uso de probiÓtico para codornas de corte coturnix ...livros01.livrosgratis.com.br/cp032873.pdf ·...
TRANSCRIPT

UNIVERSIDADE ESTADUAL DE MARINGÁ CENTRO DE CIÊNCIAS AGRÁRIAS
USO DE PROBIÓTICO PARA CODORNAS DE CORTE (Coturnix coturnix sp)
Autora: Luciana Kazue Otutumi
Orientador: Prof. Dr. Antonio Cláudio Furlan
Tese apresentada, como parte das exigências para obtenção do título de DOUTOR EM ZOOTECNIA, ao Programa de Pós-graduação em Zootecnia da Universidade Estadual de Maringá – Área de Concentração: Produção Animal.
MARINGÁ Estado do Paraná
Outubro 2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
“Aqueles que sempre têm dúvidas são
pessoas progressistas e seu futuro é
brilhante”.
Meishu-Sama

iii
A Meishu-Sama, por sempre estar presente em minha
vida, e por me dar forças nas horas mais necessárias.
Aos meus pais Youko (in memorian) e Shintaro e por mais
esta permissão.
DEDICO

iv
AGRADECIMENTOS
A Deus e a Meishu-Sama, pela permissão de concluir mais uma etapa de
minha vida, esperando sempre poder contribuir por meio da aquisição
desses novos conhecimentos, a construção de um mundo melhor;
Aos meus antepassados, em especial à minha mãe que, apesar de não
estarem diretamente presentes, contribuíram em muito para o meu
crescimento;
Ao meu pai, Shintaro, por suportar todo meu estresse durante o
desenvolvimento deste trabalho;
À Universidade Estadual de Maringá, por mais esta oportunidade;
Ao Prof. Dr. Antonio Cláudio Furlan, por mais esta orientação, paciência e
principalmente apoio e amizade, minha eterna gratidão;
Ao Prof. Dr. Elias Nunes Martins, pela amizade, incondicional paciência e
auxílio nas análises estatísticas;
À Profa. Dra. Maria Raquel Marçal Natali (DCM), pela colaboração,
amizade, incentivo e alegria em todos os momentos em que estivemos
presentes;
Ao Prof. Dr. Celso Nakamura, pela amizade e oportunidade de realização
das análises microbiológicas;

v
Ao CNPq, pela concessão da bolsa de estudos, que muito me auxiliou na
conclusão deste trabalho;
À empresa FATEC pela doação do probiótico e atenção dispendida;
À minha grande amiga, Elis Regina de Moraes Garcia e sua querida mãe
Iracema, pela amizade, apoio, incentivo, companheirismo, que foram
essenciais para mantermos sempre acesa a esperança de um final feliz;
À Ana Paula Ton e Priscila Ventura, por suas ajudas incondicionais,
amizade, minha gratidão;
Às minhas novas amizades conquistadas durante o curso: Meiby Carneiro
de Paula, Regina, Marivone, Capovilla, Ana Carolina, Andréia Fróes,
Márcia Sakamoto, Luciana Maria, Jovanir, Luís Daniel, Karina, Milene,
Elkin, Luciane, Eliane, Rafael, Fábio, Raíssa, Kelly, Alexandre e Leandro.
Às técnicas (Maria Euride e Maria dos Anjos) do Laboratório de
Histotécnica Animal/DCM/UEM, pelos ensinamentos, paciência e todos os
momentos alegres que passamos juntas;
Aos funcionários da Fazenda Experimental de Iguatemi (FEI), Célio
Passolongo e Valentim, por toda a ajuda durante o desenvolvimento do
experimento, essenciais para a conclusão;
Aos professores Luís Felipe e Cláudia e o mestrando Leandro pelo auxílio
na captura das imagens no Laboratório Zooplâncton e oportunidade de
concretizar a minha pesquisa.
E, finalmente, mas não menos importante, Carlos Gustavo Fernandes
Iwazaki, por toda sua alegria que muito me contagiou e motivou para
realmente conseguir concretizar este trabalho.

vi
BIOGRAFIA DA AUTORA
LUCIANA KAZUE OTUTUMI, filha de Shintaro Otutumi e Youko
Otutumi, nasceu em Marília, São Paulo, no dia 17 de outubro de 1976.
Em dezembro de 1999, concluiu o curso de Medicina de Veterinária pela
Universidade Estadual de Londrina.
No dia 29 de abril de 2002, adquiriu o título de Mestre em Zootecnia, área
de concentração Produção Animal, pela Universidade Estadual de Maringá,
realizando estudos na área de Nutrição de Coelhos.
No dia 23 de outubro de 2006, submeteu-se à banca para defesa de tese.

vii
ÍNDICE
Página
RESUMO............................................................................................................... 1 ABSTRACT.......................................................................................................... 3 I - INTRODUÇÃO................................................................................................ 5 CITAÇÃO BIBLIOGRÁFICA.............................................................................. 11 II - OBJETIVOS GERAIS.................................................................................... 14 III- Diferentes vias de administração de probiótico sobre o desempenho, rendimento de carcaça e população microbiana do intestino delgado de codornas de corte...................................................................................................
15 Resumo.................................................................................................................. 15 Abstract.................................................................................................................. 16 Introdução.............................................................................................................. 17 Material e Métodos................................................................................................ 19 Resultados e Discussão.......................................................................................... 25 Conclusões............................................................................................................. 34 Literatura Citada.................................................................................................... 35 IV- Efeito do probiótico sobre o desempenho, rendimento de carcaça, exigências nutricionais de proteína bruta e morfometria intestinal de codornas de corte...................................................................................................................
38 Resumo.................................................................................................................. 38 Abstract.................................................................................................................. 39 Introdução.............................................................................................................. 40 Material e Métodos................................................................................................ 42 Resultados e Discussão.......................................................................................... 47 Conclusões............................................................................................................. 67 Literatura Citada.................................................................................................... 68 V - CONCLUSÕES GERAIS............................................................................... 73

RESUMO Foram conduzidos dois experimentos para avaliar o efeito do probiótico sobre o
desempenho, rendimento de carcaça, exigências nutricionais de proteína bruta (PB),
população microbiana e morfometria do intestino delgado de codornas de corte. No
primeiro experimento, foram utilizadas duas diferentes vias de administração de
probiótico (água de bebida e ração), ou seja, avaliado o desempenho no período de um a
35 dias de idade, o rendimento de carcaça e de cortes aos 35 dias e a população
microbiana do intestino delgado aos sete e aos 14 dias de idade de codornas de corte.
Para isso, foram utilizadas 1.050 codornas distribuídas em um delineamento
inteiramente casualizado com três tratamentos e cinco repetições de 70 aves cada. Para
o rendimento de carcaça e de cortes e população microbiana do intestino delgado, foram
abatidas, respectivamente, quatro e uma ave de cada unidade experimental. As codornas
do tratamento em que se adicionou probiótico, via ração, apresentaram menor consumo
de ração (CR), em relação àquelas que receberam, via água, no período de sete a 14 dias
de idade, porém, isso não afetou o ganho de peso (GP), o que levou à melhora na
conversão alimentar (CA). Menor CR foi verificado nos períodos de 21 a 28, 28 a 35 e
um a 35 dias de idade, para as codornas dos tratamentos com adição probiótico. O
rendimento de carcaça e de cortes, a contagem total de Lactobacillus sp.,
enterobactérias e Escherichia coli não foram influenciados pela adição do probiótico.
No segundo experimento, foram utilizadas 2.304 (1.440 no período um e 864 no
período dois) codornas de corte para avaliar o desempenho e as exigências nutricionais
de PB no período de um a 35 dias de idade, o rendimento de carcaça e de cortes aos 35
dias e a morfometria da mucosa do intestino delgado aos sete, 14, 21 e 35 dias de idade.
O delineamento experimental foi inteiramente casualizado em esquema fatorial 2 x 4

2
(com e sem probiótico e quatro níveis de PB – 15, 20, 25 e 30%), com duas repetições
por tratamento, em dois períodos experimentais, totalizando 32 repetições. Para análise
morfométrica, foram abatidas duas aves de cada unidade experimental (oito aves por
tratamento). A adição do probiótico reduziu o CR e melhorou a CA na faixa etária de 1
a sete dias. De acordo com os pesos vivos médios do modelo LRP (Linear Response
Plateau), até os 14 dias, a estimativa de exigência é de 27% de PB, a partir desta idade
entre 22 e 24% de PB. A adição do probiótico não influenciou o rendimento de carcaça
e de cortes. Para o rendimento de carcaça, a exigência de PB de acordo com o modelo
quadrático, ficou estimada em 23,53%, em relação ao LRP, a estimativa reduz para
17,13% de PB. Foram estabelecidos primeiros componentes principais envolvendo as
variáveis sob estudo com a finalidade de se determinar a exigência envolvendo
simultaneamente mais de uma variável. Dentre os componentes principais estimados,
aquele envolvendo as variáveis: peso médio aos 35 dias, GP de 1 a 35 dias e CA de 1 a
28 dias foi escolhido para determinar a exigência nutricional de PB por responder por
94,61% da variação, o que resultou estimativa de exigência de 27,37% de acordo com o
modelo quadrático e 21,91% de acordo com o LRP. O probiótico não influenciou o
comprimento e a morfometria da mucosa do intestino delgado. O nível de PB da dieta
influenciou de maneira linear o comprimento do intestino delgado aos sete, 14 e 21 dias
de idade e de maneira quadrática aos 35 dias de idade. No íleo, a altura de vilo
apresentou aumento linear em função dos níveis de PB, porém a profundidade de cripta
não mostrou diferenças com o aumento dos níveis de PB. Pode-se concluir que nas
condições ambientais em que as codornas foram criadas a adição do probiótico não
influenciou o desempenho, as exigências nutricionais de PB, o rendimento de carcaça e
de cortes, a população microbiana e a morfometria intestinal.
Palavras-chave: altura vilo, desempenho, lactobacilos, população microbiana,
profundidade cripta

ABSTRACT
Two experiments were conducted to evaluate the probiotic effect on performance,
carcass yield, crude protein (CP) nutritional requirements, small intestine microbial
population and morphometry of meat quails. In the first experiment it was used two
different ways of probiotic administration (drinking water and feed), and the
performance from 1 to 35 days of age, carcass and cut yields at 35 days and small
intestine microbial population at 7 and 14 days of age of meat quails were evaluated.
Therefore, it was used 1050 quails distributed in a completely randomized design with
three treatments and five replications of seventy quails each. To carcass and cut yields
and small intestine microbial population were slaughtered respectively four and one
quail of each experimental unit. Quails from treatment that were supplemented with
probiotic in feed, had lower feed intake (FI), compared with those supplemented by
water, during the period from 7 to 14 days of age, however it did not affected the weight
gain (WG), which increased the feed conversion (FC). Lower feed intake was observed
in the periods of 21 to 28, 28 to 35 and 1 to 35 days of age for quails from probiotic
addition treatment. Carcass and cut yields, total count of Lactobacillus sp.,
enterobacters and Escherichia coli were not influenced by probiotic addition. In the
second experiment, it was used 2304 (1440 to first period and 864 to second period)
meat quails to evaluate performance and CP nutritional requirements from 1 to 35 days
of age, carcass and cut yields at 35 days and small intestine mucous morphometry at 7,
14, 21 and 35 days of age. The experimental design was completely randomized in a
factorial 2 x 4 (with and without probiotic and four CP levels – 15, 20, 25 and 30%),
with two replications by treatment, in two experimental periods, totalizing 32
replications. To morphometric analysis, two quails of each experimental unit (eight

4
quails by treatment) were slaughtered. Probiotic addition decreased FI and improved FC
of 1 to 7 days. According to mean weight of LRP (Linear Response Plateau) model, at 7
and 14 days, requirement estimative was 27% of CP and at 21, 28 and 35 days was
among 22 to 24% of CP. Probiotic addition did not influence carcass and cut yields. To
carcass yield, CP requirement according quadratic model was estimated in 23.53%,
however, in relation at LRP, the estimate decrease to 17.13% of CP. There were
established first mainly components involving the variables under study aiming to
determine the requirement involving at same time more of one variable. Among
principal components estimated, the one involving the variables medium weight at 35
days, WG of 1 to 35 days and FC of 1 to 28 days, were selected to determine CP
nutritional requirement because it was replying by 94.61% of variation, what resulted in
requirement estimative of 27.37% according quadratic model and 21.91% according
LRP. Probiotic did not influence small intestine length and mucous morphometry. Diet
CP level influenced in a linear way the small intestine length at 7, 14 and 21 days of age
and in a quadratic way at 35 days of age. In ileum, the villum height had a linear
increase in function of CP levels; however, crypt depth did not show differences with
the CP increase. It is possible to conclude that in the ambient conditions which quails
were created the probiotic addition did not influence the performance, CP nutritional
requirement, carcass and cut yields, microbial population and intestinal morphometry.
Key words: villum height, performance, lactobacillus, microbial population, crypt depth

I - INTRODUÇÃO
O crescimento da coturnicultur,a como atividade comercial no Brasil, tem
conduzido ao aumento do número de pesquisas, no intuito de melhorar a exploração
deste mercado em expansão.
Um grande crescimento na produção, particularmente de codornas de postura,
ocorreu na década de 90, por causa da mudança ocorrida no mercado atacadista
varejista, quando os ovos primordialmente comercializados “in natura” passaram
também a ser processados em indústrias beneficiadoras, ao originar os chamados ovos
descascados ou em conserva, estendendo o consumo para churrascarias, restaurantes,
bares e lanchonetes (Fujikura, 2002).
A explicação para esse crescimento se deve ao baixo custo de manutenção,
reduzido período para as aves atingirem a maturidade sexual, reduzido consumo de
alimento, grande rusticidade aliada à possibilidade da rápida reversão do capital
investido (Murakami & Ariki, 1998).
Essas características justificam, portanto, o aumento da produção nacional, em que
dados do IBGE (2006) mostram que o plantel de codornas, no Brasil, no ano de 2004,
correspondia a 6,243 milhões de aves e que houve um aumento de 29% desde o ano de
1999.
No Brasil, a produção de codornas é predominantemente voltada para a produção
de ovos e até recentemente não se praticava a criação especializada de codornas para a
produção de carne. Dessa forma, a espécie mais difundida no país é a Coturnix coturnix
japonica, linhagem de baixo peso, utilizada exclusivamente para a produção de ovos e
destinada ao abate das fêmeas em final de postura (Rezende et al., 2004).

6
Em conseqüência, as carcaças obtidas de codornas não-especializadas na produção
de carne eram muito pequenas, pesando entre 70 e 110 gramas, e a carne das fêmeas,
relativamente, dura por se tratar de aves de aproximadamente um ano de idade (Garcia,
2002), considerada inadequada para os padrões de consumo nacional.
No intuito de atender a demanda de mercado, principalmente o mercado externo,
onde o consumo da carne de codorna é bastante difundido, recentemente houve a
importação de codornas especializadas para a produção de carne (Coturnix coturnix
sp.), também conhecida como codorna européia.
As européias são maiores (peso vivo de 200 a 300 gramas), apesar de
fenotipicamente serem bastante semelhantes à Coturnix coturnix japonica, possuem
uma coloração marrom mais viva, têm temperamento nitidamente mais calmo e o peso e
o tamanho dos ovos são um pouco maiores, embora a idade de maturidade sexual seja
praticamente a mesma da codorna de postura (Rezende et al., 2004).
São escassas, entretanto, as pesquisas sobre exigências nutricionais de codornas
européias no Brasil, sendo as formulações de rações baseadas em dados estrangeiros,
pouco condizentes com as condições brasileiras, o que às vezes compromete os dados
de produtividade (Murakami, 2002); o número de pesquisas, que tem por objetivo
determinar as exigências nutricionais de codornas nas condições de Brasil, vêm
aumentando gradativamente.
Oliveira et al. (2002a), com o objetivo de estimar a exigência de proteína bruta e
energia metabolizável de codornas japonesas machos criadas para a produção de carne,
encontraram que no período total de criação (cinco a 49 dias de idade), as exigências
protéica e energética, estimada para máximo ganho de peso foram de 26% e 3.200 Kcal
EM/ Kg de ração, respectivamente.
Por outro lado, Oliveira et al. (2002b), trabalhando com níveis de proteína bruta,
que variam de 20 a 26%, verificaram que o nível de 20% mostrou-se adequado para a
produção de codornas machos e fêmeas para abate aos 49 dias.
Em pesquisa conduzida por Corrêa et al. (2004) com codornas de corte, os autores
encontraram que, para o período de sete a 14 dias, eram necessários níveis de proteína
bruta superiores a 28%, de 26% para o período de 15 a 28 dias e de 24% para o período
de 29 a 35 dias de idade, havendo, portanto, a necessidade de formular rações divididas
por fases, assim como em frangos de corte.
Com o crescimento das criações comerciais de codornas, é presumível que
aumente o número de eventuais aves portadoras de agentes infecto-contagiosos. Dessa

7
forma, o risco de ocorrência de doenças nas regiões de maior concentração de aves,
torna-se preocupação adicional para o avicultor (Oliveira, 2004).
Aliado a isso, a pouca diversidade da microflora intestinal de aves recém-nascidas,
é considerada como um fator limitante tanto para a digestão quanto à possibilidade de
colonização intestinal por patógenos entéricos (Maiorka et al., 2001).
Em condições normais, aves jovens recebem a microbiota, principalmente, das
mães; em galinhas e perus, a transferência de microrganismos é muito eficiente quando
os recém-nascidos são criados próximos aos adultos (Andreatti Filho & Sampaio, 1999).
No entanto, os sistemas modernos de produção avícola impossibilitam esse contato, e
retarda a formação de uma microbiota intestinal protetora nos animais jovens.
Nessas condições, a produção avícola com alta produtividade a baixo custo, é
quase impossível sem a utilização de aditivos alimentares ao visar à prevenção de
doenças, tais como: os antibióticos.
Em virtude do atual questionamento de seu uso, por causa da relação com a
resistência aos antibióticos usados na antibioticoterapia humana, à presença de resíduos
de antibióticos nos produtos de origem animal destinados para o consumo humano, e a
emergente exigência por parte de importadores de produtos livres de resíduos de
aditivos, houve a necessidade da busca de produtos alternativos que pudessem substituir
os antibióticos na alimentação animal, sem que isso causasse perdas na produtividade e
na qualidade do produto final.
Uma alternativa seria o uso dos probióticos, os quais são produtos constituídos por
microrganismos vivos ou suas L-forms (sem a parede celular) (Loddi et al., 2000).
O termo probiótico foi primeiramente introduzido por Lilly & Stillwell (1965)
para descrever fatores de crescimento produzidos por um microrganismo que favorece
um outro, portanto, foi usada em contraposição ao termo antibiótico.
Parker (1974) definiu os probióticos como organismos e substâncias que
contribuem para o balanço microbiano intestinal. Mais tarde, Fuller (1989) veio a
designar probióticos como produto constituído por microrganismos vivos que, uma vez
introduzidos no organismo animal, influenciam beneficamente o hospedeiro por meio
da melhoria do balanço microbiano intestinal.
Em aves, o marco do uso de probióticos foi instituído por Nurmi & Rantala
(1973). Em seus experimentos, os autores observaram que o conteúdo intestinal de aves
adultas normais, administrados, oralmente, às aves com um dia de idade, alterava sua
sensibilidade à infecção por Salmonella spp. No mesmo ano, Tortuero verificou

8
melhora na conversão alimentar e aumento no ganho de peso em aves que receberam
Lactobacillus acidophilus, que abre caminho para uma série de investigações sobre o
tema.
Trabalhos conduzidos por Andreatti Filho et al. (2000) e Oliveira et al. (2000)
também demonstraram que a administração de microrganismos da microbiota intestinal
de aves promove efeito protetor contra a colonização por Salmonella spp. Com relação à
proteção contra a colonização por Escherichia coli, Jin et al. (1996) verificaram o
mesmo efeito protetor em frangos de corte.
Os probióticos podem conter bactérias totalmente conhecidas e quantificadas ou
culturas bacterianas não-definidas, geralmente compostos por um ou mais dos seguintes
microrganismos: Lactobacillus, Streptococcus, Bifidobactéria, Bacillus ou leveduras
(Mulder, 1991); a variação da eficácia dos probióticos pode ser por causa das diferenças
nas espécies microbianas ou cepas de microrganismos utilizadas ou ainda ao método de
preparação dos produtos ofertados aos animais (Jin et al., 1998) .
Segundo Jin et al. (1997), o modo de ação dos probióticos nas aves inclui a
manutenção da microflora intestinal normal por exclusão competitiva e antagonismo;
alteração do metabolismo por aumento da atividade das enzimas digestivas e
decréscimo na atividade das enzimas bacterianas e produção de amônia; neutralização
de enterotoxinas e estimulação do sistema imune.
Edens et al. (1997) verificaram que as bactérias láticas também podem controlar
populações de bactérias patogênicas como Salmonella spp., E. coli e Enterococci, pela
produção de substâncias que inibem o crescimento bacteriano tais como: a reuterina.
Particularmente para o controle da população de E.coli, Fuller (1977) relata que a
inibição é dependente da presença do número suficiente de Lactobacillus e que a partir
de resultados de testes in vitro, parece ser necessário no mínimo 107 Unidades
Formadoras de Colônia por grama (UFC/g).
Nas galinhas, a flora bacteriana atua beneficamente no processo digestório que
conseqüentemente favorece o metabolismo dos carboidratos pela intensificação da
digestão do amido; o metabolismo das proteínas, pela recuperação do nitrogênio
endógeno e na síntese de vitaminas e digestão cecal dos componentes fibrosos (Lancini,
1994).
Pode-se perceber, em razão disso, o importante papel da microbiota intestinal na
digestão e absorção dos alimentos ingeridos pelo hospedeiro, que exerce dessa forma
influência sobre sua saúde e performance produtiva (Canalli et al., 1996).

9
Como os processos de absorção de nutrientes são totalmente dependentes dos
mecanismos que ocorrem na mucosa intestinal, a manipulação de microrganismos
probióticos dentro do trato gastrintestinal tem sido usada com o objetivo de manter ou
incrementar a flora natural, prevenir a colonização de microrganismos patogênicos e
manter a ótima utilização dos alimentos (Fuller, 1989; Vanbelle et al., 1990), em
conseqüência da proteção dos vilos intestinais e superfícies absortivas contra toxinas
produzidas por microrganismos patógenos (Dobrogsz et al., 1991; Walker & Duff,
1998).
Bradley et al. (1994) verificaram redução na profundidade de cripta ileal quando
suplementaram Saccharomyces cerevisiae var. boulardii, mas, não observaram efeito
sobre a altura dos vilos ileais. Segundo os autores, esse decréscimo na profundidade de
cripta pode indicar um decréscimo na taxa de turnover, que poderia ser resultante do
decréscimo na concentração de bactérias produtoras de metabólitos tóxicos ou a
habilidade do Saccharomyces cerevisiae var. boulardii em suprimir os efeitos destes
metabólitos tóxicos.
Pelicano et al. (2003) avaliaram a morfometria da mucosa intestinal de frangos de
corte alimentados com dietas que contém diferentes probióticos (Bacillus subtilis,
Bacillus subtilis + Bacillus licheniformis e Saccharomyces cerevisiae) na ração em
associação ou não com probiótico na água de bebida (Lactobacillus reuteri e johnsonii)
e encontraram que independente do probiótico utilizado houve maior altura e perímetro
do vilo do duodeno, mas não observada essa diferença no jejuno e íleo.
Posteriormente, Pelicano et al. (2005) encontraram maior altura de vilo do jejuno e
íleo e maior profundidade de cripta no duodeno, jejuno e íleo de frangos de corte aos 21
dias, que receberam probiótico à base de Bacillus subtilis, mostrando dessa forma
efeitos benéficos adicionais do uso desse probiótico. Todavia, nessa mesma pesquisa, o
uso do probiótico que contém Lactobacillus acidophilus e casei, Streptococcus lactis e
faecium, Bifidobacterium bifidum e Aspergillus oryzae não mostraram resultados
significativos de seu uso sobre a altura de vilo e a profundidade de cripta dos segmentos
estudados.
Em codornas, trabalhos que avaliam a utilização de probióticos são escassos, por
isso a necessidade de se determinar a eficácia de seu uso, tendo em vista, as atuais
exigências do mercado global.
No Brasil, Oliveira et al. (2004) avaliaram o efeito do uso de antibióticos e
probiótico na criação de codornas de postura nas primeiras semanas de vida, e

10
concluíram que em bom ambiente, com água e ração disponíveis e de boa qualidade, a
adição de antibióticos e probiótico não têm efeito sobre o desempenho inicial.
Tendo em vista o papel do probiótico na prevenção do desenvolvimento de
colibacilos e na inibição de enterotoxinas no sistema digestivo (Fuller, 1977), poderia
ocorrer redução da quebra da proteína a nitrogênio, aumentando a utilização de
proteínas (aminoácidos), particularmente do alimento que não o contém em quantidades
desejáveis (Mikulec et al., 1999). Dessa forma, a utilização de dietas que contenham
níveis protéicos abaixo da exigência dos animais mostraria melhores resultados com o
uso de probióticos do que dietas que o contenham em quantidades adequadas.
Evidências desse resultado foram obtidas por Kos & Wittner (1982), que não
verificaram melhoras no crescimento e na conversão alimentar de frangos em engorda
pela adição de probióticos na ração que continha todos os nutrientes em quantidades
adequadas.
De maneira similar, Mikulec et al. (1999) demonstraram a influência favorável
que os probióticos têm sobre o crescimento da massa corporal e melhora na conversão
alimentar de frangos de corte quando o nível de proteína bruta da dieta é deficiente.
O efeito sobre o crescimento da massa corporal e conversão alimentar de frangos
de corte é significativamente maior quando os níveis de metionina, cistina e lisina no
alimento estão reduzidos (Dilworth & Day, 1978).

CITAÇÃO BIBLIOGRÁFICA
ANDREATTI FILHO, R. L.; SAMPAIO, H. M. Probióticos e prebióticos: realidade na avicultura industrial moderna. Revista CRMV/SP, v.2, fasc. 2, p.59-71, 1999.
ANDREATTI FILHO, R. L.; SILVA, E. N.; RIBEIRO, A. R. et al. Use of anaerobic cecal microflora, lactose and acetic acid for the protection of broiler chicks against experimental infection with Salmonella enteritidis. Brazilian Journal of Microbiology, v. 31, n. 2, 2000.
BRADLEY, G. L.; SAVAGE, T. F.; TIMM, K. I. The effects of supplementing diets with Saccharomyces cerevisiae var. boulardii on male poult performance and ileal morphology. Poultry Science, v. 73, p. 1766-1770, 1994.
CANALLI, L. S.; FLEMMING, J. S.; MIRA, R.T. et al. Alteração da microbiota intestinal de frangos de corte pela utilização de probiótico na alimentação. Agrárias, v.15, n.1, p. 125-132, 1996.
CORRÊA, A. B.; CORRÊA, G. S. S.; SILVA, M. A. et al. Efeito de diferentes níveis de proteína bruta e energia metabolizável sobre características de codornas de corte avaliadas durante a fase de crescimento. IN: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41., 2004, Campo Grande. Anais... Campo Grande: Sociedade Brasileira de Zootecnia, [2004]. CD-ROM. Nutrição de não-ruminantes. NNR018.
DILWORTH, B.C.; DAY, E. J. Lactobacillus cultures in broiler diets. Poultry Science, v. 57, p. 1101, 1978.
DOBROGSZ, W. J.; BLACK, B. L.; CASAS, I. A. Delivery of viable Lactobacillus reuteri to the gastrointestinal tract of poultry. Poultry Science, v. 70, p. 158, 1991.
EDENS, F. W.; PARKHURST, C. R.; CASAS, I. A. et al. Principles of ex ovo competitive exclusion and in ovo administration of Lactobacillus reuteri. Poultry Science, v.76, p.179-196, 1997.
FUJIKURA, W. S. Situação e perspectivas da coturnicultura no Brasil. In: SIMPÓSIO INTERNACIONAL DE COTURNICULTURA, 1., 2002, Lavras. Anais... Lavras: Universidade Federal de Lavras, 2002. p.1.
FULLER, R. The importance of lactobacili in maintaining normal microbial balance in the crop. British Poultry Science, v. 18, p. 85-94, 1977.

12
FULLER, R. Probiotics in man and animals. Journal of Applied Bacteriology, v. 66, p. 365-378, 1989.
GARCIA, E. A. Codornas para produção de carne. In: SIMPÓSIO INTERNACIONAL DE COTURNICULTURA, 1., 2002, Lavras. Anais... Lavras: Universidade Federal de Lavras, 2002. p. 97.
INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Disponível em: http://www.sidra.ibge.gov.br/bda. Acesso em: 5 abr. 2006.
JIN, L. Z.; HO, Y. W.; ABDULLAH, N. et al. Growth performance, intestinal microbial populations, and serum cholesterol of broilers fed diets containing Lactobacillus cultures. Poultry Science, v. 77, p. 1259-1265, 1998.
JIN, L. Z.; HO, Y. W.; ABDULLAH, N. et al. Probiotics in poultry : modes of action. World’s Poultry Science Journal, v. 53, p. 351-368, dez. 1997.
JIN, L. Z.; HO, Y. W.; ABDULLAH, N. et al. Influence of dried Bacillus subtilis and Lactobacilli cultures on intestinal microflora and performance in broilers. Asian-Australian Journal Animal Science, v.9, p.397-404, 1996.
KOS, K.; WITTNER, V. Use of probiotics in the nutrition of the fattening chicks. Praxis Veterinary, v. 30, p. 283-286, 1982.
LANCINI, J. B. Fatores exógenos na função gastrointestinal. In: FUNDAÇÃO APINCO DE CIÊNCIAS E TECNOLOGIA. Fisiologia da digestão e absorção das aves. Campinas: FACTA, 1994. p. 99-126.
LIILY, D. M.; STILLWELL, R. H. Probiotics: growth promoting factors produced by microorganisms. Science, v. 147, p.747-748, 1965.
LODDI, M. M.; GONZALES, E.; TAKITA, T. S. et al. Uso de probiótico e antibiótico sobre o desempenho, o rendimento e a qualidade de carcaça de frangos de corte. Revista Brasileira de Zootecnia, v. 29, n.4, p. 1124-1131, 2000.
MAIORKA, A.; SANTIN, E.; SUGETA, S. M. et al. Utilização de prebióticos, probióticos ou simbióticos em dietas para frangos. Revista Brasileira de Ciência Avícola, v. 3, n.1, 2001.
MIKULEC, Z.; SERMAN, V.; MAS, N. et al. Effect of probiotic on production results of fattened chickens fed different quantities of protein. Veterinarski Arhiv, v. 69, n.4, p. 199-209, 1999.
MULDER, R.W.A.W. Probiotics as a tool against Salmonella contamination. World’s Poultry Science Journal, v.7, n. 3, p. 36-37, 1991.
MURAKAMI, A. E.; ARIKI, J. Produção de codornas japonesas. Jaboticabal: FUNEP, 1998, 79p.
MURAKAMI, A.E. Nutrição e alimentação de codornas japonesas em postura. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, 2002. p. 283-309.
NURMI, E.; RANTALA, M. New aspects of Salmonella infection in broiler production. Nature, v. 241, p. 210-211, 1973.

13
OLIVEIRA, B. L.; OLIVEIRA, D. D.; OLIVEIRA, A. I. G. Utilização de probióticos e antibióticos na fase inicial da criação de codornas (Coturnix coturnix japonica). In: SIMPÓSIO DE COTURNICULTURA, 2., 2004, Lavras. Anais... Lavras: Universidade Federal de Lavras, [2004]. CD-ROM.
OLIVEIRA, D.D. Sanidade de codornas. In: SIMPÓSIO INTERNACIONAL DE COTURNICULTURA, 2., 2004, Lavras. Anais... Lavras: Universidade Federal de Lavras, [2004]. CD-ROM.
OLIVEIRA, N. T. E.; SILVA, M. A.; SOARES, R. T. R. N. et al. Exigências de proteína bruta e energia metabolizável para codornas japonesas machos criadas para a produção de carne. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 54, n.2, 2002a. Disponível em: http://www.scielo.br. Acesso em: 5 abr. 2006.
OLIVEIRA, E. G.; ALMEIDA, M. I. M.; MENDES, A. A. et al. Desempenho produtivo de codornas de ambos os sexos para corte alimentadas com dietas com quatro níveis protéicos. Archives of Veterinary Science, v.7, n.2, p.75-80, 2002b.
OLIVEIRA, G. H.; BERCHIERI JUNIOR, A.; BARROW, P. A. Preventions of Salmonella infection by contact using intestinal flora of adult birds and/or a mixture of organic acids. Brazilian Journal of Microbiology, v. 31 n. 2, 2000.
PARKER, R. B. Probiotics, the other half of the antibiotic story. Animal Nutrition and Health, v. 29, p. 4-8, 1974.
PELICANO, E. R. L.; SOUZA, P. A.; SOUZA, H. B. A. et al. Intestinal mucosa development in broiler chickens fed natural growth promoters. Brazilian Journal of Poultry Science, v. 7, n. 4, p. 221-229, 2005.
PELICANO, E. R. L.; SOUZA, P. A.; SOUZA, H. B. A. et al. Morfometria e ultra-estrutura da mucosa intestinal de frangos de corte alimentados com dietas contendo diferentes probióticos. Revista Portuguesa de Ciências Veterinárias, v. 98, n. 547, p. 125-134, 2003.
REZENDE, M. J. M; FLAUZINA, L. P.; McMANUS, C. et al. Desempenho produtivo e biometria das vísceras de codornas francesas alimentadas com diferentes níveis de energia metabolizável e proteína bruta. Acta Scientiarum, v. 26, n. 3, p. 353-358, 2004.
TORTUERO, F. Influence of implantation of Lactobacillus acidophilus in chicks on the growth, feed conversion, malabsorption of fats síndrome and intestinal flora. Poultry Science, v.52, p.197-203, 1973.
VANBELLE, M.; TELLER, E.; FOCANT, M. Probiotics in animal nutrition: a review. Archives of Animal Nutrition, v. 40, p. 477-484, 1990.
WALKER, W. A.; DUFF, L. C. Diet and bacterial colonization: role of probiotics and prebiotics. Journal Nutrition Biochemical, v. 9, p. 668-675, 1998.

II - OBJETIVOS GERAIS
Avaliar o efeito da adição de probiótico à base de Lactobacillus sp. na alimentação
de codornas de corte (Coturnix coturnix sp.) sobre o desempenho, rendimento de
carcaça e de cortes, exigências nutricionais de proteína bruta, população microbiana do
intestino delgado e morfometria da mucosa intestinal.

15
III – Diferentes vias de administração de probiótico sobre o desempenho,
rendimento de carcaça e população microbiana do intestino delgado de
codornas de corte
RESUMO: O objetivo deste trabalho foi avaliar o efeito de diferentes vias de
administração de probiótico sobre o desempenho no período de um a 35 dias de idade,
rendimento de carcaça aos 35 dias de idade e a população microbiana do intestino
delgado de codornas de corte aos sete e aos 14 dias de idade. Foram utilizadas 1.050
codornas distribuídas em um delineamento inteiramente casualizado com três
tratamentos (T1 - controle, T2 -probiótico na água bebida e T3 - probiótico na ração) e
cinco repetições de 70 aves cada. O probiótico era composto por uma cultura de
Lactobacillus sp. e foi fornecido durante a primeira semana de vida. A cada sete dias, as
aves foram pesadas para avaliação do ganho de peso (GP), consumo de ração (CR),
conversão alimentar (CA) e viabilidade criatória. Aos sete aos 14 dias, uma ave de cada
repetição foi abatida para a determinação da contagem total de Lactobacillus sp.,
enterobactérias e Escherichia coli do intestino delgado de codornas de corte. Aos 35
dias, duas aves de cada repetição foram abatidas para avaliação do rendimento de
carcaça e cortes cárneos. No período de sete a 14 dias de idade, as codornas do
tratamento T3, em relação àquelas do tratamento T2, apresentaram menor CR, porém,
não afetou o GP, o que levou à melhora na CA. De 21 a 28, 28 a 35 e de 1 a 35 dias de
idade, as codornas dos tratamentos T2 e T3 apresentaram menor CR. O rendimento de
carcaça e de cortes, a contagem total de Lactobacillus sp., enterobactérias e Escherichia
coli não foram influenciados pela adição do probiótico. Conclui-se que a adição do
probiótico (Lactobacillus sp.) por sete dias não melhorou o desempenho, o rendimento
de carcaça e não alterou a flora microbiana do intestino delgado de codornas de corte.
Palavras-chave: conversão alimentar, Escherichia coli, lactobacilos

16
III – Different probiotic administration ways on performance, yield carcass
and small intestine microbial population of meat quail
ABSTRACT: The aim of this study was to evaluate the effect of probiotic
administration different ways on performance from 1 to 35 days of age, carcass yield at
35 days of age and small intestine microbial population at 7 and 14 days of age of meat
quail. A total of 1050 quails were used, distributed in a completely randomized design
with three treatments (T1 - control, T2 – probiotic in drinking water and T3 – probiotic in
ration) and five replications of seventy quail each. Probiotic was compound of
Lactobacillus sp. culture and was supplemented during first weeks of life. Each seven
days, quails were weight to evaluate weight gain (WG), feed intake (FI), feed
conversion (FC) and growth’s viability. At 7 and 14 days, one quail of each replication
was slaughtered to perform the total count of Lactobacillus sp., enterobacteria and
Escherichia coli of small intestine of meat quails. At 35 days, two quails of each
replication were slaughtered to evaluate the carcass and cut yields. From 7 to 14 days of
age, the quails of T3 treatment, compared with T2 treatment, showing lower FI, however,
did not affect the WG, which improved FC. From 21 to 28, 28 to 35 and 1 to 35 days of
age, quails from treatments T2 and T3 presented lower FI. Carcass and cut yields, total
count of Lactobacillus sp., enterobacteria and Escherichia coli were not influenced by
probiotic addition. It was possible to conclude that probiotic addition (Lactobacillus sp.)
at seven day did not improve performance, carcass yield and also did not alter small
intestine microbial flora of meat quail.
Key Words: feed conversion, Escherichia coli, lactobacillus

17
Introdução
O crescimento cada vez maior da industrialização e a necessidade de atender ao
mercado globalizado exigem dos produtores rurais a utilização de modernas tecnologias
ligadas à produção animal. Nesse contexto, a existência de um empreendimento
agropecuário lucrativo é quase impossível, sem o uso da tecnologia na busca da
eficiência produtiva, o que leva a uma dependência cada vez maior do uso nos animais
de criação de substâncias químicas como os aditivos, em especial os antimicrobianos
(Palermo-Neto, 2001).
Além disso, nos últimos anos, com a intensificação dos sistemas de produção
animal, os animais são criados em densidades cada vez mais elevadas, o que tem
aumentado os riscos de disseminação de agentes patogênicos e instalação de processos
patológicos, visto que os sistemas de desinfecção de granjas não permitem a criação de
animais em ambientes isentos de microrganismos patogênicos (Santos et al., 2003).
Dessa forma, o uso de aditivos, como os antimicrobianos promotores de crescimento,
tornou-se quase que uma obrigatoriedade.
As atuais restrições impostas pela União Européia, a exigência dos consumidores
por produtos livres de resíduos de antimicrobianos e a preocupação relacionada à
possibilidade do desenvolvimento de resistência bacteriana cruzada entre humanos e
animais têm levado os produtores e pesquisadores a buscarem alternativas aos usuais
agentes promotores do crescimento.
Entre os principais promotores de crescimento alternativos estão os probióticos,
que são definidos como organismos e/ou substâncias que contribuem para o balanço
microbiano intestinal (Parker, 1974).

18
Os probióticos, segundo Chapman (1988), agem como promotores de crescimento,
conseqüentemente melhoram a produção e o estado geral de saúde dos animais, age por
exclusão competitiva e por liberação de substâncias que desfavorecem a colonização
por bactérias indesejáveis.
Especula-se ainda que os probióticos possam produzir bacteriocinas (Fuller &
Cole, 1988), economizar energia e aumentar a disponibilidade de enzimas e vitaminas
(Fuller, 1989), o que traz benefícios ao hospedeiro.
Uma das bases teóricas que apóia o uso dos probióticos na alimentação animal é a
de que a substituição da flora intestinal normal (em particular Escherichia coli), por
outra flora, como os Lactobacillus sp., melhoraria o desempenho e aumentaria a
resistência à colonização por microrganismos patogênicos pela melhora na saúde e na
digestão dos alimentos (Dale, 1992).
Particularmente para o controle da população de Escherichia coli, Fuller (1977)
relata que tal controle é dependente da presença de número suficiente de Lactobacillus e
que a partir de resultados de testes in vitro, parece ser necessário no mínimo 107
unidades formadoras de colônia por grama (UFC/g).
A ausência de resultados benéficos com o uso do probiótico pode ser explicada
pela diferença na quantidade e nas características das cepas do microrganismo utilizado,
e o não-atendimento dos requisitos para atuar como probiótico (Loddi et al., 2000).
Resultados extremamente promissores com a adição de probióticos na ração de
frangos de corte, com melhoras no ganho de peso e/ou conversão alimentar foram
obtidos por Santoso et al. (1995), Jin et al. (1998), Cuevas et al. (2000) e Fritts et al.
(2000). Entretanto, pesquisas conduzidas por Maiorka et al. (2001), Corrêa et al. (2003)
e Pelicano et al. (2004) mostraram melhoras nos índices de conversão alimentar de
frangos de corte somente no período de 1 a 21 dias.

19
Em contraposição, trabalhos conduzidos por Henrique et al. (1998 a,b), Loddi et
al. (2000 e 2002), Pedroso et al. (2002) e Lima et al. (2003) não mostraram qualquer
benefício do uso de probióticos sobre o ganho de peso ou o consumo de ração e a
conversão alimentar em qualquer fase da criação de frangos de corte.
Oliveira et al. (2004), em codornas de postura, avaliaram o efeito da adição de
probiótico na água de bebida e ração e diferentes antibióticos via água de bebida ou
ração ou ambos, durante a primeira semana de vida, mas não encontraram diferenças
significativas entre os tratamentos, no desempenho no período de um a 21 dias.
Com relação à melhora no rendimento de carcaça e de cortes com o uso de
probiótico na alimentação de frangos de corte, os resultados disponíveis na literatura
não mostram efeitos benéficos de seu uso sobre o rendimento de carcaça, peito, pernas e
dorso (Henrique, et al. 1998 a,b; Loddi et al., 2000 e 2002; Maiorka et al., 2001, Vargas
Jr. et al., 2002) entretanto, em pesquisas desenvolvidas por Corrêa et al. (2000 e 2003),
e Pelicano et al. (2003), observou-se melhora no rendimento de pernas com a adição de
probiótico.
O objetivo do presente trabalho foi avaliar as diferentes vias de administração de
probiótico sobre o desempenho, rendimento de carcaça e população microbiana do
intestino delgado de codornas de corte.
Material e Métodos
O experimento foi conduzido no Setor de Avicultura da Fazenda Experimental de
Iguatemi (FEI) da Universidade Estadual de Maringá (UEM), no período de março a
abril de 2005, sob aprovação do Comitê de Ética em Experimentação Animal –
CEEA/UEM.

20
Foram utilizadas 1.050 codornas de corte (Coturnix coturnix sp), machos e fêmeas
de um dia de idade, por um período experimental de 35 dias, alojadas em galpão
convencional, disposto no sentido leste-oeste, com cobertura de telha francesa, piso de
concreto e paredes laterais de alvenaria com 0,40 m de altura completadas com tela de
arame até o telhado. O galpão é dividido em boxes de 3,0 m2 onde foram alojadas 70
aves por boxe, e cinco boxes para cada tratamento, totalizando 15 boxes.
Os tratamentos consistiram de um tratamento-controle –T1 (sem adição de
probiótico) e mais dois, que foram utilizadas duas diferentes vias de administração do
probiótico: água de bebida (T2) e ração (T3); o probiótico fornecido somente na primeira
semana do período experimental, em uma mesma concentração de microrganismos.
O probiótico administrado para os animais dos tratamentos T2 e T3 continha em
média 1,6 x 109 unidades formadoras de colônia por grama (UFC/g) de um “pool” de
Lactobacillus sp., administrado na dosagem de 100g/tonelada, segundo recomendações
do fabricante FATEC®.
A ração dos animais do T3 foi suplementada com probiótico em uma quantidade a
ser consumida pelos animais durante a primeira semana de vida, e posteriormente
misturada em misturador Y por um período de 15 minutos. Já, para os animais do T2, o
probiótico foi adicionado diariamente à água não-clorada.
O delineamento experimental foi inteiramente casualizado com três tratamentos e
cinco repetições.
O premix mineral vitamínico (utilizado nas rações) foi formulado para não conter
qualquer tipo de aditivo.
A ração durante todo o período experimental foi fornecida à vontade e formulada
segundo as exigências de Barral (1994) e a composição dos alimentos segundo dados
apresentados por Rostagno et al. (2005) (Tabela 1).

21
Tabela 1: Composição percentual e química da ração experimental
Table 1: Percentual and chemical composition of the experimental diets Alimentos Feeds
Composição percentual Percentual composition
Milho Corn 41,10
Farelo de soja Soybean meal 50,45
Calcário Limestone 1,05
Fosfato Bicálcico Dicalcium phosphate 1,22
Sal Comum Common salt 0,36
Premix vit.+ min.* Premix vit. + min. 0,40
Óleo de soja Vegetable oil 5,41
BHT BHT 0,01
Total Total 100
Valores calculados Calculated values Energia Metabolizável (Kcal/Kg) Metabolizable energy 3.000
Proteína Bruta (%) Crude protein 26,00
Fibra Bruta (%) Crude fiber 4,09
Cálcio (%) Calcium 0,80
Fósforo disponível (%) Available Phosphorus 0,30
Lisina Total (%) Total lysine 1,55
Met. + Cist. Total (%) Total Met + Cystine 0,83 * composição por kg do produto: Vit A, 2500.000 UI; Vit. D3, 625.000 UI; Vit. E, 3.750mg; Vit B1, 138mg; Vit. B2, 1.250mg; Vit. B6, 523mg; Vit B12, 5.000mcg; Vit. K3, 750mg; ácido fólico, 112 mg; pantotenato de cálcio 2.000 mg; niacina, 7.500 mg; metionina, 275.000 mg; colina, 87.500 mg; selênio, 63 mg; antioxidante, 2500mg; iodo, 240mg; veículo q.s.p., 1000g; zinco, 15.750mg; ferro, 12.250mg; manganês, 19.375mg; cobre, 3.900mg; cobalto, 50mg. * composition per kg: Vit A, 2500,000 UI; Vit. D3, 625,000 UI; Vit. E, 3,750mg; Vit BB1, 138mg; Vit. B2, 1,250mg; Vit. B6, 523mg; Vit B12, 5,000mcg; Vit. K3, 750mg; folic acid, 112 mg; calcium panthotenato, 2,000 mg; niacine, 7,500 mg; methionine, 275,000 mg; choline, 87,500 mg; selenium, 63 mg; antioxidant, 2,500mg; iodine, 240mg; veicul q.s.p., 1000g; zinc, 15,750mg; iron, 12,250mg; manganese, 19,375mg; copper, 3,900mg; cobalt, 50mg.
O desempenho das aves foi avaliado por meio do consumo ração, ganho de peso,
conversão alimentar e viabilidade criatória (aves vivas/total de aves*100) no período
experimental.

22
Aos 35 dias de idade foram retiradas ao acaso quatro aves de cada unidade
experimental (20 aves por tratamento) para avaliação do rendimento de carcaça e cortes
cárneos. As aves foram identificadas individualmente por meio de anilhas numeradas
colocadas nas patas e submetidas a um período de jejum alimentar de oito horas, com
água à vontade até o momento do carregamento.
Após a pesagem individual, o abate foi realizado por degola completa entre os
ossos occipital e atlas. As aves foram escaldadas por 15 a 20 segundos a uma
temperatura de 55 a 600C; e a seguir submetidas a depena manual. Em seguida, efetuou-
se a evisceração manual e a pesagem da carcaça. As partes da carcaça foram separadas e
pesadas para determinação do rendimento de peito, pernas, dorso e asa.
Para o cálculo de rendimento de carcaça, foi considerado o peso da carcaça
eviscerada, sem os pés, cabeça e gordura abdominal, em relação ao peso vivo. Para o
rendimento de cortes cárneos, foi considerado o rendimento de peito inteiro com pele,
pernas (coxa e sobrecoxa com pele), dorso e asas que foi calculado em relação ao peso
da carcaça eviscerada.
Aos sete e aos 14 dias, foi colhida uma amostra de intestino delgado de cada
unidade experimental (cinco amostras por tratamento), perfazendo um total de 30
amostras, para avaliar a influência da administração do probiótico sobre a contagem
total de Lactobacillus sp., Enterobactérias e Escherichia coli do intestino delgado de
codornas de corte.
Para isso, as aves foram abatidas por meio de degola completa entre os ossos
occipital e atlas. O material utilizado para coleta do intestino delgado foi, previamente,
esterilizado em autoclave por 30 minutos e o intestino delgado após separado, foi
pesado em placas de Petri estéreis e, em seguida, picados para acondicionamento em
tubos com tampa de rosca, contendo nove mL de solução salina estéril e pérolas de

23
vidro para auxiliar na trituração e homogeneização do material colhido para posterior
análise microbiológica. Esses tubos foram mantidos em banho de gelo até a realização
das análises no Laboratório de Microbiologia da UEM.
A contagem total de Lactobacillus sp. foi feita utilizando-se o ágar Rogosa
(Lactobacillus Selective Agar) e a contagem total de enterobactérias, o ágar EMB
(Eosin Methylene-blue Lactose Sucrose Agar), de acordo com metodologia descrita no
Manual Merck (1994).
As amostras foram diluídas até 10-4, plaqueadas em duplicata e incubadas a 37ºC
por 72 horas (Rogosa) e 24 horas (EMB). Para o ágar Rogosa, foi necessário uso da
técnica de profundidade e as placas depois de semeadas e cobertas com o restante do
ágar foram colocadas em jarras de anaerobiose. Após a incubação, as colônias foram
contadas e os resultados foram expressos em UFC/g (unidade formadora de
colônia/grama) de intestino delgado.
As colônias isoladas, que cresceram no ágar, EMB foram estocadas em ágar
nutriente e submetidas posteriormente aos testes bioquímicos, utilizando-se para isso
dos meios para identificação bacteriana da Probac® do Brasil (Enterokit B), com o
objetivo de realizar a identificação bacteriana.
Para análise estatística da contagem de Escherichia coli, no material colhido,
utilizou-se de somente duas repetições de cada tratamento.

24
A análise estatística das variáveis de desempenho e rendimento de carcaça foi
realizada levando-se em consideração o modelo estatístico:
Yij = μ + Ti + eij
Yij = variáveis estudadas obtidas com o indivíduo j, recebendo o tratamento i;
μ = constante geral;
Ti = efeito do tratamento i (sendo i1 = controle; i2 = probiótico água; i3 = probiótico
ração);
eij = erro aleatório associado a cada observação Yij.
Os dados de desempenho e rendimento de carcaça obtidos foram submetidos à
análise de variância e as médias comparadas pelo teste de Tukey em nível de 5% de
significância, utilizando o programa SAEG, desenvolvido pela Universidade Federal de
Viçosa – UFV (UFV, 1999).
A análise estatística da microbiologia foi feita por meio do método de modelos
lineares generalizados (Nelder & Wedderburn, 1972), admitindo distribuição gama com
função de ligação recíproco, e nesse caso o modelo empregado foi:
η = μ + Ti + Ij + eijk;
η = observação relativa a cada indivíduo k, recebendo o tratamento i, na idade j;
μ = constante geral;
Ti = efeito do tratamento i (sendo i1 = controle; i2 = probiótico água; i3 =
probiótico ração);
Ij = efeito da idade j (sendo j1 = 7 dias; j2 = 14 dias).
Assim, conforme McCulloch & Searle (2001), quando a distribuição foi gama, a
esperança de Y (média da variável dependente) é dada por:
Ε (Yijk) = 1/ηijk
Enquanto que para a distribuição normal é dada por:
Ε (Yijk) = ηijk

25
Resultados e Discussão
No decorrer do período experimental, a temperatura média registrada no interior
do galpão foi: máxima 300C e mínima de 240 C.
Os resultados de peso vivo (Tabela 2), nas diferentes idades, mostraram menor
peso aos 14 dias (P<0,05) para as codornas que receberam probiótico, via água de
bebida, quando comparados ao controle.
Tabela 2: Médias dos pesos vivos das codornas com 1, 7, 14, 21, 28 e 35 dias de idade, recebendo o probiótico (Lactobacillus sp.) sob duas diferentes vias de administração
Table 2: Means of live weight of quails with 1, 7, 14, 21, 28 and 35 days of age, receiving probiotic (Lactobacillus sp.) under two different ways of administration
Peso vivo (g) Live weight (g) Tratamentos
Treatments 1 dia Day 1
7 dias Day 7
14 dias Day 14
21 dias Day 21
28 dias Day 28
35 dias Day 35
Controle (T1) Control 9,37 34,31 87,16a 146,24 206,56 255,06
Probiótico na água (T2) Probiotic in Water 9,41 33,15 84,51b 143,81 203,23 249,57
Probiótico na Ração (T3) Probiotic in Feed 9,61 33,92 85,74ab 145,42 204,42 252,45
Coeficiente de variação Coefficient of variation 4,05 2,64 1,24 1,35 1,39 1,35
a,b Na mesma coluna, médias seguidas de letras diferentes, diferem pelo Teste de Tukey (P<0,05) a,b In the same column, means followed by different letters, differ by Tukey’s Test (P<0.05)
Esse resultado comprova que a adição do probiótico utilizado, via água de bebida,
não é a via mais adequada, uma vez que esses animais apresentaram maior consumo
ração (CR), no período de sete a 14 dias (Tabela 3), em comparação ao grupo que
recebeu via ração, e, portanto, era de se esperar que tivessem maior peso aos 14 dias,
fato esse que não ocorreu.
Por outro lado, quando se analisa os pesos médios aos sete, 14, 21, 28 e 35 dias de
idade, a adição do probiótico não mostrou melhoras significativas (Tabela 2), mas, com

26
exceção dos períodos de um a sete e de 14 a 21 dias, nos demais ocorreu menor CR
(Tabela 3), sem, contudo alterar o peso médio no final de cada período.
Os resultados de peso aos sete e 21 dias de idade estão de acordo com os obtidos
por Oliveira et al. (2004), que não verificaram efeito da adição de probiótico,
adicionado na primeira água de bebida, e na ração durante a primeira semana de vida,
sobre o peso das codornas nas mesmas idades. Da mesma forma, em frangos de corte,
Pedroso et al (2002) não encontraram diferenças no peso aos 21 dias quando da
suplementação de diferentes probióticos.
No período de um a sete dias de idade, período nos quais os animais dos
tratamentos T2 e T3, receberam, respectivamente, o probiótico via água de bebida e
ração, não houve diferenças (P>0,05) entre os tratamentos para as variáveis CR, ganho
de peso (GP) e conversão alimentar (CA) (Tabela 3).
Dos sete aos 14 dias de idade, os animais do T3 apresentaram significativamente
menor CR que os demais, porém, não afetou o GP, o que levou à melhora na CA
quando comparado com o tratamento T2, evidenciando efeito positivo da adição do
probiótico via ração. Cabe salientar que esta é a via recomendada pelo fabricante do
probiótico utilizado nessa pesquisa.
Esses resultados diferem dos obtidos por Maiorka et al. (2001) e Pelicano et al.
(2004), que encontraram melhoras na CA no período de um a 21 dias, no entanto, não
verificaram redução no CR de frangos de corte alimentados com dieta suplementada
com probiótico.
Por outro lado, Corrêa et al. (2003) encontraram tanto redução no CR quanto
melhora na CA de frangos de corte no período de um a 20 dias, quando se adicionou o
poliprobiótico Estibion (Lactobacillus acidophilus e casei; Streptococcus salivarium e
faecium; Bacillus subtilis e toyoi e Saccharomyces cerevisiae) à ração das aves.

27
Tabela 3: Efeito do probiótico (Lactobacillus sp.) administrado sob duas diferentes vias sobre o consumo de ração (CR), ganho de peso (GP), conversão alimentar (CA) e viabilidade (VI) de codornas de corte.
Table 3: Effect of probiotic (Lactobacillus sp.) administered under two different ways on feed intake (FI), weight gain (WG), feed conversion (FC) and viability (VI) of meat quail
TRATAMENTOS TREATMENTS Vias de administração do probiótico
Ways of probiotic administration VARIÁVEL VARIABLE
IDADE AGE Controle (T1)
Control Água (T2) Water
Ração (T3) Feed
CV %1
VC %1
1-7 39,04 38,59 38,36 3,15 7-14 95,06a 97,01a 90,72b 2,38
14-21 127,99 128,83 126,79 2,37 21-28 180,55 a 170,95 b 171,12 b 2,14 28-35 218,01 a 207,36 b 214,89 ab 2,74
CR (g) FI (g)
1-35 660,65 a 642,74 b 641,88 b 1,18 1-7 24,83 23,60 24,17 4,40
7-14 52,85 51,66 52,13 2,20 14-21 71,32 71,80 71,28 2,15 21-28 59,88 58,99 58,54 4,37 28-35 48,49 46,34 48,03 5,11
GP (g) WG (g)
1-35 257,37 252,39 254,15 1,42 1-7 1,57 1,64 1,59 3,60
7-14 1,80 ab 1,88 a 1,74 b 3,13 14-21 1,79 1,79 1,78 1,83 21-28 3,02 2,90 2,93 3,70 28-35 4,51 4,48 4,48 3,78
CA (g/g) FC (g/g)
1-35 2, 57 2,55 2,53 0,99 VI (%) VI (%) 1-35 98,286 98,286 97,714 1,823
1 Coeficiente de Variação/ 1 Coefficient of Variation a,b Na mesma linha, médias seguidas de letras diferentes, diferem pelo Teste de Tukey (P<0,05) a,b In the same line, means followed by different letters, differs by Tukey’s Test (P<0.05)
Resultados benéficos com o uso de probiótico, não somente na fase inicial de
criação, foram reportados por diferentes autores (Jin et al., 1998 e Fritts et al., 2000), os
quais obtiveram melhores pesos aos 21 e aos 42 dias e melhor conversão alimentar no
período de 21 a 42 dias, em frangos de corte que receberam probiótico.
Essas variações de resposta aos probióticos podem estar relacionadas com a
instabilidade da microbiota das aves na fase inicial, a qual está sujeita a grandes
alterações durante a primeira semana de vida das aves (Bunyan et al., 1977).

28
No período seguinte, dos 14 aos 21 dias de idade, novamente não foi verificado
diferenças entre os tratamentos para as variáveis CR, GP e CA.
Esses resultados podem refletir as boas condições de higiene em que as codornas
foram criadas, o que resultou em uma baixa condição de desafio sanitário. Nessas
condições de mínimo estresse e desafio, os probióticos podem não manifestar resultados
evidentes da eficácia de sua utilização (Fox, 1988; Dale, 1992; Maruta, 1993).
Dos 21 aos 28 e dos 28 aos 35 dias de idade, somente o CR sofreu influência dos
tratamentos, no qual os tratamentos com adição de probiótico, apresentaram menor CR.
No período total (um a 35 dias), os animais do tratamento-controle consumiram
significativamente mais ração (P<0,05) do que os tratamentos nos quais foram
adicionados probiótico por meio de água de bebida ou ração.
Dentre as várias ações benéficas do uso dos probióticos na alimentação animal,
pode-se citar o aumento da disponibilidade de aminoácidos nos locais de absorção
(Kozasa, 1989), economia de energia e aumento da disponibilidade de vitaminas e
enzimas (Fuller, 1989). Essas ações poderiam explicar o fato dos animais dos
tratamentos em que se adicionou probiótico via água de bebida ou ração, e terem menor
CR no período de um a 35 dias, sem que isso afetasse o GP e a CA.
Esses resultados diferem dos encontrados por Pelicano et al. (2004), os quais não
detectaram efeito da suplementação de probiótico, via ração, sobre o consumo de ração
de frangos de corte no período de um a 35 dias, e Loddi et al. (2000), que verificaram
um menor ganho de peso nos períodos de um a 21 e um a 42 dias quando se adicionou
probiótico na ração.
Os resultados de ganho de peso e conversão alimentar de frangos de corte obtidos
por Henrique et al. (1998 a,b), Loddi et al. (2002), Pedroso et al. (2002), Corrêa et al.
(2003), Lima et al. (2003) e Pelicano et al. (2004) corroboram àqueles que não

29
obtiveram efeito da adição do probiótico sobre esses parâmetros, no período de um a 42
dias.
Tendo em vista que a eficácia do produto é estritamente dependente da quantidade
e das características das cepas do microrganismo utilizado na elaboração do probiótico
(Tournut, 1998), fica difícil estabelecer um paralelo entre os estudos e comparar os
resultados.
A viabilidade, no período de um a 35 dias (Tabela 3), não foi influenciada pela
adição do probiótico, o que está de acordo com os resultados verificados em frangos de
corte por Pedroso et al. (2002) nos períodos de um a 21 e um a 42 dias e, Pelicano et al.
(2004), no período de um a 42 dias.
Os resultados de viabilidade obtidos, nesse experimento, denotam a boa condição
sanitária em que as aves foram criadas, já que, os animais do tratamento-controle,
mesmo não recebendo qualquer tipo de aditivo, apresentaram alta viabilidade.
A análise estatística dos resultados de rendimento de carcaça e de cortes
demonstrou não haver diferenças (P>0,05) entre os tratamentos (Tabela 4).
Tabela 4: Rendimento de carcaça e de cortes de codornas de corte, recebendo o probiótico Lactobacillus sp. sob duas diferentes vias de administração
Table 4: Carcass and cut yields of meat quails, receiving probiotic Lactobacillus sp. under two different administration ways
Tratamento Treatments
Rendimento Carcaça (%) Carcass yield
Asas (%) Wings
Dorso (%) Back
Pernas (%) Legs
Peito (%) Breast
Controle Control 70,04 6,24 24,78 23,28 45,18
Probiótico na água Probiotic in Water 70,01 6,37 25,10 24,04 43,96
Probiótico na Ração Probiotic in Feed 70,27 6,35 24,45 23,88 44,91
CV%1
CV% 1,84 4,46 4,46 3,14 1,57 1 Coeficiente de variação/ Coefficient of Variation
Resultados não diferiram pelo teste F (P>0,05)/ Results did not differ by F test (P>.05)

30
Esses resultados concordam com os obtidos por (Henrique et al., 1998 a,b; Loddi
et al 2000 e 2002; Maiorka et al., 2001 e Vargas Jr. et al., 2002), os quais não
observaram melhoras nas características de carcaça de frangos de corte com o uso do
probiótico.
Por outro lado, Corrêa et al. (2003), verificaram que frangos de corte (machos),
alimentados com poliprobiótico constituído de cepas de Lactobacillus acidophilus e
casei, Streptococcus salivarium e faecium, Bacillus subtilis e toyoi, e Saccharomyces
cerevisae, apresentaram maior rendimento de coxa em relação aos tratamentos em que
se adicionou bacitracina de zinco e probiótico, cujo ingrediente ativo eram os esporos de
Bacillus subtilis.
Resultados semelhantes, com melhoras no rendimento de pernas, foram obtidos
por Pelicano et al. (2003), em frangos de corte que receberam diferentes probióticos no
período de um a 45 dias de idade.
Não houve modificação na contagem com o aumento da idade e com a adição do
probiótico, com relação aos resultados de contagem total de Lactobacillus sp. no
intestino delgado de codornas de corte (Tabela 5).
Esses resultados são similares aos obtidos por Netherwood et al. (1999), os quais
também não verificaram alteração na contagem de Lactobacillus sp., quando adicionou
probiótico à base de Enterococcus faecium na água de bebida de galinhas.

31
Tabela 5: Contagem total de Lactobacillus sp. (UFC/g) no intestino delgado de codornas de corte aos 7 e aos 14 dias de idade, recebendo o probiótico Lactobacillus sp. sob duas diferentes vias de administração
Table 5: Total count of Lactobacillus sp. (UFC/g) in meat quails small intestine at 7 and 14 days of age, receiving probiotic Lactobacillus sp. under two different ways of administration
TRATAMENTOS Treatments
Idade (dias) Age (days)
Média Mean
Desvio padrão Standard deviation
Controle Control 15,93 x 104 25,33
Probiótico na água Probiotic in water 229,85 x 104 396,80
Probiótico na ração Probiotic in ration
7
45,24 x 104 58,22
Controle Control 333,19 x 104 570,23
Probiótico Água Probiotic in water 75,26 x 104 116,95
Probiótico ração Probiotic in ration
14
362,53 x 104 370,77 Os resultados não diferiram pelo teste Student (P>0,05)/ Results did not differ by Student test (P>0.05)
Jin et al. (1998) verificaram que, aos dez dias de idade, a população de
Lactobacillus no intestino delgado de frangos, ao receber 0,15% de uma cultura de
Lactobacillus foi significativamente maior que nas aves do grupo-controle. Mas, nessa
pesquisa a quantidade de Lactobacillus fornecida aos animais foi 15 vezes superior à
utilizada nesse experimento.
Provavelmente, os resultados obtidos nesse experimento refletem a não-
colonização do intestino delgado pelo Lactobacillus utilizado, tendo em vista que o
efeito do uso dos Lactobacillus como promotor de crescimento se deve ao fato da
bactéria ser capaz de sobreviver e colonizar o trato gastrintestinal até que suas funções
benéficas possam ser exercidas (Jin et al., 1998), fato esse que não ocorreu.
A redução na contagem total de enterobactérias em função da idade e da adição do
probiótico, também não foi verificada (Tabela 6).

32
Diferenças estatísticas entre os tratamentos não foram encontradas para a
população microbiana (Tabelas 5, 6 e 7), por causa da alta variabilidade dos dados,
apesar da metodologia de análise utilizada ser adequada a este tipo de dados.
Tabela 6: Contagem total de enterobactérias (UFC/g) no intestino delgado aos 7 e aos 14 dias de idade de codornas de corte, recebendo o probiótico Lactobacillus sp. sob duas diferentes vias
Table 6: Total count of enterobacteria (UFC/g) in small intestine at 7 and 14 days of age of meat quails, receiving probiotic Lactobacillus sp. under two different ways
TRATAMENTOS Treatments
Idade (dias) Age (days)
Média Mean
Desvio padrão Standard deviation
Controle Control 262,8 x 104a 358,53
Probiótico Água Probiotic in water 449,5 x 104a 545,27
Probiótico ração Probiotic in ration
7
170,3 x 104a 151,19
Controle Control 843,6 x 104a 1621,51
Probiótico Água Probiotic in water 107,5 x 104a 41,92
Probiótico ração Probiotic in ration
14
1244,3 x 104a 1971,98 Na mesma coluna, médias seguidas de mesma letra, não diferem pelo teste Student (P>0,05) In the same column, means followed by same letters did not differ by Student Test (P<0.05)
Jin et al. (1998) não observaram redução na população de coliformes do
intestino delgado de frangos de corte aos dez, 20, 30 e 40 dias de idade, que receberam
cultura de Lactobacillus em concentrações de 0,05; 0,10 e 0,15%. Entretanto, no ceco,
verificaram redução significativa com as três concentrações aos 10 dias; 0,05 e 0,10%
aos 20 dias e de 0,05% aos 30 dias.
Os resultados obtidos por esses autores evidenciam a diferença de
comportamento da microbiota intestinal em função da concentração do probiótico
utilizado, da porção do intestino e da idade na qual foi feita a mensuração.

33
Francis et al. (1978) verificaram, em perus, que a adição de probiótico à base de
Lactobacillus, em uma concentração de 75 mg/Kg de ração, diminui significativamente
a contagem de coliformes no ceco e intestino delgado.
Uma das bases teóricas que apóia o uso dos probióticos na alimentação animal é a
de que a substituição da flora intestinal normal (em particular Escherichia coli), por
outra flora, como os Lactobacillus sp., melhoraria o desempenho e aumentaria a
resistência à colonização por microrganismos patogênicos, pela melhora na saúde e na
digestão dos alimentos (Dale, 1992).
No presente experimento, porém, não foi verificado redução na contagem total de
Escherichia coli, em função da suplementação do probiótico (Tabela 7),
conseqüentemente, a melhora no desempenho não foi constatada, o que pode refletir a
ausência da adesão do probiótico ao epitélio intestinal e posterior colonização pela
inabilidade de utilizar os substratos disponíveis ou a sensibilidade aos agentes
antibacterianos presentes (Fuller, 1986).
Tabela 7: Contagem total de Escherichia coli (UFC/g) no intestino delgado em função
dos tratamentos e idade das codornas de corte Table 7: Total count of Escherichia coli (UFC/g) in small intestine in function of treatments
and age of meat quails Tratamentos Treatments
Média Mean
Desvio padrão Standard deviation
Controle Control
21,10 x 104ª 34,70
Probiótico Água Probiotic in water
19,22 x 104ª 22,20
Probiótico ração Probiotic in ration
51,41 x 104ª 66,59
Idade Age
Média Mean
Desvio padrão Standard deviation
7 dias 7 days
49,21 x 104ª 54,42
14 dias 14 days
11,94 x 104ª 20,15
Na mesma coluna, médias seguidas de mesma letra, não diferem pelo teste Student (P>0,05) In the same column, means followed by same letters did not differ by Student Test (P<0.05)

34
Diferentemente dos resultados obtidos nesse experimento, Jin et al. (1996) e
Edens et al. (1997) verificaram efeito protetor contra a colonização por Escherichia coli,
em frangos de corte, que receberam, respectivamente, probiótico à base de Bacillus
subtilis ou Lactobacillus sp.; e Lactobacillus reuteri.
Particularmente, para o controle da população de Escherichia coli, Fuller (1977),
relata que tal controle é dependente da presença de número suficiente de Lactobacillus e
que a partir de resultados de testes in vitro parece ser necessário, no mínimo 107UFC/g.,
o que está acima dos valores de contagem de Lactobacillus sp. encontrados no intestino
delgado de codornas de corte obtidos nessa pesquisa (Tabela 6).
Conclusões
A adição do probiótico (Lactobacillus sp.), independente da via, não melhorou o
desempenho, o rendimento de carcaça e a população microbiana do intestino delgado de
codornas de corte.

35
Literatura Citada
BARRAL, A. D. Manual de la Codorniz: cria industrial y para la caza. Lleida: Dilagro, 1994. 268p.
BUNYAN, J.; JEFRIES, L.; SAYERS, J. R. et al. Antimicrobial substances and chick growth promotion: the growth-promoting activities of antimicrobial substances, including fifty-two used either in therapy or as dietary additives. British Poultry Science, v. 18, p. 283-294, 1977.
CHAPMAN, J. D. Probiotics, acidifiers and yeast culture: a place for natural aditives in pig and poultry production. Biotechnology in the feed industry, 1988, p. 219-229.
CORRÊA, G. S. S.; GOMES, A. V. C.; CORRÊA, A. B. et al. Desempenho de frangos de corte alimentados com diferentes promotores de crescimento. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 37., 2000, Viçosa. Anais...Viçosa: Sociedade Brasileira de Zootecnia, 2000. p.37.
CORRÊA, G. S. S.; GOMES, A.V.C.; CORRÊA, A. B. et al. Efeito de antibiótico e probióticos sobre o desempenho e rendimento de carcaça de frangos de corte. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 55, n.4, p. 467-473, 2003.
CUEVAS, A. C. GONZÁLES, E. A.; HUGUENIN, M. T. C. et al. El efecto del Bacillus toyoii sobre el comportamiento productivo en pollos de engorda. Veterinária México, v.31, n.4, 2000. Disponível em: <http:// www.ejournal.unam.mx/vet_mex/vol31-04/RVM31405.pdf>. Acesso em: 10 fev. 2006.
DALE, N. Probióticos para aves. Avicultura Professional, v. 10, p. 88-89, 1992.
EDENS, F. W.; PARKHURST, C. R.; CASAS, I. A. et al. Principles of ex ovo competitive exclusion and in ovo administration of Lactobacillus reuteri. Poultry Science, v.76, p.179-196, 1997.
FOX, S. M. Probiotics: intestinal inoculants for production animals. Veterinary Medicine, v. 83, p. 806-830, 1988.
FRANCIS, C.; JANKY, D. M.; ARAFA, A. S. et al. Interrelationship of Lactobacillus and zinc bacitracin in diets of turkey poults. Poultry Science, v.57, p. 1687-1689, 1978.
FRITTS, C. A.; KERSEY, J. H.; MOTL, M. A. et al. Bacillus subtilis C-3102 (Calsporin) improves live performance and microbiological status broiler chickens. Journal of Applied Poultry Research, v. 9, n.2, p. 149-155, 2000.
FULLER, R. Probiotics: the scientific basis. London: Chapman & Hall, 1989. 307p.
FULLER, R.; COLE, C. B. The scientific basis of the probiotics concept. In: Probiotics theory and applications. Marlows: Chalcombe, 1988. p. 1-14.
FULLER, R. Probiotics. Journal of Applied Bacteriology Supplement, p. 1-7, 1986.
FULLER, R. The importance of lactobacili in maintaining normal microbial balance in the crop. British Poultry Science, v. 18, p. 85-94, 1977.

36
HENRIQUE, A. P. F.; FARIA, D. E.; FRANZOLIN, R. et al. Uso de probióticos e antibióticos como promotores de crescimento para frangos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, [1998a]. CD-ROM. Nutrição de não-ruminantes. NRU008.
HENRIQUE, A. P. F.; FARIA, D. E.; FRANZOLIN, R. et al. Efeito de ácido orgânico, probiótico e antibiótico sobre o desempenho e rendimento de carcaça de frangos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, [1998b].
CD-ROM. Nutrição de não-ruminantes. NRU009.
JIN, L. Z.; HO, Y. W.; ABDULLAH, N. et al. Influence of dried Bacillus subtilis and Lactobacilli cultures on intestinal microflora and performance in broilers. Asian-Australian Journal Animal Science, v.9, p.397-404, 1996.
JIN, L. Z.; HO, Y. W.; ABDULLAH, N. et al. Growth performance, intestinal microbial populations, and serum cholesterol of broilers fed diets containing Lactobacillus cultures. Poultry Science, v. 77, p. 1259-1265, 1998.
KOZASA, M. Probiotics for animal use in Japan. Veterinary Science Technology Off. Int. Epiz., v.8, n.2, p. 517-531, 1989.
LIMA, A. C. F.; PIZAURO JÚNIOR, J. M.; MACARI, M. et al. Efeito do uso de probiótico sobre o desempenho e atividade de enzimas digestivas de frangos de corte. Revista Brasileira de Zootecnia, v. 32, n.1, p.200-207, 2003.
LODDI, M. M.; GONZALES, E.; TAKITA, T. S. et al. Uso de probiótico e antibiótico sobre o desempenho, o rendimento e a qualidade de carcaça de frangos de corte. Revista Brasileira de Zootecnia, v. 29, n.4, p. 1124-1131, 2000.
LODDI, M. M.; TUCCI, F. M.; HANNAS, M. I. et al. Probióticos, mananoligossacarídeos + ácidos orgânicos em dietas de frangos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, [2002]. CD-ROM. Nutrição de não-ruminantes.
MAIORKA, A.; SANTIN, E.; SUGETA, S. M. et al. Utilização de prebióticos, probióticos ou simbióticos em dietas para frangos. Revista Brasileira de Ciência Avícola, v. 3, n.1, 2001. Disponível em: http://www.scielo.br . Acesso em: 5 mar. 2006.
MARUTA, K. Probióticos e seus benefícios. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1993, Santos. Anais... Santos: Fundação Apinco de Ciência e Tecnologias Avícolas, 1993. p. 203-219.
MERCK. Microbiology Manual. Darmstadt: Merck KGoA, 2000. 407p.
NETHERWOOD, T.; GILBERT, H. J.; PARKER, D. S. et al. Probiotics shown to change bacterial community structure in the avian gastrointestinal tract. Applied and Environmental Microbiology, v. 65, n. 11, p. 5134-5138, 1999.
OLIVEIRA, B. L.; OLIVEIRA, D. D.; OLIVEIRA, A. I. G. Utilização de probióticos e antibióticos na fase inicial da criação de codornas (Coturnix coturnix japonica). In: SIMPÓSIO DE COTURNICULTURA, 2., 2004, Lavras. Anais... Lavras: Universidade Federal de Lavras, [2004].CD-ROM.

37
PALERMO-NETO, J. Resíduos de antimicrobianos em alimentos. Revista CFMV, 2001, p. 65-71.
PARKER, R. B. Probiotics: the other half of the antibiotics story. Animal Nutrition and Health, v. 29, n.2, p. 4-8, 1974.
PEDROSO, A. A.; MENTEN, J. F. M.; LONGO, F. A. et al. Desempenho de frangos de corte recebendo ração suplementada com aditivos microbianos e criados em baterias ou em galpão convencional. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, [1998]. CD-ROM. Nutrição de não-ruminantes.
PELICANO, E. R. L.; SOUZA, P. A.; SOUZA, H. B. A. et al. Effect of different probiotics on broiler carcass and meat quality. Revista Brasileira de Ciência Avícola, v.5, n.3, p. 207-214, 2003.
PELICANO, E. R. L.; SOUZA, P. A.; SOUZA, H. B. A. et al. Productive traits of broiler chickens fed diets containing different growth promoters. Revista Brasileira de Ciência Avícola, v. 6, n.3, p. 177-182, 2004.
ROSTAGNO, H. S. ALBINO, L. F. T. DONZELE, J. L. et. al. Tabelas brasileiras para aves e suínos: composição de alimentos e exigências nutricionais de aves e suínos. Viçosa: UFV, 2005. 141p.
SANTOS, M. S.; FERREIRA, C.L.L.F.; GOMES, P. C. et al. Influência do fornecimento de probiótico à base de Lactobacillus sp. sobre a microbiota intestinal de leitões. Ciências Agrotecnológicas, v.27, n.6, p. 1395-1400, 2003.
SANTOSO, U.; TANAKA, K.; OHTANI, S. Effect of dried Baccilus subtilis culture on growth, body composition and hepatic lipogenic enzyme activity in female broiler chicks. British Journal of Nutrition, v. 74, n. 4, p. 523-529, 1995.
TOURNUT, J.R. Probiotics. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, 1998. p.179-199.
UNIVERSIDADE FEDERAL DE VIÇOSA. Manual de utilização do programa SAEG – Sistema para Análise Estatísticas e Genéticas. Viçosa: UFV, 1999. 59p.
VARGAS JR., J. G.; TOLEDO, R. S.; ALBINO, L. F. et al. Características de carcaça de frango de corte, submetidos a rações contendo probióticos, prebióticos e antibióticos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, [2002]. CD-ROM. Nutrição de não-ruminantes.

38
IV - Efeito do probiótico sobre o desempenho, rendimento de carcaça,
exigências nutricionais de proteína bruta e morfometria intestinal de
codornas de corte
RESUMO: Para avaliar o efeito do probiótico (Lactobacillus sp.) associado com
diferentes níveis de proteína bruta (PB) sobre o desempenho, exigências nutricionais de
PB, rendimento de carcaça e de cortes e morfometria da mucosa do intestino delgado de
codornas de corte, foram utilizadas 2.304 codornas de corte, distribuídas em um
delineamento inteiramente casualizado em esquema fatorial 2 x 4 (com e sem probiótico
e quatro níveis de PB- 15, 20, 25 e 30%), com duas repetições por tratamento, em dois
períodos experimentais, totalizando 32 unidades experimentais. A adição do probiótico
reduziu o consumo de ração e melhorou a conversão alimentar no período de um a sete
dias. De acordo com os pesos médios do modelo LRP, aos sete e aos 14 dias, as
estimativas de exigência foram próximas de 27% de PB e aos 21, 28 e 35 dias entre 22 a
24% de PB. A adição do probiótico não influenciou o rendimento de carcaça e de
cortes. Para o rendimento de carcaça, a exigência de PB de acordo com o modelo
quadrático, ficou estimada em 23,53%, mas, em relação ao LRP, a estimativa reduz para
17,13% de PB. A análise de componentes principais resultou em estimativa de
exigência igual a 27,37% de acordo com o modelo quadrático. O comprimento e a
morfometria da mucosa do intestino delgado não foram influenciados pela adição do
probiótico. O comprimento do intestino aumentou de maneira linear com o aumento dos
níveis de PB aos sete, 14 e 21 dias de idade, e a morfometria da mucosa, aumentou de
forma linear somente para altura vilo do íleo. Pode-se concluir que nas condições
ambientais em que as codornas foram criadas a adição do probiótico não influenciou o
desempenho, as exigências nutricionais de PB, o rendimento de carcaça e a morfometria
intestinal.
Palavras-chave: conversão alimentar, comprimento intestino, lactobacilos, vilosidade
intestinal

39
IV– Effect of probiotic on performance, carcass yield, crude protein
nutritional requirements and intestinal morphometry of meat quails
ABSTRACT: To evaluate the effect of probiotic (Lactobacillus sp.) associated
with different crude protein (CP) levels on performance, CP nutritional requirements,
carcass and cut yields and small intestine mucous morphometry of meat quails, a total
of 2304 meat quails were distributed in a completely randomized experimental design in
2 x 4 factorial scheme (with and without probiotic and four levels of CP – 15, 20, 25
and 30%), with two replications by treatment, in two experimental periods, totalizing 32
experimental units. Probiotic addition decreased the feed intake and improved feed
conversion at 1 to 7 days. According with mean weight of LRP model, at 7 and 14 days,
requirement estimative were of 27% of CP and to the 21, 28 and 35 days were among
22 to 24% of CP. Probiotic addition did not influence carcass and cut yields. To carcass
yield, CP requirement according with quadratic model was estimated in 23.53%,
however, in relation to LRP, the estimative decrease to 17.13% of CP. Analysis of
principal components resulted in requirement estimate equal to 27.37% according with
quadratic model. Small intestine length and mucous morphometry were not influenced
by probiotic addition. Intestine length increased in a linear way with CP levels increase
at 7, 14 and 21 days of age, and mucous morphometry increased in a linear way only to
ileum villum height. It was possible to conclude that in the evaluated ambient
conditions the probiotic addition did not influence the performance, CP nutritional
requirement, carcass yield and intestinal morphometry.
Key words: feed conversion, length intestine, lactobacillus, intestinal vilosity

40
Introdução
A qualidade da carne de codorna é conhecida desde a antiguidade, pelo seu alto
conteúdo de proteína e aminoácidos, baixa quantidade de gordura, junto a um rápido
crescimento, proporciona uma carne extraordinariamente tenra e saborosa, de
preparação gastronômica fácil e rápida, considerada uma carne superior a de outras aves
(Barral, 1994).
No Brasil, contudo, a coturnicultura de corte é uma atividade ainda pouco
difundida, o que implica em poucos conhecimentos na área de melhoramento, nutrição,
manejo e sanidade.
Particularmente, com relação à nutrição de codornas, são escassas pesquisas sobre
as exigências nutricionais de codornas que visa à produção de carne. Além disso, dos
dados disponíveis, a maioria são provenientes de literatura estrangeira, obtidas sob
condições ambientais diferentes das condições climáticas brasileiras (Oliveira et al.,
2002).
Do ponto de vista econômico, a alimentação é um fator de grande importância,
não somente porque é a principal responsável pela melhor resposta das aves, mas,
sobretudo porque representa 70% do custo da atividade (Murakami & Ariki, 1998).
Logo, as estimativas das exigências nutricionais de aves são de extrema importância
para que se formulem rações de mínimo custo ou de máximo retorno, principalmente,
no tocante aos níveis de proteína e energia, uma vez que são os componentes de maior
participação nos custos de produção.
Oliveira et al. (2002) verificaram que o nível de 26% de proteína bruta promoveu
melhor desempenho de codornas japonesas (machos) criadas para a produção de carne,
no período de cinco a 27 dias de idade, reduzindo esse nível para 18% no período de 27

41
a 38 dias. Mas, Corrêa et al. (2006) encontraram que o máximo desempenho codornas
de corte na fase de um a 21 dias foi obtido com o nível de 30% de proteína bruta.
Por outro lado, Corrêa et al. (2004a), encontraram níveis de exigência de proteína
bruta para máximo ganho de peso de codornas de corte européias, acima de 28% para o
período de sete a 14 dias, 26% para o período de 15 a 28 dias e de 24% para o período
de 29 a 35 dias.
Com relação ao rendimento de carcaça, Más et al. (2004a), avaliando diferentes
níveis de proteína na fase de um a 21 dias e 22 a 49 dias, não verificaram influência do
nível protéico da ração sobre o rendimento de carcaça de codornas de corte.
A pouca diversidade da microflora intestinal de aves recém-nascidas, além de ser
considerada como um fator limitante para a digestão, também possibilita a colonização
intestinal por patógenos entéricos, assim, os efeitos negativos desse processo foram
contornados com o uso contínuo na ração de antibióticos em doses subterapêuticas
(Maiorka et al., 2001).
Já, face as atuais proibições do uso de alguns antibióticos como promotores de
crescimento, estando inclusive em vias de discussão o total banimento de seu uso, há a
necessidade de busca de alternativas, pois a simples retirada desses produtos
provocariam grandes danos econômicos.
Pesquisas têm procurado, dessa forma, desenvolver produtos que possam ser
utilizados como substitutos aos antibióticos promotores de crescimento. Entre as
alternativas, destacam-se os probióticos, que são produtos constituídos por
microrganismos vivos que afetam beneficamente o animal hospedeiro, que promove o
equilíbrio da microbiota intestinal (Fuller, 1989).
Em pesquisa conduzida por Mikulec, et al. (1999), observou-se influência
favorável dos probióticos em dietas com deficiência protéica, esse efeito é mais

42
evidente quando há redução dos níveis de metionina, cistina, lisina e/ou triptofano
(Dilworth & Day, 1978; Kos & Wittner, 1982).
A manipulação de microrganismos probióticos dentro do trato gastrintestinal tem
sido usada com o objetivo de manter ou incrementar a flora natural, que previne a
colonização de microrganismos patogênicos e mantém a ótima utilização dos alimentos
(Fuller, 1989; Vanbelle et al., 1990), em conseqüência da proteção dos vilos intestinais
e superfícies absortivas contra toxinas produzidas por microrganismos patógenos
(Dobrogsz et al., 1991; Walker & Duff, 1998).
Pelicano et al. (2005) encontraram maior altura de vilo para o jejuno e íleo, e
maior profundidade de cripta no duodeno, jejuno e íleo de frangos que receberam
probiótico, e indicam melhoras na superfície absortiva do intestino.
O objetivo desse trabalho foi avaliar o efeito do probiótico à base de Lactobacillus
sp. em rações com diferentes níveis de proteína bruta sobre o desempenho, rendimento
de carcaça e morfometria da mucosa do intestino delgado de codornas de corte.
Material e Métodos
O experimento foi conduzido no Setor de Avicultura da Fazenda Experimental de
Iguatemi/FEI/UEM, em dois períodos (P1 - maio-junho e P2 - julho-agosto) de 2005, sob
aprovação do Comitê de Ética em Experimentação Animal – CEEA/UEM.
Foram utilizadas 2.304 (1440 P1 e 864 P2) codornas de corte (Coturnix coturnix
sp.), machos e fêmeas, no período de um a 35 dias de idade. As codornas foram alojadas
em boxes de 3,0 m2, em um galpão convencional, disposto no sentido leste-oeste, com
cobertura de telha francesa, piso de concreto e paredes laterais de alvenaria com 0,40 m
de altura completada com tela de arame até o telhado.

43
A densidade de alojamento foi de 30 aves/m2 no P1 e de 18 aves/m2 no P2.
As rações (Tabela 1) foram formuladas de acordo com as exigências propostas por
Barral (1994), e a composição dos alimentos segundo dados apresentados por Rostagno
et al. (2005) .
As rações foram calculadas de forma a serem isocalóricas e isocálcicas, com
níveis variados de proteína bruta (PB). Foram utilizados quatro níveis de proteína bruta
(PB), 15, 20, 25 e 30% e o nível de 25%, o mais próximo do recomendado por Barral
(1994), que varia entre 26 e 28% de PB.
Com o resultado da combinação probiótico e nível de proteína foram constituídos
oito tratamentos.
O delineamento experimental foi inteiramente casualizado em esquema fatorial 2
x 4 (com e sem probiótico; quatro níveis de proteína), com duas repetições por
tratamento, repetindo-se em dois períodos experimentais.
O probiótico foi administrado, via ração, durante a primeira semana experimental,
e continha em média 1,6 x 109 UFC/g de um “pool” de Lactobacillus sp., e
administrado na dosagem de 200g/tonelada de ração, ou seja, o dobro das
recomendações do fabricante FATEC®, com o objetivo de verificar melhoras no
desempenho em função do aumento na concentração de lactobacilos fornecido.
Foi avaliado o desempenho das codornas de corte por meio do consumo de ração,
ganho de peso, conversão alimentar e viabilidade criatória durante o período
experimental.

44
Tabela 1: Composição percentual e química das rações experimentais Table 1: Percentual and chemical composition of the experimental diets
Níveis de proteína (%) Protein Levels Ingredientes
Ingredients 15 20 25 30
Milho Corn 75,00 61,77 44,55 27,40
Farelo de soja Soybean meal 17,50 32,91 47,60 62,14
Farelo de trigo Wheat meal 3,84 --- --- ---
Calcário Limestone 1,10 1,04 1,05 1,07
Fosfato Bicálcico Dicalcium phosphate 1,55 1,46 1,25 1,04
Sal comum Common salt 0,40 0,40 0,40 0,40
L-lisina HCL L-lysine HCL 0,20 0,06 --- ---
Premix vit. + min. Premix vit. + min. 0,40 0,40 0,40 0,40
BHT BHT 0,01 0,01 0,01 0,01
Óleo de soja Soybean oil --- 1,96 4,77 7,60
Total 100,00 100,00 100,00 100,00 Valores calculados Calculated values EM (Kcal/Kg) ME 3.000 3.000 3.000 3.000
Proteína Bruta (%) Crude protein 15,00 20,00 25,00 30,00
Cálcio (%) Calcium 0,90 0,90 0,90 0,90
Fósforo total (%) Total phosphorus 0,60 0,60 0,60 0,60
Lisina total (%) Total lysine 0,87 1,15 1,48 1,85
Met+Cist total (%) Total Met. + Cys. 0,64 0,79 0,91 1,04 * composição por kg do produto: Vit A, 2500.000 UI; Vit. D3, 625.000 UI; Vit. E, 3.750mg; Vit B1, 138mg; Vit. B2, 1.250mg; Vit. B6, 523mg; Vit B12, 5.000mcg; Vit. K3, 750mg; ácido fólico, 112 mg; pantotenato de cálcio 2.000 mg; niacina, 7.500 mg; metionina, 275.000 mg; colina, 87.500 mg; selênio, 63 mg; antioxidante, 2500mg; iodo, 240mg; veículo q.s.p., 1000g; zinco, 15.750mg; ferro, 12.250mg; manganês, 19.375mg; cobre, 3.900mg; cobalto, 50mg. * composition per kg: Vit A, 2500,000 UI; Vit. D3, 625,000 UI; Vit. E, 3,750mg; Vit BB1, 138mg; Vit. B2, 1,250mg; Vit. B6, 523mg; Vit B12, 5,000mcg; Vit. K3, 750mg; folic acid, 112 mg; calcium panthotenato, 2,000 mg; niacine, 7,500 mg; methionine, 275,000 mg; choline, 87,500 mg; selenium, 63 mg; antioxidant, 2,500mg; iodine, 240mg; veicul q.s.p., 1000g; zinc, 15,750mg; iron, 12,250mg; manganese, 19,375mg; copper, 3,900mg; cobalt, 50mg.

45
Aos 35 dias de idade foram retiradas ao acaso cinco aves de cada unidade
experimental (20 aves por tratamento) para avaliação do rendimento de carcaça e cortes
cárneos. As aves foram identificadas individualmente por meio de anilhas numeradas
colocados nas patas e submetidas a um período de jejum alimentar de oito horas, com
água à vontade até o momento do carregamento.
Após a pesagem individual, o abate foi realizado por degola completa entre os
ossos occipital e atlas. As aves foram escaldadas por 15 a 20 segundos a uma
temperatura de 55 a 600C, e a seguir, submetidas a depena manual. Em seguida,
efetuou-se a evisceração manual e a pesagem da carcaça. As partes da carcaça foram
separadas e pesadas para determinação do rendimento de peito, pernas, dorso e asa.
Para o cálculo de rendimento de carcaça, foi considerado o peso da carcaça
eviscerada, sem os pés, cabeça e gordura abdominal, em relação ao peso vivo. Para o
rendimento de cortes cárneos, foi considerado o rendimento de peito inteiro com pele,
pernas (coxa e sobrecoxa com pele), dorso e asas que foi calculado em relação ao peso
da carcaça eviscerada.
A gordura abdominal foi constituída pelo tecido adiposo, presente ao redor da
cloaca, da bolsa cloacal, moela, proventrículo e dos músculos abdominais adjacentes,
conforme descrito por Smith (1993), e também foi calculada em relação ao peso da
carcaça eviscerada.
Para a morfometria da mucosa do intestino delgado, foram abatidas duas aves de
cada repetição (oito aves por tratamento) aos sete, 14, 21 e 35 dias de idade, por degola
completa entre os ossos occipital e atlas. Após laparatomia, o intestino delgado foi
retirado e mensurado o seu comprimento. Em seguida, duas amostras de
aproximadamente três centímetros, foram retiradas da porção inicial do duodeno e final
do íleo.

46
As amostras foram lavadas em solução salina, fixadas em solução de Bouin e,
posteriormente, desidratadas em série de concentrações crescentes de álcool,
diafanizadas em xilol, e incluídas em parafina conforme metodologia descrita por Beçak
& Paulete, (1976). Foram obtidos cortes histológicos longitudinais e semi-seriados com
sete μm de espessura, e posteriormente corados pelo método de Hematoxilina-Eosina.
Para o estudo morfométrico, as imagens foram capturadas por meio da
microscopia de luz (Olympus Bx 40), utilizando-se o sistema analisador de imagens
computadorizado (Image Proplus – versão 5.2 – Média Cibernética). Foi mensurada a
altura de 30 vilos e a profundidade de 30 criptas de cada repetição por segmento e
desses valores foi obtida a média.
A análise estatística das variáveis estudadas foi realizada, levando-se em
consideração o modelo estatístico:
Yijkl = μ + Pi + Nj + PNij + Tk + PTik + NTjk + eijkl
Yijkl = variáveis estudadas obtidas com o indivíduo k, recebendo o probiótico i,
associado com nível de proteína j;
μ = constante geral;
Pi = efeito do probiótico i (sendo i1 = ausência de probiótico; i2 = presença
probiótico);
Nj = efeito do nível de proteína j (sendo j1 = 15%; j2 = 20%; j3 = 25% e j4 = 30%);
PNij = efeito da interação probiótico e nível de proteína;
Tk = efeito do período k (sendo k1 = período 1; k2 = período 2);
PTik = efeito da interação probiótico e período;
NTjk = efeito da interação nível e período;
eijkl = erro aleatório associado a cada observação Yijkl.

47
Os graus de liberdade referentes aos níveis de proteína bruta foram desdobrados
em polinômios ortogonais, utilizando-se o programa SAEG, desenvolvido pela
Universidade Federal de Viçosa – UFV (UFV, 1999).
As estimativas da exigência de proteína bruta, levando em consideração as
características de desempenho e de carcaça, foram obtidas mediante o uso do modelo
descontínuo LRP (Linear Response Plateau), citado por Braga (1983), e/ou, o modelo
quadrático e/ou cúbico.
Adicionalmente, foram estabelecidos os primeiros componentes principais que
envolvem as variáveis sob estudo, com a finalidade de se determinar a exigência que
envolve simultaneamente mais de uma variável.
Resultados e Discussão
No decorrer do período experimental, a temperatura média registrada no interior
do galpão foi: máxima 26,50C e mínima de 22,20 C para o período 1; e máxima 22,40C e
mínima de 170 C para o período 2.
A adição do probiótico reduziu o consumo de ração (CR) e melhorou a conversão
alimentar (CA) (P<0,05) na fase de um a sete dias, entretanto, para as demais faixas,
não houve efeito da adição do probiótico sobre o CR, ganho de peso (GP) e CA de
codornas de corte (Tabela 2).
Ressalta-se que, nessa fase (um a sete dias), a dosagem do probiótico fornecido foi
o dobro do recomendado pelo fabricante, na tentativa de se melhorar os resultados de
desempenho, porém, essa tentativa, não atingiu os objetivos propostos.

48
Tabela 2: Efeito do probiótico (Lactobacillus sp.) sobre o consumo de ração (CR), ganho de peso (GP) e conversão alimentar (CA) nas fases de 1 a 7, 7 a 14, 14 a 21, 21 a 28, 28 a 35 e 1 a 35 dias de idade e viabilidade criatória (VI) de 1 a 35 dias
Table 2: Effect of probiotic (Lactobacillus sp.) on feed intake (FI), weight gain (WG) and feed conversion (FC) in the phases of 1to 7, 7 to 14, 14to 21, 21to 28, 28to 35 and 1 to 35 days of age and growth viability (VI) of 1 at 35 days
Variáveis Variables
Idade (dias) Age (days)
Probiótico Probiotic
Controle Control
CV (%)1
CV CR (g) FI (g) 34,27b 39,84a 9,20
GP (g) WG (g) 23,08 22,92 3,38
CA (g/g) FC (g/g)
1 a 7
1,53b 1,80a 10,22
CR (g) FI (g) 89,97 89,77 5,39
GP (g) WG (g) 38,90 39,23 5,11
CA (g/g) FC (g/g)
7 a 14
2,38 2,34 3,34
CR (g) FI (g) 130,32 130,96 7,67
GP (g) WG (g) 59,62 58,65 5,49
CA (g/g) FC (g/g)
14 a 21
2,26 2,30 7,96
CR (g) FI (g) 151,77 151,51 3,73
GP (g) WG (g) 48,14 48,79 6,98
CA (g/g) FC (g/g)
21 a 28
3,16 3,12 6,41
CR (g) FI (g) 209,03 210,44 4,51
GP (g) WG (g) 50,43 51,43 4,76
CA (g/g) FC (g/g)
28 a 35
4,17 4,12 5,30
CR (g) FI (g) 463,59 471,02 3,89
GP (g) WG (g) 172,03 172,23 2,46
CA (g/g) FC (g/g) 2,72 2,75 3,63
VI (g/g) VI
1 a 35
95,56 94,79 4,87 a,b Na mesma linha, médias seguidas de letras diferentes, diferem pelo Teste F (P<0,05) a,b In the same line, means followed by different letters, differs by F Test (P<0.05) 1 Coeficiente de variação 1 Coefficient of variation

49
Oliveira et al. (2004) avaliaram o efeito do uso de antibióticos e probiótico na
criação de codornas de postura (Coturnix coturnix japonica) nas primeiras semanas de
vida, e concluíram que em bom ambiente, com água e ração disponíveis e de boa
qualidade, a adição de antibióticos e probiótico não têm efeito sobre o desempenho
inicial.
A influência favorável da adição do probiótico no aumento da massa corpórea e
melhora na conversão alimentar de frangos de corte, em situações onde há quantidades
insuficientes de proteína (Mikulec, et al. 1999), não foram constatadas no presente
experimento, uma vez que não houve interação significativa entre probiótico e nível de
proteína bruta.
Em pesquisa realizada com frangos de corte por Kos & Wittner (1982), os autores
verificaram que a adição de probiótico mostrou efeito somente quando adicionado às
dietas deficientes em metionina, lisina e triptofano. Entretanto, no presente experimento
as recomendações de lisina (1,50 %) foram atendidas somente para as rações que
contém 25 e 30% de PB e de metionina + cistina (0,90 %) para o nível de 30% de PB,
de acordo com as recomendações de Barral (1994) para codornas na fase inicial de
crescimento.
Os resultados obtidos nas pesquisas desenvolvidas por Kos & Wittner (1982) e
Mikulec et al. (1999) não foram comprovadas no presente trabalho, uma vez que, as
rações deficientes em PB, e em lisina e metionina + cistina, não apresentaram melhoras
no desempenho de codornas de corte quando da administração do probiótico.
A viabilidade criatória de um a 35 dias (Tabela 2), também não foi influenciada
pela adição do probiótico, porém, houve alto percentual de aves vivas, o que pode
indicar as boas condições sanitárias em que as aves foram criadas, que resulta em uma
baixa condição de desafio sanitário.

50
Em situações de mínimo estresse e desafio, os probióticos não manifestam
resultados evidentes da eficácia de seu uso (Fox, 1988; Dale, 1992; Maruta, 1993), o
que também poderia explicar a ausência de resultados benéficos, mesmo com os níveis
nutricionais abaixo do recomendado.
O consumo de ração (CR) e o ganho de peso (GP) apresentaram comportamento
quadrático dos níveis de PB, na fase de um a sete dias de idade, que mostra o consumo
mínimo para o nível de 15,69% e maior GP para o nível de 33,74%, de acordo com a
regressão (Tabela 3).
Considerando o modelo descontínuo LRP, tal exigência ficou estimada em
22,70% e 26,95% para o CR e GP, respectivamente.
Observou-se efeito linear dos níveis de proteína sobre a CA (um a sete dias),
mostrando que à medida que se aumenta o nível de PB da dieta, há uma melhora na CA.
Para peso médio aos sete dias, as exigências determinadas, foram de 34,53 e 27,11
% de PB, respectivamente, segundo o modelo quadrático e o modelo descontínuo LRP.

Tabela 3: Equações de regressão em função dos níveis de proteína bruta para consumo de ração (CR), ganho de peso (GP), conversão alimentar (CA) e peso médio (PM) de codornas de corte e exigências nutricionais de proteína bruta considerando o modelo de regressão e o modelo descontínuo LRP (Linear Response Plateau)
Table 3: Regression equation in function of the crude protein levels to feed intake (FI), weight gain (WG), feed conversion (FC) and average weight (AW) of the meat quail and crude protein nutritition requerimenst considering regression model and descontinuous model LRP (Linear Response Plateau)
Variáveis Variables
Idade Age
Equações de regressão em função dos níveis de proteína bruta Regression equation in function of the crude protein
Regressão Regression
LRP LRP
CR (g) / FI (g) ŷ = 52,041 - 2,035x + 0,0648x2 R2 = 1,00 15,69 22,70 GP (g) / WG (g) ŷ = -20,245 + 2,976x - 0,0441x2 R2 = 1,00 33,74 26,95 CA (g/g) / FC (g/g)
1 a 7 dias 1 to 7 days ŷ = 2,481 – 0,0363x R2 = 0,95 --- ---
PM7 (g) / AW7 (g) 7 dias 7 days ŷ = - 9,72 + 2,84x -0,041x2 R2 = 1,00 34,53 27,11 CR (g) / FI (g) ŷ = 42,162 + 2,120x R2 = 0,98 --- --- GP (g) / WG (g) ŷ = -38,63 + 5,576x - 0,0889x2 R2 = 1,00 31,36 26,26 CA (g/g) / FC (g/g)
7 a 14 dias 7 to 14 days ŷ = 6,189 – 0,3075x + 0,0058x2 R2 = 1,00 26,51 21,95
PM14 (g) AW14 (g) 14 dias 14 days ŷ = - 45,326 + 8,149x -0,124x2 R2 = 1,00 32,86 26,68 CR (g) / FI (g) ŷ = 95,277 + 1,5719x --- --- GP (g) / WG (g)) ŷ = -55,800 + 9,051x - 0,165x2 R2 = 1,00 27,43 22,63 CA (g/g) / FC (g/g)
14 a 21 dias 14 to 21 days ŷ = 5,649 - 0,274x + 0,0052x2 R2 = 1,00 26,35 21,77
PM 21 (g) / AW 21 (g) 21 dias 21 days ŷ = - 100,01 + 17,12x -0,29x2 R2 = 1,00 29,78 23,56 CR (g) / FI (g) ŷ = 95,171 + 2,5098x R2 = 0,99 --- --- GP (g) / WG (g) ŷ = -7,906 + 4,486x - 0,0829x2 R2 = 0,99 27,06 21,56 CA (g/g) / FC (g/g)
21 a 28 dias 21 to 28 days ŷ = 5,582 - 0,231x + 0,0051x2 R2 = 0,81 22,65 16,75
PM 28 (g) / AW 28 (g) 28 dias 28 days ŷ = -103,79 + 21,35x -0,37x2 R2 = 1,00 29,14 23,14 CR (g) / FI (g) ŷ = 26,416 + 13,3649x – 0,2184x2 R2 = 1,00 30,60 26,29 GP (g) / WG (g) ŷ = -213,526 + 37,974x - 1,732x2 + 0,025x3 R2 = 1,00 28,29 --- CA (g/g) / FC (g/g)
28 a 35 dias 28 to 35 days ŷ = 1,991 + 0,0959x R2 = 0,91 --- ---
PM 35 (g) / AW 35 (g) 35 dias 35 days ŷ = - 66,80 + 23,04x -0,41x2 R2 = 1,00 28,06 22,23 CR (g) / FI (g)) ŷ = 57,050 + 29,687x -0,479x2 R2 = 1,00 30,99 26,45 GP (g) / WG (g) ŷ = 522,229 - 43,62x + 1,738x2 -0,0219x3 R2 = 1,00 36,24 --- CA (g/g) / FC (g/g)
1 a 35 dias 1 to 35 days ŷ = 4,440 - 0,144x + 0,00283x2 R2 = 0,97 25,44 20,26

52
Dos sete aos 14 dias de idade, o CR apresentou comportamento linear, com
aumento no consumo em função do aumento dos níveis de PB. Efeito quadrático dos
níveis de PB sobre o GP foi verificado, ao fornecer pontos de máximo peso das
codornas para dietas, contendo 31,36 % de PB. Novamente, quando se leva em
consideração o LRP, a exigência cai para 26,26 % de PB.
Para a CA, nessa fase, foi estimada exigência de PB de 26,51% e de 21,95%,
respectivamente de acordo com o modelo de regressão e LRP.
Rezende et al. (2004) não encontraram diferenças nos parâmetros: ganho de peso,
consumo de ração e conversão alimentar de codornas francesas aos 14 dias de idade ao
trabalhar com níveis de PB que variam entre 20,5 e 23,5%, o que difere dos resultados
obtidos nesse experimento, porém, estes resultados provavelmente podem ser atribuídos
aos baixos níveis de PB, à genética e à pequena variação nas dietas experimentais,
utilizada pelos autores.
Os resultados de CR e CA diferem dos resultados obtidos por Corrêa et al. (2004
a, b), em que não foi verificado efeito dos níveis de PB sobre o CR na fase de sete a 14
dias, no entanto, verificou-se efeito linear dos níveis de PB, sobre a CA de codornas de
corte. Efeito linear do nível de PB sobre o ganho de peso, foi encontrado por Corrêa et
al. (2004a) e Oliveira et al. (2002), no entanto, Corrêa et al. (2004b) observaram efeito
quadrático para as dietas com 2.900 Kcal EM/kg, cuja exigência foi de 27,40% e linear
para a dieta com 3.100 Kcal EM/Kg.
Para peso médio aos 14 dias (Tabela 3), as exigências determinadas,
respectivamente, para o modelo quadrático e descontínuo LRP, foram de 32,86 e
26,68% de PB.

53
Diferentemente dos resultados obtidos, nesse experimento, Corrêa et al. (2004b)
encontraram exigências de PB, segundo o modelo quadrático, para peso acumulado dos
sete aos 14 dias, de 26,60% PB, com dietas que contém 2.900 kcal EM/Kg.
Dos 14 aos 21 dias de idade, efeito linear dos níveis de PB foi novamente
encontrado para o CR, e quadrático para o GP e CA (Tabela 3). Esses resultados
demonstram, respectivamente, ponto de máximo GP e mínima CA para as dietas que
contém, respectivamente, 27,43 e 26,35% de PB. Com o uso do modelo descontínuo
LRP, houve redução na exigência de PB para 22,63 e 21,77%, respectivamente, para o
GP e CA.
Efeito quadrático do nível de PB sobre o CR e GP, e linear para a CA, na fase de
15 a 21 dias, foi encontrado por Corrêa et al., (2004a), com aumento do consumo até o
nível de 25,24%, e ponto de máximo peso das codornas de corte para o nível de 26,38%
de PB, o que difere dos resultados encontrados nesse trabalho.
As estimativas de exigência de PB para peso médio aos 21 dias, levando-se em
consideração o modelo quadrático e o modelo descontínuo, foram respectivamente de
29,78 e 23,56% de PB.
Dos 21 aos 28 dias de idade, as variáveis CR, GP e CA apresentaram,
respectivamente; efeito linear, quadrático e quadrático dos níveis de PB sobre essas
variáveis, com níveis de PB para máximo ganho de peso e melhor CA, de
respectivamente 27,06 e 22,65% de PB. De acordo com o modelo descontínuo LRP, a
exigência para máximo GP e melhor CA reduziu para 21,56 e 16,75%, respectivamente.
Nessa fase, a exigência para máximo ganho de peso em codornas de corte foi
superior ao encontrado por Corrêa et al. (2004a), que verificaram exigência de 26,09%
de PB. Entretanto, os mesmos autores não verificaram efeito dos níveis (22, 24, 26 e

54
28%) de PB sobre o consumo de ração na faixa etária de 22 a 28 dias, da mesma forma
que Corrêa et al. (2004b).
Para peso médio aos 28 dias, as estimativas de exigência de PB foram semelhantes
aos resultados obtidos para peso médio aos 21 dias, e obtido o nível de 29,14 e 23,14%
de PB, respectivamente, para os modelos quadrático e descontínuo.
As variáveis CR, GP na fase de 28 a 35 dias, sofreram respectivamente influência
quadrática e cúbica dos níveis de PB (Tabela 3), com máximo consumo e máximo
ganho, respectivamente, para os níveis de 30,60 e 28,29% de PB. Para a CA efeito
linear positivo foi obtido, com piora na CA com o aumento dos níveis.
Nessa fase, Corrêa et al. (2004 a, b), não encontraram efeito dos níveis de PB
sobre o consumo de ração, em contrapartida, Corrêa et al (2004a) encontraram efeito
linear para a conversão alimentar e quadrático para o ganho de peso, com estimativa de
24,05% de PB, o que é inferior ao resultado obtido nesse experimento.
Efeito linear negativo foi verificado para o GP, e linear positivo para a CA, por
Oliveira et al (2002), na fase de 27 a 38 dias, ao mostrar que nessa fase, à medida que
aumenta os níveis de PB, há um decréscimo no GP e piora na CA.
Em relação ao peso médio aos 35 dias (Tabela 3), os resultados mostraram efeito
quadrático, tendo uma exigência estimada em 28,06% de PB, mas, de acordo com o
modelo LRP essa exigência ficou estimada em 22,23%.
Segundo Kirkpinar & Oguz (1995), codornas japonesas alimentadas com 30% de
proteína bruta alcançaram maior peso vivo no 35o dia de idade, ao compararem o
desempenho de codornas alimentadas com dietas isocalóricas (2800 Kcal EM/kg),
porém com seis níveis protéicos (16, 20, 22, 25, 28 e 30%).
Na fase total (um a 35 dias), as variáveis CR, GP e CA apresentaram,
respectivamente, efeito quadrático, cúbico e quadrático dos níveis de PB sobre essas

55
características (Tabela 3), e ocorreu aumento do CR até o nível de 30,99% de PB,
máximo GP com o nível de 36,24% e melhor CA para o nível de 25,44 % de PB.
Levando-se em consideração o modelo descontínuo LRP, as estimativas de PB foram de
26,35 para o CR e de 20,26% para a CA.
Hyánová et al. (1997) verificaram que os níveis de proteína bruta (26 vs. 23,8%
inicial e 21,6 vs. 19,5% crescimento) na dieta de codornas japonesas criadas para a
produção de carne não afetaram o peso corporal, o consumo de ração e a conversão
alimentar na fase de 1 a 35 dias de idade, e que portanto, os menores níveis de proteína
bruta podem ser recomendados para uso prático. Os autores compararam somente dois
níveis de PB, o que culminou na recomendação de um baixo nível de PB.
Pode-se observar, a partir dos resultados mostrados na Tabela 3, que as
estimativas de exigência de PB variaram muito em função das diferentes variáveis
estudadas, levando-se em consideração o modelo quadrático e/ou cúbico, e o modelo
descontínuo.
A divergência entre exigências protéicas pode ser atribuída a diferenças entre
períodos, os níveis de energia metabolizável, à genética, às condições ambientais, entre
outros (Rezende et al., 2004), mas, de maneira geral, indicam a necessidade de
formulação de rações divididas por fases.
Corrêa et al. (2004a) concluíram que a exigência de proteína bruta para máximo
ganho de peso de codornas de corte européias, acima de 28% para a fase de 7 a 14 dias,
26% para a fase de 15 a 28 dias e de 24% para a fase de 29 a 35 dias.
Melhor desempenho de codornas japonesas (machos), criadas para a produção de
carne, foi encontrado por Oliveira et al. (2002), para o nível de 26% de PB, na fase de 5
a 27 dias de idade, que reduziu esse nível para 18% na fase de 27 a 38 dias.

56
Por outro lado, Más et al. (2004b) não encontraram efeito dos níveis de proteína
bruta variando de 21,5 a 26% (um a 21 dias) e de 17,5 a 22% (22 a 49 dias) sobre o
desempenho na fase de um a 35 dias de idade.
As necessidades de PB propostas por Barral et al. (1994) são divididas em duas
fases: de 26 a 28 % de PB para a fase de um a 21 dias e de 22 a 24% para a fase de 22
até o abate.
No presente trabalho, ao considerar o peso vivo médio aos sete, 14, 21, 28 e 35
dias de idade, verificou-se que houve redução das estimativas de exigência, de acordo
com o LRP. Até os 14 dias, as estimativas de exigência foram de 27% de PB e , a partir
desta idade, houve evidências de que há uma redução na exigência de proteína que pode
estar entre 22 e 24%, denotando a importância de se utilizar na alimentação de codornas
de corte, rações divididas por fases, assim como em frangos de corte.
A adição do probiótico associado com diferentes níveis de PB não influenciou o
rendimento de carcaça e de cortes (Tabela 4).
Tabela 4: Rendimento de carcaça (RCAR), rendimento peito (RPEI), rendimento asas (RASA), rendimento pernas (RPER), rendimento dorso (RDOR) e rendimento de gordura (RGOR) de codornas de corte aos 35 dias em função da presença ou não de probiótico a base de Lactobacillus sp.
Table 4: Carcass yield (CYI), breast yield (BYI), wings yield (WYI), legs yield (LYI), back yield (BAY) and fat yield (FYI) of meat quail in 35 days in function of presence or absence of probiotic formed by Lactobacillus sp.
Probiótico Probiotic
RCAR% CYI
RPEI % BYI
RASA% WYI
RPER% LYI
RDOR% BAY
RGOR% FYI
Com probiótico With probiotic 69,93 39,66 6,77 23,63 28,63 1,52
Sem probiótico Without probiotic 70,28 40,34 6,96 23,61 28,02 1,43
CV% CV% 2,80 2,49 3,05 2,12 2,57 17,31
Não-significativo pelo teste F (P>0,05) Not significant by F test (P>0.05)

57
Trabalhos conduzidos por Henrique et al. (1998 a, b), Loddi et al (2000a) e
(2002), Maiorka et al. (2001) e Vargas Jr. et al. (2002) também não mostraram melhoras
nas características de carcaça com o uso do probiótico.
Por outro lado, Corrêa et al. (2003), avaliando o poliprobiótico constituído de
cepas de Lactobacillus acidophilus e casei, Streptococcus salivarium e faecium,
Bacillus subtilis e toyoi, e Saccharomyces cerevisae e Pelicano et al. (2003), avaliando
diferentes probióticos adicionados na ração (Bacillus subtilis e/ou Bacillus
licheniformis, ou Saccharomyces cerevisiae) e/ou água de bebida (Lactobacillus reuteri
e johnsonii), verificaram, respectivamente, maior rendimento de coxa e pernas em
frangos de corte ao final do período experimental.
Para o rendimento de carcaça, a exigência de PB de acordo com o modelo
quadrático, ficou estimada em 23,53%, entretanto, em relação ao LRP, a estimativa
reduz para 17,13% de PB (Tabela 5).
Tabela 5: Equações de regressão em função dos níveis de proteína bruta (PB) para o rendimento de carcaça (RCAR), rendimento peito (RPEI), rendimento asas (RASA), rendimento pernas (RPER), rendimento dorso (RDOR) e rendimento de gordura (RGOR) de codornas de corte aos 35 dias
Table 5: Regression equations in function of crude protein (CP) levels to carcass yield (CYI), breast yield (BYI), wings yield (WYI), legs yield (LYI), back yield (BAY) and fat yield (FYI) of meat quail at 35 days
Variáveis Variables
Equações de regressão em função dos níveis de PBRegression equations in function of crude protein (CP)
levels
Regressão Regression
LRP
RCAR% CYI ŷ = 54,902 + 1,374x - 0,0292x2 R2 = 0,69 23,53 17,13
RPEI % BYI ŷ = -42,572 + 11,064x - 0,479x2 + 0,0068x3 R2 = 1,00 26,47 ---
RASA% WYI ŷ = 11,446 - 0,358x + 0,00648x2 R2 = 0,99 27,62 23,34
RPER% LYI ŷ = 24,638 - 0,0452x R2 = 0,82 --- ---
RDOR% BAY ŷ = 19,837 - 0,613x + 0,0136x2 R2 = 0,70 22,54 16,36
RGOR% FYI ŷ = -0,131 + 0,0713x R2 = 0,96 --- ---

58
Más et al (2004a), avaliando diferentes níveis de PB, variando de 21,5 a 26% no
período de um a 21 dias e de 17,5 a 22%, no período de 22 a 49 dias, não encontraram
influência do nível de PB sobre o rendimento de carcaça de codornas de corte, no
entanto, houve uma pequena variação nos níveis de PB, o que poderia explicar a
ausência do efeito, diferente dos níveis utilizados no presente trabalho.
O rendimento de peito (RPEI) apresentou comportamento cúbico, o que resultou
na exigência de 26,47% de PB, discordando dos resultados obtidos por Corrêa et al.
(2004c e 2005), nos quais não verificaram efeito dos níveis de PB (22, 24, 26 e 28%)
sobre o RPEI.
As estimativas de exigência de PB para o rendimento de asa e rendimento de
dorso, levando-se em consideração o modelo quadrático foram de respectivamente
27,62% e 22,54% de PB, e de acordo com o modelo descontínuo, de 23,34 e 16,36% de
PB, respectivamente, mostrando exigência superior para o rendimento de asa.
Para o rendimento de pernas (RPER) e de gordura (RGOR), efeito linear dos
níveis de PB foi encontrado (Tabela 5), entretanto, para o RPER verificou-se efeito
linear negativo e para o RGOR, efeito linear positivo.
Com relação ao rendimento de pernas, Corrêa et al. (2004c) encontraram efeito
quadrático do nível de PB em dietas com 2.900 Kcal EM/kg somente para os machos,
em que foi obtido máximo rendimento em dietas que contêm 24,28% de PB, e para as
fêmeas não foi verificado efeito dos níveis de PB.
O RGOR não foi afetado pelos níveis de PB (22, 24, 26 e 28%), em pesquisa
desenvolvida por Corrêa et al. (2005), o que discorda dos resultados obtidos nesse
experimento.
Wyatt et al. (1982) encontraram relação positiva e significante entre o peso
corporal e a percentagem de gordura na carcaça.

59
Por outro lado, Costa et al. (2001) verificaram maior deposição de gordura
abdominal em frangos de corte alimentados com menores níveis de PB na ração, mas,
os níveis de PB utilizados pelos autores variaram de 20 a 22,5% no período de um a 21
dias e de 17,5 a 19,5% no período de 22 a 42 dias, e as rações com menores níveis de
PB foram suplementadas com aminoácidos sintéticos (lisina, metionina e treonina).
Dentre os componentes principais estimados, aquele que envolve as variáveis:
peso médio aos 35 dias (PM35), ganho de peso de um a 35 dias (GP135) e conversão
alimentar de um a 28 (CA128) dias foi escolhido para determinar a exigência
nutricional de PB por responder por 94,61% da variação.
Ao usar esse componente principal, procedeu-se a análise da mesma forma que as
variáveis isoladamente, não observado efeito do probiótico nem da interação
probiótico*nível.
A equação ajustada para o componente principal em função do nível de PB foi ŷ =
-18,338 + 1,445x - 0,0264x2, o que resultou estimativa de exigência igual a 27,37%. A
aplicação do modelo descontínuo LRP indicou estimativa de 21,91% de PB.
Os resultados desse estudo sugerem a necessidade de formulação de rações de
codornas de corte divididas por fases, ou seja, os resultados indicam que até os 14 dias,
gira em torno de 27% e aos 21, 28 e 35 dias se encontra por volta de 22 a 24%.
O comprimento do intestino delgado (CID) não foi influenciado pela adição do
probiótico (Tabela 6), provavelmente porque não houve a colonização do intestino pela
cultura de Lactobacillus sp. contida no probiótico.
Trabalhos conduzidos por Loddi et al. (2000b) e Sato (2001) também não
apresentaram efeito significativo da suplementação do probiótico na ração de frangos de
corte sobre o comprimento do intestino.

60
Tabela 6: Comprimento do intestino delgado de codornas de corte recebendo ou não probiótico aos 7, 14, 21 e 35 dias de idade
Table 6: Small intestine length of meat quails receiving or not probiotics at 7, 14, 21 and 35 days of age
Idade (dias) Age (days)
7 14 21 35 Probiotic Probiotic
Comprimento intestino delgado (cm) Small intestine length (cm)
Com probiótico With probiotic 31,39±0,48 41,14±0,93 51,29±0,92 55,22±1,11
Sem probiótico Without probiotic 31,53±0,48 40,89±0,93 50,87±0,92 55,91±1,11
CV % VC % 6,19 9,08 7,17 7,98
Não-significativo pelo teste F (P>0,05) Not significant by F test (P>0.05)
O nível de PB da dieta influenciou de maneira linear o comprimento do intestino
delgado (CID) aos sete, 14 e 21 dias de idade e de maneira quadrática aos 35 dias de
idade (Tabela 7).
Tabela 7: Equações de regressão do comprimento do intestino delgado (CID) de
codornas de corte em função dos níveis de proteína bruta aos 7, 14, 21 e 35 dias de idade
Table 7: Regression equation of small intestine length (SIL) of meat quails in function of crude protein levels at 7, 14, 21 and 35 days of age
Valores estimados para os níveis experimentais
Estimated values to experimental levels
Níveis de PB % CP levels %
Variáveis Variables
Idade Dias Age Days
Equação de regressão Regression equation
15 20 25 30 7 ŷ = 22,953 + 0,378x R2 = 0,98 28,62 30,51 32,40 34,29
14 ŷ = 26,447 + 0,647x R2 = 0,96 36,15 39,39 42,62 45,86 21 ŷ = 41,782 + 0,413x R2 = 0,85 47,98 50,04 52,11 54,17
CID (cm) SIL (cm)
35 ŷ = 79,822 - 2,706x + 0,068x2 R2 = 0,92 54,53 52,90 54,67 59,84
Segundo Hounou et al. (2002), o comprimento do intestino correlaciona-se com o
peso e não com altura do animal, ou seja, quanto maior o ganho de peso do animal,

61
maior o comprimento do intestino. No presente experimento, a proteína exerceu efeito
quadrático sobre o ganho de peso para todas as idades, diferentemente do comprimento
do intestino.
O probiótico (Lactobacillus sp.) utilizado nesse experimento não influenciou nem
a altura de vilo (AV) nem a profundidade de cripta (PC) dos segmentos estudados
(duodeno e íleo), provavelmente por não ter ocorrido a sua colonização (Tabela 8).
Tabela 8: Altura vilo (AV) e profundidade de cripta (PC) do duodeno (duo) e íleo (ile)
de codornas de corte recebendo ou não probiótico a base de Lactobacillus sp. Table 8: Villum height (VH) and cript depth (CD) of duodenum (duo) and ileum (ile) of meat
quails receiving or not probiotic in base of Lactobacillus sp. Variáveis Variables
Com probiótico With probiotic
Sem probiótico Without probiotic
AV duo (μm) VH duo (μm)
657,23±16,44 655,05±15,96
PC duo (μm) CD duo (μm)
84,02±2,07 79,47±2,01
AV ile (μm) VH ile (μm)
354,30±7,80 346,33±7,58
PC ile (μm) CD ile (μm)
64,30±1,72 62,02±1,67
Não-significativo pelo teste F (P>0,05) Not significant by F test (P>0.05)
A maturação intestinal é fundamental para a boa performance animal no período
pós-eclosão (Sell et al., 1991) pois dela depende a digestão e absorção adequada dos
nutrientes provenientes da dieta exógena. O tamanho dos vilos está relacionado com a
perda das células (extrusão) e renovação celular (proliferação e diferenciação) pelo
epitélio da mucosa intestinal. O equilíbrio entre esses dois processos garante a
manutenção da capacidade digestiva.
As aves têm um comportamento distinto dos mamíferos, não apresentando
proliferações celulares restritas à cripta, mas também ao longo dos vilos (Uni et al.,
1998a). Em perus, as divisões mitóticas nas criptas respondem por cerca de 55% da

62
proliferação celular no intestino, a região média dos vilos por 32% e a região apical por
8 % (Applegate et al., 1994)
O uso de microrganismos probióticos na alimentação animal traz benefícios em
função da proteção dos vilos e superfícies absortivas contras toxinas produzidas por
patógenos (Dobrogsz et al., 1991; Walker & Duff, 1998). A presença de patógenos e/ou
de toxinas, por sua vez, altera a integridade do epitélio intestinal, diminui a altura do
vilo, as atividades digestivas e absortivas e, aumenta o turnover celular (Visek, 1978).
Bradley et al. (1994) também não encontraram efeito da suplementação de
Saccharomyces cerevisiae var. boulardii, sobre a altura dos vilos ileais, mas,
verificaram uma redução na profundidade de cripta, o que pode indicar, segundo os
autores, um decréscimo na taxa de turnover celular, resultante da diminuição na
concentração de bactérias produtoras de metabólitos tóxicos ou a habilidade do
Saccharomyces cerevisiae var. boulardii em suprimir os efeitos desses metabólitos.
Da mesma forma, trabalho conduzido por Loddi (1998) também não demonstrou
diferenças na morfometria dos vilos entre os animais do grupo-controle e os que
receberam o probiótico à base de Enterococcus faecium.
Pelicano et al. (2003) também não encontraram diferenças na profundidade de
cripta de frangos de corte aos 42 dias que receberam probiótico que contém Bacillus
subtilis, Bacillus subtilis e licheniformis ou Saccharomyces cerevisiae, no entanto,
verificaram maior altura de vilo do duodeno.
Posteriormente, Pelicano et al. (2005) verificaram maior profundidade de cripta
aos 21 dias de idade no duodeno e íleo de frangos de corte que receberam probiótico à
base de Bacillus subtilis, entretanto, a altura de vilo foi aumentada somente no íleo.
Esse aumento na profundidade de cripta, em contraposição à pesquisa de Bradley et al.

63
(2004) foi justificada pela alta atividade celular proliferativa, que objetiva assegurar
uma taxa de renovação celular adequada.
Dessa forma, fica claro que a ação dos probióticos tanto em características de
desempenho como histológicas do trato digestivo dependem da composição do produto
e segundo Fuller (1989) das características e sua relação com os microrganismos
presentes no trato digestivo das diferentes espécies e a partir de um determinado tempo
necessário para que o agente se estabeleça no trato digestivo e equilibre a flora
(Tournut, 1998).
Com relação aos níveis de PB avaliados, as análises morfométricas das vilosidades
e criptas do duodeno e íleo (AV e PC) de codornas de corte mostraram diferentes
comportamentos (Tabela 9).
No duodeno, tanto a AV quanto a PC não foram influenciados pelo nível de PB.
Por outro lado, no íleo, a AV apresentou aumento linear em função dos níveis de PB, e
a PC não mostrou diferenças com o aumento dos níveis de PB (Tabela 9).
Esse comportamento diferenciado está relacionado à função prioritária dos
segmentos. Em nível de duodeno, a hidrólise dos aminoácidos ainda se encontra
incompleta, logo, a absorção é mínima, além disso, nas aves, grandes quantidades de
carreadores de membrana estão localizadas no íleo, o que o torna o principal sítio de
absorção de aminoácidos (Rutz, 2002), que justifica o comportamento linear sobre a
altura da vilosidade do íleo.
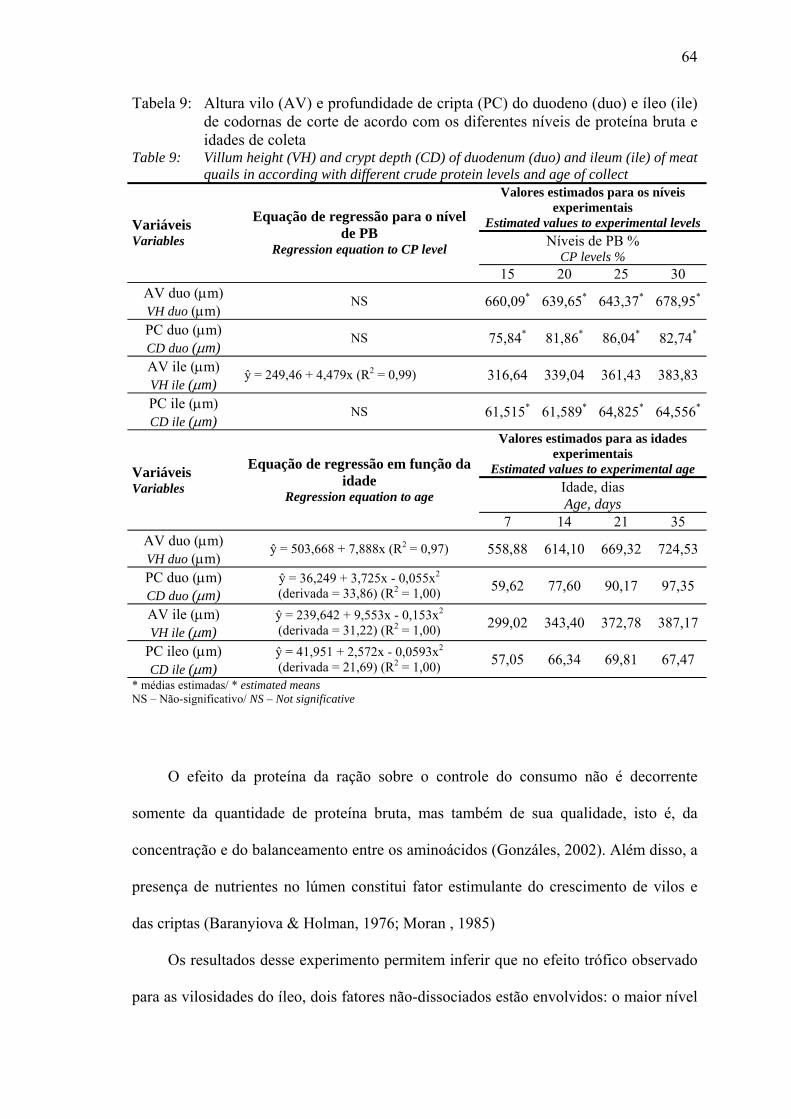
64
Tabela 9: Altura vilo (AV) e profundidade de cripta (PC) do duodeno (duo) e íleo (ile) de codornas de corte de acordo com os diferentes níveis de proteína bruta e idades de coleta
Table 9: Villum height (VH) and crypt depth (CD) of duodenum (duo) and ileum (ile) of meat quails in according with different crude protein levels and age of collect
Valores estimados para os níveis experimentais
Estimated values to experimental levelsNíveis de PB %
CP levels %
Variáveis Variables
Equação de regressão para o nível de PB
Regression equation to CP level
15 20 25 30 AV duo (μm) VH duo (μm)
NS 660,09* 639,65* 643,37* 678,95*
PC duo (μm) CD duo (μm)
NS 75,84* 81,86* 86,04* 82,74*
AV ile (μm) VH ile (μm)
ŷ = 249,46 + 4,479x (R2 = 0,99) 316,64 339,04 361,43 383,83
PC ile (μm) CD ile (μm)
NS 61,515* 61,589* 64,825* 64,556*
Valores estimados para as idades experimentais
Estimated values to experimental age Idade, dias Age, days
Variáveis Variables
Equação de regressão em função da idade
Regression equation to age
7 14 21 35 AV duo (μm) VH duo (μm)
ŷ = 503,668 + 7,888x (R2 = 0,97) 558,88 614,10 669,32 724,53
PC duo (μm) CD duo (μm)
ŷ = 36,249 + 3,725x - 0,055x2
(derivada = 33,86) (R2 = 1,00) 59,62 77,60 90,17 97,35
AV ile (μm) VH ile (μm)
ŷ = 239,642 + 9,553x - 0,153x2
(derivada = 31,22) (R2 = 1,00) 299,02 343,40 372,78 387,17
PC ileo (μm) CD ile (μm)
ŷ = 41,951 + 2,572x - 0,0593x2
(derivada = 21,69) (R2 = 1,00) 57,05 66,34 69,81 67,47
* médias estimadas/ * estimated means NS – Não-significativo/ NS – Not significative
O efeito da proteína da ração sobre o controle do consumo não é decorrente
somente da quantidade de proteína bruta, mas também de sua qualidade, isto é, da
concentração e do balanceamento entre os aminoácidos (Gonzáles, 2002). Além disso, a
presença de nutrientes no lúmen constitui fator estimulante do crescimento de vilos e
das criptas (Baranyiova & Holman, 1976; Moran , 1985)
Os resultados desse experimento permitem inferir que no efeito trófico observado
para as vilosidades do íleo, dois fatores não-dissociados estão envolvidos: o maior nível

65
de proteína na ração e o maior consumo de ração em função dos maiores níveis de
proteína.
Com relação à idade, verificou-se aumento tanto na PC quanto na AV dos
segmentos estudados (Tabela 9).
A AV do duodeno apresentou aumento linear em função da idade, já a PC do
duodeno, a AV e a PC íleo, apresentaram comportamento quadrático.
Uni et al. (1998b), ao avaliar o desenvolvimento pós-eclosão da mucosa do
intestino delgado de frangos de corte, encontraram que o desenvolvimento completo dos
vilos duodenais ocorreu até os sete dias de idade, enquanto que os vilos do jejuno e íleo
continuaram até aos 14 dias de idade, o que difere dos resultados obtidos nesse
experimento que teve até os 35 dias de idade aumento linear na AV do duodeno e
aumento na PC do duodeno, AV e PC do íleo, respectivamente, até 33,86; 31,22 e 21,69
dias de idade.
Por outro lado, Sakamoto (2005) encontrou que o desenvolvimento do duodeno de
frangos de corte completou-se aos 21 dias, enquanto que no jejuno e íleo até os 41 dias
de idade, observou-se gradual crescimento das vilosidades.
Ao comparar os dois segmentos estudados, pode-se observar maior
desenvolvimento da vilosidade do duodeno em relação ao íleo (Figura 1), o que está em
consonância com os obtidos por Uni (1999), Kondo (2003) e Sakamoto (2005).
Esse maior desenvolvimento pode ser atribuído, segundo Macari (1998), ao fato
de ser o segmento de mais rápida renovação celular e ainda por ser o primeiro segmento
do intestino delgado a receber estímulos físicos, químicos e hormonais desencadeados
pela presença da dieta no lúmen.

66
Figura 1: Fotomicrografia dos segmentos do intestino delgado de codornas de corte aos 7 dias de idade alimentados comrações contendo 15 (A e E ), 20 (B e F) , 25 (C e G) e 30% (D e H) de proteína bruta. Em A, B, C e D –duodeno; Em E, F, G e H – íleo. Barra 400 μm
Figure 1:Photomicrografy of small intestine segments of meat quails at 7 days old feed with diets containing 15 (A and E ), 20 (B and F) ,25 (C and G) and 30% (D and H) of crude protein. In A, B, C and D – duodenum; In E, F, G e H – ileum. Bar 400 μm
H
G
F
E
B
C
D
A

67
Observou-se, ainda, interação significativa entre nível e idade para a variável
profundidade de cripta do duodeno e o desdobramento dessa interação mostrou que para
o nível de 25% de proteína bruta, maior profundidade de cripta foi encontrada para as
codornas com 28,50 dias de idade, de acordo com a equação ŷ = -108,579 +14,194x - 0,249x2
(R2 =0,97).
Conclusões
Pode-se concluir que nas condições ambientais em que as codornas foram criadas
a adição de probiótico a base de Lactobacillus sp. não influenciou o desempenho, o
rendimento de carcaça, a exigência nutricional de proteína bruta e a morfometria da
mucosa intestinal de codornas de corte.
As estimativas de exigência até os 14 dias foram de 27% e, a partir desta idade,
houve evidências de que há uma redução da exigência de proteína que pode estar entre
22 e 24%.
As estimativas de exigência para o período de um a 35 dias foram de 27,37% de
proteína bruta.

68
Literatura Citada
APPLEGATE, T. J.; DIBNER, J. J.; KITCHELL, M. L., et al. Effect of turkey (Meleagridis gallopavo) breeder hen age and egg size on poult development. 2. Intetinal villus growth, enterocyte migration and proliferation of the turkey poult. Comparative Biochemistry Physiology B, v. 124, p. 381-389, 1994.
BARANYIOVA, E.; HOLMAN, J. Morphological changes in the intestinal wall in fed and fasted chickens in the first week after hatching. Acta Veterinarian Brno., v. 45, p. 151-158, 1976.
BARRAL, A. D. Manual de la Codorniz: cria industrial y para la caza. Lleida: Dilagro, 1994. 268p.
BEÇAK, W.; PAULETE, J. Técnicas de citologia e histologia. Rio de Janeiro: Livros Técnicos e Científicos, 1976. 305p.
BRADLEY, G. L.; SAVAGE, T. F.; TIMM, K. I. The effects of supplementing diets with Saccharomyces cerevisiae var. boulardii on male poult performance and ileal morphology. Poultry Science, v. 73, p. 1766-1770, 1994.
BRAGA, J. M. Avaliação da fertilidade do solo (ensaios de campo). Imprensa Universitária, n. 156, p. 15-20, 1983.
CORRÊA, G. S. S.; SILVA, M. A.; CORRÊA, A. B. et al. Exigência de proteína bruta para codornas de corte EV1 durante o período de um a 21 dias. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 43., 2006, João Pessoa, PB. Anais... João Pessoa: Sociedade Brasileira de Zootecnia, [2006]. CD-ROM.
CORRÊA, G. S. S.; SILVA, M. A.; FONTES, D. O. et al. Efeito de diferentes níveis de proteína e energia sobre o rendimento de carcaça de codornas européias. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 57, n.2, p. 266-271, 2005.
CORRÊA, A. B.; CORRÊA, G. S. S.; SILVA, M. A., et al. Efeito de diferentes níveis de proteína bruta e energia metabolizável sobre características de codornas de corte avaliadas durante a fase de crescimento. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41., 2004, Campo Grande, MS. Anais... Campo Grande: Sociedade Brasileira de Zootecnia, [2004a]. CD-ROM. Nutrição de não-ruminantes. NNR018
CORRÊA, A. B.; CORRÊA, G. S. S.; SILVA, M. A. et al. Desempenho produtivo de codornas tipo carne alimentadas com dietas contendo diferentes níveis de proteína bruta e energia metabolizável. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41., 2004, Campo Grande. Anais... Campo Grande: Sociedade Brasileira de Zootecnia, [2004b]. CD-ROM. Nutrição de não-ruminantes. NNR017.
CORRÊA, G. S. S.; SILVA, M. A.; CORRÊA, A. B. et al. Rendimento de carcaça de codornas de corte alimentadas com dietas com diferentes níveis de proteína bruta e energia metabolizável. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41., 2004, Campo Grande. Anais... Campo Grande: Sociedade Brasileira de Zootecnia, [2004c]. CD-ROM. Nutrição de não-ruminantes. NNR082.

69
CORRÊA, G. S. S.; GOMES, A.V.C.; CORRÊA, A. B. et al. Efeito de antibiótico e probióticos sobre o desempenho e rendimento de carcaça de frangos de corte. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 55, n.4, p. 467-473, 2003.
COSTA, F. G. P.; ROSTAGNO, H. S.; ALBINO, L. F. T. et al. Níveis dietéticos de proteína bruta para frangos de corte de 1 a 21 e 22 a 42 dias de idade. Revista Brasileira de Zootecnia, v. 30, n. 5, p. 1498-1505, 2001.
DALE, N. Probióticos para aves. Avicultura Professional, v. 10, p. 88-89, 1992.
DILWORTH, B. C.; DAY, E. J. Lactobacillus cultures in broiler diets (Abstract). Poultry Science, v. 57, p. 1101, 1978.
DOBROGSZ, W. J.; BLACK, B. L.; CASAS, I. A. Delivery of viable Lactobacillus reuteri to the gastrointestinal tract of poultry. Poultry Science, v. 70, p. 158, 1991.
FOX, S. M. Probiotics: intestinal inoculants for production animals. Veterinary Medicine, v. 83, p. 806-830, 1988.
FULLER, R. Probiotics: the scientific basis. London: Chapman & Hall, 1989. 307p.
GONZALES, E. Ingestão de alimentos: mecanismos regulatórios. In: MACARI, M.; FURLAN, R.L.; GONZALES, E. (Eds.) Fisiologia aviária aplicada a frangos de corte. Jaboticabal: FUNEP, 2002. p.187-199.
HENRIQUE, A. P. F.; FARIA, D. E.; FRANZOLIN, R. et al. Uso de probióticos e antibióticos como promotores de crescimento para frangos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, [1998a]. CD-ROM. Nutrição de não-ruminantes. NRU008.
HENRIQUE, A. P. F.; FARIA, D. E.; FRANZOLIN, R. et al. Efeito de ácido orgânico, probiótico e antibiótico sobre o desempenho e rendimento de carcaça de frangos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, [1998b].
CD-ROM. Nutrição de não-ruminantes. NRU009.
HOUNNOU, G.; ESTRIEUX,C.; DESME, J. et al. Anatomical study of the length of the human intestine. Surgery Radiology Anatomy, v. 24, p. 290-294, 2002.
HYÁNOVÁ, L.; DEDKOVÁ, L.; KNÍZETOVÁ, H. et al. Responses in growth, food intake and food conversion efficiency to different dietary protein concentrations in meat-type lines of Japanese quail. British Poultry Science, v. 38, p. 564-570, 1997.
KIRKPINAR, F.; OGUZ, I. Influence of various dietary protein levels on carcase composition in the male Japanese quail (Coturnix coturnix japonica). British Poultry Science, v. 36, p. 605-610, 1995.
KONDO, N. Estudo das características morfométricas de diferentes regiões do intestino delgado e índices zootécnicos em quatro linhagens de frangos de corte. 2003. 119p. Tese (Doutorado) – Faculdade de Medicina Veterinária e Zootecnia, Universidade Estadual Paulista, Botucatu, 2003.
KOS, K.; WITTNER, V. Use of probiotics in the nutrition of the fattening chicks. Praxis Veterinary, v. 30, p. 283-286, 1982.

70
LODDI, M. M.; TUCCI, F. M.; HANNAS, M. I. et al. Probióticos, mananoligossacarídeos + ácidos orgânicos em dietas de frangos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, [2002]. CD-ROM. Nutrição de não-ruminantes.
LODDI, M. M.; GONZALES, E.; TAKITA, T. S. et al. Uso de probiótico e antibiótico sobre o desempenho, o rendimento e a qualidade de carcaça de frangos de corte. Revista Brasileira de Zootecnia, v. 29, n.4, p. 1124-1131, 2000a.
LODDI, M. M.; SATO, R. N.; ARIKI, J. et al. Ação isolada ou combinada de antibiótico e probiótico como promotores de crescimento em rações iniciais de frangos de corte. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 37., 2000, Viçosa. Anais...Viçosa: Sociedade Brasileira de Zootecnia,, 2000b. p. 254.
LODDI, M. M. Aspectos produtivos e qualitativos com o uso de probióticos para frangos de corte. Botucatu: Universidade Estadual Paulista, 1998. 59f. Dissertação (Mestrado em Zootecnia) – Faculdade de Ciências Agrárias e Veterinárias, Universidade Estadual Paulista, Botucatu, 1998.
MACARI, M. Aspectos fisiológicos do sistema digestivo das aves. In: VIII SACAVET – SEMANA ACADÊMICA VETERINÁRIA, FMVZ-USP, 1998, São Paulo. Anais... São Paulo: USP, 1998. p. 4-18.
MAIORKA, A.; SANTIN, E.; SUGETA, S.M. et al. Utilização de prebióticos, probióticos ou simbióticos em dietas para frangos. Revista Brasileira de Ciência Avícola, v. 3, n.1, 2001. Disponível em: http://www.scielo.br. Acesso em: 5 mar. 2006.
MARUTA, K. Probióticos e seus benefícios. In: CONFERÊNCIA APINCO DE CIÊNCIA E TECNOLOGIA AVÍCOLAS, 1993, Santos. Anais... Santos: Fundação Apinco de Ciência e Tecnologias Avícolas, 1993. p. 203-219.
MÁS, H. A. R.; FASSANI, E. J. J.; BRITO, J. A. G. et al. Rendimento de carcaça de codornas de corte submetidas a diferentes níveis protéicos e idades de abate. In: SIMPÓSIO DE COTURNICULTURA, 2., 2004, Lavras, MG. Anais... Lavras: Universidade Federal de Lavras, [2004a]. CD-ROM.
MÁS, H.A.R.; BERTECHINI, A.G.; FASSANI, E.J. et al. Efeito da redução do nível protéico da ração sobre o desempenho de codornas de corte. In: SIMPÓSIO DE COTURNICULTURA, 2., 2004, Lavras, MG. Anais... Lavras: Universidade Federal de Lavras, [2004b]. CD-ROM.
MIKULEC, Z.; SERMAN, V.; MAS, N. et al. Effect of probiotic on production results of fattened chickens fed different quantities of protein. Veterinarski Arhiv, v. 69, n.4, p. 199-209, 1999.
MORAN, E. T. Digestion and absorption of carbohydrates in fowl and events through prenatal development. Journal of Nutrition, v. 115, p. 665-674, 1985.
MURAKAMI, A. E.; ARIKI, J. Produção de codornas japonesas. Jaboticabal: FUNEP, 1998. 79p.

71
OLIVEIRA, N. T. E.; SILVA, M. A.; SOARES, R. T. R. N. et al. Exigências de proteína bruta e energia metabolizável para codornas japonesas machos criadas para a produção de carne. Arquivo Brasileiro de Medicina Veterinária e Zootecnia, v. 54, n. 2, 2002. Disponível em: http://www.scielo.br . Acesso em: 5 abr. 2006.
OLIVEIRA, B. L., OLIVEIRA, D. D., OLIVEIRA, A. I. G. Utilização de probióticos e antibióticos na fase inicial da criação de codornas (Coturnix coturnix japonica). In: SIMPÓSIO DE COTURNICULTURA, 2004, Lavras. Anais... Lavras: [s.n.], 2004 CD-ROM.
PELICANO, E. R. L.; SOUZA, P. A.; SOUZA, H. B. A. et al. Intestinal mucosa development in broiler chickens fed natural growth promoters. Brazilian Journal of Poultry Science, v. 7, n. 4, p. 221-229, 2005.
PELICANO, E. R. L.; SOUZA, P. A.; SOUZA, H. B. A. et al. Effect of different probiotics on broiler carcass and meat quality. Revista Brasileira de Ciência Avícola, v.5, n.3, p. 207-214, 2003.
REZENDE, M. J. M.; FLAUZINA, L. P.; MCMANUS, C. et al. Desempenho produtivo e biometria das vísceras de codornas francesas alimentadas com diferentes níveis de energia metabolizável e proteína bruta. Acta Scientiarium, v. 26, n.3, p. 353-358, 2004.
ROSTAGNO, H. S.; ALBINO, L. F. T.; DONZELE, J. L. et. al. Tabelas brasileiras para aves e suínos; composição de alimentos e exigências nutricionais de aves e suínos. Viçosa: UFV, 2005. 141p.
RUTZ, F. Proteínas: digestão e absorção. In: MACARI, M.; FURLAN, R.L.; GONZALES, E. Fisiologia aviária aplicada a frangos de corte. 2. ed. Jaboticabal: FUNEP, 2002. p. 135-141.
SAKAMOTO, M. I. Influência da glutamina e vitamina “E” sobre o desempenho, resposta imunológica e morfometria intestinal de frangos de corte. Maringá: Universidade Estadual de Maringá, 2005. 70p. Dissertação (Mestrado). Universidade Estadual de Maringá- UEM, Maringá, 2005.
SATO, R. N. Ação isolada e combinada de probiótico e antibiótico como promotores de crescimento em rações iniciais de frangos de corte. Jaboticabal: Universidade Estadual Paulista, 2001. 48p. (Trabalho de Graduação) – Universidade Estadual Paulista (UNESP), 2001.
SELL, J. L.; ANGEL, C. R.; PIQUER, F. J. et al. Development patterns of selected characteristics of the gastrointestinal tract of young turkeys. Poultry Science, v. 70, p. 1200-1206, 1991.
SMITH, M. O. Parts yeld of broilers reared under cycling high temperatures. Poultry Science, v. 72, p. 1146-1150, 1993.
TOURNUT, J. R. Probiotics. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35., 1998, Botucatu. Anais... Botucatu: Sociedade Brasileira de Zootecnia, 1998. p.179-199.
UNI, Z.; PLATIN, R.; SKLAN, D. Cell proliferation in chicken intestinal epithelium occurs both in the crypt and along the villus. Journal Comparative Physiology B, v. 168, p. 241-247, 1998a.
UNI, Z., GANOT, S.; SKLAN, D. Posthatch development of mucosal function in the broiler small intestine. Poultry Science, v. 77, p. 75-82, 1998b.

72
UNI, Z. Functional development of the small intestine in domestic birds: cellular and molecular aspects. Poultry Avian Biology Reviews, v. 10, n.3, p. 167-179, 1999.
UNIVERSIDADE FEDERAL DE VIÇOSA. Manual de utilização do programa SAEG – Sistema para Análise Estatísticas e Genéticas. Viçosa: UFV, 1999. 59p.
VANBELLE, M.; TELLER, E.; FOCANT, M. Probiotics in animal nutrition: a review. Archives of Animal Nutrition, v. 40, p. 477-484, 1990.
VARGAS JR., J. G.; TOLEDO, R. S.; ALBINO, L. F. et al. Características de carcaça de frango de corte, submetidos a rações contendo probióticos, prebióticos e antibióticos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39., 2002, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, [2002]. CD-ROM. Nutrição de não-ruminantes.
VISEK, W. J. The mode of growth promotion by antibiotics. Journal of Animal Science, v. 46, p. 1447-1469, 1978.
WALKER, W. A.; DUFF, L. C. Diet and bacterial colonization: role of probiotics and prebiotics. Journal Nutrition Biochemical, v. 9, p. 668-675, 1998.
WYATT, J. F. M; SIEGEL, P. B.; CHERRY, J. A. Phenotypic relationships between adiposity, breast weight, and body weight in female Japanese quail. Poultry Science, v. 61, p. 643-646, 1982.

CONCLUSÕES GERAIS
Pode-se concluir que nas condições ambientais em que as codornas foram criadas,
a adição de probiótico não influenciou o desempenho, o rendimento de carcaça, a
exigência nutricional de proteína bruta, a população microbiana e a morfometria
intestinal de codornas de corte.
As estimativas de exigência até os 14 dias foram de 27% e, a partir desta idade,
houve evidências de que há uma redução da exigência de proteína que pode estar entre
22 e 24%.
As estimativas de exigência para o período de um a 35 dias foram de 27,37% de
proteína bruta.
Outros estudos são necessários para avaliar o uso de probióticos e estimar a
exigência de proteína bruta de codornas de corte em diferentes fases de criação.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo