universidade federal de pernambuco centro de … · 2019-10-25 · mestrado em ciências...
TRANSCRIPT

UNIVERSIDADE FEDERAL DE PERNAMBUCO CENTRO DE CIÊNCIAS BIOLÓGICAS
MESTRADO EM CIÊNCIAS BIOLÓGICAS – ÁREA DE FISIOLOGIA
Efeito da deficiência dietética em ácido ascórbico sobre
parâmetros morfológicos e funcionais da reprodução em cobaios (Cavia porcellus) machos adultos.
João Batista Ferreira Júnior
Recife

4
João Batista Ferreira Júnior
Efeito da deficiência dietética em ácido ascórbico sobre
parâmetros morfológicos e funcionais da reprodução em
cobaios (Cavia porcellus) machos adultos.
Dissertação apresentada ao Curso de
Mestrado em Ciências Biológicas - área Fisiologia da
Universidade Federal de Pernambuco, como parte
dos requisitos para a obtenção do grau de Mestre em
Ciências Biológicas - área de Fisiologia.
Orientador: Dra. Maria Inês Wanderley
Co-orientadores: Dra. Adriana da Rocha Tomé
Dra. Márcia Valéria P. Ferreira
Recife, PE-Brasil 2004.
2

5
Dados Internacionais de Catalogação na Publicação (CIP) (Setor Técnico da Biblioteca Central da Universidade de Fortaleza) ________________________________________________________________________ F383e Ferreira Júnior, João Batista. Efeito da deficiência em ácido ascórbico sobre parâmetros morfológicos e funcionais da reprodução em cobaios (Cavia porcellus) machos adultos / João Batista Ferreira Júnior. - 2004. 57f. Cópia de computador. Dissertação (mestrado) – Universidade Federal de Pernambuco, 2004. “Orientação : Profª. Drª. Maria Inês Wanderley”.
1. Ácido ascórbico - Deficiência – Função reprodutora 2. Fisiologia endócrina - Experiências. 3. Testículos – Fisiologia I. Título.
CDU 577.164.2:612.613

6

3
Dedicatória
A Deus, ser divino pai de todas as criaturas, por ter me dado forças para enfrentar este desafio. A minha esposa Jacqueline, companheira nos dias de aflição e aos meus dois filhos queridos Letícia e Davi, pela compreensão, pois sem eles não teria terminado essa dissertação.

4
Agradecimentos Ainda não havia conhecido uma pessoa, que reunisse tantas características boas. Esta pessoa é a minha querida professora Inês. Orientadora, amiga, mulher generosa, compreensiva e boa a quem devo a realização deste trabalho e todos os meus agradecimentos. Agradeço de coração a minha irmã querida professora Dra. Márcia Valeria Pitombeira Ferreira, por ter me ajudado muito na elaboração das lâminas histológicas, no estudo morfométrico e pelo suporte técnico científico. Ao meu grande amigo Reinaldo Barreto Oriá, por ter participado da elaboração do projeto de pesquisa desta dissertação, meu muito obrigado. Ao grande professor Dr. Daniel Pedro Udrisar, pelo apoio moral e pela ajuda técnica científica. Ao amigo e cunhado professor Dr. Alberto Flávio Alves Aguiar, por ter me ajudado muito na confecção das fotos e na elaboração da aula de apresentação desta dissertação e pelo apoio moral. A minha monitora Ana Patrícia Pierre Lima, pela ajuda no manejo com os cobaios e pela ajuda durante o sacrifício dos animais. Agradeço aos laboratórios de Fisiologia e Histologia da Universidade de Fortaleza (UNIFOR), pelo uso da aparelhagem técnica científica. Minha gratidão ao Departamento de Patologia da Universidade Federal do Ceará UFCE, pela ajuda na confecção das lâminas histológicas. Minha eterna gratidão ao Instituto do Câncer do Estado do Ceará, onde foram feitas as fotos e estudo morfométrico.

5
LISTA DE FIGURAS Figura 1. Biossíntese do L-ácido ascórbico em animais............................................... 12
Figura 2. Catabolismo do ácido ascórbico .................................................................... .14
Figura 3. Efeito da deficiência de AA sobre o peso corporal ....................................... 30
Figura 4. Efeito da deficiência de AA sobre a morfologia do espermatozóide ........... 33
Figura 5. Efeito da deficiência de AA sobre a estrutura do testículo ........................... 35
Figura 6. Efeito da deficiência de AA sobre a estrutura do epidídimo ........................ 36
Figura 7. Efeito da deficiência de AA sobre a produção de testosterona
estimulada pelo hCG em células intersticiais testiculares ............................ 37
Figura 8. Efeito do AA sobre a produção de testosterona em células
intersticiais testiculares.................................................................................. 39
Figura 9. Principais vias de biossíntese de testosterona ( modificado de HEDGE
e col, 1988 )......................................................................................................44

6
LISTA DE TABELAS Tabela 1 Pesos do testículo e cauda do epidídimo, nº e motilidade dos espermatozóides
e testosterona plasmática dos cobaios normais ou deficientes de AA...........31

7
SUMÁRIO Página
DEDICATÓRIA.................................................................................... 3
AGRADECIMENTOS.......................................................................... 4
LISTA DE FIGURAS........................................................................... 5
LISTA DE TABELAS........................................................................... 6
RESUMO............................................................................................ 8
ABSTRACT........................................................................................10
1. INTRODUÇÃO..............................................................................11
2. OBJETIVOS...................................................................................20
3. MATERIAL E MÉTODOS.............................................................. 21
4. RESULTADOS...............................................................................30
5. DISCUSSÃO..................................................................................40 6. CONCLUSÃO.................................................................................48
7. REFERÊNCIAS BIBLIOGRÁFICAS...............................................49

8
RESUMO
O presente trabalho estudou o efeito da deficiência em ácido ascórbico (AA) em
cobaios machos adultos, durante 21 dias, sobre o peso e a histoarquitetura (histologia e
histometria) do testículo e cauda do epidídimo, sobre a qualidade (número, motilidade e
morfologia) dos espermatozóides e sobre os níveis plasmáticos de testosterona e a resposta
secretória das células intersticiais testiculares ao estímulo gonadotrófico com hCG e ao
tratamento “in vitro” com AA. A deficiência em AA causou diminuição de 32 % no peso
corporal O peso absoluto do testículo foi reduzido em 30,5 % enquanto o seu peso relativo
não foi modificado pela deficiência em AA. O peso absoluto e relativo da cauda do
epidídimo diminuiram em 44,8 % e 25,2 %, respectivamente. A histoarquitetura dos dois
órgãos foi alterada pela carência em AA. O parênquima do testículo apresentou alterações
do epitélio germinativo dos túbulos seminíferos. A cauda do epidídimo apresentou
diminuição na altura do epitélio colunar de revestimento e do número de espermatozóides
na luz tubular. O número e motilidade dos espermatozóides diminuiram (73 % e 27,5 %,
respectivamente), como também sua morfologia foi alterada pela deficiência em AA. Os
níveis plasmáticos de testosterona foram reduzidos em 72,6 % nos animais deficientes em
AA. A produção de testosterona pelas células intersticiais testiculares estimulada com uma
dose elevada de hCG (10 mUI/ml) “in vitro” foi menor nos animais deficientes em AA. O
tratamento com uma dose alta de AA (100 µM) “in vitro” não modificou a secreção basal
de testosterona em nenhum dos grupos de animais, mas inibiu a secreção estimulada pelo

9
hCG no grupo controle, não modificando a secreção no grupo deficiente em AA. Nossos
estudos confirmam prévias referências e fornecem evidências adicionais da importância do
AA na manutenção da integridade estrutural e funcional do testículo e epidídimo. Os
resultados também indicam que o efeito primário da deficiência em AA parece estar no
testículo, reduzindo a produção de testosterona e, conseqüentemente, privando desse
hormônio os órgãos da reprodução.

10
ABSTRACT
The present work studied the effect of the deficiency in ascorbic acid (AA) in adult
male rabbits during 21 days with regard to the weight, the histoarchitecture of the testes and
the epididymis’ tail, as well as the quality of the spermatozoids, the testosterone plasmatic
levels, the secretory answer of the testis cells to the gonadotrophic stimulation with hCG
and to the treatment in vitro with AA. The deficiency in AA caused a decrease of 32 % in
the body weight. The absolute weight of the testes was reduced by 30.5 % while their
relative weight was not modified by the deficiency in AA. The absolute and relative weight
of the epididymis’ tail decreased by 44.8 % and 25.2%, respectively. The histoarchitecture
of the two organs was altered by the lack in AA. The parenchyma of the testis presented
alterations of the germinative epithelium of the seminific tube. The tail of the epidíidymis
presented decrease in the number of spermatozoids of the tubular radius. The number and
mobility of the spermatozoids decreased (73 % and 27.5 %, respectively), and their
morphology was altered by the deficiency in AA. The plasmatic testoterone levels were
reduced in 72.6 % in the in AA deficient animals. The testosterone production by the testis
cells was stimulated with a high dose of hCG (10 mUI/ml) in vitro but in the in AA
deficient animals it was smaller. The treatment with a high dose of AA (100 µM) in vitro
did not modify the basal secretion of testosterone in none of the animal groups, but it
inhibited the secretion stimulated by the hCG in the control group; the secretion in the AA
deficient group had no alterations. Our studies confirm previous evidences and supply
additional ones on the importance of AA in the maintenance of the structural and functional
integrity of the testis and epidídymis. The results also indicate that the primary effect of the
deficiency in AA seems to be in the testis, reducing the testosterone production and,
consequently, depriving the reproduction organs of that hormone.

11
1. INTRODUÇÃO
1.1 Perspectiva histórica
Os primeiros exploradores marítimos tiveram suas vidas ameaçadas nas longas
viagens, quando havia escassez de vegetais frescos e frutas. Eles desenvolviam uma
enfermidade peculiar chamada escorbuto. Os sintomas do escorbuto incluem: gengivas
inchadas e com fácil sangramento, inserção comprometida dos dentes, irritação e rigidez
das articulações, sangramento subcutâneo e cicatrização lenta dos ferimentos. Foi a partir
dos estudos do Dr. James Lind, médico da marinha Britânica, que se descobriu à
importância de frutas cítricas na prevenção e cura do escorbuto; a partir de 1792 todos os
marinheiros ingleses começaram a receber provisão diária de suco de limão para compensar
a deficiência de ácido ascórbico (Fisher e Bender, 1979). O ácido ascórbico (AA) foi
isolado primeiro de fontes naturais e estruturalmente caracterizado por Szent-Gyorgy,
Waugh e King (Naidu, 2003). Esta vitamina foi primeiro sintetizada por Haworth e Hirst
(Naidu, 2003). Atualmente o ácido ascórbico é o suplemento vitamínico mais largamente
utilizado no mundo.
1.2 Fontes de ácido ascórbico e significado evolutivo da sua biossíntese em vertebrados
terrestres
O ácido ascórbico encontra-se largamente distribuído nas frutas e vegetais frescos.
Está presente em frutas como laranja, limão, melão, mamão, uva, manga, abacaxi,

12
framboesa e cereja. Também é encontrado em vegetais de folhas verdes, tomate, brócoli,
pimenta, couveflor e abóbora. A maioria das plantas e animais sintetiza o ácido ascórbico a
partir da D-glicose ou D-galactose. A maioria dos animais produz relativamente altos níveis
de ácido ascórbico a partir da glicose no fígado (Fig. 1). Contudo, cobaios (guinea pig),
morcegos (os que se alimentam de frutas) e o homem não podem sintetizar o ácido
ascórbico devido à ausência da enzima oxidase da L-gulonolactona (LGO). Assim, nos
humanos o ácido ascórbico tem que ser suplementado através da alimentação e/ou do ácido
ascórbico sintético. Ambos os ácidos ascórbicos, natural e sintético, são quimicamente
idênticos e não há diferença na sua atividade biológica ou na sua biodisponibilidade. O
ácido ascórbico é uma molécula lábil e pode ser inativada durante o cozimento, embora
tenha a habilidade de preservar alimentos em virtude de sua propriedade redutora.
D-Glicose D-Galactose ↓
Glicose-6-fosfato ↓
Uridin-difosfoglicose ↓
Ácido uridin-difosfato glicurônico ↓
D-ácido glicurônico ↙ ↘
Lactona do ácido D-glicurônico via das pentoses fosfato ↓
γ-Lactona do ácido L-gulônico
oxidase da L-gulonolactona ↓
2-ceto-L-gulonolactona ↓
L-ácido ascórbico
Figura1. Biossíntese do L-ácido ascórbico em animais (Modificado de Naidu, 2003)

13
A expressão do gene para a LGO, a enzima terminal na via de síntese do AA em
animais, pelos primeiros vertebrados terrestres, constitui um exemplo de progresso
evolutivo. A evolução dos vertebrados do meio aquático para a atmosfera terrestre
contendo altas concentrações de oxigênio foi acompanhada pela expressão em alguns
tecidos do gene para a LGO (Chatterjee, 1973). Existem evidências indicando que a
finalidade do aparecimento da LGO foi aparentemente a de suprir os vertebrados terrestres
com quantidades adequadas de AA e assim proteger seus tecidos contra a toxicidade do
oxigênio da atmosfera terrestre (Nandi e col., 1997). Contudo, ainda não foi explicada a
causa da perda evolutiva da LGO no cobaio (guinea pig), morcegos, primatas e humanos.
Experimentos de clonagem e sequenciamento do gene da LGO de rato e seu homólogo no
cobaio indicam que a incapacidade do cobaio em sintetizar o AA deve-se a deleção de
várias regiões do gene da LGO (Nishikimi e col., 1992). Observação semelhante também
foi feita com o gene não funcional da LGO de humanos (Nishikimi e col., 1994).
1.3 Química do ácido ascórbico
L-ácido ascórbico (C6 H8 O6) é o nome trivial da vitamina C. O nome químico é 2-
oxo-L-threo-hexono-1,4-lactona-2,3-enediol. O ácido ascórbico e o ácido dehidroascórbico
são as principais formas dietéticas da vitamina C (Moser e Bendich, 1990). O palmitato do
ácido ascórbico é usado em preparações antioxidantes comerciais. Todas as formas
comerciais do ácido ascórbico, exceto o palmitato, são solúveis em água. Todas são usadas
nos alimentos como antioxidantes e estabilizadores de coloração e odor.
1.4 Catabolismo do ácido ascórbico

14
O ácido ascórbico presente nos alimentos é facilmente absorvido por transporte
ativo no intestino (Sauberlich, 1985). A maioria (80 a 90%) é absorvida quando se ingere
até 100 mg/dia, diminuindo a eficácia de sua absorção quando se ingere quantidade maior
(500 mg/dia). Baseado em estudos clínicos e epidemiológicos, foi sugerido que a ingestão
de 100 mg/dia de ácido ascórbico está associada com incidência reduzida de mortalidade
por doenças cardíacas e câncer (Carr e Frei, 1999). Contudo, o estresse, o fumo, o
alcoolismo, febre e infecção viral causam um rápido declínio do nível sanguíneo do ácido
ascórbico. O ácido ascórbico é sensível ao ar, luz, calor e facilmente destruído por
armazenamento prolongado. Os principais metabólitos do ácido ascórbico no homem são o
ácido dehidroascórbico, o ácido 2,3-dicetogulônico e o ácido oxálico (Figura 2). A
principal via de eliminação do ácido ascórbico e seus metabólitos é a urina. O ácido
ascórbico geralmente não é tóxico, mas em altas doses (2-6 g/dia) pode causar distúrbio
gastrointestinal ou diarréia (Anderson e col., 1997; Johnson, 1999).
L-ácido ascórbico ↕
Radical ascorbil → Sulfato de l-ascorbato ↓
Dehidroascorbato → CO2 ↓
2,3-ácido dicetogulônico ↓
Ácido oxálico
Figura 2. Catabolismo do ácido ascórbico (Modificado de Naidu, 2003)

15
6
1.5 Papel fisiológico do ácido ascórbico
O papel fisiológico do ácido ascórbico é largamente dependente de suas
propriedades de óxido-redução. O ácido ascórbico é um co-fator para hidroxilases e
monooxigenases envolvidas na síntese do colágeno, carnitina e neurotransmissores (Levin,
1986). O ácido ascórbico acelera reações de hidroxilação pela manutenção do centro ativo
de íons metálicos em um estado reduzido para a atividade ótima das hidroxilases e
oxigenases.
O colágeno representa cerca de um terço da proteína corporal total e constitui a
principal proteína da pele, ossos, dentes, cartilagem, tendões, vasos sanguíneos, válvulas
cardíacas, córnea e cristalino do olho. A carnitina é requerida para o transporte e
transferência de ácidos graxos na mitocôndria, onde pode ser utilizada para a produção de
energia. Além do mais, o AA atua como co-fator da dopamina-β-hidroxilase, que catalisa a
conversão do neurotransmissor dopamina para noradrenalina. Então o AA é essencial para
a síntese do colágeno, carnitina e catecolaminas (Levin, 1986). O AA também aumenta a
disponibilidade e absorção do ferro da dieta (Levin, 1986).
1.6 Ácido ascórbico e reprodução
Os efeitos diretos da deficiência em AA sobre a fertilidade de animais de laboratório
e silvestres são conhecidos há muito tempo, principalmente nos machos (Counsell e
Hornig, 1981). Baixos níveis de AA no sêmen de bovinos foram associados com baixa
performance de prenhez, enquanto cobaios escorbúticos apresentaram degeneração do

16
epitélio germinal testicular (Luck e cols., 1995). Em coelhos, o efeito das gonadotrofinas
sobre o crescimento gonadal foi aumentado pelo tratamento simultâneo com AA (Luck e
cols., 1995). Esses estudos sugerem que o AA afeta a integridade da estrutura dos túbulos
seminíferos e a função do espermatozóide (Luck e cols., 1995). No caso dos túbulos, o AA
é requerido para a secreção e manutenção das camadas de colágeno tipo IV e I, os quais
formam uma parte importante da lâmina basal (Luck, 1994). Em humanos, foi referido que
o plasma seminal contem AA, o qual é concentrado e secretado pela vesícula seminal
durante a ejaculação (Berg e cols., 1941). Em 1960, Lindahl e Kihstrom descreveram uma
substância proteinácea no plasma seminal, a qual aparece em duas formas (oxidada ou
reduzida) e que é transformada reversivelmente de uma forma para outra. A forma reduzida
tem atividade antiaglutinadora, isto é, previne o agrupamento e imobilidade dos
espermatozóides no plasma seminal. Essa forma está aderida à superfície do
espermatozóide e quando é oxidada perde essa habilidade de adesão. O AA, agente redutor
biológico, reduz a proteína oxidada (Lindahl 1966). A deficiência de AA também foi
associada com baixo número de espermatozóides, alto número de espermatozóides
anormais, reduzida motilidade e aglutinação (Dawson e cols., 1987). Os tipos de
movimento devem ser considerados ao examinar-se a motilidade individual. O movimento
normal é para frente, em linha reta. Os distúrbios do movimento padrão são muitas vezes
devidos a alterações na pressão osmótica do sêmen, levando a movimentos circulares com a
cauda curva em um círculo de raio amplo, dificilmente reconhecível, ou em um círculo tão
pequeno que a cabeça quase toca a cauda. Outra forma anormal é o movimento para trás, no
qual a cauda apresenta-se enrolada no formato de uma clave musical, de modo que repousa
acima ou próxima à cabeça (Rosemberger, 1987).

17
O tratamento dietético pode melhorar a qualidade do espermatozóide, sendo o efeito
benéfico do AA resultante da neutralização de radicais livres, os quais surgem como
conseqüência da poluição ambiental e do metabolismo celular e que poderiam causar dano
oxidativo ao DNA (Dawson e cols., 1987).
Da mesma forma que nos machos, também foi sugerido que nas fêmeas o AA atua
como regulador da fertilidade. A relação entre o AA e a função ovariana foi demonstrada
em experimentos nos quais se verificou diminuição do AA em resposta ao hormônio
luteinizante (LH) e variação da concentração de AA durante o ciclo estral (maiores níveis
no início do ciclo) (Stansfield e Flint, 1967; Mills e Schwartz, 1961; Wise, 1987). Também
foi verificado que a suplementação com AA aumenta os efeitos indutores da ovulação do
clomifeno (agonista estrogênico), por uma ação aparentemente ovariana (Igarashi, 1977).
Em contraste, o AA em altas doses parece atuar como anticoncepcional (Briggs, 1973). No
ovário o AA estimula a secreção de progesterona e ocitocina (Luck e Jungclas, 1988; Byrd
e cols., 1993) e sinergiza com neurotransmissores na estimulação da secreção hormonal
(Luck, 1990). No ovário existe uma alta taxa de remodelação tissular e de síntese do
colágeno, necessários para o crescimento do folículo, para sua reparação após ovulação e
para o desenvolvimento do corpo lúteo (Himeno e cols., 1984; Luck e Zhao, 1993); o AA é
necessário para a secreção do colágeno no líquido folicular (Christiane e cols., 1988).
Vários tecidos acumulam o AA e a sua forma oxidada, o ácido dehidroascórbico; as
concentrações mais altas ocorrem na hipófise, supra-renal e gônadas (Das e cols., 1993).
Não há diferença na concentração plasmática de ácido ascórbico entre os sexos (Wang e
cols., 1992). A alta concentração do AA nos tecidos endócrinos atesta sua importância na
síntese hormonal. A síntese de ocitocina e vasopressina na neurohipófise envolve a
amidação pós-tradução, a qual é dependente do AA (Eipper e Mains, 1991). A síntese de

18
catecolaminas também requer AA, o qual também é necessário para protegê-las da
oxidação (Roston, 1962; Levine e Morita, 1985). A esteroidogênese parece depender do
AA, principalmente nas reações de hidroxilação (Tsujl e cols., 1989; Goralczyk e cols.,
1992).
Assim, podemos dizer que o AA possui três ações biológicas de relevância para a
reprodução, cada uma dependendo de seu papel como agente redutor: ele é requerido para a
biossíntese do colágeno, dos hormônios esteróides e peptídicos e para evitar ou reduzir a
oxidação de biomoléculas, ou seja, seus efeitos podem ser explicados por mecanismos
celulares e bioquímicos semelhantes àqueles aplicados em outros tecidos (Sebrell e Harris,
1967; Millar, 1992). O entendimento mais completo da dependência da reprodução em
relação ao AA é de interesse para nós, como reguladores de nossa própria fertilidade.
1.7 Ácido ascórbico e o testículo - Cobaio (guinea pig) como modelo animal
O cobaio é um modelo animal ideal para estudar o efeito da deficiência em AA
sobre a função reprodutora porque não é capaz de sintetizar o AA, dependendo
completamente do aporte dietético dessa vitamina. Contudo, as referências sobre os efeitos
da deficiência de AA em cobaios são controversas, especialmente sobre a reprodução
masculina. Lindsay e Medes mostraram em 1926 que uma dieta deficiente em AA durante
um mês causava degeneração do epitélio germinativo de testículo de cobaios. Em 1986 os
estudos de Lindsay e Medes foram confirmados por Chinoy e cols. A lesão histológica mais
severa foi observada no testículo e cauda do epidídimo e em menor grau no vaso deferente,
próstata e vesícula seminal. A análise dos espermatozóides nas amostras de sêmen
coletadas do testículo, cauda do epidídimo e vaso deferente mostrou diminuição do seu
número e motilidade, como também um aumento das formas anormais. Como esses órgãos

19
dependem dos níveis circulantes de testosterona para a manutenção da sua integridade
estrutural e funcional, a deficiência em AA poderia causar o chamado “efeito da privação
de andrógenos” aos órgãos alvo por, provavelmente, influenciar as células de Leydig e/ou
afetar a síntese de testosterona no testículo (Chinoy e cols., 1986). Conseqüentemente, para
esclarecer o mecanismo de ação do estado de deficiência em AA sobre as funções
reprodutoras, é necessário examinar os níveis circulantes de testosterona e a produção desse
hormônio pelo testículo de cobaios deficientes.

20
2. OBJETIVOS
Os objetivos do presente estudo em cobaios machos foram:
1) Confirmar dados da literatura sobre o efeito da deficiência em AA na estrutura do
testículo e epidídimo e na qualidade dos espermatozóides (número, motilidade e
morfologia dos espermatozóides).
2) Determinar os níveis de testosterona no plasma de cobaios deficientes em AA.
3) Determinar a resposta secretória das células intersticiais testiculares de cobaios
deficientes em AA ao estímulo gonadotrófico e ao tratamento com AA “in vitro”.

21
3. MATERIAIS E MÉTODOS
3.1 Animais e dietas Foram utilizados cobaios machos adultos (Cavia porcellus), com idade entre 5 a 6
meses, pesando em torno de 350 a 450 g, fornecidos pelo HEMOCE (Hemocentro do
Ceará). Os cobaios foram mantidos em gaiolas individuais, medindo 50 x 30 x 30 cm, em
condições de temperatura variando entre 25º e 30º C e ciclo claro-escuro de 12 h cada (luz
de 5:00 às 17:00 hs). A água foi fornecida ad libitum. O grupo controle (10 animais) foi
alimentado com ração Purina para coelhos (desprovida de AA), suplementada diariamente
com 50 mg/dia de AA Shering) na água de beber e vegetal (capim) ad libitum, durante 21
dias. Essa dose de AA baseou-se no trabalho de Jager (1985). O grupo deficiente em AA
(10 animais) foi alimentado apenas com a ração Purina para coelhos durante 21 dias. A
deficiência em AA foi diagnosticada pela perda de peso corporal, perda de pelo e aparente
imobilidade no 20º dia de aplicação da dieta deficiente em AA. Os animais foram pesados a
cada 3 dias durante o período experimental.
3.2 Reagentes
Albumina sérica bovina (BSA) Fração V obtida de Miles (Naperville, IL) e de ICN
Biomedicals Inc; Ácido tricloroacético (TCA), Álcool etílico P.A., Azida de sódio,
Bicarbonato de sódio (NaHCO3), Cloreto de potássio (KCl), Cloreto de sódio (NaCl),
Sacarose obtidos de VETEC; Adenosina 5’-trifosfato (ATP), Azul tripan (direct blue

22
14), Eosina, Hematoxilina, Papanicolau, Carvão ativado, Colagenase tipo I, Dextran,
Dithithreitol (DTT), Gelatina tipo III, Histona de timo de bezerro tipo III-S, 1-(5-
Isoquinolinylsulfonyl)-2-methyl-piperazine (H-7), Inibidor da tripsina tipo 1S,
Leupeptina, 2,5-diphenyl-oxazole(PPO), Testosterona, obtidos de Sigma Chemical Co.
(St. Louis, MO); Meio de cultura 199 (Meio 199) obtido de GIBCO (Grand Island, NY);
Etanol absoluto, 2,2’-p-phenylen-bis (5-phenyloxazol) (POPOP) obtidos de MERCK;
1,2,6,7 [3H] Testosterona, obtido de Amersham International (Buckinghamshire,
England); Éter etílico P.A. obtido de Proanaysi-Isofar; Cloreto de cálcio (CaCl2), Fosfato
de potássio monobásico (KH2PO4), Fosfato de sódio dibásico (Na2HPO4.7H2O), Fosfato
de sódio monobásico (NaH2PO4.H2O), Tolueno P.A. obtidos de Reagen, Formol 10 %,
hidrato de cloral.
3.3 Procedimento de necropsia
Os animais foram anestesiados com hidrato de cloral (0,1g/100g peso corporal) por
via i.p. no 21º dia do experimento. Com os animais anestesiados, coletamos o sangue
através de punção intracardíaca, para dosagem hormonal por radioimunoensaio e retiramos
os testículos e epidídimo. Em seguida esses órgãos foram pesados e foi feita a coleta de
espermatozóides através da homogeneização da cauda do epidídimo em placa de Petri
contendo 1 ml de soro fisiológico. Os testículos foram processados para estudo histológico.
3.4 Coleta e análise dos espermatozóides
O homogeneizado da cauda do epidídimo foi utilizado para a determinação da
motilidade, concentração e morfologia dos espermatozóides. A contagem dos
espermatozóides foi realizada segundo critérios do Mies (1978), utilizando a técnica

23
empregada em hematimetria. Fizemos uma diluição de 1:10 do sêmen e introduzimos o
líquido entre a lâmina e a lamínula da câmara de Neubauer, procedendo-se ao
reconhecimento, no campo microscópico, dos espermatozóides sobre a área delimitada e
que corresponde ao mm2. Como na hematimetria, somente foram considerados os
elementos que estiveram no interior de dado quadrado e aqueles que se encontravam nas
linhas que formam o ângulo inferior esquerdo de cada quadrado a considerar. Não foram
consideradas as caudas que cruzam as linhas, mas apenas as cabeças dos espermatozóides.
A concentração espermática foi avaliada utilizando-se câmara de Neubauer de 16
quadrados centrais com altura de 0,1mm. Os quatros quadrados dos cantos, ou seja, 1/4 do
mm2, foram selecionados e procedeu-se a contagem dos espermatozóides contidos nos
mesmos. A somatória dos espermatozóides encontrados nos quatros quadrados, na diluição
de 1:10 do homogeneizado da cauda do epidídimo foi submetida aos seguintes fatores de
correção:
diluição 1:10 (x 10) Área contada 1/16 x 4 = 1/4mm2 (x 4) Altura da câmara 0,1mm (x 10) -------------- 400
A somatória dos espermatozóides presentes mos quatro quadrados foi multiplicada por 400
para a obtenção do número de espermatozóides por mm3.
A motilidade é a maneira mais fácil e mais segura de determinar a viabilidade do
espermatozóide. Ao microscópio existem dois tipos de motilidade: movimento de massa e
motilidade individual. O movimento de massa significa movimento de agregação, como
onda, em grupamentos de uma grande população de espermatozóides em ejaculados densos
e ativos. Pode-se observar melhor quando uma gota de sêmen, do tamanho de uma lentilha
é transferida em uma micropipeta esterilizada a quente para uma lâmina aquecida e

24
examinada sem aplicação de uma lamínula, por luz transmitida, em um aumento de 100
vezes, com o diafragma parcialmente fechado. Os movimentos dos espermatozóides não
obedecem a um tipo único: há elementos que se deslocam para frente, em linha reta
(movimento progressivo), enquanto que outros descrevem uma circunferência (movimento
circular) e finalmente, outros se limitam a oscilar no campo microscópico (movimento
oscilatório ou local). O movimento progressivo é fruto da rotação do espermatozóide sobre
seu eixo, aliada à movimentação propulsora da cauda, à semelhança de chicotadas (Mies,
1978). A motilidade dos espermatozóides individuais é expressa em percentagem de
espermatozóides que demonstram propulsão para diante, movimento no local e nenhum
movimento. Coletamos a amostra do homogeneizado da cauda do epidídimo com uma
micropipeta, colocamos 10 µl sobre uma lâmina de vidro limpa e desengordurada e em
seguida cobrimos com uma lamínula de 22 mm x 22mm. A preparação foi examinada com
aumento de 100 e a intensidade luminosa foi regulada de forma a apresentar claramente as
células espermáticas. A metodologia utilizada para a classificação individual dos
espermatozóides foi baseada no método de Walton (1939) abaixo resumido:
Tipo de Movimento Pontos
Todos ou quase todos os espermatozóides exibem enérgicos movimentos progressivos
5,0 (100 %)
A maioria dos espermatozóides exibem movimentos progressivos
4,0 (80 %)
Metade do movimento progressivo e oscilatório e metade imóveis
3,0 (60 %)
Só existem espermatozóides com movimento oscilatório com grande número de imóveis
2,0 (40 %)
Todos os espermatozóides são imóveis
1,0 (20 %)

25
Ausência de espermatozóides 0,0 (0 %)
Para analisar a morfologia dos espermatozóides coletamos uma amostra do
homogeneizado da cauda do epidídimo com uma micropipeta e procedemos à confecção de
um esfregaço sobre lâmina de vidro, com o auxílio de uma lâmina extensora. Os esfregaços
foram fixados e corados de acordo com a técnica de Papanicolau:
_________________________________________________________________________
Etanol 80% 10 mergulhos Etanol 70% 10 mergulhos Etanol 50% 10 mergulhos Água destilada 10 mergulhos Hematoxilina Harris 3 minutos Água corrente 3 a 5 minutos Etanol ácido 2 mergulhos Água corrente 3 a 5 minutos Solução de Scott 4 minutos Água destilada 1 mergulho Etanol 50% 10 mergulhos Etanol 70% 10 mergulhos Etanol 80% 10 mergulhos Etanol 90% 10 mergulhos Orange G6 2 minutos Etanol 95% 10 mergulhos Etanol 90% 10 mergulhos EA-50 5 minutos Etanol 95% 5 mergulhos Etanol 95% 5 mergulhos Etanol 95% 5 mergulhos Etanol 99,5% 2 minutos Xilol (3 frascos aproximadamente) 1 minuto cada
___________________________________________________________________
Em seguida foram examinados ao microscópio óptico. Durante esse procedimento foram
contados 200 espermatozóides, onde foram consideradas as seguintes alterações
morfológicas de cauda: cauda enrolada e cauda fortemente enrolada, cauda dobrada e
fortemente dobrada, cauda abaxial; e de cabeça: cabeça isolada, cabeça estreita na base,
cabeça sem capuz acrossomático.

26
3.5 Análise histológica
Fragmentos de testículo e da cauda do epidídimo foram fixados em formol
tamponado a 10 % e posteriormente desidratados em álcool, diafanizados (clareados) em
xilol e incluídos em parafina para a realização dos cortes histológicos com 5 µm de
espessura. Depois os cortes foram desparafinados, rehidratados e corados com
hematoxilina-eosina (HE). Os cortes foram examinados em microscópio óptico comum
Nikon, com objetivas de 20X, 40X, 100X e ocular de 10X. A medição da altura das células
epiteliais foi realizada com retículo quadriculado colocado na objetiva de fotografia de
microscópio Nikon. A medida do retículo foi feita com lâmina milimetrada com régua
padrão de 1 mm. Para comparação do tamanho e da altura do epitélio de revestimento da
cauda do epidídimo dos animais controle e deficiente em AA, tomou-se como referência o
aumento de 100X, onde cada quadrinho vale 0,1 do mm ou 100 micrômetros.
3.6 Preparação e incubação das células intersticiais testiculares
3.6.1 Obtenção das células intersticiais testiculares:
A obtenção das células foi realizada conforme descrito por Hedger e Eddy (1986)
com algumas modificações (Wanderley e Udrisar, 1994; Wanderley e Negro-Villar, 1996).
Os animais foram sacrificados por anestesia com hidrato de cloral (0,1g/100g peso corporal)
por via i.p. no 21º dia do experimento, e os testículos foram rapidamente removidos e
cuidadosamente descapsulados. Os testículos descapsulados foram incubados em uma
solução enzimática (2 mL/testículo) contendo colagenase (0,5 mg/mL), inibidor da tripsina
(0,2 mg/mL) e leupeptina (5 µg/mL) dissolvidos em tampão fosfato-salina (PBS) (136,9

27
mM NaCl, 2,68 mM KCl, 8,1 mM NaHPO4. 7H2O, 1,47 mM KH2PO4) contendo albumina
sérica bovina (BSA) (1 mg/mL). O pH foi ajustado para 7,4. A incubação durou 20-30
minutos em um banho-maria a 37ºC, sob agitação de 100 ciclos/minuto até a dispersão dos
túbulos em uma massa homogênea. O tecido disperso foi imediatamente diluído a 50 mL
com PBS/BSA para diminuir o efeito da enzima, os túbulos seminíferos sedimentados
durante 2 minutos e o sobrenadante foi filtrado através de malha de nylon (80 µm). O
filtrado foi centrifugado em tubos plásticos de 50 mL a 150xg, 21ºC, durante 15 minutos. O
sobrenadante foi descartado e as células sedimentadas foram ressuspendidas em meio de
cultura M199/BSA contendo NaHCO3 (2,2 mg/mL) e BSA (1 mg/mL), com o auxílio de
uma pipeta plástica até que nenhum agregado celular fosse visível. Essa suspensão foi
centrifugada como anteriormente e repetida a mesma operação duas vezes (3 lavagens). Em
seguida, as células foram contadas utilizando uma câmara de Neubauer. O número de
células a serem incubadas foi determinado previamente em 0,5 x 106 células/0,5 mL de
meio de incubação.
3.6.2 Secreção de testosterona in vitro
As preparações de células intersticiais testiculares (0,5 x 106 células/0,5 mL) foram
incubadas, em triplicata, em tubos de polipropileno 12x75mm, por 3 horas com M199,
10mUI/mL hCG ou 100µM ácido ascórbico adicionados separadamente ou 10mUI/mL
hCG + 100µM ácido ascórbico adicionados simultaneamente. As incubações foram
realizadas, logo após a obtenção da suspensão celular, a 34ºC sob atmosfera de 95% de O2 e
5% de CO2, em banho-maria tipo Dubnoff com agitação de 60 ciclos por minuto. Após o
término da incubação as células foram precipitadas por centrifugação (150xg por 15 min) a

28
4 ºC, o sobrenadante coletado e armazenado a -20ºC para posterior determinação da
testosterona por radioimunoensaio (RIA).
3.8 Viabilidade Celular
Foi utilizado o critério de exclusão do corante azul tripan (0,5%), o qual depende da
integridade da membrana celular. A leitura das células foi realizada na câmara de
Neubauer. A viabilidade celular foi expressa como a porcentagem de células não coradas
em relação ao número total de células. Essa porcentagem variou entre 90 e 95 %. Esse
estudo foi realizado após o término das incubações.
3.9 Radioimunoensaio da Testosterona
3.9.1 Testosterona no plasma:
A testosterona plasmática foi dosada utilizando-se um kit de duplo anticorpo
(Diagnostic Systems Laboratories, Inc., Texas, USA). Os coeficientes de variação intra e
inter-ensaio foram ambos 8 %.
3.9.2 Testosterona no meio de incubação (in vitro)
A testosterona foi determinada diretamente no meio de incubação (sem extração).
As amostras do meio de incubação (100 µL para os basais e 50 µL para células de Leydig
incubadas com hCG) foram incubadas com anticorpo anti-testosterona e 10.000 cpm de
[3H]-Testosterona a 4ºC, durante 20 horas, sendo o volume final da reação de 0,32 mL. O
tampão de ensaio utilizado foi o PBS - 0,1% gelatina. O anticorpo anti-testosterona foi
desenvolvido em coelhos e produzido no laboratório da Dra. Wanderley (UFPE), e sua
diluição final no ensaio foi de 1:5.000.

29
Uma mistura de carvão-dextrana na concentração de 0,625% de carvão e 0,0625%
de dextrana foi utilizada para remover a testosterona livre. Os tubos foram centrifugados a
1.300xg, à 4ºC, durante 20 minutos. O traçador ligado ao anticorpo foi lido em 0,4 mL do
sobrenadante adicionado a frascos de cintilação contendo 5 mL de solução de cintilação (3
g de PPO + 0,2 g de POPOP + 20 mL de metanol + 980 mL de tolueno). A leitura foi
realizada em um contador de cintilação líquida (Liquid Scintillation Analyzer). Foram
preparados padrões de testosterona de 8, 16, 32, 64, 128, 256 e 512 pg/20 µL e dosados em
triplicata. A testosterona das amostras foi dosada em duplicata. O coeficiente de variação
intra-ensaio foi de 8,1% e o coeficiente inter-ensaio foi de 15,1%.
3.10 Análise Estatística
Os dados experimentais foram expressos como a média ± o erro padrão da média
(média ± EPM) dos valores obtidos para cada grupo. A análise estatística dos dados foi
realizada utilizando o teste t de Student bicaudal para comparações dos valores entre os
grupos. Para a análise da testosterona no meio de incubação foram comparados os
valores basais de cada grupo com os valores estimulados (hCG, hCG+AA) e obtido um
∆ %, o qual foi utilizado para a comparação entre os grupos. Os valores com p < 0,05
foram considerados significativos.
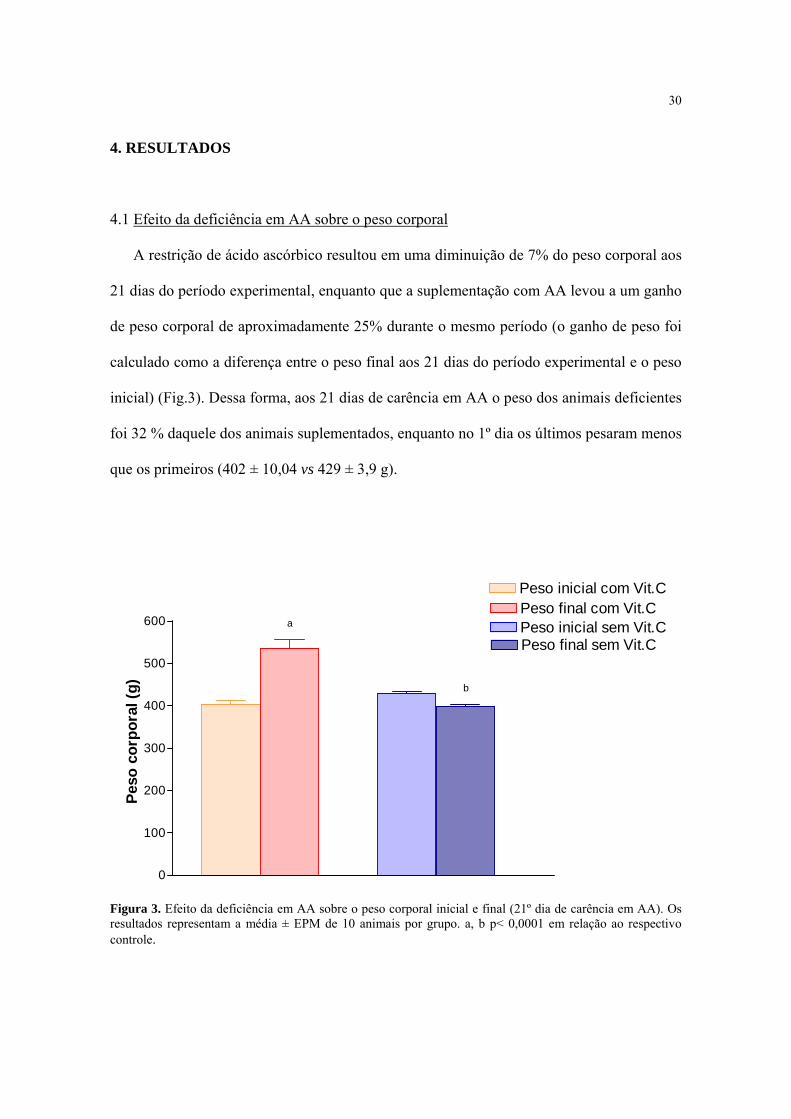
30
4. RESULTADOS
4.1 Efeito da deficiência em AA sobre o peso corporal
A restrição de ácido ascórbico resultou em uma diminuição de 7% do peso corporal aos
21 dias do período experimental, enquanto que a suplementação com AA levou a um ganho
de peso corporal de aproximadamente 25% durante o mesmo período (o ganho de peso foi
calculado como a diferença entre o peso final aos 21 dias do período experimental e o peso
inicial) (Fig.3). Dessa forma, aos 21 dias de carência em AA o peso dos animais deficientes
foi 32 % daquele dos animais suplementados, enquanto no 1º dia os últimos pesaram menos
que os primeiros (402 ± 10,04 vs 429 ± 3,9 g).
0
100
200
300
400
500
600 Peso inicial sem Vit.CPeso final sem Vit.C
Peso final com Vit.CPeso inicial com Vit.C
a
b
Peso
cor
pora
l (g)
Figura 3. Efeito da deficiência em AA sobre o peso corporal inicial e final (21º dia de carência em AA). Os resultados representam a média ± EPM de 10 animais por grupo. a, b p< 0,0001 em relação ao respectivo controle.

31
4.2- Efeito da deficiência em AA sobre o peso do testículo e da cauda do epidídimo
Os valores absolutos e relativos dos pesos do testículo e da cauda do epidídimo aos
21 dias do período experimental são mostrados na Tabela 1. Os valores médios relativos do
peso do testículo (calculados como mg/g de peso corporal) mostram que a deficiência em
AA não modificou esse parâmetro. Já o peso relativo da cauda do epidídimo diminuiu 25,2
% nos animais deficientes em AA quando comparados com o controle.
Tabela 1. Pesos absolutos (mg) e relativo (mg/g peso corporal) do testículo e cauda do epidídimo, nº e motilidade dos espermatozóides e testosterona plasmática de cobaios machos normais ou deficientes em ácido ascórbico. Os valores representam a média ± EPM de 10 animais por grupo. **p<0,01 vs controle; ***p<0,001 vs controle. Controle Deficiente em AA _______________________________________________________________________ Peso absoluto do testículo (mg) 1145,00 ± 005 795,00 ± 0,02 *** Peso relativo testículo (mg/g) 2,14 ± 0,07 1,99 ± 0,01 Peso absoluto cauda epidídimo (mg) 310,00 ± 0,03 171,00 ± 0,01*** Peso relativo cauda epidídimo (mg/g) 0,58 ± 0,02 0,43 ± 0,005** Nº espermatozóides (milhão/ml) 40,20 ± 4,00 10,80 ± 1,5*** Motilidade espermatozóide (%) 80,00 ± 5,90 58,00 ± 2,0** Testosterona plasmática (ng/ml) 2,10 ± 0,20 0,60 ± 0,1***

32
4.3 Efeito da deficiência em AA sobre o nº e motilidade dos espermatozóides
Um declínio significativo de aproximadamente 73% e 28% no número e motilidade,
respectivamente, dos espermatozóides foi observado nas cobaias deficientes em AA,
quando comparadas com as controle (Tabela 1). Os espermatozóides mostraram movimento
rotatório, motilidade muito lenta e rara progressão para diante.
4.4- Efeito da deficiência em AA sobre a concentração plasmática de testosterona
A Tabela 1 apresenta os valores médios absolutos e relativos da concentração
plasmática de testosterona nos dois grupos experimentais. A concentração de testosterona
diminuiu aproximadamente 72% nos animais deficientes em AA.
4.5 Efeito da deficiência em AA sobre a morfologia do espermatozóide
O animais com deficiência em AA apresentaram um maior número de alterações da
morfologia espermática, tanto da cabeça quanto da cauda, quando comparados com os
animais suplementados com AA (Fig. 4). As alterações morfológicas mais freqüentemente
observadas foram cabeças isoladas, estreitas na base e sem capuz acrossomático, cauda
abaxial e fortemente dobrada.
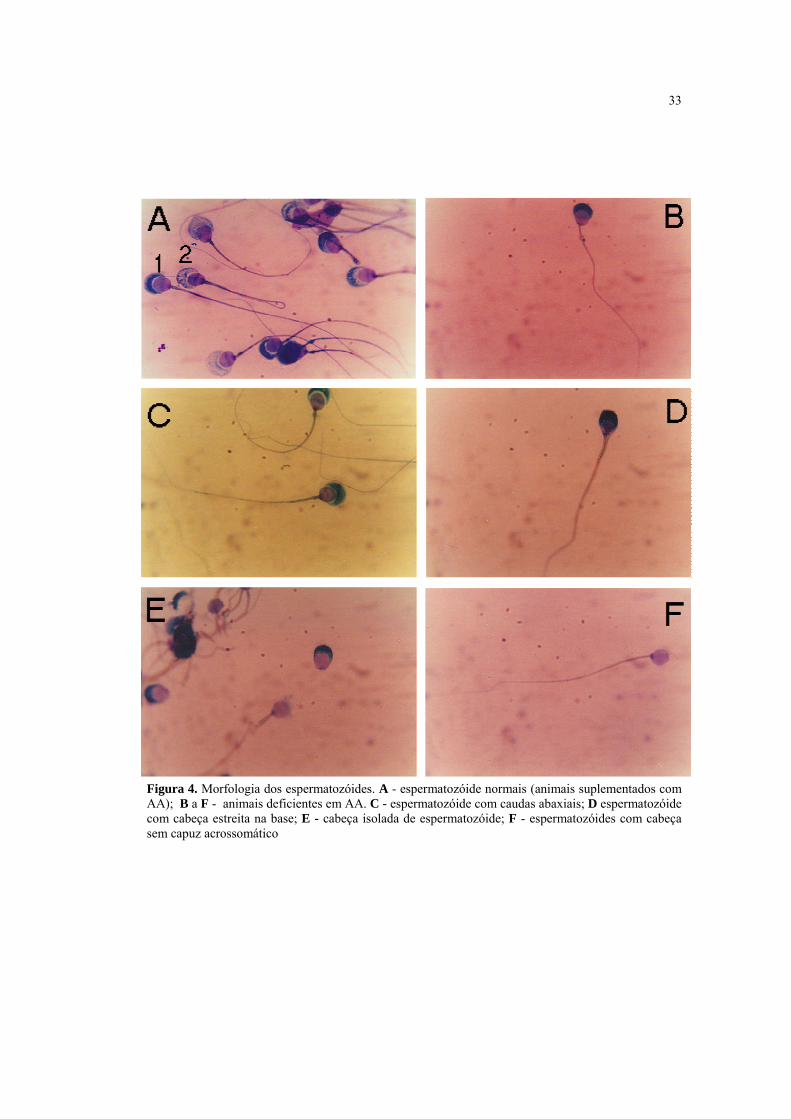
33
Figura 4. Morfologia dos espermatozóides. A - espermatozóide normais (animais suplementados com AA); B a F - animais deficientes em AA. C - espermatozóide com caudas abaxiais; D espermatozóide com cabeça estreita na base; E - cabeça isolada de espermatozóide; F - espermatozóides com cabeça sem capuz acrossomático

34
4.6 Efeito da deficiência em AA sobre a estrutura do testículo e epidídimo
Ao analisarmos (Fig.5) os cortes histológicos transversais dos testículos dos animais
deficientes em AA foi possível observar que o parênquima testicular apresentou alterações
do epitélio germinativo dos túbulos seminíferos com diminuição das camadas de
espermatogônias e dos espermatozóides, quando comparados aos animais não deficientes
em AA. Tais alterações tornaram-se mais evidentes na porção periférica do parênquima,
apresentando também desgarramento e soltura das células germinativas e áreas
vacuolizadas. As células de Leydig apresentaram-se aparentemente dentro da normalidade
histológica. Os cortes histológicos da cauda do epidídimo (Fig.6) dos animais deficientes
em AA mostraram uma aparente diminuição do nº de espermatozóides na luz tubular, o que
pode ser comprovada nos resultados da contagem de espermatozóides feita no
espermograma. Adicionalmente, houve redução na altura do epitélio colunar de
revestimento, diminuição da pseudoestratificação nuclear e áreas de redução da borda em
escova.

35
Figura 5. Histologia testicular de cobaios controle e deficientes em AA. Coloração hematoxilina eosina. Os painéis 1,3 mostram ductos seminíferos periféricos de animais controle no aumento de 20X e 40X respectivamente. Nos painéis 2,4 também no aumento de 20X e 40X respectivamente, notar a redução da celularidade germinativa nos ductos seminíferos dos animais deficientes em AA. Os painéis 5,6 e 7,8 mostram células de Leydig nos aumentos de 40X e 100X respectivamente. Notar
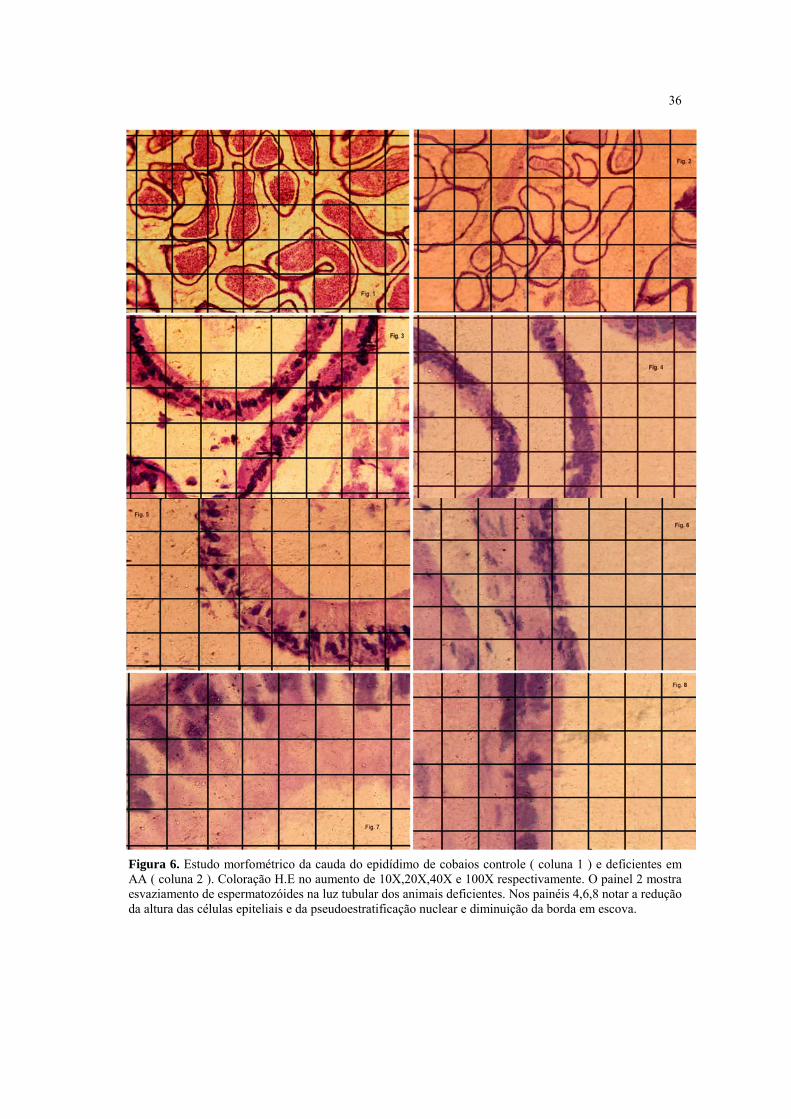
36
Figura 6. Estudo morfométrico da cauda do epidídimo de cobaios controle ( coluna 1 ) e deficientes em AA ( coluna 2 ). Coloração H.E no aumento de 10X,20X,40X e 100X respectivamente. O painel 2 mostra esvaziamento de espermatozóides na luz tubular dos animais deficientes. Nos painéis 4,6,8 notar a redução da altura das células epiteliais e da pseudoestratificação nuclear e diminuição da borda em escova.
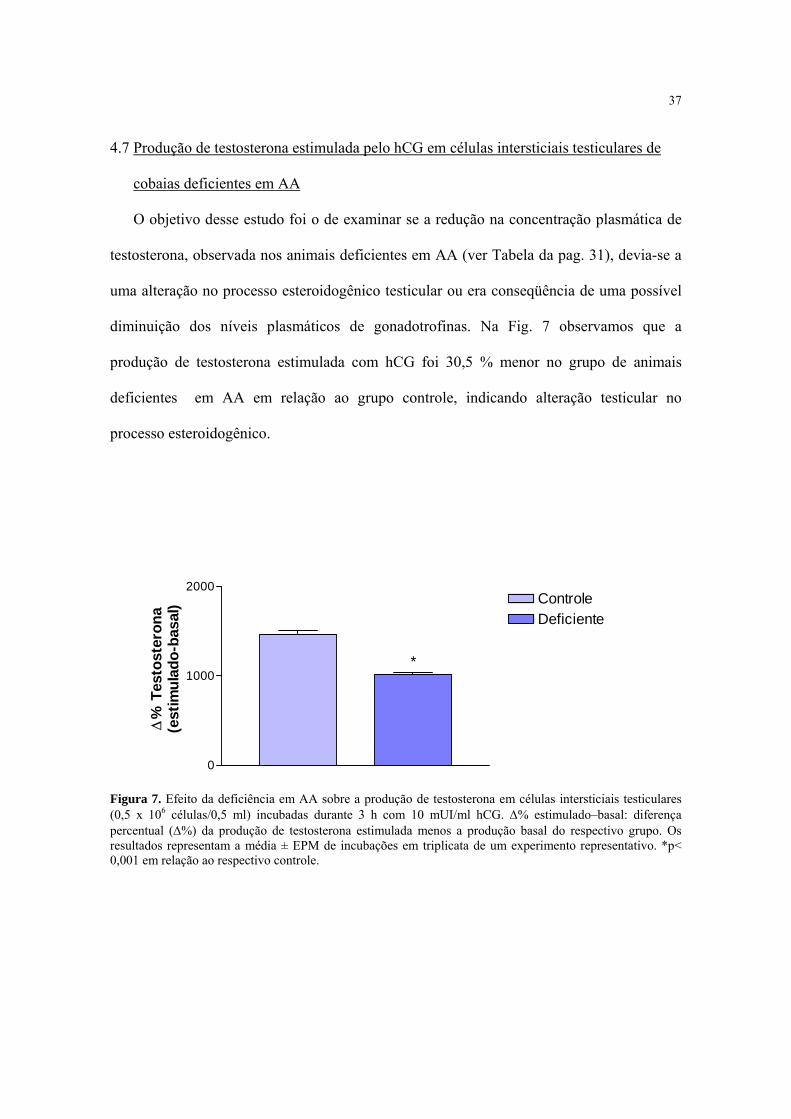
37
4.7 Produção de testosterona estimulada pelo hCG em células intersticiais testiculares de
cobaias deficientes em AA
O objetivo desse estudo foi o de examinar se a redução na concentração plasmática de
testosterona, observada nos animais deficientes em AA (ver Tabela da pag. 31), devia-se a
uma alteração no processo esteroidogênico testicular ou era conseqüência de uma possível
diminuição dos níveis plasmáticos de gonadotrofinas. Na Fig. 7 observamos que a
produção de testosterona estimulada com hCG foi 30,5 % menor no grupo de animais
deficientes em AA em relação ao grupo controle, indicando alteração testicular no
processo esteroidogênico.
0
1000
2000ControleDeficiente
*
∆%
Tes
tost
eron
a(e
stim
ulad
o-ba
sal)
Figura 7. Efeito da deficiência em AA sobre a produção de testosterona em células intersticiais testiculares (0,5 x 106 células/0,5 ml) incubadas durante 3 h com 10 mUI/ml hCG. ∆% estimulado–basal: diferença percentual (∆%) da produção de testosterona estimulada menos a produção basal do respectivo grupo. Os resultados representam a média ± EPM de incubações em triplicata de um experimento representativo. *p< 0,001 em relação ao respectivo controle.

38
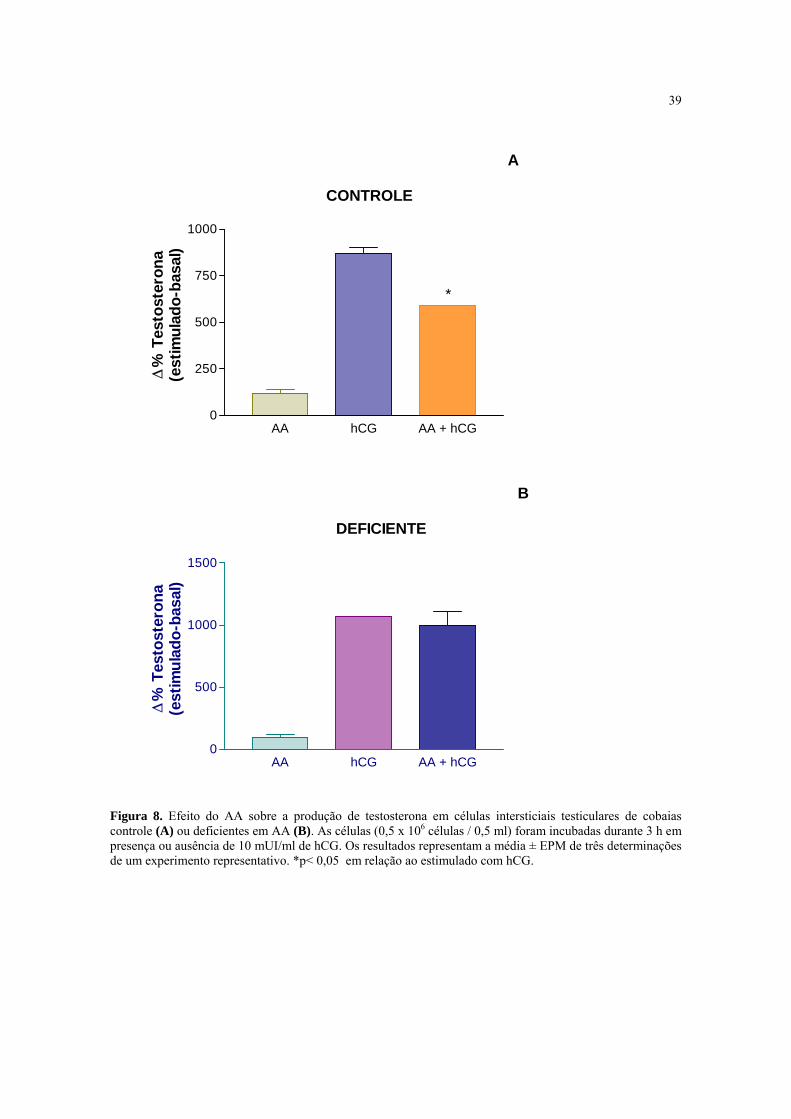
4.8 Efeito do AA sobre a produção de testosterona estimulada com hCG em células
intersticiais testiculares de cobaias controle ou deficientes em AA
O objetivo desse estudo foi o de relacionar as propriedades antioxidantes do AA com a
capacidade das células intersticiais testiculares de cobaias deficientes em AA de produzir
testosterona, um processo sensível aos danos causados pelos radicais livres originados do
O2- (Hornsby, 1980). A adição de 100 µM de AA diretamente ao meio de incubação não
modificou a produção basal de testosterona em nenhum dos dois grupos de cobaias (Fig.
8A e B). Contudo, a produção de testosterona estimulada maximamente pelo hCG foi
reduzida em 32 % no grupo controle, mas não foi modificada no grupo deficiente em AA.
39
AA hCG AA + hCG0
250
500
750
1000
∆%
Tes
tost
eron
a(e
stim
ulad
o-ba
sal)
A
CONTROLE
*
AA hCG AA + hCG0
500
1000
1500
B
DEFICIENTE
∆%
Tes
tost
eron
a(e
stim
ulad
o-ba
sal)
Figura 8. Efeito do AA sobre a produção de testosterona em células intersticiais testiculares de cobaias controle (A) ou deficientes em AA (B). As células (0,5 x 106 células / 0,5 ml) foram incubadas durante 3 h em presença ou ausência de 10 mUI/ml de hCG. Os resultados representam a média ± EPM de três determinações de um experimento representativo. *p< 0,05 em relação ao estimulado com hCG.

40
5. DISCUSSÃO
O ácido ascórbico é um componente essencial na dieta de primatas, cobaios e
morcegos. Há muitos anos ele tem sido associado com a fertilidade e parece ter um
significado evolutivo, o de proteger os tecidos contra a toxicidade do oxigênio (Nandi e
cols., 1997). Contudo, seu papel fisiológico preciso na reprodução não está completamente
esclarecido.
No presente estudo, relatamos que a deficiência em AA em cobaios machos adultos
produz alterações estruturais e funcionais nos órgãos da reprodução aos 21 dias de carência
do AA. Essas alterações consistiram em: diminuição do peso e modificação na histologia
do testículo e cauda do epidídimo, diminuição do número de espermatozóides,
malformações na cabeça e cauda dos espermatozóides e diminuição de sua motilidade,
diminuição da concentração plasmática de testosterona e da resposta secretória das células
intersticiais testiculares ao estímulo gonadotrófico em ausência ou presença de AA “in
vitro”.
Sabe-se que a deficiência em AA leva à perda de peso corporal. Estudos anteriores
(Gosiewska e cols., 1994; Peterkofsky 1991; Peterkofsky e cols., 1994) demonstraram que
durante a 3ª semana de uma dieta carente em AA, os cobaios tornam-se anoréxicos e
perdem peso. Várias conseqüências dessa deficiência de AA parecem surgir a partir do
início da anorexia e da cascata de eventos que a seguem: menor ingestão de alimentos leva
à perda de peso (Bird e cols., 1986; Peterkofsky, 1991) e a uma rápida diminuição da
concentração de insulina no plasma, a qual, por sua vez, aumenta a expressão da proteína
ligadora de fator de crescimento semelhante a insulina –1 e –2 ( IGFBP-1 e –2) no fígado e

41
aumenta os níveis de IGFBP-1 e –2 circulantes, os quais são captados pelos tecidos
(Gosiewska e Peterkofsky, 1995). Os IGFBP-1 e –2 inibem a ação do fator de crescimento
semelhante à insulina –1 (IGF-1) e então podem causar inibição da síntese de DNA, da
expressão do gene do colágeno nos ossos, pele e cartilagem (Gosiewska e cols., 1994) e da
síntese de proteoglican (Bird e cols., 1986). Contudo, nem todos os sintomas da deficiência
de AA podem ser explicados pelo efeito da perda de peso/IGFBP; estes incluem dificuldade
da cura de lesões cutâneas, deficiência de ferro e modificações na expressão de proteínas
relacionadas com o ferro (Gosiewska e cols., 1996). Nossos resultados mostraram uma
perda de peso corporal de 32 % no 21º dia (3ª semana) de carência de AA e estão em
conformidade com os resultados dos autores acima referidos.
Considerando que a perda de peso corporal acompanha a deficiência em AA, os
valores absolutos dos pesos dos órgãos estudados foram expressos em relação ao peso
corporal. Assim, foi possível observar que, se bem houve diminuição do peso absoluto do
testículo, a diferença do peso relativo desse órgão entre o grupo controle e o deficiente em
AA não foi estatisticamente significante. Contudo, houve alteração histológica e funcional
nesse órgão (diminuição da testosterona circulante e de sua produção “in vitro”). Assim, a
relação peso do órgão/peso corporal não pode ser tomada como parâmetro fisiológico que
possa ser relacionado com as diferentes disfunções observadas nos órgãos da reprodução. O
que nos leva a considerar que a deficiência em AA per se produziu alterações estruturais e
funcionais no testículo, em adição aos efeitos da desnutrição que a acompanha. Na cauda
do epidídimo a deficiência em AA causou diminuição do peso absoluto e relativo e
alteração histológica, corroborando o dito anteriormente. Chinoy e cols. (1986) também
relataram haver diminuição no peso absoluto do testículo, epidídimo e glândulas sexuais

42
acessórias em cobaios submetidos ao mesmo período de 21 dias de deficiência em AA sem,
contudo, referir-se à diminuição do peso corporal.
O presente estudo também revelou que a deficiência de AA resulta na perda da
integridade estrutural do testículo e epidídimo. As alterações observadas no testículo de
animais deficientes em AA se restringiram ao compartimento tubular, não havendo
alteração nas células de Leydig, corroborando dados de Chinoy e cols. (1983). Sabemos
que a análise histológica realizada no presente estudo é insuficiente para descartar a
possibilidade de alterações estruturais nas células de Leydig dos animais deficientes em
AA. A realização de estudos ultraestruturais quantitativos para observação do retículo
endoplasmático liso e das mitocôndrias são necessários, já que a maioria das enzimas
envolvidas na produção dos andrógenos está seqüestrada nesses compartimentos. As
modificações histológicas observadas no epidídimo dos animais deficientes em AA também
corroboram os dados de Chinoy e cols. (1983). Sabemos que esse órgão depende dos níveis
circulantes de testosterona para a manutenção de sua integridade estrutural e funcional. A
diminuição dos níveis plasmáticos de testosterona, observada nos animais deficientes em
AA, do presente estudo, suporta essa observação. Existem poucas referências sobre os
valores dos níveis plasmáticos de testosterona em cobaios normais ou escorbúticos. Os
valores de testosterona obtidos no presente estudo para os cobaios normais estão dentro dos
valores referidos na literatura (Koos Slob e cols., 1979; Gombe e cols., 1977). Com relação
aos cobaios deficientes em AA, nossos dados de diminuição dos níveis plasmáticos de
testosterona em relação aos cobaios normais, diferem dos de Gombe e cols. (1977), única
referência encontrada, os quais não observaram diferença nos níveis desse hormônio entre
os dois grupos. É possível que a diferença entre nossas observações e aquelas desses
autores seja devida às diferentes cepas dos animais utilizados. Novamente, a diminuição

43
dos níveis plasmáticos de testosterona pode ser devida tanto à carência de AA quanto à
desnutrição dela resultante ou, provavelmente, a ambas. Há referências de níveis baixos de
testosterona em cobaios desnutridos durante 23 dias a partir do desmame (Koos Slob e
cols., 1979).
Sabe-se que o comprometimento estrutural nos órgãos da reprodução dos cobaios
deficientes em AA leva ao comprometimento funcional desses órgãos (alteração do seu
meio interno e modificações do perfil dos íons metálicos e na morfologia, motilidade e
número dos espermatozóides. No presente estudo também foram observadas alterações na
morfologia, e diminuição no número e motilidade dos espermatozóides, suportando e
confirmando dados da literatura (Koehler, 1975; Chinoy e cols., 1983 e 1986).
A diminuição da concentração plasmática de testosterona associada à ausência de
alterações histológicas no compartimento das células intersticiais do testículo, observada
nos animais deficientes em AA, nos motivou a um estudo “in vitro” da secreção de
testosterona. Essa motivação deveu-se à possibilidade da existência tanto de uma
diminuição da secreção de gonadotrofina hipofisária, com a conseqüente diminuição da
resposta esteroidogênica testicular, quanto de uma alteração no processo esteroidogênico
nas células de Leydig. Para estimular a produção de testosterona utilizamos o hCG por ser,
como o LH, o principal estímulo esteroidogênico testicular. A produção de testosterona
pelo testículo é feita a partir do precursor colesterol. A maioria das reações envolvidas
nesse processo de esteroidogênese a partir do colesterol é catalizada por enzimas do
citocromo P-450, localizadas nas membranas do retículo endoplasmático e das
mitocôndrias (Hedge e col., 1988). São reações de hidroxilação em que o NADPH gerado
no ciclo das pentoses é o doador de H+. As etapas na síntese de testosterona pelo testículo
são as seguintes:

44
P-450scc 3β-ol
Colesterol → Pregnenolona → Progesterona ↓ C17α-hidroxilase 3β-ol ↓C17α-hidroxilase 17 α- hidroxipregnenolona → 17 α- hidroxiprogesterona ↓ C17-20-liase 3β-ol ↓ C17-20-liase Dehidroepiandrosterona → Androstenediona ↓ 17-cetoredutase 3β-ol ↓ 17-cetoredutase Androstenediol → Testosterona → Estradiol Aromatase
Figura 9. Principais vias para a biossíntese de testosterona no testículo (Modificado de Hedge e col., 1988)
Existem evidências que atestam a participação do AA no processo esteroidogênico,
principalmente nas reações de hidroxilação. Na adrenal o AA protege de inativação a
enzima mitocondrial 11β-hidroxilase e os sistemas enzimáticos microssomais da C17α-
hidroxilase/C17,20-liase nas células esteroidogênicas (Hornsby, 1980). Essas enzimas
podem ser destruídas por vários compostos, alguns dos quais parecendo atuar através do
mecanismo de peroxidação de lipídeos (Plaa e Witschi, 1976). Hornsby (1980) propôs que
a destruição da atividade do citocromo P-450 é conseqüência da interação dos seus (da
enzima) produtos esteróides (denominados pseudosubstratos) com a própria citocromo P-
450 para formar o complexo pseudosubstrato.P-450.O2, do qual são liberados radicais livres
derivados do oxigênio, devido à incapacidade do pseudosubstrato de ser hidroxilado. Em
estudo posterior, Hornsby e Crivello (1983) demonstraram que essa espécie reativa de
oxigênio (H2O2 e superóxido) é produzida pelo citocromo P-450 durante a catálise e sua
produção é estimulada pelos pseudosubstratos. Os radicais livres derivados do oxigênio
podem reagir com o citocromo P-450 e inativá-lo diretamente ou iniciar a peroxidação dos
lipídeos de membrana na qual o citocromo P-450 está embebido e de forma indireta inativá-

45
lo. O efeito protetor dos antioxidantes como o AA, o α-tocoferol e o dimetilsufóxido, sobre
as enzimas microssomais C17α-hidroxilase/C17,20-liase acima referidas, sugere que esse
mecanismo opere no citocromo P-450 adrenal e testicular (Hornsby, 1980; Quinn e Payne,
1985). O AA atuaria removendo os radicais livres derivados do oxigênio. Essa
susceptibilidade do citocromo P-450 aos radicais livres pode explicar a manutenção de altas
concentrações de AA e sua captação ativa na supra-renal e gônadas (Moger, 1987; Hornig,
1975). Esses tecidos mantem as condições de antioxidação para proteger o citocromo P-450
da destruição induzida pelos pseudosubstratos. Na adrenal o tratamento com o hormônio
trófico do córtex supra-renal (ACTH) aumenta a concentração dos esteróides nesse tecido,
assim como também a taxa de produção de radicais livres e a liberação de AA. Podemos
pensar que fenômeno similar ocorre nas gônadas. Dessa forma, poderíamos dizer que no
testículo a homeostase dos esteróides seria mantida quando as células de Leydig fossem
estimuladas por um pulso de hormônio luteinizante (LH), o qual levaria à indução das
enzimas P-450 e a sua degradação como resultado da estimulação da esteroidogênese. A
diminuição da resposta secretória das células intersticiais testiculares dos animais
deficientes em AA ao estímulo com hCG, observada no presente estudo, suporta a
participação do AA no processo esteroidogênico testicular. Se bem não possamos descartar
a participação da desnutrição que acompanha a deficiência em AA nesses resultados, dados
da literatura mostram que a desnutrição em ratos (estarvação durante 3 dias) diminui os
níveis plasmáticos de testosterona, mas não altera a capacidade das células de Leydig de
produzir esse hormônio sob condições basais ou estimuladas com hCG (Grizard e cols.,
1997). Esses dados reforçam a nossa sugestão da participação direta do AA na secreção
testicular de testosterona do presente estudo.

46
Por outro lado, também existem evidências de que o AA inibe muitas das reações de
hidroxilação envolvidas na biossíntese dos hormônios esteróides, incluindo a 21-
hidroxilase (Greenfield e cols., 1980; Gordon e cols., 1985), 11β-hidroxilase e 18-
hidroxilase (Kitabchi AE 1967) e a enzima de clivagem da cadeia lateral do colesterol
(Sulimovici e Boyd, 1968). No ovário de rato, o AA “in vitro” em concentrações altas inibe
a reação de clivagem da cadeia lateral do colesterol através de um efeito direto no
complexo enzimático (Sulimovici e Boyd, 1968). Essas concentrações do AA são
comparáveis àquelas observadas no ovário “em repouso”. Além do mais, redução na
concentração de AA a um nível similar àquele observado na glândula estimulada, resulta na
liberação dessa inibição. Dessa forma foi sugerido que o papel do AA pode ser o de atuar
como um agente redutor fisiológico capaz de inibir, em concentrações críticas, a cadeia de
transporte de elétrons que é parte integral das reações de hidroxilação envolvidas na
biossíntese dos hormônios esteróides (Sulimovici e Boyd, 1968; Kitabchi, 1976).
Conseqüentemente, o hormônio trófico, ao facilitar a remoção do AA pode ativar o sistema
enzimático de hidroxilação e em seguida aumentar a produção de esteróide. Essa hipótese
poderia explicar a diminuição da produção de testosterona estimulada pelo hCG observada
no presente estudo e no de Kukucka e Misra (1994), quando as células intersticiais
testiculares de cobaios controle foram incubadas com alta concentração de AA (100 µM).
Também explicaria os resultados por nós obtidos com as células intersticiais dos animais
deficientes em AA, utilizando a mesma dose de 100 µM de AA – ou seja, que, ao contrário
dos animais controle, o AA não reduziu o efeito estimulatório do hCG sobre a produção de
testosterona. Isso significaria que, para as células esteroidogênicas testiculares dos animais
deficientes, a dose de AA adicionada “in vitro” não seria uma dose alta, como o foi para os
animais controle. Podemos considerar, como na adrenal (Bjorkhem e cols., 1978) e no

47
ovário (Sulimovici e Boyd, 1968), que no testículo a concentração de AA está dentro da
faixa inibitória, levantando a possibilidade de que a inibição pelo AA tenha alguma
importância nas condições “in vivo”. O fato de que, no nosso estudo, o efeito do AA
adicionado “in vitro” sobre a produção de testosterona foi diferente nas células intersticiais
dos animais controle em relação aos deficientes em AA, está de acordo com a hipótese de
que o AA inibitório deva estar presente nas células dos animais controle. Adicionalmente, o
efeito do AA sobre a produção de testosterona nos animais deficientes atesta a participação
do AA no processo esteroidogênico testicular.
Em resumo, nossos resultados com cobaios deficientes em AA confirmam prévias
referências (dados de peso e estrutura do testículo e cauda do epidídimo e qualidade dos
espermatozóides) e fornecem evidências adicionais (níveis plasmáticos e secreção testicular
de testosterona) da importância do AA na manutenção da integridade estrutural e
fisiológica do testículo, epidídimo e espermatozóides. Os resultados também indicam que
os efeitos da deficiência em AA sobre o epidídimo e a qualidade dos espermatozóides
podem ser indiretos, conseqüentes à privação de testosterona nesse órgão alvo (nesse caso a
influência de desnutrição que acompanha a deficiência em AA também deve ser
considerada na diminuição dos níveis plasmáticos de testosterona), mas também não
descartam uma participação direta do AA no órgão. No que se refere à produção de
testosterona pelo testículo, os dados indicam a participação direta do AA nesse processo.

48
6. CONCLUSÕES 1- As alterações estruturais do testículo e epidídimo, como também do número, motilidade
e morfologia dos espermatozóides, observadas nos cobaios deficientes em AA, confirmam dados da literatura.
2- As alterações do epidídimo e espermatozóides acima referidas podem ser indiretas,
conseqüentes à privação de testosterona. 3- Os resultados da secreção de testosterona “in vitro” sugerem a participação direta do
AA na esteroidogênese, sem, contudo, descartar a influência da desnutrição e/ou alteração do eixo hipotálamo hipofisário.
4- O efeito inibitório de 100 µM de AA adicionado “in vitro”, sobre a produção de
testosterona, não foi observado nos cobaios deficientes em AA.

49
7. REFERÊNCIAS BIBLIOGRÁFICAS
ANDERSON D, PHILLIPS BJ, YU T, EDWARDS AJ, AYESH R E BUTTERWORTH
KR. The effect of vitamin C supplementation on blomakers of oxygen radical generated
damage in human volunteers with low or high cholesterol levels. Environmental
Molecular Mutagens 1997, 30:161174.
BERG OC, HUGGINS C, HODGES CV. In Effect of ascorbic acid on male fertility.
Dawson EB et al. Annals of the New York Academy of Sciences 1987, 498:312-23.
BJORKHEM I, KALLNER A, KARLMAR K-E. Effecte os ascorbic acid deficiency on
adrenal mitochondrial hydroxylations in guinea pigs. Journal of Lipid Research 1978,
19:695-704.
BRIGGS MH. Vitamin C and infertility. Lancet 1973, 2:677.
BYRD JÁ, PARDUE SL, HARGIS BM. Effect of ascorbate on luteinizing hormone
stimulated progesterone biosynthesis in chicken granulosa cells in vitro. Comparative
Biochemstry and Physiology 1993, 104A:279-281.
CARR AC E FREI B. Does vitamin C act as a pro-oxidant under physiological conditions?
FASEB Journal 1999, 13:1007-1024.

50
CHATTERJEE IB. Evolution and the biossynthesis of ascorbic acid. Science 1973,
182:1271-1272
CHINOY NJ, BUCH-NEE RP, MEHTA RR, SEETHALAKSHMI L, SHARMA JD,
CHINOY MR. Effect of vitamin C deficiency on physiology of male reproductive organs
of guinea pigs. International Journal of Fertility 1986, 31:232-39.
CHINOY MR, SHARMA JD, SANJEEVAN AG E CHINOY NJ. Structural changes in
male reproductive organs and spermatozoa of scorbutic guinea pigs. Proceedings of the
Indian National Science Academy 1983, B49:628-635.
CHRISTIANE Y, DEMOULIN A, GILLAIN D, LEROY F, LAMBOTTE R, LAPIERE
CM, NUSGENS B, FOIDART J-M. Laminin and type III procollagen peptide in human
preovulatory follicular fluid. Fertility and Sterility 1988, 50 : 48-51.
COUNSELL JN, HORNIG DH. Vitamin C (Ascorbic Acid) In Effect of ascorbic acid on
male fertility. Dawson EB et al. Annals of the New York Academy of Sciences 1987,
498:312-23.
DAS PC, DAS KP, BAGCHI K, DEY CD. Evaluation of tissue ascorbic acid status in
different hormonal states of female rat. Life Sciences 1993, 52:1493-1498.

51
DAWSON EB, HARRIS WA, RANKIN WE, CHARPENTIER LA, MCGAINTY WJ.
Effect of ascorbic acid on male fertility. Annals of the New York Academy of Sciences
1987, 498:312-323.
EIPPER BA, Mains RE. The role of ascorbate in the biosynthesis of neuroendocrine
peptides. American Journal of Clinical Nutrition 1991, 548:1153S-1156S.
GORALCZYK R, MOSER UK, MATTER U WEISER H. Regulation of steroid
hormonemetabolism requires L-ascorbic acid. Annals of the New York Academy of
Science 1992, 669:349-351.
GORDON MT, ANDERSON DC, MICHELL R, ROBERTSON WR. Some characteristics
of human adrenal microsomal 21-hydroxylase activity. Journal of Steroid Biochemstry
1985, 23: 67-72.
GREENFIELD N, PONTICORVO L, CHASALOW F, LIEBERMAN S. Activation and
inhibition of the adrenal steroid 21-hydroxylation system by cytosolic constituents:
influence of glutathione, glutathione reductase and ascorbate. Archives of Biochemstry
and Biophysic 1980, 200:232-244.
GRIZARD G, ARTONNE C, GRIZARD J, BOUCHER D. Effect of short-term starvation
on Leydig cell function in adult rats. Archives of Andrology 1997, 38:207-214.

52
HEDGE GA, COLBY HD, GOODMAN RL (eds). Fisiologia Endócrina Clínica. Rio de
Janeiro: Interlivros, 1988.
HIMENO N, KAWAMURA N, OKAMURA H, MORI T, FUKOMOTO M,
MIDORIKAWA O. Colagen synthetic activity in rabbit ovary during ovulation and its
blockage by indomethacin. Acta of Obstetrics and Gynaecology Japonica 1984,
36:1930-1934.
HORNIG D. Distribution of ascorbic acid, metabilites and analogues in man and animals.
Annals of the New York Academy of Sciences 1975, 258:103-118.
HORNSBY PJ. Regulation of cytochrome P-450-supported 11β-hydroxylation of
deoxycortisol by steroids, oxigen, and anti-oxidants in adrenocortical cell cultures. The
Journal of Biological Chemistry 1980, 255:4020-4027.
IGARASHI M. Augmentative effect of ascorbic acid upon induction of human ovulation in
Clomiphene-ineffective anovulatory women. International Journal of Fertility 1977,
22:168-173.
JAGER DH. Experimental amyotrophic lateral sclerosis in the guinea pig. Journal of
Neurological Science 1985, 67:133-142.
KITABCHI AE. Ascorbic acid in steroidogenesis. Nature 1967, 215:1385-6.

53
KUKUCKA MA MISRA HP. HPLC determination of na oxytocin-like peptide produced
by isolated guinea pig Leydig cells: stimulation by ascorbate. Archives of Andrology
1992, 29:185-190.
LEVIN M. New concepts in the biology and biochemstry of ascórbic ácid. New England
Journal of Medicine 1986, 31:892-902.
LEVINE M, MORITA K.Ascorbic acid in endocrine systems. Vitamines and Hormones
1985, 42:1-64.
Lindahl PE, Kihlstrom JE. Some factors influencing the biological activity of sperm
agglutinins. Journal of Reproduction and Fertility 1960, 1:3-22.
LINDAHL PE. Sperm agglutinating and antiagglutinating factors in normal follicular fluid
from cattle. International Journal of Fertility 1966, 11:297-305.
LINDSAY B, MEDES G. In Effect of ascorbic acid on male fertility. Dawson EB et al.
Annals of the New York Academy of Science 1987, 498:312-23.
LUCK MR, JEYASEELAN I, SCHOLES R. Ascorbic acid and Fertility. Biology of
Reproduction 1995, 52:262-266.

54
LUCK MR, JUNGCLAS B. The time course of oxytocin secretion from cultured bovine
granulosa cells, stimulated by ascorbate and catecholamines. Journal of Endocrinology
1988, 116:247-258.
LUCK MR, ZHAO Y. Identification and measurement of collagen in the bovine corpus
luteum and its relationship with ascorbic acid and tissue development. Journal of
Reproduction and Fertility 1993, 99:647-652.
LUCK MR. Cholinergic stimulation, through muscarinic receptors, of oxytocin and
progesterone secretion from bovine granulosa cells undergoing spontaneous luteinization in
serum-free culture. Endocrinology 1990, 126:1256-1263.
LUCK MR. The gonadal extra-cellular matrix. Oxford Ver Reproduction Biololy 1994,
16: 33-85.
MIES FA. Reprodução dos animais e inseminação artificial. Editora Salina, Rio Grande
do Sul, 1978.
MILLAR J. Vitamin C – the primate fertility factor? Medical Hypotheses 1992, 38:292-
295.
MILLS JH, SCHWARTZ NB. Ovarian ascorbic acid as an endogenous and exogenous
assay for cyclic proestrous LH release. Endocrinology 1961,69:844-850.

55
MOGER WH. Uptake and release of ascorbic acid by rat Leydig cells in vitro. Journal of
Andrology 1987, 8:398-402.
MOSER U E BENDICH A .Vitamin C. In: Handbook of Vitamins. Edited by Machlin LJ
e Marcel Dekker. New York, 1990, Cap. 5.
NAIDU KA. Vitamin C in human health and disease is still a mystery? An overview.
Nutrition Journal 2003, 2:7-16.
NISHIKIMI M, FUKUYAMA R, MINOSHIMA S, SHIMIZU N, YAGIK. Cloning and
chromosomal mapping of the human non-functional gene for L-gulono lactone oxidase, the
enzyme for L-ascorbic acid biosynthesis missing in man. The Journal of Biological
Chemstry 1994, 269::13685-13688.
NISHIKIMI M, KAWAI T, YAGI K. Guinea pig possess a highly mutated gene for L-
gulono lactone oxidase, the key enzyme for L-ascorbic acid biosynthesis missing in this
species. The Journal of Biological Chemstry 1992, 267::21967-21972.
PLAA GL, WITSCHI H.Chemicals, drugs, and lipid peroxidation. Annual Review of
Pharmacolgy and Toxicology 1976, 16:125-141.
QUINN PG; PAYNE AH. Steroid product-induced, oxigen-mediated damage of
microsomal cytochrome P-450 enzymes in Leydig cell cultures.The Journal of Biological
Chemistry 1985, 260:2092-2099.

56
ROSENBERG G. Exame clínico dos bovinos. Editora Guanabara Koogan, 1997.
ROSTON S. Ascorbic acid, oxygen and disappearance of adrenochrome and
noradrenochrome. Nature 1962, 194:1079-1080.
SAUBERLICH HE. Bioavailbility of vitamins. Progress in Food Nutrition Sciences
1985, 9:1-33.
SEBRELL WH, HARRIS RS. In Ascorbic acid and Fertility. Luck MR, Jeyaseelan I,
Scholes R. Biology of Reproduction 1995, 52:262-266.
STANSFIELD DA, FLINT AP. The entry of ascorbic acid into the corpus luteum in vivo
and in vitro and the effect of luteinizing hormone. Journal of Endocrinology 1967, 39:27-
35.
SULIMOVICI S E BOYD GS. The effect of ascorbic acid in vitro on the rat ovarian
cholesterol side chain cleavage enzyme system. Steroids 1968, 12:127-149.
TSUJL M, ITO Y, TERADA N, MORI H. Ovarian aromatase activity in scorbutic mutant
rats unable to synthesize ascorbic acid. Acta Endocrinologica (Copenh) 1989, 121:595-
602.

57
WALTON A The technique of artificial inseminação. Notes on artificial insemination
of sheep cattle and horses. London, 1939.
WANG Y-H, DHARIWAL KR, LEVINEM. Ascorbic acid bioavailability in humans.
Annals of the New York Academy of Science 1992, 669:383-386.

58