variabilidade genÉtica e morfolÓgica intra e ... · a base do calo onde permanece por longo...
TRANSCRIPT

PATRÍCIA LUZ RIBEIRO
VARIABILIDADE GENÉTICA E MORFOLÓGICA INTRA E
INTERPOPULACIONAL NO COMPLEXO
BULBOPHYLLUM EXALTATUM (Orchidaceae)
OCORRENTE NOS CAMPOS RUPESTRES BRASILEIROS:
IMPLICAÇÕES TAXONÔMICAS E BIOGEOGRÁFICAS
FEIRA DE SANTANA – BAHIA 2006

ii
UNIVERSIDADE ESTADUAL DE FEIRA DE SANTANA
DEPARTAMENTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
VARIABILIDADE GENÉTICA E MORFOLÓGICA INTRA E
INTERPOPULACIONAL NO COMPLEXO
BULBOPHYLLUM EXALTATUM (Orchidaceae)
OCORRENTE NOS CAMPOS RUPESTRES BRASILEIROS:
IMPLICAÇÕES TAXONÔMICAS E BIOGEOGRÁFICAS
PATRÍCIA LUZ RIBEIRO
Dissertação apresentada ao Programa de Pós-
Graduação em Botânica da Universidade
Estadual de Feira de Santana como parte dos
requisitos para a obtenção do título de Mestre em
Botânica.
ORIENTADOR: PROF. DR. EDUARDO LEITE BORBA (UFMG)
CO-ORIENTADORA: PROFA. DRA. ALESSANDRA S. SCHNADELBACH (UEFS)
FEIRA DE SANTANA – BA
2006

iii
AGRADECIMENTOS
Aos meus pais, Josemar Pires Ribeiro e Dilma Luz Ribeiro, e irmãos Adriano,
Maurício e Fernando Luz Ribeiro, pelo apoio e incentivo persistentes, que mesmo à
distância torciam e contribuíam para que chegasse aos meus objetivos. Aos meus amores,
Romualdo Júnior e João Pedro, pelo amor, pela alegria, companhia, força e principalmente,
pela paciência.
Aos companheiros de laboratório (LAMOL) que tanto ajudaram e socorreram nos
momentos de sufoco, quanto contribuíram para alegrar, divertir e enriquecer os longos
períodos de trabalho (Ana Carina, Sabrina, Lia Maria, Andrea Karla, Daiane, Adilva,
Viviane, Maria, Janaína, Alexa, Paty Cris, Paty Reyjane, Silvana, Élvia, Marlon, Jomar,
Eric, Jorge, Ricardo, Cristiano). Em especial aos amigos isoenzimáticos, Cari, Sá, Déa
Karla, Dai, Adilva, Vivi, Paty Reyjane e Marlon, hoje me divirto com as lembranças dos
nossos testes de 50 fatias, sempre atentos para a câmera secreta não flagrar as nossas
derrapadas. Ao colega e amigo Eric Smidt, por importantes críticas e contribuições ao meu
trabalho, principalmente na taxonomia. Aos demais amigos dessa caminhada, mestrandos e
doutorandos, obrigada. Para aqueles que seguirão outros caminhos, sucesso.
A Eduardo Leite Borba, meu orientador, obrigado pelos ensinamentos e
incentivos na carreira e na vida. Você foi peça chave na minha formação, sempre serei
grata por tudo que aprendi.
A Alessandra S. Schnadelbach, minha co-orientadora, obrigado pela adoção e por
importantes contribuições ao meu trabalho.
Ao professor Cássio van den Berg, pela colaboração no tratamento estatístico dos
dados morfológicos e pela confiança no meu profissionalismo.
À Universidade Estadual de Feira de Santana e ao Programa de Pós-graduação em
Botânica, pela oportunidade. À Fundação de Amparo à Pesquisa do Estado da Bahia
(FAPESB) e a CAPES pela bolsa de estudos e financiamento à pesquisa. Ao Projeto
PPBIO pelo apoio financeiro a materiais de consumo e trabalho de campo.
A todos da secretaria da Pós-graduação, Adriana, Elton e Gardênia, e ao nosso
coordenador, Luciano Paganucci de Queiroz, pela atenção e presteza durante todo período
do curso. Aos funcionários, professores e bolsistas do herbário (HUEFS) e Laboratório de
Taxonomia Vegetal (TAXON) pela colaboração e companhia.

iv
SUMÁRIO
AGRADECIMENTOS INTRODUÇÃO 1
MATERIAL E MÉTODOS 8
Populações amostradas 8
Eletroforese de aloenzimas 8
Análise dos dados 9
Análise morfométrica multivariada 11
RESULTADOS 13
Variabilidade genética intrapopulacional 13
Estruturação genética 14
Relações fenéticas 15
Análise morfométrica multivariada 17
Tabelas 20
Figuras 31
DISCUSSÃO 41
Variabilidade genética 41
Variabilidade morfológica 45
Conclusões taxonômicas 47
RESUMO 49
ABSTRACT 51
REFERÊNCIAS BIBLIOGRÁFICAS 53

1
INTRODUÇÃO
O gênero Bulbophyllum Thouars é um dos mais representativos da família
Orchidaceae, possuindo aproximadamente 1.200 espécies de distribuição pantropical, com
grande parte de seus representantes ocorrendo principalmente na Ásia e Oceania (Dressler,
1981, 1993; Vermeulen, 1991). As principais características deste gênero são
pseudobulbos formados por um único nó, inflorescência partindo da base do pseudobulbo,
presença de uma articulação entre o labelo e o pé da coluna permitindo mobilidade ao
labelo, característica importante nos diferentes mecanismos de polinização do gênero, e
dois pares de polínias desiguais sem apêndices.
O gênero Bulbophyllum apresenta miiofilia (polinização por Diptera) como
síndrome de polinização, sendo esta a segunda síndrome mais representativa em
Orchidaceae, incluindo 15 a 25% das espécies da família (van der Pijl & Dodson, 1966;
Christensen, 1994). Estudos de polinização revelaram que moscas da família Milichiidae
são os polinizadores efetivos de algumas das espécies de Bulbophyllum ocorrentes no
Brasil (Braga, 1977; Sazima, 1978; Borba & Semir, 1998b). As flores de B. weddellii
(Lindl.) Rchb.f., B. involutum Borba, Semir & F.Barros e B. ipanemense Hoehne são
polinizadas por moscas Milichiidae do gênero Pholeomyia, sendo que B. weddellii e B.
involutum compartilham as mesmas espécies de polinizadores, porém em freqüências
diferentes, onde o principal polinizador de uma espécie é o secundário da outra, enquanto
B. ipanemense é polinizado por uma espécie de Pholeomyia diferente das encontradas nas
duas espécies anteriores. Comparações entre constituintes de voláteis florais revelaram
uma maior similaridade entre B. weddellii e B. involutum do que entre B. ipanemense e
estas (Silva et al., 1999), embora B. ipanemense seja mais similar morfologicamente a B.
involutum (Borba & Semir, 1998b; Borba et al., 1999). Essas espécies ocorrem num
mesmo ambiente, campo rupestre, muitas vezes simpatricamente, possuem eventos
fenológicos sincronizados, e têm o mesmo mecanismo de polinização, no qual o inseto
pousa no labelo, atraído pelo odor ou por instinto de reprodução (oviposição), caminha até
a base do calo onde permanece por longo período se alimentando de néctar secretado neste
local (Teixeira et al., 2004) até que rajadas de vento movimentem o labelo pressionando o
inseto contra a coluna (Borba & Semir, 1998b).
Moscas normalmente apresentam um comportamento que favorece a

2
autopolinização, realizando visitas de longa duração e visitando várias flores de um mesmo
indivíduo (Borba & Semir, 1998b; Verola, 2002). Todas as três espécies são
autocompatíveis, mas precisam de um inseto vetor para realizar a polinização, sendo que a
autopolinização nestas espécies provavelmente não é freqüente na natureza devido em
parte a barreiras mecânicas como variação temporal no tamanho do polinário após sua
remoção (Borba & Semir, 1999).
Sendo as orquídeas predominantemente autocompatíveis, pode-se esperar que
espécies de orquídeas miiófilas possuam baixa variabilidade dentro das populações e
elevada diferenciação genética entre populações conspecíficas, devido à redução do fluxo
gênico, uma vez que moscas movimentam-se a distâncias muito curtas, podendo favorecer
o isolamento reprodutivo e conseqüentemente levando à diferenciação dessas populações
(Borba & Semir, 1998b; Borba et al., 2001; Farinaci, 2001). Isto poderia ajudar a explicar
o elevado número de espécies nos gêneros de orquídeas miiófilas, grande parte delas de
distribuição restrita, estando estes entre os mais numerosos da família. No entanto, em
Acianthera, outro gênero de orquídeas miiófilas, Borba et al. (2001) encontraram uma
elevada variabilidade genética dentro das populações e uma baixa a moderada
diferenciação genética entre as populações conspecíficas. Segundo os autores, a ocorrência
de auto-incompatibilidade genética, barreiras mecânicas a autopolinização e dispersão de
sementes pelo vento são fatores que provavelmente influenciam a modificação do padrão
esperado.
No Brasil, Pabst & Dungs (1975, 1977) listaram 54 espécies de Bulbophyllum,
distribuídas principalmente entre os estados do Rio de Janeiro, São Paulo e Minas Gerais.
Este número foi acrescido para cerca de 60 espécies por algumas publicações posteriores
(e.g., Borba et al., 1998; Toscano de Brito, 2000; Borba & Smidt, 2004; Fraga, 2004;
Fraga & Smidt, 2004). Para o estado da Bahia, Toscano de Brito (1995) listou a ocorrência
das primeiras três espécies para o gênero, encontradas no Pico das Almas, Chapada
Diamantina, e em um estudo recente foram citadas 13 espécies ocorrentes na Chapada
Diamantina, porção norte da Cadeia do Espinhaço (Ribeiro et al., 2005).
Pabst & Dungs (1975, 1977) distribuíram as espécies de ocorrência no Brasil em
cinco seções, Didactyle, Xiphizusa, Bulbophyllaria, Micrantha e Napellii, diferenciadas
por características tais como presença de estelídias (estruturas alongadas como braços no
ápice da coluna), protuberâncias em forma de dentes na face ventral da coluna,
comprimento e largura das sépalas, forma e posição da raque da inflorescência e número
de flores por inflorescência. Na seção Didactyle encontra-se o maior número de espécies

3
brasileiras, caracterizadas pela presença de dois dentes sob as estelídias da coluna e sépalas
quase tão largas quanto longas. Grande parte destas espécies possui morfologia muito
similar, diferenciando-se por características florais muito sutis, o que, devido à inexistência
de uma revisão taxonômica para o gênero, acarreta em determinações incertas,
sinonimizações e formação de complexos taxonômicos de difícil solução.
O complexo taxonômico aqui estudado, denominado “complexo B. exaltatum
Lindl.”, obedecendo à prioridade nomenclatural, compreende um grupo com cerca de 15
táxons da seção Didactyle ocorrentes em áreas de altitude, principalmente rupícolas em
formações de campos rupestres nas regiões Sudeste, Nordeste e Norte: B. exaltatum Lindl.,
B. geraense Rchb.f., B. gomesii Fraga, B. involutum Borba, Semir & F.Barros, B.
ipanemense Hoehne, B. longispicatum Cogn., B. machupicchuense Benn. & Cristenson, B.
meridense Rchb.f., B. roraimense Rolfe, B. sanderianum Rolfe, B. teresensis Ruschi, B.
tripetalum Lindl., B. vaughanii Brade, B. vittatum Rchb.f. & Warm., B. warmingianum
Cogn.
Os representantes deste complexo são vegetativamente muito similares, sendo
separados exclusivamente pela morfologia floral, principalmente do labelo. Bulbophyllum
ipanemense, B. vaughanii e B. warmingianum são comumente confundidos em
determinações de herbário devido à inexistência de caracteres diagnósticos consistentes.
Bulbophyllum geraense se diferiria dos primeiros apenas por uma interpretação incorreta
de sua morfologia feita por Cogniaux (1898-1902), onde ressalta que B. geraense e
também B. exaltatum possuem labelo indiviso, quando na verdade são trilobados.
Bulbophyllum machupicchuense foi descrito recentemente para o Peru (Bennett &
Christenson, 2001) sendo que suas características descritas e ilustradas são compatíveis
com os táxons citados acima, e principalmente com as populações ocorrentes no estado da
Bahia, aqui tratadas como B. exaltatum. Bulbophyllum longispicatum foi descrito por
Cogniaux (1898-1902) a partir de um material cultivado sem especificação do local de
coleta nem ilustração, porém com características florais semelhantes aos táxons já citados.
Bulbophyllum involutum era tratado anteriormente como B. warmingianum, até que através
de estudos de biologia reprodutiva e de análise de voláteis florais (Borba & Semir, 1998b,
1999; Borba et al., 1999; Silva et al., 1999) algumas populações foram segregadas em uma
espécie distinta (Borba et al., 1998). Esta espécie eventualmente hibridiza com B. weddellii
(Lindl.) Rchb.f., formando o híbrido natural B. ×cipoense Borba & Semir (Borba & Semir,
1998a; C.Azevedo et al., no prelo). Bulbophyllum roraimense e B. exaltatum são
simpátricas, diferenciadas por presença de tricomas na margem da pétala de B. exaltatum, e

4
são consideradas por Cogniaux (1898-1902) como morfologicamente próximas de B.
geraense. Estas, também compartilham o habitat com B. meridense, ocorrendo como
epífitas em áreas de altitude no estado de Roraima, na Venezuela e nas Guianas.
Bulbophyllum sanderianum é uma espécie descrita para o estado de Pernambuco e
recentemente encontrada em diferentes localidades do estado da Bahia, Alagoas e Ceará, e
morfologicamente se assemelha a B. meridense, principalmente pela forma do labelo com
lobo mediano arredondado com margem densamente longo-ciliada. Bulbophyllum
teresensis e B. gomesii são bastante semelhantes e descritas para o mesmo local, estado do
Espírito Santo, podendo ser conspecíficas, sendo a forma das sépalas e labelo relacionada
com B. tripetalum. Esta última é a espécie melhor definida morfologicamente, e a única
que a princípio não apresenta problemas taxonômicos.
Aparentemente, a maior parte dos problemas taxonômicos deste complexo ocorre
em torno de B. involutum, B. ipanemense, B. longispicatum, B. geraense e B.
warmingianum, que se distribuem nos estados de Minas Gerais, Bahia e São Paulo,
predominantemente nas áreas de campos rupestres. Observações de material de herbário
revelaram que muitos espécimes encontrados na Cadeia do Espinhaço são comumente
determinados como B. ipanemense, embora esta espécie tenha sido descrita originalmente
para o estado de São Paulo, também ocorrendo mais ao sul do estado de Minas Gerais,
desde a região de Juiz de Fora à Serra da Canastra (Farinaci, 2001), em campos rupestres
independentes do bloco constituinte da Cadeia do Espinhaço. As diferenças morfológicas
detectadas entre as populações de B. ipanemense ocorrentes no estado de Minas Gerais e
Bahia, tais como maiores dimensões do perianto e menor número de flores em antese por
inflorescência nas populações da Bahia, além de variação no padrão de coloração da flor e
forma do labelo, indicam a possibilidade destas não serem conspecíficas (Ribeiro et al.,
2005). Devido à ausência de caracteres diagnósticos claros e consistentes dos táxons deste
complexo, neste estudo utilizamos a priori a designação de B. exaltatum, nome mais
antigo, para a população do estado de Roraima, para todas as populações do estado de São
Paulo e Minas Gerais que comumente são determinadas como B. ipanemense, B.
warmingianum ou B. geraense, e também para as populações da Bahia que anteriormente
foram tratadas como B. ipanemense por Toscano de Brito (1995) e Ribeiro et al. (2005).
Diferentes populações ocorrentes no estado da Bahia apresentam ampla variação,
principalmente de tamanho e coloração da flor e da inflorescência, o que ocasiona
dificuldades na determinação entre B. involutum e B. exaltatum (Figuras 1 e 2).
Existem duas áreas principais de ocorrência dos campos rupestres no Brasil: a Cadeia

5
do Espinhaço e as serras do sul de Minas Gerais ao longo da bacia do Rio Grande. Estas
duas áreas pertencem a diferentes formações geológicas e apresentam floras distintas
(Giulietti & Pirani, 1988). A Cadeia do Espinhaço possui cerca de 1.000 km de extensão
no sentido Norte-Sul, desde a Serra de Ouro Branco no estado de Minas Gerais, até a Serra
de Jacobina ao norte do estado da Bahia, sendo a extensão Leste-Oeste variável,
aproximadamente entre 50 a 100 km (Harley, 1995). A Cadeia do Espinhaço não constitui
um maciço contínuo, possuindo algumas formações disjuntas, destacando-se a Serra do
Cabral e vários conjuntos de serras isoladas entre si, situados nas regiões de Montes
Claros, Itacambira e Grão-Mogol (Vitta, 2002). A descontinuidade das cadeias
montanhosas e dos afloramentos rochosos resulta na distribuição de muitas espécies,
principalmente as rupícolas, em populações disjuntas. Essa característica tem sido sugerida
por alguns autores como sendo responsável pela diferenciação das populações de plantas,
levando à grande diversidade e elevado grau de endemismo dos campos rupestres (Joly,
1970; Giulietti & Pirani, 1988; Borba et al., 2001; Jesus et al., 2001; Lambert et al., 2006a,
2006b), devido ao isolamento das populações. Tendo em vista que a precipitação diminui
em direção ao norte, Harley (1995) dividiu a Cadeia do Espinhaço em três setores,
considerando o tipo de vegetação. O setor Sul, ao sul de Belo Horizonte e ao redor de Ouro
Preto no estado de Minas Gerais, se localiza dentro da floresta tropical que se estende
através de uma grande área do sudeste do Brasil; o setor Central, incluindo a Serra do Cipó
e a região de Diamantina em Minas Gerais, situa-se na região dos cerrados, típica do
Planalto Central; e, o setor Norte, englobando toda a Chapada Diamantina no estado da
Bahia, é circundado pela caatinga da região Nordeste. A região de Grão Mogol, no norte de
Minas Gerais, apresenta um tipo de vegetação intermediária entre o setor Central e Norte,
devido à sua posição geográfica.
Dentre os diversos tipos vegetacionais da Cadeia do Espinhaço, os campos
rupestres dominam as regiões mais altas, geralmente acima de 1.000 m (Giulietti & Pirani,
1988; Harley, 1995). Os campos rupestres constituem uma formação ocorrente
principalmente na Cadeia do Espinhaço, nos estados de Minas Gerais e Bahia, podendo
também ser encontrada em outras localidades da porção sudoeste e sul do estado de Minas
Gerais, em Goiás e no Distrito Federal, como ilhas florísticas isoladas circundadas por
vegetação de cerrado ou caatinga (Giulietti et al, 2000; Romero, 2002). Campo rupestre é
um conjunto de comunidades predominantemente herbáceo-arbustivas, formando um rico
mosaico, floristicamente relacionadas, mas fisionomicamente distintas, em função da
topografia, natureza do substrato, profundidade do solo e do microclima (Conceição &

6
Giulietti, 2002).
Estudos de variabilidade genética no nível de populações representam uma
ferramenta importante para compreender a estrutura genética das populações e para a
taxonomia, podendo inclusive verificar os limites específicos (Solé-Cava, 1999). Esta
abordagem tem sido utilizada satisfatoriamente para estudos de populações na Cadeia do
Espinhaço pertencente a este complexo e outros grupos de Orchidaceae (Borba et al.,
2001; Farinaci, 2001; M.Azevedo et al., no prelo). Estes estudos também contribuem para
o entendimento dos padrões de distribuição das espécies e da fitogeografia da Cadeia do
Espinhaço, tendo em vista que relativamente poucos trabalhos têm abordado estas
questões. Alguns estudos de variabilidade morfológica em Orchidaceae na Cadeia do
Espinhaço também têm contribuído para compreender a variação e estabelecer
delimitações de táxons (van den Berg & Martins, 1998; Cardim et al., 2001; Borba et al.,
2002).
Borba et al. (2001) e Jesus et al. (2001), estudando espécies ocorrentes nos campos
rupestres, constataram moderada a alta diferenciação genética entre populações
conspecíficas de espécies de Acianthera (Orchidaceae) e Proteopsis argentea Mart. &
Zucc. (Asteraceae), respectivamente, indicando que a distribuição geográfica contribui
com a diferenciação genética de populações na Cadeia do Espinhaço. Estes dados
corroboram a hipótese de Giulietti & Pirani (1988), das disjunções influenciando o alto
grau de endemismo nos campos rupestres. Estudos mais recentes em outros grupos de
plantas têm fortalecido estas suposições (Machado, 2004; Lambert et al., 2006a, 2006b;
Pereira, 2006).
Em Bulbophyllum, dois trabalhos foram realizados abordando genética de
populações em campos rupestres, ambos utilizando alozimas como marcadores
moleculares. Farinaci (2001) estudou cinco populações de B. ipanemense e B. weddellii no
estado de Minas Gerais, e M.Azevedo et al. (no prelo) analisaram 12 populações de sete
espécies de Bulbophyllum: B. bidentata (Barb.Rodr.) Cogn., B. plumosum (Barb.Rodr.)
Cogn. B. insectiferum Barb.Rodr., B. regnellii Rchb.f., B. adiamantinum Brade, B.
epiphytum (Barb.Rodr.) Cogn. e B. rupicolum Barb.Rodr. nos estados de Minas Gerais e
São Paulo. Os dois trabalhos demonstraram a ocorrência de alta variabilidade genética em
todas as populações e uma baixa a moderada estruturação genética entre as populações das
espécies estudadas. Ambos estudos também apresentam padrões de variabilidade não
esperados para orquídeas miiófilas de populações disjuntas, como também encontrado por
Borba et al. (2001). A princípio, estes são os únicos trabalhos publicados abordando

7
genética de populações com orquídeas de campos rupestres até o momento.
Para este complexo de espécies “B. exaltatum” com tantas semelhanças
morfológicas, ecológicas, fenológicas e reprodutivas, um estudo de genética de populações
com um amplo número de populações, abrangendo grande parte da distribuição destas
espécies, pode nos levar a importantes considerações sobre estrutura e variabilidade
genética das espécies e manutenção desta variabilidade em ambientes como campos
rupestres. Tal abordagem é fundamental para uma melhor delimitação dos táxons
envolvidos em grupos como este, no qual apenas o exame de amostras de herbário não tem
sido suficiente para a elucidação da taxonomia do grupo.
Neste trabalho, foram utilizados estudos de genética de populações naturais
empregando marcadores aloenzimáticos para determinar a variabilidade genética dentro
das populações e o grau de diferenciação das populações de algumas espécies do gênero
Bulbophyllum pertencente ao complexo B. exaltatum, utilizando os resultados na tentativa
de delimitação dos táxons deste grupo. Adicionalmente, foram utilizados métodos de
análise multivariada para realização de estudos morfométricos em alguns indivíduos destas
populações estudadas, buscando uma delimitação mais consistente dos táxons neste
complexo. A partir destes dados, procurou-se fornecer subsídios para a definição de
padrões de distribuição das espécies estudadas do complexo B. exaltatum e contribuir para
o entendimento dos processos evolutivos e biogeográficos de espécies ocorrentes na
Cadeia do Espinhaço.

8
MATERIAL E MÉTODOS
Populações amostradas
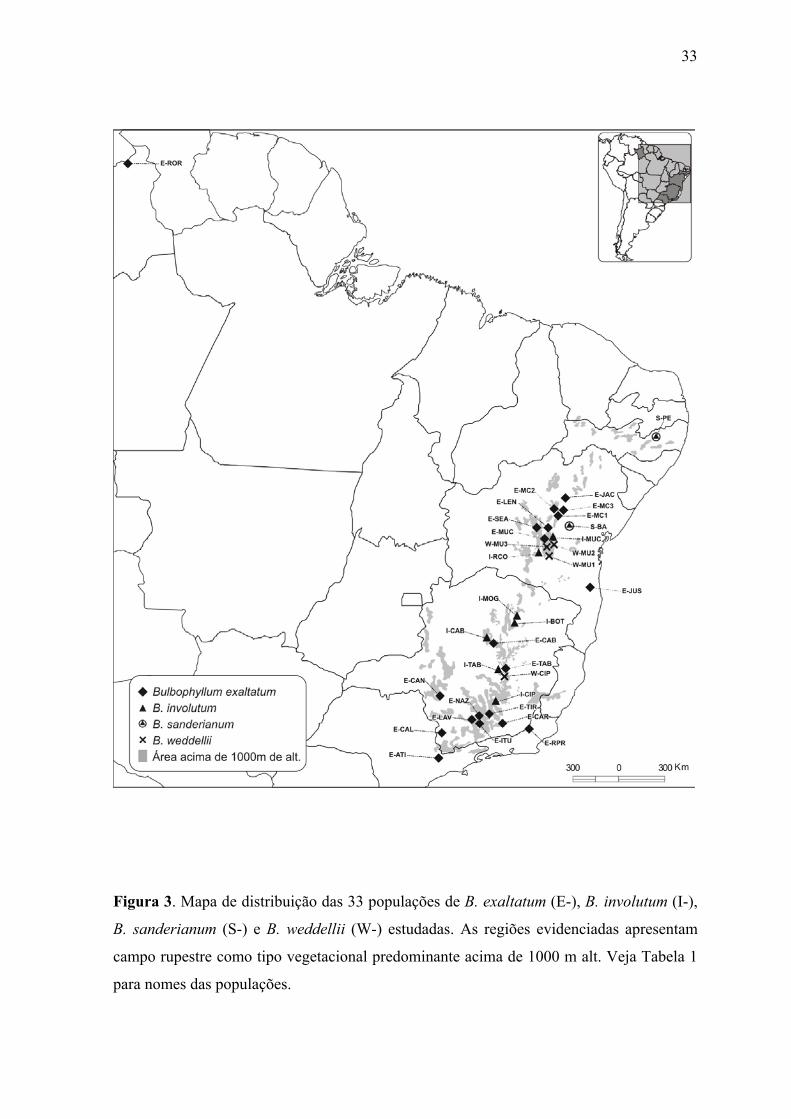
Foram coletados 601 indivíduos em 33 populações naturais distribuídas nos estados
de Roraima, Pernambuco, Bahia, Minas Gerais e São Paulo, agrupadas em três táxons, B.
exaltatum, B. involutum e B. sanderianum pertencentes ao complexo Bulbophyllum
exaltatum, e também de B. weddellii (Tabela 1; Figura 1-3). Populações desta última
espécie foram incluídas na análise devido à ocorrência de hibridação desta com uma
espécie do complexo (B. involutum) e para servir como um parâmetro para comparação de
distâncias genéticas entre espécies do grupo. Apesar de B. weddellii ter sido classificada
em uma diferente seção do gênero por Pabst & Dungs (1975), em função das suas sépalas
bem mais longas do que largas e caracteres morfológicos vegetativos. Análises
filogenéticas utilizando seqüências de DNA indicam que esta é uma espécie próxima a este
complexo, e pertencente à mesma seção do gênero (E.C.Smidt, dados não publicados).
Foram coletados indivíduos distantes entre si para evitar a coleta de clones, uma
vez que estas plantas se reproduzem vegetativamente e possuem vida longa, podendo um
único indivíduo formar um grande genet constituído por vários ramets sobre a rocha. Em
média, amostras de 20 indivíduos foram coletadas por população, dependendo do tamanho
da população, sendo que em populações pequenas este número não pode ser atingido.
Pedaços de tecido foliar foram armazenados em microtubos de 2ml em ultrafreezer a -85ºC
até a realização das corridas eletroforéticas.
Além disso, 408 indivíduos de diferentes populações deste complexo foram
plantados e estão em cultivo na casa de vegetação da Universidade Estadual de Feira de
Santana, para realização de estudos posteriores. O material testemunho das populações
estudadas foi depositado no herbário da Universidade Estadual de Feira de Santana
(HUEFS) ou no herbário da Universidade Estadual de Campinas (UEC).
Eletroforese de aloenzimas
O estudo de variabilidade genética foi realizado utilizando a técnica de eletroforese
de aloenzimas em gel de amido a 8,5%. Pequenos pedaços de folhas, congeladas no

9
momento da coleta, foram homogeneizados individualmente em 500µl do tampão de
extração [Tris 0,1M (100ml), pH 7,0, sacarose 6,846g, PVP (polivinilpirrolidona) 0,6g,
EDTA (etileno-diamina-tetraacetato) 0,029g, BSA (albumina bovina) 0,145g, DIECA
(dietil-ditiocarbonato) 0,13g, Borax (borato de sódio) 0,6g, e β-mercaptoetanol 100µl (Sun
& Garders, 1990; com modificações)]. Os extratos dos indivíduos foram absorvidos em
tiras de papel filtro (Whatman #3) medindo 0,6 × 0,3 cm e aplicados no gel. As corridas
foram realizadas até o marcador interno azul de bromofenol atingir 9 cm do ponto de
aplicação. Em todos os géis, foi colocada amostra de um indivíduo padrão proveniente da
população BELEN, para comparação da migração relativa dos alelos.
Quatro sistemas tampão foram utilizados: Sistema 1 – eletrodo: solução de histidina
0,065 M ajustada para pH 6,5 com uma solução de ácido cítrico 0,2 M; gel: mesma solução
do eletrodo diluída 1:7 (modificado a partir de Stuber et al., 1977); condição de corrida
150 V. Sistema 2 – eletrodo: solução de ácido cítrico 0,04 M ajustada para o pH 6,1 com
N-(3-aminopropil)-morfolina; gel: mesma solução do eletrodo diluída 1:20 (Clayton &
Tretiak, 1972); condição de corrida 25 mA. Sistema 3 – eletrodo: solução de hidróxido de
lítio 0,05 M, ácido bórico 0,093 M e EDTA 0,006 M, pH 8,0; gel: mesma solução do
eletrodo diluída 1:10 (modificado a partir de Ridgway et al., 1970); condição de corrida 13
mA. Sistema 4 – eletrodo: solução de ácido bórico 0,3 M, NaOH 0,06 M, pH 8,0; gel tris
0,01 M, pH 8,5 (modificado a partir de Shaw & Prasad, 1970); condição de corrida 13 mA.
Sete sistemas enzimáticos apresentaram boa resolução: Sistema tampão 1 –
isocitrato desidrogenase (IDH; EC 1.1.1.42), malato desidrogenase (MDH; EC 1.1.1.37),
leucil-aminopeptidase (LAP; EC 3.4.11.1); Sistema tampão 2 – diaforase (DIA; EC
1.8.1.4), desidrogenase do ácido 6-fosfoglucônico (6PGD; EC 1.1.1.49); Sistema tampão 3
– desidrogenase do shikimato (SKDH; EC 1.1.1.25); Sistema tampão 4 – esterase (EST;
EC 3.1.1.1.). Os procedimentos de coloração utilizados foram adaptados de Brune et al.
(1998; LAP, DIA, SKDH, EST), Corrias et al. (1991; IDH, 6PGD) e Soltis et al. (1983;
MDH), sendo que as principais modificações dizem respeito à quantidade dos
componentes utilizados. Sistemas enzimáticos que apresentaram mais de um locus foram
numerados em ordem crescente contados a partir do locus com alelos de menor
mobilidade. Os alelos foram numerados de acordo a sua mobilidade relativa a um dos
alelos do indivíduo padrão, designado como 100.
Análise de dados

10
As freqüências alélicas foram determinadas por observação direta dos padrões de
banda de homozigotos e heterozigotos corados no gel. A variabilidade genética para todas
as populações foi estimada pelos seguintes parâmetros: proporção de loci polimórfico (P;
critério 0,95), número médio de alelos por locus (A), heterozigosidade média observada
(Ho) e esperada (He). Desvios da heterozigosidade média observada em relação ao
esperado em equilíbrio de Hardy-Weinberg (HW) foram testadas utilizando χ2 com a
correção para pequenas amostras conforme Levene (1949). Foi feito um teste de
desequilíbrio de ligação utilizando o programa GENEPOP (Raymond & Rousset, 1995)
empregando 100 lotes de 1000 interações por lote e com 1000 passos de dememorização;
posteriormente, foi realizado um teste múltiplo para evitar erros do tipo 1 através do
procedimento seqüencial de Bonferroni (Rice, 1989). A partição da diversidade genética
entre populações conspecíficas foi estimada pela estatística F (FIS, coeficiente de
endogamia mede a redução do número de heterozigotos devido a cruzamentos não
aleatórios dentro de uma população; FST, mede a diferenciação entre populações; Wright,
1978). Para B. exaltatum a estrutura genética entre e dentro de regiões foi estimada pelo
indicador θ (Weir & Cockerham, 1984) calculado pelo programa TFPGA (Miller, 1997). O
fluxo gênico (Nm) entre populações foi estimado indiretamente a partir do coeficiente de
estruturação genética usando a equação de Wright (1951) modificada por Crow & Aoki
(1984): FST = 1/(1 + 4Nmα), onde α = [n/(n-1)]2, sendo n o número de populações
analisadas para cada espécie. Uma outra estimativa indireta de fluxo gênico foi feita
baseada na freqüência média de alelos exclusivos de uma população ("alelos privados";
Barton & Slatkin, 1986). Matrizes de distância genética ("unbiased genetic distance"; Nei,
1978) e de identidade genética ("unbiased genetic identity"; Nei, 1978) foram calculadas
para populações e espécies. A matriz de distância genética de Nei foi comparada com a
matriz de distância geográfica entre populações, usando o teste de Mantel com o método
de aleatorização (Monte Carlo; 1000 aleatorizações) no programa PCORD 4.10 (McCune
& Mefford, 1999), para testar correlação entre distância genética e geográfica. A distância
geográfica entre pares de populações foi calculada em quilômetros com base nas
coordenadas GPS para cada população no sistema elipsóide WGS84 utilizando o programa
INVERSE 2.0 (National Geodetic Survey, 2002). A partir da matriz de distância genética
das populações foi realizada uma análise de agrupamento utilizando o algoritmo UPGMA,
e foi também construída uma árvore utilizando o algoritmo "neighbor-joining" (NJ). Todas
as análises foram feitas utilizando o programa BIOSYS 1.0 (Swofford & Selander, 1989),
exceto para o UPGMA e NJ nas quais foi utilizado o programa MEGA 3.1 (Kumar et al.,

11
2004). O pacote PHYLIP 3.6 (Felsenstein, 1993) foi utilizado para realização de análises
de bootstrap e criação de árvores de consenso. SEQBOOT foi utilizado para obter 1000
replicações através de bootstrap convencional e aleatorização da ordem de entrada dos
dados. O arquivo de saída foi utilizado no programa GENDIST, que gerou 1000 matrizes
de distância a partir da distância de Nei (1972), sendo utilizadas no programa NEIGHBOR
para gerar 1000 árvores de NJ e 1000 de UPGMA, também com aleatorização da ordem de
entrada das populações. Os arquivos de saída de NJ e de UPGMA foram utilizados no
programa CONSENSE para criar árvores de consenso de maioria, que foram exportadas e
construídas no programa PAUP 4.0 (Swofford, 2000).
Análise morfométrica multivariada
Foram analisados 27 caracteres morfológicos florais em 77 indivíduos de B.
involutum e B. exaltatum, provenientes de 24 populações (Tabela 2). As características
foram escolhidas com base em caracteres diagnósticos das espécies do complexo e nos
caracteres utilizados em descrições florais do grupo. As peças florais foram desenhadas
com o auxílio de estereomicroscópio equipado com câmara clara. Os caracteres
quantitativos contínuos foram medidos a partir do desenho com o auxílio de régua e
transferidor.
Foi calculada uma matriz de similaridade dos indivíduos utilizando o coeficiente de
Gower (1971), devido à presença de diferentes tipos de variáveis (quantitativo, binário,
multiestado), utilizando o programa COEF do pacote FITOPAC 1.0 (Shepherd, 1995). A
partir desta matriz de similaridade (s) foi calculada uma matriz de distância (d) utilizando
dij=1-sij, e realizada uma análise de agrupamento utilizando UPGMA como algoritmo de
agrupamento no programa STATISTICA 6.0 (StatSoft, 2003).
Foi realizada uma análise de componentes principais a partir da matriz de
correlação (PCA) para examinar a estrutura geral dos dados e para detecção de ‘outliers’.
O número de eixos interpretados foi determinado pelo nível de significância obtido pelo
teste de Qui-quadrado (critério 95%). Foi também realizada uma análise de variáveis
canônicas (CVA) utilizando grupos de indivíduos pertencentes a uma mesma região
geográfica (a priori) e a um mesmo táxon (B. exaltatum ou B. involutum) como variável
categórica, devido ao baixo número de indivíduos por população, com o objetivo de
explorar a partição dos grupos pelos principais eixos canônicos e de identificar os
caracteres que mais contribuem para o padrão observado. Desta forma, foram utilizados

12
nove agrupamentos: 1- (BE)-Sul-MG (populações E-CAL, E-ITU, E-NAZ, E-LAV, E-
CAR, E-ATI, E-STL e E-SJA); 2- (BE)-Espinhaço-MG (populações E-TAB e E-SCA); 3-
(BE)-F.Doido-BA (populações E-MC2 e E-MC3); 4- grupo (BE)-Morrão-BA (população
E-MC1); 5- (BE)-Jacobina-BA (população E-JAC); 6- (BE)-Sincorá-BA (populações E-
SEA, E-LEN e E-MUC); 7- (BE)-RR (população E-ROR); 8- (BI)-MG (populações I-
MOG, I-CAB e I-TAB); 9- (BI)-BA (populações I-MUC e I-RCO). A partir da matriz de
Distância Generalizada de Mahalanobis dos centróides dos nove grupos foi também
realizada uma análise de agrupamento, utilizando UPGMA. As análises de CVA e PCA
foram realizadas considerando apenas os caracteres quantitativos contínuos, sendo
descartadas as variáveis #13, 21, 24, 25, 26 e 27, do tipo binário e multiestado não-
ordenado, que são geralmente incompatíveis com as premissas destas análises. As análises
de agrupamento, PCA e CVA foram realizadas utilizando o programa STATISTICA 6.0
(StatSoft, 2003).

13
RESULTADOS
Variabilidade genética intrapopulacional
Com os sete sistemas enzimáticos utilizados, foram obtidos nove loci com boa
resolução, que foram analisados neste estudo. Todos os loci foram polimórficos em pelo
menos uma das populações estudadas e o número de alelos em cada locus variou de três a
cinco (Tabela 3). Os loci DIA-1, DIA-2, MDH-2, EST e 6PGD foram monomórficos para
ao menos 15 populações, enquanto os demais loci foram polimórficos para a maioria das
populações.
Bulbophyllum exaltatum possui alguns alelos exclusivos: SKDH 79, SKDH 108,
IDH 90, MDH-1 108, MDH-2 120, LAP 113 e EST 92. Porém, nenhum destes alelos foi
comum a todas as populações de B. exaltatum. O alelo LAP 113, por exemplo, foi
encontrado apenas em duas populações da Bahia, em Morro do Chapéu (E-MC2) e
Jacobina (E-JAC), e o alelo EST 92 é exclusivo da população de Nazareno-MG (E-NAZ).
O alelo IDH 70 foi encontrado exclusivamente nas três populações de B. weddellii do
município de Mucugê-BA, mas em baixa freqüência. Alguns alelos são compartilhados
apenas pelas populações de B. involutum e B. exaltatum: DIA-1 90, DIA-2 95, MDH-2 90
e 110, EST 108 e 6PGD 120, sendo que este último ocorre em uma população de B.
involutum e em 10 de B. exaltatum. Nenhuma espécie apresentou alelos exclusivos fixados,
e nenhum locus diagnóstico foi encontrado.
O padrão de bandas observado neste estudo foi compatível com o de indivíduos
diplóides, não sendo evidenciadas características que suportem a presença de poliploidia
nas populações, como indivíduos com acima de três alelos por locus e/ou heterozigosidade
fixada nas populações.
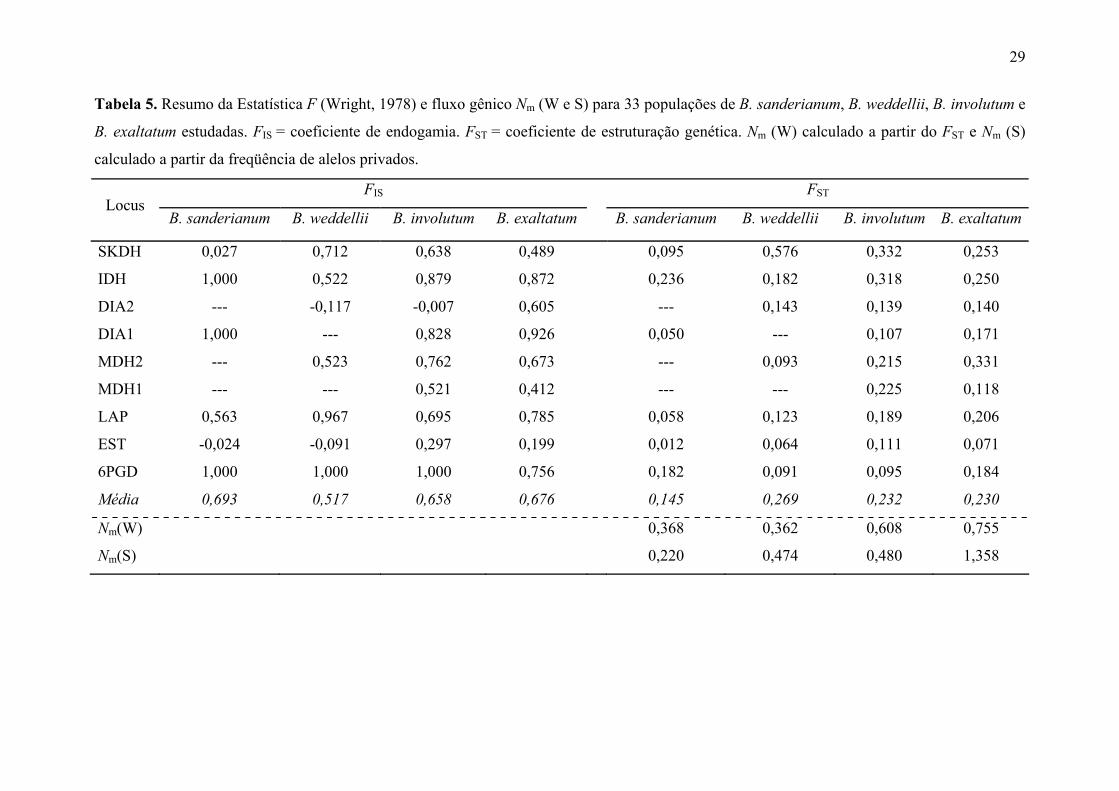
Os índices de variabilidade intrapopulacional foram, na sua maioria, elevados
(Tabela 4). O percentual de loci polimórficos (P; critério de 0,95) variou de 22,2 a 88,9%,
o número médio de alelos por locus (A) de 1,3 a 2,6 e a heterozigozidade média (He) entre
0,086 e 0,404. As populações de B. sanderianum S-PE e B. weddellii W-MU3
apresentaram os menores valores de variabilidade, enquanto os maiores valores são
encontrados nas populações E-NAZ e E-LEN de B. exaltatum e I-MOG de B. involutum.
De modo geral, as populações de B. sanderianum e B. weddellii apresentaram menores

14
valores de variabilidade genética do que as populações de B. involutum e B. exaltatum.
A variabilidade genética foi similar ao longo da área de distribuição das espécies,
com uma pequena diferença para populações isoladas geograficamente, possuindo valores
de variabilidade mais baixos, como as populações E-ATI de Atibaia-SP (He= 0,165), S-PE
de Brejo de Madre de Deus-PE (He= 0,086) e E-ROR de Pacaraima-RR (He= 0,195); e
para populações pequenas de áreas antropizadas como W-MU3 e I-MUC simpátricas de
Mucugê-BA (He= 0,098 e 0,149, respectivamente).
Todos os loci apresentaram desvios significativos em relação ao equilíbrio de HW
em mais de dez populações. Sete loci estão em equilíbrio para pelo menos uma população:
SKDH (S-PE e W-CIP), IDH (W-MU1 e W-MU3), DIA1 (I-CAB), MDH1 (E-TIR),
MDH2 (E-ITU e E-ROR), EST (S-BA, I-TAB e E-SEA) e 6PGD (E-MC2, E-MC3 e E-
ROR). Das 33 populações analisadas, 20 não possuem qualquer locus em equilíbrio. Nos
loci LAP e DIA-2, todas as populações apresentam déficit de heterozigotos. Dos nove loci
analisados, sete apresentam excesso de heterozigotos em pelo menos uma população:
SKDH, IDH, DIA1, MDH1, MDH2, EST e 6PGD. Porém, o número de loci que
apresentou excesso de heterozigotos em cada população variou de um a três loci por
população.
Foram encontrados elevados valores de FIS (Tabela 5) para todas as espécies, o que
também pode ser verificado pela grande diferença entre os valores de heterozigozidade
observada e esperada para cada população (Tabela 4).
Após correção de Bonferroni, constatou-se que para todas as populações de todas as
espécies existe desequilíbrio de ligação entre alguns dos loci relacionados. Para B.
sanderianum todos 16 testes encontrados foram significativos, com α = 0,01. Para B.
weddellii todos os 38 testes encontrados foram significativos, com α = 0,008. Em B.
involutum dos 126 testes realizados 50 foram significativos, com α = 0,00039 e, em B.
exaltatum, dos 329 testes encontrados 99 foram significativos, com α = 0,000152.
Estruturação genética
A distribuição da variação genética total foi similar em todas as espécies estudadas.
Os elevados valores médios de FST encontrados indicam alto grau de estruturação genética
para as todas as espécies (Tabela 5). Bulbophyllum sanderianum apresentou o menor FST,
sendo 14,5% da variabilidade desta espécie associada a diferenças entre populações.
Bulbophyllum weddellii apresentou o maior valor de FST (0,269), enquanto que B.

15
involutum e B. exaltatum apresentaram valores muito próximos (0,232 e 0,230
respectivamente). Em B. exaltatum, o FST reduz para 0,140 se considerarmos apenas as
populações do estado da Bahia, e praticamente se mantém (0,213) se considerarmos apenas
as populações do estado de Minas Gerais. Os valores de θ encontrados entre as duas
regiões principais de ocorrência de B. exaltatum, Bahia e Minas Gerais, revelou uma baixa
diferenciação entre estas regiões (θP = 0,058) com intervalo de confiança (IC) de -0,013 a
0,182. Valores de θ mais elevados foram encontrados entre as populações de cada região
(θS = 0,196; IC = 0,125 – 0,294).
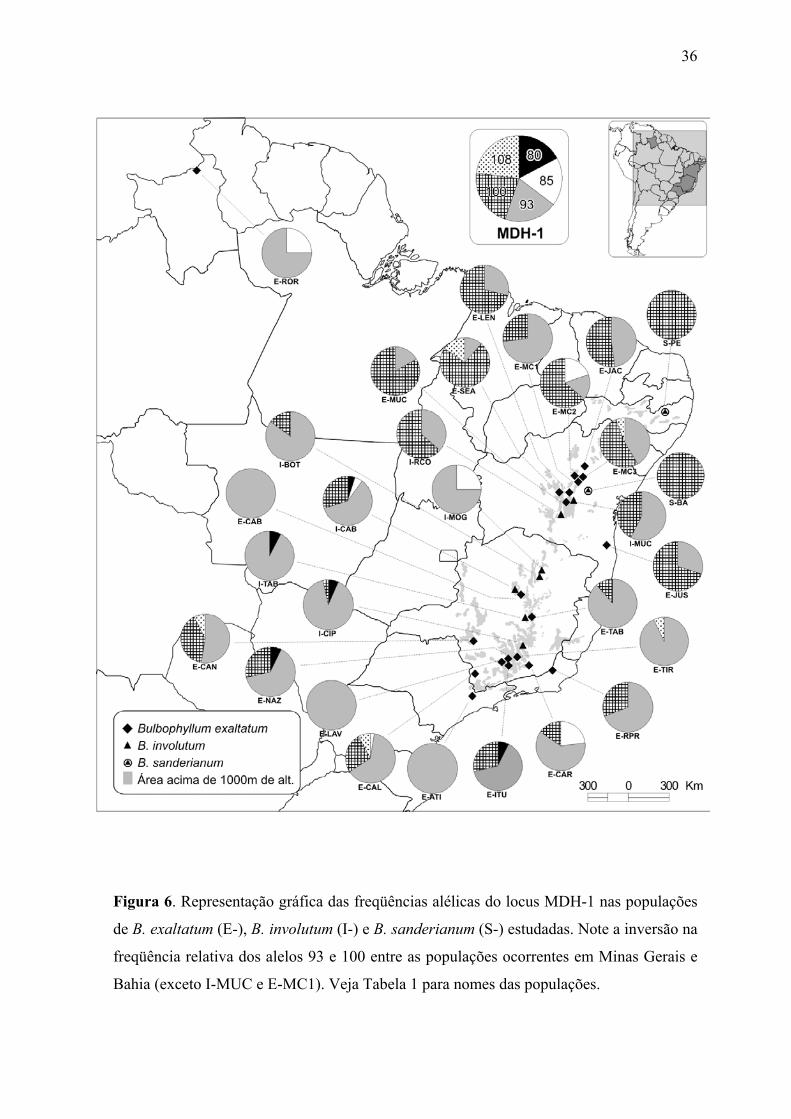
A estruturação genética encontrada em B. sanderianum está relacionada à inversão
da freqüência relativa do alelo mais comum no locus IDH entre as duas populações. Em B.
weddellii, essa inversão ocorre expressivamente no locus SKDH. Em B. involutum, além
de SKDH e IDH, o locus MDH-1, e em B. exaltatum, além dos dois primeiros, o locus
MDH-2, estão fortemente envolvidos nos elevados valores de estruturação entre as
populações destes táxons.
O fluxo gênico estimado indiretamente a partir do FST, o Nm(W) e a partir da
freqüência de alelos privados Nm(S) para B. sanderianum foi respectivamente 0,368 e
0,220, sendo a freqüência média de alelos privados 0,203; para B. weddellii o Nm(W) foi de
0,382, o Nm(S) foi de 0,474 e a freqüência média de alelos privados foi de 0,142; em B.
involutum esses valores foram respectivamente 0,608, 0,480 e 0,136; e, em B. exaltatum
foram encontrados os maiores valores de fluxo gênico, 0,755, 1,358 e 0,077,
respectivamente (Tabela 5).
Relações fenéticas
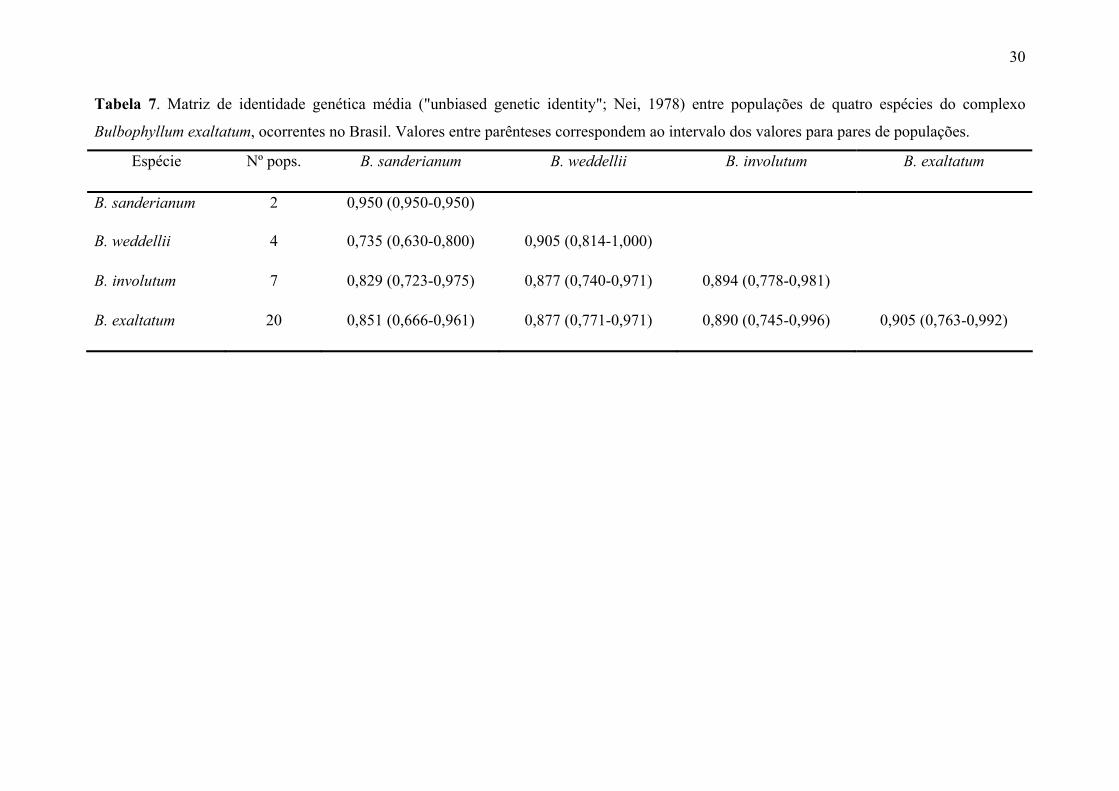
A identidade genética entre as populações conspecíficas variou de 0,76 a 1,00, com
os valores mais baixos encontrados entre populações de B. exaltatum (Tabela 6). Em vários
casos foram encontrados valores de identidade genética entre pares de populações não
conspecíficas superiores ao encontrado em pares de populações conspecíficas. A
identidade genética média entre as espécies variou de 0,735 (B. weddellii e B.
sanderianum) a 0,890 (B. exaltatum e B. involutum).
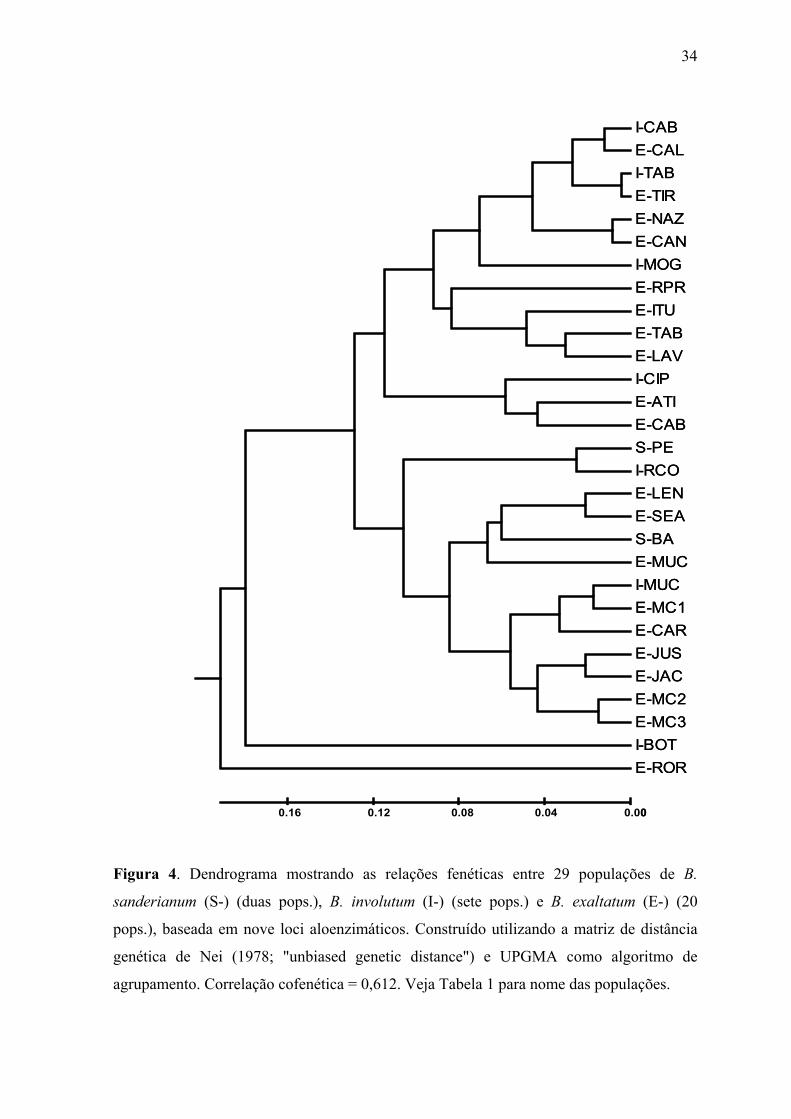
O dendrograma da análise de agrupamento (Figura 4) obtido a partir da análise
baseada na distância genética de Nei (1978), usando UPGMA como algoritmo, e excluindo
as populações de B. weddellii, revelou que nenhuma das espécies forma um grupo distinto
envolvendo exclusivamente todas as populações conspecíficas. Desta forma, não foi

16
possível a delimitação de espécies deste complexo a partir desta análise. No dendrograma,
há formação de dois grupos principais: o primeiro contém todas as populações de B.
involutum e B. exaltatum ocorrentes em Minas Gerais e São Paulo. O segundo grupo
contém as duas populações de B. sanderianum e todas as populações ocorrentes no estado
da Bahia de B. involutum e B. exaltatum, além de uma população de Minas Gerais (E-
CAR). Duas outras populações uniram-se a estes dois grupos separadamente, E-ROR,
localizada em Roraima, e I-BOT, no norte de Minas Gerais, com distâncias de 0,190 e
0,180, respectivamente. O que aparentemente distancia a população E-ROR das demais é o
monomorfismo o alelo 85 no locus SKDH, sendo este alelo raramente encontrado em
freqüência elevada nas demais populações. Na população I-BOT o responsável pela
diferenciação foi a elevada freqüência do alelo IDH 115, normalmente raro ou ausente nas
demais populações, além da fixação do alelo SKDH 95. A separação das populações de
Minas Gerais e Bahia é mantida mesmo para as populações de B. weddelli destes estados,
quando a espécie é incluída na análise.
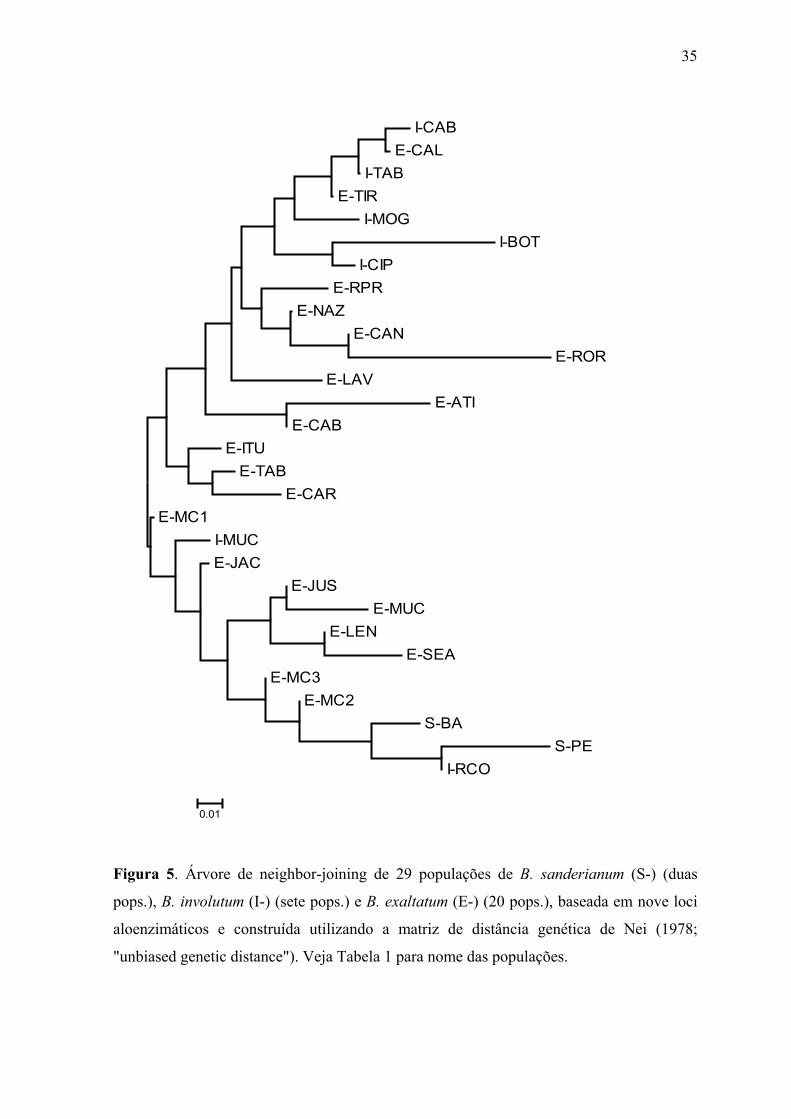
Na árvore de neighbor-joining (NJ), a separação das populações por estado é mais
patente, sendo formados dois grupos, um contendo exclusivamente as populações
ocorrentes em Minas Gerais (B. involutum e B. exaltatum) mais a população de Roraima
(E-ROR), e o outro grupo constituído por todas as populações ocorrentes na Bahia (B.
involutum, B. exaltatum e B. sanderianum) mais a população de Pernambuco S-PE (Figura
5). No primeiro grupo, todas as populações de B. involutum de Minas Gerais constituem
um subgrupo, no qual também estão incluídas as populações E-CAL e E-TIR de B.
exaltatum. No segundo grupo, as duas populações de B. involutum da Bahia não estão
proximamente agrupadas, assim como na análise de UPGMA. Porém, nenhum dos grupos
nas análises de UPGMA e NJ apresentou bootstrap (BS) superior a 50%, com todos os
ramos colapsando nas árvores de consenso estrito. As únicas exceções foram o
agrupamento formado pelas populações de B. exaltatum de Seabra (E-SEA) e Lençóis (E-
LEN) (BS 59% com algoritmo UPGMA) e o agrupamento formado pelo pelas populações
de B. exaltatum, B. involutum e B. sanderianum (BS 100%) excluindo-se as populações E-
ATI e E-CAB (algoritmo NJ) ou I-RCO e S-PE (algoritmo UPGMA).
A inversão da freqüência relativa dos alelos do locus MDH-1 provavelmente é o
principal fator responsável pela separação dos dois grandes grupos de populações, o
formado pelas populações dos estados da Bahia e Pernambuco, tendo o alelo MDH-1 100
como o mais freqüente (exceto para a população E-MC1 de Morro do Chapéu e I-MUC de
Mucugê), e outro formado pelas populações dos estados de São Paulo, Minas Gerais e

17
Roraima, onde o alelo 93 é o mais freqüente (Figura 6). O teste de diferenciação genética
FST baseado no indicador θ confirma a importante participação deste locus na estruturação
das populações por estado de ocorrência pois, em B. exaltatum o locus MDH-1 apresentou
o maior valor de estruturação genética (θP= 0,312) entre os estados da Bahia e Minas
Gerais gerando grande amplitude no intervalo de confiança (θP= -0,013 a 0,182). Porém, o
teste de Mantel indicou não haver correlação entre as distâncias genéticas e geográficas das
populações (r = -0,056; p = 0,356).
Análise morfométrica multivariada
Os intervalos dos valores encontrados nos caracteres utilizados na análise
morfométrica de B. involutum e B. exaltatum nas populações dos diferentes estados
encontram-se na Tabela 8. Na análise de agrupamento por indivíduos utilizando o
coeficiente de Gower, a grande maioria dos indivíduos constitui um grande grupo, que
pode ser subdividido em três subgrupos principais (Figura 7). O primeiro subgrupo (1) é
constituído exclusivamente por indivíduos das populações de B. exaltatum ocorrentes no
sul e na Cadeia do Espinhaço em Minas Gerais (E-CAL, E-LAV, E-ITU, E-TAB, E-SCA,
E-NAZ, E-STL, E-CAR) e em São Paulo (E-ATI). O segundo subgrupo (2) é também
formado exclusivamente por indivíduos de B. exaltatum, mas predominantemente pelas
populações ocorrentes na Bahia (E-MUC, E-JAC, E-MC1, E-MC2, E-LEN, E-SEA) e em
Roraima (E-ROR), com apenas três exceções (E-SJA, E-TAB-3 e E-SCA-2). Por outro
lado, o terceiro subgrupo (3) é constituído por indivíduos de B. exaltatum,
predominantemente de algumas populações da Bahia e raros de Minas Gerais, e a maioria
dos indivíduos de B. involutum. Três indivíduos de B. involutum (I-TAB) e um de B.
exaltatum (E-SCA), ambos de Minas Gerais, formam um pequeno grupo (4) mais
diferenciado e que se liga externamente ao grande grupo anterior.
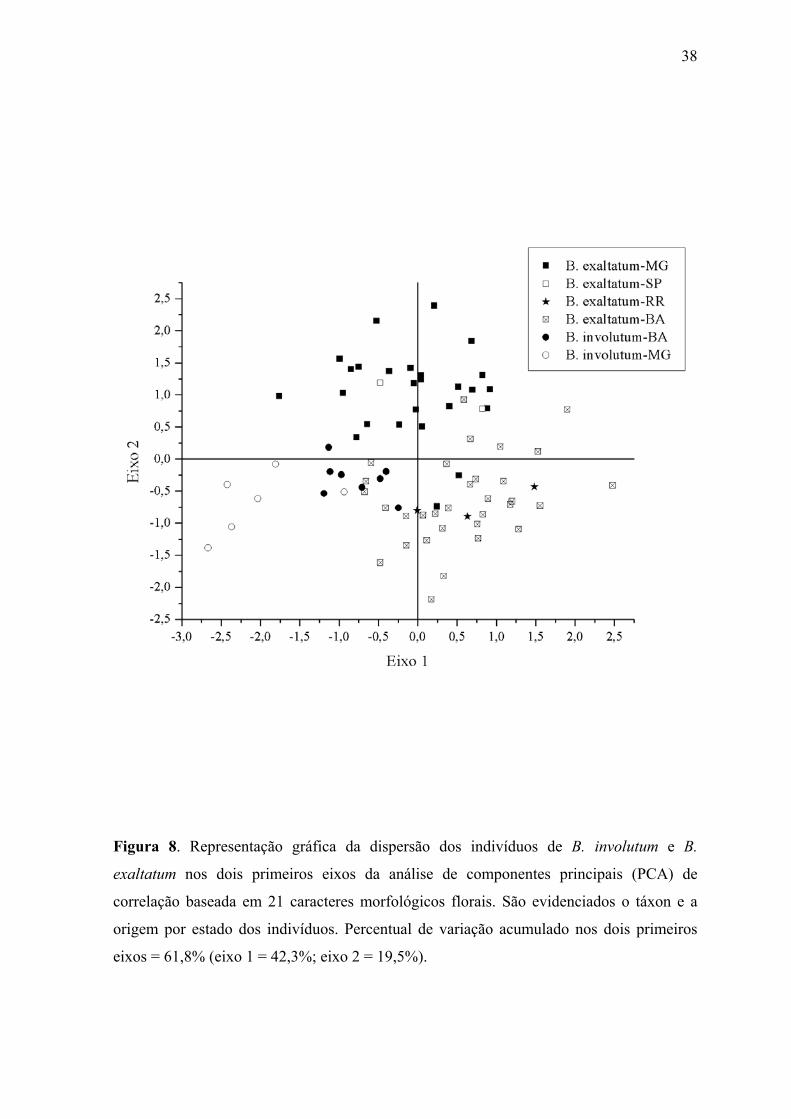
Na PCA apenas os dois primeiros eixos foram estatisticamente significativos, sendo
interpretados. O primeiro eixo acumulou 42,3% da variância, sendo explicado
principalmente pelas variáveis #14, 15, 17, 18 e 19, que correspondem a caracteres de
dimensão do labelo (Tabela 2). O segundo eixo acumulou 19,5% da variação, sendo que
variáveis de largura de sépalas e comprimento lobo mediano do labelo foram as mais
importantes na sua partição (#2, 3, 4, 7 e 16). A representação dos escores dos indivíduos
nos dois primeiros eixos revela a separação no eixo 1 das populações de B. involutum de
Minas Gerais, com exceção de um indivíduo de Grão Mogol, de todas as outras demais

18
(Figura 8). Os indivíduos de B. involutum da Bahia e o indivíduo de Grão Mogol
encontram-se em posição intermediária entre indivíduos de B. involutum de Minas Gerais e
os de B. exaltatum da Bahia, sem qualquer separação conspícua. No eixo 2, pode ser
observada uma pequena separação entre as populações de B. exaltatum dos estados de
Minas Gerais e São Paulo das populações da Bahia e Roraima, com ocorrência de algumas
sobreposições.
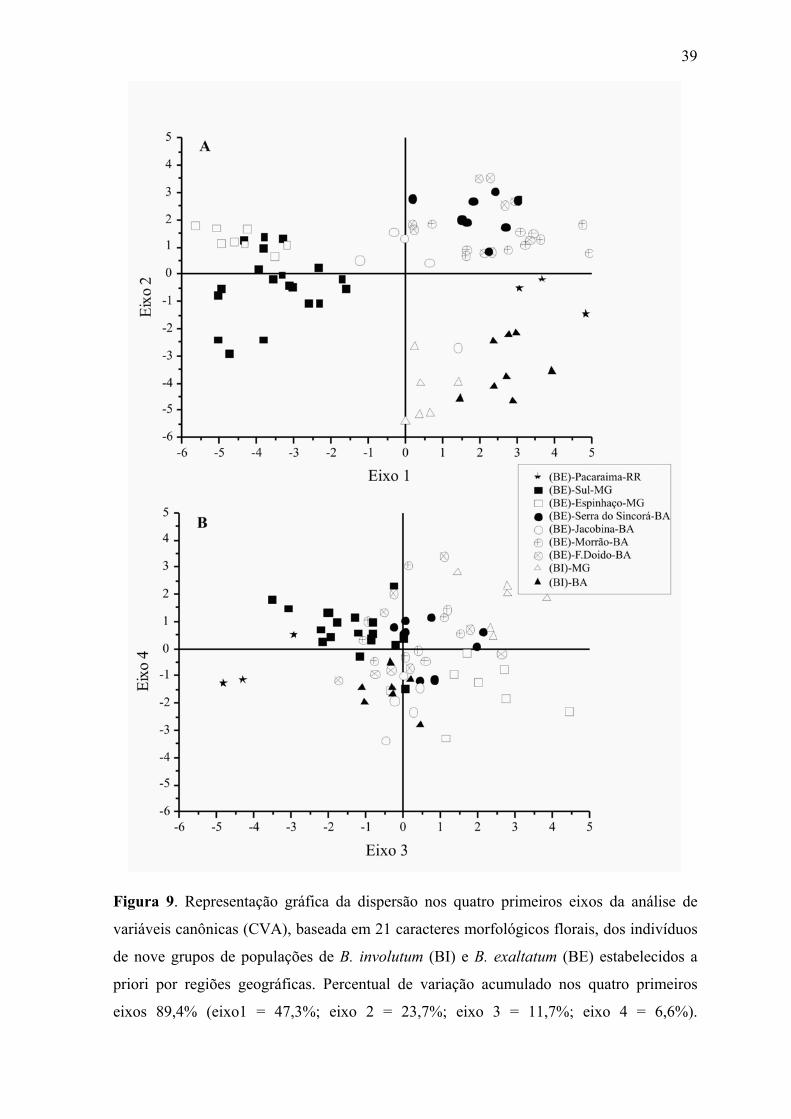
Na CVA, dos oito possíveis eixos calculados, apenas os quatros primeiros foram
estatisticamente significativos. No primeiro eixo canônico pode ser observada a separação
das populações de B. exaltatum de Minas Gerais como um grupo distinto das populações
deste táxon da Bahia e de Roraima, e também distinto de B. involutum (Figura 9A),
principalmente pelo menor tamanho do labelo e do ístimo e maior largura da sépala dorsal
e (#2, 13, 16, 20 e 22; Tabela 2). O segundo eixo separa B. exaltatum da Bahia de B.
involutum e da população de Roraima, sendo que apenas um indivíduo de Jacobina se
posiciona mais próximo ao centróide dos dois grupos de B. involutum. Os indivíduos do
grupo de Roraima encontram-se numa posição intermediária entre os B. involutum e os B.
exaltatum da Bahia. Comprimento das sépalas, ângulo do ápice da sépala dorsal,
comprimento do istmo e largura entre lobos do labelo são os caracteres que mais
contribuíram para a discriminação de grupos neste eixo (# 1, 4, 5, 15 e 20; Tabela 2). Os
eixos canônicos um e dois acumularam respectivamente 47,3 e 23,7% da variação.
O terceiro eixo canônico (Figura 9B) separa o grupo de Roraima dos B. involutum
da Bahia e este dos B. involutum de Minas Gerais. Neste mesmo eixo, pode ser observada a
separação dos dois grupos de B. exaltatum de Minas Gerais. Este eixo está
predominantemente relacionado a características de comprimento das sépalas, largura da
pétala, comprimento do lobo mediano do labelo e largura entre lobos (#1, 5, 10, 15 e 16),
todos positivamente correlacionados com o eixo. A separação entre os B. involutum da
Bahia e de Minas Gerais também pode ser percebida no quarto eixo canônico, sendo que
ângulo do ápice, comprimento e largura da pétala, comprimento do labelo e largura do lobo
mediano (# 9, 10, 12, 14 e 17) são as características que mais influenciam essa separação.
Os eixos três e quatro acumularam respectivamente 11,7 e 6,6% da variação.
Apenas os grupos formados por populações de B. exaltatum da Bahia tiveram
alguns indivíduos classificados incorretamente na análise de discriminantes. Porém, estas
classificações incorretas ocorreram entre os próprios grupos deste táxon da Bahia, exceto
um indivíduo de Jacobina, classificado como B. involutum-BA. Dos nove indivíduos do
grupo F.Doido, um foi classificado como sendo do grupo Morrão e um do grupo Serra do

19
Sincorá; dos 10 indivíduos do grupo Morrão, um foi classificado como F.Doido; do grupo
Serra do Sincorá, um indivíduo foi classificado como F.Doido.
A análise de agrupamento dos centróides dos grupos utilizando a distância de
Mahalanobis revelou a formação de quatro grupos principais (Figura 10), um contendo os
B. exaltatum de Minas Gerais, outro constituído exclusivamente por B. exaltatum de
Roraima, outro por B. exaltatum da Bahia e o último por B. involutum de Bahia e Minas
Gerais.

20
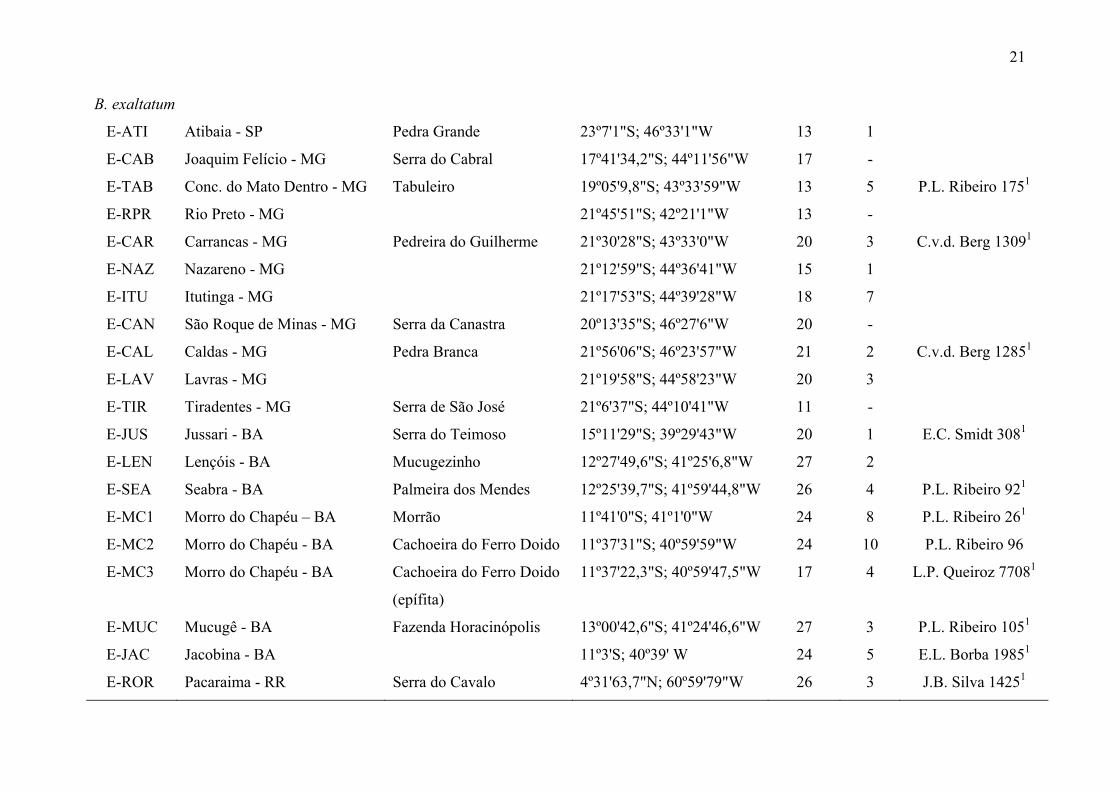
Tabela 1. Identificação, localização geográfica e número de indivíduos coletados e utilizados nas análises genéticas e morfométricas das 33
populações de B. sanderianum, B. weddellii, B. involutum e B. exaltatum estudadas. Nas análises morfométricas, adicionalmente foram utilizados
indivíduos de B. exaltatum coletados em populações em São Tomé das Letras-MG (E-STL1 indiv.), na Serra do Japi em Jundiaí-SP (E-SJA; 1
indiv.) e na Serra do Caraça em Catas Altas-MG (E-SCA; 3 indiv.). Voucher’s foram depositados no acervo dos herbários HUEFS (1) e UEC (2).
População Município Localidade Localização Nº indivíduos Voucher
genética morf.
B. sanderianum
S-PE Brejo de Madre de Deus - PE Fazenda Bituri 8º11'49,4"S; 36º24'19,3"W 12 - P.L. Ribeiro 1401
S-BA Rui Barbosa - BA Serra do Orobó 12º18'3"S; 40º28'41,9"W 24 - P.L. Ribeiro 1761
B. weddellii
W-MU1 Mucugê - BA Parque Municipal 13º43'0"S; 41º24'0"W 15 - C. Azevedo 1881
W-MU2 Mucugê - BA Serra da Tesoura 13º8'13"S; 41º20'21"W 12 - E.L. Borba 19341
W-MU3 Mucugê - BA Pousada Pé de Serra 13º0'16"S; 41º22'12"W 14 - E.L. Borba 19421
W-CIP Santana do Riacho - MG Serra do Cipó 19º14'50"S; 43º30'40"W 17 -
B. involutum
I-MUC Mucugê - BA Pousada Pé de Serra 13º0'16"S; 41º22'12"W 20 3 E.L. Borba 19431
I-RCO Rio de Contas – BA Fazenda Vacaro 13º31'55,8"S; 41º52'15,8"W 11 5
I-CAB Joaquim Felício - MG Serra do Cabral 17º41'34,25"S; 44º11'56"W 20 2
I-TAB Conc. do Mato Dentro - MG Tabuleiro 19º05'9,8"S; 43º33'59"W 13 3 P.L. Ribeiro 1741
I-MOG Grão Mogol - MG 16º33'34"S; 42º53'23"W 14 1
I-BOT Botumirim - MG 16º52'20"S; 43º0'39"W 18 -
I-CIP Santana do Riacho - MG Serra do Cipó 20º29'59"S; 43º51'28"W 15 - E.L. Borba 1502
21
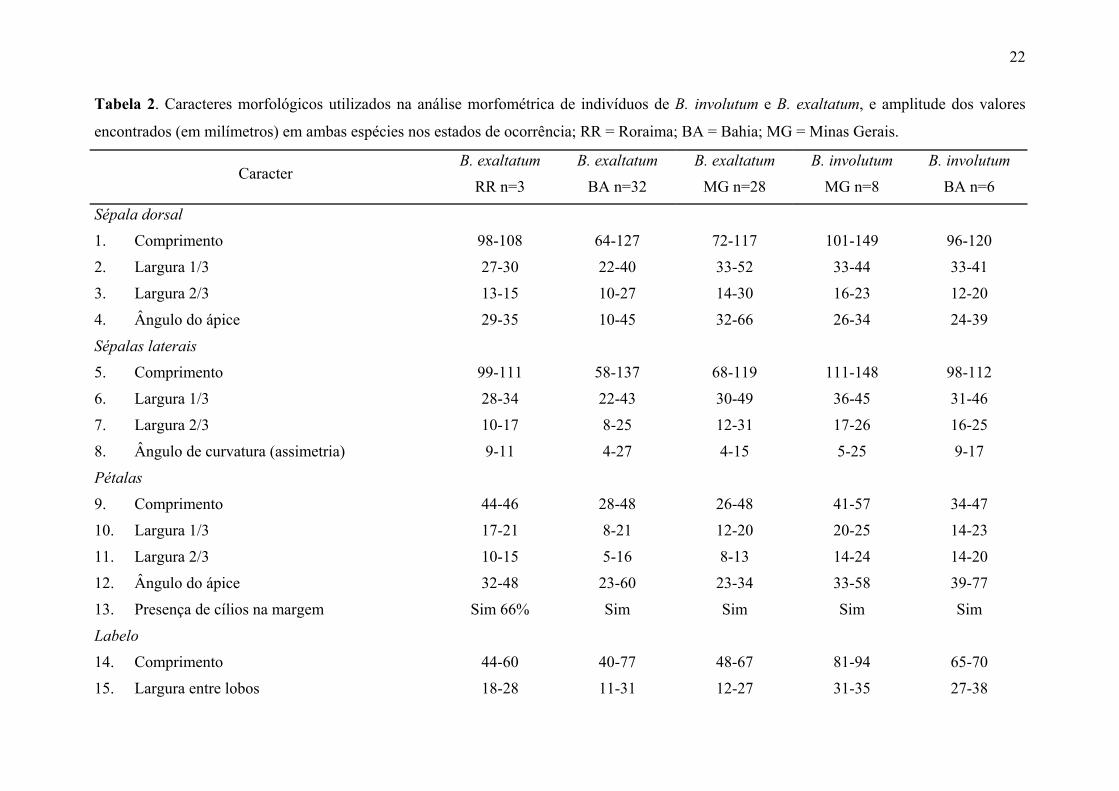
B. exaltatum
E-ATI Atibaia - SP Pedra Grande 23º7'1"S; 46º33'1"W 13 1
E-CAB Joaquim Felício - MG Serra do Cabral 17º41'34,2"S; 44º11'56"W 17 -
E-TAB Conc. do Mato Dentro - MG Tabuleiro 19º05'9,8"S; 43º33'59"W 13 5 P.L. Ribeiro 1751
E-RPR Rio Preto - MG 21º45'51"S; 42º21'1"W 13 -
E-CAR Carrancas - MG Pedreira do Guilherme 21º30'28"S; 43º33'0"W 20 3 C.v.d. Berg 13091
E-NAZ Nazareno - MG 21º12'59"S; 44º36'41"W 15 1
E-ITU Itutinga - MG 21º17'53"S; 44º39'28"W 18 7
E-CAN São Roque de Minas - MG Serra da Canastra 20º13'35"S; 46º27'6"W 20 -
E-CAL Caldas - MG Pedra Branca 21º56'06"S; 46º23'57"W 21 2 C.v.d. Berg 12851
E-LAV Lavras - MG 21º19'58"S; 44º58'23"W 20 3
E-TIR Tiradentes - MG Serra de São José 21º6'37"S; 44º10'41"W 11 -
E-JUS Jussari - BA Serra do Teimoso 15º11'29"S; 39º29'43"W 20 1 E.C. Smidt 3081
E-LEN Lençóis - BA Mucugezinho 12º27'49,6"S; 41º25'6,8"W 27 2
E-SEA Seabra - BA Palmeira dos Mendes 12º25'39,7"S; 41º59'44,8"W 26 4 P.L. Ribeiro 921
E-MC1 Morro do Chapéu – BA Morrão 11º41'0"S; 41º1'0"W 24 8 P.L. Ribeiro 261
E-MC2 Morro do Chapéu - BA Cachoeira do Ferro Doido 11º37'31"S; 40º59'59"W 24 10 P.L. Ribeiro 96
E-MC3 Morro do Chapéu - BA Cachoeira do Ferro Doido
(epífita)
11º37'22,3"S; 40º59'47,5"W 17 4 L.P. Queiroz 77081
E-MUC Mucugê - BA Fazenda Horacinópolis 13º00'42,6"S; 41º24'46,6"W 27 3 P.L. Ribeiro 1051
E-JAC Jacobina - BA 11º3'S; 40º39' W 24 5 E.L. Borba 19851
E-ROR Pacaraima - RR Serra do Cavalo 4º31'63,7"N; 60º59'79"W 26 3 J.B. Silva 14251
22
Tabela 2. Caracteres morfológicos utilizados na análise morfométrica de indivíduos de B. involutum e B. exaltatum, e amplitude dos valores
encontrados (em milímetros) em ambas espécies nos estados de ocorrência; RR = Roraima; BA = Bahia; MG = Minas Gerais.
Caracter B. exaltatum
RR n=3 B. exaltatum
BA n=32 B. exaltatum
MG n=28 B. involutum
MG n=8 B. involutum
BA n=6
Sépala dorsal 1. Comprimento 98-108 64-127 72-117 101-149 96-120
2. Largura 1/3 27-30 22-40 33-52 33-44 33-41
3. Largura 2/3 13-15 10-27 14-30 16-23 12-20
4. Ângulo do ápice 29-35 10-45 32-66 26-34 24-39
Sépalas laterais 5. Comprimento 99-111 58-137 68-119 111-148 98-112
6. Largura 1/3 28-34 22-43 30-49 36-45 31-46
7. Largura 2/3 10-17 8-25 12-31 17-26 16-25
8. Ângulo de curvatura (assimetria) 9-11 4-27 4-15 5-25 9-17
Pétalas 9. Comprimento 44-46 28-48 26-48 41-57 34-47
10. Largura 1/3 17-21 8-21 12-20 20-25 14-23
11. Largura 2/3 10-15 5-16 8-13 14-24 14-20
12. Ângulo do ápice 32-48 23-60 23-34 33-58 39-77
13. Presença de cílios na margem Sim 66% Sim Sim Sim Sim
Labelo 14. Comprimento 44-60 40-77 48-67 81-94 65-70
15. Largura entre lobos 18-28 11-31 12-27 31-35 27-38
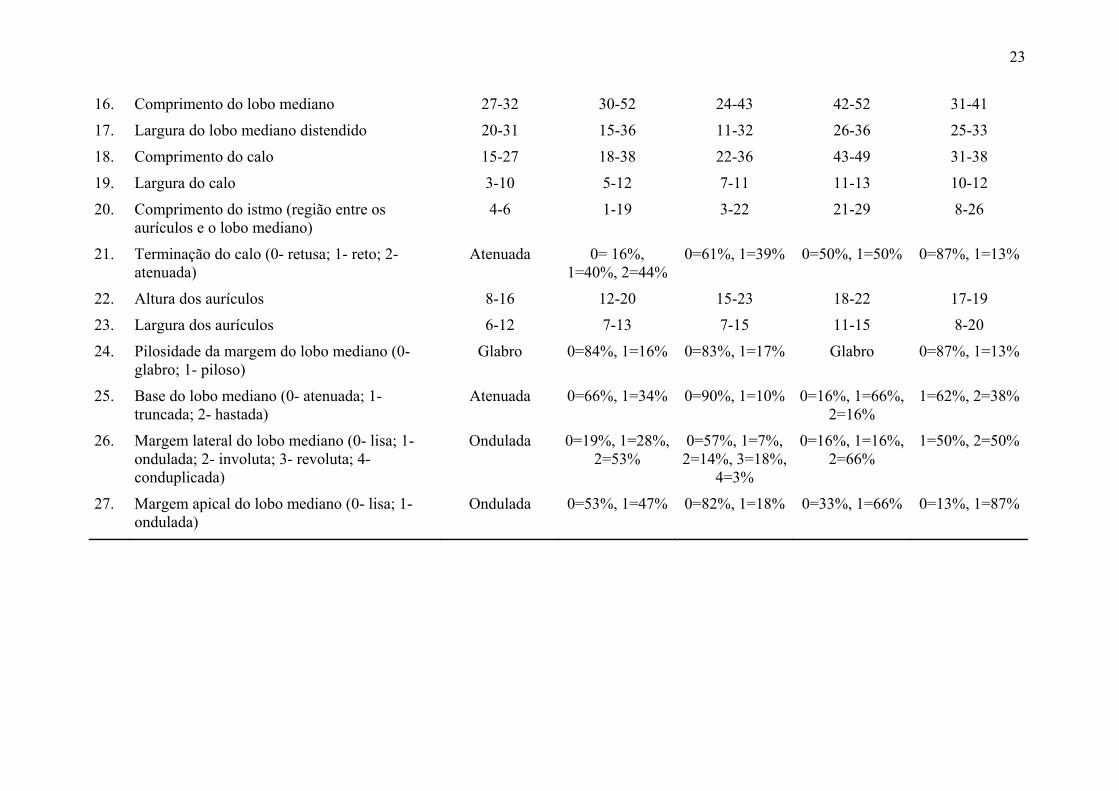
23
16. Comprimento do lobo mediano 27-32 30-52 24-43 42-52 31-41
17. Largura do lobo mediano distendido 20-31 15-36 11-32 26-36 25-33
18. Comprimento do calo 15-27 18-38 22-36 43-49 31-38
19. Largura do calo 3-10 5-12 7-11 11-13 10-12
20. Comprimento do istmo (região entre os aurículos e o lobo mediano)
4-6 1-19 3-22 21-29 8-26
21. Terminação do calo (0- retusa; 1- reto; 2- atenuada)
Atenuada 0= 16%, 1=40%, 2=44%
0=61%, 1=39% 0=50%, 1=50% 0=87%, 1=13%
22. Altura dos aurículos 8-16 12-20 15-23 18-22 17-19
23. Largura dos aurículos 6-12 7-13 7-15 11-15 8-20
24. Pilosidade da margem do lobo mediano (0- glabro; 1- piloso)
Glabro 0=84%, 1=16% 0=83%, 1=17% Glabro 0=87%, 1=13%
25. Base do lobo mediano (0- atenuada; 1- truncada; 2- hastada)
Atenuada 0=66%, 1=34% 0=90%, 1=10% 0=16%, 1=66%, 2=16%
1=62%, 2=38%
26. Margem lateral do lobo mediano (0- lisa; 1- ondulada; 2- involuta; 3- revoluta; 4- conduplicada)
Ondulada 0=19%, 1=28%, 2=53%
0=57%, 1=7%, 2=14%, 3=18%,
4=3%
0=16%, 1=16%, 2=66%
1=50%, 2=50%
27. Margem apical do lobo mediano (0- lisa; 1- ondulada)
Ondulada 0=53%, 1=47% 0=82%, 1=18% 0=33%, 1=66% 0=13%, 1=87%
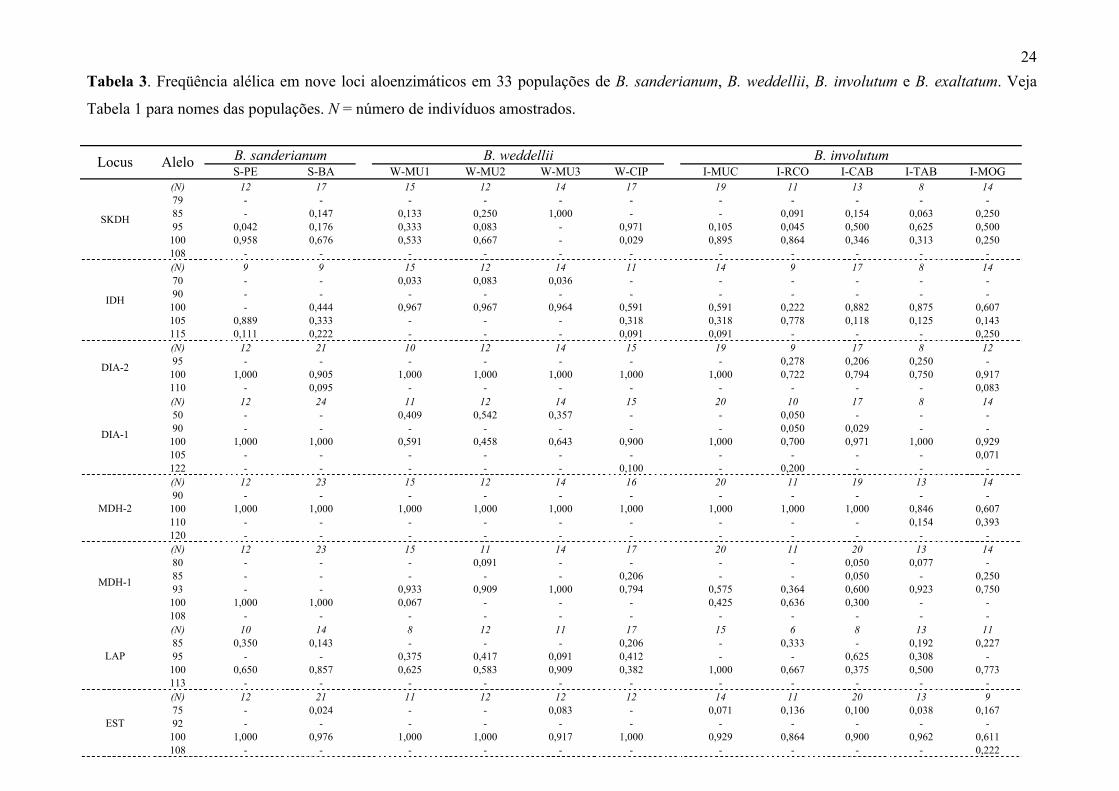
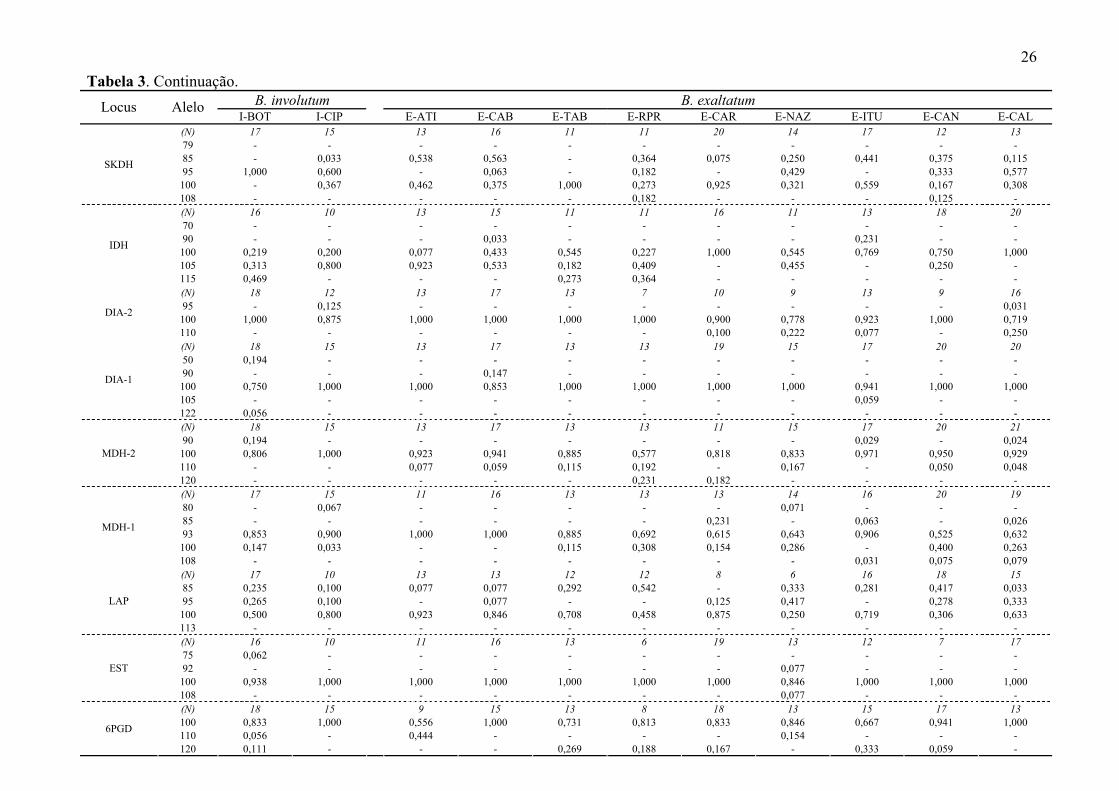
24 Tabela 3. Freqüência alélica em nove loci aloenzimáticos em 33 populações de B. sanderianum, B. weddellii, B. involutum e B. exaltatum. Veja
Tabela 1 para nomes das populações. N = número de indivíduos amostrados.
B. sanderianum B. weddellii B. involutum Locus Alelo S-PE S-BA W-MU1 W-MU2 W-MU3 W-CIP I-MUC I-RCO I-CAB I-TAB I-MOG
(N) 12 17 15 12 14 17 19 11 13 8 14 79 - - - - - - - - - - - 85 - 0,147 0,133 0,250 1,000 - - 0,091 0,154 0,063 0,250 95 0,042 0,176 0,333 0,083 - 0,971 0,105 0,045 0,500 0,625 0,500
100 0,958 0,676 0,533 0,667 - 0,029 0,895 0,864 0,346 0,313 0,250
SKDH
108 - - - - - - - - - - - (N) 9 9 15 12 14 11 14 9 17 8 14 70 - - 0,033 0,083 0,036 - - - - - - 90 - - - - - - - - - - -
100 - 0,444 0,967 0,967 0,964 0,591 0,591 0,222 0,882 0,875 0,607 105 0,889 0,333 - - - 0,318 0,318 0,778 0,118 0,125 0,143
IDH
115 0,111 0,222 - - - 0,091 0,091 - - - 0,250 (N) 12 21 10 12 14 15 19 9 17 8 12 95 - - - - - - - 0,278 0,206 0,250 -
100 1,000 0,905 1,000 1,000 1,000 1,000 1,000 0,722 0,794 0,750 0,917 DIA-2
110 - 0,095 - - - - - - - - 0,083 (N) 12 24 11 12 14 15 20 10 17 8 14 50 - - 0,409 0,542 0,357 - - 0,050 - - - 90 - - - - - - - 0,050 0,029 - -
100 1,000 1,000 0,591 0,458 0,643 0,900 1,000 0,700 0,971 1,000 0,929 105 - - - - - - - - - - 0,071
DIA-1
122 - - - - - 0,100 - 0,200 - - - (N) 12 23 15 12 14 16 20 11 19 13 14 90 - - - - - - - - - - -
100 1,000 1,000 1,000 1,000 1,000 1,000 1,000 1,000 1,000 0,846 0,607 110 - - - - - - - - - 0,154 0,393
MDH-2
120 - - - - - - - - - - - (N) 12 23 15 11 14 17 20 11 20 13 14 80 - - - 0,091 - - - - 0,050 0,077 - 85 - - - - - 0,206 - - 0,050 - 0,250 93 - - 0,933 0,909 1,000 0,794 0,575 0,364 0,600 0,923 0,750
100 1,000 1,000 0,067 - - - 0,425 0,636 0,300 - -
MDH-1
108 - - - - - - - - - - - (N) 10 14 8 12 11 17 15 6 8 13 11 85 0,350 0,143 - - - 0,206 - 0,333 - 0,192 0,227 95 - - 0,375 0,417 0,091 0,412 - - 0,625 0,308 -
100 0,650 0,857 0,625 0,583 0,909 0,382 1,000 0,667 0,375 0,500 0,773 LAP
113 - - - - - - - - - - - (N) 12 21 11 12 12 12 14 11 20 13 9 75 - 0,024 - - 0,083 - 0,071 0,136 0,100 0,038 0,167 92 - - - - - - - - - - -
100 1,000 0,976 1,000 1,000 0,917 1,000 0,929 0,864 0,900 0,962 0,611 EST
108 - - - - - - - - - - 0,222

25 (N) 10 13 5 12 14 17 16 11 17 9 12 100 1,000 0,692 1,000 1,000 1,000 0,882 1,000 1,000 1,000 0,889 1,000 110 - 0,308 - - - 0,118 - - - 0,111 - 6PGD
120 - - - - - - - - - - -
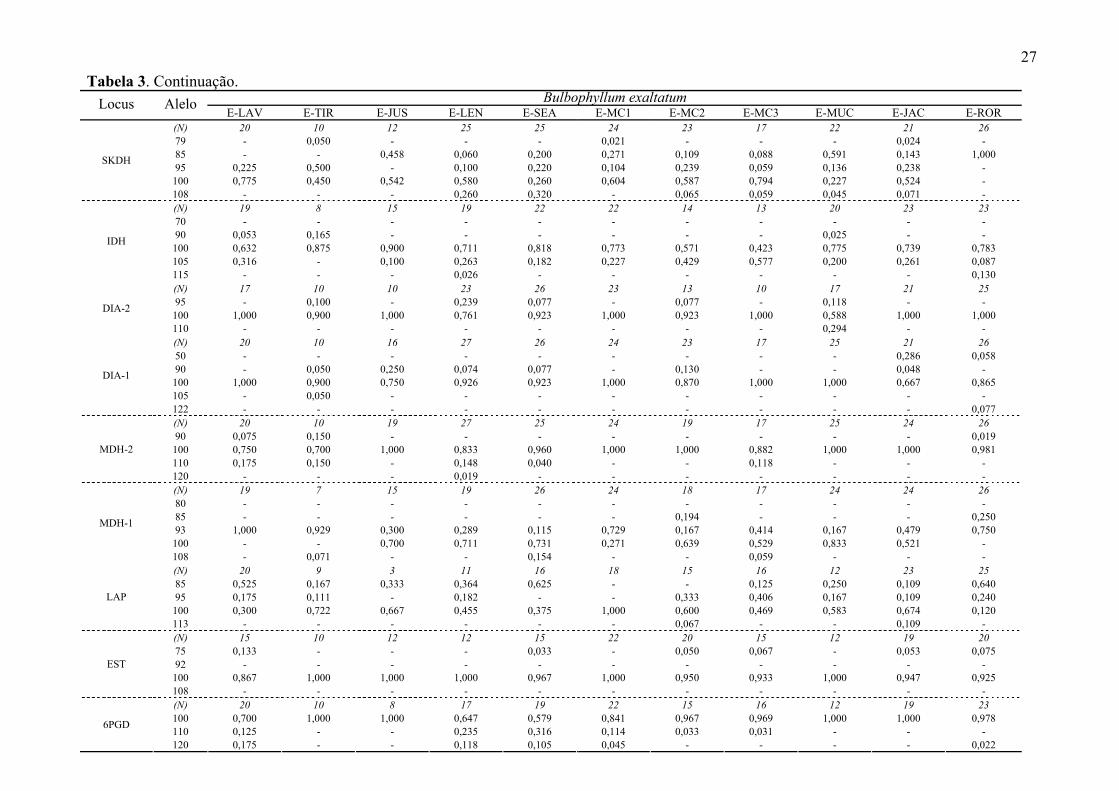
26 Tabela 3. Continuação.
B. involutum B. exaltatum Locus Alelo I-BOT I-CIP E-ATI E-CAB E-TAB E-RPR E-CAR E-NAZ E-ITU E-CAN E-CAL
(N) 17 15 13 16 11 11 20 14 17 12 13 79 - - - - - - - - - - - 85 - 0,033 0,538 0,563 - 0,364 0,075 0,250 0,441 0,375 0,115 95 1,000 0,600 - 0,063 - 0,182 - 0,429 - 0,333 0,577 100 - 0,367 0,462 0,375 1,000 0,273 0,925 0,321 0,559 0,167 0,308
SKDH
108 - - - - - 0,182 - - - 0,125 - (N) 16 10 13 15 11 11 16 11 13 18 20 70 - - - - - - - - - - - 90 - - - 0,033 - - - - 0,231 - - 100 0,219 0,200 0,077 0,433 0,545 0,227 1,000 0,545 0,769 0,750 1,000 105 0,313 0,800 0,923 0,533 0,182 0,409 - 0,455 - 0,250 -
IDH
115 0,469 - - - 0,273 0,364 - - - - - (N) 18 12 13 17 13 7 10 9 13 9 16 95 - 0,125 - - - - - - - - 0,031 100 1,000 0,875 1,000 1,000 1,000 1,000 0,900 0,778 0,923 1,000 0,719
DIA-2
110 - - - - - - 0,100 0,222 0,077 - 0,250 (N) 18 15 13 17 13 13 19 15 17 20 20 50 0,194 - - - - - - - - - - 90 - - - 0,147 - - - - - - - 100 0,750 1,000 1,000 0,853 1,000 1,000 1,000 1,000 0,941 1,000 1,000 105 - - - - - - - - 0,059 - -
DIA-1
122 0,056 - - - - - - - - - - (N) 18 15 13 17 13 13 11 15 17 20 21 90 0,194 - - - - - - - 0,029 - 0,024 100 0,806 1,000 0,923 0,941 0,885 0,577 0,818 0,833 0,971 0,950 0,929 110 - - 0,077 0,059 0,115 0,192 - 0,167 - 0,050 0,048
MDH-2
120 - - - - - 0,231 0,182 - - - - (N) 17 15 11 16 13 13 13 14 16 20 19 80 - 0,067 - - - - - 0,071 - - - 85 - - - - - - 0,231 - 0,063 - 0,026 93 0,853 0,900 1,000 1,000 0,885 0,692 0,615 0,643 0,906 0,525 0,632 100 0,147 0,033 - - 0,115 0,308 0,154 0,286 - 0,400 0,263
MDH-1
108 - - - - - - - - 0,031 0,075 0,079 (N) 17 10 13 13 12 12 8 6 16 18 15 85 0,235 0,100 0,077 0,077 0,292 0,542 - 0,333 0,281 0,417 0,033 95 0,265 0,100 - 0,077 - - 0,125 0,417 - 0,278 0,333 100 0,500 0,800 0,923 0,846 0,708 0,458 0,875 0,250 0,719 0,306 0,633
LAP
113 - - - - - - - - - - - (N) 16 10 11 16 13 6 19 13 12 7 17 75 0,062 - - - - - - - - - - 92 - - - - - - - 0,077 - - - 100 0,938 1,000 1,000 1,000 1,000 1,000 1,000 0,846 1,000 1,000 1,000
EST
108 - - - - - - - 0,077 - - - (N) 18 15 9 15 13 8 18 13 15 17 13 100 0,833 1,000 0,556 1,000 0,731 0,813 0,833 0,846 0,667 0,941 1,000 110 0,056 - 0,444 - - - - 0,154 - - - 6PGD
120 0,111 - - - 0,269 0,188 0,167 - 0,333 0,059 -
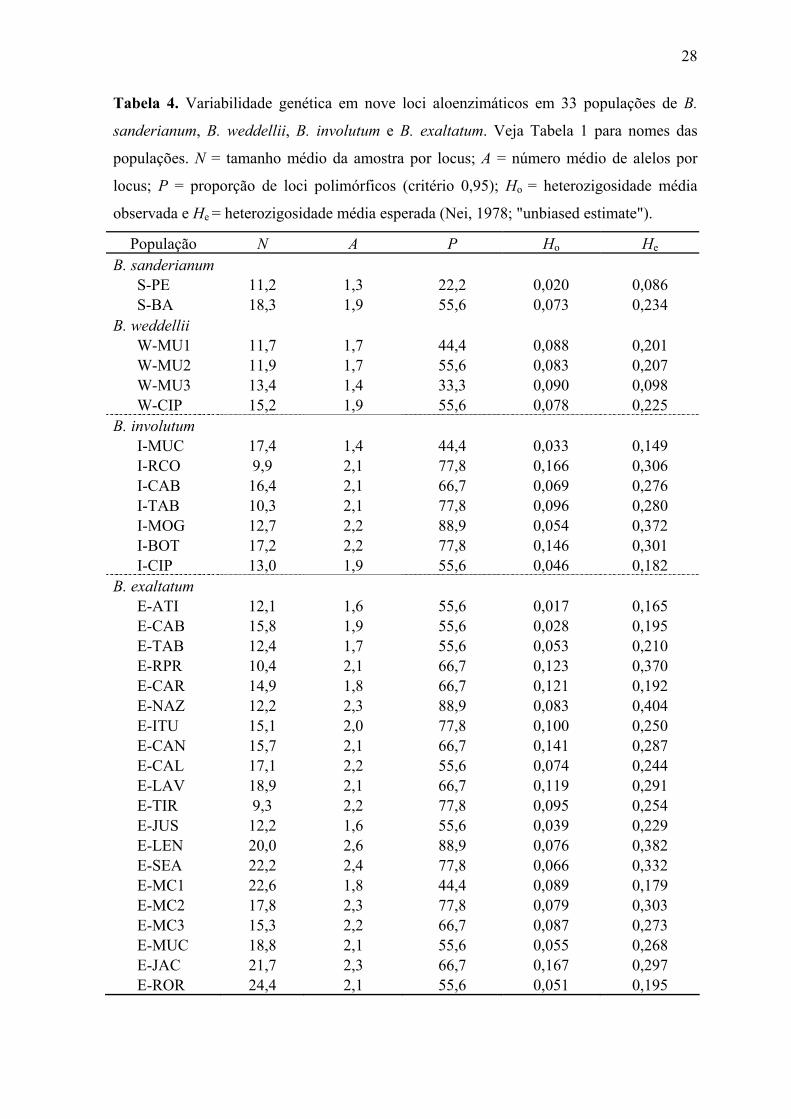
27 Tabela 3. Continuação.
Bulbophyllum exaltatum Locus Alelo E-LAV E-TIR E-JUS E-LEN E-SEA E-MC1 E-MC2 E-MC3 E-MUC E-JAC E-ROR
(N) 20 10 12 25 25 24 23 17 22 21 26 79 - 0,050 - - - 0,021 - - - 0,024 - 85 - - 0,458 0,060 0,200 0,271 0,109 0,088 0,591 0,143 1,000 95 0,225 0,500 - 0,100 0,220 0,104 0,239 0,059 0,136 0,238 - 100 0,775 0,450 0,542 0,580 0,260 0,604 0,587 0,794 0,227 0,524 -
SKDH
108 - - - 0,260 0,320 - 0,065 0,059 0,045 0,071 - (N) 19 8 15 19 22 22 14 13 20 23 23 70 - - - - - - - - - - - 90 0,053 0,165 - - - - - - 0,025 - - 100 0,632 0,875 0,900 0,711 0,818 0,773 0,571 0,423 0,775 0,739 0,783 105 0,316 - 0,100 0,263 0,182 0,227 0,429 0,577 0,200 0,261 0,087
IDH
115 - - - 0,026 - - - - - - 0,130 (N) 17 10 10 23 26 23 13 10 17 21 25 95 - 0,100 - 0,239 0,077 - 0,077 - 0,118 - - 100 1,000 0,900 1,000 0,761 0,923 1,000 0,923 1,000 0,588 1,000 1,000
DIA-2
110 - - - - - - - - 0,294 - - (N) 20 10 16 27 26 24 23 17 25 21 26 50 - - - - - - - - - 0,286 0,058 90 - 0,050 0,250 0,074 0,077 - 0,130 - - 0,048 - 100 1,000 0,900 0,750 0,926 0,923 1,000 0,870 1,000 1,000 0,667 0,865 105 - 0,050 - - - - - - - - -
DIA-1
122 - - - - - - - - - - 0,077 (N) 20 10 19 27 25 24 19 17 25 24 26 90 0,075 0,150 - - - - - - - - 0,019 100 0,750 0,700 1,000 0,833 0,960 1,000 1,000 0,882 1,000 1,000 0,981 110 0,175 0,150 - 0,148 0,040 - - 0,118 - - -
MDH-2
120 - - - 0,019 - - - - - - - (N) 19 7 15 19 26 24 18 17 24 24 26 80 - - - - - - - - - - - 85 - - - - - - 0,194 - - - 0,250 93 1,000 0,929 0,300 0,289 0,115 0,729 0,167 0,414 0,167 0,479 0,750 100 - - 0,700 0,711 0,731 0,271 0,639 0,529 0,833 0,521 -
MDH-1
108 - 0,071 - - 0,154 - - 0,059 - - - (N) 20 9 3 11 16 18 15 16 12 23 25 85 0,525 0,167 0,333 0,364 0,625 - - 0,125 0,250 0,109 0,640 95 0,175 0,111 - 0,182 - - 0,333 0,406 0,167 0,109 0,240 100 0,300 0,722 0,667 0,455 0,375 1,000 0,600 0,469 0,583 0,674 0,120
LAP
113 - - - - - - 0,067 - - 0,109 - (N) 15 10 12 12 15 22 20 15 12 19 20 75 0,133 - - - 0,033 - 0,050 0,067 - 0,053 0,075 92 - - - - - - - - - - - 100 0,867 1,000 1,000 1,000 0,967 1,000 0,950 0,933 1,000 0,947 0,925
EST
108 - - - - - - - - - - - (N) 20 10 8 17 19 22 15 16 12 19 23 100 0,700 1,000 1,000 0,647 0,579 0,841 0,967 0,969 1,000 1,000 0,978 110 0,125 - - 0,235 0,316 0,114 0,033 0,031 - - - 6PGD
120 0,175 - - 0,118 0,105 0,045 - - - - 0,022
28
Tabela 4. Variabilidade genética em nove loci aloenzimáticos em 33 populações de B.
sanderianum, B. weddellii, B. involutum e B. exaltatum. Veja Tabela 1 para nomes das
populações. N = tamanho médio da amostra por locus; A = número médio de alelos por
locus; P = proporção de loci polimórficos (critério 0,95); Ho = heterozigosidade média
observada e He = heterozigosidade média esperada (Nei, 1978; "unbiased estimate").
População N A P Ho He B. sanderianum
S-PE 11,2 1,3 22,2 0,020 0,086 S-BA 18,3 1,9 55,6 0,073 0,234
B. weddellii W-MU1 11,7 1,7 44,4 0,088 0,201 W-MU2 11,9 1,7 55,6 0,083 0,207 W-MU3 13,4 1,4 33,3 0,090 0,098 W-CIP 15,2 1,9 55,6 0,078 0,225
B. involutum I-MUC 17,4 1,4 44,4 0,033 0,149 I-RCO 9,9 2,1 77,8 0,166 0,306 I-CAB 16,4 2,1 66,7 0,069 0,276 I-TAB 10,3 2,1 77,8 0,096 0,280 I-MOG 12,7 2,2 88,9 0,054 0,372 I-BOT 17,2 2,2 77,8 0,146 0,301 I-CIP 13,0 1,9 55,6 0,046 0,182
B. exaltatum E-ATI 12,1 1,6 55,6 0,017 0,165 E-CAB 15,8 1,9 55,6 0,028 0,195 E-TAB 12,4 1,7 55,6 0,053 0,210 E-RPR 10,4 2,1 66,7 0,123 0,370 E-CAR 14,9 1,8 66,7 0,121 0,192 E-NAZ 12,2 2,3 88,9 0,083 0,404 E-ITU 15,1 2,0 77,8 0,100 0,250 E-CAN 15,7 2,1 66,7 0,141 0,287 E-CAL 17,1 2,2 55,6 0,074 0,244 E-LAV 18,9 2,1 66,7 0,119 0,291 E-TIR 9,3 2,2 77,8 0,095 0,254 E-JUS 12,2 1,6 55,6 0,039 0,229 E-LEN 20,0 2,6 88,9 0,076 0,382 E-SEA 22,2 2,4 77,8 0,066 0,332 E-MC1 22,6 1,8 44,4 0,089 0,179 E-MC2 17,8 2,3 77,8 0,079 0,303 E-MC3 15,3 2,2 66,7 0,087 0,273 E-MUC 18,8 2,1 55,6 0,055 0,268 E-JAC 21,7 2,3 66,7 0,167 0,297 E-ROR 24,4 2,1 55,6 0,051 0,195
29
Tabela 5. Resumo da Estatística F (Wright, 1978) e fluxo gênico Nm (W e S) para 33 populações de B. sanderianum, B. weddellii, B. involutum e
B. exaltatum estudadas. FIS = coeficiente de endogamia. FST = coeficiente de estruturação genética. Nm (W) calculado a partir do FST e Nm (S)
calculado a partir da freqüência de alelos privados.
FIS FST Locus
B. sanderianum B. weddellii B. involutum B. exaltatum B. sanderianum B. weddellii B. involutum B. exaltatum
SKDH 0,027 0,712 0,638 0,489 0,095 0,576 0,332 0,253
IDH 1,000 0,522 0,879 0,872 0,236 0,182 0,318 0,250
DIA2 --- -0,117 -0,007 0,605 --- 0,143 0,139 0,140
DIA1 1,000 --- 0,828 0,926 0,050 --- 0,107 0,171
MDH2 --- 0,523 0,762 0,673 --- 0,093 0,215 0,331
MDH1 --- --- 0,521 0,412 --- --- 0,225 0,118
LAP 0,563 0,967 0,695 0,785 0,058 0,123 0,189 0,206
EST -0,024 -0,091 0,297 0,199 0,012 0,064 0,111 0,071
6PGD 1,000 1,000 1,000 0,756 0,182 0,091 0,095 0,184
Média 0,693 0,517 0,658 0,676 0,145 0,269 0,232 0,230
Nm(W) 0,368 0,362 0,608 0,755
Nm(S) 0,220 0,474 0,480 1,358
30
Tabela 7. Matriz de identidade genética média ("unbiased genetic identity"; Nei, 1978) entre populações de quatro espécies do complexo
Bulbophyllum exaltatum, ocorrentes no Brasil. Valores entre parênteses correspondem ao intervalo dos valores para pares de populações.
Espécie Nº pops. B. sanderianum B. weddellii B. involutum B. exaltatum
B. sanderianum 2 0,950 (0,950-0,950)
B. weddellii 4 0,735 (0,630-0,800) 0,905 (0,814-1,000)
B. involutum 7 0,829 (0,723-0,975) 0,877 (0,740-0,971) 0,894 (0,778-0,981)
B. exaltatum 20 0,851 (0,666-0,961) 0,877 (0,771-0,971) 0,890 (0,745-0,996) 0,905 (0,763-0,992)

31
Figura 1. Flores de Bulbophyllum sanderianum de populações de Pernambuco (A) e Bahia
(B), B. weddellii de Minas Gerais (C), B. involutum da Bahia (D) e Minas Gerais (E-G) e
B. exaltatum de Minas Gerais (H-L). A, S-BA; B, S-PE; C, W-CIP; D, I-RCO; E, I-CAB;
F, I-TAB; G, I-CIP; H, E-CAB; I, E-TAB; J, E-RPR; K, E-NAZ; L, E-ITU. Veja Tabela 1
para nomes das populações.

32
Figura 2. Flores de Bulbophyllum exaltatum de populações de Minas Gerais (A-B), Bahia
(C-I) e Roraima (J-K). A, E-CAN; B, E-LAV; C, E-JUS; D, E-LEN; E e F, E-SEA; G, E-
MC2; H, E-MC3; I, E-JAC; J e K, E-ROR; L, flores de B. involutum (esquerda;
Diamantina-MG) e B. exaltatum (direita; Seabra-BA). Veja Tabela 1 para nomes das
populações.
33
Figura 3. Mapa de distribuição das 33 populações de B. exaltatum (E-), B. involutum (I-),
B. sanderianum (S-) e B. weddellii (W-) estudadas. As regiões evidenciadas apresentam
campo rupestre como tipo vegetacional predominante acima de 1000 m alt. Veja Tabela 1
para nomes das populações.
34
Figura 4. Dendrograma mostrando as relações fenéticas entre 29 populações de B.
sanderianum (S-) (duas pops.), B. involutum (I-) (sete pops.) e B. exaltatum (E-) (20
pops.), baseada em nove loci aloenzimáticos. Construído utilizando a matriz de distância
genética de Nei (1978; "unbiased genetic distance") e UPGMA como algoritmo de
agrupamento. Correlação cofenética = 0,612. Veja Tabela 1 para nome das populações.
I-CAB E-CAL I-TAB E-TIR E-NAZ E-CAN I-MOG E-RPR E-ITU E-TAB E-LAV I-CIP E-ATI E-CAB S-PE I-RCO E-LEN E-SEA S-BA E-MUC I-MUC E-MC1 E-CAR E-JUS E-JAC E-MC2 E-MC3 I-BOT E-ROR
0.000.020.040.060.080.16 0.12 0.08 0.04 0.00
I-CAB E-CAL I-TAB E-TIR E-NAZ E-CAN I-MOG E-RPR E-ITU E-TAB E-LAV I-CIP E-ATI E-CAB S-PE I-RCO E-LEN E-SEA S-BA E-MUC I-MUC E-MC1 E-CAR E-JUS E-JAC E-MC2 E-MC3 I-BOT E-ROR
0.000.020.040.060.080.16 0.12 0.08 0.04 0.00
35
Figura 5. Árvore de neighbor-joining de 29 populações de B. sanderianum (S-) (duas
pops.), B. involutum (I-) (sete pops.) e B. exaltatum (E-) (20 pops.), baseada em nove loci
aloenzimáticos e construída utilizando a matriz de distância genética de Nei (1978;
"unbiased genetic distance"). Veja Tabela 1 para nome das populações.
I-CAB E-CAL
I-TAB E-TIR
I-MOG I-BOT
I-CIP E-RPR
E-NAZ E-CAN
E-ROR E-LAV
E-ATI E-CAB
E-ITU E-TAB
E-CAR E-MC1
I-MUC E-JAC
E-JUS E-MUC
E-LEN E-SEA
E-MC3 E-MC2
S-BA S-PE
I-RCO
0.01
36
Figura 6. Representação gráfica das freqüências alélicas do locus MDH-1 nas populações
de B. exaltatum (E-), B. involutum (I-) e B. sanderianum (S-) estudadas. Note a inversão na
freqüência relativa dos alelos 93 e 100 entre as populações ocorrentes em Minas Gerais e
Bahia (exceto I-MUC e E-MC1). Veja Tabela 1 para nomes das populações.

37
Figura 7. Dendrograma mostrando as relações fenéticas entre 77 indivíduos de 24
populações de B. involutum (I-) e B. exaltatum (E-), baseado na análise morfométrica de 27
caracteres morfológicos florais. Construído utilizando o coeficiente de similaridade de
Gower (1971), com UPGMA como algoritmo de agrupamento. As setas indicam os grupos
discutidos no texto.
Gower / UPGMA
0,0 0,1 0,2 0,3 0,4
I-TAB.2I-TAB.1I-TAB.3
E-SCA.1I-CAB
I-RCO.3I-RCO.4I-RCO.2I-RCO.1I-RCO.5I-MUC.3I-MUC.1E-LEN.2E-MC2.7E-MC2.6E-MC2.5E-MC1.8E-ITU.3
E-MC2.8E-MC2.2E-MC2.1E-MC1.7E-MC1.5E-MC2.3E-SEA.4E-MC2.4
E-MC2.10E-MC1.6
E-JUSI-MUC.2E-JAC.4E-CAR.1
I-MOGI-CAB
E-CAL.2E-MC1.4E-MC1.3E-ROR.2E-ROR.3E-ROR.1E-SEA.3E-SEA.1E-JAC.2E-LEN.1E-JAC.5E-MC2.9E-SEA.2E-JAC.1
E-MUC.3E-MC1.1E-MC1.2E-SCA.2
E-SJAE-MUC.1E-JAC.3
E-MUC.2E-TAB.3E-CAR.2E-CAR.3E-LAV.3
E-STLE-LAV.2E-ITU.1E-ITU.7E-ITU.4E-ITU.5E-ITU.2E-NAZ
E-SCA.3E-TAB.4E-TAB.5E-TAB.2E-TAB.1E-ITU.6
E-ATIE-LAV.1E-CAL.1
1
2
3
4
Gower / UPGMA
0,0 0,1 0,2 0,3 0,4
I-TAB.2I-TAB.1I-TAB.3
E-SCA.1I-CAB
I-RCO.3I-RCO.4I-RCO.2I-RCO.1I-RCO.5I-MUC.3I-MUC.1E-LEN.2E-MC2.7E-MC2.6E-MC2.5E-MC1.8E-ITU.3
E-MC2.8E-MC2.2E-MC2.1E-MC1.7E-MC1.5E-MC2.3E-SEA.4E-MC2.4
E-MC2.10E-MC1.6
E-JUSI-MUC.2E-JAC.4E-CAR.1
I-MOGI-CAB
E-CAL.2E-MC1.4E-MC1.3E-ROR.2E-ROR.3E-ROR.1E-SEA.3E-SEA.1E-JAC.2E-LEN.1E-JAC.5E-MC2.9E-SEA.2E-JAC.1
E-MUC.3E-MC1.1E-MC1.2E-SCA.2
E-SJAE-MUC.1E-JAC.3
E-MUC.2E-TAB.3E-CAR.2E-CAR.3E-LAV.3
E-STLE-LAV.2E-ITU.1E-ITU.7E-ITU.4E-ITU.5E-ITU.2E-NAZ
E-SCA.3E-TAB.4E-TAB.5E-TAB.2E-TAB.1E-ITU.6
E-ATIE-LAV.1E-CAL.1
1
2
3
4
38
Figura 8. Representação gráfica da dispersão dos indivíduos de B. involutum e B.
exaltatum nos dois primeiros eixos da análise de componentes principais (PCA) de
correlação baseada em 21 caracteres morfológicos florais. São evidenciados o táxon e a
origem por estado dos indivíduos. Percentual de variação acumulado nos dois primeiros
eixos = 61,8% (eixo 1 = 42,3%; eixo 2 = 19,5%).
39
Figura 9. Representação gráfica da dispersão nos quatro primeiros eixos da análise de
variáveis canônicas (CVA), baseada em 21 caracteres morfológicos florais, dos indivíduos
de nove grupos de populações de B. involutum (BI) e B. exaltatum (BE) estabelecidos a
priori por regiões geográficas. Percentual de variação acumulado nos quatro primeiros
eixos 89,4% (eixo1 = 47,3%; eixo 2 = 23,7%; eixo 3 = 11,7%; eixo 4 = 6,6%).

40
Figura 10. Dendrograma mostrando o relacionamento fenético entre nove grupos de
populações de B. involutum (BI) e B. exaltatum (BE) estabelecidos a priori por regiões
geográficas. Construído a partir da distância generalizada de Mahalanobis entre os
centróides dos grupos e utilizando UPGMA como algoritmo de agrupamento, a partir da
análise morfométrica de 21 caracteres morfológicos florais.

41
DISCUSSÃO
Variabilidade genética
As espécies de Bulbophyllum estudadas apresentaram elevados valores de
diversidade genética na maioria das populações, sendo alguns valores similares ou ainda
maiores do que os já encontrados para Orchidaceae (Chung & Chung, 1999; Borba et al.,
2001; Chung et al., 2004; Trapnell et al., 2004). Esses valores são semelhantes na maioria
das populações e maiores do que a média dos valores referidos para espécies com
características similares: monocotiledôneas, herbáceas, dispersas pelo vento e de
reprodução sexual (Hamrick & Godt, 1990). Apesar dos elevados valores de diversidade
genética aqui observados, eles ainda são menores do que os referidos para B. ipanemense
(aqui considerado como B. exaltatum; He 0,369-0,441), e para B. weddellii (He 0,619)
detectados por Farinaci (2001). Outras espécies brasileiras do gênero não pertencentes à
seção Didactyle, B. adiamantinum, B. bidentata, B. epiphytum, B. insectiferum, B.
plumosum, B. regnellii e B. rupicolum, estudadas por M.Azevedo et al. (no prelo), também
apresentaram valores de variabilidade ainda mais elevados do que os encontrados neste
estudo (He 0,391-0,518). As populações estudadas por Farinaci (2001) e a maioria das
populações estudadas por M.Azevedo et al. (no prelo) vivem sob condições ambientais
similares, sendo rupícolas e/ou epífitas de campos rupestres, freqüentemente simpátricas
com as populações deste estudo, polinizadas por moscas e distribuídas em populações
disjuntas. No entanto, os loci mais polimórficos desses dois trabalhos não foram utilizados
neste estudo devido à falta de resolução ou definição das bandas e, possivelmente devido a
isto os valores de variabilidade encontrados tenham sido um pouco mais baixos.
A população S-PE apresentou os menores valores de variabilidade genética
detectados, que podem estar relacionados a características ecológicas desta população,
como o seu tamanho reduzido. Esta população é constituída por apenas 12 indivíduos
encontrados em um único afloramento rochoso de brejo de altitude no estado de
Pernambuco. Neste caso, esta população pode ser de colonização recente, onde a
diversidade genética reduzida seja devido ao pool gênico dos seus fundadores (efeito
fundador), ou ainda, esta população pode ter sofrido redução populacional, e a redução do
tamanho efetivo da população possibilitou a atuação da deriva reduzindo sua diversidade
genética (efeito gargalo) (Tremblay & Ackerman, 2001). Considerando a possibilidade de

42
colonização antiga desta população, o tamanho reduzido e o efeito fundador também
poderiam justificar os baixos valores de diversidade genética encontrados. Indivíduos
dessa espécie crescem sobre rochas nuas sem nenhum tipo de substrato, como também
observado na população do estado da Bahia que apresentou valores mais elevados de
variabilidade, com número maior de indivíduos, que se concentravam na face mais seca da
serra coletada. Baixa variabilidade genética também foi encontrada em uma população de
Mucugê de B. weddellii (W-MU3), com número restrito de indivíduos e localizada em uma
área antropizada, ocorrendo em simpatria com uma população de B. involutum (I-MUC)
que também apresentou baixos valores de variabilidade. A baixa variabilidade genética em
populações urbanas em relação a rurais tem sido referida para outras espécies de orquídeas,
como Epipactis helleborine (L.) Crantz. (Hollingsworth & Dickson, 1997), fortalecendo a
idéia de que a ação antrópica afeta negativamente a variabilidade genética de populações
naturais.
Todas as espécies estudadas encontram-se em populações disjuntas devido à
descontinuidade dos afloramentos rochosos e da Cadeia do Espinhaço, e algumas
populações apresentam um baixo número de indivíduos, o que favorece a ocorrência de
endogamia nessas populações. Os indivíduos também se reproduzem vegetativamente por
crescimento modular e são autocompatíveis (Borba et al., 1999). Mesmo existindo
barreiras mecânicas à autopolinização nestas espécies, esta é do tipo temporal, não
impedindo completamente a ocorrência de autopolinização, devido ao comportamento do
polinizador que permanece por longos períodos na flor, e não previne a geitonogamia entre
ramets do mesmo genet (Borba & Semir, 1999). Nesse contexto, podemos considerar que
altos valores de FIS encontrados estão relacionados a geitonogamia, como observado por
Soliva & Widmer (1999) para a orquídea Gymnadenia conopsea (L.) R. BR. e por Ehlers
& Pedersen (2000) para orquídeas do gênero Epipactis. Segundo M.Azevedo et al. (no
prelo), o fato de todas as populações não apresentarem equilíbrio de HW em pelo menos
um locus é uma evidência de que a dispersão de pólen é espacialmente restrita, o que
concorda com o comportamento dos polinizadores das espécies. Restrição na dispersão de
pólen devido ao comportamento do polinizador e o tamanho efetivo reduzido de muitas
populações são os fatores responsáveis pelo aumento da endogamia que se reflete nos
elevados valores de FIS observados.
Em plantas, a estrutura genética populacional está intimamente relacionada a
fatores como o tamanho efetivo da população, o sistema de reprodução e a dispersão do
pólen e das sementes. No gênero Bulbophyllum, os polinizadores efetivos de algumas das

43
espécies ocorrentes no Brasil são moscas da família Milichiidae (Braga, 1977; Sazima,
1978; Borba & Semir, 1998b). Estas moscas geralmente movimentam-se em distâncias
muito curtas, realizando visitas de longa duração e visitando várias flores de um mesmo
indivíduo, o que favorece a autopolinização e o isolamento de populações.
Conseqüentemente, a redução do fluxo gênico pode levar à diferenciação dessas
populações (Borba & Semir, 1998b; Borba et al., 2001; Farinaci, 2001; Verola, 2002).
Além disso, apesar do tamanho diminuto das sementes em Orchidaceae, alguns autores
demonstraram que a dispersão pelo vento nem sempre contribui para a manutenção do
fluxo gênico entre as populações, uma vez que o estabelecimento das sementes é ocasional
e específico. Em algumas espécies de orquídeas, tais como Brassavola nodosa L. (Murren
& Ellison, 1998), Cymbidium goeringii Rchb.f. (Chung et al., 1998) e Cephalanthera
longibracteata Blume (Chung et al, 2004), demonstrou-se que as sementes geralmente se
estabelecem a curtas distâncias da planta mãe. Neste contexto, e considerando os altos
valores de FIS encontrados, poderia ser esperado que as espécies de Bulbophyllum
estudadas possuam elevada estruturação genética dentro das populações.
A estruturação genética encontrada nas espécies pode ser considerada elevada
quando comparada a outros estudos realizados em Bulbophyllum (Farinaci, 2001;
M.Azevedo et al., no prelo) e em outras orquídeas (Hollingsworth & Dickson, 1997;
Sharma et al., 2000; Ehlers & Pedersen, 2000; Borba et al., 2001) e maior do que os
valores encontrados para espécies polinizadas por animais e com dispersão de sementes
pelo vento (Hamrick & Godt, 1990). A existência de estruturação nas populações de B.
sanderianum, B. weddellii e B. involutum concordam com o baixo número de migrantes
Nm(W e S) encontrados nestas espécies, sendo que altos níveis de fluxo gênico resultam em
baixos níveis de diferenciação genética entre populações.
Em B. exaltatum, apesar da alta estruturação genética, o Nm(S) também foi elevado,
sugerindo a ocorrência de fluxo gênico e concordando com a ampla distribuição e
proximidade geográfica entre as populações deste táxon. A grande redução da estruturação
genética nesta espécie quando consideradas apenas as populações do estado da Bahia pode
ser devido às populações de Minas Gerais estarem mais distantes entre si do que as
populações do estado da Bahia (isolamento por distância; veja Figura 3). Neste caso, a
dispersão de sementes pelo vento a longas distâncias deve ser a forma mais efetiva de
manutenção do fluxo gênico e, conseqüentemente, redução da diferenciação entre as
populações, uma vez que o fluxo gênico de pólen é espacialmente restrito. No entanto, a
fragmentação de habitat é um fator ainda mais expressivo para a diferenciação destas

44
populações.
As espécies estudadas são intercompatíveis (Borba et al., 1999), excluindo-se B.
sanderianum que não possui até o momento a sua biologia reprodutiva e potencialidade de
hibridação estudada. Desta forma, a existência de muitos alelos compartilhados, ocorrência
de algumas populações em simpatria e intercompatibilidade, e como hibridação natural é
conhecida no grupo (Borba & Semir, 1998a; Borba et al., 1999; C.Azevedo et al., no
prelo), possivelmente eventos de hibridação entre os táxons estudados estejam ocorrendo,
contribuindo para a manutenção da elevada identidade genética entre populações não
conspecíficas. Esta hipótese é corroborada pelo desequilíbrio de ligação constatado na
maioria dos loci de todas as populações e pelo agrupamento das populações primariamente
com base na distribuição geográfica por estados de ocorrência ao invés de por afinidade
taxonômica e morfológica. Eventos de hibridação natural são comuns em Orchidaceae
(e.g., Borba & Semir, 1998a; Nielsen, 2000; Klier et al., 1991; Barkman & Simpson, 2002)
e podem gerar um padrão reticulado de agrupamento de populações não conspecíficas,
semelhante ao aqui observado.
Porém este mesmo padrão reticulado de similaridade pode também ser gerado
simplesmente devido a uma baixa diversidade alélica detectada, com todas as populações
de todos os táxons compartilhando um estoque alélico similar, que variam de freqüência
aleatoriamente nas populações. Neste caso, a similaridade na constituição alélica seria
devido a uma diferenciação ainda incipiente entre os táxons (a priori), e não devido à
hibridação (a posteriori). Estes dois mecanismos não são mutuamente excludentes, e
sugerimos que provavelmente estejam atuando simultaneamente neste conjunto de
populações. Outra possibilidade é a limitação do marcador utilizado uma vez que a
variação de aloenzimas reflete apenas uma pequena fração de todos os eventos mutacionais
do DNA (Clegg, 1989). A natureza conservativa dos dados alozímicos, comparado a
RAPDs, ISSRs e outros marcadores baseados em DNA, poderia explicar a baixa
diversidade genética detectada no grupo estudado.
A diferenciação genética das populações de B. exaltatum ocorrentes em Minas
Gerais das populações da Bahia, conforme evidenciado nos dendrogramas e nos valores de
θ para o locus MDH-1, mesmo que com reduzida diferenciação genética média entre os
grupos (θP= 0,058), aparentemente estão relacionadas à principal disjunção da Cadeia do
Espinhaço em duas porções. A norte que constitui a Chapada Diamantina no estado da
Bahia, e a sul que compreende o Planalto de Diamantina em Minas Gerais. Esta disjunção
possui cerca de 300 km de extensão norte-sul e tem sido considerada como uma forte

45
barreira geográfica à migração de espécies de plantas, com aparentemente grande
contribuição na diversificação das plantas nos campos rupestres destas regiões (Giulietti &
Pirani, 1988; Harley, 1988). Várias outras disjunções afetando a distribuição geográfica
nestas formações têm sido freqüentemente associadas à diferenciação genética de
populações em diversos grupos de plantas (Borba et al., 2001; Jesus et al., 2001; Machado,
2004; Lambert et al., 2006a, 2006b; Pereira, 2006). Esta ainda sutil separação genética por
barreiras geográficas nos sugere o início da separação de B. exaltatum em duas linhagens
nas áreas de campos rupestres. Aparentemente, uma terceira linhagem, representada pela
população amazônica (BEROR), também está se diferenciando através de uma disjunção
maior das formações do leste com o norte do Brasil, que além de separadas por uma
distância geográfica maior também estão isoladas por formações florestais localizadas
entre elas. O fluxo gênico entre estas regiões é bastante improvável, e deriva genética pode
ser a responsável pela diferenciação observada, sendo que tal processo pode levar
rapidamente à especiação em pequenas populações isoladas (Levin, 2000). Processo
similar, porém em menor escala, foi observado para outra orquídea miiófila entre
populações da Cadeia do Espinhaço e uma população disjunta ocorrendo em afloramento
rochoso na Mata Atlântica (Borba et al., 2001).
Variabilidade morfológica
Através de observações em campo de diversas populações deste grupo, pudemos
perceber a existência de uma grande variação morfológica dentro das populações, o que é
refletido na análise de agrupamento por indivíduos. Desta forma, como esperado,
distinções claras entre populações e grupos de populações não puderam ser facilmente
encontradas, como observado nas análises de componentes principais e de variáveis
canônicas. Ainda assim, B. involutum de Minas Gerais é o grupo potencialmente mais
distinto morfologicamente, devido principalmente a maiores dimensões do labelo, embora
se assemelhando mais aos indivíduos de B. exaltatum da Bahia do que com os de Minas
Gerais.
Se considerarmos os caracteres diagnósticos de B. involutum destacados na
descrição da espécie (apenas uma flor em antese por inflorescência, flores grandes e
púrpuras, pétalas ovadas com ápice obtuso a rotundo, base do lobo mediano do labelo
truncada a subcordada e com margem involuta; Borba et al., 1998), a maioria destes
caracteres são encontradas entre os indivíduos das populações de B. exaltatum de Minas

46
Gerais e principalmente da Bahia. Além disto, a análise morfológica revelou que essas
características não são constantes nas populações, podendo variar individualmente, ou seja,
alguns indivíduos apresentam um ou mais caracteres diagnósticos dentro das populações.
Uma evidência disto foi o indivíduo da população de Grão Mogol–MG, que na análise de
PCA se posiciona entre B. involutum da Bahia. Como mencionado anteriormente, esta
região apresenta uma composição florística intermediária, devido à sua localização
geográfica intermediária entre as porções principais da Cadeia do Espinhaço da Bahia e
Minas Gerais (Harley, 1995).
As análises de dados morfológicas e a ocorrência destas duas entidades
morfológicas em sintopia em algumas localidades, com a manutenção destas diferenças,
fortalece a confirmação da existência destes dois táxons. Como ocorre em Joaquim Felício,
I-CAB e E-CAB, e Conceição do Mato Dentro, I-TAB e E-TAB, sendo os táxons
separados por eventualmente um metro de distância, e ocasionalmente observado em
outras localidades (e.g., Serra do Caraça-MG, Serra da Piedade-MG). Além disto, esta
afirmação é corroborada por estudos de biologia reprodutiva e de fitoquímica destas
espécies (Borba & Semir, 1998b; Silva et al., 1999).
Porém, a semelhança morfológica encontrada em todas as análises entre indivíduos
de B. involutum e B. exaltatum também sugere a ocorrência eventual de hibridação entre
esses táxons, principalmente nas populações do estado da Bahia. Outras populações como
E-MUC e E-JAC apresentam-se diferenciadas morfologicamente das demais devido a sua
semelhança à B. involutum, principalmente em aspectos de coloração e forma do labelo. Na
verdade, na CVA todas as populações da Bahia foram mais similares a B. involutum do que
às populações de B. exaltatum de Minas Gerais, sendo essa semelhança percebida em
campo e refletida nas determinações incorretas de material de herbário, em especial da
população de Jacobina.
Na CVA, o fato dos grupos de B. exaltatum da Bahia apresentarem classificações
incorretas entre si, fortalece a hipótese de ocorrência de fluxo gênico nas formações de
campos rupestres, mais provavelmente por sementes, entre estas populações, observado
pelo elevado Nm(S) e baixo FST no estado. Por outro lado, a população de Jussari-BA E-
JUS, que ocorre fora da Chapada Diamantina, apresenta algumas características
morfológicas qualitativas que sugerem uma diferenciação desta como: sépalas e pétalas
eretas de coloração verde, lobo mediano do labelo extenso e de cor verde (sendo
predominantemente púrpura nas outras populações). Esta é a única população de hábito
epífita que ocorre em Mata Atlântica, embora não apresente uma maior diferenciação

47
genética nem morfológica quantitativa. Situação contrária ocorreu com a população de B.
exaltatum de Roraima que se diferenciou geneticamente das demais, enquanto que
morfologicamente mostrou-se mais semelhante aos indivíduos da Bahia, se distinguindo
por comprimento das sépalas, largura da pétala e dimensões do labelo.
Todas as análises morfológicas permitem visualizar o que foi anteriormente
comentado em relação a B. exaltatum, de que as populações de Minas Gerais e da Bahia
provavelmente não constituiriam um único táxon (Ribeiro et al., 2005). No entanto,
podemos considerar um reduzido estágio de divergência morfológica entre estes grupos de
populações, refletido principalmente na maior largura da sépala e menor tamanho do labelo
e de seu ístimo, nos representantes de Minas Gerais, que talvez dificulta o reconhecimento
de duas espécies.
Conclusões taxonômicas
Das quatro espécies estudadas duas delas não possuem problemas de delimitação
específica, B. sanderianum e B. weddellii, possuindo um conjunto de caracteres
diagnósticos de fácil reconhecimento e consistentes entre os indivíduos das populações. As
outras duas, B. involutum e B. exaltatum, possuem maiores problemas taxonômicos, de
solução dificilmente consensual, mesmo com o conjunto de dados levantados neste estudo.
A ausência de loci diagnósticos e compartilhamento de maior parte do estoque alélico,
além da similaridade morfológica, sugerem ocorrência de hibridação ou baixa divergência
entre esses táxons, o que reflete na ausência de grupos constituídos apenas por populações
conspecíficas nas análises realizadas. Esses táxons possuem identidade genética que se
enquadra na faixa referida para populações conspecíficas (Crawford, 1989; van der Bank et
al., 2001). Os dados de eletroforese de isoenzimas não sustentam claramente a separação
destes dois táxons. Dados de seqüenciamento de regiões de DNA nuclear e de plastídio
indicam também a baixa divergência entre estas entidades (E.C.Smidt, dados não
publicados). Porém, a distinção morfológica e sua manutenção, mesmo quando em
simpatria e características biológicas (Borba & Semir, 1998b; Silva et al., 1999) sustentam
a separação de duas espécies distintas. O emprego de um marcador molecular com maior
variabilidade, como microsatélites, poderia ajudar a indicar se a baixa diferenciação entre
os dois táxons é devida simplesmente a uma baixa diversidade alélica detectada, com uma
diferenciação ainda incipiente em aloenzimas, ou se devido à ocorrência de hibridação. De
qualquer forma, caso exista hibridação no grupo, esta não deve ocorrer em freqüência

48
muito elevada, uma vez que a distinção morfológica nas populações simpátricas ainda é
mantida.
Com base nos dados genéticos, morfológicos e sobre a biologia das demais espécies
estudadas, podemos concluir que todas as populações consideradas como B. exaltatum
ocorrentes no estado de Minas Gerais e São Paulo devem ser tratadas taxonomicamente
como uma única entidade, não podendo ser reconhecidas mesmo categorias
infraespecíficas. Desta forma, os outros nomes comumente aplicados a estas populações,
B. warmingianum, B. ipanemense e B. geraense, deveriam ser sinonimizados sob B.
exaltatum. Por outro lado, a diferenciação apresentada pelas populações da Bahia em
relação a estas anteriores, confirma a hipótese de estarmos lidando com duas unidades
evolutivas significativamente distintas. Porém, esta separação aparentemente ainda é
incipiente, e a divisão destes dois grupos de populações em duas espécies é de pouca
aplicabilidade taxonômica prática, devido a dificuldades de diagnóstico, principalmente em
material herborizado. Uma revisão taxonômica de todo o gênero para a região Neotropical
encontra-se atualmente em fase final de desenvolvimento (E.C.Smidt, dados não
publicados). Caso a distinção morfológica aqui observada apresente-se consistente nas
demais populações dos táxons envolvidos neste complexo fora das regiões dos campos
rupestres, sugerimos o reconhecimento de duas subespécies em B. exaltatum. Desta forma,
baseado no exame do material tipo destes táxons, as populações de Bahia e da Amazônia
constituiriam a subespécie típica, e, devido à prioridade nomenclatural, as de Minas Gerais
e São Paulo deveriam ser reconhecidas como B. exaltatum subsp. geraense.

49
RESUMO
O complexo taxonômico denominado “Bulbophyllum exaltatum Lindl.”
(Orchidaceae) compreende um grupo de cerca de 15 táxons da seção Didactyle, ocorrentes
em áreas de altitude, onde a maior parte dos problemas taxonômicos ocorrem em torno de
B. involutum, B. ipanemense, B. longispicatum, B. geraense e B. warmingianum. Neste
trabalho, foram realizados estudos de genética de populações utilizando aloenzimas em 33
populações naturais para determinar a variabilidade genética dentro das populações e o
grau de diferenciação destas em algumas espécies de Bulbophyllum pertencentes a este
complexo, e empregados na delimitação de táxons no grupo. Foi também realizada análise
multivariada morfométrica em alguns indivíduos destas populações estudadas, buscando
uma delimitação mais consistente dos táxons neste complexo. A partir destes dados,
procurou-se fornecer subsídios para a definição de padrões de distribuição das espécies
estudadas do complexo B. exaltatum e contribuir para o entendimento dos processos
evolutivos e biogeográficos de espécies ocorrentes na Cadeia do Espinhaço. As populações
das espécies de Bulbophyllum estudadas, denominadas a priori como B. exaltatum, B.
involutum, B. sanderianum e B. weddellii, apresentaram elevada variabilidade genética,
sendo similares ou superiores o anteriormente reportado para Orchidaceae e espécies com
características similares. A distribuição da variação genética total foi similar em todas as
espécies estudadas. Foi detectado alto grau de estruturação genética para as espécies
estudadas indicando que o fluxo gênico é restrito, sendo este detectado somente entre as
populações de B. exaltatum, provavelmente realizado por dispersão de sementes pelo vento
a longas distâncias. A análise aloenzimática revelou que nenhuma das espécies forma um
grupo distinto envolvendo todas as populações conspecíficas, não permitindo uma
delimitação taxonômica clara das espécies deste complexo. Por outro lado, as populações
foram primariamente agrupadas com base em sua região de ocorrência por estados. Apesar
da alta variabilidade e da formação de grupos fundamentados em regiões geográficas, não
há correlação entre distância genética e geográfica entre os táxons estudados.
Possivelmente, eventos de hibridação e/ou diferenciação ainda incipiente entre os táxons
estejam contribuindo para a manutenção da elevada identidade genética entre as
populações, gerando o padrão reticulado de agrupamento de populações não conspecíficas
observado. Dados morfológicos sugerem que as populações de B. involutum de Minas
Gerais constituem um táxon distinto de B. exaltatum. Porém, as populações de ambos os

50
táxons no estado da Bahia apresentam baixa diferenciação. Dados genéticos e
morfológicos indicam que as barreiras geográficas entre Bahia, Minas Gerais e Roraima
favorecem a diferenciação genética e morfológica entre as populações destes estados. O
nome B. exaltatum deve ser aplicado a todas as populações em Minas Gerais
tradicionalmente conhecidas como B. warminginaum, B. ipanemense e B. geraense, sendo
estes sinonimizados sob o primeiro. Bulbophyllum exaltatum é uma espécie de ampla
distribuição e muito variável, mas os resultados encontrados sustentam o reconhecimento
de duas subespécies, a típica compreendendo as populações da Bahia e da Amazônia, e
outra para as populações ocorrentes em Minas Gerais e São Paulo.

51
ABSTRACT
(INFRA- AND INTERSPECIFIC GENETIC AND MORPHOLOGICAL
VARIATION IN THE COMPLEX BULBOPHYLLUM EXALTATUM
(ORCHIDACEAE) OCCURRING IN THE BRAZILIAN “CAMPOS RUPESTRES”:
IMPLICATIONS FOR TAXONOMY AND BIOGEOGRAPHY)
The taxonomic complex of Bulbophyllum exaltatum Lindl. (Orchidaceae) includes
around 15 taxa in section Didactyle which occur mostly in rocky montane areas. Most
taxonomic problems in this group consist of the delimitation of B. involutum, B.
ipanemense, B. longispicatum, B. geraense e B. wamingianum. In the present study, we
carried out population genetic studies using isozyme markers in 33 natural populations, in
order to assess the genetic variation and degree of differentiation in some species
belonging to this complex. We also performed a morphometric analysis in part of the
individuals of the genetic study, using multivariate methods as an attempt to improve
species delimitation. From these data, we tried to gather evidence for defining distribution
patterns of the B. exaltatum complex and contribute for understanding the evolutionary and
biogeographical processes of species from the Espinhaço Range, Brazil. The four species
studied, considered a priori to be B. exaltatum, B. involutum, B. sanderianum and B.
weddellii, displayed high genetic variation, the values being similar or even higher to those
previously reported for Orchidaceae and species with similar features. The distribution of
the total genetic variation was similar among all species. For all species, we detected a
high degree of genetic structure which indicates restrict gene flow. The latter was detected
only among populations of B. exaltatum, and is probably due to long-distance seed
dispersion by wind. In the results of the isozyme analysis, none of the conspecific
populations were grouped in a distinct cluster. Therefore, based on these data it is not
possible a clear taxnomic delimitation within this complex. On the other hand, the
populations clustered primarily based on the state of occurrence. Despite the high variation
and the existence of groups based on geographical regions, there is no correlation between
genetic and geographical distances among the populations. Perhaps, hybridization events
and early differentiation among the taxa contribute to keep the high genetic identity among
the populations, and thus generating the observed reticulate pattern of clustering among
different species. Morphological data suggest that the B. involutum populations in Minas

52
Gerais stand out as a distinct taxon in relation to B. exaltatum. However, the populations of
both taxa in Bahia State displayed low differentiation. Genetic and morphological data
point out that the geographical barriers among Bahia, Minas Gerais and Roraima favoured
genetic and morphological differentiation among between the populations from these
States. Bulbophyllum exaltatum is the oldest name to be applied to the populations from
Minas Gerais generally recognized as B. warminginaum, B. ipanemense and B. geraense,
requiring new synonym under the former. Bulbophyllum exaltatum is a broad, variable
species, but the results of this study give support to the recognition of two suspecies. The
typical one includes the populations from Bahia and the Amazon, wheras the second
should be established for the populations from Minas Gerais and São Paulo.

53
REFERÊNCIAS BIBLIOGRÁFICAS
AZEVEDO, C.O.; BORBA, E.L.; VAN DEN BERG, C. (no prelo). Evidence for natural
hybridization and introgression in Bulbophyllum involutum and B. weddellii (Orchidaceae)
in the Chapada Diamantina, Brazil, by using allozyme markers. Revista Brasileira de
Botânica.
AZEVEDO, M.T.A.; BORBA, E.L.; SEMIR, J.; SOLFERINI, V.N. (no prelo). Very high
genetic variability in Neotropical myophilous orchids. Botanical Journal of the Linnean
Society.
BARKMAN, T.J.; SIMPSON, B.B. 2002. Hybrid origin and parentage of Dendrochilum
acuiferum (Orchidaceae) inferred in a phylogenetic context using nuclear and plastid DNA
sequence data. Systematic Botany, 27: 209-220.
BARROS, F. 1990. Diversidade taxonômica e distribuição geográfica das Orchidaceae
brasileiras. Acta Botanica Brasilica, 4: 177-187.
BARTON, N.H.; SLATKIN, M. 1986. A quasi-equilibrium theory of the distribution of
rare alleles in a subdivided population. Heredity, 56: 409-415.
BENNETT, D. E.; CHRISTENSON, E. 2001. Bulbophyllum machupicchuense D.E.
Bennett & Christenson. In: Icones Orchidacearum Peruvianum t. 612.
BORBA, E.L.; SEMIR, J. 1998a. Bulbophyllum xcipoense (Orchidaceae), a new natural
hybrid from the Brazilian campos rupestres: description and biology. Lindleyana, 13: 113-
120.
BORBA, E.L.; SEMIR, J. 1998b Wind-assisted fly pollination on three species of
Bulbophyllum (Orchidaceae) occurring in the Brazilian campos rupestres. Lindleyana, 13:
201-218.
BORBA, E.L.; SEMIR, J.; BARROS, F. 1998. Bulbophyllum involutum Borba, Semir & F.
Barros (Orchidaceae), a new species from the Brazilian "campos rupestres". Novon, 8:
225-229.
BORBA, E.L.; SEMIR, J. 1999. Temporal variation in pollinarium size after its removal in
species of Bulbophyllum: a diferent mechanism preventing self-polination in Orchidaceae.
Plant Systematics and Evolution, 217: 197-204.
BORBA, E.L.; SHEPHERD, J.; SEMIR, J. 1999. Reprodutive systems and crossing
potential in three species of Bulbophyllum (Orchidaceae) occurring in the Brasilian

54
“campos rupestres”. Plant Systematics and Evolution, 217: 205-214.
BORBA, E.L.; FELIX, J.M.; SOLFERINI, V.N.; SEMIR, J. 2001. Fly-pollinated
Pleurothallis (Orchidaceae) species have high genetic variability: evidence from isozyme
markers. American Journal of Botany, 88: 419-428.
BORBA, E.L.; SHEPERD, G.J.; VAN DEN BERG, C.; SEMIR, J. 2002. Floral and
vegetative morphometrics in five Pleurothallis (Orchidaceae) species: correlation with
taxonomy, phylogeny, genetic variability and pollination systems. Annals of Botany, 90:
219-230.
BORBA, E.L.; SMIDT, E.C. 2004. Bulbophyllum filifolium (Orchidaceae), a new species
from southeastern Brazil. Novon, 14: 29-32.
BRAGA, P.I.S. 1977. Aspectos biológicos das Orchidaceae de uma campina na Amazônia
Central. Acta Amazônica, 7 (suppl. 2): 1-89.
BRUNE, W. ALFENAS, A.C. JUNGHANS, T.H. 1998. Identificações específicas de
enzimas em géis. In: ALFENAS, A.C. (ed.) Eletroforese de isoenzimas e proteínas afins:
fundamentos e aplicações em plantas e microorganismos. Viçosa: Universidade Federal de
Viçosa. p.: 201-218.
CARDIM, D.C.; CARLINE-GARCIA, L.A.; MONDIN, M.; MARTINS, M.; VEASEY,
E.A.; ANDO, A. 2001. Variabilidade intra-específica em cinco populações de Oncidium
varicosum Lindl. (Orchidaceae – Oncidiinae) em Minas Gerais. Revista Brasileira de
Botânica, 24 (4): 553-560.
CHRISTENSEN, D.E. 1994. Fly pollination in the Orchidaceae. In: ARDITTI, J. (ed.)
Orchid biology: reviews and perspectives VI. New York: John Wiley & Sons. p.: 415-454.
CHUNG, M.G; CHUNG, M.Y.; OH, G.S.; EPPERSON, B.K. 1998. Spatial genetic
structure in populations of Cymbidium goeringii (Orchidaceae). Genes and Genetic
Systems, 73: 281-285.
CHUNG, M.Y; CHUNG, M.G. 1999. Allozyme diversity and population structure in
Korean population of Cymbidium goeringii (Orchidaceae). Journal of Plant Research, 112:
139-144.
CHUNG, M.Y; NASON, J.D.; CHUNG, M.G. 2004. Spatial genetic structure in
populations of the terrestrial orchid Cephalanthera longibracteata (Orchidaceae).
American Journal of Botany, 91: 52-57.
CLAYTON, J.W.; TRETIAK, D.N. 1972. Amine-citrate buffers for pH control in starch
gel eletrophoresis. Journal of Fisheries Research of Board Canada, 29: 1169-1172.

55
CLEGG, M.T. 1989. Molecular diversity in plant populations. In: Brown A.H.D., Clegg
M.T., Kahler A.L., Weir (eds) Plant population genetics, breeding and genetic resources.
Sinauer Assoc, Sunderland, Massachusetts, p 98–115.
COGNIAUX, 1898-1902. Orchidaceae. In: MARTIUS, C.F.P.; EICHLER, A.G. (eds.)
Flora Brasiliensis. F. Fleischer, Lipsiae, v.3, pars 5. p.: 1-664.
CONCEIÇÃO, A.A.; GIULIETTI, A.M. 2002. Composição florística e aspectos estruturais
de campo rupestre em dois platôs do Morro do Pai Inácio, Chapada Diamantina, Bahia,
Brasil. Hoehnea, 29: 37-48.
CORRIAS, R.; ROSSI, W.; ARDUINO, P.; CIANCHI, R.; BULLINI, L. 1991. Orchis
longicornu Poiret in Sardinina: genetic, morphological and chorological data. Webbia, 45:
71-101.
CRAWFORD, D.J. 1989. Enzyme eletrophoresis and plant systematics. In: SOLTIS, E.D.;
SOLTIS, P.S. (eds.) Isozymes in plant biology. Portland: Dioscorides Press. p.: 146-164.
CROW, J.F.; AOKI, K. 1984. Group selection for a polygenic behavioral trait: estimating
the degree of population subdivision. Proceedings of the National Academy of Sciences
USA, 81: 6073-6077.
DRESSLER, R.L. 1981. The orchids: natural history and classification. Cambridge:
Harvard University Press. 332 p.
DRESSLER, R.L. 1993. Phylogeny and classification of the orchid family. Portland:
Dioscorides Press. 314 p.
EHLERS, B.K; PEDERSEN, H.A.E. 2000. Genetic variation in three species of Epipactis
(Orchidaceae): geographic scale and evolutionary inferences. Biological Journal of the
Linean Society, 69: 411-430.
FARINACI, J.S. 2001. Variabilidade genética em algumas espécies de Bulbophyllum
Thouars (Orchidaceae) de campos rupestres. Dissertação de Mestrado - Instituto de
Biologia da Universidade Estadual de Campinas. Campinas. 47 p.
FELSENSTEIN, J. 1993. PHYLIP (Phylogeny Inference Package) version 3.6. Seattle:
University of Washington, Department of Genetics.
FRAGA, C. N. 2004. Bulbophyllum boudetiana (Orchidaceae), a new species from the
Brazilian Atlantic Forest, Espírito Santo. Novon, 14: 40–42.
FRAGA, C. N.; SMIDT, E. C. 2004. Bulbophyllum arianeae (Orchidaceae), a new species
from Espirito Santo State in the Brazilian Atlantic forest. Harvard Papers in Botany, 9: 7-9.
GIULIETTI, A.M.; PIRANI, J.R. 1988. Patterns of geographic distribution of some plant
species from the Espinhaço Range, Minas Gerais and Bahia, Brazil. In: VANZOLINI,

56
P.E.; HEYER, W.R. (eds.) Proceedings of a Workshop on Neotropical Distribution
Patterns. Rio de Janeiro: Academia Brasileira de Ciências. p.: 36-69.
GIULIETTI, A.M.; HARLEY, R.M.; WANDERLEY, M.G.L.; PIRANI, J.R. 2000.
Caracterização e endemismos nos campos rupestres da Cadeia do Espinhaço. In:
CAVALCANTI, T.B.;WALTER, B.M.T. (eds.). Tópicos Atuais de Botânica. Brasília:
EMBRAPA Recursos Genéticos. p.: 311-318.
GOLDMAN, D.H.; VAN DEN BERG, C.; GRIFFITH, M.P. 2004. Morphometric
circumscription of species and infraspecific taxa in Calopogon R.Br. (Orchidaceae). Plant
Systematics and Evolution, 247: 37-60.
GOWER, J.C. 1971. A general coefficient of similarity and some one of its properties.
Biometrics, 27: 857-872.
HAMRICK, J.L.; GODT, M.J.W. 1990. Allozyme diversity in plant species. In: BROWN,
A.D.H.; CLEGG, M.T.; KAHLER, A.L.; WEIR, B.S. (eds.). Plant population genetics,
breeding and genetic resources. Sunderland: Sinauer Associates. p.: 43-63.
HARLEY, R.M. 1988. Evolution and distribution of Eriope (Labiatae) and its relatives in
Brazil. In: VANZOLINI, P.E.; HEYER, W.R. (eds.). Proceedings of a Workshop on
Neotropical Distribution Patterns. Rio de Janeiro: Academia Brasileira de Ciências. p.: 71-
120.
HARLEY, R.M. 1995. Introdução. In: STANNARD, B.L. (ed.) Flora of the Pico das
Almas, Chapada Diamantina, Bahia, Brazil. Richmond: Royal Botanic Gardens, Kew. p.:
1-40.
HOLLINGSWORTH, P.M.; DICKSON, J.H. 1997. Genetic variation in rural and urban
populations of Epipactis helleborine (L.) Crantz. (Orchidaceae) in Britain. Botanical
Journal of the Linean Society, 123: 321-331.
JESUS, F.F.; SOLFERINI, V.N.; SEMIR, J.; PRADO, P.I. 2001. Local genetic
differentiation in Proteopsis argentea (Asteraceae), a perennial herb endemic in Brazil.
Plant Systematics and Evolution, 226: 59-68.
JOLY, A.B. 1970. Conheça a vegetação brasileira. São Paulo: EDUSP. 181 p.
KLIER, K.; LEOSCHKE, M.J.; WENDEL, J.F. 1991. Hybridization and introgression in
white and yellow landyslipper orchids (Cypripedium candidum and C. pubescens). Journal
of Heredity, 82: 305-318.
KUMAR, S.; TAMURA, K.; NEI, M. 2004. MEGA3: Integrated software for molecular
evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics, 5:
150-163.

57
LAMBERT, S.M.; BORBA, E.L.; MACHADO, M.C.; ANDRADE, S.C.S. 2006a.
Allozyme diversity and morphometrics of Melocactus paucispinus (Cactaceae) and
evidence for hybridization with M. concinnus in the Chapada Diamantina, North-eastern
Brazil. Annals of Botany, 97: 389-403.
LAMBERT, S.M.; BORBA, E.L.; MACHADO, M.C. 2006b. Allozyme diversity and
morphometrics of the endangered Melocactus glaucescens (Cactaceae), and investigation
of the putative hybrid origin of Melocactus ×albicephalus (Melocactus ernestii × M.
glaucescens) in north-eastern Brazil. Plant Species Biology, 21: 93-108.
LEVENE, H. 1949. One matching problem arising in genetics. Annals of Mathematical
Statistics, 20: 91-94.
LEVIN, D.A. 2000. The origin, expansion, and demise of plant species. New York: Oxford
University Press. 240 p.
MACHADO, M.C. 2004. O gênero Discocactus Pfeiff. (Cactaceae) no estado da Bahia,
Brasil: variabilidade morfológica, variabilidade genética, taxonomia e conservação.
Dissertação de mestrado - Departamento de Ciências Biológicas da Universidade Estadual
de Feira de Santana. Feira de Santana. 113 p.
MCCUNE, B.; MEFFORD, M.J. 1999. PCOrd - Multivariate analysis of ecological data,
version 4.10. Gleneder Beach: MjM Software.
MILLER, M.P. 1997. Tools for population genetics analysis (TFPGA) 1.3: A windows
program for the analysis of allozyme and molecular population genetic data. Distribuído
pelo autor.
MURREN, C.J.; ELLISON, A.M. 1998. Seed dispersal characteristics of Brassavola
nodosa (Orchidaceae). American Journal of Botany, 85: 675-680.
NATIONAL GEODETIC SURVEY. 2002. INVERSE - Version 2.0. http://ngs.noaa.gov/.
18 de abril de 2006.
NEI, M. 1972. Genetic Distance between Populations. American Naturalist, 106: 283-292.
NEI, M. 1978. Estimation of average heterozigosity and genetic distance from a small
number of individuals. Genetics, 89: 583-590.
NIELSEN, L.R. 2000. Natural hybridization between Vanilla claviculata (W.Wright) Sw.
and V. barbellata Rchb.f. (Orchidaceae): genetic, morphological, and pollination
experimental data. Botanical Journal of the Linean Society, 133: 285-302.
PABST, G.F.J.; DUNGS, F. 1975. Orchidaceae Brasilienses, vol. 1. Hildeshein: Kurt
Schmersow. 408 p.
PABST, G.F.J.; DUNGS, F. 1977. Orchidaceae Brasilienses, vol. 2. Hildeshein: Kurt

58
Schmersow. 418 p.
PEREIRA, A.C.S. 2006. Variabilidade genética e morfológica inter e intrapopulacional de
Syngonanthus mucugensis Giul. e S. curralensis Moldenke (Eriocaulaceae) espécies
ameaçadas de extinção, Chapada Diamantina, Bahia, Brasil. Dissertação de mestrado -
Departamento de Ciências Biológicas da Universidade Estadual de Feira de Santana. Feira
de Santana. 140 p.
RAYMOND, M.; ROUSSET, F. 1995. GENEPOP (version 1-2): population genetics
software for exact tests and ecumenicism. Journal of Heredity, 86: 248-249.
RIBEIRO, P.L.; BORBA, E.L.; TOSCANO DE BRITO, A.L.V. 2005. O gênero
Bulbophyllum Thouars (Orchidaceae) na Chapada Diamantina, Bahia, Brasil. Revista
Brasileira de Botânica, 28: 423-439.
RICE, W.R. 1989. Analyzing tables of statistical tests. Evolution, 43: 223-225.
RIDGWAY, G.J.; SHERBURNE, S.W.; LEWIS, R.D. 1970. Polymorphism in the
esterases of Atlantic herring. Transactions of the American Fisheries Society, 99: 147-151.
ROMERO, R. 2002. Diversidade da flora dos campos rupestres de Goiás, sudoeste e sul de
Minas Gerais. In: ARAÚJO, E.L.; MOURA, A.N.; SAMPAIO, E.V.S.B.; GESTINARI,
L.M.S.; CARNEIRO, J.M.T. (eds.) 2002. Biodiversidade, conservação e uso sustentável da
flora do Brasil. Recife: Universidade Federal Rural de Pernambuco.
SAZIMA, M. 1978. Polinização por moscas em Bulbophyllum warmingianum Cogn.
(Orchidaceae) na Serra do Cipó, Minas Gerais, Brasil. Revista Brasileira de Botânica, 1:
133-138.
SHARMA, I.K.; CLEMENTS, M.A.; JONES, D.L. 2000. Observations of high genetic
variability in the endangered Australian terrestrial orchid Pterostylis gibosa R. Br.
(Orchidaceae). Biochemical Systematics and Ecology, 28: 651-663.
SHAW, C.R.; PRASAD, R. 1970. Starch gel eletrophoresis of enzymes – a compilation of
recipes. Biochemical Genetics, 4: 297-320.
SHEPHERD, G.J. 1995. FITOPAC 1. Campinas: Universidade Estadual de Campinas.
SILVA, U.F.; BORBA, E.L.; SEMIR, J.; MARSAIOLI, A.J. 1999. A simple solid
injection device for the analyses of Bulbophyllum (Orchidaceae) volatiles. Phytochemistry,
50: 31-34.
SOLÉ-CAVA, A.M. 1999. Biodiversidade molecular e genética da conservação. In:
MATIOLI, S.R. (ed.). Biologia molecular e evolução. Ribeirão Preto: Holos Editora. p.:
171-192.
SOLIVA, M.; WIDMER, A. 1999. Genetic and floral divergence among sympatric of

59
Gymnadenia conopsea s.l. Internatinal Journal of Plant Sciences, 160: 897-910.
SOLTIS, D.E.; HAUFLER, C.H.; DARROW, D.C.; GASTONY, G.J. 1983. Starch gel
eletrophoresis of ferns: a compilation of griding buffers, gel and eletrode buffers, and
staining schedule. American Fern Journal, 73: 9-27.
STATSOFT, INC. 2003. STATISTICA (data analysis software system), version 6. Tulsa:
StatSoft, Inc.
STUBER, C.W.; GOODMAN, M.M.; JOHNSON, F.M. 1977. Genetic control and racial
variation of β-glicosidase isozimes in maize (Zea mays L.). Biochemical Genetics, 15: 383-
394.
SUN, M.; GANDERS, F.R. 1990. Outcrossing rates and allozymes variation in rayed and
rayless morphs of Bidens pilosa. Heredity 64: 139-143.
SWOFFORD, D.L. 2000. PAUP* 4.1: phylogenetic analysis using parsimony (*and other
methods). Sunderland: Sinauer Associates.
SWOFFORD, D.L.; SELANDER, R.B. 1989. BIOSYS-1: computer program for the
analysis of alelic variation in population genetics and biochemical systematics.
Champaing: Natural History Survey.
TEIXEIRA, S.P.; BORBA, E.L.; SEMIR, J. 2004. Lip anatomy its implications for the
pollination mechanisms of Bulbophyllum species (Orchidaceae). Annals of Botany, 93:
499-505.
TOSCANO DE BRITO, A.L.V. 1995. Orchidaceae. In: STANNARD, B.L. (ed.) Flora of
the Pico das Almas, Chapada Diamantina, Bahia, Brazil. Richmond: Royal Botanic
Gardens, Kew. p.: 725-767.
TOSCANO DE BRITO, A.L.V. 2000. Two new species of Orchidaceae from Brazil.
Lindleyana, 15: 184-188.
TRAPNELL, D.W.; HAMRICK, J.L.; NASON, J.D. 2004. Three-dimensional fine-scale
structure of the neotropical epiphytic orchid, Laelia rubescens. Molecular Ecology, 13:
1111-1118.
TREMBLAY, R.L. & ACKERMAN, J.D. 2001. Gene flow and effective population size
in Lepanthes (Orchidaceae): a case for genetic drift. Biological Journal of the Linnean
Society, 72: 47-62.
VAN DEN BERG, C.; MARTINS, P.S. 1998. Biogeography of Brazilian Cattleyas:
geographic distribution, morphological variability, evolutionary and taxonomic
consequences. In: PEREIRA, C.E.B. (ed.) Proceedings of the 15th World Orchid

60
Conference (Rio de Janeiro, 1996). Naturalia Publications: Turriers, France. p.: 315-320.
VAN DER BANK, H; VAN DER BANK, M; VAN WYK, B. 2001. A review of the use of
allozyme electrophoresis in plant systematics. Biochemical Systematics and Ecology, 29:
469-483.
VAN DER PIJL, L.; DODSON, C.H. 1966. Orchid flowers: their pollination and
evolution. Coral Gables: University of Miami Press. 214 p.
VERMEULEN, J.J. 1991. Orchids of Borneo. Vol.2 - Bulbophyllum. Richmond: Royal
Botanic Gardens, Kew. 324 p.
VEROLA, C.F. 2002. Biologia floral e sistemas de reprodução em espécies de
Bulbophyllum (Orchidaceae) ocorrentes em mata de galeria, campo rupestre e floresta
estacional. Dissertação de mestrado - Instituto de Biologia da Universidade Estadual de
Campinas. Campinas. 89 p.
VITTA, F.A. 2002. Diversidade e conservação da flora dos campos rupestres da Cadeia do
Espinhaço em Minas Gerais. In: ARAÚJO, E.L.; MOURA, A. N.; SAMPAIO, E.V.S.B.;
GESTINARI, L.M.S.; CARNEIRO, J.M.T. (eds.) 2002. Biodiversidade, conservação e uso
sustentável da flora do Brasil. Recife: Universidade Federal Rural de Pernambuco
(UFRPE). p.: 90-94.
WEIR, B.S.; COCKERHAM, C.C. 1984. Estimating F-statistic for the analysis of
population-structure. Evolution, 38: 1358-1370.
WRIGHT, S. 1951. The genetical structure of populations. Annals of Eugenics, 15: 323-
354.
WRIGHT, S. 1978. Evolution and the genetics of populations. Variability within and
among natural populations. Vol. 4. Chicago: University of Chicago Press. 580 p.