dormÊncia em sementes de pupunha (baccttrriiss … nazario.pdf · 4. maturação. 5. bioquímica....
TRANSCRIPT

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS DE FLORESTAS TROPICAIS
DDOORRMMÊÊNNCCIIAA EEMM SSEEMMEENNTTEESS DDEE PPUUPPUUNNHHAA ((BBAACCTTRRIISS
GGAASSIIPPAAEESS KKUUNNTTHH)):: UUMMAA AABBOORRDDAAGGEEMM AANNAATTÔÔMMIICCAA,, HHIISSTTOOQQUUÍÍMMIICCAA EE FFIISSIIOOLLÓÓGGIICCAA
PATRÍCIA NAZÁRIO
Manaus, Amazonas Dezembro, 2011.

PATRÍCIA NAZÁRIO
DDOORRMMÊÊNNCCIIAA EEMM SSEEMMEENNTTEESS DDEE PPUUPPUUNNHHAA ((BBAACCTTRRIISS
GGAASSIIPPAAEESS KKUUNNTTHH)):: UUMMAA AABBOORRDDAAGGEEMM AANNAATTÔÔMMIICCAA,, HHIISSTTOOQQUUÍÍMMIICCAA EE FFIISSIIOOLLÓÓGGIICCAA
ORIENTADOR: SIDNEY ALBERTO DO NASCIMENTO FERREIRA Co-orientador: Eduardo Euclydes de Lima e Borges
Tese apresentada ao Instituto Nacional de Pesquisas da Amazônia como parte dos requisitos para obtenção do grau de Doutora em Ciências de Florestas Tropicais.
Manaus, Amazonas Dezembro, 2011.

i
N335 Nazário, Patrícia Dormência em sementes de pupunha (Bactris gasipaes Kunth): uma abordagem anatômica, histoquímica e fisiológica / Patrícia Nazário. --- Manaus : [s.n.], 2011. 125 f. : il. color. Tese (doutorado) --- INPA, Manaus, 2011 Orientador : Sidney Alberto do Nascimento Co-orientador : Eduardo Euclydes de Lima e Borges Área de concentração : Manejo Florestal e Silvicultura
1. Palmeira. 2. Pupunha – Sementes – Dormência. 3. Germinação. 4. Maturação. 5. Bioquímica. 6. Anatomia. I. Título.
CDD 19. ed. 584.5044
Sinopse: O trabalho visou investigar fatores relacionados à dormência em sementes de Bactris gasipaes. Foram feitos estudos anatômico e histoquímico, enfatizando-se estruturas e compostos relacionados à germinação; estudo dos compostos fenólicos em sementes de frutos em dois estádios de maturação; estudo relacionando a germinação de sementes de frutos em dois estádios de maturação com sua composição lipídica e; por fim, estabeleceu-se uma relação entre a germinação de sementes de diferentes procedências e a germinação de seus embriões in vitro. Palavras-chave: palmeira, germinação, maturação, bioquímica, anatomia

ii
Aos meus pais, Domingos e Maria Alice e ao meu amor, Giuliano,
Dedico.

iii
Agradecimentos
A Deus;
Ao Giuliano, por todo amor e dedicação;
Aos meus pais, Maria Alice e Domingos, minhas irmãs, Daniella e Juliana e
minha sobrinha, Luana, por toda força que sempre me deram;
Ao meu orientador, Dr. Sidney Alberto do Nascimento Ferreira, e meu co-
orientador, Dr. Eduardo Euclydes de Lima e Borges, pelos preciosos
ensinamentos;
Ao coordenador do Programa de Pós-Graduação em Ciências de Florestas
Tropicais, Dr. José Francisco de Carvalho Gonçalves;
Aos amigos e colegas do laboratório de sementes, Elizabeth Rebouças, Mário
Moreira Filho, Luzia Correa Dias, pelo auxílio na instação e avaliação dos
experimentos;
Aos técnicos Sr. Walderico e Sr. Diomar (Passarinho) pelo auxílio nas coletas
dos frutos;
Ao Dr. Charles Clement, pelos valioso apoio a este trabalho;
Ao Dr. Douglas Steinmacher pelas preciosas dicas e incentivo;
Ao Dr. Wander Chaves-Flores, pelo apoio durante viagem à Coari para coleta
de frutos;
Ao Sr. Wilian, pelo suporte técnico na coleta e beneficiamento dos frutos no
município de Coari;
À Marduce e à Cooperativa dos Produtores Coari/Itapeuá, por cederem
cachos de pupunha para que parte deste trabalho fosse realizada;
À Dra. Jerusa Andrade e ao MSc. Raimundo Souza pelo apoio na extração
dos lipídios e quantificação dos compostos fenólicos;

iv
À Dra. Maria Sílvia de Mendonça, responsável pelo Laboratório de Botânica
Agroflorestal (LABAF) da UFAM, pelo apoio às análises anatômicas e
histoquímicas;
Às amigas do LABAF, Maria da Glória, Ângela, Maria Elizabeth e Poliana e ao
técnico Manoel Roberto Viana pelo auxílio nas análises anatômicas e
histoquímicas;
Ao técnico do Laboratório de Sementes Florestais da Universidade Federal de
Viçosa, José Mauro Moutinho, pelo fundamental auxílio nas análises dos
ácidos graxos;
Às colegas Eliza e Glauciana pelo apoio nas análises;
À Dra. Ângela Maria Imakawa, pelas dicas e auxílio em todas as estapas da
cultura de embriões zigótocos;
À Dra. Denise Santana, pelo auxílio nos delineamentos experimentais;
Aos membros da banca avaliadora da aula de qualificação, Dr. Daniel Gentil,
Dr. Isolde Ferraz, Dra. Lucinda Garcia, Dra. Silvana Pando e Dra. Zilvanda
Melo, pelas contribuições dadas para a realização deste trabalho;
Aos amigos do INPA e do Curso de Pós-graduação em Ciências de Florestas
Tropicais;
A todas as pessoas que contribuíram direta ou indiretamente para a
realização deste trabalho, meus mais sinceros agradecimentos.

v
Sumário
INTRODUÇÃO GERAL............................................................................................... 1
OBJETIVOS ................................................................................................................ 7
Objetivo geral ............................................................................................................ 7
Objetivos específicos................................................................................................ 7
Capítulo 1 ................................................................................................................... 8
ASPECTOS ANATÔMICOS E HISTOQUÍMICOS DE SEMENTES DE PUPUNHA (Bactris gasipaes KUNTH). ....................................................................................... 9
RESUMO. .......................................................................................................................... 9
ANATOMICAL AND HISTOCHEMICAL ASPECTS OF PEACH PALM (Bactris gasipaes
KUNTH) SEEDS............................................................................................................... 10
ABSTRACT. ..................................................................................................................... 10
INTRODUÇÃO ................................................................................................................. 11
MATERIAL E MÉTODOS ................................................................................................. 13
RESULTADOS E DISCUSSÃO ........................................................................................ 14
CONCLUSÃO .................................................................................................................. 18
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 20
Capítulo 2 ................................................................................................................. 34
COMPOSTOS FENÓLICOS E DORMÊNCIA EM SEMENTES DE Bactris gasipaes KUNTH EM FUNÇÃO DA MATURAÇÃO DOS FRUTOS ........................................ 35
RESUMO. ........................................................................................................................ 35
PHENOLIC COMPOUNDS AND SEED DORMANCY IN Bactris gasipaes KUNTH AS A
FUNCTION OF FRUIT MATURITY. ................................................................................. 36
ABSTRACT ...................................................................................................................... 36
INTRODUÇÃO ................................................................................................................. 37
MATERIAL E MÉTODOS ................................................................................................. 38
Emergência de plântulas de Bactris gasipaes em função da maturação dos frutos ...... 38
Determinação do teor de compostos fenólicos em sementes de Bactris gasipaes ........ 40

vi
Efeito do extrato de sementes de Bactris gasipaes na germinação de sementes de Lactuca sativa ............................................................................................................... 42
RESULTADOS ................................................................................................................. 43
Emergência de plântulas de Bactris gasipaes em função da maturação dos frutos ...... 43
Teor de compostos fenólicos em sementes de frutos imaturos e maduros de Bactris gasipaes e sua relação com a emergência de plântulas e a dormência ........................ 44
Efeito da aplicação do extrato de sementes de frutos imaturos e maduros de Bactris gasipaes na germinação de sementes de Lactuca sativa ............................................. 46
DISCUSSÃO .................................................................................................................... 48
REFERÊNCIAS BIBLIOGRÁFICAS ................................................................................. 52
Capítulo 3 ................................................................................................................. 61
TEORES DE LIPÍDIOS E GERMINAÇÃO DE SEMENTES DE Bactris gasipaes KUNTH DE FRUTOS EM DIFERENTES ESTÁDIOS DE MATURAÇÃO ................. 62
RESUMO ......................................................................................................................... 62
Lipids and germination of seeds of Bactris gasipaes Kunth fruits in different maturation
stages .............................................................................................................................. 63
ABSTRACT ...................................................................................................................... 63
INTRODUÇÃO ................................................................................................................. 64
MATERIAL E MÉTODOS ................................................................................................. 65
Avaliação da emergência de plântulas de Bactris gasipaes a partir de sementes de frutos com diferentes estádios de maturação ................................................................ 65
Quantificação e caracterização dos lipídios .................................................................. 66
Análise estatística dos resultados ................................................................................. 68
RESULTADOS ................................................................................................................. 68
DISCUSSÃO .................................................................................................................... 71
CONCLUSÃO .................................................................................................................. 73
BIBLIOGRAFIA CITADA .................................................................................................. 75
Capítulo 4 ................................................................................................................. 80
DORMÊNCIA EMBRIONÁRIA EM SEMENTES DA PALMEIRA Bactris gasipaes KUNTH (PUPUNHA) ................................................................................................. 81
RESUMO ......................................................................................................................... 81

vii
INTRODUÇÃO ................................................................................................................. 82
MATERIAL E MÉTODOS ................................................................................................. 83
RESULTADOS ................................................................................................................. 86
DISCUSSÃO .................................................................................................................... 88
REFERÊNCIAS ................................................................................................................ 92
SÍNTESE ................................................................................................................. 102
REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................... 104

viii
Lista de Tabelas Capítulo 1 ................................................................................................................... 8
Aspectos anatômicos e histoquímicos de sementes de pupunha (Bactris gasipaes Kunth) ........................................................................................................ 8
TABELA 1. Compostos analisados, reagentes utilizados, coloração da reação positiva e
referência do teste histoquímico. ...................................................................................... 27
TABELA 2. Resultados dos testes histoquímicos aplicados em cortes histológicos de
sementes de Bactris gasipaes. ......................................................................................... 28
Capítulo 2 ................................................................................................................. 34
Compostos fenólicos e dormência em sementes de Bactris gasipaes Kunth em função da maturação dos frutos ............................................................................ 34
TABELA 1. Teor de água, massa seca, variáveis relacionadas à emergência de plântulas
e compostos fenólicos a partir de sementes de Bactris gasipaes com diferentes estádios
de maturação dos frutos. .................................................................................................. 57
TABELA 2. Correlações entre as variáveis de emergência e dormência, as características
físicas e o teor de compostos fenólicos (mg/semente) em sementes de Bactris gasipaes.
......................................................................................................................................... 58
TABELA 3. Germinação, índice de velocidade de germinação (IVG), tempo médio de
germinação (TMG), emergência, índice de velocidade de emergência (IVE) e tempo médio
de emergência (TME) de sementes de (Lactuca sativa) semeadas em substrato
umedecido com extrato fenólico de sementes de frutos de pupunha de diferentes plantas
e com diferentes estádios de maturação dos frutos. ......................................................... 59
TABELA 4. Correlação entre variáveis de germinação e de emergência de sementes de
alface submetidas ao extrato de sementes de pupunha de diferentes plantas e com
diferentes estádios de maturação dos frutos, a emergência de plântulas de pupunha a
partir dessas sementes e a concentração de compostos fenólicos nessas sementes. ..... 60
Capítulo 3 ................................................................................................................. 61
Teores de lipídios e germinação de sementes de Bactris gasipaes Kunth de frutos em diferentes estádios de maturação ........................................................ 61
TABELA 1. Características físicas, variáveis de emergência, composição de ácidos
graxos, lipídios e ponto de fusão dos lipídios em sementes de Bactris gasipaes de frutos
imaturos e maduros. ......................................................................................................... 78

ix
TABELA 2. Correlação entre variáveis de emergência, lipídios, ponto de fusão dos lipídios
e concentração de ácidos graxos em sementes de Bactris gasipaes advindas de frutos
imaturos e maduros. ......................................................................................................... 79
Capítulo 4 ................................................................................................................. 80
Dormência embrionária em sementes da palmeira Bactris gasipaes Kunth (pupunha) ................................................................................................................. 80
TABELA 1. Emergência, sementes mortas, dormentes, germinadas e não emergidas
(GNE), índice de velocidade de emergência (IVE) e tempo médio de emergência (TME) de
plântulas de Bactris gasipaes a partir de sementes de diferentes plantas. ....................... 98
TABELA 2. Emergência, sementes mortas, sementes dormentes, sementes germinadas e
não emergidas (GNE), índice de velocidade de emergência (IVE) e tempo médio de
emergência (TME), diferenciação, embriões mortos, dormentes, índice de velocidade de
diferenciação (IVD) e tempo médio de diferenciação (TMD) entre as sementes obtidas de
frutos coletados em Manaus e Coari. ............................................................................... 99
TABELA 3. Diferenciação de embriões, embriões mortos, dormentes, índice de velocidade
de diferenciação (IVD) e tempo médio de diferenciação (TMD) de embriões de Bactris
gasipaes a partir de sementes de diferentes plantas e procedências. ............................ 100

x
Lista de Figuras
Capítulo 1 ................................................................................................................... 8
Aspectos anatômicos e histoquímicos da semente de pupunha (Bactris gasipaes Kunth) ........................................................................................................ 8
FIGURA 1. Aspectos estruturais do tegumento (A, B e C), do endosperma (D) e do
embrião (E, F, G e H) de Bactris gasipaes. tg- tegumento; te- tegumento externo; ti-
tegumento interno; ct- conteúdo tanífero; en- endosperma; fv- feixe vascular; pf-
primórdios foliares; pl- plúmula; pr- primórdio da raiz. ...................................................... 29
FIGURA 2. Aspectos estruturais do embrião de Bactris gasipaes. fv- feixe vascular; co-
tecido cotiledonar. ............................................................................................................ 30
FIGURA 3. Testes histoquímicos no tegumento e no endosperma de sementes de Bactris
gasipaes detectando a presença de terpenoides (A), compostos fenólicos (B), taninos (C),
ácidos graxos (D e E), lipídios (F), lipídios ácidos e neutros (G e H). ............................... 31
FIGURA 4. Testes histoquímicos no tegumento e no endosperma de sementes de Bactris
gasipaes detectando a presença de proteínas (A e B), pectinas (C) e polissacarídeos (D).
......................................................................................................................................... 32
FIGURA 5. Testes histoquímicos no embrião de sementes de Bactris gasipaes detectando
a presença de lipídios ácidos e neutros (A), lipídios totais (B), proteínas (C) e
polissacarídeos (D). ......................................................................................................... 33
Capítulo 2 ................................................................................................................. 34
Compostos fenólicos e dormência em sementes de Bactris gasipaes Kunth em função da maturação dos frutos ............................................................................ 34
FIGURA 1. Curva de calibração do ácido tânico, com leituras de absorbância a 760 nm. 56
Capítulo 3 ................................................................................................................. 61
Teores de lipídios e germinação de sementes de Bactris gasipaes Kunth de frutos em diferentes estádios de maturação ........................................................ 61
Capítulo 4 ................................................................................................................. 80
Dormência embrionária em sementes da palmeira Bactris gasipaes Kunth (pupunha) ................................................................................................................. 80
FIGURA 1. Embrião zigótico de Bactris gasipaes cultivado in vitro apresentando emissão
de raiz. ............................................................................................................................. 97

xi
FIGURA 2. Emergência de plântulas em função da diferenciação de embriões (A); índice
de velocidade de emergência de plântulas em função do índice de velocidade de
diferenciação de embriões (B); tempo médio de emergência de plântulas em função do
tempo médio de diferenciação de embriões (C); e sementes dormentes em função dos
embriões dormentes (D) a partir de frutos Bactris gasipaes de diferentes plantas. ......... 101

xii
RESUMO
Bactris gasipaes é uma palmeira importante para as populações da Amazônia
devido, principalmente, ao valor nutricional de seus frutos. O presente trabalho está
dividido em quatro capítulos e o objetivo geral foi avaliar características anatômicas,
histoquímicas e fisiológicas e suas relações com mecanismos de dormência em
sementes de Bactris gasipaes. No primeiro, objetivou-se identificar características
anatômicas e histoquímicas de sementes de B. gasipaes associadas ao processo de
dormência e germinação. O tegumento é fino e em duas camadas, a externa mais
espessa, com células onde se verifica a presença de feixes vasculares e a interna
menos espessa, com células de formato irregular e repletas de compostos fenólicos.
O endosperma ocupa a maior parte da semente e é esbranquiçado, com células nos
formatos oblongo oboval e oval, com conteúdo lipídico. O embrião é pequeno em
relação ao tamanho da semente, cônico, indiviso e vascularizado na porção distal.
Na região proximal, em corte longitudinal, aparece a plúmula, com três primórdios
foliares. No segundo capítulo, objetivou-se quantificar compostos fenólicos em
sementes de B. gasipaes de frutos em dois estádios de maturação, correlacionando-
os com a emergência de plântulas e verificar o efeito da aplicação do extrato destas
sementes sob a germinação de sementes de Lactuca sativa. A emergência, índice
de velocidade de emergência, tempo médio de emergência, coeficiente de
uniformidade de emergência, índice de sincronização, velocidade média de
emergência, valor de emergência e coeficiente de velocidade de emergência não
variaram significativamente entre sementes obtidas de frutos imaturos e maduros.
Entretanto, as variações dessas mesmas variáveis foram maiores entre sementes de
diferentes plantas. Sementes maduras de pupunha apresentam maior concentração
de compostos fenólicos que as imaturas e o acúmulo desses compostos é um dos
fatores responsáveis pela redução da germinação em sementes de pupunha. O
extrato fenólico das sementes desta espécie foi capaz de diminuir e retardar a
germinação e emergência de sementes de alface e essa emergência apresentou
correlação significativa com a emergência de plântulas de pupunha. O teor de água
de sementes de frutos imaturos foi estatisticamente superior ao de sementes de
frutos maduros (50,4 e 46,7%, respectivamente). No terceiro capítulo, objetivou-se

xiii
avaliar a composição lipídica e o ponto de fusão dos lipídios de sementes de B.
gasipaes, bem como correlacioná-los com variáveis relacionadas à emergência de
plântulas a partir de sementes provenientes de frutos imaturos e maduros. O ponto
de fusão dos lipídios não diferiu estatisticamente entre sementes de frutos imaturos
(22,54 ºC) e maduros (22,70 ºC), porém teve alta correlação com as variáveis
relacionadas à emergência de plântulas. Os teores de ácido oleico e linoleico foram
maiores em sementes de frutos maduros (26,89 e 30,66 µg.g-1, respectivam ente)
em relação às de frutos imaturos (22,01 e 24,79 µg.g-1, respectivamente). A
concentração de ácido oleico teve correlação negativa com o tempo médio de
emergência (TME). No quarto capítulo, objetivou-se investigar a presença de
dormência embrionária em sementes de pupunha. A maioria das variáveis
relacionadas à emergência de plântulas a partir de sementes, em viveiro, e à
germinação de embriões, em meio de cultura, variou significativamente entre plantas
e procedências. Em geral, estas se relacionaram positivamente, ou seja, quanto
maior a germinação, maior a emergência. As características estruturais indicaram
que o embrião encontra-se diferenciado no momento da dispersão. A presença de
compostos fenólicos e de terpenóides no tegumento pode ser um dos fatores que
inibem a germinação de sementes de pupunha. A emergência de plântulas não
diferiu entre sementes de frutos imaturos e maduros. Sementes de frutos maduros
apresentam maior concentração de compostos fenólicos que sementes de frutos
imaturos e o acúmulo de compostos fenólicos é capaz de inibir a germinação em
sementes de pupunha. A concentração de lipídios foi maior em sementes de frutos
imaturos e sementes de frutos maduros apresentaram maiores concentrações dos
ácidos oleico e linoleico. A maior concentração de ácido oleico resultou um menor
tempo médio de emergência de plântulas. Quanto maior o ponto de fusão dos
lipídios, maior a emergência e o índice de velocidade de emergência. Evidenciou-se
que a dormência de sementes de pupunha pode ser atribuida, em parte, à
dormência embrionária.
Palavras-chave: palmeira, germinação, maturação, bioquímica, anatomia.

xiv
ABSTRACT
Bactris gasipaes is an important palm for populations in the Amazon, mainly due to
the value nutritional content of its fruit. This work is divided into four chapters and the
overall goal was evaluate anatomical, histochemical and physiological aspects and
their relationships with mechanisms of dormancy in seeds of Bactris gasipaes. In the
first, aimed identify anatomical and histochemical seed of B. gasipaes associated
with the process dormancy and germination. The integument is thin and in two layers,
the outer thicker cell where checks for the presence of vascular bundles and the
inner less thick, with irregularly shaped cells and filled with phenolic compounds. The
endosperm occupies the most of the seed and is whitish, with cells in oboval oblong
and oval, with lipid content. The embryo is small in relation to seed size, conical,
undivided and vascularized in the distal portion. In proximal region in cross section
appears plumule, with three leaf primordia. In the second chapter aimed to quantify
phenolic compounds in seeds of B. gasipaes fruit in two stages of maturation,
correlating them with the emergence of seedlings and check the effect of applying
the extract these seeds in the germination of Lactuca sativa. The emergence,
emergence rate index, mean emergence time, coefficient of uniformity emergency,
synchronization index, average speed of Emergency, emergency value and
coefficient of emergence rate did not vary significantly seeds obtained from immature
and mature fruit. However, variations of these same variables were higher among
seeds of different plants. Seeds mature peach palm have higher concentration of
phenolic compounds and the accumulation of immature compounds is one of the
factors responsible for reducing germination in seeds of peach. The phenolic extract
of seeds of this species was able to reduce and delay germination and emergence of
lettuce seeds and that Emergency correlated significantly with the emergence of
seedlings of peach palm. The water content of seeds of immature fruits was
statistically superior that of mature seeds (50.4 and 46.7%, respectively). In the third
chapter aimed to assess the lipid composition and the melting point of seed lipids of
B. gasipaes and correlate them with variables related to emergency seedlings from
seeds from fruits immature and mature. The melting point of lipids not differ
statistically between fruit seeds immature (22.54 °C) and mature (22.70 °C), but was

xv
discharged correlation with the variables related to the emergence of plantlets. The
levels of oleic and linoleic acid were higher in mature seeds (26.89 and 30.66 μg.g-1,
respective entity) in relation to fruit immature (22.01 and 24.79 μg.g-1,
respectively). The oleic acid concentration was negatively correlated with the mean
emergence time (MET). The fourth chapter aimed to investigate the presence of
numbness embryo in seeds of peach palm. Most variables related to the emergence
of seedlings from seed in the nursery, and germination of embryos in medium, varied
significantly between plants and origins. In general, these were related positively, ie
higher germination, greater the emergency. The structural characteristics indicated
that the embryo is differentiated at the time of scatter. The presence of phenolic
compounds and terpenoids in the seed coat may be one of the factors that
inhibit the germination of peach palm. The emergence seedlings did not differ
between fruit seeds immature and mature. Mature seeds have highest concentration
of phenolic compounds that seeds immature fruits and accumulation of phenolic
compounds is able to inhibit the germination of seeds of peach palm. The lipid
concentration was higher in fruit seeds immature and mature seeds showed higher
concentrations of oleic and linoleic acids. The higher concentration of oleic acid
results a lower average time of emergence. The higher the melting point of the lipids,
the greater the emergence and emergency speed index. It was evident that the seed
dormancy of peach can be attributed in part to embryo dormancy.
Key words: palm, germination, maturation, biochemistry, anatomy.

1
INTRODUÇÃO GERAL
Entre as espécies vegetais utilizadas pelo homem, diversas palmeiras
(Arecaceae) merecem destaque pelos múltiplos usos que podem apresentar, como
por exemplo, o “vinho” produzido dos frutos de açaí (Euterpe oleracea e E.
precatoria), patauá (Oenocarpus bataua) e bacaba (O. bacaba, O. mapora, O.
distichus), o palmito retirado do estipe da palmeira juçara (Euterpe edulis), da
palmeira-real australiana (Archontophoenix spp.) e da pupunha (Bactris gasipaes),
os frutos de tucumã (Astrocaryum aculeatum), pupunha, buriti (Mauritia flexuosa) e
coquinho-azedo (Butia capitata), o óleo extraídodos frutos de macaúba (Acrocomia
aculeata) e de dendê (Elaeis guiinensis), entre outros. Na Amazônia, apesar da
grande importância para as populações locais, ainda são escassos os
conhecimentos sobre o manejo e condições silviculturais adequadas para o
crescimento e desenvolvimento, desde a germinação das sementes, para a maioria
das espécies de palmeiras, incluido a pupunha.
A pupunha é cultivada, atualmente, desde Honduras, até o norte da Bolívia e
Brasil, sendo encontrada também em Trinidad, Jamaica, Porto Rico e Cuba
(Almeyda e Martin, 1980). Sua maior ocorrência é na Amazônia, principalmente na
parte ocidental, nos locais mais antigos situados próximos às margens dos principais
rios da região. É uma espécie domesticada, ocorrendo somente onde o homem a
tenha plantado e sua abundância depende de sua importância para a população
local (Clement, 1988). Na Amazônia Ocidental e no sul de América Central, por
exemplo, onde teve uma longa história de uso, pode ser muito abundante. O
processo de domesticação interferiu na tolerância ecológica ao ponto da espécie
requerer a interferência humana para se reproduzir (Clement, 1990).
A reprodução da pupunheira tem sido objeto de estudo há mais de um século
(Barbosa-Rodrigues, 1899). Porém, nos últimos 30 anos, com o aumento da
demanda pelo palmito de pupunha, mais estudos vêm sendo realizados a fim de se
obter uma melhor forma de propagação, bem como visando formas de conservação
in situ e ex situ desta espécie.
As sementes (diásporos) de pupunha apresentam forma globosa a elipsóide,
com endosperma de consistência dura, homogêneo e oleaginoso, que ocupa quase

2
todo o espaço interno da semente (Silva et al., 2006). A germinação é do tipo
adjacente ligulada, com a plântula se desenvolvendo junto à semente (Tomlinson,
1960; 1961). As sementes são recalcitrantes e perdem rapidamente a viabilidade
quando desidratadas (Mora-Urpi, 1979; Ferreira e Santos, 1992; Villalobos et al.,
1992; Carvalho e Muller, 1998; Bovi et al., 2004). A tolerância ao dessecamento
varia com o lote e com a progênie (Bovi et al., 2004).
As sementes de Bactris gasipaes germinam bem em temperaturas de 22 ºC a
30 ºC, mantendo a capacidade de germinar até 40 ºC. Temperaturas superiores a
esta são prejudiciais, pois causam a morte das sementes (Villalobos e Herrera,
1991). Em trabalhos visando acelerar a germinação por meio do aumento da
temperatura (40 ºC), não foram obtidos resultados satisfatórios (Villalobos e Herrera,
1991; Clement e Dudley, 1995). A germinação de sementes de diferentes progênies
varia de 0 a 100%, ocorrendo o processo de 38 a 133 dias (Bovi et al., 1994). A
semente apresenta dormência por períodos variáveis de um mês e meio a 14 meses
(Mora-Urpí, 1979).
A maturidade fisiológica das sementes é atingida cerca de um mês antes da
maturação do fruto; sua permanência na planta após a maturação aumenta
gradativamente a intensidade de dormência (Ferreira, 1996). Em frutos com longo
período de maturação (110 dias), a dormência pode ser reduzida antecipando-se a
colheita dos frutos bem como se retardando o processamento. O melhor resultado
foi alcançado com frutos coletados 70 dias após a antese e que passaram por um
período de 20 dias de pós-maturação (Ferreira e Ferraz, 2007). O efeito inibitório
que o endosperma exerce durante os estádios iniciais da maturação pode ser a
razão da baixa germinação das sementes, o que foi verificado em Bactris major
(Tzec-Simá et al., 2006). Sementes dessa espécie não germinaram após vários
meses, mas os resultados do teste do tetrazólio indicaram que os embriões
permaneceram viáveis. Além disso, quando os embriões foram isolados, a
germinação foi alta (90%), em meio de cultura, sem reguladores do crescimento.
Entre os substratos utilizados para a semeadura, a serragem de madeira e a
areia, ou mistura de ambos, forneceram bons resultados (Villalobos e Herrera,
1991). Para o crescimento inicial de plântulas são indicados a mistura dos substratos
terra, areia e esterco, nas mesmas proporções, e o Plantmax Hortaliças. Ótimos

3
resultados também são obtidos por meio da “semeadura” em sacos plásticos
fechados, sem substrato (Mora-Urpí, 1979). A imersão das sementes em soluções
de ácido giberélico por 1 hora, nas concentrações de 75 a 225 mg.L-1 aumenta a
germinação das sementes (acima de 40%). Sementes tratatas com etephon
(composto sintético que libera etileno), nas concentrações de 0,4 e 0,8%, por 6
horas, atingiram valores de germinação acima de 50% (Villalobos et al., 1992).
Utilizando a cultura de embriões zigóticos de pupunha, foi observado o
intumescimento do embrião e o desenvolvimento da lâmina cotiledonar após 7 dias
de inoculação. A emissão da bainha foliar, raiz primária e o completo
desenvolvimento do haustório puderam ser observados após 30 dias e aos 90 dias
foram obtidas plântulas completas. Embriões zigóticos de pupunha não apresentam
a radícula diferenciada, característica ligada à dormência embrionária (Steinmacher,
2005).
Apesar de vários trabalhos terem sido realizados visando a regularidade da
germinação e formação de plântulas e mudas em tempo reduzido, as causas da
dormência em sementes de pupunha são, até o momento, desconhecidas. A
dormência pode ser imposta pelo embrião, pelos envoltórios (tegumentos,
endosperma, etc) ou pela combinação de ambos os fatores, dependendo da espécie
(Bewley, 1997).
As sementes com dormência tegumentar têm geralmente um tegumento
impermeável ao oxigênio e, ou, à água ou resistência mecânica à protrusão da
radícula (Debeaujon et al., 2000). Porém, a dormência pode também ser causada
por algum produto químico inibidor presente na epiderme ou nas membranas
interiores adjacentes. Em circunstâncias naturais, estas sementes permanecem no
solo sem germinar até que as condições sejam favoráveis para permitir a penetração
da água, troca dos gases, ou neutralização de produtos químicos inibidores. Assim
como o tegumento, o endosperma também pode funcionar como uma barreira física
à entrada de água, barreira mecânica à protrusão da raiz primária ou conter
inibidores os que reduzem a absorção de oxigênio, afetando a germinação (Toole et
al., 1956; Baskin e Baskin, 1998).
A dormência embrionária pode ser: a) fisiológica, cujos mecanismos
responsáveis são inibidores químicos, resistência dos envoltórios ou potencial de

4
crescimento do embrião, foto equilíbrio ou fitocromo, balanço hormonal; b)
morfológica, causada pela imaturidade do embrião; e c) morfofisiológica, causada
pela dormência fisiológica de embriões imaturos (Carvalho, 2003; Baskin e Baskin,
2004). A dormência em embriões maduros pode estar localizada nos cotilédones ou
no eixo embrionário (Borghetti, 2004).
A dormência pode ser classificada, ainda em primária, quando se instala
durante a fase de desenvolvimento e maturação; e secundária, quando é instalada
após a dispersão (Cardoso, 2004). A dormência secundária pode se instalar em
sementes quiescentes ou em sementes com dormência primária.
Algumas espécies contêm inibidores químicos no fruto ou na semente que
impedem que a germinação ocorra. Entre estes inibidores, os compostos fenólicos
têm sido estudados por vários autores. A coloração marrom da testa é associada à
presença de compostos fenólicos e, consequentemente, com a germinação,
dormência e a longevidade das sementes (Debeaujon et al., 2000; Moïse et al.,
2005). Em algumas espécies, ácidos fenólicos (ácidos ferúlico, cafeico e cinâmico),
substâncias fenólicas como os polifenóis (taninos), flavonoides (quercetina)
presentes em frutos e sementes inibem a germinação das sementes (Baskin e
Baskin, 1998). Alguns autores têm observado que a presença de taninos no
tegumento controla a entrada de oxigênio no interior da semente, pois estes fixam o
O2 que a semente está absorvendo, impedindo sua chegada no interior da semente
(Dietrich, 1986), por isso, a presença desses compostos na semente pode impedir
ou retardar a germinação. Os compostos fenólicos mono-OH atuam em processos
que promovem o desenvolvimento aéreo enquanto que os derivados fenólicos bi-OH
estão envolvidos com a iniciação de raízes (George, 1996). Estas substâncias atuam
na degradação oxidativa do ácido indol acético (AIA). Por outro lado, alguns
compostos fenólicos bi-OH inibem a degradação oxidativa do AIA, tendo efeito
benéfico no enraizamento (Santiago et al., 2001).
Sementes de algumas espécies podem conter substâncias lipídicas
revestindo o tegumento, como ceras e suberinas, que impedem a entrada de água
em seu interior, impedindo a embebição do embrião (Laboriau, 1983). Alguns ácidos
graxos, presentes no interior das células do endosperma ou no embrião podem inibir
a germinação de embriões maduros (Babiano et al., 1984). Alguns tecidos de certas

5
plantas contêm ácidos graxos de cadeia curta em quantidades altas o suficiente para
afetar a dormência e o crescimento. A inibição da germinação aumenta com o
aumento da cadeia para ácidos graxos de até 9 carbonos, a partir desse número de
carbonos a inibição declina. Os ácidos graxos de 7 a 9 carbonos são mais efetivos
em reduzir a germinação, comparados aos de 6 e 10 (Ulbright et al., 1982). Em
sementes de aveia (Avena spp.), Berrie et al. (1979) verificaram que 37 dias após a
antese, as sementes de Avena sativa apresentaram 19% de germinação e 303 ng
de ácidos graxos por grão. A germinação aumentou com a maturação, assim como a
concentração os ácidos graxos. Contudo, houve diminuição do inibidor mais efetivo
(C9) da germinação a níveis insignificantes. Ácidos graxos de cadeia curta (C3 –C8)
inibiram tanto a germinação quanto o crescimento da raiz em alface, tendo a inibição
sido aumentada pelo aumento na concentração do ácido graxo. Houve também
aumento abrupto na inibição da germinação quando a cadeia foi aumentada de 7
para 8 carbonos (Ulbright et al., 1982).
Para algumas espécies de palmeiras, as causas da dormência foram
identificadas e métodos foram estabelecidos, a fim de superá-la. Sementes de Elaeis
guiinensis (dendê) tiveram aumento na porcentagem de germinação após serem
incubadas por 60 dias a 40 ºC (Martine et al., 2009). Sementes desta espécie
tiveram a performance na germinação diminuída quando o período de duração do
pré-tratamento foi aumentado para 80, 100 e 120 dias (Fondom e Etta, 2010).
Sementes de Astrocaryum aculeatum tiveram a germinação favorecida pela remoção
do endocarpo, embebição em água em temperatura ambiente por nove dias
(Ferreira e Gentil, 2006) e pela embebição em água à 40 ºC por quatro dias (Nazário
e Ferreira, 2010).
A germinação dos diásporos de macaúba (Acrocomia aculeata) é influenciada
pela presença do endocarpo e as sementes apresentam dormência fisiológica, que
pode ser superada pela remoção do tegumento opercular associada à imersão das
sementes em ácido giberélico na concentração de 2000 mg.l-1, por 24 horas,
repetindo-se o procedimento após 4 semanas (Ribeiro et al., 2011). Por outro lado, a
dormência das sementes de Butia capitata parece estar relacionada com a barreira
mecânica imposta pelos tecidos da semente que dificultam o desenvolvimento do
embrião, que pode ser superada pela retirada total do opérculo (Fior et al., 2011).

6
Sementes de Thrinax morrisii exibem dormência exógena causada exclusivamente
pelo tegumento duro, enquanto sementes de Sabal palmetto apresentam tanto
dormência exógena quanto fisiológica (Dewir et al., 2011). Para sementes de Bactris
gasipaes, as causas da dormência não foram identificadas, bem como métodos
eficientes para superá-la.
O presente trabalho foi dividido em quatro capítulos, com objetivos
específicos a cada um. No primeiro capítulo, fez-se um estudo anatômico e
histoquímicos das sementes de B. gasipaes, visando identificar estruturas e
compostos que possam atuar no mecanismo de dormência das sementes. No
segundo capítulo, estudou-se a relação entre a concentração de compostos
fenólicos nas sementes e a emergência de plântulas. No terceiro capítulo, foi feita a
caracterização do perfil de ácidos graxos presentes nas sementes e sua relação com
a emergência de plântulas. No quarto capítulo, estudou-se a dormência embrionária
nas sementes de pupunha.

7
OBJETIVOS
Objetivo geral
Avaliar características anatômicas, histoquímicas e fisiológicas e suas relações com mecanismos de dormência em sementes de Bactris gasipaes.
Objetivos específicos
Identificar características anatômicas e histoquímicas de sementes de B. gasipaes
associadas ao processo de dormência e germinação.
Quantificar compostos fenólicos em sementes de B. gasipaes de frutos em dois
estádios de maturação, correlacionando-os com a emergência de plântulas e
verificar o efeito da aplicação do extrato destas sementes sob a germinação de
sementes de Lactuca sativa;
Quantificar lipídios, ácidos graxos e o ponto de fusão dos lipídios em sementes de B.
gasipaes de frutos em diferentes estádios de maturação e correlacioná-los com a
emergência de plântulas;
Verificar se sementes de Bactris gasipaes possuem dormência embrionária.

8
Capítulo 1 __________________________________________________________________________________
Aspectos anatômicos e histoquímicos de sementes de pupunha
(Bactris gasipaes Kunth) (Elaborado de acordo com as normas da Revista Brasileira de Sementes)

9
ASPECTOS ANATÔMICOS E HISTOQUÍMICOS DE SEMENTES DE PUPUNHA (Bactris gasipaes KUNTH).
RESUMO- Bactris gasipaes (pupunha) é uma palmeira (Arecaceae) muito utilizada pelas
populações da Amazônia, cujas sementes apresentam dormência, de causa desconhecida, o
que representa dificuldade para produção uniforme de mudas. Este trabalho visou identificar
aspectos anatômicos e histoquímicos de sementes de pupunha que possam ter influência na
germinação. Foram feitos cortes histológicos de material fixado e incluso em resina, os quais
foram corados com azul de Toluidina para verificar características estruturais, avaliadas em
microscópio. Para identificação de metabólitos primários e secundários, foram feitos cortes
manuais e realizados testes histoquímicos para ácidos graxos, alcaloides, amido, compostos
fenólicos, lignina, lipídios, lipídios ácidos e neutros, pectinas, polissacarídeos, proteínas,
taninos e terpenoides. O tegumento é fino e em duas camadas, a externa mais espessa, com a
presença de feixes vasculares e a interna menos espessa, com células de formato irregular e
repletas de compostos fenólicos. O endosperma ocupa a maior parte da semente, é
esbranquiçado, com células nos formatos oblongo oboval e oval, com conteúdo de lipídios,
proteíns, pectinas e polissacarídeos. O embrião é pequeno em relação ao tamanho da semente,
cônico e vascularizado na porção distal. Na região proximal, em corte longitudinal, aparece a
plúmula, com três primórdios foliares. As células do embrião apresentam conteúdo de lipídios
ácidos, proteínas e polissacarídeos neutros na região de protrusão.
Palavras chave: palmeira, estrutura, metabólitos primários, metabólitos secundários.

10
ANATOMICAL AND HISTOCHEMICAL ASPECTS OF PEACH PALM (Bactris gasipaes
KUNTH) SEEDS
ABSTRACT- Bactris gasipaes (peach palm) is a palm (Arecaceae) widely used by people of
the Amazon, whose seeds show dormancy, cause unknown, which is difficult to produce
uniform plant. This study aimed to investigate aspects anatomical and histochemical seeds of
peach palm that may have influence on germination. Were made histological sections of fixed
material and even resin, which were stained with Toluidine blue to check the structural
characteristics and evaluated microscope. For identification of primary and secondary
metabolites cuts were made and hand made histochemical tests for fatty acids, alkalis, starch,
phenolic compounds, lignin, lipids, lipids acidic and neutral pectins, polysaccharides,
proteins, tannins and terpenoids. The seed coat is thin in two layers, external thickes with the
presence of vascular bundles and inner thinner, cells with irregularly shaped and
filled phenolic compounds. The endosperm occupies most of the seed and is white, oblong
cell formats oboval oval, with lipid, proteins, pectins polysaccharides content. The embryo
is small in relation to seed size, conical and vascularized in the distal portion. In the
region proximal, in longitudinal section, the plumule appears with three leaf primordia.
The lipid content of cells present acids, proteins andneutral polysaccharides in the region of
protrusion of the seed. The embryo cells have acid content of lipids, proteins and neutral
polysaccharides in the region of protrusion.
Key words: palm, structure, primary metabolites, secondary metabolites.

11
INTRODUÇÃO
As palmeiras pertencem à família Arecaceae, que compreende em torno de 183
gêneros e 2450 espécies, distribuídas principalmente nas regiões tropicais e subtropicais
(Johnson, 2010). A pupunheira (Bactris gasipaes) é uma espécie de palmeira de múltiplos
usos: as flores podem ser cozidas e usadas como condimento, as raizes têm ação vermicida e o
estipe fornece madeira de boa qualidade (Mora-Urpi et al., 1997; Jesus e Abreu, 2002).
Entretanto, tradicionalmente, a principal forma de utilização são os frutos cozidos, os quais
apresentam alto valor nutricional. Atualmente, a espécie tem sido plantada para produção de
palmito, pelas vantagens que apresenta em relação a outras palmeiras, como precocidade de
corte, alta produtividade, boa palatabilidade e perfilhamento abundante (Neves et al., 2005).
As sementes de várias espécies de palmeiras apresentam dormência, que pode ser
superada, em parte, utilizando-se métodos como a imersão em água, estratificação quente,
aplicação de giberelinas, entre outros (Martine et al., 2009; Fondom et al., 2010; Nazário ;
Ferreira, 2010; Ribeiro et al., 2011). Contudo, para a maior parte das espécies de palmeiras, as
causas da dormência são desconhecidas.
As sementes de Bactris gasipaes germinam de forma lenta e irregular, apresentam
graus de dormência diferenciados entre plantas e entre os estádios de maturação dos frutos
(Bovi et al., 1994; Ferreira, 2005), são recalcitrantes (Villalobos et al. 1992) e a tolerância ao
dessecamento varia com o lote e a origem (Bovi et al., 2004).
Entre os fatores que exercem influência na germinação e dormência das sementes
estão aspectos estruturais, como o número de células, o espessamento da parede celular de
seus envoltórios (Debeaujon et al., 2007) e a diferenciação e maturação do embrião (Hilhorst,
2007). A composição química das sementes também exerce influência sobre a germinação.

12
Produtos do metabolismo primário, como proteínas, lipídios e carboidratos são sintetizados
nas sementes como materiais de reserva a serem utilizados pelo embrião durante a germinação
e para a constituição de novas estruturas celulares. Contudo, entre estes compostos, podem
existir substâncias que atuam no controle da germinação, como alguns ácidos graxos nos
triacilgliceróis, e alguns açúcares (Ulbright et al., 1982; Buckeridge et al., 2004).
Os metabólitos secundários são divididos em três grandes grupos: terpenos, compostos
fenólicos e compostos nitrogenados. De uma forma geral, esses compostos são associados à
defesa nas plantas e, apesar de não apresentarem função aparente nos processos de
crescimento e desenvolvimento, alguns apresentam relação, negativa ou positiva, no processo
de germinção das sementes (Taiz e Zeiger, 2009).
O estudo anatômico em sementes pode contribuir para identificar barreiras mecânicas
e físicas à germinação e, ainda, para avaliar a diferenciação e maturação das estruturas do
embrião (Baskin et al., 2000; Cardoso, 2008). Informações sobre a estrutura de sementes de
Bactris gasipaes, tais como aspectos histoquímicos e anatômicos são importantes para o
desenvolvimento e aperfeiçoamento de técnicas de propagação in vitro, bem como para
identificar estruturas e compostos que possam impedir ou inibir a germinação de sementes e
embriões isolados.
O presente trabalho objetivou identificar aspectos anatômicos de sementes de Bactris
gasipaes e detectar a presença de substâncias ergásticas que possam influenciar na
germinação de sementes desta palmeira.

13
MATERIAL E MÉTODOS
As sementes foram coletadas em um plantio localizado no Centro de Pesquisa do
Instituto de Saúde e Biotecnologia da Universidade Federal do Amazonas (ISB/UFAM),
localizada na Estrada Coari Itapeua, no município Coari, Amazonas. O clima da região é
classificado como tropical úmido, tipo Afi pela classificação de Köppen, apresentando
precipitação média anual de 2.250 mm e temperatura média anual é de 26 ºC (Vieira e Santos,
1987). Em 14 plantas, no mesmo momento, foram coletados cachos com frutos maduros,
apresentando epicarpo completamente amarelo ou vermelho.
Após coleta, os frutos foram retirados das infrutescências, partidos um a um com
auxílio de uma faca, e retirada a semente. A limpeza consistiu da fricção das sementes sobre
uma peneira em água corrente e retirada dos resíduos remanescentes com a ponta de uma
faca. Trinta sementes de cada infrutescência foram fixadas em FAA 50%, por 72 horas e
armazenadas em álcool 70%.
Para a inclusão das sementes em historresina, estas foram fragmentadas em pedaços
menores, os quais foram desidratados em série etílica (70%-95%) e incluídos em 2-
hidroxietil-metacrilato (Historesin Leica®, preparada conforme as instruções do fabricante).
Os fragmentos foram seccionados em micrótomo rotativo (5 µm). Os cortes para análise
estrutural foram corados com azul de toluidina (O’Brien et al., 1964) e lâminas permanentes
foram montadas em Permount.
Os testes histoquímicos para os embriões foram feitos em cortes inclusos em resina,
em micrótomo rotativo, e para o tegumento e o endosperma foram confeccionados cortes
manuais com lâmina de barbear, com auxílio de um micrótomo manual, em material apenas
fixado. Os testes histoquímicos realizados constam na Tabela 1.

14
A análise do material e as fotomicrografias foram feitas em microscópio óptico, marca
Zeiss modelo Primo Star, com câmera fotográfica digital marca Canon modelo Powershot A
650 IS, acoplada.
RESULTADOS E DISCUSSÃO
As sementes de Bactris gasipaes são bitegumentadas (Figura 1-A e B). O tegumento
externo é pluriestratificado, composto por aproximadamente 10 camadas de células de
formato que varia do arredondado ao oboval. Este tecido é vascularizado (Figura 1-C). Feixes
vasculares foram observados no tegumento de Oenocarpus minor (Mendonça et al., 2008),
porém nesta espécie os feixes são mais alongados que os observados em B. gasipaes. Em
Euterpe precatoria, o tegumento, delgado, apresenta três estratos e somente no estrato externo
são observados feixes vasculares (Aguiar e Mendonça, 2003).
Foi observada a presença de terpenóides, na região do tegumento externo (Figura 3-
A). Compostos da família dos terpenos podem ter efeito de inibição da germinação com o
aumento da concentração e do tempo de exposição das sementes, conforme observado por
Dudai et al. (1999). Os terpenos presentes em folhas, flores e frutos de Tagetes minuta são
capazes de inibir a germinação de sementes de algumas espécies competidoras (López et al.,
2009). Em palmeiras, não existem estudos estabelecendo relações entre os terpenos e a
germinação.
A camada externa do tegumento é repleta de lipídios ácidos (Figura 3-H). A presença
de lipídios (Figura 3-F) no tegumento sugere que a semente seja revestida de cera ou suberina.
Estas substâncias são responsáveis pela regulação da entrada e saída de água no interior da
semente e, portanto, podem causar impedimento à germinação (Laboriau, 1983; Taiz e Zeiger,

15
2009). No teste histoquímico que utiliza sulfato azul Nilo, a coloração diferencial deve-se à
presença de duas substâncias: uma oxazona de cor vermelha reage com os lipídios neutros, ou
gorduras, e uma oxizona de cor azul que reage com os grupos carbonilos dos ácidos graxos
livres e com resíduos do ácido ortofosfórico dos fosfolipídios (Cain, 1947). A síntese de
triacilglicerol está diretamente relacionada à presença da enzima diacilglicerol aciltransferase
(DAGAT), pois quanto maior a quantidade desta enzima nos tecidos, maior a produção de
ácidos graxos (Ohlrogge e Jaworski, 1997).
A camada interna do tegumento é delgada e consiste de duas a quatro camadas de
células de formato arredondado (Figura 1-A e B). Os compostos fenólicos estão presentes em
todas as camadas do tegumento com maior intensidade nas células que compõem o tegumento
interno, evidenciado pela coloração negra para o reagente Cloreto Férrico III. (Figura 3-B). O
teste com a vanilina clorídrica confirmou que os compostos fenólicos presente na camada
interna do tegumento são taninos (Figura 3-C). A função da testa contendo inibidores
solúveis em água, como alguns compostos fenólicos, é prevenir a germinação de embriões
não dormentes até que ocorram condições ambientais favoráveis (Weber et al., 2005). Além
da prevenção da germinação, alguns compostos desta natureza, quando presentes nos
envoltórios da semente, são capazes de retardar o crescimento de plântulas (Möise et al.,
2005). A espessura fina do tegumento não oferece resistência mecânica às sementes,
possivelmente devido à presença do endocarpo duro nos frutos desta espécie. A falta de
resistência mecânica nos tegumentos das sementes de Acrocomia aculeata também foi
atribuída à presença de endocarpo esclerificado (Moura et al., 2010).
As proteínas estão presentes em todas as camadas de células do tegumento (Figura 4-
A e B). Observa-se que as pectinas, nas sementes de Bactris gasipaes estão presentes na

16
estrutura da parede celular dos tegumentos (figura 4-C). A epiderme apresenta uma camada
de células isodiamétricas, retangulares, parede periclinal alongada.
O endosperma apresenta consistência sólida, é esbranquiçado, homogêneo, composto
de células heterodimensionais nos formatos oblongo, oval e oboval, no sentido longitudinal e
arredondadas no sentido transversal, e preenche a maior parte do interior das sementes (Figura
1-D). Em sementes das palmeiras Oenocarpus minor (Mendonça et al., 2008) e Euterpe
precatoria (Aguiar e Mendonça, 2003) foi observado endosperma homogêneo, o qual também
ocupa quase todo o interior da semente. O endosperma, em monocotiledôneas, é o tecido de
reserva para o embrião durante a formação da semente, germinação e estabelecimento da
plântula (Davide e Silva, 2008). O teste para detecção de lipídios (Figura 3-F) foi positivo no
conteúdo das células do endosperma; foram observados tanto lipídios ácidos, quanto lipídios
neutros (Figura 3-G e H). Estes se encontram no interior das células organizados em corpos
oleicos; ácidos graxos livres foram observados no interior das células do endosperma (Figura
3-D e E). Observou-se uma grande concentração de corpos lipídicos no endosperma. Pode-se
dizer que este é o principal composto de reserva nas sementes desta espécie, característica
típica de palmeiras oleaginosas. Nas palmeiras Acrocomia aculeata (Moura et al., 2010) e
Euterpe edulis (Panza et al., 2004) os lipídios também aparecem estocados em corpos
lipídicos e em grande abundância. Os lipídios presentes no endosperma constituem reserva,
que será utilizada tanto na fase de germinação, quanto no desenvolvimento da plântula
(Buckeridge et al., 2004). Proteínas foram observadas nas células do endosperma (Figuras 4-
A e B) estocadas na forma de corpos proteicos. Em sementes de Euterpe oleracea, Gonçalves
et al. (2010) sugerem que a maior fonte de nitrogênio fornecido ao embrião para a germinação
venha das proteínas, que são degradadas dos tecidos de reserva. As pectinas e os
polissacarídeos estão presentes no endosperma, como conteúdo celular (Figura 4-C e D). Este

17
mesmo resultado foi observado em Acrocomia aculeata (Moura et al., 2010). Pectinas são
polissacarídeos de parede celular que tipicamente contém açúcares ácidos e neutros e,
frequentemente, contém cálcio como componente estrutural. Modificações nas pectinas
podem alterar sua conformação e ligação na parede. Quando a pectina é secretada para dentro
da parede, os grupos éster podem ser removidos da parede, o que aumenta sua capacidade de
formar um gel rígido (Taiz e Zeiger, 2009). Buckerige et al. (2000) sugerem que o galactano
presente como uma cadeia lateral da pectina seja um polissacarídeo da parede celular primária
que foi ampliado e adaptado para o armazenamento e que será mobilizado na germinação.
O embrião é lateral, cônico, coloração homogênea, composto por uma região
proximal, formada pelo eixo embrionário e região distal constituída pelo cotilédone (Figura 1-
E). Em algumas espécies de palmeiras, assim como em B. gasipaes, as regiões basal e
proximal são morfologicamente distintas. Em Oenocarpus minor a região proximal do
embrião, voltada ao poro germinativo, tem cor amarelo-pálida e região distal, amarelo clara
(Oliveira et al., 2010). Em Cocos nucifera o embrião é dividido por
uma pequena constrição na porção mediana e o eixo plúmula-raiz é encontrado na parte apical
do cotilédone (Sreekumari Kartha, 1981).
Na região proximal, em corte longitudinal, as células do embrião são ovais, a epiderme
é composta de células menores e arredondadas (Figura 1-F), o eixo embrionário tem
localização apical, constituído pelas células que formam a plúmula, composta por três
primórdios foliares (eofilo e duas bainhas cotiledonares) e pelas células precursoras da
radícula oposta a essa região (180º), caracterizadas por um aglomerado de células menores
(Figura 1-G e H). A região distal, em corte transversal, apresenta células heterodimensionais,
variando de redondas, retangulares a elipsoides (Figura 2-A e B). As células da epiderme
variam em forma e o tecido cotiledonar é formado por parênquima com muito conteúdo

18
protoplasmático, no qual são observados feixes vasculares concêntricos (Figura 2-A e B) que
vão do eixo embrionário ao extremo da região distal. A presença deste conjunto de estruturas
evidencia que no momento da dispersão o embrião apresenta-se diferenciado e maduro,
indício de que não ocorre dormência morfológica nas sementes desta espécie (Baskin ;
Baskin, 2004). Esta informação concorda com Ferreira (2005), que afirma que sementes de
Bactris gasipaes encontram-se fisiologicamente maduras aproximadamente um mês antes da
dispersão dos frutos.
Os testes histoquímicos para detecção de terpenoides, compostos fenólicos, taninos,
ácidos graxos livres, amido e lignina foram negativos. O teste para detecção de lipídios reagiu
no interior das células do embrião (Figura 5-B), os quais são encontrados na forma de corpos
oleicos (Figura 5-A). Corpos proteicos foram observados no interior das células do embrião
(Figura 5-C). De uma forma geral, a síntese de algumas proteínas é maior em sementes
dormentes do que em sementes não dormentes, apesar disso, nenhuma relação pode ser
estabelecida entre a presença ou ausência de alguma proteína e a dormência (Bewley, 1997).
Polissacarídeos foram observados apenas na zona de protrusão do botão germinativo (Figura
5-D), demonstrando que nesta região ocorre síntese de açúcares, possivelmente com a função
de fornecer energia ao início do processo germinativo.
CONCLUSÃO
O tegumento das sementes de B. gasipaes é fino e em duas camadas, a externa mais
espessa, com a presença de feixes vasculares, conteúdo de terpenóides e lipídios ácidos e a
interna menos espessa, com células de formato irregular e repletas de compostos fenólicos.

19
O endosperma ocupa a maior parte da semente, é esbranquiçado, com células nos
formatos oblongo oboval e oval, com conteúdo de lipídios, proteíns, pectinas e
polissacarídeos.
O embrião é pequeno em relação ao tamanho da semente, cônico e vascularizado na
porção distal. Na região proximal, em corte longitudinal, aparece a plúmula, com três
primórdios foliares, evidência de que o embrião no momento da dispersão encontra-se
diferenciado e maduro, indicando que não ocorre dormência morfológica nas sementes desta
espécie. As células apresentam conteúdo de lipídios ácidos, proteínas e polissacarídeos
neutros na região de protrusão do botão germinativo.
Entre os compostos identificados, os compostos fenólicos e os terpenóides são capazes
de inibir a germinação de sementes em outras espécies vegetais e as pectinas podem
influenciar a germinação devido ao enrigecimento da parece celular.

20
REFERÊNCIAS BIBLIOGRÁFICAS
AGUIAR, M.O.; MENDONÇA, M.S. Morfo-anatomia da semente de Euterpe
precatoria Mart. (Palmae). Revista brasileira de sementes. 25: 37-42. 2003.
www.scielo.br/pdf/rbs/v25n1/19628.pdf
BASKIN, J.M; BASKIN, C.C.; LI, X. Taxonomy, anatomy and evolution of physical
dormancy in seeds. Plant species biology, 15: 139-152. 2000.
http://www.aseanbiodiversity.info/Abstract/51010592.pdf
BASKIN, J.M; BASKIN, C.C. A classification system for seed dormancy. Seed science
research, 14: 1-16. 2004.
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=704528
BEWLEY, J.D. Seed dormancy and germination. The plant cell, 9: 1055-1066. 1997.
BOVI, M.L.A.; SPIERING, S.H.; MARTINS, A.L.D.; PIZZINATO, M.A.; LOURENÇÃO,
A.L.; CHAVES FLORES, W.B. Seed germination of progênies of Bactris gasipaes:
percentage, speed and duration. Acta horticulturae, 360(8):283-289. 1994.
BOVI, M.L.A.; MARTINS, C.C.; SPIERING, S.H. Desidratação de sementes de quatro lotes
de pupunheira: efeitos sobre a germinação e o vigor. Horticultura brasileira, 22(1):109–112.
2004.
BUCKERIDGE, S., M.; TINÉ, M. A. S.; SANTOS, H. P.; LIMA, D. U. Polissacarídeos de
reserva de parede celular em sementes, estrutura, metabolismo, função e aspectos ecológicos.
Revista brasileira de fisiologia vegetal, 12: 137-162. 2000.

21
BUCKERIDGE, M.S.; AIDAR, M.P.M.; SANTOS, H.P.; TINÉ, M.A.S.. Acúmulo de
Reservas. In: Ferreira, A.G.; Borghetti, F. Germinação: do básico ao aplicado. Porto
Alegre: Artmed, 2004.
CAIN, A.J. The use of Nile Blue in the examination of lipoids. Quarterly Journal of
Microscopical Science, 88: 383-392. 1947.
CARDOSO, V.J.M. Germinação e dormência. In. Kerbauy, G.B. Fisiologia vegetal. 384-407.
2 ed. Rio de Janeiro, Guanabara Koogan. 2008.
DAVIDE, A.C.; SILVA, E.A.A. Produção de sementes e mudas de espécies florestais. 1
ed. Lavras, Ed UFLA, 2008. 175 p.
DEBEAUJON, I; LEPINIEC, L; POURCEL, L; ROUTABOUL, J-M. Seed coat development
and dormancy. In: Bradford, K; Nonogaki, H. Seed development, dormancy, and
germination, 25-49, Blackwell Publishing, Oxford, United Kingdom, 2007.
DUDAI , N. ; POLJAKOF F -MAYB E R , A. ; MAYE R , A.M. ; PUTIEVSKY, E.;
LERNER, H.R. Essential oils as allelochemicals and their potential use as bioherbicides.
Journal of chemical ecology, 25: 1079-1089. 1999.
FEDER, N.; O’BRIEN, T. P. Plant microtechnique: some principles and new methods.
American journal of botany, 55 (1): 123-142. 1968.
FERREIRA, S.A.N. Pupunha: Bactris gasipaes Kunth (Arecaceae). Fascículo 5. 12 p. In:
Ferraz, I.D.K.; Camargo, J.L.C. (Eds) Manual de Sementes da Amazônia. Fascículo 5,
INPA, Manaus, AM, Brasil. 12 p. 2005. .
www.inpa.gov.br/.../fasciculo5_Bactis%20gasipaes_Pupunha-net.pdf

22
FONDOM, N.Y.; ETTA, C.E.; MIH, A.M. Breaking seed dormancy: revisiting heat-treatment
duration on germination and subsequent seedling growth of oil palm (EIaeis guineensis Jacq.)
progenies. Journal of agricultural science. 2(2):101-110. 2010.
www.ccsenet.org/journal/index.php/jas/article/download/4402/4927
GANTER, P., JOLLÉS, G. Histolgie normale et pathologique. v.1 e 2. Paris: Gauthier-
Villars, 1969, 1970.
GONÇALVES, J.F.C.; LIMA, R.B.S.; FERNANDES, A.V.; BORGES, E.E.L.;
BUCKERIDGE, M.S. Physiological and biochemical characterization of the assai palm
(Euterpe oleracea Mart.) during seed germination and seedling growth under aerobic and
anaerobic conditions. Revista árvore, 34(6).1045-1053. 2010.
www.scielo.br/pdf/rarv/v34n6/a10v34n6.pdf
HILHORST, H. W. M. Definitions and hypotheses of seed dormancy. In: Bradford, K;
Nonogaki, H. Seed development, dormancy, and germination, 50-71, Blackwell
Publishing, Oxford, United Kingdom. 2007.
JESUS, M.A.; ABREU, R.L.S. 2002. Durabilidade natural da madeira de pupunha (Bactris
gasipaes Kunth). I. Fungos. Acta amazonica, 32(4): 663-675. acta.inpa.gov.br/fasciculos/32-
4/PDF/v32n4a11.pdf
JOHANSEN, D.A. Plant microtechnique. New York: McGraw-Hill Book Co. Inc., 1940.
423p.
JOHNSON. D.V. Tropical palms. Series: Non-wood forest products 10. Food and
Agriculture Organization, Rome, . 2010. 166 p.

23
LABORIAU, L.F.G. A germinação de sementes. OEA, Washinton, 1983,174 p.
LÓPEZ, M.L. BONZANI, N.E.; ZYGADLO, J.A. Allelopathic potential of Tagetes minuta
terpenes by a chemical, anatomical and phytotoxic approach. Biochemical systematics and
ecology, 36: 882–890. 2009. http://www.biol.unlp.edu.ar/biologiavegetal/seminario4-
material4.pdf
MACE, M.E.; HOWELL, C.R. Histochemistry and identification of condensed tannin
precursor in roots of cotton seedlings. Canadian journal of botany. 52: 2423-2426, 1974.
MARTINE, B.M; LAURENT, K.K.; PIERRE, B.J., EUGÉNE, K.K.; HILAIRE, K.T.;
JUSTIN, K.Y. Effect of storage and heat treatments on the germination of oil palm (Elaeis
guineensis Jacq.) seed. African journal of agricultural research. 4 (10)- 931-937. 2009.
http://www.academicjournals.org/ajar/pdf/pdf%202009/Oct/Martine%20et%20al.pdf
MENDONÇA, M.S.; OLIVEIRA, A.B.; ARAÚJO, M.G.P. ARAÚJO, L.M. Morfo-anatomia
do fruto e semente de Oenocarpus minor Mart.(Arecaceae). Revista brasileira de sementes,
30(1): 90-95. 2008. http://www.scielo.br/pdf/rbs/v30n1/a12v30n1.pdf
MOÏSE, J.A. HAN, S; GUDYNAITE-SAVITCH, L.; JOHNSON, D.A.; MIKI, B.L.A. Seed
coats: structure, development, composition and biotechnology. In vitro cellular development
biology-plant, 41:620-644. 2005. http://www.springerlink.com/content/q56418t500843070/
MORA-URPÍ, J.; WEBER; J.C.; CLEMENT, C.R. Peach palm. Bactris gasipaes Kunth
{Pupunha}. Promoting the conservation and use of underutilized and neglected crops. 20.
Institute of Plant Genetics and Crop Plant Research - IPK, Gatersleben/ International Plant

24
Genetic Resources Institute - IPGRI, Rome, 83p. 1997.
http://www.icraf.com/downloads/publications/PDFs/b10186.pdf
MOURA, E.F.; VENTRELLA, M.C.; MOTOIKE, S.Y. Anatomy, histochemistry and
ultrastructure of seed and somatic embryo of Acrocomia aculeata (Arecaceae). Scientia
agricola, 67(4):399-407. 2010. http://www.scielo.br/pdf/sa/v67n4/v67n4a04.pdf
NAZÁRIO, P.; FERREIRA, S.A.N. Emergência de plântulas de Astrocaryum aculeatum G.
Mey. em função da temperatura e do período de embebição das sementes. Acta amazonica,
40(1):165-170. 2010. http://www.scielo.br/pdf/aa/v40n1/v40n1a21.pdf
NEVES, E.J.M. SANTOS, A.J.; LAVORANTI, A.J. MARTINS, E.G. Produção de palmito
de pupunheira sob diferentes densidades de plantio. Pesquisa florestal brasileira, 51:57-73.
2005. http://www.cnpf.embrapa.br/pfb/index.php/pfb/article/view/227/178
O’BRIEN, T. P., FEDER, N., MCCULLY, M. E. Polychromatic staining of plant cell walls
by toluidine blue O. Protoplasma, 59: 368-373, 1964.
OHLROGGE, J.B.; JAWORSKI, J.G. Regulation of fatty acid synthesis. Annual review of
plant physiology plant molecular biology, 48: 109–136. 1997.
OLIVEIRA, A.B.; MENDONCA, M.S.; ARAUJO, M.G.P. Aspectos anatômicos do embrião
e desenvolvimento inicial de Oenocarpus minor Mart: uma palmeira da Amazônia. Acta
Botânica Brasílica. 24(1): 20-24. 2010. www.scielo.br/pdf/abb/v24n1/03.pdf
PANZA, V; LÁINEZ, V; MALDONADO, S. Seed structure and histochemistry in the palm
Euterpe edulis. Botanical journal of the Linnean society, 145: 445–453. 2004.

25
PEARSE, A.G.E. Histochemistry theoretical and applied. v.2. 4.ed. Longman Group
Limited, 1980.
RIBEIRO, L.M., SOUZA, P.P., RODRIGUES JR., A.G., OLIVEIRA, T.G.S. AND GARCIA,
Q.S. Overcoming dormancy in macaw palm diaspores, a tropical species with potential for
use as bio-fuel. Seed science and technology, 39: 303-317. 2011.
SREEKUMARI KARTA, A . Embryo of coconut and its germination. Journal of plant
crops, 9: 125-127, 1981.
TAIZ, L.; ZEIGER, E. 2009. Fisiologia Vegetal. 4 ed. Porto Alegre, Artmed. 848 p.
ULBRIGHT, C.E.; PICKARD, B.G.; WARNER. J.F. Effects of short chain fatty acids on
radicle emergence and root growth in lettuce. Plant, cell and environment, 5(4): 293-301.
1982.
VIDAL, B.C. Acid glycosaminoglycans and endochondral ossification:
microespectrophotometric evaluation and macromolecular orientation. Cell and molecular
biology, 22: 45-64, 1977.
VIEIRA, L.S.; SANTOS, P.C.T.C. Amazônia: seus solos e outros recursos naturais. São
Paulo: Ceres, 416p. 1987.
VILLALOBOS, R.; HERRERA, J; GUEVARA, E. Germinación de la semilla de pejibaye
(Bactris gasipaes) II. Ruptura del reposo. Agronomía costarricense, 16(1): 61-68. 1992.
www.mag.go.cr/rev_agr/v16n01_061.pdf

26
WEBER, H.; BORISJUK, L.; WOBUS, U. Molecular physiology of legume seed
development. Annual review of plant biology, 56: 253-279. 2005.
www.annualreviews.org/doi/full/10.../annurev.arplant.56.032604.144201

27
TABELA 1. Compostos analisados, reagentes utilizados, coloração da reação positiva e
referência do teste histoquímico.
Compostos Reagente Coloração Referência
Terpenoides com
grupo carbonilo
2,4-dinitrofennilhidrazina Vermelho alaranjado Ganter e Jollès
(1969; 1970)
Compostos
fenólicos
Cloreto de ferro III Verde intenso, púrpura,
azul ou negro
Johansen
(1940)
Taninos Vanilina clorídrica Vermelho Mace e Howell
(1974)
Ácidos graxos Acetato de cobre/ácido
rubeânico
Verde-escuro Ganter e Jollès
(1970)
Lipídios Sudam IV Vermelho Pearse (1980)
Lipídios ácidos e
neutros
Sulfato azul do Nilo Lipídios neutros – rosa
Lipídios ácidos- azul
Cain (1947)
Proteínas Xilidine Ponceau Vermelho Vidal (1977)
Pectinas Vermelho de rutênio Rosa Johansen
(1940)
Polissacarídeos
neutros
PAS Rosa Feder e
O’Brien (1968)
Lignina Floroglucinol Vermelho Johansen
(1940)

28
TABELA 2. Resultados dos testes histoquímicos aplicados em cortes histológicos de
sementes de Bactris gasipaes.
Composto Tegumentos Endosperma Embrião
Terpenoides com grupo
carbonilo
+ - -
Compostos fenólicos + - -
Taninos + - -
Ácidos graxos - + -
Lipídios + + +
Lipídios ácidos e neutros + (ácidos)
- (neutros)
+ (ácidos)
+ (neutros)
+ (ácidos)
- (neutros)
Proteínas + + +
Pectinas - + +
Polissacarídeos neutros - + +
Amido - - -
Lignina - - -

29
FIGURA 1. Aspectos estruturais do tegumento (A, B e C), do endosperma (D) e do embrião
(E, F, G e H) de Bactris gasipaes. tg - tegumento; te - tegumento externo; ti -
tegumento interno; ct - conteúdo tanífero; en - endosperma; fv - feixe vascular; pf
- primórdios foliares; pl - plúmula; pr - primórdio da raiz.

30
FIGURA 2. Aspectos estruturais do embrião de Bactris gasipaes. fv - feixe vascular; co -
tecido cotiledonar.

31
FIGURA 3. Testes histoquímicos no tegumento e no endosperma de sementes de Bactris
gasipaes detectando a presença de terpenoides (A), compostos fenólicos (B),
taninos (C), ácidos graxos (D e E), lipídios (F), lipídios ácidos e neutros (G e H).

32
FIGURA 4. Testes histoquímicos no tegumento e no endosperma de sementes de Bactris
gasipaes detectando a presença de proteínas (A e B), pectinas (C) e
polissacarídeos (D).

33
FIGURA 5. Testes histoquímicos no embrião de sementes de Bactris gasipaes detectando a
presença de lipídios ácidos e neutros (A), lipídios totais (B), proteínas (C) e
polissacarídeos (D).

34
Capítulo 2 ___________________________________________________________________
Compostos fenólicos e dormência em sementes de Bactris gasipaes Kunth em função da maturação dos frutos (Elaborado de acordo com as normas da Revista Brasileira de Sementes)

35
COMPOSTOS FENÓLICOS E DORMÊNCIA EM SEMENTES DE
Bactris gasipaes KUNTH EM FUNÇÃO DA MATURAÇÃO DOS FRUTOS
RESUMO- Bactris gasipaes, espécie da família Arecaceae, conhecida vulgarmente como
pupunha, possui sementes que apresentam dormência cuja intensidade pode variar com a
planta matriz e com a maturação dos frutos. Objetivou-se, neste estudo, avaliar a emergência
de plântulas advindas de sementes de frutos em dois estádios de maturação, de diferentes
plantas, correlacionando-a com o teor de compostos fenólicos e testar o efeito do extrato
dessas sementes na germinação de sementes de Lactuca sativa. A emergência, índice de
velocidade de emergência, tempo médio de emergência, coeficiente de uniformidade de
emergência, índice de sincronização, velocidade média de emergência, valor de emergência e
coeficiente de velocidade de emergência não variaram significativamente entre sementes
obtidas de frutos imaturos e maduros. Entretanto, as variações dessas mesmas variáveis foram
maiores entre plantas. Sementes maduras de pupunha apresentam maior concentração de
compostos fenólicos que as imaturas e o acúmulo desses compostos é um dos fatores
responsáveis pela redução da germinação em sementes de pupunha. O extrato fenólico das
sementes desta espécie foi capaz de retardar e diminuir a germinação de sementes de alface e
esta apresentou correlação significativa com a emergência de plântulas de pupunha.
Termos para indexação: pupunha, emergência, inibidor da germinação.

36
PHENOLIC COMPOUNDS AND SEED DORMANCY IN Bactris gasipaes KUNTH AS A
FUNCTION OF FRUIT MATURITY.
ABSTRACT- Bactris gasipaes, species of the family Arecaceae, commonly known as peach-
palm, which has seeds present dormancy whose intensity can vary with the mother plant, and
fruit ripening. The objective of this study was to evaluate the emergence of seedlings arising
from seeds of fruits in two maturation stages in different plants, correlating it with the content
of phenolic compounds and test the effect of the extract of these seeds in the germination of
Lactuca sativa. The emergence, emergence rate index, mean emergence time, coefficient of
uniformity of emergence, synchronization index, average speed of emergency, emergency and
value of coefficient of velocity of emergence did not vary significantly from seeds obtained
from ripe and unripe fruits. However, variations of these variables were higher among plants.
Seeds mature peach palm have higher concentration of phenolic compounds in the immature
seeds and the accumulation of these compounds is one of the factors responsible for the
reduction of germination in seeds of peach-palm. The phenolic extract of the seeds of this
species was able to reduce and delay germination and emergence of lettuce seeds and that
emergency correlated significantly with the emergence of the peach palm seedlings.
Index terms: peach-palm, emergence, germination inhibitor

37
INTRODUÇÃO
As palmeiras (Arecaceae) constituem uma das mais importantes famílias de plantas
nativas utilizadas pelo homem na Amazônia, tendo o gênero Bactris como um dos mais
diversificados da bacia amazônica com, no mínimo, 30 espécies (Goulding e Smith, 2007). A
pupunha (Bactris gasipaes) é uma palmeira domesticada, portanto, sua abundância depende
de sua importância para a população local (Clement, 1988). A pupunha foi um cultivo
alimentar importante na economia de subsistência dos primeiros povos da Amazônia.
Atualmente, a economia de mercado da região demanda outros produtos, outras formas de
apresentação ou maior uniformidade de qualidade (Clement et al., 2003). A espécie tem sido
demandada principalmente para a produção de palmito, cujo mercado nacional absorve quase
a totalidade da produção, sendo o maior consumidor do mundo (Johr, 1999). Originalmente
este mercado foi ocupado pela palmeira Euterpe edulis, da Floresta Atlântica, que foi
praticamente dizimada de seu ambiente natural pelo extrativismo do palmito.
Como a maior parte das espécies de palmeiras, a principal forma de propagação da
pupunha é via sementes, que apresentam dormência, cujas causas não são ainda
compreendidas. A germinação e a dormência das sementes de pupunha são variáveis de planta
para planta. A germinação pode variar de 0 a 100% (Bovi et al., 1994) e o tempo para que a
germinação ocorra varia 90 dias a 14 meses (Mora-Urpí et al., 1997). As variações no tempo e
na velocidade de germinação estão relacionadas com a maturação fisiológica das sementes,
que é atingida cerca de um mês antes da maturação do fruto e a permanência na planta após a
maturação aumenta gradativamente a intensidade de dormência (Ferreira, 2005).
Apesar dos vários estudos realizados relacionados à germinação de sementes de
pupunha, não se sabe ao certo as causas envolvidas na dormência. Embora a dormência

38
tegumentar possa ocorrer pela presença de tegumento impermeável ao oxigênio e, ou, à água
ou resistência mecânica à protrusão da raiz primária (Debeaujon et al., 2000), a dormência
pode ser causada por algum inibidor presente no tegumento, endosperma ou mesmo no eixo
embrionário, os quais limitam a concentração de oxigênio necessária para manter o
metabolismo. Entre os inibidores da germinação conhecidos, encontram-se alguns compostos
fenólicos, como os taninos, que são capazes de retardar ou impedir a germinação (Baskin e
Baskin, 1998).
Esse estudo visou verificar a existência de relação entre a concentração de compostos
fenólicos em sementes de pupunha, de diferentes plantas, de frutos em dois estádios de
maturação com sua emergência e dormência, bem como avaliar o efeito da aplicação do
extrato das sementes sob a germinação de sementes de alface (Lactuca sativa).
MATERIAL E MÉTODOS
Emergência de plântulas de Bactris gasipaes em função da maturação dos frutos
As sementes foram coletadas em um plantio localizado no Centro de Pesquisa do
Instituto de Saúde e Biotecnologia da Universidade Federal do Amazonas (ISB/UFAM),
localizada na Estrada Coari-Itapeua, no município Coari, Amazonas. O clima da região é
classificado como tropical úmido, tipo Afi pela classificação de Köppen, apresentando
precipitação média anual de 2.250 mm e temperatura média anual de aproximadamente 26 ºC
(Vieira e Santos, 1987).
Em 14 plantas, no mesmo momento, foram coletados dois cachos de cada indivíduo
cada um com uma das seguintes características: a) Frutos imaturos: cachos com mais de 80%
dos frutos com epicarpo apresentando mudança de cor, na porção mediana, de verde para

39
amarelo ou alaranjado; b) Frutos maduros: cachos com mais de 80% dos frutos apresentando
epicarpo completamente amarelo ou vermelho.
Após coleta, os frutos foram retirados das infrutescências, partidos um a um com
auxílio de uma faca, e retirada a semente. Após retirada dos resíduos com a ponta de uma
faca, foi feita a determinação o teor de água, para cada material (sementes dos cachos das
diferentes plantas e estádios de maturação dos frutos), com duas repetições de cinco sementes,
em estufa a 105± 3 °C por 24 horas (Brasil, 2009).
As demais sementes foram colocadas em um recipiente com água, eliminando-se as
que flutuaram (Brasil, 2009). A limpeza consistiu da fricção das sementes sobre uma peneira
em água corrente e retirada dos resíduos remanescentes com a ponta de uma faca. A
semeadura foi feita em viveiro, com temperatura mínima média de 24,6 °C e máxima de 36,1
°C, em caixas plásticas contendo uma mistura de serragem e areia na proporção volumétrica
de 1:1, sendo semeadas 50 sementes para cada planta e estádio de maturação dos frutos.
Foram feitas avaliações da emergência a cada cinco dias, durante cinco meses, e o critério
utilizado foi a presença da primeira folha bífida expandida.
A partir dos dados de emergência, foram calculados: a emergência em porcentagem, o
índice de velocidade de emergência (IVE), os tempos inicial, médio e final de emergência
(TIE, TME e TFE), o coeficiente de uniformidade de emergência (CUE), a velocidade média
de emergência (VME), o valor de emergência (VE), o índice de sincronização (Z), o
coeficiente de velocidade de emergência (CVE), conforme Santana e Ranal (2004). Ao
término do experimento, foram quantificadas as sementes germinadas e não emergidas (não
apresentavam a primeira folha bífida expandida), além das sementes dormentes e mortas, pelo
teste de corte (Brasil, 2009). O delineamento estatístico utilizado foi em blocos ao acaso, com
dois tratamentos (sementes de frutos imaturos e maduros) e 14 repetições (plantas).

40
Determinação do teor de compostos fenólicos em sementes de Bactris gasipaes
Para a determinação do teor de compostos fenólicos totais, primeiramente procedeu-se
o preparo dos reagentes: a) 500 ml do reagente de Folin-Denis, adicionando-se a 375 ml de
água destilada, 50 g de tungstato de sódio dihidratado, 10 g de ácido fosfomolíbdico e 12,5 ml
de ácido fosfórico, fervendo-se a mistura por duas horas e completando-se o volume em balão
volumétrico para 500 ml (Folin e Denis, 1912); b) 1 litro de solução de carbonato de sódio 20
%, adicionando-se 200 g de carbonato de sódio anidro a 1000 ml de água, e dissolvendo-se à
temperatura de 70 °C, até que a solução se tornasse transparente (Reicher e Sierakowiski,
1981). Para a determinação da curva de calibração do ácido tânico, foram dissolvidos 100 mg
de ácido tânico p.a. em água destilada, completando-se o volume para 50 ml em balão
volumétrico e homogeneizando com movimentos de inversão do balão. Foram tomadas
alíquotas de 0,5; 1,0; 1,5; 2,0 e 2,5 ml e diluiu-se em 50 ml de água em balão volumétrico,
homogeneizando-se a mistura. Essas alíquotas corresponderam às concentrações de 20, 40,
60, 80 e 100 ųg.ml-1. Em tubos de ensaio foram colocados 1 ml de cada solução (diferentes
concentrações), 0,5 ml do reagente de Folin-Denis, 1 ml de carbonato de sódio 20%, 5,5 ml de
água destilada, agitando-se e fazendo-se a leitura em espectrofotômetro no comprimento de
onda de 760 nm. A leitura de absorbância do ácido tânico possibilitou a determinação da
equação da reta (Figura 1).
Para a determinação do teor de compostos fenólicos, foram utilizadas 30 sementes por
infrutescência (planta X estádio de maturação), as quais foram submetidas à secagem em
estufa a 30 °C por sete dias e, em seguida, trituradas em moedor portátil de café modelo MDR
301, marca Cadence e peneiradas em peneira de 2 mm. Foi feita a retirada da gordura com
solvente orgânico (hexano) pelo método a frio, em Solxhlet, durante seis horas, seguida de
evaporação total do solvente (Instituto Adolfo Lutz, 1995). A partir das sementes secas,

41
trituradas e desengorduradas foi preparado extrato alcoólico, colocando-se 1 g da amostra em
um becker e adicionando-se 20 ml de álcool metílico 50%. A mistura foi fervida em banho-
maria (90 °C) por 15 minutos, utilizando-se um funil invertido para redução da evaporação,
agitando-se ocasionalmente. A mistura foi filtrada, levada novamente ao banho-maria para
evaporação do metanol e transferida para um balão volumétrico de 50 ml, completando-se o
volume com água destilada (Goldstein e Swain, 1963). Em um tubo de ensaio foi adicionado
1,0 ml do extrato, 5,5 ml de água, 0,5 ml do reagente de Folin-Denis e 1 ml de solução de
carbonato de sódio saturado (20%) (Folin e Denis, 1912; Reicher e Sierakowiski, 1981). O
tubo foi agitado e permaneceu em repouso por 30 minutos, quando então se fez a leitura de
absorbância em espectrofotômetro (λ=760 nm)
O cálculo dos compostos fenólicos totais foi feito aplicando-se os valores de
absorbância das amostras na equação da curva de calibração do ácido tânico nas
concentrações de 20, 40, 60, 80 e 100 µg.ml-1 obtidas por meio de regressão linear simples.
Essas leituras foram realizadas a partir de amostras secas e desengorduradas, portanto, para
efeito de análise, os valores foram transformados, obtendo-se valores referentes a sementes
integrais.
O delineamento estatístico foi em blocos ao acaso, com dois tratamentos (sementes
imaturas e maduras) e 14 repetições (plantas). As leituras foram feitas em triplicata. Os dados
de concentração em mg.100g-1 e em mg/semente foram submetidos ao teste de normalidade
de e quando adequado, ao teste t a 5% de probabilidade ou ao teste U de Mann-Whitney.
Além disso, foi feito um estudo de correlação entre a concentração de compostos fenólicos
por semente e as variáveis relacionadas à emergência de plântulas (Emergência, IVE, TIE,
TME, TFE, CUE, VME, VE, Z, CVE).

42
Efeito do extrato de sementes de Bactris gasipaes na germinação de sementes de Lactuca
sativa
O extrato utilizado neste ensaio foi preparado a partir do aquecimento em banho-maria
(90 ºC), de 1 g de sementes secas e desengorduradas com 20 ml de álcool metílico 50%, por
15 minutos. Após filtragem, o extrato foi levado ao banho-maria até completa evaporação do
metanol, transferido para um balão volumétrico de 50 ml e completado o volume com água
destilada, ficando na concentração de 2 g de material seco e desengordurado para 100 ml de
água.
Em recipientes gerbox, contendo uma folha de papel filtro, foram semeadas 30
sementes de alface (Lactuca sativa). O substrato foi umedecido com o extrato de cada planta e
maturação na proporção de 2 ml do extrato para cada grama do substrato. A testemunha
consistiu em umedecer o substrato apenas com água destilada, na mesma proporção daquela
utilizada para o extrato. O teste de germinação foi conduzido em câmara de germinação à
25ºC, com fotoperíodo de 12 horas. As observações da germinação foram feitas a cada 12
horas. Dentro do ensaio foram utilizados dois critérios de avaliação: a) emissão de 2 mm de
raiz e b) formação de plântula normal (alongamento do hipocótilo com exposição dos
cotilédones). As observações foram feitas até o décimo dia e, com os dados, foram calculados
a germinação (G), o tempo médio de germinação (TMG) e o índice de velocidade de
germinação (IVG), para os dois critérios de germinação avaliados.
O delineamento experimental foi inteiramente casualizado, em esquema fatorial 2
(maturações dos frutos cujas sementes foram utilizadas no preparo do extrato) X 14 (plantas
das quais foram coletadas as sementes de pupunha utilizadas no preparo do extrato), mais um
tratamento adicional (testemunha, sem extrato), com 3 repetições de 30 sementes cada. Os
dados de germinação foram submetidos a teste de normalidade e á análise de variância,
seguida de teste de Tukey com 5% de probabilidade. Foi feito um estudo de correlação entre a

43
germinação de sementes de alface e a emergência, o índice de velocidade de emergência, o
tempo médio de emergência de plântulas de pupunha e com a concentração de compostos
fenólicos, a 5% de probabilidade.
RESULTADOS
Emergência de plântulas de Bactris gasipaes em função da maturação dos frutos
O teor de água de sementes obtidas a partir de frutos maduros (46,7%) foi
significativamente inferior ao das sementes de frutos imaturos (50,4%). A massa seca das
sementes obtidas de frutos imaturos (1,1 g), embora ligeiramente inferior à das sementes de
frutos maduros (1,3 g), não apresentou diferença significativa (Tabela 1).
A emergência de plântulas não diferiu entre sementes obtidas de frutos imaturos e
maduros. Contudo, é importante observar que o coeficiente de variação foi alto (32,1%). O
efeito da maturação não obedeceu a um único padrão para todas as plantas, sendo observados
que para algumas plantas houve maior emergência para as sementes de frutos imaturos e vice-
versa, assim como para outras as emergências de plântulas referentes a frutos imaturos e
maduros foram muito próximas. Essa grande variação também pode ser observada na variável
“sementes mortas”, que não diferiu entre sementes de frutos imaturos (26,6%) e maduros
(16,4%), com coeficiente de variação muito alto (58,5%). Da mesma forma, as sementes
dormentes não apresentaram diferenças significativas entre sementes obtidas de frutos
imaturos (5,0%) e maduros (6,0%). Esse foi resultado devido, em parte, ao coeficiente de
variação alto (95,5%), pois variou de 0 a 44% nas sementes de frutos imaturos e de 0 a 24%
nas de frutos maduros. Em decorrência, também, da grande variação observada entre as
plantas, a porcentagem de sementes germinadas e que não haviam emergido (primeira folha

44
bífida expandida) ao final do experimento não diferiu entre sementes de frutos imaturos e
maduros (8,3 e 8,7%), respectivamente.
As medidas de tempo e distribuição da emergência no tempo (Tabela 1) não
apresentaram diferenças significativas para nenhuma variáveis (IVE, TME, CUE, Z, VME,
VE e CVE), embora tenha sido observado que para algumas plantas a velocidade e a
distribuição da emergência no tempo tenha sido maior em sementes de frutos maduros e para
outras em imaturos, com alto coeficiente de variação. Essa variação alta contribuiu para que
não fossem detectadas diferenças significativas nos valores de tempo e velocidade de
emergência.
Teor de compostos fenólicos em sementes de frutos imaturos e maduros de Bactris gasipaes
e sua relação com a emergência de plântulas e a dormência
Sementes de frutos maduros apresentaram maior concentração de compostos fenólicos
por semente (1,706 mg/semente) que as de frutos imaturos (1,407 mg/semente) (Tabela 1).
Em sementes de frutos imaturos, o teor de compostos fenólicos por semente variou de 0,763
mg/semente a 2,856 mg/semente e entre sementes de frutos maduros de 0,873 a 3,458
mg/semente.
No estudo de correlação entre as variáveis de emergência e a concentração de
compostos fenólicos por semente (Tabela 2), houve correlação negativa entre a emergência e
as sementes mortas (R= -0,59), dormentes (R= -0,61), o tempo inicial de emergência (R= -
0,51) e as sementes germinadas e não emergidas (GNE; R= -0,73). A correlação foi positiva
entre a emergência e o IVE (R= 0,97) e o valor de emergência (R= 0,89). As sementes mortas
(M), observadas ao término do experimento correlacionaram-se negativamente com o IVE
(R= -0,56) e com o valor de emergência (R= -0,56) e positivamente com o teor de água das
sementes (R= 0,43). A variável sementes dormentes teve correlação negativa com o IVE (R=

45
-0,59) e com o índice de sincronização (R= -0,70) e positiva com o tempo inicial de
emergência (R= 0,60) com sementes germinadas e não emergidas (R= 0,81) e com o teor de
compostos fenólicos por semente (R= 0,47).
O IVE correlacionou-se negativamente com o TIE, com o TME e com o GNE (R= -
0,63; R= -0,45 e R= -0,72; respectivamente) e positivamente com o coeficiente de
uniformidade de emergência (R= 0,43), com o valor de emergência (R= 0,93) e com o
coeficiente de velocidade de emergência (R= 0,44). O tempo inicial de emergência
correlacionou-se negativamente com o coeficiente de uniformidade de emergência, com o
índice de sincronização, com o valor de emergência e com o coeficiente de velocidade de
emergência (R= -0,49; R= -0,43; R= -0,52 e R= -0,71; respectivamente) e positivamente com
o tempo médio, a massa seca, as sementes germinadas e não emergidas e o teor de compostos
fenólicos (R= 0,71; R= 0,46; R= 0,49 e R= 0,45; respectivamente). O tempo médio de
emergência apresentou correlação negativa com o coeficiente de uniformidade de emergência
(R= -0,61), com o valor de emergência (R= -0,47) e com o coeficiente de velocidade de
emergência (R= -0,99) e positiva com o tempo final de emergência (R= 0,60). Este se
correlacionou negativamente com o CUE e com o CVE (R= -0,69; R= -0,63;
respectivamente) e positivamente com o Z (R= 0,63).
O CUE apresentou correlação significativa positiva com o CVE (R= 0,66) e com o VE
(R= 0,58). Essas variáveis (CUE e VE) tiveram correlação positiva entre si (R=0,58) e o VE
correlacionou-se negativamente com o GNE (R= -0,64). O teor de compostos fenólicos na
semente correlacionou-se positivamente com as sementes germinadas e não emergidas e com
a massa seca das sementes (R= 0,47 e R= 0,65; respectivamente).

46
Efeito da aplicação do extrato de sementes de frutos imaturos e maduros de Bactris
gasipaes na germinação de sementes de Lactuca sativa
A germinação de sementes de alface, pelo critério de emissão de 2 mm de raiz,
submetidas ao extrato de sementes de pupunha foi inferior à da testemunha, independente da
planta ou da maturação das sementes (Tabela 3). Houve efeito de interação entre os fatores
“maturação” e “planta”. Do total das plantas, para 11 não houve diferença significativa entre a
aplicação dos extratos de sementes de frutos imaturos ou maduros. Para as plantas 9 e 11
houve maior inibição da germinação quando foi aplicado o extrato das sementes de frutos
imaturos e para a planta 10 a maior inibição da germinação deu-se com a aplicação do extrato
de sementes de frutos maduros. O mesmo comportamento foi observado para o índice de
velocidade de germinação, cujas diferenças entre os resultados de sementes de frutos maduros
e imaturos apresentaram diferenças significativas nas mesmas plantas. O tempo médio de
germinação, pelo critério de emissão da raiz, não foi diferente entre sementes submetidas ou
não à aplicação do extrato de sementes de pupunha e houve diferença significativa apenas
entre as plantas, não ocorrendo efeito de interação com a maturação das sementes.
Pelo critério de formação de plântula normal (alongamento do hipocótilo com
exposição dos cotilédones), a germinação de alface apresentou apenas diferença significativa
entre sementes submetidas ao extrato e a testemunha, não diferindo entre a maturação dos
frutos cujas sementes foram usadas para o preparo dos extratos, nem entre as plantas. O IVG
por este critério diferiu entre a testemunha e os tratamentos submetidos aos extratos e
apresentou interação entre a “maturação” e “planta”. Para duas das plantas analisadas (plantas
9 e 12), o IVG foi maior quando as sementes de alface foram submetidas ao extrato de
sementes de frutos de pupunha maduros. Com relação ao tempo médio de germinação, pelo
critério formação de plântula, não houve diferença significativa entre sementes submetidas ou
não ao extrato. Houve efeito de interação entre os fatores, sendo que para uma das plantas

47
(12) o TMG foi maior utilizando-se o extrato de sementes de frutos imaturos e para outra (14)
o TMG foi maior com extrato de sementes de frutos maduros. Para as demais plantas, não
houve diferença no TMG, pelo critério plântula, entre sementes de alface submetidas ao
extrato de sementes de frutos imaturos ou maduros.
Foi observada correlação significativa entre a germinação pelo critério de emissão de
raiz e pelo critério de formação de plântula normal de sementes de alface submetidas ao
extrato de sementes de diferentes plantas de pupunha e estádios de maturação (Tabela 4). A
germinação de sementes (emissão de raiz) correlacionou-se positivamente com: IVG-critério
raiz (R= 0,84), germinação (critério plântula - R= 0,62), IVG-critério plântula (R=0,68).
Houve correlação negativa entre a germinação pelo critério raiz e TMG pelo critério plântula.
O IVG-critério raiz correlacionou-se positivamente com a germinação (critério plântula) e o
IVG, pelo critério de formação de plântula normal (R= 0,75 e 0,84, respectivamente) e
negativamente com o TMG-critério raiz e o TMG-critério plântula, com coeficientes de
correlação R= -0,39 e R= -0,55, respectivamente. O TMG-critério raiz correlacionou-se
negativamente com o IVG-critério plântula (R= -0,39) e a germinação (critério plântula)
correlacionou-se positivamente com o IVG (critério plântula; R= 0,95). A correlação foi
significativa e positiva (R= 0,42) entre a germinação de sementes de alface, pelo critério
emissão de plântula, submetidas ao extrato de sementes de pupunha e a emergência de
plântulas de pupunha a partir dessas sementes. A emergência de plântulas de pupunha
correlacionou-se positivamente com o IVE de plântulas de pupunha (R=0,97) e este teve
correlação negativa com o TME de plântulas de pupunha (R= -0,45). A concentração de
compostos fenólicos por semente correlacionou-se positivamente apenas com a concentração
por 100 g de amostra (R= 0,69), não tendo correlação com a germinação de sementes de
alface.

48
DISCUSSÃO
Os resultados encontrados no presente estudo demonstram não haver elevação da
dormência nas sementes de Bactris gasipaes em função do avanço da idade dos frutos. Estudo
anterior sugeriu que as sementes desta espécie têm a maturação fisiológica completada cerca
de um mês antes da maturação dos frutos e apresentam dormência mais acentuada quando os
frutos estão completamente maduros (Ferreira, 2005). Contudo os resultados relacionados à
germinação, velocidade e tempo de germinação revelaram que na população em estudo,
sementes provenientes de frutos formados no mesmo período, ou seja, sujeitos às condições
climáticas semelhantes ao longo de sua maturação, e, ainda, sob as mesmas condições de
fertilidade do solo, apresentaram padrões diferenciados de germinação. Alguns trabalhos, com
outras espécies de palmeiras, mostraram haver variação entre a germinação relacionada à
maturação das sementes. Em algumas espécies, a germinação é mais elevada em sementes
imaturas, como em Livistona chinensis (Maciel, 1996). Em outras espécies, como Phoenix
canariensis, a maior germinação ocorre em sementes maduras (Pimenta et al., 2010).
De uma maneira geral, é definida como semente dormente aquela que não é capaz de
germinar em determinado período de tempo sob condições ambientais favoráveis à
germinação. Entretanto, em sementes de muitas espécies, a dormência não é um estádio de
tudo ou nada no ciclo de vida. Sementes de muitas espécies com dormência fisiológica não
profunda passam por uma série de mudanças regidas pela temperatura na sua capacidade de
responder a vários fatores entre dormência e não dormência: na maturidade a semente pode
estar em um dos estádios de dormência condicional e poderá ou não continuar dormente
(Baskin e Baskin, 2004). Durante o processo entre a dormência e a não dormência, segundo
esses autores, a faixa de temperatura em que essas sementes são capazes de germinar

49
aumenta. No presente estudo, verificou-se uma ampla variação na germinação e na velocidade
de germinação de sementes de diferentes cachos de Bactris gasipaes (alto coeficiente de
variação). Essa variação pode, em parte, ser atribuida ao período de desenvolvimento das
inflorescências, que é dividido em três fases: a) fase de crescimento lento, quando há o
desenvolvimento e diferenciação das gemas reprodutivas, as quais são controlados por um
balanço de reguladores de crescimento, luz, nutrição das plantas, umidade e genótipo, com
aumento no tamanho das gemas reprodutivas, dura cerca de dois anos; b) fase de rápido
crescimento, caracterizado pelo rompimento da bráctea externa pela bráctea peduncular, dura
cerca de dois meses; c) antese, abertura das flores e polinização (Mora-Urpi et al., 1997).
A variação no tempo de emergência entre plantas, encontrada no presente trabalho,
concorda com os resultados encontrados por Bovi et al. (1994), que verificaram diferenças no
tempo médio de emergência de sementes de diferentes progênies, variando de 38 a 133 dias.
No presente estudo, na avaliação da emergência de plântulas com a primeira folha bífida
expandida, foi também encontrada variação no tempo médio, de 79 a 118 dias.
O presente estudo revelou que houve acúmulo de compostos fenólicos no final da
maturação das sementes de pupunha. Esse resultado concorda com os encontrados por
Debaujon et al. (2000), onde sementes maduras de Arabidopsis se tornaram marrons, como
resultado da acumulação e subsequente oxidação de compostos fenólicos na camada
endotelial da testa (Moïse et al., 2005). Os compostos fenólicos geralmente são encontrados
na testa da semente (Weber et al., 2005) e existe uma correlação entre a coloração da testa e a
presença de compostos fenólicos e, consequentemente, com a germinação, dormência e a
longevidade das sementes (Debaujon et al., 2000). Porém, Henderson e Nitsch (1962)
verificaram que estes compostos fenólicos podem ter ação ativadora ou inibidora do sistema
enzimático.

50
No estudo de correlação (Tabela 2), observou-se que a emergência de plântulas foi
menor quanto maior o número de sementes mortas e dormentes. Além disso, pôde-se observar
que quanto maior o número de sementes dormentes ao término do experimento, maior o
tempo inicial de emergência, menor o índice de sincronização (indicando maior sincronia,
devido ao menor número de dias em que houve emergência) e maior o número de sementes
germinadas e não emergidas ao final do período de avaliação.
Quanto maior o teor de compostos fenólicos na semente, maior a dormência,
indicando que entre esses compostos encontram-se inibidores da germinação. Isso pode ser
confirmado observando a correlação positiva existente entre os compostos fenólicos e o
tempo inicial de emergência e com as sementes germinadas e que não emergiram, pois estas
sementes viáveis tiveram retardo na germinação. Da mesma forma, a correlação negativa
entre essa concentração e o índice de sincronização, indica que quanto mais compostos
fenólicos, mais sincrônica foi a emergência (quanto menor o índice maior a sincronia), uma
vez que esse índice demonstra quantos dos dias de observação houve germinação (Santana e
Ranal, 2004).
O extrato das sementes de pupunha a 2% foi capaz de retardar e reduzir a germinação
de sementes de alface, o que pode ser observado pela menor germinação, menor IVG, menor
emergência de plântulas e menor IVE. Essa capacidade de redução da germinação varia em
função da maturação do fruto e da planta. Embora não exista correlação entre a concentração
de compostos fenólicos no extrato e a germinação ou emergência de plântulas de alface, foi
possível observar correlação entre esta e a emergência de plântulas de pupunha. Esse
resultado demonstra que os compostos responsáveis pelo atraso na germinação em sementes
de alface, quando feita aplicação exógena, atuam também no controle da emergência de
sementes de pupunha. Segundo Bradbeer (1988) para ser considerado um inibidor endógeno,

51
deve haver correlação entre a sua concentração e o grau de dormência, o que podemos
observar nas sementes de pupunha. Ainda, segundo o autor, a aplicação exógena deve impedir
ou retardar a germinação de sementes não dormentes, o que foi observado quando se aplicou
o extrato das sementes de pupunha em sementes de alface.
No presente estudo, conclui-se que a emergência de plântulas de pupunha não é
diferente entre sementes advindas de frutos com diferentes maturações. Sementes de frutos
maduros de pupunha apresentam maior concentração de compostos fenólicos que sementes
imaturas e o acúmulo de compostos fenólicos é um dos fatores responsáveis pela inibição da
germinação em sementes de pupunha. O extrato fenólico de sementes de pupunha foi capaz de
diminuir e retardar a germinação e emergência de sementes de alface e essa emergência
apresentou correlação significativa com a emergência de plântulas de pupunha.

52
REFERÊNCIAS BIBLIOGRÁFICAS
BASKIN, C.C.; BASKIN, J.M. Seeds: ecology, biogeography, and evolution of dormancy
and germination. San Diego: Academic Press. 1998. 666p.
BASKIN, J.M; BASKIN, C.C. A classification system for seed dormancy. Seed science
research, 14: 1-16. 2004.
http://journals.cambridge.org/action/displayAbstract?fromPage=online&aid=704528
BOVI, M.L.A.; SPIERING, S.H.; MARTINS, A.L.D.; PIZZINATO, M.A.; LOURENÇÃO,
A.L.; CHAVES FLORES, W.B. Seed germination of progênies of Bactris gasipaes:
percentage, speed and duration. Acta horticulturae, 360(8):283-289. 1994.
http://www.actahort.org/books/360/360_20.htm
BRADBEER, J. W. Seed dormancy and germination. Glasgow: Blakie Son. New York,
146p. 1988.
BRASIL. MINISTÉRIO DA AGRICULTURA, PECUÁRIA E ABASTECIMENTO. Regras
para análise de sementes / Ministério da Agricultura, Pecuária e Abastecimento. Secretaria
de Defesa Agropecuária. – Brasília: Mapa/ACS, 2009. 399 p.
CLEMENT, C.R. Domestication of the pejibaye palm (Bactris gasipaes): past and present
{Domesticação da pupunha: passado e presente}. In: Balick, M.J. (ed.). The Palm - Tree of
Life. Biology, Utilization and Conservation. Advances in Economic Botany, 6:155-174. 1988.
CLEMENT, C.R.; WEBER, J.C.; COLE, D.; VAN LEEUWEN, J. A subutilização da
pupunha: lições para P&D em palmeiras amazônicas? In: 54º Congresso Nacional de
Botânica, Belém, PA. p. 212-214. 2003.

53
DEBEAUJON, I.; LÉON-KLOOSTERZIEL, K.M.; KOORNNEEF, M. Influence of the testa
on seed dormancy, germination, and longevity in Arabidopsis. Plant Physiology, 122: 403–
413. 2000.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC58877/
FERREIRA, S.A.N. Pupunha: Bactris gasipaes Kunth (Arecaceae). Fascículo 5. 12 p. In:
Ferraz, I.D.K.; Camargo, J.L.C. (Eds) Manual de Sementes da Amazônia. Fascículo 5, INPA,
Manaus, AM, Brasil. 2005. 12p.
www.inpa.gov.br/.../fasciculo5_Bactis%20gasipaes_Pupunha-net.pdf
FOLIN O. ; DENIS W. On phosphotungsticphosphomolybdic compounds as colour reagents.
Journal of Biological Chemistry. 12:239–343. 1912.
GOLDSTEIN, J.L.; SWAIN, T. Changes in tannin in ripening fruits. Biochemistry. 2:
271:383. 1963.
GOULDING, M.; SMITH, N.. Palmeiras: Sentinelas para a conservação da Amazônia.
Lima: Amazon Conservation Association-Sociedade Civil Mamirauá. 2007. 358 p.
HENDERSON, H.M.; NITSCH, J.P. Effect of certain phenolic acids on de elongation of
avena first internodes in the presence of auxins and tryptophan. Nature. 195(4843):780-782.
1962.
INSTITUTO ADOLFO LUTZ. Normas analíticas do Instituto Adolfo Lutz: Métodos
químicos e físicos para análise de alimentos. 3 ed. 1995.533p.
JOHR, J. Estadísticas del mercado interno de Brasil. In: Palmito de Pejibaye (Bactris
gasipaes Kunth): Su cultivo e industrialización. San José, CR: Editorial de la Universidad de
Costa Rica, p.220-222, 1999.
http://books.google.com/books?id=gjxTOmG2zgYC&printsec=frontcover&dq=Palmito+de+
Pejibaye+(Bactris+gasipaes+Kunth):+Su+cultivo+e+industrializaci%C3%B3n.&hl=ptBR&ei

54
=b6vNTbTqOoTfgQeSifGyDA&sa=X&oi=book_result&ct=result&resnum=1&ved=0CDkQ
6AEwAA#v=onepage&q&f=false
MACIEL, N. M. S. Efectos de la madurez y el almacenamiento del fruto, la escarificacion y el
remojo de las semillas sobre la emergencia de la palma china de abanico. Agronomia
Tropical, 46(2): 155-170. 1996.
http://sian.inia.gob.ve/repositorio/revistas_ci/Agronomia%20Tropical/at4602/arti/maciel_n.ht
m
MOÏSE, J.A. HAN, S; GUDYNAITE-SAVITCH, L.; JOHNSON, D.A.; MIKI, B.L.A. Seed
coats: structure, development, composition and biotechnology. In vitro cellular development
biology-plant, 41:620-644. 2005. http://www.springerlink.com/content/q56418t500843070/
MORA URPÍ, J., J.C. WEBER ; C.R. CLEMENT. Peach palm, Bactris gasipaes Kunth
{Pupunha}. Promoting the conservation and use of underutilized and neglected crops. 20.
Institute of Plant Genetics and Crop Plant Research - IPK, Gatersleben,
Germany/International Plant Genetic Resources Institute - IPGRI, Rome. 1997. 83 p.
PIMENTA, R.S.; LUZ, P.B.; PIVETTA,K.F.L.; CASTRO, A.; PIZETTA, P.U.C. Efeito da
maturação e temperatura na germinação de sementes de Phoenix canariensis hort. ex Chabaud
– Arecaceae. Revista Árvore, 34(1): 31-38. 2010.
www.scielo.br/pdf/rarv/v34n1/v34n1a04.pdf
REICHER, F.; SIERAKOWISKI, M.R.B.C. Determinação espectométrica de taninos pelo
reativo fosfotúngstico-fosfomolididico. Arquivo Biológico. 24(2):407-411. 1981.
SANTANA, D.G.; RANAL, M.A. Análise da. Germinação: um Enfoque Estatístico. Brasília,
Editora. Universidade de Brasília. 2004. 248p.
VIEIRA, L.S.; SANTOS, P.C.T.C. Amazônia: seus solos e outros recursos naturais. São
Paulo: Ceres, 1987. 416p.

55
WEBER, H.; BORISJUK, L.; WOBUS, U. Molecular physiology of legume seed
development. Annual review of plant biology, 56: 253-279. 2005.
www.annualreviews.org/doi/full/10.../annurev.arplant.56.032604.144201

56
FIGURA 1. Curva de calibração do ácido tânico, com leituras de absorbância a 760 nm.
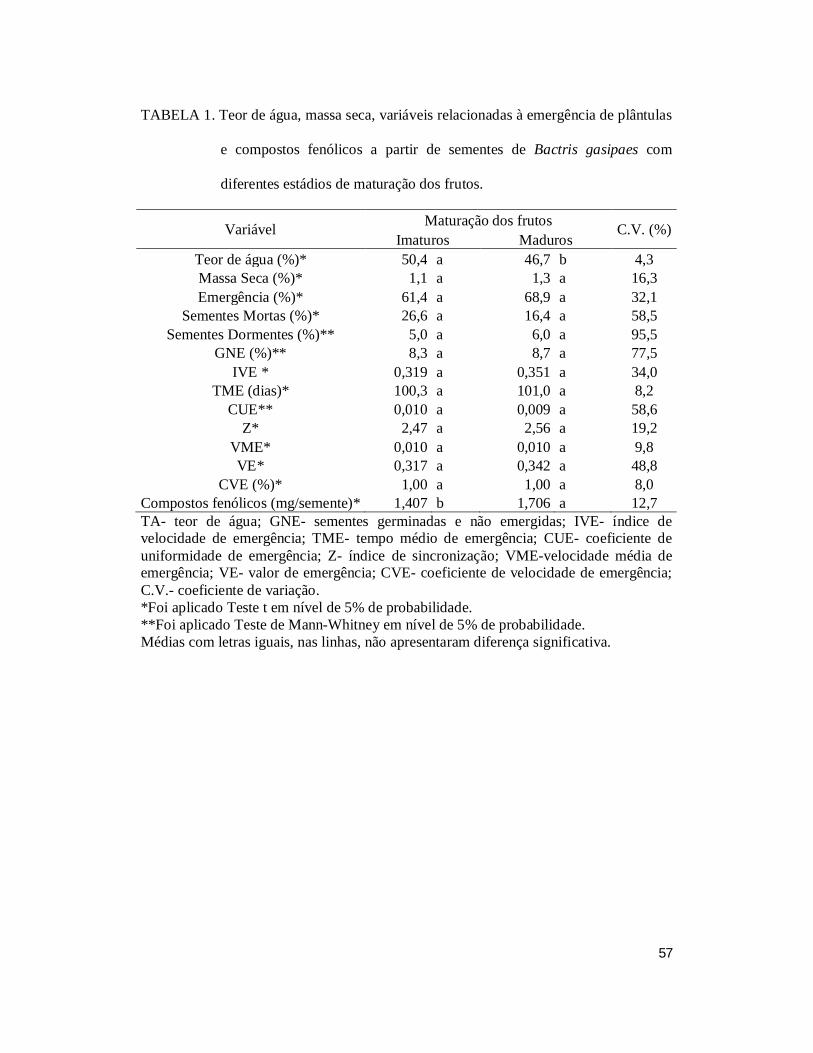
57
TABELA 1. Teor de água, massa seca, variáveis relacionadas à emergência de plântulas
e compostos fenólicos a partir de sementes de Bactris gasipaes com
diferentes estádios de maturação dos frutos.
Variável Maturação dos frutos C.V. (%) Imaturos Maduros
Teor de água (%)* 50,4 a 46,7 b 4,3 Massa Seca (%)* 1,1 a 1,3 a 16,3 Emergência (%)* 61,4 a 68,9 a 32,1
Sementes Mortas (%)* 26,6 a 16,4 a 58,5 Sementes Dormentes (%)** 5,0 a 6,0 a 95,5
GNE (%)** 8,3 a 8,7 a 77,5 IVE * 0,319 a 0,351 a 34,0
TME (dias)* 100,3 a 101,0 a 8,2 CUE** 0,010 a 0,009 a 58,6
Z* 2,47 a 2,56 a 19,2 VME* 0,010 a 0,010 a 9,8 VE* 0,317 a 0,342 a 48,8
CVE (%)* 1,00 a 1,00 a 8,0 Compostos fenólicos (mg/semente)* 1,407 b 1,706 a 12,7 TA- teor de água; GNE- sementes germinadas e não emergidas; IVE- índice de velocidade de emergência; TME- tempo médio de emergência; CUE- coeficiente de uniformidade de emergência; Z- índice de sincronização; VME-velocidade média de emergência; VE- valor de emergência; CVE- coeficiente de velocidade de emergência; C.V.- coeficiente de variação. *Foi aplicado Teste t em nível de 5% de probabilidade. **Foi aplicado Teste de Mann-Whitney em nível de 5% de probabilidade. Médias com letras iguais, nas linhas, não apresentaram diferença significativa.
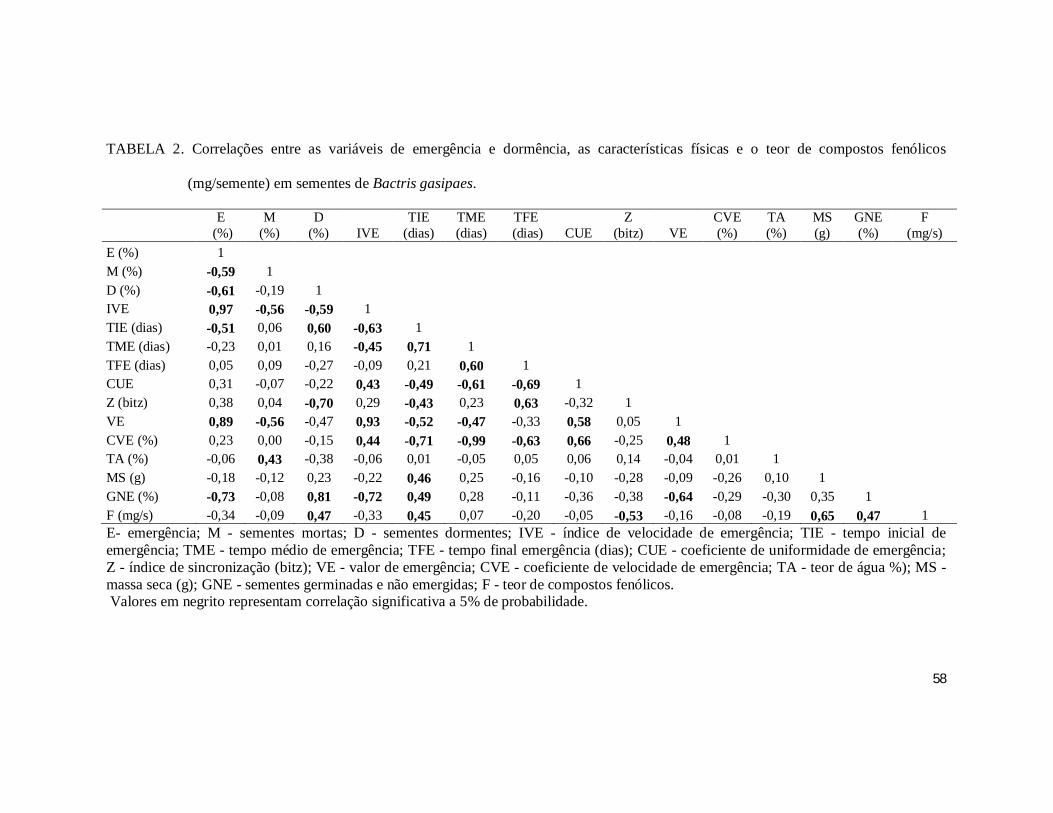
58
TABELA 2. Correlações entre as variáveis de emergência e dormência, as características físicas e o teor de compostos fenólicos
(mg/semente) em sementes de Bactris gasipaes.
E
(%) M
(%) D
(%) IVE TIE
(dias) TME (dias)
TFE (dias) CUE
Z (bitz) VE
CVE (%)
TA (%)
MS (g)
GNE (%)
F (mg/s)
E (%) 1 M (%) -0,59 1 D (%) -0,61 -0,19 1 IVE 0,97 -0,56 -0,59 1 TIE (dias) -0,51 0,06 0,60 -0,63 1 TME (dias) -0,23 0,01 0,16 -0,45 0,71 1 TFE (dias) 0,05 0,09 -0,27 -0,09 0,21 0,60 1 CUE 0,31 -0,07 -0,22 0,43 -0,49 -0,61 -0,69 1 Z (bitz) 0,38 0,04 -0,70 0,29 -0,43 0,23 0,63 -0,32 1 VE 0,89 -0,56 -0,47 0,93 -0,52 -0,47 -0,33 0,58 0,05 1 CVE (%) 0,23 0,00 -0,15 0,44 -0,71 -0,99 -0,63 0,66 -0,25 0,48 1 TA (%) -0,06 0,43 -0,38 -0,06 0,01 -0,05 0,05 0,06 0,14 -0,04 0,01 1 MS (g) -0,18 -0,12 0,23 -0,22 0,46 0,25 -0,16 -0,10 -0,28 -0,09 -0,26 0,10 1 GNE (%) -0,73 -0,08 0,81 -0,72 0,49 0,28 -0,11 -0,36 -0,38 -0,64 -0,29 -0,30 0,35 1 F (mg/s) -0,34 -0,09 0,47 -0,33 0,45 0,07 -0,20 -0,05 -0,53 -0,16 -0,08 -0,19 0,65 0,47 1 E- emergência; M - sementes mortas; D - sementes dormentes; IVE - índice de velocidade de emergência; TIE - tempo inicial de emergência; TME - tempo médio de emergência; TFE - tempo final emergência (dias); CUE - coeficiente de uniformidade de emergência; Z - índice de sincronização (bitz); VE - valor de emergência; CVE - coeficiente de velocidade de emergência; TA - teor de água %); MS - massa seca (g); GNE - sementes germinadas e não emergidas; F - teor de compostos fenólicos. Valores em negrito representam correlação significativa a 5% de probabilidade.

59
TABELA 3. Germinação, índice de velocidade de germinação (IVG), tempo médio de germinação (TMG), emergência, índice de
velocidade de emergência (IVE) e tempo médio de emergência (TME) de sementes de (Lactuca sativa) semeadas em
substrato umedecido com extrato fenólico de sementes de frutos de pupunha de diferentes plantas e com diferentes estádios de
maturação dos frutos.
Critério emissão de raiz Critério formação de plântula normal
Germinação (%) 1 IVG 1 TMG (horas) 2 Germinação (%) 2 IVG 1 TMG (horas) 1
Testemunha 71,1 * 0,416 * 68 ns 48,9 * 0,175 * 92,8 ns Plantas Imaturos Maduros Imaturos Maduros Médias Médias Imaturos Maduros Imaturos Maduros
1 60,0 a 46,7 a 0,341 a 0,252 a 71 ABC 33,9 A 0,131 a 0,093 a 100,0 a 93,8 a 2 56,7 a 63,3 a 0,344 a 0,402 a 68 ABC 36,1 A 0,133 a 0,125 a 92,0 a 92,1 a 3 54,4 a 48,9 a 0,305 a 0,247 a 77 AB 22,8 A 0,098 a 0,060 a 89,7 a 87,7 a 4 55,6 a 58,9 a 0,318 a 0,354 a 70 ABC 36,1 A 0,114 a 0,132 a 94,6 a 98,6 a 5 47,8 a 55,6 a 0,250 a 0,285 a 77 AB 32,8 A 0,114 a 0,104 a 104,4 a 97,4 a 6 56,7 a 52,2 a 0,315 a 0,265 a 74 ABC 26,1 A 0,086 a 0,085 a 94,4 a 102,9 a 7 44,4 a 46,7 a 0,235 a 0,302 a 62 BC 33,9 A 0,097 a 0,106 a 104,1 a 105,1 a 8 52,2 a 63,3 a 0,348 a 0,349 a 67 ABC 37,2 A 0,123 a 0,121 a 102,4 a 99,1 a 9 38,9 b 53,3 a 0,260 b 0,401 a 57 C 32,2 A 0,085 b 0,140 a 91,1 a 88,3 a
10 56,7 a 31,1 b 0,327 a 0,208 b 66 BC 25,0 A 0,113 a 0,071 a 82,4 a 92,4 a 11 41,1 b 65,6 a 0,224 b 0,353 a 74 ABC 27,2 A 0,060 a 0,105 a 109,9 a 101,2 a 12 45,6 a 47,8 a 0,235 a 0,277 a 68 ABC 25,0 A 0,040 b 0,104 a 133,0 a 100,1 b 13 57,8 a 55,6 a 0,344 a 0,317 a 71 ABC 34,4 A 0,113 a 0,111 a 97,6 a 96,3 a 14 46,7 a 34,4 a 0,192 a 0,162 a 85 A 23,9 A 0,066 a 0,058 a 109,1 b 137,3 a
1 Houve interação significativa entre os fatores “maturação dos frutos” e “plantas”. 2 Não houve interação significativa os os fatores “maturação dos frutos” e “plantas”. Médias com letras iguais e maiúsculas, nas colunas, não apresentaram diferença significativa. Médias com letras iguais e minúsculas, nas linhas, não apresentaram diferença significativa.

60
TABELA 4. Correlação entre variáveis de germinação e de emergência de sementes de alface submetidas ao extrato de sementes de
pupunha de diferentes plantas e com diferentes estádios de maturação dos frutos, a emergência de plântulas de pupunha a
partir dessas sementes e a concentração de compostos fenólicos nessas sementes.
Germinação de sementes de alface submetidas ao
extrato de sementes de pupunha Emergência de plântulas de
pupunha Concentração de
compostos fenólicos
Critério emissão da raiz
(A) Critério plântula
(B)
G (%)
IVG TMG (horas)
G (%)
IVG TMG (horas)
E (%)
IVE TME (dias)
Fenólicos (mg/semente)
Fenólicos (mg/100g)
Germinação de sementes de alface submetidas ao extrato de sementes de
pupunha
A
G (%) 1 IVG 0,84 1 TMG (horas) 0,11 -0,39 1
B
G (%) 0,62 0,75 -0,36 1 IVG 0,68 0,84 -0,39 0,95 1 TMG -0,40 -0,55 0,26 0,27 -0,54 1
Emergência de plântulas de pupunha
E (%) 0,02 0,08 -0,18 0,42 0,39 -0,04 1 IVE -0,02 0,06 -0,21 0,38 0,38 -0,11 0,97 1 TME (dias) 0,14 0,05 0,12 0,12 -0,04 0,35 -0,23 -0,45 1
Concentração de compostos fenólicos
Fenólicos (mg/semente) -0,00 -0,06 0,18 -0,15 -0,09 -0,19 -0,34 -0,33 0,07 1 Fenólicos (mg/100g) -0,10 -0,12 0,16 -0,14 -0,15 0,16 -0,28 -0,24 -0,09 0,69 1
A - Variáveis referentes à germinação pelo critério de emissão de 2 mm de raiz; B - Variáveis referentes à germinação pelo critério de formação de plântula normal (alongamento do hipocótilo com exposição dos cotilédones); G - Germinação; IVG - índice de velocidade de germinação, TMG - tempo médio de germinação (horas) ; E - emergência; IVE - índice de velocidade de emergência; TME - tempo médio de emergência. Valores em negrito representam correlação significativa a 5% de probabilidade.

61
Capítulo 3
___________________________________________________________________
Teores de lipídios e germinação de sementes de Bactris gasipaes
Kunth de frutos em diferentes estádios de maturação (Elaborado de acordo com as normas da Revista Acta Amazonica)

62
Teores de lipídios e germinação de sementes de Bactris gasipaes Kunth de frutos em diferentes estádios de maturação
RESUMO
Bactris gasipaes se destaca entre as palmeiras tropicais de grande importância econômica, devido ao valor nutricional de seus frutos e ao palmito de excelente qualidade. Objetivou-se avaliar a composição lipídica e o ponto de fusão dos lipídios de sementes de Bactris gasipaes, bem como correlacioná-los com variáveis relacionadas à emergência de plântulas a partir de sementes provenientes de frutos imaturos e maduros. Foram utilizadas sementes de 14 plantas a partir de cachos com frutos imaturos e maduros. Foi feita a semeadura em viveiro, utilizando-se como substrato areia e serragem na proporção volumétrica de 1:1, acompanhada da determinação do teor de água. A emergência (primeira folha bífida expandida) foi avaliada a cada cinco dias, até 150 dias. Foi feita a quantificação dos lipídios, a determinação do ponto de fusão dos lipídios e a quantificação dos ácidos graxos das sementes. O teor de água de sementes de frutos imaturos foi estatisticamente superior ao das sementes de frutos maduros (50,4 e 46,7%, respectivamente). A emergência de plântulas, o tempo médio de emergência (TME) e o índice de velocidade de emergência (IVE) não diferiram estatisticamente entre os estádios de maturação. O ponto de fusão dos lipídios não diferiu estatisticamente entre sementes de frutos imaturos (22,54 ºC) e maduros (22,70 ºC), porém teve alta correlação com as variáveis relacionadas à emergência de plântulas. Os teores de ácido oleico e linoleico foram maiores em sementes de frutos maduros (26,89 e 30,66 µg.g-1, respectivamente) em relação às de frutos imaturos (22,01 e 24,79 µg.g-1, respectivamente). A concentração de ácido oleico teve correlação negativa com o TME. Palavras-chave: pupunha, ácidos graxos, ponto de fusão.

63
Lipids and germination of seeds of Bactris gasipaes Kunth fruits in different maturation
stages
ABSTRACT
Bactris gasipaes stands out among the tropical palm trees of great economic importance due to the value nutritional content of its fruits and the heart of palm of excellent quality. The objective was to evaluate the lipid composition and the melting point of the lipids of seeds of Bactris gasipaes and correlate them with variables related to the emergence of seedlings from seeds from immature fruits and mature. Were used seeds from 14 plants from clusters with immature and mature fruits. Sowing was done in nursery, using sand as substrate and sawdust in a 1:1 volumetric ratio, together with the determination of water content.The emergence (first bifid expanded leaf) was assessed every five days, up to 150 days. Was made to quantify the lipids, the determining the melting point of lipids and quantification of fatty acids of the seeds. The moisture content from seeds of immature fruits was statistically higher than that of mature seeds (50.4 and 46.7%, respectively). The seedling emergence, the mean emergence time (MET) and emergency speed index (ESI) did not differ statistically. The point fusion of lipids did not differ statistically between seeds from immature fruits (22.54 ºC) and mature (22.70 ºC) but had a high correlation with the variables related to the emergence of seedlings. The levels of oleic and linoleic acid were higher in seeds of ripe fruits (26.89 and 30.66 μg.g-1, respectively) compared to those of immature fruits (22.01 and 24.79 μg.g-1, respectively). The concentration of oleic acid had negative correlation with MET. Keywords: peach-palm, fatty acids, melting point.

64
INTRODUÇÃO
Bactris gasipaes (Arecaceae), conhecida no Brasil como pupunha, destaca-se entre as
palmeiras tropicais de grande importância econômica (Johnson, 2010). Na Amazônia, seus
frutos são muito apreciados para o consumo humano. Estes possuem em sua composição alta
concentração de carotenoides e atividade antioxidante (Jatunov et al., 2010). A pupunha tem
sido pesquisada na América Latina focando-se o melhoramento dos frutos para consumo
humano e animal, bem como o aperfeiçoamento do cultivo para produção de palmito
(Johnson, 2010).
As sementes B. gasipaes são consideradas recalcitrantes e quando obtidas de árvores
saudáveis podem atingir mais de 80% de germinação (Clement, 2008). A protrusão da raiz
primária pode iniciar 30 dias após a semeadura (Clement, 2008) e o período de germinação
pode perdurar por 14 meses pela presença de dormência (Mora-Urpí, 1979). A maturidade
fisiológica das mesmas é atingida cerca de um mês antes da maturação do fruto e a
permanência na planta, após a maturação, pode aumentar a intensidade de dormência
(Ferreira, 2005). As causas da dormência e da grande variação no tempo necessário para que a
semente germine são, até o momento, desconhecidas.
Em algumas espécies, partes do fruto ou da semente contêm inibidores que interferem
nos processos metabólicos necessários à germinação, impedindo que ela ocorra. Em algumas
espécies, na fase de maturação, as sementes são revestidas por substâncias lipídicas, tornando-
as impermeáveis (Labouriau, 1983). Em outros casos, alguns tecidos da planta contêm ácidos
graxos de cadeia curta em quantidades altas o suficiente para afetar a dormência e o
crescimento (Berrie et al., 1979, Friedman et al., 2003).
Durante a maturação das sementes a biossíntese de lipídios varia (Voelkner e Kinney,
2001), bem como diferentes combinações de ácidos graxos saturados e insaturados resultarão

65
em diferentes pontos de fusão dos lipídios presentes na semente, pois quanto maior o tamanho
da cadeia maior o ponto de fusão e quanto maior o número de insaturações (duplas ligações)
na cadeia, menor o ponto de fusão (Linder, 2000).
Diante do exposto, o presente trabalho teve por objetivos estudar a composição
lipídica e o ponto de fusão dos lipídios de sementes de Bactris gasipaes em dois estádios
finais de maturação, bem como correlacioná-los com variáveis relacionadas à emergência de
plântulas.
MATERIAL E MÉTODOS
Avaliação da emergência de plântulas de Bactris gasipaes a partir de sementes de frutos
com diferentes estádios de maturação
As sementes utilizadas nesse estudo foram coletadas em plantio de pupunha localizado
no Centro de Pesquisa do Instituto de Saúde e Biotecnologia da Universidade Federal do
Amazonas (ISB/UFAM), localizada na Estrada Coari Itapeua, no município Coari, Amazonas.
O clima da região é classificado como tropical úmido, tipo Afi, pela classificação de Köppen,
apresentando precipitação média anual de 2.250 mm e temperatura média anual da região é
aproximadamente 26 ºC (Vieira e Santos, 1987).
Em 14 plantas, no mesmo momento, foram coletados dois cachos de cada indivíduo,
cada um com uma das seguintes características: a) Frutos imaturos: cachos com frutos com
epicarpo apresentando mudança de cor, na porção mediana, de verde para amarelo ou
alaranjado; b) Frutos maduros: cachos com frutos apresentando epicarpo completamente
amarelo ou vermelho.
Após coleta, os frutos foram retirados das infrutescências, partidos com auxílio de uma
faca e retirada a semente. A determinação do teor de água de amostra das sementes, dos

66
cachos das diferentes plantas e estádios de maturação dos frutos, foi realizada em estufa a
105± 3 °C por 24 horas, com duas repetições de cinco sementes.
As sementes foram colocadas em recipiente com água, eliminando-se as que flutuaram
(Brasil, 2009). A limpeza consistiu da fricção das sementes sobre uma peneira, com água
corrente, e retirada dos resíduos remanescentes com a ponta de uma faca. A semeadura foi
feita em viveiro, com temperatura mínima média de 24,6 °C, máxima média de 36,1 °C, em
caixas plásticas contendo uma mistura de serragem e areia na proporção volumétrica de 1:1,
sendo semeadas 50 sementes de cada planta e estádio de maturação dos frutos. Foram feitas
avaliações da emergência a cada cinco dias, durante cinco meses, e o critério utilizado foi a
presença da primeira folha bífida expandida.
A partir dos dados de emergência, foram calculados: a emergência em porcentagem, o
índice de velocidade de emergência (IVE) e o tempo médio de emergência (TME). Ao
término do experimento foram quantificadas as plântulas germinadas e que não apresentavam
a primeira folha bífida expandida (sementes germinadas e não emergidas), sementes
dormentes e mortas, identificadas pelo teste de corte (Brasil, 2009). O delineamento
estatístico utilizado foi em blocos ao acaso, com 2 tratamentos (sementes imaturas e maduras)
e 14 repetições (matrizes). Foram utilizadas 50 sementes por repetição.
Quantificação e caracterização dos lipídios
A extração e a quantificação dos lipídios foram realizadas no Laboratório de Química
e Físico-química de Alimentos da Coordenação de Tecnologia e Inovação (COTI) do Instituto
Nacional de Pesquisas da Amazônia (INPA). Foram utilizadas 30 sementes por infrutescência,
as quais foram submetidas à secagem em estufa a 35 °C, por 7 dias e, em seguida, partidas ao
meio, para retirada do embrião. As sementes sem o embrião foram trituradas em moinho,
marca Cadence, modelo MDR 301-110. Em cartuchos de papel de filtro, previamente

67
pesados, foram adicionadas aproximadamente 3 gramas de amostra e o conjunto, novamente
pesado. A extração da gordura foi feita a frio com solvente orgânico (hexano) pelo método
Soxhlet, durante 8 horas (Instituto Adolfo Lutz, 1995). Terminado o tempo de extração, os
cartuchos foram levados à estufa a 50 °C, por 12 horas, e novamente pesados. Pela diferença
entre o peso do material antes e após a extração foi determinado o teor de gordura em mg/100
g de massa seca. Foram utilizados para cada rácemo 3 repetições (cartuchos) para cada planta
e estádio de maturação.
O ponto de fusão dos lipídios (gordura) foi obtido no Laboratório de Sementes da
Universidade Federal de Viçosa (UFV). Foram utilizados tubos capilares de 10 µL,
mergulhando-os na substância fundida (gordura aquecida) até obter uma coluna de 1,5 cm. A
outra extremidade foi fechada no bico de Bünsen. Os tubos permaneceram em refrigerador
por 12 horas. Em seguida, foram presos a um termômetro com graduação de até 100 °C, por
meio de um anel de borracha, de modo que a substância no capilar ficasse junta ao bulbo do
termômetro. O conjunto capilar+termômetro foi colocado dentro de um tubo de ensaio com
água, o qual foi mergulhado em um béquer, que passou a ser aquecido lentamente, agitando-
se o tubo de ensaio. O ponto de fusão considerado referiu-se à temperatura na qual a
substância tornou-se transparente (Instituto Adolfo Lutz, 1995). Foram utilizadas três
repetições (capilar com gordura) para cada planta e maturação dos frutos.
Para a esterificação dos ácidos graxos foram pesados 20 mg das amostras de óleos em
beckers pequenos. Foram adicionados 500 µL de Tetrahidrofurano (THF) em cada repetição e
adicionado 1 mL de Metóxido de sódio (MeONa) e cada amostra foi agitada manualmente. A
solução foi transferida para tubos de ensaio (aproximadamente 15 mL) e levada a banho-
maria à 50 ºC por 10 minutos. Após, foi adicionado 50 µL de ácido acético glacial e 3 mL de
água ultra pura. A solução foi transferida para funil de separação, adicionando-se 5 mL de

68
hexano e agitando-se. A fase orgânica foi separada da fase aquosa, adicionando-se novamente
5 mL de hexano e agitando-se. As duas fases orgânicas foram misturadas em becker e
adicionado 5 g de Sulfato de sódio anidro (NaSO4), contendo 10% (0,5 g) de Carbonato de
potássio (K2CO3) para retirar o restante de água das amostras. As amostras foram filtradas em
papel filtro qualitativo sob vácuo, evaporadas em rotaevaporador, ressuspendidas em 1 mL de
hexano e transferidas para frascos de reação. Após, foram injetadas em cromatógrafo gasoso
sob as condições: Volume de injeção: 1 µL; Temperatura da coluna: 190 – 220 ºC a 2 ºC por
minuto; e temperatura do injetor: 250 ºC. Foram utilizadas três repetições para cada planta e
estádio de maturação dos frutos.
Análise estatística dos resultados
Os dados de teor de gordura, de ponto de fusão dos lipídios e de quantificação dos
ácidos graxos foram submetidos a teste de normalidade e quando adequado, à análise de
variância seguida de teste t ou às estatísticas não paramétricas correspondentes (Teste U de
Mann-Whitney).
Foi feito um estudo de correlação entre as variáveis das análises dos lipídios e as
variáveis de emergência de plântulas do material correspondente a sementes obtidas de frutos
imaturos e maduros, de cada planta.
RESULTADOS
O teor de água de sementes provenientes de frutos imaturos foi significativamente
superior ao das sementes de frutos maduros (50,4 e 46,7%, respectivamente). Contudo, a
massa seca das sementes não diferiu significativamente entre os dois estádios de maturação
dos frutos (Tabela 1). A emergência de plântulas, o índice de velocidade de emergência (IVE)

69
e o tempo médio de emergência (TME) não diferiram entre sementes de frutos imaturos e
maduros. A porcentagem de sementes que estavam mortas, ao término do experimento, não
foi diferente entre sementes de frutos imaturos e maduros. As porcentagens de sementes que
permaneceram dormentes não diferiram entre os estádios de maturação. As porcentagens de
sementes germinadas e que não haviam emergido ao término do experimento não diferiram
entre as sementes imaturas e maduras. A variação nos valores das variáveis relacionadas à
emergência de plântulas contribuiu para que não fossem detectadas diferenças entre sementes
obtidas de frutos imaturos e maduros.
A concentração dos ácidos graxos não diferiu entre sementes de frutos imaturos e
maduros para os ácidos caproico, caprílico, cáprico, láurico, mirístico, palmítico e esteárico
(Tabela 1). As concentrações dos ácidos graxos insaturados oleico (C 18:1) e linoleico (C
18:2) foram significativamente maiores em sementes de frutos maduros (26,89 µg.g-1 e 30,66
µg.g-1, respectivamente) em relação às de frutos imaturos (22,01 µg.g-1 e 24,79 µg.g-1,
respectivamente). Observou-se alto coeficiente de variação, decorrente de grande variação
entre repetições (variação entre plantas) dentro dos tratamentos.
Tanto em sementes de frutos imaturos quanto nas de frutos maduros, os ácidos graxos
que ocorreram em maior concentração foram o ácido láurico, mirístico e palmítico com
concentrações médias de 952,27, 318,72, 112,86 µg.g-1, respectivamente. O teor de lipídios
totais na semente foi significativamente maior em sementes imaturas (26,29 g.100 g-1) em
relação às maduras (24,75 g.100 g-1).
A emergência de plântulas correlacionou-se positivamente com o IVE (0,97) e
negativamente com as sementes mortas (-0,59), dormentes (-0,61) e as que haviam
germinado, porém sem apresentar a primeira folha bífida ao final das observações (-0,73)

70
(Tabela 2). Houve correlação positiva também entre a emergência de plântulas e o ponto se
fusão dos lipídios extraídos dessas sementes (0,66).
O IVE correlacionou-se negativamente com o tempo médio de emergência, com as
sementes mortas, dormentes e germinadas e não emergidas (-0,45; -0,56; -0,59; -0,72,
respectivamente) e positivamente com o ponto de fusão dos lipídios (0,70). O tempo médio de
emergência apresentou correlação significativa apenas com a concentração de ácido oleico,
sendo menor quanto maior a concentração deste ácido graxo (-0,41).
As sementes mortas correlacionaram-se negativamente com o ponto de fusão dos
lipídios (-0,59). As sementes dormentes tiveram correlação positiva com as germinadas e não
emergidas (0,81). Estas duas são medidas de dormência, visto que as sementes foram
consideradas dormentes quando o embrião permaneceu intacto ao final dos 150 dias de
avaliação e consideradas germinadas e não emergidas quando germinaram tardiamente, não
apresentando a primeira folha bífida expandida ao término do experimento.
A concentração de lipídios teve correlação negativa com o teor de ácido caprílico (-
0,43). O ácido caprílico teve correlação negativa com os teores de ácido láurico, mirístico,
palmítico e linoleico (-0,66; -0,63; -0,59 e -0,44, respectivamente). O ácido láurico apresentou
correlação positiva com os teores de ácido mirístico, palmítico e linoleico (0,93; 0,88; 0,67,
respectivamente). O ácido mirístico correlacionou-se positivamente com os teores de ácido
palmítico e linoleico (0,93 e 0,71, respectivamente). Os ácidos palmítico, esteárico e oleico
tiveram seus teores correlacionados positivamente com os de ácido linoleico (0,79; 0,49 e
0,41; respectivamente).

71
DISCUSSÃO
As variáveis relacionadas à emergência de plântulas (primeira folha bífida expandida)
não diferiram entre sementes obtidas de frutos imaturos e maduros. Contudo, foi observada,
para todas as variáveis, grande variação dentro dos tratamentos (coeficientes de variação
elevados), que pode ter contribuído para ausência de diferença entre os tratamentos. Ferreira
(1996) verificou que sementes de pupunha atingiram a maturidade fisiológica aos 80 dias após
a antese, com ao menos 50% dos frutos da infrutescência apresentando mudança de coloração
de verde para amarela ou alaranjada. Neste estudo, houve aumento da dormência primária
com o aumento da idade das sementes. Entretanto, Bactris gasipaes é uma espécie de
polinização cruzada, sendo assim, a variação na emergência entre frutos de um mesmo estádio
de maturação, de uma mesma planta pode ser, em parte, atribuída ao efeito do pólen,
fenômeno conhecido como metaxenia. Esse efeito já foi observado na palmeira Phoenix
dactylifera, onde diferentes pólens causaram variação em características como forma e
maturação do fruto (Al-Khalifah, 2006).
No presente estudo, foi verificado que entre os ácidos graxos avaliados, os
encontrados em maior concentração foram o ácido láurico, mirístico e palmítico. A
predominância desses três ácidos graxos também foi encontrada em sementes de Elaeis
guineensis (Bora et al., 2003), com percentuais de 53,2%, 19,3% e 10,4%, respectivamente.
Porém, em outro estudo com sementes de Elaeis guineensis, os três principais ácidos graxos
encontrados foram láurico, mirístico e oleico (Macaire et al., 2010). Em albúmen de Cocos
nucifera, Aroucha et al. (2005) encontraram predominância de ácido láurico, oleico e
palmítico. Em contrapartida, no óleo da semente em Phoenix dactylifera, a maior proporção é
de ácidos graxos monoinsaturados (Besbes et al., 2004).

72
No presente estudo, foram utilizadas sementes a partir de frutos em dois estádios finais
de maturação, observando-se que os frutos maduros apresentaram menor concentração de
lipídios totais em relação aos frutos imaturos. Essa redução no teor de lipídios pode ser
atribuída à ação de hidrolases e lipoxigenases, durante o amadurecimento, que degrada esses
componentes para fornecimento de energia para as células vegetais (Ingram e Bartels, 1996).
Para a maioria dos ácidos graxos avaliados, não houve diferença na concentração entre
sementes de frutos imaturos e maduros, em parte, devido à alta variação entre plantas.
Conforme Buckeridge et al. (2004), os lipídios, depositados nas sementes sob a forma de
triacilglicerídeos, apresentam variações nas concentrações de ácidos graxos entre sementes de
diferentes plantas de uma mesma espécie.
As porcentagens de sementes que permaneceram dormentes após 150 dias de
avaliação e de sementes que haviam germinado sem apresentar emergência de plântulas
representam medidas de dormência das sementes. A falta de uniformidade na germinação
pode caracterizar a presença de dormência fisiológica. Este tipo de dormência foi observado
em sementes da palmeira Sabal palmeto, nas quais o T50 (tempo para que 50% das sementes
germinem) foi reduzido tanto quando foi feita escarificação química, quanto quando foi feita a
aplicação de ácido giberélico, indicando que as sementes apresentam dormência exógena e
dormência fisiológica (Dewir et al., 2011).
A correlação negativa entre as sementes mortas e o ponto de fusão de lipídios pode
representar um indício de que a deterioração das sementes de pupunha é favorecida pela
presença de ácidos graxos insaturados. De um modo geral, quanto maior o grau de insaturação
do ácido graxo, maior a susceptibilidade à oxidação, devido à maior formação de peróxidos
(Oliveira et al., 2006).

73
A correlação positiva entre o ponto de fusão dos lipídios extraídos das sementes e o
percentual e índice de velocidade de germinação é característica de espécies de regiões
tropicais, onde a germinação das sementes é favorecida pela presença de lipídios de ponto de
fusão mais elevados, resultado de maior concentração de ácidos graxos saturados (Linder,
2000).
Verificou-se que sementes de Bactris gasipaes que apresentaram emergência de
plântulas em menor tempo médio, apresentaram maior concentração de ácido oleico. Esse
resultado assemelha-se ao encontrado em sementes de girassol (Helianthus annuus), nas quais
foi verificado que sementes de germinação rápida apresentaram maiores teores de ácido
oleico em relação às de germinação lenta (Munshi et al., 2007). O ácido oleico pode atuar
tanto como um inibidor natural da lipoxigenase, por modificar sua funcionalidade, como
quanto um substrato inadequado para esta enzima (Rodríguez-Rosales et al., 1998).
CONCLUSÃO
A emergência, o tempo médio de emergência e o índice de velocidade de emergência
de plântulas não diferiram entre sementes de frutos imaturos e maduros.
A concentração de lipídios foi maior em sementes de frutos imaturos.
Sementes de frutos maduros apresentaram maior concentração dos ácidos oleico e
linoleico.
A maior concentração de ácido oleico nas sementes de Bactris gasipaes resultou em
menor tempo médio de emergência de plântulas.

74
O ponto de fusão dos lipídios não diferiu entre sementes de frutos imaturos e maduros,
porém quanto maior o ponto de fusão dos lipídios (maior comprimento da cadeia
hidrocarbônica) maior a emergência e o índice de velocidade de emergência de plântulas.

75
BIBLIOGRAFIA CITADA
Al-Khalifah, N.S. 2006. Metaxenia : influence of pollen on the maternal tissue of fruits of two
cultivars of date palm (Phoenix dactylifera L.). Bangladesh journal of botany, 35(2):151-161.
Aroucha, E.M.M.; Magalhães, J.A.S.; Aroucha, M.C.M.; Viana, A.P.; Vianni, R. 2005
Composição lipídica do albúmen do coco anão verde e anão vermelho em diferentes estádios
de maturação. Caatinga, 18(3): 143-147.
Berrie, A.M.M.; Buller, D.; Parker, D.R. 1979. Possible role of volatile fatty acids and
abiscisic acids in the dormancy of oats. Plant physiology, 63: 758-764.
Besbes, S.; Blecker, C.; Deroanne, C. Drira, N.E.; Attia, H. 2004. Date seeds: chemical
composition and characteristic profiles of the lipid fraction. Food Chemistry, 84: 577-584.
Bora, P. S.; Rocha, R. V. M.; Narain, N.; Moreira-Monteiro, A. C.; Moreira, R. A. 2003.
Characterization of principal nutritional components of Brazilian oil palm (Elaeis guineensis)
fruits. Bioresource Technology, Fayetteville, 87: 1-5.
Brasil. 2009. Ministério da agricultura, pecuária e abastecimento. Regras para análise de
sementes / Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa
Agropecuária. – Brasília: Mapa/ACS, 399 p.
Buckeridge, M.S.; Aidar, M.P.M.; Santos, H.P.; Tiné, M.A.S. 2004. Acúmulo de Reservas.
In: Ferreira, A.G.; Borghetti, F. (orgs.) Germinação: do básico ao aplicado. Porto Alegre,
Artmed, 323 p.
Clement, C.R. 2008. Peach palm (Bactris gasipaes Kunth, Palmae). In: Janick, J.; Paull, R.E.
(eds.) The encyclopedia of fruit and nuts. pp. 93-101.

76
Dewir, Y.H.; El-Mahrouk, M.E.; Naidoo, Y. 2011. Effects of some mechanical and chemical
treatments on seed germination of Sabal palmetto and Thrinax morrisii palms. Australian
Journal of crop science, 5(3):248-253.
Ferreira, 1996. Maturação fisiológica de sementes de pupunha (Bactris gasipaes Kunth). Tese
de doutorado. Instituto Nacional de Pesquisas da Amazonia, Manaus, Amazonas. 73 p.
Ferreira, S.A.N. 2005. Pupunha: Bactris gasipaes Kunth (Arecaceae). Fascículo 5. 12 p. In:
Ferraz, I.D.K.; Camargo, J.L.C. (Eds) Manual de Sementes da Amazônia. Fascículo 5, 12p.
INPA, Manaus, AM, Brasil.
Friedman, H.; Meir, S.; Philosoph-Hads, S.; Havely, A.H. 2003. Effect of otanoic acid on
ethylene-mediated processes in Arabidopsis. Plant growth regulation, 40:239-247.
Ingram, J.; Bartels, D. 1996. The molecular basis of dehydration tolerance in plants. Annual
review of plant physiology and plant molecular biology, 47: 377-403.
Instituto Adolfo Lutz. 1995. Normas analíticas do Instituto Adolfo Lutz: Métodos químicos e
físicos para análise de alimentos. 3 ed. 533p.
Jatunov, S.; Quesada, S., D az, C.; Murillo, E. 2010. Carotenoid composition and
antioxidant activity of the raw and boiled fruit mesocarp of six varieties of Bactris
gasipaes. Archivos latinoamericanos de nutricion , 60(1): 99-104.
Johnson, D.V. 2010. Tropical palms 2010 revision. Food and agriculture organization of the
United Nations. Rome, 242 p.
Labouriau, L.G. 1983. A germinação das sementes. Washington: OEA. 174p.
Linder, C. R. 2000. Adaptive evolution of seed oils in plants: accounting for the
biogeographic distribution of saturated and unsaturated fatty acids in seed oils. The American
Naturalist. 156 (4): 442–458.

77
Macaire, W.H.; Bernard, T.; Noel, T.; Felicite, T.M.; Cesar, K.; Michel, L. Jacques, F. 2010.
Extraction of palm kernel oil in Cameroon: Effects of kernels drying on the oil quality.
Journal of food technology, 8(1): 1-7.
Mora-Urpí, J. 1979. Método practico para germinación de semillas de pejibaye. ASBANA.
3(1):14-15.
Munshi, S.K.; Sandhu, S. Sharma, S. 2007. Lipid composition in fast and slow germinating
sunflower (Helianthus annuus L.) seeds. Genetic applied to plant physiology, 33 (3-4): 235-
246.
Oliveira, D.A.; Piovesan, N.D.; José, I.C.; Barros, E.G; Dias, D.C.F; Moreira, M.A. 2006.
Lipoxigenases e teor de ácido linolênico relacionados à qualidade de sementes de soja.
Revista brasileira de sementes, 28(1): 30-35.
Rodríguez-Rosales, M.P.; Kerkeb, L.; Ferrol, N.; Donaire, J.P. 1998. Lipoxygenase activity
and lipid composition of cotyledons and oil bodies of two sunflower hybrids. Plant
physiology and biochemistry, 36(4):285-291.
Vieira, L.S.; Santos, P.C.T.C. 1987. Amazônia: seus solos e outros recursos naturais. São
Paulo: Ceres, 416p.
Voelkner,T.; Kinney, A.J. 2001. Variations in the biosynthesis of seed-starage lipids. Annual
review of plant physiology. Plant molecular biology. 52:335-361.

78
Tabela 1. Características físicas, variáveis de emergência, composição de ácidos
graxos, lipídios e ponto de fusão dos lipídios em sementes de Bactris gasipaes de frutos
imaturos e maduros.
Maturação dos frutos Coeficiente de variação (%) Imaturos Maduros Características físicas Teor de água (%)1 50,4 a 46,7 b 4,3 Massa seca (g)a1 1,1 a 1,3 a 16,3 Emergência Emergência (%)1 61,4 a 68,9 a 32,1 IVE1 0,319 a 0,351 a 34,0 TME (dias)1 100,3 a 101,0 a 8,2 Mortas (%)1 26,6 a 16,4 a 58,5 Dormentes (%)2 5,0 a 6,0 a 95,5 Germinadas e não emergidas (%)2 8,3 a 8,7 a 77,5 Ácidos graxos Capróico - 6:0 (µg.g-1)2 35,60 a 36,28 a 8,1 Caprílico - 8:0 (µg.g-1)2 52,69 a 55,33 a 13,5 Cáprico - 10:0 (µg.g-1)2 42,45 a 30,69 a 78,5 Láurico - 12;0 (µg.g-1)2 938,31 a 966,24 a 29,6 Mirístico -14:0 (µg.g-1)2 310,20 a 327,24 a 28,5 Palmítico - 16:0 (µg.g-1)2 109,92 a 115,79 a 32,9 Esteárico - 18:0 (µg.g-1)2 28,08 a 22,26 a 47,1 Oléico - 18:1 (µg.g-1)2 22,01 b 26,89 a 37,8 Linoleico - 18:2 (µg.g-1)2 24,79 b 30,66 a 63,7 Lipídios totais Ponto de fusão (°C)1 22,54 a 22,70 a 9,3 Lipídios (g.100g-1)1 26,29 a 24,75 b 17,6 a valores referentes a sementes sem o endocarpo; 1 foi aplicado Teste t ao nível de 5% de probabilidade; 2 foi aplicado Teste de Mann-Whitney ao nível de 5% de probabilidade. Médias com letras iguais, nas linhas, não apresentaram diferença significativa.

79
Tabela 2. Correlação entre variáveis de emergência, lipídios, ponto de fusão dos lipídios e concentração de ácidos graxos em sementes de
Bactris gasipaes.
Ácidos graxos Variáveis E IVE TME M D GNE L PF Caproico Caprílico Caprico Laurico Miristico Palmitico Esteárico Oleico Linoleico
E 1 IVE 0,97 1 TME -0,23 -0,45 1 M -0,59 -0,56 0,01 1 D -0,61 -0,59 0,16 -0,19 1 GNE -0,73 -0,72 0,28 -0,08 0,81 1 L -0,15 -0,11 -0,03 0,07 0,11 0,14 1 PF 0,66 0,70 -0,34 -0,59 -0,25 -0,29 -0,24 1
Áci
dos g
raxo
s
Capróico 0,05 0,00 0,20 -0,16 0,01 0,06 -0,33 0,01 1 Caprílico 0,06 -0,03 0,30 -0,05 -0,12 0,02 -0,43 -0,04 0,36 1 Caprico -0,04 -0,04 -0,04 0,16 -0,12 -0,08 -0,29 -0,15 0,24 0,28 1 Laurico 0,14 0,10 0,10 -0,15 -0,03 -0,07 0,30 0,06 -0,35 -0,66 -0,31 1 Miristico 0,10 0,07 0,06 -0,15 0,03 -0,06 0,35 -0,01 -0,36 -0,63 -0,33 0,93 1 Palmitico 0,02 -0,02 0,16 -0,07 0,06 -0,03 0,19 -0,11 -0,33 -0,59 -0,37 0,88 0,93 1 Esteárico -0,35 -0,35 0,12 0,10 0,31 0,33 -0,01 -0,19 -0,22 -0,18 -0,09 0,31 0,31 0,34 1 Oleico -0,01 0,09 -0,41 0,10 -0,04 -0,10 0,16 -0,03 -0,17 -0,12 -0,04 0,12 0,30 0,16 0,05 1 Linoleico 0,08 0,09 -0,10 -0,08 0,02 -0,10 -0,15 0,03 -0,34 -0,44 -0,28 0,67 0,71 0,79 0,49 0,41 1
Os valores em negrito são correlacionados, com um nível de significância alfa=0,05. E - emergência; IVE - índice de velocidade de emergência; TME - tempo médio de emergência; M - sementes mortas; D - sementes dormentes; GNE - sementes germinadas e não emergidas; L - lipídios; PF - ponto de fusão dos lipídios.

80
Capítulo 4
Dormência embrionária em sementes da palmeira Bactris gasipaes Kunth (pupunha) (Elaborado de acordo com as normas da Revista Seed Science Research)

81
Dormência embrionária em sementes da palmeira Bactris gasipaes Kunth
(pupunha)
Resumo
Bactris gasipaes (pupunha) é uma palmeira domesticada cujos frutos têm grande importância para a subsistência dos povos da Amazônia e seu palmito tem assumido importância econômica em outras regiões do Brasil. Neste trabalho, o objetivo foi investigar a presença de dormência embrionária em sementes de pupunha. Foram utilizadas sementes provenientes de oito plantas (infrutescências): quatro coletadas em Manaus/AM e quatro em Coari/AM. Após extração e limpeza, parte das sementes foi semeada em viveiro, em caixas plásticas contendo como substrato uma mistura de serragem e areia. Os embriões do restante das sementes foram inoculados em meio de cultura Murashige & Skoog, com metade da concentração de sais. A maioria das variáveis relacionadas à emergência de plântulas a partir de sementes integrais e à germinação de embriões variou significativamente entre plantas e procedências. Em geral, estas se relacionaram positivamente, ou seja, quanto maior a germinação, maior a emergência. Evidenciou-se que a dormência de sementes de pupunha pode ser atribuída, em parte, à dormência embrionária. Palavras-chave: Arecaceae; embrião; germinação.

82
Introdução
O gênero Bactris representa um dos mais diversificados grupos de palmeiras do
Novo Mundo, com, aproximadamente, 64 espécies. A maior diversidade de palmeiras
deste gênero está na Bacia Amazônica, com, no mínimo, 30 espécies (Goulding &
Smith, 2007). Bactris gasipaes, espécie domesticada, tem maior ocorrência na
Amazônia, principalmente na parte ocidental, nas localidades mais antigas situadas
próximas às margens dos principais rios da região e sua abundância depende de sua
importância para a população local (Clement, 2008).
A reprodução da pupunheira tem sido objeto de estudo há mais de um século
(Barbosa-Rodrigues, 1899). Porém, nos últimos 30 anos, com o aumento da demanda
pelo palmito de pupunha, mais estudos vêm sendo realizados a fim de se obter uma
melhor forma de propagação, bem como visando formas de conservação in situ e ex situ
desta espécie.
A germinação de sementes de diferentes progênies de pupunha varia de 0 a
100%, e o processo de germinação ocorre de 38 a 133 dias (Bovi et al, 1994). A
semente apresenta dormência por períodos variáveis de um mês e meio a 14 meses
(Mora-Urpí, 1979). Contudo, não são conhecidas as causas da dormência em sementes
desta espécie.
Uma semente dormente não tem a capacidade de germinar em um período
específico de tempo em uma combinação de fatores ambientais favoráveis à germinação
(Finch-Savage & Leubner-Metzger, 2006). Entre as possíveis causas de dormência das
sementes está a embrionária. Esta pode ser morfológica, quando é devida à imaturidade
dos embriões (Baskin & Baskin, 2003), ou fisiológica, quando os embriões são

83
desenvolvidos, mas mecanismos bioquímicos e fisiológicos impedem que a germinação
ocorra (Bazin et al., 2011).
A dormência das sementes e seus ”gatilhos” podem ser investigados utilizando a
cultura de embriões zigóticos (Fischer-Iglesias & Neuhaus, 2001). A cultura de
embriões zigóticos in vitro tem sido utilizada como uma técnica alternativa para a
rápida propagação vegetal. Este método vem sendo usado para a superação da
dormência de sementes, quando esta é causada pela imaturidade do embrião ou presença
de inibidores da germinação no endosperma, para estudar os aspectos nutricionais e
fisiológicos do desenvolvimento do embrião, entre outros (Hu & Ferreira, 1998). A
cultura de embriões de palmeiras permite a compreensão de aspectos fisiológicos do
desenvolvimento do embrião e da germinação das sementes (Triques et al., 1997).
No presente estudo o objetivo foi avaliar a diferenciação in vitro de embriões
zigóticos de diferentes indivíduos de pupunha, correlacionando-a com a emergência de
plântulas a partir de sementes e, assim, investigar a presença da dormência ligada ao
embrião.
Material e métodos
As sementes utilizadas no presente estudo foram obtidas de frutos maduros, de
oito infrutescências coletadas de plantas distintas: quatro coletados em um plantio
localizado no campus do INPA/V8, em Manaus/AM, com temperatura média anual de
26,7 °C e precipitação média anual de 2286,2 mm (Brasil, 1992) e quatro adquiridos
junto a produtores no município de Coari/AM, com temperatura média anual de 26 °C e
precipitação média anual de 2250 mm (Vieira & Santos, 1987).

84
Após o beneficiamento, que consistiu da extração e limpeza das sementes, parte
destas foi semeada em viveiro, em caixas plásticas contendo como substrato uma
mistura de areia e serragem na proporção volumétrica de 1:1. Concomitantemente, foi
feita a determinação do teor de água das sementes, pelo método estufa a 1053 oC,
durante 24 horas (Brasil, 2009), com duas repetições de cinco sementes.
Para investigar a presença de dormência ligada ao embrião, foi feita a cultura in
vitro dos mesmos, no Laboratório de Cultura de Tecidos Vegetais da Universidade do
Estado do Amazonas. As sementes, beneficiadas, foram extraídas do endocarpo, lavadas
com água destilada e feita a desinfestação conforme Steinmacher (2005), em câmara de
fluxo laminar, pela imersão das sementes em álcool 70%, por cinco minutos, seguida da
imersão, por 40 minutos, em hipoclorito comercial a 60%, adicionando-se uma gota de
surfactante Tween-20® para cada 100 mL desta solução. Para a remoção do excesso de
hipoclorito foram feitos três enxágues em água destilada autoclavada. Foi utilizado o
meio de cultura MS (Murashige & Skoog, 1962), com a metade da concentração dos
macro e micronutrientes, com adição de 1,5 g.L-1 de carvão ativado. A este meio foram
adicionadas 30 g L-1 de sacarose e 7 g.L-1 de Agar, com ajuste de pH para 5,8 antes da
adição do gelificante.
Tubos de ensaio contendo 20 mL do meio de cultura foram fechados com
tampas de alumínio e vedados com filme de PVC e autoclavados por 30 minutos. Os
embriões foram retirados da semente, em câmara de fluxo laminar e imediatamente
inoculados no meio de cultura. Foram inoculadas quatro repetições de 10 embriões
zigóticos por planta. As culturas foram mantidas em câmara B.O.D. à temperatura de 30
°C no escuro por 60 dias e após esse período foram transferidas para sala de

85
crescimento com fotoperíodo de 16 horas de luz, temperatura de 25±2 °C e
luminosidade de 52 µmol.m-2.s-1 de irradiância .
No estudo da emergência em viveiro o delineamento estatístico empregado foi
em inteiramente casualizado, com oito tratamentos (plantas), com quatro repetições de
25 sementes. As avaliações da emergência (emissão de plântula acima do nível do
substrato) foram feitas a cada cinco dias, durante 180 dias. Com esses dados foram
calculados a emergência em porcentagem, o índice de velocidade de emergência (IVE)
e o tempo médio de emergência (TME). Ao final do período de avaliação as sementes
remanescentes foram submetidas ao teste de corte (Brasil, 2009) e classificadas em
mortas e dormentes.
No estudo do crescimento dos embriões in vitro o delineamento estatístico
empregado foi em inteiramente casualizado, com oito tratamentos (plantas), com quatro
repetições de 10 embriões. As avaliações da diferenciação (emissão de raiz, Figura 1)
foram feitas a cada sete dias, durante 120 dias. Com esses dados foram calculados a
diferenciação da raiz do eixo embrionário, em porcentagem, o índice de velocidade de
diferenciação (IVD) e o tempo médio de diferenciação (TMD). Foram também
registrados os embriões que estavam mortos pela contaminação de fungos e bactérias ou
por oxidação, e aqueles que haviam entumescido, mas sem diferenciação da raiz. Com
estes dados, foram calculados a porcentagem de embriões mortos e dormentes.
Os dados foram submetidos aos testes de normalidade de Kolmogorov-Smirnov
e Shapiro-Wilk. Para dados com distribuição normal, esses foram submetidos à análise
de variância, ao nível de 5% de probabilidade, seguida de teste de Tukey, para
comparação das médias entre as plantas. Quando os dados não apresentaram
distribuição normal, as médias foram comparadas por meio do teste de Kruskal-Wallis.

86
Para comparação das médias entre as procedências, os dados foram submetidos ao teste
t, ao nível de 5% de probabilidade, quando apresentaram distribuição normal ou ao teste
de Mann Whitney, ao nível de 5% de probabilidade, quando não apresentaram
distribuição normal.
Foi feito um estudo de regressão, com 5 % de probabilidade, entre as variáveis
da diferenciação da raiz dos embriões in vitro (diferenciação, IVD, TMD, embriões
mortos e dormentes) e da emergência de plântulas a partir de sementes das respectivas
plantas em viveiro (emergência, IVE e TME, sementes mortas e dormentes). O
programa estatístico utilizado foi o ASSISTAT 7.6 Beta (Silva, 2011).
Resultados
A emergência de plântulas foi diferente entre as plantas, com valores variando
de 84 a 16% (Tabela 1). A porcentagem de sementes que estavam mortas ao final do
experimento, após 180 dias de semeadura, foi diferente entre as plantas e variou de
10,0% a 37,0%. As sementes que permaneceram viáveis (dormentes) ao final do
experimento variaram entre as plantas de 0,0 a 64,0%. Ao final do experimento,
verificou-se que algumas sementes haviam germinado sem ter apresentado emissão de
parte aérea, não havendo, contudo, diferença entre as plantas. O índice de velocidade de
emergência (IVE) variou entre as plantas, apresentando relação direta com a
emergência: plantas com maior emergência tiveram maior IVE e vice-versa. O tempo
médio de emergência (TME) variou entre plantas, com mínimo de 68,4 dias e máximo
de 132,7 dias.
Houve diferença em nível de 5% na emergência entre as sementes coletadas em
Manaus (57,3%) e em Coari (75,0%) (Tabela 2). Não houve diferença na porcentagem

87
de sementes mortas entre as duas procedências. Houve diferença significativa nas
sementes dormentes entre as procedências, com maior percentual na de Manaus (20%),
em relação à de Coari (5%). Não houve diferença entre procedências nas sementes
germinadas e não emergidas. O IVE foi diferente entre as procedências, com médias de
0,145 e 0,231, para as procedências Manaus e Coari, respectivamente. O tempo médio
de emergência para as sementes coletadas em Manaus foi maior que o das sementes
coletadas em Coari (110 e 92 dias, respectivamente).
A diferenciação (emissão da raiz) do eixo embrionário não foi diferente entre as
plantas, apresentando um mínimo de 37,5% e um máximo de 92,5% (Tabela 3). A
porcentagem de embriões mortos (oxidados ou contaminados) ao término do
experimento não diferiu entre as plantas e procedências. Os embriões que
permaneceram viáveis, sem oxidação (dormentes), apresentaram diferenças entre
plantas, com minimo de 0% e máximo de 30%. O índice de velocidade de diferenciação
da raiz dos embriões foi diferente entre as plantas variando de 0,136 a 0,323. O tempo
médio de diferenciação da raiz dos embriões não diferiu entre as plantas.
Não foram observadas diferenças na diferenciação da raiz dos embriões com
relação às procedências (Tabela 4). Foram observadas diferenças nos embriões
dormentes entre procedências, com média de 11,3% para a procedência Manaus e 3,1%
para a procedência Coari. Houve diferença entre o índice de velocidade de diferenciação
entre os embriões de sementes coletadas em Manaus (0,202) e em Coari (0,269). Não
houve diferença entre o tempo médio de germinação entre as procedências.
O índice de velocidade de emergência (IVE) de plântulas a partir de sementes
apresentou relação linear (R2= 0,66) significativa com o índice de velocidade de

88
diferenciação (IVD) dos embriões in vitro (Figura 2-A). O IVE estimado pela equação
da reta variou de 0,062, quando o IVD foi de 0,136, a 0,299 quando o IVD foi de 0,323.
A relação entre o tempo médio de diferenciação dos embriões in vitro e o tempo
médio emergência de plântulas a partir sementes foi linear, com coeficiente de
determinação de 0,61 (Figura 2- B). Em sementes cujos embriões tiveram tempo médio
de diferenciação da raiz de 25,6 dias, o tempo médio de emergência de plântulas foi de
83,2 dias, aumentando gradativamente, atingindo 127,0 dias quando o tempo médio de
diferenciação dos embriões foi de 49,5 dias. A porcentagem de sementes mortas ao
término do experimento não apresentou ajuste significativo em relação à porcentagem
de embriões mortos, e foi, em média, de 18,1%.
Discussão
No presente estudo, foi verificada que a emergência de plântulas de Bactris
gasipaes variou tanto entre plantas como entre procedências. Bovi et al. (1994),
trabalhando com sementes de 83 progênies de Yurimáguas, Peru, verificaram que a
variação na germinação de sementes desta espécie variou de 0% a 100%, tendo a
maioria das progênies germinação entre 60,0% e 89,9%. Esta proporção foi confirmada
para as progênies utilizadas no presente estudo. Os mesmos autores verificaram que o
IVG variou de 0,00 a 2,55 e o TMG de 38 a 133 dias. Nas progênies utilizadas no
presente estudo, o IVG e o TMG também tiveram grande variação e situaram-se dentro
destas faixas.
Sementes de palmeiras geralmente apresentam grande variação na germinação
entre progênies e procedências. Martins-Corder e Saldanha (2006) verificaram que
sementes de Euterpe edulis, de diferentes progênies, apresentaram variação na
germinação de 14% a 56%. Em sementes de Elaeis guineensis de diferentes progênies

89
submetidas aos mesmos tratamentos pré-germinativos, tiveram variação de 5,0% a
85,9% (Fondom et al., 2010). Em sementes de Euterpe edulis e de Syagrus
romanzoffiana, Bracalion et al. (2011) verificaram que a germinação e a velocidade de
germinação variaram significativavente entre áreas de coleta. Bactris gasipaes é uma
espécie de palmeira domesticada, porém, historicamente a seleção de matrizes para
coleta de sementes e propagação sempre esteve associada com a forma de utilização
pela população local, o que faz com que progênies cultivadas em diferentes regiões
tenham também diferentes características fenotípicas, como a capacidade germinativa e
a dormência das sementes.
O cultivo in vitro de embriões zigóticos em palmeiras tem sido apontada como
um método para obtenção de germinação em menor tempo (Pinheiro, 1986; Hu &
Ferreira, 1998). Contudo, no presente estudo, observou-se grande variação na
dormência embrionária entre plantas, evidenciando que a eficiência do método varia
com o genótipo.
No presente estudo, obteve-se uma alta porcentagem de diferenciação de
embriões, sem diferenças entre plantas. O tempo médio de diferenciação foi 35,2 dias,
em média, enquanto o tempo médio de emergência de plântulas foi de 100,9 dias, em
média. Embriões de Bactris major, quando isolados e cultivados in vitro iniciaram a
diferenciação após 10 dias, enquanto sementes intactas o fizeram após oito meses. Em
Desmoncus orthacanthos a redução no tempo inicial de germinação, com a utilização de
embriões in vitro, foi de dois meses para dois dias (Tzec-Sima et al., 2006).
Não foi observado o desenvolvimento completo de plântulas a partir de
embriões, em 120 dias de observação. Isso pode ser devido à concentração de sais do
meio de cultura ter sido reduzida à metade, sendo suficiente para que fosse observada

90
diferenciação da raiz, porém insuficiente para formação de uma plântula. Em alguns
casos, como em Cocos nucifera, é necessária a tranferência dos embriões para um novo
meio de cultura para que o desenvolvimento da plântula prossiga (Engelmann et al.,
2011).
Pode-se afirmar que a dormência das sementes é atribuída, em parte, à
dormência embrionária, pois sementes de germinação tardia apresentam também
embriões de diferenciação tardia. A dormência embrionária é um processo conhecido
em palmeiras. Ribeiro et al. (2011) estudando a dormência em Acrocomia aculeata,
verificaram que as sementes apresentam dormência fisiológica, associada ao embrião.
Dewir et al. (2011) demonstraram que sementes de Sabal palmetto apresentam
dormência exógena e fisiológica, enquanto em sementes de Thrinax morrisii a
dormência é totalmente imposta pela testa.
A não-diferenciação da raiz do embrião pode ser devida à imaturidade do
mesmo, ou à presença, em embriões maduros, de impedimentos metabólicos ao
alongamento (Borghetti, 2004). Sementes de algumas espécies, com embriões
pequenos, completam a fase morfogênica e tem o embrião totalmente diferenciado em
termos de estrutura, mas estes embriões não entram na fase de maturação, ou seja, eles
não se expandem e acumulam reservas. Sementes deste tipo geralmente contêm
quantidades relativamente grandes de endosperma, que muitas vezes circunda
inteiramente o embrião de pequeno porte. Nestas sementes, o embrião deve crescer
após a dispersão para que a germinação ocorra. Nesses casos a dormência é chamada
morfológica (Borghetti, 2004; Hilhorst, 2007). Em outras espécies o embrião está
maduro no momento da dispersão, mas mecanismos fisiológicos, impostos não somente
pelo embrião, mas por estruturas adjacentes, como o endosperma. Pesquisas recentes

91
têm demonstrado que diversas modalidades de dormência fisiológica são resultados da
interação entre as restrições impostas pelas estruturas que envolvem o embrião e seu
potencial de crescimento (Cardoso, 2008). A dormência, em embriões maduros, pode
ser resultado do impedimento imposto pelos cotilédones, no eixo embrionário, ou em
ambas as estruturas (Borghetti, 2004).
O presente estudo evidenciou que a dormência em sementes de Bactris gasipaes
pode ser atribuída, em parte, à dormência endógena, ou embrionária. A redução no
tempo de germinação após retirada dos envoltórios indicam que o embrião não adquiriu
crescimento potencial para suplantar as restrições impostas pelas estruturas externas.
Com base nas informações apresentadas, mais estudos devem ser realizados para
identificar se a dormência deve ser atribuída a mecanismos fisiológicos ou à
imaturidade do embrião. Estes estudos servirão de base para determinar métodos
eficientes visando à superação da dormência para uniformizar a germinação e,
consequentemente, a produção de mudas em tempo reduzido.

92
Referências
Barbosa-Rodrigues, J. (1899). As nupcias das palmeiras (primeiras notas)
[The nuptials of palm trees (first notes)]. In: Palmae Novae Paraguayensis quas
Descripsit et Iconibus Illustravit. pp. 35-51, Typographia Leuzinger, Rio de Janeiro.
Baskin, J.M.; Baskin, C.C. (2003). Classification, biogeography, and phylogenetic
relationships of seed dormancy. In: Seed conservation: turning science into practice.
(eds. R.D. Smith; J.B. Dickie; S.H. Linington, H.W. Pritchard, R.J. Probert). pp. 517-
544. The Royal Botanic Gardens, Kew. London.
Bazin, J., Batlla, D., Dussert, S., El-Maarouf-Bouteau, H.; Bailly, C. (2011) Role of
relative humidity, temperature, and water status in dormancy alleviation of sunflower
seeds during dry after-ripening. Journal of Experimental Botany, 62(2), 627–640.
Borghetti, F. (2004). Dormência embrionaria.[Embryo dormancy]. In: Germinação: Do
básico ao aplicado. (eds. A.G. Ferreira and F. Borghetti). pp. 109-123, Artmed, Porto
Alegre.
Bovi, M.L.A.; Spiering, S.H.; Martins, A.L.D.; Pizzinato, M.A.; Lourenção, A.L.;
Chaves Flores, W.B (1994). Seed germination of progênies of Bactris gasipaes:
percentage, speed and duration. Acta horticulturae, 360(8), 283-289.
Brancalion, P.H.S., Novembre, A.D.L.C. and Rodrigues, R.R. (2011). Seed
development, yield and quality of two palm species growing in different tropical forest
types in SE Brazil: implications for ecological restoration. Seed Science and
Technology, 39, 412-424.

93
Brasil. M.A.R.A. (1992). Normais climatológicas [Climatological normal] (1961-
1992). 84 p., Ministério da Agricultura e Reforma Agrária, Brasília.
Brasil. M.A.P.A. (2009) Regras para análise de sementes [Rules for seed testing], 399
p., Ministério da Agricultura, Pecuária e Abastecimento. Secretaria de Defesa
Agropecuária, Brasília.
Cardoso, V.J.M. (2008). Germinação [Germination]. In: Fisiologia vegetal (ed.
G.B.Kerbauy) 2a edição, pp. 384-407, Guanabara Koogan, Rio de Janeiro.
Clement, C.R. (2008). Peach palm (Bactris gasipaes Kunth, Palmae). In: The
encyclopedia of fruit and nuts. (eds. J. Janick and R.E. Paull) pp. 93-101. CABI,
London, United Kingdom.
Dewir, Y.H.; El-Mahrouk, M.E.; Naidoo, Y. (2011). Effects of some mechanical and
chemical treatments on seed germination of Sabal palmetto and Thrinax morrisii palms.
Australian Journal of crop science, 5(3), 248-253.
Engelmann, F.; Malaurie, B.; N’Nan, O. (2011). In vitro culture of coconut (Cocos
nucifera L.). In: Plant embryo culture: methods and protocols. (eds. T.A. Thorpe and
E.C. Yeung), pp. 63-74, Calgary, Alberta, Canada.
Finch-Savage, W.E.; Leubner-Metzger, G. (2006). Seed dormancy and the control of
germination. New Phytologist, 171, 501–523.
Fischer-Iglesias, C.; Neuhaus, G. (2001) Zygotic embryogenesis: hormonal control of
embryo development. In: Current trends in the embryology of angiosperms (eds. S.S.
Bhojwani and S.S. W.Y. Soh), pp. 223–247, Kluwer, Dordrecht.

94
Fondom, N.Y.; Etta C.E.; Mih, A.M. (2010). Breaking Seed Dormancy: Revisiting
Heat-Treatment Duration on Germination and Subsequent Seedling Growth of Oil Palm
(Elaeis guineensis Jacq.) Progenies. World Journal of Fungal and Plant Biology, 1(2),
51-58.
Goulding, M; Smith, N. (2007) Palmeiras: Sentinelas para a conservação da Amazônia
[Palms: Sentinels for Amazon conservation]. 358 p. Amazon Conservation Association-
Sociedade Civil Mamirauá, Lima.
Hilhorst, H.W.M. (2007). Definitions and hypotheses of seed dormancy. In: Seed
development, dormancy, and germination (ed. K. Bradford and H. Nonogaki), 50-71,
Blackwell Publishing, Oxford, United Kingdom.
Hu, C.Y.; Ferreira, A.G. (1998). Cultura de embriões [Embryo culture]. In: Cultura de
Tecidos e transformação genética de plantas (eds. A. C Torres; L.S. Caldas; J.A. Buso),
1: 371-394,SPI/Embrapa, Brasília.
Martins-Corder, M.P.; Saldanha, C.W. (2006). Germinação de sementes e crescimento
de plântulas de diferentes progênies de Euterpe edulis Mart. [Seed germination and
seedling growth of different progenies of Euterpe edulis Mart.]. Revista árvore, 30(5),
693-699.
Mora-Urpí, J. (1979). Método practico para germinación de semillas de pejibaye
[Practical method for germination of peach palm]. Asbana. 3(1),14-15.
Murashige, T; Skoog, F (1962). A revised medium for rapid growth and bioassays with
tobacco tissue cultures. Physiologia Plantarum, 15,473–497.

95
Pinheiro, C.U.B. (1986). Germinação de sementes de palmeiras: revisão bibliográfica
[Germination of palm trees: a literature review]. 102 p, Embrapa-UEPAE de Teresina,
Teresina.
Ribeiro, L.M., Souza, P.P., Rodrigues Jr., A.G., Oliveira, T.G.S. and Garcia, Q.S.
(2011). Overcoming dormancy in macaw palm diaspores, a tropical species with
potential for use as bio-fuel, Seed Science and Technology, 39, 303-317.
Silva, F.A.S. (2011). Assistat. Versão 7.6. Beta. (2011). Disponível em:
www.assistat.com (Acesso em: 12/08/2011/. UAEA-CTRN-UFCG, Campina Grande.
Steinmacher, D.A. 2005. Germinação in vitro, criopreservação e embriogênese
somática em pupunha [In vitro Germination, cryopreservation and somatic
embryogenesis in peach palm] Dissertação (Mestrado em Recursos Genéticos Vegetais).
124 p. UFSC, Florianópolis.
Triques, K.; Rival, A.; Beulé, T.; Puard, M.; Roy, J. Nato, A.; Lavergne, D.; Havaux,
M. Verdeil, J.L.Sangare, A.; Hamon, S (1997). Photosynthetic ability of in vitro grown
coconut (Cocus nucifera L.) plantlets derived from zigotic embryos. Plant Science, 135,
39-51.
Vieira, L.S.; Santos, P.C.T.C. (1987). Amazônia: seus solos e outros recursos naturais
[Amazon: their soils and other natural resources]. São Paulo: Ceres, 416p.
Tzec-Simá, M.A.; Orellana, R.; Robert, M.L. (2006). In vitro rescue of isolated embryos
of Bactris major Jacq. and Desmoncus orthacanthos Mart., potentially useful native

96
palms from the Yucatan Peninsula (Mexico). In vitro cellular and development biology-
Plant., 42, 54-58.
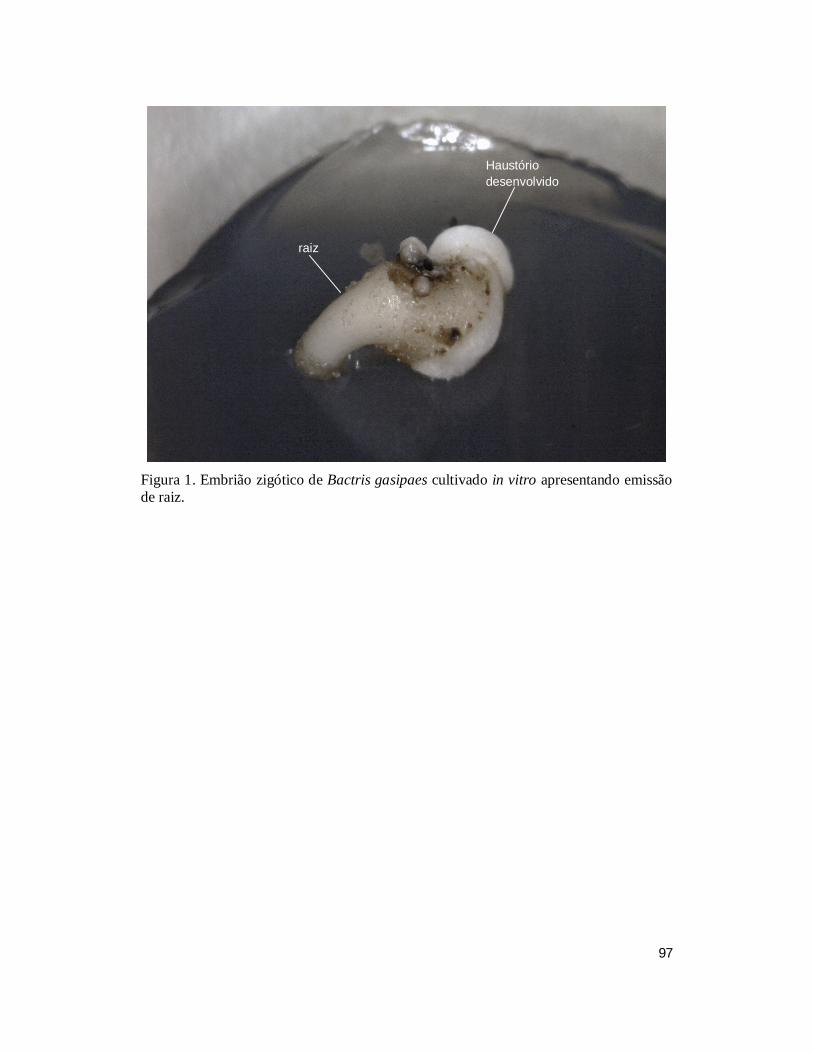
97
Figura 1. Embrião zigótico de Bactris gasipaes cultivado in vitro apresentando emissão de raiz.
raiz
Haustório desenvolvido

98
Tabela 1. Emergência, sementes mortas, dormentes, germinadas e não emergidas (GNE), índice de velocidade de emergência (IVE) e tempo médio de emergência (TME) de plântulas de Bactris gasipaes a partir de sementes de diferentes plantas.
Planta Emergência (%)2 Mortas (%)1 Dormentes (%)2 GNE (%)2
IVE 1
TME 1
L19P4 84,0 a 10,0 a 4,0 b 2,0 a 0,201 bc 108,4 d
L11P3 67,0 bc 20,0 ab 12,0 c 1,0 a 0,155 c 110,9 d
L18P3 16,0 e 17,0 ab 64,0 d 3,0 a 0,032 d 132,7 e
L15P5 62,0 d 37,0 b 0,0 a 1,0 a 0,190 bc 88,7 bc
A 82,0 a 16,0 ab 1,0 a 1,0 a 0,328 a 68,4 a
B 64,0 cd 20,0 ab 13,0 c 3,0 a 0,157 c 104,4 cd
C 70,0 bc 10,0 a 6,0 b 14,0 a 0,164 bc 112,4 d
D 84,0 a 15,0 ab 0,0 a 1,0 a 0,274 ab 80,9 ab
CV (%) 21,52 59,47 43,57 163,78 26,16 20,32 1Foi aplicado Teste de Tukey ao nível de 5% de probabilidade. 2Foi aplicado Teste de Kruskal-Wallis, ao nível de 5% de probabilidade. Médias com letras iguais, nas colunas, não apresentaram diferença significativa.

99
Tabela 2. Emergência, sementes mortas, sementes dormentes, sementes germinadas e não emergidas (GNE), índice de velocidade de emergência (IVE) e tempo médio de emergência (TME), diferenciação, embriões mortos, dormentes, índice de velocidade de diferenciação (IVD) e tempo médio de diferenciação (TMD) entre as sementes obtidas de frutos coletados em Manaus e Coari.
Sementes Procedência
Embriões Procedência
Manaus Coari Manaus Coari
Emergência (%) 2 57,3 b 75,0 a Diferenciação (%) 2 73,1 a 87,5 a
Mortas (%) 1 21,0 a 15,3 a Mortos (%) 2 15,6 a 9,4 a
Dormentes (%) 2 20,0 b 5,0 a Dormentes (%) 2 11,3 b 3,1 a
GNE (%) 2 1,8 a 4,8 a - - -
IVE 1 0,145 b 0,231 a IVD 1 0,202 b 0,269 a
TME (dias) 1 110,2 b 91,6 a TMD (dias) 2 41,8 a 28,6 a 1Foi aplicado Teste t em nível de 5% de probabilidade. 2 Foi aplicado Teste de Mann-Whitney, em nível de 5% de probabilidade. Médias com letras iguais, nas linhas, não apresentaram diferença significativa.
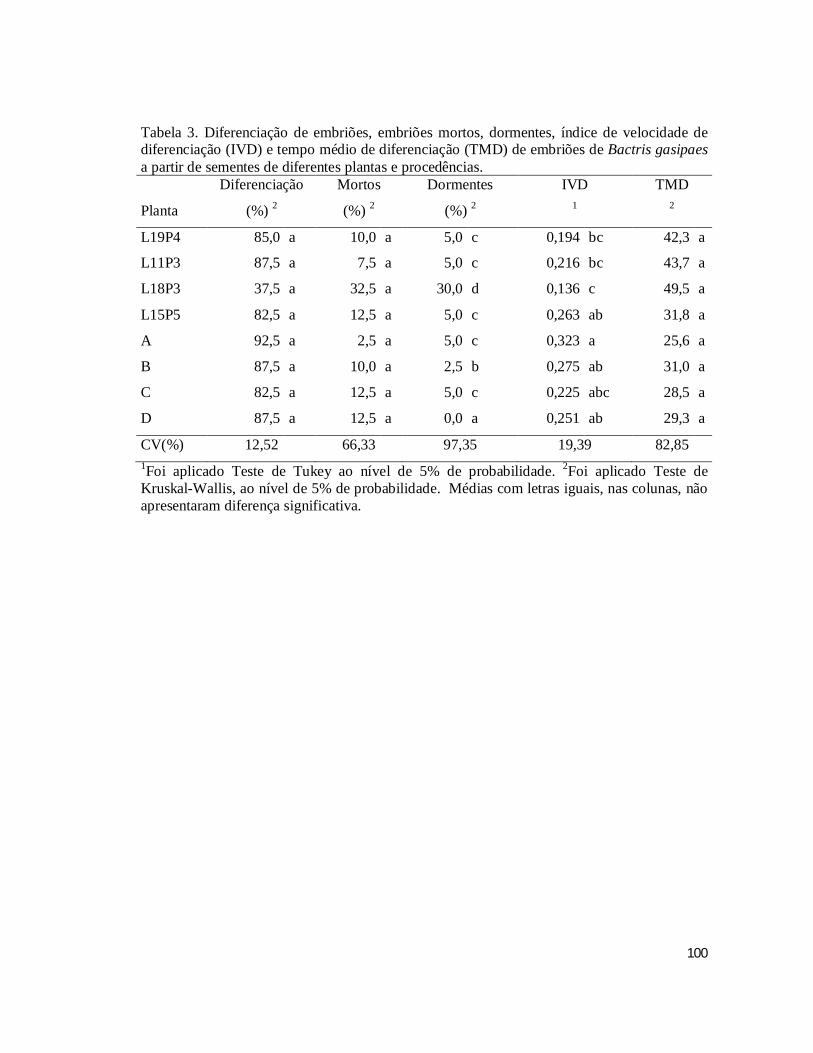
100
Tabela 3. Diferenciação de embriões, embriões mortos, dormentes, índice de velocidade de diferenciação (IVD) e tempo médio de diferenciação (TMD) de embriões de Bactris gasipaes a partir de sementes de diferentes plantas e procedências.
Planta
Diferenciação
(%) 2
Mortos
(%) 2
Dormentes
(%) 2
IVD 1
TMD 2
L19P4 85,0 a 10,0 a 5,0 c 0,194 bc 42,3 a
L11P3 87,5 a 7,5 a 5,0 c 0,216 bc 43,7 a
L18P3 37,5 a 32,5 a 30,0 d 0,136 c 49,5 a
L15P5 82,5 a 12,5 a 5,0 c 0,263 ab 31,8 a
A 92,5 a 2,5 a 5,0 c 0,323 a 25,6 a
B 87,5 a 10,0 a 2,5 b 0,275 ab 31,0 a
C 82,5 a 12,5 a 5,0 c 0,225 abc 28,5 a
D 87,5 a 12,5 a 0,0 a 0,251 ab 29,3 a
CV(%) 12,52 66,33 97,35 19,39 82,85 1Foi aplicado Teste de Tukey ao nível de 5% de probabilidade. 2Foi aplicado Teste de Kruskal-Wallis, ao nível de 5% de probabilidade. Médias com letras iguais, nas colunas, não apresentaram diferença significativa.

101
Figura 2. Índice de velocidade de emergência de plântulas em função do índice de velocidade de diferenciação de embriões (A) e tempo médio de emergência de plântulas em função do tempo médio de diferenciação de embriões (B) a partir de frutos Bactris gasipaes de diferentes plantas.

102
SÍNTESE
O presente trabalho elucidou que as sementes de Bactris gasipaes são
bitegumentadas; o tegumento consiste de um estrato interno com duas a quatro
camadas de células repletas de compostos fenólicos e o externo, com cerca de dez
camadas de células com conteúdo de lipídios e proteínas, entre as quais se
encontram feixes vasculares. Foram observados compostos fenólicos, como os
taninos, que podem atuar na inibição da germinação.
O endosperma das sementes é homogêneo, duro, esbranquiçado e ocupa
quase todo o espaço interno das sementes; suas células possuem formato oblongo,
oval e oboval, com conteúdo de lipídios ácidos e neutros, proteínas, e
polissacarídeos. Compostos lipídicos no endosperma das sementes de algumas
espécies de monocotiledôneas constituem a principal reserva que será consumida
pelo embrião durante a germinação e crescimento da plântula, porém, em algumas
espécies, a germinação pode ser inibida pela presença de determinados ácidos
graxos.
O embrião é cônico, homogêneo e pequeno, com células arredondadas, com
feixes vasculares na região distal. Na região proximal aparece a plúmula, com três
primórdios foliares e as células precursoras da radícula, oposta a esta região; o
conteúdo celular é de lipídios, polissacarídeos e proteínas. A presença da plúmula
indica que o embrião está completamente diferenciado. As células não apresentam
conteúdo fenólico.
A emergência de plântulas de pupunha não foi diferente entre sementes
advindas de frutos em dois estádios finais maturação, porém sementes de frutos
maduros de pupunha apresentam maior concentração de compostos fenólicos que
sementes de frutos imaturos e o acúmulo de compostos fenólicos é um dos fatores
responsáveis pela inibição da germinação em sementes de pupunha.
O extrato fenólico de sementes de pupunha foi capaz de diminuir e retardar a
germinação de sementes de alface e esta apresentou correlação significativa com a
emergência de plântulas de pupunha.

103
A concentração de lipídios é maior em sementes de frutos imaturos.
Sementes de frutos maduros apresentam maior concentração dos ácidos oleico e
linoleico. A maior concentração de ácido oleico nas sementes de Bactris gasipaes
resulta em menor o tempo médio de emergência de plântulas. Quanto maior o ponto
de fusão dos lipídios (maior comprimento da cadeia hidrocarbônica) maior a
emergência e o índice de velocidade de emergência de plântulas.
A dormência em sementes de Bactris gasipaes, pode ser atribuída, em parte,
à dormência endógena. A redução no tempo de germinação, após retirada dos
envoltórios, indicam que estas estruturas (endocarpo e testa) também podem
contribuir para a dormência.
Com base nas informações apresentadas, estudos futuros devem ser
voltados à identificação dos mecanismos fisiológicos da dormência. Estes estudos
servirão de base para determinar métodos eficientes visando à superação da
dormência para uniformizar a germinação e, consequentemente, a produção de
mudas de pupunha em tempo reduzido.

104
REFERÊNCIAS BIBLIOGRÁFICAS
Almeyda, N.; Martin, F.W. 1980. The pejipaye. In: Cultivation of neglected tropical
fruits with promise. USDA, pt. 8. 10p.
Babiano, M.J.; Aldasoro, J.J.; Hernándes-Nistal, J.; Rodriguez, D.; Matilla, A. 1984.
Effect of nonanoic acid and other short chain fatty acids on exchange properties
in embryonic axes of Cicer arietinum during germination. Physiologia Plantarum,
61(3): 391-395.
Baskin, C.C.; Baskin, J.M. 1998. Seeds: ecology, biogeography, and evolution of
dormancy and germination. San Diego: Academic Press. 666p.
Baskin, J.M; Baskin, C.C. 2004. A classification system for seed dormancy. Seed
science research, 14: 1-16.
Barbosa-Rodrigues, J. (1899). As nupcias das palmeiras (primeiras notas). In:
Palmae Novae Paraguayensis quas Descripsit et Iconibus Illustravit. pp. 35-51,
Typographia Leuzinger, Rio de Janeiro.
Berrie, A.M.M.; Buller, D.; Don, R. Parker. 1979. Possible role of volatile fatty acids
and abiscisic acids in the dormancy of oats. Plant physiology, 63: 758-764.
Bewley, J.D. 1997. Seed dormancy and germination. The plant cell, 9: 1055-1066.
Borghetti, F. 2004. Dormência embrionaria. In: Ferreira, A.G.; Borghetti, F.
Germinação: Do básico ao aplicado. Artmed, Porto Alegre, pp. 109-123.
Bovi, M.L.A.; Spiering, S.H.; Martins, A.L.D.; Pizzinato, M.A.; Lourenção, A.L.;
Chaves Flores, W.B. 1994. Seed germination of progênies of Bactris gasipaes:
percentage, speed and duration. Acta horticulturae, 360(8):283-289.
Bovi, M.L.A.; Spiering, S.H.; Saes, S.A. 2004. Peach palm growth and heart-of-palm
yield responses to liming. Revista brasileira de Ciência do Solo.28: 1005-1012.

105
Bovi, M.L.A.; Martins, C.C.; Spiering, S.H. 2004. Desidratação de sementes de
quatro lotes de pupunheira: efeitos sobre a germinação e o vigor. Horticultura
Brasileira, 22(1):109–112.
Cardoso, V.J.M. 2004. Dormência: estabelecimento do processo. In: Ferreira, A.G.;
Borghetti, F. Germinação: Do básico ao aplicado. Artmed, Porto Alegre, pp. 95-
108.
Carvalho, P.E.R. 2003. Espécies arbóreas brasileiras. Volume 1. 1ª edição. Embrapa
Florestas. 1040p.
Carvalho, J.E.U.; Muller, C.H. 1998. Níveis de tolerância e letal de umidade em
sementes de pupunheira, Bactris gasipaes. Revista brasileira de fruticultura.
20(3): 283-289.
Clement, C.R. 1988. Domestication of the pejibaye palm (Bactris gasipaes): past and
present. In: Balick, M.J. (ed.). The Palm - Tree of Life. Biology, Utilization and
Conservation. Advances in Economic Botany, 6:155-174.
Clement, C.R. 1990. Regeneração Natural de Pupunha (Bactris gasipaes). Acta
Amazônica. 20: 399-403.
Clement, C.R.; Dudley, N.S. 1995. Effect of botton heat and substrate on seed
germination of pejibaye (Bactris gasipaes) in Hawaii. Principes. 39(1):21-24.
Debeaujon, I. Léon-Kloosterziel, K.M., Koorneef, M. 2000. Influence of the testa on
seed dormancy, germination and longevity in Arabidopsis. Plant physiology, 122:
403-413.
Dewir, Y.H.; El-Mahrouk, M.E.; Naidoo, Y. 2011. Effects of some mechanical and
chemical treatments on seed germination of Sabal palmetto and Thrinax morrisii
palms. Australian Journal of crop science, 5(3):248-253.
Dietrich, S.M.C. 1986. Inibidores de crescimento. In: Ferri, M.G. (Coord.) Fisiologia
Vegetal. 2 ed.São Paulo: EPU, v.2, p.193-212.

106
Ferreira, S.A.N.; Santos, L.A. 1992. Viabilidade de sementes de pupunheira (Bactris
gasipaes Kunth). Acta Amazonica, 22(3): 303-307.
Ferreira, S.A.N.; Gentil, D.F.O. 2006. Extração, embebição e germinação de
sementes de tucumã (Astrocaryum aculeatum). Acta Amazonica, 36: 141-146.
Ferreira, S.A.N. 1996. Maturação fisiológica de sementes de pupunha. Tese de
doutorado. INPA/UFAM. 73p.
Ferreira, S.A.N.; Ferraz, I.D.K. 2007. Artificial maturation of pech palm (Bactris
gasipaes Kunth). In: 28th ISTA Congress/ XV Congresso Brasileiro de Sementes,
Foz do Iguaçu, p. 79.
Fior, C.S.; Rodrigues, L.R.; Leonhardt, C.; Schwarz, S.F. 2011. Superação de
dormência em sementes de Butia capitata. Ciencia rural, 41(7): 1150-1153 .
Fondom, N.Y; Etta, C.E. 2010. Breaking seed dormancy: revisiting heat-treatment
duration on germination and subsequent seedling growth of oil palm (EIaeis
guineensis Jacq.) progenies. Journal of agricultural science, 2(2):101-110.
George, E. F. 1996. Plant propagation and micropropagation: Plant propagation by
tissue culture. Exegetics. 2: 37-66.
Labouriau, L.G. 1983. A germinação das sementes. Washington: OEA. 174p.
Martine, B.M; Laurent, K.K.; Pierre, B.J., Eugéne, K.K.; Hilaire, K.T.; Justin, K.Y.
2009.Effect of storage and heat treatments on the germination of oil palm (Elaeis
guineensis Jacq.) seed. African journal of agricultural research. 4 (10)- 931-937.
Moïse, J.A.; Han, S; Gudynaite-Savitch, L.; Johnson, D.A.; Miki, B.L.A. 2005. Seed
coats: structure, development, composition and biotechnology. In vitro cellular
development biology-plant, 41:620-644.
Mora-Urpí, J. 1979. Método practico para germinación de semillas de pejibaye.
ASBANA. 3(1):14-15.

107
Nazário, P.; Ferreira, S.A.N. 2010. Emergência de plântulas de Astrocaryum
aculeatum G. May. em função da temperatura e do período de embebição das
sementes. Acta amazonica, 40(1):165-170.
Ribeiro, L.M., Souza, P.P., Rodrigues Jr., A.G., Oliveira, T.G.S. and Garcia, Q.S.
2011. Overcoming dormancy in macaw palm diaspores, a tropical species with
potential for use as bio-fuel, Seed science and technology, 39, 303-317.
Santiago, E. J. A.; Paiva, R.; Paiva, P. D. O.; Santana, J. R. F.; Gomes, G. A. C.
2001; Multiplicação: Cultura de tecidos. Paiva e Paiva, UFLA, Lavras, M.G. 5:50-
57.
Silva, V.L.; Môro, F.V.; Damião Filho, C.F. Moro, J.R.Silva, B.M.S.; Charlo, H.C.O.
2006. Morfologia e avaliação do crescimento inicial de plântulas de Bactris
gasipaes Kunth. (Arecaceae) em diferentes substratos. Revista Brasileira de
Fruticultura, 28(3): 477-480.
Steinmacher, D.A. 2005. Germinação in vitro, criopreservação e embriogênese
somática em pupunha. Dissertação (Mestrado em Recursos Genéticos Vegetais).
Florianópolis: UFSC.
Steinmacher, D.A.; Saldanha, C.W.; Clement, C.R.; Guerra, M.P. 2007a.
Cryopreservation of peach palm zygotic embryos. Cryoletters, 18(1): 13-22.
Toole, E,H.; Hendricks, S.B.; Borthwick, H.; Toole, V.K. 1956. Physiology of seed
germination. Annual Review of Plant Physiology. 7:299-324.
Tomlinson, P.B. Essays on the morphology of Palms. I Germination and the seedling.
Principes, 4(1): 56-63, 1960.
Tomlinson, P. B. Anatomy of the monocotyledons. II Palmae . Oxford:
Clarendon Press, 1961. 453p.
Triques, K.; Rival, A.; Beulé, T.; Puard, M.; Roy, J. Nato, A.; Lavergne, D.; Havaux,
M. Verdeil, J.L.Sangare, A.; Hamon, S. 1997. Photosynthetic ability of in vitro

108
grown coconut (Cocus nucifera L.) plantlets derived from zigotic embryos. Plant
Science, 135: 39-51.
Tzec-Simá, M.A.; Orellana, R.; Robert, M.L. 2006. In vitro rescue of isolated embryos
of Bactris major Jacq. and Desmoncus orthacanthos Mart., potentially useful
native palms from the Yucatan Peninsula (Mexico). In vitro cellular and
development biology- Plant.42: 54-58.
Ulbright, C.E.; Pickard, B.G.; Warner. J.F. 1982. Effects of short chain fatty acids on
radicle emergence and root growth in lettuce. Plant, cell and environment, 5(4):
293-301.
Villalobos, R.; Herrera, J. 1991. Germinacion de la semilla de pejibaye (Bactris
gasipaes). I. Efecto de la temperatura y el sustrato. Agronomia Costarricense,
15(1/2):57-62.
Villalobos, R.; Herrera, J; Guevara, E. 1992. Germinación de la semilla de pejibaye
(Bactris gasipaes) II. Ruptura del reposo. Agronomía Costarricense, 16(1): 61-68.