identificaÇÃo e caracterizaÇÃo parcial de ... - uenf.br · um ph ótimo neutro variando de 40 a...
TRANSCRIPT

IDENTIFICAÇÃO E CARACTERIZAÇÃO PARCIAL DE
PROTEASES E SEUS SUBSTRATOS DE FLUIDO
EPIDIDIMÁRIO DE Equus caballus
NATHÁLIA CURTY DE ANDRADE
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO
CAMPOS DOS GOYTACAZES-RJ
MARÇO - 2008

NATHÁLIA CURTY DE ANDRADE
IDENTIFICAÇÃO E CARACTERIZAÇÃO PARCIAL DE
PROTEASES E SEUS SUBSTRATOS DE FLUIDO
EPIDIDIMÁRIO DE Equus caballus
Dissertação apresentada ao Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte dos requisitos para a obtenção do título de Mestre em Biociências e Biotecnologia, com ênfase em Biologia Celular.
ORIENTADOR: Dr. CLAUDIO ANDRÉS RETAMAL MARTÍNEZ
CO-ORIENTADORA: Drª. MARIA LUISA LÓPEZ ALVAREZ
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
CAMPOS DOS GOYTACAZES-RJ
2008

NATHÁLIA CURTY DE ANDRADE
IDENTIFICAÇÃO E CARACTERIZAÇÃO PARCIAL DE
PROTEASES E SEUS SUBSTRATOS DE FLUIDO
EPIDIDIMÁRIO DE Equus caballus
Dissertação apresentada ao Centro de Biociências e Biotecnologia da Universidade Estadual do Norte Fluminense Darcy Ribeiro, como parte dos requisitos para a obtenção do título de Mestre em Biociências e Biotecnologia, com ênfase em Biologia Celular.
Aprovada em ____ de Março de 2008
...................................................................... Dr. Milton Masahiko Kanashiro
......................................................................
Drª. Kátia Valevski Sales Fernandes
................................................................. Dr. Marco Antônio de Oliveira
..................................................... Dr. Claudio Retamal (orientador)
.....................................................
Drª. Maria Luisa Lopez Alvarez (co-orientadora)

i
Dedico, aos meus pais: Mariléa
e Bertoni: Vocês me
transmitiram os valores mais
importantes: Amor, amizade,
carinho, compreensão, perdão,
companheirismo, honestidade e
paciência. Obrigada!!!

ii
Agradecimentos
Primeiramente a DEUS, por oferecer seu amor, por todas as bênçãos que me
concedeu e por ter me permitido chegar até aqui.
À minha família: meu pai e minha mãe, Ronaldo Carpi, meus irmãos que, em
todos os momentos, me apoiaram e me fortaleceram com muito amor e
carinho. Obrigada por existirem em minha vida!
A Marco Augusto Seabra por oferecer seu amor, paciência, apoio e carinho,
muito obrigada.
Ao professor Claudio Andrés Retamal Martínez, pela orientação, grande
amizade e confiança.
À professora Maria Luisa López, pela amizade e orientação.
À amiga Thaisa Domingos que desde o primeiro dia de aula somos como
irmãs. Muito obrigada pretinha!
Aos amigos de laboratório: Paula Amorin, Roberta Fernandes, Glauber Dias,
Joseph Albert e Renata Vasconcelos. Obrigada pela amizade,
companheirismo, pelos ensinamentos e paciência comigo!
Ao técnico do laboratório Arthur Rodrigues pela amizade e auxílio na
preparação de soluções.
Às amigas: Ana Carolina Mercadante, que me deu apoio nas horas difíceis e
pelas risadas nos momentos de descontração.
Ao pessoal do LQFPP, LBT, LBR e do CCTA pelo apoio.
À TECNORTE, FAPERJ, CNPq e UENF pelo auxílio financeiro.
E a todos que de alguma forma contribuíram para o meu trabalho e que
infelizmente não conseguiria listar a todos, muito obrigada.

iii
RESUMO
Os espermatozóides de mamíferos sofrem uma seqüência de mudanças
estruturais durante sua passagem pelo epidídimo, adquirindo
concomitantemente, um padrão motilidade maduro e capacidade fertilizante.
Algumas dessas modificações podem resultar de um processamento
proteolítico específico. Em contraste com as proteases do testículo e de células
espermáticas, as proteases epididimárias têm sido menos estudadas. Assim, o
objetivo deste trabalho foi caracterizar a atividade proteolítica no fluido
epididimário de cavalo. Amostras coletadas das diferentes regiões
epididimárias foram submetidas a eletroforese em géis de gelatina incubados
com diferentes inibidores e pHs, sob condições redutoras e não redutoras.
Tendo verificado uma migração anómala de proteases em SDS-PAGE com
gelatina, as massas moleculares aqui relatadas foram calculadas com base em
géis 2D (SDS-PAGE/SDS-PAGE 0,4% gelatina). Nossos resultados mostraram
uma distribuição regionalizada no epidídimo de metaloproteases que possuem
um pH ótimo neutro variando de 40 a 117 kDa. Na região epididimária de
cabeça proximal foram detectadas proteases de 117, 100 e 62 kDa. Na cauda
epididimária, as principais bandas de atividade foram proteínas de 117, 62, 45 e
40 kDa. O comportamento da enzima com e sem β-mercaptoetanol reflete a
existência de pontes dissulfeto e sugere a possibilidade de interações
transitórias entre proteases ou com os seus substratos/inibidores. Os
substratos endógenos foram identificados através da adição das atividades
proteolíticas isoladas de cauda a este mesmo fluido e foi observado a
diminuição da concentração das proteínas de 117, 80, 69, 57, 50, 44, 35, 68 e
60 kDa, indicando que sejam alvo da atividade proteásica. A proteína de 66
kDa se mantém o que permite descartar-la como substrato de proteases. O Rf
de algumas proteínas se altera e também aumenta a concentração de algumas
bandas (manchas) como é caso das proteínas de massas moleculares 66, 50,
49, 44, 35, 33,5 e 32,8 kDa. Analisados em conjunto, esses dados sugerem
que podemos estar frente a substratos e produtos resultantes da ação
enzimática. Isto sugere o papel biológico destas enzimas como reguladoras de
proteínas presentes em uma forma inativa e que, após seu processamento no
fluido, tornam-se ativas.

iv
ABSTRACT
Mammalian spermatozoa undergo a sequence of structural changes as
they pass through epididymis, acquiring concomitantly a mature motility pattern
and fertilizing ability. Some of these modifications would result from a specific
proteolytic processing. In contrast to the proteases of the testis and sperm cell,
epididymal proteases have been less studied. The aim of this work was to
characterize the proteolytic activities detected in stallion epididymal fluid.
Samples collected from different epididymis regions were submitted to gelatin
gel under nonreducing or reducing conditions and treated with different
protease inhibitors and pHs. Having verified an anomalous migration of
proteases in SDS-PAGE with gelatin, and molecular weights here reported were
calculated based on 2D SDS-PAGE/SDS-PAGE 0.4% gelatin gels. Our results
showed a regionalized distribution of proteolytic activity in the epididymis, which
has a neutral pH, and allowed metalloproteases detection ranging from ~40 to
117 kDa. In the proximal caput epididymidis 117, 100 and 62 kDa proteases
were detected. In the tails epididymidis the main reactive band were 117, 62, 45
and 40 kDa kDa proteins. The enzyme behavior with and without β-
mercaptoethanol likely reflects sulfhydryl cross-linking and suggests the
possibility of transient interactions, between proteases or with their
substrates/inhibitors. The endogenous substrates were identified through the
isolated proteolytic activity from tail added to this same fluid and showed that
proteins of molecular weight 117, 80, 69, 57, 50, 44, 35, 68 and 60 kDa
disappear, indicating that they are targets of protease activity. A protein of 66
kDa remains allowing discard it as a substrate for proteases. The Rf of certain
proteins changes also appear in new bands (spots) such as molecular weights
of proteins of 66, 50, 49, 44, 35, 33.5 and 32,8 kDa. Together, these data
suggest that we may be facing the substrates and products resulting from the
enzymatic action. This suggest the biological role of these enzymes, like
regulate proteins present in an inactive form and become functional after its
processing in the fluid.

v
Este trabalho foi desenvolvido no
laboratório de biologia celular e
tecidual, no setor biologia da
reprodução do centro de
biociências e biotecnologia da
Universidade Estadual do Norte
Fluminense sob orientação dos
pesquisadores Cláudio Andrés
Retamal e Maria Luisa López
Alvarez.
Apoio financeiro:
CNPq
UENF
FAPERJ
TECNORTE

vi
Lista de abreviaturas:
PH-20- Proteína com atividade hialuronidase de 30kDa.
PH30- Proteína com atividade hialuronidase de 30kDa.
HE4- Proteína relacionada à fertilida.
CRES - Cistatina relacionada com a espermatogênese epididimária.
PC4- Co-ativador transcricional p15 (PC4).
MMPs- Metaloproteases de matriz.
TIMPs- Inibidores de Metaloproteases Tecido Específicas.
BSp66- Protease de Espermatozóide Bovino de 66kDa.
MT-MMP- Metaloproteínas de membrana.
ADAM- Proteínas com um domínio desintegrina e metaloprotease.
FN- Fibronectina.
mmp- genes de MMPs.
JUN- Oncogene celular Jun.
FOS- Oncogene celular FOS.
AP1- Complexo ativador de proteínas 1.
MAPK- Proteínas quinases ativadas por mitógenos.
ERKs- Proteínas quinases reguladas por sinais extracelulares.
JNK- Proteína quinase do N-terminal de JUN ativada por estresse.
P38- Proteína quinase p38 ativada por mitógenos (MAPK).
PKC- Proteína quinase C.
EGF- Fator de crescimento epitelial.
MDC9- Metaloprotease desintegrina MDC9 (ADAM9).
BSA- Albumina sérica bovina.
SDS - Dodecil sulfato de sódio.
PAGE - Eletroforese em gel de poliacrilamida.
HCl- Ácido clorídrico.
H2O- Água.
CaCl2 - Cloreto de cálcio.
NaCl- Cloreto de sódio.
PMSF- Fenil metil sulfonil fluoreto.
EDTA - Ácido etilenodiaminotetraacético.
RNA - Ácido Desoxirribonucléico.

vii
Kr- Constante de retardação na mobilidade eletroforética.
Rf - Mobilidade relativa.
1D- Eletroforese unidimensional (primeira eletroforese).
2D- Eletroforese bidimensional (segunda eletroforese).
UV- Luz ultra-violeta
PBS- Tampão fosfato-salino.
anti-MMP- Anticorpo anti- metaloproteases de matriz.
ScPrB- Protease Schizophyllum commune B (metaloprotease).
CE9- Glicoproteína integral de membrana plasmática chamada CE9.

1
ÍNDICE
I. INTRODUÇÃO .............................................................................................................. 3
II. REVISÃO BIBLIOGRÁFICA .................................................................................... 5
1. Epidídimo e o processo de maturação espermática ....................................................... 5
2. Proteases ..................................................................................................................... 7
3. Metaloproteases de Matriz Extracelular (MMPs) ....................................................... 10
4. Maturação espermática e atividade proteolítica .......................................................... 14
III. OBJETIVOS ............................................................................................................. 16
1. Objetivo geral ............................................................................................................ 16
2. Objetivos específicos ................................................................................................. 16
IV. MATERIAIS E MÉTODOS ...................................................................................... 17
1. Obtenção das amostras .............................................................................................. 17
2. Determinação da concentração de proteínas ............................................................... 18
3. Análise do perfil eletroforético de amostras de fluido epididimário ............................ 18
4. Detecção de proteólise endógena no fluido epididimário ........................................... 18
5. Detecção de atividade metaloproteásica ..................................................................... 20
6. Ação de inibidores sobre as atividades proteolíticas ................................................... 20
7. Verificação do pH ótimo ........................................................................................... 20
8. Determinação do grau de anomalia das proteases na migração eletroforética ............. 21
9. Determinação das massas moleculares das proteases de fluido epididimário .............. 22
10. Purificação das atividades proteolíticas detectadas no fluido epididimário de cauda 24
10.1 Purificação das atividades proteolíticas a partir de géis unidimensionais ....................... 24
10.2 Purificação das atividades proteolíticas a partir de géis 2D SDS/SDS com gelatina ...... 25
10.3 Purificação das atividades proteolíticas a partir de géis nativos 10% ............................. 26
11. Detecção e identificação de substratos endógenos de fluido de cauda com atividades proteolíticas purificadas desta mesma região .................................................................... 28
12. Purificação dos substratos endógenos das atividades proteolíticas ........................... 29
12.1 Purificação dos substratos endógenos, presentes em fluido de cauda, a partir de géis nativos 10% ......................................................................................................................... 29
12.2 Purificação dos substratos endógenos, presentes em fluido de cauda, a partir de géis nativo/SDS-PAGE 12% ....................................................................................................... 31
13. Identificação dos produtos endógenos gerados pelas proteases purificadas e substratos purificados ....................................................................................................... 33
14. Western blotting .................................................................................................... 34
15. Análise densitométrica ........................................................................................... 34
V. RESULTADOS ......................................................................................................... 35
1. Análise do perfil eletroforético de amostras de fluido epididimário ............................ 35
2. Efeito da proteólise endógena no fluido epididimário ................................................ 35
3. Detecção e caracterização de proteases epididimárias ................................................ 38
3.1 Atividade proteásica no fluido epididimário, ação de inibidores sobre as atividades proteolíticas e verificação do pH ótimo ................................................................................ 38
4. Determinação da constante de retardação de proteases: eletroforeses de proteínas epididimárias em géis desnaturantes na presença de gradiente de gelatina 0 - 0,6% .......... 41
4.1 Comparação de proteínas/atividades proteásicas em géis bidimensionais SDS/SDS e SDS/gelatina 0,4% ............................................................................................................... 45
5. Detecção da atividade proteásica de fluido epididimário na ausência e presença de agente redutor .................................................................................................................. 47
6. Determinação das massas moleculares de proteases no fluido epididimário ............... 52
7. Purificação das Proteases ........................................................................................... 54

2
7.1 Purificação de proteases a partir de géis unidimensionais ............................................... 54
7.2 Purificação de proteases a partir de géis 2D SDS/SDS com gelatina ............................... 56
7.3 Purificação de proteases a partir de géis nativos 10% ..................................................... 58
8. Detecção dos substratos endógenos ........................................................................... 61
9. Purificação dos substratos endógenos das atividades proteolíticas .............................. 69
9.1 Purificação dos substratos endógenos a partir de géis nativos 10% ................................. 69
9.2 Purificação dos substratos endógenos a partir de géis nativo/SDS-PAGE 12% ............... 69
10. Identificação dos produtos endógenos gerados pelas proteases purificadas e substratos purificado ........................................................................................................ 71
10.1 Ensaio de incubação da protease P3 purificada com substrato (66kDa) purificado a partir do gel nativo 10% ................................................................................................................ 71
10.2 Ensaio de incubação das proteases P1, P2, P3 e P4 purificadas com substrato S2 purificado a partir de gel nativo/SDS 12% ............................................................................ 73
10.3 Ensaio de incubação das proteases P1, P2, P3 e P4 purificadas a partir de géis nativo 10% com os substratos S1, S2, S3 e S8 purificados a partir do gel nativo/SDS 12% ............. 76
11. Proteólise endógena ............................................................................................... 83
12. Western Blotting .................................................................................................... 86
VI. DISCUSSÃO ............................................................................................................ 87
VII. CONCLUSÕES ........................................................................................................ 96
VIII. REFERÊNCIAS BIBLIOGRAFICAS .................................................................... 97

3
I. INTRODUÇÃO
A fertilização é um processo complexo, que envolve uma cascata de eventos
ligados que progridem em forma seqüencial e altamente regulada, finalizando com a
fusão dos pró-núcleos, integração dos genomas haplóides e ativação do programa
de desenvolvimento do zigoto que dará origem a um novo indivíduo. Os
espermatozóides passam por uma série de processos preparatórios prévios à
fertilização, entre eles maturação espermática (no conduto epididimário) e
capacitação (nas vias genitais da fêmea). Somente espermatozóides maduros,
capacitados e com acrossoma intacto penetram o cumulus atingindo a zona pelúcida
do gameta homólogo (RETAMAL, 1999).
O processo de maturação espermática compreende uma série de
modificações nas características morfológicas, bioquímicas, biofísicas e funcionais
do gameta, entre elas a aquisição da motilidade progressiva unidirecional e o
potencial fertilizante. Uma expressão deste processo é a remodelação de proteínas,
glicoproteínas e lipídeos da membrana. Devido à capacidade biossintética limitada
do espermatozóide, postula-se que estas remodelações pós-testiculares estariam
fortemente influenciadas pelo microambiente epididimário (EDDY & O´BRIAN, 1994;
LÓPEZ, 1996; RETAMAL et al., 2000 a,b). Algumas destas modificações são
conseqüências de uma clivagem pré-programada e regulada de moléculas integrais,
enquanto outras podem resultar da adição ou absorção de moléculas presentes no
fluido epididimário (AMMAN, 1995; RETAMAL et al., 2000a; DIAS et al., 2004).
Estudos prévios realizados em nosso laboratório, utilizando como modelo
experimental espermatozóides de Equus caballus, têm demonstrado diferenças na
distribuição e concentração relativa de proteínas, glicoproteínas e lipídeos de
espermatozóides imaturos e maduros, obtidos das diferentes regiões do epidídimo
(LÓPEZ et al., 1987; LÓPEZ & DE SOUZA, 1991; LÓPEZ, 1996; RETAMAL, 1999;
DIAS, 2002). Essas diferenças podem estar relacionadas com a ruptura proteolítica
de componentes pré-existentes e redistribuição dos componentes clivados em
diferentes domínios da membrana (LIU et al., 2001; RUTLLANT & MEYERS, 2001).
Como exemplo, os trabalhos de PHELPS et al. (1990) e JONES et al. (1996)
demonstraram o processamento endoproteolítico das proteínas PH20 e PH30,

4
distribuídas em toda a região da cabeça nos espermatozóides testiculares, e
exclusivamente no domínio pós-acrossomal em espermatozóides epididimários.
Esses trabalhos reforçam o conceito que algumas moléculas-chave seriam
primeiramente sintetizadas como precursores e logo se ativariam - no tempo e local
apropriado - para cumprirem sua função, permitindo assim, um grau de auto-
regulação em células que estão transcricionalmente inativas (JONES et al., 1996).
Os espermatozóides são ricos em diferentes proteases, as quais se localizam,
principalmente, na vesícula acrossomal (TULSIANI et al., 1998; MÉTAYER et al.,
2002). A manutenção do equilíbrio entre proteases e seus inibidores é fundamental
na conservação da barreira hemato-testicular e no desenvolvimento do gameta.
Manter as proteases acrossomais inativas é crucial para manutenção da integridade
física desta célula. Em contraste com as proteases e seus inibidores do testículo e
da célula espermática, as proteases envolvidas na função epididimária têm sido
menos estudadas. Proteases e seus inibidores têm sido detectados no fluido
epididimário de algumas espécies (MÉTAYER et al., 2002), o que sugere um papel
similar no processo de maturação espermática e na plasticidade deste órgão, ou em
ambos.
Tendo em vista estes antecedentes, o propósito deste trabalho é caracterizar
as metaloproteases detectadas no fluido epididimário de E. caballus, bem como
identificar e caracterizar seus principais substratos endógenos. A caracterização de
proteínas comprometidas na diferenciação do gameta possibilitará uma melhor
compreensão dos processos de maturação espermática e fertilização em mamíferos.

5
II. REVISÃO BIBLIOGRÁFICA
1. Epidídimo e o processo de maturação espermática
O conhecimento a respeito das mudanças que ocorrem no espermatozóide
durante o trânsito epididimário é fundamental para compreensão dos mecanismos
envolvidos na aquisição do potencial fertilizante em mamíferos.
Convencionalmente, o epidídimo é dividido em quatro regiões
macroscopicamente bem definidas: cabeça proximal, cabeça distal, corpo e cauda
(Figura 1); porém, suas características morfológicas podem variar entre diferentes
espécies (LÓPEZ et al., 1989).
Figura 1. EPIDÍDIMO DE Eqqus caballus. 1- região de cabeça proximal. 2- região de cabeça distal. 3- região de corpo e 4- região de cauda.
O conduto epididimário oferece condições que permitem o bom
desenvolvimento do processo de maturação espermática e também a viabilidade e
transporte do gameta. Vários tipos celulares encontram-se no epitélio epididimário:
células principais, ciliadas (exclusivamente na região proximal da cabeça), basais,
apicais e leucócitos intra-epiteliais. As células principais possuem propriedades de
absorção, secreção e síntese de vários compostos do fluido epididimário (LÓPEZ et
al., 1989; FOURNIER DELPECH & THIEBAULT, 1993; RETAMAL et al., 2000b) e
apresentam diferenças regionais tanto na sua morfologia, como na sua função. Na
região proximal, há uma considerável absorção de água e fluido provenientes da
rete testis; nas regiões de cabeça e corpo os espermatozóides adquirem o potencial

6
fertilizante; e a região de cauda é o local de armazenamento dos espermatozóides
até o momento da ejaculação. Estruturalmente, o retículo endoplasmático rugoso
abundante e membranas do sistema de Golgi muito desenvolvidas refletem as
propriedades de síntese e secreção protéica destas células (ROBAIRE & HERMO,
1988; LÓPEZ et al., 1989). Na região apical, as células principais estão fortemente
unidas pela zona oclusiva que forma parte da barreira hemato-epididimária, o que
permite a manutenção de um microambiente no lúmen do conduto (LÓPEZ et al.,
1997), bem como a proteção imunológica do espermatozóide (HINTON et al., 1995).
O fluido epididimário é hiperosmótico e difere em composição do plasma
sangüíneo, do fluido testicular e do plasma seminal. Entre os constituintes orgânicos
do fluido epididimário, além de L-carnitina, mioinositol, glicerilfosforilcolina, ácido
siálico, esteróides e outros, encontram-se diferentes íons e várias proteínas como
transferrina, albumina, clusterina, imobilina, metaloproteínas e enzimas
(glicosidases, glicosiltransferases, glutationa-peroxidase, superóxido-dismutase,
transpeptidases, entre outras (SETCHELL et al., 1994). As proteínas detectadas
neste fluido podem ser de origem epididimária ou ter origem testicular, chegando via
fluido da rete testis ao lúmen deste órgão (FOUCHÉCOURT et al., 2000). A ação de
cada uma destas moléculas na fisiologia do epidídimo não está bem definida, mas
tem-se sugerido que poderiam estar envolvidas na osmoregulação, na maturação
espermática e no metabolismo de espermatozóides e células epiteliais epididimárias
(SETCHELL et al., 1994).
Além de promover o processo de maturação espermática e ser local de
armazenamento de espermatozóides até o momento da ejaculação, o epidídimo
facilita o transporte de espermatozóides, os quais se encontram em estado
quiescente por fatores do fluido luminal (ROBAIRE & HERMO, 1988). A produção e
movimento constantes do fluido proveniente dos testículos, a corrente que produzem
os cílios e as microvilosidades das células epiteliais, a reabsorção de água na região
proximal do conduto e as contrações peristálticas das células mióides e da
musculatura lisa que envolve o epitélio epididimário, facilitam o transporte dos
gametas (TURNER, 1979).
O epidídimo possui vários mecanismos de proteção ao espermatozóide, além
de barreiras estruturais como a barreira hemato-epididimária (LOPÉZ et al., 1987).
São eles: a produção de proteínas que protegem da proteólise, como a HE4 -
polipeptídio ácido rico em cisteína (10kDa), as proteínas cistatinas, relacionadas

7
com a espermatogênese epididimária (CRES - cystatin-related epididymal
spermatogenic), da superfamília de inibidores de cisteíno proteases ou as proteínas
pertencentes à família desintegrina metaloproteases (ADAM) descritas em roedores
e macacos (BLOBEL, 2000; FRAYNE et al., 1997; PRIMAKOFF & MYLES, 2000;
EVANS, 2001). Estas proteínas podem proteger os espermatozóides e/ou o epitélio
epididimário de dano proteolítico resultante da liberação prematura de enzimas
acrossomais durante o armazenamento de espermatozóides (MÉTAYER et al.,
2002). Um outro mecanismo de proteção ao gameta, constitui a presença de enzimas
antioxidantes contra espécies reativas de oxigênio e substâncias xenobióticas. Todas
essas funções epididimárias estão precisamente coordenadas e reguladas de forma a
assegurar a produção de espermatozóides viáveis e funcionais.
2. Proteases
As proteases foram inicialmente associadas ao catabolismo protéico; no
entanto, tem-se aceito que a alta especificidade de hidrólise de peptídeos pode
regular uma gama de processos biológicos (FREDERIKS & MOOK, 2004).
Proteases e seus inibidores desempenham papéis-chave em processos fisiológicos,
como migração, sinalização celular, remodelação da superfície de células e tecidos.
Estas enzimas podem pertencer a cinco diferentes famílias dependendo da natureza
do sítio ativo: 1) proteases aspárticas, como as catepsinas D e E e pepsinas; 2)
cisteíno proteases, como as catepsinas B, L, S, K e Q, calpaínas e caspases; 3)
metaloproteases, que envolvem as gelatinases, 4) serino proteases, como os
ativadores de plasminogênio, a plasmina e algumas quinases e 5) treonino
proteases, como os proteassomos (FREDERIKS & MOOK, 2004).
Um grande número de proteases e seus inibidores têm sido detectados no
testículo, órgão que experimenta importantes remodelações durante a
espermatogênese. Várias destas proteases, tais como a procatepsina L, furina tipo
PC4, ou neprilisinas estão relacionadas com processamento de proteínas
específicas (ERICKSON-LAWRENCE et al., 1991; MBIKAY et al., 1997; GHADDAR
et al., 2000), enquanto outras, como o ativador de plasminogênio e metaloproteases
da matriz (MMPs), estariam envolvidas na remodelação tecidual, migração celular ou
interações célula-célula (GUNNARSSON et al., 1999; LONGIN et al., 2001;
HOEBEN et al., 1996; ROBINSON et al., 2001; MÉTAYER et al., 2002). A presença

8
de inibidores de protease como α2-macroglobulina e cistatina (inibidor de proteína
C) e inibidores mais específicos como metaloproteinases tecido específicas (TIMPs)
tem sido descrita no testículo (CHENG et al., 1990; UHRIN et al., 2000; NOTHNICK
et al., 1998). O desequilíbrio entre estes componentes resulta em uma série de
patologias, entre elas, o câncer (LIU et al., 2001).
Os espermatozóides também são ricos em diferentes tipos de proteases
(TULSIANI et al., 1998; KOHNO et al., 1998). Uma das proteases mais bem
estudada é a acrosina, a qual apresenta um importante papel na interação do
espermatozóide com a zona pelúcida (YAMAGATA et al., 1998). Uma serino
protease com atividade do tipo tripsina, denominada BSp66 (Bovine Sperm
Protease, 66 kDa), localizada na região acrossomal tem sido observada em
espermatozóides de bovinos, roedores e humanos. Imunologicamente, BSp66 não
está relacionada com outras proteases descritas nos espermatozóides (CESARI et
al., 2004). Uma indicação da presença de cisteíno - proteases no trato reprodutor de
alguns mamíferos é a expressão de proteínas CRES, pertencentes à superfamília
das cistatinas, inibidores de cisteíno proteases (SUTTON et al., 1999). A proteína
CRES localiza-se no acrossoma e é liberada durante a reação acrossomal. A
presença desta proteína no acrossoma, sítio de alta atividade hidrolítica, sugere sua
participação na regulação do processamento de proteínas intra-acrossomais
(SYNTIN & CORNWALL, 1999). Elas são sintetizadas no epidídimo e apresentam
localizações específicas. CRES 19 e 14 kDa estão presentes em espermatozóides
obtidos do testículo e da cabeça proximal do epidídimo. Entretanto, a CRES 14 kDa
foi a forma predominante no espermatozóide epididimário maduro, obtido da região
da cauda (SYNTIN & CORNWAL, 1999). Outra função para proteases no trato
reprodutor masculino é a atividade proteásica associada com ubiquitina, um
marcador proteolítico, que foi demonstrada mediante técnicas imunológicas em
espermatozóides defeituosos no trato reprodutor masculino de homens e touros.
Também foi investigada a ubiquitinação de espermatozóides epididimários e
testiculares de garanhão. Os dados mostrados sugerem que o espermatozóide é
diferencialmente ubiquitinado durante a maturação espermática e que esta
ubiquitinação pode refletir mudanças na qualidade do sêmen (SUTOVSKY, 2003).
Diversas funções têm sido associadas a estas enzimas, como a proteólise de
moléculas do acrossoma, a participação nas cascatas de transdução de sinais que
desencadeiam a reação acrossomal, o controle destas macromoléculas, a ruptura

9
das coberturas ovocitárias e a participação como ligante na interação primária com o
ovócito (MORALES et al., 2003; SUTOVSKY, 2003; HONDA et al., 2002; HOWES &
JONES, 2002).
A maturação espermática envolve grandes mudanças em diferentes domínios
da membrana do espermatozóide, principalmente, modificações das proteínas e
glicoproteínas que fazem parte da sua composição. Algumas destas modificações
resultam de um processamento proteolítico específico, que conduz ao
desaparecimento ou a redistribuição de proteínas entre os diferentes domínios
(YANAGIMACHI, 1994; PHELPS et al., 1990). Exemplos de processamento
proteolítico são: a transformação que experimentam proteínas da família ADAMs, em
roedores e macacos (BLOBEL, 2000; FRAYNE et al., 1997; PRIMAKOFF & MYLES,
2000; EVANS, 2001), do antígeno CE9 de espermatozóides em ratos (PETRUSZAK
et al., 1991), da proteína pH20 / 2B1 (SEATON et al., 2000), de manosidase de javali
(OKAMURA et al., 1992) e da alfa D-glicosidase (DIAS et al., 2004). Algumas MMPs
e várias serino proteases com atividade gelatinolítica e inibidores de proteases têm
sido observadas no fluido testicular e epididimário de mamíferos domésticos
(MÉTAYER et al., 2002). Contudo, a presença, características e papel fisiológico
destas enzimas no epidídimo de diferentes espécies não estão bem caracterizados.

10
3. Metaloproteases de Matriz Extracelular (MMPs)
As Metaloproteinases de matriz extracelular (MMPs), coletivamente
chamadas de matrixinas, são proteinases que participam da degradação da matriz
extracelular (VISSE & NAGASE, 2003). Estas enzimas pertencem a uma família de
pelo menos 20 membros, produto de genes, homólogos ou pseudo-homólogos,
relacionados entre si (WOESSNER, 1991).
As MMPs de vertebrados podem ser classificadas em seis grupos com base
na especificidade pelo substrato, similaridade na seqüência e organização dos
domínios: colagenases (MMP1, MMP8, MMP13 e MMP18), gelatinases (MMP2 e
MMP9), estromelisinas (MMP3, MMP10 e MMP11), matrilisinas (MMP7 e MMP26) e
metaloproteínas de membrana (MMP14, MMP15, MMP16, MMP17, MMP24 e
MMP25, também conhecidas como MT1-MMP a MT6-MMP, respectivamente.) Há
também outras MMPs que possuem características especiais que impedem sua
inserção nos grupos citados (VISSE & NAGASE, 2003), tais como a metaloelastase
de macrófago (MMP12) (BELAAOUAJ et al., 1994), a estromelisina-3 (BASSET et
al., 1990), MMP-19 (PENDÁS et al.,1997) e a enamelisina (MMP20) (LLANO et al.,
1997) dentre outras. (Tabela I).
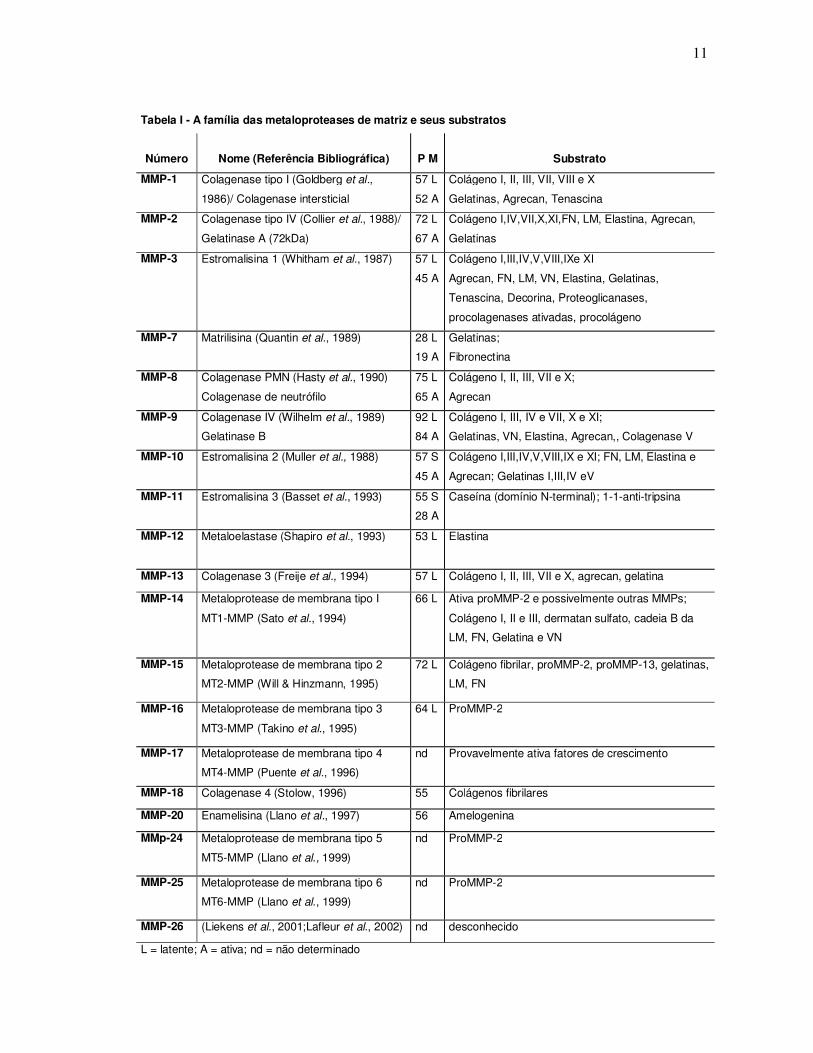
11
Tabela I - A família das metaloproteases de matriz e seus substratos
Número
Nome (Referência Bibliográfica)
P M
Substrato
MMP-1 Colagenase tipo I (Goldberg et al.,
1986)/ Colagenase intersticial
57 L
52 A
Colágeno I, II, III, VII, VIII e X
Gelatinas, Agrecan, Tenascina
MMP-2 Colagenase tipo IV (Collier et al., 1988)/
Gelatinase A (72kDa)
72 L
67 A
Colágeno I,IV,VII,X,XI,FN, LM, Elastina, Agrecan,
Gelatinas
MMP-3 Estromalisina 1 (Whitham et al., 1987)
57 L
45 A
Colágeno I,III,IV,V,VIII,IXe XI
Agrecan, FN, LM, VN, Elastina, Gelatinas,
Tenascina, Decorina, Proteoglicanases,
procolagenases ativadas, procolágeno
MMP-7 Matrilisina (Quantin et al., 1989) 28 L
19 A
Gelatinas;
Fibronectina
MMP-8 Colagenase PMN (Hasty et al., 1990)
Colagenase de neutrófilo
75 L
65 A
Colágeno I, II, III, VII e X;
Agrecan
MMP-9 Colagenase IV (Wilhelm et al., 1989)
Gelatinase B
92 L
84 A
Colágeno I, III, IV e VII, X e XI;
Gelatinas, VN, Elastina, Agrecan,, Colagenase V
MMP-10 Estromalisina 2 (Muller et al., 1988)
57 S
45 A
Colágeno I,III,IV,V,VIII,IX e XI; FN, LM, Elastina e
Agrecan; Gelatinas I,III,IV eV
MMP-11 Estromalisina 3 (Basset et al., 1993) 55 S
28 A
Caseína (domínio N-terminal); 1-1-anti-tripsina
MMP-12 Metaloelastase (Shapiro et al., 1993) 53 L
Elastina
MMP-13 Colagenase 3 (Freije et al., 1994) 57 L Colágeno I, II, III, VII e X, agrecan, gelatina
MMP-14 Metaloprotease de membrana tipo I
MT1-MMP (Sato et al., 1994)
66 L Ativa proMMP-2 e possivelmente outras MMPs;
Colágeno I, II e III, dermatan sulfato, cadeia B da
LM, FN, Gelatina e VN
MMP-15 Metaloprotease de membrana tipo 2
MT2-MMP (Will & Hinzmann, 1995)
72 L Colágeno fibrilar, proMMP-2, proMMP-13, gelatinas,
LM, FN
MMP-16 Metaloprotease de membrana tipo 3
MT3-MMP (Takino et al., 1995)
64 L ProMMP-2
MMP-17 Metaloprotease de membrana tipo 4
MT4-MMP (Puente et al., 1996)
nd Provavelmente ativa fatores de crescimento
MMP-18 Colagenase 4 (Stolow, 1996) 55 Colágenos fibrilares
MMP-20 Enamelisina (Llano et al., 1997) 56 Amelogenina
MMp-24 Metaloprotease de membrana tipo 5
MT5-MMP (Llano et al., 1999)
nd ProMMP-2
MMP-25 Metaloprotease de membrana tipo 6
MT6-MMP (Llano et al., 1999)
nd ProMMP-2
MMP-26 (Liekens et al., 2001;Lafleur et al., 2002) nd desconhecido
L = latente; A = ativa; nd = não determinado

12
Embora as MMPs sejam classificadas de acordo com o substrato pela qual
possuem afinidade, nota-se que há sobreposição entre as subclasses, significando
que as MMPs não têm necessariamente um substrato definido. Isto se torna mais
notório em processos patogênicos como a proteólise relacionada à invasão tumoral.
Além disso, as atividades das MMPs interferem umas com as outras, podendo ativar
outras proenzimas, o que sugere que a ativação controlada das mesmas pode
envolver uma cascata, englobando diferentes membros da família das MMPs
(CAWSTON, 1998).
Todas as MMPs têm algumas características em comum, incluindo, dentre
estas: 1) um peptídeo sinal que direciona o produto para secreção, 2) um domínio
pro-peptídico, 3) domínio catalítico que acomoda um átomo de Zn+2, essencial para
a manutenção de sua latência enzimática e é removido quando a enzima é ativada
(SHIMA et al., 1992). A maioria das MMPs, com exceção das MMP7, MMP26 e
MMP23, também possui um domínio C-terminal com uma seqüência de homologia a
hemopexina, que constitui um sítio de ligação para os inibidores teciduais. As
gelatinases –A e –B (MMPs 2 e 9) diferem das demais por apresentarem um
domínio semelhante a fibronectina (Fn) (Figura 2) (VISSE & NAGASE, 2003). A
gelatinase B possui ainda um domínio semelhante ao colágeno. As MT-MMPs são
caracterizadas por um domínio transmembrana que é responsável pela ancoragem
da enzima à membrana plasmática (NGUYEN et al., 2001; LIEKENS et al, 2001).
Todas as MMPs requerem um pH neutro e Ca+2 para poderem atuar e são inibidas
por uma família específica de proteínas, que se denominam inibidores tissulares de
MMP (TIMP) (MATRISIAN ,1990; NGUYEN et al., 2001). Atualmente, são
conhecidos quatro membros da família das TIMPs que são capazes de se ligar e
inibir a atividade de todos os membros da família das MMPS. Eles demonstram,
entretanto, diferenças na sua distribuição nos tecidos e na habilidade de formar
complexos com a forma inativa da MMP (MCCAWLEY & MATRISIAN, 2000).

13
Figura 2. ESTRUTURA DAS METALOPROTEINASES DE MATRIZ MMP-2 E MMP-9.
As MMPs estão amplamente distribuídas no organismo humano onde
desempenham uma série de funções fisiológicas como, por exemplo, na cicatrização
(WOLF et al., 1992), na reabsorção óssea (DELAISSÉ & VAES, 1992), na involução
mamária (TALHOUK et al., 1992) e em outras funções fisiológicas associadas a
gravidez e parto (JEFFREY, 1991). Recentemente, tem-se demonstrado que as
MMPs também estão implicadas em processos patológicos variados como na artrite
reumatóide (HARRIS, 1990), na enfermidade periodontal (PAGE, 1991), na
esclerose múltipla (CHANDLER et al., 1997), em algumas doenças cardiovasculares
(TAMARINA et al., 1997) e certas alterações hematológicas (GUEDEZ et al., 1996).
Em condições fisiológicas normais, a atividade das MMPs é regulada por
expressão gênica, por ativação da pró-enzima e por inativação da enzima ativa, por
inibidores tissulares específicos e por interação com componentes específicos da
matriz extracelular (VISSE & NAGASE, 2003).
A expressão de MMPs é induzida por fatores de crescimento, interação
célula-célula e interação célula-matriz. Muitos genes de MMPs (mmp1, mmp3,
mmp7, mmp9, mmp10, mmp12 e mmp13) são induzíveis por estímulos
extracelulares que ativam um fator de transcrição dimérico, o complexo ativador de
proteínas 1 (AP1), composto pelas proteínas JUN e FOS, que interage com o
promotor de genes MMPs e ativa sua transcrição. A indução da expressão e
atividade de AP1 é mediada por três classes de proteínas quinases ativadas por
mitógenos (MAPK), quinases reguladas por sinais extracelulares (ERKs), quinase do
N-terminal de JUN ativada por estresse (JNK) e proteína p38 (WESTERMARCK &
KAHARI, 1999). A proteína quinase C (PKC) ativa a via de sinalização de

14
ERK1/ERK2. Recentemente, tem-se mostrado que a super expressão de isoformas
de PKC resulta na ativação do promotor de MMP1. PKC também desempenha
função importante na expressão de MMP9 em células de gliomas malignos
(YABKOWITZ et al., 1999). As células endoteliais também sintetizam várias MMPS:
MMP-1, MMP-2, MMP-9 e MT1-MMP. Dentre estas a MMP-2 (gelatinase-A, 72kDa),
MMP-9 (gelatinase B, 92 kDa) e a MT1-MMP foram descritas como exercendo
importante papel na angiogênese (STETLER-STEVENSON, 1999).
4. Maturação espermática e atividade proteolítica
As mudanças observadas durante a diferenciação do gameta são de natureza
bem diversa e afetam tanto características estruturais (núcleo, acrossoma,
membranas), funcionais (padrão metabólico, grau de motilidade) como a
suscetibilidade do gameta às condições do meio. Algumas delas são provavelmente
necessárias à interação gamética, enquanto outras permitem a estabilização de
estruturas e a sobrevida do gameta no percurso pelo epidídimo (FOURNIER
DELPECH & THIEBAULT, 1993; LÓPEZ, 1996).
A qualidade do espermatozóide, em termos de mobilidade e reconhecimento do
ovócito, é fortemente dependente de diferenciações pós-testiculares que ocorrem no
epidídimo (GATTI et al., 2004). Essas modificações surgem, possivelmente, como
resultado da liberação e/ou degradação de componentes testiculares, absorção e
integração de proteínas secretadas pelo epidídimo ou por modificação de resíduos
oligossacarídicos ligados a proteínas ou lipídeos de membrana. Glicosilações,
proteólise, re-localização de componentes pré-existentes são alguns mecanismos já
descritos (DACHEUX et al., 2003; TULSIANI et al., 1995).
Há na literatura a descrição de uma série de proteínas espermáticas
proteoliticamente processadas durante o trânsito epididimário, entre elas:
hialuronidase, pH20, proteína CE9 e galactosidase, a qual existe como uma forma
de 90 kDa em espermatozóides obtidos da região de cabeça proximal e é
gradualmente convertida para uma forma majoritária de 74 kDa em espermatozóides
de cauda (CHAYKO et al., 2000). Outro exemplo de processamento proteolítico é a
fertilina. Esta é um heterodímero transmembranar com domínios pro-
metaloprotease, disintegrina, rico em cisteína e EGF, e que pertence a família ADAM

15
(YUAN et al., 1997). Esta proteína (previamente chamada PH30) sofre pelo menos
dois processamentos proteolíticos distintos: um no testículo e outro no epidídimo
(LUM & BLOBEL, 1997). Estes dados indicam que algumas proteínas podem ser
secretadas ou estarem presentes em uma pró-forma e tornam-se ativadas após o
processamento no fluido (GATTI et al., 2004). A análise da maturação intracelular e
atividade catalítica de uma desintegrina metaloprotease amplamente expressa, a
MDC9, sugere que o pro-domínio da MDC9 é removido por uma convertase tipo
furina da via secretora, depois que a proteína emerge na superfície, e que a
atividade catalítica da MDC9 pode ser inibida por inibidores de metaloprotease. A
remoção de um pró-domínio inibitório da MDC9 é um pré-requisito para a atividade
proteásica. Passos adicionais, como o de proteínas quinases C dependentes de
fosforilação, podem estar envolvidos na regulação da atividade enzimática da MDC9
(ROGHANI et al., 1999). Manter estas proteases em um estado inativo durante a
passagem do gameta pelas vias seminais é crítico para assegurar sua função
reprodutiva (UHRIN et al., 2000; ZHENG et al., 1994).

16
III. OBJETIVOS
1. Objetivo geral
Contribuir ao estudo do processo de maturação espermática de E. caballus
analisando a ocorrência de atividade metaloproteásica, bem como seus substratos e
produtos endógenos, no fluido epididimário.
2. Objetivos específicos
Detectar e caracterizar atividade de metaloproteases nos fluidos das diferentes
regiões do epidídimo de eqüinos.
Selecionar e purificar algumas metaloproteases do próprio fluido epididimário da
região de cauda.
Detectar a presença de substratos endógenos nas diferentes regiões do
epidídimo de eqüinos.
Selecionar e purificar alguns substratos endógenos das metaloproteases
purificadas.
Identificar os produtos de hidrólise mediante ensaios de metaloproteases
isoladas da região de cauda epididimária incubadas com substratos endógenos
isolados desta mesma região.

17
IV. MATERIAIS E MÉTODOS
1. Obtenção das amostras
Os epidídimos foram obtidos por castração cirúrgica de animais sadios,
sexualmente maduros, de haras da Região Norte Fluminense. As gônadas e vias
espermáticas superiores foram cuidadosamente dissecadas retirando-se o tecido
conjuntivo adjacente. Os epidídimos foram divididos em quatro regiões: (1) cabeça
proximal, (2) cabeça distal, (3) corpo e (4) cauda (Esquema 1). Cada região foi
colocada em uma placa de Petri contendo PBS, pH 7,2 e fragmentada com uma
tesoura de forma a liberar o conteúdo luminal em 5 mL de tampão na região de
cabeça proximal e em 10 mL nas demais regiões epididimárias. O conteúdo foi
filtrado em gaze dupla e centrifugado a 700 g por 30 min. a 4°C, numa centrífuga
Hitachi, para a separação do fluido epididimário dos espermatozóides. O fluido foi
novamente centrifugado, nas mesmas condições, para a remoção de células
espermáticas. As diferentes amostras foram devidamente aliquotadas, identificadas
e estocadas a -20°C, após determinação da concentração de proteínas totais.
1 2
3
4
Esquema 1: DIAGRAMA MOSTRANDO AS DIFERENTES REGIÕES DO EPIDÍDIMO: 1 – cabeça proximal, 2 – cabeça distal, 3- corpo e 4 - cauda.

18
2. Determinação da concentração de proteínas
As diferentes amostras foram devidamente identificadas e estocadas a -20°C,
após determinação da concentração de proteínas totais de cada uma delas
(BRADFORD, 1976). Na curva padrão foi utilizada albumina sérica bovina (BSA) com
concentração variando entre 0 e 20 µg, com intervalos de 2 µg. As leituras foram
realizadas em um leitor de Elisa (Lab System modelo mult-scan), a 595 nm de
comprimento de onda ou em um espectrofotômetro FEMTO 600 plus, sob mesmo
comprimento de onda.
3. Análise do perfil eletroforético de amostras de fluido epididimário
As amostras foram analisadas em SDS-PAGE 12% (LAEMMLI, 1970) (50 µg
de proteínas por poço – quantidade de proteína padronizada para a maioria dos
experimentos). Posteriormente, os géis foram corados com azul de Coomassie R-250
0,1% (v/v). As análises da mobilidade eletroforética, e a quantificação relativa das
bandas foram realizadas mediante um programa computacional "Gel Perfect" (BOZZO
& RETAMAL, 1991; RETAMAL et al., 1999). A determinação da massa molecular foi
realizada relacionando a mobilidade das proteínas das amostras em estudo, com a
mobilidade de proteínas de massa molecular conhecida, utilizando o mesmo
programa.
4. Detecção de proteólise endógena no fluido epididimário
Visando detectar proteólise endógena no fluido epididimário, amostras das
diferentes regiões epididimárias foram incubadas a 37ºC (Esquema 2A). Alíquotas de
cada amostra foram retiradas em diferentes intervalos de tempo (30 min.; 1h; 3h; 6h,
12h, 24h, 36h, 48h, 72h) e congeladas a -20ºC. Como controle uma alíquota de fluido
de cada região epididimária não foi incubada e foi mantida congelada a -20ºC. As
alíquotas foram submetidas à eletroforese em SDS-PAGE 15%, na presença de β-
mercaptoetanol 10% (v/v) para a visualização dos resultados.
Outra estratégia foi utilizada para detectar a proteólise endógena. Neste caso, a
amostra de fluido epididimário de cauda foi submetida a eletroforese em um gel de

19
poliacrilamida 10%, em condições nativas (adaptado de DAVIS, 1964). As tiras de gel
nativo foram cortadas e mantidas em tampão de amostra 2x (Tris-HCl 0,5M pH 6,8,
SDS 10% (v/v), azul de bromofenol 1%, glicerol 10% (v/v)), na temperatura ambiente,
por diferentes intervalos de tempo (24h, 48h, 72h, 96h e 120h) e congeladas a -20ºC.
Posteriormente, as tiras foram descongeladas e submetidas a uma segunda
eletroforese em géis SDS-PAGE 12% (Esquema 2B).
Esquema 2. DETECÇÃO DE PROTEÓLISE ENDÓGENA NO FLUIDO EPIDIDIMÁRIO. A- Incubação das amostras de fluido das diferentes regiões epididimárias a 37ºC; alíquotas foram retiradas em diferentes intervalos de tempo (30 min.; 1h; 3h; 6h, 12h, 24h, 36h, 48h, 72h). A visualização do experimento foi realizada em SDS-PAGE 15%. B- Esquema da incubação das tiras de gel nativo, de fluido epididimário de cauda, a temperatura ambiente, alíquotas foram retiradas em diferentes intervalos de tempo (24h, 48h, 72h, 96h e 120h).

20
5. Detecção de atividade metaloproteásica
As amostras de fluido das diferentes regiões epididimárias, de diferentes cavalos,
foram submetidas a eletroforese em SDS-PAGE 12%, 0,4% (v/v) gelatina (HUMMEL et
al., 1996, MÉTAYER et al., 2002). Os géis contendo gelatina foram incubados em Triton
X-100 2,5% (v/v) durante 3h. Posteriormente, foram lavados 3x (5 min) em H2O destilada
sob agitação e incubados em tampão de incubação (Tris 50 mM, NaCl 200 mM, CaCl2 4
mM, pH 7,5) (adaptado de MÉTAYER et al., 2002) a 37ºC por 72h e visualizados sob UV.
Em seguida, os géis foram corados com azul de Coomassie R-250 e descorados em
água destilada em ciclos de lavagem de 2 min em microondas (LG comercial) com
potência máxima. As bandas de atividade aparecem brancas em um fundo azul.
6. Ação de inibidores sobre as atividades proteolíticas
A caracterização da atividade proteolítica foi realizada mediante o uso de
diferentes inibidores de proteases. As amostras de fluido das diferentes regiões
epididimárias foram submetidas à eletroforese em SDS-PAGE 12% com gelatina
0,4% (v/v). Posteriormente, os géis foram incubados com Triton X-100 2,5% durante
3h e lavados conforme já descrito anteriormente. Cada gel foi incubado com tampão
de incubação (descrito no item 5) contendo separadamente diferentes inibidores de
proteases: PMSF 100 µM, inibidor para serino protease; EDTA 10 mM, inibidor para
metaloprotease e iodoacetamida 10 mM, inibidor para cisteíno proteases. Um gel
controle foi incubado com tampão de incubação sem inibidores (adaptado LUM &
BLOBEL, 1997).
7. Verificação do pH ótimo
A caracterização da atividade proteolítica também foi realizada mediante a
verificação das atividades a diferentes pHs. As amostras de fluido das diferentes
regiões epididimárias foram submetidas à eletroforese em SDS-PAGE 12% com
gelatina 0,4% (v/v). Os géis foram incubados com Triton X-100 2,5% durante 3h e
lavados. Cada gel foi incubado com tampão de incubação (descrito no item 5)
ajustado a diferentes pHs: 2,5; 4,5; 6,5; 7,5; 10 e 12.

21
8. Determinação do grau de anomalia das proteases na migração
eletroforética
O grau de anomalia das proteases na migração eletroforética foi detectado
através da análise das amostras em estudo por SDS-PAGE 10% na presença de um
gradiente de gelatina 0 – 0,6% (v/v). Nestes géis, uma faixa larga (1,5 cm) de SDS-
PAGE 10% sem gelatina foi polimerizada ao lado da porção do gel gradiente
(Esquema 3). As amostras das diferentes regiões epididimárias foram submetidas a
este tipo de gel em uma concentração de 100 µg (poço único) sem agente redutor.
Após a eletroforese os géis foram incubados com Triton X-100 2,5% (v/v) durante 3h,
lavados, incubados em tampão de incubação (descrito no item 5) e corados com azul
de Coomassie. Foram feitos gráficos de Ferguson para a determinação da constante
de retardação (FERGUSON, 1964). A constante de retardação (Kr) das proteases foi
determinada em um gradiente de gelatina tendo como suporte uma concentração
constante de acrilamida.
Outra abordagem para determinação do grau de anomalia na migração
eletroforética causada pela gelatina foi a sobreposição de géis 2D desnaturantes
(DIAS, 2002) com e sem gelatina 0,4% (v/v). As proteínas de fluido epididimário das
diferentes regiões foram submetidas a eletroforese em SDS-PAGE 12%. As tiras
correspondentes aos poços foram cortadas e incubadas durante 20 min. em tampão
de amostra 2x (citado no item 4) sem β-mercaptoetanol e com este agente redutor a
uma concentração de 10%. Cada tira foi então submetida a uma segunda
eletroforese (re-eletroforese) em SDS-PAGE 12% com e sem gelatina 0,4% (v/v).
Estes géis SDS/SDS e SDS/SDS com gelatina foram sobrepostos em quatro
imagens. Algumas imagens foram invertidas no programa computacional Photo
Impact. Além disso, foi feito um gráfico para ilustrar de maneira didática a variação
no coeficiente angular das retas de diferentes mobilidades relativas (Rfs) que foram
geradas pela presença de gelatina.

22
Esquema 3. DETERMINAÇÃO DO GRAU DE ANOMALIA DAS PROTEASES NA MIGRAÇÃO ELETROFORÉTICA. SDS-PAGE 10% gradiente de gelatina de 0 – 0,6% (v/v) com uma tira de SDS-PAGE 10% sem gelatina utilizado para a determinação do grau de anomalia das proteases na migração eletroforética. A seta indica o sentido crescente da concentração de gelatina.
9. Determinação das massas moleculares das proteases de fluido
epididimário
Devido à anomalia causada pela presença de gelatina nos géis (HUMMEL et
al., 1996) foram feitos géis bidimensionais (modificado de SUN & PAN, 1999; DIAS,
2002) para determinação das massas moleculares das atividades gelatinolíticas
(Esquema 4). Na primeira eletroforese (1D), as amostras de fluido epididimário das
diferentes regiões foram submetidas a SDS-PAGE 12% na ausência do polímero
gelatina (LAEMMLI, 1970). Em seguida, foram cortadas tiras do gel correspondendo
aos canais de amostra. Cada tira foi incubada durante 20 min. em tampão de amostra
2x (citado no item 4) contendo β-mercaptoetanol 10% e também sem este agente
redutor. As tiras foram submetidas a uma segunda eletroforese (2D), realizada nas
mesmas condições desnaturantes na presença de gelatina 0,4% (HUMMEL et al.,
1996; MÉTAYER et al., 2002). Os géis com gelatina foram incubados em Triton X-
100 2,5% e lavados conforme descrito anteriormente e incubados em tampão de
incubação (descrito no item 5) (adaptado de MÉTAYER et al., 2002) e visualizados
com luz ultravioleta e /ou reveladas com azul de Coomassie R-250 para sua
identificação. A determinação da massa molecular relativa das proteases foi estimada
relacionando a mobilidade das proteínas das amostras do gel 1D com a mobilidade de

23
proteínas que apresentam massa molecular conhecida, indicadas pelas atividades
proteolíticas do gel 2D.
Esquema 4. DETERMINAÇÃO DAS MASSAS MOLECULARES DAS PROTEASES NO FLUIDO EPIDIDIMÁRIO. As amostras de fluido epididimário foram submetidas à eletroforese em SDS-PAGE 12% (1D), as tiras foram submetidas a uma segunda eletroforese em SDS-PAGE 12% contendo gelatina 0,4%. PM- Padrão de Massa Molecular. As setas indicam o sentido da migração das proteínas na análise eletroforética.

24
10. Purificação das atividades proteolíticas detectadas no fluido epididimário de
cauda
10.1 Purificação das atividades proteolíticas a partir de géis
unidimensionais
As bandas com atividade proteásica foram purificadas a partir de géis
unidimensionais segundo RETAMAL et al. (1999). A atividade foi purificada de géis de
poliacrilamida 12%, SDS 0,1% (v/v) na presença de gelatina 0,4% (v/v) (Esquema 5).
As proteases foram extraídas dos locais que apresentaram atividade, seguindo como
parâmetro uma tira lateral do gel cortada e corada e também através da visualização
das mesmas sob luz UV. Em seguida, as bandas extraídas do gel foram maceradas
com 100 µl de tampão Tris HCl 5mM pH 7,4 e sonicadas, em gelo, (Sonic Dismembrator
60 Fisher Science) 20 vezes por 30 segundos, com pausa 30 de segundos, para a
extração das proteases (RETAMAL et al.,1999). As bandas extraídas, maceradas e
sonicadas foram centrifugadas a 17000 g por 30 min. (centrífuga Hermle Z 252 NK), e
os sobrenadantes coletados foram estocados em microtubos de propileno a -200C para
posterior análise.
Esquema 5. PURIFICAÇÃO DAS ATIVIDADES PROTEOLÍTICAS DETECTADAS NO FLUIDO EPIDIDIMÁRIO DE CAUDA A PARTIR DE GÉIS UNIDIMENSIONAIS. Purificação das proteases a partir de SDS-PAGE 12% com gelatina. A- Uma tira lateral do gel foi cortada, corada com Coomassie e usada como parâmetro para o corte das bandas de atividade. B- Outra abordagem para o corte das bandas de atividade foi a visualização das mesmas em luz UV.

25
10.2 Purificação das atividades proteolíticas a partir de géis 2D SDS/SDS
com gelatina
As bandas com atividade proteásica também foram purificadas a partir de géis
bidimensionais segundo RETAMAL et al. (1999), dos quais, a primeira eletroforese foi
realizada em um gel de poliacrilamida 12%, SDS 0,1% (v/v) e, a segunda eletroforese
foi realizada em um gel nas mesmas condições na presença de gelatina 0,4% (v/v)
(Esquema 6). As proteases foram extraídas do local que mostra atividade em círculos
de 1 mm de diâmetro, com o auxílio de uma ponteira de 200 µl. Os círculos cortados de
uma mesma protease foram estocados em microtubos a -200C . Em seguida, foram
macerados com 100 µl de tampão Tris, sonicados e centrifugados (conforme descrito no
item 10.1) para a extração das proteases (RETAMAL et al.,1999). Os sobrenadantes
coletados foram estocados em microtubos a -200C para posterior análise.
Esquema 6. PURIFICAÇÃO DAS ATIVIDADES PROTEOLÍTICAS DETECTADAS NO FLUIDO EPIDIDIMÁRIO DE CAUDA A PARTIR DE GÉIS 2D SDS/SDS COM GELATINA. A- Representação do gel SDS/SDS com gelatina corado e utilizado como parâmetro para o corte das bandas. B- Representação do gel não corado utilizado para a extração das bandas de atividade.

26
10.3 Purificação das atividades proteolíticas a partir de géis nativos 10%
As proteases também foram purificadas a partir de géis nativos 10% para evitar o
efeito da presença da gelatina sobre estas, depois de purificadas (Esquema 7). Para
isso foi necessário que a atividade proteásica fosse localizada em géis nativos. Assim,
foram realizados géis bidimensionais onde a primeira eletroforese foi feita em um gel
nativo e a segunda eletroforese em um gel de poliacrilamida 12%, SDS 0,1% (v/v) na
presença de gelatina 0,4% (v/v) (Esquema 7A). As proteases foram localizadas no gel
nativo 10% a partir do gel bidimensional (Esquema 7B) e assim, seguindo como
parâmetro o gel SDS bidimensional com gelatina, as bandas foram cortadas de géis
unidimensionais em condições nativas. (Esquema 7C). Em seguida, as bandas
extraídas do gel foram maceradas com 100 µl de tampão Tris (citado no item 10.1),
sonicadas e centrifugadas, conforme já descrito, para a extração das proteases. Os
sobrenadantes obtidos foram estocados em microtubos a -200C para posterior análise.

27
Esquema 7. PURIFICAÇÃO DAS ATIVIDADES PROTEOLÍTICAS DETECTADAS NO FLUIDO EPIDIDIMÁRIO DE CAUDA A PARTIR DE GÉIS NATIVOS 10%. Purificação das proteases a partir de géis nativos 10%. A- Gel 2D nativo/SDS. B- Localização das bandas de proteínas correspondentes às bandas de atividade. C- Esquema do corte das bandas de atividade a partir do gel nativo. P1-Protease 1; P2- Protease 2; P3- Protease 3 e P4- Protease 4.

28
11. Detecção e identificação de substratos endógenos de fluido de cauda com
atividades proteolíticas purificadas desta mesma região
Para identificar os substratos endógenos das proteases extraídas de fluido
epididimário de cauda estas foram incubadas com fluido epididimário da mesma região
(Esquema 8). As diferentes proteases purificadas a partir de géis com gelatina 0,4%
foram incubadas com fluido epididimário de cauda, de forma separada. Os círculos de 1
mm de diâmetro, contendo as proteases, foram misturados com 200 µl de fluido
epididimário e macerados em microtubos. Os microtubos contendo a mistura foram
mantidos a 37ºC numa estufa durante 72h. Posteriormente, os ensaios foram
visualizados em géis de poliacrilamida gradiente 5 - 15%, tanto na presença de SDS
0,1% (v/v) quanto em condições nativas (adaptado de DAVIS, 1964). Também foram
analisados em géis bidimensionais nativo/SDS (SUN & PAN, 1999). Os tubos contendo
os ensaios foram armazenados a -20ºC. O resultado foi novamente analisado em SDS-
PAGE gradiente 5 - 15%. Todos os géis foram corados com azul de Coomassie,
descorados em microondas e os possíveis substratos e produtos foram visualizados,
identificados e quantificados, através da análise no programa computacional “Gel
Perfect”.
Esquema 8. DETECÇÃO E IDENTIFICAÇÃO DE SUBSTRATOS ENDÓGENOS DE FLUIDO DE CAUDA COM ATIVIDADES PROTEOLÍTICAS PURIFICADAS DESTA MESMA REGIÃO. Incubação das amostras de fluido de cauda a 37ºC com as proteases purificadas desta mesma região. P1+F- Protease 1 + Fluido de cauda; P2+F- Protease 2 + Fluido de cauda; P3+F- Protease 3 + Fluido de cauda; P4+F- Protease 4 + Fluido de cauda; F- Fluido de cauda incubado como controle. A visualização foi realizada em géis gradiente de acrilamida com e sem SDS e géis nativo/SDS 6-15%.

29
12. Purificação dos substratos endógenos das atividades proteolíticas
12.1 Purificação dos substratos endógenos, presentes em fluido de
cauda, a partir de géis nativos 10%
Os substratos identificados através da metodologia descrita no item anterior
foram isolados de géis nativos 10%. Para tanto, os substratos foram previamente
localizados em géis bidimensionais, nas quais a primeira eletroforese foi em um gel de
poliacrilamida nativo 10% e a segunda eletroforese em um gel gradiente de
poliacrilamida 5-15%, SDS 0,1% (Esquema 9). As proteínas de fluido epididimário de
cauda foram submetidas a géis (1D) nativos 10%. Uma vez realizada a eletroforese e
antes de corar o gel, os géis foram cortados em tiras correspondentes aos canais de
amostra e, uma delas foi corada com azul de Coomassie para servir como controle.
Outra tira foi incubada em tampão de amostra 2x (descrito no item 4). Posteriormente,
esta tira foi submetida a uma segunda eletroforese (2D) em um gel gradiente de
poliacrilamida 5-15%, SDS 0,1% (v/v) (Dias, 2002). Após a eletroforese, o gel gradiente
foi corado com azul de Coomassie e o substrato pôde ser localizado no gel nativo 10%
(Esquema 9B). Onde o substrato foi localizado foram extraídas as bandas dos géis
nativos não corados, que foram armazenadas em microtubos. Em seguida, foi
adicionado tampão de incubação (descrito no item 5) às bandas em uma proporção de
1:1 e, estas foram sonicadas, em gelo, (Sonic desmembrator 60 Fisher Science) em 12
ciclos de 30s em potência máxima, a fim de extrair do gel as proteínas candidatas a
substratos. Após isto, foram centrifugadas a 17000 g durante 10 min. a 4ºC e uma
alíquota do sobrenadante, contendo o substrato purificado, foi congelada a -20ºC.
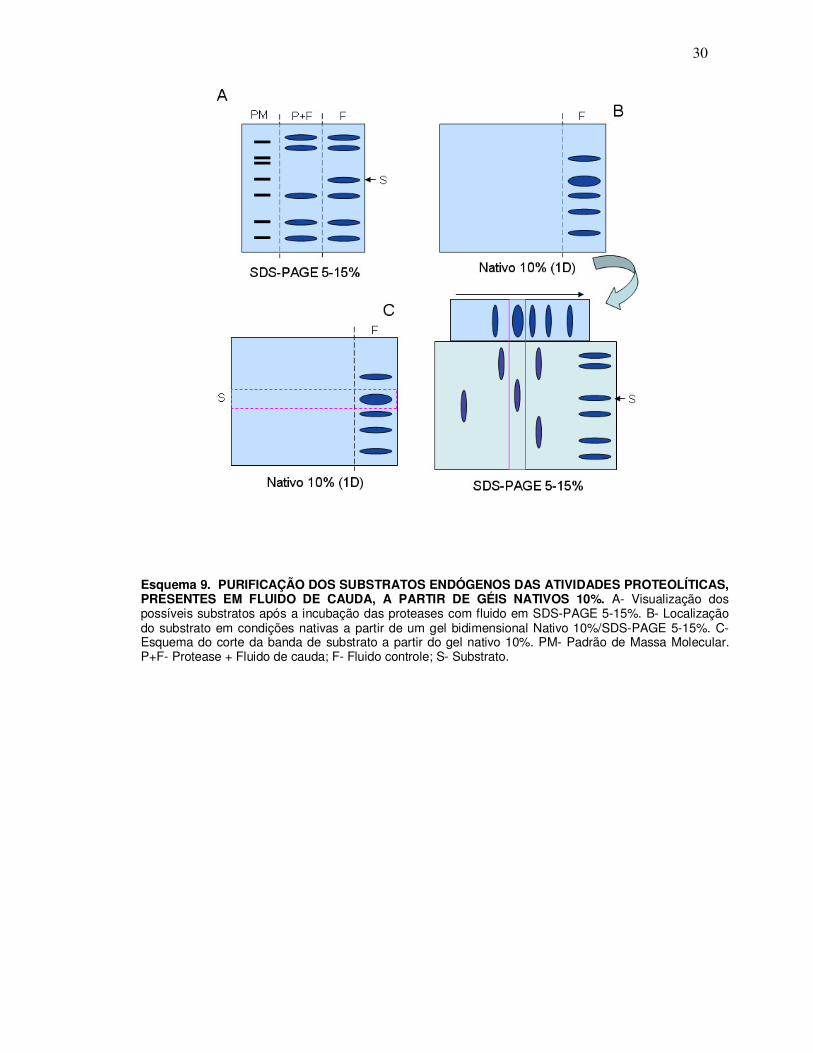
30
Esquema 9. PURIFICAÇÃO DOS SUBSTRATOS ENDÓGENOS DAS ATIVIDADES PROTEOLÍTICAS, PRESENTES EM FLUIDO DE CAUDA, A PARTIR DE GÉIS NATIVOS 10%. A- Visualização dos possíveis substratos após a incubação das proteases com fluido em SDS-PAGE 5-15%. B- Localização do substrato em condições nativas a partir de um gel bidimensional Nativo 10%/SDS-PAGE 5-15%. C- Esquema do corte da banda de substrato a partir do gel nativo 10%. PM- Padrão de Massa Molecular. P+F- Protease + Fluido de cauda; F- Fluido controle; S- Substrato.

31
12.2 Purificação dos substratos endógenos, presentes em fluido de
cauda, a partir de géis nativo/SDS-PAGE 12%
Os substratos identificados também foram isolados de géis nativos 10%/SDS-
PAGE 12% (Esquema 10). As proteínas de fluido epididimário de cauda foram
submetidas a géis (1D) nativo 10%. Uma vez realizada a eletroforese e antes de corar
o gel, os géis foram cortados em tiras correspondentes aos canais de amostra e, uma
delas, foi corada com azul de Coomassie. As outras foram incubadas em tampão de
amostra 2x (descrito no item 4) e submetidas a uma segunda eletroforese (2D) em géis
de poliacrilamida 12%, SDS 0,1% (v/v) (Dias, 2002). Foram feitos 6 géis bidimensionais
e um deles foi corado com azul de Coomassie e utilizado como padrão para o corte das
bandas de substratos dos outros géis bidimensionais não corados. Os substratos foram
visualizados e extraídos em círculos de diâmetros variados com o auxílio de diferentes
ponteiras. Os círculos cortados foram armazenados em microtubos. Ao todo foram
isolados 8 possíveis substratos. Foi adicionado aos círculos (contendo as bandas de
substratos) tampão de incubação em uma proporção de 1:1 e, em seguida, sonicados,
em gelo, (Sonic desmembrator 60 Fisher Science) em 12 ciclos de 30s em potência
máxima, a fim de extrair as proteínas do gel. Após isto, foram centrifugados a 17000 g
durante 10 min. a 4ºC e uma alíquota do sobrenadante, contendo o substrato purificado,
foi congelada a -20ºC.

32
Esquema 10. PURIFICAÇÃO DOS SUBSTRATOS ENDÓGENOS DAS ATIVIDADES PROTEOLÍTICAS, PRESENTES EM FLUIDO DE CAUDA, A PARTIR DE GÉIS NATIVO/SDS-PAGE 12%. A- Esquema do gel nativo 10% utilizado para a visualização dos possíveis substratos após a incubação das proteases com fluido. B- Esquema da visualização dos possíveis substratos no gel bidimensional nativo 10%/SDS-PAGE 5-15%. C- Esquema dos cortes das bandas de substrato a partir de géis nativo/SDS 12% não corados utilizando como parâmetro as bandas de substratos visualizadas. S- Substrato; P- Produtos; P+F- Protease + Fluido de cauda; F- Fluido controle.

33
13. Identificação dos produtos endógenos gerados pelas proteases purificadas
e substratos purificados
As proteases purificadas do fluido epididimário de cauda foram incubadas com
os substratos purificados desta mesma região visando identificar os produtos
endógenos da ação proteolítica das proteases sobre estes substratos.
A protease P3 purificada a partir de SDS-PAGE 12% com gelatina e de géis
SDS/SDS-PAGE com gelatina foi incubada com o substrato (66 kDa) purificado a partir
de géis nativos 10%. O sobrenadante contendo o substrato purificado escolhido foi
congelado a -20º C, para servir como controle. O restante do sobrenadante foi
adicionado à P3 e o ensaio foi mantido a 37 ºC, sendo retirado em diferentes intervalos
de tempo (4, 16, 25, 90 e 120h) e congelados a -20 ºC. Como outro controle, uma
alíquota do sobrenadante contendo o substrato não foi adicionada à protease, porém
também foi mantida na estufa, e retirada nos mesmos intervalos de tempo.
Posteriormente, foi realizada com estas amostras uma eletroforese em SDS-PAGE
gradiente 5 - 15%.
Também foram realizados ensaios de incubação das proteases P1, P2, P3 e P4,
purificadas a partir de SDS-PAGE 12% com gelatina uni- e bidimensionais, com o
substrato S2 purificado a partir de géis nativo/ SDS 12%. E também ensaios das
proteases P1, P2, P3 e P4 purificadas a partir de géis nativos 10% com os substratos
S1, S2, S3 e S8 purificados a partir de géis nativo/ SDS12%. Neste caso o tempo de
incubação foi fixo com 72 h de duração. Os resultados dos ensaios foram visualizados
em SDS-PAGE 12% com e sem gelatina e em géis gradiente de poliacrilamida 12 -18%
na presença de uréia gradiente 3,5 – 7,0 M e SDS 0,1% (v/v) (modificado HASHIMOTO
et al., 1983). Os géis foram corados com azul de Coomassie, descorados em
microondas. Alguns géis foram corados para prata depois de descorados.

34
14. Western blotting
As amostras de fluido das diferentes regiões epididimárias foram submetidas à
SDS-PAGE 10%. Ao término da análise, as proteínas foram eletrotransferidas a 100V,
durante 1h e 30 min, para uma matriz de nitrocelulose com poros de 0,45 µm, de acordo
com TOWBIN et al., (1979), utilizando um equipamento de transferência úmida da
BioRad e tampão fosfato 50 mM pH 7,4. Após a transferência das proteínas, a
membrana foi incubada com solução bloqueadora (5% de leite desnatado – Molico
Nestlê - em PBS) durante 1 h sob agitação a temperatura ambiente. Em seguida, a
membrana foi incubada com anticorpos policlonais anti-metaloproteases de serpente
(cedidos pelo Prof. Milton Masahiko) por 2 h a temperatura ambiente. A membrana foi
lavada três vezes em PBS Tween 0,005% (v/v). Após a lavagem, foi incubada com o
anticorpo secundário anti-IgG de camundongo conjugado com peroxidase (Southern
Biotechnology Associates, Inc.), diluído a 1:200, por 1 h. A membrana foi lavada três
vezes, nas mesmas condições descritas acima. Posteriormente, a membrana foi
revelada com DAB.
15. Análise densitométrica
A análise densitométrica dos géis foi realizada no programa computacional "Gel
Perfect" (BOZZO & RETAMAL, 1991; RETAMAL et al., 1999), após obtenção de uma
imagem cinza em formato TIFF através de um scanner comercial em 400 dpi. As
massas moleculares relativas foram obtidas por comparação da sua mobilidade
eletroforética com proteínas de massa molecular conhecida, que co-migraram no
mesmo gel. O programa calcula a mobilidade relativa (Rf) de cada banda e a área
ocupada por ela, dando também uma representação diagramática das bandas protéicas
e sua concentração relativa em relação ao total de proteínas por canal.

35
V. RESULTADOS
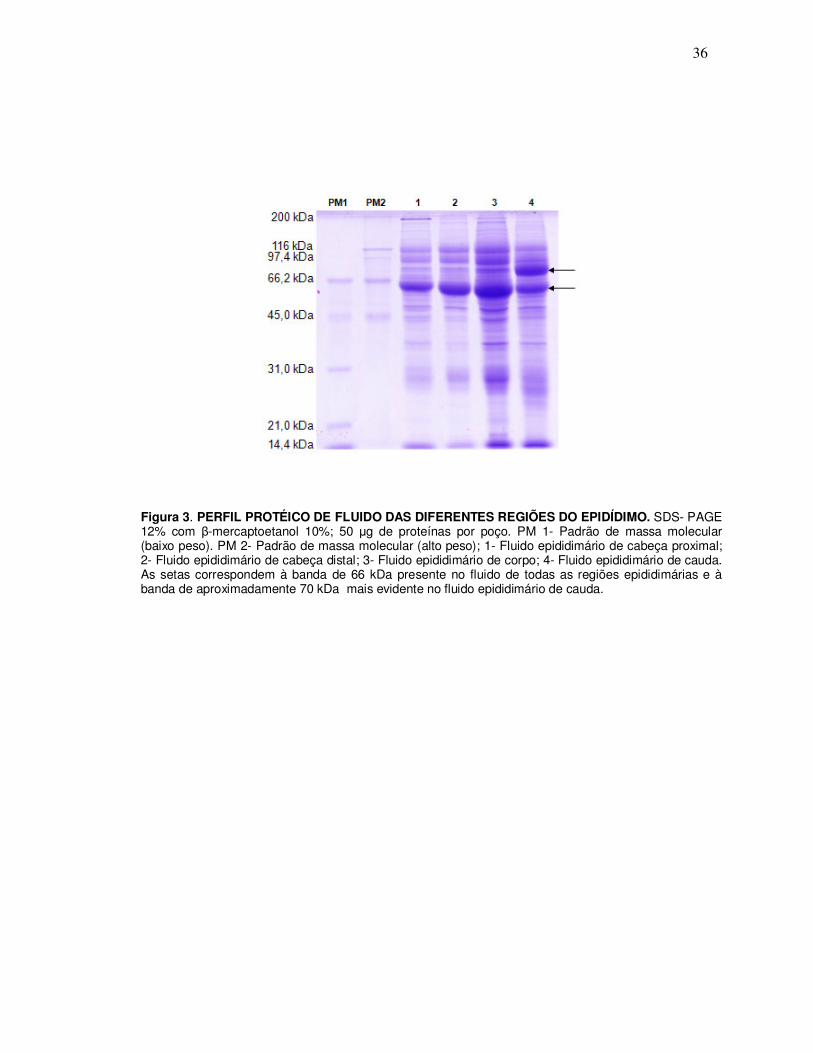
1. Análise do perfil eletroforético de amostras de fluido epididimário
Proteínas de fluido epididimário apresentaram heterogeneidade nas diferentes
regiões do conduto epididimário. Algumas proteínas da região de cauda epididimária
aparecem em menor quantidade nas regiões proximais (cabeça proximal e distal)
deste órgão. Há uma proteína de, aproximadamente 66 kDa que se distribui por todas
as regiões epididimárias, e outra, de aproximadamente 70 kDa, que é mais intensa na
região de cauda (Figura 3).
2. Efeito da proteólise endógena no fluido epididimário
A proteólise endógena no fluido epididimário foi testada através da incubação
(37ºC) de fluido de cabeça proximal, cabeça distal, corpo e cauda em diferentes
intervalos tempos. O efeito da proteólise endógena foi analisado em SDS-PAGE 15%
contendo β-mercaptoetanol 10%. Os resultados demonstraram que a concentração de
algumas proteínas aumenta e de outras diminui à medida que aumenta o tempo de
incubação (Figura 4). Este efeito é mais evidente no fluido da região de cauda e menos
pronunciado na região de cabeça proximal. Este experimento sugere fortemente a
ocorrência de atividade proteolítica endógena no fluido.
36
Figura 3. PERFIL PROTÉICO DE FLUIDO DAS DIFERENTES REGIÕES DO EPIDÍDIMO. SDS- PAGE 12% com β-mercaptoetanol 10%; 50 µg de proteínas por poço. PM 1- Padrão de massa molecular (baixo peso). PM 2- Padrão de massa molecular (alto peso); 1- Fluido epididimário de cabeça proximal; 2- Fluido epididimário de cabeça distal; 3- Fluido epididimário de corpo; 4- Fluido epididimário de cauda. As setas correspondem à banda de 66 kDa presente no fluido de todas as regiões epididimárias e à banda de aproximadamente 70 kDa mais evidente no fluido epididimário de cauda.

37
Figura 4. PROTEÓLISE ENDÓGENA NO FLUIDO EPIDIDIMÁRIO DE DIFERENTES REGIÕES DE EPIDÍDIMO. SDS-PAGE 15%; com β-mercaptoetanol 10%; 50 µg de proteínas por poço de fluido epididimário de cabeça proximal, fluido epididimário de cabeça distal, fluido epididimário de corpo e fluido epididimário de cauda previamente incubadas a 370C por diferentes intervalos tempos: 1- 0h; 2 - 0,5h; 3- 1h; 4- 3h; 5-6h; 6- 12h; 7-24h; 8-36h; 9-48h; 10-72h. As setas negras indicam as proteínas que tiveram a sua concentração diminuída e as setas vermelhas indicam as proteínas que tiveram sua concentração aumentada.

38
3. Detecção e caracterização de proteases epididimárias
3.1 Atividade proteásica no fluido epididimário, ação de inibidores sobre
as atividades proteolíticas e verificação do pH ótimo
As amostras de fluido epididimário das regiões de cabeça proximal, cabeça
distal, corpo e cauda de diferentes cavalos, foram submetidas a SDS-PAGE 12%,
gelatina 0,4% e β-mercaptoetanol 10% (Figura 5). Nossos resultados confirmaram a
presença de proteases com atividade gelatinolítica nas quatro regiões epididimárias,
com pequenas diferenças entre o perfil de atividade proteásica das amostras dos
diferentes cavalos (Figura 5A). Além disso, foram mostradas de três a cinco bandas
majoritárias de proteases com uma expressão diferencial entre as diferentes regiões
epididimárias estudadas (Figura 5B), sendo mais intensa a atividade no fluido da região
de cauda epididimária, onde foram detectadas aproximadamente cinco bandas de
atividade proteolítica (Figura 6).
A caracterização da atividade proteolítica das diferentes regiões epididimárias,
através do uso de diferentes inibidores indica que as enzimas detectadas são
metaloproteases, já que o EDTA interfere na atividade (Figura 7).
Além disso, o ensaio de incubação a diferentes pHs demonstrou que a atividade
da enzima é mantida em uma ampla faixa de pH (pH 6,5 – 10), sendo o pH ótimo em
torno de 7,5 (Figura 8).

39
Figura 5. ATIVIDADE PROTEÁSICA DE FLUIDO EPIDIDIMÁRIO ISOLADO DE DIFERENTES CAVALOS. A- SDS-PAGE 12% com gelatina 0,4%, sem β-mercaptoetanol. Cavalo A, Cavalo B, Cavalo C e Cavalo D, 50 µg de proteínas por poço de: 1- Fluido epididimário da região de cabeça proximal; 2- Fluido epididimário da região de cabeça distal; 3- Fluido epididimário de corpo; 4- Fluido epididimário de cauda. B- Análise densitográfica do gel contendo amostras das diferentes regiões epididimárias do Cavalo C.

40
Figura 6. ANÁLISE QUANTITATIVA DA ATIVIDADE PROTEOLÍTICA EM FLUIDO EPIDIDIMÁRIO. Gel SDS-PAGE 10% sem β-mercaptoetanol contendo proteínas das diferentes regiões epididimárias junto aos resultados da análise quantitativa de cada atividade. A- Fluido epididimário de cabeça proximal; B- Fluido epididimário de cabeça distal; C- Fluido epididimário de corpo e D- Fluido epididimário de cauda.
Figura 7. EFEITO DE INIBIDORES SOBRE PROTEASES DO FLUIDO DAS DIFERENTES REGIÕES EPIDIDIMÁRIAS. SDS-PAGE 10% na presença de gelatina 0,4% incubados com diferentes inibidores: EDTA 10mM, Iodoacetamida 10mM e PMSF 100µM; 1- Fluido epididimário da região de cabeça proximal; 2- Fluido epididimário da região de cabeça distal; 3- Fluido epididimário de corpo; 4- Fluido epididimário de cauda.

41
Figura 8. ENSAIO DE VERIFICAÇÃO DO PH ÓTIMO DE PROTEASES DE FLUIDO. SDS-PAGE 10% na presença de gelatina 0,4% incubados a diferentes pHs: 2,5; 4,5; 6,5; 7,5; 10 e 12. 1- Fluido epididimário da região de cabeça proximal; 2- Fluido epididimário da região de cabeça distal; 3- Fluido epididimário de corpo; 4- Fluido epididimário de cauda.
4. Determinação da constante de retardação de proteases: eletroforeses de
proteínas epididimárias em géis desnaturantes na presença de gradiente de
gelatina 0 - 0,6%
Quando as proteínas das diferentes regiões epididimárias foram submetidas a
SDS-PAGE 10% gradiente de gelatina 0 – 0,6%, com uma faixa de SDS-PAGE 10%
sem gelatina (Figura 9) foi possível determinar o grau de anomalia na migração
eletroforética causada pela presença do polímero de gelatina. A análise
densitográfica permitiu verificar a linearidade do gradiente de gelatina co-polimerizada
com acrilamida (Figura 9 B e C). Os géis gradiente de gelatina de fluido das diferentes
regiões demonstraram que há um retardo na migração das proteases à medida que a
concentração de gelatina aumenta (Figura 10). Os gráficos de Ferguson para a
determinação da constante de retardação permitiram a obtenção do Kr (constante de
retardação) de cada banda proteásica conforme indicado na Figura 11 A, B, C e D. No
gráfico se visualiza a superposição de géis em relação às suas massas moleculares e
se observa claramente o efeito de retardação na migração das proteases (Figura 11
E).

42
Figura 9. GRADIENTE DE GELATINA CO-POLIMERIZADO COM ACRILAMIDA 10%. A- Gel gradiente de gelatina (0-0,6%), acrilamida 10% com uma faixa larga de gel de acrilamida 10% sem gelatina polimerizada ao lado do gel gradiente. B e C- Gráficos obtidos da varredura da imagem com o programa computacional “Gel Perfect”, indicando a linearidade do gradiente de gelatina co-polimerizada com acrilamida.
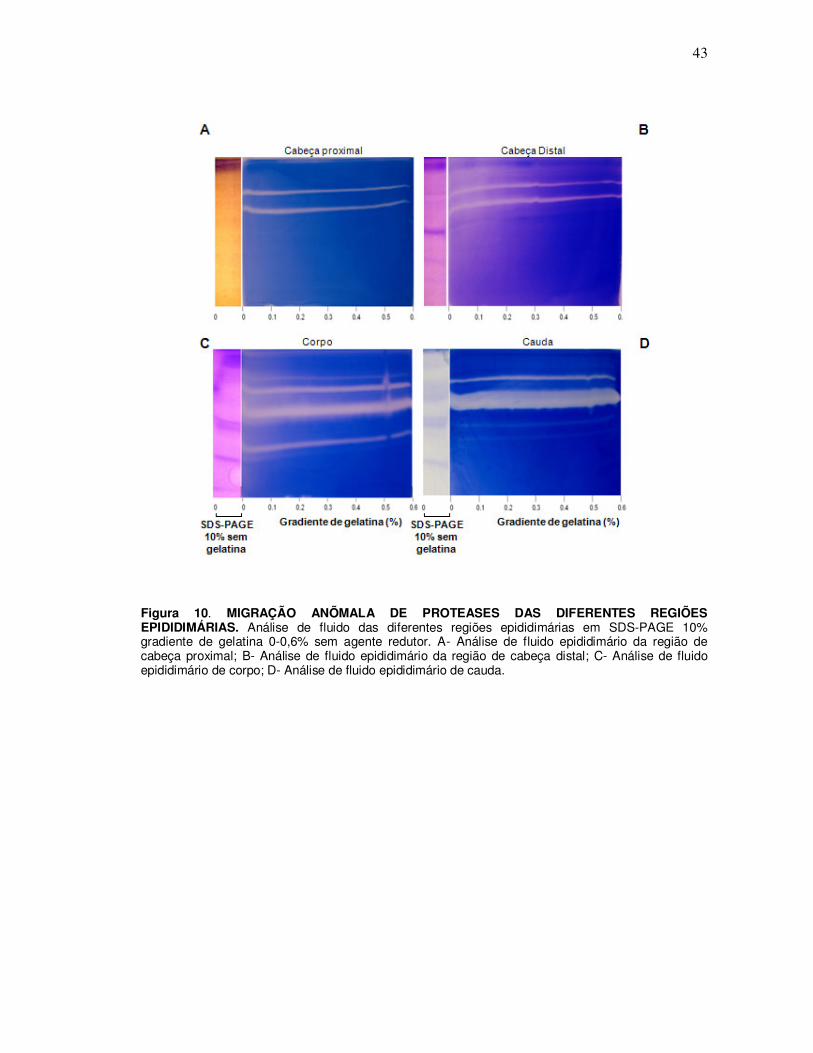
43
Figura 10. MIGRAÇÃO ANÕMALA DE PROTEASES DAS DIFERENTES REGIÕES EPIDIDIMÁRIAS. Análise de fluido das diferentes regiões epididimárias em SDS-PAGE 10% gradiente de gelatina 0-0,6% sem agente redutor. A- Análise de fluido epididimário da região de cabeça proximal; B- Análise de fluido epididimário da região de cabeça distal; C- Análise de fluido epididimário de corpo; D- Análise de fluido epididimário de cauda.

44
Figura 11. DETERMINAÇÃO DA CONSTANTE DE RETARDAÇÃO (Kr) DAS PROTEASES DE DIFERENTES REGIÕES DO EPIDÍDIMO. GRÁFICOS DE FERGUSON A-B-C-D. No gráfico (E) estão representados os Kr obtidos de cauda em relação às suas massas moleculares e se observa que existe o efeito de retardação na migração das proteases. Estes gráficos foram construídos com base na migração das proteases em géis de acrilamida 10%, gradiente de gelatina (0-0,6%).

45
4.1 Comparação de proteínas/atividades proteásicas em géis bidimensionais
SDS/SDS e SDS/gelatina 0,4%
Outra abordagem para determinação do grau de anomalia na migração das
proteases, causada pela presença de gelatina, foi a sobreposição de géis 2D
desnaturantes com e sem gelatina 0,4% das proteínas de fluido epididimário das
diferentes regiões. Estes géis (SDS/SDS e SDS/SDS com gelatina) foram sobrepostos
em imagens no programa computacional Photo Impact, onde algumas foram
invertidas (Figura 12). Nossos resultados confirmam a anomalia na migração das
proteases em géis com gelatina em relação aos géis sem gelatina. Além disso, foi feito
um gráfico para ilustrar de maneira didática a variação no coeficiente angular das retas
de diferentes Rf (Figura 12.3), onde foram utilizadas as imagens dos géis de proteínas
e proteases de fluido de cauda sobrepostas. Conforme a massa molecular das
proteínas decresce, o grau de desvio aumenta, o que significa que o polímero de
gelatina altera mais efetivamente a migração de proteínas de massas moleculares
pequenas. Esta técnica permite também localizar proteases em géis que não contêm
gelatina, com a prévia correção do valor do coeficiente angular das retas que originam
as proteínas.

46
Figura 12. MIGRAÇÃO ANÔMALA DE PROTEASES: SOBREPOSIÇÃO DE GÉIS 2D DESNATURANTES COM E SEM GELATINA 0,4%. A amostra de fluido epididimário de cauda foi analisada em géis 2D: SDS-PAGE 12% /SDS-PAGE 12% e SDS-PAGE 12% /SDS-PAGE 12% com gelatina. 1- sem β-mercaptoetanol. 2- com o agente redutor e as imagens destes géis foram sobrepostas. 1a e 2a- imagens invertidas no Photo Impact
® das imagens 1b e 2b, respectivamente. PM- Padrão de massa molecular. 3- gráfico da variação no coeficiente angular das retas de diferentes Rfs, conseqüência da presença de gelatina (amostra de fluido de cauda).

47
5. Detecção da atividade proteásica de fluido epididimário na ausência e
presença de agente redutor
A atividade proteásica de fluido de cabeça proximal (Figura 13A), cabeça
distal (Figura 13B), corpo (Figura 13C) e cauda (Figura 13D) foi detectada em géis
bidimensionais SDS-PAGE 12% / SDS-PAGE 12% com gelatina 0,4%, com e sem β-
mercaptoetanol na 2D (Figura 13). Os resultados demonstraram que o perfil
eletroforético das bandas de atividade é alterado na presença do agente redutor, o
que sugere que estas proteases possuem pontes dissulfetos (RS-SR) em sua
estrutura e podem estar formando agregados. A sobreposição das imagens dos géis
2D SDS/SDS com SDS/SDS gelatina 0,4% (com e sem agente redutor), das
proteínas das diferentes regiões epididimárias, demonstrou um retardo na migração
das proteases (migração anômala) causado pela presença do polímero de gelatina
co-polimerizado com a solução de poliacrilamida, como já observado no resultado
anterior (Figura 12).

48
Figura 13A. ANÁLISE 2D DE FLUIDO EPIDIDIMÁRIO DE CABEÇA PROXIMAL. 1- Géis 2D: SDS-PAGE 12%/ SDS-PAGE 12% sem e com β-mercaptoetanol. 2- Géis SDS-PAGE 12%/ SDS-PAGE 12% na presença de gelatina sem e com β-mercaptoetanol. 3- Sobreposição das imagens (1 e 2).
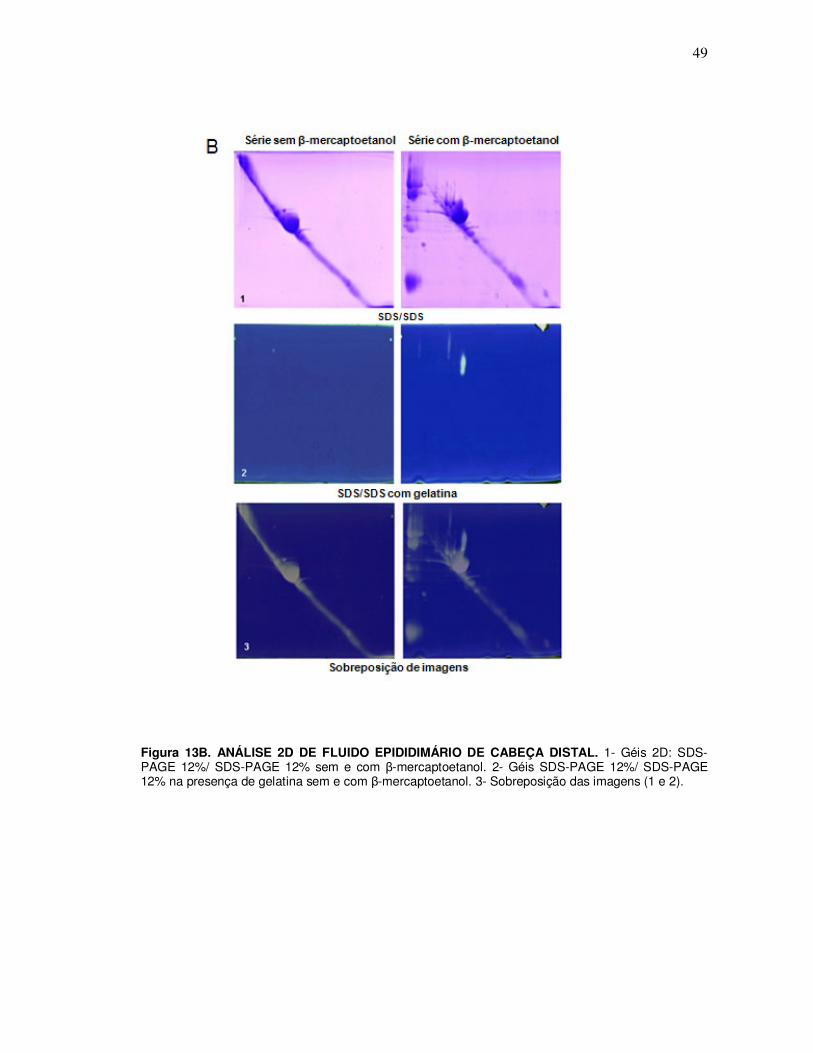
49
Figura 13B. ANÁLISE 2D DE FLUIDO EPIDIDIMÁRIO DE CABEÇA DISTAL. 1- Géis 2D: SDS-PAGE 12%/ SDS-PAGE 12% sem e com β-mercaptoetanol. 2- Géis SDS-PAGE 12%/ SDS-PAGE 12% na presença de gelatina sem e com β-mercaptoetanol. 3- Sobreposição das imagens (1 e 2).

50
Figura 13C. ANÁLISE 2D DE FLUIDO EPIDIDIMÁRIO DE CORPO. 1- Géis 2D: SDS-PAGE 12%/ SDS-PAGE 12% sem e com β-mercaptoetanol. 2-. Géis SDS-PAGE 12%/ SDS-PAGE 12% na presença de gelatina sem e com β-mercaptoetanol. 3- Sobreposição das imagens (1 e 2).

51
Figura 13D. ANÁLISE 2D DE FLUIDO EPIDIDIMÁRIO DE CAUDA. 1- Géis 2D: SDS-PAGE 12%/ SDS-PAGE 12% sem e com β-mercaptoetanol. 2- Géis SDS-PAGE 12%/ SDS-PAGE 12% na presença de gelatina sem e com β-mercaptoetanol. 3- Sobreposição das imagens (1 e 2).

52
6. Determinação das massas moleculares de proteases no fluido epididimário
Devido à anomalia na migração das proteínas em géis com gelatina em relação
a géis sem gelatina (HUMMEL et al., 1996), a determinação da massa molecular
relativa das proteases das diferentes regiões epididimárias foi realizada relacionando a
mobilidade das proteínas no gel 1D (SDS-PAGE 12%) com a mobilidade das proteases
no gel 2D (SDS-PAGE 12%, gelatina 0,4%) (Figura 14). As massas moleculares das
diferentes proteases de fluido epididimário estão resumidas na Tabela II. Neste órgão,
predomina uma protease de 45 kDa, que não é observada na cabeça proximal. Na
região de cauda, foi observada atividade em uma proteína de aproximadamente 40 kDa
que não está presente nas demais regiões do epidídimo.
Tabela II- Massas moleculares relativas das proteases de fluido das diferentes
regiões epididimárias.
Região do Epidídimo Massas moleculares relativas
Cabeça Proximal 117, 100 e 62 kDa
Cabeça Distal 117, 62 e 45 kDa
Corpo 117, 62 e 45 kDa
Cauda 117, 62, 45 e 40 kDa

53
Figura 14. DETERMINAÇÃO DAS MASSAS MOLECULARES DE PROTEASES DE FLUIDO EPIDIDIMÁRIO. Géis SDS-PAGE 12%/SDS-PAGE 12%, gelatina 0,4% com β-mercaptoetanol. A- Fluido epididimário de cabeça proximal. B- Fluido epididimário de cabeça distal. C- Fluido epididimário de corpo e D- Fluido epididimário de cauda. PM- Padrão de massa molecular.

54
7. Purificação das Proteases
7.1 Purificação de proteases a partir de géis unidimensionais
As atividades proteolíticas da região de cauda foram isoladas mediante excisão da
proteína do gel (RETAMAL et al., 1999), a partir de SDS-PAGE 12% contendo gelatina,
utilizando-se como referência para o corte das bandas tanto a visualização em uma
lâmpada UV, quanto a comparação com uma tira lateral do gel corada com azul de
Coomassie (Figura 15A). As proteases purificadas foram visualizadas em SDS-PAGE
12% com gelatina (Figura 15B). Foi possível observar que, apesar das proteases
isoladas apresentarem diferentes massas moleculares, as 4 bandas majoritárias obtidas,
passaram a apresentar o mesmo perfil de atividades (indicado pelas setas). Comparando
o SDS-PAGE 12% com o SDS-PAGE 12% com gelatina 0,4% é possível observar que as
proteases possuem uma alta atividade específica, pois, apesar de não estarem visíveis,
mesmo após o SDS-PAGE 12% ter sido corado com prata (Figura 15C), ainda assim
apresentaram atividade gelatinolítica.

55
Figura 15. PROTEASES PURIFICADAS DE FLUIDO EPIDIDIMÁRIO DE CAUDA A PARTIR DE GÉIS UNIDIMENSIONAIS. A- SDS-PAGE 12%, gelatina 0,4%, visualizado em UV e, posteriormente, corado com azul de Coomassie, utilizados como parâmetro para a excisão das proteases. B- Análise das proteases purificadas em gel SDS-PAGE 12%, gelatina 0,4%. C- Análise das proteases purificadas em gel SDS-PAGE 12%. P1- Protease 1, P2- Protease 2, P3- Protease 3, P4- Protease 4. PM- Padrão de Massa Molecular. UV- Luz ultravioleta. As setas apontam para as atividades proteolíticas.

56
7.2 Purificação de proteases a partir de géis 2D SDS/SDS com gelatina
Para obter proteases melhor purificadas, as manchas de atividade proteásica do
fluido epididimário de cauda também foram isoladas a partir de géis SDS/SDS-PAGE
12% gelatina 0,4% (Figura 16A). Após purificadas estas foram visualizadas em SDS-
PAGE 12% com gelatina. De forma semelhante aos resultados anteriores, as 4
manchas de diferentes proteases, após a re-eletroforese, apresentaram o mesmo perfil
de atividade proteásica (indicado pelas setas), ou seja, a mesma massa molecular
(Figura 16B e C). Comparando a análise das proteases purificadas em SDS-PAGE 12%
com gelatina (Figura 16B e C) e sem gelatina (Figura 16D) é possível observar,
novamente, que apesar de não estarem visíveis, mesmo após o SDS-PAGE 12% ter sido
corado com prata, ainda assim apresentaram atividade gelatinolítica.

57
Figura 16. PROTEASES PURIFICADAS DE FLUIDO EPIDIDIMÁRIO DE CAUDA A PARTIR DE GÉIS BIDIMENSIONAIS. A- Esquema da purificação das proteases de fluido epididimário de cauda, a partir de géis SDS-PAGE 12%/SDS-PAGE 12% gelatina 0,4%. Este gel 2D foi corado para servir como parâmetro para o corte das manchas de atividade. B- Análise das proteases purificadas em SDS-PAGE 12%, gelatina 0,4%. C- Imagem invertida de B. D- Análise das proteases purificadas em SDS-PAGE 12% corado com prata. P1- Protease 1, P2- Protease 2, P3- Protease, P4- Protease 4. PM- Padrão de Massa Molecular.

58
7.3 Purificação de proteases a partir de géis nativos 10%
As proteases também foram purificadas a partir de géis em condições nativas
para verificar se a gelatina poderia interferir na migração destas. Primeiramente, foi
realizada uma eletroforese (1D) em gel nativo 10% das amostras de fluido epididimário
e, em seguida, uma segunda eletroforese (2D) em gel desnaturante 12% com gelatina
0,4% (Figura 17). Observando o gel 2D foi possível verificar manchas de atividade
proteolítica de mesma massa molecular (setas vermelhas), correspondentes a
diferentes proteínas em condições nativas (vários rfs). Também foi possível observar
uma única mancha de atividade, de massa molecular maior que as demais proteases
(setas negras), correspondente a um rf em condições nativas (seta verde). Analisando a
Figura 17, é possível notar que este referido rf possui duas atividades proteásicas. Além
disso, é possível observar um aumento na intensidade das atividades nas regiões de
corpo e cauda quando comparadas à região de cabeça proximal.
Para purificar as proteases observadas na 2D estas foram correlacionadas às
proteínas no gel 1D (Figura 18A). As mesmas, após a purificação, foram analisadas em
SDS-PAGE 12% com gelatina 0,4% (Figura 18B) e os resultados demonstraram que,
apesar de terem sido purificadas de proteases de diferentes rfs em condições nativas,
estas apresentaram o mesmo perfil de atividade proteásica, mesmo rf e,
conseqüentemente, a mesma massa molecular no gel de gelatina. Além disso, foi
comparado o perfil das proteases purificadas com o perfil do SDS-PAGE 12 % sem
gelatina (Figura 18C) e foi observado que estas proteínas se encontram em baixa
concentração, conforme já observado anteriormente.

59
Figura 17. LOCALIZAÇÃO DAS PROTEASES EM GÉIS NATIVOS. Géis Nativo 10%/ SDS-PAGE 12% gelatina 0,4% utilizados para a localização das atividades proteolíticas em condições nativas. Foram localizadas as atividades de fluido das diferentes regiões epididimárias: A- Cabeça Proximal, B- Cabeça Distal, C- Corpo e D- Cauda. As setas vermelhas indicam as diferentes manchas de atividade de mesma massa molecular e a seta negra indica uma mancha de atividade de maior massa molecular, encontrada no Rf indicado pela seta verde.

60
Figura 18. PROTEASES PURIFICADAS DE FLUIDO EPIDIDIMÁRIO DE CAUDA A PARTIR DE GÉIS NATIVO 10%. A- Esquema da purificação das proteases de fluido epididimário de cauda a partir de géis Nativo 10%. Gel corado para servir como parâmetro para o corte das bandas de atividade. B- Análise das proteases purificadas em SDS-PAGE 12%, gelatina 0,4%. C- Análise das proteases purificadas em SDS-PAGE 12% corado com azul de Coomassie. P1- Protease 1, P2 – Protease 2, P3- Protease, P4- Protease 4. PM- Padrão de Massa Molecular.

61
8. Detecção dos substratos endógenos
A detecção dos substratos endógenos foi realizada mediante a incubação de
proteases isoladas com o fluido epididimário de cauda a 37 ºC por 72 h. A proteólise
resultante da incubação do próprio fluido, com as proteases isoladas a partir de géis
bidimensionais foi analisada em SDS-PAGE gradiente 5-15% (Figura 19). Foram
observadas proteínas de aproximadamente 105, 66 kDa e duas de massa molecular
entre 30 e 15 kDa que foram provavelmente proteolisadas, já que estas apresentaram
sua concentração diminuída (setas vermelhas). Também foram observadas proteínas de
aproximadamente 35 e 32 kDa, que apresentaram sua concentração aumentada, o que
sugere que sejam produtos da ação proteolítica.
A proteólise resultante da incubação do próprio fluido, com as proteases isoladas
a partir de géis unidimensionais foi analisada em SDS-PAGE gradiente 6-15% e em gel
nativo gradiente 6-15% (Figura 20). Os dados demonstram uma série de proteínas que
diminuem a concentração ou enfraquecem (setas vermelhas) e proteínas que aumentam
a concentração ou aparecem (setas roxas). Foi possível notar uma maior diferença entre
o perfil protéico, das amostras proteolisadas e não proteolisadas, em condições
desnaturantes (Figura 20A) que em condições nativas (Figura 20B).
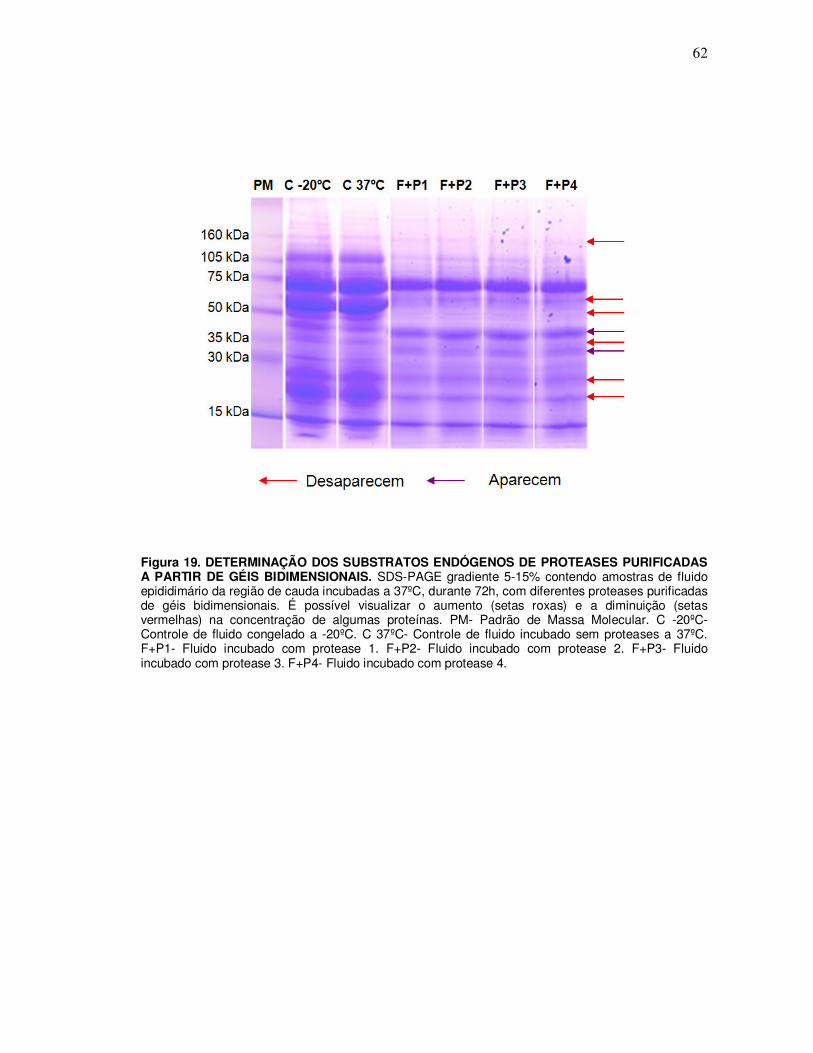
62
Figura 19. DETERMINAÇÃO DOS SUBSTRATOS ENDÓGENOS DE PROTEASES PURIFICADAS A PARTIR DE GÉIS BIDIMENSIONAIS. SDS-PAGE gradiente 5-15% contendo amostras de fluido epididimário da região de cauda incubadas a 37ºC, durante 72h, com diferentes proteases purificadas de géis bidimensionais. É possível visualizar o aumento (setas roxas) e a diminuição (setas vermelhas) na concentração de algumas proteínas. PM- Padrão de Massa Molecular. C -20ºC- Controle de fluido congelado a -20ºC. C 37ºC- Controle de fluido incubado sem proteases a 37ºC. F+P1- Fluido incubado com protease 1. F+P2- Fluido incubado com protease 2. F+P3- Fluido incubado com protease 3. F+P4- Fluido incubado com protease 4.

63
Figura 20. DETERMINAÇÃO DOS SUBSTRATOS ENDÓGENOS DE PROTEASES PURIFICADAS A PARTIR DE GÉIS UNIDIMENSIONAIS. Géis contendo amostras de fluido epididimário da região de cauda incubadas a 37ºC, durante 72h, com diferentes proteases purificadas de géis unidimensionais. É possível visualizar o aumento (setas roxas) e a diminuição (setas vermelhas) na concentração de algumas proteínas. A- SDS-PAGE gradiente 6-15%; B- Gel nativo gradiente 6-15%; A- Albumina; FF- Fluido mantido como controle no freezer -20ºC; FE- Fluido mantido como controle na estufa 37ºC. P1+F- Protease 1 + fluido epididimário de cauda. P2+F- Protease 2 + fluido epididimário de cauda. P3+F Protease 3 + fluido epididimário de cauda. P4+F- Protease 4 + fluido epididimário de cauda.
O resultado também foi visualizado em géis Nativo/SDS 6-15% (Figura 21), o
que permitiu uma visualização mais clara das proteínas que aparecem (setas roxas) e
desaparecem (setas vermelhas), e demonstra que proteínas de massa molecular 117,
80, 69, 57, 50, 44 e 35kDa desaparecem e proteínas de massa molecular 50 e 44kDa,
bem como uma mancha que engloba 32,8, 33,5 e 35kDa aparecem (Figura 22). Estes
géis foram analisados tanto em função do Rf das proteínas nas condições nativas da 1ª
Dimensão (Figura 23) quanto em função do Rf das proteínas nas condições
desnaturantes da 2ª Dimensão (Figura 24). Isto permitiu uma verificação quantitativa
das proteínas de fluido epididimário antes e após a adição de proteases.
O ensaio de incubação de proteases e fluido epididimário de cauda foi mantido a
-20ºC durante alguns dias, após ter sido visualizado nos géis das figuras 21 a 24, e foi
novamente visualizado em SDS-PAGE gradiente 6-15% (Figura 25). Foi verificado que as
proteínas apresentaram um comportamento diferenciado. Após o congelamento,
proteínas de alta massa molecular que não haviam desaparecido tiveram a sua

64
concentração diminuída, como a proteína de 66kDa. Além disso, apareceram proteínas
de massa molecular entre 31 e 21 kDa.
Figura 21. VISUALIZAÇÃO DOS SUBSTRATOS ENDÓGENOS DE PROTEASES PURIFICADAS A PARTIR DE GÉIS BIDIMENSIONAIS. Visualização do experimento realizado com proteases isoladas de géis bidimensionais SDS/SDS com gelatina. Gel SDS-PAGE 6-15%. É possível visualizar o aumento (setas roxas) e a diminuição (setas vermelhas) na concentração de algumas proteínas. PM- Padrão de Massa Molecular (alto e baixo peso). FF- Fluido mantido como controle no freezer -20ºC. FE- Fluido mantido como controle na estufa 37ºC. P1+F- Protease 1 + fluido epididimário de cauda; P2+F- Protease 2 + fluido epididimário de cauda; P3+F Protease 3 + fluido epididimário de cauda; P4+F Protease 4 + fluido epididimário de cauda;

65
Figura 22. VISUALIZAÇÃO DOS SUBSTRATOS ENDÓGENOS EM GÉIS NATIVO/SDS. Géis bidimensionais Nativo/SDS 6-15% para a análise dos ensaios de proteases isoladas a partir de géis bidimensionais SDS/SDS com gelatina. Géis controle contendo amostras de fluido epididimário de cauda incubadas a 37ºC e géis contendo amostras de proteases isoladas da região de cauda + Fluido Epididimário de Cauda. P1 + F - Protease 1 + Fluido de Cauda; P2 + F - Protease 2 + Fluido de Cauda; P3 + F - Protease 3 + Fluido de Cauda. É possível visualizar o aumento (setas roxas) e a diminuição (setas vermelhas) na concentração de algumas proteínas.
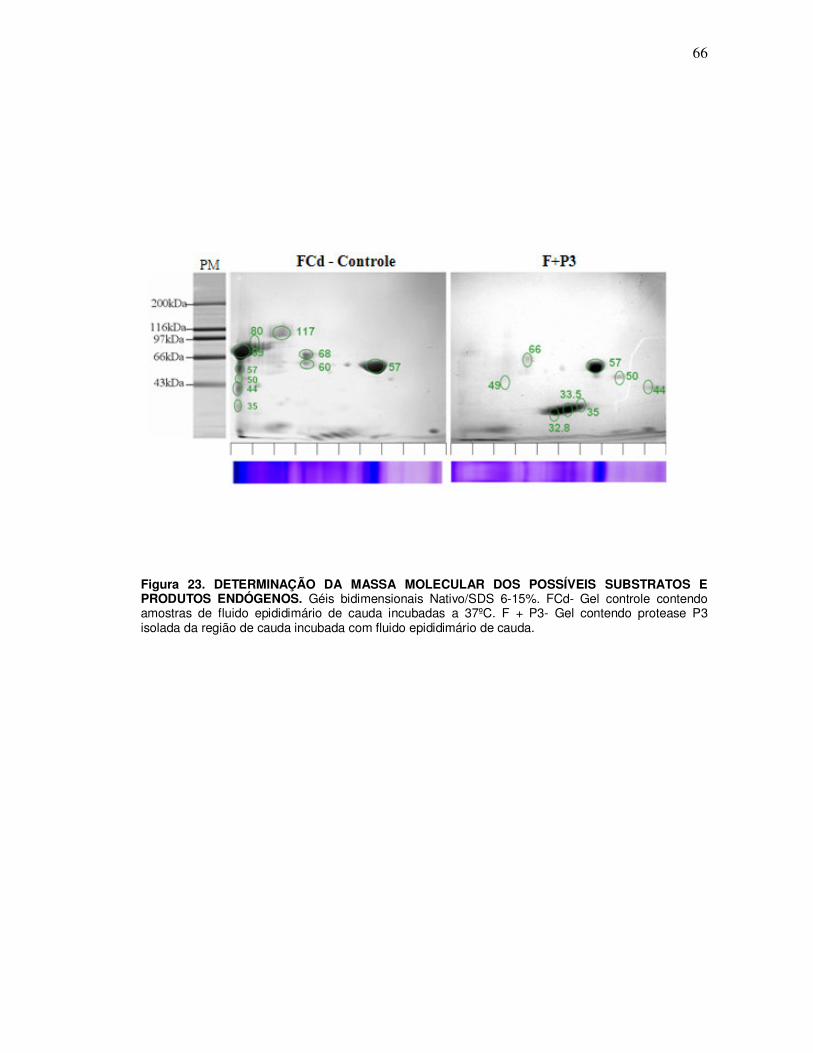
66
Figura 23. DETERMINAÇÃO DA MASSA MOLECULAR DOS POSSÍVEIS SUBSTRATOS E PRODUTOS ENDÓGENOS. Géis bidimensionais Nativo/SDS 6-15%. FCd- Gel controle contendo amostras de fluido epididimário de cauda incubadas a 37ºC. F + P3- Gel contendo protease P3 isolada da região de cauda incubada com fluido epididimário de cauda.
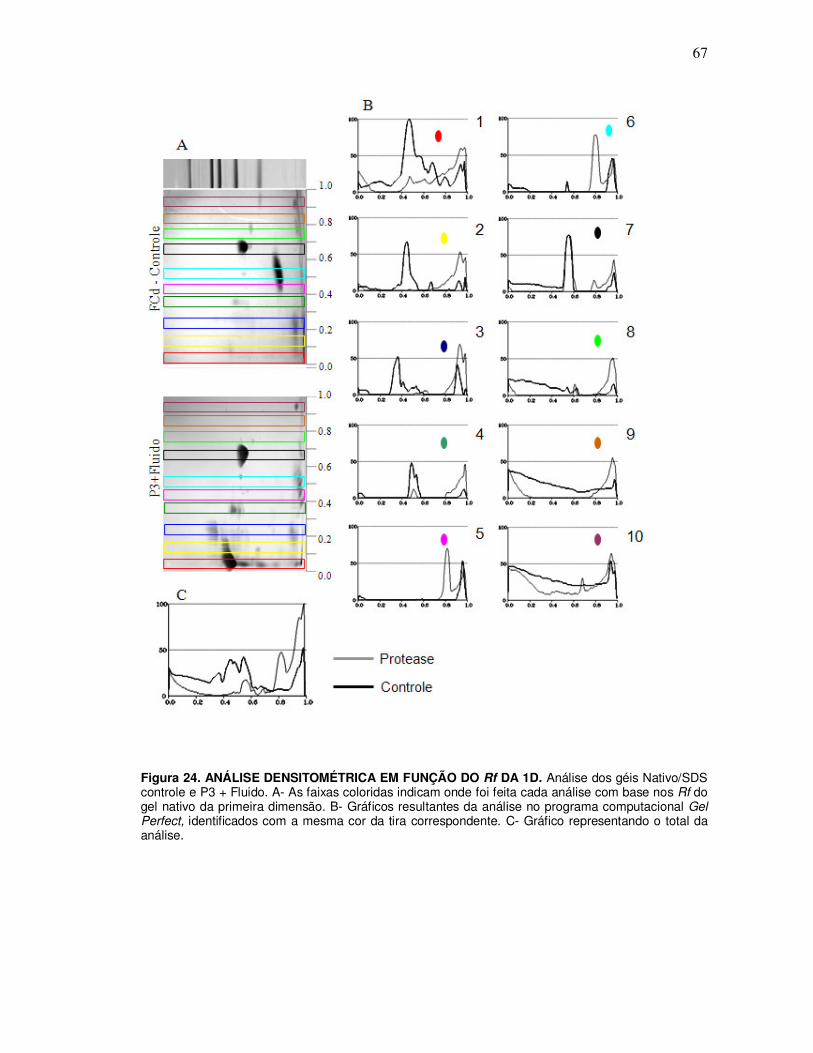
67
Figura 24. ANÁLISE DENSITOMÉTRICA EM FUNÇÃO DO Rf DA 1D. Análise dos géis Nativo/SDS controle e P3 + Fluido. A- As faixas coloridas indicam onde foi feita cada análise com base nos Rf do gel nativo da primeira dimensão. B- Gráficos resultantes da análise no programa computacional Gel Perfect, identificados com a mesma cor da tira correspondente. C- Gráfico representando o total da análise.
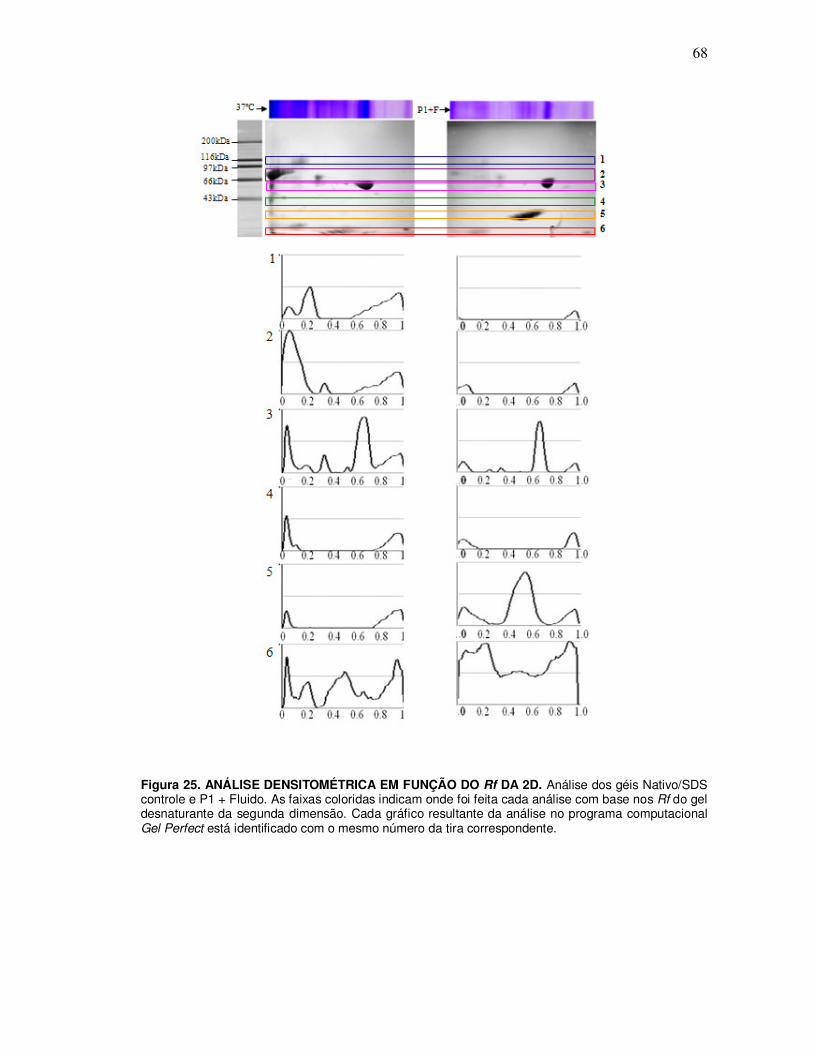
68
Figura 25. ANÁLISE DENSITOMÉTRICA EM FUNÇÃO DO Rf DA 2D. Análise dos géis Nativo/SDS controle e P1 + Fluido. As faixas coloridas indicam onde foi feita cada análise com base nos Rf do gel desnaturante da segunda dimensão. Cada gráfico resultante da análise no programa computacional Gel Perfect está identificado com o mesmo número da tira correspondente.

69
9. Purificação dos substratos endógenos das atividades proteolíticas
9.1 Purificação dos substratos endógenos a partir de géis nativos 10%
Os substratos foram isolados de géis Nativos 10% após serem localizados em
géis bidimensionais, nas quais a primeira eletroforese foi um gel de poliacrilamida nativo
10% e a segunda eletroforese um gel de poliacrilamida 5-15%, SDS 0,1%. Na Figura
26 está representado o esquema desta purificação, onde a proteína de
aproximadamente 66kDa foi purificada.
9.2 Purificação dos substratos endógenos a partir de géis nativo/SDS-
PAGE 12%
Os substratos identificados também foram isolados de géis Nativos 10%/SDS-
PAGE 12%. Ao todo foram isoladas oito proteínas candidatas a substratos,
demonstradas na Figura 27. Foi possível observar através da análise eletroforética do
substrato S2 purificado (Figura 28), que este não apresentou diferenças entre seu perfil
quando mantido na estufa a 37ºC e quando mantido no freezer -20ºC (Figura 28A). O
substrato purificado também foi analisado em SDS-PAGE 12% com gelatina 0,4% (Figura
28B) e, inesperadamente, este apresentou atividade proteolítica quando mantido a 37ºC
e a -20ºC (seta azul). No entanto, o substrato mantido a -20ºC apresentou uma segunda
banda de atividade mais intensa que no substrato mantido a 37ºC (seta negra). A seta
vermelha indica uma banda de atividade só encontrada na amostra mantida no freezer.

70
Figura 26. PURIFICAÇÃO DO SUBSTRATO A PARTIR DE GÉIS NATIVOS. A. Gel nativo 10%/SDS-PAGE 5-15% utilizado para a localização do substrato em condições nativas. A proteína da amostra de fluido epididimário de cauda de aproximadamente 66 kDa (círculo negro), foi purificada de gel nativo 10%. B. Tira corada utilizada como padrão para o corte da proteína substrato do gel nativo 1D não corado como indicado pela linha pontilhada.
Figura 27. PURIFICAÇÃO DOS SUBSTRATOS A PARTIR DE GÉIS NATIVO/SDS. Gel Nativo 10%/SDS-PAGE 12% corado com azul de Coomassie utilizado como parâmetro para a purificação dos substratos. Os substratos escolhidos para alguns ensaios estão circulados em vermelho e os substratos que não foram utilizados estão circulados em preto.

71
Figura 28. VISUALIZAÇÃO DO SUBSTRATO S2 PURIFICADO. A- Análise do substrato purificado de fluido mantido a 37ºC e a -20ºC em SDS-PAGE 12% corado com prata. B- Análise do substrato purificado mantido a 37ºC e a -20ºC em SDS-PAGE 12% com gelatina 0,4%. PM- Padrão de Massa Molecular.
10. Identificação dos produtos endógenos gerados pelas proteases purificadas
e substratos purificado
10.1 Ensaio de incubação da protease P3 purificada com substrato
(66kDa) purificado a partir do gel nativo 10%
A protease P3 isolada de géis uni- e bidimensionais com gelatina foi incubada
com o substrato (66kDa) purificado a partir de gel nativo, durante diferentes
intervalos de tempo, para verificar a proteólise deste. O resultado foi visualizado em
SDS-PAGE gradiente 5-15% (Figura 29). Foi possível verificar a diminuição gradual
de proteínas menores que 66kDa, massa aproximada do substrato purificado.
Assim, o resultado demonstra uma diminuição nas proteínas de baixo peso e não da
proteína isolada como substrato. O resultado da proteólise endógena também foi
analisado em gel gradiente de uréia e de acrilamida (Figura 30). Neste caso não foi
possível observar alterações entre o perfil protéico do substrato controle e do
substrato incubado com a protease, demonstrando que não houve proteólise
endógena.
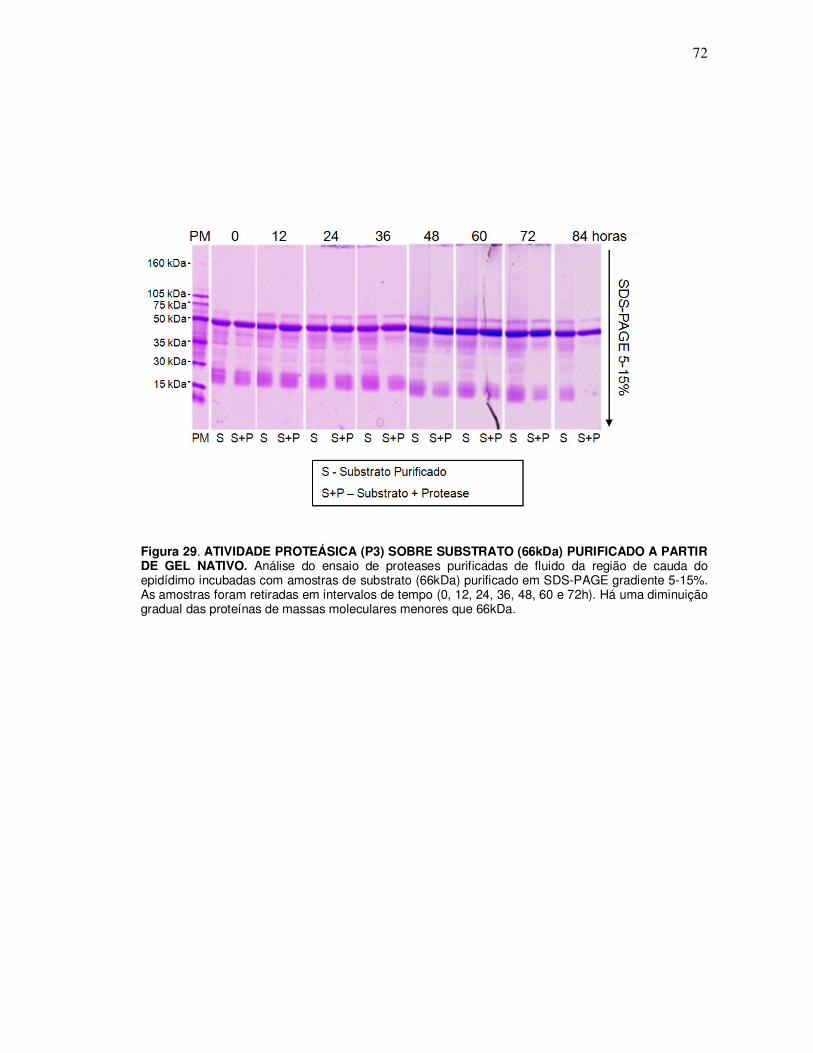
72
Figura 29. ATIVIDADE PROTEÁSICA (P3) SOBRE SUBSTRATO (66kDa) PURIFICADO A PARTIR DE GEL NATIVO. Análise do ensaio de proteases purificadas de fluido da região de cauda do epidídimo incubadas com amostras de substrato (66kDa) purificado em SDS-PAGE gradiente 5-15%. As amostras foram retiradas em intervalos de tempo (0, 12, 24, 36, 48, 60 e 72h). Há uma diminuição gradual das proteínas de massas moleculares menores que 66kDa.
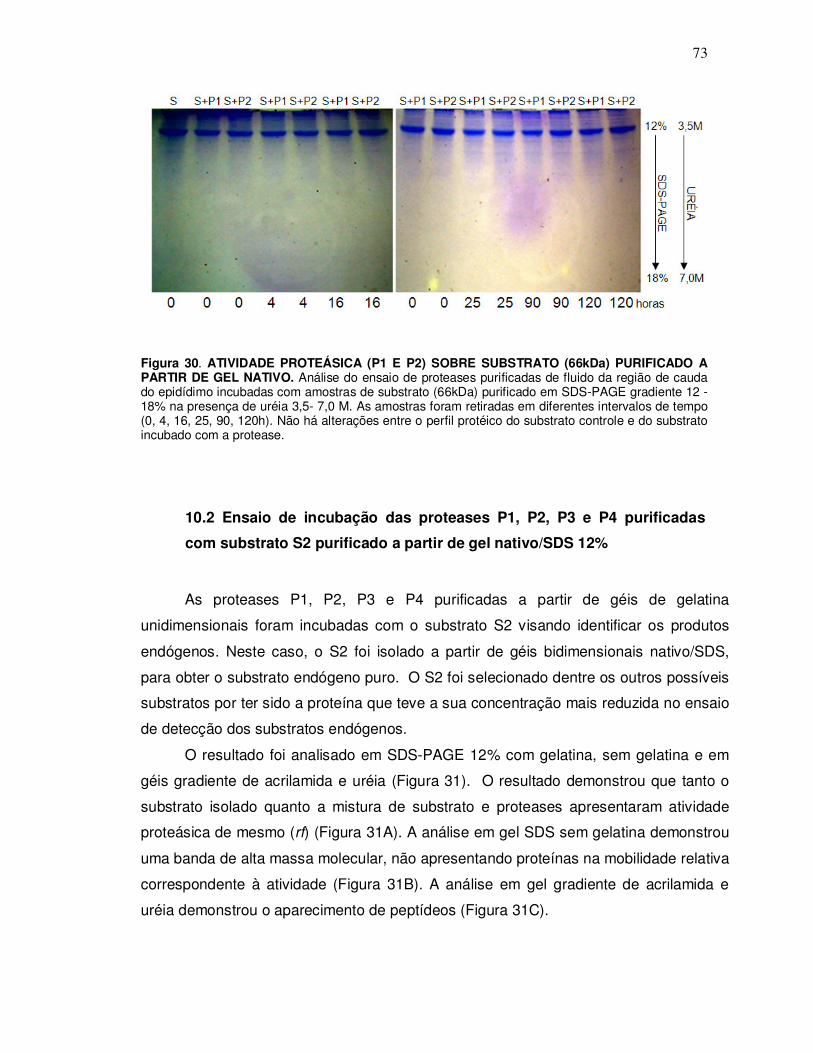
73
Figura 30. ATIVIDADE PROTEÁSICA (P1 E P2) SOBRE SUBSTRATO (66kDa) PURIFICADO A PARTIR DE GEL NATIVO. Análise do ensaio de proteases purificadas de fluido da região de cauda do epidídimo incubadas com amostras de substrato (66kDa) purificado em SDS-PAGE gradiente 12 - 18% na presença de uréia 3,5- 7,0 M. As amostras foram retiradas em diferentes intervalos de tempo (0, 4, 16, 25, 90, 120h). Não há alterações entre o perfil protéico do substrato controle e do substrato incubado com a protease.
10.2 Ensaio de incubação das proteases P1, P2, P3 e P4 purificadas
com substrato S2 purificado a partir de gel nativo/SDS 12%
As proteases P1, P2, P3 e P4 purificadas a partir de géis de gelatina
unidimensionais foram incubadas com o substrato S2 visando identificar os produtos
endógenos. Neste caso, o S2 foi isolado a partir de géis bidimensionais nativo/SDS,
para obter o substrato endógeno puro. O S2 foi selecionado dentre os outros possíveis
substratos por ter sido a proteína que teve a sua concentração mais reduzida no ensaio
de detecção dos substratos endógenos.
O resultado foi analisado em SDS-PAGE 12% com gelatina, sem gelatina e em
géis gradiente de acrilamida e uréia (Figura 31). O resultado demonstrou que tanto o
substrato isolado quanto a mistura de substrato e proteases apresentaram atividade
proteásica de mesmo (rf) (Figura 31A). A análise em gel SDS sem gelatina demonstrou
uma banda de alta massa molecular, não apresentando proteínas na mobilidade relativa
correspondente à atividade (Figura 31B). A análise em gel gradiente de acrilamida e
uréia demonstrou o aparecimento de peptídeos (Figura 31C).

74
Figura 31. ATIVIDADE PROTEÁSICA PURIFICADA A PARTIR DE GÉIS UNIDIMENSIONAIS SOBRE SUBSTRATO (S2) PURIFICADO A PARTIR DE GEL NATIVO/SDS. Análise das proteases purificadas a partir de SDS-PAGE 12% com gelatina incubadas com o substrato purificado S2, durante 72h. A- SDS-PAGE 12% gelatina 0,4%. B- SDS-PAGE 12% corado com prata. C- SDS-PAGE gradiente de acrilamida 12 - 18% na presença de uréia 3,5- 7,0 M corado com prata. SE- Substrato purificado mantido na estufa a 37ºC. P1+S- Protease 1 adicionada ao substrato, P2+S- Protease 2 adicionada ao substrato, P3+S- Protease 3 adicionada ao substrato.
As proteases isoladas de géis bidimensionais também foram incubadas com o
substrato S2 e o resultado foi analisado em SDS-PAGE 12% com gelatina e sem
gelatina (Figura 32). De forma semelhante ao resultado anterior, foi demonstrado que
tanto o substrato isolado quanto a mistura de substrato e proteases apresentam
atividade proteásica de mesmo (rf) (Figura 32A). No entanto, o número de bandas de
atividade observadas foi menor que no ensaio realizado com proteases purificadas a
partir de géis unidimensionais. A análise em gel SDS sem gelatina demonstrou uma
banda de aproximadamente 70kDa, que coincide com a banda de substrato (S2)
purificada (Figura 32B).
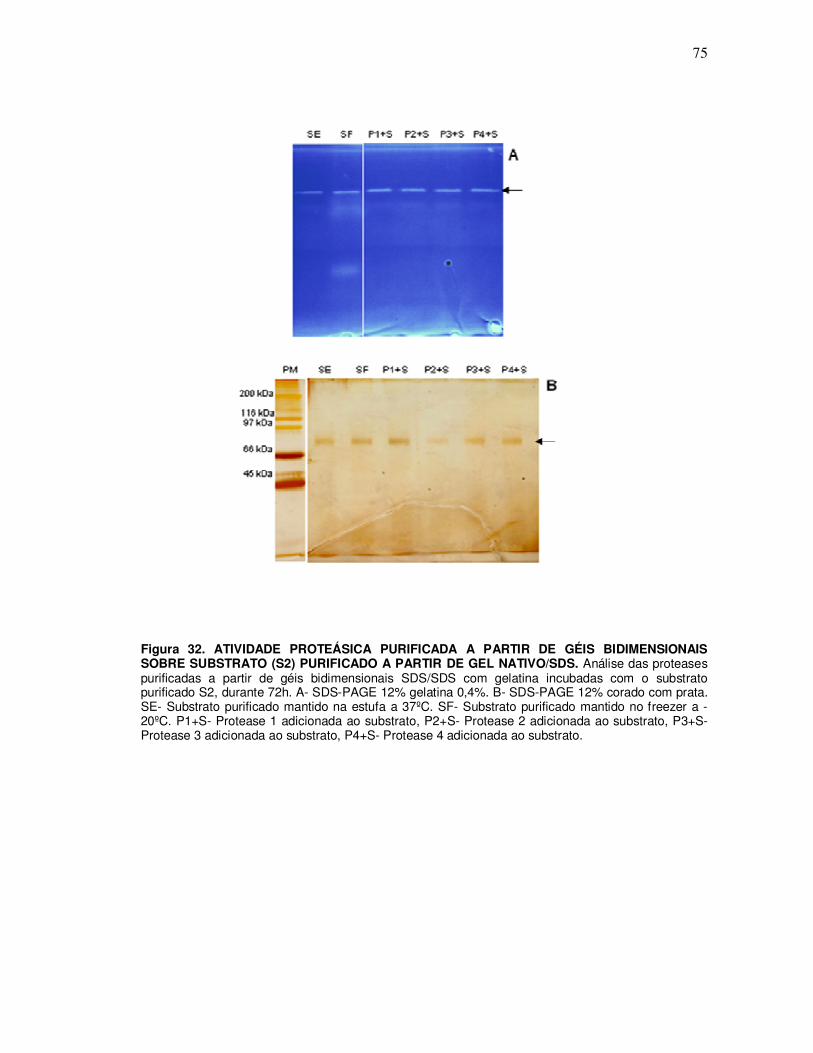
75
Figura 32. ATIVIDADE PROTEÁSICA PURIFICADA A PARTIR DE GÉIS BIDIMENSIONAIS SOBRE SUBSTRATO (S2) PURIFICADO A PARTIR DE GEL NATIVO/SDS. Análise das proteases purificadas a partir de géis bidimensionais SDS/SDS com gelatina incubadas com o substrato purificado S2, durante 72h. A- SDS-PAGE 12% gelatina 0,4%. B- SDS-PAGE 12% corado com prata. SE- Substrato purificado mantido na estufa a 37ºC. SF- Substrato purificado mantido no freezer a -20ºC. P1+S- Protease 1 adicionada ao substrato, P2+S- Protease 2 adicionada ao substrato, P3+S- Protease 3 adicionada ao substrato, P4+S- Protease 4 adicionada ao substrato.

76
10.3 Ensaio de incubação das proteases P1, P2, P3 e P4 purificadas a
partir de géis nativo 10% com os substratos S1, S2, S3 e S8 purificados
a partir do gel nativo/SDS 12%
As proteases P1, P2, P3 e P4 purificadas a partir de géis nativo 10% foram
incubadas com os substratos S1, S2, S3 e S8, visando identificar os produtos
endógenos.
O resultado foi analisado em SDS-PAGE 12% corado com prata e foram
observadas pequenas diferenças na intensidade de algumas bandas entre o perfil
protéico dos substratos isolados e o perfil protéico da mistura de substratos com
proteases (Figura 33).
O resultado também foi analisado em SDS-PAGE gradiente de acrilamida e uréia
(Figura 34) e está em concordância com as observações citadas acima. A análise em
géis SDS-PAGE 12 % com gelatina (Figura 35) reafirmou resultados anteriores de que,
tanto o substrato isolado, quanto a mistura de substratos com proteases, apresentaram
atividade proteásica de mesma massa molecular.
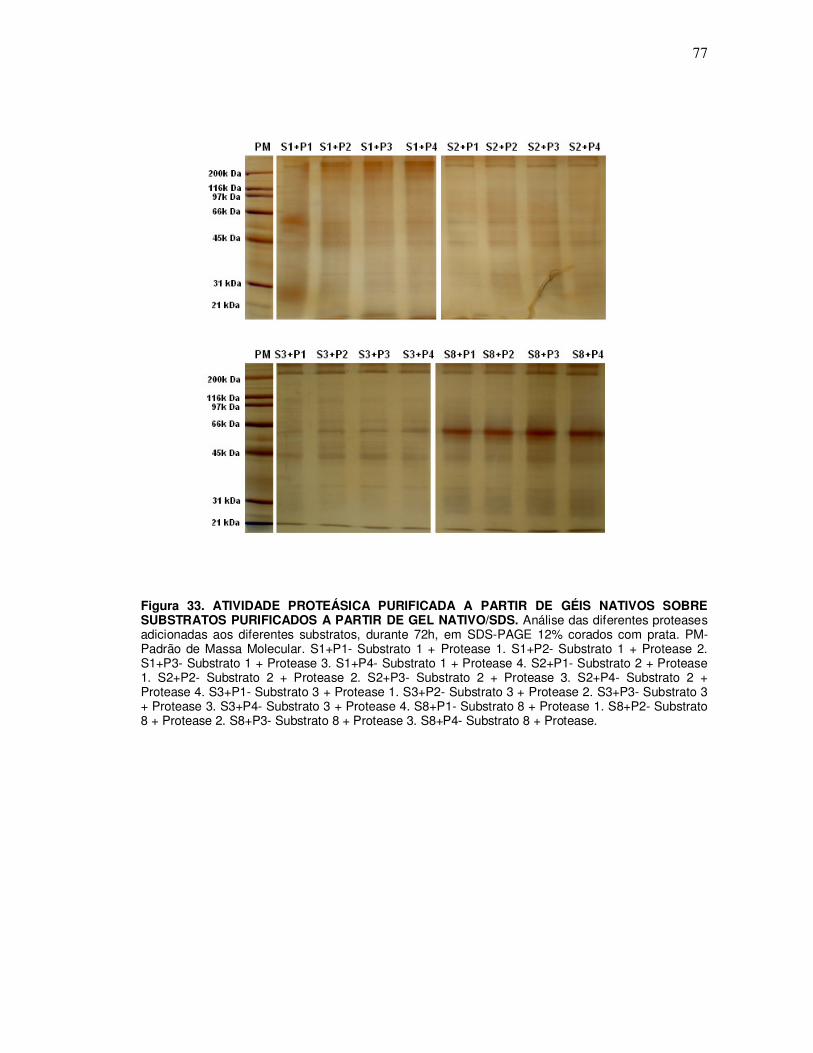
77
Figura 33. ATIVIDADE PROTEÁSICA PURIFICADA A PARTIR DE GÉIS NATIVOS SOBRE SUBSTRATOS PURIFICADOS A PARTIR DE GEL NATIVO/SDS. Análise das diferentes proteases adicionadas aos diferentes substratos, durante 72h, em SDS-PAGE 12% corados com prata. PM- Padrão de Massa Molecular. S1+P1- Substrato 1 + Protease 1. S1+P2- Substrato 1 + Protease 2. S1+P3- Substrato 1 + Protease 3. S1+P4- Substrato 1 + Protease 4. S2+P1- Substrato 2 + Protease 1. S2+P2- Substrato 2 + Protease 2. S2+P3- Substrato 2 + Protease 3. S2+P4- Substrato 2 + Protease 4. S3+P1- Substrato 3 + Protease 1. S3+P2- Substrato 3 + Protease 2. S3+P3- Substrato 3 + Protease 3. S3+P4- Substrato 3 + Protease 4. S8+P1- Substrato 8 + Protease 1. S8+P2- Substrato 8 + Protease 2. S8+P3- Substrato 8 + Protease 3. S8+P4- Substrato 8 + Protease.

78
Figura 34. ANÁLISE DOS PRODUTOS DA ATIVIDADE PROTEÁSICA PURIFICADA A PARTIR DE GÉIS NATIVOS SOBRE SUBSTRATOS PURIFICADOS A PARTIR DE GEL NATIVO/SDS. Análise das diferentes proteases adicionadas aos diferentes substratos em géis SDS gradiente de acrilamida e uréia, corados com prata. PM- Padrão de Massa Molecular. S1+P1- Substrato 1 + Protease 1. S1+P2- Substrato 1 + Protease 2. S1+P3- Substrato 1 + Protease 3. S1+P4- Substrato 1 + Protease 4. S2+P1- Substrato 2 + Protease 1. S2+P2- Substrato 2 + Protease 2. S2+P3- Substrato 2 + Protease 3. S2+P4- Substrato 2 + Protease 4. S3+P1- Substrato 3 + Protease 1. S3+P2- Substrato 3 + Protease 2. S3+P3- Substrato 3 + Protease 3. S3+P4- Substrato 3 + Protease 4. S8+P1- Substrato 8 + Protease 1. S8+P2- Substrato 8 + Protease 2. S8+P3- Substrato 8 + Protease 3. S8+P4- Substrato 8.

79
Figura 35. ANÁLISE DA ATIVIDADE PROTEÁSICA PURIFICADA SOBRE SUBSTRATOS PURIFICADOS EM GÉIS DE GELATINA. Análise das diferentes proteases adicionadas aos diferentes substratos em géis SDS-PAGE 12% gelatina 0,4%. P1- Protease 1; S1+P1- Substrato 1 + Protease 1; S1+P2- Substrato 1 + Protease 2; S1+P3- Substrato 1 + Protease 3; S1+P4- Substrato 1 + Protease 4; S1- Substrato 1; P2- Protease 2; S2+P1- Substrato 2 + Protease 1; S2+P2- Substrato 2 + Protease 2; S2+P3- Substrato 2 + Protease 3; S2+P4- Substrato 2 + Protease 4; S2- Substrato 2; P3- Protease 3; S3+P1- Substrato 3 + Protease 1; S3+P2- Substrato 3 + Protease 2; S3+P3- Substrato 3 + Protease 3; S3+P4- Substrato 3 + Protease 4; S3- Substrato 3; P4- Protease 4; S8+P1- Substrato 8 + Protease 1; S8+P2- Substrato 8 + Protease 2; S8+P3- Substrato 8 + Protease 3; S8+P4- Substrato 8 + Protease 4; S8- Substrato 8;

80
Na Figura 36 é possível observar que as amostras de proteases isoladas de géis
nativos (Figura 36A), ao serem analisadas em SDS-PAGE 12% (Figura 36B), apresentam
várias proteínas de diferentes massas moleculares, que correspondem aos agregados
protéicos presentes em uma região do gel nativo. A Tabela III demonstra a mobilidade
relativa das proteases purificadas, no gel nativo de onde foram retiradas.
Tabela III – Mobilidade relativa aproximada das proteases purificadas a partir
de géis nativos.
Protease
Purificada
Rf aproximado
P1 0 a 0,1
P2 0,1 a 0,2
P3 0,2 a 0,3
P4 0,3 a 0,4
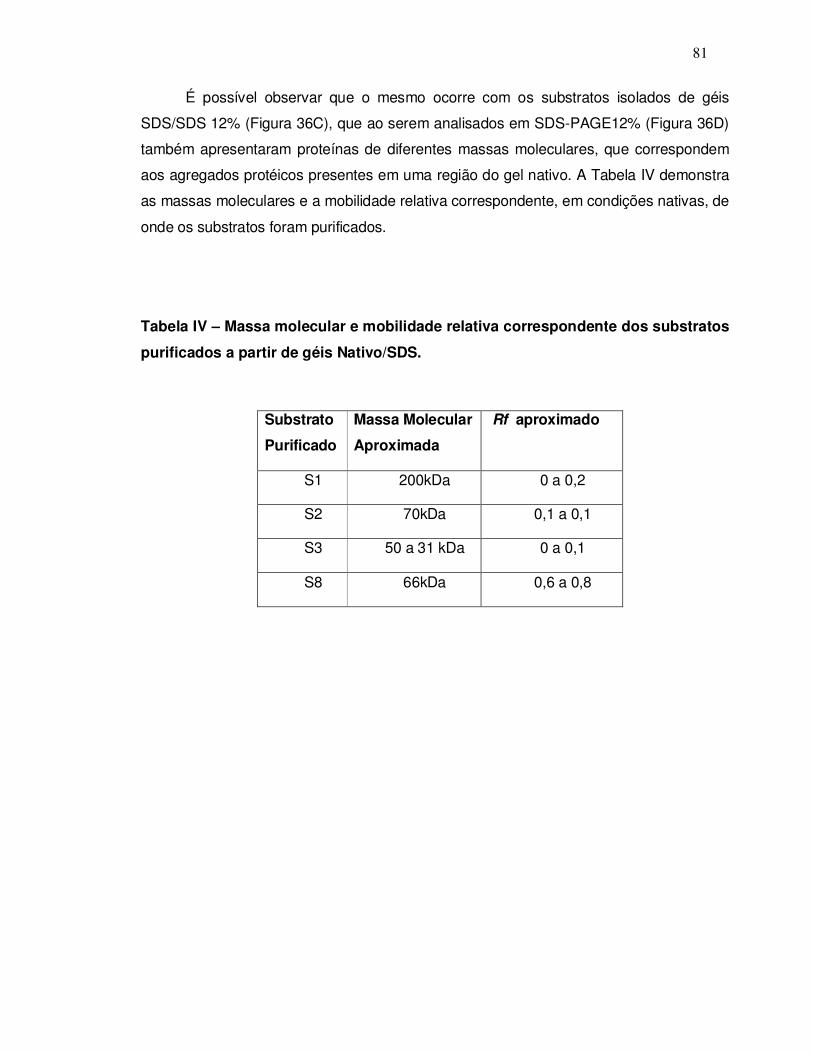
81
É possível observar que o mesmo ocorre com os substratos isolados de géis
SDS/SDS 12% (Figura 36C), que ao serem analisados em SDS-PAGE12% (Figura 36D)
também apresentaram proteínas de diferentes massas moleculares, que correspondem
aos agregados protéicos presentes em uma região do gel nativo. A Tabela IV demonstra
as massas moleculares e a mobilidade relativa correspondente, em condições nativas, de
onde os substratos foram purificados.
Tabela IV – Massa molecular e mobilidade relativa correspondente dos substratos
purificados a partir de géis Nativo/SDS.
Substrato
Purificado
Massa Molecular
Aproximada
Rf aproximado
S1 200kDa 0 a 0,2
S2 70kDa 0,1 a 0,1
S3 50 a 31 kDa 0 a 0,1
S8 66kDa 0,6 a 0,8

82
Figura 36. VISUALIZAÇÃO DAS PROTEASES E SUBSTRATOS PURIFICADOS. A- Gel Nativo 10%/SDS 12% com gelatina 0,4% demonstrando as manchas de atividade utilizadas como parâmetro para a excisão das proteínas correspondentes às proteases, a partir de gel Nativo 10%. B- Análise das proteases isoladas de gel nativo em SDS-PAGE 12% corado com prata. C- Gel Nativo 10%/SDS 12% demonstrando as proteínas isoladas candidatas a substratos. D- Análise dos substratos isolados de gel Nativo 10%/SDS-PAGE 12% em SDS-PAGE 12% corado com prata. P1- Protease 1. P2- Protease 2. P3- Protease 3. P4- Protease 4. S1- Substrato 1. S2- Substrato 2. S3- Substrato 3. S8- Substrato 8. PM- Marcador de Massa Molecular.

83
11. Proteólise endógena
Partindo do princípio que em condições nativas em um mesmo Rf, como,
por exemplo, de 0 a 0,1, existe atividade proteásica (Figura 37 A) e várias proteínas
(Figura 37B), foram utilizados géis nativos para o corte de diferentes tiras. Com base
nisto, foi realizado um ensaio de proteólise endógena com estas tiras contendo
proteínas de fluido de cauda, incubadas a diferentes tempos em temperatura ambiente
(Figura 38). Foi possível observar com a análise das proteínas presentes na tira a
(Figura 38B) que, após 72h, as proteínas de alta massa molecular ficam mais intensas,
enquanto que a proteína de 70 kDa começa a diminuir a sua concentração. Além disso,
aparecem proteínas de menor massa molecular, provavelmente produtos, de
aproximadamente 60, 40, 25 e 20kDa. A análise das proteínas presentes na tira f
(Figura 38C) demonstra que a proteína de aproximadamente 66kDa não sofre
proteólise.

84
Figura 37. VISUALIZAÇÃO DE PROTEASES E PROTEÍNAS PRESENTES EM UM MESMO Rf. A- Análise do fluido epididimário de cauda em gel Nativo 10%/SDS-PAGE 12% com gelatina. B- - Análise do fluido epididimário de cauda em gel Nativo 10%/SDS-PAGE 12%. Comparando os dois géis é possível observar que em um mesmo Rf em condições nativas (1D) corresponde a proteases (2D A) e diferentes proteínas (2D B), inclusive proteínas candidatas a substratos (circuladas em preto).
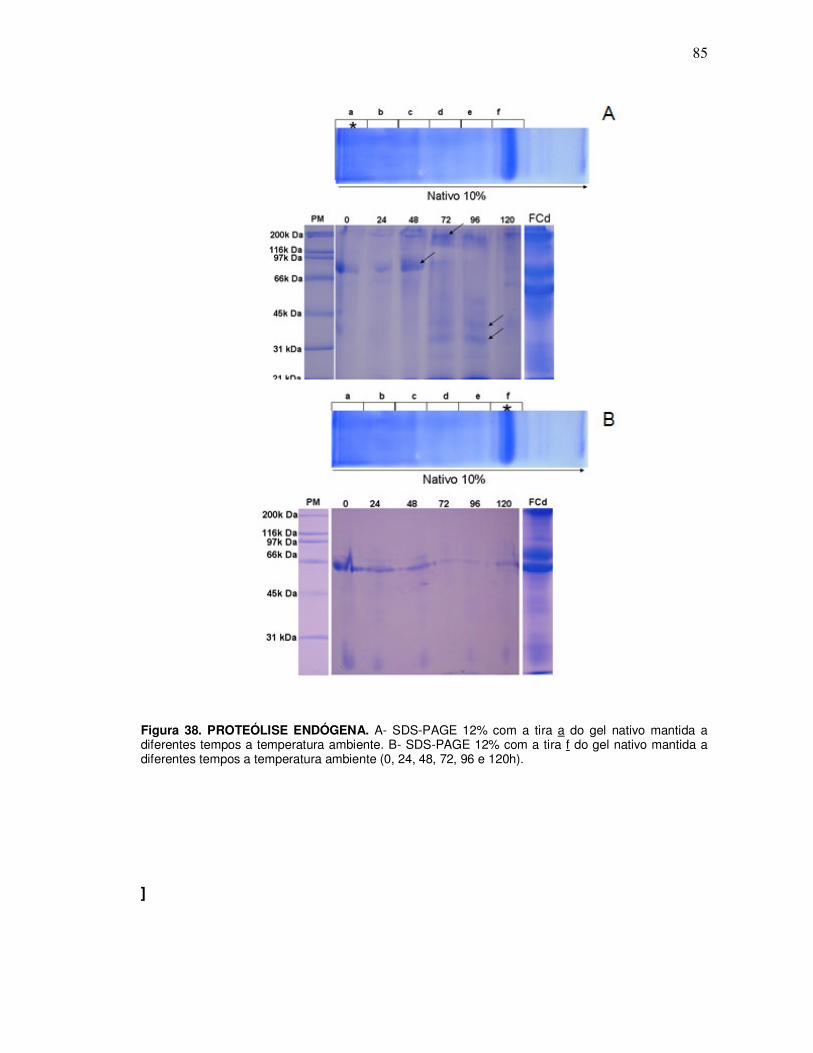
85
Figura 38. PROTEÓLISE ENDÓGENA. A- SDS-PAGE 12% com a tira a do gel nativo mantida a diferentes tempos a temperatura ambiente. B- SDS-PAGE 12% com a tira f do gel nativo mantida a diferentes tempos a temperatura ambiente (0, 24, 48, 72, 96 e 120h).
]

86
12. Western Blotting
As proteínas de fluido das regiões epididimárias analisadas em SDS-PAGE 10%
foram transferidas para a matriz de nitrocelulose e marcadas com anticorpos anti-MMP
de veneno de serpente (Figura 39). Os anticorpos reconheceram algumas proteínas
sugerindo homologia entre metaloproteases de veneno com as de fluido.
Figura 39. WESTERN BLOT. A e B- SDS-PAGE 10% contendo amostras de fluido das diferentes regiões epididimárias duplicata do gel transferido para a membrana de nitrocelulose. C- SDS-PAGE 10% contendo amostras de fluido das diferentes regiões epididimárias, transferido para a membrana de nitrocelulose. D- Membrana de nitocelulose incubada com anticorpos anti-MMP. PM- Padrão de Massa Molecular (alta massa); 1- Fluido epididimário de cabeça proximal; 2- Fluido epididimário de cabeça distal; 3- Fluido epididimário de corpo; 4- Fluido epididimário de cauda.

87
VI. DISCUSSÃO
Os espermatozóides passam por uma série de processos preparatórios prévios a
fertilização, entre eles maturação espermática (no conduto epididimário) e capacitação
(nas vias genitais da fêmea). Somente espermatozóides maduros, capacitados e com
acrossoma intacto penetram o cumulus atingindo a zona pelúcida do gameta homólogo
(FOURNIER-DELPECH & THIBAULT, 1993; LÓPEZ et al., 1987; 1989; LÓPEZ & DE
SOUZA, 1991; RETAMAL et al., 2000ab; RETAMAL, 1999). A qualidade do
espermatozóide, em termos de mobilidade e reconhecimento do ovócito, é fortemente
dependente de diferenciações pós-testiculares que ocorrem no epidídimo durante o
processo de maturação (GATTI et al., 2004).
Uma expressão do processo de maturação espermática é a profunda
remodelação dos diferentes domínios da membrana. Diversos pesquisadores têm
demonstrado modificações nas cargas de superfície, na distribuição e/ou densidade de
receptores de lectinas, nos antígenos de superfície, e no perfil protéico, glicoproteico e
lipídico de espermatozóides imaturos e maduros de mamíferos (FOURNIER-DELPECH
& THIBAULT, 1993; LÓPEZ et al., 1987; 1989; LÓPEZ & DE SOUZA, 1991; RETAMAL
et al., 2000b). Essas modificações surgem, possivelmente, como resultado da liberação
e/ou degradação de componentes testiculares, absorção e integração de proteínas
secretadas pelo epidídimo ou por modificação de resíduos oligossacarídeos ligados a
proteínas ou lipídeos de membrana. Glicosilações, proteólise, re-localização de
componentes pré-existentes são alguns mecanismos já descritos (DACHEUX et al.,
2003; TULSIANI et al., 1995).
Neste trabalho foi demonstrada a presença de proteases, mais
especificamente metaloproteinases, no fluido epididimário de eqüinos. A atividade
destas enzimas apresenta diferenças regionais na sua expressão; de forma similar
ao observado com o perfil eletroforético das proteínas obtidas de fluido coletado das
regiões de cabeça, corpo e cauda do epidídimo. A composição do fluido luminal é
fortemente modificada ao longo do conduto, provavelmente como resultado das
funções de absorção e secreção desenvolvidas pelo epitélio deste órgão
(SETCHELL et al., 1988). Tendo em consideração o menor nível de atividade
enzimática nas amostras de fluido epididimário na região de cabeça proximal, e seu
aumento nas regiões de corpo e cauda epididimária, sugere-se que estas enzimas

88
sejam secretadas e/ou ativadas no epitélio epididimário (MÉTAYER et al., 2002)
Resultados similares foram descritos por DIAS (2002) para algumas glicosidases (α-
glucosidase, α-manosidase, N-acetil glucosaminidase, entre outras. E também por
MÉTAYER et al. (2002) para proteases mediante a utilização de géis de gelatina uni
e bidimensionais. Estes autores descrevem a atividade metaloproteásica (MMPs) de
uma proteína de aproximadamente 60kDa, presente em todas as regiões
epididimárias em carneiro e cavalo, porém em javali se detecta principalmente na
região de cauda epididimária. E sugerem que as proteínas de massa molecular
maiores 54 kDa seriam proteases, entretanto, gelatinases menores seriam sensíveis
a inibidores de serino-proteases.
Os resultados aqui relatados demonstraram que as proteases
detectadas foram fortemente inibidas por inibidores de metaloproteases (EDTA). As
proteínas que contêm um domínio metaloprotease, ancorado na membrana, têm
sido relacionadas com interações célula-célula e portanto com a fertilização
(SHILLING et al., 1997; BLOBEL, 2000). Entretanto, elas foram fracamente inibidas
nos ensaios com PMSF e iodoacetamida, inibidores contra cisteíno e serino
proteases. É possível que existam proteases pertencentes a outras classes, que não
foram detectadas quer seja pelos inibidores utilizados ou por não serem gelatinases.
Neste trabalho, também foi realizado um ensaio de incubação das proteases a
diferentes pHs. Os géis de atividade foram incubados em uma faixa de pH de 2,5 a
12. Foi possível observar que as proteases de fluido epididimário possuem pH ótimo
por volta de 7,5. Sabe-se que as MMPs requerem um pH neutro e Ca+2 para
poderem atuar (MATRISIAN ,1990; NGUYEN et al., 2001). Além disso, as proteases
das diferentes regiões do epidídimo de eqüinos foram caracterizadas
qualitativamente e quantitativamente. Usando esta abordagem duas proteases foram
encontradas na região de cabeça proximal e distal, quatro proteases na região de
corpo e seis na região de cauda. Também foi demonstrado que há ligação cruzada
entre anticorpos policlonais anti-metaloproteases de serpente com as proteases de
fluido epididimário. Este resultado sugere uma homologia entre metaloproteases
destas diferentes espécies.
Há décadas atrás foi introduzido o procedimento baseado na utilização de
géis de poliacrilamida SDS contendo gelatina, caseína ou fibrina como substratos
para a localização da atividade de proteases. Estas enzimas têm a habilidade de
renaturar após a remoção do SDS e de exercer atividade proteolítica no substrato

89
co-polimerizado, e assim, serem analisadas com este método (FREDERIKS &
MOOK, 2004). Sabe-se que a presença do polímero de gelatina pode reduzir a
migração de proteínas de massa molecular conhecida de 15 a 20%. Em géis de
acrilamida a 7,5% a adição de gelatina reduz a mobilidade dos padrões 13 ± 3,4%;
em gel de acrilamida a 10% a redução é 16 ± 3,8%; e em géis 12,5%, a redução é
de 15± 2,8% (HUMMEL et al., 1996). Um retardo similar foi observado com proteínas
não proteásicas de Schizophyllum commune. As proteases migram muito mais
devagar em SDS-PAGE com gelatina do que os padrões. Para as proteases
testadas, a redução da mobilidade com a adição de gelatina em acrilamida T 10% foi
de 48% para quimiotripsina, 44% para elastase, 43% para tripsina e 41% para
papaína. (HUMMEL et al., 1996). A retardação não é constante entre proteases.
Portanto, a massa molecular das proteases não pode ser estimada precisamente em
géis SDS com gelatina. Neste trabalho a detecção da atividade proteásica foi
realizada através de géis de poliacrilamida desnaturantes. Foi observado o efeito de
retardação na migração das proteases e determinada a constante de retardação (Kr)
das mesmas das diferentes regiões do epidídimo. Foram feitos gráficos construídos
sob diferentes concentrações de gelatina e sob uma base constante de outro
polímero, a acrilamida, que permaneceu em uma concentração de 10% (v/v).
FERGUSON (1964) descreveu estes gráficos para o polímero de acrilamida e o
comportamento das proteínas neste contexto. Os géis a concentração constante de
poliacrilamida (10%) na presença de um gradiente de outro polímero (gelatina), de 0
a 0,6%, têm como propósito avaliar o efeito peneira (“sieving”) dos géis, sem alterar
a carga líquida das proteínas. Assim, tem-se sugerido várias metodologias para
determinar o tamanho das proteínas não desnaturadas utilizando esta estratégia
(FERGUSON,1964; HEDRICK & SMITH 1968; RETAMAL & BABUL, 1988;
WESTERHUIS et al., 2000).
Neste trabalho, foi utilizado um novo procedimento para determinar a massa
molecular das proteases em géis de gelatina bidimensionais SDS/SDS com gelatina,
e relacionar as massas moleculares obtidas na primeira dimensão com a posição
das proteases na segunda dimensão. Assim, foram determinadas as massas
moleculares das bandas de atividade proteásica sem o efeito anômalo causado pelo
polímero de gelatina. Na região de cabeça proximal (que em eqüinos é formada
exclusivamente por dutos eferentes) foram detectadas proteases de massas relativas
de 117, 100 e 62 kDa e nas regiões mais distais do epidídimo foram detectadas

90
proteases de massas moleculares de 117, 62 e 45 kDa. Neste órgão, predomina uma
protease de 45 kDa, que não é observada na cabeça proximal. Na região de cauda,
foi observada atividade em uma proteína de aproximadamente 40 kDa, que não se
observa em outras regiões. Estas massas moleculares diferem das calculadas por
outros autores pela sobreposição de géis unidimensionais com e sem gelatina.
Porém, o perfil eletroforético das proteases é similar.
Há dados na literatura descrevendo uma série de proteínas espermáticas
proteoliticamente processadas durante o trânsito epididimário, entre elas:
hialuronidase, pH20, proteína CE9, galactosidase (a qual existe como uma forma de
90 kDa em espermatozóides obtidos da região de cabeça proximal e é,
gradualmente, convertida para uma forma majoritária de 74 kDa em
espermatozóides de cauda) (CHAYKO et al., 2000). Outro exemplo interessante de
processamento proteolítico é a fertilina. Esta é um heterodímero transmembrana
com domínios pró- metaloprotease, disintegrina, rico em cisteína e EGF, que
pertence a família ADAMs (YUAN et al., 1997). Esta proteína (previamente chamada
PH30) sofre pelo menos dois processamentos proteolíticos distintos: primeiro, no
testículo, e outro no epidídimo (LUM & BLOBEL, 1997). Estes achados indicam que
algumas proteínas podem ser secretadas ou estarem presentes em uma pró-forma e
tornam-se ativadas após o processamento no fluido epididimário (LÓPEZ et al., 1989;
RETAMAL et al., 2000; GATTI et al., 2004, ORGEBIN-CRIST & JAHAD, 1978;
FOUCHECOURT et al., 2000). A análise da maturação intracelular e atividade
catalítica de uma desintegrina metaloprotease amplamente expressa, a MDC9,
sugere que o pró- domínio da MDC9 é removido por uma convertase tipo furina da
via secretora, depois que a proteína emerge na superfície da membrana e que a
atividade catalítica da MDC9 pode ser inibida por inibidores de metaloprotease. A
remoção de um pró-domínio inibitório da MDC9 é um pré-requisito para a atividade
proteásica. Depois do pró-domínio removido, passos adicionais, como o de
proteínas quinases C dependentes de fosforilação, podem estar envolvidos na
regulação da atividade enzimática da MDC9 (ROGHANI et al., 1999).
Sabe-se que o epidídimo modula a composição de seu conteúdo luminal pelas
funções de secreção seletiva e absorção das células epiteliais, as quais apresentam
variações regionais. Flutuações no volume do fluido, eletrólitos, proteínas e outros
componentes do fluido epididimário são presumivelmente críticas para a maturação e
sobrevida do gameta (LÓPEZ et al., 1989; RETAMAL et al., 2000; GATTI et al., 2004,

91
ORGEBIN-CRIST & JAHAD, 1978; FOUCHECOURT et al., 2000). Uma hipótese é
que além da atuação de proteases epididimárias sobre as proteínas espermáticas,
também exista uma ação proteolítica sobre as proteínas do próprio fluido. Os
resultados deste trabalho demonstraram que existe ação de proteases sobre a
fração total de proteínas de fluido da mesma região (Figuras 19, 20, 21 e 22). Estas
proteínas ativadas poderiam ter um efeito no espermatozóide ou no fluido e fazer
parte da cascata de eventos ligados ao processo de maturação. Assim as proteases
poderiam estar participando em processos de remodelação ou ativação de
componentes do fluido epididimário, remodelação e/ou redistribuição de proteínas na
superfície espermática e remoção de espermatozóides defeituosos, papéis que são
crucias na fertilização.
Uma análise proteômica parcial do fluido epididimário reflete que ambos os
processos de degradação de proteínas e adição de novos componentes ocorrem no
epidídimo (GATTI et al., 2004). Gelatinases como MMP-2 (gelatinase A, 72 kD) e
MMP-9 (gelatinase B, 92 kD) foram detectadas em zymogramas de gelatina e MMP-
7 em géis de caseína. Segundo FREDERIKS & MOOK (2004), MMPs são
secretadas das células em uma forma proteoliticamente inativa (zimogênio),
aproximadamente 10 kDa maior do que a forma ativada. A pró-forma se ativa
durante processo de desnaturação e renaturação depois da eletroforese. Ambas as
formas degradam gelatina e são detectadas em zimografias. MMPs em solução
estão frequentemente associadas com inibidores teciduais endógenos de
metaloproteases (TIMPs) (GATTI et al., 2004). Durante a eletroforese, os inibidores
são dissociados da MMP e não interferem na atividade enzimática (FREDERIKS &
MOOK, 2004) sendo mais uma indicação de que existe um processamento
proteolítico de proteínas do meio e da superfície espermática, muitas vezes sobre as
próprias proteases. A grande maioria das proteínas detectadas no epidídimo são
produtos secretados pelo epidídimo. Isto representa mais de cem proteínas diferentes,
com um alto polimorfismo (tanto na massa molecular, quanto no ponto isoelétrico)
provavelmente por glicosilações e outras modificações pós – tradução (GATTI et al.,
2004).
Neste trabalho foi demonstrada, nos experimentos de proteases isoladas
incubadas com fluido epididimário, a atividade proteásica endógena sugerindo que
as proteínas de massa molecular 117, 80, 69, 57, 50, 44 e 35kDa são substratos e
que as proteínas de massa molecular 50 e 44kDa, bem como uma mancha que

92
engloba 32,8, 33,5 e 35kDa, são produtos (Figuras 19, 20, 21 e 22). Também é
evidente que as proteases possuem um comportamento semelhante, sugerindo uma
afinidade pelo mesmo substrato. Além disso, a combinação do uso de géis Nativo
PAGE e SDS-PAGE para a análise de proteínas (géis bidimensionais) pode ser
usada para o estudo de interações proteína-proteína (SUN & PAN, 1999).
Resultados aqui relatados demonstraram que proteínas de diferentes massas
moleculares possuem o mesmo Rf em condições nativas, indicando a possibilidade
de agregados. Um exemplo é o resultado da análise das proteínas de Rf 0,1 do gel
Nativo/SDS com amostra controle (Figura 36).
Assim, tendo em vista que existe ação de proteases sobre a fração total de
proteínas de fluido da mesma região foi feito um experimento para verificar os
produtos da atividade proteásica. Para tanto, foram feitos ensaios com diferentes
proteases purificadas incubadas com diferentes substratos purificados (Figuras 29 à
35). Nestes ensaios, foi possível constatar que a proteína de aproximadamente
66kDa, não diminuiu em quantidade após sua incubação com proteases isoladas.
Além disso, com a proteína do substrato, apareceram algumas impurezas de menor
peso. Estas sim diminuíram ao longo do tempo, após a incubação com a protease
P3 (Figura 29). Uma possibilidade é que estas impurezas sejam resultantes da
proteólise da proteína de 66kDa durante o processamento desta. No entanto, uma
hipótese é a de que esta proteína esteja agregada aos inibidores de proteases
(MCCAWLEY & MATRISIAN, 2000). Esta proteína só desaparece após a incubação
de proteases com o extrato bruto de fluido, após o congelamento deste ensaio a -
200C, ou seja, após um período maior de contato entre as proteases e as proteínas
do fluido (Figura 21). Assim, esta proteína passou a ser considerada em nos
experimentos como um controle negativo. Também foram utilizadas outras proteínas
candidatas a substratos para os ensaios de detecção dos produtos. Após a
incubação destas proteínas com as diferentes proteases isoladas foi possível
observar pequenas diferenças entre o perfil eletroforético dos substratos antes e
após a incubação, demonstrando que ocorre a proteólise. No entanto, esta não foi
muito efetiva, provavelmente, pela presença de algum inibidor, quantidade de
protease ou substratos insuficientes, tempo de incubação muito curto e/ou outros
possíveis fatores.
Foram demonstradas diferentes manchas de atividades proteásicas,
tanto em géis de gelatina unidimensionais na presença de SDS quanto em géis de

93
gelatina bidimensionais (Figura 5 e 14, respectivamente), indicando proteases de
diferentes massas moleculares. Diferentemente os géis Nativo/SDS com gelatina
demonstram diferentes manchas de atividades de proteases de similar massa, ao
longo de uma faixa que corresponde ao gel nativo (Figura 17). Esta discrepância
pode ser explicada pela presença de detergente na amostra antes das eletroforeses
em géis SDS unidimensionais, que enfraquece a ligação das proteínas e,
provavelmente, dos peptídeos inibitórios ligados às proteases e/ou substratos que
estavam conformando grandes agregados protéicos, e assim, os inibidores se
desassociam das enzimas, e estas se ligam mais facilmente aos seus substratos,
interação esta mais forte do que o efeito do detergente. Ao realizar o gel
bidimensional SDS com gelatina as proteases permanecem ligadas aos seus
substratos e co-migram com estes e são visualizadas em manchas de atividades
com diferentes massas moleculares. Outro experimento que confirma esta idéia foi
quando se realizou uma eletroforese de substratos purificados em géis com gelatina
(Figuras 28 e 35), pois, os substratos isolados também apresentaram atividade
proteásica. Assim, especula-se que em géis SDS as proteases permanecem
associadas aos seus substratos apresentando uma massa molecular que é a soma
da sua própria massa com a do substrato. No entanto, quando realizado um gel
unidimensional nativo, como a amostra é preparada em condições nativas, co-
migram diversas proteínas juntas, incluindo inibidores de proteases. O perfil
eletroforético das proteases encontrado nos géis nativo/SDS com gelatina pode ser
devido a macro-complexos compostos por proteases, substratos, seus inibidores e,
possivelmente, outras proteínas que co-migram em condições nativas. E quando
estes complexos são submetidos à re-eletrofore em gel SDS, a incubação da tira do
gel nativo em tampão 2x com SDS pode não ser o suficiente para separar as
proteases dos seus inibidores a ponto destas enzimas se ligarem aos seus
substratos e co-migrarem com estes, explicando o fato de não serem observadas
diferentes massas.
Esta idéia pode ser reforçada pelos experimentos com proteases purificadas.
Estas proteases foram isoladas dos géis em condições nativa tomando como base as
manchas de atividade proteásica dos géis nativo/SDS com gelatina e, como já
mencionado, resultou em manchas de atividade proteásica de idêntica massa
molecular. Resultados similares foram encontrados por METÁYER et al. (2002) com
géis 2D. Neste caso, o autor realizou após a focalização isoelétrica das amostras de

94
fluido, géis desnaturantes com e sem gelatina. Estes, demonstraram manchas de
atividade de protease de diferentes pontos isoelétricos que, semelhantemente aos
nossos resultados, apresentam mesma massa. As proteínas correspondentes a
mancha de atividade de mesma massa foram isoladas de géis nativo e, como
esperado, ao serem analisadas em géis SDS com gelatina foi possível observar
atividades proteásicas de mesma massa molecular. No entanto, quando as atividades
proteásicas foram isoladas de géis SDS com gelatina, tanto unidimensionais quanto
bidimensionais SDS/SDS com gelatina, mesmo tendo sido retiradas de diferentes
massas, foi possível observar o mesmo resultado, ou seja, proteases de mesma
massa molecular (Figuras 15 e 16). Uma possibilidade é de que isto pudesse ser o
efeito da gelatina sobre a protease purificada, já que as proteases foram purificadas
deste tipo de gel. Para verificar se a presença da gelatina estaria influenciando no
perfil das proteases purificadas, estas foram isoladas a partir de géis nativos 10%. Os
resultados se mantiveram os mesmos, ou seja, as proteases purificadas de diferentes
Rfs do gel nativo, ao serem analisadas em géis desnaturantes, apresentaram a
mesma massa molecular (Figura 18). Provavelmente, trata-se da mesma protease
que em géis SDS com gelatina, por estarem fortemente ligadas ao substrato,
apresentam diferentes massas relativas. Mas, quando estas proteases são isoladas,
os substratos ligados se desassociam ou são metabolizados, consequentemente, as
proteases não co-migram com os substratos para diferentes massas. E além dos
substratos isolados terem apresentado atividade proteásica, como já mencionado, de
forma semelhante aos experimentos com proteases purificadas, estes substratos,
mesmo isolados de diferentes massas moleculares apresentaram atividade de mesma
massa. E esta banda de atividade foi a mesma encontrada na analise das amostras
de proteases purificadas em géis de gelatina. Ou seja, trata-se da mesma protease,
que co-migrou com diferentes substratos. O resultado do Western blot também
contribui para esta idéia (Figura 39), pois, foi possível observar um reconhecimento de
proteínas de diferentes massas do fluido epididimário pelos anticorpos anti-
metaloproteases, mostrando que existem metaloproteases de diferentes massas
moleculares. Os resultados, indicam nos experimentos com proteases purificadas e
substratos purificados demonstram que estas possuem um perfil protéico similar, o
que está de acordo com os dados discutidos anteriormente, de que as proteases
estariam co-migrando com seus substratos.

95
Foi realizado um experimento com tiras de gel nativo, contendo proteínas de
fluido de cauda, incubadas a diferentes tempos em temperatura ambiente (Figura
38). Foi possível observar, na análise da tira a, que após 72h as proteínas de alto
peso ficaram mais intensas enquanto que a banda de 70 kDa começou a
desaparecer. Além disso, apareceram proteínas de menor massa molecular,
provavelmente produtos, de aproximadamente 60, 40, 25 e 20 kDa. Isto indica que,
possivelmente, as proteínas presentes nesta tira do gel sofrem a ação de proteases
também existentes na mesma. E assim, a protease e as proteínas estariam
presentes no mesmo Rf (Figura 37). Além disso, foi possível observar na análise
desta mesma tira de gel que algumas proteínas de alto peso aumentaram a sua
concentração ao longo do tempo de incubação, indicando que os produtos da ação
proteolítica se agregam. Uma possibilidade é que a banda de 200 kDa, por exemplo,
seja um agregado formado pelas proteínas de 70, 50, 40 e 30 kDa, ou seja, um
agregado entre os substratos, proteases e produtos. Simultaneamente, conforme o
substrato é consumido, os peptídeos e as proteínas de menor peso se tornam mais
intensos, ou seja, o agregado se desfaz e tornam-se visíveis os possíveis produtos
da proteólise. Outra possibilidade é que as proteínas de 70, 60 e 25 kDa estejam se
agregando e formando a proteína de 145kDa, ou ainda as proteínas de 60 e 70 kDa
formando a proteína de aproximadamente 130 kDa. Este resultado corrobora com os
ensaios de proteólise endógena, onde o extrato bruto de fluido de cauda foi
incubado com proteases purificadas, pois, ambos demonstram que existe a ação de
proteases sobre a proteína de aproximadamente 70 kDa. Ao ser analisada a tira de
gel f, contendo a proteína de aproximadamente 66 kDa, foi demonstrado, mais uma
vez, que esta não é o substrato preferencial das proteases em estudo. No gel
Nativo/SDS com gelatina (figura 37) é evidente que no Rf da tira f não há atividade
proteásica, logo, não poderia haver proteólise das proteínas presentes nesta tira de
gel reforçando a idéia de que as proteases co-migram com seus substratos.
Assim a proteína de 70 kDa representa um substrato pela qual as proteases
possuem alta afinidade, e consequentemente co-migram com esta, enquanto que a
proteína de 66 kDa, ao contrário, corresponde a um substrato pelo qual as proteases
possuem menor afinidade, conforme discutido anteriormente. Outra possibilidade é
que existam inibidores que impedem a ação das proteases sobre esta proteína, no
entanto, esta tese não foi testada neste trabalho.

96
VII. CONCLUSÕES
- O presente trabalho demonstra a presença de atividade proteásica no fluido
epididimário de E. caballus, com diferenças regionais ao longo do conduto.
- O estudo com inibidores, indica que as proteases de fluido correspondem
majoritariamente a metaloproteases (MMPs). Na região proximal se detectaram
proteases de massas moleculares 117, 100 e 62 kDa. Na região distal predominara
aquelas de massas moleculares relativas 117, 62 e 45 kDa. Neste órgão, predomina uma
protease de 45 kDa, que não é observada na cabeça proximal. Na região de cauda, foi
observada atividade em uma proteína de aproximadamente 40 kDa. Em termos gerais,
as proteases presentes na região proximal do ducto são de maior massa molecular
relativa que aquelas detectadas na região distal.
- O estudo de incubação a diferentes pHs demonstrou que tratam-se de enzimas que
apresentam um pH ótimo neutro.
- No presente trabalho se detectou a constante de retardação das proteases, provocada
pela presença do polímero de gelatina nos géis. Assim, foi estabelecida uma abordagem
diferente para se determinar a massa molecular das proteases sem o efeito anômalo
causado pela presença de gelatina.
- O comportamento destas proteínas com β-mercaptoetanol sugere a presença de pontes
dissulfeto em sua estrutura. Estas proteínas podem estar formando agregados entre
proteases ou estar agregadas aos seus substratos.
- A caracterização das proteases após serem isoladas sugere que estas proteínas
podem estar formando agregados de proteases com proteases ou de proteases com
seus substratos.
- O presente estudo contribuiu para a compreensão da função epididimária e da
participação de proteases na função de remodelação de proteínas de fluido.

97
VIII. REFERÊNCIAS BIBLIOGRAFICAS
AMANN R.P. (1995) The vital role of the epididymis. Anais do Congresso Brasileiro
de Reprodução Animal 231-221.
BASSET P., BELLOCQ J.P., WOLF C., STOLL I., HUTIN P., LIMACHER J.M.
PODHAJCER O.L., CHENARD M.P., RIO M.C., CHAMBON P. (1990) A novel
metalloproteinase gene specifically expressed in stromal cells of breast carcinomas.
Nature 348: 699-704.
BASSET P., WOLF C., CHAMBON P. (1993) Expression of the stromelysin-3 gene in
fibroblastic cells of invasive carcinomas of the breast and other human tissues: a
review. Breast Cancer Res Treat; 24: 185-193.
BELAAOUAJ A., SHIPLEY J.M., KOBAYASHI D.K., ZIMONJIC D.B., POPESCU N.,
SILVERMAN G.A., SHAPIRO S.D. (1994) Human macrophage metalloelastase:
genomic organization, chromosomal location, gene linkage, and tissue-specific
expression. J Biol Chem 270: 14568-14575.
BLOBEL C.P. (2000) Functional processing of fertilin: evidence for a critical role of
proteolysis in sperm maturation and activation. Rev Reprod 5:75-83.
BOZZO S., RETAMAL C. (1991) Geles unidimensionales. Un nuevo método
densitométrico para computadores personales. Arch Biol Med Exp 24:181.
BRADFORD M.M. (1976) A rapid sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein dye binding. Anal Biochem
72:248-254.
CAWSTON T. (1998) Matrix metalloproteinases and TIMPs: properties and
implications for the rheumatic diseases. Mol Med Today 4:130-7.
CESARI A., CACCIATO C.S., DE CASTRO R.E., SÁNCHEZ J.J. (2004) BSp66
protease is widespread in the acrosomal region of sperm from several mammalian
species. Biochem Biophys Res Comm 324:874-877.
CHANDLER S., MILLER K.M., CLEMENTS J.M., LURY J., CORKILL D., ANTHONY
D.C.C., ADAMS S.E., GEARING A.J. (1997) Matrix metalloproteinases, tumor
necrosis factor and multiple sclerosis: an overview. J Neuroimmunol 72: 155-161.

98
CHAYKO C.A., ORGEBIN-CRIST M., SKUDLAREK M.D., TULSIANI D.R.P. (2000)
Biosynthesis, processing, and subcellular localization of rat sperm β- D-
galactosidase. Biol Reprod 63:688-696.
CHENG C.Y., GRIMA J., STAHLER M.S., GUGLIELMOTTI A., SILVESTRINI B.,
BARDIN C.W. (1990) Sertoli cell synthesizes and secretes a protease inhibitor, a2-
macroglobulin. Biochemistry 29:1063-1068.
COLLIER I.E., WILHELM S.M., EISEN A.Z., MARMER B.L., GRANT G.A., SELTZER
J.L., KRONBERGER A., HE C.S., BAUER E.A., GOLDBERG G.I. (1988) H-ras
oncogene-transformed human bronchial epithelial cells (TBE-1) secrete a single
metalloproteinase capable of degrading basement membrane colagen. J Biol Chem
263: 6579-6587.
DACHEUX J.L., GATTI J.L., DACHEUX F. (2003) Contribution of epididymal
secretory proteins for spermatozoa maturation. Microsc Res Tech 61:7-17.
DAVIS B.J. (1964) Disc electrophoresis. II. Method and application to human serum
proteins. Ann N Y Acad Sci 121:404-427.
DELAISSÉ J.M., VAES G. (1992) Mechanism of mineral solubilization and matrix
degradation in osteoclastic bone resorption. Biology and Physiology of the
Osteoclast. Boca Raton: CRC Press 289-314.
DIAS A.J.B. (2002) Detecção e caracterização de glucosidases no epidídimo e
sêmen de eqüinos. Tese de Doutorado. UENF. Campos dos Goytacazes. RJ. Brasil.
DIAS A.J.B., MAIA M.S., RETAMAL C.A., LÓPEZ M.L. (2004) Identification and
partial characterization of alfa-1,4-glucosidase activity in equine epididymal fluid.
Theriogenology 61:1545-1558.
EDDY E.M., O'BRIAN D.A. (1994) The spermatozoon. In: The Physiology of
Reproduction. 2nd ed.p.29-67.
EDMAN P., (1950) Method for determination of the aminoacid sequence in peptides.
Chem Scand 4:283-293.
ERICKSON-LAWRENCE M., ZABLUDOFF S.D., WRIGHT W.W. (1991) Cyclic
protein-2, a secretory product of rat Sertoli cells, is the proenzyme form of cathepsin
L. Mol Endocrinol 5:1789-1798.
EVANS J.P. (2001) Fertilin beta and other ADAMs as integrin ligands: insights into
cell adhesion and fertilization. Bioessays 23:628-639.
FERGUSON K.A. (1964) Starch-gel electrophoresis: Application to the classification
of pituitary proteins and polypeptides. Metabolism 13:985-1002.

99
FOUCHÉCOURT S., MÉTAYER S., LOCATELLI A., DACHEUX F., DACHEUX J.L.
(2000) Stallion epididymal fluid proteome: qualitative and quantitative
characterization secretion and dynamic changes of major proteins. Bio Reprod
62:1790-1803.
FOURNIER-DELPECH S., THIEBAULT C. (1993) Acquisition of sperm fertilizing
ability. Epididymal maturation, accessory glands and capacitation. Reprod Mammals
Man 257-278.
FRAYNE J., JURY J.A., BARKER H.L., PERRY A.C., JONES R., HALL L. (1997)
Macaque MDC family of proteins: sequence analysis, tissue distribution and
processing in the male reproductive tract. Mol Reprod Dev 48: 159-167.
FREDERIKS W.M., MOOK O.R.F. (2004) Metabolic mapping of proteinase activity
with emphasis on in situ zymography of gelatinases: Rewiew and protocols. J
Histochem & Citochem 52:711-722.
FREIJE J.M., DÍEZ-ITZA I., BALBÍN M., SÁNCHEZ L.M., BLASCO R., TOLIVIA J.,
LOPEZ-OTIN C. (1994) Molecular cloning and expression of collagenase-3, a novel
human matrix metalloproteinase produced by breast carcinomas. J Biol Chem 269:
16766-16773.
GATTI J.L., CASTELLA S., DACHEUX F., ECROYD H., MÉTAYER S., THIMON
V.,DACHEUX J.L. (2004) Post-testicular sperm environment and fertility. Animal
Reprod 82-83:321-339.
GHADDAR G., RUCHON A.F., CARPENTIER M., MARCINKIEWICZ M., SEIDAH
N.G., CRINE P., DESGROSEILLERS L., BOILEAU G. (2000) Molecular cloning and
biochemical characterization of a new mouse testis soluble-zinc-metallopeptidase of
the neprilysin family. Biochem J 347:419-429.
GOLDBERG G.I., WIHELM S.M., KRONBERGER A., BAUER E.A., GRANT G.A.,
EISEN A.R. (1986) Human fibroblast collagenase: complete primary structure and
homology to an oncogene transformation-induced rat protein. J Biol Chem 261:
6600-6605.
GUEDEZ L., LIM M.S., STETLER-STEVENSON W.G. (1996) The role of
metalloproteinases and their inhibitors in hematological disorders. Crit Rev Oncog 7:
205-225.
GUNNARSSON M., LECANDER I., ABRAHAMSSON P.A. (1999) Factors of the
plasminogen activator system in human testis, as demonstrated by in-situ
hybridization and immunohistochemistry. Mol Hum Reprod 5:934-940.

100
HARRIS E.D. (1990) Rheumatoid arthritis: pathophysiology and implications for
therapy. N Engl J Med; 322: 1277-1289.
HASHIMOTO F., HORIGOME T., KANBAYASHI M., YOSHIDA K., SUGANO H.
(1983) An improved method for separation of low-molecular-weight polypeptides by
electrophoresis in sodium dodecyl sulfate-polyacrylamide gel. Anal Biochem
129:192-9.
HASTY K.A., POURMOTABBED T.F., GOLDBERG G.I., THOMPSON J.P.,
SPINELLA D.G., STEVENS R.M., MAINARDI C.L. (1990) Human neutrophil
collagenase: a distinct gene product with homology to other matrix
metalloproteinases. J Biol Chem 265: 11421-4.
HEDRICK J.L., SMITH A.J. (1968) Size and charge isomer separation and estimation
of molecular weights of proteins by disc gel electrophoresis. Arch Biochem Biophys
126:155-64.
HINTON B.T., PALLADINO M.A., RUDOLPH D., LADUS J. (1995) The epididymis as
protector of maturing spermatozoa. Reprod Fert 7:731-745.
HOEBEN E., VAN AELST I., SWINNEN J.V., OPDENAKKER G., VERHOEVEN G.
(1996) Gelatinase A secretion and its control in peritubular and Sertoli cell cultures:
effects of hormones, second messengers and inducers of cytokine production. Mol
Cell Endocrinol 118:37-46.
HONDA A., SIRUNTAWINETI J., BABA T. (2002) Role of acrosomal matrix
proteases in sperm-zona pellucida interactions. Hum Reprod Update 8:405-12.
HOWES L., JONES R. (2002) Interactions between zona pellucida glycoproteins and
sperm proacrosin/acrosin during fertilization. J Reprod Immunol 53:181-92. Review.
HUMMEL K.H., PENHEITER A.R., GATHMAN A.C., LILLY W.W. (1996) Anomalous
estimation of protease molecular weight using gelatine-containing SDS-PAGE. Analyt
Biochem 233:140-142.
JEFFREY J.J. (1991) Collagen and collagenase: pregnancy and parturition. Semin
Perinatol 15: 118-26.
JONES R., MA A., HOU S., SHALG R., HALL L. (1996) Testicular biosynthesis and
epididymal endoproteolytic processing of rat sperm surface antigen 2B1.J Cell Sci
109:2561-2570.
KOHNO N., YAMAGATA K., YAMADA S., KASHIWABARA S., SAKAI Y., BABA T.
(1998) Two novel testicular serine proteases, TESP1 and TESP2, are present in the
mouse sperm acrosome. Biochem Biophys Res Commun 245:658–665.

101
LAEMMLI U.K. (1970) Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature Lond 227:680-685.
LAFLEUR M.A., HANDSLEY M.M., KNÄUPER V., MURPHY G., EDWARDS D.R.
(2002) Endothelial tubulogenesis within fibrin gels specifically requires the activity of
membrane-type-matrix metalloproteinases (MT-MMPs). J Cell Sci 115: 3427-38.
LLANO E., PENDÁS A.M., KNÄUPER V., SORSA T., SALO T., SALIDO E.,
MURPHY G., SIMMER J.P., BARTLETT J.D., LÓPEZ-OTÍN C. (1997) Identification
and structural and functional characterization of human enamelysin (MMP-20).
Biochemistry 36: 15101-8.
LLANO E., PENDÁS A.M., FREIJE J.P., NAKANO A., KNÄUPER V., MURPHY G.,
LÓPEZ-OTIN C. (1999) Identification and characterization of human MT5-MMP, a
new membrane-bound activator of progelatinase A overexpressed in brain tumors.
Cancer Res 59: 2570- 6.
LIEKENS S., CLERCQ E.D., NEYTS J. (2001) Angiogenesis: regulators and clinical
applications. Biochem Pharmacol 61: 253-70.
LIU Q., HAMIL K.G.P., SIVASHANMUGAM P., GROSSMAN G., SOUNDARAJAN R.,
RAO A.J., RICHARDSON R.T., ZHANG Y., O’RAND M.G., PETRUSK P., FRENCH
F.S., HALL S.H. (2001) Primate epididymis- specific proteins: characterization of
ESC42, a novel protein containing a trefoil-like motif in monkey and human
endocrinology. Copyright by The Endocrine Society 142:4529-4539.
LONGIN J., GUILLAUMOT P., CHAUVIN M.A., MORERA A. LE MAGUERESSE-
BATTISTONI B. (2001) MT1-MMP in rat testicular development and the control of
Sertoli cell proMMP-2 activation. J Cell Sci 114:2125-34.
LÓPEZ M.L., DE SOUZA W., BUSTOS E. (1987) Cytochemical analysis of the
anionic sites on the membrane of the stallion spermatozoa during the epididymal
transit. Gamete Res 18:310-332.
LÓPEZ M.L. (1996) Contribuição ao estudo do processo de maturação espermática
em eqüinos (Equus caballus). Tese de Doutorado. UFRJ. Rio de Janeiro. RJ. Brasil.
LÓPEZ M.L., DE SOUZA W. (1991) Distribution of filipin sterol complexes in the
plasma membrane of stallion spermatozoa during the epididymal maturation process.
Mol Reprod Develop 28:158-168.
LÓPEZ M.L., FUENTES P., RETAMAL C., DE SOUZA W. (1997) Regional
differentiation of the blood-epididymis barrier in stallion (Equus caballus). J
Submicrosc Cytol Pathol 29:1-6.

102
LÓPEZ M.L., GRIBBEL I., BUSTOS E. (1989) Cytochemical and ultrastructural study
of the stallion epididymis. Submicrosc Cytol Pathol 21:103-120.
LUM L., BLOBEL C.P. (1997) Evidence for distinct serine protease activities with a
potencial role in processing the sperm protein fertilin. Develop Biol 191:131- 145.
MATRISIAN L.M. (1990) Metalloproteinases and their inhibitors in matrix remodelling.
Trends Genet 6: 121-25.
MBIKAY M., TADROS H., ISHIDA N., LERNER C.P., DE LAMIRANDE E., CHEN A.,
EL-ALFY M., CLERMONT Y., SEIDAH N.G., CHRETIEN M., GAGNON C.,
SIMPSON E.M. (1997) Impaired fertility in mice deficient for the testicular germ-cell
protease PC4. Proc Natl Acad Sci 94:6842-6846.
MCCAWLEY L.J., MATRISIAN L.M. (2000) Matrix metalloproteinases: multifunctional
contributors to tumor progression. Mol Med Today 6: 149-56.
MÉTAYER S., DACHEUX F., DACHEUX J.L., GATT J.L.I. (2002) Comparison,
characterization, and identification of proteases and protease inhibitors in epididymal
fluids of domestic mammals. Matrix metalloproteinases are major fluid gelatinases.
Biol Reprod 66:1219-1229 .
MORALES P., KONG M., PIZARRO E., PASTEN C. (2003) Participation of the
sperm proteasome in human fertilization. Hum Reprod 18:1010-7.
MULLER D., QUANTIN B., GESNEL M.C., MILLON-COLLARD R., ABECASSIS J.,
BREATHNACH R. (1988) The collagenase gene family in humans consist of at least
four members. Biochem J 253: 187-92.
NGUYEN M., ARKELL J., JACKSON C.J. (2001) Human endothelial gelatinases and
angiogenesis. Int J Biochem Cell Biol 33: 960-70.
NOTHNICK W.B., SOLOWAY P.D., CURRY T.E.JR. (1998) Pattern of messenger
ribonucleic acid expression of tissue inhibitors of metalloproteinases (TIMPs) during
testicular maturation in male mice lacking a functional TIMP-1 gene. Biol Reprod
59:364–370.
OKAMURA N., DACHEUX F., VENIEN A., ONOE S., HUET J.C., DACHEUX J.L.
(1992) Localization of a maturation-dependent epididymal sperm surface antigen
recognized by a monoclonal antibody raised against a 135-kilodalton protein in
porcine epididymal fluid. Biol Reprod 47:1040-1052.
ORGEBIN-CRIST M.C., JAHAD N. (1978) The maturation of rabbit epididymal
spermatozoa in organ culture: inhibition by antiandrogens and inhibitors of
ribonucleic acid and protein synthesis 103:46-53.

103
PAGE R.C. (1991) The role of inflammatory mediators in the pathogenesis of
periodontal disease. J Period Res 26: 230-242.
PENDÁS A.M., KNÄUPER V., PUENTE X.S., LLANO E., MATTEI M.G., APTE S.,
MURPHY G., LÓPEZ-OTÍN C. (1997) Identification and characterization of a novel
human matrix metalloproteinase with unique structural characteristics chromosomal
location, and tissue distribution. J Biol Chem 272: 4281-86.
PETRUSZAK J.A., NEHME C.L., BARTLES J.R. (1991) Endoproteolytic cleavage in
the extracellular domain of the integral plasma membrane protein CE9 precedes its
redistribution from the posterior to the anterior tail of the rat spermatozoon during
epididymal maturation. J Cell Biol 114:917-927.
PHELPS B.M., KOPPEL D.E., PRIMAKOFF P., MYLES D.G. (1990) Evidence that
proteolysis of the surface is an initial step in the mechanism of formation of sperm
cell surface domains. J Cell Biol 111:1839-1847.
PRIMAKOFF P., MYLES D.G. (2000) The ADAM gene family: surface proteins with
adhesion and protease activity. Trends Genet 16:83-87.
PUENTE X.S., PENDÁS A.M., LLANO E., VELASCO G., LÓPEZ-OTÍN C. (1996)
Molecular cloning of a novel membrane-type matrix metalloproteinase from a human
breast carcinoma. Cancer Res 56: 944-49.
QUANTIN B., MURPHY G., BREATHNACH R. (1989) Pump-1 codes for a protein
with characteristics similar to those of classical collagenase family members. J
Biochem 28: 5327- 34.
RETAMAL C.A., BABUL J. (1988) Determination of the molecular weight of protein
by electrophoresis in slab gels with a transverse pore gradient of crosslinked
polyacrylamide in the absence of denaturing agents. Anal Biochem 175:544-547.
RETAMAL C.A. (1999) Caracterização e purificação de proteínas epididimárias de
eqüino (Equus caballus). Tese de Doutorado. UENF. Campos dos Goytacazes. RJ.
Brasil.
RETAMAL C.A., THIEBAUT P., ALVES E.W. (1999) Protein purification from
poliacrylamyde gels by sonication extraction. Anal Bioch 268:15-20.
RETAMAL C.A., URZÚA J., ALVES E.W., LÓPEZ M.L. (2000a) Cytochemical and
electroforetic study of the stallion epididymal glycoproteins. J. Submicrosc. Cytpl
Pathol 1:117-130.

104
RETAMAL C.A., URZÚA J., LORCA C., LÓPEZ M.L., ALVES E.W. (2000b) Changes
in the plasma membrane proteins of stallion spermatozoa during maturation in the
epididymis. J Submicrosc Cytol Pathol 2:229-239.
ROBAIRE B., HERMO L. (1988) Efferent ductis, epididymis and vas deferens:
structure, functions and their regulation. In: E. Knobil e J.D. Neill, et al., editors. The
Physiology of Reproduction. New York , Raven Press.
ROBINSON L.L., SZNAJDER N.A., RILEY S.C., ANDERSON R.A. (2001) Matrix
metalloproteinases and tissue inibidores of metalloproteinases in human fetal testis
and ovary. Mol Hum Reprod 7:641-648.
ROGHANI M., BECHERER D.J., MOSS M.L., ATHERTON R.E., ERDJUMENT-
BROMAGE H., ARRIBAS J., BLACKBURN K.R., WESKAMP G., TEMPST P.,
BLOBEL C.P. (1999) Metalloprotease-Disintegrin MDC9: Intracellular Maturation and
Catalytic Activity. J Biol Chem 274: 3531-3540.
RUDOLPH-OWEN L.A., CANNON P., MATRISIAN L.M. (1998) Overexpression of
the matrix metalloproteinase matrilysin results in premature mammary gland
differentiation and male infertility. Mol Biol Cell 9:421- 435.
RUTLLANT J., MEYERS S.A. (2001) Posttranslational processing of PH-20 during
epididymal sperm maturation in the horse. Biol Reprod 65:1324-1331.
SATO H., TAKINO T., OKADA Y., CAO J., SHINAGAWA A., YAMAMOTO E., SEIKI
M. (1994) A matrix metalloproteinase expressed on the surface of invasive tumor
cells. Nature 370: 61-5.
SEATON G.J., HALL L., JONES R. (2000) Rat sperm 2B1 glycoprotein (PH20)
contains a C-terminal sequence motif for attachment of a glycosylphosphatidylinositol
anchor. Effects of endoproteolytic cleavage on hyaluronidase activity. Biol Reprod
62:1667-1676.
SETCHELL B.P., MADDOCKS S., BROOKS D.E. (1994) Anatomy, vasculature,
inervation and fluids of the male reproductive tract. In: Physiology of Reproduction.
Eds. Raven Press, Ltda. New York.
SETCHELL B.P., POLLANEN P., ZUPP J.L. (1988) Development of the blood-testis
barrier and changes in vascular permeability at puberty in rats. J Int Androl 11:225-
33.
SHAPIRO S.D., KOBAYASHI D.K., LEY T.J. (1993) Cloning and characterization of a
unique elastolytic metalloproteinase produced by human alveolar macrophages. J
Biol Chem 268: 23824-9.

105
SHILLING F.M., KRATZSCHMAR J., CAI H., WESKAMP G., GAYKO U., LEIBOW J.,
MYLES D.G., NUCCITELLI R., BLOBEL C.P. (1997) Identification of
metalloprotease/disintegrins in Xenopus laevis testis with a potential role in
fertilization. Dev Biol 186:155-64.
SHIMA I., SASAGURI Y., KUSUKAWA J., YAMANA H., FUJITA H., KAKEGAWA T.,
MINORU MORIMATSU, M.D. (1992) Production of matrix metalloproteinase-2 and
metalloproteinase-3 related to malignant behaviour of esophageal carcinoma. Cancer
70: 2747-2753.
STETLER-STEVENSON W.G. (1999) Matrix metalloproteinases in angiogenesis: a
moving target for therapeutic intervention. J Clin Invest 103: 1237-41.
STOLOW M.A., BAUZON D.D., LI J., SEDGWICK T., LIANG V.C., SANG Q.A.
(1996) Identification and characterization of a novel collagenase in Xenopus laevis:
possible roles during frog development. Mol Biol Cell 10: 1471-1483.
SUN H.J., PAN Y.C.E. (1999) Using native gel in two-dimensional PAGE for the
detection of protePin interactions in protein extract. J Biochem Biophys Meth 39: 143-
151.
SUTOVSKY P. (2003) Ubiquitin-dependent proteolysis in mammalian
spermatogenesis, fertilization, and sperm quality control: killing three birds with one
stone. Microsc Res Tech 61:88-102. Review.
SUTTON H.G., FUSCO A., CORNWALL G.A. (1999), CRES proteins Cystatin-
Related Epididymal Spermatogenic Protein Colocalizes with Luteinizing Hormone-b
Protein in Mouse Anterior Pituitary Gonadotropes. Endocrinol 140:0013-7227/99.
SYNTIN P., CORNWALL G.A. (1999) Immunolocalization of CRES (Cystatin-Related
Epididymal Spermatogenic) Protein in the Acrosomes of Mouse Spermatozoa Biol
Reprod 60:1542–1552.
TAKINO T., SATO H., SHINAGAW A., SEIKI M. (1995) Identification of the second
membrane-type matrix metalloproteinase gene form a human placenta cDNA library.
MM-MMP form a unique membrane-type subclass in the MMP family. J Biol Chem
270: 23021-23030.
TALHOUK R.S., BISSELL M.J., WERB Z. (1992) Coordinated expression of
extracellular matrix-degrading proteinases and their inhibitors regulates mammary
epithelial function during involution. J Cell Biol 118: 1272-1282.

106
TAMARINA N.A., MCMILLAN W.D., SHIVELY V.P., PEARCE W.H. (1997)
Expression of matrix metalloproteinases and their inhibitors in aneurysms and normal
aorta. Surgery 122: 264-272.
TOWBIN H., STAEHELIN T., GORDON J. (1979) Electrophoretic transfer of proteins
from polyacrylamide gels to nitrocellulose sheets: procedure and some applications.
Proc Natl Acad Sci U S A, 76: 4350-4354.
TULSIANI D.R., ABOU-HAILA A., LOESER C.R., PEREIRA B.M. (1998) The
biological and functional significance of the sperm acrosome and acrosomal
enzymes in mammalian fertilization. Exp Cell Res 240:151-164.
TULSIANI D.R., SKUDLAREK M., ORGEBIN-CRIST M.C. (1995) Novel alpha-
Dmannosidase of rat sperm plasma membranes: characterization and potential role
in sperm -egg interactions. J Cell Biol 109:1257-67.
TURNER T.T. (1979) On the epididymis and its function. Investigative Urology. 16:
311-321.
UHRIN P., DEWERCHIN M., HILPERT M., CHRENEK P., SCHOFER C.,
ZECHMEISTER-MACHHART M., KRONKE G., VALES A., CARMELIET P., BINDER
B.R., GEIGER M. (2000) Disruption of the protein C inhibitor gene results in impaired
spermatogenesis and male infertility. J Clin Invest 06:1531-1539.
VISSE R., NAGASE H. (2003) Matrix metalloproteinases and tissue inhibitors of
metalloproteinases: structure, function, and biochemistry. Circ Res. 92:827-39.
WESTERHUIS W.H.J., STURGIS J.N., NIEDERMAN R.A. (2000) Reevaluation of
the electrophoretic migration behavior of soluble globular proteins in the native and
detergent-denatured states in polyacrylamide gels. Anal Bioch 284:143-152.
WESTERMARCK J., KAHARI V.M. (1999) Regulation of matrix mettaloproteinase
expression in tumor invasion. FASEB J 13: 781-92.
WHITHAM S.E., MURPHY G., ANGEL P., RAHMSDORF H.J., SMITH B.J., LYONS
A., HARRIS T.J., REYNOLDS J.J., HERRLICH P., DOCHERTY A.J. (1987)
Comparison of human stromelysin and collagenase by cloning and sequence
analysis. Biochem J 240: 913-16.
WILHELM S.M., COLLIER I.E., MARMER B.L., EISEN A.Z., GRANT G.A.,
GOLDBERG G.I. (1989) SV40-transformed human lung fibroblast secrete a 92-kDa
type IV collagenase which is identical to that secreted by normal human
macrophages. J Biol Chem 264: 17213-21.

107
WILL H., HINZMANN B. (1995) cDNA sequence and mRNA tissue distribution of a
novel human matrix metalloproteinase with a potential transmembrane segment. Eur
J Biochem 231: 602-8.
WOESSNER J. (1991) Matrix metalloproteinases and their inhibitors in connective
tissue remodeling. FASEB J 5: 2145-54.
WOLF C., CHENERD M.P., DURAND DE GROSSOUVRE P., BELLOCQ J.P.,
CHAMBON P., BASSET P. (1992) Breast-cancer-associated stromelysin-3 gene is
expressed in basal cell carcinoma and during cutaneous wound healing. J Invest
Dermatol 99: 870-872.
YABKOWITZ R., MEYER S., BLACK T., ELLIOTT G., MEREWETHER L.A.,
YAMANE H.K. (1999) Inflamatory cytocines and vascular endothelial growth factor
stimulate the release of soluble tie receptor from human endothelial cells via
metatalloprotease activation. Blood 93: 1969-79.
YAMAGATA K., MURAYAMA K., OKABE M., TOSHIMORI K., NAKANISHI T.,
KASHIWABARA S., BABA T. (1998) Acrosin accelerates the dispersal of sperm
acrosomal proteins during acrosome reaction. J Biol Chem 273:10470-10474.
YANAGIMACHI, R. (1994) Mammalian Fertilization. In: The Physiology of
Reproduction. 2 ed.
YUAN R., PRIMAKOFF P., MYLES D.G. (1997) A role for the disintegrin domain of
cyritestin, a sperm surface protein belonging to the ADAM family, in mouse sperm–
egg plasma membrane. J Cell Biol 137:105-112.
ZHENG X., GEIGER M., ECKE S., BIELEK E., DONNER P., EBERSPACHER U.,
SCHLEUNING W.D., BINDER B.R. (1994) Inhibition of acrosin by protein C inhibitor
and localization of protein C inhibitor to spermatozoa. Am J Physiol 267:466-472.