incidência, caracterização e estrutura genética de
TRANSCRIPT

UNIVERSIDADE FEDERAL RURAL DE PERNAMBUCO
PRÓ-REITORIA DE PESQUISA E PÓS-GRADUAÇÃO
PROGRAMA DE PÓS-GRADUAÇÃO EM FITOPATOLOGIA
Tese de Doutorado
Incidência, caracterização e estrutura genética de badnavírus nas culturas do
abacaxizeiro e da cana-de-açúcar no Nordeste do Brasil
Edlene Maria da Silva Moraes Santos
Recife – PE
2014

EDLENE MARIA DA SILVA MORAES SANTOS
INCIDÊNCIA, CARACTERIZAÇÃO E ESTRUTURA GENÉTICA DE BADNAVÍRUS NAS CULTURAS DO ABACAXIZEIRO E DA CANA-DE-
AÇÚCAR NO NORDESTE DO BRASIL
Tese apresentada ao Programa de Pós-Graduação em
Fitopatologia da Universidade Federal Rural de
Pernambuco, como parte dos requisitos para obtenção do
título de Doutor em Fitopatologia.
COMITÊ DE ORIENTAÇÃO:
Orientador: Prof. Dr. Gaus Silvestre Andrade Lima
Co-Orientador(a): Drª. Sarah Jacqueline Cavalcanti da Silva
RECIFE-PE
FEVEREIRO – 2014

Ficha catalográfica
S237i Santos, Edlene Maria da Silva Moraes Incidência, caracterização e estrutura genética de badnavírus nas culturas do abacaxizeiro e da cana-de-açúcar no Nordeste do Brasil / Edlene Maria da Silva Moraes Santos. – Recife, 2014. 78 f. :il. Orientador: Gaus Silvestre de Andrade Lima. Tese (Doutorado em Fitopatologia) – Universidade Federal Rural de Pernambuco. Departamento de Agronomia, Recife, 2014. Referências.
1. Caulimoviridae 2. Ananas comosus 3. Saccharum spp 4. Análise molecular 5. Filogenia 6. Estrutura populacional I. Lima, Gaus Silvestre de Andrade, orientador II. Título CDD 632


Aos meus pais, Geová Ferreira de Moraes e
Maria Edileuza da Silva Moraes,
Ao meu esposo, Geraldo Filho,
E a toda minha família,
Por toda força, apoio e momentos de paz e descontração.
DEDICO

AGRADECIMENTOS
A Deus, pela dádiva da vida e por sua paz que excede todo entendimento.
À Universidade Federal Rural de Pernambuco (UFRPE), por me possibilitar a
realização deste curso de doutorado.
À Universidade Federal de Alagoas (UFAL) por me proporcionar a realização dos
trabalhos de tese.
À Fundação de Amparo à Pesquisa do Estado de Alagoas (FAPEAL) pelo
financiamento da bolsa de doutorado.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) pela
concessão das bolsas de Produtividade em Pesquisa do Prof. Gaus S.A. Lima.
Ao Prof. Dr. Gaus Silvestre de Andrade Lima, pela orientação na condução deste
trabalho, pelo exemplo de profissionalismo e ética, e por todo conhecimento transmitido
durante minha formação acadêmica / profissional.
À Prof. Drª Iraildes Assunção pela amizade, colaboração e apoio.
À Drª Sarah Cavalcanti pela co-orientação, amizade e auxílio na elaboração desta
tese. Obrigada!
A todos os Professores que compõem o Programa de Pós – Graduação em
Fitopatologia da Universidade Federal Rural de Pernambuco.
Aos membros da banca examinadora, que se dispuseram a participar da avaliação
desta tese, contribuindo com sugestões para o aprimoramento do trabalho.
A todos os amigos do Laboratório de Fitovirologia Molecular do CECA/UFAL, Joyce,
Márcia, Lucas Fonseca, Renato, Jean Phellipe, Mayra, Janaíne, Aline, Lucas Jordão, Helloá,
Jackeline Laurentino, Jussara, Luis, Dayane e também aos novos membros, pelo
companheirismo e auxílio.
À Drª Liliane Dias e à futura Drª Jaqueline Figueredo, pela amizade e convivência ao
longo de nossa formação acadêmica. Sucesso!
Aos amigos da UFRPE Kátia, Bárbara, Mariote e Willie pela amizade e pelas
informações do PPGF durante minha ausência na instituição.
E a todos que contribuíram de alguma maneira para a realização deste trabalho.
Sinceros votos de gratidão a todos: sem vocês esse trabalho não seria possível. Amo
todos vocês!

vii
SUMÁRIO
SUMÁRIO.................................................................................................................. vii
RESUMO GERAL...................................................................................................... viii
GENERAL ABSTRACT............................................................................................ ix
CAPÍTULO I - INTRODUÇÃO GERAL............................................................... 01
1. Cultura do abacaxi......................................................................................... 02
2. Cultura da cana-de-açúcar............................................................................. 09
3. Família Caulimoviridae................................................................................. 15
3.1. Gênero Badnavirus........................................................................................ 17
4. Variabilidade e estrutura genética de populações de fitopatógenos.............. 20
4.1. Mutação......................................................................................................... 21
4.2. Recombinação................................................................................................ 22
REFERÊNCIAS BIBLIOGRÁFICAS....................................................................... 24
CAPÍTULO II - Incidência e caracterização molecular de badnavírus na
cultura do abacaxizeiro no Nordeste do Brasil.......................................................
35
Resumo........................................................................................................................ 36
Abstract....................................................................................................................... 37
1. Introdução...................................................................................................... 38
2. Material e Métodos........................................................................................ 39
3. Resultados e Discussão.................................................................................. 41
4. Conclusões..................................................................................................... 43
5. Referências Bibliográficas............................................................................. 43
CAPÍTULO III - Estrutura genética de populações de badnavírus que
infectam as culturas do abacaxizeiro e da cana-de-açúcar no Nordeste do
Brasil...........................................................................................................................
52
Resumo........................................................................................................................ 53
Abstract....................................................................................................................... 54
1. Introdução...................................................................................................... 55
2. Material e Métodos........................................................................................ 57
3. Resultados e Discussão.................................................................................. 58
4. Conclusões..................................................................................................... 61
5. Referências Bibliográficas............................................................................. 62
CONCLUSÕES GERAIS......................................................................................... 70

viii
RESUMO GERAL
O Brasil é o principal produtor mundial de abacaxi (Ananas comosus L. Merril) e de cana-de-
açúcar (Saccharum spp.), mas apesar da alta produção dessas culturas, ambas estão expostas a
diversos problemas fitossanitários, inclusive as viroses. Badnavírus causam doenças em
culturas economicamente importantes na maioria das regiões tropicais e subtropicais,
incluindo o Brasil. As badnaviroses do abacaxizeiro são causadas por duas espécies distintas:
Pineapple bacilliform CO virus (PBCoV) e Pineapple bacilliform ER virus (PBErV). Em
cana-de-açúcar também são descritas duas espécies: Sugarcane bacilliform IM virus
(SCBIMV) e Sugarcane bacilliform MO virus (SCBMOV). O gênero Badnavirus (família
Caulimoviridae) engloba vírus de plantas com genoma de DNA circular de fita dupla
(dsDNA) e que replicam por meio de um intermediário de RNA. Informações sobre a
variabilidade e estrutura genética de vírus de plantas com genoma de dsDNA são escassas.
Dessa forma, o objetivo do presente trabalho foi determinar a incidência de badnavírus na
cultura do abacaxi e a estrutura genética de populações de uma espécie de badnavírus que
infecta o abacaxi, PBCoV e de duas espécies que infectam a cana-de-açúcar, SCBIM e
Banana streak OL virus (BSOLV), todas provenientes da região Nordeste do Brasil. A
incidência de badnavírus foi constatada em todas as áreas de abacaxizeiros amostradas e
variou de 80 a 96,4%. Análises de comparações de sequências pareadas e filogenéticas
revelaram que todos os 83 isolados provenientes de abacaxizeiro pertencem à espécie PBCoV.
A estrutura genética das populações do PBCoV, do SCBIMV e do BSOLV, relatado
recentemente em cana-de-açúcar, foram determinadas com base no domínio que engloba a
transcriptase reversa e ribonuclease H (RT/RNaseH). Os resultados obtidos sugerem a ampla
disseminação do PBCoV no Nordeste brasileiro e registra o primeiro relato de badnavírus na
cultura do abacaxizeiro no Brasil. As análises de estrutura genética revelaram que a população
de PBCoV está estruturada com base na região geográfica. A variabilidade genética nas
populações estudadas pode ser considerada alta, com frequências de mutação na ordem de 10-
4 subs/sítio/ano para as populações de PBCoV e de 10
-3 para BSOLV e SCBIMV. Quanto
aos mecanismos evolutivos, seleção purificadora ou expansão recente da população são as
duas principais forças atuando nessas populações.
Palavras-chaves: Caulimoviridae, Ananas comosus, Saccharum spp., análise molecular,
filogenia, estrutura populacional.

ix
GENERAL ABSTRACT
Brazil is the world's leading producer of pineapple (Ananas comosus L. Merril) and sugarcane
(Saccharum spp.), but despite the high production of these crops, both are exposed to many
diseases, including viruses. Badnaviruses cause diseases in economically important crops in
most tropical and subtropical regions, including Brazil. Badnaviroses of the pineapple are
caused by two distinct species: Pineapple bacilliform CO virus (PBCoV) and Pineapple
bacilliform ER virus (PBErV). In sugarcane two species are also described: Sugarcane
bacilliform IM virus (SCBIMV) and Sugarcane bacilliform MO virus (SCBMOV). The
Badnavirus genus (family Caulimoviridae) comprises plant viruses with circular DNA
genome of double-stranded (dsDNA), which replicate by means of an RNA intermediate
virus. Information about the variability and genetic structure of plant viruses with dsDNA
genome are scarce. Thus this study aimed was determine the incidence of badnaviruses in the
pineapple crop and the genetic structure of populations of a specie of badnavirus that infects
pineapple, PBCoV and of two specie that infect sugarcane SCBIM and Banana streak OL
virus (BSOLV) all from the Northeast region of Brazil. The incidence of badnavirus was
detected in all the sampled areas and varied from 80 to 96.4%. Pairwise comparisons analysis
and phylogenetic sequences revealed that all 83 isolates from pineapple belong to the species
PBCoV. The genetic structure of populations of PBCoV, SCBIMV and of BSOLV, recently
reported in sugarcane, were determined from the domain comprising the reverse transcriptase
and ribonuclease H (RT / RNase H). The results suggest the wide spread of PBCoV in
Northeast Brazilian and records the first report of Badnavirus the pineapple crop in Brazil.
Analyses of genetic structure revealed that the population of PBCoV is structured based on
geographic region. The genetic variability at the studied populations can be considered high,
with mutation frequencies on the order of 10-4
subs/site/year for populations of PBCoV and
10-3
for BSOLV and SCBIMV. Regarding the evolutionary mechanisms purifying selection or
recent population expansion are the two main forces acting in these populations.
Keywords: Caulimoviridae, Ananas comosus, Saccharum spp., molecular analysis,
phylogeny, population structure.
CAPÍTULO I
Introdução Geral

2
INCIDÊNCIA, CARACTERIZAÇÃO E ESTRUTURA GENÉTICA DE
BADNAVÍRUS NAS CULTURAS DO ABACAXIZEIRO E DA CANA-DE-AÇÚCAR
NO NORDESTE DO BRASIL
INTRODUÇÃO GERAL
1. Cultura do abacaxi
1.1. Aspectos botânicos
O abacaxizeiro (Ananas comosus L. Merril) é uma planta originária do Brasil, de
clima tropical, herbácea e perene, da família Bromeliaceae, com caule curto e grosso, ao redor
do qual crescem folhas estreitas e resistentes, quase sempre margeadas por espinhos (SÁ,
1994). São conhecidos, aproximadamente, 50 gêneros e 2000 espécies de Bromeliaceae
(CUNHA; CABRAL, 1999). Os portugueses foram provavelmente os responsáveis pela sua
difusão nos países da África e Ásia (MARANCA, 1977).
1.2. Aspectos econômicos e produção
O abacaxi é consumido na maioria dos países e produzido fundamentalmente nos
países de clima tropical e sub-tropical (TAMAKI; CARDOSO, 1982). Segundo dados da
FAO (2011) a produção mundial foi acima de 21,8 milhões de toneladas de abacaxi, sendo o
Brasil o maior produtor, com 2,3 milhões de toneladas colhidas, o que corresponde a 10,5%
da produção no mundo, seguido pela Costa Rica que produziu 2,2 milhões de toneladas no
mesmo ano.
O IBGE (Instituto Brasileiro de Geografia e Estatística), que contabiliza a produção
em número de frutos e não em toneladas, registrou em 2012, uma produção brasileira de
abacaxi de 1.697.734 frutos. O rendimento médio da cultura alcançou a marca de 27.239
frutos por hectare e a área colhida, 60.653 ha, embora a área plantada tenha sido de 90.971 ha.
(IBGE, 2012).
O abacaxizeiro é a quinta fruteira mais cultivada no país, tendo importante papel
social, por gerar emprego e renda no meio rural (MAPA, 2010). Em geral, pequenos
agricultores cultivam de um a dez hectares em áreas arrendadas. No entanto, a produção é
prejudicada por problemas fitossanitários e deficiência nos tratos culturais, reduzindo sua
competitividade, principalmente para exportação (CUNHA, 2004).
A produção de abacaxi no país está bem distribuída em várias regiões, mas é o
Nordeste que concentra a maior produção, por possuir solos com textura média (BARREIRO

3
NETO; SANTOS, 1999), uma vez que de acordo com Gomes (2007) o abacaxizeiro não se
adapta bem a solos compactos ou impermeáveis, bem como a solos calcários.
A produção de abacaxi na região Nordeste foi de 39,9% do total nacional, seguida da
região Sudeste (34,26%). O Estado da Paraíba apresenta a maior produção desta cultura
(283,722 mil frutos), seguido de Minas Gerais, Rio de Janeiro e Rio Grande do Norte ocupam
as posições (IBGE, 2012).
Existe abacaxi plantado em todas as regiões do país. Observa-se uma tendência de
organização da produção em pólos ou núcleos especializados. Cada núcleo especializando-se
no plantio em determinada época, levando em consideração o mercado, a oferta e
principalmente o fotoperíodo. Assim, obtém-se uma oferta de abacaxi o ano todo,
provenientes de diferentes regiões tendendo a oscilações de preço cada vez menores
(CARVALHO et al., 2009).
1.3. Utilização do abacaxi
Cada planta de abacaxi produz um único fruto saboroso e de aroma intenso, que é
utilizado tanto para o consumo in natura quanto na industrialização, em diferentes formas:
pedaços em calda, suco, geleia, licor, vinho, vinagre. Como subproduto desse processo
industrial pode-se obter ainda: álcool, ácidos cítrico, málico e ascórbico; rações para animais e
a bromelina que é uma substância de alto valor medicinal, pois trata-se de uma enzima muito
utilizada como digestivo e anti-inflamatório (CARVALHO; CUNHA, 1999). Na culinária, o
suco de abacaxi é utilizado para o amaciamento de carnes. Além disso, os frutos são ótimas
fontes de vitaminas A, B e C, rico em ferro e em cálcio. Combate a hiperacidez do estômago e
quando ainda verde é diurético e vermífugo, apesar de ser também abortivo (GOMES, 2007).
1.4. Aspectos culturais
A densidade de plantio por unidade de área é um dos fatores de produção mais
importantes da cultura do abacaxi, estando diretamente relacionada ao rendimento e custo de
produção da cultura. No entanto, ainda é grande a variação das densidades de plantio nas
diversas regiões produtoras dessa fruta no mundo, sendo influenciada por fatores como
variedade usada, tipo de solo, práticas culturais e destino da produção (CUNHA; CABRAL,
1999; CUNHA et al., 1994). O cultivo seguido do abacaxizeiro num mesmo local, sem
rotação de culturas, decresce em rendimento, que pode estar ligado a problemas fitossanitários
e/ou nutricionais (SIMÃO, 1998). Segundo este mesmo autor, a temperatura favorável para o
desenvolvimento da planta situa-se entre 21ºC e 27ºC. Quando a temperatura fica abaixo de

4
20ºC, a planta entra em estado de dormência e quando se mantém acima de 32ºC, verificam-
se danos na planta, devido à transpiração e respiração.
A cultura do abacaxi é propagada vegetativamente por meio de mudas, formadas a
partir de diferentes partes vegetativas da planta, cuja qualidade depende das condições
ambientais onde são produzidas e do manejo dado à cultura. Os principais tipos de muda
convencionais são: coroa (brotação do ápice do fruto), filhote (brotação do pedúnculo, que é a
haste que sustenta o fruto), filhote-rebentão (brotação da região de inserção do pedúnculo no
caule ou talo) e rebentão (brotação do caule) apresentados na Figura 1 (REINHARDT;
CUNHA, 1999).
Figura 1. Esquema de abacaxizeiro mostrando diversos tipos de mudas convencionais.
Desenho: Almira Souza Andrade
A coroa é o tipo de muda menos usado para plantio, pois permanece nos frutos
vendidos nos mercados de frutas frescas, principalmente no Brasil, onde tal tipo de consumo
predomina. Os filhotes apresentam desenvolvimento relativamente uniforme, facilitando o
controle do florescimento e são o tipo de muda mais disponível e utilizado no Brasil,
principalmente em plantios da cultivar „Pérola‟ (MATOS et al., 2009).
Existem também as mudas não convencionais, que são aquelas produzidas a partir de
plantas que já produziram frutos, a partir de gemas axilares na base de cada folha. Mudas de
seccionamento do caule, também denominada plântula, é uma muda produzida em viveiro ou
em casa de vegetação, a partir de secções de talo ou caule de plantas cujos frutos já foram
colhidos. A principal vantagem deste tipo de muda é que as mesmas são livres de algumas
pragas e doenças, especialmente a fusariose (Fusarium subglutinans (Wollew & Reinking)

5
Nelson, Tousson & Marasas, f. sp. ananas, Ventura, Zambolim & Gilbertson). Outro tipo de
muda não convencional é a muda micropropagada. Esta muda é obtida em laboratório
mediante técnicas de cultura de tecidos vegetais. Por serem obtidas em ambiente asséptico,
apresentam excelente sanidade, entretanto seu custo ainda é muito elevado para o pequeno
produtor, o que confere à técnica um uso limitado. Por outro lado, seu elevado potencial de
multiplicação faz da micropropagação a principal tecnologia de produção de material
propagativo das novas variedades de abacaxi geradas em programas de melhoramento
genético desta cultura (MATOS et al., 2009).
1.4.1. Principais cultivares de abacaxi
Todas as cultivares de abacaxi de interesse da fruticultura pertencem à espécie Ananas
comosus (Figura 2) (PEREIRA; MELO, 2003).
A cultivar „Smooth Cayenne‟ é a mais plantada no mundo e possui muitas
características favoráveis. Estima-se que 70% da produção mundial corresponde a esta
cultivar (CUNHA; CABRAL, 1999). É uma planta robusta, de porte semi-ereto, cujas folhas
não apresentam espinhos, a não ser alguns encontrados na extremidade apical da borda da
folha. O fruto é ligeiramente cilíndrico, polpa amarela, rico em açúcar e de acidez maior do
que as outras cultivares. Essas características a tornam adequada para a industrialização e a
exportação como fruta fresca. A coroa é relativamente pequena e a planta produz poucas
mudas do tipo filhote. No entanto, é bastante suscetível ao complexo viral associado à
murcha, Pineapple mealybug wilt associated virus (PMWaV) e à fusariose (PEREIRA;
MELO, 2003). A segunda cultivar em importância para a industrialização é a „Singapore
Spanish‟, sendo amplamente cultivada na Malásia e em outros países do sul da Ásia.
(CUNHA; CABRAL, 1999).
A cultivar „Pérola‟ é de origem brasileira e é a mais plantada no país, representando
mais de 80% da área plantada (CUNHA, 2004). Seus frutos apresentam forma cônica, casca
pouco colorida, haste frutífera e folhas longas com finos espinhos. A polpa é rica em suco,
saborosa, pouco ácida e de coloração branco ou amarelo pálido (CARVALHO et al., 2009).
Apresenta tolerância ao PMWaV e é suscetível à fusariose (PEREIRA; MELO, 2003). Em
plantios comerciais do Nordeste é comum aparecer a cultivar „Jupi‟ em mistura, em lavouras
de „Pérola‟, da qual difere apenas pelo formato cilíndrico do fruto (CUNHA; CABRAL,
1999).
Apesar da ampla predominância da cultivar „Pérola‟ sobre as demais na abacaxicultura
nacional, existem outras cultivares importantes, dentre as quais a MD-2 („Gold‟), um híbrido

6
obtido da variedade „Smooth cayenne‟, que possui grande aceitação no mercado externo
(BARREIRO NETO et al., 2007). As folhas desta cultivar possuem poucos espinhos, o que
facilita o manejo da cultura. Tem ainda como vantagem uma maior durabilidade pós-colheita,
sendo adequada para exportação como fruta fresca (REINHARDT; CUNHA; MENEGUCCI,
2001).
Figura 2. Cultivares de abacaxi: a) „Smooth Cayenne‟, b)‟ Pérola‟, c) „Jup‟i, d) MD-2
(„Gold‟).
Fotos: (A, C) Domingo Haroldo Reinhardt; (B, D) Davi Theodoro Junghans
1.5. Doenças da cultura do abacaxi
O abacaxizeiro está exposto a diversos problemas que causam prejuízos variáveis a
depender da parte da planta afetada, da região produtora e da época de produção. Alguns
patógenos infectam as raízes, outros o talo e a base das folhas e outros o fruto, causando,
geralmente, danos econômicos significativos (MATOS et al., 2009).
Entre esses patógenos, aqueles que infectam as mudas revestem-se de especial
importância, haja vista que, por ser o abacaxizeiro uma cultura de propagação vegetativa, é
bastante comum a prática da compra e venda de mudas de abacaxi entre os produtores, muitas
vezes sem a fiscalização das entidades credenciadas para esta função. A movimentação de
mudas de abacaxi tem sido responsável pela dispersão de pragas e doenças, seja dentro de
uma mesma região produtora, seja de uma região para outra (MATOS et al., 2009).
A fusariose, doença de etiologia fúngica, causada por F. subglutinans f. sp. ananas, é a
mais séria ameaça à cultura do abacaxi no Brasil, onde foi encontrada pela primeira vez no
estado de São Paulo, em frutos da variedade „Smooth Cayenne‟. A fusariose pode ocasionar
perdas de até 80% na produção de frutos, além de poder infectar cerca de 40% das mudas, das
quais 20% morrem antes da produção e colheita. A maioria das variedades comerciais
cultivadas no mundo é suscetível à fusariose (SANTOS, 2010). A doença foi acidentalmente

7
introduzida em diversas regiões produtoras de abacaxi do Brasil na década de setenta através
de mudas infectadas (MATOS et al., 2009). A disseminação do patógeno pode ocorrer através
do vento ou com o auxílio de insetos, entretanto, a forma mais eficiente é por meio de mudas
infectadas (GOES, 2005).
A podridão-negra-do-fruto ou podridão-mole é a mais importante doença de pós-
colheita do fruto do abacaxizeiro, sendo causada pelo fungo Chalara paradoxa (De Seyn.)
Sacc., cujos danos podem se apresentar na haste, folhas, frutos e rebentos Presente em todas
as regiões produtoras de abacaxi do mundo, essa doença caracteriza-se pelo desenvolvimento
de uma podridão mole, aquosa, especialmente em frutos destinados ao mercado in natura.
Perdas também podem ocorrer em frutos destinados à indústria, porém, nesse caso, sua
intensidade depende de vários fatores, entre os quais o período de tempo decorrido entre a
colheita e o processamento (SANTOS, 2010).
Uma importante doença nas principais regiões produtoras de abacaxi do mundo,
principalmente em solos argilosos, úmidos e com valores de pH elevados, é causada por
Phytophthora nicotianae van Breda de Haan var. parasitica (Dastur) Waterhouse, responsável
pela podridão-do-olho. As perdas são variáveis de uma região para outra, sendo o maior
impacto econômico a morte das plantas (VENTURA; ZAMBOLIM, 2002). O patógeno está
presente em quase todas as regiões produtoras de abacaxi do Brasil, possuindo ampla gama de
hospedeiras, tais como citros, eucalipto, pera, maracujá e outras (VITALINO, 2006). Outra
espécie do gênero, P. cinnamomi Ranks é o principal agente causal da prodridão-das-raízes do
abacaxizeiro, embora, eventualmente, P. nicotianae var. parasitica e várias espécies de
Pythium também podem infectar e destruir as raízes da planta (MATOS, 1999).
A mancha-negra-do-fruto do abacaxizeiro é causada por Penicillium funiculosum
Thom. e Fusarium moniliforme Sheldon (VERZIGNASSI et al., 2009). A doença é favorecida
pelos ferimentos provocados pelo ácaro do abacaxi (Steneotarsonemus ananas Tryon) e o
ácaro alaranjado (Dolychotetranychus floridanus Banks), embora ambos não atuem como
vetores (GOES, 2005; VERZIGNASSI et al., 2009; VITALINO, 2006) e ocorre nas principais
regiões produtoras do mundo. As lesões suberificadas dos frutilhos alteram a qualidade do
produto e reduzem o aproveitamento das fatias dos frutos (VENTURA; ZAMBOLIM, 2002).
Frutos das cultivares „Smooth Cayenne‟ e „Pérola‟ não expressam sintomas externos
(MATOS; FERREIRA; CORDEIRO, 2005).
Além dessas doenças, aquelas causadas por nematóides podem provocar danos severos
na cultura do abacaxizeiro, reduzindo a eficiência na absorção dos nutrientes e,
consequentemente, provocando, além da redução do sistema radicular, clorose nas folhas, as

8
quais ficam menores e mais estreitas, bem como produção retardada e com frutos menores
(PY; LACOEUILHE; TEISSON, 1984). O abacaxizeiro é infestado por três grandes grupos
de nematóides que são agrupados em causadores de lesões radiculares (Pratylenchus
brachyurus (Godfrey) Filipjev & S. Stekhoven), o reniforme (Rotylenchus reniformis Linford
& Oliveira) e os formadores de galhas (Meloidogyne spp.) (VENTURA; ZAMBOLIM, 2002).
1.5.1. Doenças causadas por vírus
As doenças ocasionadas por vírus merecem atenção especial. Os vírus podem ser
transmitidos por insetos vetores e por meio de propagação vegetativa, o que pode ocasionar
uma ameaça à produção, tanto nas áreas onde os vírus são endêmicos, quanto naquelas livres
de vírus, mas que recebem novas mudas (FIGUEIREDO; BRIOSO, 2007).
Um fator que contribui para um baixo rendimento na produção do abacaxi é a infecção
pelo complexo viral associado à murcha, Pineapple mealybug wilt associated virus (PMWaV-
1, PMWaV-2 e PMWaV-3) pertencentes ao gênero Ampelovirus, família Closteroviridae
(GAMBLEY et al., 2008a; SETHER; HU, 2002; SETHER; MELZER; BUSTO, 2005). No
campo, estes vírus são transmitidos e disseminados pelas cochonilhas Dysmicoccus brevipes
Cockerell e D. neobrevipes Beardsley, que ocorrem associadas a formigas. Estas
desempenham um importante papel na disseminação de cochonilhas dentro e entre plantios,
pois as protegem e carregam suas ninfas de uma planta à outra (SANCHES; MATOS;
MEISSNER FILHO, 2000; SETHER; ULLMAN; HU, 1998). A murcha do abacaxizeiro foi
primeiramente relatada no Havaí em 1910 e desde então vem causando elevados prejuízos em
todo o mundo, tendo sido relatada inclusive no Brasil (GONÇALVES-GERVÁSIO; SANTA-
CECÍLIA, 2001; OLIVEIRA et al., 2013; SANCHES; MATOS; MEISSNER FILHO, 2000).
As plantas infectadas por esses vírus manifestam os sintomas de avermelhamento
foliar, bordas das folhas voltadas para baixo, apresentando seca em suas pontas. (SANCHES;
MATOS; MEISSNER FILHO, 2000). Os sintomas da murcha só surgem quando a planta
infectada pelo vírus também está sendo colonizada pelo inseto vetor (SETHER; HU, 2002).
Além dos danos diretos no vigor e produção da planta, os danos indiretos são preocupantes,
pois em alguns casos as plantas contaminadas não apresentam sintomas, dificultando a
seleção de mudas para o plantio (SANTOS; ANDRADE, 2009).
Duas espécies do gênero Badnavirus (família Caulimoviridae) também podem infectar
o abacaxizeiro: Pineapple bacilliform CO virus (PBCoV) e Pineapple bacilliform ER virus
(PBErV). Ambos são transmitidos pela cochonilha D. brevipes e, no caso do PBCoV, também
pelas cochonilhas Planococcus citri Risso (GAMBLEY et al., 2008b) e D. neobrevipes

9
Beardsley (SETHER et al., 2012). Uma associação clara entre os sintomas e a infecção por
badnavírus em abacaxi ainda não foi mostrada (GAMBLEY et al., 2008b). Os danos
econômicos ocasionados pelas badnaviroses ainda não foram estimados, mas devido a
predominância da cultura ser vegetativa e dependendo de sua incidência na lavoura, poderão
se tornar um fator limitante.
2. Cultura da cana-de-açúcar
2.1. Aspectos botânicos
A cana-de-açúcar (Saccharum spp.), é originária do sudeste Asiático com centro de
dispersão localizado entre a Nova Guiné e a Indonésia. No Brasil, foi introduzida pelos
colonizadores portugueses. Pertence à família Gramineae (Poaceae) onde as cultivares são
híbridos derivados principalmente do cruzamento entre a espécie domesticada rica em
sacarose S. officinarum e a espécie selvagem S. spontaneum. Dentro do gênero Saccharum,
ocorrem seis espécies: S. officinarum L., S. robustum Brandes e Jeswiet ex Grassl, S. barberi
Jeswiet, S. sinense Roxb., S. spontaneum L., e S. edule Hassk. (DANIELS; ROACH, 1987). A
cultura é a maior produtora de alimento energético por área (AGROLINK, 2014).
2.2. Aspectos econômicos e produção
O Brasil é o primeiro produtor desta cultura em nível mundial. A produção nacional de
cana-de-açúcar em 2013 apresentou um crescimento de 10,0% em relação a 2012, alcançando
737,9 milhões de toneladas. A área colhida apresentou um acréscimo de 4,4% em relação ao
ano anterior. Dentre os estados da federação, São Paulo é o maior produtor, responsável por
54,8% de toda produção brasileira (404,7 milhões de t), seguido por Goiás e Minas Gerais.
No Nordeste, o Estado de Alagoas ocupa a primeira posição, seguido de Pernambuco, Bahia e
Paraíba (IBGE, 2013).
As áreas em produção continuam com progressivo aumento, sendo que esse
crescimento representa a consolidação das novas variedades disponíveis no mercado, aliado
ao crescimento das áreas plantadas com variedades mais antigas (CONAB, 2012).
A cultura da cana-de-açúcar representa uma das mais importantes no cenário
socioeconômico brasileiro, por ser a principal fonte de matéria-prima utilizada pela indústria
sucroalcooleira para a produção de açúcar e álcool. O Brasil não é apenas o maior produtor de
cana, é também o primeiro do mundo na produção de açúcar e etanol, conquistando cada vez
mais o mercado externo com o uso do biocombustível como alternativa energética
(VASCONCELOS, 2002).

10
A cultura canavieira desempenha papel de destaque no agronegócio brasileiro,
representando na indústria sucroalcooleira cerca de 2% das exportações nacionais, além de
gerar quantidade significativa de empregos e contribuir de maneira efetiva para o crescimento
de mercado interno de bens de consumo. Perspectivas para o setor sucroalcooleiro são
promissoras em médio prazo, principalmente devido a quebra de subsídios do açúcar europeu
e a conquista de novos mercados consumidores (BOLOGNA-CAMPBELL, 2007).
A cadeia produtiva da cana-de-açúcar, assim como os seus produtos e subprodutos,
principalmente o açúcar e o álcool, contribuem para a distribuição de riqueza, além de ser
fonte de energia líquida e renovável propiciando a redução da poluição ambiental
(MATSUOKA; GARCIA; ARIZONO, 2005).
No sistema produtivo canavieiro brasileiro, o cultivo de variedades com boas
características agroindustriais é a melhor forma de se obter ganhos da produtividade com
baixo custo. A atuação dos programas de melhoramento genético desta cultura contribuiu
expressivamente para o desenvolvimento do setor sucroalcooleiro nacional, com a liberação
de variedades mais produtivas e mais resistentes ao ataque de pragas e doenças (BARBOSA
et al., 2003).
2.3. Utilização da cana-de-açúcar
A tendência da produção nacional é mais voltada para o etanol, que alcançou 25,4
bilhões de litros, com alta de 19,26% sobre a safra anterior. Quanto ao açúcar, foram
fabricados 34,27 milhões de toneladas, ou 0,56% a mais que na safra passada. O açúcar
continua sendo um dos principais itens dos embarques para exportação no Porto de Santos,
em São Paulo, onde foram embarcadas ao longo do ano passado 19 milhões de toneladas de
açúcar, volume 15% maior do que no ano anterior. (AGROLINK, 2014)
2.4. Melhoramento genético da cana-de-açúcar
O Programa de Melhoramento Genético da Cana-de-açúcar (PMGCA) da Rede
Interuniversitária para o Desenvolvimento do Setor Sucroalcooleiro (RIDESA) é composto
por 10 universidades: UFAL (Universidade Federal de Alagoas), UFRPE (Universidade
Federal Rural de Pernambuco), UFS (Universidade Federal de Sergipe), UFV (Universidade
Federal de Viçosa), UFRRJ (Universidade Federal Rural do Rio de Janeiro), UFSCar
(Universidade Federal de São Carlos), UFPR (Universidade Federal do Paraná), UFPI
(Universidade Federal do Piauí, UFMT(Universidade Federal do Mato Grosso) e UFG

11
(Universidade Federal de Goiás), as quais obtêm as variedades RB (República Federativa do
Brasil).
O PMGCA teve sua origem em 1971, quando foi estabecido o PLANALSUCAR-
Programa Nacional de Melhoramento da Cana-de-açúcar, que foi nomeado para atuar em
áreas de pesquisas do antigo IAA- Instituto do Açúcar e do Álcool. O programa tinha como
seu principal objetivo o de renovação da lista de variedades de cana-de-açúcar existente, entre
elas algumas importadas, as quais se consolidavam na produção sucroalcooleira do país, e que
era representada por variedades antigas que já apresentavam evidências de deterioração e que
não acompanhavam a produção por serem variedades específicas de algumas regiões
(PMGCA, 2013).
Em Alagoas, o PMGCA situa-se na Unidade Acadêmica Centro de Ciências Agrárias
(CECA) da UFAL, em parceria com o setor produtivo canavieiro. As inovações tecnológicas
desenvolvidas pelo PMGCA/CECA/UFAL e transferidas para o setor produtivo vêm
contribuindo significativamente para a elevação da produtividade e da qualidade das unidades
produtoras de açúcar, etanol e bioeletricidade (RIDESA, 2010).
Na obtenção de variedades RB, a RIDESA tem como ponto de partida um Banco de
Germoplasma da Estação de Floração e Cruzamento Serra do Ouro, situada em Murici -
Alagoas a 09º13‟S; 35º50‟W; 515 m de altitude. A área total da Estação é de 32 hectares e
apresenta pluviosidade anual média de 2.363mm, temperaturas mínima e máxima com média
de 18,2ºC e 27,9ºC, respectivamente, distando 34 km do litoral. Por sua localização e clima
privilegiados, permite o florescimento natural e profuso da cana-de-açúcar, necessário para a
realização de hibridações previamente planejadas pelos pesquisadores das Universidades que
compõem a RIDESA. Atualmente o banco de germoplasma da Serra do Ouro é composto de
2.607 genótipos, provenientes de programas nacionais e internacionais.
A estratégia básica do PMGCA-RIDESA para obtenção de novas variedades RB
baseia-se em cruzamentos de indivíduos superiores, que são realizados visando à seleção de
genótipos que apresentem características vantajosas em produtividade agroindustrial e
tolerância aos principais estresses: pragas, doenças, seca, geada, salinidade e florescimento
(RIDESA, 2010).
A adoção de tecnologias para novas variedades é o que tem contribuído para o avanço
sustentável do setor, pois ao considerar o avanço em produtividade que ocorreu com a cana-
de-açúcar nos últimos 40 anos, verificou-se um aumento em mais de 30% na produtividade
média como também a evolução significativa na qualidade da matéria-prima. Neste sentido, a

12
liberação de novas variedades disponíveis no mercado aliado ao manejo adequado pode
contribuir para elevação da produtividade com menores custos de produção. (RIDESA, 2010).
Mais importante do que o número de variedades liberadas é o nível de adoção das
mesmas pelo setor produtivo. De acordo com o censo varietal 2009, realizado pela RIDESA
considerando todas as Universidades Federais de Ensino Superior integrantes, as variedades
RB ocupam 58% dos canaviais do País, chegando a algumas regiões a representar áreas
superiores a 70%. Ao considerar os últimos censos realizados pela RIDESA verificou-se
tendência crescente na área plantada com variedades RB impulsionadas principalmente pela
liberação de novas variedades que tiveram plantio expressivo em todo território nacional.
(RIDESA, 2010).
O PMGCA/CECA/UFAL, se preocupa não só com a criação, desenvolvimento e
liberação de novas variedades de cana-de-açúcar para o mercado, mas também com a
fitossanidade das mesmas, objetivando solidificar um avanço sustentável para o setor
sucroalcooleiro, desenvolve pesquisas no banco de germoplasma da Estação de Floração e
Cruzamento Serra do Ouro para verificar possível presença de patógenos.
2.5. Doenças da cultura da cana-de-açúcar
É fato que toda essa expansão no setor sucroalcooleiro submete a cultura a uma ampla
variedade de fatores de estresse capazes de reduzir sua produtividade, sendo as doenças um
fator bastante relevante. As cinco principais doenças da cultura da cana-de-açúcar são o
raquistimo das soqueiras, a escaldadura, o carvão, o mosaico e a síndrome do amarelecimento
foliar (TOKESHI; RAGO, 2005). A cana-de-açúcar (Saccharum L. híbridos interespecíficos)
é vulnerável a diversas doenças disseminadas pelo uso de colmos contaminados, sendo que a
monocultura em extensas áreas favorece o surgimento de epidemias. A planta matriz pode se
mostrar infectada sem apresentar qualquer sintoma externo, aumentando os riscos de
disseminação de doenças para novas áreas (CHATENET et al., 2001).
O raquitismo da soqueira apresenta-se como uma das principais doenças da cana-de-
açúcar e quase todas as variedades são suscetíveis. A doença é causada por Leifsonia xyli
subsp. xyli Davis. Não existe um sintoma característico da doença, porém se observa o
crescimento desordenado dos colmos, como o próprio nome da doença retrata, além de
sintomas internos que é o desenvolvimento de uma coloração avermelhada nos feixes
vasculares. Sua transmissão se dá, por ferramentas de corte e também via plantio de toletes
contaminados (MATSUOKA, 1975). Também causada por bactérias, a escaldadura-das-
folhas tem como agente causal Xanthomonas albilineans (Ashby) Dowson. A bactéria

13
coloniza os vasos do xilema, agindo de forma sistêmica e podendo se disseminar
principalmente por meio de ferramentas de corte (facão, colhedora), por mudas infectadas,
além da gutação (TOKESHI; RAGO, 2005). Pode causar perdas de até 100%, com a queima
das folhas e apodrecimento dos colmos.
Os fungos também são causadores de algumas doenças de grande importância para a
cultura da cana. O carvão, causado por Ustilago scitaminea Syd, pertencente à classe dos
Ustomycetes, infecta a planta através dos teliósporos e coloniza preferencialmente os tecidos
meristemáticos. O principal sintoma da doença é a formação de uma estrutura chamada de
“chicote”, induzida pelo fungo, que pode chegar a até meio metro de comprimento. Sua
disseminação ocorre principalmente através do vento. Outra doença de origem fúngica que
merece destaque é a ferrugem marrom, causada pelo fungo Puccinia melanocephala Syd. &
P. Syd. Geralmente apresentam coloração avermelhada ou marrom-alaranjada nas folhas,
iniciando com pequenas manchas cloróticas que evoluem para manchas alongadas de cores
amareladas e tamanhos variáveis (MATSUOKA et al., 2005). Sua principal forma de
disseminação também se dá através do vento, que transporta os uredósporos por longas
distancias.
Nas tradicionais regiões produtoras um dos principais problemas que tem reduzido a
produtividade da cana-de-açúcar é o ataque de fitonematóides, principalmente dos gêneros
Meloidogyne e Pratylenchus (RESENDE JÚNIOR; VICENTE, 2011). Além do dano causado
pela utilização de nutrientes da planta, estes parasitos injetam toxinas no sistema radicular,
resultando em deformações nas raízes, como as galhas provocadas por Meloidogyne javanica
(Treub.) Chitwood, M. incognita (Kofoid & White) Chitwood, e extensas áreas necrosadas,
quando os nematóides presentes são Pratylenchus zeae Graham (COSTA, 2006).
2.5.1. Doenças causadas por vírus
O mosaico da cana-de-açúcar ocorre em mais de 70 países, sendo o Sugarcane mosaic
virus (SCMV) e o Sorghum mosaic virus (SrMV), pertencentes ao gênero Potyvirus, os
agentes etiológicos da doença (ALEGRIA et al., 2003). Os sintomas caracterizam-se pelo
mosaico nas folhas novas. Touceiras tem desenvolvimento retardado, podendo ter sua altura
reduzida à metade. Os sintomas mais frequentes surgem em canaviais jovens e com bom
crescimento vegetativo. Ocasionalmente em variedades extremamente suscetíveis, os colmos
podem apresentar sintomas de riscas e estrias deprimidas que podem evoluir até a necrose do
tecido sub-epidérmico. Neste caso o encurtamento dos entrenós é acentuado TOKESHI;
RAGO, 2005).

14
A síndrome do amarelecimento foliar (SAF), também conhecida por “amarelinho” foi
relatada no Brasil em 1989, mas começou a se tornar problema sério a partir do início da
década de 90 (VEGA; SCAGLIUSI; ULIAN, 1997). Plantações comerciais da variedade
SP71-6163, no Estado de São Paulo, tiveram perdas de produção de até 50%. Nesta época, as
investigações sobre a causa da doença, conduzidas no Brasil e no exterior, levantaram várias
hipóteses, entre as quais: fatores relacionados ao solo e clima ou mesmo a agentes bióticos,
como fungos, nematóides, vírus e fitoplasmas. No entanto, estudos demonstraram que o
Sugarcane Yellow Leaf Curl Virus é o agente causal do amarelinho (SCAGLIUSI;
LOCKHART, 2000).
Outros vírus vêm sendo relatados sobre a cana-de-açúcar, mas em muitos casos a
importância econômica dessas viroses ainda necessita ser determinada. Esse é o caso dos
Badnavirus, patógenos cujos relatos de ocorrência vêm aumentando na cultura em vários
países.
Em cana-de-açúcar (Saccharum spp.) foram descritas duas espécies de badnavírus:
Sugarcane bacilliform IM virus e Sugarcane bacilliform MO virus. A primeira foi descrita em
Cuba infectando S. officinarum com sintomas de estrias cloróticas ou em infecções latentes
(LOCKHART; AUTREY, 1988; RODRIGUEZ-LEMA et al., 1985). Essa espécie também já
foi relatada na Austrália, nos Estados Unidos e no Marrocos e tem como vetor a cochonilha
Saccharicoccus sacchari Cockerell (JONES; LOCKHART, 1993). Além de Saccharum spp.,
o vírus infecta plantas de bananeira (Musa acuminata Colla) em condições experimentais
(LOCKHART, 1995). O outro badnavírus que infecta a cana-de-açúcar é Sugarcane
bacilliform MO virus, relatado nos Estados Unidos sobre S. officinarum apresentando estrias
cloróticas. Ainda não se conhece o vetor desse vírus em condições naturais, sendo transmitido
apenas por propagação vegetativa, mas em condições experimentais o vírus pode infectar
bananeira (BOUHIDA; LOCKHART; OLSZEWSKI,1993).
Santos (2013) estudou a incidência de badnavírus em um dos principais Bancos de
Germoplasma de cana-de-açúcar do mundo, localizado na Estação de Floração e Cruzamento
da Cana-de-açúcar da Serra do Ouro, em Murici, Alagoas (9º13‟S; 35º50‟W), pertencente ao
PMGCA / CECA / UFAL / RIDESA e concluiu que 36,5% dos genótipos analisados estavam
infectados. Os badnavírus detectados por Santos (2013) pertencem a cinco espécies distintas,
sendo duas espécies já descritas, Banana streak OL virus (BSOLV) e Sugarcane bacilliform
IM virus (SCBIMV) e três possíveis novas espécies. Este constituiu o primeiro relato de
BSOLV em cana-de-açúcar, pois este vírus havia sido relatado apenas em plantas de banana

15
(Musa spp.) em diversos países (LOCKHART; OLSZEWSKI, 1993), inclusive no Brasil
(BRIOSO et al., 2000; LOMBARDI; HARAKAVA; COLARICCIO, 2010).
3. Família Caulimoviridae
A família Caulimoviridae engloba vírus de plantas com genoma de DNA de fita dupla
(dsDNA) classificados como pararetrovirus (vírus de DNA que utilizam a transcriptase
reversa no ciclo de replicação) e diferem dos retrovírus com base no seu genoma de DNA e na
sua integração irregular dentro do genoma do hospedeiro para a replicação (TEMIN, 1985).
Os vírus dessa família apresentam partículas não envelopadas as quais podem apresentar
morfologia isométrica (50-52 nm de diâmetro) ou baciliforme (30 nm de diâmetro e 130 -150
nm de comprimento) (GEERING; HULL, 2012).
Os vírions contêm uma única molécula de DNA fita dupla circular não covalentemente
fechada com 7,2 a 9,2 Kb (FAUQUET et al., 2005; GEERING; HULL, 2012). O genoma
possui uma região intergênica poli A, a qual pode também estar ausente, e descontinuidades
de fita simples ou “gaps” em sítios específicos de ambas as fitas (HARPER et al., 2002). O
genoma dos caulimovírus contém de um a oito ORFs (sequências de leitura aberta) e a
organização genômica é uma das principais características utilizadas para distinguir os
diferentes gêneros dessa família.
Atualmente, são reconhecidos pelo International Committee on Taxonomy of Viruses
(ICTV), sete gêneros da família Caulimoviridae: Badnavirus, Caulimovirus, Cavemovirus,
Petuvirus, Solendovirus, Soymovirus e Tungrovirus, os quais são separados de acordo com o
inseto vetor, gama de hospedeiro, organização do genoma e relacionamento filogenético. Os
membros de Caulimoviridae possuem ampla distribuição geográfica, sendo que a maioria das
espécies dos gêneros Tungrovirus e Badnavirus se localiza nas regiões tropicais e subtropicais
(GEERING; HULL, 2012).
Os hospedeiros naturais de espécies da família Caulimoviridae são Angiospermas das
classes Dicotyledonae e Monocotyledonae. Dependendo do gênero, a transmissão natural do
vírus pode ocorrer via inseto vetor ou por contato entre plantas hospedeiras, bem como por
sementes ou pelo pólen e por propagação vegetativa. A transmissão pode ser também
realizada por técnicas como a inoculação mecânica e enxertia (FAUQUET et al., 2005).
Os sintomas causados por esses vírus são variáveis e dependem da espécie viral,
hospedeira e condições climáticas. Mosaico, clareamento de nervuras, clorose entre nervuras
e estrias são os sintomas mais frequentes observados nas infecções causadas pelos diferentes
gêneros de Caulimoviridae (GEERING; HULL, 2012).

16
A replicação dos caulimovírus, em geral, envolve duas fases: transcrição de RNA a
partir do DNA viral no núcleo e transcrição reversa deste RNA para gerar dsDNA no
citoplasma. O genoma desses vírus também contêm uma sequência complementar ao
tRNAMet da planta que corresponde ao sítio de iniciação da replicação do DNA. Geralmente
este sítio está localizado dentro ou adjacente à região intergênica (região não-codificante) e é
geralmente designada como nucleotídeo 1 (DE KOCHKO et al.,1998).
Em contraste com os retrovírus, os pararetrovírus vegetais não necessitam de
integração ao genoma hospedeiro para sua replicação, portanto seu genoma não codifica a
proteína integrase. Sequências virais podem ocorrer dispersas no genoma de plantas,
originárias de uma infecção viral anterior que se fixou na linhagem germinativa. Tais
sequências são conhecidas como sequências endógenas de pararetrovirus (Endogenous
Pararetroviral Sequences - EPRVs) e são a classe mais abundante de sequências virais
endógenas em diferentes espécies de plantas (GAYRAL; ISKRA-CARUANA, 2009).
Um número cada vez maior de EPRVs em membros da família Caulimoviridae tem
sido identificado no genoma de muitas espécies de plantas (GEERING; SCHARASCHKIN;
TEYCHENEY, 2010). Todas as EPRVs descritas até o momento possuem um padrão de
arranjo similar com repetições em tandem, duplicações internas, fragmentações e inversão do
genoma viral. A maioria das EPRVs resulta em genomas virais parciais e não funcionais,
porém várias integrações contêm toda extensão do genoma viral, com sequências de leitura
aberta (ORFs) funcionais. Tais sequências podem então ser ativadas resultando na liberação
do genoma viral funcional que infectará o hospedeiro (GAYRAL; ISKRA-CARUANA,
2009). Essas sequências são integradas no genoma hospedeiro e podem dar origem a vírus
epissomais (NDOWORA et al., 1999) sendo provenientes de eventos de infecções antigas
(HARPER et al, 2005) ou sequências representativas intermediárias entre caulimovírus e
retrotransposons LTR (BOUSALEM; DOUZERY; SEAL, 2008; LLORENS et al., 2009).
Sabe-se até então que as sequências genômicas de alguns pararetrovirus vegetais são
integradas no genoma do hospedeiro, e que podem dar origem a vírus epissomais
(NDOWORA et al., 1999). Existe evidência de que algumas sequências são capazes de iniciar
uma infecção, em determinadas condições. Os melhores exemplos estudados foram para os
vírus: Banana streak OL virus (BSOLV) (NDOWORA et al., 1999), Banana streak GF virus
(BSGFV) (GAYRAL; ISKRA-CARUANA, 2009), Petunia vein clearing virus (PVCV)
(STAGINNUS; RICHERT-PÖGGELER, 2006) e Tobacco vein clearing virus (TVCV).

17
3.1. Gênero Badnavirus
O mais numeroso gênero da família Caulimoviridae é o Badnavirus, cujos membros se
apresentam como vírions baciliformes com 95-130 nm de comprimento e 24-35 nm de largura
(FAUQUET et al., 2005). O genoma de badnavírus geralmente contém uma única molécula
de dsDNA de cerca de 7200-7600 pb que forma um círculo aberto interrompido por
descontinuidades sítio-específicas e que podem conter uma região intergênica poli A
(MEDBERRY; LOCKHART; OLSZEWSKI, 1990). Os badnavírus são o segundo maior
gênero de vírus de plantas conhecido que possui um genoma de DNA (BOUHIDA;
LOCKHART; OLSZEWSKI, 1993; HAGEN et al., 1993), ficando atrás apenas do gênero
Begomovirus.
O membro tipo do gênero Badnavirus é o Commelina yellow mottle virus (CoYMV)
obtido de Commelina difusa Burm. F., em Guadaloupe (MIGLIORI; LASTRA, 1978). Todos
os badnavírus codificam para três principais ORFs (Figura 3A), e contém geralmente três
descontinuidades (gaps) em sítios específicos (BOUHIDA; LOCKHART; OLSZEWSKI,
1993; HAGEN et al., 1993; HARPER; HULL, 1998). As funções dos produtos das ORFs I e
II permanecem desconhecidas. Contudo, para o CoYMV, a proteína da ORF I está associada
com vírions imaturos, enquanto a ORF II é encontrada associada com ambos, imaturos e
maduros (CHENG; YANG; YEH, 1996). A poliproteína codificada pela ORF3 contém
domínios funcionais: rico em cisteína (CYS), motivo de ligação ao RNA (RB), aspartato
protease (PR), e replicase viral (transcriptase reversa, RT e ribonuclease, RNase H-RH)
(Figura 3B), os quais possuem domínios idênticos para os membros da família
Caulimoviridae (HARPER; HULL, 1998; LACO; BEACHY, 1994; MEDBERRY;
LOCKHART; OLSZEWSKI, 1990).

18
Figura 3A. Representação esquemática da organização genômica de Commelina yellow
mottle virus (CoYMV) membro tipo do gênero Badnavirus. O círculo completo representa o
genoma de DNA de fita dupla. As setas largas indicam a posição das ORFs 1, 2 e 3. B.
Representação linear do mapa genômico de badnavírus mostrando a identificação dos
domínios: domínios de movimento (M), motivo de ligação ao RNA (BR), região conservada
de cisteína (CYS), aspartato protease (PR), transcriptase reversa (RT), Ribonuclease H (RH)
(BRIDDON et al., 1999).
B
A
Os badnavírus são relatados infectando uma ampla gama de culturas tropicais
economicamente importantes como arroz (Oryza sativa L.) (OMURA et al., 1983), cana-de-
açúcar (Saccharum spp. L.) (LOCKHART; AUTREY, 1988), banana (Musa spp.)
(LOCKHART; OLSZEWKI, 1993), cacau (Theobroma cacao L.) (BRUNT et al., 1996),
citros (AHLAWAT et al., 1996), pimenta (Capsicum spp L.) (LOCKHART et al., 1997),
inhame (Dioscorea spp.) (PHILLIPS et al., 1999) e taro (Colocasia esculenta (L.) Schott.)
(YANG et al., 2003). A transmissão dos membros do gênero Badnavirus ocorre
principalmente por cochonilhas da espécie P. citri (Hemiptera: Pseudococcidae) e nem
sempre os vírus são detectados apenas em plantas sintomáticas, podendo ser também
detectados em plantas assintomáticas (GAUHL; PASBERG-GAUHL; HUGHES, 1997).
A identificação de badnavírus é baseada na amplificação parcial do genoma por PCR
(“Polymerase Chain Reaction” = Reação em cadeia da Polimerase). Esta técnica não é capaz
de distinguir entre sequências integradas ao genoma e cópias epissomais dos vírus na planta
hospedeira. Como consequência, o número de sequências de badnavírus tem crescido

19
rapidamente, resultando na falta de clareza sobre como determinar o estado taxonômico dos
membros desse gênero (BOUSALEM; DOUZERY; SEAL, 2008).
O genoma de diferentes badnavírus tem sido relatado como sendo altamente variável,
portanto, a maioria dos pares de primers para PCR utilizados para sua detecção foi desenhada
a partir de três regiões conservadas que ocorrem na ORF III (MEDBERRY; LOCKHART;
OLSZEWSKI, 1990), representando o domínio tRNAMet e as regiões da RNase H e RT
(GEERING et al., 2000; LOCKHART; OLSZEWSKI, 1993). Adicionalmente, para a alta
variabilidade genética de badnavírus, a descoberta da integração de Banana streak
virus (BSV) no genoma da planta, acarreta em dificuldades de testes confiáveis de diagnóstico
além de dificultar o intercambio internacional de germoplasma. A integração de partes do
genoma de BSV em espécies de Musa foi primeiramente apontada por Lafleur; Lockhart e
Olszewski (1996) através de hibridização, e demonstrado adicionalmente por técnicas
moleculares e citogenéticas por Harper et al. (1999). Geering et al. (2000) sugerem que
sequências integradas de BSV podem estar ligadas ao genoma A ou B de Musa e que a
distribuição dos integrantes de BSV podem estar restritos a determinadas espécies do
hospedeiro, pois uma infecção em banana pode ser originada a partir de sequências integradas
de badnavírus (NDOWORA et al., 1999). Cultura de tecidos e outros fatores de estresse
podem também ativar algumas dessas sequências (DAHAL et al., 2000; DALLOT et al.,
2001).
3.1.1. Outros hospedeiros de Badnavirus no Brasil
Além do abacaxizeiro, os badnavírus podem infectar uma ampla variedade de
hospedeiros, incluindo algumas culturas de grande relevância para a região Nordeste como o
inhame, a cana-de-açúcar e a bananeira.
Atualmente, apenas duas espécies de badnavírus que infectam inhame são
reconhecidas pelo ICTV, o Dioscorea bacilliform AL virus (DBALV) e Dioscorea bacilliform
SN virus (DBSNV), provenientes de D. alata L. e D. sansibarensis Pax, respectivamente
(GEERING; HULL, 2012), sendo que apenas a primeira espécie foi relatada no Brasil
(ANDRADE, 2007; GUIMARÃES, 2013; LIMA et al., 2013). Análises de sequências de
isolados do DBALV conduzidas por Lima et al. (2013) e por Guimarães, et al. (2013),
baseadas no domínio RT/RNase H revelaram uma diversidade genética relativamente baixa,
fato atribuído, em ambas as situações, a base geneticamente estreita do inhame (praticamente
uma única variedade, propagada vegetativamente). No entanto, Guimarães (2013) estudou

20
também a estrutura genética de cinco populações de DBALV concluindo que a variabilidade é
alta, semelhante àquela encontrada para vírus de ssDNA.
Estudos realizados na África e Sul do Pacífico sugerem alta diversidade de Badnavirus
em inhame e a possível presença de até doze espécies (ENI et al., 2008). O DBALV causa
distorção foliar severa e é o badnavírus mais amplamente distribuído, sendo encontrado em
infecções simples e/ou mistas com potyvírus (PIO-RIBEIRO et. al., 2005; YANG et al.,
2003).
Em bananeira são reconhecidas atualmente pelo ICTV, quatro espécies de badnavírus:
Banana streak OL virus, Banana streak GF virus, Banana streak MY virus e Banana streak
VN virus, descritos inicialmente na Nigéria, Equador, Austrália e Vietnã, respectivamente
(GEERING; HULL, 2012; HARPER; HULL, 1998; LHEUREUX et al., 2007). Os sintomas
consistem em estrias cloróticas nas folhas e redução da produção de frutos (HARPER; HULL,
1998; LOCKHART, 1995). As espécies diferem entre si com base na organização do seu
genoma (HARPER; HULL, 1998; GEERING et al., 2005; LHEUREUX et al., 2007). No
Brasil, o Banana streak virus (BSV) foi primeiramente descrito por Brioso et al. (2000) em
mudas de bananeira. Figueiredo et al. (2006a), relatou que existem no Brasil, pelo menos
quatro estirpes de BSV que podem infectar a bananeira, porém, posteriormente, essas estirpes
foram identificadas como quatro possíveis espécies de Badnavirus (FIGUEIREDO et al.,
2006b).
4. Variabilidade e estrutura genética de populações de fitopatógenos
A estrutura genética de populações de fitopatógenos refere-se à quantidade de
variabilidade genética existente e de sua distribuição dentro e entre populações
(MILGROOM, 1995). O estudo da dinâmica da variabilidade genética de populações de
fitopatógenos é necessário para conhecer como as populações evoluem (MCDONALD;
ZHAN; BURDON, 1999). Uma vez compreendidas as maneiras como populações de
fitopatógenos mantêm a variabilidade genética, é possível inferir sobre a taxa com a qual estas
populações evoluem (MCDONALD; LINDE, 2002a).
Programas de melhoramento visando resistência a patógenos podem ser orientados
pelos conhecimentos da estrutura genética de populações. McDonald e Linde (2002b)
propuseram um modelo para auxiliar na definição de estratégias de melhoramento visando
aumentar a durabilidade da resistência, que se baseia no potencial evolutivo de populações de
fitopatógenos. Por sua vez, o potencial evolutivo é determinado com base na estrutura

21
genética das populações. Em síntese, o modelo considera a quantidade e a distribuição da
variabilidade genética da população.
A durabilidade de variedades resistentes pode ser estimada com base em informações
da estrutura genética de populações de fitopatógenos (MCDONALD e LINDE, 2002a). Por
exemplo, a durabilidade do gene Xa7, que confere resistência de arroz à queima bacteriana
(Xanthomonas oryzae pv. oryzae (Ishiyama 1922) Swings et al.), foi determinada com base
em estudo da estrutura genética da população da bactéria em conjunto com estudos de
variabilidade patogênica, principalmente de atributos relacionados à adaptabilidade do
patógeno (VERA CRUZ et al., 2000).
Para compreensão da variação espacial e temporal na resistência e virulência de
patógenos, torna-se necessário combinar métodos de diferentes áreas de pesquisa como, por
exemplo, análises moleculares acopladas com análises estatísticas de genética de populações e
evolução, pois essas variações são de grande interesse para o desenvolvimento de estratégias
de manejo bem sucedidas (STUKENBROCK; MCDONALD, 2009).
Em populações de vírus o alto grau de diversidade genética é gerado por erros durante
a replicação do genoma. Os dois mecanismos de geração de erros em vírus são a mutação e
recombinação (GARCIA-ARENAL; FRAILE; MALPICA, 2003).
4.1. Mutação
A mutação é o processo pelo qual nucleotídeos que não estão presentes na fita molde
são incorporados na fita filha durante a replicação dos ácidos nucléicos (GARCIA-ARENAL;
FRAILE; MALPICA, 2003).
A taxa de mutação mede o aparecimento de mutações espontâneas, como uma função
do tempo. Taxas de mutações observadas diferem entre as espécies e também variam de
acordo com o genoma de uma dada espécie. Este parâmetro é frequentemente mensurado
como o número de substituições de nucleotídeos por base por geração. Como os vírus de
RNA são replicados por polimerases de RNA dependentes de RNA, que não possuem a
capacidade de correção de erro, eles apresentam as mais altas taxas de mutação por geração
entre 10-6
e 10-4
(DRAKE; HOLLAND, 1999). A taxa de evolução molecular para vírus de
RNA também é alta entre 10-4
e 10-2
subs/sítio/ano (JENKINS et al., 2002). Desta forma, vírus
de RNA são considerados as entidades que evoluem mais rapidamente (HOLLAND et al.,
1982).
A taxa universal de mutação para microrganismos com genoma de DNA é entre 10-10
a
10-6
mutações por base por geração, enquanto essa taxa varia em torno de um padrão médio

22
de 0,003 mutações por genoma por geração, um resultado conhecido como regra de Drake
(DRAKE, 1991). Embora essa taxa reflita estimativas obtidas para alguns vírus de DNA, a
sua universalidade é uma questão em aberto. Por exemplo, a taxa de mutação do ssDNA do
fago ΦX174 é mais alta do que a predita por esse modelo, enquanto os papilomavírus,
evoluem mais lentamente que o previsto (RANEY; DELONGCHAMP; VALENTINE,
2004). Estudos têm demonstrado que vírus de ssDNA como os parvovírus (SHACKELTON
et al., 2006) e geminivírus (DUFFY; HOLMES, 2009), podem apresentar taxas evolutivas
semelhantes as dos vírus de RNA.
As taxas de mutação correspondem à fidelidade das polimerases utilizadas na
replicação: vírus de RNA (os quais utilizam RNA polimerase dependentes de RNA, RdRps)
mutam mais rápido que os retrovírus (DNA polimerase dependente de RNA; RdDps ou
transcriptase reversa, RTs) e mais rápido que os vírus de DNA (DNA polimerase) (DUFFY;
SHACKELTON; HOLMES, 2008). DNA polimerases podem possuir domínios de correção
de erros, os quais rapidamente reduzem a taxa de mutação durante a replicação do DNA por
ao menos uma ordem de magnitude (GARCIA-DIAZ; BEBENEK, 2007). Ainda não se
conhece polimerases de RNA com capacidade de correção de erro. No caso de RTs, a
fidelidade é mais alta que a de RdRps (embora permaneça mais baixa que a de DNA
polimerases), resultando em menores taxas de mutação em retrovírus que em vírus de RNA.
Por exemplo, quando não integrados no genoma do hospedeiro, taxas 0.1-0.2 de mutações por
genoma por geração têm sido estimadas para human immunodeficiency virus type-1 (HIV-1)
(DRAKE et al., 1998), uma taxa aproximadamente cinco vezes menor do que a observada em
vírus que replicam usando RdRps (DRAKE, 1993).
Estudos com bactérias e sistemas animais indicaram que a taxa de mutação dos vírus
de dsDNA e ssDNA diferem significativamente (DUFFY; SHAKELTON; HOLMES, 2008).
As informações sobre taxas de mutação para vírus de dsDNA que infectam plantas são
escassas, sendo relatada apenas para o Cauliflower mosaic virus (CaMV), a qual estima-se
que é em torno de 10-5
subs/sítio/ano (FROISSART et al., 2005). Segundo Firth et al. (2010),
os vírus dsDNA são frequentemente descritos evoluindo através de associações co-
divergentes com seus hospedeiros, onde se espera um padrão de baixas taxas de substituição
de nucleotídeos.
4.2. Recombinação
Recombinação é o processo pelo qual segmentos de uma fita de DNA ou RNA
tornam-se incorporados na fita de um indivíduo diferente durante o mecanismo de replicação

23
(PADIDAM; BEACHY; FAUQUET; 1999). Eventos de recombinação têm sido
demonstrados estar associados com vírus aumentando sua gama de hospedeiros, (GIBBS;
WEILLER, 1999) virulência, (PITA et al., 2001; ZHOU et al., 1997) potencial evolutivo e
adaptação local (MONCI et al., 2002).
A recombinação é um evento bastante comum em vírus que infectam plantas com
genoma de ssDNA, a exemplo dos geminivírus (LEFEUVRE et al., 2007; PADIDAM;
BEACHY; FAUQUET, 1999). A elevada frequência de recombinação nesse grupo de vírus
pode ser, em parte, explicada pela ocorrência frequente de infecções mistas (DAVINO et al.,
2009; GARCÍA-ANDRES et al., 2006; PITA et al., 2001; RIBEIRO et al., 2003; SANZ et al.,
2000; TORRES-PACHECO et al., 1996), com a evidência de infecção do mesmo núcleo da
célula por mais de um begomovírus (MORILLA et al., 2004).
Estudos sugerem que eventos de recombinação contribuem para o nível de variação
genética entre os membros do gênero Caulimovirus (FROISSART et al., 2005) e Tungrovirus
(ARBOLEDA; AZZAM, 2000). Os vírus que pertencem à família Caulimoviridae (dsDNA)
são considerados objetos ideais para estudos de recombinação. Esta família de vírus de
plantas tem recebido atenção adicional desde a descoberta de que é capaz de se integrar no
genoma da planta hospedeira (SQUIRES et al., 2011). Eventos de recombinação homóloga já
foram relatados para ativação do genoma epissomal de BSV e TVCV, integrados nos
genomas de seus hospedeiros (NDOWORA et al., 1999; STAGINNUS; RICHERT-
PÖGGELER, 2006).
No entanto, a recombinação entre badnavírus de inhame em condições naturais parece
ser rara. A investigação de eventos de recombinação em espécies de badnavírus que infectam
inhame identificou apenas um único evento ocorrendo na região que codifica para a
transcriptase reversa entre as espécies tentativas DeBV A e DeBV B. Contudo, como as
análises foram realizadas para um pequeno fragmento do genoma, recombinações em outras
partes do genoma não podem ser excluídas (BOUSALEM et al., 2009).
Este trabalho teve como objetivo determinar a incidência e caracterizar os Badnavirus
detectados na cultura do abacaxi na região Nordeste do Brasil através do sequenciamento da
região RT/RNaseH e determinar a estrutura genética de populações de Badnavirus que
infectam a cultura do abacaxi e cana-de-açúcar na região Nordeste do Brasil, através da
sequência parcial da região RT/RNAseH da ORF3 do genoma viral, com a finalidade de
fornecer informações que possam contribuir para o melhor entendimento da biologia dessas
espécies e auxiliar os programas de melhoramento genético dessas culturas.

24
REFERÊNCIAS BIBLIOGRÁFICAS
AGROLINK (2014). Disponível em: <http://www.agrolink.com.br> Acesso em: 28 de jan.
2014.
AHLAWAT, Y. S.; PANT, R. P.; LOCKHART, B.E.L.; SRIVASTAVA, M.;
CHAKRABORTY, N. K.; VARMA, A. Association of a badnavirus with citrus mosaic
disease in India. Plant Disease, Saint Paul, v. 80, p. 590-592, 1996.
ALEGRIA, O.M.; ROYER, M.; BOUSALEM, M.; CHATENET, M.; PETERSCHMITT, M.;
GIRARD, J-C.; ROTT, P. Genetic diversity in the coat protein coding region of eighty-six
Sugarcane mosaic virus isolates from eight countries, particularly from Cameroom and
Congo. Archives of Virology, Austria, v. 148, p. 357-372, 2003.
ANDRADE, G. P. Diagnóstico fitossanitário da cultura do inhame (Dioscorea spp.) em
áreas produtoras do Nordeste do Brasil. 2007. 88 f. Tese (Doutorado em Fitopatologia) –
Universidade Federal Rural de Pernambuco, Recife.
ARBOLEDA, M.; AZZAM, O. Inter- and intra-site genetic diversity of natural field
populations of Rice Tungro Bacilliform Virus in the Philippines. Archives of Virology,
Austria, v. 145, n. 2, p. 275-89, 2000.
BARBOSA, G. V. S.; SOUZA, A. J. R.; ROCHA, A. M. C.; SANTOS, A. V. P.; RIBEIRO,
C. A. G.; BARRETO, E. J. S.; MOURA FILHO, G.; SOUZA, J. L.; FERREIRA, J. L. C.;
SOARES, L.; CRUZ, M. M.; FERREIRA, P. V.; SILVA, W. C. M. Três novas variedades
RB de cana-de-açúcar. Rio Largo. Boletim Técnico. n.2, 17p., 2003.
BARREIRO NETO, M.; SANTOS, E.S. Abacaxi cultura: contribuição tecnológica. João
Pessoa: EMEPA-PB, 1999. 96p. (EMEPA-PB. Documentos, 26).
BARREIRO NETO, M; LACERDA, J. F.; CARVALHO, R. A.; OLIVEIRA, E. F. Paraíba-
Rubi: cultivar de abacaxi resistente à fusariose. João Pessoa, PB: EMEPA, 2007. 4p.
BOLOGNA-CAMPBELL, I. Balanço de nitrogênio e enxofre no sistema solo-cana-de-
açúcar no ciclo de cana-planta. 2007. 112 p. Tese (Doutorado em Agronomia - Solos e
Nutrição de Plantas), Escola Superior de Agricultura Luiz de Queiroz, Universidade de São
Paulo, Piracicaba.
BOUHIDA, M. L.; LOCKHART, B. E.; OLSZEWSKI, N. E. An analysis of the complete
sequence of a sugarcane bacilliform virus genome infectious to banana and rice. Journal of
General Virology, London, v. 74, p.15-22, 1993.
BOUSALEM, M.; DOUZERY, E. J. P.; SEAL, S. E. Molecular taxonomy, phylogeny, and
evolution of plant reverse transcribing viruses (Caulimoviridae) inferred from the reverse
transcriptase sequences. Archives Virology, Austria, v. 153, p. 1085-1102, 2008.
BOUSALEM, M.; DURAND, O.; SCARCELLI, N.; LEBAS, B.S.M.; KENYON, L.;
MARCHAND, J.L.; LEFORT, F.; SEAL, S.E. Dilemmas caused by endogenous
pararetroviruses regarding the taxonomy and diagnosis of yam (Dioscorea spp.) badnaviruses:
analyses to support safe germplasm movement. Archives of Virology, Austria, v. 154, n. 2,
p. 297-314, 2009.

25
BRIDDON, R. W.; PHILLIPS, S.; BRUNT, A.; HULL, R. Analysis of the sequence of
Dioscorea Alata bacilliform virus: comparison to others members of the badnavirus group.
Virus Genes, Boston, v. 18, p. 277-283, 1999.
BRIOSO, P.S.T.; CORDEIRO, Z.J.M.; REZENDE, J.A.M.; KITAJIMA, E.W.; PIMENTEL
J.P.; FIGUEIREDO, A.R. Infecção mista em bananeiras pelos vírus do mosaico do pepino
("Cucumber mosaic virus" CMV) e da risca da bananeira ("Banana streak virus" BSV) no
Brasil. Summa Phytopathologica, Botucatu, v. 26, p. 254-257, 2000.
BRUNT A. A.; CRABTREE K.; DALLWITZ M. J.; GIBBS A. J.; WATSON L.; ZURCHER
E. J. Plant Viruses Online: Descriptions and Lists from the VIDE Database, 1996.
Disponível em: <http://biology.anu.edu.au/Groups/MES/vide/> Acesso em: 16 set. 2013.
CARVALHO V.D.; CUNHA, G.A.P. Produtos e Usos. In: CUNHA, G.A.P.; CABRAL,
J.R.S.; SOUZA, L.F.S.; O Abacaxizeiro. Cultivo, agroindústria e economia. Embrapa
Mandioca e Fruticultura (Cruz das Almas, BA). Brasília: Embrapa Comunicação para
Transferência de Tecnologia, 1999. cap. 15, p. 389-402.
CARVALHO, S.P.; PEREIRA, J.M.; BORGES, M.S.; MARIN, J.O.B. Panorama da
produção de abacaxi no Brasil e comportamento sazonal dos preços do abacaxi “pérola”
comercializados na CEASA-GO. Resumo expandido. 47º Congresso SOBER (Sociedade
Brasileira de Economia, Administração e Sociologia Rural). Porto Alegre, 2009.
CHATENET, M.; DELAGE, C.; RIPOLLES, M.; IREY, M.; LOCKHART, B. E. L.; ROTT,
P. Detection of Sugarcane yellow leaf virus in quarantine and production of virus-free
sugarcane by apical meristem culture. Plant Disease, Saint Paul, v. 85, p. 1177-1180.
2001.
CHENG, Y. H.; YANG, J. S.; YEH, S. D. Efficient transformation of papaya by coat protein
gene of papaya ringspot virus mediated by agrobacterium following liquid-phase wounding of
embryogenic tissues with caborundum. Plant Cell Reports, v. 16, p. 127-132, 1996.
CONAB 2012 (Companhia Nacional de Abastecimento). Disponível em:
<http://www.conab.gov.br > Acesso em: 30 de jan. 2013.
COSTA, F. Cana - os inimigos da produtividade. Revista Rural, v. 96, 2006. Disponível em:
<http://www.revistarural.com.br/Edicoes/2006/Artigos/rev96_cana.htm> Acesso em: 04 de
jan. 2014.
CUNHA, G. A. P.; CABRAL, J. R. S. Taxonomia, Espécies, Cultivares e Morfologia. In:
CUNHA, G. A. P.; CABRAL, J. R. S.; SOUZA, L. F. S.; O Abacaxizeiro. Cultivo,
agroindústria e economia. Embrapa Mandioca e Fruticultura (Cruz das Almas, BA).
Brasília: Embrapa Comunicação para Transferência de Tecnologia, 1999. cap. 1, p.17-51.
CUNHA, G. A. P.; MATOS, A. P.; CABRAL, J.R.S.; SOUZA, L. F. S.; SANCHES, N.F.;
REINHARDT, D.H. Abacaxi para exportação: Aspectos técnicos da produção. Brasília:
EMBRAPA/SPI, 1994. 41 p. (Série Publicações Técnicas FRUPEX, 11).
CUNHA, G.A.P. Produção integrada de abacaxi na Bahia, Paraíba e Pernambuco.
Embrapa Mandioca e Fruticultura. Cruz das Almas - BA (2004).
DAHAL G. O. R.; TENKOUANO, A.; HUGHES, J. D‟. A.; THOTTAPPILLY, G.;
VUYLSTEKE, D.; LOCKHART, E. B. L. Relationship between natural occurrence of banana

26
streak badnavirus and symptom expression, relative concentration of viral antigen, and yield
characteristics of some micropropagated Musa spp. Plant Pathology, v. 49, p. 68-79, 2000.
DALLOT, S. A. P.; RIVERA, C.; RAMIREZ, P.; CÔTE, F.; LOCKHART, B.E.L.;
CARUANA, M.L. Evidence that the proliferation stage of micropropagation procedure is
determinant in the expression of Banana streak virus integrated in the genome of FHIA 21
hybrid (Musa AAAB). Archives of Virology, Austria, v. 146, p. 2179-2190, 2001.
DANIELS, J.; ROACH, B. T. Taxonomy and evolution. p.7-84. In: HEINZ, D. J. Sugarcane
improvement through breeding. Elsevier, Amsterdam. 1987.
DAVINO, S.; NAPOLI, C.; DELLACROCE, C.; MIOZZI, L.; NORIS, E.; DAVINO, M.;
ACCOTTO, G. P. Two new natural begomovirus recombinants associated with the tomato
yellow leaf curl disease co-exist with parental viruses in tomato epidemics in Italy. Virus
Research, v. 143, n.1, p.15-23, 2009.
DE KOCHKO, A.; VERDAGUER, B.; TAYLOR, N.; CARCAMO, R.; BEACHY, R. N.;
FAUQUET, C. Cassava Vein Mosaic Virus (CsVMV), type species for a new genus of plant
double stranded DNA viruses? Archives of Virology, Austria, v. 143, p.945-962, 1998.
DRAKE, J. W. A constant rate of spontaneous mutations in DNA- based microbes.
Proceedings of the National Academy of Sciences, v. 88, p. 7160 – 7164, 1991.
DRAKE, J. W. Rates of spontaneous mutations among RNA viruses. Proceedings of the
National Academy of Sciences, v. 90, p. 4171 – 4175, 1993.
DRAKE, J. W.; HOLLAND, J. J. Mutation rates among RNA viruses. Proceedings of the
National Academy of Sciences, v. 96, n. 24, p. 13910-13913, 1999.
DRAKE, R. R.; NEAMATI, N.; HONG, H.; PILON, A. A.; SUNTHANKAR, P.; HUME, S.
D.; MILNE, G. W.; POMMIER, Y. Identification of a nucleotide binding site in HIV-1
integrase. Proceedings of the National Academy of Sciences, v. 95, n. 8, p. 4170-5, 1998.
DUFFY S.; HOLMES E.C. Validation of high rates of nucleotide substitution in
geminiviruses: phylogenetic evidence from East African cassava mosaic viruses. Journal of
General Virology, London, v. 90, p. 1539-1547, 2009.
DUFFY, S.; SHACKELTON, L. A.; HOLMES, E. C. Rates of evolutionary change in
viruses: patterns and determinants. Nature Reviews Genetics, v. 9, p. 267–276, 2008.
ENI, A. O.; , HUGHES, J. D.; ASIEDU, R.; REY, M. E. Sequence diversity among
Badnavirus isolates infecting yam (Dioscorea spp.) in Ghana, Togo, Benin and Nigeria.
Archives of Virology, Austria, v.153, p. 2263-2272, 2008.
FAO. FAO – Food and Agriculture Organization. Faostat. Disponível em:
<http://faostat3.fao.org>. Acesso em: 09 de jan. 2014.
FAUQUET, C.; MAYO, M.; MANILOFF, J.; DESSELBERGER, U.; BALL, L. Virus
taxonomy. Eight Report of the International committee on Taxonomy of Viruses.
Amsterdam. Elsevier. 2005.
FIGUEIREDO, D.V.; BRIOSO, P.S.T. PCR multiplex para a detecção de BSV e CMV em
bananeiras micropropagadas. Summa Phytopathologica, Botucatu, v. 33, n. 3, p. 229-232,
2007.

27
FIGUEIREDO, D.V.; CUNHA JUNIOR, J.O.; NOGUEIRA, M.S.R.; MONTANO, H.G.;
BRIOSO, P.S.T. Diversidade genética do Banana streak virus no Brasil. In: REUNIÃO
INTERNACIONAL DA ACORBAT, 27, 2006, Joinville. Proceedings… 2006b, v. 2, p. 757-
763.
FIGUEIREDO, D.V.; MEISSNER FILHO, P.E.; SILVA NETO, S.P.; BRIOSO, P.S.T.
Detection and analysis of Banana streak virus (BSV) sequences variability of banana from
Brazil. Summa Phytopathologica, Botucatu, v. 32, n. 2, p. 118-123, 2006a.
FIRTH, C.; KITCHEN, A.; SHAPIRO, B.; SUCHARD, M. A.; HOLMES, E. C.;
RAMBAUT, A. Using time-strutured data to estimate evolutionary rates of double-stranded
DNA viruses. Molecular Biology and Evolution, v. 27, p. 2038-2051, 2010.
FROISSART, R.; ROZE, D.; UZEST, M.; GALIBERT, L.; BLANC, S.; MICHALAKIS, Y.
Recombination every day: abundant recombination in a virus during a single multi-cellular
host infection. PLOS Biol, v. 3, n. 3, p. 89, 2005.
GAMBLEY, C. F.; GEERING, A. D. W.; STEELE, V.; THOMAS, J. E. Identification of
viral and non-viral reverse transcribing elements in pineapple (Ananas comosus), including
members of two new badnavirus species. Archives of Virology, Austria, v. 153, p. 1599–
1604, 2008b.
GAMBLEY, C. F.; STEELE, V.; GEERING, A. D. W.; THOMAS, J. E. The genetic diversity
of ampeloviruses in Australian pineapples and their association with mealybug wilt disease.
Australasian Plant Pathology, v. 37, p. 95–105, 2008a.
GARCÍA-ANDRÉS, S.; MONCI, F.; NAVAS-CASTILLO, J.; MORIONES, E. Begomovirus
genetic diversity in the native plant reservoir Solanum nigrum: Evidence for the presence of a
new virus species of recombinant nature. Virology, v. 350, n. 2, p. 433-42, 2006.
GARCIA-ARENAL, F.; FRAILE, A. E.; MALPICA, J.M. Variation and evolution of plant
virus populations. International Microbiology, v. 6, p 225-232. 2003.
GARCIA-DIAZ, M.; BEBENEK, K. Multiple functions of DNA polymerases. CRC Critical
Reviews in Plant Sciences, v. 2, p. 105-122, 2007.
GAUHL, F.; PASBERG-GAUHL,C.; HUGHES, J. D'A. First report of banana streak
badnavirus in plantain landraces in southern Cameroon, Central Africa. Plant Disease, Saint
Paul, v. 81, p. 1335-1335, 1997.
GAYRAL, P.; ISKRA-CARUANA, M. Phylogeny of Banana Streak Virus Reveals Recent
and Repetitive Endogenization in the Genome of Its Banana Host (Musa sp.). Journal of
Molecular Evolution, v. 69, p. 65-80, 2009.
GEERING, A. D. W.; HULL, R. Family Caulimoviridae. In: KING A.M.Q.; ADAMS M.J.;
CARSTENS E.B.; LEFKOWITZ, E.J. Virus Taxonomy. 9th Report of the International
Committee on Taxonomy of Viruses. London UK. Elsevier Academic Press. p. 429-443,
2012.
GEERING, A. D. W.; MCMICHAEL, L. A.; DIETZGEN, R. G.; THOMAS, J. E. Genetic
diversity among Banana streak virus isolates from Australia. Phytopathology, Saint Paul, v.
90, p. 921-927, 2000.

28
GEERING, A. D.; SCHARASCHKIN, T.; TEYCHENEY, P. Y. The classification and
nomenclature of endogenous viruses of the family Caulimoviridae. Archives of Virology,
Austria, v. 155, n. 1, p. 123-31, 2010.
GEERING, A.D.; POOGGIN, M. M.; OLSZEWSKI, N. E.; LOCKHART, B. E.; THOMAS,
J. E. Characterisation of Banana streak Mysore virus and evidence that its DNA is integrated
in the B genome of cultivated Musa. Archives of Virology, Austria, v. 150, n. 4, p. 787-796,
2005.
GIBBS, M. J; WEILLER, G. F. Evidence that a plant virus switched hosts to infect a
vertebrate and then recombined with a vertebrate-infecting virus. Proceedings of the
National Academy of Sciences, v. 96, n. 14, p. 8022-8027, 1999.
GOES, A. Doenças do Abacaxi. In: KIMATI, H.; AMORIM, L.; BERGAMIN FILHO, A.;
CAMARGO, L. E. A.; REZENDE, J. A. M. (Eds.). Manual de Fitopatologia. São Paulo:
Ceres, 2005. v.2, Doenças das Plantas Cultivadas. 4ª Ed. cap. 2, p.9-14.
GOMES, P. Fruticultura Brasileira. São Paulo, Ed. Nobel, 2007. 446p. 13ª edição.
GONÇALVES-GERVÁSIO, R. C. R.; SANTA-CECÍLIA, L. V. C. Consumo alimentar
de Chrysoperla externa sobre as diferentes fases de desenvolvimento de Dysmicoccus
brevipes, em laboratório. Pesquisa Agropecuária Brasileira, Brasília, vol. 36, n. 2, 2001.
GUIMARÃES, K. M. C. Estrutura genética de uma população do badnavirus Dioscorea
bacilliform alata virus (DBALV) que infecta inhame (Dioscorea spp.) no Nordeste do
Brasil. 2013. 47 f. Dissertação (Mestrado em Proteção de Plantas) – Universidade Federal de
Alagoas, Rio Largo.
GUIMARÃES, K. M. C.; SILVA, S. J. C.; MELO, A. M.; ASSUNÇÃO, I. P.; LIMA, G. S.
A. Diversity of badnavirus sequences infecting yam (Dioscorea cayanensis) in Alagoas,
Brazil. Resumo. In: 24 º Congresso of Virology & 8º Mercosur Meeting of Virology, v. 18,
2013, Porto Seguro. Anais… Porto Seguro: Brazilian Society for virology.
HAGEN, L. S.; JACQUEMOND, M.; LEPINGLE, A.; LOT, H.; TEPFER, M. Nucleotide
sequence and genomic organisation of cacao swollen shoot virus. Virology, v. 196, p. 619-
628, 1993.
HARPER, G.; DAHAL, G.; THOTTAPPILLY, G.; HULL, R. Detection of episomal Banana
Streak Badnavirus by IC-PCR. Journal of Virological Methods, v. 79, n. 1, p.1-8, 1999.
HARPER, G.; HART, D.; MOULT, S.; HULL, R.; GEERING, A.; THOMAS, J. The
diversity of Banana streak virus isolates in Uganda. Archives of Virology, Austria, v. 150, p.
2407-2420, 2005.
HARPER, G.; HULL, R. Cloning and sequence analysis of Banana Streak Virus DNA. Virus
Genes, v. 17, p. 271-278, 1998.
HARPER, G.; HULL, R.; LOCKHART, B.; OLSZEWSKI, N. Viral sequences integrated into
plant genomes. Annual Review of Phytopathology, Palo Alto, v. 40, p. 119-136, 2002.
HOLLAND, J. J.; SPINDLER, K.; HORODYSKI, F.; GRABAU, E.; NICHOL, S.;
VANDEPOL, S. Rapid evolution of RNA genomes. Science, v. 215, p. 1577- 1585, 1982.

29
IBGE - INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. SIDRA: Sistema
IBGE de recuperação automática. Rio de Janeiro: Instituto Brasileiro de Geografia e
Estatística, 2012. Produção agrícola Municipal. Disponível em:.
<http://www.sidra.ibge.gov.br/bda/agric/default.asp?t=2&z=t&o=11&u1=1&u2=1&u3=1&u4
=1&u5=1&u6=1> Acesso em: 27 nov. 2013
JENKINS, G. M.; RAMBAUT, A.; PYBUS, O. G.; HOLMES, E. C. Rates of molecular
evolution in RNA viruses: a quantitative phylogenetics analysis. Journal of Molecular
Evolution, v. 54, p. 152–161, 2002.
JONES, D. R.; LOCKHART, B. E. L. Banana streak disease. Musa Fact Sheet Nº 1.
International Network for Improvemente of Banana and Plantain. Montpellier, France.
1993.
LACO, G. S.; BEACHY, R. N. Rice tungro bacilliform virus encodes reverse transcriptase,
DNA polymerase and ribonuclease H activities. Proceedings of the National Academy of
Sciences USA, v. 91, p. 2654-2658, 1994.
LAFLEUR, D. A.; LOCKHART, B.E.L.; OLSZEWSKI, N.E. Portions of the banana streak
badnavirus genome are integrated in the genome of its hosts Musa spp. Phytopathology,
Saint Paul, v. 86, p. 100-101, 1996.
LEFEUVRE, P.; MARTIN, D. P.; HOAREAU, M.; NAZE, F.; DELATTE, H.; THIERRY,
M.; VARSANI, A.; BECKER, N.; REYNAUD, B.; LETT, J. M. Begomovirus 'melting pot' in
the south-west Indian Ocean islands: molecular diversity and evolution through
recombination. Journal of General Virology, London, v. 12, p. 3458-68, 2007.
LHEUREUX, F.; LABOUREAU, N.; MULLER, E.; LOCKHART, B. E.; ISKRA-
CARUANA, M. L. Molecular characterization of banana streak acuminata Vietnam virus
isolated from Musa acuminata siamea (banana cultivar). Archives of Virology, Austria, v.
152, n. 7, p. 1409-1416, 2007.
LIMA, J. S.; LIMA, A. T. M.; CASTILLO-URQUIZA, G. P.; SILVA, S. J. C.; ASSUNÇÃO,
I. P.; MICHEREFF, S. J.; ZERBINI, F. M.; LIMA, G. S. A. Variabilidade genética de
isolados de badnavírus infectando inhame (Dioscorea spp.) no Nordeste do Brasil. Tropical
Plant Pathology, Brasília, v. 38, n. 4, p. 349-353, 2013.
LLORENS, C.; MUNOZ-POMER, A.; BERNARD, L.; BOTELLA, H.; MOYA, A. Network
dynamics of eukaryotic LTR retroelements beyond phylogenetic trees. Biology Direct, v. 4,
p.41, 2009.
LOCKHART, B. E. L. Banana Streak Badnavirus infection in Musa: Epidemiology, diagnosis
and control. Food and Fertilizer Technology Center. Tech. Bull. v. 143, 1995.
LOCKHART, B. E. L.; AUTREY, L. J. C. Occurrence in sugarcane ofa bacilliform virus
related serologically to banana streak virus. Plant Disease, Saint Paul, v. 72, p. 230-233,
1988.
LOCKHART, B. E. L.; KIRATIYA-ANGUL, K.; SILVA, P. De; OLSZEWSKI, N.E.;
LOCKHART, N.; DEEMA, N.; SANGALANG, J. Identification of Piper yellow mottle virus,
a mealybug transmitted badnavirus infecting Piper spp. Southeast Asia. European of Plant
Pathology, v. 103, p. 303-311, 1997.

30
LOCKHART, B. E. L.; OLSZEWSKI, N.E. Serological and genomic heterogeneity of banana
streak badnavirus: implications for virus detection in Musa germplasm, Breeding Banana and
Plantain for Resistance to Diseases and Pests. CIRAD/INIBAP, p. 105-113, 1993.
LOMBARDI, R.; HARAKAVA, R.; COLARICCIO, A. Clonagem e purificação de
fragmento da proteína capsidial de Banana streak OL vírus. Pesquisa Agropecuária
Brasileira, Brasília, v. 45, n. 8, p. 811-817, 2010.
MAPA – Ministério da Agricultura, Pecuária e Abastecimento. Portaria Nº 187, de 5 de julho
de 2010. Diário Oficial da União. Brasília – DF, 06 de julho de 2010 – Seção 1. Disponível
em: <http://www.agricultura.gov.br> Acesso em: 12 nov. 2013.
MARANCA, G. Fruticultura comercial: mamão, goiaba e abacaxi. São Paulo, Nobel,
1977. 118p.
MATOS, A. P. Doenças e seu controle. In: CUNHA, G.A.P.; CABRAL, J.R.S.; SOUZA,
L.F.S.; O Abacaxizeiro. Cultivo, agroindústria e economia. Embrapa Mandioca e
Fruticultura (Cruz das Almas, BA). Brasília: Embrapa Comunicação para Transferência de
Tecnologia, 1999. cap. 11, p. 269-305.
MATOS, A. P.; FERREIRA, D. M. V.; CORDEIRO, Z. J. M. Doenças do abacaxizeiro.
Informe Agropecuário, Belo Horizonte, v. 26, n. 228, p. 7-11. 2005.
MATOS, A. P.; REINHARDT, D. H.; SANCHES, N. F.; SOUZA, L. F. S.; TEIXEIRA, F.
A.; ELIAS JÚNIOR, J.; GOMES, D. C. Produção de Mudas Sadias de Abacaxi. Circular
Técnica 89, EMBRAPA mandioca e fruticultura tropical, 2009.
MATSUOKA, S. Disseminação de controle do raquitismo da soqueira da cana de açúcar.
Summa Phytopathologica, Botucatu, v. 1, n. 4, p. 245-257, 1975.
MATSUOKA, S.; GARCIA, A. A. F.; ARIZONO, H. Melhoramento da cana-de-açúcar. p.
205-251. In: BORÉM, A. Melhoramento de espécies cultivadas. 2.ed. Editora UFV, Viçosa,
2005. 969 p.
MCDONALD, B. A.; LINDE, C. Pathogen population genetics, evolutionary potential, and
durable resistance. Annual Review of Phytopathology, Palo Alto, v. 40, p. 349-379, 2002a.
MCDONALD, B. A.; LINDE, C. The population genetics of plant pathogens and breeding
strategies for durable resistance. Euphytica, New York, v. 124, p. 163–180, 2002b.
MCDONALD, B. A.; ZHAN, J.; BURDON, J. J. Genetic structure of Rhynchosporium
secalis in Australia. Phytopathology, Saint Paul, v. 89, p. 639-45, 1999.
MEDBERRY, S. L.; LOCKHART, B. E. L.; OLSZEWSKI, N. E. Properties of Commelina
yellow mottle virus complete DNA sequence, genomic discontinuities and transcript suggest
that it is a pararetrovirus. Nucleic Acids Research, v. 18, p.5505-5513, 1990.
MIGLIORI, A.; LASTRA, R. Etude de vi rus present chez Commelina diffusa Burm. en
Guadeloupe. Annals of Phytopathology, Palo Alto, v. 10, p.467-477, 1978.
MILGROOM, M. G. Analysis of population structure in fungal plant pathogens. In: Leslie, J.
F. e Frederiksen, R. A. (Eds.) Disease analysis through genetics and biotechnology. Iowa
State University, Ames. p. 213-229, 1995.

31
MONCI, F.; SÁNCHEZ-CAMPOS, S.; NAVAS-CASTILLO, J.; MORIONES, E. A natural
recombinant between the geminiviruses Tomato yellow leaf curl Sardinia virus and Tomato
yellow leaf curl virus exhibits a novel pathogenic phenotype and is becoming prevalent in
Spanish populations. Virology, New York, v. 303, n. 2, p. 317-26, 2002.
MORILLA, G.; KRENZ, B.; JESKE, H.; BEJARANO, E. R.; WEGE, C. Tête à tête of
tomato yellow leaf curl virus and tomato yellow leaf curl sardinia virus in single nuclei.
Journal of Virology, Washington, v. 78, p. 10715-10723, 2004.
NDOWORA, T. D. G.; LAFLEUR, D.; HARPER, G.; HULL, R.; OLSZEWSKI, N.E.;
LOCKHART, B.E.L. Evidence that badnavirus infection in Musa can originate from
integrated pararetroviral sequences. Virology, New York, v. 255, p.214-220, 1999.
OLIVEIRA, J. P. S.; SANTOS, K. C.; JUNGHANS, D. T.; ABREU, E. F. M.; ANDRADE,
E. C. Prevalência do Pineapple Mealybug Wilt-Associated Virus (PMWaV) nas principais
regiões produtoras de abacaxi no Brasil. Resumo. In: JORNADA CIENTÍFICA EMBRAPA
MANDIOCA E FRUTICULTURA TROPICAL, 7, 2013, Cruz das Almas. Anais... Cruz das
Almas: Embrapa Mandioca e Fruticultura, 2013. Publicação online. Disponível em:
<http://ainfo.cnptia.embrapa.br/digital/bitstream/item/89345/1/Prevalencia-do-Pineapple-151-
13-JPaulo-Davi.pdf> Acesso em: 06 jan. 2014.
OMURA, T.; MINOBE, Y.; KIMURA, I.; HIBINO, H.; TSUCHIZAKI, T.; SAITO, Y.
Improved purification procedure and RNA segments of rice ragged stunt virus. Annals of the
Phytopathological Society of Japan, v. 49, p. 670-675, 1983.
PADIDAM, M.; BEACHY, R.N.; FAUQUET, C.M. A phage single-stranded DNA (ssDNA)
binding protein complements ssDNA accumulation of a geminivirus and interferes with viral
movement. Journal of Virology, v. 73, p. 1609-1616, 1999.
PEREIRA, P.C.; MELO, B. Cultura do Abacaxizeiro. 2003. Disponível em:
<http://www.fruticultura.iciag.ufu.br/abacaxi.html> Acesso em: 17 out. 2013.
PHILLIPS, S.; BRIDDON, R. W.; BRUNT, A.; HULL, R. The Partial Characterization of a
Badnavirus Infecting the Greater Asiatic or Water Yam (Dioscorea alata). Journal of
Phytopatholology, Washington, v. 147, p. 265-269, 1999.
PIO-RIBEIRO, G. MELO-FILHO, P. A.; KITAJIMA, E. W.; LIMA, S. K.; ANDRADE, G.
P.; DOMINGOS, C. A. Detecção de potyvírus em inhame em áreas produtoras de
Pernambuco e obtenção de matrizes in vitro. Fitopatologia Brasileira, Brasília, v. 30, p. 183,
2005.
PITA, J. S.; FONDONG, V. N.; SANGARÉ, A.; OTIM-NAPE, G. W.; OGWAL, S.;
FAUQUET, C. M. Recombination, pseudorecombination and synergism of geminiviruses are
determinant keys to the epidemic of severe cassava mosaic disease in Uganda. Journal of
General Virology, London, v. 82, p. 655-665, 2001.
PMGCA (Programa de Melhoramento Genético da Cana-de-açúcar). Disponível em:
<http://pmgca.dbv.cca.ufscar.br/htm/pmg/histor.php> Acesso em: 12 dez. 2013.
PY, C.; LACOEUILHE,J.J.; TEISSON, C. L‟Ananas sa culture, ses produits. Paris: G.M.
Maisoneuve et Larose, 1984, 562p.
RANEY, J. L.; DELONGCHAMP, R. R.; VALENTINE, C. R. Spontaneous mutant
frequency and mutation spectrum for gene A of phiX174 grown in E. coli. Environmental
and Molecular Mutagenesis, v. 44, p. 119–127, 2004.

32
REINHARDT, D.H.R.C.; CUNHA, G.A.P. Métodos de Propagação. In: CUNHA, G.A.P.;
CABRAL, J.R.S.; SOUZA, L.F.S.; O Abacaxizeiro. Cultivo, agroindústria e economia.
Embrapa Mandioca e Fruticultura (Cruz das Almas, BA). Brasília: Embrapa Comunicação
para Transferência de Tecnologia, 1999. cap.5, p. 105-138.
REINHARDT, D.H.R.C.; CUNHA, G.A.P.; MENEGUCCI, J.L.P. Indução Floral. In:
REINHARDT, D.H.R.C.; SOUZA, L.F da S.; CABRAL, J.R.S. Abacaxi irrigado em
condições semi-áridas. 1ª Edição. Embrapa Mandioca e Fruticultura. Cruz das Almas - BA
(2001).
RESENDE JÚNIOR, J. C.; VICENTE, C. B. Controle de Pratylenchus zeae Graham em
cana-de-açúcar: aplicação de nematicidas, indutores de resistência e rotação de cultura.
Enciclopédia Biosfera, Centro Científico Conhecer - Goiânia, v.7, n. 12, p. 1-8, 2011.
RIBEIRO, S. G.; AMBROZEVÍCIUS, L. P.; AVILA, A. C.; BEZERRA, I. C.;
CALEGARIO, R. F.; FERNANDES, J. J.; LIMA, M. F.; MELLO, R. N.; ROCHA, H.;
ZERBINI, F. M. Distribution and genetic diversity of tomato-infecting begomoviruses in
Brazil. Archives of Virology, Austria, v. 148, p. 281-295, 2003.
RIDESA (2010) Rede Interuniversitária para o Desenvolvimento do Setor Sucroalcooleiro.
Catálogo nacional de variedades “RB” de cana-de-acúcar. Curitiba, 2010. 136 p.
RODRIGUEZ-LEMA, E.; RODRIGUEZ, D.; FERNANDEZ, E.; ACEVEDO, R. ; LOPEZ,
D. Reporte de un nuevo vírus de la cana de azucar. Ciencias de la Agricultura, México, v.
23, p. 130, 1985.
SÁ, L.F. Cultura do abacaxi. Goiânia, Empresa de Assistência Técnica e Extensão Rural,
EMATER-GO, 1994. 18p. (BOLETIM TÉCNICO, 1).
SANCHES, N. F.; MATOS, A. P.; MEISSNER FILHO, P. E. Murcha associada à cochonilha.
In: REINHARDT, D. H., SOUZA, L. F. DA S., CABRAL, J. R. S. (Org.). Abacaxi -
Produção: aspectos técnicos. Cruz das Almas: Embrapa Mandioca e Fruticultura; Brasília,
DF: Embrapa Comunicação para Transferência de Tecnologia, 2000. p. 62-65.
SANTOS, J. M. S. Incidência e caracterização de badnavírus no banco de germoplasma
de cana-de-açúcar (Saccharum spp.) na Serra do Ouro, Murici-AL. 2013. 54 f.
Dissertação (Mestrado em Proteção de Plantas) - Universidade Federal de Alagoas, Rio
Largo.
SANTOS, K. C.; ANDRADE, E. C. Detecção do vírus que causa a murcha do abacaxi por
meio de RT-PCR, utilizando primers específicos e degenerados. In: JORNADA
CIENTÍFICA EMBRAPA MANDIOCA E FRUTICULTURA TROPICAL, 3, 2009,
Cruz das Almas. Anais... Cruz das Almas: Embrapa Mandioca e Fruticultura Tropical, 2009.
1 CD-ROM.
SANTOS, M.C.P.B. Manual de identificação de pragas, doenças e deficiências
nutricionais na cultura do abacaxi. Embrapa Mandioca e Fruticultura. Cruz das Almas –
BA, 2010.
SANZ, A. I.; FRAILE, A.; GARCÍA-ARENAL, F.; ZHOU, X.; ROBINSON, D. J.;
KHALID; S.; BUTT, T.; HARRISON, B. D. Multiple infection, recombination and genome
relationships among begomovirus isolates found in cotton and other plants in Pakistan.
Journal of General Virology, London, v. 81, p. 1839-1849, 2000.

33
SCAGLIUSI, S. M.; LOCKHART, B. E. Transmission, characterization and serology of
sugarcane yellow leaf luteovirus. Phytopathology, Saint Paul, v.90, p.120-124, 2000.
SETHER, D. M.; MELZER, M. J.; BORTH, W. B.; HU, J. S. Pineapple bacilliform CO
virus: Diversity, Detection, Distribution, and Transmission. Plant Disease, Saint Paul, v. 96,
n. 12, p. 1798-1884, 2012.
SETHER, D. M.; MELZER, M. J.; BUSTO, J. Diversity and mealybug transmissibility of
ampeloviruses in pineapple. Plant Disease, Saint Paul, v. 89, p. 450-456, 2005.
SETHER, D. M.; ULLMAN, D. E.; HU, J. S. Transmission of pineapple mealybug wilt–
associated virus by two species of mealybug (Dysmicoccus spp.). Phytopathology, Saint
Paul, v. 88, p.1224-1230, 1998.
SETHER, D.M.; HU, J.S. Closterovirus infection and mealybug exposure are necessary for
the development of mealybug wilt of pineapple disease. Phytopathology, Saint Paul, v. 92, n.
9, p. 928-935, 2002.
SHACKELTON, L. A.; RAMBAUT, A.; PYBUS, O. G.; HOLMES, E. C. JC virus evolution
and its association with human populations. Journal of Virology, v. 80, p. 9928-9933, 2006.
SIMÃO, S. Tratado de Fruticultura. FEALQ: Piracicaba, 1998. 760p.
SQUIRES, J.; GILLESPIE, T.; SCHOELZ, J.E; PALUKAITIS, P. Excision and episomal
replication of cauliflower mosaic virus integrated into a plant genome. Plant Physiology, v.
155, p. 1908-1919, 2011.
STAGINNUS, C.; RICHERT-PÖGGELER, K. R. Endogenous pararetroviruses: twofaced
travelers in the plant genome. Trends in Plant Sciences, v. 11, p. 485-491, 2006.
STUKENBROCK, E. H.; MCDONALD, B. A. Population genetics of fungal and Oomycete
effectors involved in gene-for-gene interactions. Molecular Plant-Microbe Interactions, v.
22, p. 371–380, 2009.
TAMAKI, T.; CARDOSO, J. L. Aspectos comerciais e econômicos da cultura do
abacaxizeiro no Brasil. In: Anais do 1 ° Simpósio Brasileiro de Abacaxicultura. Jaboticabal,
p.359, 1982.
TEMIN, H. M. Reverse transcription in the eukaryotic genome: Retroviruses,
pararetroviruses, retrotransposons, and retrotranscripts. Molecular Biology and Evolution, v.
2, p. 455-468, 1985.
TOKESHI, H.; RAGO, A. Doenças da cana-de-açúcar. In: KIMATI, H.; AMORIM, L.;
BERGAMIN FILHO, A.; CAMARGO, L. E. A.; REZENDE, J. A. M. (Eds.). Manual de
Fitopatologia. São Paulo: Ceres, 2005. v.2, Doenças das Plantas Cultivadas. 4ª Ed., cap.21,
p.185-196.
TORRES-PACHECO, I.; GARZÓN-TIZNADO, J. A.; BROWN, J. K., BECERRA-FLORA,
A.; RIVERA-BUSTAMANTE, R. F. Detection and distribution of geminiviruses in Mexico
and the Southern United States. Phytopathology, Saint Paul , v. 86, p. 1186-1192, 1996.
VASCONCELOS, J. N. Derivados da cana-de-açúcar. In: STAB: açúcar, álcool e
subprodutos, v. 20, n. 3, p. 16-18, 2002.

34
VEGA, J.; SCAGLIUSI, S. M. M.; ULIAN, E. C. Sugarcane yellow leaf disease in Brazil:
evidence of association with a luteovirus. Plant Disease, Saint Paul, v.81, p.21-26, 1997.
VENTURA, J. A.; ZAMBOLIM, L. Controle das doenças do abacaxizeiro. In: ZAMBOLIM,
L.; VALE, F. X. R.; MONTEIRO, A. J. A.; COSTA, H. Controle de doenças de
plantas: fruteiras. Viçosa: UFV, 2002. p. 445-510.
VERA CRUZ, C. M.; BAI, J.; OÑA, I.; LEUNG, H.; NELSON, R. J.; MEW; T.; LEACH, J.
E. Predicting durability of a disease resistance gene based on assessment of the fitness loss
and epidemiological consequences of avirulence gene mutation. Proceedings of the National
Academy of Sciences USA, v. 97, p. 13500-13505, 2000.
VERZIGNASSI, J. R.; MATOS, A. P.; SANTOS, M. F.; POLTRONIERI, L. S.;
BENCHIMOL, R. L.; SANCHES, N. F. Mancha negra do abacaxi no Pará. Summa
Phytopathologica, Botucatu, v. 35, n. 1, p. 76, 2009.
VITALINO, R.C. Recomendação técnica do cultivo do abacaxi irrigado no Distrito
Federal. Planaltina – DF, 2006.
YANG, I. C.; HAFNER, G.J.; DALE, J.L.; HARDING, R. M. Genomic characterisation of
taro bacilliform virus. Archives of Virology, Austria, v. 148, p. 937-949, 2003.
ZHOU, X.; LIU, Y.; CALVERT, L.; MUNOZ, C.; OTIM-NAPE, G. W.; ROBINSON, D. J.;
HARRISON, B. D. Evidence that DNA-A of a geminivirus associated with severe cassava
mosaic disease in Uganda has arisen by interspecific recombination. Journal of General
Virology, London, v. 78, p. 2101-2111, 1997.

CAPÍTULO II
Incidência e caracterização molecular de badnavírus na cultura do abacaxizeiro
no Nordeste do Brasil

36
Incidência e caracterização molecular de badnavírus na cultura do abacaxizeiro no
Nordeste do Brasil
Edlene M. S. M. Santos1; Sarah J. C. Silva
2; Jaqueline F. O. Costa
2; Iraildes P. Assunção
2;
Gaus S. A. Lima1,2
.
RESUMO
O Brasil é o principal produtor mundial de abacaxi (Ananas comosus L. Merril), tendo a
região Nordeste destaque como maior produtora no âmbito nacional. O abacaxizeiro está
exposto a diversos problemas fitossanitários, inclusive as viroses. As badnaviroses do
abacaxizeiro são causadas por duas espécies distintas: Pineapple bacilliform CO virus
(PBCoV) e Pineapple bacilliform ER virus (PBErV). O objetivo do presente trabalho foi
determinar a incidência de badnavírus na cultura do abacaxi no Nordeste do Brasil utilizando
técnicas moleculares através de PCR e caracterizá-los mediante sequenciamento da região
RT/RNaseH. Amostras foliares foram coletadas nos Estados de Alagoas, Maranhão, Paraíba e
Pernambuco e submetidas à extração de DNA. A incidência de badnavírus foi constatada em
todas as áreas amostradas e variou de 80 a 96,4%. Análises de comparações de sequências
pareadas revelaram que todas as sequências obtidas neste trabalho apresentaram identidade
superior a 80% com a sequência da espécie PBCoV (EU377664), oriunda da Austrália,
corroborando integralmente as análises filogenéticas. Estes resultados sugerem a ampla
disseminação do PBCoV no Nordeste brasileiro e registra o primeiro relato de badnavírus na
cultura do abacaxizeiro no Brasil.
Palavras-chave:1 Caulimoviridae, Ananas comosus, análise molecular.
1Departamento de Agronomia/Universidade Federal Rural de Pernambuco (UFRPE), 52171-900, Recife, PE,
Brasil. 2Centro de Ciências Agrárias/ Universidade Federal de Alagoas (UFAL), 57100-000, Rio Largo, AL, Brasil.
Autor para correspondência: Gaus S. A. Lima. e-mail: [email protected]

37
Incidence and molecular characterization of badnavirus in culture of pineapple in
Northeast of the Brazil.
ABSTRACT
Brazil is the world's leading producer of pineapple (Ananas comosus L. Merril), having the
Northeast highlighted as major producing nationally. The pineapple plant is exposed to
various problems, including viruses. Badnaviroses of the pineapple are caused by two
distinct species: Pineapple bacilliform CO virus (PBCoV) and Pineapple bacilliform ER virus
(PBErV). The aim of this work was determine the incidence of Badnavirus in the pineapple
crop in Northeast of the Brazil using molecular techniques by PCR and characterize them by
sequencing of the region RT/RNaseH. Leaf samples were collected in the states of Alagoas,
Maranhão, Paraíba and Pernambuco and subjected to DNA extraction. The incidence of
badnavirus was detected in all the sampled areas and varied from 80 to 96.4%. Analyzes
comparing paired sequences revealed that all sequences obtained in this study showed identity
greater than 80% to the sequence of the specie PBCoV (EU377664), coming from Australia,
fully corroborating the phylogenetic analyzes. These results suggest the wide spread of
PBCoV in Northeast Brazilian and records the first report of Badnavirus the pineapple crop in
Brazil.
Key words: Caulimoviridae, Ananas comosus, molecular analysis.

38
1. INTRODUÇÃO
O abacaxizeiro (Ananas comosus L. Merril) é uma planta monocotiledônea, herbácea,
perene, pertencente à família Bromeliaceae, que apresenta aproximadamente 50 gêneros e
2000 espécies. Do ponto de vista econômico, o gênero Ananas é o mais importante desta
família, pois nele estão incluídos os abacaxis (CUNHA & CABRAL, 1999).
Segundo dados da FAO, em 2011, a produção mundial foi de 21,8 milhões de
toneladas de abacaxi. O Brasil é o maior produtor, com 2,3 milhões de toneladas, o que
corresponde a 10,5% da produção no mundo, sendo a cultivar „Pérola‟ a mais plantada no país
(CARVALHO et al., 2009). A região Nordeste destaca-se com 39,9% do total da produção do
país, e o Estado da Paraíba como maior produtor nacional (IBGE, 2013). Apesar do grande
volume de abacaxi produzido no Brasil e no mundo, o abacaxizeiro está exposto a diversos
problemas fitossanitários que causam, geralmente, danos econômicos significativos (MATOS
et al., 2009), incluindo as viroses.
Dentre as viroses de ocorrência na cultura se destacam a murcha do abacaxizeiro,
causada pelo Pineapple mealybug wilt associated virus (PMWaV-1, PMWaV-2) (SETHER &
HU, 2002) e PMWaV-3 (GAMBLEY et al., 2008a), pertencentes ao gênero Ampelovirus,
família Closteroviridae e as badnaviroses, causadas por Pineapple bacilliform CO virus
(PBCoV) e Pineapple bacilliform ER virus (PBErV), do gênero Badnavirus, família
Caulimoviridae (GAMBLEY et al., 2008b). O PMWaV-1, 2 e 3 tem ampla distribuição,
sendo relatados inclusive no Brasil (OLIVEIRA et al., 2013), enquanto PBCoV e PBErV
foram relatados apenas na Austrália (WAKMAN et al., 1995), na China (WU et al., 2010) e
no Havaí (SETHER & HU, 2002). O PMWaV é transmitido e disseminado pelas cochonilhas
Dysmicoccus brevipes Cockerell e D. neobrevipes Beardsley e causam sintomas de
avermelhamento foliar, as bordas das folhas ficam voltadas para baixo e ocorre seca de suas
pontas (SANCHES et al., 2000). Os dois badnavírus que ocorrem na cultura (PBCoV e
PBErV) são transmitidos pela cochonilha D. brevipes e o PBCoV também por Planococcus
citri Risso (GAMBLEY et al., 2008b) e D. neobrevipes (SETHER et al., 2012). Uma
associação clara entre os sintomas e a infecção por badnavírus em abacaxi ainda não foi
mostrada (GAMBLEY et al., 2008b). No entanto, como o abacaxizeiro é uma planta de
propagação predominantemente vegetativa, a doença pode ser disseminada por meio de
mudas infectadas, mesmo na ausência dos insetos vetores.
De um modo geral os membros da família Caulimoviridae possuem ampla distribuição
geográfica. No entanto, a maioria das espécies dos gêneros Tungrovirus e Badnavirus se
localiza nas regiões tropicais e subtropicais (GEERING & HULL, 2012). Esta família engloba

39
vírus de plantas com genoma de DNA de fita dupla (dsDNA) classificados como
pararetrovirus, ou seja, vírus de DNA que utilizam a transcriptase reversa no ciclo de
replicação (TEMIN, 1985). Os vírus dessa família apresentam partículas não envelopadas as
quais podem apresentar morfologia isométrica (50-52 nm de diâmetro) ou baciliforme (30 nm
de diâmetro e 130 -150 nm de comprimento) (GEERING & HULL, 2012). Essa família tem
recebido maior atenção desde a década passada devido à descoberta de sequências endógenas
pararetrovirais (EPRVs). Acredita-se que EPRVs integram-se por recombinação ilegítima no
genoma do hospedeiro, e sua presença não está necessariamente associada com a infecção
(GEERING et al., 2005; HARPER et al., 1999; NDOWORA et al., 1999).
Os membros do gênero Badnavirus são caracterizados por partículas baciliformes não
envelopadas contendo um genoma circular de DNA de dupla fita, com 7.0–7.6 kb
(FAUQUET et al., 2005). São relatados infectando uma ampla gama de culturas tropicais
economicamente importantes como arroz (Oryza sativa L.) (OMURA et al., 1983), cana-de-
açúcar (Saccharum officinarum L.) (LOCKHART & AUTREY, 1988), banana (Musa spp.)
(LOCKHART & OLSZEWKI, 1993), cacau (Theobroma cacao L.) (BRUNT et al., 1996),
citros (Citrus spp. L.) (AHLAWAT et al., 1996), espécies de pimenta (LOCKHART et al.,
1997), inhame (Dioscorea spp.) (PHILLIPS et al., 1999), taro (Colocasia esculenta (L.)
Schott.) (YANG et al., 2003) e abacaxi (A. comosus) (GAMBLEY et al., 2008b).
Diante do exposto, o objetivo do presente trabalho foi determinar a incidência de
Badnavirus na cultura do abacaxi na região Nordeste do Brasil através de PCR e caracterizá-
los por meio do sequenciamento da região que engloba a transcriptase reversa e ribonuclease
H (RT/RNaseH).
2. MATERIAL E MÉTODOS
Foram coletadas amostras foliares de abacaxizeiros provenientes de cinco áreas de
cultivo localizadas nos estados de Alagoas, Paraíba, Pernambuco e Maranhão (Tabela 1).
Algumas amostras foram coletadas de plantas apresentando sintomas de estrias cloróticas e
outras de plantas assintomáticas.
Para a extração do DNA total foram utilizados de 100 a 200 mg de tecido foliar
provenientes de folhas jovens de abacaxizeiro, seguindo o protocolo CTAB (cationic
hexadecyl trimethyl ammonium bromide) de acordo com FERREIRA & GRATTAPAGLIA
(1998). O DNA total serviu de molde para as reações de amplificação por PCR de fragmentos
de aproximadamente 580 pb. O par de oligonucleotídeos utilizados neste trabalho, BadnaFP
(5‟-ATGCCITTYGGIITIAARAAYGCICC-3‟) e BadnaRP (5‟-

40
CCAYTTRCAIACISCICCCCAICC-3‟) foi baseado no domínio RT/RNaseH da ORF 3 do
genoma de vários badnavírus já descritos (YANG et al., 2003).
As reações foram realizadas inicialmente em volume total de 15 μL para avaliar a
presença ou ausência de badnavírus. As amostras positivas foram então submetidas a PCR em
volume total de 60 μL. As condições de amplificação foram: desnaturação inicial a 94°C por
4 min, seguida de 94°C por 30 s, 50°C por 30 s e 72°C por 30 s. Os ciclos foram repetidos 35
vezes exceto o passo inicial e a extensão final a 72°C por 10 min. Após a amplificação, uma
alíquota dos produtos da PCR foi aplicada em gel de agarose a 1,2% e submetidos à
eletroforese em tampão TAE (Tris-Ácido acético, EDTA 0,5M pH 8,0) por aproximadamente
uma hora. Em seguida, o gel foi corado com brometo de etídeo e visualizado sob luz
ultravioleta. Os produtos de PCR foram purificados usando o kit GFX ™ PCR DNA and Gel
Band Purification Kit (GE Healthcare) e enviados para sequenciamento na Macrogen Inc.
(Seul, Coréia do Sul).
As sequências obtidas foram submetidas ao algoritmo BLASTn (Nucleotide Basic
Local Alignment Search Tool) (ALTSCHUL et al., 1990) e, posteriormente, editadas usando o
programa The Staden Package, ver 1.7 (STADEN, 1998) para gerar a sequência consenso de
cada isolado.
Comparações pareadas entre as sequências obtidas nesse trabalho e outras espécies de
Caulimoviridae disponíveis no GenBank (Tabela 2) foram geradas utilizando-se a ferramenta
Species Demarcation Tool, v. 1.0 (SDT) (MUHIRE et al., 2013) e estimada a percentagem de
identidade das sequências de nucleotídeos entre os isolados. De acordo com os critérios do
Comitê Internacional de Taxonomia Viral (ICTV) para delimitação de espécies dentro do
gênero Badnavirus, as diferenças entre as sequências na região RT/RNaseH não devem ser
maiores que 20%, pois acima desse valor serão considerados vírus de espécies distintas
(GEERING & HULL, 2012).
As sequências foram alinhadas utilizando o software MUSCLE (EDGAR, 2004),
disponível no pacote computacional MEGA 5 (Molecular Evolutionary Genetics Analysis)
(TAMURA et al., 2011) e submetidas à análise filogenética de Máxima Verossimilhança
(MV) (RIDLEY, 2006) utilizando o modelo de substituição de nucleotídeos General Time
Reversible com distribuição gama (GTR+G). A confiabilidade da árvore gerada foi obtida a
partir de 2000 repetições bootstrap. A sequência outgroup consistiu da espécie Rice tungro
bacilliform virus (RTBV), a qual pertence ao gênero Tungrovirus, família Caulimoviridae
(Tabela 2).

41
3. RESULTADOS E DISCUSSÃO
Considerando todas as áreas, um total de 277 amostras foram analisadas, sendo 244
amostras positivas, o que correspondeu a uma incidência média de 88,1%. O número de
amostras de cada local de coleta e sua respectiva incidência de badnavírus é apresentado na
Tabela 3. A técnica de PCR vem sendo amplamente utilizada para a detecção de membros do
gênero Badnavirus, inclusive para análises de variabilidade genética e caracterização de
espécies desse gênero, mesmo não discriminando sequências integradas (endógenas) ao
genoma das sequências epissomais (DELANOY et al., 2003).
Na Austrália, THOMSON et al. (1996) utilizaram a técnica de PCR para detecção de
Pineapple bacilliform virus (PBV) em extratos de plantas de abacaxi infectadas e foi possível
constatar a presença do PBV em todas as áreas de plantio da costa leste da Austrália. Os
autores concluíram que a sequência de nucleotídeos obtida foi distinta de outros badnavírus já
descritos. Recentemente, SETHER et al. (2012) utilizaram PCR e relataram uma provável
nova espécie designada PBCoV-HI1 e nove variantes (A-I) do PBCoV no Havaí. Esses nove
variantes não podem ser considerados como novas espécies porque compartilham >92/95%
(nucleotídeos/aminoácidos) de identidade com os PBCoVs já relatados no Hawaí.
A forma endógena do vírus é de extrema importância na detecção de viroses e no que
se refere ao controle de doenças, pois vários fatores têm sido descritos como desencadeantes
de doenças devido à geração de formas epissomais dos vírus a partir das formas endógenas
(DAHAL et al., 2000; HARPER et al., 1999), apesar desse processo ser ainda pouco
estudado, já é sabido que ele de fato ocorre (GAYRAL & ISKRA-CARUANA, 2009). Certas
condições de estresse como a cultura de tecidos, hibridação e eventos de recombinação que
ocorrem nas sequências endógenas, podem levar à reconstituição do genoma viral na forma
ativada e, assim, resultar em infecções epissomais (CÔTE et al., 2010; DALLOT et al., 2001;
NDOWORA et al., 1999). Todas as sequências utilizadas neste trabalho foram distintas entre
si. Essa alta variabilidade observada indica que as mesmas são provavelmente sequências
epissomais, pois todas as EPRVs (Endogenous Pararetroviral Sequences) descritas até o
momento possuem um padrão de arranjo similar com repetições em tandem, duplicações
internas, fragmentações e inversão do genoma viral (GAYRAL & ISKRA-CARUANA,
2009).
Dentre as amostras positivas para badnavírus, foram sequenciadas vinte de cada um
dos cinco locais de coleta, totalizando sequências de 100 amostras, no entanto foi possível
montar contigs de boa qualidade apenas para 83 sequências.

42
As análises preliminares utilizando o algorítmo BLAST e comparações de sequências
pareadas com SDT revelaram que todas as sequências obtidas neste trabalho apresentaram
identidade superiores a 80% com a sequência da espécie Pineapple bacilliform comosus virus
(PBCoV) depositada no GenBank com o número de acesso EU377664 (oriunda da Austrália),
o que indica que todas as sequências caracterizadas nesse estudo correspondem a isolados do
PBCoV (Figura 1).
Os relacionamentos filogenéticos analisados com base nas sequências de nucleotídeos
do domínio RT/RNAseH da ORF3 dos badnavírus, revelaram que os isolados sequenciados
neste estudo foram posicionados junto à sequência de PBCoV da Austrália obtida do
GenBank (Figura 2). Essa análise indicou que os 83 isolados utilizados neste trabalho
formaram um ramo monofilético (Grupo I) incluindo a espécie PBCoV, as quais possuem um
ancestral comum com a espécie PBErV (Figura 2). O subgrupo 1 contém isolados
provenientes de todos os municípios amostrados, enquanto o subgrupo 2 mostrou uma
tendência a agrupar por região geográfica, incluindo a maioria dos isolados provenientes dos
municípios de Arapiraca e Coruripe, ambos pertencentes ao Estado de Alagoas. O grupo II
inclui as outras espécies pertencentes ao gênero Badnavirus (Figura 2).
A proximidade geográfica entre o município de Arapiraca e o Povoado Pindorama
(Coruripe), que distam em apenas 66 Km, é a provável causa para os isolados oriundos destes
locais agruparem entre si, pois essa condição facilita a troca de mudas de abacaxizeiro entre
produtores. Estes resultados diferem dos obtidos por GUIMARÃES (2013) ao realizar
análises filogenéticas baseadas no domínio RT/RNAseH de badnavírus que infectam
Dioscorea spp. no Nordeste do Brasil, onde os isolados virais não agruparam de acordo com a
sua origem geográfica.
As análises pareadas corroboraram integralmente com os resultados obtidos na
filogenia, ou seja, o grupo I da árvore filogenética incluiu apenas os isolados obtidos neste
trabalho junto com a sequência do PBCoV da Austrália e que na análise par a par
compartilharam acima de 80% de identidade de sequência, o que de acordo com o ICTV é um
critério decisivo para demarcação de espécies do gênero Badnavirus (GEERING & HULL,
2012; HULL et al., 2005).
Os badnavírus nem sempre são detectados em plantas sintomáticas e podem ser
também detectados em plantas assintomáticas (GAUHL et al., 1997). A ausência de sintomas
em plantas de abacaxizeiro infectadas por badnavírus impossibilita a seleção de mudas sadias
para o plantio e comercialização, acarretando na maior disseminação / introdução do vírus em
outras áreas de cultivo. Além disso, os sintomas causados por badnavírus em plantas de

43
abacaxizeiro ainda não estão claramente determinados (GAMBLEY et al., 2008b; SETHER et
al., 2012).
A transmissão de PBCoV por cochonilhas foi observada na Austrália com incidência
de infecção de 20% e 10% para D. brevipes e P. citri, respectivamente (GAMBLEY et al.,
2008b). Posteriormente, no Havaí, SETHER et al. (2012) observaram alta incidência de
PBCoV (80%) após o contato com a cochonilha D. neobrevipes. A ocorrência da espécie D.
brevipes é relatada em todos os países onde o abacaxi é cultivado e em todos os Estados
brasileiros (LACERDA et al., 2009). A espécie P. citri apresenta uma ampla distribuição
geográfica, ocorrendo em regiões tropicais, subtropicais e temperadas (CORREIA et al.,
2008) e D. neobrevipes já foi relatada em várias fruteiras cultivadas no Brasil (COSTA,
2002). Contudo, a taxa de transmissão de badnavírus por cochonilha é geralmente considerada
baixa, sendo o maior risco de disseminação do vírus causado pelo uso de material propagativo
infectado (LOCKHART & OLSZEWSKI, 1993). Desta forma, acredita-se que a propagação
vegetativa do abacaxizeiro seja um dos principais fatores que contribuem para a prevalência
de PBCoV na região Nordeste do Brasil.
4. CONCLUSÕES
Este constitui o primeiro registro na cultura do abacaxizeiro de infecção por
Badnavirus no Brasil. A presença de Pineapple bacilliform CO virus em Alagoas, Maranhão,
Paraíba e Pernambuco, sugere uma ampla distribuição e prevalência desta espécie na região
Nordeste do Brasil devido à proximidade geográfica das áreas de cultivo avaliadas e ausência
de barreiras fitossanitárias, permitindo assim a prática comum de troca de mudas infectadas
entre as mesmas.
5. REFERÊNCIAS BIBLIOGRÁFICAS
AHLAWAT, Y. S. et al. Association of a badnavirus with citrus mosaic disease in India.
Plant Disease, v.80, p.590-592, 1996.
ALTSCHUL, S. F. et al. Basic local alignment search tool. Journal of Molecular Biology,
v.215, p.403-410, 1990.
BRUNT, A. A. et al. Plant Viruses Online: Descriptions and Lists from the VIDE Database,
1996. Disponível em: <http://biology.anu.edu.au/Groups/MES/vide/> Acesso em: 16 set.
2013.
CARVALHO, S.P. et al. Panorama da produção de abacaxi no Brasil e comportamento
sazonal dos preços do abacaxi “pérola” comercializados na CEASA-GO. In: 47º Congresso
SOBER (Sociedade Brasileira de Economia, Administração e Sociologia Rural), 2009, Porto
Alegre, Resumo expandido.

44
CORREA, L.R.B. et al. Desenvolvimento da cochonilha-branca Planococcus citri (Risso,
1813) (Hemiptera: Pseudococcidae) em frutíferas. Arquivos do Instituto Biológico, São
Paulo, v.75, p.239-242, 2008.
COSTA, C.L. Inter-relações dos insetos vetores com vírus de plantas frutíferas cultivadas no
Brasil. In: ZAMBOLIM, L. (Ed.) Manejo Integrado – Fruteiras Tropicais. Doenças e
Pragas. Viçosa, UFV, 2002. p. 105-153.
CÔTE, F. X. et al. Micropropagation by tissue culture triggers differential expression of infectious
endogenous Banana streak virus sequences (eBSV) present in the B genome of natural and
synthetic interspecific banana plantains. Molecular Plant Pathology, v.11, p.137-144, 2010.
CUNHA, G.A.P.; CABRAL, J.R.S. Taxonomia, Espécies, Cultivares e Morfologia. In:
CUNHA, G.A.P. et al. O Abacaxizeiro. Cultivo, agroindústria e economia. Embrapa
Mandioca e Fruticultura (Cruz das Almas, BA). Brasília: Embrapa Comunicação para
Transferência de Tecnologia, 1999. cap. 1, p.17-51.
DAHAL, G. O. R. et al. Relationship between natural occurrence of banana streak badnavirus
and symptom expression, relative concentration of viral antigen, and yield characteristics of
some micropropagated Musa spp. Plant Pathology, v.49, p.68-79, 2000.
DALLOT, S. A. P. et al. Evidence that the proliferation stage of micropropagation procedure is
determinant in the expression of Banana streak virus integrated in the genome of FHIA 21 hybrid
(Musa AAAB). Archives of Virology, v.146, p. 2179-2190, 2001.
DELANOY, M. et al. Development of realtime PCR for the rapid detection of episomal
Banana streak virus (BSV). Plant Disease, v.87, p.33-8, 2003.
EDGAR, R.C. MUSCLE: A multiple sequence alignment method with reduced time and
space complexity. BMC Bioinformatics, v.5, p.1-19, 2004.
FAO - FAO – Food and Agriculture Organization. Faostat. Disponível em:
<http://faostat3.fao.org>. Acesso em: 09 de jan. 2014.
FAUQUET, C. et al. Virus taxonomy. Eight Report of the International committee on
Taxonomy of Viruses. Amsterdam. Elsevier. 2005.
FERREIRA, M. E.; GRATTAPAGLIA, D. Introdução ao uso de marcadores moleculares
em análise genética. 3. ed. Brasília: Embrapa – CENARGEN. 1998. 220p.
GAMBLEY, C. F. et al. Identification of viral and non-viral reverse transcribing elements in
pineapple (Ananas comosus), including members of two new badnavirus species. Archives of
Virology, v.153, p.1599–1604, 2008b.
GAMBLEY, C. F. et al. The genetic diversity of ampeloviruses in Australian pineapples and
their association with mealybug wilt disease. Australasian Plant Pathology, v.37, p.95–105,
2008a. Disponível em: www.publish.csiro.au/journals/app
GAUHL, F. et al. First report of banana streak badnavirus in plantain landraces in southern
Cameroon, Central Africa. Plant Disease, v.81, p.1335-1335, 1997.
GAYRAL, P.; ISKRA-CARUANA, M. Phylogeny of Banana Streak Virus Reveals Recent
and Repetitive Endogenization in the Genome of Its Banana Host (Musa sp.). Journal of
Molecular Evolution, v.69, p.65-80, 2009.

45
GEERING, A. D. W.; HULL, R. Family Caulimoviridae. In: KING A.M.Q.; ADAMS M.J.;
CARSTENS E.B.; LEFKOWITZ, E.J. Virus Taxonomy. 9th Report of the International
Committee on Taxonomy of Viruses. London UK. Elsevier Academic Press. p. 429-443,
2012.
GEERING, A.D.W. et al. Banana contains a diverse array of endogenous badnaviruses.
Journal of General Virology, v.86, p.511-520, 2005.
GUIMARÃES, K. M. C. Estrutura genética de uma população do badnavirus Dioscorea
bacilliform alata virus (DBALV) que infecta inhame (Dioscorea spp.) no Nordeste do
Brasil. 2013. 47 f. Dissertação (Mestrado em Proteção de Plantas) – Universidade Federal de
Alagoas.
HARPER, G.O. et al. Integration of banana streak badnavirus into the Musa genome:
molecular and cytogenetic evidence. Virology, v.255, p.207- 219, 1999.
HULL, R. et al. Family Caulimoviridae. Elsevier, p. 385-396, 2005.
IBGE. INSTITUTO BRASILEIRO DE GEOGRAFIA E ESTATÍSTICA. Levantamento
sistemático da produção agrícola. Rio de Janeiro, v. 26, n. 1, p.1-83, 2013. On-Line.
Disponível em: <http://www.sidra.ibge.gov.br> Acesso em: 16 jan. 2014.
LACERDA, J. T. et al. Cochonilha Dysmicoccus brevipes: a praga cosmopolita da
abacaxicultura. Tecnol. & Ciên. Agropec., v.3, p.15-21, 2009.
LOCKHART, B. E. L. et al. Identification of Piper yellow mottle virus, a mealybug
transmitted badnavirus infecting Piper spp. Southeast Asia. European of Plant Pathology,
v.103, p.303-311, 1997.
LOCKHART, B. E. L.; AUTREY, L. J. C. Occurrence in sugarcane of a bacilliform virus
related serologically to banana streak virus. Plant Disease, v.72, p.230-233, 1988.
LOCKHART, B. E. L.; OLSZEWSKI, N.E. Serological and genomic heterogeneity of banana
streak badnavirus: implications for virus detection in Musa germplasm, Breeding Banana and
Plantain for Resistance to Diseases and Pests. CIRAD/INIBAP, p.105-113, 1993.
MATOS, A.P. et al. Produção de Mudas Sadias de Abacaxi. Brasília: EMBRAPA
mandioca e fruticultura tropical, 2009. (Circular Técnica 89).
MUHIRE, B. et al. A genome-wide pairwise-identity-based proposal for the classification of
viruses in the genus Mastrevirus (family Geminiviridae). Archives of virology, v. 158,
p.1411-1424, 2013. Disponível em: <http://www.ncbi.nlm.nih.gov/pubmed/23340592>.
Acesso em 28 jul. 2013.
NDOWORA, T. et al. Evidence that badnavirus infection in Musa can originate from
integrated pararetroviral sequences. Virology, v.255, p.214-220, 1999.
OLIVEIRA, J. P. S. et al. Prevalência do Pineapple Mealybug Wilt-Associated Virus
(PMWaV) nas principais regiões produtoras de abacaxi no Brasil. Resumo. In: JORNADA
CIENTÍFICA EMBRAPA MANDIOCA E FRUTICULTURA TROPICAL, 7, 2013, Cruz das
Almas. Anais... Cruz das Almas: Embrapa Mandioca e Fruticultura, 2013. Publicação online.
Disponível em: <http://ainfo.cnptia.embrapa.br/digital/bitstream/item/89345/1/Prevalencia-
do-Pineapple-151-13-JPaulo-Davi.pdf> Acesso em: 06 jan. 2014.

46
OMURA, T. et al. Improved purification procedure and RNA segments of rice ragged stunt
virus. Annals of the Phytopathological Society of Japan, v.49, p.670-675, 1983.
PHILLIPS, S. et al. The Partial Characterization of a Badnavirus Infecting the Greater Asiatic
or Water Yam (Dioscorea alata). Journal of Phytopatholology, v.147, p.265-269, 1999.
RIDLEY, M. Evolução. 3ª Ed. São Paulo. Artmed, 2006.
SANCHES, N. F. et al. Murcha associada à cochonilha. In: REINHARDT, D. H., SOUZA, L.
F. DA S., CABRAL, J. R. S. (Org.). Abacaxi - Produção: aspectos técnicos. Cruz das
Almas: Embrapa Mandioca e Fruticultura; Brasília, DF: Embrapa Comunicação para
Transferência de Tecnologia, 2000. p.62-65.
SETHER, D. M.; HU, J. S. Closterovirus Infection and Mealybug Exposure Are Necessary
for the Development of Mealybug Wilt of Pineapple Disease. Phytopathology, v.92, p.928-
935, 2002.
SETHER, D. M. et al. Pineapple bacilliform CO virus: Diversity, Detection, Distribution, and
Transmission. Plant Disease, v.96, p.1798-1804, 2012. Disponível em:
<http://dx.doi.org/10.1094/PDIS-08-11-0718-RE> Acesso em: 30 jan. 2014.
STADEN, R. et al. The Staden package, 1998. In MISENER, S.; KRAWETZ, S.A.
(eds). Bioinformatics methods and protocols Humana, p.115-130, 1998.
TAMURA, K. et al. MEGA 5: Molecular evolutionary genetics analysis using maximum
likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and
Evolution, v.28, p.2731–2739, 2011.
TEMIN, H. M. Reverse transcription in the eukaryotic genome: Retroviruses,
pararetroviruses, retrotransposons, and retrotranscripts. Molecular Biology and Evolution,
v.2, p.455-468, 1985.
THOMSON, K. G. et al. Detection of pineapple bacilliform virus using the polymerase chain
reaction. Annals of Applied Biology, v.129, p.57-69, 1996.
WAKMAN, W. et al. Presence of a clostero-like virus and a bacilliform virus in pineapple
plants in Australia. Australian Journal of Agricultural Research, v.46, p.947-958, 1995.
WU, L. et al. Sequencing and Analysis of the Complete Genomic Sequence of Pineapple
bacilliform comosus virus. Scientia Agricultura Sinica, v.43, p.1969-1976, 2010.
YANG, I.C. et al. Sequence diversity of South Pacific isolates of Taro bacilloform virus and
the development of a PCR-based diagnostic test. Archives of Virology, v.148, p.1957-1968,
2003.

47
Tabela 1. Procedência, cultivar, número e código das amostras de abacaxi coletadas na região
Nordeste do Brasil. Ano de coletas: 2013
Áreas de coleta Latitude Longitude Cultivar Amostras
coletadas
Código das
Amostras
Arapiraca (AL) 09°45'09'' 36°39'40'' „Jupi‟ 60 A1A a A60A
Pedras de Fogo (PB) 07°24'07'' 35°06'59'' „Pérola‟ 60 A1PF a A60PF
São Domingos do
Maranhão (MA) 05°34'33'' 44°23'07'' „Pérola‟ 56 A1SD a A56SD
Coruripe* (AL) 10°07'32'' 36°10'32'' „Pérola‟ 51 A1Pi a A51Pi
Pombos (PE) 08°08'29'' 35°23'45'' „Pérola‟ 60 A1Po a A60Po
* As amostras do município de Coruripe foram coletadas no Povoado Pindorama.
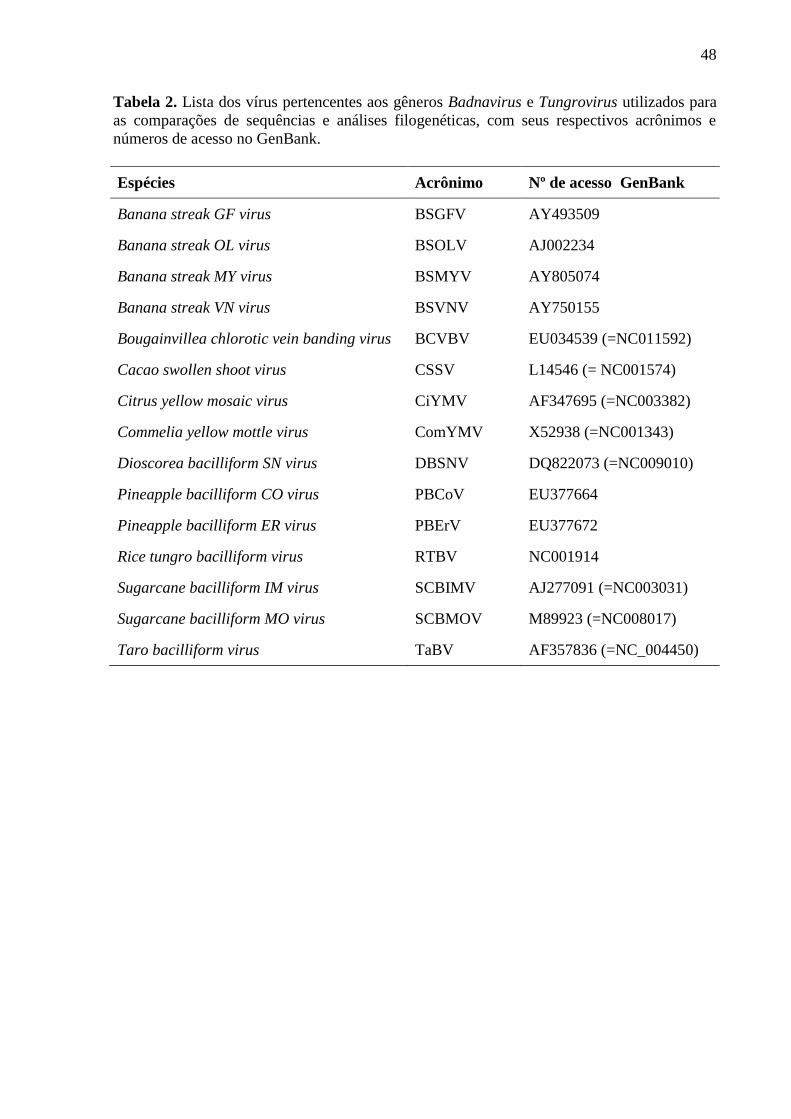
48
Tabela 2. Lista dos vírus pertencentes aos gêneros Badnavirus e Tungrovirus utilizados para
as comparações de sequências e análises filogenéticas, com seus respectivos acrônimos e
números de acesso no GenBank.
Espécies Acrônimo Nº de acesso GenBank
Banana streak GF virus BSGFV AY493509
Banana streak OL virus BSOLV AJ002234
Banana streak MY virus BSMYV AY805074
Banana streak VN virus BSVNV AY750155
Bougainvillea chlorotic vein banding virus BCVBV EU034539 (=NC011592)
Cacao swollen shoot virus CSSV L14546 (= NC001574)
Citrus yellow mosaic virus CiYMV AF347695 (=NC003382)
Commelia yellow mottle virus ComYMV X52938 (=NC001343)
Dioscorea bacilliform SN virus DBSNV DQ822073 (=NC009010)
Pineapple bacilliform CO virus PBCoV EU377664
Pineapple bacilliform ER virus PBErV EU377672
Rice tungro bacilliform virus RTBV NC001914
Sugarcane bacilliform IM virus SCBIMV AJ277091 (=NC003031)
Sugarcane bacilliform MO virus SCBMOV M89923 (=NC008017)
Taro bacilliform virus TaBV AF357836 (=NC_004450)
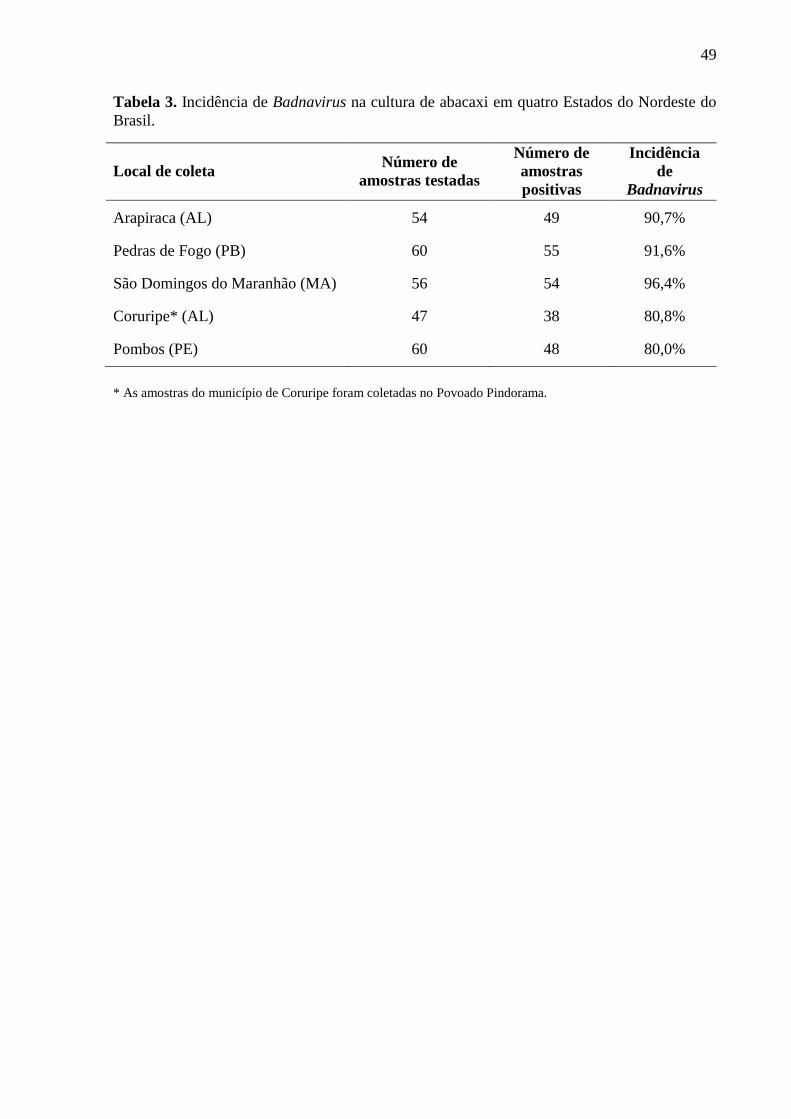
49
Tabela 3. Incidência de Badnavirus na cultura de abacaxi em quatro Estados do Nordeste do
Brasil.
Local de coleta Número de
amostras testadas
Número de
amostras
positivas
Incidência
de
Badnavirus
Arapiraca (AL) 54 49 90,7%
Pedras de Fogo (PB) 60 55 91,6%
São Domingos do Maranhão (MA) 56 54 96,4%
Coruripe* (AL) 47 38 80,8%
Pombos (PE) 60 48 80,0%
* As amostras do município de Coruripe foram coletadas no Povoado Pindorama.

50
Figura 1. Matriz bidimensional representando a percentagem de identidade de comparações
pareadas de sequências nucleotídicas da região RT/RNaseH dos isolados avaliados neste
trabalho e outras espécies do gênero Badnavirus disponíveis no GenBank e uma espécie do
gênero Tungrovirus.

51
Figura 2. Árvore filogenética de Máxima Verossimilhança baseada em sequências da região
RT/RNaseH de isolados de Pineapple bacilliform CO virus obtidas neste trabalho e outras
espécies de Badnavirus disponíveis no GenBank. O Rice tungro bacilliform virus (RTBV),
gênero Tungrovirus, foi utilizado como outgroup. Valores de suporte de bootstrap de 2000
repetições são apresentados nos ramos.
Sub
grup
o 1
Sub
grup
o 2
Gru
po I
Gru
po I
I

CAPÍTULO III
Estrutura genética de populações de badnavírus que infectam as culturas do
abacaxizeiro e da cana-de-açúcar no Nordeste do Brasil

53
Estrutura genética de populações de badnavírus que infectam as culturas do
abacaxizeiro e da cana-de-açúcar no Nordeste do Brasil
Edlene M. S. M. Santos1; Sarah J. C. Silva
2; Joyce S. Lima
2; Iraildes P. Assunção
2; Gaus
S. A. Lima1,2
.
RESUMO
O gênero Badnavirus (família Caulimoviridae) engloba vírus de plantas com genoma de DNA
circular de fita dupla (dsDNA) e que replicam por meio de um intermediário de RNA.
Badnavírus causam doenças em culturas economicamente importantes na maioria das regiões
tropicais e subtropicais do mundo, incluindo o Brasil. Informações sobre a variabilidade e
estrutura genética de vírus de plantas com genoma de dsDNA são escassas. Dessa maneira, o
presente trabalho objetivou determinar a estrutura genética de populações de uma espécie de
badnavírus que infecta o abacaxizeiro (Pineapple bacilliform CO virus (PBCoV)) e de duas
espécies que infectam a cana-de-açúcar (Banana streak OL virus (BSOLV) e Sugarcane
bacilliform IM virus (SCBIMV)), todas provenientes da região Nordeste do Brasil. A
estrutura genética das populações foi determinada com base no domínio que engloba a
transcriptase reversa e ribonuclease H (RT/RNaseH) e revelaram que a população de PBCoV
está estruturada com base na região geográfica de origem. A variabilidade genética nas
populações estudadas pode ser considerada alta, com frequências de mutação na ordem de 10-
4 subs/sítio/ano para as populações de PBCoV e de 10
-3 para BSOLV e SCBIMV. Quanto
aos mecanismos evolutivos, seleção purificadora ou expansão recente da população são as
duas principais forças atuando nessas populações.
Palavras-chave:2 Caulimoviridae, estrutura populacional, Ananas comosus, Saccharum spp.
1Departamento de Agronomia/Universidade Federal Rural de Pernambuco (UFRPE), 52171-900, Recife, PE,
Brasil. 2Centro de Ciências Agrárias/ Universidade Federal de Alagoas (UFAL), 57100-000, Rio Largo, AL, Brasil.
Autor para correspondência: Gaus S. A. Lima. e-mail: [email protected]

54
Genetic structure of badnavirus populations that infect the cultures of pineapple and
sugarcane in Northeastern Brazil
Edlene M. S. M. Santos1; Sarah J. C. Silva
2; Joyce S. Lima
2; Iraildes P. Assunção
2; Gaus
S. A. Lima1,2
.
ABSTRACT
The Badnavirus genus (family Caulimoviridae) comprises plant viruses with circular DNA
genome of double-stranded (dsDNA), which replicate by means of an RNA intermediate.
Badnaviruses cause diseases in economically important crops in most tropical and subtropical
regions of The World, including Brazil. Information about the variability and genetic structure
of plant viruses with dsDNA genome are scarce. Thus, this study aimed to determine the
genetic structure of populations of a specie of badnavirus that infects pineapple (Pineapple
bacilliform CO virus (PBCoV)) and two specie that infect sugarcane (Banana streak OL virus
(BSOLV) and Sugarcane bacilliform IM virus (SCBIMV)) all from the Northeast region of
Brazil. Pairwise comparisons analysis and phylogenetic sequences revealed that all 83 isolates
from pineapple belong to the species Pineapple bacilliform virus CO (PBCoV). The genetic
structure was determined from the domain comprising the reverse transcriptase and
ribonuclease H (RT / RNase H) and revealed that the population of PBCoV is structured
based on geographic region of origin. The genetic variability at the studied populations can be
considered high, with mutation frequencies on the order of 10-4
subs/site/year for populations
of PBCoV and 10-3
for BSOLV and SCBIMV. Regarding the evolutionary mechanisms,
purifying selection or recent population expansion are the two main forces acting in these
populations.
Key words: Caulimoviridae, population structure, Ananas comosus, Saccharum spp.

55
1. INTRODUÇÃO
A família Caulimoviridae engloba vírus de plantas com genoma de DNA circular de
fita dupla (dsDNA) classificados como pararetrovirus, ou seja, vírus de DNA que utilizam a
transcriptase reversa no ciclo de replicação. Apresentam partículas não envelopadas as quais
podem ter morfologia isométrica ou baciliforme. Atualmente, são reconhecidos pelo ICTV
(International Committee on Taxonomy of Viruses), sete gêneros na família: Badnavirus,
Caulimovirus, Cavemovirus, Petuvirus, Solendovirus, Soymovirus e Tungrovirus, os quais são
classificados de acordo com o inseto vetor, gama de hospedeiros, organização do genoma e
relacionamento filogenético (GEERING & HULL, 2012).
O mais numeroso gênero da família Caulimoviridae é Badnavirus, cujos membros se
apresentam como vírions baciliformes com 95-130 nm de comprimento e 24-35 nm de largura
(FAUQUET et al., 2005). O genoma dos badnavírus geralmente contém uma única molécula
de dsDNA de cerca de 7200-7600 pb que forma um círculo aberto interrompido por
descontinuidades sítio-específicas e que podem conter uma região intergênica poli A
(MEDBERRY et al., 1990). Atualmente, o gênero Badnavirus possui 25 espécies
reconhecidas pelo ICTV, sendo o segundo maior gênero de vírus de plantas conhecido com
genoma de DNA (BOUHIDA et al., 1993; HAGEN et al., 1993), ficando atrás apenas dos
begomovírus.
O genoma de todos os badnavírus possui três ORF‟s, dando origem a três proteínas,
P1, P2 e P3. A função de P1 é desconhecida, enquanto P2 é uma proteína associada ao vírion.
A poliproteína codificada pela ORF3 contém quatro domínios funcionais: rico em cisteína
(CYS), motivo de ligação ao RNA (RB), aspartato protease (PR), replicase viral (transcriptase
reversa – RT e ribonuclease – RNase H-RH), os quais são conservados para os membros da
família (MEDBERRY et al., 1990; LACO & BEACHY, 1994; HARPER & HULL, 1998;
GEERING & HULL, 2012).
Os badnavírus são transmitidos de forma semi-persistente por cochonilhas (GEERING
& HULL, 2012) para uma ampla gama de culturas tropicais economicamente importantes,
incluindo cana-de-açúcar (Saccharum spp. L.) (LOCKHART & AUTREY, 1988), inhame
(Dioscorea spp.) (PHILLIPS et al., 1999) e abacaxi (Ananas comosus) (GAMBLEY et al.,
2008). Nem sempre os vírus são detectados em plantas sintomáticas, podendo ser também
detectados em plantas assintomáticas (GAUHL et al., 1997; GEERING & HULL, 2012).
Atualmente, são reconhecidas pelo ICTV duas espécies de badnavírus que infectam
abacaxi: Pineapple bacilliform CO virus (PBCoV) e Pineapple bacilliform ER virus (PBErV),
originalmente associadas a A. comosus var. comosus e A. comosus var. erectifolius,

56
respectivamente (GAMBLEY et al., 2008; GEERING & HULL, 2012). Para a cana-de-açúcar
também são descritas duas espécies de badnavírus: Sugarcane bacilliform IM virus
(SCBIMV) e Sugarcane bacilliform MO virus (SCBMOV), ambas infectando S. officinarum
(BOUHIDA et al., 1993; LOCKHART & AUTREY, 1988; RODRIGUEZ-LEMA et al.,
1985).
No Brasil o registro de DBALV em inhame (D. cayanensis e D. alata) é bem
documentado (ANDRADE, 2007; LIMA et al., 2013), porém os registros de incidência de
badnavírus em cana-de-açúcar e em abacaxi são recentes. Em cana-de-açúcar, dados
preliminares apontam para a ocorrência de pelo menos cinco espécies distintas, sendo SBIMV
e Banana streak OL virus (BSOLV) as de maior prevalência (SANTOS, 2013). Já para o
abacaxi apenas o PBCoV foi constatado em amostras provenientes de quatro estados do
Nordeste (SANTOS, 2014). Todos esses relatos se restringiram a registrar a incidência e
realizar a caracterização parcial dos vírus.
O Brasil é o maior produtor mundial de abacaxi e de cana-de-açúcar (FAO, 2012).
Apesar da importância destas culturas para o país, há uma escassez de estudos sobre
variabilidade genética de badnavírus na região Nordeste, dificultando a adoção de medidas de
controle que possam auxiliar programas de melhoramento no desenvolvimento de variedades
resistentes e a troca de material nacional e internacional de boa qualidade.
O abacaxizeiro pertence à família Bromeliaceae, gênero Ananas, sendo uma planta de
clima tropical, herbácea e perene, com folhas estreitas e resistentes, quase sempre margeadas por
espinhos (SÁ, 1994). A cultura do abacaxi é propagada vegetativamente por meio de mudas,
formadas a partir de diferentes partes vegetativas da planta, cuja qualidade depende das condições
ambientais onde são produzidas e do manejo dado à cultura (REINHARDT & CUNHA, 1999).
A cana-de-açúcar pertence à família Gramineae (Poaceae), gênero Saccharum, onde as
cultivares são híbridos derivados principalmente do cruzamento entre a espécie domesticada
S. officinarum, rica em sacarose, e a espécie selvagem S. spontaneum, do cruzamento com
híbridos naturais (DANIELS & ROACH, 1987).
No sistema produtivo canavieiro brasileiro, o cultivo de variedades com boas
características agroindustriais é a melhor forma de se obter ganhos da produtividade com
baixo custo. A atuação dos programas de melhoramento genético desta cultura contribui
expressivamente para o desenvolvimento do setor sucroalcooleiro nacional, com a liberação
de variedades mais produtivas e mais resistentes ao ataque de pragas e doenças (BARBOSA
et al., 2003).
O estudo da dinâmica da variabilidade genética de populações de fitopatógenos é
necessário para conhecer como as populações evoluem (MCDONALD et al., 1999). A

57
estrutura genética de populações está relacionada à quantidade de variabilidade genética
existente e de sua distribuição dentro e entre populações (MILGROOM, 1995). Populações de
vírus apresentam um alto grau de diversidade genética a qual é gerada por erros durante a
replicação do genoma (GARCIA-ARENAL, 2003). No entanto, informações sobre a
variabilidade e estrutura genética de vírus de plantas com genoma de dsDNA são escassas.
O objetivo do presente trabalho foi determinar a estrutura genética das populações de
badnavírus que infectam as culturas do abacaxizeiro e da cana-de-açúcar na região Nordeste
do Brasil através da análise da sequência parcial da região RT/RNAseH da ORF3 do genoma
viral, e assim, fornecer informações que possam contribuir para o melhor entendimento da
biologia dessas espécies e auxiliar os programas de melhoramento genético existentes.
2. MATERIAL E MÉTODOS
2.1. Alinhamentos das sequências de badnavírus
Alinhamentos utilizando o software MUSCLE (EDGAR, 2004), disponível no pacote
computacional MEGA 5 (Molecular Evolutionary Genetics Analysis) (TAMURA et al.,
2011), foram realizados com 83 sequências de PBCoV obtidas de abacaxi provenientes de
cultivos comerciais de quatro estados do Nordeste do Brasil (SANTOS, 2014) e com 17
sequências de BSOLV e 15 de SCBIMV obtidas de badnavírus de cana-de-açúcar
provenientes do banco de germoplasma da Estação de Floração e Cruzamento da Cana-de-
açúcar da Serra do Ouro, em Murici, Alagoas (9º13‟S; 35º50‟W), pertencente ao Programa de
Melhoramento Genético da cana-de-açúcar (PMGCA) do Centro de Ciências Agrárias
(CECA) / Universidade Federal de Alagoas (UFAL) / Rede Interinstitucional de
Desenvolvimento do Setor Sucroalcooleiro (RIDESA) (SANTOS, 2013). As sequências
alinhadas foram utilizadas para análise da estrutura genética de populações de badnavírus.
2.2. Descritores gerais da estrutura genética de populações virais
Os principais descritores de variabilidade molecular foram estimados para as
populações virais incluindo número total de sítios segregantes (s), número total de mutações
(Eta), número médio de diferenças de nucleotídeos entre sequências (k), diversidade de
nucleotídeos (π), frequência de mutações, número de haplótipos (h), diversidade haplotípica
(Hd). Estas análises foram realizadas com o programa DnaSP v. 5 (ROZAS et al., 2009).
Adicionalmente, foi calculado o índice de fixação F de Wright (WEIR, 1996) utilizado
para detecção de subdivisão da população de badnavírus por região geográfica na cultura de
abacaxi. Ainda de acordo com esse autor, valores de Fst de 0 a 0,05 indicam pouca

58
diferenciação genética; de 0,05 a 0,15 moderada diferenciação; de 0,15 a 0,25, grande
diferenciação; e > 0,25, alta diferenciação.
2.3. Parametrização de mecanismos evolutivos
Três tipos de testes de neutralidade foram empregados para testar a hipótese de
ocorrência de seleção em populações utilizando o programa DnaSP (ROZAS et al., 2009):
Tajima‟s D, Fu & Li‟s D* e F*. O teste de Tajima é utilizado para testar a hipótese de que
todas as mutações são seletivamente neutras e leva em consideração a diversidade de
sequência, sendo que a base deste teste é que os sítios segregantes e o número médio de
diferenças par a par divergem significativamente quando sob seleção. Valores negativos de
Tajima D representam um excesso de polimorfismo de baixa frequência relacionado ao
esperado, indicando expansão no tamanho da população (por exemplo, após um efeito
gargalo) e/ou seleção purificadora, enquanto que valores positivos de Tajima D representam
baixos níveis de polimorfismos de baixa e alta frequência, indicando decréscimo no tamanho
da população e/ou seleção balanceadora (TAJIMA, 1989). Já os testes Fu & Li D e F
baseiam-se na frequência de mutações, de forma que o teste D avalia o excesso de mutações
recentes e o teste F avalia excesso de mutações antigas (FU & LI, 1993). Para comprovar a
hipótese de neutralidade o valor esperado para os testes Fu & Li D e F é zero, e ambos os
desvios positivos ou negativos são característicos de forças demográficas e/ou eventos
seletivos semelhantes aos encontrados para o teste Tajima D, atuando na população (BISWAS
& AKEY, 2006).
Para detectar sítios de aminoácidos sob seleção positiva ou negativa e as taxas de
substituições sinônimas e não-sinônimas (dN/dS), foi analisado o domínio da Transcriptase
Reversa usando o método o single-likelihood ancestor counting (SLAC) implementado no
servidor DataMonkey (DELPORT et al., 2010). A razão dN/dS é esperada exceder uma
unidade quando a seleção natural promove mudanças na sequência de proteínas (seleção
diversificadora), enquanto numa razão mais baixa que uma unidade é esperada se a seleção
natural suprime mudanças na proteína (seleção purificadora ou negativa) (YANG &
BIELAWSKI, 2000). O esperado para regiões neutras é que acumulem ambos os tipos de
substituições (sinônimas e não sinônimas) em uma taxa similar (HUGHES & NEI, 1988).
3. RESULTADOS E DISCUSSÃO
3.1. Descritores gerais da estrutura genética de populações virais

59
A subdivisão das populações de PBCoV foi estimada utilizando os testes estatísticos
Fst e Nst. As análises de diferenciação confirmaram subdivisão por região geográfica para o
PBCoV (Tabela 1).
O índice de fixação F de Wright é uma medida da proporção de variação genética total
contida numa subpopulação em relação à variação genética total. Os valores podem variar
entre 0 e 1 e valores Fst > 0,05 (5%) sugerem um certo grau de diferenciação entre as
subpopulações (WRIGHT, 1951). As populações de PBCoV provenientes de Arapiraca e
Coruripe apresentaram a menor diferenciação genética entre si (0,085), enquanto as de
Arapiraca e Pombos apresentaram a mais alta diferenciação genética (0,529), como mostra a
Tabela 1, indicando que há diferenciação entre as populações e que, portanto, as mesmas não
podem ser tratadas como uma única população. Este resultado corrobora com as análises
filogenéticas realizadas por SANTOS (2014), onde a maioria dos isolados de PBCoV
provenientes dos municípios de Arapiraca e Coruripe, ambos no Estado de Alagoas,
compartilha o mesmo clado na árvore de Máxima Verossimilhança (MV). No entanto,
análises de filogeografia de badnavírus que infectam banana, cana-de-açúcar, citros e inhame
demonstram que estes tendem a agrupar, na maioria dos casos com seus hospedeiros, em
detrimento da região geográfica (BOUSALEM et al., 2009; VARMA, 2012). Este resultado
se deve, provavelmente, à propagação vegetativa dessas espécies que permite a troca de
material propagativo infectado entre áreas de cultivo, aliada à baixa taxa de transmissão do
vírus pelas cochonilhas.
As análises da estrutura das populações de PBCoV, BSOLV e SCBIMV obtidas nesse
estudo foram comparadas com uma população de DBALV, proveniente de inhame da região
Nordeste do Brasil (GUIMARÃES, 2013). Observou-se que o PBCoV e DBALV
apresentaram um alto grau de variabilidade genética, porém menor que os vírus obtidos de
cana-de-açúcar. Por exemplo, os valores para diversidade nucleotídica são 0,0350 para
PBCoV, 0,0733 para BSOLV, 0,1516 para SCBIMV e 0,0534 para DBALV (Tabela 2). A
alta variabilidade observada para badnavírus que infectam abacaxi, cana-de-açúcar e inhame
no Nordeste do Brasil é consistente com aquela encontrada para badnavírus de banana
(JAMES et al., 2011), cacau (MULLER & SACKEY, 2005), cana-de-açúcar (MULLER et al.,
2011), inhame (LIMA et al., 2013) e taro (YANG et al., 2003).
A frequência de mutação determinada para população total e para todas as
subpopulações do PBCoV foi da ordem de 10-4
substituições/sítio/ano, assim como
encontrada para DBALV (GUIMARÃES, 2013). No entanto, as populações de BSOLV e
SCBIMV apresentaram frequência de mutação na ordem de 10-3
substituições/sítio/ano, sendo

60
esse resultado uma possível consequência da introdução de genótipos de cana-de-açúcar
infectados com badnavírus procedentes de diversos países, e em diferentes períodos, no
Banco de germoplasma da Estação de Floração e Cruzamentos da Serra do Ouro (Tabela 2).
Vírus de dsDNA frequentemente evoluem através de associações de co-divergência
com seus hospedeiros, onde um padrão de baixas taxas de substituição de nucleotídeos são
esperadas (FIRTH et al., 2010). Taxas de evolução molecular estimada para herpes simplex
virus, HSV (3,5x10-8
) e human papiloma virus, HPV (4,5x10-7
) são baixas, reforçando a
hipótese de co-divergência com populações humanas (ONG et al., 1993; SAKAOKA et al.,
1994). No entanto, as frequências de mutação encontradas tanto para as populações de
PBCoV, BSOLV e SCBIMV obtidas neste estudo, como para o DBALV são similares
àquelas encontradas para vírus de RNA e ssDNA. Estes resultados corroboram as altas
frequências de mutação, também na ordem de 10-4
subs/sítio/ano determinadas para duck
hepatites B virus (DHBV), hepatites B virus, (HBV), os quais infectam aves e humanos; e
Cauliflower mosaic virus (CaMV) que infecta plantas, os quais também possuem genoma de
dsDNA e replicam via um intermediário de RNA utilizando uma transcriptase reversa
(YASAKA et al., 2014; ZHOU & HOLMES, 2007).
A alta variabilidade genética de badnavírus tem sido atribuída à replicação inacurada
por transcrição reversa presente em Caulimoviridae (BOUSALEM et al., 2009). Embora não
seja conhecida a medida precisa da fidelidade da transcriptase reversa (RT) codificada pelos
vírus que pertencem às famílias Caulimoviridae e Hepadnaviridae, acredita-se que as altas
taxas de mutação encontradas para esses vírus se deve à perda da atividade de correção de
erros de suas RTs. Transcriptases reversas são conhecidas por serem altamente tendenciosas à
produzirem erros em todos os retrovírus e outros retroelementos para os quais a taxa de
fidelidade foi estimada (DRAKE et al., 1998; SVAROVSKAIA et al., 2003).
3.2. Parametrização de mecanismos evolutivos
Testes de neutralidade foram usados para avaliar os tipos de seleção que estão atuando
sobre a sequência codificadora do domínio RT da ORF III de BSOLV, PBCoV e SCBIMV.
Valores negativos para Tajima D, Fu & Li D e Fu & Li F foram obtidos para as populações de
BSOLV e PBCoV, porém estes não foram suportados estatisticamente a p < 0.10. A
população de SCBIMV apresentou valores positivos nos testes de Fu & Li D e F (Tabela 3).
Os valores negativos encontrados para os testes Tajima D, Fu & Li D e Fu & Li F
sinalizam que o domínio da transcriptase reversa das populações de BSOLV e PBCoV estão
sob seleção purificadora ou expansão demográfica recente. Enquanto que a população de

61
SCIMV apresentou valores negativos para o teste Tajima D, indicando possível ocorrência de
seleção purificadora ou expansão demográfica recente. No entanto, os valores estimados para
os testes Fu & Li D e F, foram positivos.
O método SLAC detectou valores de dN/dS < 1, e 47, 34 e 19 sítios foram encontrados
envolvidos sob seleção negativa versus nenhum sítio envolvido sob seleção positiva em
PBCoV, BSOLV e SCBIMV, respectivamente. Estes resultados são um indício de seleção
purificadora (negativa) agindo nesta população (Tabela 4), corroborando os resultados
encontrados para os testes Tajima D, Fu & Li D e Fu & Li F.
Durante o ciclo de infecção viral, vários fatores podem impor seleção purificadora na
população. A biologia do vetor e seus hábitos alimentares criam pontos de estrangulamento
para a manutenção das características estruturais e funcionais das proteínas virais. A interação
com fatores do hospedeiro também pode impor seleção purificadora (GARCÍA-ARENAL et
al., 2001). A presença de seleção purificadora no domínio RT/RNAse H, enfatiza a função
crítica realizada por esta região e seu requerimento para sobrevivência de PBCoV, BSOLV e
SCBIMV, e portanto limitado número de substituições não sinônimas de nucleotídeos era
esperado. Seleção purificadora e expansão populacional também foram as principais forças
evolutivas atuando sobre a população de DBALV ocorrendo no nordeste brasileiro
(GUIMARÃES, 2013). Mecanismos evolutivos como efeito gargalo induzidos por
populações locais do vetor ou hospedeiros, efeito fundador e eventos de mudança de
hospedeira tem, provavelmente, contribuído para variabilidade genética de badnavírus de
inhame (GARCIA-ARENAL et al., 2001; BOUSALEM et al., 2009). Já para populações de
CaMV, mutação, recombinação e fluxo gênico são as principais forças evolutivas (YASAKA
et al., 2014). Contudo, a ocorrência de mutações não é suficiente para explicar completamente
a variabilidade genética de PBCoV, BSOLV e SCBIMV, reforçando a possível influência de
outras forças evolutivas como migração e recombinação atuando sobre estas populações.
4. CONCLUSÕES
A estrutura genética das populações de Pineapple bacilliform CO virus, Banana streak
OL virus e Sugarcane bacilliform IM virus é provavelmente consequência de diferentes
mecanismos os quais incluem seleção purificadora, expansão geográfica e adaptação
evolutiva dos vírus a diferentes populações de hospedeiras e vetor. A variabilidade genética é
alta e reflete a capacidade dessas populações evoluírem rapidamente, principalmente em
resposta a variações das populações locais de hospedeiras, sobretudo para BSOLV e
SCBIMV. Portanto, deve-se considerar a variabilidade genética dessas três espécies em casos

62
de implantação de medidas de controle baseadas em resistência e métodos diagnósticos
baseados em sequência de DNA, uma vez que ambos os métodos poderiam falhar devido a
um longo tempo de exposição contínua as rápidas taxas de mutação desta população.
5. REFERÊNCIAS BIBLIOGRÁFICAS
ANDRADE, G.P. Diagnóstico fitossanitário da cultura do inhame (Dioscorea spp.) em
áreas produtoras do Nordeste do Brasil. 2007. 88 f. Tese (Doutorado em Fitopatologia) –
Universidade Federal Rural de Pernambuco, Recife.
BARBOSA, G. et al. Três novas variedades RB de cana-de-açúcar. Rio Largo, 2003, 17p.
(BOLETIM TÉCNICO, 1).
BISWAS, S.; AKEY, J.M. Genomic insights into positive selection. Trends in Genetics,
v.22, p.437- 446, 2006.
BOUHIDA, M.L. et al. An analysis of the complete sequence of a sugarcane bacilliform virus
genome infectious to banana and rice. Journal of General Virology, v.74, p.15-22, 1993.
BOUSALEM, M. et al. Dilemmas caused by endogenous pararetroviruses regarding the
taxonomy and diagnosis of yam (Dioscorea spp.) badnaviruses: analyses to support safe
germplasm movement. Archives of Virology, v.154, p.297-314, 2009.
DANIELS, J.; ROACH, B.T. Taxonomy and Evolution. In: HEINZ, D.J. (ed.). Sugarcane
improvement through breeding. Elsevier, Amsterdam, 1987. p.7-84.
DELPORT, W. et al. Datamonkey 2010: a suite of phylogenetic analysis tools for
evolutionary biology. Bioinformatic, v.19, p.2455-7, 2010. Acesso em: 10 jan. 2014.
DRAKE, R.R.et al. Identification of a nucleotide binding site in HIV-1 integrase.
Proceedings of the National Academy of Sciences, v.95, p.4170-4175, 1998.
EDGAR, R.C. MUSCLE: A multiple sequence alignment method with reduced time and
space complexity. BMC Bioinformatics, v.5, p.1-19. 2004.
FAO. FAO – Food and Agriculture Organization. Faostat. Disponível em:
<http://faostat3.fao.org>. Acesso em: 09 de jan. 2014.
FAUQUET, C. et al. Virus taxonomy. Eight Report of the International committee on
Taxonomy of Viruses. Amsterdam. Elsevier. 2005.
FIRTH, C. et al. Using time-strutured data to estimate evolutionary rates of double-stranded
DNA viruses. Molecular Biology and Evolution, v.27, p.2038-2051, 2010.
FU, Y.; Li, W. H. Statistical tests of neutrality of mutations. Genetics, v.133, p.693-709,
1993.
GAMBLEY, C.F. et al. Identification of viral and non-viral reverse transcribing elements in
pineapple (Ananas comosus), including members of two new badnavirus species. Archives of
Virology, v.153, p.1599–1604, 2008.

63
GARCIA-ARENAL, F. et al. Variability and genetic structure of plant virus populations.
Annual Reviews of Phytopathology, v.39, p.157–186, 2001.
GARCIA-ARENAL, F. et al. Variation and evolution of plant virus populations.
International Microbiology, v.6, p.225-232. 2003.
GAUHL, F. et al. First report of banana streak badnavirus in plantain landraces in southern
Cameroon, Central Africa. Plant Disease, v.81, p.1335-1335, 1997.
GEERING, A.D.W.; HULL, R. Family Caulimoviridae. In: KING A.M.Q.; ADAMS M.J.;
CARSTENS E.B.; LEFKOWITZ, E.J. Virus Taxonomy. 9th Report of the International
Committee on Taxonomy of Viruses. London UK. Elsevier Academic Press. p.429-443,
2012.
GUIMARÃES, K.M. C. Estrutura genética de uma população do badnavirus Dioscorea
bacilliform alata virus (DBALV) que infecta inhame (Dioscorea spp.) no Nordeste do
Brasil. 2013. 47 f. Dissertação (Mestrado em Proteção de Plantas) – Universidade Federal de
Alagoas.
HAGEN, L.S. et al. Nucleotide sequence and genomic organisation of cacao swollen shoot
virus. Virology, v.196, p.619-628, 1993.
HARPER, G.; HULL, R. Cloning and sequence analysis of Banana Streak Virus DNA. Virus
Genes, v.17, p.271-278, 1998.
HUGHES, A.L.; NEI, M. Pattern of nucleotide substitution at major histocompatibility
complex class I loci reveals overdominant selection. Nature, v.335, p.167–170, 1988.
JAMES, A.P. et al. Development of a Novel Rolling - Circle Amplification Technique to
Detect Banana streak virus that also Discriminates Between Integrated and Episomal Virus
Sequences Plant Disease, v.95, p.57-62, 2011.
LACO, G.S.; BEACHY, R.N. Rice tungro bacilliform virus encodes reverse transcriptase,
DNA polymerase and ribonuclease H activities. Proceedings of the National Academy of
Sciences USA, v.91, p.2654-2658, 1994.
LIMA, J.S. et al. Variabilidade genética de isolados de badnavírus infectando inhame
(Dioscorea spp.) no Nordeste do Brasil. Tropical Plant Pathology, v.38, p.349-353, 2013.
LOCKHART, B.E.L.; AUTREY, L.J. C. Occurrence in sugarcane ofa bacilliform virus
related serologically to banana streak virus. Plant Disease v.72, p.230-233, 1988.
MCDONALD, B. A. et al. Genetic structure of Rhynchosporium secalis in Australia.
Phytopathology, v.89, p.639-45, 1999.
MEDBERRY, S.L. et al. Properties of Commelina yellow mottle virus complete DNA
sequence, genomic discontinuities and transcript suggest that it is a pararetrovirus. Nucleic
Acids Research, v.18, p.5505-5513, 1990.
MILGROOM, M.G. Analysis of population structure in fungal plant pathogens. In: Leslie, J.
F. e Frederiksen, R. A. (Eds.) Disease analysis through genetics and biotechnology. Iowa
State University, Ames. p.213-229, 1995.
MULLER, E.; SACKEY, S. Molecular variability analysis of five new complete cocoa
swollen shoot virus genomic sequences. Archives of Virology, v.150, p.53-66, 2005.

64
MULLER, E. et al. High molecular variability of sugarcane bacilliform viruses in
Guadeloupe implying the existence of at least three new species. Virus Research, v.160,
p.414 - 419, 2011.
ONG, C.K. et al. Evolution of human papillomavirus type 18: an ancient phylogenetic root in
Africa and intratype diversity reflect coevolution with human ethnic groups. Journal of
Virology, v.67, p.6424–6431, 1993.
PHILLIPS, S. et al. The Partial Characterization of a Badnavirus Infecting the Greater Asiatic
or Water Yam (Dioscorea alata). Journal of Phytopatholology, v.147, p.265-269, 1999.
REINHARDT, D.H.R.C.; CUNHA, G.A.P. Métodos de Propagação. In: CUNHA, G.A.P. et
al. O Abacaxizeiro. Cultivo, agroindústria e economia. Embrapa Mandioca e Fruticultura
(Cruz das Almas, BA). Brasília: Embrapa Comunicação para Transferência de Tecnologia,
1999. cap.5, p.105-138.
RODRIGUEZ-LEMA, E. et al. Reporte de un nuevo vírus de la cana de azucar. Ciencias de
la Agricultura, México, v. 23, p. 130, 1985.
ROZAS, et al. DnaSP v5: A software for comprehensive analysis of DNA polymorphism
data. Bioinformatics, v.25, p.1451-1452, 2009.
SÁ, L.F. Cultura do abacaxi. Goiânia, Empresa de Assistência Técnica e Extensão Rural,
EMATER-GO, 1994. 18p. (BOLETIM TÉCNICO, 1).
SAKAOKA, H. et al. Quantitative analysis of genomic polymorphism of herpes simplex virus
type 1 strains from six countries: studies of molecular evolution and molecular epidemiology
of the virus. Journal of Geneneral Virology, v.75, p.513–527, 1994.
SANTOS, E.M.S.M. Incidência, caracterização e estrutura genética de badnavírus nas
culturas do abacaxizeiro e da cana-de-açúcar no Nordeste do Brasil. 2014. 78 f. Tese
(Doutorado em Fitopatologia) – Universidade Federal Rural de Pernambuco.
SANTOS, J.M.S. Incidência e caracterização de badnavírus no banco de germoplasma
de cana-de-açúcar (Saccharum spp.) na Serra do Ouro, Murici-AL. 2013. 54 f.
Dissertação (Mestrado em Proteção de Plantas) - Universidade Federal de Alagoas.
STADEN, R. et al. The Staden package, 1998. In MISENER, S.; KRAWETZ, S.A.
(eds). Bioinformatics methods and protocols Humana, New York, p.115-130, 1998.
SVAROVSKAIA, E.S. et al. Retroviral mutation rates and reverse transcriptase fidelity.
Fronties in Bioscience, v.8, p.117–134, 2003.
TAJIMA, F. Statistical method for testing the neutral mutation hypothesis by DNA
polymorphism. Genetics, v.123, p.585-595. 1989.
TAMURA, K. et al. MEGA 5: Molecular evolutionary genetics analysis using maximum
likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and
Evolution, v.28, p.2731 – 2739, 2011.
VARMA, A.; PRAVEEN, S. Phylogeographic evolution of plant virus. In: SHARMA, V. P.
Nature at work ongoing saga of evolution. India: Sringer, 2010. p.75–92.
WEIR, B.S. Genetic data analysis II: methods for discrete population genetic data. Sinauer
Associated Inc., Sunderland, MA, 1996.

65
WRIGHT, S. The genetical structure of populations. Annals of Eugenics, v.15, p. 23-354,
1951.
YANG, I.C. et al. Sequence diversity of South Pacific isolates of Taro bacilloform virus and
the development of a PCR-based diagnostic test. Archives of Virology, v.148, p.1957-1968,
2003.
YANG, Z.; BIELAWSKI, J.P. Statistical methods for detecting molecular adaptation. Trends
in Ecology and Evolution, v.15, p.496-503, 2000.
YASAKA, R., et al. The Temporal Evolution and Global Spread of Cauliflower mosaic virus,
a Plant Pararetrovirus. PLOS One, v.9, p.1–12, 2014.
ZHOU, Y.; HOLMES, E.C. Bayesian estimates of the evolutionary rate and age of hepatitis
B virus. Journal of Molecular Evolution, v.65, p.197–205, 2007.
ZHOU, X. et al. Evidence that DNA-A of a geminivirus associated with severe cassava
mosaic disease in Uganda has arisen by interspecific recombination. Journal of General
Virology, v.78, p.2101-2111, 1997.

66
Tabela 1. Resultados de testes de subdivisão na população de Pineapple bacilliform CO virus
do Nordeste do Brasil, de acordo com a origem geográfica.
Valores de 0 a 0,05 indicam baixa diferenciação genética; valores de 0,05 a 0,15 indicam moderada
diferenciação; valores de 0,15 a 0,25 indicam grande diferenciação e valores > 0,25 indicam alta diferenciação.
População Nst Fst
Arapiraca / Coruripe 0,086 0,085
Arapiraca / Pedras de Fogo 0,386 0,382
Arapiraca / Pombos 0,536 0,529
Arapiraca / São Domingos do Maranhão 0,511 0,505
Coruripe / Pedras de Fogo 0,292 0,288
Coruripe / Pombos 0,370 0,365
Coruripe / São Domingos do Maranhão 0,367 0,362
Pedras de Fogo / Pombos 0,117 0,116
Pedras de Fogo / São Domingos do Maranhão 0,115 0,114
Pombos / São Domingos do Maranhão 0,122 0,121
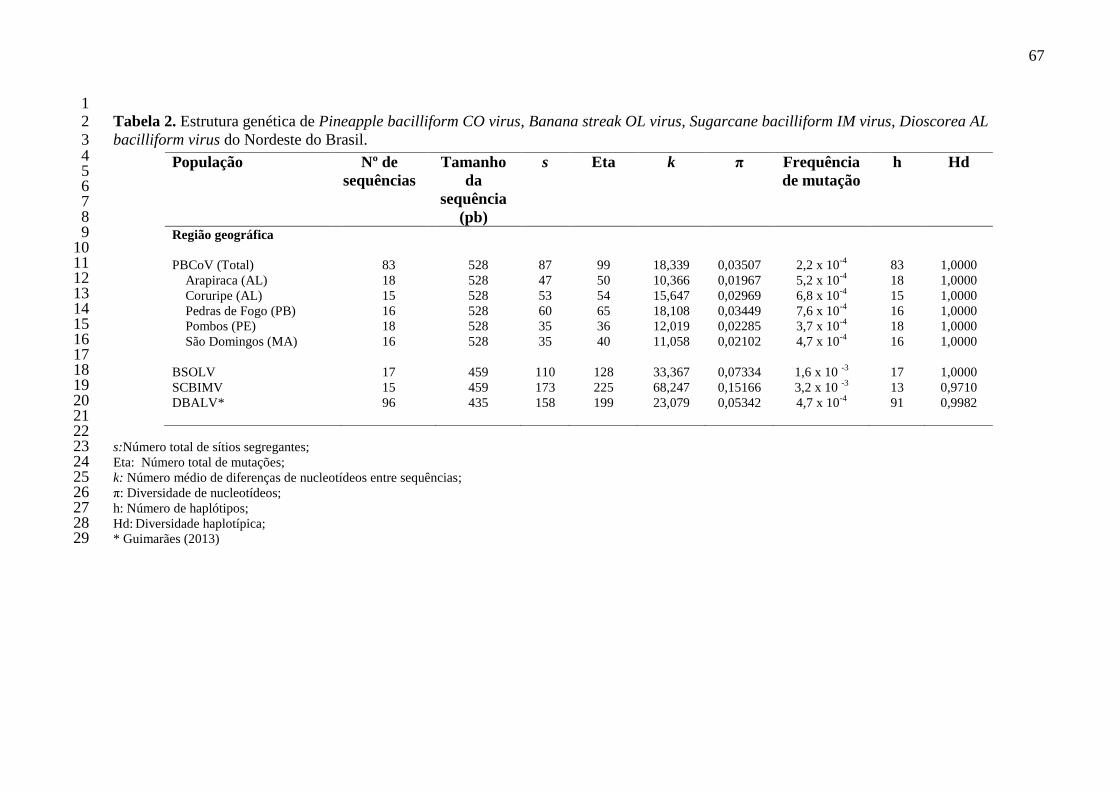
67
1 Tabela 2. Estrutura genética de Pineapple bacilliform CO virus, Banana streak OL virus, Sugarcane bacilliform IM virus, Dioscorea AL 2
bacilliform virus do Nordeste do Brasil. 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 s:Número total de sítios segregantes; 23 Eta: Número total de mutações; 24 k: Número médio de diferenças de nucleotídeos entre sequências; 25 π: Diversidade de nucleotídeos; 26 h: Número de haplótipos; 27 Hd:
Diversidade haplotípica; 28
* Guimarães (2013) 29
População Nº de
sequências
Tamanho
da
sequência
(pb)
s Eta k π Frequência
de mutação
h Hd
Região geográfica
PBCoV (Total)
Arapiraca (AL)
Coruripe (AL)
Pedras de Fogo (PB)
Pombos (PE)
São Domingos (MA)
BSOLV
SCBIMV
DBALV*
83
18
15
16
18
16
17
15
96
528
528
528
528
528
528
459
459
435
87
47
53
60
35
35
110
173
158
99
50
54
65
36
40
128
225
199
18,339
10,366
15,647
18,108
12,019
11,058
33,367
68,247
23,079
0,03507
0,01967
0,02969
0,03449
0,02285
0,02102
0,07334
0,15166
0,05342
2,2 x 10-4
5,2 x 10-4
6,8 x 10-4
7,6 x 10-4
3,7 x 10-4
4,7 x 10-4
1,6 x 10 -3
3,2 x 10 -3
4,7 x 10-4
83
18
15
16
18
16
17
13
91
1,0000
1,0000
1,0000
1,0000
1,0000
1,0000
1,0000
0,9710
0,9982

68
Tabela 3. Resultado dos diferentes testes de neutralidade realizados para o domínio RT da
ORF III das populações de Pineapple bacilliform CO virus (PBCoV), Banana streak OL virus
(BSOLV) e Sugarcane bacilliform IM virus (SCBIMV) do Nordeste do Brasil.
População Tajima D Fu & Li D Fu & Li F
PBCoV -0,25350 (ns) -0,10663 (ns) -0,20023 (ns)
BSOLV -0,50708 (ns) -0,12028 (ns) -0,26959 (ns)
SCBIMV -0,06078 (ns) 0,47013 (ns) 0,37019 (ns)
ns, valores não-significativos p < 0.10

69
Tabela 4. Valores médios de taxas de substituições não sinônimas e sinônimas (dN/dS) e
sítios selecionados positivamente e negativamente em populações de badnavírus usando o
método SLAC.
População dN/dS Selecionados
Positivamente Selecionados Negativamente
PBCoV 0.0423 -
1, 5, 12, 17, 19, 24, 27, 34, 36, 38, 49, 62,
67, 69, 72, 74, 76, 77, 78, 81, 85, 86, 98,
101, 102, 105, 108, 110, 112, 117, 119,
120, 122, 124, 128, 136, 142, 144, 147,
153, 154, 159, 169, 170, 171, 172, 173
BSOLV 0.0673 -
1, 5, 9, 18, 20, 23, 30, 32, 35, 37, 42, 44,
45, 47, 59, 61, 70, 71, 75, 81, 82, 83, 89,
90, 91, 107, 122, 129, 131, 135, 136, 137,
138, 151
SCBIMV 0.1535 - 1, 5, 8, 9, 13, 31, 33, 35, 36, 47, 69, 73, 79,
101, 113, 130, 133, 144, 146

70
CONCLUSÕES GERAIS
Este constitui o primeiro registro de infecção por Badnavirus na cultura do
abacaxizeiro no país;
A presença de Pineapple bacilliform CO virus em Alagoas, Maranhão, Paraíba e
Pernambuco, sugere uma ampla distribuição e prevalência dessa espécie em todas as
áreas amostradas da região Nordeste do Brasil;
A estrutura genética das populações de Pineapple bacilliform CO virus, Banana streak
OL virus e Sugarcane bacilliform IM virus é provavelmente consequência de
diferentes mecanismos os quais incluem seleção purificadora, expansão geográfica e
adaptação evolutiva dos vírus a diferentes populações de hospedeiras e vetor;
A variabilidade genética é alta e reflete a capacidade dessas populações evoluir
rapidamente principalmente em resposta a variações das populações locais de
hospedeiras, sobretudo para Banana streak OL virus e Sugarcane bacilliform IM virus.