respostas da fotossÍntese À variaÇÃo da temperatura...
TRANSCRIPT

DISSERTAÇÃO
RESPOSTAS DA FOTOSSÍNTESE À VARIAÇÃO DA TEMPERATURA DO AR E DAS RAÍZES EM
MUDAS DE LARANJEIRA ‘VALÊNCIA’
Carlos Marcos Alves dos Santos
Campinas, SP 2010

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA TROPICAL E SUBTROPICAL
RESPOSTAS DA FOTOSSÍNTESE À VARIAÇÃO DA TEMPERATURA DO AR E DAS RAÍZES EM MUDAS
DE LARANJEIRAS ‘VALÊNCIA’
CARLOS MARCOS ALVES DOS SANTOS
Orientador: Eduardo Caruso Machado
Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical, Área de Concentração em Tecnologia da Produção Agrícola
Campinas, SP Abril de 2010

S237r Santos, Carlos Marcos Alves dos Respostas da fotossíntese à variação da temperatura do ar e das raízes em mudas de laranjeiras ‘Valência’ / Carlos Marcos Alves dos Santos. Campinas, 2010. 51 fls. Orientador: Eduardo Caruso Machado Dissertação (Mestrado em Concentração em Tecnologia da Produção Agrícola) – Instituto Agronômico
1. Citros – fotoinibição 2. Citros - carboidratos, 3. Citros - trocas gasosas. I. Machado, Eduardo Caruso II. Título
CDD 634.3

ii

iii
A Deus,
A minha mãe Maria do Carmo Alves dos Santos
e ao meu pai Manoel Ramalho dos Santos
DEDICO
A todos os meus melhores amigos,
incluindo aqueles que não fazem
parte do cotidiano de trabalho, mas
que estão presentes no meu coração
OFEREÇO

iv
AGRADECIMENTOS
- Aos meus familiares, minha tia Maria Marta Alves e meu tio Julio Fernandes Ciriaco e
minhas primas Emília, Ivete e Geralda por estarem presentes nas minhas conquistas e
me proporcionarem apoio e tranqüilidade no decorrer desta etapa da minha vida;
- Ao Instituto Agronômico de Campinas e ao programa de pós-graduação, que
proporcionaram condições para a realização deste trabalho;
- Ao meu orientador, Dr Eduardo Caruso Machado pelo apoio, ensinamentos, paciência,
companheirismo e exemplo de profissionalismo, caráter e sabedoria;
- Aos pesquisadores componentes da Banca examinadora: Dra. Norma Magalhães
Erismann, Dr. João Domingos Rodrigues pela presença, apoio e sugestões para
melhoria do trabalho;
- Ao Conselho Nacional de Desenvolvimento Científico e tecnológico (CNPq) pela
bolsa de estudos;
- Aos amigos que auxiliaram na execução dos trabalhos, em especial José Rodrigues
(Zé), Severino Nogueira e Daniela Fávero São Pedro Machado pelo apoio nos
momentos em que mais precisei, foram amigos e grandes colaboradores no projeto;
- Aos amigos de pós-graduação: Thiago, Leandro, Verônica, Paulo, Ricardo, Cíntia,
Daniele, Karina, Cristina e Ana Carolina pelo auxílio, conversas, companheirismo e
amizade no decorrer do curso. Sucesso para todos vocês;
- Aos amigos que me incentivaram direta ou indiretamente na entrada do curso de pós-
graduação, em especial Gilberto Braz e André Luís Resende;
- Aos amigos do alojamento do IAC: Gustavo Becari, João Francisco da Capadócia,
Eduardo Cajuru, Anderson da Cruz (Fogo) e João Carlos pela amizade e convivência
durante o curso, que me servirão de aprendizado para o resto da minha vida;
- À Universidade Federal Rural do Rio de Janeiro pela minha formação;
- À todos os professores e funcionários da PG – IAC, em especial Adilza, Célia e Beth
pela amizade, conselhos e atenção durante o curso;

v
SUMÁRIO LISTA DE TABELAS...................................................................................................... vi LISTA DE FIGURAS....................................................................................................... vii LISTA DE ABREVIATURAS E SÍMBOLOS................................................................ ix RESUMO........................................................................................................................... xi ABSTRACT....................................................................................................................... xiii 1 INTRODUÇÃO.............................................................................................................. 1 2 REVISÃO DA LITERATURA...................................................................................... 4 3 MATERIAL E MÉTODOS............................................................................................ 11 3.1 Material Vegetal.......................................................................................................... 11 3.2 Procedimento Experimental........................................................................................ 11 3.3 Trocas gasosas e atividade fotoquímica...................................................................... 13 3.4 Conteúdo de carboidratos nas folhas, caules e raízes................................................. 13 3.5 Delineamento estatístico............................................................................................. 16 4 RESULTADOS E DISCUSSÃO................................................................................... 16 4.1 Biometria..................................................................................................................... 16 4.2 Trocas gasosas............................................................................................................. 16 4.3 Assimilação diurna de CO2......................................................................................... 26 4.4 Atividade fotoquímica................................................................................................. 28 4.5 Variação do conteúdo de carboidratos........................................................................ 34 4.6 Relações entre a fotossíntese e o teor de carboidrato na folha.................................... 37 5 CONCLUSÕES.............................................................................................................. 41 6 REFERÊNCIAS BIBLIOGRÁFICAS........................................................................... 42

vi
LISTA DE TABELAS
Tabela 1 Tratamentos aplicados com quatro repetições em laranjeira ‘Valência’ sobre limoeiro ‘Cravo’ dentro de câmara de crescimento. Foi feita aclimatação das plantas por um dia nas seguintes condições: fotoperíodo de 12h, DFFFA = 900 mol m-2 s-
1 e umidade do ar de 60/65% (dia/noite), temperatura noturna = 20 oC, temperatura diurna = 28 oC e temperatura das raízes = 26 oC. Os tratamentos foram aplicados por um período de seis dias consecutivos, variando apenas as temperaturas, conforme descrito na tabela..............................................................................................
12
Tabela 2 Área foliar específica (AFE) em laranjeira ‘Valência’ sobre limoeiro ‘Cravo’, após 6 dias nos seguintes tratamentos térmicos: T1 = 28, 20, 26 oC; T2R = 28, 20, 10 oC; T3N = 28, 10, 26 oC; T4NR = 28, 10, 10 oC ; T5DN = 17, 10, 26 oC; T6DNR = 17, 10, 10 oC, na parte aérea no período diurno e noturno e na parte radicular, respectivamente...................................................................................
16

vii
LISTA DE FIGURAS
Figura 1 Fotografia ilustrando a disposição das plantas dentro da câmara de crescimento durante o período experimental.......................................
14
Figura 2 Características biométricas (em A) de laranjeira ‘Valência’ sobre
limoeiro ‘Cravo’, utilizadas no experimento e e distribuição relativa de fitomassa (R/T, C/T e F/T em B). R, C, F e T representam respectivamente fitomassa seca das raízes, caules, folhas e total. As barras indicam desvio padrão da média referente a 4 repetições e em B indicam desvio padrão da média em 24 repetições...........................
16
Figura 3 Variação diurna da assimilação de CO2 (A em A), da condutância estomática (gs em B), da concentração interna de CO2 (Ci em C) e da eficiência aparente de caboxilação (A/Ci em D) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas a seis regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TR = 28, TN = 10, TR = 26 oC; T4NR, TR = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições.....................
18
Figura 4 Relação entre assimilação de CO2 (A) com a condutância estomática (gs em A) e com a eficiência aparente de carboxilação (A/Ci em B) e relação entre a eficiência aparente de carboxilação com gs (em C) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições...............................................................................................
25
Figura 5 Assimilação diurna de CO2 (Ai) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições.....................
27

viii
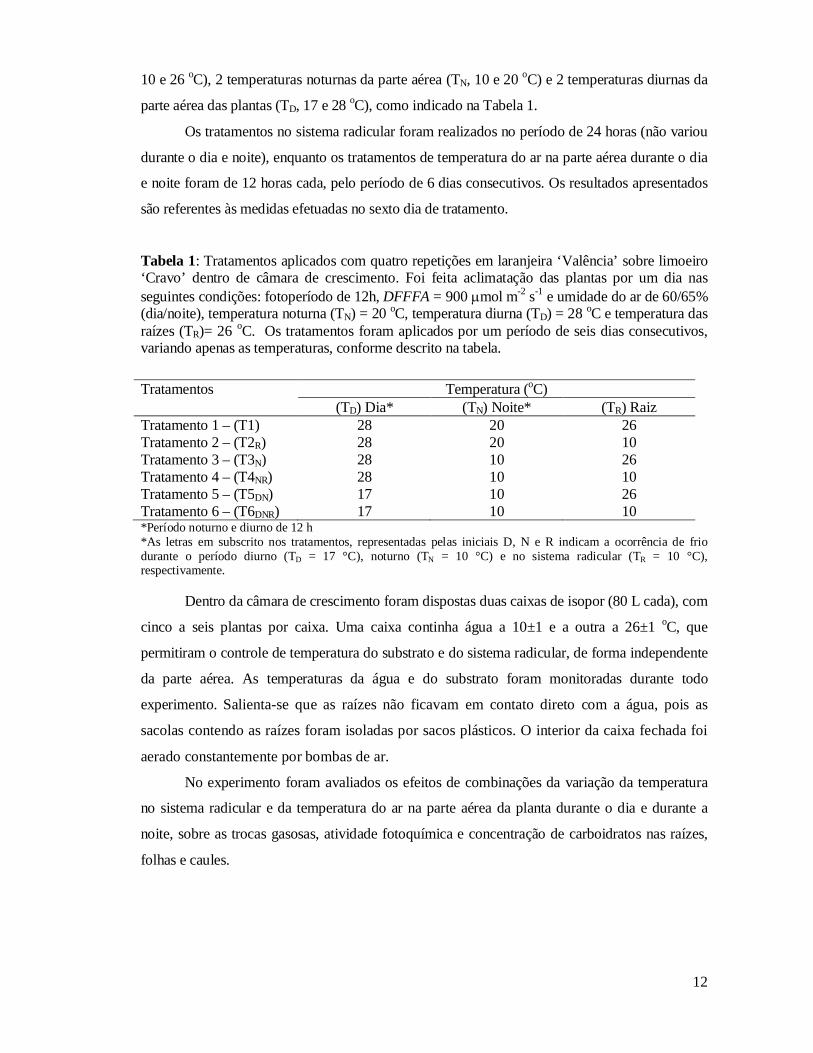
Figura 6 Fluorescência basal (Fo em A, fluorescência máxima (Fm em B) e
eficiência quântica máxima do FSII (Fv/Fm em C) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições....................
30
Figura 7 Conteúdo de açúcares solúveis, de sacarose, de amido e carboidratos
totais em folhas, caules e raízes em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições....................
34
Figura 8 Assimilação máxima (Amax em A) e diurna de CO2 (Ai em B) em função do conteúdo de açúcares solúveis totais e de amido em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC.TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições..............................................................................................
37

ix
LISTA DE ABREVIATURAS E SÍMBOLOS
A Assimilação de CO2 (µmol m-2 s-1) ABA Ácido abscísico
Ai Assimilação diária de CO2 (mmol m-2 d-1) Amax Assimilação máxima de CO2 no decorrer do dia
Amin Assimilação mínima de CO2 no decorrer do dia A/Ci Eficiência aparente de carboxilação (mol m-2 s-1 Pa-1) AFE Área foliar específica (m2 kg-1)
AM Amido [g Glu (kg MS)-1]
AS Açúcares solúveis [g Glu (kg MS)-1] CT Carboidratos totais [g Glu (kg MS)-1]
ATP Adenosina trifosfato Ci Concentração intercelular de CO2 (µmol mol-1)
Cc Concentração de CO2 no cloroplasto DPV Déficit de pressão de vapor do ar (kPa)
DPVar-folha Diferença de pressão de vapor entre o ar e a folha DFFFA Densidade de fluxo de fótons fotossinteticamente ativos (µmol m-2 s-1)
E Transpiração (mmol m-2 s-1) EF Exportação de fotoassimilados (g CH2O m-2)
ETR Transporte aparente de elétrons (µmol m-2 s-1) FSI Fotossistema I
FSII Fotossistema II Fm Fluorescência máxima em tecidos foliares adaptados ao escuro
Fo Fluorescência basal em tecidos foliares adaptados ao escuro Fv Fluorescência variável no escuro
Fv/Fm Eficiência quântica potencial do fotossistema II gs Condutância estomática (mol m-2 s-1)
Glu Glicose
gm Condutância do mesófilo (mol m-2s-1)
Jmax Taxa máxima de regeneração da RuBP ( mol m-2 s-1)
KL Condutividade hidráulica (mmol H2O m-2 s-1
MPa-1)
MS Massa seca NADPH Nicotinamida adenina dinucleotídeo de piridina fosfato reduzida
QA Quinona aceptora de elétrons

x
Rubisco Ribulose-1,5-bifosfato carboxilase/oxigenase
RuBP Ribulose-1,5-bifosfato Tar Temperatura do ar
TD Temperatura diurna TN Temperatura noturna
TR Temperatura da raiz T1 Controle
T2R Frio somente no sistema radicular
T3N Frio somente durante o período noturno
T4NR Frio durante o período noturno e no sistema radicular T5DN Frio durante o período diurno e noturno
T6DNR Frio durante o período diurno, noturno e no sistema radicular Vc,max Taxa máxima de carboxilação da RuBP ( mol m-2s-1)
Ψ Potencial da água na folha

xi
Respostas da fotossíntese à variação da temperatura do ar e das raízes em mudas de laranjeira ‘Valência’
RESUMO A fotossíntese é um processo regulado por fatores fisiológicos e ambientais, sendo a
temperatura um elemento importante na interação planta-ambiente. O objetivo deste estudo
foi avaliar os efeitos da variação da temperatura noturna, diurna e das raízes sobre a
fotossíntese e sua interação com a dinâmica de carboidratos em mudas de laranjeira
‘Valência’ enxertada em limoeiro ‘Cravo’. A nossa hipótese de trabalho é que a variação
sazonal da fotossíntese está relacionada com a variação da temperatura do ar e das raízes e
com a demanda de fotoassimilados para o crescimento. A pesquisa foi realizada em câmara de
crescimento com mudas de sete meses de idade. O delineamento experimental foi em parcelas
subdivididas com quatro repetições. As variáveis medidas foram submetidas à análise de
variância e as médias comparadas pelo teste de Tukey com 0,05 de probabilidade. As plantas
foram submetidas a seis regimes térmicos, por seis dias consecutivos. Os tratamentos foram: T1
onde TD=28, TN=20, TR=26 oC; T2R onde: TD=28, TN=20, TR=10 oC; T3N onde: TD=28, TN=10,
TR=26 oC; T4NR onde: TD=28, TN=10, TR=10 oC; T5DN onde: TD=17, TN=10, TR=26 oC; e T6DNR
onde: TD=17, TN=10, TR=10 oC. Os símbolos TD, TN e TR, representam respectivamente,
temperatura diurna, noturna e das raízes. Foram avaliados a assimilação de CO2 (A), a
condutância estomática (gs), a eficiência aparente de carboxilação (A/Ci, sendo Ci concentração
intercelular de CO2), a eficiência quântica potencial do fotossistema II (Fv/Fm) e o teores de
carboidratos nas folhas, raízes e caules. O tratamento referência (T1) apresentou o melhor
desempenho da fotossíntese, visto que as temperaturas adotadas são as ideais para fotossíntese de
laranjeiras. Os maiores valores de A, gs, A/Ci e Ai foram verificados nas plantas com temperatura
das raízes mais elevadas (TR de 26 °C em T1, T3N e T5DN), independentemente da temperatura
da parte aérea. Nos tratamentos sob baixa temperatura nas raízes (T2R, T4NR e T6DNR, TR de 10
°C), mesmo aqueles com a parte aérea sob condições ideais (28/20 °C diurna/noturna em T2R), a
produção fotossintética foi aproximadamente a metade em relação à média dos tratamentos com
temperatura das raízes a 26 °C e frio noturno ou diurno na parte aérea. O decréscimo da
fotossíntese em todos os tratamentos, em relação ao T1 foi devido à redução tanto de gs como
em A/Ci, ou seja, ocorreram limitações de origem estomática e bioquímica. Com exceção de
T6DNR, nos regimes térmicos em que as plantas foram expostas a baixas temperaturas, ocorreu
maior acúmulo de amido nos caules e raízes, apesar de apresentarem menor Ai em relação a

xii
T1, indicando que o balanço entre produção e utilização de fotoassimilados determina o
acúmulo de reservas. Não se verificou relação entre A e o teor de carboidratos foliares. As
baixas temperaturas do ar durante o dia e a noite causaram queda em Fv/Fm. A fotoinibição nos
tratamentos de frio noturno foram discretas, porém a fotoinibição foi acentuada e crônica nas
plantas submetidas ao frio diurno.
Palavras-Chave: Citrus sinensis, fotoinibição, carboidratos, relações fonte:dreno, trocas
gasosas.

xiii
Photosynthesis responses to air and root temperature in young ‘Valencia’ orange plants
ABSTRACT
The photosynthesis is a process regulated by physiological and environmental factors, being
the temperature an important element in plant-environment interaction. The aim of this study
was to evaluate the effects of night, diurnal and root temperature variations on photosynthesis
and its interaction with the carbohydrates dynamics in ‘Valência’ orange plants grafted on
‘Rangpur lime’. Our hypothesis is that the seasonal variation of photosynthesis is related to
the air and the root temperature variation and to the photoassimilates demand for growth. The
research was conducted in a growth chamber with seven-month old plants. The experiment
was arranged in a split-plot design, with four replication. The measured variables were
submitted to variance analysis and the averages were compared by the Tukey test at 0,05 of
probability. The plants were submitted to six thermal regimes, for six consecutive days. The
treatments were: T1 where TD=28, TN=20, TR=26 oC; T2R where: TD=28, TN=20, TR=10 oC;
T3N where: TD=28, TN=10, TR=26 oC; T4NR where: TD=28, TN=10, TR=10 oC; T5DN where:
TD=17, TN=10, TR=26 oC; and T6DNR where: TD=17, TN=10, TR=10 oC. The symbols TD, TN
and TR, represent, temperature of the day, the night and the roots, respectively. The CO2
assimilation (A), stomatal conductance (gs), the apparent carboxylation efficiency (A/Ci,
being Ci intercellular CO2 concentration), the potential quantum yield of photosystem II
(Fv/Fm) and the carbohydrates content in leaves, roots and stems were evaluated. The
reference treatment (T1) presented the best photosynthetic performance, because the adopted
temperatures are ideal for orange trees photosynthesis. The highest values of A, gs, A/Ci and
Ai were verified in plants under the highest root temperature (TR of 26 °C in T1, T3N and
T5DN), independenty of the aerial part temperature. Under the lowest root temperature
treatments (T2R, T4NR and T6DNR, TR of 10 °C), even those with the aerial part under ideal
conditions (28/20 °C diurnal/nocturnal, T2R), the photosynthetic production was
approximately one half of the average of the treatments under root temperature of 26 °C and
the aerial part under low nocturnal or diurnal temperatures. The photosynthesis decrease in all
treatments, in relation to T1, was due to a reduction in gs as well as in A/Ci, in other words,
stomatal and biochemical limitations. In exception to T6DNR, those plants exposed in a low
temperature conditions presented higher starch content in stalks and roots, even though a
value reduced in Ai in relation to T1 treatment. These response suggest that the balance
between produce and consume to photoassimilates can to determine pool storage. The

xiv
relationship between A and leaf carbohydrate content was not verified. The low air
temperatures during the day and the night caused a decrease in Fv/Fm. The photoinhibition in
treatments under low night temperature was discreet, however photoinhibition was
accentuated and chronicle in plants submitted to diurnal cold.
Key words: Citrus sinensis, photoinhibition, carbohydrates, source-sink relationships, gas
exchanges.

1
1 INTRODUÇÃO
A citricultura brasileira é responsável por aproximadamente 37 e 51% da produção
mundial de frutas frescas e suco concentrado de laranja, respectivamente (FNP
CONSULTORIA & COMÉRCIO, 2006). O Estado de São Paulo é o principal produtor
nacional, sendo responsável por aproximadamente 85% da produção. Entretanto a
produtividade média dos pomares paulistas é baixa, em torno de 2 caixas/planta/ano, quando
comparada à média da Flórida nos Estados Unidos, ao redor de 6 caixas/planta/ano
(MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO, 2009), o que
prejudica sua competitividade no mercado mundial. No entanto, é importante observar que o
Brasil tem alto potencial para aumentar a produtividade e assim continuar competindo pelo
comércio externo de suco concentrado.
A produtividade de laranjais depende da interação dos fatores ambientais com a
fotossíntese, o crescimento das copas, a indução de floração, a fixação e crescimento dos
frutos (GOLDSCHMIDT & KOCH, 1996). O conhecimento básico das interações entre tais
fatores possibilita o manejo da cultura de forma a obter-se maior produtividade dos pomares.
A fotossíntese é o processo responsável pelo acúmulo de todo material orgânico nas plantas e
sendo assim, qualquer fator interferente sobre ela afetará a produtividade.
As laranjeiras são plantas sempre-verdes, sendo ativas fotossinteticamente durante todo
o ano, apresentando baixa atividade fotossintética, atingindo em São Paulo valores máximos
entre 8 e 13 mol m-2 s-1 na primavera e verão, respectivamente, e valores duas a três vezes
mais baixos no inverno (HABERMANN & RODRIGUES, 2009; RIBEIRO, 2006; RIBEIRO
& MACHADO, 2007; SYVERTSEN & LLOYD, 1994). Portanto, a fotossíntese é submetida
a intensa variação das condições ambientais em função das estações do ano, o que afeta sua
eficiência (HABERMANN & RODRIGUES, 2009; MACHADO et al., 2002; RIBEIRO,
2006; RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2009 a,b). De fato, a fotossíntese em
laranjeiras é afetada pela disponibilidade hídrica (ERISMANN et al., 2008; MAGALHÃES
FILHO et al., 2008; MEDINA et al., 1999), pela variação da temperatura do ar
(HABERMANN & RODRIGUES, 2009; MACHADO, 2009; MACHADO et al., 2005;
MAGALHÃES FILHO et al., 2009; MEDINA et al., 2003; RAMOS, 2009; RIBEIRO &
MACHADO, 2007; RIBEIRO et al., 2009a, b), pelo déficit de pressão de vapor do ar
(HABERMANN et al., 2003; KHAIRI & HALL, 1976a; RIBEIRO et al., 2009a, b); pela

2
intensidade de radiação solar (MACHADO et al., 2005; MEDINA et al., 2002; RIBEIRO,
2006) e pela demanda sazonal de fotoassimilados e relação fonte-dreno (IGLESIAS et al.,
2002; MACHADO et al., 2002; RAMOS, 2009; RIBEIRO, 2006; RIBEIRO & MACHADO,
2007).
Praticamente em todas as regiões do Estado de São Paulo encontram-se pomares de
citros. No Estado de São Paulo, o tipo climático predominante é o tropical de altitude ou
mesotérmico úmido existindo um período úmido e quente que corresponde ao verão
(novembro a março) e outra estação ou período seco e frio (inverno) que normalmente
abrange os meses de junho a agosto (CAMARGO et al., 1974). Entre os fatores ambientais
que regulam o desenvolvimento da laranjeira a temperatura, fator climático incontrolável, tem
papel decisivo sobre a fotossíntese, afetando o crescimento vegetativo (HALLGREE et al.,
1991; KHAIRI & HALL, 1976a, b; PRADO et al., 2007; RAMOS, 2009) e a floração e
produção dos frutos (HABERMANN & RODRIGUES, 2009; PRADO et al., 2007).
A região norte do Estado, caracteriza-se por apresentar temperaturas mais elevadas em
relação a região sul (RIBEIRO et al., 2006; ROLIM et al., 2007). Independentemente da
região do Estado, no verão e na primavera as temperaturas do ar e do solo são mais elevadas
e, em geral, apresentam temperaturas mais elevadas no período diurno. No inverno observa-se
a predominância de noites frias seguidas de dias claros que podem ou não ser quente. A
assimilação de CO2 (A) e o crescimento durante o inverno são significativamente menores que
na primavera e o verão (HABERMANN & RODRIGUES, 2009; MACHADO et al., 2002;
PRADO et al., 2007; RIBEIRO et al., 2009a, b). A queda na assimilação de CO2 no inverno
ocorre mesmo em dias com temperatura diurna do ar variando entre 20 e 26 °C, ou seja,
próxima à faixa considerada ideal para citros que está entre 25 e 30 oC (GUO et al., 2006;
MACHADO et al., 2005; RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2009a, b). Assim,
a redução de A no inverno em relação ao verão, não pode ser atribuída à temperatura diurna,
mas a baixa temperatura noturna do ar e do solo (MACHADO, 2009; MACHADO et al.,
2009; MAGALHÃES FILHO et al., 2009; RIBEIRO et al., 2009a, b). Durante os meses de
inverno se observa com freqüência, além de noites frias, também dias com temperaturas
baixas. Nesse caso, é possível que a queda de A seja ainda maior (DAY et al., 1991;
DELUCIA et al., 1991; MEDINA, 2002).
Além do abaixamento da temperatura do ar no inverno, também ocorre queda na
temperatura do solo, e conseqüentemente, das raízes (PEREIRA et al., 2002; RIBEIRO,
2006). A redução da fotossíntese durante o inverno pode ser induzida por noites frias e pelo
abaixamento da temperatura do sistema radicular e, além de ocorrer em laranjeiras

3
(HABERMANN & RODRIGUES, 2009) também ocorre em outras espécies sempre verdes,
como em cafeeiro (BAUER et al., 1985) e mangueira (ALLEN & ORT, 2001).
Também há a possibilidade de que a redução de A no inverno esteja relacionada ao
aparente repouso vegetativo devido a diminuição da temperatura e ainda com a diminuição do
comprimento do dia (MACHADO et al., 2002; RIBEIRO, 2006; SYVERSTSEN & LLOID,
1994). Nessa época, a baixa temperatura do ar (<13oC) causa redução do metabolismo da
parte aérea, ocasionando uma aparente paralisação do crescimento (KHAIRI & HALL, 1976a;
REUTHER, 1973, RIBEIRO, 2006). SPIEGEL-ROY & GOLDSCHMIDT (1996) relataram
que o crescimento radicular pode ocorrer de forma ininterrupta, variando, entretanto, a
velocidade de crescimento em função da disponibilidade de água, aeração e temperatura, que
deve ser superior a 13 oC e inferior a 36 oC (BEVINGTON & CASTLE, 1985).
A queda na intensidade de crescimento diminui a demanda por fotoassimilados.
Porém, devido a contínua produção fotossintética em plantas sempre verdes, mesmo que em
menor intensidade, pode haver acúmulo de carboidratos na folha (GOLDSCHMIDT &
KOCH, 1996). De fato, observam-se teores altos de amido no fim do inverno e começo da
primavera, com posterior decréscimo em virtude da sua remobilização durante o fluxo
vegetativo e reprodutivo primaveril (JONES & STEINACKER, 1951; PRADO et al., 2007;
RIBEIRO, 2006). O acúmulo de carboidratos nas folhas pode causar decréscimo na
fotossíntese (AZCON-BIETO, 1983; GOLDSHMIDT & HUBER, 1992; IGLESIAS et al.,
2002). No entanto, também há relatos segundo os quais a fotossíntese pode ser regulada pela
dinâmica de carboidratos, em função da demanda, e não somente pelo conteúdo de
carboidratos (RAMOS, 2009; RIBEIRO et al., 2005; RIBEIRO & MACHADO, 2007;
RIBEIRO et al., 20101).
O conhecimento de como o ambiente modifica a fisiologia da planta é fundamental na
atual conjuntura da agricultura nacional, uma vez que se busca o aumento da eficiência na
produção sem incrementos no uso de insumos e energia. Nesta pesquisa tem-se como hipótese
que a variação sazonal da fotossíntese (trocas gasosas e reações fotoquímicas) está
relacionada com a variação da temperatura do ar (dia e noite) e das raízes e com a demanda de
fotoassimilados para o crescimento. Para testar essas hipóteses os objetivos desta pesquisa
foram analisar os efeitos da variação das temperaturas noturna, diurna e das raízes em mudas
de laranjeira ‘Valência’: 1) sobre os processos difusivos, bioquímicos e fotoquímicos da
fotossíntese e 2) sobre a dinâmica de carboidratos na planta.
1 RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G.; OLIVEIRA, R.F. Evidences towards the regulation of photosynthesis by carbohydrates dynamics in citrus leaves (submitted), 2010.

4
2 REVISÃO DE LITERATURA
A produção agrícola é, em essência, o manejo da fotossíntese, portanto é desejável ter-
se o conhecimento das respostas desse processo às variações das condições do ambiente. Tais
conhecimentos possibilitam-nos manejar alguns fatores ambientais (água, nutrição, solo) e
estruturais da planta (espaçamento, arquitetura, arranjo espacial das plantas) visando
condições mais propícias para que os mecanismos fisiológicos ocorram com máxima
eficiência. A produtividade pode ser avaliada pela relação entre produção e utilização de
carboidratos, sendo que a fotossíntese e o teor de reservas podem afetar a fertilização, a
fixação e o crescimento dos frutos. A produtividade de um pomar resulta de uma complexa
cadeia de eventos, envolvendo os efeitos do clima sobre a produção fotossintética, o
crescimento da copa, a indução e a intensidade de florescimento, a fixação dos frutos e a
massa final de frutos maduros colhidos, além da eficiência do uso da água e de nutrientes
(GOLDSCHMIDT, 1999).
Os elementos orgânicos são acumulados a partir da fotossíntese, que representa a fonte
primária de carboidratos para todos os processos de crescimento e qualquer fator biótico ou
abiótico que a afetar afetará também a produção. Obviamente a fotossíntese ocorre somente
durante a presença de luz, mas os processos de crescimento e de manutenção da planta
ocorrem tanto à luz como em sua ausência. Os fotoassimilados são disponíveis à noite porque
a assimilação de CO2 (A) durante o dia, em condições adequadas, é suficiente para atender a
demanda imediata e ainda acumular reservas, principalmente como amido na folha, as quais
podem ser remobilizadas à noite (RIBEIRO et al., 2010; SMITH & STITT, 2007). Em
herbáceas quase todo amido pode ser consumido durante à noite. No entanto, em citros essa
variação diurna não é tão acentuada, ou seja parte significativa do amido acumulado
permanece na folha (GOMES et al., 2003; RIBEIRO et al., 2010). Em qualquer fase do
desenvolvimento, quando a demanda por carboidratos for menor que a produção de
fotoassimilados, o excesso é armazenado em folhas, ramos e principalmente raízes, e quando
a demanda for maior que a produção de fotoassimilados pelas folhas, as reservas podem ser
remobilizadas para órgãos em crescimento (GOLDSCHMIDT, 1999, GOLDSCHIMIDT &
COLOMB, 1982; MAGALHÃES FILHO et al., 2008). Há relação entre a produção
fotossintética, o acúmulo de reservas e a remobilização das reservas durante o decorrer de um
ano (MEDINA, 2002; RIBEIRO, 2006; GOLDSCHIMIDT & COLOMB, 1982; PRADO et

5
al., 2007), ficando evidente que a queda da fotossíntese em qualquer fase do desenvolvimento
implica em prejuízo dos processos relacionados ao crescimento e acúmulo de matéria seca e
de reservas na planta.
O clima de São Paulo caracteriza-se por verão úmido, com níveis de energia solar típico
do trópico e um período de inverno seco com temperaturas amenas e precipitação
pluviométrica reduzida. Na transição do verão para o inverno, a temperatura do ar,
comprimento do dia e intensidade de fluxo de energia radiante decaem, tendo como
consequência a queda da temperatura do solo. O inverno caracteriza-se por noites frias e dias
mais curtos em relação ao verão. Após noites frias podem ocorrer dias relativamente mais
quentes ou mais frios. De julho a agosto existem condições térmicas e hídricas que
condicionam o repouso vegetativo e em setembro condições favoráveis para o florescimento
em laranjeiras. Na primavera, após a antese, nas fases iniciais de crescimento do fruto e da
área foliar, a intensidade de radiação solar é mais elevada, os dias são mais longos, as
temperaturas mais altas e, geralmente, há boa disponibilidade de água. O crescimento dos
frutos depende do suprimento de substrato fotossintetizado sendo, portanto, favorecido por
condições adequadas e altos valores de fotossíntese. No verão, as condições climáticas no
Estado de São Paulo favorecem a produção de fotoassimilados, que mantém o fornecimento
de carboidratos para o crescimento da planta e acúmulo de reservas, fornecendo substrato e
água para o crescimento dos frutos (GOLDSCHIMIDT & KOCH, 1996; SYVERSTSEN &
LLOYDE, 1994).
As principais áreas produtoras de citros do Estado de São Paulo encontram-se em
regiões onde a temperatura do ar atinge valores inferiores a 15 °C no inverno e o total anual
de precipitação varia entre 1200 e 1500 mm (DAVIES, 1997). De acordo com valores limites
de temperatura do ar e deficiência hídrica, CAMARGO et al. (1974) elaboraram o
zoneamento agroclimático para os citros no Estado de São Paulo, tendo sido estabelecida a
temperatura média anual de 17 °C como limite, abaixo da qual seria inapropriado o cultivo
dos citros.
A valores máximos de fotossíntese em laranjeiras (Citros sinensis (L.) Osbeck) ocorrem
em temperaturas do ar em torno de 25 a 30 oC (GUO et al., 2006; HABERMANN &
RODRIGUES, 2009; MACHADO et al., 2005; RIBEIRO, 2006). Sob condições naturais,
sem deficiência hídrica no solo e com densidade de fluxo de fótons fotossinteticamente ativos
(DFFFA) saturante, A máximo ocorre ao redor das 9:30 h, decrescendo posteriormente, com o
aumento da temperatura e do déficit de pressão de vapor do ar (DPV) (HABERMANN &
RODRIGUES, 2009, MACHADO et al., 2002; MEDINA et al., 1999; RIBEIRO et al., 2009a,

6
b). A redução de A com o aumento do DPV e da temperatura está relacionada com a queda da
condutância estomática e da condutância do mesófilo, respectivamente (KHAIRI & HALL,
1976a; LLOYD et al., 1992; MACHADO, 2009; MACHADO et al., 1994; MAGALHÃES
FILHO et al., 2009; MEDINA et al.,1998, 1999; RIBEIRO et al., 2009a, b).
A fotossíntese e as reservas acumuladas em folhas, ramos e raízes constituem as fontes
de carboidratos para a formação de novos ramos, raízes, flores e frutos. As laranjeiras mantêm
suas folhas sempre-verdes, ou seja, são ativas fotossinteticamente durante todo ano. Assim
sendo, esse processo é submetido a grande variação das condições do ambiente (MACHADO
et al., 2002, RIBEIRO, 2006; RIBEIRO & MACHADO, 2007), tendo assim desenvolvido
mecanismos para tolerar estresse hídrico e nutricional (GOLDSCHIMIDT, 1999). A laranjeira
apresenta baixa taxa de fotossíntese (GOLDSCHMIDT & KOCH, 1996; MACHADO et al.,
2002) devida à alta resistência interna à difusão de CO2 (ERISMANN et al, 2008;
MACHADO, 2009; MAGALHÃES FILHO et al., 2009; SYVERTSEN & LLOYD, 1994).
Que está relacionada às características anatômicas e bioquímicas, associadas com a
longevidade das folhas, variando com as condições ambientais, com a disponibilidade de
nutrientes e com a tolerância à seca e ao frio (MACHADO, 2009; WARREN & ADAMS,
2006). No decorrer de um ano, as variações nas condições ambientais (precipitação, DPV,
umidade do solo, radiação solar) e nos estádios de desenvolvimento da planta afetam a
eficiência fotossintética. A fotossíntese máxima em laranjeiras nas condições do Estado de
São Paulo ocorre no verão quente e úmido, decaindo progressivamente até alcançar valores
mínimos nos meses de inverno, frios e secos (MACHADO et al., 2002; MEDINA, 2002;
RIBEIRO, 2006). A produção fotossintética diária, em um dia claro de verão, chega a ser 2,9
vezes maior que no inverno (MACHADO et al., 2002; RIBEIRO et al., 2009a; RIBEIRO &
MACHADO, 2007). Esses valores foram observados em plantas sob irrigação, estando eles
relacionados às variações na temperatura e no DPV, ao comprimento do dia e à fase de
desenvolvimento da planta (MACHADO et al., 2002) porém, há dúvidas sobre os
mecanismos envolvidos no controle da fotossíntese.
BEVINGTON & CASTLE (1985) observaram que a variação do crescimento anual da
parte aérea e das raízes de laranjeiras está relacionada com a variação da temperatura do ar e
do solo. KHAIRI & HALL (1976b), mostraram que em temperatura do ar igual a do solo,
variando em torno de 24°C e 31ºC, há em laranjeiras indução (doce e azeda) a uma maior
brotação e crescimento do sistema radicular, em comparação a temperaturas de 15°C e 20°C.
A temperatura mínima para o desenvolvimento da raiz de mudas de citros é de 12 ºC, a
ótima de 26 °C e a máxima de 37 °C (KRIEDEMANN & BARRS, 1981). O crescimento das

7
raízes tem inicio na primavera, quando a temperatura supera os 13ºC (SPIEGEL-ROY &
GOLDSCHMIDT, 1996). Até 19 ºC, a taxa de crescimento radicular é pequena, acelerando-se
daí até 30 ºC, tanto para as raízes pioneiras como para as fibrosas (BEVINGTON &
CASTLE, 1985; DAVIES & ALBRIGO, 1994; KRIEDEMANN & BARRS, 1981;
SPIEGEL-ROL & GOLDSCHMIDT, 1996). Temperaturas acima de 36 °C inibem o
crescimento radicular (CASTLE, 1980).
Em laranjeiras no campo, a maior concentração das raízes localiza-se até 50 cm de
profundidade no perfil do solo e a temperatura média pode variar desde valores acima de 30 oC no verão, até valores abaixo de 10 oC, no inverno. Na superfície do solo, até 5 cm de
profundidade, a amplitude de variação é maior, ou seja, entre 5 até 45 oC (RIBEIRO, 2006).
A distribuição temporal e espacial da temperatura do solo, em um laranjal, varia entre
9 e 28 oC do inverno ao verão, nas profundidades entre 5 e 40 cm (RIBEIRO, 2006). A
variação do crescimento anual da parte aérea e das raízes em citros está relacionada com essa
variação da temperatura do ar e do solo (BEVINGTON & CASTLE, 1985). A temperatura
adequada do solo incrementa a divisão celular, o que resulta em maior crescimento das raízes,
e afeta a condutividade hidráulica e a permeabilidade de membranas (DODD et al., 2000).
Nos meses de inverno, em plantas sob irrigação, a menor taxa de fotossíntese pode
estar relacionada tanto com a queda na demanda por fotoassimilados, devido à menor taxa de
crescimento, como com o abaixamento da temperatura (BEVINGTON & CASTLE, 1985;
KHAIRI et al., 1976b; MACHADO et al., 2002; RIBEIRO & MACHADO, 2007).
A queda da fotossíntese durante o inverno é causada pela associação de noites e solo
frios, mesmo quando as condições de variação de temperatura durante o dia não são limitantes
(RIBEIRO et al., 2009b). A ocorrência somente de noites frias ou de solos com baixas
temperaturas (MACHADO, 2009; MAGALHÃES FILHO et al., 2009) também afetam
negativamente a fotossíntese. A baixa temperatura causa decréscimo na condutância
estomática (gs) e na atividade de carboxilação da ribulose-1,5-bisfosfato
carboxilase/oxigenase (Rubisco) e regeneração da ribulose-1,5-bisfosfato (RuBP)
(MACHADO, 2009; MAGALHÃES FILHO et al., 2009; RIBEIRO & MACHADO, 2007;
RIBEIRO et al., 2009a,b). A queda de gs pode estar relacionada com a queda da atividade
fotossintética em si, com conseqüente aumento da concentração intercelular de CO2 (Ci),
causando fechamento parcial dos estômatos. Alternativamente o estômato em si poderia ser o
alvo inicial do resfriamento e seu fechamento causar redução em Ci, acarretando a queda na
fotossíntese (ALLEN et al., 2000; ALLEN & ORT, 2001).

8
Há trabalhos que demonstram os efeitos isolados da baixa temperatura noturna e do
solo sobre a fotossíntese em citros e outras espécies arbóreas, como mangueira e cafeeiro
(ALLEN et al., 2001; BAUER et al., 1985; JOLY & HAHN, 1991; MACHADO, 2009;
MAGALHÃES FILHO, 2009).
MACHADO (2009) verificou que com a ocorrência de frio noturno a assimilação de
CO2
(A), a condutância estomática (gs), a condutância do mesofilo (gm) e a transpiração (E)
decrescem em laranjeiras. Sob baixa temperatura há ainda redução da concentração de CO2 no
cloroplasto (Cc), do transporte máximo de elétrons (Jmax) e da eficiência máxima de
carboxilação da Rubisco (Vc,max) além de aumento da concentração de amido na folha,
causada pela menor remobilização no período noturno, pois o frio noturno estaria diminuindo,
ou mesmo inibindo a atividade das enzimas de mobilização de amido. Ocorre ainda
fotoinibição caracterizada pela queda da eficiência quântica potencial do fotossistema II e da
eficiência quântica efetiva, porém sem serem as causas da queda em A. Efeitos semelhantes
foram descritos para outras espécies arbóreas submetidas ao frio noturno, tais como
mangueira, cafeeiro e cacaueiro (ALLEN et al., 2001; BAUER et al., 1985; JOLY & HAHN,
1991).
MAGALHÃES FILHO et al. (2009) estudaram o efeito de três temperaturas do solo
sobre a fotossíntese em laranjeiras: 10, 20 e 30 oC. O resfriamento das raízes de 20 para 10 oC
reduziu significativamente A, gs e gm, o potencial da água na folha (), a condutância
hidráulica (KL), Vc,max e Jmax. O aumento da temperatura radicular para 30 oC promoveu
aumento significativo em A, gs, gm,, KL, Jmax em relação ao tratamento referência a 20 oC.
A condutância estomática decresceu, em função da queda em e KL. O incremento em A nas
raízes a 30 oC ocorre principalmente em resposta ao aumento de Jmax e em menor grau a gs e
gm e a eficiência fotoquímica da fotossíntese. O aumento da temperatura do solo para 30 °C
aumentou KL e manteve os tecidos foliares mais hidratados, elevando gs e gm, permitindo
maior fluxo de CO2 para o cloroplasto e aumento da regeneração da RuBP. Segundo
GEORGE et al. (2002) as médias de potenciais hídricos no xilema da folha em árvores de
carambola (Averrhoa carambola L.) são mais negativos em árvores crescendo em
temperaturas da raiz de 5, 10 ou 38°C, em comparação àquelas temperaturas da raiz de 20 ou
25°C. As médias de A aumentam consistentemente para árvores crescendo em temperaturas
da raiz de 20 ou 25°C, comparado a todos os outros tratamentos. A faixa de temperatura
considerada não prejudicial para plantas de carambola se situou entre 10 e 38ºC, e a faixa
ótima entre 20 e 25°C. Exposição de mudas de carambola a baixa (< 10ºC) ou alta (38°C)

9
temperatura da raiz causou estresse hídrico indicado pela redução no potencial da água das
folhas. Tendências similares no potencial da água também foram observadas por ISMAIL
(1994) em mudas de carambola com seis meses de idade, apresentado estresse hídrico quando
submetidas à baixa temperatura do sistema radicular. Estudos de potencial da água
conduzidos com outras espécies lenhosas tropicais e subtropicais apresentaram respostas
similares quanto ao efeito da baixa temperatura da raiz.
Durante o inverno, o crescimento das laranjeiras diminui acentuadamente devido às
baixas temperaturas (KHAIRI & HALL,1976b; RAMOS, 2009; REUTHER, 1973),
evidenciando o papel regulatório da temperatura do ar em relação ao crescimento dos citros.
Porém, há atividade fotossintética durante o inverno, mesmo que reduzida, podendo ainda
ocorrer acúmulo de reservas nas folhas (fotoassimilados) e assim causar a inibição de A
(AZCON-BIETO, 1983, IGLESIAS et al., 2002). De fato, há acúmulo de carboidratos nas
folhas no período de inverno, diminuindo na primavera, devido à alta demanda em
consequência do florescimento, pegamento e desenvolvimento inicial dos frutos (PRADO,
2006; PRADO et al., 2007; RIBEIRO, 2006).
O carbono fixado fotossinteticamente é direcionado para a síntese de amido no
cloroplasto ou de sacarose no citoplasma celular (PAUL & FOYER, 2001), sendo a rota
preferencial dependente da demanda de carboidratos pela planta. Assim, quando o
crescimento é reduzido, a via preferencial é a síntese de amido, que fica armazenado nos
cloroplastos. Quando o crescimento é intenso, a rota preferencial é a síntese de sacarose, que é
distribuída para toda planta. O amido pode ser reutilizado, caso a atividade metabólica fique
novamente intensa. Interessante notar que, em citros ocorre acúmulo de reservas em todos os
órgãos (GOLDSCHMIDT & GOLOMB, 1982; GOLDSCHMIDT & KOCH, 1996;
MAGALHÃES FILHO et al., 2008), e da mesma forma que o amido armazenado nas folhas,
podem ser remobilizados e utilizados em outros órgãos em crescimento (GOLDSCHMIDT &
GOLOMB, 1982; MAGALHÃES FILHO et al., 2008). A queda da fotossíntese devido ao
aumento do teor de carboidratos na folha esta relacionada com o decréscimo da reciclagem de
Pi no cloroplasto, causando queda na síntese de ATP e NADPH e na regeneração de RuBP
(PAUL & FOYER, 2001). No entanto, alguns autores destacam (GOLDSCHIMIDT &
KOCH, 1996; RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2010) que esse controle da
fotossíntese devido ao acúmulo de carboidratos nas folhas de citros não é evidente sob
condições naturais e pode ser complementar a um mecanismo dinâmico da relação fonte-
dreno. A inibição da fotossíntese pelo acúmulo de carboidratos é evidenciado em experimento
em que se manipula artificialmente a relação fonte:dreno (GOLDSHIMIDT & HUBER, 1992;

10
GOLDSCHIMIDT & KOCH, 1996; IGLESIAS et al., 2002; SYVERTSEN, 1994). Porém, se
observou, em laranjeira sem a manipulação da relação fonte:dreno, a ocorrência de altos
valores de A em presença de altos teores de carboidratos (RIBEIRO & MACHADO, 2007;
RAMOS, 2009; RIBEIRO et al., 2010), sugerindo que A está mais relacionado à dinâmica
diária de produção e de consumo de carboidratos na folha e não propriamente ao teor de
reservas (RIBEIRO et al., 2005; RIBEIRO & MACHADO, 2007; RAMOS, 2009; RIBEIRO
et al., 2010). No verão, tanto a produção como o consumo e o teor de carboidratos são mais
elevados que no inverno, sendo A mais relacionado com a exportação de carboidratos da folha
(RIBEIRO et al., 2005; RIBEIRO & MACHADO, 2007; RAMOS, 2009; RIBEIRO et al.,
2010).
A variação da temperatura do solo também induz respostas específicas sobre a parte
aérea da planta, incluindo mudanças morfológicas (STONEMAN & DELL, 1993) e em gs e A
(BOUCHER et al., 2001; DAY et al.,1991; DELUCIA et al., 1991; OJEDA et al., 2004;
STRAND et al., 2002; XU & HUANG, 2000). De acordo com LEE & HE (2001), quando se
observa o efeito das temperaturas altas sobre a fotossíntese, a queda de A com a elevação da
temperatura acima da faixa fisiológica é causada pela inativação da atividade da Rubisco.
HUREWITZ & JANES (1987) mostraram que a ativação da Rubisco decresce quando a
temperatura da zona radicular aumenta.
A baixa temperatura do solo pode afetar a fotossíntese de várias formas. Sob alta
radiação pode ocorrer fotoinibição (DELUCIA et al., 1991), talvez devido à queda da
absorção de água e favorecimento ao fechamento parcial dos estômatos. DAY et al. (1991)
observaram em Pinus, que sob temperatura baixa no sistema radicular há fechamento parcial
dos estômatos, causando queda da fotossíntese. Outros autores por sua vez, observaram
também em plantas sempre verdes, que sob baixa temperatura a queda da fotossíntese estava
relacionada em parte a fatores estomáticos e em parte a fatores bioquímicos (MAGALHÃES
FILHO et al., 2009; STRAND et al., 2003). Assim, a queda da fotossíntese pode ser devida
tanto a fatores relacionados ao estômato, como a fatores bioquímicos e fotoquímicos
(MACHADO et al., 2002).
A associação de baixa temperatura do ar durante o período noturno e diurno com baixa
temperatura do solo poderia acarretar maiores quedas na fotossíntese, pois pode ocorrer
também fotoinibição ou, em casos mais severos, fotooxidação (DELUCIA et al., 1991; LONG
et al., 1994), porém ainda não se tem informações sobre os efeitos dessa condição sobre a
fotossíntese da laranjeira.

11
3 MATERIAL E MÉTODOS
3.1 Material vegetal
O experimento foi conduzido com mudas de laranjeira ‘Valência’ [Citrus sinensis (L.)
Osbeck] sobre limoeiro ´Cravo’ (Citrus limonia Osbeck), com sete meses de idade, contidas
em sacolas plásticas com capacidade para 7 L preenchidas com substrato composto de 95% de
casca de Pinus e 5% de vermiculita.
A nutrição das plantas foi realizada utilizando-se duas soluções-estoque (A e B). A
solução A foi preparada com a adição dos seguintes compostos em 1 L de água destilada: 80 g
de nitrato de cálcio, 41,5 g de nitrato de magnésio, 0,23 g de sulfato de manganês, 0,133 g de
sulfato de zinco, 1,54 g de ferro-EDTA e 5,6 mL de cobre-EDTA. A solução B foi preparada
com a adição dos seguintes compostos em 1 L de água destilada: 36 g de nitrato de potássio,
8,0 g de fosfato monoamônio, 12,0 g de sulfato de potássio e 1 mL de molibdato de sódio.
Aplicou-se duas vezes por semana 200 mL de solução nutritiva diluída (10 mL de cada
solução estoque por litro de água).
3.2 Procedimento experimental
A condução do experimento (aplicação dos tratamentos e medidas) foi feita com as
plantas dentro de câmara de crescimento (PGR14, Conviron, Canadá) o que possibilitou o
controle de umidade e temperatura do ar, fotoperíodo e densidade de fluxo de fótons
fotossinteticamente ativos (DFFFA) (Figura 1).
As mudas de laranjeiras foram mantidas em casa de vegetação até o momento do
experimento, quando foram transferidas para a câmara de crescimento onde permaneceram por
um dia para aclimatação nas seguintes condições: fotoperíodo de 12 h, temperatura do ar
28±1/20±1 °C (dia/noite), temperatura das raízes de 26±1 °C, umidade relativa de 60/65%
(dia/noite) com déficit de pressão de vapor (DPV) mantida ao redor de 1 kPa, DFFFA de 900
mol m-2 s-1. Essas condições são consideradas ótimas para a fotossíntese dos citros
(MACHADO et al., 2005; RIBEIRO et al., 2004). Excetuando-se a temperatura, as demais
condições ambientais na câmara de crescimento foram mantidas constantes durante todo período
experimental que teve duração de 7 dias. Após o primeiro dia nas condições descritas, as plantas
foram submetidas a seis tratamentos resultantes da combinação de 2 temperaturas das raízes (TR,
12
10 e 26 oC), 2 temperaturas noturnas da parte aérea (TN, 10 e 20 oC) e 2 temperaturas diurnas da
parte aérea das plantas (TD, 17 e 28 oC), como indicado na Tabela 1.
Os tratamentos no sistema radicular foram realizados no período de 24 horas (não variou
durante o dia e noite), enquanto os tratamentos de temperatura do ar na parte aérea durante o dia
e noite foram de 12 horas cada, pelo período de 6 dias consecutivos. Os resultados apresentados
são referentes às medidas efetuadas no sexto dia de tratamento.
Tabela 1: Tratamentos aplicados com quatro repetições em laranjeira ‘Valência’ sobre limoeiro ‘Cravo’ dentro de câmara de crescimento. Foi feita aclimatação das plantas por um dia nas seguintes condições: fotoperíodo de 12h, DFFFA = 900 mol m-2 s-1 e umidade do ar de 60/65% (dia/noite), temperatura noturna (TN) = 20 oC, temperatura diurna (TD) = 28 oC e temperatura das raízes (TR)= 26 oC. Os tratamentos foram aplicados por um período de seis dias consecutivos, variando apenas as temperaturas, conforme descrito na tabela.
*Período noturno e diurno de 12 h *As letras em subscrito nos tratamentos, representadas pelas iniciais D, N e R indicam a ocorrência de frio durante o período diurno (TD = 17 °C), noturno (TN = 10 °C) e no sistema radicular (TR = 10 °C), respectivamente.
Dentro da câmara de crescimento foram dispostas duas caixas de isopor (80 L cada), com
cinco a seis plantas por caixa. Uma caixa continha água a 10±1 e a outra a 26±1 oC, que
permitiram o controle de temperatura do substrato e do sistema radicular, de forma independente
da parte aérea. As temperaturas da água e do substrato foram monitoradas durante todo
experimento. Salienta-se que as raízes não ficavam em contato direto com a água, pois as
sacolas contendo as raízes foram isoladas por sacos plásticos. O interior da caixa fechada foi
aerado constantemente por bombas de ar.
No experimento foram avaliados os efeitos de combinações da variação da temperatura
no sistema radicular e da temperatura do ar na parte aérea da planta durante o dia e durante a
noite, sobre as trocas gasosas, atividade fotoquímica e concentração de carboidratos nas raízes,
folhas e caules.
Tratamentos Temperatura (oC) (TD) Dia* (TN) Noite* (TR) Raiz Tratamento 1 – (T1) 28 20 26 Tratamento 2 – (T2R) 28 20 10 Tratamento 3 – (T3N) 28 10 26 Tratamento 4 – (T4NR) 28 10 10 Tratamento 5 – (T5DN) 17 10 26 Tratamento 6 – (T6DNR) 17 10 10

13
3.3 Trocas gasosas e atividade fotoquímica
As medidas de assimilação de CO2 (A, mol m-2 s-1); de transpiração foliar (E, mmol
m-2 s-1); de condutância estomática (gs, mol m-2 s-1) e da concentração intercelular de CO2 (Ci,
mol mol-1) foram efetuadas com analisador portátil de fotossíntese por radiação infra-
vermelha (Licor 6400, Lincoln, NE, USA) em folhas totalmente expandidas, com
aproximadamente dois meses de idade. As condições dentro da câmara de fotossíntese do
analisador portátil de fotossíntese foram as mesmas que as condições dentro da câmara de
crescimento em cada etapa do experimento. A diferença de pressão de vapor entre o ar e a folha
(DPVar-folha) foi mantida em todos tratamentos ao redor de 1,5 kPa. As medidas de trocas gasosas
foram efetuadas entre 7:00 e 18:00 h, em intervalos de 2 horas no 6o dia de tratamento, iniciando-
as 1 hora após o início do período iluminado.
A partir das variáveis de trocas gasosas calculou-se a eficiência aparente de carboxilação
(A/Ci, mol m-2 s-1 Pa-1) (FLEXAS et al., 2001; MACHADO et al., 2005). Calculou-se a
assimilação diurna de CO2 (Ai, mmol m-2 d-1) a partir da integração da curva diurna de A por 12
horas, correspondente ao período iluminado com DFFFA de 900 mol m-2 s-1, no 6º dia de
tratamento.
Medidas da emissão de fluorescência da clorofila a foram realizadas com fluorômetro
modulado (PAM-2000, Walz, Alemanha) pelo método do pulso de saturação, (nm, e
densidade de fluxo de fótons de ~ 10.000 mol fótons m-2 s-1) (MAXWELL & JOHNSON,
2000). A fluorescência basal (Fo) e máxima (Fm) foram medidas em tecidos foliares
adaptados ao escuro, às 6:00 h, antes do início do período de luz e às 12:00 e 18:00 h, após 30
min de adaptação ao escuro, utilizando-se clipe foliar específico. Estas variáveis foram
utilizadas para calcular a eficiência quântica máxima, Fv/Fm, onde Fv = Fm-Fo.
3.4 Conteúdo de carboidratos nas folhas, caules e raízes
As folhas coletadas para quantificação de carboidratos foram similares em idade e
posição no dossel àquelas avaliadas em relação às trocas gasosas e fluorescência da clorofila.
As amostras de folhas foram coletadas às 6:00 horas no 7° dia. A coleta de caules e raízes foi
feita no início da manhã do 7° dia. As amostras de caule, folhas e raízes foram imersas em
CO2 sólido (-70ºC) imediatamente após a coleta e mantidas em freezer (-20ºC).
Para obtenção da massa seca o material foi seco em estufa (modelo MAO32, Marconi)
com circulação forçada de ar à temperatura de 50 ºC por 4 dias ou até massa constante. As

14
amostras foram então maceradas em almofariz e armazenadas em frascos de vidro até que as
análises de açúcares fossem realizadas.
Nas determinações do conteúdo de carboidratos (g de Glu kg-1 de massa seca) foram
considerados os açúcares solúveis (AS), sacarose (Sac), amido (AM) e carboidratos totais
(CT), sendo CT=AS+AM. As frações de AS e Sac foram determinadas em amostras de
aproximadamente 75 mg de massa seca (MS) e as frações de AM em amostras de 10 mg.
Figura 1. Fotografia ilustrando a disposição das plantas dentro da câmara de crescimento durante o período experimental
O extrato para quantificação de sacarose e açúcares solúveis totais foi obtido a partir
de extração pelo método do MCW, descrito por BIELESK & TURNER (1966), onde foram
transferidos para um tubo de rosca contendo 75 mg de amostra, 3 mL de MCW (metanol,
clorofórmio e água na proporção 12:5:3 v/v) que ficaram sob refrigeração por 2 dias. Após a
separação de fases obtida pela adição de clorofórmio e água, coletou-se o sobrenadante que

15
em seguida foi concentrado em banho-maria (50 °C), para evaporação do metanol e resíduos
de clorofórmio. Os extratos foram então armazenados em freezer (-20 °C) até o momento da
quantificação.
Para a quantificação de amido utilizou-se o método descrito por AMARAL et al.
(2007) para obtenção do extrato e determinação enzimática de seu conteúdo. Em eppendorf
contendo 10 mg de amostra, foram realizadas quatro extrações com 500 µL de etanol 80 % a
80 °C (em banho-maria) por 20 minutos, para retirada de açúcares, pigmentos, fenóis e outras
substâncias. O precipitado foi então seco a temperatura ambiente até completa evaporação do
resíduo de etanol. Em seguida foram adicionados 500 µL (120U mL-1) de α-amilase (EC
3.2.1.1) termoestável de Bacillus licheniformis (cód. E-ANAAM, MEGAZYME, Irlanda),
diluída em tampão MOPS 10 mM e pH 6,5. As amostras foram incubadas a 75 °C (em banho-
maria) por 30 minutos. Este passo foi repetido por mais uma vez, totalizando 120 unidades de
enzima. Em seguida adicionou-se 500 µL (30U mL-1) de amiloglucosidase (EC 3.2.1.3) de
Aspergillus Níger (cód. E-AMGPU, MEGAZYME, Irlanda) em tampão acetato de sódio 100
mM e pH 4,5. As amostras foram incubadas a 50 °C (em banho-maria) por 30 minutos. Este
passo foi repetido por mais uma vez, totalizando 30 unidades de enzima. Foram acrescentados
então, 100 µL de ácido perclórico (HClO4) 0,8 M para parar a reação e precipitar proteínas.
Para dosagem de amido procedeu-se da seguinte forma: em eppendorf contendo 20 µL de
extrato, foi adicionado 30 µL de água destilada e 750 µL do reagente glicose PAP Liquiform
(Labtest Diagnóstica S.A.). A mistura foi levada ao banho-maria a 30 °C por 15 minutos. O
teor de glicose foi determinado em leitor de microplacas de ELISA (modelo EL307C, Bio-
Tek Instruments, Winooski, Vermont) em comprimento de onda de 490 nm. As concentrações
de glicose foram calculadas a partir de uma curva-padrão obtida com leituras de soluções
contendo 5, 10, 15, 25 e 30 µg de glicose. Como branco, colocou-se água no lugar da amostra.
Determinou-se a concentração de sacarose pelo método descrito por HANDEL (1968)
e a dosagem foi feita pelo método do fenol-sulfúrico. Em tubo de ensaio adicionou-se 10 µL
de extrato, 490 µL de água destilada e 500 µL de solução de hidróxido de potássio (KOH) 30
%. Os tubos foram vedados e levados ao banho-maria 100 °C por 10 minutos. Em seguida
adicionou-se 500 µL de solução de fenol 5 % e 2 mL de ácido sulfúrico (H2SO4) concentrado.
Agitou-se bem e após resfriar, fez-se a leitura de absorbância em espectrofotômetro
(Micronal, modelo B342II) a 490 nm. As concentrações de sacarose foram calculadas a partir
de uma curva-padrão obtida com leituras de soluções contendo 5, 10, 20, 30, 40 e 50 µg de
sacarose. Como branco, colocou-se água destilada no lugar da amostra.

16
A concentração de açúcares solúveis totais foi determinada pelo método do fenol-
sulfúrico (DUBOIS et al., 1956). Em tubo de ensaio adicionou-se 10 µL de extrato e 490 µL
de água destilada, 500µL de solução de fenol 5 % e 2 mL de ácido sulfúrico (H2SO4)
concentrado. Agitou-se bem e após resfriar, fez-se a leitura de absorbância em
espectrofotômetro (Micronal, modelo B342II) a 490nm. As concentrações de açúcares
solúveis foram calculadas a partir de uma curva-padrão obtida com leituras de soluções
contendo 5, 10, 20, 30, 40 e 50 µg de glicose. Como branco, colocou-se água no lugar da
amostra.
3.6 Delineamento experimental
O delineamento experimental foi em parcelas subdivididas (split-plot) com quatro
repetições, tendo os tratamentos (6 combinações de temperaturas) como parcelas e o tempo (6
horários de avaliação) como subparcelas. As variáveis medidas foram submetidas à análise de
variância (ANOVA) e as médias comparadas pelo teste de Tukey com 0,05 de probabilidade.
4 RESULTADOS E DISCUSSÃO
4.1 Biometria
As plantas utilizadas no experimento foram semelhantes não havendo diferenças
significativas nas dimensões de suas folhas, caules e raízes (Figura 2A). Consequentemente, a
distribuição porcentual de fitomassa de cada órgão também não apresentou diferenças
importantes, apresentando cerca de 63% de raízes, 18% de caule e 19% de folhas. Observou-
se que nessa fase, houve uma tendência de estabelecer um sistema radicular bem
desenvolvido em relação à planta. As plantas também apresentaram um padrão similar quanto
ao número de folhas e área foliar específica (AFE, Tabela 2).
4.2 Trocas gasosas
Os cursos diários da assimilação de CO2 (A), da condutância estomática (gs), da
concentração intercelular de CO2 (Ci) e da eficiência aparente de carboxilação (A/Ci) em
mudas de laranjeira ‘Valência’ submetidas a seis regimes térmicos são mostrados na Figura 3.
Fica evidenciada a influência do regime térmico sobre as trocas gasosas em laranjeiras,
havendo diferenças significativas (P<0,05) entre eles.

17
Figura 2 – Características biométricas (em A) de laranjeira ‘Valência’ sobre limoeiro ‘Cravo’, utilizadas no experimento e distribuição relativa de fitomassa (R/T, C/T e F/T em B). R, C, F e T representam respectivamente fitomassa seca das raízes, caules, folhas e total. Em A, barras indicam desvio padrão da média referente a 4 repetições e em B, indicam desvio padrão da média em 24 repetições. Tabela 2. Área foliar específica (AFE) em laranjeira ‘Valência’ sobre limoeiro ‘Cravo’, após 6 dias nos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R , TD = 28, TN = 20, TR = 10 oC; T3N, TR = 28, TN = 10, TR = 26 oC; T4NR, TR = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes.
Ao longo do dia, plantas do tratamento T1, cujas temperaturas do ar diurna (TD) e
noturna (TN) e do sistema radicular (TR) são consideradas as ideais para laranjeira, 28, 20 e 26 oC, respectivamente (MACHADO et al., 2005; MAGALHÃES FILHO, 2009; RIBEIRO,
2006; SYVERTSEN & LLOYD, 1994) apresentaram os maiores valores de A durante todo
período de medidas em relação aos outros tratamentos (Figura 3 A).
Tratamento AFE (m2/kg) T1 7,5±0,3 T2R 7,9±0,4 T3N 7,7±1,8 T4NR 8,8±1,1 T5DN 9,1±0,3 T6DNR 8,9±0,5
0
5
10
15
20
25
30
Fitomassa de órgão/fitomassa total
B
Fito
mas
sa s
eca
(g p
lant
a-1)
Tratamento
Raiz Caule Folha TotalA
T1 T2R T3N T4NR T5DN T6DNR
0
10
20
30
40
50
60
70
F/TC/T
Porc
enta
gem
(%)
R/T

18
As medidas foram feitas em folhas jovens e maduras, apresentando-se dentro dos
padrões superiores de fotossíntese de plantas sadias e bem nutridas (MACHADO et al., 2005;
RAMOS, 2009; RIBEIRO, 2006). Apesar das plantas terem sido mantidas em condições
ambientais constantes, houve tendência de A ser máximo (AmaxT1 = 9,6 mol m-2 s-1) nos
primeiros horários de medida, para diminuir após às 15:00 h, atingindo um mínimo ao redor
das 18:00 (AminT1 = 4,3 mol m-2 s-1), em resposta à queda de gs e de A/Ci. Esse padrão de
variação havia sido relatado anteriormente por outros autores em laranjeiras também sob
Figura 3 – Variação diurna da assimilação de CO2 (A em A), da condutância estomática (gs em B), da concentração intercelular de CO2 (Ci em C) e da eficiência aparente de caboxilação (A/Ci em D) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas a seis regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TR = 28, TN = 10, TR = 26 oC; T4NR, TR = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes (24 h).
6 8 10 12 14 16 180,0
0,1
0,2
0,3
0,4
6 8 10 12 14 16 18
-4
-2
0
2
4
6
8
10
12
14
6 8 10 12 14 16 180,00
0,05
0,10
0,15
0,20
6 8 10 12 14 16 18200
250
300
350
Hora
A/C
i (m
ol m
-2 s
-1 P
a-1)
D
A (
mol
m-2 s
-1)
T1 T2R T3
N
T4NR T5DN T6DNR
A B
gs (m
ol m
-2 s
-1)
C
Ci (m
ol C
O2 m
ol-1)
Hora

19
condições ambientais constantes (RAMOS, 2009; RIBEIRO et al., 2009b), sugerindo a
existência de algum tipo de regulação endógena causando queda em A (RAMOS, 2009;
RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2009b).
No tratamento T2R, cuja diferença em relação a T1 é a baixa temperatura das raízes
(TR=10 oC), mostrou um padrão diário semelhante, porém em patamares bem menores (Figura
3A). Assim, AmaxT2R = 5,2 µmol m-2 s-1, ao redor das 8:00h e AminT2R = 2,3µmol m-2 s-1, no
final da tarde. Em T2R a baixa temperatura das raízes causou queda significativa de A. No
tratamento T3N, em que a diferença em relação a T1 foi a baixa temperatura noturna (TN=10 oC) o padrão de resposta foi diferente de T1 e T2R, ou seja os menores valores de A ocorreram
nas primeiras horas da manhã (AminT3N = 4,3 µmol m-2 s-1), para em seguida com o
aquecimento do ar durante o dia (TD=28 oC) aumentar gradativamente até um máximo ao
redor das 12:00 h. Interessante notar que após este máximo, A seguiu a mesma tendência de
T1 e T2R, diminuindo até o final da tarde (Figura 3A).
Em T4NR em que houve a combinação de baixa temperatura noturna do ar e das raízes
(TR=10 e TN=10 oC), A foi baixo no início da manhã (AmaxT4NR = 3,1 µmol m-2 s-1) mantendo-
se praticamente constante até o final da tarde, quando seguiu a tendência dos outros
tratamentos decrescendo a um mínimo ao redor de AminT4NR = 2,5 mol m-2 s-1 (Figura 3 A).
Os tratamentos T5DN e T6DNR mantiveram o mesmo padrão de T4NR, mantendo valores de A
praticamente constantes durante o dia e abaixando no final da tarde. Assim, além dos valores
absolutos de A serem diferentes conforme o tratamento térmico, os padrões de resposta da
fotossíntese durante o dia também variaram em relação ao tratamento referência, T1.
Na Figura 3 também se observa com clareza que além das variações significativas nas
respostas de A houve variações em gs, Ci e em A/Ci com o regime térmico. Para ocorrer a
fixação do CO2 em moléculas de carboidratos no ciclo de Calvin pela ação da enzima ribulose-
1,5-bisfosfato (RuBP) carboxilase/oxigenase (Rubisco) o CO2 movimenta-se, desde a atmosfera
ao redor das folhas, através da camada limite e do ostíolo dos estômatos para a cavidade
subestomática e daí para o sítio de carboxilação dentro do estroma, através do mesofilo da folha.
Esse processo pode ser afetado por fatores que provocam limitação difusiva, e/ou bioquímica e
fotoquímica. A primeira diz respeito ao fato de a fotossíntese ser limitada devido à menor
abertura estomática, ocasionando diminuição do fluxo de CO2 entre a atmosfera e o mesófilo
foliar. Outras limitações se referem a danos que podem ocorrer nas reações bioquímicas e/ou
fotoquímicas. Essas últimas envolvem a absorção de energia radiante, hidrólise da água e
liberação de O2, com posterior produção de moléculas ricas em energia química (ATP) e com

20
poder redutor (NADPH), as quais serão utilizadas nas reações bioquímicas que fixam o CO2 em
moléculas de carboidratos. A influência de gs na difusão de CO2 e conseqüentemente em A já
é bem conhecida em laranjeiras crescidas em campo ou em condições protegidas (MEDINA
et al., 1998, 1999; RIBEIRO, 2006; RIBEIRO & MACHADO, 2007) ou em ambientes
controlados (MACHADO et al., 2005; RIBEIRO et al., 2004). Da mesma forma, observa-se
variação de A em função de variações nas eficiências de carboxilação e das atividades
fotoquímicas. A condutância estomática varia em função de fatores ambientais como
temperatura do ar e do solo, temperatura noturna, umidade do solo, turgescência e potencial
da água da folha, DPVar-folha, DFFFA e fatores relacionados com a própria planta (ALLEN et
al., 2000; ALLEN & ORT, 2001; BAUER et al., 1985; ERISMANN et al., 2008; FENELL &
MARKHART, 1998; HABERMANN et al., 2003; MACHADO et al., 2002, 2005; RIBEIRO
& MACHADO, 2007; RIBEIRO et al., 2009a,b).
Observa-se na Figura 3 que a queda de A em relação a T1 esta sempre relacionada com
a queda em gs e em A/Ci. Ci manteve-se aproximadamente igual em todos os tratamentos,
exceto em T6DNR no qual foi maior, possivelmente devido a danos mais acentuados no
aparelho fotossintetizador, como sugere a queda acentuada em A/Ci e em Fv/Fm, como será
discutido posteriormente. No entanto, as causas responsáveis pelas quedas em gs e A/Ci são
diferentes em função das diferentes formas de ocorrência de baixas temperaturas.
Os maiores valores de A (P<0,05) ocorreram nas plantas com temperatura radicular de
26 °C, independentemente da temperatura da parte aérea, ou seja, nos tratamentos T1, T3N e
T5DN. A influência positiva da maior temperatura radicular (26 °C) sobre A pode ser
confirmada em T5DN em que as temperaturas diurna/noturna do ar foram baixas, 17 e 10 °C.
Nessa condição os valores de A foram superiores aos encontrados nas plantas em que as
temperaturas da parte aérea estavam em condições ótimas (28/20 °C dia/noite), mas com
raízes a 10 °C (tratamento T2R), que apresentaram redução de A superior aos tratamentos T3N
e T5DN (Figura 3A). Sob temperatura radicular mais elevada nesses tratamentos, observam-se
em concordância, maiores gs e A/Ci, o que também foi relatado por outros autores em
laranjeiras (MAGALHÃES FILHO et al., 2009; RIBEIRO et al., 2009b).
A temperatura do solo influencia fortemente o crescimento da parte aérea em várias
espécies e parece ser mais importante do que a temperatura do ar no controle do crescimento
da planta (XU & HANG, 2000). Temperaturas mais elevadas, dentro da faixa fisiológica,
favorecem o metabolismo celular aumentando a velocidade das reações químicas.
Metabolismo mais acelerado requer reposição mais rápida de substratos para a continuidade
das reações e, por isso, alterações fisiológicas nas plantas devem ocorrer para atender a

21
demanda por esses substratos. Em citros o crescimento das raízes aumenta com a temperatura
na faixa entre 13 e 27 °C (BEVINGTON & CASTLE, 1985). Maior crescimento requer maior
fornecimento de carboidratos, estimulando a produção fotossintética (PAUL & FOYER,
2001). Assim, o maior valor de A nas plantas com temperatura do sistema radicular adequada
pode ser em resposta a maior demanda relativa das raízes. Note-se, que nessa fase do
desenvolvimento, as raízes representam aproximadamente 60% da fitomassa seca total da
planta (Figura 2B). A maior temperatura das raízes proporciona a maior eficiência aparente de
carboxilação da Rubisco (Figura 3D). MAGALHÃES FILHO et al. (2009), observaram
também em laranjeiras que Vc,max e Jmax foram significativamente mais altos em plantas
cujas raízes estavam a 30 °C em comparação com 20 e 10 oC.
No geral, sinais hidráulicos e/ou químicos estão envolvidos no decréscimo de gs.
Quando as raízes estão em condições adequadas de temperatura, a condutividade hidráulica é
elevada (KL; HUBBARD et al., 2001), permitindo absorção adequada de água, o que implica
também em maior abertura estomática e portanto de gs, como se verifica na Figura 3B, ou
seja, maiores valores de A estão relacionados com gs. Sob condições de temperatura radicular
baixa, os teores de ABA aumentam e os de citocinina decrescem (WAN et al., 2004; ZHANG
et al., 2008). No entanto, a queda em gs em função da ocorrência de frio nas diferentes partes
da planta ou períodos do dia (diurno ou noturno) deve-se a efeitos diferentes, como será
discutido à frente. Mudanças no potencial da água na folha são sinais hidráulicos que podem
afetar a abertura estomática. Haja vista que a baixa temperatura do solo causa um aumento na
viscosidade da água devido ao aumento da força de ligação de hidrogênio, reduzindo KL da
planta, ou seja, reduz o fluxo através dos vasos do xilema (KRAMER, 1983). Existem canais
de proteínas intrínsecas, denominados aquaporinas, que estão presentes na membrana
plasmática. Esses canais facilitam a absorção de água do solo e mediam a regulação da
condutividade hidráulica. Aquaporinas são encontradas nas células da epiderme e ponta da
raiz, e tecidos próximos do xilema da raiz (BOHNERT et al.,1995). A atividade desses canais
de proteínas, responsáveis pelo aumento/redução da resistência ao fluxo da água na planta,
parece comandar a captação de água do solo (TOURNAIRE-ROUX et al., 2003). A atividade
das aquaporinas é afetada pela temperatura (BERNACHI et al., 2002). De acordo com
TOURNAIRE-ROUX et al. (2003), a redução do pH da seiva pode bloquear a atividade
dessas vesículas reduzindo a capacidade de absorção de água e consequentemente o KL. Em
estudo realizado por eles, a redução do pH do citossol, promovido pela baixa disponibilidade
de O2 nas raízes, reduziu drasticamente o KL reduzindo a absorção de água devido à mudança
da permeabilidade das membranas, relacionada às aquaporinas. Possivelmente essa é a primeira

22
resposta ao estresse nas raízes (TOURNAIRE-ROUX et al., 2003). A baixa temperatura na parte
aérea pode incrementar a cavitação dos vasos do xilema reduzindo KL (SPERRY et al., 1988).
Em videira, FLEXAS et al. (1999) observaram que a queda de A em função do decréscimo em gS
estaria relacionada com as propriedades hidráulicas do xilema, causando queda do conteúdo
relativo de água nas folhas.
Assim, o resfriamento das raízes a 10 °C, possivelmente reduziu a atividade das
aquaporinas, a absorção de água pelas raízes e a hidratação das folhas, o que causou diminuição
em gS (FLEXAS et al., 2008). Entretanto, a manutenção da temperatura das raízes a 26 °C
(dentro da faixa ideal) aumenta a atividade das aquaporinas, e mantém elevada a absorção de
água e o fluxo de CO2 no mesofilo foliar (BERNACCHI et al. 2002).
Quanto aos sinais químicos podemos citar a modificação na composição da seiva
(variação da concentração de íons na seiva do xilema), mudanças no pH (WAN et al., 2004),
alteração dos teores de citocinina e do conteúdo de ABA (DAVIES & ZHANG, 1991;
TARDIEU & SIMONNEAU, 1998; GOMES et al., 2003). A presença de sinais químicos foi
sugerida por MEDINA & MACHADO (1998) em mudas de laranjeiras ‘Valência’ submetidas à
deficiência hídrica. Observou-se que logo nos primeiros dias após a suspensão da irrigação
houve decréscimo de gs, mesmo antes de reduzir a hidratação das folhas, ou seja, gS respondeu
mais rapidamente à queda da umidade do solo do que ao potencial da água na folha. Esse fato
está coerente com a hipótese de que há uma comunicação química entre a raiz e a parte aérea,
provocando o movimento estomático, conforme sugerem vários autores (SCHURR et al., 1992;
TARDIEU et al., 1992). Sugere-se que esse sinal seja intermediado por substâncias de
crescimento, possivelmente o ABA, produzido nas raízes (TARDIEU et al., 1992). Sob baixa
temperatura, a composição e o pH da seiva no xilema mudam e ocorre aumento no teor de
ABA e decréscimo do teor de citocinina nas folhas, causando queda em gs (NADA et al.,
2003; VELESOVA et al., 2005; WAN et al., 2004; ZHANG et al., 2008; ZHOU et al., 2007).
O decréscimo do teor de citocinina na folha é devido à queda do seu fluxo a partir da raiz e ao
aumento da atividade de oxidase de citocinina na folha (VELESOVA et al., 2005). ZHOU et
al. (2007) também relataram aumento do teor de ABA e queda em citocinina em raízes
submetidas ao frio. ZHOU et al. (2007) observaram que o frio nas raízes causa queda no teor
de citocinina, que pode ser um dos fatores fisiológicos que explicariam parcialmente os
decréscimos da eficiência de carboxilação, do conteúdo e da atividade de Rubisco. Citocinina
pode aumentar o teor de mRNA relacionado a síntese e atividade da Rubisco e FBPase
(DAVIS & ZHANG, 1991).

23
Segundo XU & HUANG (2000), a temperatura do solo em faixas ótimas mesmo
quando a parte aérea é exposta a temperaturas supra-ótimas proporciona aumento de A no
dossel, reduz o consumo de carbono através da respiração, aumenta a alocação de carbono
para raiz elevando o acúmulo de carboidratos em duas cultivares de grama (Agrostis palustris
Huds).
No tratamento T3N (TD=28, TN=10 e TR=26 °C) o resfriamento noturno causou queda
em A, também nesse caso em consonância com a queda em gs e em A/Ci (Figura 3A, B e D).
Interessante notar que em laranjeira a queda em A devido ao frio noturno ocorreu principalmente
nas primeiras horas da manhã (Figura 3A e B), conforme também verificou MACHADO (2009).
No entanto, mangueiras resfriadas durante a noite apresentaram decréscimo em A somente após
o meio-dia (ALLEN et al., 2000; ALLEN & ORT, 2001), mostrando haver diferenças de
respostas conforme a espécie estudada.
A ocorrência de frio noturno (T3N e T4NR) pode afetar diretamente a fisiologia do
estômato ou ainda a fotossíntese devido a inibição direta pelo frio, causando aumento em Ci e
fechamento parcial dos estômatos (ALLEN & ORT, 2001). No entanto, não ocorreu variação
em Ci (Figura 3C), permanecendo aproximadamente igual no período de medidas (Figura
3C). Apesar da queda de A em T3N, Ci não aumentou em função da queda do consumo de
CO2, pois gs também decaiu. Qualquer que seja o efeito do frio sobre os estômatos, o fato é
que o fator difusivo foi uma das causas da queda de A. Também fica evidente que a queda da
razão A/Ci é uma das causas da queda em A em T3N (Figura 3D). Isso pode ser confirmado
pelos decréscimos acentuados tanto de A como de gs nas plantas submetidas ao resfriamento.
A redução de gs foi mais intensa nos tratamentos em que a raiz foi resfriada a 10 °C,
como mostram os dados de trocas gasosas (Figura 3B). Todavia o menor gs não foi o
principal fator determinante para o decréscimo de A, haja vista que os valores de Ci variando
entre 190 e 270 mol m-2 s-1 não devem limitar a fotossíntese (Figura 3C, RIBEIRO, 2006).
Verificou-se também que o decréscimo de A em função do resfriamento (TD, TN e TR)
estava sempre relacionado com a queda na eficiência aparente de carboxilação da Rubisco,
dado por A/Ci (figura 3D). As reduções mais acentuadas na razão A/Ci ocorreram também nos
tratamentos em que houve resfriamento das raízes. Assim a queda de A devido ao
resfriamento também apresentou um componente metabólico, além do difusivo, conforme
também observaram outros autores (MACHADO, 2009; MAGALHÃES FILHO, 2009;
RIBEIRO et al, 2009b).
Em T4NR em que houve a combinação de baixa temperatura no ar à noite e das raízes
(TD=28, TN=10 e TR=10 oC), foi observado o segundo menor valor de assimilação de CO2,

24
tendo um efeito cumulativo tanto da noite como do sistema radicular sob baixa temperatura.
Os efeitos sobre gs foram mais acentuados que nos tratamentos T2R, T3N e T5DN. As
explicações e análises dos efeitos da baixa temperatura são os mesmos apresentados
anteriormente.
Em relação ao T5DN (TD=17, TN=10 e TR=26 oC), A foi menor somente em relação a
T1 e T3N. Esse tratamento pode ser comparado diretamente com o tratamento T3N (TD=28,
TN=10 e TR=26 oC), sendo a diferença entre os dois devido provavelmente à ocorrência de frio
diurno. A baixa temperatura diurna age diretamente sobre a fisiologia dos estômatos afetando
gs. Observa-se que gs decresceu em função da ocorrência de frio diurno e noturno, sendo mais
afetado do que quando ocorreu somente frio noturno em T3N. Isso mostra efeito cumulativo
dos períodos noturno e diurno. Também a baixa temperatura diurna causou queda na razão
A/Ci, sugerindo efeitos sobre o metabolismo. De fato, o frio diminui a atividade das enzimas
do ciclo de Calvin, afetando a ativação da Rubisco e das enzimas relacionadas com o ciclo
redutivo das pentoses e, portanto, com a regeneração de RuBP (ALLEN & ORT, 2001;
HÄLLGREN et al., 1991; RIBEIRO et al., 2009b). Assim, também em T5DN A decresceu em
consonância a gs e A/Ci.
O tratamento T6DNR (TD=17, TN=10 e TR=10 °C) foi o mais drástico, sendo uma
combinação de frio tanto de dia como de noite e no sistema radicular, tal como num dia típico
de inverno. Dessa forma, todos os efeitos discutidos nos outros tratamentos ocorreram aqui,
mas de maneira mais acentuada. Somente em T6DNR ocorreu um aumento significativo em Ci
em relação aos demais tratamentos. Esse aumento em Ci pode estar relacionado, em parte,
com a grande queda que ocorreu em A/Ci (Figura 3D). Outro aspecto que pode ter contribuído
para o aumento de Ci seria uma possível queda no valor da condutância do mesofilo (gm).
Durante a fotossíntese o CO2 se move da atmosfera para o interior da cavidade subestomática
através da abertura estomática e daí para o cloroplasto através do mesofilo, caracterizando gm.
A condutância do mesofilo varia em função das condições do ambiente (BERNACCHI et al.,
2002; FLEXAS et al., 2008), sendo que a variação da temperatura noturna na parte aérea e/ou
nas raízes afeta negativamente gm (MACHADO 2009; MAGALHÃES FILHO et al., 2009).
Um baixo valor de gm em T6DNR diminuiria o movimento de CO2 no sentido do cloroplasto
causando um aumento em Ci.
A Figura 4A mostra a relação entre A e gs considerando todos os tratamentos. No
início da curva o aumento de A é linear em função da abertura estomática, atingindo um platô,
em que o aumento de gs não significa novos incrementos de A. Em condições ambientais não
limitantes, a estabilização dos valores de A com o aumento de gs ocorre devido à limitação

25
imposta por reações enzimáticas, como por exemplo, a capacidade limitada de regenerar a
RuBP (LARCHER, 2000). Já nos tratamentos onde o frio foi imposto na parte aérea durante
Figura 4 – Relação entre assimilação de CO2 (A) e condutância estomática (gs em A) e eficiência aparente de carboxilação (A/Ci em B) e relação entre A/Ci e gs (em C) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes (24 h). Barras indicam desvio padrão da média referente a 4 repetições.
0,03 0,06 0,09 0,12 0,15 0,18 0,210
2
4
6
8
10
0,0 0,1 0,2 0,3 0,40
2
4
6
8
10
0,03 0,06 0,09 0,12 0,15 0,18 0,210,0
0,1
0,2
0,3
0,4
0,5
A=Amax[1-exp(gs-0,01)]R=0,94 0,312
A (
mol
m-2 s
-1)
gs (mol m-2 s-1)
T1 T2R T3N T4NR T5DN T6DNR
A
A (
mol
m-2 s
-1)
A/Ci (mol m-2 s-1 Pa-1)
A=0,0697+24,59*(A/Ci)R=0,99 P<0,0001
B
C
A/C
i (m
ol m
-2 s
-1 P
a-1)
gs (mol m-2 s-1)

26
o dia ou a noite, ou ainda, no sistema radicular, implicando em queda de gs, A diminui,
mostrando que a diminuição do fluxo de CO2 para o interior da folha limita a fotossíntese. Na
figura 4A fica evidente que o frio limita a fotossíntese devido a fatores difusivos, como já foi
relatado por MACHADO, 2009; MACHADO et al., 2009; MAGALHÃES FILHO, 2009;
RIBEIRO et al., 2009a,b.
Nos seis tratamentos térmicos houve correlação positiva significativa (p<0,05) entre A
e A/Ci (Figura 4B). Os maiores valores de A/Ci e de A correspondem ao tratamento T1. Nos
demais tratamentos onde pelo menos alguma situação era sob temperatura baixa, a razão A/Ci
sofria de alguma forma queda na eficiência metabólica. Nos tratamentos térmicos a 26 °C no
sistema radicular, A manteve-se em um patamar superior quando comparados aos tratamentos
de 10 °C na raiz.
O padrão de resposta induzido pelos tratamentos térmicos na relação A x A/Ci sugere
novamente que além da regulação estomática e fotoquímica (discutida no item 4.4, Atividade
fotoquímica), a fotossíntese é limitada bioquimicamente pela atividade da Rubisco, enzima
que representa cerca de 19-25% da proteína solúvel total em citros (VU & YELENOSKY,
1988a). A incorporação de CO2 atmosférico mediado pela atividade carboxilase da Rubisco é
uma das principais reações bioquímicas responsáveis pela fotossíntese (BERRY &
BJORKMAN, 1980; HALLGREN et al., 1991), havendo distinção entre o efeito da baixa
temperatura nas diferentes partes da planta ou períodos do dia (diurno ou noturno).
A Figura 4C mostra a relação entre a razão A/Ci e gs. Verifica-se que inicialmente o
aumento de A/Ci é praticamente linear com o aumento de gs, sugerindo que a eficiência
aparente de carboxilação é limitada também pelo fluxo de CO2 para o interior da folha, ou
seja, limitação pelo substrato (MACHADO et al., 2005). Quanto maior a limitação de gs em
função do tratamento térmico menor A/Ci, indicando que o efeito da baixa temperatura é
devido à interação de fatores difusivos e metabólicos.
4.3 Assimilação diurna de CO2
A assimilação diurna de CO2 (Ai), isto é, o somatório de todo CO2 assimilado em um
dia, mostra com clareza as diferenças entre as respostas da fotossíntese em função dos
regimes térmicos. A maior produção diurna foi do tratamento T1 (367,0±15,7 mmol m-2 dia-
1), visto que as condições ambientais foram as ideais para a fotossíntese de laranjeiras
(MACHADO et al., 2005), seguido por T3N (263,5±21,3 mmol m-2 dia-1 ou AiT1=1,39*AiT3N),
T5DN (215,5±12,1 mmol m-2 dia-1 ou AiT1=1,70*AiT5DN), T2R (146,3±5,8 mmol m-2 dia-1 ou
AiT1=2,50*AiT2R), T4NR (115,3±2,2 mmol m-2 dia-1 ou AiT1=3,18*AiT4NR) e T6DNR (85,6±5,8

27
mmol m-2 dia-1 ou AiT1=4,31*AiT6DNR). Essa forma de expressar a fotossíntese, como a
integração diurna, evidencia a importância do regime térmico para a produção da planta.
Interessante notar que a ocorrência de frio noturno, comum mesmo fora do inverno, quando
há entrada de frentes frias, pode causar prejuízos consideráveis na produção fotossintética,
mesmo se os dias subseqüentes apresentarem temperaturas não limitantes, como se apresenta
no tratamento T3N.
A ocorrência de baixa temperatura no período diurno e noturno e temperatura alta das
raízes é possível ocorrer na transição de verão para outono (RIBEIRO, 2006), quando ocorre
queda da temperatura do ar, mas ainda não houve tempo para a temperatura do solo baixar.
Essa situação foi simulada pelo tratamento T5DN. Observou-se em T5DN queda de 40% em Ai
em relação a T1.
Também é possível a situação em que o solo apresenta baixa temperatura e dias
ensolarados com temperatura elevada. Essa situação pode ocorrer no fim de inverno, quando a
temperatura do ar está se elevando, mas ainda o solo não foi aquecido. Essas defasagens de
temperaturas são possíveis devido ao efeito ‘tampão’ do solo, não ocorrendo variação da
temperatura tão rápida quanto a do ar (PEREIRA et al., 2000). Essa situação foi simulada pelo
tratamento T2R, em que a temperatura das raízes foi de 10 oC.
Figura 5 - Assimilação diurna de CO2 (Ai) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições.
0
50
100
150
200
250
300
350
400
Ai (
mm
ol m
-2 s
-1)
Tratamento T1 T2
R T3
N T4
NR T5
DN T6
DNR

28
Por fim, a situação mais drástica foi apresentada pelo tratamento T6DNR, em que a
temperatura do ar diurna e noturna e a das raízes foram baixas, num típico dia de inverno no
Estado de São Paulo, em que a queda na produção fotossintética foi muito pronunciada,
representando apenas 23% em relação a T1 (verão), como também foi observado por
RIBEIRO et al. (2009b), comparando Ai de plantas sob condições naturais no verão e no
inverno.
Essa forma de expressar a fotossíntese (Ai) evidencia mais ainda a importância da
ocorrência da baixa temperatura das raízes na determinação das respostas da fotossíntese. Na
média os tratamentos que tiveram baixa temperatura nas raízes (T2R, T4NR e T6DNR) a
produção fotossintética foi aproximadamente a metade em relação a media dos tratamentos
com raízes com temperatura de 26 oC e frio noturno ou diurno (T3N e T5DN).
4.4 Atividade fotoquímica
A atividade fotoquímica envolve a absorção de luz pelos pigmentos no complexo
coletor de luz (light-havesting complex, LHC) e a transferência dessa energia para o centro de
reação do fotossistema II (FSII) e desse para o fotossistema I (FSI) por meio de uma cadeia de
transportadores de elétrons. Durante o transporte linear de elétrons há formação de ATP e
NADPH que são consumidos na redução do CO2, do nitrogênio, do enxofre e na
fotorrespiração (HUNER et al., 1998). As reações fotoquímicas primárias de FSII (absorção e
excitação dos centros de reação da clorofila) são mais rápidas que as reações bioquímicas, e a
exposição da planta a energia que excede àquela requerida nas reações da fotossíntese resulta
em um desbalanço que geralmente causa fotoinibição (HUNER et al., 1998; LONG et al.,
1994). A fotoinibição é o termo utilizado para descrever a redução da capacidade
fotossintética induzida pela exposição a luz visível. A fotoinibição da fotossíntese é sinônimo
de danos ao FSII, mas é importante reconhecer que danos ao FSII por excesso de energia de
excitação somente será fotoinibição, por definição, se ocorrer decréscimo nos valores de
fotossíntese. As condições ambientais que podem levar ao excesso de energia de excitação
são alta irradiância, déficit hídrico, alta temperatura, baixa temperatura, entre outros.
Funcionalmente, a consequência da fotoinibição é a redução da eficiência quântica máxima de
conversão do CO2 absorvido e decréscimo da convexidade da curva de resposta da
fotossíntese à luz, sendo que a exposição excessiva a luz por tempo prolongado pode também
causar decréscimo da fotossíntese máxima sob luz saturante (LONG et al., 1994). Todos os

29
fótons absorvidos pelos pigmentos fotossintéticos excitam as clorofilas, porém quando o fluxo
de fótons excede a capacidade de utilização, pode resultar em danos às membranas
fotossintéticas, se não for dissipado convenientemente. Para evitar esses danos, as plantas
apresentam mecanismos de defesa para dissipar o excesso de energia. Quando tais
mecanismos não são suficientes ocorre a fotoinibição da fotossíntese, devido ao desbalanço
entre a taxa de dano e de reparo do FSII. Os danos ao FSII estão relacionados com o dano à
proteína D1 integrante do complexo protéico do centro de reação. A perda fotoinibitória de
D1 somente inibe a fotossíntese se a taxa de reparo for menor que a taxa de dano (LONG et
al., 1994; CRITCHEY, 1998; HUNER et al., 1998). Há dois tipos de fotoinibição: a dinâmica
e a crônica (OSMOND, 1994). A fotoinibição dinâmica é transiente e está associada
principalmente com variações no ciclo da xantofila (DEMMING-ADAMS & ADAMS III,
1992). A fotoinibição crônica está mais relacionada com os danos à proteína D1 (LONG et
al., 1994; CRITCHEY, 1998; HUNER et al., 1998). Na fotoinibição dinâmica a eficiência
quântica decresce, mas a assimilação máxima de CO2 permanece inalterada. A fotoinibição
dinâmica é causada pelo desvio da energia luminosa absorvida em direção à dissipação do
calor, por isso, o decréscimo na eficiência quântica. As pesquisas anteriores sobre fotoinibição
interpretavam todos os decréscimos de eficiência quântica como dano ao aparato
fotossintético. Hoje, reconhece-se que o decréscimo de curto prazo na eficiência quântica
parece refletir mecanismos protetores, enquanto a fotoinibição crônica representa o dano
presente ao cloroplasto, resultante da luz excessiva ou de falha dos mecanismos protetores.
Além disso, a exposição da planta a condições fotoinibitórias pode resultar na formação de
espécies reativas de oxigênio devido a redução do O2 em ânion super-óxido (1O2-) ou pela
transferência de energia de exitação da clorofila em estado triplet para o O2 formando
oxigênio singlet (1O2) (ASADA, 2000). O 1O2- é muito reativo, e uma vez formado reage
rapidamente com lipídios, pigmentos e proteínas que se encontram próximas a seu sítio de
produção, especialmente na região do FSII (NIYOGI,1999). Os efeitos mais nocivos do 1O2-
reativo são degradação de clorofila e outros pigmentos fotossintéticos, além da destruição da
proteína D1 do FSII e formação de radicais orgânicos (livres) nas membranas dos lipídios. A
perdurar a fotoinibição, podem ocorrer perdas importantes na produtividade das plantas
(LONG et al., 1994). A técnica usada para quantificação da fotoinibição é a medida do
rendimento quântico pela fluorescência da clorofila. A fotoinibição usualmente é medida pela
razão Fv/Fm, que indica a máxima eficiência pela qual a luz absorvida pelo FSII é utilizada
para a redução da quinona A (QA). A eficiência quântica potencial (Fv/Fm) é uma variável

30
que se correlaciona com o número de complexos funcionais do FSII (CRITCHLEY, 1998). A
ocorrência de estresses bióticos e abióticos causa queda em Fv/Fm.
A eficiência quântica potencial do FSII apresentou respostas características em função
do regime térmico a que as plantas foram submetidas (Figura 5). No tratamento T1 Fv/Fm
apresentou valores ao redor de 0,78±0,02, após 12 h de escuro, indicando que o aparato
fotoquímico das folhas estava em bom estado fisiológico e não apresentava sintomas de
fotoinibição (BJÖRKMAN & DEMMIG, 1987; CRITCHLEY, 1998; LONG et al., 1994).
Esse resultado era esperado, visto que as condições impostas no tratamento T1 correspondem
às ideais para laranjeiras, e coincidiu com os valores já observados por outros autores
(MACHADO, 2009; RIBEIRO et al., 2009a,b). Nesse mesmo tratamento os valores de Fv/Fm
foram 0,73 e 0,70, respectivamente, ao redor das 12:00 e 18:00 h. Possivelmente a queda em
Fv/Fm nesses horários foi devida ao maior período de iluminação prévio e, no fim do dia, a
queda que ocorreu em A (Figura 6).
Quando a energia absorvida é maior do que a capacidade de utilização pelas reações
fotoquímicas e/ou bioquímicas da fotossíntese ocorre fotoinibição (BARBER &
ANDERSSON, 1992; LONG et al., 1994; OSMOND, 1994), indicada por Fv/Fm menor que
0,725 (CRITCHLEY, 1998). A queda na razão Fv/Fm usualmente reflete a alteração da taxa
de transporte de elétrons por meio do FSII para o FSI (BERTANI et al., 2005).
A redução de Fv/Fm em T1 (às 12:00 e 18:00 h) foi transiente, visto que os valores
após o período noturno são recuperados, caracterizando fotoinibição dinâmica (CRITCHLEY,
1998; OSMOND, 1994). Esse tipo de fotoinibição ocorre em folhas expostas e reflete redução
da eficiência quântica da fotossíntese sem, no entanto, afetar a capacidade de fixação de CO2
em DFFFA saturante (OSMOND, 1994), e por ser reversível não ocasiona danos ao aparato
fotoquímico (CRITCHLEY, 1998). ÖQUIST et al. (1992) sugerem que a fotoinibição,
conseqüência do funcionamento do FSII sob exposição prolongada à luz, representa uma
retro-regulação estável e a longo prazo, da atividade fotoquímica, que substitui parte da
regulação freqüentemente exercida pelo gradiente de pH. OSMOND & GRACE (1995) citam
que redução de Fv/Fm é essencial para preservar a funcionalidade fotossintética em alto
DFFFA. Logo, pode-se sugerir a função protetora para a fotoinibição ocorrida nos horários
das 12:00 e 18:00 h (Figura 6), quando houve decréscimo de A devido ao aumento das
limitações estomáticas e não estomáticas, após as 10:00 h (Figura 3).
Os tratamentos T2R, T3N e T4NR apresentaram valores de Fv/Fm nos 3 horários de
medida ao redor de 0,7, indicando uma discreta fotoinibição (CRITCHLEY, 1998), que no
entanto não deve ter influenciado A. Logo, pode-se presumir que os tecidos fotossintetizantes

31
estavam submetidos a excesso de energia radiante (SCHREIBER & BILGER, 1987). Nesses
tratamentos (T2R, T3N e T4NR), a temperatura no período diurno foi de 28 oC, ou seja, ideal
para a fotossíntese, porém em cada um deles ou a temperatura noturna (T3N) ou a temperatura
das raízes (T2R) ou ambas estavam em condições estressantes (T4NR).
De acordo com FLEXAS et al. (2001) a redução de Fv/Fm devido a baixas
temperaturas é uma conseqüência da baixa fixação de CO2, indicando a retro-regulação da
atividade fotoquímica para manter um suprimento adequado de elétrons e ao mesmo tempo
evitar quantidade excessiva de energia radiante nos fotossistemas, capaz de levar à
fotoinibição crônica e fotooxidação. Assim parece que nos tratamentos T2R, T3N e T4NR a
queda de Fv/Fm foi devida a queda na fixação de CO2 e teve função protetora do sistema de
transporte de elétrons.
Nos tratamentos T5DN (TD=17, TN=10, TR=26 oC) e T6DNR (TD=17, TN=10, TR=10 oC),
as temperaturas no período diurno foram menores do que nos outros tratamentos. Em ambos
os tratamentos houve queda acentuada em Fv/Fm, ou seja de 0,78, observado em T1, para
0,63 e 0,51, respectivamente, em T5DN e T6DNR, caracterizando fotoinibição crônica visto que
os valores foram baixos após o período noturno de 12 h, não voltando a apresentar
funcionamento normal da atividade fotoquímica (Figura 6). A queda em Fv/Fm nesses dois
tratamentos foi relacionada com a queda em Fm (Figura 6B), indicando inativação
fotoquímica do FSII pelo efeito sobre o centro de reação. Fo foi semelhante em todos os
tratamentos.
No tratamento T2R (TD=28, TN=20, TR=10 oC), apesar da queda significativa de A
(Figura 3), a redução em Fv/Fm (0,72) foi relativamente baixa em relação a T1 (0,78). Assim,
possivelmente a baixa fotoinibição, não foi a causa principal da queda de A. Provavelmente o
mecanismo de decréscimo de A quando o sistema radicular esta sob condições de baixa
temperatura esteja mais relacionado com a significativa queda em gs e em A/Ci (Figura 3).
MAGALHÃES FILHO et al. (2009) também observaram que a queda de A em laranjeira com
o sistema radicular submetido a baixa temperatura (10 oC) foi relacionada tanto com aspectos
difusivos como metabólicos, ou seja, queda acentuada em gs e na eficiência de carboxilação
(Vc,max) e taxa de regeneração de RuBP (Jmax). DELUCIA et al. (1991) relataram queda em
Fv/Fm devido a baixa temperatura no sistema radicular (1 oC) em Pinus silvestris L., mas
encontraram evidências indicando que a queda em A foi associada a danos no metabolismo
fotossintético.
32
Figura 6 – Fluorescência basal (Fo em A, fluorescência máxima (Fm em B) e eficiência quântica máxima do FSII (Fv/Fm em C) em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes (24 h). Barras indicam desvio padrão da média referente a 4 repetições.
6 12 180
100
200
300
400
500
Hora
A
Fo
T1 T2R T3N
T4NR
T5DN
T6DNR
6 12 180,3
0,4
0,5
0,6
0,7
0,8C
Fv/F
m
Hora
6 12 180
200
400
600
800
1000
1200
1400
Hora
B
Fm

33
No tratamento T3N (TD=28, TN=10, TR=26 oC) em que o fator estressante foi a baixa
temperatura noturna, também a queda em Fv/Fm foi baixa, ou seja, de 0,78 em T1 para 0,7
em T3N. Como no caso anterior (T2R) possivelmente a queda em Fv/Fm foi uma conseqüência
da redução em A e não a causa. MACHADO (2009) observou, também em laranjeira
‘Valência’ sobre ‘Cravo’, que a queda em Fv/Fm somente ocorreu após a terceira noite
submetida ao tratamento de frio noturno, enquanto a redução em A ocorreu já após a primeira
noite de tratamento. Resultados semelhantes foram relatados também para mangueira, em que
a redução de Fv/Fm não foi a causa da redução em A após frio noturno (ALLEN et al., 2000).
Também nesse caso, a queda em A foi devida a causas difusivas e metabólicas, ou seja, houve
grande queda nos valores de gs e nos valores de A/Ci, em conformidade com o que foi
relatado por MACHADO (2009).
No tratamento T4NR (TD=28, TN=10, TR=10 oC) em que houve combinação de frio nas
raízes e frio noturno, apesar da redução em A ter sido maior que em T2R e T3N, Fv/Fm foi
semelhante, indicando também que a queda de A, possivelmente foi a causa da queda em
Fv/Fm e não conseqüência (Figuras 3 e 6). RIBEIRO et al. (2009a) observaram queda de
aproximadamente 70% em A em plantas submetidas a baixa temperatura noturna e nas raízes,
contudo sem queda significativa em Fv/Fm. Conseqüentemente, esses autores sugeriram que
as causas da queda em A com ocorrência de frio noturno e nas raízes concomitantemente,
foram relacionadas a fatores difusivos e metabólicos expressos respectivamente, pela queda
em gs, Vc,max e Jmax.
Vários estudos têm demonstrado que o excesso de radiação induz, à partir de
violaxantina, a formação de zeaxantina e anteroxantina, as quais estão correlacionadas com
aumento de extinção não fotoquímica da fluorescência e queda de Fv/Fm (LONG et al.,
1994). Durante a manifestação do ciclo das xantofilas há dissipação de energia, sendo assim, a
queda em Fv/Fm pode ser interpretada como um mecanismo de fotoproteção.
Já foi discutido que a queda de A em T5DN (TD=17, TN=10, TR=26 oC) pode ser
comparada diretamente com o T3N (TD=28, TN=10, TR=26 oC), tratamento do qual difere
apenas pela temperatura no período diurno. Assim, a queda de A em T5DN em relação a T3N
esta relacionada com a menor temperatura no período diurno. Verificou-se que tal diferença
foi devida a valores relativamente menores de gs e A/Ci em T5DN em relação a T3N. É
interessante notar, quando se considera a fotossíntese diurna, que a diferença entre Ai em T3N
e T5DN (Figura 5) não foi tão acentuada (AiT3 = 1,23*AiT5), porém ocorreu efeito acentuado da

34
temperatura diurna sobre o aparato fotossintético, demonstrado pela queda de Fv/Fm em T5DN
(Figuras 3 e 6C).
O tratamento T6DNR (TD=17, TN=10, TR=10 oC) pode ser diretamente comparado com
o T4NR (TD=28, TN=10, TR=10 oC), sendo a maior queda em T6DNR causada pelos menores
valores em gs e A/Ci (Figura 3), sendo a relação entre T4NR e T6DNR descrita por AiT4 =
1,35*AiT6. Obviamente na comparação entre T5DN e T6DNR (AiT5 = 2,52*AiT6), a diferença esta
relacionada com a baixa temperatura no sistema radicular.
A principal causa da queda de Fv/Fm em T5DN e T6DNR deve estar relacionada com a
baixa temperatura no período diurno. É conhecida a ocorrência de fotoinibição sob condições
de baixa temperatura em folhas, associada com alta radiação (ALLEN & ORT, 2002;
BERTAMINI et al., 2005; LONG et al., 1994), efeito esse caracterizado pela redução em
Fv/Fm (LONG et al., 1994). Baixa temperatura durante o dia causa redução da atividade das
reações bioquímicas (Figura 3D), limitando a utilização de energia (ATP e NADPH) na
fixação de carbono pelo ciclo de Calvin e do fosfoglicolato (ALLEN & ORT, 2002). Baixo
dreno para a absorção da energia de excitação, combinado com baixa eficiência de síntese da
proteína D1 danificada, aumenta o potencial de dano oxidativo do FSII (LONG et al., 1994).
Também a baixa temperatura foliar reduz significativamente a fluidez de membranas,
prejudicando a difusão de metabólicos envolvidos na fotossíntese (ALLEN & ORT, 2002).
4.5 Variação do conteúdo de carboidratos
Nos citros, a sacarose é a principal forma pela qual os fotoassimilados são
transportados pelo floema. O amido é uma reserva estável e insolúvel de carboidratos,
presente em quase todas as plantas. Na folha, tanto o amido como a sacarose são sintetizados
a partir da triose-fosfato gerada pelo ciclo de Calvin. As sínteses de sacarose e de amido
reações competitivas, sendo as concentrações relativas de ortofosfato (Pi) e triose-fosfato os
principais fatores que controlam se o carbono fixado fotossinteticamente será
compartimentado como amido no cloroplasto ou como sacarose no citossol, sendo a primeira
via acionada preferencialmente quando o crescimento é reduzido, ao passo que a segunda via
é mais ativa quando há drenos ativos. Uma baixa concentração de Pi no citossol limita a
exportação de triose-fosfato do cloroplasto por intermédio do transportador, promovendo,
assim, a síntese de amido. Por outro lado, uma abundância de Pi no citosol inibe a síntese de
amido dentro do cloroplasto e estimula a exportação de triose-fosfato para o citossol, onde é
convertida em sacarose (PAUL & FOYER, 2001).

35
Figura 7 – Conteúdo de açúcares solúveis, de sacarose, de amido e carboidratos totais em folhas, caules e raízes em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC. TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições.
0
30
60
90
120
150
T1 T2R T3
N T4
NR T5
DN T6
DNR
Folha
g G
lu (k
g M
S)-1
0
20
40
60
80
100
g G
lu (k
g M
S)-1
Caule
Açúcar solúvel Sacarose Amido Carboidrato total
0
20
40
60
80
100T1 T2
R T3
N T4
NR T5
DN T6
DNR
T1 T2R T3
N T4
NR T5
DN T6
DNR
g G
lu (k
g M
S)-1
Tratamento
Raiz

36
Os carboidratos exportados da fonte, como sacarose, para qualquer órgão dreno são
utilizados pela respiração e para o próprio crescimento do dreno. Quando a quantidade de
sacarose que chega ao órgão dreno é maior que a utilizada, há acúmulo de reservas,
principalmente como amido no próprio órgão dreno ou de armazenamento. Assim, o balanço
de reservas em uma folha difere do acúmulo de reservas nos órgãos-dreno. A dinâmica de
produção, exportação, acúmulo/armazenamento e utilização de fotoassimilados é complexa,
visto que envolve vários aspectos metabólicos e suas interações com o ambiente.
Nas folhas o teor de açúcares solúveis foi semelhante em todos os tratamentos, porém
houve diferenças quando considerados os teores de sacarose e de amido, em função do
tratamento térmico (Figura 7). Em T1 os teores de sacarose e amido foram relativamente
baixos, ainda que a produção fotossintética tenha sido alta (Figura 5).
Nas folhas, o resfriamento somente da parte aérea durante a noite, ou em conjunto com
o resfriamento das raízes nos tratamentos T3N (TD=28, TN=10, TR=26 °C) e T4NR (TD=28,
TN=10, TR=10 °C), respectivamente, promoveu aumento no conteúdo de amido e sacarose em
relação ao tratamento referência (Figura 7). A maior concentração de sacarose pode estar
relacionada ao transporte menos intenso das folhas para outras partes da planta, em função da
diminuição do metabolismo causado pela baixa temperatura noturna (FOYER, 1988).
MACHADO (2009) observou resposta semelhante também em laranjeira ‘Valência’ sobre
‘Cravo’. O amido normalmente é hidrolisado à noite para atender a demanda de órgãos-dreno
em crescimento (RIBEIRO et al., 2010). MACHADO (2009) sugeriu que a causa de aumento
do amido devido à baixa temperatura noturna poderia estar relacionado com: 1) inibição ou
diminuição da atividade das enzimas de mobilização de amido (LEEGOOD & EDWARD,
1996) e/ou 2) diminuição da demanda por carboidratos devido a possível queda no
crescimento (IGLESIAS et al., 2002; RIBEIRO & MACHADO, 2007). Porém, é importante
lembrar que a produção de fotoassimilados (Ai) em T3N e, principalmente, em T4NR, foi mais
baixa que em T1 (Figura 5), mesmo que os teores de sacarose e de amido tenham sido mais
baixos em T1. Em T3N e T4NR, o teor de amido nos caules e nas raízes foi mais alto que em
T1. No regime térmico de frio somente na parte aérea (noturno e diurno), representado
pelo tratamento T5DN (TD=17, TN=10, TR=26 °C) os teores de carboidratos nas folhas foram
semelhantes aos do T1, mas ocorreu maior acúmulo (p<0,05) de amido na raiz em T5DN em
relação a T1 (Figura 7C). Também se deve considerar que Ai em T5DN foi somente menor que
em T1 e T3N. Assim, o acúmulo de amido nas raízes em T5DN pode ser justificado pelo fato
de que sob baixa temperatura na parte aérea, a demanda de carboidrato pelas brotações e

37
folhas em crescimento possivelmente foi pequena, e o excedente pode ter migrado para as
raízes. Sob temperatura aérea diurna e noturna de 17 e 10 oC, respectivamente, o crescimento
é baixo (BEVINGTON & CASTLE, 1985; GOLDSCHMIDT & GOLOMB; 1982; PRADO et
al., 2007; RIBEIRO, 2006)
Os teores de carboidratos nas folhas em T2R foram semelhantes à T1, embora Ai tenha
sido bem menor em T2R (Figuras 5, 7 e 8). Mas, tanto no caule como na raiz os teores de
amido foram maiores que em T1. Isso implica em que a produção de fotoassimilados foi
maior que a utilização. O crescimento radicular ocorre de forma ininterrupta, variando apenas
sua velocidade em função da disponibilidade de água, aeração e temperatura, que deve ser
superior a 13 °C e inferior a 36 °C (CASTLE, 1980; SPIECEL-ROY & GOLDSCHMIDT,
1996). Dessa forma, quando a raiz, que nessa fase representa 60% da fitomassa da planta
(Figura 2B), foi submetida a uma temperatura de 10 °C, a demanda por carboidratos
possivelmente se tornou baixa devido à inibição do crescimento. Mesmo sob baixa produção
fotossintética deve ter ocorrido excesso de fotoassimilados, que proporcionaram aumento das
reservas em caules e raízes em T2R.
Em T6DNR, os teores de carboidratos em todos os órgãos (folhas, caules e raízes) foram
semelhantes aos encontrados em T1, apesar de neste tratamento Ai (Figura 5) ter sido
aproximadamente 23% de T1. Portanto, se no tratamento T1, Ai era alto para atender a
demanda de carboidratos sem que houvesse excesso, em T6DNR, Ai foi muito baixo, mas,
devido a baixa temperatura em todos os órgãos, sua demanda também foi baixa, não havendo
acúmulos de amido em caules e raízes.
Em relação aos tratamentos T2R, T3N, T4NR e T5DN, apesar de apresentarem menores
produções de fotoassimilados em relação a T1, ocorreu maior acúmulo de amido nos caules e
raízes, indicando que o balanço entre o que chega aos drenos e o que é utilizado na respiração
e crescimento é que determina o acúmulo e/ou a utilização das reservas.
4.6 Relações entre a fotossíntese e o teor de carboidratos nas folhas O crescimento de uma planta está relacionado com a produção de fotoassimilados na
fonte e com a forma como eles são distribuídos e utilizados nos diversos órgãos-dreno. Por
outro lado, as atividades dos drenos afetam a eficiência fotossintética e a forma com que
fotossintatos são distribuídos. Dessa forma há inter-relação entre esses processos, que são
dependentes do estádio fenológico das plantas e afetados pelo ambiente (DICKINSON, 1991;
GOLDSCHMIDT & KOCH, 1996; PAUL & FOYER, 2001).

38
Em todos os tratamentos em que houve queda em A e Ai em relação ao tratamento
referência, T1, a causa foi a ocorrência de temperaturas baixas nas raízes, ou na parte aérea
durante o período diurno ou noturno ou ainda a combinação entre eles. Geralmente, a baixa
temperatura limita tanto o crescimento como a fotossíntese (BOESE & HUNER, 1990). As
causas relativas a baixa temperatura para queda em A foram discutidas nos itens anteriores.
No entanto, uma das causas relatadas na literatura para queda em A sob baixa temperatura é a
restrição à atividade de crescimento dos drenos (BOESE & HUNER, 1990). O carbono fixado
fotossinteticamente é direcionado para a síntese de amido no cloroplasto ou de sacarose no
citoplasma celular (PAUL & FOYER, 2001), sendo a rota preferencial dependente da
demanda de carboidratos pela planta. Assim, quando o crescimento é reduzido, a via
preferencial é síntese de amido, que fica armazenado nos cloroplastos. Quando o crescimento
é intenso, a rota preferencial é a síntese de sacarose, que é distribuída para toda planta. O
amido pode ser reutilizado, caso a atividade metabólica torne-se novamente intensa. A queda
da fotossíntese, devido ao aumento do teor de carboidratos na folha, está relacionada com o
decréscimo da reciclagem de Pi no cloroplasto, causando queda na síntese de ATP e NADPH
e na regeneração de RuBP (PAUL & FOYER, 2001). No entanto, no caso presente não
ocorreu esse tipo de resposta, ou seja, queda de A com o acúmulo de amido ou de carboidratos
totais nas folhas (Figura 8). Por outro lado, tanto Amax como Ai não se correlacionaram com
o conteúdo de amido e/ou teor de carboidratos totais na folha (Figura 8A e B). Por exemplo,
os teores de amido (41,1±7,3 g Glu kg-1 folha) e de carboidratos totais (97,1±3,4 g Glu kg-1
folha) no tratamento T1, que apresentou mais altas Amax e Ai, foram maiores que os teores de
amido (24,2±7,8 g CH2O kg-1) e carboidratos totais (76,0±12,6 g CH2O kg-1) no tratamento
T6DNR, que apresentou os valores mais baixos de Amax e Ai (Figura 8A e B). No entanto,
também em citros, IGLESIAS et al. (2002) de forma contrária ao que observamos, sugeriram
que a elevada concentração de açúcares, per se, induz a inibição da fotossíntese. Os trabalhos
que indicam a inibição ou estímulo da fotossíntese de citros pelo conteúdo de carboidratos
foram conduzidos com a imposição de fatores exógenos que alteravam a relação fonte-dreno
por meio de modificações em um determinado órgão da planta, tais como o anelamento, a
poda, o suprimento externo de sacarose e o desbaste de frutos (IGLESIAS et al., 2002;
SYVERTSEN, 1994). No entanto, DICKSON (1991) cita que o controle da alocação de
carbono, da síntese de sacarose e amido e da translocação de açúcares deve ser estudado em
tecidos e plantas intactas, ou seja, em condições mais próximas das naturais, uma vez que os
artifícios utilizados para modificar as relações fonte-dreno podem afetar as resposta das
plantas de forma descontrolada.

39
Alguns autores destacam (GOLDSCHIMIDT & KOCH, 1996; RAMOS, 2009;
RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2010) que o controle da fotossíntese devido
ao acúmulo de carboidratos nas folhas de citros não é evidente sob condições naturais e pode
ser complementar a um mecanismo dinâmico da relação fonte-dreno (RIBEIRO et al, 2010).
A inibição da fotossíntese pelo acúmulo de carboidratos é evidenciada em experimento em
que se manipula artificialmente a relação fonte:dreno (GOLDSHIMIDT & HUBER, 1992;
GOLDSCHIMIDT & KOCH, 1996; IGLESIAS et al., 2002; SYVERTSEN, 1994). Porém,
observou-se, em laranjeira, sem a manipulação artificial da relação fonte:dreno, a ocorrência
de altos valores de A em presença de altos teores de carboidratos (RIBEIRO & MACHADO,
2007; RAMOS, 2009, RIBEIRO et al., 2010), sugerindo que A está mais relacionada à
dinâmica diária de produção e de consumo de carboidratos na folha e não propriamente ao
maior teor de reservas (RIBEIRO et al., 2005; RIBEIRO & MACHADO, 2007; RAMOS,
2009; RIBEIRO et al., 2010 ).
Recentemente RIBEIRO et al. (2010), analisaram comparativamente a relação entre
exportação de fotoassimilados, conteúdo de carboidratos e A em laranjeiras durante o verão e
inverno. As condições ambientais no verão e no inverno induzem diferentes intensidades de
crescimento de forma natural. Esses autores observaram relação negativa entre teor de amido
e A em tecidos foliares submetidos a diferentes períodos de ausência de luz (12, 48 e 96 h).
Independentemente da estação de crescimento, quanto maior o período em que a folha
permanecia no escuro, mais baixas as reservas (consumidas no escuro) e, consequentemente,
no período subseqüente de luz, maior a taxa de fotossíntese. Ou seja, foi observada relação
negativa entre teor de carboidratos e fotossíntese. No entanto, quando as mesmas plantas não
foram submetidas à manipulação artificial da relação fonte:dreno os autores observaram, que
durante o verão, tanto A como a exportação de fotoassimilados foliares e o teor de
carboidratos eram bem mais altos que no inverno e que A era relacionado positivamente com
a exportação de fotoassimilados foliares, mas não com o teor de carboidratos. RAMOS
(2009), de forma semelhante observou correlação entre exportação de fotoassimilados e A em
plantas submetidas a diferentes regimes térmicos, porém, não encontrou relação entre A e teor
de carboidratos. No experimento aqui apresentado, nossos resultados estão de acordo com os
observados por RIBEIRO et al. (2010) e RAMOS (2009), em relação à ausência de correlação
entre o teor de amido e/ou de carboidratos totais e fotossíntese.
Apesar da hipótese de que a fotossíntese pode ser controlada pelo teor de assimilados
na folha fonte ser largamente aceita, tal mecanismo de regulação nem sempre ocorre sob
condições naturais e é sujeito a contestações (PAUL & FOYER, 2001; GOLDSHIMIDT &
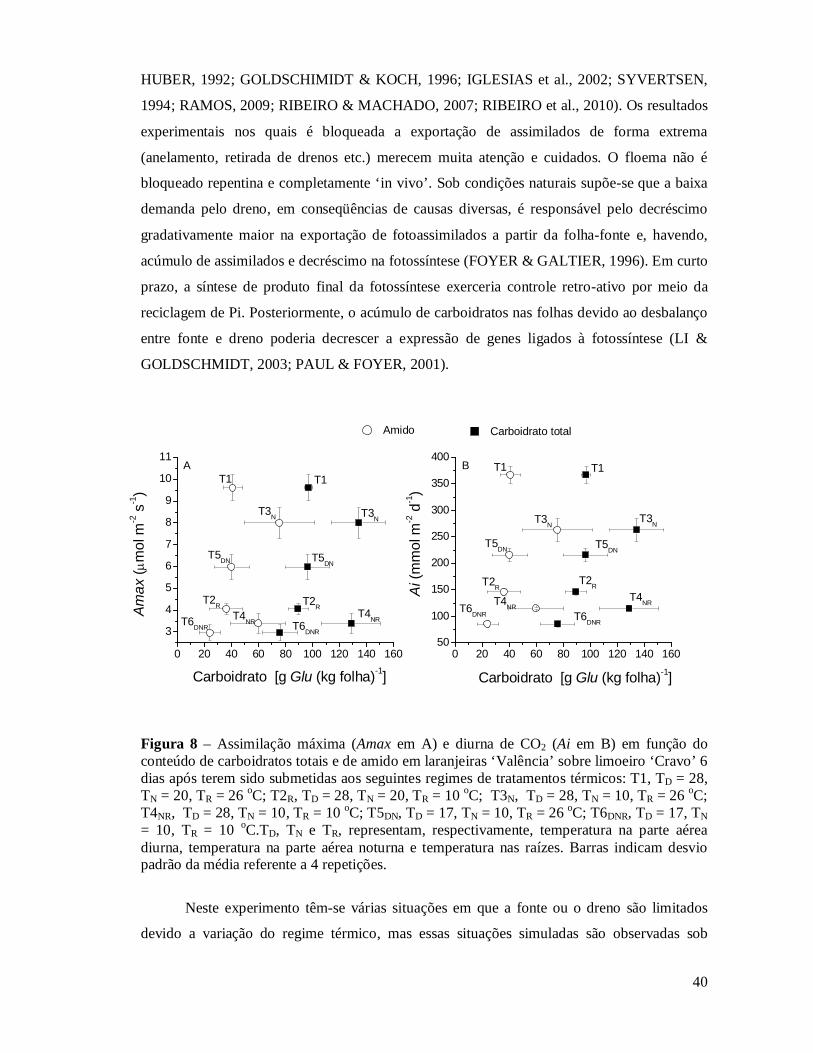
40
HUBER, 1992; GOLDSCHIMIDT & KOCH, 1996; IGLESIAS et al., 2002; SYVERTSEN,
1994; RAMOS, 2009; RIBEIRO & MACHADO, 2007; RIBEIRO et al., 2010). Os resultados
experimentais nos quais é bloqueada a exportação de assimilados de forma extrema
(anelamento, retirada de drenos etc.) merecem muita atenção e cuidados. O floema não é
bloqueado repentina e completamente ‘in vivo’. Sob condições naturais supõe-se que a baixa
demanda pelo dreno, em conseqüências de causas diversas, é responsável pelo decréscimo
gradativamente maior na exportação de fotoassimilados a partir da folha-fonte e, havendo,
acúmulo de assimilados e decréscimo na fotossíntese (FOYER & GALTIER, 1996). Em curto
prazo, a síntese de produto final da fotossíntese exerceria controle retro-ativo por meio da
reciclagem de Pi. Posteriormente, o acúmulo de carboidratos nas folhas devido ao desbalanço
entre fonte e dreno poderia decrescer a expressão de genes ligados à fotossíntese (LI &
GOLDSCHMIDT, 2003; PAUL & FOYER, 2001).
Figura 8 – Assimilação máxima (Amax em A) e diurna de CO2 (Ai em B) em função do conteúdo de carboidratos totais e de amido em laranjeiras ‘Valência’ sobre limoeiro ‘Cravo’ 6 dias após terem sido submetidas aos seguintes regimes de tratamentos térmicos: T1, TD = 28, TN = 20, TR = 26 oC; T2R, TD = 28, TN = 20, TR = 10 oC; T3N, TD = 28, TN = 10, TR = 26 oC; T4NR, TD = 28, TN = 10, TR = 10 oC; T5DN, TD = 17, TN = 10, TR = 26 oC; T6DNR, TD = 17, TN = 10, TR = 10 oC.TD, TN e TR, representam, respectivamente, temperatura na parte aérea diurna, temperatura na parte aérea noturna e temperatura nas raízes. Barras indicam desvio padrão da média referente a 4 repetições.
Neste experimento têm-se várias situações em que a fonte ou o dreno são limitados
devido a variação do regime térmico, mas essas situações simuladas são observadas sob
0 20 40 60 80 100 120 140 160
3
4
5
6
7
8
9
10
11
0 20 40 60 80 100 120 140 16050
100
150
200
250
300
350
400
T6DNR
T4NR
T2R
T5DN
T3N
Am
ax (
mol
m-2 s
-1)
T6DNR
T2R
T4NR
T5DN
T3N
T1T1
Amido
A
Carboidrato total
T6DNR
T6DNR
T4NRT4NR
T2RT2R
T5DNT5DN
T3NT3N
Carboidrato [g Glu (kg folha)-1]
T1
Carboidrato [g Glu (kg folha)-1]
T1B
Ai (
mm
ol m
-2 d
-1)

41
condições naturais, principalmente em culturas que permanecem no campo por vários anos
seguidos, como é o caso de laranjeiras. Assim, as interações fonte:dreno em citros estariam
baseadas na dinâmica de carboidratos mais do que no teor de carboidratos per se, conforme
sugeriram RIBEIRO et al. (2010).
5 CONCLUSÕES
1) Em laranjeiras o regime de temperatura influencia a fotossíntese, agindo sobre
fatores difusivos, metabólicos e fotoquímicos. Também influencia as relações fonte:dreno e o
armazenamento de reservas nos vários órgãos da planta.
2) A ocorrência de baixa temperatura da parte aérea (diurna e/ou noturna) e/ou das
raízes, causa queda na atividade fotossintética, afetando a difusão e a eficiência aparente de
carboxilação (A/Ci). A baixa temperatura no período diurno causa fotoinibição crônica,
evidenciada pela queda na eficiência quântica potencial (Fv/Fm).
3) A baixa temperatura das raízes é mais prejudicial ao processo fotossintético quando
comparada à ocorrência de baixa temperatura na parte aérea, no período diurno e/ou noturno.

42
6 REFERÊNCIAS BIBLIOGRÁFICAS
ALLEN, D.J.; ORT, D.R. Impacts of chilling temperatures on photosynthesis in warm-climate plants. Trends in Plant Science, London, v. 6, p. 36-42, 2001. ALLEN, D.J.; RATNER, K.; GILLER, Y.E.; GUSSAKOVSKY, E.E.; SHAHAK, Y.; ORT, D.R. An overnight chill induces a delayed inhibition of photosynthesis at midday in mango (Manguifera indica L.). Journal of Experimental Botany, Oxford, v. 51, p. 1893-1902, 2000. AMARAL, L.I.V.; GASPAR, M.; COSTA, P.M.F.; AIDAR, M.P.M.; BUCKERIDGE, M.S. Novo método enzimático rápido e sensível de extração e dosagem de amido em materiais vegetais. Hoehnea, São Paulo v. 34, p. 425-431, 2007. ASADA, K. The water-water cycle as alternative photon and electron sinks. Philosophical Transactions of the Royal Society of London Series B – Biological Sciences, London, v. 355, p. 1419-1431, 2000. AZCÓN-BIETO, J. Inhibition of photosynthesis by carbohydrates in wheat leaves. Plant Physiology, Rockville, v. 73, p. 681-686, 1983. BARBER, J.; ANDERSSON, B. Too much of a good thing: light can be bad for photosynthesis. Trends in Biochemical Sciences, London, v. 17, p. 61-66, 1992. BAUER, H.; WIERER, W.H.; HATHEWAY, H.; LARCHER, W. Photosynthesis of Coffea arabica after chilling. Physiologya Plantarum, Copenhagen, v. 64, p. 449-454, 1985. BERNACCHI, C.J.; PORTIS, A.R.; NAKANO, H.; VON CAEMMERER, S.; LONG, S.P. Temperature response of mesophyll conductance. Implications for the determination of Rubisco enzyme kinetics an for limitations to photosynthesis in vivo. Plant Physiology, Rockville, v. 130, p. 1992-1998, 2002. BERRY, J.; BJÖRKMAN, O. Photosynthetic response and adaptation to temperature in higher plants. Annual Review of Plant Physiology, Palo Alto, v. 31, p. 491-543, 1980. BERTAMINI, K. MTHUCHELIAN, K.; RUBINIGG, M. ZORER, R. NEDUNCHEZHIAN, N. Low-night temperature (LNT) induce changes of photosynthesis in grapevine (Vitisvinifera L.) plants. Plant Physiology and Biochemistry, Paris, v. 43, p. 693-699, 2005. BEVINGTON, B.K.; CASTLE, W.S. Annual root growth pattern of young citrus trees in relation to shoot growth, soil temperature, and soil water content. Journal of American Society of Horticultural Science, Alexandria, v.110, p. 840-845, 1985. BIELESKI, R.L.; TURNER, A. Separation and estimation of amino acids in crude plant extracts by thin-layer electrophoresis and chromatography. Analytical biochemistry, Amsterdam, v. 17, p. 278-293, 1966.

43
BJÖRKMAN, O.; BADGER, M.R.; ARMOND, P.A. Response and adaptation of photosynthesis to high temperatures. In: TURNER, N.C.; KRAMER, P.J. Adaptation of plants to water and high temperature stress. New York: John Wiley, 1980. chap. 15, p. 233-249. BJÖRKMAN, O.; DEMMIG, B. Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77K among vascular plants of diverse origins. Planta, New York, v. 170, p. 489-504, 1987. BOESE, S.R.; HUNER, N.P.A. Effect of growth temperature and temperature shifts on spinach morphology and photosynthesis. Plant Physiology, Rockville, v. 94, p. 1830-1836, 1990. BOHNERT, H.J; NELSON, D.E.; JENSEN, R.G. Adaptations to environmentalstresses. Plant, Cell and Environment, Oxford, v.7, p. 1099-1111, 1995. BOUCHER, J.F.; BERNIER, P.Y.; MUNSON, A.D. Radiation and soil temperatureinteractions on the growth and physiology of eastern white pine (Pinus strobus L.) seedlings. Plant and Soil, Dordrecht, v.236, p. 165-174, 2001. CAMARGO, A.P.; PINTO, H.S.; BRUNINI, O.; PEDRO JUNIOR, M.J.; ORTOLANI, A.A.; ALFONSI, R.R. Clima do Estado de São Paulo. In: SÃO PAULO (Estado). Secretaria de Agricultura. Zoneamento agrícola do Estado de São Paulo. São Paulo, 1974. v. 1, cap. 2, p. 51-88). CASTLE, W.S. Citrus root systems: their structure, function, growth and relationship totree performance. Proceeding of International Society of Citriculture, Griffith, p. 62-69, 1980. CRITCHLEY, C. Photoinhibition. In: RAGHAVENDRA, A.S. Photosynthesis: a comprehensive treatise. Cambridge: Cambridge University Press. 1998. chap. 20, p. 264-272. DAVIES, F.S.; ALBRIGO, L.G. Citrus. Wallingford: CAB International, 1994. 254 p. DAVIES, W.J.; ZHANG, J. Root signals and the regulation of growth and development of plants in drying soils. Annual Review of Plant Physiology and Plant Molecular Biology, Palo Alto, v. 42, p. 55-76, 1991. DAY, T.A.; HECKATHORN, S.A.; DELUCIA, E.H. Limitations of photosynthesis in Pinus taeda L. (loblolly-pine) at low soil temperatures. Plant Physiology, Rockville, v.96, p. 1246-1254, 1991. DELUCIA, E.H.; DAY, T.A.; OQUIST, G. The potential for photoinhibition of pinus sylvestris.Seedlings exposed to high light and low soil-temperature. Journal of Experimental Botany, Oxford, v.42, p. 611-617, 1991. DEMMIG-ADAMS, B.; ADAMS III, W.W. Photoprotection and other responses of plants to high light stress. Annual Review of Plant Physiology and Plant Molecular Biology, Palo Alto, v. 43, p. 599-626, 1992.

44
DODD, I.C.; HE, J.; TURNBULL, C.G.N.; LEE, S.K.; CRITCHLEY, C. The influence of supra-optimal root-zone temperatures on growth and stomatal conductance in Capsicum annuum L. Journal of Experimental Botany, Oxford, v.51, p. 239–248, 2000. DUBOIS, M.; GILLES, K.A.; HAMILTON, J.K.; REBERS, P.A.; SMITH, F. Colorimetric method for determination of sugars and related substances. Analytical Chemistry, Washington, v. 28, n.30, p. 350-356, 1956. ERISMANN, N.M.; MACHADO, E.C.; TUCCI, M.L.S. Photosynthetic limitation by CO2 diffusion in drought stressed orange leaves on three rootstocks. Photosynthesis Research, Dordrecht, v.96, p. 163-172, 2008. FENNELL, A.; MARKHART, A.H. Rapid acclimation of root hydraulic conductivity to low temperature. Journal of Experimental Botany, Oxford, v.49, p. 879-884, 1998. FLEXAS, J.; BADGER, M.; CHOW, W.S.; MEDRANO, H.; OSMOND, C.B. Analysis of the relative increase in photosynthetic O2 uptake when photosynthesis in grapevine leaves is inhibited following low night temperatures and/or water stress. Plant Physiology, Rockville, v. 121, p. 675-684, 1999. FLEXAS, J.; GULIAS, J.; JONASSON, S.; MEDRANO, H.; MUS, M. Seasonal patterns and control of gas exchange in local populations of the Mediterranean evergreen shrub Pistacia lentiscus L. Acta Oecologica, Paris, v. 22, p. 33-43, 2001. FLEXAS,J.; RIBAS-CARBÓ, M.; DIAZ-ESPEJO, A.; GALMÉS, J.;MEDRANO,H. Mesophyll conductance to CO2: current knowledge and future prospects. Plant, Cell and Environment , Oxford, v. 31, p. 602–621, 2008. FNP CONSULTORIA & COMÉRCIO. Agrianual 2006: anuário da agricultura brasileira. 11. ed. São Paulo, 2006, 536p. FOYER, C.H. Feedback inhibition of photosynthesis through source-sink regulation in leaves. Plant Physiology and Biochemistry, Paris, v.26, p. 483-492, 1988. FOYER, H.F.; GALTIER, N. Source-sink interation and communicatio in leaves. In: ZAMSKI, E.; SCHAFFER, A.A. Photoassimilate distribution in plants and crops: source-sink relationships. New York: Marcel Dekker, 1996. chap. 34, p. 797-823. GENTY, B.; BRIANTAIS, J.-M.; BAKER, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochimica et Biophysica Acta, Amsterdam, v. 990, p. 87-92, 1989. GEORGE, H.L..; DAVIES, F.S.; CRANE, J.H.; SCHAFFER, B. Root temperature effects on ‘Arkin’ carambola (Averrhoa carambola L.) trees. I. Leaf gas exchange and water relations. Scientia Horticulturae, Amsterdam v. ,96 p. 53-65, 2002. GOLDSCHMIDT, E.E.; GOLOMB, A. The carbohydrate balance of alternate-bearing citrus trees and the significance of reserves for flowering and fruiting. Journal of American Society of Hortultural Science, Alexandria, v.107, p. 06-208, 1982.

45
GOLDSCHMIDT, E. E. Carbohydrate supply as a critical factor for citrus fruit development and productivity. HortScience, Alexandria, v. 34, p. 1020-1024, 1999. GOLDSCHMIDT, E.E.; HUBER, S.C. Regulation of photosynthesis by end-product accumulation in leaves of plants storing starch, sucrose, and hexose sugars. Plant Physiology, Rockville, v. 99, p.1443-1448, 1992. GOLDSCHMIDT, E.E.; KOCH, K.E. Citrus. In: ZAMSKI, E.; SCHAFFER, A.A. (Eds.) Photoassimilate distribution in plants and crops. Source-sink relationships. Marcel Dekker, Inc. New York, 1996. p. 797-823. GOMES, M.M.A.; LAGÔA, A.M.M.A.; MACHADO, E.C.; MEDINA, C.L.; MACHADO, M.A. Gas exchanges and carbohydrate metabolism in orange trees with citrus variegated chlorosis. Brazilian Journal of Plant Physiology, Brasília, v. 15, p. 25-31, 2003 GUO, Y.P.; ZHOU, H.F.; ZHANG, L.C. Photosynthetic characteristics and protective mechanisms against photooxidation during high temperature stress in two citrus species. Scientia Horticulturae, Amsterdam, v.108, p. 260-267, 2006. HABERMANN, G; MACHADO, E. C. ; RODRIGUES, J D ; MEDINA, C L . Gas exchange rates at different vapor pressure deficits and water relations of ‘Pera’ sweet orange plants with citrus variegated chlorosis (CVC). Scientia Horticulturae, Amsterdam, v. 98, p. 233-245, 2003. HABERMANN, G.; RODRIGUES, J.D. Leaf gas exchange and fruit yield in sweet orange trees as affected by Citrus Variegated Chlorosis and environmental conditions. Scientia Horticulturae, Amsterdam v. 122, p, 69-76, 2009. HÄLLGREN, J.-E.; STRAND, M.; LUNDMARK, T. Temperature stress. In: RAGHAVENDRA, A.S. Physiology of trees. New York: John Wiley, 1991. chap. 13, p. 301-335. HANDEL, E. V. Direct microdetermination of sucrose. Analytical Biochemistry, Amsterdam, v. 22, p. 280-283, 1968. HE, J.; LEE, S. K. Relationship among photosynthesis, ribulose-1,5-bisphosphate carboxylase (Rubisco) and water relations of the subtropical vegetable Chinese broccoli grown in the tropics by manipulation of root-zone temperature. Environmental and Experimental Botany, Oxford, v.46, p.119-128, 2001. HUBBARD, R.M.; RYAN, M.G.; STILLER, V.; SPERRY, J.S. Stomatal conductance and photosynthesis vary linearly with plant hydraulic conductance in ponderosa pine. Plant, Cell and Environment, Oxford, v.24, p. 113-121, 2001. HUNER, N.P.A.; ÖQUIST, G.; SARHAN, F. Energy balance and acclimation to light and cold. Trends in Plant Science, London, v. 3, p. 224-230, 1998.

46
HUREWITZ, J.; JANES, H.W. The relationship between the activity and the activation state of the RuBP carboxylase and carbon exchange rate as affected by sink and developmental changes. Photosynthesis Research, Dordrecht, v.12, p. 105-117, 1987. IGLESEAS, D.J.; LLISO, I.; TADEO, F.R.; TALON, M. Regulation of photosynthesis throught source-sink imbalance in citrus is mediated by carboydrate content in leaves. Physiologia Plantarun, Copenhagen, v. 116, p. 563-572, 2002. ISMAIL, M. R.; BURRAGE, S. W.; TARMIZI, H.,AZIZ, M. A. Growth, plant water relations, photosynthesis rate and accumulation of proline in young carambola plants in relation to water stress. Scientia Horticulturae, Amsterdam, v.60, p.101.114, 1994. JOLY, R.J.; D.T. HAHN. Net CO2 assimilation of cacao seedlings following dark chilling. Tree Physiology, Victoria, v.9, p.415-424, 1991. JONES, W.W.; STEINACKER, M.L. Seasonal changes in concentration of sugars and starch in leaves and twigs of citrus trees. Proceedings of the American Society for Horticultural Science, Geneva, v. 58, p. 1-4, 1951. KALT-TORRES, W.; KERR, P.S.; USUDA, H.; HUBER, S.C. Diurnal changes in maize leaf photosynthesis. 1. Carbon exchange rate, assimilate export rate, and enzyme activities. Plant Physiology, Rockville, v. 83, p. 283-288, 1987. KHAIRI, M.M.; HALL, A.E. Effects of air and soil t on vegetative growth of citrus. Journal of American Society for Horticultural Science, Alexandria , v. 101, p. 337-341, 1976a. KHAIRI, M.M.; HALL, A.E. Temperature and humidity effects on net photosynthesis and transpiration of citrus. Physiologia Plantarum, Copenhagen, v. 36, p. 29-34, 1976b. KRAMER, P.J. Water relations of plants. New York: Academic Press, 1983. 489 p. KRIEDEMANN, P.E.; BARRS, H.D. Citrus orchards. In: KOZLOWSKI, T.T. Water deficits and plant growth. New York: Academic Press, 1981. p. 325-417. LARCHER, W. Ecofisiologia vegetal. São Carlos: RIMA Artes & Textos, 2000. 531 p. LI, C.Y.; WEISS, D.; GOLDSCHMIDT, E.E. Effects of carbohydrate starvation on gene expression in citrus root. Planta, New York, v. 217, p. 11-20, 2003. LLOYD, J.; SYVERTSEN, J.P.; KRIEDEMANN, P.E.; FARQUHAR, G.D. Low conductances for CO2 diffusion from stomata to the sites of carboxylation in leaves of woody species. Plant, Cell and Environment, Oxford, v. 15, p. 873-899, 1992. LONG, S.P.; HUMPHRIES, S.; FALKOWSKI, P.G. Photoinhibition of photosynthesis in nature. Annual. Review of. Plant Physiology and Plant Mololecular Biology, Palo Alto, v.45, p: 633-661, 1994. MACHADO, D.F.S.P. A resposta da fotossíntese à baixa temperatura noturna em laranjeira ‘Valência’ é dependente do porta-enxerto. 58 f. 2009 - Dissertação (Mestrado), Instituto Agronômico, Campinas.

47
MACHADO, D.F.S.P; MACHADO, E.C.; MACHADO, R.S; RIBEIRO, R.V. Efeito da baixa temperatura noturna e do porta-enxerto na variação diurna das trocas gasosas e na atividade fotoquímica de laranjeira ‘Valência’. Revista Brasileira de fruticultura, Jabuticabal, v. 32, 2010 (no prelo). MACHADO, E.C.; QUAGGIO, J.A.; LAGÔA, A.M.M.A.; TICELLI, M.; FURLANI, P.R. Trocas gasosas e relações hídricas de laranjeiras com clorose variegada dos citros. Revista Brasileira de Fisiologia Vegetal, Londrina, v. 6, p. 53-57, 1994. MACHADO, E.C.; MEDINA, C.L.; GOMES, M.M.A.; HABERMANN, G. Variação sazonal da fotossíntese, condutância estomática e potencial da água na folha de laranjeira ‘Valência’. Scientia Agrícola, Piracicaba, v. 59, p. 53-58, 2002. MACHADO, E.C.; SCHMIDT, P.T.; MEDINA, C.L.; RIBEIRO, R.V. Respostas da fotossíntese a fatores ambientais em três espécies de citros. Pesquisa Agropecuária Brasileira, Brasília, v. 40, p. 1161-1170, 2005. MAGALHÃES FILHO, J.R.; AMARAL, L.R.; MACHADO, D.F.S.P.; MEDINA, C.L.; MACHADO, E.C. Deficiciência hídrica, trocas gasosas e crescimento de raízes em laranjeira 'Valência' sobre dois tipos de porta-enxerto. Bragantia, Campinas, v. 67, p. 693-699, 2008. MAGALHÃES FILHO, J,R. Resposta da fotossíntese à variação da temperatura no sistema radicular em laranjeira ‘Valência’. 2009. 50 f. Dissertação (Mestrado) - Instituto Agronômico, Campinas. MAGALHÃES FILHO, J.R.; MACHADO, E.C.; MACHADO, D.F.S.P.; RAMOS, R.A.; RIBEIRO, R.V. Variação da temperatura do substrato e fotossíntese em mudas de laranjeira 'Valência'. Pesquisa Agropecuária Brasileira, Brasília, v. 44, p. 1118-1126, 2009. MAXWELL, K.; JOHNSON, G.N. Chlorophyll fluorescence - a practical guide. Journal of Experimental Botany, Oxford, v. 51, p. 659-668, 2000. MEDINA, C.L.; MACHADO, E.C.; PINTO, J.M. Fotossíntese de laranjeira ‘Valencia’ enxertada sobre quatro porta-enxertos e submetida à deficiência hídrica. Bragantia, Campinas, v. 57, p. 1-14, 1998. MEDINA, C. L.; MACHADO, E. C.; GOMES, M. M. A. Condutância estomática, transpiração e fotossíntese em laranjeira ‘Valencia’ sob deficiência hídrica. Revista Brasileira de Fisiologia Vegetal, Londrina, v. 11, p. 29-34, 1999. MEDINA, C.L.; MACHADO, E.C. Trocas gasosas e relações hídricas em laranjeira ‘Valência’ enxertada sobre limoeiro ‘Cravo’ e ‘Trifoliata’ e submetida à deficiência hídrica. Bragantia, Campinas, v.57, p. 15-22, 1999. MEDINA, C.L. Fotossíntese, relações hídricas e alterações bioquímicas em laranjeira ‘Pêra’ com CVC e submetida à deficiência hídrica. Campinas, 2002. 147f. Tese (Doutorado) - Universidade Estadual de Campinas.

48
MEDINA, C.L.; SOUZA, R.P.; MACHADO, E.C.; RIBEIRO, R.V.; SILVA, J.A.B. Photosynthetic response of citrus grown under reflective aluminized polypropylene shading nets. Scientia Horticulturae, Amsterdam, v. 96, p. 115-125, 2002. MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO. Culturas. Brasil: Produtividade média de lavouras temporárias e permanentes. Disponível em: <http://www.agricultura.gov.br/>. Acesso em: 28 jan. 2009. NADA, K.; HE, L. XIONG; TACHIBANA, S. Impaired photosynthesis in Cucumber (Cucumis sativus L.) by hight root-zone temperature involves ABA – induced stomatal closure and reduction in ribulose-1,5-bisphosphate carboxylase/oxygenase activity. Journal of the Japanese Society for Horticultural Science, Tokyo, v.72, p. 504-510, 2003. NIYOGI, K.K. Photoprotection Revisited: Genetic and Molecular Approaches. Plant Physiology and Plant Mololecular Biology, California, v. 333, p. 50-59, 1999. OJEDA, M.; SCHAFFER, B.; DAVIES, F.S. Soil temperature, physiology, and growth of containerized Annona species. Scientia Horticulturae, Amsterdam, v.102, p. 243-255, 2004. ÖQUIST, G.; CHOW, W.S.; ANDERSON, J.M. Photoinhibition of photosynthesis represents a mechanism for the long-term regulation of photosystem II. Planta, New York, v. 186, p. 450-460, 1992. OSMOND, C.B. What is photoinhibition? Some insights from comparisons of sun and shade plants. In: BAKER, N.R.; BOWYER, J.R. Photoinhibition and photosynthesis: from molecular mechanisms to the field. Oxford: Bios Scientific Publishers, 1994. chap. 1, p.1-24. OSMOND, C.B.; GRACE, S.C. Perspectives on photoinhibition and photorespiration in the field: quintessential inefficiencies of the light and dark reactions of photosynthesis? Journal of Experimental Botany, Oxford, v. 46, p. 1351-1362, 1995. PAUL, M.J.; FOYER, C.H. . Sink regulation of photosynthesis. . Journal of Expermental Botany, Oxford, v. 52, p. 1383-1400, 2001. PEREIRA, A.R.; ANGELOCCI, L.R.; SENTELHAS, P.C. Agrometeorologia: fundamentos e aplicações práticas. Guaíba: Agropecuária, 2002. 478 p. PRADO, A.K.S. Florescimento, frutificação e teores de carboidratos em laranjeiras ‘Valência’ com diferentes níveis de produtividade e submetidas à irrigação. 2006. 46 f. Dissertação (Mestrado em Agricultura Tropical e Subtropical) – Instituto Agronômico, Campinas, 2006. PRADO, A.K.S.; MACHADO, E. C.; MEDINA, C.L.; MACHADO, D.F.S.P.; MAZZAFERA, P.. Florescimento e frutificação em laranjeiras 'Valência' com diferentes cargas de frutos e submetidas ou não à irrigação. Bragantia, Campinas, v. 66, p. 173-182, 2007. RAMOS, R.A. Regime térmico e relação fonte-dreno em mudas de laranjeiras: dinâmica de carboidratos, fotossíntese e crescimento. 2009. 52 f. Dissertação (Mestrado em Agricultura Tropical e Subtropical) - Instituto Agronômico, Campinas.

49
REUTHER, W. Climate and citrus behavior. In: The Citrus Industry. Riverside: University of California, 1973. v. 3, chap. 9, p. 280-337. RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G.; OLIVEIRA, R.F. Variação sazonal e diária do conteúdo de carboidratos em folhas de laranjeira ‘Valência’. In: Congresso Brasileiro de Fisiologia Vegetal, 10.; Congresso Latino Americano de Fisiologia Vegetal, 12., 2005, Recife. Resumos expandidos. CD-ROM. RIBEIRO, R.V. Variação sazonal da fotossíntese e relações hídricas de laranjeira ‘Valência’. 2006. 157f. Tese (Doutorado). Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba. RIBEIRO, R.V.; MACHADO, E. C.; BRUNINI, O. Ocorrência de condições ambientais para a indução do florescimento de laranjeiras no Estado de São Paulo. Revista Brasileira Fruticultura, Jaboticabal, v.28, p .247-253, 2006. RIBEIRO, R.V.; MACHADO, E.C. Some aspects of citrus ecophysiology in subtropical climates: re-visiting photosynthesis under natural conditions. Brazilian Journal of Plant Physiology, Londrina, v.19, p. 393-411, 2007. RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G.; OLIVEIRA, R.F. Photosynthesis and water relations of well-watered orange plants as affected by winter and summer conditions. Photosynthetica, Pragha, v.49, 2009a. RIBEIRO, R.V.; MACHADO, E.C.; SANTOS, M.G.; OLIVEIRA, R.F. Seasonal and diurnal changes in photosynthetic limitation of young sweet orange trees. Environmental and Experimental Botany, Oxford, v.66, p. 203-211, 2009b. ROLIM, G.S.; CAMARGO, M.B.P; LANIA, D.G.; MORAES, J.F.L. Classificação climática de Köppen e de Thornthwaite e sua aplicabilidade na determinação de zonas agroclimáticas para o estado de são Paulo. Bragantia, Campinas, v.66, p. 711-720, 2007. SCHREIBER, U.; BILGER, W. Rapid assessment of stress effects on plant leaves by chlorophyll fluorescence measurements. In: TENHUNEN, J.D.; CATARINO, F.M.; LANGE, O.L.; OECHEL, W.C. Plant response to stress. Berlin: Springer Verlag, 1987. p. 27-53. (NATO ASI Series G: Ecological Sciences, 15). SCHREIBER, U.; BILGER, W.; HORMANN, H.; NEUBAUER, C. Chlorophyll fluorescence as a diagnostic tool: basics and some aspects of practical relevance. In: RAGHAVENDRA, A.S. Photosynthesis: a comprehensive treatise. Cambridge: Cambridge University Press. 1998. chap. 24, p. 320-336. SCHURR, U.; GOLLAN, T. & SHULZE, E.D. Stomatal response to drying soil in reaction to changes in xylem sap composition of Helianthus annuus. II. Stomatal sensitivity to abscisic acid imported from xylem sap. Plant, Cell and Environment, Baltimore, 15:561-567, 1992. SMITH, A.M.; STITT, M. Coordination of carbon supply and plant growth. Plant, Cell and Environment, Oxford, v.30, p. 1126-1149, 2007.

50
SPERRY, J.S.; DONNELLY, J.R.; TYREE, M.T. Seasonal occurrence of xylem embolism in sugar maple (Acer saccharum). American Journal of Botany, Saint Louis, v. 75, p. 1212-1218, 1988. SPIEGEL-ROY, P.; GOLDSCHMIDT, E.E. Biology of citrus. Cambridge: Cambridge University Press, 1996. 230 p. STONEMAN, G.L.; DELL, B. Growth of Eucalyptus marginata (Jarrah) seedlings in agreenhouse in response to shade an soil temperature. Tree Physiology, Victoria, v.13,p. 239–252, 1993. STRAND, M.; LUNDMARK, T.; SODERBERGH, I.; MELLANDER, P.E. Impacts of seasonal air and soil temperatures on photosynthesis in Scots pine trees. TreePhysiology, Victoria, v.22, p. 839-847, 2002. SYVERTSEN, J.P. Partial shoot removal increases net CO2 assimilation and alters water relations of Citrus seedlings. Tree Physiology, Victoria, v. 14, p. 497-508, 1994. SYVERTSEN, J.P.; LLOYD, J. Citrus. In: SCHAFFER, B.; ANDERSEN, P.C. Handbook of environmental physiology of fruit crops: sub-tropical and tropical crops. Boca Raton: CRC Press, 1994. v. 2, chap. 4, p.65-99. SYVERSTSEN, J.P.; GOÑI, C.; OTELO, A. Fruits load and canopy shading affest leaf characteristics and net gas exchange of ‘Spring’ navel orange trees. Tree Physiology, Victoria, v.23, p. 899-906, 2003. TARDIEU, F.; ZHANG, J.; KATERJI, N.; BETHENOD, O., PALMER, S. & DAVIES, W.J. Xylem ABA controls the stomatal conductance of field-grow maize as a function of the predraw leaf water potential. Plant, Cell and Environment, Baltimore, 15:193-197, 1992. TARDIEU, F.; SIMONNEAU T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours. Journal of Experimental Botany, Oxfod, v.49, p.419–432, 1998. TOURNAIRE-ROUX, C.; SUTKA, M.; JAVOT, H.; GOUT, E.; GERBEAU, P.; LUU, D-T.; BLIGNY, R.; MAUREL, C. Cytosolic pH regulates root water transport during anoxic stress through gating of aquaporins. Nature, London, v.425, p. 393-397, 2003. VESELOVA, S.V.; FARHUTDINOV, R.G.; VESELOV, S.Y.; KUDOYAROVA, G.R.; VESELOV, D.S.; HARTUNG, W. The effect of root cooling on hormone content, leaf conductance and root hydraulic conductivity of durum wheat seedlings (Triticum durum L.). Journal of Plant Physiology, Stuttgart, v.162, p. 21-16, 2005. VU, J.C.V.; YELENOSKY, G. Water deficit and associated changes in some photosynthetic parameters in leaves of ‘Valencia’ orange (Citrus sinensis Osbeck). Plant Physiology, Rockville, v. 88, p. 375-378, 1988. WAN, X.; LANDHÄUSSER, S.M.; ZWIAZEK, J.J.; LIEFFERS, V.J. Stomatal conductance and xylem sap properties of aspen (Populus tremuloides) in response to low soil temperature. Physiologia Plantarum, Copenhagen, v. 122, p. 79-85, 2004.

51
WARREN, C.R.; ADAMS, M.A. Internal conductance does not scale with photosynthetic capacity: implications for carbon isotope discrimination and the economics of water and nitrogen use in photosynthesis. Plant, Cell and Environment, Oxford, v. 29, p. 192-201, 2006. XU, Q.; HUANG, B. Effects of differential air and soil temperature on carbohydrate metabolism in creeping bentgrass. Crop Science, Columbia, v. 40, p. 368-374, 2000. ZHANG, Y.P.; QIAO, Y.X; ZHANG, Y.L.; ZHOU, Y.H.; YU, Y.Q. Effects of root temperature on leaf gas exchange and xylem sap abscisic acid concentrations in six Curcubitaceae species. Photosynthetica, Praga, v. 46, p. 356-362, 2008. ZHOU, Y.; HUANG, L.; ZHANG, Y, SHI, K.; YU, J.; NOGUÉS, S. Chill-induced decrease in capacity of RuBP carboxylation and associated H
2O
2 accumulation in cucumber leaves are
alleviated by grafting onto figleaf gourd. Annals of Botany, Oxford, v. 100, p. 839-848, 2007.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo