trends in Biochemistry Science, v. 20, p. 500-506,
1995.
ROSQVIST, R.; MAGNUSSON, K. E.; WOLF-WATZ, H. Target cell contact triggers
expression and polarized transfer of Yersinia YopE cytotoxin into mammalian cells.
EMBO Journal, v. 13, p. 964-972, 1994.
ROSS, W.; GOSINK, K. K.; SALOMON, J.; IGARASHI, K.; ZOU, C.; ISHIHAMA,
A.; SEVERINOV, K.; GOURSE, R. L. A third recognition element in bacterial
promoters: DNA binding by the α subunit of RNA polymerase. Science, v. 262, p.
1407-1413, 1993.
SARKER, M. R.; SORY, M. P.; BOYD, A. P.; IRIARTE, M.; CORNELIS, G. R. LcrG
is required for efficient translocation of Yersinia Yop effector proteins into eukaryotic
cells. Infection Immunology, v. 66, p. 2976-2979, 1998.
SKRZYPEK, E.; STRALEY, S. C. LcrG, a secreted protein involved in negative
regulation of the low-calcium response in Yersinia pestis. Journal of Bacteriology, v.
175, p. 3520-3528, 1993.
SMYTH, C.P.; LUNDBACK, T.; RENZONI, D.; SILIGARDI, G.; BEAVIL, R.;
LAYTON, M.; SIDEBOTHAM, J. M.; HINTON, J. C.; DRISCOLL, P. C.; HIGGINS,
39

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
C. F.; LADBURY, J. E. Oligomerization of the chromatin-structuring protein H-NS.
Molecular Microbiology, v. 36, n. 4, p. 962-972, 2000.
SNYDER, L.; CHAMPNESS, W. Molecular genetics of bacteria. ASM Press, p. 265-
284, 1997.
SORY, M. P.; BOLAND, A.; LAMBERMONT, I.; CORNELIS, G. R. Identification of
the YopE and YopH domains required for secretion and internalization into the cytosol
of macrophages, using the cyaA gene fusion approach. Proceedings of the National
Academy of Science USA, v. 92, p. 11998-12002, 1995.
SUH, S. J.; SILO-SUH, L.; WOODS, D. E.; HASSETT, D. J.; WEST, S. E.; OHMAN,
D. E. Effect of rpoS mutation on the stress response and expression of virulence factors
in Pseudomonas aeruginosa. Journal of Bacteriology, v. 181, n. 13, p. 3890-3897,
1999.
THOMAS, N. A.; FINLAY, B. B. Pathogens: bacterial needles ruled to length and
specificity. Current Biology, v. 14, n. 5, p. R192-194, 2004.
VALLANCE, B. A.; FINLAY, B. B. Exploitation of host cells by enteropathogenic
Escherichia coli. Proceedings of the National Academy of Science USA, v. 97, n. 16,
p. 8799-8806, 2000.
VICENTE, M.; KUSHNER, S. R.; GARRIDO, T.; ALDEA, M. The role of the
'gearbox' in the transcription of essential genes. Molecular Microbiology, v. 5, n. 9, p.
2085-2091, 1991.
VILAR, J. M. G.; GUET, C. C.; LEIBLER, S. Modeling network dynamics: the lac
operon, a case study. The Journal of Cell Biology, v. 161, n. 3, p. 471-476, 2003.
WAGNER, R. Trascription regulation in prokaryotes. Oxford University Press, 2000.
40

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
WATTIAU, P.; BERNIER, B.; DESLEE, P.; MICHIELS, T.; CORNELIS, G. R.
Individual chaperones required for Yop secretion by Yersinia. Proceedings of the
National Academy of Science USA, v. 91, p. 10493-10497, 1994.
WATTIAU, P.; CORNELIS, G. R. Identification of DNA sequences recognized by
VirF, the transcriptional activator of the Yersinia yop regulon. Journal of Bacteriology,
v. 176, n. 13, p. 3878-3884, 1994.
WATTIAU, P.; CORNELIS, G. R. SycE, a chaperone-like protein of Yersinia
enterocolitica involved in the secretion of YopE. Molecular Microbiology, v. 8, p.
123-131, 1993.
WHO. Human plague in 2002 and 2003. Weekly Epidemiological Record, v. 79, n.
33, p. 301-308, 2004.
WILLIAMS, R. M.; RIMSKY, S. Molecular aspects of the E. coli nucleoid protein, H-
NS: a central controller of gene regulatory networks. FEMS Microbiology Letters, v.
156, n. 2, p. 175-185, 1997.
WISE, A.; BREMS, R.; RAMAKRISHNAN, V.; VILLAREJO, M. Sequences in the -
35 region of Escherichia coli rpoS-dependent genes promote transcription by E sigma
S. Journal of Bacteriology, v. 178, n. 10, p. 2785-2793, 1996.
WOLFF, C.; NISAN, I.; HANSKI, E.; FRANKEL, G.; ROSENSHINE, I. Protein
translocation into host epithelial cells by infecting enteropathogenic Escherichia coli.
Molecular Microbiology, v. 28, p. 143-155, 1998.
YAMADA, H.; MURAMATSU, S.; MIZUNO, T. An Escherichia coli protein that
preferentially binds to sharply curved DNA. Journal of Biochemistry, v. 108, p. 420-
425, 1990.
ZHANG, G.; DARST, S. A. Structure of the Escherichia coli RNA polymerase alpha
subunit amino-terminal domain. Science, v. 281, n. 5374, p. 262-266, 1998.
41

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
JUSTIFICATIVA
42

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Bactérias com um sistema de secreção do tipo III possuem um mecanismo de
defesa que permite a introdução direta de toxinas no interior de macrófagos e outras
células. Dessa forma, a utilização dessas toxinas fusionadas a proteínas imunogênicas
de parasitas como a leishmania, poderia ser capaz de desencadear uma resposta imune
específica contra os mesmos. Assim, o uso de bactérias enteropatogênicas como
carreadores para apresentação de antígenos ao sistema imune, poderia servir de
instrumento para a produção de uma vacina, no mínimo, protetora. Entretanto, para que
isso ocorra de maneira bem sucedida, é importante estudar os mecanismos que regulam
a expressão dessas toxinas e seu comportamento in vitro e in vivo, como também obter
um promotor forte que possa ser ativado pelas próprias condições de virulência. Estudos
também são necessários para identificar novos alvos que possam ser utilizados no
desenvolvimento de novos testes diagnósticos.
43

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
OBJETIVOS
44

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Objetivo Geral
Investigar a possibilidade de utilizar modelos bacterianos como
vetores para a introdução de antígenos imunogenicamente importantes em
animais de laboratório, visando a indução de resposta imunológica
específica contra estes antígenos. Verificar se a proteína YopH
recombinante é reconhecida por soros de pacientes suspeitos de peste e
identificar novos alvos para diagnóstico.
Objetivos Específicos
• Analisar o promotor da proteína YopH quanto a presença de seqüências
regulatórias.
• Demonstrar o reconhecimento das proteínas de fusão, pelo anticorpo
anti-YopH.
• Analisar a expressão da YopH, na presença e ausência do seu promotor,
em diferentes plasmídeos.
• Transformar diferentes culturas de Escherichia coli com plasmídeos
contendo a proteína YopH e analisar a sua expressão, determinando os
fatores que influenciam a sua expressão.
• Avaliar o potencial da YopH no diagnóstico da peste (infecção pela Y.
pestis).
45

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
ARTIGOS
46

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
ARTIGO I
Positioning of the Yersinia enterocolitica yopH promoter after
the Escherichia coli lac promoter induces a strong and
regulated expression of YopH in E. coli.
Patrícia R. O. Haver1, Suzana M. R. Costa2, Bruno L. A. da Silva3, Mariana
M. S. C. Chagas3, Alzira M. P. de Almeida4, Osvaldo P. de Melo Neto3
1. Doutorado em Ciências Biológicas, CCB/UFPE 2. Mestrado em Genética, Departamento de Genética, CCB/UFPE 3. Departamento de Microbiologia, Centro de Pesquisas Aggeu Magalhães, FIOCRUZ/MS, Recife, PE, Brasil. 4. Laboratório de Peste, Departamento de Microbiologia, Centro de Pesquisas Aggeu Magalhães, FIOCRUZ/MS, Recife, PE, Brasil.
Palavras chave: YopH, Yersinia, HNS
Manuscrito a ser submetido para publicação no:
FEMS Microbiology Letters
47

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Positioning of the Yersinia enterocolitica yopH promoter after the
Escherichia coli lac promoter induces a strong and regulated
expression of YopH in E. coli.
Patrícia R. O. Haver, Suzana M. R. Costa, Bruno L. A. da Silva, Mariana
M. S. C. Chagas, Alzira M. P. de Almeida and Osvaldo P. de Melo Neto.
Abstract
The tyrosine phosphatase YopH is a virulence factor produced by pathogenic Yersinia
and required for its survival within the mammalian host. Expression of YopH is tightly
regulated at the transcriptional level and is dependent on the transcriptional activator
VirF, which binds to the yopH promoter. In Escherichia coli VirF is absent and the
yopH promoter has a low residual activity. Here we describe a synergistic effect
between the yoph and lac promoters in E. coli, observed when the full length Y.
enterocolitica yopH gene was cloned downstream of the lac promoter in a pUC derived
plasmid. A very strong expression of YopH was achieved even in conditions when the
lac promoter would be inactive. Similar levels of expression were also seen for various
YopH fusion proteins. Optimal YopH synthesis required an increase in temperature or
in the culture growth phase and was observed under different plasmid backgrounds. It
was also enhanced in E. coli strains defective for the histone like protein H-NS, which
presumably may bind to a strong DNA curve predicted within the yopH promoter.
48

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
1. Introduction
Pathogenic Yersinia spp. manage to survive inside their mammalian hosts through their
ability to avoid the antimicrobial activities of professional phagocytes. This is
accomplished through the secretion of different virulence proteins encoded in a 65-
70Kb plasmid called pYV (for reviews see (Cornelis, 1998; Cornelis et al., 1998; Bleves
and Cornelis, 2000). Foremost amongst these protein are those called Yops, many of
which are secreted into the cytoplasm of target cells through a type III secretion system
(Cheng and Schneewind, 2000; Hueck, 1998) and are required to disarm the host
immune response. The yop genes have been shown to be contained in different operons
and their expression is regulated at the level of transcription by temperature as well as
by extracelular Ca2+ concentration.
Many of the virulence factors required for colonisation and survival of
pathogenic bacteria in mammalian hosts have their expression activated upon an
increase in temperature. In Yersinia the temperature-dependent transcriptional activation
of many genes encoding virulence factors, including the Yop proteins, is dependent on
the product of the regulatory gene, virF (Cornelis et al., 1987; Lambert de Rouvroit et
al., 1992) or lcrF (Hoe et al., 1992). The expression of virF is itself thermoregulated and
involves a negatively acting chromosomal factor, YmoA (Cornelis et al., 1991). In a Y.
enterocolitica ymoA mutant, virF expression is derepressed, although it is still enhanced
by increased temperature (Cornelis, 1993). Expression of the gene coding for the
tirosine phosphatase YopH, involved in preventing bacterial phagocytosis by
macrophages (Bliska et al., 1991), also requires VirF which binds to a sequence -64 to -
34 nucleotides upstream of the yopH gene transcription start site (Lambert de Rouvroit
et al., 1992). However, expression of yopH becomes independent of VirF at 37oC in the
ymoA mutant, although it still remains thermoinducible (Cornelis, 1993). Constitutive
expression of VirF at 25oC leads to yopH expression only in the absence of YmoA
(Cornelis, 1993; Lambert de Rouvroit et al., 1992). Both YmoA, because of its histone-
like structure, and temperature are likely to influence DNA conformation, and changes
in DNA superhelicity have been shown to coincide with virF transcription and Yop
expression (Rohde et al., 1994).
Here we report the construction of a double promoter consisting of the
Escherichia coli lac promoter and the Y. enterocolitica yopH promoter (comprising 199
nucleotides upstream of the predicted yopH transcription start site), which mediates a
very a strong expression of YopH in E. coli. The expression of YopH in this
49

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
heterologous system is regulated both by temperature and the culture growth phase.
Other results, coupled with computer curvature predictions which indicates the
existence of multiple curves in the yopH promoter (Rohde et al., 1999), also suggest a
role for the histone-like protein H-NS in regulating the promoter activity possibly
through binding to the curved promoter DNA.
2. Materials and Methods
2.1. Cloning of the Y. enterocolitica yopH gene.
The full length yopH gene from Y. enterocolitica O:3 Ye35, including its promoter, was
amplified from total genomic DNA using the following primers: P1 (5`CCC GAG CTC
GTA TTA CCA TCT GTT CC3`) designed to anneal to a segment starting about 199
nucleotides upstream of the predicted transcription start site and including a Sac I
restriction site (underlined); and P4 (5`CCC CTG CAG ATA AAC CTC AAC3`) which
anneals downstream of the transcription terminator sequence and includes a Pst I
restriction site. The PCR fragment was cloned between the Sac I and Pst I sites of the
cloning vector pTZ18R (GenBank accession: L08956). Transformation was carried out
using E. coli DH5α. Transformed cells were plated at either 37 or 24oC. No
recombinant clones were obtained from the ligations after incubating the transformation
plates at 37oC overnight. However upon incubation at the lower temperature for 36
hours clones were observed which, upon DNA preparation from liquid LB cultures, also
grown at 24oC, yielded recombinant plasmids containing the expected 1.8 Kb insert
(pTZ/YopH). Restriction enzyme digests and sequencing of selected clones confirmed
the identity of the insert. All later cloning steps were performed by growing the cells at
24oC.
2.2. Construction of the Y. enterocolitica yopH fusions.
To obtain the pTZ/YopH-HSP83 construct the pTZ/YopH plasmid was first digested
with Xba I, end-filled and ligated to an end-filled BamH I fragment encoding the N-
terminal half (amino acids 1-303) of the 83 kDa Heat Shock Protein (HSP83) from
Leishmania braziliensis. This cloning strategy was designed so as to maintain the same
reading frame between the YopH N-terminus and the HSP83 fragment. This strategy
maintained the two BamH I sites flanking the HSP83 insert while deleting the Xba I
50

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
site. Translation was predicted to terminate immediately after the HSP83 portion of the
fusion protein.
Alternatively, to obtain the pTZ/YopH-GP63 plasmid, the 680 bp Sac I/BamH I
fragment from pTZ/YopH-HSP83, encoding the YopH N-terminus, was subcloned into
the same sites of the original pTZ18R vector previously modified by the cloning of a
Xba I/Pst I PCR fragment encoding the remaining segment of the yopH gene. This latter
fragment was amplified using the primers P3 (encompassing the original Xba I site, 5’GCG TTC ACA TCT AGA CCC3’) and P4 (described above), and contained a stop
codon at position 708 of the second yopH fragment. A 500 bp PCR fragment consisting
of the central region of a gene encoding a gp63 cysteine proteinase from L. braziliensis
was subsequently cloned into the BamH I site of the resulting plasmid generating the
pTZ/YopH-GP63 construct. This PCR fragment corresponds to amino acids 168-333
from the described L. guyanensis GP63 homologue (Steinkraus and Langer, 1992;
Steinkraus et al., 1993). The resulting plasmid allowed the expression of a fusion
protein containing the 168 amino acids fragment of the GP63 protein positioned
between the first 155 amino acids of the YopH N-terminal end and followed by the 166
amino acids of the YopH central region.
2.3. Protein expression analysis.
DH5α cells containing the recombinant plasmids were maintained by growth in LB
medium at 25oC prior to storage in glycerol at –80oC. For the YopH expression
experiments aliquots of frozen cells were transferred to fresh medium and allowed to
grow at 25oC until reaching stationary phase. They were then diluted to O.D. 0.01 and
allowed to grow for various time periods and ODs at either 25 or 37oC. The cells were
then collected by centrifugation and ressuspended in sample buffer for SDS-PAGE.
Samples were analysed through a 15% gel stained with Coomassie Blue. Cells
transformed with the pTZ18R derived plasmids were always grown in the presence of
ampicillin at 50 µg/ml. For the cells transformed with both the pTZ/YopH and pREP4
(QIAGEN) plasmids Kanamycin at 20 µg/ml was also added to the growth media. In the
experiments with the various hha and hns deletion strains supplementary antibiotics
were added depending on the strain as follows: YK4122H (hha deletion) – Kanamycin
at 20 µg/ml; YK4124 (hns deletion) – Hygromycin at 100 µg/ml; YK4124H (hha/hns
double deletion) – Both Kanamycin at 20 µg/ml and Hygromycin at 100 µg/ml. For the
IPTG induction experiments the DH5α cells were allowed to grow at 37oC until
51

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
reaching OD 0.5 when IPTG was added to 0.1 mM. Subsequently the cells were
harvested at various ODs and processed as before to analyse YopH expression.
2.4. Antibody production and western-blots.
DH5α cells transformed with the pTZ/YopH plasmid were grown in large scale as
described above, ressuspended in SDS-sample buffer and the protein extract ran in a
preparative SDS gel. The putative YopH protein band was then excised from the gel and
used to immunise rabbits in order to produce serum against the protein. The antibodies
were kept at 4oC with 0.03% sodium azide and used in a dilution of 1/90000. Western
blots were performed with the Immobilon-P PVDF membrane (Millipore), using as
second antibody peroxidase conjugated goat anti-rabbit IgG serum (Jackson
ImmunoResearch Laboratories) diluted 1/5000. The reactions were detected by
enhanced chemiluminescence (ECL). Densitometric scanning of the various results was
performed with the Kodak 1D Image Analysis Software, version 3.5 for Windows.
3. Results
Analysis of the expression of YopH and YopH fusion proteins in E. coli.
This work started with the goal of cloning the Yersinia enterocolitica yopH gene in
order to express YopH fusions proteins in this bacteria and analyse their secretion
through the type III secretion system. As cloning vector we used the plasmid pTZ18R, a
pUC18 derived plasmid modified by the insertion of the T7 promoter upstream of the
multiple cloning sites. The yopH gene was cloned in the same orientation as the lac
promoter, between the vector’s unique Sac I/Pst I sites, leading to the introduction of a
stop codon within the LacZ ORF. Figure 1 shows the sequence comprising the 500 nt
upstream of the YopH coding sequence including the lac promoter, the truncated LacZ
sequence and the 191 nt remaining between the two ORFs including the yopH promoter.
During the initial cloning stages we did not expect to see much expression of the YopH
protein in Escherichia coli due to the lack of the transcriptional activator VirF in this
bacteria. A minimal residual expression due to leakage of the lac promoter could be
expected but we did not anticipate it to be significant. However, from the beginning we
noticed a strong inhibition in the growth of E. coli transformed with the plasmid
pTZ/YopH, and its derivatives, when grown in solid media plates at 37oC. This
behaviour, also seen to a minor extent in liquid LB media as well, was not observed
when cells were grown at 24 or 30oC. This temperature dependent restriction of cell
52

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
growth at 37oC was reminiscent of the behaviour of wild type Yersinia and led us to
investigate the possibility of YopH being differentially expressed at this temperature in
E. coli transformed with the pTZ/YopH plasmid.
First total protein extracts from wild type E. coli DH5α as well as bacteria
transformed with either the original pTZ18R vector or the pTZ/YopH construct were
analysed through SDS-PAGE after growth at 37oC. A very strong band of 52kDa was
observed in extracts from the bacteria transformed with plasmid pTZ/YopH (Figure
2A). This protein was by far the strongest one being synthesised by the bacteria and it
was absent from the control DH5α or the cells transformed with the vector alone. A
similar result was seen when E. coli BL21 or YK4122 cells were used (not shown). Next
rabbit antisera was raised against the 52kDa band and assayed in western blots against
extracts from cells transformed with either pTZ18R or the pTZ/YopH construct. The
serum obtained proved to be specific for the 52kDa protein with a very high titer and
minimum cross-reaction against E. coli proteins (Figure 2B). The 52kDa protein could
not be detected in the control cells transformed with the original pTZ18R vector.
We then investigated whether the expression of fusion proteins consisting of
parts of YopH plus fragments of heterologous eukaryotic proteins would be expressed at
similar levels as wild type YopH in E. coli. Two different constructs were made, both
making use of an internal Xba I site present in the yopH gene so that both resulting
proteins contain the first 154 aminoacids of the YopH N-terminus and are under the
control of the same transcription regulatory elements as the wild type YopH. For the
first protein the YopH N-terminus is followed by the N-terminal half of the L.
braziliensis heat shock protein HSP83 (Skeiky et al., 1995) and a stop codon.
Alternatively, a fragment of a L. braziliensis GP63 protein was also cloned at the same
site but this time half of the remaining YopH C-terminus was also left in frame after the
GP63 segment (Figure 2C). E. coli DH5α transformed with the resulting plasmids
(pTZ/YopH-HSP83 and pTZ/YopH-GP63) were incubated in conditions described
above for YopH expression. SDS-PAGE and immunoblotting analysis showed the
induction of the predicted chimeric proteins and confirmed their identity since they
reacted with the polyclonal serum directed against the 52kDa protein (reinforcing its
identity as YopH - Figure 2D). The HSP83 fusion protein was also confirmed by a
second serum directed against a L. braziliensis protein fraction known to contain HSP83
(data not shown). The expression levels for the YopH-GP63 protein was equivalent to
53

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
that of wild type YopH but it was not possible to make a similar comparison with
YopH-HSP83 since the antisera seems to have a stronger preference for the YopH C-
terminus, absent from this protein.
Both the lacZ and yopH promoters contribute to the strong expression of YopH in
E. coli.
In order to specifically investigate the role of the lacZ promoter in the strong expression
of the YopH protein in E. coli we set out to compare its expression in the presence or
absence of the lacZ inducer IPTG. Since the high copy number of the pTZ18R plasmid
might prevent an adequate regulation of the lacZ promoter in wild type DH5α cells we
co-transformed bacteria previously transformed with the pTZ/YopH construct with a
second plasmid, pREP4, which allows for the constitutive expression of the LacIq
repressor at high levels. Since it contains a p15A origin of replication it is compatible
with the ColE1 origin of replication present in pTZ18R. Figure 3A compares the YopH
expression levels of E. coli cells grown in the presence and absence of the pREP4
plasmid before and after induction with IPTG at 37oC. In the absence of the pREP4
plasmid no effect could be observed by the addition of IPTG and high levels of
expression of YopH could be seen throughout. Even in the presence of excess LacIq
repressor high levels of YopH could be detected although addition of IPTG did increase
its expression to a certain extent. Since expression by the lacZ promoter would be
tightly regulated and responsive to IPTG at least in the presence of the LacIq repressor
we conclude then that the lacZ promoter is not the sole responsible for the strong YopH
expression observed in our experiments.
To analyse the role of the yopH promoter in the YopH expression, the yopH
gene was subcloned into another cloning vector, pGEM3zf+, which is similar to
pTZ18R in many aspects such as size, copy number and the availability of multiple
cloning sites within the LacZ coding frame. The major difference is that the multiple
cloning site is positioned in a reverse orientation regarding the lacZ promoter so we did
not expect any influence from this promoter in YopH transcription. The same sites used
for pTZ/YopH plasmid were used to generate the pGEM/YopH construct. E. coli
DH5α cells transformed with this constructed were then grown and analysed for YopH
expression using the same conditions as described above (Figure 3B). Here a very
significant decrease in the expression of YopH is observed. The yopH promoter is active
on its own but is not capable of supporting the strong levels of expression seen in the
54

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
pTZ18R construct. Similar results were observed when the yopH gene was subcloned
into two other vectors, again with opposing orientations regarding the lacZ promoter,
pK19 and pSU2718 (Conchas and Carniel, 1990). The pK19 plasmid is another pUC
derived plasmid conferring resistance to Kanamycin which contains the lacZ gene plus
multiple cloning sites but with its promoter in the opposite orientation regarding the
cloning of the yopH gene. Expression of YopH is similar to that observed for the
pGEM/YopH construct (Figure 3B). In contrast the pSU2718 vector is derived from
pACYC184, contains a chloramphenicol resistance gene and similar lacZ sequences and
restriction sites. Cloning of yopH in this vector (as for pTZ18R) leads to the yopH being
positioned downstream of the lacZ promoter and induces high levels of expression of
the resulting protein (data not shown). In summary our results indicate then that the
presence of both the lacZ and yopH promoters is required for the elevated levels of
YopH and that a synergism may be occurring which enhances expression.
YopH expression in E. coli is regulated both by temperature as well as the cell
culture density.
As described above, we had previously noticed a temperature dependent inhibition of
growth in cells transformed with pTZ/YopH, presumably due to the expression of the
YopH protein at 37oC but not at lower temperatures. Since E. coli lacks the transcription
regulation machinery present in Yersinia which, in part at least, is required for the
temperature regulation of YopH expression, we set out to understand to what extend this
expression is regulated by temperature in E. coli transformed with the pTZ18R derived
constructs. First, DH5α cells maintained at 24oC were diluted to OD 0.01 and allowed
to grow for several hours either at 24 or 37oC. At different time periods, but the same
optical density, these cells were then harvested and equal load of cellular extract applied
to SDS-PAGE and analysed by Coomassie Blue staining and western-blotting (Figure
4A). When the cells are grown exponentially at 24oC, up to early stationary phase, the
levels of YopH expression are very low (cell densities ranging from 0.2 to 1). However
when they are allowed to grow for significant periods after reaching stationary phase (as
in an overnight growth) the levels of YopH increase several fold and it is easily detected
even in Coomassie Blue stained gels. The same cells grown at 37oC produce a very
different behaviour. At very low optical densities (OD 0.2) the YopH protein is hardly
visible, as seen at 24oC, however soon after a dramatic increase in YopH expression is
seen which is maintained throughout the growth curve. The protein is easily visible in
55

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
the Coomassie Blue gels as well as through the Western-blot assays. Figure 4B shows a
graphic analysis of the blots performed after the densitometric scanning of the YopH
band. The difference in expression levels at 24oC and 37oC is clearly seen as well as the
importance of the cell culture density for YopH expression.
YopH expression is enhanced in a E. coli strain lacking the histone like protein H-
NS.
The protein product of the gene hha is a histone-like protein which shares significant
homology with Yersinia YmoA and can substitute for YmoA in a Yersinia ymoA mutant
((Mikulskis and Cornelis, 1994). Similarly expression of YmoA in a E. coli hha mutant
restores regulation mediated by Hha of α-Haemolisin expression (Balsalobre et al.,
1996). Hha interacts directly with the chromatin associated protein H-NS and this
interaction may be required for it to exert its function in regulating transcription of the
α-Haemolisin gene (Nieto et al., 2002; Madrid et al., 2002). H-NS is a very abundant
histone-like protein which binds preferentially to curved DNA and is involved in DNA
structuring as well as regulating transcription (Owen-Hughes et al., 1992; Atlung and
Ingmer, 1997; Rimsky, 2004). Due to the known effect of YmoA in Yop expression in
Yersinia, we decided to investigate the expression of YopH induced by the pTZ/YopH
plasmid in E. coli mutants lacking either Hha, H-NS or both. E. coli K5 strains with
deletions in the hha (YK4122H), hns (YK4124), or hha/hns (YK4124H) genes (Nieto et
al., 1997) were transformed with plasmid pTZ/YopH and assayed for induction of
YopH at 24oC or 37oC and comparing different cell culture densities.
First the parental strain YK4122 was analysed at the same conditions performed
as described in Figure 4. The expression profiles for YK4122 were very similar to those
obtained for the DH5α cells with very little YopH expression at the lower temperature
except after reaching stationary phase and strong expression in general at 37oC with the
only exception being very early in the growth curve (Figure 4 – WT). Similar profiles
were observed in cells lacking the hha gene the suggesting that the protein Hha is not
involved in the regulation of transcription of the yoph gene (Figure 4 – Hha). In
contrast, in strains where the expression of H-NS is abolished, either in the presence or
absence of Hha, a very significant increase in YopH expression is observed (Figure 4 –
H-NS and Hha/H-NS). This increase is more noticeable at 24oC, during the culture´s
various growth phases, but also at 37oC at low OD values. Strikingly, at latter stages of
growth at 37oC, the levels of YopH are reduced and nearly disappear during late
56

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
stationary phase. This reduction in YopH levels are accompanied by a very reduced
growth rate which led us to speculate whether the lack of YopH is due to low
transcription or to loss of the pTZ/YopH plasmid.
To sort out what was happening aliquots from both the parental strain YK4122
as well as the hns deletion strain YK4124 grown at 24oC or 37oC cells were taken at
early log growth phase (OD 0.2) or late stationary phase (after overnight growth) and
the numbered of viable cells containing the pTZ/YopH plasmid quantitated by plating
serial dilutions in LB amp plates. Table 1 summarizes the results obtained which show
that when the YK4124 cells were grown overnight at 37oC a very significant loss of the
pTZ/YopH plasmid occurred, presumably because the very high expression of YopH
prevented the cellular growth and only cells which had lost the plasmid survived. In
contrast in the parental strain YK4122 at 37oC the high levels of expression of YopH
were still compatible with cellular survival. We conclude then that the presence of H-
NS is required to repress expression of YopH, predominantly at 24oC and during the
early phases of culture growth.
Curvature predictions of the YopH promoter.
Plasmid pYV has been shown to contain multiple curvatures along its sequence (Rohde
et al., 1999) and curved DNA is known to be associated with events of transcriptional
regulation dependent on temperature. In the case of the Shigella virF promoter
repression is achieved by binding of H-NS to the curved DNA in the promoter at
temperatures below 32oC, preventing expression. Temperature shift to 37oC induces a
conformational change in the DNA which allows transcription of the promoter (Tobe et
al., 1993; Tobe et al., 1995). With the use of the program WEDGE 430 (Figure 6) we
attempted to identify any sequences within the yopH promoter which might produce
curves in the DNA. The profile obtained using Trifonov´s co-ordinates (Bolshoy et al.,
1991) imply a significant curvature within this promoter, similar in extent to that
observed in the Shigella virF promoter which is known to be regulated by the binding of
H-NS (Falconi et al., 1998b).
4. Discussion
Expression of Yop fusion proteins in E. coli using the original Yop promoters
have been analysed with YopE and YopH fused to β-Galactosidase. Thermal induction
of Y. pestis YopE/β-Gal expression in E. coli is strictly dependent on the presence of a
57

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
plasmid containing the lcrF locus (Hoe et al., 1992). Y. enterocolitica YopH/β-Gal
fusion protein was expressed at a very low level in E. coli but was seen to have a slight
increase in expression at 37oC, as compared to 25oC, in the absence of VirF (Cornelis et
al., 1987). In both cases expression was assayed by dosage of β-Gal activity and both
papers agreed that thermal regulation of Yop expression in E. coli wasn´t effective in
the absence of VirF (Cornelis et al., 1987; Hoe et al., 1992; Michiels and Cornelis,
1988), and overall promoter strength was weak.
VirF is a transcriptional activator, member of the AraC family of transcriptional
regulators (Lambert de Rouvroit et al., 1992). The VirF binding motif has been
postulated as degenerate half sites observed in the different promoters regulated by
VirF, with the sequence TTTTaGYcTtTat as one half site. We searched the E. coli
genome database for possible homologues of VirF which could recognize the YopH
promoter and mediate VirF transcriptional activation. Although several proteins
belonging to the AraC family can be identified, including many with yet unassigned
functions, no homologues with greater than 35% overall identity could be seen using the
complete VirF ORF. Therefore, VirF-related proteins are not likely to be required for
the transcriptional activation of YopH expression at 37oC seen in our system.
Temperature dependent alterations in structured (curved) DNA are recognised as
important contributors in the regulation of expression of several virulence proteins
(Perez-Martin and de, V, 1997). Temperature shifts from 25 to 37oC have been shown
to abrogate curves in the DNA and therefore relieve transcriptional repression mediated
by histone-like proteins such as H-NS, which binds preferentially to curved DNA
(Falconi et al., 1998a; Owen-Hughes et al., 1992). Thermo-regulation of virulence
factors in Yersinia is sensitive to changes in DNA supercoiling (Rohde et al., 1994), and
multiple DNA curves have been detected in Yersinia pYV plasmid which are sensitive
to temperature changes and might be involved in the transcriptional regulation of
virulence factors (Rohde et al., 1999). The prediction of a very strong curve in the
YopH promoter fragment used in this study is in agreement with this hypothesis and
suggests that the curved DNA might be responsible for the thermoregulation of YopH
transcription in E. coli. As to protein factors which interact with the curved DNA, aside
from H-NS, another histone-like protein involved in temperature mediated
transcriptional regulation of virulence factors in E. coli (Mourino et al., 1996) is Hha
which has also been shown to be capable of having a synergistic effect with H-NS in
regulating gene expression (Nieto et al., 1997). Our results confirm that H-NS might be
58

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
involved in preventing promoter activity at 25oC in E. coli with or without additional
factors such as Hha.
Our results indicate that protein factors present in both E. coli and Yersinia are
involved in the temperature regulation of YopH expression and that the role of Yersinia
specific factors might be to create another level of regulation superimposed upon the
basic transcriptional regulation mediated by temperature.
Acknowledgements. We would like to thank Cristina Madrid and Dr. Antonio
Juarez for the E. coli strains lacking the hha and hns genes and Gioacchino Micheli for
access to the WEDGE 430 software. SMRC was funded by the Brazilian funding
agency CAPES. BLAS and PROH were recipients of scholarships given by FACEPE.
Part of this work was funded by a PAPES II Grant from FIOCRUZ.
Reference List
Atlung,T. and Ingmer,H. (1997). H-NS: a modulator of environmentally regulated gene expression. Mol. Microbiol. 24, 7-17.
Balsalobre,C., Juarez,A., Madrid,C., Mourino,M., Prenafeta,A., and Munoa,F.J. (1996). Complementation of the hha mutation in Escherichia coli by the ymoA gene from Yersinia enterocolitica: Dependence on the gene dosage. Microbiology-Uk 142, 1841-1846.
Bleves,S. and Cornelis,G.R. (2000). How to survive in the host: the Yersinia lesson. Microbes Infect. 2, 1451-1460.
Bliska,J.B., Guan,K.L., Dixon,J.E., and Falkow,S. (1991). Tyrosine Phosphate Hydrolysis of Host Proteins by An Essential Yersinia-Virulence Determinant. Proceedings of the National Academy of Sciences of the United States of America 88, 1187-1191.
Bolshoy,A., Mcnamara,P., Harrington,R.E., and Trifonov,E.N. (1991). Curved Dna Without A-A - Experimental Estimation of All 16 Dna Wedge Angles. Proceedings of the National Academy of Sciences of the United States of America 88, 2312-2316.
Cheng,L.W. and Schneewind,O. (2000). Type III machines of Gram-negative bacteria: delivering the goods. Trends Microbiol. 8, 214-220.
Conchas,R.F. and Carniel,E. (1990). A Highly Efficient Electroporation System for Transformation of Yersinia. Gene 87, 133-137.
Cornelis,G., Vanootegem,J.C., and Sluiters,C. (1987). Transcription of the yop regulon from Y. enterocolitica requires trans acting pYV and chromosomal genes. Microb. Pathog. 2, 367-379.
59

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Cornelis,G.R. (1998). The Yersinia Yop virulon, a bacterial system to subvert cells of the primary host defense. Folia Microbiologica 43, 253-261.
Cornelis,G.R. (1993). Role of the transcription activator virF and the histone-like protein YmoA in the thermoregulation of virulence functions in yersiniae. Zentralbl. Bakteriol. 278, 149-164.
Cornelis,G.R., Boland,A., Boyd,A.P., Geuijen,C., Iriarte,M., Neyt,C., Sory,M.P., and Stainier,I. (1998). The virulence plasmid of Yersinia, an antihost genome. Microbiology and Molecular Biology Reviews 62, 1315-+.
Cornelis,G.R., Sluiters,C., Delor,I., Geib,D., Kaniga,K., Lambert de Rouvroit,C., Sory,M.P., Vanooteghem,J.C., and Michiels,T. (1991). ymoA, a Yersinia enterocolitica chromosomal gene modulating the expression of virulence functions. Mol. Microbiol. 5, 1023-1034.
Falconi,M., Colonna,B., Prosseda,G., Micheli,G., and Gualerzi,C.O. (1998c). Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. EMBO J. 17, 7033-7043.
Falconi,M., Colonna,B., Prosseda,G., Micheli,G., and Gualerzi,C.O. (1998b). Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. Embo Journal 17, 7033-7043.
Falconi,M., Colonna,B., Prosseda,G., Micheli,G., and Gualerzi,C.O. (1998a). Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. Embo Journal 17, 7033-7043.
Hoe,N.P., Minion,F.C., and Goguen,J.D. (1992). Temperature Sensing in Yersinia-Pestis - Regulation of Yope Transcription by Lcrf. Journal of Bacteriology 174, 4275-4286.
Hueck,C.J. (1998). Type III protein secretion systems in bacterial pathogens of animals and plants. Microbiology and Molecular Biology Reviews 62, 379-+.
Lambert de Rouvroit,C., Sluiters,C., and Cornelis,G.R. (1992). Role of the transcriptional activator, VirF, and temperature in the expression of the pYV plasmid genes of Yersinia enterocolitica. Mol. Microbiol. 6, 395-409.
Madrid,C., Nieto,J.M., and Juarez,A. (2002). Role of the Hha/YmoA family of proteins in the thermoregulation of the expression of virulence factors. Int. J. Med. Microbiol. 291, 425-432.
Michiels,T. and Cornelis,G. (1988). Nucleotide sequence and transcription analysis of yop51 from Yersinia enterocolitica W22703. Microb. Pathog. 5, 449-459.
Mikulskis,A.V. and Cornelis,G.R. (1994). A new class of proteins regulating gene expression in enterobacteria. Mol. Microbiol. 11, 77-86.
60

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Mourino,M., Madrid,C., Balsalobre,C., Prenafeta,A., Munoa,F., Blanco,J., Blanco,M., Blanco,J.E., and Juarez,A. (1996). The Hha protein as a modulator of expression of virulence factors in Escherichia coli. Infection and Immunity 64, 2881-2884.
Nieto,J.M., Madrid,C., Miquelay,E., Parra,J.L., Rodriguez,S., and Juarez,A. (2002). Evidence for direct protein-protein interaction between members of the enterobacterial Hha/YmoA and H-NS families of proteins. J. Bacteriol. 184, 629-635.
Nieto,J.M., Mourino,M., Balsalobre,C., Madrid,C., Prenafeta,A., Munoa,F.J., and Juarez,A. (1997). Construction of a double hha hns mutant of Escherichia coli: effect on DNA supercoiling and alpha-haemolysin production. Fems Microbiology Letters 155, 39-44.
Owen-Hughes,T.A., Pavitt,G.D., Santos,D.S., Sidebotham,J.M., Hulton,C.S., Hinton,J.C., and Higgins,C.F. (1992). The chromatin-associated protein H-NS interacts with curved DNA to influence DNA topology and gene expression. Cell 71, 255-265.
Perez-Martin,J. and de,L., V (1997). Clues and consequences of DNA bending in transcription. Annu. Rev. Microbiol. 51, 593-628.
Rimsky,S. (2004). Structure of the histone-like protein H-NS and its role in regulation and genome superstructure. Curr. Opin. Microbiol. 7, 109-114.
Rohde,J.R., Fox,J.M., and Minnich,S.A. (1994). Thermoregulation in Yersinia-Enterocolitica Is Coincident with Changes in DNA Supercoiling. Molecular Microbiology 12, 187-199.
Rohde,J.R., Luan,X.S., Rohde,H., Fox,J.M., and Minnich,S.A. (1999). The Yersinia enterocolitica pYV virulence plasmid contains multiple intrinsic DNA bends which melt at 37 degrees C. Journal of Bacteriology 181, 4198-4204.
Skeiky,Y.A., Benson,D.R., Guderian,J.A., Whittle,J.A., Bacelar,O., Carvalho,E.M., and Reed,S.G. (1995). Immune responses of leishmaniasis patients to heat shock proteins of Leishmania species and humans. Infect. Immun. 63, 4105-4114.
Steinkraus,H.B., Greer,J.M., Stephenson,D.C., and Langer,P.J. (1993). Sequence heterogeneity and polymorphic gene arrangements of the Leishmania guyanensis gp63 genes. Mol. Biochem. Parasitol. 62, 173-185.
Steinkraus,H.B. and Langer,P.J. (1992). The protein sequence predicted from a Leishmania guyanensis gp63 major surface glycoprotein gene is divergent as compared with other Leishmania species. Mol. Biochem. Parasitol. 52, 141-144.
Tobe,T., Yoshikawa,M., Mizuno,T., and Sasakawa,C. (1993). Transcriptional Control of the Invasion Regulatory Gene Virb of Shigella-Flexneri - Activation by Virf and Repression by H-Ns. Journal of Bacteriology 175, 6142-6149.
Tobe,T., Yoshikawa,M., and Sasakawa,C. (1995). Thermoregulation of Virb Transcription in Shigella-Flexneri by Sensing of Changes in Local Dna Superhelicity. Journal of Bacteriology 177, 1094-1097.
61

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figure 1. Scheme of the construct used in this study. (A) Nucleotide sequence near the
beginning of the yoph gene in the pTZ/YopH plasmid. Shown are the 401 nt upstream
of the translation start codon for YopH as well as the sequence encoding the first 33
amino acids. Sequences derived from the pTZ18R vector, including the lac promoter are
in italic. Capital letters indicate sites of curved DNA. (B) Schematic description of the
sequences shown in A plus the full length YopH orf and neighbouring sequences.
Relevant restriction sites are shown.
62

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figure 2. Expression of plasmid pTZ/YopH in E. coli. Control E. coli DH5α cells as
wells as transformants containing the wild type pTZ18R or the pTZ/YopH plasmids
were grown in liquid LB medium supplemented with Ampicillin at 37°C until reaching
OD 0.7. The arrow shows the YopH protein. (A) SDS-PAGE of total protein extracts
from control E. coli DH5α cells as wells as transformants containing the original
pTZ18R vector or its pTZ/YopH derivative (B) SDS-PAGE and western-blot of
selected samples from A with the antibody against the 52kDa band. (C) Scheme of the
constructs used in this study. (D) SDS-PAGE and western-blot with the YopH-GP63
and YopH-HSP83 fusions.
63

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figure 3. Comparison of the expression levels of the yoph gene under the control of
different promoters. (A) E. coli DH5α cells transformed with the pTZ/YopH plasmid, in
the presence or absence of pREP4, were grown in liquid LB medium supplemented with
Ampicillin, or Ampicillin/Kanamycin, at 37°C until reaching OD 0.5. Cultures were
then divided into 4 aliquots which differed as to the presence or not of IPTG and the
O.D. at which the cells were harvested (0.5 or 1.0 as indicated). The samples were the
analysed through Western-blotts using the antibody directed against YopH. (B) E. coli
DH5α cells transformed with different plasmid vectors containing the yoph gene, as
described in figure 2C, were analysed as in A in the absence of IPTG.
64

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figure 4. SDS-PAGE and Western-blots showing the different levels of expression of
the YopH and fusion proteins at different temperatures and O.D.s. (A) SDS-PAGE and
Western-blot using the anti-YopH antisera with the samples shown in A. ON stands for
overnight. (B) Analysis produced by the densitometric scanning of the YopH bands in
the Western-blot shown in A. The values shown in the graph are arbitrary units where
the value obtained for the O.D. of the ON sample at 24oC was defined as 1.
65

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
66

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figure 6. yopH promoter sequence used in this work as well as the predicted curvature
profile obtained using the "Wedge" software. A. Wedge software profile for the curving
of the DNA sequence shown in figure 1 (nucleotides -109 to +41 of the YopH
promoter). Shown are the VirF binding sites I (blue) and II (red), as well as the
transcription start site (blue arrow). B. Same as A but using the Shigella VirF promoter
sequence (-289 to +67), showing the binding sites for the H-NS protein (red and pink
lines for sites I and II respectively - based on (Falconi et al., 1998c).
67

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
68

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
ARTIGO II
Co-expressão das proteínas SycH e YopH de Yersinia enterocolitica.
Thiago de Salazar e Fernandes1, Mariana Guerra1, Patrícia R. de O. Haver2,
Suzana M. R. Costa3, Bruno L. A. Silva1, Alzira M. P. Almeida4 e Osvaldo
P. de Melo Neto1
1. Departamento de Microbiologia, Centro de Pesquisas Aggeu Magalhães, FIOCRUZ/MS, Recife, PE, Brasil. 2. Doutorado em Ciências Biológicas, CCB/UFPE, Recife, PE, Brasil 3. Mestrado em Genética, Departamento de Genética, CCB/UFPE 4. Laboratório de Peste, Departamento de Microbiologia, Centro de Pesquisas Aggeu Magalhães, FIOCRUZ/MS, Recife, PE, Brasil.
Palavras chave: YopH, Yersinia, AraC, EPEC
Manuscrito a ser submetido para publicação na
Genetics and Molecular biology.
69

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Co-expressão das proteínas SycH e YopH de Yersinia enterocolitica
Thiago de Salazar e Fernandes1, Mariana Guerra, Patrícia Haver2, Suzana Costa, Bruno
Silva, Alzira Almeida3 e Osvaldo de Melo Neto3
1 Universidade Federal de Pernambuco, Depto. de Energia Nuclear, Pernambuco, Brasil 2 Universidade Federal de Pernambuco, Centro de Ciências Biológicas, Pernambuco, Brasil 3 Centro de Pesquisas Aggeu Magalhães, Depto. de Microbiologia, Pernambuco, Brasil Resumo A proteína YopH é uma tirosina fosfatase que é secretada/translocada pelo sistema de secreção tipo III, presente em bactérias do gênero Yersinia. Para uma eficiente secreção, ela necessita da chaperona SycH, que previne sua degradação e a mantém em conformação apropriada. Neste trabalho, foi amplificada a sequência codificadora da SycH, que foi clonada em plasmídios da série pBAD. Foi realizada a expressão dos plasmídios pBAD18/SycH e pBAD33/SycH, e sua co-expressão com o plasmídio pTZ/YopH, em células de Escherichia coli enteropatogênica (EPEC) também dotadas de sistema de secreção tipo III. A adição de arabinose e glicose, no meio de cultura, provocou uma redução na expressão da YopH, que também foi observada na ausência dos plasmídios pBAD. O plasmídio pTZ/YopH contém o promotor yoph, que apresenta os sítios de ligação para a proteína VirF e regiões curvas de DNA. VirF age como um regulador transcricional em Yersinia e pertence a família AraC/Xyls. Em E. coli, a YopH é regulada positivamente pela proteína H-NS, temperatura e fase de crescimento. O mecanismo responsável pela diminuição da expressão da YopH, na presença de arabinose e glicose permanece indefinido. Futuros estudos são necessários para elucidar todos os fatores envolvidos na transcrição da YopH. Palavras chave: YopH, Yersinia, AraC, EPEC
Endereço de correspondência para Dr. Osvaldo P. de Melo Neto, CPqAM/FIOCRUZ/MS. Campus da
UFPE, Cidade Universitária, 50670-420. Recife, PE, Brasil. Tel: +55 81 2101 2636; fax:+ 55 81 3453
2449. E-mail: [email protected].
70

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Introdução Bactérias do gênero Yersinia e Escherichia coli enteropatogênica (EPEC),
utilizam o sistema de secreção tipo III (TTSS) para injetar proteínas no interior de
macrófagos e leucócitos polimorfonucleares (PMNs), como mecanismo de defesa contra
a resposta imune (Hueck, 1998).
O gênero Yersinia possui 3 espécies patogênicas para humanos e outros animais:
Y. pestis, o agente da peste; e a Y. enterocolitica e Y. pseudotuberculosis,
enteropatógenos responsáveis pela yersiniose. Para sobreviver e se multiplicar nos
tecidos linfóides, estas bactérias necessitam de um plasmídio de virulência chamado
pYV, que codifica as proteínas responsáveis pelo TTS, as proteínas Yops (Yersinia
Outer Proteins) e suas chaperonas Sycs (Specific Yop Chaperone). Entre as principais
Yops, encontra-se a YopH, uma tirosina fosfatase que participa do bloqueio da
fagocitose e indução da apoptose. Para a sua secreção, é necessária uma chaperona
específica, chamada SycH, no citoplasma bacteriano, que previne sua degradação e a
mantém em conformação apropriada para a secreção (Cornelis, 2002a, 2002b; Cornelis
et al., 1998; Perry and Fetherston, 1997).
A patogenicidade de E. coli enteropatogênica é conferida pela presença de uma
ilha de patogenicidade, formada pela região LEE (locus for enterocyte effacement).
LEE é responsável pela codificação dos componentes do sistema de secreção tipo III, as
proteínas Esps (EPEC-secreted proteins) e suas chaperonas. As Esps são fatores
similares as Yops, que promovem rearranjos no citoesqueleto da célula do hospedeiro e
induzem a apoptose (Groisman, 2001; Wolff e cols., 1998).
A regulação da transcrição é um dos principais mecanismos de controle da
expressão gênica em procariotas e alterações na estrutura do DNA podem modular a
ativação ou repressão de genes. Em Yersiniae, a expressão das Yops é ativada por
temperatura (37oC) e controlada pela proteína VirF, que é codificada no pYV. VirF
pertence à família de reguladores AraC/Xyls, com o qual compartilha 23% de
homologia na região C-terminal. A proteína H-NS apresenta uma grande homologia
com as histonas eucarióticas, e é responsável pela regulação da proteína YopH, em E.
coli. Em EPEC, essa proteína interage com sequências regulatórias e influencia a
expressão dos genes codificados pela região LEE (MacLeod and Johnson, 2001;
Cornelis et al., 1998; Puente et al., 1996; Lambert de Rouvroit et al., 1992; Cornelis et
al., 1989; Haver, dados não publicados).
71

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Neste trabalho, foi amplificada a sequência codificadora da SycH, que foi
clonada em plasmídios da série pBAD. Foi realizada a expressão dos plasmídios
pBAD18/SycH e pBAD33/SycH, e sua co-expressão com o plasmídio pTZ/YopH, que
contém o promotor yoph. A expressão dessas proteínas foi analisada em culturas de E.
coli enteropatogênicas.
Material e Métodos Culturas bacterianas
Para as preparações de DNA, foi utilizada a linhagem E. coli DH5α e para as
análises de expressão, E. coli TG2 e E. coli enteropatogênica. As bactérias foram
crescidas em meio de cultura LB (Luria Bertani) e MGM (Meio Glicerol Mínimo),
suplementados ou não com antibióticos nas seguintes concentrações (µg/ml):
Ampicilina (Amp) 100, Kanamicina (Km) 20 e Cloranfenicol (Cm) 30.
Plasmídios
Foi utilizado o plasmídio pTZ/YopH, contendo o promotor yoph e a seqüência
codificadora para a proteína YopH (Costa, 1999). As clonagens da proteína SycH foram
realizadas nos plasmídios pTZ18R (GenBank accession: L08956), pBAD18 e pBAD33
(Guzman e cols., 1995).
Clonagem do gene sych, de Y. enterocolitica
A seqüência codificante para o gene sych foi amplificada a partir do DNA total
de Y. enterocolitica O:3 Ye35. Foram utilizados os seguintes oligonucleotídeos: P 3116
(5´GGTCTAGATTCTAATTTTAAAGGGTCAG3´), desenhado para anelar antes da
sequência Shine Delgarno e contendo um sítio de restrição Xba I; e P3117
(5’GGCTGCAGATTAACAAAAATATAAACAAC3’), desenhado para anelar após a
sequência codificadora e contendo um sítio de restrição Pst I. O fragmento de PCR foi
clonado no plasmídio pTZ18R (GenBank accession: L08956), entre os sítios Xba I e Pst
I. A transformação foi realizada em E. coli DH5α, segundo Sambrook et al. (2001).
Preparações de DNA a partir de culturas crescidas a 37oC, em meio LB, revelaram
plasmídios recombinantes contendo um fragmento de 530 pb (pTZ/SycH). Digestão por
enzimas de restrição e sequenciamento de clones selecionados confirmaram a identidade
do fragmento.
72

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Subclonagem da proteína SycH nos plasmídios da série pBAD
O plasmídio contendo o gene codificante para a proteína SycH foi digerido a
partir do plasmídio pTZ/SycH, utilizando as enzimas de restrição Xba I e Pst I. O
fragmento foi purificado, utilizando iodeto de sódio (NaI) e sílica, e subclonado para os
plasmídios pBAD18 e pBAD33. A clonagem foi realizada por um período de 15 horas,
a 16°C, com a enzima T4 DNA Ligase (GIBCO BRL).
Expressão protéica
Para a expressão da proteína SycH, as culturas de E. coli foram incubadas a
37ºC, em 3ml de meio glicerol mínimo acrescido de 0,2% de glicose, por 16 horas.
Foram transferidos 100 µl da cultura para 5ml do mesmo meio e o crescimento
bacteriano foi monitorado até atingir uma DO (densidade ótica) de 0,5 (comprimento de
onda 600nm). Após a coleta da amostra controle, a cultura foi centrifugada e
ressuspendida em meio acrescido de 0,05% de arabinose. A cultura foi colocada a 37oC
e as demais alíquotas foram retiradas após 24 e 48 horas. No meio LB, as culturas foram
crescidas até atingir uma densidade ótica igual a 0,5; e foi então retirada uma alíquota
controle. A arabinose (0,05%) foi adicionada como indutora da expressão; e a glicose
(0,2%) como repressora.
A expressão da proteína YopH foi realizada em culturas incubadas a 24ºC até
atingirem uma DO igual a 0,5. Foi retirada uma alíquota controle. As culturas foram
então incubadas a 37oC e as alíquotas retiradas após 02 e 04 horas de indução.
Na co-expressão das proteínas SycH e YopH, as culturas foram incubadas a
24ºC, até atingir uma DO igual a 0,5. Foi retirada uma alíquota controle. A arabinose foi
adicionada em três concentrações (0,02%, 0,2% e 2%). As culturas foram então
colocadas a 37oC, e as amostras foram coletadas no início do crescimento e após 4
horas.
Para a preparação das amostras, foi utilizado o tampão de Laemmli (duas vezes
concentrado) acrescido de β-mercaptoetanol (50µl/ml), segundo Laemli (1970).
Análise da expressão protéica
As amostras obtidas foram separadas em gel de poliacrilamida 15% e 20%, em
condições desnaturantes (SDS-PAGE). Os géis foram corados com azul de Coomassie e
depois submetidos a uma solução descorante (25% de metanol, 7% de ácido acético e
H2O).
73

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Para o Western-blot, as proteínas foram transferidas para membrana de
nitrocelulose PVDF (“Polyvinylidene Difluoride Membrane” – Immobilon-P –
Milipore), em sistema semi-seco. O anticorpo anti-YopH foi utilizado nas
concentrações de 1:15.000 e 1:90.000. A anti-IgG de coelho conjugado a peroxidase
(Jackson Immunoresearch laboratories INC) foi diluído 1/5.000. A revelação foi feita
por quimioluminescência (ECL) , sendo utilizada uma solução com 1,2mM de Luminol,
0,4mM de Iodofenol diluído em DMSO e H2O2 (10 volumes) a 0,03%. A membrana foi
incubada por 1 minuto e exposta a filme de auto-radiografia por 5 a 10 minutos.
Resultados Expressão da proteína SycH
Para expressar a proteína SycH, culturas bacterianas de E. coli TG2 foram
transformadas com os plasmídios pBAD18/SycH e pBAD33/SycH, em meio MGM
contendo arabinose a 0,05%. Este meio de cultura foi escolhido por ser um meio pobre
em glicose, pois este açúcar exerce um estímulo repressor sobre o promotor PBAD.
Entretanto, na presença de arabinose, este promotor é estimulado e ocorre a transcrição
do gene sycH. A análise das amostras separadas por SDS/PAGE 17,5%, revelou o
surgimento de uma proteína de aproximadamente 17 kDa, correspondente ao tamanho
esperado da proteína SycH. A expressão ocorreu apenas na presença de arabinose,
sendo mais forte após 24 horas de crescimento (Figura 1).
Para as análises de expressão, foi escolhida a bactéria E. coli EPEC por
apresentar o TTSS e poder ser utilizada, no futuro, para analisar a secreção da proteína
YopH, na presença de SycH. Culturas de E. coli EPEC foram transformadas com o
plasmídio pBAD33/SycH e crescidas em meio LB, que permite um crescimento
bacteriano mais rápido. A análise das amostras em SDS/PAGE 20% revelou a presença
da proteína SycH apenas no meio contendo arabinose a 0,05%. A proteína não foi
expressa na presença de glicose (dados não mostrados).
Expressão da proteína YopH
Para avaliar a expressão da proteína YopH em culturas de E. coli EPEC,
bactérias transformadas com o plasmídio pTZ/YopH foram incubadas a 37oC. As
amostras foram separadas por eletroforese e submetidas a Western-blot, para o
reconhecimento pelo anticorpo específico anti-YopH. Foi observada uma banda de
74

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
52kDa correspondendo à presença da YopH, que apresentou uma expressão intensa a
37oC (Figura 2).
Co-expressão das proteínas SycH e YopH
Com a finalidade de analisar a influência da SycH na secreção da YopH,
culturas de EPEC foram co-transformadas com os plasmídios pTZ/YopH e
pBAD33/SycH. Para que os dois plasmídios fossem mantidos na bactéria, eles
apresentavam diferentes origens de replicação e conferiam resistências a antibióticos
diferentes. Culturas bacterianas sem o plasmídio pBAD33/SycH e sem adição de
arabinose foram utilizadas como controle. As amostras foram separadas por SDS/PAGE
15% e submetidas a Western-blot. Nas culturas contendo apenas o plasmídio
pTZ/YopH, foi observada uma forte expressão da YopH, que também foi observada na
ausência de arabinose. Entretanto, nas culturas contendo os dois plasmídios, observou-
se uma redução da expressão da YopH, proporcional a concentração de arabinose, sendo
maior a 2% (Figura 3).
Para verificar se a diminuição da expressão da YopH estava relacionada com a
presença da proteína SycH, culturas de EPEC foram transformadas apenas com o
plasmídio pTZ/YopH. Foram utilizadas arabinose 2%, glicose 2% e IPTG 0,1mM; além
de um grupo controle. As amostras foram analisadas por eletroforese e Western-blot. A
redução nos níveis de expressão da YopH foi observada na presença de arabinose e
glicose, não sendo observada na presença de IPTG (Figura 4).
Discussão O sistema de secreção tipo III é utilizado por muitas bactérias Gram negativas,
sendo um dos mecanismos mais sofisticados para a translocação de proteínas através de
membranas lipídicas. As proteínas chaperonas são essenciais para a secreção e sua
ausência resulta em secreção deficiente da proteína cognata (Feldman and Cornelis,
2003). Em estudos com a proteína YopH, em E. coli, foi observado que a expressão
desta proteína é regulada pela temperatura, como ocorre em Yersiniae, devido à
presença do promotor yoph. Análise da sequência promotora revelou a presença de
regiões curvas de DNA e dos sítios para a ligação da proteína VirF, que regula a
expressão das Yops, em yersínia (Haver, dados não publicados).
75

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Para estudar a expressão da proteína SycH, foram utilizados plasmídios da série
pBAD por ser um sistema que pode ser ligado e desligado de maneira rápida e eficiente,
além da expressão poder ser modulada através de diferentes concentrações de arabinose
(Guzman et al., 1995). Entretanto, a co-expressão desta proteína com a YopH, produziu
uma diminuição na expressão da YopH na presença de arabinose. Esta redução também
foi observada na ausência do plasmídio pBAD33/SycH, indicando que a proteína YopH
não estava sendo secretada.
A utilização destes vetores se mostrou eficiente para a expressão da proteína
SycH isoladamente, porém o mesmo não ocorreu na sua co-expressão com a proteína
YopH, já que o fator de transcrição que ativa a expressão da SycH reprime a expressão
da YopH. Para futuros estudos sobre a secreção, terá que ser utilizado um vetor de
expressão diferente que não influencie a expressão da YopH. Uma outra possibilidade é
clonar os dois genes no mesmo plasmídio, sob o controle de um promotor que permita a
expressão das duas proteínas.
Referências Cornelis GR (2002a). The Yersinia Ysc-Yop 'type III' weaponry. Nat Rev Mol Cell Biol
3 (10): 742-752.
Cornelis GR (2002b). Yersinia type III secretion: send in the effectors. J Cell Biol 158
(3): 401–408.
Cornelis GR, Boland A, Boyd AP, Geuijen C, Iriarte M, Neyt C, Sory MP and Stainier
I. (1998). The virulence plasmid of Yersinia, an antihost genome. Microbiol. Mol
Biol Rev 62: 1315-1352.
Cornelis GR, Sluiters C, Lambert de Rouvroit C and Michiels T (1989). Homology
between VirF, the transcriptional activator of the Yersinia virulence regulon, and
AraC, the Escherichia coli arabinose operon regulator. J Bacteriol 171: 254-262.
Costa SMR (1999). Análise da expressão de proteínas fusionadas ao fator de virulência
YopH de Yersinia enterocolitica sob o controle do promotor yopH em Escherichia
coli. Tese de mestrado UFPE.
Feldman MF and Cornelis GR (2003). The multitalented type III chapeones: all you can
do with 15 kDa. FEMS Microbiol Lett 219: 151-158.
Groisman EA (2001). Principles of bacterial pathogenesis. Academic Press.
76

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Guzman LM, Belin D, Carson MJ and Beckwith J (1995). Tight regulation, modulation,
and high-level expression by vectors containing the arabinose PBAD promoter. J
Bacteriol 177 (14): 4121-4130.
Hueck CJ (1998). Type III protein secretion systems in bacterial pathogens of animals
and plants. Microbiol. Mol. Biol. Rev. 62: 379-433.
Laemmli UK (1970). Cleavage of structural proteins during the assembly of the head of
the bacteriophag T4. Nature 227: 680-685.
Lambert de Rouvroit C; Sluiters C; Cornelis GR. Role of the transcriptional activator
VirF and temperature in the expression of the pYV plasmid genes of Yersinia
enterocolitica. Mol. Microbiol. 6: 379-388, 1992.
MacLeod SM; Johnson RC. Control of transcription by nucleoid proteins. Curr. Op.
Microbiol. 4: 152-159, 2001.
Perry RD; Fetherston JD. Yersinia pestis-etiologic agent of plague. Clin. Microbiol.
Rev. 10: 35-66, 1997.
Puente JL; Bieber D; Ramer SW; Murray W; Schoolnik GK. The bundle-forming pili of
enteropathogenic Escherichia coli: transcriptional regulation by environmental
signals. Mol. Microbiol. 20 (1): 87-100, 1996.
Wolff C; Nisan I; Hanski E; Frankel G; Rosenshine I. Protein translocation into host
epithelial cells by infecting enteropathogenic Escherichia coli. Mol. Microbiol. 28:
143-155, 1998.
77

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 1 - SDS/PAGE 17,5% mostrando a síntese da proteína SycH (seta), em culturas
de E. coli TG2 transformadas com os plasmídios pBAD18/SycH (A) e pBAD33/SycH
(B). C-controle.
78

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 2 - Western-blot mostrando a síntese da proteína YopH a 37oC(seta), em
culturas de E. coli EPEC, transformadas com o plasmídio pTZ/YopH. C-controle, 24oC.
79

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 3 - Co-expressão das proteínas YopH (seta) e SycH (asterico). A- SDS/PAGE,
B- Western-blot.
80

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Figura 4 - Expressão da proteína YopH, em EPEC, na presença de glicose 2%,
arabinose 2% e IPTG 0,1mM. C-controle.
81

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
ARTIGO III
Avaliação da YopH recombinante no diagnóstico sorológico
da peste
Marina M. V. Petribú1, Patrícia R. O. Haver2, S. Santos, A. I. Aragão3,
Alzira M. P. de Almeida1
1. Laboratório de Peste, Departamento de Microbiologia, Centro de Pesquisas Aggeu Magalhães, FIOCRUZ/MS, Recife, PE, Brasil. 2. Doutorado em Ciências Biológicas, CCB/UFPE, Recife, PE, Brasil 3. Secretaria Estadual de Saúde, CE, Brasil
Palavras chave: Peste, Diagnóstico, F1, YopH, Yersinia pestis
Manuscrito a ser submetido para publicação na:
Memórias do Instituto Osvaldo Cruz
82

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Avaliação da YopH recombinante no diagnóstico sorológico da peste
MMV Petribú, PO Haver, S Santos, AI Aragão, AMP de Almeida*
Departamento de Microbiologia, Centro de Pesquisas Aggeu Magalhães-Fiocruz,
Campus da UFPE, Cidade Universitária, 50670-420 Recife, PE, Brasil
A morbimortalidade por peste entrou em declínio nos focos brasileiros a partir
da década de 70. No decorrer dos últimos anos, a hipótese diagnóstica tem sido
aventada e as amostras coletadas são testadas pela prova de hemaglutinação (HA) para
detecção de anticorpos contra o antígeno F1 da Yersinia pestis e, apesar de sua
fundamentação clínico-epidemiológica, os casos têm sido descartados por serem
reiteradamente não reagentes (HA-). A transcendência da peste justifica estudar se tais
resultados decorrem da falta de resposta à F1, constituindo uma situação de risco pela
ocorrência de falso-negativos, e se outras proteínas da Y. pestis são reconhecidas pelos
soros reagentes e não reagentes por HA, bem como identificar novos alvos para
diagnóstico da peste. Assim sendo, soros humanos de pacientes suspeitos de peste, soros
de coelho anti-YopH e soros de coelhos imunizados com cepas brasileiras e estrangeiras
de Y. pestis foram analisados por western-blot utilizando a proteína YopH
recombinante, a proteína F1 purificada, e proteínas totais de cepas de Y. pestis de
diversas origens e de uma cepa de Y. pseudotuberculosis. Nas análises por western-blot
os padrões de reconhecimento foram muito variados com os diversos soros. A F1 e a
YopH não foram reconhecidas pelos soros humanos HA- e nenhuma proteína
majoritária, comum a todos os soros humanos e coelhos, foi identificada, o que permite
concluir que os casos suspeitos devem ser submetidos a uma avaliação clínica mais
rigorosa, aprofundando a investigação epidemiológica em busca de outras etiologias.
Palavras-chaves: Yersinia pestis, peste, F1, YopH, diagnóstico
Endereço para correspondência: Dra Alzira de Almeida. CPqAM/FIOCRUZ/MS.
Campus da UFPE, Cidade Universitária, 50670-420. Recife, PE, Brasil. Tel: 55 81
33012634 Fax: 55 81 3453 2449 e-mail: [email protected]
83

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
A peste, infecção pela Yersinia pestis, mantem-se em vários focos naturais na
África, Ásia e Américas e atualmente é considerada uma doença reemergente pela
Organização Mundial da Saúde (WHO 2004, Higgins 2004), constituindo um problema
de Saúde Pública. A epidemiologia da peste é bastante complexa e, apesar dos avanços
científicos e tecnológicos, a sua erradicação ainda não é exeqüível (Gage & Kosoy
2004). A elevada letalidade e potencial de epidemização justificam a sua inclusão na
Classe I do Regulamento Sanitário Internacional vigente, que exige notificação
compulsória de toda atividade pestosa e manutenção de vigilância permanente nos focos
e locais por onde a infecção possa ser introduzida a partir de focos ativos.
Vários fatores de virulência da Y. pestis são codificados em genes localizados no
cromossomo e nos três plasmídeos prototípicos: pPst, pFra e pYV. O plasmídeo pPst,
específico da Y. pestis, codifica para uma protease (Pla) ou ativador do plasminogênio.
No plasmídeo pFra, também específico, estão localizados os genes que codificam para
uma proteína de envoltório, a fração antigênica F1, e a toxina murina (Perry &
Fetherston 1997). A F1 é imunogênica para homens e animais, o que fundamenta a sua
utilização na maioria dos testes diagnósticos da peste (Chu 2000). A toxina murina
(Ymt) é uma exotoxina que parece desempenhar um importante papel na colonização da
Y. pestis no estômago das pulgas (Hinnebush 2002). Payne et al. (1956) observaram que
ela é menos imunogênica que a F1 e o seu emprego no diagnóstico sorológico da peste é
irrelevante. O plasmídio pYV é indispensável à virulência da bactéria e também é
encontrado nas outras yersínias patogênicas: Y. pseudotuberculosis e Y. enterocolitica.
O pYV codifica para um complexo sistema de secreção tipo III, responsável pela síntese
de várias proteínas envolvidas na neutralização das defesas dos hospedeiros (Cornelis
2002) e cujo uso no diagnóstico da peste limita-se a uns poucos testes (Mazza et al.
1985, Gomes-Solecki et al. 2005).
A prova da hemaglutinação (HA) com hemácias de carneiro, para detecção de
anticorpos contra o antígeno F1 (WHO 1970), é utilizada há várias décadas no
diagnóstico e vigilância da peste humana e em animais nos focos brasileiros. As
amostras de soro pareadas, colhidas na fase aguda e na convalescença, devem mostrar
uma diferença de quatro títulos ou mais. Na maioria dos pacientes a soroconversão
ocorre dentro de uma a duas semanas após o início dos sintomas, mas em alguns
acontece mais precocemente, até cinco dias, e noutros ela não ocorre (Almeida et al.
1981, 1989, 1995, Aragão et al. 2002).
84

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Nos focos, caso não se disponha de pessoal qualificado para desenvolver a
investigação epidemiológica, quaisquer linfoadenopatias podem conduzir a um
diagnóstico equivocado, o que determinaria a negatividade dos exames. A plasticidade
do genoma da Y. pestis, com perda de plasmídeos, de segmentos cromossômicos e
inserção de seqüências (Perry & Fetherston 1997), por sua vez, permite supor que a falta
de resposta de casos poderia decorrer de infecções por cepas atípicas não produtoras de
F1, o que também implicaria na ocorrência de casos falso-negativos. Cumpre, pois,
identificar novos alvos, o que possibilitará o desenvolvimento de uma nova geração de
técnicas diagnósticas.
O objetivo deste trabalho é verificar se outras proteínas da Y. pestis são
reconhecidas pelos soros reagentes e não reagentes nos testes de detecção de anticorpos
anti-F1, e identificar outros alvos para o desenvolvimento de novos testes de
diagnóstico, confirmando ou descartando os suspeitos.
MATERIAIS E MÉTODOS
O estudo envolveu cinco cepas de Y. pestis (P. Exu 769, P. PB 881, P. CE 882,
A1122 e EV76) da coleção de culturas do Departamento de Microbiologia do Centro de
Pesquisas Aggeu Magalhães (CPqAM) (Almeida et al. 1985, 1989) e uma Y.
pseudotuberculosis (IP 32950); Escherichia coli DH5α transformada com o plasmídeo
PTZ/YopH (Costa 1999); antígeno F1 produzido no Serviço de Referência em Peste
(SRP) do CPqAM segundo descrito em Chu (2000); soros de coelhos imunizados com
F1 (anti-F1), com Y. pseudotuberculosis IP32950 (C/YP 2950) e com as cepas de Y.
pestis (C/P. Exu 769, C/P. PB 881, C/P.CE 882, C/YP A1122, C/YP EV76) produzidos
no SRP segundo descrito por Bahmanyar & Cavanaugh (1976); soro de coelho anti-
YopH (Costa 1999), soro normal de coelho (SNC), 34 soros humanos (04 HA+) da
soroteca do SRP, obtidos pelas equipes do serviço de vigilância e controle da peste no
Brasil (Almeida et al. 1995, FUNASA 2002).
Os soros foram analisados por HA segundo WHO (1970) e por western-blot com
proteínas totais das culturas bacterianas e com o antígeno F1, segundo protocolo
descrito por Chu (2000).
Para avaliar o potencial da proteína YopH recombinante no diagnóstico da peste,
membranas contendo proteínas totais da cultura E. coli DH5α transformada com o
plasmídeo PTZ/YopH foram testadas com soro de coelho anti-YopH, soros de coelhos
85

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
imunizados com cepas de Y. pestis originadas de três focos pestosos brasileiros (P. Exu
769, P. PB 881 e P. CE 882) e duas cepas de referência estrangeiras (A1122 e EV76) e
soros humanos com suspeita clínica de peste (HA-).
Buscando identificar outras proteínas de interesse para o diagnóstico da peste, soros
de coelhos imunizados com cepas brasileiras e estrangeiras de Y. pestis foram testados
com o antígeno F1 e proteínas totais das mesmas cepas.
RESULTADOS E DISCUSSÃO
Nos focos brasileiros numerosos casos humanos diagnosticados clinicamente
como peste benigna ou ambulatorial resultam negativos pela HA. Estes resultados
poderiam ser atribuídos a uma baixa sensibilidade da HA ou falta de resposta ao
antígeno F1. Nesse contexto, dois aspectos devem ser ressaltados: a qualidade
discutível do diagnóstico clínico e a progressiva redução da coleta de amostras para
exames bacteriológicos observada nos últimos 30 anos, em decorrência das vantagens
oferecidas pelos exames sorológicos, desconsiderando-se procedimentos eficazes que
poderiam subsidiar a investigação, inclusive no que concerne à avaliação das rotinas e
de novos ensaios (Almeida et al. 1981, Aragão et al. 2002).
A qualidade da assistência, por desconhecimento da doença, implica
diagnósticos equivocados e supressão de componentes da investigação, exigindo a
pesquisa de novas técnicas que reduzam ao mínimo possível a ocorrência de falso-
negativos. Assim sendo, para verificar se as hipóteses de baixa sensibilidade da HA e
da ausência de resposta à F1 justificariam os resultados obtidos, amostras negativas por
HA foram reexaminadas por Dot-Elisa (Almeida & Ferreira 1992). Saliente-se que as
doenças elencadas no diagnóstico diferencial da peste e a coleta inadequada foram
consideradas e que, apesar da melhor sensibilidade do teste, todas as amostras também
resultaram negativas (Aragão et al. 2002, Sousa et al. 1995).
A identificação de novos alvos para diagnóstico e o desenvolvimento de testes
dirigidos a outros antígenos da Y. pestis contribuiria para solucionar um eventual
problema de resultados falso-negativos com os testes tradicionais de detecção de
anticorpos anti-F1.
A YopH inicialmente denominada Yop51 em função do seu peso molecular
(Bolin & Wolf-Watz 1988) desempenha importante papel na resistência bacteriana ao
sistema imune do hospedeiro (Cornelis 2002). A proteína YopH recombinante expressa
86

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
em E. coli DH5α mostrou-se imunogênica em coelhos (Costa 1999). Anticorpos contra
Yops (YopD, YopE) foram reconhecidas por soros de convalescentes de peste e Yops
produzidas por qualquer das três yersinías patogênicas são reconhecidas por soros de
animais imunizados com qualquer uma delas mostrando a similaridade intraespecífica
dessas proteínas (Bolin et al. 1985, Straley & Bowmer 1986, Wolf-Watz et al. 1985).
Nos ensaios de western-blot realizados com as proteínas totais de E. coli
transformada com o plasmídio pTZ/YopH houve reconhecimento de uma banda
correspondendo ao tamanho da proteína YopH (~52 kDa) apenas com o soro anti-YopH
(Fig. 3). Nos soros humanos houve reconhecimento de outras proteínas, mas não da
YopH. Apesar da similaridade das Yops das yersinías a YopH recombinante não foi
reconhecida pelos soros dos coelhos imunizados com cepas de Y. pestis. É possível que
a falta de sinal no western-blot com os soros dos coelhos anti-Y. pestis seja devido à
baixa quantidade de anticorpos anti-YopH produzida. Uma banda de ~65 kDa foi
reconhecida pelos soros humanos e dos coelhos exceto pelo soro normal de coelho. Esta
banda é provavelmente uma proteína da E. coli comum às outras bactérias da família
Enterobacteriaceae.
Nos ensaios de western-blot com antígeno F1 e proteinas totais de Y. pestis
realizados com soros de coelhos imunizados com cepas brasileiras e estrangeiras de Y.
pestis todos os soros reconheceram uma banda correspondente ao tamanho da proteína
F1 (17 kDa). Nos extratos de proteínas totais além dessa banda várias outras proteínas
foram reconhecidas pelos soros. Os padrões de reconhecimento dessas proteínas nas
diferentes cepas de Y. pestis foram muito variados e nenhuma proteína majoritária foi
reconhecida por todos os soros, salvo a F1 (Fig. 1).
Um soro de coelho anti-F1 testado por western-blot com o antígeno F1 e
proteínas totais das cepas de Y. pestis reconheceu tanto a banda da proteína F1
purificada quanto uma banda de igual tamanho nos extratos de proteínas totais de todas
as cepas (dados não mostrados) refletindo a similaridade da F1 produzida pelas cepas de
diferentes focos do Brasil e outros paises. Um soro de coelho anti-Y. pseudotuberculosis
IP32950 não mostrou reação com a membrana contendo F1 o que era previsível, pois a
bactéria não sintetiza a proteína F1 e, conseqüentemente, o soro do coelho infectado por
tal bactéria não deve conter anticorpos contra este antígeno (Fig. 1).
Dos soros humanos testados com a F1 purificada apenas os soros positivos por
HA+ reconheceram a proteína F1. Esses mesmos soros também reconheceram a F1 nos
extratos de proteínas totais das cepas de Y. pestis (Fig. 2). Os soros HA- não
87

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
reconheceram a proteína F1. Outras proteínas foram reconhecidas, porém os padrões
foram muito variados com os diversos soros e as diferentes cepas (Fig. 4 e 5).
A análise desses resultados permite concluir que as investigações
epidemiológicas realizadas devem ser desenvolvidas por equipes altamente qualificadas,
pela necessidade de uma avaliação criteriosa dos casos suspeitos, com rigorosa coleta de
espécimes para exames bacteriológicos e sorológicos, valorizando-se devidamente a
epidemiologia e nosologia regionais, assim como outros recursos diagnósticos, o que
permitirá descartar ou reforçar oportunamente a hipótese aventada, permitindo o pronto
desencadeamento das ações de controle, enquanto se aguarda a confirmação
laboratorial. A HA deve continuar sendo utilizada como padrão, mas esta confirmação
tende a se estear em técnicas moleculares, sensíveis e específicas, mesmo trabalhando-
se com bactérias inviáveis contidas em amostras putrefeitas, mumificadas e / ou
contaminadas, que devem ser aperfeiçoadas, visando à identificação de novos alvos e
técnicas que possam ser executadas nas diversas condições de uma investigação.
REFERÊNCIAS
Almeida CR, Almeida, AMP, Vieira JB, Guida U, Butler T 1981. Plague in Brazil
during two years of bacteriological and serological surveillance. Wld Hlth Org Bull
59: 591-597.
Almeida AMP, Brasil DP, Carvalho FG, Almeida CR 1985. Isolamento de Yersinia
pestis nos focos pestosos do nordeste do Brasil no período de 1966 a 1982. Rev Inst
Med Trop São Paulo 27: 207-218.
Almeida AMP, Brasil DP, Leal NC, Melo MEB, Rego RVB, Almeida CR 1989.
Estudos bacteriológicos e sorológicos de um surto de peste no Estado da Paraíba,
Brasil. Mem Inst Oswaldo Cruz 84: 249-256, 1989.
Almeida AMP, Ferreira LCS 1992. Evaluation of three sorological tests for the
detection of human plague in Northeast Brazil. Mem Inst Oswaldo Cruz 87: 87-92.
Almeida AMP, Leal NC, Carvalho FG, Dantas-Sobrinho J, Almeida CR 1995. Plague
surveillance in Brazil: 1983–1992. Rev Inst Med Trop São Paulo 37: 511-516.
Aragão AI, Seoane AC, Leal NC, Almeida AMP 2002. Vigilância da peste no Estado
do Ceará: 1990-1999. Rev Soc Bras Med Trop 35: 143-148.
Bahmanyar M, Cavanaugh DC 1976. Plague Manual, WHO, Geneva.
88

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Bölin I, Portnoy DA, Wolf-Watz 1985. Expression of the temperature-inducible outer
membrane proteins of Yersiniae. Infect Immun 48: 234-240.
Bölin I, Wolf-Watz 1988. The plasmid-encoded Yop2 protein of Yersinia
pseudotuberculosis is a virulence determinant regulated by calcium and
temperature at the level of transcription. Mol Microbiol 2: 237-245.
Chu M 2000. Laboratory Manual of Plague Diagnostic Tests, CDC/WHO, Geneve,
129pp.
Cornelis GR 2002. Molecular and cell biology aspects of plague. Proc Natl Acad Sci
97: 8778-8783.
Costa SMR 1999. Análise da expressão de proteinas fusionadas ao fator de virulência
YopH de Yersinia enterocolitica sob o controle do promotor yopH em E. coli. Tese
de Mestrado UFPE.
FUNASA 2002. Peste, in: Guia de Vigilância Epidemiológica 5a ed. Vol. II: 641-652.
Gage KL, Kosoy MY 2004. Natural History of Plague: Perspectives from More than a
Century of Research. Ann Rev Entomol 50: 505-528.
Gomes-Solecki MJ, Savitt AG, Rowehl R, Glass JD, Bliska JB, Dattv 2005. LcrV
capture enzyme-linked immunosorbent assay for detection of Yersinia pestis from
human samples. Clin Diagn Lab Immunol 12: 339-346.
Higgins R 2004. Emerging or re-emerging bacterial zoonotic diseases: bartonellosis,
leptospirosis, Lyme borreliosis, plague. Rev sci tech Off int Epiz 23: 569-581.
Hinnebush BJ, Rudolph AE, Cherepanov P, Dixon JE, Schwan TG, Forsberg A 2002.
Role of Yersinia murine toxin in survival of Yersinia pestis in the midgut of the flea
vector. Science 296: 733-735.
Mazza G, Karu AE, Kingsbury DT 1985. Immune response to plasmid- and
chromosome-encoded Yersinia antigens. Infect Immun 48: 676-685.
Payne FE, Smadel JE, Courdurier J 1956. Immunologic studies on persons residing in
a plague area. Infect Immun 77: 24-33.
Perry RD, Fetherston JD 1997. Yersinia pestis – Etiologic Agent of Plague. Clin
Microbiol Rev 10: 35-66.
Sousa AQ, Parise ME, Pompeu ML, Coelho-Filho JM, Vasconcelos IAB, Lima JWO,
Oliveira EG, Vasconcelos AW, David JR, Maguire JH 1995. Bubonic
leishmaniasis: a common manifestation of Leishmania (Viannia) brasiliensis
infection in Ceara, Brazil. Am J Trop Med Hyg 53: 380-385.
89

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Straley S, Bowmer 1986. Virulence genes regulated at the transcriptional level by Ca2+
in Yersinia pestis include structural genes for outer membrane proteins. Infect
Immun 51: 445-454.
WHO 1970. Technical Report Series, WHO Expert Committee on Plague, Fourth
Report. Wld Hlth Org Bull 447: 23-25.
WHO 2004. Human plague in 2002 and 2003. Wkly Epidemiol Rec 79: 301-308.
Wolf-Watz H, Portnoy DA, Bölin I, falkow 1985. Transfer of the virulence plasmid of
Yersinia pestis to Yersinia pseudotuberculosis. Infect Immun 48: 241-243.
90

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Fig. 1: Western-blot com soro de coelho anti-A1122 e a proteína F1 purificada (linha 1),
proteínas totais da cepa P. CE 882 (linha 2); soro de coelho anti-Y. pseudotuberculosis
IP 32950 e a proteína F1 (linha 3).
91

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Fig. 2: Western-blot com proteínas totais da cepa P. PB 881 e soros humanos HA+.
Linhas 1: 76 Ya BA (61) 1982, 2: 24/12081, 3: 04/80 Ipubi, 4: 248515 Humano CE.
92

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Fig. 3: Proteínas totais de E. coli contendo o plasmídio pTZ/YopH separadas por
SDS/PAGE e coradas pelo Azul de Coomassie (A); Western-blot com soro de coelho
anti-YopH (B).
93

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Fig. 4: Western-blot com proteínas totais de E. coli contendo o plasmídio pTZ/YopH e
soros de coelho. Linhas 1: Soro anti-YopH; 2: Soro normal de coelho; 3: Soro C/YP
2950; 4: C/YP A1122; 5: C/YP EV76; 6: C/P. PB 881; 7: C/P. CE 882; 8: C/P. Exu
769.
94

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Fig. 5: Western-blot das proteínas totais de E. coli contendo o plasmídio pTZ/YopH e
soros de coelho anti-YopH e soros humanos HA-. Linhas 1: Soro anti-YopH; 2:
318233H; 3: 318235H; 4: 318222H; 5: 318221H.
95

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
CONCLUSÕES
A partir da análise dos resultados obtidos neste estudo, podemos
concluir que:
• Em E. coli, a expressão da proteína YopH é regulada pela
temperatura (37oC) e fase de crescimento, sendo maior na fase
estacionária.
• A proteína H-NS está envolvida na atividade promotora,
funcionando como reguladora da expressão da YopH, pelo menos
em E. coli.
• O mecanismo responsável pela diminuição da expressão da YopH,
na presença de glicose e arabinose permanece indefinido.
• Não houve reconhecimento da proteína YopH pelos soros de
pacientes suspeitos de peste e coelhos imunizados com Y. pestis.
96

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
ANEXOS
97

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
BASIC INFORMATION
Genetics and Molecular Biology ISSN 1415-4757 (formerly named Revista Brasileira de Genética/Brazilian Journal of Genetics - ISSN 0100-8455) is published quarterly by the Sociedade Brasileira de Genética (Brazilian Society of Genetics).
The Journal considers contributions that present the results of original research in genetics, evolution and related scientific disciplines.
Genetics and Molecular Biology begins with vol. 21, issue 1, of March 1998, following the sequence of numbering of its predecessor, which was published from 1978 to 1997, V. 1 to V. 20.
The journal's short title is Genet. Mol. Biol., which should be used in bibliographies, footnotes and bibliographical references and strips.
INDEXED IN
Genetics and Molecular Biology is abstracted or indexed in:
• ISI Science Citation Index • Biological Abstracts • Excerpta Medica • Genetics Abstracts • Animal Breeding Abstracts • Plant Breeding Abstracts • Chemical Abstracts • Referativnyi Zhurnal (Abstracts Journal, Russia) • Periódica (México)
COPYRIGHT / PHOTOCOPYING
Articles may be reproduced without specific permission from this journal in the following cases:a) single copies for the purpose of private study or research only;b) multiple copies for the use in a non-profit basis in teaching.
98

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
I NSTRUCTIONS TO AUTHORS
Scope and policy Submission of papers
SCOPE AND POLICY
Genetics and Molecular Biology (formerly named Revista Brasileira deGenética/Brazilian Journal of Genetics - ISSN 0100-8455) is publishedquarterly by the Sociedade Brasileira de Genética (Brazilian Society ofGenetics).
The Journal considers contributions that present the results of originalresearch in genetics, evolution and related scientific disciplines.
Although Genetics and Molecular Biology is an official publication of theBrazilian Society of Genetics, contributors are not required to be members ofthe Society.
It is a fundamental condition that submitted manuscripts have not been andwill not be published elsewhere. With the acceptance of a manuscript forpublication, the publishers acquire full and exclusive copyright for alllanguages and countries.
Manuscripts considered in conformity with the scope of the journal as judgedby the Editor in conjunction with the Editorial Board are reviewed by theAssociate Editors and two or more external reviewers. Acceptance by theEditor is based on the quality of the work as substantial contribution to thefield and on the overall presentation of the manuscript .
SUBMISSION OF PAPERS
1. Manuscripts should be submitted to:
Fábio de Melo Sene, Editor-in-Chief Genetics and Molecular Biology Rua Capitão Adelmio Norberto da Silva, 736 14025-670 Ribeirão Preto, SP - Brasil
2. A submission package sent to the Editorial Office must contain:
a) A cover letter signed by all authors stating that they have approved the submission of the manuscript and that the findings have not been published or are not under
99

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
consideration for publication elsewhere;
b) A copy of the manuscript, including original figures.
c) A cop of any unpublished or in-press companion articles referred to in the submission.
d) A copy of the text, tables and figures on a disk. Be sure that the disk is adequately protected. Formats for text are Word or RTF, in Windows platform. Images in TIFF or JPEG formats should be sent in separate files (For Figures, see detailed instructions in 3.1.g). Disk must be labeled with the first author's last name, platform and software. (See detailed instructions below). Failure to adhere to these guidelines can delay the handling of your contribution, and manuscripts may be returned before being reviewed.
3. Categories of Contribution:
3.1. Research Articles
Manuscripts must be written in English in double-spaced, 12-pointtype throughout, including the References Cited section, appendices,tables and legends; printed on one side only of A4 paper with 2.5 cmmargins; marked with consecutive page numbers, beginning with thecover page.
The following elements must start on a new page and be ordered asthey are listed below:
a) The title page must contain: a concise and informative title; theauthors' names (first name at full length); the authors' institutionalaffiliation, including department, institution, city, state or provinceand country; different affiliations indicated with superscript numbers;a short running title of about 35 characters, including spaces; up tofive key words; the corresponding author's name, postal address,phone and fax numbers and email address. The corresponding authoris the person responsible for checking the page proofs, arranging forthe payment of color illustrations and author's alteration charges.
b) The Abstract must be a single paragraph that does not exceed200 words and summarizes the main results and conclusions of thestudy. It should not contain references.
c) The text must be as succinct as possible. Text citation s: articlesshould be referred to by authors' surnames and date of publication;citations with two authors must include both names; in citations withthree or more authors, name the first author and use “et al”. Onlyarticles that are published or in press should be cited. In the case ofpersonal communications or unpublished results, all contributorsmust be listed by initials and last name (“et al” should not be used).Number s: In the text, numbers nine or less must be written outexcept as part of a date, a fraction or decimal, a percentage, or a unitof measurement. Use Arabic numerals for numbers larger than nine.Avoid starting a sentence with a number. Binomial Name s: Latin
100

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
names of genera, species and intraspecific taxa in the text must beprinted in italics; names of orders and families should be in the Title.
The text includes the following elements:
Introduction – Description of the background that led to the study.
Material (or Subjects) and Methods – Details relevant to the conductof the study. Statistical methods should be explained at the end ofthis section.
Results – Undue repetition in text and tables should be avoided.Comment on significance of results is appropriate but broaderdiscussion should be part of the Discussion section.
Discussion – The findings of the study should be placed in context ofrelevant published data. Ideas presented in other publications shouldnot be discussed solely to make an exhaustive presentation.
Some manuscripts may require different formats appropriate to theircontent.
d) The Acknowledgments must be a single paragraph thatimmediately follows the discussion and includes references to grantsupport.
e) The References Section: citations must be orderedalphabetically by the first author; only articles that are published orin press should be included; personal communications must be citedwithin the text; journal titles must be abbreviated according toMedline (http://www.ncbi.nlm.nih.gov/entrez/jrbrowser.cgi).
Sample journal article citatio n:
Breuer ME and Pavan C (1955) Behaviour of polytene chromosomesof Rhynchosciara angelae at different stages of larval development.Chromosoma 7:371-386.
Bertollo LAC, Takahashi CS and Moreira-Filho O (1978)Cytotaxonomic consideration on Hoplias lacerdae (Pisces,Erythrinidae). Rev Bras Genet 1:103-120.
Sample book citation:
Salzano FM and Freire-Maia N (1967) Populações Brasileiras.Companhia Editora Nacional and EDUSP, São Paulo, 178 pp.
Dobzhansky T (1951) Genetics and Origin of Species. 3rd edition.Columbia University Press, New York , 364 pp.
Sample chapter-in-book citation:
Carvalho A, Monaco LC and Krug CA (1966) Melhoramento genéticodas plantas e sua repercussão econômica. In: Pavan C and da CunhaAB (eds) Elementos de Genética. 2nd ed. EDUSP and Companhia
101

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Editora Nacional, São Paulo, pp 587-653.
Sample abstracts in meeting citation
Basile R (1973) Cromossomos Politênicos em células nutritivas deovócitos de ovário atrofiado de Rhyncosciar a. Ciênc e Cult 25(suppl): 248. XXV Reunião Anual da SBPC, Rio de Janeiro, Brazil.
Sample Thesis/Dissertation citation
Frota-Pessoa O (1953) Revision of the Tripunctata group ofDrosophila with description of fifteen new species. PhD Thesis,Universidade do Brasil, Rio de Janeiro .
Sample Electronic Article citation:
Simin K, Wu H, Lu L, Pinkel D, Albertson D, Cardiff RD, Van Dyke T(2004) pRb Inactivation in Mammary Cells Reveals CommonMechanisms for Tumor Initiation and Progression in DivergentEpithelia. Plos Biol 2: 194-205. http://www.plosbiology.org/.
Sample Electronic Database citation:
Online Mendelian Inheritance in Man (OMIM),http://www.ncbi.nlm.nih.gov/OMIM
f) Tables each table must start on a new page. A concise title shouldbe provided above the table. Tables must be numbered consecutivelyin Arabic numerals. Each column must have a title in the box head.Footnotes typed directly below the table should be indicated inlowercase superscript numbers.
g) Figures must be numbered consecutively in Arabic numerals.Legends should be typed on a separate sheet. A set of originalillustrations of the highest quality must be provided in glossy paper.If you have created figures electronically submit them also as hardcopies. Scanned figures should not be submitted. Images should bein TIFF or JPEG format and provided in separate files. Figures in Wordformat cannot be published. Journal quality reproduction will requiregrayscale and color at resolution yielding 300 dpi. Authors shouldsubmit bitmapped line art at resolution yielding 600–1200 dpi. Theseresolutions refer to the output size of the file; if it is anticipated thatimages will be enlarged or reduced, the resolutions should beadjusted accordingly. Identify each illustration by affixing on the backa label containing: the number of the figure, the name of the firstauthor and an arrow indicating top of illustration. Illustrationssupplied on disks must follow instructions in item 2 (Submissionpackage). Color illustration can be accepted, but authors are asked todefray the cost. For costs of color figures, check with the EditorialOffice.
h) Nomenclature: current standard international nomenclatureshould be adhered to.
i) Sequences may appear in text or in figure. DNA, RNA and protein
102

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
sequences equal to or greater than 50 units must be entered intopublic databases. The accession number must be provided andreleased to the general public together with publication of the article.Long sequences requiring more than two pages to reproduce will notbe published unless the Editorial decision is that the publication isnecessary. Complete mtDNA sequence will not be published.
j) Data acces s: reference should be made to availability of detaileddata and materials used for reported studies.
k) Ethical issues: Reports of experiments on live vertebrates mustinclude a brief statement that the work was approved by theinstitutional review board. For experiments involving human subjects,authors must also include a statement that informed consent wasobtained from all subjects. If photos or any other identifiable data areincluded, a copy of the signed consent must accompany themanuscript.
3.2 Short Communications present brief observations that do notwarrant full-length articles. They should not be consideredpreliminary communications. They should be 15 or fewer typed pagesin double spaced 12-point type, including literature cited. They shouldinclude an Abstract no longer than five percent of the paper's lengthand no further subdivision with introduction, material and methods,results and discussion in a single section. Up to two tables and twofigures may be submitted. The title page and reference sectionformat is that of full-length article.
3.3 Letters to the Editor relate or respond to recent publisheditems in the journal. Discussions of political, social and ethical issuesof interest to geneticists are also welcome in this form.
3.4 Review Articles are welcome.
3.5 Book Review s: publishers are invited to submit books onGenetics, Evolution and related disciplines, for review in the journal.Aspiring reviewers may propose writing a review.
3.6 History, Story and Memories: accounts on historical aspects ofGenetics relating to Brazil .
4. Proofs: Page proofs will be sent to the corresponding author.Changes made to page proofs, apart from printer's errors, will becharged to the authors. Notes added in proof require Editorialapproval.
5. Reprints are free of charge and provided as a pdf-file.
103

Haver, P. R. O.
ANÁLISE DA REGULAÇÃO TRANSCRICIO
As Memórias do Instituto Oswaldo Cruz são uma revista multidisciplinar que publica pesquisas originais relativas aos campos da medicina tropical (incluindo patologia, epidemiologia de campo e estudos clínicos), parasitologia médica e veterinária (protozoologia, helmintologia, entomologia e malacologia) e microbiologia médica (virologia, bacteriologia e micologia). A revista aceita, especialmente, pesquisas básicas e aplicadas em bioquímica, imunologia, biologia molecular e celular, fisiologia, farmacologia e genética relacionada a essas áreas. Comunicações breves são também consideradas. Artigos de revisão só quando solicitados. Ocasionalmente, trabalhos apresentados em simpósios ou congressos são aparecem como suplementos.
Os artigos apresentados devem ser escritos preferencialmente em inglês. Quando neste idioma, devem ser checados por alguém que tenha o inglês como primeira língua e que, preferencialmente, seja um cientista da área.
A submissão de um manuscrito às Memórias requer que este não tenha sido publicado anteriormente (exceto na forma de resumo) e que não esteja sendo considerado para publicação por outra revista. A veracidade das informações e das citações bibliográficas é de responsabilidade exclusiva dos autores.
Os manuscritos serão analisados por pelo menos dois pareceristas; a aprovação dos trabalhos será baseada no conteúdo científico e na apresentação.
Todo o material deve ser enviado para a Produção Editorial, Memórias do Instituto Oswaldo Cruz, Av. Brasil 4365, Pavilhão Mourisco, sala 308, 21045-900 Rio de Janeiro, RJ, Brasil.
Os manuscritos que não estiverem de acordo com estas instruções serão imediatamente devolvidos.
Ao encaminhar um manuscrito para a revista, os autores devem estar cientes de que, se aprovado para publicação, o copyright do artigo, incluindo os direitos de reprodução em todas as mídias e formatos, deverá ser concedido exclusivamente para as Memórias. A revista não recusará as solicitações legítimas dos autores para reproduzir seus trabalhos.
Favor providenciar e checar cada item abaixo antes de submeter seu
NAL...
104

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
105
manuscrito para as Memórias do Instituto Oswaldo Cruz:
• Carta de submissão do trabalho, assinada por todos os autores, especificando o autor de contato, bem como endereço, telefone, fax e e-mail. • Quatro cópias completas do artigo, incluindo as ilustrações e um disquete contendo o texto, tabelas, gráficos e fotografias digitalizadas. • O manuscrito (incluindo tabelas e referências) deve ser preparado em um software para edição de textos, em espaço duplo, fonte 12, impresso em papel padrão e paginado. As margens devem ser de pelo menos 3 cm. • A seqüência do artigo deve ser: título resumido (com até 40 caracteres - letras e espaços), título (com até 250 caracteres), autores (sem títulos ou graduação), afiliação institucional (endereço completo somente do autor correspondente), resumo, palavras-chave, notas de rodapé indicando a fonte de financiamento ou mudanças de endereço, introdução, material e métodos, resultados, discussão, agradecimentos (os mínimos necessários), referências, tabelas (fora do texto e com título), e figuras (com legendas em folha separada). • Só as referências citadas no texto devem aparecer nas lista e devem seguir o estilo do Index Medicus. Se a referência for de artigo ainda não publicado, mas já aceito, deverá ser apresentada carta da revista que publicará o manuscrito ou de outros autores autorizando a referida citação.
Para maiores informações sobre o formato e o estilo da revista, favor consultar um número recente da Revista ou entrar em contato com a Editoria Científica pelo telefone (+55-21-2598.4335), fax (+55-21-2561.1442 / 2280-5048), ou e-mail ([email protected] / [email protected])

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Format / Style
The manuscript should be prepared using standard word processing software and should be printed (font size 12) double-spaced throughout the text, figure captions, and references, with margins of at least 3 cm. The manuscript should be arranged in the following order: running title, title, authors' names, institutional affiliations, summary, key words, introduction, materials and methods, results, discussion, acknowledgements, and references. Sponsorships and corresponding author (mentioning fax number and e-mail address) should be as a footnote on the first page. Summary: up to 200 words (100 words in case of short communications). It should emphasize new and important aspects of the study or observations. Key words: 3-6 items must be provided. Terms from the Medical Subject Headings (Mesh) list of Index Medicus should be used. Introduction: should set the purpose of the study, give a brief summary (not a review) of previous relevant works, and state what new advance has been made in the investigation. It should not include data or conclusions from the work being reported. Materials and Methods: should briefly give clear and sufficient information to permit the study to be repeated by others. Standard techniques need only be referenced. Ethics: when reporting experiments on human subjects, indicate whether the procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (institutional or regional) and with the Helsinki Declaration of 1975, as revised in 1983. When reporting experiments on animals, indicate whether the institution's or a national research council's guide for, or any national law on the care and use of laboratory animals was followed. Results: should be a concise account of the new information discovered, with the least personal judgement. Do not repeat in text all the data in the tables and illustrations. Discussion: should be limited to the significance of the new information and relate the new findings to existing knowledge. Only unavoidable citations should be included. Acknowledgements: should be short and concise, and restricted to those absolutely necessary. References: must be accurate. Only citations that appear in the text should be referenced. Unpublished papers, unless accepted for publication, should not be cited. Work accepted for publication should be referred to as "in press" and a letter of acceptance of the journal must be provided. Unpublished data should only be cited in the text as
106

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
"unpublished observations", and a letter of permission from the author must be provided. The references at the end of the paper should be arranged in alphabetic order according to the surname of the first author. The titles of journals should be abbreviated according to the style used in the Index Medicus. Consult www.nlm.nih.gov/serials/lii.html. - In the text use authors' surname and date: Lutz (1910) or (Lutz 1910). With two authors it is (Lutz & Neiva 1912) or Lutz and Neiva (1912). When there are more than two authors, only the first is mentioned: Lutz et al. (1910) or (Lutz et al. 1910). - At the end of the paper use the following styles: Journal article Chagas C, Villela E 1922. Forma cardiaca da tripanosomiase americana. Mem Inst Oswaldo Cruz 14: 15-61. Book or Thesis Morel CM 1983. Genes and Antigens of Parasites. A Laboratory Manual, 2nd ed., Fundação Oswaldo Cruz, Rio de Janeiro, xxii + 580 pp. Chapter in book Cruz OG 1911. The prophylaxis of malaria in central and southern Brasil. In R Ross, The Prevention of Malaria, John Murray, London, p. 390-398.
Illustrations: Figures and tables must be understandable without reference to the text. - Figures should be mounted on a manuscript-size sheet. Photographs must be sharply focused, well contrasted, black and white glossy prints. Photographs and line drawings must be marked on the back with the author's name, the figure number and an arrow pointing to the top. If mounted onto a plate, the figures should be numbered consecutively with Arabic numbers. Magnification must be indicated by a line or bar in the figure, and referenced, if necessary in the caption (e.g., bar = 1 mm). Plates and line figures should either fit one column (8 cm) or the full width (16.5 cm) of the page and should be shorter than the page length to allow inclusion of the legend. Legends must be provided on a separate sheet. Letters and numbers on figures should be of a legible size upon reduction or printing. Collour illustrations can only be accepted if the authors defray the cost. However, a colour photograph illustrates the cover of each issue of the Journal and authors are invited to submit illustrations with legends from their manuscript for consideration for the cover at no charge. - Tables should supplement, not duplicate, the text and should be numbered with Roman numerals. A short descriptive title should appear above each table, with any explanations or footnotes (identified with a, b, c, etc.) below.
Short communications should communicate rapidly single results or techniques. They should occupy no more than four printed pages including figures and/or tables. They should not contain excessive references. References should be cited at the end of the paper using the
107

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
same format as in full papers. A brief summary and three key words must be provided.
Alternative format: manuscripts may be submitted following the "Uniform Requirements for Manuscripts Submitted to Biomedical Journals" produced by the International Committee of Medical Journal Editors also known as the Vancouver Style. In this case, authors should follow the guidelines in the fifth edition (Annals of Internal Medicine 1997; 126: 36-47, or at the website http://www.acponline.org/journals/resource/unifreqr/htm) and will be responsible for modifying the manuscript where it differs from the instructions given here, if the manuscript is accepted for publication. Authors should also follow the Uniform Requirements for any guidelines that are omitted in these Instructions.
Once a paper is accepted for publication, the authors must provide: • a diskette containing the text of the final approved version of the manuscript (including tables and graphics) in Word or Word Perfect for Windows format (Macintosh formats should be converted); • an affidavit, provided by the Editorial Office, signed by all authors. Authors from different countries or institutions may sign in different sheets containing the same basic statement; • a copyright assignment form, provided by the Editorial Office, signed by the corresponding author. Page charges: there will be no page charges. Proofs: one set of page proofs will be supplied for the author to check for typesetting accuracy, to be returned by the stipulated date. No changes to the original manuscript will be allowed at this stage. Offprints: authors will receive 30 free offprints. An order form will be sent to the author enabling further offprints to be ordered at prices listed on the form.
108

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
Checklist for manuscripts to be submitted
Please check every item below before submitting your manuscript to Memórias do Instituto Oswaldo Cruz.
• Please provide a cover letter, signed by all authors, with your submission specifying the corresponding author as well as an address, telephone and facsimile number, and e-mail. • Submit four printed copies of your manuscript, each with a complete set of original illustrations, and a diskette containing the text, tables, graphics and digitalized photographs. • The entire manuscript (including tables and references) must be typed double-spaced, font size 12, and printed on standard-sized paper. The left and right margins must be at least 3 cm. • Please number pages beginning with the title page. • The title page must include a running headline of not more than 40 characters, a title of no more than three printed lines (250 letters and spaces), authors (no titles or degrees), institutional affiliations, complete address of author to whom correspondence must be addressed, and footnotes indicating sources of financial support and changes of address. • The order of appearance of material in all manuscripts should be as follows: running headline, title, authors, institutional affiliations, abstract, key words, footnotes, introduction, materials and methods, results, discussion, acknowledgements, references, tables, legends for figures, and figures. • References must be cited on the line of the text and in parentheses, e.g., (Chagas 1909). References not cited in the text cannot appear in the reference section. Reference citations must follow the format established by the "Index" (see examples in Instructions to Authors). • If you reference your own unpublished work (i.e., an "in press" article) you must enclose an acceptance letter from the journal. • If you cite unpublished data that are not your own, you must provide a letter of permission from the author of that publication. • Please provide three glossy or laser-produced prints of each figure. Label all figures with the first author's name and figure number (place typed label on back of figure). Provide a figure legend for each figure on a separate page at the end of the manuscript. • Tables must be on a separate page at the end of the manuscript. A short descriptive title must be provided for each table.
109

Haver, P. R. O. ANÁLISE DA REGULAÇÃO TRANSCRICIONAL...
110