universidade tecnolÓgica federal do paranÁ...
TRANSCRIPT

UNIVERSIDADE TECNOLÓGICA FEDERAL DO PARANÁ
PPGTA – PROGRAMA DE PÓS-GRADUAÇÃO EM TECNOLOGIA DE
ALIMENTOS
LARISSA CRISTINA COSTA
CARACTERIZAÇÃO PARCIAL DE TERMORESISTÊNCIA E ESTABILIDADE
A COMPOSTOS QUÍMICOS DE BACTERIOCINAS PRODUZIDAS POR
Enterococcus sp
DISSERTAÇÃO
CAMPO MOURÃO
2017

LARISSA CRISTINA COSTA
CARACTERIZAÇÃO PARCIAL DE TERMORESISTÊNCIA E ESTABILIDADE
A COMPOSTOS QUÍMICOS DE BACTERIOCINAS PRODUZIDAS POR
Enterococcus sp
Dissertação apresentada ao programa de Pós
Graduação em Tecnologia de Alimentos da
Universidade Tecnológica Federal do Paraná,
como parte dos requisitos para obtenção do
título de mestre em Tecnologia de Alimentos.
Orientadora: Profª. Dra. Luciana Furlaneto Maia.
CAMPO MOURÃO
2017

Dados Internacionais de Catalogação na Publicação
C837c
Costa, Larissa Cristina
Caracterização parcial de termoresistencia e estabilidade a compostos químicos de bacteriocinas produzidas por Enterococcus sp / Larissa Cristina Costa – 2017.
63 f. : il. ; 30 cm. Orientadora: Prof.ª Dra. Luciana Furlaneto Maia Dissertação (Mestrado) – Universidade Tecnológica Federal do
Paraná. Programa de Pós-Graduação em Tecnologia de Alimentos. Medianeira, 2017.
Inclui bibliografias. 1. Produtos Quimícos. 2. Inibição. 3. Tratamentos térmicos. 4.
Alimentos – Dissertações. I. Maia, L, orient. II. Universidade Tecnológica Federal do Paraná. Programa de Pós-Graduação em Tecnologia de Alimentos. III. Título.
CDD: 664
Biblioteca Câmpus Medianeira Fernanda Cristina Gazolla Bem dos Santos CRB: 9-1735

TERMO DE APROVAÇÃO
CARACTERIZAÇÃO PARCIAL DE TERMORESISTÊNCIA E ESTABILIDADE
A COMPOSTOS QUÍMICOS DE BACTERIOCINAS PRODUZIDAS POR
Enterococcus sp
POR
LARISSA CRISTINA COSTA
Essa dissertação foi apresentada às quatorze horas, do dia vinte de fevereiro de
dois mil e dezessete, como requisito parcial para obtenção do titulo de Mestre em
Tecnologia de Alimentos, Linha de Pesquisa: Processos Tecnológicos na Indústria
de Alimentos, no Programa de Pós-graduação em Tecnologia de Alimentos, da
Universidade Tecnológica Federal do Paraná. A candidata foi arguida pela Banca
Examinadora composta pelos professores abaixo assinados. Após a deliberação, a
Banca Examinadora considerou o trabalho APROVADO.
__________________________________________________
Profª. Draª. Luciana Furlaneto-Maia (Orientadora – PPGTA)
__________________________________________________
Profª Draª. Marcia Cristina Furlaneto (Membro Externo – UEL)
__________________________________________________
Profª. Draª. Giselle Apª Nobre Costa (Membro Externo – UNOPAR)
Ministério da Educação
Universidade Tecnológica Federal do Paraná
Programa de Pós-Graduação em Tecnologia de Alimentos

Dedico esse trabalho à Deus,
minha mãe, minhas irmãs
e meus sobrinhos.

AGRADECIMENTOS
Quero agradecer primeiramente a Deus pelo dom da vida.
Agradeço a minha mãe, guerreira que me deu tanto apoio quando mais precisei que
me incentivou nas horas difíceis de desânimo e cansaço.
À as minhas irmãs pelo apoio que me deram e, toda ajuda oferecida.
Aos meus professores por todo conhecimento adquirido durante o mestrado.
Aos novos amigos que fiz durante essa fase da minha vida.
À professora Dra Marcia Cristina Furlaneto por disponibilizar seu Laboratório de
pesquisa para realização deste estudo bem como a sua equipe de pesquisa que me
deu auxílio quando precisei.
À Marcia Terra por todo apoio e ajuda na realização deste trabalho.
À Katia Real Rocha que me ajudou com as análises e me fez companhia até nos
final de semana no laboratório. Minha imensa gratidão.
À minha orientadora Luciana Furlaneto Maia por ter aceitado a me orientar e me dar
todo suporto durante a realização da pesquisa. Obrigada por todo conhecimento
adquirido, orientação e paciência.
A todos que direta ou indiretamente fizeram parte desta fase em minha vida, o meu
muito obrigado.

“O fruto de um trabalho de amor atinge sua plenitude na colheita, e esta chega
sempre no seu tempo certo”
Autor desconhecido

RESUMO
COSTA, Larissa Cristina. Caracterização parcial de Termoresistência e estabilidade
a compostos químicos de bacteriocinas produzidas por Enterococcus sp. 2017. 63 f.
Dissertação – Programa de Pós-Graduação em Tecnologia de Alimentos,
Universidade Tecnológica Federal do Paraná. Campo Mourão, 2017.
As bacteriocinas possuem amplo espectro de atividade antimicrobiana sobre
diversas bactérias patogênicas e deteriorantes de alimentos. Diversos gêneros
bacterianos produzem enterocinas, dentre estes, o gênero Enterococcus sp.
Portanto, o presente trabalho teve como objetivo estudar a produção de enterocinas
produzidas por linhagens de Enterococcus faecium, nos pHs 4.5, 5.0, 5.5, 6.0 e 6.5
onde o meio de cultura foi suplementado com lactose, manose, manitol, glicerol,
tween® 20 e tween® 80 com diferentes concentrações. Os testes de antagonismo
foram realizados contra a bactéria Listeria innocua CLIP 12612. Diante dos melhores
parâmetros para a produção de enterocina, estas foram testadas quanto a
estabilidade diante da ação dos produtos químicos SDS, EDTA, Tween® 20,
Tween® 80 e NaCl, temperaturas de 40, 50, 60, 70, 80, 90, 100°C por 30 minutos e
121°C por 15 minutos, diversos pHs 3, 5, 7 e 9, adsorção e sobre sua ação
bactericida e/ou bacteriostática. Os resultados obtidos mostram que, os pHs ótimo
de produção foram entre 6,0 e 6,5 e as suplementações com maior atividade de
inibição foram lactose 20 g/L, glicerol 20 g/L, tween®20 5,0g/L. Estes suplementos
também foram acrescidos juntos no meio MRS sendo denominado de MIX. As
enterocinas permaneceram estáveis aos pHs analisados e mantiveram sua atividade
inibitória após tratamento térmico até 100°C por 30 minutos. Porém, houve uma
redução na atividade das enterocinas quando submetida a 121°C por 15 minutos.
Dentre os produtos químicos avaliados, o SDS e EDTA foram os que apresentaram
maior atividade sendo que a enterocina produzida com o Mix foi a que apresentou
maior halo de inibição. Também foi evidenciado efeito bactericida e/ou
bacteriostática e a adsorção de maior porcentagem ocorreu entre 60-65% entre
ambas enterocinas produzidas. Os resultados obtidos indicam que, as enterocinas
produzidas pelas linhagens de Enterococcus faecium possuem potencial de
aplicabilidade em alimentos como bioconservadores.
PALAVRA CHAVE: Bioconservantes, Inibição, Listeria innocua, Produtos químicos,
Tratamento térmico.

ABSCTRACT
COSTA, Larissa Cristina. Characterization partial of thermoresistance and stability of
compounds chemicals of bacteriocins produced by Enterococcus sp. 2017. 63 f.
Dissertação – Programa de Pós-Graduação em Tecnologia de Alimentos,
Universidade Tecnológica Federal do Paraná. Campo Mourão, 2017.
Bacteriocins has a broad spectrum of antimicrobial activity on various pathogenic and
deteriorating bacterium in food. Several bacterial genera produce enterocins,
between them, the genus Enterococcus sp. The objective of this study was to study
the production of enterocins produced by Enterococcus faecium in the pH 4.5, 5.0,
5.5, 6.0 and 6.5 where the culture medium was supplemented with lactose,
mannose, mannitol, glycerol, tween® 20 and tween 80 with different concentrations.
The antagonistic tests used bacterium Listeria innocua CLIP 12612. About the best
parameters for the production of enterocin, these were tested for stability against the
action of the chemicals SDS, EDTA, Tween® 20, Tween® 80 and NaCl,
Temperatures of 40, 50, 60, 70, 80, 90, 100°C for 30 minutes and 121°C for 15
minutes, various pHs 3, 5, 7 and 9, adsorption and on their bactericidal and/or
bacteriostatic action. The results showed that the pH great of production was
between 6.0 and 6.5, and the supplements with larger inhibition activity were lactose
20 g/L, glycerol 20 g/L and tween®20 5.0 g/L. These supplements also were added
in the medium MRS, referred to as MIX. The enterocins remained stable at the
analyzed pHs and maintained their inhibitory activity after heat treatment at 100°C for
30 minutes. However, had reduction in the activity of the enterocins when submitted
to 121°C for 15 minutes. Among the chemicals evaluated, the SDS and EDTA were
the ones that presented the highest activity, and the enterocins produced with the
Mix showed the highest inhibition halo. It was also evidenced effect bactericidal
and/or bacteriostatic, and the highest percentage adsorption occurred between 60-
65% between enterocins produced. The results indicate that the enterocins produced
by Enterococcus faecium strains have potential applicability in foods as
bioconservatives.
KEYWORDS: Bioconservatives, Inhibition, Listeria innocua, Chemical products, Heat
treatment.

LISTA DE FIGURAS
Figura 1 – Classificação cronológica das bacteriocinas ...................................... 19
Figura 2 – Biossíntese e transporte de bacteriocina ............................................ 21
Figura 3 – Mecanismos de ação bacteriocinas. Bacteriocinas classe I: inibição da
síntese de peptidoglicano. Bacteriocinas classe II: formação de pores na membrana
celular .............. .................................................................................................... 22
ARTIGO
Figura 1: Resultado do halo de inibição em meio MRS suplementado com lactose
20,0 g/L, glicerol 20,0 g/L e tween 20 5,0 g/L, pelos isolados Efm20 e Efm22 .... 50
Figura 2: Resultado do efeito do pH sobre as enterocinas obtidas pelos isolados
Efm20 (A) e Efm22 (B) ........................................................................................ 51
Figura 3: Halos de inibição formados com enterocina produzida pelos isolados Efm
20 e Efm 22 .......................................................................................................... 52
Figura 4: Atividade residual do efeito da temperatura na estabilidade de enterocina
produzidas pelos isolados Efm20 (A) e Efm22 (B) (2).........................................52
Figura 5: Resultado do efeito dos produtos químicos na estabilidade das enterocina
dos isolados Efm20 (A) e Efm22 (B) .................................................................... 53
Figura 6: Resultado do efeito bactericida contra L. innocua das enterocinas
produzidas pelos isolados Efm20 e Efm22 em meio MRS suplementado ........... 53

LISTAS DE TABELAS
Tabela 1. Tampões utilizados na estabilidade das enterocina ............................. 28
ARTIGO
Tabela 1: Mensuração do efeito do substrato em meio MRS na produção de
enterocina por isolados de Enterococcus sp ........................................................ 50
Tabela 2: Adsorção de enterocinas produzidas pelos isolados Efm20 e Efm22..54

LISTA DE ABREVIATURAS E SIGLAS
AU: Unidade arbitrária de atividade;
BAL: Bactérias ácidas láticas;
BHI: Meio Brain Heart Infusion;
CFS: Sobrenadante livre de células;
EDTA: Ácido etilenodiamino tetra-acético;
FDA: Food and Drug Administration (Administração de comidas e remédios);
GRAS: Generally Recognized as Safe (Geralmente reconhecido como seguro);
g/L; Gramas por litro;
KDa: Unidade de massa atômica;
MRS: Meio Man Rogosa
NaCl: Cloreto e sódio
NaOH: Hidróxido de sódio
PBS: Tampão fosfato
PCR: Reação em cadeia polimerase
pH: Potencial de hidrogênio iônico
rpm: Rotações por minuto
UFC: Unidade formadora de colônia
TRIS: Trisaminometano
v/v: Volume por volume

SUMÁRIO
1. INTRODUÇÃO ............................................................................................................. 13
2. OBJETIVO GERAL E ESPECÍFICO ............................................................................ 15
2.2 Objetivo Geral ........................................................................................................................... 15
2.2 Objetivos Específicos .............................................................................................................. 15
3. REVISÃO BIBLIOGRÁFICA ........................................................................................ 16
3.1 Gênero Enterococcus sp ................................................................................................... 16
3.2 Bacteriocinas ............................................................................................................................ 17
3.2.1 Biossíntese e Transporte ............................................................................................... 20
3.2.2 Mecanismo de ação ......................................................................................................... 21
3.2.3 Fatores que afetam a produção e ação das bacteriocinas ........................................ 23
3.3 Biopreservação ................................................................................................................... 24
4 METODOLOGIA ........................................................................................................... 26
4.1 Material Biológico ..................................................................................................................... 26
4.2 Ensaio da ação antagônica da enterocina ........................................................................... 26
4.3 Efeito do pH na produção de bacteriocinas ......................................................................... 27
4.4 Efeito das suplementações no meio MRS na produção de bacteriocinas...................... 27
4.5 Efeito do pH na estabilidade das bacteriocinas .................................................................. 28
4.6 Efeito da temperatura na estabilidade das bacteriocinas ................................................. 29
4.7 Efeito da estabilidade das bacteriocinas na presença de SDS, tween® 20, tween® 80,
EDTA e NaCl ................................................................................................................................... 29
4.8 Determinação do efeito bactericida/bacteriostático das bacteriocinas ........................... 30
4.9 Adsorção de bacteriocinas nas células alvo ........................................................................ 30
4.10 Análise de dados ................................................................................................................... 31
5. REFERÊNCIAS ............................................................................................................ 32
ARTIGO ............................................................................................................................... 43
INTRODUÇÃO ................................................................................................................................ 44
MATERIAIS E MÉTODOS ............................................................................................................ 45
RESULTADOS ................................................................................................................................ 49
DISCUSSÃO ................................................................................................................................... 54
CONCLUSÃO ................................................................................................................................. 57
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................................ 57

13
1. INTRODUÇÃO
Atualmente, existem diversas tecnologias de preservação e conservação de
alimentos. Contudo, há inúmeros estudos para o desenvolvimento de tecnologias
e/ou métodos de conservação para que, juntamente com as tecnologias existentes,
possam oferecer alimentos seguros aos consumidores (SCHULZ; BONELLI;
BATISTA, 2005; VÁSQUEZ; SUÁREZ; ZAPATA, 2009).
As bacteriocinas são peptídeos antimicrobianos que possuem ação bactericida
e/ou bacteriostática sobre diversos microrganismos. São sintetizadas nos
ribossomos das células e são liberados no meio extracelular. Há um grande
interesse pela indústria de alimentos em substituir conservantes químicos pelas
bacteriocinas, visto que estas não promovem nenhuma alteração sensorial e
nutricional no produto (NASCIMENTO; MORENO; KUAYE, 2008).
Segundo Cleveland et al. (2001), as bactérias gram-positivas produtoras de
bacteriocinas são Staphylococcus aureus, S. epidermidis, Bacillus subtilis, Bacillus
cereus e Enterecoccus sp; já as bactérias gram-negativas produtoras são
Escherichia coli, Klebsiella oxytoca e Pseudomonas sp.
As bacteriocinas produzidas pelas espécies de Enterococcus são chamadas de
enterocinas. Enterococcus são bactérias que colonizam o trato gastrointestinal de
seres humanos e de animais, solos e em água contaminada. Esse grupo de
bactérias possui resistência ao estresse químico e físico e conseguem sobreviver
fora de seu habitat por um longo período de tempo (KUHN et al., 2000).
Enterococcus são cocos gram-positivos, não esporulados, podem ocorrer isolados,
em pares ou pequenas cadeias. São anaeróbios e, de seu metabolismo, o ácido
lático é o principal produto.
Segundo Schulz et al. (2003), o mecanismo de ação das bacteriocinas ocorre
de duas formas, em que pode promover efeito letal bactericida associada ou não
com uma lise celular, ou ainda, ter efeito bacteriostático, que causa a inibição da
multiplicação de microrganismos. Inicialmente, o modo de ação das bacteriocinas
ocorre com a sua ligação às bactérias alvo, onde ocorrerá inserção das

14
bacteriocinas na membrana que provocará modificações. Tais modificações levam à
formação de poros na membrana citoplasmática, fazendo com que o meio
intracelular seja liberado para fora, ou altera a força próton-motora que é utilizada
para a produção de energia e para a síntese de proteínas, tornando inviável a
manutenção da célula alvo (MARTINS et al., 2006; DELBONI, 2009).
Existem alguns fatores que influenciam na produção de bacteriocinas como
condições de incubação, de temperatura, pH e tempo de incubação (CARRIM.,
2005). O pesquisador também afirma que, a composição do meio de crescimento
com uma fonte rica de nitrogênio, fosfato, carbono e substâncias surfactantes ou até
mesmo o aumento destes nutrientes favorece a produção destes compostos.
Trabalho similar foi realizado por Sakira, Lipton e Aishwaryz (2010), que avaliaram a
produção de bacteriocinas por Lactobacillus rhamnosus GP1 isolados em diversas
condições de cultivo no meio MRS, e concluiu que há influência da composição do
meio na produção do composto antimicrobiano.
Dos compostos que podem ser adicionados ao meio de cultura, o extrato de
levedura tem a capacidade de promover o crescimento celular, pois é capaz de
fornecer nitrogênio e vitaminas importantes. Já substâncias surfactantes como
tween® 20 e tween® 80 influenciam positivamente a produção de bacteriocinas, pois
interagem com a membrana celular fazendo com que ocorra uma maior adsorção ou
até mesmo remoção da camada externa lipídica promovendo uma ação bactericida
(BALLONGUE, 2004; SCHEPERS; THIBAULT; LACROIX, 2002; COLLADO et al.,
2005).
Diante do exposto, o objetivo deste trabalho foi avaliar as enterocinas
produzidas pelas espécies de Enterococcus faecium bem como avaliar as melhores
condições que influenciam sua atividade para que se torne uma alternativa na
aplicação em alimentos como conservantes naturais.

15
2. OBJETIVO GERAL E ESPECÍFICO
2.2 Objetivo Geral
Avaliar a estabilidade das enterocinas produzidas por cepas de Enterococcus
faecium diante de várias variáveis bem como analisar as melhores condições que
influenciam a produção destes compostos antimicrobianos.
2.2 Objetivos Específicos
Verificar a influência da produção de enterocinas nos pHs 4.5, 5.0, 5.5,
6.0, 6.5;
Verificar a influência da adição de nutrientes no meio de cultura como
lactose, glicerol, manitol, manose, extrato de levedura, tween® 20 e tween® 80, para
produção de enterocinas;
Avaliar a estabilidade da enterocina frente à estabilidade térmica sob as
temperaturas de 40, 50, 60, 70, 80, 90 e 100°C por 30 minutos e a 121°C por 15
minutos;
Avaliar a estabilidade das enterocinas nos pHs 3, 5, 7 e 9;
Avaliar a estabilidade das enterocinas sob ação de produtos químicos
como o NaCl, SDS, EDTA, Tween® 20, Tween® 80;
Avaliar efeito bactericida ou bacteriostático das enterocinas;
Avaliar a adsorção das enterocinas pelas células alvo em estudo.

16
3. REVISÃO BIBLIOGRÁFICA
3.1 Gênero Enterococcus sp
O gênero Enterococcus constitui-se de bactérias cocos, gram positivos, não
formador de esporos, anaeróbios facultativos que vivem em colônias ou isolados
(DOMIG; MAYER; KNEIFEL., 2003; FOULQUIÉ-MORENO; SARANTINOPOULOS;
TSAKALIDOU, 2006; VAN DEN BERGHE; DE WINTER; DE VUYST., 2006).
Habitam o trato gastrointestinal de humanos e animais, solos, água e alimentos.
Entre suas características, encontram-se a adaptabilidade e resistência em diversos
ambientes (REFFUVEILLE et al., 2011).
As espécies de Enterococcus têm despertado atenção nos últimos anos, pois
muitas das espécies bacterianas estão mostrando-se resistentes aos antibióticos
sendo consideradas como patógenos emergentes (NES; DIEP; IKE, 2014). Devido a
esta resistência aos antibióticos, principalmente com a vancomicina, espécies de E.
faecium e E.faecalis são responsáveis por 38 a 75% das infecções hospitalares
(WILLEMS et al., 2011).
A resistência a antibióticos pode ocorrer por fatores intrínsecos ou por agentes
antimicrobianos como os glicopeptídeos, p-lactamas e as fluoroquinolonas (ARIAS;
CONTRERAS; MURRAY, 2010). Conforme Kuhn et al. (2003), a resistência dos
Enterococcus de origem animal deve-se ao uso de drogas que são utilizadas nas
rações animais e tal resistência também se propagou na população humana visto
que tais bactérias estão presentes em alimentos, principalmente em carnes.
São conhecidas cerca de 37 espécies de enterococos, dentre eles encontram-
se as espécies E. aquimarinus, E. asini, E. avium, E. caccae, E. canintestini, E.
canis, E. casseliflavus, E. cecorum, E. columbae, E. devriesei, E. dispar, E. durans,
E. faecalis, E. faecium, E. gallinarum, E. gilvus, E. haemoperoxidus, E.
hermanniensis, E. hirae, E. italicus, E. malodoratus, E. moraviensis, E. munditi, E.
pallens, E. phoeniculicola, E. pseudoavium, E. raffinosus, E. ratti, E. saccharolyticus,
E. silesiacus, E. sulfureus, E. termitis e E. villorum (KOHLER, 2007). Porém, nem
todas as espécies são patogênicas para o homem.

17
As espécies de Enterococcus faecalis e Enterococcus faecium foram
responsáveis por aproximadamente 12% das infecções hospitalares notificados nos
EUA (SIEVERT et al., 2013). Em animais e humanos estão relacionadas com
infecções urinárias, infecções de feridas, meningites, endocardites, bacteremia e
septicemia (CAUWERTS et al., 2007; GAMA, 2008; KENSE; LANDMAN, 2011; ZOU
et al., 2011).
E. faecium é considerado um patógeno multirresistente o qual possui
resistência, por exemplo, à vancomicina onde se faz necessário o melhoramento ou
terapias novas para combater tal patógeno (BOUCHER et al., 2009; ARIAS;
CONTRERAS; MURRAY, 2010). A espécie E. faecium é comumente isolada de
produtos cárneos processados e em carnes suínas e, a espécie de E. faecalis
encontra-se em carne bovina e suína (FRANZ; HOLZAPFEL; STILES, 1999).
A resistência destas bactérias sobre diversos fatores como pH, temperatura,
adaptação a diversos substratos e ao uso de aditivos químicos faz com que estas
bactérias se adaptem ao meio e sobrevivam durante o processamento de alguns
tipos de alimentos como o leite e a carne, entre outros (FURLANETO-MAIA et al.,
2014). Ainda, quando adicionadas intencionalmente, algumas espécies de
Enterococcus têm a vantagem de contribuir com as propriedades sensoriais do
produto e, apresenta-se como biopreservantes com ação sobre outros patógenos.
Estudos com a utilização de espécies de Enterococcus, em especial a E.
faecium, destaca-se pelo seu potencial probiótico em alimentos, a qual é capaz de
reduzir o colesterol LDL e, tem a capacidade no aumento da absorção intestinal de
nutrientes (LUND; EDLUND, 2001).
3.2 Bacteriocinas
Segundo Lee e Kim (2011), as bacteriocinas constituem-se de um grupo de
defesa que age especificamente contra as bactérias, eliminando-as e/ou inibindo-as.
Através de suas diversas estruturas, as bacteriocinas possuem diferentes
mecanismos de ação com diversas aplicações, dentre elas as terapêuticas.

18
São sintetizadas nos ribossomos das células que após atingirem seu grau de
maturidade são liberados no meio extracelular com atividades bactericida ou
bacteriostática sobre diversos patógenos. Geralmente são compostas por 20 a 60
aminoácidos as quais possuem atividade antimicrobiana associada à tolerância
ácida e a termoestabilidade (CASTILHO et al., 2013; EIJSINK et al., 2002; TAMIME,
2005).
As bacteriocinas têm despertado grande interesse na indústria de alimentos
visto que, através de seu papel como biopreservantes, podem eliminar bactérias
patogênicas e deteriorantes. No entanto, tais antimicrobianos sofrem a interferência
de alguns fatores para a sua produção como: ambiente inadequado para expressão,
redução da produção da enzima, antagonismo pela microbiota, interação entre
bacteriocina e as macromoléculas formando complexos não ativos e
desenvolvimento de organismos resistentes às bacteriocinas (CUI et al., 2012).
Cada bacteriocina possui características de acordo com as bactérias
produtoras que muitas vezes são imune às suas próprias bacteriocinas, porém sua
atividade antimicrobiana varia de acordo com a bactéria alvo podendo apresentar
uma ação reduzida contra as bactérias relacionadas e/ou possuir maior ação contra
bactérias de outros gêneros (COTTER; ROSS; HILL, 2013; DIGAITENE et al., 2012;
MILLS et al., 2011).
As bacteriocinas diferem em si em relação à massa molecular, modificação
pós-tradução, presença de aminoácidos modificados, estrutura química e modo de
ação (COTTER; ROSS; HILL, 2013). Até o ano de 2015, um total de 185
bacteriocinas foram identificadas, porém apenas 53% destas foram sequenciadas.
No decorrer dos anos, as classificações das bacteriocinas sofreram
modificações como mostra a figura 1, sendo que, a mais recente classificação é
proposta por Cotter, Ross e Hill (2013), onde há separação entre as bacteriocinas
produzidas por bactérias gram-positivas e gram-negativas. Nessa nova classificação,
foram excluídas as classes III e IV as quais foram relatadas por pesquisadores
anteriormente.

19
Figura 1: Classificação cronológica das bacteriocinas. Fonte: OGAKI, M. B.; FURLANETO, M. C.; MAIA, L. F., 2015.
As classes dos lantibióticos vêm despertando interesse pelo fato de que, a
nisina, uma bacteriocina comercializada em vários países, inclusive no Brasil,
pertence a essa classe. Produzida a partir de Lactococcus lactis, a nisina pertence à
classe I dos lantibióticos onde as moléculas caracterizam-se por serem flexíveis e
agir na despolarização de membranas (MOLLOY et al., 2014). Já a classe II são
formadas por peptídeos não modificados com massa molecular inferior a 10 kDa,
estáveis ao calor e é a classe que possui maior atividade antimicrobiana
principalmente sobre o gênero Listeria spp (JERONYMO, 2013).
Gratia
Principio V
Jacob et al.,
Sugerem termo bacteriocinas
Klaenhammer
Propõe classificação:
I – lantibióticos II - Peptídeos não modificados III- Proteínas grandes termo-lábeis IV-complexos de bacteriocinas
NES et al.,
Exclusão da classe IV
VAN BELKUM e Stiles
Classe II subdividida com base no numero
de resíduos e cisteínas
Cotter et al.,
Eliminaram classe IIc, III e IV; Reestruturaram a classe II.
I – Lantibióticos (11 subclasses) II – Peptídeos não modificados IIa – tipo pediocina IIb – dois peptídeos IIc – peptídeos cíclicos IId – Grupo heterogêneo
Cotter et al.,
I – Peptídeos modificados (11 subclasses) II–Peptídeos não modificados / cíclicos IIa – Tipo Pediocina IIb – dois peptídeos IIc – peptídeos cíclicos IId – os sem modificações, lineares, tipo não pediocina, ou bacteriocinas de único peptídeo IId – Contem serina na região C-terminal, modificação pós-traducional tipo sideróforo.
Heng e Tagg
I - Lantibióticos Ia - Linear Ib - Globular Ic - multi-componentes II - Peptídeos não modificados IIa -Tipo pediocina IIb - Grupo heterogêneo IIc - multi-componentes III - peptídeos grandes IIIa - bacteriocliticos IIIb - não líticos IV - Peptídeos cíclicos
1925
1953
1993
1996 2000
2005
2006
2013

20
De acordo com Cui e colaboradores (2012), a produção de bacteriocinas sofre
influência do estado fisiológico da bactéria produtora, pois é ela que determinará o
crescimento bacteriano. Fatores como: composição do meio de cultura de produção,
o uso de suplementações ao meio, temperatura e pH também podem influenciar a
produção de bacteriocinas (DE VUYST; LEROY, 2007).
As bacteriocinas produzidas pelo gênero Enterococcus são denominadas de
enterocinas. Elas possuem grande espectro de inibição contra bactérias do gênero
Listeria spp (PAPAGIANNI, 2003). Possuem estabilidade a altas temperaturas e a
uma ampla faixa de pH.
Muitas enterocinas foram identificadas e purificadas, porém como há uma
necessidade da caracterização química e também molecular destes compostos, os
impedem de ser utilizadas como biopreservantes em alimentos (OHMOMO et al.,
2000).
3.2.1 Biossíntese e Transporte
Segundo Lee e Kim (2011), a síntese das bacteriocinas é composta por um
agrupamento de genes que dentre eles são responsáveis pelas estruturas de
transporte, imunidade, regulação e processamento. Após sofrerem modificações,
são enzimaticamente clivadas para remoção da sequência sinal e são transportadas
para o meio extracelular, já como uma bacteriocina madura.
A formação de uma bacteriocina madura, inicia-se com peptídeos prematuros
que irão ligar-se a uma chaperona Sec B que esta localizada no citoplasma. A
mesma o transportará a outra chaperona, denominada de Sec A. Por sua vez, a
chaperona Sec A irá acionar a hidrólise de ATP e através de um regulador
intramolecular o IRA 1, irá interagir com o pré-peptídeo prematuro auxiliando na
translocação pela região Sec YEG. Nesta mesma região, descrita anteriormente, irá
passar o pré-peptídeo prematuro onde ocorrerá a formação da bacteriocina madura
(FELTCHER; BRAUNSTEIN, 2012; LYCKLAMA; DRIESSEN, 2012). A ilustração
pode ser verificada na figura 2.

21
Figura 2: Biossíntese e transporte de bacteriocina. Fonte: FELTCHER; BRAUNSTEIN, 2012; LYCKLAMA; DRIESSEN, 2012.
As proteínas translocase são responsáveis pelo transporte dos pré-peptídeos
no interior da membrana plasmática para o meio extracelular onde através da
hidrólise de ATP e por força próton motora, a bacteriocina madura é acionada para o
exterior da célula (FELTCHER; BRAUNSTEIN, 2012).
3.2.2 Mecanismo de ação
Muitos mecanismos de ação das bacteriocinas têm sido propostos nos últimos
anos. Para Chen e Hoover (2003), e Sabo et al. (2014), a ação depende muito da
espécie bacteriana produtora, bem como das condições de crescimento entre outros
fatores.
A ação de mecanismo das bacteriocinas pode ocorrer de duas formas:
bactericida ocasionando a lise ou não celular com a inibição da síntese de alguns
compostos importantes para a sobrevivência da célula ou ocasionando ação
bacteriostática aonde irá apenas inibir a multiplicação celular (CINTAS et al., 2001;
DEVLIEGHERE; VERMEIREN; DEBEVERE, 2004; SABO et al., 2014).
O mecanismo de ação das bacteriocinas ocorre em 2 modelos como mostra a
figura 3. No primeiro modelo, a da classe I, ocorre o envelope celular. Neste modelo,
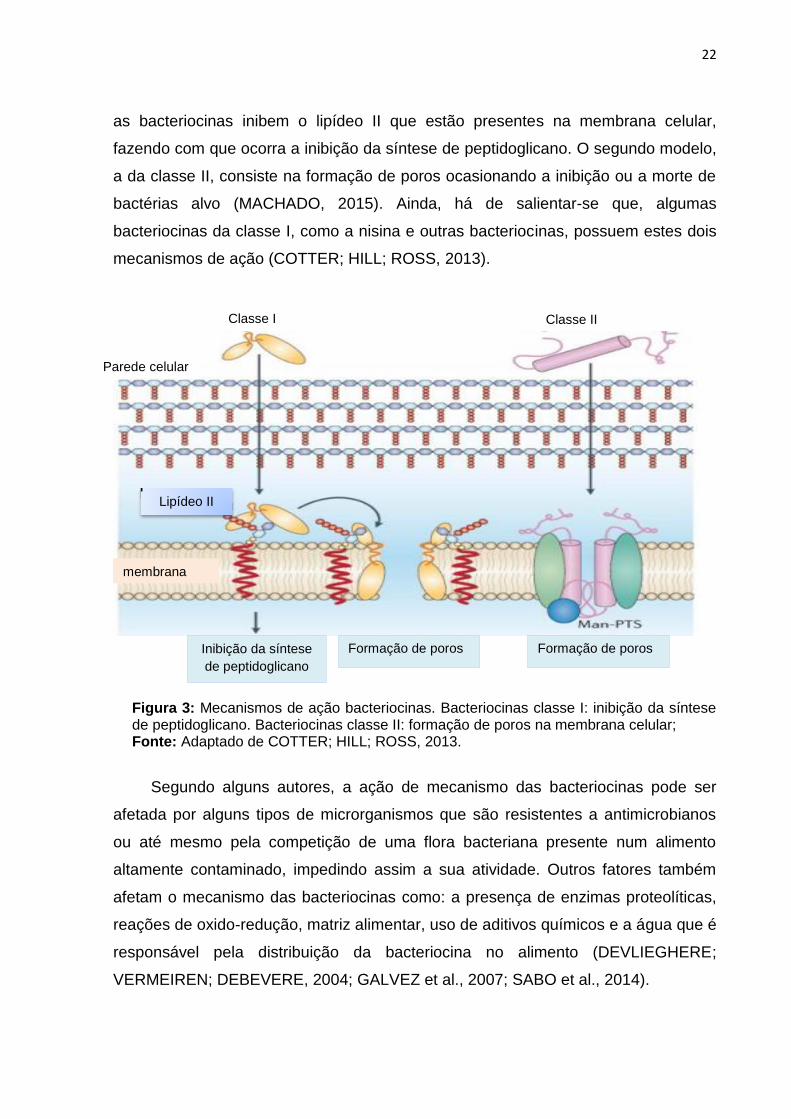
22
as bacteriocinas inibem o lipídeo II que estão presentes na membrana celular,
fazendo com que ocorra a inibição da síntese de peptidoglicano. O segundo modelo,
a da classe II, consiste na formação de poros ocasionando a inibição ou a morte de
bactérias alvo (MACHADO, 2015). Ainda, há de salientar-se que, algumas
bacteriocinas da classe I, como a nisina e outras bacteriocinas, possuem estes dois
mecanismos de ação (COTTER; HILL; ROSS, 2013).
Figura 3: Mecanismos de ação bacteriocinas. Bacteriocinas classe I: inibição da síntese de peptidoglicano. Bacteriocinas classe II: formação de poros na membrana celular; Fonte: Adaptado de COTTER; HILL; ROSS, 2013.
Segundo alguns autores, a ação de mecanismo das bacteriocinas pode ser
afetada por alguns tipos de microrganismos que são resistentes a antimicrobianos
ou até mesmo pela competição de uma flora bacteriana presente num alimento
altamente contaminado, impedindo assim a sua atividade. Outros fatores também
afetam o mecanismo das bacteriocinas como: a presença de enzimas proteolíticas,
reações de oxido-redução, matriz alimentar, uso de aditivos químicos e a água que é
responsável pela distribuição da bacteriocina no alimento (DEVLIEGHERE;
VERMEIREN; DEBEVERE, 2004; GALVEZ et al., 2007; SABO et al., 2014).
Inibição da síntese
de peptidoglicano
Formação de poros Formação de poros
Classe I Classe II
Parede celular
Lipídeo II
membrana

23
Ainda há de salientar-se que, as bactérias gram-negativas são resistentes a
ação das bacteriocinas, pois estas bactérias possuem em sua estrutura celular uma
barreira adicional impermeável não permitindo a passagem de moléculas de
tamanho médio de 3 KDa, por exemplo (ABEE; KROCKEL; HILL, 1995).
3.2.3 Fatores que afetam a produção e ação das bacteriocinas
Durante a produção de bacteriocinas, fatores como crescimento e atividade
fisiológica da bactéria produtora, é responsável pela quantidade do antimicrobiano
produzido (LEROY; DE VUYST, 2003; YANG; RAY, 1994) o qual ainda pode ser
influenciado pela composição do meio de cultura, adição de suplementos, pH e
temperatura (YANG; RAY, 1994).
Estudo realizado por Kawai et al. (2001), obtiveram que a produção da
enterocina AS-48 se estabilizou em pH 6,55 com meio de cultura suplementado com
1% de glicose. Audisío, Oliver e Apella (2001), ao suplementarem o meio de cultura
com frutooligassacarídeos como fonte de carbono, observaram que a adição de
nutrientes foi determinante para a produção de bacteriocinas. De acordo com Van
Reenen et al. (2003), o caldo MRS suplementado com peptona bacteriológica,
triptona e extrato de levedura foram essenciais para a produção de plantaricina 423.
Para aplicação em alimentos, as bacteriocinas devem englobar diversos fatores
como: possuir atividade contra microrganismos patogênicos e deteriorantes, ser
termotolerantes, possuir atividade bactericida, não ser tóxicas entre outras (GALVEZ
et al., 2007).
Estudo com a bacteriocina Nisina produzida por L. lactis subsp. Lactis WNC20
manteve-se estável em pH de 3 a 5 porém a pH neutro (≥7) houve perda total de sua
atividade, estes quando incubados a 121ºC por 15 minutos (NOONPAKDEE et al.,
2003). Outro estudo realizado com L. sakei R1333 manteve atividade nos pH 2 a 12
quando incubados a 30°C por 30 minutos (TODOROV et al., 2011).

24
3.3 Biopreservação
Os alimentos são perecíveis por natureza própria e, portanto necessitam de
uma proteção contra ação de microrganismos patogênicos e deteriorantes, as quais
podem surgir em alguma fase do beneficiamento do alimento, seja ele no
processamento, armazenamento ou na sua distribuição, com o objetivo de assegurar
sua qualidade durante seu tempo de prateleira (HOLLEY; PATEL, 2005).
A maior preocupação da indústria de alimentos e dos complexos circuitos de
distribuição é fazer com que os alimentos tenham um maior tempo de prateleira
mantendo suas características sensoriais e nutricionais originais (SOUSA et al.,
2013).
Neste aspecto, a indústria de alimentos utiliza-se de vários métodos de
conservação para prevenir e inibir os patogênicos indesejáveis. Dentre os métodos
destacam-se o tratamento térmico, congelamento, secagem, a cura, processos de
conserva, embalagens (a vácuo, atmosfera modificada, inteligentes), uso da
fermentação e dos conservantes (GALVEZ et al., 2007; GRAM et al., 2002).
Por outro lado, os consumidores estão cada vez mais preocupados com a
saúde e a qualidade de vida. Com isso, cada vez mais é constante a procura de
alimentos com propriedades benéficas, entre elas, nutricionais e terapêuticas,
levando em conta a segurança alimentar da mesma e, a forma com que o alimento
foi processado (DIGAITIENE et al., 2012; MILLS et al., 2011).
Esta demanda por alimentos seguros faz com que as indústrias de alimentos
procurem alternativas seguras para atender estes requisitos, para que juntamente
com as tecnologias já existentes, possam oferecer alimentos de qualidade do ponto
de vista da segurança alimentar (SHULZ et al., 2003). Portanto, a bioconservação
mostra-se como uma alternativa promissora.
A bioconservação é a técnica que faz uso de microrganismos que estão
naturalmente inseridas aos alimentos e/ou adicionadas intencionalmente para inibir
microrganismos deteriorantes ou patogênicos. É uma alternativa para prolongar o
tempo de prateleira do produto bem como reforçar a segurança microbiológica por

25
meio de uma microbiota natural (VASQUEZ et al., 2009) bem como fazer com que
haja uma redução do uso de aditivos nos alimentos (NASCIMENTO, 2007).
Grande parte dos estudos voltados para a bioconservação ou biopreservação
de alimentos volta-se para a produção de bacteriocinas bem como a sua detecção,
produção, isolamento, mecanismo de ação, caracterização bioquímica, propriedades
antimicrobianas e etc (VAZQUEZ et al., 2009). Atualmente, existem muitos estudos
com bacteriocinas e por isso existe uma ampla coleção de bacteriocinas para uso
alimentar. Porém, comercialmente, apenas três bacteriocinas estão disponíveis
sendo elas a nisina produzida por Lactococcus lactis, a ALTA 2351® e ALTA 2341®,
ambas produzidas por Pediococcus spp. e usadas em processos fermentativos
(MILLS et al., 2011; SABO et al., 2014).
Conforme Mills et al. (2011), a aplicação de bacteriocinas na alimentação deve
respeitar três critérios sendo eles a segurança, a qualidade e nutrição/saúde. Por ser
de origem proteica e de baixa massa molecular, as bacteriocinas são reconhecidas
como seguros (GRAS), que não causam efeitos colaterais, e são considerados
seguros para consumo humano, pois acabam sendo inativadas pelas enzimas
digestivas no estômago (SHARMA et al., 2011). De acordo com o Food and Drug
Administration (FDA), os antimicrobianos naturais aplicados em alimentos devem ser
produzidos por microrganismos seguros (GRAS).
As bacteriocinas podem ser introduzidas aos alimentos pela adição direta ou
através da inoculação de microrganismos produtores, sendo que é necessário que
haja condições favoráveis para a produção do composto. A adição direta da
bacteriocina nos alimentos, pode ocorrer de duas formas: (I) pela adição direta ao
alimento sendo ela purificada ou parcialmente purificada e, (II) pela forma de um
ingrediente concentrado reconhecido como uma cultura starter. Ambas as
aplicações em alimentos exigem legislação e a sua aprovação como conservante
alimentar (CARVALHO, 2016).
Verificou-se que, a aplicação de bacteriocinas possui maior espectro de ação
quando utilizada em sinergismo com outros agentes antimicrobianos como NaCl,
ácidos orgânicos, agentes quelantes, óleos essenciais ou até na presença de outras

26
bacteriocinas (GALVEZ et al., 2007; MARTIN-VISSCHER et al., 2011; MILLS et al.,
2011).
Nos estudos realizados por Mills et al. (2011), os autores apresentaram
diversas aplicações das bacteriocinas em alimentos ondem enfatizam o uso destes
na promoção da saúde, na redução dos riscos de doenças vasculares, e até mesmo
na prevenção de doenças cancerígenas. Ainda, afirmam que, tais compostos
antimicrobianos têm efeitos semelhantes em relação a alguns fármacos.
O uso de bacteriocinas também pode ser aplicado em películas para uso
alimentar (MILLS et al., 2011) ou no desenvolvimento de embalagens bioativas
(GALVEZ et al., 2007) onde estas irão interagir na conservação do alimento
inativando microrganismos. Trata-se de um potencial conservante de alimentos.
Portanto, o uso das bacteriocinas na conservação de alimentos oferece vantagens
que satisfazem a indústria alimentar assim como o consumidor que procura um
alimento saudável.
4 METODOLOGIA
4.1 Material Biológico
Neste estudo, utilizou-se isolados de E. faecium provenientes de queijo, sendo
designados de Efm20 e Efm22, previamente analisados por Furlaneto-Maia et al.,
(2014) e Ogaki et al. (2015). As culturas encontravam-se estocadas em freezer e
foram re-hidratadas em 5 mL de caldo MRS por 10 minutos à temperatura ambiente,
seguido de incubação a 37ºC por 24 horas. Listeria innocua CLIP 12612 foi utilizado
como bactéria indicadora.
4.2 Ensaio da ação antagônica da enterocina
O estudo da ação antagônica de uma enterocina tem um papel relevante
quando se pretende avaliar uma possível aplicação de agentes antimicrobianos ou
da bactéria como agente probiótico, cultura starter ou simplesmente como inibidor de

27
patógenos, quando adicionados em um alimento (GUERREIRO et al., 2014; KOCH
et al., 2010).
O ensaio antagônico da enterocina produzida em diversas condições seguiu a
metodologia de difusão em poços. Para tanto, 20 mL de ágar soft semi-sólido 0,8%
(m/v), contendo a bactéria indicadora L. innocua na concentração final
correspondente à escala 4 de McFarland (12x108 células/mL) foi transferido para
uma placa de Petri e, após a solidificação foram feitos 3 poços de aproximadamente
5mm de diâmetro com auxílio de ponteiras estéreis. Foram adicionados 45μL do
sobrenadante livre de células (CFS) neutralizado nos poços. As placas foram
incubadas a 37°C por 24h e posteriormente os halos de inibição foram mensurados
em milímetros com auxílio de uma régua.
4.3 Efeito do pH na produção de bacteriocinas
A análise do efeito do pH seguiu a metodologia descrita por Furtado et al.,
(2014). Para tanto, as células provenientes de um cultivo em caldo MRS a 37°C por
18 horas, foram centrifugadas e o sedimento foi lavado duas vezes em solução
salina 0,85% estéril. A suspensão resultante foi ajustada para o valor 0,5 da escala
de McFarland (1,5x108 células/mL). Uma alíquota de 100 µL foi inoculada em 10 mL
de caldo MRS com pH ajustado para: 4.5, 5.0, 5.5, 6.0 e 6.5 e foram incubados a
37°C por 24 horas em estufa sob agitação. O pH foi ajustado com NaOH ou HCl
1Mol/L. Após incubação, os tubos que obtiverem crescimento celular foram
centrifugados a 10000 rpm por 15 minutos e o sobrenadante livre de célula (CFS) foi
esterilizado por filtração em membrana com poros de 0,22 μm com baixa capacidade
de ligação de proteínas (Millipore®). O pH do CFS foi ajustado em 6,5 para eliminar a
inibição causada pela presença de ácido lático.
4.4 Efeito das suplementações no meio MRS na produção de
bacteriocinas
O melhor resultado obtido na análise de pH foi submetido à análise de
diferentes componentes do meio de cultivo, seguindo a metodologia de Furtado et al.

28
(2014). Foram testadas diferentes formulações, acrescidas ao caldo MRS: lactose
(5,0 g/L; 10,0 g/L; 20,0 g/L; 30,0 g/L; 50,0 g/L), manose (20,0 g/L), manitol (20,0 g/L),
extrato de levedura (20,0 g/L), glicerol (2,0 g/L; 5,0 g/L; 8,0 g/L; 10,0 g/L; 20,0 g/L),
Tween® 20 (2,0 g/L; 5,0 g/L; 10,0 g/L), Tween® 80 (2,0 g/L; 5,0 g/L; 10,0 g/L) e foram
incubados a 37°C por 24 horas. Após incubação, os tubos que obtiverem
crescimento celular foram centrifugados (10000 rpm / 15 min) e o sobrenadante livre
de célula (CFS) foi neutralizado com NaOH 0,1 Mol/Le esterilizado por filtração em
membrana com poros de 0,22 μm.
Os experimentos cujos resultados apresentaram-se maiores halos de inibição
na ação antagônica das enterocinas foram testados para as demais variáveis a fim
de avaliar sua eficiência diante da célula alvo.
4.5 Efeito do pH na estabilidade das bacteriocinas
Como as bacteriocinas são constituídas de proteínas, a modificação do pH do
meio onde se encontra, exerce grande influência na sua estabilidade e atividade,
pois pode ocorrer interações entre as cadeias dos aminoácidos que por
consequência afetará a estrutura e a estabilidade da conformação tridimensional, a
qual é fundamental para a sua atividade (CHEN; HOOVER, 2003; MATAGARAS et
al., 2003; VAN REENEN et al., 2003).
A estabilidade das enterocinas foi testada em pH 3 a 9, utilizando os tampões
conforme descrito na tabela 1. Para tanto, 200 µL de CFS foram acrescidos de 200
µL de tampão.
Tabela 1. Tampões utilizados na estabilidade das enterocina. pH Tampão Concentração (M)
3 Glicina 0,25
5 Citrato de sódio 0,25
7 Fosfato de sódio 0,25
9 Tris 0,25
Após incubação foi realizado o teste de ação antagônica contra a bactéria
indicadora L. innocua. As placas foram incubadas em estufa a 37°C por 18 horas e o

29
halo de inibição foi mensurado em milímetros com auxilio de uma régua. Como
controle, foi adicionado aos poços somente os tampões de pH, a fim de verificar se
este provocaria inibição da bactéria indicadora.
4.6 Efeito da temperatura na estabilidade das bacteriocinas
Para avaliar a estabilidade das bacteriocinas ao tratamento térmico foram
utilizadas alíquotas de CFS (1 mL) e foram incubados em banho Maria a
temperaturas constantes de 40°C, 50°C, 60°C, 70°C, 80°C, 90°C e 100°C por 30
minutos e a 121°C na autoclave por 15 minutos. Após o tratamento térmico as
alíquotas foram submetidas ao teste de ação antagônica contra a bactéria
indicadora. A atividade antimicrobiana residual de cada temperatura também foi
avaliada. Sobrenadantes sem tratamento térmico foram aplicados nos testes de
difusão em poços e os resultados foram aplicados na Equação 1.
Atividade Residual (%) =
(Equação 1)
Onde:
Ht = halo do sobrenadante após tratamento térmico (mm);
Hc = halo do controle (mm);
t = tamanho do poço
4.7 Efeito da estabilidade das bacteriocinas na presença de SDS, tween®
20, tween® 80, EDTA e NaCl
Para a avaliação da estabilidade diante dos produtos químicos, foram
adicionados 10 mg/mL de tween® 20, tween® 80, NaCl, EDTA e SDS aos CFS
contendo a enterocina. Após, 20 mL de ágar semi-sólido 0,8% (m/v), contendo a
bactéria indicadora Listeria innocua na concentração final correspondente à escala 4
de McFarland (12x108 células/mL) foi transferido para uma placa de Petri e, após a
solidificação foram feitos 3 poços de aproximadamente 5mm de diâmetro com
auxílio de ponteiras estéreis. Foram adicionados 45μL do CFS neutralizado nos
poços.

30
4.8 Determinação do efeito bactericida/bacteriostático das bacteriocinas
Para a determinação do efeito bactericida e/ou bacteriostático das enterocinas,
L. innocua foi cultivada em meio BHI a 37°C por 24h, em leve agitação.
Posteriormente, 100 µL da suspensão celular e 100 µL do CFS foram adicionadas
em poços de microplaca de titulação. A leitura em espectrofotômetro (DO660nm) e a
determinação da UFC foram feitas a cada duas horas até 8 horas de incubação.
4.9 Adsorção de bacteriocinas nas células alvo
Dá-se grande importância ao conhecimento do grau de adsorção das
bacteriocinas às células alvos visto que é necessário para compreensão da
sensibilidade e mecanismo de ação das bacteriocinas (YILDIRIM; AVSAR;
YILDIRIM, 2002).
O teste de adsorção foi realizado segundo método de Todorov (2008). As
linhagens foram cultivadas por 18 h em meio caldo BHI a 37°C e após centrifugadas
(10000 x g) por 15 minutos. Em seguida, as células foram lavadas duas vezes com
tampão fosfato 5 M estéril pH 6,5 e foram ressuspendidas no volume original do
mesmo tampão. Posteriormente, foi feita 8 diluições seriadas da solução as quais
foram submetidas ao teste de ação antagônica. Os halos de inibição foram
mensurados em mm e para obter a adsorção de cada diluição foi aplicado a
Equação 2.
(Equação 2)
Onde:
UA/mL1: atividade da bacteriocinas após tratamento;
UA/mL0: atividade original do sobrenadante contendo a bacteriocinas (antes do
tratamento).

31
4.10 Análise de dados
Todos os experimentos foram realizados em triplicata e com 2 repetições.
Análise de variância (ANOVA) foi aplicado para verificar diferenças significativas (p <
0.05) entre os valores obtidos em cada experimento, utilizando Teste de Tukey com
auxílio do Programa Assistat 7.7 Beta.

32
5. REFERÊNCIAS
ABEE, T.; KROCKEL, L.; HILL, C.: Bacteriocins: modes of action and potentials in
food preservation and control of food poisoning. International Journal of Food
Microbiology, v. 28, n. 2, p. 169-185, 1995.
ARIAS, C. A.; CONTRERAS, G.; MURRAY, B. E.: Management of multidrug‐
resistant enterococcal infections. Clinical Microbiology and Infection, v. 16, n. 6, p.
555-562, 2010.
AUDISIO, M. C.; OLIVER, G.; APELLA, M. C.: Effect of different complex carbon
sources on growth and bacteriocin synthesis of Enterococcus faecium. International
Journal Of Food Microbiology, v. 63, n. 3, p. 235-241, 2001.
BALLONGUE, J. E. A. N.: Bifidobacteria and probiotic action. Food Science And
Technology-New York-Marcel Dekker, v. 139, p. 67-124, 2004.
BOUCHER, H. W.; TALBOT, G. H.; BRADLEY, J. S.; EDWARDS, J. E.; GILBERT,
D.; RICE, L. B.; BARTLETT, J.: Bad bugs, no drugs: no ESKAPE! An update from the
Infectious Diseases Society of America. Clinical Infectious Diseases, v. 48, n. 1, p.
1-12, 2009.
CAUWERTS, K.; DECOSTERE, A.; DE GRAEF, E. M.; HAESEBROUCK, F.;
PASMANS, F.: High prevalence of tetracycline resistance in Enterococcus isolates
from broilers carrying the erm (B) gene. Avian Pathology, v. 36, n. 5, p. 395-399,
2007.
CARRIM, A. J. I.: Bioprospecção de microrganismos endofíticos com atividade
enzimática e bacteriocinogênica em isolados de Jacaranda decurrens Cham
(Carobinha do campo). Dissertação de mestrado do Programa de Pós-graduação
em Microbiologia Ambiental, Universidade Federal de Goiás, 2005.

33
CARVALHO, M. J. C.: Isolamento e caracterização de bacteriocinas com
potencial interesse na área alimentar. Dissertação de mestrado em gestão da
qualidade e segurança alimentar, Instituto politécnico de Viana do Castelo, 2016.
CASTILHO, F. M.; BALCIUNAS, E. M.; CONVERTI, A.; COTTER, P.; OLIVEIRA, R.:
Bacteriocin prodution by Bifidobacterium spp. A review. Biotechnology Advances,
v.31, n.4, p. 482-488, 2013.
CLEVELAND, J.; MONTVILLE, T. J.; NES, I. F.; CHIKINDAS, M. L.: Bacteriocins:
safe, natural antimicrobials for food preservation. International Journal of Food
Microbiology, v. 71, n. 1, p. 1-20, 2001.
CHEN, H.; HOOVER, D. G.: Bacteriocins and their food
applications. Comprehensive Reviews in Food Science and Food Safety, v. 2, n.
3, p. 82-100, 2003.
CINTAS, L. M.; CASAUS, M. P.; HERRANZ, C.; NES, I. F.; HERNÁNDEZ, P. E.:
Review: bacteriocins of lactic acid bacteria. Food Science and Technology
International, v. 7, n. 4, p. 281-305, 2001.
COLLADO, M. C.; GONZALEZ, A.; GONZALEZ, R.; HERNANDEZ, M.; FERRUS, M.
A.; SANZ, Y.: Antimicrobial peptides are among the antagonistic metabolites
produced by Bifidobacterium against Helicobacter pylori. International Journal of
Antimicrobial Agents, v. 25, n. 5, p. 385-391, 2005.
COTTER, P. D.; HILL, C.; ROSS, R. P.: Bacteriocins: developing innate immunity for
food. Nature Reviews Microbiology, v. 3, n. 10, p. 777-788, 2013.
CUI, Y.; ZHANG, C.; WANG, Y.; SHI, J.; ZHANG, L.; DING, Z.; CUI, H.: Class IIa
bacteriocins: diversity and new developments. International Journal Of Molecular
Sciences, v. 13, n. 12, p. 16668-16707, 2012.

34
DE VUYST, L.; LEROY, F.: Bacteriocins from lactic acid bacteria: production,
purification, and food applications. Journal of Molecular Microbiology and
Biotechnology. V. 13, n. 4, p. 194–199, 2007.
DELBONI, R. R.: Dinâmica populacional de microrganismos e a conservação de
alimentos. Dissertação de mestrado do Programa de Pós-graduação em Estatística
e Computação Científica da Universidade Estadual de Campinas – UNICAMP.
Campinas/SP, 2009.
DEVLIEGHERE, F.; VERMEIREN, L.; DEBEVERE, J.: New preservation
technologies: Possibilities and limitations. International Dairy Journal, v. 14, n. 4,
p. 273-285, 2004.
DIGAITIENE, A.; HANSEN, A. S.; JUODEIKIENE, G.; EIDUKONYTE, D.;
JOSEPHSEN, J.: Lactic acid bacteria isolated from rye sourdoughs produce
bacteriocin‐like inhibitory substances active against Bacillus subtilis and
fungi. Journal of Applied Microbiology, v. 112, n. 4, p. 732-742, 2012.
DOMIG, K. J.; MAYER, H. K.; KNEIFEL, W.: Methods used for the isolation,
enumeration, characterisation and identification of Enterococcus spp.: 2. Pheno-and
genotypic criteria. International Journal of Food Microbiology, v. 88, n. 2, p. 165-
188, 2003.
EIJSINK, V. G.; AXELSSON, L.; DIEP, D. B.; HAVARSTEIN, L. S.; HOLO, H.; NES,
I. F.: Production of class II bacteriocins by lactic acid bacteria; an example of
biological warfare and communication. Antonie Van Leeuwenhoek, v. 81, n. 1, p.
639-654, 2002.
FELTCHER, M. E.; BRAUNSTEIN, M.: Emerging themes in SecA2-mediated protein
export. Nature Reviews Microbiology, London, v. 10, n. 11, p. 779-789, 2012.

35
FOULQUIÉ-MORENO M.R.; SARANTINOPOULOS P.; TSAKALIDOU E.: The role
and application of enterococci in food and health. International Journal of Food
Microbiology, v.106, n.1, p.1-24, 2006.
FRANZ, C. M.; HOLZAPFEL, W. H.; STILES, M. E.: Enterococci at the crossroads of
food safety?. International Journal of Food Microbiology, v. 47, n. 1, p. 1-24,
1999.
FURTADO, D. N; TODOROV, S. D; LANDGRAF, M; DESTRO, M. T; FRANCO, B.
D.: Bacteriocinogenic Lactococcus lactis subsp. lactis DF04Mi isolated from goat
milk: Characterization of the bacteriocin. Brazilian Journal of Microbiology , v 45,
n. 4, p. 1541-1550, 2014.
FURLANETO-MAIA, L.; ROCHA, K. R.; HENRIQUE, F. C.; GIAZZI, A.;
FURLANETO, M. C.: Antimicrobial resistance in Enterococcus sp isolated from soft
cheese in Southern Brazil. Advances in Microbiology, v. 2014, 2014.
FURLANETO-MAIA, L.; OGAKI, M. B.; BRANDALIZE, C. C.; ROCHA, K. R.;
FURLANETO, M. C.: Enterococcus sp. em alimentos: Paradigmas. Tópicos
especiais em Microbiologia. Programa de Pós-graduação em Microbiologia.
Universidade Estadual de Londrina –PR. 1º ed, p. 71-90, Londrina – PR, 2015.
GALVEZ, A.; ABRIOUEL, H.; LÓPEZ, R. L.; OMAR, N. B.: Bacteriocin-based
strategies for food biopreservation. International Journal Of Food Microbiology, v.
120, n. 1, p. 51-70, 2007.
GAMA, B. A.: Análise da resistência antimicrobiana e de genes de virulência de
Enterococcus spp. Tese de Doutorado do Programa de Pós-graduação em
Biologia Celular e Molecular pela Universidade Federal Do Rio Grande Do Sul. Porto
Alegre, RS. 2008.
GUERREIRO, J.; MONTEIRO, V.; RAMOS, C.; DE MELO FRANCO, B. D. G.;
MARTINEZ, R. C. R.; TODOROV, S. D.; FERNANDES, P.: Lactobacillus pentosus
B231 isolated from a Probiotics and Antimicrobial Proteins Portuguese PDO

36
cheese: production and partial characterization of its bacteriocin. , v. 6, n. 2, p. 95-
104, 2014.
GRAM, L.; RAVN, L.; RASCH, M.; BRUHN, J. B.; CHRISTENSEN, A. B.; GIVSKOV,
M.: Food spoilage—interactions between food spoilage bacteria. International
Journal of Food Microbiology, v. 78, n. 1, p. 79-97, 2002.
HOLLEY, R. A.; PATEL, D.: Improvement in shelf-life and safety of perishable foods
by plant essential oils and smoke antimicrobials. Food Microbiology, v. 22, n. 4, p.
273-292, 2005.
JERONYMO, A. B. O. Avaliação do potencial probiótico de bactérias
acidoláticas produtoras de substância antimicrobiana isoladas de mussarela
de búfala. Dissertação de mestrado. Universidade Estadual Paulista Júlio de
Mesquita Filho. Instituto de Biociências, Letras e Ciências Exatas de São José do
Rio Preto, 2013.
KAWAI, Y.; ISHII, Y.; UEMURA, K.; KITAZAWA, H.; SAITO, T.; ITOH, T.:
Lactobacillus reuteri LA6 and Lactobacillus gasseri LA39 isolated from faeces of the
same human infant produce identical cyclic bacteriocin. Food Microbiology, v. 18, n.
4, p. 407-415, 2001.
KENSE, M. J.; LANDMAN, W. J. M.: Enterococcus cecorum infections in broiler
breeders and their offspring: molecular epidemiology. Avian Pathology, v. 40, n. 6,
p. 603-612, 2011.
KOHLER, W.: The present state of species within the genera Streptococcus and
Enterococcus. International Journal of Medical Microbiology, v. 297, n. 3, p. 133-
150, 2007.
KOCH, J.; DWORAK, R.; PRAGER, R.; BECKER, B.; BROCKMANN, S.; WICKE, A.;
STARK, K.: Large listerios is outbreak linked to cheese made from pasteurized milk,
Germany, 2006–2007. Food borne Pathogens and Disease, v. 7, n. 12, p. 1581-
1584, 2010.

37
KUHN, I.; IVERSEN, A.; BURMAN, L. G.; OLSSON-LILJEQUIST, B.; FRANKLIN,
A.; FINN, M.; MOLLBY, R.: Epidemiology and ecology of enterococci, with special
reference to antibiotic resistant strain, in animals, humans and the environment.
Example of an ongoing project within de European research programmer.
International Journal of Antimicrobial Agents, v. 14, n. 4, p. 337-342, 2000.
KUHN, I.; IVERSEN, A.; BURMAN, L. G.; OLSSON-LILJEQUIST, B.; FRANKLIN, A.;
FINN, M.; TAYLOR, H.: Comparison of enterococcal populations in animals, humans,
and the environmental European study. International Journal of Food
Microbiology, v. 88, n. 2, p. 133-145, 2003.
LEE, H. j.; KIM, A.: Review: Lantibiotics, Class I bacteriocins from the genus
Bacillus. Journal of Microbiology and Biotechnology, v. 21, n. 3, p. 229-235,
2011.
LEROY, F.; DE VUYST, L.: A combined model to predict the functionality of the
bacteriocin-producing Lactobacillus sakei strain CTC 494. Applied And
Environmental Microbiology, v. 69, n. 2, p. 1093-1099, 2003.
LUND, B.; EDLUND, C.: Probiotic Enterococcus faecium strain is a possible recipient
of the vanA gene cluster. Clinical Infectious Diseases, v. 32, n. 9, p. 1384-1385,
2001.
LYCKLAMA, J. A.; DRIESSEN, A. J. M.: The bacterial Sec-translocase: Structure
and mechanism. Philosophical Transactions of the Royal Society B, London, v.
367, p. 1016-1028, 2012.
MACHADO, S. I. G.: Estudo de atividades antimicrobianas em bactérias do
género Bacillus isoladas nos Açores: otimização das condições de produção,
identificação e caracterização de bacteriocinas. Dissertação de Mestrado do
Programa de Pós-graduação em Ciências Biomédicas da Universidade de Açores.
Departamento de Biologia, Ponta Delgada, 2015.

38
MARTINS, A. D. O.; MENDONÇA, R. C. S.; SILVA, D. L.; RAMOS, M. S.; MARTINS,
M. C.; DONZELE, J. L.; ANDRADE, N. J.: Resistência de bactérias lácticas, isoladas
de fezes de suínos e sua capacidade antagônica frente a microrganismos
indicadores. Revista de Ciências Agro Veterinárias. Lages, v.5, n.1, p. 53-59,
2006.
MARTIN-VISSCHER, L. A.; YOGANATHAN, S.; SIT, C. S.; LOHANS, C. T.;
VEDERAS, J. C.: The activity of bacteriocins from Carnobacterium maltaromaticum
UAL307 against Gram-negative bacteria in combination with EDTA treatment. FEMS
Microbiology Letters, v. 317, n. 2, p. 152-159, 2011.
MATARAGAS, M.; METAXOPOULOS, J.; GALIOTOU, M.; DROSINOS, E. H.:
Influence of pH and temperature on growth and bacteriocin production by
Leuconostoc mesenteroides L124 and Lactobacillus curvatus L442. Meat Science,
v. 64, n. 3, p. 265-271, 2003.
MILLS, S.; STANTON, C.; HILL, C.; ROSS, R. P.: New developments and
applications of bacteriocins and peptides in foods. Annual Review of Food Science
and Technology, v. 2, p. 299-329, 2011.
MOLLOY, E. M.; FIELD, D.; COTTER, P. D.; HILL, C.; ROSS, R. P.: Saturation
mutagenesis of lysine 12 leads to the identification of derivatives of nisin A with
enhanced antimicrobial activity. Plos One, v. 8, n. 3, 2014.
NASCIMENTO, M. S.: Caracterização da atividade antimicrobiana e tecnológica
de três culturas bacteriocinogenicas e a avaliação de sua eficiência no
controle de Listeria monocytogenes, Staphylococcus aureus em queijo minas
frescal. Tese de doutorado em tecnologia de alimentos da Faculdade de engenharia
de Alimentos, Universidade Estadual de Campinas, Campinas, 2007.
NASCIMENTO, M. S.; MORENO, I.; KUAYE, A. Y.: Bacteriocinas em alimentos: uma
revisão. Brazilian Journal of Food Technology, v.11, n.2, p.120-127, 2008.

39
NES I. F.; DIEP D. B.; IKE, Y.: Enterococcal bacteriocins and antimicrobial
proteins that contribute to niche control. 2014.
Disponível em: https://www.ncbi.nlm.nih.gov/books/NBK190428/?report=reader
Acesso em: 09/11/2016.
NOONPAKDEE, W.; SANTIVARANGKNA, C.; JUMRIANGRIT, P.; SONOMOTO, K.;
PANYIM, S.: Isolation of nisin-producing Lactococcus lactis WNC 20 strain from
nham, a traditional Thai fermented sausage. International Journal of Food
Microbiology, v. 81, n. 2, p. 137-145, 2003.
OGAKI, M. B; FURLANETO, M. C.; MAIA, L. F.: Review: General aspects of
bacteriocins. Brazilian Journal of Food Technology, v. 18, n. 4, p. 267-276, 2015.
OHMOMO, S.; MURATA, S.; KATAYAMA, N.; NITISINPRASART, S.; KOBAYASHI,
M.; NAKAJIMA, T.; NAKANISHI, K.: Purification and some characteristics of
enterocin ON‐157, a bacteriocin produced by Enterococcus faecium NIAI
157. Journal of Applied Microbiology, v. 88, n. 1, p. 81-89, 2000.
PAPAGIANNI, M.: Ribosomally synthesized peptides with antimicrobial properties:
biosynthesis, structure, function, and applications. Biotechnology Advances, v. 21,
n. 6, p. 465-499, 2003.
REFFUVEILLE, F.; LENEVEU, C.; CHEVALIER, S.; AUFFRAY, Y.; RINCÉ, A.:
Lipoproteins of Enterococcus faecalis: bioinformatic identification, expression
analysis and relation to virulence. Microbiology, v. 157, n. 11, p. 3001-3013, 2011.
SABO, S.; VITOLO, M.; GONZÁLEZ, J. M. D.; DE SOUZA OLIVEIRA, R. P.:
Overview of Lactobacillus plantarum as a promising bacteriocin producer among
lactic acid bacteria. Food Research International, v. 64, p. 527-536, 2014.

40
SARIKA, A.R.; LIPTON, A.P.; AISHWARYZ, M. S.: Bacteriocin Production by a New
Isolate of Lactobacillus rhamnosus GP1 under Different Culture Conditions. Advance
Journal of Food Science and Technology, v.2, n.5, p. 291-297, 2010.
SIEVERT, D. M.; RICKS, P.; EDWARDS, J. R.; SCHNEIDER, A.; PATEL, J.;
SRINIVASAN, A.; FRIDKIN, S.: Antimicrobial-resistant pathogens associated with
healthcare-associated infections summary of data reported to the National
Healthcare Safety Network at the Centers for Disease Control and Prevention, 2009–
2010. Infection Control & Hospital Epidemiology, v. 34, n. 01, p. 1-14, 2013.
SCHULZ, D.; PEREIRA, M. A.; BONELLI, R. R.; NUNES, M. M.; BATISTA, C. R. V.:
Bacteriocinas: Mecanismo de ação e uso na conservação de alimentos. Alimentos e
Nutrição. Araraquara, v.14, n.2, p.229-235, 2003.
SCHULZ, D.; BONELLI, R. R.; BATISTA, C. R. V.: Bacteriocinas e enzimas
produzidas por Bacillus spp. para conservação e processamento de alimentos.
Alimentos e Nutrição. V.16, n.4, p. 403-411, 2005.
SCHEPERS, A. W.; THIBAULT, J.; LACROIX, C.: Lactobacillus helveticus
crescimento e produção de ácido lático durante culturas lote controlado por pH em
meio de soro de leite extrato permeado / levedura. Parte I. A análise cinética fator
múltipla. A Enzima e Tecnologia Microbiana, 30 v., n. 2, p. 176-186, 2002.
SHARMA, N.; KAPOOR, R.; GAUTAM, N.; KUMARI, R.: Purification and
characterization of bacteriocin produced by Bacillus subtilis R75 isolated from
fermented chunks of mung bean (Phaseolus radiatus). Food Technology and
Biotechnology, v. 49, n. 2, p. 169, 2011.
SOUSA, L. C. F. S.; DA SILVA SOUSA, J.; BORGES, M. D. G. B.; MACHADO, A. V.;
DA SILVA, M. J. S.; FERREIRA, R. T. F. V.; SALGADO, A. B.: Tecnologia de
embalagens e conservação de alimentos quanto aos aspectos físico, químico e
microbiológico. Agropecuária Científica no Semiárido, v. 8, n. 1, p. 19-28, 2013.

41
TAMINE, A. Y.: Probiotic dairy products. Dairy Sciense and Technology. n.1, p. 39-
62. 2005.
TODOROV, S. D.: Bacteriocin production by Lactobacillus plantarum AMA-K isolated
from Amasi, a Zimbabwean fermented milk product and study of the adsorption of
bacteriocin AMA-K to Listeria sp. Brazilian Journal of Microbiology, v. 39, n. 1, p.
178-187, 2008.
TODOROV, S. D.; RACHMAN, C.; FOURRIER, A.; DICKS, L. M.; VAN REENEN, C.
A.; PRÉVOST, H.; DOUSSET, X.: Characterization of a bacteriocin produced by
Lactobacillus sakei R1333 isolated from smoked salmon. Anaerobe, v. 17, n. 1, p.
23-31, 2011.
VAN DEN BERGHE E.; DE WINTER T.; DE VUYST, L.: Enterocin A production by
Enterococcus faecium FAIR-E 406 is characterised by a temperature- and pH-
dependent switch-off mechanism when growth is limited due to nutrient depletion.
International Journal of Food Microbiology, v.107, p. 159-170, 2006.
VAN REENEN, C. A.; CHIKINDAS, M. L.; VAN ZYL, W. H.; DICKS, L. M. T.:
Characterization and heterologous expression of a class IIa bacteriocin, plantaricin
423 from Lactobacillus plantarum 423, in Saccharomyces cerevisiae. International
Journal Of Food Microbiology, v. 81, n. 1, p. 29-40, 2003.
VÁSQUEZ, S. M.; SUÁREZ, H.; ZAPATA, S.: Utilización de sustancias
antimicrobianas producidas por bacterias acido lácticas en la conservación de la
carne. Revista Chilena de Nutrición, v. 36, n. 1, p. 64-71, 2009.
YANG, R.; RAY, B.: Factors influencing production of bacteriocins by lactic acid
bacteria. Food Microbiology, v. 11, n. 4, p. 281-291, 1994.
YILDIRIM, Z.; AVS, Y. K.; YILDIRIM, M.: Factors affecting the adsorption of
buchnericin LB, a bacteriocin produced by Lactocobacillus buchneri. Microbiological
Research, v. 157, n. 2, p. 103-107, 2002.

42
WILLEMS, R. J.; HANAGE, W. P.; BESSEN, D. E.; FEIL, E. J.: Population biology of
Gram-positive pathogens: high-risk clones for dissemination of antibiotic
resistance. FEMS Microbiology Reviews, v. 35, n. 5, p. 872-900, 2011.
ZOU, L. K.; WANG, H. N.; ZENG, B.; LI, J. N.; LI, X. T.; ZHANG, A. Y.; XIA, Q. Q.:
Erythromycin resistance and virulence genes in Enterococcus faecalis from swine in
China. New Microbiologica, v. 34, n. 1, p. 73-80, 2011.

43
ARTIGO
ESTUDO DA ESTABILIDADE DE ENTEROCINAS PRODUZIDAS POR
Enterococcus faecium
RESUMO: As enterocinas possuem atividade antimicrobiana sobre diversas
bactérias patogênicas e deteriorantes em alimentos. O objetivo do trabalho foi
estudar a produção de enterocinas por Enterococcus faecium e avaliar sua
estabilidade sob diversas variáveis. Para tanto, testou-se os pHs 4.5, 5.0, 5.5, 6.0 e
6.5 e o meio de cultivo MRS suplementado com lactose, manose, manitol, glicerol,
tween 20 e tween 80. A bactéria indicadora utilizada nos testes de antagonismo foi
Listeria innocua CLIP 12612. Os pHs ótimo de produção foram entre 6,0 e 6,5 e as
suplementações que obtiveram maior inibição foram lactose 20 g/L, glicerol 20 g/L,
tween 20 5,0 g/L, obtendo o mix que é a mistura dos nutrientes. Nenhum dos pHs
testados interferiu na redução da ação das enterocinas e a estabilidade térmica
variou conforme o isolado e a suplementação utilizada. Em relação à estabilidade a
diferentes produtos químicos o Tween 20 e 80, de ambos isolados, apresentou
redução da ação. Ainda, observamos que o mix de suplementos apresentou maior
adsorção e que o efeito bactericida foi observado com 2 horas de incubação. Os
resultados obtidos indicam que, as enterocinas que foram produzidas possui
potencial de aplicabilidade em alimentos como bioconservadores.
Palavras chaves: Bioconservantes, Inibição, Listeria innocua, Produtos químicos,
Tratamento térmico.
ABSTRACT: Enterocins has antimicrobial activity on several pathogenic and
deteriorating bacterium in food. The objective of this work was to study the
production of enterocins by E. faecium and evaluate your stability about different
variable. Therefore, production in various pH 4.5, 5.0, 5.5, 6.0 and 6.5, and medium
culture was tested for lactose, mannose, mannitol, glycerol, tween 20 and tween 80
at different concentrations using a Listeria innocua bacterium indicator. The best pHs
produced was 6.0 and 6.5, and as supplements that obtained the highest inhibition
for lactose 20 g/L, glycerol 20 g/L, tween® 20 5.0 g/L, obtaining the mix which is a

44
mixture of Nutrients. None of the pHs tested interfered in the reduction of the action
of the enterocines and a thermal stability varied according to the isolate and a
supplementation used. In relation to the stability and different chemical products,
Tween 20 and Tween 80, of both isolates, obtained reduction of the action. Still,
observed that the set of supplements for the one that presented greater adsorption
and that the bactericidal effect was observed as soon as the first two hours of
incubation. The results obtained indicate that, as enterocins that produce has
potential of application in food as bioconservatives.
Keywords: Bioconservantes, Inhibition, Listeria innocua, Chemical products,
Thermal treatment.
INTRODUÇÃO
As pesquisas sobre metabólitos antimicrobianos produzidos por bactérias ácido
láticas (BAL) tiveram um avanço nas últimas décadas. Isto ocorreu devido à
emergência de microrganismos potencialmente patogênicos em alimentos e na
crescente tendência da indústria de alimentos em substituir os aditivos químicos
pelos naturais (YANG, 2014).
Bacteriocinas são peptídeos antimicrobianos, produzidos por BAL e que
apresentam atividade antagônica contra diversas bactérias (DE VUYST; LEROY,
2007). Várias bacteriocinas produzidas por BAL têm sido isoladas de leite e
produtos lácteos (FRANCO et al., 2013; SCHIRRU et al., 2012). Estas bacteriocinas
apresentam um amplo espectro de atividade, modo de ação, origem genética,
massa molecular e propriedades bioquímicas (ALVAREZ-SIEIRO et al., 2016).
Entre as BAL, o gênero Enterococcus (especialmente Enterococcus faecium e
Enterococcus faecalis) é amplamente distribuído no ambiente e, principalmente, em
produtos alimentícios (GIRAFFA, 2003), sendo comuns as espécies de
Enterococcus produzirem múltiplas bacteriocinas (enterocinas) (NES; DIEP; HOLO,
2007). Enterococcus produtores de enterocinas já foram extensivamente estudados
e isolados de diversas fontes, com o propósito de aplicação como cultura

45
preservativa em alimentos e controle de patógenos (BAYOUB et al., 2011; HADJI-
SFAXI et al., 2011).
A ação protetiva de enterocinas já foi descrita em diversos alimentos, como a
enterocina CCM 4231 em produtos lácteos e salame fermentado (LAUKOVA et al.,
1999), enterocinas RZS C13 e CCM 4231 em salsichas (CALLEWAERT et al.,
2000), em presunto suíno cozido (AYMERICH et al., 2002), enterocina AS-48 usada
em produtos lácteos (MUNOZ et al., 2007), e a enterocina B a qual protege os
alimentos contra bactérias contaminantes (AYMERICH et al., 2002).
As enterocinas já foram bem descritas quanto seu efeito antilisterial
(CALLEWAERT; HUGAS; DE VUYST, 2000) além de diversas bactérias esporuladas
e Gram negativas (GARCIA et al., 2004). No entanto, o isolamento e caracterização
de novos compostos antimicrobianos produzidos por microrganismos isolados de
alimentos indica utilização industrial mais efetiva (LAUKOVA et al., 1999).
Experimentalmente e industrialmente têm sido demonstrado que as bacteriocinas
protegem os produtos alimentares contra contaminantes alimentares frequentes, tais
como Listeria e Bacillus (McAULIFFE et al., 1999).
Embora várias enterocinas já tenham sido descritas, neste estudo, avaliou-se
alguns constituintes do meio de cultura e condições de cultivo para averiguar a
melhor produção de enterocinas por isolados de E. faecium.
Diante do exposto, o objetivo do trabalho foi avaliar a produção de enterocinas
pelas linhagens de Enterococcus faecalis bem como analisar as melhores condições
que influenciam a produção de enterocinas para que se torne uma alternativa na
aplicação em alimentos como conservantes naturais.
MATERIAIS E MÉTODOS
Linhagens
Utilizou-se isolados de E. faecium provenientes de queijo, sendo designados de
Efm20 e Efm22, previamente analisados por Furlaneto-Maia et al. (2014), e Ogaki et
al. (2016). As culturas estavam estocadas em freezer e foram re-hidratadas em 5 mL

46
de caldo MRS por 10 minutos em temperatura ambiente, seguido de incubação a
37ºC por 24 horas. Listeria innocua CLIP 12612 foi utilizado como bactéria
indicadora.
Ensaio da ação antagônica
O ensaio antagônico seguiu a metodologia de difusão em poços. Para tanto, 20
mL de ágar soft semi-sólido 0,8% (m/v), contendo a bactéria indicadora L. innocua
na concentração final correspondente à escala 4 de McFarland (12x108 células/mL)
foi transferido para uma placa de Petri e, após a solidificação foram feitos 3 poços de
aproximadamente 5mm de diâmetro com auxílio de ponteiras estéreis. Foram
adicionados 45μL do sobrenadante livre de células (CFS) neutralizado nos poços. As
placas foram incubadas a 37°C por 24h e posteriormente os halos de inibição foram
mensurados em milímetros com auxílio de uma régua.
Efeito do pH e suplementação do meio na produção de enterocina
Para averiguar o efeito de diferentes pHs na produção de enterocina, seguiu a
metodologia descrita por Furtado et al. (2014), com modificações. Uma alíquota de
100 µL de cada inóculo (concentração celular final de 1,5x108 células/mL) foi
inoculada em 10 mL de caldo MRS com pH ajustado para: 4,5, 5,0, 5,5, 6,0 e 6,5 e
foram incubados a 37°C por 24 horas, sob agitação. Após realizou o ensaio da ação
antagônica.
O efeito da suplementação do meio MRS seguiu a metodologia descrita por
Furtado et al. (2014). Foram testadas diferentes formulações, acrescidas ao caldo
MRS: lactose (5,0 g/L; 10,0 g/L; 20,0 g/L; 30,0 g/L; 50,0 g/L), manose (20,0 g/L),
manitol (20,0 g/L), extrato de levedura (20,0 g/L), glicerol (2,0 g/L; 5,0 g/L; 8,0 g/L;
10,0 g/L; 20,0 g/L), Tween® 20 (2,0 g/L; 5,0 g/L; 10,0 g/L), Tween® 80 (2,0 g/L; 5,0
g/L; 10,0 g/L) (Tabela 1) e foram incubados a 37°C por 24 horas.
Após incubação em todos os tratamentos, as células foram recuperadas por
centrifugação a 10000 rpm por 15 minutos e o sobrenadante livre de célula (CFS) foi

47
esterilizado por filtração em membrana com poros de 0,22 μm com baixa capacidade
de ligação de proteínas (Millipore®). O pH do CFS foi ajustado para 6,5 e tratado
com catalase para evitar a presença de H2O2 (OGAKI et al., 2016) e foi realizado o
teste de ação antagônica.
Os suplementos que obtiverem o maior efeito na produção de enterocinas
foram testadas frente a diversas variáveis a fim de avaliar sua eficiência diante da
célula alvo.
Efeito do pH e temperatura na estabilidade das enterocinas da enterocina
produzida
A estabilidade das enterocinas foi testada em pH 3 a 9, utilizando os tampões
glicina 25 mM (pH 3), citrato de sódio 25 mM (pH 5), fosfato de sódio 25 mM (pH 7,2)
e tris 25 mM (pH 9).
Para o teste de estabilidade à temperatura, um volume de 1 mL de CFS foi
incubados em temperaturas constantes de 40°C, 50°C, 60°C, 70°C, 80°C, 90°C e
100°C por 30 minutos. Uma alíquota também foi submetida a 121°C a 1 atm
(autoclavagem) por 15 minutos. A atividade antimicrobiana residual de cada
temperatura foi avaliada de acordo com protocolo descrito por Cladera-Oliveira
(2004).
Após obtenção do CFS foi realizado o ensaio antagônico contra a bactéria
indicadora L. innocua. Como controle, foi adicionado aos poços somente os tampões
de pH, a fim de verificar se este provocaria inibição da bactéria indicadora.
Efeito do tween® 20, tween® 80, NaCl, EDTA e SDS na estabilidade da
enterocina produzida
A avaliação da estabilidade da ação da enterocina frente aos compostos
químicos tween® 20, tween® 80, NaCl, EDTA e SDS seguiu protocolo descrito por
Todorov et al., (2008) com modificações. Foi adicionado ao CFS 10 mg/mL de cada

48
composto, seguido de 30 minutos de incubação a 37°C. Em seguida, procedeu-se a
análise da atividade antagônica.
Determinação do efeito bactericida e/ou bacteriostático das bacteriocinas
Para a determinação do efeito bactericida e/ou bacteriostática das enterocinas
sobre a bactéria indicadora L. innocua, um volume de 100 µL da suspensão celular
foi adicionada em microplaca de titulação e adicionados 100 µL do CFS. A leitura em
espectrofotômetro (DO660 nm) e a determinação da UFC foram feitas a cada duas
horas até 8 horas de incubação.
Adsorção de bacteriocinas pelas células alvo
O teste de adsorção foi realizado segundo método descrito por Todorov (2008).
As linhagens foram cultivadas por 18 h em meio caldo BHI a 37°C e após
centrifugado (10000 x g) por 15 minutos. Em seguida, as células foram lavadas duas
vezes com tampão fosfato 5 M estéril pH 6,5 e foram ressuspendidas no volume
original do mesmo tampão. Após, foi feita 8 diluições seriadas da solução as quais
foram submetidas ao teste de antagonismo. O halo de inibição foi mensurado em
mm e para obter a adsorção de cada diluição foi aplicado a Equação 1.
(Equação 1)
Onde:
UA/mL1 : atividade da bacteriocinas após tratamento;
UA/mL0: atividade original do sobrenadante contendo a bacteriocinas (antes do
tratamento).
Análise estatística
Todos os experimentos foram realizados em triplicata e com 2 repetições.
Análise de variância (ANOVA) foi aplicado para verificar diferenças significativas (p <
0.05) entre os valores obtidos em cada experimento, utilizando Teste de Tukey com
auxílio do Programa Assistat 7.7 Beta.

49
RESULTADOS
O efeito antagônico de enterocinas, produzidas por enterococos provenientes
de alimentos lácteos, sobre bactérias patogênicas e/ou deteriorantes pode ser um
potencial candidato na bioconservação de alimentos. Neste estudo, foram avaliados
os isolados E. faecium denominados de Efm20 Efm22, provenientes de queijo. Os
parâmetros de tempo e temperatura de incubação foram mantidos em todos os
tratamentos. Todos os dados foram apresentados com duas ou três repetições.
Para determinação do melhor pH para produção de enterocinas, o crescimento
dos isolados ocorreu na presença do caldo MRS em diferentes valores de pH. Os
resultados obtidos mostraram que não houve diferença significativa (p<0,05) entre
os isolados na produção de enterocinas nos pHs 6.0 e 6.5. Contudo, estes valores
de pH foram estatisticamente melhores para a produção de enterocinas, quando
comparado com os pHs 5.0 e 5.5. No pH 4.5 não foi detectada ação antagônica
contra a bactéria teste. A partir destes resultados, elencamos o pH 6.5 para os
ensaios posteriores.
Nos testes realizados com diversos componentes e concentrações do meio,
observou-se que a suplementação de lactose na concentração 20,0 g/L, glicerol 20,0
g/L e tween® 20 5,0 g/L, quando avaliados separadamente, foram os que mais
influenciaram positivamente a produção da enterocina (Tabela 1) em ambos
isolados. Em contrapartida, as demais suplementações com extrato de levedura,
manitol, manose e tween® 80, não alteraram a produção de enterocinas,
apresentando produção similar ao controle (meio MRS), não havendo diferença
significativa.
As análises de produção de enterocina também foi produzida em meio MRS
contendo lactose 20,0 g/L, glicerol 20,0 g/L e tween® 20 5,0 g/L, e denominamos de
mix. Esse mix apresentou elevada produção de enterocinas quando comparado com
o meio MRS.

50
Tabela 1: Mensuração do efeito do substrato em meio MRS na produção de enterocina por isolados de Enterococcus sp.
Componente Concentração
Medida do halo em mm
Efm20 Efm22
Lactose 5,0 g/L 10,6 ± 0,2b 11,1 ± 0,0b
Lactose 10,0 g/L 10,6 ± 0,5b 11,3 ± 0,5b
Lactose 20,0 g/L 13,1 ± 0,0a 13,3 ± 0,5a
Lactose 30,0 g/L 10,5 ± 0,7b 11,1 ± 0,0b
Lactose 50,0 g/L 10,5 ± 0,7b 11,1 ± 0,0b
Extrato de Levedura 20,0 g/L 10,3 ± 0,5b 11,5 ± 0,7b
Manose 20,0 g/L 10,3 ± 0,5b 11,1 ± 0,0b
Manitol 20,0 g/L 10,5 ± 0,7b 11,1 ± 0,7b
Glicerol 2,0 g/L 10,8 ± 0,2b 11,5 ± 0,5b
Glicerol 5,0 g/L 10,8 ± 0,7b 11,6 ± 0,5b
Glicerol 8,0 g/L 10,8 ± 0,2b 11,3 ± 0,5b
Glicerol 10,0 g/L 10,3 ± 0,5b 11,1 ± 0,2b
Glicerol 20,0 g/L 13,0 ± 1,0a 13,2 ± 0,4a
Tween® 20 2,0 g/L 10,5 ± 0,5b 11,1 ± 0,2b
Tween® 20 5,0 g/L 12,8 ± 0,0a 13,2 ± 0,3a
Tween® 20 10,0 g/L 10,5 ± 0,5b 11,3 ± 0,5b
Tween® 80 2,0 g/L 10,8 ± 0,0b 11,6 ± 0,7b
Tween® 80 5,0 g/L 10,5 ± 0,8b 11,6 ± 0,3b
Tween® 80 10,0 g/L 10,8 ± 0,0b 11,1 ± 0,0b
*mix - 12,3 ± 0,5a 13,0 ± 1,0a a, b
Letras diferentes na mesma coluna, indicam diferença significativa (p<0,05). *Mix: Mistura de substrato (lactose 20 g/L; glicerol 20 g/L; Tween® 20 g/L).
De posse dos valores de pH e as suplementações que forneceram os
melhores resultados para produção de enterocinas (Figura 1), estas foram testadas
sobre diversas variáveis para avaliar sua viabilidade.
Figura 1: Resultado do halo de inibição em meio MRS suplementado com lactose 20,0 g/L, glicerol 20,0 g/L e tween® 20 5,0 g/L, pelos isolados Efm20 e Efm22.
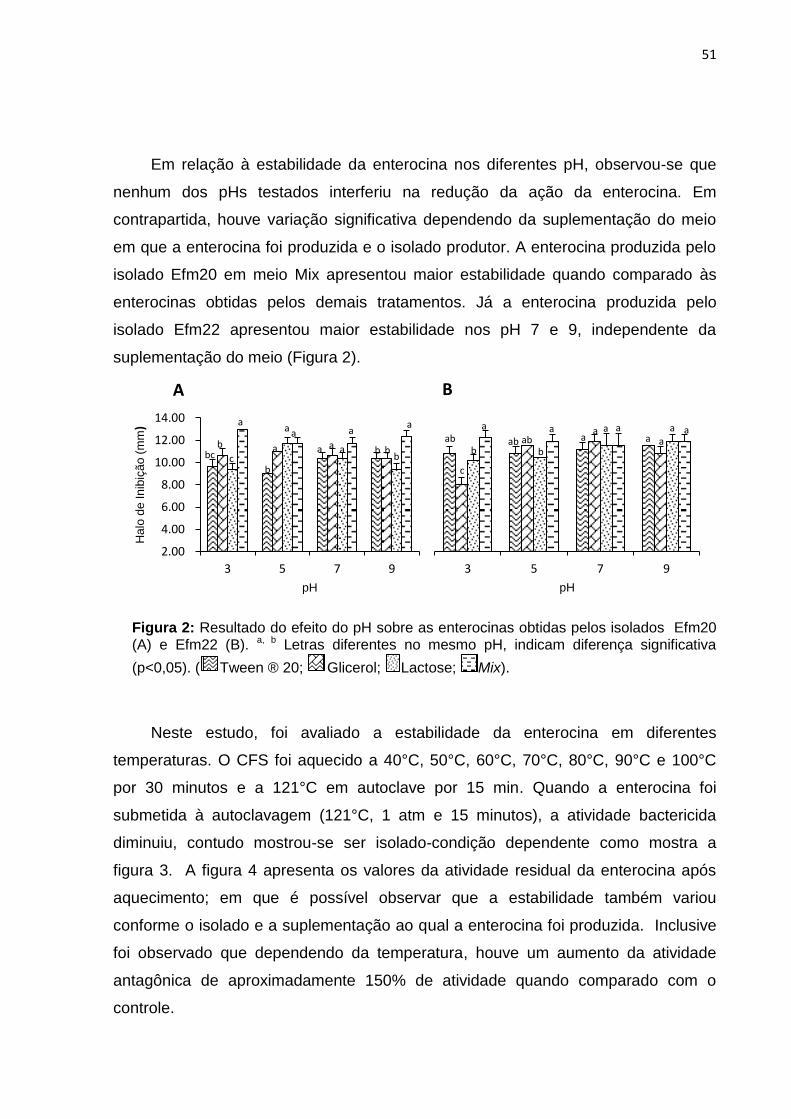
51
Em relação à estabilidade da enterocina nos diferentes pH, observou-se que
nenhum dos pHs testados interferiu na redução da ação da enterocina. Em
contrapartida, houve variação significativa dependendo da suplementação do meio
em que a enterocina foi produzida e o isolado produtor. A enterocina produzida pelo
isolado Efm20 em meio Mix apresentou maior estabilidade quando comparado às
enterocinas obtidas pelos demais tratamentos. Já a enterocina produzida pelo
isolado Efm22 apresentou maior estabilidade nos pH 7 e 9, independente da
suplementação do meio (Figura 2).
Figura 2: Resultado do efeito do pH sobre as enterocinas obtidas pelos isolados Efm20 (A) e Efm22 (B). a, b Letras diferentes no mesmo pH, indicam diferença significativa
(p<0,05). ( Tween ® 20; Glicerol; Lactose; Mix).
Neste estudo, foi avaliado a estabilidade da enterocina em diferentes
temperaturas. O CFS foi aquecido a 40°C, 50°C, 60°C, 70°C, 80°C, 90°C e 100°C
por 30 minutos e a 121°C em autoclave por 15 min. Quando a enterocina foi
submetida à autoclavagem (121°C, 1 atm e 15 minutos), a atividade bactericida
diminuiu, contudo mostrou-se ser isolado-condição dependente como mostra a
figura 3. A figura 4 apresenta os valores da atividade residual da enterocina após
aquecimento; em que é possível observar que a estabilidade também variou
conforme o isolado e a suplementação ao qual a enterocina foi produzida. Inclusive
foi observado que dependendo da temperatura, houve um aumento da atividade
antagônica de aproximadamente 150% de atividade quando comparado com o
controle.
A B
2.00
4.00
6.00
8.00
10.00
12.00
14.00
3 5 7 9
Ha
lo d
e In
ibiç
ão
(m
m)
pH
b
c
a
b
a
a a
a a a
a
b b b
a
bc
3 5 7 9
pH
c
b
a
ab ab b
a
a
a
a
a
a
a
a
a
ab

52
Figura 3: Halos de inibição formados com enterocina produzida pelos isolados Efm20 e Efm 22, nas temperaturas 40°C, 60°C, 100°C e 121°C respectivamente.
Figura 4: Atividade residual do efeito da temperatura na estabilidade de enterocina produzidas pelos isolados Efm20 (B) e Efm 22 (C) (2). ■ Glicerol 20,0 g/L; ▲tween 5,0 g/L; ♦lactose 30,0 g/L;●mix.
A estabilidade das enterocinas frente aos produtos químicos tween® 20,
tween® 80, NaCl, EDTA e SDS foi avaliado, visto que vários agentes químicos
podem estar presentes no processamento de alimentos, e observamos que a
estabilidade das enterocinas aos diferentes produtos é isolado/condição
dependente, como pode ser visto na Figura 5. Embora a presença do tween® 20 e
80 tenha apresentado maior redução da ação das enterocinas, estas não foram
afetadas ao nível de perda total de ação. Não foi observada inibição pelos produtos
químicos nos testes controle, sendo assim, a inibição somente ocorreu devido à
ação das enterocinas.
30.00
50.00
70.00
90.00
110.00
130.00
150.00
40 50 60 70 80 90 100 121
Ativid
ad
e R
esid
ua
l (%
)
Temperatura (°C)
30.00
50.00
70.00
90.00
110.00
130.00
150.00
40 50 60 70 80 90 100 121
Ativid
ad
e R
esid
ua
l (%
)
Temperatura (°C)
A B

53
Figura 5: Resultado do efeito dos produtos químicos na estabilidade das enterocina dos Isolados Efm20 (A) e Efm22 (B). a, b Letras diferentes no mesmo produto químico, indicam
diferença significativa (p<0,05). Tween® 20; Glicerol; Lactose; Mix.
O efeito bactericida e/ou bacteriostática dos CFS provenientes dos isolados
Efm20 e Efm22 foi observado já nas 2 primeiras horas de incubação. Após a
redução máxima, o crescimento de L. innocua manteve-se praticamente inalterado
dependendo da enterocina utilizada, enquanto que para outras, as células
sobreviventes retomaram seu crescimento (Figura 6). Concomitante houve também
a redução na medida da absorbância, o que revela a lise de células de L. innocua,
resultando então numa ação bactericida e bacteriostática das enterocinas.
Figura 6: Resultado do efeito bactericida e/ou bacteriostática contra L. innocua das
enterocinas produzidas pelos isolados Efm20 (A) e Efm22 (B) em meio MRS suplementado.
■ Lactose 20,0 g/L; ▲Glicerol 5,0 g/L; ♦ Listeria innocua ;● Tween 20 20,0 g/L; *Mix.
EDTA
a
c
b
ab ab
bc
bc
a
b
c bc
a
b b b
a
Tween NaCl Tween 80
c
a
b
ab
Tween 20 5.00
7.00
9.00
11.00
13.00
15.00
17.00
19.00
21.00
23.00
Halo
de Inib
ição (
mm
)
EDTA Tween 80 SDS NaCl
c
b
a ab
c
a
b
a
c
ab
c
bc
a
b b
a
b
ab
bc
a
Tween 20
A B
A B
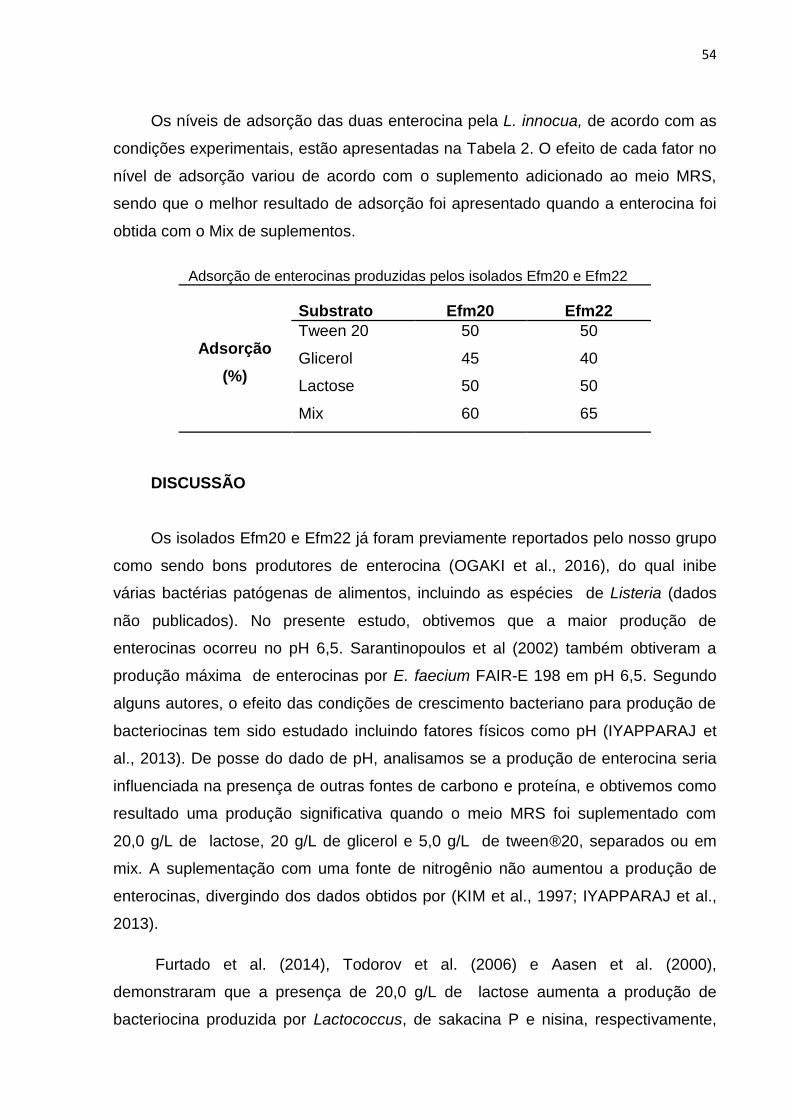
54
Os níveis de adsorção das duas enterocina pela L. innocua, de acordo com as
condições experimentais, estão apresentadas na Tabela 2. O efeito de cada fator no
nível de adsorção variou de acordo com o suplemento adicionado ao meio MRS,
sendo que o melhor resultado de adsorção foi apresentado quando a enterocina foi
obtida com o Mix de suplementos.
Adsorção de enterocinas produzidas pelos isolados Efm20 e Efm22
Adsorção
(%)
Substrato Efm20 Efm22
Tween 20 50 50
Glicerol 45 40
Lactose 50 50
Mix 60 65
DISCUSSÃO
Os isolados Efm20 e Efm22 já foram previamente reportados pelo nosso grupo
como sendo bons produtores de enterocina (OGAKI et al., 2016), do qual inibe
várias bactérias patógenas de alimentos, incluindo as espécies de Listeria (dados
não publicados). No presente estudo, obtivemos que a maior produção de
enterocinas ocorreu no pH 6,5. Sarantinopoulos et al (2002) também obtiveram a
produção máxima de enterocinas por E. faecium FAIR-E 198 em pH 6,5. Segundo
alguns autores, o efeito das condições de crescimento bacteriano para produção de
bacteriocinas tem sido estudado incluindo fatores físicos como pH (IYAPPARAJ et
al., 2013). De posse do dado de pH, analisamos se a produção de enterocina seria
influenciada na presença de outras fontes de carbono e proteína, e obtivemos como
resultado uma produção significativa quando o meio MRS foi suplementado com
20,0 g/L de lactose, 20 g/L de glicerol e 5,0 g/L de tween®20, separados ou em
mix. A suplementação com uma fonte de nitrogênio não aumentou a produção de
enterocinas, divergindo dos dados obtidos por (KIM et al., 1997; IYAPPARAJ et al.,
2013).
Furtado et al. (2014), Todorov et al. (2006) e Aasen et al. (2000),
demonstraram que a presença de 20,0 g/L de lactose aumenta a produção de
bacteriocina produzida por Lactococcus, de sakacina P e nisina, respectivamente,

55
corroborando com nossos resultados. Todorov e Dicks (2009) afirmam que a
produção de enterocina está relacionado com substrato e consequentemente com o
crescimento celular; embora nosso estudo não focou no desenvolvimento da cultura,
porém deduzimos que algumas quantidades especificas de substrato possa ter
contribuído pelo desenvolvimento da cultura.
Ainda, ambos isolados possuem 3 genes produtores de enterocinas, sendo
entA, entB e entP (Ogaki et al., 2015), estes podem ser expressos de maneira
diferente em cada tratamento realizado.
Posteriormente, avaliamos a estabilidade da enterocina formada em diferentes
temperaturas, pH e presença de detergente e sal. Verificamos que a variação de pH
não interferiu na ação da enterocina, já a ação da temperatura afetou de forma
variável, sendo que CFS provenientes de algumas condições apresentaram
atividades maiores após o aquecimento. Independente dos resultados, os dados do
presente trabalho, revelaram que as enterocinas testadas foram termo-estáveis.
Diversos autores relataram a característica termoestável de enterocinas produzidas
por Enterococcus (NES et al., 1996; MORENO et al., 2002; SABIA et al., 2002;
KANG; e LEE, 2005) e por E. faecium (TODOROV et al., 2010; BELGACEM et al.,
2010; CAMPOS et al., 2006; GHRAIRI et al., 2008). Bacteriocinas da classe II
apresentam em sua estrutura uma ponte dissulfeto entre duas cisteínas, que
desempenha importante papel na atividade antimicrobiana e faz com que seja
resistente a temperaturas elevadas (FIMLAND et al., 2005; DRIDER et al., 2006;
RICHARD et al., 2006).
Todorov e Dicks (2009) e Moreno et al. (2002), afirmam que a capacidade das
bacteriocinas suportarem elevadas temperaturas é desejável pois permite sua para
que haja sua aplicação destes em alimentos, uma vez que não sofrerão impacto
quando submetidas à etapa de tratamento térmico, mantendo suas propriedades
antimicrobianas. Resultado interessante obtido em nosso estudo foi que em
determinadas temperaturas a atividade antagônica foi maior quando comparado com
o controle, sugerindo que a temperatura elevada ativa a ação da enterocina.
Em relação à estabilidade em pHs variados, estudo realizado por Rodriguez et
al. (2000) mostraram que E. faecium TAB 7 não foram inativadas nos pH 6, 7 e 9

56
exceto em pH 2,0. Semelhante à temperatura, é desejável que as enterocinas
apresentem atividade antimicrobiana em diversos pH, permitindo sua utilização em
diversos alimentos, principalmente em alimentos com pH baixo (FRANZ;
SCHILLINGE; HOLZAPFEL, 1996).
A presença dos produtos químicos SDS, EDTA, tween® 20, tween®80 e NaCl
afetaram a ação da enterocina de maneira isolado-condição dependente, contudo,
não anularam o efeito antilisterial da enterocina. Os tensoativos tween®20 e
tween®80 foram os que mais afetaram negativamente a ação antagônica da
enterocina. Embora, a presença do tween possa alterar a tensão superficial da
célula produtora de enterocina, facilitando a liberação desta para o meio extracelular
(VERELLEN et al., 1998), de alguma maneira estes interferiram na ação desta,
contra a bactéria indicadora. Estes resultados corroboram com diversos estudos em
que confirmaram que a presença desses reagentes afeta a ação de bacteriocinas de
diferentes maneiras, dependendo dos isolados testados (FURTADO et al., 2014;
STROMPFOVÁ; LAUKOVA, 2007). É de conhecimento que bacteriocinas
produzidas por diferentes bactérias podem diferir na sua natureza química
(MOJGANI; HUSSAINI; VASEJI, 2015).
Segundo estudo realizado por Von Mollendorff et al. (2007), foi detectada a
produção de enterocinas pelo isolado E. faecalis HV219 na presença de diversos
compostos químicos, como triton, b-mercapto-etanol, etanol, metanol, clorofórmio,
NaCl, KCl, KH2PO4, K2HPO4, MgCl2 e acetato de sódio.
O efeito bactericida e/ou bacteriostática dos CFS provenientes de diferentes
substratos foi confirmado neste estudo, mostrando sua eficiência nas primeiras
horas de incubação. O crescimento de L. innocua com CFS ocorreu de maneira
não expressiva nas horas subsequentes, pois as células sobreviventes retomaram
seu crescimento. Uma hipótese a este fato foi a adição de concentrações
insuficientes de enterocina para a inibição total desse microrganismo. Conforme
Galvez et al. (2007), para que haja efeito inibidor de microrganismos em alimentos é
necessário adicionar concentrações bem mais elevadas de bacteriocinas em relação
a quantidade utilizada para obter efeito no meio de cultura. O modo de ação das
bacteriocinas é dependente de muitos fatores, dentre os quais a sua concentração.

57
A inibição de células persiste enquanto houver bacteriocina ativa remanescente no
meio de crescimento.
Em relação à adsorção da enterocina pela célula alvo, este trabalho mostrou
que a porcentual maior de adsorção ocorreu quando a enterocina foi produzida no
meio Mix, chegando a 60% da enterocina Efm20 e 65% para Efm22. O teste de
adsorção demonstra a capacidade da enterocina atuar na membrana da célula alvo
ocasionando a lise celular (YILDIRIM; WINTERS; JOHNSON, 1999). Segundo
Cleveland et al. (2001), o alvo principal das enterocinas é a membrana
citoplasmática da célula alvo, onde formam poros na membrana fazendo com que
ocorra o esgotamento do meio intracelular e do gradiente de pH resultando na morte
celular.
CONCLUSÃO
A condição de crescimento da bactéria é fundamental para a produção de
enterocinas. A suplementação do meio MRS com lactose, glicerol e tween® 20
mostrou ser ideal para a produção de enterocinas pelos isolados Efm20 e Efm22.
Ainda, a enterocina produzida por estes isolados mostrou efeito antilisterial, além de
serem termoestáveis e estáveis em ampla faixa de pH e concentração de NaCl.
Estes parâmetros são fundamentais para a otimização e produção de bacteriocinas
como agentes de biopreservação para a indústria de alimentos e bacteriocinas
produzidas por bactérias ácidas láticas isoladas de produtos lácteos são bons
candidatos para serem estudados como bioconservadores, principalmente por
estarem adaptados às condições ambientais desses alimentos.
REFERÊNCIAS BIBLIOGRÁFICAS
AASEN, I. M.; MORETRO, T.; KATLA, T.; AXELSSON, L.; STORRO, I. Influence.
INFLUENCE of complex nutrients, temperature and pH on bacteriocin production by
Lactobacillus sakei CCUG 42687. Applied Microbiology and Biotechnology, v. 53,
n. 2, p. 159-166, 2000.

58
ALVAREZ-SIEIRO, P.; MONTALBÁN-LÓPEZ, M.; MU, D.; KUIPERS, O. P.:
Bacteriocins of lactic acid bacteria: extending the family. Applied microbiology and
biotechnology, v. 100, n. 7, p. 2939-2951, 2016.
AYMERICH, M. T.; GARRIGA, M.; COSTA, S.; MONFORT, J. M.; HUGAS, M.:..:
Prevention of ropiness in cooked pork by bacteriocinogenic cultures. International
Dairy Journal, v. 12, n. 2/3, p. 239-246, 2002.
BAYOUB, K.; MARDAD, I.; AMMAR, E.; SERRANO, A.; SOUKRI, A.: Isolation and
purification of two bacteriocins 3D produced by Enterococcus faecium with inhibitory
activity against Listeria monocytogenes. Current Microbiology, v. 62, n. 2, p. 479-
485, 2011.
BELGACEM, Z. B.; ABRIOUEL, H.; OMAR, N. B.; LUCAS, R.; MARTÍNEZ-
CANAMERO, M., GÁLVEZ, A.; MANAI, M. Antimicrobial activity, safety aspects, and
some technological properties of bacteriocinogenic Enterococcus faecium from
artisanal Tunisian fermented meat. Food Control, v. 21, n. 4, p. 462-470, 2010.
CALLEWAERT, R.; HUGAS, M.; DE VUYST, L.: Competitiveness and bacteriocin
production of Enterococci in the production of Spanish-style dry fermented sausages.
International Journal of Food Microbiology, v. 57, n. 1/2, p. 33-42, 2000.
CAMPOS, C. A.; RODRÍGUEZ, Ó.; CALO-MATA, P.; PRADO, M.; BARROS-
VELÁZQUEZ, J. Preliminary characterization of bacteriocins from Lactococcus lactis,
Enterococcus faecium and Enterococcus mundtii strains isolated from turbot (Psetta
maxima). Food Research International, v. 39, n. 3, p. 356-364, 2006.
CHEN, H.; HOOVER, D. G.: Bacteriocins and their food
applications. Comprehensive Reviews In Food Science And Food Safety, v. 2, n.
3, p. 82-100, 2003.

59
CLADERA-OLIVEIRA, F. Produção, caracterização, purificação parcial, e
aplicação de um peptídeo antimicrobiano produzido por Bacillus licheniformis
P40. 2004. 118f. Dissertação de (Mestrado do) – Programa de Pós Graduação em
Microbiologia Agrícola e do Ambiente, Faculdade de Agronomia, Universidade
Federal do Rio Grande do Sul, Porto Alegre, 2004.
CLEVELAND, J.; MONTVILLE, T. J.; NES, I. F.; CHIKINDAS, M. L.: Bacteriocins:
safe, natural antimicrobials for food preservation. International Journal of Food
Microbiology, v. 71, n. 1, p. 1-20, 2001.
DE VUYST, L.; E LEROY, F. Bacteriocins from lactic acid bacteria: production,
purification, and food applications. Journal of Molecular Microbiology and
Biotechnology, vol. 13, n. 4, p. 194–199, 2007.
DRIDER, D.; FIMLAND, G.; HÉCHARD, Y.; MCMULLEN, L. M.; PRÉVOST, H. The
continuing story of class IIa bacteriocins. Microbiology and Molecular Biology
Reviews, v. 70, n. 2, p. 564-582, 2006.
FRANZ, C. M. A. P.; SCHILLINGER, U.; HOLZAPFEL, W. H. Production and
characterization of enterocin 900, a bacteriocin produced by Enterococcus faecium
BFE 900 from black olives. International Journal of Food Microbiology, v. 29, n. 2,
p. 255-270, 1996.
FIMLAND, G.; JOHNSEN, L.; DALHUS, B.; NISSEN‐MEYER, J. PEDIOCIN‐like
antimicrobial peptides (class IIa bacteriocins) and their immunity proteins:
biosynthesis, structure, and mode of action. Journal of Peptide Science, v. 11, n.
11, p. 688-696, 2005.
FRANCO, B. D. G.; BALCIUNAS, E. M.; MARTINEZ, F. A. C.; TODOROV, S. D.;
CONVERTI, A.; DE SOUZA OLIVEIRA, R. P. Novel biotechnological applications of
bacteriocins: a review. Food Control, v. 32, n. 1, p. 134-142, 2013.
FURTADO, D. N.; TODOROV, S. D.; LANDGRAF, M.; DESTRO, M. T.; FRANCO, B.
D. Bacteriocinogenic Lactococcus lactis subsp. lactis DF04Mi isolated from goat milk:

60
Characterization of the bacteriocin. Brazilian Journal of Microbiology , v 45, n. 4,
p. 1541-1550, 2014.
FURLANETO-MAIA, L.; ROCHA, K. R.; HENRIQUE, F. C.; GIAZZI, A.;
FURLANETO, M. C.: Antimicrobial resistance in Enterococcus sp isolated from soft
cheese in Southern Brazil. Advances in Microbiology, v. 2014, 2014.
GÁLVEZ, A.; ABRIOUEL, H.; LÓPEZ, R. L.; OMAR, N. B.: Bacteriocin-based
strategies for food biopreservation. International Journal Of Food Microbiology, v.
120, n. 1, p. 51-70, 2007.
GARCÍA, M.; LUCAS, R.; ABRIOUEL, H.; OMAR, N. B.; PÉREZ, R.; GRANDE, M.;
GÁLVEZ, A.: Antimicrobial activity of enterocin EJ97 against ‘Bacillus
macroides/Bacillus maroccanus’ isolated from zucchini purée. Journal of applied
microbiology, v. 97, n. 4, p. 731-737, 2004.
GIRAFFA, G. Functionality of enterococci in dairy products. International Journal of
Food Microbiology, v. 88, n. 2, p. 215-222, 2003.
GHRAIRI, T.; FRERE, J.; BERJEAUD, J. M.; MANAI, M. Purification and
characterisation of bacteriocins produced by Enterococcus faecium from Tunisian
rigouta cheese. Food Control, v. 19, n. 2, p. 162-169, 2008.
HADJI-SFAXI, I.; EL-GHAISH, S.; AHMADOVA, A.; BATDORJ, B.; LE BLAY-
LALIBERTÉ, G.; BARBIER, G.; CHOBERT, J. M.: Antimicrobial activity and safety of
use of Enterococcus faecium PC4. 1 isolated from Mongol yogurt. Food Control, v.
22, n. 12, p. 2020-2027, 2011.
IYAPPARAJ, P.; MARUTHIAH, T.; RAMASUBBURAYAN, R.; PRAKASH, S.;
KUMAR, C.; IMMANUEL, G.; PALAVESAM, A.: Optimization of bacteriocin
production by Lactobacillus sp. MSU3IR against shrimp bacterial pathogens. Aquatic
Biosystems, v. 9, n. 1, p. 12, 2013.

61
KANG, J. H.; LEE, M. S. Characterization of a bacteriocin produced by Enterococcus
faecium GM‐1 isolated from an infant. Journal of Applied Microbiology, v. 98, n. 5,
p. 1169-1176, 2005.
KIM, W. S.; HALL, R. J.; DUNN, N. W.: The effect of nisin concentration and nutrient
depletion on nisin production of Lactococcus lactis. Applied Microbiology and
Biotechnology, v. 48, n. 4, p. 449-453, 1997.
LAUKOVA, A.; CZIKKOVÁ, S.; LACZKOVÁ, S.; TUREK, P. USE of enterocin CCM
4231 to control Listeria monocytogenes in experimentally contaminated dry
fermented Hornad salami. International Journal of Food Microbiology, v. 52, n. 1,
p. 115-119, 1999.
MCAULIFFE, O.; ROSS, R. PAUL; HILL, COLIN. Lantibiotics: structure, biosynthesis
and mode of action. FEMS Microbiology Reviews, v. 25285-308, 1999.
MOJGANI, N.; HUSSAINI, F.; VASEJI, N. Characterization of indigenous
Lactobacillus strains for probiotic properties. Jundishapur Journal of Microbiology,
v. 8, n. 2, 2015.
MORENO, M. R. F.; LEISNER, J. J.; TEE, L. K.; LEY, C.; RADU, S.; RUSUL, G.; DE
VUYST, L. Microbial analysis of Malaysian tempeh, and characterization of two
bacteriocins produced by isolates of Enterococcus faecium. Journal of Applied
Microbiology, v. 92, n. 1, p. 147-157, 2002.
MUNOZ, A.; ANANOU, S.; GÁLVEZ, A.; MARTÍNEZ-BUENO, M.; RODRÍGUEZ, A.;
MAQUEDA, M.; Valdivia, E. Inhibition of Staphylococcus aureus in dairy products by
enterocin AS-48 produced in situ and ex situ: bactericidal synergism with
heat. International Dairy Journal, v. 17, n. 7, p. 760-769, 2007.
NES, I. F.; DIEP, D. B.; HÅVARSTEIN, L. S.; BRURBERG, M. B.; EIJSINK, V.;
HOLO, H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie Van
Leeuwenhoek, v. 70, n. 2/-/4, p. 113-128, 1996.

62
NES, I. F.; DIEP, D. B.; HOLO, H. Bacteriocin diversity in Streptococcus and
Enterococcus. Journal of Bacteriology, v. 189, n. 4, p. 1189-1198, 2007.
OGAKI, M. B.; FURLANETO, M. C.; MAIA, L. F. Review: General aspects of
bacteriocins. Brazilian Journal of Food Technology, v. 18, n. 4, p. 267-276, 2015.
OGAKI, M. B.; ROCHA, K. R.; TERRA, M. R.; FURLANETO, M. C.; MAIA, L. F.:
Screening of the Enterocin-Encoding Genes and Antimicrobial Activity in
Enterococcus Species. Journal of microbiology and biotechnology, v. 26, n. 6, p.
1026-1034, 2016.
RICHARD, C.; CANON, R.; NAGHMOUCHI, K .; BERTRAND, D.; PREVOST, H
.; DRIDER, D. Evidências sobre correlação entre o número de dissulfeto de ponte e
toxicidade da classe bacteriocinas IIa . Food Microbiol, v. 23, p.175-183, 2006.
RODRÍGUEZ, E.; GONZÀLEZ, B.; GAYA, P.; NUNEZNUÑEZNUNEZ, M.; MEDINA,
M. Diversity of bacteriocins produced by lactic acid bacteria isolated from raw milk.
International Dairy Journal, v. 10, n. 1/2, p. 7-15, 2000.
SABIA, C.; MANICARDI, G.; MESSI, P.; DE NIEDERHÄUSERN, S.; BONDI, M.
Enterocin 416K1, an antilisterial bacteriocin produced by Enterococcus casseliflavus
IM 416K1 isolated from Italian sausages. International Journal of Food
Microbiology, v. 75, n. 1, p. 163-170, 2002.
SARANTINOPOULOS, P.; KALANTZOPOULOS, G.; TSAKALIDOU, E. Effect of
Enterococcus faecium on microbiological, physicochemical and sensory
characteristics of Greek Feta cheese. International Journal of Food Microbiology,
v. 76, n. 1, p. 93-105, 2002.
SCHIRRU, S.; TODOROV, S. D.; FAVARO, L.; MANGIA, N. P.; BASAGLIA, M.;
CASELLA, S.; DEIANA, P.LAUKOVA.: Sardinian goat’s milk as source of

63
bacteriocinogenic potential protective cultures. Food Control, v. 25, n. 1, p. 309-320,
2012.
STROMPFOVA, V.; LAUKOVA, A. In vitro study on bacteriocin production of
Enterococci associated with chickens. Anaerobe, v. 13, n. 5, p. 228-237, 2007.
TODOROV, S. D.; DICKS, L. M. Screening for bacteriocin-producing lactic acid
bacteria from boza, a traditional cereal beverage from Bulgaria: Comparison of the
bacteriocins. Process Biochemistry, London, v. 41, n. 1, p. 11-19, 2006.
TODOROV, S. D. Bacteriocin production by Lactobacillus plantarum AMA-K isolated
from Amasi, a Zimbabwean fermented milk product and study of the adsorption of
bacteriocin AMA-K to Listeria sp. Brazilian Journal of Microbiology, v. 39, n. 1, p.
178-187, 2008.
TODOROV, S. D.; DICKS, L. M. T. Bacteriocin production by Pediococcus
pentosaceus isolated from marula (Scerocarya birrea). International Journal of
Food Microbiology, v. 132, n. 2, p. 117-126, 2009.
TODOROV, S. D.; FRANCO, B. D. G. D. M.:.: Lactobacillus plantarum:
Characterization of the species and application in food production. Food Reviews
International, v. 26, n. 3, p. 205-229, 2010.
VAN REENEN, C. A.; CHIKINDAS, M. L.; VAN ZYL, W. H.; DICKS, L. M. T.:
Characterization and heterologous expression of a class IIa bacteriocin, plantaricin
423 from Lactobacillus plantarum 423, in Saccharomyces cerevisiae. International
Journal Of Food Microbiology, v. 81, n. 1, p. 29-40, 2003.
VERELLEN, T. L.; BRUGGEMAN, G.; VAN REENEN, C. A.; DICKS, L. M.;
VANDAMME, E. J.: Fermentation optimization of plantaricin 423, a bacteriocin
produced by Lactobacillus plantarum 423. Journal of Fermentation and
Bioengineering, v. 86, n. 2, p. 174-179, 1998.

64
VON MOLLENDORFF, J. W.; TODOROV, S. D.; DICKS, L. M. T.: Factors Affecting
the Adsorption of Bacteriocins to Lactobacillus sakei and Enterococcus sp. Applied
Biochemistry And Biotechnology, v. 142, n. 2, p. 209-220, 2007.
YANG, S.: Antibacterial activities of bacteriocins:application in foods and
pharmaceuticals – review article. Frontiers In Microbiology, 26 may, 2014.
YILDIRIM, Z.; WINTERS, D. K.; JOHNSON, M. G.:.: Purification, amino acid
sequence and mode of action of bifidocin B produced by Bifidobacterium bifidum
NCFB 1454. Journal Of Applied Microbiology, v. 86, n. 1, p. 45-54, 1999.