universidade federal do cearÁ centro de ciÊncias ... · 1.4 produção de lectinas vegetais em...
TRANSCRIPT

UNIVERSIDADE FEDERAL DO CEARÁ
CENTRO DE CIÊNCIAS
DEPARTAMENTO DE BIOQUÍMICA E BIOLOGIA MOLECULAR
CURSO DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
CLONAGEM E EXPRESSÃO DE UMA LECTINA DE Artocarpus incisa L.
(FRUTALINA) EM Escherichia coli
DENISE ROCHA NEPOMUCENO
FORTALEZA
2008

UNIVERSIDADE FEDERAL DO CEARÁ
CENTRO DE CIÊNCIAS
DEPARTAMENTO DE BIOQUÍMICA E BIOLOGIA MOLECULAR
CURSO DE PÓS-GRADUAÇÃO EM BIOQUÍMICA
CLONAGEM E EXPRESSÃO DE UMA LECTINA DE Artocarpus incisa L.
(FRUTALINA) EM Escherichia coli
DENISE ROCHA NEPOMUCENO
Dissertação submetida à coordenação do curso de Pós-graduação em Bioquímica, da Universidade Federal do Ceará, como requisito para obtenção do grau de mestre em Bioquímica, sob a orientação do prof. Dr. Renato de Azevedo Moreira.
FORTALEZA
2008

CLONAGEM E EXPRESSÃO DE UMA LECTINA DE Artocarpus incisa L.
(FRUTALINA) EM Escherichia coli
Dissertação submetida à coordenação do curso de Pós-graduação em Bioquímica, da Universidade Federal do Ceará, como requisito para obtenção do grau de mestre em Bioquímica.
Dissertação aprovada em: _____ / _____ / _____
Denise Rocha Nepomuceno
Prof. Dr. Renato de Azevedo Moreira (Orientador)
Universidade Federal do Ceará – UFC
Profa. Dra. Cristina Paiva da Silveira Carvalho (Co-Orientadora)
Universidade Federal do Ceará – UFC
Profa. Dra. Maria Izabel Florindo Guedes
Universidade Estadual do Ceará – UECE

Ó profundidade da riqueza, tanto da sabedoria
como do conhecimento de Deus! Quão
insondáveis são os seus juízos, e quão
inescrutáveis, os seus caminhos! Quem, pois,
conheceu a mente do Senhor? Ou quem primeiro
deu a Ele para que lhe venha a ser restituído?
porque dEle, e por meio dEle, e para Ele são todas
as coisas. A Ele, pois, toda a glória eternamente.
Romanos 11:33-36

Aos meus pais
Luís e Helena,
aos meus irmãos
Eduardo e Liliane
e ao meu marido
Hélcio
Dedico

AGRADECIMENTOS
Ao Prof. Dr. Renato de Azevedo Moreira pela orientação e pela oportunidade que me deu ao
me receber no Laboratório de Lectinas e Glicoconjugados 1.
À Profa. Dra. Cristina Paiva da Silveira Carvalho pela orientação, pela paciência, pela
dedicação e pela amizade.
À professora Dra. Maria Izabel Florindo Guedes, pela disponibilidade em avaliar o trabalho e
compor a banca examinadora.
Ao Prof. Dr. Thalles Barbosa Grangeiro pela orientação e pela indispensável contribuição
para a realização deste trabalho.
Ao Prof. Dr. Humberto Muniz Pereira pela ajuda na realização de parte dos experimentos.
À amiga Tuana pela indispensável ajuda na elaboração dos modelos.
Ao Prof. Dr. Francisco de Assis Paiva Campos por permitir a utilização do Laboratório de
Biologia Molecular de Plantas para a realização de experimentos.
A todos os professores do Departamento de Bioquímica e Biologia Molecular.
A todos os integrantes e ex-integrantes do Laboratório de Citogenética e Genética Molecular,
Prof. Dr. Thalles Barbosa Grangeiro, Profa. MS Maria Aparecida Alves, Adriana, Bruno,
Cândida, Camila, Júlio, Marina, Nathália, Patrícia, Sandra, Tuana e Wal. Agradeço pela
contribuição na realização deste trabalho, pelas palavras de incentivo e pela amizade.
Ao Wagner, pela alegria e disposição em ajudar.
Ao Hélio, pela ajuda em parte dos experimentos.
Às amigas inseparáveis Camila, Clarissa, Ivana, Juliana e Luana. Agradeço pela amizade,
pelo carinho e pelos momentos de descontração que sempre serão lembrados.
Aos colegas de pós-graduação, pela amizade.
Às grandes amigas Aline, Carol, Deiviene, Denise, Elane, Karinny, Monique e “Patrícias”,
por todo apoio, carinho e amizade durante os muitos anos de convivência.
Aos meus queridos pais, Luís e Helena, pela forma tão especial que me educaram e me
incentivaram.

Aos meus irmãos, Eduardo e Liliane, pela amizade, amor e incentivo.
Ao meu amado Hélcio, por seu meu maior incentivador. Agradeço por toda paciência e
companheirismo.

ESTE TRABALHO FOI REALIZADO GRAÇAS AO AUXÍLIO DAS SEGUINTES
INSTITUIÇÕES:
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)
Fundação Cearense de Amparo à Pesquisa e Tecnologia do Estado do
Ceará (FUNCAP)
Ministério da Ciência e Tecnologia (MCT)
Programa Nacional de Cooperação Acadêmica (PROCAD)
Laboratório de Genética Molecular, Departamento de Biologia, Universidade Federal do
Ceará
Laboratório de Biotecnologia Vegetal, Departamento de Bioquímica e Biologia
Molecular, Universidade Federal do Ceará
Laboratório de Bioquímica, Centro de Ciências da Saúde (CCS), Universidade de
Fortaleza
Departamento de Bioquímica e Biologia Molecular, Centro de Ciências, Universidade Federal
do Ceará

SUMÁRIO
Lista de Figuras ..............................................................................................................................xi
Lista de Quadros ..........................................................................................................................xiii
Resumo .........................................................................................................................................xiv
Abstract..........................................................................................................................................xv
1. Introdução....................................................................................................................................1
1.1 Lectinas de origem vegetal ...............................................................................................1
1.2 Aplicações das lectinas no diagnóstico e terapia do câncer .............................................4
1.3 Lectinas Jacalina-relacionadas .........................................................................................5
1.3.1 Modificações Pós-traducionais e Estrutura Tridimensional ..................................6
1.3.2 Lectina de Artocarpus incisa (Frutalina).................................................................7
1.4 Produção de lectinas vegetais em Escherichia coli ..........................................................9
2. Objetivos....................................................................................................................................13
2.1 Objetivo geral .................................................................................................................13
2.2 Objetivos específicos......................................................................................................13
3. Material......................................................................................................................................14
3.1 Material vegetal ..............................................................................................................14
3.2 Bactérias .........................................................................................................................14
3.3 Enzimas ..........................................................................................................................14
3.4 Plasmídeos ......................................................................................................................14
3.5 Reagentes e outros materiais ..........................................................................................15
4. Métodos .....................................................................................................................................16
4.1 Extração de RNA total de sementes de Artocarpus incisa .............................................16
4.2 Amplificação do cDNA da frutalina por RT- PCR ........................................................19
4.3 Preparação de células eletrocompetentes de Escherichia coli........................................19
4.4 Clonagem do cDNA da frutalina em células de E. coli DH10B ....................................21
4.5 Isolamento de plasmídeos recombinantes de E. coli ......................................................22
4.6 Seqüenciamento de DNA ...............................................................................................24
4.7 Comparação do cDNA da frutalina obtidos de diferentes fases de maturação do fruto
de A. incisa ...........................................................................................................................25
4.7.1 Modelagem por homologia ..................................................................................25
4.8 Expressão da rFrutalina em células de E. coli Origami (DE3).......................................28
4.8.1 Subclonagem da seqüência codificadora da frutalina ..........................................28

4.8.2 Transformação de células de E. coli Origami(DE3) com pET-15b::frutalinaE4 .32
4.8.3 Indução da expressão da rFrutalina em E. coli Origami (DE3).............................32
4.8.4 Purificação dos corpos de inclusão ......................................................................33
4.9 Eletroforese em gel de poliacrilamida em presença de SDS e β-mercaptoetanol
(PAGE-SDS) ........................................................................................................................33
4.10 Imunodetecção da rFrutalina imobilizada em membrana (Western Blotting) .............34
5. Resultados e Discussão..............................................................................................................37
5.1 Obtenção do cDNA da frutalina .....................................................................................37
5.2 Clonagem do cDNA da frutalina ....................................................................................37
5.3 Seqüenciamento de DNA ...............................................................................................40
5.4 Modelagem por homologia.............................................................................................46
5.5 Expressão da rFrutalina em E. coli .................................................................................57
5.5.1 Subclonagem da seqüência codificadora da frutalina ..........................................57
5.5.2 Transformação de células de E. coli Origami (DE3) com pET-15b::FTLE4........59
5.5.3 Indução da expressão da rFrutalina em células de E. coli .....................................63
6. Conclusões.................................................................................................................................73
7. Referências Bibliográficas.........................................................................................................74

LISTA DE FIGURAS
Figura 1 - Frutos de Artocarpus incisa em três diferentes estágios de maturação..................17
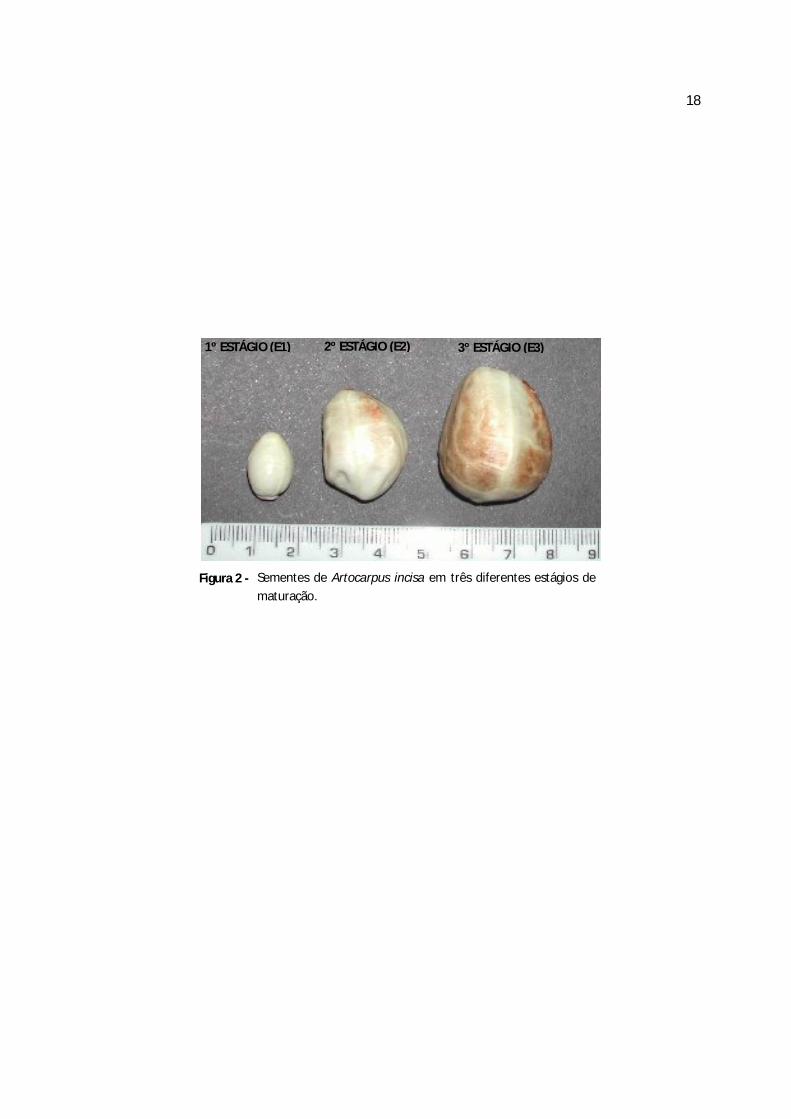
Figura 2 - Sementes de Artocarpus incisa em três diferentes estágios de maturação.............18
Figura 3 - Eletroforese em gel de agarose 1,2% do RNA total extraído de sementes de A.
incisa em diferentes estágios de maturação..............................................................................38
Figura 4 - Eletroforese em gel de agarose 1,0% dos produtos de RT-PCR, com iniciadores
específicos para a seqüência codificadora da frutalina, a partir do RNA total extraído de
sementes de Artocarpus incisa.................................................................................................39
Figura 5 - Colônias brancas e azuis em meio LB ágar............................................................41
Figura 6 - Eletroforese em gel de agarose 1,0% dos produtos de PCR, com iniciadores específicos para a seqüência codificadora da frutalina, a partir dos plasmídeos pGEM-T::frutalina...............................................................................................................................42
Figura 7 - Alinhamento das seqüências de nucleotídeos dos clones obtidos.........................44
Figura 8 - Alinhamento da seqüência de aminoácidos correspondente a seqüência da frutalina nos clones obtidos....................................................................................................................45
Figura 9 - Estrutura tridimensional em formato “cartoon” do clone E2.1 da frutalina e da jacalina......................................................................................................................................63
Figura 10 - Estrutura tridimensional em formato “ribbon” da jacalina...................................64
Figura 11 - Estrutura tridimensional em formato “ribbon” dos clones E2.1; E2.2, E3.1, E4.1; E4.2; E4.3 e E4.4 da frutalina, e dajacalina......................................................................................................................................65
Figura 12 - Estrutura tridimensional em formato “ribbon” clone E2.1...................................67
Figura 13 - Estrutura tridimensional em formato “ribbon” do clone E2.2, E3.1, E4.1...........68
Figura 14 - Estrutura tridimensional em formato “ribbon” do clone E4.2..............................69
Figura 15 - Estrutura tridimensional em formato “ribbon” do clone E4.3..............................70
Figura 16 - Estrutura tridimensional em formato “ribbon” do clone E4.4..............................71
Figura 16 - Eletroforese em gel de agarose 0,8% dos produtos da digestão de plasmídeos pGEM-T::frutalina purificados de E.coli com as enzimas de restrição BamHI e XhoI..........................................................................................................................................47
Figura 17 - Eletroforese em gel de agarose 0,8% dos produtos da digestão de plasmídeos pET15b::frutalina purificados de E.coli com as enzimas de restrição XhoI ou BamHI......................................................................................................................................49

Figura 18 - Eletroforese em gel de agarose 0,8% dos produtos da digestão de plasmídeos pET15b::frutalina purificados de E.coli com as enzimas de restrição XhoI e BamHI......................................................................................................................................50
Figura 19 - Eletroforese em gel de agarose 1,0% dos produtos de PCR, com iniciadores específicos para a seqüência codificadora da frutalina, a partir dos plasmídeos pET15b íntegro e pET15b::frutalinaE4.............................................................................................................51
Figura 21 - Eletroforese em gel de poliacrilamida 17,5% das células e do sobrenadante da cultura de E. coli transformada com pET-15b::frutalinaE4 e induzidas com IPTG 1 mM por 10 h...........................................................................................................................................53
Figura 22 - Eletroforese em gel de poliacrilamida 17,5% das células de E. coli transformadas com o pET-15b::frutalinaE4, induzidas (IPTG+) e não-induzidas (IPTG-) por 10 h...........................................................................................................................................54
Figura 23 - Eletroforese em gel de poliacrilamida 17,5% das células de E. coli transformadas com o vetor pET15b íntegro e pET-15b::frutalinaE4, induzidas (IPTG+) e não-induzidas (IPTG-) por 10 h......................................................................................................................55
Figura 24 - Imunodetecção da rFrutalina a partir do extrato total de células da cultura de E. coli transformadas com o pET-15b::frutalinaE4......................................................................57
Figura 25 - Eletroforese em gel de poliacrilamida 17,5% do extrato total solúvel e insolúvel de células de E. coli transformadas com o pET-15b::frutalinaE4........................................................................................................................59

LISTA DE QUADROS
Quadro 1 - Oligonucleotídeos específicos para a seqüência codificadora da frutalina com sítios de restrição para clonagem no vetor pGEM-T ...............................................................20
Quadro 2 - Oligonucleotídeos específicos para a seqüência codificadora da frutalina com
sítios de restrição para clonagem no vetor pET15b..................................................................27
Quadro 3 - Protocolo para confecção dos géis de poliacrilamida...........................................31
Quadro 4 - Relação entre o número de clones de cada estágio de maturação e a quantidade de
modelos gerados ......................................................................................................................34

RESUMO
As lectinas constituem ferramentas biotecnológicas de fundamental importância em
estudos histoquímicos e citoquímicos para a detecção de glicoconjugados nos tecidos. Entre
estas aplicações, destacam-se a identificação de receptores de membrana e a detecção de
estruturas características de neoplasias. A Frutalina, uma lectina -D-galactose-ligante
encontrada em sementes de Artocarpus incisa, já foi utilizada na detecção histoquímica de
lesões malignas de mama e tireóide. No presente trabalho foi realizada a clonagem e
expressão do gene da frutalina em células de Escherichia coli Origami (DE3) e foi
determinada a estrutura tridimensional dessa proteína através da modelagem por homologia.
A clonagem do gene da frutalina foi realizada a partir do produto amplificado da RT-PCR de
sementes em diferentes estágios de maturação dos frutos de A. incisa. Foram obtidos 12
clones, dos quais 5 foram seqüenciados. A lectina recombinante foi expressa como cadeia
única, formada pela cadeia beta com 20 resíduos, ligada a um peptídeo de ligação com quatro
resíduos, e a cadeia alfa com 133 resíduos. A expressão da lectina foi indicada pelo
aparecimento de uma banda protéica com massa molecular aparente de 19,2 kDa, superior a
da lectina nativa (15 kDa), após a indução com IPTG 1mM. A lectina recombinante foi
mantida exclusivamente em corpos de inclusão e foi reconhecida imunologicamente por
anticorpos policlonais anti-Frutalina. Os modelos gerados para os clones obtidos indicam que
as mutações ocorridas não alteram os sítios de ligação a carboidratos das moléculas. As
mutações nos clones obtidos sugerem a ocorrência de isoformas dessa proteína.

ABSTRACT
The lectins constitute biotechnological tools of fundamental importance in
hystochemical and cytochemical studies for the detection of tissue glycoconjugates. Among
their applications are the identification of membrane receptors and the detection of neoplastic
characteristic structures. Frutalin, an -D-galactose-binding lectin from Artocarpus incise
seeds, has already been used in the hystochemical detection of thyreoid and breast neoplasia.
In this work was made a study of the cloning and expression of the frutalin gene in
Escherichia coli Origami (DE3) cells and was determined the three-dimensional structure of
this protein through homology modeling. The frutalin gene cloning was accomplished from
the product on RT-PCR with seeds in distinct stages of development of the Artocarpus incisa
fruits. Twelve clones were obtained, from which five were sequenced. The frutalin gene was
expressed in E. coli using the pET15b expression vector. A recombinant lectin (rFrutalin) was
expressed by growing the bacteria in the presence of isopropyl -D-thiogalactopyranoside
1mM. All the recombinant lectin was found in an insoluble aggregated form as inclusion
bodies. The recombinant lectin had a higher molecular mass (19,2 kDa) than the native lectin
(15 kDa) as estimated by SDS-PAGE and Western blot analyses, showing that the
recombinant single chain Frutalin is not processed in E. coli cells. The models generated for
the clones obtained indicate that the mutations do not change the molecules active sites.
Mutations in the clones obtained suggest the occurrence of isoforms of this protein.

1
1. INTRODUÇÃO
1.1. Lectinas de origem vegetal
Lectinas são proteínas ou glicoproteínas que apresentam a propriedade comum de
reconhecimento específico e ligação reversível a resíduos de carboidratos, sem alterar a
estrutura química dos mesmos. A capacidade da lectina de se ligar especificamente a mono ou
oligossacarídeos confere a este grupo de proteínas uma grande diversidade de atividades
biológicas , como a capacidade de aglutinar células e da estimulação mitogênica em linfócitos
(CARRINGTON et al.,1985; SHARON; LIS; 2004).
Com base em suas estruturas moleculares, as lectinas são divididas em quatro
grandes grupos: merolectinas, hololectinas, quimerolectinas e superlectinas (VAN DAMME et
al.,1998a).
As merolectinas são proteínas formadas por uma única cadeia polipeptídica,
possuindo apenas um sitio de ligação a carboidratos. Desta forma, são incapazes de precipitar
glicoconjugados ou aglutinar células. A heveína e as proteínas monoméricas de orquídeas que
ligam manose são exemplos de lectinas pertencentes a este grupo. As hololectinas
compreendem proteínas que apresentam dois ou mais sítios de ligação a carboidratos, os quais
são idênticos ou homólogos e ligam o mesmo açúcar ou outro estruturalmente similar. Como
as hololectinas têm múltiplos sítios ligantes a carboidratos são capazes de aglutinar células e
de precipitar glicoconjugados. A maioria das lectinas de plantas se enquadra no subgrupo das
hololectinas. As quimerolectinas são proteínas de fusão compostas por um domínio ligante a
carboidrato e outro domínio não relacionado. Este domínio pode ter uma atividade catalítica
bem definida, mas que atue independentemente do domínio ligante a carboidrato. As
quitinases de plantas pertencentes à classe 1 são exemplos desse grupo. As superlectinas são
um tipo especial de quimerolectinas. Elas são proteínas de fusão com dois domínios ligantes a
carboidratos que são estruturalmente diferentes e reconhecem açúcares não relacionados
(VAN DAMME et al.,1998a). A superlectina TxLC-I isolada de bulbos de tulipa é exemplo de
lectina pertencente a este grupo (CAMMUE et al.,1986).

2
Monteiro-Moreira (2002) sugeriu um novo grupo de lectinas denominado de
multilectinas, devido à frutalina também interagir, embora fracamente, com -D-manose. Este
grupo foi definido como proteínas constituídas de domínios ligantes a carboidratos que são
idênticos ou homólogos, entretanto reconhecem açúcares estruturalmente não relacionados, ou
seja, são lectinas de especificidade múltipla. As lectinas galactose-ligante de Artocarpus
integrifolia (jacalina) e de A. incisa (frutalina) são exemplos desse grupo. No entanto, na
classificação de Peumans e Van Damme (1995a) a frutalina é definida como uma hololectina,
pois se trata de uma lectina que tem dois sítios de ligação a carboidratos.
As lectinas têm sido encontradas em quase todos os organismos, incluindo plantas,
vertebrados, invertebrados e bactérias (MEJÍA; PRISECARU, 2005). Nas plantas, as lectinas
são especialmente abundantes nas sementes (CARLINI; GROSSI-DE SÁ, 2002). Nas
sementes de Leguminosas, por exemplo, as lectinas chegam a constituir 10 % do total das
proteínas (ETZLER, 1986). As lectinas estão localizadas principalmente nos corpos protéicos,
tendo sido encontradas também no citoplasma e no espaço intercelular (RÜDIGER; GABIUS,
2001).
As lectinas vegetais podem ser classificadas em sete famílias de acordo com Van
Damme et al. (1998): lectinas de leguminosas; lectinas de monocotiledôneas ligantes a
manose; lectinas com domínio heveínico; proteínas inativadoras de ribossomos (RIPs) tipo 2;
lectinas relacionadas à jacalina; amarantina e lectinas de floema de Curcubitaceae.
O maior grupo de lectinas vegetais já isoladas e estudadas são as de leguminosas.
Estas proteínas, em geral, são sintetizadas como protômeros de aproximadamente 30 kDa,
com uma seqüência sinalizadora de 20 resíduos de aminoácidos precedendo a cadeia de lectina
madura, que guia seu transporte ao retículo endosplamático. Essas cadeias de 30 kDa podem
se combinar em dímeros, ou tetrâmeros (VAN DAMME et al.,1998).
Apesar de diversas lectinas de plantas terem sido estudadas em grandes detalhes, o
papel fisiológico destas proteínas ainda é pouco conhecido (VAN DAMME et al., 2004).
Algumas funções biológicas exógenas, a exemplo da proteção contra insetos e fungos, e
endógenas, como a interação com proteínas de armazenamento ou enzimas, têm sido
encontradas para as lectinas de plantas (RÜDIGER; GABIUS, 2001).

3
Algumas evidências têm sustentado a idéia de que quando as plantas são
estimuladas por fatores bióticos abióticos específicos, elas respondem por meio da expressão
de lectinas de plantas nucleares ou citoplasmáticas. A localização e a regulação da expressão
destas lectinas indicam que as mesmas estão envolvidas em interações proteína-carboidrato
endógenas específicas (VAN DAMME et al., 2004).
Várias lectinas de plantas, principalmente das leguminosas, são conhecidas por
conseguirem estabelecer uma relação simbiótica com bactérias do solo pertencentes ao gênero
Rhizobium ou de outros microrganismos capazes de fixar o nitrogênio atmosférico, fazendo
com que estas plantas fiquem independentes de suprimento externo de nitrogênio (RÜDIGER,
1984).
As lectinas que se acumulam em grandes quantidades, provavelmente combinam
um papel relacionado à defesa contra invertebrados fitófagos e animais herbívoros com a
função de armazenamento (VAN DAMME et al.,1998).
Propõe-se também que, devido à ampla distribuição em vegetais, lectinas podem
estar envolvidas em papéis de defesa dos vegetais contra ataques de fungos. A ligação das
mesmas nos constituintes da parece celular do fungo pode interferir no seu crescimento
(RÜDIGER; GABIUS, 2001).
Estudos realizados com a lectina de germe de trigo e a de batata mostraram que as
mesmas ativaram uma fosfatase endógena de suas respectivas plantas, sugerindo que as
lectinas podem não apenas se ligar a enzimas, mas também modificar sua atividade
(LORENC-KUBIS et al.,1981; WIERZBA-ARABSKA et al.,1987 citado por RÜDIGER;
GABIUS, 2001).
Keyaerts e colaboradores (2007) estudaram a atividade antiviral de lectinas de
plantas com diferentes especificidades contra o coronavirus da síndrome respiratória aguda
grave (SARS-CoV) e o vírus da peritonite infecciosa felina (FIPV). A maior atividade anti-
coronavirus foi detectada predominantemente entre as lectinas manose-ligantes.
As lectinas são usadas como ferramentas em diversas áreas como bioquímica,
biologia celular, medicina e áreas relacionadas. A aplicação das lectinas vai desde a

4
purificação e caracterização de glicoconjugados de superfície celular; análise dos mecanismos
envolvidos na glicosilação até o emprego na medicina, como na tipagem sanguínea e definição
de secretores; no diagnóstico e terapia na pesquisa sobre o câncer (RÜDIGER; GABIUS,
2001; VAN DAMME et al.,1998)
1.2. Aplicações das lectinas no diagnótico e terapia do câncer
A especificidade das lectinas por carboidratos faz com que estas proteínas
sejam ferramentas úteis para estudos glicobiológicos em biomedicina e na pesquisa sobre
câncer. Essa especificidade por carboidratos permite a caracterização funcional e fenotípica de
glicoproteínas de membranas expressas em células cancerosas (MEJÍA; PRISECARU, 2005).
Os primeiros relatos referentes ao reconhecimento de células cancerosas pelas
lectinas foram feitos na década de 1960 por Aub e colaboradores (1963, 1965). Os autores
sugeriram que a diferença entre as células normais e as células cancerosas residia na sua
superfície, ou seja, que alterações nas propriedades da superfície celular habilitariam as células
cancerosas a continuar se multiplicando e se desprender do seu sítio primário, se espalhar pelo
corpo, alojar-se em um novo meio e se desenvolver.
Diversos estudos têm sugerido uma forte relação entre certos padrões de ligação
das lectinas e seu comportamento biológico em vários tumores. Fik e colaboradores (2001)
descreveram o efeito das lectinas em células normais e cancerosas in vitro. Wang e
colaboradores (2000) estudaram os efeitos de lectinas com diferentes especificidades de
ligação a carboidratos em hepatoma humano (H3B), coriocarcinoma humano, melanoma e
osteosarcoma de rato. Nesse estudo, a lectina de Pleurotus ostreatus foi mais eficiente na
inibição do tumor de sarcoma S-180 do que de hepatoma H-22, e apresentou um efeito
significante no tempo de sobrevivência dos ratos. O mecanismo do efeito antitumoral pode ser
devido à melhora no sistema imunológico do hospedeiro (MEJÍA; PRISECARU, 2005).
A lectina de Vicia faba (VFA) estimulou a diferenciação morfológica e reduziu o
fenótipo maligno de células cancerosas de cólon (JORDINSON et al.,1999). Em outro estudo,

5
a aglutinina do germe de trigo (WGA) apresentou efeito inibitório em células tumorais
pancreática de ratos (MIKKAT et al.,2001)
No estudo do câncer humano, as lectinas vegetais têm sido utilizadas como
ferramentas para revelar alterações na glicosilação de glicoconjugados. A importância deste
reconhecimento específico se deve ao fato de que a maioria destes glicoconjugados estão
relacionados ao comportamento tumoral quanto à invasividade e ao processo de metástase
(BROCKHAUSEN, 1999).
As lectinas de plantas têm apresentado atividade quimiopreventiva para o câncer
in vitro e in vivo, e em estudos de casos humanos. Estas proteínas estão sendo empregadas
como agentes terapêuticos em estudos de tratamento de câncer (MEJÍA; PRISECARU, 2005).
1.3. Lectinas Jacalina-relacionadas
A lectina D-galactose-ligante de sementes de jaca (Artocarpus integrifolia),
conhecida como jacalina, foi purificada por Moreira (1977). O termo “lectinas jacalina
relacionadas” é usado como coletivo para todas as lectinas que possuem relação estrutural e
evolucionária com a jacalina (VAN DAMME et al.,1998).
A família das lectinas jacalina relacionadas são dividas em dois subgrupos: as
lectinas manose-ligantes (mJRLs) e as galactose-ligantes (gJRLs). Os membros desta família
apresentam diferenças em suas estruturas moleculares, especificidades por carboidratos e
localização celular. Embora o subgrupo das galactose-ligantes tenha sido encontrada apenas na
família Moraceae, as manose-ligantes são amplamente distribuídas no reino das plantas
(NAKAMURA-TSURUTA et al.,2008).
As lectinas de Moraceae, até agora caracterizadas, são compostas por quatro
protômeros idênticos consistindo de uma cadeia alfa e uma cadeia beta pequena. Ambas as
cadeias são derivadas de um único precursor e não são unidas por pontes dissulfeto. Cada
protômero contém um único sitio ligante de carboidrato. A lectina galactose ligante de
Artocarpus integrifolia (jacalina) apresenta quatro cadeias alfas e quatro betas com 133 e 20
resíduos, respectivamente (YOUNG et al.,1991).

6
Lectinas muitos semelhantes à jacalina também têm sido isoladas de sementes de
Artocarpus incisa (frutalina) (MOREIRA et al.,1998), A. altilis, A. champeden, A. integer, A.
lakocha, A. tonkinensis (BLASCO et al.,1996).
1.3.1. Modificações Pós-traducionais e Estrutura Tridimensional
A Jacalina é sintetizada como uma pré-pro-proteína, consistindo de um peptídeo
sinal seguido por um pro-peptídeo de 39 resíduos de aminoácidos, a cadeia beta com 20
resíduos, ligada a um peptídeo de ligação com quatro resíduos, e a cadeia alfa com 133
resíduos. É provável que a jacalina seja sintetizada no retículo endoplasmático e processada
co-traducionalmente pela remoção do peptídeo sinal. O pro-peptídeo resultante é transportado
para os corpos protéicos. Durante ou depois do transporte, o pro-peptídeo é clivado em três
diferentes sítios, resultando na remoção do pro-peptídeo N-terminal e da excisão do peptídeo
de ligação. As cadeias alfa e beta permanecem unidas por meio de ligações não-covalentes e
formam o protômero da jacalina (YANG et al.,1993).
A estrutura tridimensional da jacalina foi resolvida por análises de difração de
raios-X (SANKARANARAYANAN et al., 1996). Cada protômero consiste de um β-prisma
triplo simétrico, constituído a partir de três folhas β de quatro fitas. Das doze fitas, onze são
formadas pela cadeia alfa e apenas uma pela cadeia beta. Quatro subunidades estão associadas
não covalentemente formando uma estrutura tetramérica. As cadeias betas ocupam a região
central do tetrâmero e desempenham um papel crucial na associação das quatro subunidades.
Cada protômero possui um único sitio ligante de carboidrato formado por Gly1,
Tyr122, Trp123 e Asp125 da cadeia alfa que cria um sistema complexo de nove ligações de
hidrogênio com O3, O4, O5 e O6 da galactose. A liberação da Gly1 da cadeia alfa, após o
processamento proteolítico da pro-jacalina em cadeias alfa e beta, é essencial para a formação
do sítio ligante de galactose e requer a correta excisão do peptídeo de ligação (HOULÈS
ASTOUL et al., 2002).
A especificidade da jacalina não é restrita a galactose, mas se estende a epímeros
como glucose e manose. As principais diferenças estruturais entre as lectinas galactose ligante,

7
como a jacalina, e as manose ligante consistem no tamanho e na conformação do sítio ligante
de carboidrato. Devido à clivagem proteolítica do peptídeo de ligação entre as cadeias alfa e
beta, o tamanho do sítio ligante de carboidrato da jacalina é mais estendido do que de seus
homólogos manose específicos que permanecem com seus protômeros intactos (HOULÈS
ASTOUL et al., 2002).
Análises estruturais demonstraram que o tamanho relativamente grande do sítio
ligante de carboidrato permite que a jacalina acomode monossacarídeos com diferentes
conformações hidroxil e mostraram evidências de que a estrutura β-prisma da jacalina é
suficientemente flexível para conferir diferentes especificidades com carboidratos para uma
única lectina (BOURNE; HOULÈS ASTOUL, 2002).
1.3.2 Lectina de Artocarpus incisa (Frutalina)
Artocarpus incisa L. (Moraceae), conhecida popularmente como fruta-pão é uma
espécie arbórea que chega a medir 15 m de altura e cujo centro de origem se encontra na Ásia.
No Brasil, a espécie é cultivada do Pará a São Paulo (BRAGA, 1976).
A lectina encontrada em sementes de Artocarpus incisa L. (frutalina) foi isolada e
caracterizada por Moreira et al. (1998), por cromatografia de afinidade em coluna de
galactomanana de Adenanthera pavonina. A frutalina (FTL) é uma glicoproteína, -D-
galactose ligante, que possui 2,1% de carboidratos em sua estrutura e apresenta elevados
teores de aminoácidos ácidos, hidroxilados e hidrofóbicos e baixo teor de aminoácidos
sulfurados (MONTEIRO, 1998; MONTEIRO-MOREIRA, 2002).
A frutalina apresenta duas bandas protéicas com massas moleculares aparentes de 12 e
15 kDa quando submetida à eletroforese em gel de poliacrilamida sob condições desnaturantes
(PAGE-SDS), na presença e ausência de -mercaptoetanol, evidenciando assim que esta lectina
não apresenta pontes dissulfeto (MONTEIRO-MOREIRA, 2002).
A estrutura primária da frutalina é composta pela cadeia alfa com 133 resíduos de
aminoácidos e pela cadeia beta com 20 resíduos. A estrutura secundária desta lectina é formada
predominantemente por folhas β antiparalelas. A frutalina é uma proteína oligomérica,

8
encontrando-se como tetrâmero apenas em pH alcalino, com massa molecular aparente de 60 kDa.
Alinhando as seqüências de aminoácidos da frutalina com outras lectinas isoladas de moráceas
observou-se que a frutalina apresenta em torno de 40% de identidade com a KM+ (lectina
manose-ligante de Artocarpus integrifolia), 76% com a lectina de Maclura pomifera e 98%
com a jacalina (lectina galactose-ligante de A. integrifolia). Como a estrutura primária da
frutalina é muito semelhante a da jacalina, é bastante razoável se pensar que a estrutura
tridimensional da frutalina seja bem próxima a da jacalina (MONTEIRO-MOREIRA, 2002).
Monteiro-Moreira (2002) estudou o efeito do pH na estrutura quaternária da
frutalina e sua estabilização pelo açúcar ligante. Na ausência do ligante, a lectina apresentou-
se na forma tetramérica a pH 10,0, com Mr de 60 kDa. O mesmo não ocorreu em valores de
pH mais baixos (8,0 e 6,0), onde a lectina apresentou massa molecular aparente que
corresponde à forma dimérica da lectina; a mesma se encontrou sob a forma monomérica a pH
2,6 e 4,0. No entanto, quando este experimento foi realizado na presença de D-galactose 0,2
M, um comportamento diferente foi verificado. Em valores de pH 2,6 e 4,0, observou-se uma
estabilização da estrutura quaternária da frutalina, uma vez que foi verificado um equilíbrio
entre as formas dimérica e tetramérica. Em valores de pH 6,0 e 8,0, a lectina apresenta-se na
forma tetramérica, enquanto que na ausência do açúcar a mesma permanece na forma
dimérica. Em pH 10,0 a frutalina apresentou massa molecular aparente ligeiramente maior que
a Mr do tetrâmero, e este comportamento foi independente da presença de açúcares.
É evidente que a presença do açúcar contribui na estabilização da estrutura
quaternária da frutalina quando esta é submetida a diferentes valores de pH, fato evidenciado
por Campana e colaboradores (2002), quando realizaram estudos de desenovelamento e
renovelamento da frutalina e mostraram a necessidade da presença da D-galactose para que a
lectina, após passar por um processo de desnaturação, fosse renaturada.
Diversas atividades biológicas da frutalina foram descritas. Devido a sua
especificidade anomérica por -D-galactose, esta lectina mostrou-se eficaz na identificação de
lesões malignas de mama (FERREIRA, 2001). A marcação em tecidos neoplásicos foi
predominantemente membranar, apresentando menor intensidade no citoplasma. Não houve
marcação no tecido normal ou estroma. A marcação de carcinoma in situ foi intensa e houve

9
aumento progressivo da intensidade da marcação com a frutalina das lesões menos agressivas
para as mais agressivas.
Milhome (2003) avaliou o uso da frutalina no estudo de lesões benignas e
malignas de tireóide humana. A frutalina marcou intensamente a membrana de carcinoma
papilar e houve marcação citoplasmática de moderada a forte. Não houve marcação
membranar de estroma ou células normais, nem de lesões benignas.
A ligação da frutalina a receptores da superfície celular elicita uma série de
respostas biológicas, tais como indução da migração de neutrófilos na cavidade pleural de
ratos, indução da quimiotaxia e reorganização do citoesqueleto de actina em neutrófilos
humano (BRANDO-LIMA et al., 2005), ativação mitogênica de linfócitos humanos
(BRANDO-LIMA et al., 2006). Estes efeitos foram inibidos quando realizados na presença de
galactose, sugerindo que o reconhecimento seja carboidrato específico.
1.4. Produção de lectinas vegetais em Escherichia coli
A dificuldade de separação das isoformas das lectinas, por técnicas convencionais,
tem feito com que as propriedades funcionais e atividades biológicas das lectinas sejam
definidas por uma mistura de proteínas. As diferenças das seqüências entre as isoformas
podem causar um efeito significante na atividade biológica da molécula. Os sistemas
heterólogos produzem proteínas com uma seqüência de aminoácidos definida e permitem que
a relação entre seqüência e função da proteína possa ser explorada (LUO et al., 2005).
A produção de proteínas recombinantes envolve a clonagem de um determinado
gene em um vetor de expressão sob o controle de um promotor induzível. Os fatores que
influenciam o nível de expressão incluem a estabilidade e a eficiência do mRNA, o correto
enovelamento da proteína, a degradação por proteases e a toxicidade da proteína
(SCHUMANN et al., 2004).
A bactéria Gram-negativa E. coli tem sido o sistema biológico procariótico mais
empregado para a produção de proteínas heterólogas. A escolha por esse sistema resulta do
fato de ser um organismo de bioquímica, fisiologia e genética bem caracterizadas (CHOI;

10
LEE, 2004; MAKRIDES et al., 1996 citado por CHOI et al, 2006). Esse sistema de expressão
também apresenta uma rápida taxa de multiplicação, capacidade para fermentação contínua e
um baixo custo (YIN et al., 2007).
Até 2003, aproximadamente 80% das proteínas usadas para resolver estruturas
tridimensionais do PDB (Protein Data Bank) foram preparadas em E. coli de forma
recombinante (SORENSEN; MORTENSEN, 2005).
As proteínas recombinantes de eucariotos produzidas em E. coli são
caracteristicamente destituídas de modificações pós-traducionais, tais como pontes dissulfeto
(CHOI et al., 2006). Por ser o processamento pós-traducional de uma pré-pro-proteína um
evento fundamental para a produção da proteína biologicamente ativa, os mecanismos de
processamento pós-traducional de proteínas devem ser compreendidos em detalhes. A criação
de sistemas heterólogos para a produção de lectinas recombinantes tem contribuído de forma
efetiva com o esclarecimento da biossíntese e processamento dessas proteínas (STREICHER;
SHARON, 2003).
Muitas lectinas vegetais recombinantes têm sido produzidas em Escherichia coli a
exemplo das lectinas: PSL de Pisum sativum (STUBBS et al., 1986; PRASTHOFER et al.,
1989), PHA de Phaseolus vulgaris (HOFFMAN; DONALDSON, 1987, ConA de C.
ensiformis (YAMAUCHI; MINAMIKAWA, 1990; MIN; JONES, 1992), LBL de Phaseolus
lunatus (JORDAN; GOLDSTEIN, 1994), PNA de Arachis hypogea (RODRIGUEZ-
ARANGO et al.,1992; SHARMA; SUROLIA, 1994), GS-II de Griffonia simplicifolia (ZHU
et al.,1996 citado por STREICHER; SHARON, 2003), SBA de Glycine max (ADAR et
al.,1997), RBL de Robinia pseudocacia (NISHIGUCHI et al., 1997 citado por STREICHER;
SHARON, 2003), MLI de Viscum album (ECK et al.,1999), ConBr de C. brasiliensis
(NOGUEIRA et al.,2002), jacalina de Artocarpus integrifolia (SAHASRABUDDHE et al.,
2004), AHA de Arisaema heterophyllum (ZHAO et al., 2006), GNA de Galanthus nivalis
(LUO et al., 2005), KM+ de Artocarpus integrifolia (SILVA et al., 2005).
Na produção de lectinas recombinantes de leguminosas, o sistema E. coli tem sido
o mais empregado. Diversas lectinas de leguminosas já foram expressas nesse sistema.
Nogueira et al.,(2002) expressaram e purificaram a lectina de Canavalia brasiliensis em E.
coli. A produção da pré-pro-ConBr em células de E. coli (BL21 DE3) não resultou no

11
processamento da lectina na sua forma madura. Mesmo não havendo processamento, a ConBr
recombinante produzida em E. coli se mostrou capaz de induzir a migração de neutrófilos para
a cavidade peritoneal de ratos e causar degranulação de mastócitos peritoniais murinos, com
conseqüente liberação de histamina. No entanto, a intensidade das respostas obtidas com a
proteína recombinante foi inferior às repostas apresentadas pela ConBr nativa (NOGUEIRA et
al., 2002). Para a ConA de Canavalia ensiformis expressa em E. coli, nenhum processamento
foi observado, mas a proteína foi expressa de maneira estável (MIN; JONES, 1992).
O número de lectinas jacalina-relacionadas expressas em E. coli é bem menor que
das lectinas de leguminosas. A maioria destas lectinas recombinantes produzidas em E. coli
são manose-ligantes.
A lectina manose/glucose ligante de Castanea crenata (CCA) expressa em E. coli
apresentou o espectro de dicroísmo circular e a capacidade de ligação à carboidratos
semelhante a da lectina nativa (nCCA). A CCA recombinante foi produzida em uma escala de
4,2 mg/L de cultura (NAKAMURA et al., 2002).
Tateno et al.,(2003) clonaram e expressaram um novo membro das lectinas
jacalina-relacionadas de rizomas da samambaia Phlebodium aureum. A lectina recombinante
apresentou atividade biológica similar à da proteína nativa.
A lectina galactose ligante de Artocarpus integrifolia (Jacalina) foi expressa em E.
coli visando a comparação em alguns aspectos, como: afinidade por açúcar e enovelamento,
com a proteína nativa. A Jacalina recombinante não sofreu processamento proteolítico. A
mesma apresentou uma menor afinidade por açúcar, mas seu enovelamento foi semelhante ao
da proteína nativa (SAHASRABUDDHE et al., 2004).
Silva et al., (2005) clonaram e expressaram a lectina manose ligante de Artocarpus
integrifolia (KM+) em E. coli. Para avaliar a expressão da lectina recombinante, a mesma foi
produzida como sua forma nativa (rKM+) e em fusão com a proteína glutationa S-transferase
(GST). A expressão da rGST-KM+ resultou em um aumento significativo no rendimento da
proteína recombinante quando comparada a rKM+. Nas mesmas condições de crescimento, o
rendimento da rGST-KM+ foi cerca de 40 vezes maior que o da rKM+, após 4 horas de
indução. Contudo, a proteína de fusão se acumulou exclusivamente em corpos de inclusão.

12
A rápida produção de proteínas recombinantes pode ocasionar a formação de
agregados insolúveis conhecidos como corpos de inclusão (BETTS; KING, 1999 citado por
SCHUMANN et al., 2004). Estas partículas esféricas são claramente separadas do citoplasma
e resultam de uma falha do sistema de reparo ou remoção de proteínas desenoveladas ou
enoveladas incorretamente (SCHUMANN et al., 2004). Para a obtenção de proteínas solúveis,
faz-se necessária a aplicação de métodos apropriados de solubilização e renaturação das
proteínas recombinantes (MIDDELBERG, 2002). Vários relatos demonstraram que lectinas
sintetizadas em E. coli são funcionais após passarem por um tratamento de solubilização e
renaturação (SHARMA; SUROLIA, 1994; SCHROEDER; RAIKHEL, 1992; ARANGO et
al., 1992).
Segundo Haraguchi et al. (2006), a lectina jacalina-relacionada de Cycas revoluta
(CRLL) foi expressa em E. coli de forma ativa. A lectina recombinante (rCRLL) se acumulou
nos corpos de inclusão, por isso passou por um tratamento de solubilização e renaturação para
que pudesse ser purificada. O rendimento foi de 15 mg da rCRLL para 800 mL da cultura. A
atividade específica da rCRLL foi bastante similar da proteína nativa (nCRLL).
O fator de agregação da -glucosidase de milho (BGAF), uma lectina quimérica
jacalina-relacionada, foi expresso em E. coli e suas propriedades foram comparadas com a
lectina nativa. A lectina recombinante (rBGAF) mostrou atividade hemaglutinante e afinidade
por carboidratos semelhantes às da proteína nativa (KITTUR, et al., 2007).
Apesar das lectinas recombinantes expressas em E. coli não sofrerem os
processamentos que ocorrem quando da sua biossíntese nas células vegetais, elas apresentam,
via de regra, atividades biológicas semelhantes às exibidas pelas lectinas nativas, como
capacidade de aglutinar hemácias, interação com carboidratos e atividade inseticida (ZHU et
al., 1996).

13
2. OBJETIVOS
2.1. Objetivo geral
O presente trabalho teve como objetivo comparar a seqüência codificadora da
lectina de Artocarpus incisa L. (FTL), isolada de sementes em diferentes estágios de
maturação, e expressar um dos clones seqüenciados em células de Escherichia coli.
2.2. Objetivos específicos
- Clonar a seqüência codificadora da frutalina a partir do cDNA de sementes em diferentes estágios de maturação;
- Seqüenciar os clones obtidos e avaliar as mutações entre os mesmos;
- Determinar a estrutura tridimensional dos clones obtidos através da modelagem por homologia. Analisar os modelos e observar a ocorrência de mutações no sítio ligante de carboidrato;
- Induzir a expressão da frutalina em células de E. coli;
- Avaliar a expressão da frutalina recombinante por meio de ensaio de imunodetecção.

14
3. MATERIAL
3.1. Material vegetal
Frutos de três diferentes estágios de maturação (Estágio 1, Estágio 2 e Estágio 3)
foram coletados de uma planta adulta da espécie A. incisa no município de Maranguape (CE).
3.2. Bactérias
As cepas de E. coli TOP10 e DH10B foram utilizadas nos experimentos de
clonagem do cDNA da frutalina. A cepa recombinante de E. coli Origami pET::FTL foi
utilizada como hospedeiro de expressão da lectina recombinante de A. incisa.
3.3. Enzimas
As enzimas de restrição XhoI (10 U/L), BamHI (10 U/L) e HindIII (10 U/L),
acompanhadas dos seus respectivos tampões de reação 10X, foram adquiridas da Gibco BRF-
Life Technologies. A DNA ligase T4 foi obtida da Promega. Ribonuclease I “A” de pâncreas
bovino acompanhada de seu tampão foi obtida da Invitrogen Life Technologies (USA). Taq
DNA polimerase foi produzida e purificada no Laboratório de Genética Molecular – UFC.
3.4. Plasmídeos
Os plasmídeos pGEM-T easy vector (Promega) e pET15b (EMD/Novagen,
Germany) foram utilizados como vetor de clonagem e de expressão, respectivamente. O
plasmídeo recombinante pCR4-TOPO contendo o cDNA da frutalina de outro estágio de

15
maturação, denominado de Estágio 4, utilizado nos experimentos de expressão foi cedido por
Felix (2008).
3.5. Reagentes e outros materiais
Oligonucleotídeos iniciadores complementares à seqüência codificadora da
frutalina foram sintetizados pela Integrated DNA Technologies (USA). O kit de purificação de
DNA (GFX DNA) foi adquirido da Amersham Biosciences. O Kit para a reação de RT-PCR
(Access RT-PCR System) foi obtido da Promega.
O marcador de proteínas (BenchMark Pré-Stained Protein Ladder) (Invitrogen) foi
utilizado nas corridas eletroforéticas. Membranas de nitrocelulose HybondTM C Extra
(Amersham Biosciences Corp., USA) e papel de filtro Whatman 3 MM foram utilizados nos
experimentos de Western Blotting . Leite desnatado Molico® obtido comercialmente foi
utilizado nos experimentos de imunodetecção.
O anticorpo primário de coelho contra a lectina nativa de A. incisa (frutalina) foi
cedido por Felix (2008).
Os demais reagentes foram todos de grau analítico.

16
4. MÉTODOS
4.1. Extração de RNA total de sementes de Artocarpus incisa
RNA total foi isolado de sementes de A. incisa (fruta-pão) coletadas em três
estágios de amadurecimento diferentes (Estágio 1, Estágio 2 e Estágio 3), de acordo com a
metodologia baseada no uso do reagente Cloreto de Lítio, como descrito por Chang et
al.,(1993). As Figuras 1 e 2 apresentam os frutos e as sementes de A. incisa em três diferentes
estágios de maturação. Uma semente de cada estágio foi macerada na presença de nitrogênio
líquido, até a obtenção de um pó fino, que foi transferido para um tubo contendo 20 mL de
tampão CTAB pré-aquecido a 65 ºC e mantidos por 50 minutos. Em seguida, foi adicionado 1
volume de clorofórmio/álcool isoamílico (24:1) e após 15 minutos de incubação a temperatura
ambiente, sob agitação, as amostras foram centrifugadas a 3.500 x g, por 20 minutos. Após a
transferência do sobrenadante para um novo tubo contendo 15 mL de clorofórmio, as amostras
permaneceram sob agitação por 5 minutos e foram centrifugadas a 3.500 x g, por 20 minutos.
O sobrenadante (cerca de 20 mL) foi coletado em tubos de centrífuga estéreis onde foi feita a
adição de 6 mL de cloreto de lítio 10,0 M para a precipitação do RNA. As amostras foram
mantidas a 4 ºC por 18 horas.
Após a centrifugação das amostras por 45 minutos a 3.500 x g, o tubo foi invertido
e o precipitado foi lavado duas vezes com 7 mL de etanol 70%, sendo centrifugado após cada
lavagem por 10 minutos a 3.500 x g. Em seguida, após evaporação do etanol, o precipitado foi
ressuspenso em 200 L de água estéril tratada com DEPC (dietilpirocarbonato), inibidor de
RNAse. As amostras foram armazenadas a -80 ºC. Todas as soluções utilizadas para extração
de RNA foram preparadas com água ultrapura estéril tratada com DEPC.
Após a extração, foi feita a análise quantitativa do RNA total por meio de
espectrofotometria (DO260). Para análise qualitativa, uma alíquota de 5,0 L foi misturada com
2,0 L do tampão da amostra (azul de bromofenol 0,25%, glicerol 30% em TE pH 8,0) e então
aplicada em gel de agarose 1%. A eletroforese foi realizada a 100 V com o gel submerso em
tampão TBE (Tris-Borato 45 mM, EDTA 1,0 mM, pH 8,0) em cuba horizontal.

17
1º ESTÁGIO (E1)
2º ESTÁGIO (E2)
3º ESTÁGIO (E3)
Frutos de Artocarpus incisa em três diferentes estágios de maturação. Barra: 2,65 cm.
Figura 1 -
18
1º ESTÁGIO (E1) 2º ESTÁGIO (E2) 3º ESTÁGIO (E3)
Sementes de Artocarpus incisa em três diferentes estágios de maturação.
Figura 2 -

19
O gel corado com brometo de etídio (0,5 g/mL) foi visualizado no transluminador AFL
(Pharmacia Biotech) com luz UV.
4.2. Amplificação do cDNA da frutalina por RT- PCR
A reação de RT-PCR (Reação em cadeia da polimerase com transcrição reversa)
para a amplificação e obtenção do cDNA da frutalina dos três diferentes estágios de maturação
foi realizada utilizando o kit Access RT-PCR System (Promega), usando como substrato o
RNA total isolado de sementes. A quantidade de RNA utilizada como molde para a reação foi
de 1000 ng. Os oligonucleotídeos específicos para a seqüência codificadora da frutalina
utilizados na reação estão descritos no Quadro 1. Os ciclos de temperatura foram realizados
em aparelho termociclador PTC 200 (DNA Thermal Cycler, MJ Research) e os parâmetros
utilizados foram: 45 ºC por 45 minutos e 94 ºC por 2 minutos, seguido por mais 40 ciclos de
94 ºC por 30 segundos (etapa de desnaturação), 60 C por um minuto (etapa de anelamento) e
68 ºC por 2 minutos (etapa de extensão). Terminados esses ciclos, foi feita uma extensão a 68
C por 7 minutos.
Após a amplificação do cDNA, foi realizada uma eletroforese em gel de agarose
1%, contendo brometo de etídio 0,5 g/mL.
O cDNA da frutalina de outro diferente estágio de maturação, denominado de
Estágio 4, foi previamente clonado no vetor pCR4-TOPO e cedido por Felix (2008) para
estudos de expressão do gene e modelagem da proteína.
4.3. Preparação de células eletrocompetentes de Escherichia coli
Células eletrocompetentes de E. coli DH10B e TOP10 foram utilizadas nos
experimentos de clonagem e E. coli Origami (DE3) na expressão do gene. Inicialmente, uma
colônia isolada de E. coli foi inoculada em 10 mL de meio líquido 2xYT (triptona 1,6%,

20
Iniciadores
N-terminal (5’–3’) CATATGAACGAACAAAGCGGGA
C-terminal (3’–5’) GGATCCTCAAAGGGACAAGTACAT
Oligonucleotídeos específicos para a seqüência codificadora da frutalina com sítios de
restrição para as enzimas NdeI e BamHI para clonagem no vetor pGEM-T
Os sítios de restrição para as enzimas NdeI e BamHI estão evidenciados em itálico, enquanto a seqüência correspondente a seqüência codificadora da frutalina está em negrito. Os códons de iniciação (ATG) e terminação (TCA) estão sublinhados.
Quadro 1-

21
extrato de levedura 1%, NaCl 0,5%, pH 7,0) contendo 30 g/mL de estreptomicina em
erlenmeyer de 50 mL e a cultura foi incubada por 18 horas sob agitação constante de 250 rpm
a 37 C.
Ao final da incubação, 5 mL da cultura foram transferidos para 500 mL de meio
2xYT contendo estreptomicina (30 g/mL). A cultura foi mantida sob agitação constante de
250 rpm, a 37 ºC, até atingir uma DO600 igual a 0,6. Em seguida, a suspensão de células foi
mantida no gelo por 30 minutos e então, centrifugada a 10.000 x g, 4 ºC por 5 minutos. O
precipitado celular foi ressuspenso em 20 mL de solução resfriada de glicerol 10% estéril. A
suspensão foi novamente centrifugada e a lavagem com glicerol foi realizada duas vezes. Após
as lavagens, as células foram ressuspensas em 2 mL de meio GYT (glicerol 10%, extrato de
levedura 0,125%, triptona 0,25%) resfriado. Ao final do processo, alíquotas de 50 L de
células eletrocompetentes foram transferidas para microtubos estéreis de 1,5 mL e
armazenadas a -80 ºC.
4.4. Clonagem do cDNA da frutalina em células de Escherichia coli DH10B
O cDNA da frutalina dos estágios 2 e 3 amplificados na reação de RT-PCR foram
inseridos no vetor de clonagem pGEM-T Easy Vector (Promega) de acordo com as
especificações do fabricante. Os plasmídeos transformados com o cDNA da frutalina do
estágio 2 foram denominados de pGEM-T::FTL-E2 e os transformados com o cDNA da
frutalina do estágio 3 foi denominado de pGEM-T::FTL-E3. A reação de ligação entre o
inserto e o vetor consistiu na mistura dos seguintes componentes: 3,0 L do produto da RT-
PCR, 5,0 L do tampão de ligação 2x, 1,0 L do vetor (50 ng/L), 1,0 L da T4 DNA ligase
(3 U/L). A reação foi mantida a 4 C por quinze horas em termociclador PTC 200 (DNA
Thermal Cycler, MJ Research). Em seguida, o produto da reação de ligação foi utilizado para
transformar células eletrocompetentes de E. coli.
A transformação foi realizada por eletroporação e consistiu na mistura de 10 L da
reação de ligação com 50 L de células eletrocompetentes de E. coli DH10B em uma cubeta
de eletroporação (Eppendorf), com espaço interno de 4 mm, que foi submetida a um pulso de

22
2,5 kV no eletroporador Eppendorf, modelo 2510. No tratamento controle, as células foram
eletroporadas na ausência do plasmídeo, sendo o volume completado com água ultrapura
estéril. Imediatamente após o pulso, foi adicionado, a cada cubeta, 1,0 mL de meio SOC
(triptona 2%, extrato de levedura 0,5%, NaCl 10 mM, KCl 2,5 mM, MgCl 10 mM, glucose 20
mM, pH 7,0), pré-aquecido a 37 ºC. A suspensão foi transferida para um tubo de 15 mL e
incubada sob agitação constante de 150 rpm a 37 C por 1 hora. Após a incubação, uma
alíquota de 100 L da suspensão de células eletroporadas foi transferida para placas de Petri
contendo meio LB ágar (triptona 1%, extrato de levedura 0,5%, NaCl 1%, ágar 1,6%, pH 7,5)
suplementado com carbenicilina 100 g/mL, estreptomicina 30 g/mL, X-Gal 80 g/mL e
IPTG 0,5 mM. A suspensão foi espalhada sobre o meio de cultura com o auxílio de uma alça
de Drigalski. As placas foram mantidas na estufa a 37 C por 16 horas e avaliadas de acordo
com a coloração das colônias: brancas (transformada com o plasmídeo recombinante) ou azuis
(transformadas com o plasmídeo íntegro).
Doze colônias brancas foram selecionadas e inoculadas em 5 mL de meio LB
(triptona 1%, extrato de levedura 0,5%, NaCl 1%, pH 7,5) suplementado com carbenicilina
100 g/mL e estreptomicina 30 g/mL. As culturas foram mantidas por 16 horas a 37 C sob
agitação constante de 200 rpm. Em seguida, as células foram coletadas e os plasmídeos foram
isolados com o auxílio do kit Wizard Sv Plasmid DNA Purification System (Promega, USA) de
acordo com as especificações do fabricante. Os plasmídeos purificados foram analisados por
meio de uma reação de amplificação por PCR com os iniciadores específicos para a seqüência
codificadora da frutalina.
4.5. Isolamento de plasmídeos recombinantes de E. coli
As células de E.coli DH10B eletroporadas com os plasmídeos transformados
(pGEM-T::FTL-E2 e pGEM-T::FTL-E3) tiveram seus plasmídeos isolados com o auxílio do
kit Wizard Sv Plasmid DNA Purification System (Promega, USA) de acordo com as
especificações do fabricante. Como outro método alternativo de purificação, os plasmídeos
foram isolados pelo método de lise alcalina, como descrito por SAMBROOK e colaboradores
(1989). Foram expandidas doze colônias selecionadas, aleatoriamente, da cultura de E. coli

23
transformadas com pGEM-T::FTL-E2 e pGEM-T::FTL-E3 em erlenmeyers de 50 mL
contendo 5 mL de meio LB suplementado com 100 g/mL de carbenicilina e 30 g/mL de
estreptomicina. As culturas foram mantidas sob agitação constante de 200 rpm a 37 C, por
aproximadamente 18 horas. Após a incubação das células, uma alíquota de 1,5 mL de cada
amostra foi transferida para tubos de microcentrífuga e procedeu-se com a extração de
plasmídeos.
Após a centrifugação, por cinco minutos, a 12.000 rpm (MiniSpin-Eppendorf), as
células foram coletadas e ressuspensas em 100 L de GET (glicose 50 mM, Tris-HCl 25 mM,
EDTA 10 mM, pH 8,0). Em seguida foi feita a adição da solução de lise (SDS 10%, NaOH
4,0 M) e após três minutos foi adicionada a solução de neutralização (acetato de potássio 3,0
M, ácido acético glacial 5,0 M). As amostras foram mantidas por cinco minutos no gelo e
depois centrifugadas por cinco minutos a 12.000 rpm. O sobrenadante foi coletado em
microtubos estéreis onde foi feita a adição de dois terços do volume de isopropanol 100%.
Após uma hora, as amostras foram centrifugadas e o sobrenadante descartado. Após lavagem
com etanol 70%, os plasmídeos foram ressuspensos em 50 L de água ultrapura estéril
acrescidos de 2,0 L de RNAse (1,0 mg/mL).
A presença e qualidade dos plasmídeos foi verificada por meio de eletroforese em
gel de agarose 0,8% contendo EtBr 0,5 g/mL. Uma vez constatada a eficiência do processo
de extração dos plasmídeos, estes foram submetidos a uma reação de amplificação por PCR
utilizando-se os iniciadores específicos para a seqüência codificadora da frutalina, segundo os
seguintes parâmetros: para uma reação de 10 L, foram adicionados 1,0 L do tampão 10x
(Tris-HCl 100 mM, pH 9,0; MgCl2 15 mM e KCl 500 mM), 2,0 L de dNTPs (1,0 mM), 1,0
L de cada iniciador (5 M) e 0,5 L de Taq polimerase (5,0 U/L). A quantidade de DNA
utilizada como molde para a reação foi de 100 ng. Como controle, a reação de amplificação foi
realizada sem a adição do DNA molde, sendo o volume da reação completado com água.
A reação de amplificação por PCR do gene da frutalina foi realizada no
termociclador PTC 200 (DNA Thermal Cycler, MJ Research) e consistiu inicialmente de uma
etapa de desnaturação a 95 C por cinco minutos, seguido de uma etapa de anelamento a 60 C
por dois minutos e uma etapa de extensão a 72 C por dois minutos. Esse ciclo foi seguido
por mais 30 ciclos de 94 C por um minuto, 60 C por um minuto e 72 C por dois minutos.

24
Ao final da reação, uma alíquota de 5 L foi misturada com 2,0 L do tampão da amostra
(azul de bromofenol 0,25%, glicerol 30% em TE, pH 8,0) e então aplicada em gel de agarose
1,0%. A eletroforese foi realizada a 100 V com o gel submerso em tampão TBE (Tris-Borato
45 mM, EDTA 1,0 mM, pH 8,0) em cuba horizontal. O gel corado com brometo de etídio (0,5
g/mL) foi visualizado no transluminador AFL (Pharmacia Biotech) com luz UV. Após a
confirmação da transformação das células de E. coli com a seqüência codificadora da
frutalina, os clones foram mantidos em ágar estoque em camada alta e em glicerol a -80 C.
4.6. Sequenciamento de DNA
A partir dos plasmídeos pGEM-T transformados com seqüência codificadora da
frutalina dos estágios de maturação 2 e 3, os clones obtidos foram seqüenciados. A reação de
sequenciamento foi preparada com o auxilio do kit DYEnamic ET Dye terminator kit (GE
Healthcare, USA) para o seqüenciador MegaBACE 1000, de acordo com o descrito o manual
do fabricante. Na reação foram utilizados oligonucleotídeos específicos para o plasmídeo
pGEM-T Easy Vector. Foram usados como molde 350 ng dos plasmídeos purificados. A
reação foi realizada no termociclador PTC 200 (DNA Thermal Cycler, MJ Research) e
consistiu inicialmente de uma etapa de desnaturação a 95 ºC por 20 segundos, seguidos de
uma etapa de anelamento a 50 ºC por 15 segundos e uma etapa de extensão a 60 ºC por 1
minuto. Esse ciclo foi repetido por 25 vezes.
Ao término da reação, os fragmentos foram precipitados e purificados por
centrifugação com 1/10 do volume de acetato de amônio 7,5 M e etanol absoluto (2,5
volumes). Em seguida, foram lavados com etanol 70% para remoção de resíduos que
poderiam afetar a qualidade das seqüências.
Após o sequenciamento em MegaBACE 1000 (GE Healthcare, USA), as
diferenças obtidas de todos os clones foram analisadas com o auxilio do pacote
Phred/Phrap/Consed (EWING et al.,1998; GORDON et al.,1998) para a obtenção das
seqüências consenso dos clones. As seqüências dos diferentes clones foram alinhadas com o
auxílio do programa Clustal W (HIGGINS et al.,1994) e as mutações foram observadas.

25
4.7. Comparação do cDNA da frutalina obtidos de diferentes fases de maturação do fruto
de A. incisa
4.7.1. Modelagem por homologia
A técnica de modelagem por homologia foi utilizada para a previsão da possível
estrutura tridimensional de sete clones obtidos. Os clones foram escolhidos do 2º e 3º estágio
de maturação (E2 e E3) que foram seqüenciados, juntamente com seis clones de outro estágio
de maturação (E4), que foram seqüenciados previamente e os dados foram cedidos por Felix
(2008). Os clones obtidos foram analisados e comparados quanto à presença de mutações entre
os resíduos nos diferentes estágios de maturação.
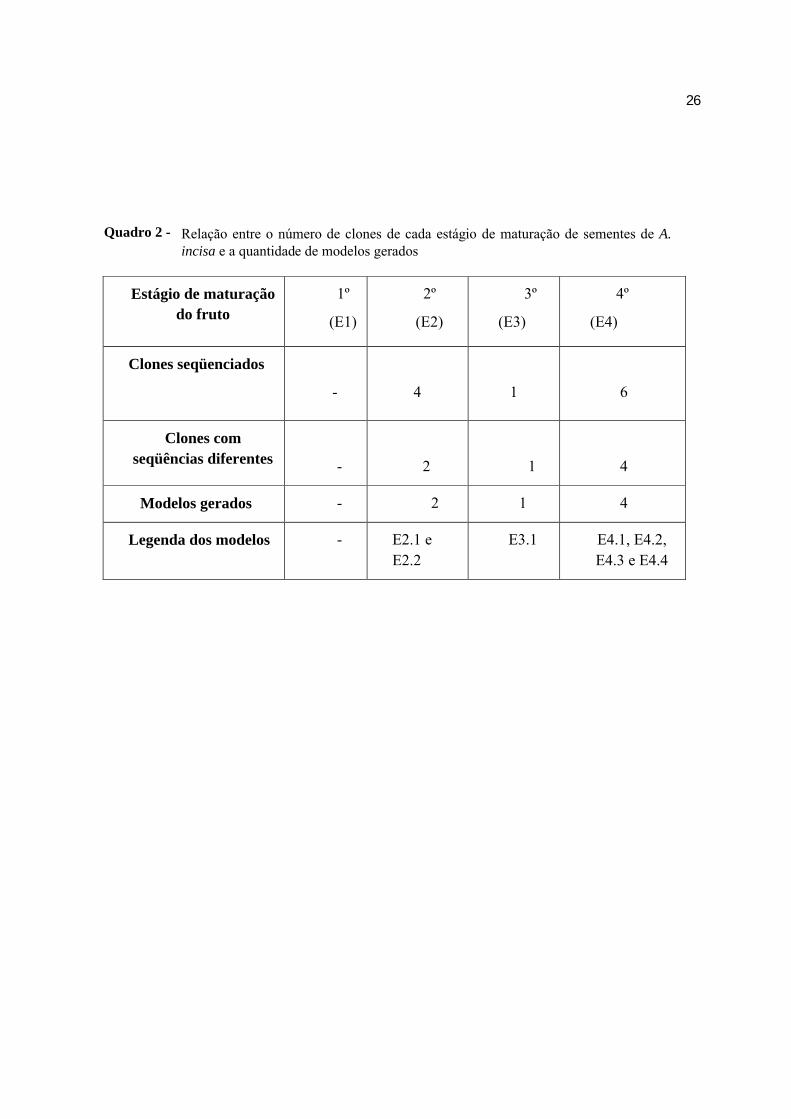
O número de clones com seqüências de qualidade obtidas para cada um dos
estágios de maturação e o número de modelos gerados estão descritos no Quadro 2. Os clones
2.2, 3.1 e 4.1 possuem a mesma seqüência, o que gerou o mesmo modelo para esses três
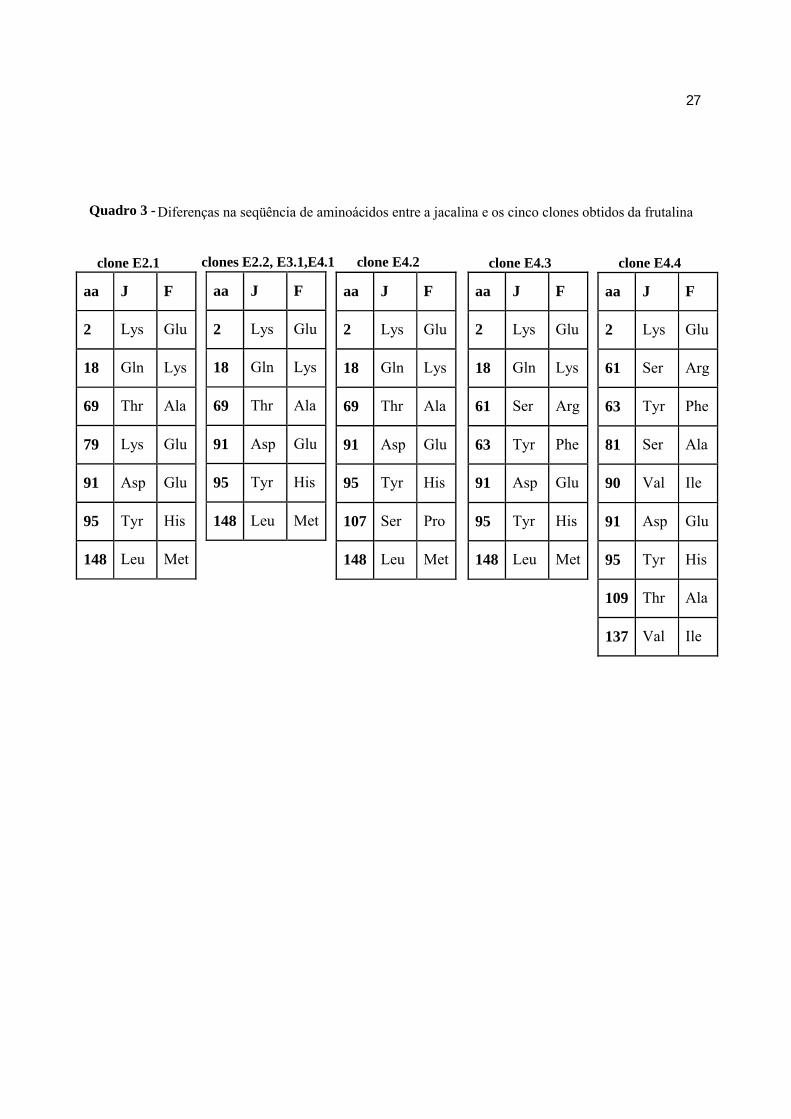
clones. As diferenças observadas entre os cinco clones obtidos estão descritas no Quadro 3.
A escolha da proteína molde para a modelagem da frutalina foi realizada de acordo
com o critério padrão de preferência pela estrutura disponível de maior grau de identidade.
Para tal, a seqüência de aminoácidos deduzida da frutalina foi comparada ao banco de dados
de estruturas resolvidas (Protein Data Bank – PDB) através das ferramentas de busca e
alinhamento BLASTp (www.ncbi.nlm.nih.gov). Dessa forma, a estrutura que apresentou
maior identidade com a proteína de interesse – 98% - foi a lectina galactose ligante de
Artocarpus integrifolia (jacalina) com o código de acesso 1KU8, resolvida por
Sankaranarayanan et al.,(1996).
Para cada um dos cinco clones foram gerados cem modelos utilizando o programa
Modeller (SALI; BLUNDELL, 1993). O método utilizado foi a randomização inicial das
coordenadas do modelo e sua conseqüente otimização. As coordenadas cartesianas foram
deslocadas até 4.0 Å, e selecionou-se o grau máximo de otimização para os modelos. Os
critérios de seleção da escolha do melhor modelo para cada caso foram a função energética
gerada pelo programa e a qualidade do mapa de Ramachandran (RAMACHANDRAN et
al.,1963) gerado pelo programa Procheck (LASKOWSKI et al.,1993).
26
Estágio de maturação do fruto
1º
(E1)
2º
(E2)
3º
(E3)
4º
(E4)
Clones seqüenciados
- 4 1 6
Clones com seqüências diferentes - 2 1 4
Modelos gerados - 2 1 4
Legenda dos modelos - E2.1 e E2.2
E3.1 E4.1, E4.2, E4.3 e E4.4
Relação entre o número de clones de cada estágio de maturação de sementes de A. incisa e a quantidade de modelos gerados
Quadro 2 -
27
aa J F
2 Lys Glu
18 Gln Lys
69 Thr Ala
79 Lys Glu
91 Asp Glu
95 Tyr His
148 Leu Met
aa J F
2 Lys Glu
18 Gln Lys
69 Thr Ala
91 Asp Glu
95 Tyr His
148 Leu Met
aa J F
2 Lys Glu
18 Gln Lys
69 Thr Ala
91 Asp Glu
95 Tyr His
107 Ser Pro
148 Leu Met
aa J F
2 Lys Glu
18 Gln Lys
61 Ser Arg
63 Tyr Phe
91 Asp Glu
95 Tyr His
148 Leu Met
aa J F
2 Lys Glu
61 Ser Arg
63 Tyr Phe
81 Ser Ala
90 Val Ile
91 Asp Glu
95 Tyr His
109 Thr Ala
137 Val Ile
Quadro 3 - Diferenças na seqüência de aminoácidos entre a jacalina e os cinco clones obtidos da frutalina
clone E2.1 clones E2.2, E3.1,E4.1 clone E4.2 clone E4.3 clone E4.4

28
Dentre os dez modelos com as menores energias geradas, selecionou-se aquele
com o melhor mapa de Ramachandran (sendo escolhido o de energia mais baixa no caso de
haver mais de um modelo com qualidades iguais de resíduos fora das regiões favoráveis).
O mapa de Ramachandran mostra a distribuição dos ângulos torsionais φ e ψ para
cada aminoácido. Há regiões energeticamente favoráveis para cada resíduo, e a presença de
um aminoácido fora dessas regiões indica um erro ou algum fenômeno não convencional
naquele local, como uma forte interação causando o deslocamento.
Outras informações a respeito da qualidade dos modelos foram verificadas
utilizando os programas Whatcheck (HOOFT et al.,1996) e Verify3D (BOWIE, et al.,1991). O
Verify3D julga a verossimilhança da vizinhança de cada aminoácido através de três
propriedades: a área exposta ao solvente, a fração da área da cadeia lateral coberta por átomos
polares e a estrutura secundária local. A combinação desses três parâmetros permite colocar o
resíduo em uma de dezoito regiões. Para cada tipo de resíduo há escore que reflete a sua
compatibilidade com tal região (definida previamente por análise estatística de proteínas de
estrutura conhecida).
O software Whatcheck verifica uma larga gama de possíveis erros em estruturas
tridimensionais modeladas ou experimentais, como atribuições não convencionais de célula
unitária, valores do coeficiente de Matthews (MATTHEWS, 1968) fora da faixa considerada
aceitável, erros de nomenclatura, ausência de átomos, ângulos e distâncias de ligação,
planaridade e ângulos torsionais de cadeias laterais, choques entre átomos, posicionamento de
moléculas de água, pontes de hidrogênio, etc.
A análise dos modelos escolhidos foi feita com o uso do software Pymol
(DELANO, 2002), responsável pela visualização das ilustrações dos modelos.
4.8. Expressão da rFrutalina em células de E. coli Origami (DE3)
4.8.1. Subclonagem da seqüência codificadora da frutalina
A seqüência codificadora da frutalina de outro diferente estágio de maturação,
denominado de Estágio 4, foi previamente clonada no vetor pCR4-TOPO e cedida por Felix

29
(2008) para estudos de expressão da proteína. Essa seqüência foi subclonada no vetor pGEM-
T. Para tanto, uma extração de plasmídeos foi realizada a partir das culturas E.coli Mach 1-T1.
Os plasmídeos pCR4-TOPO::FTL-E4 (plasmídeos transformados com a seqüência
codificadora da frutalina do estágio 4) foram submetidos a uma reação de amplificação por
PCR com iniciadores específicos para o inserto da frutalina. Os iniciadores foram desenhados
com sítios de restrição para XhoI e BamHI (Quadro 4). Os parâmetros utilizados foram: para
uma reação de 10 L, foram adicionados 1,0 L do tampão 10x (Tris-HCl 100 mM, pH 9,0;
MgCl2 15 mM e KCl 500 mM), 2,0 L de dNTPs (1,0 mM), 1,0 L de cada iniciador (5 M)
e 0,5 L de Taq polimerase (5,0 U/L). A quantidade de DNA utilizada como molde para a
reação foi de 100 ng.
A reação de amplificação por PCR da seqüência codificadora da frutalina foi
realizada no termociclador PTC 200 (DNA Thermal Cycler, MJ Research) e consistiu
inicialmente de uma etapa de desnaturação a 95 C por cinco minutos, seguido de uma etapa
de anelamento a 60 C por dois minutos e uma etapa de extensão a 72 C por dois minutos.
Esse ciclo foi seguido por mais 30 ciclos de 94 C por um minuto, 60 C por um minuto e 72
C por dois minutos. O produto amplificado foi ligado ao plasmídeo pGEM-T Easy Vector
(Promega) de acordo com as especificações do fabricante. A reação de ligação entre o inserto
e o vetor consistiu na mistura dos seguintes componentes: 3,0 L do produto da PCR, 5,0 L
do tampão de ligação 2x, 1,0 L do vetor (50 ng/L), 1,0 L da T4 DNA ligase (3 U/L). A
reação foi mantida a 4 C por 15 horas em termociclador PTC 200 (DNA Thermal Cycler, MJ
Research). Em seguida, o produto da reação de ligação foi utilizado para transformar células
eletrocompetentes de E. coli. A transformação foi realizada por eletroporação de acordo com o
protocolo descrito no item 5.4.
Vinte colônias brancas foram selecionadas e inoculadas em 5 mL de meio LB
suplementado com carbenicilina 100 g/mL e estreptomicina 30 g/mL. As culturas foram
mantidas por 16 horas a 37 C sob agitação constante de 200 rpm. Em seguida, as células
foram coletadas e os plasmídeos foram isolados pelo método de lise alcalina, como descrito
por SAMBROOK e col. (1989).

30
Iniciadores
N-terminal (5’–3’) CCGCTCGAGATGAACGAACAAAGCG
C-terminal (3’–5’) CGGGATCCTCAAAGGGACAAGTAC
Oligonucleotídeos específicos para a seqüência codificadora da frutalina com sítios de
restrição para as enzimas XhoI e BamHI para clonagem no vetor pET15b
Os sítios de restrição para as enzimas XhoI e BamHI estão evidenciados em itálico, enquanto a seqüência correspondente ao gene da frutalina está em negrito. Os códons de iniciação (ATG) e terminação (TCA) estão sublinhados.
Quadro 4 -

31
Os plasmídeos recombinantes (pGEM-T::FTL-E4) obtidos pelo método de lise
alcalina foram submetidos a uma reação de digestão com as enzimas XhoI e BamHI. Foram
usados cerca de 1g dos plasmídeos na digestão.
Uma primeira reação de 20 L consistiu em: 10 L de pGEM-T::FTL-E4, 2,0 L
do tampão React 2 (Tris-HCl 50 mM, pH 8,0, MgCl2 10 mM, NaCl 100 mM), 2,0 L de XhoI
(10 U/L) e 6,0 L de água. A reação foi mantida a 37 C por três horas. Após este período
foi feita a adição de 2,0 L da enzima BamHI (10 U/L), 1,5 L de NaCl 1,0 M, 2,5 L de
BSA 10X e 4,0 L de água. Em seguida, os produtos da digestão foram aplicados em gel de
agarose 0,6 %.
A banda referente ao inserto foi cortada do gel e purificada com o Kit GFX gel
band purification system (Invitrogen), de acordo com as especificações do fabricante. O
plasmídeo pET15b (Invitrogen) foi submetido à mesma reação de digestão descrita acima.
Após a quantificação dos produtos purificados do gel de agarose, a reação de
ligação entre o inserto e o vetor foi feita com a enzima T4 DNA ligase (Promega), nas razões
3:1, 5:1, 8:1, obedecendo a seguinte fórmula:
vetor (ng) x tamanho do inserto (pb) x razão inserto (pb)
tamanho do inserto(pb)
A reação foi mantida a 4 C por aproximadamente dezoito horas. Em seguida, o
produto da reação de ligação foi utilizado para transformar células eletrocompetentes de E.
coli TOP10. As células transformadas foram incubadas por 1 hora sob agitação a uma
temperatura de 37 C. Em seguida, uma alíquota de 100 L das células foi transferida para
uma placa contendo meio LB ágar suplementado com carbenicilina 100 g/mL e
estreptomicina 30 g/mL.
As placas foram mantidas na estufa a 37 C por aproximadamente 16 horas e
avaliadas quanto ao aparecimento de colônias. Foram selecionadas doze colônias que foram
inoculadas em erlenmeyers de 50 mL contendo 5,0 mL de meio LB suplementado com
carbenicilina 100 g/mL e estreptomicina 30 g/mL. As culturas foram incubadas a 37 C sob
agitação constante (200 rpm) por 16 horas. Em seguida, os plasmídeos foram purificados e a

32
transformação foi confirmada através da reação de digestão com a enzima de restrição HindIII.
Os clones confirmados pela digestão foram mantidos em estoque em glicerol (-80 C).
4.8.2. Transformação de células de E. coli Origami (DE3) com pET-15b::FTL-E4
Uma alíquota de 3 L dos plasmídeos recombinantes pET-15b::FTL-E4
purificados foi usada para transformar 50 L de células eletrocompetentes de E. coli Origami
(DE3) como descrito anteriormente. A transformação foi confirmada através de uma reação de
amplificação por PCR com os iniciadores específicos para o inserto da frutalina. Os clones
confirmados por PCR foram mantidos em estoque em glicerol (-80 C).
4.8.3. Indução da expressão da rFrutalina em E. coli Origami (DE3)
Um clone de E. coli Origami (DE3) transformado com a seqüência codificadora da
frutalina foi selecionado para a indução da expressão.
Uma colônia de E. coli Origami (DE3) do clone selecionado foi inoculada em um
erlenmeyer de 50 mL, contendo 5 mL de meio LB com carbenicilina 100 g/mL, canamicina
15 g/mL e tetraciclina 12,5 g/mL e a suspensão, ou pré-inóculo, foi mantida sob agitação
constante (180 rpm) a 37 °C por aproximadamente 16 horas. Uma alíquota de 2 mL do pré-
inóculo foi transferida para um erlenmeyer de 1000 mL, contendo 100 mL de meio LB com
carbenicilina 100 g/mL, canamicina 15 g/mL e tetraciclina 12,5 g/mL e a suspensão foi
incubada sob agitação constante (200 rpm) a 37 °C até atingir uma DO600 entre 0,4 e 0,6,
quando então foi adicionado IPTG para uma concentração final de 1 mM. A cultura foi
incubada nas mesmas condições por 10 horas.
Alíquotas de 1,5 mL foram coletadas em zero, 2, 4, 6, 8 e 10 horas após o início da
indução e transferidas para microtubos estéreis de 1,5 mL. Após centrifugação por 10 minutos
a 13.000 rpm (MiniSpin - Eppendorf), o sobrenadante foi transferido para microtubos de 1,5
mL. Tanto o precipitado de células como o sobrenadante foram armazenados a -4 °C.

33
O restante da cultura, cerca de 90 mL, foi centrifugado a 7.000 x g, por 20 minutos
a 4 °C para a purificação dos corpos de inclusão.
Também foi realizada uma indução controle com uma cultura de E. coli Origami
(DE3) contendo o plasmídeo pET15b íntegro. A indução do clone transformado e a indução
controle foram feitas na presença e ausência de IPTG.
4.8.4. Purificação dos corpos de inclusão
As células bacterianas obtidas como descrito no item 4.8.3 foram ressuspensas em
9 mL de tampão TES tampão TES (Tris-HCl 50 mM, EDTA 2 mM, NaCl 0,15 M, pH 8,0)
adicionado de 900 L de Triton X-100 1% e lisozima 100 g/mL. A mistura foi incubada a 30
°C por 15-30 minutos até que ocorresse a lise das células, evidenciada pelo desenvolvimento
da viscosidade.
A digestão do DNA genômico da bactéria foi realizada pela adição de DNAse I
para uma concentração final de 20 g/g de células. A mistura foi novamente incubada a 30 °C
por 1 hora, até a perda da viscosidade, e os corpos de inclusão coletados por centrifugação a
13.000 rpm por 40 minutos (MiniSpin-Eppendorf).
4.9. Eletroforese em gel de poliacrilamida em presença de SDS e -mercaptoetanol
(PAGE-SDS)
Os experimentos de SDS-PAGE com -mercaptoetanol foram conduzidos de
acordo com o protocolo descrito por LAEMMLI (1970), adaptado para géis montados em
placas de vidro.
O precipitado de células obtido em 1,5 mL da cultura, descrito no item 5.7.3, foi
submetido a tratamento químico com a adição de 50 µL de tampão de amostra SDS-PAGE 1X
(Tris-HCl 62,5 mM pH 6,8, SDS 2% (m/v), glicerol 10%, -mercaptoetanol 5% (v/v), azul de
bromofenol 0,001%). A 150 µL do sobrenadante, foram adicionados 50 µL do tampão de
amostra SDS-PAGE 4X. Ambos foram submetidos a um tratamento térmico a 95 °C por 10

34
minutos. As amostras foram então mantidas a -4 °C até o momento das análises. Os corpos de
inclusão também foram submetidos a este mesmo tratamento.
Após ser descongelada, uma alíquota de 10 µL das amostras foi utilizada para a
realização das corridas eletroforéticas, usando-se o sistema Dual Gel Caster (Hoefer, São
Francisco), sendo a concentração do gel de separação 17,5% e o gel de concentração 3,5%
(Quadro 5). Como controle, 5 L do marcador (BenchMark Pré-Stained Protein Ladder)
(Invitrogen) foi utilizado. As corridas eletroforéticas foram conduzidas sob voltagem
constante de 100 V em tampão de corrida (Tris-HCl 0,025 M, glicina 0,192 M, SDS 0,1%
(m/v), pH 8,3)
Após a corrida, as bandas protéicas foram visualizadas através da coloração com
Comassie Briliant Blue R-250 0,1% em metanol 30% e ácido acético 10% (v/v) por 1 hora, e
posterior descoloração do gel através de uma solução de isopropanol 12,5% e ácido acético
10% (v/v).
4.10. Imunodetecção da rFrutalina imobilizada em membrana (Western blotting)
Ao final da corrida eletroforética, os géis, as membranas de nitrocelulose
(HybondTM C Extra) e papéis de fitro (Whatman) foram mergulhados em solução de
transferência (Tris-HCl 25 mM pH 8,3, glicina 185 mM, metanol 20 %, SDS 0,01 %) por dez
minutos. A montagem do “sanduíche” foi feita de acordo com as instruções do fabricante do
sistema.
A eletrotransferência foi realizada num sistema vertical (Mini V8 Life
Technologies Gibco BRL) contendo a solução de transferência. A voltagem foi mantida
constante (100 V) por uma fonte Pharmacia LBK GPS 200/400 e o tempo de transferência foi
de duas horas. Todo o processo foi realizado sob agitação constante, mantendo-se a cuba em
banho de gelo.
Para o bloqueio dos sítios ativos livres, as membranas foram mantidas em solução
de lavagem (Tris-HCl 50 mM pH 7,4, NaCl 0,15 M, Tween-20 0,05%, glucose 0,1 M)
contendo leite desnatado (Molico®) por aproximadamente 17 horas a 4 °C. Após este período,

35
Gel de separação 17,5% Gel de concentração 3,5%
Acrilamida:bisacrilamida (30:0,8 %) 5,8 mL 625 µL
Tris-HCl 3 M pH 8,8 1,25 mL -
Tris-HCl 0,5 M pH 6,8 - 1,25 mL
SDS 10 % (m/v) 0,1 mL 50 µL
TEMED 5 µL 5 µL
Persulfato de amônio 1,5 mg/mL 0,5 mL 250 µL
Água destilada 2,31 mL 2,82 mL
Volume final 10 mL 5,0 mL
Quadro 5 - Protocolo para confecção dos géis de poliacrilamida.

36
as membranas foram novamente imersas no mesmo tampão de lavagem contendo leite
desnatado 5% e anticorpo primário de coelho anti-FTL 1:300 (v/v). As membranas ficaram em
contato com o anticorpo primário por duas horas sob agitação constante, à temperatura
ambiente, quando foram submetidas a cinco lavagens de dez minutos com a mesma solução,
para que fosse eliminado o excesso de anticorpo primário não ligado às proteínas retidas na
membrana.
Em seguida, foi adicionado o anticorpo secundário (anticorpo anti-coelho
complexado a fosfatase alcalina) à solução de lavagem em uma diluição de 1:5.000, onde as
membranas permaneceram por duas horas, sob agitação constante. Após um ciclo de mais
cinco lavagens de dez minutos, foi feita a revelação utilizando o 5-bromo-4-cloro-3-indol
fosfato e o nitro azul tetrazólio (BCIP/NBT) por 10 minutos na ausência da luz. A fosfatase
alcalina reage com BCIP/NBT para formar um precipitado púrpura insolúvel. A reação foi
interrompida com a lavagem em água destilada. Após secagem à temperatura ambiente as
membranas foram guardadas envolvidas em papel alumínio.

37
5. RESULTADOS E DISCUSSÃO
5.1. Obtenção do cDNA da frutalina
RNA total extraído de sementes coletadas de frutos de A. incisa, em três diferentes
estágios de maturação, foi usado na reação de RT-PCR (Figura 3).
A reação de RT-PCR usando oligonucleotídeos específicos para a seqüência
codificadora da frutalina resultou na amplificação do cDNA da frutalina nos estágios 2 e 3
(Figura 4). Uma banda com aproximadamente 500 pb foi observada, resultado coerente com o
esperado, pois a seqüência codificadora possui 459 pb. A ausência de amplificação do cDNA
da frutalina no estágio 1, provavelmente foi devido ao fruto ser imaturo e ainda não está
havendo a expressão do gene. Um aumento na intensidade da banda foi detectado entre os
estágios 2 e 3. Esse resultado sugere que o amadurecimento do fruto aumenta a expressão do
gene da frutalina.
5.2. Clonagem do cDNA da frutalina
O cDNA da frutalina dos estágios 2 e 3 de maturação amplificados na reação de
RT-PCR foram inseridos no vetor de clonagem pGEM-T Easy Vector (pGEM-T::FTL-E2 e
pGEM-T::FTL-E3). A seleção de colônias de E. coli transformadas com os plasmídeos
pGEM-T::FTL- E2 e pGEM-T::FTL-E3 em meio LB suplementado com carbenicilina 100
g/mL, estreptomicina 30 g/mL, X-Gal 80 g/mL e IPTG 0,5 mM baseou-se no fato destes
plasmídeos apresentarem a seqüência do gene LacZ que codifica para a enzima -
galactosidase. Essa enzima hidroliza X-Gal (5-bromo-4-cloro-3-indoil--D-
galactopiranosídeo) em galactose e 4-cloro-3-bromo-indigo, que forma um precipitado de cor
azul. Como o sítio múltiplo de clonagem (local de inserção do fragmento de DNA exógeno)
do plasmídeo pGEM-T ocupa a região do gene que codifica a porção N-terminal da -
galactosidase, e o fragmento de DNA clonado foi inserido nesta região, as bactérias com

38
Eletroforese em gel de agarose 1,2% do RNA total extraído de sementes de Artocarpus incisa nos estágios de maturação 1 (poço 2), 2 (poço 3) e 3 (poço 4).Foram aplicados 5 L nos poços 2 a 4. O poço 1 contém 250 ng do marcador X174 RF DNA/Hae III Fragments (Invitrogen).
pb
1353 1078 872 603
310
1 2 3 4
Figura 3 -
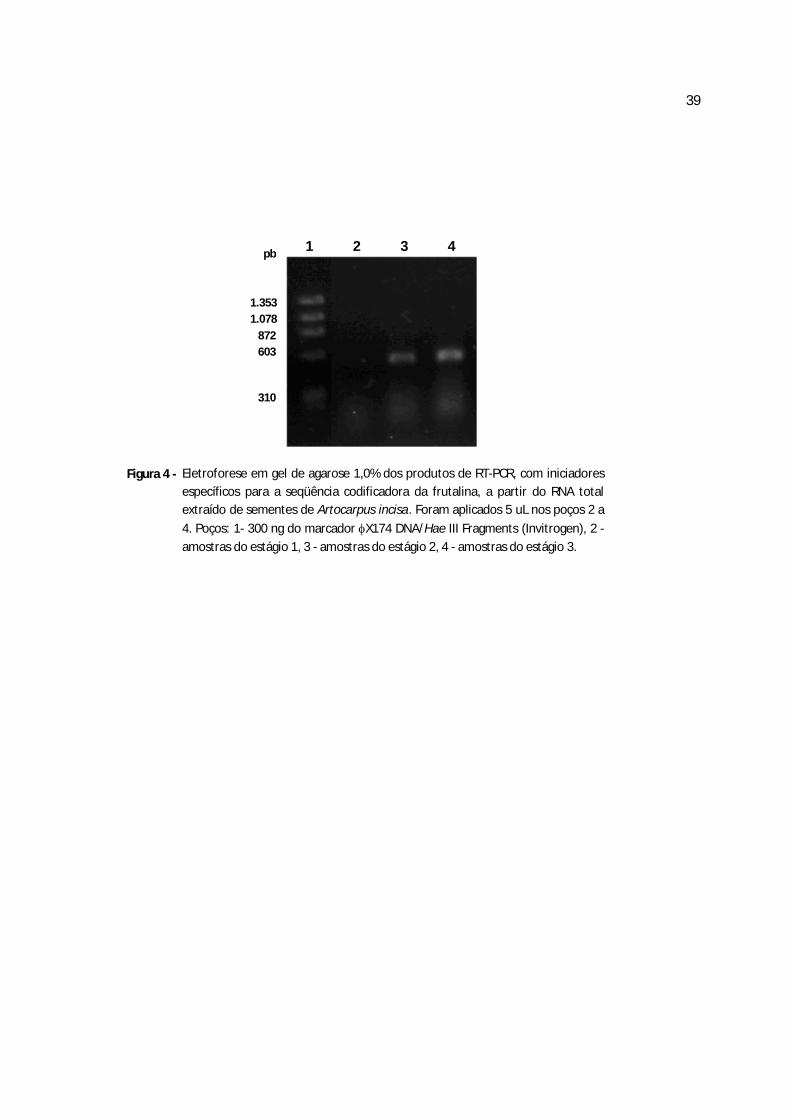
39
1 2 3 4 pb
1.3531.078 872 603
310
Eletroforese em gel de agarose 1,0% dos produtos de RT-PCR, com iniciadores específicos para a seqüência codificadora da frutalina, a partir do RNA total extraído de sementes de Artocarpus incisa. Foram aplicados 5 uL nos poços 2 a 4. Poços: 1- 300 ng do marcador X174 DNA/Hae III Fragments (Invitrogen), 2 -amostras do estágio 1, 3 - amostras do estágio 2, 4 - amostras do estágio 3.
Figura 4 -

40
plasmídeos recombinantes, incapazes de produzir a -galactosidase funcional, formaram
colônias brancas, permitindo assim uma seleção inicial dos clones recombinantes. A
visualização das colônias foi feita após 24 horas do inóculo. No tratamento controle não foram
observadas colônias em meio seletivo. A figura 5 apresenta uma placa contendo colônias
brancas (transformadas com o plasmídeo recombinante) e azuis (transformadas com o
plasmídeo íntegro).
Doze colônias brancas de E. coli transformadas com os vetores de clonagem
pGEM-T::FTL-E2 e pGEM-T::FTL-E3 foram selecionadas para a extração dos plasmídeos e
confirmação da transformação. A Reação de PCR resultou na amplificação do gene da
frutalina em dez clones E2 (E2.2, E2.3, E2.4, E2.5, E2.6, E2.7, E2.8, E2.9, E2.11 e E2.12) e
dois clones E3 (E3.5, e E3.6). A eletroforese do fragmento amplificado revelou a presença de
uma banda específica com o tamanho esperado, cerca de 459 pb, confirmando a eficiência da
transformação de 12 clones de E. coli DH10B (Figura 6).
Os transformantes confirmados por PCR foram usados no preparo de culturas
estoques de E. coli em glicerol mantidas a -80 C e de culturas em ágar em camada alta
mantidas a 4 C.
5.3. Sequenciamento de DNA
Os doze clones que tiveram sua transformação confirmada por PCR foram
seqüenciados. A seqüência de nucleotídeos de cinco clones foi obtida. Os clones E2.3, E2.4,
E2.6, E2.8, E2.9, E2.12 e E3.6 não apresentaram seqüências de qualidade para se obter uma
seqüência consenso. A Figura 7 mostra o alinhamento das seqüências de nucleotídeos dos
clones E2.2, E2.5, E2.7, E2.11, e E3.5, juntamente com seis clones do estágio 4 de maturação
(E4), que foram seqüenciados e seus dados cedidos por Felix (2008). A seqüência de
aminoácidos correspondente a seqüência da frutalina nos clones está representada na Figura 8.

41
Colônias brancas e azuis em meio LB ágar (suplementado com carbenicilina 100 g/mL, estreptomicina 30 g/mL, X-Gal 80 g/mL e IPTG 0,5 mM) após 16 horas do inóculo de células eletrocompetentes de Escherichia coli DH10B eletroporadas com o vetor pGEM-T::FTL.
Figura 5 -

42
E2 E3
Eletroforese em gel de agarose 1,0% dos produtos de PCR, com iniciadores específicos para a seqüência codificadora da frutalina, a partir dos plasmídeos pGEM-T::FTL. Os poços 2 a 13 contêm 5 uL dos produtos amplificados do gene da frutalina de 12 clones do 2° estágio (E2). Os poços 15 a 26 contêm 5 uL dos produtos amplificados do gene da frutalina de 12 clones do 3° estágio (E3). Os poços 1 e 14 contêm 300 ng do marcador X174 DNA/Hae III Fragments (Invitrogen).
Figura 6 -
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 pb
1.3531.078 872 603
310

43
cloneE2.2 1 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE2.5 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE2.7 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE2.11 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE3.5 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE4.1 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE4.2 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE4.3 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCcloneE4.4 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCCAAGTAAGCcloneE4.5 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCCAAGTAAGCcloneE4.6 AACGAACAAAGCGGGAAGAGCCAGACTGTAATAGTAGGACCATGGGGAGCCAAAGTAAGCARPJACA AACAAACAAAGCGGGAAGAGCCAGACCGTAATAGTAGGACCATGGGGAGCCCAAGTAAGC *** ********************** ************************ ********
cloneE2.2 61 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE2.5 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE2.7 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE2.11 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE3.5 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE4.1 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE4.2 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE4.3 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE4.4 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE4.5 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACcloneE4.6 ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAACARPJACA ACTAGCTCCAATGGTAAAGCTTTTGATGACGGTGCATTCACCGGAATCAGAGAAATCAAC ************************************************************
cloneE2.2 121 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE2.5 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE2.7 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE2.11 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE3.5 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE4.1 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE4.2 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATTTACGACTTGAATGGAcloneE4.3 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAcloneE4.4 CTCTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATTTACGACTTGAATGGAcloneE4.5 CTCTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATTTACGACTTGAATGGAcloneE4.6 CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATCTACGACTTGAATGGAARPJACA CTTTCATATAATAAGGAGACCGCCATTGGGGACTTCCAAGTTATTTACGACTTGAATGGA ** ***************************************** ***************
cloneE2.2 181 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGGAGATTcloneE2.5 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE2.7 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE2.11 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE3.5 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE4.1 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE4.2 AGGCCATTTGTTGGACAAAATCATACTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE4.3 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE4.4 AGGCCATTTGTTGGACAAAATCATACTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE4.5 AGGCCTTTTGTTGGACAAAATCATACTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTcloneE4.6 TCGCCATATGTTGGACAAAATCATGCTAGTTTTATAAAAGGCTTCACACCAGTGAAGATTARPJACA TCGCCATATGTAGGACAAAATCATACAAGTTTTATAAAAGGCTTCACACCAGTGAAGATT *** * *** ************ * *************************** *****

44
cloneE2.2 241 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE2.5 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE2.7 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE2.11 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE3.5 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE4.1 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE4.2 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE4.3 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTcloneE4.4 GCTCTAGACTTTCCAAGCGAGTATATTATAGAAGTGAGCGGGCACACTGGTAAAGTGAGTcloneE4.5 GCTCTAGACTTTCCAAGCGAGTATATTATAGAAGTGAGCGGGCACACTGGTAAAGTGAGTcloneE4.6 TCTCTAGACTTTCCAAGCGAGTATATAGTAGAAGTGAGCGGACACACTGGTAAAGTGAGTARPJACA TCTCTAGACTTTCCAAGCGAGTATATAGTAGATGTAAGCGGGTACACTGGTAAGGTGAGC ************************* **** ** ***** ********** *****
cloneE2.2 301 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE2.5 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE2.7 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE2.11 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE3.5 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE4.1 GGGTATGTAGTAGTACGCCCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE4.2 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE4.3 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATcloneE4.4 GGGTATGTAGTAGTACGCTCTTTGGCATTCAAGACTAATAAGAAAACTTATGGACCATATcloneE4.5 GGGTATGTAGTAGTACGCTCTTTGGCATTCAAGACTAATAAGAAAACTTATGGACCATATcloneE4.6 GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAAAAAACCTATGGACCATATARPJACA GGGTATGTAGTAGTACGCTCTTTGACATTCAAGACTAATAAGAAAACCTATGGACCATAT ****************** ***** **************** ***** ************
cloneE2.2 361 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE2.5 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE2.7 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE2.11 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE3.5 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE4.1 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE4.2 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE4.3 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCcloneE4.4 GGAGTTACAAGCGGCACACCTTTCAATCTCCCAATCGAAAATGGCTTAATTGTTGGATTCcloneE4.5 GGAGTTACAAGCGGCACACCTTTCAATCTCCCAATCGAAAATGGCTTAATTGTTGGATTCcloneE4.6 GGAGTTACAAGCGGCACACCTTTCAACCTTCCAATCGAGAATGGCTTAGTTGTTGGATTCARPJACA GGAGTTACAAGTGGTACACCTTTCAATCTCCCAATCGAAAATGGCTTAGTTGTTGGATTC *********** ** *********** ** ******** ********* ***********
cloneE2.2 421 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE2.5 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE2.7 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE2.11 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE3.5 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE4.1 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE4.2 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE4.3 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTcloneE4.4 AAAGGAAGTATCGGCTACTGGTTGGACTACTTTAGTATGTACTTGTCCCTTcloneE4.5 AAAGGAAGTATCGGCTACTGGTTGGACTACTTTAGTATGTACTTGTCCCTTcloneE4.6 AAAGGAAGTATCGGCTACTGGATGGACTACTTTAGCATGTACTTGTCCCTTARPJACA AAAGGAAGTATCGGCTACTGGTTGGACTACTTTAGTATGTACTTGTCCCTT ********************* ************* ***************
Alinhamento das seqüências de nucleotídeos dos clones pGEM-T::FTL-E2, pGEM-T::FTL-E3 e pCR4-TOPO::FTL-E4 obtidos. ARPJACA representa a seqüência da jacalina. O primeiro códon transcrito está marcado de amarelo.
Figura 7 -

45
cloneE2.2 1 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE2.5 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE2.7 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE2.11 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE3.5 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE4.1 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE4.2 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE4.3 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE4.4 NEQSGKSQTVIVGPWGAQVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE4.5 NEQSGKSQTVIVGPWGAQVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGcloneE4.6 NEQSGKSQTVIVGPWGAKVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNGARPJACA NKQSGKSQTVIVGPWGAQVSTSSNGKAFDDGAFTGIREINLSYNKETAIGDFQVIYDLNG *:***************:******************************************
cloneE2.2 61 SPYVGQNHASFIKGFTPVEISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE2.5 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE2.7 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE2.11 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE3.5 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE4.1 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRPLTFKTNKKTYGPYcloneE4.2 RPFVGQNHTSFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE4.3 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYcloneE4.4 RPFVGQNHTSFIKGFTPVKIALDFPSEYIIEVSGHTGKVSGYVVVRSLAFKTNKKTYGPYcloneE4.5 RPFVGQNHTSFIKGFTPVKIALDFPSEYIIEVSGHTGKVSGYVVVRSLAFKTNKKTYGPYcloneE4.6 SPYVGQNHASFIKGFTPVKISLDFPSEYIVEVSGHTGKVSGYVVVRSLTFKTNKKTYGPYARPJACA SPYVGQNHTSFIKGFTPVKISLDFPSEYIVDVSGYTGKVSGYVVVRSLTFKTNKKTYGPY *:*****:*********:*:********::***:***********.*:***********
cloneE2.2 121 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE2.5 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE2.7 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE2.11 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE3.5 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE4.1 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE4.2 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE4.3 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLcloneE4.4 GVTSGTPFNLPIENGLIVGFKGSIGYWLDYFSMYLSLcloneE4.5 GVTSGTPFNLPIENGLIVGFKGSIGYWLDYFSMYLSLcloneE4.6 GVTSGTPFNLPIENGLVVGFKGSIGYWMDYFSMYLSLARPJACA GVTSGTPFNLPIENGLVVGFKGSIGYWLDYFSMYLSL ****************:**********:*********
Alinhamento da seqüência de aminoácidos correspondente a seqüência da frutalina nos clones pGEM-T::FTL-E2, pGEM-T::FTL-E3 e pCR4-TOPO::FTL-E4 obtidos. ARPJACA representa a seqüência da jacalina. Os resíduos marcados de: (cinza) – cadeia beta; (vermelho) – peptídeo de ligação; (amarelo) – fazem parte do sítio de ligação a carboidratos. Após o peptídeo de ligação, os demais resíduos constituem a cadeia alfa.
Figura 8 -

46
5.4. Modelagem por homologia
Foram obtidos cinco modelos diferentes da possível estrutura tridimensional da
frutalina por meio da modelagem por homologia. Dos sete clones com seqüências de
qualidade obtidos, três possuem a mesma seqüência (clone 2.2, clone 3.1 e clone 4.1), o que
gerou o mesmo modelo para esses três clones. Foram gerados modelos dos três diferentes
estágios de maturação:
1- clone E2.1 (modelo obtido do 2º estágio de maturação);
2- clone E2.2, clone E3.1, clone E4.1 (modelo obtido do 2º, 3° e 4° estágio de
maturação);
3- clone E4.2 (modelo obtido do 4º estágio de maturação);
4- clone E4.3 (modelo obtido do 4º estágio de maturação);
5- clone E4.4 (modelo obtido do 4º estágio de maturação);
A proteína molde escolhida para a modelagem da frutalina foi realizada de acordo
com o critério padrão de preferência pela estrutura disponível de maior grau de identidade.
Para tal, a seqüência de aminoácidos deduzida da frutalina foi comparada ao banco de dados
de estruturas resolvidas (Protein Data bank – pdb). A lectina galactose ligante de Artocarpus
integrifolia (jacalina) foi a que apresentou uma maior identidade com a frutalina (98%), por
isso foi utilizada como proteína molde na obtenção dos modelos da estrutura tridimensional da
frutalina.
A ferramenta mais bem sucedida de predição de estruturas tridimensionais de
proteínas é a modelagem por homologia, também conhecida como modelagem comparativa.
Esta abordagem baseia-se em alguns padrões gerais que têm sido observados, em nível
molecular, no processo de evolução biológica: (1) homologia entre seqüências de aminoácidos
implica em semelhança estrutural e funcional; (2) proteínas homólogas apresentam regiões
internas conservadas (principalmente constituídas de elementos de estrutura secundária:
hélices- e fitas-); (3) as principais diferenças estruturais entre proteínas homólogas ocorrem
nas regiões externas, constituídas principalmente por alças (“loops”), que ligam os elementos
externos de estruturas secundárias (FILHO; ALENCASTRO, 2003).

47
A Figura 9 apresenta os modelos gerados da jacalina e da frutalina (clone E2.1)
evidenciando as quatro subunidades que formam o tetrâmero. A principal diferença entre os
modelos está nas alças da região central, pois o peptídeo de ligação de quatro resíduos faz
parte desta região. Esta diferença foi devido à estrutura tridimensional da jacalina ter sido
obtida com a proteína nativa, sem o peptídeo de ligação, e o modelo gerado da frutalina possui
esse peptídeo.
A estrutura tridimensional da jacalina foi resolvida por análises de difração de
raios-X (SANKARANARAYANAN et al.,1996). Cada protômero consiste de um β-prisma
triplo simétrico, constituído a partir de três folhas β de quatro fitas. Das doze fitas, onze são
formadas pela cadeia alfa e apenas uma pela cadeia beta. Quatro subunidades estão associadas
não covalentemente formando uma estrutura tetramérica. As cadeias betas ocupam a região
central do tetrâmero e desempenham um papel crucial na associação das quatro subunidades.
Os resíduos que fazem parte do sítio de ligação a carboidratos na jacalina estão
evidenciados na Figura 10. Cada protômero possui um único sitio ligante de carboidrato
formado por Gly1, Tyr122, Trp123 e Asp125 da cadeia alfa que cria um sistema complexo de
nove ligações de hidrogênio com O3, O4, O5 e O6 da galactose (HOULÈS ASTOUL et al.,
2002). Nos clones obtidos para a frutalina, não foi observada nenhuma mutação nestes
resíduos, indicando que os mesmos permanecem conservados nesta proteína.
A Figura 11 apresenta um alinhamento das estruturas tridimensionais da jacalina
com os cinco modelos dos clones obtidos. Devido ao alto grau de identidade entre estas duas
proteínas, as estruturas tridimensionais das mesmas são bastante semelhantes.
As estruturas dos modelos gerados para todos os clones da frutalina apresentaram
diferenças em alguns resíduos em comparação com a jacalina. Em todos os clones obtidos, o
2º resíduo da cadeia beta é glutamato, enquanto que na jacalina é uma lisina. Essa diferença
deve acarrretar alguma modificação estrutural entre as duas proteínas, pois estes dois resíduos
possuem características bioquímicas diferentes, o glutamato tem o grupo lateral ácido e a
lisina tem o grupo lateral básico. As outras diferenças são: na posição 91, um glutamato nos
clones da frutalina por um aspartato na jacalina, e na posição 95, uma histidina nos clones da
frutalina para uma tirosina na jacalina. No primeiro caso, glutamato e aspartato possuem
características bioquímicas semelhantes.
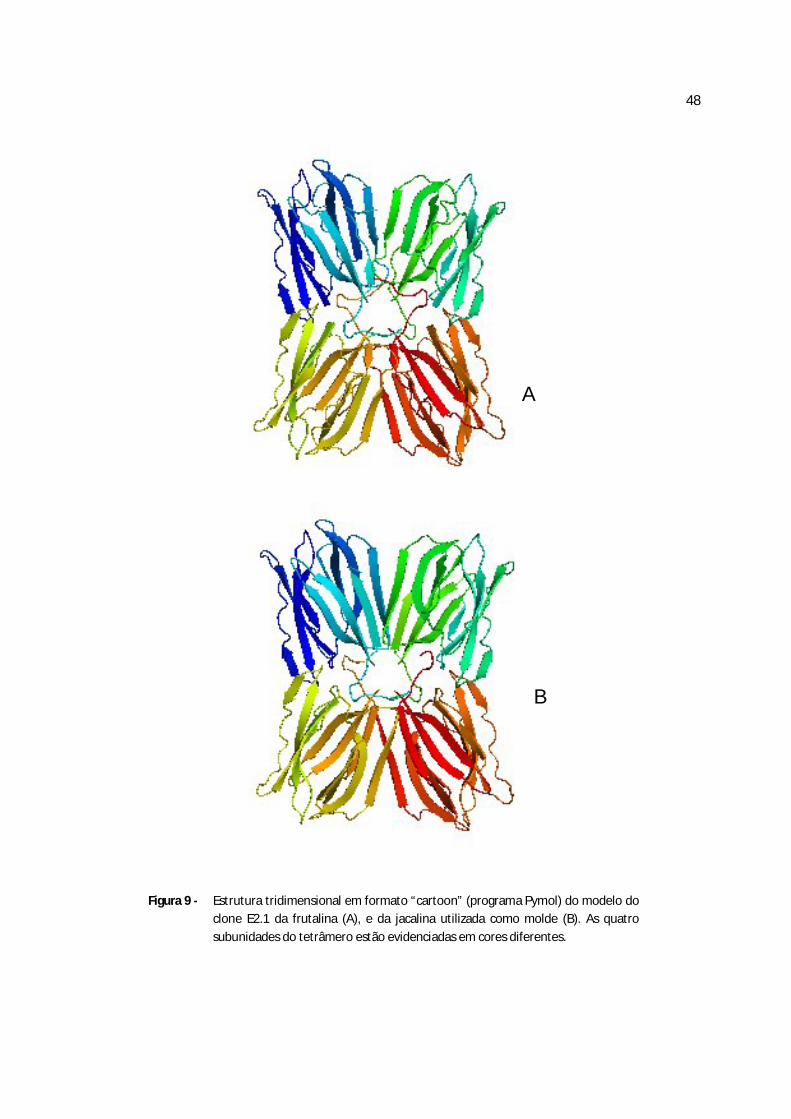
48
A
B
Estrutura tridimensional em formato “cartoon” (programa Pymol) do modelo do clone E2.1 da frutalina (A), e da jacalina utilizada como molde (B). As quatro subunidades do tetrâmero estão evidenciadas em cores diferentes.
Figura 9 -

49
Estrutura tridimensional em formato “ribbon” (programa Pymol) do modelo da jacalina, utilizada como molde. Os resíduos que fazem parte do sítio de ligação a carboidratos estão identificados em uma das subunidades.
Figura 10 -
Tyr 122
Gly 1Asp 125
Trp 123

50
Estrutura tridimensional em formato “ribbon” (programa Pymol) dos modelos dos clones E2.1(azul); E2.2, E3.1, E4.1 (rosa), E4.2 (amarelo); E4.3 (vermelho) e E4.4 (laranja), da frutalina, e da jacalina, utilizada como molde para gerar os modelos (verde).
Figura 11 -

51
Como estas diferenças nos resíduos ocorreram em todos os clones obtidos, esse
resultado sugere que estes três resíduos representam diferenças entre as duas proteínas
analisadas.
Para analisar as mutações ocorridas, foram observadas as diferenças de resíduos
entre os clones da frutalina obtidos. Nas Figuras 12 a 16 estão evidenciados os resíduos
diferentes entre os cinco clones. Ao analisar as figuras de cada clone, podemos observar que
na posição 18 apenas o clone E4.4 possui um glutamina, enquanto que os outros clones
possuem uma lisina. Os clones E4.3 e E4.4 possuem na posição 61 uma serina e os outro
clones uma arginina, e na posição 63 uma tirosina e os outros uma fenilalanina. Essa última
mutação evolutiva provavelmente não acarretou alterações na atividade lectínica, pois ambos
os aminoácidos apresentam o anel aromático na sua cadeia lateral, preservando suas
propriedades energéticas da região do sítio ativo.
Os clones E2.1; E2.2, E3.1, E4.1 e E4.2 possuem na posição 69 uma treonina,
enquanto os outros, uma alanina. Para a posição 79, apenas o clone E2.1 possui um glutamato
ao invés de uma lisina, presente nos outros clones. Essa diferença deve causar alguma
modificação estrutural neste clone, pois estes dois resíduos possuem características
bioquímicas diferentes, o glutamato tem o grupo lateral ácido e a lisina tem o grupo lateral
básico.
Algumas mutações foram observadas apenas no clone E4.4, como: troca de uma
serina por uma alanina na posição 81, troca de uma valina por uma isoleucina na posição 90,
troca de uma treonina por uma alanina na posição 109, troca de uma valina por uma isoleucina
na posição 137 e troca de uma metionina por uma leucina na posição 148.
Mutações de resíduos com grupos de cadeias laterais semelhantes devem acarretar
uma menor influência na estrutura e na atividade das proteínas do que mutações de resíduos
com características bioquímicas diferentes, pois as mesmas influenciam diretamente nos tipos
de interações que serão formadas.
O clone E4.2 possui uma mutação na posição 107, que é troca de uma serina por
uma prolina. O grupo amino secundário (imino) dos resíduos de prolina é mantido em uma
conformação rígida que leva à redução na flexibilidade estrutural de regiões polipeptídicas que
contêm prolina (NELSON; COX, 2002).
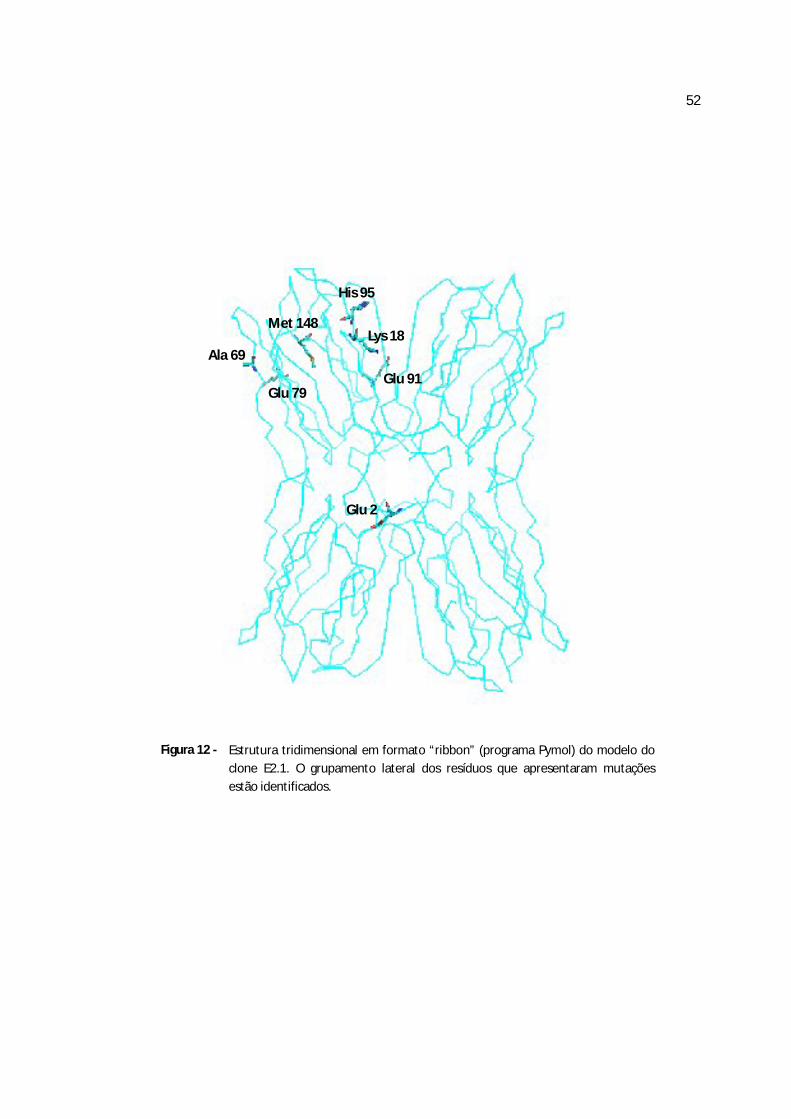
52
Estrutura tridimensional em formato “ribbon” (programa Pymol) do modelo do clone E2.1. O grupamento lateral dos resíduos que apresentaram mutações estão identificados.
Figura 12 -
Glu 2
Lys 18Ala 69
Glu 91Glu 79
Met 148
His 95
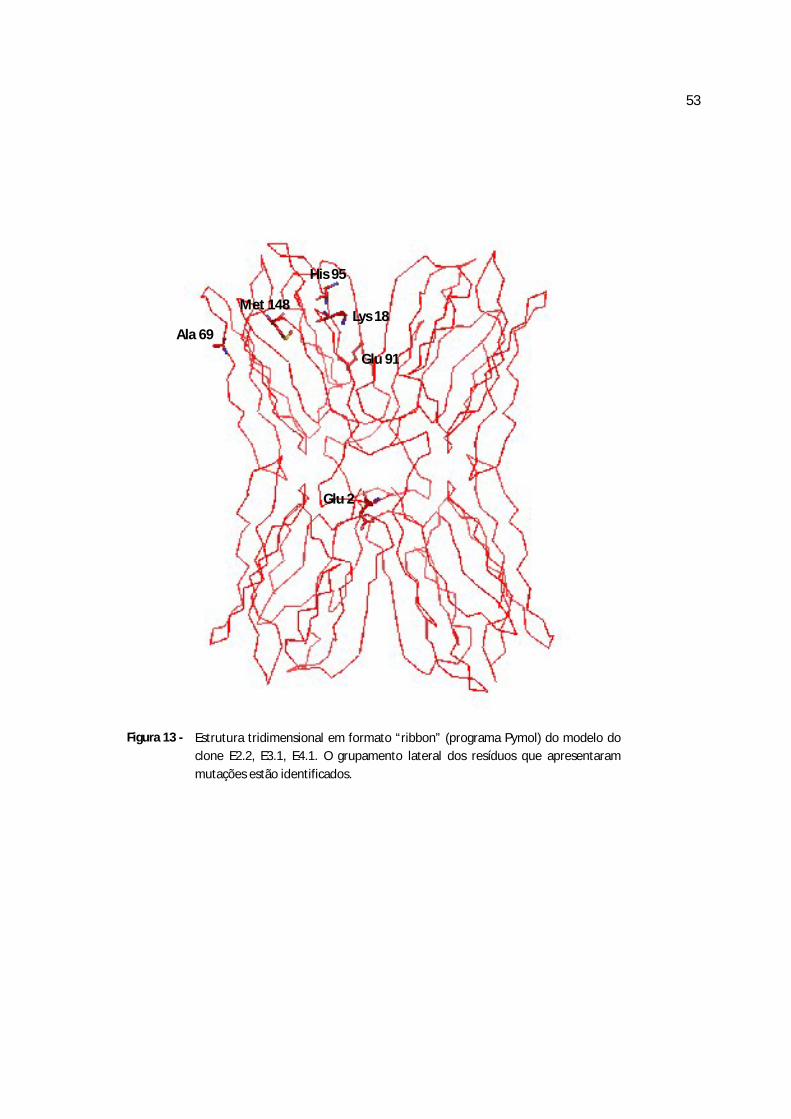
53
Estrutura tridimensional em formato “ribbon” (programa Pymol) do modelo do clone E2.2, E3.1, E4.1. O grupamento lateral dos resíduos que apresentaram mutações estão identificados.
Figura 13 -
Glu 2
Lys 18Ala 69
Glu 91
Met 148
His 95
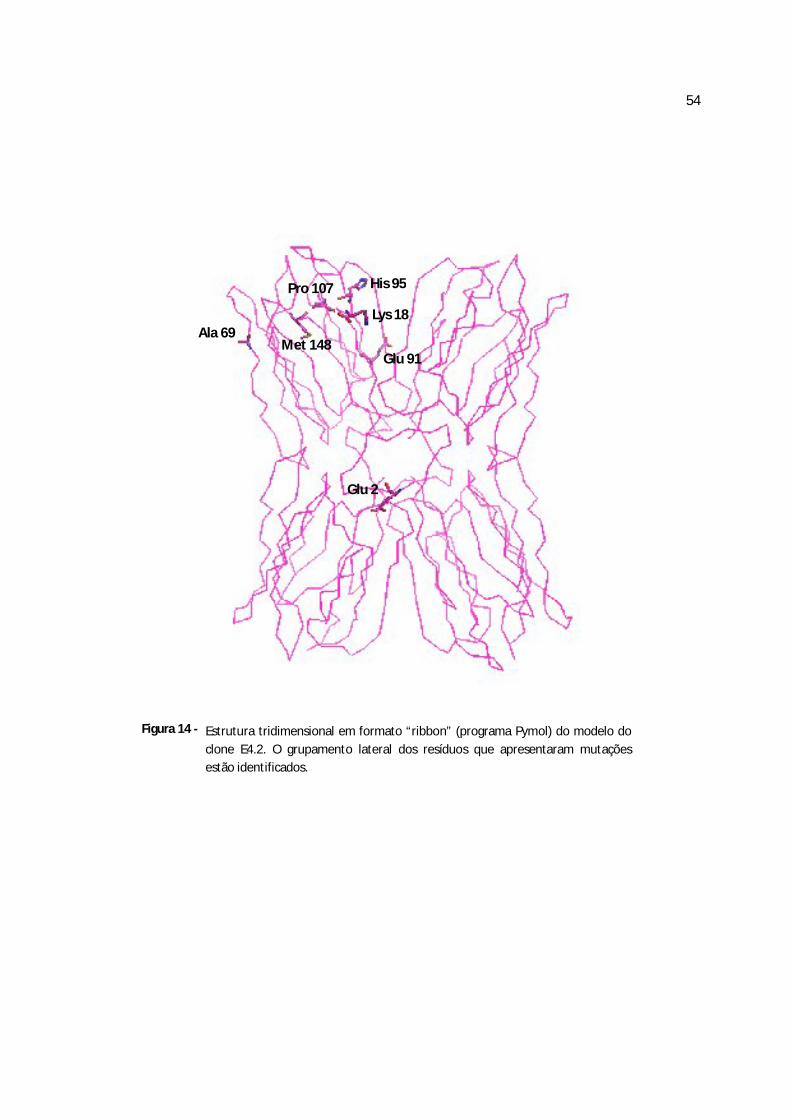
54
Estrutura tridimensional em formato “ribbon” (programa Pymol) do modelo do clone E4.2. O grupamento lateral dos resíduos que apresentaram mutações estão identificados.
Figura 14 -
Glu 2
Lys 18Ala 69
Glu 91Met 148
His 95Pro 107

55
Estrutura tridimensional em formato “ribbon” (programa Pymol) do modelo do clone E4.3. O grupamento lateral dos resíduos que apresentaram mutações estão identificados.
Figura 15 -
Glu 2
Lys 18
Arg 61
Glu 91
Phe 63
Met 148
His 95
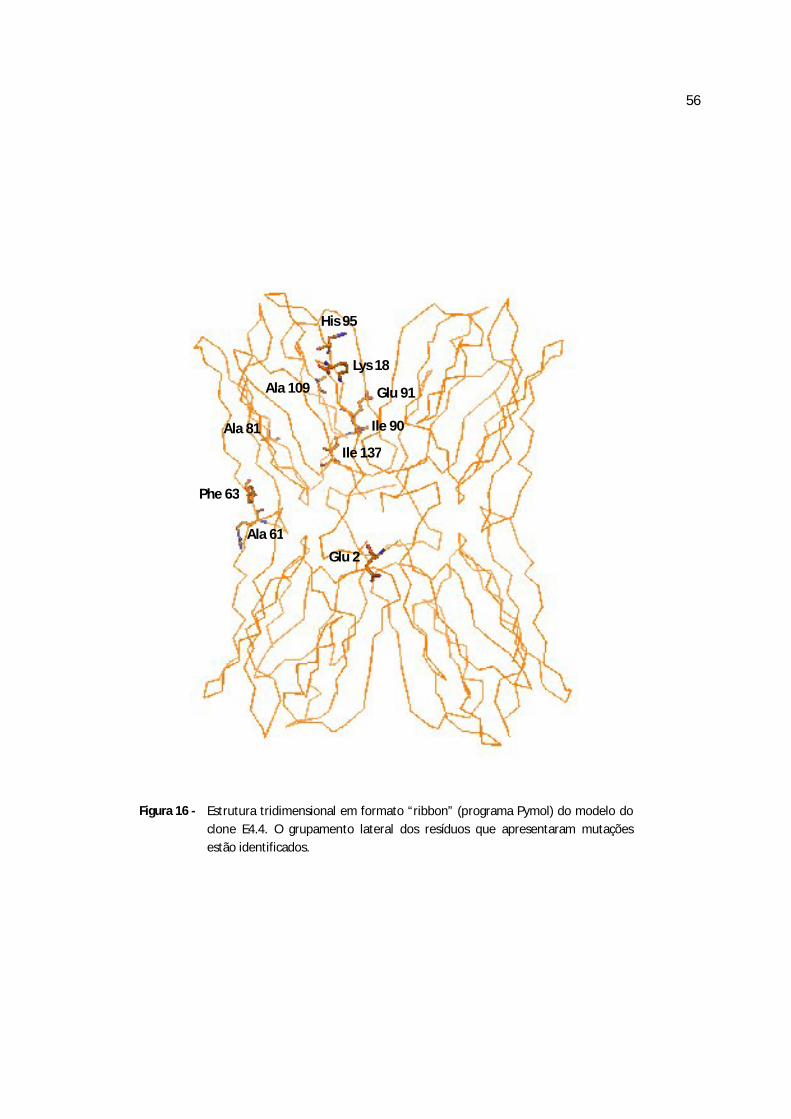
56
Estrutura tridimensional em formato “ribbon” (programa Pymol) do modelo do clone E4.4. O grupamento lateral dos resíduos que apresentaram mutações estão identificados.
Figura 16 -
Glu 2
Ala 61
Glu 91
Phe 63
Ala 81
His 95
Ala 109
Ile 137
Ile 90
Lys 18

57
A expressão da seqüência codificadora da frutalina foi realizada com este clone. A influência
desta mutação na atividade da proteína poderá ser avaliada após a purificação da frutalina
recombinante.
As diferenças funcionais entre membros de uma mesma família protéica são, em
geral, conseqüência de diferenças estruturais na superfície externa das proteínas. Estas
diferenças provêm de substituições, eliminações e inserções de resíduos nas cadeias de
proteínas homólogas, principalmente nas alças, as regiões mais expostas da proteína. Por
serem regiões estruturalmente variáveis, as alças geralmente determinam a especificidade das
proteínas, contribuindo para a estrutura dos sítios de ligação (FILHO; ALENCASTRO, 2003).
Ao analisar os clones da frutalina, podemos observar que algumas mutações se
repetem entre os clones. Esse resultado pode ser uma evidência da existência de isoformas da
frutalina nativa.
5.4. Expressão da rFrutalina em E. coli
5.4.1. Subclonagem da seqüência codificadora da frutalina
A seqüência codificadora da frutalina do estágio 4 de maturação (E4), previamente
clonada no vetor pCR4-TOPO, foi subclonada no vetor pGEM-T.
Para tanto, uma extração de plasmídeos foi realizada a partir das culturas E.coli Mach
1-T1. Os plasmídeos pCR4-TOPO::FTL-E4 foram submetidos a uma reação de amplificação
por PCR com iniciadores desenhados com sítio de restrição para XhoI e BamHI.
O plasmídeo recombinante pGEM-T::FTL-E4 foi utilizado para transformar células
eletrocompetentes de E. coli TOP10. Doze colônias de E. coli transformadas com os vetores
de clonagem pGEM-T::FTL-E4 foram selecionadas para a extração de plasmídeos e
confirmação da transformação. A reação de digestão com as enzimas de restrição BamHI e
XhoI realizada com três clones resultou na presença de duas bandas com o tamanho esperado,
uma com cerca de 3.600 pb, correspondente ao vetor pGEM-T, e outra com cerca de 459 pb,
correspondente ao inserto, para dois dos três clones selecionados (Figura 17). Essa eletroforese
confirmou a eficiência da transformação de dois clones. A banda referente ao inserto foi

58
Eletroforese em gel de agarose 0,8% dos produtos da digestão de plasmídeos pGEM-T::FTL-E4purificados de Escherichia coli TOP10 com as enzimas de restrição BamHI e XhoI. Os poços 2 a 4 contêm 10 L das reações dos clones 4, 11 e 12, respectivamente. O poço 1 contém 250 ng do marcador 250 ng do marcador DNA/Hind III Fragments (Invitrogen). O poço 5 contém 250 ng do marcador X174 DNA/Hae III Fragments (Invitrogen). As setas indicam as bandas referentes ao plasmídeo e ao inserto da frutalina.
Figura 17 -
1.3531.078 872 603
310
pb
23.130 9.416 6.557 4.361
2.322 2.027
1 2 3 4 5
3600 pb (pGEM-T)
459 pb

59
cortada do gel, purificada e ligada ao vetor de expressão pET15b.
O plasmídeo recombinante pET15b::FTL-E4 foi utilizado para transformar células
eletrocompetentes de E. coli TOP10. Doze colônias de E. coli TOP10 transformadas com os
vetores de expressão pET15b::FTL-E4 foram selecionadas para a extração de plasmídeos. A
transformação foi confirmada por meio de três reações de digestão, a primeira com a enzima
de restrição XhoI, a segunda com a enzima de restrição BamHI e a terceira com essas duas
enzimas juntas (Figuras 18 e 19). A digestão do pET15b íntegro foi realizada como controle
da reação.
A reação de digestão com as duas enzimas juntas foi realizada com dois clones e
resultou na presença de duas bandas com o tamanho esperado, uma com cerca de 5.700 pb,
correspondente ao plasmídeo pET15b, e outra com cerca de 459 pb, correspondente ao inserto,
para um dos dois clones selecionados. A eficiência da transformação de um clone foi
confirmada na eletroforese.
5.4.2 Transformação de células de E. coli Origami (DE3) com pET-15b::FTL-E4
O plasmídeo recombinante pET-15b::FTL-E4 foi utilizado para transformar
células eletrocompetentes de E. coli Origami (DE3). Dez colônias de E. coli transformadas
com o plasmídeo pET15b íntegro e pET-15b::FTL-E4 foram selecionadas para a extração de
plasmídeos e confirmação da transformação.
A Reação de PCR resultou na amplificação do gene da frutalina em oito clones. A
eletroforese do fragmento amplificado revelou a presença de uma banda específica com o
tamanho esperado, cerca de 459 pb, confirmando a eficiência da transformação de oito clones
de E. coli Origami (DE3) (Figura 20). O plasmídeo pET15b íntegro foi utilizado como
controle na reação. Os clones confirmados por PCR foram mantidos em estoque em glicerol (-
80 C).
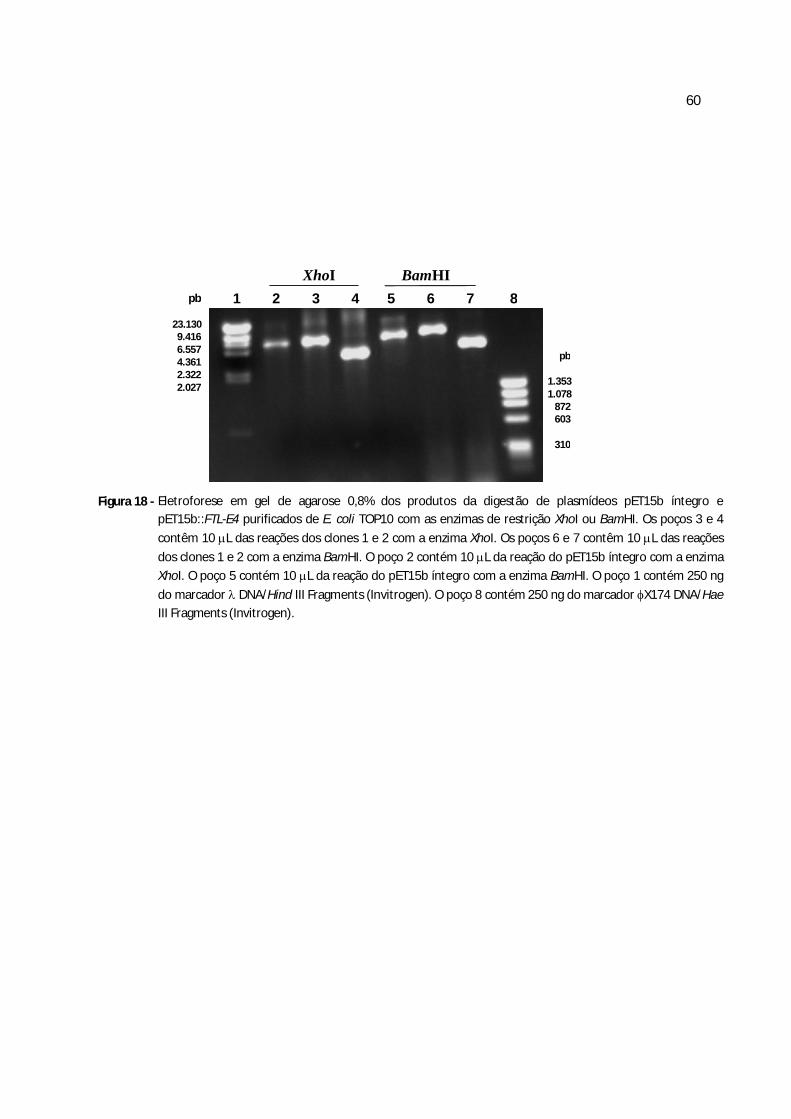
60
Eletroforese em gel de agarose 0,8% dos produtos da digestão de plasmídeos pET15b íntegro e pET15b::FTL-E4 purificados de E. coli TOP10 com as enzimas de restrição XhoI ou BamHI. Os poços 3 e 4 contêm 10 L das reações dos clones 1 e 2 com a enzima XhoI. Os poços 6 e 7 contêm 10 L das reações dos clones 1 e 2 com a enzima BamHI. O poço 2 contém 10 L da reação do pET15b íntegro com a enzima XhoI. O poço 5 contém 10 L da reação do pET15b íntegro com a enzima BamHI. O poço 1 contém 250 ng do marcador DNA/Hind III Fragments (Invitrogen). O poço 8 contém 250 ng do marcador X174 DNA/Hae III Fragments (Invitrogen).
1 2 3 4 5 6 7 8
pb
1.3531.078 872 603
310
pb
23.130 9.416 6.557 4.361 2.322 2.027
Figura 18 -
XhoI BamHI
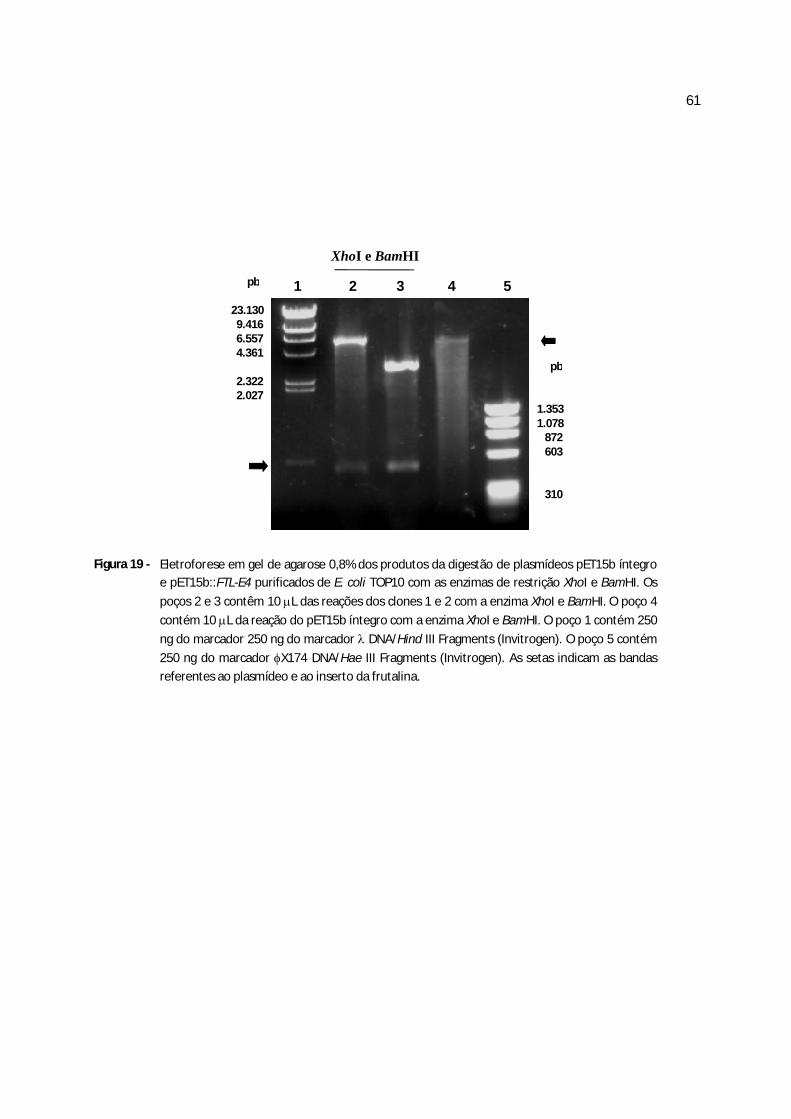
61
Eletroforese em gel de agarose 0,8% dos produtos da digestão de plasmídeos pET15b íntegroe pET15b::FTL-E4 purificados de E. coli TOP10 com as enzimas de restrição XhoI e BamHI. Os poços 2 e 3 contêm 10 L das reações dos clones 1 e 2 com a enzima XhoI e BamHI. O poço 4 contém 10 L da reação do pET15b íntegro com a enzima XhoI e BamHI. O poço 1 contém 250 ng do marcador 250 ng do marcador DNA/Hind III Fragments (Invitrogen). O poço 5 contém 250 ng do marcador X174 DNA/Hae III Fragments (Invitrogen). As setas indicam as bandas referentes ao plasmídeo e ao inserto da frutalina.
Figura 19 -
1 2 3 4 5
pb
1.3531.078 872 603
310
pb
23.130 9.416 6.557 4.361
2.322 2.027
XhoI e BamHI

62
Eletroforese em gel de agarose 1,0% dos produtos de PCR, com iniciadores específicos para o gene da frutalina, a partir dos plasmídeos pET15b íntegro e pET15b::FTL-E4. Os poços 2 e 3 contêm 5 uL dos produtos amplificados a partir do pET15b íntegro (controle). Os poços 4 a 12 contêm 5 uL dos produtos amplificados do gene da frutalina dos clones 1 a 8. O poço 13 contém 5 uL do controle negativo da reação. O poço 1 contém 300 ng do marcador X174 DNA/Hae III Fragments (Invitrogen).
Figura 20 -
pb
1.3531.078 872 603
1 2 3 4 5 6 7 8 9 10 11 12 13

63
5.4.3. Indução da expressão da rFrutalina em células de Escherichia coli
A expressão da Frutalina recombinante foi induzida pela adição de IPTG 1 mM
durante 10 horas. Foram realizadas quatro induções de 100 mL. A indução realizada com as
células de E. coli Origami (DE3) transformadas com o plasmídeo recombinante pET-
15b::FTL-E4 foi feita na presença e ausência de IPTG. A indução controle, com células de E.
coli Origami (DE3) transformadas com o plasmídeo pET-15b íntegro, também foi realizada na
presença e ausência de IPTG.
A Figura 21 apresenta a eletroforese em gel de poliacrilamida 17,5% do extrato
total de células e do sobrenadante da cultura de E. coli Origami (DE3) transformadas com o
plasmídeo recombinante pET-15b::FTL-E4 e induzidas com IPTG 1 mM durante 10 horas.
Nenhuma banda foi detectada no sobrenadante da cultura de E. coli, o que está de acordo com
o esperado, já que a Frutalina não possui nenhum sinal de secreção.
Uma proteína com massa molecular aparente de 19,2 kDa foi detectada a partir de
duas horas após a indução. A maior expressão da proteína foi atingida após oito horas da
indução das células com IPTG 1 mM, e a partir de dez horas foi observado um decréscimo na
intensidade da banda detectada. Esse polipeptídeo com massa molecular aparente superior a da
lectina nativa (15 kDa) foi detectado apenas na indução com as células de E. coli Origami
(DE3) transformadas com o plasmídeo recombinante pET-15b::FTL-E4 feita na presença de
IPTG. Na indução controle, realizada na ausência de IPTG no meio, nenhuma banda foi
detectada (Figura 22). A Figura 23 apresenta a eletroforese em gel de poliacrilamida 17,5%
das quatro induções realizadas.
Sahasrabuddhe e colaboradores, (2004), expressaram a lectina galactose ligante de
A. integrifolia (jacalina) em E. coli como cadeia única, formada pela cadeia beta com 20
resíduos, ligada a um peptídeo de ligação com quatro resíduos, e a cadeia alfa com 133
resíduos. A jacalina é uma glicoproteína e sua expressão como cadeia única foi o primeiro
relato da contribuição do peptídeo de ligação (T-S-S-N) no sítio ligante da jacalina. A proteína
recombinante de 19 kDa foi expressa com massa molecular superior a da lectina nativa (15
kDa). A jacalina recombinante não sofreu processamento proteolítico.

64
M 0h 2h 4h 6h 8h 10h 0h 2h 4h 6h 8h 10h
Eletroforese em gel de poliacrilamida 17,5 % em condições desnaturantes, na presença de SDS, do extrato total de células e do sobrenadante da cultura de E. coli Origami (DE3) transformadas com o plasmídeo recombinante pET-15b::FTL e induzidas com IPTG 1 mM durante 10 horas. A seta indica a banda referente à frutalina recombinante. Linha M: marcador; Linhas 0h, 2h, 4h, 8h e 10h representam os tempos de coleta da indução.
25.9 kDa
19.4 kDa
14.8 kDa
Figura 21 -
37.1 kDa
CÉLULAS MEIO

65
M 0h 2h 4h 6h 8h 10h 0h 2h 4h 6h 8h 10h
IPTG - IPTG +
Figura 22 - Eletroforese em gel de poliacrilamida 17,5 % em condições desnaturantes, na presença de SDS, do extrato total de células de E. coli Origami (DE3) transformadas com o plasmídeo recombinante pET-15b::FTL, induzidas (IPTG+) e não-induzidas (IPTG-) por 10 horas. A seta indica a banda referente à frutalina recombinante. Linha M: marcador; Linhas 0h, 2h, 4h, 8h e 10h representam os tempos de coleta da indução.
37.1 kDa
25.9 kDa
19.4 kDa
14.8 kDa
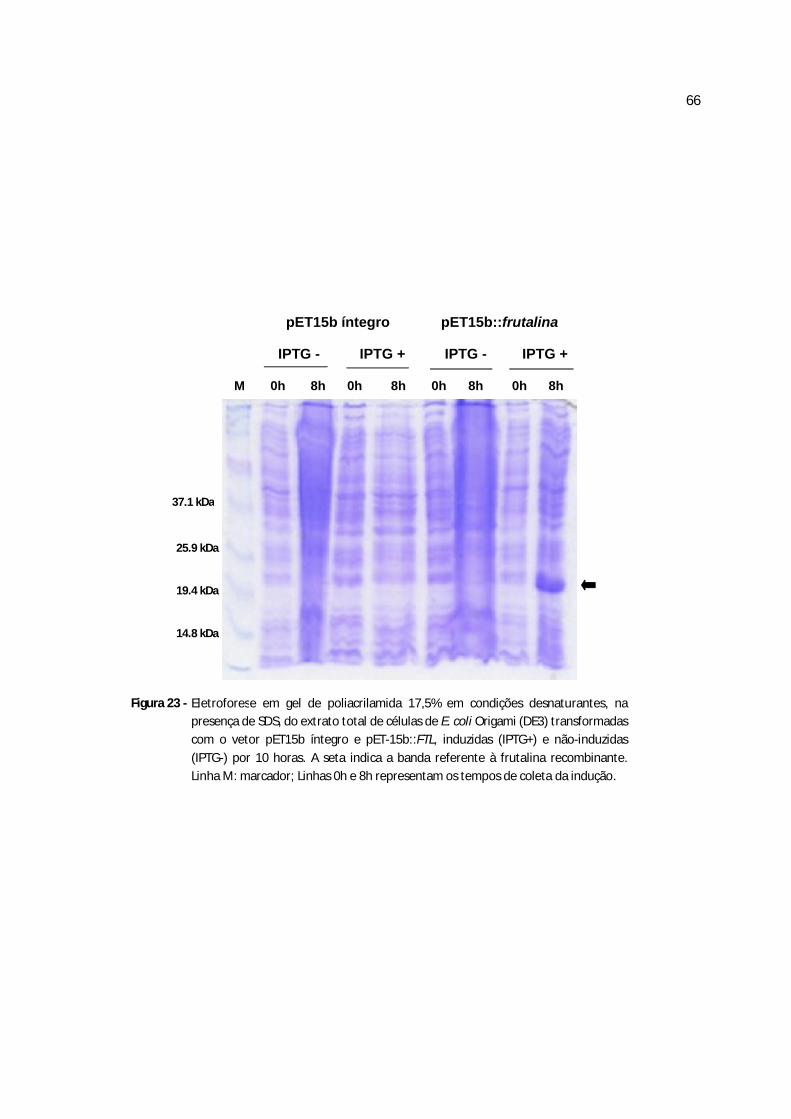
66
pET15b íntegro pET15b::frutalina
IPTG - IPTG + IPTG - IPTG +
M 0h 8h 0h 8h 0h 8h 0h 8h
Figura 23 - Eletroforese em gel de poliacrilamida 17,5% em condições desnaturantes, na presença de SDS, do extrato total de células de E. coli Origami (DE3) transformadas com o vetor pET15b íntegro e pET-15b::FTL, induzidas (IPTG+) e não-induzidas (IPTG-) por 10 horas. A seta indica a banda referente à frutalina recombinante. Linha M: marcador; Linhas 0h e 8h representam os tempos de coleta da indução.
37.1 kDa
25.9 kDa
19.4 kDa
14.8 kDa

67
Houlès Astoul e colaboradores (2002) sugerem que a clivagem proteolítica do
peptídeo de ligação é fundamental para a formação do sítio ligante da jacalina. Contudo, a
jacalina recombinante expressa com o peptídeo de ligação intacto apresentou maior
especificidade por galactose do que por manose, similarmente a jacalina nativa. Esses
resultados sugerem que a ligação da jacalina com monossacarídeos não é dependente da
excisão do tetrapeptídeo. Contudo, esse evento deve ser requerido para aumentar a afinidade,
como sugerido por Vijayan e colaboradores, (1999) (citado por SAHASRABUDDHE et al.,
2004).
A frutalina nativa apresenta duas bandas protéicas com massas moleculares aparentes
de 12 e 15 kDa quando submetida à eletroforese em gel de poliacrilamida sob condições
desnaturantes (PAGE-SDS). Entretanto, no plasmídeo pET15b, a proteína recombinante é
produzida na forma de uma proteína de fusão com um peptídeo N-terminal que contém na sua
estrutura uma seqüência de seis resíduos de Histidina consecutivos (cauda de polihistidina). A
presença desse peptídeo rico em Histidina causa um retardamento na migração eletroforética da
proteína induzida, acarretando um aumento de 2 kDa em sua massa molecular aparente. A
diferença da massa da frutalina recombinante (19,2 kDa) expressa em E. coli e massa da lectina
nativa (12 e 15 kDa) deve ser atribuída também a ausência do processamento pós-traducional de
clivagem do peptídeo de ligação.
Levando em consideração esses fatos, o valor de 19,2 kDa da banda induzida é
aproximadamente a massa molecular aparente esperada para a frutalina recombinante.
Em análises por “Western blotting”, a banda de 19,2 kDa induzida pelo IPTG nas
células recombinantes de E. coli foi reconhecida especificamente por anticorpos policlonais de
coelho contra a frutalina nativa (anti-FTL) isolada de sementes maduras de A.incisa. Nenhuma
banda protéica foi reconhecida no lisado das bactérias não induzidas (Figura 24). Esses
resultados confirmam a identidade da proteína sintetizada em E. coli em resposta ao IPTG
como sendo a frutalina.
O fracionamento das proteínas citoplasmáticas do lisado celular após a indução
com IPTG e análise por PAGE-SDS revelou que praticamente toda proteína induzida
permanece na fração insolúvel na forma de corpos de inclusão (Figura 25).
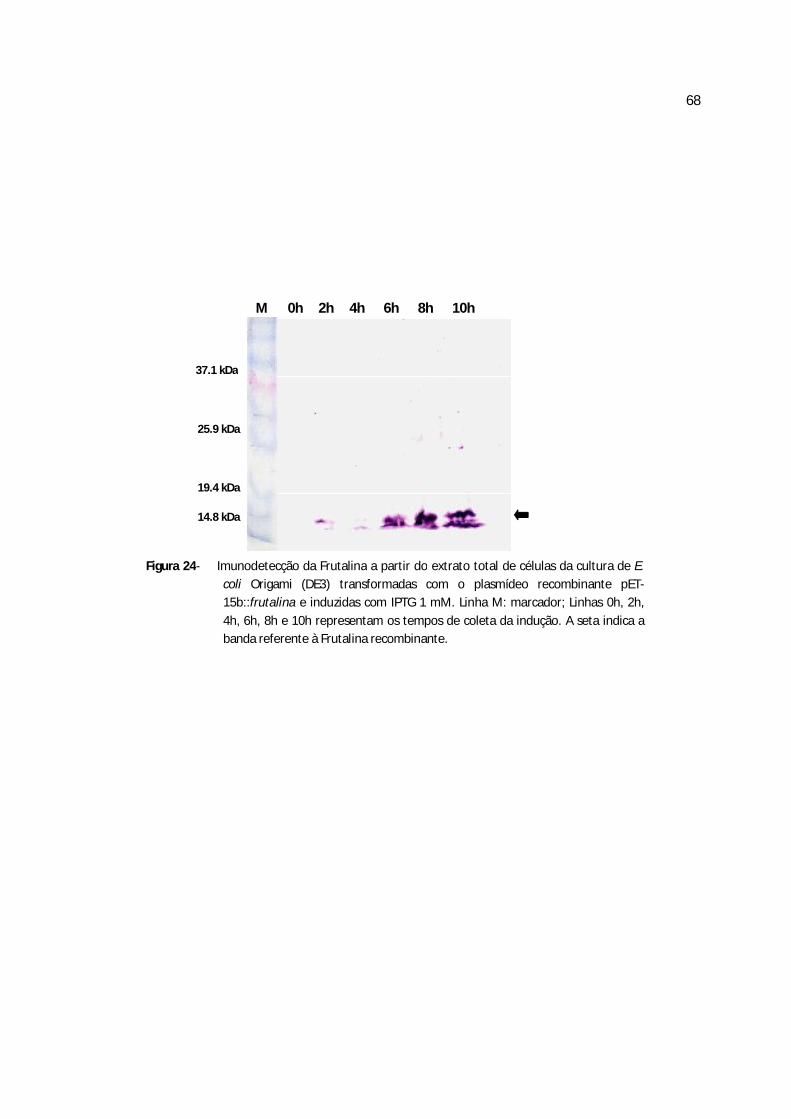
68
Figura 24- Imunodetecção da Frutalina a partir do extrato total de células da cultura de E. coli Origami (DE3) transformadas com o plasmídeo recombinante pET-15b::frutalina e induzidas com IPTG 1 mM. Linha M: marcador; Linhas 0h, 2h, 4h, 6h, 8h e 10h representam os tempos de coleta da indução. A seta indica a banda referente à Frutalina recombinante.
M 0h 2h 4h 6h 8h 10h
37.1 kDa
25.9 kDa
19.4 kDa
14.8 kDa

69
37.1 kDa
19.4 kDa
25.9 kDa
M 1 2 3
Figura 25 - Eletroforese em gel de poliacrilamida 17,5 % em condições desnaturantes, na presença de SDS, das amostras de proteínas obtidas a partir do extrato total solúvel e insolúvel (corpos de inclusão) de células de E. coli Origami (DE3) transformadas com o plasmídeo pET-15b::frutalina induzidas com IPTG 1mM. A seta indica a banda referente à frutalina recombinante. Linha M: marcador; Linhas 1 e 2: extrato total insolúvel; Linha 3: extrato total solúvel.

70
A frutalina recombinante expressa em E. coli foi mantida exclusivamente na forma
de corpos de inclusão insolúveis no interior bacteriano, à semelhança do relatado para a
maioria das lectinas vegetais produzidas nesse procarioto. A formação de corpos de inclusão
resulta de uma falha do sistema de reparo ou remoção de proteínas desenoveladas ou
enoveladas incorretamente (SCHUMANN et al., 2004). Essa formação parece ser uma
propriedade natural, pois mesmo para proteínas nativas da bactéria, quando expressas em altos
níveis, os corpos de inclusão também são produzidos (SHARMA, 1986). Esses agregados não
são formados pela proteína recombinante pura, mas contêm várias impurezas como proteínas
do hospedeiro, componentes ribossomais e DNA plasmidial (SCHUMANN et al., 2004).
Da mesma maneira, várias lectinas vegetais expressas em E. coli foram produzidas
exclusivamente na forma insolúvel no interior bacteriano.
A lectina da leguminosa Canavalia brasiliensis (ConBr) expressa em E. coli foi
mantida exclusivamente em corpos de inclusão no citoplasma bacteriano, atingindo um
rendimento de 15 a 30 mg por litro de cultura. Este rendimento foi bastante elevado, quando
comparado ao obtido com outras proteínas expressas também em E. coli, como a pré-pro-
ConA de C. ensiformis (MIN; JONES, 1992), com um rendimento de 0,5 a 1,3 mg por litro, e
a pro-ConA (MIN et al.,1992), com um rendimento de 0,4 mg por litro. Mesmo não havendo
processamento da pré-pro-ConBr, a mesma se mostrou capaz de induzir a migração de
neutrófilos para a cavidade peritoneal de ratos e causar degranulação de mastócitos peritoniais
murinos, com conseqüente liberação de histamina. No entanto, a intensidade das respostas
obtidas com a proteína recombinante foi inferior às repostas apresentadas pela ConBr nativa
(NOGUEIRA et al.,2002).
A maioria das lectinas jacalina-relacionadas expressas em E. coli são manose-
ligantes. Como exemplo da lectina de Castanea crenata (CCA) que foi expressa por
Nakamura et al.,(2002). A lectina recombinante (rCCA) se acumulou nos corpos de inclusão,
por isso passou por um tratamento de solubilização com uréia 8 M e renaturação para que sua
atividade biológica pudesse ser recuperada.
O rendimento obtido foi de 4,2 mg/L de cultura. A rCCA apresentou o espectro de
dicroísmo circular similar ao da lectina nativa, indicando que as duas foram enoveladas de

71
forma semelhante e adotaram a mesma estrutura. A capacidade de ligação a carboidratos
também foi similar à da lectina nativa (nCCA). Os resultados indicaram que a rCCA obtida
era equivalente à lectina nativa.
A lectina manose ligante de Artocarpus integrifolia (KM+) foi expressa em E.
coli na sua forma nativa (rKM+) e em fusão com a proteína GST (rGST-KM+). A produção da
proteína de fusão resultou em um aumento significante no rendimento da proteína
recombinante quando comparada a rKM+. Nas mesmas condições de crescimento, o
rendimento da rGST-KM+ foi cerca de 40 vezes maior que o da rKM+, após 4 horas de
indução. A proteína de fusão se acumulou completamente nos corpos de inclusão, entretanto, o
rendimento conseguido foi muito baixo, de 1,5 mg por litro (SILVA et al.,2005).
Haraguchi et al.,(2006) expressaram a lectina jacalina-relacionada de Cycas
revoluta (CRLL) em E. coli de forma ativa. A lectina recombinante (rCRLL) também se
acumulou nos corpos de inclusão e assim como outras lectinas, precisou passar por um
tratamento de solubilização com uréia 8 M e renaturação para que pudesse ser purificada. A
rCRLL purificada apresentou massa molecular aparente de 31 kDa, similarmente a proteína
nativa (nCRLL), e foi reconhecida especificamente em experimentos de imunodetecção. O
rendimento da rCRLL foi de 15 mg da rCRLL para 800 mL da cultura. Em análises de
inibição da hemaglutinação, a proteína recombinante obteve os mesmos resultados que a
proteína nativa.
A formação de corpos de inclusão na produção de proteínas recombinantes em E.
coli é o principal obstáculo para a obtenção de polipeptídeos biologicamente ativos
(VILLAVERDE et al., 2003). A principal desvantagem é o requerimento de tratamentos de
solubilização e renaturação para a obtenção da proteína solúvel. Algumas estratégias podem
ser realizadas para minimizar a formação destes agregados insolúveis, como: vetor com baixo
número de cópias, promotor fraco, indução em baixas temperaturas, co-expressão de
chaperonas moleculares (SCHUMANN et al.,2004).
A recuperação de proteínas biologicamente ativas de corpos de inclusão também
tem algumas vantagens. Os corpos de inclusão podem ser acumulados no citoplasma em níveis
muito mais elevados que as proteínas solúveis. Os mesmos são resistentes a proteólise por
proteases de E. coli, permitindo um alto rendimento da proteína recombinante. Esses

72
agregados não têm atividade biológica, facilitando a produção de proteínas que normalmente
seriam tóxicas em E. coli (CHOI et al, 2006).
Após a purificação da Frutalina recombinante, será possível a realização de
estudos da atividade biológica em comparação com a da lectina nativa, assim como o
seqüenciamento N-terminal e análises da estrutura secundária.

73
6. CONCLUSÕES
A Frutalina recombinante foi expressa em Escherichia coli com massa molecular
aparente superior a da nativa e foi mantida exclusivamente em corpos de inclusão insolúveis
no interior bacteriano.
Os sítios de ligação a carboidratos não foram alterados pelas mutações ocorridas
nos clones da frutalina. As mutações pontuais nos clones sugerem a existência de isoformas
dessa proteína.

74
7. REFERÊNCIAS BIBLIOGRÁFICAS
ADAR, R., STREICHER, H., ROZENBLATT, S., SHARON, N. Synthesis of soybean
agglutinin in bacterial and mammalia cells. European Journal of Biochemistry. v.249 n.3, p.
684-689, 1997.
ARANGO, R.; ADAR, R. ROZENBLAT; SHARON, N. Expression of Erythrina
corallodendron lectin in Escherichia coli. European Journal Biochemistry. v. 205, p. 575-
581, 1992.
AUB, J. C., TIESLAU, C.; LANKESTER, A. Reactions of normal and tumor cell to enzymes.
I. Wheat-germ lipase and associated mucopolysaccharides. Proc. Natl. Acad. Sci. USA. v. 50,
p. 613-619, 1963.
AUB, J. C., SANFORD, B. H.; COTE, M. N. Studies on reactivity of tumor and normal cells
to a wheat germ agglutinin. Proc. Natl. Acad. Sci. USA. v. 54, p. 396-399, 1965.
BLASCO, E.; NGOG, L. D.; AUCOUTURIER, P.; PREUD’HOMME, J. L. Mitogenic
activity of new lectins from seeds of wild Artocarpus species from Vietnam. C.R. Acad. Sci.
Paris. v. 319, p. 405-405, 1996.
BOURNE, Y.; HOULÈS ASTOUL, C. Structural basis for the unusual carbohydrate-binding
specificity of jacalin towards galactose and mannose. Biochem. J. v. 364, p. 173-180, 2002.
BOWIE, J. U.; LÜTHY, R.; EISENBERG, D. A method to identify protein sequences that
fold into a known three-dimensional structure. Science. v. 253, p. 164-170, 1991.
BRAGA, R. Plantas do Nordeste, especialmente do Ceará. Fortaleza (Reproduzido na Col.
Mossoroense, em 3o ed., v. XLII, Mossoró, 1976). 1953.
BRANDO-LIMA, A. C.; SALDANHA-GAMA, R. F.; HENRIQUES, M.G.M.O.;
MONTEIRO-MOREIRA, A. C. O.; MOREIRA, R. A.; BARJA-FIDALGO, C. Frutalin, a
galactose-binding lectin, induces chemotaxis and rearrangement of actin cytoskeleton in
human neutrophils: Involvement of tyrosine kinase and phosphoinositide 3-kinase. Toxicology
and Applied Pharmacology, v. 208, p. 145-154, 2005.

75
BRANDO-LIMA, A. C.; SALDANHA-GAMA, R. F.; PEREIRA, C. R.; VILLELA, C. G.;
SAMPAIO, A. L. F.; MONTEIRO-MOREIRA, A. C. O.; HENRIQUES, M. G. M. O.;
MOREIRA, R. A.; BARJA-FIDALGO, C. Involvement of phosphatidylinositol-3 kinase–Akt
and nuclear factor kappa-B pathways in the effect of frutalin on human
lymphocyte. International Immunopharmacology. v. 6, p. 465-472, 2006.
BROCKHAUSEN, I. Pathways of O-glican biosinthesis in cancer cells. Biochimica et
Biophysica Acta- General Subjects, v. 1473, n. 1, p. 67-95, 1999.
CARLINI, C. R.; GROSSI-DE SÁ, M. F. Plant toxic proteins with insecticidal properties. A
review on their potentialities as bioinsecticides. Toxicon, v. 40, p. 1515-1539, 2002.
CAMPANA, P. T.; MORAES, D. I.; MONTEIRO-MOREIRA, A. C. O.; BELTRAMINI, L.
M. Unfolding and refolding studies of frutalin, a tetrameric D-galactose binding lectin. Eur. J.
Biochem. 268, 753 – 758, 2002.
CAMMUE, B. P. A.; PEETERS, B.; PEUMANS, W. J. A new lectin from tulip (Tulipa)
bulbs. Plant. V. 169, n. 4, p. 583-588, 1986.
CARRINGTON, D.M.; AUFFRET, A.; HANKE, D.E. Polypeptide ligation occurs during
post-translational modification of concanavalin A. Nature, vol. 313, p. 64-67, 1985.
CHANG, S.; PURYEAR, J.; CAIRNEY, J. A simple and efficient method for isolating RNA
from pine trees. Plant Molecular Biology. v. 11, p. 113-116, 1993.
CHOI, J. H.; KEUM, K. C.; LEE, S. Y. Production of recombinant proteins by high cell
density culture of Escherichia coli. Chemical Engineering Science, v. 61, p. 876-885, 2006.
DELANO, W. L. The PYMOL. Molecular Graphics System. Delano Scientific, San Carlos,
CA, USA, 2002.
ECK, J., LANGER, MÖCKEL, B, BAUR,A., ROTHE, M., ZINKE, H., LENTZEN, H.
Cloning of the mistletoe lectin gene and characterization of the recombinant A-chain.
European Journal of Biochemistry. v. 264, p.775-784, 1999.
ELTZER, M.E. Distribution and function of plant lectins. In: LIENER, I.E.; SHARON, N.;
GOLDSTEIN, I.J. (Eds.) The Lectins: Properties, Functions, and Applications in Biology
and Medicine. Academic Press, Inc. 1986, 600 p.

76
EWING, B.; HILLIER, L.; WENDL, M.; GREEN, P. Basecalling of automated sequencer
traces using phred. I. Accuracy assessment. Genome Research. v. 8, p. 175-185, 1998.
FELIX, W. P. Produção Heteróloga de Frutalina em Pichia pastoris. Fortaleza, 2008. Tese
de Doutorado. Departamento de Bioquímica e Biologia Molecular, Universidade Federal do
Ceará, ? p.
FERREIRA, M. V. P. Frutalina, Lectina -D-Galactose Ligante de Artocarpus incisa L.
um estudo com Câncer de Mama. Fortaleza, 2001. Tese de Doutorado. Departamento de
Bioquímica e Biologia Molecular, Universidade Federal do Ceará, 145 p.
FIK, E.; WOLUN-CHOLEWA, M.; KISTOWSKA, M.; WARCHOL, J. B; GOZDZICKA-
JOZEFIAK, A. Effect of lectin from Chelidonium majus L. on normal and cancer cells in
culture. Folia Histochem. Cytobiol. v. 39, n. 2, p. 215-216, 2001.
FILHO, O. A. S.; ALENCASTRO, R. B. Modelagem de proteínas por homologia. Química
Nova. v. 26, n. 2, p. 253-259, 2003.
GORDON, D.; ABAJIAN, C.; GREEN, P. Consed: a graphical tool for sequence finishing.
Genome Research. v. 8, p. 195-202, 1998.
HARAGUCHI, T.; NOMURA, K.; YAGI, F. Cloning and expression of a mannose-binding
Jacalin-related lectin from leaves of Japanese Cycad (Cycas revoluta Thunb.). Biosci.
Biotechnol. Biochem., v. 70, n. 9, p. 2222-2229, 2006.
HIGGINS, D. G; THOMPSON, J. D; GIBSON, T. J; Clustal W: improving the sensitivity of
progressive multiple sequence alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res. v. 22, p. 4673-4680, 1994.
HOFFMAN, L., DONALDSON, D. Synthesis of mitogenic phytohemmaglutinin-L in
Escherichia coli. Bio/Technol. vol 5, p.157-160, 1987.
HOOFT, R. W. W.; VRIEND, G.; SANDER, C.; ABOLA, E. E. Erros in protein structures.
Nature. v. 381, p. 83-85, 1996.
HOULÈS ASTOUL, C.; PEUMANS, W. J.; VAN DAMME, E. J. M.; BARRE, A.;
BOURNE, Y.; ROUGÉ, P. The size, shape and specificity of the sugar-binding site of the

77
jacalin-related is profoundly affected by the proteolytic cleavage of the subunits. Biochem. J.,
v. 367, p. 817-824, 2002.
JORDAN, E. T., GOLDSTEIN, I.J. The sequence of a second member of the Lima Bean lectin
gene family and the expression and characterization of recombinant lectin in Escherichia coli.
The Journal of Biological Chemistry. v. 269, n.10, p.7674-1681, 1994.
JORDISON, M.; EL-HARIRY, I.; CALNAN, D.; CALAM, J.; PIGNATELLI, M. Vicia faba
agglutinin, the lectin present in broad beans, stimulates differentiation of undifferentiated
colon cancer cells. Gut. v. 44, p.709-714, 1999.
KEYAERTS, E.; VIJGEN, L.; PANNECOUQUE, C.; VAN DAMME, E. J. M.; PEUMANS,
W. J.; EGBERINK, H.; ALZARINI, J.; RANST, M. V. Plant lectins are potent inhibitors of
coronaviruses by interfering with two targets in the viral replication cycle. Antiviral
Research, v. 75, p. 179-187, 2007.
KITTUR, F. S.; LALGONDAR, M.; YU, H. Y.; BEVAN, D. R.; ESEN, A. Maize β-
glucosidase- aggregating factor is a polyspecific jacalin-related chimeric lectin, and its lectin
domain is responsible for β-glucosidase aggregation. The Journal of Biological Chemistry.
v. 282, n. 10, p. 7299-7311, 2007.
LASKOWSKI, R. A.; MACARTHUR, M. W.; MOSS, D. S; THORNTON, J. M.
PROCHECK: a program to check the stereochemical quality of protein structures. Journal of
Applied Crystallography. v. 26, p. 283-291, 1993.
LAEMMLI, U.K. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature, v. 227, p. 680-685, 1970.
LUO, S.; ZHANGSUN, D.; TANG, K. Functional GNA expressed in Escherichia coli with
high efficiency and its effect on Ceratovacuna lanigera Zehntner. Applied Genetics and
Molecular Biotechnology, v. 69, p. 184-191, 2005.
MATTHEWS, B. W. Solvent content of protein crystals. Journal of Molecular Biology. v.
33, p. 491-497, 1968.
MEJÍA, E. G.; PRISECARU, V, I. Lectins as bioactive plant proteins: a potencial in câncer
treatment. Critical Reviews in Food Science and Nutrition. v. 45, p. 425-445, 2005.

78
MIDDELBERG, A. P. J. Preparative protein refolding. Trends in Biotechnology. vol. 20, n
10, p. 437- 443, 2002.
MIKKAT, U.; DAMM, I.; KIRCHHOFF, F.; ALBRECHT, E.; NEBE, B.; JONAS, L. Effects
of lectins on CCk-8-stimulated enzyme secretion and differentiation of the rat pancreatic cell
line AR42J. Pancreas. v. 23, n. 4, p. 368-374, 2001.
MILHOME, M. V. L. Frutalina, Lectina -D-Galactose Ligante de Artocarpus incisa L.,
no Estudo do Câncer de Tireóide Humana Análise Comparativa com a Galectina-3.
Fortaleza, 2003. Dissertação de Mestrado. Departamento de Bioquímica e Biologia Molecular,
Universidade Federal do Ceará, 69 p.
MIN, W., JONES, D.H. Stability and detection of recombinant pre-pro-concanavalin A after
cytoplasmic expression in Escherichia coli. FEBS Letters. v. 301, n.3, p. 315-318, 1992.
MONTEIRO, A. C. O. Características químicas, físico-químicas e estruturais da
frutalina: uma lectina D-galactose ligante de Artocarpus incisa. Fortaleza, 1998.
Dissertação de Mestrado. Departamento de Bioquímica e Biologia Molecular, Universidade
Federal do Ceará.
MONTEIRO-MOREIRA, A. C. O. Caracterização Estrutural de três Lectinas
Apresentando Especificidades Distintas, Presentes em Sementes de Fruta-Pão (Artocapus
incisa). Fortaleza, 2002. Tese de Doutorado. Departamento de Bioquímica e Biologia
Molecular, Universidade Federal do Ceará, 164 p.
MOREIRA, R. A. Purification of a lectin from jackfruit (Artocarpus integrifolia) seeds. Plant
Physiology, v. 59, p. 68, 1977.
MOREIRA, R. A., CASTELO-BRANCO, C.C., MONTEIRO, A.C.O., TAVARES, R.O.,
BELTRAMINI, L.M. Isolation and characterization of a lectin from Artocarpus incisa L.
seeds. Phytochemistry, v. 47, n. 7, p. 1183-1188, 1998.
NAKAMURA, S.; IKEGAMI, A.; MATSUMURA, Y.; NAKANISHI, T.; NOMURA, K.
Molecular cloning and expression of the mannose/glucose specific lectin from Castanea
crenata cotyledons. J. Biochem. v. 131, p. 241-246, 2002.
NAKAMURA-TSURUTA, S.; UCHIYAMA, N.; PEUMANS, W. J.; VAN DAMME, E. J.
M.; TOTANI, K.; YUKISHIGE, I.; HIRABAYASHI, J. Analysis of the sugar-biding

79
specificity of mannose-binding-type Jacalin-related lectins by frontal affinity chromatography
– an approach to functional classification. FEBS Journal, v.1, p.1227-1239, 2008.
NELSON, D. L.; COX, M. M. Lehninger Principios de Bioquímica. – Tradução: SIMÕES,
A. A.; LODI, W. R. N. 3ª edição. Editora Sarvier. 2002. 975 p., São Paulo.
NOGUEIRA, N.A.P.; GRANGEIRO, M.B.; CUNHA, R.M.; ALVES, M.A.O.; TEIXEIRA,
E.H.; BARRAL-NETO, M.; CALVETE, J.J.; CAVADA, D.S.; GRANGEIRO, T.B.
Expression and purification of the recombinant ConBr (Canavalia brasiliensis lectin)
produced in Escherichia coli cells. Proteins and peptide letters, v.9, n.1, p.59-66, 2002.
PEUMANS, W. J.; VAN DAMME, E. J. M. The role of lectins in plant defence. Histoch.
Journal, v.27, p.253-271, 1995.
PRASTHOFER, T., PHILLIPS, S.R., SUDDATH, F.L., ENGLER, J.A . Design, expression
and crystallization of recombinant lectin from the Garden pea (Pisum sativum) The Journal
of Biological Chemistry. v.264, n.12, p. 6793-6796, 1989.
RAMACHANDRAN, G. N.; RAMAKRISHNAN, C.; SASISEKHARAN, V. Stereochemistry
of polypeptide chain configurations. Journal of Molecular Biology. v. 7, p. 95-99, 1963.
RODRIGUEZ-ARANGO, E. ARANGO, R., ADAR, R, GALILI, G, SAHRON, N. Cloning,
sequence analysis and expression in Escherichia coli of the cDNA encoding a precursor of
peanut agglutinin. FEBS Letters v.307, n.2, p.185-189, 1992.
RÜDIGER, H. On the physiology role of plant lectins. BioScience, v. 34, p. 95-99, 1984.
RÜDIGER, H.; GABIUS, H. J. Plant lectins: Occurrence, biochemistry, functions and
applications. Glycoconjugate Journal, v.18, p. 589-613, 2001.
SAHASRABUDDHE, A. A.; GAIKWAD, S. M.; KRISHNASASTRY, M. V.; ISLAM
KHAN, M. Studies on recombinant single chain Jacalin lectin reveal reduced affinity for
saccharides despite normal folding like native Jacalin. Protein Science, v. 13, p. 3264-3273,
2004.
SALI, A.; BLUNDELL, T. L. Comparative Protein Modelling by Satisfaction of Spatial
Restraints. Journal of Molecular Biology. v. 234, p.779-815, 1993.

80
SAMBROOK, J.; FRITSCH, E.F.; MANIATIS, T. Molecular Cloning – A Laboratory
Manual. Second Edition. Cold Spring Harbor Laboratory Press, 1989.
SANKARANARAYANAN, R.; SEKAR, K.; BANERJEE, R.; SHARMA, V.; SUROLIA, A.;
VIJAYAN, M. A novel mode of carbohydrate recognition in jacalin, a Moraceae plant lectin
with a -prisma fold. Nature Structural Biology, v. 3, p. 596-603, 1996.
SCHROEDER, M.R., RAIKHEL, N.V. Isolation and characterization of pro-barley lectin
expressed in Escherichia coli. Protein Expression and Purification. v. 3, p. 508-511, 1992.
SCHUMANN, W.; FERREIRA, L. C. S. Production of recombinant proteins in Escherichia
coli. Genetics and Molecular Biology, v. 27, n. 3, p. 442-453, 2004.
SHARMA, S. K. On the recovery of genetically engineered proteins from Escherichia coli.
Separation Science and Technology. v. 21, n. 8, p. 701-726, 1986.
SHARMA, V., SUROLIA, A. Cloning by genomic PCR and production of peanut agglutinin
in Escherichia coli. Gene. v.148, p.299-304, 1994.
SHARON, N.; LIS, H. History of lectins: from hemagglutinins to biological recognition
molecules. Glycobiology. v. 14, p. 53-62, 2004.
SILVA, L. L. P.; MOLFETTA-MACHADO, J. B.; PANUNTO-CASTELO, A.; DENECKE,
J.; GOLDMAN, G. H.; ROQUE-BARREIRA, M.; GOLDMAN, M. H. S. cDNA cloning and
functional expression of KM+, the mannose-binding lectin from Artocarpus integrifolia seeds.
Biochimica et Biophysica Acta. v. 1726, p. 251-260, 2005.
SORENSEN, H. P.; MORTENSEN, K. K. Advanced genetic strategies for recombinant
protein expression in Escherichia coli. Journal of Biotechnology, v. 115, p. 113-128, 2005.
STREICHER, H, SHARON, N. Recombinant plant lectins and their mutants. Methods in
Enzimology, vol 363, p.47-77, 2003.
STUBBS, M.E., CARVER, J.P., DUNN, R.J. Production of pea lectin in Escherichia coli.
The Journal of Biological Chemistry. vol. 261, n.14, p. 6141-6144, 1986.
TATENO, H.; WINTER, H. C.; PETRYNIAK, J.; GOLDSTEIN, I. J. Purification,
characterization, molecular cloning, and expression of novel members of jacalin-related lectins

81
from rhizomes of the true fern Phlebodium aureum (L) J. Smith (Polypodiaceae). The
Journal of Biological Chemistry. v. 278, n. 13, p. 10891-10899, 2003.
VAN DAMME, E. J. M.; PEUMANS, W. J.; BARRE, A.; ROUGÉ, P. Plant lectins: a
composite of several distinct families of structurally and evolutionary related proteins with
diverse biological roles. Critical Reviews in Plant Sciences, v. 17, n. 6, p. 575-692, 1998.
VAN DAMME, E. J. M.; BARRE, A.; ROUGÉ, P; PEUMANS, W. J. Cytoplasmic/nuclear
plant lectins: a new story. Trends in Plant Science. v. 9, n. 10, p. 484-489, 2004.
VILLAVERDE, A.; CARRIÓ, M. M. Protein aggregation in recombinant bacteria: biological
role of inclusion bodies. Biotechnology Letters. v. 25, p. 1385-1395, 2003.
WANG, H.; NG, T. B.; OOI, V. E.; LIU, W. K. Effects of lectins with different carbohydrate-
binding specifities on hepatoma, choriocarcinoma, melanoma and osteosarcoma cell lines. Int.
J. Biochem. Cell. Biol. v. 32, n. 3, p. 365-372, 2000.
YAMAUCHI, D.; MINAMIKAWA, T. Structure or the gene encoding concanavalin A from
Canavalia gladiata and its expression in Escherichia coli. FEBS Letters, vol. 260, n.1, p.
127-130, 1990.
YANG, H.; CZAPLA, T. H. Isolation and Characterization of cDNA Clones Encoding Jacalin
Isolectins. The Journal of Biological Chemistry, 268(8), 5905 - 5910, 1993.
YIN, J.; LI, G.; REN, X.; HERRLER, G. Select what you need: A comparative evaluation of
the advantages and limitations of frequently used expression systems for foreign genes.
Journal of Biotechnology, v. 127, p. 335-347, 2007.
YOUNG, N.M.; JOHNSTON, R.A.Z.; WATSON, D.C. The amino acid sequences of jacalin
and the Maclura pomifera agglutinin. FEBS Letters, v. 282, n. 2, p. 382-384, 1991.
ZHAO, X.; CHEN, Z.; LIN, J.; KONG, W.; SUN, X.; TANG, K. Expression and purification
of Arisaema heterophyllum agglutinin in Escherichia coli. Journal of Plant Physiology, v.
163, p. 206-212, 2006.
ZHU, K.; HUESING, J. E.; SHARE, R, E.; BRESSAN, R. A.; HASEGAWA, P. M.;
MURDOCK, L. An insecticidal N-acetylglucosamine-specific lectin gene from Griffonia
simplicifolia (Leguminosae). Plant Physiol. v. 110, n. 1, p. 195-202, 1996.

82