anais - ufrgs.br · corretamente balanceadas após o parto é o fator unificador de sucesso durante...
TRANSCRIPT

ANAIS
EDITORES
Félix H. D. González
Ana C. R. Berreta
Anne R. Guadagnin
Faculdade de Veterinária
Universidade Federal do Rio Grande do Sul
Porto Alegre - RS, Brasil
2014


EDITORES
Félix Gonzalez
Faculdade de Veterinária – Universidade Federal do Rio Grande do
Sul
Av. Bento Gonçalves 9090. Porto Alegre - RS 91.540-000
Ana Claudia Ramos Berreta Faculdade de Veterinária – Universidade Federal do Rio Grande do
Sul
Av. Bento Gonçalves 9090. Porto Alegre - RS 91.540-000
Anne Rosi Guadagnin Faculdade de Veterinária – Universidade Federal do Rio Grande do
Sul
Av. Bento Gonçalves 9090. Porto Alegre - RS 91.540-000


COMISSÃO ORGANIZADORA
Prof. Dr. Félix Hilario Diaz González
Ana Claudia Ramos Berreta
Angélica Petersen Dias
Anne Rosi Guadagnin
Mariana Soares da Silva
Rafael Marques Gomes


Catalogação na fonte: Ana Vera Finardi Rodrigues – CRB 10/884
Copyright 2014 by Félix H.D. González.
Todos os direitos reservados. Não é permitida a reprodução total ou parcial
desta publicação sem a autorização escrita e prévia dos editores.
S612 Simpósio Nacional da Vaca Leiteira (1. : 2014 : Porto Alegre).
Anais do 1º Simpósio Nacional da Vaca Leiteira / Editores:
Félix H. D. González, Ana C. R. Berreta, Anne R. Guadagnin. –
Porto Alegre, 2014.
239 p. ; il.
1. Medicina veterinária : vacas leiteiras I. González, Félix
H. D. II. Berreta, Ana C. R. III. Guadagnin, Anne R.
CDD 636.2


AUTORES CONTRIBUINTES
Bolívar Nóbrega de Faria
Recursos Humanos no Agronegócio Ltda - Belo Horizonte
Carlos Bondan
Serviço de Análises de Rebanhos Leiteiros – Universidade de Passo
Fundo
Enrico Ortolani
Faculdade de Medicina Veterinária e Zootecnia– Universidade de São
Paulo
Felipe Cardoso
Department of Animal Science – University of Illinois
José Luiz Vasconcelos
Departamento de Produção Animal – Universidade Estadual Paulista
Júlio de Mesquita Filho
Manoel Francisco de Sá Filho
Faculdade de Medicina Veterinária e Zootecnia – Universidade de São
Paulo
Marcelo Cecim
Departamento de Clínica de Grandes Animais, Centro de Ciências
Rurais – Universidade Federal de Santa Maria

Márcio Nunes Correa
Faculdade de Veterinária – Universidade Federal de Pelotas
Marcos Neves Pereira
Departamento de Zootecnia - Universidade Federal de Lavras
Vivian Fischer Departamento de Zootecnia – Universidade Federal do Rio Grande do
Sul

APRESENTAÇÃO
A presente publicação reúne as palestras proferidas durante o
1º Simpósio Nacional da Vaca Leiteira, que o Laboratório de Análises
Clínicas Veterinárias, da Faculdade de Veterinária da Universidade
Federal do Rio Grande do Sul, organizou por iniciativa de um grupo
de alunos de Medicina Veterinária desta Universidade formandos em
2015/1.
O rebanho leiteiro do Rio Grande do Sul está entre os mais
produtivos e de maior taxa de crescimento na indústria leiteira do
Brasil. O potencial de crescimento corre em paralelo com as maiores
expectativas de demanda regional e nacional.
Os médicos veterinários que militam na pecuária de leite
devem ser cientes de que, os trabalhos, tanto de campo, como de
pesquisa estão permeados pela multidisciplinariedade, que envolve
conceitos de nutrição, metabolismo, clínica, reprodução, medicina
preventiva, produção e manejo alimentar.
No evento testemunhado pelo presente material, foram
convidados a participar palestrantes de várias Universidades do Brasil
e dos Estados Unidos, engajados no estudo da clínica, a nutrição e a
reprodução da vaca leiteira de alta produção. Trata-se de três áreas
responsáveis por mais de 80% dos limitantes na produção de gado
leiteiro. Os aspectos clínicos tratados aqui enfocam basicamente os
transtornos metabólicos e as doenças do periparto. O propósito do

evento é iniciar uma sequência de eventos que permitam a reciclagem
de conhecimentos em buiatria leiteira, de forma bianual, permitindo a
atualização aos profissionais que cada vez exigem mais atividades de
profundização.
Nosso mais sincero agradecimento aos professores e
pesquisadores que atenderam ao nosso chamado. Especiais
agradecimentos também às empresas e instituições que se vincularam
e apoiaram este evento, Laboratório BRAVET, Bayer Animal Care,
Ibasa/Importadora Bagé, KetoVet, Kera e Revista Balde Branco, bem
como a Universidade UniRitter (Laureate International Universities).
O simpósio contou com o apoio institucional da Fundação de Apoio a
Pesquisa do Rio Grande do Sul (FAPERGS) e da Pró-reitoria de
Extensão da Universidade Federal do Rio Grande do Sul.
Os editores
Porto Alegre, novembro de 2014.

SUMÁRIO
1 MANEJO NUTRICIONAL PRÉ E PÓS-PARTO PARA
OTIMIZAR A FERTILIDADE EM VACAS DE LEITE 15
Felipe C. Cardoso
2 SUPLEMENTAÇÃO DE METIONINA PARA
MELHORAR A PRODUTIVIDADE DA VACA
LEITEIRA. ......................................................................... 34
Marcos Neves Pereira
3 COMO A DIETA PODE AFETAR A COMPOSIÇÃO
DO LEITE. ......................................................................... 52
Bolívar Nóbrega de Faria
4 CONTROLE LEITEIRO .................................................. 91
Carlos Bondan
5 TRANSTORNOS METABÓLICOS DA VACA
LEITEIRA NO PERÍODO DE TRANSIÇÃO ................ 107
Enrico Lippi Ortolani
6 TRANSTORNOS NO PERÍODO DE TRANSIÇÃO DA
VACA LEITEIRA COM ÊNFASE NAS
ENFERMIDADES UTERINAS E DA GLÂNDULA
MAMÁRIA ......................................................................... 127
Márcio Nunes Côrrea

7 BIOTÉCNICAS DA REPRODUÇÃO PARA
MELHORAR A FERTILIDADE DA VACA
LEITEIRA. ......................................................................... 152
Manoel Francisco de Sá Filho
8 O QUE DEVO SABER PARA DEFINIR O
PROTOCOLO DE IATF A SER UTILIZADO? ............ 189
José Luiz Moraes Vasconcelos
9 ESTRESSE CALÓRICO EM VACAS LEITEIRAS:
EFEITO SOBRE O METABOLISMO E A
QUALIDADE DO LEITE ................................................. 206
Vivian Fischer
10 BEM-ESTAR DE VACAS LEITEIRAS: MELHOR
PRODUÇÃO COM A VACA FELIZ .............................. 230
Marcelo da Silva Cecim

15
1. MANEJO NUTRICIONAL PRÉ E PÓS-PARTO PARA
OTIMIZAR A FERTILIDADE EM VACAS DE LEITE1.
Felipe C. Cardoso
1.1 Introdução
O sucesso reprodutivo é crítico para o sucesso econômico e
para a sustentabilidade de fazendas de leite em qualquer sistema de
produção. O sucesso reprodutivo depende de uma série de eventos
fisiológicos coordenados, incluindo retorno da ciclicidade ovariana
após o parto, desenvolvimento e ovulação de um oócito viável,
fertilização, involução uterina, desenvolvimento e implantação
embrionária e permanência da prenhez até a maturação fetal (Butler,
2003; Garnsworthy et al., 2008). A formulação de dietas e o manejo
da alimentação durante o período seco, período do periparto e período
imediato após o parto podem facilitar ou interromper diversos dos
passos citados acima antes da prenhez ser estabelecida e mantida
(Butler, 2003; Garnsworthy et al., 2008; Thatcher et al., 2011). As
taxas de concepção para vacas de alta produção que estão ciclando é
alta (> 70%) mas muitas vacas (> 50%) apresentam morte embrionária
1 Cardoso, F. C. Manejo nutricional pré e pós-parto para otimizar a
fertilidade em vacas de leite. Anais. I Simpósio Nacional da Vaca Leiteira.
Porto Alegre: Universidade Federal do Rio Grande do Sul. 2014. 239 p.

16
precoce e falham em manter a prenhez (Diskin and Morris, 2008). Os
maiores pontos de ênfase relacionando nutrição e reprodução em
vacas de leite são a severidade e a duração do estado de nutrientes
negativo, o chamado balanço energético negativo (BEN), que no
começo da lactação é fortemente influenciado pelo manejo nutricional
e do ambiente durante o período de transição desde o final da prenhez
até o início da lactação.
O período de 6 a 8 semanas em torno do parto é conhecido
como o período de transição ou peripartum, crítico para determinar o
bem-estar e a lucratividade de vacas individualmente durante a
lactação seguinte (Drackley et al., 2005). Programas de manejo do
período de transição que não são adequados resultam em alta
prevalência de doenças da produção (i.e. deslocamento de abomaso)
em rebanhos leiteiros (Mulligan & Doherty, 2008). Distúrbios no
metabolismo durante o período de transição podem ter influência
direta ou indireta na fertilidade e transições difíceis têm impacto
negativo posteriormente na reprodução (Chapinal et al., 2012). Uma
grande variedade de estratégias nutricionais para facilitar as
adaptações metabólicas e fisiológicas da gestação até o início da
lactação têm sido propostas (Friggens et al., 2004; Roche et al., 2013).
Os objetivos deste artigo são de revisar as causas do BEN e o
impacto em distúrbios metabólicos, de relacionar como estes
distúrbios influenciam a reprodução e de avaliar como diferentes
estratégias nutricionais antes e depois do parto podem afetar o BEN e
a adequação dos nutrientes. O tema central do artigo é que o manejo

17
para promover maior ingestão de matéria seca (IMS) de dietas
corretamente balanceadas após o parto é o fator unificador de sucesso
durante o período de transição para otimizar a reprodução.
1.2 Fertilidade, Produção de Leite e Doenças do Periparto
Uma informação que é aceita por todos é a de que a fertilidade
de vacas de leite modernas está piorando, especialmente para a
genética Holstein, pelo menos em parte pela consequência indesejada
de seleção continuada para alta produção de leite. Esta informação foi
desafiada recentemente (LeBlanc, 2010; Bello et al., 2012). Existe
uma grande distribuição do sucesso reprodutivo tanto dentro do
rebanho como entre rebanhos. Por exemplo, entre cinco rebanhos na
California totalizando 6.396 vacas, encontrou-se que vacas que
estavam no mais baixo quartil para produção de leite nos primeiros 90
dias após o parto (32,1 kg/dia) tiveram menor probabilidade de retorno
ao ciclo estral aos 65 dias pós-parto quando comparado a vacas no 2º
quartil (39,1 kg/dia), no 3º quartil (43,6 kg/dia) ou no 4º quartil (50,0
kg/dia), de forma que a produção de leite não afetou a taxa de prenhez
(Santos et al., 2009). Mudanças de manejo e sistemas de manejo
inadequados potencialmente são mais limitantes para a fertilidade de
vacas de leite modernas do que a sua genética per se.
Vacas de leite são suscetíveis a doenças da produção durante
o periparto e início da lactação (Mulligan et al., 2006; Ingvartsen &
Moyes, 2013; Roche et al., 2013). Existe pouca evidência de que a
produção de leite per se contribua para maior ocorrência de doenças.

18
Entretanto, o pico de ocorrência de doenças (logo após o parto)
corresponde com o momento de maior BEN, com o pico de
concentração sanguínea de ácidos graxos não esterificados (AGNE) e
com a maior aceleração na produção de leite (Ingvartsen et al., 2003).
O pico na produção de leite ocorre muitas semanas após. Doenças
associadas com maior BEN no pós-parto estão também relacionadas
com subótima performance reprodutiva, incluindo fígado gorduroso
(Rukkwamsuk et al., 1999; Jorritsma et al., 2003) e cetose (Walsh et
al., 2007; McArt et al., 2012). Vacas que perderam mais de 1 unidade
(escala de 1-5) no escore de condição corporal (ECC) apresentaram
maior incidência de metrite, retenção de placenta e doenças
metabólicas (deslocamento de abomaso, hipocalcemia e cetose) e
também maior intervalo do parto até a primeira cobertura do que vacas
que perderam menos de 1 unidade durante o período de transição.
Indicadores do BEN são altamente correlacionados com perda
de produção de leite, aumento da prevalência de doenças e redução na
fertilidade (Ospina et al., 2010; Chapinal et al., 2012). Entretanto, o
grau no qual o BEN é o causador de doenças do periparto e não
somente um fenômeno associado, precisa ser examinado criticamente
(Roche et al., 2013). Por exemplo, a resposta inflamatória pode
reduzir a IMS em vacas no período de transição, causando alterações
no metabolismo e predispor vacas a um maior BEN ou maior
prevalência de doenças (Bertoni et al., 2008; Graugnard et al., 2012,
2013; Ingvartsen & Moyes, 2013).

19
Aumento na concentração de AGNE no pós-parto imediato e
redução na concentração de glicose estão fortemente associados com
a taxa de prenhez à primeira cobertura em um programa de
inseminação artificial em tempo fixo (IATF) (Garverick et al., 2013).
Apesar da concentração de AGNE não ter sido diferente entre vacas
que ovularam ou não antes do programa de IATF, a taxa de prenhez
reduziu com a maior concentração de AGNE e aumentou com a maior
concentração de glicose 3 dias após o parto (Garverick et al., 2013).
McArt et al. (2012) concluíram que vacas com cetose subclínica
diagnosticada entre 3 a 7 dias pós-parto tiveram 0,7 chances de serem
classificadas como prenhes ao primeiro serviço e 4,5 vezes mais
chances de serem removidas do rebanho nos primeiros 30 dias após o
parto quando comparadas a vacas que desenvolveram cetose 8 dias
após o parto ou mais tardiamente. Vacas que conseguem se adaptar à
subsequente lactação (Jorritsma et al., 2003) e conseguem evitar
problemas metabólicos (Ingvartsen et al., 2003) ou desbalanço
fisiológico (Ingvartsen and Moyes, 2013) são capazes de suportar alta
produção de leite e reproduzir com sucesso.

20
1.3 Balanço Energético Negativo (BEN) e Fertilidade Subótima
As vacas não têm a capacidade de consumir nutrientes
energéticos através de IMS voluntária após o parto para suprir os
requerimentos para produção de leite. Consequentemente, o BEN
ocorre por um período de dias a semanas durante o começo da
lactação. Apesar de estudos não terem demostrado uma forte relação
entre o grau de BEN e a fertilidade, o tempo para o valor mínimo do
BEN e a direção e/ou grau de mudança no BEN parecem ser fortes
indicadores (Whitaker et al., 1993; Butler, 2003; Reist et al., 2003).
Os mecanismos potencialmente envolvidos nos efeitos danosos do
BEN na reprodução incluem: (1) lento retorno da ciclicidade ovariana,
(2) impacto na qualidade, viabilidade, ou função do ovócito ou do
corpo lúteo, e (3) desenvolvimento de lipidose hepática.
Em geral, vacas que ovulam mais cedo depois do parto tem
maior fertilidade (Butler, 2003; Bossaert et al., 2008; Galvão et al.,
2010). A produção de estrógeno pelo folículo dominante, a
recuperação da secreção e pulsatilidade de LH, e a resposta do ovário
ao LH vão determinar o sucesso da ovulação. O estado do BEN no
pós-parto é negativamente associado com a performance reprodutiva
em parte porque interrompe aqueles 3 fatores (Butler, 2003). A
insulina é o hormônio que normalmente reflete o estado energético e
a adequação nutricional à dieta, podendo ser a primeira ligação entre
os sistemas metabólico e reprodutivo. Baixas concentrações de
insulina e IGF-I (insulin-like growth factor 1) foram associadas à
baixa qualidade do corpo lúteo e do ovócito (Jorritsma et al., 2003).

21
A lipidose hepática está negativamente associada com a
fertilidade (Rukkwamsuk et al., 1999), o que pode ser um efeito
indireto do excessivo BEN em vacas. Entretanto, talvez existam
efeitos negativos diretos da infiltração hepática por lipídeos e a
reprodução. Um grande número de funções vitais do fígado é
negativamente afetado por cetose e infiltração hepática de lipídeos no
pós-parto imediato (Loor et al., 2007). Um exemplo é o aumento da
taxa de apoptose, identificada pela análise do fígado pela técnica de
micro arranjo (Loor et al., 2007) e mais recentemente por métodos
funcionais diretos (Tharwat et al., 2012).
1.4 Manejo nutricional para controlar o BEN e otimizar a
fertilidade.
O balanço energético negativo no pós-parto inicia com a
síntese de leite, entretanto, a severidade do BEN em vacas é
fracamente associada com a produção de leite ou a secreção energética
de leite. O grau e duração do BEN está fortemente associado com IMS
(Zurek et al., 1995; Drackley et al., 2005). Consequentemente,
alimentação e estratégias de manejo para vacas no período seco,
durante o período de transição ou no pós-parto imediato devem buscar
fornecer os nutrientes necessários mas também promover o apetite e a
vigorosa IMS após o parto (Grummer et al., 2004).
Alguns fatores importantes a serem considerados para
promover o apetite e a alta IMS após o parto são: (1) reduzir fatores
estressantes externos e manter as vacas confortáveis, (2) evitar escore

22
de condição corporal (ECC) excessivamente alto ou baixo, (3)
prevenir o consumo exagerado de energia durante o período seco, (4)
assegurar a ingestão de forragens com alto valor de fibra efetiva e
evitar a ingestão excessiva de carboidratos rapidamente fermentáveis
na dieta após o parto, (5) reduzir a diferença cátion-ânion da dieta
(DCAD) antes do parto mas aumentar na dieta da vaca durante o pós-
parto imediato, e (6) garantir suporte de nutrientes para o sistema
imunológico. Estratégias nutricionais para auxiliar vacas em fazer
estas adaptações foram revisados (Friggens et al., 2004; Beever, 2006;
Ingvartsen, 2006; Drackley & Dann, 2008; Lean et al., 2013a, b;
Roche et al., 2013).
Bach et al. (2008) concluíram que o manejo não-nutricional
representou mais de 50% da variação média de produção de leite (20,6
a 33,8 kg/dia) entre 47 rebanhos que consumiam exatamente a mesma
dieta. Emmanuel et al. (2007, 2008) demonstraram que quantidades
crescentes de grão de cevada aumentaram as concentrações de
endotoxina no fluído ruminal e que, em situações de baixo pH no
rúmen, aumentou a absorção ruminal de endotoxinas. Fatores
estressantes não-nutricionais podem reduzir a IMS e predispor vacas
a problemas de saúde no pós-parto. Vacas que desenvolveram metrite
(Hammon et al., 2006; Huzzy et al., 2007) ou cetose (Goldhawk et al.,
2009) após o parto tiverem menor IMS ou comportamento de consumo
reduzido antes do parto.
Enquanto a utilização de dietas steam-up ou close-up antes do
parto, recomendadas por muitos anos, existe na literatura uma falta de

23
fatores positivos associados ao seu uso com a saúde, a produção, ou a
reprodução de vacas. A estratégia mais simples e o princípio
nutricional mais facilmente defendido para a alimentação de vacas nos
períodos seco e de transição é o de alimentar vacas para atender mas
não exceder os requerimentos nutricionais (Drackley & Dann, 2008).
Diversos trabalhos do nosso grupo de pesquisa demonstraram que
dietas com energia controlada durante o período seco contribuem para
um período de transição de maior sucesso (Grum et al., 1996; Dann et
al., 2005, 2006; Douglas et al., 2006; Janovick et al., 2011; Graugnard
et al., 2012, 2013; Ji et al., 2012). Nossos estudos indicam que o
consumo prolongado de energia durante o período seco pode reduzir
a IMS após o parto (Douglas et al., 2006; Dann et al., 2006; Janovick
& Drackley, 2010, 2014). O consumo excessivo de energia resulta em
resposta negativa de indicadores metabólicos como altas
concentrações de AGNE e beta-hidroxibutirato (BHB) no sangue e
mais triglicerídeos no fígado após o parto (Douglas et al., 2006;
Janovick et al., 2011). Alterações celulares e de genes no fígado (Loor
et al., 2006, 2007) e no tecido adiposo (Ji et al., 2012) potencialmente
podem explicar muitas das alterações que ocorrem na vaca. O
consumo excessivo de energia no pré-parto aumenta o “maquinário”
enzimático no tecido adiposo para mobilização de triglicerídeos após
o parto, com alterações na transcrição de genes que levam à redução
na lipogênese, aumento da lipólise e redução na capacidade da insulina
de inibir lipólise (Ji et al., 2012). Controlar o consumo de energia no
período seco também aumentou a função de neutrófilos no pós-parto

24
(Graugnard et al., 2012) ocasionando melhoria na função do sistema
imune.
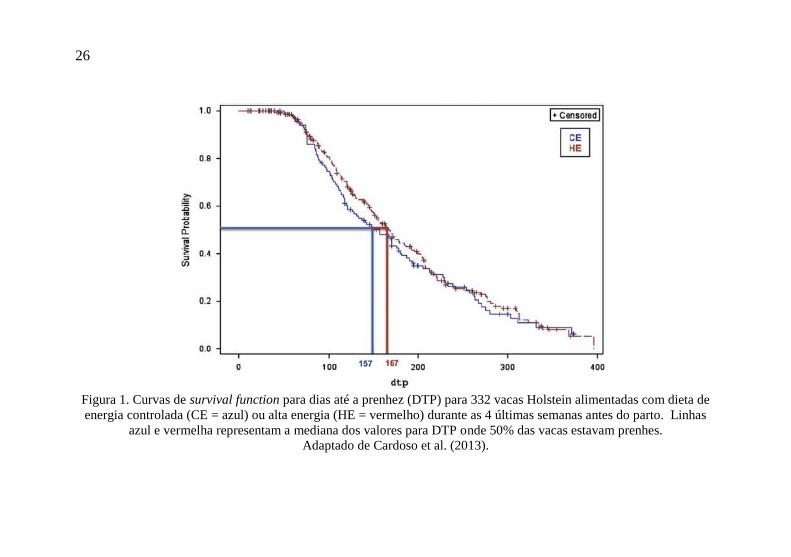
Cardoso et al. (2013) analisaram as associações entre o regime
energético de alimentação no pré-parto e a performance reprodutiva
de vacas de raça Holandesa. Dados individuais de 408 vacas foram
utilizados. Tratamentos nutricionais foram divididos em vacas
recebendo dieta com energia controlada (CE; NEL = 13,7 Mcal/dia)
ou vacas recebendo dieta com alta energia (HE; NEL = 22,1 Mcal/dia)
durante o período seco distante (FO, 60 até 28 dias antes do parto) ou
período seco próximo (CU, 28 dias antes do parto até o parto). O
modelo COX revelou uma diferença estatística em dias até a prenhez
(DTP) entre HE e CE durante CU (mediana = 167 e 157 dias, hazard
ratio = 0,696; Figura 1). Vacas que foram alimentadas com dietas HE
durante CU perderam mais ECC (escala 1–5) durante as primeiras 6
semanas pós-parto do que vacas alimentadas CE (−0,43 e −0,30,
respectivamente). Vacas que foram alimentadas com HE durante o
período seco tiveram mais chances de apresentarem deslocamento de
abomaso ou cetose quando comparadas a vacas que receberam CE.
1.5 Conclusão
Formulação e alimentação de dietas apropriadas, que limitam
o consumo total de energia para suprir, e não exceder, os
requerimentos de vacas durante o pré-parto podem auxiliar a aliviar o
BEN após o parto. Efeitos destas dietas em indicadores de
metabolismo para saúde são geralmente positivos, sugerindo o

25
potencial para redução das doenças do periparto e melhora da
fertilidade. Estratégias para a formulação de dietas que estimulem a
IMS e diminuam o BEN no pós-parto imediato são menos
pesquisadas. Entretanto, o balanço entre fibra fisicamente efetiva e
fermentação do amido nestas dietas é crítico.
26
Figura 1. Curvas de survival function para dias até a prenhez (DTP) para 332 vacas Holstein alimentadas com dieta de
energia controlada (CE = azul) ou alta energia (HE = vermelho) durante as 4 últimas semanas antes do parto. Linhas
azul e vermelha representam a mediana dos valores para DTP onde 50% das vacas estavam prenhes.
Adaptado de Cardoso et al. (2013).

27
REFERÊNCIAS BIBLIOGRÁFICAS
BACH A, VALLS N, SOLANS A and TORRENT T 2008. Association
between nondietary factors and dairy herd performance. Journal of Dairy
Science 91, 3529-3267.
BEEVER DE 2006. The impact of controlled nutrition during the dry
period on dairy cow health, fertility and performance. Animal
Reproduction Science 96, 212-226.
BELLO NM, STEVENSON JS and TEMPELMAN RJ 2012. Invited
review: Milk production and reproductive performance: Modern
interdisciplinary insights into an enduring axiom. Journal of Dairy
Science 95, 5461-5475.
BERTONI G, TREVISI E, HAN X and BIONAZ M 2008. Effects of
inflammatory conditions on liver activity in puerperium period and
consequences for performance in dairy cows. Journal of Dairy Science
91, 3300-3310.
BOSSAERT P, LEROY JLMR, DE VLIEGHER S and OPSOMER G 2008.
Interrelations between glucose-induced insulin response, metabolic
indicators, and time of first ovulation in high-yielding dairy cows. Journal
of Dairy Science 91, 3363-3371.
BUTLER WR 2003. Energy balance relationships with follicular
development, ovulation and fertility in postpartum dairy cows. Livestock
Production Science 83, 211-218.
CARDOSO FC, LEBLANC SJ, MURPHY MR and DRACKLEY JK 2013.
Prepartum nutritional strategy affects reproductive performance in dairy
cows. Journal of Dairy Science 96, 5859-5871.
CHAPINAL N, LEBLANC SJ, CARSON ME, LESLIE KE, GODDEN S,
CAPEL M, SANTOS JE, OVERTON MW and DUFFIELD TF. 2012.
Herd-level associations of serum metabolites in the transition period with
disease, milk production, and early lactation reproductive performance.
Journal of Dairy Science 95, 5676-5682.
DANN HM, LITHERLAND NB, UNDERWOOD JP, BIONAZ M,
D’ANGELO A, MCFADDEN JW and DRACKLEY JK 2006. Diets during

28
far-off and close-up dry periods affect periparturient metabolism and
lactation in multiparous cows. Journal of Dairy Science 89, 3563-3577.
DANN HM, MORIN DE, MURPHY MR, BOLLERO GA and
DRACKLEY JK 2005. Prepartum intake, postpartum induction of ketosis,
and periparturient disorders affect the metabolic status of dairy cows.
Journal of Dairy Science 88, 3249-3264.
DISKIN MG and MORRIS DG 2008. Embryonic and early foetal losses in
cattle and other ruminants. Reproduction in Domestic Animals 43 (Suppl.
2), 260-267.
DOUGLAS GN, OVERTON TR, BATEMAN HG, DANN HM and
DRACKLEY JK 2006. Prepartal plane of nutrition, regardless of dietary
energy source, affects periparturient metabolism and dry matter intake in
Holstein cows. Journal of Dairy Science 89, 2141-2157.
DRACKLEY, JK and FC CARDOSO 2014. Prepartum and postpartum
nutritional management to optimize fertility in high-yielding dairy cows in
confined TMR systems. Animal, 8:S1, 5-14.
DRACKLEY JK and DANN HM 2008. A scientific approach to feeding
dry cows. In Recent advances in animal nutrition – 2007. (eds PC
Garnsworthy and J Wiseman), pp. 43-74. Nottingham University Press,
Nottingham, UK.
DRACKLEY JK, DANN HM, DOUGLAS GN, JANOVICK GURETZKY
NA, LITHERLAND NB, UNDERWOOD JP and LOOR JJ 2005.
Physiological and pathological adaptations in dairy cows that may increase
susceptibility to periparturient diseases and disorders. Italian Journal of
Animal Science 4, 323-344.
EMMANUEL DG, DUNN SM and AMETAJ BN 2008. Feeding high
proportions of barley grain stimulates an inflammatory response in dairy
cows. Journal of Dairy Science 91, 606-614.
EMMANUEL DG, MADSEN KL, CHURCHILL TA, DUNN SM and
AMETAJ BN 2007. Acidosis and lipopolysaccharide from Escherichia coli
B:055 cause hyperpermeability of rumen and colon tissues. Journal of
Dairy Science 90, 5552-5557.

29
FRIGGENS NC, ANDERSEN JB, LARSEN T, AAES O, and
DEWHURST RJ 2004. Priming the dairy cow for lactation: a review of dry
cow feeding strategies. Animal Research 53, 453–473.
GALVÃO KN, FRAJBLAT M, BUTLER WR, BRITTIN SB, GUARD CL
and GILBERT RO 2010. Effect of early postpartum ovulation on fertility
in dairy cows. Reproduction in Domestic Animals 45, e207-e211.
GARNSWORTHY PC, SINCLAIR KD and WEBB R 2008. Integration of
physiological mechanisms that influence fertility in dairy cows. Animal 2,
1144-1152.
GARVERICK HA, HARRIS MN, VOGEL-BLUEL R, SAMPSON JD,
BADER J, LAMBERSON WR, SPAIN JN, LUCY MC and
YOUNGQUIST RS 2013. Concentrations of nonesterified fatty acids and
glucose in blood of periparturient dairy cows are indicative of pregnancy
success at first insemination. Journal of Dairy Science 96, 181-188.
GOLDHAWK C, CHAPINAL N, VEIRA DM, WEARY DM and VON
KEYSERLINGK MA 2009. Prepartum feeding behavior is an early
indicator of subclinical ketosis. Journal of Dairy Science 92, 4971-4977.
GRAUGNARD DE, BIONAZ M, TREVISI E, MOYES KM, SALAK-
JOHNSON JL, WALLACE RL, DRACKLEY JK, BERTONI G and LOOR
JJ 2012. Blood immunometabolic indices and polymorphonuclear leukocyt
e function in peripartum dairy cows are altered by level of dietary energy
prepartum. Journal of Dairy Science 95, 1749-1758.
GRAUGNARD DE, MOYES KM, TREVISI E, KHAN MJ, KEISLER D,
DRACKLEY JK, BERTONI G and LOOR JJ 2013. Liver lipid content and
inflammometabolic indices in peripartal dairy cows are altered in response
to prepartal energy intake and postpartal intramammary inflammatory
challenge. Journal of Dairy Science 96, 918-935.
GRUM DE, DRACKLEY JK, YOUNKER RS, LACOUNT DW and
VEENHUIZEN JJ 1996. Nutrition during the dry period and hepatic lipid
metabolism of periparturient dairy cows. Journal of Dairy Science 79,
1850-1864.

30
GRUMMER RR, MASHEK DG and HAYIRLI A 2004. Dry matter intake
and energy balance in the transition period. Veterinary Clinics North
America Food Animal Practice 20, 447-470.
HAMMON DS, EVJEN IM, DHIMAN TR, GOFF JP and WALTERS JL
2006. Neutrophil function and energy status in Holstein cows with uterine
health disorders. Veterinary Immunology and Immunopathology 113,
21-29.
HUZZEY JM, VEIRA DM, WEARY DM and VON KEYSERLINGK MA
2007. Prepartum behavior and dry matter intake identify dairy cows at risk
for metritis. Journal of Dairy Science 90, 3220-3233.
INGVARTSEN KL 2006. Feeding- and management-related diseases in
the transition cow. Physiological adaptations around calving and strategies
to reduce feeding-related diseases. Animal Feed Science and Technology
126, 175-213.
INGVARTSEN KL, DEWHURST RJ and FRIGGENS NC 2003. On the
relationship between lactational performance and health: is it yield or
metabolic imbalance that cause production diseases in dairy cattle? A
position paper. Livestock Production Science 83, 277–308.
INGVARTSEN KL and MOYES K 2013. Nutrition, immune function and
health of dairy cattle. Animal 7 (Suppl. 1), 112-122.
JANOVICK NA and DRACKLEY JK 2010. Prepartum dietary
management of energy intake affects postpartum intake and lactation
performance by primiparous and multiparous Holstein cows. Journal of
Dairy Science 93, 3086–3102.
JANOVICK NA, BOISCLAIR YR and DRACKLEY JK 2011. Prepartum
dietary energy intake affects metabolism and health during the
periparturient period in primiparous and multiparous Holstein cows.
Journal of Dairy Science 94, 1385-1400.
JI P, OSORIO JS, DRACKLEY JK and LOOR JJ 2012. Overfeeding a
moderate energy diet prepartum does not impair bovine subcutaneous
adipose tissue insulin transduction and induces marked changes in peripartal
gene network expression. Journal of Dairy Science 95, 4333-4351.

31
JORRITSMA R, WENSING T, KRUIP TAM, VOS PLAM and
NOORDHUIZEN JPTM 2003. Metabolic changes in early lactation and
impaired reproductive performance in dairy cows. Veterinary Research
34, 11-26.
LEAN IJ, VAN SAUN R and DEGARIS PJ 2013a. Energy and protein
nutrition management of transition dairy cows. Veterinary Clinics North
America Food Animal Practice 29, 337-366.
LEAN IJ, VAN SAUN R and DEGARIS PJ 2013b. Mineral and
antioxidant management of transition dairy cows. Veterinary Clinics North
America Food Animal Practice 29, 367-386.
LEBLANC, S 2010. Assessing the association of the level of milk
production with reproductive performance in dairy cattle. Journal of
Reproduction and Development 56(Suppl.), S1-S7.
LOOR JJ, DANN HM, JANOVICK GURETZKY NA, EVERTS RE,
OLIVEIRA R, GREEN CA, LITHERLAND NB, RODRIGUEZ-ZAS SL,
LEWIN HA and DRACKLEY JK 2006. Plane of nutrition prepartum alters
hepatic gene expression and function in dairy cows as assessed by
longitudinal transcript and metabolic profiling. Physiological Genomics
27, 29-41.
LOOR JJ, EVERTS RE, BIONAZ M, DANN HM, MORIN DE,
OLIVEIRA R, RODRIGUEZ-ZAS SL, DRACKLEY JK and LEWIN HA
2007. Nutrition-induced ketosis alters metabolic and signaling gene
networks in liver of periparturient dairy cows. Physiological Genomics 32,
105-116.
MCART JA, NYDAM DV and OETZEL GR 2012. Epidemiology of
subclinical ketosis in early lactation dairy cattle. Journal of Dairy Science
95, 5056-5066.
MULLIGAN FJ and DOHERTY ML 2008. Production diseases of the
transition cow. Veterinary Journal 176, 3-9.
MULLIGAN FJ, O’GRADY L, RICE DA and DOHERTY ML. 2006. A
herd health approach to dairy cow nutrition and production diseases of the
transition cow. Animal Reproduction Science 96, 331-353.

32
OSPINA PA, NYDAM DV, STOKOL T and OVERTON TR 2010.
Association between the proportion of sampled transition cows with
increased nonesterified fatty acids and β-hydroxybutyrate and disease
incidence, pregnancy rate, and milk production at the herd level. Journal of
Dairy Science 93, 3595-3601.
REIST M, ERDIN DK, VON EUW D, TSCHÜMPERLIN KM,
LEUENBERGER H, HAMMON HM, MOREL C, PHILIPONA C,
ZBINDEN Y, KÜNZI N and BLUM JW 2003. Postpartum reproductive
function: association with energy, metabolic and endocrine status in high
yielding dairy cows. Theriogenology 59, 1707-1723.
ROCHE JR, BELL AW, OVERTON TR and LOOR JJ 2013. Nutritional
management of the transition cow in the 21st century – a paradigm shift in
thinking. Animal Production Science 53, 1000-1023.
RUKKWAMSUK T, WENSING T and KRUIP TAM 1999. Relationship
between triacylglycerol concentration in the liver and first ovulation in
postpartum dairy cows. Theriogenology 51, 1133-1142.
SANTOS JEP, RUTIGLIANO HM and SA FILHO MF 2009. Risk factors
for resumption of postpartum estrous cycles and embryonic survival in
lactating dairy cows. Animal Reproduction Science 110, 207-221.
THARWAT M, TAKAMIZAWA A, HOSAKA YZ, ENDOH D and
OIKAWA S. 2012. Hepatocyte apoptosis in dairy cattle during the
transition period. Canadian Journal of Veterinary Research 76, 241-247.
THATCHER W, SANTOS JEP and STAPLES CR 2011. Dietary
manipulations to improve embryonic survival in cattle. Theriogenology
76, 1619-1631.
WALSH RB, WALTON JS, KELTON DF, LEBLANC SJ, LESLIE KE and
DUFFIELD TF 2007. The effect of subclinical ketosis in early lactation on
reproductive performance of postpartum dairy cows. Journal of Dairy
Science 90, 2788-2796.
WHITAKER DA, SMITH EJ, DA ROSA GO and KELLY JM 1993. Some
effects of nutrition and management on the fertility of dairy cattle.
Veterinary Record 133, 61-64.

33
ZUREK E, FOXCROFT GR and KENNELLY JJ 1995. Metabolic status
and interval to first ovulation in postpartum dairy cows. Journal of Dairy
Science 78, 1909-1920.

34
2. SUPLEMENTAÇÃO DE METIONINA PARA MELHORAR
A PRODUTIVIDADE DA VACA LEITEIRA2
Marcos Neves Pereira
2.1 Introdução
A formulação de dietas para gado leiteiro considerando a
exigência nutricional por aminoácidos (AA) é uma ferramenta para
reduzir o teor de proteína bruta (PB) na dieta e para melhorar o
desempenho animal. Dietas com baixa PB e adequadas em AA são
desejáveis, pois aumentam a eficiência de uso do N alimentar (relação
entre o N secretado no leite e o N consumido), reduzem a perda
ruminal de amônia e o impacto ambiental negativo da produção
animal, e podem reduzir o custo alimentar do rebanho e aumentar o
desempenho leiteiro, especialmente a secreção de proteína no leite
(Schwab, 2010). A suplementação com metionina (Met) em torno do
parto também pode atuar positivamente sobre a saúde e a longevidade
de vacas leiteiras (Osorio et al., 2013).
2 Pereira, M.N. Suplementação de metionina para melhorar a produtividade
da vaca leiteira Anais. I Simpósio Nacional da Vaca Leiteira. Porto Alegre:
Universidade Federal do Rio Grande do Sul. 2014. 239 p.

35
Mais de 700 AA existem na natureza (Wu, 2013) mas apenas
20 constituem as proteínas presentes nos animais. Dentre estes 20 AA,
10 são considerados nutricionalmente essenciais, ou indispensáveis,
devendo ser supridos pela dieta. A síntese de proteína é um evento
determinado geneticamente, sendo que cada proteína é formada por
uma sequência única e imutável de AA. Especialmente importante na
pecuária leiteira é a síntese de caseína pela glândula mamária. Além
da função na síntese de proteína, que afeta todo o metabolismo em
células vivas (e.g. enzimas são proteínas), os AA livres também
regulam vários processos fisiológicos, incluindo a resposta imune. Os
AA também são usados para a síntese de vários compostos contendo
N, como hormônios, nucleotídeos (RNA e DNA), neurotransmissores,
etc. Os aminoácidos “não-proteicos” são encontrados nos fluídos
fisiológicos e são determinantes do metabolismo, e portanto da saúde
e produção de vacas leiteiras. Exemplos destes AA são a
homocisteína, a glutationa e a taurina, todos sintetizados a partir de
Met.
Modelos nutricionais utilizados para a formulação de dietas
estimam o desaparecimento da proteína e de AA do lúmen intestinal
(metabolizáveis) e assumem que estes são utilizados com eficiência de
conversão fixa para suportar funções metabólicas (INRA, 1989; NRC,
2001; Fox et al., 2004). Mesmo sabendo que os procedimentos
matemáticos contidos nestes modelos são insuficientes para descrever
a complexa biologia de ruminantes (Doepel et al, 2004; Hanigan et al,
2006; Lapierre et al., 2006; Lapierre et al, 2012), estes têm sido

36
considerados como de acurácia suficiente para direcionar a
formulação de dietas para rebanhos leiteiros (Pacheco et al., 2012).
Segundo predições destes modelos, a Met é o aminoácido mais
limitante do desempenho leiteiro em dietas formuladas com grão e
forragem de milho e soja e/ou concentrados proteicos de origem
animal (exceto de peixe).
O objetivo deste artigo é discutir de forma prática a
formulação de dietas para vacas leiteiras por AA, enfatizando a
necessidade da suplementação com Met em dietas baseadas em milho
e soja, típicas dos sistemas de produção de leite do Brasil. O modelo
do NRC (2001) será adotado para exemplificar os conceitos, por ser o
modelo nutricional oficial dos EUA, pela sua larga adoção pela
indústria leiteira e por possibilitar o acesso gratuito dos usuários
(https://nanp-nrsp-9.org/nrc-dairy-model/).
2.2 Aspectos Básicos da Nutrição Proteica de Ruminantes
Em ruminantes, os AA absorvidos no intestino podem ter
origem na proteína microbiana sintetizada no rúmen, na proteína
dietética não degrada no rúmen (Proteína Não-Degradável no Rúmen,
PNDR) e na proteína endógena (NRC, 2001). A proteína microbiana
é a maior fonte de AA para absorção no intestino delgado,
especialmente em dietas formuladas com baixo teor proteico. Em
dietas contendo baixo teor de PB, a contribuição dos AA microbianos
para o total de AA absorvidos (metabolizáveis) é maior que em dietas
formuladas com teor excessivo de proteína e de PNDR. Maximizar a

37
função ruminal é o primeiro passo para obter sucesso na formulação
de dietas por AA e obter alto desempenho animal em dietas com baixa
inclusão de concentrados proteicos. Manter teores dietéticos
adequados de carboidratos fibrosos e não-fibrosos e o suprimento
adequado de amônia, aminoácidos e peptídeos para o rúmen (Proteína
Degradável no Rúmen, PDR) são necessários para obter alta eficiência
de síntese de proteína microbiana (g de microrganismos / g de matéria
orgânica fermentada no rúmen). Compreender a demanda por AA dos
microrganismos ruminais é uma fronteira a ser explorada na nutrição
proteica de precisão para vacas leiteiras (Kajikawa et al., 2002).
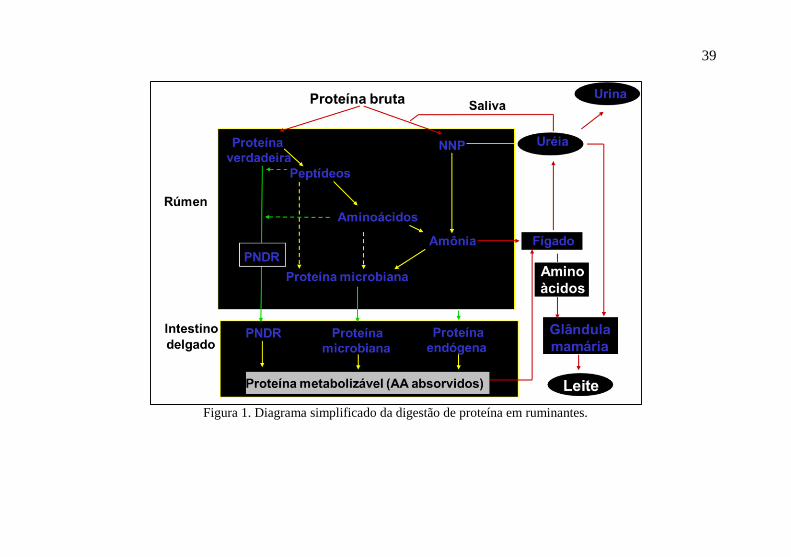
A Figura 1 ilustra de forma simplificada a digestão de proteína
em ruminantes. Parte da proteína verdadeira na dieta e o nitrogênio
não-proteico da dieta ou reciclado do sangue para o rúmen são
degradados no rúmen e o N é incorporado à proteína microbiana ou
absorvido como amônia para o sangue. Em condições normais (fluxo
não excessivo de amônia do rúmen para o sangue), a amônia será
convertida em ureia pelo fígado, sendo esta reciclada para o rúmen ou
excretada pela urina e pelo leite (N-Ureico no Leite, NUL). A perda
urinária de amônia para o ambiente representa um poluente e pode
contribuir para o aquecimento global, sendo uma ineficiência dos
ruminantes. A taxa de crescimento microbiano no rúmen é maior
quando ocorre aporte adequado de carboidratos fermentáveis,
simultaneamente ao suprimento dietético de fibra efetiva em teor
adequado para controlar a ocorrência de acidose ruminal. Alta taxa
(velocidade) de síntese microbiana e/ou aporte ruminal de PDR não

38
excessivo podem reduzir a perda ruminal de amônia, o que é desejável
nutricionalmente e ambientalmente.
39
Figura 1. Diagrama simplificado da digestão de proteína em ruminantes.

40
Parte da proteína verdadeira da dieta pode passar pelo rúmen
em forma não degradável (PNDR). A digestibilidade intestinal da
PNDR difere entre as fontes proteicas, bem como a composição em
AA essenciais da PNDR digerida no intestino (NRC, 2001). Fontes
proteicas de alto valor biológico fornecem AA que são limitantes na
proteína microbiana, relativamente à exigência de AA para síntese de
proteína no corpo (incluindo a glândula mamária). Animais requerem
AA, não PB. O perfil de aminoácidos da proteína microbiana (% de
AA essenciais na PB) é bom, relativamente ao perfil de AA da proteína
do leite. Farelo de soja tem alto teor de Lisina (Lis) na PNDR digerida
no intestino e baixo teor de Met, enquanto proteína oriunda de milho
(glúten) é rica em Met, mas é pobre em Lis. A proteína microbiana
sintetizada no rúmen é deficiente em Histidina (His), relativamente à
proteína do leite.
2.3 Etapas na Formulação de Dietas por Aminoácidos
2.3.1 Fornecer carboidratos de forma adequada.
A proteína microbiana sintetizada no rúmen tem bom perfil de
AA e boa digestibilidade intestinal. Para maximizar a síntese ruminal
de proteína microbiana deve-se fornecer forragens de alta
digestibilidade e com adequado tamanho de partícula e grãos
adequadamente processados, provendo uma mistura de fontes de
carboidratos fermentáveis e fibra fisicamente efetiva que maximize a

41
função (saúde) ruminal, sendo capaz de propiciar alto consumo de
matéria seca e alto fluxo de proteína microbiana para o duodeno. A
nutrição de proteína se inicia com bom domínio e compreensão da
nutrição de carboidratos. Uma dieta adequadamente formulada em
carboidratos resultará em alta secreção de gordura e de proteína no
leite
2.3.2 Atender a demanda de proteína do rúmen.
A exigência de proteína do rúmen (PDR) é diretamente
proporcional à disponibilidade de matéria orgânica fermentável no
rúmen. Em maior consumo de matéria orgânica fermentável, maior é
a demanda ruminal por PDR, já que mais proteína microbiana será
sintetizada. Excesso de PDR relativamente à demanda para
crescimento microbiano deve ser evitado, pois aumentará a perda
ruminal de amônia e reduzirá a proporção do N na dieta incorporado
na proteína do leite. O modelo do NRC (2001) prediz o balanço de
PDR no rúmen (Figura 2). O balanço representa o suprimento dietético
de PDR subtraído da exigência de PDR dos microrganismos do rúmen.
Na metodologia do NRC 2001, a exigência de PDR do rúmen é
diretamente proporcional ao consumo de NDT (energia), quando o
rúmen está em balanço positivo de PDR (N não é limitante do
crescimento microbiano). A meta na formulação é manter o balanço
de PDR levemente positivo, indicando que a exigência ruminal por
AA, peptídeos e amônia está sendo atendida, mas sem excesso
relativamente à disponibilidade de energia para crescimento

42
microbiano. Excesso de N no rúmen pode resultar em excesso de ureia
no leite e no sangue, o que tem implicações negativas para a indústria
(menor proporção de proteína verdadeira na proteína bruta do leite) e
pode afetar negativamente a eficiência reprodutiva de rebanhos
leiteiros.
O teor de NUL do rebanho é uma ferramenta para monitorar
o balanço entre o teor dietético de PDR e o suprimento de carboidratos
fermentáveis no rúmen. O teor de NUL do rebanho deve ser avaliado,
com o intuito de fazer ajustes no suprimento dietético de PDR e na
disponibilidade de carboidratos fermentáveis, adequando a predição
do modelo à realidade dos ingredientes da dieta e dos animais sendo
alimentados. Norte-americanos têm recomendado teores de NUL
entre 8 e 10 mg/dL, dificilmente obtiveis no Brasil, provavelmente em
decorrência da baixa degradabilidade ruminal do amido no milho com
textura dura do endosperma (milho flint) cultivado no país. Teores de
NUL entre 11 e 14 mg/dL são considerados baixos, e desejáveis, em
nossos rebanhos leiteiros. Teor muito baixo de NUL sugere que
deficiência de PDR pode estar limitando a síntese de proteína
microbiana no rúmen, enquanto teor excessivamente alto representa
alta perda ruminal de N dietético na forma de amônia.

43
Figura 2. Estimativas do modelo do NRC (2001) para o balanço de PDR no
rúmen e PNDR, fluxos de proteína metabolizável, e teores de PDR e PNDR
da dieta.
2.3.3 Atender a demanda de proteína metabolizável e energia do grupo
de animais.
A proteína metabolizável (PM) representa a PB (N x 6,25) que
desapareceu do lúmen intestinal e que é assumida como disponível
para os tecidos do animal. A PM representa a mistura de todos os AA
digeridos no intestino, não sendo uma medida precisa da exigência
nutricional por AA essenciais (Arriola et al., 2014). Uma regra prática,

44
seria formular a dieta considerando o consumo de matéria seca
estimado pela média do grupo de animais (peso vivo e produção de
leite médios), e ter a estimativa da produção de leite permissível por
PM equivalente à produção de leite média + 1 desvio padrão (Figura
3). Esta é uma sugestão que agrega alguma margem de segurança ao
nutricionista, apesar de alguns nutricionistas usarem o leite médio para
estimar o leite permissível por PM, em vez do leite médio + 1 desvio
padrão. Dietas adequadamente balanceadas em proteína e energia têm
diferença entre a produção de leite permissível por PM de no máximo
2 kg da produção de leite permissível por energia líquida (NEl).

45
Figura 3. Estimativas do modelo do NRC (2001) para produção de leite
permissível por proteína metabolizável (PM) e por energia líquida (NEl) e
do balanço de PM e energia.

46
2.3.4 Atender à exigência de lisina na proteína metabolizável.
Em dietas com baixo teor proteico formuladas para atender a
demanda por PM, mas sem serem excessivas em balanço de PDR,
atender a demanda de Lis como % da PM é um desafio, especialmente
no Brasil, onde o uso de concentrados proteicos de origem animal é
proibido (e.g. farinha de sangue). Mesmo em dietas com farelo de soja
como concentrado proteico majoritário (alimento rico em Lis), o teor
de Lis na PM pode ser abaixo do recomendado (Figura 4). Segundo o
modelo do NRC (2001), o teor de Lis na PM deve ser ao redor de
6,83% (Whitehouse et al., 2013). Atingir esta recomendação pode ser
difícil na prática, a experiência sugere que almejar teores de Lis ao
redor de 97% (6,6% da PM) a 95% (6,5% da PM) deste valor parece
ser adequado. Nestes casos, pode ser requerida a suplementação de Lis
através de suplementos de baixa degradação ruminal (Lis encapsulada
ou parcialmente protegida da degradação ruminal).
2.3.5 Atender à exigência de metionina na proteína metabolizável.
Uma fonte de Met protegida da degradação ruminal ou um
precursor de Met de baixa metabolização no rúmen (éster isopropílico
do ácido 2-hidróxi-4-metiltio-butírico, HMBi) deve ser acrescida à
dieta para resultar em relação entre Lis e Met na PM de 3,00 (2,28%
de Met na PM, caso tenha 6,83% de Lis na PM), a recomendação para
o modelo do NRC 2001 (Whitehouse et al., 2013). Estimativas do
fluxo de AA essenciais digestíveis são fornecidas pelo modelo (Figura

47
4), e devem ser usadas para monitorar a adequacidade da dieta quanto
aos teores de Met e Lis na PM. Existem sugestões que His pode ser o
próximo AA limitante da produção de leite em dietas formuladas com
baixo teor de PB (Lee et al., 2012). Isto ocorre porque nestes casos a
dependência de AA microbianos para o fluxo de PM para o animal é
proporcionalmente maior que em dietas com excesso de proteína, e
His é um AA cujo teor na proteína microbiana é baixo relativamente
ao teor na proteína do leite. Como regra prática, tem sido sugerido que
o teor de His na PM deveria ser mantido no mínimo 0,1 unidades
percentuais acima do teor de Met, apesar desta recomendação não ter
forte suporte experimental.

48
Figura 4. Estimativas do modelo do NRC (2001) para o fluxo de AA essenciais digestíveis e teor de AA na proteína
metabolizável.

49
2.4 Conclusão
Vacas leiteiras exigem AA, não proteína bruta. Modelos
nutricionais são uma ferramenta para formular dietas por AA, criando
oportunidade para aumentar a eficiência de utilização do N dietético e
reduzir o custo de concentrados proteicos da dieta, sem penalizar, ou
mesmo aumentando, o desempenho leiteiro. O AA mais limitante em
dietas baseadas em milho e soja, típicas do Brasil, é a Met. Atender à
exigência nutricional por este AA requer suplementação,
maximização da função ruminal e suprimento adequado de Lis na PM.
REFERÊNCIAS BIBLIOGRÁFICAS
ARRIOLA APELO, S.I.; KNAPP J.R.; HANIGAN M.D. Current
representation and future trends of predicting amino acid utilization in the
lactating dairy cow. J. Dairy Sci., 97: 4000-4017, 2014.
DOEPEL, L.; PACHECO D.; KENNELLY J.J.; HANIGAN M.D.; LÓPEZ
I.F.; LAPIERRE H.. Milk protein synthesis as a function of amino acid
supply. J. Dairy Sci., 87: 1279-1297, 2004.
FOX, D.G.; TEDESCHI L.O.; TYLUTKI T.P.; RUSSELL J.B.; VAN
AMBURGH M.E.; CHASE L.E.; PELL A.N.; T.R. Overton. The Cornell Net
Carbohydrate and Protein System model for evaluating herd nutrition and
nutrient excretion. Anim. Feed Sci. Technol., 112: 29-78, 2004.
HANIGAN, M.D., BATEMAN H.G., FADEL J.G., MCNAMARA J.P..
Metabolic models of ruminant metabolism: recent improvements and current
status. J. Dairy Sci., 89(E.Suppl.): E52-E54, 2006.
INRA. Ruminant nutrition: Recommended allowances and feed tables.
INRA Editions, Paris, França. 1989

50
KAJIKAWA, H.; MITSUMORI M.; OHMOMO S.. Stimulatory and
inhibitory effects of protein amino acids on growth rate and efficiency of
mixed ruminal bacteria. J. Dairy Sci., 85: 2015-2022, 2002.
LAPIERRE, H.; PACHECO D.; BERTHIAUME R.; OUELLET D.R.;
SCHWAB C.G.; DUBREUIL P.; HOLTROP G.; LOBLEY G.E.. What is the
true supply of amino acids for a dairy cow? J. Dairy Sci., 89(E-Suppl.): E1-
E14, 2006.
LAPIERRE, H.; LOBLEY G.E.; DOEPEL L.; RAGGIO G.; RULQUIN H.;
LEMOSQUET S.. Triennial Lactation Symposium: Mammary metabolism
of amino acids in dairy cows. J. Anim. Sci., 90: 1708-1721, 2012.
LEE, C.; HRISTOV A.N.; CASSIDY T.W.; HEYLER K.S.; LAPIERRE H.;
VARGA G.A.; DE VETH M.J.; PATTON R.A.. Rumen-protected lysine,
methionine, and histidine increase milk protein yield in dairy cows fed a
metabolizable protein-deficient diet. J. Dairy Sci., 95: 6042-6056, 2012.
National Research Council. Nutrient Requirements of Dairy Cattle. 7th
rev. ed. Natl. Acad. Sci., Washington, EUA. 2001
OSORIO, J.S.; JI P.; DRACKLEY J.K.; LUCHINI D.; LOOR J.J..
Supplemental Smartamine M or MetaSmart during the transition period
benefits postpartal cow performance and neutrophil function. J. Dairy Sci.,
96: 6248-6263, 2013.
PACHECO, D.; PATTON R.A.; PARYS C.; LAPIERRE H.. Ability of
commercially available dairy ration programs to predict duodenal flows of
protein and essential amino acids in dairy cows. J. Dairy Sci., 95: 937-963,
2012.
SCHWAB, C.G. Balancing diets for amino acids: nutritional,
environmental and financial implications. Pages 1-13 In: Proc. Tri-State
Dairy Nutr. Conf., Fort Wayne, EUA. The Ohio State University, Columbus,
2010.
WHITEHOUSE, N.L.; SCHWAB C.G.; TYLUTKI T.; SLOAN B.K..
Optimal lysine and methionine concentrations for milk protein production as
determined with the latest versions of Dairy NRC 2001 and AMTS.Cattle. J.
Dairy Sci., 93 (Suppl.1): 253, 2013.

51
WU, G. Amino acids - Biochemistry and Nutrition. CRC Press, Boca
Raton, EUA. 2013.

52
3. COMO A DIETA PODE AFETAR A COMPOSIÇÃO DO
LEITE.3
Bolivar Nóbrega de Faria
3.1 Introdução
A composição e a porcentagem de proteína do leite de vacas
têm recebido grande interesse nos últimos anos, principalmente por
influir diretamente no rendimento industrial. Uma queda na
concentração de proteína de 3,55% para 2,90% representa a
necessidade de aproximadamente 1.580 litros de leite adicionais para
se produzir uma tonelada de queijo do tipo Cheddar.
Os resultados da alteração no teor de proteína do leite através
da manipulação da dieta giram em torno de 0,6 unidades percentuais
(sendo mais prováveis 0,1 a 0,2 unidades). É modesto, se comparado
ao potencial de alteração do teor de gordura do leite, que gira em torno
de 3,0 unidades percentuais. Porém, à medida que se aumenta o teor
de proteína do leite, geralmente também aumenta a produção de leite,
o que já não ocorre com a gordura (Waldner et al., 2004).
3 Faria, B.N. Como a dieta pode afetar a composição do leite. Anais. I
Simpósio Nacional da Vaca Leiteira. Porto Alegre: Universidade Federal do
Rio Grande do Sul. 2014. 239 p.

53
Além da nutrição, vários aspectos como estação do ano,
composição racial, estádio de lactação e doenças podem afetar o teor
de proteína do leite. A sazonalidade relaciona-se com a temperatura
ambiente e disponibilidade de alimentos. Altas temperaturas e
umidade reduzem o teor de proteína e a produção de leite, pois o
animal reduz o consumo de alimento. Entre as raças leiteiras, a
Holandesa possui a menor porcentagem de proteína no leite enquanto
a Jersey é a que possui a maior. A mudança na composição do leite
utilizando a seleção de animais é lenta, e não produz resultados
imediatos, pois a herdabilidade das características é baixa.
Os níveis de gordura do leite podem ser mais influenciados
pela dieta. Maiores níveis de gordura no leite também têm sido
buscados pelos laticínios, uma vez que aumenta do rendimento de
alguns produtos nobres, como alguns tipos de queijo. Com isso, novas
formas de pagamento têm sido propostas se valorizando cada vez mais
os constituintes do leite. Além disso, concentrações baixas de gordura
do leite têm sido relacionadas direta ou indiretamente a várias doenças
metabólicas, como o deslocamento de abomaso, laminite e,
principalmente, a acidose ruminal subaguda. Dessa forma, saber como
a dieta afetas o perfil dos constituintes do leite pode ser uma boa forma
de se monitorar ou estimar a saúde dos animais.
A compreensão da composição do leite pode ser interessante
ao produtor que precisa planejar a lactação da vaca para maximizar os
lucros. Isso envolve a compreensão do efeito da alimentação, do
manejo reprodutivo e da genética sobre a lactação. O conhecimento da
54
composição do leite também é importante para a indústria
processadora, que depende da manipulação das suas características
físicas e químicas para a elaboração de diferentes produtos lácteos,
assim como para assegurar a sua qualidade.
O objetivo desta revisão é abordar os fatores nutricionais da
vaca que interferem na concentração de alguns componentes do leite.
3.2 Síntese dos Carboidratos do Leite
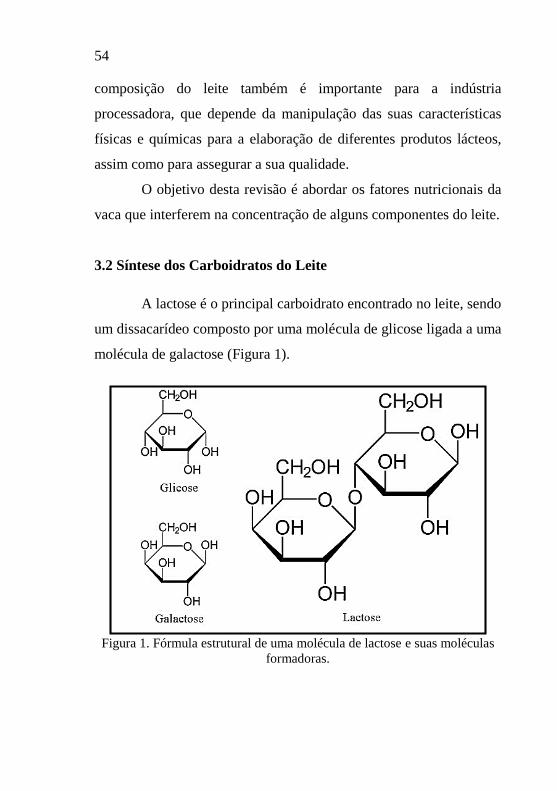
A lactose é o principal carboidrato encontrado no leite, sendo
um dissacarídeo composto por uma molécula de glicose ligada a uma
molécula de galactose (Figura 1).
Figura 1. Fórmula estrutural de uma molécula de lactose e suas moléculas
formadoras.

55
A lactose possui um papel muito importante na síntese do
leite. Sua função está relacionada com a manutenção da osmolaridade
do leite e nos processos de produção e secreção (Larson, 1995), ou
seja, é o grande responsável por atrair a água para o leite (cerca de
50% da pressão osmótica para a síntese do leite), sendo o principal
responsável pelo volume do leite produzido (Muhlbach, 2003). Cada
grama de lactose do leite arrasta aproximadamente 10 vezes o mesmo
peso em água. Devido à estreita relação entre a síntese de lactose e a
quantidade de água atraída para o leite, a lactose é o componente
menos variável do leite (Tabela 1).
Tabela 1. Variações nos constituintes de 257.540 amostras de leite nos
estados de São Paulo, Paraná e Santa Catarina (Ribas et al., 2004).
Constituinte do
leite
Concentração média
(%)
Desvio
padrão
Lactose 4,55 0,19
Proteína 3,24 0,24
Gordura 3,69 0,62
Sólidos totais 12,32 0,79
Segundo Hurley (2004) a lactose é relativamente insensível às
mudanças na dieta das vacas, no entanto, vacas subnutridas
apresentam redução na produção de leite e na percentagem de lactose,
sendo que estes sintomas são revertidos quando dietas adequadas são
fornecidas.

56
3.3 Compostos Nitrogenados do Leite
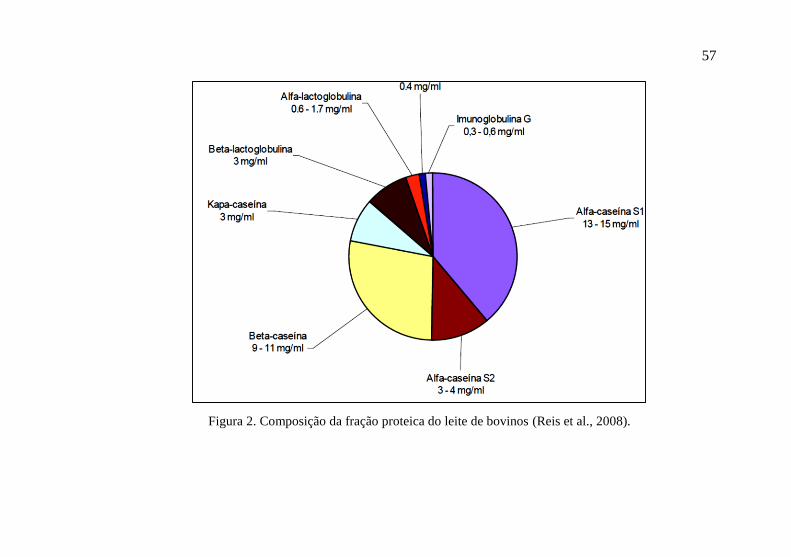
Como principais representantes das proteínas do leite estão as
caseínas (α, β, κ), e as lactoglobulinas (α e β), que representam mais
de 90% da proteína total do leite, além de albumina sérica e
imunoglobulina G (Figura 2). A caseína constitui de 76 a 86% da
proteína presente no leite, sendo o componente determinante do
rendimento industrial na produção de queijo. É sintetizada pelas
células secretoras, como resultado da expressão de quatro genes (dois
genes para α-caseína, s1 e s2; um para β-caseína e um para κ-caseína),
sendo secretada na forma de micelas, que são grupamentos de várias
moléculas de caseína ligadas a íons como o fosfato e o cálcio, e se
encontram em suspensão no leite (De Peters & Cant, 1992).
57
Figura 2. Composição da fração proteica do leite de bovinos (Reis et al., 2008).

58
Ao contrário das caseínas, existem proteínas que se encontram
em solução no leite, sendo chamadas de proteínas do soro do leite.
Estas proteínas possuem duas fontes principais, podendo ser
produzidas na glândula mamária, como a α-lactoalbumina e β-
lactoglobulina, ou podem passar diretamente do sangue para a
glândula mamária, como a albumina sérica e as imunoglobulinas (De
Peters & Ferguson, 1992). O leite possui ainda uma fração de
nitrogênio-não proteico (NNP), constituindo cerca de 5% da proteína
bruta do leite, composta principalmente de ureia (aproximadamente
48%) e em menor quantidade de creatinina, amônia e outros
compostos nitrogenados (De Peters & Ferguson, 1992) (Tabela 2).
Tabela 2. Fração de nitrogênio-não proteico do leite bovino (Bremel, 1995)
Componentes Concentração
(mg N/dL)
Aminoácidos 3,7
Ureia 8,4
Creatina 1,2
Creatinina 0,2
Ácido úrico 0,8
Ácido orótico 1,2
Carnitina e acetil-carnitina 0,2
3.4 Gordura do Leite
A gordura é o principal componente energético do leite, sendo
responsável pelas propriedades físicas, características industriais,
qualidades organolépticas do leite e seus derivados e, por isso,

59
possuem um importante valor econômico (Bauman e Griinari, 2003).
Segundo Riel (1991), o tipo de gordura predominante no leite de vaca,
com 97% a 99% dos lipídeos totais são os triglicerídeos. Os restantes
são os fosfolipídeos e esteróis, especialmente o colesterol. Os
triglicerídeos são constituídos principalmente de ácidos graxos, sendo
na gordura do leite constituídos em sua maioria de ácidos graxos de
cadeia saturada (Tabela 3). A gordura é o constituinte do leite mais
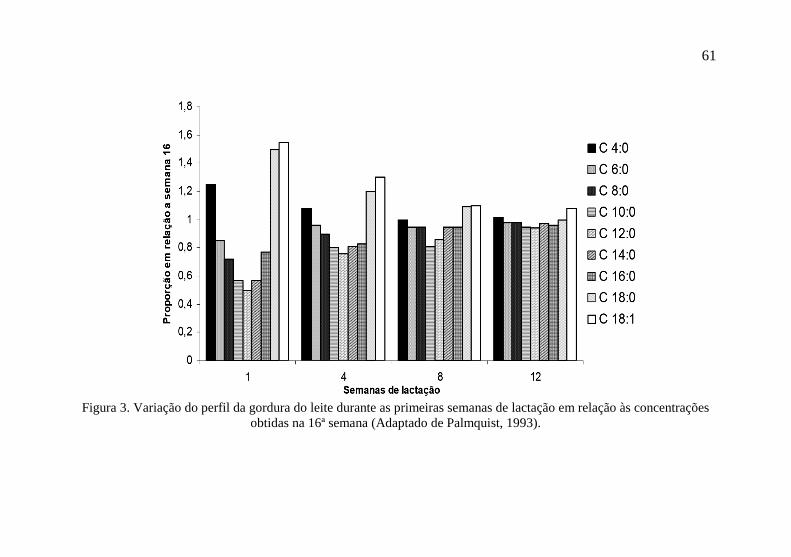
sensível às variações, sendo possível observar variações não somente
em relação à raça, mas também ao longo da lactação (Figura 3).
Tabela 3. Composição de gordura do leite in natura ou em pó (Adaptado de
Jenkins e McGuire, 2006).
Constituinte do leite In Natura Em Pó
Água 88,32 2,47
Gordura 3,25 26,71
Ácidos graxos
(% do total)
Saturados 64,90 66,10
Monoinsaturados 28,30 31,30
Poli-insaturados 6,80 2,60
Ultimamente várias pesquisas têm despertado interesse para o
perfil de lipídeos do leite. Isso está mais relacionado aos seus efeitos
na saúde humana, como é o interesse nas concentrações de ácido
linoleico conjugado (CLA) e não somente em relação aos seus efeitos
sobre os bovinos. Os CLA são compostos naturalmente encontrados
na gordura de ruminantes que exercem efeitos benéficos à saúde
humana. Dentre os benefícios destacam-se efeitos anticarcinogênicos,

60
alterações na partição de nutrientes e no metabolismo de lipídeos,
propriedades antidiabéticas e antiaterogênicas, modulação da resposta
imune, dentre outras (Bauman et al., 2001). Portanto, seria desejável
que as concentrações do CLA no leite fossem aumentadas, já que são
a maior fonte de CLA na dieta de humanos (Bauman et al., 2000).
61
Figura 3. Variação do perfil da gordura do leite durante as primeiras semanas de lactação em relação às concentrações
obtidas na 16ª semana (Adaptado de Palmquist, 1993).

62
Com relação aos ruminantes, o foco tem sido dado a dois
isômeros específicos, o cis-9 trans-11 e o trans-10 cis-12 (Figura 4).
O primeiro devido ao seu papel como um anticarcinogênico e o
segundo, devido aos seus efeitos sobre o metabolismo de lipídeos
(Bauman et al., 2001).
Figura 4. Estrutura química do ácido linoleico e dos isômeros de CLA
C18:2 cis-9, trans-11 e C18:2 trans-10, cis-12 (Bauman et al., 2001).
3.5 Síntese de Carboidratos do Leite
A produção de lactose no úbere está relacionada à síntese de
propionato no rúmen e disponibilidade de produtos gliconeogênicos.
Quanto maior a disponibilidade de glicose para a glândula mamária,
maior será a possibilidade de produção de lactose (Figura 5) e,
consequentemente, de produção de leite.

63
UTP + glicose 1-P UDP-glicose + P-P (1)
UDP-glicose UDP-galactose (2)
UDP-galactose + glicose lactose + UDP (3)
Figura 5. Via metabólica de síntese de glicose na glândula mamária
(Larson, 1995).
UTP= uridina trifosfato; UDP= uridina difosfato; UDP-glicose= uridina
difosforil glicose; UDP-galactose= uridina difosforil galactose; P-P=
pirofosfato; Enzimas: (1) uridina difosforil glicose pirofosfarilase; (2)
uridina difosforil galactose-4-epimerase; e (3) lactose sintetase (composta
por galactosil transferase e α-lactoalbumina).
A glicose que o bovino necessita como precursor para a
síntese de lactose na glândula mamária é sintetizada no fígado via
gliconeogênese, principalmente a partir do ácido propiônico,
sintetizado via degradação ruminal dos carboidratos oriundos da dieta.
Outra fonte importante de substrato para a gliconeogênese são os
aminoácidos, oriundos da dieta ou de mobilização corporal,
principalmente no período pós-parto e/ou balanço energético negativo
(Figura 6). Após a absorção pelas células secretoras da glândula
mamária, cerca de 60 a 70% da glicose é destinada para a síntese de
lactose. No entanto, a glicose também tem outras funções importantes
na síntese dos constituintes do leite, sendo um importante estimulador
para a síntese de proteica, substrato para síntese de glicerol e da
gordura do leite.
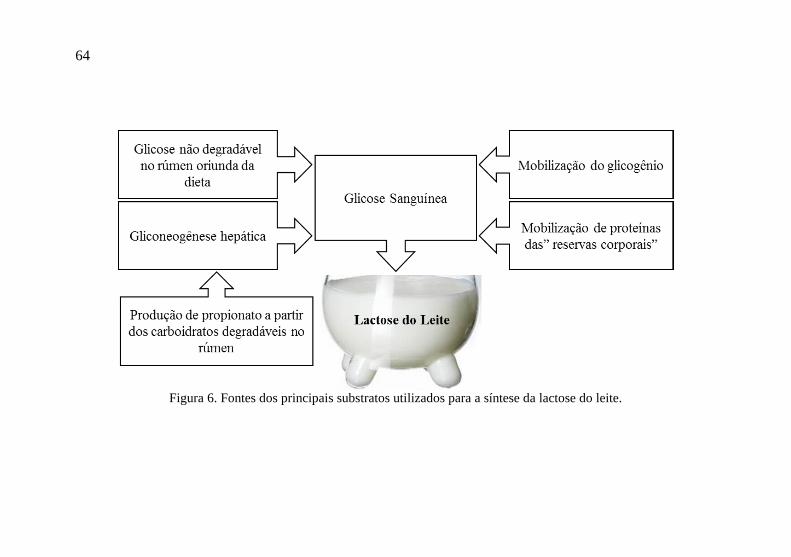
64
Figura 6. Fontes dos principais substratos utilizados para a síntese da lactose do leite.

65
3.6 Síntese de Compostos Nitrogenados do Leite
As proteínas do leite (caseínas e lactoglobulinas) são
sintetizadas nas células secretoras da glândula mamária a partir de
aminoácidos provenientes do sangue ou transportados do sangue
(Figura 7). No entanto, alguns aminoácidos podem ser sintetizados na
própria glândula mamária através da utilização de precursores como a
glicose e outros aminoácidos. A síntese de proteína nas células
secretoras ocorre no retículo endoplasmático rugoso. A estrutura da
proteína é pré-determinada pela informação genética contida no DNA
presente no núcleo da célula. O DNA serve de molde para a síntese de
uma fita de RNA que contém a informação sobre a sequência de
aminoácidos para as proteínas. Vacúolos presentes no citoplasma são
formados para transporte da proteína sintetizada até o lúmen.
Algumas proteínas presentes no leite não são sintetizadas na
glândula mamária e são transportadas pelo sangue até entrarem no
lúmen alveolar. A albumina encontrada no leite é produzida no fígado
e sua concentração no leite reflete a concentração no sangue. As
imunoglobulinas são transportadas para o leite via sangue e vasos
linfáticos de origem do baço e linfonodos. Linfócitos B produtores de
anticorpos podem se alojar na glândula mamária para auxiliar na
síntese de imunoglobulinas do colostro (Larson, 1995).
Outro composto nitrogenado importante do leite, a ureia, não
é produzido diretamente pela glândula mamária. O nível de nitrogênio
ureico no leite (MUN) está negativamente associado à eficiência da

66
utilização do nitrogênio da dieta para síntese proteica do leite. Como
a ureia se difunde livremente entre os fluidos do organismo animal,
sua concentração na glândula mamária reflete a concentração no
sangue (Nousiainen et al., 2004). Qualquer fator que interfira na
eficiência da utilização da amônia presente no rúmen pelos
microrganismos ruminais irá resultar em aumento da quantidade de
ureia no sangue. A baixa eficiência é parcialmente devido à perda de
nitrogênio amoniacal no rúmen. Estima-se que entre 40 a 80% do
nitrogênio bacteriano tem origem na amônia ruminal (Hristov e
Broderick, 1996), e que a concentração ruminal de amônia é
inversamente relacionada à disponibilidade de carboidratos.
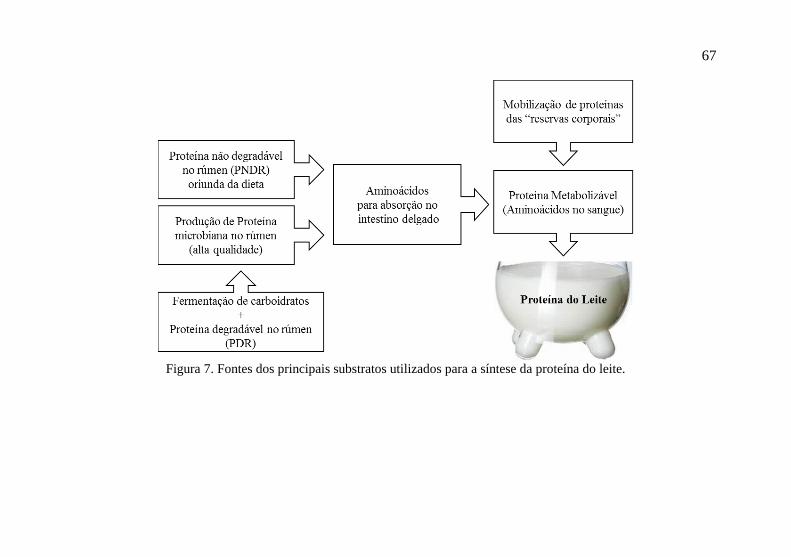
67
Figura 7. Fontes dos principais substratos utilizados para a síntese da proteína do leite.

68
3.7 Síntese de Gordura do Leite
Segundo Harding (1995), a gordura do leite é sintetizada nas
células epiteliais alveolares, mais especificamente nas áreas do
citoplasma ocupadas pelo retículo endoplasmático rugoso. Os
principais precursores da gordura do leite são ácidos graxos voláteis
(AGV), o acetato e o butirato, originados na fermentação ruminal de
forragens e outros alimentos ricos em fibra, e os ácidos graxos não
esterificados da corrente sanguínea. Esses substratos irão dar origem
aos ácidos graxos, que posteriormente formarão os triglicerídeos do
leite (Figura 8).
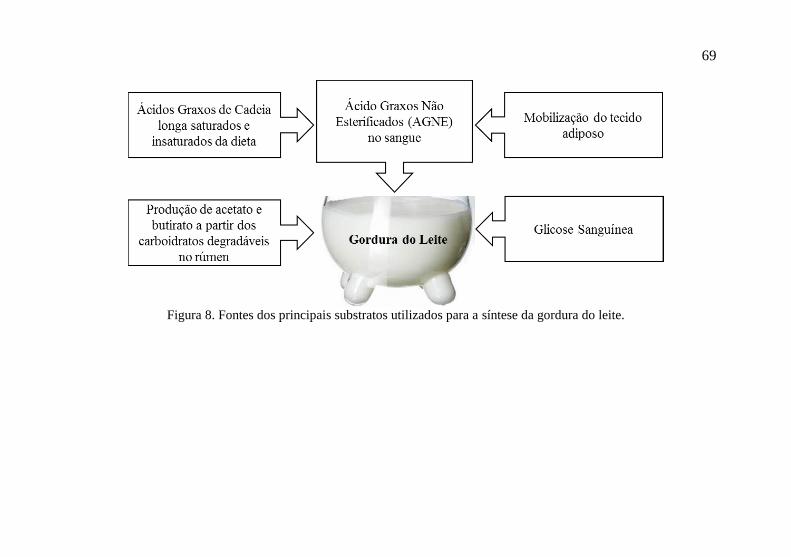
69
Figura 8. Fontes dos principais substratos utilizados para a síntese da gordura do leite.

70
Ácidos graxos de cadeia curta, contendo entre 4 e 8 carbonos,
e de cadeia média, com 10 a 14 carbonos, provem quase que
exclusivamente da síntese “de novo”. Já os ácidos graxos de cadeia
longa, constituídos por cadeias superiores a 16 carbonos, são
derivados da captação direta dos lipídios da corrente sanguínea pela
glândula mamária (Bauman e Griinari, 2003). Os ácidos graxos com
16 carbonos podem ser obtidos através das duas fontes. Cerca de
metade dos ácidos graxos do leite são sintetizados pela própria
glândula mamária, a partir da síntese “de novo” tendo como principal
fonte de carbono, principalmente, o acetato produzido na fermentação
ruminal (Bauman e Griinari, 2003).
Outra fonte de carbonos para a síntese “de novo” na glândula
mamária é o β-hidroxibutirato produzido pelo epitélio ruminal, a partir
do butirato. Os ácidos graxos pré-formados captados pela glândula
mamária e diretamente usados para a síntese de gordura do leite são
derivados das lipoproteínas circulantes e dos ácidos graxos não
esterificados originários da absorção dos lipídios no trato
gastrointestinal e da mobilização de reservas corporais,
respectivamente. A lipólise (mobilização de gordura corporal) é
responsável por menos de 10% dos ácidos graxos da gordura do leite,
sendo a maior parte dos ácidos graxos de cadeia longa do leite
proveniente da absorção intestinal (Bauman e Griinari, 2003).
As gorduras de origem vegetal da dieta das vacas leiteiras são
altamente insaturadas, desta forma, essas gorduras quando ingeridas

71
sofrem no rúmen a biohidrogenação, antes de serem absorvidas pela
corrente sanguínea na forma de triglicerídeos ligados a lipoproteínas.
Esses triglicerídeos são transportados pelo sangue até a glândula
mamária, onde sofrem a quebra em subunidades de glicerol e ácidos
graxos livres que podem, então, ser absorvidos pelas células da
glândula mamária.
3.8 Fatores nutricionais que interferem nos compostos
nitrogenados do leite.
De forma geral, as estratégias nutricionais que visam o
aumento da proteína do leite devem ter como princípio o maior
suprimento de aminoácidos e energia (glicose) para a glândula
mamária. A utilização de aminoácidos pela glândula mamária é
dependente de uma série de fatores como, irrigação sanguínea da
glândula mamária, concentração arterial de aminoácidos, eficiência no
transporte e regulação intracelular das vias metabólicas (Bequette et
al. 1998). A eficiência na conversão de aminoácidos em proteína é
baixa e inconstante (27 a 42%), e varia de acordo com o fornecimento
de aminoácidos metabolizáveis (Doeppel et al., 2004). Segundo
Murphy e O’Mara, (1993), a limitação da produção de proteína pela
glândula mamária é devida principalmente aos aminoácidos
essenciais, como a metionina, lisina e histidina. De forma geral, lisina
e metionina são consideradas aminoácidos limitantes (Schwab et al.,
1992).

72
Vários experimentos têm sido realizados na tentativa de se
corrigir a relação lisina/metionina através da suplementação desses
aminoácidos na forma protegida, como hidroxi-análogos ou
precursores. No entanto, os resultados quanto aos amentos das
concentrações de proteína no leite são variáveis (Figura 9), uma vez
que está resposta depende de outras variáveis importantes como dieta
base, ingestão de matéria seca, níveis energéticos e proteicos da dieta,
quantidade de fibra fisicamente efetiva, além do potencial genético do
animal.
O aumento no fornecimento de aminoácidos para a glândula
mamária pode ser obtido através do aumento na quantidade de
aminoácidos que chegam ao intestino delgado, ou seja, proteína
microbiana e proteína degradável no rúmen (PNDR). Com o maior
suprimento de aminoácidos absorvidos e disponíveis, além de um
melhor perfil de aminoácidos não haveria ou diminuiria a limitação
dos principais aminoácidos essenciais para a síntese proteica (Murphy
e O’Mara, 1993). Além disso, os aminoácidos resultantes da
mobilização proteica endógena também colaborariam para esse
melhor e maior pool.

73
Figura 9. Variação da proteína do leite (%) de vários experimentos realizados com diferentes dietas, fontes e
concentrações proteicas (Jenkins & McGuire, 2006).
2.6 2.7 2.8 2.9 3 3.1 3.2 3.3 3.4
Met (16,1)
Met (18,9)
F. Soja + Met (15,6)
Met + Lys (19,5)
Met (19,5)
Glutem Milho + Met (16,2)
Met (19,5)
F. Soja Tostado + Met (15,0)
Milho moído (15,7)
F. Soja Tostado (15,0)
Soja espandida + Met (15,7)
Soja espandida (15,7)
Controle (16,1)
Controle (18,8)
Controle (15,6)
Controle (19,5)
Proteína do Leite (%)

74
Em dietas a base de silagem de milho e/ou suplementadas com
milho, o aminoácido mais limitante tende a ser a lisina. Em dietas ricas
em soja e outras leguminosas pode haver limitação de metionina, por
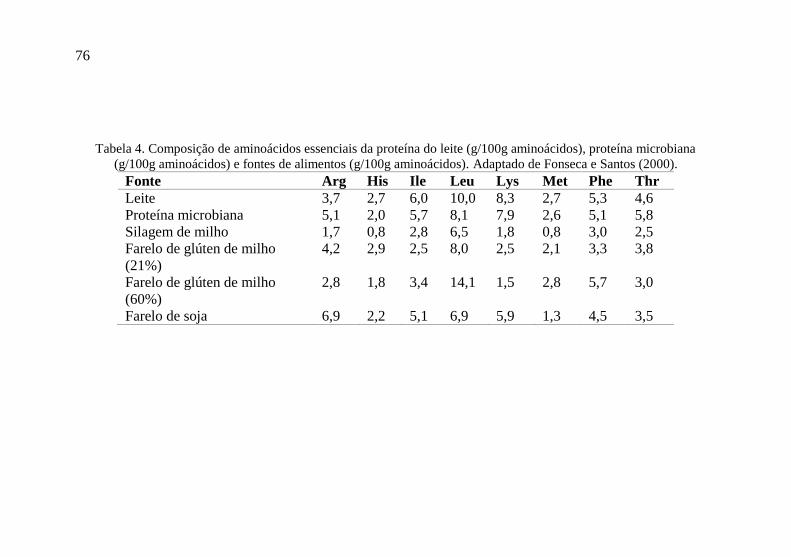
haver grande suprimento de lisina (Tabela 4). A utilização de
aminoácidos protegidos, em especial lisina e metionina, tem
apresentado resultados modestos (Misciatteli et al., 2003). No entanto,
há aumentos significativos na eficiência de absorção dos aminoácidos
em função da energia metabolizável da dieta. Mais estudos são
necessários para verificar outros aminoácidos possivelmente
limitantes. Além disso, o alto custo de suplementação dos
aminoácidos protegidos tem limitado seu uso nos países onde estes
suplementos estão disponíveis.
O teor de proteína bruta da dieta possui efeito muito pequeno
sobre o teor proteico do leite (De Peters e Ferguson, 1992), relatou
aumento de 0,02% para cada 1% de aumento na proteína da dieta. No
entanto, o fornecimento de dietas com deficiências de proteína pode
reduzir a concentração deste nutriente em 0,1 a 0,2 unidades
percentuais (Sutton, 1989), além de limitar a produção de leite. Pode-
se dizer que a variação no teor proteico da dieta afeta muito mais a
produção de leite do que sua composição.
A proteína bruta da dieta não está correlacionada (p> 0,25)
com a porcentagem de proteína no leite, mas fracamente
correlacionada com a produção de proteína no leite. Por outro lado, a

75
proteína do leite aumenta linearmente com o teor de PNDR na dieta
(NRC, 2001).
76
Tabela 4. Composição de aminoácidos essenciais da proteína do leite (g/100g aminoácidos), proteína microbiana
(g/100g aminoácidos) e fontes de alimentos (g/100g aminoácidos). Adaptado de Fonseca e Santos (2000).
Fonte Arg His Ile Leu Lys Met Phe Thr
Leite 3,7 2,7 6,0 10,0 8,3 2,7 5,3 4,6
Proteína microbiana 5,1 2,0 5,7 8,1 7,9 2,6 5,1 5,8
Silagem de milho 1,7 0,8 2,8 6,5 1,8 0,8 3,0 2,5
Farelo de glúten de milho
(21%)
4,2 2,9 2,5 8,0 2,5 2,1 3,3 3,8
Farelo de glúten de milho
(60%)
2,8 1,8 3,4 14,1 1,5 2,8 5,7 3,0
Farelo de soja 6,9 2,2 5,1 6,9 5,9 1,3 4,5 3,5

77
No entanto, é muito difícil separar o efeito proveniente do
aumento do teor da proteína da dieta e o aumento no teor de energia
ingerido pelo animal, tendo em vista que o teor de proteína da dieta
aumenta a eficiência microbiana, influenciando positivamente a
ingestão de matéria seca e a digestibilidade da dieta. A relação entre o
aumento na ingestão de energia e o aumento no teor de proteína do
leite é mais importante que a relação entre o teor de proteína ingerido
na dieta e o aumento no teor de proteína do leite (Griinari et al., 1997).
O aumento da ingestão de energia por vacas em lactação aumenta a
concentração de proteína no leite. Estas mudanças na ingestão de
energia são geralmente, obtidas por aumento da relação
concentrado/volumoso, por mudanças na fonte de carboidratos ou por
utilização de grãos processados.
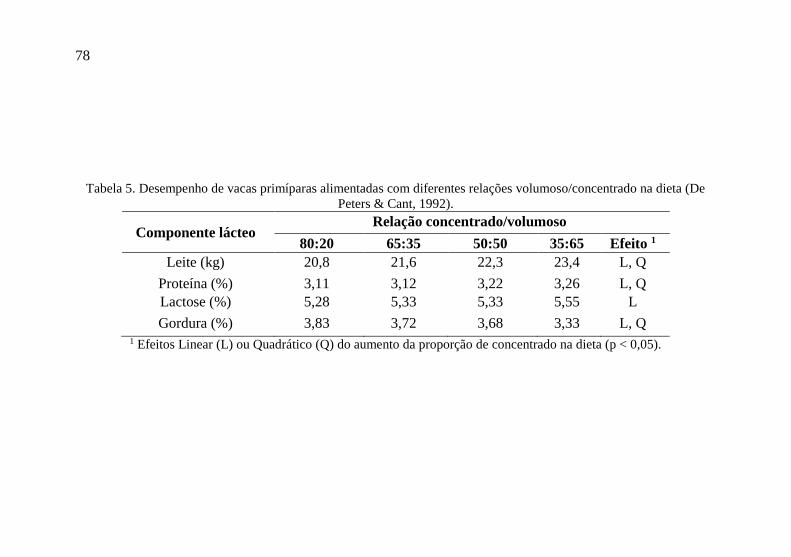
78
Tabela 5. Desempenho de vacas primíparas alimentadas com diferentes relações volumoso/concentrado na dieta (De
Peters & Cant, 1992).
Componente lácteo Relação concentrado/volumoso
80:20 65:35 50:50 35:65 Efeito 1
Leite (kg) 20,8 21,6 22,3 23,4 L, Q
Proteína (%) 3,11 3,12 3,22 3,26 L, Q
Lactose (%) 5,28 5,33 5,33 5,55 L
Gordura (%) 3,83 3,72 3,68 3,33 L, Q 1 Efeitos Linear (L) ou Quadrático (Q) do aumento da proporção de concentrado na dieta (p < 0,05).

79
Há limites para a elevação da produção de proteína do leite,
com o aumento de concentrados na dieta, que seria no ponto em que o
pH cai sensivelmente e reduz a população celulolítica do rúmen.
Dietas com elevados teores energéticos costumam reduzir a gordura e
aumentar a proteína do leite. Pode-se aumentar potencialmente o
fornecimento ou perfil de aminoácidos disponíveis no intestino através
da utilização de maiores quantidades de proteína não degradável nu
rúmen. No entanto, só haverá benefício quando a fonte de PNDR
completar positivamente a proteína microbiana e a composição da
proteína dos demais alimentos, uma vez que as fontes de PNDR
normalmente substituem as fontes de degradação ruminal na dieta, que
estimulam a fermentação microbiana (Wright et al., 1998). A
subnutrição e dietas com baixos teores de proteína diminuem a
porcentagem de proteína no leite, sendo que este quadro pode ser
revertido por meio da adição de proteína extra, quando a proteína for
o limitante.
3.9 Fatores nutricionais que afetam a gordura do leite.
O conteúdo e composição da gordura do leite podem ser
fortemente afetados pela dieta. Em ruminantes, a composição dos
ácidos graxos da dieta não é fortemente refletida na composição dos
ácidos graxos da gordura do leite. Isso ocorre devido à presença do
rúmen, onde os constituintes lipídicos da dieta são alterados pelo
metabolismo microbiano (Bauman e Griinari, 2003).

80
Os triglicerídeos não são fermentados no rúmen, portanto não
são fonte de energia para os microrganismos ruminais. Ao atingirem
o rúmen, as gorduras podem passar por duas transformações pelos
microrganismos ruminais: lipólise e biohidrogenação. A lipólise é um
processo que libera ácidos graxos livres no rúmen a partir de lipídeos
esterificados das plantas e, depois passam por um processo de
biohidrogenação (Jenkins, 1993). A taxa de lipólise varia de acordo
com a quantidade e a composição dos ácidos graxos da gordura
fornecida na dieta (NRC, 2001). A biohidrogenação é uma forma de
proteção, uma vez que os triglicerídeos insaturados possuem certa
toxicidade aos microrganismos ruminais. Esse processo consiste em
desfazer as duplas ligações dos triglicerídeos insaturados e acrescentar
um átomo de hidrogênio, formando uma ligação simples com o
carbono (Figura 10). Isso é nada mais que um processo de saturação
do triglicerídeo.

81
Figura 10. Esquema da biohidrogenação ruminal.
No processo de biohidrogenação, enzimas microbianas
saturam o ácido linoleico (C18:2) adicionando hidrogênio nas duplas
ligações até que a molécula seja totalmente saturada e transformada a
ácido esteárico. No processo de formação do ácido esteárico, produtos
intermediários são formados, como os ácidos trans 18:1 e ácidos
linoleicos conjugados (CLA). Esses intermediários passam do rúmen
ao intestino onde são absorvidos. Os maiores substratos da
biohidrogenação ruminal são o ácido linoleico e linolênico e a taxa de
biohidrogenação dos ácidos graxos é mais alta à medida que o grau de
insaturação aumenta. Para a maioria das dietas a taxa de

82
biohidrogenação do ácido linoleico e linolênico é de 70-95% e 85-
100%, respectivamente (Doreau & Ferlay, 1994; Beam et al., 2000).
Quando dietas ricas em concentrado são fornecidas, a taxa de
hidrogenação é reduzida, o que pode ser atribuído à inibição da lipólise
em pH ruminal baixo provocado por essas dietas (Van Nevel &
Demeyer, 1995; Van Nevel & Demeyer, 1996b). A hidrogenação
também é afetada quando uma quantidade excessiva de lipídios não
protegidos está presente na dieta.
Dois importantes intermediários no processo de
biohidrogenação são o trans-11 18:1 (ácido trans-vaccênico)
formados a partir de ácido linoleico e ácido linolênico e cis-9, trans-
11 ácido linoleico conjugado (CLA) formado na biohidrogenação do
ácido linoleico. Esses intermediários estão presentes em quantidades
apreciáveis na gordura dos ruminantes, em uma relação de 3:1 (CLA:
trans-vaccênico), mas no rúmen CLA cis-9, trans-11 é apenas um
intermediário transitório, enquanto o trans-vaccênico é que se acumula
no rúmen.
Em condições de ácidos graxos insaturados em excesso no
rúmen ou pH ruminal baixo, alguns isômeros trans de ácidos graxos
podem ser formados. Esses ácidos graxos estão associados com
depressão da gordura do leite (Griinari et al., 1998; Baumgard et al.,
2000; Bauman & Griinari, 2003). Portanto, dependendo de quanto os
ácidos graxos saem do rúmen na forma saturada, como ácido esteárico,
ou na forma dos intermediários insaturados influenciará em quanto de
energia será disponibilizada para absorção e os efeitos dos ácidos

83
graxos na gordura do leite. Dessa forma, a dieta e alterações no
ambiente ruminal podem mudar as vias da biohidrogenação resultando
em mudanças nos ácidos graxos intermediários.
Outro aspecto importante que deve ser levado em
consideração na dieta é a quantidade e efetividade da fibra utilizada, o
que reflete diretamente na condição ruminal e gordura do leite (Tabela
6). A fibra efetiva atua estimulando a ruminação e produção de saliva,
o que mantém o pH em níveis favoráveis para digestão da fibra,
resultando em maior disponibilidade de ácido acético, o principal
precursor da gordura no leite.
84
Tabela 6. Ácidos graxos voláteis no rúmen e depressão da gordura do leite em vacas alimentadas com dieta de baixa
fibra, composta por alta quantidade de grãos e pouca forragem (Bauman & Griinari, 2003).
Dietas
Controle Baixa fibra
Leite (kg/dia) 19,1 20,9
Conteúdo de gordura, % 3,6 1,7*
Produção de gordura, g/dia 683 363*
Ácidos graxos voláteis no rúmen (% molar)
Acetato 67 46*
Propionato 21 46*
Butirato 11 9
Proporção acetato: propionato 3,2 1,0*
Produção no rúmen (moles/dia)
Acetato 29,4 28,1
Propionato 13,3 31,0*
Taxa de entrada no corpo (mg/min kg.75)
β-hidroxibutirato 3,40 4,43 * p< 0,05

85
Dietas com grandes quantidades de carboidratos prontamente
fermentáveis e reduzida quantidade de fibra, ou dietas com quantidade
adequada de fibra, mas com fibra efetiva física, possuem pouca
habilidade de manutenção do funcionamento normal do rúmen,
podendo assim, levar a depressão na gordura do leite (Bauman e
Griinari, 2003).
A inclusão de lipídeos na dieta pode promover alterações
sobre a gordura do leite, o que mais uma vez é influenciado pelas
alterações no ambiente ruminal (Palmquist et al, 1993). Os efeitos
observados, quando da adição de gordura à dieta, foram dependentes
do tipo e quantidade da gordura fornecida. O fornecimento de
quantidades moderadas ou elevadas de gordura insaturada reduz de
maneira significativa o teor de gordura do leite, podendo esta queda
ser de até uma unidade percentual.
Griinari et al. (1998), confirmaram que dietas pobres em fibra
e ricas em ácidos graxos insaturados (óleo de milho) aumentam o
conteúdo de C18:1 trans. Segundo estes mesmos autores, este tipo de
dieta está associado a uma diminuição significativa na produção e
conteúdo da gordura do leite, bem como na concentração de ácido
esteárico, e menores valores ruminais de pH. Isso provavelmente
indica que houve uma redução das etapas finais da biohidrogenação
ruminal. O aumento da concentração intestinal de trans-vaccênico
diminui a capacidade lipogênica da glândula mamária (taxas de
incorporação de acetato em ácidos graxos) e a expressão de genes de
enzimas relacionadas ao transporte de ácidos graxos circulantes,

86
diminui a síntese “de novo” de ácidos graxos, a dessaturação de ácidos
graxos e a formação de triglicerídeos. O mecanismo que
provavelmente os esses ácidos graxos insaturados diminuem a
produção de gordura do leite envolve a redução na expressão do
mRNA de enzimas chave associadas à síntese de gordura do leite
(Baumgard et al., 2002). Kalscheur et al, (1997) observaram que vacas
alimentadas com dietas ricas em concentrado apresentaram
diminuição da gordura do leite, acompanhada por aumento no
conteúdo de C 18:1 trans.
Certos aspectos relacionados a formas de manejo de
alimentação também alteram a ingestão de fibra e, consequentemente,
a gordura do leite. Pode-se destacar entre as deficiências no manejo a
baixa quantidade e qualidade de volumoso, sendo muitas vezes
compensados pela substituição por subprodutos fibrosos e
concentrados; cochos mal dimensionados, o que estimula a
competição entre os animais, instalações inadequadas quanto ao
conforto animal, proporcionando menores tempos de ruminação e
elevação do estresse.
3.10 Considerações Finais
O aumento no teor de proteína do leite é interessante para as
indústrias processadoras, pois permite maior rendimento industrial,
além da menor quantidade de água a ser transportada e removida no
beneficiamento. No entanto, para o produtor o aumento no teor de
proteína do leite só trará benefícios com a implementação de um

87
sistema de pagamento por componentes, onde o pagamento pela
produção extra de proteína cobre os custos de produção. No entanto, a
opção por aumentar o teor de proteína do leite através da manipulação
nutricional, deve sempre avaliar a alternativa mais rentável disponível
antes de proceder às alterações.
REFERÊNCIAS BIBLIOGRÁFICAS
BAUMAN, D. E. et al. Update on conjugated linoleic acids (CLA).
Proceedings of Cornell Nutrition Conference, p.170-177, 2001.
BAUMAN, D. E.; et al. Technical note: production of butter with enhanced
conjugated linoleic acid for use in biomedical studies with animal models.
J. Dairy Sci., v. 83, n. 11, p. 2422-2425, 2000.
BAUMAN, D. E.; GRIINARI, J. K. Nutritional regulation of milk fat
synthesis. Annu. Rev. Nutr., v. 23, p. 203-227, 2003.
BAUMGARD, L. H. et al. Trans-10, cis-12 conjugated linoleic acid
decreases lipogenic rates and expression of genes involved in milk lipid
synthesis in dairy cows. J. Dairy Sci., v. 85, p. 2155-2163, 2002.
BAUMGARD, L.H., CORL, B.A., DWYER, D.A., SAEBO, A. BAUMAN,
D.E. Identification of the conjugated linoleic acid isomer that inhibits milk
fat synthesis. Am. J. Physiol., v. 278, p. R179-184, 2000.
BEAM, T. M., JENKINS, T. C., MOATE, P. J., KOHN, R. A.,
PALMQUIST, D. L... Effects of amount and source of fat on the rates of
lipolysis and biohydrogenation of fatty acids in ruminal contents. J. Dairy
Sci. v. 83, p. 2564-2573, 2000.
BECKETT, S. et al. Effects of monensin on the reproduction, health, and
milk production of dairy cows J. Dairy Sci., v. 81, p. 1563-1573, 1998.
BEQUETTE, B. J. et al. Current concepts of amino acids and protein
metabolism in the

88
DE PETERS, E. J.; CANT, J. P. Nutritional factors influencing the nitrogen
composition of bovine milk: a review. J. Dairy Sci. v. 75, p. 2043-2070,
1992.
DE PETERS, E. J.; FERGUNSON, J. D. Non nitrogen and protein
distribution in the milk of cows. J. Dairy Sci. v. 75, p. 3192-3209, 1992.
DOEPEL, L. et al. Milk protein synthesis as a function of amino acid
supply. J. Dairy Sci. v. 87, p. 1279-1297, 2004.
DOREAU, M., FERLAY, A. Digestion and utilization of fatty-acids by
ruminants. Anim. Feed Sci. Technol., v. 45, p. 379-396, 1994.
FONSECA, L. F. L; SANTOS, M. V. Conceitos básicos sobre composição
de leite e métodos utilizados. 1° Curso on line sobre qualidade do leite.
Instituto Fernando Costa, Milkpoint. 2000.
GRIINARI, J. M. et al. Trans-Octadecenoic acids and milk fat depression in
lactating dairy cows J. Dairy Sci., v. 81, p. 1251-1261, 1998.
GRIINARI, J. M. MCGUIRE, M. A. DWYER, D. A. BAUMAN, D. E.,
BARBANO, D. M. HOUSE, W. A. The role of insulin in the regulation of
milk protein synthesis in dairy cows. J. Dairy Sci. v. 80, p. 2361-2371,
1997.
GRIINARI, J.M., DWYER, D.A., MCGUIRE, M.A., BAUMAN, D.E.,
PALMQUIST, D.L., Nurmela, K.V.V. Transoctadecenoic acids and milk fat
depression in lactating dairy cows. J. Dairy Sci., v. 81, p. 1251-1261, 1998.
HARDING, F. Milk quality. 1 ed., Glasgow: Blackie, p. 166, 1995.
HRISTOV, A.; BRODERICK, G. A. Synthesis of microbial protein in
ruminally cannulated cows fed alfafa silage, alfafa hey or corn silage. J.
Dairy Sci., v. 79, p. 1627-1637, 1996.
HURLEY, W. L. Nutritional factors affecting milk yield and
composition. University of Illinois. 2004.
JENKINS, T. C., MCGUIRE, M. A. Major Advances in Nutrition: Impact
on Milk Composition, J. Dairy Sci., v. 89, p. 1302–1310, 2006.
JENKINS, T.C. Lipid metabolism in the rumen. J. Dairy Sci. v. 76, p.
3851-3863, 1993.

89
KALSCHEUR, K. F. B. B. et al. effect of dietary forage concentration and
buffer addition on duodenal flow of trans C 18:1 fatty acids and milk fat
production in dairy cows. J. Dairy Sci., v. 80, p. 2104-2114, 1997.
KIRCHGESSNER M. et al. Fütterung und Milchzusammensetzung,
BLV, 292 p., 1965.
LARSON, B. L. Biosynthesis & cellular secretion of milk. In: LARSON,
B. L. Lactation. Cap. 4. p.129-163. Iowa State University Press. 1995.
MISCIATTELLI, L. et al. Milk production, nutrient utilization, and
endocrine responses to increased postruminal lysine and meyhionine supply
in dairy cows. J. Dairy Sci. v. 86, p 275286, 2003.
MÜHLBACH, P. R. F. Nutrição da vaca em lactação e a qualidade do
leite. In: SIMPÓSIO DE BOVINOCULTURA DE LEITE, 2003, Chapecó.
Socied. Catarinense de Méd. Vet. –Núcleo Oeste.
MURPHY, J. J.; O’MARA, F. Nutritional manipulation of milk protein
concentration and its impact on dairy industry. Lives. Produc. Sci. v. 35, p
117-134, 1993.
NATIONAL RESEARCH COUNCIL (NRC). Nutrient requirement of
dairy cattle. Washington DC. National Academy Press. 2001.
NOUSIAINEN, J. et al. Evaluation of milk urea as a diagnostic of protein
feeding. J. Dairy Sci. v. 87, p. 386-398. 2004.
NOUSIAINEN, J., SHINGFIELD, K. J., HUHTANEN, P. Evaluation of
Milk Urea Nitrogen as a Diagnostic of Protein Feeding. J. Dairy Sci. v. 87,
p. 386–398, 2004.
PALMQUIST, D. L. et al. Adsa foundation symposium: milk fat synthesis
and modification. J. Dairy Sci. v. 76, p. 1753-1771, 1993.
REIS, R. B., GLÓRIA, J. R., VIEIRA, L. R., FARIA, B. N. Manipulação
da composição do leite pela nutrição da vaca. In Anais do Simpósio
Mineiro de Nutrição, UFMG, 2008.
RIBAS, N.P., HARTMANN, W., MONARDES, H. G., ANDRADE, U. V
.C. Sólidos Totais do Leite em Amostras de Tanque nos Estados do Paraná,
Santa Catarina e São Paulo. R. Bras. Zootec., v.33, n.6, p.2343-2350, 2004
(Supl. 3)

90
RIEL, R. Composicion y estructura fisico-quimica de la leche. Ed.
Acribia S.A.: Espanha, pp. 1-54, 1991.
SCHWAB, C.G., BOZAK, C.K., WHITEHOUSE, N.L., MESBAH,
M.M.A. Amino acid limitation and flow to the duodenum at four stages of
lactation. I. Sequence of lysine and methionine limitation. J. Dairy Sci., v.
75, p. 3486–3502, 1992.
SUTTON, J. D. Altering milk composition by feeding. J. Dairy Sci. v. 72,
p. 2801-2814, 1989.
VAN NEVEL, C. J., DEMEYER, D. I. Influence of pH on lipolysis and
biohydrogenation of soybean oil by rumen contents in vitro. Reprod. Nutr.
Dev., v.36, p. 53-63, 1996
VAN NEVEL, C. J., DEMEYER, D. I. Lipolysis and biohydrogenation of
soybean oil in the rumen in vitro: Inhibition by antimicrobials. J. Dairy
Sci., v. 78, p. 2797-2806, 1995.
WALDNER, D. N. et al. Managing milk composition: normal sources of
variation. OSU: Extension facts. 2004.
WRIGHT, T. C. et al. Effects of rúmen-undegradable protein and feed
intake on nitrogen balance and milk production in dairy cows. J. Dairy Sci.
v. 81, p 784-793, 1998.

91
4. CONTROLE LEITEIRO4
Carlos Bondan
4.1 Introdução
O controle leiteiro é uma ferramenta de mensuração mensal
da produção, dos eventos e da qualidade do leite de cada vaca do
rebanho em um determinado período de tempo. A interpretação destas
informações fornece subsídios para as decisões de manejo nutricional,
sanidade da glândula mamária, reprodução, descartes, acasalamentos
e qualidade do leite. Permite ainda, manter registros vitalícios,
valorizando o plantel através das informações de lactações encerradas,
avaliações genéticas e rastreabilidade.
Os benefícios do controle leiteiro podem ser comprovados
pela média de produção dos rebanhos que utilizam esta ferramenta.
Enquanto a produção média brasileira em 2013 foi de 4,39 L/vaca/dia
e a gaúcha de 7,96 L/vaca/dia, os rebanhos sob controle leiteiro
apresentaram médias de 26,8 ± 8,63 L/vaca/dia com teores de sólidos
totais de 12,2 ± 0,95, demonstrando que a profissionalização é o
caminho para o sucesso na atividade.
4 Bondan, C. Controle leiteiro. Anais. I Simpósio Nacional da Vaca Leiteira.
Porto Alegre: Universidade Federal do Rio Grande do Sul. 2014. 239 p.

92
Este texto tem o objetivo de esclarecer a metodologia utilizada para a
operacionalização do controle leiteiro, desde o cadastro das
propriedades e dos animais, até a coleta mensal das informações
individuais de cada vaca e a interpretação dos resultados. Porém, estas
informações não terminam aqui, pois há um aperfeiçoamento contínuo
do controle leiteiro conforme as demandas dos produtores e da
assistência técnica.
4.2 Como iniciar o controle leiteiro.
Primeiramente o produtor ou técnico deve solicitar ao SARLE
pelo e-mail: [email protected] ou pelo telefone: (54) 3316-8191/8194 uma
ficha de cadastro do cliente e as fichas de cadastro dos animais.

93
4.2.1 Cadastro de Cliente
DADOS DO CRIADOR
Rebanho nº Região
Produtor
CPF Fone Fax
Endereço
Cidade UF CEP
DADOS DO CONTROLADOR
Controlador
Endereço
Cidade UF CEP
Fone Cel.
E-Mail:
DADOS PARA COBRANÇA DAS ANÁLISES
Nome destino da cobrança
Endereço
Cidade UF CEP
Fone Cel.
E-mail:
CGC ou CPF Número de animais:

94
4.2.2 Cadastro dos Animais
FICHA INICIAL DE IDENTIFICAÇÃO DOS ANIMAIS
SCL Nº_________
Vaca: (nome por extenso)
N° do brinco e/ou apelido:
RAÇA: GS: Nº REGISTRO: DATA NASCIMENTO:
NOME DO PAI:
RAÇA: GS: Nº REGISTRO:
NOME DA MÃE:
RAÇA: GS: Nº REGISTRO:
DADOS REPRODUTIVOS
DATA DE PARIÇÃO: ORDEM DE LACTAÇÃO:
DATA ÚLTIMA COBERTURA: REPRODUTOR:
BEZERRO STFC*
PROPRIETÁRIO:
Depois de realizado o cadastro no SARLE o produtor receberá
um relatório de campo, o qual deverá ser preenchido no dia da
realização do controle.
95
Exemplo de Relatório de Campo

96
4.2.3.Legendas
1. Nome do produtor
2. Data do controle
3. Número do cadastro do rebanho no SARLE (código)
4. Cidade onde se encontra a propriedade
5. Assinatura do proprietário/responsável
6. Número do registro genealógico e nº do animal no cadastro
do SARLE
7. Data do nascimento, último parto e último serviço
8. Produção de leite do controle leiteiro realizado no mês
anterior
9. Número da amostra e apelido do animal (o nº da amostra é
correspondente ao apelido do animal). No exemplo: a
amostra 1 corresponde ao animal de apelido 547
10. Campo para preenchimento das pesagens de leite
correspondentes as ordenhas: manhã, tarde e noite
11. Códigos de lactação: são códigos de explicação da lactação
que serão utilizados para anotar eventos ocorridos no animal
no dia do controle como, por exemplo, mastite, cio, doenças
e será usado também para o envio de informações de
descarte (eliminação) e venda de animais
12. Evento: São códigos de eventos reprodutivos acontecidos no
mês anterior do controle atual como, por exemplo, data de
parto, coberturas e data de secagem
13. Data (dia/mês) é a data em que ocorreram os eventos
reprodutivos. Exemplo: O animal pariu, com isto teve o
código 2 (parto) e em seguida a data do evento “parto”
14. Reprodutor: campo a ser preenchido com o número do
registro do reprodutor que a vaca foi inseminada
15. Bezerro: são informações referentes ao parto

97
4.2.4 Tabela de Códigos de Explicação de Lactação
CÓDIGOS DOS
FATORES QUE
AFETAM O
CONTROLE
CÓDIGOS DE
DESCARTE
(ELIMINAÇÃO)
CÓDIGOS DE
CAUSAS DE
MORTE
CÓDIGOS DE
VENDA
11-aborto 40-baixa produção 60-choque
elétrico, raio
70-para rebanho
dentro do estado
12-amamentando 41-baixa gordura 61-deslocamento
do abomaso
71-para rebanho
fora do estado
13-cetose 42-causa
desconhecida
62-doença 72-em leilão
14-cio 43-deslocamento
do abomaso
63-intoxicação
15-corpo estranho 44-esgotamento do
úbere e problemas
na ordenha
64-febre do leite
16-deslocamento
de abomaso
45-febre do leite 65-ferimento
17-diarréia 46-ferimento 66-idade
avançada
18-em exposição 47-idade avançada 67-timpanismo
19-febre do leite 48-mastite 68-outros
20-ferimento do
úbere
49-ordenha lenta
21-lactação
induzida
50-outras causas
22-mastite 51-outras doenças
23-metrite 52-problemas de
pernas e pés
24-nervosismo 53-problemas
reprodutivos
25-ordenha
perdida
54-temperamento
nervoso

98
26-outros
ferimentos
55-açougue
27-outros
problemas de
saúde
28-podridão no
casco
29-amostra de leite
perdida
30-recém parida
31-sem apetite
32-timpanismo
33-tratado com
ocitocina
34-amostras com sangue
35-sem amostra
36-coagulada
37-sem conservante
4.2.5 Tabela de Códigos dos Eventos Reprodutivos
Código Descrição
1 Em Lactação
2 Parto
3 Secagem
4 Cobertura
8 Aborto

99
4.2.6 Códigos de Bezerros
Sexo (S) Tamanho (T) Facilidade de parto (F) Condição (C)
1-macho
2-fêmea
3-
gêmeos
1-pequeno
2-médio
3-grande
1-parto sem auxílio
2-parto com tração fácil
3-parto com tração
difícil
4-parto com cirurgia
5-parto com má
apresentação
1-bezerro vivo
2-bezerro
morto
4.2.7 Observações
Não esquecer de preencher e assinar a ficha de controle com
a data de coleta, informações da ordenha e dos eventos
ocorridos durante o mês
No caso de cadastro de novos animais no controle, preencher
obrigatoriamente a ficha inicial de identificação dos animais
A assinatura do proprietário e/ou controlador são campos
obrigatórios

100
4.3 Como realizar as coletas.
4.3.1 Material necessário:
Relatório de campo
Frascos para análise de CCS e composição (contendo
conservante Bronopol) fornecidos pelo SARLE
Material para a pesagem do leite conforme o sistema de
ordenha
4.3.2 Execução
Pesagem/volume do leite:
O leite individual de cada animal deverá ser pesado ou o
volume de cada animal deverá ser registrado conforme as
características da ordenhadeira e transcrito para o relatório de campo.
Balde ao pé: O leite poderá ser pesado ou seu volume medido
em balde transparente com escalas. A espuma deve ser descontada.
Canalizado: O leite deverá ser medido em amostradores
próprios para ordenhadeiras canalizadas.
Coleta da amostra:
A amostra deverá ser coletada em frasco específico, fornecido
pelo SARLE contendo conservante (Bronopol) O frasco deverá ser
identificado na tampa com o número da ordem amostral fornecida pelo
SARLE
Homogeneização do leite antes da coleta da amostra

101
Balde ao pé: o leite deverá ser passado de um balde para outro
por 3 (três) vezes consecutivas.
Canalizado: agitar o leite no amostrador utilizando fluxo de ar
durante um segundo por litro de leite, e no mínimo 10 segundos.
Como coletar a amostra:
Balde ao pé: utilizar uma concha para a coleta, a concha deve
ser ambientada mergulhando-a no mínimo três vezes na massa de leite,
coletar o volume respeitando as marcas de cada ordenha. Nunca
encher o frasco totalmente.
Canalizado: abrir o registro do medidor e coletar o volume
respeitando as marcas de cada ordenha. Nunca encher o frasco
totalmente.
Após 20 minutos da coleta da amostra os frascos deverão ser
homogeneizados para dissolução do conservante.
Caixa de amostras:
Ordenar os frascos na caixa na mesma sequência do relatório
deixando os novos animais por último (Figura 1).

102
1 2 3 4 5 6
7 8 9 10 11 12
13 14 15 16 17 18
19 20 21 22 23 24
25 26 27 28 29 30
Figura 1. Sequência na qual os frascos devem ser acondicionados na caixa,
conforme consta no relatório de campo.
Enviar na caixa o relatório de campo preenchido.
Preencher sobre a tampa da caixa o nome do produtor (número
do rebanho no SARLE), número das caixas caso haja mais de uma e o
nome do interessado (Figura 2).
Independentemente do tipo de transporte, as amostras deverão
ser entregues diretamente no SARLE de 2ª a 6ª das 08h às 20h e aos
sábados das 08h às 12h, em até 72h após a coleta.

103
CAIXA: 01 de 03
INTERESSADO: CEPAGRO
ou REBANHO 45
Figura 2. Marcação da caixa para envio ao laboratório.
4.4 Resultados
Os resultados são encaminhados via e-mail, ou poderão ser
acessados pelo site www.sarle.upf.br. O produtor deverá solicitar o
login e a senha ao SARLE pelo e-mail [email protected].
São encaminhados 6 relatórios mensais:
a) Relatório R1- Coleta
Este relatório deverá ser preenchido com as pesagens/volume
do leite de cada animal e os eventos, deverá acompanhar a caixa com
as amostras de leite.

104
b) Relatório R2- Informações dos últimos 12 meses
Neste relatório constam as informações dos resultados das
análises de cada animal (% gordura, proteína, produção e CCS), bem
como as médias individuais e do rebanho, referente ao último controle
e aos 12 meses anteriores.
c) Relatório R2.2- CCS
Este relatório informa os resultados das análises do leite para
composição (% gordura, proteína, lactose, sólidos), contagem de
células somáticas (CCS) e a pesagem/volume do leite de cada animal.
Neste relatório constam ainda as seguintes informações de cada
animal: % da CCS que cada animal representa no tanque e o impacto
de CCS na média geral do tanque.
d) Relatório R3.1- Produção
Apresenta dados de produção individual dos animais, com
projeção de produção para 305 dias, projeção para a idade adulta e a
média do rebanho.
e) Relatório R3.2- Desempenho reprodutivo
Neste relatório constam todas os informes reprodutivos (parto,
idade, dias em lactação, 1º serviço, número de coberturas, projeção
intervalo entre partos, dias em aberto, próximo parto, data de secagem
e dias secos) de cada animal em individual e uma média do rebanho.

105
f) Relatório GR- Gráficos
Apresenta gráficos de desempenho do rebanho.
i. Avaliação do potencial de produção de leite em
função da proteína
ii. Avaliação da dieta: risco de acidose/cetose
iii. Adequação de fibra e energia da dieta
iv. Dispersão da ureia
v. Produção de leite individual
vi. Distribuição das matrizes em lactação CCS atual
x CCS anterior em escore linear
vii. Avaliação do coeficiente G:P (gordura: proteína)
4.5 Problemas e Procedimentos
Perda de uma das pesagens: deixar o espaço correspondente
em branco, sem estimar valores.
Perda de parte do leite de uma amostra: desprezar a quantidade
restante e deixar o espaço correspondente em branco, sem estimar
valores.
Marcação de produção de leite no campo de outra vaca: se não
for possível identificar a tempo, deixar o espaço correspondente em
branco, sem estimar valores.
Perda de amostra total de leite: colocar o código de perda de
amostra na coluna correspondente e manter o frasco na caixa de
amostras em seu respectivo lugar.

106
Perda de uma das amostras de leite: colocar o código de perda
de amostra na coluna correspondente e manter o frasco na caixa de
amostras em seu respectivo lugar.
Animais doentes: se o animal estiver em ordenha, realizar o
controle normal e anotar o código de explicação da lactação; se não
estiver em ordenha, anotar o código de explicação da lactação e deixar
os espaços de produção em branco. O frasco de amostra do leite deve
permanecer em seu lugar na caixa. Em hipótese alguma estimar
produção.
Animal fora da propriedade no dia do controle: deixar os
espaços de produção em branco, e o frasco de amostra de leite em seu
lugar correspondente.
4.6 Contatos
SERVIÇO DE ANÁLISES DE REBANHOS LEITEIROS (SARLE)
UNIVERSIDADE DE PASSO FUNDO - Campus I
BR 285 km 171
Caixa postal 611
CEP 99001-970
Passo Fundo-RS
Fone: (54)- 3316-8191/8194
E-mail: [email protected]

107
5. TRANSTORNOS METABÓLICOS DA VACA LEITEIRA NO
PERÍODO DE TRANSIÇÃO5
Enrico Lippi Ortolani
5.1 Introdução
Define-se período de transição como as quatro últimas e
primeiras semanas em relação ao parto. Nesse período a vaca leiteira
está sujeita a ser acometida por várias enfermidades, com destaque às
de origem metabólica (Herdt, 2013). Várias condições predisponentes
facilitam o surgimento dessas enfermidades. No período pré-parto
citam-se os seguintes fatores: (i) a grande mobilização de nutrientes
para o crescimento fetal, colostrogênese e o desenvolvimento
mamário; (ii) ajustes metabólicos necessários para favorecer a
mobilização de gordura e os teores de cálcio ionizável; e (iii) a
representativa diminuição na ingestão de matéria seca. No período
pós-parto, embora ocorra uma lenta recuperação no apetite, o animal
deve subsistir a uma rápida mobilização e perda de nutrientes para
atender a crescente produção leiteira, em especial no concernente a
gliconeogênese (Herdt, 2013).
5 Ortolani, E. Transtornos metabólicos da vaca leiteira no período de
transição. Anais. I Simpósio Nacional da Vaca Leiteira. Porto Alegre:
Universidade Federal do Rio Grande do Sul. 2014. 239 p.

108
Dentre as enfermidades metabólicas que se manifestam no
período de transição, destacam-se as seguintes: hipocalcemia
acompanhada ou não de síndrome do animal caído; cetose com
presença ou não de esteatose hepática; edema mamário; deslocamento
do abomaso e hipomagnesemia (Herdt, 2013). Considerando a
frequência e a relevância em nosso meio, serão destacadas duas
enfermidades: a hipocalcemia da vaca parturiente e a cetose da vaca
leiteira.
5.2 Hipocalcemia da Vaca Parturiente
5.2.1 Definição e Sinonímias
Define-se a hipocalcemia como a dificuldade das vacas
leiteiras em manter a homeostase do cálcio livre plasmático,
momentos antes ou em seguida ao parto, gerando inicialmente quadro
de tetania, seguido de paresia e paralisia, que se não foram tratadas
levam frequentemente o animal à morte (Ortolani, 1995b). A
hipocalcemia tem as seguintes sinonímias: paresia da parturiente,
febre vitular ou febre do leite. Porém, essas duas últimas
denominações são incorretas, pois as vacas não têm síndrome febre e
numa das fases clínicas podem até apresentar hipotermia.

109
5.2.2 Aspectos Epidemiológicos
O primeiro estudo epidemiológico no Brasil detectou uma
incidência de 4,25% em rebanhos leiteiros do Vale do Paraíba (SP).
Contudo, deve-se mencionar que os rebanhos retinham vacas além da
6ª gestação, e que a partir da 3ª lactação ocorreu aumento exponencial
na incidência, atingindo na 12ª lactação 31% (Ortolani, 1995a). A
letalidade foi alta (12,8%), em especial em vacas velhas ou tratadas
em fases adiantadas da doença. Outros estudos nacionais identificaram
incidências de 1,8% e 5,2% (Corassin, 2004; Coelho, 2004). Vacas
das raças Jersey podem ser mais acometidas que outras raças,
ocorrendo o mesmo em vacas gordas (> 3,5 de condição corporal com
escores entre 1-5) no dia do parto em relação às magras (Radostits et
al., 2007).
De acordo com dados nacionais, a hipocalcemia é
inicialmente notada nas primeiras 24 horas pós-parto (64%),
reduzindo-se no segundo dia (31%) e diminuindo muito no 3º dia a
seguir (5%) (Ortolani, 1995 a). Porém, também é detectada em
algumas vacas nos últimos dias anteriores ao parto (Radostits et al.,
2007).
5.2.3 Prejuízos Econômicos
Essa enfermidade provoca grandes prejuízos econômicos,
pois além de diminuir a produção leiteira (até 80 L), nas primeiras seis
semanas de lactação, causa mortalidade (Ortolani, 1995a) e

110
predispõem a chance do surgimento de outras doenças do período de
transição, na seguinte ordem: cetose (23,5x), distocia (7,3x quando
ocorre no período pré-parto), mamite (5,4x) , metrite (4,7x), retenção
de secundinas (4,3x), menor eficiência reprodutiva (4,2 x) e
deslocamento do abomaso à esquerda (3x) (Corassin, 2004; Herdt,
2013). A hipocalcemia é, depois da distocia, a segunda principal causa
de decúbito patológico (doença da vaca caída), sendo responsável pelo
surgimento de 38% destes casos (Herdt, 2013).
5.2.4 Metabolismo do Cálcio na Vaca Leiteira
No plasma se encontram duas frações principais de cálcio
(Ca): o cálcio livre (Ca2+) ou ionizável (48%) e o cálcio ligado às
proteínas (52%). Quadros de hipocalcemia só ocorrem quando a
fração de cálcio ionizável se reduz abaixo dos 50%. Os teores de cálcio
ionizável são finamente mantidos dentro de valores específicos (1,0 a
1,25 mmol/L) por vários mecanismos hormonais. Na hipocalcemia a
paratireoide produz o hormônio da paratireoide (PTH), que estimula a
hidroxilação da molécula 25-hidroxicolecalciferol nos rins, formando
a vitamina D3, muito mais potente que o composto anterior. A
vitamina D3 aumenta a absorção intestinal de Ca pela síntese de
proteínas específicas para carrear Ca dos intestinos. Porém, há
necessidade dessa vitamina se ligar a receptores específicos e aptos
nos enterócitos. Além disso, a vitamina D3 aumenta a ressorção de Ca
nos ossos e a reabsorção nos túbulos renais. O PTH pode per se ter
uma ação semelhante à vitamina D3 nos ossos e rins. Na hipercalcemia

111
entra em ação a calcitonina, produzida pela tireoide, que excreta Ca
pela urina e promove deposição de cálcio nos ossos (Ortolani, 1995
b).
5.2.5 Patogenia da Hipocalcemia
A hipocalcemia é fruto do déficit entre a demanda de Ca
dispendido e a quantidade deste elemento obtido na absorção intestinal
e reabsorção óssea no período do periparto. A maior perda de Ca é
para a produção de colostro, que contém o dobro deste elemento (2,2
g/L) em comparação ao leite normal. Vacas entre a 3ª e 6ª lactações
atingem o ápice da produção de colostro. Vacas boas produtoras
podem perder muito mais Ca pelo leite (23 g/dia) que para formação
do feto no final da gestação (11 g/dia). Superordenha do colostro nos
primeiros dois dias de lactação podem desencadear quadros de
hipocalcemia.
Toda vaca no dia do parto apresenta uma diminuta
hipocalcemia. Para reduzir este déficit de Ca tanto os processos de
ressorção óssea como de absorção intestinal necessitam estar bastante
ativos. Porém, exatamente nestes dias existe uma crise na obtenção de
cálcio. Quanto mais velha for a vaca menor é a capacidade de
mobilizar Ca dos ossos devido ao reduzido número de receptores para
a vitamina D3 nos enterócitos. Vacas Jersey também têm o mesmo
problema em relação ao número de receptores. Vacas que, em
sucessivas lactações, têm hipocalcemia apresentam dificuldade de
aumentar os receptores para vitamina D3 antes do parto (Goff, 2008).

112
Dietas muito ricas em Ca, P, Na e K no pré-parto interferem na
produção de vitamina D3, ocorrendo o contrário em relação às dietas
pobres em Ca. Vacas que recebem no final da gestação dietas ricas em
sais catiônicos (Na+; K+) apresentam pH sanguíneo superior a 7,35, o
que parece interferir negativamente na ligação do PTH aos seus
receptores nos ossos, diminuindo a ressorção óssea, e nos rins
interferindo na reabsorção tubular de cálcio (Herdt, 2000; Goff, 2008,
DeGaris e Lean, 2008).
5.2.6 Quadro Clínico
A hipocalcemia pode apresentar três estágios clínicos, de
acordo com o teor de Ca livre no plasma (Ortolani, 1995b). No
primeiro estágio, o animal mantém a consciência, mas apresenta os
seguintes sinais nervosos: excitamento, tremores de cabeça e tetania
com hipersensibilidade. Manifesta ranger de dentes e protrusão de
língua, rigidez dos membros e aspecto de cavalete, podendo ficar em
decúbito lateral. A temperatura retal pode se elevar até os 41º C. Esse
estágio dura no máximo 8 h.
No segundo estágio, a consciência fica deprimida e o animal
se torna sonolento, a tetania desaparece e há paresia muscular, as
pupilas se dilatam e ocorre insensibilidade cutânea. O esfíncter anal se
relaxa e as fezes se tornam secas. E comum a atitude de “auto
auscultação”. As vacas apresentam hipotermia, esfriamento de
extremidade e a pele e muflo ficam secos. Chama a atenção a
taquicardia com hipofonese (Barreto Jr. et al., 2011). O pulso pode

113
ficar imperceptível, o movimento de rúmen desaparece e sobrevêm o
meteorismo gasoso e é comum a instalação da síndrome do animal
caído. A temperatura retal diminuí sensivelmente apresentando
quando de hipotermia.
No terceiro estágio, há perda de consciência, flacidez de
musculatura e instalação de quadro comatoso, com hipotermia
(<36ºC). A taquicardia é destacada (> 130 bat./min). A ausência de
tratamento provoca iminente morte por falência cardiorrespiratória.
Em condições nacionais os quadros de hipocalcemia são
acompanhados de hipofosfatemia e normomagnesemia (Ortolani,
1995a).
5.2. 7 Tratamento
Deve ser realizado com infusão lenta (IV) de soluções
contendo cálcio. A reversão do quadro está intimamente ligada ao grau
de hipocalcemia. Geralmente, o déficit de Ca plasmático é na ordem
de 6 g e tratamentos com essa quantidade resolvem 70% dos casos,
porém parte deles necessitam quantidades superiores (Ortolani,
1995a). Doses excessivas de cálcio provocam recorrência da
hipocalcemia horas após a terapia, devido à liberação de calcitonina.
Intoxicações iatrogênicas pelo Ca podem ocorrer matando o animal
por parada cardíaca, mas podem ser revertidas com uso de sulfato de
atropina (10 mg/animal). Vacas em decúbito respondem bem ao
tratamento com doses adicionais (9 g) de fósforo (IV).

114
5.2.8 Prevenção
A prevenção deve ser realizada quando a incidência for
superior a 1%. Isso é feito manipulando a dieta nos últimos 20 dias
pré-parto. Deve-se evitar o oferecimento dietético excessivo de
potássio e fósforo, que devem ficar restritos a 15 e 35 g/vaca/dia,
respectivamente. Por outro lado, estimula-se a adição de cloreto e
enxofre na dieta para gerar um balanço ligeiramente aniônico. As
dietas usuais a base de volumosos, concentrados e silagem têm mais
cátions que ânions, contendo uma diferença cátion-ânion da dieta
(DCAD) em média de +100 a +300 mEq/kg MS. Para tornar a dieta
aniônica existem sais comerciais específicos ricos em cloreto
(NH4+Cl) e enxofre (SO4Ca.2H20), os quais são suplementados no
final de gestação, em torno de 120 a 200 g/vaca/dia, suprimindo-os no
dia do parto. Essa quantidade proporciona que o balanço permaneça
entre -150 a 0 mEq/kg MS, onde se obtêm os melhores resultados na
prevenção. A constatação que a dieta aniônica está atuando é a queda
no pH urinário, que deve-se situar-se em torno de 6,0 em vacas de raça
Holandesa e entre 5,5 e 6,0 em vacas Jersey (Goff, 2008; DeGaris e
Lean, 2008).
A dieta aniônica provoca uma ligeira acidose metabólica
sistêmica que por seu turno aumenta a produção de vitamina D3 e de
PTH, aumentando a ação dos osteoclastos na ressorção óssea, na
reabsorção urinária e na absorção intestinal de Ca. Essa acidose
provoca também melhor aderência do PTH aos seus receptores ósseos
e renais. Recomenda-se ainda na dieta pré-parto que a quantidades de

115
Ca não ultrapasse 70 g/vaca/dia e ofereçam entre 40 a 50 g de Mg
(Ortolani, 1995b; Goff, 2008; DeGaris e Lean. 2008).
5.3 Cetose da Vaca Leiteira
5.3.1 Definição
A cetose ocorre no início da lactação, devido a um déficit no
metabolismo energético, em especial na gliconeogênese, causado pela
alta demanda de nutrientes no pico da lactação podendo levar a um
quadro de hipoglicemia e acetonemia. Isso provoca diminuições na
produção leiteira e no apetite e o surgimento de sinais nervosos.
5.3.2 Aspectos Epidemiológicos
Teoricamente, quanto mais alta for a produção leiteira maior
é risco do surgimento de cetose. Vários trabalhos nacionais já
relataram a presença de cetose em nossos rebanhos, com uma
incidência acumulada de 13 % a 24 % (Coelho, 2004; Corassin; 2004;
Garcia, 2010). Ela é mais frequente em vacas no decorrer da 2ª a 6ª
lactações, com maior incidência na 3ª e 4ª lactações, quando a
produção leiteira é mais alta. A cetose tipo I é mais frequente em vacas
que pariram em condições corporais menores (< 3,5), enquanto na
cetose tipo II ocorre o contrário (> 3,5); a cetose tipo III (secundária)
pode acometer fêmeas com diferentes condições corporais. O pico de

116
ocorrência é mais tardio no tipo I (3ª semana pós-parto) do que no tipo
II (1ª semana), podendo ser variado no tipo III.
5.3.3 Prejuízos Econômicos
Estudo nacional constatou que vacas cetóticas têm uma
redução láctea durante a lactação de 427 L, ou seja, 4,11% inferior a
vacas hígidas (Corassin, 2004). A cetose aumenta o surgimento de
risco de doenças na seguinte ordem: 6,17x de deslocamento do
abomaso à esquerda e 3,4x para metrite e mamite. Por outro lado,
aumentam o risco de cetose as seguintes enfermidades: hipocalcemia
23,6x; retenção de placenta 3x e pneumonia 4,75 x (Corassin, 2004;
Herdt, 2013; Ortolani, 2014).
5.3.4 Metabolismo da Glicose e Geração de Corpos Cetônicos na Vaca
Nos monogástricos, cerca de 60% da glicose sistêmica é
proveniente da absorção dos carboidratos dietéticos, porém nos
ruminantes estes são fermentados no rúmen. Assim, os ruminantes têm
que gerar glicose quase que unicamente pelo processo de
gliconeogênese hepática. Cerca de 60% da fonte de glicose é derivada
do propionato produzido no rúmen, 30% dos aminoácidos, 5% do
lactato e 5% do glicerol. Os requerimentos de glicose são variáveis de
acordo com a fase produtiva. Uma vaca prenhe no final de gestação
necessita de 500 g de glicose/dia, porém no pico da lactação esta

117
quantidade mais que triplica numa fêmea com produção acima de 20
L de leite (Herdt, 1988).
Os ruminantes têm pequeno estoque de glicogênio hepático,
sendo os depósitos de gordura a principal forma de energia
armazenada no organismo. A mobilização dessas gorduras é feita pela
ação das lipases. No superávit energético as gorduras são estocadas,
ocorrendo o inverso em casos de déficit. Assim, a alta secreção de
insulina bloqueia a ação das lipases, porém o glucagon, o hormônio
do crescimento, a adrenalina, o balanço energético negativo (BEN), o
estresse e o jejum fortemente estimulam as lipases, que mobilizam os
triglicerídeos e os transformam em glicerol e ácidos graxos livres,
também denominados de ácidos graxos não esterificados (AGNEs).
Esses últimos são carreados aos hepatócitos para serem oxidados ou
esterificados.
No déficit energético os AGNEs são grandemente oxidados
gerando energia e acetilcoenzima A (acetil-CoA). Em condições
normais, esse composto se combina com o oxalacetato formando o
citrato, que entra no ciclo de Krebs, gerando ATP e podendo formar
glicose. Porém, quando a produção de acetil-CoA for superior ao
oxalacetato disponível ele é transformado em acetoacetato e este em
acetona e β-hidroxibutirato (βHB), conhecidos como corpos
cetônicos. Isso ocorre abundantemente na cetose, devido à grande
oxidação dos AGNEs. Excesso de corpos cetônicos provocam redução
da disponibilidade de glicose para a glândula mamária, menor
proliferação de linfócitos e decréscimo de atividade dos neutrófilos.

118
Conjuntamente com os altos AGNEs os corpos cetônicos deprimem o
apetite e aumentam o grau de resistência insulínica. O βHB pode ser
transformado, em 5% das vacas cetóticas em álcool isopropílico, que
provoca o surgimento de vários sintomas nervosos.
Os AGNEs podem também ser esterificados no citosol dos
hepatócitos, se unindo ao glicerol e sendo envelopados por colesterol,
fosfolipídeos e proteína formando lipoproteína (LDH-VLDL) para ser
utilizada nos tecidos extra-hepáticos. Porém, se a quantidade de
AGNEs no citosol for muito grande superior a capacidade da
esterificação, as moléculas de AGNEs se condensam e são depositadas
em forma de gordura provocando quadro de esteatose hepática,
comum na cetose tipo II.
O principal fator predisponente para o surgimento da cetose é
a depressão e lenta recuperação do apetite a partir do parto,
acompanhado do pico da lactação nesse período quando a mobilização
de nutrientes é máxima. Assim, a cetose prevalece quando o BEN é
muito significativo, quer seja por desbalanço nutricional ou por menor
apetite, ou pelo fato que as vacas gordas apresentam menor depressão
no apetite pós-parto e maior mobilização de gorduras que vacas
magras (Herdt, 1988; 2000; 2013).

119
5.3.5 Categorias de Cetose
Várias classificações de cetose foram propostas até hoje.
Classicamente, classificava-se como primária ou secundária (Herdt,
2000). A primária causada pela falta de carboidrato na dieta no pós-
parto para prover de glicose e manter a alta produção de leite e a
secundária, determinada pelo menor apetite causado por outra doença
concorrente.
Nova proposta foi feita por Holtenius e Holtenius (1996),
comparando-se a cetose à diabetes mellitus humana. A cetose tipo I
ocorre no pico da lactação sem outra doença concorrente e com
esteatose hepática, caracterizada por hipoglicemia e hipoinsulinemia
em vacas que receberam dietas pobres em energia no pós-parto, e que
parem com condição corporal abaixo de 3,5 semelhante ao descrito na
forma primária clássica.
A cetose tipo II se manifesta nos primeiros 15 dias pós-parto,
caracterizada por hiperinsulinemia, hiperglicemia e alta resistência
insulínica (estado que a insulina diminui a sua ação biológica nos
tecidos insulino-dependentes: por exemplo, musculatura e gordura),
em vacas superalimentadas no pré-parto e que pariram com alta
condição corporal (> 3,5), frequentemente com doenças concorrentes
e esteatose hepática. Contudo, estudos posteriores verificaram que a
hiperinsulinemia e hiperglicemia eram pouco frequentes e que muitos
quadros inflamatórios poderiam incrementar a resistência insulínica
(Herdt, 2013).

120
Uma classificação errônea divide a cetose em clínica e
“subclínica” de acordo com os teores de corpos cetônicos e a presença
ou não de sintomas clínicos (Herdt, 2013) ignorando que o diagnóstico
clínico é firmado pela presença de sinais clínicos e/ou exame
laboratorial positivo. Além disso, a presença de sinais clínicos está
ligada a capacidade das vacas tolerarem corpos cetônicos, pois
algumas se mantem assintomáticas com alta acetonemia, e outras
manifestam a doença com teores menores (Herdt, 2000).
Assim, é lógico classificar a cetose em três formas. A cetose
tipo I causada por privação alimentar no pós-parto; a cetose tipo II
gerada por excesso de alimentação no pré-parto; e a cetose tipo III, ou
secundária, em vacas bem manejadas dieteticamente no pré e pós-
parto, mas acometidas por enfermidades concorrentes após o parto.
5.3.6 Quadro Clínico
Embora etiologicamente existam três tipos de cetose, quanto
ao quadro clínico pode-se dividir em duas formas: a típica, presente
em 95% dos casos, e a nervosa ou bizarra. A forma típica tem uma
evolução clínica média de 16 dias no tipo I e de 8 dias no tipo 2. Em
ambos os casos, é marcada por um tripé de sintomas: hipogalaxia,
hipofagia e sintomas nervosos. A hipogalaxia gira em torno de 30%,
mas pode atingir até 80%. A hipofagia chega a 50%, sendo mais
destacada nos casos tipo II. O apetite pode ser seletivo e o animal
priva-se de consumir primeiro o concentrado, depois o volumoso,
podendo desenvolver pica, ingerindo cama, areia ou terra. Isso leva a

121
uma perda evidente de peso corpóreo atingindo até 70 kg no primeiro
mês pós-parto. Os animais podem apresentar sonolência, olhar fixo,
ataxia, pequenos cambaleios e cegueira parcial. As grandes funções
podem estar modificadas devido aos quadros concorrentes. Devido à
diminuição do apetite pode ocorrer afundamento na fossa paralombar
esquerda em 70% dos casos e constipação e fezes ressecadas em 50%.
Muitas vacas com cetose tipo I e III apresentam pelame eriçado. Cerca
de 50% das fêmeas manifestam odor cetótico pelo ar expirado.
O quadro de cetose nervosa ou bizarra surge subitamente. São
comuns os sintomas de hiperestesia, delírio, agressividade, intensa
sialorreia, amaurose levando a ataxia, cambaleios e quedas, e apoio da
cabeça em obstáculos. Caso esses animais não sejam devidamente
tratados podem sucumbir após o surgimento de quadro depressivo
(Radostits et al., 2007; Herdt, 2013).
5.3.7 Diagnóstico
Como os sintomas nem sempre são tão evidentes assim, é
fundamental o diagnóstico confirmativo da enfermidade. Nova
ferramenta foi desenvolvida, disponível e barata em nosso meio, que
permite o diagnóstico rápido ao “pé-do-animal” por meio da
determinação de βHB em sangue total mensurado em tiras reativas,
por química seca (Precision Xtra), o qual apresenta alta sensibilidade
e especificidade (Araújo, 2014). Confirma-se o diagnóstico com
valores iguais ou superiores a 1,4 mmol/L de βHB, e suspeito entre
1,2 e 1,3 mmol/L. No rebanho pode ser realizada essa prova de rotina

122
para o diagnóstico de cetose, estabelecendo exames na cetose tipo I no
15º, 21º e 28º, e na cetose tipo II no 4º, 7º e 12º dias pós-parto.
5.3.8 Tratamento
O tratamento deve atingir as seguintes metas: diminuições da
lipólise e da cetogênese, e aumentos da glicemia e do consumo de
matéria seca. Geralmente, 50% das vacas não tratadas apresentam
recuperação espontânea lenta. O arsenal de tratamento conta com
diferentes tipos de medicamentos, a base de glicose (1.500 mL solução
20% IV); precursores de glicose (propileno-glicol 300 mL/dia por 5
ou mais dias ou glicerol/glicerina 300 mL/dia por quatro dias via oral);
corticosteroides (dose única de 20 mg de dexametasona, 1 g de
cortisona ou 5 mg de flumetasona); e medicamentos contendo fósforo
e vitamina B12 (Butafosfan 25 mL/dia, correspondente a 0,43 g de P e
1.250 µg de vitamina B12/dose), por 3 dias consecutivos (Herdt, 2013).
Os tratamentos devem ser priorizados de acordo com as
condições clínicas, assim: a glicose deve ser administrada na cetose
tipos I e III e nas formas bizarras; precursores de glicose em todos os
tipos de cetose; corticosteroides na cetose tipo II, devendo ser evitado
seu uso em animais com quadros inflamatórios; e os suplementos de
fósforo e B12 na cetose tipo I. Nos quadros mais rebeldes de cetose
tipo II (glicemia > 2,2 mmol/L; βHB > 2,4 mmol/L e alta resistência
insulínica) deve-se empregar glicose, os precursores e principalmente
corticosteroides (Gordon, 2013).

123
5.3.9 Prevenção
Deve-se iniciar na 8ª semana pré-parto com a oferta de
quantidades adequadas de nutrientes. Daí até o parto a ingestão de
matéria seca (MS) reduz-se sensivelmente (2% do peso vivo a 1,5%
em kg de MS; ex. vaca de 500 kg passa da ingestão de 10 kg MS/dia
para 7,5 kg), principalmente nas últimas três semanas. Isso deve ser
compensado com o aumento na densidade energética nessas semanas
finais. Da 8ª à 4ª semana pré-parto as vacas devem receber dieta com
12 % de proteína bruta (PB), 50% de NDT e 40% de Fibra Detergente
Neutro (FDN), passando a oferecer, em seguida, 15% de PB, 60% de
NDT e 35% de FDN. Do ponto de vista prático, no primeiro período
a dieta deve conter 10% de concentrados energéticos e 90% de
volumosos (50% de silagem e 40% de capins tenros e feno),
recebendo, em seguida, 30% de concentrados e 70% de volumosos.
Durante as últimas oito semanas pré-parto o ganho de peso
diário deve atingir 0,2 kg/dia fazendo com que a condição corporal
permaneça entre 3,0 e 3,25. No primeiro mês pós-parto a dieta deve
ser fornecida com 19% de PB, 73% de NDT e 30% de FDN,
mantendo-se a relação volumoso: concentrado em 60:40. Nos meses
posteriores a dieta conterá 18% de PB, 60% de NDT e 32 % de FDN.
Do ponto de vista prático, a quantidade de concentrados energéticos
oferecido nos primeiros três meses pós-parto é baseado na produção
láctea, ou seja: vacas produzindo mais de 27 L recebem 1 kg de
concentrado/2 L de leite; com produção de 18 a 27 L recebem 1 kg de

124
concentrado/2,5 L de leite e com produção entre 13 e 18 L recebem 1
kg de concentrado/3 L CE leite.
A suplementação da dieta com 335 g/vaca/dia de monensina
no último mês de gestação e no decorrer do pós-parto aumentou a
gliconeogênese, diminuiu os teores de βHB e aumentou a produção
láctea. O combate aos vários tipos de estresse às vacas no período de
transição é essencial, para não provocar reduções na ingestão da
matéria seca e aumento na incidência de cetose. A prevenção das
várias enfermidades presentes no pós-parto é de grande importância
para diminuir o risco de ocorrência de cetose (Herdt, 2013).
REFERÊNCIAS BIBLIOGRÁFICAS
ARAÚJO, C.A.S.C. Perfil metabólico e hormonal de ovelhas
superalimentadas submetidas a hipercetonemia e hiperlipidemia, e
tratadas com somatotropina recombinante bovina (r-bST). 114p. Tese
(Doutorado). Faculdade de Medicina Veterinária e Zootecnia, Universidade
de São Paulo, São Paulo, 2014.
BARRETO JÚNIOR, R.A.; Minervino, A.H.H.; Rodrigues, F.A.M.L.;
Meira Júnior, E.B.S.; Lima, A.S.; Mori, C.S.; Barros, I.O.; Ortolani, E.L.
Avaliação do quadro clínico e perfil bioquímico de bovinos durante a
indução e tratamento da hipocalcemia. Brazilian Journal Veterinary
Research Animal Science, v. 48, p.192-199, 2011.
COELHO, K.O. Impacto de eventos ocorridos antes e após o parto sobre
o desempenho produtivo e reprodutivo na lactação atual e na posterior
de vacas Holandesas. 70p. Dissertação (Mestrado), Escola Superior de
Agricultura “Luis de Queiroz”, Universidade de São Paulo, Piracicaba,
2004.

125
CORASSIN, C.H. Determinação e avaliação de fatores que afetam a
produtividade de vacas leiteiras: Aspectos sanitários e reprodutivos.
113p. Tese (Doutorado), Escola Superior de Agricultura “Luiz de Queiroz”,
Universidade de São Paulo, Piracicaba, 2004.
DEGARIS, P.J.; Lean, I.J. Milk fever in dairy cows: a review of
pathophysiology and control principals. The Veterinary Journal, v. 176,
p.58-69, 2008.
GARCIA, A.M.B. Avaliação metabólica de vacas leiteiras submetidas a
diferentes estratégias de prevenção do balanço energético negativo do
pós-parto. 83p. Dissertação (Mestrado), Faculdade de Veterinária,
Universidade Federal do Rio Grande do Sul, Porto Alegre, 2010.
GOFF, J.P. The monitoring, prevention and treatment of milk fever and
subclinical hypocalcemia in dairy cows. The Veterinary Journal, v.176,
p.50-57, 2008.
GORDON, J. Factors risks and treatment for ketosis in lactating dairy
cows. 162 p. Tese (Doutorado), Universidade de Guelph, Guelph, 2013.
HERDT, T.H. Metabolic diseases of ruminant livestock. Veterinary
Clinics North America – Food Animal Practice, v.4, p. 213 - 439, 1988.
HERDT, T.H. Metabolic disorders of ruminants. Veterinary Clinics
North America. – Food Animal. Practice, v.16, p.215-408, 2000.
HERDT, T.H. Metabolic diseases of dairy cows. Veterinary Clinics North
America. – Food Animal Practice, v. 29, p.267-468, 2013.
HOLTENIUS, P.; HOLTENIUS, K. New aspects of ketone bodies in
energy metabolism of dairy cows: a review. Journal Veterinary Medicine
A, v.43, p. 579-587, 1996.
ORTOLANI, E.L. Aspectos clínicos, epidemiológicos e terapêuticos da
hipocalcemia de vacas leiteiras. Arquivo Brasileiro de Medicina
Veterinária e Zootecnia, v.47, p. 799-808, 1995a.
ORTOLANI, E.L. Hipocalcemia da vaca parturiente. Cadernos Técnicos
Escola Veterinária UFMG, v.14: p. 59-71, 1995b.

126
RADOSTITS, O.M.; GAY, C.C.; HINCHCLIFF, K.W.; CONSTABLE, P.J.
Veterinary Medicine: A textbook of the diseases of cattle, horses, sheep,
pigs and goats. 10 ed. Philadelphia, Saunders Elsevier, 2007, p. 2156.

127
6. TRANSTORNOS METABÓLICOS NO PERÍODO DE
TRANSIÇÃO DA VACA LEITEIRA COM ÊNFASE NAS
ENFERMIDADES UTERINAS E DA GLÂNDULA MAMÁRIA6
Marcio Nunes Corrêa
Andressa Stein Maffi
Beatriz Riet-Correa
Ismael Mateus Cavazini
Marcelo Moreira Antunes
Viviane R. Rabassa
Cássio C. Brauner
6.1 Introdução
O período de transição em vacas leiteiras compreende as três
semanas que antecedem e as três que sucedem o parto (Goff & Horst,
1997). Doenças como a hipocalcemia, metrite, cetose, deslocamento
de abomaso e mastite geralmente ocorrem neste período (Ziguer et al.
2007). As doenças do periparto de vacas leiteiras constituem um
complexo, de modo que uma condição patológica predispõe a
ocorrência de outra. Por exemplo, uma vaca que apresenta
6Correa, M.N. et al. Transtornos metabólicos no período de transição da vaca
leiteira com ênfase na enfermidades uterinas e da glândula mamária. Anais. I
Simpósio Nacional da Vaca Leiteira. Porto Alegre: Universidade Federal do
Rio Grande do Sul. 2014. 239 p.

128
hipocalcemia é 4 vezes mais susceptível à ocorrência de retenção de
placenta e 16,4 vezes mais susceptível à ocorrência de cetose.
O início da produção de leite impõe grandes desafios para os
mecanismos responsáveis pela homeostase energética, proteica e
mineral da vaca. Durante o início da lactação, fatores homeorréticos
são responsáveis pelo direcionamento de grande quantidade de energia
para a glândula mamária e síntese de leite. Nesta fase, a vaca acaba
mobilizando reservas corporais e passa a enfrentar um estado de
balanço energético negativo (BEN). Neste período, as vacas sofrem
uma diminuição da resposta imunológica. Este estado de
imunossupressão em que o animal se encontra deve-se à diminuição
da atividade dos neutrófilos e dos linfócitos (Kehrli et al. 1989),
decorrentes das alterações do padrão de consumo alimentar e das
alterações hormonais e metabólicas a que a vaca leiteira está sujeita
durante o periparto. Todos esses fatores predispõem a um aumento
acentuado do risco de desenvolver doenças que influenciam
negativamente a resposta produtiva e reprodutiva (Youngquist &
Threlfall, 2007). Assim, o BEN, a inflamação e a resposta imune estão
diretamente relacionados, podendo atuar como causa ou consequência
de enfermidades comuns no periparto.
No periparto, a concentração de cálcio associa-se com o
decréscimo na contração muscular e com a imunocompetência. O
cálcio é necessário para a realização de várias funções vitais, na síntese
de leite, na contração muscular e desempenhando funções como
segundo mensageiro ou como cofator em diversas vias metabólicas

129
intracelulares (Bruno, 2010). Durante o período seco, as necessidades
de cálcio da vaca são baixas (manutenção e desenvolvimento fetal),
ocorrendo incremento acentuado após o parto para a síntese de leite.
Entretanto, os mecanismos de controle do organismo não conseguem
mobilizar tamanha exigência, levando a quadros de hipocalcemia
subclínica e clínica.
A hipocalcemia na forma subclínica contribui para a
diminuição da contração da musculatura lisa, da motilidade do rúmen
e abomaso, redução da ingestão de matéria seca e da produção de leite
no início da lactação, assim como predispõe ao desenvolvimento de
outras doenças. Esta desordem atinge cerca de 20 a 40% de vacas
leiteiras no período pós-parto, e pode persistir em até cerca de 22%
dos animais até os 10 dias pós-parto (Charbonneau et al. 2008). A
diminuição das reservas de cálcio intracelular no sistema imune
prejudica de forma direta a resposta celular a um estímulo de ativação,
contribuindo para um estado de supressão imunitária. Diversos autores
indicam que cortisol, hormônio do crescimento (GH) e fator de
crescimento semelhante a insulina (IGF) alteram a proliferação de
linfócitos. Também, níveis elevados de ácidos graxos não
esterificados (AGNE) e de beta- hidroxibutirato (BHB) são sugeridos
como agentes imunossupressores. Estudos in vitro demonstram que a
presença desses metabólitos em níveis elevados reduz a atividade de
polimorfonucleares e torna os animais mais suscetíveis a doenças
uterinas e a mastite.

130
6.2 Metabolismo e Relação com Enfermidades Uterinas
Os casos de retenção de placenta, metrite e endometrite clínica
e subclínica afetam a regeneração do endométrio e o retorno a
ciclicidade, resultando em atraso na primeira inseminação, aumento
no número de inseminações por concepção, aumento no intervalo
entre partos e redução da taxa de parição (Hussain & Daniel, 1991).
Os tratamentos dessas enfermidades resultam no aumento dos custos
de produção.
Dentre os fatores decisivos para o sucesso de uma nova
concepção no pós-parto recente de vacas, estão o retorno à ciclicidade
ovariana e o término da involução uterina de forma fisiológica. Muitos
são os fatores que influenciam o desempenho reprodutivo de bovinos
de leite, como a nutrição, o manejo, a detecção e sincronização de cio,
e a correta inseminação (Garnsworthy et al. 2008; Schefers et al.
2010). Além disso, algumas alterações ocorridas durante o pré-parto,
como a redução de ingestão de matéria seca e a maior demanda de
energia, podem comprometer o estado nutricional do animal e o seu
sistema imunológico, facilitando a ocorrência de doenças uterinas,
afetando o desempenho reprodutivo (Sordillo, 2009).
Antes do parto o lúmen uterino é estéril, ocorrendo a
contaminação do útero por micro-organismos durante o parto, os quais
são eliminados durante o processo de involução uterina. Quadros de
infecção uterina ocorrem quando os mecanismos de defesa do
organismo não conseguem debelar a infecção havendo um desbalanço
entre os mecanismos de defesa local e as bactérias patogênicas.

131
Animais que tiveram quadros de retenção de placenta, distocias, partos
gemelares, abortos e curtos períodos de gestação, têm mais risco ao
estabelecimento destas infecções (Sheldon et al. 2006; Benzaquen et
al. 2007; Bell & Roberts, 2007). O estabelecimento, a gravidade e a
persistência dos diferentes tipos de infecção são influenciados
basicamente pela condição do ambiente uterino, por fatores genéticos,
além da imunidade inata e adquirida (Willians et al. 2007). A
expressão dos sinais clínicos depende da interação entre resposta
imunológica, quantidade e patogenicidade dos agentes microbianos
(Azawi, 2008).
6.3 Retenção de Placenta
A retenção de placenta (RP) caracteriza-se pela falta de
expulsão da placenta no período de 12 horas após o parto, sendo
associado mais frequentemente com abortos, distocias e gestações
gemelares, mas apresentando também fatores de risco como idade
avançada, estresse, falhas de manejo, hipocalcemia e cetose,
deficiências de vitaminas A e E, deficiências minerais como iodo e
selênio, intoxicações, distúrbios hormonais, doenças como brucelose,
leptospirose e rinotraqueite infecciosa bovina. A incidência de RP
varia entre rebanhos, sendo de 3 a 27%, podendo chegar a 46% em
partos gemelares. Casos não complicados não têm efeito significativo
sobre a fertilidade e sobre o intervalo parto-concepção, porém podem
apresentar como sequela a ocorrência de metrite. Os prejuízos
causados pela retenção placentária podem ultrapassar 150 reais (em

132
torno de U$68,00) por caso tratado, e são devidos a perdas de leite,
que alcançam 250 L em média (levando em conta o leite descartado
pelo uso de antibióticos mais o que a vaca deixou de produzir pela
enfermidade), gastos com tratamento e mão de obra, atraso de 15 dias
para concepção, descarte de 6% dos animais acometidos pela doença,
além da mortalidade que apresenta valores médios de 1,5% dos casos
(Corrêa et al. 2010).
Além das consequências diretas da RP sobre a produção de
leite, também é um fator de risco para outras desordens reprodutivas e
doenças como a cetose, laminite e mastite. Como consequência, a RP
afeta negativamente o desempenho reprodutivo, podendo ser
observado o aumento no intervalo entre o parto e a primeira ovulação,
redução da taxa de concepção ao primeiro serviço e menor taxa de
prenhez (Fourichon et al. 1999; Bell & Roberts, 2007; Tillard et al.
2008; Sartori et al. 2013). Relatos de ocorrência de RP em vários
países incluem os Estados Unidos com 7,8% dos animais, Coréia com
18,3% e Alemanha com 27,8% (Drillich et al. 2006).
Estudos indicam a relação de RP com desbalanços de cálcio e
fósforo (Pelissier, 1976). Vacas hipocalcêmicas são mais pré-
dispostas à ocorrência de RP, por via indireta, devido ao maior risco
de partos distócicos. Ademais, a imunossupressão gerada pelos baixos
níveis de cálcio, associada a quadros de RP, predispõem estes animais
a um quadro clínico de endometrite.
Segundo Gunnink (1984), o sistema imune identifica a
placenta como um corpo estranho. Em seu estudo, avaliando nível de

133
fagocitose dos leucócitos, observou que vacas com RP tiveram menor
capacidade fagocítica comparada à de vacas saudáveis. Esta menor
atividade de leucócitos foi observada antes, durante e após o parto em
outros estudos (Gunnink, 1984a, 1984b, 1984c). Vacas com redução
da atividade de leucócitos apresentam redução na capacidade de
ingestão de matéria seca duas semanas antes e duas semanas após o
parto (Romaniukowa, 1984). No estudo realizado por Kimura et al.
(2002), vacas com RP apresentavam baixa função de neutrófilos desde
o pré-parto até duas semanas pós-parto.
Vacas com níveis elevados de AGNE, indicando alta
lipomobilização, têm alteração tanto da função dos linfócitos quanto
da proliferação e secreção de imunoglobulina M e de interferon
(Lacetera et al. 2004), afetando também a capacidade oxidativa e
viabilidade dos polimorfonucleares (Scalia et al. 2006).
Concentrações plasmáticas de AGNE pré e pós-parto acima de 300 e
600 µmol/L, respectivamente, são consideradas fatores de risco para
RP e metrite (Ospina et al. 2010). A RP é um dos principais fatores
predisponentes da metrite, sendo que 20 a 25% das vacas com RP
desenvolvem metrite (Dubuc et al. 2011).

134
6.4 Metrite
Os eventos do pós-parto, juntamente com vários mecanismos
de defesa do útero, facilitam a eliminação da flora bacteriana
indesejável (Senger, 2005). As contrações do miométrio permitem a
eliminação de grande parte dos lóquios. O retorno à ciclicidade
ovariana (estro) é o fator natural mais importante, assim como o
aumento das concentrações de estradiol, que favorece a migração de
leucócitos para o trato reprodutivo. A secreção ovárica de estrogênio
antes da primeira ovulação promove uma maior resistência uterina a
infecções, possibilitando o retorno precoce à atividade reprodutiva
(Hafez & Hafez, 2000). Porém, casos de lipidose hepática têm sido
associados com uma extensão no período anovulatório no pós-parto
(Reist et al. 2000), reduzindo o desempenho reprodutivo de vacas
leiteiras em lactação (Jorritsma et al. 2000), e favorecendo a doenças
uterinas, como a metrite.
A metrite séptica no pós-parto de vacas leiteiras caracteriza-
se por toxemia grave e abundante corrimento uterino com odor
pútrido, acompanhado ou não de retenção de placenta, é uma
inflamação que atinge todas as camadas do útero. A sua etiologia é
multifatorial, admitindo-se uma combinação de fatores, como
retenção de placenta, insuficiente involução uterina pós-parto e
infecção uterina ascendente. A ocorrência é maior no período de 2 a 4
dias pós-parto, tendo como fatores predisponentes a ocorrência de
distocias, retenção de placenta, obesidade e debilidade orgânica.

135
A metrite puerperal aguda pode apresentar incidência variável
entre 11 e 36%. As perdas econômicas ocorrem de forma indireta,
como a redução na produção de 266 L de leite até o dia 119 de
lactação, incluindo o leite descartado pelo tratamento. Além disso,
prejuízos com o aumento de 7% nas taxas de descarte e redução da
vida útil da vaca em 6 a 8 meses. A metrite ainda altera a involução
uterina e reduz o desenvolvimento folicular pós-parto, elevando o
intervalo parto-concepção e comprometendo os resultados da
inseminação artificial (Corrêa et al. 2010).
A contaminação bacteriana do útero no pós-parto é comum,
requerendo uma resposta imune de forma rápida das células de defesa,
principalmente neutrófilos. O recrutamento de neutrófilos é iniciado
por meio de citocinas pró-inflamatórias, geradas a partir do
endométrio, no momento de reconhecimento das bactérias. Produção
de baixos níveis de citocinas pró-inflamatórias, incluindo o fator
necrose tumoral α (TNFa), IL-1β e IL-6, tem sido associada com o
desenvolvimento de metrite (Galvão et al. 2011). Vacas com atividade
de leucócitos reduzidos apresentam maior propensão a apresentarem
quadros de metrite e mastite no pós-parto (Cai et al. 1994). Além disso,
a redução na ingestão de matéria seca que se inicia no pré-parto e se
estende até o pós-parto imediato, associada à grande demanda
metabólica, gera um déficit negativo de energia e, conforme abordado
anteriormente, vacas com níveis elevados de AGNE apresentam
alteração da capacidade dos linfócitos de proliferar e secretar
imunoglobulina M e interferon (Lacetera et al. 2004), afetando

136
também a capacidade oxidativa e viabilidade dos polimorfonucleares
(Scalia et al. 2006). Vacas que perderam entre 1 e 1,5 pontos de
condição corporal entre o período seco e o pós-parto apresentam um
balanço energético negativo mais intenso e maior tempo para
recuperação da condição corporal, entre a primeira e quarta semana de
lactação, do que as vacas que perderam 0,75 pontos de ECC. Esses
animais apresentam maior ocorrência de metrite (62% x 27%) e
doenças metabólicas, como cetose, deslocamento de abomaso e
hipocalcemia (23% x 2%). Os animais com maior perda de ECC
apresentaram 103 dias em aberto, enquanto os animais com perda
moderada de ECC tiveram 87 dias em aberto (Kim & Suh, 2003).
Atualmente, vem-se estudando alguns marcadores que
indicam o risco do animal apresentar metrite antes da ocorrência de
sinais clínicos. O quadro clínico de metrite é marcado por alterações
em proteínas hepáticas, conhecidas como proteínas de fase aguda,
sendo a haptoglobina uma proteína de fase aguda positiva e a
paraoxanase e a albumina proteínas de fase aguda negativa. Um estudo
demonstrou aumento dos níveis de haptoglobina em torno do parto,
em vacas com metrite (Huzzey et al. 2009). Redução nos níveis de
albumina indicam alterações hepáticas e está relacionado com
incidência de doenças uterinas (Bertoni et al. 2008; Burke et al. 2010).
No estudo realizado por Schneider et al. (2013) pode-se observar que
vacas com metrite apresentavam redução na atividade de paraoxanase
no dia 7 pré-parto e de albumina aos 21 dias antes do parto. Neste
estudo também foi possível observar uma maior taxa de prenhez dos

137
animais saudáveis aos 150 dias pós-parto, comparado aos animais com
metrite.
6.5 Endometrite
A endometrite é uma enfermidade caracterizada por uma
inflamação do endométrio e sua ocorrência se dá a partir dos 21 dias
pós-parto, não estando associada com envolvimento sistêmico. Esta
enfermidade tem sido subdividida em endometrite clínica e
endometrite subclínica, sendo a endometrite clínica caracterizada pela
presença de conteúdo purulento ou mucopurulento dos 21 aos 26 dias
pós-parto. O quadro de endometrite subclínica caracteriza-se pela
presença de mais de 18% de polimorfonucleares na citologia uterina
no período de 21 aos 33 dias pós-parto, ou mais de 10% de
polimorfonucleares dos 34 aos 47 dias pós-parto. Vacas com quadros
de endometrite subclínica não apresentam descarga de conteúdo
uterino, porém, apresentam efeito negativo na reprodução (Sheldon et
al. 2006).
Um estudo recente realizado por Krause et al. (2014)
demonstrou que vacas com ciclos estrais normais tiveram menores
níveis de células polimorfonucleares (26,3%) quando comparado a
vacas anovulatórias (53,4%). Estudos realizados por Sheldon et al.
(2008, 2009) demostraram que em torno de 40% dos animais de alta
produção, mantidos em sistemas intensivos, podem apresentar metrite
nas três primeiras semanas após o parto, com persistência da
enfermidade na forma de endometrite clínica em até 20% dos animais.

138
Assim como na retenção de placenta e na metrite, os casos de
endometrite estão associados com uma redução da função imune. Kim
et al. (2005) observaram que vacas com endometrite apresentaram
linfócitos com capacidade fagocítica reduzida já no pré- parto.
Há uma relação positiva entre endometrite, BEN
(caracterizado por AGNE e BHB elevados) e quadros de cetose
(Hammon et al. 2006; Galvão et al. 2010). Demonstrou-se maior
incidência de endometrite em animais com esteatose hepática (Fronk
et al. 1980) ou com acetonemia durante o puerpério recente
(Markusfeld, 1985).
Outro fator a ser levado em consideração é a maior tendência
que animais com sobrepeso no período seco têm de desencadearem
quadros mais intensos de BEN e cetose no pós-parto, por apresentarem
maior perda de condição corporal e terem um maior atraso na
recuperação de condição corporal. É sabido que animais com
sobrepeso no pré-parto tem maior tendência à hiperlipomobilização no
pós-parto, o que leva a quadros mais intensos de BEN e por
consequência acentuação da imunossupressão. Vacas com elevada
condição corporal (acima de 3,5) no pré-parto apresentam uma
redução no metabolismo hepático, resultando em uma menor atividade
oxidativa quando desafiadas no pós-parto. A concentração elevada de
AGNE no fígado leva também ao aumento de proteínas de fase aguda,
principalmente haptoglobina, sendo possível detectar este perfil
durante o pré-parto. Sua detecção pode ser um bom indicador no
diagnóstico precoce de enfermidades do peri-parto, uma vez que as

139
vacas com endometrite apresentam níveis mais elevados desta
proteína já no pré-parto (Schneider et al. 2013).
6.6 Metabolismo e Relação com a Mastite
Mastite é a inflamação da glândula mamária que afeta uma
elevada proporção de vacas (Seegers et al. 2003). Economicamente, é
considerada a principal doença do gado leiteiro, uma vez que
apresenta elevada prevalência e os custos com perdas e tratamentos
são geralmente elevados (Seegers et al. 2003). Animais acometidos
por mastite apresentam alterações na glândula mamária (Schwegler et
al. 2013), como edema, vermelhidão, dor; alteração no aspecto e na
composição do leite, como o aumento da contagem de células
somáticas (Hortet & Seegers, 1998), além de apresentarem um queda
crônica na produção (Bar et al. 2007). Dependendo da severidade da
mastite, alterações sistêmicas também podem estar presentes e
incluem a diminuição da ingestão de matéria seca e alterações dos
padrões fisiológicos durante o exame clínico. Em casos mais graves
há severa desidratação e endotoxemia. Além disso, a mastite clínica
altera as concentrações metabólicas sanguíneas, modificando assim o
perfil hormonal da vaca, prejudicando sua fertilidade (Hockett et al.
2000; Lavon et al. 2010).
Dependendo do agente etiológico e das outras condições
envolvidas, pode ocasionar a morte do animal. Não é incomum a
mastite ser um dos principais, senão o principal motivo de descarte de
vacas leiteiras nos rebanhos. Grande parte do que determina o animal

140
a desenvolver ou não a doença está relacionada com a capacidade do
agente causador em vencer as defesas inatas e adquiridas da glândula
mamária e se multiplicar, assim como a capacidade do animal em
responder aos estímulos patogênicos e desenvolver uma resposta
imune de forma rápida e que seja eficaz.
Como já citado, durante o periparto, além do BEN, as vacas
passam por uma fase de imunossupressão, que pode ser definida como
a baixa capacidade do organismo em formar uma resposta imune
rápida e eficaz para combater uma infecção. Durante esta fase, os
animais estão mais propensos ao surgimento de doenças infecciosas,
entre elas a mastite. Observa-se que aproximadamente 25% das
mastites ocorrem durante as primeiras duas semanas de lactação e que
a maioria são causadas por agentes oportunistas, o que indica
influência da condição imune nesta fase.
Com o objetivo de avaliar se o BEN é o principal causador
desses distúrbios, pesquisadores induziram restrição alimentar em
vacas no meio da lactação e procederam com infusões intramamárias
de endotoxinas, observando que o BEN neste momento não afetou
significativamente a expressão de moléculas de adesão sob a
superfície de leucócitos bovinos (Perkins et al. 2001; Moyes et al.
2009). Em outro estudo, inocularam quantidades semelhantes de
Escherichia coli em grupos de vacas no início ou meio da lactação,
identificando que as vacas periparturientes tiveram crescimento
bacteriano mais rápido, infecção mais severa e maior temperatura
corporal do que as vacas no meio da lactação (Shuster et al. 1996).

141
As principais células envolvidas no sistema imune da glândula
mamária são os neutrófilos e os linfócitos. Os neutrófilos são um dos
principais tipos celulares do sistema imune, pois são a primeira linha
de defesa, respondendo de forma rápida (em minutos) e não precisam
de contato prévio com o agente. Uma de suas principais funções é a
fagocitose. Os linfócitos representam uma segunda linha de defesa,
muito importante pelo fato da memória imunológica. O
estabelecimento de uma infecção em qualquer órgão ou tecido é
dependente de um balaço delicado entre os mecanismos de defesa
nativos do organismo e a capacidade da bactéria de resistir a condições
desfavoráveis de crescimento. Quando há desequilíbrio, como no caso
da imunossupressão, a doença ocorre.
Com o objetivo de avaliar o efeito da produção de leite sobre
a imunossupressão durante o periparto, foram utilizadas um grupo de
vacas com glândula mamária (inteiras) e outro com vacas
mastectomizadas, nas quais se avaliou a capacidade dos neutrófilos
em eliminar micro-organismos (atividade da mieloperoxidade). As
vacas mastectomizadas tiveram uma rápida recuperação da habilidade
fagocitária dos neutrófilos, enquanto que nos animais inteiros foram
necessárias cerca de três semanas após o parto para haver essa
completa recuperação (Kimura et al. 1999). Em virtude desse fato,
alguns trabalhos vêm investigando componentes metabólicos
individuais associados ao BEN. A cetose é a elevação dos corpos
cetônicos na corrente sanguínea, urina ou leite, associado a um
declínio na glicose sanguínea. Os corpos cetônicos são resultado da

142
excessiva mobilização lipídica que ocorre nas vacas em BEN e da
incapacidade do fígado em metabolizar o excesso de ácidos graxos
(González et al. 2000). Os corpos cetônicos são utilizados como
energia alternativa por diversos tecidos. Entretanto, muitas das células
imunes são afetadas negativamente pelos níveis de metabólitos típicos
de um ambiente cetônico (baixa glicose, elevação nos corpos
cetônicos e presença de AGNE).
Dessa forma, diversos trabalhos vêm demonstrando que vacas
cetônicas apresentam quadros mais severos de mastite do que vacas
saudáveis (Oltenacu & Ekesbo, 1994; Kremer et al. 1993). Linfócitos
de vacas com cetose clínica e subclínica, cultivados in vitro,
responderam menos a desafios patogênicos, quando comparados com
linfócitos de vacas saudáveis (Kandefer & Szerszen et al. 1992).
Ademais, houve uma diminuição na expressão de moléculas de
superfície associadas com a função de neutrófilos sanguíneos (Zerbe
et al. 2000).
A excessiva mobilização lipídica no período de transição leva
a um acúmulo de AGNE na corrente sanguínea, o que causa efeitos
maléficos sobre a função das células imunes. Estudos in vitro
demonstraram a diminuição da função, viabilidade e proliferação das
células mononucleares quando em contanto com AGNE (Scalia et al.
2006; Ster et al. 2012). Portanto, níveis elevados de AGNE na
proximidade do parto são preditivos para a subsequente mastite
(Moyes et al. 2009; Schwegler et al. 2013), além de outras desordens

143
como o deslocamento de abomaso, cetose clínica, metrite e retenção
de placenta (Ospina et al. 2010).
Além dos metabólicos envolvidos diretamente com o BEN
(AGNE, corpos cetônicos) sabe-se que o cálcio também exerce um
papel importante sobre a ocorrência das mastites. A função principal
do cálcio está relacionada com a contração muscular. Dessa forma, em
vacas hipocalcêmicas o canal do teto da glândula mamária pode ficar
aberto e favorecer a penetração de patógenos ambientais para o
interior da luz do órgão. Além disso, vacas caídas e que ficam com os
tetos expostos ao solo por longos períodos também tem chances
maiores de desenvolver mastites. A hipocalcemia também age como
mais um fator estressante para a vaca durante o pré-parto, já que os
animais com hipocalcemia subclínica apresentam níveis mais
elevados nas concentrações plasmáticas de cortisol, quando
comparados com vacas saudáveis, prejudicando ainda mais o
organismo já imunossuprimido (Horst & Jorgensen, 1982). Ademais,
o cálcio também exerce influência sobre a imunocompetência celular,
uma vez que é importante na sinalização intracelular para a ativação
das células imunes (Kimura et al. 2006). Apesar dos maiores prejuízos
serem observados com a forma clínica da doença, é importante
ressaltar que grande parte das vacas permanecem com hipocalcemia
subclínica durante as primeiras semanas após o parto, influenciando a
função imune (Goff et al. 1996). Foi demonstrado que as vacas com
hipocalcemia clínica foram 8,1 vezes mais propensas a desenvolverem
mastite do que as vacas saudáveis (Curtis et al. 1983).

144
6.7 Considerações Finais
O período de transição influencia a vaca leiteira em nível
produtivo e reprodutivo no decorrer da lactação, sendo por isso
considerado um momento crucial no ciclo de produção da vaca. A
ocorrência de mastite e enfermidades uterinas são reconhecidas como
os principais problemas na sanidade dos rebanhos leiteiros e o
metabolismo está diretamente relacionado com a ocorrência dessas
enfermidades, uma vez que fatores como o BEN, corpos cetônicos e
hipocalcemia influenciam diretamente a resposta imune dessas vacas
nesse período.
REFERÊNCIAS BIBLIOGRÁFICAS
AZAWI, O.I. Postpartum uterine infection in cattle. Animal Reproduction
Science, v. 105, p. 187-208, 2008.
BAR, D.; GRÖHN, Y.T.; BENNETT, G. et al. Effect of repeated episodes
of generic clinical mastitis on milk yield in dairy cows. Journal of Dairy
Science, v. 90, p. 4643-4653, 2007.
BELL, M. J & ROBERTS, D. J. 2007. The impact of uterine infection on a
dairy cow’s performance. Theriogenology 68: 1074-1079.
BENZAQUEN, M. E., RISCO, C. A., ARCHBALD, L.F. et al. 2007. Rectal
temperature, calving-related factors, and the incidence of puerperal metritis
in postpartum dairy cows. Journal of Dairy Science 90: 2804-2814.
BERTONI, G., TREVISI, E., HAN, X. et al. 2008. Effects of inflammatory
conditions on liver activity in puerperium period and consequences for
performance in dairy cows. Journal of Dairy Science 91(9): 3300-3310.

145
BRUNO, R. G. S. 2010. Nutrition and reproduction in modern dairy cows.
Mid-south ruminant nutrition conference, Texas, USA, 51-56.
BURKE, C. R., MEIER, S., MCDOUGALL, S. et al. 2010. Relationships
between endometritis and metabolic state during the transition period in
pasture-grazed dairy cows. Journal of Dairy Science 93(11): 5363-5373.
CAI T. Q., WESTON, P. G., LUND, L. A. et al. 1994. Association between
neutrophil functions and periparturient disorders in cows. American
Journal of Veterinary Research 55: 934-943.
CHARBONNEAU, E., CHOUINARD, P. Y., TREMBLAY, G. F. et al.
2008. Hay to reduce dietary cation-anion difference of dairy cows. Journal
of Dairy Science 91: 1585-1596.
CORRÊA, M. N., GONZÁLEZ, F. H. D. and DA SILVA, S. C. 2010.
Transtornos metabólicos nos animais domésticos. Editora e gráfica
universitária PREC-UFPel, Pelotas, RS, ed. 1.
CURTIS, C. R., ERB, H. N., SNIFFEN, C. J. et al. 1983. Association of
parturient hypocalcemia with eight periparturient disorders in Holstein
cows. Journal of the American Medical Association 183(5): 559-561.
DRILLICH, M. M., MAHLSTEDT, U., REICHERT, B. A. et al. 2006.
Strategies to improve the therapy of retained fetal membranes in dairy cows.
Journal of Dairy Science 89: 627-635.
DUBUC, J., DUFFIELD, T. F., LESLIE, K. E. et al. 2011. Randomized
clinical trial of antibiotic and prostaglandin treatments for uterine health and
reproductive performance in dairy cows. Journal of Dairy Science 94:
1325-1338.
FOURICHON, C., SEEGERS, H., BAREILLE, N. et al. 1999. Effects of
disease on milk production in the dairy cow: a review. Preventive
Veterinary Medicine 1: 1-35.
FRONK, T. J., SCHULTZ, L. H. and HARDIE, A. R. 1980. Effect of
period overconditioning on subsequent metabolic disorders and
performance of dairy cows. Journal of Dairy Science 63: 1080-1090.
GALVÃO, K. N., FLAMINIO, M. J., BRITTIN, S. B. et al. 2010.
Association between uterine disease and indicators of neutrophil and

146
systemic energy status in lactating Holstein cows. Journal of Dairy
Science 93: 2926-2937.
Galvão, K. N., Santos, N. R., Galvão, J. S. et al. 2011. Association between
endometritis and endometrial cytokine expression in postpartum Holstein
cows. Theriogenology 76: 290-299.
GARNSWORTHY, P. C., LOCK, A., MANN, G. E. et al. 2008. Nutrition,
metabolism and fertility in dairy cows: Dietary fat content and ovarian
function. Journal of Dairy Science 91: 3824-3833.
GOFF, J. P., HORST, R. P., JARDON, P. W. et al. 1996. Field trials of an
oral calcium propionate paste as an AID to prevent milk fever in
periparturient dairy cows. Journal of Dairy Science 79: 378-383.
GOFF, J. P. & HORST, R. L. 1997. Physiological changes at parturition
and their relationship to metabolic disorders. Journal of Dairy Science 80:
1260-1268.
GONZÁLEZ, F., BARCELLOS, J., PATIÑO, H. 2000 Perfil metabólico
em ruminantes: seu uso em nutrição e doenças nutricionais. Editora
UFRGS, Porto Alegre, RS.
GUNNINK, J. W. 1984a. Pre-partum leucocytic activity and retained
placenta. Veterinary Quaterly 6(2): 52-54.
GUNNINK, J. W. 1984b. Post-partum leucocytic activity and its
relationship to caesarian section and retained placenta. Veterinary
Quaterly 6(2): 55-57.
GUNNINK, J. W. 1984c. Retained placenta and leukocytic activity.
Veterinary Quaterly 6(2): 49-51.
HAFEZ, E. S. E. & B. HAFEZ. 2000. Reproduction in Farm Animals.
Lea and Febiger, Philadelphia, USA. ed. 7.
HAMMON, D. S., EVJEN, I. M., DHIMAN, T. R., GOFF, J. P. et al. 2006.
Neutrophil function and energy status in Holstein cows with uterine health
disorders. Veterinary Immunology and Immunopathology 113: 21-29.
HOCKETT, M. E., HOPKINS, F. M., LEWIS, M. J. et al. 2000. Endocrine
profile of dairy cows following experimentally induced clinical mastitis
during early lactation. Animal Reproduction Science 58: 241-251.

147
HORST, R. L. & JORGENSEN, N. A. 1982. Elevated plasma cortisol
during induced and spontaneous hypocalcemia in ruminants. Journal of
Dairy Science 65: 23-32.
HORTET, P. & SEEGERS, H. 1998. Review - Loss in milk yield and
related composition changes resulting from clinical mastitis in dairy cows.
Preventive Veterinary Medicine 37: 1-20.
HUSSAIN, A. M. & DANIEL, R. C. W. 1991. Bovine endometrites:
current and future alternative therapy. Journal of the Veterinary Medicine
38: 641-651.
HUZZEY, J. M., DUFFIELD, T. F., LEBLANC, S. J. et al. 2009. Short
communication: Haptoglobin as an early indicator of metritis. Journal of
Dairy Science 92(2): 621-625.
JORRITSMA, R., JORRITSMA, H., SCHUKKEN, Y. H. et al. 2000.
Relationships between fatty liver and fertility and some periparturient
diseases in commercial Dutch dairy herds. Theriogenology 54: 1065-1074.
KANDEFER-SZERSZEN, M., FILAR, J., SZUSTER-CIESIELSKA, A. et
al. 1992. Suppression of interferon response of bovine leukocytes during
clinical and subclinical ketosis in lactating cows. Dtsch. Tierarztl.
Wochenschr. 99: 440-443.
KEHRLI, M. E. J. R., NONNECKE, B. J. and ROTH, J. A. 1989.
Alterations in bovine peripheral blood lymphocyte function during the
periparturient period. American Journal of Veterinary Research 50: 215-
220.
KIM I. H., NA, K. J. and YANG M. P. 2005. Immune responses during the
peripartum period in dairy cows with postpartum endometritis. Journal of
Reproduction and Development 51: 757-764.
KIM, I. H & SUH, G. H. 2003. Effect of the amount of body condition loss
from the dry to near calving periods on the subsequent body condition
change, occurrence of postpartum diseases, metabolic parameters and
reproductive performance in Holstein dairy cows. Theriogenology 60:
1445-1456.

148
KIMURA, K., GOFF, J. P. and KEHRLI, M. E. 1999. Effects of the
presence of the mammary gland on expression of neutrophil adhesion
molecules and myeloperoxidase activity in periparturient dairy cows.
Journal of Dairy Cows 82: 2385-2392.
KIMURA, K., GOFF, J. P., KEHRLI, M. E. Jr. et al. 2002. Decreased
neutrophil function as a cause of retained placenta in dairy cattle. Journal
of Dairy Science 85: 544-550.
KIMURA, K., REINHARDT, T. A. and GOFF, J. P. 2006. Parturition and
hypocalcemia blunts calcium signals in immune cells of dairy cattle.
Journal of Dairy Science. 89: 2588-2595.
KRAUSE, A. R. T., PFEIFER, L. F. M., MONTAGNER, P. et al. 2014.
Associations between resumption of postpartum ovarian activity, uterine
health and concentrations of metabolites and acute phase proteins during the
transition periods in Holstein cows. Animal Reproduction Science 145: 8-
14.
KREMER, W. D., NOORDHUIZEN-STASSEN, E. N., GROMMERS, F. J.
et al. 1993. Severity of experimental Escherichia coli mastitis in ketonemic
and nonketonemic dairy cows. Journal of Dairy Science 76: 3428-3436.
LACETERA, N., SCALIA, D., FRANCE, O. et al. 2004. Effects of
nonesterified fatty acids on lymphocyte function in dairy heifers. Journal
of Dairy Science 87: 1012-1014.
LAVON, Y., LEITNER, G., VOET, H. et al. 2010. Naturally occurring
mastitis effects on timing of ovulation, steroid and gonadotrophic hormone
concentrations, and follicular and luteal growth in cows. Journal of Dairy
Science 93: 911-921.
MARKUSFELD O. 1985. Relationship between overfeeding, metritis and
ketosis in high yielding dairy cows. Veterinary Record 116: 489-491.
MOYES, K. M., DRACKLEY, J. K., SALAK-JOHSON, J. L. et al. 2009.
Dietary-induced negative energy balance has minimal effects on innate
immunity during a Streptococcus uberis mastitis challenge in dairy cows
during midlactation. Journal of Dairy Science 92: 4301-4316.

149
OLTENACU, P. A. & EKESBO, I. 1994. Epidemiological study of clinical
mastitis in dairy cattle. Veterinary Research 25: 208-212.
OSPINA, P. A., NYDAM, D. V., STOKOL, T. et al. 2010. Evaluation of
nonesterified fatty acids and beta-hydroxybutyrate in transition dairy cattle
in the northeastern United States: Critical thresholds for prediction of
clinical diseases. Journal of Dairy Science 93: 546-554.
PELISSIER, C. L. 1976. Dary cattle breeding problems and their
consequences. Theriogenology 6: 575-583.
PERKINS, K. H., VANDEHAAR, M. J., BURTON, J. L. et al. 2001.
Negative energy balance does not decrease expression of leukocyte
adhesion or antigen-presenting molecules in cattle. Journal of Dairy
Science 84: 421-428.
REIST, M., KOLLER, A., BUSATO, A. et al. 2000. First ovulation and
ketone body status in the early postpartum period of dairy cows.
Theriogenology 54: 685-701.
ROMANIUKOWA, K. 1984. Phagocytosis in the uterus. In: Proceedings
of the 10th International Congress on Animal Reproduction and AI.
Urbana, Champaign, IV: XI, 30–33.
SARTORI, R., GUARDIEIRO, M. M., SURJUS, R. S. et al. 2013.
Metabolic hormones and reproductive function in cattle. Animal
Reproduction 10: 199-205.
SCALIA, D., LACETERA, N., BERNABUCCI, U. et al. 2006. In vitro
effects of nonesterified fatty acids on bovine neutrophils oxidative burst and
viability. Journal of Dairy Science 89: 147-154.
SCHEFERS J. M., WEIGEL, K. A., RAWSON, C. L. et al. 2010.
Management practices associated with conception rate and service rate of
lactating Holstein cows in large, commercial dairy herds. Journal of Dairy
Science 93(4): 1459-1467.
SCHNEIDER A., CORRÊA, M. N. and BUTLER, W. R. 2013. Short
communication: Acute phase proteins in Holstein cows diagnosed with
uterine infection. Research in Veterinary Science 95: 269-271.

150
SCHWEGLER, E., SCHNEIDER, A., MONTAGNER, P. et al. 2013.
Predictive value of prepartum serum metabolites for incidence of clinical
and subclinical mastitis in grazing primiparous Holstein cows. Tropical
Animal Health and Production 45: 1549-1555.
SEEGERS, H., FOURICHON, C., BEAUDEAU, F. 2003. Production
effects related to mastitis and mastitis economics in dairy cattle herds.
Veterinary Research 34: 475-491.
SENGER, P. L. 2005. Pathways to pregnancy and parturition. Current
conceptions Inc., Pullan, WA, USA, ed. 2.
SHELDON, I. M., CRONIN, J., GOETZE, L. et al. 2009. Defining
postpartum uterine disease and the mechanisms of infection and immunity
in the female reproductive tract in cattle. Biology of Reproduction 81:
1025-1032.
SHELDON, I. M., LEWIS, G. S., LEBLANC, S. et al. 2006. Defining
postpartum uterine disease in cattle. Theriogenology 65(8): 1516-1530.
SHELDON, I M., WILLIAMS, E. J., MILLER, A. N. et al. 2008. Uterine
diseases in cattle after parturition. Veterinary Journal 176(1): 115-121.
SHUSTER, D. E., LEE, E. K. and KEHRLI, M. E. Jr. 1996. Bacterial
growth, inflammatory cytokine production, and neutrophil recruitment
during coliform mastitis in cows within ten days after calving, compared
with cows at midlactation. American Journal of Veterinary Research 57:
1569-1575.
SORDILLO, L. M. 2009. Current concepts on immunity and mastitis.
WCDS Advances in Dairy Technology 21: 111-119.
STER, C., LOISELLE, M. C. and LACASSE, P. 2012. Effect of
postcalving serum nonesterified fatty acids concentration on the
functionality of bovine immune cells. Journal of Dairy Science 95: 708-
717.
TILLARD, E., HUMBLOT, P., FAYE, B. et al. 2008. Postcalving factors
affecting conception risk in Holstein dairy cows in tropical and sub-tropical
conditions. Theriogenology 69: 443-457.

151
WILLIAMS, E. J., FISHER, D. P., NOAKES, D. E. et al. 2007. The
relationship between uterine pathogen growth density and ovarian function
in the postpartum dairy cow. Theriogenology 68: 549-559.
YOUNGQUIST, R. S. & THRELFALL, W. R. 2007. Current therapy in
large animal theriogenology. WB Saunders, USA, ed. 2.
ZERBE, H., SCHNEIDER, N. SCHNEIDER, W. et al. 2000. Altered
functional and immunophenotypical properties of neutrophilic granulocytes
in postpartum cows associated with fatty liver. Theriogenology 54(5): 771-
786.
ZIGUER, E. A., GOULART, M. A., JÚNIOR, J. A. S. F. et al. 2007.
Alterações clínicas em bovinos de leite na região centro-norte do Rio
Grande do Sul. A Hora Veterinária 156: 1-4.

152
7. BIOTÉCNICAS DA REPRODUÇÃO PARA MELHORAR A
FERTILIDADE DA VACA LEITEIRA7
Manoel F. Sá Filho
Laís M. Vieira
Bruna M. Guerreiro
Carlos A. Rodrigues
Pietro S. Baruselli
7.1Resumo
A inseminação artificial (IA) e a transferência de embrião
(TE) são relevantes tecnologias para aumentar a produtividade em
rebanhos especializados na produção de leite. Os programas de
sincronização da ovulação para inseminação artificial em tempo fixo
(IATF) ou a transferência de embrião em tempo fixo (TETF) são
ferramentas que permitem aumentar o número de fêmeas inseminadas
ou aptas a receberem um embrião, eliminando a necessidade de
detecção de estro, aumentando a eficiência reprodutiva dos rebanhos.
Atualmente, os protocolos de sincronização da ovulação estão
estabelecidos e apresentam resultados satisfatórios e previsíveis
quando aplicados em propriedades com adequado manejo nutricional
e sanitário. Vários estudos foram realizados para estabelecer
7 Sá Filho, M.F. et al. Biotécnicas da reprodução para melhorar a fertilidade
da vaca leiteira. Anais. I Simpósio Nacional da Vaca Leiteira. Porto Alegre:
Universidade Federal do Rio Grande do Sul. 2014. 239 p.

153
programas reprodutivos aplicados ao manejo das propriedades
comerciais com os protocolos de IATF e de TETF existentes. Tais
programas visam facilitar e intensificar a aplicação dessas
biotecnologias, bem como adequá-los aos objetivos específicos de
cada fazenda. Atualmente, dentre as formas de intensificação do uso
da IA/IATF e/ou TE/TETF, o estabelecimento de programas de
ressincronização das fêmeas não gestantes após o primeiro serviço
merecem destaque. Os programas de ressincronização eliminam a
necessidade de observação de estro (facilitam o manejo e aumentam a
taxa de serviço), reduzem o intervalo entre o parto e a concepção
(aumentam a eficiência reprodutiva) e aumentam o número de
produtos oriundos de IA/TE (incremento do ganho genético),
determinando maior lucratividade às propriedades produtoras de leite.
Em resumo, o aprimoramento das técnicas de IATF e produção in vitro
de embriões (TE/TETF) associadas aos programas de
ressincronização são ferramentas eficientes para o alcance máximo da
eficiência reprodutiva dos rebanhos de forma organizada, pré-
determinada e previsível, ou seja, com elevada precisão. Portanto, uma
vez estabelecido o objetivo da atividade rural, esses programas são
incorporados ao sistema de produção e de gestão, aumentando a
produtividade e a rentabilidade da propriedade.

154
7.2 Introdução
A produtividade em fazendas de leite está diretamente
vinculada à eficiência reprodutiva do rebanho. De maneira resumida,
a eficiência reprodutiva pode ser definida como a habilidade de fazer
a vaca se tornar gestante rapidamente após o parto (para obtenção de
intervalo entre partos próximos a 12 meses a vaca deve conceber entre
75 e 85 dias pós-parto) e com o menor número de coberturas possível.
A inseminação artificial (IA) é a biotécnica mais utilizada em todo o
mundo para difundir material genético superior nos rebanhos bovinos.
Apesar do Brasil ainda empregar pouco essa biotecnologia, verifica-
se um considerável aumento no percentual de matrizes bovinas
inseminadas, passando de cerca de 5% em 2002 para
aproximadamente 10% no ano de 2012 (Baruselli et al., 2012b). Com
a intensificação do uso da IA, o país vem apresentando satisfatório
avanço no melhoramento genético do rebanho, pelo incremento do
número de bezerros nascidos de touros geneticamente superiores.
Assim, o uso de biotecnologias da reprodução, tais como a IA, visando
o aumento no ganho genético do rebanho pode proporcionar
significativo aumento da produtividade e da rentabilidade da
agropecuária.
Verifica-se que o crescimento da utilização da IA no país está
associada ao maior emprego dos programas reprodutivos que utilizam
a técnica da inseminação artificial em tempo fixo (IATF). Em 2013,
segundo dados levantados pelo nosso grupo de pesquisa, estima-se que
8.750.000 sincronizações para IATF foram comercializadas no Brasil

155
(estimativa levando em consideração a venda de produtos para
sincronização). A Associação Brasileira de Inseminação Artificial
(ASBIA, 2014) reportou a comercialização de 13.024.033 de doses de
sêmen no mesmo ano. Dessa forma, estima-se que 67% das
inseminações artificiais realizadas no Brasil empregam técnicas de
sincronização para inseminação artificial em tempo fixo.
A IATF baseia-se na utilização de fármacos comercialmente
disponíveis para sincronizar o ciclo estral e a ovulação de vacas e
novilhas, viabilizando a IA em momentos pré-determinados, sem a
necessidade da prévia detecção do estro. Atualmente existem diversos
protocolos de IATF comercialmente disponíveis em todo o mundo. No
entanto, os protocolos comumente utilizados na América do Sul são a
base de progesterona (P4) e estradiol (E2) (e.g. dia 0 = E2+P4; dia 8 ou
9 = remoção da P4+E2 + prostaglandina; dia 10 = IATF 48 a 54 h após
a remoção da P4). A técnica de IATF já está bem estabelecida e os
resultados são satisfatórios e previsíveis. Esta biotecnologia é capaz
de contornar, com relativo sucesso, os principais entraves que
reduzem a eficiência reprodutiva em fêmeas submetidas a programas
de IA. Entre as dificuldades, pode-se citar a dificuldade de mão de
obra qualificada no campo e falhas na detecção de estro.
Após vários estudos para ajustar tecnicamente os protocolos
de IATF, surgiu a necessidade de adequar a IATF na rotina de manejo
das propriedades comerciais brasileiras. Tais ajustes visam
principalmente, facilitar e intensificar a utilização dessa ferramenta,
bem como adequar os programas reprodutivos aos objetivos

156
específicos de cada fazenda. Dentre as formas de intensificação do uso
da IATF, o estabelecimento de programas de ressincronização das
fêmeas não gestantes após a primeira IATF merece destaque. Esse tipo
de programa elimina a necessidade de observação de estro e,
principalmente, aumenta o número de produtos oriundos de IA, que
determinam aumento de produtividade e maior lucratividade à
atividade.
Além do emprego da IA para o melhoramento genético, a
utilização da transferência de embrião (TE) tem apresentado
expressivo aumento no Brasil, passando de 200.000 embriões
produzidos em 2004 para 350.000 em 2011 (Viana, 2012). Na
atualidade, a Sociedade Brasileira de Tecnologia de Embriões (SBTE)
estima que mais de 400.000 embriões bovinos são produzidos por ano
no Brasil. A TE potencializa o ganho genético por multiplicar tanto
touros quanto doadoras geneticamente superiores. Os protocolos para
TE em tempo fixo (TETF) em receptoras de embriões também foram
intensamente pesquisados e atualmente estão estabelecidos para o
emprego nas propriedades rurais (Baruselli et al., 2010). Semelhante
à IATF, os protocolos de TETF empregam E2+P4 no dia 0; remoção
da P4+E2 + prostaglandina + gonadotrofina coriônica equina (eCG) no
dia 8; TETF no dia 17. Os requisitos necessários para a utilização da
TE em larga escala são semelhantes aos observados para IA. Além
disso, devido à dificuldade na criopreservação de embriões produzidos
in vitro (90,7% do total de embriões produzidos no Brasil), na
atualidade os programas de TE estão limitados à utilização de

157
embriões não criopreservados. Nesse sentido, vários grupos de
pesquisa estão trabalhando para estabelecer um protocolo de
criopreservação de embriões produzidos in vitro para impulsionar a
utilização da TE em larga escala. Adicionalmente, visando agregar
vantagens de ambas as biotecnologias (IA e TE), pesquisas têm sido
desenvolvidas para avaliar a eficiência reprodutiva e o incremento
genético quando são associadas em fazendas comerciais produtoras de
leite.
Portanto, o presente manuscrito tem como objetivo discorrer
sobre o emprego de diferentes programas reprodutivos que visam a
intensificação do emprego da IATF e da TETF, bem como a
associação dessas biotecnologias em fazendas comerciais. Serão
abordadas, principalmente, as técnicas que apresentam impacto
significativo na eficiência reprodutiva de fêmeas leiteiras.
7.3 Inseminação Artificial em Tempo Fixo (IATF)
Durante as últimas décadas, a seleção de fêmeas bovinas
geneticamente superiores para a produção de leite tem sido
correlacionada com a reduzida fertilidade dos rebanhos leiteiros. Além
dos fatores relacionados à alta produção de leite, o estresse térmico
também tem sido relatado como contribuinte das alterações
fisiológicas responsáveis pela menor fertilidade de vacas de leite
(Lopez et al., 2004, Wiltbank et al., 2006). No entanto,
reconhecidamente, a redução do desempenho reprodutivo de vacas de
leite de alta produção foi modificada e ocorreu uma tendência de

158
melhora nos últimos anos (Norman et al., 2009). Alguns pesquisadores
correlacionam a melhor eficiência reprodutiva com o emprego
estratégico e sistemático da IATF em larga escala nos rebanhos
(Norman et al., 2009).
Essa técnica é uma importante ferramenta para minimizar
falhas reprodutivas (e.g. baixa taxa de detecção de estro) observadas
no sistema de produção de leite, seja devido às falhas na detecção do
estro ou ao anestro pós-parto. Dados da literatura relatam semelhante
taxa de concepção (Cerri et al., 2004) e menor intervalo parto-
concepção (87,4 ± 1,8 vs. 94,6 ± 1,8 dias, P= 0,005; Teixeira, 2010)
quando as vacas de leite são submetidas à IATF comparado ao manejo
de IA após detecção de estro. Em resumo, o uso de IATF pode
melhorar os resultados reprodutivos e consequentemente, a
viabilidade da propriedade leiteira, especialmente quando a taxa de
detecção de estro é baixa (Giordano et al., 2011, Giordano et al., 2012,
Ribeiro et al., 2012, Bisinotto et al., 2013, Galvão et al., 2013).
Visando a intensificação do uso da IATF, novas abordagens
têm sido feitas em relação à ressincronização da ovulação para as
demais IATFs nos animais que não se tornaram gestantes nas
inseminações anteriores (Cavalieri et al., 2007). Os programas de
ressincronização em fêmeas de leite também têm dois possíveis
momentos para serem iniciados: (1) antes do diagnóstico de gestação
(ressincronizar as fêmeas gestantes ou não gestantes à IATF prévia), e
(2) no momento do diagnóstico de gestação (ressincronizar somente
as vacas não gestantes). Estudo realizado em vacas de leite

159
demonstrou semelhante taxa de prenhez após ressincronização 25 dias
(antes do diagnóstico de gestação) ou 32 dias (no momento do
diagnóstico de gestação) após IATF prévia (Thompson et al., 2010).
Apesar da similar taxa de prenhez, quando a ressincronização foi
realizada antes do diagnóstico de gestação (25 dias após IATF prévia)
o intervalo parto-concepção reduziu 3,2 dias comparado com a
ressincronização realizada no momento do diagnóstico de gestação
(Thompson et al., 2010).
Outros estudos foram realizados na tentativa de acelerar ainda
mais a ressincronização após a IATF de vacas de leite. Vieira et al.
(2014b) avaliaram a eficiência reprodutiva de vacas em lactação
submetidas à ressincronização 13 dias (dia 13) após IATF
(Experimento 1) e verificaram o efeito do tratamento com benzoato de
estradiol (1,5 mg) na sincronização da emergência da nova onda
folicular e na atividade do corpo lúteo (Experimento 2). No
Experimento 1, um total de 183 vacas em lactação (25,2 ± 8,8 L
leite/dia) foi distribuído em dois grupos: Controle (n=93) e
Ressincronização (n=90). Posteriormente, a primeira IATF as fêmeas
do grupo Controle foram inseminadas após a detecção de estro entre
os dias 18 e 25 pós IATF. As fêmeas do grupo Ressincronização foram
ressincronizadas com a inserção de um dispositivo de P4 e 1,5 mg de
benzoato de estradiol no dia 13. No dia 21, os dispositivos de P4 foram
removidos. As fêmeas do grupo Ressincronização classificadas como
não gestantes baseado na vascularização do corpo lúteo (ultrassom
modo doppler) ou ausência de corpo lúteo receberam 1 mg de

160
cipionato de estradiol, 0,150 mg de PGF2α e foram inseminadas 48 h
após. Nesse estudo, as taxas de corpo lúteo com vascularização no dia
21 (P= 0,003) e taxa de prenhez aos 30 dias após a 1ª IATF (P= 0,02)
foram menores nas fêmeas do grupo Ressincronização (40,0%, 36/90
e 14,9%, 13/87) em relação às do grupo Controle (62,4%, 58/93 e
32,6%, 28/86; respectivamente, Figura 1). No entanto, não houve
diferença (P= 0,98) na taxa de prenhez obtida após a 2ª IA entre o
grupo Controle (25,0%; 3/12) e Ressincronização (22,6%, 12/53).
Ainda, Vieira et al. (2014b) verificaram em outro experimento que
apesar do grupo Ressincronização apresentar a emergência da nova
onda folicular 3,2 ± 1,4 dias após tratamento com benzoato de
estradiol e apresentar semelhante taxa de vascularização do corpo
lúteo nos dias 13 (Controle: 87,8% e Ressincronização: 84,4%) e 15
(Controle: 90,6% e Ressincronização: 86,9%), a vascularização do
corpo lúteo do grupo Ressincronização foi reduzida a partir do dia 17
(dia 17: 71,9%, dia 19: 51,9% e dia 21: 40,6%) em relação ao grupo
Controle (dia 17: 89,4%; dia 19: 88,9% e dia 21: 76,1%; Figura 2).
Esses dados são indicativos de que o tratamento com 1,5 mg de
benzoato de estradiol 13 dias após IATF prévia induziu a regressão do
corpo lúteo e diminuiu a taxa de concepção à IATF realizada
previamente, inviabilizando o emprego dessa metodologia de
ressincronização.

161
Figura 1. Taxa de corpos lúteos (CL) vascularizados no dia 21 pós IATF
prévia e taxa de prenhez à primeira e segunda IATF em animais
ressincronizados no dia 13 com 1,5 mg de benzoato de estradiol e
dispositivo de progesterona ou sem tratamento após primeira IATF
(Controle). Adaptado de Vieira et al. (2014b).
Figura 2. Taxa de vascularização do CL após IATF (dia 0) em animais
ressincronizados no dia 13 com 1,5 mg de benzoato de estradiol e
dispositivo de progesterona ou sem posterior tratamento (Controle).
Adaptado de Vieira et al. (2014b).

162
Dessa forma, para melhor compreensão da eficiência de
diferentes manejos de ressincronização, foram simulados três
programas reprodutivos: IA 12 h após observação de estro (grupo
Estro) com taxa de serviço de 50% para cada 21 dias, IATF com
ressincronização a cada 25 (grupo Ressinc 25 d) ou 32 dias (grupo
Ressinc 32 d), com o objetivo de estimar o intervalo parto-concepção,
intervalo entre partos e taxa de fêmeas gestantes aos 250 dias pós-
parto. Para os diferentes manejos foram estabelecidas as seguintes
taxas de concepção: até o terceiro serviço semelhante a 30%, após o
terceiro serviço 20% e perda gestacional entre 30 e 60 dias de 15%.
Diferentemente da ressincronização em bovinos de corte que tem
início 22 dias após a IATF prévia, em bovinos de leite sugere-se
intervalo de 25 dias para o início da ressincronização para que o
diagnóstico de gestação, os tratamentos de sincronização e a IATF
sejam realizados sempre nos mesmos dias da semana (considerando
visita semanal do veterinário no mesmo dia da semana). Diante disso,
pesquisas recentes mostraram a possibilidade de utilização de GnRH
ou benzoato de estradiol no início do protocolo de ressincronização 25
dias após IATF com taxas de concepção semelhantes entre os
fármacos.
O manejo reprodutivo com IATF associada às consecutivas
ressincronizações (25 ou 32 dias após IATF prévia) reduziu o intervalo
parto-concepção (grupo Ressinc 25d: 146,3 dias, grupo Ressinc 32d:
151 dias e grupo Estro: 155,4 dias), intervalo entre partos (Ressinc
25d: 14,1 meses, Ressinc 32d: 14,3 meses e Estro: 14,4 meses) e o

163
percentual de fêmeas gestantes 250 dias pós-parto (Ressinc 25d:
75,8%, Ressinc 32d: 73,9% e Estro: 69,3% dias; Figura 3). Portanto,
com o estabelecimento do manejo reprodutivo conforme a
disponibilidade do médico veterinário e principalmente, de acordo
com o objetivo da propriedade é possível a obtenção de índices
zootécnicos que favoreçam a produtividade e a rentabilidade da
propriedade de leite.
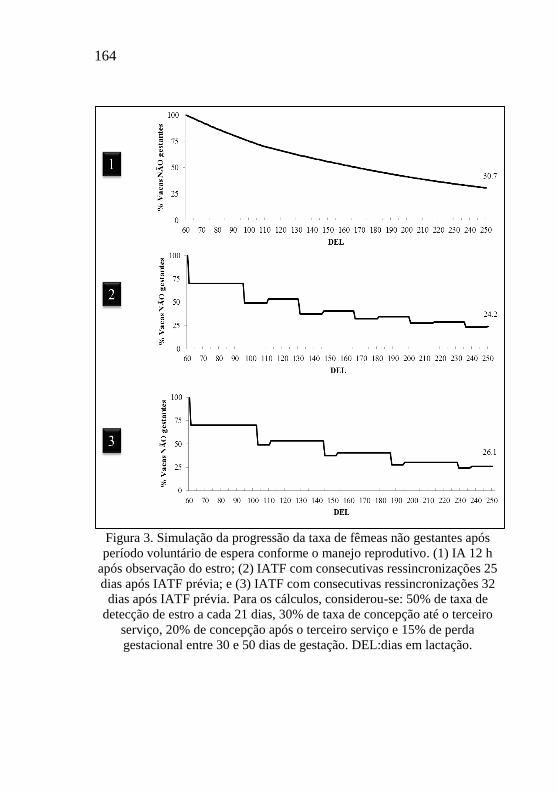
164
Figura 3. Simulação da progressão da taxa de fêmeas não gestantes após
período voluntário de espera conforme o manejo reprodutivo. (1) IA 12 h
após observação do estro; (2) IATF com consecutivas ressincronizações 25
dias após IATF prévia; e (3) IATF com consecutivas ressincronizações 32
dias após IATF prévia. Para os cálculos, considerou-se: 50% de taxa de
detecção de estro a cada 21 dias, 30% de taxa de concepção até o terceiro
serviço, 20% de concepção após o terceiro serviço e 15% de perda
gestacional entre 30 e 50 dias de gestação. DEL:dias em lactação.

165
7.4 Transferência de Embrião (TE)
A utilização da TE em vacas de leite não é apenas uma
ferramenta que acelera o ganho genético, mas também uma
biotecnologia que melhora a eficiência reprodutiva dos rebanhos. Com
o emprego da TE é possível minimizar os efeitos deletérios do estresse
térmico (Rodrigues et al., 2007b) e aumentar os índices reprodutivos
de vacas repetidoras de serviço (fêmeas que não possuem
comprometimento do trato reprodutivo, porém recebem três ou mais
inseminações e não se tornam gestantes; Rodrigues et al., 2007a).
Ainda, visto que com o manejo da TE é possível ressincronizar as
fêmeas que não apresentam corpo lúteo (CL) no momento da TE,
existe a possibilidade de diminuir o intervalo entre serviços e aumentar
a probabilidade da fêmea se tornar gestante antecipadamente no
período pós-parto.
O efeito prejudicial do estresse térmico na fertilidade de vacas
da raça Holandesa já está bem estabelecido. É um fator extrínseco que
compromete negativamente a eficiência reprodutiva (Hansen et al.,
2001) e provoca consideráveis perdas econômicas nos rebanhos
leiteiros (Rensis e Scaramuzzi, 2003). Pesquisas relatam que vacas em
lactação apresentam menor capacidade de termorregulação durante o
período quente do ano comparado às novilhas (Sartori et al., 2002,
Ferreira et al., 2011). No entanto, em ambas as categorias tanto os
ovócitos (Ferreira et al., 2011), quanto em embriões em estágios
iniciais de desenvolvimento (Ealy et al., 1993, Hansen e Aréchiga,
1999, Hansen et al., 2001, Al-Katanani e Hansen, 2002) são

166
comprometidos pelo estresse térmico. Em temperaturas elevadas, o
processo de maturação do ovócito é interrompido e,
consequentemente, a capacidade de fecundação é prejudicada
(Hansen, 2009). Além disso, embriões sobre estresse térmico
apresentam comprometimento no desenvolvimento inicial,
apresentando bloqueio em 8 a 16 células por incapacidade de realizar
a ativação de seu genoma durante o desenvolvimento inicial (Hansen,
2009). Desta forma, visto que os embriões bovinos são transferidos
para o útero da receptora entre o estágio de mórula e blastocisto (> 32
células) e os principais efeitos deletérios do estresse térmico ocorrem
no início do desenvolvimento embrionário, a TE tem sido utilizada
como estratégia reprodutiva para atenuar os efeitos prejudiciais do
estresse térmico na fertilidade (Putney et al., 1989, Drost et al., 1999,
Hansen e Aréchiga, 1999, Al-Katanani e Hansen, 2002, Rodrigues et
al., 2004a).
Na tentativa de avaliar o desempenho reprodutivo de vacas
leiteiras brasileiras após IA ou TE ao longo do ano, dados
retrospectivos com considerável número de animais foram analisados
pelo nosso grupo de pesquisa (Rodrigues et al., 2007b, Freitas et al.,
2010). Na primeira análise, 12.875 IA e 4.822 TE foram avaliadas no
período de 2001 a 2006 na Fazenda Agrindus em Descalvado, SP
(Rodrigues et al., 2007b). Os dados foram indicativos de que durante
os meses mais quentes do ano a taxa de concepção reduziu quando a
IA foi utilizada. No entanto, quando a TE foi empregada nesse mesmo
período, não houve redução na taxa de concepção conforme a época

167
do ano, sendo superior em fêmeas submetidas a TE (41,9%)
comparada às vacas inseminadas artificialmente (28,5%; Figura 4).
Uma segunda análise foi realizada com os dados da mesma
fazenda, porém, utilizando vacas da raça Holandesa repetidoras de
serviço (> 4 serviços). Assim como no estudo anterior, foi observado
que a TE apresentou taxas de gestação constantes ao longo do ano,
sendo sempre superior a TE (41,7%, 1.609/3.858) comparado com a
IA (17,9%, 1.019/5.693). Esses estudos são indicativos de que o
manejo com TE pode ser uma alternativa eficaz para alcançar índices
reprodutivos satisfatórios em fêmeas durante os períodos de estresse
térmico e em fêmeas repetidoras de serviço (Rodrigues et al., 2007a).
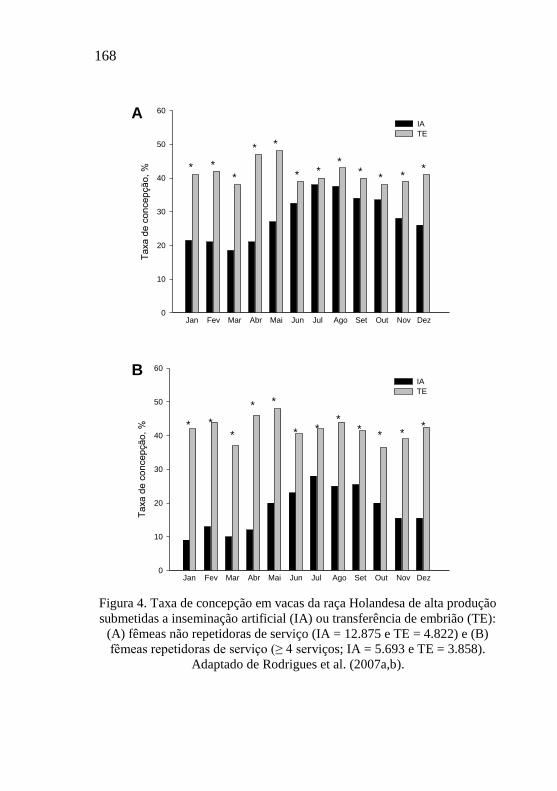
168
Taxa d
e c
oncepção, %
0
10
20
30
40
50
60
IA
TE
* **
* *
* **
** *
*
Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
A
Taxa d
e c
on
cep
ção
, %
0
10
20
30
40
50
60
IA
TE
* **
* *
* **
** *
*
Jan Fev Mar Abr Mai Jun Jul Ago Set Out Nov Dez
B
Figura 4. Taxa de concepção em vacas da raça Holandesa de alta produção
submetidas a inseminação artificial (IA) ou transferência de embrião (TE):
(A) fêmeas não repetidoras de serviço (IA = 12.875 e TE = 4.822) e (B)
fêmeas repetidoras de serviço (≥ 4 serviços; IA = 5.693 e TE = 3.858).
Adaptado de Rodrigues et al. (2007a,b).

169
Assim como para os programas de IA, a utilização da TE em
larga escala constatou o estabelecimento de protocolos para
transferência de embriões em tempo fixo (TETF), que eliminou a
necessidade de detecção de estro e aumentou a eficiência reprodutiva
das receptoras de embrião (Bó et al., 2002, Baruselli et al., 2010,
Baruselli et al., 2011). Estudos realizados pelo nosso grupo de
pesquisa nas condições brasileiras de manejo (Rodrigues et al. 2010)
demonstraram que a utilização da TETF resultou em maior taxa de
aproveitamento (P< 0,0001) e superior taxa de prenhez (P= 0,001)
comparado aos animais submetidos ao tratamento com prostaglandina
(i.e., apenas receptoras com CL) e observação de estro seguida de TE
(Figura 5). Diferentemente da TE após a observação do estro, o
emprego da TETF aumentou o número de receptoras (vacas da raça
Holandesa em lactação) aptas à transferência de embrião. No grupo de
animais que receberam TETF, receptoras sem CL apresentaram a
mesma taxa de prenhez a TETF que receptoras com CL.
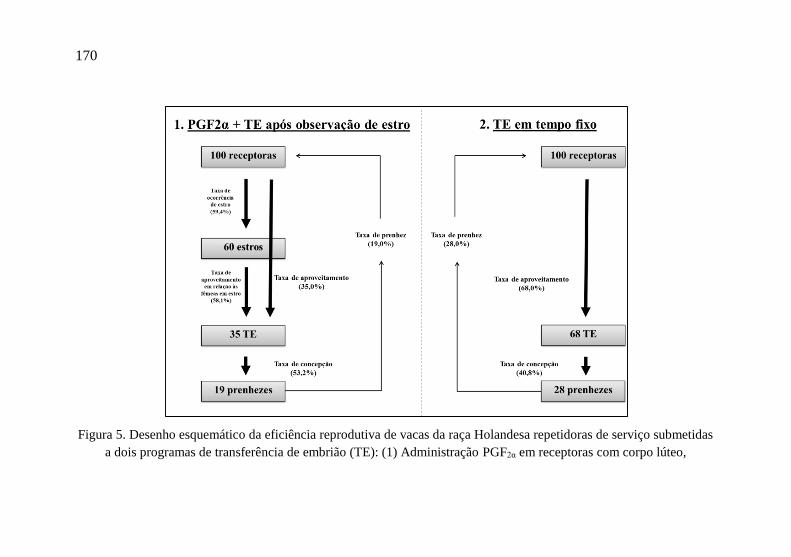
170
Figura 5. Desenho esquemático da eficiência reprodutiva de vacas da raça Holandesa repetidoras de serviço submetidas
a dois programas de transferência de embrião (TE): (1) Administração PGF2α em receptoras com corpo lúteo,

171
(cont.) observação visual de estro 1 a 5 dias após tratamentos e TE 6 a 8 dias após observação de estro e (2) TE em
tempo fixo (dia 0: TETF; implante auricular de norgestomet + 2 mg de benzoato de estradiol; dia 8: remoção do
implante + 400 UI de eCG + 1,0 mg de cipionato de estradiol + PGF2α; dia 17: transferência de embrião em tempo fixo).
Taxa de ocorrência de estro: número de animais observados em estro em relação ao número de animais tratados com
PGF2α; Taxa de aproveitamento em relação às fêmeas em estro: número de animais aptos a receber um embrião em
relação ao número de animais observados em estro; Taxa de aproveitamento: número de animais aptos a receber um
embrião em relação ao número de animais tratados (PGF2α ou protocolos de TETF); Taxa de concepção: número de
animais gestantes em relação ao número de animais que receberam a TE; Taxa de prenhez: número de animais gestantes
em relação ao número de animais tratados. Adaptado de Rodrigues et al. (2010).

172
Vale ressaltar que a obtenção de resultados satisfatórios em
manejos que envolvem a TE é dependente de diversos fatores, dentre
eles, o congelamento do embrião. Em um estudo retrospectivo com
vacas da raça Holandesa criadas em regiões tropicais, nosso grupo de
pesquisa avaliou a influência do congelamento do embrião produzido
in vivo no estabelecimento da gestação (Rodrigues et al., 2007c).
Nessa análise, verificou-se taxas superiores em receptoras que
receberam um embrião fresco (43,9%, n = 2.634) comparado com as
que receberam um embrião previamente congelado (39,5%, n =
2.237). Resultados semelhantes (27,2% para embriões fresco vs.
24,1% para embriões congelados) foram obtidos recentemente por
Vieira et al. (2014a), sugerindo que o uso de embriões produzidos in
vivo frescos aumentam os índices reprodutivos de receptoras da raça
Holandesa em programas de TE.
7.4.1 Transferência de embriões produzidos in vitro.
Dentre as biotecnologias da reprodução existentes, a produção
in vitro de embriões (PIVE) pode ser considerada uma importante
ferramenta para promover aumento no ganho genético nos rebanhos
leiteiros. Entretanto, diferentemente dos resultados obtidos em fêmeas
Bos indicus (Pontes et al., 2010), a PIVE em vacas leiteiras de alta
produção (Bos taurus) ainda necessita de aprimoramentos. Além da
variação individual existente na resposta entre as doadoras aos
programas (Merton et al., 2003, Pontes et al., 2009), têm sido relatadas
diferenças nas taxas de blastocistos produzidos entre as categorias de

173
fêmeas existentes nas propriedades (Ferreira et al., 2011, Guerreiro et
al., 2014b).
A menor resposta de vacas Bos taurus nos programas de OPU-
PIVE (ovum pick-up 8 -produção in vitro de embriões) pode ser
relacionada à menor população ovariana de folículos antrais quando
comparado a Bos indicus (Batista et al., 2014) além de diversos
aspectos adicionais relacionados principalmente a menor competência
oocitária (Ambrose et al., 1999, Rodrigues et al., 2004b, Block et al.,
2010, Baruselli et al., 2012a). Neste sentido, a tentativa de melhorar
os resultados na PIVE em vacas leiteiras, a utilização de protocolos
superestimulatórios em doadoas de ovócitos adultas utilizando FSH
previamente à OPU-PIVE pode ser considerada como uma alternativa
(Goodhand et al., 1999, Sendag et al., 2008). Vieira et al. (2014b)
reportaram aumento na PIVE em vacas lactantes (VL) e não lactantes
(VNL) da raça Holandesa após tratamento de superestimulação (VL:
controle 1,0 ± 0,4; FSH 1,5 ± 0,5; VNL: controle 2,7 ± 0,6; FSH 4,4 ±
0,8; P= 0,01).
A fase do desenvolvimento folicular em que a punção
folicular (OPU) é realizada tem sido associada ao sucesso na PIVE
(Pavlok et al., 1992, Blondin e Sirard, 1995, Fair et al., 1995,
Hagemann et al., 1999, Hendriksen et al., 2004). A aquisição do
potencial de desenvolvimento oocitário tem sido associada com o
8 OPU: Ovum Pick-Up pode ser traduzido como aspiração folicular
ovariana.

174
crescimento folicular, ou seja, o potencial de desenvolvimento eleva-
se à medida que o diâmetro folicular aumenta e aproxima-se do pico
de LH (Lonergan et al., 1994, Arlotto et al., 1996, Sirard et al., 2006,
Caixeta et al., 2009, Sirard, 2011, Sirard, 2012). Mourot et al. (2006)
relataram as diferenças nos perfis de expressão gênica de ovócitos
oriundos de folículos com diferentes tamanhos, por exemplo, os níveis
de mRNA mais elevados para PSMB2, SKIIP, CDC5L, RGS16, e
PRDX1 em ovócitos de folículos mais do que 8 mm. Além disso, Chu
et al. (2012) demonstraram aumento da expressão de PTTG1, BTG4,
Papola e genes LEO1 em ovócitos de doadoras superovuladas quando
comparado a doadoras não tratadas. Considerando-se que esses genes
estão relacionados com a transcrição e a regulação do ciclo celular, os
autores sugerem que a estimulação com FSH exógeno poderia permitir
a acumulação de mais mRNA, o que resultaria no aumento da
qualidade dos ovócitos. Portanto, o uso de protocolos de sincronização
da onda folicular e superestimulação antes da OPU é uma estratégia
importante para melhorar a eficiência dos programas de OPU-PIVE
em bovinos de leite.
No entanto, além da variação individual existente na resposta
entre as doadoras aos programas de PIVE (Merton et al., 2003, Pontes
et al., 2009), tem sido relatadas diferenças nas taxas de blastocistos
entre as categorias de fêmeas existentes nas propriedades (Ferreira et
al., 2011, Guerreiro et al., 2014b{Ferreira, 2011 #886). Dessa forma,
na tentativa de verificar o impacto da categoria animal na eficiência
dos programas de OPU-PIVE, Guerreiro et al. (2014b) relataram que

175
quando se utiliza novilhas pré-púberes, da raça Holandesa, de 8-10
meses de idade, obtem-se menor número de embriões por sessão de
OPU-PIVE (novilhas pré-púberes: 0,5 ± 0,2a; novilhas púberes: 1,0 ±
0,5b; vacas lactantes: 1,1 ± 0,6b; vacas não lactantes: 3,9 ± 0,6c;
P<0,0001). Porém, novilhas púberes apresentaram produção de
embriões semelhante àquela obtida em fêmeas em lactação durante a
época não quente do ano. Essa baixa taxa de produção de embriões
observada em fêmeas pré-púberes pode ser decorrentes do número de
genes diferencialmente expressos entre doadoras jovens e adultas
(Dorji et al., 2012). Ainda, a baixa competência oocitária observada
em fêmeas pré-púberes pode ser associada ao grande número de
células apoptóticas presentes nos blastocistos produzidos por essa
categoria (Zaraza et al., 2010). Dessa forma, novilhas pré-púberes da
raça Holandesa ainda apresentam menor eficiência quando submetidas
à programas de OPU-PIVE, portanto, novos estudos devem ser
desenvolvidos na tentativa de melhorar tais resultados.
Ainda, a necessidade de aprimorar a técnica para aplicação em
animais jovens tem sido cada vez mais requerida devido ao emprego
da avaliação genômica, ferramenta capaz de identificar,
precocemente, indivíduos com maior potencial genético. A avaliação
genômica aumenta a confiabilidade da seleção genética tradicional
(provas zootécnicas) e diferenças esperadas na progênie,
principalmente quando estas são aplicadas em animais jovens
(Coutinho et al., 2010). Portanto, a intensificação da PIVE em fêmeas
jovens e geneticamente superiroes viabiliza a obtenção de

176
descendentes antes das mesmas atingirem a puberdade, aumentando o
potencial reprodutivo da fêmea, bem como reduzindo o intervalo de
gerações de maneira significativa.
Fêmeas pré-puberes apresentam elevada população folicular
(Erickson, 1966, Revel et al., 1995) o que determina elevado interesse
na utilização desta categoria animal em programas de OPU-PIVE.
Entretanto, estudos relataram que ovócitos de animais jovens possuem
menor eficiência para PIVE quando comparado a animais adultos
(Khatir et al., 1996, Presicce et al., 1997, Majerus et al., 1999, Palma
et al., 2001). Alternativamente, existem relatos de aumento na
eficiência da produção de embriões quando se utilizou oócitos de
bezerras previamente superestimuladas com gonadotrofinas exógenas
(Armstrong et al., 1994, Armstrong et al., 1997). No Brasil, estudos
recentes avaliaram a PIVE em bezerras da raça Holandesa submetidas
a protocolo superestimulatório utilizando progesterona durante os sete
dias que entecederam a OPU e 140 mg de FSH administrado em quatro
doses (40, 40, 30 e 30 mg), sendo a primeira, 48h antes do processo
de OPU (Guerreiro et al, dados ainda não publicados). Um total de 32
fêmeas foi distribuído em quatro grupos experimentais: novilhas
púberes sem tratamento prévio (NOV, controle positivo; n=8);
bezerras de 2-4 meses de idade sem tratamento prévio (BEZ, controle
negativo; n=8); bezerras de 2-4 meses de idade com progesterona
(BEZ-P4, n=8) e bezerras de 2-4 meses de idade com progesterona e
140mg de FSH (BEZ-P4FSH, n=8). Verificou-se que bezerras
superesestimuladas com FSH obtivieram maior número de folículos

177
aspirados (BEZ-P4FSH: 58.6a vs. NOV: 22.3b vs. BEZ: 23.1ab vs.
BEZ-P4: 25.6ab; P=0.03), oócitos totais (BEZ-P4FSH: 29.6a vs.
NOV: 9.4b vs. BEZ: 11.9b vs. BEZ-P4: 11.4b; P=0.006) e oócitos
viáveis (BEZ-P4FSH: 17.0a vs. NOV: 5.0b vs. BEZ: 2.6b vs. BEZ-
P4: 4.1b; P=0.004) por sessão de aspiração quando comparado aos
demais grupos. Porém, estudos adicionais são necessários para
melhorar a compreensão e os resultados obtidos nos programas de
OPU-PIVE nesta categoria animal.
O número de embriões produzidos por sessão de aspiração é
fundamental para viabilidade econômica dos programas reprodutivos
dentro dos rebanhos leiteiros. Estudos sugerem que tanto fêmeas Bos
taurus quanto Bos indicus que possuem elevada população de
folículos antrais (PFA) presentes nos ovários produzem mais embriões
por sessão de aspiração quando comparadas àquelas com baixa
população (Pontes et al., 2010, Guerreiro et al., 2014a). Entretanto,
estudos retrospectivos mostraram que apesar da quantidade de oócitos
recuperados estar diretamente relacionda com o número de embriões
produzidos por sessão de OPU em doadoras da raça Holandesa, essa
relação não foi confirmada quando avaliou-se o estabelecimento
gestacional dos embriões conforme número de oócitos recuperados
(Bragança et al., 2014). A taxa de concepção foi semelhante (P=0,66)
após transferência de embriões provenientes de doadoras com alto
(43,5%), médio (46,2%) e baixo (38,3%) número de oócitos
recuperados após a OPU. Dessa forma, estudos adicionais necessitam
ser desenvolvimentos para esclarecer e confirmar tal associação.

178
O real sentido prático do uso de PIVE está estreitamente
relacionado com a geração de descendentes em quantidades e custos
satisfatórios. Ainda existem poucos estudos que relataram a prenhez
por TE (P/TE) em seus resultados. Estudos recentes compararam a
eficiência da taxa de concepção de embriões produzidos in vivo com
embriões produzidos in vitro em receptoras da raça Holandesa em
lactação (Rodrigues et al., 2014). Verificou-se menor taxa de
concepção para os embriões produzidos in vitro tanto aos 30 (43,8%,
273/624 vs. 26,5%, 82/309; P <0,0001), como aos 60 dias de gestação
(34,4%, 213/620 vs. 20,4%, 63/309; P <0,0001). Entretanto, a perda
gestacional entre 30 dias e 60 dias foi semelhante entre os grupos (in
vivo: 20,8%, 56/269 vs. in vitro: 23,2%, 19/82; P = 0,79). Diante do
potencial da técnica, vários estudos estão sendo realizados em todo o
mundo com o objetivo de melhorar a quantidade e qualidade dos
embriões produzidos in vitro para aumentar a eficiência reprodutiva
dos programas que utilizam essa biotecnologia.
7.5. Conclusão
O emprego das técnicas de reprodução (IA e TE) para
multiplicar indivíduos superiores resulta em significativo avanço
genético dos rebanhos. No entanto, a aplicação da IA e TE nas
propriedades rurais ainda apresenta limitações, principalmente devido
às dificuldades de execução do manejo e da disponibilidade de mão de
obra qualificada. Dessa forma, o desenvolvimento e aprimoramento
das técnicas de sincronização para IATF e programas de

179
ressincronização e TE são ferramentas eficientes para alcançar elevada
eficiência reprodutiva de forma organizada e com alta precisão.
Portanto, uma vez estabelecido o objetivo da atividade rural, esses
programas são incorporados ao sistema de produção e de gestão,
aumentando a produtividade e a rentabilidade da propriedade.
REFERÊNCIAS BIBLIOGRÁFICAS
AL-KATANANI, Y. M.; HANSEN, P.J. Induced thermotolerance in bovine
two-cell embryos and the role of heat shock protein 70 in embryonic
development. Molecular Reproduction and Development, v. 62, p. 174-
180, 2002.
AMBROSE, J. D., DROST M., MONSON R. L., RUTLEDGE J. J.,
LEIBFRIED-RUTLEDGE M. L., THATCHER M. J., KASSA T., BINELLI
M., HANSEN P. J., CHENOWETH P. J., and THATCHER W. W.. 1999.
Efficacy of Timed Embryo Transfer with Fresh and Frozen In Vitro
Produced Embryos to Increase Pregnancy Rates in Heat-Stressed Dairy
Cattle1. J. Dairy Sci. 82(11):2369-2376.
ARLOTTO, T., SCHWARTZ J.-L., FIRST N. L., and LEIBFRIED-
RUTLEDGE M. L.. 1996. Aspects of follicle and oocyte stage that affect in
vitro maturation and development of bovine oocytes. Theriogenology
45:943-956.
ARMSTRONG, D. T., B. J. IRVINE, C. R. EARL, D. MCLEAN, and R. F.
SEAMARK. 1994. Gonadotropin stimulation regimens for follicular
aspiration and in vitro embryo production from calf oocytes.
Theriogenology 42(7):1227-1236.
ARMSTRONG, D. T., KOTARAS P. J., and EARL C. R.. 1997. Advances
in production of embryos <i>in vitro</i> from juvenile and prepubertal
oocytes from the calf and lamb. Reproduction, Fertility and Development
9(3):333-340.

180
BARUSELLI, P., VIEIRA L. M., GUERREIRO B. M., SOARES J.,
OLIVEIRA T., and FREITAS B.. 2014. Relationship between antral follicle
population and pregnancy results following fixed-time AI in Nelore (bos
indicus) cows. Anim. Reprod. Sci. 11(3):390.
BARUSELLI, P. S., FERREIRA R. M., FILHO M. F. S., NASSER L. F. T.,
RODRIGUES C. A., and BÓ G. A.. 2010. Bovine embryo transfer recipient
synchronisation and management in tropical environments. Reproduction,
Fertility and Development 22(1):67-74.
BARUSELLI, P. S., FERREIRA R. M., SALES J. N. S., GIMENES L. U.,
SÁ FILHO M. F., MARTINS C. M., RODRIGUES C. A., and BÓ G. A..
2011. Timed embryo transfer programs for management of donor and
recipient cattle. Theriogenology 76(9):1583-1593.
BARUSELLI, P. S., SÁ FILHO M. F., FERREIRA R. M., SALES J. N. S.,
GIMENES L. U., VIEIRA L. M., MENDANHA M. F., and BÓ G. A..
2012a. Manipulation of Follicle Development to Ensure Optimal Oocyte
Quality and Conception Rates in Cattle. Reprod. Domest. Anim. 47:134-
141.
BARUSELLI, P. S., SALES J. N. S., SALA R. V., VIEIRA L. M., and SÁ
FILHO M. F.. 2012b. History, evolution and perspectives of timed artificial
insemination programs in Brazil. Animal Reproduction 9:139-152.
BISINOTTO, R. S., GRECO L. F., RIBEIRO E. S., MARTINEZ N., LIMA
F. S., STAPLES C. R., THATCHER W. W., and SANTOS J. E. P.. 2013.
Influences of nutrition and metabolism on fertility of dairy cows. Animal
Reproduction 9(3):260-272.
BLOCK, J., BONILLA L., and HANSEN P. J.. 2010. Efficacy of in vitro
embryo transfer in lactating dairy cows using fresh or vitrified embryos
produced in a novel embryo culture medium1. J. Dairy Sci. 93(11):5234-
5242.
BLONDIN, P. and SIRARD M. A.. 1995. Oocyte and follicular
morphology as determining characteristics for developmental competence
in bovine oocytes. Molecular Reproduction and Development (41):54-62.
BÓ, G. A., BARUSELLI P. S., MORENO D., CUTAIA L., CACCIA M.,
TRÍBULO R.,. TRÍBULO H, and MAPLETOFT R. J.. 2002. The control of

181
follicular wave development for self-appointed embryo transfer programs in
cattle. Theriogenology 57(1):53-72.
BRAGANÇA, L., MARQUES K., VIEIRA L., SILVEIRA C., PICADO L.,
MONTEIRO B., SÁ FILHO M., and B. PS. 2014. The number of oocytes
recovered from Holstein donors has no effect on the pregnancy
establishment of in vitro embryos programs. Anim. Reprod. Sci. 11(3):433.
CAIXETA, E. S., RIPAMONTE P., FRANCO M. M., JUNIOR J. B., and
DODE M. A. N.. 2009. Effect of follicle size on mRNA expression in
cumulus cells and oocytes of Bos indicus: an approach to identify marker
genes for developmental competence. Reproduction, Fertility and
Development 21(5):655-664.
CAVALIERI, J., HEPWORTH G., SMART V. M., RYAN M., and
MACMILLAN K. L.. 2007. Reproductive performance of lactating dairy
cows and heifers resynchronized for a second insemination with an
intravaginal progesterone-releasing device for 7 or 8d with estradiol
benzoate injected at the time of device insertion and 24h after removal.
Theriogenology 67(4):824-834.
CERRI, R. L. A., SANTOS J. E. P., JUCHEM S. O., GALVÃO K. N.,
andCHEBEL R. C.. 2004. Timed artificial insemination with estradiol
cypionate or insemination at estrus in high-producing dairy cows. J. Dairy
Sci. 87(11):3704-3715.
CHU, T., DUFORT I., and SIRARD M. A.. 2012. Effect of ovarian
stimulation on oocyte gene expression in cattle. Theriogenology
77(9):1928-1938.
COUTINHO, L. L., ROSÁRIO M. F., and JORGE E. C.. 2010.
Biotecnologia animal. Estudos Avançados 24:123-147.
DORJI, Y. OHKUBO, K. MIYOSHI, and YOSHIDA M.. 2012. Gene
expression profile differences in embryos derived from prepubertal and
adult Japanese Black cattle during in vitro development. Reprod Fertil Dev
24(2):370-381.
DROST, M., AMBROSE J. D., THATCHER M. J., CANTRELL C. K.,
WOLFSDORF K. E., HASLER J. F., and THATCHER W. W.. 1999.

182
Conception rates after artificial insemination or embryo transfer in lactating
dairy cows during summer in florida. Theriogenology 52(7):1161-1167.
EALY, A., DROST M., and HANSEN P.. 1993. Developmental changes in
embryonic resistance to adverse effects of maternal heat stress in cows. J
Dairy Sci 76:2899 - 2905.
ERICKSON, B. H. 1966. Development and Senescence of the Postnatal
Bovine Ovary. J. Anim. Sci. 25(3):800-805.
FAIR, T., HYTTEL P., and GREVE T.. 1995. Bovine oocyte diameter in
relation to maturational competence and transcriptional activity. Mol
Reprod Dev 42(4):437-442.
FERREIRA, R. M., AYRES H., CHIARATTI M. R., FERRAZ M. L.,
ARAÚJO A. B., RODRIGUES C. A., WATANABE Y. F., VIREQUE A.
A., JOAQUIM D. C., SMITH L. C., MEIRELLES F. V., and BARUSELLI
P. S.. 2011. The low fertility of repeat-breeder cows during summer heat
stress is related to a low oocyte competence to develop into blastocysts. J.
Dairy Sci. 94(5):2383-2392.
FREITAS, B. G., RODRIGUES C. A., SALES J. N. S., TEIXEIRA A. A.,
FERREIRA R. M., AYRES H., RANIERI A. L., and BARUSELLI P. S..
2010. Perda embrionária (entre 30 e 60 dias) à inseminação artificial e à
transferência de embrião de vacas holandesas de alta produção. in Proc.
XXIV Reunião Anual da Sociedade Brasileira de Tecnologia de
Embriões (SBTE), Porto de Galinhas.
GALVÃO, K. N., FEDERICO P., DE VRIES A., and SCHUENEMANN
G. M.. 2013. Economic comparison of reproductive programs for dairy
herds using estrus detection, timed artificial insemination, or a combination.
Journal of Dairy Science 96(4):2681-2693.
GIORDANO, J. O., FRICKE P. M., WILTBANK M. C., and CABRERA
V. E.. 2011. An economic decision-making support system for selection of
reproductive management programs on dairy farms. Journal of Dairy
Science 94(12):6216-6232.
GIORDANO, J. O., KALANTARI A. S., FRICKE P. M., WILTBANK M.
C., and CABRERA V. E.. 2012. A daily herd Markov-chain model to study
the reproductive and economic impact of reproductive programs combining

183
timed artificial insemination and estrus detection. Journal of Dairy Science
95(9):5442-5460.
GOODHAND, K. L., WATT R. G., STAINES M. E., HUTCHINSON J. S.
M., and BROADBENT P. J.. 1999. In vivo oocyte recovery and in vitro
embryo production from bovine donors aspirated at different frequencies or
following FSH treatment. Theriogenology 51(5):951-961.
GUERREIRO, B. M., BATISTA E. O. S., VIEIRA L. M., SÁ FILHO M.
F., RODRIGUES C. A., CASTRO NETTO A., SILVEIRA C. R. A.,
BAYEUX B. M., DIAS E. A. R., MONTEIRO F. M., ACCORSI M.,
LOPES R. N. V. R., and BARUSELLI P. S.. 2014a. Plasma anti-mullerian
hormone: an endocrine marker for in vitro embryo production from Bos
taurus and Bos indicus donors. Domest. Anim. Endocrinol. 49(0):96-104.
GUERREIRO, B. M., RODRIGUES C. A., CASTRO NETTO A.,
SILVEIRA C. R. A., VIEIRA L. M., OLIVEIRA R. C., FREITAS B. G.,
and BARUSELLI P. S.. 2014b. Prepubertal Holstein have a low efficiency
when submitted to ovum pick-up and in vitro embryo production. Anim.
Reprod. Sci. 11(3):405.
HAGEMANN, L. J., BEAUMONT S. E., BERG M., DONNISON M. J.,
LEDGARD A., PETERSON A. J., SCHURMANN A., andTERVIT H. R..
1999. Development during single IVP of bovine oocytes from dissected
follicles: interactive effects of estrous cycle stage, follicle size and atresia.
Molecular Reproduction and Development 53(4):451-458.
HANSEN, P. J. 2009. Effects of heat stress on mammalian reproduction.
Philosophical Transaction of the Royal Society of London B Biology
Society 364:3341-3350.
HANSEN, P. J. and ARÉCHIGA C. F.. 1999. Strategies for managing
reproduction in the heat-stressed dairy cow. Journal of Animal Science
77(2):36-50.
HANSEN, P. J., DROST M., RIVERA R. M., PAULA-LOPES F. F., AI-
KATANANIT Y. M., KRININGER III C. E., andCHASE JR C. C.. 2001.
Adverse impact of heat stress on embryo production: causes and strategies
for mitigation. Theriogenology 55(1):91-103.

184
HENDRIKSEN, P. J., STEENWEG W. N., HARKEMA J. C., MERTON J.
S., BEVERS M. M., VOS P. L., and DIELEMAN S. J.. 2004. Effect of
different stages of the follicular wave on in vitro developmental competence
of bovine oocytes. Theriogenology 61(5):909-920.
KHATIR, H., LONERGAN P., and CAROLAN C.. 1996. Prepubertal
bovine oocyte: a negative model for studying oocyte developmental
competence. . Molecular Reproduction and Development 45:231-239.
LONERGAN, P., MONAGHAN P., RIZOS D., BOLAND M. P., and
GORDON I.. 1994. Effect of follicle size on bovine oocyte quality and
developmental competence following maturation, fertilization, and culture
in vitro. Mol Reprod Dev 37(1):48-53.
LOPEZ, H., SATTER L. D., and WILTBANK M. C.. 2004. Relationship
between level of milk production and estrous behavior of lactating dairy
cows. Anim. Reprod. Sci. 81(3-4):209-223.
MAJERUS, V., DE ROOVER R., ETIENNE D., KAIDI S., MASSIP A.,
DESSY F., and DONNAY I.. 1999. Embryo production by ovum pick up in
unstimulated calves before and after puberty. Theriogenology 52(7):1169-
1179.
MERTON, J. S., DE ROOS A. P., MULLAART E., de RUIGH L., KAAL
L., VOS P. L., and DIELEMAN S. J.. 2003. Factors affecting oocyte quality
and quantity in commercial application of embryo technologies in the cattle
breeding industry. Theriogenology 59(2):651-674.
MOUROT, M., DUFORT I., GRAVEL C., ALGRIANY O., DIELEMAN
S., and SIRARD M.-A.. 2006. The influence of follicle size, FSH-enriched
maturation medium, and early cleavage on bovine oocyte maternal mRNA
levels. Molecular Reproduction and Development 73(11):1367-1379.
NORMAN, H. D., WRIGHT J. R., HUBBARD S. M., MILLER R. H., and
HUTCHISON J. L.. 2009. Reproductive status of Holstein and Jersey cows
in the United States. J. Dairy Sci. 92(7):3517-3528.
PALMA, G. A., TORTONESE D. J., and SINOWATZ F. 2001.
Developmental Capacity In Vitro of Prepubertal Oocytes. Anatomia,
Histologia, Embryologia 30(5):295-300.

185
PAVLOK, A., LUCAS-HAHN A., and NIEMANN H.. 1992. Fertilization
and developmental competence of bovine oocytes derived from different
categories of antral follicles. Molecular Reproduction and Development
31:63-67.
PONTES, J. H. F., NONATO-JUNIOR I., SANCHES B. V., ERENO-
JUNIOR J. C., UVO S., BARREIROS T. R. R., OLIVEIRA J. A.,
HASLER J. F., and SENEDA M. M.. 2009. Comparison of embryo yield
and pregnancy rate between in vivo and in vitro methods in the same Nelore
(Bos indicus) donor cows. Theriogenology 71(4):690-697.
PONTES, J. H. F., SILVA K. C. F., BASSO A. C., RIGO A. G.,
FERREIRA C. R., SANTOS G. M. G., SANCHES B. V., PORCIONATO
J. P. F., VIEIRA P. H. S., FAIFER F. S., STERZA F. A. M., SCHENK J.
L., and SENEDA M. M.. 2010. Large-scale in vitro embryo production and
pregnancy rates from Bos taurus, Bos indicus, and indicus-taurus dairy
cows using sexed sperm. Theriogenology 74(8):1349-1355.
PRESICCE, G. A., JIANG S., SIMKIN M., ZHANG L., LOONEY C. R.,
GODKE R. A., and YANG X.. 1997. Age and hormonal dependence of
acquisition of oocyte competence for embryogenesis in prepubertal calves.
Biology of Reproduction 56(2):386-392.
PUTNEY, D. G., DROST' M., and THATCHER W. W.. 1989. Influence of
summer heat stress on pregnancy rates of lactating dairy cattle following
embryo transfer or artificial insemination. Theriogenology 31(4):765–778.
RENSIS, F. D. and SCARAMUZZI R. J.. 2003. Heat stress and seasonal
effects on reproduction in the dairy cow—a review. Theriogenology
60(6):1139-1151.
REVEL, F., MERMILLOD P., PEYNOT N., RENARD J. P., and
HEYMAN Y.. 1995. Low developmental capacity of in vitro matured and
fertilized oocytes from calves compared with that of cows. Journal of
Reproduction and Fertility 103(1):115-120.
RIBEIRO, E. S., BISINOTTO R. S., FAVORETO M. G., MARTINS L. T.,
CERRI R. L. A., SILVESTRE F. T., GRECO L. F., THATCHER W. W.,
and SANTOS J. E. P.. 2012. Fertility in dairy cows following
presynchronization and administering twice the luteolytic dose of

186
prostaglandin F2α as one or two injections in the 5-day timed artificial
insemination protocol. Theriogenology 78(2):273-284.
RODRIGUES, C. A., AYRES H., FERREIRA R. M., TEIXEIRA A. A.,
MANCILHA R. F., OLIVEIRA M. E. F., SOUZA A. H., and BARUSELLI
P. S. 2007a. Comparison of pregnancy rates after artificial insemination or
embryo transfer in high producing repeat breeder holstein cows. Acta
Scientiae Veterinariae 35(3):1255.
RODRIGUES, C. A., AYRES H., FERREIRA R. M., TEIXEIRA A. A.,
MANCILHA R. F., OLIVEIRA M. E. F., SOUZA A. H., and BARUSELLI
P. S. 2007b. Conception rate after artificial insemination and embryo
transfer in high Producing holstein cows. Acta Scientiae Veterinariae
35(3):1254.
RODRIGUES, C. A., AYRES H., FERREIRA R. M., TEIXEIRA A. A.,
MANCILHA R. F., OLIVEIRA M. E. F., SOUZA A. H., and BARUSELLI
P. S. 2007c. Efeito da sincronia entre a doadora e a receptora na taxa de
concepção após transferêcnia de embriões frescos e criopreservados em
vacas holandesas de alta produção. Acta Scientiae Veterinariae
35(3):1257.
RODRIGUES, C. A., AYRES H., REIS E. L., NICHI M., BO G. A., and
BARUSELLI P. S.. 2004a. Artificial insemination and embryo transfer
pregnancy rates In high production holstein (repeat breeder) breedings.
Acta Scientiae Veterinariae 32:219.
RODRIGUES, C. A., AYRES H., REIS E. L., NICHI M., BÓ G. A., and
BARUSELLI P. S.. 2004b. Taxa de concepção de embriões frescos e
criopreservados transferidos em vacas holandesas de alta produção. Acta
Scientiae Veterinariae 32:159.
RODRIGUES, C. A., SILVA P. R. L., CASTRO A., SILVEIRA C. R. A.,
VIEIRA L. M., GIMENES L. U., and BARUSELLI P. S.. 2014. Pregnancy
establishment in Holstein cows after transferring in vivo or in vitro
produced embryo. Animal Reproduction.
RODRIGUES, C. A., TEIXEIRA A. A., FERREIRA R. M., AYRES H.,
MANCILHA R. F., SOUZA A. H., and BARUSELLI P. S.. 2010. Effect of
fixed-time embryo transfer on reproductive efficiency in high-producing

187
repeat-breeder Holstein cows. Animal Reproduction Science 118(2-
4):110-117.
SARTORI, R., SARTOR-BERGFELT R., MERTENS S. A., GUENTHER
J. N., PARRISH J. J., and WILTBANK M. C.. 2002. Fertilization and early
embryonic development in heifers and lactating cows in summer and
lactating and dry cows in winter. Journal of Dairy Science 85:2803-2812.
SENDAG, S., CETIN Y., ALAN M., HADELER K.-G., and NIEMANN
H.. 2008. Effects of eCG and FSH on ovarian response, recovery rate and
number and quality of oocytes obtained by ovum pick-up in Holstein cows.
Anim. Reprod. Sci. 106(1):208-214.
SIRARD, M.-A. 2011. Follicle environment and quality of in vitro matured
oocytes. Journal of Assisted Reproduction and Genetics 28(6):483-488.
SIRARD, M.-A., RICHARD F., BLONDIN P., and ROBERT C.. 2006.
Contribution of the oocyte to embryo quality. Theriogenology 65(1):126-
136.
SIRARD, M. A. 2012. Factors Affecting Oocyte and Embryo
Transcriptomes. Reprod. Domest. Anim. 47:148-155.
TEIXEIRA, A. A. 2010. Impacto da inseminação artificial em tempo
fixo na eficiência reprodutiva de vacas de leite de alta produção. Page
69 in Reprodução Animal. Vol. Mestrado. Faculdade de Medicina
Veterinária e Zootecnia, Universidade de São Paulo.
THOMPSON, I. M., CERRI R. L., KIM I. H., GREEN J. A., SANTOS J.
E., and THATCHER W. W.. 2010. Effects of resynchronization programs
on pregnancy per artificial insemination, progesterone, and pregnancy-
associated glycoproteins in plasma of lactating dairy cows. Journal of
Dairy Science 93(9):4006-4018.
VIANA, J. H. M. 2012. Levantamento estatístico da produção de embriões
bovinos no Brasil em 2011: mudanças e tendências futuras. Page 32 in O
embrião. 51 ed.
VIEIRA, L. M., RODRIGUES C. A., MENDANHA M. F., SÁ FILHO M.
F., SALES J. N. S., SOUZA A. H., SANTOS J. E. P., and BARUSELLI P.
S.. 2014a. Donor category and seasonal climate associated with embryo

188
production and survival in multiple ovulation and embryo transfer programs
in Holstein cattle. Theriogenology 82(2):204-212.
VIEIRA, L. M., SÁ FILHO M. F., PUGLIESI G., GUERREIRO B. M.,
CRISTALDO M. A., BATISTA E. O. S., FREITAS B. G., CARVALHO F.
J., GUIMARÃES L. H. C., and BARUSELLI P. S.. 2014b.
Resynchronization in dairy cows 13 days after tai followed by pregnancy
diagnosis based on corpus luteum vascularization by color doppler. Animal
Reproduction (accepted).
WILTBANK, M., LOPEZ H., SARTORI R., SANGSRITAVONG S., and
GÜMEN A.. 2006. Changes in reproductive physiology of lactating dairy
cows due to elevated steroid metabolism. Theriogenology 65(1):17-29.
ZARAZA, J., OROPEZA A., VELAZQUEZ M. A., KORSAWE K.,
HERRMANN D., CARNWATH J. W., and NIEMANN H.. 2010.
Developmental competence and mRNA expression of preimplantation in
vitro–produced embryos from prepubertal and postpubertal cattle and their
relationship with apoptosis after intraovarian administration of IGF-1.
Theriogenology 74(1):75-89.

189
8. O QUE DEVO SABER PARA DEFINIR O PROTOCOLO DE
IATF A SER UTILIZADO?9
Marcos H.C. Pereira
José Luiz .M. Vasconcelos
8.1 Introdução
Protocolos para sincronização do ciclo estral têm sido
utilizados no manejo reprodutivo de vacas em lactação e envolvem a
administração sequencial de hormônios para sincronizar o momento
da ovulação e permitir a ovulação de um ovócito fértil e inseminação
(IA) em momento pré-determinado. As taxas de ovulações
sincronizadas são superiores a 80% [1] e o maior desafio é aumentar
a prenhez e a manutenção da gestação. A expressão de estro em
protocolos de sincronização da ovulação pode ser um bom preditor da
fertilidade por estar associado à maior sincronização [1], melhor
ambiente hormonal pré e pós-IA [1], menor incidência de ciclos curtos
[2] e por aumentar a manutenção da gestação [1, 3, 4].
Vacas em lactação têm alto metabolismo dos hormônios
esteroides [5] com menor concentração de estrógeno (E2) próximo ao
9 Pereira, M.H.C.; Vasconcelos, J.L.M. O que devo saber para definir o
protocolo de IATF a ser utilizado? Anais. I Simpósio Nacional da Vaca
Leiteira. Porto Alegre: Universidade Federal do Rio Grande do Sul. 2014.
239 p.

190
estro ou ovulação [5-7]. Concentração sérica de E2 tem correlação
positiva (0,57) com a proporção de vacas apresentando
comportamento de estro e intensidade de estro e correlação negativa
entre produção de leite e duração do estro (-0,45) [8]. Estratégias para
aumentar E2 e expressão de estro em protocolos têm sido avaliadas em
estudos, como: a adição de E2 em protocolos Ovsynch [9], tratamento
com E2 para induzir ovulação [3, 10], aumento do intervalo entre
prostaglandina (PGF) e momento da IATF [11] e aumento da duração
do protocolo [1].
Uma sequência de experimentos foi realizada para avaliar se
vacas em lactação que apresentam estro em protocolos a base de E2/P4
tem maior prenhez e menor perda de gestação.
8.2 Materiais e Métodos
Esta analise retrospectiva foi realizada a partir de projetos
desenvolvidos em 13 fazendas no Brasil, de janeiro de 2010 a janeiro
de 2014. Foram utilizadas 7.433 vacas em lactação que ovularam ao
seguinte protocolo de IATF: dispositivo intravaginal de P4 contendo
1,9 g de P4 (CIDR, Zoetis) novo, utilizado previamente por 9 ou 18
dias e 2,0 mg (i.m.) de benzoato de estradiol (BE, 2,0 mL de Estrogin,
Farmavet) no dia -11, 25 mg (i.m.) de dinoprost tromethamine (PGF;
5,0 mL de Lutalyse, Zoetis) no dia -4, retirada do CIDR e 1,0 mg (i.m.)
de cipionato de estradiol (ECP 0,5 mL, Zoetis) no dia -2, e IATF no
dia 0 ou TET no dia 7. No dia -4 as vacas foram marcadas na base da

191
cauda com bastão de cera (tail-chalk) e vacas em que o tail-chalk foi
removido completamente no dia 0 foram consideradas em estro.
Os ovários foram avaliados por ultrassonografia para avaliar
o diâmetro do maior folículo presente no dia 0 e para avaliar a presença
de corpo lúteo (CL) no dia 7. Vacas com CL no dia 7 foram
consideradas sincronizadas ao protocolo e apenas estes animais foram
utilizados nesta análise.
As variáveis binomiais foram analisadas utilizando
procedimento GLMMIX do SAS (SAS Institute Inc.) com fazendas
como efeito aleatório e outras variáveis incluídas no modelo quando
apropriado (P< 0,10). O procedimento GLM foi utilizado para
determinar se cada medida individual influenciou na prenhez de forma
linear, quadrática ou cúbica. O procedimento LOGISTIC foi utilizado
para terminar os valores de intercept e slope(s) e construir as curvas
utilizando os valores mínimos e máximos para cada medida
individual. Os resultados são mostrados como média dos quadrados
mínimos. Foi considerado significativo quando P≤ 0,05, e tendência
quando P< 0,10.
8.3 Resultados
A Tabela 1 mostra que, no diagnóstico de gestação aos 32 e
aos 60 dias, vacas que expressaram estro tiveram maior prenhez por
IA (P/IA) e prenhez por transferência de embriões (P/TE) e menor
perda de gestação. Não houve interação entre técnica reprodutiva e
expressão de estro na prenhez e na perda de gestação.

192
Tabela 1. Efeito da expressão de estro na prenhez (%) por IA, prenhez (%) por TE e perda de gestação em vacas em
lactação sincronizadas ao protocolo.
IATF TETF P
Sem estro Estro Sem estro Estro IAxTE Estro
Prenhez
32 d 25,5 (222/846) 38,9 (1785/4584) 32,7 (193/606) 46,2 (645/1397) 0,01 < 0,01
60 d 20,1 (179/846) 33,3 (1530/4584) 25,1 (150/606) 37,5 (525/1397) 0,06 < 0,01
Perda de gestação 20,1 (43/222) 14,4 (255/1785) 22,7 (43/193) 18,6 (120/645) 0,21 0,01 IATF: inseminação artificial em tempo fixo; TETF: transferência de embrião em tempo fixo
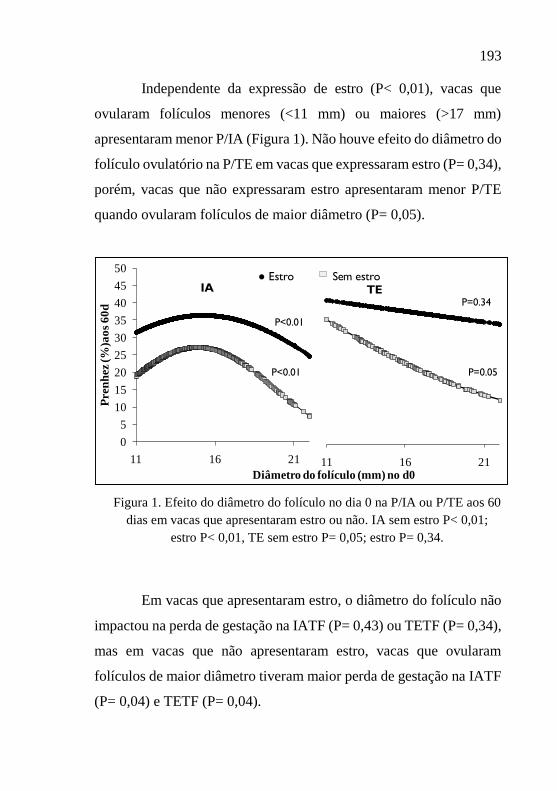
193
Independente da expressão de estro (P< 0,01), vacas que
ovularam folículos menores (<11 mm) ou maiores (>17 mm)
apresentaram menor P/IA (Figura 1). Não houve efeito do diâmetro do
folículo ovulatório na P/TE em vacas que expressaram estro (P= 0,34),
porém, vacas que não expressaram estro apresentaram menor P/TE
quando ovularam folículos de maior diâmetro (P= 0,05).
Figura 1. Efeito do diâmetro do folículo no dia 0 na P/IA ou P/TE aos 60
dias em vacas que apresentaram estro ou não. IA sem estro P< 0,01;
estro P< 0,01, TE sem estro P= 0,05; estro P= 0,34.
Em vacas que apresentaram estro, o diâmetro do folículo não
impactou na perda de gestação na IATF (P= 0,43) ou TETF (P= 0,34),
mas em vacas que não apresentaram estro, vacas que ovularam
folículos de maior diâmetro tiveram maior perda de gestação na IATF
(P= 0,04) e TETF (P= 0,04).
0
5
10
15
20
25
30
35
40
45
50
11 16 21
Pren
hez
(%)a
os
60
d
Diâmetro do folículo (mm) no d0
P<0.01
IA Estro Sem estro
P<0.01
11 16 21
P=0.34
P=0.05
TE

194
Figura 2. Efeito do diâmetro do folículo no dia 0 na perda de gestação entre
32 e 60 dias na IA e TE em vacas que apresentaram estro ou não. IA sem
estro P= 0,04; estro P= 0,43, TE sem estro P= 0,04; estro P= 0,34.
Houve efeito positivo da concentração de P4 no dia 7 na P/IA
em vacas que expressaram estro (P= 0,01) ou não (P= 0,02). Não
houve efeito da P4 no dia 7 na P/TE (sem estro P= 0,76; estro P= 0,52).
0
10
20
30
40
50
60
11 16 21
Perd
a d
e g
esta
çã
o (%
)
Diâmetro do folículo (mm) no d0
IA Estro Sem estro
P=0.43
P=0.04
11 16 21
P=0.34
P=0.04
TE

195
Figura 3. Efeito da concentração de P4 no dia 7 na P/IA e P/TE aos 60 dias
em vacas que apresentaram estro ou não. IA sem estro P= 0,02; estro P=
0,01, TE sem estro P= 0,52; estro P= 0,76.
A expressão de estro em protocolos de IATF e TETF está
associada a aumento na fertilidade. Em protocolos de IATF, a
otimização do diâmetro do folículo ovulatório, aumento da P4 no dia
7 após IA e expressão de estro está associada a maior fertilidade.
Entretanto na TETF, a associação de fertilidade com diâmetro do
folículo ovulatório ou P4 no dia 7 é menos evidente e parece estar
principalmente relacionada à expressão de estro.
8.3.1 Protocolos podem aumentar a detecção de estro e a concepção?
Realizou-se estudo para avaliar se em protocolos de
sincronização da ovulação, o momento da aplicação da PGF interfere
na concepção de vacas Holandesas em lactação submetidas à
inseminação artificial em tempo fixo (IATF) ou transferência de
0
5
10
15
20
25
30
35
40
45
50
0 1 2 3 4 5
Pren
hez (%
) a
os
60
d
Concentração de Progesterona (ng/mL) no d7
P=0.02
IA
Estro Sem estro
P=0.01
0 1 2 3 4 5
P=0.76
P=0.52
TE

196
embrião em tempo fixo (TETF). Em cada fazenda (n=10), vacas
(n=1085; DEL 205 ± 136; produzindo 34 ± 11 kg/leite/dia) foram
distribuídas aleatoriamente para receber a aplicação de PGF no dia 7
ou 8 do protocolo (dia 0 BE + CIDR; PGF dia 7 ou 8; dia 8 ECP e
retira CIDR).
Melhores taxas de concepção foram observadas no grupo de
animais que receberam a PGF no dia 7, na IATF e na TETF, em
relação ao grupo que recebeu a PGF no dia 8 (Tabela 2). Resultados
mostram a importância de antecipar a aplicação da PGF e que os
benefícios podem estar relacionados tanto como no transporte de
gametas, fertilização, desenvolvimento embrionário inicial, bem
como mudanças no ambiente uterino e hormonal.

197
Tabela 2. Concepção por IA ou por TE nos dias 28 e 60 de gestação e perda de gestação, em vacas recebendo PGF nos
dias 7 ou 8 do protocolo de sincronização
IATF TETF P
PGF d7 PGF d8 PGF d7 PGF d8 d7 x d8
Concepção 28 d 32,9 (238) 20,6 (168) 47,0 (243) 40,7(244) < 0,01
Concepção 60 d 30,0 (238) 19,2 (168) 37,9 (243) 33,5 (244) < 0,01
Perda 28 – 60 d 7,7 (87) 5,5 (42) 20,4 (116) 19,4 (100) 0,68

198
Em outro estudo aumentou-se o intervalo entre a PGF e a
IATF. Vacas Holandesas em lactação ciclando foram dividas
aleatoriamente para receber os protocolos: (8 d) d0 BE + CIDR; PGF
7 ou 8; d8 ECP e retira CIDR; ou (9d) d0 BE + CIDR; PGF 7; d9 ECP
e retira CIDR. Neste estudo (Tabela 3) o protocolo 9 d aumentou a
expressão de cio, mas não houve efeito do protocolo na concepção.
Entretanto o protocolo com maior intervalo entre a PGF e a IA resultou
em menores taxas de perda de gestação. Os resultados deste trabalho
indicam que melhoras na fertilidade pela menor taxa de perda de
gestação podem estar relacionadas à otimização do folículo
ovulatório, aumento da expressão de estro e otimização do proestro.

199
Tabela 3. Efeitos da duração do protocolo na expressão de cio,
sincronização e concepção.
Duração do protocolo
8 d 9 d P
Expressão de cio¹ 63,4
(240/385)
73,0
(269/374)
< 0,01
Sincronização² 92,8
(352/379)
91,5
(339/370)
0,52
Concepção¹
Aos 32 d 45,0
(175/385)
43,9
(166/374)
0,77
Aos 60d 38,1
(150/385)
40,4
(154/374)
0,52
Perda de gestação 14,7 (25/175) 7,6
(12/166)
0,04
Concepção²
Aos 32 d 48,1
(170/352)
47,9
(163/339)
0,96
Aos 60d 40,5
(145/352)
43,9
(151/339)
0,37
Perda de gestação 15,2 (25/170) 7,8
(12/163)
0,03
¹Todas vacas inseminadas. ² Vacas sincronizadas (CL 7d após IA)
8.3.2 Aumento da P4 em protocolos de IATF também pode aumentar
a concepção?
Um estudo foi realizado para avaliar se a fertilidade de vacas
em lactação submetidas a protocolos a base de E2 e P4 poderiam
aumentar através da combinação de GnRH com benzoato de estradiol

200
no início do protocolo, pelo aumento da concentração de P4 durante o
período de desenvolvimento do folículo. Além disso, foi avaliada a
utilização de duas doses de PGF ao final do protocolo.
Vacas Holandesas em lactação (n=1.808) foram divididas
aleatoriamente durante 2 estações do ano (quente ou fria) para receber
um dos seguintes tratamentos:
I. Controle: CIDR + 2 mg de benzoato de estradiol no
dia -11, PGF no dia -4, retirada do CIDR + 1,0 mg de
cipionato de estradiol no dia -2, e IATF no dia 0
II. 2PGF: igual ao controle, com adição de uma segunda
dose de PGF no dia -2
III. GnRH: igual ao 2PGF com adição de 100 µg de
GnRH no dia -11.
A estação do ano teve grande efeito na fertilidade das vacas,
sendo que durante a estação fria foi observada melhor P/IA (Tabela
4). O protocolo com GnRH aumentou a percentagem de vacas com
CL no momento da PGF (Controle= 56,9%; 2PGF= 55,8%; GnRH=
70,5%) e P4 na PGF (Controle= 3,28 ± 0,22; 2PGF= 3,35 ± 0,22;
GnRH= 3,70 ± 0,21 ng/mL).
O protocolo GnRH aumentou a P/IA aos 32 e 60 após IATF
em relação ao controle, sendo que o protocolo 2PGF apresentou
resultados intermediários (Tabela 4). O efeito positivo do protocolo
GnRH foi detectado apenas na estação fria do ano. O uso de GnRH
combinado com benzoato de estradiol no início de protocolos a base

201
de E2/P4 aumentou a fertilidade, particularmente durante a estação fria
do ano.

202
Tabela 4. Efeitos do protocolo de IA na fertilidade de vacas em lactação
Critérios analisados Protocolo
P Controle 2PGF GnRH
P/IA 32 d todas vacas
Fria 41,0 (116/283) 44,2 (125/283) 50,9 (148/291) 0,05
Quente 19,0 (61/321) 21,8 (71/326) 23,4 (71/304) 0,40
P < 0,01 < 0,01 < 0,01
Combinados 30,0 (177/604) 33,2 (196/609) 37,3 (219/595) 0,02
P/IA 60 d todas vacas
Fria 32,9 (93/283) 36,4 (103/283) 41,6 (121/291) 0,09
Quente 16,2 (52/321) 18,7 (61/326) 19,1 (58/304) 0,59
P < 0,01 < 0,01 < 0,01
Combinados 25,1 (145/604) 28,0 (164/609) 31,0 (179/595) 0,06
P/IA 32 d vacas sincronizadas
Fria 46,6 (110/236) 49,0 (120/245) 55,7 (142/255) 0,11
Quente 22,5 (52/231) 26,0 (60/231) 27,4 (61/223) 0,47
P < 0,01 < 0,01 < 0,01
Combinados 34,7 (162/467) 37,8 (180/476) 42,5 (203/478) 0,05
P/IA 60 d vacas sincronizadas
Fria 37,7 (89/236) 40,4 (99/245) 45,5 (116/255) 0,20
Quente 19,5 (45/231) 21,7 (50/231) 21,5 (48/223) 0,81
P < 0,01 < 0,01 < 0,01
Combinados 28,7 (134/467) 31,3 (149/476) 34,3 (164/478) 0,18
.

203
8.4 Conclusões
A concentração de P4 durante o desenvolvimento do folículo
ovulatório e a expressão de estro em protocolos de IATF são fatores
importantes para se obter melhore índices de P/IA.
REFERÊNCIAS BIBLIOGRÁFICAS
PEREIRA MH, RODRIGUES AD, DE CARVALHO RJ, WILTBANK
MC, VASCONCELOS JL, Increasing length of an estradiol and
progesterone timed artificial insemination protocol decreases pregnancy
losses in lactating dairy cows, J Dairy Sci, 2014,
SÁ FILHO OG, THATCHER WW, VASCONCELOS JL, Effect of
progesterone and/or estradiol treatments prior to induction of ovulation on
subsequent luteal lifespan in anestrous Nelore cows, Anim Reprod Sci,
2009;112:95-106,
CERRI RL, SANTOS JE, JUCHEM SO, GALVÃO KN, CHEBEL RC,
Timed artificial insemination with estradiol cypionate or insemination at
estrus in high-producing dairy cows, J Dairy Sci, 2004;87:3704-15,
PEREIRA MHC, RODRIGUES ADP, MARTINS T, OLIVEIRA WVC,
SILVEIRA PSA, WILTBANK MC, et al, Timed Artificial Insemination
Programs During Summer in Lactating Dairy Cows: Comparison of the 5-d
Cosynch protocol with an Estrogen/Progesterone-based protocol, J Dairy
Sci (Submitted), 2013,
VASCONCELOS JL, SANGSRITAVONG S, TSAI SJ, WILTBANK MC,
Acute reduction in serum progesterone concentrations after feed intake in
dairy cows, Theriogenology, 2003;60:795-807,
SANGSRITAVONG S, COMBS DK, SARTORI R, ARMENTANO LE,
WILTBANK MC, High feed intake increases liver blood flow and
metabolism of progesterone and estradiol-17beta in dairy cattle, J Dairy
Sci, 2002;85:2831-42,

204
WILTBANK M, LOPEZ H, SARTORI R, SANGSRITAVONG S,
GÜMEN A, Changes in reproductive physiology of lactating dairy cows
due to elevated steroid metabolism, Theriogenology, 2006;65:17-29,
LOPEZ H, SATTER LD, WILTBANK MC, Relationship between level of
milk production and estrous behavior of lactating dairy cows, Anim
Reprod Sci, 2004;81:209-23,
SOUZA AH, GÜMEN A, SILVA EP, CUNHA AP, GUENTHEr JN, PETO
CM, et al, Supplementation with estradiol-17beta before the last
gonadotropin-releasing hormone injection of the Ovsynch protocol in
lactating dairy cows, J Dairy Sci, 2007;90:4623-34,
PANCARCI SM, JORDAN ER, RISCO CA, SCHOUTEN MJ, LOPES FL,
MOREIRA F, et al, Use of estradiol cypionate in a presynchronized timed
artificial insemination program for lactating dairy cattle, J Dairy Sci,
2002;85:122-31,
PEREIRA MH, SANCHES CP, GUIDA TG, RODRIGUES AD, ARAGON
FL, VERAS MB, et al, Timing of prostaglandin F2α treatment in an
estrogen-based protocol for timed artificial insemination or timed embryo
transfer in lactating dairy cows, J Dairy Sci, 2013;96:2837-46,
GALVÃO KN, SANTOS JE, JUCHEM SO, CERRI RL, COSCIONI AC,
VILLASEÑOR M, Effect of addition of a progesterone intravaginal insert
to a timed insemination protocol using estradiol cypionate on ovulation rate,
pregnancy rate, and late embryonic loss in lactating dairy cows, J Anim
Sci, 2004;82:3508-17,
HILLEGASS J, LIMA FS, SÁ FILHO MF, SANTOS JE, Effect of time of
artificial insemination and supplemental estradiol on reproduction of
lactating dairy cows, J Dairy Sci, 2008;91:4226-37,
SOUZA AH, SILVA EP, CUNHA AP, GÜMEN A, AYRES H,
BRUSVEEN DJ, et al, Ultrasonographic evaluation of endometrial
thickness near timed AI as a predictor of fertility in high-producing dairy
cows, Theriogenology, 2011;75:722-33,
MILLER BG, MOORE NW, MURPHY L, STONE GM, Early pregnancy
in the ewe: effects of oestradiol and progesterone on uterine metabolism and
on embryo survival, Aust J Biol Sci, 1977;30:279-88,

205
BARTOL FF, THATCHER WW, LEWIS GS, BLISS EL, DROST M,
BAZER FW, Effect of estradiol-17beta on PGF and total protein content in
bovine uterine flushings and peripheral plasma concentration of 13, 14-
dihydro-15-keto-PGF(2alpha), Theriogenology, 1981;15:345-58,
BUHI WC, Characterization and biological roles of oviduct-specific,
oestrogen-dependent glycoprotein, Reproduction, 2002;123:355-62,
ROBERTS C, PERRY G, MINTEN M, ROBERTS A, MACNEIL M,
GEARY T, Effects of preovulatory estradiol concentration on embryo
survival and pregnancy establishment in beef cows, Western Section of
Animal Science Proceedings 2012,

206
9. ESTRESSE CALÓRICO EM VACAS LEITEIRAS: EFEITO
SOBRE O METABOLISMO E A QUALIDADE DO LEITE10
Vivian Fischer
Marcelo Tempel Stumpf
Alexandre Sausenbach Abreu
Elissa Forgiarini Vizzotto
9.1 Cenário – por que o estresse térmico é importante?
Cenários mais pessimistas apontam para um inequívoco
aumento nas médias de temperatura na Terra em até 4°C até 2100
(IPCC, 2007). Os sistemas de produção animal, ao estarem inseridos
e serem diretamente dependentes das condições naturais vigentes,
serão afetados por tais modificações.
As condições climáticas afetam e desafiam o animal a se
adaptar às condições ambientais para manter a homeostase e o bem-
estar. Nos trópicos e em condições de pastagem, o gado é exposto ao
sol, além de outros agentes meteorológicos como umidade, chuva e
10 Fischer, V.; Stumpf, M.T.; Abreu, A.S.; Vizzotto, E.F. Estresse calórico
em vacas leiteiras: efeito sobre o metabolismo e a qualidade do leite. Anais.
I Simpósio Nacional da Vaca Leiteira. Porto Alegre: Universidade Federal
do Rio Grande do Sul. 2014. 239 p.

207
vento durante várias horas por dia. A intensificação da atividade
pecuária, reduzindo o espaço por animal e modificando o ambiente
pode dificultar ou mesmo impedir que animais encontrem alternativas
para se adaptarem a essas condições, tornando-os mais suscetíveis ao
estresse, resultando em mudanças fisiológicas que diminuem o seu
desempenho produtivo (Deitenbach et al ., 2008).
Podem se esperar efeitos diretos em animais expostos a altas
temperaturas, tais como alterações comportamentais, como o aumento
na ocorrência de interações agonísticas (Vizzotto, 2014), maior tempo
em ócio, redução tempo de pastejo diurno e aumento no noturno,
maior ingestão de água (West, 2003; Fischer et al., 2014); alterações
fisiológicas, como aumento nas frequências respiratória e cardíaca
(Mitlöhner et al., 2001; Dalcin, 2013), escore de ofegação e
temperatura retal (McManus et al., 2009; Stumpf, 2014; Fischer ET
AL., 2014) e redução no consumo de alimento (West, 1994; Rhoads
et al., 2009).
Mecanismos homeorréticos são acionados de forma a tornar o
animal capaz de se adaptar e se manter em um equilíbrio dinâmico
com as novas contingências impostas. O animal necessita dissipar
calor ao meio em busca de manter sua homeostase, ou seja, manter a
temperatura interna dentro dos níveis normais de 38 a 39,5˚C (Stober,
1993). Mecanismos de dissipação de calor requerem energia, a qual
pode provir da sua partição em detrimento da glândula mamária
através de alterações hormonais acionadas e controladas pelo eixo
hipotálamo-hipófise-adrenal (Baumgard & Rhoads, 2013).

208
Os efeitos negativos do estresse térmico são mais evidentes em
animais com elevada produção devido à maior geração de calor
metabólico (Kadzere et al., 2002). Animais zebuínos, normalmente
mais tolerantes ao calor, devido às suas adaptações de forma corporal,
superfície de pele, número de glândulas sudoríparas e características
do pelame, sofrem mais intensamente à medida que a sua
produtividade aumenta. Entre as estratégias dos animais para
minimizarem o estresse térmico, se destacam a redução da carga
térmica, redução da produção de calor metabólico e aumento da
dissipação de calor (Berman, 2012).
Na zona de conforto térmico, em torno de 75% da perda de calor
ocorre por radiação, convecção e/ou condução. No entanto, quando a
temperatura ambiente excede a temperatura crítica superior, o
gradiente de temperatura torna-se pequeno para que ocorra
resfriamento por esses meios. Inicia-se então a termólise,
especialmente por mecanismos evaporativos para manter o balanço
térmico e a evaporação, por meio da sudorese e/ou respiração,
tornando-se a principal via de dissipação de calor, responsável por
80% da perda de calor corporal. Em condições de umidade relativa do
ar aumentada, há dificuldade na evaporação e o ambiente torna-se
mais estressante para o animal (Collier et al., 2006).
Animais mantidos em ambientes com temperaturas acima da
temperatura crítica superior podem se considerar em estresse térmico,
já que dependem de mecanismos de dissipação de calor. A mediação
de tais mecanismos se dá pelo eixo hipotálamo-hipófise-adrenal, o

209
qual é abastecido de informações provenientes das unidades
termoreceptoras e termosensitivas no sistema nervoso central (Baker,
1989). Uma das primeiras respostas do animal ao ambiente estressante
é a redução no consumo de alimentos (Castanheira, 2009), na busca
de reduzir a produção de calor proveniente de processos digestivos
(calor metabólico) e maior consumo de água, para repor as perdas
promovidas pela evaporação. Estima-se aumento de consumo de 1,2
kg de água para cada 1˚C de aumento na temperatura ambiental
mínima (West, 2003). Em adição, os animais aumentam suas
exigências de mantença, necessárias para suportar as mudanças
fisiológicas para dissipação de calor: ofegacão, suor, aumento nas
reações químicas, síntese e mantença da funcionalidade das proteínas
de choque térmico (Tomanek et al., 2010), e aumento na atividade
celular induzida pelos níveis elevados de adrenalina (Gaffin &
Hubbard, 1996). De acordo com o NRC (2001), estresse médio a
severo pode elevar entre 7 a 25% tais exigências, embora Fox &
Tyluki (1998) sugiram aumentos de até 30%. Ocorre vasodilatação das
vias sanguíneas e aumento do fluxo de sangue para as zonas
periféricas do animal, para que ocorra resfriamento por contato com o
ar mais frio. A elevação na frequência cardíaca promove aumento
nesse fluxo sanguíneo, auxiliando o processo. Na busca de elevar as
perdas de calor por evaporação, os animais elevam também a
frequência respiratória, podendo prevenir o aumento da temperatura
retal até índices de temperatura e umidade (ITU) de 80 (Silanikove et
al., 2000). O aumento nas perdas de gás carbônico em virtude do

210
aumento da frequência respiratória altera a proporção entre as
concentrações de CO2:HCO3 sanguíneos, a qual deve se encontrar por
volta de 20:1, e é o principal sistema de tamponamento do sangue
(Kadzere et al., 2002). Para compensar essas perdas, o CO2 diminui,
levando a um quadro de alcalose respiratória e aumento do pH
sanguíneo. A redução no teor de CO2 força os rins a excretarem maior
quantidade de bicarbonato (tamponante), na tentativa de manter a
proporção de 20:1 estável (Beatty et al., 2006). Além disso, a reação
CO2 ↔ HCO3 se desloca para a esquerda, para compensar as perdas
de gás carbônico. O menor teor de bicarbonato promove uma
subsequente redução no pH sanguíneo, provocando acidose
metabólica compensatória.
Mecanismos de sobrevivência sobrepõem aqueles relacionados
à produção de leite. Os efeitos aditivos do menor consumo de
alimentos, maiores exigências de energia para mantença, possíveis
distúrbios metabólicos e maior destinação de energia aos processos de
perda de calor geram as reduções nos níveis produtivos dos animais.

211
9.2 Efeitos sobre produção de leite
Até recentemente se creditava a redução na produção de leite à
diminuição no consumo de alimento (Baccari, 2001). A redução da
produção leiteira varia entre 20 e 40% em animais severamente
estressados (West, 2003; Abreu et al., 2011). Segundo Wheelock et
al. (2010), 40 a 50% dessa redução se deve ao menor consumo de
alimentos (efeito indireto), sendo o restante em virtude de outros
mecanismos induzidos pelo estresse (efeitos diretos). Além do menor
consumo, ocorre redução de até 35% na eficiência de utilização de
energia para fins produtivos (McDowell et al., 1976).
Silanikove et al. (2009) sugerem outro mecanismo de ação para
explicar a redução na produção leiteira. Sob estresse térmico agudo,
ocorre aumento na concentração de um peptídeo derivado da clivagem
da β-caseína pela plasmina, o qual bloqueia os canais de potássio da
membrana apical das células epiteliais mamárias, promovendo
despolarização potencial da membrana, reduzindo o metabolismo
celular, sua atividade secretória e, por conseguinte, diminuindo a
produção láctea. Os efeitos negativos do estresse térmico sobre a
produção leiteira (Figura 1) dependem do nível de produção antes do
estresse (quanto maior a produtividade, maior a redução na produção)
e do estádio de produção (vacas no terço médio reduzem mais a
produção do que vacas ao início ou final da lactação (Bernabucci et
al., 2010).
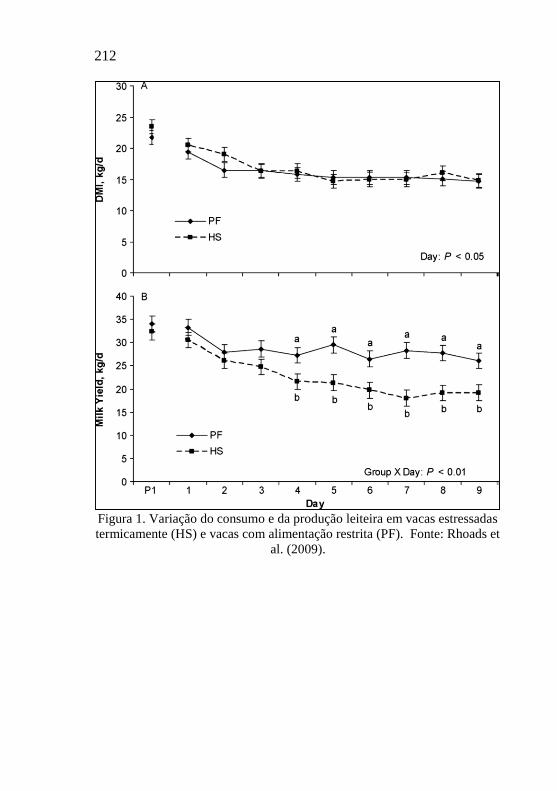
212
Figura 1. Variação do consumo e da produção leiteira em vacas estressadas
termicamente (HS) e vacas com alimentação restrita (PF). Fonte: Rhoads et
al. (2009).

213
9.3 Efeitos do estresse calórico sobre a composição do leite –
características químicas
Vacas mantidas em ambientes com índice de temperatura e
umidade (ITU) superiores a 72, produziram leite com menores teores
de proteína, alfa e beta caseínas. Todavia, os efeitos sobre os teores de
gordura foram contraditórios (Bernabucci et al., 2002). A principal
causa da redução nos teores de proteína nesse estudo foi a redução nos
níveis de αs e β-caseínas, provavelmente em virtude do menor aporte
de energia e proteína. Essas caseínas são ricas em grupos fosfato e são
os componentes acídicos das micelas (Schmidt, 1980).
Reduções no teor de proteína no leite foram descritos em
bovinos por Rhoads et al. (2009), embora não tenham detectado
diferenças nos teores de gordura e lactose (Figura 2). Ao contrário do
relatado por Abreu et al. (2011) os quais verificaram que vacas
leiteiras sob estresse térmico severo sem acesso a sombra produziram
leite com menores teores de proteína (2,75 vs 2,99%), porém maiores
teores de gordura (3,61 vs 3,40%) e nitrogênio ureico (23,70 vs 18,81
mg/dL), não encontrando diferença para os teores de lactose (4,37 vs
4,34).
Segundo Sevi & Caroprese (2012), a exposição ao sol em altas
temperaturas pode promover aumento na permeabilidade dos
capilares, elevando a quantidade de enzimas proteolíticas no leite, de
forma a reduzir a concentração proteica. Em adição, a redução na
oferta e consumo de pasto nos meses mais quentes, bem como o menor
aporte de energia e nitrogênio, contribuem para os valores

214
encontrados. O citado aumento na permeabilidade dos capilares
também promove aumento nas concentrações de enzimas lipolíticas,
resultando em decréscimo na síntese e alteração no perfil lipídico do
leite (Sevi & Caroprese, 2012). Esses autores observam que a menor
taxa de passagem no rúmen aumenta o tempo de exposição da digesta
ao processo de biohidrogenação, de forma que os ácidos graxos
insaturados são convertidos em saturados e têm seus teores reduzidos
no leite. O estresse térmico e a redução na qualidade da forragem em
virtude da maior atividade metabólica vegetal atuam em conjunto
diminuindo o consumo (Renna et al., 2011) e a produção de ácido
acético no rúmen, principal precursor da gordura láctea.
Os efeitos negativos do estresse térmico sobre os níveis de
lactose no leite são decorrentes do uso da glicose como fonte de
energia ao animal, reduzindo o aporte à glândula mamária e posterior
síntese desse componente, além do efeito direto da queda no consumo
de alimentos. Alterações na concentração de lactose, por sua vez, são
mais difíceis de ocorrer, já que sua queda é acompanhada por menores
níveis de produção de leite.
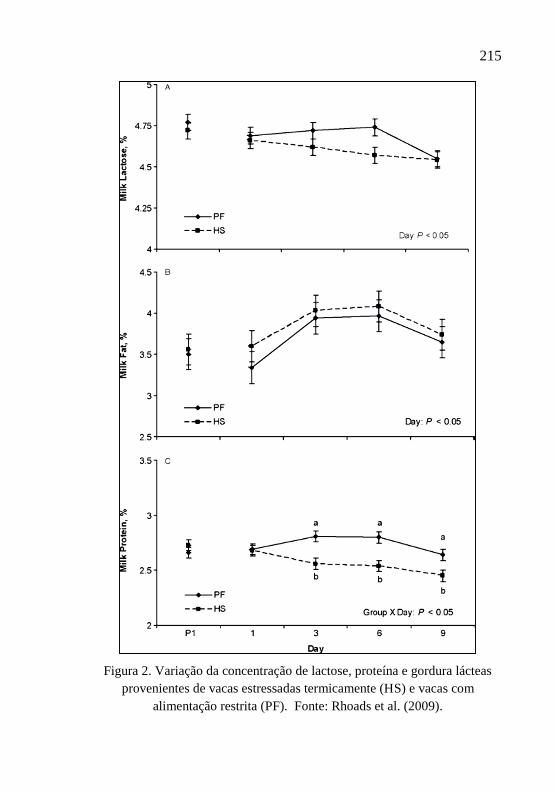
215
Figura 2. Variação da concentração de lactose, proteína e gordura lácteas
provenientes de vacas estressadas termicamente (HS) e vacas com
alimentação restrita (PF). Fonte: Rhoads et al. (2009).

216
9.4 Efeitos do estresse calórico sobre a composição do leite –
características físicas
Em trabalhos realizados durante o verão (Mariani et al., 1994;
Bernabucci & Calamari, 1998; Calamari & Mariani, 1999), foram
verificados valores de pH mais alto e de acidez titulável mais baixa
em animais em estresse térmico. No entanto, Abreu et al. (2011)
relataram menores valores de pH, crioscopia e densidade e maior valor
de acidez titulável em vacas estressadas termicamente sem acesso a
sombra em relação àquelas com acesso a sombra. Também, os autores
encontraram que vacas estressadas termicamente produziram leite
com menor estabilidade (testes do álcool e do tempo de coagulação
sob temperatura de 144°C, Figura 3).
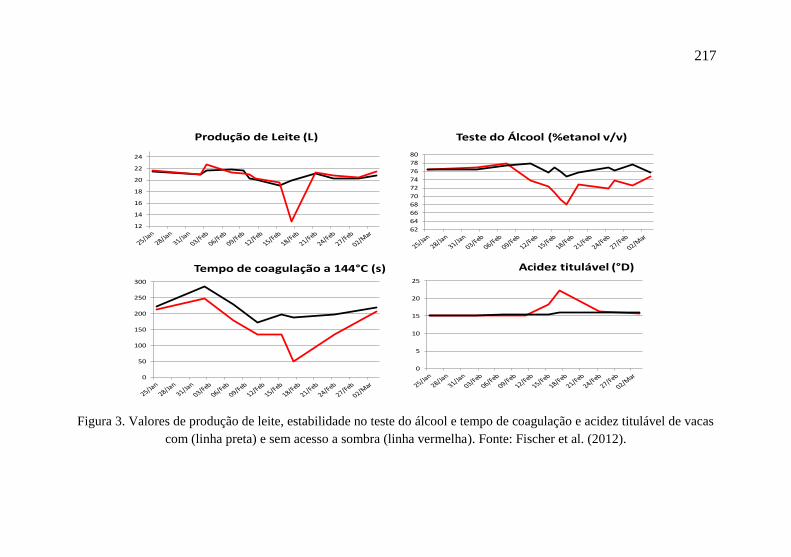
217
12
14
16
18
20
22
24
Produção de Leite (L)
62
64
66
68
70
72
74
76
78
80
Teste do Álcool (%etanol v/v)
0
50
100
150
200
250
300
Tempo de coagulação a 144°C (s)
0
5
10
15
20
25
Acidez titulável (°D)
Figura 3. Valores de produção de leite, estabilidade no teste do álcool e tempo de coagulação e acidez titulável de vacas
com (linha preta) e sem acesso a sombra (linha vermelha). Fonte: Fischer et al. (2012).

218
Os efeitos do estresse térmico sobre a estabilidade do leite no
teste do álcool podem ser relacionados à redução no consumo de
alimentos dos animais. De fato, a restrição de consumo per se resultou
em redução na estabilidade do leite (Zanela et al., 2006; Abreu et al.,
2011; Stumpf et al., 2013) e épocas do ano de menor oferta de
alimentos promovem maior incidência de leite instável (Ponce &
Hernández, 2001; Marques, 2004).
Ademais, distúrbios metabólicos provenientes do estresse
térmico, ao alterar de forma significativa as condições fisiológicas do
animal, apresentam grande potencial de reduzir a estabilidade do leite,
conforme relatado por Marques et al. (2011) ao induzirem acidose
metabólica através do fornecimento de sal aniônico em bovinos
leiteiros.
9.5 Efeitos do estresse calórico sobre o metabolismo
O estresse térmico provoca mudanças na homeostase e na
homeorrese das vacas lactantes. Ocorrem alterações no perfil
hormonal caracterizadas principalmente por um declínio e aumento,
respectivamente, naqueles com efeitos anabolizantes e catabolizantes,
alterando o metabolismo pós-absortivo da energia, lipídeos e
proteínas, com prejuízos à função hepática e aumento do estresse
oxidativo. Ocorre prejuízo à resposta imunológica e ao desempenho
reprodutivo. Os efeitos sobre o metabolismo podem ser quantificados
pela mensuração das variáveis fisiológicas, tais como a temperatura
corporal, frequência respiratória, concentrações hormonais e

219
equilíbrio ácido-base e hidroeletrolítico (Bernabucci et al., 2010).
Assim, ocorre diminuição do hormônio do crescimento, catecolaminas
e glicocorticoides. Este estado endócrino reduz os níveis circulantes
de tiroxina (T4) e triiodotironina (T3), e, em consequência, a
diminuição da taxa metabólica basal e da produção de calor (Johnson,
1980; Yousef, 1987).
Vacas em balanço energético negativo decorrente de estresse
térmico, apesar de ingerirem menos, não apresentam reduções na
secreção e na sensitividade à insulina (Wheelock et al., 2010) e podem
ter seus níveis de somatotropina reduzidos (Li et al., 2006). Dessa
forma, as vacas se tornam metabolicamente inflexíveis, uma vez que
não podem oxidar ácidos graxos para gerar energia, pois esse processo
contribui ao incremento de calor metabólico (Baumgard & Rhoads,
2013). Não se detectam elevações em ácidos graxos não esterificados
no sangue (Shwartz et al., 2009) e os animais passam a ser
dependentes de glicose como fonte de energia. Como consequência, o
aporte de glicose à glândula mamária diminui, com consequente
redução da síntese de lactose, o que diminui a produção de leite.
Vacas sem acesso a sombra, mesmo em condição de estresse
térmico leve, apresentaram maiores valores de Na, mas valores
menores de hematócrito, PCO2, TCO2, HCO3, excesso de base, e
tenderam a apresentar hemoglobina baixa (Abreu, comunicação
pessoal). Foram observados maiores concentrações de ureia,
creatinina e albumina no sangue de vacas severamente estressadas
pelo calor (Abreu, comunicação pessoal). Devido às maiores perdas

220
renais de HCO3- e perda de saliva, pela redução da ruminação nos
animais ofegantes no calor, as vacas estressadas pelo calor são mais
propensas a sofrer acidose ruminal e metabólica.
9.6 Estresse térmico em fases não produtivas (crescimento de
novilhas e período seco de vacas)
Vacas que sofreram estresse térmico durante o período seco
apresentaram menor produção leiteira durante 280 dias na lactação
subsequente (28,9 vs 33,9 kg/dia), possivelmente devido à menor
proliferação celular observada no pré-parto, não sendo observados
efeitos sobre a apoptose (Tao et al., 2011). Também foi observado
menor concentração de proteína láctea (3,01 vs 2,87%). O estresse
térmico durante o período seco reduz o consumo e debilita a
imunidade dos animais (Tal et al., 2013).
Em comparação com as vacas estressadas termicamente durante
o período seco, os animais que ficaram sob resfriamento (ventiladores
e aspersores) ganharam mais peso antes do parto, mas perderam mais
peso e condição corporal no início da lactação. As vacas com
resfriamento produziram mais leite do que vacas mantidas sob estresse
térmico (34,0 vs 27,7 kg/dia), mas os tratamentos não afetaram a
composição do leite (Figura 4). Os tratamentos não influenciaram a
insulina circulante e os metabólitos pré-parto, mas vacas sob
resfriamento apresentaram menor concentração plasmática de glicose,
aumento dos ácidos graxos livres, e tenderam a ter menor
concentração de insulina no pós-parto em comparação com vacas
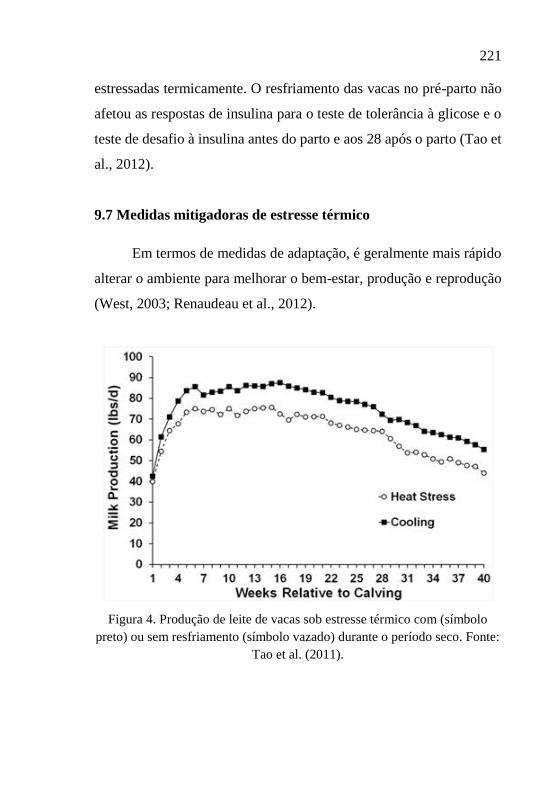
221
estressadas termicamente. O resfriamento das vacas no pré-parto não
afetou as respostas de insulina para o teste de tolerância à glicose e o
teste de desafio à insulina antes do parto e aos 28 após o parto (Tao et
al., 2012).
9.7 Medidas mitigadoras de estresse térmico
Em termos de medidas de adaptação, é geralmente mais rápido
alterar o ambiente para melhorar o bem-estar, produção e reprodução
(West, 2003; Renaudeau et al., 2012).
Figura 4. Produção de leite de vacas sob estresse térmico com (símbolo
preto) ou sem resfriamento (símbolo vazado) durante o período seco. Fonte:
Tao et al. (2011).

222
Raças de origem europeia sofrem mais o estresse térmico
devido à sua alta produtividade, reduzindo o seu limiar de conforto
térmico (Silva et al., 2002). Por isso, os criadores brasileiros têm
procurado combinar as características desejáveis das raças europeias
e zebuínas, através da produção de animais mestiços, geralmente
usando Holandês e Gir (Girolando). O rebanho nacional é composto
por aproximadamente 95% de animais mestiços (Martinez &
Verneque, 2001), representando cerca de 70% da produção de leite
(Alvim et al., 2005).
As raças diferentes reagem de forma diferente ao estresse
térmico (McManus et al., 2009). Além disso, os avanços na produção
animais em áreas como a nutrição e a reprodução resultaram em
aumento significativo na produção de leite, o que resultou em uma
maior taxa de calor metabólico e, portanto, aumentando a necessidade
de dissipação de calor (Silanikove et al, 2000;. Baumgard et al ., 2007).
O cruzamento entre as raças Holandês e Gir é usado há décadas
para obter animais mais tolerantes ao calor. A comparação entre
animais pertencentes aos grupos genéticos Holandês, 50% Holandês e
50% Gir e 75% holandês e 25% Gir, mostrou que as vacas 50% H 50%
G apresentaram menores frequências respiratórias e cardíacas,
temperatura retal e escore de ofegação comparadas ao grupo 100%
Holandês, enquanto o grupo 75% H 25% G apresentou valores
intermediários (Stumpf, 2014).
Criar animais em um ambiente com conforto para proporcionar
bem-estar, pode melhorar o desempenho produtivo do animal.

223
Portanto, para minimizar os efeitos nocivos do clima sobre os animais
em países tropicais é importante para mitigar o efeito negativo das
variáveis climáticas consideradas responsáveis pelo estresse por calor.
Nos locais, caracterizado pelo clima subtropical de altitude com
umidade elevada, a velocidade do vento contribui para a dissipação de
calor dos processos de animais por convecção, mas a alta umidade
reduz a capacidade de dissipação de calor do corpo, por meio de
evaporação (Azevedo et al, 2009). O resfriamento evaporativo é a
forma principal de perda de calor disponível para os animais
homeotérmicos quando a temperatura ambiente é superior à
temperatura do corpo, e este processo é mais eficiente quando a
umidade relativa é baixa (Robinson, 2004).
O uso de sombra serve para atenuar o efeito da radiação solar,
diminuindo a temperatura do corpo (Mitlohner et al., 2001; Kendall et
al., 2006; Tucker et al., 2007; Fischer et al., 2014). No entanto, deve-
se prover área em torno de 9 m2 por vaca para evitar competição entre
os animais pela sombra (Schutz et al., 2010).
O provimento de instalações como galpões dotados de
aspersores na linha do cocho aliado a ventiladores, tipo fixo ou tipo
móvel, ou galpões com sistema de resfriamento no piso ou nas camas,
são ferramentas nos rebanhos comerciais dos países desenvolvidos.
Existem diferenças entre os sistemas quanto à capacidade de reduzir a
temperatura corporal e a frequência respiratória, aumentar o tempo em
que a vaca fica deitada e aumentar o consumo de alimentos. O
provimento de conforto térmico durante o período seco exerce efeitos

224
positivos sobre o sistema imune, desenvolvimento placentário e
desenvolvimento do terneiro, com reflexos positivos sobre a produção
de leite na lactação subsequente (Tao et al., 2013).
REFERÊNCIAS BIBLIOGRÁFICAS
ABREU, A.S.; FISCHER, V.; KOLLING, G. et al. Estresse calórico
induzido por privação de acesso à sombra em vacas Holandesas reduz a
produção leiteira e a estabilidade térmica do leite. Proceedings. 2ª
Conferência Internacional de Leche Inestable, Colonia del Sacramento,
Uruguay, 2011.
ALVIM, M.J.; PACIULLO, D.S.C.; CARVALHO, M.M. et al. Sistema de
produção de leite com recria de novilhas em sistemas silvipastoris.
2005. EMBRAPA.
AZEVÊDO, D.M.R; ALVES A.A. Bioclimatologia aplicada à produção
de bovinos leiteiros nos trópicos. Embrapa Meio-Norte, 83 p. Teresina.
2009.
BAKER, M. A. Effect of dehydration and rehydration on thermoregulatory
sweating in goats. Journal of Physiology, v. 417, p. 421 – 435, 1989.
BAUMGARD, L.H.; RHOADS, R.P. Effects of heat stress on
postabsorptive metabolism and energetics. Annu. Rev. Anim. Biosci. v.1, p.
311-333, 2013.
BEATTY, D. T. et al. Physiological responses of Bos taurus and Bos
indicus cattle to prolonged, continuous heat and humidity. J. Animal Sci.,
v. 84, p. 972-985, 2006.
BERMAN, A. From heat tolerance to heat stress relief: An evolution of
notions in animal farming. In: Environmental Physiology of Livestock. Ed.
Collier, R.J. & Collier, J.L. 2012. p.1-16.

225
BERNABUCCI, U.; LACETERA, N.; RONCHI, B.; NARDONE, A.
Effects of the hot season on milk protein fractions in Holstein cows. Anim.
Res. v. 51, p. 25-33, 2002.
BERMANBUCCI, U.; LACETERA, N.; BAUMGARD, L.H.; RHOADS,
R.P.; RONCHI, B.; NARDONE, A. Metabolic and hormonal acclimation to
heat stress in domesticated ruminants. Animal, v. 4, p. 1167–1183, 2010.
CASTANHEIRA, M. Análise multivariada de características que
influenciam a tolerância ao calor de equinos, ovinos e bovinos. 2009.
107 f. Tese (Doutorado em Ciência Animal). Universidade Federal de
Goiás, Goiânia, 2009.
CERUTTI, W.G.; BERMUDES, R.F.; VIÉGAS, J.; MARTINS, C.M.M.
Physiological and productive responses of Holstein cows in fosterage
submitted or not to shading and aspersion in premilking. Rev. Bras. Saúde
Prod. Anim., v. 14, p. 406-412, 2013.
COLLIER, R.J.; DAHL, G.E.; VAN BAALE, M.J. Major advances
associated with environmental effects on dairy cattle. J. Dairy Sci., v. 89, p.
1244-1253, 2006.
COSTA, M.J.R. Ambiência na produção de bovinos de corte a pasto.
Anais de Etologia, v. 18, p. 26-42, 2000.
COSTA, M.J.R.; CROMBERG, V.U. Alguns aspectos a serem
considerados para melhorar o bem-estar de animais em sistemas de pastejo
rotacionado. In: Peixoto, A.M.; Moura, J.C.; Faria, V.P. Fundamentos do
pastejo rotacionado. Piracicaba: FEALQ, 1997, p. 327.
DALCIN, V.C. Parâmetros fisiológicos em bovinos leiteiros submetidos ao
estresse térmico. Dissertação (mestrado). PPG Zootecnia, UFRGS. Porto
Alegre, 2013. 31 p.
DIETENBACH, A.; FLORIANI, G.S.; DUBOIS, J.C.L et al. Manual
agroflorestal para a Mata Atlântica. Ministério do Desenvolvimento
Agrário. Brasília, 2008.
DRACKLEY, J.K. Biology of dairy cows during the transition period: The
final frontier? J. Dairy Sci., v. 82, p. 2259–2273, 1999.

226
FISCHER, V.; VIZZOTTO, E.F.; ABREU, A.S.; THALER, A.; STUMPF,
M.T.; WERNCKE, D.; SCHMIDT, F.A. Behavior of lactating dairy cows
under mild and severe heat stress with free access or not to shade. In: Joint
Annual Meeting, Kansas, MI, July 2014, Proceedings….
FOX, D.G.; TYLUTKI, T.P. Accounting for the effects of environment on
the nutrient requirements of dairy cattle. J. Dairy Sci., v. 81, p. 3085-
3089, 1998.
GAFFIN, S.L.; HUBBARD, R. Experimental approaches to therapy and
prophylaxis for heat stress and heatstroke. Wilderness & Environmental
Medicine, v. 7, p. 312-334, 1996.
Intergovernmental Panel on Climate Change (IPCC). 4th Assessment
Report, 2007. Disponível em: <http://
www.ipcc.ch/publications_and_data/publications_and_data_reports.shtml#2
>. Acesso em: 25 março, 2013.
KADZERE, C.T. et al. Heat stress in lactating dairy cows: a review.
Livestock Production Science, v. 77, p. 59-91, 2002.
LI, G.; ALI, I.; CURRIE, R.W. Insulin induces myocardial protection and
Hsp70 localization to plasma membranes in rat hearts. American Journal
of Physiology. Heart and Circulatory Physiology, v. 291, p. 1709-1721,
2006.
MCDOWELL, R.E. et al. Effects of climate on performance of Holsteins in
first lactation. J. Dairy Sci., v. 59, p. 965-973, 1976.
MCMANUS, C.; PALUDO, G.R.; LOUVANDINI, H.; GUGEL, R.;
SASAKI, L.C.B.; PAIVAS, R. Heat tolerance in Brazilian sheep:
physiological and blood parameters. Tropical Animal Health and
Production, v. 41, p. 95–101, 2009.
MARQUES, L.T.; FISCHER, V.; ZANELA, M.B. et al. Produção leiteira,
composição do leite e perfil bioquímico sanguíneo de vacas lactantes sob

227
suplementação com sal aniônico. Rev. Bras. Zoot. v. 40, p. 1088-1094,
2011.
MITLÖHNER, F.M. et al. Shade and water misting effects on behavior,
physiology, performance, and carcass traits of heat-stressed feedlot cattle. J.
Anim. Sci., v. 79, p. 2327–2335, 2001.
National Research Council. Nutrient Requirement of Dairy Cattle. 7th
ed. Washington: National Academic Press, 2001. 408p.
SILANIKOVE, N. Effects of heat stress on the welfare of extensively
managed domestic ruminants. Livestock Production Science, v. 67, p. 1-
18, 2000.
SILANIKOVE, N.; SHAPIRO, F.; SHINDER, D. Acute heat stress brings
down milk secretion in dairy cows by up-regulating the activity of milk-
borne negative feedback regulatory system. BMC Physiology, v. 9, p. 13-
22, 2009.
RENNA, M.; LUSSIANA, C.; MALFATTO, V.; MIMOSI, A.;
BATTAGLINI, L.M. Effect of exposure to heat stress conditions on milk
yield and quality of dairy cows grazing on Alpine pasture. Proceedings, 9th
European IFSA Symposium, Vienna, Austria, July 2011.
RENAUDEAU, D.; COLLIN, A.; YAHAV, S.; de BASILIO, V.;
GOURDINE, J.L.; COLLIER, R.J. Adaptation to hot climate and strategies
to alleviate heat stress in livestock production. Animal, v. 6, p. 707-728,
2012.
RHOADS, M.L.; RHOADS, R.P.; VAN BAALE, M.J. et al. Effects of heat
stress and plane of nutrition on lactating Holstein cows: I. Production,
metabolism and aspects of circulating somatotropin. J. Dairy Sci., v. 92, p.
1986–1997, 2009.
ROBINSON, N.E. Homeostase e Termorregulação. In: Cunningham, J.G.
Tratado de Fisiologia Veterinária. 3ª ed. Rio de Janeiro: Guanabara
Koogan, 2004. p. 550-561.
SEVI, A.; ANNICHIARICO, G.; ALBENZIO, M. et al. Effects of solar
radiation and feeding time on behavior, immune response and production of

228
lactating ewes under high ambient temperature. J. Dairy Sci., v. 84, p. 629-
640, 2001.
STOBER, M. Identificação, anamnese, regras básicas da técnica do exame
clínico geral. In: Rosemberg, G. Exame clínico dos bovinos. Rio de
Janeiro: Guanabara Koogan, 1993. p. 419.
STUMPF, M.T.; FISCHER, V.; MCMANUS, C.M. et al. Severe feed
restriction increases permeability of mammary gland cell tight junctions and
reduces ethanol stability of milk. Animal, v. 7, p. 1137-1142, 2013.
TAO, S.; BUBOLZ, J.W.; AMARAL, B.C.; THOMPSON, I.M.; HAYEN,
M.J.; JOHNSON, S.E.; DAHL, G.E.. Effect of heat stress during the dry
period on mammary gland development. J. Dairy Sci., v. 94, p. 5976–5986,
2011.
TAO, S.; THOMPSON, I.M.; MONTEIRO, A.P.; HAYEN, M.J.; YOUNG,
L.J.; DAHL, G.E. Effect of cooling heat-stressed dairy cows during the dry
period on insulin response. J. Dairy Sci., v. 95, p. 5035-5046, 2012.
TAO, S.; DAHL, G.E. Invited review: Heat stress effects during late
gestation on dry cows and their calves. J. Dairy Sci., v. 96, p. 4079-4093,
2013.
TOMANEK, L. Variation in the heat shock response and its implication for
predicting the effect of global climate change on species’ biogeographical
distribution ranges and metabolic costs. Journal of Experimental Biology,
v. 213, p. 971-979, 2010.
VIZZOTTO, E.F. Comportamento animal e atributos fisiológicos de
vacas leiteiras submetidas a ambientes com e sem sombreamento
durante a estação quente. Dissertação (mestrado). PPG Zootecnia,
UFRGS, Porto Alegre, 2014.
WEST, J.W. Interactions of energy and bovine somatotropin with heat
stress. J. Dairy Sci., v. 77, p. 2091–2102, 1994.
WEST, J.W. Physiological effects of heat stress on production and
reproduction. In: Tri-State Dairy Nutrition Conference, Fort Wayne.
Proceedings, 2002.

229
WEST, J.W. Effects of heat-stress on production in dairy cattle. J. Dairy
Sci., v. 86, p. 2131-2144, 2003.
WHEELOCK, J.B.; RHOADS, R.P.; VAN BAALE, M.J.; SANDERS,
S.R., BAUMGARD, L.H. Effects of heat stress on energetic metabolism in
lactating Holstein cows. J. Dairy Sci., v. 93

230
10. MONITORAMENTO DO BEM-ESTAR ANIMAL COMO
FERRAMENTA DE DIAGNÓSTICO PRECOCE E
MANUTENÇÃO DA SAÚDE PRODUTIVA EM REBANHOS
LEITEIROS11
Marcelo Cecim
Não há muito tempo atrás falar na preocupação com o
conforto e o bem-estar de animais de produção era um assunto que
gerava risadas e desdém, pois acreditava-se que para um animal ser
lucrativo precisava apenas de boa sanidade, boa genética e boa
nutrição. Hoje depois de décadas de seleção, chegamos a um modelo
de vaca leiteira que, apesar de produzir mais leite, tem uma expectativa
de vida cada vez menor e uma crescente infertilidade. Em termos de
evolução das espécies, uma raça que vive e se reproduz menos está
fadada a extinção. No entanto, insiste-se em chamar este fenômeno de
"melhoramento genético".
É necessário entender que à medida que optamos por animais
maiores e mais produtivos, suas exigências serão maiores também.
11 Cecim, M. Monitoramento do bem-estar animal como ferramenta de
diagnóstico precoce e manutenção da saúde produtiva em rebanhos leiteiros.
Anais. I Simpósio Nacional da Vaca Leiteira. Porto Alegre: Universidade
Federal do Rio Grande do Sul. 2014. 239 p.

231
Muito trabalho tem sido direcionado no sentido de reconhecer as
demandas nutricionais crescentes nestes animais para poder produzir
mais. Infelizmente, isso não tem sido suficiente para mantê-las
felizes. Ainda entendemos muito pouco de necessidades sociais, de
adaptação a técnicas de manejo e até mesmo da expectativa individual
de cada vaca. Em relação a isso, sempre vale lembrar o comentário de
Louis Patenaude, proprietário da recordista Gillette Smurf, uma vaca
canadense que viveu 16 anos e produziu mais de 225.000 kg de leite,
quem disse: "Ela foi uma vaca que nunca precisou ser mimada".
Longevidade e fertilidade são indiscutivelmente a opinião das vacas
sobre a sua qualidade de vida. Em nossa realidade, o valor econômico
da longevidade é enorme. A maior parte das fazendas leiteiras no sul
do Brasil está tentando aumentar o plantel, e cada descarte
involuntário atrasa o processo. Em rebanhos estáveis, cada descarte
involuntário precoce, representa uma novilha que o produtor precisa
comprar de si mesmo. Com um custo de recria da novilha oscilando
entre R$ 850,00 e R$ 1.300,00 e um valor de mercado de venda desta
novilha entre R$ 4.500,00 e R$ 6.000,00, cada vez que isso acontece
o produtor faz uma operação comercial com algo próximo a 400% de
prejuízo. Em um rebanho em crescimento como o nosso, a venda de
novilhas rende muito mais que a produção de leite. Além disso, com
um descarte menor, as novilhas selecionadas terão sem dúvida melhor
mérito genético. Também a lucratividade individual de uma vaca
aumenta com a idade, pois não só sua produção bruta aumenta até a
quarta ou quinta lactação, mas quanto maior o número de lactações

232
menor será o custo proporcional da recria deste animal. O resultado
final é que animais com baixa vida produtiva, dificilmente serão
lucrativos, mesmo que produzam muito leite. Aí temos o impacto do
bem-estar sobre a própria sustentabilidade do negócio.
Qual a expectativa de vida produtiva das vacas leiteiras ao
redor do mundo? Estima-se que esteja ao redor de 4,7 lactações no
Reino Unido e 5,3 na Nova Zelândia. Um estudo avaliou 69 milhões
de lactações de vacas de raça Holandesa em diferentes regiões dos
EUA. A expectativa de vida produtiva variou de 2,73 lactações na
região nordeste até 1,94 lactações na região sudeste. Este resultado
assustador indica que se não fosse o advento da sexagem, em breve
haveria rebanhos em que cada vaca produziria menos de uma filha, e
a raça entraria em extinção. Qual a situação no sul do Brasil, onde a
linhagem canadense é a mais utilizada? Estes dados não existem. No
entanto é bastante comum encontrarmos rebanhos onde 30 ou até 35%
dos animais em lactação são vacas de primeira cria. Nestes casos, a
expectativa de vida é menor que 3 lactações e é óbvio que a vida dos
animais nestes rebanhos não é boa.
Existem diferentes formas para a vaca externar a sua
insatisfação com o sistema, as mais leves estão relacionadas a quadros
depressivos e incluem comer menos, ficar menos tempo deitada, estar
mais tempo em pé, ter menor tempo de ruminação, ter cios mais curtos,
etc. Tais sinais, como são de difícil observação acabam passando
despercebidos. Ou seja, como elas não são escutadas, os transtornos
tentam falar mais alto, deslocando abomaso, fazendo cetose, metrite

233
ou uma mastite séria. Quando nada funciona e elas de fato desistem,
já que vaca não foge, a solução é morrer. Um produtor ou técnico
quando afirma que vai descartar esta ou aquela vaca por problema de
casco ou úbere, está no mínimo tendo uma atitude covarde, jogando a
culpa na vaca. Na verdade é a vaca que está dizendo ao dono que não
quer mais viver ali, ou seja, ela o está descartando.
Hoje, na medicina humana, entendemos que prevenção e
diagnóstico precoce são as melhores estratégias de lidar com os nossos
problemas de saúde. Em medicina de produção leiteira temos a mesma
situação, apenas chamamos de ambiência e monitoramento.
Ambiência são todos aqueles fatores pelos quais permitimos que as
vacas expressem seu comportamento normal, ou seja, ambiência são
as ferramentas do bem-estar.
Se uma vaca pudesse de fato escolher o que fazer durante o
dia, o que ela faria? A distribuição do tempo das atividades é uma
ferramenta básica na análise do bem-estar. Mais especificamente, uma
vaca gosta de deitar em cama seca por 12 a 14 horas por dia, a maior
parte deste tempo ruminando. Em torno de 5 horas são dedicadas à
alimentação; 2 a 3 horas elas passam bebendo caminhando e
socializando; em torno de uma hora ruminando em pé; até 3 horas por
dia ela aceita estar presa ou na ordenha.
Aí iniciam os problemas: vacas pastando em piquetes com
baixa oferta precisam comer por cinco horas ou mais, vacas com calor
deitam por menos tempo, duas ordenhas diárias mais o tempo de
espera normalmente passam de 4 horas, falta de cama seca aumenta o

234
ócio em pé. Este “tempo extra” acaba sendo retirado do tempo
deitada, e ruminando, que é a grande expressão de conforto da vaca.
Todos estes fatos comuns no dia a dia de um rebanho são sinais de que
falta conforto e bem-estar para as vacas. No final das contas, prover
conforto para um rebanho leiteiro é um conjunto de atitudes,
instalações e manejo que permitam que o animal expresse o
comportamento normal, em outras palavras, que façam o que uma
vaca feliz gostaria de fazer.
Reconhecer bem-estar de um rebanho leiteiro é algo que pode
ser feito por qualquer um sem nenhum equipamento especial, apenas
precisa saber o que, quando e como observar, quantificar os resultados
e anotar. Para isso foram criadas classificações que são conhecidas
como escores de monitoramento, usando principalmente os escores
de: consumo, preenchimento de rúmen, uso do tempo, ambiência,
interação social, conforto térmico, consistência e composição de fezes,
locomoção etc. Estes comportamentos representam a opinião da vaca
sobre as coisas que lhe são ofertadas. A análise conjunta de todos os
escores representa a análise do próprio sistema produtivo, mas, sob a
ótica da vaca. Isto pode ser usado com muita eficiência para a
composição de um mapa de pontos de riscos e seguranças do sistema,
como uma análise FOFA (pontos fortes e oportunidades, pontos fracos
e ameaças). Esta é a estratégia básica das companhias de seguro na
definição de risco do cliente e do prêmio a ser pago. O problema inicia
com o tempo que tudo isso consome, e, com a capacidade do avaliador
em entender a inter-relação dos escores e formular soluções práticas e

235
viáveis. É humanamente impossível acompanhar e anotar atividade de
um rebanho 24 horas por dia. Todo tratador experiente sabe quando
uma vaca não está bem, muito embora ela não esteja doente.
Infelizmente esta informação, embora valiosa, é tardia. Se a vaca é
reconhecida como não estando bem, é porque ela não comeu, está
deprimida, deu menos leite, andou mais devagar etc. Tudo isso na
verdade são sinais que a vaca “está mal” e o diagnóstico já é tardio,
em pelo menos 24 horas. Aí entram os sistemas de monitoramento de
comportamento. São ferramentas pelas quais a vaca pode dizer que
não se sente bem, 1 ou 2 dias antes de deixar de comer, 3 dias antes de
diminuir a produção e 4 ou 5 dias antes de ficar doente, permitindo
desta forma uma ação proativa e não reativa ao problema. Entendemos
que muito embora as alterações de comportamento não digam qual é
exatamente o problema da vaca, elas avisam ao tratador que este
animal requer uma olhada especial, ou seja, elas permitem um
diagnóstico precoce.
O uso do tempo e do consumo de alimento hoje são
considerados os principais indicadores de que um animal está bem.
Portanto, cada vez que uma vaca altera estes parâmetros do seu
comportamento diário, sabemos que alguma coisa não vai bem, e que
ela merece uma olhada mais próxima. Esta é, sem dúvida, é a melhor
medida preventiva que podemos usar. Os pedômetros são usados
essencialmente na identificação do cio. As coleiras que associam
deslocamento e ruminação tornam esta identificação bem mais
precisa. A coleira brasileira C-Tech, recentemente lançada no

236
mercado, é a primeira a conter algoritmos relacionados ao conforto,
além da detecção de cio. Este sistema é provido de um acelerômetro
que detecta a posição da cabeça da vaca com 40 leituras por segundo,
daí são formados pacotes de comportamento a cada hora, onde são
identificados minutos em: atividade (caminhando, comendo,
montando bebendo), minutos em ruminação e minutos em ócio. Estes
pacotes de dados são enviados automaticamente, 2 ou 3 vezes ao dia,
para um programa que compara o comportamento de cada vaca com
ela mesma nos 15 dias anteriores e também com o resto do rebanho.
Cada vez que uma vaca aumenta seu tempo de atividade, diminuindo
a ruminação e o ócio, o programa reconhece como cio. Por outro lado,
quando existe um aumento no ócio com redução da ruminação e
também da atividade, o sistema entende como alteração de
comportamento e o animal deve ser examinado. Ao final da ordenha,
duas listas são geradas contendo os brincos dos animais alterados. Se
houver internet na fazenda, as listas podem ser automaticamente
enviadas por SMS para o celular do veterinário, por exemplo. Também
o comportamento individual e do lote a cada hora pode ser
acompanhado de qualquer lugar pelo gerente ou proprietário. É
importante salientar, que este equipamento à exceção de cio, não gera
diagnósticos, mas reconhece que algo mudou para pior ou melhor no
dia de uma vaca em particular, ou no seu lote.
Classicamente, um bovino dedica seu dia a 3 atividades
principais, de 8 horas cada: ruminação, deslocamento e ócio. Hoje
entende-se que a ruminação é uma variável mais sensível que o

237
consumo, pois ruminando menos, a taxa de esvaziamento cai e no dia
seguinte cai o consumo. O tempo de ruminação pode aumentar em
consequência do aumento do conforto, do consumo total e de fibra
longa na dieta. Vai diminuir no estresse térmico, nos quadros de dor,
na redução de consumo ou simplesmente na falta de conforto. Os
bovinos preferem ruminar deitados, portanto a qualidade de cama
também afeta a ruminação.
As primeiras observações de campo em rebanhos que utilizam
esta tecnologia mostram informações surpreendentes, como por
exemplo, a alta frequência de cios curtos (5 ou 6 horas). A queda de
minutos de ruminação é normal no dia do parto (até 50%), porém,
vacas que fazem uma boa transição, retornam aos 400-500 minutos de
ruminação por dia, já no quinto dia pós-parto. As vacas que
apresentarão mastite, metrite ou cetose até o dia 21, mostram uma
ascendência da curva de ruminação bem mais lenta. As vacas que
deslocam abomaso entre o dia 14 e 21 mostram baixo tempo de
ruminação entre os dias 0 e 7 depois do parto.
Portanto, estamos aprendendo a escutar a opinião das vacas,
principalmente durante uma fase crítica que é a transição. Os próximos
estudos devem direcionar-se no acúmulo destes dados e a construção
de uma base dados comportamentais das vacas em sistemas produtivos
no Brasil. Isto permitirá a criação de um programa de predição de
desafios e doenças, que levará ao diagnóstico precoce e, por fim,
poderá diminuir o descarte involuntário através do entendimento das
necessidades individuais da vaca.

238
REFERÊNCIAS BIBLIOGRÁFICAS
CARAVIELLO, DZ et al. (2006) Survey on management practices on
reproductive performance of dairy cattle on large US commercial farms. J.
Dairy Sci 89:4723-4735.
COOK, N. (2007) The Dual Roles of Cow Comfort in the ‘Get Lame –
Stay Lame’ Hypothesis Sixth International Dairy Housing Conference
Proceedings16-18 June 2007, (Minneapolis, Minnesota, USA)
COOK, N; NORLUND, K.(2009) The influence of the environment on
dairy cow behavior, claw health and herd lameness dynamics. The
veterinary journal 179 (3):360-369
HALEY, DB; RUSHEN J.Behavioural indicators of cow comfort: activity
and resting behaviour of dairy cows in two types of housing. (2000)
Canadian Journal of Animal Science: 80(2): 257-263.
HARE, E; NORMAN, HD; WRIGHT, JR. (2006). Survival Rates and
Productive Herd Life of Dairy Catt in the United States J. Dairy Sci.
89:3713–3720.
WEARY, D.M. & TUCKER, C. 2003. The science of cow comfort.
Proceedings of the Joint Meeting of the Ontario Agri Business Association
and the Ontario Association of Bovine Practitioners, Guelph, Ont.,
April,2003.

239