estudos de estrutura e função de aldeído desidrogenases...
TRANSCRIPT

Estudos de estrutura e função de Aldeído Desidrogenases
envolvidas em sinalização e detoxificação
Bolsista Fábio Neves do Amaral
Estudante de Biomedicina da Universidade Federal de Pernambuco
Orientador
Prof. Dr. José Xavier Neto/LNBio

Estudos de estrutura e função de Aldeído Desidrogenases
envolvidas em sinalização e detoxificação
Bolsista Fábio Neves do Amaral
Estudante de Biomedicina da Universidade Federal de Pernambuco
Orientador
Prof. Dr. José Xavier Neto/LNBio
2011

Agradecimentos
Agradeço primeiramente a Deus, que meus deu as forças necessárias para
sempre seguir em frente em qualquer situação.
Agradeço a todos que me incentivaram a vir para o 20° Programa Bolsas
de Verão, principalmente meus pais, amigos mais próximos, esposa e parentes.
Agradeço aos organizadores do programa pelo excelente trabalho e a
ótima oportunidade que aqui tivemos.
Agradeço a todos que me ajudaram no desenvolvimento deste trabalho,
principalmente a Andrea Balan, Hozana Castillo, Vanessa Rodrigues e Tereza
Cristina, que me acompanharam de perto em minhas atividades e me trouxeram
muito conhecimento.
Agradeço ao Prof. Dr. José Xavier Neto, pela sua orientação neste trabalho
e por tudo que me foi ensinado.
Por fim, dedico este trabalho ao meu filho, Daniel Lira, que mesmo em
minha ausência durante este tempo de trabalho, com certeza desenvolveu um
aprendizado muito maior em relação a sua vida.

Sumário
AGRADECIMENTOS ............................................................................................................................ 3
RESUMO ............................................................................................................................................ 5
1. INTRODUÇÃO ............................................................................................................................ 6
1.1 ÁCIDO RETINÓICO ........................................................................................................................ 6
1.2 ÁCIDO RETINÓICO NO DESENVOLVIMENTO CARDÍACO ......................................................................... 7
1.3 ÁCIDO RETINÓICO NO DESENVOLVIMENTO NEURAL ............................................................................ 7
1.4 ALDHS ...................................................................................................................................... 7
2. HIPÓTESE ................................................................................................................................. 11
3. OBJETIVOS ............................................................................................................................... 13
3.1 OBJETIVOS ESPECÍFICOS .............................................................................................................. 13
4. METODOLOGIA ........................................................................................................................ 14
4.1 SUBCLONAGEM DOS GENES BFALDH1A, CIALDH1D E BFALDH1E ................................................... 14
4.2 EXPRESSÃO DE CIALDH2 ............................................................................................................ 18
5. RESULTADOS E DISCUSSÃO ...................................................................................................... 21
5.1 SUBCLONAGEM ......................................................................................................................... 21
5.2 EXPRESSÃO ............................................................................................................................... 22
6. PERSPECTIVAS ......................................................................................................................... 23
7. REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................................ 24

Resumo
As Aldeído Desidrogenases (ALDHs) formam uma superfamília de
proteínas que catalisa a oxidação de uma grande variedade de aldeídos. Muitas
destas proteínas apresentam papéis distintos nos organismos, como exemplo,
ALDH1 e ALDH2. A ALDH1 desempenha um papel fundamental no
desenvolvimento dos sistemas orgânicos de eucariotos, transformando o
retinaldeído em ácido retinóico, que atua como sinalizador celular, enquanto a
ALDH2 degrada pequenos aldeídos tóxicos, tendo assim um forte papel na
detoxificação e proteção.
Estudos mostram que as preferências dessas ALDHs pelos seus substratos
dependem do tamanho do canal de acesso ao sítio catalítico, que varia com a
modificação de três aminoácidos chave. Alguns organismos apresentam
variantes da ALDH1, com estrutura e padrões de expressão semelhantes aos da
ALDH2, sugerindo que ao longo da evolução estas proteínas adquiriram uma
nova estrutura/função.
Para testar a hipótese que a modificação dos aminoácidos chave pode
reprogramar a função de ALDHs envolvidas em sinalização e detoxificação, o
objetivo desse trabalho foi estabelecer a estrutura e função das ALDH1 e 2 e suas
variantes em Branchiostoma floridae (bf) e Ciona intestinalis (ci) e compará-las
com a estrutura e função da ALDH2 destes organismos.
Para isso, foram clonados os genes bfALDH1A, ciALDH1D e bfALDH1E,
enquanto paralelamente foi testada a metodologia de expressão da ciALDH2,
estabelecida com temperatura de 28°C com 6 horas de indução. Esses dados
darão base para a avaliação das ALDHs em relação a sua estrutura/função.

1. Introdução
1.1 Ácido retinóico
O desenvolvimento em boa parte dos eucariotos depende de complexos
sistemas de sinalização celular, que desempenham um papel crucial no
desenvolvimento embrionário. Dentro desse contexto, está a via de sinalização
da qual faz parte o ácido retinóico (Figura 1).
O ácido retinóico (RA) é um derivado da vitamina A e está relacionado
com o desenvolvimento embrionário neural e cardíaco, assim como de outros
tecidos.1, 2
Figura 1 - Cascata de sinalização do Ácido Retinóico (RA). Apenas os principais atuantes desta
via estão demonstrados. Enzimas relacionadas à síntese e degradação de RA estão destacadas nas
caixas pretas (adaptado de CAMPO-PAYSAA et al., 2008).
O papel do RA na biologia do desenvolvimento vem sido amplamente
estudado em modelos animais e supõe-se que ele apresente diferentes
mecanismos de ação entre grupos como invertebrados e vertebrados.2, 4

1.2 Ácido retinóico no desenvolvimento cardíaco
Indícios mostram que a proliferação dos cardiomiócitos e a formação do
coração a nível embrionário é regulada pelo RA, e alterações nos níveis deste
sinalizador podem causar mudanças drásticas no padrão de formação do tecido
cardíaco.5, 6, 7
Um dos pontos principais é a influencia da ALDH1A2 (RALDH2) no
controle dos níveis de RA, uma vez que a alteração nos níveis ou atividade da
RALDH2 causa uma mudança nos níveis de RA circulante, resultando numa
variação inversamente proporcional do numero de cardiomiócitos. Além disso,
os níveis de RA são importantes em outros fatores do desenvolvimento cardíaco,
como a diferenciação do miocárdio e a maturação e crescimento ventricular. 5, 6, 7
1.3 Ácido retinóico no desenvolvimento neural
Há uma possível correlação entre os níveis de ácido retinóico e o
desenvolvimento embrionário neural de alguns grupos de cordados, como os
cefalocordados. Alguns autores também afirmam haver uma relação entre a
sinalização exercida pelo RA e a diferenciação da notocorda, a transição de
cordados invertebrados para os vertebrados na evolução.4
Portanto, entendendo a correlação das funções metabólicas desses
organismos cordados invertebrados com os vertebrados e como se desenvolveu
o processo de sinalização por RA, pode-se obter alguma informação que aponte
as peças chaves deste processo evolutivo.4, 2
1.4 ALDHs
Durante o processo de degradação da vitamina A até o RA em
vertebrados, a etapa de oxidação do retinaldeído a ácido retinóico é catalisada
por uma enzima chamada aldeído desidrogenase 1A (ALDH1A) ou retinaldeído
desidrogenase (RALDH). Existe uma grande variedade destas enzimas, que
formam uma superfamília que se estende aos mais variados organismos. A
estrutura da RALDH é extremamente importante em relação a sua função nestes
e em outros organismos.3, 7, 8, 9, 10
As ALDHs catalisam a oxidação do retinaldeído utilizando o NADP agente
oxidante. Organizam-se em tetrâmeros e sua estrutura monomérica apresenta

um canal de acesso ao sítio catalítico na porção globular livre e uma região de
oligomerização, responsável pela formação do tetrâmero, com quatro porções
globulares livres. (figura 2)8, 9
Figura 2 – Modelo de um monômero da ALDH (a) e sua forma tetramérica (b).Procurar
referencia.
Pesquisas indicam que existem aminoácidos que são responsáveis pela
especificidade das ALDHs ao aldeído que elas catalisam, modificando o tamanho
do canal de acesso ao sítio catalítico, restringindo assim a passagem de grandes
aldeídos em proteínas que tenham esse canal reduzido, como o RA em ALDH2, e
também a permanência de pequenos aldeídos em ALDHs de canal mais largo,
como acetaldeído em ALDH1.9
Desses aminoácidos, três (a.a. 124, a.a.459, a.a. 303) se destacam por ter
um papel mais importante na regulação do volume desse túnel. Basicamente, o
tamanho desses aminoácidos define o tamanho dessa passagem, com os
aminoácidos de cadeia lateral longa obstruindo o canal, no caso da ALDH2, e os
aminoácidos de cadeia curta mantendo-os livres e abertos, como na ALDH1
(figura 3).9
a b

Figura 3 - Estrutura dos tuneis de entrada ao sítio catalítico em ALDH1 e ALDH2, comparados a
dois aldeídos de diferentes tamanhos (Adaptado de SOBREIRA, T. J. P. et al. - 2010).
A modificação no tipo de aldeído catalisado em função da diferença nestes
aminoácidos ocasiona em funções distintas para essas proteínas. A ALDH1, com
seu papel na sinalização celular, e a ALDH2, no papel de detoxificação de
pequenos aldeídos tóxicos.9
Mutações no aminoácido Lys504 por Glu504 em ALDH2 (mutante
ALDH2*2) em humanos, comuns em populações do leste asiático, estão
correlacionadas com o aumento do risco de câncer estomacal quando associados
ao alcoolismo, pois a capacidade de degradar os aldeídos tóxicos gerados na
ingestão alcoólica fica reduzida. O estudo destas mutações mostrou que a
molécula Alda-1 funciona como uma chaperona (Figura 4), retomando a
atividade destas ALDH2*2, com uma eficiência semelhante às formas nativas.
Essa descoberta e o entendimento deste mecanismo de ação podem auxiliar no
desenvolvimento de novos fármacos que recuperem a atividade de enzimas
mutantes.3, 8

Figura 4– Modelo de um potencial mecanismo de ativação da ALDH2*2 através de Alda-1
(PEREZ-MILLER, S. et al. – 2010)
As ALDHs são uma família com sequencias de aminoácidos muito
conservada, sugerindo que tenham uma origem comum. Considerando esse fato,
possivelmente houve uma adaptação estrutural entre essas ADLHs, sugerindo
assim uma reprogramação de suas funções originais.9 Analisando as implicações
práticas, é possível que mutagenizando esses genes nas regiões correspondentes
aos três aminoácidos chave principais supracitados, haverá uma reprogramação
da função baseada nesta modificação estrutural induzida. Ou seja, podemos
induzir mutações no gene da ALDH2, recuperando a função da ALDH1 e vice-
versa.
Em alguns cordados, foram encontrados homólogos da ALDH1, mas
apresentaram características semelhantes a ALDH2 em relação ao padrão de
expressão, que é focalizado nas ALDH1 e disperso nos tecidos nas ALDH2. Elas
apresentam variações em relação ao tamanho do canal, como intermediários
entre as estruturas da ALDH1 e ALDH2 (Figura 5).9

Figura 5 - Filogenia (A) estrutura do canal (B) e expressão (C) de ALDH1/2 em anfioxo
(SOBREIRA, T.J.P. et al. 2011).
2. Hipótese
As ALDHs apresentam um padrão de catálise semelhante, modificado
apenas pelo tipo de aldeído a ser catalisado, utilizando como parâmetro o
tamanho destes. Essa seleção ocorre pelo tamanho do canal de entrada para o
sítio catalítico, que é determinado por alguns aminoácidos chave, dos quais três
são mais importantes. A ALDH1 apresenta subtipos em organismos como Ciona
intestinalis (ci)e Branchiostoma floridae(bf) que se assemelham com a ALDH2 em
alguns aspectos. Estes dados levam a hipótese de que a ALDH2 pode ter uma
origem comum com a ALDH1, sendo que a modificação desses aminoácidos
chave pode ter reprogramado a função desta enzima. Os subtipos da ALDH1 nos
organismos acima podem conter alguma informação sobre essa possível
reprogramação, mas ainda são necessáriosestudos sobre suas estruturas e
funções.

Considerando estes fatos, supõe-se que a modificação dos três principais
aminoácidos relacionados com o padrão de tamanho dos canais de acesso ao
sítio catalítico da ALDHs pode reprogramar as enzimas entre as funções
sinalização e detoxificação, sendo que os subtipos encontrados em C. intestinalis
e B. floridae podem ter relação com essas modificações e ajudem a esclarecer a
evolução das funções dessas ALDHs.

3. Objetivos
Para testar a hipótese que a modificação dos aminoácidos chave pode
reprogramar a função de ALDHs envolvidas em sinalização e detoxificação, o
objetivo desse trabalho é estabelecer a estrutura e função da ALDH2 e das
variantes de ALDH1 em Branchiostoma floridae e Ciona intestinalis e compará-las
com a estrutura e função da ALDH2 destes organismos.
3.1 Objetivos específicos
Sub-clonar os genes ALDH1 de Branchiostoma floridae e Ciona intestinalis;
Expressar, extrair e purificar essas proteínas, assim como as ALDH2 de
Branchiostoma floridae e Ciona intestinalis.

4. Metodologia
4.1 Subclonagem dos genes bfALDH1A, ciALDH1D e bfALDH1E
4.1.1 Vetor de clonagem
O vetor utilizado para realizar a subclonagem foi o pET28a-SUMO (Figura
6), previamente construído utilizando a tecnologia SUMO. Para isso, o vetor
pET28a é clonado com o gene da SUMO, que facilita a expressão, solubilização e
purificação, segundo fabricantes. 11
pET28a SUMO
5632 bp
N-6His
Kan
T7 promoter
lacI
SUM O
C-6His
End
BamH I ( 5092)
ClaI (912)
EcoRI (5098)
HindIII (5117 )
Nco I (47 31)
SmaI (7 31)
XmaI (7 29)
SapI (1920)
NotI (5124)
SacI (5108)
SalI (5111)
XhoI (5132)
NdeI (47 91)
NheI (47 96)
AvaI (7 29)
AvaI (5132)
ApaLI (1489)
ApaLI (1989)
ApaLI (3924)
Figura 6 – Mapa do pET28a-SUMO, com o gene de resistência a kanamicina.

4.1.2 Amplificação dos genes para clonagem
Para amplificar os genes bfALDH1A, ciALDH1D e bfALDH1E, previamente
clonados em vetores comerciais, foram desenhados oligonucleotídeos
direcionais contendo em sua região 5´ o sítio de restrição para a enzima BamHI e
oligonucleotídeos reverso contendo na sua região 3´ o sítio de restrição para a
enzima XhoI. Os genes bfALDH1A, ciALDH1D e bfALDH1E foram amplificados
através de reação em cadeia da polimerase (PCR), utilizando os
oligonucleotídeos abaixo:
Tabela 1 – Oligonucleotídeos utilizados na clonagem dos genes bfALDH1A, ciALDH1D e
bfALDH1E. As regiões destacadas correspondem aos sítios para as enzimas de restrição
correspondentes (BamHI e XhoI)

Foram realizados testes para a melhor temperatura de anelamento, onde
as temperaturas obtidas ideais foram de 60°C para bfALDH1E e ciALDH1D e de
57,5°C para bfALDH1A (Tabela 2).
Desnaturação Anelamento Extensão
bfALDH1A 95°C - 1 minuto 57,5°C - 30 seg. 72°C – 2 minutos
ciALDH1D 95°C - 1 minuto 60°C - 30 seg. 72°C – 2 minutos
bfALDH1E 95°C - 1 minuto 60°C - 30 seg. 72°C – 2 minutos
Tabela 2 – Temperaturas utilizadas na amplificação dos genes bfALDH1A, ciALDHD e bfALDH1E
4.1.3 Gel de agarose
As amplificações foram confirmadas por eletroforese em gel de agarose
1% em tampão TAE.
4.1.4 Purificação dos produtos de PCR
Os produtos de PCR foram purificados usando o illustra GFX™ PCR DNA
and Gel Band Purification Kit, seguindo o protocolo do fabricante.
4.1.5 Digestão enzimática
Foi executada a digestão dos genes amplificados e do vetor pET28a-SUMO
utilizando as enzimas de restrição BamHI e XhoI.
4.1.6 Inativação das enzimas de restrição
As enzimas de restrição foram inativadas por choque térmico a 80°C,
durante 20 minutos.
4.1.7 Ligação dos insertos
Os insertos foram ligados utilizando-se a enzima T4 ligase e as
quantidades de DNA utilizadas foram estabelecidas com a seguinte formula:

A quantidade inicial de vetor era 100ng, mas a proporção inserto/vetor
foi aumentada 5 vezes para melhorar a eficiência da ligação. Esta foi executada a
temperatura ambiente por 1h.
4.1.8 Transfecção dos vetores clonados para células competentes
Os vetores foram transfectados em linhagem de E. coli DH5a
termocompetentes, utilizando o seguinte protocolo:
Incubar 100uL de bactérias competentes adicionadas de 100 ηg de DNA
em gelo por 30 minutos;
Executar choque térmico, 42°C, 45 segundos;
Incubar mais 3 minutos no gelo e adicionar 900 µl de meio LB sem
antibiótico.
Deixar em shaker a 37°C, 200 rpm por 1h.
Centrifugar a 11000 rpm, 30 segundos;
Retirar 900 μL do sobrenadante e ressuspender o pellet no restante;
Inocular em meio ágar-LB adicionado de kanamicina numa concentração
final de 50 ηg/μL, utilizando alça de drigalski;
Incubar em estufa a 37°C, overnight.
4.1.9 Inóculo
Retirar a placa e inocular cada colônia isolada em 3 mL de meio LB
adicionado de kanamicina a 50 ηg/μL, agitando em shaker a 37°C, 200 rpm,
overnight.

4.1.10 Extração do plasmídeo
Após a incubação, foi utilizado para extrair os plasmídeos o kit comercial
Invisorb® Spin Plasmid Mini Two, sendo o DNA plasmidial extraído quantificado
posteriormente no NanoDrop™ 2000c.
4.1.11 Screening de clonagem
O screening de clonagem foi realizado através de digestão enzimática,
utilizando as enzimas de restrição BamHI e XhoI, seguido de eletroforese em gel
de agarose 1%.
4.2 Expressão de ciALDH2
4.2.1 Linhagem de expressão e tecnologia pRARE
A linhagem utilizada para expressão foi a E. coli BL21 pRARE (resistente a
cloranfenicol), que auxilia no melhor desempenho em relação à expressão do
gene.
4.2.2 Transfecção
O gene da ciALDH2 clonado em vetor pET28a-SUMO foi transfectado
através de eletroporação. Posteriormente, foi adicionado de 1 mL de LB sem
antibiótico, incubando em shaker a 37°C, 200rpm, 1h, para a recuperação do
dano causado as células na eletroporação.
A seguir, 200 μL do conteúdo foi retirado e plaqueado com auxilio de alça
de drigalski, em meio LB adicionado com cloranfenicol (34 ηg/μL) e kanamicina
(50 ηg/μL), incubado a 37°C, overnight.
4.2.3 Pré-indução
Após o crescimento, uma colônia isolada é retirada e inoculada em 10 ml
de LB adicionado de cloranfenicol (34 ηg/μL) e kanamicina (50 ηg/μL),
incubados em shaker a 37°C, overnight.

4.2.4 Testes de indução da expressão
Após a realização do pré-indução, foram realizados testes de indução,
para encontrar a melhor relação entre tempo e temperatura de indução.
Para isso, são inoculados 50 μL da pré-indução em 50 mL de meio LB
adicionado de cloranfenicol (34 ηg/μL) e kanamicina (50 ηg/μL). Estes são
incubados em shaker a 37° até atingirem uma densidade optica de 0,5 a 0,6 em
600 ηm .
Nessa etapa, uma alíquota de 1 mL de cada inóculo é retirado, como uma
referência de tempo 0 (T0) da quantidade basal de proteína do gene alvo que
pode ter sido expressa. Essa alíquota é centrifugada a 14000 rpm por 5 minutos
e o sobrenadante descartado. O pellet é guardado a -20 para posterior análise.
Após a retirada da alíquota, 0,1 mM de IPTG é adicionado, e os inóculos
são distribuídos em shakers com as condições abaixo:
Temperatura (°C) Tempo de indução (Horas) RPM
1 37 2 200
2 28 2 200
3 28 6 200
4 18 Overnight (16-18) 200
Passado o tempo de indução, uma alíquota de 1 mL é (T2) retirada como
referência da quantidade de proteína que foi expressa em relação a T0,
centrifugada a 14000 rpm por 5 minutos, o sobrenadante descartado e o pellet
estocado a -20°C. Posteriormente, o inóculo é centrifugado e o pellet é estocado a
-20°C.
4.2.5 Análise da expressão
São adicionados 50 μL de tampão de amostra aos pellets T0 e T2. Em
seguida, foi feita uma eletroforese em SDS-PAGE 12%, onde o padrão de bandas
indica se houve expressão.

4.2.6 Extração
Após a confirmação da extração, foi executada a extração das proteínas do
pellet da indução, seguindo o protocolo abaixo:
Ressuspender pellet em 5mL de tampão A (Tris-EDTA);
Adicionar 25 μL de lisozima (10 mg/mL);
Incubar 30 minutos no gelo;
Sonicar
Centrifugar em microcentrifuga, 10 minutos, 4°, 14000 Rpm
Retirar e guardar o sobrenadante (E1);
Adicionar 0,5 mL de tampão B (tampão A + 2% de Triton X-100);
Sonicar;
Repetir passo de centrifugação;
Retirar o sobrenadante e guardar (E2);
Ressuspender o pellet em 50 μL de tampão A.
4.2.7 Teste de solubilidade
Realizar eletroforese em SDS-PAGE 12% dos extratos 1 (E1), 2 (E2) e
pellet, para verificar se a proteína se encontra em fração solúvel.
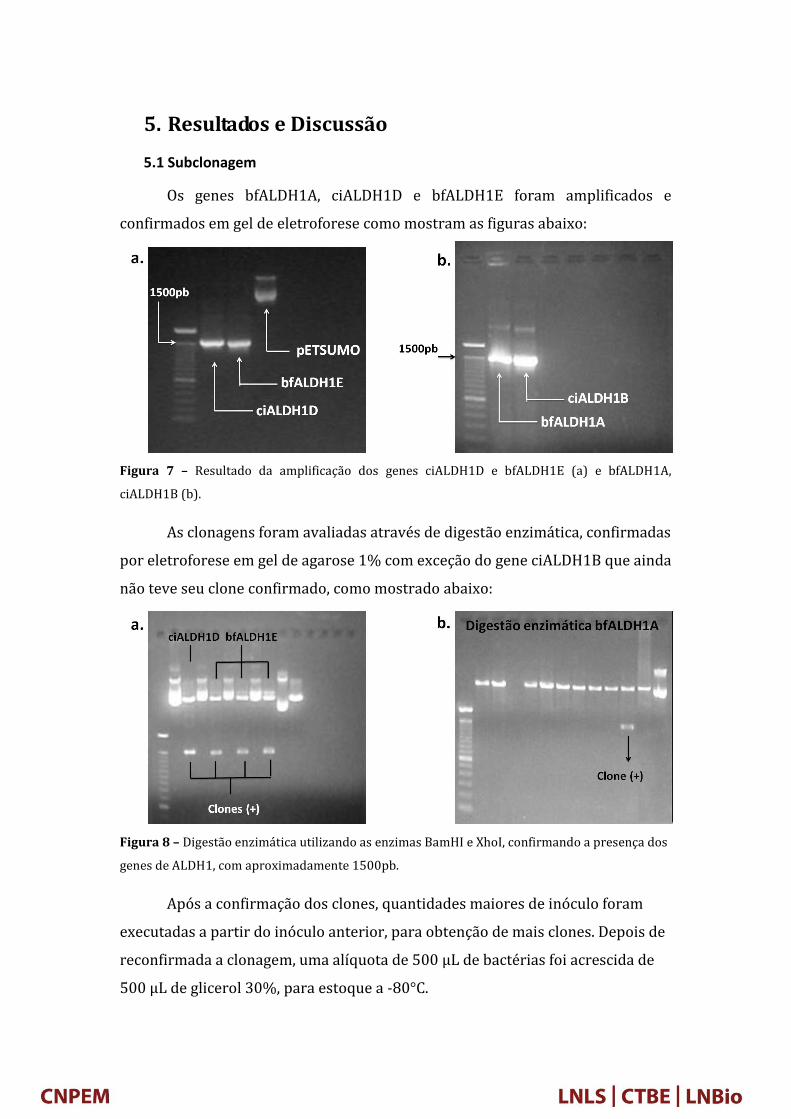
5. Resultados e Discussão
5.1 Subclonagem
Os genes bfALDH1A, ciALDH1D e bfALDH1E foram amplificados e
confirmados em gel de eletroforese como mostram as figuras abaixo:
Figura 7 – Resultado da amplificação dos genes ciALDH1D e bfALDH1E (a) e bfALDH1A,
ciALDH1B (b).
As clonagens foram avaliadas através de digestão enzimática, confirmadas
por eletroforese em gel de agarose 1% com exceção do gene ciALDH1B que ainda
não teve seu clone confirmado, como mostrado abaixo:
Figura 8 – Digestão enzimática utilizando as enzimas BamHI e XhoI, confirmando a presença dos
genes de ALDH1, com aproximadamente 1500pb.
Após a confirmação dos clones, quantidades maiores de inóculo foram
executadas a partir do inóculo anterior, para obtenção de mais clones. Depois de
reconfirmada a clonagem, uma alíquota de 500 μL de bactérias foi acrescida de
500 μL de glicerol 30%, para estoque a -80°C.

5.2 Expressão
O gene ciALDH2 foi avaliado quanto à expressão e solubilidade, como
indicam as figuras abaixo:
Figura 9 – Teste de expressão (a) e teste de indução (b) de ciALDH2, onde os quadrados em
vermelho mostram a proteína expressa.
Como se pode observar, a melhor condição de expressão foi a de 28°C
durante 6h de indução. Também houve uma boa expressão na condição 18°C
overnight (dados não mostrados), mas a primeira opção foi a que se apresentou
com tempo de execução mais aproveitável e maior quantidade de proteína
expressa.
Nos testes de solubilidade, foi visualizada a proteína obtida na fase
solúvel. Este dado é importante, uma vez que é necessária a produção solúvel de
proteína para realização de ensaios enzimáticos.

6. Perspectivas
Estabelecidos os parâmetros acima descritos, espera-se dar base para o
prosseguimento do projeto. Com as clonagens preparadas, a próxima etapa será
a preparação dos mutantes, seguindo os mesmos protocolos.
Além disso, conseguindo expressar todas estas proteínas, ensaios
enzimáticos e testes de cristalização podem ser realizados ao surgir das etapas
realizadas.
Com isso, as perspectivas para a continuação desse trabalho são:
Estabelecer a atividade dessas proteínas frente à retinaldeído e pequenos
aldeídos;
Reprogramar as funções de sinalização e detoxificação das ALDHs através
de mutações sítio dirigidas;
Estabelecer a estrutura e função destas proteínas;
Assim realizados, estes dados auxiliarão na compreensão dos mecanismos
que implicam na sinalização celular e na evolução desse sistema.

7. Referências bibliográficas
1. Balmer, J.E. and R. Blomhoff, Gene expression regulation by retinoic
acid. Journal of Lipid Research, 2002. 43(11): p. 1773-1808.
2. Campo-Paysaa, F., et al., Retinoic acid signaling in development:
Tissue-specific functions and evolutionary origins. genesis, 2008. 46(11): p.
640-656.
3. Chen, C.-H., et al., Activation of Aldehyde Dehydrogenase-2 Reduces
Ischemic Damage to the Heart.Science, 2008. 321(5895): p. 1493-1495.
4. Ferdinand Marlétaz, L.Z.H., Vincent Laudet, and Michael Schubert,
Retinoic acid signaling and the evolution of chordates.International Journal
of Biological Sciences, 2006. 2((2)): p. 9.
5. Keegan, B.R., et al., Retinoic Acid Signaling Restricts the Cardiac
Progenitor Pool.Science, 2005. 307(5707): p. 247-249.
6. Moss, J.B., et al., Dynamic Patterns of Retinoic Acid Synthesis and
Response in the Developing Mammalian Heart.Developmental Biology,
1998. 199(1): p. 55-71.
7. Pavan, M., et al., ALDH1A2 (RALDH2) genetic variation in human
congenital heart disease.BMC Medical Genetics, 2009. 10(1): p. 113.
8. Perez-Miller, S., et al., Alda-1 is an agonist and chemical chaperone
for the common human aldehyde dehydrogenase 2 variant.Nat Struct Mol
Biol, 2010. 17(2): p. 159-164.
9. Sobreira, T.J.P., et al., Structural shifts of aldehyde dehydrogenase
enzymes were instrumental for the early evolution of retinoid-dependent
axial patterning in metazoans.Proceedings of the National Academy of
Sciences, 2011. 108(1): p. 226-231.
10. Yoshida, A., et al., Human aldehyde dehydrogenase gene family.
European Journal of Biochemistry, 1998. 251(3): p. 549-557.

11. Champion™ pET SUMO Protein Expression System. Catalog guide. Manual
part no. 25-0709, Invitrogen life technologies corporation ®