diversidade e ecologia do fitoplâncton ... -...
TRANSCRIPT

Universidade Federal de Minas Gerais
Instituto de Ciências Biológicas
Programa de Pós-Graduação em Ecologia, Conservação e Manejo de Vida Silvestre
Tese apresentada ao Curso de Ecologia, Conservação e Manejo da Vida Silvestre da Universidade Federal de Minas Gerais, como requisito parcial à obtenção do título de Doutor em Ecologia.
Orientador: Prof. Dr. Francisco Antônio Rodrigues Barbosa
Cristiane Freitas de Azevedo Barros
Belo Horizonte
2010
Diversidade e ecologia do fitoplâncton em 18 lagoas naturais
do médio Rio Doce

II
À minha família, com todo meu amor
e minha gratidão

III
Agradecimentos
Agradeço ao orientador e amigo, Professor Francisco Barbosa, por me acolher na limnologia desde os primeiros passos, sempre acompanhando e incentivando meu crescimento.
Ao pessoal dos laboratórios de Limnologia e Ecologia do zooplâncton, pelo apoio e agradável convivência ao longo desses anos. Em especial à Professora Paulina, Rosinha, Marcelo e Graça, que me acompanham desde sempre e por estarem sempre tão disponíveis para ajudar.
Aos amigos do laboratório de Ecologia do Zooplâncton, que me acolheram sempre com tanto carinho, me fazendo sentir parte da equipe, esquecendo, em vários momentos, que eu era uma das “meninas do fito”.
Aos amigos que me ajudaram nas coletas (as que deram e as que não deram certo): Filipe, Atenágoras, Sofia, Akemi, Diego, Lu Brandão, Sidmar, Nelson e Thiago. Agradeço imensamente pela boa vontade e paciência de todos vocês com os contratempos e estresses de campo. O projeto não teria saído do papel sem vocês! Espero que, como eu, considerem que passamos semanas divertidas, apesar da correria e do cansaço.
Aos que ajudaram com as análises de laboratório, Simone, Marcelo, Diego, Pedro, Raquel, Akemi, Atenágoras, Débora, pela disponibilidade, dedicação e comprometimento, e por abrirem mão do descanso em alguns fins de semana.
Ao Diego Pujoni, com quem tive a imensa sorte e a felicidade de dividir este trabalho. Muitíssimo obrigada por aceitar trabalhar com as amostras de zooplâncton, mesmo tendo a metade do tempo que eu tive, por ter tido a paciência e de me ajudar (e me ensinar) com as análises estatísticas, mesmo na correria para cumprir seus próprios prazos.
Ao Nelson Mello, pela ajuda com a CCA. À Maria Bêtania Gonçalves Souza e ao Professor José Fernandes Bezerra-Neto pelas valiosas discussões e pela revisão dos textos. À Professora Judit Padisák pelas importantes sugestões para o último capítulo. Ao Luiz, pela ajuda na formatação da tese.
Às “meninas do fito” de todas as gerações: Vi, a primeira a chegar, e a que me ajudou a descobrir e me apaixonar pelas algas. Às segunda e terceira gerações, que já se misturam na minha cabeça: Bê, Erikita, Paula, João, Fábio, Maíra, que tornaram o trabalho no laboratório a melhor parte do dia. Nossas discussões científicas me acrescentaram muito! Às mais novas: Paulinha e Aline (empatia desde o primeiro segundo!), obrigada pela amizade, pelo apoio, pelo empenho e pela ajuda na identificação das espécies, na organização das amostras e planilhas e, por assumirem várias vezes funções do laboratório atribuídas a mim, para que eu pudesse ficar “por conta” da tese.
Aos amigos para todo o sempre: Ana Letícia, Bê, Érika, Flávio, Renata, Raquel Simone e Vi, por me apoiarem nas decisões loucas, me acudirem nos momentos de desespero, ouvirem meus desabafos e sempre me inspirarem a

IV
ser uma pessoa melhor. A vida, com certeza, não teria a mesma graça e leveza sem vocês por perto.
A minha família e ao Luiz, um muito obrigada infinito por representarem só coisas boas na minha vida: apoio, amor, carinho, incentivo... Pela paciência com a minha falta de paciência nos últimos meses. Sei que vocês também sofreram pra este trabalho sair... com o “está acabando” que não virava “acabou” nunca!...Tenham a certeza de que contribuíram demais pra essa tese acontecer! Amo vocês demais!
Aos professores Carlos Bicudo, Célia Sant’Anna, Vera Werner e Thelma Ludwig, pela ajuda na identificação das espécies.
Ao Instituto de Ciências Biológicas e ao Curso de Pós-Graduação em Ecologia, Conservação e Manejo de Vida Silvestre da UFMG, pela infra-estrutura e apoio durante a realização desse trabalho. Em especial, ao Fred, Cris e Mary, pela ajuda com as questões burocráticas.
Ao Conselho Nacional de Pesquisa (MCT-CNPq) pelo financiamento do projeto através do Programa de Pesquisas Ecológicas de Longa Duração (PELD-Site 4).
À Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), pela bolsa de estudos concedida.
Ao Instituto Estadual de Florestas de Minas Gerais (IEF-MG), pela licença de coleta, e à administração do Parque Estadual do Rio Doce (PERD) pelo apoio logístico e disponibilização dos alojamentos e mateiros, fundamentais para a realização deste estudo. Obrigada também aos Srs. Tomé e Valtinho, que nos acompanharam, sempre que possível, às lagoas de acesso mais complicado.
À Companhia Agrícola Florestal – Santa Bárbara (Grupo Arcelor) pela autorização para coleta nas lagoas localizadas dentro das áreas da empresa.
À Polícia Ambiental de MG, especialmente ao pelotão do PERD, na pessoa do Sr. Eduardo Dutra, pelo acompanhamento durante a escolha das lagoas e primeiras amostragens.

V
SUMÁRIO
ÍNDICE DE TABELAS ...................................................................................................................... VI
ÍNDICE DE FIGURAS ...................................................................................................................... VII
RESUMO GERAL ............................................................................................................................... 1
GENERAL ABSTRACT ...................................................................................................................... 2
INTRODUÇÃO GERAL ...................................................................................................................... 3
ÁREA DE ESTUDOS .......................................................................................................................... 8
CAPÍTULO 1 ...................................................................................................................................... 15
PADRÕES REGIONAIS E VARIAÇÃO ESPACIAL DA DIVERSIDADE FITOPLANCTÔNICA
NO SISTEMA LACUSTRE DO MÉDIO RIO DOCE, MINAS GERAIS......................................... 15
Resumo ........................................................................................................................................... 16
Abstract ........................................................................................................................................... 17
Introdução ....................................................................................................................................... 18
Materiais e métodos ..................................................................................................................... 20
Resultados ...................................................................................................................................... 25
Discussão ....................................................................................................................................... 34
CAPÍTULO 2 ...................................................................................................................................... 49
ATENUAÇÃO DA RADIAÇÃO VISÍVEL EM 16 LAGOAS NATURAIS DO MÉDIO RIO DOCE, MINAS GERAIS: FATORES DETERMINANTES E IMPLICAÇÕES PARA O
FITOPLÂNCTON ................................................................................................................................ 49
Resumo ........................................................................................................................................... 50
Introdução ....................................................................................................................................... 52
Área de estudo .............................................................................................................................. 54
Metodologia .................................................................................................................................... 55
Resultados ...................................................................................................................................... 57
Discussão ....................................................................................................................................... 64
CAPÍTULO 3 ...................................................................................................................................... 70
RESPOSTA DOS GRUPOS FUNCIONAIS FITOPLANCTÔNICOS AOS GRADIENTES
AMBIENTAIS NO SISTEMA LACUSTRE DO MÉDIO RIO DOCE, MG ..................................... 70
Resumo ........................................................................................................................................... 71
Abstract ........................................................................................................................................... 72
Introdução ....................................................................................................................................... 73
Materiais e métodos ..................................................................................................................... 75
Resultados ...................................................................................................................................... 79
Discussão ....................................................................................................................................... 95
DISCUSSÃO GERAL ..................................................................................................................... 102
REFERÊNCIAS BIBLIOGRÁFICAS ............................................................................................. 104

VI
ÍNDICE DE TABELAS
TABELA 1 - AMBIENTES ESTUDADOS NO BRASIL ATÉ 2009 UTILIZANDO A ABORDAGEM DOS GRUPOS
FUNCIONAIS PARA ANÁLISE DO FITOPLÂNCTON. .......................................................................... 6
TABELA 2 - COORDENADAS GEOGRÁFICAS, ALTITUDE, PERÍMETRO, ÍNDICE DE DESENVOLVIMENTO DE
MARGEM (DL) E ÁREA DAS LAGOAS ESTUDADAS. ..................................................................... 11
TABELA 1. 1 - COORDENADAS GEOGRÁFICAS, ALTITUDE, ÍNDICE DE DESENVOLVIMENTO DE MARGEM
(DL), ÁREA DA LAGOA E ÁREA DA BACIA DE DRENAGEM DAS 18 LAGOAS AMOSTRADAS ENTRE
AGOSTO DE 2007 E MAIO DE 2008 NO PARQUE ESTADUAL DO RIO DOCE E ÁREA DE ENTORNO. . 21
TABELA 1. 2 - MÉDIAS E COEFICIENTES DE VARIAÇÃO DAS VARIÁVEIS LIMNOLÓGICAS MEDIDAS NA SUB-SUPERFÍCIE DE CADA LAGOA AO LONGO DOS QUATRO PERÍODOS AMOSTRADOS ......................... 26
TABELA 1. 3 - CORRELAÇÕES ENTRE AS CARACTERÍSTICAS MORFOLÓGICAS E LIMNOLÓGICAS E A
RIQUEZA DE ESPÉCIES FITOPLANCTÔNICAS PARA O CONJUNTO DE 18 LAGOAS AMOSTRADAS ENTRE
AGOSTO DE 2007 E MAIO DE 2008 NO SISTEMA LACUSTRE DO MÉDIO RIO DOCE. VALORES EM
NEGRITO INDICAM CORRELAÇÕES SIGNIFICATIVAS (P<0,05). .................................................... 29
TABELA 1. 4 - MATRIZ DE SIMILARIDADE (ÍNDICE DE JACCARD) OBTIDA PARA AS 18 LAGOAS
AMOSTRADAS NO PERD ENTRE AGOSTO DE 2007 E MAIO DE 2008 .......................................... 31
TABELA 2. 1 - ÁREA E PROFUNDIDADE DAS 16 LAGOAS AMOSTRADAS. .............................................. 57
TABELA 2. 2 - RESULTADOS OBTIDOS PARA REGRESSÃO LINEAR SIMPLES DA PROFUNDIDADE TOTAL NO
PONTO DE COLETA E OS TEORES DE FÓSFORO TOTAL (PTOTAL) E MATÉRIA ORGÂNICA DISSOLVIDA
COLORIDA (MODC) E O COEFICIENTE DE ATENUAÇÃO DA LUZ K0 .............................................. 59
TABELA 2. 3 - RESULTADOS OBTIDOS PARA ANÁLISE DE REGRESSÃO MÚLTIPLA DO K0 UTILIZANDO
BIOMASSA DO FITOPLÂNCTON, MATÉRIA ORGÂNICA DISSOLVIDA COLORIDA E CONCENTRAÇÃO DE
FÓSFORO TOTAL COMO VARIÁVEIS INDEPENDENTES ................................................................. 59
TABELA 2. 4 - PRINCIPAIS ESPÉCIES E PORCENTAGEM DA BIOMASSA REPRESENTADA EM CADA LAGOA
DURANTE AS QUATRO CAMPANHAS DE AMOSTRAGEM. .............................................................. 63
TABELA 3. 1 - MÍNIMOS E MÁXIMOS OBTIDOS PARA AS PRINCIPAIS VARIÁVEIS AMBIENTAIS DE 18 LAGOAS
NO PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO DURANTE AS AMOSTRAGENS
REALIZADAS NOS PERÍODOS DE CHUVAS (NOV/2007 E FEV/2008) E SECA (AGO/2007 E
MAIO/2008). COMPARANDO-SE OS PERÍODOS EM CADA LAGOA, OS VALORES EM NEGRITO FORAM
OS MAIS ELEVADOS (TESTE DE MANN-WHITNEY; P<0,05) ........................................................ 81
TABELA 3. 2 - CLASSIFICAÇÃO DE 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E SEU
ENTORNO EM 5 TIPOS, IDENTIFICADOS COM BASE NA PROFUNDIDADE TOTAL, COEFICIENTE DE
ATENUAÇÃO ESCALAR DA LUZ E TROFIA (AMOSTRAGENS REALIZADAS ENTRE AGOSTO DE 2007 E
MAIO DE 2008) ...................................................................................................................... 85
TABELA 3. 3 - ESPÉCIES FITOPLANCTÔNICAS DESCRITORAS (>5% DA BIOMASSA TOTAL), CLASSES
TAXONÔMICAS E GRUPOS FUNCIONAIS (GF) DE 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE
E NO SEU ENTORNO (AMOSTRAGENS REALIZADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008). .... 87
TABELA 3. 4 - CONTRIBUIÇÃO DAS PRINCIPAIS ESPÉCIES E GRUPOS FUNCIONAIS FITOPLANCTÔNICOS
MAIS FREQUENTES PARA A BIOMASSA NAS 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO, DURANTE PERÍODOS DE CHUVAS (NOVEMBRO DE 2007 E FEVEREIRO DE
2008) E SECA (AGOSTO DE 2007 E MAIO DE 2008). ................................................................. 92
TABELA 3. 5 - CORRELAÇÕES SIGNIFICATIVAS (P<0,05) ENTRE AS VARIÁVEIS ABIÓTICAS, A DENSIDADE
DE ZOOPLÂNCTON E A BIOMASSA DOS PRINCIPAIS GRUPOS FUNCIONAIS DO FITOPLÂNCTON EM 18

VII
LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO (AMOSTRAGENS
REALIZADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008) ........................................................... 93
TABELA 3. 6 - COEFICIENTES DE CORRELAÇÃO CANÔNICA (INTRA-SET) DOS VETORES AMBIENTAIS E
BIÓTICOS COM OS EIXOS 1 E 2 PARA OS GRUPOS FUNCIONAIS FITOPLANCTÔNICOS OBSERVADOS
EM 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO (AMOSTRAGENS
REALIZADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008) ........................................................... 94
Índice de Figuras
FIGURA 1 - SISTEMA LACUSTRE DO MÉDIO RIO DOCE, EVIDENCIANDO OS LIMITES DO PARQUE ESTADUAL
DO RIO DOCE, AS LAGOAS AMOSTRADAS (1 A18) E OS PONTOS DE COLETA (+). IMAGEM CEDIDA
PELO INSTITUTO ESTADUAL DE FLORESTAS DE MINAS GERAIS – IEF/MG (MODIFICADA)............ 10
FIGURA 2 - IMAGEM DE SATÉLITE DA ÁREA DE ESTUDO, COM DESTAQUE PARA AS LAGOAS CARIOCA, DOM HELVÉCIO, GAMBÁ E GAMBAZINHO, LOCALIZADAS NO PARQUE ESTADUAL DO RIO DOCE. FONTE: WWW.EARTH.GOOGLE.COM ........................................................................................ 12
FIGURA 3 - IMAGEM DE SATÉLITE DA ÁREA DE ESTUDO, COM DESTAQUE PARA AS LAGOAS ANÍBAL, PATOS E SANTA HELENA, LOCALIZADAS NO PARQUE ESTADUAL DO RIO DOCE. FONTE: WWW.EARTH.GOOGLE.COM .................................................................................................... 12
FIGURA 4 - IMAGEM DE SATÉLITE DA ÁREA DE ESTUDO, COM DESTAQUE PARA AS LAGOAS CENTRAL, LOCALIZADA NO PARQUE ESTADUAL DO RIO DOCE, E PIMENTA, LOCALIZADA NA ÁREA DE
ENTORNO. FONTE: WWW.EARTH.GOOGLE.COM ........................................................................ 13
FIGURA 5 - IMAGEM DE SATÉLITE DA ÁREA DE ESTUDO, COM DESTAQUE PARA AS AMARELA, ÁGUAS
CLARAS E PALMEIRINHA, LOCALIZADAS NA ÁREA DE ENTORNO DO PARQUE ESTADUAL DO RIO
DOCE. FONTE: WWW.EARTH.GOOGLE.COM .............................................................................. 13
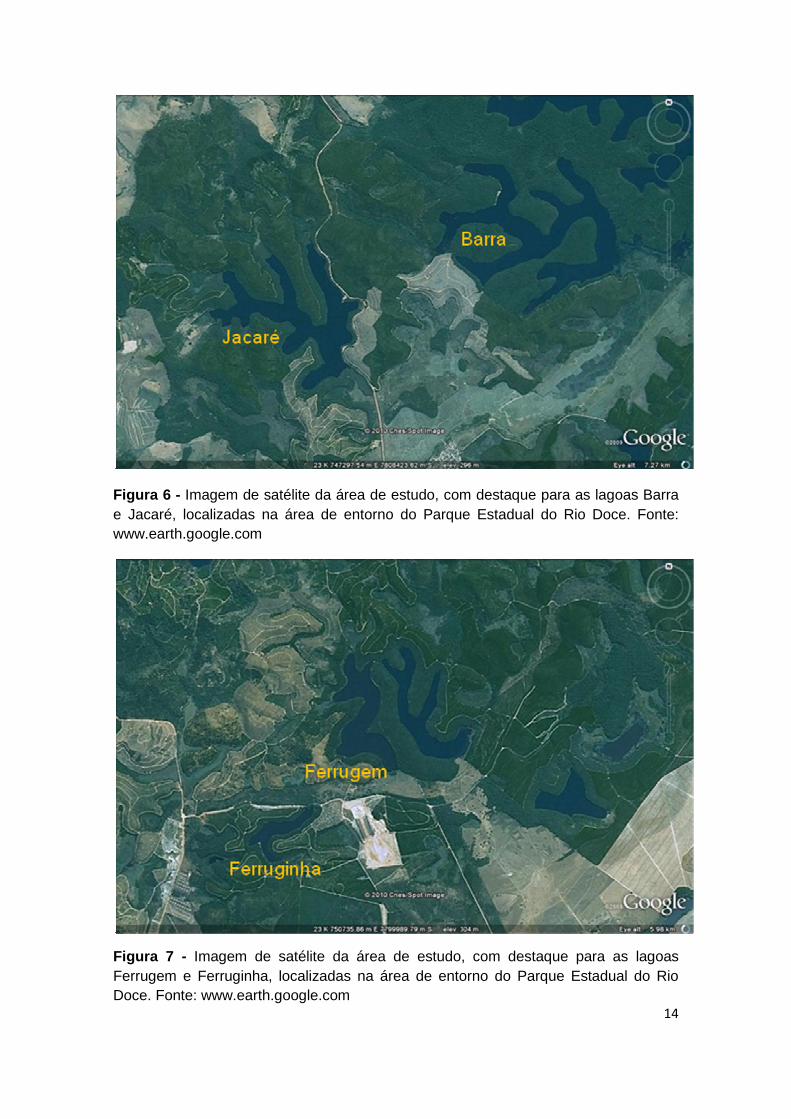
FIGURA 6 - IMAGEM DE SATÉLITE DA ÁREA DE ESTUDO, COM DESTAQUE PARA AS LAGOAS BARRA E
JACARÉ, LOCALIZADAS NA ÁREA DE ENTORNO DO PARQUE ESTADUAL DO RIO DOCE. FONTE: WWW.EARTH.GOOGLE.COM .................................................................................................... 14
FIGURA 7 - IMAGEM DE SATÉLITE DA ÁREA DE ESTUDO, COM DESTAQUE PARA AS LAGOAS FERRUGEM E
FERRUGINHA, LOCALIZADAS NA ÁREA DE ENTORNO DO PARQUE ESTADUAL DO RIO DOCE. FONTE: WWW.EARTH.GOOGLE.COM .................................................................................................... 14
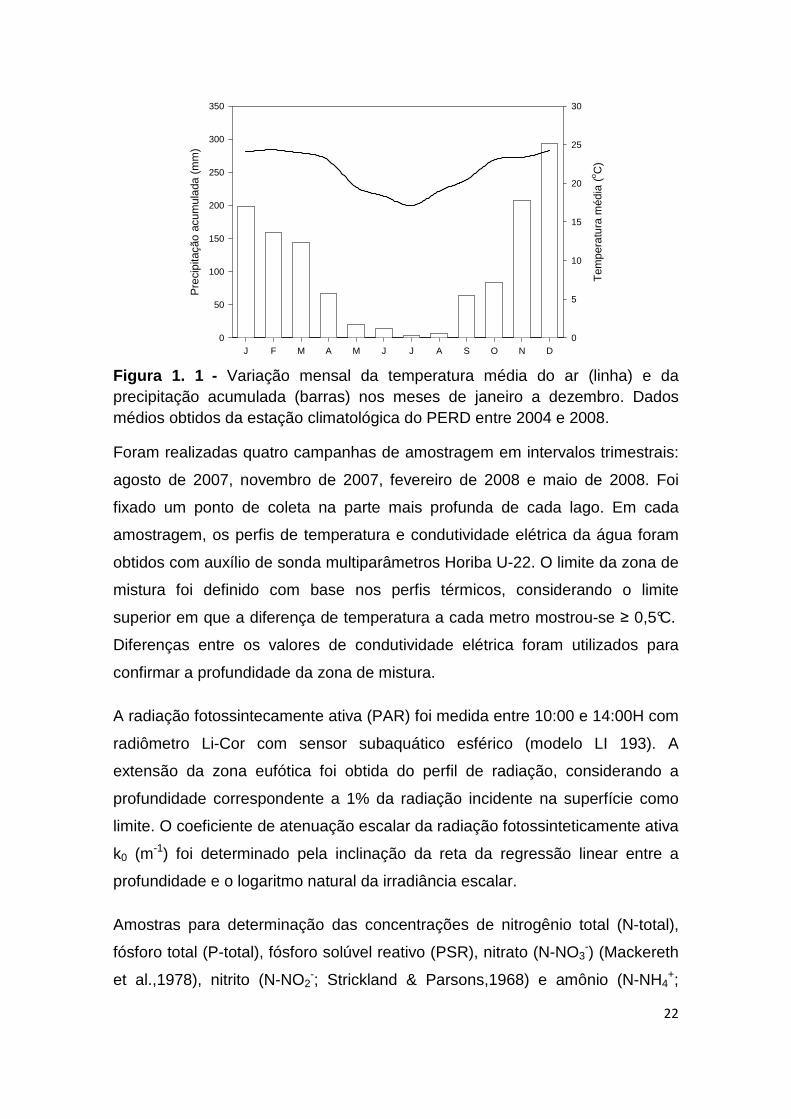
FIGURA 1. 1 - VARIAÇÃO MENSAL DA TEMPERATURA MÉDIA DO AR (LINHA) E DA PRECIPITAÇÃO
ACUMULADA (BARRAS) NOS MESES DE JANEIRO A DEZEMBRO. DADOS MÉDIOS OBTIDOS DA
ESTAÇÃO CLIMATOLÓGICA DO PERD ENTRE 2004 E 2008. ...................................................... 22
FIGURA 1. 2 - CURVA DE ACUMULAÇÃO DE ESPÉCIES REGISTRADA PARA AS 18 LAGOAS AMOSTRADAS
ENTRE AGOSTO DE 2007 E MAIO DE 2008 NO SISTEMA LACUSTRE DO MÉDIO RIO DOCE. ............ 27
FIGURA 1. 3 - TOTAL DE ESPÉCIES E CONTRIBUIÇÃO DE CADA CLASSE TAXÔMICA PARA AS 18 LAGOAS
AMOSTRADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008 NO SISTEMA LACUSTRE DO MÉDIO RIO
DOCE. ................................................................................................................................... 28
FIGURA 1. 4 - VARIAÇÃO NA RIQUEZA TOTAL (A) E NO NÚMERO DE ESPÉCIES EXCLUSIVAS (B) DAS
LAGOAS LOCALIZADAS NO PERD E EM SUA ÁREA DE ENTORNO. EM CADA CAIXA, A LINHA
REPRESENTA A MEDIANA E A CAIXA E O WHISKER REPRESENTAM 75 E 90% DOS DADOS, RESPECTIVAMENTE. ............................................................................................................... 29

VIII
FIGURA 1. 5 - CLUSTER OBTIDO PARA AS 18 LAGOAS AMOSTRADAS NO PERD, CONSIDERANDO A MÉDIA
DAS VARIÁVEIS AMBIENTAIS OBTIDA PARA AS QUATRO COLETAS REALIZADAS ENTRE AGOSTO DE
2007 E MAIO DE 2008. OS CÓDIGOS DAS LAGOAS SÃO OS MESMOS DA TABELA 1.4. .................. 31
FIGURA 1. 6 - CLUSTER OBTIDO PARA AS 18 LAGOAS AMOSTRADAS NO PERD, CONSIDERANDO A
PRESENÇA E AUSÊNCIA DAS ESPÉCIES, ENTRE AGOSTO DE 2007 E MAIO DE 2008. OS CÓDIGOS
DAS LAGOAS SÃO OS MESMOS DA TABELA 1.4. ........................................................................ 32
FIGURA 1. 7 - VARIAÇÃO NA BIOMASSA TOTAL E NOS VALORES DO ÍNDICE DE SHANNON (H’) E DE
PIEOLOU (E) NAS 18 LAGOAS AMOSTRADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008. EM CADA
CAIXA, A LINHA REPRESENTA A MEDIANA E A CAIXA E O WHISKER REPRESENTAM 75 E 90% DOS
DADOS, RESPECTIVAMENTE. ÁGUAS CLARAS (AC), ALMÉCEGA (AL), AMARELA (AM), ANÍBAL
(AN), BARRA (BA), CARIOCA (CA), CENTRAL (CE), DOM HELVÉCIO (DH), FERRUGEM (FE), FERRUGINHA (FN), GAMBÁ (GA), GAMBAZINHO (GN), JACARÉ (JA), PALMEIRINHA (PA), PATOS
(PT), PIMENTA (PI), SANTA HELENA (SH) E VERDE (VE). ........................................................ 33
FIGURA 2. 1 - VARIAÇÃO MENSAL DA TEMPERATURA MÉDIA DO AR (LINHA) E DA PRECIPITAÇÃO
ACUMULADA (BARRAS) NOS MESES DE JANEIRO A DEZEMBRO. DADOS MÉDIOS OBTIDOS DA
ESTAÇÃO CLIMATOLÓGICA DO PERD ENTRE 2004 E 2008. ...................................................... 54
FIGURA 2. 2 – MÉDIA E DESVIO PADRÃO OBSERVADOS PARA O COEFICIENTE DE ATENUAÇÃO DA
RADIAÇÃO FOTOSSINTETICAMENTE ATIVA (K0), DA CONCENTRAÇÃO DE MATÉRIA ORGÂNICA
DISSOLVIDA COLORIDA (A250), DO TEOR DE FÓSFORO TOTAL (PTOTAL), FÓSFORO SOLÚVEL REATIVO
(PSR), E NITROGÊNIO INORGÂNICO DISSOLVIDO (DIN) NAS LAGOAS DURANTE OS PERÍODOS DE
SECA E CHUVA. ...................................................................................................................... 58
FIGURA 2. 3 - CONTRIBUIÇÃO DAS CLASSES TAXONÔMICAS PARA O TOTAL DE ESPÉCIES REGISTRADO EM
CADA LAGOA AMOSTRADA ENTRE AGOSTO DE 2007 E MAIO DE 2008......................................... 60
FIGURA 2. 4 - VARIAÇÃO DA RIQUEZA EM ESPÉCIES DE BACILLARYOPHYCEAE NAS LAGOAS COM
COEFICIENTES DE ATENUAÇÃO ESCALAR (K0) MENORES (ÁGUAS CLARAS) E MAIORES QUE 0,8
(ÁGUAS ESCURAS). EM CADA CAIXA, O PONTO REPRESENTA A MEDIANA E OS LIMITES INFERIOR E
SUPERIOR COMPREENDEM OS PERCENTIS 25 E 75% DOS DADOS, RESPECTIVAMENTE. .............. 60
FIGURA 2. 5 - VARIAÇÃO DA BIOMASSA TOTAL, DAS CLASSES CYANOBACTERIA E CRYPTOPHYCEAE E DA
ABUNDÂNCIA RELATIVA (%) DE ZYGNEMATOPHYCEAE NAS LAGOAS COM COEFICIENTE DE
ATENUAÇÃO ESCALAR DA LUZ K0 <0,8 (ÁGUAS CLARAS) E >0,8 (ÁGUAS ESCURAS). EM CADA
CAIXA, O PONTO REPRESENTA A MEDIANA E OS LIMITES INFERIOR E SUPERIOR COMPREENDEM OS
PERCENTIS 25 E 75%, RESPECTIVAMENTE. OS CÍRCULOS REPRESENTAM OS OUTLIERS. LETRAS
DIFERENTES INDICAM DIFERENÇAS SIGNIFICATIVAS (P<0,05). ................................................... 61
FIGURA 3. 1 - LOCALIZAÇÃO DAS 18 LAGOAS ESTUDADAS NO PARQUE ESTADUAL DO RIO DOCE/PERD
(MG) E NO SEU ENTORNO E DO PONTO EM QUE FORAM REALIZADAS AS COLETAS LIMNOLÓGICAS
(+). FONTE: INSTITUTO MINEIRO DE GESTÃO DAS ÁGUAS (IGAM-MG). A LINHA CONTÍNUA INDICA
OS LIMITES DO PERD. ........................................................................................................... 76
FIGURA 3. 2 - VARIAÇÃO MENSAL DA TEMPERATURA MÉDIA DO AR (LINHA) E DA PRECIPITAÇÃO
ACUMULADA (BARRAS) NOS MESES DE JANEIRO A DEZEMBRO. DADOS MÉDIOS OBTIDOS DA
ESTAÇÃO CLIMATOLÓGICA DO PARQUE ESTADUAL DO RIO DOCE (MG), ENTRE 2004 E 2008. ... 77
FIGURA 3. 3 - PERFIS DE TEMPERATURA DA COLUNA D’ÁGUA OBTIDOS EM 18 LAGOAS DO PARQUE
ESTADUAL DO RIO DOCE E DO SEU ENTORNO, AMOSTRADAS ENTRE AGOSTO DE 2007 E MAIO DE
2008. .................................................................................................................................... 82
FIGURA 3. 4 - DIAGRAMA DE ORDENAÇÃO DA CORRELAÇÃO NA ANÁLISE DE COMPONENTES PRINCIPAIS
PARA AS AMOSTRAS LIMNOLÓGICAS DE 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E
NO SEU ENTORNO (AMOSTRAGENS REALIZADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008).

IX
VARIÁVEIS DESCRITORAS: ÍNDICE DE ESTADO TRÓFICO (IET), PROFUNDIDADE (PROF), COEFICIENTE DE ATENUAÇÃO DA LUZ (KO), EXTENSÃO DA ZONA EUFÓTICA (ZEU), EXTENSÃO DA
ZONA DE MISTURA (ZMIX), TEMPERATURA DA ÁGUA (TEMP) E CONCENTRAÇÃO DE SILICATO (SIO4). ÁGUAS CLARAS (AC), ALMÉCEGA (AL), GAMBÁ (GA), GAMBAZINHO (GN), PALMEIRINHA (PA), SANTA HELENA (SH), ANÍBAL (AN), BARRA (BA), CARIOCA (CA), JACARÉ (JA), PATOS (PT), CENTRAL (CE), FERRUGINHA (FN), PIMENTA (PI), AMARELA (AM) E FERRUGEM (FE).OS
NÚMEROS 1 A 4 INDICAM A PRIMEIRA À QUARTA COLETA. .......................................................... 84
FIGURA 3. 5 - VARIAÇÃO ESPACIAL E TEMPORAL DA BIOMASSA FITOPLANCTÔNICA EM 18 LAGOAS NO
PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO CLASSIFICADAS EM CINCO TIPOS
IDENTIFICADOS COM BASE NA PROFUNDIDADE TOTAL, COEFICIENTE DE ATENUAÇÃO ESCALAR DA
LUZ E TROFIA (AMOSTRAGENS REALIZADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008). A LINHA
HORIZONTAL DENTRO DAS CAIXAS REPRESENTA A MEDIANA E OS LIMITES DA CAIXA E A LINHA
VERICAL ENGLOBAM 75% E 90% DOS DADOS, RESPECTIVAMENTE. DOM HELVÉCIO (DH), VERDE
(VE), ÁGUAS CLARAS (AC), ALMÉCEGA (AL), GAMBÁ (GA), GAMBAZINHO (GN), PALMEIRINHA
(PA), SANTA HELENA (SH), ANÍBAL (AN), BARRA (BA), CARIOCA (CA), JACARÉ (JA), PATOS
(PT), CENTRAL (CE), FERRUGINHA (FN), PIMENTA (PI), AMARELA (AM) E FERRUGEM (FE). .... 86
FIGURA 3. 6 - MÉDIA E DESVIO-PADRÃO DA BIOMASSA FITOPLANCTÔNICA DE 18 LAGOAS NO PARQUE
ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO, CLASSIFICADAS EM CINCO TIPOS
IDENTIFICADOS COM BASE NA PROFUNDIDADE TOTAL, COEFICIENTE DE ATENUAÇÃO ESCALAR DA
LUZ E TROFIA (AMOSTRAGENS REALIZADAS ENTRE AGOSTO DE 2007 E MAIO DE 2008). LETRAS
DIFERENTES INDICAM DIFERENÇAS SIGNIFACATIVAMENTE DIFERENTES (P<0,05). ...................... 88
FIGURA 3. 7 - MÉDIA E DESVIO PADRÃO DA BIOMASSA FITOPLANCTÔNICA DOS PRINCIPAIS GRUPOS
FUNCIONAIS DE 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU ENTORNO
CLASSIFICADAS EM CINCO TIPOS IDENTIFICADOS COM BASE NA PROFUNDIDADE TOTAL, COEFICIENTE DE ATENUAÇÃO ESCALAR DA LUZ E TROFIA (AMOSTRAGENS REALIZADAS ENTRE
AGOSTO DE 2007 E MAIO DE 2008). ........................................................................................ 90
FIGURA 3. 8 - CONTRIBUIÇÃO MÉDIA DOS PRINCIPAIS GRUPOS FUNCIONAIS PARA A BIOMASSA
FITOPLANCTÔNICA DE 18 LAGOAS NO PARQUE ESTADUAL DO RIO DOCE (MG) E NO SEU
ENTORNO, CLASSIFICADAS EM CINCO TIPOS, IDENTIFICADOS COM BASE NA PROFUNDIDADE TOTAL, COEFICIENTE DE ATENUAÇÃO ESCALAR DA LUZ E TROFIA, DURANTE PERÍODOS DE CHUVAS (C, NOVEMBRO DE 2007 E FEVEREIRO DE 2008) E SECA (S, AGOSTO DE 2007 E MAIO DE 2008). .... 91
FIGURA 3. 9 - DIAGRAMA DE ORDENAÇÃO DA ACC DOS GRUPOS FUNCIONAIS E SETE VARIÁVEIS
DESCRITORAS CONSIDERANDO AS 18 LAGOAS AMOSTRADAS EM QUATRO CAMPANHAS ENTRE
AGOSTO DE 2007 E MAIO DE 2008 NO MÉDIO RIO DOCE (N=72). .............................................. 95

1
RESUMO GERAL
Os ecossistemas naturais são dinâmicos e sua diversidade de espécies é
controlada por mecanismos múltiplos. O conhecimento dos padrões existentes
é essencial para a definição dos métodos de monitoramento e estratégias de
conservação. Por serem entidades ecológicas bem delimitadas na paisagem,
lagoas constituem bons modelos para o estudo da riqueza de espécies em
relação aos gradiente ambientais. O presente estudo foi desenvolvido em 18
lagoas naturais do sistema lacustre do médio Rio Doce, localizado na região
sudeste do Brasil, e teve como objetivos avaliar: 1) a riqueza de espécies e a
composição fitoplanctônica desses ambientes, bem como seus padrões e
fatores determinantes, 2) a variação espacial e temporal do coeficiente de
absorção da radiação fotossinteticamente ativa (k0) bem como sua relação com
os teores de matéria orgânica dissolvida colorida (MODC), fósforo total e
composição e biomassa do fitoplâncton e 3) a variação espacial e temporal dos
grupos funcionais fitoplanctônicos e sua relação com os fatores ambientais. As
diversidades local (97 a 169 espécies) e regional (490 espécies) mostraram-se
elevadas e não foram diretamente afetadas pela área e forma de cada lagoa. A
similaridade florística foi baixa, acarretando em elevada diversidade beta (356
espécies). Os valores de k0 variaram espacial e sazonalmente, em função das
variações na profundidade da zona de mistura, nas concentrações de P-total e
MODC e na biomassa fitoplanctônica. Lagoas mais claras apresentaram maior
riqueza de diatomáceas e maior contribuição de desmídias para a biomassa
total. Lagoas mais escuras apresentaram maior biomassa total, composta
principalmente por cianofíceas e de criptofíceas. Os resultados sugerem não
haver limitação por luz nas lagoas estudadas. Lagoas mais claras e
oligotróficas foram dominadas pelos grupos funcionais NA e A, enquanto
grupos descritos como tolerantes à baixa luminosidade (SN, S1, MP, Y, X1, J,
Lo, X2) dominaram nas lagoas mais escuras e meso-eutróficas. A distribuição
dos grupos funcionais foi determinada principalmente pelas condições de
transparência da água, profundidade e trofia das lagoas estudadas e reforça
sua eficiência como indicadores das condições ambientais também em lagoas
naturais tropicais.

2
GENERAL ABSTRACT
Natural ecosystems are dynamic and their diversity of species is controlled by
multiple mechanisms. Knowledge of existing patterns is essential to the
definition of methods for monitoring and conservation strategies. Because they
are well-defined ecological entities in the landscape, lakes are good models for
the study of species richness in relation to environmental gradient. This study
was conducted in 18 natural lake of the middle Rio Doce lake district, located in
southeastern Brazil, and aimed to evaluate: 1) species richness and
composition of phytoplankton in these environments, as well as its patterns and
determinants, 2) the spatial and temporal variation of the absorption coefficient
of photosynthetically active radiation (k0) and their relation with the
concentration of colored dissolved organic matter (CDOM), total phosphorus
and composition and biomass of phytoplankton and 3) the spatial and temporal
of phytoplankton functional groups and their relationship to environmental
factors. The local diversity (97-169 species) and regional (490 species) were
high and were not directly affected by area and shape of each pond. Floristic
similarity was low, resulting in high beta diversity (356 species). The values of
k0 varied spatially and seasonally, depending on the variations in the depth of
mixing zone concentrations of total P and CDOM and phytoplankton biomass.
Clearer lakes have greater richness of diatoms and desmids greatest
contribution to total biomass. Darker lakes had higher total biomass, composed
mainly of cyanophytes and cryptophytes. The results suggest no limitation by
light in the lakes studied. More clear and oligotrophic lakes were dominated by
functional groups NA and A, while the groups described as tolerant to low light
(SN, S1, MP, Y, X1, J Lo, X2) dominated in ponds darker and meso-eutrophic.
The distribution of functional groups was determined mainly by conditions of
water transparency, depth and trophic status of the lakes studied and enhances
its efficiency as indicators of environmental conditions also in natural tropical
lakes.

3
INTRODUÇÃO GERAL
Lagoas são ambientes muito valorizados pelas qualidades recreativas,
estéticas e paisagísticas e por serem reservatórios naturais de água, recurso
imprescindível para a manutenção da vida (Garn et al., 2003). Apesar dessa
importância, pouco se tem feito em direção à conservação desses
ecossistemas, de modo que, juntamente com rios e áreas alagadas, estão
entre os ecossistemas mais ameaçados do mundo (Abell et al., 2008).
A condição de uma lagoa em um dado momento é o resultado da interação de
vários fatores, incluindo bacia de drenagem, clima, geologia, influência
antrópica e morfometria. Assim, a expansão dos dados disponíveis sobre esse
tipo de ecossistema possibilita aos limnólogos o entendimento dos problemas
em uma lagoa em particular e podem ser usados na previsão de respostas a
variações ambientais futuras (Garn et al., 2003). No entanto, estudos em
ecossistemas de água doce são muito menos frequentes que os de
ecossistemas marinhos (Carneiro et al., 2008), especialmente nas regiões
tropicais, onde informações fundamentais sobre diversidade, ciclagem de
nutrientes, propagação das variações ambientais através da cadeia alimentar e
diferenças nas estratégias adaptativas dos organismos em relação aos
ambientes temperados ainda são escassas (Lewis Jr., 2000).
As principais ameaças às lagoas são a super-exploração de água, a poluição e
a introdução de espécies invasoras (Revenga et al., 2005). Levantamentos
taxonômicos e avaliações das alterações na organização estrutural e funcional
das comunidades ou ecossistemas podem ser utilizados como medidas do
impacto das atividades humanas, possibilitando uma estimativa da perda de
diversidade de espécies e dos efeitos sinergéticos das diferentes alterações
antropogênicas na bacia hidrográfica (Buss et al., 2003).
Os ecossistemas naturais são dinâmicos e sua diversidade de espécies é
controlada por mecanismos múltiplos (Flöder & Burns, 2005). Assim, um bom
conhecimento dos padrões existentes é essencial para a definição dos
métodos de monitoramento e estratégias de conservação (Declerck et al.,
2005). Por se tratarem de entidades ecológicas bem delimitadas na paisagem,

4
lagoas constituem bons modelos para o estudo da riqueza de espécies em
relação aos gradiente ambientais (Dodson et al., 2000). Ainda assim, estudos
dessa natureza são sub-representados em comparação a ambientes terrestres
(Waide et al., 1999).
A ecologia do fitoplâncton está intimamente relacionada às características
dominantes, como estrutura física do ambiente e disponibilidade de recursos, e
à freqüência e escala em que estas características são alteradas (Reynolds,
1993). Devido a essa estreita relação, o fitoplâncton é considerado um
componente-chave na biocenose de lagoas e reservatórios (Sarmento &
Descy, 2008).
A luz, especialmente, a radiação fotossinteticamente ativa, está entre os
recursos essenciais ao desenvolvimento do fitoplâncton, sendo determinante
na estrutura. Nessa comunidade, é reconhecida uma série de características e
estratégias que permitem a captura eficiente e a utilização das diferentes
intensidades e domínios espectrais da luz (Litchman & Klausmeier, 2008). Por
se tratar de um recurso, sua utilização por um indivíduo reduz sua
disponibilidade para os demais indivíduos da mesma espécie e para as
espécies competidoras. Dessa forma, a tolerância a baixas intensidades
luminosas pode ser vista como uma importante vantagem competitiva. Essa
tolerância pode ser garantida por atributos associados à capacidade de
absorção da luz, como qualidade e quantidade dos pigmentos
fotossintetizantes, ou à capacidade do organismo regular sua posição para se
manter em local de luminosidade favorável, por meio de produção de
mucilagem, óleos, presença de flagelos ou aerótopos (Reynolds, 1997a).
As mudanças na comunidade fitoplanctônica em função das variações
ambientais são tradicionalmente verificadas através das variações na biomassa
e na composição em termos de grandes grupos taxonômicos e espécies
dominantes. No entanto, é muito difícil prever a ocorrência da maioria das
espécies, e as condições que favorecem certos grupos taxonômicos
freqüentemente se sobrepõem (Costa et al., 2009). Considerando as diferenças
nas demandas e formas de aquisição dos recursos essenciais observadas

5
entre os organismos fitoplanctônicos (Falkowski et al., 2004), tem-se assumido
que o entendimento e o manejo dos ambientes aquáticos se tornam mais fáceis
quando as espécies são agrupadas conforme a semelhança de características
ou comportamentos (Salmaso & Padisák, 2007).
A primeira abordagem considerando as características das espécies para
prever a composição da comunidade ao longo de gradientes ambientais foi
feita por Margalef em 1978 (Litchman & Klausmeier, 2008). Mais recentemente,
Reynolds (1997a) sugeriu grupos funcionais que poderiam co-dominar ou
dominar, conforme as condições ambientais predominantes. Esse autor e seus
colaboradores (Reynolds et al., 2002; Kruk et al., 2002; Padisák et al., 2003a,
Padisák et al., 2009) construíram uma extensa lista de grupos morfo-funcionais,
considerando as demandas para as combinações de propriedades físicas,
químicas e biológicas da lagoa (profundidade da zona de mistura, luz,
temperatura, disponibilidade de fósforo, sílica, carbono e pressão de
herbivoria).
Atualmente, estão descritos 38 grupos funcionais identificados por códigos alfa-
numéricos (Padisák et al., 2009). Tais grupos foram formados de acordo com
as características adaptativas coincidentes, considerando as sensibilidades e
as tolerâncias das espécies (Reynolds, 2006). No Brasil, os estudos utilizando
essa abordagem tiveram início ainda nos anos 90 (Huszar & Reynolds, 1997) e
mostraram-se importantes na década atual. No entanto, a maioria deles se
concentrou em reservatórios e pouca informação está disponível para
ambientes naturais (Tabela 1 ).
Formado a partir de sedimentação das drenagens originais do rio Doce e de
seus tributários (Pflug, 1969 apud De Meis & Tundisi, 1997), o sistema lacustre
do médio Rio Doce é um dos grandes sistemas de água doce do Brasil. A
importância biológica desta área úmida, constituída por cerca de 150 lagoas, foi
reconhecida recentemente em nível internacional, quando se tornou 11º sítio
brasileiro a integrar a lista de Ramsar (RAMSAR, 2010).

6
Tabela 1 - Ambientes estudados no Brasil até 2009 utilizando a abordagem dos grupos funcionais para análise do fitoplâncton.
Ecossistemas estudados Referências Artificiais Açude Namorados (PB) Araújo & Barbosa, 2009 Açude Taperoá (PB) Araújo & Barbosa, 2009 Açude Soledade (PB) Araújo & Barbosa, 2009 Reservatório Faxinal (RS) Becker et al. 2008, 2009a, 2009b Lago Monte Alegre (SP) Rangel et al., 2009 Lago das Garças (SP) Crossetti & Bicudo, 2005, 2008a, 2008b
Fonseca & Bicudo, 2008 Huszar et al., 2000 Lopes et al., 2005
Reservatório Billings (SP) Gemelgo et al., 2009 Reservatório Guarapiranga (SP) Gemelgo et al., 2009 Reservatório Algodões (PE) Huszar et al., 2000 Reservatório Chapéu (PE) Huszar et al., 2000 Reservatório Ingazeiras (PE) Huszar et al., 2000 Reservatório Mandaú (PE) Moura et al., 2007 Reservatório Poço da Cruz (PE) Huszar et al., 2000 Reservatório Pão de Açúcar (PE) Huszar et al., 2000 Reservatório Tapacurá (PE) Bouvy et al., 2003 Reservatório Capivari (PR) Borges et al., 2008 Reservatório Foz do Areia (PR) Silva et al., 2005 Reservatório Salto Caxias (PR) Silva et al., 2005 Reservatório Salto Osório (PR) Silva et al., 2005 Reservatório Salto Santiago (PR) Silva et al., 2005 Reservatório Segredo (PR) Borges et al., 2008
Silva et al., 2005 Reservatório Juturnaíba (RJ) Huszar et al., 2000
Marinho & Huszar, 2002 Reservatório Duas Bocas (ES) Delazari-Barroso et al., 2009 Naturais Estuário do rio Paraíba do Sul (RJ) Costa et al., 2009 Lagoa Comprida (RJ) Alves-de-Souza et al., 2006 Lagoa Barra (RJ) Huszar et al., 2000 Lago Batata (AM) Huszar & Reynolds, 1997
Melo & Huszar, 2000 Lago Dom Helvécio (MG) Souza et al., 2008 Lagoa Ventura (MS) Bovo-Scomparin & Train 2008 Rio Paraibuna (MG) Soares et al., 2007 Rio Pomba (MG) Lago Tigres (GO) Nabout & Nogueira, 2007 Lagoas de inundação do rio Araguaia (GO/ MT)
Nabout et al., 2006 Nabout et al., 2009

7
As pesquisas limnológicas no médio Rio Doce foram iniciadas na década de
1970 com os estudos pioneiros de Tundisi e colabores (Tundisi et al., 1978;
CETEC, 1981). A partir de então, diversos aspectos da limnologia das lagoas
que formam esse sistema foram investigados (ex. Barbosa & Tundisi, 1980;
Henry & Barbosa, 1989). Os estudos realizados através de cooperação com
centros de pesquisas japoneses (Tundisi & Saijo, 1997a) possibilitaram uma
intensificação do conhecimento dos aspectos geológicos, morfológicos, físicos,
químicos e biológicos das lagoas da região. No entanto, ainda havia a carência
de informações sobre a diversidade da região e a dinâmica das comunidades.
O Programa de Pesquisas Ecológicas de Longa Duração (PELD/UFMG)
contribuiu para o preenchimento dessa lacuna, permitindo um olhar
aprofundado para a compreensão de fenômenos em diferentes escalas de
tempo e espaço (Souza, 2005; Garcia, 2007; Petrucio & Barbosa, 2004;
Petrucio et al., 2006, Bezerra-Neto et al., 2006; Maia-Barbosa et al., 2006),
bem como o monitoramento de mudanças na qualidade das águas frente a
diferentes impactos decorrentes de ações antrópicas e propostas de ações
mitigadoras destes impactos (Barbosa et al., 2004).
A heterogeneidade ambiental observada nas lagoas do médio Rio Doce,
resultante do processo de formação geológica, da evolução diferenciada dos
processos físicos (De Meis & Tundisi, 1997) e do grau de impacto antrópico em
cada lagoa, sugere a ocorrência de elevada diversidade de organismos
aquáticos.
O presente estudo foi desenvolvido no âmbito do Programa de Pesquisas
Ecológicas de Longa Duração – PELD/MCT-CNPq/UFMG. Trata-se de um
esforço concentrado para ampliar o conhecimento sobre a diversidade e a
ecologia da comunidade fitoplanctônica nesse sistema lacustre ao comparar
um número maior de ambientes. Para tanto, focou-se nos seguintes objetivos:
1. Avaliar a diversidade fitoplanctônica em 18 lagoas do sistema lacustre
do médio Rio Doce;
2. Verificar a ocorrência de padrões no que se refere à estrutura do
fitoplâncton, em termos de riqueza e composição;

8
3. Identificar os grupos funcionais descritores do fitoplâncton nas lagoas
estudadas;
4. Identificar os principais fatores ambientais determinando a composição e
estrutura do fitoplâncton nas lagoas estudadas.
O estudo foi dividido em três capítulos, assim descritos: I) apresenta a
diversidade de espécies nos níveis local e regional; II) avalia o efeito das
variações na atenuação da radiação incidente observada entre as lagoas sobre
os grupos taxonômicos e III) variações espaciais e temporais dos grupos
funcionais, apontando seus fatores determinantes.
ÁREA DE ESTUDOS
A bacia do Rio Doce está situada na região leste do Brasil e abrange uma área
de 83.400 km2 distribuídos entre os estados de Minas Gerais (86%) e Espírito
Santo (14%) (Marques & Barbosa, 2002). Dois sistemas lacustres são
observados ao longo dessa bacia: um no médio curso do Rio Doce, com 158
lagoas (Castellano-Solá & Barbosa, 2003) e outro no baixo curso, com
aproximadamente 70 lagoas (Cavati & Fernandes, 2008).
Aproximadamente 1/3 das lagoas do sistema lacustre do médio Rio Doce está
protegido dentro do Parque Estadual do Rio Doce (PERD, 19º29’24”-
19º48’18”S, 42º28’18”-42º38’30”W), criado em 1939 e que constitui o maior
remanescente contínuo de Mata Atlântica do estado de Minas Gerais (35.976
ha). As demais lagoas localizam-se em área de entorno do PERD e tiveram
sua bacia de drenagem alterada principalmente pela substituição da vegetação
nativa por plantações de eucalipto e pela ocupação humana.
Para este estudo, foram selecionadas 18 lagoas, sendo 8 localizadas dentro
dos limites do PERD e 10 em sua área de entorno. A escolha das lagoas
amostradas baseou-se na inclusão das maiores diferenças fisiográficas das
mesmas bem como na viabilidade de acesso, particularmente no período de
chuvas. A localização dos ambientes e dos pontos de coleta podem ser

9
observadas na Figura 1 . As coordenadas geográficas, bem como algumas
características morfométricas de cada ambiente encontram-se na Tabela 2 .
A região, conhecida como Vale do Aço, é um dos importantes pólos
siderúrgicos do estado. Nove municípios ocupam o entorno do PERD, sendo a
maioria deles com baixas populações. A maior parte da população se
concentra nos municípios que formam a região metropolitana do Vale do Aço:
Timóteo, Coronel Fabriciano e Ipatinga, que, juntos, abrigam 381.000
habitantes e onde o grau de urbanização chega a 99%. Em Dionísio e
Marliéria, importantes municípios da área de abrangência do PERD, o grau de
urbanização é de 55% e 22%, respectivamente (IEF, 2002).
O clima da região é definido como tropical semi-úmido, com 4-5 meses de seca
e características mesotérmicas (Nimer, 1989), com temperatura em torno de
25ºC. Segundo Tundisi (1997), a precipitação máxima ocorre no mês de
dezembro (350 mm) e a mínima, em julho ou agosto (10 mm).
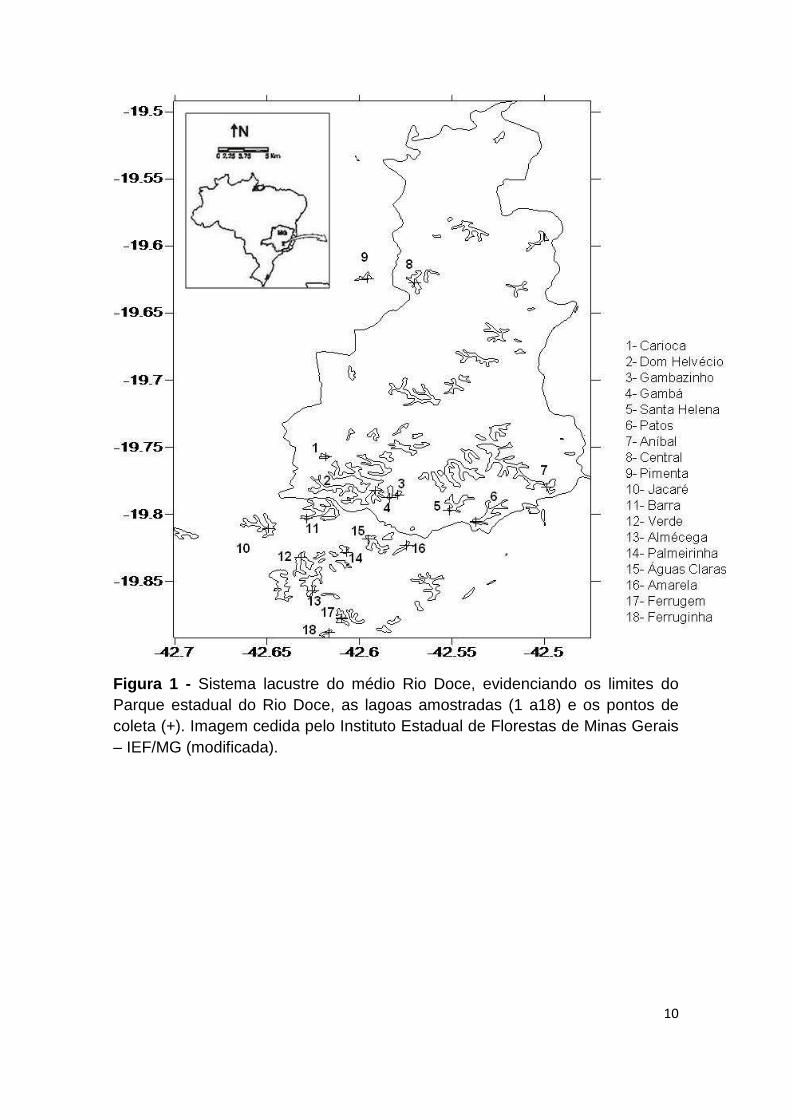
10
Figura 1 - Sistema lacustre do médio Rio Doce, evidenciando os limites do Parque estadual do Rio Doce, as lagoas amostradas (1 a18) e os pontos de coleta (+). Imagem cedida pelo Instituto Estadual de Florestas de Minas Gerais – IEF/MG (modificada).
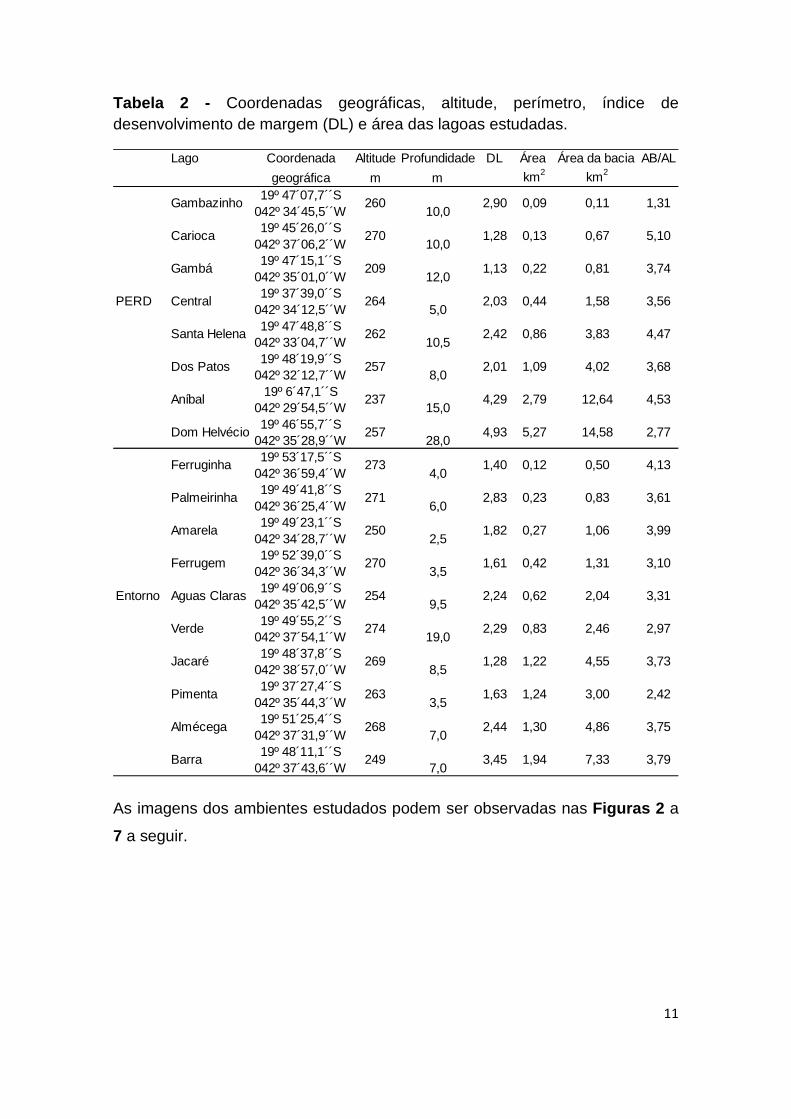
11
Tabela 2 - Coordenadas geográficas, altitude, perímetro, índice de desenvolvimento de margem (DL) e área das lagoas estudadas.
Lago Coordenada Altitude Profundidade DL Área Área da bacia AB/AL
geográfica m m km2 km2
Gambazinho19º 47´07,7´´S
042º 34´45,5´´W260
10,02,90 0,09 0,11 1,31
Carioca19º 45´26,0´´S
042º 37´06,2´´W270
10,01,28 0,13 0,67 5,10
Gambá19º 47´15,1´´S
042º 35´01,0´´W209
12,01,13 0,22 0,81 3,74
PERD Central19º 37´39,0´´S
042º 34´12,5´´W264
5,02,03 0,44 1,58 3,56
Santa Helena19º 47´48,8´´S
042º 33´04,7´´W262
10,52,42 0,86 3,83 4,47
Dos Patos19º 48´19,9´´S
042º 32´12,7´´W257
8,02,01 1,09 4,02 3,68
Aníbal19º 6´47,1´´S
042º 29´54,5´´W237
15,04,29 2,79 12,64 4,53
Dom Helvécio19º 46´55,7´´S
042º 35´28,9´´W257
28,04,93 5,27 14,58 2,77
Ferruginha19º 53´17,5´´S
042º 36´59,4´´W273
4,01,40 0,12 0,50 4,13
Palmeirinha19º 49´41,8´´S
042º 36´25,4´´W271
6,02,83 0,23 0,83 3,61
Amarela19º 49´23,1´´S
042º 34´28,7´´W250
2,51,82 0,27 1,06 3,99
Ferrugem19º 52´39,0´´S
042º 36´34,3´´W270
3,51,61 0,42 1,31 3,10
Entorno Aguas Claras19º 49´06,9´´S
042º 35´42,5´´W254
9,52,24 0,62 2,04 3,31
Verde19º 49´55,2´´S
042º 37´54,1´´W274
19,02,29 0,83 2,46 2,97
Jacaré19º 48´37,8´´S
042º 38´57,0´´W269
8,51,28 1,22 4,55 3,73
Pimenta19º 37´27,4´´S
042º 35´44,3´´W263
3,51,63 1,24 3,00 2,42
Almécega19º 51´25,4´´S
042º 37´31,9´´W268
7,02,44 1,30 4,86 3,75
Barra19º 48´11,1´´S
042º 37´43,6´´W249
7,03,45 1,94 7,33 3,79
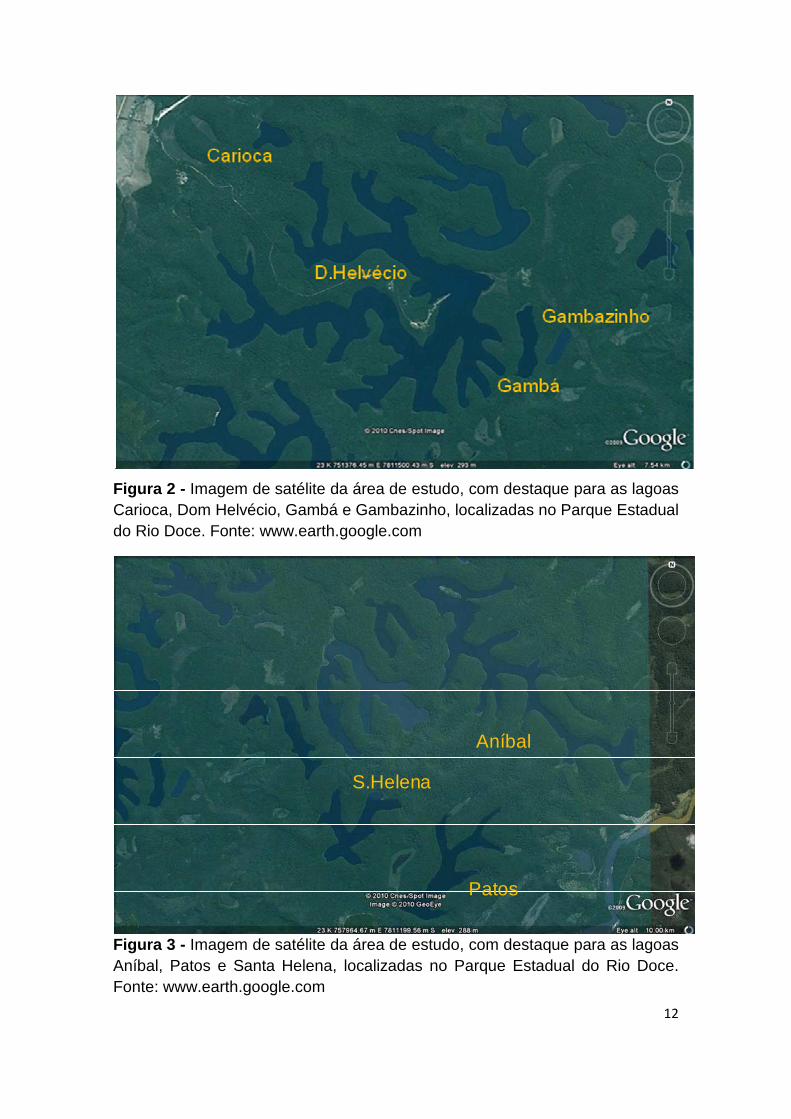
As imagens dos ambientes estudados podem ser observadas nas Figuras 2 a
7 a seguir.
12
Figura 2 - Imagem de satélite da área de estudo, com destaque para as lagoas Carioca, Dom Helvécio, Gambá e Gambazinho, localizadas no Parque Estadual do Rio Doce. Fonte: www.earth.google.com
Aníbal
Patos
S.Helena
Figura 3 - Imagem de satélite da área de estudo, com destaque para as lagoas Aníbal, Patos e Santa Helena, localizadas no Parque Estadual do Rio Doce. Fonte: www.earth.google.com
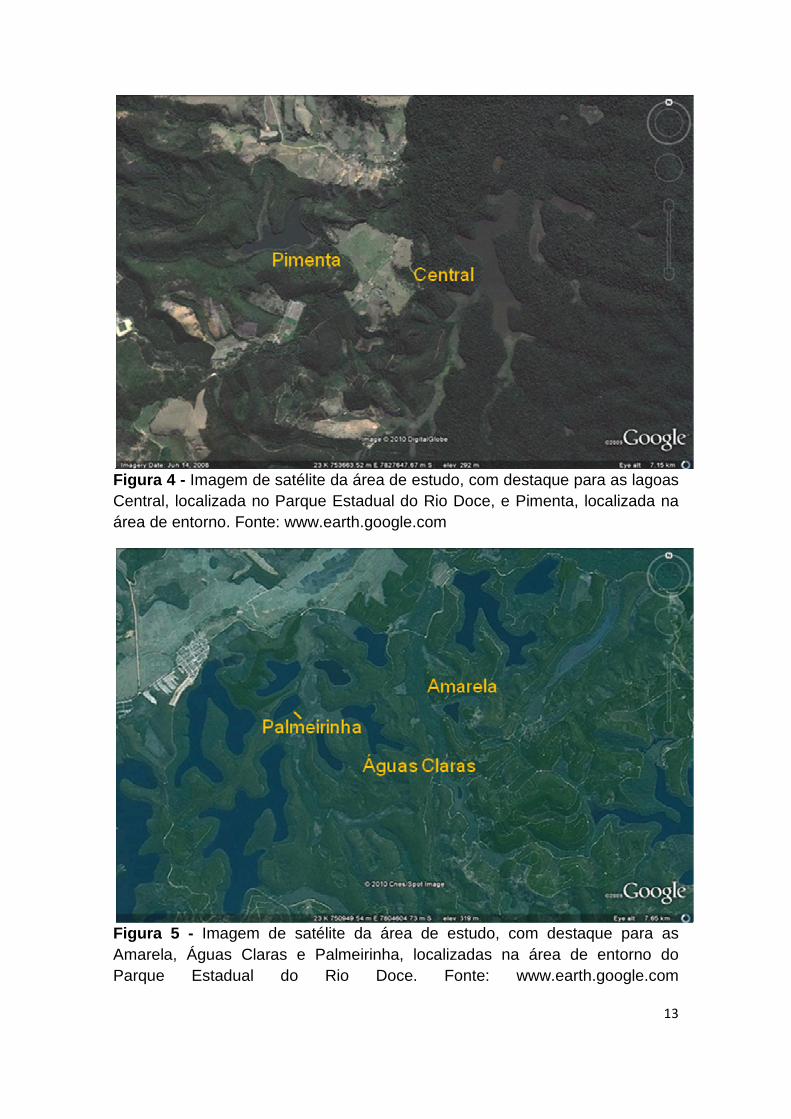
13
Figura 4 - Imagem de satélite da área de estudo, com destaque para as lagoas Central, localizada no Parque Estadual do Rio Doce, e Pimenta, localizada na área de entorno. Fonte: www.earth.google.com
Figura 5 - Imagem de satélite da área de estudo, com destaque para as Amarela, Águas Claras e Palmeirinha, localizadas na área de entorno do Parque Estadual do Rio Doce. Fonte: www.earth.google.com
14
Figura 6 - Imagem de satélite da área de estudo, com destaque para as lagoas Barra e Jacaré, localizadas na área de entorno do Parque Estadual do Rio Doce. Fonte: www.earth.google.com
Figura 7 - Imagem de satélite da área de estudo, com destaque para as lagoas Ferrugem e Ferruginha, localizadas na área de entorno do Parque Estadual do Rio Doce. Fonte: www.earth.google.com

15
CAPÍTULO 1
PADRÕES REGIONAIS E VARIAÇÃO ESPACIAL DA DIVERSIDAD E FITOPLANCTÔNICA NO SISTEMA LACUSTRE DO MÉDIO RIO DO CE,
MINAS GERAIS

16
Resumo
Padrões regionais e variação espacial da diversidad e fitoplanctônica no
sistema lacustre do médio Rio Doce, Minas Gerais
A riqueza de espécies e a composição taxonômica representam os principais
componentes da diversidade biológica e podem ser consideradas maneiras
simples de quantificar a complexidade em uma dada região. O presente estudo
apresenta a compoisção do florística de 18 lagoas do sistema lacustre do
médio Rio Doce, considerando: 1) sua diversidade alfa, gama e beta; 2) a
similaridade entre as lagoas estudadas e sua relação com a distância
geográfica e 3) a influência de fatores como morfometria, características físicas
e químicas da água e grau de impacto antrópico sobre a riqueza de espécies.
Foram estudadas 18 lagoas, oito localizadas no Parque Estadual do Rio Doce,
PERD, e dez em seu entorno. O trabalho resultou no aumento de 54% da
riqueza fitoplanctônica conhecida para a região, totalizando 490 espécies. A
diversidade local mostrou-se elevada em todas as lagoas (97 a 169 espécies) e
esteve inversamente correlacionada à profundidade da zona de mistura e à
razão N:P. O tamanho e a forma das lagoas parecem não interferir diretamente
na riqueza de espécies. A similaridade entre as lagoas estudadas foi baixa
(<0,522), acarretando em elevada diversidade beta (356 espécies). A
heterogeneidade ambiental do sistema lacustre do médio Rio Doce confirmou-
se pelas diferenças observadas nas características físicas e químicas e na
composição do fitoplâncton. A hipótese de que a proteção do PERD garante
maior riqueza de espécies fitoplanctônicas foi refutada e a composição florística
em cada lagoa parece resultar da interação de diversos fatores, inclusive de
processos geológicos e históricos. Assim, o sistema lacustre como um todo
exibe alto valor para conservação, devendo ser considerada a possibilidade de
conservação do sistema lacustre como um todo, com a proposição de
estratégias específicas para os ambientes da área de entorno.
Palavras-Chave: Médio Rio Doce, composição do fitoplâncton, desmídias

17
Abstract
Regional patterns and spatial variation of phytopla nkton diversity in the
the middle Rio Doce lake system, Minas Gerais
Species richness and taxonomic composition represent the main components of
biological diversity and can be considered simple ways to quantify complexity in
a given region. This study aims to contribute to the floristic knowledge of the
middle Rio Doce lake system estimating: 1) alpha, beta and gamma diversity, 2)
lakes similarity and their relation to geographic distance and 3) the influence of
morphology, physical and chemical features and human impact degree on
species richness. We studied 18 lakes, 8 located in the Rio Doce State Park,
RDSP, and 10 in their surroundings. An increase of 54% of phytoplankton
diversity known for the region was verified, totaling 490 species. Local diversity
was high (97-169 species) and was inversely correlated to the depth of the
mixing zone and N:P ratio. The size and shape of the lakes do not appear to
directly interfere in species richness. The similarity between the lakes studied
was small (<0.522), resulting in high beta diversity (356 species). The
heterogeneity of the lake system of the middle Rio Doce was confirmed by
differences in and chemical physical characteristics and phytoplankton
composition. The idea that the protection of RDSP ensures greater species
richness was refuted. The floristic composition resulting in each lake appears to
result from the interaction of several factors, including historical and geological
processes. Thus, the lake system as a whole exhibits high conservation value
and should be considered to conserve the lake system as a whole, with the
proposition of specific strategies for the environments of the surrounding area.
Key-Words: Middle Rio Doce, phytoplankton composition, desmids

18
Introdução
A riqueza de espécies e a composição taxonômica representam os principais
componentes da diversidade biológica e são importantes características dos
ecossistemas. O conhecimento de seus padrões é considerado essencial para
a formulação de hipóteses sobre os fatores controladores e suas
conseqüências em um ecossistema e para a proposição de estratégias de
monitoramento e conservação da biodiversidade (Downing & Leibold, 2002;
Declerck et al., 2005; Nogueira et al., 2008).
A riqueza de espécies pode ser considerada a medida mais simples para
expressar e quantificar a complexidade em uma dada região (Nabout et al.,
2007), uma vez é influenciada por vários fatores, como padrões de mistura da
coluna d’água, disponibilidade de luz e nutrientes, temperatura da água e
herbivoria (Reynolds, 1987).
A diversidade local, ou diversidade alfa (α) é o total de espécies existente em
cada habitat, enquanto a diversidade regional, ou diversidade gama (γ), é o
total de espécies observado em um conjunto de habitats (Magurran, 2004). O
termo diversidade beta (β) foi inicialmente introduzido por Whittaker em 1960
para descrever as mudanças na composição de espécies ao longo de
gradientes de altitude e umidade através de diferenças nas taxas de espécies
ganhas e perdidas, mas tem sido considerado hoje como a diferença
taxonômica entre as amostras, independentemente se elas ocorrem ao longo
de um gradiente ambiental (Veech et al., 2002).
A bacia do Rio Doce está situada na região sudeste do Brasil e abrange uma
área de 83.400 km2 distribuídos entre os estados de Minas Gerais (86%) e
Espírito Santo (14%) (Marques & Barbosa, 2002). Dois sistemas lacustres são
observados ao longo dessa bacia: um no médio curso do Rio Doce, com cerca
de 150 lagoas (Castellano-Solá & Barbosa, 2003) e outro no baixo curso, com
cerca de 70 lagoas (Cavati & Fernandes, 2008).
Como parte do sistema lacustre do médio Rio Doce, merecem destaque as
cerca de 50 lagoas localizadas dentro dos limites do Parque Estadual do Rio

19
Doce, considerada o maior remanescente contínuo de Mata Atlântica do estado
de Minas Gerais (35.976 ha), enquanto as demais se localizam em áreas
sujeitas a impactos variados, com destaque para as atividades de plantação de
eucalipto. A heterogeneidade ambiental observada nas lagoas do médio Rio
Doce, resultante do processo de formação geológica, da evolução diferenciada
dos processos físicos (De Meis & Tundisi, 1997) e do grau de impacto
antrópico em cada lagoa, sugere a ocorrência de elevada diversidade de
organismos aquáticos.
Embora os estudos limnológicos no médio Rio Doce tenham se iniciado na
década de 70 (Barbosa & Tundisi, 1980), aqueles abrangendo o fitoplâncton
consideraram uma (Hino et al., 1986; Barbosa, 2004, Barros et al., 2006; Souza
et al., 2008) ou duas lagoas (Barbosa, 2009) e enfocara essencialmente as
lagoas Dom Helvécio e Carioca. Um número maior de ambientes foi
considerado por Tundisi e colaboradores, que estudaram 15 lagoas (Tundisi et
al., 1989) e, mais recentemente, por Maia-Barbosa e colaboradores, que
abordaram a diversidade planctônica em sete lagoas (Maia-Barbosa et al.,
2006). Assim, a carência de informações sobre a diversidade fitoplanctônica
nesse sistema lacustre, evidenciada por Barbosa et al. (1994), ainda
permanece.
O presente estudo visa contribuir ao conhecimento florístico do sistema
lacustre do médio Rio Doce, avaliando: 1) sua diversidade alfa, gama e beta; 2)
a similaridade entre as lagoas estudadas e sua relação com a distância
geográfica e 3) a existência de padrões e a influência de fatores como
morfometria, características físicas e químicas da água e grau de impacto
antrópico sobre a riqueza de espécies.
Foram testadas as predições das seguintes hipóteses:
I. Em função da diversidade ambiental, o sistema lacustre do médio Rio Doce
apresenta elevada diversidade nos níveis alfa, beta e gama
i.elevada riqueza de espécies é observada em cada ambiente e no
conjunto de lagoas estudadas;

20
ii. a similaridade florística entre as lagoas estudadas é baixa;
II. A riqueza de espécies está diretamente relacionada à área e forma das
lagoas
i. lagoas maiores e/ou mais dendríticas apresentam maior riqueza de
espécies;
III. A distância entre as lagoas interfere na similaridade florística
i. lagoas mais próximas geograficamente apresentam composição do
fitoplâncton mais similar que lagoas mais distantes;
IV. O grau de preservação da vegetação nativa no entorno das lagoas
influencia as características limnológicas e, consequentemente, a riqueza e
composição e diversidade da comunidade fitoplanctônica.
i. As lagoas localizadas no PERD apresentam maior riqueza de
espécies, maior contribuição de zygnemaphyceae e maiores
valores de diversidade e equitabilidade que aquelas localizadas
em área de plantação de eucalipto.
Materiais e métodos
Foram estudadas 18 lagoas do sistema lacustre do médio Rio Doce, sendo oito
localizadas na área do PERD e dez em seu entorno. As coordenadas
geográficas e as características morfométricas de cada ambiente são
fornecidas na Tabela 1.1.
O clima da região é definido como tropical semi-úmido, com quatro a cinco
meses de seca e características mesotérmicas (Nimer, 1989) e temperaturas
em torno de 25ºC. O período chuvoso se estende de outubro a março e o
período de seca, de abril a setembro (Figura 1.1 ).

21
Tabela 1. 1 - Coordenadas geográficas, altitude, índice de desenvolvimento de margem (DL), área da lagoa e área da bacia de drenagem das 18 lagoas amostradas entre agosto de 2007 e maio de 2008 no Parque Estadual do Rio Doce e área de entorno.
Lago Coordenada Altitude Profundidade DL Área Área da bacia AB/AL
geográfica m m km2 km2
Gambazinho19º 47´07,7´´S
042º 34´45,5´´W260
10,02,90 0,09 0,11 1,31
Carioca19º 45´26,0´´S
042º 37´06,2´´W270
10,01,28 0,13 0,67 5,10
Gambá19º 47´15,1´´S
042º 35´01,0´´W209
12,01,13 0,22 0,81 3,74
PERD Central19º 37´39,0´´S
042º 34´12,5´´W264
5,02,03 0,44 1,58 3,56
Santa Helena19º 47´48,8´´S
042º 33´04,7´´W262
10,52,42 0,86 3,83 4,47
Dos Patos19º 48´19,9´´S
042º 32´12,7´´W257
8,02,01 1,09 4,02 3,68
Aníbal19º 6´47,1´´S
042º 29´54,5´´W237
15,04,29 2,79 12,64 4,53
Dom Helvécio19º 46´55,7´´S
042º 35´28,9´´W257
28,04,93 5,27 14,58 2,77
Ferruginha19º 53´17,5´´S
042º 36´59,4´´W273
4,01,40 0,12 0,50 4,13
Palmeirinha19º 49´41,8´´S
042º 36´25,4´´W271
6,02,83 0,23 0,83 3,61
Amarela19º 49´23,1´´S
042º 34´28,7´´W250
2,51,82 0,27 1,06 3,99
Ferrugem19º 52´39,0´´S
042º 36´34,3´´W270
3,51,61 0,42 1,31 3,10
Entorno Aguas Claras19º 49´06,9´´S
042º 35´42,5´´W254
9,52,24 0,62 2,04 3,31
Verde19º 49´55,2´´S
042º 37´54,1´´W274
19,02,29 0,83 2,46 2,97
Jacaré19º 48´37,8´´S
042º 38´57,0´´W269
8,51,28 1,22 4,55 3,73
Pimenta19º 37´27,4´´S
042º 35´44,3´´W263
3,51,63 1,24 3,00 2,42
Almécega19º 51´25,4´´S
042º 37´31,9´´W268
7,02,44 1,30 4,86 3,75
Barra19º 48´11,1´´S
042º 37´43,6´´W249
7,03,45 1,94 7,33 3,79
22
J F M A M J J A S O N D
Pre
cipi
taçã
o ac
umul
ada
(mm
)
0
50
100
150
200
250
300
350
Tem
pera
tura
méd
ia (
o C)
0
5
10
15
20
25
30
Figura 1. 1 - Variação mensal da temperatura média do ar (linha) e da precipitação acumulada (barras) nos meses de janeiro a dezembro. Dados médios obtidos da estação climatológica do PERD entre 2004 e 2008.
Foram realizadas quatro campanhas de amostragem em intervalos trimestrais:
agosto de 2007, novembro de 2007, fevereiro de 2008 e maio de 2008. Foi
fixado um ponto de coleta na parte mais profunda de cada lago. Em cada
amostragem, os perfis de temperatura e condutividade elétrica da água foram
obtidos com auxílio de sonda multiparâmetros Horiba U-22. O limite da zona de
mistura foi definido com base nos perfis térmicos, considerando o limite
superior em que a diferença de temperatura a cada metro mostrou-se ≥ 0,5°C.
Diferenças entre os valores de condutividade elétrica foram utilizados para
confirmar a profundidade da zona de mistura.
A radiação fotossintecamente ativa (PAR) foi medida entre 10:00 e 14:00H com
radiômetro Li-Cor com sensor subaquático esférico (modelo LI 193). A
extensão da zona eufótica foi obtida do perfil de radiação, considerando a
profundidade correspondente a 1% da radiação incidente na superfície como
limite. O coeficiente de atenuação escalar da radiação fotossinteticamente ativa
k0 (m-1) foi determinado pela inclinação da reta da regressão linear entre a
profundidade e o logaritmo natural da irradiância escalar.
Amostras para determinação das concentrações de nitrogênio total (N-total),
fósforo total (P-total), fósforo solúvel reativo (PSR), nitrato (N-NO3-) (Mackereth
et al.,1978), nitrito (N-NO2-; Strickland & Parsons,1968) e amônio (N-NH4
+;

23
Koroleff, 1976) foram coletadas na sub-superfície de cada lagoa e congeladas
para análise em laboratório.
As amostras para análise qualitativa (riqueza de espécies) foram coletadas
através de arrastos verticais e horizontais sucessivos com rede de 20µm de
abertura de malha e fixadas com solução de formalina a 4%. A identificação
das espécies foi feita sob microscópio óptico em aumentos de 400 e 1000
vezes. Foram analisadas oito lâminas por amostra, totalizando 32 lâminas por
lagoa. O sistema de classificação adotado para as classes taxonômicas foi o de
Round (1983). Os organismos foram identificados até o menor nível
taxonômico possível, com auxílio de bibliografia específica: Föster (1969 e
1974), Prescott et al. (1975; 1977; 1981 e 1982), Komárek & Fott (1983),
Sant’Anna (1984), Komárek & Anagnostidis (1989 e 1999), Menezes et al.
(1995), Bicudo & Menezes (2006).
As amostras para análise quantitativa do fitoplâncton foram coletadas com
garrafa de van Dorn em três profundidades, sendo a primeira a sub-superfície
das lagoas e as demais, definidas com o auxílio do disco de Secchi (Cole,
1983). Para tanto, assumiu-se que a profundidade do desaparecimento visual
do disco de Secchi representa cerca de 10% da radiação solar incidente na
superfície e que, multiplicada por 3 resulta na profundidade correspondente a
1% da radiação que atinge a superfície e que delimita o limite inferior da zona
eufótica (Wetzel & Likens 1991). As amostras foram fixadas com lugol acético
e armazenadas ao abrigo da luz. As contagens foram realizadas sob
microscópio invertido pelo método de Utermöhl (1958). Foram contados até
400 indivíduos da espécie mais abundante, garantindo erro de ±10% para um
intervalo de confiança de 95% (Lund et al., 1958). Os cálculos de densidade
foram feitos conforme Villafañe & Reid (1995). O biovolume específico foi
obtido associando as unidades contadas (células/colônias) a formas
geométricas (Hillebrand et al., 1999; Sun & Liu, 2003) e multiplicando-se a
densidade da população pelo volume médio obtido. Foram consideradas as
espécies que contribuíram, em média, com pelo menos 1% da densidade total
de cada lagoa, para as quais foram medidos 20 indivíduos. A densidade das
algas e cianobactérias foi considerada igual a 1 g.cm-3. A diversidade de

24
espécies foi calculada com base na biomassa, através do índice de Shannon-
Wiener e a equitabilidade, segundo o índice de Pielou (Magurran, 2004).
A similaridade florística entre as lagoas foi medida através do índice de
similariadade de Jaccard. Análises de agrupamento hierárquico (cluster)
considerando a distância de jaccard e utilizando-se o método de ligação Ward
foram realizadas para avaliar a similaridade das lagoas em relação às variáveis
ambientais e à composição do fitoplâncton.
A riqueza de espécies em cada lagoa (diversidade alfa) foi avaliada com base
no número de espécies encontradas ao longo do estudo considerando os
dados provenientes das análises qualitativas e quantitativas. A riqueza de
espécies para a região amostrada (diversidade gama) foi estimada através do
índice de extrapolação Jackknife de 1ª ordem (Nabout et al., 2007) utilizando-
se o software Stimate S (Colwell, 2006). A diversidade beta, que avalia o
quanto a diversidade alfa média é excedida pela diversidade regional, foi
estimada pelos índices β-1 (Harrison et al., 1992) e Whittaker (βw; sensu
Wilson & Shmida, 1984), que consideram o total de espécies encontrado no
conjunto de lagoas (S), e a riqueza média por lagoa (α médio) e, no primeiro
caso, o número de lagoas amostradas (N): β-1=[(S/αmédio)-1] / [N-1] x 100 e
βw=S/αmédio-1. A diversidade beta também foi estimada pela diferença entre a
riqueza media de espécies das lagoas e a diversidade gama (total de espécies
no conjunto de lagoas), conforme sugerido por Crist et al. (2003): β = γ – α
médio.
A normalidade dos dados foi testada através do teste de Komolgorov-Smirnov.
Uma vez que os dados obtidos não apresentaram a normalidade e
homocedasticidade necessárias para a aplicação de testes paramétricos, as
diferenças nos parâmetros físicos e químicos e na riqueza, biomassa,
diversiadae e equitabilidade do fitoplâncton entre as lagoas foram verificadas
através de testes de Kruskal-Wallis e Mann-Whitney. As relações entre as
variáveis físicas, químicas e biológicas foram avaliadas através de correlações
de Spearman. Foi utilizado o software Past versão 1.9 (Hammer et al., 2001).

25
Resultados
Os valores médios e os coeficientes de variação obtidos ao longo das quatro
amostragens realizadas em cada lagoa para variáveis limnológicas são
apresentados na Tabela 1.2. Diferenças entre as profundidades total (H=66,7;
p=0,000), da zona de mistura (H=44,5; p=0,003) e da zona eufótica (H=53,1;
p=000), coeficiente de atenuação da luz (H=55,7; p=0,000), teores de fósforo
total (H=34,4; p=0,008), nitrogênio total (H=33,9; p=0,009), condutividade
elétrica (H=34,0; p=0,008) foram verificadas entre as lagoas amostradas,
enquanto a razão Zeu:Zmis, a temperatura da água e os teores de nitrogênio e
fósforo dissolvidos não diferirem entre as mesmas.
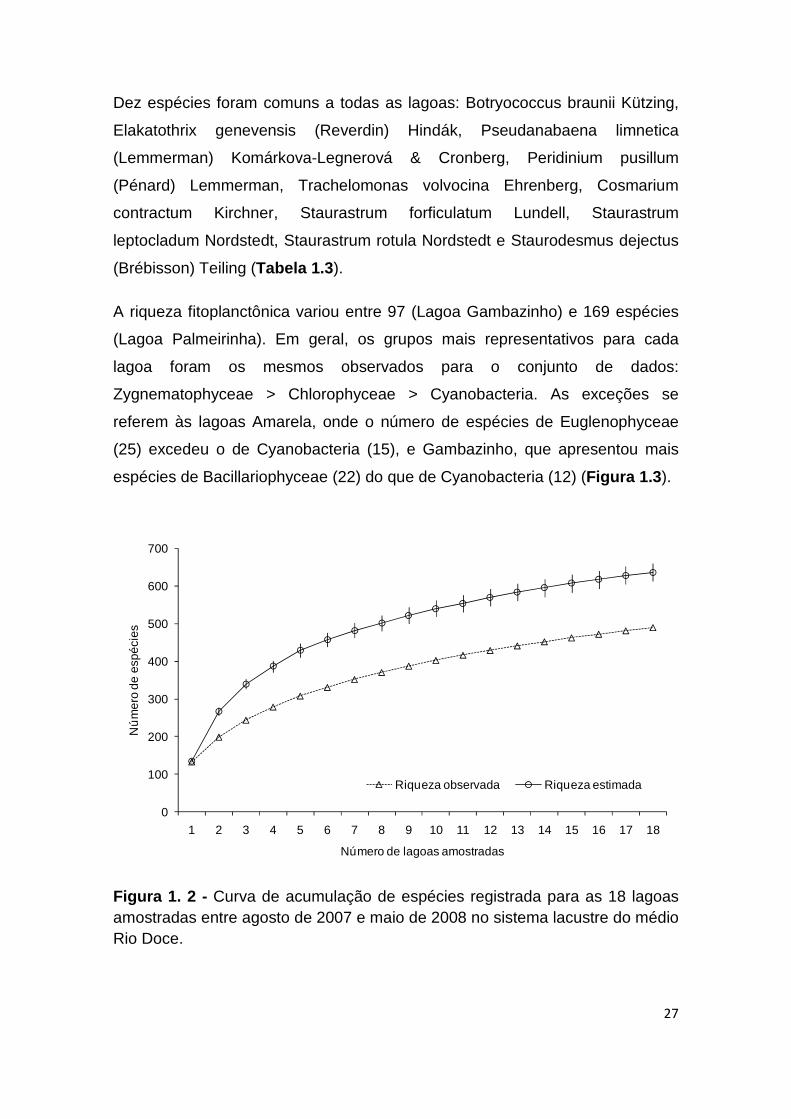
Foi registrado um total de 490 taxa (Figura 1.2 ), valor correspondente a 77%
da riqueza esperada, estimada pelo índice de extrapolação de riqueza
(Jackknife 1 = 636). As espécies registradas distribuíram-se entre as classes
Zygnematophyceae (171), Cyanobacteria (102), Chlorophyceae (98),
Bacillariophyceae (45), Euglenophyceae (42), Dinophyceae (10),
Xanthophyceae (8), Chrysophyceae (6), Cryptophyceae (6) e
Oedogoniophyceae (2). Quase metade dos taxa (47%) mostraram-se raros,
ocorrendo em uma ou duas lagoas, enquanto 90 deles podem ser
considerados de frequência importante no sistema lacustre do médio Rio Doce,
ocorrendo em pelo menos 10 das 18 lagoas amostradas.
26
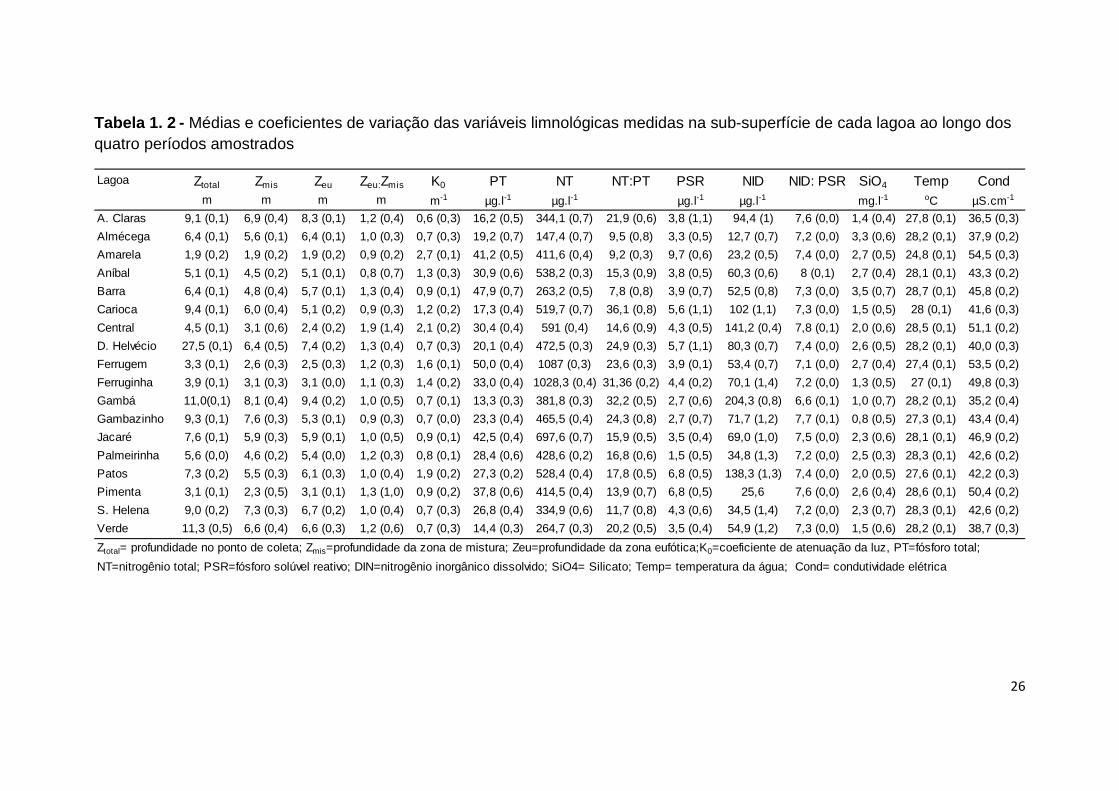
Tabela 1. 2 - Médias e coeficientes de variação das variáveis limnológicas medidas na sub-superfície de cada lagoa ao longo dos quatro períodos amostrados
Lagoa Ztotal Zmis Zeu Zeu:Zmis K0 PT NT NT:PT PSR NID NID: PSR SiO4 Temp Condm m m m m-1 µg.l-1 µg.l-1 µg.l-1 µg.l-1 mg.l-1 oC µS.cm-1
A. Claras 9,1 (0,1) 6,9 (0,4) 8,3 (0,1) 1,2 (0,4) 0,6 (0,3) 16,2 (0,5) 344,1 (0,7) 21,9 (0,6) 3,8 (1,1) 94,4 (1) 7,6 (0,0) 1,4 (0,4) 27,8 (0,1) 36,5 (0,3)
Almécega 6,4 (0,1) 5,6 (0,1) 6,4 (0,1) 1,0 (0,3) 0,7 (0,3) 19,2 (0,7) 147,4 (0,7) 9,5 (0,8) 3,3 (0,5) 12,7 (0,7) 7,2 (0,0) 3,3 (0,6) 28,2 (0,1) 37,9 (0,2)
Amarela 1,9 (0,2) 1,9 (0,2) 1,9 (0,2) 0,9 (0,2) 2,7 (0,1) 41,2 (0,5) 411,6 (0,4) 9,2 (0,3) 9,7 (0,6) 23,2 (0,5) 7,4 (0,0) 2,7 (0,5) 24,8 (0,1) 54,5 (0,3)
Aníbal 5,1 (0,1) 4,5 (0,2) 5,1 (0,1) 0,8 (0,7) 1,3 (0,3) 30,9 (0,6) 538,2 (0,3) 15,3 (0,9) 3,8 (0,5) 60,3 (0,6) 8 (0,1) 2,7 (0,4) 28,1 (0,1) 43,3 (0,2)
Barra 6,4 (0,1) 4,8 (0,4) 5,7 (0,1) 1,3 (0,4) 0,9 (0,1) 47,9 (0,7) 263,2 (0,5) 7,8 (0,8) 3,9 (0,7) 52,5 (0,8) 7,3 (0,0) 3,5 (0,7) 28,7 (0,1) 45,8 (0,2)
Carioca 9,4 (0,1) 6,0 (0,4) 5,1 (0,2) 0,9 (0,3) 1,2 (0,2) 17,3 (0,4) 519,7 (0,7) 36,1 (0,8) 5,6 (1,1) 102 (1,1) 7,3 (0,0) 1,5 (0,5) 28 (0,1) 41,6 (0,3)
Central 4,5 (0,1) 3,1 (0,6) 2,4 (0,2) 1,9 (1,4) 2,1 (0,2) 30,4 (0,4) 591 (0,4) 14,6 (0,9) 4,3 (0,5) 141,2 (0,4) 7,8 (0,1) 2,0 (0,6) 28,5 (0,1) 51,1 (0,2)
D. Helvécio 27,5 (0,1) 6,4 (0,5) 7,4 (0,2) 1,3 (0,4) 0,7 (0,3) 20,1 (0,4) 472,5 (0,3) 24,9 (0,3) 5,7 (1,1) 80,3 (0,7) 7,4 (0,0) 2,6 (0,5) 28,2 (0,1) 40,0 (0,3)
Ferrugem 3,3 (0,1) 2,6 (0,3) 2,5 (0,3) 1,2 (0,3) 1,6 (0,1) 50,0 (0,4) 1087 (0,3) 23,6 (0,3) 3,9 (0,1) 53,4 (0,7) 7,1 (0,0) 2,7 (0,4) 27,4 (0,1) 53,5 (0,2)
Ferruginha 3,9 (0,1) 3,1 (0,3) 3,1 (0,0) 1,1 (0,3) 1,4 (0,2) 33,0 (0,4) 1028,3 (0,4) 31,36 (0,2) 4,4 (0,2) 70,1 (1,4) 7,2 (0,0) 1,3 (0,5) 27 (0,1) 49,8 (0,3)
Gambá 11,0(0,1) 8,1 (0,4) 9,4 (0,2) 1,0 (0,5) 0,7 (0,1) 13,3 (0,3) 381,8 (0,3) 32,2 (0,5) 2,7 (0,6) 204,3 (0,8) 6,6 (0,1) 1,0 (0,7) 28,2 (0,1) 35,2 (0,4)
Gambazinho 9,3 (0,1) 7,6 (0,3) 5,3 (0,1) 0,9 (0,3) 0,7 (0,0) 23,3 (0,4) 465,5 (0,4) 24,3 (0,8) 2,7 (0,7) 71,7 (1,2) 7,7 (0,1) 0,8 (0,5) 27,3 (0,1) 43,4 (0,4)
Jacaré 7,6 (0,1) 5,9 (0,3) 5,9 (0,1) 1,0 (0,5) 0,9 (0,1) 42,5 (0,4) 697,6 (0,7) 15,9 (0,5) 3,5 (0,4) 69,0 (1,0) 7,5 (0,0) 2,3 (0,6) 28,1 (0,1) 46,9 (0,2)
Palmeirinha 5,6 (0,0) 4,6 (0,2) 5,4 (0,0) 1,2 (0,3) 0,8 (0,1) 28,4 (0,6) 428,6 (0,2) 16,8 (0,6) 1,5 (0,5) 34,8 (1,3) 7,2 (0,0) 2,5 (0,3) 28,3 (0,1) 42,6 (0,2)
Patos 7,3 (0,2) 5,5 (0,3) 6,1 (0,3) 1,0 (0,4) 1,9 (0,2) 27,3 (0,2) 528,4 (0,4) 17,8 (0,5) 6,8 (0,5) 138,3 (1,3) 7,4 (0,0) 2,0 (0,5) 27,6 (0,1) 42,2 (0,3)
Pimenta 3,1 (0,1) 2,3 (0,5) 3,1 (0,1) 1,3 (1,0) 0,9 (0,2) 37,8 (0,6) 414,5 (0,4) 13,9 (0,7) 6,8 (0,5) 25,6 7,6 (0,0) 2,6 (0,4) 28,6 (0,1) 50,4 (0,2)
S. Helena 9,0 (0,2) 7,3 (0,3) 6,7 (0,2) 1,0 (0,4) 0,7 (0,3) 26,8 (0,4) 334,9 (0,6) 11,7 (0,8) 4,3 (0,6) 34,5 (1,4) 7,2 (0,0) 2,3 (0,7) 28,3 (0,1) 42,6 (0,2)
Verde 11,3 (0,5) 6,6 (0,4) 6,6 (0,3) 1,2 (0,6) 0,7 (0,3) 14,4 (0,3) 264,7 (0,3) 20,2 (0,5) 3,5 (0,4) 54,9 (1,2) 7,3 (0,0) 1,5 (0,6) 28,2 (0,1) 38,7 (0,3)
Ztotal= profundidade no ponto de coleta; Zmis=profundidade da zona de mistura; Zeu=profundidade da zona eufótica;K0=coeficiente de atenuação da luz, PT=fósforo total;
NT=nitrogênio total; PSR=fósforo solúvel reativo; DIN=nitrogênio inorgânico dissolvido; SiO4= Silicato; Temp= temperatura da água; Cond= condutividade elétrica
27
Dez espécies foram comuns a todas as lagoas: Botryococcus braunii Kützing,
Elakatothrix genevensis (Reverdin) Hindák, Pseudanabaena limnetica
(Lemmerman) Komárkova-Legnerová & Cronberg, Peridinium pusillum
(Pénard) Lemmerman, Trachelomonas volvocina Ehrenberg, Cosmarium
contractum Kirchner, Staurastrum forficulatum Lundell, Staurastrum
leptocladum Nordstedt, Staurastrum rotula Nordstedt e Staurodesmus dejectus
(Brébisson) Teiling (Tabela 1.3 ).
A riqueza fitoplanctônica variou entre 97 (Lagoa Gambazinho) e 169 espécies
(Lagoa Palmeirinha). Em geral, os grupos mais representativos para cada
lagoa foram os mesmos observados para o conjunto de dados:
Zygnematophyceae > Chlorophyceae > Cyanobacteria. As exceções se
referem às lagoas Amarela, onde o número de espécies de Euglenophyceae
(25) excedeu o de Cyanobacteria (15), e Gambazinho, que apresentou mais
espécies de Bacillariophyceae (22) do que de Cyanobacteria (12) (Figura 1.3 ).
0
100
200
300
400
500
600
700
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Nú
mer
o de
esp
écie
s
Número de lagoas amostradas
Riqueza observada Riqueza estimada
Figura 1. 2 - Curva de acumulação de espécies registrada para as 18 lagoas amostradas entre agosto de 2007 e maio de 2008 no sistema lacustre do médio Rio Doce.

28
0
20
40
60
80
100
120
140
160
180
An
íbal
Car
ioca
Cen
tral
D.H
elvé
cio
Gam
bá
Gam
bazi
nh
o
Pat
os
S. H
elen
a
A. C
lara
s
Alm
éceg
a
Am
arel
a
Bar
ra
Fer
ruge
m
Fer
rugi
nha
Jaca
ré
Pal
mei
r
Pim
enta
Ver
de
Riq
ueza
de
espé
cies
Bacillariophyceae Chlorophyceae Chrysophyceae Cryptophyceae Cyanobacteria
Dinophyceae Euglenophyceae Oedogoniophyceae Xanthophyceae Zygnemaphyceae
Figura 1. 3 - Total de espécies e contribuição de cada classe taxômica para as 18 lagoas amostradas entre agosto de 2007 e maio de 2008 no sistema lacustre do médio Rio Doce.
As lagoas localizadas na área de entorno do PERD apresentaram riqueza
média de 144 espécies/lagoa, maior que aquelas localizadas dentro da
Unidade de Conservação, com média de 122 espécies/lagoa (U= 13,0;
p=0,018). O conjunto de lagoas localizadas na área de entorno também
apresentou maior número de espécies exclusivas (148) que o conjunto
localizado dentro do PERD (77 espécies). No entanto, os valores médios da
riqueza se mostraram muito próximos (7,6 e 9 espécies/lagoa,
respectivamente) e as diferenças observadas não foram significativas (U= 38,0;
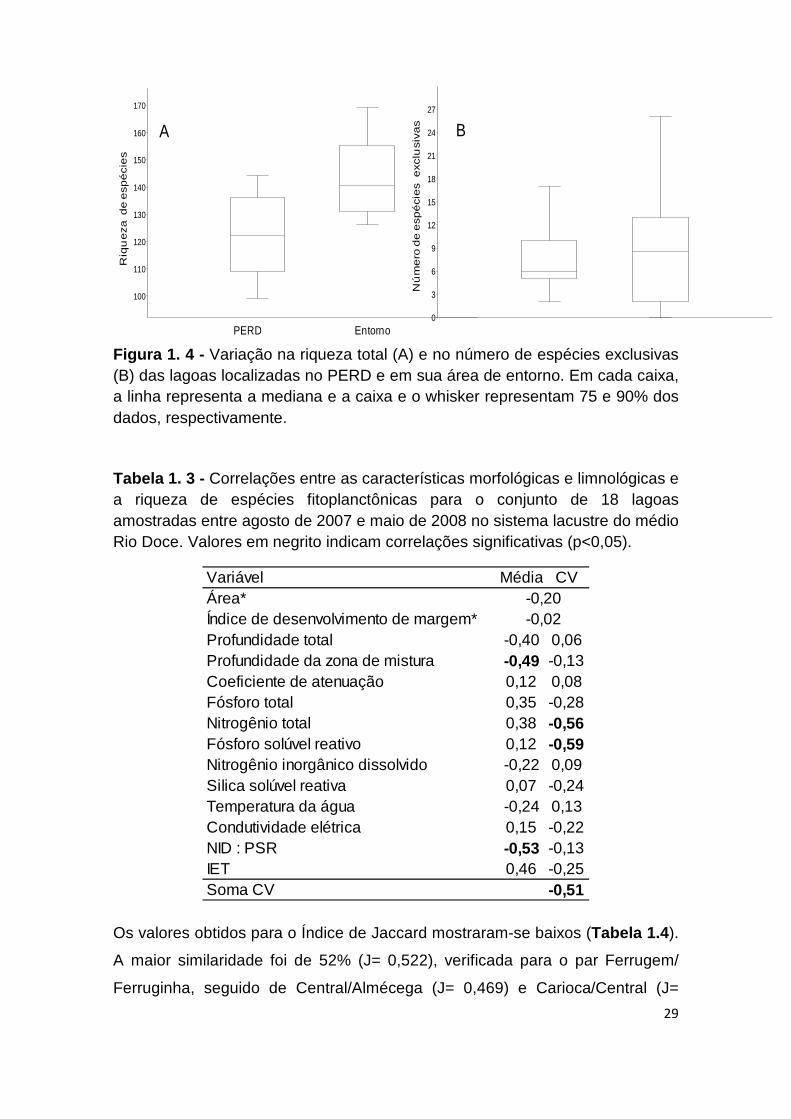
p= 0,894; Figura 1.4 ). A lagoa Amarela foi a que apresentou o maior número
de taxa exclusivos (25), seguida das lagoas Gambazinho (18) e Águas Claras
(15). As menores ocorrências de espécies exclusivas foram observadas nas
lagoas Barra (0), Ferrugem e Ferruginha (2 espécies cada).
Os resultados das correlações entre os valores médios e os coeficientes de
variação das variáveis ambientais medidas e a riqueza de espécies encontram-
se na Tabela 1.3 . A riqueza de espécies mostrou-se inversamente
correlacionada à profundidade da zona de mistura e à razão entre as formas
inorgânicas dissolvidas de nitrogênio e fósforo, bem como ao coeficiente de
variação do nitrogênio total e fósforo solúvel reativo.
29
100
110
120
130
140
150
160
170
Riq
ue
za
de
esp
écie
s
PERD Entorno0
3
6
9
12
15
18
21
24
27
Nú
me
ro d
e e
sp
écie
s e
xclu
siv
as
A B
Figura 1. 4 - Variação na riqueza total (A) e no número de espécies exclusivas (B) das lagoas localizadas no PERD e em sua área de entorno. Em cada caixa, a linha representa a mediana e a caixa e o whisker representam 75 e 90% dos dados, respectivamente.
Tabela 1. 3 - Correlações entre as características morfológicas e limnológicas e a riqueza de espécies fitoplanctônicas para o conjunto de 18 lagoas amostradas entre agosto de 2007 e maio de 2008 no sistema lacustre do médio Rio Doce. Valores em negrito indicam correlações significativas (p<0,05).
Variável Média CVÁrea*Índice de desenvolvimento de margem*Profundidade total -0,40 0,06Profundidade da zona de mistura -0,49 -0,13Coeficiente de atenuação 0,12 0,08Fósforo total 0,35 -0,28Nitrogênio total 0,38 -0,56Fósforo solúvel reativo 0,12 -0,59Nitrogênio inorgânico dissolvido -0,22 0,09Silica solúvel reativa 0,07 -0,24Temperatura da água -0,24 0,13Condutividade elétrica 0,15 -0,22NID : PSR -0,53 -0,13IET 0,46 -0,25Soma CV -0,51
-0,20-0,02
Os valores obtidos para o Índice de Jaccard mostraram-se baixos (Tabela 1.4 ).
A maior similaridade foi de 52% (J= 0,522), verificada para o par Ferrugem/
Ferruginha, seguido de Central/Almécega (J= 0,469) e Carioca/Central (J=

30
0,449). A lagoa Amarela apresentou os menores valores para esse índice (J=
0,142 a 0,286).
As análises de cluster evidenciaram a formação de quatro agrupamentos em
relação às variáveis ambientais ( Figura 1.5 ): as lagoas Carioca, Gambazinho,
Águas claras, Verde, Gambá e Patos foram as mais similares, seguidas do
grupo Barra,Santa Helena,Almécega, Palmeirinha e Gambazinho. As lagoas
maiores e mais profundas, Aníbal e Dom Helvécio, se isolaram em um grupo à
parte, e também se observou o agrupamento das mais rasas e escuras
(Ferrugem, Ferruginha, Central, Pimenta e Amarela).
Cinco agrupamentos foram observados em relação à composição do
fitoplâncton (Figura 1.6 ): 1) lagoas Aníbal, Carioca, Pimenta, Patos, Barra,
Dom Helvécio e Santa Helena; 2) Central, Almécega, Ferrugem e Ferruginha;
3) Águas Claras, Palmeirinha, Jacaré e Verde; 4) Gambá e Gambazinho e 5)
lagoa Amarela. No entanto, um baixo coeficiente cofenético (0,33) foi
observado. A similaridade na composição fitoplanctônica das lagoas não se
mostrou correlacionada à distância entre as mesmas (r = 0,03; p > 0,05).
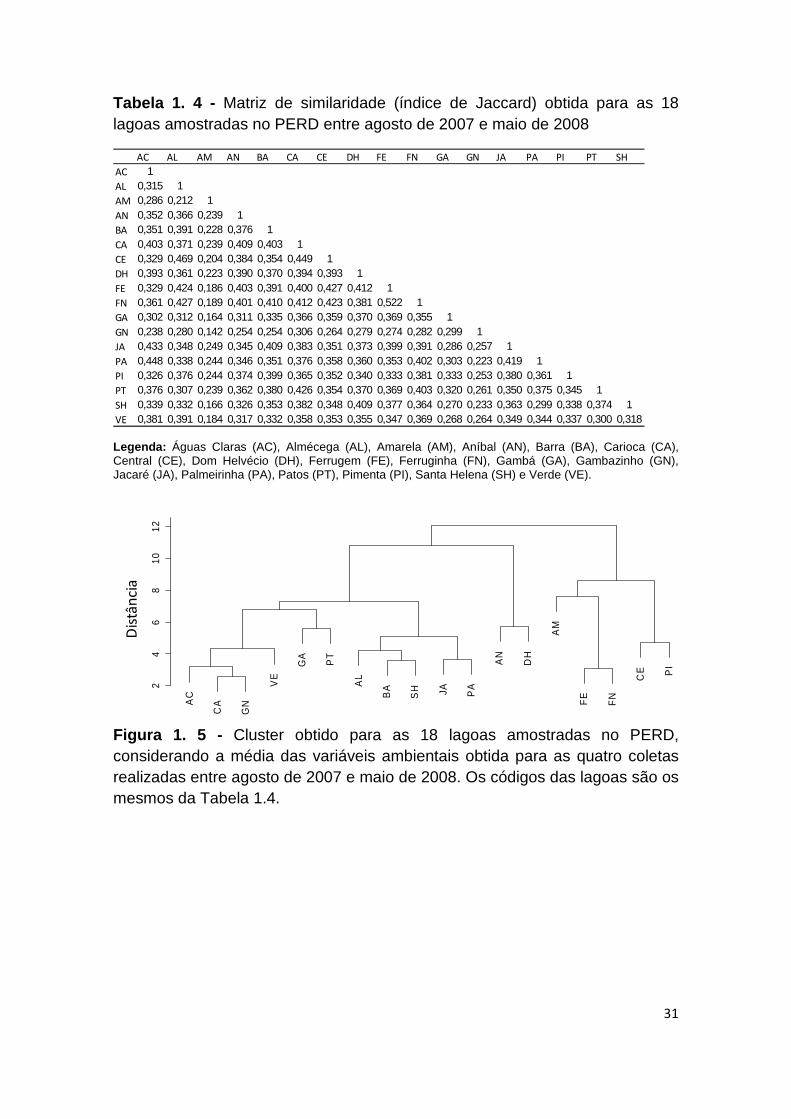
31
Tabela 1. 4 - Matriz de similaridade (índice de Jaccard) obtida para as 18 lagoas amostradas no PERD entre agosto de 2007 e maio de 2008
AC AL AM AN BA CA CE DH FE FN GA GN JA PA PI PT SH
AC 1AL 0,315 1AM 0,286 0,212 1AN 0,352 0,366 0,239 1BA 0,351 0,391 0,228 0,376 1CA 0,403 0,371 0,239 0,409 0,403 1CE 0,329 0,469 0,204 0,384 0,354 0,449 1DH 0,393 0,361 0,223 0,390 0,370 0,394 0,393 1FE 0,329 0,424 0,186 0,403 0,391 0,400 0,427 0,412 1FN 0,361 0,427 0,189 0,401 0,410 0,412 0,423 0,381 0,522 1GA 0,302 0,312 0,164 0,311 0,335 0,366 0,359 0,370 0,369 0,355 1GN 0,238 0,280 0,142 0,254 0,254 0,306 0,264 0,279 0,274 0,282 0,299 1JA 0,433 0,348 0,249 0,345 0,409 0,383 0,351 0,373 0,399 0,391 0,286 0,257 1PA 0,448 0,338 0,244 0,346 0,351 0,376 0,358 0,360 0,353 0,402 0,303 0,223 0,419 1PI 0,326 0,376 0,244 0,374 0,399 0,365 0,352 0,340 0,333 0,381 0,333 0,253 0,380 0,361 1PT 0,376 0,307 0,239 0,362 0,380 0,426 0,354 0,370 0,369 0,403 0,320 0,261 0,350 0,375 0,345 1SH 0,339 0,332 0,166 0,326 0,353 0,382 0,348 0,409 0,377 0,364 0,270 0,233 0,363 0,299 0,338 0,374 1VE 0,381 0,391 0,184 0,317 0,332 0,358 0,353 0,355 0,347 0,369 0,268 0,264 0,349 0,344 0,337 0,300 0,318
Legenda: Águas Claras (AC), Almécega (AL), Amarela (AM), Aníbal (AN), Barra (BA), Carioca (CA), Central (CE), Dom Helvécio (DH), Ferrugem (FE), Ferruginha (FN), Gambá (GA), Gambazinho (GN), Jacaré (JA), Palmeirinha (PA), Patos (PT), Pimenta (PI), Santa Helena (SH) e Verde (VE).
AC
CA
GN
VE
GA
PT
AL
BA
SH JA PA
AN
DH
AM
FE
FN
CE PI
24
68
1012
Dis
tân
cia
Figura 1. 5 - Cluster obtido para as 18 lagoas amostradas no PERD, considerando a média das variáveis ambientais obtida para as quatro coletas realizadas entre agosto de 2007 e maio de 2008. Os códigos das lagoas são os mesmos da Tabela 1.4.

32
AN
CA PI
PT
BA
DH
SH
CE AL
FE FN
AC PA
JA
VE
GA
GN
AM
0.4
0.5
0.6
0.7
0.8
0.9
D en drogram o f agnes(x = paCr is.jac, d iss = T , meth o d = "ward ")
Hei
ght
AN
CA PIPT BA
DH SH
CE AL
FE FN
AC PAJA
VEGA GN
AM
0,9
0,8
0,7
0,6
0,5
0,4
Distância
Figura 1. 6 - Cluster obtido para as 18 lagoas amostradas no PERD, considerando a presença e ausência das espécies, entre agosto de 2007 e maio de 2008. Os códigos das lagoas são os mesmos da Tabela 1.4.
Embora a maior parte das amostras (74%) tenha apresentado biomassa
fitoplanctônica menor que 2,0 mg.L-1, diferenças entre as lagoas foram
observadas em relação a este parâmetro (H=44,7; p=0,0003). Valores
extremamente elevados foram registrados em agosto de 2007 nas lagoas Barra
(13,2 mg.L-1) Aníbal (12,1 mg.L-1), Central (7,7 mg.L-1) e Ferruginha (5,2 mg.L-
1), enquanto as menores biomassas foram registradas nas lagoas Amarela
(0,09 mg.L-1), Águas Claras (0,28 mg.L-1) e Gambá (0,34 mg.L-1). A diversidade
calculada através do índice de Shannon-Wiener apresentou maior amplitude de
variação e mostrou-se diferente entre as lagoas estudadas (H=28,5; p= 0,039).
As maiores medianas foram obtidas para as lagoas Verde (2,12 bits.mg-1),
Patos (2,07 bits.mg-1), Santa Helena (1,84 bits.mg-1), Pimenta (1,82 bits.mg-1) e
Carioca (1,81 bits.mg-1) (Figura 1.7 ). As lagoas Ferrugem (J=0,53), Jacaré
(J=0,53), Amarela (J=0,53), Ferruginha(J=0,53), Almécega (J=0,53) e Patos
(J=0,53) apresentaram as menores medianas do índice de equitabilidade
(Pielou). Alta relação entre o valor do índice de diversidade e o índice de
equitabilidade mostrou-se elevada (R2= 0,82), enquanto a relação do primeiro
com a riqueza de taxa em cada amostra foi baixa (R2= 0,14).
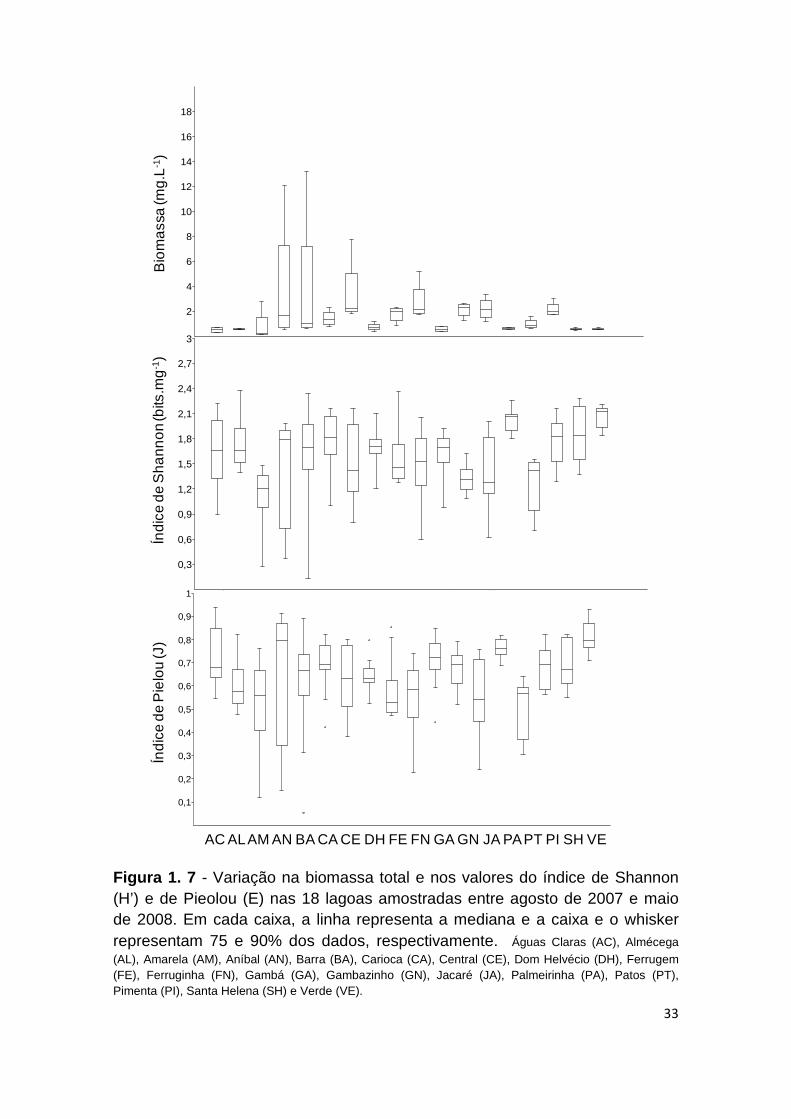
33
AC AL
AM
AN
BA
CA
GA JA PA
PT
2
4
6
8
10
12
14
16
18
Bio
ma
ssa
(mg.
L-1
)
A B C D E F G H I J K L M N O P Q R
0,3
0,6
0,9
1,2
1,5
1,8
2,1
2,4
2,7
3
Índi
ce d
e S
hann
on
(bits
.mg
-1)
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Índ
ice
de
Pie
lou
(J)
AC AL AM AN BA CA CE DH FE FN GA GN JA PA PT PI SH VE
Figura 1. 7 - Variação na biomassa total e nos valores do índice de Shannon (H’) e de Pieolou (E) nas 18 lagoas amostradas entre agosto de 2007 e maio de 2008. Em cada caixa, a linha representa a mediana e a caixa e o whisker representam 75 e 90% dos dados, respectivamente. Águas Claras (AC), Almécega (AL), Amarela (AM), Aníbal (AN), Barra (BA), Carioca (CA), Central (CE), Dom Helvécio (DH), Ferrugem (FE), Ferruginha (FN), Gambá (GA), Gambazinho (GN), Jacaré (JA), Palmeirinha (PA), Patos (PT), Pimenta (PI), Santa Helena (SH) e Verde (VE).

34
Os valores da diversidade beta obtidos através do índice β-1 foi 15,64 e,
através do índice Whittaker (βW), 2,66. A diferença entre a diversidade gama e
a alfa média, também considerada uma estimativa de diversidade beta, foi de
356 espécies.
Discussão
A heterogeneidade ambiental do sistema lacustre do médio Rio Doce foi
confirmada através das diferenças observadas nas características físicas e
químicas e na composição do fitoplâncton das lagoas.
Embora ainda pouco empregados para o fitoplâncton (Nabout et al., 2007;
Nogueira et al., 2008), os índices de extrapolação de riqueza, como o
Jackknife, podem ser uma importante ferramenta para se avaliar a
expressividade do esforço amostral. No presente estudo, a realação entre a
riqueza observada e a estimada pelo índice de extrapolação indicou que a
metodologia utilizada foi apropriada para o levantamento da diversidade.
A diversidade fitoplanctônica conecida para o sistema lacustre do médio Rio
Doce foi ampliada: Maia-Barbosa et al. (2006) registraram 267 espécies ao
estudarem sete lagoas desse sistema. A diversidade regional (γ) foi elevada
em 54% com a expansão do número de ambientes estudados, reforçando a
dissimilaridade ambiental e biótica entre as lagoas.
O predomínio de desmídias, especialmente dos gêneros Cosmarium,
Staurastrum e Staurodesmus, no sistema lacustre do médio Rio Doce foi
salientado por Reynolds (1997), que associou a dominância do grupo à
oligotrofia e às condições de preservação das lagoas. A atelomixia também tem
sido apontada como um fator chave para o predomínio das desmídias nesse
sistema lacustre (Barbosa & Padisák, 2002; Souza et al. 2008). Tal padrão de
estratificação térmica, observado em várias lagoas, permite que espécies
desse grupo, consideradas de densidade específica relativamente elevada
(Padisák et al., 2003b), permaneçam nas camadas superiores da coluna
d’água. O destaque das euglenofíceas na lagoa Amarela, também levantado
por Reynolds (1997), está associado ao seu avançado estágio de eutrofização,

35
já bastante raso e com a maior parte de seu espelho d’água coberto por
macrófitas, especialmente dos gêneros Nymphaea, Utricularia e Eleocharis.
A diversidade de espécies em um dado momento é proporcional ao número de
recursos limitantes (Tilman, 1982), de modo que as flutuações ambientais,
percebidas como distúrbios, são capazes de promover o aumento da
diversidade (Hutchinson, 1961). Além disso, os ambientes naturais são
dinâmicos e vários mecanismos atuam na regulação da diversidade de
espécies (Flöder & Burns, 2005) e fatores como a quebra da estratificação
térmica e variações nos níveis de transparência e turbidez da água podem ser
caracterizados como distúrbios nos ambientes aquáticos, afetando as
condições de equilíbrio entre as espécies (Figueredo & Giani, 2001). A
substituição da Mata Atlântica por silvicultura pode ter aumentado a
susceptibilidade das lagoas do entorno do PERD às variações ambientais. A
riqueza de espécies mais elevada no conjunto de lagoas da área de entorno do
PERD sugere uma expressão da hipótese do distúrbio intermediário (Connell,
1978), que prevê que distúrbios com frequência e intensidade intermediárias
têm um efeito positivo sobre a diversidade. No entanto, a diversidade local (α)
pode ser considerada elevada em todos os ambientes e os menores valores
foram obtidos em duas lagoas localizadas dentro do PERD (Gambazinho, com
97 espécies, e Gambá, com 99 espécies), sugerindo que os impactos
antrópicos sobre os ambientes estudados podem ser considerados pouco
severos mesmo na área de entorno. As correlações indicaram maior riqueza de
espécies em condições de menor zona de mistura e menor variação no teor de
nitrogênio total e no conjunto de variáveis avaliadas, condições ambientais
associadas aos ambientes mais rasos considerados nesse estudo.
A extensão da zona de mistura e a razão NID: PSR, bem como as variações
temporais nos teores destes nutrientes parecem ser determinantes para a
riqueza fitoplanctônica das lagoas estudadas. No entanto, o baixo número de
correlações entre a riqueza de espécies e as variáveis ambientais, a despeito
da confirmação das diferenças entre as lagoas, sugere maior importância da
interação entre fatores do que de variáveis isoladas para a definição da
diversidade local. Reynolds (1997) salienta que a sazonalidade relativamente

36
fraca (comparada a ambientes temperados), combinada ao isolamento
geográfico, à extensiva cobertura vegetal das bacias hidrográficas e à ausência
de poluição nesses ambientes, caracteriza uma situação de pobreza de
distúrbios nesse sistema lacustre. No entanto, dados publicados (Barros et al.
2006; Souza et al., 2008) mostram que embora menos marcante nos trópicos,
a sazonalidade exerce um efeito importante sobre a dinâmica do fitoplâncton
na lagoa Dom Helvécio, controlando os padrões de mistura da coluna d’água e
determinando sua dinâmica biológica. Este efeito se evidencia no presente
estudo através da variação temporal observada para a biomassa
fitoplanctônica e para os valores do índice de Shannon em cada lagoa.
Flöder & Sommer (1999) e Passarge et al. (2006) salientam que a variabilidade
temporal pode ser mais importante que a heterogeneidade espacial para a
manutenção da diversidade, especialmente em lagos sem gradientes
ambientais proeminentes. Com exceção das lagoas Amarela e Ferrugem, as
demais lagoas estudadas são classificadas como oligo (11 lagoas) ou
mesotróficas (5 lagoas), evidenciando um fraco gradiente de estado trófico.
Adicionalmente, variáveis descritas como importantes para o desenvolvimento
do fitoplâncton (teores de fósforo solúvel reativo (PSR), nitrogênio inorgânico
dissolvido (NID), temperatura, razões PSR:NID e Zeu:Zmix), não diferiram
entre as lagoas, considerando o conjunto das quatro amostragens,
evidenciando que as variações climáticas podem ser importantes na
manutenção da diversidade. Além disto, o tamanho das lagoas, descrito na
literatura como uma importante variável para a explicação da variação da
diversidade em lagoas (Allen et al., 1999), também não apresentou relação
com a diversidade alfa no presente estudo.
As diferenças nas características limnológicas são também evidenciadas pela
baixa similaridade florística, refletidas pela heterogeneidade ambiental no
sistema lacustre. Um terço das espécies registradas foi restrita a somente uma
das lagoas estudaddas e poucas espécies (10, ou seja, 2%) foram comuns às
18 estudadas. Embora tenha apresentado a menor diversidade alfa, na lagoa
Gambazinho foi registrada a segunda maior incidência de espécies exclusivas
(18), ficando atrás apenas da lagoa Amarela, com 25 taxa exclusivos. Em

37
ambos, a elevada ocorrência de taxa exclusivos indica a contribuição da região
litorânea, pois, no caso da lagoa Gambazinho, os principais representantes
foram diatomáceas da ordem Pennales (7) e grandes desmídias (5). As
desmídias de maior ramanho também se destacaram na lagoa Amarela (12),
juntamente com as euglenofíceas (5). Esses dois ambientes apresentam
características que os destacam dos demais estudados: o elevado teor de
matéria orgânica na lagoa Amarela, em decorrência do avançado estágio de
eutrofização, e o padrão polimítico de estratificação térmica da lagoa
Gambazinho, enquanto o padrão dominante no sistema lacustre do médio Rio
Doce é monomítico-quente. Tais características parecem permitido o registro
de um número maior de espécies típicas do perifiton nas amostras do
fitoplâncton.
As análises de agrupamento evidenciaram pouca importânica da distância
geográfica para a similaridade entre as lagoas tanto em relação às
características ambientais quanto em relação à composição fitoflorística. Com
exceção das lagoas mais próximas entre si, Gambá e Gambazinho (500m de
distância), Ferrugem e Ferruginha (1.200m de distância), e Águas Claras e
Palmeirinha (1.500m de distância), em ambos os casos, os grupos são
formados por lagoas distantes umas das outras, inclusive mesclando
ambientes localizados dentro do PERD e na área de entorno. Esse resultado
aponta que as características limnológicas observadas em cada lagoa
independem de sua localização. Assim os fatores locais parecem assumir
maior importância que os fatores regionais na seleção das espécies que
compõem o fitoplâncton de cada uma das lagoas estudadas.
Os valores do índice de diversidade podem ser considerados baixos,
comparando-se a valores obtidos para ambientes impactados, como o
reservatório da Pampulha (Figueredo & Giani, 2001). Os máximos de
diversidade coincidiram com os máximos de equitabilidade e verificou-se uma
forte relação entre os valores do índice de Shannon-Wiener e os do índice de
Pielou. Esses resultados mostram que a baixa equitabilidade foi responsável
pelos reduzidos valores do índice de diversidade, uma vez que a maioria das

38
lagoas mostrou dominância de alguma espécie durante as quatro amostragens
realizadas.
Embora a diversidade beta ofereça informações importantes para a escolha de
áreas a serem preservadas (Ward et al., 1999), apenas três trabalhos
realizados no Brasil consideraram este parâmetro em estudos limnológicos:
Huszar et al., (1990), Nabout et al. (2007) e Nogueira et al., (2008). Os valores
obtidos no presente estudo para os índices de diversidade beta (β-1=15,9 e
βW=2,96) são considerados baixos se comparados àqueles obtidos por Nabout
et al. (2007) para o conjunto de 21 lagoas da bacia do Rio Araguaia (β-1=30 a
46), por Nogueira et al. (2008) para o conjunto de 4 lagoas artificiais localizadas
no município de Goiânia (β-1 > 40) e por Huszar et al. (1990) para as 18 lagoas
estudadas no sistema lacustre do baixo Rio Doce (βW=7,5). Tal fato pode ser
explicado pela ocorrência de elevada diversidade gama nas lagoas do presente
estudo, uma vez que ambos os índices levam em consideração a riqueza
média de espécies do conjunto de lagoas. Nos três casos citados, as
diferenças na diversidade alfa entre os ambientes estudados foram mais
expressivas e a riqueza de espécies registrada em cada local foi menor do que
o observado no presente estudo. Veech et al. (2002), no entanto, sugerem que
se considere a diversidade gama como a soma e não o produto, das
diversidades alfa e beta, mantendo as propostas originais de MacArthur e
colaboradores e Levins (MacArthur et al., 1966; Levins, 1968 apud Veech et al,
2002). Assim, tem-se maior importância para as diferenças na composição de
espécies entre os ambientes amostrados, mantendo-se a proposta original do
termo diversidade beta. Utilizando-se desta abordagem, a diversidade beta
obtida através da diferença entre a diversidade gama (regional) e a diversidade
alfa (local) média encontradas neste estudo pode ser considerada elevada,
uma vez que o valor obtido (356 espécies) corresponde a 72% do total de
espécies registradas.
Os resultados obtidos contribuem para informações sobre o conhecimento da
diversidade fitoplanctônica do sistema lacustre do médio Rio Doce. Reportando
às hipóteses testadas, foram evidenciados elevados níveis de diversidade alfa
e gama e a baixa similaridade entre os ambientes (= alta diversidade beta,

39
sensu Veech et al, 2002). No entanto, a área, forma ou a distância geográfica,
por si só, não definem as espécies presentes em cada lagoa ou os padrões da
distribuição de espécies. Assim, parece não haver um único fator determinando
os padrões de riqueza nesse sistema lacustre e a composição florística
resultante em cada ambiente provavelmente resulta da interação de diversos
fatores. Adicionalmente, a hipótese de que a proteção do PERD garante maior
riqueza de espécies foi refutada. Coesel (2001) propõe que a diversidade e a
raridade são importantes critérios para se avaliar o valor de conservação de
uma dada região. Deve-se considerar, portanto, a possibilidade de
conservação do sistema lacustre como um todo, com a proposição de
estratégias específicas para os ambientes da área de entorno, para a
manutenção da diversidade fitoplanctônica em nível regional.
40
Apêndice - Taxa observados nas 18 lagoas amostradas sistema lacustre do médio Rio Doce e número de lagoas (n) em que ocorreram. Taxon n Taxon n Bacillariophyceae Urosolenia cf eriensis (H. L.) Round & Crawf 2 Synedra acus Kützing 16 Pennales NI11 1 Cyclotella sp 14 Caloneis cf 1 Gomphonema gracile Ehrenberg 14 Encyonema sp5 1 Eunotia lineolata Husted 13 Encyonema sp6 1 Stenopterobia curvula (Wm. Smith) Krammer 13 Eunotia sp6 1 Urosolenia cf longiseta (Zacharias) Edlung & Stoermer 13 Frustulia sp1 1 Encyonema sp1 9 Gomphonema subtile Ehrenberg 1 Navicula sp 8 Hantzschia amphioxys (Ehrenberg) Grunow 1 Eunotia sp1 7 Neidium sp 1 Aulacoseira sp1 6 Nitzschia sp 1 Pinnularia viridis (Nitzsch) Ehrenberg 5 Pinnularia sp2 1 Pennales NI5 4 Pinnularia sp3 1 Encyonema sp2 4 Pinnularia sp4 1 Eunotia sp4 3 Pinnularia sp5 1 Eunotia sp5 3 Pinnularia sp6 1 Frustulia crassinervia (Brébisson) Costa 3 Chlorophyceae Frustulia krammeri Lange-Bertalot e Metzeltin 3 Botryococcus braunii Kützing 18 Pennales NI3 2 Elakatothrix genevensis (Reverdin) Hindák 18 Pennales NI6 2 Chlorella sp 17 Aulacoseira ambigua (Grunow) Simonsen 2 Closteriopsis sp1 17 Aulacoseira granulata (Ehrenberg) Simonsen 2 Tetraedron caudatum (Corda) Hansgirg 17 Aulacoseira sp2 2 Monoraphidium sp 16 Encyonema sp3 2 Botryococcus terribilis J. Komárek & P. Marvan 15 Encyonema sp4 2 Eutetramorus planctonicus (Korsikov) Bourrely 15 Eunotia sp2 2 Monoraphidium contortum (Thur.) Kom.-Legn 15 Melosira varians Agardh 2 Nephrocytium agardhianum Nägeli 15 Stauroneis phoenicenteron (Nitzsch) Ehrenberg 2 Oocystis sp4 15 Surirella linearis W. Smith 2 Botryococcus protuberans W. & G.S. West 14 Surirella sp1 2 Chlorophyceae NI1 14

41
Apêndice - Continuação.
Taxon n Taxon n Crucigenia tetrapedia (Kirchner) W.& G.S. West 14 Botryococcus NI4 4 Kirchneriella lunaris (Kirchner) K. Möbius 14 Chlamydomonas sp 4 Oocystis lacustris Chodat 14 Coelastrum microporum Nägeli 4 Oocystis sp3 14 Crucigenia cf fenestrata (Schimidle) Shimidle 4 Scenedesmus bijugus (Turpin) Kutzing 14 Desmodesmus denticulatus (Lagerheim) An et al. 4 Dictyosphaerium pulchellum Wood 13 Dictyosphaerium erenbergianum Nägeli 4 Ankistrodesmus fusiformis Corda 12 Eudorina elegans Ehrenberg 4 Tetraedron minimum (A. Braun) 12 Kirchneriella cf obesa (W.West) Schmidle 4 Crucigeniella retangularis (Nägeli) Kom. 11 Oocystis cf solitaria Wittrock 4 Ankistrodesmus sp2 10 Ankistrodesmus densus Korsikov 3 Coelastrum reticulatum (Dang.) Senn 10 Pediastrum duplex Meyen 3 Desmodesmus quadricauda (Turp.) Bréb. 10 Radiococcus cf 3 Coelastrum pulchrum Schimidle 9 Scenedesmus acunae Comas 3 Coelastrum sphaericum Nägeli 9 Chlorotetraedron incus (Teiling) G.M. Smithi 3 Desmodesmus armatus var. bicaudatus (Guglielmetti) Hegewald 9 Chlamydomonadaceae NI2 2 Keratococcus sp 9 Chlorococcales NI1 2 Actinastrum aciculare Playf 8 Dimorphococcus cf lunatus A. Braun 2 Scenedesmus acuminatus (Lagerheim) Chodat 8 Kirchneriella roselata Hindák 2 Ankyra judayi (G.M. Smithi) Fott 7 Oocystis cf nephrocytioides Fott & Cado 2 Closteriopsis sp2 7 Oocystis sp6 2 Coelastrum cambricum Arch. 7 Pediastrum tetras (Ehrenberg) Ralfs 2 Scenedesmus bijugus var disciformis (Chod.) Leite 7 Planctonema cf 2 Tetrallantos lagerheimii Teiling 7 Scenedesmus acuminatus f. maximus Uherk. 2 Pediastrum privum (Printz) Hegew 6 Desmodesmus spinosus Chodat 2 Quadrigula closterioides (Bohlin) Printz 6 Selenastrum sp1 2 Golenkinia radiata Chodat 5 Tetranephris brasiliense Leite & C.Bicudo 2 Oocystis sp1 5 Tetranephris cf 2 Scenedesmus arcuatus Lemmerman 5 Volvocales NI 2 Sphaerocystis sp1 5 Actinastrum hantszchii Lagerheim 1 Ankistrodesmus spiralis (Turner) Lemmermann 4 Ankistrodesmus falcatus (Corda) Ralfs 1
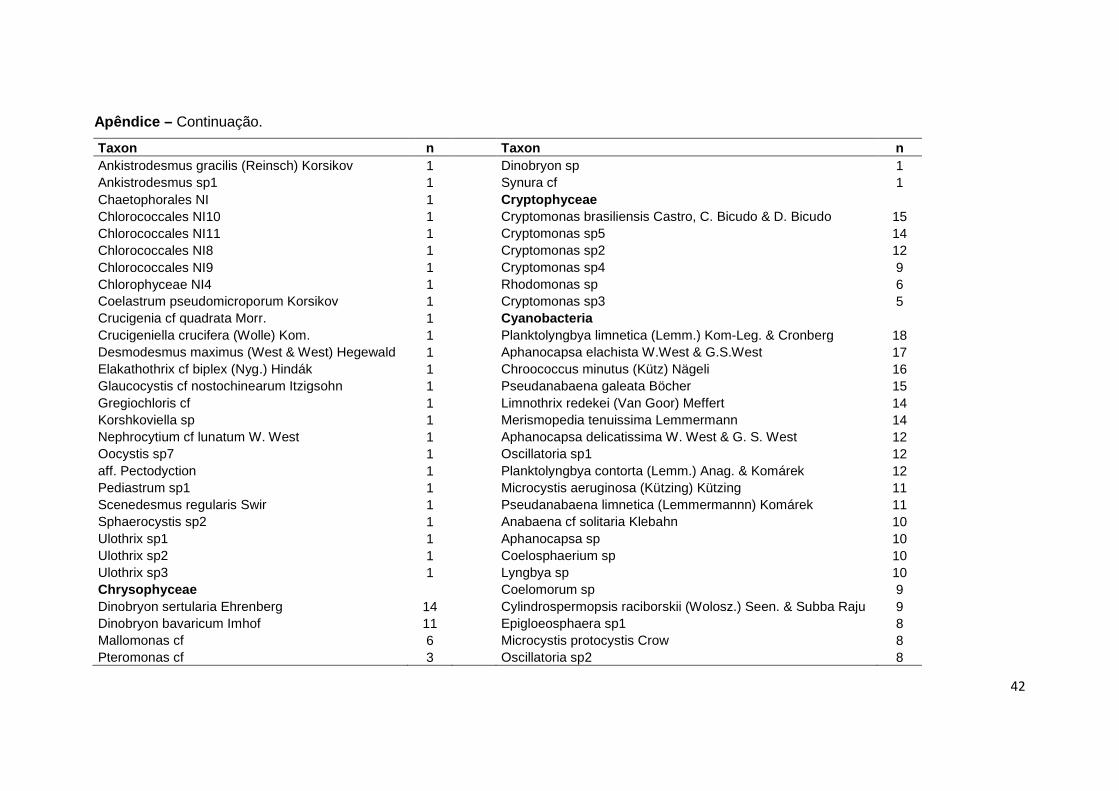
42
Apêndice – Continuação.
Taxon n Taxon n Ankistrodesmus gracilis (Reinsch) Korsikov 1 Dinobryon sp 1 Ankistrodesmus sp1 1 Synura cf 1 Chaetophorales NI 1 Cryptophyceae Chlorococcales NI10 1 Cryptomonas brasiliensis Castro, C. Bicudo & D. Bicudo 15 Chlorococcales NI11 1 Cryptomonas sp5 14 Chlorococcales NI8 1 Cryptomonas sp2 12 Chlorococcales NI9 1 Cryptomonas sp4 9 Chlorophyceae NI4 1 Rhodomonas sp 6 Coelastrum pseudomicroporum Korsikov 1 Cryptomonas sp3 5 Crucigenia cf quadrata Morr. 1 Cyanobacteria Crucigeniella crucifera (Wolle) Kom. 1 Planktolyngbya limnetica (Lemm.) Kom-Leg. & Cronberg 18 Desmodesmus maximus (West & West) Hegewald 1 Aphanocapsa elachista W.West & G.S.West 17 Elakathothrix cf biplex (Nyg.) Hindák 1 Chroococcus minutus (Kütz) Nägeli 16 Glaucocystis cf nostochinearum Itzigsohn 1 Pseudanabaena galeata Böcher 15 Gregiochloris cf 1 Limnothrix redekei (Van Goor) Meffert 14 Korshkoviella sp 1 Merismopedia tenuissima Lemmermann 14 Nephrocytium cf lunatum W. West 1 Aphanocapsa delicatissima W. West & G. S. West 12 Oocystis sp7 1 Oscillatoria sp1 12 aff. Pectodyction 1 Planktolyngbya contorta (Lemm.) Anag. & Komárek 12 Pediastrum sp1 1 Microcystis aeruginosa (Kützing) Kützing 11 Scenedesmus regularis Swir 1 Pseudanabaena limnetica (Lemmermannn) Komárek 11 Sphaerocystis sp2 1 Anabaena cf solitaria Klebahn 10 Ulothrix sp1 1 Aphanocapsa sp 10 Ulothrix sp2 1 Coelosphaerium sp 10 Ulothrix sp3 1 Lyngbya sp 10 Chrysophyceae Coelomorum sp 9 Dinobryon sertularia Ehrenberg 14 Cylindrospermopsis raciborskii (Wolosz.) Seen. & Subba Raju 9 Dinobryon bavaricum Imhof 11 Epigloeosphaera sp1 8 Mallomonas cf 6 Microcystis protocystis Crow 8 Pteromonas cf 3 Oscillatoria sp2 8
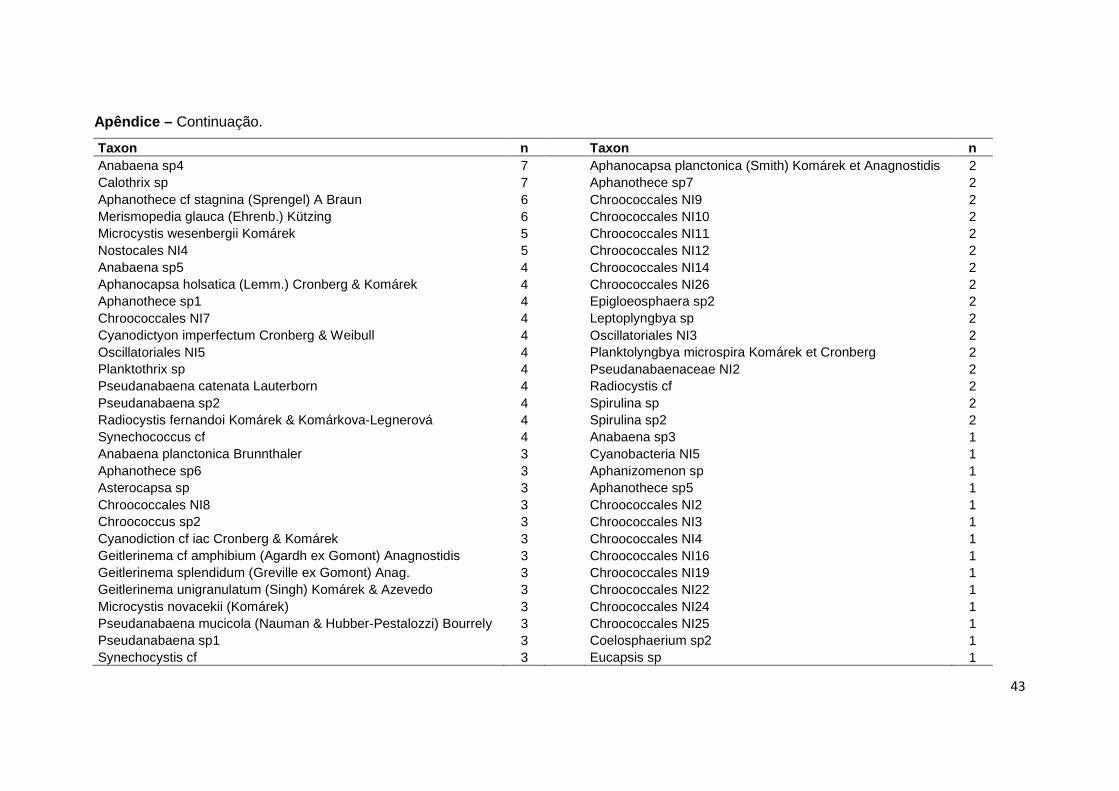
43
Apêndice – Continuação.
Taxon n Taxon n Anabaena sp4 7 Aphanocapsa planctonica (Smith) Komárek et Anagnostidis 2 Calothrix sp 7 Aphanothece sp7 2 Aphanothece cf stagnina (Sprengel) A Braun 6 Chroococcales NI9 2 Merismopedia glauca (Ehrenb.) Kützing 6 Chroococcales NI10 2 Microcystis wesenbergii Komárek 5 Chroococcales NI11 2 Nostocales NI4 5 Chroococcales NI12 2 Anabaena sp5 4 Chroococcales NI14 2 Aphanocapsa holsatica (Lemm.) Cronberg & Komárek 4 Chroococcales NI26 2 Aphanothece sp1 4 Epigloeosphaera sp2 2 Chroococcales NI7 4 Leptoplyngbya sp 2 Cyanodictyon imperfectum Cronberg & Weibull 4 Oscillatoriales NI3 2 Oscillatoriales NI5 4 Planktolyngbya microspira Komárek et Cronberg 2 Planktothrix sp 4 Pseudanabaenaceae NI2 2 Pseudanabaena catenata Lauterborn 4 Radiocystis cf 2 Pseudanabaena sp2 4 Spirulina sp 2 Radiocystis fernandoi Komárek & Komárkova-Legnerová 4 Spirulina sp2 2 Synechococcus cf 4 Anabaena sp3 1 Anabaena planctonica Brunnthaler 3 Cyanobacteria NI5 1 Aphanothece sp6 3 Aphanizomenon sp 1 Asterocapsa sp 3 Aphanothece sp5 1 Chroococcales NI8 3 Chroococcales NI2 1 Chroococcus sp2 3 Chroococcales NI3 1 Cyanodiction cf iac Cronberg & Komárek 3 Chroococcales NI4 1 Geitlerinema cf amphibium (Agardh ex Gomont) Anagnostidis 3 Chroococcales NI16 1 Geitlerinema splendidum (Greville ex Gomont) Anag. 3 Chroococcales NI19 1 Geitlerinema unigranulatum (Singh) Komárek & Azevedo 3 Chroococcales NI22 1 Microcystis novacekii (Komárek) 3 Chroococcales NI24 1 Pseudanabaena mucicola (Nauman & Hubber-Pestalozzi) Bourrely 3 Chroococcales NI25 1 Pseudanabaena sp1 3 Coelosphaerium sp2 1 Synechocystis cf 3 Eucapsis sp 1

44
Apêndice – Continuação.
Taxon n Taxon n Johannesbaptistia sp 1 Dinophyceae NI2 1 Microcrocis cf 1 Peridinium sp2 1 Nostoc sp1 1 Peridinium sp5 1 Nostoc sp2 1 Euglenophyceae Nostoc sp3 1 Trachelomonas volvocina Ehrenberg 18 Nostocales NI5 1 Phacus longicauda (Ehrenberg) Dujardin 10 Nostocales NI6 1 Phacus raciborskii Drez 10 Nostocales NI7 1 Trachelomonas armata (Ehrenberg) Stein 8 Nostocales NI8 1 Euglenales NI1 7 Nostocales NI9 1 Phacus hamatus Pochmann 7 Oscillatoriales NI6 1 Euglenophyceae Planktolyngbya sp2 1 Trachelomonas volvocina Ehrenberg 18 Porphyrosiohon cf 1 Phacus longicauda (Ehrenberg) Dujardin 10 Pseudanabaenaceae NI 1 Phacus raciborskii Drez 10 Pseudanabaenaceae NI4 1 Trachelomonas armata (Ehrenberg) Stein 8 Pseudanabaenaceae NI6 1 Euglenales NI1 7 Rabdoderma cf 1 Phacus hamatus Pochmann 7 Raphidiopsis cf 1 Euglena oxyuris Schmarda 6 Snowella cf 1 Lepocinclis sp1 6 Sphaerocavum brasiliense M. T. P. Azevedo & Sant’Anna 1 Trachelomonas cf hispida (Perty) Stein em. Defl. var duplex Defl. 6 Stigonematales NI 1 Trachelomonas lacustris Drezepolski 6 Stigonematales NI2 1 Trachelomonas sp2 6 Dinophyceae Euglena acus Ehremberg 5 Peridinium pusillum (Pénard) Lemmerman 18 Euglena ehrenbergii Klebs 4 Gymnodynium sp 16 Euglenales NI7 4 Peridinium baliense Lindemann 16 Phacus sp3 4 Peridinium cf africanum Lemmermann 10 Phacus cf suecicus 3 Dinophyceae NI1 9 Phacus onyx Pochmann 3 Peridinium cf volzii Lemmerman 4 Trachelomonas cf mirabilis Woronichin 3 Peridinium sp3 3 Trachelomonas cf zingeri Roll 3

45
Apêndice – Continuação.
Taxon n Taxon n Trachelomonas similis Stokes 3 Tetraplekton sp2 7 Lepocinclis fusiformis (Carter) Lemm. emend.Conr. 2 Centritactus sp 6 Phacus sp5 2 Isthmochlorum gracile (Reinsch) Skuja 5 Trachelomonas megalacantha Cunha 2 Tetraplekton torsum 5 Trachelomonas sp1 2 Pseudostaurastrum sp 3 Trachelomonas sp3 2 Zygnematophyceae Euglenales NI2 1 Cosmarium contractum Kirchner 18 Euglenales NI3 1 Staurastrum forficulatum Lundell 18 Euglenales NI4 1 Staurastrum leptocladum Nordstedt 18 Lepocinclis cf ovum (Ehrenberg) Lemmermann 1 Staurastrum rotula Nordstedt 18 Lepocinclis sp2 1 Staurodesmus dejectus (Brébisson) Teiling 18 Lepocinclis sp3 1 Cosmarium asphaerosporum 17 Lepocinclis sp4 1 Teilingia granulata (Roy & Bisset) Bourrely 17 Phacus orbicularis Hübner emend. Zakry´s & Kosmala 1 Cosmarium moniliforme West & West 16 Rhabdomonas sp 1 Cosmarium pseudoconnatum Nordst. 16 Strombomonas sp 1 Staurastrum ionatum Wolle 16 Strombomonas sp1 1 Staurastrum laeve Ralfs 16 Trachelomonas cf magdaleniana Deflandre 1 Staurastrum trifidum Nordstedt 16 Trachelomonas cf oblonga Lemmermann 1 Staurastrum sp5 16 Trachelomonas hispida (Perty) Stein emend. Deflandre 1 Staurodesmus crassus (West & West) Florin 16 Trachelomonas sp5 1 Staurodesmus jaculiferus (W. West) Teiling 16 Trachelomonas sp6 1 Staurastrum smithii (G.M. Smith) Teiling 15 Trachelomonas sp8 1 Staurodesmus sp3 15 Oedogoniophyceae Staurastrum setigerum Gleve 13 Oedogoniophyceae NI 1 Staurodesmus convergens (Ehr. Ex Rafs) Teiling 13 Oedogonium sp 9 Staurodesmus cuspidatus (Brébisson) Teiling 13 Xanthophyceae Staurastrum tetracerum (Kützing) Ralfs ex Ralfs 12 Tetraplekton sp1 12 Staurodesmus subulatus (Kützing) Croasdale 12 Isthmochloron lobulatum (Nägeli) Skuja 10 Cosmarium bioculatum Brébisson 11 Tetraplekton cf bourrelyi 7 Cosmarium depressum (Nageli) Lundell 11

46
Apêndice – Continuação.
Taxon n Taxon n Spondylosium panduriforme (Heimerl) Teiling 11 Staurastrum sp4 5 Staurastrum chaetoceras (Schr.) G.M.Smith 11 Staurastrum sp7 5 Cosmarium monomazum P.Lundell 10 Staurastrum sp8 5 Cosmarium quadrum P.Lundell 10 Staurastrum sp15 5 Micrasterias truncata (Corda) Bréb. ex Ralfs 10 Staurastrum sp18 5 Staurastrum cf muticum (Brébisson) Ralfs 10 Staurastrum sp24 5 Staurastrum cerastes Lundell 9 Staurodesmus convergens (Ehr. Ex Rafs) Teiling var.2 5 Staurastrum manfeldtii Delponte 9 Staurodesmus sp6 5 Staurastrum sp6 9 Cosmarium cf conspersum Ralfs 4 Cosmarium pyramidatum Brébisson in Ralfs 8 Cosmarium portianum Archer 4 Sphaerozosma sp 8 Haplotaenium minutum (Ralfs) Delponte 4 Staurastrum grallatorium Nordstedt 8 Micrasterias pinnatifida (Kütz.) Ralfs 4 Staurastrum sp26 8 Octacanthium mucronulatum (Nordstedt) 4 Staurastrum sp27 8 Staurastrum teliferum Ralfs 4 Staurodesmus sp5 8 Staurodesmus incus (Brébisson) Teiling 4 Closterium gracile Brebisson ex Ralfs 7 Staurodesmus sp7 4 Staurastrum leptacanthum Nordstedt 7 Closterium kuetzingii Brebisson 3 Staurastrum taylorii Grönblad 7 Cosmarium sp20 3 Staurastrum teliferum Ralfs var.2 7 Cosmarium sp23 3 Staurodesmus lobatus (Borgesen) Bourrely 7 Cosmarium sp4 3 Bourrellyodesmus jolyanus C. Bicudo & Azevedo 6 Cosmarium sp7 3 Closterium cf setaceum Ehrenberg 6 Cosmarium sp8 3 Cosmarium sp16 6 Euastrum cf abruprum Nordstedt 3 Hyaloteca sp 6 Gonatozygon brebissonii De Bary 3 Staurodesmus o'mearii (Archer) Teiling 6 Pleurotaenium sp2 3 Mougeotia sp1 5 Staurastrum orbiculare (Ehrenberg) Ralfs, Brit. 3 Staurastrum depressiceps Scott et Grönblad 5 Staurastrum quadrangulare (Brébisson) 3 Staurastrum minnesotense Wolle 5 Staurastrum tentaculiferum Borge 3 Staurastrum nudibrachiatum Borge 5 Staurastrum sp22 3 Staurastrum wolleanum Butler apud Wolle 5 Staurastrum sp46 3
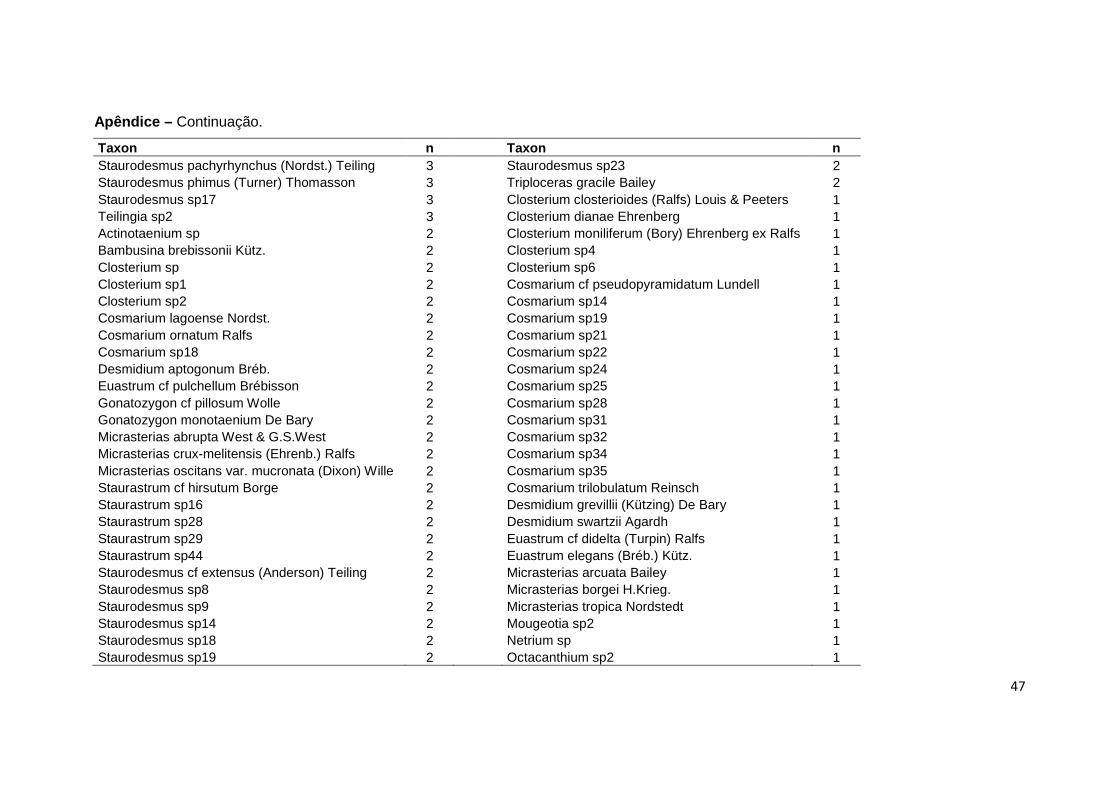
47
Apêndice – Continuação.
Taxon n Taxon n Staurodesmus pachyrhynchus (Nordst.) Teiling 3 Staurodesmus sp23 2 Staurodesmus phimus (Turner) Thomasson 3 Triploceras gracile Bailey 2 Staurodesmus sp17 3 Closterium closterioides (Ralfs) Louis & Peeters 1 Teilingia sp2 3 Closterium dianae Ehrenberg 1 Actinotaenium sp 2 Closterium moniliferum (Bory) Ehrenberg ex Ralfs 1 Bambusina brebissonii Kütz. 2 Closterium sp4 1 Closterium sp 2 Closterium sp6 1 Closterium sp1 2 Cosmarium cf pseudopyramidatum Lundell 1 Closterium sp2 2 Cosmarium sp14 1 Cosmarium lagoense Nordst. 2 Cosmarium sp19 1 Cosmarium ornatum Ralfs 2 Cosmarium sp21 1 Cosmarium sp18 2 Cosmarium sp22 1 Desmidium aptogonum Bréb. 2 Cosmarium sp24 1 Euastrum cf pulchellum Brébisson 2 Cosmarium sp25 1 Gonatozygon cf pillosum Wolle 2 Cosmarium sp28 1 Gonatozygon monotaenium De Bary 2 Cosmarium sp31 1 Micrasterias abrupta West & G.S.West 2 Cosmarium sp32 1 Micrasterias crux-melitensis (Ehrenb.) Ralfs 2 Cosmarium sp34 1 Micrasterias oscitans var. mucronata (Dixon) Wille 2 Cosmarium sp35 1 Staurastrum cf hirsutum Borge 2 Cosmarium trilobulatum Reinsch 1 Staurastrum sp16 2 Desmidium grevillii (Kützing) De Bary 1 Staurastrum sp28 2 Desmidium swartzii Agardh 1 Staurastrum sp29 2 Euastrum cf didelta (Turpin) Ralfs 1 Staurastrum sp44 2 Euastrum elegans (Bréb.) Kütz. 1 Staurodesmus cf extensus (Anderson) Teiling 2 Micrasterias arcuata Bailey 1 Staurodesmus sp8 2 Micrasterias borgei H.Krieg. 1 Staurodesmus sp9 2 Micrasterias tropica Nordstedt 1 Staurodesmus sp14 2 Mougeotia sp2 1 Staurodesmus sp18 2 Netrium sp 1 Staurodesmus sp19 2 Octacanthium sp2 1

48
Apêndice- continuação
Taxon n Onychonema laeve Nordstedt 1
Pleurotaenium cf baculoides (Skuja) Krieger 1 Sirogonium cf 1 Spirogyra sp1 1 Spirogyra sp2 1 Spirogyra sp3 1 Spondylosium planum (Wolle) West &
G.S.West 1 Staurastrum brasiliense Nordstedt 1 Staurastrum sexangulare Bulnheim
(Rabenhorst) 1 Staurastrum sp17 1 Staurastrum sp25 1 Staurastrum sp30 1 Staurastrum sp35 1 Staurastrum sp38 1 Staurastrum sp39 1 Staurastrum sp40 1 Staurastrum sp41 1 Staurastrum sp42 1 Staurastrum sp45 1
Staurodesmus sp13 1 Staurodesmus sp20 1 Staurodesmus sp22 1 Staurodesmus sp24 1 Staurodesmus sp25 1 Teilingia sp1 1 Xanthidium regulare Nordst. 1 aff. Zygnema sp 1

49
CAPÍTULO 2
ATENUAÇÃO DA RADIAÇÃO VISÍVEL EM 16 LAGOAS NATURAIS DO MÉDIO RIO DOCE, MINAS GERAIS: FATORES DETERMINANTES E IMP LICAÇÕES
PARA O FITOPLÂNCTON

50
Resumo
Atenuação da radiação visível em 16 lagoas naturais do médio Rio Doce, Minas
Gerais: fatores determinantes e implicações para o fitoplâncton
Este estudo apresenta a variação espacial e temporal do coeficiente de absorção da
radiação fotossinteticamente ativa (k0) em 16 lagoas do sistema lacustre do médio
Rio Doce, bem como sua relação com os teores de matéria orgânica dissolvida
colorida (MODC), fósforo total e composição e biomassa do fitoplâncton. Os valores
de k0 (0,4 a 2,93 m-1) variaram espacial e sazonalmente, em função das variações
na profundidade da zona de mistura, nas concentrações de P-total e matéria
orgânica dissolvida colorida (MODC) e na biomassa fitoplanctônica. Lagoas mais
claras (k0 < 0,8 m-1) apresentaram maior riqueza de diatomáceas e maior
contribuição de desmídias para a biomassa total. As lagoas mais escuras (k0 > 0,8)
apresentaram maior biomassa total, composto principalmente por cianofíceas e de
criptofíceas. A sazonalidade da atenuação da luz foi determinada pelos padrões de
estratificação térmica, reforçando a importância do padrão monomítico-quente nesse
sistema lacustre. A relação positiva ente os valores de k0 e a biomassa
fitoplanctônica sugere que mesmo nas lagoas com maiores valores de k0 não houve
limitação do desenvolvimento dessa comunidade. Embora mesmo os valores de k0
observados nas lagoas de águas escuras não possam ser considerados elevados se
comparados a outros ambientes húmicos, a influência do grau de atenuação da luz
sobre o fitoplâncton foi percebida pelas diferenças na composição da comunidade
fitoplanctônica, com o favorecimento de espécies mixotróficas, tolerantes à baixa
disponibilidade de luz ou oportunistas.
Palavras-Chave: atenuação da PAR, fitoplâncton, MODC

51
Abstract
Attenuation of visible radiation in 16 natural lake s of the middle Rio Doce,
Minas Gerais: determinants and implications for phy toplankton
This study highlights the spatial and temporal variation of absorption coefficient of
photosynthetically active radiation (k0) in 16 lakes of the lacustrine system of the
middle Rio Doce and its relation to the concentration of colored dissolved organic
carbon, CDOC, total phosphorus and phytoplankton composition and biomass. k0
values (0.4 to 2.93 m-1) varied spatially and seasonally, related to variations in the
mixing zone depth, total-P and CDOM concentrations and phytoplankton biomass.
Lighter lakes (k0 <0.8 m-1) showed greater richness of diatoms, and greater
contributions of desmids to the total biomass. Darker lakes (k0> 0.8) had higher total
and cyanobacteria and cryptophytes biomass. The seasonality of the light attenuation
was determined by thermal stratification patterns, reinforcing the importance of warm-
monimitico pattern for the system. The positive relationship between k0 values and
phytoplankton biomass suggests that even in lakes with higher CDOM levels, the k0
valaues were not sufficient to limit the development of planktonic algae. Even though
the values of k0 observed in darker lakes can not be considered high compared to
other humic environments, the influence of light attenuation on phytoplankton was
perceived by the differences in the composition of the phytoplankton community, with
the encouragement of mixotrophic, low-light tolerant or opportunistic species.
Key-words: PAR atenuation, phytoplankton, CDOM

52
Introdução
A radiação solar tem uma importância fundamental no fluxo de energia de
ecossistemas aquáticos. Fatores como posição do sol, nebulosidade e concentração
de carbono orgânico dissolvido e material particulado influenciam a profundidade de
penetração da radiação solar (Kostoglidis et al., 2005), afetando a absorção e
dissipação na forma de calor e determinando a estrutura térmica, a ciclagem de
nutrientes e a distribuição dos gases dissolvidos e dos organismos nos ambientes
aquáticos (Wetzel, 2001).
A disponibilidade de luz, especialmente a radiação fotossinteticamente ativa (PAR),
está entre os principais fatores que controlam a composição e a biomassa do
fitoplâncton. As diferenças temporais e espaciais na intensidade e no espectro de
luz possibilitam a sucessão fitoplanctônica, uma vez que a presença ou ausência de
atributos fisiológicos que permitam crescer sob uma dada condição luminosa podem
determinar o sucesso ou fracasso de uma espécie em se estabelecer em um
ambiente (Litchman & Klausmeier, 2008).
A quantificação da profundidade da zona eufótica e a influência dos diferentes
fatores sobre a atenuação da radiação fotossinteticamente ativa fornecem
informações importantes acerca do clima de luz para o fitoplâncton (Foden et al.,
2008). Compostos orgânicos, especialmente a matéria orgânica dissolvida colorida
(MODC) são conhecidos por seu importante papel na atenuação da luz em lagoas. A
concentração de MODC influencia a penetração de luz nos sistemas (Devol et al.,
1984; Bezerra-Neto et al., 2006; Karlsson et al., 2009), alterando sua qualidade e
quantidade, uma vez que absorve especialmente a porção azul da faixa do visível
(400-700nm) do espectro eletromagnético (Branco & Kremer, 2005). Ambientes com
águas escuras devido a elevados teores de MODC são também conhecidos como
ambientes húmicos e têm uma rápida atenuação da radiação fotossinteticamente
ativa (Jones, 1992).
Devido aos efeitos da MODC sobre a disponibilidade e qualidade da luz, ambientes
húmicos são associados a baixas biomassa e produtividade fitoplanctônicas (Jones,
1992) bem como baixa riqueza de espécies (Alves-de-Souza et al., 2006),
normalmente com elevada contribuição de organismos mixotróficos, como

53
crisofíceas e dinoflagelados (Jansson et al., 1996; Jones 2000; Drakare et al., 2002).
No entanto, a maior parte dos estudos que contemplam o fitoplâncton de sistemas
húmicos foi realizada em clima temperado e pouco se conhece ainda sobre a
relação do teor de MODC e o fitoplâncton em ambientes tropicais. No Brasil, foram
estudados alguns ambientes de águas escuras na Amazônia e na região de restinga
do Estado do Rio de Janeiro (Pålsson & Granéli, 2004; Alves-de-Souza et al., 2006).
O sistema lacustre do médio Rio Doce está entre os mais importantes sistemas de
lagoas naturais do Brasil, com cerca de 150 lagoas que se diferenciam em relação
as características morfométricas e limnológicas. Uma particularidade marcante
observada neste sistema é a ocorrência de um gradiente de atenuação da luz, com
a existência de lagoas com elevada transparência lado a lado com lagoas com
elevada turbidez. Tal gradiente possibilita o desenvolvimento de estudos sobre as
variações espaciais e temporais do coeficiente de atenuação da luz nos quais as
interferências decorrentes de diferenças geográficas e geomorfológicas são
minimizadas. Além disso, embora as lagoas do médio Rio Doce venham sendo
estudadas desde a década de 70 (Tundisi & Saijo, 1997b), existe ainda uma
carência de informações sobre os padrões de atenuação da luz e suas
conseqüências para o fitoplâncton nesses ambientes.
Até o presente, apenas o estudo de Bezerra-Neto et al. (2006) considerou a
variação da atenuação da radiação fotossintéticamente ativa em quatro lagoas do
sistema, sem contudo considerar a influência sobre a composição e distribuição das
espécies fitoplanctônicas. Na expectativa de ampliar tais estudos e a compreensão
dos fatores responsáveis pela atenuação da luz visível no sistema lacustre do médio
Rio doce, o presente estudo apresenta a variação espacial e temporal do coeficiente
de absorção da radiação fotossinteticamente ativa (k0) em 16 lagoas do médio Rio
Doce, bem como a relação com os teores de MODC, fósforo total e composição e
biomassa do fitoplâncton.

54
Área de estudo
O sistema lacustre do médio Rio Doce é formado por cerca de 150 lagoas, das quais
50 estão protegidas dentro dos limites do Parque Estadual do Rio Doce-PERD
(19º29’S; 42º28’W). Esta Unidade de conservação, o maior remanescente de Mata
Atlântica do Estado de Minas Gerais (35.976 ha), tem seu entorno ocupado
principalmente por plantações de Eucaliptus. O clima da região é definido como
tropical semi-úmido, com 4-5 meses de seca e características mesotérmicas (Nimer,
1989), com temperatura em torno de 25ºC. O período chuvoso se estende de
outubro a março e o período de seca, de abril a setembro (Figura 2.1 ).
J F M A M J J A S O N D
Pre
cipi
taçã
o ac
umul
ada
(mm
)
0
50
100
150
200
250
300
350
Tem
pera
tura
méd
ia (
o C)
0
5
10
15
20
25
30
Figura 2. 1 - Variação mensal da temperatura média do ar (linha) e da precipitação acumulada (barras) nos meses de janeiro a dezembro. Dados médios obtidos da estação climatológica do PERD entre 2004 e 2008.
Para este estudo, foram selecionadas 16 lagoas, sendo sete localizadas dentro dos
limites do PERD e nove, em sua área de entorno. A área e a profundidade de cada
lagoa podem ser observadas na Tabela 2.1 .

55
Metodologia
Foram realizadas quatro campanhas de amostragem com intervalos trimestrais:
agosto de 2007, novembro de 2007, fevereiro de 2008 e maio de 2008.
Os perfis de temperatura e condutividade elétrica da água foram determinados com
auxílio de sonda multiparâmetros Horiba U-22. O limite da zona de mistura foi
definido com base nos perfis térmicos, considerando o limite superior em que a
diferença de temperatura a cada metro mostrou-se igual ou superior a 0,5 °C.
Diferenças entre os valores de condutividade elétrica foram utilizados para confirmar
a profundidade definida para a zona de mistura.
A radiação fotossintecamente ativa foi medida com radiômetro Li-Cor com sensor
quantum subaquático esférico (modelo LI 193). O coeficiente de atenuação escalar
da radiação fotossinteticamente ativa k0 (m-1) foi determinado pela inclinação da reta
da regressão linear entre a profundidade e o logaritmo natural da irradiância escalar.
Todas as medidas foram tomadas entre 10:00 e 14:00H.
Amostras para determinação dos teores de matéria orgânica dissolvida colorida
(MODC) foram coletadas a 0,5m da superfície. As amostras foram filtradas em filtro
GF/F, Whatman e estocadas no escuro a 4°C até o mom ento da análise. O teor de
MODC foi estimado através de espectrofotometria, considerando a absorbância
obtida a 250nm em cubetas de quartzo de 1cm e utilizando água Milli-Q como
referência (Tranvik & Bertilsson, 2001). Os valores de absorbância a 250nm foram
convertidos no coeficiente de absorção através da equação (Kirk, 1994):
aMODC (m-1) = 2,303 A/r
onde: A= absorbância a 250 nm
r=caminho óptico da cubeta (mm)
Amostras para análise da concentração do fósforo total (P-total), fósforo solúvel
reativo (PSR) e nitrogênio inorgânico dissolvido (NID) e contagem do fitoplâncton
foram coletadas com garrafa de van Dorn na sub-superfície, na profundidade do
desaparecimento visual do disco de Secchi, que corresponde a cerca de10% da

56
radiação incidente na superfície e na profundidade correspondente a 1% da radiação
solar incidente na superfície de cada lagoa (profundidade do Secchi x 3). Para
determinação das concentrações de fósforo total, fosfóro solúvel reativo, nitrogênio
inorgânico dissolvido (nitrato-N, nitrito-N e amônio-N) e silicato foram utilizadas as
metodologias propostas por Golterman & Climo, 1967; Koroleff, 1976 e Mackereth
et al., 1978.
Para análise qualitativa do fitoplâncton, as amostras foram coletadas através de
arrastos verticais e horizontais sucessivos com rede de 20µm de abertura de malha.
A identificação das espécies foi feita sob microscópio óptico em aumentos de 400 e
1000 vezes. As contagens do fitoplâncton foram realizadas sob microscópio invertido
Zeiss após sedimentação em câmaras de 50mL (Utermöhl, 1958). Os organismos
foram contados até que se atingisse um total de 400 indivíduos da espécie mais
abundante, garantindo erro de ±10% para um intervalo de confiança de 95% (Lund
et al., 1958). Os cálculos de densidade foram feitos conforme Villafañe & Reid
(1995).
O biovolume específico foi calculado associando cada espécie a uma forma
geométrica conhecida (Hillebrand et al., 1999; Sun & Liu, 2003) e multiplicando-se a
densidade da população pelo volume médio obtido. Em geral, foram realizadas 20
medições para os taxa que contribuíram em média com pelo menos 1% da
densidade total de cada lagoa considerado. Assumiu-se peso fresco como unidade
de massa, considerando 1 mm3.L-1 = 1 mg.L-1 (Wetzel & Likens 2000).
A importância relativa da concentração de P-total, do fitoplâncton (biomassa) e do
teor de MODC (A250nm) sobre o coeficiente de atenuação escalar da radiação
fotossinteticamente ativa (k0) foram avaliadas através de regressões lineares
múltiplas, após transformação dos dados (log x+1). As diferenças na biomassa,
composição e estrutura do fitoplâncton entre os ambientes foram verificadas através
do teste de Kruskal-Wallis.

57
Resultados
A Tabela 2.1 apresenta os valores de Ptotal, MODC e k0 observada nos ambientes
estudados durante os períodos de seca e chuvas. A concentração de P-total variou
entre lagoas e amostragens, sendo o menor valor (7,2 µg.l-1) observado na lagoa
Águas Claras em agosto de 2007 (seca) e o maior (78,8 µg.l-1) na lagoa Barra em
fevereiro de 2008 (chuvas). O coeficiente da MODC (A250) variou entre 1,9 e 105,5
m-1, com valores significativamente mais elevados nas lagoas Central, Pimenta,
Ferrugem, Ferruginha e Patos (KW-H= 5,33; p<0,05). As lagoas Central, Ferrugem,
Ferruginha, Patos, Carioca, Barra e Pimenta apresentaram os maiores valores para
o coeficiente de atenuação da luz (k0 > 0,8), que variou de 0,4 a 2,93 m-1 (KW-H=
5,33; p<0,05). Foi observada uma tendência a maiores valores de k0 durante a
estação seca na maioria das lagoas. Os teores de Ptotal, MODC e os valores de k0
mostraram-se positivamente relacionados à profundidade de mistura (Tabela 2.2 ).
Tabela 2. 1 - Área e profundidade das 16 lagoas amostradas.
Área Prof
(Km2) (m)
A. Claras 0,617 9,5Jacaré 0,086 8,5Gambá 0,122 12,0Carioca 0,131 10,0Gambazinho 0,216 10,0Patos 0,230 8,0Ferruginha 0,422 4,0Central 0,443 5,0Verde 0,826 19,0S.Helena 0,858 10,5Ferrugem 1,091 3,5Palmeirinha 1,220 6,0Pimenta 1,241 3,5Almécega 1,298 7,0Barra 1,936 7,0D. Helvécio 5,268 28,0

58
Figura 2. 2 – Média e desvio padrão observados para o coeficiente de atenuação da radiação fotossinteticamente ativa (k0), da concentração de matéria orgânica dissolvida colorida (A250), do teor de fósforo total (Ptotal), fósforo solúvel reativo (PSR), e nitrogênio inorgânico dissolvido (DIN) nas lagoas durante os períodos de seca e chuva.

59
Tabela 2. 2 - Resultados obtidos para regressão linear simples da profundidade total no ponto de coleta e os teores de fósforo total (Ptotal) e matéria orgânica dissolvida colorida (MODC) e o coeficiente de atenuação da luz k0
Variável R2 F p
P-total 0,386 8,82 0,010
MODC 0,455 11,69 0,004
k0 0,297 5,93 0,029
Os resultados da regressão múltipla de k0 versus Ptotal, MODC e biomassa do
fitoplâncton podem ser observados na Tabela 2.3 . A importância de cada uma
dessas variáveis no coeficiente de atenuação da luz variou sazonalmente. Durante o
período de seca, 85% da variação de k0 foi explicada pelos teores de MODC e de
fósforo total, enquanto a biomassa do fitoplâncton e os teores de MODC explicaram
87% da variação nos valores de k0 no período de chuvas.
Tabela 2. 3 - Resultados obtidos para análise de regressão múltipla do k0 utilizando biomassa do fitoplâncton, matéria orgânica dissolvida colorida e concentração de fósforo total como variáveis independentes
Período Modelo R2 ajustado p
seca + chuva log k0= 0,045 + 0,306 log Biov + 0,502 log MODC 0,80 0,000000
seca k0= 0,0004 + 0,637 log MODC + 0,335 log PT 0,85 0,000007
chuva log k0= 0,01 + 0,462 log Biov + 0,502 log MODC 0,87 0,000003
Foram registrados 490 taxa fitoplanctônicos nos ambientes estudados, sendo
Zygnematophyceae (171), Cyanobacteria (102) e Chorophyceae (98) as classes
mais ricas em espécies. A contribuição de cada classe taxonômica nas lagoas
amostradas pode ser observada na Figura 2.3 . As classes mais representativas em
cada lagoa foram as mesmas observadas para o conjunto de lagoas. No entanto,
registrou-se maior número de espécies de diatomáceas (Bacillariophyceae) nas
lagoas com coeficiente de atenuação da luz menor que 0,8 m-1 (Figura 2.4 ).
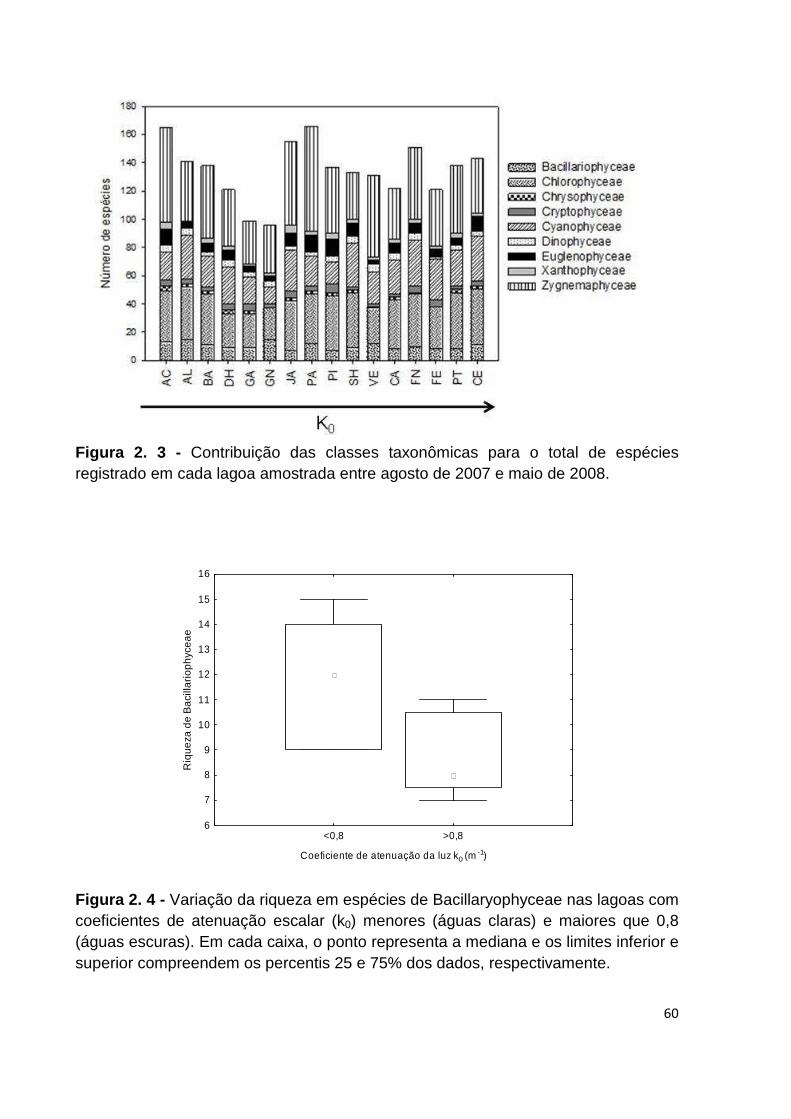
60
Figura 2. 3 - Contribuição das classes taxonômicas para o total de espécies registrado em cada lagoa amostrada entre agosto de 2007 e maio de 2008.
<0,8 >0,8
Coeficiente de atenuação da luz k0 (m-1)
6
7
8
9
10
11
12
13
14
15
16
Riq
ueza
de
Ba
cilla
riop
hyc
ea
e
Figura 2. 4 - Variação da riqueza em espécies de Bacillaryophyceae nas lagoas com coeficientes de atenuação escalar (k0) menores (águas claras) e maiores que 0,8 (águas escuras). Em cada caixa, o ponto representa a mediana e os limites inferior e superior compreendem os percentis 25 e 75% dos dados, respectivamente.
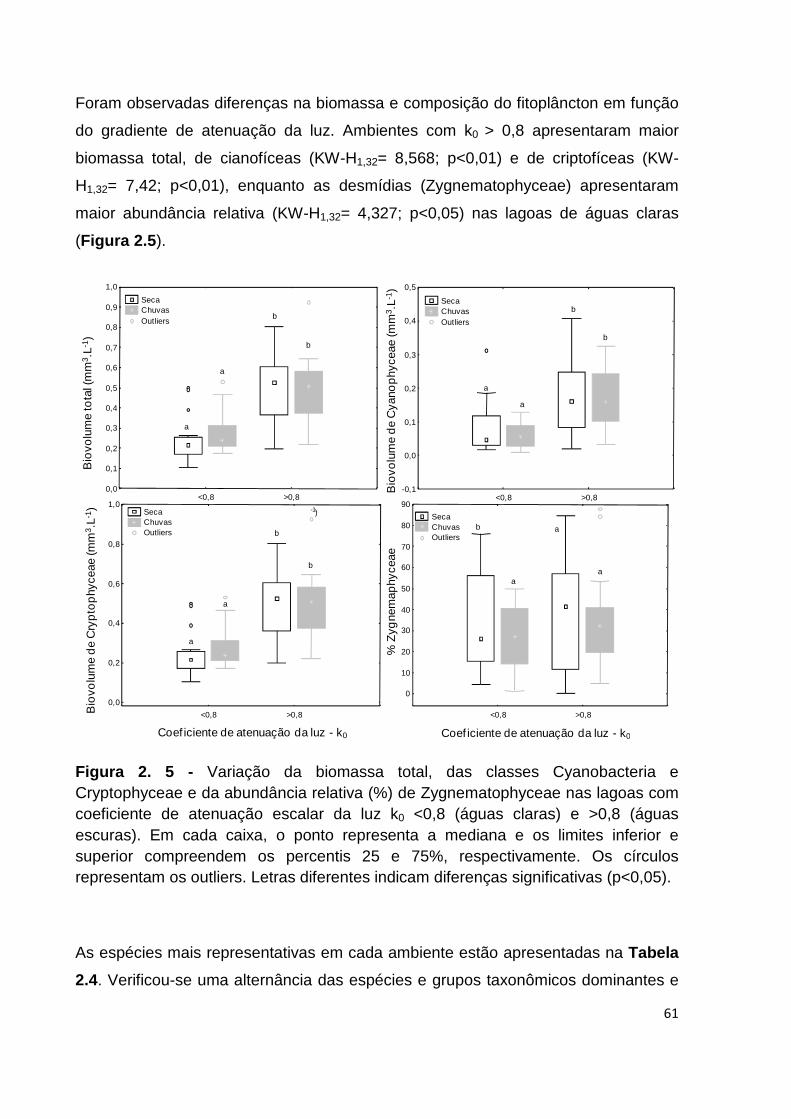
61
Foram observadas diferenças na biomassa e composição do fitoplâncton em função
do gradiente de atenuação da luz. Ambientes com k0 > 0,8 apresentaram maior
biomassa total, de cianofíceas (KW-H1,32= 8,568; p<0,01) e de criptofíceas (KW-
H1,32= 7,42; p<0,01), enquanto as desmídias (Zygnematophyceae) apresentaram
maior abundância relativa (KW-H1,32= 4,327; p<0,05) nas lagoas de águas claras
(Figura 2.5 ).
<0,8 >0,8
Coef iciente de atenuação da luz - k0
0,0
0,2
0,4
0,6
0,8
1,0
Bio
volu
me
de
Cry
pto
phy
ceae
(mm
3 .L-1
) SecaChuvasOutliers
a
b
b
a
<0,8 >0,8
-1)
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1,0
SecaChuvasOutliers
a
b
b
a
Bio
volu
me
tota
l (m
m3 .
L-1 )
<0,8 >0,8
Coeficiente de atenuação da luz k0 (m-1)
-0,1
0,0
0,1
0,2
0,3
0,4
0,5
SecaChuvasOutliers
a
b
b
a
Bio
volu
me
de
Cya
nop
hyce
ae (m
m3 .
L-1 )
<0,8 >0,8
0
10
20
30
40
50
60
70
80
90
SecaChuvasOutliers
a
b
a
a
%Z
ygne
map
hyce
ae
Coef iciente de atenuação da luz - k0
Figura 2. 5 - Variação da biomassa total, das classes Cyanobacteria e Cryptophyceae e da abundância relativa (%) de Zygnematophyceae nas lagoas com coeficiente de atenuação escalar da luz k0 <0,8 (águas claras) e >0,8 (águas escuras). Em cada caixa, o ponto representa a mediana e os limites inferior e superior compreendem os percentis 25 e 75%, respectivamente. Os círculos representam os outliers. Letras diferentes indicam diferenças significativas (p<0,05).
As espécies mais representativas em cada ambiente estão apresentadas na Tabela
2.4. Verificou-se uma alternância das espécies e grupos taxonômicos dominantes e

62
sub-dominantes na maioria das lagoas com k0 < 0,8, com maior biomassa relativa de
desmídias, especialmente Cosmarium bioculatum Brébisson ex Ralfs, Staurodesmus
crassus (West et G.S. West) Florin e Cosmarium moniliforme Turpin ex Ralfs,
durante o período de chuvas. Nas lagoas com k0 > 0,8 m-1 observou-se maior
constância entre as quatro campanhas de amostragem no que se refere à
composição do fitoplâncton. Nas lagoas com k0 > 0,8 m-1 as cianobactérias
Limnothrix redekeii (van Goor) Meffert, Cylindrospermopsis raciborskii (Wollosz.)
Seenayya & Subba Raju e Planktolyngbya limnetica (Lemmermann) Komárková-
Legnerová & Cronberg e P. contorta (Lemmermann) Anagnostidis et Komárek se
destacaram, juntamente com os dinoflagelados Gymnodynium sp e Peridinium
pusillum (Penard) Lemmerman e clorofíceas (Botryococcus braunii Kützing, Chlorella
spp, Monoraphidium contortum (Thuret) Komárkova-Legnerová nas lagoas escuras e
nas amostragens realizadas no período de seca nas lagoas de águas claras.
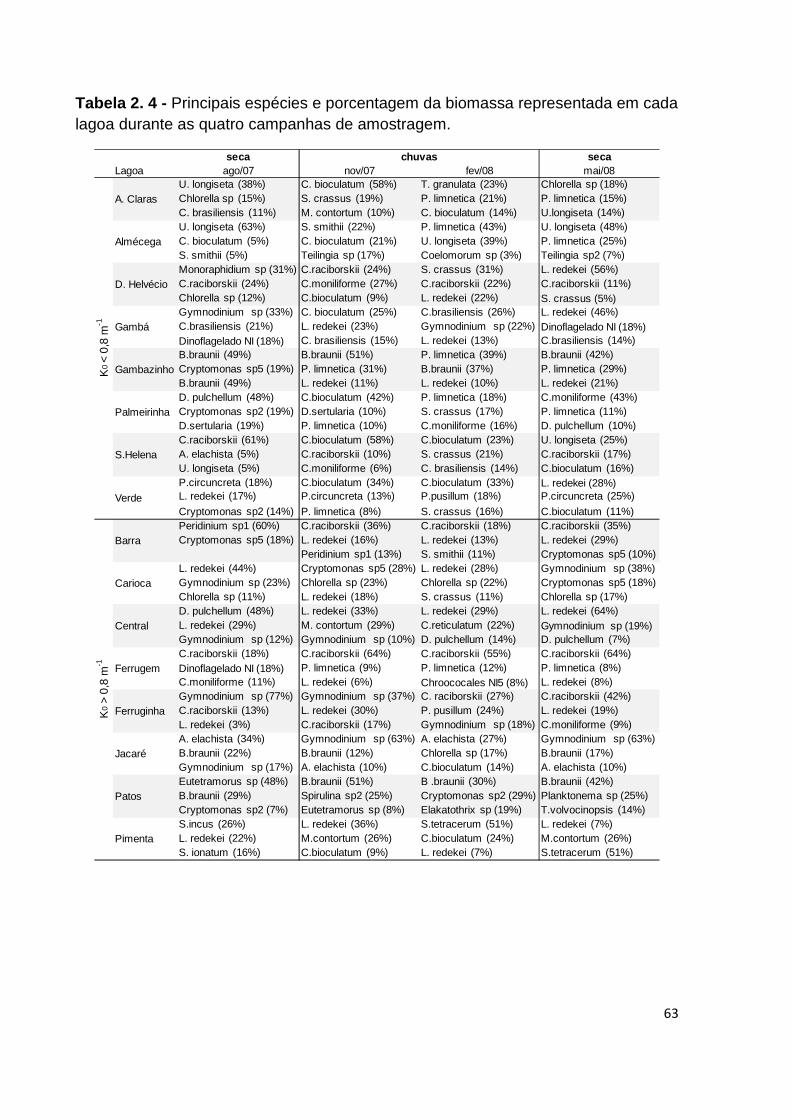
63
Tabela 2. 4 - Principais espécies e porcentagem da biomassa representada em cada lagoa durante as quatro campanhas de amostragem.
seca secaLagoa ago/07 nov/07 fev/08 mai/08
U. longiseta (38%) C. bioculatum (58%) T. granulata (23%) Chlorella sp (18%)A. Claras Chlorella sp (15%) S. crassus (19%) P. limnetica (21%) P. limnetica (15%)
C. brasiliensis (11%) M. contortum (10%) C. bioculatum (14%) U.longiseta (14%)U. longiseta (63%) S. smithii (22%) P. limnetica (43%) U. longiseta (48%)
Almécega C. bioculatum (5%) C. bioculatum (21%) U. longiseta (39%) P. limnetica (25%)S. smithii (5%) Teilingia sp (17%) Coelomorum sp (3%) Teilingia sp2 (7%)Monoraphidium sp (31%) C.raciborskii (24%) S. crassus (31%) L. redekei (56%)
D. Helvécio C.raciborskii (24%) C.moniliforme (27%) C.raciborskii (22%) C.raciborskii (11%)Chlorella sp (12%) C.bioculatum (9%) L. redekei (22%) S. crassus (5%)Gymnodinium sp (33%) C. bioculatum (25%) C.brasiliensis (26%) L. redekei (46%)C.brasiliensis (21%) L. redekei (23%) Gymnodinium sp (22%) Dinoflagelado NI (18%)Dinoflagelado NI (18%) C. brasiliensis (15%) L. redekei (13%) C.brasiliensis (14%)B.braunii (49%) B.braunii (51%) P. limnetica (39%) B.braunii (42%)
Gambazinho Cryptomonas sp5 (19%) P. limnetica (31%) B.braunii (37%) P. limnetica (29%)B.braunii (49%) L. redekei (11%) L. redekei (10%) L. redekei (21%)D. pulchellum (48%) C.bioculatum (42%) P. limnetica (18%) C.moniliforme (43%)
Palmeirinha Cryptomonas sp2 (19%) D.sertularia (10%) S. crassus (17%) P. limnetica (11%)D.sertularia (19%) P. limnetica (10%) C.moniliforme (16%) D. pulchellum (10%)C.raciborskii (61%) C.bioculatum (58%) C.bioculatum (23%) U. longiseta (25%)
S.Helena A. elachista (5%) C.raciborskii (10%) S. crassus (21%) C.raciborskii (17%)U. longiseta (5%) C.moniliforme (6%) C. brasiliensis (14%) C.bioculatum (16%)P.circuncreta (18%) C.bioculatum (34%) C.bioculatum (33%) L. redekei (28%)
Verde L. redekei (17%) P.circuncreta (13%) P.pusillum (18%) P.circuncreta (25%)
Cryptomonas sp2 (14%) P. limnetica (8%) S. crassus (16%) C.bioculatum (11%)Peridinium sp1 (60%) C.raciborskii (36%) C.raciborskii (18%) C.raciborskii (35%)
Barra Cryptomonas sp5 (18%) L. redekei (16%) L. redekei (13%) L. redekei (29%)Peridinium sp1 (13%) S. smithii (11%) Cryptomonas sp5 (10%)
L. redekei (44%) Cryptomonas sp5 (28%) L. redekei (28%) Gymnodinium sp (38%)Carioca Gymnodinium sp (23%) Chlorella sp (23%) Chlorella sp (22%) Cryptomonas sp5 (18%)
Chlorella sp (11%) L. redekei (18%) S. crassus (11%) Chlorella sp (17%)D. pulchellum (48%) L. redekei (33%) L. redekei (29%) L. redekei (64%)
Central L. redekei (29%) M. contortum (29%) C.reticulatum (22%) Gymnodinium sp (19%)Gymnodinium sp (12%) Gymnodinium sp (10%) D. pulchellum (14%) D. pulchellum (7%)C.raciborskii (18%) C.raciborskii (64%) C.raciborskii (55%) C.raciborskii (64%)
Ferrugem Dinoflagelado NI (18%) P. limnetica (9%) P. limnetica (12%) P. limnetica (8%)C.moniliforme (11%) L. redekei (6%) Chroococales NI5 (8%) L. redekei (8%)Gymnodinium sp (77%) Gymnodinium sp (37%) C. raciborskii (27%) C.raciborskii (42%)
Ferruginha C.raciborskii (13%) L. redekei (30%) P. pusillum (24%) L. redekei (19%)L. redekei (3%) C.raciborskii (17%) Gymnodinium sp (18%) C.moniliforme (9%)A. elachista (34%) Gymnodinium sp (63%) A. elachista (27%) Gymnodinium sp (63%)
Jacaré B.braunii (22%) B.braunii (12%) Chlorella sp (17%) B.braunii (17%)Gymnodinium sp (17%) A. elachista (10%) C.bioculatum (14%) A. elachista (10%)Eutetramorus sp (48%) B.braunii (51%) B .braunii (30%) B.braunii (42%)
Patos B.braunii (29%) Spirulina sp2 (25%) Cryptomonas sp2 (29%) Planktonema sp (25%)Cryptomonas sp2 (7%) Eutetramorus sp (8%) Elakatothrix sp (19%) T.volvocinopsis (14%)S.incus (26%) L. redekei (36%) S.tetracerum (51%) L. redekei (7%)
Pimenta L. redekei (22%) M.contortum (26%) C.bioculatum (24%) M.contortum (26%)S. ionatum (16%) C.bioculatum (9%) L. redekei (7%) S.tetracerum (51%)
chuvas
K0
< 0,
8 m
-1K
0 >
0,8
m-1
Gambá

64
Discussão
Os resultados apontam claras diferenças espaciais e temporais em relação à
atenuação da radiação fotossinteticamente ativa, os fatores determinantes e suas
conseqüências para o fitoplâncton no conjunto das lagoas analisadas contribuindo
com informações significativas sobre limnologia desse importante complexo de
lacustre.
O valor de k0 = 0,8 m-1 foi considerado como o limite do coeficiente de atenuação da
luz que separa condições de elevada (k0<0,8) e reduzida (k0>0,8) penetração da
radiação luminosa. Esse valor foi utilizado por Pfannkuche (2002) em ambientes da
plataforma continental da Nova Zelândia e por Foden et al. (2008) em corpos d’água
do Reino Unido, que registraram valores acima daquele limite em estuários e águas
costeiras de elevada turbidez.
Aplicando este critério aos ambientes estudados, as lagoas Águas Claras,
Almécega, Gambá, Gambazinho, Palmeirinha, Santa Helena e Verde podem ser
consideradas como de “águas claras”, uma vez que os valores de k0 mantiveram-se
inferiores a 0,8 m-1 em todas ou três das quatro amostragens realizadas. Por sua
vez, as lagoas Barra, Carioca, Central, Ferrugem, Ferruginha, Jacaré e Patos podem
ser consideradas ambientes de águas escuras, com valores de k0 > 0,8 m-1 em
quase todas as amostragens.
A maioria das lagoas apresentou redução nos valores de k0 durante o período de
chuvas. A sazonalidade da atenuação da luz visível foi mais marcante na lagoa
Central (80%), mas também foi expressiva nas lagoas Verde (28%), Santa Helena
(24%) e Dom Helvécio (23%). Uma redução em menor escala (12 – 19%) dos
valores de k0 também foi registrada na estação chuvosa nas lagoas Palmeirinha,
Carioca, Gambazinho e Pimenta. Esses resultados corroboram aqueles obtidos por
Bezerra-Neto et al. (2006), que observaram redução de 26% nos valores para k0
para a lagoa Carioca e 37% para a lagoa Dom Helvécio durante o período de chuvas
de 2004/2005. Tal sazonalidade, observada tanto nas lagoas de águas claras quanto
nas de águas escuras, está provavelmente associada aos padrões de estratificação
térmica, uma vez que a circulação da coluna d’água que ocorre no período de seca

65
pode proporcionar a suspensão de matéria orgânica particulada do sedimento,
conforme sugerido por Calijuri et al. (1997).
As relações significativas entre a profundidade e os valores de Ptotal, A250 e k0
observados em cada lagoa demonstram a importância da morfometria para as
características limnológicas dos ambientes e reforçam o papel determinante das
condições de mistura da coluna d’água sobre a atenuação da luz (Reynolds, 1994).
O maior biomassa fitoplanctônico nos ambientes com maior coeficiente de
atenuação de luz vai de encontro ao sugerido por Devol et al. (1984), que
observaram baixas densidades nos ambientes de águas escuras da Amazônia, por
Carpenter et al. (1998), que apontam efeitos negativos da concentração do carbono
orgânico dissolvido (COD) sobre a concentração de clorofila-a e sobre a
produtividade primária do fitoplâncton em lagos do Wisconsin, e por Alves-de-Souza
et al., 2006, que também registraram baixa biomassa do fitoplâncton (max. 0,5 mg.L-
1) na lagoa Comprida, Rio de Janeiro, que tem elevadas concentrações de
compostos húmicos.
Alguns autores, por sua vez, sugerem que concentrações intermediárias de COD
podem ser mais favoráveis ao crescimento do fitoplâncton do que baixas
concentrações, pois reduzem os efeitos deletérios da radiação UV (Aitkenhead-
Peterson et al., 2003). Os efeitos da MODC sobre o fitoplâncton são geralmente
associados a limitação por luz. No entanto, as lagoas que apresentaram maior
atenuação da luz apresentaram maior biomassa fitoplanctônica, sugerindo que
mesmo nas lagoas com maiores valores de k0 os teores de MODC não foram
suficientes para limitar o desenvolvimento dessa comunidade. Jones (1992) sugeriu
que um coeficiente de atenuação de 1,33m-1 não representou uma atenuação
expressiva da radiação incidente na superfície de lagos no Canadá. Com exceção
das lagoas Central e Patos, os ambientes estudados mostraram valores de k0
abaixo, ou ligeiramente superiores a 1,33m-1, reforçando a hipótese que mesmo os
mais escuros dentre os ambientes estudados não tiveram sua produção
fitoplanctônica limitada, sobretudo, se considerarmos que a insolação é mais
elevada na região tropical.

66
A relação direta observada entre a biomassa fitoplanctônica e o coeficiente de
atenuação da radiação fotossinteticamentem ativa também evidencia a contribuição
do fitoplâncton para essa atenuação, uma vez que o fitoplâncton está entre os
componentes que afetam as propriedades ópticas inerentes a cada corpo de água
(Mobley 1994, apud Green et al., 2003). Os resultados das regressões múltiplas
realizadas reforçam esta hipótese, uma vez que a biomassa fitoplanctônica aparece
entre os fatores responsáveis pelas variações da atenuação da luz nas lagoas
estudadas.
A importância da atenuação da radiação fotossinteticamente ativa para o fitoplâncton
é evidenciada quando se compara a composição do fitoplâncton entre lagoas de
águas claras e águas escuras. A maior importância das desmídias
(Zygnematophyceae) para a biomassa fitoplanctônica nas lagoas com baixo
coeficiente de atenuação da luz (“águas claras”) durante o período chuvoso
corrobora estudos já realizados, que apontam o padrão de estratificação térmica
predominante nas lagoas da região, monomítico-quente, como fator determinante
para a abundância desses organismos. A importância da atelomixia para a
manutenção dessas algas que possuem baixa resistência à sedimentação (Padisák
et al., 2003b) nas camadas superiores da coluna d’água foi evidenciada por
Reynolds (1997) e Barbosa & Padisák (2002).
A dependência desses organismos de maior disponibilidade de luz, já reportada por
Souza et al. (2008) em estudos realizados na lagoa Dom Helvécio, é corroborada
pela ausência de variação sazonal significativa nas lagoas com k0 >0,8m-1 (“águas
escuras”), apesar de esses ambientes apresentarem o mesmo padrão monomítico-
quente. Tal resultado sugere que, apesar de também ter sido observada uma
redução dos valores de k0 durante o período de chuvas nesses ambientes, a
radiação fotossinteticamente ativa ainda manteve níveis insuficientes para o
desenvolvimento mais expressivo das desmídias. A diatomácea Urosolenia longiseta
(Zacharias) M. B. Edlund & Stoermer também aparece entre as espécies dominantes
das lagoas Águas Claras, Almécega e Santa Helena, todas com baixos valores de k0
(< 0,8m-1), e é da mesma forma associada a ambientes de águas claras e pobres em
nutrientes (Padisák et al., 2009).

67
Dentre os ambientes com águas claras, a lagoa Gambazinho foi o único no qual as
desmídias não estão listadas entre as espécies dominantes ou subdominantes em
nenhuma das amostragens, embora Zygnematophyceae também tenha se mostrado
a classe com maior riqueza taxonômica. A ausência de uma biomassa expressiva de
desmídias na lagoa Gambazinho pode estar relacionada ao padrão de estratificação
térmica. Embora tenha se mostrado estratificada durante as amostragens realizadas
no período de chuvas, a estratificação térmica é um evento pouco freqüente nessa
lagoa (dados não publicados), que foge ao padrão monomítico-quente confirmado
para as demais lagoas estudadas. Assim, a constante mistura da coluna d’água
pode estar atuando como fator limitante ao desenvolvimento das desmídias.
Em lagoas húmicas, a mixotrofia pode ser vantajosa em relação à fagotrofia
obrigatória, pois permite algum uso da luz disponível, e também em relação à
fototrofia obrigatória, por possibilitar a utilização de uma fonte extra para obtenção
de carbono e nutrientes (Jones, 2000). A frequência dos dinoflagelados
Gymnodinium sp e Peridinium pusillum entre as espécies dominantes foi maior nas
lagoas com maiores coeficientes de atenuação da luz (k0 > 0,8m-1). Esses
organismos são mixotróficos (Jansson et al., 1996) e sua relação com elevadas
concentrações de Ptotal e MODC já havia sido verificada no lago GroßeFuchskuhle,
na Alemanha (Hehmann et al., 2001) e no lago Örträsket na Suécia (Drakare et al.,
2002). Hehmann et al. (2001) atribuem o sucesso dos dinoflagelados à capacidade
de realizar heterotrofismo, se alimentando de bactérias, e ao comportamento
migratório que possibilita maior acesso às regiões com maior disponibilidade de luz
e/ou bactérias (Findlay, 1999).
A maior biomassa das classes Cryptophyceae, representada por diferentes espécies
de Cryptomonas, e Cyanobacteria, representada por C. raciborskii, L. redekeii, A.
elechista e Planktolyngbya spp., nas lagoas com k0 > 0,8m-1 corrobora a hipótese de
que as condições de luz nesses ambientes estão limitando a fotossíntese e
selecionando as espécies do fitoplâncton. As Cryptomonas podem ser consideradas
mixotróficas (Jones, 2000) ou potencialmente mixotróficas (Jansson et al 1996) e a
presença de flagelos as permite migrar na coluna d’água. Dentre as cianobactérias,
a migração vertical é viabilizada pela presença de aerótopos e mucilagem (Bold &
Wyne, 1985). Izaguirre et al. (2004) e Rodrigues & Pizarro (2007) também

68
registraram elevadas percentagens desses grupos taxonômicos na Laguna Grande,
um ambiente húmico localizado em área de estuário na Argentina e atribuíram tal
dominância ao prevalecimento da radiação na faixa do vermelho devido à absorção
dos menores comprimentos de onda pela MODC em elevadas concentrações.
Nesses ambientes com menor disponibilidade de luz, clorofíceas como Botryococcus
braunii, capaz de controlar sua posição na coluna d’água devido à produção de
gotículas de óleo e Monoraphidium contortum e Chlorella sp., considerados
organismos oportunistas (Rodrigo et al., 2009), também se destacaram entre as
espécies dominantes.
Ao compararem dados de lagoas temperadas e tropicais com elevados teores de
MODC, Pålsson & Granéli (2004) encontraram baixa contribuição de gêneros
mixotróficos na biomassa fitoplanctônica dos ambientes tropicais. No entanto, os
próprios autores ressaltaram duas hipóteses para a ocorrência de tal resultado: a
amostragem não refletir a situação predominante, já que havia sido realizada em
uma única ocasião, e a contribuição de tais organismos poderia ser mais expressiva
em outras épocas do ano; ou a trofia e as elevadas temperaturas registradas nos
trópicos teriam favorecido a dominância de cianobactérias nos ambientes estudados
(lagoas Imboassica, Comprida e Cabiúnas – Brasil).
A ocorrência de Dinobryon sertularia Ehrenberg, considerada mixotrófica, como
espécie subdominante na lagoa Palmeirinha, com baixo coeficiente de atenuação da
luz, refuta a hipótese de favorecimento das Chrysophyceae nos ambientes mais
turvos. Apesar de poder contribuir significativamente para a biomassa fitoplanctônica
em ambientes húmicos (Alves-de-Souza et al. 2006), a dominância do gênero
Dinobryon em ambientes com elevada disponibilidade de luz já foi observada em
estudos anteriores (Jansson et al., 1996). Isaksson et al. (1999) sugerem que a
mixotrofia não é uma estratégia restrita a baixa disponibilidade de luz e que, em
situação de limitação por fósforo, pode ser particularmente eficiente se alimentar de
bactérias ricas neste nutriente. Outros autores destacam que a mixotrofia pode ser
útil tanto em condições de reduzida disponibilidade de luz (Pålsson & Granéli, 2004),
quanto em condições de elevada disponibilidade de luz e limitação por nutrientes
(Arenowski et al., 1995).

69
Os resultados apresentados mostraram que existem variações no que se refere à
radiação fotossinteticamente ativa nas lagoas do médio Rio Doce e que tais
diferenças estão associadas à profundidade, ao clima, à trofia, indicada pelos teores
de Ptotal e biomassa do fitoplâncton e à concentração de MODC. Embora nem
mesmo os valores de k0 observados nas lagoas de águas escuras possam ser
considerados elevados se comparados a outros ambientes húmicos, a influência do
grau de atenuação da luz sobre o fitoplâncton é percebida pelas diferenças na
composição da comunidade fitoplanctônica, com o favorecimento de espécies
mixotróficas, tolerantes à baixa disponibilidade de luz ou oportunistas. Esses
resultados contribuem de maneira significativa para o entendimento da relação entre
as propriedades ópticas e a comunidade fitoplanctônica no sistema lacustre do
médio Rio Doce.

70
CAPÍTULO 3
RESPOSTA DOS GRUPOS FUNCIONAIS FITOPLANCTÔNICOS AOS GRADIENTES AMBIENTAIS NO SISTEMA LACUSTRE DO MÉDIO RIO DOCE,
MG

71
Resumo
Resposta dos grupos funcionais fitoplanctônicos aos gradientes ambientais
no sistema lacustre do médio Rio Doce, MG
O presente artigo teve como objetivos avaliar a variação espacial e temporal dos
grupos funcionais fitoplanctônicos em 18 lagoas que integram o sistema lacustre do
médio Rio Doce e identificar os principais fatores que regulam essa variação. A
transparência da água, a profundidade e o índice de estado trófico foram as
principais variáveis definindo as condições ambientais nas lagoas estudadas e foram
utilizadas para separá-las em cinco tipos: 1) profundas, claras e oligotróficas; 2) de
profundidade intermediária, claras e oligotróficas; 3) de profundidade intermediária,
escuras e mesotróficas; 4) rasas, escuras e mesotróficas e 5) rasas, escuras e
eutróficas. Uma tendência ao aumento da biomassa total ao longo desse gradiente
ambiental foi verificada. Foram identificadas 51 espécies descritoras, representando
18 grupos funcionais. Os grupos funcionais F, Lo, NA, S1, SN, X1, X2 e Y foram os
mais frequentes nas lagoas estudadas. Em geral, as lagoas mais claras e
oligotróficas, tipos 1 e 2, foram dominadas pelos grupos NA e A, enquanto grupos
descritos como tolerantes à baixa luminosidade, dominaram o fitoplâncton das
lagoas tipo 3 (SN, S1,MP, Y, X1, K e F), tipo 4 ( F, S1, Y, J, SN e Lo) e tipo 5 (F, Y,
X2, SN, S1, Lo). Os resultados obtidos evidenciaram uma grande concordância
entre os grupos funcionais dominantes e as condições ambientais reforçando a
eficiência dos grupos funcionais como indicadores das condições ambientais
também em lagoas naturais tropicais. Como mostrado para outros sistemas, não
somente a disponibilidade de recursos (luz e nutrientes) ou a estrutura física
(estratificação térmica) direcionaram a comunidade fitoplanctônica e sim a interação
entre esses fatores.
Palavras-chave: Grupos funcionais fitoplanctônicos lagos naturais

72
Abstract
Response of phytoplankton functional groups to envi ronmental gradients in
the lacustrine system of the middle Rio Doce, MG
This study aimed at evaluating the spatial and temporal variation of phytoplankton
functional groups in 18 lakes of the middle Rio Doce lake district and identify the key
factors governing this variation. Water transparency, depth and trophic state index
were the main variables defining the environmental conditions in studied lakes and
were used to separate them into five types: 1) deep, clear and oligotrophics, 2)
intermediate depth, clear and oligotrophics, 3) intermediate depth, dark and
mesotrophic, 4) shallow, dark and mesotrophic and 5) shallow, dark and well
nourished. Despite low biomass in all lakes, a tendency to higher values along this
environmental gradient was verified. Fifty-one descriptions species (> 5% of total
biomass) were identified and accounted for 18 functional groups. Groups F, Lo, NA,
S1, SN, X1, X2 and Y were the most frequent. In general, the more clear and
oligotrophic lakes (types 1 and 2) were dominated by groups NA and A. Groups
described as tolerant to low light, dominated the phytoplankton in lakes type 3 (SN,
S1, MP, Y, X1, K and F), type 4 (F, S1, Y, J, NS, and Lo) and type 5 (F, Y, X2, SN,
S1, Lo). The results showed close agreement between the dominant functional
groups and environmental conditions, improving the efficiency of the functional
groups as indicators of environmental conditions also in natural tropical lagoons. As
shown for others systems not only the resources (light and nutrients) availability or
the physical structure (thermal stratification) directed the phytoplankton community
and yes, interaction among them.
Key-words: Phytoplankton functional groups, tropical lakes

73
Introdução
A importância dos processos físicos e químicos para a composição e sucessão de
espécies em lagoas é bem clara (Huszar & Caraco, 1998; Reynolds, 2006).
Reorganizações na composição e estrutura do fitoplâncton ao longo do tempo são
frequentes e vistas como essenciais para o metabolismo dos ecossistemas
aquáticos (Calijuri et al., 2002).
Considerando a capacidade de dispersão relativamente elevada das espécies que
compõem o fitoplâncton, a composição e estrutura dessa comunidade são moldadas
por fatores locais (Beisner et al., 2006), especialmente pelas variações no regime de
luz e disponibilidade de nutrientes (Reynolds, 1984). As características morfológicas
das lagoas, associadas aos diferentes tipos, frequência e intensidade dos impactos
antrópicos (Hajnal & Padisák, 2008), aos ciclos sazonais e à combinação das
variáveis ambientais, criam uma arena competitiva onde as espécies mais
adaptadas à situação prevalente em cada período se sobressaem (Padisák et al.,
2010). O resultado dessa competição por recursos, influenciado também pelas
respostas às interações bióticas como herbivoria e parasitismo (Padisák et al.,
2006), pode ser percebido em nível florístico ou estrutural, através de mudanças na
distribuição da biomassa entre classes de tamanho (Naselli-Flores et al., 2007), ou
grupos funcionais (Reynolds et al., 2002). Tais mudanças podem ser entendidas
como respostas populacionais, uma vez que as diferentes espécies possuem uma
variedade de estratégias adaptativas, com atributos morfológicos (ex. tamanho,
relação volume/superfície), fisiológicos (ex. produção de vacúolos gasosos e
mucilagem, afinidade por diferentes formas de nitrogênio) e comportamentais (ex.
migração vertical) que as permitem utilizar os recursos de maneira diferenciada
(Reynolds, 1997; Crossetti & Bicudo, 2005).
A utilização dos atributos morfo-funcionais como ferramenta para estudos da
dinâmica do fitoplâncton vem sendo bastante incentivada (Huszar & Caraco, 1998;
Kruk et al., 2002; Reynolds et al., 2002; Huszar et al., 2003; Salmaso & Padisák,
2007) e tem substituído, ou ao menos complementado, as abordagens baseadas na
biomassa total e variações nas classes taxonômicas.

74
Com base nesses atributos, Reynolds (1997) sugeriu grupos funcionais que
poderiam co-dominar ou dominar conforme as condições ambientais predominantes.
Diversas contribuições foram dadas à proposição original (Kruk et al., 2002;
Reynolds et al. 2002; Padisák et al., 2003a; Reynolds, 2006; Padisák et al., 2009) e
hoje estão descritos 38 grupos funcionais identificados por códigos alfa-numéricos
(Padisák et al., 2009). Tais grupos, geralmente polifiléticos, foram formados de
acordo com as características adaptativas coincidentes, considerando as
sensibilidades e as tolerâncias das espécies (Reynolds, 2006).
Embora proposta inicialmente para lagoas temperadas, os grupos funcionais têm
sido utilizados também no entendimento da dinâmica do fitoplâncton em ambientes
subtropicais (Kruk et al., 2002; Bonilla et al., 2005; Borges et al 2008; Becker et al.,
2009a; 2009b; Devercelli 2010) e tropicais (Marinho & Huszar, 2002; Nabout et al.,
2006; Souza et al., 2008; Costa et al.; 2009; Delazari-Barroso et al., 2009).
No Brasil, a maioria dos estudos foram realizados em reservatórios e poucos
abrangendo ambientes naturais como lagoas costeiras (Alves-de Souza et al., 2006),
lagoas de inundação (Nabout et al., 2006; Bovo-Scomparin & Train, 2008), estuários
(Costa et al., 2009) e lagos isolados (Nabout & Nogueira, 2007; Bovo-Scomparin &
train, 2008; Souza et al., 2008).
Os grupos funcionais têm se mostrado mais bem relacionados às variações
ambientais que espécies ou grupos taxonômicos, de modo que sua identificação e
uso podem ser ferramentas importantes na análise da comunidade fitoplanctônica,
sobretudo quando se trata da comparação de diferentes ambientes (Salmaso &
Padisák, 2007).
O trecho médio da bacia do Rio Doce é uma região peculiar e extremamente
importante. Além de abrigar o maior remanescente de Mata Atlântica do Estado de
Minas Gerais, comporta um sistema de lagoas formado por cerca de 150 ambientes
que variam em relação às suas características físicas (área, forma e profundidade),
químicas (cor da água e disponibilidade de nutrientes) e biológicas (ocupação por
macrófitas) e são sujeitos a diferentes tipos e intensidades de impactos antrópicos
(introdução de espécies exóticas, preservação da área de entorno, uso para
recreação).

75
Estudos comparando a dinâmica biológica nessas lagoas são escassos,
especialmente no que concerne à comunidade fitoplanctônica, sendo a maioria deles
concentrados nas lagoas Dom Helvécio e Carioca.
O presente estudo avalia a variação espacial e temporal do fitoplâncton em 18
lagoas que integram o sistema lacustre do médio Rio Doce e identifica os principais
fatores que regulam essa variação. Duas hipóteses foram testadas: (i) os grupos
funcionais respondem às variações temporais e espaciais das condições ambientais
observadas nesse sistema lacustre; (ii) os grupos funcionais fornecem uma
descrição adequada das características limnológicas predominantes em cada lagoa.
Materiais e métodos
O sistema lacustre do médio Rio Doce é formado por cerca de 150 lagoas, dos quais
aproximadamente 50 estão protegidos dentro dos limites do Parque Estadual do Rio
Doce/ PERD (19º29’S - 42º28’W), o maior remanescente de Mata Atlântica do
Estado de Minas Gerais (35.976 ha), cujo entorno é ocupado principalmente por
plantações de Eucaliptus. Dezoito lagoas foram consideradas para este estudo,
sendo oito localizadas dentro dos limites do PERD e dez localizadas em sua área de
entorno (Figura 3.1 ).
O clima da região é definido como tropical semi-úmido, com quatro a cinco meses de
seca e características mesotérmicas (Nimer, 1989), com temperatura em torno de
25ºC. Dados da estação climatológica instalada dentro do PERD obtidos entre 2004
e 2008 mostram que o período mais quente e chuvoso se estende de outubro a
março e o período de seca, de abril a setembro (Figura 3.2 ).
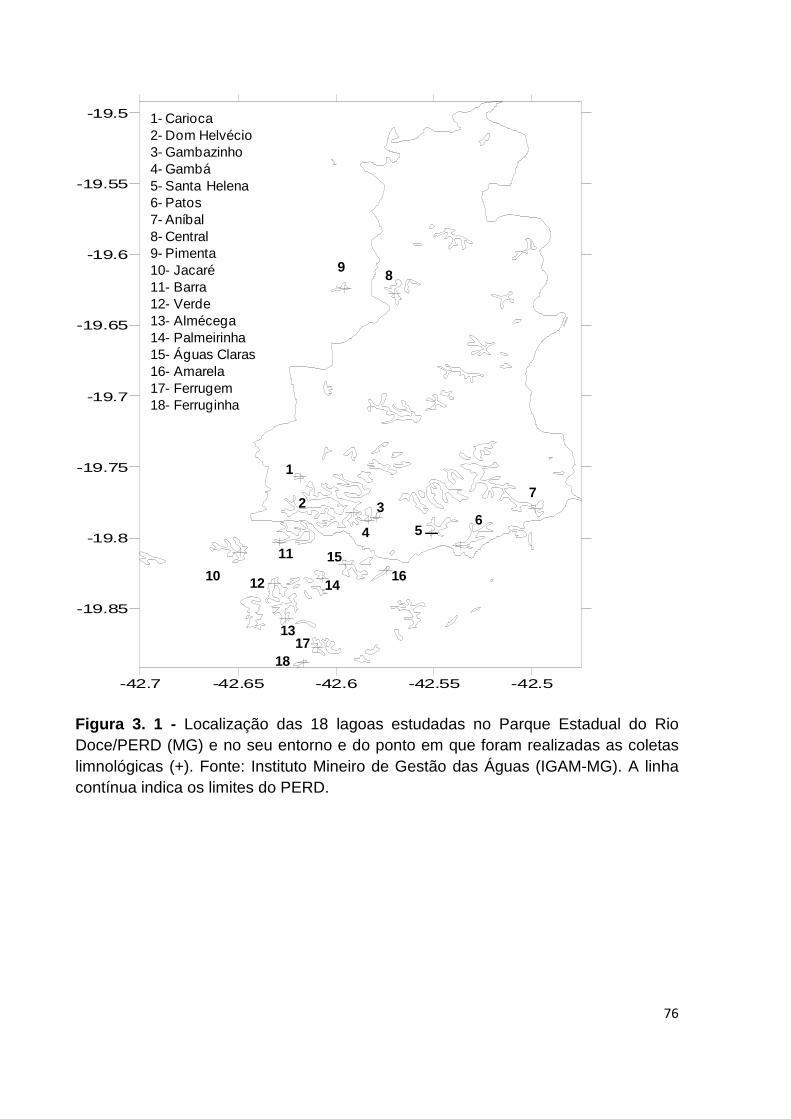
76
-42.7 -42.65 -42.6 -42.55 -42.5
-19.85
-19.8
-19.75
-19.7
-19.65
-19.6
-19.55
-19.5
1
32
4 56
7
89
10
11
12 14
1516
1713
18
1- Carioca2- Dom Helvécio3- Gambazinho4- Gambá5- Santa Helena6- Patos7- Aníbal8- Central9- Pimenta10- Jacaré11- Barra12- Verde13- Almécega14- Palmeirinha15- Águas Claras16- Amarela17- Ferrugem18- Ferruginha
Figura 3. 1 - Localização das 18 lagoas estudadas no Parque Estadual do Rio Doce/PERD (MG) e no seu entorno e do ponto em que foram realizadas as coletas limnológicas (+). Fonte: Instituto Mineiro de Gestão das Águas (IGAM-MG). A linha contínua indica os limites do PERD.
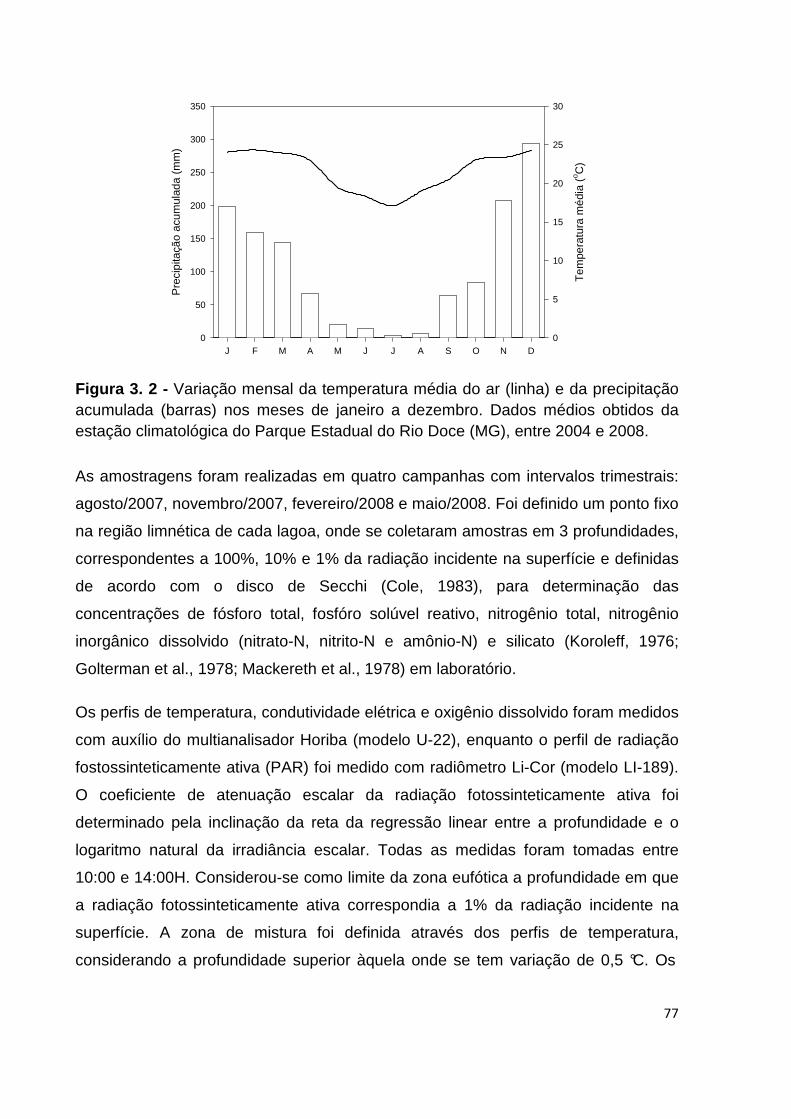
77
J F M A M J J A S O N D
Pre
cipi
taçã
o ac
umul
ada
(mm
)
0
50
100
150
200
250
300
350
Tem
pera
tura
méd
ia (
o C)
0
5
10
15
20
25
30
Figura 3. 2 - Variação mensal da temperatura média do ar (linha) e da precipitação acumulada (barras) nos meses de janeiro a dezembro. Dados médios obtidos da estação climatológica do Parque Estadual do Rio Doce (MG), entre 2004 e 2008.
As amostragens foram realizadas em quatro campanhas com intervalos trimestrais:
agosto/2007, novembro/2007, fevereiro/2008 e maio/2008. Foi definido um ponto fixo
na região limnética de cada lagoa, onde se coletaram amostras em 3 profundidades,
correspondentes a 100%, 10% e 1% da radiação incidente na superfície e definidas
de acordo com o disco de Secchi (Cole, 1983), para determinação das
concentrações de fósforo total, fosfóro solúvel reativo, nitrogênio total, nitrogênio
inorgânico dissolvido (nitrato-N, nitrito-N e amônio-N) e silicato (Koroleff, 1976;
Golterman et al., 1978; Mackereth et al., 1978) em laboratório.
Os perfis de temperatura, condutividade elétrica e oxigênio dissolvido foram medidos
com auxílio do multianalisador Horiba (modelo U-22), enquanto o perfil de radiação
fostossinteticamente ativa (PAR) foi medido com radiômetro Li-Cor (modelo LI-189).
O coeficiente de atenuação escalar da radiação fotossinteticamente ativa foi
determinado pela inclinação da reta da regressão linear entre a profundidade e o
logaritmo natural da irradiância escalar. Todas as medidas foram tomadas entre
10:00 e 14:00H. Considerou-se como limite da zona eufótica a profundidade em que
a radiação fotossinteticamente ativa correspondia a 1% da radiação incidente na
superfície. A zona de mistura foi definida através dos perfis de temperatura,
considerando a profundidade superior àquela onde se tem variação de 0,5 °C. Os

78
perfis de condutividade elétrica foram utilizados para confirmar a extensão da zona
de mistura.
O estado trófico das lagoas foi definido de acordo com o índice de Calrson (Calrson,
1977) adotando as modificações sugeridas por Toledo et al. (1983) e Lamparelli
(2004), considerando os dados de profundidade de desaparecimento visual do disco
de Secchi e concentração de fósforo total.
Amostras para análise quantitativa do fitoplâncton foram coletadas nas três
profundidades definidas pelo disco de Secchi. A densidade fitoplanctônica foi
estimada em campos aleatórios, utilizando o método de Utermöhl (1958). A
contagem dos indivíduos foi realizada até que fossem quantificados 400 indivíduos
da espécie mais freqüente (Lund et al. 1958). A biomassa específica foi estimada
associando cada indivíduo a uma forma geométrica conhecida (Hillebrand et al.,
1999; Sun & Liu, 2003) e multiplicando-se a densidade da população pelo volume
médio obtido. Para o cálculo do volume celular foram medidos pelo menos 20
indivíduos, exceto no caso de espécies raras. Assumiu-se o peso fresco como
unidade de massa, considerando 1 mm3.L-1 = 1 mg.L-1 (Wetzel & Likens, 2000).
As amostras de zooplâncton foram coletadas nos mesmos períodos e pontos de
amostragem e processadas pelo laboratório de Ecologia do Zooplâncton,
ICB/UFMG. Para análise qualitativa, as amostras foram coletadas através de
arrastos verticais e horizontais sucessivos com rede de 68µm de abertura de malha.
Para as análises quantitativas, com o auxílio de uma bomba hidráulica, filtraram-se
aproximadamente 200 litros de água em rede de 68µm de abertura de malha em
cada uma das profundidades correspondentes a 100%, 10% e 1% da radiação solar
incidente na superfície da lagoa. As contagens foram realizadas sob microscópio
óptico em aumento de 100x, utilizando-se câmaras de Sedgwich-Rafter, até o
mínimo de 200 indivíduos da espécie dominante.
As espécies que contribuíram com pelo menos 5% da biomassa total em pelo menos
uma amostra foram consideradas descritoras e agrupadas em grupos funcionais,
definidos conforme Reynolds et al. (2002) e Padisák et al. (2003a, 2006 e 2009).
Para cada coleta, considerou-se a biomassa média dos grupos funcionais nas três

79
profundidades, a fim de se reduzir a variabilidade espacial entre as amostras de uma
mesma lagoa.
A normalidade dos dados foi verificada pelo teste de Kolmogorov–Smirnov. Como os
dados não apresentaram distribuição normal, os testes de Kruskal-Wallis e o pós-
teste de Dunn foram utilizados para comparar lagoas e períodos. A ordenação das
amostras em função das variáveis ambientais foi avaliada através de Análise de
Componentes Principais (ACP), utilizando a matriz de correlação. Foi utilizado o
Programa PAST 1.9 (Hammer et al., 2001). As relações entre os grupos funcionais e
as variáveis bióticas e abióticas foram avaliadas através de correlações de
Spearman e de Análise de Correspondência Canônica (ACC; Ter Braak, 1986). Na
ACC, testou-se a hipótese nula de ausência de relação entre as matrizes principal
(grupos funcionais do fitoplâncton) e secundária (abiótica e composta pela
comunidade zooplanctônica) através do teste de Monte Carlo, considerando 1000
repetições aleatórias. Todos os dados foram transformados previamente (logaritmo
na base 10 do valor adicionado a uma unidade; Log(x+1)) e a ACP foi realizada com o
programa PC-ORD (McCune & Mefford, 1997).
Resultados
Variáveis ambientais
A maioria dos ambientes apresentou coluna d’água termicamente estratificada
durante as coletas realizadas em novembro de 2007 e fevereiro de 2008 (períodos
de chuvas) e desestratificação da coluna d’água em agosto de 2007 e maio de 2008
(períodos de seca). As exceções foram a lagoa Dom Helvécio, que se manteve
estratificada durante as quatro campanhas de amostragem, Verde, onde a
circulação completa da coluna d’água foi verificada somente em agosto de 2007 e
Amarela e Ferruginha, nas quais a coluna d’água mostrou-se homogênea durante
todo o período de estudo (Figura 3.3 ).
As lagoas diferiram em relação às principais variáveis ambientais medidas tanto na
estação seca quanto na estação chuvosa e também foram registradas diferenças
entre os períodos de seca e chuvas em cada lagoa (Tabela 3.1 ). Maiores
temperaturas da água foram observadas no período de chuva em todas as lagoas. A

80
extensão da zona de mistura e o coeficiente de atenuação escalar da luz mostraram-
se mais elevados na estação seca na maioria das lagoas em que se observou
variação sazonal destas variáveis.

81
Tabela 3. 1 - Mínimos e máximos obtidos para as principais variáveis ambientais de 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno durante as amostragens realizadas nos períodos de chuvas (nov/2007 e fev/2008) e seca (ago/2007 e maio/2008). Comparando-se os períodos em cada lagoa, os valores em negrito foram os mais elevados (Teste de Mann-Whitney; p<0,05)
Lagoa Periodo P-total PSR N-total SiO4 NID Temp Zeu Zmix ko
µg.L-1 µg.L-1 µg.L-1 µg.L-1 µg.L-1 oC m m m-1
A.Claras Chuvas 10,8-56,5 0,8-4,3 53,2-1211,8 0,7-1,7 9,6-15,3 26,2-31,2 6,8-7,8 4,0-4,5 0,5-0,7
Seca 6,4-31,5 0,6-9,4 330,2-412,8 0,6-1,9 73,7-194,2 23,2-26,8 9,0-9,5 9,5-9,5 0,4-0,7
Almécega Chuvas 11,9-40,1 2,4-7,2 37,3-166,5 0,7-5,4 1,6-30,7 28,5-31,0 6,0-7,0 5,0-5,0 0,5-0,8
Seca 9,3-15,7 1,7-7,3 121,1-283,2 0,7-5,3 15,6-23,0 24,4-27,3 6,0-6,5 6,0-6,5 0,5-0,9
Amarela Chuvas 26,4-174,9 6,2-8,3 231,2-836,5 3,2-4,4 35,7-74 26,5-27,3 1,5-1,8 1,5-1,8 2,2-2,9
Seca 31,8-65,5 2,6-10,0 220,5-410,5 1,2-5,7 13,5-25,8 22,0-22,4 1,8-2,4 1,8-2,5 2,7-3,0
Aníbal Chuvas 20,3-58,1 1,9-6,3 431,1-1038,4 1,5-3,8 30,8-60,2 28,6-30,8 4,5-6,0 4,0-4,0 1,0
Seca 18,6-28,0 1,7-4,5 398,6-905,5 1,0-3,8 104,7-119,6 23,3-26,2 4,5-5,5 4,5-5,5 0,8-1,5
Barra Chuvas 28,7-184,5 2,2-7,3 242,2-611,5 2,1-7,0 4,2-168,5 25,6-31,5 5,4-6,5 3,0-4,0 0,8-1,0
Seca 22,0-44,9 1,2-7,4 122,1-334 0,8-2,4 30,4-176,3 23,2-26,8 5,0-6,0 5,0-7,0 0,8-1,0
Carioca Chuvas 10,9-70,6 2,4-6,5 274,6-567,8 1,9-2,4 19,1-65,4 23,5-31,0 5,4-6,3 4,0-6,0 0,8-1,4
Seca 12,4-34,6 1,9-3,9 201,4-990,1 0,7-1,2 73,2-278,0 21,6-27,9 4,2-4,5 5,0-9,0 1,2-1,4
Central Chuvas 20,7-44,3 3,0-7,5 833,7-981,2 1,0-3,7 70,3-175,4 27,2-30,6 2,7-3,0 0,5-3,0 1,7-1,8
Seca 22-43,5 1,9-3,6 321,9-811,9 1,0-3,5 123,1-213,3 21,8-27,6 1,8-2,3 4,5-4,5 2,4-2,7
D.Helvécio Chuvas 12,6-53,8 0,3-4,6 422,4-678,4 3,2-3,7 24-77,3 26,9-30,9 6,0-8,7 5,0-7,5 0,5-0,6
Seca 17,4-27,4 1,8-9,4 308-494,2 0,9-2,4 21,8-227,6 23,2-27,5 6,0-9,0 3,0-10,0 0,7-0,9
Ferrugem Chuvas 43,1-78,8 3,4-4,6 686,3-1470,4 2,9-5,0 95,4-153,6 28,2-31,0 2,0-2,0 2,0-2,0 1,6-1,7
Seca 21,5-56,1 3,8-5,0 700,4-1489,7 0,7-2,9 29,6-47,1 23,6-25,5 2,7-3,5 3,0-3,5 1,5-1,5
Ferruginha Chuvas 25,4-55,0 2,1-5,0 824,1-1329,4 1,4-1,9 82,5-185,1 28,2-31,0 3,0-3,3 2,5-4,0 1,2-1,6
Seca 20,0-55,6 4,2-8,2 501,2-1117,4 0,5-1,1 10,4-24,5 22,3-24,4 3,0-3,0 2,0-4,0 1,2-1,7
Gambá Chuvas 9,2-38,7 0,9-4,1 231,5-631,5 0,9-1,7 - 23,9-31,0 10,2-11,3 5,5-6,0 0,7-0,7
Seca 11,5-20,1 2,1-4,8 353,9-612,8 0,2-1,6 87,8-320,9 23,2-26,9 6,8-9,5 9,5-11,5 0,7-0,8
Gambazinho Chuvas 21,6-35,2 0,8-4,3 387,4-614,4 0,4-1,3 23,2-268,0 26-30,1 5,7-6,0 5,0-6,5 0,7-1,0
Seca 14,0-23,1 2,4-5,6 240-780,5 0,3-0,8 20,8-218,1 23,4-26 4,5-5,1 9,0-10,0 0,7-0,8
Jacaré Chuvas 44,0-87,2 1,1-6,7 273,5-1310,2 1,3-4,1 16,1-186 27-31,7 5,1-6,0 3,5-5,0 0,8-0,9
Seca 7,2-32,0 3,8-6,1 171,4-753,2 0,9-3,4 36,2-246,9 22,9-26,8 6,0-6,3 7,0-8,0 0,9-1,0
Palmeirinha Chuvas 50,2-82,0 0,9-5,9 468,9-1998,7 2,2-3,6 4,8-5,3 27,4-31,5 5,4-5,5 3,5-4,0 0,8-0,8
Seca 3,1-41,0 0,5-4,1 369,9-691,8 0,7-2,9 14,4-94,3 23,3-27,1 5,25-5,5 5,5-5,5 0,7-0,9
Patos Chuvas 31,6-211,2 2,1-8,4 281,7-742,4 1,4-3,2 3,3-78,9 24,6-30,4 7,2-7,5 4,0-5,0 0,8-1,0
Seca 12,6-27,4 5,3-9,7 389,2-647,1 1,0-1,7 157,2-412,9 22,6-26,2 4,5-5,0 5,0-8,0 0,8-1,1
Pimenta Chuvas 11,2-67,8 5,0-7,5 471,2-736,9 2,7-3,7 - 28,0-31,5 3,0-3,0 1,0-1,5 1,4-2,1
Seca 15432,0 2,9-6,1 261,3-419,8 0,9-2,8 23,7-25,6 22,9-27,7 3,0-3,5 3,0-3,5 1,6
S.Helena Chuvas 38,9-194,9 2,4-9,5 194,2-599,4 1,1-4,9 4,8-33,2 27,7-31,5 6-8,3,0 6,0-6,0 0,6-0,6
Seca 15,3-44,8 1,5-6,4 209,3-336,9 0,5-1,2 17,4-164,9 23,4-27,2 5,4-7,0 7,0-10,0 0,6-1,0
Verde Chuvas 11,6-20,6 2,7-5,6 152,4-377,0 1,7-2,3 8,7-21,7 26,1-30,8 6,8-8,7 4,0-5,0 0,5-0,6
Seca 9,7-18,8 1,8-5,7 219,9-346,9 0,5-2,6 4,7-132,8 23,6-28 4,5-6,3 7,5-10,0 0,7-0,9
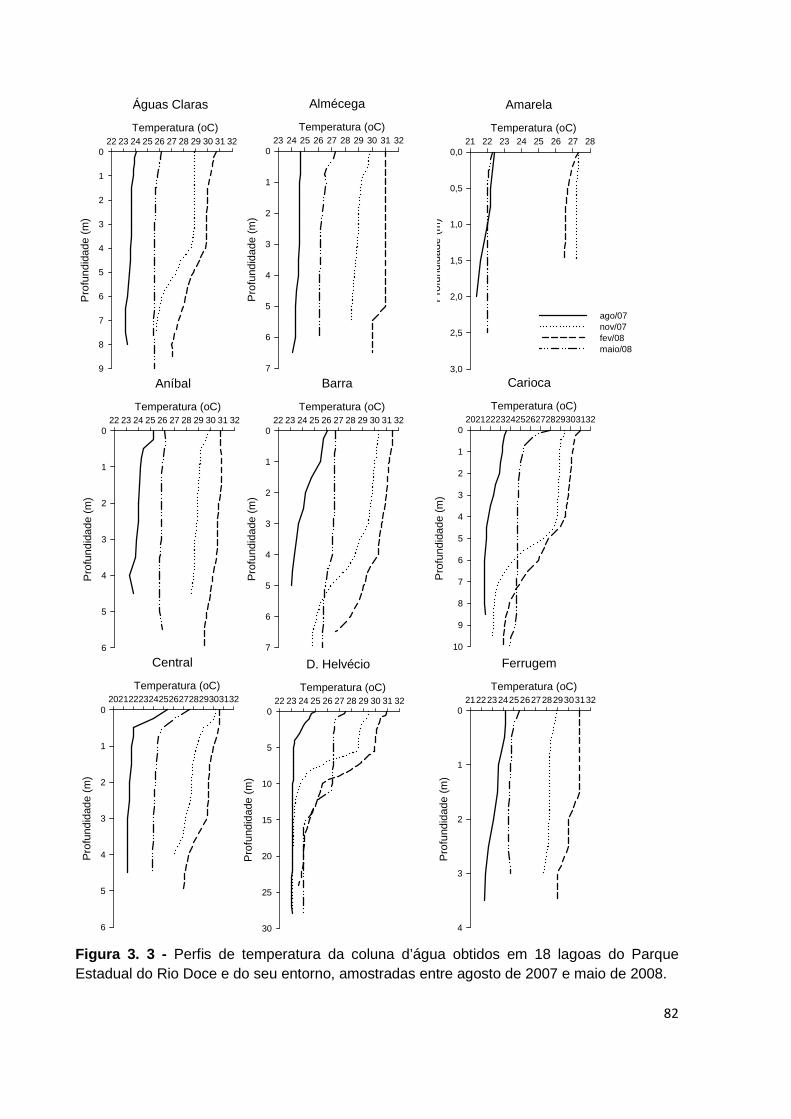
82
Amarela
Temperatura (oC)21 22 23 24 25 26 27 28
Pro
fund
idad
e (m
)
0,0
0,5
1,0
1,5
2,0
2,5
3,0
Central
Temperatura (oC)20212223242526272829303132
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
D. Helvécio
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
5
10
15
20
25
30
ago/07nov/07fev/08maio/08
Ferrugem
Temperatura (oC)212223242526272829303132
Pro
fund
idad
e (m
)
0
1
2
3
4
Águas Claras
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
8
9
Almécega
Temperatura (oC)23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
Aníbal
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
Barra
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
Carioca
Temperatura (oC)20212223242526272829303132
Pro
fund
idad
e (m
)0
1
2
3
4
5
6
7
8
9
10
Figura 3. 3 - Perfis de temperatura da coluna d’água obtidos em 18 lagoas do Parque Estadual do Rio Doce e do seu entorno, amostradas entre agosto de 2007 e maio de 2008.

83
Ferruginha
Temperatura (oC)212223242526272829303132
Pro
fund
idad
e (m
)
0
1
2
3
4
Gambá
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
8
9
10
11
12
Gambazinho
Temperatura (oC)22 23 24 25 26 27 28 29 30 31
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
8
9
10
Jacaré
Temperatura (oC)222324252627282930313233
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
8
9
10
Palmeirinha
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
Patos
Temperatura (oC)22 23 24 25 26 27 28 29 30 31
Pro
fund
idad
e (m
)0
1
2
3
4
5
6
7
8
Pimenta
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
ago/07nov/07fev/08maio/08
Sta. Helena
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0
1
2
3
4
5
6
7
8
9
10
Verde
Temperatura (oC)22 23 24 25 26 27 28 29 30 31 32
Pro
fund
idad
e (m
)
0123456789
10111213141516171819
Figura 3.3 (continuação). Perfis de temperatura da coluna d’água obtidos nas 18 lagoas do Parque Estadual do Rio Doce e do seu entorno, amostradas entre agosto de 2007 e maio de 2008.
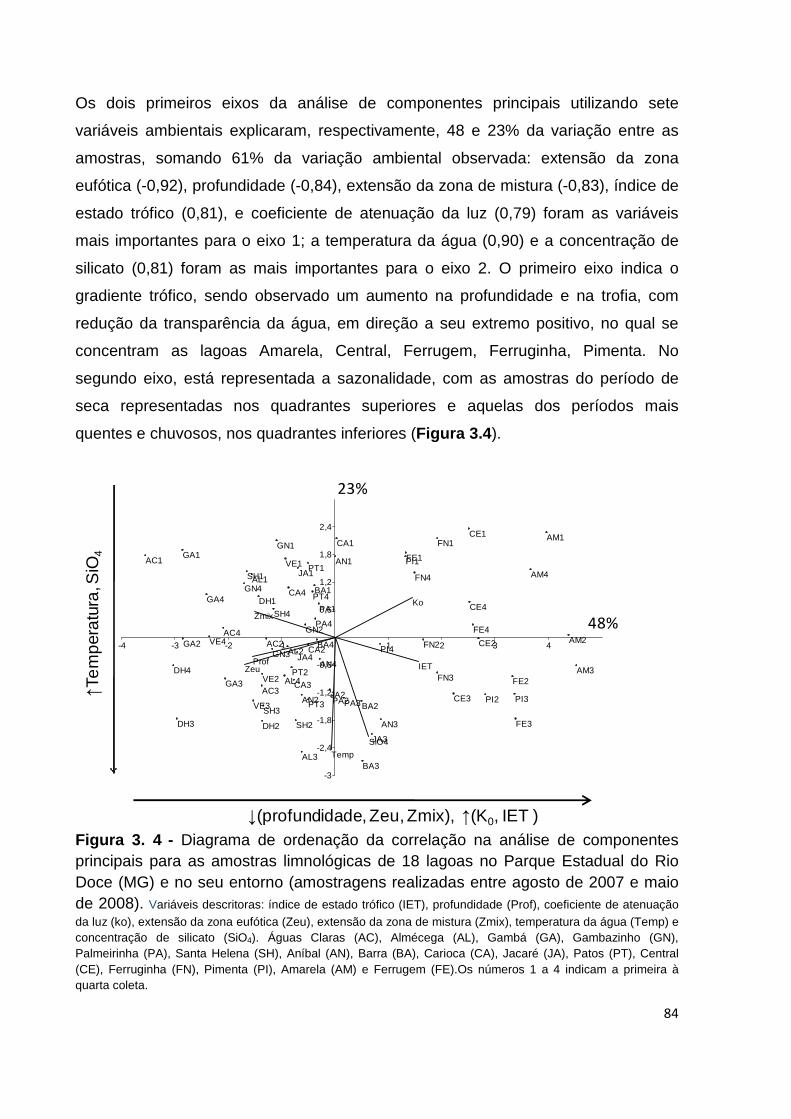
84
Os dois primeiros eixos da análise de componentes principais utilizando sete
variáveis ambientais explicaram, respectivamente, 48 e 23% da variação entre as
amostras, somando 61% da variação ambiental observada: extensão da zona
eufótica (-0,92), profundidade (-0,84), extensão da zona de mistura (-0,83), índice de
estado trófico (0,81), e coeficiente de atenuação da luz (0,79) foram as variáveis
mais importantes para o eixo 1; a temperatura da água (0,90) e a concentração de
silicato (0,81) foram as mais importantes para o eixo 2. O primeiro eixo indica o
gradiente trófico, sendo observado um aumento na profundidade e na trofia, com
redução da transparência da água, em direção a seu extremo positivo, no qual se
concentram as lagoas Amarela, Central, Ferrugem, Ferruginha, Pimenta. No
segundo eixo, está representada a sazonalidade, com as amostras do período de
seca representadas nos quadrantes superiores e aquelas dos períodos mais
quentes e chuvosos, nos quadrantes inferiores (Figura 3.4 ).
SiO4
Temp
IETProf
Ko
Zeu
Zmix
AC1
AC2
AC3
AC4
AL1
AL2
AL3
AL4
AM1
AM2
AM3
AM4
AN1
AN2
AN3
AN4
BA1
BA2
BA3
BA4
CA1
CA2
CA3
CA4
CE1
CE2
CE3
CE4DH1
DH2DH3
DH4
FE1
FE2
FE3
FE4
FN1
FN2
FN3
FN4
GA1
GA2
GA3
GA4
GN1
GN2
GN3
GN4
JA1
JA2
JA3
JA4
PA1
PA2PA3
PA4
PT1
PT2
PT3
PT4
PI1
PI2 PI3
PI4
SH1
SH2
SH3
SH4
VE1
VE2
VE3
VE4-4 -3 -2 -1 1 2 3 4
-3
-2,4
-1,8
-1,2
-0,6
0,6
1,2
1,8
2,4
↓(profundidade, Zeu, Zmix), ↑(K0, IET )
↑Te
mpe
ratu
ra, S
iO4
48%
23%
Figura 3. 4 - Diagrama de ordenação da correlação na análise de componentes principais para as amostras limnológicas de 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno (amostragens realizadas entre agosto de 2007 e maio de 2008). Variáveis descritoras: índice de estado trófico (IET), profundidade (Prof), coeficiente de atenuação da luz (ko), extensão da zona eufótica (Zeu), extensão da zona de mistura (Zmix), temperatura da água (Temp) e concentração de silicato (SiO4). Águas Claras (AC), Almécega (AL), Gambá (GA), Gambazinho (GN), Palmeirinha (PA), Santa Helena (SH), Aníbal (AN), Barra (BA), Carioca (CA), Jacaré (JA), Patos (PT), Central (CE), Ferruginha (FN), Pimenta (PI), Amarela (AM) e Ferrugem (FE).Os números 1 a 4 indicam a primeira à quarta coleta.

85
A transparência da água, a profundidade e o índice de estado trófico foram os
principais fatores definindo as condições ambientais nas lagoas estudadas. Com
base nestas variáveis, as lagoas foram separadas em cinco tipos: 1) profundas,
claras e oligotróficas; 2) de profundidade intermediária, claras e oligotróficas; 3) de
profundidade intermediária, escuras e mesotróficas; 4) rasas, escuras e mesotróficas
e 5) rasas, escuras e eutróficas (Tabela 3.2 ).
Tabela 3. 2 - Classificação de 18 lagoas no Parque Estadual do Rio Doce (MG) e seu entorno em 5 tipos, identificados com base na profundidade total, coeficiente de atenuação escalar da luz e trofia (amostragens realizadas entre agosto de 2007 e maio de 2008)
Tipo Profundidade k0 Trofia Lagoas
m m-1
1 >19 <0,7 oligotróficas Dom Helvécio (DH), Verde (VE)
2 6 - 12 <0,9 oligotróficas Águas Clara (AC), Almécega (AL), Gambá (GA),
Gambazinho (GN), Palmeirinha (PA), Santa Helena (SH)
3 7-10 >0,9 mesotróficas Aníbal (AN), Barra (BA), Carioca (CA), Jacaré (JA), Patos (PT)
4 <5 >1 mesotróficas Central (CE), Ferruginha (FN), Pimenta (PI)
5 <4 >1,5 eutróficas Amarela (AM), Ferrugem (FE)
Dinâmica do fitoplâncton
Foram identificadas 490 espécies no fitoplâncton, distribuídas em 10 classes
taxonômicas e 29 grupos funcionais. Cinquenta e uma dessas espécies foram
consideradas descritoras e representaram 18 grupos funcionais (Tabela 3.3 ).
Em relação à biomassa total, as medianas variaram entre 0,2 e 3,3 mg.L-1. Menores
valores foram observados nas lagoas Amarela, Águas Claras, Santa Helena,
Almécega, Palmeirinha, Verde e Dom Helvécio (Figura 3.5 ). Com exceção da lagoa
Amarela, cuja biomassa fitoplanctônica alcançou 3,0 mg.L-1 nas amostragens de
novembro de 2007 devido ao predomínio de Botryococcus braunii Kützing e Dom
Helvécio, que alcançou 1,3 mg.L-1 em fevereiro de 2008, devido ao aumento de
Cylindrospermopsis raciborskii (Woloszynska) Seenayya & Subba Raju, estes
ambientes mantiveram biomassas inferiores a 1,0 mg.L-1 nos quatro períodos de
amostragem.

86
As maiores biomassas foram registradas nas lagoas Ferrugem (0,8 a 2,7 mg.L-1;
mediana = 2,2 mg.L-1), Jacaré (1,2 a 2,9 mg.L-1; mediana = 2,5 mg.L-1), Ferruginha
(2,4 a 7,4 mg.L-1; mediana = 3,0 mg.L-1) e Central (2,2 a 7,1 mg.L-1; mediana = 3,4
mg.L-1). Os picos de biomassa na lagoa Ferruginha, devido a Dictyosphaerium
pulchellum Wood (48% da biomassa total) e Limnothrix redekei (Van Goor) Meffert
(29 % da biomassa total) e na lagoa Central, devido a Gymnodinium sp (77% da
biomassa total), ocorreram nas amostragens de agosto de 2007.
Diferenças entre os períodos de seca e chuva fora observadas para as lagoas
Águas Claras (U=0,0; Z=-2,88; p=0,004), Amarela (U=0,00; Z=-2,61; p=0,009) e
Dom Helvécio (U=0,00; Z=-2,56; p=0,010), que mostraram maior biomassa no
período chuvoso, e Aníbal, onde a biomassa foi mais elevada durante a seca
(U=0,00; Z=2,40; p=0,016).
DH
VE
AC AL
GA
GN
PA
SH
AN
BA
CA JA PT
CE
FN PI
AM FE0
1
2
3
4
5
6
7
8
9
DH VE AC AL GA GN PA SH AN BA CA JA PT CE FN PI AM FE
Tipo 1 Tipo 2 Tipo 3 Tipo 4 Tipo 5
Bio
mas
sa fi
topl
anct
ônic
a (m
g.L-1
)
Figura 3. 5 - Variação espacial e temporal da biomassa fitoplanctônica em 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno classificadas em cinco tipos identificados com base na profundidade total, coeficiente de atenuação escalar da luz e trofia (amostragens realizadas entre agosto de 2007 e maio de 2008). A linha horizontal dentro das caixas representa a mediana e os limites da caixa e a linha verical englobam 75% e 90% dos dados, respectivamente. Dom Helvécio (DH), Verde (VE),
Águas Claras (AC), Almécega (AL), Gambá (GA), Gambazinho (GN), Palmeirinha (PA), Santa Helena (SH), Aníbal (AN), Barra (BA), Carioca (CA), Jacaré (JA), Patos (PT), Central (CE), Ferruginha (FN), Pimenta (PI), Amarela (AM) e Ferrugem (FE).

87
Tabela 3. 3 - Espécies fitoplanctônicas descritoras (>5% da biomassa total), classes taxonômicas e grupos funcionais (GF) de 18 lagoas no Parque Estadual do Rio Doce e no seu entorno (amostragens realizadas entre agosto de 2007 e maio de 2008).
Espécie Classe taxonômica GFUrosolenia longiseta (Zacharias) Edlung & Stoermer Bacillariophyceae ACyclotella sp Bacillariophyceae BDinobryon bavaricum Imhof Chrysophyceae EDinobryon sertularia Ehrenberg Chrysophyceae EBotryococcus braunii Kützing Chlorophyceae FDictyosphaerium pulchellum Wood Chlorophyceae FElakatothrix genevensis (Reverdin) Hindák Chlorophyceae FEutetramorus planctonicus (Korsikov) Bourrely Chlorophyceae FOocystis cf solitaria Wittrock Chlorophyceae FOocystis sp Chlorophyceae FCoelastrum pseudomicroporum Korsikov Chlorophyceae JCoelastrum pulchrum Schimidle Chlorophyceae JCoelastrum reticulatum (Dangeard) Senn Chlorophyceae JCoelastrum sphaericum Nägeli Chlorophyceae JCrucigenia tetrapedia (Kirchner) West & West Chlorophyceae JScenedesmus bijugus (Turpin) Kutzing Chlorophyceae JAphanocapsa elachista West & West Cyanobacteria KAphanocapsa sp Cyanobacteria KPeridinium pusillum (Pénard) Lemmermann Dinophyceae Lo
Peridinium baliense Lindemann Dinophyceae Lo
Peridinium sp (~34um) Dinophyceae Lo
Navicula sp1 Bacillariophyceae MPPseudanabaena galeata Böcher Cyanobacteria MPCosmarium asphaerosporum Nordstedt Zygnemaphyceae NA
Cosmarium bioculatum Brébisson Zygnemaphyceae NA
Cosmarium moniliforme West & West Zygnemaphyceae NA
Staurodesmus crassus (West & West) Florin Zygnemaphyceae NA
Staurodesmus dejectus (Brébisson) Teiling Zygnemaphyceae NA
Staurodesmus incus (Brébisson) Teiling Zygnemaphyceae NA
Teilingia granulata (Roy & Bisset) Bourrely Zygnemaphyceae NA
Teilingia sp2 Zygnemaphyceae NA
Staurastrum ionatum Wolle Zygnemaphyceae NA
Staurastrum smithii (G.M. Smith) Teiling Zygnemaphyceae NA
Staurastrum tetracerum (Kützing) Ralfs ex Ralfs Zygnemaphyceae NA
Limnothrix redekei (Van Goor) Meffert Cyanobacteria S1Planktolyngbya contorta (Lemmermann) Anagnostidis & Komárek Cyanobacteria S1Planktolyngbya limnetica (Lemmermann) Komárek Cyanobacteria S1Spirulina sp Cyanobacteria S2Cylindrospermopsis raciborskii (Wolos.) Seenayya & Subba Raju Cyanobacteria SNPlanktonema sp Chlorophyceae TTrachelomonas volvocina Ehrenberg Euglenophyceae W2Chlorella sp (~5um) Chlorophyceae X1Monoraphidium contortum (Thur.) Komárkova-Legnerová Chlorophyceae X1Monoraphidium sp Chlorophyceae X1Cryptomonas brasiliensis Castro, C. Bicudo & D. Bicudo Cryptophyceae X2Cryptomonas marsonii Skuja Cryptophyceae X2Cryptomonas sp5 (~13um) Cryptophyceae X2Chlorella sp1 Chlorophyceae X3Cryptomonas sp3 (~18um) Cryptophyceae YCryptomonas sp4 (~22um) Cryptophyceae YGymnodynium sp Dinophyceae YDinoflagelado NI (~8um) Dinophyceae Y
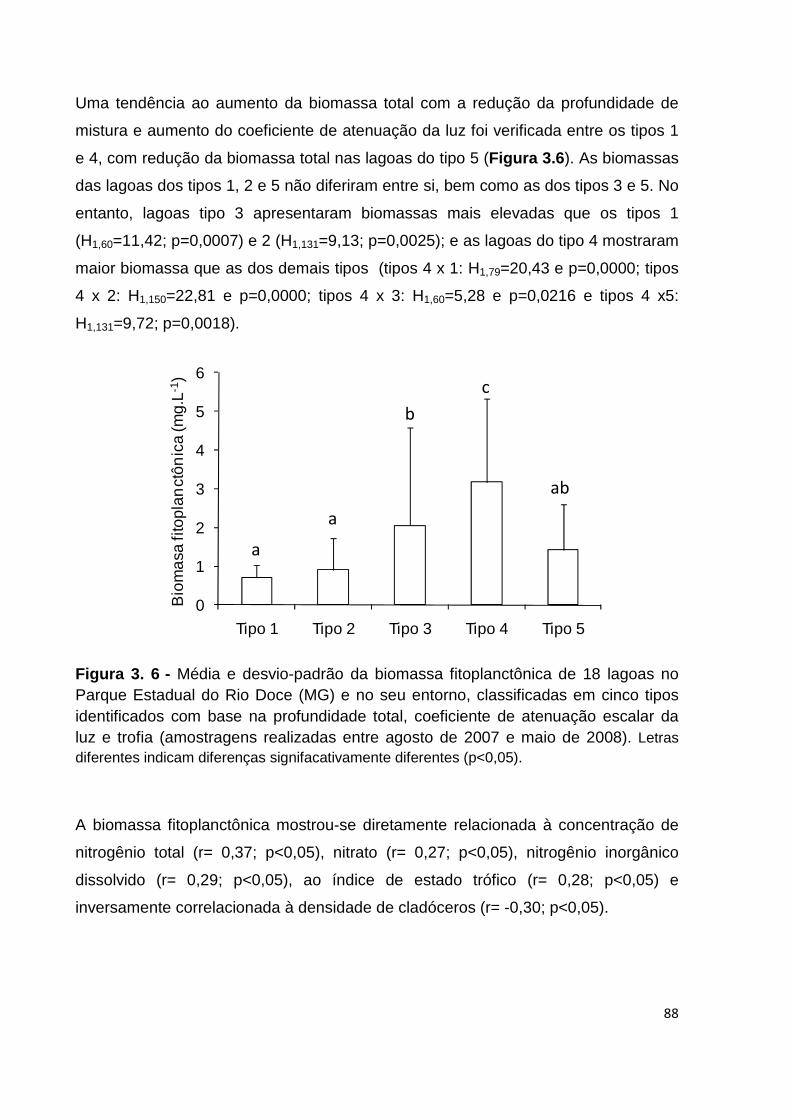
88
Uma tendência ao aumento da biomassa total com a redução da profundidade de
mistura e aumento do coeficiente de atenuação da luz foi verificada entre os tipos 1
e 4, com redução da biomassa total nas lagoas do tipo 5 (Figura 3.6 ). As biomassas
das lagoas dos tipos 1, 2 e 5 não diferiram entre si, bem como as dos tipos 3 e 5. No
entanto, lagoas tipo 3 apresentaram biomassas mais elevadas que os tipos 1
(H1,60=11,42; p=0,0007) e 2 (H1,131=9,13; p=0,0025); e as lagoas do tipo 4 mostraram
maior biomassa que as dos demais tipos (tipos 4 x 1: H1,79=20,43 e p=0,0000; tipos
4 x 2: H1,150=22,81 e p=0,0000; tipos 4 x 3: H1,60=5,28 e p=0,0216 e tipos 4 x5:
H1,131=9,72; p=0,0018).
0
1
2
3
4
5
6
Tipo 1 Tipo 2 Tipo 3 Tipo 4 Tipo 5
Bio
mas
a fit
opla
nct
ônic
a (m
g.L-1
)
b
a
a
ab
c
Figura 3. 6 - Média e desvio-padrão da biomassa fitoplanctônica de 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno, classificadas em cinco tipos identificados com base na profundidade total, coeficiente de atenuação escalar da luz e trofia (amostragens realizadas entre agosto de 2007 e maio de 2008). Letras diferentes indicam diferenças signifacativamente diferentes (p<0,05).
A biomassa fitoplanctônica mostrou-se diretamente relacionada à concentração de
nitrogênio total (r= 0,37; p<0,05), nitrato (r= 0,27; p<0,05), nitrogênio inorgânico
dissolvido (r= 0,29; p<0,05), ao índice de estado trófico (r= 0,28; p<0,05) e
inversamente correlacionada à densidade de cladóceros (r= -0,30; p<0,05).

89
Oito grupos funcionais (GF) foram frequentes nas lagoas estudadas: F, Lo, NA, S1,
SN, X1, X2 e Y, e se alternaram entre as campanhas de amostragem,
especialmente entre os períodos de seca e chuvas. Destaca-se o predomínio do GF
“NA”, representado principalmente pelas espécies Cosmarium bioculatum,
Cosmarium moniliforme, Staurodesmus crassus, Staurastrum chaetoceras e
Staurastrum smithii, durante o período chuvoso na maioria das lagoas amostradas
(Tabela 3.5 ). Representantes deste grupo não foram listados entre as espécies
descritoras das lagoas Gambazinho, Patos, Central e Amarela. O GF “A”,
representado por Urosolenia longiseta, ocorreu apenas nas lagoas Águas Claras e
Almécega, enquanto os GFs “T” (Planctonema sp) e “S2” (Spirulina sp) estiveram
representados apenas na lagoa dos Patos.
A biomassa do GF F (Botryococcus braunii e Dictyosphaerium pulchellum) aumentou
com o aumento da trofia e redução da profundidade (H4,210=32,29; p=0,0000). Maior
biomassa do GF NA (Cosmarium spp; Teilingia spp; Stautodesmus crassus) foi
observada nas lagoas tipo 1 e 4 (H4,210=19,78; p=0,0006), enquanto GF Y (grandes
Cryptomonas; Gymnodinium sp; H4,210=60,57; p=0,0000) e GF Lo (Peridinium spp)
apresentaram biomassas mais elevadas (H4,210=38,87; p=0,0000) nas lagoas tipo 3 e
4. O GF X2 (pequenas Cryptomonas spp) também se destacou nas lagoas tipo 3
(H4,210=51,52; p=0,0000) e GF S1 (Limnothrix redekei e Planktolyngbya limnetica) e
GF X1, nas lagoas tipo 4. O GF SN (Cylindrospermopsis raciborskii) apresentou
elevada biomassa nas lagoas tipo 5 (Figura 3.7 ). Representantes do GF J
(Coelastrum reticulatum e Crucigenia tetrapedia) encontram-se entre as principais
espécies das lagoas tipo 4 (Tabela 3.4 ).
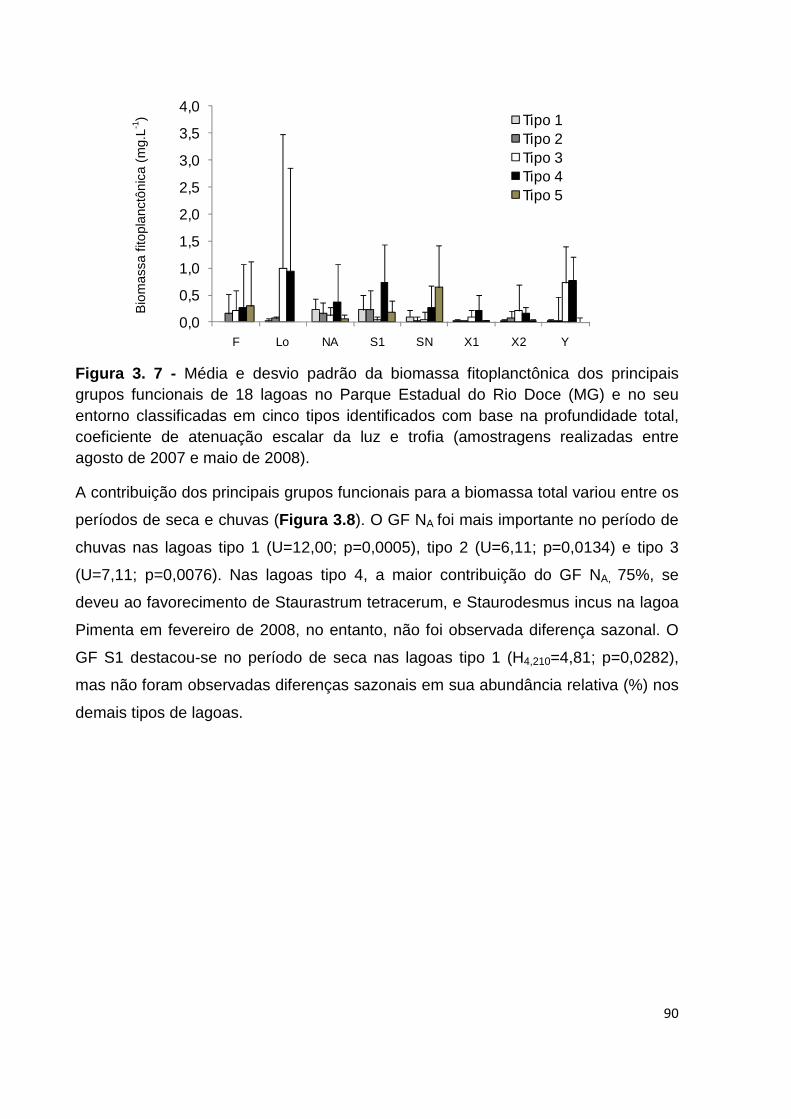
90
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
F Lo NA S1 SN X1 X2 Y
Tipo 1Tipo 2Tipo 3Tipo 4Tipo 5
Figura 3. 7 - Média e desvio padrão da biomassa fitoplanctônica dos principais grupos funcionais de 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno classificadas em cinco tipos identificados com base na profundidade total, coeficiente de atenuação escalar da luz e trofia (amostragens realizadas entre agosto de 2007 e maio de 2008).
A contribuição dos principais grupos funcionais para a biomassa total variou entre os
períodos de seca e chuvas (Figura 3.8 ). O GF NA foi mais importante no período de
chuvas nas lagoas tipo 1 (U=12,00; p=0,0005), tipo 2 (U=6,11; p=0,0134) e tipo 3
(U=7,11; p=0,0076). Nas lagoas tipo 4, a maior contribuição do GF NA, 75%, se
deveu ao favorecimento de Staurastrum tetracerum, e Staurodesmus incus na lagoa
Pimenta em fevereiro de 2008, no entanto, não foi observada diferença sazonal. O
GF S1 destacou-se no período de seca nas lagoas tipo 1 (H4,210=4,81; p=0,0282),
mas não foram observadas diferenças sazonais em sua abundância relativa (%) nos
demais tipos de lagoas.
Bio
mas
sa fi
topl
anct
ônic
a (m
g.L-1
)
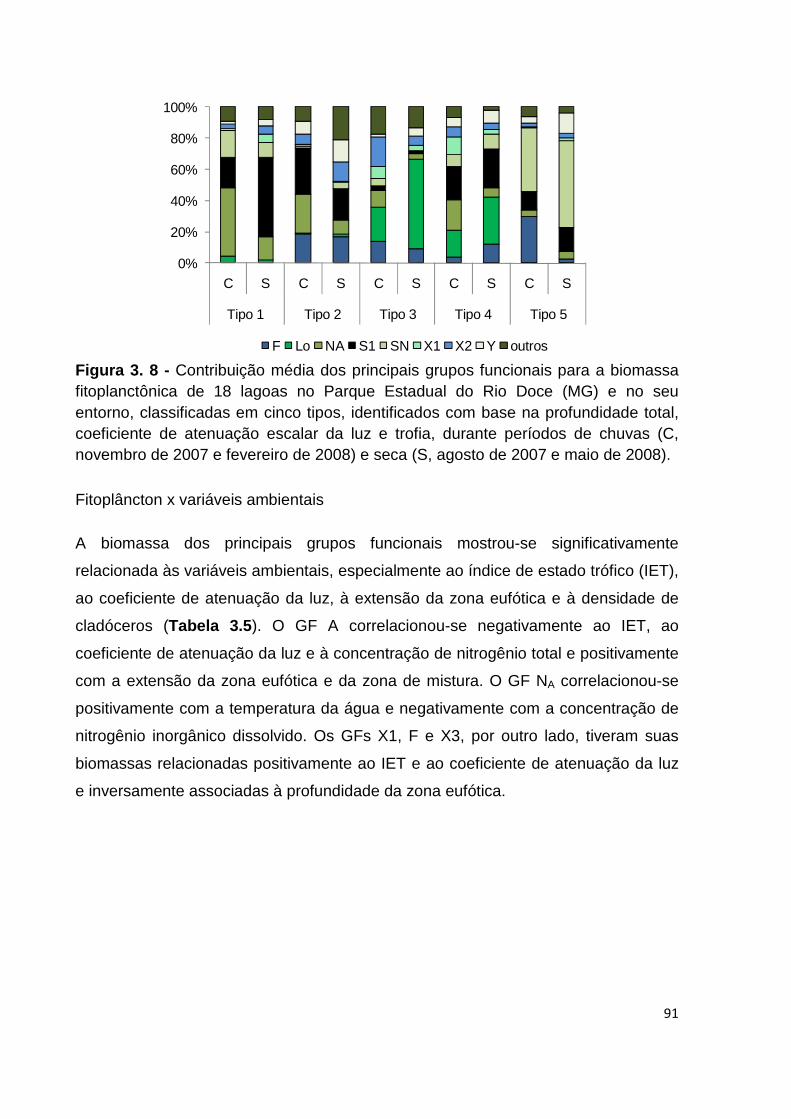
91
0%
20%
40%
60%
80%
100%
C S C S C S C S C S
Tipo 1 Tipo 2 Tipo 3 Tipo 4 Tipo 5
F Lo NA S1 SN X1 X2 Y outros
Figura 3. 8 - Contribuição média dos principais grupos funcionais para a biomassa fitoplanctônica de 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno, classificadas em cinco tipos, identificados com base na profundidade total, coeficiente de atenuação escalar da luz e trofia, durante períodos de chuvas (C, novembro de 2007 e fevereiro de 2008) e seca (S, agosto de 2007 e maio de 2008).
Fitoplâncton x variáveis ambientais
A biomassa dos principais grupos funcionais mostrou-se significativamente
relacionada às variáveis ambientais, especialmente ao índice de estado trófico (IET),
ao coeficiente de atenuação da luz, à extensão da zona eufótica e à densidade de
cladóceros (Tabela 3.5 ). O GF A correlacionou-se negativamente ao IET, ao
coeficiente de atenuação da luz e à concentração de nitrogênio total e positivamente
com a extensão da zona eufótica e da zona de mistura. O GF NA correlacionou-se
positivamente com a temperatura da água e negativamente com a concentração de
nitrogênio inorgânico dissolvido. Os GFs X1, F e X3, por outro lado, tiveram suas
biomassas relacionadas positivamente ao IET e ao coeficiente de atenuação da luz
e inversamente associadas à profundidade da zona eufótica.

92
Tabela 3. 4 - Contribuição das principais espécies e grupos funcionais fitoplanctônicos mais frequentes para a biomassa nas 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno, durante períodos de chuvas (novembro de 2007 e fevereiro de 2008) e seca (agosto de 2007 e maio de 2008).
ago/07 nov/07 fev/08 mai/08Lagoa GF Representantes GF Representantes GF Representantes Representantes
X1 Monoraphidium sp (31%) NA C.moniliforme (27%) NA S. crassus (31%) S1 L. redekei (56%)
D. Helvécio SN C.raciborskii (24%) C.bioculatum (9%) SN C.raciborskii (22%) SN C.raciborskii (11%)
X3 Chlorella sp (12%) SN C.raciborskii (24%) S1 L. redekei (22%) NA S. crassus (5%)
S1 P.contorta (18%) NA C.bioculatum (34%) NA C.bioculatum (33%) S1 L. redekei (28%)
Verde L. redekei (17%) S1 P.contorta (13%) S. crassus (16%) P.contorta (25%)
X2 Cryptomonas sp2 (14%) P. limnetica (8%) Lo P.pusillum (18%) NA C.bioculatum (11%)
A U. longiseta (38%) NA C. bioculatum (58%) NA T. granulata (23%) X3 Chlorella sp (18%)
A.Claras X3 Chlorella sp (15%) S. crassus (19%) C. bioculatum (14%) S1 P. limnetica (15%)X2 C. brasiliensis (11%) X1 M. contortum (10%) S1 P. limnetica (21%) A U.longiseta (14%)
A U. longiseta (63%) NA S. smithii (22%) S1 P. limnetica (43%) A U. longiseta (48%)
Almécega NA C. bioculatum (5%) C. bioculatum (21%) A U. longiseta (39%) S1 P. limnetica (25%)
S. smithii (5%) Teilingia sp (17%) NA Teilingia sp2 (7%)
Y Gymnodinium sp (33%) NA C. bioculatum (25%) X2 C.brasiliense (26%) S1 L. redekei (46%)
Gambá Dinoflagelado NI (15%) S1 L. redekei (23%) Y Gymnodinium sp (22%) Y Dinoflagelado NI (17%)X2 C.brasiliense (21%) X2 C. brasiliensis (15%) S1 L. redekei (13%) X2 C.brasiliense (14%)F B.braunii (49%) F B.braunii (51%) S1 P. limnetica (39%) F B.braunii (42%)
Gambazinho X2 Cryptomonas sp5 (19%) S1 P. limnetica (31%) L. redekei (10%) S1 P. limnetica (29%)L. redekei (11%) F B.braunii (37%) L. redekei (21%)
F D. pulchellum (48%) NA C.bioculatum (42%) NA S. crassus (17%) NA C.moniliforme (43%)
Palmeirinha X2 Cryptomonas sp2 (19%) E D.sertularia (10%) C.moniliforme (16%) S1 P. limnetica (11%)E D.sertularia (19%) S1 P. limnetica (10%) S1 P. limnetica (18%) F D. pulchellum (10%)
SN C.raciborskii (61%) NA C.bioculatum (58%) NA C.bioculatum (23%) A U. longiseta (25%)
S. Helena K A. elachista (5%) C.moniliforme (6%) S. crassus (21%) SN C.raciborskii (17%)
A U. longiseta (5%) SN C.raciborskii (10%) X2 C. brasiliensis (14%) NA C.bioculatum (16%)
Lo Peridinium sp3 (90%) Lo Peridinium sp3 (26%) NA C. bioculatum (25%) NA C. bioculatum (35%)
Aníbal F O. cf solitaria (5%) NA C. bioculatum (22%) X2 Cryptomonas sp5 (24%) S. dejectus (13%)
X2 Cryptomonas sp5 (3%) X2 Cryptomonas sp5 (17%) S1 P. limnetica (17%) S. ionatum (12%)Lo Peridinium sp1 (60%) SN C.raciborskii (36%) SN C.raciborskii (18%) SN C.raciborskii (35%)
Barra X2 Cryptomonas sp5 (18%) S1 L. redekei (16%) S1 L. redekei (13%) S1 L. redekei (29%)
Lo Peridinium sp1 (13%) NA S. smithii (11%) X2 Cryptomonas sp5 (10%)
MP P.galeata (44%) X2 Cryptomonas sp5 (28%) MP P.galeata (28%) Y Gymnodinium sp (38%)Carioca Y Gymnodinium sp (23%) X1 Chlorella sp (23%) X1 Chlorella sp (22%) X2 Cryptomonas sp5 (18%)
X1 Chlorella sp (11%) MP P.galeata (18%) NA S. crassus (11%) X1 Chlorella sp (17%)
K A. elachista (34%) Y Gymnodinium sp (63%) K A. elachista (27%) Y Gymnodinium sp (63%)Jacaré F B.braunii (22%) F B.braunii (12%) X3 Chlorella sp (17%) F B.braunii (17%)
Y Gymnodinium sp (17%) K A. elachista (10%) NA C.bioculatum (14%) K A. elachista (10%)
F E.planctonicus (48%) F B.braunii (51%) F B.braunii (30%) F B.braunii (42%)Patos B.braunii (29%) E.planctonicus (8%) Elakatothrix sp (19%) T Planctonema sp (25%)
X2 C. marsonii (7%) S2 Spirulina sp2 (25%) X2 C. marsonii (29%) W2 T.volvocinopsis (14%)F D. pulchellum (48%) S1 L. redekei (33%) S1 L. redekei (29%) S1 L. redekei (64%)
Central S1 L. redekei (29%) X1 M. contortum (29%) J C.reticulatum (22%) Y Gymnodinium sp (19%)Y Gymnodinium sp (12%) Y Gymnodinium sp (10%) F D. pulchellum (14%) F D. pulchellum (7%)Y Gymnodinium sp (77%) Y Gymnodinium sp (37%) SN C.raciborskii (27%) SN C.raciborskii (42%)
Ferruginha SN C.raciborskii (13%) S1 L. redekei (30%) Lo P.pusillum (24%) S1 L. redekei (19%)
S1 L. redekei (3%) SN C.raciborskii (17%) Y Gymnodinium sp (18%) NA C.moniliforme (9%)
NA S.incus (26%) S1 L. redekei (36%) NA S.tetracerum (51%) S1 L. redekei (47%)
Pimenta S. ionatum (16%) X1 M.contortum (26%) C.bioculatum (24%) X1 M.contortum (13%)
S1 L. redekei (22%) C.bioculatum (9%) S1 L. redekei (7%) NA S.tetracerum (7%)
F B.braunii (50%) F B.braunii (92%) F B.braunii (64%) Y Cryptomonas sp4 (41%)Amarela Y Cryptomonas sp4 (20%) J C. tetrapedia (4%) Y Cryptomonas sp4 (18%) X2 Cryptomonas sp2 (29%)
X2 Cryptomonas sp2 (15%) X2 Cryptomonas sp2 (13%) F B.braunii (19%)SN C.raciborskii (18%) SN C.raciborskii (64%) SN C.raciborskii (55%) SN C.raciborskii (64%)
Ferrugem Y Dinoflagelado NI (18%) S1 P. limnetica (9%) S1 P. limnetica (12%) S1 P. limnetica (8%)
NA C.moniliforme (11%) L. redekei (6%) L. redekei (8%)
Seca Chuvas Seca
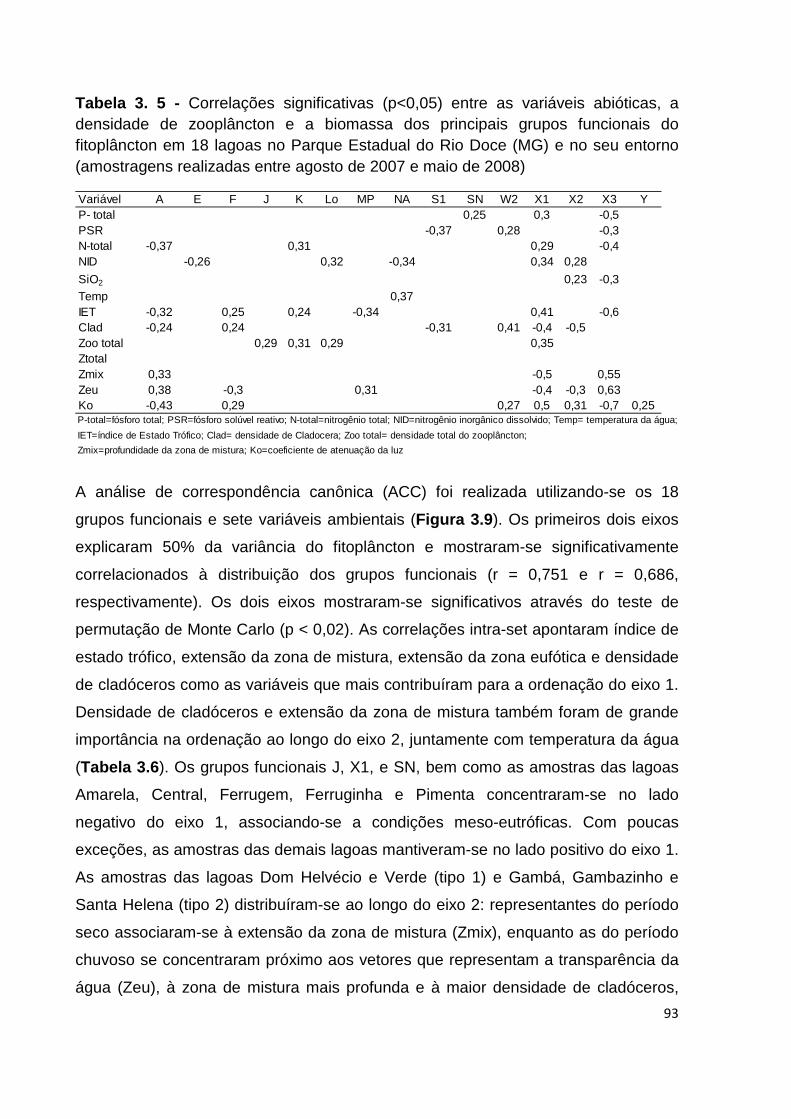
93
Tabela 3. 5 - Correlações significativas (p<0,05) entre as variáveis abióticas, a densidade de zooplâncton e a biomassa dos principais grupos funcionais do fitoplâncton em 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno (amostragens realizadas entre agosto de 2007 e maio de 2008)
Variável A E F J K Lo MP NA S1 SN W2 X1 X2 X3 YP- total 0,25 0,3 -0,5PSR -0,37 0,28 -0,3N-total -0,37 0,31 0,29 -0,4NID -0,26 0,32 -0,34 0,34 0,28
SiO2 0,23 -0,3Temp 0,37IET -0,32 0,25 0,24 -0,34 0,41 -0,6Clad -0,24 0,24 -0,31 0,41 -0,4 -0,5Zoo total 0,29 0,31 0,29 0,35ZtotalZmix 0,33 -0,5 0,55Zeu 0,38 -0,3 0,31 -0,4 -0,3 0,63Ko -0,43 0,29 0,27 0,5 0,31 -0,7 0,25P-total=fósforo total; PSR=fósforo solúvel reativo; N-total=nitrogênio total; NID=nitrogênio inorgânico dissolvido; Temp= temperatura da água;
IET=índice de Estado Trófico; Clad= densidade de Cladocera; Zoo total= densidade total do zooplâncton;
Zmix=profundidade da zona de mistura; Ko=coeficiente de atenuação da luz
A análise de correspondência canônica (ACC) foi realizada utilizando-se os 18
grupos funcionais e sete variáveis ambientais (Figura 3.9 ). Os primeiros dois eixos
explicaram 50% da variância do fitoplâncton e mostraram-se significativamente
correlacionados à distribuição dos grupos funcionais (r = 0,751 e r = 0,686,
respectivamente). Os dois eixos mostraram-se significativos através do teste de
permutação de Monte Carlo (p < 0,02). As correlações intra-set apontaram índice de
estado trófico, extensão da zona de mistura, extensão da zona eufótica e densidade
de cladóceros como as variáveis que mais contribuíram para a ordenação do eixo 1.
Densidade de cladóceros e extensão da zona de mistura também foram de grande
importância na ordenação ao longo do eixo 2, juntamente com temperatura da água
(Tabela 3.6 ). Os grupos funcionais J, X1, e SN, bem como as amostras das lagoas
Amarela, Central, Ferrugem, Ferruginha e Pimenta concentraram-se no lado
negativo do eixo 1, associando-se a condições meso-eutróficas. Com poucas
exceções, as amostras das demais lagoas mantiveram-se no lado positivo do eixo 1.
As amostras das lagoas Dom Helvécio e Verde (tipo 1) e Gambá, Gambazinho e
Santa Helena (tipo 2) distribuíram-se ao longo do eixo 2: representantes do período
seco associaram-se à extensão da zona de mistura (Zmix), enquanto as do período
chuvoso se concentraram próximo aos vetores que representam a transparência da
água (Zeu), à zona de mistura mais profunda e à maior densidade de cladóceros,
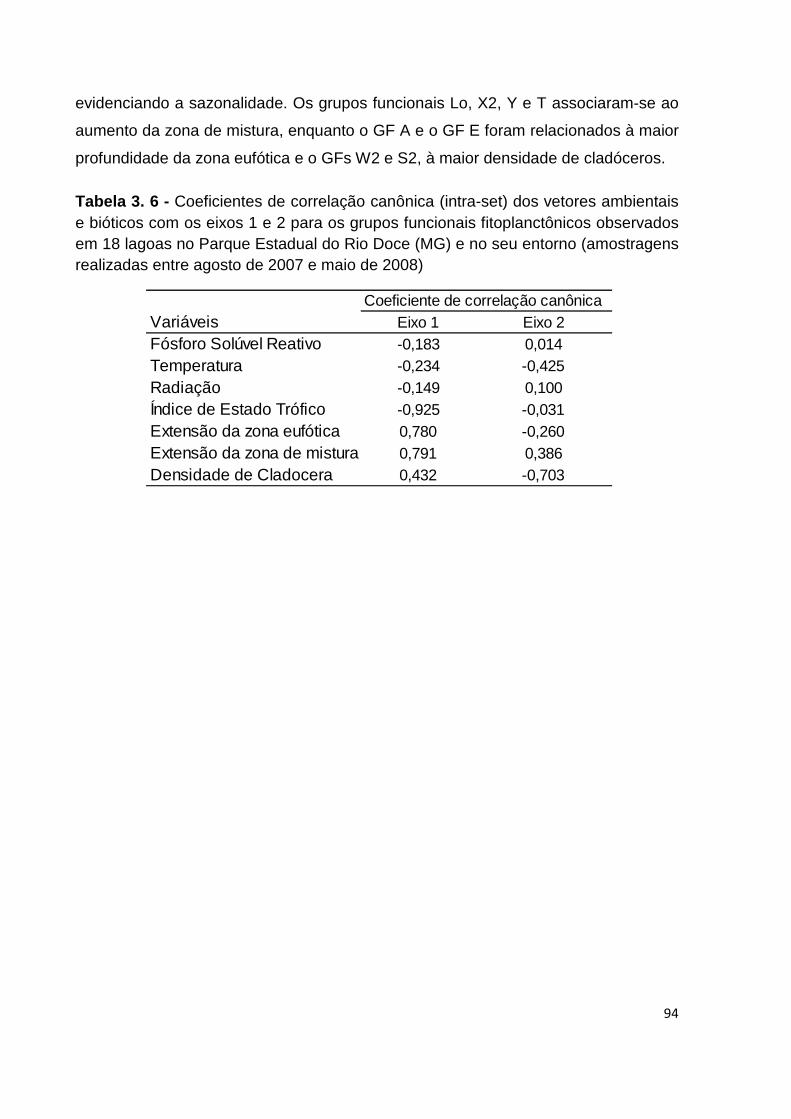
94
evidenciando a sazonalidade. Os grupos funcionais Lo, X2, Y e T associaram-se ao
aumento da zona de mistura, enquanto o GF A e o GF E foram relacionados à maior
profundidade da zona eufótica e o GFs W2 e S2, à maior densidade de cladóceros.
Tabela 3. 6 - Coeficientes de correlação canônica (intra-set) dos vetores ambientais e bióticos com os eixos 1 e 2 para os grupos funcionais fitoplanctônicos observados em 18 lagoas no Parque Estadual do Rio Doce (MG) e no seu entorno (amostragens realizadas entre agosto de 2007 e maio de 2008)
Coeficiente de correlação canônicaVariáveis Eixo 1 Eixo 2Fósforo Solúvel Reativo -0,183 0,014Temperatura -0,234 -0,425Radiação -0,149 0,100Índice de Estado Trófico -0,925 -0,031Extensão da zona eufótica 0,780 -0,260Extensão da zona de mistura 0,791 0,386Densidade de Cladocera 0,432 -0,703
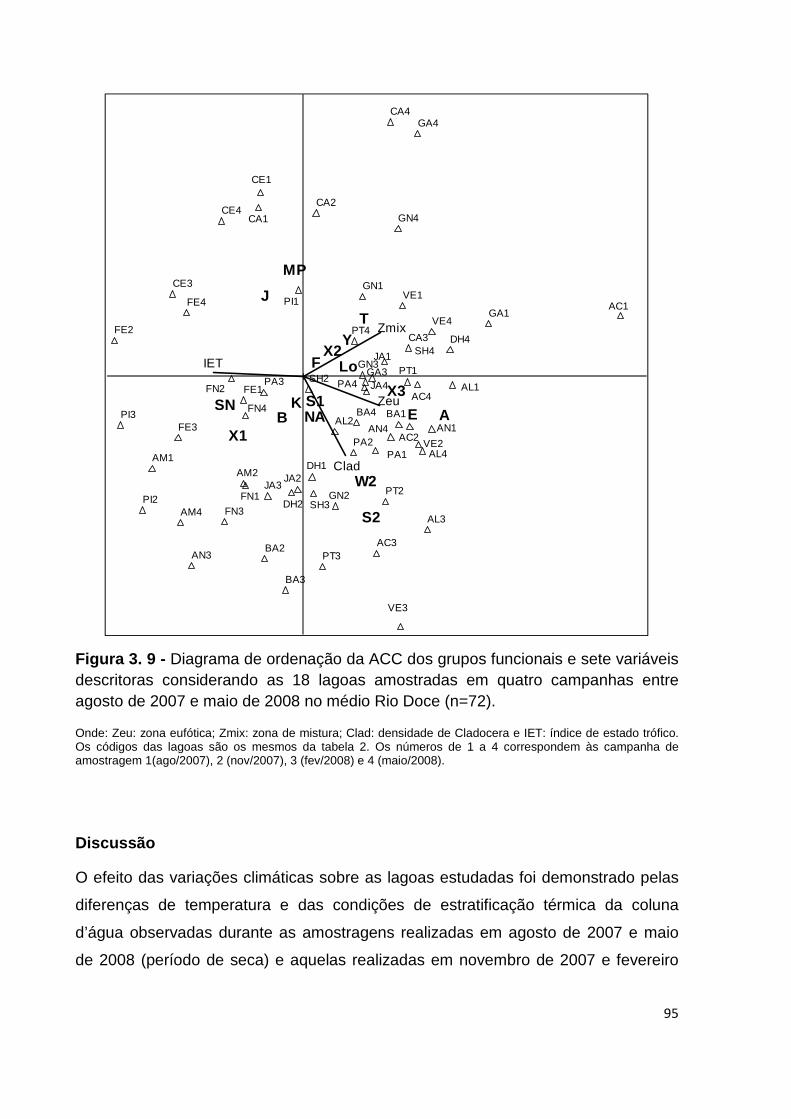
95
AC1
AC2
AC3
AC4AL1
AL2
AL3
AL4AM1
AM2
AM4
AN1
AN3
AN4
BA1
BA2
BA3
BA4
CA1
CA2
CA3
CA4
CE1
CE3
CE4
DH1
DH2
DH4
FE1
FE2
FE3
FE4
FN1
FN2
FN3
FN4
GA1
GA3
GA4
GN1
GN2
GN3
GN4
JA1
JA2JA3
JA4
PA1PA2
PA3 PA4PT1
PT2
PT3
PT4
PI1
PI2
PI3
SH2
SH3
SH4
VE1
VE2
VE3
VE4
AB E
F
J
K
Lo
MP
NAS1
S2
SN
T
W2
X1
X2
X3
Y
IET
Zeu
Zmix
Clad
Figura 3. 9 - Diagrama de ordenação da ACC dos grupos funcionais e sete variáveis descritoras considerando as 18 lagoas amostradas em quatro campanhas entre agosto de 2007 e maio de 2008 no médio Rio Doce (n=72).
Onde: Zeu: zona eufótica; Zmix: zona de mistura; Clad: densidade de Cladocera e IET: índice de estado trófico. Os códigos das lagoas são os mesmos da tabela 2. Os números de 1 a 4 correspondem às campanha de amostragem 1(ago/2007), 2 (nov/2007), 3 (fev/2008) e 4 (maio/2008).
Discussão
O efeito das variações climáticas sobre as lagoas estudadas foi demonstrado pelas
diferenças de temperatura e das condições de estratificação térmica da coluna
d’água observadas durante as amostragens realizadas em agosto de 2007 e maio
de 2008 (período de seca) e aquelas realizadas em novembro de 2007 e fevereiro

96
de 2008 (período de chuvas). A influência dos processos físicos na dinâmica do
fitoplâncton nas lagoas estudadas ficou evidente.
Embora a biomassa fitoplanctônica tenha aumentado ao longo do gradiente
profundidade total-turbidez-trofia, a maioria das amostras mostraram valores
inferiores a 3 mg.L-1, limite estabelecido por Vollenweider (1968) para ambientes
oligotróficos. A frequência de valores superiores a este limite, no entanto, aumentou
ao longo do gradiente, tendo ocorrido em 0, 2, 17 e 35% das amostras nas lagoas
tipo 1, 2, 3 e 4, respectivamente e decaindo para 10% das amostras nas lagoas tipo
5.
Picos de biomassa em níveis considerados eutróficos (>5 mg.L-1) foram registrados
nas lagoas Carioca, Aníbal, Central (~7 mg.L-1) e Ferruginha (~10 mg.L-1). Estes
resultados corroboram parcialmente aqueles obtidos para o índice de estado trófico
(IET) criado por Calrson e modificado por Toledo et al. (1983), que apontaram
condições oligotróficas nas lagoas do tipo 1 e 2, mesotróficas nas lagoas tipo 3 e 4 e
eutróficas nas lagoas tipo 5. As duas lagoas consideradas eutróficas apresentaram
situações opostas no que se refere à biomassa: enquanto na lagoa Amarela, a
biomassa fitoplanctônica manteve-se inferior a 0,3 mg.L-1, na maioria das amostras,
na Ferrugem, predominaram biomassas superiores a 2 mg.L-1. Tal diferença deve-se
ao fato da lagoa Amarela ter a maior parte do espelho d’água coberta por Nymphaea
sp. Conforme apontam Meerhoff et al. (2003) e Pinto et al. (2007), macrófitas
flutuantes podem inibir o desenvolvimento do fitoplâncton ao reduzir drasticamente a
penetração da radiação solar e os elevados valores de k0 obtidos para os períodos
de chuvas e seca confirmam a limitação por luz nessa lagoa. Na lagoa Ferrugem, as
maiores biomassas se deveram ao predomínio da cianobactéria filamentosa
Cylindrospermopsis raciborskii (GF SN), tolerante a baixas luminosidades (Bouvy et
al., 1999).
Mesmo as lagoas classificadas como eutróficas segundo o índice de Toledo et al.
1983), apresentaram biomassas fitoplanctônicas compatíveis com o nível oligotrófico
(sensu Vollenveider,1968) na maior parte do tempo. Naselli-Flores et al. (2007)
sugerem que a limitação por luz é a principal força moldando a comunidade
fitoplanctônica nos ambientes eutróficos enquanto a capacidade de absorver

97
nutrientes é mais relevante nos ambientes oligo- ou mesotróficos. De acordo com
Reynolds (1999), a concentração mínima de fósforo solúvel reativo necessária para
sustentar a taxa de crescimento do fitoplâncton é 3-6 µg.L-1. Com exceção de
Amarela, Pimenta e Patos, os teores de fósforo solúvel reativo observados nas
lagoas estudadas foram inferiores a 6 µg.L-1, indicando que as baixas concentrações
desse nutriente podem limitar o crescimento do fitoplâncton, como já sugerido por
Henry et al. (1997) e Barros et al. (2006) para o lago Dom Helvécio. Os elevados
coeficientes de atenuação da luz registrados para as duas lagoas eutróficas também
corroboram a colocação de Naselli-Flores et al. (2007).
Embora mais divulgado para ambientes grandes e profundos, o padrão monomítico-
quente prevalece nas lagoas tropicais como um todo (Townsend, 2006), onde a
estratificação térmica tende a ser mais longa e menos estável que aquela observada
para as maiores latitudes (Lewis, 2000). Com exceção de Amarela e Ferruginha, as
lagoas estudadas se mostraram estratificadas durante os períodos mais quentes
(novembro e fevereiro), evidenciando a importância das variações temporais da
temperatura para a estabilidade da coluna d’água também para ambientes rasos
(Barbosa & Padisák, 2002; Padisák & Reynolds, 2003). A seleção de espécies
depende, dentre outros fatores, das condições locais (Costa et al. 2009) e o
movimento da coluna d’água é considerado uma das principais variáveis controlando
a dinâmica do fitoplâncton, uma vez que afeta a disponibilidade dos recursos
essenciais para seu crescimento (Reynolds, 2006). Essa importância foi constatada
no presente estudo através das variações temporais observadas para a estrutura do
fitoplâncton em cada lagoa.
Somente em 4 das 18 lagoas estudados não se observou a presença de
representantes do grupo funcional NA entre as espécies descritoras (>5% da
biomassa total). Tal fato corrobora a importância do padrão monomítico-quente, e da
atelomixia em especial, neste sistema lacustre, conforme sugerido por Barbosa &
Padisák (2002), Souza et al. (2008) e Barbosa (2004; 2009) após estudos realizados
nas lagoas Carioca e Dom Helvécio. Esse grupo predominou especialmente nas
lagoas oligotróficas, profundas e claras (tipos 1 e 2) e durante o período de chuvas,
quando a coluna d’água esteve estratificada. Dentre os ambientes com estas
características, a lagoa Gambazinho foi o único em que espécies do grupo NA não

98
figuraram entre as descritoras. Apesar de apresentar-se estratificada durante as
amostragens realizadas em novembro de 2007 e fevereiro de 2008, o
monitoramento mensal realizado através do programa de Pesquisas Ecológicas de
longa Duração (PELD/UFMG) permitiu a identificação do padrão de mistura
polimítico nesta lagoa, condição que desfavorece o desenvolvimento das desmídias
(Barbosa & Padisák, 2002).
A importante contribuição do grupo funcional NA para o fitoplâncton das lagoas
Aníbal (tipo 3), Pimenta (tipo 4) e Ferrugem (tipo 5), por outro lado, demonstra a
capacidade dos organismos deste grupo em tolerar condições de menor
luminosidade. No entanto, ressalta-se que o enquadramento da lagoa Aníbal no tipo
3 deve-se ao fato do ponto de coleta localizar-se próximo à margem. A profundidade
máxima relatada em estudos anteriores foi 16 m (Calijuri et al. 1997), enquanto o
ponto de coleta apresentou somente 6 m. Assim, se a coleta fosse realizada em seu
ponto mais profundo, provavelmente esse ambiente estaria enquadrado no tipo 1.
No caso da lagoa Pimenta, Staurastrum ionatum e Staurastrum tetracerum foram
importantes espécies representando o GF NA. Experimentos realizados por
Spijkerman & Coesel (1996) indicaram que espécies desse gênero necessitam de
maior disponibilidade de fósforo que pequenas espécies de Cosmarium e este
nutriente manteve maiores concentrações nessa lagoa, especialmente durante as
amostragens de fevereiro, quando S. tetracerum representou 51% da biomassa
fitoplanctônica.
Condições de turbidez elevada são descritas como favorecedoras do grupo funcional
S1 (Reynolds et al., 2002). Membros desse grupo foram importantes na maioria das
lagoas estudadas, com as maiores biomassas associadas a estados tróficos
intermediários, mas baixas concentrações de fósforo solúvel reativo. A dominância
ou co-dominância em lagoas oligotróficas, como Dom Helvécio, Gambá e
Gambazinho, parecem desvincular a ocorrência desse grupo a ambientes eutróficos
(Krasznai et al., 2010). No entanto, seus representantes se concentraram nas
camadas mais profundas dessas lagoas, confirmando a adaptação a condições de
baixa luminosidade.

99
O grupo funcional SN (Cylindrospermopsis raciborskii), também tolerante à baixa
luminosidade (Reynolds et. al. 2002), destacou-se nas lagoas estudadas. Maiores
biomassas foram favorecidas pelo aumento da trofia, tendo sido registradas nas
lagoas Ferrugem (2 mg.L-1) e Ferruginha (1 mg.L-1). No entanto, ainda que com
baixas biomassas (< 0,5 mg.L-1), essa espécie co-dominou nas lagoas Dom Helvécio
e Santa Helena no período de seca. C. raciborskii é reconhecida como espécie
invasora (Padisák, 1997; Pivato et al. 2006) e seu maior desenvolvimento nesses
ambientes oligotróficos coincidiu com os períodos de maior atenuação da radiação
fotossinteticamente ativa (>k0) e pode ter sido favorecido pelas elevadas
temperaturas que predominam na região (Bouvy et al., 2000). Diversos trabalhos
(ex. Borics et al., 2000; Istvánovics et al., 2000; Huszar et al., 2000) descrevem a
estabilidade da coluna d’água como um importante fator beneficiando o
desenvolvimento dessa cianobactéria. No entanto, seu predomínio na lagoa
Ferruginha, que não apresentou estratificação térmica e sua maior
representatividade nas lagoas Dom Helvécio e Santa Helena em condições de
homogeneidade da coluna d’água, indicam sua capacidade de adaptação a
diferentes condições ambientais.
Os grupos funcionais A e E foram favorecidos em condições de elevada
transparência da água, ocorrendo somente nas lagoas tipo 2. Urosolenia longiseta
(GF A), mostrou-se importante nas lagoas Águas Claras, Almécega e Santa Helena,
oligotróficas e claras, como a lagoa Palmeirinha, onde Dinobryon bavaricum (E) teve
uma contribuição expressiva para a biomassa total. O menor tamanho (0,2 km2) e
profundidade (6 m) da lagoa Palmeirinha, podem ter favorecido o GF E, associado a
ambientes mais rasos e menores (Padisák et al., 2009).
Botryococcus braunii (F) e espécies grandes (Y) e pequenas (X2) de Cryptomonas
se alternaram na lagoa Amarela. Flöder et al. (2002) sugerem que a limitação por luz
seleciona espécies que toleram baixa intensidade luminosa. Espécies desses grupos
são adaptadas a viver em uma variedade de condições ambientais e toleram baixa
luminosidade (Reynolds et al,. 2002). A dominância de Cryptomonas já foi reportada
em ambientes húmicos como a lagoa Comprida (Alves-de-Souza et al., 2006) e tem-
se sugerido que a capacidade de mixotrofia (Jones, 2000) e de migração vertical,
possibilitada pelo flagelo (Jansson et al., 1996), são características que aumentam o

100
potencial competitivo do gênero em condições de limitação por luz. A dominância de
espécies desses grupos apenas em maio de 2008 pode ser explicada pela
sensibilidade à predação, uma vez que as densidades de zooplâncton total e de
cladóceros foram menores nessa ocasião. Ressalta-se ainda que Trachelomonas
volvocina (GF W2), também associada a condições eutróficas, esteve entre as
espécies descritoras dessa lagoa.
Representantes do GF F ainda se destacaram nas lagoas Jacaré, Central (B.
braunii) e Patos (B. braunii, Eutetramorus planctonicus), também pouco
transparentes. A tolerância de B. braunii à elevada turbidez (Costa et al., 2009) é
conferida pela capacidade de se manter na camada superficial devido à produção de
mucilagem e óleo e pode ter contribuído para seu estabelecimento nesses
ambientes mesotróficos. B. braunii também foi importante para a biomassa total na
lagoa Gambazinho, enquanto Dictyosphaerium pulchellum representou o mesmo
grupo na lagoa Palmeirinha. Ambas as lagoas são claras (Ko<0,8m-1), tipo de habitat
que favorece o desenvolvimento desse grupo funcional (Reynolds et al., 2002).
A ocorrência de Spirulina sp (GF S2) e Planctonema sp (GF T) entre as espécies
dominantes (> 50% da biomassa total) e Trachelomonas volvocina (GF W2) entre as
descritoras (> 5% da biomassa total) na lagoa dos Patos sugere condições de
mistura frequente da coluna d’água nesse ambiente, que tem extensos bancos de
macrófitas (Eleocharis e Nymphaea). Representantes dos grupos funcionais J, B e
W2, que também toleram baixa luminosidade, estão entre as espécies descritoras
das lagoas Central (Coelastrum reticulatum, GF J), Barra (Cyclotella sp, GF B),
Amarela e Patos (em ambas, T. volvocina, GF W2).
Estudos sobre a dinâmica do fitoplâncton têm mostrado uma forte relação entre as
condições ambientais e os grupos funcionais (ex. Crossetti & Bicudo 2008a; Bovo-
Scomparin & Train, 2008). O presente estudo corrobora essa afirmação ao
evidenciar uma grande concordância entre os grupos funcionais dominantes e as
condições ambientais. Em geral, as lagoas mais claras e oligotróficas, tipos 1 e 2,
foram dominadas pelos grupos NA e A. Grupos descritos como tolerantes à baixa
luminosidade dominaram o fitoplâncton das lagoas tipo 3 (SN, S1,MP, Y, X1, K e F),
tipo 4 ( F, S1, Y, J, SN e Lo) e tipo 5 (F, Y, X2, SN, S1, Lo). O predomínio de

101
espécies dos grupos S1 e F na lagoa Gambazinho (clara e oligotrófica) e a
importância das desmídias (GF NA) na lagoa Pimenta (escura e meso-trófica)
evidenciam ainda a importância dos padrões de mistura da coluna d’água e da
sazonalidade nesse sistema lacustre.
Como mostrado para outros sistemas (Becker et al, 2009b), não somente a
disponibilidade de recursos (luz e nutrientes) ou a estrutura física (estratificação
térmica) direcionaram a comunidade fitoplanctônica, e sim a sua interação. As
hipóteses de que os grupos funcionais respondem às variações temporais e
espaciais das condições ambientais observadas no sistema lacustre do médio Rio
Doce e de que essa abordagem fornece uma boa descrição das características
limnológicas predominantes em cada lagoa foram corroboradas, reforçando,
portanto, a eficiência dos grupos funcionais como indicadores das condições
ambientais também em lagoas naturais tropicais.

102
DISCUSSÃO GERAL
O sistema lacustre do médio Rio Doce comporta um conjunto de ecossistemas
aquáticos tropicais únicos e importantes e deve ser conservado em sua totalidade, o
que torna imprescindível o conhecimento e a manutenção dos processos inerentes
ao mesmo (Tundisi & Saijo, 1997). O presente estudo reforça a importância da
conservação dessa região, já apontada para outros grupos de organismos
(Drummond et al., 2005), também para a manutenção da vida aquática ao evidenciar
elevados níveis de diversidade fitoplanctônica em níveis locais e regional. A raridade
das espécies e a baixa similaridade entre as lagoas também chamaram a atenção, e
podem ser considerados critérios que reforçam o valor de conservação da região
(Coesel, 2001).
Embora a substituição da Mata Atlântica por silvicultura esteja, juntamente com a
introdução de espécies exóticas de peixes (piranha, Pygocentrus nattereri, e
tucunaré, Cichla cf. ocellaris), entre os principais impactos antrópicos descritos para
a região estudada, não se detectaram diferenças na riqueza de espécies ou índices
de diversidade e equitabilidade do fitoplâncton entre os ambientes localizados dentro
ou fora da Unidade de Conservação, sugerindo um estado ecológico similar. O
predomínio de desmídias, tanto em relação à riqueza quanto à abundância, além de
comprovar a importância da atelomixia, é outro forte indício das boas condições
ecológicas desses ambientes, uma vez que a maioria das espécies desse grupo é
restrita a habitats oligo- ou mesotróficos (Coesel, 1996).
Crossetti & Bicudo (2008b) salientam que as diferenças fisiológicas e morfológicas
de cada grupo funcional garantem uma heterogeneidade de atributos extremamente
necessários a seu sucesso ecológico, o que torna essa abordagem uma ferramenta
útil no monitoramento e manejo dos ecossistemas aquáticos. Vinte e nove grupos
funcionais estiveram representados nos ambientes estudados, sendo 18
representativos em termos de biomassa. Os diferentes nichos representados em
cada um deles se complementam em relação à utilização de um dado recurso
(Dukes, 2001) e podem estar conferindo uma maior resistência à invasão e maior
resiliência frente a estresses como a introdução de espécies exóticas e alterações
ambientais (Scheffer et al., 2001; Vinebrooke et al., 2004). No entanto, a ocorrência

103
de Cylindrospermopsis raciborskii em metade das lagoas estudadas e sua
dominância em algumas delas, por sua vez, alertam para a fragilidade e
susceptibilidade dessa comunidade a espécies invasoras.
Clima, profundidade, estado trófico e concentração de MODC apareceram como os
principais fatores ambientais direcionando a dinâmica do fitoplâncton neste sistema
lacustre. No entanto, essas variáveis não atuaram de maneira isolada e exerceram
um efeito sinérgico sobre o fitoplâncton ao interferirem nos processos de penetração
de luz, estratificação térmica e disponibilidade de nutrientes. Nesse contexto, deve-
se salientar que, especialmente nas lagoas do entorno do PERD, muito das
mudanças ambientais são direcionadas por ações humanas (manejo das plantações
de eucalipto e exploração turística) e a combinação de impactos como a introdução
de peixes exóticos, o aumento do aporte de nutrientes e a erosão do solo, podem
levar à perda de diversidade funcional e ao estabelecimento de estados não
desejados e não-reversíveis (Folke et al., 2004). Dessa forma, há que se enfocar
estratégias que considerem a possibilidade de conservação do sistema lacustre
como um todo, com a proposição de ações específicas para os ambientes da área
de entorno, para a manutenção da diversidade fitoplanctônica em nível regional. A
medida mais urgente é a regeneração do entorno das lagoas, respeitando o limite
mínimo de 50m estabelecido para Áreas de Preservação Permanentes (APPs) em
lagoas naturais localizadas em áreas rurais (CONAMA, 2002). Na maioria das
lagoas estudadas, a essa medida está sendo atendida e o plantio de eucalipto nas
margens foi abandonado, permitindo a regeneração da vegetação nativa.

104
Referências Bibliográficas
Abell, R., Thieme, M.L., Revenga, C., Bryer, M., Kottelat, M., Bogutskaya, N., Coad, B., Mandrak, N., Contreras-Balderas, S., Bussing, W., Stiassny, M.L., Skelton, P. Allen, G.R., Unmack, P., Naseka, A., Ng, R., Sindorf, N., Robertson, J., Armijo, E., Higgins, V.J., Heibel, T.J., Wikramanayake, E., Olson, D., López, H.L., Reis, R.E., Lundberg, J.G., Sabaj-Pérez, M.H. & Petry, P. 2008. Freshwater Ecoregions of the World: A New Map of Biogeographic Units for Freshwater Biodiversity Conservation. BioScience, 58:403–414.
Aitkenhead-Peterson, J.A., McDowell, W.H. & Neff, J.C. 2003. Sources, production and Regulation of allochthonous dissolved organic matter inputs to surface waters. In: Findley, S.E.G. & Sinsabaugh, R.L. (eds). Aquatic Ecosystems: Interactivity of Dissolved Organic Matter. Elsevier Science. USA. p. 25–70.
Allen, A.P., Whittier, T.R., Kaufmann, P.R., Larsen, D.P., O’Connor, R.J., Hughes, R.M., Stemberger, R.S., Dixit, S.S., Brinkhurst, R.O., Herlihy, A.T. & Paulsen, S.G. 1999. Concordance of taxonomic richness patterns across multiple assemblages in lakes of the northeastern United States. Canadian Journal of Fisheries and Aquatic Sciences, 56:739–747.
Alves-de-Souza, C., Menezes, M. & Huszar, V. 2006. Phytoplankton composition and functional groups in a tropical humic coastal lagoon, Brazil. Acta Botanica Brasilica, 20:701–708.
Araújo, G.J.M. & Barbosa, J.E.L. 2009. Grupos funcionais da comunidade fitoplanctônica de três ecossistemas aquáticos do trópico semi-árido, Brasil. Anais do IX Congresso de Ecologia do Brasil:1–4.
Arenowski, A.L., Lim, E.L. & Caron, D.A. 1995. Mixotrophic nanoplankton in oligotrophic surface waters of the Sargasso Sea may employ phagotrophy to obtain major nutrients. Journal of Plankton Research, 17:801–820.
Barbosa, F.A.R. & Padisák, J.G. 2002. The forgotten lake stratification pattern: atelomixis, and its ecological importance. International Association of Theoretical and Applied Limnology, 28:1385–1395.
Barbosa, F.A.R. & Tundisi, J.G. 1980. Primary production of phytoplankton and environmental characteristics of a shallow Quarternary lake at Eastern Brasil. Archiv fur Hydrobiologie, 90:139 –161.
Barbosa, F.A.R., Godinho, A.L., Gentil, J.G. & Pinto-Coelho, R.M. 1994. Limnologia e Ictiologia. In: Instituto Estadual de Florestas/ MG (Ed.). Anais do Workshop sobre pesquisas prioritárias para o Parque Estadual do Rio Doce. Marliéria, MG. p. 17–23.
Barbosa, F.A.R., Scarano, F.R., Sabará, M.G. & Esteves, F.A. 2004. Brazilian LTER: Ecosystem and biodiversity information in support of decision-making. Environmental Monitoring and Assessment, Dordrecht - The Netherlands, 90: 121–133.

105
Barbosa, L.G. 2004. Variações diurnas na composição, estrutura e biomassa da comunidade fitoplanctônica da lagoa Carioca, Parque Estadual do Rio Doce, Minas Gerais Brasil. Dissertação de mestrado. Universidade Federal de Minas Gerais.
Barbosa, L.G. 2009. Estrutura, dinâmica e adaptações de comunidades de Desmidias (Zygnemaphyceae) no período de cinco anos em dois lagos naturais (Parque Estadual do Rio Doce, MG): tendências em lago raso e profundo. Tese de Doutorado. Universidade Federal de Minas Gerais.
Barros, C.F.A., Souza, M.B.G. & Barbosa, F.A.R. 2006. Seasonal forces driving phytoplankton size structure in a tropical deep lake (Dom Helvécio Lake, South-East Brasil). Acta Limnologica Brasiliensis. 18:55–66.
Becker, V., Cardoso, L. & Huszar, V.L.M. 2009a. Diel variation of phytoplankton functional groups in a subtropical reservoir in southern Brazil during an autumnal stratification period. Aquatic Ecology, 43:285–293.
Becker, V., Huszar, V.L.M. & Crossetti, L.O. 2009b. Responses of phytoplankton functional groups to the mixing regime in a deep subtropical reservoir. Hydrobiologia, 628:137–151.
Becker, V., Huszar, V.L.M., Naselli-Flores, L. & Padisák, J. 2008. Phytoplankton equilibrium phases during thermal stratification in a deep subtropical water supply reservoir. Freshwater Biology, 53:952–963.
Beisner, B., Peres-Neto, P., Lindstro, E., Barnett, B. & Longhi, M. 2006. The role of environmental and spatial processes in structuring lake communities from bacteria to fish. Ecology, 87:2985–2991.
Bezerra-Neto, J.F., Barbosa, P.M. & Barbosa, F.A.R. 2006. Temporal variation of attenuation of the visible radiation in four lakes of Parque Estadual do Rio Doce (PERD), Minas Gerais, Brasil. Acta Limnologica Brasiliensis, 18:39–46.
Bicudo, C.E.M. & Menezes, M. 2006. Gêneros de Algas de Águas Continentais do Brasil: Chave para Identificação e Descrições. 2ª ed. Rima. São Carlos. 502pp.
Bold, H.C. & Wyne, M.J. 1985. Introduction to the Algae. 2ª Ed. Prentice-Hall. USA. 720pp.
Bonilla, S., Conde, D., Aubriot, L. & Perez, M.D. 2005. Influence of hydrology on phytoplankton species composition and life strategies in a subtropical coastal lagoon periodically connected with the Atlantic Ocean. Estuaries, 28:884–895.
Borges, P.A.F., Train, S. & Rodrigues, L.C. 2008. Spatial and temporal variation of phytoplankton in two subtropical Brazilian reservoirs. Hydrobiologia, 607:63–74.
Borics, G., Grigorsky, I., Szabó, S. & Padisák, J. 2000. Phytoplankton associations in a small hypertrophic fishpond in East Hungay during a change from bottom-up to top-down control. Hydrobiologia, 424:79–90.

106
Bouvy, M., Falcão, D., Marinho, M., Pagano, M. & Moura, A. 2000. Occurence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquatic Microbial Ecology, 23:13-27.
Bouvy, M., Molica, R.O.S., Marinho, M.M. & Beker, B. 1999. Dynamic of a toxic cyanobacterial bloom (Cylindrospermopsis raciborskii) in a shallow reservoir in the semi-arid region of northeast Brazil. Aquatic Microbial Ecology, 20:285–297.
Bouvy, M., Nascimento, S.M., Molica, R.J.R. & Ferreira, A. 2003. Limnological features in Tapacurá reservoir (northeast Brazil) during a severe drought. Hydrobiologia, 493:115–130.
Bovo-Scomparin, V.M. & Train, S. 2008. Long-term variability of the phytoplankton community in an isolated floodplain lake of the Ivinhema River State Park, Brazil. Hydrobiologia, 610:331–344.
Branco, A.B. & Kremer, J.N. 2005. The Relative Importance of Chlorophyll and Colored Dissolved Organic Matter (CDOM) to the Prediction of the Diffuse Attenuation Coefficient in Shallow Estuaries. Estuaries, 28:643–652.
Buss, D.F., Baptista, D.F. & Nessimian, J.L. 2003. Bases conceituais para a aplicação de biomonitoramento em programas de avaliação da qualidade da água de rios. Caderno de Saúde Pública, 19:465–473.
Calijuri, M.C., dos Santos, A.C.A. & Jati, S. 2002.Temporal changes in the phytoplankton community structure in a tropical and eutrophic reservoir (Barra Bonita, SP - Brazil). Journal of Plankton Research, 24:617–624.
Calijuri, M.C., Tundisi, J.G., Henry, R., Ibañez, M.S.R., Matsumura-Tundisi, T. & Rocha, O. 1997. Changes in light attenuation in fifteen lakes and their relationship with chlorophyll and particulate material during summer and winter. In: Tundisi, J.G. & Saijo, Y. (eds). Limnological Studies on the Rio Doce Valley Lakes, Brazil. Brazilian Academy of sciences, University of São Paulo, School Engineering at S. Carlos and Center for Water Resources and Applied Ecology, São Paulo, p. 83–93.
Calrson, R.E. 1977. A trophic state index for lakes. Limnology and Oceanography, 22:361–369.
Carneiro, F.M., Nabout, J.C. & Bini, L.M. 2008. Trends in the scientific literature on phytoplankton. Limnology, 9:153–158.
Carpenter, S.R., Cole, J.J., Kitchel, J.F. & Pace, M.L. 1998. Impact of dissolved organic carbon, phosphorus and grazing on phytoplankton biomass and production in experimental lakes. Limnology and Oceanography, 43:73–80.
Castellano-Solá, M. E. & F. A. R. Barbosa, 2003. Sistema lacustre do médio rio Doce, MG: caracterização morfológica de 158 lagos tropicais utilizando Sistema de Informações Geográficas. In IX Congresso Brasileiro de Limnologia: Água-rompendo fronteiras entre ciência, educação e cidadania, Juiz de Fora.

107
Cavati, B. & Fernandes, V.O. 2008. Algas perifíticas em dois ambientes do baixo rio Doce (lagoa Juparanã e rio Pequeno – Linhares, Estado do Espírito Santo, Brasil): variação espacial e temporal. Acta Scientiarium, 30:439–448.
CETEC. 1981. Pragramas de pesquisas ecológicas no Parque Estadual do Rio Doce. Fundação Centro Tecnológico de Minas Gerasi. 1º Vol. 264p. Relatório Final não publicado.
Coesel, P.F.M. 1996. Biogeography of desmids. Hydrobiologia, 336: 41–53.
Coesel, P.F.M. 2001. A method for quantifying conservation value in lentic freshwater habitats using desmids as indicator organisms. Biodiversity and Conservation, 10:177–187.
Cole, G.A. 1983. Textbook of Limnology. The C.V. Mosby Company, St. Louis. 401pp.
Colwell, R.K. 2006. Estimate S - Statistical estimation of species richness and shared species from samples. Version 8. University of Connecticut. Disponível em: http://purl.oclc.org/estimates.
CONAMA, Resolução Nº 303 de 20 de março de 2002. Dispões sobre parâmetros, definições e limites de Áreas de Preservação Permanente.
Connell, J. 1978. Diversity in tropical rain forests and coral reefs. Science, 199:1304–310.
Costa, L.S., Huszar, V.L.M. & Ovale, A.R. 2009. Phytoplankton Functional Groups in a Tropical Estuary: Hydrological Control and Nutrient Limitation. Estuaries and Coasts, 32:508–521.
Crist, T.O., Veech, J.A., Gering, J.C. & Summerville, K.S. 2003. Partitioning Species Diversity across Landscapes and Regions: A Hierarchical Analysis of α, β, and γ Diversity. The American Naturalist, 162:734–743.
Crossetti, L.O. & Bicudo, C.E.M. 2005. Structural and functional phytoplankton responses to nutrient impoverishment in mesocosms placed in a shallow eutrophic reservoir (Garça Pond), São Paulo, Brazil. Hydrobiologia, 541:71–85.
Crossetti, L.O. & Bicudo, C.E.M. 2008a. Phytoplankton as a monitoring tool in a tropical urban shallow reservoir (Garças Pond): The assemblage index application. Hydrobiologia, 610:161–173.
Crossetti, L.O. & Bicudo, C.E.M. 2008b. Adaptations in phytoplankton life strategies to imposed change in a shallow urban tropical eutrophic reservoir, Garças Reservoir, over 8 years. Hydrobiologia, 614:91–105.
De Meis, M.R.M. & Tundisi, J.G. 1997. Geomorphological and Limnological process as a basis for lake typology. The middle Rio Doce lake system. In: Tundisi, J.G. &

108
Saijo, Y. (eds), Limnological Studies in the Rio Doce Valley Lakes. Brazilian Academy of Sciences, São Carlos: p. 25–50.
Declerck, S., Vandekerkhove, J., Johansson, L., Muylaert, K., Conde-Porcuna, J. M., Van Der Gucht, K., Pérez-Martinéz, C., Lauridsen, T., Schwenk, K., Zwart, G., Rommens, W., López-Ramos, J., Jeppesen, E., Vyverman, W., Brendonck, L. & De Meester, L. 2005. Multi-group biodiversity in shallow lakes along gradients of phosphorus and water plant cover. Ecology, 86:1905–1915.
Delazari-Barroso, A., Barroso, G.F., Huszar, V.L.M. & Azevedo, S.M.F.O. 2009. Physical regimes and nutrient limitation affecting phytoplankton growth in a meso-eutrophic water supply reservoir in southeastern Brazil. Lakes & Reservoirs: Research and Management, 14:269–278.
Devercelli, M. 2010. Changes in phytoplankton morpho-functional groups induced by extreme hydroclimatic events in the Middle Parana´ River (Argentina). Hydrobiologia, 639:5–19.
Devol, A.H., Dos Santos, A., Forsberg, B.R. & Zaret, T.M. 1984. Nutrient addition experiments in Lago Jacaretinga, Central Amazon, Brazil: 2. The effect of humic and fulvic acids. Hydrobiologia, 109:97–103.
Dodson S.I., Arnott, S.E. & Cottingham, K.L. 2000. The relationship in lake communities between primary productivity and species richness. Ecology, 81:2662–2679.
Downing, A.L. & Leibold, M.A. 2002. Ecosystem consequences of species richness and composition in pond food webs. Nature 416:837–841.
Drakare, S., Blomqvist, P., Bergström, A.M. & Jansson, M. 2002. Primary production and phytoplankton composition in relation to DOC input and bacterioplankton production in humic Lake Örträsket. Freshwater Biology, 47:41–52.
Drummond et al., 2005. Biodiversidade em Minas Gerais: um atlas para sua conservação. 2. ed. Fundação Biodiversitas. Belo Horizonte. 222 p.
Dukes, J.S. 2001. Biodiversity and invasibility in grassland microcosms. Oecologia, 126: 563–568.
Falkowski, P.G., Katz, M.E., Knoll, A.H., Quigg, A. & Raven, J.A. 2004. The evolution of modern eukaryotic phytoplankton. Science, 305:354–360.
Figueredo, C.C. & Giani, A. 2001. Seasonal variation in the diversity and species richness of phytoplankton in a tropical eutrophic reservoir. Hydrobiologia, 445:165–174.
Findlay, D.L., Hecky, R.E., Kasian, S.E.M., Stainton, M.P., Hendzel, L.L. & Schindler, E.U. 1999. Effects on phytoplankton of nutrients added in conjunction with acidification. Freshwater Biology, 41:131–145.

109
Flöder, S. & Burns, C.W. 2005. The influence of fluctuating light on diversity and species number of nutrient-limited phytoplankton. Journal of Phycology, 41:950–956.
Flöder, S. & Sommer, U. 1999. Diversity in Planktonic Communities: An Experimental Test of the Intermediate Disturbance Hypothesis. Limnology and Oceanography, 44:1114–1119.
Flöder, S., Urabe, J. & Kawabata, Z. 2002. The influence of fluctuating light intensities on species composition and diversity of natural phytoplankton communities. Oecologia, 133:395–401.
Foden, J., Sivyer, D.B., Mills, D.K. & Devlin, M.J. 2008. Spatial and temporal distribution of chromophoric dissolved organic matter (CDOM) fluorescence and its contribution to light attenuation in UK waterbodies. Estuarine, Coastal and Shelf Science, 79:707–717.
Folke, C., Carpenter, S. Walker, B., Scheffer, M., Elmqvist, T., Gunderson, L. & Holling, C.S. 2004. Regime shifts, resilience, and biodiversity in ecosystem management. Annual Reviews of Ecology, Evolution and Systematics, 35:557–81.
Fonseca, B.M. & Bicudo, C.E.M. 2008. Phytoplankton seasonal variation in a shallow stratified eutrophic reservoir (Garças Pond, Brazil). Hydrobiologia, 600:267–282.
Förster, K. 1969. Amazonische Desmidieen. 1. Areal Santarém. Amazoniana, 2:5–232.
Förster, K. 1974. Amazonische Desmidieen. 2. Areal Maués-n Abacaxis. Amazoniana, 5:135–242.
Garcia, F.C. 2007. Estudos Ecológicos da cianobactéria Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju no Lago Dom Helvécio, Parque Estadual do Rio Doce, MG, Brasil. Tese de Doutorado
Garn, H.S., Elder, J.F. & Robertson, D.M. 2003. Why Study Lakes? An Overview of USGS Lake Studies in Wisconsin. USGS Science for a changing world, fact sheet 063-03:1–8.
Gemelgo, M.C.P., Mucci, J.L.N. & Navas-Pereira, D. 2009. Population dynamics: seasonal variation of phytoplankton functional groups in Brazilian reservoirs (Billings and Guarapiranga, São Paulo). Brazilian Journal of Biology, 69:1001–1013.
Golterman, H.L., Clymo, R.S. & Ohnstad, M.A.M. 1978. Methods for physical and chemical analysis of freshwaters. Blackwell Scientific Publications, Oxford. 213pp.
Green, R. E., Sosik, H.M., Olson, R.J. Contributions of phytoplankton and other particles to inherent optical properties in New England continental shelf waters. Limnology and Oceanography, 48(6): 2377–2391.
Hajnal, E. & Padisák, J. 2008. Analysis of long-term ecological status of Lake Balaton based on the ALMOBAL phytoplankton database. Hydrobiologia, 599:227–237.

110
Hammer, O., Harper, D.A.T. & Ryan, P.D. 2001. Palenontological statistics software package for education and data analysis. Paleontologia electronic, 4. 9 pp.
Harrison, S., Ross, S.J. & Lawton, J.H. 1992. Beta diversity on geographic gradients in Britain. Journal of Animal Ecology, 62:151–158.
Hehmann, A., Krienitz, L. & Koschel, R. 2001. Long-term phytoplankton changes in an artificially divided, top-down manipulated humic lake. Hydrobiologia, 448:83–96.
Henry, R. & Barbosa, F.A.R. 1989. On the thermal structure, heat content and stability in two lakes of the Rio Doce Valley Park (Minas Gerais, Brazil). Hydrobiologia, 171:189–199.
Henry, R., Tundisi, J.G., Calijuri, M.C., Ibañez, M.S.R., Mitamura, O., Matsumura-Tundisi, T., Rocha, O., Espindola, E.L.G. & Gullén, S.M. 1997. Addition of phosphate and ammonia and their effects on primary productivity by phytoplankton of Lake Dom Helvécio in experiments using large enclosures. In: Tundisi, J.G. & Saijo, Y. (eds.) Limnological Studies in the Rio Doce Valley Lakes. Brazilian Academy of Sciences, São Carlos. p.391–397.
Hillebrand, H., Dürselen, C., Kirschtel, D., Pollingher U. & Zohary, T. 1999. Biovolume calculation for pelagic and benthic microalgae. Journal of Phycology, 35:403–424.
Hino, K., Tundisi, J.G. & Reynolds, C.S. 1986. Vertical distribution of phytoplankton in a stratified lake (Lago Dom Helvécio, Southeastern Brazil), with special reference to the metalimnion. Japanese Journal of Limnology, 47:239–246.
Huszar, V.L.M. & Caraco, N. 1998. The relationship between phytoplankton composition and physical–chemical variables: a comparison of taxonomic and morphological–functional descriptors in six temperate lakes. Freshwater Biology, 40:679–696.
Huszar, V.L.M. & Reynolds, C.S. 1997. Phytoplankton periodicity and sequences of dominance in on Amazonian flood-plain lake (Lago Batata, Pará, Brazil): responses to gradual environmental change. Hydrobiologia, 346:169–181.
Huszar, V.L.M., Kruk, C. & Caraco, N. 2003. Steady-state assemblages of phytoplankton in four temperate lakes (NE USA). Hydrobiologia, 502:97–109
Huszar, V.L.M., Silva, L.H.S. & Esteves, F.A. 1990. Estrutura das comunidades fitoplanctônicas de 18 lagoas da região do Baixo Rio Doce, Linhares, Espírito Santo, Brasil. Revista Brasileira de Biologia, 50:585–598.
Huszar, V.L.M., Silva, L.H.S., Marinho M., Domingos P. &. Sant’Anna, C.L. 2000. Cyanoprokariote assemblages in eight productive tropical Brazilian waters. Hydrobiologia, 424:67–77.
Hutchinson, G. E. 1961. The paradox of plankton. American Naturalist, 95:137–147.

111
Instituto Estadual de Florestas / IEF-MG. 2002. Plano de Manejo do Parque Estadual do Rio Doce. Disponível em: http://www.ief.mg.gov.br/component/content/306?task=view
Isaksson, A., Bergstrom, A. K., Blomqvist & P. & Jansson, A. 1999. Bacterial grazing by phagotrophic phytoflagellates in a deep humic lake in notherm Sweden. Journal of Planton Research, 21:247–268.
Istvánovics, V., Shafik, H.M., Présing, M. & Szilveszter, C. 2000. Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanobacteria) in throughflow cultures. Freshwater Biology, 43:257–275.
Izaguirre, I., O’Farrell, I., Unrein, F., Sinistro, R., Dos Santos Afonso, M. & Tell, G. 2004. Algal assemblages and limnological features across a wetland, from a shallow lake to the relictual oxbow lakes (Lower Parana´ River, South America). Hydrobiologia, 511:25–36.
Jaccard, P. 1908. Nouvelles recherches sur la distribution florale. Bulletin de la Societé Vanddoise des Sciences Natureles, 44:223–270.
Jansson, M., Blomqvist, P., Jonsson, A. & Bergström, A.K. 1996. Nutrient limitation of bacterioplankton, autotrophic and mixotrophic phytoplankton, and heterotrophic nanoflagellates in Lake Örträsket. Limnology and Oceanography, 41:1552–1559.
Jones, R.I. 1992. The influence of humic substances on lacustrine planktonic food chains. Hydrobiologia, 229:73–91.
Jones, R.I. 2000. Mixotrophy in planktonic protists: an overview. Freshwater Biology, 45:219–226.
Karlsson, J., Byström, P., Ask, J., Ask, P., Persson, L. & Jansson, M. 2009. Light limitation of nutrient-poor lake ecosystems. Nature, 460:506–510.
Kindt, R. & Coe, R. 2005. Tree diversity analysis. A manual and software for common statistical methods for ecological and biodiversity studies. World Agroforestry Centre (ICRAF), Nairobi.
Kirk, J.T.O. 1994. Light and photosynthesis in aquatic ecosystems. 2ª Ed. Cambridge University Press. Cambridge. 509pp.
Komárek, J. & Anagnostidis, K. 1989. Modern aprroach to the classification system of cyanophytes, 4: Nostocales. Algological Studies, 56:247–345.
Komárek, J. & Anagnostidis, K. 1999. Cyanoprokaryota I. Teil Chroococcales. In: Ettl, H., Gärtner, G., Heynig, H. & Mollenhauer, D. (eds.). Süsswasserflora von Mitteleuropa 19. G. Fischer, Verlag, p. 1–548.
Komárek, J. & Fott, B. 1983. Chlorophyceae (Grünalgen), Ordiniung: Chlorococcales. In: Huber-Pestalozzi, G., Heynig, H. & Mollenhauer, D. (eds.). Das Phytoplankton des

112
Sübwassers: Systematik und Biologie. E. Schwiezerbat‘sche Verlagsbuchlandlung, Stuttgart. 1044pp.
Koroleff, F. 1976. Determination of ammonia. In Grasshoff, K. (ed.), Methods of seawater analysis. Verlag Chemie, Weinheim, p. 126–133.
Kostoglidis, A., Pattiaratchi, C.B. & Hamilton, D.P. 2005. CDOM and its contribution to the underwater light climate of a shallow, microtidal estuary in south-western Australia. Estuarine, Coastal and Shelf Science, 63:469–477.
Krasznai, E., Borics, G., Várbíró, G, Abonyi, A, Padisák, J, Deák, C. & Tóthmérész, B. 2010. Characteristics of the pelagic phytoplankton in shallow oxbows. Hydrobiologia, 639:173–184.
Kruk, C., Mazzeo, N., Lacerot, G. & Reynolds, C.S. 2002. Classification schemes for phytoplankton: a local validation of a functional approach to the analysis of species temporal replacement. Journal of Plankton Research, 24:901–912.
Lamparelli, M.C. 2004. Grau de trofia em corpos d’água do estado de São Paulo: avaliação dos métodos de monitoramento – São Paulo – Tese de (Doutorado) – Instituto de Biociências – USP.
Lewis, W.M. 2000. Basis for the protection and management of tropical lakes. Lakes & Reservoirs: Research and Management, 5:35–48.
Litchman, E. & Klausmeier, C.A. 2008. Trait-based community ecology of phytoplankton. Annual Reviews of Ecology, Evolution and Systematics, 39:615–639.
Lopes, M.R.M., Bicudo, C.E.M. & Ferragut, M.C. 2005. Short term spatial and temporal variation of phytoplankton in a shallow tropical oligotrophic reservoir, southeast Brazil. Hydrobiologia, 542:235–247.
Lund, J.W.G., Kipling, C. & LeCren, E.D. 1958. The inverted microscope method of estimating algal number and the statistical basis of estimation by counting. Hydrobiologia, 11:143–170.
MacArthur, R.H., Recher, H. & Cody, M. 1966. On the relation between habitat selection and species diversity. American Naturalist, 100:319–332.
Mackereth, F.J.H., Heron, J. & Talling, J.F. 1978. Water analysis and some revised methods for limnologists. New York, Freshwater Biological Association, 36. 120pp.
Maechler, M., Rousseeuw, P., Struyf, A. & Hubert, M. 2005. Cluster Analysis Basics and Extensions; unpublished.
Magurran, A.F. 2004. Measuring Biological diversity. Blackwell, Oxford. 256 pp.
Maia-Barbosa, P.M., Barros, C.F.A, Souza, M.B.G, Faria, V.R., Barbosa, L.G, Brito, S.L., Souza, R.A., Rietzler, A., Eskinazi Sant’Anna, E. & Barbosa, F.A.R. 2006. The

113
middle Rio Doce lakes, southeast Brazil: biodiversity and its controlling forces at local and regional scales. Verhandlungen des Internationalen Verein Limnologie, 29:1–5.
Marinho, M.M. & Huszar, V.L.M. 2002. Nutrient availability and physical conditions as controlling factors of phytoplankton composition and biomass in a tropical reservoir (Southeastern Brazil). Archiv für Hydrobiologie, 153:443–468.
Marques, M.M. & Barbosa, F.A.R. 2002. Áreas prioritárias para a conservação da diversidade aquática no trecho médio da bacia do Rio Doce, MG. Naturalia, 27:211–229.
McCune, B. & Mefford, M. J. 1997. Multivariate Analysis of Ecological Data. Version 3.0. MjM software, Gleneden Beach, Oregon.
Meerhoff, M., Mazzeo, N., Moss, B. & Rodriguez Gallego, L. 2003. The structuring role of free floating versus submerged plants in a subtropical shallow lake. Aquatic Ecology, 37:377–391.
Melo, S. & Huszar, V.L.M. 2000. Phytoplankton in an Amazonian floo-plain lake (Lago Batata, Brasil): Diel variation and species strategies. Journal of Plankton Research, 22:63–76.
Menezes, M., Nascimento, E.P. & Fonseca, C.G. 1995. Flora dos Estados de Goiás e Tocantins - Criptógamos volume 4: Euglenophyceae. Editora UFG. Goiânia. 77pp.
Moura, A.N., Bittencourt-Oliveira, M.C., Dantas, E.W. & Arruda Neto, J.D.T. 2007. Phytoplanktonic associations: A tool to understanding dominance events in a tropical Brazilian reservoir. Acta Botanica Brasiliensis, 21:641–648.
Nabout, J.C. & Nogueira, I. 2007. Spatial and temporal dynamics of phytoplankton functional group in a blocked valley (Brazil). Acta Limnologica Brasiliensis, 19:305–314.
Nabout, J.C., Nogueira, I.S. & Oliveira, L.G. 2006. Phytoplankton community of floodplain lakes of the Araguaia River, Brazil, in the rainy and dry seasons. Journal of Plankton Research, 28:181–193.
Nabout, J.C., Nogueira, I.S. & Oliveira, L.G. 2009. Estrutura de populações de fitoflagelados nas lagoas de inundação do rio Araguaia, Brasil. Acta Botanica Brasilica, 23:67–72.
Nabout, J.C., Nogueira, I.S., Oliveira, L.G. & Morais, R.R. 2007. Phytoplankton diversity (alpha, beta, and gamma) from the Araguaia River tropical floodplain lakes (central Brazil). Hydrobiologia, 557:455–461.
Naselli-Flores, L., Padisák, J. & Albay, M. 2007. Shape and size in phytoplankton ecology: do they matter? Hydrobiologia, 578:157–161.
Nimer, E. 1989. Climatologia do Brasil. IBGE. 421 pp.

114
Nogueira, I.S., Nabout, J.C., Oliveira, J.E. & Silva, K.D. 2008. Diversidade (alfa, beta e gama) da comunidade fitoplanctônica de quatro lagos artificiais urbanos do município de Goiânia, GO. Hoehnea, 35:219–233.
Padisák, J. & Reynolds, C.S. 2003. Shallow lakes: the absolute, the relative, the functional and the pragmatic. Hydrobiologia, 506–509:1–11.
Padisák, J. 1997. Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: worldwide distribution and review of its ecology. Archiv für Hydrobiologie, 4:563–593.
Padisák, J., Borics, G., Fehér, G., Grigoszrsky, I., Oldal, I., Schimdt, A. & Zámbóné-Doma, Z. 2003a. Dominant species, functional assemblages and frequency of equilibrium phases in late summer phytoplankton assemblages in Hungarian small shallow lakes. Hydrobiologia, 502:157–168.
Padisák, J., Soróczki-Pintér, E. & ReznerSinking, Z. 2003b. Properties of some phytoplankton shapes and the relation of form resistance to morphological diversity of plankton – an experimental study. Hydrobiologia, 500:243–257.
Padisák, J., Borics, G., Grigorszky, I. & Soróczki-Pintér, E. 2006. Use of phytoplankton assemblages for monitoring ecological status of lakes within the Water Framework Directive: the assemblage index. Hydrobiologia, 553:1–14.
Padisák, J., Crossetti, L.O. & Naselli-Flores, L. 2009. Use and misuse in the application of the phytoplankton functional classification: a critical review with updates. Hydrobiologia, 621:1–19.
Padisák, J., Hajnal, E., Naselli-Flores, L., Dokulil, M.T., Nõges, P & Zohary, T. 2010. Convergence and divergence in organization of phytoplankton communities under various regimes of physical and biological control. Hydrobiologia, 639:205–220.
Pålsson, C. & Granéli, W. 2004. Nutrient limitation of autotrophic and mixotrophic phytoplankton in a temperate and tropical humic lake gradient. Journal of Plankton Research, 26:1005–1014.
Passarge, J., Hol, S., Escher, M. & Huisman, J. 2006. Competition for nutrients and light: stable coexistence, alternative stable states, or competitive exclusion? Ecological Monographs, 76:57–72.
Petrucio, M.M. & Barbosa, F.A.R. 2004. Diel variations of phytoplankton and bacterioplankton production rates in four tropical lakes in the middle Rio Doce basin (Southeastern Brazil). Hydrobiologia, 513:71 –76.
Petrucio, M.M., Barbosa, F.A.R. & Furtado, A.L.S. 2006. Bacterioplankton and phytoplankton production in seven lakes in the middle Rio Doce, south-east Brazil. Limnologica, 36:192-203.
Pfannkuche, J. 2002. Optical properties of Otago shelf waters: South Island New Zealand. Estuarine, Coastal and Shelf Science, 55:613–627.

115
Pinto, P.T., Allende, L. & O’farrel, I. 2007. Influence of free-floating plants on the structure of a natural phytoplankton assemblage: an experimental approach. Journal of Plankton Research, 29:47–56.
Pivato, B.M., Train, S. & Rodrigues, L.C. 2006. Dinâmica nictemeral das assembléias fitoplanctônicas em um reservatório tropical (reservatório de Corumbá, Estado de Goiás, Brasil), em dois períodos do ciclo hidrológico. Acta Scientiarium, 28:19–29.
Prescott, G.W., Bicudo, C.E.M. & Vinard, W.C. 1982. A Synopsis of North American Desmids Part II. Desmidiaceae: Placodermae Section 4. University of Nebraska Press. USA. 699pp.
Prescott, G.W., Croasdale, H.T. & Vinard, W.C. 1975. A Synopsis of North American Desmids Part II. Desmidiaceae: Placodermae Section 1. University of Nebraska Press. USA. 275pp.
Prescott, G.W., Croasdale, H.T. & Vinard, W.C. 1977. A Synopsis of North American Desmids Part II. Desmidiaceae: Placodermae Section 2. University of Nebraska Press. USA. 413pp.
Prescott, G.W., Croasdale, H.T., Vinard, W.C., & Bicudo, C.E.M. 1981. A Synopsis of North American Desmids Part II. Desmidiaceae: Placodermae Section 3. University of Nebraska Press. USA. 720pp.
RAMSAR. 2010. The RAMSAR List of Wetlands of International Importance. Disponível em http://www.ramsar.org/pdf/sitelist_order.pdf
Rangel, L.M., Silva, L.H.S., Arcifa, M.S. & Perticarrari, A. 2009. Driving forces of the diel distribution of phytoplankton functional groups in a shallow tropical lake (Lake Monte Alegre, Southeast Brazil). Brazilian Journal of Biology, 69:75–85.
Revenga, C., Campbell, I., Abell, R., de Villiers, P. & Bryer, M. 2005. Prospects for monitoring freshwater ecosystems towards the 2010 targets. Philosophical Transactions of the Royal Society B, 360:397–413.
Reynolds, C.S. 1984. Phytoplankton periodicity: the interactions of form, function and environmental variability. Freshwater Biology, 14:111–142.
Reynolds, C.S. 1987. The response of phytoplankton communities to change lake environments. Schweizerische Zeitschrift fur Hydrologie, 49:220–236.
Reynolds, C.S. 1993. Scales of disturbance and their role in plankton ecology. Hydrobiologia, 249:157–171.
Reynolds, C.S. 1994. The long, the short and the stalled: on the attributes of phytoplankton selected by physical mixing in lakes and rivers. Hydrlobiologia, 289:9– 21.
Reynolds, C.S. 1997a. Vegetation Processes in the Pelagic. A Model for Ecosystem Theory. The International Ecology Institute, Oldendorf. 371pp.

116
Reynolds, C.S. 1997b. On the vertical distribution of phytoplankton in the middle Rio Doce Valley lakes. In: Tundisi, J.G. & Saijo, Y. (eds). Limnological Studies on the Rio Doce Valley Lakes, Brazil. Brazilian Academy of sciences, University of São Paulo, School Engineering at S. Carlos and Center for Water Resources and Applied Ecology, São Paulo, p. 265–274.
Reynolds, C.S. 1999. Non-determinism to Probability, or N:P in the community ecology of phytoplankton. Arch. Hydrobiol., 146:23–35.
Reynolds, C.S. 2006. The ecology of phytoplankton. Cambridge: Cambridge University Press. 552pp.
Reynolds, C.S., Huszar, V.L.M., Kruk, C., Naselli-Flores, L. & Melo, S. 2002. Towards a functional classification of the freshwater phytoplankton. Journal of Plankton Research, 24:417–428.
Rodrigo, M.A., Rojo, C., Segura, M. & Larrosa, J. 2009 Mechanisms of microalgae selection during the assembly of a planktonic community. Aquatic Ecology, 43:61–72.
Rodrigues, P. & Pizarro, H. 2007. Phytoplankton productivity in a highly colored shallow lake of a South American floodplain. Wetlands, 27:1153–1160.
Round, F. 1983. Biologia das Algas. 2a. edição. Ed. Guanabara Dois. Rio de Janeiro. 263pp.
Salmaso, N. & Padisák. J. 2007. Morpho-functional groups and phytoplankton development in two deep lakes (Lake Garda, Italy and Lake Stechlin, Germany). Hydrobiologia, 578:97–112.
Sant'Anna, C.L. 1984. Chlorococcales (Chlorophyceae) do Estado de São Paulo, Brasil. Bibliotheca Phycologica. J.Cramer. Germany. 348 pp.
Sarmento, H. & Descy, J.P. 2008. Use of marker pigments and functional groups for assessing the status of phytoplankton assemblages in lakes. Journal of Applied Phycology, 20:1001–1011.
Scheffer, M., Carpenter, S., Foley, J.A., Folke, C. & Walker, B. 2001. Catastrophic shifts in ecosystems. Nature, 413: 591–596.
Shannon, C.E. & Weaver, W. 1963. The mathematical theory of communication. Illinois University Press. Urbana. 144 pp.
Silva, C.A., Train, S. & Rodrigues, L.C. 2005. Phytoplankton assemblages in a Brazilian subtropical cascading reservoir system. Hydrobiologia, 537:99–109.
Soares, M.C.S., Huszar, V.L.M. & Roland, F. 2007. Phytoplankton dynamics in two tropical rivers with different degrees of human impact (Southeast Brazil). River Research and Applications, 23:698–714.

117
Souza, M.B.G. 2005. Flutuação temporal e avaliação do potencial invasor de Cylindrospermopsis raciborskii (Cyanoprokariota, Nostocales) em lagos do trecho médio da bacia do Rio Doce, MG. Dissertação de mestrado. Universidade Federal de Minas Gerais.
Souza, M.B.G., Barros, C.F.A., Barbosa F., Hajinal, E. & Padisák, J. 2008. Role of atelomixis in phytoplankton assemblages’ replacement in Dom Helvécio Lake, South-East Brazil. Hydrobiologia, 607:211–224.
Spijkerman, E. & Coesel, P.F.M. 1996. Competition for phosphorus among planktonic desmid species in continuous-flow culture. Journal of Phycology, 32:939–948.
Strickland, J.D.H. & Parsons, T.R. 1968. Determination of reactive nitrite. In: A practical Handbook of seawater analysis. Fisheries Research Board of Canada, Bulletin, 167:71–75.
Sun, J. & Liu, D. 2003. Geometric models for calculating cell biovolume and surface area for phytoplankton. Journal of Plankton Research, 25:1331–1346.
Ter Braak, C.J.F. 1986. Canonical correspondence analysis: A new eigenvector technique for multivariate direct gradient analysis. Ecology, 67:1167–1179.
Tilman, D. 1982. Resource competition and community structure. Princeton University Press. Princeton. 296 pp.
Toledo, A., Talarico, M., Chinez, S.J. & Agudo, E.G. 1983. A aplicação de modelos simplificados para a avaliação do processo de eutrofização em lagos e reservatórios tropicais. Poster presented in the Congresso Brasileiro de Engenharia Sanitária e Ambiental, 12, Camboriú. Anais. p. 1–34.
Townsend, S.A. 2006. Hydraulic phases, persistent stratification and phytoplankton in a tropical floodplain lake (Mary River. Northern Australia). Hydrobiologia, 556:163–179.
Tranvik, L.J & Bertilsson, S. 2001. Contrasting effects of solar UV radiation on dissolved organic sources for bacterial growth. Ecology Letters, 4:458–463.
Tundisi, J.G., Matsumura-Tundisi, T., Barbosa, F.A., Gentil, J.G., Rugani, C.A., Forte-Pontes, M.C., Aleixo, R.C., Okano, W.Y. & Santos, L.C. 1978. Estudos Limnológicos no sistema de lagos do Parque Florestal do Rio Doce, MG. Convênio CETEC-UFSCar. 45p.
Tundisi, J.G., Matsumura-Tundisi, T., Fukumura, H., Mitamura, O., Guillén, S.M., Henry, R., Rocha, O., Calijuri, M.C., Ibañez, M.S.R, Espíndola, E.L.G., Govoni, S.H. 1989. Limnology of fifteen lakes in the middle Rio Doce: results of a winter survey. In: Saijo, Y. & Tundisi, J.G. (Eds.). Limnological studies in Rio Doce lakes, Brazil. Water Research Institute, Nagoya. p. 139–159.

118
Tundisi, J.G. 1997. Climate. In: Tundisi, J.G. & Saijo, Y. (eds). Limnological Studies on the Rio Doce Valley Lakes, Brazil. Brazilian Academy of sciences, University of São Paulo, School Engineering at S. Carlos and Center for Water Resources and Applied Ecology, São Paulo, p. 7–11.
Tundisi, J.G. & Saijo, Y. 1997a. Historical rewiew of international research in the Rio Doce lakes and Brazil/Japan cooperation in the field of limnology. In: Tundisi, J.G. & Saijo, Y. (eds). Limnological Studies on the Rio Doce Valley Lakes, Brazil. Brazilian Academy of sciences, University of São Paulo, School Engineering at S. Carlos and Center for Water Resources and Applied Ecology, São Paulo, p. 3–4.
Tundisi, J.G. & Saijo, Y. 1997b. Limnological Studies in the Rio Doce Valley Lakes, Brazil. Brazil. Brazilian Academy of sciences, University of São Paulo, School Engineering at S. Carlos and Center for Water Resources and Applied Ecology, São Paulo. 513pp.
Utermöhl, H. 1958. Zur Vervollkomnung der quantitativen Phytoplankton-Methodik. Verhandlungen Internationale Vereinigung für Theoretische und Angewandte Limnologie, 9:1–38.
Veech, J.A., Summerville, K.S., Crist, T.O. & Gering, J.C. 2002. The additive partitioning of species diversity: recent revival of an old idea. Oikos, 99:3–9.
Villafañe, V.E. & Reid, F.M.H. 1995. Métodos de microscopia para la cuatificacion del fitoplancton. In: Aveal, K., Ferrario, M.E., Oliveira, E.C. & Sar, E. (Eds.) Manual de métodos ficológicos. Universidad de Concepción, Chile. P. 169–185.
Vinebrooke, R.D., Cottingham, K.L., Norberg,J., Scheffer, M., Dodson, S.I., Maberly, S.C. & Sommer, U. 2004. Impacts of multiple stressors on biodiversity and ecosystem functioning: the role of species co-tolerance. Oikos, 104:451–457.
Vollenweider, R.A. 1968. Scientific fundamentals of the eutrophication of lakes and flowing water, with particular reference to phosphorus and nitrogen as factors in eutrophication.Technical Report. OECD Paris, DAS/CSI/58-27, 1–159p.
Waide, R.B., Willig, M.R., Steiner, C.F., Mittelbach, G., Gough, L., Dodson, S.I., Juday, G.P. & Parmenter, R. 1999. The relationship between productivity and species richness. Annual Review of Ecology and Systematics, 30:257–300.
Ward, J.H. 1963. Hierarchical grouping to optimize an objective function. Journal of the American Statistic Association, 58:236–244.
Ward, J.V., Tockner, K. & Schiemer, F. 1999. Biodiversity of floodplain river ecosystems: ecotones and connectivity. Regulation River: Reservoir & Management, 11:105–119.
Wetzel, R.G. & Likens, G.E. 2000. Limnological Analysis. 2.ed. Spring-Verlag. New York. 429 pp.

119
Wetzel, R.G. 2001. Limnology – Lake and River Ecosystems. 3ª Ed. Academic Press. San Diego, USA. 1006 pp.
Wilson, M.V. & Shmida, A. 1984. Measuring Beta Diversity with Presence-Absence Data. Journal of Ecology, 72:1055–1064.