distribuiÇÃo de galhas de neopelma baccharidis...
TRANSCRIPT

DISTRIBUIÇÃO DE GALHAS DENEOPELMA BACCHARIDIS (HOMOPTERA: PSYLLIDAE)
EM BACCHARIS DRACUNCULIFOLIA (ASTERACEAE)
ANGELA CHRISTINA F. LARA e G. WILSON FERNANDES
Ecologia Evolutiva de Herbívoros Tropicais. Departamento de Biologia Geral, Caixa Postal 486,ICB/Universidade Federal de Minas Gerais -30161-970 Belo Horizonte. MG
(Com 4 figuras)
RESUMO
Neopelma,baccharidis (Homoptera: Psyllidae) induz galhas em folhas de Eaccharis dracun-culifolia (Asteraceae). No presente trabalho descrevemos relações existentes entre a abundân-
cia de galhas, tamanho da planta hospedeira e o impacto do inseto galhador no número de
folhas e tamanho dos ramos. O estudo foi realizado no Campus da Universidade Federal de
Minas Gerais, Belo Horizonte, MG. Cem arbustos de E. dracunculifolia foram medidos (al-
tura em centímetros) e tiveram seus números de galhas contados. Cinco pares de ramos vizin-
hos galhados e não galhados foram coletados de 43 dos cem arbustos, dos quais obteve-se seu
tamanho (cm) e número de folhas (jovens e adultas).
O número de galhas correlacionou-se positiva, embora fracamente, com o aumento da planta
hospedeira (y = -16.3 + 0.0304X, r2 = 0.11, p < 0.001). Ramos galhados (8.30 :t 0.44 cm, n =
135) foram menores do que ramos não galhados (9.94 :t 0.46 cm, n = 140) (t = 2.58, p <
0.01). O número de folhas maduras foi menor em ramos galhados (14.1 :t 1.3, n = 135) do
que em ramos não galhados (22.6 :t 1.4, n = 140) (t = 4.39, p < 0.001). O número de folhas
jovens foi também inferior nos ramos galhados (8.5 :t 1.1, n = 135) em relação aos ramos não
galhados (11.8 :t 1.3, n = 140) (t = 2.01, p < 0.05). Os dados obtidos indicam que o impacto
das galhas ocasiona uma redução no tamanho dos ramos atacados e em seu número de folhas.
Palavras-chave: Eaccharis dracunculifolia, galhas de insetos, herbivoria, Neopelma baccha-
ridis, padrões de distribuição.
ABSTRACT
Distribution of Galls of Neopelma baccharidis (Homoptera: Psy"idae) on
Baccharis dracunculifolia (Asteraceae)
Neopelma baccharidis (Homoptera: Psyllidae) induce ga11s on Baccharis dracunculifolia(Asteraceae) leaves. In this study we present data on the relationship between ga11 abundanceand plant height, and on the impact of the galling insect on the number of leaves and shoot
Recebido em 18 de março de 1991Aceito em 26 de março de 1993
Distribuído em 30 de novembro de 1994
Rev. Brasil. Biol., 54 (4):661-668

662 ANOELA CHRISTINA F. LARA e O. WILSON FERNANDF~"
length. The study was conducted at the Campus of the Universidade Federal de Minas Gerais,Belo Horizonte, MG. In a hundred shrubs of B. dracunculifolia we measured plant height(cm) and gall abundance. Five pars of neighbor galled and ungalled stems were collectedfrom 43 shrubs. From these, we measured stem length and number of leaves (young and ma-ture leaves).
Eleven percent of the variation in gall density was explained by variation in plant height (y =-16.3 + 0.0304X, r2 = 0.11, p < 0.001). There was a statisticaly significant difference betweenthe average length of galled (8.30 :t 0.44 cm, n = 135) and ungalled stems (9.94 :t 0.46 cm, n= 140) (t = 2.58, p < 0.01). There were fewer mature leaves on galled (14.1 :t 1.3 cm, n =
135) than on ungalled stems (22.6 :t 1.4 cm, n = 140) (t = 4.39, p < 0.001). There were also
fewer young leaves on galled (8.5 :t 1.1, n = 135) than on ungalled (11.8 :t 1.3, n = 140) stems(t = 2.01, p < 0.05). Gall induction causes a significant reduction in stem length and in the
number of leaves supported by the stem.
Key words: Baccharis dracunculifolia, distribution pattems, herbivory, insect galls, Neopelmabaccharidis.
INTRODUÇÃO idade (Karban, 1987), fenologia (Feeny, 1976;
Dirzo, 1984; Aide, 1988; Kearsley e Whitham,1989), defesas induzidas (Haukioja, 1982), com-postos secundários (Janzen, 1979), característicasmorfológicas como pubescência (Levin, 1973),esclerofilia (Coley, 1983) e hipersensibilidade dotecido do hospedeiro (Fernandes, 1990), isolada-mente ou em conjunto.
Tempo e energia são investidos por insetosherbívoros na seleção de micro-habitats em seushospedeiros. Em galhadores, a escolha do sítio deoviposição pela fêmea influencia a sobrevivênciada prole, uma vez que os estágios imaturos têmseus movimentos restritos aos limites da galha
(Fernandes, 1990; Price e! al., 1990). Assim espe-ra-se que a distribuição de galhas varie dentro eentre hospedeiros devido à mortalidade diferencial(Fernandes and Price, 1991) bem como à seleçãopela fêmea (Fernandes, 1992).
A ocorrência de galhas induz mudançasmorfofisiológicas no tecido "parasitado" e em ór-gãos e tecidos adjacentes (veja Fernandes, 1987).{}alhas fUncionam como drenadores de subst[fi-cias nutricionais de seus hospedeiros, pelo blo-queio do fluxo normal de recursos e/ou pela
mobilização (drenagem) ativa destes das partes vi-zinhas à galha (Stinner and Abrahamson, 1979;Abrahamson and McCrea, 1986). Isso possibilitao uso de algumas espécies de insetos galhadoresno controle biológico de plantas invasoras (Fer-
nandes, 1987; Dennill and Moran, 1989; Dennilland {}ordon, 1990).
Plantas são recursos heterogêneos para inse-tos herbívoros. O conjunto de aspectos particula-res de cada espécie, como tamanho, hábito, formade crescimento e variedade das formas aéreas, ouseja sua arquitetura (Lawton, 1978, 1983), defineo grau de complexidade estrutural da espécie (Ju-niper and Southwood, 1986). Tal complexidadeinfluencia a riqueza e abundância de insetos herbí-voros associados (Lawton and Sch[õder, 1977;Lawton and Price, 1979; Strong et al., 1984). As-sim sendo, árvores teriam uma maior complexida-de estrutural, abrigando uma fauna mais rica eabundante de insetos em relação a arbustos e er-vas. Entretanto, Leather (1986) e Fernandes andPrice ( 1988, 1991) têm apresentado algumas exce-ções a este padrão.
A heterogeneidade de plantas se expressa aníveis geográfico, populacional e individual. Ateoria do crescimento modular (Harper, 1977)postula serem as plantas mosaicos de genótipos efenótipos que seriam gerados principalmente apartir de mutações somáticas ocorridas em tecidomeristemático. Assim sendo, plantas poderiamapresentar resistência diferencial ao ataque de her-bívoros (Whitham and Slobodchikoff, 1981) e ariqueza e abundância das espécies de herbívorospoderia ser influenciada pela dinâmica e modo decrescimento dessa população de módulos (Whit-ham et al., 1984).
A distribuição de insetos herbívoros pode,também, ser influenciada por parâmetros como
Rev. Brasil. Biol.. 54 (4J:66J--.6/íR

DISTRIBUIÇÃO DE GALHAS DE NEOPELMA BACCHARIDIS... 663
ProcedimentosOs cem primeiros arbustos encontrados na
área de estudo foram marcados, medidos (altura) etiveram seus números totais de galhas contados. Aaltura das plantas amostradas foi tomada comomedida indireta da quantidade de recurso (brotosfoliares para oviposição) disponível ao galhador(veja Lawton, 1983). Ramos foram amostradosem 43 dos 100 indivíduos. Coletou-se 5 pares deramos vizinhos (5 galhados e 5 não galhados) aoredor de cada um dos 43 arbustos (veja Price etal., 1990); suas galhas e folhas j~vens e madurasforam contadas. As galhas foram abertas para con-tagem do número de ninfas por galha e observa-ção de possíveis parasitas e predadores.
Para análise dos dados, utilizou-se regressÕ-es lineares, comparações entre as inclinações e in-terceptos de regressões e testes t (Zar, 1984). Asmédias das medidas tomadas são apresentadas e
seguidas pelo seu erro padrão (X I EP).
RESULTADOS
o gênero Eaccharis (Asteraceae) apresentadistribuição cosmopolita, podendo ser encontradona Oceania, América do Sul e América do Norte(Palmer, 1987). Existem diversos estudos sobre afauna de insetos associada a Eaccharis, objetivan-do o conhecimento de herbívoros potenciais parao seu controle populacional (e.g. Tilden, 1961;Kraft and Denno, 1982; Boldt and Robbins, 1987;Palmer, 1987). Embora esses levantamentos já te-nham sido feitos na América do Norte, não há da-dos recentes publicados sobre a fauna de insetosherbívoros associados a Eaccharis no Brasil.
Em observações casuais em uma populaçãode E. dracunculifolia no Campus da UniversidadeFederal de Minas Gerais, os herbívoros mais fre-qUentes foram insetos galhadores [nove espéciesdistintas e não identificadas de Cecidomyiidae(Diptera) e uma espécie de Psyllidae, Neopelmabaccharidis (Homoptera: Psyllidae)]. Neopelmabaccharidis é restrita à Região Neotropical (Buck-hardt, 1987) e sua biologia e história natural são,em grande parte, desconhecidas. Todos os hospe-deiros de N. baccharidis conhecidos pertencem aogênero Eaccharis (veja Buckhardt, 1987). De ummodo geral, os psilídeos são hospedeiros específi-cos a nível de fam1lia, gênero e espécie (Buck-hardt, 1988). ' ,
"O presente estudo aborda as seguintes ques-
tões sobre a distribuição de galhas de N. bacchari-dis em E. dracunculifolia: a) quais as relaçõesexistentes entre abundância de galhas e tamanhoda planta hospedeira? b) como varia o número defolhas maduras e jovens e o tamanho dos ramosgalhados e não galhados?
MATERIAL E MÉTODOS
Área de estudo
O estudo foi realizado entre agosto e dezem-bro de 1990, no Campus da Universidade Federalde Minas Gerais, Belo Horizonte, MG, a uma alti-
tude de 805 m (19°30'8 e 44°W). A vegetação doCampus apresenta-se bastante heterogênea devidoa intervenções antr6picas, com espécies nativas decerrado, mata e brejo, juntamente com espéciesintroduzidas (veja Ferrari, 1977). A área de coletaapresenta-se em sucessão secundária com predo-mínio de R. dracunculifolia, duas outras espéciesde Raccharis, gramíneas e leguminosas de porteherbáceo e arbustivo (Ferrari, 1977).
Aspectos da história natural e biologia do
galhadorAs galhas de N. baccharidis são foliares,
verdes, glabras, fusiformes, uniloculares e comuma ninfa por galha. As fêmeas ovipõem na ner-vura principal das folhas jovens, onde inicialmen-te o tecido foliar sofre um intumescimento(hipertrofia e/ou hiperplasia), dobrando-se sobre simesmo e formando uma cápsula fusiforme (Fig.I). As paredes da galha são suculentas durantetodo o processo de desenvolvimento do psilídeo.Mesmo após a eclosão do galhador, as galhas per-manecem aderidas ao ramo, sofrendo um processogradual de dessecamento, assumindo uma aparên-
.,cla lenhosa. E bastante comum encontrar-se ga-lhas remanescentes de gerações passadas e galhasem diversas fases de desenvolvimento em umamesma planta.
O principal parasita do galhador foi uma es-pécie não identificada de Eulophidae (Hymenop-tera) (Patrício Fidalgo, com. pess.).
Distribuição e impacto das galhas
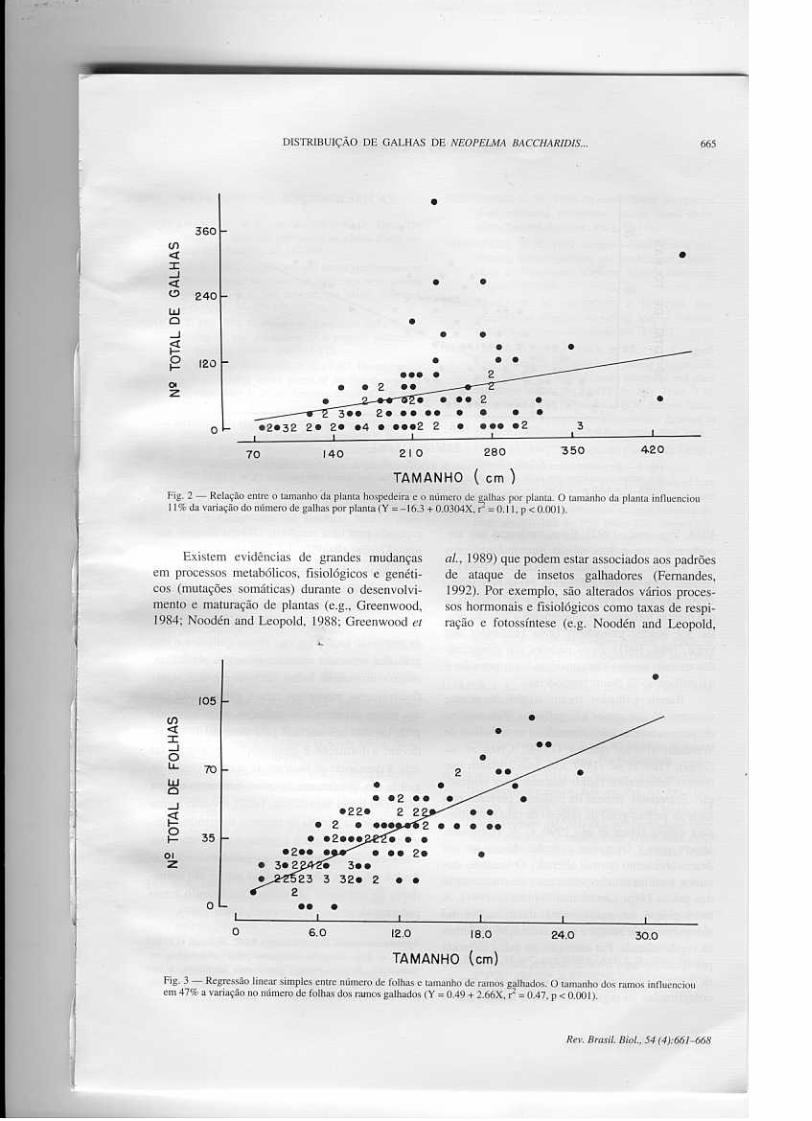
O número de galhas aumentou com a alturada planta hospedeira (y = -16.3 + 0.0304X, ~ =
0.11, p < 0.001, Fig. 2).
Ramos galhados foram significativamente
mais curtos do que ramos não galhados. O com-
Rev. Brasil. Biol.. 54 (4):661-668

664 ANOELA CHRIS11NA F. LARA e O. WILSON FERNANDES
~
~
~
~::==.
~
-E=:::~
1
,
,é'c,.
~
BA c D
Fig. I -Padrão de desenvolvimento das galhas foliares de Neopelma baccharidis em Baccharis dracunculifolia. a) seta indicaponto de inserção do ovo do psilídeo; h) galha em fase inicial de desenvolvimento. Entumescido das lâminas foIiares a partir danervura principal; c) galha madura; d) deiscência da galha.
primento médio de ramos galhados foi 8.30 cm
(:t 0.44, n = 135) e de não galhados foi 9.94 cm
(:t 0.46, n = 140) (t = 2.58, p < 0.01).
O número total de folhas por ramo variou
positivamente com o aumento no comprimento doramo nas categorias "galhado" (y = 0.49 + 2.66X,r2 = 0.47, p < 0.001, Fig. 3), e "não galhado" (y =
3.73 + 3.08X, ~ = 0.64, p < 0.001, Fig~4). Com-
parações entre as inclinações e interceptos dessas
duas regressões mostraram que elas foram signifi-
cativamente diferentes (Tangentes: t = 4.5334, gl= 270, p < 0.0001; Interceptos: t = 4.0885, gl =
271, p < 0.0001).
O número de folhas maduras e jovens foi
significativamente menor em ramos galhados. Onúmero médio de folhas maduras em ramos galha-
dos foi 14.1 (:t 1.3, n = 135) e 22.6 em não galha-
dos (:t 1.4, n = 140) (t = 4.39, p < 0.001). O
número médio de folhas jovens em ramos galha-
dos foi 8.5 (:t 1.1, n = 135) e 11.8 em ramos não
galhados (:t 1.3, n = 140) (t = 2.01, p < 0.05).
ridis. A sincronização entre fenologia da plantahospedeira e insetos galhadores está documentadana literatura (Mani, 1964; Rohfritsh and Shorthou-se, 1982; Weis et al., 1988). Galhas de N. baccha-ridis podem ser multivoltinas, hipótese essa
sugerida pela sobreposição de gerações de galhas.Embora alguns trabalhos já tenham abordadoquestões referentes a estratégias de univoltinismoe multivoltinismo em insetos galhadores (e.g.,
Mani, 1964; Yukawa, 1987), os mecanismos eprocessos envolvidos não foram totalmente escla-recidos. Os estudos de Chen and Appleby (1984)sobre a biologia de Taxodiomyia cupressiananassa(Diptera: Cecidomyiidae) em ciprestes reforçam ahipótese do multivoltinismo de galhas em plantas
que apresentam crescimento durante todo o ano(veja Weis et al., 1988).
Lawton (1983) argumenta que hospedeirosmaiores concentram densidade e riqueza mais al-tas de insetos herbívoros. Embora o número degalhas tenha aumentado com o tamanho do hospe-deiro, apenas 11% da variação na abundância degalhas foi correlacionada com a altura da planta.Outros fatores como fenologia, idade, diferenças
morfofisiológicas entre os sexos da planta e com-postos secundários devem ser levados em consi-deração na análise deste resultado, pois 89% davariação do número de galhas não foram devida-mente explicados.
DISCUSSÃO
As fêmeas de N. baccharidis ovipõem emfolhas jovens. A produção de brotos foliares de E.dracunculifolia propícios à oviposição pelo insetogalhador durante todo o ano pode ser relevante nageração dos padrões de distribuição de N. baccha-
Rev. Brasil. Biol.. 54 (4):661-668

666 ANOELA CHRISTINA F. LARA e O. WILSON FERNANDES
.
. .75(/)<t:1:-Jo~
LLJo
-J
~o1-01Z
~~~
8 8. . .
350 .2 .
...
...2 ..
25
..-
...3 .
.2 2..224 .2. ..
6 oL. .223.4..2.2.. ..2
1 I I I I I
0.0 5,0 10.0 15.0 20.0 25.0
TAMANHO ( cm )
Fig. 4 -Regressão linear simples entre número de folhas e tamanho de ramos não galhados. O tamanhodos ramos influenciou em 64% a variação no número de folhas dos ramos não galhados (y = 3.73 +
23.08X, r= 0.64, p < 0.001). ,
1988; Fay et al., 1992). Estas mudanças são im-portantes para o sucesso de insetos herbívoros(e.g.. Karban, 1987; Larsson and Ohmart, 1988) ecruciais para insetos galhadores (veja Femandes,1990; Fay et al., 1992). Variações de resistênciadas plantas influenciam diretamente a seleção des-tas por insetos galhadores (veja FerDandes and
&Price, 1988, 1991). Estes padrões são subestima-dos quando apenas são considerados o tamanho ea distribuição da planta hospedeira.
Ramos galhados foram significativamentemenores do que ramos não galhados. Este resulta-do parece, a princípio, contradizer os trabalhos deWhitham (1978), Ahman (1984), Craig et al.(1989), Price et al. (1987 a,b), que afirmam quefêmeas selecionam ramos maiores para oviposi-ção. Entretanto, fêmeas de psilídeos parecem nãofugir ao padrão geral de seleção de ramos maiorespara ovipor (Price et al., 1990, G.W. Femandes,observ. pess.). Os ramos galhados devem ter seudesenvolvimento normal alterado. O tamanho dosramos é influenciado pelo custo de manutençãodas galhas (veja Larson and Whitham, 1991). Amorfogênese da galha induz modificações noelongamento de ramos e na proliferação de folhasna região afetada. Por exemplo, na galha induzidapor Rhabdophaga strobiloides em Salix, o númerode folhas presentes em uma secção de 1 cm decomprimento na região afetada é o mesmo que o
esperado para uma secção de 150 cm de ramo nor-
mal (Ahman, 1984).
-.~..~ ,,~..."~~~ e não galhados apresentaram-se
ambas positivas. Entretanto, as diferenças estatís-
ticas entre as inclinações e interceptos destas retas
de regressão indicaram que ramos galhados e não
galhados suportam números diferentes de folhas,sendo o número de folhas maduras e jovens signi-
ficativamente menor em ramos galhados do que
em ramos vizinhos não galhados. Este resultado
devido à forn1ação e -
seja, .Tecidos de
galhas são, geralmente, ricos em nutrientes e água
(Rohfritsch and Shorthouse, 1982), levando a uma
diminuição na parcela de recursos destinados a re-
giões e órgãos vizinhos à galha. A diferença entre
o número de folhas em ramos galhados e não ga-
lhados reforça a idéia de que as galhas são drena-
dores de nutrientes do ramo, funcionando como
parasitas de plantas (Price et al., 1986, 1987).
Agradecimentos -Agradecemos a J.E.C. Figueira, H.R. pi-menta e dois revisores anônimos pelos comentários nomanuscrito. Patrício Fidalgo gentilmente identificou o eu-lofídeo parasita. Também agradecemos ao CNPq (processo
300.728/91-3) pela bolsa de pesquisa a GWF.
Rev. Brasil. Biol.. 54 (4J:661-668

DISTRIBUICÃO DE G, ,H, 'EOPELMA BACCHARlDl,Ç 66~
REFERÊNCIAS BmLIOGRÁFICAS FERNANDES, G. W., 1990, Hypersensitivity: a neglected
plant resistance mechanism against insect herbi-
vores. Envjron. Entomol., 19: 1173-1182.
FERNANDES, G. W., 1992, Adaptive distribution of ga11-
forming insects: patterns and mechanisms. Disser-
tação de Doutorado, Northern Arizona University,
F1agstaff, Arizona.
FERNANDES, G. W. and PRICE, P. W., 1988, Bio-
geographica1 gradients in ga11ing species richness:tests ofhypotheses. Oecologja, 76: 161-167.
FERNANDES, G. W. and PRICE, P. W., 1991, Compari-
sons of tropica1 and temperate ga11ing species rich-
ness: the role of environmenta1 harshness and plant
nutritiona1 status. pp. 91-115. ln: P.W. Price, T.M.
Lewinsohn, G.W. Fernandes e W.W. Benson (eds.).
Plant-Anjmallnteractions: Evolutjonary Ecology jn
Tropjcal and Temperate Regjons. Wiley, New York.
FERRARI, J. M., 1977, Vegetação do Campus da UFMG.
Oréades, 6: 3-5.
FRANKIE, G. I. and MORGAN, D. L., 1984, Role of host
plant and parasites in regulating insect herbivore
abundance, with an emphasis on ga11-inducing in-sects. pp. 101-140. ln: P.W. Price, C.N. Slobodchik-
off e W.S. Gaud (eds.). A New Ecology: Novel
Approaches to lnteractjve Systems. Wiley, New
York.
GREENWOOD, M. S., 1984, Phase change in loblolly
pine: shoot development as a function of age. Plant
Physjol., 61: 518-522.
GREENWOOD, M. S., HOPPER, C. A. and HUTCHIN-
SON, K. W., 1989, Maturation in larch. I. Effect ofage on shoot growth, foliar characteristics, and DNA
methylation. Plant Physjol., 90: 406-412.
HARPER, J. L., 1977, Populatjon Bjology of Plants. Aca-
dernic, New York.
HAUKIOJA, E., 1982, Inducible defenses of white birch toa geometric defo1iator, Epjrrita autumnata. pp. 199-
203. ln: Proc 5th lnt. Symp. Insect-Plant Relatjon-
shjps. Pudoc, Wageningen, The Netherlands.
HODKlNSON, I. D., 1974, The biology of the Psylloidea
(Homoptera): a review. Bull. ent. Res., 64: 325-339.
JANZEN, D. H., 1979, New horizons in the biology of
plant defenses. pp. 331-350. ln: Herbjvores: thejr
lnteractjon wjth Secondary Plant Metaboljtes. G.A.
Rosentha1 e D.H. Janzen (eds.). Academic, New
York.
JUNIPER, B. and SOUTHWOOD, T. R. E., 1986, lnsects
and the Plant Surface. Edward Arnold, London.
KARBAN, R., 1987, Herbivory dependent on plant age: ahypothesis based on acquired resistance. Ojkos, 43:
336-338.
KEARSLEY, M. J. C. and WHITHAM, T. G., 1989, Devel-
opmenta1 changes in resistance to herbivory: impli-cations for individuais and populations. Ecology, 70:422-434.
ABRAHAMSON, W. G. and McCREA, K. D., 1986, Theimpact of gal1s and gal1makers on p1ants. Proc. En-tomol. Soc. Wash., 88: 364-367.
AHMAN, I., 1984, Oviposition and 1arval performance ofRhabdophaga tenninalis on Salix ssp. with specialconsideration to bud size of host p1ants. Entomol.Exp. Appl., 35: 129-136.
AIDE, T. M., 1988, Herbivory as a selective agent on thetiming of leaf production in a tropical understorycommunity. Nature, 336: 574-575.
BOLDT, P. E. and ROBBINS, T. O., 1987, Phytophagousand pollinating insect fauna of Baccharis neglecta
.(Compositae) in Texas. Environ. Entomol., 16: 887-895.
BUCKHARDT, D., 1987, Jumping plant 1ice (Homoptera:Psylloidea) of the temperate Neotropical Region.
.Part 1: Psyllidae (subfami1ies Aphalarinae, Rhino-colinae and Aphalaroidinae). Zool.. J. Linn. Soc., 89:299-392.
BUCKHARDT, D., 1988, Jumping p1ant lice (Homoptera:Psylloidea) of the temperate Neotropical Region.
.Part 3: Calophyidae and Triozidae. Zool. J. Linn., Soc., 92: 115-191.
CHEN, D. and APPLEBY, J. E., 1984, Biology of the cy-
press twig gall rnidge, Taxodiomyia cupres-siananassa (Diptera: Cecidomyiidae), in centrallllinois. Ann. Entomol. Soc. Am., 77: 203-207.
COLEY, P. D., 1983, Herbivory and defensive charac-teristics of trees speciesin a low1and tropical forest.Ecol. Monog1:, 53: 209-Z34.
CRAIG, T. P., ITAMI, J. K. and PRICE, P. W., 1989,
Strong relationships between oviposition preferenceand larval performance in a shoot gal1ing sawfly.
Ecology, 70: 1691-1699.
DENNILL, G. B. and GORDON, A. J., 1990, C1imate-re-lated differences in the efficacy ofthe Australian gall
wasp (Hymenoptera- Pteromalidae) re1eased for thecontrol of Acacia longifolia in South Africa. Envi-ron. Entomol.. 19: 130-136.
DENNILL, G. B. and MORAN, V. C., 1989, On insect-plant associations in agricu1ture and the se1ection ofagents for weed biocontro1. Ann. Appl. Biol., 114:157-166.
DIRZO, R., 1984, Herbivory: A phytocentric overview. pp.141-165. ln: R. Dirzo, J. Sarukhán, (eds.). Plant
,.,.,..Population Ecology. Sinauer, Sunder1and, Massa-chussetts.
FAY, P. A., HARTNETT, D. C. and KNAPP, A. K., 1992,
Increased photosynthesis and water potentials in Sil-
phium integrifolium gal1ed by cynipid wasps. Oe-cologia (in press).
FEENY, P., 1976, P1ant apparency and chernical defense.Rec. Adv. Phytochem., 10: 1-40.
FERNANDES, G. W., 1987, Gal1-forrning insects: theireconornic importance and contro1. Rev. bras. Ento-mol.. 31. ~7<)-~<)R
Rev. Brasil. Bia 54 (4):661-668

fifiR ANGELA CHRISTINA F. ,ARA e O. WILSON FERNANDES
KRAFf, S. K. and DENNO, R. F., 1982, Feeding responsesof adapted insects to the defensive properties of Bac-charjs haljmifolja L. (Compositae). Oecologja, 52:156-163.
LARSON, K. C. and WHITHAM, T. G., 1991, Manipu1a-tion of food resource by a gal1-forming aphid: the
physio1ogy of sink-source interactions. Oecologja,88: 15-21.
LARSSON, S. and OHMART, C. P., 1988, Leaf age and1arval performance of the 1eaf beetle Paropsjsatomaria. Ecol. Entomol., 13: 19-24.
LAWTON, J. H., 1978, Host-p1ant influences on insect di-versity: the effects of space and time. Symp. R. Ento-mol. Soc. London, 9: 105-125.
LAWTON, J. H., 1983, P1ant architecture and diversity ofphytophagous insects. Ann. Rev. Entomol., 28: 23-29.
LAWTON, J. H. and PRlCE, P. W., 1979, Species richnessof parasites on hosts: agromyzid flies on British Um-be1iferae. J. Anjm. Ecol., 48: 619-637.
LAWTON, J. H. and SCHRÔDER, D., 1977, Effects ofp1ant type, size of geographical range and taxonomiciso1ation on number of insect species associated withBritish p1ants. Nature, 256: 137-140.
LAWTON, J. H. and SCHRÔDER, D., 1978, Some obser-vations on the structure of phytophagous insect com-munities: the implications for bio1ogica1 contro1. pp.57-73. ln: Proc. lVth. lnt. Symp. Bj()l. Cont. Weeds,University of Florida, Gainsville, Florida.
LEATHER, S. R., 1986, Insect species richness of the Brit-ish Rosaceae: the importance of host range, plant ar-chitecture, age of establishment, taxonomic iso1ationand species-area re1ationships. J. Anjm. Ecol., 55:841-860.
LEVIN, D., 1973, The role of trichomes in p1ant defense.Quart. Rev. Bjol., 48: 3-15.
MANI, M. S., 1964, The Ecology of Plant Galls. Dr. Junk,The Hague, The Netherlands.
NOODÉN, L. D. and LEOPOLD, A. C., 1988, Senescenceand Agjng jn Plants. Acadernic, New York.
PALMER, W. A., 1987, The phytophagous insect fauna as-sociated with Baccharis haljmifolja L. and B. ne-glecta Britton in Texas, Louisiana and northemMexico. Proc. Entomol. Soc. Wash., 89: 185-199.
PRlCE, P. W., COBB, N., CRAIG, T. P., FERNANDES, G.W., ITAMI, J. K., MOPPER, S. and PRESZLER, R.W., 1990, Insect herbivore population dynafuics ontrees and shrubs: new approaches relevant to latent
and eruptive species and life table development. pp.1-38. In: E. A. Bernays, (ed.). Insect-Plant Interac-tion. Vol. II. CRC, Boca Raton, Florida.
PRICE, P. W., ROININEN, .H. and TAHVANAINEN, J.,1987a, Plant age and attack by the bud galler, Euuramucronata. Oecologia, 73: 334-337.
PRICE, P. W., ROININEN, H. and TAHVANAINEN, J.,1987b, Why does the bud-galling sawfly, Euura mu-cronata, attack long shoots? Oecologia, 74: 1-6.
PRICE, P. W., WARING, G. L. and FERNANDES, G. W.,
1986, Hypotheses on the adaptive nature of ga11s.Proc. Entomol. Soc. Wash., 88: 361-363.
ROHFRITSCH, O. and SHORTHOUSE, J. D.,. 1982, In-sect ga11s, pp. 132-151. In: G. Kahl e J.S. Schel1(eds.). Molecular Biology of Plant Tumors. Aca-dernic, New York.
STINNER, B. R. and ABRAHAMSON, W. G., 1979, Ener-getics of the Solidago canadensis stem gal1 insect-
parasitoid guild interaction. Ecology, 60: 918-926.
STRONG, D. R., LAWTON, J. H. and SOUTHWOOD, T.R. E., 1984, Insects on Plants: Community Patternsand Mechanisms. Blackwel1, Oxford.
TILDEN, J. W., 1961, The insect associates of Baccharispilularis De Condol1e. Microentomol., 16: 149-188.
WEIS, A. E., WALTON, R. and CREGO, C. L., 1988, Re-active plant tissue sites and the population biology ofga11 makers. Ann. Rev. Entomol., 33: 467-486.
WHITHAM, T. G., 1978, Habitat selection by Pemphigusaphids in response to resource lirnitation and compe-tition. Ecology, 59: 1164-1176.
WHITHAM, T. G. and SLOBODCHIKOFF, C. N., 1981,Evolution by individua1, plant-herbivore interac-tions, and mosaics of genetic variability: the adap-tive significance of somatic mutation in plants.Oecologia, 49: 287-292.
WHITHAM, T. G., WILLIAMS, A. G. and ROBINSON,A. M., 1984, The variation principIe: individualplants as tempora1 and spatial mosaics of resistanceto rapidly evolving pest. pp. 16-51. In: P. W. Price,C. N. Slobodchikoff e W. S. Gaud, (eds.), A NewEcology: Novel Approaches to Interactive Systems.Wiley, New York.
YUKAWA, J., 1987, Life history strategies of univoltinega1l-making Cecidomyiidae (Diptera) in Japan. Phy-
tophaga, 1: 121-139.
ZAR, J. H., 1984, Bioestatistical Analysis. Prentice-Ha1I,Englewood Cliffs, New Jersey.
Rev. Brasil. Biol.. 54 (4):661-668