04 - função tubular
DESCRIPTION
ÂTRANSCRIPT

Capítulo
4Função Tubular
Antonio Carlos Seguro, Lúcia H. Kudo e Claudia M. de B. Helou
INTRODUÇÃO
TRANSPORTE ATRAVÉS DA MEMBRANA EPITELIAL
PROCESSOS REGULADORES DE TRANSPORTE
TRANSPORTE AO LONGO DO NÉFRON
Túbulo proximal
Alça de Henle
características de transporte de suas duas membranas:apical e basolateral (Fig. 4.2).
A membrana apical ou luminal, que está em contatodireto com o fluido tubular, apresenta diferentes canaisiônicos, carregadores, trocadores e co-transportadores, deacordo com as necessidades de transporte do segmento,além de bombas de transporte ativo, como a H�-ATPase.
A membrana basolateral é a que está em contato com oespaço intercelular e o capilar peritubular. Além de canaise outros tipos de transportes facilitados, a membrana ba-solateral apresenta uma densidade variável de bombas,que utilizam a energia liberada pela hidrólise do ATP paratransportar ativamente o Na� para fora e o K� para o inte-rior da célula (Fig. 4.3). Essas bombas são na verdade en-zimas transportadoras e são denominadas de Na�,K�-ATPases. Em condições normais as Na�,K�-ATPases distri-buem-se apenas na face basolateral das células tubularesrenais. Como esta enzima necessita de ATP, a sua distri-buição nos segmentos do néfron é diretamente proporcio-nal aos segmentos que possuem maior quantidade de mi-tocôndrias. Portanto, o túbulo contornado proximal e aporção espessa ascendente da alça de Henle são os segmen-tos do néfron que apresentam maior distribuição quanti-tativa da Na�,K�-ATPase.
A maior parte do transporte de solutos e de água noepitélio renal é realizada pela via transcelular, ou seja, atra-vés da célula. Mas o fluido e os solutos podem atingir o
Túbulo contornado distal
Túbulo de conexão
Ducto coletor
BIBLIOGRAFIA SELECIONADA
ENDEREÇOS RELEVANTES NA INTERNET
INTRODUÇÃO
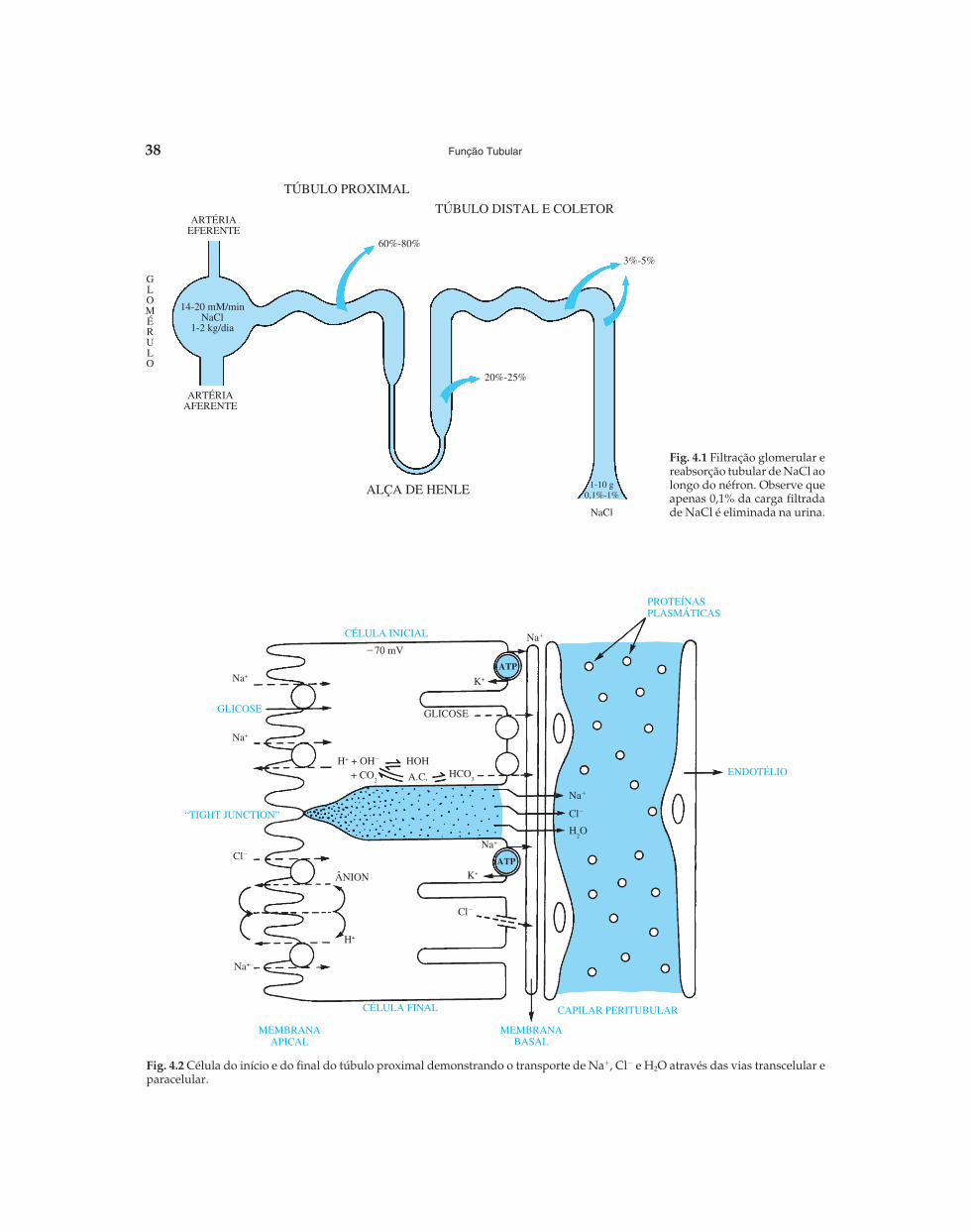
O néfron é a unidade funcional do rim e é constituídopelo glomérulo e 14 segmentos tubulares. O trabalho demilhões de néfrons resulta na formação da urina. Cerca de25% do plasma que atinge o rim são ultrafiltrados pelos glo-mérulos, levando à formação de 100 a 120 ml/min deultrafiltrado em média no homem. Entretanto, apenas 1,2%desse volume é eliminado, e o restante reabsorvido da luztubular para o espaço peritubular (Fig. 4.1).
Ao lado deste intenso processo de reabsorção temosoutro, não menos importante, o de secreção tubular. Este secaracteriza pelo transporte de substâncias do espaço peri-tubular (vasos e interstício) para a luz tubular. Este pro-cesso permite a excreção pela urina de substâncias que nãopassaram pela barreira dos capilares glomerulares, comomacromoléculas ou partículas ligadas a proteínas.
Portanto, a formação da urina resulta de três processos:
1. Filtração glomerular2. Reabsorção tubular3. Secreção tubular
O túbulo renal é formado por uma parede de epitéliosimples, ou seja, uma única camada de células que repou-sa sobre a membrana basal birrefringente. As células epi-teliais renais são ditas polarizadas devido às diferentes
38 Função Tubular
GLOMÉRULO
ARTÉRIAEFERENTE
14-20 mM/minNaCl
1-2 kg/dia
ARTÉRIAAFERENTE
TÚBULO DISTAL E COLETOR
60%-80%
TÚBULO PROXIMAL
3%-5%
20%-25%
ALÇA DE HENLE 1-10 g0,1%-1%
NaCl
Fig. 4.1 Filtração glomerular ereabsorção tubular de NaCl aolongo do néfron. Observe queapenas 0,1% da carga filtradade NaCl é eliminada na urina.
Fig. 4.2 Célula do início e do final do túbulo proximal demonstrando o transporte de Na�, Cl� e H2O através das vias transcelular eparacelular.
PROTEÍNASPLASMÁTICAS
Na�
Na�
Cl�
H2O
CAPILAR PERITUBULAR
K+
ATP
GLICOSE
CÉLULA INICIAL
�70 mV
GLICOSE
H+ + OH�
+ CO2
HOH
A.C. HCO3
“TIGHT JUNCTION”
Cl�
ATP
K+
Cl�
CÉLULA FINAL
MEMBRANAAPICAL
MEMBRANABASAL
Na+
Na+
Na+
H+
ÂNION
ENDOTÉLIO
Na+

capítulo 4 39
capilar pela via paracelular, que é através das junções estrei-tas (tight junctions) e do espaço intercelular, portanto, omovimento é realizado pela face lateral das células. Asjunções estreitas variam de morfologia e de componentesdependendo do segmento, e por isso são denominadasatualmente de complexos juncionais. É através da alta ou da
baixa condutância dos complexos juncionais que se deter-mina a resistência ao movimento molecular pela via para-celular em muitas células. Pode-se citar como exemplo otúbulo contornado proximal, que é considerado como seg-mento do néfron cujo epitélio é de vazamento devido à altacondutância do complexo juncional (Fig. 4.2). O contrárioé observado no ducto coletor medular interno, onde ascélulas epiteliais são fortemente aderidas devido à presençade complexos juncionais de baixa condutância, além dedesmossomos.
TRANSPORTE ATRAVÉS DAMEMBRANA EPITELIAL
O transporte de uma substância através de uma mem-brana epitelial pode ser feito por:
1. Mecanismo passivo2. Mecanismo ativo
Nos processos de transporte passivo, o movimentotransepitelial (reabsorção ou secreção) se faz sem gasto deenergia, obedecendo às forças físicas como gradiente quí-mico (reabsorção de uréia), pressão hidrostática (filtraçãoglomerular), gradiente elétrico (reabsorção de cloretos notúbulo proximal) ou pela diferença de potencial eletroquí-mico ocorrido pelo transporte de algum íon, ou então pelaforça física resultante do movimento do arrasto do solvente(solvent drag). O transporte passivo pode ser então por sim-ples difusão ou por difusão facilitada através de poros, carre-gadores ou canais existentes na membrana.
O processo de difusão simples através do epitélio ocor-re com muitas substâncias ao longo do néfron, caracteri-zando-se pela migração transmembrana de uma substân-cia apenas sob a ação do gradiente químico, elétrico ouentão de pH. Neste caso a quantidade transportada depen-derá apenas do gradiente existente e da maior ou menorpermeabilidade da membrana em relação à substância a sertransportada.
Com relação ao solvente como a água, que também éreabsorvida em muitos segmentos do néfron, a difusãopassiva se dá no túbulo renal por osmose, isto é, a água semovimenta do meio menos concentrado (com menor os-molalidade) para o mais concentrado (com maior osmola-lidade). O coeficiente de reflexão do soluto, que pode va-riar de zero a um, é que determina o movimento da águaatravés da membrana. Quanto maior o coeficiente de re-flexão, maior a capacidade do soluto de produzir um mo-vimento de água através da membrana. Isto é, o soluto quepossui alto coeficiente de reflexão exerce maior pressãoosmótica para um mesmo gradiente de concentração. Aosmose determina a reabsorção de 99% da água filtradapelo glomérulo, e é este tipo de transporte que permite aformação de urina concentrada (alta osmolalidade).
Fig. 4.3 Estrutura da Na�-K�-ATPase. (A) A bomba pode ser umheterodímero �, �. A subunidade � contém os sítios de ligaçãopara Na� (1), para ATP (4), para fosforilação (5), para K� (2) e paraouabaína (3). (B) O painel inferior mostra a subunidade � atra-vessando a membrana sete a oito vezes. A subunidade �, que églicosilada em sua porção extracelular, atravessa somente umavez a membrana. A função da subunidade � não é conhecida, masela é indispensável para o completo funcionamento da Na�-K�-ATPase.
A
B
2K+
�Ouabaína
Citoplasma
3 Na+
Mg ATP Mg ADP + Pi
subunidade � subunidade �
32
1
4 5
C
C
2 3
N N1
4 5
C
?
?
�
c
a
b
Pontos-chave:
• A formação da urina se deve à filtraçãoglomerular e ao trabalho do epitélio tubularem processos de reabsorção e secreção
• O transporte tubular se faz pelas viastranscelular e paracelular através doscomplexos juncionais
• O gradiente eletroquímico gerado pelaNa�,K�-ATPase inserida na membranabasolateral é o responsável por diversostransportes que ocorrem na membranaluminal

40 Função Tubular
O gradiente gerado por pH também pode induzir difu-são passiva de uma substância pela membrana epitelial.Provavelmente devido à natureza hidrofóbica da membra-na celular, formas não-ionizadas de ácidos e bases fracaspenetram mais rapidamente do que formas ionizadas.Considerando que em muitos segmentos do néfron o pHdo fluido tubular difere do existente no espaço peritubu-lar, a geração de um gradiente de pH favorece a difusãode ácidos e bases fracas pelo epitélio. Se o pH do fluidotubular for mais ácido, como ocorre normalmente, o gra-diente resultante favorecerá a reabsorção de ácidos fracosdo lúmen para o espaço peritubular. Mesmo que a concen-tração do ácido fraco seja idêntica nos dois lados do epité-lio, o baixo pH luminal favorecerá a não-dissociação doácido e portanto a sua difusão do espaço luminal para operitubular. Entretanto, se o pH luminal for mais elevadoque o do espaço peritubular, a dissociação do ácido seráfavorecida, resultando em menor reabsorção, por ser estaforma menos permeável (Fig. 4.4).
O inverso ocorre com bases fracas. A acidificação do flui-do tubular aumenta a dissociação de bases fracas, dificul-tando então a sua difusão do lúmen para o espaço peritu-bular (Fig. 4.4).
Em resumo, a evidência de transporte passivo origina-se de duas observações básicas: 1a) desaparecimento dotransporte quando se abole ou anula o gradiente elétricoe/ou químico; 2a) quando o uso de inibidores metabólicosnão altera o transporte da substância em estudo.
No caso de transporte ativo, a reabsorção ou a secreçãode uma determinada substância se faz contra gradienteelétrico, químico ou ambos, e por conseguinte é feita à custade energia. No transporte ativo temos uma dependênciaimediata do metabolismo celular, e a inibição deste deter-mina a parada do transporte.
Os transportadores que utilizam diretamente a energialiberada pela hidrólise do ATP são considerados como ele-mentos de transporte ativo primário e são chamados de bom-bas. Na verdade, as bombas são enzimas que possuem umsítio de ligação para o ATP e por isso são também conheci-das como ATPases. A fosforilação destas enzimas permiteque íons sejam transportados contra gradientes químicose/ou elétricos (Fig. 4.3). Um bom exemplo é a Ca��-ATPa-se, que ativamente transporta o Ca�� do intracelular, cujaconcentração é de 100 a 150 nM, para o interstício, onde aconcentração deste íon é aproximadamente 6.000 a 10.000vezes maior (1 mM).
A energia liberada por uma ATPase para o transportede um íon pode induzir um gradiente eletroquímico quefacilita o movimento desse íon a favor do gradiente gera-do. A este transporte iônico pode-se acoplar um outro so-luto que poderá ser na mesma direção, co-transporte, ou emsentido oposto, antiporte. Por isso, este transporte acopla-do é tido como transporte secundariamente ativo (Fig. 4.2).
Como exemplo de co-transporte secundariamente ati-vo podemos citar o de Na�-glicose que existe na face lu-minal das células do túbulo proximal. As Na�,K�-ATPasespresentes na face basolateral dessas células geram um gra-diente eletroquímico que facilita a entrada de Na� pela faceluminal (Fig. 4.2). Esta entrada pode ser através de umaproteína transportadora que possui sítios específicos paraNa� e para glicose (Fig. 4.5). Primeiro, o Na� se liga ao seurespectivo sítio e produz uma alteração na conformaçãoprotéica do carregador, expondo o sítio para a ligação daglicose. Essa segunda ligação (glicose e receptor) provocauma nova alteração na estrutura da proteína, permitindoque tanto o Na� quanto a glicose atravessem a membrana.Portanto, Na� e glicose passam pela membrana lipoprotéi-ca utilizando a energia liberada pela Na�,K�-ATPase. Aflorizina pode inibir este co-transporte, competindo coma glicose pelo mesmo sítio de ligação no carregador. A li-gação da florizina ao sítio não promove a segunda altera-ção na proteína carregadora, impedindo então o co-trans-porte Na�-glicose (Fig. 4.5).
Em muitos segmentos do néfron a secreção de H� ocor-re através do transportador Na�-H�. Este sistema trocador deíons é também secundariamente ativo, pois a secreção deH� para a luz tubular é feita acoplada a um movimentocontrário de Na�. O Na� movimenta-se da luz para o in-tracelular a favor de gradiente eletroquímico gerado pelaatividade da Na�,K�-ATPase (Fig. 4.2).
Convém também citar um tipo especial de transporteativo, que é a endocitose. Macromoléculas são reabsorvidasatravés do seu envolvimento pela membrana apical, resul-tando em invaginações e formação de vacúolos. Quandoo conteúdo dos vacúolos é de substâncias sólidas, esseprocesso recebe o nome de fagocitose, e quando o vacúoloé formado por fluido, a denominação é de pinocitose. Nocitoplasma, o material fagocitado pode sofrer ações de di-gestão. A extrusão do conteúdo vacuolar para o extracelu-
A
HA HA H� A�
B� OH�
H�
A
pH 5,5 pH 7,4PERITUBULAR
LUZ TUBULAR
BOH BOH
pH 8,5
B�
OH�
pH 7,4PERITUBULARLUZ TUBULAR
B
Fig. 4.4 Difusão transtubular à custa de um gradiente de pH.Esquema A: reabsorção de um ácido fraco (HA) e ausência dereabsorção de base fraca (BOH) em virtude de o pH do fluidotubular ser inferior ao peritubular. Esquema B: reabsorção de umabase fraca (BOH) e não-reabsorção de ácido fraco decorrente deum pH urinário alcalino.

capítulo 4 41
lar recebe o nome de exocitose e consiste na fusão da mem-brana vacuolar à membrana basolateral da célula e conse-qüente extrusão do conteúdo do vacúolo para o espaçoextracelular.
Nos túbulos renais o transporte de macromoléculas érepresentado principalmente pela reabsorção de proteínasfiltradas pelo glomérulo, que ocorre logo no primeiro seg-mento do néfron, túbulo contornado proximal.
Pontos-chave:
• Transporte passivo: difusão, difusãofacilitada, “solvent-drag”
• O transporte ativo é realizado por ATPases,enzimas que hidrolisam o ATP
• O gradiente eletroquímico gerado pelasATPases pode permitir o transportesecundário de outros íons
PROCESSOS REGULADORES DETRANSPORTE
Didaticamente podemos dividir os processos regulado-res de transporte em: fatores cinéticos, endocitoses-exocitoses e segundos mensageiros.
Os fatores cinéticos modulam a velocidade de transportealterando a concentração de solutos. O transporte de uma
substância pode ser saturável ou insaturável, independentede ele ser ativo ou passivo.
Um transporte é classificado como saturável quando aquantidade da substância transportada na unidade de tem-po aumenta até um certo limite, acima do qual o aumentoda substância a ser transportada não mais incrementa otransporte, pois alcançou o transporte máximo, Tm. Portan-to, quando se atinge o Tm de uma substância, nem a adi-ção de energia, no caso de transporte ativo, nem o aumen-to do gradiente químico e/ou elétrico, no caso de transpor-te passivo, aumenta o transporte.
A existência de um transporte máximo saturável podeser decorrente de vários mecanismos:
1. Existência de um carregador auxiliando no transporte.Então, o Tm da substância a ser transportada é deter-minado pela quantidade de carregadores existentes, ou,então, se o sítio de ligação a uma determinada substân-cia apresenta afinidade a uma outra, resultando em umprocesso de competição. A galactose por exemplo com-pete com a glicose pelos mesmos receptores da proteí-na carregadora presente no túbulo contornado proxi-mal.
2. Limite de energia para transporte ativo. Por exemplo, oTm de glicose pode ser diminuído pela presença detransporte de fosfato que compete pela energia libera-da pela Na�,K�-ATPase.
3. Limite do gradiente eletroquímico gerado pelo transpor-te ativo. Assim, uma substância ou íon sendo transpor-tado da luz tubular para o espaço peritubular por um
Na�
Na�
Na�
FLORIZINA
TIRO-SINA
LISINA
GLICOSE
Na�
GLICOSE OUFLORIZINA
FLORIZINA
GLICOSE
Na�
Fig. 4.5 Representação esquemática do co-transporte Na�-glicose. Os sítios de ligação de sódio e glicose na proteína transportadoralocalizam-se no lado externo da membrana celular. A ligação do sódio causa alteração estrutural na enzima transportadora, resul-tando na exposição do sítio de ligação à glicose. A interação glicose e receptor induz uma segunda alteração estrutural que permitea passagem do Na� e da glicose para o interior da célula. A florizina pode competir com a glicose pelo receptor. Entretanto, a ligaçãoflorizina-receptor não induz alteração estrutural, impedindo então que tanto florizina quanto Na� sejam transportados para o intra-celular.

42 Função Tubular
mecanismo ativo diminuiria progressivamente sua con-centração luminal, aumentando-a no espaço peritubu-lar se esses fluidos não fossem removidos. Este aumen-to de concentração no espaço peritubular e o gradienteelétrico criado pelo transporte favorecem a volta desteíon ou da substância para a luz tubular, anulando o tra-balho ativo efetuado.
O processo de endocitose-exocitose é considerado comoregulador de transporte, pois em condições de repouso ostransportadores podem estar seqüestrados em vesículaslogo abaixo da membrana apical. É necessário um estímu-lo apropriado para que ocorra a inserção dessas proteínasformando evaginações na face luminal da membrana. Oaumento de inserções dessas proteínas favorece o transpor-te da substância em questão. Como exemplos podemoscitar a secreção de H� e o fluxo de água induzido pela va-sopressina. No caso da secreção de H�, a acidificação dacélula é o estímulo para a inserção na borda luminal dasvesículas que contêm as H�-ATPases. No caso do transpor-te de água estimulado pela vasopressina, os canais de água(aquaporinas) são ancorados à membrana através da ge-ração de AMP cíclico e portanto com a utilização de umsegundo mensageiro. A exocitose por sua vez requer a açãointegrada do citoesqueleto celular. Assim, um estímulo in-duz o aumento de circulação de vesículas ativando tantoa endocitose quanto a exocitose.
A regulação de transporte através da ação de segundosmensageiros vem sendo amplamente estudada, principal-mente nos últimos anos. Entre eles podemos citar a gera-ção do AMP e GMP cíclicos e a variação da concentraçãodo Ca�� livre intracelular ([Ca��i]), que podem modulardiretamente as proteínas transportadoras ou afetar a aber-tura de um canal iônico.
Na regulação de transporte existe ainda o fenômeno deadaptação ao longo do tempo. O melhor exemplo é o da esti-mulação da reabsorção de Na� no ducto coletor induzidopela aldosterona. Este hormônio estimula a produção deproteínas que ativam os canais de Na+ existentes na mem-brana luminal, como também aumenta a síntese de Na�,K�-ATPase. Este mineralocorticóide também favorece a inser-
ção e a ativação desta bomba na membrana basolateral. Acapacidade metabólica da célula também é influenciadapela mediação da aldosterona a nível de mitocôndria eportanto pela produção de ATP (Fig. 4.3). Assim, a aldos-terona é um agonista que participa da adaptação da célulado ducto coletor para aumentar o transporte de Na� nestesegmento do néfron.
TRANSPORTE AO LONGO DONÉFRON
Túbulo ProximalO túbulo proximal, segmento que segue imediatamen-
te o glomérulo, é responsável pela reabsorção da maiorparte das substâncias que são filtradas pelo glomérulo. Porisso, este segmento do néfron desempenha importantepapel no controle da eliminação de diversas substâncias.Assim, pequenas alterações na intensidade de reabsorçãoao nível do túbulo proximal podem causar variações sig-nificantes na excreção urinária de uma dada substância.
O túbulo proximal é constituído por três segmentos. Osdois primeiros, que são denominados de S1 e S2, correspon-dem à parte convoluta do túbulo e a eles se segue umaporção retificada, S3, conhecida também como pars recta. Amaior parte de água, sódio e cloro filtrados pelo gloméru-lo (60% a 70% da carga filtrada) é reabsorvida pelo túbuloproximal (Fig. 4.1).
A análise da composição química do fluido obtido dotúbulo proximal mostra que a concentração de Na� perma-nece idêntica à do plasma (�140 mEq/L), assim como aosmolaridade. Estes dados indicam, então, que a reabsor-ção do Na� nesta região do néfron é acompanhada pelamesma proporção de água, portanto, uma reabsorção isotô-nica.
Como já foi referido em parágrafos anteriores, a entra-da do Na� pela membrana apical das células do túbuloproximal ocorre através de mecanismos passivos a favorde um gradiente eletroquímico gerado pelas Na�,K�-ATPases presentes na membrana basolateral. Na verdade,esses mecanismos são secundariamente ativos, pois utili-zam a energia liberada pela quebra do ATP. A entrada desódio na célula se faz através de dois mecanismos:
1. co-transporte que pode ser com a glicose, com o fosfatoinorgânico, com os aminoácidos, com os sulfatos ouentão com os outros ácidos orgânicos (Fig. 4.2). Este sis-tema ocorre principalmente nos segmentos S1 e S2 e éatravés de um processo de difusão facilitada que essassubstâncias saem passivamente da célula pela membra-na basolateral;
2. trocador Na�-H�. Através da quebra da molécula daágua o íon H+ é liberado e secretado para a luz tubularatravés de uma troca com o Na�. A hidroxila, por sua
Pontos-chave:
• Certos transportadores como o da glicosesão saturáveis. Portanto, atingem umtransporte máximo (Tm)
• O processo de endocitose permite estocardentro das células ATPases e outrasproteínas, como por exemplo asaquaporinas. O inverso, a exocitose, permitea inserção dessas proteínas na membranacelular em condições de estímulo

capítulo 4 43
vez, em presença da anidrase carbônica, reage com oCO2 formando o HCO3
� que sai da célula pela membra-na basolateral por um co-transporte ligado ao Na� naproporção de 1 cátion para 3 ânions (Fig. 4.2).
No início do túbulo proximal, o gradiente elétrico entrea luz tubular e o espaço peritubular é da ordem de �2 a�4 mV, lúmen negativo (Fig. 4.6). Estes dados sugerem quea reabsorção de Na� se faz contra gradiente elétrico. Omovimento de cargas positivas devido à ação das Na�,K�-ATPases existentes na face basolateral das células seriaresponsável por essa diferença de potencial transtubular.Entretanto, nos segmentos finais do túbulo proximal ondepraticamente toda a glicose, o fosfato e os aminoácidosforam reabsorvidos, a diferença de potencial transtubularpassa a ser de �1 a �2 mV, lúmen positivo (Fig. 4.6). Istoé explicado pela difusão de íons cloro, cuja concentraçãoaumenta progressivamente ao longo do túbulo proximal.No início do túbulo proximal, a reabsorção de sódio é pre-ferencialmente acompanhada pela reabsorção do bicarbo-nato. Dessa maneira, a concentração de cloro na luz tubu-lar aumenta progressivamente ao longo deste túbulo, atin-gindo a concentração de 135 mEq/L no segmento S3, valoreste superior à do plasma e à do espaço peritubular, que éde 105 a 110 mEq/L, como está ilustrado na Fig. 4.6.
A reabsorção de cloro se faz tanto pela via paracelularquanto pela transcelular. Neste último caso, o cloro entrapela membrana apical através de um trocador de Cl� aco-plado a outro ânion, e através de gradiente eletroquímicofavorável, o cloro se difunde pela membrana basolateralda célula. Em conseqüência à difusão passiva dos íons Cl�,o gradiente elétrico é gerado com lúmen positivo, favore-cendo portanto a reabsorção passiva de cátions como Na�,K� e Ca�� neste segmento do néfron.
Outro importante íon reabsorvido pelo túbulo proximalé o potássio. Este íon utiliza principalmente a via parace-
lular e mecanismos passivos. O fato de a água ser ampla-mente reabsorvida ao longo do néfron induz um aumentona concentração de potássio na luz tubular, criando-se en-tão um gradiente químico que facilita a sua reabsorção.Além desse mecanismo, também se tem sugerido a possi-bilidade de o K� ser reabsorvido neste segmento por umtransporte ativo. Experimentos inibindo a reabsorção deNa� com acetazolamida (inibidor da anidrase carbônica)mostraram que a concentração de potássio no fluido tubu-lar diminui, atingindo valores inferiores aos observados noespaço peritubular e plasma, indicando que a reabsorçãode potássio no túbulo contornado proximal envolve tam-bém um mecanismo ativo de transporte.
O transporte de água através do túbulo proximal se faztanto pela via transcelular quanto paracelular devido aogradiente de pressão osmótica existente entre o fluido tu-bular e o espaço peritubular. Apesar do baixo gradienteosmótico, de 2 a 5 mOsm/kg H2O, ele é suficiente parainduzir a reabsorção da água, uma vez que as membranasapical, basolateral e complexo juncional das células do tú-bulo proximal são muito permeáveis a este solvente. Porisso, como já foi referido anteriormente, este epitélio é con-siderado como de vazamento.
Nas porções iniciais do túbulo proximal essa ligeira hi-pertonicidade do fluido peritubular em relação ao lúmené induzida pela reabsorção de Na� acoplada ao HCO3
� ouao co-transporte com outros solutos como a glicose. Nametade final deste túbulo, embora a concentração luminalde Cl� (�135 mEq/L) seja maior que a do espaço peritu-bular, a reabsorção da água também é feita por osmose,uma vez que o sódio, o bicarbonato e os outros solutos queforam reabsorvidos na porção inicial geram um gradienteosmótico maior que o Cl�.
A intensa reabsorção de Na� e água ao longo do túbulocontornado proximal forma o gradiente químico que fa-
Fig. 4.6 Transporte de água e solutos ao longo do túbulo contornado proximal.
TÚBULO CONTORNADO PROXIMALA. EFERENTE
GLOMÉRULO
Cl� 110
Na+
A. AFERENTE
INICIAL
�4 mV
HCO�3
FOSFATO
GLICOSE
AMINOÁCIDOS
FINAL
+ 1,0 mV
CAPILARPERITUBULAR
Cl� 135
Cl�
Na+
H2O
Ca2+

44 Função Tubular
vorece a reabsorção passiva de outras substâncias perme-áveis a este epitélio, como a uréia, o ácido úrico e os íonsK� e Cl�. Desta maneira, a diminuição na reabsorção pro-ximal de Na� acarreta também a diminuição da reabsor-ção desses outros solutos. O transporte de Ca�� e Mg�� émodulado por fatores hormonais, mas existem evidênciasde que também está relacionado com o transporte ativo deNa�. O fosfato também é intensamente reabsorvido, prin-cipalmente nas porções iniciais do túbulo contornado pro-ximal. Este transporte diminui com a redução na quanti-dade de Na� reabsorvida e com o aumento da concentra-ção de paratormônio através do estímulo da adenilciclase.
Ainda em relação ao transporte de Na� no túbulo pro-ximal, é importante descrever a teoria do balanço gloméru-lo-tubular. Verifica-se que frente a variações fisiológicas dafiltração glomerular ocorrem alterações paralelas da reab-sorção de Na� no túbulo proximal, de modo que perma-nece constante a quantidade do íon reabsorvido em rela-ção à sua carga filtrada, ou seja, a fração de reabsorção de Na�
mantém-se inalterada. O balanço glomérulo-tubular é de-corrente pelo menos em grande parte das variações daconcentração de proteínas nos capilares, pressão oncótica,que ocorre durante as alterações da filtração glomerular,como mostra a Fig. 4.7. Quanto à finalidade da existênciado balanço glomérulo-tubular, acredita-se que esse proces-so, juntamente com o feedback túbulo-glomerular, que serádescrito adiante, constituem os dois mecanismos pelosquais o rim impede a perda de sódio durante variações fi-siológicas da carga filtrada de sódio devido a alterações dafiltração glomerular.
Quanto às proteínas, que eventualmente escapam noprocesso de ultrafiltração glomerular, são reabsorvidas atra-vés de mecanismo de endocitose já descrito anteriormente.
A pars recta ou segmento S3 do túbulo proximal se iniciano córtex renal a partir da última alça da parte convoluta ese dirige em linha reta para a medula terminando ao nívelde medula externa. Na microscopia óptica as células des-se segmento são semelhantes às da parte convoluta. Entre-tanto, os estudos de microscopia eletrônica revelam que apars recta é constituída por células epiteliais retangularescom grande quantidade de mitocôndrias junto à membra-na peritubular, mas com menor número de invaginaçõesna membrana basolateral.
Quanto à fisiologia da pars recta, a reabsorção de sódiotambém se faz à custa da geração de gradiente eletroquími-co induzido pelas Na�-K�-ATPases presentes na membra-na basolateral. O gradiente elétrico e químico criado pelotransporte de Na� é que determina a reabsorção passiva deCl�, cuja concentração é elevada neste segmento. A reabsor-ção de Na� também é do tipo isotônica, pois a mesma quan-tidade de água acompanha este cátion (Fig. 4.8).
Apesar de a pars recta dos néfrons superficiais possuirum comprimento de 5 mm, a quantidade reabsorvida deNa�Cl� e água é apenas em torno de 5 a 10% da carga fil-trada, e portanto significativamente menor do que nasporções convolutas.
Entretanto, analisando a capacidade de secreção de áci-dos orgânicos, verifica-se que a pars recta tem maior capa-cidade em secretar ácido úrico, para-amino-hipurato eoutros ácidos que os segmentos S1 e S2. O transporte des-
Fig. 4.7 Mecanismos que impediriam a perda de NaCl: balançoglomérulo-tubular e feedback túbulo-glomerular.
BALANÇO
GLOMÉRULO-TUBULAR
70%
14 mEq/min
Na+
A. AFERENTE
MÁCULA
DENSA
10%
Na+ 0,9 mEq/min
Cl� 15%
3-5%
“FEEDBACK”TÚBULO-GLOMERULAR
2,8mEq/min
Fig. 4.8 Processos de reabsorção e secreção na pars recta do túbu-lo proximal.
GLO
MÉ
RU
LO
T.C. Proximal
Na+Cl�H
2O
Ác. Orgânicos
K+
UréiaNa+Cl�H
2O
Ác. Orgânicos
PARS
RECTA
A. AFERENTE

capítulo 4 45
ses ácidos orgânicos é mediado por carregadores e portantopor mecanismo saturável. Do ponto de vista clínico e far-macológico, a alta capacidade do segmento S3 em secretarácidos orgânicos constitui uma via importante de excreçãode muitos medicamentos como a aspirina, antibióticos ediuréticos.
Uma outra função muito importante atribuída ao seg-mento S3 é a sua capacidade de secretar K� e uréia. Portan-to, a pars recta participa dos mecanismos de concentraçãourinária como elemento integrante no sistema de contra-corrente.
Pontos-chave:
• O túbulo proximal é responsável pelareabsorção isotônica de 60 a 70% da cargafiltrada de Na�Cl� e água
• O sódio é reabsorvido na membranaluminal através de diferentes mecanismos:trocador Na�-H�, co-transporte com glicose,fosfato e aminoácido
• O bicarbonato é preferencialmentereabsorvido nos segmentos S1 e S2
• Na pars recta (segmento S3) ocorrereabsorção preferencial de Cl� e secreção deácidos orgânicos
Alça de HenleA alça de Henle é dividida em porção fina descendente,
porção fina ascendente, porção espessa ascendente medular eporção espessa ascendente cortical.
A porção fina descendente é altamente permeável àágua e pouco permeável a solutos. Aproximadamente 20%da água filtrada é reabsorvida neste segmento. A diferen-ça de potencial transtubular é próxima a zero com lúmennegativo (�2 a �4 mV).
O segmento que se segue à porção fina descendente daalça de Henle é a curvatura. Esta porção do néfron é mui-to utilizada pelos micropuncionadores para o estudo dafunção dos néfrons justamedulares.
A porção fina ascendente da alça de Henle apresentacomo característica ser impermeável à água mas permeá-vel a Cl� e a Na�, que são reabsorvidos por um processopassivo na sua maior parte.
A porção espessa ascendente da alça de Henle que tam-bém é impermeável à água é responsável pela reabsorçãode 25% da carga filtrada de sódio. A Na�, K�-ATPase pre-sente na membrana basolateral gera um gradiente eletro-químico que favorece a entrada do Na� pela membranaapical através de um co-transporte Na�-K�-2Cl (Fig. 4.9).
Existem indícios de que o co-transporte Na�-K�-2Cl�
obedece a uma seqüência de ligações iônicas que se suce-
dem resultando em alterações na estrutura do co-transpor-tador para poder permitir as uniões seguintes. Primeiro éo Na� que se liga, seguindo-se um íon Cl� e em terceirolugar o K�, e só então é que se liga o segundo Cl�. A furo-semida e a bumetanida podem inibir este sistema de co-transporte ao se ligarem no lugar do segundo Cl� na últi-ma etapa.
Uma vez no intracelular, o Na� é ativamente transporta-do para o interstício através da ação da Na�-K�-ATPase namembrana basal, mas o K� e o Cl� são transportados passi-
Fig. 4.9 Célula da porção espessa ascendente da alça de Henlemostrando o co-transporte Na�-K�-2Cl� e o contratransporteNa�-H� na membrana luminal. Os íons Na� são ativamente trans-portados através da membrana basal pela Na�-K�-ATPase e osíons K� e Cl� saem passivamente da célula através de canais.Outro dado importante a assinalar é o potencial positivo do flui-do tubular em relação ao peritubular.
PORÇÃO ESPESSA ASCENDENTE DA ALÇA DE HENLE
LÚMEN+3 a +10 mV
CÉLULA�70 mV
PERITUBULAR0 mV
1 Na+
2 Cl�
1 K+
Na+
Na+
K++ CO
2
H+ + OH HOHA.C.
HCO�3
K+
K+
Cl�
Na+ e
OUTROSCÁTIONS
ATP
Pontos-chave:
• A porção fina descendente é permeável àágua e muito pouco a solutos
• 25% da carga filtrada de Na�Cl� éreabsorvida nas porções ascendentes da alçade Henle
• Presença do co-transportador Na�-K�-2Cl�,sensível ao furosemide, na membranaluminal da porção espessa ascendente. Esteco-transportador é elemento muitoimportante nos mecanismos deconcentração e diluição urinárias
• Ca2� e Mg2� são reabsorvidos pela viaparacelular

46 Função Tubular
vamente. O K� retorna ao lúmen através de um canal espe-cífico (pertencente à família ROMK) na membrana apical eo Cl� sai da célula pela membrana basal através de um ca-nal específico a este ânion. A saída de carga positiva para olúmen e de uma carga negativa para o interstício gera umpotencial positivo luminal de cerca de �7 mV. Esta diferen-ça de potencial permite que o Na+ e outros cátions como opróprio K�, Ca�� e Mg�� sejam reabsorvidos passivamentepelos espaços intercelulares laterais, como ilustra a Fig. 4.9.
No segmento cortical da porção espessa ascendente daalça de Henle é descrita também a secreção de H� atravésdo trocador Na�-H� presente na membrana luminal; cer-ca de 10% da carga filtrada de bicarbonato são reabsorvi-dos neste segmento.
Túbulo Contornado Distal
O túbulo contornado distal é a continuidade do segmen-to cortical da porção espessa ascendente da alça de Henlese estendendo da mácula densa até a região de transiçãocom o ducto coletor. Este segmento do néfron também éimpermeável à água e apresenta características especiaisquanto ao transporte de sódio e cloro. Através de um co-transporte com o Cl�, o Na� é transportado pela membra-na luminal de maneira passiva. Este co-transporte pode serinibido por tiazídicos e é secundariamente ativo à ação daNa�-K�-ATPase que transporta ativamente o Na+ pelamembrana basal, mas o Cl� sai do interior da célula atra-vés de mecanismo passivo por canal específico (Fig. 4.10).
A teoria do feedback túbulo-glomerular relaciona a quan-tidade de Na� que chega aos segmentos distais do néfron
e a regulação da filtração glomerular. De acordo com estateoria, a quantidade de Na� ao atingir o início do túbulodistal sensibiliza a mácula densa, que por sua vez ativamecanismos efetores que irão modular a resistência dosvasos pré-glomerulares. Portanto, se grande quantidade deNa� atinge o início do túbulo distal, a renina é liberada,induzindo vasoconstrição da arteríola aferente com con-seqüente redução do fluxo sanguíneo renal, pressão glo-merular e filtração glomerular (Fig. 4.7).
Ponto-chave:
• Presença do co-transportador Na�-Cl�
sensível a tiazídicos na membrana luminal
Túbulo de ConexãoA região de transição entre o túbulo contornado distal
e o ducto coletor pode ser abrupta ou gradual, e como nestelocal vários túbulos distais se reúnem para formar o ductocoletor, esse segmento do néfron recebe então o nome detúbulo de conexão. Ele é constituído por dois tipos de célu-las: as do túbulo de conexão e as intercaladas. As célulasdo túbulo de conexão apresentam características morfoló-gicas compatíveis com a transição entre as células do dis-tal e as principais do ducto coletor. A principal funçãodessas células está relacionada à secreção de potássio, queé em parte regulada por mineralocorticóides. As célulasintercaladas, por sua vez, desempenham importante papelnos mecanismos de secreção de H�, que será descrito emdetalhes no parágrafo referente ao túbulo coletor.
Neste segmento do néfron, o sódio pode ser reabsorvi-do através de um co-transporte acoplado ao Cl� semelhan-te ao acima descrito nas células do túbulo contornado dis-tal. Aliás, estudos em coelhos onde o túbulo de conexão ébem evidente sugerem que o co-transporte Na�-Cl� sensí-vel a tiazídico ocorre apenas neste segmento. O Na� podetambém ser transportado para o intracelular através decanais sensíveis ao amiloride e do trocador Na�-H� aco-plado a um trocador Cl�-HCO3
�. A ação ativa da Na�,K�-ATPase presente na membrana basal também é respon-sável pela saída do Na� da célula (Fig. 4.11).
É importante ressaltar que o túbulo de conexão e o cole-tor cortical são os únicos segmentos do néfron onde o bicar-
Fig. 4.10 Célula do túbulo distal inicial mostrando o transportede Na� acoplado a Cl� sensível ao tiazídico na membrana lumi-nal secundariamente ativo à ação de Na�-K�-ATPase da mem-brana basal.
TÚBULO DISTAL INICIAL
LÚMENNEGATIVO
CÉLULA�70 mV
PERITUBULAR0 mV
Na+
ATP
Cl�
INIBIÇÃOPELOTIAZÍDICO
K+
Na+
Cl�
Pontos-chave:
• Constituído por células de transição entre asdo túbulo distal e do ducto coletor
• Presença do trocador Cl�-HCO3� na
membrana luminal pode permitir a secreçãode bicarbonato em certas condições dealcalemia

capítulo 4 47
bonato pode ser secretado para a luz tubular utilizando paratal o trocador Cl�-HCO3
� presente na membrana apical.Também é descrita a presença de uma ATPase na face
luminal das células intercaladas neste segmento, denomi-nada H�,K�-ATPase. Esta enzima seria estimulada peladepleção de K�.
Ducto ColetorO ducto coletor é dividido em cortical, medular exter-
no e medular interno, apresentando dois tipos de células:as principais e as intercaladas.
As células principais caracterizam-se na microscopia ele-trônica por apresentarem um cílio central. O sódio é reabsor-vido nestas células por mecanismo passivo através de canaisna membrana luminal sensíveis ao amiloride ou trianterene,denominados ENaC. É também através da Na�,K�-ATPaseque o transporte ativo de Na� gera potencial negativo nolúmen na ordem de �30 mV no coletor cortical (Fig. 4.12).
A concentração de K� no intracelular das células princi-pais do ducto coletor é elevada devido à alta atividade dasNa�,K�-ATPases presentes na membrana basal. É através decanais específicos (ROMK) tanto na membrana apical quan-to na basolateral que o K� é transportado passivamente parafora da célula (Fig. 4.12). A secreção de K� está diretamenterelacionada à diferença de potencial gerada pela quantida-de de Na� reabsorvida.
Tanto a reabsorção de sódio quanto a secreção de potás-sio nas células principais do ducto coletor são moduladaspela aldosterona. Este mineralocorticóide entra no citoplas-ma da célula induzindo através de RNAm a síntese de pro-teínas pelo núcleo, que aumentam o número de canais de
sódio da membrana apical, a densidade de Na�,K�-ATPa-ses da membrana basal e por fim estimulam a produção deATP pelas mitocôndrias, resultando em aumento na ativi-dade das Na�,K�-ATPases. A espironolactona interfere como sítio citoplasmático da aldosterona impedindo a produ-ção do RNAm, e dessa maneira a reabsorção de Na� e a se-creção de K� ficam prejudicadas.
As células intercaladas caracterizam-se por serem célu-las escuras devido à presença de grânulos em seu citoplas-ma. São descritos atualmente dois tipos de células interca-ladas: as � e as �. Elas estão relacionadas ao transporte ati-vo de H� através de H�-ATPases. Estas se localizam namembrana luminal nas células do tipo � e na membranabasal nas do tipo �. No caso das células intercaladas do tipoα a secreção luminal de H� está acoplada ao sistema troca-dor Cl�-HCO3
� na membrana basal. O inverso é observadonas células intercaladas do tipo �, onde o H� é transporta-do pela H�-ATPase agora localizada na membrana basal dacélula e o sistema trocador Cl�-HCO3
� tem localização namembrana apical. Acredita-se que as condições ácido-bási-cas determinam a quantidade de células � ou �. Na acidosepredominam as células do tipo � e na alcalose, as do tipo �.
Como já foi referido no túbulo de conexão, uma ATPa-se relacionada à reabsorção de K� está presente nas célu-las intercaladas do ducto coletor. A H�,K�-ATPase é maisabundante nos segmentos corticais e diminui à medida quese aproxima da papila. Ainda é controverso na literaturase alterações ácido-básicas modulam a atividade dessaenzima. Há indícios de que no ducto coletor medular in-terno outros cátions possam ocupar o lugar do H� naH�,K�-ATPase, e por isso ela tem sido denominada comoX�,K�-ATPase, como o amônio (NH4
�). O ducto coletor medular interno é dividido em três seg-
mentos devido à sua heterogeneidade morfológica e fun-cional: IMCD1, IMCD2 e IMCD3. As células que compõemo IMCD1 são muito semelhantes às do ducto coletor me-dular externo, estando presentes as células do tipo princi-pal e cerca de 10% de intercaladas. Entretanto, as porçõesIMCD2 e IMCD3 parecem representar um segmento distin-to. Estudos recentes têm considerado que o ducto coletormedular interno apresenta dois segmentos funcionalmentedistintos: a porção inicial que corresponde ao IMCD1 e aporção distal com os segmentos IMCD2 e IMCD3.
Este último segmento do néfron tem importante papelna regulação final da composição da urina pelo ajuste dareabsorção de sódio, potássio, uréia e água. Convém sali-entar que o ducto coletor medular interno é o único seg-mento do néfron que possui sítio de ação aos peptídiosatriais natriuréticos, e também existem evidências da pre-sença do co-transporte Na�-K�-2Cl�.
O transporte de água no túbulo distal final e ducto co-letor varia com a concentração plasmática do hormônio an-tidiurético, HAD, que altera a permeabilidade destes seg-mentos à água. A ação do hormônio antidiurético tornatambém o epitélio do coletor medular permeável à uréia.
Fig. 4.11 Célula do túbulo de conexão mostrando na membranaluminal os canais de Na� sensíveis ao amiloride, o co-transporteNa�-Cl� e os co-transportes Na�-H� e Cl�-HCO3
� secundariamen-te ativos à Na�-K�-ATPase da membrana basal.
TÚBULO DE CONEXÃO
LÚMEN�5 mV
CÉLULA�85 mV
Na+
Na+INIBIÇÃOPELO
AMILORIDE Na+
Cl�
K+
ATP
KCl KCl
Cl� Na+
PERITUBULAR0 mV
H+
HCO�3

48 Função Tubular
Esta permeabilidade aumenta em direção à papila, promo-vendo a hipertonicidade do interstício. Esta propriedade,que é importante na determinação da osmolaridade daurina, será discutida com mais detalhes em outro capítuloreferente a mecanismos de concentração urinária.
Antes de finalizar este capítulo sobre a função tubular,é importante salientar dois aspectos da função renal:
1.º) A descrição da função tubular foi feita considerandoo rim como constituído por uma população homogênea denéfrons; entretanto, sabemos que existem diferenças mor-fológicas e funcionais entre os néfrons justamedulares (cór-tex profundo) e os néfrons superficiais. Entre as diferençasdeve-se destacar a maior capacidade dos néfrons justame-dulares de variar a excreção de Na�Cl� frente a variaçõesdo volume extracelular. Em condições de depleção intensado volume extracelular, observa-se uma maior reabsorçãode Na�Cl� pelos néfrons justamedulares e, em condições deexpansão, também são os néfrons profundos os que apre-sentam a maior capacidade de excreção de Na�Cl�.
2.º) A função renal foi apresentada como simplesmenteum processo de filtração glomerular, reabsorção e secreção
Fig. 4.12 Representação dos três tipos de células do ducto coletor: intercalada α, intercalada � e principal. Observa-se que na inter-calada α a inserção da H�-ATPase é na membrana luminal, e do contratransporte Cl�-HCO3
� , na membrana basolateral, o que favo-rece a secreção de ácidos. O inverso é observado nas células intercaladas �. Na face luminal das células principais o Na� é reabsor-vido através de canais sensíveis ao amiloride. O K� é secretado passivamente para a luz tubular. Ambos os transportes são decor-rentes do gradiente eletroquímico gerado pela Na�-K�-ATPase na membrana basal.
CÉLULA �
Cl�
PERITUBULAR
ATPCl�
CÉLULA � CÉLULA PRINCIPAL
ATP
PERITUBULAR
Cl� Cl�
H+
Cl�
Cl�
H+
K+
Na+
K+ K+
Na+
K+
ATP
PERITUBULAR
LUMINAL LUMINAL
Cl�
HCO�3
Cl�
HCO�3
tubular que permite a manutenção do balanço de sódio,potássio, hidrogênio e água; entretanto, o rim tem outrasimportantes funções do ponto de vista metabólico, como,por exemplo: no metabolismo de hidratos de carbono pelasíntese de glicose que ocorre no córtex renal e pela inativa-ção de insulina e glucagon, diminuindo a meia-vida desseshormônios; no metabolismo ósseo pela regulação da excre-ção de cálcio, fósforo, ativação de vitamina D e inativaçãode paratormônio. Não devemos esquecer também o impor-tante papel do rim na regulação da pressão arterial.
BIBLIOGRAFIA SELECIONADA
KRIZ, W. and BANKIR, L. A standard nomenclature for structures of thekidney. American Journal of Physiology, 254(23):F1-F8, 1988.
MOE, O.W.; BERRY, C.A. and RECTOR JR, F.C. Renal transport ofglucose, amino acids, sodium, chloride, and water. In: Brenner, B.M.and Rector, F.C., Jr. (eds.) The Kidney, 6th ed. W.B. Saunders Company,Philadelphia, pp. 375-415, 2000.
SEGURO, A.C.; MAGALDI, A.J.B.; HELOU, C.M.B.; MALNIC, G. e ZATZ,R. Processamento de água e eletrólitos pelos túbulos renais. In Zatz, R.(ed.) Fisiopatologia Renal, 1.ª edição, Atheneu, pp. 71-96, 2000.
STOKES, J.B. Principles of epithelial transport. In: Narins, R.G. (ed.)Maxwel & Kleeman’s Clinical Disorders of Fluid and Electrolyte Metabo-lism, 5th ed. McGraw-Hill, Inc. New York, pp. 21-44, 1994.
VALTIN, H. and SCHAFER, J.A. Tubular reabsorption. In: Valtin, H. andSchafer, J.A. (eds.) Renal Function, 3rd ed. Little, Brown and Company,pp. 62-82, 1995.
VALTIN, H. and SCHAFER, J.A. Tubular secretion. In: Valtin, H. andSchafer, J.A. (eds.) Renal Function, 3rd ed. Little, Brown and Company,pp. 84-93, 1995.
ENDEREÇOS RELEVANTES NA INTERNET
http://www.hdcn.comhttp://www.nephron.comhttp://www.renalnet.org
Pontos-chave:
• As células principais são responsáveis pelareabsorção de sódio e secreção de potássio,sendo estes processos modulados pelaaldosterona
• As células intercaladas são células escurasresponsáveis pela acidificação urinária
• O ducto coletor medular interno é a porçãofinal do néfron, onde ocorrem os ajustesfinais para a formação da urina
LUMINAL