treinamento fÍsico antes e durante a gestaÇÃo e … · 2019-10-25 · submetido ao corpo...
TRANSCRIPT

FILIPPE FALCÃO TEBAS OLIVEIRA
TREINAMENTO FÍSICO ANTES E DURANTE A GESTAÇÃO
E DESNUTRIÇÃO PERINATAL: RESPOSTAS ADAPTATIVAS
NA PROLE DE RATOS
RECIFE
2011

FILIPPE FALCÃO TEBAS OLIVEIRA
TREINAMENTO FÍSICO ANTES E DURANTE A GESTAÇÃO
E DESNUTRIÇÃO PERINATAL: RESPOSTAS ADAPTATIVAS
NA PROLE DE RATOS
Dissertação apresentada ao Programa de
Pós-Graduação em Nutrição do Centro
de Ciências da Saúde da Universidade
Federal de Pernambuco, para obtenção
do título de Mestre em Nutrição.
Orientadora: Profa. Dr
a. Carol Virgínia
Góis Leandro.
Co-orientadora: Profa. Dr
a. Cláudia
Jacques Lagranha.
RECIFE
2011

Oliveira, Filippe Falcão Tebas
Treinamento físico antes e durante a gestação e desnutrição perinatal: respostas adaptativas na prole de ratos / Filippe Falcão Tebas Oliveira. – Recife: O
Autor, 2011. 113 folhas: il., fig. quadro; 30 cm.
Orientador: Carol Virgínia Góis Leandro
Dissertação (mestrado) – Universidade Federal de Pernambuco. CCS. Nutrição, 2011.
Inclui bibliografia e anexos.
1. Desnutrição. 2. Atividade física. 3. Gestação. 4. Desenvolvimento sensório-motor. 5.
Histologia muscular. I. Guedes, Rubens Carlos Araújo. II.Título.
UFPE
616.39 CDD (20.ed.) CCS2011-161


Aos meus pais, Edésio e Derli (in memoriam), e minha “mãe do
coração”, Tânia (in memoriam). A base de tudo, meus primeiros professores e
suas lições eternas.
À Camila, minha doce companheira com quem compartilho tudo! E
à nossa pequena Beatriz, onde em cada abraço, sorriso e palavra sua, o amor
parece fluir... e assim, a nossa felicidade se satisfaz. Amo muito vocês!

AGRADECIMENTOS ESPECIAIS (PARTE 1)
Às minhas orientadoras:
Carol,
Só tenho a agradecer e muito, por toda formação acadêmica, profissional e moral que você tem
assumido comigo. Saber dizer um “não” (ou um precisa melhorar) na hora certa é tão importante quanto
um “sim” (ou um parabéns). Geralmente se associa isso aos pais e mães para com seus filhos. E você,
querida Carol, além desta característica mostra muito carinho, zelo, confiança e outras qualidades
“maternas” quando se trata de seus orientandos. Obrigado por me aceitar, me ensinar e por estar ao meu
lado, desde o início. Tenha certeza que dedico a você respeito e admiração imensuráveis!
Cláudia,
A simplicidade e a atenção com que você lida com as situações e com as pessoas, te põe em
destaque naturalmente. Basta conversar um minuto com aqueles que lhe conhecem (nem que seja um
pouquinho só) para que sua identidade seja revelada: uma pessoa animada, carinhosa, dedicada, efetiva,
enfim... Essas (e outras, que acredito que ainda não tive tempo de conhecer) são as qualidades que tornam
você especial. Obrigado por caminhar comigo nessa estrada!
Ao professor Raul,
Ah! Se eu conseguisse escrever aqui o obrigado do tamanho que realmente quero expressar...
Desde o início quando você me instigou (na sua sala, quando Carol me apresentou, lembra?!), eu não fui
mais a mesma pessoa. Gostaria, portanto, de lhe dizer meus sinceros agradecimentos por todas as
oportunidades cedidas, pela atenção dispendida, pelos conselhos e o cuidado dedicado.
Tenham certeza que vocês ganharam um fiel
“escudeiro” ao longo dessa caminhada.

AGRADECIMENTOS ESPECIAIS (PARTE 2)
Ao meu irmão Will e minha irmã Cris, tenho carinho incondicional por vocês!
A Marco, afinal tudo começou com você. Sou eternamente grato a você por abrir meus olhos e
estimular sempre o conhecimento! “Mais um dia (ou, se me permite), uma fase, ganha com honestidade!”.
À Ana Elisa, por toda sua paciência e dedicação e conhecimento compartilhados comigo.
A Adriano, Antônio e Diogo, devo muitos obrigados vocês: pela sinceridade, pelo
companheirismo... e sobretudo, pela amizade! E, como “quem avisa amigo é”: Cuidado , isso vai dá . . . . . . .
A todos os integrantes de nosso grupo NNI-DOHaD, pelas significativas contribuições nas
discussões científicas e, porque não, pelos momentos de (des)concentração!
À Rafaela e Amanda, sem a ajuda de vocês certamente o caminho seria muito mais árduo. Muito
obrigado!
Ao Dr. França e “Seu” Paulino (in memoriam), pelas sábias orientações no dia-a-dia.
Ao prof. Dr. Rubem e ao Departamento de Anatomia (UFPE), pelo espaço cedido.
A Neci e Cecília por toda atenção.
Aos colegas do mestrado, funcionários e professores do Departamento e da Pós-Graduação em
Nutrição da UFPE.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela concessão da
bolsa de estudo.

RESUMO
O objetivo desta dissertação foi avaliar os efeitos da desnutrição protéica e/ou do treinamento
físico no período perinatal sobre o desenvolvimento do sistema nervoso, indicadores de
crescimento somático e bioquímicos, e a histologia das fibras musculares esqueléticas da
prole de ratos. Ratas Wistar realizaram treinamento físico moderado (5 dias/semana e
60min/dia, a 65% VO2max) antes e leve (5 dias/semana e 20min/dia, a 40% VO2max) durante a
gestação. A manipulação nutricional ocorreu na gestação e lactação, com dieta a base de
caseína 17% (controle e treinada) ou caseína 8% (desnutrida e treinada desnutrida). As
avaliações de maturação reflexa e características físicas, indicadores murinométricos e
bioquímicos e histologia das fibras musculares foram realizadas nos filhotes, os quais foram
divididos de acordo com a manipulação materna: controle (CF,), treinado (TF), desnutrido (DF)
e treinado desnutrido (TDF). A partir do desmame, os filhotes foram mantidos com dieta
padrão. O grupo DF apresentou como resposta adaptativa imediata o atraso na maturação dos
reflexos e de características físicas, além de déficit de crescimento somático comparado ao
grupo CF. Com relação ao contraste gerado pela resposta adaptativa preditiva e o ambiente
nutricional ao longo da vida, o grupo DF exibiu maiores valores de circunferência abdominal,
glicemia e colesterolemia e uma histologia muscular direcionada às fibras glicolíticas,
comparado ao grupo CF. O treinamento físico durante a gestação per si, pouco modificou os
parâmetros avaliados. Todavia, quando associado à desnutrição perinatal, o treinamento físico
foi capaz de atenuar e/ou normalizar a maioria dos parâmetros avaliados na prole ao longo da
vida. Nossos dados demonstram que o treinamento físico durante a gestação possa ter um
papel fundamental nas respostas adaptativas imediatas e preditivas.
Palavras-chave: Desnutrição; Atividade Física; Gestação; Desenvolvimento sensório-motor;
Histologia muscular; Ratos.

ABSTRACT
The aim of this study was to evaluate the effects of undernutrition and/or physical training in
the perinatal period on the development of the nervous system, somatic growth and
biochemical indicators and the histology of skeletal muscle fibers of offspring rats. Female
rats were submitted to a moderate (5 d/week and 60min/d at 65% VO2max) physical training
before and light (5 d/week and 20min/d, up to 40% VO2max) during pregnancy. The nutritional
manipulation was carried out during pregnancy and lactation with either a low-protein diet
17% casein (control and trained groups) or 8% casein (undernourished and undernourished
trained groups). The evaluations of reflex maturation and physical characteristics,
murinometrics and biochemical indicators and histology of muscle fibers were performed in
pups, which were divided according to maternal manipulation: control (CP), trained (TP),
undernourished (UNP) and trained undernourished (T+UNP). At weaning, pups were
maintained on standard diet. The UNP group showed as an immediate adaptive response, a
delay in the maturation of reflexes and physical characteristics, and a lower somatic growth
when compared to the CP group. In terms of the contrast generated by the predictive adaptive
response and nutritional environment throughout life, the UNP group showed a high values of
waist circumference, blood glucose and cholesterol and muscle histology toward to glycolytic
fibers, when compared to CP group. Physical training during pregnancy itself had non
pronounceable changes in the evaluated parameters. However, when associated with perinatal
undernutrition, physical training was able to attenuate and/or normalize most of the
parameters in the offspring throughout life. We demonstrated that physical training during
pregnancy may have a role in both predictive and immediate adaptive responses.
Keywords: Undernutrition, Physical Activity, Pregnancy, sensory-motor development,
muscle histology; Rats.

SUMÁRIO
1. APRESENTAÇÃO .......................................................................................................... 9
2. REVISÃO DA LITERATURA
Nutrição, atividade física e a origem desenvolvimentista da saúde e da doença ........... 11
3. OBJETIVOS .................................................................................................................. 34
4. HIPÓTESES .................................................................................................................. 35
5. MÉTODOS
5.1 Animais .......................................................................................................... 36
5.2 Protocolo experimental de treinamento físico das mães ................................ 36
5.3 Manipulação da dieta das mães ...................................................................... 39
5.4 Consumo alimentar ...................................................................................... 40
5.5 Avaliação do peso corporal, do ganho de peso e da circunferência
abdominal .................................................................................................... 40
5.6 Avaliações murinométricas .......................................................................... 41
5.7 Avaliação da maturação de características físicas .......................................... 42
5.8 Avaliação da ontogênese dos reflexos ............................................................ 42
5.9 Avaliação da glicemia e colesterolemia dos filhotes ...................................... 43
5.10 Preparação dos músculos soleus e extensor longo dos dedos ...................... 43
5.11 Análise histológica ....................................................................................... 44
5.12 Análise estatística ......................................................................................... 45
6. RESULTADOS
Maternal low-protein diet-induced delayed reflex ontogeny is attenuated by moderate
physical training during gestation in rats ....................................................................... 46
Efeitos do treinamento físico durante a gestação sobre a evolução ponderal, glicemia
e colesterolemia de ratos adultos submetidos à desnutrição perinatal ........................... 64
Treinamento físico e desnutrição perinatal: efeitos sobre a histologia de fibras
musculares esqueléticas em ratos adultos ...................................................................... 86
7. CONCLUSÕES ............................................................................................................... 106
REFERÊNCIAS .................................................................................................................. 108
ANEXOS ........................................................................................................................... 109

9
1. APRESENTAÇÃO
Embora vários estudos associem a pandemia da obesidade, diabetes e síndrome
metabólica ao estilo de vida atual do indivíduo, uma nova explicação à luz da origem
desenvolvimentista da saúde e da doença vem se consolidando. De acordo com essa hipótese,
o organismo em desenvolvimento é capaz de interagir com ambiente atual gerando uma
resposta adaptativa imediata e/ou preditiva (curto e longo prazo, respectivamente). Um
estímulo investigado é a desnutrição perinatal, e suas consequências tanto em curto quanto em
longo prazo têm sido observadas. A resposta em curto prazo pode acarretar consequências
negativas, mas garantem a sobrevivência do indivíduo até a idade reprodutiva. Enquanto que
na resposta em longo prazo, o grau de diferença entre a previsão realizada no início da vida e
o ambiente ao longo do seu desenvolvimento podem influenciar o risco de desenvolver
doenças metabólicas.
De fato, alguns estudos têm observado os efeitos da desnutrição perinatal em curto-
prazo, no desenvolvimento do sistema nervoso e crescimento somático. Em longo prazo, as
alterações fenotípicas em animais cujas mães foram desnutridas parecem ser constantes. A
desnutrição perinatal causa em ratos adultos um perfil semelhante ao da obesidade, da
diabetes tipo II e da síndrome metabólica. Ademais, há relação entre as doenças metabólicas e
o fenótipo do sistema muscular esquelético de animais previamente desnutridos. Acredita-se
que o metabolismo no músculo tenha papel fundamental no risco de desenvolvimento das
doenças metabólicas.
Neste estudo, propomos que a atividade física regular materna possa agir como
intervenção precoce para combater os efeitos em curto e/ou em longo prazo da desnutrição
perinatal. Tendo em consideração o anteriormente exposto, este estudo tem com objetivo
principal: analisar as repercussões do treinamento físico associado à desnutrição perinatal
sobre o desenvolvimento sensório-motor e a proporção de fibras musculares esqueléticas na
prole de ratos adultos. Nossas hipóteses são (1) a desnutrição durante a gestação e lactação
retarda o desenvolvimento sensório-motor e aumenta o percentual de fibras musculares
esqueléticas do tipo IIb e diminui do tipo I da prole adulta; (2) o treinamento físico durante a
gestação, por sua vez, reverte os efeitos da desnutrição perinatal no desenvolvimento
sensório-motor e no fenótipo de fibras musculares da prole adulta.
Esta dissertação gerou quatro artigos científicos, um foi submetido, um foi publicado,
um foi aceito para publicação, e o último será enviado após suas correções. O primeiro artigo,

10
intitulado “Nutrição, atividade física e a origem desenvolvimentista da saúde e da doença”, foi
submetido ao corpo editorial do Jornal de Pediatria. O objetivo deste artigo de revisão foi
discutir (1) as bases teóricas da origem desenvolvimentista da saúde e da doença, e (2) as
repercussões na prole do fator nutricional e da atividade física materna.
Para testar a resposta adaptativa imediata à desnutrição e/ou o treinamento físico na
gestação, o segundo artigo, intitulado: “Maternal low-protein diet-induced delayed reflex
ontogeny is attenuated by moderate physical training during gestation in rats”, foi
confeccionado. Este artigo original foi aceito para publicação pelo British Journal of
Nutrition. O objetivo deste trabalho foi avaliar os efeitos do treinamento físico durante a
gestação sobre a ontogênese reflexa em ratos neonatos provindos de mães que foram
desnutridas na gestação e lactação. Com este artigo, mostramos as respostas adaptativas
imediatas geradas pela desnutrição e pelo treinamento físico na gestação.
O terceiro artigo, intitulado: Efeitos do treinamento físico durante a gestação sobre a
evolução ponderal, glicemia e colesterolemia de ratos adultos submetidos à desnutrição
perinatal, foi aceito para publicação na Revista Brasileira de Medicina do Esporte. Neste
artigo nós avaliamos os efeitos do treinamento físico durante a gestação na evolução ponderal,
circunferência abdominal, glicemia e colesterolemia de filhotes adultos submetidos à
desnutrição perinatal. Foi demonstrado que o treinamento físico durante a gestação atenua os
efeitos da desnutrição perinatal sobre alguns indicadores murinométricos e bioquímicos nos
filhotes adultos.
O último artigo original resultante desta dissertação encontra-se em processo de
construção. O título (temporário) do manuscrito é “Treinamento físico moderado na gestação
aumenta o percentual de fibras oxidativas em filhotes adultos submetidos à desnutrição
perinatal”. Neste artigo foi analisada a histologia das fibras musculares esqueléticas em ratos
adultos, cujas mães foram desnutridas durante a gestação e lactação e/ou submetidas a um
protocolo de treinamento físico.

11
2. REVISÃO DA LITERATURA – ARTIGO DE REVISÃO
ARTIGO DE REVISÃO – SUBMETIDO AO CONSELHO EDITORIAL DO JORNAL DE PEDIATRIA
TÍTULO: Nutrição, atividade física e a origem desenvolvimentista da saúde e da doença
TÍTULO ABREVIADO: ATIVIDADE FÍSICA E RESPOSTAS ADAPTATIVAS
Nome dos autores Titulação Endereço eletrônico Currículo
Lattes
Contribuição específica
Filippe Falcão-Tebas Graduado [email protected] Sim Consulta às bases de dados, discussão
teórica, escrita e revisão final
Raul Manhães-de-
Castro
Doutor [email protected] Sim Discussão teórica e revisão final do
texto
Carol G. Leandro Doutora [email protected] Sim Discussão teórica, escrita e revisão
final
DECLARAÇÃO DE CONFLITO DE INTERESSE
Nada a declarar.
DEFINIÇÃO DE INSTITUIÇÃO
Departamento de Nutrição, Universidade Federal de Pernambuco.
Núcleo de Educação Física e Ciências do Esporte, Centro Acadêmico de Vitória,
Universidade Federal de Pernambuco.

12
AUTOR RESPONSÁVEL PELA CORRESPONDÊNCIA E PELOS CONTATOS PRÉ-
PUBLICAÇÃO
Filippe Falcão-Tebas
Rua Prof. Moraes Rego, 1235 – CEP: 50670-901 – Departamento de Nutrição – Cidade
Universitária – Recife, PE – Brasil.
Programa de Pós-graduação em Nutrição – UFPE
Telefone: (00 55 81) 21268463
E-mail: [email protected]
FONTE FINANCIADORA OU FORNECEDORA DE EQUIPAMENTO E
MATERIAIS
Não é o caso.
CONTAGEM TOTAL DAS PALAVRAS DO TEXTO
4.446 palavras.
CONTAGEM TOTAL DAS PALAVRAS DO RESUMO
224 palavras (excluindo-se os termos de indexação)
NÚMERO DE TABELAS E FIGURAS
Tabelas – 0 (zero).
Figuras – 1 (um).

13
RESUMO
Objetivo: Discutir (1) as bases teóricas da origem desenvolvimentista da saúde e da doença, e
(2) as repercussões na prole do fator nutricional e da atividade física materna.
Fontes dos dados: Foram utilizadas as bases de dados do Medline/PubMed e SciELO,
restringindo-se a publicações de 2000 a 2011. Os termos de indexação utilizados foram:
programming, adaptive responses, perinatal, undernutrition, gestation, physical activity,
physical exercise.
Síntese dos dados: A hipótese da origem desenvolvimentista da saúde e da doença propõe
explicar as associações entre o estado de saúde ao longo da vida e o ambiente durante o
crescimento e desenvolvimento do indivíduo. A nutrição é um fator capaz de induzir um
contraste em diferentes fases da vida. As consequências causadas pela desnutrição perinatal
seguida da normalização ou do excesso de ingestão de nutrientes, estão associadas à
obesidade e ao diabetes mellitus tipo dois. Procurando conter a incidência destas doenças, a
investigação de medidas de intervenção assume importância. Tem sido relatado que a
atividade física durante a gestação tem efeitos benéficos em ambos os períodos fetal e
neonatal.
Conclusões: Os mecanismos subjacentes da origem desenvolvimentista da saúde e da doença
parecem envolver respostas homeostáticas e epigenéticas em curto e longo prazo,
respectivamente. A atividade física durante a gestação é capaz de aumentar o aporte de
nutrientes e oxigênio no ambiente feto-placentário e promover uma resposta adaptativa
imediata favorável.
Palavras-chave: Desnutrição; Atividade Física; Plasticidade no Desenvolvimento; Respostas
Adaptativas.

14
TITLE: Nutrition, physical activity and developmental origins of health and disease
SHORT TITLE: PHYSICAL ACTIVITY AND ADAPTIVE RESPONSES
ABSTRACT
Objectives: To discuss: (1) the theoretical basis of the developmental origin of health and
disease, and (2) the repercussions of a nutritional factor and maternal physical activity
Sources: For this review, it was used the databases Medline/PubMed, SciELO, with
publications since 2000 to 2011. The index terms were: programming, adaptive responses,
perinatal, undernutrition, gestation, physical activity, physical exercise.
Summary of the findings: The proposal of Developmental Origin of Heath and Disease is to
explain relationship between the status of adult health with the critical period of growth and
development. Nutrition plays an important role in this context once it can induce a mismatch
during different periods of life. In addition the long-last effects of perinatal undernutrition are
related to earlier appearance of obesity and diabetes mellitus type 2. Different procedures of
intervention have been tested, for example the habitual physical activity during gestation.
Physical activity during gestation is associated to beneficial effects for both mother and fetus.
Conclusions: In conclusion, underlying mechanisms of developmental origin of health and
disease involve homeostatic and epigenetic responses in short and long-last, respectively.
Maternal physical activity is able to enhance the amount of nutrients and oxygen into the fetal
environment and promotes an positive adaptive immediate response.
Keywords: Undernutrition; Physical Activity; Developmental Plasticity; Adaptive Responses

15
INTRODUÇÃO
A alta prevalência de doenças crônicas não transmissíveis como obesidade,
hipertensão e diabetes tem sido estudada em uma nova óptica, a “origem desenvolvimentista
da saúde e da doença” ou DOHaD1, 2
. Neste sentido, estudos epidemiológicos e experimentais
têm relatado que o aparecimento de doenças da vida adulta tem uma associação com as fases
iniciais do desenvolvimento que incluem a gestação, lactação e primeira infância 2. Por
exemplo, a obesidade em homens adultos foi associada com a exposição à fome materna
durante a gestação3. Em animais adultos expostos à desnutrição protéica na gestação, foi
observado um maior risco de desenvolver diabetes tipo dois4.
Um dos focos de estudo da DOHaD envolve o contraste entre os ambientes pré e pós-
natal, que pode induzir respostas em curto e/ou em longo prazo no organismo em
desenvolvimento. As respostas em curto prazo são chamadas de “respostas adaptativas
imediatas”, enquanto que as respostas em longo-prazo são chamadas de “respostas adaptativas
preditivas”5, 6
. Tais respostas podem ser observadas no âmbito nutricional uma vez que a
desnutrição perinatal pode induzir em curto prazo: menor crescimento somático, atraso na
maturação reflexa e desregulação de parâmetros metabólicos em animais com até 30 dias de
vida7, 8
. Já os estudos voltados às respostas em longo prazo demonstram que a desnutrição
perinatal seguida da normalização ou do excesso de nutrientes ingeridos na vida adulta pode
elevar as chances de desenvolver as doenças crônicas não transmissíveis4, 9, 10
. Desse modo, a
investigação de um fator ambiental que possa atuar protegendo o organismo em
desenvolvimento é de interesse.
Tem sido observado que a atividade física durante a gestação pode ter um efeito
benéfico para o organismo em desenvolvimento11
. Mulheres que praticam exercício físico
antes e durante a gestação têm uma melhor interação materno-fetal e funcionalidade
placentária, resultando em uma melhor oferta de oxigênio e nutrientes12, 13
. Contudo, estes

16
efeitos são diretamente relacionados à magnitude do esforço e o período de início do
programa de atividade física. Alguns estudos têm mostrado que intervir com o treinamento
físico de baixa intensidade (~40% VO2max) na gestação pode ser benéfico sobretudo na prole
exposta à desnutrição, normalizando a taxa de crescimento e o desenvolvimento sensório-
motor após o nascimento14, 15
. Diante do exposto, sugerimos que a atividade física durante a
gestação é um importante fator modulador que pode ser considerado no âmbito da origem
desenvolvimentista da saúde e da doença. Portanto, o objetivo desta revisão é discutir (1) as
bases teóricas da origem desenvolvimentista da saúde e da doença, e (2) as repercussões do
fator nutricional e da atividade física materna durante a gestação.
Para compor a literatura citada, foram consultadas as bases de dados
MedLine/PubMed (National Library of Medicine, EUA) e Scientific Electronic Library
Online (SciELO), utilizando-se os seguintes termos de indexação: programming, adaptive
responses, perinatal, undernutrition, gestation, physical activity, physical exercise. Foram
selecionados os artigos publicados de 2000 a 2011, além de estudos clássicos, de acordo com
a relevância do trabalho para os tópicos abordados.
ORIGEM DESENVOLVIMENTISTA DA SAÚDE E DA DOENÇA E A NUTRIÇÃO PERINATAL
A hipótese da origem desenvolvimentista da saúde e da doença (DOHaD) propõe
explicar os fenômenos biológicos que relacionam o fenótipo do adulto ao seu ambiente no
início da vida16
. Na conjectura proposta, se introduz a ideia de uma plasticidade relacionada
ao desenvolvimento (developmental plasticity) ou fenótipo, em períodos com acentuada
proliferação e diferenciação celular. A plasticidade do desenvolvimento, ou plasticidade
fenotípica, reflete a capacidade de um único genótipo produzir formas alternativas de
estrutura, estados fisiológicos ou comportamentais em resposta às condições ambientais1, 16, 17
.

17
A hipótese da DOHaD surgiu, em grande parte, de estudos epidemiológicos retrospectivos,
observando estímulos/insultos nestes períodos de alta plasticidade17
.
Os primeiros trabalhos relacionando a vida perinatal e as consequências na vida adulta
surgiram no início do século passado. A partir de 1911, no condado de Hertfordshire, no
Reino Unido, todos os nascimentos foram assistidos e o peso ao nascer registrado18, 19
. A
seguir, o estado de saúde ou doença destes indivíduos na vida adulta foi associado ao período
perinatal. Foi observado em 10.636 homens com até 65 anos de idade, que a taxa de
incidência de mortes por doenças coronárias era inversamente proporcional ao peso ao
nascer18
. Mais adiante, entre 1944-1945, durante a segunda guerra mundial, a Holanda sofreu
embargo do transporte de alimentos imposto pelas forças de ocupação alemã (período
chamado de “Fome Holandesa” - Dutch Famine). Nesta ocasião, a população sofreu uma
restrição alimentar em torno de 50% (~1400 para 670 kcal diárias), inclusive entre as
gestantes3, 20
. Esta restrição na alimentação materna resultou em filhos nascidos com baixo
peso21
. Além disso, a exposição à restrição alimentar na primeira metade da vida intrauterina,
foi associada com uma maior prevalência de obesidade3 e doenças coronarianas
22 em homens
aos 19 anos. O modelo da DOHaD portanto, assume que consequências agudas e duradouras
podem ser observadas.
De fato, um dos conceitos da DOHaD reflete a ideia que o feto pode ser capaz de
desenvolver uma resposta adaptativa imediata (RAI; em curto prazo) e/ou uma resposta
adaptativa preditiva (RAP; em longo prazo) em decorrência de um estímulo ambiental5, 23
. As
RAIs são modificações do organismo em desenvolvimento, geralmente associadas a uma
vantagem adaptativa, para regular sua resposta fisiológica para o ambiente
intrauterino/perinatal atual17, 24
. Estas respostas, por exemplo, homeostáticas que incluem
alterações no metabolismo e/ou no fluxo sanguíneo, aumentam as chances de sobrevivência
em curto prazo17
. Uma RAI metabólica do feto à desnutrição aguda é uma redução do

18
anabolismo, e caso a desnutrição seja mantida, a RAI fetal observada é um crescimento mais
lento de órgãos25
. Em resposta à desnutrição materna, o feto tende a proteger órgãos vitais,
como o encéfalo e o coração, pela redistribuição do fluxo sanguíneo em seu favor26
. Com um
menor fluxo sanguíneo para os demais órgãos fetais, há uma menor transferência materno-
fetal de aminoácidos e de glicose via placenta, diminuindo sua disponibilidade27
.
Consequentemente, as concentrações fetais de glicose, de insulina, do fator de crescimento
semelhante à insulina (IGF-1) diminuem26
. É de ressalva que estes efeitos são transitórios e
podem ou não comprometer o crescimento fetal para garantir sua sobrevivência24
.
Outra resposta que pode ser observada em relação à desnutrição intrauterina/perinatal
é a RAP que surge primariamente assumindo vantagens adaptativas futuras para o organismo
em desenvolvimento1, 5
. Ao interagir com o milieu intrauterino (dependente do ambiente
materno) e perinatal, o feto tem a capacidade de prever e adaptar-se a este mesmo ambiente ao
longo de sua vida pós-natal, de modo dependente ou não de uma RAI5, 16, 17, 24
. Se a dedução
fetal contrastar com o ambiente pós-natal, ou seja, o ambiente intrauterino/perinatal não for
mantido, haverá um maior risco para o desenvolvimento de doenças não-
transmissíveis/metabólicas na vida adulta, como a obesidade e a diabetes mellitus tipo dois5,
24. Consequentemente, a DOHaD considera que haja uma zona de menor risco de desenvolver
doenças metabólicas e se o ambiente ao longo da vida se encontrar fora deste limite tanto
superior quanto inferior, o risco de desenvolver doenças aumenta. Assim, o grau de
concordância ou contraste (match or mismatch) entre o ambiente previsto no período de alta
plasticidade e o ambiente real em que o indivíduo vive quando adulto irá modular seu
fenótipo6, 16, 28
.
O papel da nutrição perinatal

19
As repercussões do contraste provocado pela dieta inadequada/insuficiente no período
de alta plasticidade seguida de uma dieta adequada/excedente ao longo da vida podem ser
entendidas a partir da transição nutricional observada em alguns países29
. A transição
nutricional é caracterizada pelo rápido declínio da prevalência de desnutrição na população
infantil e concomitante aumento da prevalência de sobrepeso/obesidade em adultos29, 30
. Há
aproximadamente 30 anos, a China introduziu reformas na estrutura de sua economia rural e
obteve um aumento rápido da economia no país, logo, a transição alimentar ocorreu de forma
acentuada31
. Um estudo prospectivo, com um total de aproximadamente 16.000 crianças e
adultos na China, mostrou que o sobrepeso e a obesidade têm aumentado rapidamente nas
ultimas três décadas31
. No Brasil, a transição nutricional também tem sido documentada30
.
Estudos epidemiológicos relatam a tendência contínua de redução da desnutrição infantil no
Brasil, concomitante ao aumento do excesso de peso e da obesidade da população em
diferentes fases da vida30, 32
. Assim, os estudos com a transição nutricional parecem assumir
um papel importante no âmbito da DOHaD em humanos.
Outras situações como a imigração entre países socioeconomicamente diferentes
envolvem mudanças culturais na dieta, na saúde e no comportamento. Por exemplo, há cerca
de 40 anos, a prevalência de diabetes mellitus tipo dois em Israel era 0,06% e, após as
primeiras imigrações iemenitas para Israel, a prevalência passou a ser aproximadamente 12%,
25 anos depois33
. Outro estudo similar foi realizado com 158 etíopes (homens e mulheres),
com aproximadamente 30 anos de idade, que transitaram para Israel refugiando-se da fome
severa34
. Embora todos os imigrantes fossem magros, eles revelaram grandes mudanças nos
hábitos alimentares desde a sua chegada em Israel e, segundo o teste oral de tolerância à
glicose, ~25% tornaram-se diabéticos não insulino-dependentes em 4 anos ou menos34
. Na
Suécia, um estudo observou 30.705 homens e mulheres, com faixa etária entre 35-64 anos,
dos quais 25% eram estrangeiros35
. A maior prevalência de diabetes mellitus tipo dois foi

20
encontrada em imigrantes de origem não europeia (5,82%), enquanto que a prevalência nos
imigrantes de origem europeia (1,96%) e suecos (1,80%) foram iguais35
.
As interações entre o ambiente pré e pós-natal não são facilmente perceptíveis a partir
de estudos em humanos, devido à falta de controle da composição e quantidade da dieta ou o
grau da desnutrição nas mães e filhos. Assim, estudos com modelos animais assumem grande
importância na investigação das respostas adaptativas imediatas e preditivas.
Modelos experimentais
Os estudos com animais utilizando manipulações dietéticas durante o período perinatal
têm fornecido maior plausibilidade biológica para as respostas adaptativas tanto imediatas
quanto preditivas. Modelos experimentais bem estabelecidos submetem ratos a dietas
hipoprotéicas (5 – 9% caseína) ou restrição alimentar (30 – 70% de redução da dieta oferecida
ao controle) durante a gestação e/ou lactação4, 7, 36
. Relativamente à RAI, ratas submetidas à
dieta hipoprotéica de 8% durante a gestação tiveram neonatos com menor peso comparados
aos controles (20% caseína)4, 37
. A desnutrição perinatal na prole de ratos causa atraso na
maturação reflexa e retardo no crescimento somático durante a lactação8, 38
, além de
alterações no perfil das fibras musculares esqueléticas e cardíacas39
. Ademais, em
camundongos alimentados durante a lactação, com restrição de 70% da dieta, foram
observados maiores concentrações de colesterol e triglicerídeos aos 22 dias de idade7.
As RAPs por sua vez, ocorrem de acordo com o contraste entre o ambiente nutricional
pré e pós-natal, o qual aumenta as possibilidades de desenvolver doenças metabólicas. Ratos
expostos à desnutrição protéica (8% caseína) durante a gestação e lactação, e à dieta padrão
de manutenção ao longo da vida apresentaram menor peso corporal absoluto até os 180 dias40
.
Porém aos 240 dias de vida o peso dos animais desnutridos foi igual ao peso dos controles,
por um mecanismo independente do comportamento alimentar40
. Corroborando estes

21
resultados, em outro estudo com mesmo desenho experimental, os animais apresentaram
maior risco de desenvolver resistência à insulina com ~450 dias de idade devido a menor
expressão de componentes da via de sinalização da insulina no músculo e no adipócito4. Em
uma prole exposta à desnutrição intrauterina, foi observada uma maior concentração sérica de
ácidos graxos livres, além da menor expressão de enzimas-chave da via de sinalização da
insulina (AKT-1 e PKC-) aos 120 dias41
. Ratos adultos cujas mães foram submetidas à
restrição alimentar (70% da dieta ofertada às controles) apresentaram maior gordura
suprarrenal relativa ao peso corporal, maiores concentrações de leptina, insulina e glicose
séricas, considerados parte dos indicadores do fenótipo da obesidade9. Embora esses fatos
venham sendo amplamente observados, suas bases teóricas necessitam ser mais exploradas.
Mecanismos propostos
O mecanismo subjacente à RAI parece envolver uma menor disponibilidade de
substratos e de oxigênio ao feto via placenta42
. Ovelhas alimentadas com ~30% da dieta
ofertada ao grupo controle durante a gestação apresentaram uma diminuição no fluxo
sanguíneo uterino43
. Em cobaias (Cavia porcellus) alimentadas com 70% da dieta do grupo
controle até a metade da gestação, foi observado aumento na espessura da barreira placentária
e diminuição no tamanho relativo da rede placentária44
, dificultando o aporte de substratos
para o feto. Adicionalmente, ratas expostas a restrição calórica de 50% da dieta na última
semana de gestação tiveram menor expressão de transportadores de glicose tipo 3 (GLUT3)
na placenta45
. Com um menor número de transportadores, a disponibilidade de substrato ao
feto é menor, afetando seu crescimento e desenvolvimento. A desnutrição (80% da dieta
ofertada ao controle) em ratas, também diminuiu o conteúdo de glicogênio na placenta, no 16º
dia de gestação46
. Com relação à dieta hipoproteica (8% caseína) materna, foi observado um
aumento no tamanho da placenta47
. Este aumento foi associado a um maior número de

22
estruturas relacionadas à função placentária, como a área total de interface materno-fetal e o
volume não-labiríntico relativo ao peso fetal47
. Esses mecanismos podem estar relacionados
às respostas adaptativas imediatas sofridas pelos filhotes devido ao aporte inadequado de
substratos.
Em relação às RAPs, o mecanismo sugerido envolve a epigenética, onde um fator
externo pode modular a estrutura do DNA sem mudança na sua sequência28
. Estes
mecanismos incluem, mas não são limitados à, metilação e acetilação do DNA, e
modificações nas histonas48
. Assim, os estímulos ambientais são capazes de conduzir o
organismo ao seu ambiente esperado na vida adulta além do que seria possível através do
genótipo herdado16
. Em um estudo com ratos desnutridos (30% da dieta ofertada ao controle)
durante a gestação, o fenótipo e a expressão de genes diretamente relacionados às vias
metabólicas dos carboidratos, lipídios e proteínas em diferentes tecidos foram avaliados10
. A
prole cuja mãe foi desnutrida apresentou aos 55 dias de vida, um menor peso corporal e um
percentual de gordura corporal igual ao grupo controle. No músculo esquelético e no tecido
adiposo branco da prole nessa idade, não foram encontradas alterações na expressão gênica
entre os grupos10
, entretanto, foi observado que o fígado apresentou 249 genes expressos
irregularmente (143 mais expressos e 106 menos expressos) relacionados à glicogênese,
glicogenólise e glicólise. Ainda neste estudo, o fenótipo do grupo desnutrido aos 110 dias de
vida foi modificado, incluindo: maior percentual de gordura corporal, maiores concentrações
de leptina, insulina e peptídeo-C comparado ao controle10
. Estes resultados suportam o
modelo das respostas adaptativas preditivas sugerido pela DOHaD16
.
Com o intuito de amenizar os efeitos deletérios tardios relacionados à desnutrição
perinatal, a busca por uma intervenção assume importância. Neste sentido, nosso grupo tem
questionado a cerca da influência do estilo de vida materno, no âmbito da atividade física,
durante períodos de alta plasticidade49
. A atividade física durante a gestação é uma

23
intervenção capaz de atenuar o ganho de peso excessivo, melhorar a função placentária e
cardiovascular50
. Esta intervenção durante a gestação origina efeitos concomitantes ao feto, os
quais parecem gerar uma RAI e modular a trajetória normal de desenvolvimento.
ATIVIDADE FÍSICA DURANTE A GESTAÇÃO E SEU PAPEL NA DOHAD
Atividade física regular tem sido associada com um risco reduzido de morbidade e
mortalidade, melhora de funções cardiorrespiratórias, metabólicas e psicológicas. Durante a
gestação, os efeitos benéficos podem ser observados a depender da magnitude do exercício e
o nível de aptidão física da mãe49
. Em 2002, o American College of Obstetricians and
Gynecologists51
publicou um guia para exercício físico durante a gestação. Neste guia,
mulheres gestantes poderiam se exercitar 30 minutos ou mais (intensidade moderada) todos os
dias da semana desde que não apresentassem complicações médicas ou obstétricas.
Entretanto, este guia não especificou o que significa “exercício moderado” ou a quantidade de
calorias gastas por semana na realização de atividade física regular. Recentemente, um novo
posicionamento oficial com as recomendações foi sugerido com base no dispêndio energético
(expresso em taxa de equivalente metabólico, MET)52
. Neste guia, foi sugerida a realização de
atividade física de forma a aumentar o dispêndio energético entre 16 - 28 METs/hora/semana,
ou um exercício de intensidade moderada (≥ 60% da frequência cardíaca de reserva durante a
gestação ou até 60% do consumo máximo de oxigênio, VO2max)52
. Para realizar uma atividade
de 28 METs hora/semana, as gestantes devem caminhar a 3.2 km/hora, por 11.2 horas por
semana (2.5 METs, intensidade leve, ≤ 40% do VO2max).
A atividade física regular de intensidade leve parece resultar em benefícios para a mãe
e o feto. Na mãe, tem sido observado uma melhor função cardiovascular, ganho de peso
limitado, diminuição do desconforto músculo-esquelético, menor incidência de câimbras
musculares e edema de membros inferiores, estabilidade no humor, redução de diabetes

24
mellitus gestacional e a hipertensão gestacional53
. Para o feto, foi observada uma diminuição
da massa gorda, melhor tolerância ao estresse e avançada maturação neurocomportamental53
.
Em modelos animais, o nosso grupo tem observado que ratas gestantes treinadas em esteira (5
dias/semana, progressiva diminuição da duração 50 – 20 minutos/dia, a 40% do VO2max)
apresentaram um menor ganho de peso corporal e um aumento no consumo de oxigênio de
repouso (VO2 de repouso)14
. Nos filhotes durante a lactação, foi observada uma melhora na
ontogênese reflexa e na maturação física15, 54
.
Respostas adaptativas imediatas à atividade física materna e à desnutrição
As repercussões da atividade física regular podem ser entendidas no contexto das
respostas preditivas adaptativas imediatas sugeridas pela DOHaD55
. Um dos trabalhos
pioneiros foi uma coorte realizada na década de 90 que incluiu 800 gestantes praticantes de
atividade física com intensidade leve/moderada, praticantes com intensidade alta e não
praticantes56
. Foi verificado que os recém-nascidos de mães que se exercitaram com
intensidade leve/moderada durante a gestação apresentaram peso normal56
. Na Índia, um
questionário de atividade física foi estruturado para classificar 797 mulheres de acordo com
seu nível de atividade física. Neste estudo, foi observado que um nível de atividade física
intensa regular (ambiente doméstico e rural) repercutiu negativamente no peso placentário,
peso ao nascer, circunferências da cabeça e do braço do neonato57
. Resultado similar em
relação ao peso ao nascer foi observado em um ensaio clínico envolvendo 46 gestantes
exercitadas (60 minutos/dia, 5dias/semana durante 20 semanas, diminuindo gradualmente
para 20 minutos/dia, 5dias/semana por 24 semanas)13
.
É de ressalva, contudo, que o balanço energético durante a gestação é um fator
importante que afeta a relação entre nutrição materna e o peso ao nascer49
. Mulheres
subnutridas de comunidades rurais dos países em desenvolvimento têm cargas altas de

25
atividade física (trabalho agrícola e atividades domésticas) e seus filhos apresentam baixo peso
ao nascer57
. Extrema carga de trabalho também tem sido associada ao aumento da taxa de
aborto e bebês prematuros57
. Por outro lado, atividade física de baixa intensidade e realizada
sistematicamente está associada ao aumento do peso ao nascer mesmo em mulheres que
passaram por privação dietética58
.
Estudos têm utilizado o rato como modelo experimental para avaliar os efeitos
moduladores da atividade física em animais submetidos à desnutrição perinatal15, 54
. Ratos
cujas mães foram desnutridas (caseína 8%) durante a gestação e lactação e/ou treinadas (50, 30
e 20min/dia na 1ª, 2ª e 3ª semana respectivamente; 5 dias/semana, 4 semanas antes e 3 semanas
durante a gestação, com intensidade de ~40% do VO2max), foram avaliados quanto à
parâmetros murinométricos e sensório-motores15
. Observamos que a desnutrição perinatal
atrasa o crescimento somático e a maturação dos reflexos em animais. Todavia, filhotes de
mães submetidas à desnutrição associada ao treinamento físico, tiveram o peso corporal, o
comprimento nasoanal e a maturação de reflexos normalizados em relação aos seus controles
apenas desnutridos15
. Utilizando o mesmo desenho experimental, observamos também que o
treinamento físico na gestação atenuou os efeitos da desnutrição relativamente ao peso ao
nascer, eixo latero-lateral e ântero-posterior do crânio54
.
Os mecanismos subjacentes parecem se fundamentar em alterações na comunicação
materno-fetal via placenta58, 59
. Gestantes exercitadas (20 min/dia, 3-5 dias/semana, com
intensidade a 55-60% VO2max) tiveram maior eficiência em interagir com o feto devido ao
maior volume funcional placentário12
. Estima-se que o volume placentário de sangue seja
maior na 20ª e 40ª semana de gestação em mulheres exercitadas (60% do VO2max) comparadas
as não exercitadas60
. Outro mecanismo proposto é que o treinamento físico de intensidade
leve/moderada aumenta o consumo de oxigênio de repouso mesmo diante da desnutrição14
.
Sugere-se então, que uma maior oferta de oxigênio e nutrientes possa ser destinada ao feto,

26
podendo modular positivamente seu crescimento e desenvolvimento mesmo diante de uma
menor demanda de nutrientes. Logo, é possível que a atividade física durante a gestação reduza
as chances de uma RAI inapropriada/incompatível com seu ambiente pós-natal caso este seja
diferente do ambiente intrauterino, reduzindo assim os riscos à saúde (Figura 1).
Respostas adaptativas preditivas à atividade física materna e à desnutrição
Com relação às respostas adaptativas preditivas à atividade física e à desnutrição, os
estudos ainda são escassos. Todavia, este é um tópico de particular importância uma vez que
mulheres gestantes subnutridas ou com excesso de peso apresentam um alto nível de atividade
física por questões sociais57
e culturais61
. Não há estudos com humanos que relatam o efeito
em longo-prazo da atividade física materna associado à desnutrição/sobrenutrição na prole
adulta. Em animais, os efeitos da RAP têm sido estudados recentemente pelo nosso grupo14,
15. Amorim et al (2009) observaram um menor catch up de crescimento em filhotes adultos
provindos de mães desnutridas/treinadas quando comparados aos seus pares apenas
desnutridos. Ademais, aos 90 dias de idade, estes filhotes apresentaram um ganho de peso
normal e indicadores de crescimento e desenvolvimento similares aos seus controles.
Utilizando o mesmo modelo experimental, nós observamos que estes filhotes aos 30, 60 e 90
dias de idade apresentaram padrões normais relativamente à atividade locomotora no teste de
“open field” (tempo de parada, potência, velocidade de reação, distância percorrida e número
de paradas) [dados não publicados].
Recentemente, Falcão-Tebas et al. (2011) observaram que filhotes adultos (270 dias
de idade) provindos de mães desnutridas/treinadas apresentaram um perfil normal de gordura
abdominal, colesterolemia e glicemia (após jejum de 6 horas). Estes resultados apontam para
uma RAP positiva do treinamento físico de intensidade leve/moderada em animais propensos
a desenvolver um padrão metabólico comprometedor à sua saúde. Os mecanismos subjacentes

27
ainda permanecem a ser estabelecidos, mas acredita-se que fatores epigenéticos possam
regular os efeitos benéficos do exercício físico.
De fato, a atividade física voluntária pode regular a expressão de genes via
mecanismos epigenéticos. Por exemplo, tem sido encontrado que o exercício estimula a
desmetilação do DNA de fatores neurotróficos do encéfalo (BDNF-IV) e eleva os níveis de
proteínas envolvidas na metilação e na expressão de RNAm, e aumenta a acetilação de
histonas62
. Caso estas alterações ocorram durante o período crítico do desenvolvimento, a
atividade física assume importância fundamental no controle da transcrição gênica no
contexto dos efeitos da desnutrição perinatal e da plasticidade. De acordo com os avanços dos
estudos sobre a origem desenvolvimentista da saúde e da doença e os fatores epigenômicos, o
estudo da atividade física durante a gestação abre novas possibilidades terapêuticas de baixo
custo para atenuar os efeitos de desordens nutricionais precoces.
CONCLUSÕES
A atividade física durante a gestação está sendo estudada quanto ao seu papel na
origem desenvolvimentista da saúde e da doença. Tem sido observado que esta intervenção
com exercício físico precoce, realizada em baixa a moderada intensidade, é capaz de aumentar
o aporte de nutrientes e oxigênio no ambiente feto-placentário. Nesse sentido, é provável que
a atividade física durante a gestação promova uma resposta adaptativa imediata favorável.
Sugerimos que a atividade física materna também possa modular a resposta adaptativa
preditiva ao longo da vida via mecanismos epigenéticos. Ademais, tanto a resposta adaptativa
imediata quanto a resposta adaptativa preditiva geradas pela atividade física na gestação
parecem ser mais expressivas se associadas à desnutrição perinatal, modulando o fenótipo
adulto e diminuindo o risco de desenvolver doenças.

28
AGRADECIMENTOS
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
REFERÊNCIAS
1. Gluckman PD, Hanson MA. Developmental plasticity and human disease: research
directions. J Intern Med. 2007 May;261(5):461-71.
2. McMillen IC, Robinson JS. Developmental origins of the metabolic syndrome:
prediction, plasticity, and programming. Physiol Rev. 2005 Apr;85(2):571-633.
3. Ravelli GP, Stein ZA, Susser MW. Obesity in young men after famine exposure in
utero and early infancy. N Engl J Med. 1976 Aug 12;295(7):349-53.
4. Ozanne SE, Jensen CB, Tingey KJ, Storgaard H, Madsbad S, Vaag AA. Low
birthweight is associated with specific changes in muscle insulin-signalling protein
expression. Diabetologia. 2005 Mar;48(3):547-52.
5. Gluckman PD, Hanson MA. The developmental origins of the metabolic syndrome.
Trends Endocrinol Metab. 2004 May-Jun;15(4):183-7.
6. Godfrey KM, Gluckman PD, Hanson MA. Developmental origins of metabolic
disease: life course and intergenerational perspectives. Trends Endocrinol Metab. 2010
Apr;21(4):199-205.
7. Kumon M, Yamamoto K, Takahashi A, Wada K, Wada E. Maternal dietary restriction
during lactation influences postnatal growth and behavior in the offspring of mice.
Neurochem Int. 2010 Aug;57(1):43-50.
8. Barros KM, Manhaes-De-Castro R, Lopes-De-Souza S, Matos RJ, Deiro TC, Cabral-
Filho JE, et al. A regional model (Northeastern Brazil) of induced mal-nutrition delays
ontogeny of reflexes and locomotor activity in rats. Nutr Neurosci. 2006 Feb-Apr;9(1-2):99-
104.
9. Thompson NM, Norman AM, Donkin SS, Shankar RR, Vickers MH, Miles JL, et al.
Prenatal and postnatal pathways to obesity: different underlying mechanisms, different
metabolic outcomes. Endocrinology. 2007 May;148(5):2345-54.
10. Morris TJ, Vickers M, Gluckman P, Gilmour S, Affara N. Transcriptional profiling of
rats subjected to gestational undernourishment: implications for the developmental variations
in metabolic traits. PLoS One. 2009;4(9):e7271.
11. Hopkins SA, Baldi JC, Cutfield WS, McCowan L, Hofman PL. Exercise training in
pregnancy reduces offspring size without changes in maternal insulin sensitivity. J Clin
Endocrinol Metab. 2010 May;95(5):2080-8.
12. Clapp JF, 3rd, Kim H, Burciu B, Lopez B. Beginning regular exercise in early
pregnancy: effect on fetoplacental growth. Am J Obstet Gynecol. 2000 Dec;183(6):1484-8.
13. Clapp JF, 3rd, Kim H, Burciu B, Schmidt S, Petry K, Lopez B. Continuing regular
exercise during pregnancy: effect of exercise volume on fetoplacental growth. Am J Obstet
Gynecol. 2002 Jan;186(1):142-7.
14. Amorim MF, dos Santos JA, Hirabara SM, Nascimento E, de Souza SL, de Castro
RM, et al. Can physical exercise during gestation attenuate the effects of a maternal perinatal
low-protein diet on oxygen consumption in rats? Exp Physiol. 2009 Aug;94(8):906-13.
15. Falcão-Tebas F, Bento-Santos A, Fidalgo MA, Almeida MB, Santos JA, Lopes de
Souza S, et al. Maternal low protein diet-induced delayed reflex ontogeny is attenuated by
moderate physical training during gestation in rats. British Journal of Nutrition. 2011 in press.

29
16. Bateson P, Barker D, Clutton-Brock T, Deb D, D'Udine B, Foley RA, et al.
Developmental plasticity and human health. Nature. 2004 Jul 22;430(6998):419-21.
17. Gluckman PD, Hanson MA. Living with the past: evolution, development, and
patterns of disease. Science. 2004 Sep 17;305(5691):1733-6.
18. Barker DJ, Winter PD, Osmond C, Margetts B, Simmonds SJ. Weight in infancy and
death from ischaemic heart disease. Lancet. 1989 Sep 9;2(8663):577-80.
19. Barker DJ. Early growth and cardiovascular disease. Arch Dis Child. 1999
Apr;80(4):305-7.
20. Lumey LH. Compensatory placental growth after restricted maternal nutrition in early
pregnancy. Placenta. 1998 Jan;19(1):105-11.
21. Lumey LH, Stein AD, Ravelli AC. Timing of prenatal starvation in women and birth
weight in their first and second born offspring: the Dutch Famine Birth Cohort study. Eur J
Obstet Gynecol Reprod Biol. 1995 Jul;61(1):23-30.
22. Roseboom TJ, van der Meulen JH, Osmond C, Barker DJ, Ravelli AC, Schroeder-
Tanka JM, et al. Coronary heart disease after prenatal exposure to the Dutch famine, 1944-45.
Heart. 2000 Dec;84(6):595-8.
23. Silveira PP, Portella AK, Goldani MZ, Barbieri MA. Origens desenvolvimentistas da
saúde e da doença (DOHaD). Jornal de Pediatria. 2007;83:494-504.
24. Gluckman PD, Hanson MA, Beedle AS. Early life events and their consequences for
later disease: a life history and evolutionary perspective. Am J Hum Biol. 2007 Jan-
Feb;19(1):1-19.
25. Harding JE, Johnston BM. Nutrition and fetal growth. Reprod Fertil Dev.
1995;7(3):539-47.
26. Barker DJ. In utero programming of chronic disease. Clin Sci (Lond). 1998
Aug;95(2):115-28.
27. Oliver MH, Harding JE, Breier BH, Evans PC, Gluckman PD. Glucose but not a
mixed amino acid infusion regulates plasma insulin-like growth factor-I concentrations in
fetal sheep. Pediatr Res. 1993 Jul;34(1):62-5.
28. Bruce KD, Hanson MA. The developmental origins, mechanisms, and implications of
metabolic syndrome. J Nutr. 2010 Mar;140(3):648-52.
29. Chan JC, Malik V, Jia W, Kadowaki T, Yajnik CS, Yoon KH, et al. Diabetes in Asia:
epidemiology, risk factors, and pathophysiology. JAMA. 2009 May 27;301(20):2129-40.
30. Batista Filho M, Rissin A. [Nutritional transition in Brazil: geographic and temporal
trends]. Cad Saude Publica. 2003;19 Suppl 1:S181-91.
31. Zhai F, Wang H, Du S, He Y, Wang Z, Ge K, et al. Prospective study on nutrition
transition in China. Nutr Rev. 2009 May;67 Suppl 1:S56-61.
32. Pinheiro ARdO, Freitas SFTd, Corso ACT. Uma abordagem epidemiológica da
obesidade. Revista de Nutrição. 2004;17:523-33.
33. Trostler N. Health risks of immigration: the Yemenite and Ethiopian cases in Israel.
Biomed Pharmacother. 1997;51(8):352-9.
34. Cohen MP, Stern E, Rusecki Y, Zeidler A. High prevalence of diabetes in young adult
Ethiopian immigrants to Israel. Diabetes. 1988 Jun;37(6):824-8.
35. Wandell PE, Gafvels C. High prevalence of diabetes among immigrants from non-
European countries in Sweden. Prim Care Diabetes. 2007 Feb;1(1):13-6.
36. Langley-Evans SC, Daniel ZC, Wells CA, Ryan KJ, Plant R, Welham SJ. Protein
restriction in the pregnant mouse modifies fetal growth and pulmonary development: role of
fetal exposure to {beta}-hydroxybutyrate. Exp Physiol. 2011 Feb;96(2):203-15.
37. Fernandez-Twinn DS, Ozanne SE, Ekizoglou S, Doherty C, James L, Gusterson B, et
al. The maternal endocrine environment in the low-protein model of intra-uterine growth
restriction. Br J Nutr. 2003 Oct;90(4):815-22.

30
38. Zhang Y, Li N, Yang J, Zhang T, Yang Z. Effects of maternal food restriction on
physical growth and neurobehavior in newborn Wistar rats. Brain Res Bull. 2010 Aug
30;83(1-2):1-8.
39. Toscano AE, Manhaes-de-Castro R, Canon F. Effect of a low-protein diet during
pregnancy on skeletal muscle mechanical properties of offspring rats. Nutrition. 2008
Mar;24(3):270-8, Toscano AE, Amorim MA, de Carvalho Filho EV, Aragao Rda S, Cabral-
Filho JE, de Moraes SR, et al. Do malnutrition and fluoxetine neonatal treatment program
alterations in heart morphology? Life Sci. 2008 May 23;82(21-22):1131-6.
40. Orozco-Solis R, Lopes de Souza S, Barbosa Matos RJ, Grit I, Le Bloch J, Nguyen P,
et al. Perinatal undernutrition-induced obesity is independent of the developmental
programming of feeding. Physiol Behav. 2009 Mar 2;96(3):481-92.
41. Camm EJ, Martin-Gronert MS, Wright NL, Hansell JA, Ozanne SE, Giussani DA.
Prenatal hypoxia independent of undernutrition promotes molecular markers of insulin
resistance in adult offspring. FASEB J. 2010 Oct 5.
42. Fowden AL, Ward JW, Wooding FP, Forhead AJ, Constancia M. Programming
placental nutrient transport capacity. J Physiol. 2006 Apr 1;572(Pt 1):5-15.
43. Chandler KD, Leury BJ, Bird AR, Bell AW. Effects of undernutrition and exercise
during late pregnancy on uterine, fetal and uteroplacental metabolism in the ewe. Br J Nutr.
1985 May;53(3):625-35.
44. Roberts CT, Sohlstrom A, Kind KL, Earl RA, Khong TY, Robinson JS, et al. Maternal
food restriction reduces the exchange surface area and increases the barrier thickness of the
placenta in the guinea-pig. Placenta. 2001 Feb-Mar;22(2-3):177-85.
45. Lesage J, Hahn D, Leonhardt M, Blondeau B, Breant B, Dupouy JP. Maternal
undernutrition during late gestation-induced intrauterine growth restriction in the rat is
associated with impaired placental GLUT3 expression, but does not correlate with
endogenous corticosterone levels. J Endocrinol. 2002 Jul;174(1):37-43.
46. Coan PM, Vaughan OR, Sekita Y, Finn SL, Burton GJ, Constancia M, et al.
Adaptations in placental phenotype support fetal growth during undernutrition of pregnant
mice. J Physiol. 2010 Feb 1;588(Pt 3):527-38.
47. Doherty CB, Lewis RM, Sharkey A, Burton GJ. Placental composition and surface
area but not vascularization are altered by maternal protein restriction in the rat. Placenta.
2003 Jan;24(1):34-8.
48. Hahn M, Dambacher S, Schotta G. Heterochromatin dysregulation in human diseases.
J Appl Physiol. 2010 Jul;109(1):232-42.
49. Leandro CG, Amorim MF, Hirabara SM, Curi R, Castro RMd. Pode a atividade física
materna modular a programação fetal induzida pela nutrição? Revista de Nutrição.
2009;22:559-69.
50. Artal R, O'Toole M. Guidelines of the American College of Obstetricians and
Gynecologists for exercise during pregnancy and the postpartum period. Br J Sports Med.
2003 Feb;37(1):6-12; discussion
51. ACOG committee opinion. Exercise during pregnancy and the postpartum period.
Number 267, January 2002. American College of Obstetricians and Gynecologists. Int J
Gynaecol Obstet. 2002 Apr;77(1):79-81.
52. Zavorsky GS, Longo LD. Exercise guidelines in pregnancy: new perspectives. Sports
Med. 2011 May 1;41(5):345-60.
53. Melzer K, Schutz Y, Boulvain M, Kayser B. Physical activity and pregnancy:
cardiovascular adaptations, recommendations and pregnancy outcomes. Sports Med. 2010 Jun
1;40(6):493-507.

31
54. Fidalgo M, Macêdo ÉMCd, Dantas R, Foerster P, Neves C, Manhães-de-Castro R, et
al. Efeito do treinamento físico e da desnutrição durante a gestação sobre os eixos cranianos
de ratos neonatos. Revista Brasileira de Medicina do Esporte. 2010;16:441-4.
55. Rosa BV, Firth EC, Blair HT, Vickers MH, Morel PC. Voluntary exercise in pregnant
rats positively influences fetal growth without initiating a maternal physiological stress
response. Am J Physiol Regul Integr Comp Physiol. 2011 May;300(5):R1134-41.
56. Hatch MC, Shu XO, McLean DE, Levin B, Begg M, Reuss L, et al. Maternal exercise
during pregnancy, physical fitness, and fetal growth. Am J Epidemiol. 1993 May
15;137(10):1105-14.
57. Rao S, Kanade A, Margetts BM, Yajnik CS, Lubree H, Rege S, et al. Maternal activity
in relation to birth size in rural India. The Pune Maternal Nutrition Study. Eur J Clin Nutr.
2003 Apr;57(4):531-42.
58. Clapp JF. Effects of Diet and Exercise on Insulin Resistance during Pregnancy. Metab
Syndr Relat Disord. 2006 Summer;4(2):84-90.
59. Clapp JF, 3rd. The effects of maternal exercise on fetal oxygenation and feto-placental
growth. Eur J Obstet Gynecol Reprod Biol. 2003 Sep 22;110 Suppl 1:S80-5.
60. Thomas DM, Clapp JF, Shernce S. A foetal energy balance equation based on
maternal exercise and diet. J R Soc Interface. 2008 Apr 6;5(21):449-55.
61. Davies GA, Maxwell C, McLeod L, Gagnon R, Basso M, Bos H, et al. Obesity in
pregnancy. J Obstet Gynaecol Can. 2010 Feb;32(2):165-73.
62. Gomez-Pinilla F, Zhuang Y, Feng J, Ying Z, Fan G. Exercise impacts brain-derived
neurotrophic factor plasticity by engaging mechanisms of epigenetic regulation. Eur J
Neurosci. 2011 Feb;33(3):383-90.

32
PÁGINA DE FIGURAS
Figura 1.

33
PÁGINA DE LEGENDA DAS FIGURAS
Figura 1. Modelo teórico referente ao papel da atividade física durante a gestação
beneficiando o indivíduo ao longo da vida que previu no período intrauterino, um ambiente
nutricional restrito. Os efeitos em curto prazo da atividade física durante a gestação para o
feto são conhecidos (seta cinza-escuro). Entretanto, sugerimos que a atividade física materna
possa exercer efeitos benéficos ao feto previamente desnutrido em longo prazo através de
mecanismos epigenéticos (seta cinza-claro com contorno pontilhado).

34
3. OBJETIVOS
Geral
Avaliar os efeitos da desnutrição protéica perinatal e/ou do treinamento físico antes e
durante a gestação sobre o desenvolvimento do sistema nervoso, indicadores de crescimento
somático e bioquímicos, e a proporção das fibras musculares esqueléticas da prole de ratos.
Específicos
Em ratas nutridas ou desnutridas durante a gestação e lactação, submetidas ou não ao
protocolo de treinamento físico antes e durante a gestação, avaliar:
- o ganho de peso corporal
- o consumo alimentar
Nos filhotes, analisar:
- o número e o peso dos filhotes ao nascer;
- a maturação de características físicas e dos reflexos;
- o peso corporal, o ganho de peso e indicadores murinométricos;
- a glicemia e colesterolemia aos 270 dias de idade;
- o peso, comprimento e largura dos músculos soleus e extensor longo dos dedos
(EDL);
- a proporção de fibras oxidativas (tipo I) e glicolíticas (tipo IIa e IIb) nos músculos
soleus e EDL.

35
4. HIPÓTESES
A desnutrição perinatal causa, em curto prazo, retardo na maturação das características
físicas e na ontogênese dos reflexos e induz um catch up de crescimento da prole.
A desnutrição materna durante a gestação diminui, em longo prazo, a proporção de
fibras tipo I do músculo soleus e do tipo IIa do músculo EDL em ratos com 270 dias
de idade.
O treinamento físico antes e durante a gestação atenua os efeitos da desnutrição
perinatal em curto prazo. Da mesma forma, reduz os efeitos em longo prazo, gerados
pela incompatibilidade nutricional pré e pós-natal.

36
5. MÉTODOS
5.1 Animais
Este estudo foi aprovado pelo comitê de ética em estudos com animais do Centro de
Ciências Biológicas da UFPE (protocolo nº. 23076.049077/2010-80). A manipulação e os
cuidados com os animais seguiram as recomendações do Comitê Brasileiro de
Experimentação Animal (COBEA).
Foram utilizadas ratas da linhagem Wistar, provenientes do biotério de criação do
departamento de Nutrição (UFPE). Os animais foram mantidos em biotério de
experimentação, com temperatura de 22°C 2, ciclo claro-escuro invertido de 12/12 horas e
livre acesso à água e dieta padrão do biotério (52% carboidratos, 21% proteínas, 4% lipídeos
– Nuvilab CR1-Nuvital®, Curitiba, Paraná, Brasil). Toda manipulação foi realizada nas
gestantes, enquanto que os filhotes foram apenas mantidos para avaliação posterior (Figura 1).
Figura 1. Desenho experimental geral do estudo.
5.2 Protocolo experimental de treinamento físico das mães
As ratas foram divididas, aleatoriamente, em dois grupos: treinada (T, n=6) e não-
treinada (NT, n=6). O grupo T realizou uma corrida de baixa intensidade (0,3 km.h-1
, durante
10 minutos) em esteira, durante 3 dias consecutivos, para adaptação ao equipamento. A

37
seguir, foram submetidas a um programa de treinamento físico moderado de acordo com o
protocolo de Amorim, et al. (2009). O protocolo experimental de treinamento consistiu, 5 dias
por semana e 60 minutos por dia, durante 7 semanas. A intensidade utilizada foi de
aproximadamente 50% da velocidade máxima atingida em teste de desempenho máximo e
70% do VO2max (Quadro 1) (Amorim et al., 2009). O grupo NT permaneceu em suas
respectivas gaiolas, próximo à esteira do treinamento, simulando o estresse sonoro e o
induzido pela manipulação das gaiolas.
Após quatro semanas de treinamento físico, as ratas dos dois grupos, foram acasaladas
(machos reprodutores, n=6; 2:1 na relação fêmea e macho). Quando diagnosticado como
gestantes, através do exame de esfregaço vaginal para a presença de espermatozoides
(Marcondes et al., 2002), o protocolo de treinamento foi modificado para as ratas do grupo T.
A velocidade e duração dos estágios foram progressivamente diminuídas, caracterizando um
treinamento de intensidade leve, com aproximadamente 25% da velocidade máxima atingida
em teste de desempenho máximo e 30% do VO2max (Quadro 2). Na lactação não houve
treinamento físico, apenas a manipulação dietética. Foram mantidos 6 ratos por ninhada, e
cada ninhada, foram utilizados (aleatoriamente) 3 filhotes machos. Os filhotes foram
divididos em 4 grupos, de acordo com a manipulação das mães: Não-Treinado (NTF; n=9);
Treinado (TF; n=9); Desnutrido (DF; n=9); Treinado Desnutrido (TDF; n=9).

38
Semanas Velocidade
(km.h-1)
% de esforço relativo ao TDM
(Velocidade máxima
atingida = 1.8 km.h-1)
Duração
(min)
Duração Total
do Treino (min)
1ª Semana
(adaptação)
0.3 16.7 5
20 0.4 22.2 5
0.5 27.8 5
0.3 16.7 5
2ª semana
0.4 22.2 5
50 0.5 27.8 10
0.6 33.3 30
0.4 44.4 5
3ª semana
0.4 22.2 5
60
0.5 27.8 10
0.6 33.3 10
0.8 44.4 30
0.4 22.2 5
4ª semana
0.5 27.8 5
60
0.6 33.3 10
0.8 44.4 10
0.9 50.0 30
0.5 27.8 5
Quadro 1. Caracterização do protocolo experimental de treino de acordo com a velocidade e
a duração de cada sessão para cada uma das quatro semanas de treino (Amorim et al, 2009).

39
Semana de
gestação
Velocidade
(km.h-1)
% de esforço relativo ao TDM
(Velocidade máxima
atingida = 1.8 km.h-1)
Duração
(min)
Duração Total
do Treino (min)
1ª semana
0.4 22.2 5
50
0.5 27.8 10
0.6 33.3 10
0.8 44.4 20
0.4 22.2 5
2ª semana
0.4 22.2 5
30 0.5 27.8 10
0.6 33.3 10
0.4 22.2 5
3ª semana
0.3 16.7 5
20 0.4 22.2 5
0.5 27.8 5
0.3 16.7 5
Quadro 2. Caracterização do protocolo experimental de treino de acordo com a velocidade e
a duração de cada sessão para cada uma das semanas de treino durante a gestação (Amorim et
al, 2009).
5.3 Manipulação da dieta das mães
A manipulação nutricional ocorreu durante a gestação e lactação. Imediatamente após
o diagnóstico de prenhez, metade das ratas de cada grupo (T e NT) foi submetida à dieta
hipoprotéica (dieta a base de caseína à 8%). As demais ratas receberam dieta normoprotéica
(dieta a base de caseína à 17%). As dietas confeccionadas foram isocalóricas com alteração
apenas no conteúdo de proteína (Tabela 1).

40
Tabela 1. Composição centesimal da dieta à base de caseína(*).
Ingredientes Quantidade por Kg de ração
CASEÍNA a 8% CASEÍNA a 17%
Caseína 79,30g 179,30g
Mix de vitamina 10,00g 10,00g
Mix de sais minerais 35,00g 35,00g
Celulose 50,00g 50,00g
Óleo de soja 70ml 70ml
Bitartarato de colina 2,50 g 2,50 g
DL-metionina 3,00g 3,00g
Amido 750,20g 650,20g
(*) Fonte: (Reeves et al., 1993)
5.4 Consumo alimentar
O consumo alimentar (g) das ratas foi obtido pela subtração do peso da dieta ofertada
e o peso da dieta restante após 24h, de acordo com a seguinte fórmula: Consumo alimentar =
Peso da ração (g) – Peso da ração do dia anterior (g) (Lopes de Souza et al., 2008).
5.5 Avaliação do peso corporal, do ganho de peso e da circunferência abdominal
O peso corporal dos animais foi avaliado diariamente antes, durante e após a gestação.
A partir do desmame, o peso foi aferido semanalmente através de uma balança eletrônica
digital – Marte, modelo S-1000, com capacidade máxima de 1000g e sensibilidade de 0,01g.
O percentual de ganho de peso corporal foi calculado tendo como base o peso do 1° dia de
avaliação, segundo a fórmula: % ganho de peso=[Peso do dia (g) x 100/ Peso do 1° dia (g)]
– 100 (Bayol, et al, 2004).

41
A circunferência abdominal (cm) foi determinada através da maior circunferência
entre a borda superior da crista ilíaca e a última costela (Novelli et al., 2007).
5.6 Avaliações Murinométricas
Cada animal foi avaliado diariamente do 10º ao 17º dia pós-natal, a partir das
15h30mim. Avaliamos às seguintes medidas (Fox, 1965; Silva et al., 2005):
Eixo Látero-lateral do Crânio (ELLC) – Este eixo foi considerado tendo como
referência a linha perpendicular ao eixo longitudinal do crânio, dividindo os pavilhões
auriculares ao meio. O pesquisador continha o animal com uma das mãos, tendo a cabeça
deste entre os dedos indicador e polegar. Assim, com auxílio do paquímetro digital (Stainless
Hardened®), procedia-se a medida, em mm, do eixo látero-lateral do crânio.
Eixo Ântero-posterior do Crânio (EAPC) – Foi aceito como referência, a linha
média que vai da extremidade do focinho até o ponto de interseção com outra linha
perpendicular imaginária. Essa última passa tangencialmente às extremidades posteriores dos
pavilhões auriculares. O pesquisador continha o rato com uma das mãos, mantendo a cabeça
do animal entre os dedos indicador e polegar. Procedia-se à medida com auxílio do
paquímetro digital.
Eixo Longitudinal do Corpo (EL) – O eixo longitudinal foi medido, contendo
delicadamente o corpo do animal com os dedos de forma que o corpo fosse mantido sob os
dedos anular e médio e a cauda sob o indicador, causando uma rápida imobilização. Dessa
forma com ajuda de uma caneta, os pontos entre o focinho e a base da cauda foram marcados
e a distância entre estes, efetuada com paquímetro digital.
Comprimento da Cauda (CC) – O animal foi contido delicadamente com uma das
mãos do pesquisador. Em seguida, encostava-se a região anal do animal na borda de uma
mesa lisa e plana. Por sobre a mesa, a cauda do animal era delicadamente mantida bem
estirada. Com uma caneta, foi feita uma marca na mesa, coincidente com a extremidade da

42
cauda. Media-se então a distância em mm, entre as duas extremidades, com a ajuda de um
paquímetro digital.
5.7 Avaliação da maturação de características físicas
Cada animal dos diferentes grupos foi analisado diariamente, do 10º ao 17º dia pós-
natal, quanto à maturação dos seguintes caracteres físicos: Abertura do conduto auditivo
(ACA) e abertura dos olhos (AO) (Smart, Dobbing, 1971). Quando constatamos que o animal
já havia adquirido as características físicas por três dias consecutivos, o primeiro foi
considerado o dia de maturação. Estas características físicas foram observadas da seguinte
forma:
ACA - Foi registrado no dia em que os dois orifícios auriculares, primitivamente
obliterados, abriam-se.
AO - Foi registrado o dia em que ambos os olhos se encontravam abertos e com a
presença de movimento palpebral.
5.8 Avaliação da ontogênese dos reflexos
Foram analisados diariamente, em cada animal, do 10º ao 17º dia pós-natal, o
desenvolvimento dos seguintes reflexos: Colocação pelas Vibrissas (CV), Geotaxia Negativa
(GN), Resposta ao Susto (RS) e Reação de Aceleração (RA) (Smart, Dobbing, 1971). Para
cada um dos reflexos, foi registrado o dia da sua consolidação, após três dias consecutivos de
confirmação. Procedeu-se às avaliações do desenvolvimento sensório-motor (reflexos),
utilizando-se instrumentos elaborados ou existentes no laboratório, conforme descrição
abaixo:

43
CV - O rato foi suspenso pela cauda de tal forma, que suas vibrissas tocassem
levemente a borda de uma mesa. A resposta foi considerada positiva, quando o animal, no
tempo máximo de 10s, colocasse as patas anteriores sobre a mesa tentando caminhar.
GN - O animal foi colocado ao centro de uma rampa de 45º de inclinação forrada com
material antiderrapante (papel crepom), com a cabeça no sentido descendente. A resposta
reflexa foi considerada positiva quando o animal, em um período máximo de 10s, o animal se
voltasse completamente, girando o corpo aproximadamente 130° e posicionando a cabeça em
sentido ascendente.
RS - O rato foi submetido a um estampido agudo, produzido pela percussão de um
bastão metálico sobre um recipiente também metálico. A resposta foi considerada positiva
quando ocorresse uma simultânea retração e imobilização rápida e involuntária do corpo do
animal, característica do susto.
RA - Segurando-se o rato pelas quatro patas, com o dorso voltado para baixo, a uma
altura de 30 cm (uma régua de 30 cm, perpendicular ao plano serviu como guia), foi
observada sua queda livre sobre um leito de espuma sintética (30 x 12 cm). A resposta foi
considerada positiva, quando o animal girava completamente o corpo, voltando o ventre para
baixo, caindo na superfície apoiado sobre as quatro patas.
5.9 Avaliação da Glicemia e Colesterolemia dos Filhotes
Após jejum de 6 horas, os animais sofreram um corte na extremidade da cauda para
coleta do sangue. A glicose e o colesterol séricos (mg.dL-1) dos filhotes aos 270 dias de idade
foram avaliados. As concentrações de glicose sanguínea foram identificadas pelo método
enzimático colorimétrico da glicose-oxidase/peroxidase e a leitura através de glicosímetro
(Accu-Chek Performa®, Roche Diagnostics). A quantificação da colesterolemia foi realizada
por meio método da colesterol-oxidase e analisada por fotometria, sendo os valores calculados
através do monitor (Accutrend Colesterol®, Roche Diagnostics).

44
5.10 Preparação dos músculos soleus e extensor longo dos dedos (EDL)
Após seis horas de jejum, os animais foram decapitados aos 270 dias de idade, sem
aplicação de anestésico. A partir de uma incisão na face lateral da pata posterior, os músculos
soleus e o extensor longo dos dedos (EDL) foram dissecados e imersos em N-Hexano
(C6H14). Esse método de congelamento evita a formação de cristais de água no fragmento do
músculo, além de fixar sem desnaturar as enzimas a serem caracterizadas. Nessas condições
os músculos congelados foram estocados e conservados em tubos armazenados em freezer a -
80º C para posterior processamento (Toscano et al., 2008).
5.11 Análise histológica
Para obtenção de coloração característica da atividade ATPasica da miosina foi
utilizado o método proposto por Brooke e Kaiser (1970). Os músculos foram mantidos em
suporte com a utilização do TissueTek® O.C.T Compound (Sakura Finetek USA, Inc.,
Torrance, CA 9050 U.S.A.), que foi utilizado após a fixação do tecido e antes da criosecção e
foram mantidos a -20±2 ºC. Para as secções transversais dos músculos (10µm) foi utilizado
um criostato (LEICA modelo CM1100, Nussloch - Alemanha). Os cortes foram fixados em
lâminas e ao atingirem a temperatura ambiente foi iniciada a coloração (Pestronk et al., 1992).
Os cortes transversais dos músculos foram pré-incubados em temperatura ambiente
por 10 minutos em 400 mL de solução contendo 280 mL de acido acético a 0,2M (12,7 mL de
acido acético e 1000 mL de água destilada) e 120 mL de acetato de sódio a 0,2M (27,2g
acetato de sódio) cujo pH foi ajustado para 4,3 ou 4,55. Em seguida, as lâminas foram lavadas
em água destilada e incubadas a 37°C em uma solução, contendo ATP (pH 9,4) durante 40
minutos. Após o período de incubação, as lâminas foram lavadas em água destilada e
mergulhadas em solução de cloreto de cobalto a 2% durante 5 minutos, para revelação em
solução de sulfeto de amônio a 5%. Após a revelação, os fragmentos de músculos foram
desidratados em uma bateria crescente de álcoois (70° a 100°), por fim foram imersas duas
vezes em tolueno. Após a secagem, as lâminas foram montadas utilizando New Entellan®
Merck (Brooke and Kaiser, 1970).

45
Os diferentes tipos de fibras musculares presentes nos cortes histológicos foram
analisados. As fibras musculares foram classificadas de acordo com a intensidade de reação
da ATPase após a pré-incubação em meio ácido (pH 4,3 e 4,55), nos três tipos principais: I
(fibras lentas oxidativas, escuras), IIa (fibras rápidas oxidativas-glicolíticas, clara) e IIb (fibras
rápidas glicolíticas, coloração intermediária) (Brooke and Kaiser, 1970). A composição dos
tipos de fibras do músculo foi determinada contando-se aproximadamente 1500 fibras
igualmente distribuídas em campos sobre a amostra. Os campos microscópicos foram obtidos
através de microscópio óptico OLYMPUS modelo U-CMAD-2 (objetiva 4 X) acoplado a um
programa para captação de imagens (TV TUNER APPLICATION – TelSignal Company
Limited, Taiwan). Para contagem das células foi utilizado o software MESURIN PRO versão
3.4 (Jean-François Madre-Amiens, França). A contagem das fibras foi realizada em duplicata
e de forma cega.
5.12 Análise estatística
As variáveis contínuas foram testadas, segundo a normalidade da distribuição. Os
dados que se mostraram dentro da distribuição gaussiana foram expressos em média e erro
padrão da média (EPM). Na comparação entre as médias, foi usado o ANOVA (one-way ou
two-way) seguido de Tukey ou de Bonferroni como testes a posteriori, a depender da análise
(com um ou dois fatores). A associação entre as variáveis foi avaliada pelo teste de correlação
de Pearson. O nível de significância adotado foi de 5% em todos os casos. A construção do
banco de dados e as análises estatísticas foram desenvolvidas no programa Excel (versão
2007, Microsoft, USA) e Graphpad Prisma 5® (GraphPad Software Inc., La Jolla, CA, USA),
respectivamente.

46
6. RESULTADOS – ARTIGOS ORIGINAIS
Title: Maternal low protein diet-induced delayed reflex ontogeny is attenuated by moderate
physical training during gestation in rats
Running title: Physical training and undernutrition
Authors:
Filippe Falcão-Tebasa, Adriano Bento-Santos
b, Marco Antônio Fidalgo Amorim
c, Marcelus
Brito de Almeidac, José Antônio dos Santos
b, Sandra Lopes de Souza
a,b, Raul Manhães-de-
Castroa,b
, Carol Góis Leandroa,c
a Department of Nutrition, Federal University of Pernambuco, Brazil.
b Department of Neuropsychiatry and Behavioral Science, Federal University of Pernambuco,
Brazil.
c Department of Physical Education and Sports Science, CAV, Federal University of
Pernambuco, Brazil.
Mailing address:
Filippe Falcão-Tebas
Rua Prof. Moraes Rego, 1235 – CEP: 50670-901 – Departamento de Nutrição - Cidade
Universitária - Recife, PE – Brasil.
Telephone: (00 55 81) 21268463. Fax: (00 55 81) 21268473
E-mail: [email protected]

47
Abstract
We evaluated the effects of moderate to low-intensity physical training during gestation on
reflex ontogeny in neonate rats whose mothers were undernourished. Virgin female Wistar
rats were divided into four groups as follows: untrained (NT, n =7); trained (T, n =7);
untrained with a low-protein diet (NT+LP, n =7); and trained with a low-protein diet (T+LP, n
=4). Trained rats were subjected to a protocol of moderate physical training on a treadmill
over a period of 4 weeks (5 days week−1
and 60 min day−1
, at 65% of VO2max). After
confirming the pregnancy, the intensity and duration of the exercise was reduced. Low-
protein groups were provided with an 8% casein diet, and controls were provided with a 17%
casein diet. Their respective offspring were evaluated (during the 10th
to 17th day of postnatal
life) in terms of physical feature maturation, somatic growth, and reflex ontogeny. Pups born
to mothers provided with the low-protein diet during gestation and lactation showed delayed
physical feature and reflex maturation and a deficit in somatic growth when compared with
controls. However, most of these deficiencies were attenuated in pups of undernourished
mothers undergoing training. In conclusion, physical training during gestation attenuates the
effects of perinatal undernutrition on some patterns of maturation in the central nervous
system during development.
Key-words: Brain; Pregnancy; Exercise; Diet, Low-Protein; Rats, Wistar

48
1. Introduction
It has been well established that insults during the critical period of development of the
nervous system, such as malnutrition, can have noticeable impacts on the development of
brain structure and function, resulting in developmental dysfunctions and diseases in later
life(1)
. A perinatal low-protein diet is associated with impaired cerebral vascularization, and
reduced dendrite numbers in rat offspring(2)
. Human infants born with fetal intrauterine
growth restriction (IUGR) have reduced hippocampal volume, which has been associated with
functional behavioral differences(3)
.
The reflex-maturation pattern, “a behavior elicited by exactly prescribed stimulation”,
is used to demonstrate an appropriate maturation of the central nervous system (CNS) during
development(4)
. Using this model, our previous study demonstrated a delay in the maturation
of reflexes, as well as deficits in locomotion patterns caused by postnatal malnutrition in
neonatal rats(5)
. In addition, reductions in protein or caloric intake during the perinatal period
may influence feeding behavior and locomotor activity in adult rats(5-6)
. Such structural and
functional changes in the brains of offspring may be due to changes in cerebral metabolism,
reduced nutrients in placental space or reduced oxygen consumption by dams(7)
.
Recently, it has been recognized that an active maternal lifestyle, including regular to
moderate physical activity, improves aerobic fitness and the maternal-fetal physiological
reserve by enhancing nutrient and oxygen availability to the fetus(8)
. In the nervous system,
maternal exercise during pregnancy has beneficial effects on brain functions and structures of
both mother and offspring(9)
. In rats, exercise during pregnancy increases the expression of
brain-derived neurotrophic factor (BDNF) hippocampal mRNA and spatial learning in rat
pups(10)
. Maternal treadmill running during pregnancy (30 min at a mild-intensity, once a day
until delivery) enhanced hippocampal cell survival and improved the short-term memory
capability of pups of rats as compared to control group(11)
. A human study showed that

49
mothers who trained during gestation showed better self-esteem and a lower incidence of
depression during pregnancy, and their offspring showed fewer signs of stress during delivery
and were in better general condition (as evidenced by higher Apgar scores)(9)
.
In our previous study, we demonstrated that in rats, a controlled moderate to low-
intensity exercise before and during gestation (5 days week-1
, 60 min day-1
, 40% - to 70% of
VO2max) attenuated the impact of the low-protein diet by improving the mothers’ resting
oxygen consumption(12)
. In the present study, the impact of the moderate to low protocol of
physical training during gestation on the reflex ontogeny in neonates whose mothers were
undernourished was evaluated. Our hypothesis was that exercise-induced protective changes
in the brain during gestation attenuate the impact of a low-protein diet on somatic
development and reflex ontogeny in the offspring.
2. Methods
The experimental protocol was approved by the Ethical Committee of the Biological
Sciences Center (protocol n° 23076.049077/2010-80), Federal University of Pernambuco,
Brazil, and we followed the Guidelines for the Care and Use of Laboratory Animals(13)
.
2.1 Animals
Sixty-day old virgin female albino Wistar rats (Rattus novergicus) were obtained from
the Department of Nutrition, Federal University of Pernambuco, Brazil and were maintained
at a room temperature of 22 ± 1oC with a controlled light–dark cycle (dark 06.00–18.00
hours). Standard laboratory chow (52% carbohydrate, 21% protein, and 4% lipids - Nuvilab
CR1-Nuvital
, Curitiba, Parana, Brazil) and water were given ad libitum. Animals were

50
randomly divided into two groups: untrained rats (NT, n = 14) and trained rats (T, n = 11).
Trained rats were submitted to a training program of moderate running over a period of 4
weeks (5 days week–1
and 60 min day–1
) on a treadmill (Insight Equipments, EP-131®
, São
Paulo, Brazil) at a controlled intensity based on their VO2max(12)
. After a 4-week training
period, the rats were mated (2 females for 1 male). The day on which spermatozoa were
present in a vaginal smear was designated as the day of conception, day 0 of pregnancy.
Pregnant rats were then transferred to individual cages. Half of the rats of each group received
a 17% casein diet and the other half received an 8% casein isocaloric diet (low-protein group,
LP) ad libitum (Table 1). Thus, two more groups were formed as follows: untrained (NT, n
=7); trained (T, n =7); untrained with low-protein diet (NT+LP, n =7); and trained with low-
protein diet (T+LP, n =4). The exercise program was maintained during gestation, with a
progressive reduction of intensity until day 19 of gestation(12)
. During the suckling period, the
offspring were maintained as litters of six pups, and their mothers continued to be provided
with the respective diet of either 8% casein or 17% casein. The litters of six pups represent the
sample that was evaluated: (NTL, n =7); trained (TL, n =7); untrained with low-protein diet
(NT+LPL, n =7); and trained with low-protein diet (T+LPL, n =4). The evaluation of physical
characteristics, growth, and reflex ontogeny of pups was performed between days 10 and 21
of the suckling period. The collection of physical characteristics and behavioral data was
performed under blind conditions.
2.2 Protocol of physical training
The protocol of physical training was performed according to Amorim et al. (2009).
Rats ran on a treadmill during the four weeks prior to pregnancy (5 days week–1
and 60 min
day–1
, at 65% VO2max). The protocol was divided into four progressive stages in each session:

51
(i) warm-up (5 minutes); (ii) intermediary (10 minutes); (iii) training (30 minutes), and (iv)
cool-down (5 minutes) periods. The percentage of VO2max during the sessions of training was
kept around 55 – 65%(12)
. During pregnancy, rats ran at a progressively lower intensity of
effort (40% VO2max, 5 days week–1
and 20 min day–1
) until the 19th
day of gestation. There
was no physical training during the lactation period.
2.3 Mother´s body weight and food intake
Mother´s body weight was recorded daily, and daily food consumption was
determined by the difference between the amount of food provided at the onset of the dark
cycle (06.00 hours) and the amount of food remaining 24 h later(6)
. Body and food weights
were recorded with a Marte Scale (AS-1000) with a 0·01-g accuracy.
2.4 Physical feature maturation of the offspring
An internal auditory conduit opening of both ears and eyes, (i.e., when any visible
break in the covering membrane of both eyes) was observed. The features were evaluated
daily between 13.00 and 15.00 hours by the method of Smart and Dobbing (1971) during the
suckling period, until maturation. The maturation age of a particular feature was defined as
the day it was first observed.
2.5 Somatic growth
Somatic growth was assessed in terms of body weight, tail length, and mediolateral
and anteroposterior head axis measurements performed from the 10th
to 17th postnatal day

52
between 13.00 and 15.00 hours as follows: body weight of the pups was recorded daily
throughout the experiment with a Marte scale with 100 mg precision. Body weight gain was
calculated as follows: Percentage weight gain = [body weight (g) × 100/birth weight (g)] −
100. Changes in body weight (Δ body weight) was calculated by the gain of grams per day (g
day−1
)(14)
. Changes in body length (Δ body length) was calculated by the gain of millimeters
per day (mm day−1
)(14)
. Tail length (distance from tail tip to tail base), length of the
laterolateral skull axis (distance between the ear holes) and length of the anteroposterior axis
of the head (distance between snout and head-neck articulation) were measured with a digital
caliper (Starrett®, Series 799, São Paulo, Brazil) with a 0·01-mm precision.
2.6 Reflex testing
Daily examinations evaluating a spectrum of reflexologic tests (Table 2)(15)
as
indicators of neurologic development were conducted from the 10th
to the 17th
postnatal day.
Reflex tests were conducted between 11.00 and 13.00 hours. The time of the appearance of
each reflex was defined as the first day of its occurrence during a period of three consecutive
days.
2.7 Statistical analyses
Body weight and food intake of mothers are presented as means ± S.E.M. Each litter
of six pups was considered one sample, and statistical analyses were performed by using the
mean values of each litter. For statistical analysis, data were analyzed by two-way repeated
measures ANOVA, with mothers’ diet (NT, NT + LP) and physical training (T, T + LP) as
factors. Bonferroni’s post hoc test was used. For reflex ontogeny and physical characteristics,
data are presented as median ± minimal and maximal values. Kruskal-wallis’ test followed by

53
Dunn´s test (reflexes) was applied for multiple comparisons. Significance was set at P <0·05.
Data analysis was performed using the statistical program Graphpad Prism 5® (GraphPad
Software Inc., La Jolla, CA, USA).
3. Results
Gain of body mass was lower in the second and last third of gestation in the groups
submitted to a low-protein diet, whereas physical training had no effect in the gain of body
mass (NT, 33.9±2.4; T, 35.9±1.5; NT+LP, 26.9±1.2; and T+LP, 28.8±2.1; P <0.05; Figure
1A). Data were adjusted for the number of pups born to each dam [NT, 10.0 (9–12); T, 9.5
(9–13); NT+LP, 9.5 (8–10); and T+LP, 10.0 (8–10); values expressed as median (minimum
and maximum)]. Pearson’s correlation coefficient between number of pups and body weight
gain of the mother was not significant (r2 = 0.29). Daily food intake during gestation and the
first week of lactation was not different among groups (Figure 1B). On the second week of
lactation, rats of trained groups ingested more food, whereas groups provided with the low-
protein diet ingested less food daily when compared to controls (Figure 1B).
Litters from NT+LP mothers showed a deficit in the somatic growth indicators when
compared with litters from NT control group (Table 3). When compared to NT+LPL group,
T+LPL showed higher values of growth rate, tail length, laterolateral skull axis and
anteroposterior axis of the head (Table 3).
During the lactation period (from 10th
to 17th
days of life), pups in each litter were
analyzed in terms of the development of physical characteristics and reflex ontogeny (Table
4). Physical features of pups from trained mothers (TL) did not change, while undernourished
pups appeared two days later as compared to control pups. For the time of “eyes opening”,
T+LPL group was not different when compared to NTL group. Similarly, the reflex ontogeny

54
of pups from trained mothers did not change, while animals from mothers provided a low-
protein diet during gestation and lactation showed a delayed maturation of reflexes when
compared with the control (NTL) group, with the exception of the auditory-startle response
(Table 4). In comparison with the NT+LPL group, animals from trained and undernourished
mothers (T+LPL) showed an early maturation of the vibrissa-placing, cliff-avoidance and
negative-geotaxis (Table 4).
4. Discussion
Several studies on critical periods in brain development and the vulnerability of the
developing brain to undernutrition have been previously performed(16,15,17-19,5)
. Undernutrition
at the time of critical development appears to depress the rate of all brain growth events(17)
. In
fact, our data showed that physical features, somatic growth, and ontogeny reflexes are
severely affected in pups from undernourished mothers. However, these undernutrition-
induced alterations were less pronounced in pups whose mothers were submitted to physical
training during gestation.
Maternal weight gain is the standard measure used to assess pregnancy outcome, and it
also allows examination of the extent to which an undernourished dam is able to change body
composition to maintain lactational performance(20)
. In the present study, patterns of response
in terms of body weight gain and food intake by undernourished mothers were similar to
those of the previous studies(21,20,22)
. During lactation, undernourished mothers showed a
reduced food intake, whereas trained mothers showed an increase in the food intake. Because
physical training can induce an increase in maternal lean body mass, the dietary low-protein-
induced catabolism can be attenuated by a mechanism that includes an increase in availability
of protein(8)
. In addition, maternal physical exercise seems to have a positive impact on the

55
maternal protein reserve, thereby improving the female’s ability to sustain lactation, with
more protein available for mobilization allowing an increased food intake and improved
lactational performance(23)
.
To minimize the effects of stress from mother/pups separation, which itself has a
significant impact on behavioral responses(24)
, pups were evaluated in a shortened range of
time because most reflexes mature after the 10th
day of life(4-5)
. Our data showed that pups
from undernourished mothers were smaller than those of controls. Furthermore, the
dimensions of the head were reduced. Maternal protein restriction is associated with lower
stores of maternal nutrients and, subsequently, less transfer of nutrients to the offspring,
which is related to reduce postnatal growth(25)
. Intrauterine malnutrition also has long-term
effects on brain cell number and cell size, probably via alteration in the fetal mitotic rate(18)
.
However, moderate exercise during gestation may have beneficial effects on indicators of
somatic growth in the offspring from undernourished mothers by a mechanism that could
include increasing uterine blood flow(26)
, redistributing blood flow, changes in the production
of fetal and placental hormones that control growth(27)
and a higher resting oxygen
consumption(12)
.
The repercussions of perinatal malnutrition on the development of the structural,
neurochemical and functional integrity of the nervous cells are well known(28)
. In the present
study, there was a delay in the appearance of physical features and most reflexes, with the
exception of the auditory-startle response. In a previous study using a multideficient diet
(regional diet basic, 8% protein) during lactation, it was observed that reflexes – including
free-fall righting, negative-geotaxis, cliff-avoidance, and vibrissa-placing – showed the same
range of delayed days(5)
. Although the different undernutrition models and the period of time
(gestation, lactation, or both) differed by experimental designs, one can be certain that the
normal development of the brain is strictly related to balanced nutritional intake.

56
Maternal physical training was able to attenuate the delayed reflex ontogeny-induced
by undernutrition. The physiological mechanism can be associated with an increase of uterine
blood flow, which occurs during moderate-intensity exercise, as well as with the expanded
blood volume, and increased resting oxygen consumption found in response to regular
exercise(8,12)
. The present study suggests that exercise should be considered as a therapeutic
means of countering the effects of maternal undernutrition, and it may provide a useful
strategy for enhancing the neuronal activity of children born to mothers who experience a
restricted diet during pregnancy.
Acknowledgements
The authors declare no conflicts of interest. This research was supported by CNPq and
CAPES. The authors’ contributions are as follows: M.A.F.A. and C.G.L. performed the study
design. F.F.T., A.B.S. and J.A.S. performed the experiments and data collection. F.F.T. and
C.G.L. performed statistical analyses and interpretation of data. F.F.T. and C.G.L. wrote the
manuscript. All authors were responsible for critical revisions to the manuscript and approval
of the final version. The authors are indebted to Lucia Pires and Edeones França for technical
assistance.
References
1. Dauncey MJ & Bicknell RJ (1999) Nutrition and neurodevelopment: mechanisms of
developmental dysfunction and disease in later life. Nutr Res Rev 12, 231-253.
2. Bennis-Taleb N, Remacle C, Hoet JJ et al. (1999) A low-protein isocaloric diet during
gestation affects brain development and alters permanently cerebral cortex blood vessels in rat
offspring. J Nutr 129, 1613-1619.

57
3. Lodygensky GA, Seghier ML, Warfield SK et al. (2008) Intrauterine growth restriction
affects the preterm infant's hippocampus. Pediatr Res 63, 438-443.
4. Smart JL & Dobbing J (1971) Vulnerability of developing brain. II. Effects of early
nutritional deprivation on reflex ontogeny and development of behaviour in the rat. Brain Res
28, 85-95.
5. Barros KM, Manhaes-De-Castro R, Lopes-De-Souza S et al. (2006) A regional model
(Northeastern Brazil) of induced mal-nutrition delays ontogeny of reflexes and locomotor
activity in rats. Nutr Neurosci 9, 99-104.
6. Lopes de Souza S, Orozco-Solis R, Grit I et al. (2008) Perinatal protein restriction reduces
the inhibitory action of serotonin on food intake. Eur J Neurosci 27, 1400-1408.
7. Gallagher EA, Newman JP, Green LR et al. (2005) The effect of low protein diet in
pregnancy on the development of brain metabolism in rat offspring. J Physiol 568, 553-558.
8. Clapp JF, 3rd (2003) The effects of maternal exercise on fetal oxygenation and feto-
placental growth. Eur J Obstet Gynecol Reprod Biol 110 Suppl 1, S80-85.
9. Wolfe LA & Weissgerber TL (2003) Clinical physiology of exercise in pregnancy: a
literature review. J Obstet Gynaecol Can 25, 473-483.
10. Parnpiansil P, Jutapakdeegul N, Chentanez T et al. (2003) Exercise during pregnancy
increases hippocampal brain-derived neurotrophic factor mRNA expression and spatial
learning in neonatal rat pup. Neurosci Lett 352, 45-48.
11. Kim H, Lee SH, Kim SS et al. (2007) The influence of maternal treadmill running during
pregnancy on short-term memory and hippocampal cell survival in rat pups. Int J Dev
Neurosci 25, 243-249.
12. Amorim MF, dos Santos JA, Hirabara SM et al. (2009) Can physical exercise during
gestation attenuate the effects of a maternal perinatal low-protein diet on oxygen consumption
in rats? Exp Physiol 94, 906-913.
13. Bayne K (1996) Revised Guide for the Care and Use of Laboratory Animals available.
American Physiological Society. Physiologist 39, 199, 208-111.
14. Bayol S, Jones D, Goldspink G et al. (2004) The influence of undernutrition during
gestation on skeletal muscle cellularity and on the expression of genes that control muscle
growth. Br J Nutr 91, 331-339.
15. Dobbing J & Hopewell JW (1971) Permanent deficit of neurones in cerebral and
cerebellar cortex following early mild undernutrition. Arch Dis Child 46, 736-737.
16. Dobbing J (1964) The Influence of Early Nutrition on the Development and Myelination
of the Brain. Proc R Soc Lond B Biol Sci 159, 503-509.

58
17. Smart JL (1981) Under nutrition during early life and its effects on animal development
and behaviour. Neuropharmacology 20, 1251-1252.
18. Morgane PJ, Austin-LaFrance R, Bronzino J et al. (1993) Prenatal malnutrition and
development of the brain. Neurosci Biobehav Rev 17, 91-128.
19. Barreto Medeiros JM, Cabral Filho JE, De Souza SL et al. (2002) Early malnourished rats
are not affected by anorexia induced by a selective serotonin reuptake inhibitor in adult life.
Nutr Neurosci 5, 211-214.
20. Pine AP, Jessop NS, Allan GF et al. (1994) Maternal protein reserves and their influence
on lactational performance in rats. 4. Tissue protein synthesis and turnover associated with
mobilization of maternal protein. Br J Nutr 72, 831-844.
21. Zamenhof S (1981) Maternal nutrient storage and efficiency in production of fetal brain
tissue in rats. Biol Neonate 39, 45-51.
22. Moita L, Lustosa MF, Tobias Silva AT et al. (2011) Moderate Physical Training
Attenuates the Effects of Perinatal Undernutrition on the Morphometry of the Splenic
Lymphoid Follicles in Endotoxemic Adult Rats. Neuroimmunomodulation 18, 103-110.
23. Chandler KD & Bell AW (1981) Effects of maternal exercise on fetal and maternal
respiration and nutrient metabolism in the pregnant ewe. J Dev Physiol 3, 161-176.
24. Dauge V (2003) [Neurobiological impact of separating mothers from newborns in
rodents]. Med Sci (Paris) 19, 607-611.
25. Passos MC, Toste FP, Dutra SC et al. (2009) Role of neonatal hyperleptinaemia on serum
adiponectin and suppressor of cytokine signalling-3 expression in young rats. Br J Nutr 101,
250-256.
26. May LE, Glaros A, Yeh HW et al. (2010) Aerobic exercise during pregnancy influences
fetal cardiac autonomic control of heart rate and heart rate variability. Early Hum Dev 86,
213-217.
27. Clapp JF, 3rd (2008) Long-term outcome after exercising throughout pregnancy: fitness
and cardiovascular risk. Am J Obstet Gynecol 199, 489 e481-486.
28. Deiro TC, Manhaes-de-Castro R, Cabral-Filho JE et al. (2006) Sertraline delays the
somatic growth and reflex ontogeny in neonate rats. Physiol Behav 87, 338-344.

59
Table 1. Composition of the diets (control 17% and low-protein 8%).
Ingredients Amount for 1 Kg of diet
Low-protein Control
Casein 79,3 g 179,3 g
Vitamin Mixa 10 g 10 g
Mineral Mixtureb
35 g 35 g
Cellulose 50 g 50 g
Bitartarato of choline 2,5 g 2,5 g
DL-Methionine 3,0 g 3,0 g
Soya oil 70 ml 70 ml
Cornstarch 750,2 g 650,2 g
a Vitamin mixture contained the following (mg/kg of diet): retinol, 12; cholecalciferol, 0.125;
thiamine, 40; riboflavin, 30; pantothenic acid, 140; pyridoxine, 20; inositol, 300;
cyanocobalamin, 0.1; menadione, 80; nicotinic acid, 200; choline, 2720; folic acid, 10; p-
aminobenzoic acid, 100; biotin, 0.6.
b Mineral mixture contained the following (mg/kg of diet): CaHPO4, 17200; KCI, 4000; NaCl,
4000; MgO, 420; MgSO4, 2000; Fe2O2, 120; FeSO4·7H2O, 200; trace elements, 400
[MnSO4·H2O, 98; CuSO4·5H2O, 20; ZnSO4·7H2O, 80; CoSO4·7H2O, 0.16; KI, 0.32;
sufficient starch to bring to 40 g (per kg of diet)].

60
Table 2. Description of the tests reflexes.
Reflex Stimulus Response
Free-fall righting
(acceleration
righting)
Rat held by the paws, back
downwards, is dropped
from 30cm on a cotton
wool pad
Turns body in mid-air, to land on
all fours. All legs must be free of
body on landing.
Negative-
Geotaxis
Rat placed with head
downwards, on a 45° slope
Turns to face up the slope, at least
130°, in 10s.
Cliff-avoidance
Rat put on edge of bench,
with nose and forefeet just
over edge
Withdrawal of head and both
forefeet from edge, moving away
from “cliff”, in 10 s
Auditory-startle
response
Sudden sound stimulus by
percussion with a metallic
stick in a metal surface
Body retraction, with a transitory
immobility. The stimulus was
given twice in each test, with an
interval of 1 min.
Vibrissa-placing
Rat held by the tail, head
facing an edge of bench,
vibrissa just touching
vertical surface.
Lifts head and extends forepaws
in direction of the bench, making
oriented “walking” movements to
go far from the edge, in 10 s.
According to Smart and Dobbing (1971).

61
Table 3. Indicators of somatic growth at 17th day of life of litters (6 pups per litters).
Groups
NTL TL NT + LPL T + LPL
Mean
(S.E.M.)
Mean
(S.E.M.)
Mean
(S.E.M.)
Mean
(S.E.M.)
Body weight (g) 39.0 (1.1) 33.9 (0.9) 19.5 (0.7) a 19.9 (1.0)
a
Body length (mm) 104.9 (1.3) 107.5 (0.9) 81.0 (1.2) a 86.0 (1.2)
a
Δ body weight (g.day-1
) 2.8 (0.9) 2.7 (0.4) 1.1 (0.1) a 1.4 (0.01)
a,b
Δ body length (mm.day-1
) 30.1 (1.5) 29.7 (1.4) 25.3 (2.2) 19.9 (2.6) a
Tail length (mm) 47.8 (0.4) 44.2 (1.2) 37.0 (0.6) a 41.0 (0.6)
a,b
Laterolateral skull axis
(mm) 16.7 (0.06) 16.1 (0.09) 14.3 (0.1)
a 15.1 (0.2)
a,b
Anteroposterior axis of
the head (mm) 34.3 (0.09) 34.1 (0.1) 30.9 (0.4)
a 31.4 (0.1)
a,b
During gestation, the dams were submitted to physical training and fed a low-protein diet.
During lactation, dams remain receiving a low-protein diet. The pups into each litter were
evaluated from 10th
to 17th
during lactation. Groups: non-trained (NTL, n=7); trained (TL,
n=7); non-trained + low protein diet (NT + LPL, n=7) and trained + low protein diet (T + LPL,
n=4). The values are presented as mean (S.E.M.) a p<0.05 vs. NTL, and
b p<0.05 vs. NT +
LPL using two-way ANOVA, and Bonferroni´s post hoc test.

62
Table 4. Development of physical features and the reflexes in rats.
Groups
NTL TL NT + LPL T + LPL
Age (days)
Median
(Min-Max)
Age (days)
Median
(Min-Max)
Age (days)
Median
(Min-Max)
Age (days)
Median
(Min-Max)
Physical features
Auditory conduit
opening 12.0 (12-14) 13.0 (12-14) 14.0 (13-15)
a 14.0 (13-15)
a
Eyes opening 13.0 (12-14) 13.0 (12-14) 15.0 (14-16) a 14.0 (13-15)
Reflexes
Vibrissa-placing 12.0 (11-13) 12.0 (11-13) 16.0 (15-17) a 14.0 (13-16)
a,b
Cliff-avoidance 10.0 (10-11) 10.0 (10-11) 12.0 (11-13) a 10.0 (10-12)
b
Free-fall righting 12.0 (11-13) 13.0 (11-15) 17.0 (16-18) a 15.0 (14-16)
a
Auditory-startle
response 15.0 (14-16) 14.0 (13-15) 15.0 (14-16) 15.0 (14-16)
Negative-Geotaxis 13.0 (12-14) 14.0 (12-15) 17.0 (16-18) a 14.5 (14-16)
a,b
During gestation, the dams were submitted to physical training and fed a low-protein diet.
During lactation, dams remain receiving a low-protein diet. The pups into each litter were
evaluated from 10th
to 17th
during lactation. Groups: non-trained (NTL, n=7); trained (TL,
n=7); non-trained + low protein diet (NT + LPL, n=7) and trained + low protein diet (T + LPL,
n=4). The values are presented as median followed by the minimum and maximum values (in
parentheses). a p<0.05 vs. NTL and
b p<0.05 vs. NT+LPL using Kruskal-wallis test; Dunn´s
test.

63
1 2 30
10
20
30
40
NT T
NT + LP T+LP
Weeks of gestation
*
*
st nd rd
A
% g
ain
of body w
eig
ht
during g
esta
tion
1 2 3 1 2
0
10
20
30
40
NT
T
NT + LP
T+LP
st nd rd
B
st nd
Weeks of gestation Weeks of lactation
Delivery
*
*
Daily
food in
take (
g/d
ay)
Figure 1. Gain of body weight during gestation and food intake during gestation and lactation
by untrained (NT, n = 7), trained (T, n = 7), untrained + low protein (NT+LP, n = 7) and
trained + low protein dams (T+LP, n = 4). (A) Percentage of body weight gain in each third of
gestation, relative to the body mass on the first day of pregnancy, and (B) daily food intake
expressed in g/day. The values are presented as means ± S.E.M. ∗ p<0.05 vs NT group using
two-way ANOVA and Bonferroni’s post hoc test.

64
Título: Efeitos do treinamento físico durante a gestação sobre a evolução ponderal,
glicemia e colesterolemia de ratos adultos submetidos à desnutrição perinatal.
Autores: Filippe FALCÃO-TEBAS1, Amanda Thereza TOBIAS1, Adriano BENTO-
SANTOS2, José Antônio dos SANTOS2, Diogo Antônio Alves de VASCONCELOS1,
Marco Antônio FIDALGO3, Raul MANHÃES-DE-CASTRO1, Carol Góis LEANDRO3.
1Departamento de Nutrição – Universidade Federal de Pernambuco, Recife,
Pernambuco.
2Departamento de Neuropsiquiatria e Ciências do Comportamento – Universidade
Federal de Pernambuco, Recife, Pernambuco.
3Núcleo de Educação Física e Ciências do Esporte – CAV – Universidade Federal de
Pernambuco, Vitória de Santo Antão, Pernambuco.
Endereço para correspondência:
Filippe Falcão-Tebas
Rua Prof. Moraes Rego, 1235 – CEP: 50670-901 – Departamento de Nutrição –
Cidade Universitária – Recife, PE – Brasil.
Telefone: (00 55 81) 21268463
E-mail: [email protected]

65
Resumo
A incompatibilidade entre a desnutrição perinatal e uma nutrição adequada durante o desenvolvimento aumenta o risco de aparecimento precoce de doenças não transmissíveis na vida adulta. Todavia, acredita-se que a atividade física materna possa atenuar estas consequências. O presente estudo teve como objetivo avaliar os efeitos do treinamento físico durante a gestação na evolução ponderal, circunferência abdominal, glicemia e colesterolemia de filhotes adultos submetidos à desnutrição perinatal. Ratas Wistar (n=12) foram divididas em quatro grupos: controle (C, n=3), treinada (T, n=3), desnutrida (D, n=3) e treinada desnutrida (T+D, n=3). Durante a gestação e lactação, os grupos D e T+D receberam dieta baixa em proteína (8% caseína) e os grupos C e T receberam dieta normoprotéica (caseína a 17%). O protocolo de treinamento físico moderado foi realizado em esteira ergométrica (5 dias/semana, 60 min/dia, a 65% do VO2max) e iniciou 4 semanas antes da gestação. Na gestação, a duração e a intensidade do treinamento foram reduzidas (5 dias/semana, 20 min/dia, a 30% do VO2max) até o 19º dia pré-natal. Após o desmame, os filhotes (CF=9, TF=9, DF=7, T+DF=9) receberam dieta padrão de biotério e foram avaliados aos 270 dias de idade. A circunferência abdominal (CA) foi avaliada relativa ao peso corporal. Para avaliação da glicemia e colesterolemia foi utilizado o método enzimático colorimétrico da glicose-oxidase/peroxidase e da colesterol-oxidase, respectivamente. Ratos do grupo DF apresentaram um maior ganho de peso corporal ao longo do crescimento, maiores valores de CA, glicemia e colesterolemia quando comparados ao grupo CF. Para o grupo T+DF, o ganho de peso foi atenuado, e a CA, a glicemia e a colesterolemia foram normalizados (p<0,05). Esses resultados demonstram que o treinamento físico durante a gestação atenua os efeitos da desnutrição perinatal sobre alguns indicadores murinométricos e bioquímicos nos filhotes adultos. PALAVRAS-CHAVE: Plasticidade fenotípica; Dieta com baixa proteína; Exercício Físico

66
Title: Effects of physical training during pregnancy on body weight gain, glycaemia
and cholesterolemia of adult rats submitted to a perinatal undernutrition Abstract The incompatibility of perinatal undernutrition and adequate nutrition during development increases the risk of early onset of non-communicable diseases in adulthood. However, it has been considered that maternal physical activity may attenuate these effects. This study aimed to evaluate the effects of physical training during pregnancy on body weight gain, waist circumference, glycaemia and cholesterolemia in adult offspring submitted to a perinatal undernutrition. Female Wistar rats (n = 12) were divided into four groups: control (C, n=3), trained (T, n=3), undernourished (D, n=3) undernourished and trained (T+D, n=3). During gestation and lactation, D and T+D groups were fed a low protein diet (8% casein) and C and T groups fed a normal protein diet (casein 17%). The protocol of moderate physical training was performed on a treadmill (5 days/week, 60 min/day, at 65% of VO2max) and began 4 weeks before pregnancy. At pregnancy, the duration and intensity of training were reduced (5 days/week, 20 min/day, at 30% VO2max) until the 19th day prenatal. At weaning, male pups (CP=9, TP=9, DP=7, T+DP=9) received standard diet and evaluations took place at 270 days old. The abdominal circumference (AC) was evaluated relative to body weight. Enzymatic colorimetric method glucose-oxidase/peroxidase and cholesterol-oxidase was used to evaluate fasting glycaemia and cholesterolemia, respectively. Rats from DP group showed a high body weight gain during growth, values of CA, glycaemia and cholesterolemia when compared to CP. For the group T+DP, body weight gain was attenuated, and the CA, glycaemia and cholesterolemia were normalized (p<0.05). These results demonstrate that physical training during pregnancy reduces the effects of perinatal undernutrition on some murinometric and biochemical indicators of adult offspring. KEYWORDS: Phenotypic plasticity; Low-Protein Diet; Physical Exercise

67
INTRODUÇÃO
O crescimento e o desenvolvimento do feto dependem de fatores genéticos,
hormonais, placentários, do milieu materno, e do suprimento adequado de oxigênio
e nutrientes(1). Particularmente o aporte inadequado de nutrientes durante a vida
fetal tem sido associado ao baixo peso ao nascer e déficits durante o crescimento e
maturação(2). Da mesma forma, a incompatibilidade entre a desnutrição durante o
período perinatal e a nutrição ao longo da vida pode estar relacionada ao
aparecimento de doenças metabólicas(2). A desnutrição perinatal influencia
negativamente o desenvolvimento do sistema nervoso, causa atraso na ontogênese
reflexa em ratos lactantes e altera o comportamento alimentar na vida adulta (3, 4). Há
ainda alterações na morfologia do músculo cardíaco, atraso na aquisição dos
padrões normais da atividade locomotora e deficiência nas propriedades contráteis e
elásticas do músculo esquelético de ratos adultos(3-7).
O termo “programação” ou “plasticidade fenotípica” tem sido utilizado para
explicar que durante a ontogênese, o desenvolvimento de cada órgão ou sistema
passa por uma janela crítica de sensibilidade ou plasticidade, onde fatores
ambientais podem gerar ajustes no fenótipo que permanecem ao longo da vida (8). A
atividade física materna induz adaptações fisiológicas durante a gestação
envolvendo o crescimento feto-placentário e o aumento da disponibilidade de
nutrientes e oxigênio para o feto(9, 10). Entretanto, tais efeitos sobre a oxigenação e o
crescimento feto-placentário estão diretamente correlacionados com o nível de
aptidão física da mãe e em que momento durante a gestação o programa de
exercício é realizado(11).
De acordo com o American College of Obstetricians and Gynecologists(13),
mulheres com gestação de baixo-risco podem praticar exercício físico moderado (até

68
70% do VO2max) e leve (até 40% do VO2max) por cerca de 30 minutos por dia, todos
os dias da semana. Contudo, ainda é difícil estabelecer recomendações de
percentual de esforço físico durante a gestação uma vez que a intensidade, o tipo e
a duração do exercício físico são determinantes para as adaptações fisiológicas na
mãe e as repercussões no filho(12). Estudos epidemiológicos realizados em uma
comunidade rural da Índia com mulheres gestantes demonstraram uma relação
inversa entre a intensidade do esforço e o peso ao nascer dos filhos(14, 15). Em
animais, ratas treinadas antes da gestação (cinco dias/semana, 60 min/dia, a 65%
do VO2max) com diminuição progressiva do esforço durante a gestação (cinco
dias/semana, 30 min/dia, a 40% do VO2max) apresentaram uma menor queda no
consumo de oxigênio de repouso(16). Este efeito foi determinante para atenuar os
efeitos da desnutrição protéica perinatal relativamente à maturação do sistema
nervoso e a taxa de crescimento somático dos filhotes(16, 17).
Os estudos que associam o treinamento físico durante a gestação com a
hipótese da plasticidade fenotípica ainda são escassos. Da mesma forma que ainda
sabe-se pouco sobre as repercussões em longo-prazo de um programa de
treinamento físico durante a gestação. Assim, o presente estudo teve como objetivo
avaliar os efeitos do treinamento físico durante a gestação na evolução ponderal,
circunferência abdominal, glicemia e colesterolemia de filhotes adultos submetidos à
desnutrição perinatal. Nossa hipótese é que o treinamento físico durante a gestação
atenua os efeitos da programação induzida pela desnutrição protéica perinatal.
MATERIAIS E MÉTODOS
Animais

69
Este estudo foi aprovado pelo comitê de ética em estudos com animais do
Centro de Ciências Biológicas da UFPE (protocolo nº. 23076.049077/2010-80). A
manipulação e os cuidados com os animais seguiram as recomendações do Comitê
Brasileiro de Experimentação Animal (COBEA).
Foram utilizadas 12 ratas albinas (60 dias de idade) da linhagem Wistar, com
peso corporal entre 180 ± 11 g, obtidas da colônia do Departamento de Nutrição da
Universidade Federal de Pernambuco. As ratas foram mantidas em biotério de
experimentação em condições padronizadas de temperatura de 23°C 1 e
iluminação claro das 18:00 às 06:00 h, com livre acesso à água e alimentação
padrão do biotério (52% carboidratos, 21% proteínas, 4% lipídeos – Nuvilab CR1-
Nuvital®, Curitiba, Paraná, Brasil). As ratas foram divididas em dois grupos
experimentais: controle (C, n=6) e treinada (T, n=6). O grupo T realizou um
programa de treinamento físico moderado em esteira (EP-131®, Insight Equipments,
SP, Brasil)(16) (Tabela 1). Após as quatro semanas de treinamento físico, as ratas
dos dois grupos, foram postas para acasalar (duas fêmeas para um macho). O
diagnóstico da gestação foi realizado através do exame de esfregaço vaginal para a
presença de espermatozóides(18). Detectada a gestação, metade das ratas de cada
grupo (C e T) foi submetida à dieta hipoprotéica (caseína a 8%), enquanto que as
demais ratas receberam dieta normoprotéica (caseína a 17%). As dietas
confeccionadas foram isocalóricas com alteração apenas no conteúdo de
proteína(19). Sendo formados então os seguintes grupos: controle (C, n=3, 17%
caseína), treinada (T, n=3, 17% caseína), desnutrida (D, n=3, 8% caseína) e
treinada desnutrida (T+D, n=3, 8% caseína). O programa de treinamento físico foi
mantido ao longo da gestação, até o 19º dia, com diminuição progressiva da
intensidade e da duração das sessões (Tabela 2). Durante todo o experimento, o

70
peso corporal das mães foi acompanhado semanalmente. Na lactação, as mães
permaneceram recebendo a dieta experimental à base de caseína e as ninhadas
foram ajustadas para seis filhotes por lactente. Após o desmame (aos 22 dias de
vida dos filhotes), foram utilizados aleatoriamente três filhotes machos, de cada
ninhada. Os filhotes foram alimentados com a dieta padrão do biotério e divididos
em quatro grupos de acordo com a manipulação das suas respectivas mães: filhotes
de mães controle (CF, n=9), filhotes de mães treinadas (TF, n=9), filhotes de mães
desnutridas (DF, n=7) e filhotes de mães treinadas e desnutridas (T+DF, n=9).
Protocolo de Treinamento físico
Os animais foram previamente adaptados ao biotério de ciclo invertido
(durante 15 dias) e à esteira ergométrica por um período de três dias (10 min/dia,
com velocidade de 0.3 km.h-1). O protocolo de treinamento físico moderado utilizado
consistiu de quatro semanas de treinamento, cinco dias por semana, 60 minutos por
dia a 65% do VO2max(16). Na primeira semana houve a adaptação ao treinamento
físico, que consistiu em sessões de 20 minutos, divididos em quatro estágios de
cinco minutos, durante cinco dias. Após a adaptação, o protocolo foi dividido em
estágios progressivos em cada sessão: (1) aquecimento (5 minutos); (2) zona
intermediária (10 minutos); (3) zona de treinamento (30 minutos), e (4) período final
(5 minutos) (Tabela 1).
No período gestacional, ocorreu a diminuição progressiva na intensidade e na
duração do treinamento, que consistiu de três semanas de treinamento, cinco dias
por semana, reduzindo para 20 minutos por dia a 30% do VO2max (Tabela 2).
TABELA 1.

71
Avaliação do peso corporal e do ganho de peso
O peso corporal, expresso em gramas, foi avaliado semanalmente nas mães
e mensalmente nos filhotes através de uma balança eletrônica digital – Marte,
modelo S-1000, com capacidade máxima de 1000g e sensibilidade de 0,01g. O
percentual de ganho de peso corporal foi calculado tendo como base o peso do
desmame, segundo a fórmula: % ganho de peso=[Peso do dia (g) x 100/ Peso do
desmame (g)] – 100 (20). A circunferência abdominal foi determinada através da
maior circunferência entre a borda superior da crista ilíaca e a última costela, e
expressa em centímetros(21).
Avaliação da Glicemia e Colesterolemia dos Filhotes
Após jejum de 6 horas, os animais sofreram um corte na extremidade da
cauda para coleta do sangue. A glicose e o colesterol séricos (mg.dL-1) dos filhotes
aos 270 dias de idade foram avaliados. As concentrações de glicose sanguínea
foram identificadas pelo método enzimático colorimétrico da glicose-
oxidase/peroxidase e a leitura através de glicosímetro (Accu Chek Performa®, Roche
Diagnostics). A quantificação da colesterolemia foi realizada por meio método da
colesterol-oxidase e analisada por fotometria, sendo os valores calculados através
do monitor (Accutrend Colesterol®, Roche Diagnostics).
Análise estatística
Inicialmente, todas as variáveis foram submetidas ao teste de normalidade
(Kolmogorov-Smirnov). Ao revelarem normalidade e homogeneidade de variâncias,
TABELA 2.

72
para comparação de parâmetros das ratas durante o período pré-gestacional foi
utilizado o t-test Student. Para a análise entre os grupos durante a gestação, o peso
dos filhotes ao nascer e durante o seu crescimento e a avaliação dos parâmetros
bioquímicos dos animais na idade adulta foi utilizado o ANOVA two-way seguido do
teste post-hoc de Bonferroni. Os valores estão expressos em média e erro padrão
da média (EPM). O nível de significância foi mantido em 5% (p<0,05) em todos os
casos. Toda análise estatística foi realizada utilizando o programa GraphPad Prism®
(GraphPad Software Inc., La Jolla, CA, USA; versão 5.0 para Windows).
RESULTADOS
As ratas submetidas ao protocolo de treinamento físico moderado
apresentaram valores menores de ganho de peso corporal no período pré-
gestacional a partir da terceira semana (Tabela 3).
Durante a gestação, os grupos foram subdivididos de acordo com a dieta
oferecida às mães. As mães que receberam dieta hipoprotéica apresentaram um
menor ganho de peso corporal a partir da 2ª semana de gestação. As mães
treinadas que foram desnutridas apresentaram valores menores de ganho de peso
corporal, na última semana de gestação (Tabela 4).
O valor médio do peso ao nascer dos filhotes de mães desnutridas foi menor
quando comparados ao controle (CF = 6,1 ± 0,3; DF = 4,9 ± 0,3; TF = 7,1 ± 0,4; T +
DF = 5,9 ± 0,2). O crescimento dos filhotes foi acompanhado desde o desmame até
TABELA 3.
TABELA 4.

73
os 270 dias de idade. Os animais provindos de mães desnutridas e treinadas
desnutridas apresentaram valores inferiores de peso corporal ao longo do
crescimento (Figura 1A). Contudo, o ganho de peso relativo ao desmame dos
animais desnutridos foi maior a partir do 30º dia de vida quando comparado ao
controle (Figura 1B). Embora os animais provindos de mães treinadas desnutridas
tenham apresentado maior ganho de peso corporal ao longo do crescimento, estes
valores foram inferiores ao grupo desnutrido (Figura 1B).
Os valores médios da circunferência abdominal relativa ao peso corporal dos
filhotes de mães desnutridas foram maiores comparados ao controle. No grupo de
animais provindos de mães treinadas desnutridas não houve diferença em relação
ao controle e foi menor com relação ao grupo desnutrido (Figura 2).
A glicemia e a colesterolemia dos filhotes na idade adulta que foram
submetidos à desnutrição perinatal foi maior em comparação ao grupo controle
(Figura 3). Os animais provindos de mães treinadas desnutridas não apresentaram
diferença em relação ao controle e tiveram valores médios inferiores ao grupo
desnutrido (Figura 3).
DISCUSSÃO
O objetivo do presente estudo foi avaliar os efeitos do treinamento físico
moderado no ganho de peso corporal das mães submetidas ou não à desnutrição
FIGURA 1.
FIGURA 2.
FIGURA 3.

74
perinatal, bem como as repercussões ocorridas nos filhotes com idade de 270 dias.
Confirmando nossa hipótese, o treinamento físico materno durante a gestação
atenuou os efeitos deletérios da desnutrição perinatal na evolução ponderal e em
indicadores murinométricos e bioquímicos em ratos adultos.
Em relação às mães desnutridas foi observado um menor ganho de peso
corporal e seus filhotes nasceram com baixo peso, corroborando com estudos
prévios(20, 22, 23). Os filhotes de mães desnutridas permaneceram com uma menor
trajetória de crescimento até a idade adulta mesmo quando a dieta equilibrada foi
oferecida. É interessante observar que o ganho de peso corporal (expresso em
percentual a partir do desmame) foi maior nos filhotes de mães desnutridas. Este
fenômeno é chamado de “catch up” de crescimento, onde o organismo compensa
em longo-prazo uma taxa lenta de crescimento por déficit de proteínas na dieta
durante os períodos iniciais da vida(24). Este crescimento acelerado pós-natal tem
sido associado com o aumento: do percentual do tecido adiposo, da pressão arterial
e, do risco de desenvolver intolerância a glicose, em longo prazo(25).
Por outro lado, as alterações no “catch up” dos animais desnutridos foram
atenuadas nos filhotes de mães treinadas desnutridas, demonstrando que o
treinamento físico pode atenuar os efeitos da desnutrição hipoprotéica. No presente
estudo, as ratas iniciaram o protocolo de treinamento físico quatro semanas antes da
gravidez, a fim de normalizar os parâmetros fisiológicos em resposta ao estresse
agudo de uma sessão de exercício. Ademais, exercícios físicos de intensidade leve
ao longo da gestação, similares ao utilizado em nosso trabalho, têm sido
recomendados, pois além de contribuir com a manutenção do ganho de peso,
aumentam a taxa de crescimento feto-placentário e o peso ao nascer(11). Os
mecanismos subjacentes parecem incluir: (a) o aumento do fluxo de sangue uterino;

75
(b) redistribuição de fluxo sanguíneo; (c) alterações na produção fetal e placentária
de hormônios que controlam o crescimento e, (d) uma menor diminuição do
consumo de oxigênio de repouso em resposta ao treinamento físico(9, 16, 26). Estes
parâmetros fisiológicos podem estar contribuindo para uma melhor resposta
adaptativa preditiva (um modelo teórico que dá suporte a hipótese da plasticidade
fenotípica) gerada pelo estímulo precoce(8). Esta resposta tem sido associada a
mecanismos epigenéticos, onde um fator externo pode modular a estrutura do DNA
sem mudança na sua sequência(2, 8). Em ratos expostos à desnutrição intrauterina
foram observados maiores concentrações de leptina, insulina e peptídeo-C,
indicadores da síndrome metabólica na idade adulta, devido a alterações no
genótipo os animais(27). Assim, tem sido relatado que estímulos ambientais precoces
podem modular o fenótipo ao longo da vida.
A circunferência abdominal tem sido utilizada como um indicador de
obesidade em ratos(21). Em estudos populacionais, a circunferência abdominal tem
sido utilizada como um indicador de risco ao aparecimento de doenças cárdio-
vasculares e obesidade em adultos. Em acordo com a hipótese da plasticidade
fenotípica, os animais submetidos à desnutrição perinatal apresentaram maiores
valores de circunferência abdominal e os nossos resultados são similares aos
estudos prévios(28, 29). Em ratos, tem sido demonstrado que filhotes recém-nascidos
provindos de mães desnutridas (50% de restrição da dieta recebida pelo controle ad
libitum) apresentaram, aos nove meses de idade, um aumento na expressão de
fatores de transcrição lipogênicos e adipogênicos, levando a hipertrofia dos
adipócitos(30). Por outro lado, nos animais provindos de mães treinadas desnutridas,
estes resultados foram atenuados. É provável que o menor “catch up” de

76
crescimento observado nestes animais possam justificar uma menor circunferência
abdominal.
O aumento da circunferência abdominal pode também indicar uma maior
concentração plasmática de triglicerídeos, ácidos graxos e colesterol, que são
indicadores de risco aterogênico(31). Nossos resultados demonstram que filhos de
mães desnutridas apresentaram um aumento nas concentrações séricas de glicose
e colesterol quando adultos. Estes efeitos têm sido verificados em estudos prévios e
são indicadores clássicos do início da resistência periférica à insulina e dislipidemias
associados com a origem desenvolvimentista da síndrome metabólica(32). Contudo,
os animais provindos de mães treinadas desnutridas não apresentaram este quadro
de hiperglicemia e hipercolesterolemia.
Diante disto, podemos sugerir alguns mecanismos que podem dar suporte
teórico a ação atenuante do treinamento físico: aumento no volume placentário,
aumento do fluxo de sangue para a placenta após o exercício, aumento na
passagem de nutrientes e oxigênio para o feto, aumento na massa magra corporal
da mãe(33); e aumento no consumo de oxigênio de repouso(16). É de ressalva que
todos estes efeitos são diretamente relacionados à magnitude do esforço. No
presente estudo, foi utilizado um protocolo de treino padronizado a partir das
medidas diretas de consumo de oxigênio. A intensidade e duração foram
controladas de forma que as sessões de treinamento fossem de intensidade
moderada e leve(16).
Em conclusão, o treinamento físico moderado antes e durante a gestação
atenuou os efeitos da desnutrição perinatal sobre o crescimento somático e os níveis
séricos de glicose e colesterol nos filhotes adultos. Os nossos dados corroboram os
estudos que testam a hipótese da plasticidade fenotípica e, abre um cenário para os

77
efeitos de um estímulo ambiental positivo sendo capaz de atenuar os efeitos da
desnutrição.
AGRADECIMENTOS
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e
à Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco
(FACEPE) por tornarem possível a realização deste estudo.
REFERÊNCIAS
1. Harding JE. The nutritional basis of the fetal origins of adult disease. Int J
Epidemiol. 2001 Feb;30(1):15-23.
2. Bateson P, Barker D, Clutton-Brock T, Deb D, D'Udine B, Foley RA, et al.
Developmental plasticity and human health. Nature. 2004 Jul 22;430(6998):419-21.
3. Barros KM, Manhaes-De-Castro R, Lopes-De-Souza S, Matos RJ, Deiro TC,
Cabral-Filho JE, et al. A regional model (Northeastern Brazil) of induced mal-nutrition
delays ontogeny of reflexes and locomotor activity in rats. Nutr Neurosci. 2006 Feb-
Apr;9(1-2):99-104.
4. Lopes de Souza S, Orozco-Solis R, Grit I, Manhaes de Castro R, Bolanos-
Jimenez F. Perinatal protein restriction reduces the inhibitory action of serotonin on
food intake. Eur J Neurosci. 2008 Mar;27(6):1400-8.
5. Toscano AE, Amorim MA, de Carvalho Filho EV, Aragao Rda S, Cabral-Filho
JE, de Moraes SR, et al. Do malnutrition and fluoxetine neonatal treatment program
alterations in heart morphology? Life Sci. 2008 May 23;82(21-22):1131-6.

78
6. Toscano AE, Manhaes-de-Castro R, Canon F. Effect of a low-protein diet
during pregnancy on skeletal muscle mechanical properties of offspring rats.
Nutrition. 2008 Mar;24(3):270-8.
7. Orozco-Solis R, Lopes de Souza S, Barbosa Matos RJ, Grit I, Le Bloch J,
Nguyen P, et al. Perinatal undernutrition-induced obesity is independent of the
developmental programming of feeding. Physiol Behav. 2009 Mar 2;96(3):481-92.
8. Gluckman PD, Hanson MA. Developmental plasticity and human disease:
research directions. J Intern Med. 2007 May;261(5):461-71.
9. Clapp JF, 3rd, Schmidt S, Paranjape A, Lopez B. Maternal insulin-like growth
factor-I levels (IGF-I) reflect placental mass and neonatal fat mass. American journal
of obstetrics and gynecology. 2004 Mar;190(3):730-6.
10. Haakstad LA, Voldner N, Henriksen T, Bo K. Physical activity level and weight
gain in a cohort of pregnant Norwegian women. Acta Obstet Gynecol Scand.
2007;86(5):559-64.
11. Clapp JF, 3rd. Long-term outcome after exercising throughout pregnancy:
fitness and cardiovascular risk. Am J Obstet Gynecol. 2008 Nov;199(5):489 e1-6.
12. Leandro CG, Amorim MF, Hirabara SM, Curi R, Castro RMd. Can maternal
physical activity modulate the nutrition-induced fetal programming? Revista de
Nutrição. 2009;22(4):559-69.
13. ACOG Committee opinion. Number 267, January 2002: exercise during
pregnancy and the postpartum period. Obstet Gynecol. 2002 Jan;99(1):171-3.
14. Dwarkanath P, Muthayya S, Vaz M, Thomas T, Mhaskar A, Mhaskar R, et al.
The relationship between maternal physical activity during pregnancy and birth
weight. Asia Pac J Clin Nutr. 2007;16(4):704-10.

79
15. Rao S, Kanade A, Margetts BM, Yajnik CS, Lubree H, Rege S, et al. Maternal
activity in relation to birth size in rural India. The Pune Maternal Nutrition Study. Eur J
Clin Nutr. 2003 Apr;57(4):531-42.
16. Amorim MF, dos Santos JA, Hirabara SM, Nascimento E, de Souza SL, de
Castro RM, et al. Can physical exercise during gestation attenuate the effects of a
maternal perinatal low-protein diet on oxygen consumption in rats? Exp Physiol. 2009
Aug;94(8):906-13.
17. Falcão-Tebas F, Bento-Santos A, Fidalgo MA, Almeida MB, Santos JA, Lopes
de Souza S, et al. Maternal low protein diet-induced delayed reflex ontogeny is
attenuated by moderate physical training during gestation in rats. British Journal of
Nutrition. 2011 in press.
18. Marcondes FK, Bianchi FJ, Tanno AP. Determination of the estrous cycle
phases of rats: some helpful considerations. Braz J Biol. 2002 Nov;62(4A):609-14.
19. Reeves PG, Nielsen FH, Fahey GC, Jr. AIN-93 purified diets for laboratory
rodents: final report of the American Institute of Nutrition ad hoc writing committee on
the reformulation of the AIN-76A rodent diet. J Nutr. 1993 Nov;123(11):1939-51.
20. Bayol S, Jones D, Goldspink G, Stickland NC. The influence of undernutrition
during gestation on skeletal muscle cellularity and on the expression of genes that
control muscle growth. Br J Nutr. 2004 Mar;91(3):331-9.
21. Novelli EL, Diniz YS, Galhardi CM, Ebaid GM, Rodrigues HG, Mani F, et al.
Anthropometrical parameters and markers of obesity in rats. Lab Anim. 2007
Jan;41(1):111-9.
22. Lucas A, Baker BA, Desai M, Hales CN. Nutrition in pregnant or lactating rats
programs lipid metabolism in the offspring. Br J Nutr. 1996 Oct;76(4):605-12.

80
23. Ozanne SE, Hales CN. The long-term consequences of intra-uterine protein
malnutrition for glucose metabolism. Proc Nutr Soc. 1999 Aug;58(3):615-9.
24. Barker DJ, Eriksson JG, Forsen T, Osmond C. Fetal origins of adult disease:
strength of effects and biological basis. Int J Epidemiol. 2002 Dec;31(6):1235-9.
25. Hales CN, Ozanne SE. The dangerous road of catch-up growth. J Physiol.
2003 Feb 15;547(Pt 1):5-10.
26. Clapp JF, 3rd, Kim H, Burciu B, Schmidt S, Petry K, Lopez B. Continuing
regular exercise during pregnancy: effect of exercise volume on fetoplacental growth.
American journal of obstetrics and gynecology. 2002 Jan;186(1):142-7.
27. Morris TJ, Vickers M, Gluckman P, Gilmour S, Affara N. Transcriptional
profiling of rats subjected to gestational undernourishment: implications for the
developmental variations in metabolic traits. PLoS One. 2009;4(9):e7271.
28. Ozanne SE, Hales CN. Lifespan: catch-up growth and obesity in male mice.
Nature. 2004 Jan 29;427(6973):411-2.
29. Vuguin PM. Animal models for small for gestational age and fetal
programming of adult disease. Horm Res. 2007;68(3):113-23.
30. Ozanne SE, Hales CN. Early programming of glucose-insulin metabolism.
Trends Endocrinol Metab. 2002 Nov;13(9):368-73.
31. Taylor PD, Poston L. Developmental programming of obesity in mammals.
Exp Physiol. 2007 Mar;92(2):287-98.
32. Ozanne SE. Metabolic programming in animals. Br Med Bull. 2001;60:143-52.
33. Thomas DM, Clapp JF, Shernce S. A foetal energy balance equation based on
maternal exercise and diet. J R Soc Interface. 2008 Apr 6;5(21):449-55.

81
Tabela I. Programa de treinamento físico para ratas de acordo com a
velocidade, duração e intensidade de cada sessão durante o período de quatro
semanas no período pré-gestacional(16).
Semanas Velocidade
(km.h-1) % do VO2max
Duração de cada estágio
(min)
Duração total de cada sessão (min)
1a semana (adaptação)
0.3 36.1 2.7 5
20 0.4 38.7 2.9 5
0.5 37.8 1.9 5
0.3 35.7 3.1 5
2ª semana
0.4 42.5 1.7 5
50 0.5 47.8 3.1 10
0.6 57.3 5.0 30
0.4 54.4 2.4 5
3ª semana
0.4 32.4 2.0 5
60
0.5 41.8 3.0 10
0.6 51.1 2.8 10
0.8 64.0 3.3 30
0.4 57.2 3.4 5
4ª semana
0.5 26.8 1.7 5
60
0.6 43.4 3.9 10
0.8 49.1 5.1 10
0.9 65.3 4.7 30
0.5 57.8 3.9 5

82
Tabela 2. Programa de treinamento físico para ratas de acordo com a velocidade,
duração e intensidade de cada sessão durante o período de três semanas, durante
a gestação(16).
Semanas Velocidade
(km.h-1) % do VO2max
Duração de cada estágio
(min)
Duração total de cada sessão
(min)
1ª semana
0.4 52.9 3.4 5
50
0.5 57.8 4.4 10
0.6 63.1 1.4 10
0.8 66.4 4.9 20
0.4 62.0 2.6 5
2ª semana
0.4 42.0 1.3 5
30 0.5 47.8 5.1 10
0.6 43.5 5.7 10
0.4 42.1 3.2 5
3ª semana
0.3 36.7 1.8 5
20 0.4 32.2 2.9 5
0.5 32.8 1.7 5
0.3 29.9 2.6 5

83
Tabela 3. Valores médios ± EPM do ganho de peso corporal das ratas submetidas a
um protocolo de treinamento físico moderado no período pré-gestacional.
Ganho de peso corporal pré-gestacional
Grupos
Controle Treinada
2a semana 9,9 ± 0,9 9,5 ± 0,7 3a semana 13,0 ± 1,1 11,2 ± 0,9* 4a semana 17,0 ± 2,3 13,0 ± 1,1*
*p< 0,05 vs controle, t-test student.
Tabela 4. Valores médios ± EPM do ganho de peso corporal das ratas submetidas a
um protocolo de treinamento físico moderado no período pré-gestacional e leve
durante a gestação e/ou à uma dieta hipoprotéica durante a gestação.
Ganho de peso corporal gestacional
Grupos
Controle Desnutrida Treinada Treinada + Desnutrida
1a semana 4,02 ± 1,1 3,9 ± 0,9 4,2 ± 0,9 3,9 ± 1,0 2a semana 16,0 ± 2,1 14,2 ± 3,0* 17,8 ± 2,4 15,1 ± 2,9 3a semana 36,9 ± 5,3 30,1 ± 5,1* 38,2 ± 5,5 31,5 ± 6,1*
*p< 0,05 vs controle, ANOVA two-way seguido do teste de Bonferroni.

84
Desmame 30 60 90 120 150 180 210 240 2700
100
200
300
400
500CF
TF
DF
T + DF
*
* * * * * *
Pe
so
(g
)
*
* * * * * *
A
Idade (dias)
30 60 90 120 150 180 210 240 2700
500
1000
1500
2000CF
TF
DF
T + DF
*
*
** * * * *
#
#
##
# # # #
*
Ga
nh
o d
e p
eso
em
pe
rce
ntu
al
B
Idade (dias)
Figura 1. Peso corporal expresso em gramas (A) e percentual do ganho de peso
semanal dos 30 aos 270 dias em relação ao desmame (B) de ratos. Grupos: CF
(n=9); TF (n=9); DF (n=7) e T+DF (n=9). Os dados estão representados em média ±
EPM. * p<0,05 vs CF; # p<0,05 vs DF. Anova two-way seguida do teste de
Bonferroni.
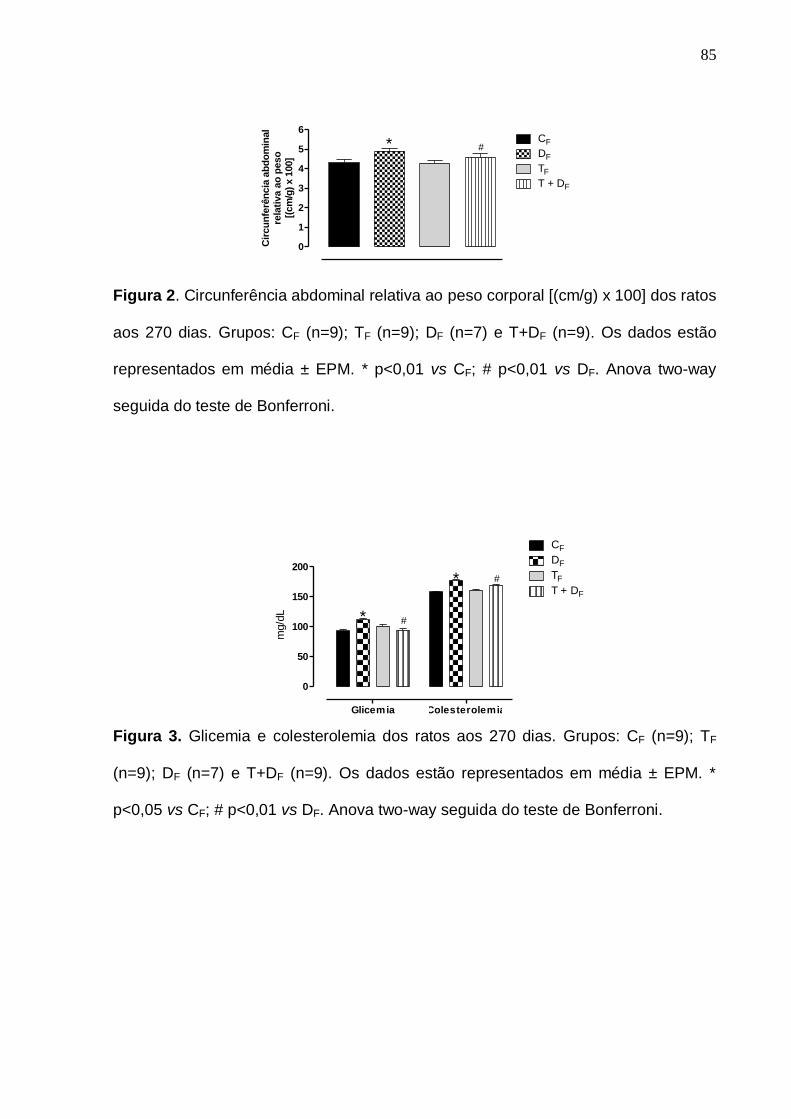
85
Glicemia Colesterolemia
0
50
100
150
200
CF
TF
DF
T + DF
mg/d
L *
* #
#
0
1
2
3
4
5
6CF
TF
DF
T + DF
* #
Cir
cu
nfe
rên
cia
ab
do
min
al
rela
tiva a
o p
eso
[(cm
/g)
x 1
00]
Figura 2. Circunferência abdominal relativa ao peso corporal [(cm/g) x 100] dos ratos
aos 270 dias. Grupos: CF (n=9); TF (n=9); DF (n=7) e T+DF (n=9). Os dados estão
representados em média ± EPM. * p<0,01 vs CF; # p<0,01 vs DF. Anova two-way
seguida do teste de Bonferroni.
Figura 3. Glicemia e colesterolemia dos ratos aos 270 dias. Grupos: CF (n=9); TF
(n=9); DF (n=7) e T+DF (n=9). Os dados estão representados em média ± EPM. *
p<0,05 vs CF; # p<0,01 vs DF. Anova two-way seguida do teste de Bonferroni.

86
TREINAMENTO FÍSICO MODERADO NA GESTAÇÃO AUMENTA O PERCENTUAL DE FIBRAS
OXIDATIVAS EM FILHOTES ADULTOS SUBMETIDOS À DESNUTRIÇÃO PERINATAL
TÍTULO CURTO: Resposta preditiva adaptativa e o fenótipo de fibras musculares esqueléticas
Filippe Falcão-Tebas1, Rafaela Araújo Dias da Silva
1, Adriano Bento-Santos
2, José Antônio
dos Santos2, Cláudia Jacques Lagranha
3, Ana Elisa Toscano
3, Carol Góis Leandro
3
1Departamento de Nutrição – Universidade Federal de Pernambuco, Recife, Pernambuco.
2Departamento de Neuropsiquiatria e Ciências do Comportamento – Universidade Federal de
Pernambuco, Recife, Pernambuco.
3Núcleo de Educação Física e Ciências do Esporte – CAV – Universidade Federal de
Pernambuco, Vitória de Santo Antão, Pernambuco.
Endereço para correspondência:
Filippe Falcão-Tebas
Rua Prof. Moraes Rego, 1235 – CEP: 50670-901 – Departamento de Nutrição – Cidade
Universitária – Recife, PE – Brasil.
Telefone: (00 55 81) 21268463
E-mail: [email protected]

87
RESUMO
O objetivo do estudo foi analisar o efeito do treinamento físico durante a gestação na
proporção de fibras musculares esqueléticas de ratos adultos submetidos à desnutrição
perinatal. Ratas Wistar (n=12) foram divididas em dois grupos (controle, C; n=6 e treinada, T;
n=6). O protocolo de treinamento físico moderado em esteira ergométrica foi iniciado 4
semanas antes da gestação (5 dias/semana, 60 min/dia, a 65% do VO2max). Na gestação, a
duração e a intensidade do treinamento foram reduzidas (5 dias/semana, 20 min/dia, a 30% do
VO2max) até o 19º dia pré-natal. Durante a gestação e lactação, os grupos C e T receberam
dieta normoprotéica (caseína a 17%) e os grupos desnutrida (D, n=3) e treinada desnutrida
(TD, n=3) receberam dieta hipoprotéica (8% caseína). Após o desmame, os filhotes foram
divididos de acordo com a manipulação materna (CF=7, TF=7, DF=7, TDF=7) e receberam
dieta padrão de biotério. Aos 270 dias de idade, os filhotes foram avaliados quanto ao: peso e
ganho de peso corporal (expresso em percentual), além do peso, comprimento e largura dos
músculo soleus e extensor longo dos dedos (EDL). A proporção de fibras em cada músculo
foi avaliada pela técnica da atividade ATPase da miosina. O efeito da desnutrição perinatal foi
observado tanto no peso corporal ao longo da vida e no ganho de peso corporal aos 90 dias.
Este efeito foi atenuado nos animais do grupo TDF (p < 0,05). O grupo DF apresentou
diminuição no percentual de fibras tipo I e um aumento na proporção de fibras tipo IIb no
soleus, embora não tenha ocorrido alterações na proporção de fibras do músculo EDL. Nos
animais do grupo TDF, os efeitos da desnutrição materna foram normalizados (p < 0,05).
Nossos resultados confirmam a associação entre desnutrição perinatal e as alterações no
fenótipo do músculo esquelético na prole adulta, enquanto o treinamento físico durante a
gestação atenua estes efeitos.
PALAVRAS-CHAVE: Atividade física, desnutrição, histologia, músculo esquelético

88
ABSTRACT
The purpose of this study was to analyze the histology of skeletal muscle fibers in adult rats
whose mothers were undernourished during pregnancy and lactation and/or subjected to
physical training protocol. Female Wistar rats (n = 12) submitted to a protocol of moderate
physical training on a treadmill (5 days week-1
, 60 min day-1
, 65% of VO2max) initiate 4 weeks
before gestation. During pregnancy, the duration and intensity of training were reduced (5
days week-1
, 20 min day-1
at 30% VO2max) until the 19-day pre-natal care. The nutritional
manipulation occurred during pregnancy and lactation with 17% casein diet for the control
and trained groups, and 8% casein diet for the undernourished and trained undernourished
groups. After weaning, the pups were divided according to maternal manipulation (NTp, n =
7, pups from untrained mothers; Tp, n = 7, pups from trained mothers; NT+LPp, n = 7, pups
from undernourished mothers; and T-LPp, n = 7, pups from trained undernourished mothers)
and received standard diet until 270 days old. Were evaluated on pups: body weight and
weight gain (expressed in percentage) and with respect to muscle: weight, length and width as
well as analysis of soleus and extensor digitorum longus (EDL) muscles fibers by the
technique of myosin ATPase activity. The effect of perinatal undernutrition was observed
both in body weight throughout life and in body weight gain at 90 days, which was reduced
by T+LPP (p<0.05). Relatively to the analysis of the muscle fiber proportions at 270 days, the
NT+LPP group presented an increase in the IIb fibers in the soleus, whereas there was no
effect in EDL fibers. However, in T+LPP these effects of maternal undernutrition were
normalized (p<0.03). Our results confirm the association between perinatal undernutrition and
changes in skeletal muscle phenotype in the adult offspring, while physical training during
pregnancy attenuates these consequences.
KEYWORDS: Physical activity, undernutrition, gestation, histology, skeletal muscle

89
INTRODUÇÃO
A origem desenvolvimentista da saúde e da doença (DOHaD) propõe que o risco de
desenvolver doenças crônicas ao longo da vida é diretamente proporcional ao grau de
incompatibilidade entre os ambientes pré e pós-natal (Gluckman e Hanson, 2004; Gluckman,
Hanson et al., 2005). Estudos epidemiológicos e modelos experimentais têm observado as
consequências devido ao contraste entre o período perinatal e a idade adulta, sobretudo, no
âmbito nutricional (Mcmillen e Robinson, 2005). De fato, a desnutrição durante os períodos
de alta plasticidade é capaz de gerar respostas adaptativas preditivas (RAP, em longo prazo),
revelando a capacidade fetal de prever e se adaptar a um ambiente nutricional inadequado ao
longo de sua vida (Gluckman, Hanson et al., 2005). Ratos desnutridos na gestação (30% da
dieta ofertada ao controle), mas normonutridos no período pós-natal apresentaram aumento no
índice de massa corporal, percentual de gordura e, concentrações plasmáticas de leptina,
insulina e glicose (Thompson, Norman et al., 2007). Tem sido especulado que o tecido
muscular esquelético esteja relacionado ao aparecimento dessas doenças crônicas não-
transmissíveis devido ao envolvimento tanto no movimento corporal quanto no metabolismo
de proteínas, lipídios e carboidratos (Brameld, 2004).
Estudos relatam que a desnutrição perinatal pode afetar os principais tipos de fibras
musculares (I, oxidativas; IIa, glicolíticas-oxidativas; IIb, predominantemente glicolíticas)
(Bayol, Jones et al., 2004; Mallinson, Sculley et al., 2007). Aos 90 dias de idade, ratos
desnutridos (7,8% caseína) durante a gestação apresentam maior percentual de fibras tipo IIa
no músculo soleus, concomitante a uma menor proporção das fibras tipo I e IIa e maior das
fibras IIb, no músculo extensor longo dos dedos (EDL) (Toscano, Manhaes-De-Castro et al.,
2008). Em humanos, o baixo peso ao nascer foi associado com um aumento de 66% de fibras
do tipo IIb e uma diminuição de 22% de fibras IIa no músculo vasto lateral de jovens aos 19
anos (Jensen, Storgaard et al., 2007). Ademais, é provável que a musculatura esquelética atue

90
na patogênese da resistência à insulina, uma vez que indivíduos obesos e diabéticos
apresentam maior quantidade de fibras glicolíticas e menor percentual de fibras oxidativas
(Tanner, Barakat et al., 2002). Assim, o fenótipo do tecido muscular parece estar envolvido
nas modificações promovidas pela RAP e, portanto, assume importância para a DOHaD.
A busca por um meio de intervenção capaz de atenuar os efeitos tardios da desnutrição
perinatal, tem revelado um modulador potencial das respostas adaptativas, a atividade física
durante a gestação. O exercício físico antes e durante a gestação otimiza a disponibilidade de
oxigênio e nutrientes, devido a uma melhor interação materno-fetal e melhor funcionalidade
placentária (Clapp, Kim et al., 2000; Clapp, Kim et al., 2002). Ademais, ratas treinadas antes
da gestação (cinco dias/semana, 60 min/dia, a 65% do VO2max) com diminuição progressiva
do esforço durante a gestação (cinco dias/semana, 30 min/dia, a 40% do VO2max)
apresentaram uma menor queda no consumo de oxigênio de repouso comparadas às não-
treinadas (Amorim, Dos Santos et al., 2009). A atividade física materna de baixa intensidade
(até 30% do consumo máximo de oxigênio, VO2max) e realizada sistematicamente está
associada ao aumento do peso ao nascer (Barakat, Lucia et al., 2009). Ratos provindos de
mães desnutridas (8% caseína) e treinadas (20 min/dia, 5 dias/semana, a ~30% do VO2max) na
gestação tiveram o peso ao nascer, indicadores de crescimento e o desenvolvimento sensório-
motor normalizados comparado ao seu controle (Fidalgo, Macêdo et al., 2010; Falcão-Tebas,
Bento-Santos et al., 2011). Estas respostas adaptativas imediatas (RAI, em curto prazo)
aumentam as chances de sobrevivência (Gluckman e Hanson, 2004) e sugerem que a
atividade física materna possa influenciar também a RAP. De fato, temos observado que ratos
cujas mães foram desnutridas e treinadas apresentaram um ganho de peso e perfil de gordura
abdominal normal, além da colesterolemia e glicemia similares aos seus controles (Amorim,
Dos Santos et al., 2009; Falcão-Tebas, Tobias et al., 2011).

91
Diante do exposto, o presente estudo teve como objetivo analisar o efeito de um
protocolo de treinamento físico durante a gestação sobre a proporção dos tipos de fibras dos
músculos soleus e EDL de ratos adultos submetidos à desnutrição perinatal. Nossas hipóteses
são (1) a desnutrição perinatal diminui a proporção de fibras oxidativas (tipo I) e aumenta a
proporção de fibras glicolíticas (tipo IIa e IIb); enquanto que (2) o treinamento físico durante
a gestação atenua os impactos da desnutrição na proporção de fibras oxidativas, afetando
minimamente a proporção de fibras glicolíticas na prole adulta.
MATERIAIS E MÉTODOS
Animais e manipulações experimentais
Este estudo foi aprovado pelo comitê de ética em estudos com animais do Centro de
Ciências Biológicas da Universidade Federal de Pernambuco (UFPE) (protocolo nº.
23076.049077/2010-80) e realizado no Departamento de Nutrição da UFPE. A manipulação e
os cuidados com os animais seguiram as recomendações do Comitê Brasileiro de
Experimentação Animal (COBEA).
As ratas-matrizes (n=12; peso corporal entre 180 ± 11 g) foram mantidas em biotério
de experimentação com temperatura controlada (23°C 1) e iluminação claro das 18:00 às
06:00 h, com água e alimentação padrão (52% carboidratos, 21% proteínas, 4% lipídeos –
Nuvilab CR1-Nuvital®, Curitiba, Paraná, Brasil) ad libitum. Metade das ratas (n=6) foram
submetidas a um protocolo de treinamento físico 4h após o início da fase escura do ciclo, de
intensidade moderada (5 dias/semana, 60 min/dia, a 65% do VO2max) durante 4 semanas
(Amorim, Dos Santos et al., 2009). Ao término deste período, as ratas foram acasaladas (na
proporção 2:1, fêmeas e macho) e, confirmada a prenhez, a intensidade e duração do
treinamento físico foram reduzidas (5 dias/semana, 20 min/dia, a 30% do VO2max) até o 19º

92
dia pré-natal (Amorim, Dos Santos et al., 2009). A manipulação nutricional teve início desde
a gestação até a lactação, onde foi ofertada uma dieta a base de caseína 17% para os grupos
controle (n=3) e treinada (n=3), ou caseína 8% para os grupos desnutrida (n=3) e treinada
desnutrida (n=3) (Reeves, Nielsen et al., 1993). As dietas confeccionadas foram isocalóricas
com alteração apenas no conteúdo de proteína (Reeves, Nielsen et al., 1993). As ninhadas
foram ajustadas para seis filhotes por lactente. Após o desmame (22 dias), foram utilizados
aleatoriamente três filhotes machos, de cada ninhada para compor os grupos experimentais.
Assim, as avaliações foram realizadas nos filhotes, os quais foram divididos em quatro
grupos, de acordo com a manipulação realizada em suas mães: controle (CF, n=9), treinado
(TF, n=9), desnutrido (DF, n=7) e treinado desnutrido (TDF, n=9). Os filhotes foram mantidos
do desmame (22 dias) até a vida adulta (270 dias) com dieta padrão.
Parâmetros avaliados
Indicadores murinométricos
A mensuração do peso corporal foi realizada diariamente a partir do décimo dia da
lactação e semanalmente até o 270º dia. Foi utilizada uma balança eletrônica digital, marca
Marte (modelo ASF11), capacidade máxima 500g e mínima 0,002g. O percentual de ganho de
peso corporal foi calculado tendo como base o peso do desmame, segundo a fórmula: %
ganho de peso=[Peso do dia (g) x 100 / Peso no 21º dia (g)] – 100 (Bayol, Jones et al., 2004).
O comprimento nasoanal (CNA) foi mensurado durante a lactação, com as medidas
registradas em papel milimetrado.
Análise histológica do músculo esquelético
Aos 270 dias, os animais foram eutanasiados por decapitação e tiveram os músculos
soleus e EDL cuidadosamente dissecados e pesados rapidamente. A seguir, foram imersos em

93
n-hexano (C6H14) a baixa temperatura, prevenindo a formação de cristais de água no
fragmento do músculo, além de fixar sem desnaturar as enzimas a serem estudadas.
Finalmente, foram estocados a -82ºC até a análise.
No criostato (Jung Frigocut 2800E-Leica) mantido a -20±1°C, os músculos foram
medidos quanto ao comprimento (distância entre os tendões de origem e inserção do músculo)
e a largura (no ponto médio do ventre do músculo) com um paquímetro digital Starrett®
(Series 799, São Paulo, Brasil). Logo após, foram obtidas secções transversas (10μm) fixados
a um suporte com a ajuda de TissueTek® O.C.T Compound (Sakura Finetek USA, Inc.,
Torrance, CA 9050 U.S.A.). Os cortes foram fixados em lâminas e ao atingirem a temperatura
ambiente foi iniciada a coloração. A fim de se obter uma coloração característica da atividade
ATPase da miosina foi utilizado o método proposto por Brooke e Kaiser (1970). Essa técnica
permite obter uma coloração característica da atividade enzimática específica através de
reações físico-químicas, realizadas em etapas sequenciais. As fibras musculares foram
classificadas de acordo com a intensidade de reação à ATPase miofibrilar cálcio-ativada
(mATPase) após a pré-incubação em meio ácido (pH 4,3 e 4,55), nos três tipos principais: I
(fibras lentas oxidativas), IIa (fibras rápidas oxidativas-glicolítica) e IIb (fibras rápidas
glicolíticas) (Figura 1) (Brooke e Kaiser, 1970). Após a secagem, as lâminas foram montadas
utilizando New Entellan® Merck. Os campos microscópicos foram obtidos através de
microscópio óptico OLYMPUS modelo U-CMAD-2 (objetiva 4 X) acoplado a um programa
para captação de imagens (TV TUNER APPLICATION – TelSignal Company Limited,
Taiwan). Finalmente, as fibras foram contadas utilizando o software MESURIN PRO (Jean-
François Madre-Amiens, França, versão 3.4 para Windows) e os valores referentes aos
diferentes tipos de fibras foram expressos em percentual do número total. A contagem das
fibras foi realizada em duplicata e de forma cega.

94
Análise estatística
Os valores estão expressos em média e erro padrão da média (EPM). Para comparação
entre grupos, em relação às medidas dos músculos e aos tipos de fibras musculares, foi
utilizado o teste de variância ANOVA Two-way seguido do teste de Bonferroni. Para o testes
ANOVA two-way foram utilizados como fatores a dieta e o treinamento físico. O nível de
significância foi mantido em 5% (p<0,05) em todos os casos. A construção do banco de dados
e as análises estatísticas foram desenvolvidas no programa Excel®
(Microsoft, USA, versão
2007) e GraphPad Prism® (GraphPad Software Inc., La Jolla, CA, USA; versão 5.0 para
Windows), respectivamente.
RESULTADOS
O treinamento físico antes e durante a gestação per si, não alterou o peso corporal no
10º e 21º dia de vida em relação ao controle (p>0,05). Os filhotes provindos de mães
alimentadas com dieta hipoprotéica, independente do treinamento físico materno, tiveram
menor peso corporal em relação aos seus respectivos controles, no 10º (CF = 22,63 ± 0,37; TF
= 22,36 ± 0,39; DF = 11,88 ± 0,18; TDF = 12,20 ± 0,44; p<0,001) e no 21º dia de vida (CF=
49,23 ± 0,65; TF = 46,68 ± 0,80; DF = 18,43 ± 0,59; TDF = 23,83 ± 0,76; p<0,001). Com
relação ao CNA, o grupo TF não apresentou diferença em relação ao CF tanto no 10º (CF =
81,65 ± 0,25; TF = 83,45 ± 0,66; DF = 66,77 ± 0,57; TDF = 70,96 ± 0,44; p<0,01) quanto no
21º dia pós-natal. Novamente, o efeito da desnutrição perinatal foi demonstrado ao 10º e 21º
dia de vida, diminuindo o CNA quando comparado aos seus controles (p<0,05). Entretanto,
aos 21 dias de idade, o grupo TDF apresentou maior CNA comparado ao grupo DF (CF =
124,10 ± 4,76; TF = 123,76 ± 6,44; DF = 88,07 ± 3,06; TDF = 93,79 ± 4.43; p<0,05).
O grupo TF não apresentou o peso ou ganho de peso corporal alterado em relação ao
controle, desde o desmame até o 270º dia de vida (p>0,05). Os grupos desnutridos (DF e TDF)

95
tiveram menor peso corporal quando comparados ao grupo controle desde o desmame até o
final do experimento (p<0,05) (Tabela 1). É de ressalva que, embora o ganho que peso
corporal do 30º ao 90º dia pós-natal em relação ao desmame tenha permanecido maior
comparado ao grupo CF, o grupo TDF obteve menores valores comparado ao grupo DF
(p<0,01) (Tabela 1). Os valores de peso corporal e de peso, comprimento e largura dos
músculos EDL e soleus aos 270 dias de idade estão expressos na tabela 1.
Na avaliação da proporção dos tipos de fibras do músculo EDL, não houve diferente
entre os grupos tanto no percentual de fibras do tipo I quanto do tipo II (p>0,05) (Figura 2A).
Em relação aos subtipos, os animais do grupo TF quando comparados ao grupo CF
apresentaram maior percentual de fibras do tipo IIa (TF = 77,32 ± 0,09; CF = 67,72 ± 0,21;
p<0,05) e menor percentual de fibras do tipo IIb (TF = 18,19 ± 0,09; CF = 25,78 ± 0,21;
p<0,05) (Figura 2B). No músculo EDL, o efeito apenas da desnutrição não foi observado em
nenhum pH, em relação ao grupo controle (p>0,05) (Figura 2A e 2B). Na comparação com o
grupo DF, os animais cujas mães foram treinadas e desnutridas apresentaram maiores valores
percentuais de fibras IIa (DF = 65,26 ± 1,45; TDF = 73,54 ± 1,74; p<0,05) e menores valores
percentuais do tipo IIb no músculo EDL (DF = 28,32 ± 1,45; TDF = 22,14 ± 1,74; p<0,05)
(Figura 2B). Ademais, não foi observada diferença entre os grupos TDF e CF (p>0,05).
Os valores médios do percentual de fibras oxidativas no músculo soleus do grupo TF
foi maior quando comparado ao grupo CF (CF = 72,13 ± 0,01; TF = 91,16 ± 0,02; p<0,01),
simultaneamente houve uma diminuição no percentual das fibras glicolíticas (p<0,01; Figura
2C). A desnutrição perinatal causou uma diminuição da proporção das fibras do tipo I e
concomitante aumento das fibras do tipo II no músculo soleus comparado ao controle
(p<0,05) (Figura 2C). Todavia, no grupo TDF houve maior predomínio das fibras oxidativas
(DF = 69,41 ± 0,05; TDF = 79,55 ± 0,29; p<0,01) e menor percentual das fibras glicolíticas
com relação ao grupo DF (Figura 2C). Em relação aos subtipos de fibras musculares do

96
músculo soleus, o grupo TF apresentou menor proporção de fibras tipo IIa (TF = 10,02 ± 0,09;
CF = 23,22 ± 0,23; p<0,01) mas não houve diferença nas fibras IIb quando comparados ao CF
(p>0,05) (Figura 2D). O grupo DF apresentou menores valores percentuais de fibras do tipo
IIa e do tipo IIb quando comparado ao grupo CF (p<0,05) (Figura 2D). Entretanto, o grupo
TDF apresentou menores valores percentuais tanto nas fibras oxidativas e glicolíticas (IIa) (DF
= 20,80 ± 0,01; TDF = 13,63 ± 1,21; p<0,03) quanto das fibras predominantemente glicolíticas
(IIb) quando comparado ao CF e DF (DF = 9,79 ± 0,01; TDF = 6,81 ± 1,21; p<0,05) (Figura
2D).
DISCUSSÃO
Nos períodos de alta plasticidade, caracterizados pela acentuada proliferação e
diferenciação celular, o organismo em desenvolvimento é capaz de produzir formas
alternativas de estrutura, estados fisiológicos ou comportamentais em resposta à desnutrição
(Barker, Eriksson et al., 2002; Barker, 2007). Nossos resultados corroboram com estudos
prévios, onde a desnutrição perinatal causou retardo no crescimento durante a lactação
(Toscano, Manhaes-De-Castro et al., 2008; Falcão-Tebas, Bento-Santos et al., 2011).
Segundo a hipótese da origem desenvolvimentista da saúde e da doença (DOHaD), esta
resposta pode ser esperada visto que a desnutrição atua como estímulo ambiental e é capaz de
gerar uma RAI (Gluckman, Hanson et al., 2005). Estas respostas, por exemplo,
homeostáticas, alteram o metabolismo, e aumentam as chances de sobrevivência em curto
prazo (Gluckman e Hanson, 2004). Uma resposta metabólica à desnutrição é o catabolismo,
que pode promover um crescimento lento de estruturas e órgãos, atingindo assim o peso
corporal (Harding e Johnston, 1995).
Por outro lado, um dos benefícios do treinamento físico antes e durante a gestação é o
aumento da massa magra da mãe e, portanto, a indução do catabolismo pela dieta hipoprotéica

97
pode ser atenuada devido ao aumento da disponibilidade de proteínas (Clapp, 2003). De fato,
os nossos dados demonstram que embora os animais provindos de mães treinadas e
desnutridas apresentem menor peso comparado ao controle ao final da lactação, estes valores
tendem a ser maiores que os animais cujas mães foram desnutridas.
A desnutrição precoce está relacionada a um déficit no crescimento embora tenha sido
relatada uma compensação até a idade adulta (Eriksson, Forsen et al., 2001; Barker, 2007).
Em nossos resultados observamos que o grupo DF apresentou menor peso corporal comparado
ao grupo CF desde o 10º até o 270º dia, mesmo quando a dieta padrão (normoprotéica) foi
oferecida. Entretanto, ao analisar o ganho de peso corporal (um indicador da velocidade de
crescimento, expresso em percentual a partir do desmame), o grupo DF apresentou os
primeiros indícios de uma RAP devido à dieta hipoprotéica perinatal, estabelecendo maiores
valores tanto aos 60 quanto aos 90 dias de vida. Nessa velocidade ou “catch up” de
crescimento, o organismo procura compensar ao longo da vida, o crescimento lento por déficit
de proteínas na dieta durante os períodos iniciais de seu desenvolvimento (Barker, Eriksson et
al., 2002). Portanto, no modelo da DOHaD, onde o risco de desenvolver síndrome metabólica
na idade adulta é diretamente proporcional à incompatibilidade entre os ambientes
nutricionais pré e pós-natal, o rápido catch up de crescimento tem sido considerado (Eriksson,
Forsen et al., 2001; Barker, Eriksson et al., 2002). Por outro lado, exercícios físicos de
intensidade leve (~40% VO2max) durante a gestação aumentam a taxa de crescimento feto-
placentário e o peso ao nascer (Clapp, Kim et al., 2000; Clapp, Kim et al., 2002; Clapp,
2003), indicando benefícios imediatos ao feto. No presente estudo embora os animais TDF
tenham apresentado maior catch up em relação ao controle, sugerindo que os efeitos gerados
pela desnutrição não foram totalmente normalizados, esses resultados foram menores que o
exibido pelo grupo DF. Os efeitos da desnutrição perinatal associados ao rápido crescimento

98
pós-natal parecem estar envolvidos com as características exibidas pelo individuo quando
adulto, inclusive no fenótipo muscular (Bayol, Jones et al., 2004).
Estudos mostram que uma dieta baixa em proteína durante a gestação e lactação atinge
o período de alta plasticidade do tecido muscular. Em roedores, a embriogênese de fibras
primárias e secundárias do músculo soleus ocorre no período gestacional e vai até o início da
vida pós-natal (Brameld, 2004). Nossos resultados demonstram que o peso muscular relativo
ao peso corporal dos músculos soleus e EDL dos grupos TF e DF não diferiram do grupo CF.
Esses resultados conflitam com achados prévios onde a desnutrição perinatal diminuiu o peso
do músculo soleus e EDL (Toscano, Manhaes-De-Castro et al., 2008). É provável que a data
de avaliação (25º e 90º dias) e o período da desnutrição (apenas na gestação) tenham
influenciado a diferença entre nossos resultados em animais desnutridos durante gestação e
lactação, aos 270 dias de idade. Entretanto, na comparação entre os animais TDF e os DF,
houve aumento do peso muscular, conferindo o efeito do treinamento físico materno. Isso
sugere que o treinamento físico durante a gestação pode atuar no peso relativo do músculo
frente ao insulto nutricional precoce.
Ainda com relação ao fenótipo muscular, o percentual de fibras na prole adulta, do
músculo EDL não sofreu influência da desnutrição perinatal. Em estudos prévios utilizando
desnutrição intrauterina (30% da dieta ofertada ao controle) não foi observado diferença na
proporção de fibras do músculo gastrocnêmio superficial, descrito como glicolítico,
semelhante ao EDL (Huber, Miles et al., 2009). Embora o músculo EDL tenha apresentado
indiferença nos subtipos de fibras musculares, tem sido relatado que a desnutrição pode afetar
o metabolismo de músculos predominantemente glicolíticos (Huber, Miles et al., 2009). As
fibras do tipo I exibem uma maior sensibilidade à insulina quando comparadas ao tipo IIa
(He, Watkins et al., 2001). O tipo IIb é menos dependente de sinalização de insulina
(Nyholm, Qu et al., 1997) e, portanto, um aumento destes tipos de fibras era esperado.

99
Embora não tenhamos observado este aumento, outros estudos referem que os efeitos da
desnutrição podem ser expressos tanto nas análises histológicas quanto no metabolismo
muscular (Huber, Miles et al., 2009). Ratos cujas mães foram desnutridas [modelos de
restrição calórica (30%) ou dieta hipoprotéica (8% caseína)] na gestação apresentaram menor
conteúdo de transportadores de glicose (GLUT4, na musculatura esquelética) e de proteínas
intermediárias da via de sinalização da insulina (PCK-zeta, p85 e p110-beta) (Ozanne, Jensen
et al., 2005; Huber, Miles et al., 2009). O treinamento físico materno per si ou associado à
desnutrição, foi capaz de aumentar o percentual de fibras IIa e IIb, comparado aos seus
respectivos controles. De acordo com as bases teóricas do DOHaD (Gluckman e Hanson,
2004; Gluckman, Hanson et al., 2005), é provável que o treinamento físico atue como um
estímulo precoce positivo podendo diminuir o risco induzido pelo contraste nutricional ao
longo da vida.
Alguns mecanismos podem ser sugeridos: (1) aumento do fluxo sanguíneo feto-
placentário; (2) aumento na passagem de nutrientes e oxigênio para o feto; e (3) aumento do
consumo de oxigênio no repouso (Clapp, 2003; Thomas, Clapp et al., 2008; Amorim, Dos
Santos et al., 2009). Desta forma, o milieu intrauterino foi alterado e uma resposta positiva na
mãe pode ter repercutido diretamente no desenvolvimento do feto. Como consequência, a
RAP do organismo em desenvolvimento, pode não seguir a mesma resposta dos animais cujas
mães foram apenas desnutridas e, portanto, o grau de incompatibilidade pré e pós-natal no
âmbito nutricional seria menor, diminuindo o risco de desenvolver doenças metabólicas ao
longo da vida.
CONSIDERAÇÕES FINAIS
Os efeitos negativos do contraste gerado pela RAP devido a uma desnutrição perinatal
e ao meio ambiente nutricional ao longo da vida estão bem estabelecidos na histologia do

100
músculo esquelético. Sugerimos que o treinamento físico durante a gravidez pode funcionar
como um estímulo capaz de gerar uma RAP de forma a não seguir a mesma resposta dos
animais desnutridos. Portanto, o grau de incompatibilidade em relação à nutrição pré e pós-
natal deve ser mais baixo, reduzindo os efeitos negativos sobre o fenótipo muscular.
AGRADECIMENTOS
Ao CNPq e a FACEPE pelo apoio financeiro. Os autores declaram que não há conflitos de
interesse.
REFERÊNCIAS
AMORIM, M. F. et al. Can physical exercise during gestation attenuate the effects of a maternal
perinatal low-protein diet on oxygen consumption in rats? Exp Physiol [S.I.], v. 94, n. 8, p. 906-13,
Aug 2009.
BARAKAT, R. et al. Resistance exercise training during pregnancy and newborn's birth size: a
randomised controlled trial. Int J Obes (Lond) [S.I.], v. 33, n. 9, p. 1048-57, Sep 2009.
BARKER, D. J. The origins of the developmental origins theory. J Intern Med [S.I.], v. 261, n. 5, p.
412-7, May 2007.
BARKER, D. J. et al. Fetal origins of adult disease: strength of effects and biological basis. Int J
Epidemiol [S.I.], v. 31, n. 6, p. 1235-9, Dec 2002.
BAYOL, S. et al. The influence of undernutrition during gestation on skeletal muscle cellularity and
on the expression of genes that control muscle growth. Br J Nutr [S.I.], v. 91, n. 3, p. 331-9, Mar 2004.
BIESWAL, F. et al. The importance of catch-up growth after early malnutrition for the programming
of obesity in male rat. Obesity (Silver Spring) [S.I.], v. 14, n. 8, p. 1330-43, Aug 2006.
BOL, V. V. et al. Postnatal catch-up growth after fetal protein restriction programs proliferation of rat
preadipocytes. Obesity (Silver Spring) [S.I.], v. 16, n. 12, p. 2760-3, Dec 2008.
BRAMELD, J. M. The influence of undernutrition on skeletal muscle development. Br J Nutr [S.I.], v. 91, n. 3, p. 327-8, Mar 2004.
BROOKE, M. H.; KAISER, K. K. Three "myosin adenosine triphosphatase" systems: the nature of their pH lability and sulfhydryl dependence. J Histochem Cytochem [S.I.], v. 18, n. 9, p. 670-2, Sep
1970.
CLAPP, J. F., 3RD. The effects of maternal exercise on fetal oxygenation and feto-placental growth.
Eur J Obstet Gynecol Reprod Biol [S.I.], v. 110 Suppl 1, p. S80-5, Sep 22 2003.
CLAPP, J. F., 3RD et al. Beginning regular exercise in early pregnancy: effect on fetoplacental
growth. Am J Obstet Gynecol [S.I.], v. 183, n. 6, p. 1484-8, Dec 2000.

101
CLAPP, J. F., 3RD et al. Continuing regular exercise during pregnancy: effect of exercise volume on
fetoplacental growth. Am J Obstet Gynecol [S.I.], v. 186, n. 1, p. 142-7, Jan 2002.
ERIKSSON, J. et al. Size at birth, childhood growth and obesity in adult life. Int J Obes Relat Metab
Disord [S.I.], v. 25, n. 5, p. 735-40, May 2001.
FALCÃO-TEBAS, F. et al. Maternal low protein diet-induced delayed reflex ontogeny is attenuated by moderate physical training during gestation in rats. British Journal of Nutrition [S.I.], in press
2011.
FALCÃO-TEBAS, F. et al. Efeitos do treinamento físico durante a gestação sobre a evolução
ponderal, glicemia e colesterolemia de ratos adultos submetidos à desnutrição perinatal. Rev Brasileira
de Medicina do Esporte [S.I.], v. in press, 2011.
FIDALGO, M. et al. Efeito do treinamento físico e da desnutrição durante a gestação sobre os eixos
cranianos de ratos neonatos. Revista Brasileira de Medicina do Esporte [S.I.], v. 16, p. 441-444, 2010.
GLUCKMAN, P. D.; HANSON, M. A. Living with the past: evolution, development, and patterns of
disease. Science [S.I.], v. 305, n. 5691, p. 1733-6, Sep 17 2004.
GLUCKMAN, P. D. et al. Predictive adaptive responses and human evolution. TRENDS in Ecology
and Evolution [S.I.], v. 20, p. 527 - 533, 2005.
HARDING, J. E.; JOHNSTON, B. M. Nutrition and fetal growth. Reprod Fertil Dev [S.I.], v. 7, n. 3,
p. 539-47, 1995.
HE, J. et al. Skeletal muscle lipid content and oxidative enzyme activity in relation to muscle fiber
type in type 2 diabetes and obesity. Diabetes [S.I.], v. 50, n. 4, p. 817-23, Apr 2001.
HUBER, K. et al. Prenatally induced changes in muscle structure and metabolic function facilitate
exercise-induced obesity prevention. Endocrinology [S.I.], v. 150, n. 9, p. 4135-44, Sep 2009.
JENSEN, C. B. et al. Altered skeletal muscle fiber composition and size precede whole-body insulin
resistance in young men with low birth weight. J Clin Endocrinol Metab [S.I.], v. 92, n. 4, p. 1530-4,
Apr 2007.
MALLINSON, J. E. et al. Fetal exposure to a maternal low-protein diet during mid-gestation results in
muscle-specific effects on fibre type composition in young rats. Br J Nutr [S.I.], v. 98, n. 2, p. 292-9, Aug 2007.
MCMILLEN, I. C.; ROBINSON, J. S. Developmental origins of the metabolic syndrome: prediction, plasticity, and programming. Physiol Rev [S.I.], v. 85, n. 2, p. 571-633, Apr 2005.
NYHOLM, B. et al. Evidence of an increased number of type IIb muscle fibers in insulin-resistant
first-degree relatives of patients with NIDDM. Diabetes [S.I.], v. 46, n. 11, p. 1822-8, Nov 1997.
OZANNE, S. E. et al. Low birthweight is associated with specific changes in muscle insulin-signalling
protein expression. Diabetologia [S.I.], v. 48, n. 3, p. 547-52, Mar 2005.
REEVES, P. G. et al. AIN-93 purified diets for laboratory rodents: final report of the American
Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J Nutr [S.I.], v. 123, n. 11, p. 1939-51, Nov 1993.
TANNER, C. J. et al. Muscle fiber type is associated with obesity and weight loss. Am J Physiol Endocrinol Metab [S.I.], v. 282, n. 6, p. E1191-6, Jun 2002.

102
THOMAS, D. M. et al. A foetal energy balance equation based on maternal exercise and diet. J R Soc
Interface [S.I.], v. 5, n. 21, p. 449-55, Apr 6 2008.
THOMPSON, N. M. et al. Prenatal and postnatal pathways to obesity: different underlying
mechanisms, different metabolic outcomes. Endocrinology [S.I.], v. 148, n. 5, p. 2345-54, May 2007.
TOSCANO, A. E. et al. Effect of a low-protein diet during pregnancy on skeletal muscle mechanical properties of offspring rats. Nutrition [S.I.], v. 24, n. 3, p. 270-8, Mar 2008.

103
Figura 1. Ilustração de cortes transversais dos músculos EDL (A, pH = 4,3 e B, pH = 4,55) e
soleus (C, pH = 4,3 e D, pH = 4,55). Os quadros estão 4 ou 10 x aumentados, utilizados para
análise dos tipos de fibras. Setas vermelhas indicam fibras do tipo I; setas amarelas, fibras do
tipo II; e sua subdivisão está identificada em setas verdes, fibras do tipo IIa; setas azuis, fibras
do tipo IIb.

104
Tabela 1. Valores médios ± EPM do peso corporal e do peso, comprimento e largura dos
músculos soleus e EDL, de filhotes adultos provindos de mães controle (CF), treinadas (TF),
desnutridas (DF) ou treinadas e desnutridas (TDF). N=9 em todos os grupos.
GRUPOS
CF
(média ± EPM)
TF
(média ± EPM)
DF
(média ± EPM)
TDF
(média ± EPM)
Peso corporal aos
270 dias (g) 459,22 9,88
440,11 12,82
354,36a 10,10
367,56a 10,19
% do ganho de
peso corporal até
90 dias
429,51 8,92
475,09 19,91
1167,58a 9,43
1038,26ab 6,83
Músculo EDL
Peso (mg) 261,40 25,60 272,80 26,20 249,20 20,70 284,00 15,10
Peso do
músculo*100/peso
corporal (mg/g)
57,50 4,40
62,10 7,00
68,60 2,20
81,10a 4,80
Comprimento
(mm) 33,90 0,54
35,12 0,57
31,19 1,19
32,56 1,05
Largura (mm) 3,80 0,21 3,58 0,09 4,03 0,33 3,96 0,43
Músculo soleus
Peso (mg) 302,40 23,2 290,20 10,8 265,30 23,4 306,40 33,5
Peso do
músculo*100/peso
corporal (mg/g)
63,90 3,40
65,70 3,00
74,40 7,00
87,90ab 10,02
Comprimento
(mm) 31,13 0,96
33,81 0,79
30,54 0,32
32,93 0,06
Largura (mm) 3,83 0,22 4,18 0,04 3,52 0,03 3,27 0,08
a p< 0,05 vs CF,
b p< 0,05 vs DF. ANOVA two-way seguido do teste de Bonferroni.

105
Type I Type II0
20
40
60
80
100 a
a
b
b
a
a
Soleus (pH = 4,3)
% f
ibers
Type I Type IIa Type IIb0
20
40
60
80
100
aab
a
a
a
aab
ab
Soleus (pH = 4,55)
% f
ibers
CP TP DP TDP
A B
Type I Type II0
20
40
60
80
100
EDL (pH = 4,3)%
fib
ers
Type I Type IIa Type IIb0
20
40
60
80
100
a
a b
b
EDL (pH = 4,55)
% f
ibers
C D
Figura 2. Valores percentuais a partir da contagem total de fibras nos cortes (média ± EPM).
Músculo soleus em pH 4,3 (A) e 4,55 (B) e extensor longo dos dedos (EDL) em pH 4,3 (C) e
4,55 (D). Controle (CF, n=7), treinados (TF, n=6), desnutridos (DF, n=7), e treinados
desnutridos (TDF, n=7). Teste ANOVA two-way seguido do teste de Bonferroni. a p<0.05 vs
CF. b p<0.05 vs grupo DF.

106
7. CONCLUSÕES
Uma das respostas adaptativas imediatas à desnutrição perinatal é o retardo no
desenvolvimento de características físicas e na maturação reflexa dos ratos. Ratificando os
efeitos em curto prazo, o crescimento pós-natal acelerado (catch up) está aumentado em ratos
provindos de mães desnutridas na gestação e lactação. Em relação à resposta adaptativa
preditiva, a desnutrição perinatal seguida da alimentação normal ao longo da vida induz
alterações no fenótipo do rato na idade adulta, aumentando: a circunferência abdominal, as
concentrações séricas de glicose e colesterol. Ademais, as consequências em longo prazo
devido à incompatibilidade gerada pela desnutrição perinatal e a alimentação normal ao longo
da vida diminuem a proporção de fibras oxidativas no músculo soleus.
O treinamento físico durante a gestação, por sua vez, parece ser capaz de induz tanto
respostas adaptativas imediatas quanto preditivas. Com esta intervenção precoce, foi possível
modular os efeitos da desnutrição perinatal na maturação de características físicas e na
maturação reflexa em ratos. Adicionalmente, o catch up de crescimento, embora ainda não
tenha sido normalizado, é atenuado devido ao treinamento físico leve no período pré-natal.
Relativamente às consequências em longo prazo, o treinamento físico na gestação reverte o
perfil característico à obesidade em ratos adultos provindo de mães desnutridas. Ademais, o
fenótipo da musculatura esquelética responde ao treinamento físico na gestação. Um
direcionamento ao perfil oxidativo das fibras foi observado mesmo perante a
incompatibilidade gerada pela desnutrição perinatal e a alimentação normal ao longo da vida.
Assim, o treinamento físico durante a gestação é capaz de atenuar as consequências devido as
respostas adaptativas imediatas e preditivas à desnutrição perinatal.

107
PERSPECTIVAS
Em ratos provindos de mães que foram submetidas ao treinamento físico antes e
durante a gestação e à desnutrição durante a gestação e lactação, avaliar:
a atividade de enzimas relacionadas ao metabolismo oxidativo e glicolítico em músculos de
contração rápida e lenta de ratos aos, 270 dias de idade;
a expressão de proteínas-chave envolvidas na cascata de sinalização intracelular da insulina
em músculos de contração rápida e lenta em ratos jovens e adultos;
a concentração sérica de insulina e leptina em ratos adultos.

108
REFERÊNCIAS
Amorim MF, dos Santos JA, Hirabara SM, Nascimento E, de Souza SL, de Castro RM, Curi
R, Leandro CG. 2009. Can physical exercise during gestation attenuate the effects of a
maternal perinatal low-protein diet on oxygen consumption in rats? Exp Physiol 94(8):906-
913.
Brooke MH, Kaiser KK. 1970. Three "myosin adenosine triphosphatase" systems: the nature
of their pH lability and sulfhydryl dependence. J Histochem Cytochem 18(9):670-672.
Fox WM. 1965. Reflex-ontogeny and behavioural development of the mouse. Anim Behav
13(2):234-241.
Lopes de Souza S, Orozco-Solis R, Grit I, Manhaes de Castro R, Bolanos-Jimenez F. 2008.
Perinatal protein restriction reduces the inhibitory action of serotonin on food intake. Eur J
Neurosci 27(6):1400-1408.
Marcondes FK, Bianchi FJ, Tanno AP. 2002. Determination of the estrous cycle phases of
rats: some helpful considerations. Braz J Biol 62(4A):609-614.
Novelli EL, Diniz YS, Galhardi CM, Ebaid GM, Rodrigues HG, Mani F, Fernandes AA,
Cicogna AC, Novelli Filho JL. 2007. Anthropometrical parameters and markers of obesity in
rats. Lab Anim 41(1):111-119.
Pestronk GJ, Kaiser KK, Brooke MH. 1992. ATPase stain in muscle histochemistry. Muscle
Nerve 15(2):258.
Reeves PG, Nielsen FH, Fahey GC, Jr. 1993. AIN-93 purified diets for laboratory rodents:
final report of the American Institute of Nutrition ad hoc writing committee on the
reformulation of the AIN-76A rodent diet. J Nutr 123(11):1939-1951.
Silva HJd, Barros KMFTd, Cunha DAd, Santos FdAdS, Soares EB, Silva CNdSd, Moraes
SRAd, Castro RMd. 2005. Malnutrition and Somatic Development: Application of Protocol
of Mensuration to Evaluation of Indicators of Wistar Rats. International Journal of
Morphology 24:469-474.
Toscano AE, Manhaes-de-Castro R, Canon F. 2008. Effect of a low-protein diet during
pregnancy on skeletal muscle mechanical properties of offspring rats. Nutrition 24(3):270-
278.

109
ANEXOS
Anexo A Parecer do Comitê de Ética em Pesquisa.
Anexo B Comprovante de aceite do manuscrito “Maternal low protein diet-induced
delayed reflex ontogeny is attenuated by moderate physical training during
gestation in rats” no periódico British Journal of Nutrition.
Anexo C Comprovante de aceite do manuscrito “Efeitos do treinamento físico durante a
gestação sobre a evolução ponderal, glicemia e colesterolemia de ratos adultos
submetidos à desnutrição perinatal” no periódico Revista Brasileira de
Medicina do Esporte.
Anexo D Comprovante de encaminhamento do manuscrito “Nutrição, atividade física e
a origem desenvolvimentista da saúde e da doença” no periódico Jornal de
Pediatria.

110
ANEXO A – Parecer do Comitê de Ética em Pesquisa

111
ANEXO B - Comprovante de aceite do manuscrito “Maternal low protein diet-induced
delayed reflex ontogeny is attenuated by moderate physical training during gestation in rats”
no periódico British Journal of Nutrition.
- -
From: [email protected]
Sent: Thursday, April 28, 2011 6:33:28 AM
Mr. Filippe Falcão-Tebas Federal University of Pernambuco Departament of Nutrition Av. Prof. Moraes Rego, 1235 - Cidade Universitária Recife, Pernambuco 50670-901 Brazil 27th Apr 2011 Dear Mr. Falcão-Tebas Paper No. BJN-2010-016235R: "Maternal low protein diet-induced delayed reflex ontogeny is attenuated by moderate physical training during gestation in rats". We have now been able to assess the revised version of your paper, and I am very pleased to tell you that it has been accepted for publication in the British Journal of Nutrition. We will now process the manuscript for publication. Emma Pearce, Production Editor at Cambridge University Press, will be contacting you regarding your proofs in due course ([email protected]). Please let her know if your contact details change or you have any specific issues regarding your accepted paper. Please download and return a completed 'Licence to Publish' form (signed by all authors) by post, fax or email to: Publications Office, British Journal of Nutrition, The Nutrition Society, 10 Cambridge Court, 210 Shepherds Bush Road, London W6 7NJ, UK [fax +44 (0)20 7602 1756, email: [email protected]]. Please ensure that your names are clearly printed next to the signatures. Licence to Publish Form: http://bjn.msubmit.net/cgi-bin/main.plex?el=A7P4FSx3B3Tkz4G6E9owIaCoc77XfsXKsK1Tx1gZ Please note that due to the high level of manuscripts processed by BJN we are unable to respond to individual requests for confirmation of receipt of licence to publish forms. You will only be notified if your form has not been received. We are sorry for any inconvenience this may cause. Please contact the Editor-in-Chief ([email protected]) if you are planning to issue a press release for this article. Thank you for submitting your interesting study to the British Journal of Nutrition. Yours sincerely I.S. Wood Deputy Editor

112
ANEXO C - Comprovante de aceite do manuscrito “Efeitos do treinamento físico durante a
gestação sobre a evolução ponderal, glicemia e colesterolemia de ratos adultos submetidos à
desnutrição perinatal” na Revista Brasileira de Medicina do Esporte.
[RBME] Decisão editorial -
From: Sistema SciELO de Publicação ([email protected])
Sent: Monday, June 13, 2011 4:44:30 PM
To: Senhor Filippe Falcão-Tebas ([email protected])
Prezado(a) Doutor(a) Senhor Filippe Falcão-Tebas,
Informamos a V.Sa. a conclusão da avaliação do manuscrito "Efeito do
treinamento físico moderado durante a gestação sobre parâmetros
somáticos de ratos adultos submetidos à desnutrição perinatal" foi
aprovado para publicação na Revista Brasileira de Medicina do Esporte.
Pedimos sua atenção aos comentários dos revisores do artigo de sua
autoria, submetido para apresentação no site Scielo. Necessitamos que as
adequações solicitadas pelos revisores sejam providenciadas e anexadas no
Sistema de Submissão Online pelo link versão do autor ou como documento
suplementar.
O artigo será encaminhado para publicação após verificarmos as
correções efetuadas.
Estamos enviando abaixo o parecer dos avaliadores para apreciação.
Avaliadores:
Apesar do estudo estar bem estruturado, os autores discutem timidamente os
resultados encontrados. É necessário uma discussão mais ampla sobre a
repercussão dos principais achados do estudo frente a investigações
anteriores, indicando claramente a aplicação prática e as informações
adicionais estabelecidas. Também é necessário que os autores discutam as
principais limitações do estudo e analisem o impacto dessas sobre os
resultados encontrados.
Colocamo-nos à disposição para suas observações.
Atenciosamente
Revista Brasileira de Medicina do Esporte RBME
Fernanda Colmatti/Arthur T. Assis
Atha Comunicação e Editora
Tel/Fax:55-11-5579-5308
Revista Brasileira de Medicina do Esporte
http://submission.scielo.br/index.php/rbme

113
ANEXO D - Comprovante de encaminhamento do manuscrito “Nutrição, atividade física e a
origem desenvolvimentista da saúde e da doença” ao conselho editorial do periódico Jornal de
Pediatria.
From: Filippe Falcão ([email protected])
Sent: Saturday, June 25, 2011 9:37:05 PM
1 attachment (29.5 KB)
Download as zip
Prezados Editores do Jornal de Pediatria, Segue em anexo uma carta-proposta para submissão de um artigo de revisão intitulado: Nutrição, atividade física e a origem desenvolvimentista da saúde e da doença. Filippe Falcão-Tebas Raul Manhães-de-Castro Carol Góis Leandro Os autores declaram a originalidade do artigo e garantem que nunca foi publicado e, caso venha a ser aceito pelo Jornal de Pediatria, não será publicado em outra revista. Ademais, atestamos que o presente artigo não foi enviado a outra revista e não o será enquanto sua publicação estiver sendo considerada pelo Jornal de Pediatria. Todos os autores participaram da concepção do trabalho, da análise e interpretação dos dados e de sua redação e/ou revisão crítica além de lerem e aprovarem a versão final. Por fim, reconhecemos que a Sociedade Brasileira de Pediatria passa a ter os direitos autorais, caso o artigo venha a ser publicado. Sem mais, aguardamos resposta em relação à proposta do artigo de revisão. Agradecemos antecipadamente a atenção, Filippe Falcão-Tebas Raul Manhães-de-Castro Carol Góis Leandro