universidade federal de sÃo joÃo del-rei ufsj … · avaliaÇÃo da modulaÇÃo do perfil...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE SÃO JOÃO DEL-REI – UFSJ CAMPUS CENTRO-OESTE DONA LINDU – CCO
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
LÍLIAN NARA DAVID E SILVA
AVALIAÇÃO DA MODULAÇÃO DO PERFIL LIPÍDICO E ATIVIDADE DA Na,K-
ATPase DE CÉLULAS TUMORAIS TRATADAS COM GLICOSÍDEOS
CARDÍACOS
Divinópolis-MG
Fevereiro-2016

UNIVERSIDADE FEDERAL DE SÃO JOÃO DEL-REI – UFSJ CAMPUS CENTRO-OESTE DONA LINDU – CCO
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DA SAÚDE
LÍLIAN NARA DAVID E SILVA
AVALIAÇÃO DA MODULAÇÃO DO PERFIL LIPÍDICO E ATIVIDADE DA Na,K-
ATPase DE CÉLULAS TUMORAIS TRATADAS COM GLICOSÍDEOS
CARDÍACOS
Orientador: Leandro Augusto de Oliveira Barbosa
Co-orientadora: Hérica de Lima Santos
Divinópolis- MG
Fevereiro- 2016
Dissertação apresentada ao Programa de Pós Graduação em Ciências da Saúde, da Universidade Federal de São João Del Rei, como requisito parcial para a obtenção do
grau de Mestre.

ii
À minha mãe pelas orações, apoio, incentivo e
por ser minha fonte inesgotável de amor.
Ao meu marido, pelo imenso carinho e
companheirismo.

iii
AGRADECIMENTOS
Primeiramente à Deus, toda gratidão! Dele vem toda a sabedoria, força e luz.
Gratidão também pelas pessoas que colocou em meu caminho e que foram
importantes para que eu chegasse até aqui.
À minha mãe Heloísa, meu maior presente Divino, minha base e meu
principal exemplo de ser humano. Obrigada por me ensinar a lutar com fé, amor e
dedicação. Eu não seria nada sem você!
Ao meu pai Wagner, que ainda tão distante, se fez presente em alguns
momentos.
Aos meus irmãos Letícia, Guilherme e Karol; e meu sobrinho-afilhado, por
estarem sempre ao meu lado e por alegrarem minha vida. Obrigada por confiarem
em mim. Por vocês o amor mais puro e verdadeiro que existe!
Ao meu marido Adriano, por ser tão companheiro e amigo. Por cada palavra
de incentivo e carinho, pelo abraço apertado e sorriso de todos os dias. Por ouvir
inúmeras apresentações de trabalho, lamentações e alegrias e por me tranquilizar
sempre. Essa conquista também é por você!
Às minhas queridas avós Conceição e Ana, e tios, em especial Maria, Ulisses,
André e Vitor, pelas orações em minha intenção. Obrigada pela torcida!
Aos meus amigos Ana Elisa e Douglas, pelos bons encontros. Esses
momentos de descontração foram muito importantes pra mim! À minha amiga
querida, Fernanda, que mesmo há tantos quilômetros de distância está sempre
presente. É muito bom ter amigos, com quem posso contar. A amizade é um bem
muito valioso!
Ao meu orientador, Leandro, pela orientação do trabalho e por transmitir tanto
conhecimento. Pela oportunidade de aprender um pouco mais a cada dia. Obrigada
pela atenção mesmo quando esteve longe e pelo exemplo de profissionalismo. À
você minha admiração e eterna gratidão!
À minha co-orientadora, Hérica, por ser tão atenciosa e solícita. Obrigada pela
orientação durante todo o projeto, principalmente enquanto o Leandro esteve longe.

iv
Nossas conversas foram muito valiosas para a construção do trabalho. Muito
obrigada!
À professora Vanessa, sempre sorridente, pelo carinho e apoio que também
foram fundamentais para a concretização desse projeto.
À equipe do laboratório de Bioquímica Celular, em especial Grazi, por não ser
simplesmente a técnica do laboratório, por ser nossa amiga. Obrigada por todo seu
cuidado com o laboratório e conosco! Aos alunos de iniciação científica, mestrado e
doutorado. Aqui, agradeço com carinho à Duane (minha duplinha) e Marco Túlio por
terem me ensinado uma das técnicas tão necessárias para a realização desse
trabalho, o Western Blotting. Agradeço com o mesmo afeto à Letícia, por tudo que
sei sobre as técnicas da cultura celular. Obrigada por me ensinar com tanta
paciência e pela sua amizade! Aos amigos, ‘Cris’ e ‘Isma’, meus companheiros não
só de laboratório, mas também graduação e mestrado, pelos muitos e proveitosos
dias de estudos, trabalhos e pelas tantas discussões e padronização de protocolos.
Pelos tantos anos de convivência e principalmente pela amizade! Ao casal amigo,
Luana e Marco Túlio, pelos bons encontros ‘regados’ por boa comida e boa
conversa, muitas vezes aliviaram o meu coração. Aos amigos que fiz aqui, meu
sincero obrigado!
Ao professor José Villar por ceder os compostos que utilizei no trabalho e ao
professor Fabio Vieira por ceder gentilmente espaço e materiais necessários em
alguns experimentos. À Fundação Ezequiel Dias (FUNED-BH) pela doação de
composto e principalmente pela doação das células HeLa.
As professoras Luciana Maria Silva e Nayara Delgado A. Bordoleto, por terem
aceitado o convite para compor a minha banca de defesa de dissertação.
À UFSJ e ao Programa de Pós-graduação em Ciências da Saúde, pelo apoio
institucional para a realização do mestrado. À CAPES, pelo suporte financeiro para
realização deste trabalho.

v
“Depois de termos conseguido subir a uma grande montanha, só descobrimos que
existem ainda mais grandes montanhas para subir.”
Nelson Mandela

vi
RESUMO
Os esteroides cardiotônicos, entre eles a digoxina, tem se mostrado como potenciais
fármacos antitumorais. A 21-benzilideno digoxina (21-BD) é um derivado sintético da
digoxina que apresenta um grupo estireno adicional em sua estrutura. Essa
alteração na estrutura tem mostrado efeitos interessantes e diferentes dos
observados com a digoxina. O objetivo do trabalho foi avaliar possíveis alterações
na atividade da Na,K-ATPase, composição lipídica e estado redox de células HeLa
tratadas com digoxina e 21-BD. A atividade da Na,K-ATPase e a expressão da
subunidade α1 foi avaliada nas células tratadas por 48 horas com 21-BD. A 21-BD
apresentou um efeito bifásico na atividade da Na,K-ATPase, aumentando em 5µM e
diminuindo em 50µM, entretanto, a expressão proteica não foi alterada. A modulação
do perfil lipídico foi avaliado na célula e na membrana celular. Após o tratamento das
células com as drogas, foi realizado um homogenato total e preparação de
membrana. As determinações do conteúdo de colesterol e fosfolipídios das células
foram realizadas nos dois tipos de amostra. A digoxina diminuiu os níveis de
colesterol e fosfolipídios totais das células e não alterou o conteúdo desses lipídios
na membrana. A 21-BD não alterou os níveis de colesterol total e diminuiu o
conteúdo de fosfolipídios das células, porém na membrana foi observado o aumento
desses lipídios. A diminuição da atividade da Na,K-ATPase parece estar
correlacionada com a diminuição da fluidez da membrana celular. Para avaliar o
estado redox das células após o tratamento com as drogas, foi avaliada a
peroxidação lipídica, a atividade catalítica da superóxido dismutase, catalase e o
conteúdo de glutationa reduzida (GSH). A peroxidação lipídica e a atividade da
superóxido dismutase foram diminuídas após o tratamento com digoxina e 21-BD.
Os níveis de GSH e atividade da catalase permaneceram inalterados nas células.
Os esteroides cardiotônicos, 21-BD e digoxina, apresentaram resultados
interessantes sobre a modulação da composição lipídica e estado redox das células.
As alterações observadas podem fazer parte do mecanismo de ação dos esteroides
cardiotônicos.
Palavras- Chave: esteroides cardiotônicos, Na,K-ATPase, estado redox, perfil
lipídico

vii
ABSTRAT
Cardiotonic steroids, including digoxin, has proven as potential antitumor drugs. 21-
benzylidene digoxin (21-BD) is a synthetic derivative of digoxin and presents an
additional styrene group in its structure. This structure modification has shown
interesting and different effects of the observed with digoxin. The objective of the
work was to evaluate possible changes in Na,K-ATPase activity, lipid composition
and redox state on in HeLa cells treated with digoxin and 21-BD . The 21-BD showed
a biphasic effect on Na,K-ATPase activity, increasing in 5 µM and decreasing in 50
µM, however, the protein expression was not altered. Modulation of lipid profile was
evaluated in cell and cell plasma membrane. After cell treatment, it was carried out a
total homogenate and a membrane preparation. Cell cholesterol and phospholipids
contents were assessed in both samples. Digoxin decreased cholesterol levels, and
total phospholipids of the cell, but did not alter phospholipids content in plasma
membrane. The 21-BD did not change total cholesterol and decreased phospholipid
content of the cells, but in the membrane was observed the increase of these lipids.
The decrease in Na, K-ATPase activity appears to be correlate with the decrease in
cell membrane fluidity. To evaluate the redox state of cells after treatment with the
drugs, the lipid peroxidation, the catalytic activity of superoxide dismutase, catalase
and reduced glutathione (GSH) content was evaluated. Lipid peroxidation and activity
of superoxide dismutase were decreased after treatment with digoxin and 21-BD.
The GSH levels and activity of catalase remained unchanged in cells. The
cardiotonic steroids, digoxin, and 21-BD, showed interesting results On lipid profile
modulation and cell redox state. The observed changes coud be part of the
mechanism of action of cardiotonic steroids.
Key-words: Cardiotonic steroids, Na,K-ATPase, redox state, lipid profile

viii
SUMÁRIO
RESUMO .................................................................................................................... vi
ABSTRACT ............................................................................................................... vii
LISTA DE FIGURAS.................................................................................................... x
I) INTRODUÇÃO ....................................................................................................... 12
I.1) REVISÃO DA LITERATURA ............................................................................... 13
I.1.1) Câncer .............................................................................................................. 13
I.1.1) Células de carcinoma de colo uterino (Células HeLa) ........................... 14
I.1.2) Esteroides cardiotônicos .................................................................................. 16
I.1.2.1) 21-benzilideno digoxina (21-BD) ......................................................... 20
I.1.3) Na,K-ATPase .................................................................................................. 21
I.1.4) Membrana Celular ........................................................................................... 26
I.1.4.1) Interação lipídio-proteina.................................................................... 29
I.1.4.2) Esteroides cardiotônicos e modulação do perfil lipídico......................30
I.1.5) Estresse oxidativo e peroxidação lipídica ........................................................ 31
I. 2) JUSTIFICATIVA E OBJETIVOS ......................................................................... 33
I.2.1) Justificativa ...................................................................................................... 33
I.2.2) Objetivo Geral ................................................................................................. 34
I.2.3) Objetivos específicos ...................................................................................... 34
II) MATERIAL E MÉTODOS ...................................................................................... 35
II.1) Cultura de células .............................................................................................. 35

ix
II.2) Tratamento com esteroides cardiotônicos ......................................................... 35
II.3) Preparação do homogenato total ....................................................................... 36
II.4) Preparação de membrana plasmática ............................................................... 36
II.5) Preparação da membrana livre de hemoglobina (Ghost) .................................. 36
II.6) Dosagem da atividade ATPásica de Na,K-ATPase............................................ 37
II.7) Dosagem de proteínas totais ............................................................................. 38
II.8) Extração de lipídios ............................................................................................ 38
II.9) Dosagem do conteúdo de fosfolipídios totais ..................................................... 38
II.10) Dosagem do conteúdo de colesterol total ........................................................ 39
II.11) Determinação de substâncias reativas ao ácido tiobarbitúrico (TBARS) ......... 39
II.12) Dosagem da atividade das enzimas antioxidantes........................................... 40
II.13) Determinação da atividade enzimática da Superóxido Dismutase (SOD) ........ 40
II.14) Determinação da atividade enzimática da catalase (CAT) .............................. 40
II.15) Determinação do conteúdo de Glutationa Reduzida (GSH) ............................. 41
II.16) Análises de expressão proteica por Western Blotting ..................................... 41
II.17) Análise Estatística ............................................................................................ 42
III. RESULTADOS E DISCUSSÃO ............................................................................ 43
IV. CONSIDERAÇÕES FINAIS ................................................................................ 57
V. REFERÊNCIAS BIBLIOGRÁFICAS ...................................................................... 58

x
LISTA DE FIGURAS
Figura 1: Estrutura química geral de esteroides cardiotônicos. ................................ 17
Figura 2: Mecanismo de ação de esteroides cardiotônicos. ..................................... 18
Figura 3: Estrutura química da digoxina e da 21-benzilideno digoxina. .................... 21
Figura 4: Representação esquemática da estrutura da Na,K-ATPase . ................... 23
Figura 5: Representação esquemática do ciclo de reação da Na,K-ATPase. .......... 25
Figura 6: Modelo do mosaico fluido. ......................................................................... 27
Figura 7: Esquema de integração das enzimas antioxidantes ................................ 33
Figura 8: Atividade da Na,K-ATPase de células Hela tratadas por 48h com 21-BD 43
Figura 9: Expressão da subunidade α1 da Na,K-ATPase da membrana plasmática
de células HeLa tratadas por 48 horas com 21-BD .................................................. 45
Figura 10: Porcentagem de colesterol total do homogenato e da membrana
plasmática de células HeLa tratadas por 48 horas com digoxina ............................. 46
Figura 11: Porcentagem de colesterol total do homogentato e da membrana
plasmática de células HeLa tratadas por 48 horas com 21-BD. ................................ 47
Figura 12: Porcentagem de fosfolipídios totais do homogento e da membrana
plasmática de células HeLa tratadas por 48 horas com digoxina ............................. 47
Figura 13: Porcentagem de fosfolipídios totais do homogento e da membrana
plasmática de células HeLa tratadas por 48 horas com 21-BD.. ............................... 48
Figura 14: Porcentagem de colesterol total e de fosfolipídios totais de membrana de
eritrócitos incubadas com digoxina e 21-BD por 1 hora. ......................................... 49
Figura 15: Determinação da peroxidação lipídica de células HeLa tratadas por 48
horas e por 1 hora com 21-BD e digoxina ................................................................ 52
Figura 16: Atividade catalítica da superóxido dismutase de células HeLa tratadas
por 1 hora com digoxina e 21-BD ............................................................................. 54
Figura 17: Atividade catalítica da catalase de células HeLa tratadas por 1 hora com
digoxina e 21-BD. ...................................................................................................... 54

xi
Figura 18: Níveis de GSH de células HeLa tratadas por 1 hora com digoxina e 21-
BD ............................................................................................................................ 55

12
I. INTRODUÇÃO
Os esteroides cardiotônicos, inibidores da Na,K-ATPase, são fármacos
utilizados há muitos anos para tratar a insuficiência cardíaca congestiva e
atualmente tem apresentado interessantes efeitos antitumorais frente a diversas
linhagens celulares. No entanto, o mecanismo de ação do efeito antitumoral exercido
por esses fármacos não está claramente elucidado. Alguns efeitos causados pelos
esteroides cardiotônicos como, inibição da Na,K-ATPase, alterações da fluidez de
membrana e o aumento da geração de espécies reativas de oxigênio (EROS)
parecem estar envolvidos no mecanismo de ação pelo qual esses fármacos exercem
seus efeitos citotóxicos (CERELLA; DICATO; DIEDERICH, 2013; MIJATOVIC; KISS,
2013).
Devido à estrutura química dos esteroides cardiotônicos, eles podem interagir
com a membrana e alterar sua fluidez e microviscosidade, como foi demonstrado em
células tratadas com oleandrina. A modulação do perfil lipídico da membrana celular
pode alterar a funcionalidade de proteínas de membrana e por consequência
interferir em processos celulares fundamentais (RAGHAVENDRA; SREENIVASAN;
MANNA, 2007).
Efeitos como carbonilação de proteínas (JORNADA et al., 2011) e
peroxidação lipídica (BROCARDO et al., 2010) são consequências do estresse
oxidativo observadas após o tratamento com esses fármacos. O tratamento de
células com esteroides cardiotônicos induz o estresse oxidativo, por aumentar a
produção de EROS e modular o sistema de defesa antioxidante, como foi
demonstrado por NEWMAN e colaboradores (2006), onde o tratamento de células
BRO (melanoma humano) com oleandrina induziu uma diminuição dos níveis de
glutationa reduzida (GSH) (NEWMAN et al., 2006). Além disso, tem sido
demonstrado que EROS podem ativar vias de sinalização celular que induzem as
células à apoptose ou autofagia (NEWMAN et al., 2006; XIE et al., 2011).
É importante ressaltar que EROS podem inativar a Na,K-ATPase por levar à
modificações na estrutura da enzima (YAN et al., 2013), ou indiretamente por causar
danos na membrana celular através da peroxidação lipídica. A peroxidação lipídica
pode alterar a fluidez de membrana que está diretamente correlacionada com
alterações na atividade da Na,K-ATPase (RODRIGO et al., 2007)

13
I.1) REVISÃO DA LITERATURA
I.1.1) Câncer
Câncer é um termo utilizado para denominar um conjunto de doenças onde as
células apresentam um crescimento anormal, acelerado, e que são capazes de
invadir outros tecidos, propagando-se para outras partes do corpo em um processo
conhecido como metástase. O crescimento desordenado das células pode ter várias
causas externas e internas como, o ambiente e alterações no material genético. As
alterações no material genético são as principais causas das replicações infinitas
que conferem a imortalidade das células, além disso, elas se tornam resistentes aos
supressores de crescimento e aos mecanismos de morte celular. As células tumorais
passam também a induzir a formação de novos vasos, processo conhecido como
angiogênese, o que facilita a invasão das células em outros tecidos (HANAHAN;
WEINBERG, 2011; INCA, 2015).
Além disso, o metabolismo de células tumorais pode se diferenciar das
células normais. Devido a proliferação acelerada das células tumorais, elas
necessitam de uma maior disponibilidade de blocos de construção para a síntese de
proteínas, lipídios, nucleotídeos, etc. O metabolismo de lipídios se encontra alterado
no câncer e parece estar envolvido nos mecanismos da tumorigênese. As células
tumorais frequentemente apresentam níveis de expressão anormais das enzimas
envolvidas na biossíntese do colesterol, em especial, um aumento da expressão e
da atividade de HMG-CoA redutase (3-hidroxi-3-metil-glutaril-CoA redutase). Por
essa razão, o conteúdo de colesterol geralmente é maior em tecidos tumorais do
que em tecidos normais (CASEY et al., 2015; FABIANI, 2015). A composição dos
fosfolipídios de membrana também é diferente em algumas células tumorais em
relação às células normais, sendo que os diferentes tipos de fosfolipídios se
encontram em níveis aumentados no câncer. As modificações na composição de
fosfolipídios poderiam ser usados como mais um biomarcador relacionado ao câncer
(CÍFKOVÁ et al., 2014).
A alteração do sistema de enzimas antioxidantes e o aumento da produção de
espécies reativas de oxigênio (EROS) é outra diferença encontrada em células
tumorais. As EROS contribuem para o desenvolvimento do câncer através da
mutagênese e efeitos no microambiente tumoral, além disso, os oncogenes e genes
supressores tumorais podem ser regulados por EROS. As células tumorais

14
apresentam níveis mais elevados de EROS e alterações no sistema de defesa
antioxidante, que podem ser encontradas em níveis menores ou maiores
dependendo do tipo celular. O aumento da atividade das enzimas antioxidantes em
células tumorais pode ser entendido como uma forma de adaptação, para que
possam sobreviver mesmo com altos níveis de EROS. Esse processo de adaptação
ao estresse oxidativo pode contribuir para progressão do câncer, invasão e
resistência a medicamentos (BULDAK et al., 2014; NOGUEIRA; HAY, 2013).
O câncer acomete milhões de pessoas no mundo todo. A incidência de câncer
em 2012 foi de 14 milhões, sendo 8,2 milhões de mortes em todo o mundo. A
estimativa de novos casos de câncer para 2015 chega a 15,2 milhões e 8,8 milhões
de mortes. O câncer de colo uterino é o terceiro tipo de câncer mais comum entre as
mulheres, correspondendo a aproximadamente 4% dos casos de câncer no mundo
todo (GLOBOCAN, 2012).
Devido ao crescente número de novos casos e por ser uma doença que
apresenta diversas causas e efeitos, a busca e o desenvolvimento de novos
fármacos, vem sendo um intenso alvo de estudos para encontrar terapias eficientes
contra o câncer.
I.1.1.1) As células de carcinoma de colo uterino (Células HeLa)
A linhagem celular de adenocarcinoma cervical (HeLa) , foi a primeira
linhagem de células humana mantida em cultura. Essas células foram obtidas de
tecido proveniente da biopsia do tumor de colo uterino de uma jovem afroamericana
de aproximadamente 31 anos, chamada Henrietta Lacks, por essa razão foram
nomeadas "HeLa" (sigla formada pela primeira sílaba do primeiro e do segundo
nome). No entanto, por uma questão de confidencialidade, foi dito o doador ser
Helen Lane, ou Helen Larson (R.MASTERS, 2002).
Em 1951, Henrietta Lacks se apresentou à ginecologia do Hospital de Johns
Hopkins em Baltimore, com suspeita de câncer de colo uterino. Os tecidos obtidos
da biopsia foram enviados ao departamento de patologia da universidade Johns
Hopkins e o câncer foi confirmado. George Gey, pesquisador na universidade,
retirou uma amostra do tecido enviado para biopsia e levou para seu laboratório,
onde trabalhava com o isolamento e manutenção de células em cultura obtidas a
partir de tecidos normais e tumorais. Ao contrário de outras células, as células HeLa

15
cresceram em tubo de cultivo de forma acelerada, em um meio composto por
extrato de embrião bovino e soro do cordão de placenta humana em tipo de cultura
utilizando a técnica . Era a primeira vez que se conseguia cultivar células humanas
“imortalizadas”, elas estavam sempre em divisão. Henrietta Lacks faleceu no mesmo
ano em que iniciou os tratamentos, porém suas células continuam sendo
comercializadas até hoje (GEY; COFFMAN; KUBICEK, 1952; LUCEY; NELSON-
REES; HUTCHINS, 2009). O cultivo celular da HeLa representou um grande
benefício para a área médica e biológica, pois permitiu estudos sobre os processos
celulares e desenvolvimento de fármacos. Um dos primeiros usos dessa linhagem
celular foi para o desenvolvimento da vacina contra o vírus da poliomielite
(SCHERER; SYVERTON; GEY, 1953).
As células HeLa possuem morfologia de células epiteliais, são células
aderentes e se proliferam rapidamente (o tempo de replicação é de
aproximadamente 24 horas), tais características a tornam um bom modelo celular à
ser empregado em diversas linhas de pesquisa. A capacidade de proliferação
dessas células tem sido atribuída à atividade da telomerase, que faz com que a
célula possa se dividir infinitamente sem perder a estabilidade do genoma
(BLACKBURN, ELIZABETH H. GREIDER; SZOSTAK, 2009; BRYAN et al., 1998).
Também foi demonstrado que as células HeLa possuem o vírus do papiloma
humano (HPV) 18 e desde então têm-se associado o HPV com adenocarcinomas
agressivos. Além da atividade da telomerase, o fato de as células HeLa possuírem o
HPV-18 também pode explicar o crescimento acelerado das células em cultura,
observado primeiramente por Gey (YEE et al., 1985). As pesquisas sobre a
presença do vírus HPV em células HeLa desempenharam um papel fundamental,
pois permitiram correlacionar o vírus do papiloma humano com a ocorrência do
câncer cervical (HAUSEN, 2008).
O fator limitante de se trabalhar com esse tipo celular é a facilidade dessa
linhagem em contaminar outras linhagens celulares. Porém, por muitos anos esse
problema não era conhecido e as células HeLa que eram comercializadas e
cultivadas em laboratórios do mundo todo haviam contaminado outras linhagens
celulares (LUCEY; NELSON-REES; HUTCHINS, 2009). No entanto, mesmo com
essa limitação as células HeLa ainda são bastante utilizadas como modelo de
células tumorais em estudos de citotoxicidade e mecanismo de ação de novos
fármacos.

16
Os esteroides carditônicos são fármacos já utilizados há alguns anos no
tratamento da insuficiência cardíaca congestiva e têm também apresentado efeito
antitumoral frente a algumas linhagens tumorais. As células HeLa tem sido utilizadas
nesses estudos e elas são sensíveis ao efeito citotóxico de digitalis (Gupta et al.,
1989). O efeito antiproliferativo e apoptótico da ouabaína e digoxina, foram
demonstrados em células HeLa e a citotoxicidade desses compostos tem sido
correlacionada à eventos envolvendo a Na,K-ATPase expressa nesse tipo celular,
como inibição da enzima e desequilibrio iônico ou ativação de vias de sinalização
desencadeada pela ligação das drogas na enzima (RAMIREZ-ORTEGA et al.,
2006). Outros estudos tem demonstrado o efeito citótóxico da ouabaína (ALONSO et
al., 2013), digoxina e derivados sintéticos da digoxina (ALVES et al., 2015; ROCHA
et al., 2014) em células HeLa. Contudo, essa linhagem celular tem sido empregada
como modelo na investigação do efeito antiproliferativo e do mecanismo de ação de
esteroides cardiotônicos.
I.1.2) Esteroides cardiotônicos
Os esteroides cardiotônicos têm sido utilizados por mais de 200 anos para
tratar a insuficiência cardíaca congestiva. Em 1785, o médico britânico William
Withering descreveu as propriedades da dedaleira (Digitalis purpurea), durante seus
estudos sobre o uso medicinal da planta para o tratamento de pacientes com
hidropsia (WILKINS; KENDALL; WADE, 1985; WITHERING, 1785). Anos mais tarde,
John Ferriar propôs a ação do extrato da planta no tecido cardíaco (FERRIAR,
1799).
Essa classe de compostos pode ser obtida através do metabolismo
secundário de algumas espécies de plantas e também podem ser encontrados em
algumas espécies de animais. A planta mais conhecida por conter esses compostos
é a dedaleira (Digitalis lanata e Digitalis purpurea), por isso os esteróides
cardiotônicos são comumente chamados de digitálicos cardíacos. Os esteróides
cardiotônicos mais encontrados são originados das plantas pertencentes as famílias
Scrophulariaceae (digoxina e digitoxina) e Apocynaceae (ouabaína e oleandrina). A
digoxina, digitoxina, ouabaína e oleandrina são isolados de plantas como, Digitalis
lanata, Digitalis purpurea, Strofanthus gratus e Nerium oleander, respectivamente.

17
Outras famílias de plantas como Asclepiadacea e Ranunculacea também contém
esteroides cardiotônicos (ALEVIZOPOULOS et al., 2014; MELERO; MEDARDE;
SAN FELICIANO, 2000).
Em animais, esses compostos são obtidos principalmente das secreções das
glândulas da pele de sapos do gênero Bufo. Exemplos de esteroides cardiotônicos,
mais precisamente bufadenolídeos, encontrados nos venenos de sapos das
espécies Bufo gargarizans e Bufo marinus, são a bufalina e marinobufagenina
respectivamente (PRASSAS; DIAMANDIS, 2008; STEYN; VAN HEERDEN, 1998).
Os esteroides cardiotônicos também estão presentes em tecidos e fluidos corporais
de mamíferos, como por exemplo, cérebro, glândulas adrenais, coração, plasma,
fluido cerebroespinal e urina (BAGROV; SHAPIRO; FEDOROVA, 2009).
A estrutura dos esteroides cardiotônicos é composta por um núcleo esteroidal
ligado à um anel lactônico na posição C17, o que determina se o composto é um
cardenolídeo (anel lactônico de cinco membros), ou bufadenolídeo (anel lactônico de
seis membros). A estrutura pode ou não conter uma ou mais unidades de açúcar na
posição C3 do núcleo esteroidal e predominantemente em posição β,
caracterizando-os em glicosídeos (glicosídeos cardíacos) ou genina. Os açúcares
mais comumente encontrados são a ramnose, glicose, digitalose e digitoxose
(MIJATOVIC; KISS, 2013; MIJATOVIC et al., 2007). A estrutura química geral dos
esteroides cardiotônicos está representada na figura 1.
Figura 1: Estrutura química geral de esteroides cardiotônicos (Adaptado de
MIJATOVIC; KISS, 2013).

18
O uso de esteroides cardiotônicos para tratar a insuficiência cardíaca, deve-se
ao seu efeito inotrópico positivo causado pela inibição parcial da Na,K-ATPase.
Esses compostos têm a capacidade de se ligarem a uma alça extracelular na
subunidade catalítica da enzima e, por consequência, inibir sua atividade. Em doses
terapêuticas, a inibição parcial da Na,K-ATPase provoca um aumento da
concentração de Na+ intracelular. O desbalanço de Na+/K+ causa inibição do
transporte de Ca2+ pelo trocador Na+/Ca2+ levando ao acúmulo de Ca2+ no interior da
célula. O aumento dos níveis de Ca2+ no interior da célula favorece a ativação das
proteínas miosina II e actina, aumentando a contração muscular (figura 2). No
entanto, em doses mais altas, os altos níveis de Na+ e Ca2+ intracelulares promovem
efeitos tóxicos demonstrados pelos sintomas de arritmias cardíacas (STUCKY;
GOLDBERGER, 2015; THOMAS; GRAY; ANDREWS, 1990).
Figura 2: Mecanismo de ação de esteroides cardiotônicos (Adaptado de
KLABUNDE, 2015).
Observações importantes feitas por Stenkvist e colaboradores sobre a
utilização de esteroides cardiotônicos e a inibição de células tumorais fizeram com
que crescesse o interesse dos pesquisadores por esses fármacos. Em uma série de
trabalhos eles demonstraram o potencial antitumoral desses fármacos. Foi
demonstrado que a capacidade proliferativa de células de câncer de mama era
menor em pacientes que faziam uso de esteroides cardiotônicos e que as células
tumorais das pacientes possuíam alterações em sua morfologia, apresentando maior
característica benigna do que as células cancerígenas obtidas de pacientes controle,

19
que não estavam sob uso desses fármacos (STENKVIST, 1980; STENKVIST et al.,
1979). Em outro estudo foi relatado que mulheres que foram mastectomizadas e que
não faziam uso de digitalina apresentaram uma taxa de recorrência 9,6 vezes maior
do que mulheres que faziam uso desse medicamento (STENKVIST, 1982). Além
disso, observaram que os pacientes que faziam uso de esteroides cardiotônicos
raramente morriam de câncer (STENKIVIST, 2001). Desde então, interessantes
efeitos antitumorais têm sido observados e apontado que além do uso para o
tratamento de insuficiência cardíaca, os esteroides cardiotônicos podem ser
potentes fármacos para o tratamento do câncer.
O efeito antitumoral dos esteroides cardiotônicos já foi demonstrado para uma
série de linhagens tumorais como células T Jurkat e células B Daudi humanas
(HAUX, 1999), TK-10 (adenocarcinoma renal), MCF-7 (adenocarcinoma de mama),
UACC-62 (melanona maligno), e K-562 (células leucêmicas) (LÓPEZ-LÁZARO et al.,
2005) e células de linfoma (U937) (CUOZZO et al., 2012).
A digoxina, usada para tratar insuficiência cardíaca, também tem apresentado
resultados relevantes como antitumoral. O efeito antiproliferativo e pró-apoptótico
induzido pela digoxina, foi observado em diversas linhagens de células tumorais
como, células de câncer de mama (MCF-7) (BIELAWSKI; WINNICKA; BIELAWSKA,
2006; LÓPEZ-LÁZARO et al., 2005), próstata (PC-3) (MCCONKEY et al., 2000),
pulmão (H69 AR), leucemia (CCRF-CEM), renal (ACHN), mieloma (8226-S, 8226-
LR5, 8226-DOX-40) (JOHANSSON et al., 2001) e células de carcinoma oral
escamoso (SCC-25) (CHAVES et al., 2014).
O mecanismo de ação pelo qual os esteroides cardiotônicos exercem seus
efeitos antitumorais tem sido alvo de estudo de muitos pesquisadores. Possíveis
mecanismos de ação têm sido apontados, como por exemplo, alteração da
homeostase de Na+, K+, e Ca2+, por inibição da Na,K-ATPase, aumento da geração
de EROS (NEWMAN et al., 2006), alterações na fluidez de membrana (MANNA,
SUNIL K.; SREENIVASAN, YASHIN; SARKAR, 2006), inibição da topoisomerase II
(BIELAWSKI; WINNICKA; BIELAWSKA, 2006), entre outros (CERELLA; DICATO;
DIEDERICH, 2013). No entanto, o mecanismo de ação exato dos esteroides
cardiotônicos ainda não foi completamente elucidado.

20
I.1.2.1) 21-benzilideno digoxina (21-BD)
A 21-benzilideno digoxina é um derivado sintético da digoxina que apresenta
em sua estrutura um grupo estireno adicional no carbono C21 do anel da lactona
(Figura 3). A adição do grupo estireno no anel lactônico, fornece a 21-BD
características diferentes das observadas para outros esteroides carditônicos,
provavelmente porque o grupo aromático adicional pode promover um impedimento
estérico no sítio de ligação da Na,K-ATPase. Por essa razão a Na,K-ATPase possui
uma menor afinidade por 21-BD, em relação a digoxina e ouabaína. Foi
demonstrado que além da Na,K-ATPase, a 21-BD pode ter outros alvos celulares
como Pdr5p, uma proteína da família de transportadores ABC de Saccharomyces
cerevisiae, que teve sua atividade inibida por 21-BD e não pela digoxina. Esses
dados sugerem que 21-BD poderia atuar como um medicamento para reverter o
fenótipo de tumores resistentes a múltiplas drogas (ROCHA et al., 2014).
Avaliando o potencial antitumoral da 21-BD em células de carcinoma de colo
uterino e carcinoma de cólon (HeLa e RKO) verificou-se que ela apresenta uma
citotoxicidade menor (IC50= 56,16 μM ±8,12 μM) em relação a digoxina (IC50=2,2 μM
±0,8 μM) e que não apresentou toxicidade para as células normais de rim canino
(MDCK). A 21-BD induziu a morte de células de ovário de hamister chinês (CHO-K1)
e células HeLa, por apoptose. Mesmo apresentando uma baixa citotoxicidade, a 21-
BD apresentou resultados interessantes que motivaram a continuação dos estudos
com essa molécula. A 21-BD modulou as junções oclusivas aumentando a
expressão de claudina-4 e ZO-1, e inibindo a expressão de claudina-2. Dessa forma
a 21-BD pode fortalecer as junções oclusivas, podendo regular características
importantes na metástase como: migração, transformação, proliferação e invasão
celular (ROCHA et al., 2014).

21
Figura 3: Estrutura química da digoxina e da 21-benzilideno digoxina (Adaptado de
ROCHA et al., 2014)
I.1.3) Na,K-ATPase
A Na,K-ATPase, ou bomba de sódio e potássio, foi descrita em 1957 por Jens
Skou em experimentos com nervo de caranguejo, onde foi demonstrado a existência
de uma adenosina trifosfatase na membrana que estaria envolvida na extrusão de
íons sódio (SKOU, 1957). Através das observações de Skou, sabe-se hoje que a
enzima atua acoplando a energia obtida a partir da hidrólise do ATP, para
transportar íons Na+ para fora da célula e íons K+ para o interior da célula.
Essa enzima está presente em todas as células de animais e o gradiente de
Na+ gerado, através do transporte de Na+/K+, dirige muitos processos por meio de
co-transportadores (ex: sódio/glicose) e trocadores (Na+/Ca2+). Este gradiente
conduz o transporte de aminoácidos e vitaminas para dentro da célula. Além disso, o
gradiente eletroquímico fornece energia para muitos processos fisiológicos
essenciais como, excitabilidade elétrica, transmissão nervosa e contração muscular
(HORISBERGER, 2004).
A Na,K-ATPase (figura 4) é encontrada na maioria das células como um
heterodímero composto por duas subunidades, α e β . A subunidade catalítica, α,

22
que tem uma massa molecular de aproximadamente 112 kDa e apresenta 10
segmentos transmembrânicos onde estão os sítios de ligação para os íons sódio,
potássio, ATP e para os esteroides cardiotônicos, como por exemplo, ouabaína que
é um inibidor específico dessa enzima (BLANCO; MERCER, 1998; LINGREL;
KUNTZWEILER, 1994). São descritas 4 isoformas da subunidade α (α1, α2, α3
(SHULL; GREEB; LINGREL, 1986) e α4 (BLANCO et al., 1999; SHAMRAJ;
LINGREL, 1994). A subunidade α1 é encontrada na maioria dos tecidos, a α2 é
encontrada no músculo esquelético, cérebro e coração, α3 é essencialmente de
tecido cardíaco e neural (LINGREL; KUNTZWEILER, 1994) e a subunidade α4 é
encontrada em testículos (BLANCO et al., 1999; SHAMRAJ; LINGREL, 1994).
A subunidade glicosilada, β, regula a atividade catalítica da enzima. Ela
apresenta um único segmento transmembrânico de massa molecular que varia de
40 a 60kDa dependendo do grau de glicosilação (BLANCO; MERCER, 1998;
KAPLAN, 2002). Existem 3 isoformas da subunidade β : β1, β2 e β3 (MALIK et al.,
1996; MARTIN-VASALLO et al., 1989). A subunidade β1 é amplamente distribuída
entre os tecidos, a β2 é encontrada principalmente em tecido nervoso e a
subunidade β3 está presente na retina, fígado, testículos e pulmão (BLANCO;
MERCER, 1998). As possíveis combinações de α e β resultam em enzimas
cataliticamente funcionais nas células.
Em alguns tipos celulares como, células renais, é encontrada uma terceira
subunidade, a subunidade γ, que foi recentemente identificada. Essa subunidade
apresenta um caráter hidrofóbico e faz parte de um grupo de proteínas com resíduo
conservado FXYD composto por sete proteínas pequenas, possuindo uma massa
molecular entre 7,5 e 14kDa. A subunidade γ parece estar envolvida na modulação
da atividade da enzima (BLANCO; MERCER, 1998).

23
Figura 4: Representação esquemática da estrutura da Na,K-ATPase (Adaptado de
BAGROV; SHAPIRO; FEDOROVA, 2009)
A Na,K-ATPase é classificada como uma ATPase do tipo P, uma classe de
enzimas responsáveis pelo transporte de íons através da membrana, cujo nome
deriva do fato de que um intermediário fosforilado é formado em um resíduo de
aspartato durante a catálise. As ATPases do tipo P contêm cinco domínios
funcionais estruturalmente distintos. Três são domínios citoplasmáticos (um atuador,
A; um de ligação à nucleotídeos, N; um de fosforilação, P) e dois domínios de
membrana (transporte, T; apoio específico, S). Apresenta também um domínio
regulamentar (R) que está situado na região N-terminal ou C-terminal (ou ambos).
Durante cada ciclo catalítico, o domínio P é fosforilado em um resíduo de aspartato
pelo domínio N e subsequentemente desfosforilado pelo domínio A. Desse modo, o
domínio N opera como uma proteína quinase, o domínio A como uma fosfatase e o
domínio P como substrato tanto para as proteínas quinase e fosfatase (PALMGREN;
NISSEN, 2011; PEDERSEN; CARAFOLI, 1987).

24
O mecanismo de reação da Na,K-ATPase, baseia-se no modelo descrito por
Albers-Post, que fundamenta-se na mudança de afinidade da enzima pelos íons Na+
e K+ e na mudança conformacional da mesma (ALBERS; FAHN; KOVAL, 1963;
POST; SEN; ROSENTHAL, 1965). Na etapa inicial a enzima encontra-se no estado
E1, que possui afinidade pelo Na+ citoplasmático. Nessa etapa, a enzima apresenta
um ATP previamente ligado e por afinidade três íons Na+ ligam-se à ela no lado
citoplasmático, o que provoca uma mudança conformacional, fazendo com que o
resíduo de aspartato da enzima seja fosforilado pelo ATP presente no sítio N. A
mudança conformacional para o estado E2 faz com que a enzima perca a afinidade
por Na+ e tenha maior afinidade pelos íons K+. Os sítios de ligação ao sódio são
expostos para a superfície extracelular e nessa conformação os três íons Na+ são
liberados e dois íons K+ extracelulares ligam-se à enzima. A ligação dos íons K+ faz
com que a enzima mude novamente sua conformação o que resulta na
desfosforilação do resíduo de aspartato, pelo domínio A. O domínio N apresenta-se
agora acessível ao nucleotídeo intracelular e a ligação do ATP promove a mudança
do estado E2 para a conformação E1. Na conformação E1 a enzima possui baixa
afinidade para K+ e a mudança conformacional resulta na abertura do canal
intracelular e a subsequente liberação de K+ para o lado citoplasmático. O esquema
do ciclo de reação está representado na figura 5 (KÜHLBRANDT, 2004;
HORISBERGER, 2004; KAPLAN, 2002).

25
Figura 5: Representação esquemática do ciclo de reação da Na,K-ATPase
(Adaptado de KAPLAN, 2002).
.
Além da funcionalidade no transporte de íons e ser extremamente importante
para a homeostase iônica, evidências sugerem que a Na,K-ATPase pode ter função
como receptor e que a ligação de esteroides cardiotônicos na enzima ativa vias de
sinalização intracelular que estão envolvidas na regulação da expressão gênica,
síntese de proteínas e proliferação celular (CHEN et al., 2006; XIE; ASKARI, 2002).
Considerando que a Na,K-ATPase é importante para diversas funções
celulares, a modulação da atividade da enzima pode resultar em danos para a
célula. A atividade da Na,K-ATPase pode ser inibida diretamente pelos esteroides
cardiotônicos ou indiretamente por alterações na fluidez da membrana plasmática
(RAUCHOVÁ; DRAHOTA; KOUDELOVÁ, 1999).

26
I.1.4) Membrana Celular
A membrana celular é formada por uma dupla camada de moléculas
anfifílicas, os lipídios, onde as porções hidrofóbicas se associam excluindo as
interações com a água e as porções hidrofílicas interagem com o ambiente aquoso.
Os lipídios que compõe a membrana são o colesterol e os fosfolipídios como,
fosfatidilserina, fosfatidiletanolamina, fosfatidilinositol, fosfatidilcolina e
esfingomielina. Além de lipídios, as membranas biológicas, apresentam também
proteínas e glicoproteínas. As proteínas presentes na membrana podem estar
inseridas (integrais de membrana), periféricas ou associadas à membrana. As
proteínas integrais são ligadas à membrana por interações hidrofóbicas, enquanto
que proteínas periféricas são mantidas na membrana por interações eletrostáticas.
Os componentes proteicos e lipídicos específicos e as proporções de proteínas para
lipídios variam de acordo com o tipo celular (MARQUARDT; GEIER; PABST, 2015;
NELSON; COX, 2014; NICOLSON, 2014).
O modelo atualmente aceito para representar a membrana plasmática é o que
foi proposto por Singer e Nicolson em 1972. O modelo do mosaico fluido (Figura 6)
apresentado descreve a dinâmica e a estrutura da membrana e foi assim
denominado pela disposição das proteínas e lipídios e pelo fato de que os lipídios
podem se movimentar pela camada e permitem que as proteínas também alterem
sua posição (SINGER; NICOLSON, 1972).

27
Figura 6: Modelo do mosaico fluido (Adaptado de PIETZSCH, 2004).
A membrana da célula é assimétrica, apresentando uma distribuição de
diferentes lipídios, proteínas e glicoproteínas entre as camadas internas e externas.
A camada interna apresenta fosfatidilserina, fosfatidilinositol e fosfatidiletanolamina e
a membrana externa apresenta fosfatidilcolina e esfingomielina. Assim, alterações
nessa distribuição dos fosfolipídios entre a bicamada podem sinalizar diversos
processos como agregação, adesão e apoptose (MARQUARDT; GEIER; PABST,
2015; NICOLSON, 2014).
A membrana plasmática apresenta diferentes fases, ou diferentes estados de
organização e fluidez. Na fase de líquido desordenado, os lipídios estão
desordenados e a membrana possui uma maior fluidez o que permite uma rápida
difusão lateral dos fosfolipídios no plano da camada lipídica. Na fase líquido-
ordenado a movimentação dos lipídios é mais restrita. A fluidez da membrana
plasmática depende da composição lipídica e da temperatura. A presença do
colesterol diminui a fluidez, pois sua estrutura rígida devido ao núcleo esteroidal
restringe a movimentação dos fosfolipídios. O conteúdo de ácidos graxos saturados

28
também pode diminuir a fluidez da membrana, pois as cadeias de ácidos graxos
saturados se acondicionam bem em um conjunto paracristalino (empacotado). No
entanto, as insaturações fornecem ‘dobras’ na cadeia dos ácidos graxos insaturados
e interferem no acondicionamento (empacotamento) aumentando a fluidez da
membrana (NELSON; COX, 2014; VOET; VOET; PRATT, 2008).
As membranas celulares representam importantes barreiras, atuando como
filtros que podem transmitir seletivamente sinais e substâncias do meio externo ou
de células adjacentes para o interior de uma célula. Íons, hormônios, citocinas,
fatores de crescimento, entre outros devem primeiro interagir com os lipídios e
proteínas da membrana para serem internalizados ou iniciar os processos de
sinalização (NELSON; COX, 2014; VOET; VOET; PRATT, 2008).
A primeira estrutura de contato de células normais e cancerosas é a
membrana plasmática. As membranas celulares são essenciais em muitos
processos bioquímicos e em particular, estão intensamente envolvidas na metástase
do câncer. O processo de invasão e metástase requer uma interação complexa
entre as células malignas e as células normais e estas interações são mediadas
principalmente pela membrana das células (NICOLSON, 2015).
As células cancerígenas, em contraste com células normais, apresentam um
metabolismo acelerado e usam intermediários metabólicos principalmente para
processos anabólicos em vez de para a produção de ATP. O crescimento das
células é dependente da síntese de lipídios (FABIANI, 2015) e a composição lipídica
da membrana determina as propriedades bioquímicas e biofísicas dos processos
celulares. A biossíntese dos lipídios tem se tornado um alvo interessante na
bioquímica do câncer devido ao elevado grau da dependência dessas biomoléculas,
em especial do colesterol, para a célula tumoral (GORIN; GABITOVA;
ASTSATUROV, 2012)
A modulação da atividade das proteínas envolvidas na síntese de lipídios, ou
a modulação direta da composição do colesterol e fosfolipídios das membranas
celulares pode causar um efeito intenso sobre o tráfico vesicular, sinalização e
atividade de enzimas integrais com potenciais implicações para a terapia do câncer.

29
I.1.4.1) Interação lipídio-proteína
A composição lipídica e a fluidez da bicamada podem modular a atividade de
proteínas associadas a membrana, como, a Na,K-ATPase (DE LIMA SANTOS et al.,
2005). Além disso, a cristalografia de raio-x da estrutura da proteína revelou a
ligação de fosfolipídios e colesterol em sítios específicos da enzima, e que a
atividade catalítica também é dependente dessas interações específicas entre
lipídios e proteína (CORNELIUS et al., 2015; HABECK et al., 2015). Foi
demonstrado que a enzima possui três sítios específicos de ligações aos
fosfolipídios e colesterol. Um sítio (A) onde ocorre a interação da fosfatidilserina e
colesterol, estabilizando a atividade da enzima; um sítio (B) de interação com
fosfatidilcolina e fosfatidiletanolamina poliinsaturadas que estimulam a atividade
catalítica e o sítio (C) onde a interação com o colesterol e fosfatidilcolina causam a
inibição da atividade enzimática (CORNELIUS et al., 2015).
A estabilização das proteínas integrais depende da interação entre elas e os
lipídios da membrana celular. A primeira camada de lipídios que revestem a
superfície das proteínas integrais de membrana é denominada de lipídios anulares e
as interações entre esses lipídios e as proteínas são estabilizadas por interações
polares entre o grupo cabeça polar dos fosfolipídios e aminoácidos específicos da
proteína e por interações apolares entre a cadeia de ácidos graxos e o domínio
transmembranar. Existem também os lipídios não anulares, que não circundam a
proteína, mas que também contribuem para a estabilização desses na membrana,
através de interações hidrofóbicas. A composição dos lipídios anulares da Na,K-
ATPase são principalmente fosfatidilcolina, fosfatidilserina e colesterol
(CONTRERAS; ERNST; WIELAND, 2011; CORNELIUS et al., 2015).
Geralmente, os modelos utilizados para se estudar a relação entre a
composição lipídica e a atividade da Na,K-ATPase são os lipossomos. Nesses
modelos a reconstituição da enzima é feita em lipossomos de composição lipídica
definida a fim de entender a influência e as interações entre os lipídios e a proteína.
Os estudos têm demonstrado que o conteúdo de colesterol, comprimento da cadeia
de ácidos graxos dos fosfolipídios, saturações e tipo de fosfolipídios (grupo polar)
influenciam na atividade e na estabilidade da Na,K-ATPase (DE LIMA SANTOS et
al., 2005). Cornelius e colaboradores (2001 e 2003) demonstraram que a
composição lipídica da membrana plasmática é provavelmente muito importante na

30
regulação da atividade da Na,K-ATPase e que o mecanismo de reação da enzima é
modificada através da interação proteína-lipídio (CORNELIUS; TURNER;
CHRISTENSEN, 2003; CORNELIUS, 2001).
I.1.4.2) Esteroides Cardiotônicos e Modulação do perfil lipídico
Existem poucos relatos na literatura mostrando o efeito do tratamento com
esteroides cardiotônicos sobre o conteúdo de lipídios e a modulação da fluidez da
membrana celular. Manna e colaboradores (2006) demonstraram que o tratamento
com oleandrina causou alteração da fluidez e da microviscosidade da membrana
plasmática em diferentes linhagens celulares. Os autores sugeriram que devido a
estrutura química da oleandrina, por conter um núcleo esteroidal como outros
esteroides cardiotônicos, ela poderia interagir diretamente com os lipídios da
membrana e assim alterar a fluidez. Em células, como macrófagos e neutrófilos, foi
observado que a oleandrina impede a ligação de interleucina-8 (IL-8) ao receptor (IL-
8R). Além desse ligante, a oleadrina afeta a ligação de fator de crescimento
epidermal (EGF) e fator de crescimento nervoso (NGF) em seus respectivos
receptores. A diminuição da afinidade e ligação dos ligantes em seus respectivos
receptores pode ter sido causada pela alteração da fluidez de membrana que
consequentemente pode levar a uma alteração na conformação dos receptores
(MANNA, SUNIL K.; SREENIVASAN, YASHIN; SARKAR, 2006).
O mesmo grupo de pesquisadores demonstrou que a oleandrina induz
apoptose em diferentes linhagens tumorais humanas (células T Jurkat; linfoma -U-
937 e HL-60; colo uterino -Hela; mama -MCF-7), porém células de murinos foram
insensíveis à esse esteroide cardiotônico. Dessa forma, sugeriram que as diferentes
respostas dessas linhagens celulares frente ao tratamento com oleandrina seria pela
interação da droga com a membrana de células humanas e não com células
murinas. Os autores sugerem ainda que a ligação da oleandrina pode ser afetada
pela composição lipídica da membrana plasmática, uma vez que a composição
lipídica da membrana das células humanas pode ser diferente das células murinas
(RAGHAVENDRA; SREENIVASAN; MANNA, 2007).
Além dos estudos com oleandrina, existe um relato na literatura
demonstrando que a ouabaína alterou a fluidez da membrana plasmática de células

31
de câncer de próstata. Os pesquisadores demonstraram alterações nos níveis de
esfingomielina e fosfatidilinositol, que são lipídios importantes na sinalização celular
(GASPER; VANDENBUSSCHE; GOORMAGHTIGH, 2011).
I.1.5) Estresse oxidativo e peroxidação lipídica
As espécies reativas de oxigênio (EROS) são moléculas que apresentam um
par de elétrons desemparelhados e por isso são instáveis e extremamente reativas.
São produzidos na mitocôndria durante a respiração celular pelos complexos da
cadeia de transporte de elétrons. São moléculas importantes para diversas funções
celulares, porém o excesso pode levar ao estresse oxidativo que é definido como um
desequilíbrio entre moléculas oxidantes (EROS) e antioxidantes, onde os primeiros
estão em nível mais elevado do que o segundo, resultando em danos moleculares e
celulares (LICHTENBERG; PINCHUK, 2015).
EROS como, radicais hidroxila (OH-), peróxido de hidrogênio (H2O2) e radical
superóxido (O2) podem atacar as macromoléculas das células como proteínas, DNA
e lipídios, formando um novo radical que pode se propagar em uma reação em
cadeia. O ataque às proteínas pode resultar em modificações em sua estrutura e
alteração ou perda da função; o ataque ao DNA pode causar mutações e o ataque
aos lipídios de membrana pode causar uma variedade de modificações funcionais
(LICHTENBERG; PINCHUK, 2015).
A peroxidação lipídica é uma das consequências mais estudadas da ação de
EROS na estrutura e função da membrana. A membrana celular contem fosfolipídios
com a cadeia de ácidos graxos insaturados e a peroxidação ocorre quando as
EROS atacam as insaturações desses ácidos graxos formando radicais
hidroperóxidos que propagam a reação (MAFRA, 1999). Um dos produtos formados
na peroxidação lipídica é o malondialdeído, que é utilizado como biomarcador desse
processo oxidativo (ABDALLA; DE SENA, 2008). A reação em cadeia causa lesões
na membrana que contribuem para a perda de funções celulares através da
alteração na composição lipídica, fluidez e inativação de proteínas de membrana.
A atividade de enzimas integrais da membrana, como a Na,K-ATPase, podem
ser inibidas em consequência do estresse oxidativo. Diretamente, a atividade pode
ser inibida pelo ataque de EROS à proteína, causando modificações em sua

32
estrutura como carbonilação ou indiretamente, por alterações na fluidez da
membrana, provocada pela peroxidação lipídica (RAUCHOVÁ; DRAHOTA;
KOUDELOVÁ, 1999).
A redução da atividade da Na,K-ATPase foi observada em pacientes
hipertensos e os dados indicam uma correlação entre a diminuição da atividade da
enzima e o aumento da peroxidação lipídica, consequência do estresse oxidativo
que é um fato central desse processo patológico. A peroxidação lipídica provocou
mudanças na composição dos ácidos graxos, aumentando o conteúdo de ácidos
graxos saturados em relação aos ácidos graxos mono e poliinsaturados. A diferença
na composição lipídica da membrana pode resultar na diminuição da fluidez da
membrana e ,consequentemente, na diminuição da atividade da Na,K-ATPase
(RODRIGO et al., 2007). Em um modelo animal de isquemia cerebral, também foi
demonstrado o aumento da peroxidação lipídica e a diminuição da atividade da
Na,K-ATPase. Nesse estudo o uso do antioxidante resveratrol, reverteu a inibição da
Na,K-ATPase observada em ratos com isquemia, sugerindo que a atividade da
enzima é inibida sob o estresse oxidativo (SIMÃO et al., 2011).
As células possuem mecanismos de defesa que as protegem contra os danos
provocados por EROS. Uma das principais barreiras das células contra o ataque de
EROS à membrana celular é a glutationa reduzida (GSH) e as principais enzimas
antioxidantes presentes nas células são a catalase (CAT), superóxido dismutase
(SOD), glutationa peroxidase (GPx) e glutationa redutase (GR) (ARMSTRONG,
2011).
A GSH é um tripeptídeo com grupos tiol (-SH), um bom nucleófilo, o que a
torna um potente agente redutor. Ela remove as EROS e lipidioperóxidos em uma
reação catalisada pela glutationa peroxidase (GPx), convertendo H2O2 em H2O e os
lipidioperóxidos em seus respectivos alcoóis, resultando na formação de glutationa
oxidada (GSSH). A GSSH é novamente convertida à GSH pela glutationa redutase
(GR). A superóxido-dismutase é uma metaloproteína e se classifica de acordo com o
metal presente em sua estrutura, que pode ser cobre (CuSOD), zinco (ZnSOD), ferro
(FeSOD) ou manganês (MnSOD). MnSOD é encontrada na matriz mitocondrial e
CuSOD e ZnSOD são encontrada no espaço intermembranas da mitocôndria e no
citosol. A superóxido dismutase faz a conversão do radical superóxido em peróxido
de hidrogênio que é decomposto pela catalase. A catalase está localizada em
peroxisomos e ela decompõe o peróxido de hidrogênio em água e oxigênio

33
(ARMSTRONG, 2011). As enzimas antioxidantes estão integradas em um sistema
eficiente de defesa contra os danos celulares (Figura 7).
Figura 7: Esquema de integração das enzimas antioxidantes. SOD: Superóxido
dismutase; CAT: catalase; GPx: glutationa peroxidase; GSH: glutationa reduzida;
GSSG: glutationa oxidada; GRD: glutationa redutase; NADPH: Fosfato de
dinucleotído de nicotinamida e adenina.
I.2) JUSTIFICATIVA E OBJETIVOS
I.2.1) Justificativa
O câncer está entre as principais causas de morte no mundo todo
(GLOBOCAN, 2012) e por essa razão, tem sido crescente a busca por
medicamentos eficazes contra essa doença. Os esteroides cardiotônicos, entre eles
a digoxina, têm mostrado efeitos antitumorais interessantes. No entanto, o
mecanismo de ação pelo qual eles exercem o efeito citotóxico não está totalmente
compreendido.
A 21-benzilideno digoxina é um esteroide cardiotônico, mais precisamente um
glicosídeo cardíaco, que foi recentemente sintetizado a partir da estrutura da
digoxina. Apesar de essa molécula apresentar uma menor citotoxicidade em células
tumorais, em relação à digoxina, ela tem apresentado efeitos relevantes e diferentes

34
em relação aos efeitos já descritos por outros esteroides cardiotônicos, como por
exemplo a inibição da proteína Pdr5p e fortalecimento das junções oclusivas que
foram observado após o tratamento com 21-BD e não com digoxina (ROCHA et al.,
2014). Tais observações motivam a continuação dos estudos sobre os efeitos
causados nas células após o tratamento com essa molécula.
Existem poucos registros na literatura mostrando o efeito dos esteroides
cardiotônicos sobre a modulação do perfil lipídico e do estado redox de células
tumorais. Devido a importância do conteúdo lipídico da membrana celular para as
funções intracelulares e tendo indícios de que a alteração do conteúdo lipídico pode
estar envolvida com processos característicos de células malignas (FABIANI, 2015)
faz-se necessário analisar a modulação, causada por esses fármacos, no conteúdo
dos lipídios em células tumorais. As células tumorais apresentam altos níveis de
EROS e alterações da atividade das enzimas antioxidantes, como forma de
adaptação ao estresse oxidativo. O aumento da geração de EROS poderia induzir
danos nas células e até a morte celular (BULDAK et al., 2014).
Tendo em vista a importância da integridade da membrana e a manutenção
do equilíbrio redox para a funcionalidade adequada da célula, é importante investigar
se a modulação do perfil lipídico e do estado redox das células HeLa podem estar
incluídos no mecanismo de ação dos efeitos antitumorais apresentados pelos
esteroides cardiotônicos, digoxina e 21-BD.
I.2.2) Objetivo geral
Avaliar o efeito dos esteroides cardiotônicos, digoxina e 21-BD, sobre a
atividade da Na,K-ATPase, modulação do perfil lipídico e estado redox de células de
carcinoma de colo uterino (HeLa).
I.2.3) Objetivos específicos
Avaliar a atividade da Na,K-ATPase e a expressão da subunidade α1 nas
células de carcinoma de colo uterino, após o tratamento com 21-BD;
Analisar o conteúdo de fosfolipídios totais e colesterol total nas células e na
membrana plasmática, após o tratamento com digoxina e 21-BD;

35
Avaliar a formação de malondialdeído (MDA), produto da peroxidação lipídica,
após o tratamento das células com os glicosídeos cardíacos;
Determinar a atividade da catalase e superóxido dismutase; e os níveis de
glutationa reduzida após o tratamento das células com os glicosídeos
cardíacos;
II. MATERIAL E MÉTODOS
II.1) Cultura de células
A linhagem de células HeLa (carcinoma de colo uterino, ATCC CCL2) foram
gentilmente cedidas pela Fundação Ezequiel Dias (FUNED) de Belo Horizonte, MG.
As células foram cultivadas em estufa a 37ºC, atmosfera de 5% de CO2 e em meio
DMEM estéril suplementados com 10% de soro fetal bovino e 0,1% de gentamicina.
O meio de cultura foi trocado a cada dois dias e as células lavadas com tampão PBS
estéril [pH=7,4] (NaCl 137 mM, KCl 2,7 mM e PO43- 10 mM). Para o repique celular,
o meio foi descartado, as células lavadas com PBS e incubadas por 3 minutos com
tripsina 0,025%. Em seguida, as células foram ressuspendidas em meio DMEM e
uma alíquota foi retirada para contagem das células em câmara de Neubauer. A
concentração de células adequada para cada experimento foi adicionada em
garrafas ou placas de cultivo.
II.2) Tratamento com esteroides cardiotônicos
A digoxina foi obtida por doação da Fundação Ezequiel Dias (FUNED) de
Belo Horizonte, MG. A 21-benzilideno digoxina, derivado da digoxina, foi
recentemente sintetizada no laboratório de Síntese Orgânica da Universidade
Federal de São João del-Rei (ROCHA et al., 2014). As células foram tratadas com
50 nM, 150 nM e 1 µM de digoxina e 5 µM e 50 µM 21-BD. As soluções estoque
foram preparadas diluindo-se as drogas em 100% de DMSO (dimetilsulfóxido). Em
seguida, a solução de digoxina foi diluída em tampão PBS autoclavado e a solução
de 21-BD foi diluída em água MiliQ ultrapura autoclavada. Para evitar o efeito

36
citotóxico do DMSO, as drogas foram diluídas à uma concentração que continha no
máximo 1% de DMSO. As soluções preparadas foram mantidas a -20°C.
II.3) Preparação do homogenato total
Um volume da suspensão correspondente a 5x105 células foram plaqueadas
em placa de seis poços e cultivadas até atingirem 80% de confluência, quando
foram tratadas com as concentrações estabelecidas anteriormente, por 48 horas.
Em seguida, as células foram retiradas da placa com o auxílio de rupper em 500 µL
de tampão (Tris-HCl 6 mM, imidazol 20 mM, sacarose 0,25 M, SDS 0,01%, EDTA
0,1 M e inibidor de protease PMSF 2 mM) e sonicadas por três ciclos de 15
segundos e 45% de potência. As amostras foram mantidas em -20°C.
II.4) Preparação de membrana plasmática
Uma alíquota correspondente a 2,25x106 células foram colocadas em garrafas
de 75 cm2 e cultivadas até atingirem 80% de confluência. Em seguida as células
foram tratadas por 48 horas com diferentes concentrações de 21-BD e digoxina.
Após o tratamento as células foram lavadas com PBS e removidas com auxílio do
rupper em 3mL de tampão (Tris-HCl 6 mM, imidazol 20 mM, sacarose 0,25 M, SDS
0,01%, EDTA 0,1 M e inibidor de protease PMSF 2mM). As amostras foram
homogeneizadas em homogeinizador de tecidos tipo potter por 20 vezes e
posteriormente centrifugadas em ultracentrífuga rotor WTi 45 por 20 minutos a
10.000g. O pellet foi descartado e o sobrenadante centrifugado novamente a
70.000g. O pellet obtido após a segunda centrifugação foi ressuspendido em 300 µL
do tampão. As amostras foram armazenadas em freezer a -20°C.
II.5) Preparação da membrana livre de hemoglobina (Ghost)
A preparação da membrana plasmática de eritrócitos humanos (ghost) foi
realizada pela metodologia descrita por REGA e colaboradores (1979). As hemácias

37
foram separadas do plasma e dos leucócitos por centrifugação a 6000g por 17
minutos a 4°C. Em seguida, foram resuspensas em tampão (Tris-HCl 20 mM, pH
7,4, KCl 130 mM e inibidor de protease PMSF 0,6 mg/ml) e centrifugadas a 6000g
por 17 minutos, a 4°C. O pellet foi submetido a congelamento em nitrogênio líquido e
descongelado à temperatura ambiente, sendo em seguida resuspenso em tampão
(Hepes 5 mM, pH 7,4 EDTA 1mM e PMSF 0,6 mg/mL) e centrifugado novamente a
8000g por 17 minutos a 4°C. Esta etapa foi repetida 4 vezes, sendo a última
lavagem feita na ausência de PMSF e de EDTA. Após cada lavagem com tampão as
amostras eram homogeneizadas em homogeneizador de tecidos tipo Potter e após
cada centrifugação o sobrenadante foi descartado. Em seguida o pellet obtido foi
ressuspenso em tampão (Hepes pH 7,4) e centrifugado 8000g por 17 minutos a 4°C.
Novamente o sobrenadante foi descartado e o pellet ressuspendido em tampão
(Tris-HCl 10 mM, pH 7,4, KCl 130 mM, MgCl2 0,5 mM e CaCl2 50 μM) e centrifugado
a 8000g por 17 minutos, a 4ºC. O pellet obtido corresponde a fração de membranas
de eritrócitos livre de hemoglobina, e foi resuspenso no último tampão e estocado
em freezer -80ºC.
II.6) Dosagem da atividade ATPásica de Na,K-ATPase
A atividade ATPásica foi determinada pela quantificação do fosfato inorgânico
(Pi) liberado pela hidrólise de ATP pela Na,K-ATPase, usando o método
colorimétrico de FISKE e SUBBAROW (1925). A atividade catalítica foi realizada em
meio reacional contendo: Hepes 50 mM, NaCl 120 mM, KCl 20 mM, MgCl2 2 mM.
Esse experimento foi realizado em placa de ELISA de 96 poços. Foi adicionado em
cada poço 15 µg de proteína, 25 µL de meio reacional e a reação foi iniciada com a
adição de 3 mM de ATP. O experimento foi realizado sem e com o inibidor da
enzima, ouabaína 1 mM. A placa foi incubada em estufa a 37ºC por 1 hora,
marcando a partir do primeiro poço onde se iniciou a reação. Após 1 hora a placa é
retirada da estufa e adicionado 100 µL de SDS 1% sem fosfato (dodecil sulfato de
sódio) em cada poço, para parar a reação, seguindo a ordem em que a reação foi
iniciada. Posteriormente, foi adicionado 100 µL da solução de molibdato de
amônio/reagente de Fiske na proporção 5:1. Após a adição da solução esperou-se
15 minutos e em seguida realizou-se a leitura da placa de ELISA em
espectrofotômetro, em um comprimento de onda de 660 nm.

38
II.7) Dosagem de proteínas totais
A dosagem de proteína foi realizada pelo método colorimétrico de
BRADFORD (1976) em placa de 96 poços. As amostras foram diluídas em 50x e em
seguida foram adicionados 40 µL dessas diluições nos poços. Posteriormente foi
adicionado 200 µL de reagente de Bradford e a placa foi deixada em repouso por 15
minutos. As leituras foram realizadas em espectrofotômetro no comprimento de onda
de 595 nm. Foi ultilizada uma curva padrão variando-se as concentrações de
albumina.
II.8) Extração de lipídios
As frações de membrana de células HeLa, de eritrócitos e o homogenato total
de células foram submetidas à extração por solvente pelo método de Folch adaptado
(FOLCH; LESS; STANLEY, 1987). Foi adicionado às amostras 5 mL de
clorofórmio/metanol (2:1), deixando-as em agitação por 1 hora a temperatura
ambiente. Em seguida, foi adicionado 1 mL de NaCl 0,9% e agitadas em vórtex por
alguns segundos. As amostras foram centrifugadas a 670g por 20 minutos e
deixadas em repouso por 30 minutos, para melhorar a separação de fases. Após a
separação de fases, foi adicionado à fase orgânica metanol/água (1:1) e à fase
aquosa foi adicionado clorofórmio. As amostras foram agitadas novamente em
vórtex por alguns segundos e deixadas em repouso na geladeira até o dia seguinte.
As fases foram separadas e a fase orgânica foi concentrada em rotaevaporador e
ressuspendidas em clorofórmio. Os extratos foram estocados em freezer -20ºC.
II.9) Dosagem do conteúdo de fosfolipídios totais
A quantificação de fosfolipídios totais foi realizada utilizando-se o
procedimento descrito por CHEN e colaboradores (1956). Uma alíquota de 40 µL
das amostras dos extratos lipídicos foi seca em nitrogênio e digerida com 0,5 mL de
ácido nítrico 65 % (v/v) à temperatura de 120ºC. Em seguida, foi adicionada à
amostra 0,5 mL de água destilada e 1,0 mL do reagente de Chen. As amostras

39
foram levadas ao banho-maria a 45ºC por 20 minutos. O reagente de Chen
complexa-se com o fosfato restante da digestão da cadeia de ácidos graxos dos
fosfolipídios resultando em um produto de coloração azulada. As amostras então
são submetidas à análise espectrofotométrica em comprimento de onda de 820 nm.
Como padrão, foi utilizada uma solução de dihidrogênio fosfato de sódio 1,0 µM.
II.10) Dosagem do conteúdo de colesterol total
A determinação do conteúdo de colesterol foi realizada pelo método descrito
por HIGGINS (1987), baseado na complexação do cloreto férrico (FeCl3.6H2O ) com
o colesterol em meio ácido. Uma alíquota de 40 µL das amostras de extrato de
lipídico foi seca em nitrogênio, e em seguida foi adicionado 0,75 mL de ácido acético
glacial e 0,5 mL do Reagente B. A amostra foi mantida em repouso por 10 minutos à
temperatura ambiente e quantificada em espectrofotômetro em um comprimento de
onda de 550 nm. O reagente B foi preparado pela diluição de 4 mL do Reagente A
em 46 mL de ácido sulfúrico. Para a preparação do reagente A, foi realizada a
dissolução de 2,5 g de FeCl3.6H2O em 100 mL de ácido ortofosfórico 85 %. As
determinações foram feitas em triplicata e a quantidade foi calculada com relação a
uma curva padrão. A curva padrão foi feita através de uma solução padrão de
colesterol 1 mg/mL.
II.11) Determinação de substâncias reativas ao ácido tiobarbitúrico (TBARS)
Uma quantidade de 7,5x105 células HeLa foi adicionada em garrafas de
25cm2 e cultivadas até atingirem confluência. Em seguida as células foram tratadas
com digoxina e 21-BD por 1 hora e por 48 horas. Após o tratamento as células foram
removidas com rupper, ressuspendidas em 0,5 mL de água ultrapura e sonicadas
por 5 ciclos de 20 segundos em 35% de potência. Foram utilizados 150 µL das
amostras para a dosagem de TBARS. Foi adicionado 1 mL de solução (15% TCA
(ácido tricloro acético), 0,8% TBA (ácido tiobarbitúrico), 0,25 M HCl) às amostras e
em seguida elas foram aquecidas em banho-maria à uma temperatura de 95ºC por
15 minutos. Posteriormente foram deixadas em repouso a temperatura ambiente, até

40
esfriarem. As leituras foram realizadas em espectrofotômetro em comprimento de
onda de 532 nm. Foi utilizado como padrão o malondialdeído (MDA).
II.12) Dosagem da atividade das enzimas antioxidantes
Uma alíquota correspondente a 7,5x105 células foi adicionada em garrafas de
25 cm2. As células foram tratadas por 1 hora com as concentrações pré-definidas de
digoxina e 21-BD e em seguida, foram lavadas com PBS e removidas com o auxílio
de rupper. Posteriormente as amostras foram sonicadas por 3 ciclos de 20 segundos
a 20% de potência e centrifugadas a 13.000g por 10 minutos. O pellet foi descartado
e o sobrenadante foi utilizado nas análises do conteúdo de glutationa reduzida,
atividade enzimática da superóxido dismutase e da catalase.
II.13) Determinação da atividade enzimática da Superóxido Dismutase (SOD)
A determinação da atividade da SOD foi determinada pelo método de Misra e
Fridovich (1972). Esse método se baseia na capacidade da SOD em inibir a
autoxidação da epinefrina à adrenocromo (MISRA; FRIDOVICH, 1972).
Primeiramente realizou-se a leitura do meio reacional (tampão glicina 50 mM pH 10
e 25 µL de epinefrina (60 mM pH 2)), o que corresponde a 100% da reação. Em
seguida foram adicionados 10, 20 e 30 µL de cada amostra ao meio e a reação foi
acompanhada por dois minutos em cada situação. As leituras foram realizadas em
480 nm. A atividade enzimática foi expressa em unidades da enzima necessárias
para inibir 50% da velocidade de formação de adenocromo. Os dados foram
plotados em unidade de enzima/µg de proteína
II.14) Determinação da atividade enzimática da catalase (CAT)
A atividade enzimática da catalase foi determinada pelo método de Aebi
(1984) que avalia a decomposição do peróxido de hidrogênio. O espectrofotômetro
foi zerado com o meio de reação (tampão fosfato 50 µM pH 7,4 e peróxido de

41
hidrogênio), para cada amostra foi utilizada uma alíquota 50 µL e adicionada ao
meio de reação. A decomposição do peróxido foi acompanhada por 1 minuto,
anotando-se a absorbância a cada 15 segundos. As leituras foram realizadas em
240 nm. Uma unidade de enzima foi determinada como sendo 1µmol de H202
decomposto por minuto. Os gráficos foram plotados em razão de Unidade de
enzima/µg de proteína (AEBI, 1984).
II.15) Determinação do conteúdo de Glutationa Reduzida (GSH)
A dosagem do conteúdo de GSH consiste na reação do grupamento sulfidrila
da GSH com o reagente DTNB (ácido 5,5'-ditio-bis-2- nitrobenzóico), formando o
TNB ou (ácido 5-tio-2-nitrobenzóico) de coloração amarela, ue pode ser quantificado
em espectrofotometro em 412 nm. A formação de TNB é diretamente proporcional à
concentração de GSH na amostra. A concentração de GSH foi avaliada nas
amostras de sobrenadante em placa de 96 poços. Uma alíquota de 50 μL das
amostras foi adicionada à 225 μL de meio reacional (tampão fosfato 0,1 M pH 8,0 e
EDTA1 mM), em seguida foi adicionado DNTB 10 mM. A placa foi deixada em
repouso por 15 minutos em temperatura ambiente e quantificada em
espectrofotômetro em comprimento de onda de 412 nm. Como padrão, foi utilizada
uma solução de cisteína 1,5mM (BEUTLER; DURON; KELLY, 1963; JOLLOW,
1974).
II.16) Análises de expressão proteica por Western Blotting
As amostras de preparação de membrana foram diluídas em água MiliQ
ultrapura e tampão de amostra (Tris-HCl 0,125mM [pH=6,8], Azul de Bromofenol
0,004%, 2-Mercaptoetanol 10%, Glicerol 20%, e SDS 4%) para uma concentração
de 20µg de proteína. Posteriormente, as amostras foram corridas em um gel de
poliacrilamida 10% (Sodium dodecyl sulfate-polyacrylamide gel electrophoresis -
SDS-PAGE) (Gel de corrida – Tris 1,5M [pH=8,8], Acrilamida/bis-acrilamida
30,8%/2,7%, SDS 10%, Persulfato de Amônio 10% e TEMED [N,N,N’,N’-
tetramethylethylenediamine]; Gel de empacotamento – Tris 0,5M [pH=6,8],

42
Acrilamida/bis-acrilamida 30,8%/2,7%, SDS 10%, Persulfato de Amônio 10% e
TEMED; Tampão de corrida – Tris-base 25mM, Glicina 192mM e SDS 0,1%). Após a
corrida das amostras, foi realizada a transferência das proteínas do gel de
poliacrilamida para uma membrana de nitrocelulose por 2 horas em tampão de
transferência –Tris-base 25mM, Glicina 192mM e Metanol 20%. A eficiência da
transferência foi avaliada corando-se o gel de poliacrilamida com solução de Azul de
Coomassie Brilhante G-250 e a membrana de nitrocelulose com solução de
vermelho Ponceau Xilidina. Para bloquear ligações inespecíficas dos anticorpos
utilizados, a membrana foi bloqueada por 1 hora com leite desnatado 5% diluído em
T-TBS (Tris-Base 100mM, NaCl 0,9% e Tween 0,1%). Em seguida, a membrana foi
lavada com T-TBS por 5 minutos e por duas vezes e incubada overnight com
anticorpo primário anti-α1 (Santa Cruz – sc-21712) diluído em T-TBS 1:1000. No dia
seguinte, a membrana foi lavada três vezes por 10 minutos e incubada por 2 horas
com anticorpo secundário goat anti-mouse IgG-HRP (Santa Cruz – sc-2005) diluído
1:2000 em T-TBS. Posteriormente, a membrana foi lavada duas vezes por 5 minutos
e revelada por método de quimioluminescência utilizando uma mistura equimolar de
duas soluções (Solução 1: Tris 100mM [pH=8,5], Luminol 2,5mM e ácido p-cumárico
0,396mM; Solução 2: Tris 100mM [pH=8,5] e H2O2 0,06%). Após a exposição à
mistura, a membrana foi revelada em filme radiográfico (Medical X-ray Film -
KODAK). Os filmes radiográficos foram digitalizados e as bandas correspondentes a
cada amostra foi quantificada por densitometria utilizando-se o programa Image J
1.47.
II.17) Análise Estatística
Foi utilizado o teste One-way ANOVA (Analysis of Variance) para comparação
de mais de duas médias, seguida do teste de Tukey (Tukey’s post-hoc test) para
identificação das médias com diferença estatisticamente significantes. Todas as
análises e gráficos foram feitos utilizando-se o programa GraphPad Prism 5.

43
III. RESULTADOS E DISCUSSÃO
Os esteroides cardiotônicos são inibidores clássicos da Na,K-ATPase. Em um
trabalho recente do nosso gupo foi demostrado a inibição da enzima pela digoxina
nas concentrações testadas (50 nM, 150 nM e 1 µM) (PESSOA, 2014), dessa forma,
como já é conhecido o efeito da digoxina sobre a atividade da Na,K-ATPase, no
presente trabalho avaliamos a atividade da enzima após o tratamento somente com
21-BD.
Após o tratamento das células por 48 horas com 21-BD a atividade catalítica
foi mensurada. Foi observado que o tratamento com 5 µM ocasionou um aumento
de 34% da atividade da Na,K-ATPase, enquanto que, o tratamento com 50 µM
diminuiu em 64% a atividade da enzima. O perfil apresentado na Figura 8 sugere um
efeito bifásico da 21-BD, sendo que a menor concentração da droga estimulou a
atividade da Na,K-ATPase e a maior concentração inibiu a atividade da enzima.
Figura 8: Atividade da Na,K-ATPase de células Hela tratadas por 48h com 21-BD.
*Diferença significativa em relação ao grupo controle (p< 0,05); n=3.
O grupo estireno adicional na molécula da 21-BD pode promover um
impedimento estérico no sítio de ligação da Na,K-ATPase, além disso, essa

44
modificação proporciona efeitos diferentes em relação aos observados com a
digoxina. Nosso grupo demonstrou através de modelagem molecular que a 21-BD
se liga à Na,K-ATPase com uma afinidade similar a ouabaína e digoxina. Porém
experimentos cinéticos mostraram que a 21-BD foi capaz de inibir a atividade da
enzima com isoforma α1, expressa em células de inseto Sf9 e em preparações de
membrana de rim de ratos, somente em concentrações elevadas. E não causou a
inibição da enzima de hemisfério cerebral de ratos, que possui em sua maioria as
isoformas α2 e α3. Tais resultados sugerem uma menor afinidade da enzima pela 21-
BD em relação à ouabaína. Além disso o tratamento de células HeLa por 48 horas
com 10µM de 21-BD causou um aumento da atividade da Na,K-ATPase (ROCHA et
al., 2014).
Já foi relatado o aumento da atividade da Na,K-ATPase após o tratamento
com esteróides cardiotônicos, por exemplo digoxina e ouabaína, porém em doses
mais baixas (em concentrações nanomolares) (GAO et al., 2002; OSELKIN; TIAN;
BERGOLD, 2010) do que a menor dose testada de 21-BD. Sabe-se também que
essas doses podem alterar a expressão das subunidades da Na,K-ATPase. Em
miocárdio ventricular de ratos normais a ouabaína diminuiu a expressão da
subunidade α1 e aumentou a expressão de α3, a digoxina também aumentou a
expressão de α3 e a expressão de α1 permaneceu inalterada nesse tratamento. A
expressão de α2 permaneceu inalterada em ambos os tratamentos. (WANG; YUAN;
LU, 2001). Tian e colabores (2009) demonstraram o aumento da expressão da
subunidade α1 em células normais de rim de porco (LLC-PK1) e a diminuição da
expressão em células tumorais de câncer de mama humano (BT-20) e células de
câncer de próstata humano (DU145), sugerindo que a regulação da expressão seja
dependente do tipo celular (TIAN et al., 2009).
O tratamento com 50 µM de 21-BD aumentou os níveis de mRNA de Na,K-
ATPase em células HeLa e aumentou a expressão proteica em células normais de
rim canino (MDCK) (ROCHA et al., 2014). Para comprender se a 21-BD altera a
atividade da enzima por modular a expressão proteica, foi realizado um westen
blotting das frações de membrana das células após 48 horas de tratamento.
Não foram observadas diferenças significativas na expressão da subunidade
α1 da Na,K-ATPase na membrana plasmática das células HeLa após o tratamento
(Figura 9). Esse dado sugere que a atividade da Na,K-ATPase pode estar sendo
modulada por outro fator que não seja a expressão proteica, pois mesmo com os

45
níveis de expressão de Na,K-ATPase inalterados na membrana celular, a atividade
da enzima foi alterada nas células HeLa.
Figura 9: Expressão da subunidade α1 da Na,K-ATPase da membrana plasmática
de células HeLa tratadas por 48 horas com 21-BD.
A fluidez da membrana plasmática pode modular a atividade de enzimas
integrais de membrana, como a Na,K-ATPase. Tendo em vista que a alteração da
fluidez de membrana é um dos efeitos provocados nas células pelos esteroides
cardiotônicos (RAGHAVENDRA; SREENIVASAN; MANNA, 2007), foi avaliada a
composição lipídica das células HeLa após o tratamento por 48 horas com 21-BD e
com digoxina.
As dosagens lipídicas das células foram observadas em dois tipos de
amostras. Primeiramente foi avaliado o conteúdo lipídico total da célula, para isso foi
realizado um homogenato total e foi também realizado a preparação de membrana
das células, para avaliar a modulação dos lipídios presentes na membrana.

46
O tratamento com doses mais baixas de digoxina não causou efeito
significativo no conteúdo de colesterol, porém a concentração de 1 µM causou uma
diminuição de 30% do colesterol total das células. Não houve alterações no
conteúdo do colesterol da membrana plasmática em nenhuma das concentrações de
digoxina testadas (Figura 10).
Figura 10: Porcentagem de colesterol total do homogenato (A) e da membrana
plasmática (B) de células HeLa tratadas por 48 horas com digoxina. * Diferença
significativa em relação ao grupo controle (p< 0,05); n=3.
A 21-BD não alterou significativamente o conteúdo de colesterol total das
células, no entanto, aumentou em duas vezes os níveis desse lipídio na membrana
plasmática das células que foram tratadas na concentração de 50 µM (Figura 11).
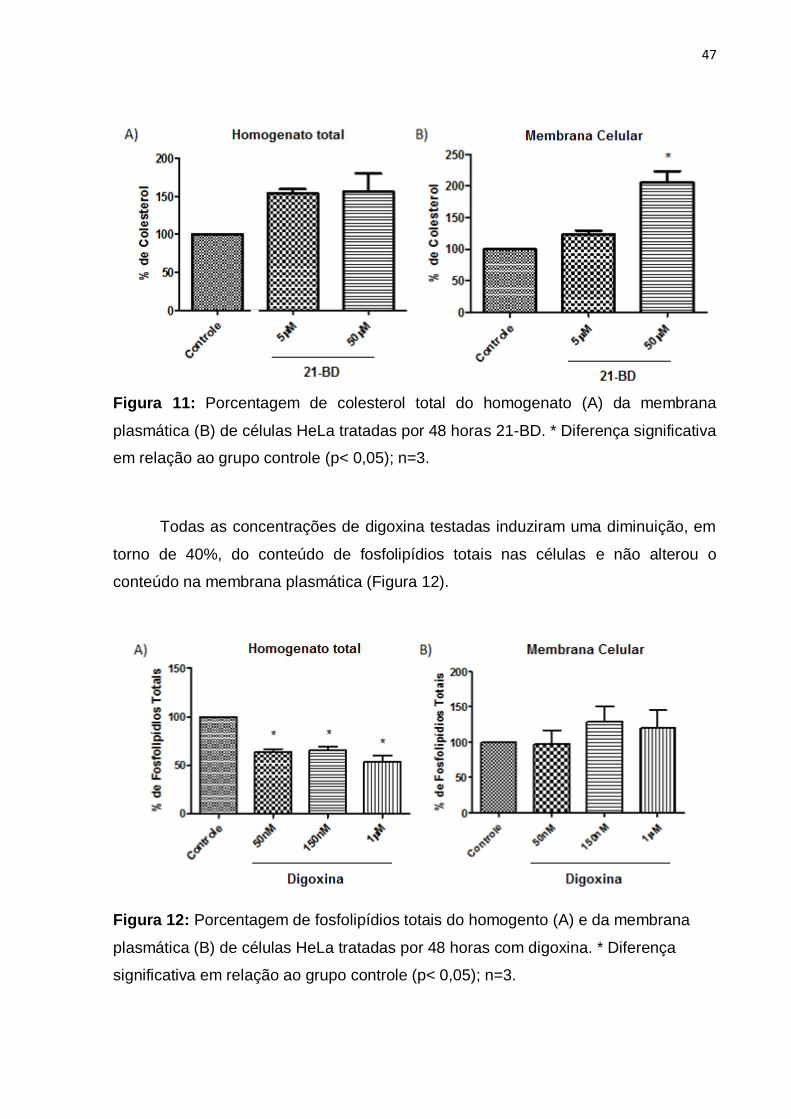
47
Figura 11: Porcentagem de colesterol total do homogenato (A) da membrana
plasmática (B) de células HeLa tratadas por 48 horas 21-BD. * Diferença significativa
em relação ao grupo controle (p< 0,05); n=3.
Todas as concentrações de digoxina testadas induziram uma diminuição, em
torno de 40%, do conteúdo de fosfolipídios totais nas células e não alterou o
conteúdo na membrana plasmática (Figura 12).
Figura 12: Porcentagem de fosfolipídios totais do homogento (A) e da membrana
plasmática (B) de células HeLa tratadas por 48 horas com digoxina. * Diferença
significativa em relação ao grupo controle (p< 0,05); n=3.

48
A concentração de 50 µM de 21-BD causou uma diminuição em torno de
40% dos fosfolipídios totais nas células e um aumento de 50% do conteúdo de
fosfolipídios totais da membrana plasmática (Figura 13). Esse resultado indica que a
21-BD pode estar causando a translocação de fosfolipídios totais da célula para a
membrana.
Figura 13: Porcentagem de fosfolipídios totais do homogento (A) e da membrana
plasmática (B) de células HeLa tratadas por 48 horas com 21-BD. * Diferença
significativa em relação ao grupo controle (p< 0,05); n=3.
Sabe-se que os esteróides cardiotônicos podem interagir com a membrana
através da interação com os lipídios e assim exercerem seus efeitos nas células.
Com o objetivo de verificar se os esteróides utilizados conseguem modular os
lipídios por agirem diretamente na membrana das células, foram utilizadas frações
de membrana plasmática de eritrócitos e essas foram incubadas com as maiores
concentrações de digoxina e 21-BD.
Não foi observado efeito significativo dos esteróides sobre o perfil lipídico da
membrana de eritrócitos (Figura 14), sugerindo que não seja uma modulação lipídica
direta causada pela droga na membrana, mas possivelmente a alteração observada
no conteúdo dos lipídios na célula e na membrana celular seja provocado por
alguma resposta da própria célula por exemplo, uma alteração no metabolismo dos
lipídios.

49
Figura 14: Porcentagem de colesterol total (A) e de fosfolipídios totais (B) de
membrana de eritrócitos incubadas com digoxina e 21-BD por 1 hora.
O primeiro estudo que avaliou o efeito do tratamento com esteroides
cardiotônicos no metabolismo de colesterol em células demonstrou que a digoxina e
ouabaína estimularam a síntese de colesterol em diferentes tipos celulares (células
hepáticas - HepG2; células de câncer de cólon - HT29; células de leucemia - THP-1
e cardiomioblasto H9c2). As duas drogas regularam a etapa limitante da síntese de
colesterol nessas células. Os dados mostram que ambas aumentaram a síntese de
colesterol, por aumentar a atividade e a expressão da HMG-CoA redutase, de
maneira dose e tempo-dependente. O aumento da expressão e atividade da enzima
é provocado pela ativação do complexo SRBP2/SCAP. Os autores sugerem que
ouabaína e digoxina podem se ligar ao domínio de ligação a esterol na SCAP,
impedindo que o colesterol se ligue e por essa razão mantenha o complexo
SRBP2/SCAP ativo (CAMPIA et al., 2009). No entanto, em nosso modelo de estudo,
células HeLa, a digoxina causou a diminuição do conteúdo de colesterol total
enquanto que a 21-BD aumentou os níveis desse lipídio nas células.
Nossos dados mostram que a 21-BD, causou efeitos interessantes sobre o
conteúdo de colesterol e fosfolipídios totais, aumentando o conteúdo desses lipídios,
na membrana das células HeLa. As mudanças na composição de fosfolipídios e do
conteúdo de colesterol alteram a fluidez da membrana plasmática das células

50
podendo alterar as funções celulares que dependem da membrana (BLOJ et al.,
1973).
Há muitos anos tem se investigado se existe uma correlação entre a fluidez
de membrana e a atividade da Na,K-ATPase. Como essa proteína é integral de
membrana e são necessárias interações entre seus resíduos hidrofóbicos e os
lipídios da bicamada, as alterações no conteúdo de fosfolipídios e colesterol
poderiam alterar a fluidez da membrana e interferir na atividade catalítica de tais
enzimas. Muitos desses estudos têm apontado que a atividade da Na,K-ATPase é
modulada por seu microambiente lipídico ( CORNELIUS et al., 2015; RODRIGO et
al., 2014; YONEDA et al., 2014; DE LIMA SANTOS et al., 2005; DIOMEDE et al.,
1992; TANAKA et al., 1988; YEAGLE, 1983;KIMELBERG, 1975; BLOJ et al., 1973).
Muitos dos trabalhos encontrados na literatura apontam uma correlação direta
entre a diminuição da fluidez da membrana e a inibição da atividade da Na,K-
ATPase. Em 1975, Kimelberg demonstrou que o aumento do colesterol diminuia a
atividade da Na,K-ATPase purificada de rim de coelho, solubilizada em lipossomos
(KIMELBERG, 1975). Em um outro estudo com Na,K-ATPase reconstituída em
lipossomos, foi demonstrado que a quantidade de colesterol afeta diretamente a
atividade da atividade enzimática. A incorporação do colesterol no modelo da
bicamada lipídica pode alterar o estado de líquido-cristalino por uma fase com um
grau intermediário de organização. O efeito inibidor do colesterol é provavelmente
causado por uma redução do movimento molecular ou limitação da liberdade
conformacional da enzima na bicamada. Isso porque a alteração na composição
lipídica, pelo aumento do colesterol, provoca uma diminuição da fluidez de
membrana que restringe as alterações conformacionais da enzima prejudicando a
atividade catalítica (YONEDA et al., 2014).
Tanaka e colaboradores (1988) demonstraram a diminuição da atividade da
enzima assim como, a diminuição da fluidez da membrana durante o processo de
diferenciação de células epidérmicas de porquinho da índia (TANAKA et al., 1988). A
diminuição da fluidez da membrana e a redução da atividade da Na,K-ATPase
também foi demonstrada em células leucêmicas humanas, onde as células que
apresentavam um maior conteúdo de colesterol exibiram uma menor atividade da
Na,K-ATPase (células HL60 suplementadas com colesterol em relação as células
HL60), e que células que apresentavam um menor conteúdo de colesterol exibiram
uma maior atividade enzimática (K562 depletadas de colesterol e células K562)

51
(DIOMEDE et al., 1992). Além disso, a diminuição da fluidez de membrana causada
pelo aumento de ácidos graxos saturados em relação aos ácidos graxos
insaturados, provocou a inibição da Na,K-ATPase na membrana de eritrócitos de
pacientes hipertensos (RODRIGO et al., 2014).
No presente trabalho, o tratamento com 50 µM de 21-BD causou o aumento
do conteúdo de colesterol e fosfolipídios totais da membrana plasmática das células
HeLa. Essa mesma concentração diminuiu significativamente a atividade da Na,K-
ATPase nessas células. Esses resultados em conjunto, sugerem que a diminuição
da atividade da enzima esteja correlacionada com a possível diminuição da fluidez
da membrana celular, causada pelo aumento do conteúdo de colesterol e
fosfolipídios (especialmente se forem fosfolipídios de cadeia saturada), uma vez que
a Na,K-ATPase possui baixa afinidade para 21-BD e que não foi encontrado
alteração na expressão proteica da subunidade α1. Os mecanismos moleculares
pelos quais a composição lipídica da membrana influencia na atividade da Na,K-
ATPase provavelmente envolve alterações conformacionais da proteína, devido ao
estado físico dos lipídios que se encontram aumentados levarem à restrições do
movimento molecular necessário para o ciclo de reação.
Tem sido demonstrado que o aumento de colesterol, aumenta a espessura da
bicamada lipídica, o que pode deslocar o equilíbrio conformacional da Na,K-ATPase
de E2P/E1P para E1P e de E2/E1 para E1 (CORNELIUS et al., 2015). Além disso,
as alterações estruturais podem resultar em um aumento da exposição do sítio de
ligação ao esteroide cardiotônico e por consequência, aumentarem a sensibilidade
ao ligante (RODRIGO et al., 2014).
A fluidez e integridade da membrana celular podem ser comprometidas por
ataque de EROS aos lipídios da membrana, levando à peroxidação lipídica. Alguns
estudos demonstram que o tratamento com esteroides cardiotônicos causam um
aumento na produção de EROS em diferentes linhagens celulares, podendo causar
danos as células e atuar como segundo mensageiros em eventos de sinalização
celular, que desencadeiam processos como apoptose e autofagia. O tratamento de
células de osteosarcoma (MG-63) com bufalina causou um aumento da geração de
EROS de maneira dose dependente e induziu a apoptose das células por aumentar
a permeabilidade mitocondrial e permitir a ativação de caspases (WANG; BI, 2014).
Em células de câncer de cólon (HT-29 e Caco-2) a bufalina causou autofagia das
células através da geração de EROS via JNK (XIE et al., 2011). A digoxina e

52
ouabaína induziram o aumento de EROS, mediada por Src, em linhagens de células
não pequenas de pulmão (A549 e H460) e também induziram a morte celular por
autofagia (WANG et al., 2014). Em um outro estudo, foi demonstrado que a
ouabaína aumentou a produção de EROS e a concentração de Ca2+ em células de
linfoma (U937), o que causou a morte celular (CUOZZO et al., 2012).
Tendo em vista alguns estudos demonstrando que alguns esteroides
cardiotônicos induzem o estresse oxidativo, e que por essa razão podem causar
danos celulares como, a peroxidação lipídica; foi avaliado o conteúdo de
malondialdeído (MDA), atividade de enzimas antioxidantes (CAT e SOD) e os níveis
de GSH após o tratamento 21-BD e digoxina. Tais parâmetros podem ser utilizados
como biomarcadores do estresse oxidativo.
O tratamento com digoxina nas concentração de 150 nM por 48 horas e, 1
μM de digoxina e 50 μM de 21-BD por 1 e 48 horas causaram uma redução da
peroxidação lipídica da membrana celular, representada pela diminuição de
aproximadamente 30% dos níveis de MDA (Figura 15).
Figura 15: Determinação da peroxidação lipídica de células HeLa tratadas por 48
horas (A) e por 1 hora (B) com digoxina e 21-BD. Determinação foi realizada pela
quantificação de MDA produzido em homogenato total de células. *Diferença
significativa em relação ao grupo controle (p< 0,05); n=3.

53
A maioria dos estudos encontrados sobre os efeitos do estresse oxidativo
causado pelos esteroides cardiotônicos são realizados em animais. A administração
de ouabaína in vivo causou o aumento de EROS, peroxidação lipídica e do nível de
proteínas carboniladas no cérebro como um todo (BRÜNING et al., 2012) e em
algumas regiões específicas (RIEGEL et al., 2010) como prefrontal (JORNADA et al.,
2011), córtex cerebral e hipocampo (BROCARDO et al., 2010; SOUZA et al., 2014).
Também foi observado o aumento de EROS seguido do aumento da peroxidação
lipídica e diminuição dos níveis de GSH, em homogenato de fígado que foram
incubados com digoxina. Tais observações foram revertidas quando as amostras
foram tratadas com antioxidantes em conjunto com a droga, demonstrando que a
peroxidação lipídica foi consequência do estresse oxidativo provocado pelo
tratamento com os esteroides cardiotônicos (ROY; SAHA; SENGUPTA, 2001).
O pré-tratamento de células de linfoma (U937) por 1 hora com oleandrina
diminuiu a produção de EROS e diminuiu também a peroxidação lipídica induzida
por TNF (fator de necrose tumoral), o que os autores relacionaram com o bloqueio
da ativação de NF-κB (fator nuclear kappa B) por TNF causada pela oleandrina
(MANNA et al., 2000). Porém, em células HeLa, a oleandrina aumentou a produção
de EROS e a peroxidação lipídica consideravelmente. E o pré-tratamento com
oleandrina por 4 horas potencializou o efeito da ceramida aumentando a geração de
EROS e a peroxidação lipídica nesse tipo celular (SREENIVASAN; SARKAR;
MANNA, 2003).
No presente trabalho, a peroxidação lipídica diminuída após o tratamento com
21-BD, pode ser explicada pelo aumento do colesterol e fosfolipídios totais na
membrana celular. A peroxidação lipídica ocorre principalmente em ácidos graxos
insaturados, pode ser que a 21-BD aumente a composição lipídica de ácidos graxos
saturados em relação aos insaturados que são mais susceptíveis ao ataque de
EROS. Embora a digoxina não tenha modulado o conteúdo de colesterol e
fosfolipídios totais, pode haver alterações em de fosfolipídios específicos que
conduzam essa diminuição observada na peroxidação lipídica da membrana.
Após o tratamento com as drogas, as enzimas antioxidantes sofreram
alterações em sua atividade. A atividade da superóxido dismutase (Figura 16) foi
reduzida significativamente, enquanto que os níveis da catalase (Figura 17) e da
GSH (Figura 18) se mantiveram.

54
Contr
ole
Dig
oxina
50nM
Dig
oxina
150n
M M
Dig
oxina
1M
21-B
D 5
M
21-B
D 5
0
0
10
20
30
40
50
* * * **
U/
g d
e p
rote
ína
Figura 16: Atividade catalítica da superóxido dismutase de células HeLa tratadas
por 1 hora com digoxina e 21-BD. *Diferença significativa em relação ao grupo
controle (p< 0,05); n=3.
Contr
ole
Dig
oxina
50nM
Dig
oxina
150n
M M
Dig
oxina
1M
21-B
D 5
M
21-B
D 5
0
0.00
0.02
0.04
0.06
U/
g d
e p
rote
ína
Figura 17: Atividade catalítica da catalase de células HeLa tratadas por 1 hora com
digoxina e 21-BD. *Diferença significativa em relação ao grupo controle (p< 0,05);
n=3.

55
Contr
ole
Dig
oxina
50nM
Dig
oxna
150n
M M
Dig
oxina
1M
21-B
D 5
M
21-B
D 5
0
0.00
0.05
0.10
0.15
nm
ol
GS
H/
g p
rote
ína
Figura 18: Níveis de GSH de células HeLa tratadas por 1 hora com digoxina e 21-
BD. *Diferença significativa em relação ao grupo controle (p< 0,05); n=3.
Os relatos encontrados na literatura demonstram que a atividade das enzimas
antioxidantes são alteradas em resposta ao aumento de EROS após o tratamento
com esteroides cardiotônicos. A atividade da glutationa redutase, glutationa
peroxidase e catalase foram reduzidas em algumas regiões específicas do cérebro
(BROCARDO et al., 2010; BRÜNING et al., 2012; JORNADA et al., 2011; RIEGEL et
al., 2010; SOUZA et al., 2014). O desequilíbrio entre a produção de EROS e a
atividade das enzimas antioxidantes, é a causa dos danos observados em lipídios e
proteínas.
Em células de melanoma humano, a oleandrina aumentou a produção de
EROS, especificamente de radicais superóxido. O aumento dos níveis de EROS
causou a diminuição dos níveis de GSH, levando ao aumento do estresse oxidativo,
e induziu a morte celular (NEWMAN et al., 2006). No presente trabalho, os níveis de
GSH permaneceram praticamente inalterados. Além do aumento dos lipídios da
membrana, que podem ser a causa da diminuição da peroxidação lipídica discutido
anteriormente, nós podemos inferir que os níveis de GSH continuariam protegendo a
membrana do ataque de EROS e por essa razão, os níveis de MDA foram
reduzidos.

56
Alguns tipos de células tumorais possuem níveis aumentados de EROS
(BULDAK et al., 2014) e que o tratamento com esteroides cardiotônicos
potencializam a produção dessas moléculas reativas, principalmente radicais
superóxido (NEWMAN et al., 2006). O aumento de EROS pode levar ao aumento da
atividade das enzimas antioxidantes como uma forma de proteger as células aos
danos celulares, ou também podem inibir a atividade dessas enzimas. A inibição das
enzimas antioxidantes causada por EROS pode ser provocada pelo ataque das
próprias moléculas reativas à estrutura da proteína. Por exemplo, o aumento de
peróxido de hidrogênio ou íons superóxido induzido pelos esteroides cardiotônicos
podem resultar na inibição da atividade da superóxido dismutase por oxidar resíduos
de aminoácidos no sítio de ligação ao substrato. A diminuição da atividade da
superóxido dismutase resulta em uma menor formação de moléculas de peróxido de
hidrogênio, devido a menor decomposição dos íons superóxido pela enzima
(ARMSTRONG, 2011; CHAUDHARY; PARVEZ, 2012). A atividade da catalase se
manteve inalterada, assim como os níveis de GSH, podemos inferir que esses dados
sejam devido a inibição da superóxido dismutase e consequente diminuição de
peróxidos de hidrogênio.
As modificações oxidativas nas proteínas, causadas por EROS como,
carbonilação em alguns resíduos de aminoácidos podem resultar na diminuição da
atividade de outras enzimas além das antioxidantes, por exemplo a Na,K-ATPase.
Os resíduos de lisina, argenina, treonina e prolina das subunidades da enzima
podem ser carbonilados por EROS. Além disso, os resíduos de cisteínas podem
sofrer glutationilação. Foi demonstrado em células LLC-PK1, que o aumento da
geração de EROS causou a carbonilação reversível da subunidade α1 da Na,K-
ATPase, que resultou na inibição do fluxo de Na+. Tais modificações são
responsáveis pela perca da atividade catalítica, pois podem estabilizar a enzima na
conformação E2 ou bloqueiam a ligação do ATP. Além de levar a inibição da
enzima, o aumento de EROS e o estresse oxidativo podem levar a enzima à
degradação (YAN et al., 2013).
A diminuição da atividade da Na,K-ATPase, observada quando as células
foram tratadas com 50 µM de 21-BD, também pode estar correlacionada com o
aumento da produção de EROS induzida por esse esteroide cardiotônico.

57
IV. CONSIDERAÇÕES FINAIS
Os efeitos dos esteroides cardiotônicos em células tumorais não são
totalmente conhecidos, são poucos os estudos existentes na literatura em relação
aos efeitos desses fármacos sob a modulação do perfil lipídico e o estado redox de
células tumorais.
A 21-BD, derivado sintético da digoxina, tem apresentado efeitos
interessantes em células tumorais. Foi demonstrado um apresentou um efeito
bifásico na atividade da Na,K-ATPase das células após o tratamento, onde a droga
foi capaz de inibir a atividade da enzima somente na maior concentração testada.
Nossos resultados em conjunto indicam que a inibição da atividade da Na,K-ATPase
está correlacionada com a diminuição da fluidez da membrana e com o possível
aumento de EROS, uma vez que a expressão da subunidade α1 não sofreu
alterações e que a enzima possui baixa afinidade pela 21-BD.
Tanto a digoxina, quanto a 21-BD provocaram alterações no conteúdo celular
de colesterol e fosfolipídios totais, no entanto efeitos na composição lipídica da
membrana foram observados somente em células tratadas com 21-BD. As
alterações na composição lipídica da membrana podem comprometer eventos
fundamentais para a sobrevivência celular, como alterações conformacionais das
proteínas integrais de membrana e de receptores, podendo comprometer vias de
sinalização celular.
As duas drogas inibiram a atividade da superóxido dismutase, possivelmente
por causa do aumento da produção de EROS induzido pelo tratamento. No entanto
a atividade da catalase e os níveis de GSH não foram alterados, o que pode explicar
a diminuição da peroxidação lipídica observada após os tratamentos.
Portanto, a modulação lipídica e a alteração do estado redox das células
HeLa tratadas com 21-BD e digoxina, são efeitos que provavelmente façam parte do
complexo mecanismo de ação dos fármacos dessa classe.

58
V. REFERÊNCIAS BIBLIOGRÁFICAS
ABDALLA, D. S. P.; DE SENA, K. C. M. Biomarcadores de peroxidação lipídica na aterosclerose. Revista de Nutricao, v. 21, n. 6, p. 749–756, 2008.
AEBI, H. Oxygen Radicals in Biological Systems. Methods in Enzymology, v. 105,
n. 1947, p. 121–126, 1984.
ALBERS, R. W.; FAHN, S.; KOVAL, G. J. The Role Of Sodium Ions In The Activation Of Electrophorus Eletric Organ Adenosine Triphosphatase. Proceedings of the National Academy of Sciences of the United States of America, v. 50, p. 474–
481, 1963.
ALEVIZOPOULOS, K. et al. Na+/K+ ATPase Inhibitors in Cancer. Current Drug Targets, v. 15, p. 988–1000, 2014.
ALONSO, E. et al. Nanomolar ouabain elicits apoptosis through a direct action on HeLa cell mitochondria. Steroids, v. 78, n. 11, p. 1110–1118, 2013.
ALVES, S. L. G. et al. γ-Benzylidene digoxin derivatives synthesis and molecular modeling: Evaluation of anticancer and the Na,K-ATPase activity effect. Bioorganic & Medicinal Chemistry, v. 23, n. 15, p. 4397–4404, 2015.
ARMSTRONG, D. Oxidative Stress in Applied Basic Research and Clinical Practice. [s.l: s.n.].
BAGROV, A. Y.; SHAPIRO, J. I.; FEDOROVA, O. V. Endogenous Cardiotonic Steroids : Physiology , Pharmacology , and Novel Therapeutic Targets. Pharmacological Reviews, v. 61, n. 1, p. 9–38, 2009.
BEUTLER, E. .; DURON, O. .; KELLY, B. M. Improved method for the determination of blood glutathione. J Lab Clin Med, v. 61, p. 882–888, 1963.
BIELAWSKI, K.; WINNICKA, K.; BIELAWSKA, A. Inhibition of DNA topoisomerases I and II, and growth inhibition of breast cancer MCF-7 cells by ouabain, digoxin and proscillaridin A. Biological & pharmaceutical bulletin, v. 29, n. 7, p. 1493–1497,
2006.
BLACKBURN, ELIZABETH H. GREIDER, C. W.; SZOSTAK, J. W. Nobel Prize.
Disponível em: <http://www.nobelprize.org/nobel_prizes/medicine/laureates/2009/>.
BLANCO, G. et al. Functional characterization of a testes-specific alpha-subunit isoform of the sodium/potassium adenosinetriphosphatase. Biochemistry, v. 38, n. 41, p. 13661–13669, 1999.
BLANCO, G.; MERCER, R. W. Isozymes os the Na,K-ATPase: heterogeneity in structure, diversity in function. Review Literature And Arts Of The Americas, v. 11,
n. 6, p. 633–650, 1998.
BLOJ, B. et al. Membrane fluidity, cholesterol and allosteric transitions of membrane bound Mg2-ATPase, Na,K-ATPase e acetylcholinesterase from rat erytrhocytes. v.

59
38, n. 1, p. 101–105, 1973.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry, v. 72, p. 248–254, 1976.
BROCARDO, P. S. et al. Folic acid administration prevents ouabain-induced hyperlocomotion and alterations in oxidative stress markers in the rat brain. Bipolar Disorders, v. 12, n. 4, p. 414–424, 2010.
BRÜNING, C. A. et al. Diphenyl diselenide ameliorates behavioral and oxidative parameters in an animal model of mania induced by ouabain. Progress in Neuro-Psychopharmacology and Biological Psychiatry, v. 38, n. 2, p. 168–174, 2012.
BRYAN, T. M. et al. Telomere length dynamics in telomerase-positive immortal human cell populations. Experimental cell research, v. 239, n. 2, p. 370–8, 1998.
BULDAK, R. J. et al. Significance of selected antioxidant enzymes in cancer cell progression. Pol J Pathol, v. 65, n. 3, p. 167–175, 2014.
CAMPIA, I. et al. Digoxin and ouabain increase the synthesis of cholesterol in human liver cells. Cellular and Molecular Life Sciences, v. 66, n. 9, p. 1580–1594, 2009.
CASEY, S. C. et al. Cancer prevention and therapy through the modulation of the tumor. Seminars in Cancer Biology, 2015.
CERELLA, C.; DICATO, M.; DIEDERICH, M. Assembling the puzzle of anti-cancer mechanisms triggered by cardiac glycosides. Mitochondrion, v. 13, n. 3, p. 225–234, 2013.
CHAUDHARY, S.; PARVEZ, S. An in vitro approach to assess the neurotoxicity of valproic acid induced oxidative stress in cerebellum and cerebral cortex of young rats. Neuroscience, v. 225, p. 258–268, 2012.
CHAVES, A. L. et al. Effects of digoxin and Na, K-ATPase immunoexpression on human oral squamous carcinomas. Anticancer Res, v. 34, p. 5397–403, 2014.
CHEN, J. Q. et al. Sodium/potasium ATPase (Na+, K+-ATPase) and ouabain/related cardiac glycosides: A new paradigm for development of anti- breast cancer drugs? Breast Cancer Research and Treatment, v. 96, n. 1, p. 1–15, 2006.
CHEN, P. S. et al. Microdetermination of phosphorus. Analytical chemistry, v. 28, p. 1756–1758, 1956.
CÍFKOVÁ, E. et al. Determination of lipidomic differences between human breast cancer and surrounding normal tissues using HILIC-HPLC/ESI-MS and multivariate data analysis. Analytical and Bioanalytical Chemistry, v. 407, n. 3, p. 991–1002, 2014.
CONTRERAS, F.; ERNST, A. M.; WIELAND, F. Specificity of Intramembrane Protein − Lipid Interactions. Cold Spring Harbor Perspectives in Biology, v. 3, p. 1–18,
2011.

60
CORNELIUS, F. Modulation of Na , K-ATPase and Na-ATPase Activity by Phospholipids and. Biochemistry, v. 40, n. 15, p. 8842–8851, 2001.
CORNELIUS, F. et al. General and specific lipid–protein interactions in Na,K-ATPase. Biochimica et Biophysica Acta (BBA) - Biomembranes, v. 1848, n. 9, p. 1729–1743, 2015.
CORNELIUS, F.; TURNER, N.; CHRISTENSEN, H. R. Z. Modulation of Na,K-ATPase by phospholipids and cholesterol. II. Steady-state and presteady-state kinetics. Biochemistry, v. 42, n. 28, p. 8541–8549, 2003.
CUOZZO, F. et al. Pro-death and pro-survival properties of ouabain in U937 lymphoma derived cells. Journal of Experimental & Clinical Cancer Research, v. 31, n. 1, p. 95, 2012.
DE LIMA SANTOS, H. et al. Na,K-ATPase reconstituted in liposomes: Effects of lipid composition on hydrolytic activity and enzyme orientation. Colloids and Surfaces B: Biointerfaces, v. 41, n. 4, p. 239–248, 2005.
DIOMEDE, L. et al. Modulations of ATPase activity by cholesterol and synthetic ether lipids in leukemic cells. v. 43, n. 4, 1992.
FABIANI, E. DE. Targeting lipid metabolism to starve and stop cancer cells. European Journal of Lipid Science and Technology, v. 117, n. 7, p. 905–907, 2015.
FERRIAR, J. An Essay On The Medical Properties Of The Digitalis purpurea Or Foxglove. In: Medical Histories And Reflections. First Amer ed. Manchester:
Sowler and Russell, 1799. p. 253–289.
FISKE, C. H.; SUBBAROW, Y. The colorimetric determination of phosphorus. The Journal of Biological Chemistry, v. 66, p. 375–400, 1925.
FOLCH, J.; LESS, M.; STANLEY, G. H. S. A simple method for the isolation and purification of total lipides from animal tissues. Journal of Biological Chemistry, v. 226, p. 497–509, 1987.
GAO, J. et al. Isoform-specific stimulation of cardiac Na/K pumps by nanomolar concentrations of glycosides. J Gen Physiol, v. 119, n. 4, p. 297–312, 2002.
GASPER, R.; VANDENBUSSCHE, G.; GOORMAGHTIGH, E. Ouabain-induced modifications of prostate cancer cell lipidome investigated with mass spectrometry and FTIR spectroscopy. Biochimica et Biophysica Acta - Biomembranes, v. 1808, n. 3, p. 597–605, 2011.
GEY, G. O.; COFFMAN, W. D.; KUBICEK, M. T. Tissue culture studies of the proliferative capacity of cervical carcinoma and normal epithelium. Cancer Res., v.
12, p. 264–265, 1952.
GLOBOCAN. Cancer Incidence, Mortality and Prevalence Worldwide. Disponível
em: <http://globocan.iarc.fr/old/burden.asp?selection_pop=224900&Text-p=World&selection_cancer=4162&Text-

61
c=Cervix+uteri&pYear=3&type=0&window=1&submit= Execute>.
GORIN, A.; GABITOVA, L.; ASTSATUROV, I. Regulation of cholesterol biosynthesis and cancer signaling. Current Opinion in Pharmacology, v. 12, n. 6, p. 710–716,
2012.
HABECK, M. et al. Stimulation , Inhibition , or Stabilization of Na , K-ATPase Caused by Specific Lipid Interactions at Distinct Sites. The Journal of Biological Chemistry, v. 290, n. 8, p. 4829–4842, 2015.
HANAHAN, D.; WEINBERG, R. A. Hallmarks of cancer: The next generation. Cell, v. 144, n. 5, p. 646–674, 2011.
HAUSEN, H. ZUR. Nobel Prize. Disponível em: <http://www.nobelprize.org/nobel_prizes/medicine/laureates/2008/hausen-facts.html>.
HAUX, J. Digitoxin is a potential anticancer agent for several types of cancer. Medical hypotheses, v. 53, n. 6, p. 543–548, 1999.
HIGGINS, G. In biological Membranes. [s.l.] Pratical Approach Series, 1987.
HORISBERGER, J.-D. Recent insights into the structure and mechanism of the sodium pump. Physiology (Bethesda, Md.), v. 19, p. 377–387, 2004.
INCA. O que é o câncer? Disponível em: <http://www1.inca.gov.br/conteudo_view.asp?id=322>.
JOHANSSON, S. et al. Cytotoxicity of digitoxin and related cardiac glycosides in human tumor cells. Anti-cancer drugs, v. 12, n. 5, p. 475–483, 2001.
JOLLOW, D. J. ET AL. Bromobenzene-induced liver necrosis. Protective role of glutathione and evidence for 3,4-bromobenzene oxide as the hepatotoxic metabolite. Pharmacology, v. 11, p. 151–169, 1974.
JORNADA, L. K. et al. Lithium and valproate modulate antioxidant enzymes and prevent ouabain-induced oxidative damage in an animal model of mania. Journal of Psychiatric Research, v. 45, n. 2, p. 162–168, 2011.
KAPLAN, J. H. Biochemistry of Na,K-ATPase. Annual Review of Biochemistry, v. 71, n. 1, p. 511–535, 2002.
KIMELBERG, H. K. Activity due to lipid fluidity. Biochimica et Biophysica Acta, v. 413, p. 143–156, 1975.
KLABUNDE, R. E. Cardiac Glycosides (Digitalis Compounds). Disponível em: <http://www.cvpharmacology.com/cardiostimulatory/digitalis>. Acesso em: 4 ago. 2015.
KÜHLBRANDT, W. Biology, structure and mechanism of P-type ATPases. Nature Reviews Molecular Cell Biology, v. 5, n. 4, p. 282–295, 2004.

62
LI, Z.; XIE, Z. The Na/K-ATPase/Src complex and cardiotonic steroid-activated protein kinase cascades. Pflugers Archiv European Journal of Physiology, v. 457, n. 3, p. 635–644, 2009.
LICHTENBERG, D.; PINCHUK, I. Oxidative stress, the term and the concept. Biochemical and Biophysical Research Communications, v. 461, n. 3, p. 441–
444, 2015.
LINGREL, J. B.; KUNTZWEILER, T. Na,K-ATPase Minireview. The Journal of Biological Chemistry, v. 269, p. 19659–19662, 1994.
LÓPEZ-LÁZARO, M. et al. Digitoxin inhibits the growth of cancer cell lines at concentrations commonly found in cardiac patients. Journal of Natural Products, v. 68, n. 11, p. 1642–1645, 2005.
LUCEY, B. P.; NELSON-REES, W. A.; HUTCHINS, G. M. Henrietta Lacks, HeLa cells, and cell culture contamination. Archives of Pathology and Laboratory Medicine, v. 133, n. 9, p. 1463–1467, 2009.
MAFRA, D. Peroxidação Lipídica Em Pacientes Com Insuficiência Renal Crônica Lipid. Rev. Nutr., v. 12, n. 2, p. 205–212, 1999.
MALIK, N. et al. Identification of the mammalian Na,K-ATPase 3 subunit. The Journal of biological chemistry, v. 271, n. 37, p. 22754–8, 1996.
MANNA, S. K. et al. Oleandrin Suppresses Activation of Nuclear Transcription Factor- KB , Activator Protein-1, and c-Jun NH2-Terminal Kinase. Cancer Research, v. 60, p. 3838–3847, 2000.
MANNA, SUNIL K.; SREENIVASAN, YASHIN; SARKAR, A. Cardiac Glycoside Inhibits IL-8-Induced Biological Responses by Downregulating IL-8 Receptors Through Altering Membrane Fluidity. Journal of cellular physiology, v. 207, n. 2, p. 195–207, 2006.
MARQUARDT, D.; GEIER, B.; PABST, G. Asymmetric Lipid Membranes: Towards More Realistic Model Systems. Membranes, v. 5, n. 2, p. 180–196, 2015.
MARTIN-VASALLO, P. et al. Identification of a putative isoform of the Na,K-ATPase beta subunit. Primary structure and tissue-specific expression. J Biol Chem, v. 264,
n. 8, p. 4613–4618, 1989.
MCCONKEY, D. J. et al. Cardiac Glycosides Stimulate Ca 2 + Increases and Apoptosis in Androgen-independent , Metastatic Human Prostate Adenocarcinoma Cells Cardiac Glycosides Stimulate Ca 2 ؉ Increases and Apoptosis in Androgen-. Cancer Research, v. 60, n. 713, p. 3807–3812, 2000.
MELERO, C. P.; MEDARDE, M.; SAN FELICIANO, A. A short review on cardiotonic steroids and their aminoguanidine analogues. Molecules, v. 5, n. 1, p. 51–81, 2000.
MIJATOVIC, T. et al. Cardiotonic steroids on the road to anti-cancer therapy. Biochimica et Biophysica Acta - Reviews on Cancer, v. 1776, n. 1, p. 32–57, 2007.

63
MIJATOVIC, T.; KISS, R. Cardiotonic steroids-mediated Na+/K+-ATPase targeting could circumvent various chemoresistance pathways. Planta Medica, v. 79, n. 3-4, p. 189–198, 2013.
MISRA, H. P.; FRIDOVICH, I. The Role of Superoxide Anion in the Autoxidation of Epinephrine and a Simple Assay for Superoxide Dismutase The Role of Superoxide Anion in the Epinephrine and a Simple Assay for Superoxide Dismutase. J. Biol. Chem, v. 247, n. 10, p. 3170–3175, 1972.
NELSON, D. L.; COX, M. M. Princípios de Bioquímica. 4. ed. [s.l.] Sarvier, 2014.
NEWMAN, R. A. et al. Oleandrin-mediated oxidative stress in human melanoma cells. Journal Experimental Therapeutics and Oncology, v. 5, p. 167–181, 2006.
NICOLSON, G. L. Biochimica et Biophysica Acta The Fluid — Mosaic Model of Membrane Structure : Still relevant to understanding the structure , function and dynamics of biological membranes after more than 40 years. BBA - Biomembranes,
v. 1838, n. 6, p. 1451–1466, 2014.
NICOLSON, G. L. Cell Membrane Fluid-Mosaic Structure and Cancer Metastasis. Cancer Research, p. 1169–1177, 2015.
NOGUEIRA, V.; HAY, N. Molecular Pathways: Reactive Oxygen Species Homeostasis in Cancer Cells and Implications for Cancer Therapy. Clinical Cancer Research, v. 19, n. 16, p. 4309–4314, 2013.
OSELKIN, M.; TIAN, D.; BERGOLD, P. J. Low-dose cardiotonic steroids increase sodium–potassium ATPase activity that protects hippocampal slice cultures from experimental ischemia. Neuroscience Letters, v. 473, n. 2, p. 67–71, 2010.
PALMGREN, M. G.; NISSEN, P. P-type ATPases. Annual review of biophysics, v.
40, p. 243–266, 2011.
PEDERSEN, P. L.; CARAFOLI, E. Ion motive ATPases. I. Ubiquity, properties, and significance to cell function. Trends in Biochemical Sciences, v. 12, p. 146–150, 1987.
PESSOA, M. T. C. Mecanismo de ação da 21-Benzilideno digoxina em células tumorais. [s.l.] Universidade Federal de São João del Rei, 2014.
PIETZSCH, P. Mind the membrane. Nature, p. 1–4, 2004.
POST, R. L.; SEN, A. K.; ROSENTHAL, A. S. A Phosphorylated Intermediate in Adenosine Triphosphate- dependent Sodium and Potassium Transport across Kidney Membranes. The Journal of biological chemistry, v. 240, n. 3, p. 1437–
1445, 1965.
PRASSAS, I.; DIAMANDIS, E. P. Novel therapeutic applications of cardiac glycosides. Nature reviews. Drug discovery, v. 7, n. 11, p. 926–935, 2008.
R.MASTERS, J. HeLa cells 50 years on: the good, the bad and the ugly. Nat rev Cancer, v. 2, n. April, p. 311–315, 2002.

64
RAGHAVENDRA, P. B.; SREENIVASAN, Y.; MANNA, S. K. Oleandrin induces apoptosis in human, but not in murine cells: Dephosphorylation of Akt, expression of FasL, and alteration of membrane fluidity. Molecular Immunology, v. 44, n. 9, p.
2292–2302, 2007.
RAMIREZ-ORTEGA, M. et al. Proliferation and apoptosis of HeLa cells induced by in vitro stimulation with digitalis. European Journal of Pharmacology, v. 534, n. 1-3, p. 71–76, 2006.
RAUCHOVÁ, H.; DRAHOTA, Z.; KOUDELOVÁ, J. The role of membrane fluidity changes and thiobarbituric acid-reactive substances production in the inibition of cerebral cortex Na,K-ATPase activity. Physiol. Res., v. 48, p. 73–78, 1999.
RIEGEL, R. E. et al. Intracerebroventricular ouabain administration induces oxidative stress in the rat brain. International Journal of Developmental Neuroscience, v. 28, n. 3, p. 233–237, 2010.
ROCHA, S. C. et al. 21-Benzylidene Digoxin: A Proapoptotic Cardenolide of Cancer Cells That Up-Regulates Na,K-ATPase and Epithelial Tight Junctions. PLoS ONE, v.
9, n. 10, p. e108776, 2014.
RODRIGO, R. et al. Relationship between (Na + K)-ATPase activity, lipid peroxidation and fatty acid profile in erythrocytes of hypertensive and normotensive subjects. Molecular and Cellular Biochemistry, v. 303, n. 1-2, p. 73–81, 2007.
RODRIGO, R. et al. Modulation of (Na,K)-ATPase activity by membrane fatty acid composition: therapeutic implications in human hypertension. Clinical & Experimental Hypertension, v. 36, n. 1, p. 17–26, 2014.
ROY, K. D. K.; SAHA, A.; SENGUPTA, C. Evaluation of alpha-tocoferol, probucol and ascorbic acid as supressors of digoxin induced lipid peroxidation. Acta Poloniac Pharmaceutica-Drug Researc, v. 58, p. 391–400, 2001.
SCHERER, W. F.; SYVERTON, J. T.; GEY, G. O. Studies on the propagation in vitro of poliomyelitis viruses. IV. Viral multiplication in a stable strain of human malignant epithelial cells (strain HeLa) derived from an epidermoid carcinoma of the cervix. The Journal of experimental medicine, v. 97, n. 5, p. 695–710, 1953.
SHAMRAJ, O. I.; LINGREL, J. B. A putative fourth Na+,K(+)-ATPase alpha-subunit gene is expressed in testis. Proceedings of the National Academy of Sciences of the United States of America, v. 91, n. 26, p. 12952–6, 1994.
SHULL, G. E.; GREEB, J.; LINGREL, J. B. Molecular Cloning of Three Distinct Forms of the Na,K-ATPase &-Subunit from Rat Brain. Biochemistry, v. 25, n. 25, p. 8125–32, 1986.
SIES, H. Oxidative stress: a concept in redox biology and medicine. Redox Biology, v. 4, p. 180–183, 2015.
SIMÃO, F. et al. Resveratrol prevents oxidative stress and inhibition of Na+K+-ATPase activity induced by transient global cerebral ischemia in rats. The Journal of Nutritional Biochemistry, v. 22, n. 10, p. 921–928, 2011.

65
SINGER, S. J.; NICOLSON, G. L. The fluid mosaic model of the structure of cell membranes. Science, v. 175, p. 720–731, 1972.
SKOU, J. C. H. R. The influence of some cations on an adenosine triphosphatase from peripheral nerves. Biochimica et Biophysica Acta, v. 23, p. 394–401, 1957.
SOUZA, L. C. et al. The protective effect of melatonin against brain oxidative stress and hyperlocomotion in a rat model of mania induced by ouabain. Behavioural Brain Research, v. 271, p. 316–324, 2014.
SREENIVASAN, Y.; SARKAR, A.; MANNA, S. K. Oleandrin suppresses activation of nuclear transcription factor- k B and activator protein-1 and potentiates apoptosis induced by ceramide. Biochemical Pharmacology, v. 66, p. 2223–2239, 2003.
STENKIVIST, B. Cardenolides and cancer. Anti-Cancer Drugs, v. 12, p. 635–636,
2001.
STENKVIST, B. ET AL. Evidence of a modifying influence of heart glucosides on the development of breast cancer. Analytical And Quantitative Cytology And Histology, v. 1, p. 49–54, 1980.
STENKVIST, B. ET AL. Cardiac glycosides and breast cancer, revisited. New England Journal Of Medicine, v. 306, p. 484, 1982.
STENKVIST, B. et al. Cardiac Glycosides And Breast Cancer. Lancet, v. 1, p. 563, 1979.
STEYN, P. S.; VAN HEERDEN, F. R. Bufadienolides of plant and animal origin. Natural product reports, v. 15, n. 4, p. 397–413, 1998.
STUCKY, M. A; GOLDBERGER, Z. D. Digoxin: its role in contemporary medicine. Postgraduate Medical Journal, v. 91, p. 514–518, 2015.
TANAKA, T. et al. Changes of membrane fluidity and Na+,K+-ATPase activity during cellular differentiation in the guinea pig epidermis. Archives for dermatological research. Archiv fur dermatologische Forschung, v. 280, n. 1, p. 29–32, 1988.
THOMAS, R.; GRAY, P.; ANDREWS, J. Digitalis: Its Mode of Action, Receptor, and Structure–Activity Relationships. In: ACADEMIC PRESS (Ed.). . Advances In Drug Research. London: Bernard Testa, 1990. p. 313–575.
TIAN, J. et al. Changes in Sodium Pump Expression Dictate the Effects of Ouabain on Cell Growth. Journal of Biological Chemistry, v. 284, n. 22, p. 14921–14929,
2009.
VOET, D.; VOET, J. G.; PRATT, C. W. Fundamentos de Bioquímica. [s.l.] Artmed,
2008.
WANG, D.; BI, Z. Bufalin inhibited the growth of human osteosarcoma MG-63 cells via down-regulation of Bcl-2/Bax and triggering of the mitochondrial pathway. Tumor Biology, v. 35, n. 5, p. 4885–4890, 2014.

66
WANG, H.; YUAN, W.; LU, Z. Effects of ouabain and digoxin on gene expression of sodium pump alpha-subunit isoforms in rat myocardium. Chinese Medical Journal, v. 114, n. 10, p. 1055–9, 2001.
WANG, Y. et al. Src mediates extracellular signal-regulated kinase 1/2 activation and autophagic cell death induced by cardiac glycosides in human non-small cell lung cancer cell lines. Molecular Carcinogenesis, v. 34, n. October 2013, p. 26–34, 2014.
WILKINS, M. R.; KENDALL, M. J.; WADE, O. L. William Withering and digitalis, 1785 to 1985. British medical journal (Clinical research ed.), v. 290, n. 6461, p. 7–8,
1985.
WITHERING, W. An Account Of The Foxglove, And Some Of Its Medical Uses: With Practical Remarks On Dropsy, And Other Diseases. Birmingham, U. K.: M. Swinney, 1785.
XIE, C.-M. et al. Bufalin induces autophagy-mediated cell death in human colon cancer cells through reactive oxygen species generation and JNK activation. Free radical biology & medicine, v. 51, n. 7, p. 1365–1375, 2011.
XIE, Z.; ASKARI, A. Na+/K+-ATPase as a signal transducer. European Journal of Biochemistry, v. 269, n. 10, p. 2434–2439, 2002.
YAN, Y. et al. Involvement of reactive oxygen species in a feed-forward mechanism of Na/K-ATPase-mediated signaling transduction. Journal of Biological Chemistry, v. 288, n. 47, p. 34249–34258, 2013.
YEAGLE, P. L. Cholesterol modulation of Na,K-ATPase ATP hidrolyzing activity in the human erythrocyte. v. 727, p. 39–44, 1983.
YEE, C. et al. Presence and expression of human papillomavirus sequences in human cervical carcinoma cell lines. The American journal of pathology, v. 119, n.
3, p. 361–366, 1985.
YONEDA, J. S. et al. Na,K-ATPase reconstituted in ternary liposome: The presence of cholesterol affects protein activity and thermal stability. Archives of Biochemistry and Biophysics, v. 564, p. 136–141, 2014.