variabilidade genÉtica inter e intrapopulacional de
TRANSCRIPT

GABRIEL DE MENEZES YAZBECK
VARIABILIDADE GENÉTICA INTER E INTRAPOPULACIONAL DE AKODON CURSOR (RODENTIA: SIGMODONTINAE) EM
AMBIENTES FRAGMENTADOS
BELO HORIZONTE DEPARTAMENTO DE BIOLOGIA GERAL INSTITUTO DE CIÊNCIAS BIOLÓGICAS
2002
Dissertação apresentada ao Curso de Pós Graduação em Genética, do Departamento de Biologia Geral do Instituto de Ciências Biológicas, da Universidade Federal de Minas Gerais, como requisito parcial à obtenção do título de Mestre em Genética. Orientadora: Profa Dra Maria Bernadete Lovato.

2
043 Yazbeck, Gabriel de Menezes. Y935v Variabilidade genética inter e intrapopulacional de
Akodon cursor (Rodentia: Sigmodontinae) em ambientes fragmentados. / Gabriel de Menezes Yazbeck. – Belo Horizonte : Universidade Federal de Minas Gerais / Instituto de Ciências Biológicas, 2002.
VIII, 67 f.: il. graf., tab.
Dissertação (mestrado) – Universidade Federal de Minas Gerais, Departamento de Biologia Geral.
1. Genética de populações – Teses. 2. Akodon – Teses. 3. Marcadores genéticos – Teses. 4. Fragmentação florestal. I. Título. II. Universidade Federal de Minas Gerais. Departamento de Biologia Geral.

3
AGRADECIMENTOS
Este trabalho foi uma conquista, dura, tortuosa, porém muito gratificante. Qualquer
lista de agradecimento que eu fizer não conseguirá abranger todas as pessoas que, de uma
forma ou de outra, fizeram sua contribuição presente ao longo destes anos, desde as mais
simples até as mais decisivas. Para você que ficou de fora e me ajudou neste caminho,
minhas desculpas e meus agradecimentos. Todavia não poderia de deixar registrado minha
imensa gratidão a um grupo (grande) de pessoas que tornaram esse estudo viável: à minha
orientadora, Bernadete, por ter aceitado o desafio e acreditado no trabalho mesmo nos
momentos críticos, com paciência e perseverança, e pelos ensinamentos a mim
proporcionados. Ao amigo Adriano por participar de forma decisiva na elaboração e
execução do projeto e pelas infindáveis discussões e esclarecimentos nas análises, que
muito me incentivaram. À amiga Rosângela pelo treinamento na parte laboratorial, pelo
incentivo e, principalmente, pela participação fundamental na execução do trabalho, o que
tornou possível o término do mesmo. À Dolores pelo auxílio no trabalho laboratorial e por
proporcionar um ambiente de trabalho providencial. À Marlene pelo contínuo apoio no
trabalho de bancada. Aos demais amigos do Laboratório de Genética de Populações,
Renata, Maíra, Luciene, Reinaldo, Valéria, Carol, Daiane e Juliano pelo ótimo convívio e
constante ajuda nos detalhes do cotidiano e, também, à Daniela pela imensa paciência para
explicar algumas análises e pelas constantes contribuições. Às inúmeras pessoas que se
envolveram nas coletas de campo e obtenção das amostras biológicas, principalmente Pablo
(além das discussões e ótimos momentos), Léo Vieira (também por todo ensinamentos
práticos no campo), Rafael Goreti, e os demais alunos do Lab. de Ecologia Quantitativa
(UFV). Ao Cláudio Nessralla, aos estagiários do Lab. de Mastozoologia (UFMG), Maria
Olímpia (importante material bibliográfico e identificações de animais), Bárbara, Camila,
Heitor, Fernando, também à Débora. Aos irmãos Marco Aurélio e Eduardo Sábato e,
também à Mônica Fonseca. À Sônia Talamoni pelas amostras da Mutuquinha, que apesar
de não terem sido utilizadas, me foram gentilmente cedidas. Ao Fabrício Santos pela ajuda
e sugestões, e aos seus alunos, principalmente ao Rodrigo Redondo pelo auxílio com o
protocolo de extração de DNA. Aos professores Lucio Campos e Paulo De Marco, pelos
incentivos e exemplos de conduta e profissionalismo. Aos amigos “viçosenses” André
Irsigler (pela sugestão decisiva na PCR), Samuel (pela ajuda com os programas de análise
populacional, sugestões e discussões) e Vander. Também ao Eduardo Almeida, pela ajuda

4
no laboratório, pelas discussões e ensinamentos musicais. À Roberta e Teofânia (René
Rachou), pelo auxílio e prestatividade. À Lucia , que mesmo do outro lado do mundo me
incentiva e me serve de exemplo de vida. Às várias pessoas, funcionários, alunos e
professores da UFMG que me ajudaram: às secretárias Marina e Regina; às professoras
Cleusa, Mônica, Marisa, Maria Raquel, Adlane e Andréa, pelo auxílio no dia a dia e aos
professores Edmar Chartone, Fernando da Silveira e Francisco Barbosa, por gentilmente
ceder equipamentos de laboratório ou veículos para a realização de algumas coletas. Aos
colegas Lino e Cláudia Carvalho. Ao Lab. de Genética Microorganismos e seus estagiários
(sempre prestativos), e Lab. de Genética Bioquímica (a seus estagiários e principalmente à
Neusa, pelo uso da Polaroid), ao pessoal do Geoprocessamento e da Matozoologia de
primatas André Hirsch e Helena Charllote (ambos pela ajuda no importantíssimo trabalho
de determinação dos tamanhos e distância de fragmentos) e seus estagiários Leandro e
Natália, aos primatólogos Luiz e Cláudio P. Nogueira (in memorian), ao Alexsander
Azevedo por providenciar a caracterização da Faz. Corredor e a Érica Maria. Também
gostaria de agradecer aos vários profissionais espalhados pelo Brasil e pelo mundo: George
Dergan e aos funcionários do Museu de Zoologia João Moojen (UFV), à YatitoYonenaga-
Yassuda, Lena Geise e ao Fernando Fernadez (pela presteza e rapidez de ambos no envio
de seus valiosos trabalhos) e ao Mark Miller pelo auxílio com seu programa de análise
populacional e no esclarecimento de dúvidas. Gostaria de agradecer também às empresas
de consultoria SETE e Brandt, às empresas Valourec-Mannesmann (Guilherme Freitas e
Devanei) e MBR (Miguel Andrade, José Maurício Ramos e Seu Antônio) e à COPASA,
por permitirem a realização das coletas em suas propriedades. Ao IBAMA pela concessão
da licença de coleta. À CAPES pela imprescindível bolsa de estudos. A São Judas Tadeu,
Hermeto Pascoal e Frank Zappa (in memorian). Finalmente (porém não menos importante),
gostaria de agradecer às pessoas com as quais convivi em Belo Horizonte (Andrei, Natalie,
Júlio, Sandra, Laura, Samuel, Junior, Elder, Lorenzo, Betânia e à Solange), à minha tia
Stella e minha prima Sílvia e aos meus amigos Bilunga, David, Viti e Kiti. Gostaria de
agradecer especialmente a minha linda namorada Carolina e, acima de tudo, a meus pais
Fuad e Lola e aos meus irmãos, por todo o amor, educação, compreensão e conforto que
eles me proporcionam e a quem dedico este trabalho....

5
SUMÁRIO LISTA DE FIGURAS............................................................................................................vi LISTA DE TABELAS..........................................................................................................vii RESUMO...............................................................................................................................ix ABSTRACT............................................................................................................................x
1 INTRODUÇÃO..................................................................................................................11 1.1. Considerações gerais....................................................................................................11 1.2. Efeitos genéticos da diminuição do tamanho populacional.........................................14 1.3. Variação genética versus variação morfológica...........................................................18 1.4. A espécie analisada......................................................................................................18 1.5. A técnica do DNA Polimórfico Amplificado Aleatoriamente (RAPD).......................21
2 OBJETIVOS.....................................................................................................................23 3 MATERIAIS E MÉTODOS............................................................................................24
3.1. Localidades de coleta..................................................................................................24 3.2. Captura, obtenção das amostras de tecido e medidas morfológicas...........................27 3.3. Extração de DNA e Reação em Cadeia da Polimerase (PCR)....................................28 3.4. Seleção de iniciadores, bandas e indivíduos...............................................................29 3.5. Análises estatísticas.....................................................................................................31
4 RESULTADOS.................................................................................................................34 4.1. Padrão geral de variação dos marcadores RAPD........................................................34 4.2. Variação intrapopulacional.........................................................................................38 4.3. Estrutura de populações..............................................................................................40 4.4. Relação da variabilidade genética com tamanho de fragmento..................................45 4.5. Relação da distância geográfica com a divergência genética.....................................47 4.6. Relação da variação morfológica com a variação genética intrapopulacional............47
5 DISCUSSÃO.....................................................................................................................51 5.1. Marcadores RAPD......................................................................................................51 5.2. Variabilidade intrapopulacional..................................................................................52 5.3. Estrutura Genética de Populações...............................................................................56 5.4. Relação da variabilidade genética com tamanho de fragmento..................................61 5.5. Relação distância geográfica com a divergência genética..........................................62 5.6. Relação da variação morfológica com a variação genética intrapopulacional............62 5.7. Considerações Finais...................................................................................................63
6 CONCLUSÕES................................................................................................................65 7 BIBLIOGRAFIA..............................................................................................................66

6
LISTA DE FIGURAS
1 Mapa com as localizações das populações amostradas no estado de Minas Gerais e
do Espírito Santo.......................................................................................................26 2 Perfil de amplificação pelo iniciador OPA-2. As canaletas assinaladas com as letras
“M” contêm padrões de comprimentos de bandas. Os indivíduos numerados de 1 a 8 são da população ES e de 9 a 14 da população Paraíso, sendo o indivíduo 5 um exemplar de Thaptomys e o 15 um exemplar de Akodon serrensis..........................34
3 Número de marcadores polimórficos encontrado em cada população, pelo método
jackknife de reamostragem. As barras verticais representam os intervalos de confiança (95%,).......................................................................................................36
4 Estimativas do número de bandas polimórficas feita através de Jackknife,
considerando-se desde apenas um indivíduo na amostra até os tamanhos amostrais de cada população. Os valores médios representam o número de bandas em cada população e as barras verticais os intervalos de confiança (95%)............................37
5 Análise de Componentes Principais (PCA) com o conjunto total de bandas RAPD
das seis populações estudadas. Cada eixo representa uma combinação das bandas
que melhor explicam as diferenças entre os indivíduos............................................43
6 Fenograma de distância genética de Nei (1972), pelo método UPGMA. Os números
próximos às bifurcações indicam os valores bootstrap (1000 réplicas)...................45
7 Regressão entre a variação molecular estimada pela AMOVA e a área (valores log
transformados na base dois) dos fragmentos amostrados (resultados não
significativos)............................................................................................................46
8 Regressão entre a variação molecular estimada pelo índice de diversidade de
Shannon (Ho) e a área (valores log transformados na base dois) dos fragmentos
amostrados (resultados não significativos)...............................................................47

7
LISTA DE TABELAS
1 Localidades de coletas, tamanhos amostrais, área dos fragmentos, posição relativa do local de captura nos fragmentos e localização geográfica...................................26
2 Iniciadores usados na análise dos 58 indivíduos de Akodon cursor, número e tamanho dos marcadores e número de fenótipos RAPD encontrados.....................35
3 Medidas de variação intrapopulacional e classificação das populações em ordem decrescente de variação.............................................................................................38
4 Estimativas do índice de diversidade fenotípica de Shannon (Ho) e partição da variabilidade dentro (Hpop/Hsp) e entre ((Hsp-Hpop)/Hsp) as seis populações de Akodon
cursor para os cinco iniciadores................................................................................39
5 Análise de variância das distâncias euclidianas entre pares de indivíduos...............40
6 Valores de φst por pares de populações. Todos valores de φst entre populações são
estatisticamente significativos com P<0,001............................................................41
7 Matriz de probabilidades combinadas sobre o total dos 55 marcadores RAPD para os pares de populações, segundo teste exato de Fischer para diferenças nas freqüências de marcadores........................................................................................41
8 Cargas dos Fatores (extração dos componentes principais). Aquelas maiores que 0,7 estão em negrito........................................................................................................44
9 Médias morfológicasdas seis populações analisadas de A. cursor. DP=Desvio Padrão........................................................................................................................48
10 Resultados das comparações de variação das características morfológicas entre as populações. O valor de P com um asterisco (*) é estatisticamente significativo, ao nível de 5%................................................................................................................49
11 Resultados das regressões entre variação morfológica do tarso (DP=desvio padrão) e variação genética (AMOVA e índice de Shannon, Ho)..........................................49

8
12 Sumário das análises de variâncias para as variáveis morfológicas. Valores significativos de P, ao nível de 1%, estão assinalados com dois asteriscos (**). SQ=Soma de Quadrados, QM=Quadrado Médio e GL=Graus de Liberdade..........40

9
RESUMO
A destruição de habitats naturais e a fragmentação das paisagens estão entre
os principais fatores que influenciam na extinção de espécies silvestres. As ameaças
da fragmentação estão relacionadas principalmente à diminuição dos tamanhos
populacionais e suas conseqüências como, estocasticidade ambiental, deriva
genética e depressão endogâmica. Neste estudo foram analisados aspectos da
estrutura genética de seis populações (n=58) fragmentadas de uma espécie de
pequeno mamífero da fauna brasileira, Akodon cursor, um roedor encontrado na
Mata Atlântica, campos abertos associados a matas e matas de galeria no Cerrado e
que possui alta variabilidade cariotípica (2n=14,15 e 16). A diversidade e estrutura
populacional foram analisadas através de marcadores moleculares do tipo RAPD
(DNA polimórfico amplificado aleatoriamente), utilizado-se cinco iniciadores
arbitrários, totalizando 55 bandas, das quais 41 foram polimórficas. Não houve
relação significativa entre tamanho de fragmentos e variabilidade genética
intrapopulacional. Os níveis de variação genética parecem ser mais fortemente
influenciados pela presença de ambientes propícios acessíveis nas proximidades das
populações do que pelo tamanho do fragmento florestal à qual as mesmas se
encontram associadas. Os níveis de variabilidade genética das populações não
foram relacionados com a quantidade de variação morfológica em quatro caracteres
(comprimentos do corpo, cauda, orelha e tarso). Estimou-se que 44% da
variabilidade genética está distribuída entre as populações, através da AMOVA
(análise de variância molecular) e do índice de diversidade fenotípica de Shannon.
As populações não seguiram o modelo de isolamento pela distância e se mostraram
altamente divergentes geneticamente, provavelmente devido a um considerável
isolamento entre as mesmas e em função da movimentação limitada desses
roedores. A fim de se manter variabilidade genética em A. cursor é necessário se
conservar o maior número possível de populações diferentes. Estudos associando
variação molecular e citogenética em A. cursor, além de estudos demográficos são
necessários para uma melhor compreensão dos mecanismos que mantém variação
genética nessa espécie.

10
ABSTRACT
Landscape fragmentation and habitat destruction are amongst the main factors leading
wild species extinction. Fragmentation treats are related mainly to the lowering of
population size and its consequences such as, environmental stocasticity, genetic drift
and endogamic depression. In this study genetic structure aspects were accessed from
six fragmented populations (n=58) of a Brazilian wild small mammal species, Akodon
cursor, a rodent inhabiting the Brazilian Atlantic rain forest, open fields associated
with woods and gallery forests in the Brazilian Cerrado, which displays a high level
of cariotipic polymorphism, (2n=14, 15 e 16). Population structure and diversity were
analyzed by Random Amplified Polymorphic DNA (RAPD), using five arbitrary
primers, making a total of 55 scored bands, from which 41 were polymorphic. There
was no significative relation between fragment size and intrapopulational diversity.
Genetic variation levels seems to more strongly influenced by the presence of
accessible suitable habitats nearby the fragments associated with the population.
Genetic variation levels were not related to the amount of morphological variation for
four traits (lengths of body, tail, ear and hind foot). 44% of genetic variability was
estimated to be distributed between populations by the Molecular Analysis of
Variance (AMOVA) and Shannon’s phenotypic diversity index. Populations did not
show congruence with the isolation by distance model and were genetically highly
divergent, probably due to a considerable isolation among them and to the very
limited movement of this rodent species. In order to maintain genetic variability in A.
cursor it is necessary to conservate as much different populations as possible. Studies
associating citogenetics and molecular variations in A. cursor, besides demographics
studies are necessary to a better understanding of the mechanisms which maintain
genetic variation in this species.

11
1) INTRODUÇÃO
1.1) Considerações Gerais
A taxa de desflorestamento nos trópicos sofreu um grande aumento na década
passada, seguindo uma tendência contínua do século XX (Dean, 1995). No sudeste
brasileiro, os desmatamentos de áreas florestais foram impulsionados principalmente pela
extração de madeira para obtenção de carvão vegetal, para emprego de áreas na pecuária e
agricultura e para o assentamento da crescente população humana.
A destruição e fragmentação de habitats são consideradas as maiores causas da
extinção de espécies (Pimm e Raven, 2000; Clarke e O’Dwer, 2000; Lawrence e Cochrane,
2001), e possivelmente estejamos sendo responsáveis por mais um evento de extinção em
massa (Lande, 1988; Brito e Fernandez, 2000).
Desde o início do processo de ocupação européia no Brasil o impacto da exploração
antrópica intensiva vem causando danos acentuados aos ambientes naturais. Inicialmente a
perturbação ocorreu com a retirada seletiva de espécies arbóreas na Mata Atlântica para a
obtenção de madeiras altamente valorizadas no velho mundo. Posteriormente a mineração
se tornou a principal atividade econômica no Brasil colônia, com considerável impacto
ambiental. Já nos últimos séculos, um desmatamento mais extenso se intensificou, com a
ascensão da agropecuária, o que resultou na destruição de áreas de mata para serem
utilizadas como lavouras, principalmente de café ou cana de açúcar e mais tarde como áreas
de pastagem para a criação extensiva de gado. Porém no último século o processo de
desmatamento atingiu taxas altíssimas, devido à demanda de energia nas indústrias
(extração de carvão) e ao desenvolvimento urbano, principalmente no sudeste. A população
brasileira cresceu muito (passando de aproximadamente 17,4 milhões em 1900, para cerca
de 100 milhões em 1980, e no ano de 2000 ultrapassou a marca de 166 milhões - IBGE,
2000).
O resultado atual dessa seqüência de eventos é uma paisagem completamente
fragmentada, restando, por exemplo, aproximadamente 7,5% da cobertura vegetal original
da Mata Atlântica que correspondia a mais de 1,2 milhões de km2 continuamente
distribuídos (Myers et al, 2000).

12
O processo de fragmentação antropogênica das paisagens naturais leva a
modificações na forma, tamanho e distribuição espacial dos habitats originais das espécies
silvestres. Conseqüentemente, populações de distribuição contínua, submetidas à
fragmentação têm importantes parâmetros demográfico-populacionais afetados (e.g. taxas
de extinções locais, padrões e taxas de dispersão de indivíduos entre populações e tamanhos
populacionais locais - Gibbs, 2001 - em função de alterações abióticas e por impactos
causados nas relações estabelecidas por uma dada espécie em sua comunidade, como por
exemplo, na dinâmica entre presas-predadores - Schneider, 2001).
Os efeitos da fragmentação na diminuição dos tamanhos populacionais certamente
têm conseqüências diferentes entre espécies, dependendo de suas características como,
distribuição geográfica, tamanho corporal, nível trófico, limites de tolerância, capacidade
de dispersão e sistema de acasalamento. Por exemplo, muitas espécies apresentam uma
distribuição geográfica descontínua e, portanto, podem não ser fortemente afetadas pela
destruição da vegetação, considerando-se uma escala geográfica maior. Por outro lado,
animais de grande tamanho corporal devem sofrer rapidamente os efeitos da redução de seu
habitat, devido aos seus requerimentos energéticos acentuados, que podem ser satisfeitos
somente se existir uma determinada quantidade de recursos alimentares, o que muitas vezes
deve estar correlacionado com a área.
Populações fragmentadas ou de pequenos tamanhos ficam sujeitas a efeitos
genéticos e ecológicos aleatórios (estocasticidade ambiental, demográfica e deriva genética
- Simberloff, 1988), além de efeitos determinísticos (efeito Allee, efeito de borda e
depressão endogâmica - Lande, 1999). Os principais efeitos não genéticos da fragmentação
e da redução populacional são:
i) A estocasticidade demográfica, que consiste na mudança aleatória nos
tamanhos populacionais e nos parâmetros como taxas de nascimento e
morte, que podem inviabilizar a persistência de uma população isolada.
Esses efeitos, geralmente, são determinados por flutuações ambientais
(Simberloff, 1988; Figurny-Puchalska et al, 2000; Mack, 2000), como
mudanças climáticas, ocorrência de destruição por fogo, introdução de
doenças, competidores ou predadores, etc.
ii) O efeito Allee consiste na perda de valor adaptativo dos indivíduos de uma
população reduzida, pela insuficiência de estabelecimentos de interações

13
sociais entre seus componentes. Muitas espécies dependem de cooperação
social para defesa, forrageamento ou aninhamento, migração e outras
características de sua história de vida, o que se torna comprometido com a
redução dos tamanhos populacionais, afetando diretamente animais que
têm forte organização social. Populações pequenas e esparsas de espécies
não-sociais também sofrem efeitos negativos devido à maior dificuldade
de encontros entre potenciais parceiros reprodutivos (Lande, 1999;
Grevstad, 1999). O efeito Allee tende a acelerar a diminuição
populacional, quando ultrapassado um limite inferior de numero de
indivíduos. Os benefícios da presença de indivíduos da mesma espécie
têm conquistado cada vez mais importância em considerações
conservacionistas (Stephens e Suterland, 1999).
iii) O efeito de borda é um fenômeno que acontece em função da diminuição
da área florestal, que altera as características físicas e biológicas de regiões
da periferia de fragmentos, denominadas bordas, que anteriormente
ocupavam áreas mais centrais do habitat, tornando-as semelhantes às áreas
de transição com outros habitats (antropogênicos ou não) como, por
exemplo, Mata Atlântica-campo de gramíneas ou mata de Cerrado-
cultivos de eucalipto. O efeito de borda influencia fortemente as
comunidades de invertebrados (Foggo et al., 2001) e de vertebrados,
incluindo pequenos mamíferos (Stevens e Husband, 1995; Malcon, 1995
apud Malcom, 1998; Lynam e Billick, 1999). As alterações abióticas
como, por exemplo, níveis de insolação, umidade do solo, variação
térmica, etc., se estendem até o interior dos fragmentos, alterando, por
exemplo, a dinâmica térmica dentro da mata, em caso de limites entre
florestas/campos abertos (Malcom, 1998), além de facilitar a incursão de
espécies exóticas (Lande, 1999; Pivello et al. 1999). Certas espécies são
resilientes, isto é, suportam bem o impacto do efeito de borda, ou até
mesmo podem ser favorecidas em tais ambientes, devido às suas
características biológicas, tornando-se mais abundantes do que no interior
da mata, modificando a estrutura local da comunidade (Elliot et al., 1999;
Foggo et al., 2001).

14
Os Modelos de Análise de Viabilidade Populacional (PVA) devem incorporar todos
esses fatores e mais uma série de características da biologia básica da espécie estudada a
fim de possuírem um poder preditivo efetivo (Lindenmayer et al., 2000; Lindenmayer e
Lacy, 2002) e para serem úteis nas tomadas de decisões em conservação (Lande, 1988), o
que justifica estudos sobre características biológicas gerais das espécies nativas, como
história de vida, estruturas demográfica, genética e social, aspectos do comportamento, etc.
Autores como Soulé, 1980, Lande 1999 e Gibbs 2001 consideram a estocasticidade
demográfica a maior responsável pela extinção de populações demasiadamente pequenas.
Quando o tamanho populacional é suficiente para “tamponar” esse efeito, mas não para
evitar a deriva genética, importantes fenômenos genéticos tendem a se manifestar (Lande
1988; Frankham, 1996; Lande,1999). As conseqüências genéticas da fragmentação de
habitats nas populações naturais constituem uma preocupação importante na biologia da
conservação. Esses efeitos podem ser decisivos em levar à extinção local uma população
reduzida (Mills e Smouse, 1994; Frankham, 1995; Tanaka, 1997; Saccheri et al., 1998).
1.2) Efeitos genéticos da diminuição do tamanho populacional
A primeira conseqüência da diminuição do tamanho populacional é uma influência
grande do acaso na determinação da variabilidade dentro da população. A deriva genética
leva à depauperação da variabilidade genética dentro das populações isoladas (aumento da
homozigosidade), em função de uma grande proporção de alelos que se fixam, devido ao
desvio aleatório das freqüências alélicas entre gerações (perda da diversidade total). O
resultado da subdivisão de populações contínuas é o aumento da divergência genética entre
populações isoladas, isto é, o aumento da variação atribuída a diferenças entre populações
(Wright 1943).
A taxa de perda de variabilidade genética neutra exclusivamente devido à deriva é
inversamente proporcional ao tamanho da população e depende do número de gerações que
a população se encontrou retraída em seu menor tamanho efetivo (Amos e Balmford,
2001). O número efetivo equivale ao número de indivíduos de uma população ideal (onde
não há seleção, migração e onde os acasalamentos são ao acaso) que tem a mesma
magnitude de perda de variabilidade genética observada em uma dada população real (Harlt
e Clark, 1997). O efeito nocivo dessa perda é atribuído à redução na capacidade de

15
responder de forma adequada a desafios evolutivos. Essa proposição, no entanto, baseia-se
nas prerrogativas que a seleção natural atua predominantemente de forma balanceadora
(favorecendo a heterozigosidade através dos loci de um organismo), ou que a retenção de
variabilidade genética é vantajosa em ambientes heterogêneos espacial e temporalmente, e
que a perda passiva da variabilidade neutra acontece de forma semelhante à variabilidade
seletivamente importante à espécie (Amos e Harwood, 1998; Amos e Balmford, 2001).
Outra conseqüência do tamanho populacional reduzido é o aumento da endogamia,
que leva a efeitos detrimentais nos indivíduos, fenômeno denominado depressão
endogâmica (Harlt e Clark, 1997). Esse tem sido um dos principais enfoques das discussões
em genética da conservação recentemente (Hedrick e Kalinowski, 2000; Amos e Balmford,
2001; Keller e Waller, 2002). Com a ocorrência intensa de cruzamentos endogâmicos, há
depreciação do sucesso reprodutivo médio da população (Crnokrak e Roff, 1999; Keller e
Waller, 2002) e aumento nos riscos de extinção (Finke e Jetschke, 1998). Os efeitos da
depressão endogâmica são expressos de várias formas, como na redução da fecundidade, da
sobrevivência juvenil e do tempo de vida (Frankham e Ralls, 1998), aumento na
susceptibilidade a patologias e parasitismo (Liersch e Scimid-Hempel, 1997; Edwards e
Hedrick, 1998), aumento da assimetria flutuante (Waldmann, 2001 e Winding et al. 2001),
queda do desempenho na predação (De Clercq et al, 1998). A depressão endogâmica tem
sido considerada como principal responsável pela extinção local de algumas populações de
insetos lepidópteros (Saccheri et al., 1998; Nieminen et al., 2001). A endogamia leva a um
decréscimo do valor adaptativo de populações de cativeiro (Meagher et al., 1999;
Kalinowski e Hedrick, 2001) e silvestres de espécies naturalmente não endogâmicas (Keller
e Waller, 2002) e na literatura há experimentos bem conhecidos onde se demonstra o
aumento das extinções de populações endogâmicas de camundongos e moscas-da-fruta em
laboratórios (Frankham e Ralls, 1998).
Jiménez e colaboradores (1994) submeteram a programas de cruzamento, em
laboratório, roedores capturados no campo. Posteriormente os indivíduos das gerações
resultantes foram introduzidos e monitorados em ambientes naturais. Esses estudos
mostraram que as linhagens endogâmicas tiveram maior perda de massa corporal, maior
mortalidade e menor sucesso reprodutivo do que as linhagens exogâmicas (de
acasalamentos não relacionados). Tais evidências são consideradas contundentes em
demonstrar os efeitos nocivos da endogamia nesses roedores. Tem sido sugerido que a

16
depressão endogâmica em populações naturais seja bem mais severa do que em animais
experimentais (Jiménez et al., 1994; Weller e Keller, 2002) e que seus efeitos sejam
dependentes das pressões competitivas entre os indivíduos (Meagher et al., 1999).
O mecanismo pelo qual cruzamentos aparentados são prejudiciais à população é a
exposição da carga genética, que leva à redução da adaptabilidade média da população em
decorrência do aumento da proporção de genótipos homozigotos de menor adaptabilidade
(Futuyma, 1993). Os efeitos da depressão endogâmica são sentidos no momento (ou logo
após) da redução do tamanho populacional (Amos, 1999; Crnokrak et al., 1999). Tem sido
descrita, entretanto, a manutenção de populações naturais bem monitoradas com alto
coeficiente de endogamia como, por exemplo, em algumas espécies de lobos (Wauters et
al., 1994; Kalinowski et al 1999, Crnokrak et al., 1999). Groon e Preuninger (2000)
argumentam que os níveis de depressão endogâmica variam entre espécies, mas que mesmo
quando estes não parecem ser importantes ameaças para a persistência de uma população,
devem ser monitorados em populações fragmentadas. Quando um gargalo populacional
(redução súbita do número efetivo de indivíduos que contribuem na formação das próximas
gerações) severo ocorre, em poucas gerações muitos indivíduos se tornam parentes
próximos. Quando os cruzamentos endogâmicos predominarem, os alelos letais serão
eliminados, por seleção natural, em poucas gerações nessa população (Reed e Bryant,
2001), o que explicaria, em parte, a “saúde” genética de tais populações naturais
endogâmicas (para evidências contrárias ver Frankham et al., 2001; Groon e Pruniger,
2000). No entanto, alelos não-letais e/ou de efeitos quase aditivos tenderiam a se fixar em
populações de tamanho demasiadamente diminuído, por não estarem sujeitas à eliminação
rigorosa dos alelos letais, diminuindo a adaptabilidade média da população (Lande, 1999;
Sari et al., 2001; Amos e Balmford, 2001).
Outros fatores, além dos tamanhos populacionais que determinam os padrões de
distribuição da variabilidade genética dentro e entre populações (essa distribuição espacial
da variação é chamada estrutura genética da população), são as migrações, os sistemas de
acasalamento e as pressões seletivas sofridas pelos indivíduos de cada população (Slatkin,
1994, Via, 1994). As migrações são um importante fator na determinação da variabilidade
genética e consistem na saída (emigração) e entrada (imigração) de indivíduos em
populações estabelecidas ou em áreas adequadas não ocupadas. Na biologia da
conservação, tem sido atribuída uma atenção especial ao papel das migrações e seus efeitos

17
são a uniformização genética entre diferentes populações e o aumento da variabilidade
dentro das populações que recebem novos indivíduos (Harlt e Clark, 1997).
Evolutivamente, as migrações podem agir de maneira oposta à seleção natural, por diminuir
a freqüência de alelos de maior valor seletivo dos loci relacionados com adaptações locais
(maior adaptabilidade dos indivíduos nascidos em um dado ambiente em relação aos
imigrantes, nesse local), instaurando uma carga migratória na população (Lenormand,
2002). Já em populações ameaçadas por efeitos genéticos prejudiciais decorrentes do
isolamento e diminuição populacional, atribui-se uma grande importância à chegada de
indivíduos externos à população na restauração da diversidade genética intrapopulacional: o
chamado efeito resgate (Richards 2000; Ingvarsson, 2001).
O comportamento reprodutivo da espécie é outro determinante da estrutura genética
das populações (Antonovics, 1994; Sun 1995). Em animais vertebrados os sistemas de
acasalamento mais freqüentes são poliginia (um macho acasala com várias fêmeas),
monogamia (um macho tem apenas uma parceira), poliandria (a fêmea tem vários parceiros
machos) e a promiscuidade (sem associação entre pares), podendo ainda ocorrrer espécies
com sistemas mistos de acasalamento (Groon e Pruniger, 2000). Nesses sistemas as
contribuições genéticas podem ser assimétricas em relação ao sexo e indivíduos da
população. Os próprios parâmetros da estrutura demográfica das populações, como razão
sexual, estrutura etária, tempo máximo de vida, influenciam a sua estrutura genética
(Slatkin, 1994).
Finalmente, se atribui uma grande importância na determinação da estrutura
genética das populações fragmentadas à dinâmica de meta-populações, isto é, ao conjunto
de populações em uma determinada região, que trocam indivíduos entre si e permitem a re-
colonização de áreas onde há ocorrência de extinções locais (Simberloff, 1988). O
intercâmbio recorrente de indivíduos em uma localidade pode manter uma menor
diferenciação genética entre populações de uma região, atuando em direção oposta a deriva
genética (Aars e Ims, 2000; Antolin et al., 2001) e permite a retomada das áreas que sofrem
eventos pontuais de extinção.

18
1.3) Variação genética versus variação morfológica
Na biologia da conservação uma das preocupações acerca dos níveis de
variabilidade genética se refere aos seus efeitos em características fenotípicas
ecologicamente importantes, pois essas atuam diretamente nas adaptações locais (Suggs et
al.,1997). Alguns componentes dos fenótipos são combinações de características
morfológicas interligadas, e a forma como cada característica relevante à sobrevivência
responde à seleção natural depende de como esta varia entre indivíduos e de como ela se
correlaciona com outras características seletivamente importantes em um mesmo indivíduo
(Hartt e Haeffner, 1998). Muitas características morfológicas certamente têm uma
importância ecológica e podem estar relacionadas à capacidade de disputar territórios,
parceiros sexuais e evitar predação, como já foi demonstrada, por exemplo, em peixes
salmonídeos (Hard et al.1999). Porém, em recente meta-análise, Reed e Frankham (2001)
mostraram que a variação genética e fenotípica de caracteres quantitativos são muito
fracamente correlacionadas com a variação de marcadores moleculares. Por exemplo, não
se encontrou relação entre variação fenotípica e em características ecológicas com a
variabilidade molecular estimada por RAPD em uma espécie da árvore mexicana Mailcara
zapota (Heaton et al. 1999). Em uma revisão recente, Mackay e Latta (2002) propõem que
cautela deve ser tomada na associação da influência da diversidade de marcadores
moleculares nas características quantitativas, e que deve haver discernimento entre variação
genética obtida através de marcadores moleculares (potencialmente neutros) e variação
genética em características quantitativas.
1.4) A espécie analisada
Akodon cursor (Winge, 1887) é um pequeno roedor sigmodontíneo da fauna
silvestre, encontrado no sul, sudeste e nordeste brasileiros (Cristoff, 1997), de tamanho
(comprimento do corpo mais comprimento da cauda) entre 10 e 25 cm e peso variando de
20 a 90 g (Emmons 1997). Segundo descrição de Emmons (1997), os indivíduos das
espécies do gênero Akodon ocorrentes na Mata Atlântica têm as partes superiores
uniformemente amarronzadas a marrom-oliváceo, ventre cinza escuro, pelo macio, orelhas
arredondadas e pequenas, focinho ligeiramente pontiagudo, bigodes finos e curtos e cauda

19
geralmente menor que o corpo. Sua ocorrência é principalmente na Floresta Atlântica densa
(floresta ombrófila densa) (Cristoff, 1997), em campos abertos associados a matas
(Emmons, 1997) e matas de galeria no Cerrado. Esta espécie não se encontra em ambientes
urbanizados, como muitas espécies de roedores nativos, porém, pode ser associada a
ambientes agro-florestais. Tem sido descrita como insetívoro-omnívora uma vez que sua
dieta parece ter uma importante parcela constituída por insetos, mas também se alimenta de
frutos, sementes, partes vegetativas de gramíneas e outros invertebrados como anelídeos, e
são principalmente terrestre com alguma habilidade escansorial (Fonseca et al., 1988;
Fonseca et al. 1997). Sua reprodução parece ser sazonal, com picos de densidade
populacional nos períodos de menor pluviosidade do ano (Fonseca et al. 1988), e a espécie
tem sido reportada durante todo o ano em estudos de longo prazo (Cerqueira et al. 1993).
Em um estudo feito em um campo de regeneração natural, Yazbeck e De Marco (dados não
publicados) encontraram evidências de que ocorre poliginia na população observada,
conforme inferências feitas em mapas sobrepostos das áreas de vida de machos e fêmeas.
Isso porque a área de vida de machos é claramente maior que área de vida de fêmeas, o que
é compatível com a explicação de que machos podem assim acasalar-se com um maior
número de fêmeas (Antolin et al., 2001). Maiores áreas de vida em machos de A. cursor
também foram observadas por Gentile e colaboradores (1997).
Estudos da variabilidade em A. cursor foram realizados através do seqüênciamento
do gene mitocondrial para citocromo b (Geise et al, 2001) e sugerem uma alta estruturação
genética, mesmo para distâncias razoavelmente curtas. No entanto, a maior parte dos
trabalhos com marcadores genéticos nesses animais tem o intuito de resolver questões de
filogenia e biogeografia (por exemplo, Smith e Patton, 1991; Smith et al., 1992; Smith e
Patton, 1993; Rieger, 1995; Smith et al., 2001).
O gênero Akodon está inserido na tribo Akodontini, sendo um dos maiores gêneros
de mamíferos nos neotrópicos, com cerca de 43 espécies (Emmons, 1997), sendo que novas
espécies continuam sendo descritas (Silva et al., 1998; Cristoff et al., 2000). Há uma pobre
compreensão sobre a sua sistemática, devido à documentação incompleta de espécimes e da
descrição de caracteres taxonomicamente importantes (Cristoff, 1997). A classificação de
suas espécies se baseia principalmente na morfologia crânio-dentária, porém depende
fortemente da verificação dos complementos cromossômicos, uma vez que espécies
diferentes dentro desse gênero, muitas vezes, são demasiadamente semelhantes do ponto de

20
vista morfológico. Os tipos e números cariotípicos nessas espécies são extremamente
variáveis (Liascovich et al., 1989; Geise, 1995; Fagundes et al., 1997; Silva et al., 1998;
Fagundes et al., 1998).O gênero possui o menor número diplóide conhecido em roedores
(2n=10-9) em uma espécie recém descoberta (Silva et al., 1998), ao mesmo tempo em que
outra espécie, Akodon nigita, possui 2n=52 (Liascovich et al., 1989; Fagundes e Yonenaga-
Yassuda, 1998). Há a ocorrência de espécies com cromossomos supranumerários (ou B), ou
com eventuais mosaicismos cariotípicos (um mesmo indivíduo com mais de um cariótipo),
como 2n=10-9, ou 2n=14-13 (Palhares e colaboradores, dados não publicados) e fêmeas
XO (Yonenaga,1975). Também já foram descritas reversões no sistema de determinação
sexual (ocorrem fêmeas XY) em certos indivíduos de diferentes espécies do gênero
(Koekstra e Edwards, 2000).
As formas do gênero com ocorrência na Mata Atlântica e matas de galeria do
Cerrado são praticamente indistinguíveis por características morfológicas e padrão de
pelagem, constituindo espécies crípticas (Fagundes e Yonenaga-Yassuda, 1998). Estas
formas são, algumas vezes, indiscriminadamente chamadas A. cursor, A. montensis, A.
“arviculoides” e A. aff cursor, podendo ser separadas através do cariótipo, e integram o
chamado grupo de espécies Akodon cursor (=complexo A. cursor). Relações filogenéticas
dentro desse grupo têm sido estabelecidas por meio da análise de marcadores
isoenzimáticos (Rieger et al, 1995) e seqüenciamento de segmentos do DNA mitocondrial
(Geise et al., 2001). As formas de menor número cromossômico são consideradas as mais
derivadas (Geise et al. 1998).
A espécie Akodon cursor, anteriormente também referida como uma sub-espécie de
Akodon arviculoides (Yonenaga,1972; Yonenaga et al., 1975) tem um cariótipo ao qual se
atribui a maior variabilidade conhecida em mamíferos com 2n=14, 15 e 16 (Fagundes, et al,
1998; Cristoff, 1997). Essa variação no cariótipo se deve a complexos rearranjos
(fusão/fissão), que acontecem no primeiro par do complemento, além do fato de que
existem polimorfismos cromossômicos envolvendo fusões/fissões e inversões pericêntricas
que resultam na manutenção de pares heterozigotos, envolvendo os pares 2, 3 e 5
(Yonenaga,1972; Fagundes et al, 1997; Cristoff, 1997). Essa diversidade cromossômica
está refletida na descrição de pelo menos 31 cariótipos distintos relatados na literatura para
essa espécie (Cristoff et al., 2000; Fagundes, et al, 1998), de 81 tipos esperados (Yonenaga-
Yassuda, com. pess.). Esses polimorfismos estão distribuídos entre e dentro de populações,

21
podendo haver fixação para algum tipo em certas localidades. A forma 2n=16 é quase que
ausente no sudeste brasileiro e prevalente nos animais do nordeste, sendo referida como A.
aff cursor (Rieger et al. 1995; Cristoff, 1997; Geise et al. 1998). Em Minas Gerais a
predominância é dos tipos de 2n=14-15. A localidade tipo de A. cursor é Lagoa Santa, MG,
onde animais recentemente coletados têm sido descritos como portadores do cariótipo
2n=14 (Cristoff, 1997).
1.5) A técnica do DNA Polimórfico Amplificado Aleatoriamente (RAPD)
Informações sobre a estrutura a genética das populações podem ser obtidas por meio
de emprego de marcadores moleculares (DNA) ou enzimáticos (Cavalli-Sforza, 1999).
Atualmente, a análise da variabilidade genética em populações naturais é realizada
principalmente através da utilização de marcadores moleculares de DNA (Haig, 1998,
Amos, 1998). A técnica de PCR (Reação em Cadeia da Polimerase) abriu um novo capítulo
na avaliação da quantidade de variação molecular em populações naturais (Lewin, 1999). A
PCR permite que segmentos específicos de DNA sejam amplificados, gerando milhões de
cópias dessa seqüência, a partir de quantidades ínfimas de DNA molde com a utilização de
um par de iniciadores (ou primers) que flanqueiam a seqüência-alvo, utilizando-se uma
enzima termoestável. A técnica de RAPD (Random Amplified Polymorphic DNA)
(Williams et al., 1990) é uma variação da técnica de PCR classificada como AP-PCR
(Arbitrary Primer), que consiste na amplificação de fragmentos de DNA genômico total,
com o emprego de iniciadores aleatórios, geralmente de dez bases (oligonucleotídeos), em
condições de baixa estringência. Os segmentos amplificados são tipicamente menores que
4000 pares de base, são determinados pela presença de sítios de anelamento
complementares aos iniciadores, em direções opostas, dentro de um intervalo de
comprimento, no genoma analisado. O emprego de diferentes iniciadores gera um conjunto
de marcadores moleculares que formam um perfil RAPD nos indivíduo analisado. A
análise simultânea de vários marcadores torna a técnica adequada a amostras pequenas de
indivíduos (Stweart e Excoffier, 1996). Esses marcadores são considerados de natureza
dominante, já que só se pode avaliar, em um gel de eletroforese, a presença (homozigoto
dominante e heterozigoto) ou ausência (homozigoto recessivo) de uma determinada banda.
Os marcadores RAPD são considerados com os segmentos amplificados distribuídos ao

22
acaso pelo genoma. Essa metodologia é de grande valor e aplicabilidade, já que não há pré-
requisitos de conhecimento prévio do genoma da espécie a se estudar, além do relativo
baixo custo, simplicidade e por requerer quantidades mínimas de DNA para a realização
das análises (Ferreira e Grattapaglia, 1998).
A técnica vem sendo extensivamente utilizada em estudos de populações de plantas
(Huff et al., 1993; Lu et al., 1997; Fahima et al., 1999; Heaton et al. 1999; Lacerda et al.,
2000), sendo também empregada em estudos de estrutura genética de populações de
vertebrados (e.g. Haig et al., 1994; Gibbs et al., 1994; Nausser et al., 1996; Mockford et
al., 1999). Para se utilizar essa ferramenta molecular, no entanto, os procedimentos
laboratoriais devem ser otimizados para a realização consistente das reações, de forma a se
alcançar boa repetibilidade dos fragmentos (Ferreira e Grattapaglia, 1998).

23
2) OBJETIVOS
Considerando os aspectos anteriormente mencionados, neste trabalho a técnica de
RAPD foi utilizada para um estudo genético-populacional em uma espécie de roedor nativo
Akodon cursor (Winge 1887; Rodentia: Sigmodontinae), na tentativa de elucidar aspectos
da estrutura genética de populações e dos possíveis impactos da fragmentação na
variabilidade desse animal. Será testado se existe relação entre os níveis de variabilidade
genética e os tamanhos de fragmentos. Esse trabalho tem os seguintes objetivos específicos:
i. Estimar a variação genética dentro e entre diferentes populações de Akodon cursor.
ii. Analisar a relação entre a variabilidade intrapopulacional com os tamanhos dos
fragmentos.
iii. Comparar a variação genética entre diferentes populações.
iv. Analisar a relação entre a variação genética estimada através de marcadores RAPD e a
variabilidade em certos caracteres morfométricos.
v. Testar a existência de correlação entre as distâncias genéticas e geográficas das
diferentes populações.

24
3) MATERIAIS E MÉTODOS
3.1) Localidades de coleta
Um total de seis áreas em cinco municípios foi amostrado: UFV e Mata do Paraíso
em Viçosa, MG; Corredor em Bocaiúva, MG; Jambreiro em Nova Lima, MG; Domingos
Martins, ES e Pedra Negra na região de Lavras, MG. O número de indivíduos por área
amostrada e suas localizações geográficas estão mostrados na Tabela 1. As áreas de coleta
(Figura 1) estavam dentro da região de ocorrência da Mata Atlântica, excetuando-se a
população Corredor, que constitui-se de uma mata de galeria no Cerrado, ambos biomas
considerados prioritários para a conservação (Myers et al., 2000). A seguir será apresentada
uma breve descrição de cada área e da respectiva coleta:
• UFV: Essa área se encontra dentro do campus da Universidade Federal de
Viçosa em MG. Os indivíduos dessa população foram capturados por meio de
uma grade (ver adiante) de amostra em um campo em regeneração (essa área
havia sido completamente destruída por um incêndio logo antes das coletas, que
se estenderam de 2000 a 2001), adjacente a uma área de fragmento de mata em
topo de morro com cerca de 11 ha, situado na proximidade de vários outros
fragmentos de tamanhos variáveis. A área apresenta uma declividade leve a
moderada. Os indivíduos foram capturados próximo à borda do fragmento em
uma área dominada por capim gordura (Melinis minultiflora). Foram amostrados
14 indivíduos, dos quais seis eram fêmeas e os seis outros eram machos.
• Corredor: Situada em Bocaiúva na Fazenda da Valourec-Mannesmann que está
localizada dentro de uma região de ocorrência do Cerrado. Nesse fragmento se
encontra uma mata de galeria com características muito semelhantes às da Mata
Atlântica, conhecido no local como “Lapa D’água”. Essa mata está situada
dentro de uma porção maior de mata de Cerrado totalizando, junto com áreas de
cascalheira e campo de gramíneas, uma área de 542,3ha inserida em uma
paisagem totalmente dominada pelo cultivo de eucaliptos. A mata de galeria
possui declividade suave a média. Os animais foram capturados em um
transecto de captura que cruzava um corpo d’água presente no local. Os 12

25
indivíduos dessa população foram capturados em diferentes campanhas de
captura, entre 1998 e 2001, sendo seis machos e seis fêmeas.
• Mata do Paraíso (Paraíso): Também nos limites do município de Viçosa, essa
área da mesma forma da UFV se apresenta em um campo de M. minultiflora,
adjacente à borda do fragmento florestal. Esse fragmento possui uma área de
194 ha, porém considerando-se os fragmentos intimamente “conectados” a esse,
a área efetiva passa a ser de 384 ha (Ribon, 1998). Os indivíduos foram
amostrados entre 1999 e 2001, em diferentes campanhas. A grade de captura se
encontra em uma suave encosta sobre uma barragem artificial de água. Cinco
indivíduos eram machos e quatro eram fêmeas.
• Jambreiro: O fragmento do Jambreiro é o maior dentre os aqui analisados e
está situado em uma APA (Área de Proteção Ambiental) da MBR no município
de Nova Lima, na região metropolitana de Belo Horizonte. As armadilhas foram
dispostas no interior da mata, ao longo de uma das trilhas de educação ambiental
do CEA-MBR e em um transecto contíguo, na mata, ao logo de uma suave
encosta. Há a presença de um corpo d’água que corta a trilha e o local possui
uma declividade moderada. Nove indivíduos foram amostrados para análise
molecular, no intervalo de um ano, entre 2000 e 2001. Seis indivíduos eram
fêmeas e apenas três eram machos.
• ES: O transecto de coleta estava situado na borda externa de um fragmento na
encosta e topo de um morro, em um campo de gramíneas próximo a um pequeno
corpo d’água. O local possui relevo ondulado e está dentro do município de
Domingues Martins, ES, apesar de ser mais próximo a Venda Nova dos
Imigrantes, ES. A localidade não é distante do Parque Estadual da Pedra Azul
em ES e se encontra cercado de vários outros fragmentos de tamanhos e formas
semelhantes, principalmente nos topos de morro. Os sete indivíduos foram
coletados em uma única sessão de capturas no final de 2000, sendo três fêmeas e
quatro machos.
• Lavras: Local de relevo plano, contou com um transecto que cortava o interior
do fragmento de mata, onde há a presença de um corpo d’água. A localidade é
denominada Pedra Negra, próxima ao município de Lavras, MG. Os sete
indivíduos foram amostrados em duas sessões de coletas realizadas com menos

26
de dois meses de intervalo no final de 2001. Novamente, aqui há uma paisagem
fragmentada com a presença de vários outros fragmentos próximos ao
amostrado. Foram analisados quatro fêmeas e três machos desse local.
Tabela 1. Localidades de coletas, tamanhos amostrais, área dos fragmentos, posição relativa
do local de captura nos fragmentos e localização geográfica.
Coordenadas Geográficas Localidade N
Área
(ha)
Posição do Transecto
ou da Grade de coleta Longitude Latitude
UFV 14 10, 7 Borda 42º52'55''W 20º45'14''S
Corredor 12 542,3 Interior do fragmento 43º53'34''W 17º22'52''S
Paraíso 9 384 Borda 42º51'W 20º45'S
Jambreiro 9 912 Interior do fragmento 43º50'48''W 19º59'08''S
ES 7 16 Borda 41º08'03''W 20º20'23''S
Lavras 7 54,4 Interior do fragmento 44º59'59''W 21º14'47''S
Figura 1. Mapa com as localizações aproximadas das populações amostradas no
estado de Minas Gerais e do Espírito Santo.

27
3.2) Captura, obtenção das amostras de tecido e medidas morfológicas
Os animais foram capturados por meio de armadilhas para animais vivos, do tipo
Shermann e Tomahawk, dispostas no esquema de transecto (linha reta), com armadilhas
distanciadas cerca de 30 m, ou de grade (linhas e colunas ortogonais), com as armadilhas
separadas de 10 a 25m, em sessões de captura de no mínimo cinco noites, em campanhas de
esforços amostrais variáveis (mínimo de 100 armadilhas-noites, máximo de 420
armadilhas-noites; o número de armadilhas-noites é igual ao número de armadilhas abertas
no local vezes o número de noites de captura naquela sessão).
A metodologia padrão utilizada foi a de captura-marcação-recaptura apenas para
individualização dos animais. Em todas as localidades espécimes-testemunho foram
removidos do campo para identificação e deposição em coleções de referência (Museu de
Zoologia João Moogen, na Universidade Federal de Viçosa, MG, Museu Nacional, da
Universidade Federal do Rio de Janeiro, RJ e na coleção de referência da Universidade
Federal de Minas Gerais, MG). A totalidade dos indivíduos capturados na população ES foi
removida.
Os animais liberados no campo foram medidos para quatro variáveis morfológicas
externas (comprimento do corpo, comprimento da cauda, comprimento da orelha e
comprimento da pata traseira), pesados, marcados por meio de anilhamento numerado na
orelha, ou por amputação de falanges distais das patas dianteiras (unidades) e traseiras
(dezenas). Foi verificado o sexo do animal e finalmente, foi colhida uma pequena porção
distal da cauda (cerca de 1cm), que foi fixada no ato da coleta com etanol 70%, em um
recipiente estéril e devidamente identificado. Os ferimentos causados foram higienizados
com tintura de iodo e os animais foram soltos nos pontos de coleta. Recapturas posteriores
mostraram que as feridas, de maneira geral, eram rapidamente cicatrizadas entre recapturas.
O instrumento utilizado para o corte da porção da cauda (um aparador de unhas de uso
veterinário) foi limpo primeiramente com solução fisiológica e algodão e novamente com
etanol, entre coletas de amostras, a fim de se evitar contaminação cruzada entre indivíduos.
Animais sacrificados no laboratório tiveram porções do fígado, na maioria das vezes, ou
coração, fixadas em etanol 70%. Para a análise de reprodutibilidade de resultados (PCR)
entre diferentes tecidos, foram obtidas amostras de dois indivíduos (fígado, coração,
músculo esquelético e cauda).

28
3.3) Extração de DNA e Reação em Cadeia da Polimerase (PCR)
O DNA foi extraído utilizado-se um protocolo de fenol-clorofórmio-álcool
isoamílico com as seguintes etapas gerais:
1. As amostras de tecido foram cuidadosamente picotadas com auxílio de uma
lâmina de bisturi limpa e em seguida foram adicionadas ao tampão de
extração, TrisNH4Cl (37o C). Foi realizada uma nova limpeza do material
com solução salina (NaCl) a 0,85%.
2. Para o processo de lise e desproteinização inicial o material foi suspenso em
high-TE pH 8,0 e foram adicionadas solução de lise de Madisen (Tris HCL
0,1 M/EDTA 40mM/SDS 0,2%/NaCl 1M; 50o C) e Proteinase K (10 µg/µl).
3. O próximo passo foi o da desproteinização do DNA realizado com fenol-
cloroformio-álcool isoamílico (25:24:1). Esse passo é de fundamental
importância, pois determina a o quanto o DNA vai estar isento de
contaminantes como, por exemplo, proteínas.
4. Para a precipitação dos ácidos nucléicos utilizou-se acetato de sódio (3M) e
isopropanol gelado. Os tubos foram incubados no freezer para finalizar a
precipitação do DNA e centrifugados para a formação do pellet, que por sua
vez foi re-suspenso em Low-TE (pH 8,0). Finalmente, os tubos foram
incubados em banho-maria a 37o C e homogeneizados antes de serem
estocados em freezer convencional (-20o).
5. A quantificação foi feita visualmente em gel de agarose 1%, corado com
brometo de etídeo (1 ηg/µl) e as amostras foram diluídas em TE para atingir
uma concentração de cerca de 5 ηg/µl.
Os ensaios PCR foram feitos por um protocolo encontrado em Ferreira e
Grattapaglia (1998), com modificações. O volume final de cada reação foi 16 µl: 10 mM de
Tris-HCl pH 8,0 e 50 mM de KCl (tampão), 2 mM de Mg Cl2++, 1 µM de cada um dos
quatro dNTPs diferentes, 1 unidade de Taq polimerase (Phoneutria), 0,25 µM de iniciador
(oligonucleotídeo de 10 bases aleatórias, Operon Technologies) e de 1 a 5 ηg/µl de DNA
do organismo estudado. Os tubos foram selados com uma pequena gota de óleo mineral,
para impedir evaporação durante o aquecimento na amplificação. Reações onde o DNA foi

29
substituído por água serviram de controle negativo, para assegurar a não contaminação das
reações.
Os ciclos de amplificação foram os seguintes: 1) desnaturação inicial a 95o C
durante 1 min, seguida de 35 ciclos de desnaturação, anelamento do iniciador e
polimerização (94o C por 10 seg, 36o C por 1min e 72o C por 2min, respectivamente) e
extensão final a 72o C por 7 min. As amplificações foram feitas em termocicladores do
modelo PTC-100 (M.J. Research).
A visualização dos resultados da amplificação foi feita aplicando-se a totalidade do
produto de amplificação (corado com tampão de corrida contendo azul de bromo fenol e
sacarose) em gel de agarose a 1% submerso em TBE (89 mM de Tris-HCl pH 8,0; 89mM
de ácido bórico e 2 mM de EDTA), submetido a um campo elétrico de 100 V durante três
horas e meia. Depois desse período o gel foi corado em uma cuba contendo TBE 1x e
brometo de etídeo (1 ηg/µl), de 20 a 30 minutos. O gel foi então lavado com água para
eliminar-se o excesso de brometo de etídeo por mais vinte minutos e então fotografado sob
luz ultra-violeta em película polaróide monocromática.
Devido aos tamanhos populacionais reduzidos, mais de uma população pôde ser
visualizada em um mesmo gel, para um mesmo iniciador. Indivíduos previamente
amplificados foram repetidos em outras reações e aplicados em géis de outras populações,
para servirem de controle positivo, para verificar a reprodutibilidade de bandas entre
reações e facilitar a leitura dos géis. Em todos os géis foram incluídos padrões de tamanho
de fragmentos moleculares (DNA-Ladder 100 bp, Life Technologies), para se determinar os
tamanhos aproximados dos fragmentos amplificados.
3.4) Seleção de iniciadores, bandas e indivíduos
Um total de 56 iniciadores de 10 bases (Operon Technologies, 20 do conjunto OPA,
20 do OPB, 8 do OPC e 8 do OPL) foram testados em quatro indivíduos de populações
diferentes, para se determinar quais resultariam em amplificações. Os iniciadores que
produziram um bandeamento nítido tiveram seus géis fotografados. As reações para esses
iniciadores foram refeitas em dias diferentes e aqueles que repetiram amplificações foram
fotografadas e comparadas com os primeiros resultados. Apenas os iniciadores que
produziram padrões fortemente recorrentes entre as reações e onde podiam ser observados

30
polimorfismos foram selecionados para este estudo (OP-A2, OP-A4, OP-A7, OP-C16 e
OP-L1).
A técnica de RAPD, para ser aplicada a estudos populacionais, depende de uma
criteriosa seleção dos fragmentos amplificados (Cooper, 2000) e muitas vezes comparações
entre diferentes estudos têm seu poder de comparação restrito devido à subjetividade na
escolha dos marcadores (Sunnucks, 2001). A seleção dos marcadores foi feita
considerando-se os critérios apresentados em Lacerda, 2000 e Cooper, 2000. Foram
consideradas apenas bandas nítidas, facilmente distinguíveis de outras bandas, e de
reprodutibilidade constatada entre diferentes reações. Bandas que não se reproduziram em
diferentes dias e bandas muito claras ou indistinguíveis não foram consideradas como
marcadores. Como mais de um tipo de tecido foi utilizado como fonte de DNA, reações de
PCR com DNA proveniente de cauda, fígado, coração e músculo de um mesmo animal
foram realizadas, utilizando-se dois iniciadores para averiguar a repetição dos resultados
entre diferentes tecidos. Esses ensaios piloto demonstram não haver diferenças de padrão
de bandeamento entre tecidos alternativos de um mesmo indivíduo.
Além de Akodon cursor, indivíduos de outras três espécies de roedores akontotíneos
tiveram seus DNAs amplificados com os iniciadores aqui usados. A primeira foi Tapthomys
(ES), um pequeno roedor de um gênero que já foi considerado subgênero de Akodon (Smith
et al., 1993; Cristoff, 1997) muito semelhante a um jovem Akodon cursor, exceto pela sua
curta cauda. Outras duas foram Bolomys lasiurus (UFV) e Akodon serrensis (coletado na
Serra do Brigadeiro, MG, próximo ao município de Viçosa), espécies muito semelhante a
A. cursor, porém discerníveis. Todas as espécies apresentaram uma diferença clara nos
padrões de bandeamento dos indivíduos de A. cursor. Isso serviu de controle para se excluir
indivíduos de classificação duvidosa, já que a falta dos dados de cariotipagem dos
indivíduos analisados pode levantar dúvidas sobre identificação dos mesmos (Yonenaga-
Yassuda, com. pess.). Akodon montensis é morfologicamente indistinguível de A. cursor
(espécies crípticas), excetuando-se por uma abordagem multi-variada (Cristoff, 1997). No
estado do Rio de Janeiro as duas espécies se segregam segundo um gradiente de altitude,
sendo que A. montensis só ocorre a partir dos 800m de altitude (Geise, 1995). Não houve
um indivíduo de A. montensis cariotipado, disponível para comparação por RAPD, neste
trabalho. Indivíduos de A. cursor, que apresentaram um DNA com baixa qualidade de

31
amplificação, foram eliminados da amostra. Desta forma o tamanho total da amostra foi de
58 indivíduos.
3.5) Análises estatísticas
Para análises estatísticas foi utilizado o programa Statistica 5.0. Para análises
populacionais, utilizou-se o programa TFPGA. Para a Análise de variância molecular foi
utilizado o programa AMOVA 1.55.
A partir da análise cuidadosa das fotografias dos géis foi construída uma matriz
fenotípica composta de “0” e “1”, sendo “0” a ausência da banda e “1” a presença. Foi
realizado um procedimento jackknife de reamostragem sobre o número de bandas
amostradas em cada população. Uma comparação entre esses números foi realizada por
meio de intervalos de confiança calculados em função dos erros padrão associados às
médias jackknife de cada população. Os valores jackknife também foram recalculados para
sub-amostras aleatórias de cada população (N-1, N-2, ... 1; onde N é o tamanho de cada
população) e plotados em um gráfico de acumulação para avaliar a representatividade da
amostra em relação ao número de bandas amostradas.
Na determinação da variabilidade intrapopulacional, foram consideradas as
proporções de marcadores RAPD polimórficos nas populações, a variância
intrapopulacional calculada pela Análise de Variância Molecular (AMOVA) e o índice de
diversidade de Shannon (considerando-se as bandas como espécies). O teste exato de Fisher
foi realizado para se comparar as freqüências de presença/ausência das bandas.
A proporção de marcadores polimórficos foi avaliada em todo o conjunto amostral,
sendo consideradas polimórficas as bandas que estivessem presentes ou ausentes em pelo
menos um indivíduo. Foi calculada a freqüência de cada marcador e determinada a
percentagem de marcadores polimórficos dentro de cada população.
A AMOVA (Excoffier et al., 1992; Stewart e Excoffier, 1996) consiste em uma
análise de variância tradicional realizada com os quadrados das distâncias euclidianas (que
aqui é igual ao número de diferenças de estados no conjunto de marcadores) entre todos os
pares de indivíduos, obtida a partir da matriz original de 0 e 1 de marcadores RAPD de
todos os indivíduos associados a suas respectivas populações. Dessa forma a AMOVA
determina a quantidade de variação entre as populações, de forma similar ao Fst de Wright,

32
chamada de φst e testa sua significância contra a hipótese nula de ausência de estruturação
(panmixia), por procedimentos de permutação aleatória entre indivíduos e populações. Por
meio dessa análise também se extrai a quantidade de variação dentro de populações. A
AMOVA gera valores de φst entre pares de populações, permitindo inferir-se sobre a
divergência genética entre as mesmas. Para saber se as populações variam da mesma forma,
a heterogeneidade de variâncias (heterocedasticidade) entre as populações foi verificada
pelo teste de Bartlett, sendo sua significância verificada com a realização de 1000
permutações.
O índice de Shannon deriva da teoria da informação (Pielou, 1975) e sua aplicação
prática remonta à segunda guerra mundial, quando essa metodologia era aplicada para se
avaliar a quantidade de informação em mensagens secretas codificadas e assim decidir
quais mensagens se estudar primeiro. Seu uso tem sido amplamente difundido no estudo de
comunidades como índice de diversidade de espécies. O índice foi aplicado pela primeira
vez em estudos genético-populacionais, por Richard Lewontin em 1972. Aqui ele é usado
de uma forma análoga, considerando-se diversidade de marcadores, seguindo os
procedimentos sugeridos por Yeh, 1995, utilizando simultaneamente a freqüência de
presença e de ausência das bandas. Esse índice parece adequado para estudos de
diversidade genética, uma vez que seu comportamento é parecido com o da
heterozigosidade esperada. O cálculo do índice de diversidade fenotípica de Shannon (Ho)
foi calculado da seguinte forma: Ho= (-Σpilog2pi)/n; onde pi é a freqüência da presença ou a
ausência da banda i, e n é o número de bandas consideradas. O procedimento foi calculado
para o conjunto de marcadores obtidos com cada iniciador separadamente e para o total de
marcadores. O valor do índice dentro de cada população foi obtido por uma média
ponderada (pelo número de marcadores de cada iniciador) e foi chamado de Hpop. Uma
medida da proporção da variação dentro de cada população foi feita com a razão entre o
índice atribuído à média das populações e o índice calculado para a espécie (Hpop/Hsp). O
índice de diversidade da espécie (Hsp) foi calculado desconsiderando as populações. Com
essas medidas foi possível calcular a partição dessa variação entre as populações de
maneira similar ao Fst: (Hsp -Hpop)/ Hsp. (Wachira et al., 1995; Wolff et al., 1997, Lacerda,
2000).
Uma análise de componentes principais (PCA) foi feita com o intuito de se observar
se há congruência entre os resultados da AMOVA e a distribuição dos dados segundo os

33
dois principais eixos que melhor explicam a variação total dos dados. Essa análise é feita a
partir de uma matriz de correlação gerada pelos dados de presença/ausência das bandas
polimórficas analisadas.
Para se avaliar diferenças entre populações um procedimento Monte Carlo usando o
algoritmo de cadeias de Markov (Raymond e Rousset, 1995) foi realizado a fim de se obter
aproximações ótimas da probabilidade exata da diferença observada na freqüência de
marcadores entre populações (teste exato de Fisher), utilizando os dados de
presença/ausência de bandas em cada indivíduo. As probabilidades foram determinadas
entre a média dos loci para cada par de populações e para o conjunto total das mesmas.
Também foi determinada a distância de genética de Nei (1972) entre as populações. Para
esse cálculo foi assumido o equilíbrio de Hardy-Weinberg, e as freqüências alélicas foram
estimadas pelo método sugerido por Lynch e Milligan, (1994). Esses valores foram
utilizados para a construção de um fenograma de uma análise de agrupamento de médias
pareadas não ponderadas, ou UPGMA (Backeljau et al., 1996), e os valores bootstrap
(Felsenstein, 1986), para cada nó da árvore foram calculadas a partir de 1000 réplicas.
A constatação da relação entre variabilidade genética com o tamanho de fragmento
foi realizada através de regressão linear entre log da área e duas estimativas de variação
intrapopulacionais: as da AMOVA e do índice de Shannon. Também foi testado se existe
correlação entre as matrizes de distância genética (distância genética de Nei (1972) e φst) e
de distância geográfica entre os pares de localidades, por meio do teste de Mantel (Sokal,
1979), para verificar se há um aumento da divergência genética em função da distância
geográfica.
Finalmente as medidas morfológicas externas tomadas das populações foram
submetidas a uma análise de variância (ANOVA) e a estatística de teste de Bartlett para
heterocedasticidade: medidas que eventualmente apresentassem diferenças entre médias e
heterocedasticidades significativas tinham a ANOVA repetida com valores da variável log
transformadas. As características que apresentaram heterogeneidade de variâncias tiveram
seus desvios-padrão submetidos a uma regressão com os valores de variação
intrapopulacional da AMOVA e do índice de Shannon, independentemente de diferenças
das médias entre populações.

34
4) RESULTADOS
4.1) Padrão geral de variação dos marcadores RAPD
Dentre os 56 iniciadores testados, cinco foram utilizados neste estudo, OPA-2,
OPA-4, OPA-7, OPC-16 e OPL-1. A partir desse conjunto um total de 55 fragmentos
RAPD foi selecionado e identificado, com base nos critérios de nitidez, reprodutibilidade e
distinção em relação a outras bandas. Os nomes dos marcadores foram dados pela letra do
conjunto ao qual o primer pertencia seguido do seu tamanho aproximado em pares de base.
As bandas variaram em comprimento dentro de um intervalo de aproximadamente 390pb-
2050pb (Tabela 2). A Figura 2 mostra um exemplo de visualização de fragmentos RAPD
gerados pelo primer OPA-2.
Figura 2. Perfil de amplificação pelo iniciador OPA-2. As canaletas assinaladas com as
letras “M” contêm padrões de comprimentos de bandas. Os indivíduos numerados de 1 a 8
são da população ES e de 9 a 14 da população Paraíso, sendo o indivíduo 5 um exemplar de
Thaptomys e o 15 um exemplar de Akodon serrensis.
Cada iniciador produziu entre quatro a 14 bandas, com uma média de 11 bandas por
iniciador (Tabela 2). No total de marcadores gerados por cada iniciador, pode-se notar uma
acentuada variação na capacidade de escrutinar fenótipos RAPD exclusivos nos 58
indivíduos analisados (um perfil único para cada indivíduo), sendo que apenas 3 fenótipos
diferentes são evidenciados com o uso do iniciador OP-A4, até 47 perfis RAPD distintos no
iniciador OPA-7. Portanto, nenhum iniciador pôde sozinho detectar um perfil genético
único para cada indivíduo analisado (Tabela 2).

35
Tabela 2. Iniciadores usados na análise dos 58 indivíduos de Akodon cursor, número e
tamanho dos marcadores e número de fenótipos RAPD encontrados.
Iniciador Seqüência (5’-3’) Número de bandas
selecionadas (polimórficas/monomórficas)
No de perfis RAPD
distintos gerados
Tamanho de fragmentos
amplificados (pb)
OPA-2 TGCCGAGCTG 9 (4/5) 11 600-1700 OPA-4 AATCGGGCTG 4 (2/2) 03 420-1200 OPA-7 GAAACGGGTG 14 (13/1) 47 600-1600 OPC-16 CACACTCCAG 14 (13/1) 41 500-2050 OPL-1 GGCATGACCT 14 (9/5) 35 390-1400
Conjunto dos cinco iniciadores 55 (41/14) 58 390-2050
Os números de marcadores RAPD polimórficos das populações foram comparados
(Figura 3). Populações com intervalos de confiança que estão sobrepostos à média de outras
populações não são estatisticamente diferentes ao nível de 5%. Pode-se evidenciar que a
população do ES apresenta um número superior de marcadores polimórficos, seguida da
população do Jambreiro. Mata do Paraíso e Lavras são iguais entre si e iguais a ambas,
UFV e Corredor, porém as duas últimas são diferentes entre si em relação à esse parâmetro
(Figura 3). A população do ES tem o menor número de alelos nulos fixados (variando de 6
a 12, média de 9,6). A freqüência de bandas exclusivas nas populações (i.e. presentes em
todos os indivíduos de uma população e ausentes das demais populações) foi muito baixa,
havendo apenas um marcador (1,8 % de todos os marcadores) que era exclusivo da
população do Jambreiro.
Também foi verificada a eficiência do tamanho das amostras populacionais em
relação ao número de bandas polimórficas consideradas nesse estudo, por meio de um
procedimento jackknife. Para todas populações o número de indivíduos coletados amostra
eficientemente o número de bandas polimórficas selecionadas para a análise, conforme
sugere a disposição gráfica dos valores jackknife nos gráficos abaixo (Figura 4). Essa
disposição é análoga à “curva do coletor de espécies” em uma comunidade e sugere que o
aumento do tamanho da amostra não aumentará o número de marcadores encontrados na
população.

36
Núm
ero
de m
arca
dore
s po
limór
ficos
30
32
34
36
38
40
42
UFV Corredor Paraíso Jambreiro ES Lavras
Figura 3. Número de marcadores polimórficos encontrado em cada população, pelo método jackknife de reamostragem. As barras verticais representam os intervalos de confiança (95%,).

37
Número de indivíduos na amostra
Núm
ero
de
mar
cado
res
polim
órfic
os
0
10
20
30
40
50
60
70
1 3 5 7 9 11 13 15 17 19
UFV Número de indivíduos na amostra
Núm
ero
de
mar
cado
res
polim
órfic
os
0
10
20
30
40
50
60
70
1 3 5 7 9 11 13 15 17 19
Corredor
Número de indivíduos na amostra
Núm
ero
de m
arca
dore
s po
limór
ficos
0
10
20
30
40
50
60
70
1 3 5 7 9 11 13 15 17 19
Paraíso Número de indivíduos na amostra
Núm
ero
de m
arca
dore
s po
limór
ficos
0
10
20
30
40
50
60
70
1 3 5 7 9 11 13 15 17 19
Jambreiro
Número de indivíduos na amostra
Núm
ero
de m
arca
dore
s po
limór
ficos
0
10
20
30
40
50
60
70
1 3 5 7 9 11 13 15 17 19
ES Número de indivíduos na amostra
Núm
ero
de m
arca
dore
s po
limór
ficos
-10
0
10
20
30
40
50
60
70
0 2 4 6 8 10 12 14 16 18 20
Lavras
Figura 4. Estimativas do número de bandas polimórficas feita através de jackknife,
considerando-se desde apenas um indivíduo na amostra até os tamanhos amostrais totais em
cada população. Os valores médios representam o número de bandas em cada população e
as barras verticais os intervalos de confiança (95%).

38
4.2) Variação intrapopulacional
A Tabela 3 apresenta medidas de variabilidade intrapopulacional estimadas por
métodos independentes, por meio da variação obtida pela AMOVA, pelo índice de
diversidade de Shannon e pela percentagem de marcadores polimórficos. Os níveis de
variação genética intrapopulacional foram relativamente concordantes entre os três métodos
aqui empregados: em todos a população UFV apresentou a maior variabilidade e Jambreiro
a menor. As posições relativas de ordem decrescente nos níveis de variação dentro das
populações foram quase que inteiramente coincidentes entre a AMOVA e o índice de
Shannon, índices que consideram as freqüências de marcadores na população, havendo
apenas a troca de lugares entre Corredor e Lavras nas 2a e 3a posições (Tabela 3). Na
percentagem de loci polimórficos há um empate entre Paraíso, ES e Lavras, populações que
nas outras metodologias mostram níveis intermediários e relativamente próximos de
variabilidade (Tabela 3). Esses resultados mostram que a população do Jambreiro tem uma
variabilidade reduzida em relação às outras populações.
Tabela 3. Medidas de variação intrapopulacional e classificação das populações em ordem
decrescente de variação.
AMOVA Shannon Percentagem de marcadores
polimórficos População
�2 dentro Ordem (Hpop) Ordem % Ordem
UFV 5,39 1o 0,384 1o 45,4% 1o
Corredor 5,11 3o 0,362 2o 43,6% 2o
Paraíso 4,83 5o 0,331 5o 40,0% 3o
Jambreiro 3,30 6o 0,240 6o 34,5% 4o
ES 5,04 4o 0,333 4o 40,0% 3o
Lavras 5,14 2o 0,338 3o 40,0% 3o

39
Tabela 4. Estimativas do índice de diversidade fenotípica de Shannon (Ho) e partição da variabilidade dentro (Hpop/Hsp) e entre ((Hsp-
Hpop)/Hsp) as seis populações de Akodon cursor para os cinco iniciadores.
Iniciadores UFV Corredor Paraíso Jambreiro ES Lavras Hpop Hsp Hpop/Hsp (Hsp-Hpop)/Hsp
OPA-2 0,179 0,400 0,212 0,170 0,241 0,271 0,247 0,358 0,691 0,309
OPA-4 0,093 0 0,191 0 0,246 0,148 0,100 0,446 0,224 0,776
OPA-7 0,401 0,429 0,328 0,373 0,481 0,366 0,397 0,728 0,544 0,456
OPC-16 0,528 0,427 0,374 0,388 0,391 0,236 0,410 0,782 0,524 0,476
OPL-1 0,437 0,309 0,407 0,072 0,211 0,509 0,331 0,539 0,614 0,386
Média geral 0,384 0,362 0,331 0,240 0,333 0,338 0,331 0,613 0,557 0,443

40
A Tabela 4 mostra que existe grande variação entre iniciadores na capacidade de
estimar isoladamente os níveis de variação intrapopulacionais. O iniciador OPA-4, por
exemplo, não indica variação dentro de algumas populações com o índice de Shannon, ao
passo que outros iniciadores são capazes de detectar variação intrapopulacional
considerável nessas mesmas populações.
A ausência de relação entre o número de bandas amostradas por população e os
níveis de variação intrapopulacionais, além da baixa freqüência de marcadores exclusivos
dentro de populações indicam que as diferenças encontradas entre populações devem ser
explicadas principalmente pelas freqüências das bandas RAPD.
4.3) Estrutura de populações
A análise de variância molecular (AMOVA) mostrou que aproximadamente 44% da
variação total se encontra distribuída entre as 6 populações estudadas (φst=0,441) e que
cerca de 56% é devida a diferença entre indivíduos dentro de populações (Tabela 5). O teste
de homogeneidade de variâncias de Bartlett, feito através da relocação aleatória de
indivíduos através das populações, mostrou que a probabilidade de se encontrar pelo acaso
uma heterocedasticidade maior do que a observada é pouco maior que 2%, depois de 1000
permutações (X2=0,597; gl=5; p=0,0239), indicando que as populações apresentam
diferentes níveis de variação genética.
Tabela 5. Análise de variância das distâncias euclidianas entre pares de indivíduos.
Fonte de
Variação GL SQ QM
Componentes
da Variância
Percentagem
do Total Valor de P
Entre Populações 5 206.801 41.360 3.829 44.08% <0.001
Dentro de
Populações 52 252.646 4.859 4.858 55.92% -
φst=0,441

41
Na Tabela 4 são apresentados os resultados das medidas análogas ao Fst obtidos
com os índices de Shannon. A análise de partição da variação usando o índice de
diversidade de Shannon foi extremamente congruente com as estimativas da AMOVA. A
proporção da variação total atribuída à variação dentro das populações foi
aproximadamente 0,56 (Hpop/Hsp) e, portanto, a proporção devida à variação entre
populações ((Hsp-Hpop)/Hsp=0,443) foi 0,44. Esses valores, no entanto, apresentaram
acentuada variação entre iniciadores (Tabela 4), o que ressalta a importância de se utilizar
vários iniciadores para estudos de diversidade e estrutura genética de populações.
Os valores de φst entre pares de populações foram moderados a muito elevados
(0,116 a 0,622) e são apresentados na Tabela 6. Curiosamente, o valor mais baixo
encontrado (0,116) foi entre as populações ES e UFV, sendo que estas estão distantes uma
da outra 187 km, ao passo que UFV e Paraíso se separam por apenas 15 km e apresentaram
um valor de φst semelhante, igual a 0,14. De acordo com esses valores de pares de φst as
populações de Lavras e do Jambreiro são as mais divergentes das demais. A média de
variação dos valores φst de Lavras com as demais populações foi igual a 0,53 e da
população do Jambreiro com as demais foi de 0,59, enquanto que a média dos outros
valores entre pares de φst foi inferior, igual a 0,20.
Tabela 6. Valores de φst por pares de populações. Todos valores de φst entre populações são
estatisticamente significativos com P<0,001
UFV Corredor Paraíso Jambreiro ES Lavras
UFV -
Corredor 0.2475 -
Paraíso 0.1499 0.2406 -
Jambreiro 0.5839 0.5715 0.6120 -
ES 0.1160 0.2407 0.2287 0.6221 -
Lavras 0.5557 0.4853 0.5412 0.5413 0.5534 -

42
O teste exato de Fisher encontrou diferenças nas freqüências dos marcadores RAPD
entre populações, considerando-se todos os 55 marcadores (teste de probabilidade
combinada de Fisher: X2= 211,3020; gl=82; pgeral < 0,0001). Três pares de populações,
entretanto, não apresentaram diferenças significativas nas suas freqüências de marcadores
(Tabela 7), apesar da presença de estruturação significativa, em relação a todos os pares de
populações segundo a AMOVA. Notadamente esses foram os pares de populações que
exibiram os menores valores de φst, UFV-Paraíso, UFV-ES e Paraíso-ES (Tabela 6).
Tabela 7. Matriz de probabilidades combinadas sobre o total dos 55 marcadores RAPD para
os pares de populações, segundo teste exato de Fischer para diferenças nas freqüências de
marcadores.
UFV Corredor Paraíso Jambreiro ES Lavras
UFV -
Corredor 0,0000 -
Paraíso 0,1326 0,0024 -
Jambreiro 0,0000 0,0000 0,0000 -
ES 0,5267 0,0035 0,1110 0,0000 -
Lavras 0,0000 0,0000 0,0000 0,0000 0,0000 -
As diferenças entre indivíduos e populações podem ser visualizadas através do
gráfico dos dois componentes que melhor explicam a variação dos dados, na análise de
componentes principais (PCA) (Figura 5 e Tabela 8). Nessa análise os componentes 1 e 2
explicam apenas 24 e 13% da variação total e apresentam auto-valores de 9.89 e 5,44,
respectivamente. A disposição dos dados no gráfico mostra a clara distinção das populações
de Lavras e do Jambreiro em relação ao segundo fator e a separação destas das demais, em
relação ao primeiro fator. Não se observa, por outro lado, uma separação nítida entre as
outras quatro populações, ES, UFV, Corredor e Paraíso. Portanto, os resultados da análise
de PCA concordam com os valores de φst obtidos através da AMOVA. Os marcadores que
mais influenciaram na separação dos indivíduos e populações (cargas > 0,7) no fator 1
foram dois do iniciador OPA-4, um do OPA-7, três do OPC-16 e um do OPL-1. Em relação
ao fator 2 contribuíram principalmente três marcadores do iniciador OPA-7 (Tabela 8).

43
.
FATOR 1
FA
TO
R 2
-3
-2
-1
0
1
2
-1,5 -1,0 -0,5 0,0 0,5 1,0 1,5 2,0 2,5
UFVCorredorParaísoJambreiroESLavras
Figura 5. Análise de Componentes Principais (PCA) com o conjunto total de bandas RAPD das seis populações estudadas.
Cada eixo representa uma combinação das bandas que melhor explicam as diferenças entre os indivíduos

44
Tabela 8. Cargas dos Fatores (extração dos componentes principais). Aquelas maiores que
0,7 estão em negrito.
Marcadores Fator 1 Fator 2
A2-990 0,338 -0,319
A2-1050 0,104 -0,509
A2-1320 -0,651 -0,002
A4-1170 -0,934 0,0582
A4-1200 0,779 -0,006
A7-600 0,673 0710
A7-690 0,723 -0,541
A7-720 -0,365 0,2586
A7-800 -0,109 0,2450
A7-900 0,3593 0,2668
A7-920 -0,409 -0,776
A7-1000 -0,406 -0,272
A7-1020 -0,334 -0,331
A7-1100 -0,127 -0,408
A7-1200 0,116 -0,332
A7-1400 -0,253 -0,728
A7-1500 -0,104 -0,036
A7-1600 0,293 -0,541
C16-500 0,417 0,454
C16-550 0,815 -0,035
C16-700 -0,971 0,007
C16-720 0,078 -0,317
C16-850 0,943 0,028
C16-1000 0,463 -0,476
C16-1100 -0,092 -0,610
C16-1150 -0,275 0,107
C16-1500 -0,378 -0,493
C16-1550 0,390 -0,356
C16-1950 0,649 -0,246
C16-2000 -0,583 -0,281
C16-2050 -0,225 0,003
L1-450 -0,971 0,007
L1-490 0,485 0,580
L1-600 -0,045 -0,403
L1-710 0,354 -0,498
L1-850 -0,506 0,307
L1-950 -0,111 -0,007
L1-1100 -0,119 -0,567
L1-1200 -0,388 0,190
L1-1400 -0,224 0,087
Variação explicada 90,890 50,443
Proporção Total 0,241 0,132

45
Um fenograma foi realizado por UPGMA, com as matrizes binárias das seis
populações, através da distância de Nei (1972) (Figura 6). A árvore resultante foi refeita por
bootstrap (1000 réplicas). Nenhuma árvore resultante teve entroncamentos com “empates”.
Nessa figura pode-se observar que as populações mais próximas geograficamente (UFV e
Paraíso) agora se agrupam mais fortemente (exibem menor distância genética) e essas se
agrupam com o Corredor e o ES. As populações do Jambreiro e de Lavras permanecem as
mais divergentes de todas.
Figura 6. Fenograma de distância genética de Nei (1972), pelo método UPGMA. Os
números próximos às bifurcações indicam os valores bootstrap (1000 réplicas).
4.4) Relação da variabilidade genética com tamanho de fragmento
A fim de se determinar se há uma relação da área do fragmento com a variabilidade
genética intrapopulacional foram realizadas duas regressões, uma com a variância estimada
pela AMOVA e a outra com o índice de Shannon de diversidade fenotípica (Ho). Nenhuma
das duas análises resultou em regressões estatisticamente significativas (R2= 0,46; P=0,13;
F(1,4)= 3,44 e R2=0,37; F(1,4)=2,34 e P=0,20, respectivamente). De fato, Jambreiro, o maior
fragmento (912 ha) apresentou uma variação genética bem inferior às demais populações e
ES, um dos menores fragmentos, apresentou variação genética semelhante às populações de
Lavras e Paraíso, fragmentos com áreas de 54 e 384 ha, respectivamente. Esses resultados
sugerem que fragmentos relativamente pequenos podem manter variação genética de

46
Akodon cursor e que outros fatores, como por exemplo a migração, devem ser mais
importantes na determinação da quantidade de variação mantida em suas populações. Os
gráficos da análise de regressão podem ser visualizados nas Figuras 7 e 8.
Log 2 Área
Var
iânc
ia (
AM
OV
A)
UFV
Corredor
Mata do Paraíso
Jambreiro
ESLavras
3,0
3,4
3,8
4,2
4,6
5,0
5,4
5,8
2 3 4 5 6 7 8 9 10 11
Figura 7. Regressão entre a variação molecular estimada pela AMOVA e a área (valores
log transformados na base dois) dos fragmentos amostrados (resultados não significativos).

47
Log 2 ÁREA
Índi
ce d
e S
hann
onUFV
Corredor
Mata do Paraíso
Jambreiro
ESLavras
0,22
0,24
0,26
0,28
0,30
0,32
0,34
0,36
0,38
0,40
2 3 4 5 6 7 8 9 10 11
Figura 8. Regressão entre a variação molecular estimada pelo índice de diversidade de
Shannon (Ho) e a área (valores log transformados na base dois) dos fragmentos amostrados
(resultados não significativos).
4.5) Relação da distância geográfica com a divergência genética
Com o objetivo de verificar se a estrutura das populações está de acordo com o
modelo de isolamento por distância (Wright, 1943) foi utilizado o teste de Mantel (Sokal,
1979). As correlações entre os pares de distâncias geográficas e genéticas (distância
genética de Nei (1972) e valores de φst) foram testadas e em nenhuma das duas
comparações se encontrou significância (r = -0,0428 e p(z-aleatório < z observado) = 0,52 e r =
0.00029 e p(z-aleatótio < z observado)= 0,50, respectivamente). Portanto, as populações mais
distantes geograficamente não são mais divergentes entre si do que as populações mais
próximas.
4.6) Relação da variação morfológica com a variação genética
intrapopulacional
Foram tomadas medidas morfológicas externas comumente utilizadas nos estudos
de campo em pequenos mamíferos: comprimento do corpo (cujas médias populacionais

48
variaram de 94,44mm a 115,71mm), comprimento da cauda (com médias variando de
83,12mm a 89,90mm), comprimento da orelha, com médias variando de 15,00mm a
19,40mm e comprimento da pata traseira, vulgarmente chamada “tarso”, com médias
variando de 22,00mm a 25,40mm (Tabela 9). Essas diferenças entre populações, entretanto,
só foram significativas para a variável comprimento da orelha (Tabela 12). Não houve
dimorfismo sexual para as características morfológicas analisadas, conforme apontado por
comparação de intervalos de confiança, a 5% de probabilidade (análises não apresentadas).
Tabela 9. Médias morfológicasdas seis populações analisadas de A. cursor. DP=Desvio
Padrão.
Corpo (mm) Cauda (mm) Tarso (mm) Orelha (mm) População
Média DP Média DP Média DP Média DP
UFV 102,12 9,41 83,12 11,70 22,00 1,85 16,87 0,99
Corredor 96,25 12,17 85,83 7,120 24,41 1,56 17,00 2,00
Paraíso 115,71 25,00 88,28 14,55 23,85 2,19 15,57 1,71
Jambreiro 94,44 10,69 88,33 11,57 23,55 3,04 15,00 3,09
ES 105,28 13,23 86,14 11,82 23,06 2,47 16,14 1,46
Lavras 104,00 11,37 89,80 6,572 25,40 0,547 19,40 1,14
Geral 101,85 15,16 86,55 10,29 23,66 2,26 16,62 2,13
N Total 48 45 49 45
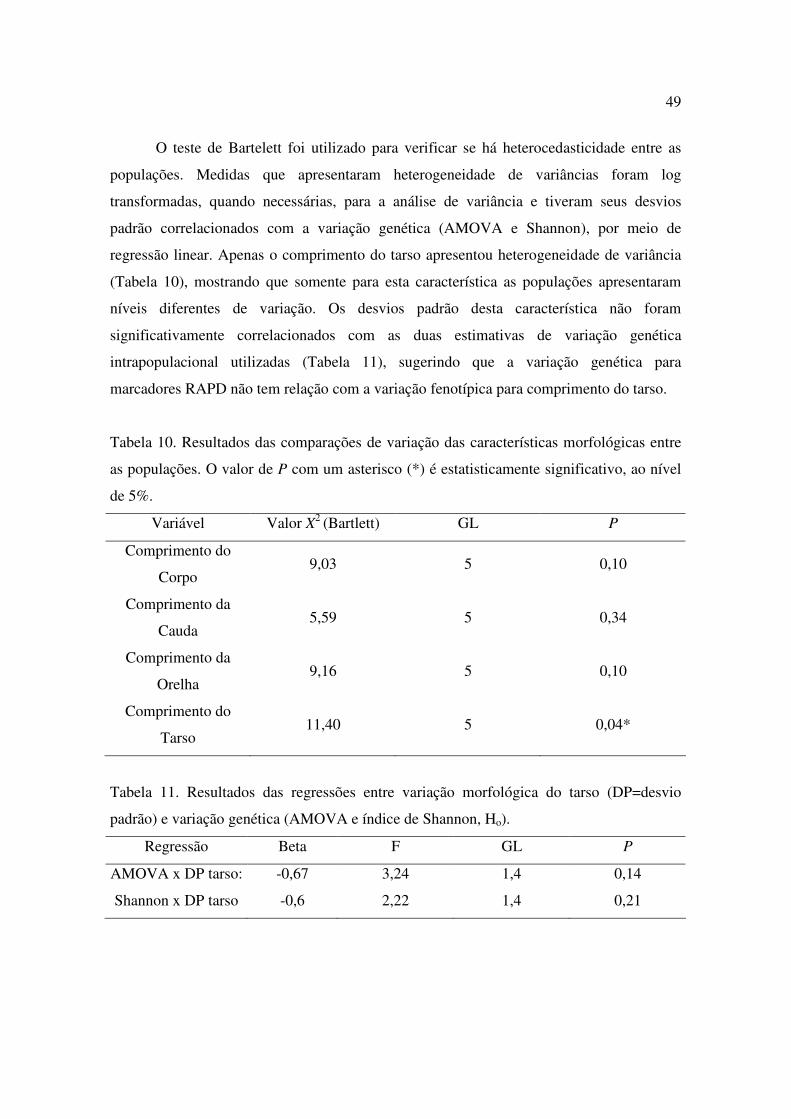
49
O teste de Bartelett foi utilizado para verificar se há heterocedasticidade entre as
populações. Medidas que apresentaram heterogeneidade de variâncias foram log
transformadas, quando necessárias, para a análise de variância e tiveram seus desvios
padrão correlacionados com a variação genética (AMOVA e Shannon), por meio de
regressão linear. Apenas o comprimento do tarso apresentou heterogeneidade de variância
(Tabela 10), mostrando que somente para esta característica as populações apresentaram
níveis diferentes de variação. Os desvios padrão desta característica não foram
significativamente correlacionados com as duas estimativas de variação genética
intrapopulacional utilizadas (Tabela 11), sugerindo que a variação genética para
marcadores RAPD não tem relação com a variação fenotípica para comprimento do tarso.
Tabela 10. Resultados das comparações de variação das características morfológicas entre
as populações. O valor de P com um asterisco (*) é estatisticamente significativo, ao nível
de 5%.
Variável Valor X2 (Bartlett) GL P
Comprimento do
Corpo 9,03 5 0,10
Comprimento da
Cauda 5,59 5 0,34
Comprimento da
Orelha 9,16 5 0,10
Comprimento do
Tarso 11,40 5 0,04*
Tabela 11. Resultados das regressões entre variação morfológica do tarso (DP=desvio
padrão) e variação genética (AMOVA e índice de Shannon, Ho).
Regressão Beta F GL P
AMOVA x DP tarso: -0,67 3,24 1,4 0,14
Shannon x DP tarso -0,6 2,22 1,4 0,21
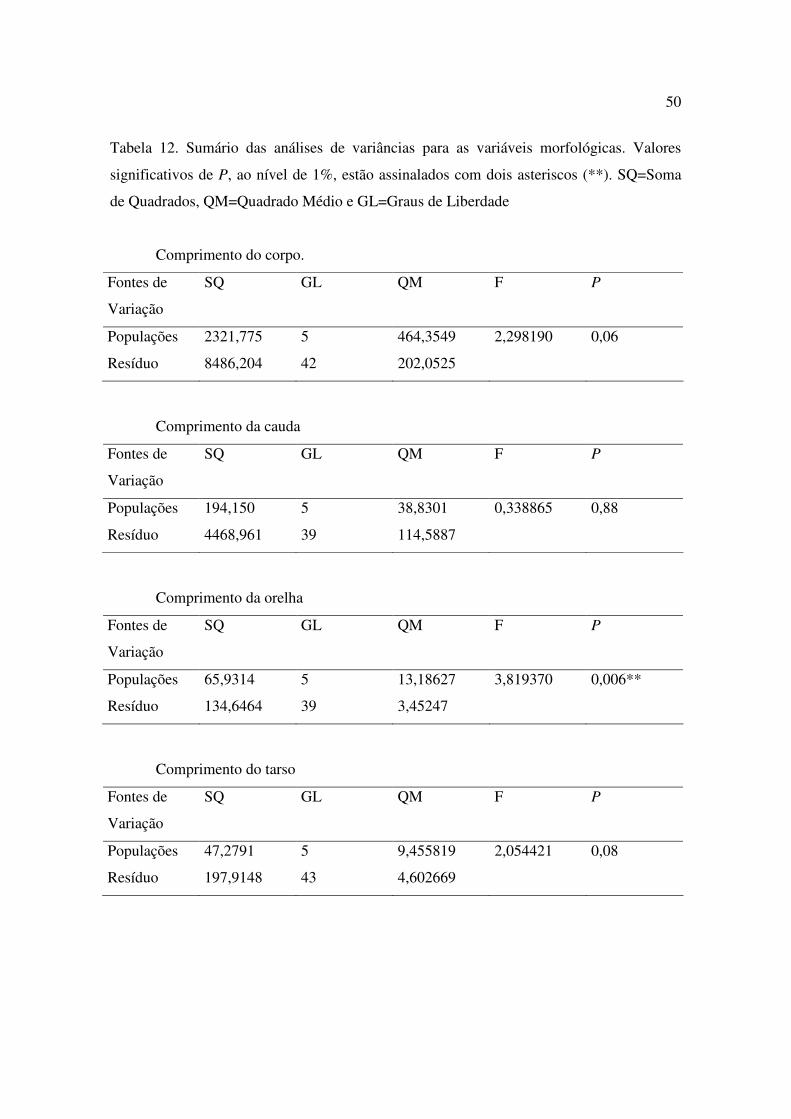
50
Tabela 12. Sumário das análises de variâncias para as variáveis morfológicas. Valores
significativos de P, ao nível de 1%, estão assinalados com dois asteriscos (**). SQ=Soma
de Quadrados, QM=Quadrado Médio e GL=Graus de Liberdade
Comprimento do corpo.
Fontes de
Variação
SQ GL QM F P
Populações 2321,775 5 464,3549 2,298190 0,06
Resíduo 8486,204 42 202,0525
Comprimento da cauda
Fontes de
Variação
SQ GL QM F P
Populações 194,150 5 38,8301 0,338865 0,88
Resíduo 4468,961 39 114,5887
Comprimento da orelha
Fontes de
Variação
SQ GL QM F P
Populações 65,9314 5 13,18627 3,819370 0,006**
Resíduo 134,6464 39 3,45247
Comprimento do tarso
Fontes de
Variação
SQ GL QM F P
Populações 47,2791 5 9,455819 2,054421 0,08
Resíduo 197,9148 43 4,602669

51
5) DISCUSSÃO
5.1) Marcadores RAPD
Neste trabalho, aspectos da genética populacional em Akodon cursor foram
analisados por meio de marcadores RAPD. O método de extração de DNA aqui empregado
resultou em amostras passíveis de boas amplificações com os iniciadores selecionados,
resultando na produção de marcadores de repetibilidade constatável, criteriosamente
escolhidos. A possibilidade da ocorrência de mais de uma espécie na amostra foi controlada
com a eliminação de indivíduos com perfis de amplificação muito díspares dos demais,
visto que A. cursor é morfologicamente indistinguível de A. montensis (Cristoff, 1997),
porém não o é do ponto de vista genético (Rieger et al., 1995), e é muito parecido com
outras espécies do gênero Akodon e mesmo com espécies de outros gêneros, como Bolomys
lasiurus (Fagaundes e Yonenaga-Yassuda, 1998). As motivações para o uso de tais
marcadores foram: i) a possibilidade de se estudar muitos marcadores genéticos
simultaneamente em um intervalo de tempo relativamente curto, com gastos relativamente
baixos, ii) com uma amostra limitada de indivíduos (Stewart e Excoffier, 1996), iii) devido
à sua capacidade de discernir entre espécies próximas e, até mesmo, linhagens dentro de
espécies (Morgan-Richards, 1998; Landry e Lapointe, 1999; Mischra et al., 2002) e, iv)
principalmente, por não depender de caracterização prévia de partes do genoma da espécie
(Ferreira e Grattapaglia, 1997).
O número de marcadores considerados foi igual a 55, sendo que 41 se mostraram
polimórficos. Em uma espécie de pequeno mamífero australiano, o marsupial Isodon
obdeus estudos com RAPD revelaram 39 marcadores polimórficos em três inicadores
(Cooper, 2000), resultado semelhante ao aqui descrito. Em trabalhos empregando
marcadores RAPD na espécie norte-americana de roedor Peromyscus maniculatus, Landry
e Lapointe (1999) selecionaram 45 marcadores (três iniciadores) e Vucetich et al (2001),
selecionaram 12 bandas polimórficas altamente reprodutíveis. Já na espécie de pinipédio
Phoca vitulina, porém, com 25 iniciadores analisados foi encontrada variação em apenas
nove bandas (Kappe et al., 1995). Vucetich e seus colaboradores (2001) argumentam que a
falta de compreensão da influência do número de bandas no poder estatístico, não justifica

52
afirmar que o número reduzido de bandas implique necessariamente em baixo poder de
análise, e que o uso de marcadores de repetibilidade verificável somado à obtenção de
valores significativos de probabilidade, permitem a interpretação parcimoniosa de que os
resultados encontrados devem refletir padrões subjacentes (Vucetich et al., 2001). O fato de
que o número de bandas amostradas em todas populações de A. cursor parece atingir um
limite abaixo do número de indivíduos em cada população, verificado pelo jackknife,
sugere que os tamanhos amostrais são suficientes em relação a essa variável. Segundo Nei
(1978), as variâncias amostrais de estimativas genético-populacionais são mais fortemente
afetadas por um reduzido número de loci do que pelo número de indivíduos. O
comportamento multiplex do RAPD (i.e. a obtenção de vários marcadores em um único
ensaio PCR) o torna particularmente interessante para estudos com pequenos mamíferos,
onde os sucessos de captura são tipicamente baixos. O número de bandas consideradas em
estudos que usam essa metodologia irá variar muito em função do número de inicadores
triados e dos critérios de seleção de marcadores, o que costuma em implicar em limitações
para a comparação de resultados entre diferentes estudos com esse marcador (Sunnucks,
2001). Em uma revisão sobre uso de RAPDs em estudos genético-populacionais de plantas,
Nybom e Bartish (2000) mostraram que apenas estudos com menos de cinco indivíduos por
população apresentaram alterações importantes nas estimativas de diversidade entre
populações.
5.2) Variabilidade intrapopulacional
Nos estudos de Biologia de Populações, a mensuração da variabilidade genética tem
sido empregada como meio de acessar-se a “saúde” genética de uma população (Fristch et
al., 1993), e essa tem sido encarada como variável central para a Biologia da Conservação
(Lande, 1988; Amos, 1999; Lande 1999; Amos e Balmford, 2001). O nível de variabilidade
genética exibida por uma população qualquer é o saldo entre eventos que geram e mantêm
variação genética e eventos que levam à perda de variabilidade (Amos e Harwood, 1998).
A mutação é a fonte primordial de variação e é um processo que acontece desvinculado das
pressões ambientais. Assim também o é o fenômeno da recombinação, outro promotor da
variabilidade. A entrada de novos indivíduos em uma população (imigração) é outro fator

53
que pode contribuir para o aumento da variabilidade genética, pela introdução de novos
alelos anteriormente ausentes ali, ou pelo aumento da freqüência de um alelo que antes da
imigração era raro. Alguns tipos de seleção natural, como a seleção a favor do heterozigoto
(Futuyma, 1993), seleção dependente da freqüência (Futuyma, 1993) e a seleção em
ambiente heterogêneo (Hedrick, 2000), também podem contribuir para a manutenção da
variabilidade no(s) locus(i) em questão. Por outro lado, geralmente, a seleção natural para
adaptações locais desgasta os níveis de variabilidade (Frankhan, 1996). Os efeitos da perda
de variabilidade são sentidos pela população ao longo de várias gerações (Harlt, 1988 apud
Amos, 1999) e são justamente relativas as deficiências adaptativas às possíveis situações
ambientais adversas, como a falta de variabilidade em um locus para resistência a uma
determinada patologia (Sommers et al., 2002).
Em todas as três mensurações da variabilidade genética dentro de cada uma das
populações analisadas (AMOVA, índice de diversidade de Shannon e percentagem de
marcadores polimórficos), a população do Jambreiro, em Nova Lima, MG, foi a que
apresentou a menor variabilidade. Na hipótese de haver um número suficientemente grande
de indivíduos nessa população para evitar os efeitos da deriva genética (Lande, 1988), uma
explicação plausível seria a ocorrência de adaptações locais, em conjunto com uma taxa
baixa de ingresso de indivíduos nessa população. Isto porque, para muitas destas
adaptações, a seleção natural tende a erodir a variabilidade, regulando a freqüência dos
alelos envolvidos nessas características, e a não entrada de novos indivíduos na população
evitaria o efeito de carga migratória, que causaria aumento da variabilidade
intrapopulacional, mas atuaria exercendo uma depressão exogâmica na população
(Lenormand, 2002). A provável neutralidade dos marcadores RAPD, entretanto, torna essa
explicação menos atraente. A baixa variabilidade genética encontrada na população
Jambreiro poderia ser devida a um tamanho populacional reduzido durante várias gerações
e/ou a maior dificuldade de entrada de migrantes nessa população, sob a ótica da variação
RAPD ser predominantemente neutra. O fragmento do Jambreiro está situado dentro da
região metropolitana de Belo Horizonte, tendo em grande parte de seu perímetro áreas de
mineração da MBR e áreas de ocupação antrópica. Esse entorno, potencialmente, pode
impedir ou dificultar as migrações. Portanto, parece que esta população está mais isolada de
outros fragmentos do que as outras populações aqui analisadas. De fato, a grande

54
divergência entre essa população e as demais, estimada pelos valores de φst e pela análise
de componentes principais favorecem essa hipótese. Apesar de aspectos da demografia de
Akodon cursor não terem sido aqui analisados, isso não impede de se fazer uma constatação
sobre a aparente baixa densidade populacional dessa espécie na mata do Jambreiro: no
intervalo de um ano de coleta capturou-se quase que o mesmo número de indivíduos em
apenas uma e duas sessões de captura, no ES e em Lavras, respectivamente (resultados não
apresentados). Populações de pequenos mamíferos, notadamente de alguns roedores,
experimentam ciclos de alternância de densidades populacionais, algumas vezes muito
complexos, com padrões pluri-anuais (Fonseca, com. pess.), e A. cursor poderia apresentar
estes padrões, alternando bruscamente os tamanhos populacionais. Populações com
variação no tamanho populacional entre gerações têm um tamanho efetivo que é a média
harmônica dos tamanhos de cada geração, o que significa que o tamanho efetivo médio será
mais próximo do menor tamanho populacional (Harlt e Clark, 1997). Como os efeitos de
deriva genética são proporcionais ao tamanho efetivo, a ocorrência dessas flutuações
populacionais pode estar levando à perda de variabilidade nessa população, que não seria
restaurada devido ao seu provável isolamento.
Os efeitos da endogamia devido ao pequeno tamanho efetivo populacional poderiam
ser minimizados pelos mecanismos de prevenção de endogamia (i.e. mecanismos que
permitem indivíduos proximamente aparentados de se reconhecerem e conseqüentemente
evitar acasalar-se (Pusey e Wolf, 1996)) e que operam em algumas espécies de roedores.
Em várias espécies de animais têm sido documentados mecanismos, em diferentes níveis,
pelos quais, indivíduos acabam evitando acasalamentos endogâmicos. Uma forma de se
evitar acasalamentos aparentados em pequenos mamíferos é dispersar do local de
nascimento em idade reprodutiva. A mata do Jambreiro tem uma dimensão que permite
facilmente que os indivíduos se distribuam separadamente dentro da área. Em
camundongos de laboratório foi demonstrada a capacidade de reconhecimento de parentes
por meio semio-químicos secretados na urina, o que talvez impeça o cruzamento entre esses
indivíduos (Pusey e Wolf, 1996).
Assim como aconteceu com as medidas de variabilidade intrapopulacional na
população do Jambreiro, os resultados da alta variabilidade na população da UFV foram
concordantes entre diferentes métodos. A área da UFV está situada em um local com vários

55
fragmentos de mata proximamente localizados e muitas vezes separados por capim, ou com
pequenos obstáculos facilmente transponíveis por roedores, como estradas de terra estreitas
(observação pessoal). Uma particularidade dessa área, em relação a todas as outras é o fato
desse local se tratar de um campo de gramíneas, adjacente a um fragmento e de ter sido
completamente removido pelo fogo, deixando o solo nu (Goretti e De Marco, dados não-
publicados). O fato desta área ter sido destruída em um incêndio há dois anos e ter sido
gradualmente modificada pela sucessão pode ter induzido um efeito de “dreno” na área: a
partir do momento em que as condições do local se tornaram favoráveis à colonização por
Akodon cursor, o local pode ter se comportado como um ponto de convergência de
estabelecimentos bem sucedidos, de indivíduos oriundos de diferentes “fontes” adjacentes.
Isso pode ter ocasionado uma contribuição efetiva de indivíduos geneticamente diversos na
formação dessa população, em contraste com populações já estabelecidas. Em algumas
populações de pequenos mamíferos, a entrada de novos indivíduos sofre uma maior
resistência exercida pela competição direta com os animais residentes (Fernadez et al,
1999), como por exemplo, por parceiros reprodutivos, espaço para estabelecimento de área
de vida, território, e sítios de aninhamento. Poderia se esperar, portanto, que o conjunto
genético-populacional de uma população já estabelecida conte, possivelmente, com uma
contribuição genética diversa diminuída em função da maior restrição à entrada de novos
indivíduos. Essa é uma possível explicação para a relativa alta variabilidade dentro da
população da UFV, acima do nível observado em outras localidades aqui amostradas, com
a mesma característica de estarem diretamente acessíveis para indivíduos migrantes de
diferentes localidades, que é o caso de três fragmentos analisados, Paraíso, Lavras e ES.
Essas três populações apresentam níveis de variações genética semelhantes e bem
superiores à do Jambreiro. Essas localidades estão inseridas em paisagens altamente
fragmentadas com a ocorrência de diversos fragmentos de vários tamanhos, muitas vezes
separados por campos de gramíneas. Os campos de gramíneas associados a fragmentos são
ambientes propícios à espécie aqui estudada (Pires et al., 2001, Paglia et al, 1995,
Fernadez, comunicação pessoal, observação pessoal). Em estudo de movimentação de
espécies de pequenos mamíferos entre fragmentos proximamente localizados, e separados
por matriz campo aberto de gramíneas, (realizado nas chamadas “Ilhas dos Barbados”, na
reserva biológica Poço das Antas, RJ), Pires e colaboradores (2002), mostraram que

56
existem diferenças nos padrões de movimentação fragmento-fragmento/fragmento-matriz,
entre espécies, mesmo que proximamente relacionadas. No intervalo de tempo
compreendido pelo estudo evidenciou-se apenas a movimentação de uma fêmea adulta de
A. cursor entre fragmentos, separados pela distância mínima de 675m. Entretanto foi
observado um maior número de indivíduos, machos e fêmeas adultos, se locomovendo
entre fragmentos florestais e áreas de coleta situadas na matriz de gramíneas. As conclusões
desse trabalho são que essa espécie, provavelmente exibe as características de uma única
grande população na área considerada, uma vez que essa matriz presente no local não
restringe a dispersão de Akodon cursor entre os fragmentos, apesar da natureza
relativamente sedentária dessa espécie (Gentile et al., 2000; Pires et al.,2002; Yazbeck e De
Marco, dados não publicados).
A população do Corredor também apresentou um nível relativamente alto de
variação genética. A mata de galeria em Bocaiúva, onde essa população foi coletada, está
inserida dentro de uma reserva de cerrado, cercada por extensas plantações de eucaliptos.
Em um estudo de comunidade de pequenos mamíferos, realizado em matriz de eucaliptal,
com presença de sub-bosque (Fonseca, 1997), não foram encontradas diferenças na
composição da comunidade de roedores entre fragmento de mata e a matriz, sugerindo que
as espécies estudadas não seriam restringidas diante da existência de tais alterações
antropogênicas no entorno de fragmentos. Dessa forma, é plausível assumir que os
eucaliptais da fazenda Corredor não impeçam o processo migratório de A. cursor nessa
área, permitindo o intercâmbio de indivíduos com populações locais, mantendo o nível da
variabilidade intrapopulacional.
5.3) Estrutura Genética de Populações
Os resultados da estrutura de populações em A. cursor aqui estudadas, mostram que
há uma subdivisão e diferenciação marcante entre as populações, corroborada por métodos
independentes, a AMOVA que estimou um valor de φst = 0,441 e o índice de Shannon cujo
valor de (Hsp-Hpop)/Hsp=0,443. Variações intrapopulacionais superiores às
interpopulacionais são comumente encontradas na literatura, usando-se marcadores RAPD
em mamíferos (e.g., Antolin et al. 2001, Cooper, 2000; Landry e Lapointe,1999).

57
Estimativas de Fst muito altas, porém, são mais inconspícuas, embora ocorram entre
populações fortemente isoladas (Vucetich et al., 2001). Uma hipótese a ser levantada em
relação à esse resultado é se ele não é o reflexo dos tamanhos amostrais reduzidos. Nybom
e Bartish (2000) argumentam que em plantas não se encontram influências fortes de
número de indivíduos nas estimativas de variabilidade interpopulacionais, a partir de 10
indivíduos, com marcadores RAPD. Isso sugere que possivelmente os resultados aqui
obtidos não sejam artefatos causados pelo tamanho amostral.
Os resultados de estruturação de populações devem refletir, entre outras variáveis, a
amplitude espacial de abrangência dos indivíduos amostrados. Apesar de não ter havido
uma correlação significativa entre os pares de distâncias geográficas e de distância
genética, nesse trabalho as distâncias geográficas abrangidas são, em média, muito maiores
do que a capacidade de dispersão a longas distâncias para estes roedores. Considerando que
a área de vida de Akodon cursor é bem limitada (Cerqueira et al. 2000, Yazbeck e De
Marco, dados não publicados), e sua capacidade de dispersão é obviamente restrita em
relação a animais alados, estruturação e diferenciação genética grande, na escala espacial
deste trabalho, são esperadas (Ditchfield, 2000). Geise e colaboradores (2001), também
evidenciaram uma estruturação genética bem acentuada em populações de Akodon cursor, e
atribuíram esse fato à baixa vagilidade dessa espécie. A falta de movimentação expressiva
desses animais poderia estar proporcionando o distanciamento genético das populações
estudadas, mesmo quando se considerando as duas localidades mais próximas (UFV-
Paraíso), pela deficiência de migrantes entre as localidades, fator que pode interromper a
continuidade genética entre populações (Antolin et al., 2001). Os menores valores de φst
entre pares de populações foram entre as populações da UFV e ES, seguido de UFV e Mata
do Paraíso. As duas últimas localidades são as mais próximas dentro deste estudo, estando
separadas por cerca de 15 km, no entanto apresentam uma estruturação significativa
(φst=0,1499) pela AMOVA, mas não pelo teste exato de Fisher. Isso pode ser o reflexo de
um fluxo gênico, que poderia estar ocorrendo de forma indireta entre essas localidades, por
exemplo, por sistema de alpondras, onde indivíduos não se dispersam diretamente de uma
população para outra, mas estabelecem fluxo gênico por meio de dispersão de indivíduos
entre populações adjacentes, intermediárias às áreas consideradas (Futuyma, 1993). Em um
estudo com as formas do gênero de Akodontíneos Oxymycterus, através da análise de

58
caracteres morfométricos e por seqüênciamento do citocromo-b, Gonçalves (2001) concluiu
que animais do estado de Espírito Santo, coletados na mesma localidade da população de
Akodon cursor aqui analisadas, apresentavam menor divergência com uma forma ocorrente
em Viçosa do que com as amostras de outras localidades estudadas.
As médias dos valores de pares de φst mostram claramente o maior distanciamento
genético das populações do Jambreiro (0,59) e Lavras (0,53), em relação às demais. O
isolamento relativo das populações entre si e a manutenção de baixos tamanhos
populacionais, poderiam estar levando a uma diferenciação dessas populações em virtude
da deriva genética e falta de migração entre as mesmas. Essa explicação parece se aplicar
mais rigorosamente à população do Jambreiro, que se encontra inserida dentro de uma área
majoritariamente urbana e de atividade mineradora, que ocasionaria maior isolamento a
este fragmento. A falta de conhecimentos detalhados sobre a ecologia deste animal
dificulta o esclarecimento de potenciais barreiras que estariam levando ao provável maior
isolamento da população de Lavras, em relação às outras populações.
A fixação de tipos cariotípicos distintos entre localidades poderia refletir na elevada
diferenciação observada entre as populações, uma vez que os polimorfismos RAPD podem
ser marcadores diretos de certos tipos cromossômicos (Tosta et al., 2000). A espécie A.
cursor tem o maior polimorfismo cariotípico já descrito para mamíferos, tendo esses
polimorfismos distribuídos entre, e até mesmo, dentro de populações. Dado a presença de
vários pares heterozigotos no complemento, e a variação numérica encontrada nesses
animais, existe um total de 81 tipos citótipos esperados. Desses, apenas 31 são
caracterizados na literatura (Cristoff, 1997). As formas alternativas de cariótipos podem ser
fixadas em localidades distintas, o que poderia estar contribuindo para um aumento na
estimativa de φst entre as populações estudadas, apesar da falta de estudos citogenéticos dos
indivíduos analisados.
Uma outra hipótese para explicar os valores elevados de variabilidade
interpopulacional é a possibilidade de ocorrência de mais de uma espécie entre os
indivíduos considerados nas análises. Essa hipótese é verificável se for possível obter
indivíduos cariotipados das diversas localidades estudadas, o que não ocorreu nesse
trabalho. A. cursor tiveram seus perfis de amplificação RAPD comparados com
akodontíneos disponíveis no estudo e a literatura foi consultada para verificação de

59
diagnósticos de animais descritos como 2n=14, para as localidades de Viçosa (Geise et al.,
1998; Gonçalves, comunicação pessoal) e do ES (Cristoff, 1997), além de Lagoa Santa,
localidade próxima a Nova Lima (Jambreiro). Além disso, um animal da amostra do ES
teve parte de seu DNA mitocondrial seqüenciado, e diretamente relacionado, em
comparação com seqüências depositadas no GeneBank, à A. cursor (Gonçalves,
comunicação pessoal).
O maior distanciamento das populações de Lavras e do Jambreiro, pode ser
corroborado tanto pela análise de componentes principais (PCA) (Figura 5), quanto pelo
dendrograma do UPGMA realizado com a distância genética de Nei (1972) (Figura 6). Na
hipótese da presença de mais de uma espécie na amostra, essas duas populações poderiam
formar um grupo distinto. Na PCA, o conjunto de variáveis genéticas estudadas (os 41
marcadores RAPD polimórficos) é reduzido a duas novas variáveis que melhor explicam a
variação entre observações. Dessa forma pode-se verificar os marcadores que mais
influenciaram na diferenciação das populações. Observando-se a tabela de cargas de
fatores, que são correlações (variando de –1 a +1), entre cada banda e cada fator (a nova
“variável resumida”), pode-se verificar se é a presença ou a ausência de determinada banda
(observando-se os sinais positivo e negativo, respectivamente) que mais fortemente
contribuem para as diferenças entre indivíduos e populações. No gráfico da PCA (Figura 5)
as populações de Lavras e do Jambreiro se separaram das demais populações em relação ao
primeiro fator, e entre si em relação ao segundo fator. Uma análise cuidadosa dos
marcadores de maior carga de fator (módulo maior que 0,7) paralelamente a uma análise
citogenética, talvez pudesse revelar marcadores específicos relacionados a certos tipos
cromossômicos. Futuros trabalhos nessa linha ajudariam a elucidar a relação da
variabilidade cariotípica e a variabilidade molecular nesta espécie.
O teste exato de Fisher tem sido mais freqüentemente aplicado a marcadores
genotípicos como RFLP, VNTRs (Guo e Thompson, 1992). Aqui sua aplicação se dá por
meio da comparação da freqüência de marcadores RAPD binários, entre as populações por
aproximação Monte-Carlo e informa se existe diferença de freqüência da presença/ausência
através do conjunto de marcadores polimórficos (Raymond e Rosset, 1995). Os resultados
das análises aqui realizadas mostram que existem diferenças significativas entre todos os
pares de populações (o que é de se esperar a priori, considerando-se os resultados da

60
AMOVA), com exceção dos pares UFV-Paraíso (separadas por cerca de apenas 15 km) e
UFV-ES e ES-Paraíso (as populações que exibiram o menor valor de φst entre pares de
populações). Os resultados do Teste Exato reforçam os resultados da AMOVA, por não
mostrar diferenças nas freqüências de marcadores, justamente entre os pares de populações
que exibiram diferenciação baixa a moderada (critérios de Wright), na AMOVA. O
resultado do Teste Exato foi, no entanto, feito através de todos os marcadores polimórficos
e não implica que existam sub-conjuntos de marcadores com diferenças estatísticas em suas
freqüências entre esses pares de populações. Essas diferenças podem não ser o bastante
para indicar diferenças no Teste Exato, porém, determinar as diferenças significativas no
AMOVA.
A distância genética de Nei (1972) separa as populações de Lavras e Jambreiro das
demais, com suporte bootstrap de100%, e mais uma vez confirma um maior distanciamento
genético dessas populações em relação às outras. Dos demais agrupamentos, o que engloba
a população do Corredor e as duas populações de Viçosa é o que tem menor suporte
bootstrap (55,4%). Nessa análise, as duas populações de Viçosa se agregam mais
fortemente do que nas estimativas de φst entre pares de populações. É pertinente lembrar
que a distância genética de Nei (1972) depende das estimativas das freqüências alélicas,
que são realizadas assumindo-se o equilíbrio de Hardy-Weinberg nas populações
amostradas, o que pode não estar ocorrendo em algumas ou mesmo em todas as
populações. Cristoff (1997) constatou a existência de equilíbrio de Hardy-Weinberg para
pares de cromossomos heterozigotos em algumas populações de A. cursor no estado de São
Paulo, o que não torna esse pressuposto assumido tão improvável. Esses resultados
sugerem que, de fato as duas populações de Viçosas devem manter, de alguma forma, um
certo nível de fluxo gênico. Isso é plausível, dado que as duas áreas se separam
principalmente por uma paisagem de mosaico de fragmentos florestais/campos de
gramíneas e propriedades rurais, ambientes propícios ou ao menos toleráveis para essa
espécie de roedor (Pires et al. 2002).

61
5.4) Relação da variabilidade genética com tamanho de fragmento
A falta de correlação entre duas medidas de variabilidade intrapopulacional e as
medidas logaritmizadas das áreas dos fragmentos, aqui estudados (Figuras 7 e 8), pode ser
um indicativo da maior importância da distribuição espacial das populações de A. cursor
em fragmentos/matrizes próximos, capazes de efetivarem migrações, na determinação dos
níveis intrapopulacionais de variabilidade, do que o tamanho de fragmento florestal,
isoladamente. Outros fatores poderiam estar mascarando uma eventual correlação entre
essas variáveis, como por exemplo, a qualidade ambiental das diferentes localidades para
essa espécie, ou a estrutura das comunidades de pequenos mamíferos em cada local. A
presença de certas espécies que poderiam competir diretamente com o roedor aqui
estudado, poderia estar mantendo as populações dos mesmos em baixas densidades
demográficas, por exemplo, na Mata do Jambreiro, uma vez que o estabelecimento de
espécies mais competitivas poderia influir na capacidade suporte de A. cursor (Goretti e De
Marco, dados não publicados). Essa explicação seria compatível com o cenário da área em
regeneração da UFV, onde espécies com melhores características colonizadoras (maior taxa
intrínseca de crescimento, hábitos de dieta generalista, resistência a variações térmicas
acentuadas, etc), que parece ser o caso de A. cursor, poderiam manter maiores densidades
populacionais, antes do avanço da sucessão ecológica, quando as condições passam a ser
favoráveis a melhores competidores e promovem a diminuição ou mesmo a exclusão de
espécies de estágios mais precoces da sucessão.
A abordagem demográfica possivelmente seria mais apropriada para se avaliar o
efeito da fragmentação na variação genética, do que tamanhos de fragmentos. Alguns
trabalhos (e.g Wauters et al., 1994) têm encontrado uma forte relação entre tamanhos
populacionais e a variação genética, enquanto outros encontram essa relação ou não
dependendo dos marcadores usados (e.g. Loeschcke et al. 1994; Pertoldi et al., 2001). A
falta de informações adicionais sobre estrutura espacial e demográfica em A. cursor e as
poucas publicações discutindo seus padrões de dispersão dificultam a elaboração de
hipóteses explicativas para os resultados discutidos nesse trabalho.

62
5.5) Relação distância geográfica com a divergência genética
Neste trabalho não foram encontradas correlações significativas entre medidas de
divergência genética interpopulacional e distâncias geográficas . Esses resultados estão de
acordo com aqueles descritos no estudo realizado por Geise e colaboradores (2001), com
haplótipos de seqüências do gene mitocondrial para citocromo-b, onde se mostra que o
padrão espacial de diferenciação genética em Akodon cursor não segue uma tendência de
isolamento por distância. No trabalho desses autores, animais de localidades mais
próximas, nem sempre evidentemente isoladas por barreiras geográficas, apresentaram
diferenças maiores na seqüência de DNA do que animais mais distantes geograficamente.
O mesmo padrão não foi observado na espécie irmã de Akodon cursor, A. montensis, que
mostrou uma relação maior entre estruturação genética e distância geográfica. Esses autores
atribuem essas diferenças possivelmente a diferenças no status da distribuição geográfica
das espécies: A. cursor estaria sujeito à expansão geográfica, enquanto que as populações
de A. montensis teriam sua distribuição geográfica mais coincidente com a área de
distribuição original das espécies Akodontíneas ancestrais, seriam mais antigas e estáveis, e
estariam, portanto, sujeitas ao isolamento por distância, por exibirem equilíbrio fluxo
gênico/deriva genética (Geise et al., 2001). Os resultados aqui obtidos com A. cursor estão
de acordo com a hipótese proposta por Geise e colaboradores (2001).
5.6) Relação da variação morfológica com a variação genética
intrapopulacional
As variáveis morfológicas foram analisadas por meio de ANOVA, para detectar
diferenças entre populações e o teste de Bartlett foi aplicado para averiguar se as
populações variavam de forma diferente. As análises foram realizadas com todos
indivíduos em conjunto, uma vez que nenhuma variável morfológica apresentou
dimorfismo sexual. A única variável que apresentou heterocedasticidade (comprimento do
tarso) teve seus desvios padrão submetidos a regressão com duas estimativas de
variabilidade intrapopulacional, porém em nenhuma das duas houve uma correlação
significativa. A ausência de associação entre as variações morfológicas e genéticas é um
fato conspícuo em Akodon cursor, e mesmo a nível supra-específico. Tanto assim o é que

63
A. cursor possui uma espécie irmã morfologicamente indistinguível (Akodon montensis),
que possui um número de cromossomos muito maior que o seu. Apesar da grande variação
de cariótipo encontrada entre e dentro das populações de A. cursor, geralmente não existe
diferenciação morfológica entre diferentes populações (Cristoff, 1997). As características
morfológicas (caracteres quantitativos) são influenciadas por vários loci gênicos e, também,
pelas condições ambientais do desenvolvimento dos indivíduos, como por exemplo,
quantidade de recurso disponível no ambiente. O nível de polimorfismo molecular aqui
encontrado e a virtual falta de diferenciação morfológica nessa espécie são fatos curiosos, e
estudos da herdabilidade dessas características seriam necessários para ajudar na
compreensão dessa aparente falta de associação. Na espécie de pequeno mamífero
australiano Isodon obedeus, foi demonstrado com sucesso a associação entre variação
molecular, por RAPD, e diferenças morfológicas associadas a tipos de ambiente ocupados
pela espécie. No entanto, outros autores (Reed e Frankham, 2001) demonstram com dados
publicados na literatura, que a variação de caracteres quantitativos, principalmente daquelas
cujas importâncias são evidentes na ecologia e na história de vida, não se correlaciona com
a variabilidade de marcadores moleculares de forma expressiva.
5.7) Considerações Finais
Os resultados aqui apresentados mostram que a espécie de roedor da fauna
brasileira, Akodon cursor é um animal peculiar do ponto de vista genético-populacional,
uma vez que apresenta uma marcante estruturação genética, superior às encontradas na
literatura para outros pequenos mamíferos. Essa espécie, que é associada a florestas
ombrófilas de encostas e matas de galeria do cerrado no sudeste brasileiro parece sofrer
com a redução de áreas florestais, pelo grau de isolamento destes fragmentos. Alguns
resultados aqui discutidos sugerem que certas perturbações ambientais poderiam estar
promovendo, pelo menos momentaneamente, o aumento da variabilidade local, sob a ótica
dessa espécie ser uma colonizadora competente em relação às abundantes espécies de
roedores desses ambientes.
Os estudos citogenéticos de indivíduos, em conjunto com estudos moleculares,
poderiam abrir novos horizontes na compreensão da relação dos diversos cariótipos

64
encontrados na espécie com a diversidade genética. Da mesma forma, a elucidação mais
profunda de aspectos da ecologia populacional em A. cursor, poderia esclarecer quais são
os fatores preponderantes na manutenção da variabilidade nesse roedor. O entendimento de
tais determinantes poderia lançar uma luz nos modelos de conservação de populações
naturais, possivelmente em outras espécies cujas variáveis ecológicas tenham aspectos em
comum com esse pequeno mamífero.
Do ponto de vista da conservação da diversidade genética nessa espécie, a maneira
mais efetiva de se manter a variabilidade seria com a conservação de populações diversas,
através do raio de distribuição geográfica da mesma, uma vez que grande parte desta
variação está distribuída entre populações distintas. A melhor maneira de se fazer isso seria
a manutenção de populações uniformemente distribuídas, accessíveis a migração. A
manutenção dos fragmentos restantes de Mata Atlântica e meios de viabilizar a dispersão
(que em Akodon cursor, não implica necessariamente na existência de corredores
ecológicos, sendo suficiente a presença de campos de gramíneas entre fragmentos), para
essa espécie parece ser de fundamental importância na manutenção da variabilidade dentro
de suas populações.

65
6 CONCLUSÕES
Os estudos genético-populacionais aqui realizados com as populações da espécie de roedor
silvestre Akodon cursor, por meio de marcadores moleculares do tipo RAPD, permitem
chegar às seguintes conclusões:
• As populações apresentaram uma marcante estruturação, com grande parte da
variação genética total (44%) sendo distribuída entre populações.
• A população do Jambreiro apresenta variabilidade intrapopulacional reduzida em
relação às outras populações e elevada divergência das mesmas, possivelmente
devido a um maior isolamento dessa área, que se encontra inserida dentro de uma
paisagem altamente urbanizada e impactada por atividades de mineração.
• A população da UFV tem a maior variabilidade genética intrapopulacional, dentre
as populações analisadas e essa alta variabilidade pode ser devido a uma recente
colonização da área por indivíduos de diferentes populações próximas.
• O tamanho de fragmento florestal não parece ser o fator preponderante na
determinação da variabilidade genética em Akodon cursor. O fator mais importante
parece ser a capacidade das populações de receberem indivíduos de áreas diversas.
• Para se conservar a variabilidade genética nessa espécie a melhor estratégia seria a
conservação do maior número de populações distintas, dado que grande parte da
variabilidade genética está distribuída entre as populações.
• A variabilidade molecular estimada nesse trabalho pode estar diretamente
relacionada ao grande polimorfismo cariotípico dessa espécie. Estudos utilizando
marcadores moleculares, como o RAPD, realizados concomitantemente com
estudos citogenéticos são desejáveis para o esclarecimento dessa eventual relação.

66
7 BIBLIOGRAFIA Aars, J. e Ims, R.A. (2000). Population dynamic and genetic consequences of spatial
density-dependent dispersal in patchy populations. American Naturalist. 155(2): 252-265.
Amos,W. (1999). Two problems with the measurement of genetic diversity and genetic
distance. in Genetics and extinction of species. Princeton, West Sussex: 75-100. ______; e Balmford, A. (2001). When does conservation genetics matters? Heredity 87.
257-265.
______; e Harwood, J. (1998). Factors affecting levels of genetic diversity in natural populations. Philosophical transaction: Royal Society of London. 353: 177-186.
Antolin, M.F., Van Horne, B., Berger Jr., M.D., Holloway, A.K., Roach, J.L. e Weeks, Jr., R.D. (2001). Effective population size and the genetic structure of a piute ground squirrel (Spermophilus mollis) population. Canadian Journal of Zoology 79: 26-34.
Antonovics, J. (1994). Ecological genetics of metapopulations: the Silene-Ustilago plant-pathogen system. in Ecological Genetics. Princeton, West Sussex: 146-170.
Backeljau T., De Bruyn, L., De Wolf, H., Jordaens, K., Van Dongen S. e Winnepenninckx, B. (1996). Multiple UPGMA and neighbor-joining trees, and the performance of some computer packages. Molecular biology and evolution 13(2): 309-313.
Brito, B. e Fernandez, F.A.S. (2000). Dealing with extinction is forever: understanding
the risks faced by small populations. Ciência e Cultura 52(3). 161-169. Cavalli-Sforza, L.L. (1988). The DNA revolution in population genetics. Trends in
Genetics. 14(2): 60-65. Cerqueira, R., Gentile, R., Fernandez, F.A.S. e D'Andrea, P.S. (1993). A five-year
population study of an assemblage of small mammals in southeastern Brazil. Mammalia, 57:5 07-517.
Clarke, M.G. e O’Dwyer, C. (2000). Genetic variability and population structure of the
endangered olden sun moth, Synemon plana. Conservation Biology 92. 371-381.

67
Cooper, M.L. (2000). Random amplified polymorphic DNA analysis of southern brown bandicoot (Isoodon obesulus) populations in Western Australia reveals genetic differentiation related to environmental variables. Molecular Ecology 9: 469-479.
Cristoff, A.U. (1997). Contribuição à sistemática das espécies do gênero Akodon
(Rodentia: Sigmodontinae) do leste do Brasil: estudos anatômicos, citogenéticos e de distribuição geográfica. Tese de Doutorado, São Paulo, Departamento de Biologia, Instituto de Biociências, USP: 163pp.
______; Fagundes, V.; Sbalqueiro, I.J.; Mattevi, M.S. e Yonenaga-Yassuda, Y. (2000).
Description of a new species of Akodon (Rodentia: Sigmodontinae) from southern of Brazil. Journal of Mammalogy 81(3): 838-851.
Crnokrak, P.e. Roff, D.A. (1999). Inbreeding depression in the wild. Heredity 83: 260-
270. De Clercq, P., Vandewalle, M. e Tirry, L. (1998). Impact of inbreeding on performance
of the predator Podisus maculiventris. BioControl 43. 299-310. Dean, W., (1995). With broadaxe and firebrand: the destruction of the Brazilian Atlantic
forest. University of Califormia Press, Berkley, 197p. Ditchfield, A.D. (2000). The comparative phylogeography of neotropical mammals:
patterns of intraspecific mitochondrial DNA variation among bats, contrasted with nonvolant small mammals. Molecular Ecology 9: 1307-1318.
Edwards, S.V. e Hedrick, P.W. (1998). Evolution and ecology of MHC molecules: from
genomics to sexual selection. Trends in Ecology and Evolution. 13(8): 305-311. Elliot, K.J., Hendrick, L.R., Major, A.E., Vose, J.M. e Swank, W.T. (1999). Vegetation
dynamics after a prescribed fire in the southern Appalachians. Forest Ecology and
Management. 114. 199-213. Emmons, L.H. (1998) Neotropical rainforest mammals. A field guide. 2nd ed. Chicago,
Chicago. 307pp. Excoffier, L., Smouse, P.E. e Quattro, J.M. (1992). Analysis of molecular variance
inferred from metric distances among haplotypes: applications to human mitichondrial DNA restriction data. Genetics 131: 479-491.
Fagundes, V., Christoff, A.U. e Yonenaga-Yassuda, Y. (1998) Extraordinary
chromosomal polymorphism with 28 different karyotypes in the neotropical species Akodon cursor (Muridae, Sigmodontinae), one of the smallest diploid number in rodents (2n=16, 15 and 14). Hereditas 129: 263-274.
______; Vianna-Morgante, A.M. e Yonenaga-Yassuda, Y. (1997). Telomeric sequences
localization and G-banding patterns in the identification of a polymorphic

68
chromosomal rearrangement in the rodent Akodon cursor (2n=14, 15 and 16). Chromosome Research 5: 228-232.
______; e Yonenaga-Yassuda, Y. (1998) Evolutionary conservation of whole
homeologous chromosome arms in the akodont rodents Bolomys and Akodon: maintenance of interstitial telomeric segments (ITBs) in recent event of centric fusion. Chromossome Research 6: 643-648.
Fahima, T., Sun, G.L., Beharav, A., Krigman, T., Beiles, A. e Nevo, E. (1999). RAPD
polymorphism of wild emmer wheat populations, in Israel. Theoretical and Applied
Genetics 98: 434-447. Felsenstein, J. (1986). Confidence limits on phylogenies: an approach using the bootstrap.
Evolution 39(4): 783-791.
Fernandez, F.A.S., Dunstone, N. e Evans, P.R. (1999). Density-dependence in habitat utilization by wood mice in Sitka spruce sucessional mosaic: the roles of immigration, emigration, and variation among local parameters. Canadian Journal
of Zoology 77: 397-405. Ferreira, D. e Grattapaglia, M.E. (1998). Introdução ao uso de marcadores moleculares
em análise genética. 3a. ed. Embrapa,-Cenargen, Brasília: 220pp.
Figurny-Puchalska, M., Gadeberg, R.M.E. e Boomsma, J.J. (2000). Comparison of genetic population structure of the large blue butterflies Maculinea nausithous and M. teleius. Biodiversity and Conservation. 9. 419–432.
Finke, E. e Jetschke, G. (1998). How inbreeding and outbreeding influence the risk of
extinction : a genetically explicit model. Mathematical Biosciences 156: 309-314. Foggo, A., Ozanne, C.M.P., Speight, M.R e Hambler, C. (2001). Edge effects and
tropical forest canopy invertebrates. Plant Ecology 153. 347–359. Fonseca, G.A.B. (1988). Biology and natural history of Brazilian Atlantic Forest small
mammals. Bulletin of the Florida State Museum, Biological Sciences, 34: 99-152. ______; Herrmann, G., Leite, Y.L.R. e Mittermeier, R.A. (1997) Lista anotada de
mamíferos do Brasil. Occassional Papers in Conservation Biology.
Fonseca, M.T. (1997). A estrutura da comunidade de pequenos mamíferos em um
fragmento de Mata Atlântica e monocultura de eucalipto: a importância da matriz de habitat. Dissertação de Mestrado. Belo Horizonte, Departamento de Biologia geral, Instituto de ciências biológicas, UFMG. 52pp.
Frankham, R. (1995). Inbreeding and extinction: a threshold effect. Conservation
Biology.9. 792-799.

69
______; (1996). Relationship of genetic variation to population size in Wildlife. Conservation Biology 10(6). 1500-1508.
______; Gilligan,D.M., Morris, D. e Briscoe, D.A. (2001). Inbreeding and extinction:
effects of purging. Conservation Genetics 2: 279–285. ______; e Ralls, K. (1998). Inbreeding leads to extinction. Nature 392: 441-442.
Fritsch, P., Hanson, M.A., Spore, C.D., Pack, P.E. e Rieseberg, R.H. (1993). Constancy of RAPD primer amplification strength among distantly related taxa of flowering plants. Plant Molecular Biology Reporter 11: 10-20.
Futuyma, D.J. (1997). Evolutionary biology. 3rd ed. Sinauer Associates Inc. Sunderland.
751pp. Geise, L. (1995). Os roedores sigmodontinae (Rodentia, Muridae) do estado do Rio de
Janeiro. Sistemática, citogenética, distribuição e variação geográfica. Dissertação de Mestrado, Rio de Janeiro, Departamento de Genética, Instituto de Biologia, UFRJ: 389pp.
______; Canavez, F.C. e Seuánez, H.N. (1998). Comparative karyology in Akodon
(Rodentia, Sigmodontinae) from southeastern Brazil. Journal of Heredity 89: 158-163.
______; Smith, M.F. Patton, J.L. (2001). Diversification in the genus Akodon (Rodentia:
Sigmodontinae) in southeastern South America: mitochondrial DNA sequence analysis. Journal of Mammalogy 82(1). 92-101.
Gentile, R., D’Andrea, P.S. e Cerqueira, R. (1997). Home ranges in Philander frenata
and Akodon cursor in the Brazilian Restinga (coastal shurbland). Mastozoologia
Neotropical. 42(2): 105-112. Gibbs, J.P. (2001). Demography versus habitat fragmentation as determinants of genetic
variation in wild populations. Biological Conservation 100. 15-20. Gibbs, H.L, Prior, K.A. e Weatherhead, P.J. (1994). Genetic analysis of populations of
threatened snake species using RAPD markers. Molecular Ecology 3: 329-337. Gonçalves, P.R. (2001). Identificação e alocação taxonômica de duas forma simpátridas de
Oxymycterus (Rodentia, Sigmodontinae) do sudeste do Brasil: uma avaliação de hipóteses dobre a diversidade do gênero. Dissertação de Mestrado. Rio de Janeiro, Museu Nacional, UFRJ. 135pp.
Grevstad, F.S. (1999). Experimental invasions using biological control introductions: the
influence of release size on the chance of population establishment. Biological
Invasion. 1: 313-323.

70
Groon, M.J. e Preuninger, T.E. (2000). Population type can influence the magnitude of
inbreeding depression in Clarckia coccina. Evolutionary Ecology. 14: 155-180. Guo, S.W. e Thompson, E.A. (1992). Performing the exact test of Hardy-Weinberg
proportions for multiple alleles. Biometrics 48(2): 361-372. Haig, S.M. (1998). Molecular contributions to conservation. Ecology 79: 413-425. ______; Rhymer, J.M. e Heckel, D.G. (1994). Population differentiation in randomly
amplified polymorphic DNA of red-cockaded woodpeckers Picoides borealis. Molecular Ecology 3: 581-595.
Hard, J.J., Winans, G.A. e Richardson, J.C. (1999). Phenotypic and genetic architecture
of juvenile morphometry in chinook salmon.. The Journal of Heredity 90(6): 597-606.
Harlt, D.L. e Clark, A.G. (1997). Principles of population genetics. 3rd ed. Sinauer
Associates Inc., Sunderland: 542pp. Hartt, L. e Haeffner, J.W. (1998). How phenotypic variation and life history trait
correlation enhance mean fitness in prey populations. Theoretical Population
Biology 54: 50-61. Heaton, H.J., Whitkus, R. e Gómez-Pompa, A. (1999). Extreme ecological and
phenotypic differences in the tropical tree chicozapote (Manilkara zapota) are not matched by genetic divergence: a RAPD analysis. Molecular Ecology 8: 627-632.
Hedrick, P.W. (2001). Conservation Genetics: where are we now? Trends in Ecology and
Evolution 16(11): 629-636. ______; (2000). Genetics of populations. 2nd ed. Jones & Bartlett, Sudbury. 553pp. ______; e Kalinowski, S.T. (2000). Inbreeding depression in conservation biology.
Annual Review of Ecology and Systematics 31: 139–62. Hoestra, H.E. e Edwards, E.V. (2000). Multiple origins of XY female mice (genus
Akodon): phylogenetic and chromosomal evidence. Proceedings of the Royal
Society of London, Biological Sciences 267: 1825-1831. Huff, D.R., Peakall,R. e Smouse, P.E. (1993). RAPD variation within and among natural
populations of outcrossing buffalograss. Theoretical and Applied Genetics 86: 927-934.
IBGE (2000). Instituto Brasileiro de Geografia e Estatística, Rio de Janeiro, RJ,
<http://www.ibge.gov.br>.

71
Ingvarsson, P.K. (2001) Restoration of genetic variation lost - the genetic rescue hypothesis. Trends in Ecology and Evolution 16(2): 62-63.
Jiménez, J.A., Hughes, K.A., Alaks, G., Graham, L. e Lacy, R.C. (1994) An
Experimental study of inbreeding depression in a natural habitat. Science 266: 271-273.
Kalinowski, S. T., Hedrick, P. W. e Miller, P. S. (1999). No evidence of inbreeding
depression in Mexican and red wolves. Conservation Biology 13: 1371–1377. Kappe, A.L., Van De Zande, L., Vedder, E.J., Bijlsma, R. e Van Delden, W. (1995)
Genetic Variation in Phoca virtulina revealed by DNA fingerprints and RAPDs. Heredity 74: 647-653.
Keller, L.F. e Waller, D.M. (2002). Inbreeding effects in wild populations. Trends in
Ecology and Evolution. 17(5): 230-241. Lacerda, D.R. (2000). Diversidade e estrutura genética de populações de Plathymenia
reticulata e P. foliolosa (Vinhático do campo e Vinhático da mata) e diferenciação molecular nas duas espécies. Dissertação de Mestrado. Belo Horizonte, Departamento de biologia geral, Instituto de ciências biológicas, UFMG. 69pp.
______; Acedo, M.D.P., Lemos Filho, J.P. e Lovato, M.B. (2001) Genetic diversity and
structure of natural populations of Plathymenia reticulata a tropical tree from the Brazilian Cerrado. Molecular Ecology. 10(5): 1143-1152.
Lande, R. (1988). Genetics and demography in biological conservation. Science 241:
1455-1460. ______; (1999). Extinction risks from anthropogenic, ecological and Genetic Factors. in
Genetics and extinction of species. 1 ed. Princeton, West Sussex: 1-22. Landry, P.A., e Lapointe, F.J. (1999). The genetic heterogeneity of deer mouse
populations (Peromyscus maniculatus) in an insular landscape. Researches in
Population Ecology. 41: 263-268. Lawrence, W.F. e Cochrane, M.A. (2001). Synergistic effects in fragmented landscapes.
Conservation Biology 15(6): 1488-1489. Lenormand, T. (2002). Gene flow and the limits to natural selection. Trends in Ecology
and Evolution 17(4): 183-189. Lewin, R. (1999). Patterns in Evolution: the new molecular view. 1st ed. Scientific
American Library, New York. 245pp. Lewontin, R.C. (1972) The apportionnmt of human diversity. Evolutionary Biology 6:
381-398.

72
Liascovich, R.C. e Reig, O.A. (1989). Low chromossomal number in Akodon cursor
montensis Thomas, and karyologic confirmation of Akodon serrensis Thomas in Missiones Argentina. Journal of Mammalogy 70(2): 391-395.
Liersch, S. e Schmid-Hempel, P. (1998). Genetic variation within social insect colonies
reduces parasite load. Proceedings of the Royal Society of London, Biological
Sciences 265: 221-225. Lindenmayer D.B. e Lacy, R.C. (2002). Small mammals, habitat patches and PVA
models: a field test of model predictive ability Biological Conservation 103(3): 247-265.
______; Lacy, R.C. e Pope. M.L. (2000). Testing a Population Viability Analysis model.
Ecological Applications 10: 580-597. Loeschcke, V., Tomiuk, J. e Jain, S.K., (1994). Conservation Genetics. 1st ed. Birkhäuser
Verlag, Berlin. 438p. Lu, M.Z., Wang,X.R.N e Nszmit, A.E. (1997). Molecular properties of RAPD in Pinus
silvestris and their implications for the genetic analysis. Forest Genetics 4: 227-234. Lynam, A.J. e Billick, I. (1999). Differential responses of small mammals to
fragmentation in a Thailand tropical forest. Biological Conservation. 91: 191-200. Lynch, M. e Milligan, B.G. (1994). Analysis of population genetic structure with RAPD
markers. Molecular Ecology 3: 91-99. Mack, R.N. (2000). Cultivation fosters plant naturalization by reducing environmental
stochasticity. Biological Invasion. 2: 111-122. Malcon, J.R. (1998). A model of conductive heat flow in forest edges and fragmented
landscapes. Climatic Change. 39: 487-502. McKay, J.K. e Latta, R.G. (2002). Adaptive population divergence: markers, QTL
and traits. Trends in Ecology and Evolution. 17(6): 285-291. Meagher, S., Penn, D.J. e Potts, W.K. (2000). Male–male competition magnifies
inbreeding depression in wild house mice. Proceedings of the National Academy of
Science 97(7): 3324-3329. Mills, S.L. e Smouse, P. (1994). Demographic consequences of inbreeding in remnant
populations. American Naturalist. 144: 412-431. Mishra, M., Dubey, N., Totey, S.M., Bhat, K.V., Babu, S., Awasthi-Kalia, M. e Anand,
R.K. (2002). Phylogenetic relationships and genetic polymorphisms in wild Indian mice. Biomolecular Engineering 18: 281–288.

73
Mockford, S.W., Snyder, M. e Herman, T.B. (1999) A preliminary examination of genetic variation in a peripheral population of Blandings turtle, Emydoidea
blandingii. Molecular Ecology 8(2): 323-326.
Morgan-Richards, M e Wolff, K. (1999). Genetic structure and differentiation of Plantago major reveals a pair of sympatric sister species. Molecular Ecology 8: 1027-1036.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., Fonseca, G.A.B. e Kent, J. (2000).
Biodiversity hotspots for conservation priorities. Nature 403: 853-858. Nausser, J.A., Goto, R.M., Ledig, D.B., Fleischer, R.C. e Miller, M.M. (1996). RAPD
analysis reveals low genetic variability in the endangered light-footed clapper rail. Molecular Ecology 5: 463-472.
Nei, M. (1972). Genetic distance between populations. American naturalist 106(949): 283-
292.
______; (1978). Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89: 583-590.
Nieminen, M., Singer, M.C. Fortelius, W., Schöps e Hanski, I. (2001). Experimental
Confirmation that Inbreeding Depression Increases Extinction Risk in Butterfly Populations. American Naturalist 157(2): 237-244.
Nybon, H. e Bartish, I.V. (2000). Effects of life history traits and sampling strategies on
genetic diversity estimates obtained with RAPD markers in plants. Perspectives in
plant ecology, evolution and systematics 3(2): 93-114. Pertoldi, C., Hansen, M.M., Loeschcke, V., Madsen, A.B., Jacobsen, L. e Baagoe, H.
(2001). Genetic consequences of population decline in the European otter (Lutra
lutra): an assessment of microsatellite DNA variation in Danish otters from1883 to 1993. Proceedings of the Royal Society of London, Biological Sciences 268: 1775-1781.
Pielou, E.C. (1975). Ecological Diversity. 1st ed. Willey-Interscience, New York. 165pp. Pimm, S.L. e Raven, P. (2000). Extinction by numbers. Nature 403: 842-843. Pires, A.S., Lira, P.K., Fernandez, F.A.S., Schittini, G.M. e Oliveira, L.C. (2002).
Frequency of movements of small mammals among Atlantic Coastal forest fragments in Brazil. Biological Conservation 108: 229-237.

74
Pivello, V.R., Shida, C.N. e Meirelles, S.T. (1999). Alien grasses in Brazilian savannas: a threat to the biodiversity. Biodiversity and Conservation. 8: 1281–1294.
Pusey, A. e Wolf, M. (1996). Inbreeding avoidance in animals. Trends in Ecology and
Evolution 11(5): 201-206. Raymond, M. e Rousset, F. (1995). An exact test for population differentiation. Evolution
49(6): 1280-1283. Reed, D.H. e Bryant, E.H. (2001). Fitness, genetic load and purging in experimental
populations of the housefly. Conservation Genetics 2: 57–62. ______; e Frankham, R. (2001). How closely are correlated molecular and quantitative
measures of genetic variation? A meta-analysis. Evolution 55(6): 1095-1103. Ribon, R. (1998). Fatores que influenciam a distribuição da avifauna em fragmentos de
Mata Atlântica nas montanhas em Minas Gerais. Dissertação de Mestrado, Minas Gerais, Departamento de Biologia Geral, Instituto de Ciências Biológicas, UFMG: 90pp
Richards, C.M. (2000). Inbreeding depression and genetic rescue in a plant
metapopulation. American Naturalist 155(3): 383-394. Rieger, T.T., Langguth, A. e Waimer, T.A. (1995). Allozymic characterization and
evolutionary relationships in the Brazilian Akodon cursor species group (Rodentia: Cricetidae). Biochemical Genetics 33(9/10): 283-295.
Saccheri, I., Kuussaari, M., Kankare, M., Vikman, P., Fortelius, W. e Hanski, I.
(1998). Inbreeding and extinction in a butterfly metapopulation. Nature. 392: 491-494.
Sari, H., Wilhelm F., Robert, O., Mikko, M., Niklas, W., Saccheri, I., Singer, M. e
Ilkka. (2001) Inbreeding depression and the maintenance of genetic load in Melitaea cinxia metapopulations. Conservation Genetics 2(4):325-335.
Schneider, M.F. (2001). Habitat loss, fragmentation and predator: spatial implications for
prey conservation. Journal of applied ecology 38: 720–735. Silva, M.J.J. e Yonenaga-Yassuda, Y. (1998) Karyotype and chromosomal polymorphism
of an undescribed Akodon from central Brazil, a species with the lowest known diploid chromosome number in rodents. Cytogentics and Cell Genetics 81: 46-50.
Simberloff, D. (1988). The contribution of population and community biology to
conservation science. Annual Review of Ecology and Systematics. 19: 473-511. Slatkin, M. (1994). Gene flow and population structure. in Ecological Genetics. Princeton,
West Sussex: 1-17.

75
Smith, M.F. e Patton, J.L. (1991) variation in mitochondrial cytochrome b sequence in
natural populations of South American akodontine rodents (Muridae: Sigmodontinae). Molecular Biology and Evolution 8(1): 85-103.
______; Thomas, W.K. e Patton,J.L (1992). Mitochondrial DNA-like sequence in the
nuclear genome of an akodontine rodent. Molecular Biology and Evolution 9(2): 204-215.
______; e Patton,J.L. (1993). The diversification of South American murid rodents:
evidence from the mitochondrial DNA sequence data for the Akodontine tribe. Biological Journal of Linnean Society 50: 149-177.
______; Kelt, D.A. e Patton,J.L. (2001). Testing the model of diversification in mice in
Chile and Argentina. Molecular Ecology 10: 397-405. Sokal, R. (1979). Testing statistical significance of geographic variation patterns.
Systematic Zoology 28: 227-232. Stephens, P.A. e Sutherland, W.J. (1999). Consequences of the Allee effect for behavior,
ecology and conservation. Trends in Ecology and Evolution. 14(10): 401-405. Stevens, S.M. e Husband, T.P. (1998). The influence of edge on small mammals:
evidence from Brazilian Atlantic forest fragments. Biological Conservation. 85: 1-8. Stewart Jr., C.N. e Excoffier, L. (1996). Assessing population genetic structure and
variability with RAPD data: application to Vaccinum macrocarpon. Journal of
Evolutionary Biology 9: 153-171. Sugg, D.W., Cheeser, R.K. e Long, J.C. (1997). Assessment of genetic information in
morphometric traits: geographic patterns and evolutionary interpretation. Journal of
Mammalogy 78(2): 405-416. Sun, M. (1996). Effects of population size, mating system, and evolutionary origin on
genetic diversity in Spiranthes snensis and S. hongkongensis. Conservation Biology 10: 785-795.
Sunnucks, P. (2000). Efficient genetic markers for population biology. Trends in Ecology
and Evolution 15(5): 199-203. Tanaka, Y. (1997). Extinction of populations due to inbreeding depression with
demographic disturbance. Researches in Population Ecology. 39: 57-66. Tosta, V. C. Waldschimit, A. M. Good-God, P. I. V. Pompolo. S. Barros. E. G.
Campos. L.A.O. (2000). Uso de marcadores moleculares para estudo de cromossomos supranumerários.em Partamona helleri (Hymenoptera, Apidae, Meliponinae). Genetics and Molecular Biology 22 (supplement): 182.

76
Via, S. (1994). Population structure and local adaptation in a clonal herbivore. in
Ecological Genetics. Princeton, West Sussex: 58-85. Vucetich, L.M., Vucetich, J.A., Joshi, C.P., Waite, T.A. e Peterson, R.O. (2001).
Genetic (RAPD) diversity in Peromyscus maniculatus populations in a naturally fragmented landscape. Molecular Ecology 10: 35-40.
Wachira, F.N., Waugh, R., Hackett, C.A. e Powell, W. (1995). Detection of genetic
diversity in tea (Camellia sinensis) using RAPD markers. Genome 38: 201-210. Waldmann, P. (2001) The effect of inbreeding on fluctuating asymmetry in Scabiosa
canescens (Dipsacaceae). Evolutionary Ecology 15: 117-127.
Wauters, L., Hutchinson, I., Parkin,D.T. e Dhondt, A.A.. (1994). The effects of habitat fragmentation on demography and on the loss of genetic variation in the red squirrel. Proceedings of the Royal Society of London, Biological
Sciences 255:107-111.
Williams, J.G.K., Kubelik, A.R. Livak, K.J. Rafalski, J.A. e Tigey, S.V. (1990). DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Research 18: 6531-6535.
Windig, J.J., Rintäki; P.T¨, Cassel, A. e Nylin, S. (2001). How useful is fluctuating
asymmetry in conservation biology: Asymmetry in rare and abundant in Coenonympha butterflies. Journal of Insect Conservation 4: 253-261.
Winge, H. (1887). Jordfundne og nulevende Gnavere (Rodentia) fra Lagoa Santa, Minas
Geraes, Brasilien. E Museo Lundii, 1: 1-200. Wolff, K., El-Akkad, S. e Abbott, R.J. (1997). Population substructure in Alkanna
orientalis in the Sinai desert, in relation to its pollinators behavior. Molecular
Ecology 6: 365-372. Wright, S. (1943). Isolation by distance. Genetics. 28: 114-138. Yonenaga, Y. (1972). Chromosomal polymorphism in the rodent Akodon arviculoides (2n
= 14) resulting from two pericentric inversions. Cytogenetics 11: 488-499. ______; Kasahara, S., Almeida, E.J.C. e Peracchi, A.L. (1975). Chromosomal banding
patterns in Akodon arviculoides (2n = 14), Akodon sp (2n = 24 and 25) and male hybrids with 19 chromosomes. Cytogenetics and Cell Genetics 15: 388-399.

77
Yeh, F.C., Chong, D.K.X. e Yang, R.C. (1995). RAPD variation within and among natural populations of trembling aspen (Populus tremuloides). From Alberta. Journal of Heredity 86: 454-460.