função de proteínas: mioglobina e...
TRANSCRIPT

Aula de Bioquímica I
Tema:
Função de Proteínas:
Mioglobina e Hemoglobina
Prof. Dr. Júlio César BorgesDepto. de Química e Física Molecular – DQFM
Instituto de Química de São Carlos – IQSC
Universidade de São Paulo – USP
E-mail: [email protected]

Proteínas
Funções
Transporte e Estoque;
Catálise enzimática;
Movimento;
Suporte mecânico;
Proteção imune;
Sinalização intra e extracelular;
etc.
Variedade Funcional ���� Diferentes Estruturas Tridimensionais
Estrutura 3D ���� Estrutura Estável em função do tempo
Estabilidade versus flexibilidade
Função versus Conformação.

Como as Proteínas Funcionam
Grande maioria → interação com outras moléculas
Interações reversíveis → ligante (qualquer tipo de molécula, inclusive proteína);***Natureza transitória das interações proteína-ligante***
↓VIDA
↓RESPOSTA RÁPIDA E REVERSÍVEL A MUDANÇAS AMBIENTAIS E CONDIÇÕES
METABÓLICAS
SÍTIO DE LIGAÇÃO
→ Interação específica
→ Acoplada a uma mudança conformacional → ENCAIXE INDUZIDO
*ENZIMAS: Substratos; Sítio catalítico ou Sítio ativo.

Introdução���� Organismos aeróbicos ���� Necessitam de suprimento de O2 constante
�Organismos multicelulares vertebrados
- Desenvolvimento de sistema circulatório
- Desenvolveram proteínas de transporte de O2
1 – Mioglobina ���� músculo
- mantém suprimento local de O2
2 – Hemoglobina ���� sangue ���� hemácias
- Transporta O2 dos pulmões para os tecidos
- Transporta CO2 e H3O+ dos tecidos para os pulmões
Mioglobina versus Hemoglobina
- São similares na Estrutura Primária, Secundária e Terciária
- Ligam O2 de maneira muito similar
- Em condições similares aos tecidos, a Hemoglobina está 7% saturada com O2 enquanto que a Mioglobina apenas 90% saturada
- Hemoglobina liga O2 de maneira cooperativa e é uma proteína alostérica
- Ambas são importantes proteínas-modelo de estudo Bioquímico
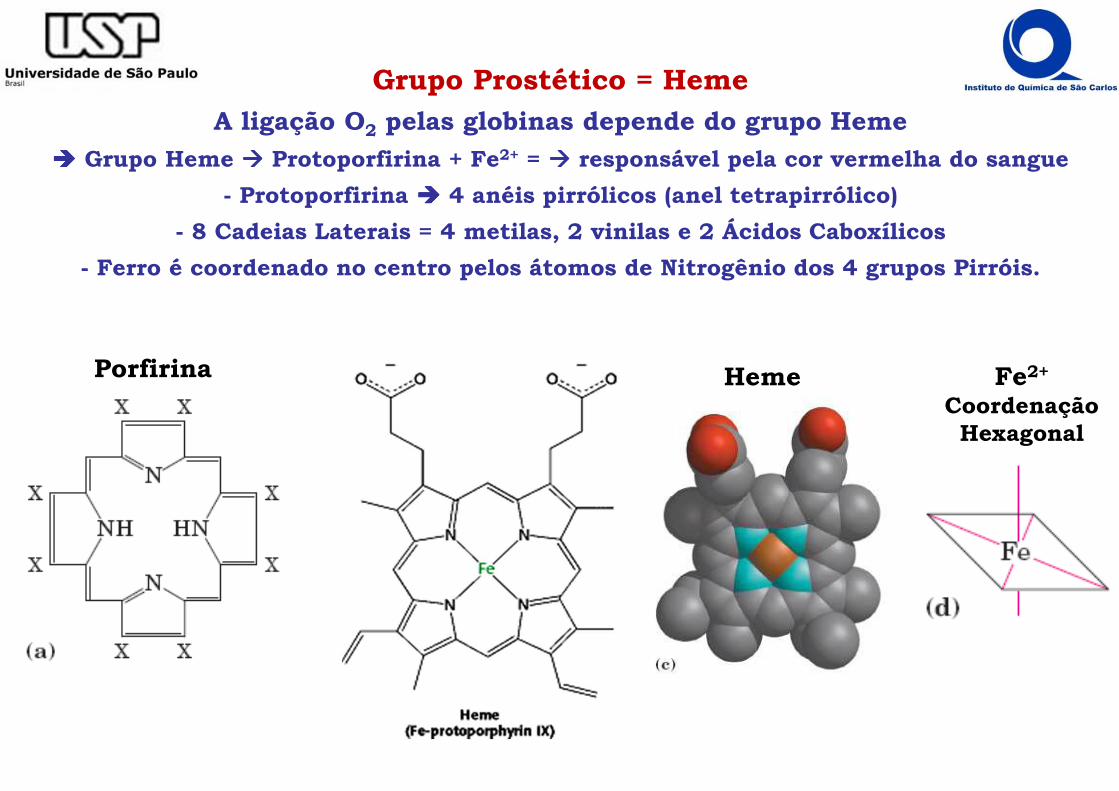
Grupo Prostético = HemeA ligação O2 pelas globinas depende do grupo Heme
���� Grupo Heme ���� Protoporfirina + Fe2+ = ���� responsável pela cor vermelha do sangue
- Protoporfirina ���� 4 anéis pirrólicos (anel tetrapirrólico)
- 8 Cadeias Laterais = 4 metilas, 2 vinilas e 2 Ácidos Caboxílicos
- Ferro é coordenado no centro pelos átomos de Nitrogênio dos 4 grupos Pirróis.
Porfirina Heme Fe2+
Coordenação Hexagonal

HolomioglobinaGrupo heme está ligado à apoproteína:
1) pelo ocupação do Quinto ponto de coordenação do Fe2+ pela His proximal
2) Interações Hidrofóbicas do anel tetrapirrólico
- Sexto ponto de coordenação do Fe2+ é ocupado pelo O2
- Na forma deoxi, a His Proximal retira o Fe2+ do plano central da porfirina

HolomioglobinaGrupo heme permite monitoramento:

Holomioglobina���� 1º Proteína a ter a sua estrutura 3D resolvida
- Estrutura em Hélices Alfa - Possui 153 resíduos de AA
- Total de 8 hélices anfipáticas – nomeadas de A a H
���� Grupo Heme está ligado entre as hélices E e F
- Hélice alfa F contém a His Proximal (Chamada de His F8)
- Hélice alfa E contém a His Distal (Chamada de His E7)

Holomioglobina���� Grupo Heme está ligado entre as hélices E e F
- Hélice alfa F contém a His Proximal (Chamada de His F8)
- Hélice alfa E contém a His Distal (Chamada de His E7)
���� A ligação O2–Fe2+ gera a transferência parcial de 1 elétron do Fe2+ para o O2���� forma o íon superóxido
O2-
���� A His E7 estabiliza o O2ligado ao Fe2+ por uma
ligação de H;
���� Evita a formação do íon superóxido;
- O componente protéico estabiliza a reatividade do
O2

Interação Proteína-Ligante
pode ser quantificada
Proteínas são capazes de interagir especificamente com ligantes
A interação Proteína P com o Ligante L obedece as leis da Termodinâmica
Questões apresentadas
1) Qual é o máximo número de moles de ligante por proteína?
2) Qual é a constante de equilíbrio deste ligante por cada um dos sítios de ligação?
3) A constante de equilíbrio do L por cada um dos sítios é independente da ocupação dos demais?
4) A constante de equilíbrio do L é modificada pela presença de um segundo ligante?
P + L PLk1
k2

Interação Proteína-Ligante
pode ser quantificadaA interação Proteína P com o Ligante L obedece as leis da Termodinâmica
k1 e k2 são as constantes de velocidade de associação e dissociação
KA e KD são as constantes de equilíbrio no sentido de associação e dissociação
KA é dado em (mol/L)-1 (M-1)���� Quanto maior o valor de KA maior a afinidade da interação PL
KD é dado em mol/L (M)���� Quanto menor o valor de KD maior a afinidade da interação PL
� Quando a [L] é >>> [P] podemos considerar que a formação do PL não altera a [L]
� A fração de Sítios de ligação na P frente ao L pode ser descrita pela seguinte equação
P + L PLk1
k2 ADAeq K
KLP
PL
k
kKK
1
]][[
][
2
1 =∴===
][][
][
sDisponívei Sítios de Total
Ocupados Sítios
PPL
PL
+==θ

Interação Proteína-Ligante
pode ser quantificadaΘ pode ser monitorado e seu valor em função da [L] resulta numa hiperbólica retangular
][][
][
PPL
PL
+=θ
]][[][]][[
][LPKPL
LP
PLK AA =∴=
D
A
A
A
A
A
KL
L
KL
L
LK
LK
PLPK
LPK
+=
+=
+=
+=
][][
1][
][1][
][][]][[
]][[θ
Dividindo ambos os
por [P]
Dividindo ambos por
KA
DA
KK
=1
Quando [L] = KD ���� [L]onde metade dos sítios
de ligação estão ocupados pelo Ligante
���� Para 90% de saturação [L] = 9 x KD
Substituindo [PL]

Interação Mioglobina-O2
A interação entre a mioglobina e O2 segue as leis da termodinâmica mesmo considerando o Oxigênio como gás
- Adequação experimental e matemática é necessária
DKOO
θ+
=][
][
2
2
Se P50 = [O2] na qual metade dos sítios estão ocupados
Quando pO2 for equivalente à metade do sinal máximo temos KD e assim podemos
definir P50
DKpOpO
θ+
=2
2
O2 liga fortemente à mioglobina com um P50 de
0,26 kPa - Condição na qual metade dos sítios estão ocupados
A [O2] em solução depende diretamente da pressão de O2 na
atmosfera que pode ser controlada.
502
2
PpOpO
θ+
=
DKP =50

Estrutura da Hemoglobina���� A Hemoglobina é um heterotetrâmero
- Duas cadeias Alfa (141 resíduos) e duas cadeias Beta (146 resíduos)
- Possui estrutura em hélice alfa
���� Hemoglobina A (HbA) = par de dímeros α1β1 e α2β2
- A interface α1β1 e α2β2 apresenta ~30 contatos
- A interface α1β2 e α2β1 apresenta ~19 contatos
���� Maioria hidrofóbicos e alguns hidrofílicos ���� Importantes para regulação
- Grupos Heme estão bem separados (24-40 Å)

Estrutura da Hemoglobina� Cadeia Alfa e Beta da HbA apresentam ~25% de identidade com a Mioglobina
���� Alta identidade estrutural
- Apesar da baixa identidade sequencial
- Identidade estrutural é mais conservada
���� Mesma forma de ligar grupo Heme
-Possui His Proximal e Distal

Estrutura da Hemoglobina���� A HbA liga O2 de maneira cooperativa – curva Sigmoidal
���� A ligação de O2 em uma cadeia da HbA aumenta a ligação de O2 nas demais cadeias
���� A dissociação de O2 em uma cadeia da HbA facilita a dissociação de O2 nas demais cadeias

Estrutura da Hemoglobina���� A ligação de O2 com HbA é >10 x mais fraca do que com a Mioglobina
���� A baixa afinidade da HbA por O2 está relacionada com a função desta proteína
- Captar O2 nos pulmões (“alta” pO2) e liberar nos tecidos (“baixa” pO2)
���� A alta afinidade da Mioglobina por O2 está relacionada com a função: Armazenar O2

Cooperatividade� A Alosteria – “outra forma” – está envolvida na cooperatividade
Sistema de variação de afinidade/atividade frente à modulação.
���� Envolve a participação de um Modulador
- Modulador ���� Ativador ou inibidor
- Modulação homotrópica: Ligante e modulador iguais
- Modulação heterotrópica: Ligante e modulador diferentes
���� Uma curva de ligação sigmoidal é um sinal de ligação cooperativa.

Cooperatividade� A ligação cooperativa de O2 na HbA foi analisada por Archibald Hill (1910)
P + nL PLn
k1
k2
nn
Aeq LPPL
kk
KK]][[][
2
1 ===
D
n
KL
θnθ ][=− DKnLn
θnθ
log]log[log −=
−�Equação de Hill
1][][+
=n
An
nAn
LK
LKnθ
1) Reorganizando2) Dividindo ambos por KA
3) Substituindo por KD
� Curva de Hill ���� log [θ/(1-θ)] versus log[L] onde a inclinação = n = nH
Aplicando Log

Cooperatividade� A ligação cooperativa de O2 na HbA foi analisada por Archibald Hill (1910)
( ) ( )502 log0loglog Pnpnθn
θ −=
−
�nH não representa o número de sítios de ligação, mas o grau de interação
entre eles: o grau de cooperatividade
� nH representa a cooperatividadeentre os sítios de ligação
� nH < n

Ligação de O2 na Hemoglobina� Em exercício, o sangue arterial, após uma passagem pelo pulmão, apresenta ~98% de
saturação da HbA com O2
���� Após passar pelos tecidos, o sangue venoso apresenta ~32% de saturação de O2
- Portanto, ~66% dos pontos de ligação contribuíram para o transporte de O2
� A ação cooperativa da HbA favorece a DESCARGA adequada de O2
- Nestas condições, a Mioglobina permaneceria com 91% de saturação ���� 7% de transporte
���� A HbA transporta ~10 x mais O2 do que a Mioglobina

O2
A cooperatividade da HbA� A ligação de O2 na HbA resulta em mudanças conformacionais na tetrâmero de HbA-O2
- Um dímero Alfa-Beta sofre rotação de 15º em relação ao outro
���� A HbA existe em duas conformações principais:
1) T (Tenso) ���� Estado deoxigenado
2) R (Relaxado) ���� Estado Oxigenado
- O Estado R apresenta maior afinidade por O2 do que o estado T
- Na presença de O2, o estado R é mais estável

A cooperatividade da HbA� Estado T apresenta um intricada rede de pares iônicos na interface α1β2 e α2β1
���� A ligação de O2 altera a estrutura do grupo Heme que perturba a rede de pares iônicos
- Mudanças no ambiente do grupo
Heme são transmitidas para as outras cadeias
do oligômero

A cooperatividade da HbA� Estado T apresenta um intricada rede de pares iônicos na interface α1β2 e α2β1
���� A ligação de O2 altera a estrutura do grupo Heme que perturba a rede de pares iônicos

A cooperatividade da HbA� A HbA pura no estado T é muito instável
- Liga avidamente O2 em baixas [O2] e libera apenas 8% de O2 em condições fisiológicas
���� A transição T ���� R adequada depende de um ligante – o 2,3-Bisfosfoglierato – 2,3-BPG
���� A [2,3-BPG] nas hemácias é aumentada na altitude.
- Adaptação em ~ 24 horas
- Permite menor afinidade entre HbA-O2
- A HbA liga menos O2,
MAS descarga de O2 nos tecidos é MAIOR apesar da
menor pO2 atmosférica

A cooperatividade da HbA� Efeito do 2,3-Bisfosfoglierato – 2,3-BPG na p50

A cooperatividade da HbA
O 2,3-BPG:
1) Liga-se no centro do Tetrâmero da HbA;
2) É liberado após transição T ���� R;
3) Age mantendo HbA no Estado T;
4) É um Efetor Alostérico.

A cooperatividade da HbA
A transição T ���� R expulsa o BPG do sítio de ligação
Estado REstado T

A Hemoglobina Fetal- A Hb fetal contem duas cadeia Alfa e duas cadeias Gama
���� A cadeia Gama é 72% idêntica à cadeia Beta na seqüência de aminoácidos
- A modificação Beta-His143 �������� Gama-Ser143
- Modifica o sítio de interação do 2,3-BPG com a Hb
Consequências:
- Redução da afinidade da Hb Fetal por 2,3-BGP
- Aumento da afinidade da Hb Fetal por O2
���� Permite que a Hb Fetal receba eficientemente o O2 transportado pela HbA
maternal
- O feto não “experimenta” pO2 atmosférica
Outros genes
���� Existem outras 3 cadeias de hemoglobina expressas em diferentes estágios do
desenvolvimento
�Existem ainda: Neuroglobina e citoglobina
(+ similares à mioglobina)

Integração das trocas gasosas� O CO2 é transportado como HCO3
- (maior parte)
� A Hidratação do CO2 gera H+

A HbA transporta H+ e CO2
dos tecidos para o pulmão� A HbA é sensível aos níveis de H+ e CO2 nos tecidos
- H+ e CO2 são efetores alostéricos da HbA ���� aumentam a liberação de O2 para os tecidos
� O estado T é estabilizado pela redução do pH
- Resultado: descarga de O2 nos tecidos
���� É chamado de Efeito Bohr (Christian Bohr, pai de Niels Bohr)
� O CO2 é hidratado pela enzima Anidrase Carbônica nas hemácias
���� A afinidade da HbA por O2diminui com a redução do pH

A HbA liga H+ e CO2
���� A HbA transporta H+ dos tecidos para o pulmão
No capilar sanguíneo (tecidos)
���� A Hidratação do CO2 é
catalisada pela Anidrasecarbônica
�O próton liberado pela hidratação do CO2 liga-se à cadeia alfa e beta da HbA estabilizando uma ponte salina
���� estabilização do estado T
� Permite o transporte de próton para os pulmões
- 80% do transporte
Ponte salina na interface α2β1

A HbA liga H+ e CO2
���� A HbA transporta CO2 dos tecidos para o pulmão
���� O CO2 liga-se à aminas terminais
- ~15-20% do transporte ligado à HbA
- Restante é transportado na forma de HCO3- solúvel
� Forma grupos carbamato ���� carga negativa
���� Estabiliza estado T por favorecer interações eletrostáticas na interface cadeia Alfa-Beta da HbA
Descarga de O2 pela HbA66%77%88%

Fatores influentes na HbA
Efeitos na curva de saturação

Mutações na HbA� Os genes da Hb sofrem mutações que alteram a cadeia protéica
- Podem prejudicar a afinidade da Hb pelo O2
- Originam as Hemoglobinopatias – são as anemias hereditárias
- ~ 500 tipos de mutações identificadas na população humana
- Muitas mutações são inócuas
- O Surgimento da anemia depende se o indivíduo é heterozigoto ou homozigoto
���� Anemia falciforme
- devido à mutação D6V na cadeia Beta
- Forma a Hemoglobina S
- Forma um contato hidrofóbico com a F85 e V88 entre as cadeias beta
inter-tetrâmeros

Mutações na HbA� As pontes hidrofóbicas inter-cadeias propicia a fibrilação

Mutações na HbA� As pontes hidrofóbicas inter-cadeias precipitam a HbS ���� não liga O2

Mutações na HbA�A HbS forma fibras que deformam a estrutura das hemácias
� HbS aumenta resistência à malária ���� parasita vive dentro das hemácias
-possível fator seletivo positivo da HbS ���� por isto foi “selecionada” e persiste presente