josÉ luiz de barros araÚjo -...
TRANSCRIPT

ALGUNS MOLUSCOS TERRESTRES COMO HOSPEDEIROS INTERMEDIÁRIOS DE
PARASITOS DE ANIMAIS DOMÉSTICOS, NO BRASIL: ESTUDOS SOBRE
ANATOMIA, SISTEMÁTICA E PARTICIPAÇÃO EM HELMINTOSES
JOSÉ LUIZ DE BARROS ARAÚJO
1982

JOSÉ LUZ DE BARROS ARAÚJO
ALGUNS MOLUSCOS TERRESTRES COMO HOSPEDEIROS INTERMEDIÁRIOS DE
PARASITOS DE ANIMAIS DOMÉSTICOS, NO BRASIL: ESTUDOS SOBRE
ANATOMIA, SISTEMÁTICA E PARTICIPAÇÃO EM HELMINTOSES
Tese apresentada no Decanato de Pesquisa
e Pós-Graduação da Universidade Federal
Rural do Rio de Janeiro, para habilita-
ção ao grau de Doutor em Ciências, na
área de Parasitologia Veterinária.
ORIENTADOR:
MICHAEL ROBIN HONER
ITAGUAÍ
RIO DE JANEIRO - BRASIL
1982

iii.
AGRADECIMENTOS
Ao Dr. HUGO DE SOUZA LOPES pelo apoio e sugestões que
sempre nos ofereceu.
Ao Prof. ARNALDO CAMPOS DOS SANTOS COELHO pelo auxí-
lio nos trabalhos de revisão bibliográfica.
Aos Profs. MICHAEL ROBIN HONER, CARLOS WILSON GOMES
LOPES e CARLOS LUIZ MASSARD pelas sugestões apresentadas.
Ao Sr. OSWALDO DUARTE GONÇALVES pelo auxílio na revi-
são do texto.
Às Sras. MARCIA DA SILVA RICARDO DE AGUIAR e MARIA AU-
XILIADORA VIEIRA BRITO pelos trabalhos datilográficos iniciais
e ao Sr. GILMAR FERREIRA VITA pelo trabalho definitivo.
Ao CNPq pelo auxílio financeiro para a confecção do
trabalho.

BIOGRAFIA
JOSÉ LUIZ DE BARROS ARAÚJO, filho de Eduardo Maurício
de Araújo e Anna de Barros Araújo, nasceu em Rio das Flores,
Estado do Rio de Janeiro, a 28 de janeiro de 1937, formou-se em
Veterinária pela então Escola Nacional de Veterinária, da Uni-
versidade Rural do Brasil, em 1962. Após cursar a Escola de
Pós-Graduação desta Universidade, obteve o títuto de "Magister
Scientiae" em 1971.
Em 1963 ingressou na carreira de magistério, galgando
os vários níveis desde Auxiliar de Ensino até ao de Professor
Adjunto, na Área de Parasitologia do Departamento de Biologia
Animal do Instituto de Biologia. Além dos encargos referentes
ao ensino de graduação, é também vinculado à lecionação das
disciplinas de Malacologia e de Helmintologia do Curso de
Pós-Graduação em Parasitologia.
Ainda como acadêmico, iniciou-se na pesquisa sob a
orientação do Prof. Hugo de Souza Lopes, estudando os moluscos.
Para o desenvolvimento destes estudos, foi bolsista do Institu-
to de Economia Rural, do Instituto Oswaldo Cruz e do Conselho

v.
Nacional de Desenvolvimento Científico e Tecnológico. Em todas
estas instituições, procurou manter a linha de estudos traçada
inicialmente, apenas acrescentando mais tarde a participação
dos moluscos nos ciclos evolutivos de parasitos, devido à sua
formação voltada para a Parasitologia. No momento participa do
projeto "Parasitismo em bovinos de leite no Estado do Rio de
Janeiro: Epidemiologia, importância econômica e controle". Par-
ticipa também de outros projetos em que alguns parasitos em es-
tudo não usam necessariamente os moluscos como hospedeiros in-
termediários, como o de "Levantamento e identificação das para-
sitoses de ruminantes e suínos no Território Federal do Amapá,
interferentes no processo de produção. Importância como zoono-
ses", no qual atua como responsável.
Publicou vários trabalhos sobre moluscos, helmintos e
relações entre eles.

ÍNDICE
1. INTRODUÇÃO
2. REVISÃO DA LITERATURA
3. MATERIAL E MÉTODOS
Origem e manuseio do material estudado
Estudos morfológicos
Estudos bionômicos
4. RESULTADOS
Observações bionômicas
L e p t i n a r i a u n i l a m e l l a t a
S u b u l i n a o c t o n a
Bradybaena s i m i l a r i s
Sistemática e aspectos morfológicos dos moluscos
L e p t i n a r i a u n i l a m e l l a t a (Orbigny, 1835)
Concha
Câmara paleal
Sistema genital
S u b u l i n a o c t o n a (Brugière, 1789)
Concha
Páginas
1
3
15
15
16
18
20
20
20
21
22
22
31
31
32
32
38
38

vii.
Páginas
39
40
40
45
45
46
45
47
48
56
60
86
91
Rádula
Mandíbula
Sistema genital
Bradybaena s i m i l a r i s (Férussac, 1821)
Concha
Rádula
Câmara Paleal
Mandíbula
Aparelho genital
Os parasitos de interesse
Distribuição geográfica e relações dos moluscos com
parasitos dos animais domésticos, no Brasil
5. DISCUSSÃO E CONCLUSÕES
6. REFERÊNCIAS BIBLIOGRÁFICAS

RESUMO
Foram estudadas as espécies de moluscos terrestres
Leptinaria unilamellata (Orbigny, 1835), Subulina octona (Bru-
guière, 1972) e Bradybaena similaris (Férussac, 1821), que
tuam ou têm possibilidade de atuarem como hospedeiros inter-
mediários, dentre outros, de parasitos de animais domésticos
no Brasil. São abordados os aspectos anatômicos das conchas
e parte moles (sistema genital), mais seguros para a caracte-
rização das espécies, como também aspectos bionômicos e os
hábitos destas espécies, e as circunstâncias e oportunidades
em que se fazem as infestações dos hospedeiros definitivo e
intermediário.

SUMMARY
The terrestrial molluscs Leptinaria unilamellata (Or-
bigny, 1835), Subulina octona (Bruguière, 1789) and Bradybaena
similaris (Férussac, 1821) were studied, as the actual or
possible intermediate hosts of parasites of domestic animals
in Brazil. Those aspects of the morphology of the shell and
internal organs (genital system) are emphasized, which can be
most safely used for the identification of the species. Also
given are aspects of the biometry and ecology anda survey of
the circumstances and opportunities for the infection of the
definitive and intermediate hosts.

1. INTRODUÇÃO
Muitas espécies de moluscos são conhecidas como hos-
pedeiras intermediárias de parasitos dos animais domésticos e
do homem.
Trabalhos que estudem a participação destes moluscos
no ciclo evolutivo de parasitos devem, necessariamente, desen-
volver-se por etapas, a fim de que seus objetivos sejam alcan-
çados. A nosso ver, é fundamental que partam de seguras deter-
minações das espécies envolvidas. No que se refere aos parasi-
tos dos animais domésticos, poucas controvérsias ainda persis-
tem, porém, quanto aos moluscos, inúmeros aspectos são ainda
pouco conhecidos, principalmente em relação às espécies cosmo-
politas. Muitas destas espécies são apontadas na literatura co-
mo vetoras de parasitos sem, contudo, se ter a certeza de sua
validade e, conseqüentemente, de sua participação, pois as de-
terminações desses vetores são, às vezes, imprecisas, porque ba-
seadas apenas nas conchas e desprovidas, portanto, de dados mor-
fológicos capazes de elucidar as dúvidas que, não raro, surgem
e que se agravam pelo fato de muitas espécies exibirem compor-

2.
tamentos diferentes em regiões diferentes. Ainda sob o mesmo
ponto de vista médico-veterinário, outro aspecto também relati-
vo ao ciclo evolutivo desses moluscos é o das suas exigências
ambientais, uma vez que em seu ambiente estão certamente inse-
ridos os parasitos que deles dependem para desenvolver seus
próprios ciclos, freqüentemente até à forma infectante para o
hospedeiro definitivo.
Este trabalho teve por objetivo o estudo dos aspectos
do ciclo evolutivo e da anatomia das partes moles de algumas
espécies de moluscos terrestres e a avaliação, quanto à obriga-
toriedade, das relações dessas espécies com parasitos de ani-
mais domésticos no Brasil.

2. REVISÃO DA LITERATURA
CHEMNITZ (1786) refere pela primeira vez a Helix octo-
na apresentando uma figura.
FÉRUSSAC & DESHAYES (1820/1851) fez comentários sobre
a grande distribuição de Helix similaris, além de uma descri-
ção conquiliológica.
LAMARCK (1822) fez apenas uma breve descrição conqui-
liológica, descrevendo Bulimus octonus como uma concha alonga-
da, com 10 linhas (voltas?).
D'ORBIGNY (1835) descreveu Bulimus octonus apontando
as figuras de CHEMNITZ como as que correspondiam à sua espé-
cie. Tal fato deixou dúvidas que foram apontadas por D'ORBIGNY
(in La Sagra) quanto à abertura da concha.
D'ORBIGNY (1847) fez uma descrição da concha de Buli-
mus unilamellatus que a nosso ver não coincide com a espécie,
pois afirma que a volta corporal ocupa a metade do comprimento
total da concha. Indica na sua sinonímia Helix unilamellata de
D'ORBIGNY (1835).
REEVE (1849), descreve as conchas de Achatina octona

4.
e Achatina lamellata, assinalando as particularidades importan-
tes. Da última, atribui a espécie a POTIEZ & MICHAND, frizando
ser muito semelhante a Achatina funcki Pfeiffer, 1847.
LA SAGRA (1853) fez comentários acerca dos caracteres
das conchas de Achatina octona, comparando-os com CHEMNITZ
(1786), contudo as figuras não coincidem com suas indicações.
REEVE (1854) fez uma curta descrição da concha de He-
lix similaris.
FISCHER & CROSSE (1878) fizeram uma descrição simples
dos detalhes das conchas de Subulina octona ao lado de uma dis-
tribuição geográfica na qual é citada para o Brasil, assinalan-
do sua ocorrência nos estados do Ceará e Bahia.
VON MARTENS (1890/1901) considerou na família Stenogy-
ridae os gêneros Subulina, onde inclui entre outras espécies
Subulina octona e Leptinaria, não considerando Leptinaria uni-
lamellata. Em todas as espécies apresentou apenas os caracte-
res das conchas.
TRYON & PILSBRY (1894) conceituaram a família Frutici-
colidae, redescrevendo-a e assinalando a distribuição geográfi-
ca das espécies.
TRYON & PILSBRY (1906) estudaram os caracteres morfo-
lógicos, especialmente das conchas dos Subulinidae. Caracteri-
zaram os gêneros Subulina e Leptinaria, incluindo as espécies
de interesse para o Brasil e a distribuição geográfica por con-
tinentes.
BAKER (1913) relaciona acompanhando uma descrição con-

5.
quiliológica, várias espécies de moluscos colecionados durante
uma expedição efetuada no Brasil.
BAKER (1931) apresentou, como resultado de uma expedi-
ção realizada no Brasil, nas regiões Norte, Nordeste e Centro-
Oeste, uma lista de espécies de moluscos com descrições conqui-
liológicas e indicação dos locais onde foram coletados os es-
pécimens.
TRAVASSOS (1928) estudou vários trematódeos da super-
família Fascioloidea, dentre os quais Harmostomum mazzantii
(= Brachylaemus mazzantii) de pombo doméstico.
THIELE (1931) classificou os moluscos a partir dos
grandes grupos, caracterizando-os até o nível de subgênero.
PILSBRY (1933) relatou os resultados de uma expedição
realizada ao Estado de Mato Grosso em 1931, incluindo os acha-
dos de Bulimulus tenuissimus e Leptinaria lamllata concentrica.
SANTOS (1934), com material procedente do Estado do
Rio de Janeiro, descreveu Tamerlania bragai, ao mesmo tempo em
que apresentou uma chave para determinação dos gêneros da famí-
lia Eucotylidae e relatou aspectos da ação patológica do para-
sito sobre o hospedeiro.
DOLLFUS (1935a), em estudo sobre alguns Brachylaemus
da fauna francesa, apresentou uma lista da distribuição por
hospedeiro na qual aparecem algumas espécies também brasilei-
ras.
DOLLFUS (1935b,c,d,e) catalogou os Stylommatophora que
abrigam formas larvais, as quais descreveu resumidamente.

6.
HOBMAIER & HOBMAIER (1935) estudaram os hospedeiros in-
termediários de Aelurostrongylus abstrusus. Entre os vários mo-
luscos nos quais conseguiram as etapas evolutivas do parasito,
encontram-se Agriolimax agrestis e Helix aspersa, referidos pa-
ra o Brasil. Consideraram estas duas espécies de importância se-
cundária no ciclo daquele parasito.
PINTO (1938), em importante obra que versa sobre espé-
cies de helmintos de interesse médico-veterinário, com alguns
dados acerca dos seus ciclos evolutivos, ressaltou nos casos de-
vidos os moluscos.
PENNER (1939) descreveu Tamerlania melospizae, apre-
sentando também notas sobre o gênero.
ALICATA (1940) estudou o ciclo evolutivo da Posthar-
mostomum gallinum, apontando Bradybaena similares como primei-
ro hospedeiro intermediário e Subulina octona como segundo.
MORRETES (1943), em sua contribuição ao conhecimento
da fauna brasileira de moluscos, divulgou os resultados do es-
tudo de uma pequena coleção de moluscos obtida em uma excur-
são, assinalando vários pontos de coleta das espécies encontra
das, entre elas
MALDONADO (1945a), estudando Tamerlania bragai, esta-
beleceu o ciclo desta espécie, apontando um molusco terrestre,
Subulina octona, como hospedeiro intermediário. Ambos, molusco
e parasito, são extremamente comuns no Brasil.
MALDONADO (1945b), em estudo sobre o ciclo evolutivo
de Platynosomum fastosum, apontou Subulina octona como primei-

7.
ro hospedeiro intermediário, frizando também a participação des-
te molusco no ciclo evolutivo de outros parasitos (trematódeos,
cestódeos e nematódeos).
SCOTT (1945) descreveu, com boas ilustrações, várias
espécies de moluscos que ocorrem em Tilcara, na Argentina. Des-
tas, algumas ocorrem também no Brasil.
MORRETES (1949) relacionou, dentro da sistemática, to-
das as espécies brasileiras de moluscos, incluindo as de inte-
resse médico-veterinário; este levantamento foi complementado
por OLIVEIRA & CASTRO (1979).
GERICHTER (1949) estudou a morfologia de Ael. abstru-
sus do pulmão de Felídeos na Palestina. Referindo-se ao ciclo
evolutivo desta espécie, assinala Agriolimax sp. e Limax flavus
entre outros, como hospedeiro intermediário.
HAAS (1949), em trabalho sobre moluscos aquáticos e
terrestres provenientes do Peru, listou Leptinaria lamellata,
de ocorrência também no Brasil.
FREITAS (1951), em revisão da família Eucotylidae, in-
cluiu as espécies parasitas de aves brasileiras.
JAECKEL (1952), relacionando os moluscos colecionados
no período 1934-38 no Nordeste Brasileiro e depositados no Mu-
seu de Berlin, grupou as espécies em algumas classes como Ter-
restres, Univalvos de água doce, Bivalvos de água doce e Lito-
râneos, ou então, em relação às condições de umidade, em Higró-
filos, Mesófilos e Xerófilos. Tais classificações não distin-
guem as posições sistemáticas dentro dos grandes grupos.

8.
HAAS (1952) faz uma distribuição geográfica de espé-
cies de moluscos sul americanos, recebidas do Brasil. Algumas
delas tem uma breve descrição conquiliológica.
BONDESEN (1954), estudando várias espécies de molus-
cos terrestres desprovidos de concha, caracterizou-as pelo as-
pecto externo, levando em conta também seus habitats.
HAAS (1955) incluiu, em trabalho sobre uma pequena co-
leção de conchas provenientes da América do Sul, entre outras
espécies, Bradybaena similaris, provenientes de São Paulo, Brasil.
VICENTE (1955) estudou formas intermediárias de várias
espécies de Brachylaemus encontradas em moluscos terrestres.
Destes, Agriolimax reticulatus é uma espécie citada para o Bra-
sil e considerada uma das mais importantes como hospedeiro in-
termediário de uma das espécies de Brachylaemus estudadas.
CAMARGO (1956), trabalhando com moluscos nocivos às
plantas cultivadas no Rio Grande do Sul, incluiu espécies de
grande ocorrência, umas somente naquele Estado e outras em vá-
rios Estados do Brasil.
VENMANS & FRÖMMING (1957) estudaram a anatomia e a bio-
logia de Subulina kassaiana, a qual mostra muitas semelhanças
com Subulina octona em vários de seus aspectos.
BUCKUP & BUCKUP (1957) confirmaram a presença de Helix
aspersa no Brasil, relacionando em seu catálogo espécimens cap-
turados no Rio Grande do Sul.
ROSE (1957a) fez importantes observações sobre a bio-
nomia do primeiro estádi o de vida livre de Muellerius capilla-

9.
ris, assinalando a possibilidade de penetração na massa pedio-
sa de Agriolimax reticulatus que, por ter atividade em grande
parte do ano, daria maior oportunidade ao parasito.
ROSE (1975b) estudou os estágios larvais de Muellerius
capillaris em Agriolimax agrestis e Agriolimax reticulatus. Ava-
liou a influência da temperatura sobre o tempo para alcançar a
fase infectante, o tempo de sobrevivência no hospedeiro inter-
mediário, as diferenças entre as fases infectantes e pré-infectante
sugerida por outro autor, além de outros importantes detalhes.
GETZ (1959) estudou, comparativamente, aspectos ecoló-
gicos de três espécies de moluscos desprovidos de concha: Arion
circunscriptus, Deroceras reticulatum e Deroceras laeve.
BURCH (1960) trabalhou com alguns moluscos para os
quais havia prescrição de quarentena nos EUA; entre eles figu-
ram espécies das quais existem registros de ocorrência no Bra-
sil. Fez também uma breve exposição da distribuição geográfica
mundial daquelas espécies.
ARAÚJO, REZENDE & RODRIGUES (1960) estudaram a anato-
mia de Bulimulus tenuissimus nos detalhes macroscópicos e his-
tológicos, apontanto os principais caracteres da espécie.
HAAS (1962), em estudo sobre a fauna de Curação e ou-
tras ilhas do Caribe, incluiu Subulina octona e Leptinaria la-
mellata entre os Subulinidae.
LAPAGE (1962), em obra sobre helmintologia e entomolo-
gia veterinárias, relacionou os parasitos em cujos ciclos se in-
cluem moluscos terrestres, assinalando as espécies destes.

10.
ASH (1962) estudando os helmintos parasitos de cães e
gatos no Hawaii, refere a Subulina octona e Eulota similaris co-
mo hospedeiros intermediários de Ael. abstrusus e Platynosomum
fastosum. Para este trematódeo a comprovação foi feita de for-
ma experimental.
FIGUEIRAS (1963) enumerou 51 espécies e subespécies de
moluscos do Uruguai, dentre as quais várias ocorrem também no Brasil.
BASH (1965) estudou a etapa final do desenvolvimento
de Eurytrema pancreaticum, concluindo pela existência de um se-
gundo hospedeiro intermediário, após o desenvolvimento das eta-
pas iniciais em um molusco.
FRANCO (1965) assinalou, no Estado do Pará, a presença
de Paratanaisia bragai em Melleagris gallopavo e em Odontopho-
rus capueira capueira.
SOULSBY (1965), entre as várias espécies que entram
na composição da obra, refere-se também àquelas que ocorrem no
Brasil, relacionando-as aos moluscos nos casos devidos.
LANZIERI (1966a) estudou o aparelho genital de Succinea
meridionalis em seus aspectos microanatômicos.
LANZIERI (1966a) estudou os aspectos morfoestruturais
de Subulina octona, consistindo este o trabalho anatômico mais
importante sobre a espécie.
LANZIERI (1966b) estudou espécimens provenientes da
Ilha da Trindade, concluindo pela descrição de duas novas espé-
cies, uma no gênero Succinea e outra no gênero Oxiloma, assina-
lado pela primeira vez na América do Sul.

11
STEPHENSON & JNUTSON (1966) apresentaram um resumo dos
diferentes estudos de invertebrados associados a moluscos sem
concha, entre os quais se encontram, de interesse para o Bra-
sil, espécies pertencentes à família Limacidae.
HYMAN (1967) relatou as relações biológicas dos molus-
cos com outros animais, incluindo algumas espécies de interes-
se veterinário direto e de ocorrência no Brasil.
SOULSBY (1968), em sua obra sobre os helmintos, artró-
podos e protozoários dos animais domésticos, assinala os para-
sitos e os moluscos que têm relações entre si.
GUILHON & AFHGAHI (1969) estudaram a evolução larvar
de Angiostrongylus vasorum no organismo de diversas espécies de
moluscos terrestres, entre os quais incluíram Helix aspersa.
TRAVASSOS, FREITAS & KOHN (1969) fizeram um estudo mor-
fológico completo acerca dos trematódeos do Brasil, separando
as espécies a partir dos grandes grupos. Tais espécies foram tam-
bém separadas pelos seus hospedeiros, igualmente classificados
a partir dos grandes grupos.
COSTA & FREITAS (1970) listaram as espécies de helmin-
tos parasitos de animais domésticos no Brasil. Tais espécies fo-
ram classificadas dentro de suas famílias e separadas por hos-
pedeiro.
ROSEN, ASH & WALLACE (1970) estudaram o ciclo evoluti-
vo de Angiostrongylus vasorum, afirmando que Biomphalaria gla-
brata foi um excelente hospedeiro intermediário, e que vários
moluscos terrestres, como Bradybaena similaris e Subulina octo-

12.
na, que também ocorrem no Brasil, foram igualmente positivos
para a infecção.
SCHELL (1970), em obra sobre os trematódeos, incluiu
os de interesse para o Brasil como parasitos de animais domés-
ticos. De modo resumido, refere os ciclos evolutivos de al-
guns, assinalando moluscos e parasitos de ocorrência também
no Brasil.
GRISI (1971) assinalou a ocorrência de Angyocaulus
raillieti (Travassos, 1927) em Canis familiaris no Brasil.
Propôs, na oportunidade, que a primeira citação de Angiostron-
gylus vasorum (Baillet, 1866), feita por GONÇALVES (1961) no
Rio Grande do Sul, fosse revista, pois acreditava tratar-se
da espécie por ele estudada. Além disso, julga que a ocorrên-
cia de Angiostrongylus vasorum no Brasil ainda carece de con-
firmação.
COSTA & FREITAS (1972) descreveram, pela primeira
vez, Eucotyle freitasi das vias urinárias de pato doméstico.
FERNANDEZ (1973) elaborou um catálogo da malacofauna
argentina, no qual várias espécies e gêneros, referidos para
aquele país, não o são para o Estado brasileiro limítrofe, de
monstrando a importância do desenvolvimento de estudos sobre
o assunto.
MALEK & CHENG (1974) caracterizaram os grandes gru-
pos de moluscos, incluindo em cada um deles as espécies de im-
portância médica e econômica. Na sua forma de participação, en-
contram-se várias citações importantes para o Brasil, muitas

13
vezes sem esta referência.
KOHN & FERNANDES (1975) estudaram as espécies do gêne-
ro Echinostoma descrita por ADOLPHO LUTZ em 1924, incluindo en-
tre elas as de aves domésticas.
PITONI, VEITENHEIMER & MANSUR (1976), em trabalho so-
bre coleta, preparação e conservação de moluscos, assinalaram
a presença de várias espécies de interesse médico-veterinário,
fazendo uma breve caracterização de cada uma delas.
DUARTE (1977) descreveu o ciclo de Postharmostomum
gallinum no Brasil, constituindo este o trabalho mais completo
sobre a espécie; além de Bradybaena similaris e Subulina oc-
tona, comuns no Brasil, outras espécies de moluscos, ainda não
identificadas como hospedeiros intermediárias de qualquer para-
sito, nele tiveram comprovada sua ocorrência.
SEIXAS (1977) estudou duas espécies de Limacidae da
fauna de Portugal, caracterizando-as pela sua anatomia inter-
na. Fez importante comentário sobre a separação de Agriolimax
agrestis (L.) e Agriolimax laevis (Müller).
BOFFI (1979), estudando os moluscos brasileiros de in-
teresse médico e econômico, colocou as espécies de interesse
veterinário, em parte, como pragas da agricultura, e em parte,
como moluscos hospedeiros intermediários de parasitos do ho-
mem (que também o são de alguns dos animais domésticos), fazen-
do um arranjo sistemático dentro de conceitos atualizados. Nes-
se estudo, são mencionadas espécies que, em nossa conceitua-
ção, atuam de diferentes maneiras como hospedeiras intermediá-

14.
rias de parasitos de animais domésticos, sendo algumas já com-
provadas no Brasil.
BREURE (1979) fez importante trabalho sobre os Bulimu-
lidae (Bulimulinae), listando as espécies com suas distribui-
ções geográficas.
OLIVEIRA, REZENDE & CASTRO (1981) catalogaram os mo-
luscos pertencentes à Universidade Federal de Juiz de Fora, in-
cluindo várias espécies de interesse médico-veterinário e in-
formando sobre suas procedências.
BUSCH (1982) em breve comunicação comenta e atualiza
a sistemática de moluscos de significação para quarentena nos
Estados Unidos, baseando-se em seu trabalho de 1960.

3. MATERIAL E MÉTODOS
O estudo foi conduzido na Universidade Federal Rural
do Rio de Janeiro, (UFRRJ), no Distrito de Seropédica, Itaguaí,
Estado do Rio de Janeiro, para onde era transportado o mate-
rial coletado.
Origem e manuseio do material estudado
Para o estudo morfológico dos moluscos, foram utiliza-
dos espécimens capturados em vários habitats terrestres nos
Estados do Rio de Janeiro (municípios de Valença, Rio das Flo-
res, Três Rios, Paraíba do Sul, Itaguaí), de São Paulo (municí-
pios de Santos e Piquete) e do Rio Grande do Sul (município de
Porto Alegre) e Território Federal do Amapá (município de Ma-
capá).
Os moluscos terrestres foram capturados nos locais de
suas preferências (sob pedras, em madeira em decomposição, so-
bre plantas de jardim, em fendas junto a construções, sob fe-
zes dessecadas nas pastagens, em locais lamacentos, etc). Em

16.
maioria, as espécies ocorriam com maior freqüência após perío-
dos de chuvas intensas. Outras espécies, ao contrário, eram
mais freqüentes nos períodos mais secos do ano, mesmo com tem-
peratura mais baixa. Os espécimens coletados eram transporta-
dos para o laboratório em frascos de vidro com tampa perfura-
da que permitiam uma pequena aeração. No laboratório, eram se-
parados em lotes de acordo com a finalidade. Alguns eram des-
tinados ao estabelecimento de colônias, sendo colocados em cai-
xas onde se procurava reproduzir, tanto quanto possível, seu ha-
bitat. Tais moluscos eram utilizados em estudos morfológicos
posteriores, observações bionômieas (alimentação, reprodução,
etc.) ou infecções experimentais com parasitos para avaliação
de sua susceptibilidade. Este material, quando fixado, era de-
positado na Coleção de Moluscos Hospedeiros Intermediários (Col.
Mol. HOSD. Int.), na Área de Parasitologia do Departamento de
Biologia Animal, do Instituto de Biologia da UFRRJ.
Estudos morfológicos
Para os estudos morfológicos, o material, uma vez no
laboratório, era dissecado, procurando-se isolar as partes mais
importantes para a determinação das espécies, no caso, a con-
cha, o sistema digestivo (para a retirada da rádula e mandíbu-
la) e o sistema genital.
Os espécimens vivos, providos de concha, eram coloca-
dos em água e aquecidos a 60°C para sacrifício, sendo retira-

17.
das as partes moles da concha. Os moluscos sem concha eram co-
locados em solução fisiológica, ocorrendo em poucos momentos a
morte, e neste mesmo líquido eram dissecados. Outras vezes os
moluscos eram dissecados em álcool glicerinado, quando se uti-
lizava este líquido para sua conservação por ocasião da cole-
ta. Quando, da dissecação, um detalhe da anatomia topográfica
era julgado importante, executavam-se desenhos em câmara cla-
ra.
Concluídas as dissecações e separadas as partes de in-
teresse, procedia-se do seguinte modo: o bulbo da rádula era
fervido em potassa a 10% para o isolamento de rádula e mandí-
bula. As rádulas eram coradas em uma mistura de Vermelho Von-
go e Orange G após desidratação na série de álcoois xilol. Em
seguida, eram montadas em bálsamo-do-canadá entre lâmina e la-
mínula. O sistema genital era desidratado na série de álcoois
e posteriormente corado em carmin clorídrico, clarificado em
creosoto, onde era conservado e estudado.
Para a distribuição geográfica dos moluscos e dos pa-
rasitos no Brasil, além das comprovações pessoais foram consul-
tados: ARAÚJO, RESENDE & RODRIGUES (1960), BAKER (1931), BARTH
(1957), BARRETO & MIES FILHO (1942), BOFFI (1979), BRANT & COS-
TA (1963), BUCKUP & BUCKUP (1957), CAMARGO (1956), CORREIA (1952,
1965), COSTA (1965), COSTA & FREITAS (1970, 1972), DUARTE (1977),
FRANCO (1965), FREITAS (1951), GETZ (1959), GRISI (1971), HAAS
(1952), JAECKEL (1952), KOHN & FERNANDES (1975), LANZIERI (1965,
1966a e 1966b) MORRETES (1949), OLIVEIRA, RESENDE & CASTRO (1981),

18.
PILSBRY (1933), PITONI, VEITENHEIMER & MANSUR (1976), SANTOS
(1934), TEIXEIRA DE FREITAS, KOHN & IBANEZ (1967), TRAVASSOS
(1927, 1928).
Estudos bionômicos
Algumas observações acerca do comportamento das espé-
cies foram feitas nos seus habitats, onde se procurava acompa-
nhar a atividade destas espécies em vários momentos, quando as
circunstâncias se modificavam, Outras vezes, alguns dados fo-
ram obtidos em criadouros, onde os moluscos eram mantidos e ob-
servados. Estes criadouros constaram de caixas pláticas, fe-
chadas com tampa telada para permitir aeração do ambiente. O
fundo era coberto por uma camada de terra, sempre rica em maté-
ria orgânica sobre a qual se colocava pedras, pedaços de madei-
ta em decomposição, sendo permanentemente mantido um certo
grau de umidade. A alimentação dos moluscos constava de vege-
tais folhosos (principalmente alface) e de ração para aves le-
vemente umedecida e colocada em pequenas placas de Petri. vi-
sando a conhecer as relações de algumas espécies com parasi-
tos, foram feitos ensaios com o intuito apenas de se averiguar
a susceptibilidade, quando era pesquisado o aparecimento de
formas intermediárias dos parasitos utilizados. Estes ensaios
constavam de infecções de moluscos com os parasitos em estudo.
Colocavam-se ovos já embrionados nos alimentos que lhes desti-
naram. Passados cerca de 7 dias, eram sacrificados e pesquisa-

19.
das as formas intermediárias dos parasitos, principalmente na
glândula digestiva.
O estudo da participação dos moluscos no ciclo de pa-
rasitos torna necessária a conceituação de alguns termos para
deixar mais claras as nuances de que se revestem as várias si-
tuações. Uma relação é considerada "obrigatória" quando as eta-
pas evolutivas de determinado parasito, só são possíveis em de-
terminada espécie de molusco. Podem ser incluídas nesse concei-
to aquelas espécies que poderiam ser chamadas eficientes, in-
dependentemente do tempo em que decorram as formas evolutivas.
Assim, para um mesmo parasito, poderá haver espécies em que o
tempo de decorrência no molusco e o número de formas resultan-
tes diferente não impediria sua inclusão no grupo. Uma relação
é considerada "facultativa" quando várias espécies de moluscos
podem abrigar uma mesma etapa evolutiva de um parasito. É "po-
tencial" quando ocorrem, no Brasil, espécies de moluscos que,
em outros países, são comprovadas como hospedeiras intermediá-
rias de parasitos, ou quando, ocorrendo ambos, hospedeiro in-
termediário e parasito, ainda não tenha sido realizado, em nos
so país, estudo que estabeleça relação entre ambos. É evidente
que, com base nestes conceitos, uma mesma espécie poderá apre-
sentar-se em situações diferentes.

4. RESULTADOS
Observações bionômicas
O estudo foi limitado às três espécies de maior ocor-
rência, cujo comportamento é redumidamente apresentado a se-
guir.
Leptinaria unilamellata (Orbigny, 1835)
É uma espécie ainda pouco estudade em quase todos os
seus aspectos. Alguns autores, como PILSBRY (1906), questionam
sua colocação na sistemática, comparando-a com alguns grupos
próximos e analisando suas características conquiliológicas.
encontrada em locais semelhantes àqueles frequentados por Subu-
lina octona, porém, suas colônias são sempre compostas de pe-
queno número de indivíduos. No Brasil, são freqüentemente en-
contradas em convivência com algumas espécies do gênero Opeas.
Citações de sua ocorrência em nosso pais são apenas as de PILS-
BRY (1906) e DUARTE (1977).

21.
Sua participação no ciclo evolutivo de parasitos se
restringe, até o momento, a apenas um trematódeo, do qual pode
servir de segundo hospedeiro intermediário facultativo.
Subulina octona (Bruguière, 1789)
espécie cosmopolita, ocorrendo preferentemente nos
meses chuvosos e quentes do ano. Abriga-se no solo, onde é en-
contrada refugiada, em certos momentos não propícios à sua ati-
vidade, sob pedras, fragmentos de madeira, etc. Tanto a campo
como em laboratório, tem sido comprovada, na sua alimentação,
uma predileção por vegetais tenros ou facilmente fragmentá-
veis. Suas posturas são sempre constituídas por pequenos gru-
pos de ovos esféricos porém com dois achatamentos paralelos,
colocados em fendas no solo. Durante seu período de atividade
reprodutiva, os ovos são vistos por transparência da concha,
sempre em número de dois ou três, ocupando as últimas porções
do sistema genital, prestes a serem postos. Freqüentam muito
as instalações de animais, onde são encontrados refugiados nos
bordos das construções, em locais para onde possam escoar águas
capazes de conduzir ovos e deste modo contaminar vegetais que
lhes sirvam de alimento.
A revisão da bibliografia mundial não forneceu dados
acerca desta espécie. Somente foram encontrados trabalhos sobre
sua morfologia conquiliológica e citações em ciclos evolutivos
de parasitos, é frequentemente citada como envolvida no ciclo

22.
evolutivo de nematódeos, trematódeos e cestódeos.
Bradybaena similaris (Férussac, 1821)
uma espécie encontrada com muita frequência em nos-
so meio, especialmente, como S. octona, nos meses quentes e chu-
vosos do ano. Habitualmente, é encontrada em hortas e jardins,
sobre as plantas, e pode danificar severamente os vegetais. Em
seu período de atividade, é sempre encontrada acima do solo,
nos caules das plantas, sobre detritos, etc. As oportunidades
de captura que oferece aos seus predadores parecem ser maiores
que as oferecidas por outras espécies. Suas posturas são sem-
pre colocadas em fendas do solo ou detritos diversos. Seu pe-
ríodo de maior atividade é curto, coincidindo com os dias chu-
vosos ou posteriormente a eles, quando ainda grande quantida-
de de água permanece retida no solo; refugia-se bem mais rapi-
damente que as demais, quando se inicia o período de estia-
gem. É uma espécie relativamente resistente à dessecação, su-
portando longos dias sem alimentação. Tem vários inimigos na-
turais principalmente as aves, pelas suas dimensões e oportu-
nidade de captura que lhes oferece.
Sistemática e aspectos morfológicos
Dentro dos vários grupos de moluscos de interesse do
ponto de vista médico-veterinário, que atuam como hospedeiros

23.
intermediários, tem destaque os Pulmonata, vindo em seguida,
com menor evidência, os Prosobranchia. Era um sentido mais am-
plo, poderiam também ser incluídos, em menor escala, os Opis-
tobranchia. Mas, é nos Pulmonata, nas suas duas ordens, que en-
contramos o maior interesse nesses estudos para o Brasil. Sen-
do nosso objetivo o estudo de moluscos de hábitos terrestres,
nossa atenção se volta para os moluscos de Ordem Stylommatopho-
ra.
As famílias a ela pertencentes e já assinaladas no Bra-
sil, nas quais são incluídas espécies de alguma forma importan-
tes, estão nos Sigmurethra e nos Heterurethra. Nos Sigmurethra
encontramos Subulinidae, Zonitidae, Limacidae, Bulimulidae, Bra-
dybaenidae e Helicidae. De interesse indireto, devido aos para-
sitos que ocorrem no Brasil e que se servem de espécies a elas
pertencentes, mas que ainda não tem representantes assinalados
no Brasil, podem ser enumerados Cochlicopidae, Vertiginidae e
Enidae, nos Orturethra e Arionidae e Phylomicidae, nos Sigmure-
thra. Nos Heterurethra, Succineidae tem importância pelo fato
de ter sido assinalado, em algumas oportunidades, o gênero Suc-
cinea como hospedeiro intermediário de alguns parasitos em ou-
tros países. Conforme nosso critério, tal fato nos leva à ci-
tação das espécies que aqui existem até o momento.
Subulinidae tem três representantes de interesse para
o Brasil: Subulina octona (Bruguière, 1789), Leptinaria unila-
mellata (Orbigny, 1835) e Rumina decollata (L.). A primeira
constitui um hospedeiro obrigatório de parasitos também assina-

24.
lados no Brasil, porém, sem seus ciclos realizados em nosso
país. Tem participação facultativa ou provável em outros ca-
sos. A segunda tem participação apenas de uma forma faculta-
tiva e a terceira é um hospedeiro potencial.
Em Zonitidae, apenas Zonitoides arboreus (Say, 1816),
pode ser considerado um hospedeiro potencial.
Em Limacidae foram assinalados: Limax maximus L., 1758,
Limax flavus L., 1758, Limax valentianus (Férussac, 1821) e De-
roceras reticulatum (Muller, 1774) (= Agriolimax reticulatus).
Ambos os gêneros são referidos na literatura como hospedeiros
intermediários de parasitos que ocorrem em nosso país, sem ha-
ver, contudo, indicação de algum trabalho que estabeleça rela-
ções entre ambos.
Bulimulidae é uma família largamente representada no
Brasil. Contudo, apenas Bulimulus tenuissimus (Orbigny, 1835)
tem participação facultativa no ciclo de parasitos.
Bradybaenidae tem como representante de interesse Bra-
dybaena similaris. Sendo uma espécie cosmopolita, sua distri-
buição no Brasil é, também, ampla. Sua participação no ciclo
de parasitos foi comprovada, sendo obrigatória em alguns ca-
sos. Em outros, embora não comprovada, certamente acontecerá o
mesmo quando forem efetuados os respectivos estudos.
Helicidae é representada no Brasil apenas por Helix as-
persa Muller, 1774, constituindo um hospedeiro potencial.
As espécies de moluscos de interesse como hospedeiros
intermediários de parasitos dos animais domésticos assinaladas

25.
no Brasil ou que atuam desta forma em outros países, mas que
têm relação com parasitos também assinalados no Brasil, orga-
nizam-se dentro da sistemática nos seguintes grupos:
Classe Gastropoda
Subclasse Pulmonata
Ordem Stylommatophora
Subordem Orthurethra
Família Cochlicopidae
Gênero Cochlicopa (Férussac, 1821) Risso, 1826
(= Cionella Jeffreys, 1829)
Cionella lubrica (Müller, 1774)
Família Vertiginidae
Gênero Abida Leach in Turton, 1831
(= Torquilla Studer, 1820 part)
Torquilla frumentum (Draparnaud, 1801)
Família Enidae
Gênero Chondrula Beck, 1837
Chondrula tridens (Muller, 1774)
Gênero Zebrina Held, 1837
Zebrina detrita (Muller, 1774)
Zebrina hohenhackeri (Krynicki, 1837)
Gênero Ena (Leach in) Turton, 1831
Ena obscura (Muller, 1774)
Subordem Heterurethra
Família Succineidae
Gênero Succinea Draparnaud, 1801

26.
*Succinea meridionalis Orbigny, 1837
Succinea meridionalis: Lanzieri, 1965
*Succinea lopesi Lanzieri, 1966
*Succinea pusilla Pfeiffer, 1849
Succinea pusilla: Jaeckel, 1952
Subordem Sigmurethra
Família Arionidae
Gênero Arion Férussac, 1819
Arion circuscriptus Johnston, 1828
Arion empiricorum Férussac, 1819
Arion hortensis Férussac, 1819
Arion intermedius Normand, 1852
Família Zonitidae
Gênero Philomycus (Rafinesque, 1820) Férussac, 1821
Philomycus bilineatus?
Família Zonitidae
Gênero Zonitoides Lehmann, 1862
*Zonitoides arboreus (Say, 1816)
Zonitoides nitidus (Muller, 1774)
Gênero Oxychilus Fitzinger, 1833
Oxychilus derbentinus Andrz.?
(= Helicella ericetorum Muller, 1774?)
Família Limacidae
Gênero Limax Linnaeus, 1758
Limax flavus Linnaeus, 1758
Limax maximus Linnaeus, 1758

27.
*Limax valentianus (Férussac, 1821)
Limax cinereus?
Gênero Agriolimax Mörch, 1865
*Agriolimax agrestis Linnaeus, 1758
Gênero Deroceras Rafinesque, 1820
*Deroceras reticulatum (Muller, 1774)
Gênero Lehmania
Lehmania marginata (Muller, 1774)
Família Subulinidae
Gênero Leptinaria Beck, 1837
*Leptinaria unilamellata (Orbigny, 1835)
Gênero Rumina Risso, 1826
*Rumina decollata Linnaeus, 1758
Gênero Subulina Beck, 1837
*Subulina octona (Brugière, 1789)
Família Clausiliidae
Gênero Armenica O. Boettger, 1877
Armenica brunnea (Anton, 1839)
Família Bulimulidae
Gênero Bulimulus Leach, 1814
*Bulimulus tenuissimus (Orbigny, 1835)
Bulimulus tenuissimus: Araújo et al., 1960
Família Polygyridae
Gênero Praticolella Martens, 1892
Praticolella griseola (Pfeiffer, 1841)
Família Bradybaenidae

28.
Gênero Cathaica Mollendorf, 1884
Cathaica ravida sieboldtiana (Benson, 1842)
Gênero Fruticicola Held, 1837
(= Eulota Hartmann, 1843)
Eulota lantzi?
Gênero Euhadra Pilsbry, 1890
Euhadra sp.
Gênero Bradybaena Beck, 1837
*Bradybaena similaris (Fêrussac, 1821)
Bradybaena arcasiana Crosse e Debeaux.?
Bradybaena fragilis (Pfeiffer, 1839)
Bradybaena maacki (Gerstfeldt, 1859)
Bradybaena middendorfi (Gerstfeldt, 1859)
Bradybaena selskii (Gerstfeldt, 1859)
Família Helminthoglyptidae
Gênero Epiphragmophora Doring, 1873
*Epiphragmophora bernardius Ihering, 1900
Gênero Helminthoglypta Ancey, 1887
Helminthoglypta arrosa (Gould, 1855)
Helminthoglypta nickliana (Lea, 1838)
Helminthoglypta californiensis (Lea, 1838)
Família Helicidae
Gênero Helicella Férussac, 1821
Helicella acuta (Muller, 1774)
Helicella candidula (Studer, 1820)
Helicella uniforciola?

29.
Helicella crenimargo Krynicki, 1833
Helicella rugosiuscula (Michaud, 1831)
Gênero Cochlicella Risso, 1826
Cochlicella acuta (Muller, 1774)
Gênero Monacha Fitzinger, 1833
Monacha sp.
Gênero Zenobila Gude & B.B. Woodward, 1921
Zenobiela rubiginosa (Gould, 1846)
Gênero Fruticampylea Kobelt, 1871
Fruticampylea nazzanensis (Krynicki, 1837)
Gênero Helicigona (Férussac, 1821) Risso, 1826
Subgênero Arianta (Leach, 1320) Turton, 1831
Arianta arbustorum (Linnaeus, 1758)
Gênero Euomphalia Westerlund, 1389
Euomphalia s electa (Klika, ?)
Euomphalia ravergiensis (Férussac, 1835)
Euomphalia strigella (Draparnaud, 1801)
Gênero Theba Risso, 1826
Theba cartusiana (Muller, 1774)
Theba fruticola (Krynicki, 1833)
Gênero Cepaea Held, 1837
Cepaea nemoralis (Linnaeus, 1758)
Gênero Helix Linnaeus, 1758
*Helix aspersa (Muller, 1774).

30.
EXPLICAÇÃO DAS FIGURAS
bm: bainha muscular do pênis
cd: canal deferente
cp: complexo peniano
co: coração
cr: complexo do receptáculo
cs: canal da espermateca
em: embrião
ep: epifalus
es: esôfago
fa: falus
ga: glândula de albumina
gd: glândula digestiva
gm: glândulas multífidas
mc: músculo columelar
mr: músculo retrator do pênis
op: ovispermoduto
ov: oviduto
ot: ovotestis
pr: próstata
rm: rim
rt: feto
sd: saco do dardo
sp: espermateca
ur: ureter
us: ureter secundário
ut: útero
vs: vesícula seminal

31.
Leptinaria unilamellata (Orbigny, 1835)
(Figs. 1 a 11)
Helix unilamellata Orbigny, 1835: 9.
Bulimus unilamellatus Orbigny, 1835: 257.
Achatina lamellata Potiez e Michaud, 1838: 128, pl. 11,
f. 7-8.
Tornatelina ferussaci Pfeiffer, 1848: 394.
Achatina lamellata Reevie, 1849: sp. 97, pl. XVIII.
Tornatellina unilamellata Pfeiffer, 1859: 649-650.
Leptinaria lamellata Pilsbry, 1906: 288-289. pl. 42,
figs. 39-40, pl. 43, fig. 50.
Leptinaria unilamellata Wenz, 1960: 348, fig. 1280.
Leptinaria (Leptinaria) lamellata Haas, 1962: 52-53,
pl. VII, figs. F-G.
Leptinaria lamellata Duarte, 1977: ii, 28.
CONCHA (Figs. 1 e 2). É pequena, com 4,5 mm e cerca de
5 a 6,5 voltas de espira, com a superfície mostrando as linhas
de crescimento não muito marcadas, suturas bem evidentes, mas
não muito profundas. Volta corporal com comprimento equivalente
a pouco mais que as demais. Abertura ovalada com peristoma cor-
tante, bordo não refletido e desprovido de dentes ou lamelas no
bordo externo. Bordo columelar com uma lamela lisa, triangular,
baixa e projetada para a abertura. Bordo parietal com uma lame-
la transversal à abertura e situada a meia distância entre tér-
mino da lamela columelar e a inserção da última volta.

32.
CÂMARA PALEAL (Fig. 4). Apresenta poucos vasos, desta-
cando-se apenas a veia pulmonar, que recebe algumas afluentes.
Sua origem junto ao pneumóstomo é pouco marcada. O rim é o ór-
gão mais característico para a espécie, nesta região do molus-
co. Sua forma é alongada e estreita no sentido transversal à câ-
mara paleal. Do lado de sua extremidade columelar, para o ure-
ter primário, que se coloca paralelamente a toda a extensão do
rim. A curvatura se faz logo em seguida à outra extremidade do
rim, caminhando o ureter paralelamente ao reto.
SISTEMA GENITAL (Fig. 6). O ovotestis é formado por um
número pequeno de grupamentos de folículos, que estão profunda
mente incluídos na glândula digestiva. Os canais excretores de
cada folículo desembocam no canal coletor de ovotestis, que per-
corre parte da face côncava da glândula digestiva até a sua de
sembocadura na vesícula seminal. A vesícula seminal (Fig. 7) é
mais ou menos curta, sinuosa e apoiada em toda sua extensão na
glândula digestiva. Seu término junto ao receptáculo seminal é
bem mais delgado, tornando estes órgãos pouco perceptíveis, por
introduzirem-se parcialmente na glândula de albumina. A glându-
la de albumina (Fig. 7) é curvada, acompanhando a curvatura da
volta da concha, com a extremidade distal mais delgada e com
duas dobras fortes, limitando a face côncava.
OVISPERMODUTO. O útero (Fig. 6) é constituído de pre-
gueamento baixo, tornando-se ligeiramente mais forte no senti-
do distal. Quando em fase de atividade reprodutiva intensa, dis-
tende-se muito, deixando de um lado o pregueamento uterino e

33.
do outro a próstata, ambos como se estivessem colados ao ór-
gão. Os ovos (Fig. 8) são de envoltório transparente deixando
ver no seu interior os moluscos jovens. Geralmente são encon-
trados de 2 a 3 ovos "in utero", deixando ver nitidamente os
de desenvolvimento mais avançado ocupando uma posição mais dis-
tal no órgão. O oviduto (Fig. 6) é longo, de luz mais alarga-
da em sua porção distal, onde se vê a origem do canal da esper-
mateca, (Fig. 3) na mesma altura da parte distal do complexo
peniano. Em conseqüência, o átrio genital é amplo e bem desen-
volvido. O complexo peniano (Fig. 9 a 11) é pouco evidente em
relação à parte feminina. É muito simples, sem uma distinção
marcada entre as regiões que o compõem. A região que correspon-
de ao falus é mais ampla, sem distinguir-se riqueza de tecido
glandular. A região epifálica é longa e delgada, constituída
praticamente por um tubo contínuo, exibindo modificações mar-
cadas somente em sua parte proximal. Esta parte é constituída
por uma dilatação pouco extensa, que corresponde internamente
também a uma modificação pronunciada de sua estrutura, consti-
tuída de vilosidades dispostas no sentido transversal. A re-
gião que poderia corresponder ao flagelo é inconspícua e cons-
tituída apenas por estreito e curto espaço colocado em posição
proximal à anteriormente descrita, limitado por células coloca-
das em posição oblíqua, e envolvida amplamente pela inserção
do músculo retrator do pênis. O músculo retrator do pênis (Figs.
10 e 11) tem origem no músculo columelar como uma faixa que se
destaca. É muito longo, apoiando-se em seu trajeto sobre o bul-

34.
bo da rádula, contornando-o parcialmente em sua parte superior,
passando por baixo do tentáculo esquerdo e inserindo-se termi-
nalmente no complexo peniano, envolvendo-o amplamente. O canal
deferente (Figs. 6, 10 e 11) é a parte mais marcante do siste-
ma masculino, iniciando-se na porção distal da próstata, onde
também termina o pregueamento uterino, prende-se ao oviduto ao
cruzá-lo dirigindo-se para a porção distal do falus, aumenta de
calibre ao mesmo tempo que se torna muito enovelado, mantendo-
se preso por tecido conjuntivo frouxo, junto ao complexo penia-
no, com sua terminação delgada, como no início. Em seguida ca-
minha paralelamente à região epifálica por uma distância bem lon-
ga, correspondendo à parte mais delgado do órgão, desembocando
em uma posição proximal ao término do órgão em uma porção mais
alargada.

35.
Leptinaria unilamellata (Orbigny, 1835): Flg. 1: Concha(abertura); Fig. 2: Concha; Fig. 3: Cana] da espermate-ca e espermateca; Fig. 4: Porção proximal da câmara pe-leal; Fig. 5: Porção média do ovispermoduto.

36.
Leptinaria unilamellata (Orbigny, 1835): Fig. 6:
Aparelho genital (porções média e distal).

37.
Leptinaria unilamellata (Orbigny, 1835): Fig. 7: Glândula de albumina evesícula seminal; Fig. 8: Embrião retirado do útero; Fig. 9: Complexopeniano (porção proximal); Fig. 10: Complexo peniano; Fig. 11. Complexopeniano (porção proximal).

38.
Subulina octona (Bruguière, 1789)
(Figs. 12 a 21)
Helix octona Indiae occidentalis, Chemnitz, 1786: 190,
pl. 136, f. 1264.
Bulimus octonus Bruguière, 1789: 325.
Bulimus octonus Lamarck, 1822 2a. part.: 124, fig. 27.
Achatina octona Orbigny, in La Sagra, 1853: 168, 169
pl. 11, f. 4-6 in La Sagra, 1853.
Achatina octona Reeve, 1849. Achatina sp., 84 pl.
XVII.
Subulina octona Fisher e Crosse, 1878: 639, pl. XXV,
figs. 15 e 15a.
Subulina octona Von Martens, 1890/1901: 298-300.
Subulina octona Tryon e Pilsbry, 1906: 72-74, 222-224,
pl. 12, figs. 8-9, pl. 39, figs. 28, 37,
39 e 40.
C.
A-F.
Subulina octona Burch, J.B., 1960: 29, pl. VI, fig.
Subulina octona Hass, 1962: 49-52, pl. VIII, figs.
Subulina octona Lanzieri, 1966: 1-29, 32-44, figs. 2-
13.
CONCHA (Figs. 15 e 16. É pequena, medindo aproximada-
mente 13mm x 4 mm, com a espira muito longa, graças ao número
de voltas que apresenta (8). A amplitude das voltas aumenta pro-

39.
gressivamente até a volta corporal. Não apresenta nehuma orna-
mentação característica, evidenciando de forma não muito marca-
da, as linhas de crescimento. A abertura é de forma ovalada sem
dentes ou lamelas e com o peristoma cortante. O bordo colume-
lar apresenta uma leve projeção alongada no sentido do eixo,
mais alargada em sua metade. As suturas são evidentes, porém
não muito profundas.
RÁDULA (Fig. 12). É uma lâmina muito delgada, com 20
dentes de cada lado, mais desenvolvidos e mostrando particula-
ridades apreciáveis somente até a quinta fileira. Daí em dian-
te os dentes são muito pequenos e reduzem-se progressivamente
até a margem, onde são rudimentares. A margem anterior do den-
te recobre a posterior do dente anterior da mesma fileira. O
dente central é muito reduzido e sem características marcan-
tes. Tem a forma retangular, com uma leve protuberância cen-
tral, no mesmo sentido do eixo maior do dente. Pelas pequenas
dimensões deste dente, a fileira central mostra-se como uma es-
treita linha em toda a extensão da rádula. O 1º dente é quase
simétrico, apenas com o ângulo posterior externo mais agudo. A
cúspide central é desenvolvida e com duas cúspides menores co-
locadas interna e externamente a ela. Estas características se
repetem até o 4º dente. O 5º dente é um pouco menor e provido
de duas pequenas cúspides. Do 6º dente até a margem, os dentes
não têm características marcantes, aparecendo então discreta-
mente, nos primeiros que se seguem, um espessamento um pouco
maior da margem anterior.

40.
MANDÍBULA (Fig. 14). É do tipo odontognato, com pla-
cas de forma aproximadamente retangular, cujos bordos anterio-
res são levemente arredondados e mais estreitos que o restante
do corpo da mandíbula. A metade anterior tem os bordos late-
rais marcados, articulando-se com as placas contíguas, porém
com muito pouca quitinização, o que lhe dá a flacidez necessá-
ria para a mobilidade.
OVOS (Fiqs. 13 e 13a). São esféricos, de côr esbran-
quiçada, medindo aproximadamente 2 mm x 1,6 mm, devido aos dois
achatamentos paralelos que possuem.
SISTEMA GENITAL. Inicia-se pelo ovotestis aproximada-
mente a partir da 3a. volta da espira, sendo em seu conjunto,
um sistema bastante longo, decorrente da forma da concha e ocu-
pando sempre uma posição próxima do eixo columelar. O ovotes-
tis é formado por um pequeno grupo de folículos pouco indivi-
dualizados. Seus canais vão ter ao canal coletor do ovotestis
(Fig. 21) de extensão mais ou menos retilínea, apoiado na glân-
dula de albumina e fracamente preso a ela por tecido conjunti-
vo frouxo. Bruscamente modifica sua forma, tornando-se um tubo
fortemente enrolado, de calibre maior, sem contudo, modificar
sua direção, e apresentando importantes diferenças histológi-
cas das regiões vizinhas (Lanzieri, 1966). O canal hemafrodita
tem aspecto semelhante ao canal coletor do ovotestis, porém, é
mais curto e ligeiramente mais espesso. Sua desembocadura se
faz no receptáculo seminal, que se coloca na face côncava da
glândula de albumina em pouco mais próximo da extremidade pro-

41.
ximal desta. Glândula de albumina. De forma achatada acompanhan-
do a volta da espira e em grande parte colocando-se entre esta
e a glândula digestiva, variando sua forma, dimensões e aspecto
geral de acordo com a atividade reprodutiva do animal. O ovis-
permoduto, e também um órgão alongado decorrente da forma geral
do molusco. Contudo, em face de raramente, em exemplares adul-
tos, não serem encontrados ovos "in utero" o órgão destende-se
muito, modificando-se a posição do preguiamento uterino, tornan-
do-o pouco mais baixo, bem como modificando a próstata, em um
órgão laminar, com aparência mais curta que o que se contata com
o útero em outras condições. O oviduto (Fig. 17) e longo, tendo
junto ao canal deferente (Figs. 17, 18 e 20) que é preso em gran-
de parte de sua extensão por tecido conjuntivo frouxo. Este ca-
nal abandona este trajeto a alguma distância do átrio genital
para adotar a mesma posição junto ao complexo peniano. Desembo-
ca no complexo peniano, sem demonstrar externamente modifica-
ções importantes.
CANAL DA ESPERMATECA (Fig. 19). É um curto e calibro-
so canal que tem sua origem bastante distante do átrio genital,
deixando com isso o duto vaginal longo. Sua luz é estreita em
toda sua extensão, mesmo nas proximidades da espermateca, onde
se reduzo calibre do canal. Apoia-se ao longo do ovispermodu-
to, ficando a espermateca sobre as últimas dobras do útero, nas
proximidades da origem do canal deferente.
COMPLEXO PENIANO (Figs. 17, 18 e 20). Muito longo e
delgado, tendo esta característica na maior parte de sua vida,

42.
com diferença bem marcada para as demais partes do sistema ge-
nital. A região fálica representa a maior parte do órgão, isto
porque, nas preparações efetuadas não demonstrou diferenças im-
portantes que a diferenciariam da região epifálica, se existis
se esta, como por exemplo a presença do tecido glandular. Não
possue bainha muscular envolvendo-a. O que se nota de forma
mais evidente, é um maior calibre até o Ponto onde se insere
uma faixa muscular originada de uma faixa do músculo colume-
lar. O músculo retrator do pênis tem inserção terminal no fla-
gelo e é relativamente longo. O flagelo e curto, terminando em
fundo cego, possue uma luz não muito evidente nas preparações
habituais. Seu calibre pode apresentar-se diferente em relação
ao falus, conforme o estado fisiológico do órgão no momento.

43.
Subulina octona (Bruguière, 1972): Fig. 12: Rádula;Fig. 13 e 13a: ovo; Fig. 14: Mandíbula; Fig. 15.Concha; Fig. 16. Concha (abertura}.

44.
Subulina octona (Bruguière, 1792): Fig. 17: Aparelhogenital (porção distal, destacando-se o complexo peni-ano); Fig. 18: Complexo peniano; Fig. 19: Canal daespermateca e espermateca; Fig. 20: Complexo peniano(porção proximal); Fig. 21: Vesícula seminal.

45.
Bradybaena similaris (Férussac, 1821)
(Figs. 22 a 31)
Helix similaris Férussac e Deshayes, 1820/1851: 171,
pl. 25B, f. 1-4, pl. 27-A, f. 1-5.
Helix similaris Reeve, 1854, sp. 149, pl. XXXIV, figs.
a, b.
Helix similaris Tryon e Pilsbry. 1887: 205-206, pl.
46, figs. 27-30, pl. 47, figs. 33-37.
Eulota similaris Tryon e Pilsbry, 1894: 205.
Bradybaena similaris Thiele, 1931: 691.
Bradybaena similaris Malek e Cheng, 1974: 80, fig.
3-17, 0.
Bradybaena similaris Pitoni e cols., 1976: 27, fig. 2.
CONCHA (Fig. 22). De aspecto geral heliciforme, com pa-
rede espessa e forte, é constituída de 5 a 6 voltas de espira,
mostra suturas evidentes porém não muito profundas, acompanha-
das no seu limite superior por uma linha de coloração castanha
que se continua na volta corporal até o bordo da abertura e é
visível também pela face interna da concha. As linhas de cres-
cimento são mais pronunciadas na volta corporal, dando um as-
pecto levemente rugoso à superfície da concha. O umbigo e aber-
to, com os bordos abrindo-se amplamente, sem contudo ser muito
profundo. A abertura da concha é semilunar, com os bordos re-
fletidos largamente, não sendo guarnecida por lamelas ou dentes.
CÂMARA PAELAL (Fig. 26). É ampla, tendo maiores dimen-

46.
sões em sua porção anterior, onde os vasos são mais numerosos.
Nesta parte, encontram-se os vasos formadores de veia pulmonar
e que praticamente existem no seu lado direito. Do lado esquer-
do encontram-se vasos mais longos, mas que se dirigem para uma
veia marginal que corre paralela ao colar do manto. A veia pul-
monar, em sua metade final, é mais ou menos retilínea e cami-
nha paralelamente à face esquerda do rim até desembocar no co-
ração. A parte posterior do lado esquerdo da câmara paleal pra-
ticamente não tem vasos, apenas se observando o esboço de al-
guns nas proximidades do limite columelar. Os vasos do lado di-
reito são curtos, sempre em posição transversal ao eixo longi-
tudinal da câmara, e distribuem-se de modo mais ou menos uni-
forme até à curvatura do ureter. O rim é um órgão alongado e
estreito que se projeta no sentido anterior na câmara paleal,
alcançando sua metade, e situado um pouco para a direita des-
ta. O ureter faz uma curva curta e caminha paralelamente ao re-
to até à abertura excretora junto ao pneumóstomo.
RÁDULA (Fig. 23). Tem 32 dentes de cada lado da filei-
ra central. São dentes na maioria monocúspides e bem evidentes
até aproximadamente à altura do 15º, a partir de onde de forma
progressiva, vão tendo reduzido seu tamanho e menos evidentes
suas peculiaridades. O dente central é simétrico com sua largu-
ra posterior, que é pouco maior que a anterior, que é mais
delgada que o restante do dente. A cúspide central é cuneifor-
me, acompanhando a forma de sua base, e termina em um Ângulo
fortemente agudo. Os dentes de 1 a 8 têm forma geral muito se-

47.
melhante. São dentes retangulares com sua parte anterior leve-
mente voltada para a face lateral do órgão. Com isto, o ângulo
anterior externo do cente é mais agudo que o seu oposto. A cús-
pide central é também aguda como a do dente central, originan-
do-se de uma base aproximadamente lasangular em que o ângulo mais
anterior representa a projeção anterior da base do cúspide. A
partir do 4º dente, a cúspide se situa levemente mais para a
face lateral interna do dente. Do 9º ao 15º dente começam a apa-
recer leves modificações no padrão geral da forma. O ângulo ex-
terno é mais agudo, embora mais curta a saliência no 15º dente.
A cúspide central é também mais curta e colocada, como foi di-
to anteriormente, mais próximo da face interna. Na base da cús-
pide central começa também, a partir do 11º dente, uma curta
projeção no lado anterior externo. Do 16 dente em diante, a di-
minuição progressiva do tamanho até ao 32º, bem como uma modi-
ficação em suas peculiaridades no sentido de redução, conduza
praticamente dentes redimentares na margem do órgão.
MANDíBULA (Fig. 24). É um órgão curvado e colocado na
parte superior da entrada da cavidade bucal. Seu bordo anterior
é mais quitinizado, apresentando-se como uma linha sinuosa. Tais
sinuosidades representam o limite anterior de placas cujos bor-
dos externos são mais quitinizados. São presentes três placas
de cada lado de uma crista central fortemente quitinizada, que
possui duas projeções agudas na sua parte posterior. Projeções
como estas são vistas também no ângulo posterior externo de ca-
da placa.

48.
APARELHO GENITAL. Ovotestis. É um órgão formado por
três ácinos, e estes, por numerosos folículos, implantados na
face columelar da glândula digestiva, mais ou menos na altura
da terceira volta da espira. De cada um dos ácinos parte um fi-
no canal que desemboca em um canal comum. Este canal, apoiado
em seu início na glândula digestiva, modifica-se para formar
a vesícula seminal. Esta acompanha a volta da espira, sendo for
mada por um canal que tem maior calibre em sua parte média e,
em todo o trajeto é sinuoso, com exceção do seu início, logo
após o canal comum do ovotestis, e do seu término, no complexo
do receptáculo. Complexo do receptáculo (Fig. 28). É um órgão
muito delgado, situado nas proximidades da junção da glândula
de albumina com o ovispermoduto. Em seu interior contém finos
túbulos alongados no sentido longitudinal do órgão, que é pre-
so à glândula de albumina por tecido conjuntivo frouxo. Apenas
em sua base faz conexão com a vesícula seminal por um fino canal
com a glândula. Ovispermoduto. O útero é formado por um pre-
gueamento nítido, sem ser muito forte, cujo término é mais bai-
xo que suas primeiras porções junto à glândula de albumina. A
próstata é uma lâmina estreita, cujo comprimento não correspon-
de ao do útero, distinguindo-se claramente em seu término a for-
mação de canal deferente. Glândula de albumina (Fig. 28). Ocu-
pando grande parte da extensão da glândula digestiva, coloca-
se entre esta e a concha. Seu limite distal pode alcançar gran-
de parte da curvatura da glândula digestiva. Seu limite proxi-
mal é nas proximidades do fundo da cavidade paleal, junto à cur-

49.
vatura do ureter secundário. Seu desenvolvimento e coloração pa-
recem estar em função da atividade do sistema reprodutor. Sua
forma é sempre alongada, acompanhando a volta da espira. Ovidu-
to (Fig. 30). É um órgão simples, curto, com luz relativamente
ampla, porém sem peculiaridades. Canal de espermateca, (Figs.
27, 30 e 31). É um canal espesso, de luz ampla, preso ao ovidu-
to por frouxo tecido conjuntivo e também ao ovispermoduto, até
desembocar na espermateca, que se situa na altura do inicio do
pregueamento uterino. Saco do dardo (Fig. 27). Órgão volumoso,
em forma de cunha, com sua base largamente implantada na porção
terminal do trato genital. Glândulas multífidas (Fig. 27). Cons-
tituídas por pequenos tubos alongados de fundo cego, cuja de-
sembocadura se faz na parte distal do saco do dardo. São em nú-
mero de duas, que se dicotomizam algumas vezes. As ramificações
apoiam-se na curvatura do saco de dardo, mantendo-se presas Dor
frágil tecido conjuntivo. Complexo peniano (Fig. 29). É a es-
trutura que mais se destaca dentre as demais por suas peculia-
ridades. É um órgão fortemente musculoso, com sua porção dis-
tal correspondente ao falus envolvida por uma bainha muscular
bem individualizada, porém não muito desenvolvida. Sua luz não
é ampla, porém mostra paredes espessas, demonstrando riqueza de
tecido glandular. Da bainha muscular parte um pequeno ligamen-
to que também o canal deferente junto ao órgão. De sua termina-
ção parte outro ligamento que termina nas imediações do ponto
onde se observa uma modificação na estrutura do complexo, carac-
terizada por uma constrição, e onde também existe uma modifica-

50.
ção parte outro ligamento que termina nas imediações do ponto
onde se observa uma modificação na estrutura do complexo, carac-
terizada por uma constrição, e onde também existe uma modifica-
ção da luz do órgão. O complexo peniano diminui gradativamen-
te de calibre, sendo esta diminuição um pouco mais brusca nas
proximidades de sua parte média, onde a luz se torna mais estrei-
ta e o tecido glandular é escasso. A metade final do órgão é re-
lativamente longa e, em posição terminal, desemboca o canal de-
ferente, no mesmo ponto onde termina a segunda faixa ligamen-
tosa. O músculo retrator do pênis é uma lâmina muscular bem for
te que se origina no início do terço distal do complexo penia-
no. Canal deferente (Fig. 27 e 29). É um delgado canal, com dis-
posição sinuosa, especialmente entre o epifalus, sua desemboca-
dura, e o fino ligamento que o prende ao bordo da bainha museu-
lar. No restante do trajeto, tem contiguidade com o saco do dar-
do e as glândulas multífidas e o oviduto, cruzando este órgão
nas proximidades da origem do canal da espermateca.

51.
Bradybaena similaris (Férussac, 1821): Fig. 22: Concha

52.
Bradybaena similaris (Ferussac, 1821); Fig. 23: Rádula;Fig. 24: Mandíbula; Fig. 25: Massa visceral (porção pro-ximal), destacando-se o ovotestis e vesícula seminal.

53.

54.
Bradybaena similaris (Férussac, 1821); Fig. 27: Apa-relho genital (porções média e distal); Fig. 28: Com-plexo do receptáculo e vesícula seminal.

55.
Bradybaena similaris (Férussac, 1821): Fig. 29:Complexo peniano; Fig. 30: Oviduto e início docanal da espermateca; Fig. 31: Ovispermoduto.

56.
Os parasitos de interesse
Os parasitos que ocorrem no Brasil e que têm seu ci-
clo evolutivo parcialmente desenvolvido, de forma obrigatória
ou facultativa, nos moluscos terrestres, ou que, por suas afi-
nidades, devem ter fases de seus ciclos neste grupo de molus-
cos estão relacionados a seguir.
Classe Trematoda
Ordem Digenea
Subordem Prosostomata
Família Dicrocoelidae Odhner, 1911
Gênero Dicrocoelium Dujardin, 1845
Dicrocolium dendriticum (Rudolphi, 1819)
Gênero Eurytrema Looss, 1907
Eurytrema coelomaticum (Giard e Billet, 1892)
Looss, 1907
Gênero Platynosomum Looss, 1907
Platinosomum illiciens (Braun, 1901) Kossak, 1910
Família Echinostomatidae Poche, 1926
Gênero Echinostoma Rudolphi, 1809
Echinostoma revolutum (Froelich, 1802) Looss, 1899
Família Brachylamidae
Gênero Brachylaemus Dujardin, 1843
Brachylaemus mazzamatii (Travassos, 1927)
Brachylaemus suis Balozet, 1936

57.
Gênero Postharmostomum Witenberg, 1923
Postharmostomum gallinum Witenberg, 1923
sin. Postharmostomum commutatum (Diesing, 1858)
Postharmostomum fleuryi (Fonseca, 1939)
Brachylaemus fleuryi Fonseca, 1939
Família Eucotylidae
Gênero Eucotyle Cohn, 1904
Eucotyle freitasi Costa e Freitas, 1972
Gênero Paratanaisia Freitas, 1959
Paratanaisia bragai (Santos, 1934) Freitas, 1959
sin. Tamerlania bragai Santos, 1934
Tanaisia bragai Santos, 1934
Classe Nematoda
Superfamília Strongyloidea
Família Protostrongylidae
Gênero Protostrongylus Kamensky, 1905
Protostrongylus rufescens (Leuckart, 1865)
Gênero Cystocaulus Schulz, Orloff e Kutass, 1933
Cystocaulus ocreatus (Railliet e Henry, 1907)
sin. Protostrongylus ocreatus Railliet e Henry, 1907
Cystocaulus nigrescens (Jerke, 1911)
Gênero Muellerius Cameron, 1927
Muellerius minutissimus (Mégnin, 1878)
sin. Muellerius capilaris (Muller, 1889)

58.
Gênero Aclurostrongylus Cameron, 1927
Aclurostrongylus abstrusus (Railliet, 1898)
Gênero Bronchostrongylus Cameron, 1931
Bronchostrongylus s ubcrenatus (Railliet e Henry, 1913)
Gênero Angiostrongylus Kamensky, 1905
Angiostrongylus vasorum (Baillet, 1866)
Angiostrongylus cantonensis Chen, 1935
Gênero Filaroides v. Beneden, 1858
Filaroides rostratus Cerichter, 1949
Filaroides martis (Werner, 1782)
Classe Cestoda
Superfamília Taenioidea
Família Davaineidae Fuhrmann, 1907
Gênero Davainea Blanchard, 1981
Davainea proglottina (Davaine, 1860)
Gênero Raillietina Fuhrmann, 1920
Rallietina bonini (Mégnin, 1899)

59.
EXPLICAÇÃO DO TEXTO E DOS QUADROS 2 A 8
+ : Presente
- : Ausente ou ainda não identificada a espécie
* : Espécies assinaladas no Brasil
: Relação obrigatória estabelecida
--------------- : Relação facultativa estabelecida
Indicações superiores: Brasil
Indicações inferiores: Outros países

60.
DISTRIBUIÇÃO GEOGRÁFICA E RELAÇÕES DOS MOLUSCOS COM PARASITOS
DOS ANIMAIS DOMÉSTICOS, NO BRASIL
As espécies de moluscos e de parasitos ou somente uma
delas, citadas para o Brasil, situam-se dentro de várias alter-
nativas, de acordo também com suas relações estabelecidas ou
não em nosso país, levando-nos ao interesse no desenvolvimen-
to de estudos nos casos apropriados. Podemos abranger estas
relações do seguinte modo:

QUADRO I.
DISTRIBUIÇÃO GEOGRÁFICA DOS MOLUSCOS
E DOS PARASITOS A ELES RELACIONADOS, DOS ANIMAIS DOMÉSTICOS, NO BRASIL

QUADRO l.
Conti nuação

OUADRO 1.
Continuação

QUADRO 1.
Continuação

QUADRO 1.
Continuação

QUADRO 2.
ESPÉCIES DE MOLUSCOS E DE PARASITOS QUE OCORREM NO BRASIL
E EM OUTROS PAÍSES, COM RELAÇÕES ENTRE AMBOS CONHECIDAS EM AMBAS AS DISTRIBUIÇÕES

QUADRO 3.
ESPÉCIES DE MOLUSCOS E DE PARASITOS QUE OCORREM NO BRASIL
E EM OUTROS PAíSES, COM RELAÇÕES ENTRE AMBOS CONHECIDAS SOMENTE NESTES ÚLTIMOS

QUADRO 3.
Continuação

QUADRO 3.
Continuação

QUADRO 3.
Continuação

QUADRO 3.
Continuação

QUADRO 4.
ESPÉCIES DE MOLUSCOS E DE PARASITOS QUE OCORREM NO BRASIL
E EM OUTROS PAÍSES, COM RELAÇÕES ENTRE AMBOS CONHECIDAS SOMENTE NO BRASIL

QUADRO 5.
ESPÉCIES DE MOLUSCOS DE OCORRÊNCIA NO
BRASIL E EM OUTROS PAÍSES, CUJAS RELAÇÕES COM PARASITOS SÃO CONHECIDAS
SOMENTE NESTES ÚLTIMOS; AS ESPÉCIES DE PARASITOS NÃO ESTÃO AINDA ASSINALADAS NO BRASIL

QUADRO 5.
Continuação
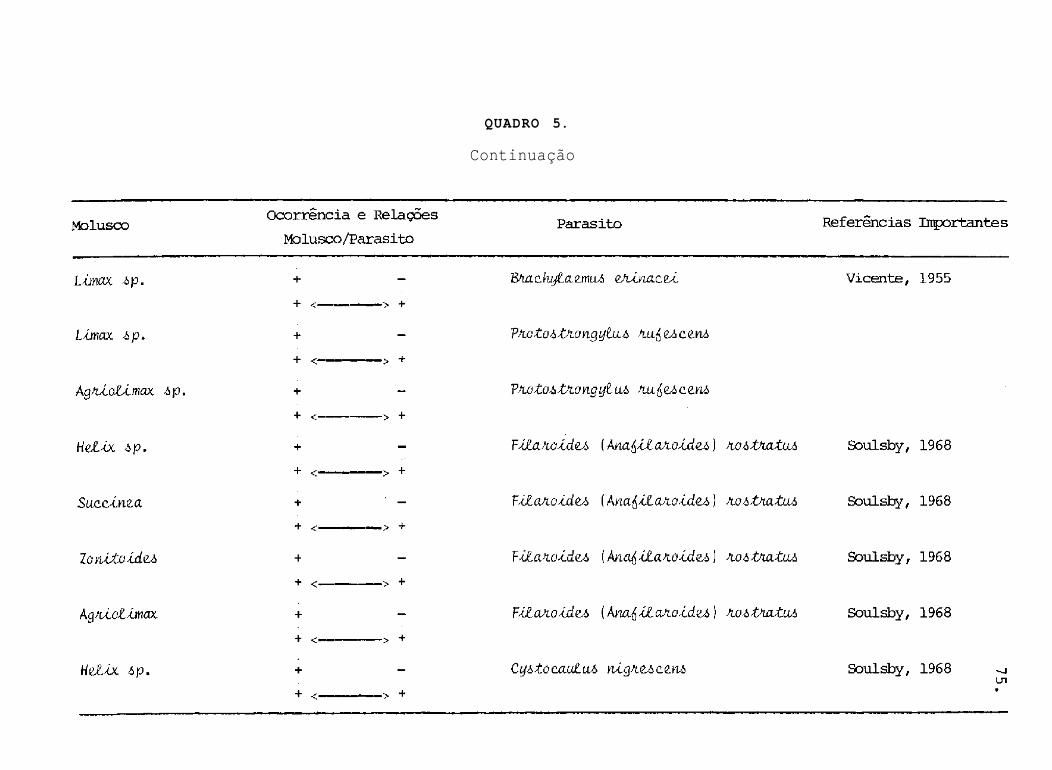
QUADRO 5.
Continuação

QUADRO 6.
ESPÉCIES DE MOLUSCOS AINDA NÃO ASSINALADOS NO BRASIL, MAS QUE
SÃO RELACIONADAS, EM OUTROS PAÍSES, A PARASITOS QUE TAMBÉM OCORREM NO PAÍS

QUADRO 6.
Continuação

QUADRO 6.
Continuação

QUADRO 6.
Continuação

QUADRO 6.
Continuação

QUADRO 6.
Continuação

QUADRO 6.
Continuação

QUADRO 6.
Continuação

QUADRO 7.
ESPÉCIES DE PARASITOS QUE OCORREM NO BRASIL E EM OUTROS PAÍSES
E QUE CERTAMENTE SÃO RELACIONADAS A MOLUSCOS CUJAS ESPÉCIES AINDA NÃO FORAM DETERMINADAS

QUADRO 8.
ESPÉCIES DE PARASITOS QUE OCORREM SOMENTE NO BRASIL E QUE
CERTAMENTE SÃO RELACIONADAS A MOLUSCOS, CUJAS ESPÉCIES AINDA NÃO FORAM DETERMINADAS

5. DISCUSSÃO E CONCLUSÕES
A posição sistemática das espécies envolvidas nos ci-
clos evolutivos de parasitos no Brasil, pôde ser avaliada de
maneira mais precisa através do estudo da anatomia visceral.
No caso dos Subulinidae, muitas espécies são citadas para o Bra-
sil, nos géneros Subulina, Leptinaria, Opeas, Megaspira,
Obeliscus, etc. cuja morfologia das conchas é, em alguns casos
muito semelhante, e podem ser encontradas nos mesmos habitats.
Subulina octona sendo uma espécie cosmopolita, é citada em vá-
rios estados do Brasil. Suas determinações foram sempre basea-
das nas conchas, e por elas foram descritas algumas varieda-
des. O estudo morfológico das partes moles mais importantes foi
realizado por LANZIERI (1966) dando ênfase aos aspectos histo-
lógicos. Graças a estes estudos puderam ser confirmados deta-
lhes anatômicos, como a ausência de epifalus e bainha muscular
e a presença de uma faixa muscular inserida no falus, que acre-
ditamos com função possivelmente, também retratora. Quanto a
Leptinaria unilamellata, os detalhes da anatomia visceral são
agora conhecidos e podem assim serem comparados com as demais

87.
espécie do gênero, bem como dos gêneros afins.
No caso dos Bradybaenidae, encontramos Bradybaena si-
milaris, que possui muitos caracteres comuns à família onde era
considerada anteriormente Fruticicolidae. Alguns caracteres, en-
tre outros, puderam ser evidenciados, no sentido do conhecimen-
to da fisiologia do sistema genital, como a disposição do ca-
nal diferente do complexo peniano.
Assim, estes estudos nos levam ao melhor conhecimento
das espécies e, naturalmente à maior segurança de suas posi-
ções na sistemática, mostrando que alguma modificação, se ne-
cessária, deve proceder a um estudo comparativo das partes mo-
les, envolvendo outras espécies afins.
Outras espécies como Limax flavus, Limax maximus, Li-
max valentianus, Deroceras reticulatum, Agriolimax agrestis,
Succinea meridionalis, Succinea lopesi, Succinea pusilla, Zoni-
toides aeboreus, Bulimulus tenuissimus, Epiphragmophora bernar-
dius. Helix aspersa citadas para o Brasil, devem merecer tam-
bém estudos em seus vários aspectos, já que são espécies tam-
bém envolvidas, ou, possivelmente envolvidas, em ciclos de pa-
rasitos no Brasil.
Os parasitos de animais domésticos que, de forma obri-
gatória ou não, tem os moluscos como hospedeiros intermediá-
rios, deles dependendo, portanto, para desenvolver seus ci-
clos, restringem-se aos trematodeos digenéticos (principalmen-
te), aos nematódeos em menor escala, e eventualmente, os cestó-
deos. Dentro da conceituação que se dá ao trabalho, os primei-

88.
ros, sempre e algumas espécies dos demais têm os moluscos como
hospedeiros obrigatórios. Ainda levando-se em conta a concei-
tuação geral do assunto, observa-se que, para alguns grupos de
parasitos, determinadas espécies de moluscos assumem maior in-
portância porque nelas tais parasitos se multiplicam intensa-
mente enquanto que em outras eles apenas se transformam, atin-
gindo estágios mais avançados e capazes de dar seqüência do ci-
clo quando levadas as formas infectantes aos hospedeiros defi-
nitivos. Desta análise, conclui-se que os moluscos terrestres
são muito importantes, especialmente quando os parasitos neles
se multiplicam, e, em particular, os hospedeiros intermediá-
rios facultativos, com cuja participação as chances dos parasi-
tos são ampliadas com maiores possibilidades de atingir os hos-
pedeiros definitivos, como também de se proteger de condições
adversas para uma determinada espécie somente.
Estes estudos permitiram esclarecer o que ocorre com
várias espécies de moluscos, em conseqüência dos sinônimos usa-
dos em inúmeras ocasiões, quando citado o hospedeiro interme-
diário de determinado parasito, ou com relação aos próprios pa-
rasitos, embora nestes com menor freqüência. Evidenciaram tam-
bém que d-terminados parasitos, portadores de afinidades evolu-
tivas, têm hospedeiros com esta mesma característica, mostran-
do as espécies eficientes vários aspectos comuns com as obriga-
tórias, especialmente com relação ao seu habitat, embora even-
tualmente apareçam exceções. Deste modo, pôde-se verificar que:
Leptinaria unilamellata é uma espécie encontrada com

89.
muita frequência nos mesmos habitats que Subulina octona, embo-
ra com populações compostas de menor número de indivíduos que
esta, e outras vezes como populações isoladas, compostas de um
número não muito grande de indivíduos; sua importância, até o
momento, está ligada à participação no ciclo evolutivo de Pos-
tharmostomum gallinum das aves, como mais uma alternativa para
o parasito, abrigando a forma infectante, constituindo-se no
segundo hospedeiro intermediário, facultativo.
Subulina octona, como a espécie anterior, é extrema-
mente freqüente em nosso meio; tendo atividade muito intensa
em seu período favorável, constitui presa fácil para os preda-
dores, especialmente aves, dando excelentes oportunidades aos
parasitos que hospeda para neles desenvolverem seus ciclos; sua
participação é obrigatório no caso de Platynosomun illiciens das
vias biliares do gato, constituindo-se no seu primeiro hospe-
deiro intermediário; ainda de forma obrigatória, participa do
ciclo de Paratanaisia bragai (SANTOS, 1934) das aves, abrigan-
do inclusive a forma infectante para o hospedeiro definitivo;
no ciclo evolutivo de Postharmostomum gallinum é, no Brasil, um
hospedeiro facultativo, uma vez que outras espécies de molus-
cos podem abrigar a forma infectante para as aves; tal fato não
foi considerado do mesmo modo por ALICATA (1940), já que somen-
te ela foi encontrada abrigando a metacercária; também é cita-
da abrigando facultativamente a forma infectante (cisticercói-
de) de Davainea proglottina das aves, como também Aclurostron-
gylus abstrusus do gato, que pode evoluir até à forma infectan-

90.
te (larva de 3º estádio).
Bradybaena similaris é extremamente freqüente no Bra-
sil; especialmente nos períodos mais adequados à sua ativida-
de, prolífera e está presente em grande quantidade; suas rela-
ções com parasitos, conhecidas no Brasil, são somente com Pos-
tharmostomum gallinum das aves (DUARTE, 1977), do qual conside-
ramos um hospedeiro obrigatório, já que as primeiras fases do
ciclo se dão somente neste molusco e as demais são facultativa
mente passadas em outras espécies; ainda duas outras espécies
de parasitos são apontadas pela literatura, como relacionadas
a ela: Eurytrema coelomaticum, cos canais pancreáticos de bovi-
nos, para o qual é o primeiro hospedeiro intermediário, o que
também interpretamos como uma relação obrigatória, já que se
trata da única espécie citada pela literatura, e Aclurostrongylus
abstrusus que é um nematódeo que se serviria facultativamente
dela para desenvolver seu ciclo até larva de 3º estágio, uma
vez que outras espécies poderiam atuar de maneira análoga.

6. REFERÊNCIAS BIBLIOGRÁFICAS
ASH, L.R. 1962. Helminth parasites of dogs and cats in Hawaii.
J. Parasitol. 48: 63-65.
ADBOU, A. 1956. Life cycle of Davainea proglottina. J. Helmin-
thol. 30: 189-202.
AGASSIZ, L. 1807/1873. Nomenclator zoologicus, continens nomina
systematica generum Animalium tan viventium quan fossilium.
v. 1. Mammalia-Mollusca. v.3. Index Universalis.
ALICATA, J.E. 1940. The life cicle of Postharmostomum gallinum
the cecal fluke of poultry. J. Parasitol. 26(2): 135-143.
ALICATA, J.E. 1964. Parasitic infections of man and animals in
Hawaii Hawaii Agricultural Experiment Station University of
Hawaii Tech. Bul. 61: 1-138.
ARAÚJO, J.L. de B.; REZENDE, H.E.B. & RODRIGUES, P.A.F. 1960.
Sobre "Bulimulus tenuissimus" (Orbigny, 1835) (Gastro-
poda, Pulmonata). Rev. Bras. Biol. 20(1): 33-42, figs.
1-23.

92.
BALOZET, L. 1937. Cycle evolutif de Brachylaemus suis. C. R.
Acad. Sci. Paris 204: 622-624.
BECKLUND, W.W. 1964. Revised check list of internal and external
parasites of domestic animals in the United States and Pos-
sessions and in Canadá. Amer. J. Vet. Res. 25: 1380-1416.
BACKER, F. 1913. The land and fresh-water mollusks os the Stan-
ford Expedition to Brazil. Proc. Acad. Nat. Sci. Philad.:
618-672, pls. XXI-XXVII.
BARRETO, J.F. & MIES, Fº A. 1942. Primeiras observações sobre
a presença de Tamerlania bragai (Violantino dos Santos, 1934)
nos rins de Meleagris gallopavo domestica. Serviço de Informação
Agrícola, Ministério da Agricultura, Rio de Janeiro, 3 pp.
BASH, P.F. 1965. Completion of the life cycle of Eurytrema pan
creaticum (Trematoda; Dicrocoelidae). J. Parasitol. 51(3):
350-355.
BOFFI, A.V. 1979. Moluscos brasileiros de interesse médico e
econômico. 376 pp., 105 Ests., 162 figs. Hucitec Ed. Rio de
Janeiro.
BONDENSEN, P. 1954. Slugs (Snails without a House). Natur. og
Museum. 3(2): 1-9, 3 figs., 1 pl.
BRANT, P.C. & COSTA, A.S. 1963. Distribuição geográfica da Hi-
datidose, Cisticercose, Eurytrematose, Dictyocaulose e Fas-

93.
ciolose bovinas no Estado de Minas Gerais. Arq. Esc. Vet.
Belo Horizonte. 15: 47-59.
BREURE, A.S.H. 1979. Systematics, Phylogeny and Zoogeography
of Bulimulidae (Bulimulinae) (Mollusca). Zool. Verh. 186:
1-215, 182 figs., 3 pls.
BRUGIÈRE, J.G. 1789/1792. Encyclopedie Méthodique. Histoire
Naturelle des Vers. Paris et Liège. T. I part. 1 (1789): 1-
344. T. II part. 2 (1792): 345-758.
BUCKUP, L. & BUCKUP, E.H. 1957. Catálogo dos moluscos do Museu
Rio-Grandense de Ciências Naturais. Zoologia. (1): 1-40.
BURCH, J.B. 1960. Some snails and slugs of Quarantine signi-
ficance to the United States. Sterkiana. 2: 13-53.
CAMARGO, O.R. 1956. Caracóis nocivos às plantas cultivadas
no Rio Grande do Sul. Sec. de Estado dos Negócios da Agri-
cultura, Industria e Comércio. Série C. Porto Alegre.
CAMERON, T.W.M. 1931. Experimental infection with. Dicrocoelium
dendriticum. J. Helm. 9: 41-44.
CHEMNITZ, J.H. 1786 in MARTINI, F.H.W. & CHEMNITZ, J.H. Neus
Systematisches Conchylien Cabinet, 9, 151 + XXVI + 194 pp,
pls. 103-136. Nurnberg.
COSTA, H.M.A. & FREITAS, M.G. 1972. Eucotyle freitasi sp. n. do
pato doméstico. Rev. Bras. Biol. 32(3): 391-393.

94.
COSTA, H.M.A. & FREITAS, M.G. 1970. Lista dos helmintos para-
sitos dos animais domésticos no Brasil. Arq. Esc. Vet.
XXII: 33-94.
COSTA, H.M.A. 1965. Alguns aspectos sobre helmintos parasitos
de Sus domesticus Linnaeus, 1758, procedentes do Estado da
Bahia, Brasil. Arq. Esc. Vet. Belo Horizonte. 17: 11-44.
CORREIA, O. 1952. Incidência helmíntica em suínos, equinos,
ovinos e bovinos no Rio Grande do Sul. Mundo Agrícola São
Paulo. 1(1): 66-67.
CORREIA, O. 1965. Incidência da Hidatidose, Fasciolose, Ste-
phanurose e Cisticercose nos rebanhos riograndenses. Rev.
Fac. Agron. Vet. Porto Alegre. 7(2): 137-146.
CRAGG, J.B.; FOSTER, R. & VINCENT, M. 1957. Larval Trematodes
(Brachylaemidae) from the slugs Milax sowerby (Férussac),
Agriolimax reticulatus (Muller) and Arion l usitanicus Ma-
bille. Parasitology. 47.
DOLLFUS, R.P. 1935. Les Distomes des Stylommatophores
terrestres (excl. Succineidae). Catalogue par hôtes
et resumé des descriptions. Ann. Parasitol. XIII(2):
176-188.
DOLLFUS, R.P. 1935. Sur quelques Brachylaemus de la faune
française récoltés principalement a Richelieu (Indre-et-
Loire). Ann. Parasit. Hum. Comp. XIII(1): 52-79.

95.
DOLLFUS, R.P. 1935. Les Distomes des Stylolmnatophores terres-
tres (excl. Succineidae). Catalogue par hôtes et resumé des
descriptions. Ann. Parasit. XIII(5): 445-485.
DOLLFUS, R.P. 1935. Les Distomes des Stylommatophores terres-
tres (excl. Succineidae). Catalogue par hôtes et resumé des
descriptions. Ann. Parasit. XIII(4): 369-385.
DOLLFUS, R.P. 1935. Les Distomes des Stylommatophores terres-
tres (excl. Suecineidae). Catalogue par hôtes et resumé des
descriptions. Ann. Parasit. XIII(3): 259-278.
DUARTE, M.J.F. 1977. O ciclo evolutivo de Postharmostomum
gallinum Witenberg, 1923, no Estado do Rio de Janeiro.
U.F.R.R.J. Tese. 69 pp., 17 figs. Mimeografado.
EUZÉBY, J. 1964. Les zoonoses helminthiques 389 pp., 154 figs.
Vigot Frères Ed. Paris.
FERNANDEZ, D. 1973. Catálogo de la Malacofauna terrestre argen-
tina. Comission de Investigaciones Cientificas de la Provin-
cia de Buenos Aires. Monografias 4. 197 pp, 29 figs.
FÉRUSSAC, D. & DESHAYES, G.P. 1819/1851. Histoire Naturelle
génerale et particulière des Mollusques. Test 1, VIII + 402
pp., 2, XVI, (1), 184 pp.; (2), 260 pp.; Atlas 1, 24 pp.,
69 pls. Paris.
FIGUEIRAS, A. 1963. Ennumeracion sistematica de los moluscos
terrestres del Uruguay. Com. Soc. Malac. Uauguay. 1(4): 79-96.

96.
FISHER, P. & CROSSE, H. 1878. Mission Scientifique au Mexique
et dans l'Amérique Centrale. Recherches zoologiques, pp.
153-699.
FOSTER, R. 1958. The effects of trematode metacercariae (Bra-
chylaemidae) on the slugs Milax sowerbii and Agriolimax re-
ticulatus. Parasitology. 48.
FRANCO, S.R.N.S. 1965. Nota sobre trematódeos Eucotylideos (Tre-
matoda, Eucotylidae). Mem. Inst. Oswaldo Cruz. 63 (único):
59-65, 7 figs., 1 quadro.
FREITAS, J.F.T. 1951. Revisão da família Eucotylidae Skrja-
bin, 1924 (Trematoda). Mem. Inst. Oswaldo Cruz. 49: 33-271,
74 Ests. 223 figs.
FRÖMMING, E. 1956. Experimentelle Untersuchungen über den Ein-
flub von Umweltfaktoren auf das Gedeihen der Lungeschnecke
Rumina decollata (L.). Zool. Jahr. Band 84, Heft 6: 577-602.
GERICHTER, C.B. 1948. Observations on the life history of lung
nematodes using snails as intermediate hosts. Amer. J. Vet.
Res. 9: 109-112.
GERICHTER, C.B. 1949. Studies on the nematodes parasitic in
the lungs of Felidae in Palestine. Parasitology. 39: 251-
254.
GERMAIN, L. 1930. Faune de France. Mollusques terrestres et
fluviatiles, 1ª part.: 1-477, pls. I-XXVI.

97.
GETZ, L.L. 1959. Notes on the ecology of slugs: Arion circuns-
criptus, Deroceras reticlatum and D. laeve. Am. Med. Nat.
61(2): 485-498.
GRISI, L. 1971. Ocorrência de Angyocaulus railfieti (Travas-
sos, 1927) comb. n. em Canis familiaris (Nematoda, Protos-
trongylidae).
GUILHON, J. & AFGHAHI, A. 1969. Evolution larvaire d'Angyos-
trongylus vasorum (Baillet, 1866) dans l'organisme de di-
verses especes de Mollusques terrestres. C. R. Acad. Sci.
Paris. 268: 434-436.
HAAS, F. 1949. Land and fresh-water mollsks from Perú. Field.
Zool. 31(28): 235-250, figs. 50-59.
HAAS, F. 1952. South American non-marine shells: further remarks
and descriptions. Fild. Zool. 34(9): 107-132, figs. 14-26.
HAAS, F. 1955. On some collections of in land shells from South
America. Field. Zool. 34/35: 361-387.
HAAS, F. 1962. Caribbean Land Molluscs: Subulinidae and Olea-
cinidae, Studies on the fauna of Curação and other Caribbean
Islands XIII (58): 49-60, pls. VII-XI. The Hague Martius
Nijhoff.
HOBMAIER, M. & HOBMAIER, A. 1935. Intermediate hosts of Aele-
rostrongylus abstrusus of the cat. Proc. Soc. Expl. Biol.
Med. 32: 1641-1647.

98.
HYMAN, L.H. 1967. The Invertebrates vol VI Mollusca I - VII
+ 1 - 792 pp, 249 figs.
JAECKEL, J. 1952. Short review of the land and fresh-water
mollusks of the North-East States of Brazil. Dusenia. 3: 1-
10.
KLEWER, H.L. 1958. The indicence of helminth lung para-
sites of Lynx rufus (Shreber) and the life cicle of
Anafilaroides rostratus (Gerichter, 1949). J. Parasit. 44
(supp.): 29.
KOHN, A. & FERNANDES, B.M.M. 1975. Sobre as espécies do gênero
Echinostoma Rudolphi, 1809 descritas por Adolpho Lutz em
1924. Mem. Inst. Oswaldo Cruz. 73(1-2): 77-89, Est. I-X,
figs. 1-41.
LAMARCK, J.B.P.A. 1819/1822. Histoire Naturelle des Animaux
sans vertèbres 6(1) (1819), VI + 343 pp, (2) (1922), 232 pp.
Paris.
LANZIERI, P.D. 1965. Sobre a cópula e a constituição do apare-
lho genital de Succinea meridionalis Orbigny, 1846 (Gastro-
poda, Pulmonata, Succineidae Mem. Inst. Oswaldo Cruz. 63
(único): 207-228, 115 figs.
LANZIERI, P.D. 1966a. A família Succineidae da ilha da Trin-
dade, Costa do Brasil (Gastropoda, Pulmonata). Pap. Av. Depto.
Zool. 19: 169-183, 79 figs.

99.
LANZIERI, P.D. 1966b. Alguns aspectos morfo-estruturais do apa-
relho genital de Subulina octona (Bruguière, 1792) (Gastropo-
da, Pulmonata, Subulinidae). Tese. 44 pp., 13 figs. Mimeogra-
fado.
LAPAGE, G. 1962. Veterinary Helminthology and Entomology. 600
pp., 26 pls. The Williams and Wilkins Company. Baltimore.
LA SAGRA, M.R. de. 1953. Histoire Physique, Politique et Natu-
relle de l'Ile de Cuba. Mollusques (par Alcide D'Orbigny). T.
I 1841: 1-208. T. II 1842: 209-264.
MALDONADO, J.F. 1945a. The life cicle of Tamerlania bragai
Santos, 1934 (Eucotylidae) a Kidney fluke of domestic pi-
geons. J. Parasitol. 31(5): 306-314.
MALDONADO, J.F. 1945b. Ciclo vital e biologia del Platynoso-
muro fastosum Kossak, 1910 (Trematoda: Dicrocoelidae). 3. Publ.
Hlth. Trop. Med. 20(17): 34-60.
MALEK, E.A. & CHENG, T.C. 1974. Medical and Economic Mala-
cology. X + 398 pp., Academic Press Ed.
MARTENS VON, E. 1890/1901. Biologia Centrali-Americana. Land
and Freshwater mollusca: I-XXVIII + 1-705.
MORRETES, F.L. 1943. Contribuição ao estudo da fauna brasilei-
ra de moluscos. Pap. Av. Depto. Zool. III(7): 111-126.
MORRETES, F.L. 1949. Ensaio de Catálago dos Moluscos do Bra-

100
sil. Arq. Mus. Par. VII: 5-216.
NADITKO, N.V. 1973. Development of Eurytrema pancreaticum (Jan-
son, 1889) (Trematoda: Dicrocoelidae) in the Primorsk terri-
tory. Parazitologiya. 7: 408-417.
NEVEU-LEMAIRE, M. 1936. Traité d'Helminthologie Médicale et
Veterinaire. 1514 pp., 787 figs. Vigot-Frères Ed. Paris.
OLIVEIRA, M.P. & CASTRO, G.A. 1979. Adenda ao Ensaio de Ca-
tálogo de Moluscos do Brasil e de Frederico Lange de Morre-
tes. Bol. Inst. Cienc. Biol. e Geoc. 26:8 pp.
OLIVEIRA, M.P.; REZENDE, G.J.R. & CASTRO, G.A. 1981. Catálogo
dos moluscos da Universidade Federal de Juiz de Fora. 520
pp. Universidade Federal de Juiz de Fora Ed.
OLSEN, O.W. 1974. Parasitologia Animal II. Platelmintos, Acan-
tocefalos y Nematelmintos. 719 pp. Aedos Ed. Barcelona.
ORBIGNY, A. D. 1835/1846. Yoyage dans l'Amerique Meridiona-
le. Paris. Mollusques 5(3): 185-376 (1837): 489-758 (1846).
PFEIFFER, L. 1848. Monographia Heliceorum Viventium. II: 1-
594.
PFEIFFER, L. 1859. Monographia Heliceorum Viventium. IV: 1-
920.
PENNER, L.R. 1939. Tamerlania melospizae n. sp. (Trematoda, Eucoti-
lidae) with notes on the genus. J. parazitol. XXV(5): 421-424.

101.
PILSBRY, H.A. 1933. Zoolofical results of the Matto Grosso ex-
pedition to Brazil in 1931. II - Mollusca. Proc. Acad. Nat.
Sci. Philad. LXXXV: 67-76.
PINTO, C. 1938. Zooparasitos de interesse médico e veteriná-
rio. 376 p. Pimenta de Melo Ed.
PITONI, V.L.L.; VEITENHEIMER, I.L. & MANSUR, M.C.D. 1976. Mo-
luscos do Rio Grande do Sul: coleta, preparação e conser-
vação, lheringia Div. 5: 25-68, 61 figs.
REEVE, L.A. 1849. Conchologia Iconica. Illustrations of the
shells of molluscous animals. Vol. V.
REEVE, L.A. 1854. Conchologia Iconica. Vol. VII, 1495 spp,
pls. I-CCX.
ROSE, J.H. 1957. Observations on the larval stages of Muelle-
rius capillaris within intermediate hosts Agriolimax agres-
tis e Agriolimax reticulatus. J. Helmint. XXXI (1-2): 1-16.
ROSEN, L.; ASH, L.A. & WALLACE, G.D. 1970. Life history of
the canine lungworm Angyostrongylus vasorum (Baillet). Am
J. Vet. Res. 31(1): 131-143.
SANTOS, V. 1934. Monostomose renal das aves domésticas. Rev.
Dep. Nac. Prod. An. I(1): 203-211, 4 figs.
SCHELL, S.C. 1970. The Trematodes. W. M. C. Brown Company
Publishers I -VII + 1 - 3-5 pp. 685 figs.

102.
SCOTT, M.I.H. 1945. Faunula malacologica de Tilcara. Rev. Mus.
de la Plata (n.s.) Sec. Zool. IV: 195-211, 6 figs., 1 lam.
SEIXAS, M.M.P. 1977. Duas novas espécies de Limacidae (Gas-
tropoda, Pulmonata), novas para a fauna portuguesa. Bol.
Soc. Port. Cienc. Nat. 17: 57-61, 2 figs., 1 mapa.
SOULSBY, E.J.L. 1965. Text-book of veterinary clinical Para-
sitology I Helminths I - XXXI + l-ll20 pp., 214 figs.
SOULSBY, E.J.L. 1968. Helminths, Arthropods, and Protozoa
of domesticated animals (Monning) 824 pp., 288 figs., XXXVIII
pls. The Williams and Wilkins Co. Baltimore.
STEPHENSON, J.W. & KNUTSON, L.V. 1966. A resumé of recent
studies of invertebrates associated with slugs. J. Econ.
Entomol. 59(2): 356-360.
TEIXEIRA DE FREITAS, J.F.; KOHN, A. & IBANEZ, N. 1967. Sobre
as espécies de Brachylaems (Brachylaemus) Dujardin, 1843 (Tre-
matoda). Mem. Inst. Oswaldo Cruz. 65(1): 41-61, 16 figs.
THIELE, J. 1931. Handbuch der Systematischen Weichtierkunde
1: VI + 1-778, 783 figs. Jena.
THOMPSON, F.G. 1957. A collection of land and fresh-water
molluscs from Tabasco, México. Nautilus. 70(3): 97-102.
TRAVASSOS, L. 1927. Nematódeos novos. Bol. Biol. 6: 52-61, 6
figs.

103.
TRAVASSOS, L., 1928. Fauna helmintológica de Mato Grosso. Mem.
Inst. Oswaldo Cruz. XXI (2): 309-372, pl. 42-54.
TRAVASSOS, L.; FREITAS, J.F.T. & KOHN, A. 1969. Trematódeos do
Brasil. Mem. Inst. Oswaldo Cruz. 67 (único): 1-886, figs. 1-
577.
TRYON, G.W. & PILSBRY. 1894. Manual of Conchology. IX: XLVIII
+ 1 - 366, 71 pls.
TRYON, G.W. & PILSBRY, H.A. 1906. Manual of Conchology. XVIII:
I -VII + 1 - 357, 51 pls.
VENMANS, L.A.W.C. & FRÖMMING, E. 1957. Notes on the anatomy and
biology of Subulina kassaiana Rochebrune & Germain. Basteria.
21(1-2): 14-28.
VICENTE, F.S. 1955. Brachylaemus en infestacion experimental
y natural. Rev. Iber. Parasit. 15: 301-320.
YMAGUTI, S. 1958. Systema Helminthum, I. The Digenetic trema-
todes of vertebrates. Part I: XI+979 pp. Part II: 980-1232,
VAN VOLKEMBERG, H.L. 1939. An annotated check list of the pa-
rasites of animals in Puerto Rico. Puerto Rico Exper. Sta.
Circ. 22: 12.
VAN VOLKEMBERG, H.L. 1938. Check list of Parasites found among
principal domestic animals in Puerto Rico. Proc. Helminthol.
Soc. Washington. 5: 7-8.

104.
1445-1575, 106 pls., 1302 figs. Interscience Publishers, Inc.
Ed. New York.
ZILCH, A. 1960 in WENZ, W. Handbuch der Paläozoologie, Gastro-
poda, Euthyneura, 6 (2) (1959-1960): XII+1 - 834,2515 figs.
Berlin.