estrutura da assemblÉia de peixes de - livros grátislivros01.livrosgratis.com.br/cp062157.pdf ·...
TRANSCRIPT

UNIVERSIDADE FEDERAL DA PARAÍBA CENTRO DE CIÊNCIAS EXATAS E DA NATUREZA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS ÁREA DE CONCENTRAÇÃO: ZOOLOGIA
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE
POÇAS DE MARÉ DO ARQUIPÉLAGO DE
FERNANDO DE NORONHA – PE, BRASIL, A PARTIR
DE MÉTODOS NÃO DESTRUTIVOS
LEANDRO PERRIER DE FARIA VALENTIM
João Pessoa
2008

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

LEANDRO PERRIER DE FARIA VALENTIM ESTRUTURA DA ASSEMBLEIA DE PEIXES DE POÇAS DE MARÉ DO ARQUIPÉLAGO DE FERNANDO DE NORONHA – PE, BRASIL A PARTIR DE MÉTODOS NÃO DESTRUTIVOS
Dissertação apresentada ao Programa de
Pós-graduação em Ciências Biológicas
(área de concentração: Zoologia) da
Universidade Federal Paraíba.
ORIENTADOR: PROF. DR. RICARDO DE SOUZA ROSA.
JOÃO PESSOA
2008

ii
LEANDRO PERRIER DE FARIA VALENTIM
ESTRUTURA DA ASSEMBLEIA DE PEIXES DE POÇAS DE MARÉ DO ARQUIPÉLAGO DE FERNANDO DE NORONHA – PE, BRASIL A PARTIR DE MÉTODOS NÃO
DESTRUTIVOS
BANCA EXAMINADORA
________________________________________________
Prof. Dr. Ricardo de Souza Rosa (Orientador) Departamento de Sistemática e Ecologia – Universidade Federal da Paraíba
________________________________________________
Prof. Dr. Sergio Ricardo Floeter Departamento de Ecologia e Zoologia – Universidade Federal de Santa Catarina
________________________________________________
Profa. Dra. Ierecê Maria de Lucena Rosa Departamento de Sistemática e Ecologia – Universidade Federal da Paraíba
________________________________________________
Prof. Dr. Robson Tamar (Suplente) Departamento de Sistemática e Ecologia – Universidade Federal da Paraíba

“É preciso força para sonhar e perceber
que a estrada vai além do que se vê”
Marcelo Camelo
iii

AGRADECIMENTOS
Mãe, pai, por todo investimento em minha formação, pelo choro da ida e o
sorriso da volta pra casa, pelo apoio e amor incondicional, as palavras sempre
parecerão pouco. Muito obrigado!
Ao meu orientador, Prof. Dr. Ricardo Rosa, pela oportunidade de realizar
este trabalho sob sua orientação, pela confiança e pelos dois anos de
aprendizado;
Aos Professores, Coordenadores e Funcionários do PPGCB/Zoologia –
UFPB, principalmente os professores Celso, Ierecê e Robson, pelo apoio,
sugestões, e a ótima convivência durante o curso; a CAPES pela bolsa de
mestrado e apoio financeiro ao projeto via PPG;
Ao Dr. Sergio Floeter por aceitar o convite para compor a banca e suas
contribuições para melhoria do texto e das análises;
Aos relatores do projeto, Drs. Cassiano Monteiro Neto e Francisca Edna de
Andrade Cunha, pelas sugestões e dicas valiosas;
Ao IBAMA de Brasília, em nome de Adriana Carvalhal Fonseca, pela
emissão da licença para esta pesquisa; ao IBAMA de Noronha (Instituto Chico
Mendes), especialmente ao Sr. Wilson e ao Chefe da APA, Thiago, pelo apoio
logístico à equipe; à Administração do Distrito Estadual de Fernando de Noronha,
pelo apoio ao projeto;
Aos companheiros de campo, Camilla, Letícia e Wander, pelo empenho,
dedicação e seriedade nas coletas, mas também a amizade e descontração das
horas vagas;
Aos Drs. Sergio Lourenço e Elisabete Barbarino, pelo espaço no laboratório
e equipamentos utilizados na aferição de salinidade das amostras coletadas;
Ao Tamar de Fernando de Noronha, especialmente ao vizinho Rafael, que
quebrou muitos galhos, sempre procurou ajudar e pelos almoços de domingo e a
amizade de sempre;
Jesus, Val, Maneco, a galera do Tamar e da Águas Claras e todas as
novas amizades feitas durante a estadia em Noronha; a todos os conhecidos e
“anônimos” que ajudaram com as fundamentais caronas, os passeios de barco e
bate-papos ocasionais;
iv

Carlos, Flávia, Maíse, Paloma, e todos os amigos e colegas da Pós pela
amizade e companheirismo durante esses dois anos. Allan e Martina pelo abrigo
durante a seleção, a companhia em casa e a ajuda nas análises estatísticas.
Amanda, José Luis e Gledson, companheiros amigos da nova casa, mesmo que
por pouco tempo;
Thiago, Zé, Bob e os “velhos” e queridos amigos do Rio (e de Niterói,
claro!), pelas inúmeras despedidas e os braços sempre abertos a cada retorno. O
apoio de vocês foi fundamental nesse período longe de casa!
A Tamara, por ter se aventurado nessa comigo e, embora não tenha dado
certo, ter continuado me apoiando da melhor maneira possível. Obrigado por
todas as lições de amizade, solidariedade, entrega, amor. Espero poder retribuir
tudo que você fez por mim e o quanto cresci como pessoa só por estar ao seu
lado e ter seu amor. Te amo!
v

SUMÁRIO LISTA DE FIGURAS .............................................................................................. vii
LISTA DE TABELAS .............................................................................................. ix
RESUMO ............................................................................................................ xi
ABSTRACT .......................................................................................................... xii
1. INTRODUÇÃO ................................................................................................. 1
2. OBJETIVOS ................................................................................................... 8
3. ÁREA DE ESTUDO .......................................................................................... 9
4. MATERIAL E MÉTODOS ................................................................................... 12
4.1. CARACTERIZAÇÃO DAS ÁREAS DE ESTUDO ................................................ 14
4.2. PROCEDIMENTOS DE CAMPO .................................................................... 19
4.3. ANÁLISE DOS DADOS ............................................................................... 22
5. RESULTADOS ................................................................................................ 25
6. DISCUSSÃO ................................................................................................... 38
7. CONCLUSÃO ................................................................................................. 48
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 50
vi

LISTA DE FIGURAS
Figura 1 Poça de maré no Arquipélago de Fernando de Noronha, PE, Brasil (foto: Leandro Valentim). ................................................................................................. 1
Figura 2 Mapas mostrando a América do Sul e a localização do Arquipélago de Fernando de Noronha na região nordeste do Brasil e, em detalhe, com as áreas do Parque Nacional Marinho e da Área de Proteção Ambiental demarcadas (Modificado de IBAMA, 2008)................................................................................. 9
Figura 3 Perfil da montanha submarina em cujo topo encontra-se o Arquipélago de Fernando de Noronha, indicando os principais pontos emersos e submersos (modificado de Teixeira et al., 2003). ................................................................... 10
Figura 4 Localização dos pontos de coleta no Arquipélago de Fernando de Noronha: 1 – Atalaia; 2 – Baía dos Porcos; 3 – Buraco da Raquel; 4 – Enseada das Caieiras; 5 – Enseada dos Abreus; 6 – Ponta do Air France; 7 – Porto de Santo Antônio; 8 – Praia do Bode; 9 – Praia do Boldró; 10 – Praia do Meio (Fonte: GoogleTM Earth).................................................................................................... 13
Figura 5 Frasco de vidro âmbar com 20 ml de óleo de cravo da marca K-dent, fabricado pela Quimidrol. ..................................................................................... 19
Figura 6 Procedimentos de campo: a medida de comprimento de poça de maré; tomada com trena de 30 m, graduada em centímetros; b medida de profundidade tomada com régua de 50 cm graduada em milímetros; c medida da distância da poça de maré até o mar, medida com a trena de 30 m; d medida do comprimento total de Holocentrus adscensionis, com a régua de 50 cm (fotos: Leandro Valentim). ............................................................................................................. 21
Figura 7 Recipiente utilizado como aquário de recuperação para os peixes capturados nas poças de maré do Arquipélago de Fernando de Noronha, com capacidade de 52 l e aerado constantemente por bombas de oxigênio portáteis (foto: Wander Godinho)........................................................................................ 21
Figura 8a Abundância relativa das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. b Abundância relativa das famílias. Ver Tabela III para as espécies e Tabela IV para as famílias englobadas pela categoria “Outras”. ............................................................................................... 26
Figura 9 Gráficos MDS em relação à composição de espécies das poças de maré do Arquipélago de Fernando de Noronha. a Por status de conservação. b Por categoria de fundo dominante. ............................................................................. 29
Figura 10 Gráficos de abundância relativa das categorias ecológicas determinadas para as espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. De cima para baixo: em relação à residência nas poças de maré; em relação ao hábito; em relação à alimentação....................................... 32
vii

Figura 11 Número de indivíduos (N) capturados nas poças de maré do Arquipélago de Fernando de Noronha por classe de tamanho. ........................... 33
Figura 12 Distribuição da abundância de espécies por classe de tamanho (cm). a Áreas do mar de fora (protegidas) e b áreas do mar de dentro (livres)................ 35
Figura 13 Percentual de indivíduos por classe de tamanho determinada para as espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. As espécies Acyrtus pauciradiatus (n = 1), Chaetodon ocellatus (n = 1), Dactyloscopus cf. tridigitatus (n = 3), Enneanectes altivelis (n = 1) e Haemulon parra (n = 1) não foram representadas devido ao baixo número de indivíduos capturados............................................................................................................ 35
Figura 14 Mediana, intervalo no qual se encontram 50% dos dados (25-75%) e mínimo e máximo do comprimento total dos indivíduos capturados nas poças de maré de áreas livres (mar de dentro) e protegidas (mar de fora) do Arquipélago de Fernando de Noronha. ......................................................................................... 36
Figura 15 Mediana, intervalo no qual se encontram 50% dos dados (25-75%) e mínimo e máximo do tempo de indução a anestesia das espécies que primeiro apresentaram sintomas de anestesia ao óleo de cravo nas poças de maré do Arquipélago de Fernando de Noronha. Absa – Abudefduf saxatilis (n = 15); Baso – Bathygobius soporator (n = 6); Emsi – Emblemariopsis signifera (n = 1); Envo – Entomacrodus vomerinus (n = 21); Masp – Malacoctenus sp. (n = 3); Optr – Ophioblennius trinitatis (n = 3); Scsp – Scartella sp. (n = 1); e Thno – Thalassoma noronhanum (n = 2).............................................................................................. 37
Figura 16 Correlações significativas para p < 0,05. a Tempo de indução a anestesia das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha pelo volume da poça. b Tempo de indução a anestesia pela área da poça. ....................................................................................................... 37
viii

LISTA DE TABELAS
Tabela I Locais escolhidos, localização no arquipélago e status da Unidade de Conservação (UC) dos pontos de trabalho. ......................................................... 12
Tabela II Descritores estatísticos dos dados abióticos coletados, dos tempos de indução a anestesia, coleta e total de trabalho e da mortalidade registrada em cada poça de maré do Arquipélago de Fernando de Noronha............................. 26
Tabela III Relação das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha, indicando o número de indivíduos capturados (n, total 3551), a abundância relativa (%), a freqüência de ocorrência (%) e a incidência de mortalidade (%). ................................................................................................... 27
Tabela IV Relação das famílias representadas nas poças de maré do Arquipélago de Fernando de Noronha, indicando o número de indivíduos capturados (n, total 3551), a abundância relativa (%), a freqüência de ocorrência (%), o número de mortes e a incidência de mortalidade (%) em relação ao total capturado. ........... 28
Tabela V Número de espécies (S) e de indivíduos (N) e índices de equitabilidade (J’, Pielou), diversidade (H’, Shannon) e riqueza (d, Margalef) por local amostrado no Arquipélago de Fernando de Noronha, destacando os valores para as áreas livres e protegidas. ............................................................................................... 29
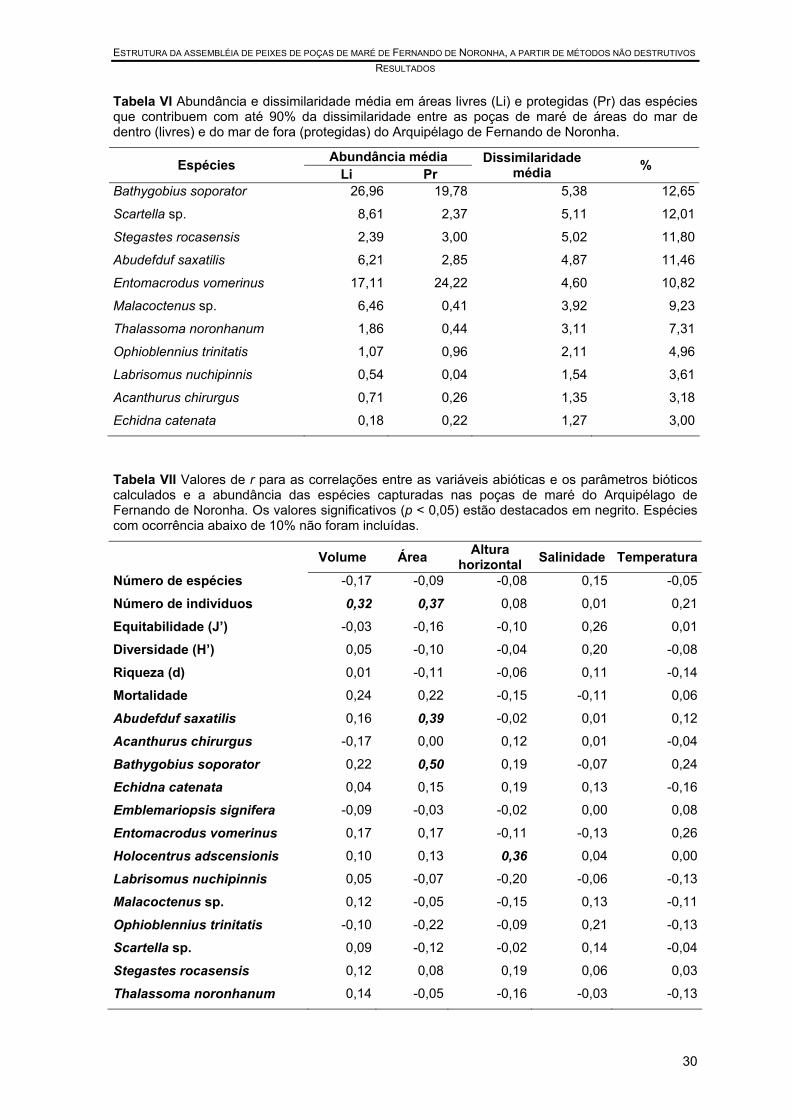
Tabela VI Abundância e dissimilaridade média em áreas livres (Li) e protegidas (Pr) das espécies que contribuem com até 90% da dissimilaridade entre as poças de maré de áreas do mar de dentro (livres) e do mar de fora (protegidas) do Arquipélago de Fernando de Noronha. ................................................................ 30
Tabela VII Valores de r para as correlações entre as variáveis abióticas e os parâmetros bióticos calculados e a abundância das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. Os valores significativos (p < 0,05) estão destacados. Espécies com ocorrência abaixo de 10% não foram incluídas. .............................................................................................................. 30
Tabela VIII Classificação das espécies nas categorias ecológicas determinadas.............................................................................................................................. 31
Tabela IX Número de indivíduos por local de coleta, destacando o total de áreas livres (Li) e protegidas (Pr), para cada categoria ecológica determinada. Bd – Praia do Bode; Bo – Praia do Boldró; Me – Praia do Meio; Pó – Baía dos Porcos; Pt – Porto de Sto. Antônio; Ab – Enseada dos Abreus; AF – Ponta do Air France; At – Atalaia; Ca – Enseada das Caieiras; Ra – Buraco da Raquel. ..................... 32
Tabela X Número de indivíduos em cada classe de tamanho determinada e tamanho médio de primeira maturação (Lm) para as espécies capturadas nas poças de maré de áreas livres (Li) e protegidas (Pr) do Arquipélago de Fernando de Noronha........................................................................................................... 34
ix

Tabela XI Total de espécies de peixes recifais registradas e número e percentual (%) das espécies observadas na zona entre-marés em diferentes países tropicais.............................................................................................................................. 39
Tabela XII – Características das poças de maré amostradas no Arquipélago de Fernando de Noronha no presente estudo. Ab – Enseada dos Abreus; AF – Ponta do Air France; At – Atalaia; Bd – Praia do Bode; Bo – Praia do Boldró; Ca – Enseada das Caieiras; Me – Praia do Meio; Po – Baía dos Porcos; Pt – Porto de Sto. Antônio; Ra – Buraco da Raquel. Absa – Abudefduf saxatilis; Baso – Bathygobius soporator; Emsi – Emblemariopsis signifera; Envo – Entomacrodus vomerinus; Masp – Malacoctenus sp. ; Optr – Ophioblennius trinitatis; Thno – Thalassoma noronhanum..................................................................................... 58
x

RESUMO
Os peixes que utilizam a zona entre-marés, incluindo as poças, o fazem por
diferentes períodos de tempo ou etapas da vida. A composição e os padrões de
diversidade, distribuição e abundância de espécies de peixes em poças de maré
foram estudadas em diversas áreas, sobretudo de regiões temperadas, sendo
escassos os trabalhos na região tropical. Sendo assim, este trabalho tem por
objetivo identificar as espécies de peixes que ocorrem nas poças de maré do
Arquipélago de Fernando de Noronha e determinar a estrutura da assembléia de
peixes de tais ambientes. Foi utilizada uma solução de 20 mg/l de óleo de cravo
como anestésico para facilitar a captura dos indivíduos, que foram identificados e
tiveram o comprimento total medido. Dez pontos foram selecionados e 55 poças
de maré amostradas no total. Parâmetros ambientais, como temperatura,
salinidade, volume da poça e altura horizontal foram medidos. Um total de 3551
indivíduos de 21 espécies e quatorze famílias foi capturado. As famílias
Blenniidae e Gobiidae foram as mais abundantes, representando quase 80% do
total. Outras 16 espécies de dez famílias foram observadas em censos visuais e
observações livres em poças maiores. A mortalidade foi baixa (5,49%), sendo o
maior índice verificado para Bathygobius soporator (46,15% do total de mortes), a
espécie mais abundante. Não foram verificadas correlações fortes entre os
parâmetros ambientais e a composição das poças, porém o aumento do número
de indivíduos foi proporcional ao volume e área das poças. A análise das
categorias ecológicas revelou a dominância de espécies residentes, crípticas e
herbívoras, sendo esta última não significativa. Indivíduos na faixa de 2,0 – 5,9 cm
de comprimento total foram os mais comuns (68,80%), e as poças do mar de
dentro abrigam indivíduos menores que as do mar de fora. O óleo de cravo
mostrou-se um anestésico eficiente, com baixos tempos de indução à anestesia
(105,83±30,98 s) e baixa mortalidade. Os resultados mostram que apenas duas
espécies dominam as poças de maré do Arquipélago, sendo ambas as espécies
características da zona entre-marés, com adaptações para a vida neste ambiente.
A diversidade encontrada neste estudo é comparável a outros trabalhos
realizados no Brasil, porém baixa para áreas tropicais. Isto se deve principalmente
a baixa diversidade local e ao volume reduzido das poças amostradas.
xi

xii
ABSTRACT
The fishes which inhabit the intertidal zone, including tidepools, do so for different
periods of time or life stages. The fish species composition, diversity, distribution
and abundance of rocky tidepools have been studied in several tempered areas,
whilst works in the tropical region are scarce. This work aims to identify the
tidepool fish species of the Fernando de Noronha Archipelago and to determine its
fish assemblage structure. A 20 mg/l clove oil solution was used as anaesthetic to
capture the individuals. Each one was identified to species level and had its total
length measured. Ten locals were selected and 55 tidepools were sampled.
Abiotic parameters, such as temperature, salinity, volume of the tidepool and
horizontal height were measured. A total of 3551 individuals of 21 species and
fourteen families were captured. The families Blenniidae and Gobiidae were the
most abundant, representing almost 80% of the total. Other 16 species of ten
families were observed in visual censuses from inside and outside bigger
tidepools. Mortality was low (5,49%) and the highest index verified for Bathygobius
soporator (46,15% of total deaths), the most abundant species. Strong correlations
between the abiotic parameters and the tidepools composition were not verified;
however the increment of the number of individuals was proportional to the volume
and area of the pools. The ecological categories analysis disclosed the dominance
of resident, cryptic and herbivorous species. Individuals ranging from 2,0-5,9 cm
total length were the most common (68,80%), and the “mar de fora” tidepool fishes
were bigger than the “mar de dentro” ones. The clove oil revealed an efficient
anaesthetic action showing low anesthesia induction times (105,83±30,98 s) and
mortality. The results show that only two species dominate the Fernando de
Noronha Archipelago tidepools and both are characteristic intertidal species,
showing adaptations for life in this environment. The diversity found in this study is
comparable to other works carried out in Brazil, however low for tropical areas.
This is mainly due to the low local diversity and to the reduced volume of the pools
sampled.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
1. INTRODUÇÃO
A zona entre-marés certamente possui a menor área dentre todos os
ambientes marinhos e, por ser mais acessível ao homem, uma vez que não são
necessários equipamentos especiais para o seu estudo, este talvez seja o habitat
marinho melhor conhecido. Apesar da sua pequena extensão, apresenta a maior
variação de fatores abióticos dentre todas as regiões marinhas, tais como maré,
temperatura, ação de ondas e salinidade, e, por isso, possui também uma imensa
variedade de espécies, com diferentes adaptações, cuja diversidade pode ser
comparada àquela da zona sublitoral (Nybakken & Bertness, 2004).
A ação contínua das ondas, ventos, marés e dos organismos que habitam
a zona entre-marés torna bastante irregular e complexa a superfície destes
ambientes, o que colabora com a elevada diversidade. Tal irregularidade também
favorece a formação de “poças de maré” (Figura 1), que são depressões e
cavidades onde a água do mar fica represada durante a maré baixa, sem
comunicação direta com o mar (Zander et al., 1999).
Figura 1 Poça de maré no Arquipélago de Fernando de Noronha, PE, Brasil (foto: Leandro Valentim).
1

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
A presença de poças de maré com diferenças no tamanho, volume,
profundidade, distância da linha de baixamar, ação das ondas e padrão de
drenagem, é característica de muitos costões rochosos, recifes de coral e arenito
e bancos de areia. O ambiente físico é altamente regulado pelas marés e suas
variações são menores em relação às áreas expostas vizinhas, uma vez que a
água está presente durante todo o ciclo de maré. Entretanto, todas as variáveis
abióticas oscilam diariamente e sazonalmente, podendo formar gradientes
verticais, com a profundidade, e horizontais, ao longo da poça (Metaxas &
Scheilbling, 1993; Horn et al., 1999; Nybakken & Bertness, 2004).
Considerando que cada poça de maré possui características específicas de
volume, profundidade e outros parâmetros físicos, dificilmente existirão duas
idênticas entre si, o que torna cada poça de maré única, resultando em grandes
variações na estrutura de suas comunidades. Devido a isto, a replicação de
estudos em diferentes poças de maré deve ser feita com cautela (Metaxas &
Scheilbling, 1993).
Os três principais fatores físicos sujeitos à variação nas poças de maré
são: (1) temperatura, que oscila muito pouco e quase sempre lentamente nos
oceanos, porém, durante a maré baixa, a temperatura da água de uma poça de
maré pode mudar rapidamente e atingir níveis letais, além disso, durante a maré
cheia, com a entrada da água oceânica, a temperatura de toda a poça irá mudar
novamente, ainda mais rapidamente; (2) salinidade, um fator praticamente estável
em águas oceânicas, pode elevar-se rapidamente durante a maré baixa em uma
poça de maré exposta ao calor intenso. Ao contrário, chuvas fortes podem
diminuir a salinidade destes corpos d'água, podendo atingir os limites de
tolerância ecológica dos organismos que ali vivem; e (3) concentração de
oxigênio, que está intimamente relacionada à temperatura, de maneira que o
aumento da temperatura da água diminui a concentração de oxigênio no meio. Se
a concentração de organismos no local for grande, este fator pode tornar-se
problemático (Metaxas & Scheilbling, 1993; Zander et al., 1999; Nybakken &
Bertness, 2004).
Os organismos que habitam as poças de maré são similares aos que
habitam o substrato exposto adjacente, porém diferenças relativas à abundância
destes podem ser observadas. Isto ocorre porque as espécies devem estar
adaptadas às oscilações dos fatores já citados (Nybakken & Bertness, 2004).
2

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
Muitas algas, invertebrados e, principalmente, peixes se aproveitam das poças de
maré para estender sua distribuição na zona entre-marés além dos limites
possíveis sem as poças (Metaxas & Scheilbling, 1993). Apesar das grandes
variações as quais estão sujeitas, as assembléias de peixes de poças de maré
apresentam marcada estabilidade e resiliência, dando suporte às evidências de
que processos determinísticos são predominantes nestes ambientes (Almada &
Faria, 2004). De maneira geral, o número de espécies e de organismos é maior em
poças de maré mais próximas a zona sublitoral (Zander et al., 1999) e nas de
maior tamanho (Mahon & Mahon, 1994). As poças de maré das regiões tropicais
também apresentam maior diversidade, quando comparadas as de regiões
temperadas, porém este fato pode estar associado ao tamanho das poças
estudadas, por serem estas maiores nos trópicos (Barreiros et al., 2004).
Variações sazonais na estrutura das comunidades também estão associadas a
gradientes latitudinais (Gibson & Yoshiyama, 1999).
Os peixes que utilizam a zona entre-marés, incluindo as poças, o fazem por
diferentes períodos de tempo ou etapas da vida. Num extremo estão as espécies
que lá vivem por todo o ciclo de vida (residentes “típicas”). Estas são pequenas,
crípticas e possuem diversas adaptações fisiológicas, morfológicas, e
comportamentais para a vida neste ambiente. No outro extremo estão as espécies
que entram apenas na maré alta, por curtos períodos, podendo ficar presas na
maré baixa, (“casuais” ou “acidentais”) e não apresentam adaptações especiais
(Horn et al., 1999). Nos diferentes trabalhos, diversas classificações foram
utilizadas para caracterizar as espécies encontradas (Crabtree & Dean, 1982;
Mahon & Mahon, 1994; Rosa et al., 1997; Barreiros et al., 2004). A principal crítica
a tais classificações é que elas são muito subjetivas, uma vez que cada autor
utiliza critérios diferentes para defini-las, deixando uma lacuna entre os dois
extremos reconhecidos e tornando impossível a comparação de resultados
(Gibson & Yoshiyama, 1999; Castellanos-Galindo et al., 2005).
De qualquer maneira, as espécies que passam todo seu ciclo de vida nas
poças de maré (residentes “típicas”) são apontadas como as mais abundantes na
maioria dos estudos (Mahon & Mahon, 1994; Rosa et al., 1997; Griffiths, 2003;
Barreiros et al., 2004; Castellanos-Galindo et al., 2005; Cunha, 2005). A grande
importância das poças para a reprodução e crescimento dos jovens destas
3

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
espécies já foi demonstrada em alguns estudos (Crabtree & Dean, 1982;
Rangueley & Kramer, 1995), porém muito pouco se sabe a respeito da ecologia
ou da composição básica das espécies de peixes que habitam a zona entre-
marés (Griffiths, 2003). As poças são também importantes áreas de criadouro
para espécies da zona sublitoral (Lardner et al., 1993; Mahon & Mahon, 1994).
A composição da ictiofauna e os padrões de diversidade, distribuição e
abundância de espécies de peixes em poças de maré foram estudadas em
diversas áreas, sobretudo de regiões temperadas e subtropicais, como a costa da
Califórnia (Yoshiyama et al., 1986), New South Wales (Lardner et al., 1993),
Santa Catarina (Barreiros et al., 2004) e outros. De acordo com Metaxas &
Scheibling (1993), a zonação em peixes não foi quantitativamente documentada,
mas qualitativamente ocorre um declínio no número de espécies à medida que
aumenta a distância da linha de maré.
Em regiões tropicais, poucos trabalhos foram realizados, sendo a maioria
na costa oeste do Pacífico (Castellanos-Galindo et al., 2005). Mahon & Mahon
(1994) e Castellanos-Galindo et al. (2005) trabalharam com a estrutura de
comunidade dos peixes de poças de maré de Barbados e Colômbia,
respectivamente. No Brasil, estudos em poças de maré são escassos (Barreiros
et al., 2004). Almeida (1973) amostrou a ictiofauna de poças de maré da região de
Salvador, Bahia. Rosa et al. (1997) estudaram a diversidade da ictiofauna das
poças de maré e Cunha (2005) fez um estudo comparativo de metodologias de
amostragem não-destrutivas, ambos em João Pessoa – PB. Mais recentemente,
Godinho (2007) e Cunha et al. (2007) verificaram as relações entre fatores
bióticos e abióticos e as variações espaço-temporais de poças de maré no litoral
do Ceará.
Diversas técnicas têm sido utilizadas para caracterizar qualitativa e
quantitativamente as assembléias de peixes em recifes e outros ambientes.
Gibson (1999) classifica tais técnicas em observações diretas e indiretas,
incluindo censo visual, vídeo, telemetria e diversas técnicas de captura, tanto
manuais, como químicas, ou através de redes, armadilhas e outros. Podem ainda
ser divididas em destrutivas, que implicam na remoção dos peixes do ambiente
com o uso de ictiotóxicos, redes, armadilhas e outros; e não-destrutivas,
representadas principalmente por censos visuais e, mais recentemente, pelo uso
de anestésicos (Ackerman & Bellwood, 2000). Nos últimos anos, muitos trabalhos
4

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
foram publicados discutindo a eficácia dos métodos utilizados e testando novas
possibilidades (Munday & Wilson, 1997; Ackerman & Bellwood, 2000; Griffiths,
2000; Willis, 2001; Ackerman & Bellwood, 2002).
Ainda que tenha sido a base dos estudos ecológicos de peixes recifais e
que seja uma técnica não-destrutiva, o censo visual é duramente criticado,
principalmente para estudos quantitativos, pois já há muito se sabe que é uma
técnica que subestima a diversidade, principalmente com relação às espécies
pequenas e/ou crípticas (Gibson, 1999; Ackerman & Bellwood, 2000; Willis, 2001).
Amostragens com técnicas visuais são mais úteis para obter medidas de
abundância relativa (Gibson, 1999).
A amostragem com auxílio de produtos químicos também apresenta suas
limitações. A rotenona, um ictiotóxico eficaz e não-seletivo, que causa a morte de
peixes por asfixia (Mascaro et al., 1998), é extremamente destrutivo, e por esta
razão, é um método muitas vezes inaceitável para coleta de peixes recifais
(Ackerman & Bellwood, 2002), e seu uso para pesca é proibido em todo o
território nacional (Lei de Crimes Ambientais, No. 9.605/98). Foi também
considerado como perigoso para a saúde do pesquisador (Betarbet et al., 2000).
Outros produtos químicos, usados como anestésicos (quinaldina, MS-222),
mostraram-se letais para algumas espécies e ainda extremamente caros, além de
também colocarem em risco a saúde do pesquisador (Griffiths, 2000).
Existe um crescente interesse no uso do óleo de cravo como anestésico
para peixes, e este produto tem sido considerado o mais adequado para o estudo
da ictiofauna de poças de maré (Griffiths, 2000). Estudos comparativos indicaram
os seguintes aspectos positivos de seu uso: indução a anestesia rápida e mais
tranqüila e tempos mais longos de recuperação; efetivo em concentrações
inferiores a de outros produtos; uso seguro, inclusive tornando-se uma alternativa
aos ictiocidas à base de rotenona por apresentar a mesma eficiência que este
produto como método de amostragem (Munday & Wilson, 1997; Griffiths, 2000;
Ackerman & Bellwood, 2002). Cunha & Rosa (2006) investigaram a eficiência do
óleo de cravo em um grupo de sete espécies de peixes tropicais do litoral do
nordeste do Brasil em experimentos em laboratório, concluindo que baixas
concentrações são capazes de produzir calma indução a anestesia com baixa
mortalidade.
5

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
Recentemente, Cunha (2005) observou quase duas vezes mais espécies e
indivíduos, a maioria deles críptica, utilizando óleo de cravo em comparação ao
censo visual em poças de maré, com mortalidade média de apenas 6,81% dos
indivíduos amostrados por poça de maré.
Além disso, o uso do óleo de cravo é aprovado para aqüicultura na
Austrália, Nova Zelândia e outros países da Oceania sem necessidade de
depuração para o consumo humano (Kildea et al., 2004) e considerado como
aditivo alimentar pela Food and Drug Administration (FDA, EUA) (Anderson et al.,
1997).
Muitos outros trabalhos têm demonstrado a eficiência do óleo de cravo
para a manipulação de indivíduos criados em cultivos (Soto & Burhanuddin, 1995;
Cho & Heath, 2000; Walsh & Pease, 2002; Iversen et al., 2003; Inoue et al., 2005;
Mylonasa et al., 2005) e ressaltando os relativamente pequenos impactos
fisiológicos de seu uso (Tort et al., 2002; Wagner et al., 2002; Pirhonen &
Schreck, 2003; Small, 2003; Cooke et al., 2004; Davis & Griffin, 2004; Holloway et
al., 2004; Kildea et al., 2004; King V et al., 2005), sendo atualmente uma
importante ferramenta para a piscicultura marinha e de água doce.
Alguns trabalhos recentes têm verificado a ação do óleo de cravo sobre
outros organismos. Venarsky & Wilhelm (2006) testaram a ação do anestésico
sobre duas espécies de anfípodas de água doce norte-americanos, Gammarus
minus e G. acherondytes, encontrando concentrações para uso seguro em
campo. Frisch et al. (2007) estudaram os efeitos do óleo de cravo sobre o coral
Pocillopora damicornis, do Recife da Grande Barreira, Austrália, e concluíram que
quantidades limitadas da solução não são capazes de prejudicar este coral.
O óleo de cravo é um líquido marrom escuro, resultado da destilação das
folhas e flores, incluindo talos, das árvores de cravo (Eugenia aromatica), sendo a
substância ativa o eugenol, com concentração que varia de 70 a 90% da
composição total do óleo essencial do cravo (Keene et al., 1998). Há séculos vem
sendo utilizado no tratamento de diversas dores no Sudeste asiático (Soto &
Burhanuddin, 1995), tendo ampla aplicação mundial na odontologia (Curtis, 1990).
Devido às características de sua formação vulcânica recente, o
Arquipélago de Fernando de Noronha apresenta uma costa bastante acidentada
(Ferreira et al., 1990), o que favorece a formação de inúmeras poças de maré.
Apesar de estar bastante afastado da costa do Brasil – cerca de 450 km da
6

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS INTRODUÇÃO
7
cidade de Natal, RN – e ser protegido pelo IBAMA (Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis), o fluxo turístico é grande,
gerando, potencialmente, diversos impactos sobre o ambiente.
Uma vez que a zona entre-marés faz parte da área mais utilizada pela
maioria dos banhistas, turistas e muitos pescadores, o impacto sobre as
comunidades desta região pode ser considerável, principalmente nas poças de
maré, que são muito atraentes por serem verdadeiras “piscinas naturais”. Desta
forma, considerando a grande importância das poças para as diversas espécies
que as ocupam, tornam-se necessários estudos que caracterizem este ambiente,
para que estes, associados a programas de educação junto aos freqüentadores,
possam servir de base para planos de conservação e manejo cada vez mais
eficazes.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS OBJETIVOS
2. OBJETIVOS
Objetivo geral
• Identificar as espécies de peixes que ocorrem nas poças de maré do
Arquipélago de Fernando de Noronha e determinar a estrutura da
assembléia de peixes de tais ambientes.
Objetivos específicos
• Identificar as espécies que ocorrem em poças de maré do Arquipélago de
Fernando de Noronha;
• medir a abundância, a diversidade e a riqueza de tais espécies;
• analisar a influência de fatores abióticos na composição da ictiofauna das
poças;
• determinar a estrutura da assembléia de peixes de poças de maré a partir
de categorias ecológicas;
• comparar tal estrutura em áreas protegidas/controladas e de livre acesso.
• determinar as espécies mais rapidamente induzidas a anestesia pelo óleo
de cravo;
• determinar o tempo de indução à anestesia de tais espécies.
8

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS ÁREA DE ESTUDO
3. ÁREA DE ESTUDO
O Arquipélago de Fernando de Noronha está localizado no oceano
Atlântico Sul (3°45' - 3°56' S e 32°20' - 32°30' W), a 345 km do Cabo de São
Roque (RN) e 545 km da costa de Recife (PE) (Figura 2). De origem vulcânica,
possui 21 ilhas, ilhotas e rochedos, sendo Fernando de Noronha a maior ilha e a
única habitada, com 18,4 km2 de área (91% do total), maior eixo com cerca de 10
km, largura máxima de 3,5 km e perímetro de 60 km. Trata-se do topo de um dos
cones vulcânicos da Cordilheira Dorsal Meso-atlântica, cuja base de 74 km de
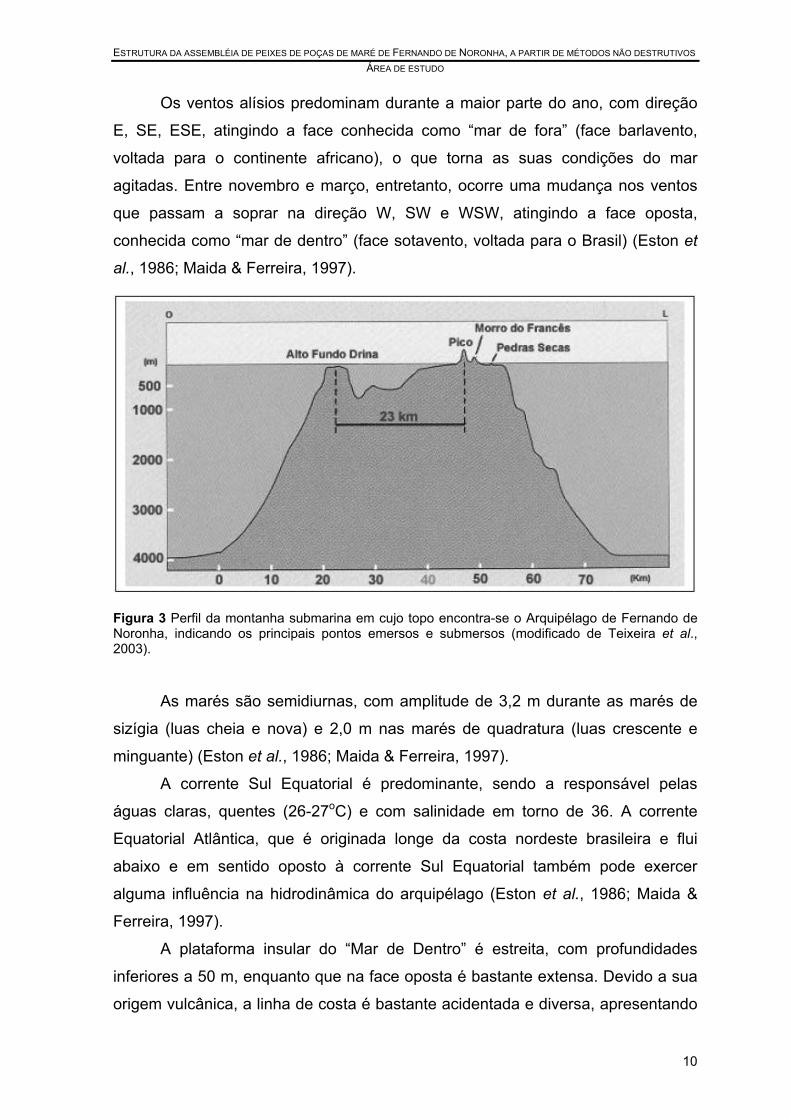
diâmetro se encontra a cerca de 4000 m de profundidade (Figura 3) (Teixeira et
al., 2003; Ibama, 2008).
Figura 2 Mapas mostrando a América do Sul e a localização do Arquipélago de Fernando de Noronha na região nordeste do Brasil e, em detalhe, com as áreas do Parque Nacional Marinho e da Área de Proteção Ambiental demarcadas (Modificado de IBAMA, 2008).
O clima do arquipélago é tropical oceânico, com duas estações bem
definidas: a estação chuvosa, de fevereiro/março a julho/agosto, e a seca, de
agosto/setembro a janeiro/fevereiro. Médias anuais de pluviosidade e temperatura
chegam aos 1300 mm e 25°C, respectivamente (Ferreira et al., 1990; Soto, 2001).
9
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS ÁREA DE ESTUDO
Os ventos alísios predominam durante a maior parte do ano, com direção
E, SE, ESE, atingindo a face conhecida como “mar de fora” (face barlavento,
voltada para o continente africano), o que torna as suas condições do mar
agitadas. Entre novembro e março, entretanto, ocorre uma mudança nos ventos
que passam a soprar na direção W, SW e WSW, atingindo a face oposta,
conhecida como “mar de dentro” (face sotavento, voltada para o Brasil) (Eston et
al., 1986; Maida & Ferreira, 1997).
Figura 3 Perfil da montanha submarina em cujo topo encontra-se o Arquipélago de Fernando de Noronha, indicando os principais pontos emersos e submersos (modificado de Teixeira et al., 2003).
As marés são semidiurnas, com amplitude de 3,2 m durante as marés de
sizígia (luas cheia e nova) e 2,0 m nas marés de quadratura (luas crescente e
minguante) (Eston et al., 1986; Maida & Ferreira, 1997).
A corrente Sul Equatorial é predominante, sendo a responsável pelas
águas claras, quentes (26-27oC) e com salinidade em torno de 36. A corrente
Equatorial Atlântica, que é originada longe da costa nordeste brasileira e flui
abaixo e em sentido oposto à corrente Sul Equatorial também pode exercer
alguma influência na hidrodinâmica do arquipélago (Eston et al., 1986; Maida &
Ferreira, 1997).
A plataforma insular do “Mar de Dentro” é estreita, com profundidades
inferiores a 50 m, enquanto que na face oposta é bastante extensa. Devido a sua
origem vulcânica, a linha de costa é bastante acidentada e diversa, apresentando
10

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS ÁREA DE ESTUDO
11
paredões íngremes, pontas, reentrâncias, platôs, costões rochosos e praias de
seixos rolados ou de areia. Os costões rochosos situados no “mar de fora” sofrem
a ação das ondas durante a maior parte do ano. A principal característica desta
face do arquipélago é a presença de extensas barreiras de recifes formadas
principalmente por concreções de algas calcárias. Estes recifes desenvolvem-se
contra a arrebentação das ondas podendo atingir até quatro metros de espessura,
além de formar plataformas, franjas e terraços, que delimitam piscinas e lagunas
(Laborel & Kempf, 1966; Eston et al., 1986; Maida & Ferreira, 1997).
Por ser um arquipélago oceânico, a peculiaridade de sua fauna marinha
aliada a beleza de suas paisagens naturais fez com que, em 1980, a IUCN (União
Internacional para a Conservação da Natureza e dos Recursos Naturais),
juntamente com a UNEP (Programa das Nações Unidas para o Meio Ambiente) e
o WWF, considerasse o arquipélago como “área de alta prioridade para a
conservação” (Mitraud, 2001). Em 1988, o Governo Federal implantou o Parque
Nacional Marinho e a Área de Proteção Ambiental (Figura 2), limitando os
impactos da ação antropogênica no arquipélago, até então desordenada, irrestrita
e predatória. A área do Parque abriga cerca de 70% da costa do arquipélago e
inclui áreas de acesso restrito e outras que são fechadas para visitação e uso
público (Ibama, 2008). Isto possibilita o equilíbrio ecológico observado, tornando
Fernando de Noronha um excelente local para o estudo de espécies marinhas em
ambiente natural.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
4. MATERIAL E MÉTODOS
Para determinar qualitativamente as espécies de peixes que ocorrem nas
poças de maré do Arquipélago de Fernando de Noronha foram selecionados dez
pontos de trabalho ao redor da ilha principal (Figura 4), abrangendo locais da
Área de Proteção Ambiental (APA), abertos à visitação, e do Parque Nacional
Marinho (PARNAMAR), cuja visitação é proibida, controlada, ou livre, conforme
indica a Tabela I. Todas as áreas de visitação livre encontram-se no “mar de
dentro” e são praias arenosas limitadas por costões rochosos, enquanto as áreas
protegidas encontram-se no “mar de fora”, sendo predominantemente costões
rochosos. A descrição mais detalhada dos pontos encontra-se no próximo tópico
desta seção.
Os locais foram escolhidos principalmente devido à presença de poças de
maré próprias ao trabalho proposto e também pela facilidade de acesso, pois o
deslocamento nem sempre é fácil em Fernando de Noronha.
Tabela I Locais escolhidos, localização no arquipélago e status da Unidade de Conservação (UC) dos pontos de trabalho.
Ponto Localização* Face do arquipélago UC Tipo de
visitação Atalaia 03º51’29,7” S / 32º24’31,2” W Mar de fora PARNAMAR Controlada
Baía dos Porcos 03º51’02,5” S / 32º26’28,9” W Mar de dentro PARNAMAR Livre
Buraco da Raquel 03º50’03,9” S / 32º23’51,6” W Mar de fora PARNAMAR Proibida
Enseada das Caieiras 03º50’24,4” S / 32º23’52,1” W Mar de fora PARNAMAR Controlada
Enseada dos Abreus 03º51’47,5” S / 32º24’51,9” W Mar de fora PARNAMAR Controlada
Ponta do Air France 03º49’47,8” S / 32º23’55,6” W Mar de fora PARNAMAR Controlada
Porto de Sto. Antonio 03º50’03,5” S / 32º24’04,7” W Mar de dentro APA Livre
Praia do Bode 03º50’52,0” S / 32º26’06,8” W Mar de dentro APA Livre
Praia do Boldró 03º50’43,5” S / 32º25’50,2” W Mar de dentro APA Livre
Praia do Meio 03º50’22,0” S / 32º24’52,0” W Mar de dentro APA Livre
*Obtida através do software GoogleTM Earth
Os seguintes critérios foram utilizados para a escolha das poças:
• Isolamento – foi dada preferência às poças mais isoladas, com o mínimo
de conexão com poças adjacentes ou com o mar;
• Volume – devido à limitação de pessoas na equipe (quatro) e da
quantidade disponível do anestésico, poças pequenas e médias e/ou
12

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
relativamente rasas foram escolhidas, sempre visualmente, não sendo
estabelecido um volume ou área máxima.
• Outros – tipos de fundo, distância da linha de maré e presença de espécies
ainda não amostradas.
As coletas foram realizadas entre os dias 19 de maio e 27 de junho de
2007, sempre durante o dia, iniciando-se entre 07h25 e 16h57, de acordo com o
horário da maré baixa diurna, cujo nível variou de 0,2 a 0,8 m nos dias de
amostragem. Cada coleta foi realizada em uma poça de maré diferente, não
havendo repetição de poças já amostradas. Foram realizadas tantas amostragens
quantas fossem possíveis a cada dia, variando de uma a três poças de maré. Em
alguns dias, a maré mais baixa ocorreu em horários que não permitiram a
execução do trabalho, e em uma oportunidade o mar muito agitado impossibilitou
a coleta de dados.
Figura 4 Localização dos pontos de coleta no Arquipélago de Fernando de Noronha: 1 – Atalaia; 2 – Baía dos Porcos; 3 – Buraco da Raquel; 4 – Enseada das Caieiras; 5 – Enseada dos Abreus; 6 – Ponta do Air France; 7 – Porto de Santo Antônio; 8 – Praia do Bode; 9 – Praia do Boldró; 10 – Praia do Meio (Fonte: GoogleTM Earth).
13

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
4.1. CARACTERIZAÇÃO DOS LOCAIS DE ESTUDO
1. Atalaia 03º51’29.7” S / 32º24’31.2” W
Apresenta uma praia arenosa,
protegida por crista recifal que forma
uma grande poça de maré durante a
maré baixa. A visitação nesta poça é
controlada e limitada a 100 pessoas
por dia, em grupos de até 25, sempre
com a presença de um fiscal do
IBAMA. A esquerda da praia estende-
se uma longa faixa de grandes rochas soltas, enquanto do lado direito o limite é
um costão rochoso plano, alagado na preamar, e que na baixa-mar forma várias
poças de maré menores que a principal. Tais poças possuem fundo rochoso e
grande abundância de algas filamentosas e corais. A visitação nesta parte do
Atalaia é proibida. Foto: Leandro Valentim
2. Baía dos Porcos 03º51’02,5” S / 32º26’28,9” W
Baía com praia de fundo
arenoso e seixos rolados, exibindo
grandes lajes de rocha vulcânica que
afloram na superfície. É limitada a
esquerda e a direita por costas
rochosas, onde se observam diversas
poças de maré, principalmente no
lado direito. Tais poças são rasas (até
40 cm de profundidade) e apresentam
fundo rochoso e de areia. Toda a baía está incluída no PARNAMAR, entretanto a
visitação é livre, exceto por uma poça de maré localizada no costão mais ao sul,
com 3 m de profundidade, considerada como importante local de reprodução e
berçário de várias espécies de peixes e invertebrados. Foto: Leandro Valentim
14

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
3. Buraco da Raquel 03º50’03,9” S / 32º23’51,6 ”W
Costa rochosa plana e extensa,
onde se localiza o monumento natural
que dá nome ao lugar. Devido ao
fundo irregular, são formadas diversas
poças de maré rasas durante a baixa-
mar, onde também são observados
vários corais. Uma franja recifal
protege a maior parte do lugar,
formando um alagado que atinge até 6 m de profundidade e abriga neonatos e
adultos de tubarão-limão (Negaprion brevirostis), que entram através de falhas
naturais existentes na franja. Existe ainda uma pequena baía onde, durante a
maré baixa, formam-se três grandes poças de maré e diversas outras menores.
Considerada área intangível do PARNAMAR, não sendo permitido nenhum tipo
de atividade. Foto: Leandro Valentim
4. Enseada das Caieiras 03º50’24,4” S / 32º23’52,1” W
Apresenta uma faixa de areia
de cerca de 200 m limitada por
extensos costões rochosos planos e
de fundo irregular em ambos os lados.
Exposta diretamente as ondas do mar
de fora, a longa faixa do costão é
coberta por corais e algas e apresenta
diversas poças de maré de vários
tamanhos e profundidades durante a baixa-mar. A partir desta enseada é feita
uma trilha até o Atalaia, obrigatoriamente guiada por fiscal credenciado pelo
IBAMA. A visitação é permitida, porém a utilização das poças de maré ao longo
da trilha é proibida. Foto: Camilla Fahning
15

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
5. Enseada dos Abreus 03º51’47,5” S / 32º24’51,9” W
Local com o mais difícil acesso
dentre os pontos amostrados, recebe
visitação apenas de grupos
acompanhados de guia credenciado
pelo IBAMA, entretanto a visitação é
muito pequena. Localizado entre a
Baía de Sueste e o Atalaia, é uma
grande costa rochosa, que recebe
diretamente as ondas do mar de fora. Relativamente plano, porém bastante
irregular, durante a baixa-mar possibilita a formação de diversas poças de maré
com diferentes tamanhos ao longo de todo o costão, embora este não seja muito
largo. Presença de algas arribadas, como sargaço, que ficam presas nas poças e
também corais nos locais mais abrigados. Foto: Letícia Duran
6. Ponta do Air France 03º49’47,8” S / 32º23’55,6” W
Localizada no extremo norte da
ilha principal, onde ocorre o encontro
simbólico dos mares de dentro e de
fora, é ponto de visitação turística,
embora esteja incluído no
PARNAMAR, e único acesso por terra
à ilha de São José, que só pode ser
visitada por terra durante a maré baixa. Marcada por uma costa rochosa larga,
plana e baixa por toda sua extensão, onde durante a maré baixa se formam
inúmeras poças de maré rasas (até 40 cm) e algumas maiores e mais profundas,
sendo o ponto com maior número de poças de todos amostrados. Neste ponto
são observados diversos ouriços brancos (Tripneustes ventricosus), cujo recente
crescimento populacional pode estar causando impactos sobre a fauna local,
principalmente na face voltada para o mar de dentro. Foto: Leandro Valentim
16

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
7. Porto de Santo Antônio 03º50’03,5” S / 32º24’04,7” W
Praia arenosa utilizada
historicamente para o embarque e
desembarque de pessoas e
mercadorias devido ao acesso fácil e as
águas relativamente tranqüilas durante
a maior parte do ano. Recentemente foi
construído um molhe protegendo parte
da praia para facilitar tais operações,
inclusive em dias de ondas grandes. É a praia mais ao norte do mar de dentro e
limite entre a APA e o PARNAMAR, estando inclusa na primeira. Uma grande laje
estende-se por boa parte da praia ao sul do molhe, desde a zona entre-marés até
cerca de 10 m de profundidade. Durante a maré baixa, formam-se diversas poças
de maré, de tamanho e profundidade variáveis. A direita do molhe existe apenas
uma pequena faixa de areia e um costão rochoso que se estende até a Ponta do
Air France, tendo as mesmas características descritas para este local. Foto: Leandro Valentim
8. Praia do Bode 03º50’52,0” S / 32º26’06,8” W
Praia arenosa que se estende
pelas praias da Quixabinha e da
Cacimba do Padre, mais ao sul, todas
na área da APA, e é limitada por um
pequeno e estreito costão rochoso
mais ao norte. Nos primeiros metros,
este costão é mais largo e são
observadas poças de maré com fundo
arenoso e rochoso, que estão diretamente sujeitas a ação das ondas. Durante o
período de novembro a abril predominam ondas grandes, de até 4 m, enquanto
no restante do ano, o mar é bem mais calmo. Neste último período, boa parte das
poças de maré é completamente coberta pela areia. Essa mudança muitas vezes
é drástica, podendo mudar completamente o perfil da praia de um dia para o
outro, como foi observado durante o período de estudo. Foto: Leandro Valentim
17

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
9. Praia do Boldró 03º50’43,5” S / 32º25’50,2” W
Praia arenosa, localizada na
APA, possui uma laje principal
composta por algas calcárias,
gastrópodes vermetídeos e outros
organismos incrustantes que se
estende por praticamente metade da
praia. Durante a maré baixa, parte da
laje fica exposta, formando diversas poças de maré. Limitada por costas
rochosas, à direita pela base do Morro do Pico, também nesses locais são
formadas poças de maré na baixa-mar, com fundo de areia e rocha e
profundidade máxima de 60 cm. Na laje central, onde foi instalado o sistema de
captação de água para dessalinização, formam-se poças de maré mais profundas
e complexas que as da praia e dos costões. O perfil da praia é modificado ao
longo do ano devido à ação das ondas, como na Praia do Bode. Foto: Leandro Valentim
10. Praia do Meio 03º50’22,0” S / 32º24’52,0” W
Praia arenosa localizada na
APA, entre as praias da Conceição,
mais ao sul, e do Cachorro, mais ao
norte, e separada de ambas por
costões rochosos de extensão média,
porém bastante irregulares,
principalmente ao sul. Ainda entre as
praias do Meio e da Conceição está a Ilha do Morro de Fora, onde se localiza o
monumento natural da Pedra do Pião. Tanto neste local, quanto no costão
rochoso, formam-se diversas poças de maré de fundo rochoso que são marcadas
pela presença de algas variadas. Foto: Leandro Valentim
18

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
4.2. PROCEDIMENTOS DE CAMPO
O óleo de cravo foi utilizado como anestésico para facilitar a captura dos
peixes em cada poça. O produto, da marca K-dent, fabricado pela Quimidrol
(Figura 5), foi adquirido em lojas de material odontológico, em frascos de 20 ml.
Como a densidade do óleo de cravo é aproximadamente a mesma da água (1
g/ml) (Soto & Burhanuddin, 1995), assume-se que cada frasco comporta 20 g do
óleo.
A solução de óleo de cravo utilizada para anestesia foi preparada com base
em trabalhos anteriores (Griffiths, 2000; Cunha & Rosa, 2006). Foi utilizada uma
concentração de 20 mg/l, seguindo recomendações de Cunha & Rosa (2006) para
uma amostragem eficiente e com baixa mortalidade, porém é recomendável a
realização de testes com diferentes concentrações do óleo de cravo, pois o
produto tem ação espécie-específica e os resultados podem variar de região para
região (Cunha & Rosa, 2006). O volume necessário para atingir essa
concentração foi dissolvido em cinco partes (1:5) de etanol 41% antes da
aplicação nas poças.
Figura 5 Frasco de vidro âmbar com 20 ml de óleo de cravo da marca K-dent, fabricado pela Quimidrol.
Todas as poças de maré amostradas foram medidas (Figura 6a-c)
utilizando uma trena de 30 m graduada em centímetros, obtendo-se os valores de
comprimento máximo (C, paralelo à costa), largura máxima (L, perpendicular à
19

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
costa), área (A = C x L), profundidade máxima (P), volume (V = A x 0,5P) e
distância até o mar (altura horizontal). Também foi medida a temperatura, com
termômetro de mercúrio em escala de 0,5oC, e coletada água para aferição da
salinidade, feita posteriormente com refratômetro, em laboratório. O tipo de fundo
foi caracterizado a partir da categoria dominante presente.
A partir do volume estimado da poça, foi preparada a quantidade
necessária de solução de óleo de cravo, utilizando um Becker de 50 ml graduado
a cada 5 ml. A solução foi distribuída uniformemente ao longo da poça e a água
agitada para acelerar a mistura.
Para calcular o tempo de indução à anestesia, o cronômetro era iniciado
juntamente com a aplicação da solução. Foi considerado anestesiado o primeiro
indivíduo a atingir o estágio “3” de anestesia, segundo Keene et al. (1998),
caracterizado pela perda parcial do tônus muscular, natação errática, aumento da
taxa opercular e reação apenas a fortes estímulos táteis e vibracionais.
Após a identificação do primeiro individuo anestesiado, foi anotada a
espécie e o tempo de indução, sendo iniciada a captura através de puçás,
procurando remover todos os indivíduos sem mortalidade. Os indivíduos
capturados eram transferidos para um recipiente com aproximadamente sete litros
de água do mar, sob aeração constante a partir de uma bomba de ar portátil. Foi
feita a identificação visual sempre que possível e medido o comprimento total
utilizando uma régua graduada em milímetros (Figura 6d). Os indivíduos
identificados e medidos eram então transferidos para um recipiente maior, com
cerca de 25 l de água do mar e também com aeração constante, para
recuperação (Figura 7).
Os indivíduos que não foram identificados visualmente em campo foram
fotografados para identificação utilizando literatura especializada (Humann &
Deloach, 2002; Reefnet, 2003) ou com a ajuda de especialistas.
Os tempos de coleta – tempo decorrido da aplicação do anestésico até a
captura do último indivíduo – e total de trabalho foram anotados. Após o término
da identificação e medição dos indivíduos, todos que exibiam comportamento
normal foram considerados recuperados e liberados em poças adjacentes. No
caso de óbitos, os indivíduos foram quantificados e medidos mais uma vez para
que fossem identificados na planilha de campo. A resposta de outros organismos
à ação do óleo de cravo também foi observada.
20

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
Figura 6 Procedimentos de campo: a medida de comprimento de poça de maré; tomada com trena de 30 m, graduada em centímetros; b medida de profundidade tomada com régua de 50 cm graduada em milímetros; c medida da distância da poça de maré até o mar, medida com a trena de 30 m; d medida do comprimento total de Holocentrus adscensionis, com a régua de 50 cm (fotos: Leandro Valentim).
Figura 7 Recipiente utilizado como aquário de recuperação para os peixes capturados nas poças de maré do Arquipélago de Fernando de Noronha, com capacidade de 52 l e aerado constantemente por bombas de oxigênio portáteis (foto: Wander Godinho).
21

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
Poças de maré maiores, nas quais a amostragem seria difícil com o óleo de
cravo, foram observadas para complementar qualitativamente a ocorrência de
espécies não amostradas. Foram realizados censos visuais com duração de 10 a
30 minutos, de acordo com o tamanho da poça, em cinco poças de maré,
utilizando técnicas de mergulho livre, com máscara e snorkel. Também foram
realizadas observações livres de fora da água em diversas outras poças.
4.3. ANÁLISE DOS DADOS
Para determinar as espécies mais abundantes nas poças de maré de
Fernando de Noronha, foi calculada a abundância relativa de cada uma, como
uma fração do numero de indivíduos capturados da espécie “x” pelo total
capturado, expresso em porcentagem. O mesmo foi feito para as famílias. Para
calcular a freqüência de ocorrência de cada espécie, foi dividido o número de
poças na qual a espécie “x” foi encontrada pelo total de poças amostradas.
Para calcular a diversidade foi utilizado o Índice de Shannon (H’),
comumente empregado em trabalhos de ecologia de comunidades, através do
programa Primer 5 for Windows. No mesmo programa foram calculados o Índice
de Pielou (J’), referente à equitabilidade, e a riqueza de Margalef (d).
Ainda utilizando o programa Primer, foi feita a análise SIMPER, para
verificar a dissimilaridade entre as áreas, destacando as espécies que mais
contribuem para tal dissimilaridade. A análise gráfica MDS (Non-metric Multi-
Dimensional Scaling), também baseada nos dados de diversidade, foi realizada
visando verificar agrupamentos entre as amostras. Para esta análise foi feita uma
matriz de similaridade de Bray-Curtis entre as poças, padronizada e com
transformação de log(x+1). O gráfico foi gerado a partir de dez reentradas e
seleção de fatores para agrupamento, no caso o status de conservação das áreas
e o tipo de fundo predominante em cada poça de maré.
Os dados coletados não passaram nos testes de normalidade de Shapiro-
Wilk e homogeneidade de Levene, com nível de significância de 95%. Portanto,
foram realizadas análises estatísticas não-paramétricas. Para verificar diferenças
significativas entre duas variáveis independentes (mar de dentro e mar de fora,
por exemplo) foi aplicado o teste “U” de Mann-Whitney, enquanto as análises que
envolviam mais de duas variáveis independentes foram realizadas com os testes
22

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
de análise de variância (ANOVA) de Kruskal-Wallis. O nível de significância
utilizado foi de 95%, e os testes rodados no programa Statistica 7 (Statsoft, 2004).
Para verificar possíveis relações entre os parâmetros ambientais e
biológicos das poças de maré foram feitas correlações de Spearman entre as
variáveis abióticas e os índices calculados, com nível de significância de 95%,
utilizando o programa Statistica 7 (Statsoft, 2004).
A divisão e classificação em categorias ecológicas foram baseadas em
Gibson & Yoshiyama (1999), em relação ao tempo de residência nas poças de
maré, Floeter et al. (2004) e Froese & Pauly (2008), em relação a alimentação, e
Cunha (2005) em relação ao hábito, da seguinte maneira:
⇒ Residência
Residente – espécies que durante todo o ciclo de vida ocupam a
região entre-marés, incluindo as poças de maré;
Secundária – espécies que ocupam a região entre-marés durante
algum estágio do ciclo de vida, mais comumente nas fases iniciais;
Ocasional – espécies que transitam pela região entre-marés por
motivos variados, e acabam presas nas poças de maré durante a
baixa-mar.
⇒ Alimentação
Carnívora – espécies que se alimentam de peixes, crustáceos,
moluscos e outros animais;
Herbívora – espécies que se alimentam de algas e plâncton;
Onívora – espécies que se alimentam de animais e/ou algas e ainda
de detritos.
⇒ Hábito
Críptica – espécies que apresentam características como:
comportamento secretivo, coloração críptica e/ou pequeno porte;
Sedentária – espécies que ocupam uma área relativamente
pequena, deslocando-se pouco;
Vagante – espécies de natação constante, que se deslocam por
grandes áreas.
23

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS MATERIAL E MÉTODOS
24
A Tabela VIII lista as categorias na qual cada espécie foi classificada. Foi
calculada a abundância relativa de cada categoria e as análises entre as áreas
livres e protegidas foram feitas conforme descrito anteriormente.
Os indivíduos capturados e medidos nas poças de maré foram agrupados
em seis classes de tamanho de 2 cm: < 2,0 cm; 2,0 – 3,9 cm; 4,0 – 5,9 cm; 6,0 –
7,9 cm; 8,0 – 9,9 cm; e > 9,9 cm. O mesmo foi feito para cada espécie, a fim de
determinar as classes de comprimentos totais mais freqüentes.
A freqüência na qual cada espécie foi a primeira a exibir os efeitos da
anestesia pelo óleo de cravo e o tempo médio de indução a anestesia de tais
espécies também foram analisadas. Correlações de Spearman entre o tempo de
indução a anestesia e as variáveis ambientais foram calculadas no programa
Statistica 7 (Statsoft, 2004), a fim de determinar fatores importantes que podem
influenciar no tempo de ação do anestésico.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
5. RESULTADOS
Um total de 55 poças de maré foi amostrado nos dez pontos selecionados
no Arquipélago de Fernando de Noronha, distribuídos de acordo com a Tabela I.
O volume das poças amostradas variou de 63 – 2864 l (0,063 – 2,864 m3), e a
área de 0,73 – 19,44 m2, A temperatura da água oscilou entre 26 – 34oC,
enquanto a salinidade ficou entre 34 – 42, sendo que em duas poças foram
registrados os valores 10 e 11 (Tabela II). O teste U de Mann-Whitney revelou
diferença significativa (Z = 2,00; p = 0,04) entre a altura horizontal das poças do
mar de dentro (8,95±8,82 m) e do mar de fora (13,00±13,11 m), sendo estas mais
distantes da linha de maré. Em relação ao tipo de fundo, o mais comum foi o
rochoso (69,09%), com (41,82%) ou sem cobertura algal (27,27%). Outros tipos
de fundo das poças amostradas foram areia (16,36%), coral (1,82%), crista algal
(3,64%) e “Mix” (9,09%), no qual não foi observada categoria dominante dentre
duas ou mais das citadas acima. As características detalhadas de cada poça de
maré estão indicadas no Anexo (Tabela XII).
Foram capturados 3551 indivíduos nas 55 poças de maré amostradas, com
média de 64,56 (±51,45) indivíduos por poça, representando 21 espécies de
quatorze famílias, conforme mostrado na Tabela III. As espécies mais abundantes
foram Bathygobius soporator e Entomacrodus vomerinus, com 36,30% e 31,91%,
respectivamente. As demais espécies somadas tiveram abundância de 31,91%,
sendo a mais representativa Scartella sp., com 8,59% (Figura 8a e Tabela III). Ao
considerar as famílias registradas, a mais abundante foi a Blenniidae (42,07%),
representada por três espécies, seguida por Gobiidae (36,30%), representada por
B. soporator, e Pomacentridae (11,24%), representada por duas espécies (Figura
8b e Tabela IV).
Em relação à ocorrência, as espécies mais freqüentes foram E. vomerinus
(98,18%), ausente em apenas uma das poças de maré amostradas, B. soporator
(90,91%) e Abudefduf saxatilis (78,18%) [Tabela III]. Tais espécies são
representantes das famílias mais freqüentes, Blenniidae (98,18%), Gobiidae
(90,91%) e Pomacentridae (89,09%) [Tabela IV].
25
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
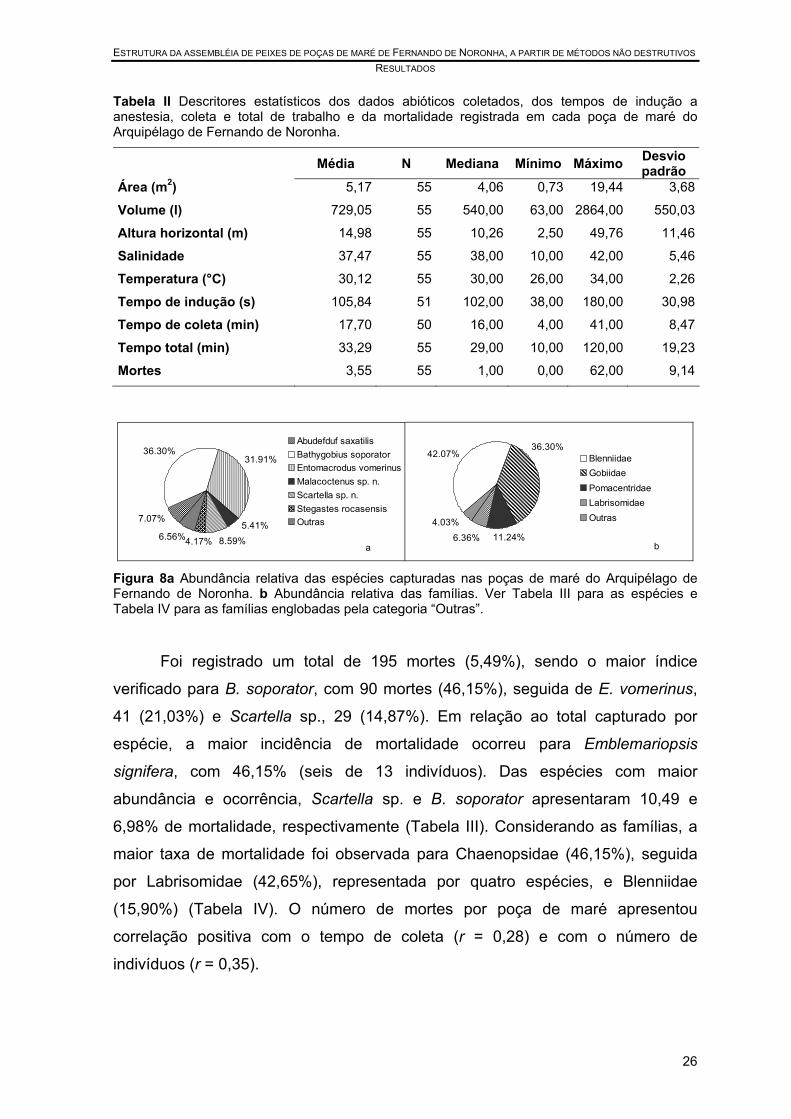
Tabela II Descritores estatísticos dos dados abióticos coletados, dos tempos de indução a anestesia, coleta e total de trabalho e da mortalidade registrada em cada poça de maré do Arquipélago de Fernando de Noronha.
Média N Mediana Mínimo Máximo Desvio padrão
Área (m2) 5,17 55 4,06 0,73 19,44 3,68
Volume (l) 729,05 55 540,00 63,00 2864,00 550,03
Altura horizontal (m) 14,98 55 10,26 2,50 49,76 11,46
Salinidade 37,47 55 38,00 10,00 42,00 5,46
Temperatura (°C) 30,12 55 30,00 26,00 34,00 2,26
Tempo de indução (s) 105,84 51 102,00 38,00 180,00 30,98
Tempo de coleta (min) 17,70 50 16,00 4,00 41,00 8,47
Tempo total (min) 33,29 55 29,00 10,00 120,00 19,23
Mortes 3,55 55 1,00 0,00 62,00 9,14
a
31.91%36.30%
4.17% 8.59%
5.41%7.07%
6.56%
Abudefduf saxatilisBathygobius soporatorEntomacrodus vomerinusMalacoctenus sp. n.Scartella sp. n.Stegastes rocasensisOutras
b
42.07%
11.24%4.03%
6.36%
36.30%BlenniidaeGobiidaePomacentridaeLabrisomidaeOutras
Figura 8a Abundância relativa das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. b Abundância relativa das famílias. Ver Tabela III para as espécies e Tabela IV para as famílias englobadas pela categoria “Outras”.
Foi registrado um total de 195 mortes (5,49%), sendo o maior índice
verificado para B. soporator, com 90 mortes (46,15%), seguida de E. vomerinus,
41 (21,03%) e Scartella sp., 29 (14,87%). Em relação ao total capturado por
espécie, a maior incidência de mortalidade ocorreu para Emblemariopsis
signifera, com 46,15% (seis de 13 indivíduos). Das espécies com maior
abundância e ocorrência, Scartella sp. e B. soporator apresentaram 10,49 e
6,98% de mortalidade, respectivamente (Tabela III). Considerando as famílias, a
maior taxa de mortalidade foi observada para Chaenopsidae (46,15%), seguida
por Labrisomidae (42,65%), representada por quatro espécies, e Blenniidae
(15,90%) (Tabela IV). O número de mortes por poça de maré apresentou
correlação positiva com o tempo de coleta (r = 0,28) e com o número de
indivíduos (r = 0,35).
26

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
Tabela III Relação das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha, indicando o número de indivíduos capturados (n, total 3551), a abundância relativa (%), a freqüência de ocorrência (%) e a incidência de mortalidade (%).
Família Espécie n Abundância Relativa Ocorrência Mortalidade
Acanthuridae Acanthurus chirurgus 27 0,76 12,73 0,00
Blenniidae Entomacrodus vomerinus 1133 31,91 98,18 3,62
Ophioblennius trinitatis 56 1,58 23,64 1,79
Scartella sp.* 305 8,59 32,73 10,49
Chaenopsidae Emblemariopsis signifera 13 0,37 10,91 46,15
Chaetodontidae Chaetodon ocellatus 1 0,03 1,82 0,00
Dactyloscopidae Dactyloscopus cf. tridigitatus 3 0,08 1,82 0,00
Gobiesocidae Acyrtus pauciradiatus 1 0,03 1,82 0,00
Tomicodon sp.* 4 0,11 3,64 0,00
Gobiidae Bathygobius soporator 1289 36,30 90,91 6,98
Haemulidae Haemulon parra 1 0,03 1,82 0,00
Holocentridae Holocentrus adscensionis 17 0,48 10,91 0,00
Labridae Thalassoma noronhanum 64 1,80 32,73 4,69
Labrisomidae Labrisomus kalisherae 7 0,20 3,64 14,29
Labrisomus nuchipinnis 16 0,45 14,55 12,50
Malacoctenus sp.* 192 5,41 29,09 6,77
Starksia sp. 11 0,31 9,09 9,09
Muraenidae Echidna catenata 11 0,31 12,73 0,00
Pomacentridae Abudefduf saxatilis 251 7,07 78,18 2,79
Stegastes rocasensis 148 4,17 56,36 0,68
Tripterygiidae Enneanectes altivelis 1 0,03 1,82 0,00
*Espécies em descrição por outros autores.
Os censos visuais realizados em poças de maré maiores e as observações
livres adicionaram dezesseis espécies e dez famílias cuja ocorrência não foi
reportada nas amostras com óleo de cravo, aumentando para 37 o total de
espécies e para 24 o total de famílias. São elas: Platybelone argalus (Belonidae);
Caranx aff. crysos e C. latus (Carangidae); Negaprion brevirostris
(Carcharhinidae), Haemulon chrysargyreum (Haemulidae); Hemiramphus
brasiliensis (Hemiramphidae), Myripristis jacobus (Holocentridae); Halichoeres
radiatus (Labridae); Aluterus scriptus (Monacanthidae), Mulloidichthys martinicus
e Pseudupeneus maculatus (Mullidae); Muraena pavonina (Muraenidae);
Pomacanthus paru (Pomacanthidae), Sparisoma amplum e S. axillare (Scaridae);
e Cephalopholis fulva (Serranidae).
27

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
Tabela IV Relação das famílias representadas nas poças de maré do Arquipélago de Fernando de Noronha, indicando o número de indivíduos capturados (n, total 3551), a abundância relativa (%), a freqüência de ocorrência (%), o número de mortes e a incidência de mortalidade (%) em relação ao total capturado.
Família n Abundância Relativa Ocorrência Mortes Mortalidade
Acanthuridae 27 0,76 12,73 0 0,00
Blenniidae 1494 42,07 98,18 71 4,75
Chaenopsidae 13 0,37 10,91 6 46,15
Chaetodontidae 1 0,03 1,82 0 0,00
Dactyloscopidae 3 0,08 1,82 0 0,00
Gobiesocidae 5 0,14 5,45 0 0,00
Gobiidae 1289 36,30 90,91 90 6,98
Haemulidae 1 0,03 1,82 0 0,00
Holocentridae 17 0,48 10,91 0 0,00
Labridae 64 1,80 32,73 3 4,69
Labrisomidae 226 6,36 34,55 17 7,52
Muraenidae 11 0,31 12,73 0 0,00
Pomacentridae 399 11,24 89,09 8 2,01
Tripterygiidae 1 0,03 1,82 0 0,00
O número de indivíduos e de espécies e os índices de equitabilidade de
Pielou (J’), diversidade de Shanon (H’) e riqueza de Margalef (d) para cada local
amostrado são apresentados na Tabela V. Não houve diferença significativa entre
os locais para nenhum dos parâmetros citados. Também não foi verificada
diferença significativa entre as áreas livres e protegidas, embora os valores dos
parâmetros para as áreas livres tenham sido maiores que os das áreas
protegidas.
A análise SIMPER revelou uma dissimilaridade média de 42,52% em
relação à composição das espécies de áreas do mar de dentro (livres) e do mar
de fora (protegida). As espécies que mais contribuíram para tal dissimilaridade
foram B. soporator (12,65%), Scartella sp. (12,01%) e Stegastes rocasensis
(11,80%), sendo que onze espécies foram responsáveis por 90% da
dissimilaridade (Tabela VI). A abundância das espécies Malacoctenus sp. (Z = -
2,57 e p = 0,01) e Scartella sp. (Z = -2,35 e p = 0,01) foi significativamente maior
em áreas do mar de dentro. Já os gráficos do MDS não mostraram agrupamentos
claros por status da área, nem pelo tipo de fundo predominante das poças, com
stress de 0,18 para a configuração em 2d (Figura 9).
28

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
Tabela V Número de espécies (S) e de indivíduos (N) e índices de equitabilidade (J’, Pielou), diversidade (H’, Shannon) e riqueza (d, Margalef) por local amostrado no Arquipélago de Fernando de Noronha, destacando os valores para as áreas livres e protegidas.
Local Número de poças S N J' H' D
Baía dos Porcos 2 6 122 0,57 0,92 0,97
Porto de Sto. Antônio 7 16 589 0,74 1,38 1,45
Praia do Bode 4 3 218 0,61 0,59 0,38
Praia do Boldró 8 14 631 0,71 1,10 1,05
Praia do Meio 7 13 489 0,72 1,36 1,36
Livre 28 20 2049 0,69 1,15 1,13
Atalaia 4 9 117 0,71 1,02 0,99
Buraco da Raquel 7 10 434 0,69 1,01 0,86
Enseada das Caieiras 6 10 365 0,67 1,06 1,11
Enseada dos Abreus 5 10 162 0,73 1,07 1,03
Ponta do Air France 5 10 424 0,65 1,11 1,08
Protegido 27 15 1502 0,69 1,05 1,01
Livre
Protegidoa
Livre
Protegido
Livre
Protegidoa
“Mix”
Areia
Coral
Crista algal
Rocha
Rocha + algab
“Mix”
Areia
Coral
Crista algal
Rocha
Rocha + alga
“Mix”
Areia
Coral
Crista algal
Rocha
Rocha + alga
“Mix”
Areia
Coral
Crista algal
Rocha
Rocha + algab
Figura 9 Gráficos MDS em relação à composição de espécies das poças de maré do Arquipélago de Fernando de Noronha. a Por status de conservação. b Por categoria de fundo dominante. “Mix” – duas ou mais das categorias listadas presentes, sem dominância clara.
As correlações entre a área da poça de maré e as abundâncias de A.
saxatilis e B. soporator e o número de indivíduos, e entre o volume da poça de
maré e o número de indivíduos foram significativas para p < 0,05, Desta forma,
verificou-se um aumento do número de indivíduos (r = 0,37), da abundância de A.
saxatilis (r = 0,39) e B. soporator (r = 0,50) com o aumento da área da poça de
maré, e um aumento do número de indivíduos com aumento do volume da poça (r
= 0,32). Também foi significativa a correlação entre a abundância de Holocentrus
adscensionis e a altura horizontal (r = 0,36). Os valores de r de todas as
correlações estão na Tabela VII.
29
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
Tabela VI Abundância e dissimilaridade média em áreas livres (Li) e protegidas (Pr) das espécies que contribuem com até 90% da dissimilaridade entre as poças de maré de áreas do mar de dentro (livres) e do mar de fora (protegidas) do Arquipélago de Fernando de Noronha.
Abundância média Espécies Li Pr Dissimilaridade
média %
Bathygobius soporator 26,96 19,78 5,38 12,65
Scartella sp. 8,61 2,37 5,11 12,01
Stegastes rocasensis 2,39 3,00 5,02 11,80
Abudefduf saxatilis 6,21 2,85 4,87 11,46
Entomacrodus vomerinus 17,11 24,22 4,60 10,82
Malacoctenus sp. 6,46 0,41 3,92 9,23
Thalassoma noronhanum 1,86 0,44 3,11 7,31
Ophioblennius trinitatis 1,07 0,96 2,11 4,96
Labrisomus nuchipinnis 0,54 0,04 1,54 3,61
Acanthurus chirurgus 0,71 0,26 1,35 3,18
Echidna catenata 0,18 0,22 1,27 3,00
Tabela VII Valores de r para as correlações entre as variáveis abióticas e os parâmetros bióticos calculados e a abundância das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. Os valores significativos (p < 0,05) estão destacados em negrito. Espécies com ocorrência abaixo de 10% não foram incluídas.
Volume Área Altura horizontal Salinidade Temperatura
Número de espécies -0,17 -0,09 -0,08 0,15 -0,05
Número de indivíduos 0,32 0,37 0,08 0,01 0,21
Equitabilidade (J’) -0,03 -0,16 -0,10 0,26 0,01
Diversidade (H’) 0,05 -0,10 -0,04 0,20 -0,08
Riqueza (d) 0,01 -0,11 -0,06 0,11 -0,14
Mortalidade 0,24 0,22 -0,15 -0,11 0,06
Abudefduf saxatilis 0,16 0,39 -0,02 0,01 0,12
Acanthurus chirurgus -0,17 0,00 0,12 0,01 -0,04
Bathygobius soporator 0,22 0,50 0,19 -0,07 0,24
Echidna catenata 0,04 0,15 0,19 0,13 -0,16
Emblemariopsis signifera -0,09 -0,03 -0,02 0,00 0,08
Entomacrodus vomerinus 0,17 0,17 -0,11 -0,13 0,26
Holocentrus adscensionis 0,10 0,13 0,36 0,04 0,00
Labrisomus nuchipinnis 0,05 -0,07 -0,20 -0,06 -0,13
Malacoctenus sp. 0,12 -0,05 -0,15 0,13 -0,11
Ophioblennius trinitatis -0,10 -0,22 -0,09 0,21 -0,13
Scartella sp. 0,09 -0,12 -0,02 0,14 -0,04
Stegastes rocasensis 0,12 0,08 0,19 0,06 0,03
Thalassoma noronhanum 0,14 -0,05 -0,16 -0,03 -0,13
30

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
Tabela VIII Classificação das espécies nas categorias ecológicas determinadas.
Categoria ecológica Espécie Residência Alimentação Hábito
Abudefduf saxatilis Secundário Onívoro Sedentário
Acanthurus chirurgus Secundário Herbívoro Vagante
Acyrtus pauciradiatus Ocasional Carnívoro Críptico
Bathygobius soporator Residente Carnívoro Críptico
Chaetodon ocellatus Ocasional Onívoro Vagante
Dactyloscopus cf. tridigitatus Ocasional Carnívoro Críptico
Echidna catenata Ocasional Carnívoro Críptico
Emblemariopsis signifera Ocasional Carnívoro Críptico
Enneanectes altivelis Ocasional Carnívoro Críptico
Entomacrodus vomerinus Residente Herbívoro Críptico
Haemulon parra Secundário Carnívoro Sedentário
Holocentrus adscensionis Ocasional Carnívoro Sedentário
Labrisomus kalisherae Ocasional Carnívoro Críptico
Labrisomus nuchipinnis Ocasional Carnívoro Críptico
Malacoctenus sp. Ocasional Carnívoro Críptico
Ophioblennius trinitatis Ocasional Herbívoro Críptico
Scartella sp. Residente Herbívoro Críptico
Starksia sp. Ocasional Carnívoro Críptico
Stegastes rocasensis Ocasional Herbívoro Sedentário
Thalassoma noronhanum Ocasional Herbívoro Vagante
Tomicodon sp. Residente Carnívoro Críptico
A análise das categorias ecológicas revelou a dominância de espécies
residentes, crípticas e herbívoras (Figura 10 e Tabela VIII, para classificação das
espécies). A abundância de cada categoria foi significativamente diferente
(residência: H = 87,75 e p < 0,01; hábito: H = 122,44 e p < 0,01; trófico: H = 73,60
e p < 0,01). Como análises não-paramétricas não podem ser testadas
posteriormente para se verificar onde tais diferenças ocorrem, foi aplicada
também a ANOVA unifatorial paramétrica, que mostrou os mesmos resultados da
Kruskal-Wallis, inclusive com valores de p < 0,01, O teste post-hoc de Tukey
revelou que a abundância das espécies residentes e crípticas é significativamente
maior, porém a das herbívoras não, estando a diferença da categoria trófica na
menor abundância de onívoros. Comparando áreas livres e protegidas, o padrão
se manteve, exceto para as categorias tróficas, nas quais se observou maior
31
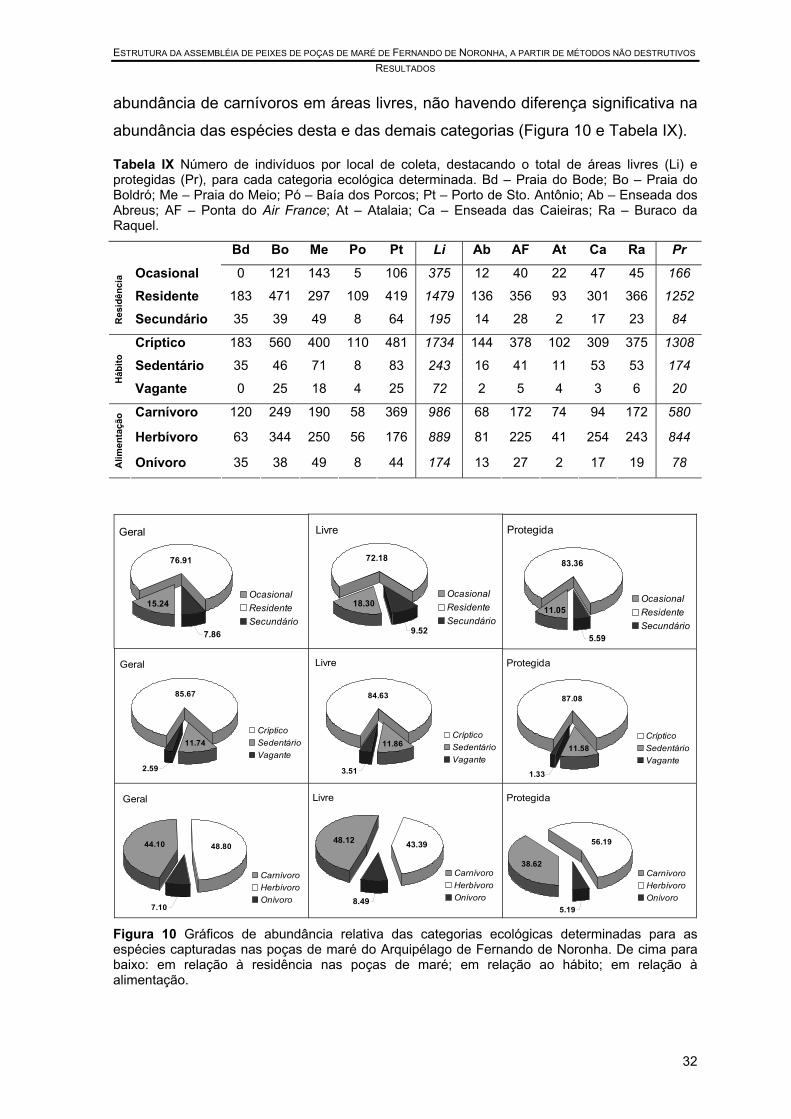
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
abundância de carnívoros em áreas livres, não havendo diferença significativa na
abundância das espécies desta e das demais categorias (Figura 10 e Tabela IX).
Tabela IX Número de indivíduos por local de coleta, destacando o total de áreas livres (Li) e protegidas (Pr), para cada categoria ecológica determinada. Bd – Praia do Bode; Bo – Praia do Boldró; Me – Praia do Meio; Pó – Baía dos Porcos; Pt – Porto de Sto. Antônio; Ab – Enseada dos Abreus; AF – Ponta do Air France; At – Atalaia; Ca – Enseada das Caieiras; Ra – Buraco da Raquel.
Bd Bo Me Po Pt Li Ab AF At Ca Ra Pr
Ocasional 0 121 143 5 106 375 12 40 22 47 45 166
Residente 183 471 297 109 419 1479 136 356 93 301 366 1252
Res
idên
cia
Secundário 35 39 49 8 64 195 14 28 2 17 23 84
Críptico 183 560 400 110 481 1734 144 378 102 309 375 1308
Sedentário 35 46 71 8 83 243 16 41 11 53 53 174
Háb
ito
Vagante 0 25 18 4 25 72 2 5 4 3 6 20
Carnívoro 120 249 190 58 369 986 68 172 74 94 172 580
Herbívoro 63 344 250 56 176 889 81 225 41 254 243 844
Alim
enta
ção
Onívoro 35 38 49 8 44 174 13 27 2 17 19 78
Geral
15.24
76.91
7.86
OcasionalResidenteSecundário
Livre
9.52
18.30
72.18
OcasionalResidenteSecundário
Protegida
5.59
11.05
83.36
OcasionalResidenteSecundário
Geral
85.67
11.74
2.59
CrípticoSedentárioVagante
Livre
84.63
11.86
3.51
CrípticoSedentárioVagante
Protegida
87.08
11.58
1.33
CrípticoSedentárioVagante
Geral
7.10
44.10 48.80
CarnívoroHerbívoroOnívoro
Livre
8.49
48.12 43.39
CarnívoroHerbívoroOnívoro
Protegida
38.62
5.19
56.19
CarnívoroHerbívoroOnívoro
Figura 10 Gráficos de abundância relativa das categorias ecológicas determinadas para as espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. De cima para baixo: em relação à residência nas poças de maré; em relação ao hábito; em relação à alimentação.
32

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
O menor indivíduo capturado foi da espécie B. soporator e mediu 0,6 cm de
comprimento total, enquanto o maior indivíduo mediu 62,0 cm, da espécie
Echidna catenata. Excluindo esta espécie – uma moréia, cujos indivíduos
apresentaram os maiores comprimentos totais – o maior indivíduo é também da
espécie B. soporator, com 14,4 cm de comprimento total.
Foram determinadas seis classes de tamanho com intervalos de 2,0 cm,
sendo a distribuição dos indivíduos capturados por classe de tamanho mostrada
na Figura 11. Foi observada uma maior ocorrência de indivíduos (68,80%) na
faixa de 2,0-5,9 cm de comprimento total, sendo que as classes de tamanho 2,0-
3,9 cm (37,54%) e 4,0-5,9 cm (31,26%) foram significativamente mais abundantes
que as demais, embora não significativamente diferentes entre si.
Também houve diferença significativa entre a abundância das classes de
tamanho do mar de dentro (livre) e do mar de fora (protegido) quando analisadas
separadamente e em comparação. Nas áreas livres, a classe de tamanho 2,0-3,9
cm foi significativamente mais abundante que as demais, enquanto que em áreas
protegidas a classe de tamanho 4,0-5,9 cm foi a mais abundante. Já as classes
de tamanho < 2,0 cm e 2,0-3,9 cm foram significativamente mais abundantes no
mar de dentro (Z = -2,81; p < 0,01 e Z = -2,22; p = 0,03, respectivamente), em
relação ao mar de fora (Figura 12).
0
200
400
600
800
1000
1200
1400
< 2.2 2.0 - 3.9 4.0 - 5.9 6.0 - 7.9 8.0 - 9.9 > 9.9
Classe de tamanho (cm)
N
Figura 11 Número de indivíduos (N) capturados nas poças de maré do Arquipélago de Fernando de Noronha por classe de tamanho.
33
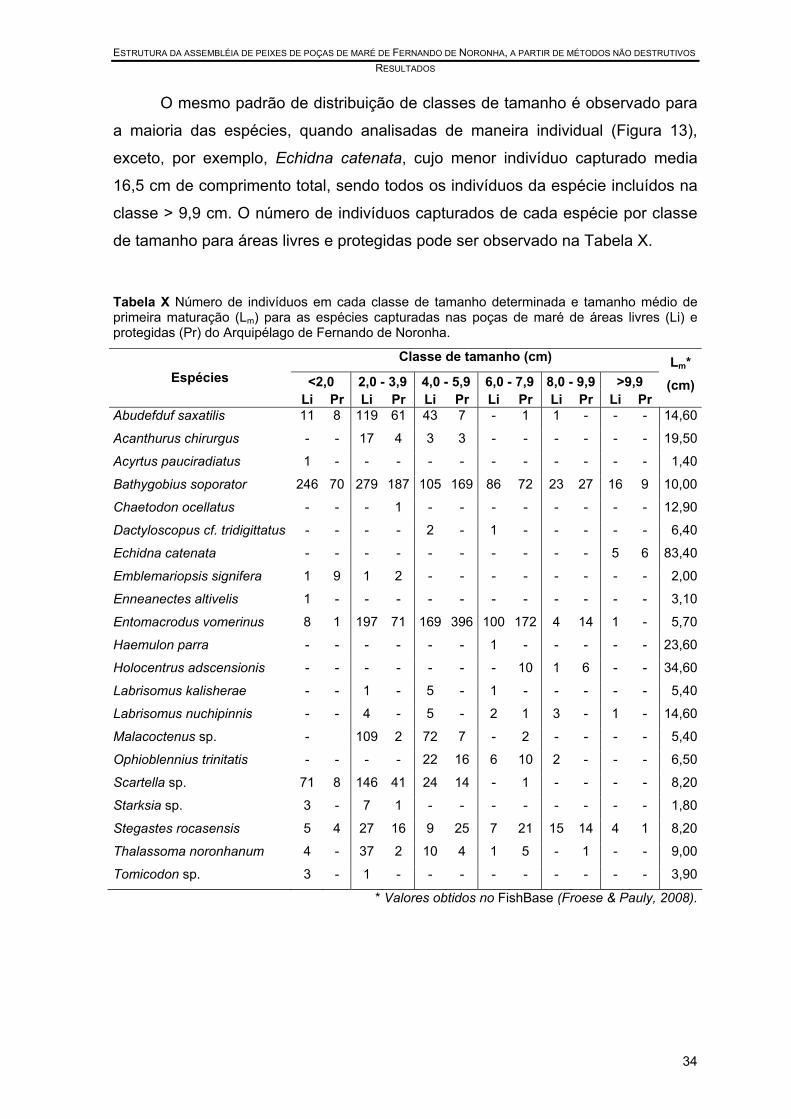
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
O mesmo padrão de distribuição de classes de tamanho é observado para
a maioria das espécies, quando analisadas de maneira individual (Figura 13),
exceto, por exemplo, Echidna catenata, cujo menor indivíduo capturado media
16,5 cm de comprimento total, sendo todos os indivíduos da espécie incluídos na
classe > 9,9 cm. O número de indivíduos capturados de cada espécie por classe
de tamanho para áreas livres e protegidas pode ser observado na Tabela X.
Tabela X Número de indivíduos em cada classe de tamanho determinada e tamanho médio de primeira maturação (Lm) para as espécies capturadas nas poças de maré de áreas livres (Li) e protegidas (Pr) do Arquipélago de Fernando de Noronha.
Classe de tamanho (cm)
<2,0 2,0 - 3,9 4,0 - 5,9 6,0 - 7,9 8,0 - 9,9 >9,9 Espécies Li Pr Li Pr Li Pr Li Pr Li Pr Li Pr
Lm* (cm)
Abudefduf saxatilis 11 8 119 61 43 7 - 1 1 - - - 14,60
Acanthurus chirurgus - - 17 4 3 3 - - - - - - 19,50
Acyrtus pauciradiatus 1 - - - - - - - - - - - 1,40
Bathygobius soporator 246 70 279 187 105 169 86 72 23 27 16 9 10,00
Chaetodon ocellatus - - - 1 - - - - - - - - 12,90
Dactyloscopus cf. tridigittatus - - - - 2 - 1 - - - - - 6,40
Echidna catenata - - - - - - - - - - 5 6 83,40
Emblemariopsis signifera 1 9 1 2 - - - - - - - - 2,00
Enneanectes altivelis 1 - - - - - - - - - - - 3,10
Entomacrodus vomerinus 8 1 197 71 169 396 100 172 4 14 1 - 5,70
Haemulon parra - - - - - - 1 - - - - - 23,60
Holocentrus adscensionis - - - - - - - 10 1 6 - - 34,60
Labrisomus kalisherae - - 1 - 5 - 1 - - - - - 5,40
Labrisomus nuchipinnis - - 4 - 5 - 2 1 3 - 1 - 14,60
Malacoctenus sp. - 109 2 72 7 - 2 - - - - 5,40
Ophioblennius trinitatis - - - - 22 16 6 10 2 - - - 6,50
Scartella sp. 71 8 146 41 24 14 - 1 - - - - 8,20
Starksia sp. 3 - 7 1 - - - - - - - - 1,80
Stegastes rocasensis 5 4 27 16 9 25 7 21 15 14 4 1 8,20
Thalassoma noronhanum 4 - 37 2 10 4 1 5 - 1 - - 9,00
Tomicodon sp. 3 - 1 - - - - - - - - - 3,90
* Valores obtidos no FishBase (Froese & Pauly, 2008).
34

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
Mediana 25%-75% Mín-Máx
< 2.
0
2.0
- 3.9
4.0
- 5.9
6.0
- 7.9
8.0
- 9.9
> 9.
9Classe de tamanho
0
10
20
30
40
50
60
70
80A
bund
ânci
a
a
Mediana 25%-75% Mín-Máx
< 2.
0
2.0
- 3.9
4.0
- 5.9
6.0
- 7.9
8.0
- 9.9
> 9.
9
Classe de tamanho
0
20
40
60
80
100
120
140
160
180
b Figura 12 Distribuição da abundância de espécies por classe de tamanho (cm). a Áreas do mar de fora (protegidas) e b áreas do mar de dentro (livres).
0
20
40
60
80
100
0.0 - 1.9 2.0 - 3.9 4.0 - 5.9 6.0 - 7.9 8.0 - 9.9 >9.9
%
Abudefduf saxatilis Bathygobius soporator
Entomacrodus vomerinus Scartella sp. n.
0
20
40
60
80
100
0.0 - 1.9 2.0 - 3.9 4.0 - 5.9 6.0 - 7.9 8.0 - 9.9 >9.9
Ophioblennius trinitatis Malacoctenus sp. n.Stegastes rocasensis Thalassoma noronhanum
0
20
40
60
80
100
0.0 - 1.9 2.0 - 3.9 4.0 - 5.9 6.0 - 7.9 8.0 - 9.9 >9.9
Classe de tamanho (cm)
%
Echidna catenata Emblemariopsis signiferaHolocentrus adcensionis Acanthurus chirurgus
0
20
40
60
80
100
0.0 - 1.9 2.0 - 3.9 4.0 - 5.9 6.0 - 7.9 8.0 - 9.9 >9.9
Classe de tamanho (cm)
Labrisomus nuchipinnis Labrisomus kalisheraeStarksia sp. Tomicodon sp. n.
Figura 13 Percentual de indivíduos por classe de tamanho determinada para as espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha. As espécies Acyrtus pauciradiatus (n = 1), Chaetodon ocellatus (n = 1), Dactyloscopus cf. tridigitatus (n = 3), Enneanectes altivelis (n = 1) e Haemulon parra (n = 1) não foram representadas devido ao baixo número de indivíduos capturados.
O Teste U de Mann-Whitney revelou diferenças significativas entre as
áreas livres e protegidas em relação ao comprimento dos indivíduos capturados
(Z = 17,59 e p < 0,01) [Figura 14], sendo os maiores valores observados para as
35

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
áreas protegidas. Indivíduos das espécies Bathygobius soporator (Z = 8,65 e p <
0,01), Entomacrodus vomerinus (Z = 9,51 e p < 0,01), Malacoctenus sp. (Z = 3,67
e p < 0,01), Scartella sp. (Z = 3,30 e p < 0,01) e Thalassoma noronhanum (Z =
3,86 e p < 0,01) também seguiram esse padrão, enquanto os indivíduos de
Abudefduf saxatilis (Z = -2,68 e p = 0,01) foram maiores no mar de dentro (áreas
livres).
A Tabela XII, no anexo, mostra a primeira espécie afetada pelo óleo de
cravo e o tempo de indução a anestesia em cada uma das poças amostradas,
exceto em três delas, quando não foi possível observar com certeza a primeira
espécie afetada, e uma outra poça na qual o tempo de indução não foi registrado.
Oito espécies se incluem nesse grupo, as mais comuns foram Entomacrodus
vomerinus (40,38%) e Abudefduf saxatilis (28,85%). O tempo médio de indução
foi de 105,83 s (±30,98 s), sendo o mais rápido 38 s, para Bathygobius soporator
e o mais longo 180 s, para Thalassoma noronhanum (Tabela II). A Figura 15
mostra a mediana dos tempos de indução à anestesia de cada espécie.
As correlações entre o tempo de indução a anestesia e as variáveis
abióticas foram significativas para o volume (r = 0,33) e a área (r = 0,36) das
poças de maré (Figura 16).
Mediana 25%-75% Mín-Máx Protegido Livre
Status
0
3
6
57
60
63
Com
prim
ento
tota
l (cm
)
Figura 14 Mediana, intervalo no qual se encontram 50% dos dados (25-75%) e mínimo e máximo do comprimento total dos indivíduos capturados nas poças de maré de áreas livres (mar de dentro) e protegidas (mar de fora) do Arquipélago de Fernando de Noronha.
36

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS RESULTADOS
37
Mediana 25%-75% Min-Máx
Abs
a
Bas
o
Em
si
Env
o
Mas
p
Opt
r
Scs
p
Thno
Espécies
20
40
60
80
100
120
140
160
180
200
Tem
po (s
)
Figura 15 Mediana, intervalo no qual se encontram 50% dos dados (25-75%) e mínimo e máximo do tempo de indução a anestesia das espécies que primeiro apresentaram sintomas de anestesia ao óleo de cravo nas poças de maré do Arquipélago de Fernando de Noronha. Absa – Abudefduf saxatilis (n = 15); Baso – Bathygobius soporator (n = 6); Emsi – Emblemariopsis signifera (n = 1); Envo – Entomacrodus vomerinus (n = 21); Masp – Malacoctenus sp. (n = 3); Optr – Ophioblennius trinitatis (n = 3); Scsp – Scartella sp. (n = 1); e Thno – Thalassoma noronhanum (n = 2).
Volume = 129.02 + 5.9475 * Tempor = .33495
0 40 80 120 160 200Tempo de indução (s)
0
500
1000
1500
2000
2500
3000
Vol
ume
(l)
a
Área = .68831 + .04315 * Tempo r = .35719
0 40 80 120 160 200Tempo de indução (s)
0
4
8
12
16
20
Áre
a (m
2 )
b Figura 16 Correlações significativas para p < 0,05. a Tempo de indução a anestesia das espécies capturadas nas poças de maré do Arquipélago de Fernando de Noronha pelo volume da poça. b Tempo de indução a anestesia pela área da poça.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
6. DISCUSSÃO
A maioria dos trabalhos acerca da ictiofauna de poças de maré foi
realizada em áreas temperadas e subtropicais (Thomson & Lehner, 1976;
Yoshiyama, 1981; Grossman, 1982; Moring, 1986; Bennett, 1987; Varas & Ojeda,
1990; Santos et al., 1994; Underwood & Skilleter, 1996; Pfister, 1998; Griffiths,
2003; Barreiros et al., 2004). Trabalhos em poças de maré da região tropical
ainda são escassos, principalmente no Brasil. Estudos realizados em Taiwan
(Chang et al., 1977; Lee, 1980) e Barbados (Mahon & Mahon, 1994) registraram a
maior diversidade de espécies, chegando a 194, porém em poças de maré com
comunicação com a zona sublitoral (Chang et al., 1977) [Tabela XI]. A grande
diversidade da ictiofauna observada nesses locais possibilita essa grande
diversidade na zona entre-marés e nas poças, especificamente. Quando
comparadas a outras regiões do planeta, a proporção de espécies que ocorrem
em poças de marés em relação à ictiofauna local é maior nas regiões citadas
(Tabela XI).
No Brasil, a diversidade de espécies é bem menor. Entretanto os trabalhos
registraram relativamente um alto número de espécies nas poças de maré
amostradas. Rosa et al. (1997) coletaram 44 espécies em duas poças de maré do
Cabo Branco, em João Pessoa (PB), o que representa cerca de 12% da
diversidade de peixes recifais no país, totalizando 3143 indivíduos ao longo de um
ano. Mais recentemente, Cunha & Rosa (2005), também em João Pessoa,
coletaram 716 indivíduos de 16 espécies em apenas onze poças de maré e, no
Ceará, Cunha et al. (2007) registraram 4750 indivíduos de 26 espécies, que
representam 7% da diversidade da ictiofauna recifal do Brasil.
Neste trabalho, os 3551 indivíduos de 21 espécies seguem o padrão de
abundância e diversidade da região, sendo baixa em comparação com outros
locais da região tropical. A baixa diversidade da ictiofauna do Arquipélago e o
volume reduzido das poças amostradas são os principais responsáveis por estes
resultados. Entretanto, somando-se as espécies observadas em mergulhos livres
e de fora das poças, as 37 espécies registradas representam quase metade do
número total observado por Ilarri (2008) – 91 espécies – em censos visuais
realizados no Arquipélago, 22% do total registrado por Soto (2001) [168 espécies]
38
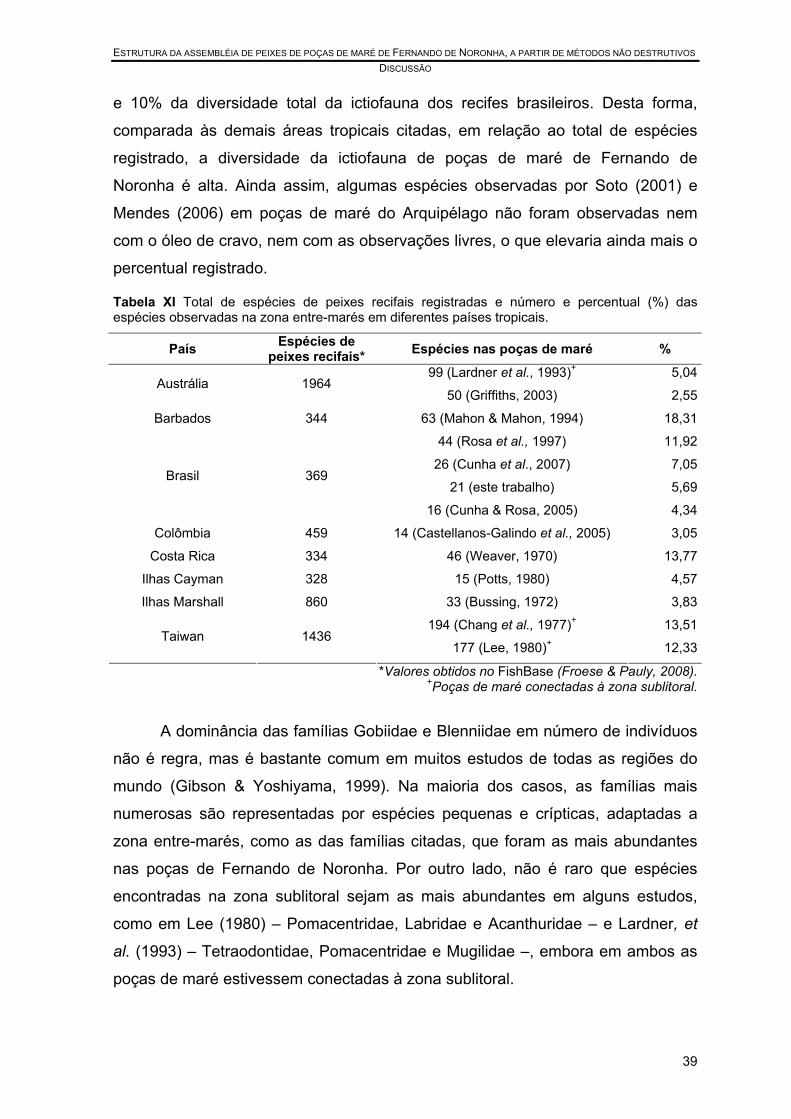
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
e 10% da diversidade total da ictiofauna dos recifes brasileiros. Desta forma,
comparada às demais áreas tropicais citadas, em relação ao total de espécies
registrado, a diversidade da ictiofauna de poças de maré de Fernando de
Noronha é alta. Ainda assim, algumas espécies observadas por Soto (2001) e
Mendes (2006) em poças de maré do Arquipélago não foram observadas nem
com o óleo de cravo, nem com as observações livres, o que elevaria ainda mais o
percentual registrado.
Tabela XI Total de espécies de peixes recifais registradas e número e percentual (%) das espécies observadas na zona entre-marés em diferentes países tropicais.
País Espécies de peixes recifais* Espécies nas poças de maré %
99 (Lardner et al., 1993)+ 5,04Austrália 1964
50 (Griffiths, 2003) 2,55
Barbados 344 63 (Mahon & Mahon, 1994) 18,31
44 (Rosa et al., 1997) 11,92
26 (Cunha et al., 2007) 7,05
21 (este trabalho) 5,69Brasil 369
16 (Cunha & Rosa, 2005) 4,34
Colômbia 459 14 (Castellanos-Galindo et al., 2005) 3,05
Costa Rica 334 46 (Weaver, 1970) 13,77
Ilhas Cayman 328 15 (Potts, 1980) 4,57
Ilhas Marshall 860 33 (Bussing, 1972) 3,83
194 (Chang et al., 1977)+ 13,51Taiwan 1436
177 (Lee, 1980)+ 12,33
*Valores obtidos no FishBase (Froese & Pauly, 2008). +Poças de maré conectadas à zona sublitoral.
A dominância das famílias Gobiidae e Blenniidae em número de indivíduos
não é regra, mas é bastante comum em muitos estudos de todas as regiões do
mundo (Gibson & Yoshiyama, 1999). Na maioria dos casos, as famílias mais
numerosas são representadas por espécies pequenas e crípticas, adaptadas a
zona entre-marés, como as das famílias citadas, que foram as mais abundantes
nas poças de Fernando de Noronha. Por outro lado, não é raro que espécies
encontradas na zona sublitoral sejam as mais abundantes em alguns estudos,
como em Lee (1980) – Pomacentridae, Labridae e Acanthuridae – e Lardner, et
al. (1993) – Tetraodontidae, Pomacentridae e Mugilidae –, embora em ambos as
poças de maré estivessem conectadas à zona sublitoral.
39

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
O amplo domínio das espécies Bathygobius soporator e Entomacrodus
vomerinus, espécies residentes típicas, de comportamento críptico e adaptadas a
zona entre-marés, alavancou a maior abundância das espécies classificadas
como residentes e também as crípticas. Apenas essas duas espécies
representaram quase 70% de todos os indivíduos capturados. Resultado
semelhante foi observado nos trabalhos nos quais as espécies residentes foram
as mais abundantes. Em Griffiths (2003) e Castellanos-Galindo, et al. (2005), as
espécies B. cocosensis (no primeiro) e B. ramosus (no segundo) foram as mais
abundantes, com 34 e 55% do total, respectivamente. B. soporator representou
36% da abundância total das poças do Arquipélago de Fernando de Noronha.
Estes dados reforçam a importância das poças de maré como abrigo específico
de algumas espécies, tornando fundamental a conservação e o manejo adequado
nesses ambientes.
As espécies consideradas residentes – neste trabalho foram quatro – foram
amplamente mais abundantes que as secundárias e ocasionais. A comparação
com outros estudos é difícil, pois muitos autores utilizaram classificações
subjetivas (Gibson & Yoshiyama, 1999), entretanto é comum que as espécies
residentes sejam poucas, mas muito numerosas. Esse padrão é observado nos
trabalhos de Mahon & Mahon (1994), Rosa, et al. (1997) Griffiths (2003),
Barreiros, et al. (2004) e Castellanos-Galindo, et al. (2005), por exemplo, que
incluem áreas tropicais e temperadas. Alguns trabalhos, como Lee (1980),
registraram maior abundância de espécies secundárias. Mahon & Mahon (1994)
sugerem que esse fato é devido a coletas repetidas nas mesmas poças de maré,
o que aumenta possibilidade de captura de espécies ocasionais, e também ao
tamanho da poça, que quanto maior, se torna mais propícia a espécies da zona
sublitoral.
O mesmo foi observado para as categorias comportamentais. As espécies
de comportamento críptico foram amplamente abundantes, muito devido a grande
abundância das duas espécies citadas. Embora as espécies residentes sejam, na
maioria dos casos, crípticas, as poças de maré abrigam ainda outras espécies de
comportamento críptico que habitam áreas rasas e podem acabar presas com a
descida da maré (Gibson & Yoshiyama, 1999), tornando a dominância desta
categoria ainda mais ampla, como foi observado. Esse é um resultado esperado,
40

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
pois uma vez que as poças de maré são ambientes restritos, espécies que não
têm a capacidade de “se esconder”, ficam muito expostas a possíveis predadores.
Em relação à categoria trófica foi observado um grande equilíbrio entre
carnívoros e herbívoros. Isso se deve basicamente à elevada abundância de B.
soporator, classificado como carnívoro, e E. vomerinus, classificado como
herbívoro. Como a abundância das outras espécies foi muito baixa e a dessas
duas semelhante, de maneira geral, não foram observadas diferenças
significativas entre essas categorias, apenas os onívoros foram muito menos
abundantes. Não é comum que trabalhos com peixes de poças de maré analisem
a estrutura trófica, entretanto na zona entre-marés de costas rochosas a maioria
das espécies é carnívora e se alimenta inteiramente de invertebrados bênticos
(Gibson & Yoshiyama, 1999). Por outro lado, é esperado que a estrutura trófica
varie de acordo com a estrutura etária das espécies que habitam a zona entre-
marés em cada local, uma vez que muitas espécies modificam completamente
sua dieta ao longo da vida (Horn, 1983). Embora não fosse encontrada uma
relação significativa, a presença e abundância de Entomacrodus vomerinus, o
principal herbívoro capturado neste trabalho, parece estar associada à presença
de algas nas poças, enquanto Bathygobius soporator parece preferir poças com
fundo rochoso.
Mendes (2006) observou indivíduos de B. soporator investindo contra
indivíduos de E. vomerinus. Durante as coletas, esse tipo de interação foi também
observada: dois indivíduos de B. soporator de 9,7 e 10,0 cm de comprimento total
predaram dois indivíduos de E. vomerinus com 6,5 e 5,9 cm de comprimento total,
respectivamente, provavelmente aproveitando-se do estado de anestesia das
presas, uma vez que o primeiro foi visto ainda na poça, após a aplicação do
anestésico, e ou outro foi visto no aquário de recuperação. Um outro E. vomerinus
com 8,2 cm de comprimento total foi predado por B. soporator de 14,4 cm e ainda
em uma outra poça de maré, a mesma interação foi observada antes da aplicação
do anestésico. Outras interações observadas incluem predação de B. soporator
(2,6 cm) por outro B. soporator (6,5 cm), por um polvo (Octopus sp.) e por
paguros, e predação de Ophioblennius trinitatis por anêmona.
O volume das poças amostradas neste trabalho é comparável com as
amostradas por Castellanos-Galindo, et al. (2005) na costa da Colômbia, cujo
valor variou entre 87-1350 l, sendo também poças de maré relativamente
41

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
pequenas e rasas. O número de espécies encontrado foi baixo (14) para uma
área tropical, entretanto nas poças de maré de Fernando de Noronha também
foram capturadas poucas espécies, devido ao volume reduzido. Pelo menos
outras 16 espécies foram observadas em poças de maré maiores, nas quais não
era apropriada a amostragem com o óleo de cravo. Além disso, Prochazka &
Griffiths (1992) e Mahon & Mahon (1994) verificaram um aumento significativo no
número de indivíduos e espécies com o aumento do volume da poça, o que não
foi claramente observado na costa colombiana, provavelmente por causa do
volume semelhante da maioria das poças de maré amostradas. Neste trabalho, tal
correlação foi também significativa, embora não tenha apresentado uma relação
muito forte.
Nenhum parâmetro ambiental medido teve forte correlação com os dados
bióticos, embora algumas correlações tenham sido significativas. O trabalho de
Castellanos-Galindo, et al. (2005) também não registrou relações claras entre os
fatores físicos e biológicos e os autores atribuem essa ausência de relação ao
volume das poças amostradas, que assim como neste trabalho, foi reduzido.
Todavia, além do já citado volume, no Arquipélago de Fernando de Noronha
também a área da poça de maré está diretamente relacionada ao número de
indivíduos, tanto de maneira geral, quanto específica, para Bathygobius soporator
e Abudefduf saxatilis. Como a área está relacionada diretamente com o volume,
este era um resultado esperado, mesmo considerando a pouca variação no
tamanho das poças amostradas.
A outra correlação significativa neste trabalho – altura horizontal e
abundância de Holocentrus adscensionis – provavelmente se deve a algum outro
aspecto das poças nas quais a espécie foi capturada, como por exemplo, a
complexidade do fundo e a presença de cavidades e abrigos, pois esta é uma
espécie de hábitos noturnos (Humann & Deloach, 2002) e mantém-se abrigada no
horário das coletas. Além disso, é uma espécie comum na zona sublitoral e sua
abundância deveria diminuir com o aumento da distância da poça de maré ao
mar. Possivelmente, as poucas poças nas quais a espécie foi capturada têm
contato diário com o mar, apesar da grande distância de algumas delas.
A classificação dos indivíduos capturados em classes de tamanho revelou
a dominância de indivíduos pequenos nas poças de maré do Arquipélago de
Fernando de Noronha. Este é o padrão observado para a zona entre-marés, já
42

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
que as espécies residentes são, em sua maioria, pequenas e crípticas (Gibson &
Yoshiyama, 1999), e, como já visto, geralmente muito numerosas. O volume
reduzido das poças também pode influenciar, uma vez que indivíduos maiores
foram observados nas poças de maré amostradas a partir de mergulhos e
observações livres, até mesmo um filhote de tubarão-limão (Negaprion
brevirostris), com cerca de 70 cm, avistado na poça principal do Atalaia, o que é
relativamente comum em outras poças do Arquipélago (Garla, 2003).
Mahon & Mahon (1994) observaram um aumento do tamanho médio dos
indivíduos de várias espécies com o aumento do volume das poças de maré e
sugerem que isto pode ser devido ao aumento da vulnerabilidade a predação, ou
a ação física de correntes ou ondas em poças menores. No Arquipélago de
Fernando de Noronha, além das espécies residentes e crípticas, os demais
indivíduos eram, em sua maioria, jovens de espécies encontradas adultas em
recifes e costões. Isto indica que tais espécies procuram águas rasas e protegidas
até atingirem o tamanho que lhes permite sobreviver no ambiente recifal, ficando
ocasionalmente presas nas poças de maré. Sale (1978) propõe que estes
indivíduos aproveitam os espaços que se tornam disponíveis nos recifes ao longo
do ano, sendo a zona entre-marés – consequentemente, as poças – um
suprimento contínuo de recrutas e importante na regulação das assembléias de
peixes recifais (Ayal & Safriel, 1982). Por outro lado, existe a possibilidade de
estes jovens representarem um excesso nos ambientes recifais, terminando por
morrer na zona entre-marés (Smith & Tyler, 1975).
A exceção à regra de comprimentos totais reduzidos foi a moréia Echidna
catenata, que apesar de ser encontrada até 12 m de profundidade, utiliza a zona
entre-marés para se alimentar na descida da maré, durante o dia (Sazima &
Sazima, 2004). Como apresenta corpo estreito e alongado, é capaz de entrar em
fendas e alcançar locais que outros peixes maiores não são capazes. Entretanto
todos os indivíduos capturados nas poças de maré do Arquipélago de Fernando
de Noronha são também jovens, considerando o Lm de 83 cm (Froese & Pauly,
2008).
Esses dados, somados ao fato de que a maioria dos indivíduos de quinze
das 21 espécies capturadas pode ser considerada jovem segundo o Lm
apresentado em Froese & Pauly (2008), reforçam a importância das poças de
maré como berçário de espécies recifais do sublitoral. Tais espécies procuram
43

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
com sucesso águas mais rasas durante as primeiras fases de desenvolvimento,
pois na zona entre-marés, principalmente nas poças, o número de predadores –
espécies piscívoras – é muito reduzido, o que aumenta as chances dos jovens
atingirem a fase adulta, quando voltam a ocupar os recifes e costões adjacentes.
Além disso, a captura de indivíduos jovens e adultos de espécies residentes
mostra que as poças são áreas importantes para a reprodução e todo o ciclo de
vida destas espécies, adaptadas para a vida na zona entre-marés.
O tamanho dos indivíduos e a altura horizontal das poças foram os únicos
parâmetros significativamente diferentes entre áreas do mar de dentro (livres) e
do mar de fora (protegidas). Como a face do Arquipélago conhecida como mar de
fora é caracterizada por costões rochosos, em sua maioria, largos e extensos,
entretanto planos e provavelmente alagados na maioria das marés, a formação de
poças de maré nos locais amostrados é favorecida, mesmo em pontos afastados
mais de 50m da linha de maré. Já os locais amostrados no mar de dentro são
costões mais estreitos que delimitam praias, alguns relativamente elevados em
relação ao nível do mar, ou ainda cristas recifais e algais formadas na zona entre-
marés das praias, o que limita um pouco a distância da linha de maré.
Os indivíduos capturados no mar de fora (áreas protegidas) foram
significativamente maiores que os do mar de dentro. A classe de tamanho mais
abundante em cada local também foi significativamente diferente, sendo classes
de tamanho menores mais abundantes no mar de dentro. Essa diferença se dá
porque os indivíduos das espécies Malacoctenus sp. e Scartella sp. foram
significativamente mais abundantes no mar de dentro além de significativamente
menores que os do mar de fora, assim como os indivíduos das espécies
Bathygobius soporator, Entomacrodus vomerinus e Thalassoma noronhanum.
Essa diferença talvez seja explicada pela maior proximidade ao mar das poças de
maré do mar de dentro, estando mais sujeitas as ações das ondas e correntes, o
que corrobora a hipótese de Mahon & Mahon (1994) acerca da influência desses
fatores físicos em poças de maré menores, favorecendo a presença indivíduos de
menor porte.
Apenas duas espécies apresentaram abundância significativamente
diferente entre o mar de dentro e o mar de fora, entretanto a dissimilaridade de
42,52% entre essas áreas pode ser considerada alta. Nenhuma espécie
apresentou participação muito elevada na dissimilaridade e dentre as espécies
44

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
com diferença significativa, Scartella sp. foi a que mais contribuiu, sendo a
segunda mais relevante dentre todas. Todavia não foram observadas relações
claras para essas suaves diferenças na abundância de cada espécie, podendo
estas ser atribuídas a uma variação normal, que pode ser minimizada com o
aumento do esforço amostral.
Embora as áreas do mar de fora coincidam com áreas do PARNAMAR,
onde a visitação é proibida ou controlada, não eram esperadas diferenças em
relação à composição da assembléia de peixes entre as áreas. A carga turística
no Arquipélago só recentemente aumentou e de maneira relativamente ordenada,
visto que as Unidades de Conservação foram criadas há vinte anos, com medidas
visando à limitação de visitantes e de uso das áreas protegidas (Ferreira et al.,
1990; Mitraud, 2001). Contudo o número de visitantes e moradores tem
aumentado bastante nos últimos anos, o que é preocupante, já que a fiscalização
e a infra-estrutura local não têm acompanhado tal crescimento (Carvalho, 1999).
Sendo o turismo uma atividade potencialmente poluidora e transformadora de
espaços naturais devido a todas as transformações sociais que provoca (Ortega
et al., 1992), é importante considerar possíveis impactos que podem estar em
curso, principalmente nas áreas do mar de dentro de visitação livre, inseridas na
APA.
O uso do óleo de cravo para amostrar a ictiofauna de poças de maré tem
se mostrado extremamente eficiente em diversos trabalhos (Griffiths, 2000;
Ackerman & Bellwood, 2002; Griffiths, 2003; Castellanos-Galindo et al., 2005;
Cunha, 2005), embora seja recomendado para poças menores (Griffiths, 2000).
Mesmo sendo um anestésico que permite a manipulação dos indivíduos
capturados – como é feito na aqüicultura –, a maioria dos trabalhos envolveu
coleta. Cunha (2005), assim como neste trabalho, devolveu os indivíduos ao
ambiente após a captura e obtenção dos dados, registrando uma mortalidade em
média de 6,81% dos indivíduos por poça de maré. Griffiths (2000) não registrou
mortalidade.
Neste trabalho, somente 5,49% dos indivíduos capturados morreram,
sendo a maior incidência para Bathygobius soporator, que também foi a espécie
mais abundante. Das 21 espécies capturadas, nove não apresentaram
mortalidade e em apenas quatro, essa foi superior a 10%. A alta mortalidade
registrada para Emblemariopsis signifera (46,15%) está relacionada ao tamanho
45

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
reduzido dos indivíduos e seu comportamento críptico. Na maioria das vezes em
que foi capturada, a espécie foi uma das últimas a ser encontrada. Não foi
observada nenhuma relação entre a mortalidade das espécies e parâmetros
abióticos.
A maior dificuldade da amostragem com óleo de cravo é o fato de muitos
indivíduos não saírem logo de seus abrigos, ou ficarem presos sob rochas e
algas, dificultando a coleta e aumentando o tempo de exposição ao anestésico.
Em Fernando de Noronha, poucos indivíduos procuravam a superfície durante a
anestesia, repousando sobre o fundo à medida que o estágio anestésico
avançava. Devido a isso, foi observada uma relação direta entre a mortalidade e o
tempo de coleta e o número de indivíduos nas poças de maré, sendo que o
número de indivíduos também influencia o tempo de coleta, embora não tenha
sido encontrada nenhuma relação direta, estatisticamente.
O tempo médio de indução da primeira espécie afetada nas poças foi bem
próximo ao observado por Cunha & Rosa (2006) em laboratório, para a
concentração de 20 mg/l. A rápida indução a anestesia é desejada, porém é
também a responsável pela morte das espécies mais sensíveis. Embora os
tempos de coleta tenham sido relativamente curtos – média de 17,7±8,47 min – a
ação prolongada do anestésico culmina na parada respiratória, levando os
indivíduos ao óbito (Keene et al., 1998). A área e o volume das poças de maré
foram os únicos parâmetros correlacionados significativamente com o tempo de
ação do anestésico, sendo este maior com o aumento dos parâmetros. Isso se
deve a difusão mais lenta do anestésico em volumes maiores de água, ainda que
fosse priorizada uma distribuição homogênea ao longo da poça.
Não foi observado um padrão claro de sensibilidade das espécies. Exemplo
disso é que na poça de maré na qual foi registrado o menor tempo de indução –
38 s – a espécie anestesiada foi Bathygobius soporator, enquanto que a de maior
tempo – 180 s – registrou Thalassoma noronhanum como primeira espécie
anestesiada. Ou seja, a sensibilidade dos indivíduos a ação do óleo de cravo
provavelmente depende de muitos fatores em ambientes naturais, já que
indivíduos de uma mesma espécie podem levar de 38 a mais de 180 s para
serem induzidos a anestesia.
Uma outra dificuldade observada em campo são as fugas e invasões
devido a conexões e proximidade a poças adjacentes. Em muitos casos foram
46

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS DISCUSSÃO
47
observados indivíduos anestesiados em outras poças próximas, sendo impossível
saber com certeza se o anestésico vazou, ou se os indivíduos fugiram da poça
amostrada. Da mesma forma, alguns indivíduos foram avistados em locais bem a
mostra muito tempo após o fim da coleta, caracterizando uma possível invasão.
Entretanto, quando a coleta ainda estava em curso, não era possível avaliar com
segurança se o individuo se encontrava ou não na poça no início da coleta. De
qualquer forma, como sempre se tratavam de espécies abundantes na poça em
questão, especificamente B. soporator e Entomacrodus vomerinus,
estatisticamente este fato não foi relevante.
Foi possível observar em algumas poças amostradas a mudança de
comportamento de alguns outros animais. Corais, crustáceos, moluscos e
equinodermas parecem ser também afetados pelo óleo de cravo, embora sejam
aparentemente mais resistentes que os peixes. Alguns corais da espécie
Zoanthus sociatus se abriam durante a coleta, com os tentáculos visivelmente
relaxados. Já os crustáceos, como siris, procuravam deixar a poça; paguros não
se mostraram afetados e aproveitavam para predar pequenos peixes
anestesiados; pequenos camarões e lagostins foram capturados em avançado
grau de anestesia, alguns já aparentemente mortos. Pequenos moluscos
indicavam efeito anestésico ao relaxar completamente o pé e ficando incapazes
de se fixar às rochas; já os polvos, mesmo sob efeito anestésico, procuravam
abrigos e soltavam jatos de tinta. Os ouriços não mostraram nenhum tipo de efeito
anestésico, entretanto alguns ofiúros pequenos foram coletados em estágio
avançado de anestesia.
Trabalhos como os de Venarsky & Wilhelm (2006), com anfípodas, Seol, et
al. (2007), com uma espécie de polvo e, principalmente, Frisch et al. (2007), com
uma espécie de coral são necessários para se conhecer cada vez melhor os
efeitos do óleo de cravo sobre outros animais e as concentrações não letais, para
que a coleta de dados seja cada vez mais segura e menos impactante para as
comunidades como um todo.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
7. CONCLUSÃO Foram capturadas 21 espécies de quatorze famílias nas poças de maré do
Arquipélago de Fernando de Noronha. As famílias mais especiosas são
Labrisomidae, quatro espécies, Blenniidae, três espécies, e Pomacentridae e
Gobiesocidae, duas espécies. Outras 16 espécies de dez famílias foram
observadas em poças maiores a partir de censo visual com mergulho livre e
observações livres de fora das poças.
As espécies mais abundantes nas poças de maré do Arquipélago são
Bathygobius soporator e Entomacrodus vomerinus, que juntas representam quase
70% da abundância total dos 3551 indivíduos capturados.
Em relação às categorias ecológicas, as dominantes nas poças de maré de
Fernando de Noronha são espécies residentes e crípticas, com abundância
significativamente superior às demais. A categoria trófica não apresentou
dominância clara, porém a maioria é de herbívoros, o que não é comum para a
zona entre-marés, normalmente dominada por espécies carnívoras.
Não foram registradas relações claras entre fatores físicos e a composição
da assembléia de peixes das poças de maré. Assim como descrito na literatura,
no Arquipélago de Fernando de Noronha o número de indivíduos tende a ser
maior em poças de maré maiores e de maior volume.
Não foram também registradas diferenças em relação à composição das
assembléias de peixes de poças de maré do mar de dentro (áreas livres) e do mar
de fora (áreas protegidas). Apenas a abundância de duas espécies foi
significativamente maior no mar de dentro, o que pode estar relacionado ao
esforço amostral, já que não foram identificados fatores que possam estar
relacionados a essa diferença. A coleta de dados mais precisos acerca da
complexidade e da composição do fundo pode trazer algumas respostas.
O óleo de cravo se mostrou um anestésico eficiente para auxiliar na
captura de peixes em poças de maré pequenas e rasas, apresentando rápida
indução à anestesia dos indivíduos e tempos de recuperação suficientes para a
obtenção de dados biológicos. Além disso, foi observada uma baixa mortalidade,
o que é desejável em estudos ecológicos, principalmente em Unidades de
Conservação. Apesar das dificuldades citadas neste trabalho, da ação observada
48

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
49
em alguns outros organismos e da ação desconhecida na maioria dos demais, o
óleo de cravo é uma alternativa eficiente aos ictiotóxicos historicamente usados,
já que apresenta a mesma, ou melhor, eficiência, sem causar grandes prejuízos
aos indivíduos capturados e a comunidade, em geral.

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
REFERÊNCIAS BIBLIOGRÁFICAS
ACKERMAN, J. L. & BELLWOOD, D. R. 2000. Reef fish assemblages: a re-evaluation using enclosed rotenone stations. Marine Ecology Progress Series, 206: 227-11.
ACKERMAN, J. L. & BELLWOOD, D. R. 2002. Comparative efficiency of clove oil and rotenone for sampling tropical reef fish assemblages. Journal of Fish Biology, 60: 893-901.
ALMADA, V. C. & FARIA, C. 2004. Temporal variation of rocky intertidal resident fish assemblages – patterns and possible mechanisms with a note on sampling protocols. Reviews in Fish Biology and Fisheries, 14: 239-250.
ALMEIDA, V. G. 1973. New records of tidepool fishes from Brazil. Papéis Avulsos de Zoologia, 26 (14): 187-191.
ANDERSON, W. G.; MCKINLEY, R. S. & COLAVECCHIA, M. 1997. The use of clove oil as an anesthetic for rainbow trout and its effects on swimming performance. North American Journal of Fisheries Management, 17: 301–307.
AYAL, Y. & SAFRIEL, U. N. 1982. Species diversity of coral reef - a note on the role of predation and of adjacent habitats. Bulletin of Marine Science, 32: 787-790.
BARREIROS, J. P.; BERTONCINI, A.; MACHADO, L.; HOSTIM-SILVA, M. & SANTOS, R. S. 2004. Diversity and seasonal changes in the ichthyofauna of rocky tidal pools from Praia Vermelha and São Roque, Santa Catarina. Brazilian Archives of Biology and Technology, 47 (2): 291-299.
BENNETT, B. A. 1987. The rock-pool fish community of Koppie Alleen and an assessment of the importance of Cape rock-pools as nurseries for juvenile fish. South African Journal of Zoology, 22: 25-32.
BETARBET, R. T.; SHERER, G. & MACKENZIE, M. 2000. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nature Neuroscience, 3: 1301–1306.
BUSSING, W. A. 1972. Recolonization of a population of supratidal fishes at Eniwetok Atoll, Marshall Islands. Atoll Research Bulletin, 154: 1-7.
CARVALHO, L. F. A. 1999. Impactos do turismo no Arquipélago de Fernando de Noronha : um estudo no caminho do desenvolvimento sustentável. Graduação em Turismo, Departamento de Relações Públicas, Propaganda e Turismo, Universidade de São Paulo, São Paulo. 126pp.
50

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
CASTELLANOS-GALINDO, G. A.; GIRALDO, A. & RUBIO, E. A. 2005. Community structure of an assemblage of tidepool fishes on a tropical eastern Pacific rocky shore, Colombia. Journal of Fish Biology, 67: 392-17.
CHANG, K.-H.; LEE, S.-C. & WU, W.-L. 1977. Fishes of reef limestone platform at Maopitou, Taiwan: diversity, and abundance. Bulletin of the Institute of Zoology Academia Sínica, 16: 9-21.
CHO, G. K. & HEATH, D. D. 2000. Comparison of tricaine methanesulphonate (MS222) and clove oil anesthesia effects on the physiology of the juvenile chinook salmon Oncorhynchus tshwaytshca (Walbaum). Aquaculture Research, 31: 537-10.
COOKE, S. J.; SUSKIB, C. D.; OSTRANDA, K. G.; TUFTSB, B. L. & WAHL, D. H. 2004. Behavioral and physiological assessment of low concentrations of clove oil anaesthetic for handling and transporting largemouth bass (Micropterus salmoides). Aquaculture, 234: 509-21.
CRABTREE, R. E. & DEAN, J. M. 1982. The structure of two South Caroline estuarine tidal pool fish assemblages. Estuaries, 5 (1): 2-9.
CUNHA, F. E. A. 2005. Estudo comparativo de metodologias de amostragens não destrutivas em peixes recifais no nordeste brasileiro. Doutorado em Ciências Biológicas - Zoologia, Departamento de Sistemática e Ecologia, Universidade Federal da Paraíba, João Pessoa. 95pp.
CUNHA, F. E. A.; MONTEIRO-NETO, C. & NOTTINGHAN, M. C. 2007. Temporal and spatial variations in tidepool fish assemblages of the northeast coast of Brazil. Biota Neotropica, 7 (1): 111-118.
CUNHA, F. E. A. & ROSA, I. L. 2006. Anaesthetic effects of clove oil on seven species of tropical reef teleosts. Journal of Fish Biology, 69: 1504-1512.
CURTIS, E. K. 1990. In pursuit of palliation: oil of cloves in the art of dentistry. Bulletin of the History of Dentistry, 38: 9-14.
DAVIS, K. B. & GRIFFIN, B. R. 2004. Physiological responses of hybrid striped bass under sedation by several anesthetics. Aquaculture, 233: 531-18.
ESTON, V. R.; MIGOTTO, A. E.; OLIVEIRA-FILHO, E. C.; RODRIGUES, S. A. & FREITAS, J. C. 1986. Vertical distribution of benthic marine organisms on rocky coasts of the Fernando de Noronha Arquipelago (Brazil). Boletim do Instituto Oceanográfico, 34: 37-53.
FERREIRA, L. M.; JESUS, F. & SILVA, H. A. 1990. Plano de Manejo do Parque Nacional Marinho de Fernando de Noronha, IBAMA/FUNATURA, Brasília, 253pp.
51

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
FLOETER, S. R.; FERREIRA, C. E. L.; DOMINICI-AROSEMENA, A. & ZALMON, I. R. 2004. Latitudinal gradients in Atlantic reef fish communities: trophic structure and spatial use patterns. Journal of Fish Biology, 64: 1680-1699.
FRISCH, A. J.; ULSTRUP, K. E. & HOBBS, J. P. A. 2007. The effects of clove oil on coral: an experimental evaluation using Pocillopora damicornis (Linnaeus). Journal of Experimental Marine Biology and Ecology, 345: 101-109.
FROESE, R. & PAULY, D. 2008. FishBase. www.fishbase.org, 10/2007.
GARLA, R. C. 2003. Ecologia e conservação dos tubarões do Arquipélago de Fernando de Noronha, com ênfase no tubarão-cabeça-de-cesto Carcharhinus perezi (Poey, 1876) (Carcharhiniformes, Carcharhinidae). Doutorado em Ciências Biológicas (Zoologia), Instituto de Biociências, Universidade Estadual Paulista, Rio Claro. 170pp.
GIBSON, R. N. 1999. Methods for Studying Intertidal Fishes. In: Intertidal Fishes - Life In Two Worlds, (HORN, M. H.; MARTIN, K. L. & CHOTKOWSKI, M. A., eds). Academic Press, San Diego, 7-25.
GIBSON, R. N. & YOSHIYAMA, R. M. 1999. Intertidal Fish Communities. In: Intertidal Fishes - Life In Two Worlds, (HORN, M. H.; MARTIN, K. L. & CHOTKOWSKI, M. A., eds). Academic Press, San Diego, 264-296.
GODINHO, W. O. 2007. Influência de fatores bióticos e abióticos na ictiofauna de poças de maré nas praias de Iparana, Pacheco e Flecheiras, Ceará. Graduação em Engenharia de Pesca, Departamento de Engenharia de Pesca, Universidade Federal do Ceará, Fortaleza. 41pp.
GRIFFITHS, S. P. 2000. The use of clove oil as an anaesthetic and method for sampling intertidal rockpool fishes. Journal of Fish Biology, 57: 1453-1464.
GRIFFITHS, S. P. 2003. Rockpool ichthyofaunas of temperate Australia: species composition, residency and biogeographic patterns. Estuarine, Coastal and Shelf Science, 58: 173-14.
GROSSMAN, G. D. 1982. Dynamics and organization of a rocky intertidal fish assemblage: the persistence and resilience of taxocene structure. American Naturalist, 119: 611-637.
HOLLOWAY, A. C.; KEENE, J. L.; NOAKES, D. G. & MOCCIA, R. D. 2004. Effects of clove oil and MS-222 on blood hormone profiles in rainbow trout Oncorhynchus mykiss, Walbaum. Aquaculture Research, 35: 1025-6.
HORN, M. H. 1983. Optimal diets in complex environments: feeding strategies of two herbivorous fishes from a temperate rocky intertidal zone. Oecologia (Berlin), 58: 345-350.
52

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
HORN, M. H.; MARTIN, K. L. & CHOTKOWSKI, M. A. 1999. Introduction. In: Intertidal Fishes - Life In Two Worlds, (HORN, M. H.; MARTIN, K. L. & CHOTKOWSKI, M. A., eds). Academic Press, San Diego, 1-6.
HUMANN, P. & DELOACH, N. 2002. Reef Fish Identification - Florida, Caribbean, Bahamas, 3rded. New World Publications, Jacksonville, 512pp.
IBAMA. 2008. Parque Nacional Marinho de Fernando de Noronha. www.ibama.gov.br, 01/2008.
ILARRI, M. I. 2008. Comparação da assembléia de peixes recifais de áreas de proteção integral e uso sustentável no Arquipélago de Fernando de Noronha-PE. Mestrado em Ciências Biológicas (Zoologia), Departamento de Sistemática e Ecologia, Universidade Federal da Paraíba, João Pessoa. 82pp.
INOUE, L. A. K. A.; AFONSO, L. O. B.; IWAMA, G. K. & MORAES, G. 2005. Effects of clove oil on the stress response of matrinxã (Brycon cephalus) subjected to transport. Acta Amazonica, 35 (2): 289-296.
IVERSEN, M.; FINSTADA, B.; MCKINLEYC, R. S. & ELIASSEN, R. A. 2003. The efficacy of metomidate, clove oil, Aqui-STM and Benzoak® as anaesthetics in Atlantic salmon (Salmo salar L.) smolts, and their potential stress-reducing capacity. Aquaculture, 221: 549-18.
KEENE, J. L.; NOAKES, D. L. G.; MOCCIA, R. D. & SOTO, C. G. 1998. The efficacy of clove oil as an anesthetic for rainbow trout, Oncorhynchus mykiss (Walbaum). Aquaculture Research, 29: 89-101.
KILDEA, M. A.; ALLANB, G. L. & KEARNEY, R. E. 2004. Accumulation and clearance of the anaesthetics clove oil and AQUI-STM from the edible tissue of silver perch (Bidyanus bidyanus). Aquaculture, 232: 265-13.
KING V, W.; HOOPER, B.; HILLSGROVE, S.; BENTON, C. & BERLINSKY, D. L. 2005. The use of clove oil, metomidate, tricaine methanesulphonate and 2-phenoxyethanol for inducing anaesthesia and their effect on the cortisol stress response in black sea bass (Centropristis striata L.). Aquaculture Research, 36: 1442-8.
LABOREL, J. L. & KEMPF, M. 1966. Formação de vermetos e algas calcárias nas costas do Brasil. Trabalhos do Instituto Oceanográfico, Universidade Federal de Pernambuco, Recife, 7/8: 33-50.
LARDNER, R.; IVANTSOFF, W. & CROWLEY, L. E. L. M. 1993. Recolonization by fishes of a rocky intertidal pool following repeated defaunation. Australian Zoology, 29: 85-92.
LEE, S.-L. 1980. Intertidal fishes of the rocky pool at Lanyu (Botel Tobago), Taiwan. Bulletin of the Institute of Zoology Academia Sínica, 19: 1-13.
53

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
MAHON, S. & MAHON, S. D. 1994. Structure and resilience of a tidepool fish assemblage at Barbados. Environmental Biology of Fishes, 41: 171-190.
MAIDA, M. & FERREIRA, B. P. 1997. Coral reefs of Brazil: an overview. Proceedings of the 8th International Coral Reef Symposium, 1: 263-274.
MASCARO, U. C. P.; RODRIGUES, L. A.; BASTOS, J. K.; SANTOS, E. & CHAVES DA COSTA, J. P. 1998. Valores de DL50 em peixes e no rato tratados com pó de raízes de Derris spp e suas implicações ecotoxicológicas. Pesquisa Veterinária Brasileira, 18 (2): 53-4.
MENDES, L. F. 2006. História natural dos amborés e peixes-macaco (Actinopterygii, Blennioidei, Gobioidei) do Parque Nacional Marinho do Arquipélago de Fernando de Noronha, sob um enfoque comportamental. Revista Brasileira de Zoologia, 23 (3): 817-823.
METAXAS, A. & SCHEILBLING, R. E. 1993. Community structure and organization of tidepools. Marine Ecology Progress Series, 98: 187-198.
MITRAUD, S. F. 2001. Uso Recreativo no Parque Nacional Marinho de Fernando de Noronha: um Exemplo de Planejamento e Implementação, WWF-Brasil, Brasília, 100pp.
MORING, J. R. 1986. Seasonal presence of tidepool fish species in a rocky intertidal zone of northern California, USA. Hydrobiologia, 134: 21-27.
MUNDAY, P. L. & WILSON, S. K. 1997. Comparative efficacy of clove oil and other chemicals in anaesthetization of Pomacentrus amboinensis, a coral reef fish. Journal of Fish Biology, 51: 931-938.
MYLONASA, C. C.; CARDINALETTIA, G.; SIGELAKIA, I. & POLZONETTI-MAGNI, A. 2005. Comparative efficacy of clove oil and 2-phenoxyethanol as anesthetics in the aquaculture of European sea bass (Dicentrarchus labrax) and gilthead sea bream (Sparus aurata) at different temperatures. Aquaculture, 246: 467-15.
NYBAKKEN, J. W. & BERTNESS, M. D. 2004. Marine Biology - An Ecological Approach, 6thed. Benjamin Cummings, San Francisco, 579pp.
ORTEGA, R.; RIBEIRO, W. & WINTHER, J. R. 1992. Planejamento ambiental e desenvolvimento do ecoturismo. Turismo em Análise, 3 (1).
PFISTER, C. A. 1998. Extinction, colonization, and species occupancy in tidepool fishes. Oecologia, 114: 118-9.
PIRHONEN, J. & SCHRECK, C. B. 2003. Effects of anaesthesia with MS-222, clove oil and CO2 on feed intake and plasma cortisol in steelhead trout (Oncorhynchus mykiss). Aquaculture, 220: 507-8.
54

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
POTTS, G. W. 1980. The littoral fishes of Little Cayman (West Indies). Atoll Research Bulletin, 241: 43-52.
PROCHAZKA, K. & GRIFFITHS, C. L. 1992. The intertidal fish fauna of the west coast of South Africa - species, community and biogeographic patterns. South African Journal of Zoology, 27: 115-120.
RANGUELEY, R. W. & KRAMER, D. L. 1995. Tidal effects on habitat selection and aggregation by juvenile pollock Polachius virens in the rocky intertidal zone. Marine Ecology Progress Series, 126 (1-3): 19-29.
REEFNET. 2003. Fishes of the Caribbean and Adjacent Waters, 3rded. ReefNet Inc. Mississauga.
ROSA, R. S.; ROSA, I. L. & ROCHA, L. A. 1997. Diversidade da ictiofauna de poças de maré da praia do Cabo Branco, João Pessoa, Paraíba, Brasil. Revista Brasileira de Zoologia, 14 (1): 201-212.
SALE, P. F. 1978. Coexistence of coral reef fishes - a lottery for living space. Environmental Biology of Fishes, 3: 85-102.
SANTOS, R. S.; NASH, R. D. M. & HAWKINS, S. J. 1994. Fish assemblages on intertidal shores of the island of Faial, Azores. Arquipélago-Life Marine Sciences, 12A: 87-100.
SAZIMA, I. & SAZIMA, C. 2004. Daytime hunting behaviour of Echidna catenata (Muraenidae): why chain morays foraging at ebb tide have no followers. aqua, Journal of Ichthyology and Aquatic Biology, 8 (1): 1-8.
SEOL, D. W.; LEE, J.; IM, S. Y. & PARK, I. S. 2007. Clove oil as an anaesthetic for common octopus (Octopus minor, Sasaki). Aquaculture Research, 38: 45-49.
SMALL, B. C. 2003. Anesthetic efficacy of metomidate and comparison of plasma cortisol responses to tricaine methanesulfonate, quinaldine and clove oil anesthetized channel catfish Ictalurus punctatus. Aquaculture, 218: 177-9.
SMITH, C. L. & TYLER, J. C. 1975. Succession and stability in fish communities of dome-shaped patch reefs in the West Indies. American Museum Novitates, 2572: 1-18.
SOTO, C. G. & BURHANUDDIN. 1995. Clove oil as a fish anaesthetic for measuring length and weight of rabbitfish (Siganus lineatus). Aquaculture, 136: 149-152.
SOTO, J. M. R. 2001. Peixes do Arquipélago de Fernando de Noronha. Mare Magnum, 1 (2): 147-169.
55

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
STATSOFT. 2004. Statistica - Data Analysis Software System, 7ed. www.statsoft.com.
TEIXEIRA, W.; CORDANI, U. G. & MENOR, E. A. 2003. Caminhos do Tempo Geológico. In: Arquipélago de Fernando de Noronha – o Paraíso do Vulcão, (LINSKER, R., ed). Terra Virgem Editora, São Paulo, 26-63.
THOMSON, D. A. & LEHNER, C. E. 1976. Resilience of rocky intertidal fish community in a physically unstable environment. Journal of Experimental Marine Biology and Ecology, 22: 1-29.
TORT, L.; PUIGCERVER, M.; CRESPO, S. & PADROS, F. 2002. Cortisol and haematological response in sea bream and trout subjected to the anesthetics clove oil and 2-phenoxyethanol. Aquaculture Research, 33: 907-4.
UNDERWOOD, A. J. & SKILLETER, G. A. 1996. Effects of patch-size on the structure of assemblages in rock pools. Journal of Experimental Marine Biology and Ecology, 197: 63-26.
VARAS, E. & OJEDA, F. P. 1990. Intertidal fish assemblages of the central Chilean coast: diversity, abundance and trophic patterns. Revista de Biologia Marina, 25: 59-70.
VENARSKY, M. P. & WILHELM, F. M. 2006. Use of clove oil to anaesthetize freshwater amphipods. Hydrobiologia, 568: 425-8.
WAGNER, E.; ARNDTA, R. & HILTON, B. 2002. Physiological stress responses, egg survival and sperm motility for rainbow trout broodstock anesthetized with clove oil, tricaine methanesulfonate or carbon dioxide. Aquaculture, 211: 353-14.
WALSH, T. C. & PEASE, B. C. 2002. The use of clove oil as an anesthetic for the longfinned eel, Anguilla reinhardtii (Steindachner). Aquaculture Research, 33: 627-9.
WEAVER, P. L. 1970. Species diversity and ecology of tidepool fishes in three Pacific costal areas of Costa Rica. Revista de Biología Tropical, 17: 165-185.
WILLIS, T. J. 2001. Visual census methods underestimate density and diversity of cryptic reef fishes. Journal of Fish Biology, 59: 1408-4.
YOSHIYAMA, R. M. 1981. Distribution and abundance patterns of rocky intertidal fishes in central California. Environmental Biology of Fishes, 6: 315-332.
YOSHIYAMA, R. M.; SASSAMAN, C. & LEA, R. N. 1986. Rocky intertidal communities of California: temporal and spatial variation. Environmental Biology of Fishes, 20: 203-218.
56

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS REFERÊNCIAS BIBLIOGRÁFICAS
57
ZANDER, C. D.; NIEDER, J. & MARTIN, K. L. 1999. Vertical Distribution Patterns. In: Intertidal Fishes - Life In Two Worlds, (HORN, M. H.; MARTIN, K. L. & CHOTKOWSKI, M. A., eds). Academic Press, San Diego, 26-53.
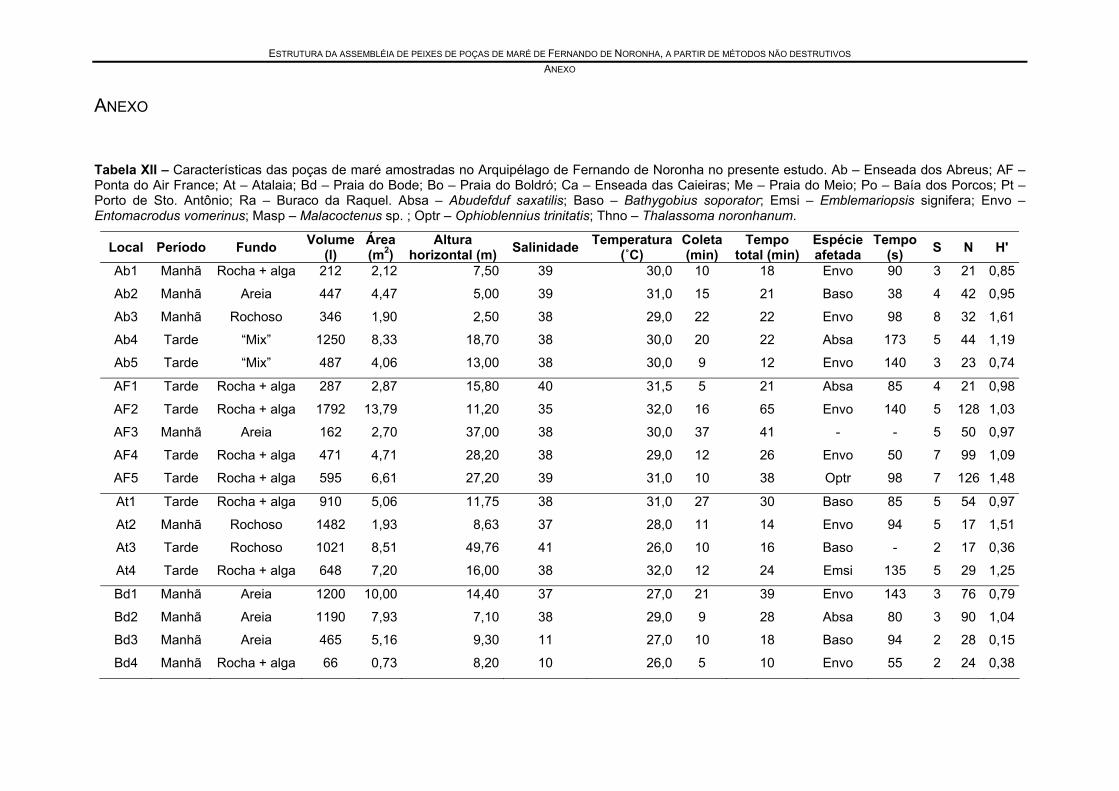
ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS ANEXO
ANEXO
Tabela XII – Características das poças de maré amostradas no Arquipélago de Fernando de Noronha no presente estudo. Ab – Enseada dos Abreus; AF – Ponta do Air France; At – Atalaia; Bd – Praia do Bode; Bo – Praia do Boldró; Ca – Enseada das Caieiras; Me – Praia do Meio; Po – Baía dos Porcos; Pt – Porto de Sto. Antônio; Ra – Buraco da Raquel. Absa – Abudefduf saxatilis; Baso – Bathygobius soporator; Emsi – Emblemariopsis signifera; Envo – Entomacrodus vomerinus; Masp – Malacoctenus sp. ; Optr – Ophioblennius trinitatis; Thno – Thalassoma noronhanum.
Local Período Fundo Volume (l)
Área (m2)
Altura horizontal (m) Salinidade Temperatura
(˚C) Coleta (min)
Tempo total (min)
Espécie afetada
Tempo (s) S N H'
Ab1 Manhã Rocha + alga 212 2,12 7,50 39 30,0 10 18 Envo 90 3 21 0,85
Ab2 Manhã Areia 447 4,47 5,00 39 31,0 15 21 Baso 38 4 42 0,95
Ab3 Manhã Rochoso 346 1,90 2,50 38 29,0 22 22 Envo 98 8 32 1,61
Ab4 Tarde “Mix” 1250 8,33 18,70 38 30,0 20 22 Absa 173 5 44 1,19
Ab5 Tarde “Mix” 487 4,06 13,00 38 30,0 9 12 Envo 140 3 23 0,74
AF1 Tarde Rocha + alga 287 2,87 15,80 40 31,5 5 21 Absa 85 4 21 0,98
AF2 Tarde Rocha + alga 1792 13,79 11,20 35 32,0 16 65 Envo 140 5 128 1,03
AF3 Manhã Areia 162 2,70 37,00 38 30,0 37 41 - - 5 50 0,97
AF4 Tarde Rocha + alga 471 4,71 28,20 38 29,0 12 26 Envo 50 7 99 1,09
AF5 Tarde Rocha + alga 595 6,61 27,20 39 31,0 10 38 Optr 98 7 126 1,48
At1 Tarde Rocha + alga 910 5,06 11,75 38 31,0 27 30 Baso 85 5 54 0,97
At2 Manhã Rochoso 1482 1,93 8,63 37 28,0 11 14 Envo 94 5 17 1,51
At3 Tarde Rochoso 1021 8,51 49,76 41 26,0 10 16 Baso - 2 17 0,36
At4 Tarde Rocha + alga 648 7,20 16,00 38 32,0 12 24 Emsi 135 5 29 1,25
Bd1 Manhã Areia 1200 10,00 14,40 37 27,0 21 39 Envo 143 3 76 0,79
Bd2 Manhã Areia 1190 7,93 7,10 38 29,0 9 28 Absa 80 3 90 1,04
Bd3 Manhã Areia 465 5,16 9,30 11 27,0 10 18 Baso 94 2 28 0,15
Bd4 Manhã Rocha + alga 66 0,73 8,20 10 26,0 5 10 Envo 55 2 24 0,38

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS ANEXO
Local Período Fundo Volume (l)
Área (m2)
Altura horizontal (m) Salinidade Temperatura
(˚C) Coleta (min)
Tempo total (min)
Sp afetada
Tempo (s) S N H'
Bo1 Manhã Areia 1250 3,91 9,00 38 28,5 32 68 Envo 145 4 67 1,10
Bo2 Manhã Rocha + alga 217 2,71 8,05 39 30,0 13 34 Absa 86 5 48 0,95
Bo3 Manhã Rocha + alga 167 2,09 10,60 40 31,0 17 17 - - 2 9 0,64
Bo4 Manhã Rocha + alga 406 2,71 40,00 40 33,0 22 67 Absa 120 5 97 1,19
Bo5 Tarde Rocha + alga 950 4,32 35,30 39 32,0 17 32 Envo 102 5 92 1,04
Bo6 Manhã Areia 700 2,80 6,52 34 27,5 9 19 Envo 100 3 23 0,69
Bo7 Manhã Crista algal 540 6,00 15,20 38 29,5 27 68 Thno 180 9 131 1,58
Bo8 Manhã Areia 573 1,79 10,20 40 30,0 33 70 Masp 73 12 164 1,57
Ca1 Tarde Rochoso 63 2,75 40,00 40 31,0 - 25 - - 4 22 1,21
Ca2 Tarde Rochoso 1250 5,68 40,00 38 32,0 35 50 Envo 135 4 102 0,86
Ca3 Manhã Rocha + alga 389 3,89 7,35 38 30,5 - 33 Masp 115 7 62 1,28
Ca4 Tarde “Mix” 418 2,79 9,50 39 31,0 - 40 Envo 100 9 80 0,93
Ca5 Tarde Rocha + alga 164 2,05 10,26 38 34,0 - 12 Envo 126 3 31 0,80
Ca6 Manhã Rocha + alga 414 5,17 10,20 39 26,0 29 30 Absa 109 6 68 1,26
Me1 Tarde Rochoso 1354 8,20 3,40 39 30,0 21 37 Baso 85 7 69 1,53
Me2 Tarde “Mix” 263 1,75 6,25 39 28,0 18 24 Thno 102 8 45 1,79
Me3 Tarde Rocha + alga 1170 4,68 7,10 38 32,0 12 15 Envo 135 4 26 0,69
Me4 Tarde Rochoso 739 5,28 7,30 39 34,0 15 28 Baso 105 5 41 1,06
Me5 Tarde Rochoso 1662 8,30 8,25 42 31,0 25 53 Absa 70 8 194 1,67
Me6 Manhã Rocha + alga 336 3,36 14,70 37 26,0 12 28 Absa 135 5 34 1,05
Me7 Manhã Rocha + alga 1856 9,28 8,90 37 27,0 23 42 Absa 134 9 80 1,73
Po1 Tarde Rochoso 2864 17,90 14,00 38 31,0 25 41 Absa 120 5 66 0,85
Po2 Tarde Rochoso 1182 6,22 5,30 38 28,5 9 37 Envo 116 5 56 0,99

ESTRUTURA DA ASSEMBLÉIA DE PEIXES DE POÇAS DE MARÉ DE FERNANDO DE NORONHA, A PARTIR DE MÉTODOS NÃO DESTRUTIVOS ANEXO
Local Período Fundo Volume (l)
Área (m2)
Altura horizontal (m) Salinidade Temperatura
(˚C) Coleta (min)
Tempo total (min)
Sp afetada
Tempo (s) S N H'
Pt1 Tarde Rochoso 347 3,47 3,00 39 32,0 18 35 Absa 80 6 62 1,23
Pt2 Manhã Rocha + alga 1166 19,44 16,80 38 33,0 41 120 Envo 138 9 315 0,95
Pt3 Tarde Rochoso 168 2,80 5,00 39 34,0 11 29 Absa 100 4 57 1,12
Pt4 Tarde Rochoso 559 6,21 26,00 40 34,0 12 22 Absa 85 4 31 0,92
Pt5 Manhã Areia 232 2,58 17,63 40 29,0 25 42 Masp 119 10 43 1,94
Pt6 Manhã Crista algal 453 2,52 5,85 39 29,0 21 29 Optr 90 8 39 1,79
Pt7 Tarde Rocha + alga 584 3,25 11,20 38 30,0 20 21 Optr 73 7 42 1,71
Ra1 Tarde Rochoso 429 3,90 4,73 35 32,0 16 31 Envo 115 4 84 0,93
Ra2 Tarde Rocha + alga 943 7,86 6,90 40 33,0 - 33 Absa 109 3 74 0,82
Ra3 Manhã “Mix” 870 3,62 8,50 39 30,0 12 18 Envo 150 4 30 0,67
Ra4 Tarde Coral 157 1,57 15,70 39 31,0 17 48 Scsp 40 5 109 1,12
Ra5 Tarde Rochoso 1239 6,20 40,00 40 33,0 11 20 Envo 135 4 51 1,29
Ra6 Manhã Rocha + alga 518 3,69 22,12 38 26,0 14 23 Envo 80 6 39 1,41
Ra7 Manhã Rocha + alga 475 5,28 21,90 38 27,5 13 25 Absa 93 5 47 0,85

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo