determinação de carga fúngica em órgãos de camundongos ... · universidade de sÃo paulo...
TRANSCRIPT

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE FILOSOFIA, CIÊNCIAS E LETRAS DE RIBEIRÃO PRETO
Departamento de Biologia
Determinação de carga fúngica em órgãos de
camundongos infectados com Paracoccidioides
brasiliensis por reação em cadeia da polimerase
quantitativa.
Marcelo Vieira Costa
Monografia apresentada à Faculdade de
Filosofia, Ciências e Letras de Ribeirão
Preto da Universidade de São Paulo
como trabalho de Conclusão de Curso
para obtenção do título de Bacharel em
Ciências Biológicas.
Ribeirão Preto - SP
– 2015 –

UNIVERSIDADE DE SÃO PAULO
FACULDADE DE FILOSOFIA, CIÊNCIAS E LETRAS DE RIBEIRÃO PRETO
Departamento de Biologia
Determinação de carga fúngica em órgãos de
camundongos infectados com Paracoccidioides
brasiliensis por reação em cadeia da polimerase
quantitativa.
Marcelo Vieira Costa
Monografia apresentada à Faculdade de
Filosofia, Ciências e Letras de Ribeirão
Preto da Universidade de São Paulo
como trabalho de Conclusão de Curso
para obtenção do título de Bacharel em
Ciências Biológicas.
Orientador: Prof. Dr.
Ademilson Panunto Castelo
Ribeirão Preto - SP
– 2015 –

Autorizo a reprodução total ou parcial deste trabalho, por qualquer meio convencional ou eletrônico, para fins de estudo ou pesquisa, desde que citada a fonte.
FICHA CATALOGRÁFICA
Costa, Marcelo Vieira Determinação de carga fúngica em órgãos de camundongos
infectados com Paracoccidioides brasiliensis por reação em cadeia da polimerase quantitativa / Marcelo Vieira Costa; Orientador Ademilson Panunto Castelo. Ribeirão Preto, 2015.
34p.:il.; 30cm. Monografia (TCC) Faculdade de Filosofia, Ciências e Letras
de Ribeirão Preto da Universidade de São Paulo, 2015. 1.Reação em cadeia da polimerase. 2.PCR em tempo real. 3.
Paracoccidioidomicose experimental. 4. Fungo. 5.Infecção

Trabalho realizado no Laboratório de Imunologia, Departamento de Biologia da
Faculdade de Filosofia Ciências e Letras de Ribeirão Preto – Universidade de São
Paulo, com o auxílio financeiro do CNPq (2175)

Dedicatória
Aos meus pais, Marcelo e Elizeth, pelo
carinho e apoio incondicionais, aos meus
avós, Luís, Antônio, Heda e Olivina, pelos
ensinamentos e amor à família, e à minha
namorada Priscila, pelo companheirismo e
por permitir momentos tão incríveis

Agradecimentos
Ao meu orientador Prof. Dr. Ademilson Panunto-Castelo, pelos ensinamentos, apoio,
amizade, paciência e humanidade, que foram elementos cruciais no desenvolvimento deste
trabalho
À Profa Dra. Maria Helena Goldman e à Profa Dra. Arlete Ap. Martins Coelho-Castelo,
pelo espaço cedido e por permitir o uso de equipamentos de seus laboratórios, contribuindo
muito para este trabalho
Aos meus pais, Marcelo e Elizeth, por serem minha fonte de admiração e inspiração, pelo
apoio e incentivo, pela dedicação e trabalho, por serem exemplo de caráter e, por fim, pelo amor
e carinho que foram tão importantes na minha formação
Ao meu avô Luís pelas pescarias, momentos hilários e esforço em reunir a família em sua casa,
que foi palco de momentos únicos. À minha avó Heda, pelas horas no piano, conversas sobre
literatura, esforço para tornar esse mundo um lugar melhor para todos. Ao meu avô Antônio,
pelas mais animadas rodas de violão, visitas ao aeroporto e oficina e divagações sobre teoria da
relatividade, astronomia, existencialismo e assuntos tão fascinantes quanto possível. À minha
avó Olivina, pelo carinho, cuidado, preocupação, bondade e comida mais gostosa que já
experimentei
À minha namorada Priscila, pela cumplicidade, simplicidade, sinceridade, carinho, incentivo,
apoio, ajuda no laboratório, pelo amor ao próximo e aos animais e, principalmente, por trazer
esperança num momento tão conturbado quanto o momento em que nos conhecemos
À amiga Taíse, pelos ensinamentos, pela ajuda com o trabalho e estudo, pelas conversas
sinceras e até broncas, sendo minha irmã dentro e fora do laboratório
Ao amigo Fabrício e Gabriela, por toda ajuda, paciência, pelos ensinamentos no laboratório e
horas de descontração
Aos amigos do grupo dois, Adriano, Assyr, Igor Alves, Igor Caetano e Murilo, pela amizade
sincera, por fazerem da vida acadêmica um período hilário, pelo apoio dentro e fora da academia
e por marcarem uma época
Aos amigos de Ribeirão Preto, Alexandre, Arthur, Christiano, Gustavo, José Humberto,
Júlio, Rodrigo Ramos, pelos momentos de lazer e risadas nos fins de semana

Aos amigos da Quadragésima Sétima Turma de Ciências Biológicas da Universidade de
São Paulo de Ribeirão Preto, pelo acolhimento e pelas horas de trabalho e descontração
Aos familiares e amigos que apoiaram de alguma forma o desenvolvimento deste trabalho
Aos técnicos Wander Ribeiro, Andréa Carla Quiapim da Silva, Susie Keiko Teixeira
Rocha, pelo auxílio prestado
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), pelo auxílio
financeiro

RESUMO
COSTA, M. V. Determinação da carga fúngica em órgãos de camundongos infectados com
Paracoccidioides brasiliensis por reação em cadeia da polimerase quantitativa. 2015. 34 f.
Monografia (Trabalho de Conclusão de Curso). Faculdade de Filosofia, Ciências e Letras de
Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto.
Espécies de fungos dimórficos do gênero Paracoccidioides causam a paracoccidioidomicose
(PCM), uma micose sistêmica que é endêmica na América Latina, sendo um problema de saúde
pública no Brasil, que abriga 80% dos casos. A infecção ocorre pela a inalação de propágulos
micelianos, que se transformam em leveduras, a 37oC, e induzem uma inflamação crônico-
granulomatosa, sendo o pulmão o órgão mais afetado. Uma parte significativa dos
conhecimentos sobre a PCM advém de estudos em modelo experimental em camundongos. Por
desenvolver uma patologia similar à PCM humana, o modelo murino é amplamente usado,
possibilitando o desenvolvimento de diversos protocolos experimentais, tais como os de
vacinas e terapias. Embora o método clássico para quantificação da carga fúngica por unidades
formadoras de colônias (UFC) em cultura microbiana seja amplamente usado nos estudos de
PCM experimental, em outros modelos de infecção se verifica uma forte tendência à inclusão
de métodos moleculares para complementar ou substituir os métodos clássicos, tal como a
reação em cadeia da polimerase quantitativa (qPCR). Este trabalho visou a padronização da
metodologia de qPCR para a quantificação de DNA genômico de P. brasiliensis obtido do
pulmão de camundongos infectados, através do gene de uma glicoproteína gênero-específica
de 43kDa (gp43). Para tanto, grupos de camundongos infectados tiveram seus pulmões
extraídos, macerados e submetidos à PCR, nested-PCR e qPCR. O baixo nível de detecção de
amplicons em todas as situações ensaiadas sugeriu que um baixo número de cópias do gene
alvo era encontrado no DNA extraído diretamente do macerado de pulmão. Buscamos, então,
isolar os fungos dos pulmões antes da extração de DNA, pelo método fenol:clorofórmio, através
da lise das células murinas com água, filtração para eliminação de matriz extracelular e fibras
do órgão, seguido por lavagem das leveduras. Com esse protocolo de isolamento de fungo do
órgão foi possível estabelecer um coeficiente de determinação muito alto (R2 ~ 0,9) entre o
número de cópias do gene de gp43 e o número de UFCs obtidos do pulmão. Esses resultados
demonstram que essa técnica é robusta e relativamente rápida e fácil de ser realizada, sendo
aplicável aos modelos de PCM experimental murina.
Palavras-chave: Reação em cadeia da polimerase. PCR em tempo real. Paracoccidioidomicose
experimental. Fungo. Infecção.

ABSTRACT
COSTA, M. V. Determination of fungal burden in the organs from Paracoccidioides
brasiliensis-infected mice by quantitative polymerase chain reaction. 2015. X f. Monografia
(Trabalho de Conclusão de Curso). Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto,
Universidade de São Paulo, Ribeirão Preto.
Species of dimorphic fungi of the genus Paracoccidioides cause the paracoccidioidomycosis
(PCM), a systemic mycosis that is endemic in Latin America. PCM is a public health problem
in Brazil, which has 80% of all PCM cases in the world. The infection occurs by inhalation of
mycelial propagules. At 37°C, the mycelial form is tran sformed in yeast, which induce a
chronic-granulomatous inflammation, mainly in the lungs. Because mouse develops pathology
similar to human PCM, this murine model is widely used and responsible for significant part of
the knowledge about PCM. Therefore, the model enables the development of various
experimental protocols, such as those related to vaccines and therapies. Although the classical
method for evaluation of fungal burden by quantification of colony forming units (CFU) in
microbial culture is widely used in studies of experimental PCM, currently, in other models of
infection, there is a strong tendency for the inclusion of molecular methods to complement or
replace the traditional methodology, such as the quantitative polymerase chain reaction (qPCR).
In this study, we aim to standardize a qPCR to quantify the genus-specific gene of the
glycoprotein of 43kDa (gp43) from the genomic DNA of P. brasiliensis in the lungs of infected
mice. So mice infected with yeast had their lungs extracted, homogenized and submitted to
PCR, nested-PCR and qPCR. The low level detection of amplicons in all tested situations
suggested that there was a low number of copies target gene in the genomic DNA extracted
directly from lung homogenates. Then we sought to isolate fungi from the lungs before DNA
extraction with phenol:chloroform through the lysis of murine cells with water, filtration to
remove extracellular matrix and organ fibers, and washing of yeast. This protocol of yeast
isolation allowed establishing a very high correlation coefficient (R2 ~ 0.9) between the number
of copies of the gp43 gene and the number of CFU obtained from the lungs. These results
demonstrate that this technique is robust and relatively quick and easy to perform, and applies
to PCM murine experimental models.
Keywords: Polymerase chain reaction. Real-time PCR. Experimental paracoccidioidomycosis.
Fungus. Infection.

SUMÁRIO
1. INTRODUÇÃO ............................................................................................................................. 1 1.1. Paracoccidioidomicose e seu agente etiológico ................................................................... 1 1.2. PCR ....................................................................................................................................................... 5 1.3. Gp43 ..................................................................................................................................................... 7 1.4. Quantificação de fungos dos órgãos de animais infectados ............................................ 8
2. OBJETIVOS ................................................................................................................................... 9
3. MATERIAL E MÉTODOS ........................................................................................................... 9 3.1. Animais ............................................................................................................................................... 9 3.2. Infecção dos camundongos com leveduras de P. brasiliensis ......................................... 9 3.3. Isolamento de leveduras de P. brasiliensis a partir do pulmão ................................... 10 3.4. Extração de DNA genômico de P. brasiliensis ..................................................................... 10 3.5. Eletroforese ..................................................................................................................................... 11
3.5.1. Gel de agarose ........................................................................................................................................ 11 3.5.2. Gel de poliacrilamida ........................................................................................................................... 11
3.6. PCR ..................................................................................................................................................... 11 3.6.1. Convencional: ......................................................................................................................................... 11 3.6.2. qPCR ........................................................................................................................................................... 12
3.7. Cálculo do número de cópias .................................................................................................... 13
4. Resultados ................................................................................................................................ 13 4.1. Padronização dos iniciadores e teste de especificidade da PCR .................................. 13 4.2. Correlação entre quantidade de DNA genômico de P. brasiliensis e Cq .................... 15 4.3. qPCR de amostras de DNA extraídas de pulmão de camundongos infectados com
P. brasiliensis .............................................................................................................................................. 16 4.4. qPCR de camundongos infectados com quantidades diferentes de leveduras de P. brasiliensis ................................................................................................................................................... 17 4.5. qPCR de camundongos infectados utilizando iniciadores de -actina fúngica....... 18 4.6. Nested-qPCR .................................................................................................................................... 21 4.7. Extração de DNA de leveduras isoladas do pulmão de camundongos infectados por de P. brasiliensis ................................................................................................................................ 24 4.8. qPCR de DNA de leveduras de camundongos infectados com P. brasiliensis .......... 25 4.9. qPCR de camundongos infectados com P. brasiliensis ..................................................... 27
5. Discussão .................................................................................................................................. 28
6. Conclusão ................................................................................................................................. 31
7. Referências .............................................................................................................................. 32

1
1. INTRODUÇÃO
1.1. Paracoccidioidomicose e seu agente etiológico
A paracoccidioidomicose (PCM) é uma micose sistêmica causada por fungos do gênero
Paracoccidioides, tais como P. brasiliensis e P. lutzi (Matute et al., 2006; Carrero et al., 2008;
Theodoro et al., 2008; Teixeira et al., 2009). Similarmente a outros fungos dimórficos, tais
como Histoplasma capsulatum, Blastomyces dermatitidis, Coccidioides spp., fungos do gênero
Paracoccidioides pertencem ao filo Ascomycota, classe Euromycetes, ordem Onygenales e
família Ajellomycetaceae (Onygenaceae). O dimorfismo desses fungos está condicionado à
temperatura, uma vez que crescem como micélio à temperatura de 25oC e leveduras em
temperaturas entre 35 e 37oC (Bagagli et al., 2008).
Figura 1. Microscopia óptica da forma miceliana (A) e leveduriforme (B) de P. brasiliensis
Apesar de a PCM ser endêmica em uma extensa área da América Latina (Brummer et
al., 1993), com um número relativamente alto de casos na Venezuela, Colômbia e norte da
Argentina, certamente, o problema é mais sério no Brasil, por abrigar cerca de 80% dos casos
(Brummer et al., 1993; Sifuentes-Osornio et al., 2012). A PCM é encontrada em todas as
regiões do território brasileiro, muito embora, no Nordeste, tenham sido relatados apenas casos
esporádicos da doença. A PCM representa um importante problema de saúde pública devido ao
seu alto potencial incapacitante, que está correlacionado ao comprometimento da função
pulmonar e à extensão das áreas endêmicas de valor econômico-produtivo (Shikanai-Yasuda et
al., 2006). Outro dado que chama atenção é taxa de mortalidade que supera a da leishmaniose,
2
sendo a PCM a oitava causa mais comum de morte entre as doenças infecciosas crônicas e
recorrentes (Coutinho et al., 2002). Os dados de mortalidade e diagnóstico positivo são
subestimados, uma vez que a PCM não e uma doença de notificação compulsoria. Assim, não
há dados precisos sobre a incidência no Brasil. Além disso, muitos dos afetados fazem parte de
uma população que trabalha no campo, particularmente desprivilegiados economicamente. Isso
se deve, em parte, à falha na mobilização de recursos e investimentos para tal segmento social
no setor da saúde (Coutinho et al., 2002; Ministério da Saúde, 2009).
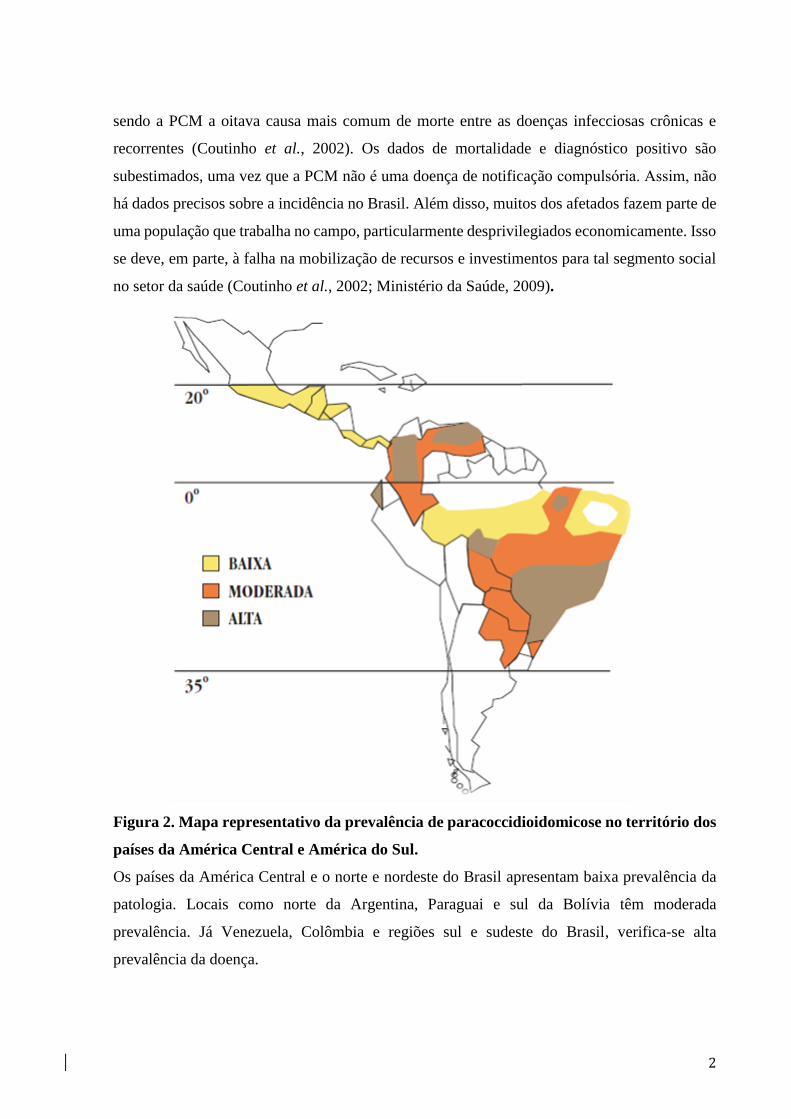
Figura 2. Mapa representativo da prevalência de paracoccidioidomicose no território dos
países da América Central e América do Sul.
Os países da América Central e o norte e nordeste do Brasil apresentam baixa prevalência da
patologia. Locais como norte da Argentina, Paraguai e sul da Bolívia têm moderada
prevalência. Já Venezuela, Colômbia e regiões sul e sudeste do Brasil, verifica-se alta
prevalência da doença.

3
A ecologia de P. brasiliensis ainda não é muito bem elucidada. Porém alguns estudos
mostram que o fungo é capaz de se desenvolver no solo. Através de análises moleculares, foi
possível detectar esses fungos em tatu, preá, guaxinim, porco espinho e furão, sendo esses
apontados como os prováveis hospedeiros silvestres de Paracoccidoides (Richini-Pereira et al.,
2008). Apesar dos esforços, o isolamento do fungo a partir do solo limita-se a algumas poucas
tentativas. Essa esporádica observação da ocorrência natural do fungo no ambiente torna difícil
a realização de inferências acerca do real micronicho ocupado. Uma série de estudos analisou
variáveis ecológicas atreladas à dados epidemiológicos de pacientes com PCM ou de pessoas
simplesmente infectadas com o fungo (teste de pele com paracoccidioidina). Conforme
revisado por Restrepo et al. (2001), existe um padrão de características ecológicas associadas à
ocorrência de infecção por Paracoccidoides. Dentre estas, verifica-se ocorrência de exposição
ao fungo em regiões desmatadas, próximas a cursos d’água, de oscilação térmica anual entre
17 e 24oC, localizadas em altitudes entre 47 e 1.300m, pluviosidade moderada, de cultivo de
tabaco e café. A verificação do fungo em tatus da espécie Dasypus novemcinctus mostrou
correlação com a observação de casos de PCM em humanos em certos estados do Brasil como
Pará, São Paulo e Minas Gerais (Restrepo et al., 2001).
A infecção por Paracoccidioides se inicia pela inalação de propágulos infectantes
(conídios ou fragmentos micelianos) da forma saprofítica do fungo, que cresce na forma
filamentosa à temperatura ambiente. Uma vez que o conídio se encontre no trato respiratório
do animal, acontece a modificação para levedura que é a forma patogênica do fungo.
Curiosamente, a infecção é debelada quando não há a termo-conversão dos fungos de forma
filamentosa para a de leveduras. Esse fenômeno foi bem relatado quando se estudou a
resistência à infecção que as mulheres têm após a adolescência, pois estrógenos agem no fungo
na forma filamentosa e impedem que se transforme em leveduras. Esse é o motivo pelo qual a
razão entre pacientes com PCM é de 10 a 15 homens para cada mulher infectada (Restrepo et
al., 1984; Borges-Walmsley et al., 2002). Ao menos nos pacientes, a imunidade inata é incapaz
de restringir o crescimento fúngico. As primeiras células a capturar os fungos são macrófagos
residentes, no entanto, a parede das leveduras de Paracoccidioides é composta principalmente
por -glucanas que não é substrato para enzimas macrofágicas (San-Blas e San-Blas, 1977;
Brummer et al., 1993; Borges-Walmsley et al., 2002). As leveduras se replicam e geram uma
inflamação crônica. A geração de resposta imunológica celular é um fenômeno importante para
evitar a disseminação do fungo, uma vez que, a formação de granulomas, estruturas esféricas
bem organizadas e formadas por macrófagos e linfócitos, que circundam o foco infeccioso,

4
impedem sua dispersão (De Brito e Franco, 1994). A depender de fatores do hospedeiro, tais
como resposta imunológica, background genético, idade, e do fungo, como virulência, tamanho
do inóculo, etc, a infecção por Paracoccidioides pode variar de assintomática a uma infecção
sistêmica (Borges-Walmsley et al., 2002; Franco et al., 1987).
Na maioria dos casos, a PCM afeta primariamente os pulmões, porém, em alguns casos
pode se disseminar para outros órgãos (Franco, 1987; Brummer et al., 1993). As formas clínicas
da PCM são decorrentes, em grande parte, do tipo de resposta imunológica do paciente. Muitos
indivíduos infectados por Paracoccidioides desenvolvem a infecção assintomática, mostrando
um certo grau de resistência ao fungo. Entretanto, outros indivíduos são susceptíveis e
desenvolvem a PCM (Franco, 1987; Borges-Walmsley et al., 2002). Quando a resposta
imunológica é menos eficiente, há um pior prognóstico e recorrências mais frequentes da
doença, gerando doença disseminada, como ocorre nas formas agudas e subagudas e na forma
crônica grave (Fortes et al., 2011). Mais de 90% dos pacientes com PCM apresentam a forma
crônica da doença. Esta caracteriza-se por progressão lenta e silenciosa, podendo levar anos para
ser diagnosticada. Tal forma é marcada por envolvimento pulmonar, muco-cutâneo e das adrenais.
A forma aguda da PCM é uma forma clínica de evolução mais rápida, na qual há acometimento de
indivíduos jovens, com envolvimento de linfonodos, fígado, baço, ossos, glândulas supra-renais
e medula óssea (Franco, 1987; Shikanai-Yasuda et al., 2006).
Grande parte do conhecimento sobre imunidade à Paracoccidioides são decorrentes de
estudos em animais. Através de modelo experimental de infecção por P. brasiliensis em
camundongos susceptíveis (B10/A) e resistentes (A/Sn) se evidenciou que macrófagos de
animais susceptíveis secretam altos níveis de TGF- e baixos níveis de TNF-, enquanto
linfócitos T CD4 secretam baixas concentrações de IFN- e altas de IL-5 e IL-10. Já nos
camundongos resistentes, constatou-se perfil oposto de secreção de citocinas por linfócitos T
CD4, com produções precoce de IFN- e IL-2 e tardia de IL-4 e IL-5. Desse modo, consolidou-
se a ideia de que a resistência à infecção por Paracoccidioides esteja atrelada à produção de
citocinas por linfócitos T CD4 do tipo helper (Th) 1, qual seja, altos níveis de TNF- e IFN-.
Por outro lado, a pequena produção de TNF- e IFN-, associada à produção de IL-5, IL-10 e
TGF- (linfócitos Th2), ocorre na forma aguda e subaguda da PCM, como a que acomete os
animais susceptíveis (Calich e Kashino, 1998; Kashino et al., 2000). Alguns estudos têm
demonstrado de forma clara a participação de TNF- e IFN- no desenvolvimento de uma
resposta imunitária protetora à infecção experimental por P. brasiliensis (Cano et al., 1998;
Souto et al., 2000; Calvi et al., 2003).

5
A reação granulomatosa na PCM humana e experimental desempenha um papel
importante na defesa do hospedeiro contra o fungo. A IL-10 e o NO parecem ser importantes
na regulação da formação do granuloma (Diniz et al., 2001). Em pacientes, a PCM, em sua
forma aguda, apresenta padrão da resposta inflamatória granulomatosa difusa, que está
associada à supressão de células T (Diniz et al., 1999). Nesses casos, o mecanismo pelo qual o
fungo evade da resposta imunológica do hospedeiro e contribui para a forma disseminada da
PCM está associado à produção de citocinas anti-inflamatórias, tais como IL-10 e TGF-β, nos
linfonodos dos pacientes (Neworal et al., 2003). Além disso, há necessidade de uma cinética de
produção de citocinas, e se o fungo induzir, precocemente, a síntese de citocinas com atividade
supressora e anti-inflamatória, tais como TGF-β e IL-10, resultará na supressão da resposta dos
macrófagos, permitindo a instalação e reprodução do fungo nos tecidos e sua disseminação para
vários órgãos (Benard, 2008).
Em termos de padrões de resposta imunológica e regulação na PCM em humanos, tem-
se evidenciado que pacientes com a doença ativa tem a supressão da resposta celular
caracterizada pela diminuição da síntese de citocinas Th1, tais como, IL-2 e IFN- e aumento
nos níveis de IL-4, IL-10 e IL-5 da resposta Th2, que não protege o hospedeiro (Benard et al.,
2001). Indivíduos com PCM que vivem em áreas endêmicas e que não desenvolvem a doença
mostram um padrão de resposta Th1 evitando a replicação fúngica e mantendo um balanço
entre o parasita e o hospedeiro, enquanto que os pacientes com a forma aguda e subaguda
desenvolvem a resposta Th2. Já em doentes com a forma crônica de gravidade moderada
apresentam uma resposta imunológica intermediária entre os padrões Th1 e Th2 (Mamoni;
Blotta, 2005). Recentemente, um outro padrão de resposta inflamatória tem sido caracterizado
(Th17), sendo estabelecido que uma doença mais branda está associada com um padrão misto
de resposta entre Th1 e Th17 (CASTRO et al., 2013).
1.2. PCR
A reação em cadeia da polimerase (PCR – do inglês polymerase chain reaction) é uma
técnica de biologia molecular que se baseia na amplificação numérica de um fragmento de DNA
de interesse. A partir dessa técnica, amplifica-se em progressão geométrica uma sequência de
DNA de modo a se obter muitas cópias de um molde (template) a partir de uma mistura
complexa de DNA. As raízes da estória da técnica PCR moderna se iniciam com a descoberta
da enzima Taq DNA polimerase, que atua e mantem sua atividade em altas temperaturas. Tal
enzima foi purificada a partir da bactéria extremófila Thermus aquaticus, o que foi responsável

6
por grande avanço da técnica. A reação se inicia com a desnaturação (ou separação) da dupla
fita do fragmento a ser sintetizado, normalmente a 94oC. Em seguida, diminui-se a temperatura
para ocorrer o anelamento de duas sequências oligonucleotídicas curtas de DNA, conhecidas
como iniciadores (primers), complementares ao fragmento de interesse. Esses iniciadores
anelam em sequências complementares à sua sequência presentes na fita senso e antissenso do
molde, delimitando assim, os limites do produto da reação (amplicon). Subsequentemente,
normalmente a 72oC, a DNA polimerase reconhece as regiões aneladas com os iniciadores
(dupla fita de DNA) e dá início à polimerização nucleotídica (extensão) no sentido 5’→3’, o
que resultará numa fita de DNA idêntica ao fragmento flanqueado pelos iniciadores. Tais etapas
descritas são marcadas por ciclagem das temperaturas de desnaturação, anelamento e extensão.
O resultado da reação é um aumento geométrico do número de moléculas de DNA da sequência
de interesse de acordo com o número de ciclos realizados. As aplicações desta técnica de
replicação in vitro são numerosas e diversas, tangendo: diagnóstico microbiológico,
identificação de patógenos, detecção de DNA com baixo número de cópias, detecção de
mutações e polimorfismos, análise filogenética, dentre muitas outras. Pode-se dizer que o
desenvolvimento e aperfeiçoamento da técnica de PCR contribuíram de forma impactante para
pesquisa clínica, forense, estudos evolucionários, projeto genoma, biotecnologia, patologia de
planta, etc (Bustin, 2010).
Uma das variações da técnica de PCR consiste na quantificação dos amplicons obtidos
em cada ciclo da reação, sendo a técnica conhecida como PCR quantitativa (qPCR - quantitative
polimerase chain reaction). Tal técnica faz uso moléculas fluorescentes que reportam um
aumento de sinal de fluorescência que é emitida, sendo correlacionado com aumento na
quantidade de DNA da reação. Inicialmente, a fluorescência se mantem a níveis basais, de modo
que o acúmulo de amplicons não é detectável. Na fase exponencial, a quantidade de produtos
de PCR, aproximadamente, duplica a cada ciclo. Eventualmente, o produto amplificado atinge
um nível de fluorescência cujo sinal é detectável. O número do ciclo em que tal evento ocorre
é chamado de Ciclo de Quantificação (Cq – do inglês quantification cycle) (Bustin et al. 2009).
Com o prosseguimento da reação, os componentes da reação, são consumidos ao ponto de não
haver mais amplificação considerável. Essa fase é conhecida como platô não exponencial. Uma
vez que os valores de Cq são medidos numa fase em que os reagentes não são limitantes, tem-
se que a fase exponencial é adequada para se inferir a quantidade inicial de DNA presente numa
reação de qPCR. O Cq da reação é determinado pela quantidade de molde de DNA inicial
presente na amostra, alta quantidades de DNA molde necessitam de menos ciclos para acumular

7
produto suficiente para emitir um sinal de fluorescência acima do nível basal, sendo que
pequenas quantidades necessitarão de mais ciclos (Bustin, 2010).
Verifica-se o uso de dois tipos principais de qPCR: um que utiliza fluoróforos
intercalantes de DNA dupla fita, tais como SYBR green I e brometo de etídio, e outra com
sondas que são hidrolisadas. No caso dos fluoróforos intercalantes, após o desenvolvimento do
SYBR green I e de outras cianinas, abandonou-se o uso de brometo de etídio. SYBR green I e
outras cianinas não inibem a PCR e tem uma maior sensibilidade quando comparados ao
brometo de etídeo. Além disso, brometo de etídeo tem potencial carcinogênico e um alto nível
de toxicidade. Uma das desvantagens dos fluoróforos intercalantes é que tais corantes se ligam
ao DNA de dupla fita independentemente da sequência, ou seja, de modo inespecífico e caso
haja amplificações inespecíficas, estas emitirão fluorescência da mesma forma que o produto
desejado (Kubista et al., 2005). Nas qPCRs com sondas que são hidrolisadas, utiliza-se
oligonucleotídeos marcados por um fluoroforo no terminal 5’ e um quencher no terminal 3’. A
sonda deve ser desenhada para que se ligue numa região interna ao par de iniciadores que
flanqueia a região a ser amplificada na reação. Enquanto a sonda está intacta, a proximidade
entre quencher e fluoróforo decresce significativamente a emissão de fluorescência do
fluoróforo. Na etapa de extensão, enquanto a DNA polimerase adiciona nucleotídeos ao longo
da fita molde, a enzima cliva a sonda devido a sua atividade de exonuclease 5’-3’. Tal evento
precede o distanciamento entre fluoróforo e quencher. Dessa forma, o fluoróforo emitirá sinal
de fluorescência que será lido pelo aparelho a cada ciclo. A especificidade dessa PCR reside no
fato de que apenas será emitido sinal caso a sonda se ligue em sua região complementar de
interesse (Arya et al. 2005). A especificidade e sensibilidade inerentes à técnica de qPCR,
tornam essa técnica muito atrativa para análise quantitativa e qualitativa.
1.3. Gp43
As sequências alvo mais frequentemente usadas para detecção molecular de P.
brasiliensis por PCR são as do gene da gp43 e de DNAs ribossômicos do fungo, por serem
específicas para esse gênero de microrganismo. Ensaios de PCR têm sido usados
experimentalmente para detectar P. brasiliensis no soro e tecido de camundongos infectados,
em solos contaminados e em amostras ambientais (Koishi et al). Popi et al. (2002) mostraram
que a molécula gp43 está envolvida com a inibição de fagocitose de P. brasiliensis e da
liberação de NO por macrófagos. Além de ser uma glicoproteína predominante em P.
brasiliensis, a notoriedade dessa glicoproteína se deve principalmente ao fato de ser
reconhecida por soro de 100% dos pacientes com PCM (Travassos et al., 1995), sendo também

8
aplicada ao teste de DTH em substituição a paracoccidioidina (Saraiva et al., 1996).
Incontestavelmente, a gp43 é a mais estudada dentre as glicoproteínas de Paracoccidioides,
isso graças a sua gênero-especificidade, o que faz dessa molécula um alvo para abordagens
moleculares de detecção. Com isso, busca-se evitar reações cruzadas entre formas de vida com
proximidade filogenética. O gene que codifica a glicoproteína de 43kDa de P. brasiliensis é
consensualmente considerado um bom marcador molecular para identificação molecular desse
fungo por PCR.
1.4. Quantificação de fungos dos órgãos de animais infectados
Atualmente, na maioria dos grupos de pesquisa que estudam PCM experimental, a
técnica laboratorial utilizada para quantificação de Paracoccidioides oriundos dos órgãos de
animais infectados tem sido a contagem de unidades formadoras de colônia (UFC) de
macerados de órgãos plaqueados em meio de cultura. Como descrito na literatura, a análise da
carga fúngica por quantificação de UFC é um parâmetro confiável para analisar o curso de uma
infecção, progressão ou cura, discriminando animais resistentes de susceptíveis (Clemons et
al., 2005). A queda do número de colônias recuperadas dos pulmões de animais infectados é
um traço reconhecido de resistência a P. brasiliensis, como previamente verificado (Singer-
Vermes et al., 1993; Souto et al., 2000; Nascimento et al., 2002; De Mattos Grosso et al., 2003;
Pina et al., 2004).
Nosso grupo tem estudado o efeito de adjuvantes em camundongos infectados por 20
dias com P. brasiliensis. Mostramos o efeito benéfico em animais infectados e tratados com
adjuvante completo de Freund (CFA) (De Oliveira et al., 2008), que apresentaram uma incrível
diminuição da carga fúngica, do número de granulomas e do infiltrado inflamatório quando
comparado com animais infectados e não tratados com CFA (De Oliveira et al., 2008). Os
estudos têm se expandido para adjuvantes liberados para uso em humanos, como o monofosforil
lipídeo A (MPLA), que apresentou efeito tão bom quanto o CFA (Taise N. Landgraf,
comunicação pessoal). Temos analisado a carga fúngica dos animais através da quantificação
de UFC dos órgãos, principalmente do pulmão, assim como fazem os grupos de pesquisa que
estudam PCM experimental. Apesar dessa técnica ser amplamente usada, essa apresenta certas
limitações, tais como (1) o crescimento do fungo é relativamente lento – 7 a 14 dias para formar
colônias, (2) o consumo de recursos materiais é muito grande por conta dos vários passos
envolvidos na técnica e, de certa forma o mais preocupante, (3) as diversas etapas envolvidas
no método aumentam as chances de contaminação das amostras e/ou intercorrências, que
podem retardar ainda mais a obtenção de resultados claros e fidedignos. Tais limitações do

9
método de quantificação por UFC justifica a nossa motivação em padronizar um método de
quantificação mais rápido e prático.
2. OBJETIVOS
O presente projeto visa padronizar um método de determinação da carga fúngica em
pulmão de animais infectados com P. brasiliensis através de qPCR e comparar com o método
tradicional, qual seja, contagem de UFC.
3. MATERIAL E MÉTODOS
3.1. Animais
Foram utilizados camundongos machos da linhagem BALB/c, de 6 a 8 semanas de
idade. Este projeto consta de parte do projeto de doutorado da estudante Taise N. Landgraf, do
Programa de Pós-Graduação em Imunologia Básica e Aplicada da Faculdade de Medicina de
Ribeirão Preto-USP (FMRP-USP), aprovado pelo Comitê de Ética em Experimentação Animal
(CETEA) da FMRP-USP, protocolo No. 105/2007.
3.2. Infecção dos camundongos com leveduras de P. brasiliensis
Os camundongos foram infectados, por via endovenosa, com 100µL de uma suspensão
de 107 leveduras de P. brasiliensis por mililitro. Em alguns experimentos, os camundongos
foram infectados com suspensão de 106 ou 105 leveduras/mL. Após 20 dias, os animais foram
divididos em grupos e injetados subcutaneamente com 100µL de adjuvante completo de Freund
(CFA – Sigma-Aldrich. St. Louis, EUA) emulsificado em PBS (1:1) ou 20µg monofosforil
lipídeo A (MPLA – Novagen, Madison, EUA) em 100µL de PBS. Alguns animais foram usados
como controle e, para tanto, não foram tratados. Decorridos 30 dias do tratamento, os animais
foram mortos e tiveram os pulmões assepticamente retirados e pesados.
Um dos pulmões de cada animal foi transferido para um tubo de ensaio de vidro
(1590mm) contendo 1mL de PBS estéril e macerado em homogeneizador de tecidos (Ultra-
Turrax T25 Basic, IKA Works, Inc., Wilmington, EUA). Para determinar a recuperação de
fungos, primeiramente, diluiu-se o macerado a 1:10 em PBS estéril, adicionando-se 50µL de
macerado em 450µL de PBS estéril. Logo após, foram semeados 100µL dessa diluição, em

10
duplicata, em placas de Petri contendo meio BHI-ágar enriquecido com 4% de soro bovino fetal
inativado (Hyclone, Utah, EUA) e 100mg/mL de ampicilina (Invitrogen, Carlsbad, EUA). As
placas foram mantidas a 37oC por até 14 dias. A recuperação de fungos nos órgãos foi avaliada
a partir da contagem de UFC aos 7 dias de cultivo, sendo determinado o número de UFC por
grama de órgão (UFC/g).
3.3. Isolamento de leveduras de P. brasiliensis a partir do pulmão
O primeiro passo para isolamento da levedura foi a lise com solução hipotônica das
células dos camundongos contidas no macerado. Para tanto, o restante do macerado de pulmão
em PBS estéril (950 µL), obtido como descrito no item anterior, foi adicionado a 20mL de água
destilada gelada estéril. Para eliminação das fibras da matriz extracelular do pulmão do
macerado, passou-se o macerado diluído por uma peneira fina de poliéster (tecido organza),
com o auxílio do embolo emborrachado de uma seringa. O filtrado foi centrifugado a 700 g,
por 5 minutos, a 4oC. Após descartar o sobrenadante, o sedimento foi lavado com 20mL de
água ultrapura gelada estéril por centrifugação, a 700 g, por 5 minutos, a 4oC. O sedimento
de levedura foi ressuspendido em 500µL de água ultrapura gelada estéril.
3.4. Extração de DNA genômico de P. brasiliensis
DNA genômico de leveduras de P. brasiliensis foi extraído com protocolo
convencional. Macerados de pulmão dos animais, obtidos como descrito acima, ou leveduras
da cepa virulenta Pb18 de P. brasiliensis cultivada em meio F12 Nutrient Mixture (Life
Technologies, Carlsbad, EUA), por 7 dias, a 37°C, foram ressuspendidos em 200μL de solução
de lise (Triton X-100 a 2%, SDS a 1%, NaCl a 100mM e Tris-HCl a 10mM, EDTA a 1mM, pH
8,0), seguido pela adição de 200μL de fenol:cloroformio (1:1) e 200μL de perolas de vidro
(Sigma-Aldrich, St. Louis, EUA). Os tubos contendo as amostras foram agitados vigorosamente
(agitadores do tipo vórtex – Phoenix, Araraquara, SP), por 3 minutos, e submetidos a
centrifugação, a 700 g, por 5 minutos, a 4oC. As fases aquosas foram submetidas à
precipitação com etanol a 100% e os precipitados (DNA) ressupendidos em tampão TE (Tris-
HCl a 10mM contendo EDTA a 1mM, pH 8,0).

11
3.5. Eletroforese
3.5.1. Gel de agarose
Às amostras de DNA foi adicionada uma solução aquosa contendo azul de bromofenol
a 0,25% e glicerol a 60%, na razão de razão 5:1, sendo as preparações aplicadas nos géis de
agarose. Esses géis foram preparados fundindo-se agarose em tampão TAE (Tris-acetato a
40mM e EDTA a 1mM, pH 8,0), em concentrações de 0,8 e 1%, sendo adicionado SYBR Safe
DNA Gel Stain (Thermo-Fisher Scientific, Waltham, EUA) antes da gelificação. As corridas
foram feitas em uma cuba horizontal Sub-Cell GT (Bio-Rad Laboratories, Hercules, EUA) em
tampão contendo TAE com uma tensão constante de 80V. As amostras de DNA corridas no gel
foram visualizadas em transiluminador de luz azul (Safe Imager 2.0 Blue Light
Transilluminator, Thermo-Fisher Scientific).
3.5.2. Gel de poliacrilamida
Algumas análises eletroforéticas em gel de poliacrilamida a 6,5% (PAGE) foram
realizadas em sistema Mini-Protean Tetra (Bio-Rad Laboratories). Às amostras de DNA foi
adicionado tampão de amostra (azul de bromofenol a 0,25%, glicerol a 50%, SDS a 10% e Tris-
HCl a 0,25M, pH 8,0), na razão de 4:1, sendo as preparações aplicadas no gel. As corridas
eletroforéticas foram feitas em tampão TAE, em uma tensão constante de 100V (80-120mA).
Os géis foram corados com prata, conforme descrito por Blum et al. (1987).
3.6. PCR
3.6.1. Convencional:
As PCRs convencionais foram realizadas para padronização dos reagentes da reação,
em um aparelho C1000 ThermoCycler (Bio-Rad). Como molde das reações foram usadas
amostras de DNA obtidas de leveduras de cultura ou de pulmão dos animais infectados ou DNA
plasmidial (pTE28a) contendo o gene completo de gp43 (pET28a-GP43). Essa construção
plasmidial foi gentilmente cedida pelo Dr. Fabrício Freitas Fernandes, Pós-Doutorando do
Departamento de Biologia Celular e Molecular e Bioagentes Patogênicos da Faculdade de
Medicina de Ribeirão Preto-USP. A reação foi realizada com 200ng de DNA ou diluições na
base 10 dessa amostra, diluído em tampão de PCR, 10mM de uma mistura dNTP, 50mM de
MgCl2, 10µM de cada iniciador (Tabela 1), 0,2U de Taq DNA polimerase (Thermo-Fisher
Scientific) e água ultrapura, totalizando uma reação de 25µL. As amostras foram submetidas
inicialmente à temperatura de 94°C, por 2 minutos, para desnaturação do DNA. Seguiu-se 35
ciclos, sendo que cada ciclo consistiu de desnaturação por 30 segundos a 94°C, anelamento dos

12
iniciadores (tabela 1), primariamente testado em três diferentes temperaturas (64, 61 e 58°C),
durante 30 segundos, e extensão por 30 segundos, a 72°C. No último ciclo, as amostras foram
mantidas a 72oC, por 5 minutos.
3.6.2. qPCR
a) Fluoróforos intercalantes de DNA dupla fita: Esse tipo de qPCR foi realizada com
Platinum Quantitative PCR SuperMix-UDG with ROX (Thermo-Fisher Scientific). O DNA foi
adicionado a 12,5µL do SYBR Green qPCR SuperMix-UDG with ROX, 10mM de cada iniciador
(tabela 1) e água ultrapura, totalizando uma reação de 25µL. Foi usada inicialmente uma
temperatura de 94°C, por 3 minutos, para desnaturação do DNA. Seguiu-se 35 ciclos, sendo
que cada ciclo consistiu de desnaturação por 30 segundos, a 94°C, anelamento dos iniciadores
(tabela 1), primariamente testado em três diferentes temperaturas (64°C, 61°C e 58°C), durante
30 segundos, e extensão por 30 segundos, a 72°C.
Nos ensaios de nested-PCR, amplicons obtidos de uma PCR convencional (subitem
3.6.1) foram submetidos a reação descrita acima. No entanto, foram usados, nessa segunda
etapa (qPCR), iniciadores que anelam a sequências internas desses amplicons da PCR
convencional.
b) TaqMan: As qPCRs TaqMan foram feitas com quantidades variáveis de molde
(DNA), 12,5µL do Advanced Master Mix, 10mM de cada iniciador (descrito na tabela 1 como
Gp43-137, Applied Biosystem, Foster City, EUA), 10µM da sonda (6-FAM-5’-
TTAGGACCTTCACCATTGACCAGCAC-3’ MGB/NFQ – Applied Biosystem) e água
ultrapura, totalizando uma reação de 25µL. Foi usada inicialmente uma temperatura de 94°C,
por 3 minutos, para desnaturação do DNA. Seguiu-se 35 ciclos, sendo que cada ciclo consistiu
de desnaturação por 30 segundos, a 94°C, anelamento dos iniciadores, primariamente testado
em três diferentes temperaturas (64°C, 61°C e 58°C), durante 30 segundos, e extensão a 72°C,
por 30 segundos.
As leituras de emissão de fluorescência das qPCRs com SYBR green e TaqMan foram
feitas em um aparelho CFX96 Touch Real-Time PCR Detection System (Bio-Rad).

13
3.7. Cálculo do número de cópias
Para se chegar ao número de cópias do plasmídeo, a partir da quantidade de DNA
plasmidial conhecida, foi utilizado o programa Calculator for Determining the Number of
Copies of a Template encontrado no endereço web http://cels.uri.edu/gsc/cndna.html.
Tabela 1. Pares de iniciadores (primers) usados nas reações de PCR e/ou qPCR.
Nome Tamanho1 D/R2 Sequência (5’ 3’)
Gp43-137 137 D TCGACACATACCACAACCAAG
R CACTCTTTCACAATCAACGGC
Gp43a 148 D AACTAGAATATCTCACTCCCAGTCC
R AATCCGTTACGCCAATCGTA
Gp43b 354 D AACTAGAATATCTCACTCCCAGTCC
R TGTAGACGTTCTTGTATGTCTTGGG
Gp43 (gene
completo) 1234
D CGAATTCGCAGGATCAGCAATATAT
R CTCGAGCCTGCATCCACCATACTTCC
-actina 187 D GGATGAGGAGATGGATTATGG
R GAAACACTCGACGCACACGAC
1 Tamanho do material amplificado dado em pares de base (pb). 2 D – direto (forward), R – reverso (reverse).
4. Resultados
4.1. Padronização dos iniciadores e teste de especificidade da PCR
Para padronizar as condições da qPCR, primeiramente, realizou-se PCRs convencionais
com amostras de DNA de leveduras de P. brasiliensis extraídas com o protocolo tradicional
(fenol:clorofórmio). Obteve-se bons resultados com os iniciadores Gp43a e Gp43b nas três
temperaturas de anelamento (64°C, 61°C e 58°C) (Figura 1). Em relação aos pares de
iniciadores específicos para -actina de P. brasiliensis, a reação mostrou um resultado melhor
com a temperatura de anelamento de 58oC (Figura 3). Optou-se pelo iniciador Gp43a e a
temperatura de anelamento de 64ºC (Figura 3), na qual há uma condição de maior estringência
e uma banda visualmente mais fluorescente e espessa na análise da eletroforese em gel de
agarose, podendo-se inferir uma maior quantidade de material amplificado.

14
Figura 3. Padronização da PCR. DNA de leveduras de P. brasiliensis foi submetido à PCR
com quatro pares de iniciadores diferentes: -actina e dois iniciadores para gp43 (tabela 1).
Foram testadas três temperaturas de anelamento: 64°C, 61°C e 58°C. Os produtos da PCR
foram analisados em gel de agarose a 1%. O tamanho molecular em pares de bases dos
amplicons está mostrado à esquerda da figura. O DNA foi marcado com SYBR green e
visualizado em transiluminador de lâmpada azul.
Para confirmar que o par de iniciadores GP43a amplificavam apenas o segmento gênico
de gp43, foram realizadas reações de PCR com amostras de DNA extraídas do pulmão de
camundongos infectados com P. brasiliensis. Como mostrado na figura 4, todas as amostras
obtidas de pulmão de camundongos infectados apresentaram apenas uma banda de

15
aproximadamente 148pb. Não houve detecção de bandas referentes à amplificação de DNA em
pulmão de um camundongo não infectado (dado não mostrado). Esses resultados mostraram
que o par de iniciadores GP43a possibilitava a detecção de P. brasiliensis em amostras contendo
DNA de camundongo, conferindo relativa especificidade da reação.
Figura 4. PCR realizado a partir do DNA de macerado de pulmão de camundongos
infectados com 106 leveduras de P. brasiliensis. As reações foram realizadas com par de
iniciadores gp43a, que amplifica 148pb do gene gp43. Os produtos da PCR foram analisados
em gel de agarose a 1%. /HindIII é a raia contendo marcadores de pares de bases (fago lambda
digerido com a enzima de restrição HindIII). O DNA foi marcado com SYBR green e
visualizado em transiluminador de lâmpada azul.
4.2. Correlação entre quantidade de DNA genômico de P. brasiliensis e Cq
Com intuito de determinar se havia correlação entre a quantidade de DNA de fungo e
amplificação do gene de gp43, foram realizadas qPCRs com amostras de DNA genômico
extraído de leveduras de P. brasiliensis obtidas de cultura, em quatro massas diferentes, quais
sejam 200, 20, 2 e 0,2ng, e par de iniciador GP43a. Como esperado, observou-se uma correlação
inversa entre as concentrações de DNA com o Cq de cada amostra, sendo determinado o Cq
como o ponto de intersecção entre a linha de corte e a fase exponencial da curva de amplificação
da amostra (Figura 5), ou seja, quanto maior a quantidade de molde, mais precoce será o
aparecimento da fase exponencial da reação, enquanto quantidades menores de DNA
determinarão a fase exponencial mais tardia nos ciclos da reação.

16
Figura 5. Reta de correlação entre quantidade de DNA genômico de P. brasiliensis e Cq.
Quatro doses de DNA genômico de P. brasiliensis (ng) foram submetidas à qPCR usando o par
de iniciadores gp43a, que amplifica 148pb do gene gp43. Cq (ciclo de quantificação) consiste
no número de ciclos onde há intersecção da linha de corte na fase exponencial da curva de
amplificação de cada amostra (número de ciclos). A reação foi realizada no aparelho CFX96
(Bio-Rad).
4.3. qPCR de amostras de DNA extraídas de pulmão de camundongos infectados com
P. brasiliensis
Para determinar se amplicons poderiam ser detectados por qPCR de amostras de tecido,
DNA de pulmões de animais não infectados e infectados com 106 leveduras de P. brasiliensis
foram amplificados com o par de iniciadores GP43a. Observou-se que as amostras de pulmões
oriundos de três animais infectados com P. brasiliensis foram amplificadas. Embora estivesse
fora do limite de detecção da qPCR (Figura 6), esses dados foram contrastantes em relação às
amostras oriundas de animais não infectados, uma vez que não detectamos curva de
amplificação nessas amostras (dados não mostrados).

17
Figura 6. qPCR de DNA extraído do pulmão de camundongos infectados com P.
brasiliensis. DNA (200ng) extraído de macerado de pulmão de camundongos infectados com
106 leveduras de P. brasiliensis foi submetido à qPCR usando o par de iniciadores gp43a, que
amplifica 148pb do gene gp43. Cq (ciclo de quantificação) foi determinado pela intersecção
entre a linha de corte e a curva de amplificação de cada amostra. A reação foi realizada no
aparelho CFX96 (Bio-Rad).
4.4. qPCR de camundongos infectados com quantidades diferentes de leveduras de P.
brasiliensis
Para verificar se haveria correlação entre a carga fúngica e Cq, foram realizadas qPCRs
de amostras de DNA obtidas de pulmões de animais infectados por 50 dias com 104, 105 e 106
leveduras de P. brasiliensis, utilizando-se o par de iniciadores Gp43a. Apesar de haver
diferenças nos inóculos, na ordem de 10 e 100 vezes, não foi constatada correlação nos Cqs de
tais amostras (Figura 7). Além disso, pode-se observar que todas as amostras se encontram
numa faixa de Cq entre 30,9 e 33,4, mostrando, assim, um intervalo relativamente restrito
quando confrontando com o número de leveduras usadas para a infecção.

18
Figura 7. Correlação entre Cq e quantidade de DNA expressa em femtogramas (fg).
Duzentos nanogramas de DNA extraídos de amostras de pulmão de camundongos infectados
com 106, 105 ou 104 leveduras de P. brasiliensis foram submetidos à qPCR utilizando o par de
iniciadores gp43a. Conversão de Cq em quantidade de DNA foi determinado por uma equação
da reta obtida pela regressão linear dos dados de Cq e quantidade de DNA plasmideal contendo
o gene de gp43 (pET28a-GP43). Cq (ciclo de quantificação) foi determinado pela intersecção
entre a linha de corte e a curva de amplificação de cada amostra. A reação foi realizada no
aparelho CFX96 (Bio-Rad).
4.5. qPCR de camundongos infectados utilizando iniciadores de -actina fúngica
Para avaliar se outro par de iniciadores para um novo alvo poderia mostrar maior
sensibilidade nas amostras de camundongos infectados com P. brasiliensis, utilizou-se um par
de iniciadores para a -actina fúngica. Primeiramente, foi construída uma curva com seis
amostras de DNA genômico de leveduras de cultura, correlacionando-se Cq e quantidade de
DNA, a qual revelou um coeficiente de determinação (R²) muito bom (0,99) (Figura 8).
Posteriormente, foram usadas amostras de camundongos infectados e tratados com CFA ou
MPLA, e como controle, camundongos apenas infectados. Quando analisadas as amostras,
verificou-se uma correlação baixa entre quantidade de DNA (ng) e contagem de UFC (Tabela
2 e Figura 9). Além disso, a maior parte das amostras apresentou uma quantidade do molde
muito pequena. Em muitas das amostras não foi observado curva de amplificação exponencial
ou mesmo não foi possível contar suas UFC devido a questões metodológicas. Tomados juntos,

19
esses resultados mostraram que a técnica realizada dessa forma não poderia ser usada para
substituição da determinação de carga fúngica por contagem de UFC.
Figura 8. Curva padrão construída a partir dos Cqs de amostras de DNA genômico de P.
brasiliensis diluído na base 10 (20.000, 2.000, 200, 20, 2 e 0,2pg de DNA). Os iniciadores
utilizados amplificam a sequência de β-actina fúngica. Cq (ciclo de quantificação) foi
determinado pela intersecção entre a linha de corte e a curva de amplificação de cada amostra.
A reação foi realizada no aparelho CFX96 (Bio-Rad). Log Starting Quantity se refere ao
logaritmo do número de cópias (ou log concentração relativa). Apenas as amostras padrões
(DNA diluído na base 10) foram plotadas neste gráfico. A qPCR de cada amostra foi realizada
em duplicata.

20
Tabela 2. Correlação entre quantificação relativa de DNA e UFC/g de pulmão.
Tratamento1 Camundongo # UFC2 DNA (ng)3
Nenhum
1 60.000 0
2 120.000 1
3 816.666 6,7
4 102.000 0
MPLA
1 0 1,4
2 0 0,7
3 60.000 1,9
4 290.000 9,7
5 5.000 1,3
CFA
1 730.000 1
2 0 0,2
3 0 0,007
4 32.000 0
5 25.000 0
6 120.000 5,9
7 0 1,1
1Camundongos foram infectados com leveduras de P. brasiliensis e tratados com CFA ou
MPLA. Como controle foram usados camundongos apenas infectados. Trinta dias após os
tratamentos, todos os animais foram mortos e o macerado de seus pulmões plaqueados para
obtenção de 2unidades formadora de colônia (carga fúngica) e DNA. 3Quantidade de DNA
relativo à curva padrão de amplificação de DNA genômico de P. brasiliensis (Figura 8) com os
pares de iniciadores específicos para -actina.

21
Figura 9. Gráfico de correlação entre quantificação relativa de DNA e UFC/g
de pulmão. Resultados plotados a partir dos dados mostrados na tabela 2.
4.6. Nested-qPCR
O fato de a sensibilidade das qPCRs que foram testadas acima estar muito baixa,
motivou-nos a avaliar amostras pré-amplificadas com par de iniciadores que geraria um
amplicon maior, seguindo de uma qPCR desse amplicon com iniciadores internos, técnica
conhecida com nested-PCR. Inicialmente, foram feitas PCRs em plasmídeos contendo como
inserto o gene completo de gp43 (pET28a-GP43) e DNA genômico de P. brasiliensis. As
análises dos amplicons feitas em gel de agarose a 0,8% possibilitaram a visualização de
amplicons a partir de 2pg do DNA plasmidial (figura 10A). Já com o DNA genômico, houve
necessidade de uma quantidade de DNA 1.000 vezes maior do que DNA plasmidial para ocorrer
amplificação similar, ou seja 2ng de DNA genômico (Figura 10B).

22
Figura 10. PCRs de (A) DNA plasmidial e (B) genômico em diferentes concentrações. PCR
realizado a partir de (A) diferentes quantidades de plasmídeo clonado com a sequência completa
de Gp43 (pET28a-GP43) e de (B) DNA genômico de P. brasiliensis. As reações foram
realizadas com par que amplificam a sequência completa de Gp43. Os produtos da PCR foram
analisados em gel de agarose a 1%. /HindIII é a raia contendo marcadores de pares de bases
(fago lambda digerido com a enzima de restrição HindIII). O DNA foi marcado com SYBR
green e visualizado em transiluminador de lâmpada azul.
Para determinar se amplicons poderiam ser detectados por PCR de amostras de tecido,
DNA de pulmões de camundongos infectados ou não com P. brasiliensis foram amplificados
com o par de iniciadores que amplificam o gene completo de gp43. Como observado na figura
11, houve amplificação da amostra contendo 200ng de DNA de pulmão de camundongo
infectado, mas não com a dos camundongos infectados e tratados com o adjuvante MPLA,
indicando que a terapia com esse o adjuvante funcionou, diminuindo a carga fúngica, que foi
vista em termos de UFC (dados não mostrados). Nas amostras dos controles não foram
detectadas bandas, ou seja, não houve reações inespecíficas.

23
Figura 11. PCR realizado a partir do DNA de macerado de pulmão de camundongos. Três
animais foram infectados com leveduras de P. brasiliensis: um animal não tratado (raia 1), e
dois animais tratados com MPLA (raias 2 e 3). Um animal não infectado foi usado como
controle (Controle-1). Br indica ausência de DNA molde na reação. As reações foram realizadas
com par de iniciadores Gp43 gene completo, que amplifica 1.234pb do gene gp43. Os produtos
da PCR foram analisados em gel de agarose a 1%. /HindIII é a raia contendo marcadores de
pares de bases (fago lambda digerido com a enzima de restrição HindIII). O DNA foi marcado
com SYBR green.
Em uma tentativa de se melhorar a reação foram testados iniciadores em duas
concentrações diferentes. Nas reações com 2ng de DNA, tanto genômico quanto plasmidial,
nota-se que as reações com 10pmol de iniciadores foi mais eficiente (Figura 12). Quando se fez
a qPCR dessas e de outras amostras, observou-se um problema com a reação, qual seja, algumas
amostras que seriam controles negativos também amplificaram (dados não mostrados). Esses
resultados inviabilizaram a continuação dos testes com nested-PCR.

24
Figura 12. Nested-PCR realizado a partir de produto das PCRs com DNA plasmidial e
genômico. Para a diluição de 2ng foi realizada a tentativa de padronizar uma melhor
concentração de iniciadores nas reações. Para tanto, levou-se 5 e 10pmol dos iniciadores por
reação de tal diluição. Os produtos da PCR foram analisados em PAGE a 6,5%. O DNA foi
corado com prata (Blum et al., 1987).
4.7. Extração de DNA de leveduras isoladas do pulmão de camundongos infectados
por de P. brasiliensis
O principal problema de nossas abordagens até esse ponto do trabalho se relacionava
com a pequena quantidade de DNA fúngico nas amostras extraídas de tecido de animal
infectado. Assim, surgiu a ideia de se fazer um enriquecimento das células fúngicas presentes
no tecido pulmonar antes de se extrair o DNA. Para tanto, pulmões dos animais infectados
foram macerados em água para lise das células do camundongo. A amostra resultante foi
filtrada em organza, para eliminação de material fibroso da matriz extracelular, e lavada por
centrifugação do material (modificação da técnica de descrito por Marty et al., 2013). Pela
resistência da parede fúngica ao choque osmótico, foi possível obter leveduras sem
contaminação por células dos camundongos (dados não mostrados).
O protocolo de extração de DNA das leveduras isoladas do pulmão foi feito com
fenol:clorofórmio. Como observado na figura 13, as amostras extraídas revelaram boa
qualidade. Embora as bandas indiquem uma extração de baixo rendimento, verifica-se que tais
amostras apresentaram boa integridade.

25
Figura 13. Extração de DNA de leveduras isoladas de pulmão de camundongos infectados
com P. brasiliensis. Os animais foram mortos no dia 70 após a infecção e tiveram os pulmões
retirados e macerados. DNA foi extraído com fenol:clorofórmio. Foi corrido um gel de agarose
a 0,8%. λ/HindIII e a raia contendo marcadores de pares de bases (fago lambda digerido com a
enzima de restrição HindIII). O DNA foi marcado com SYBR green e visualizado em
transiluminador de lâmpada azul.
4.8. qPCR de DNA de leveduras de camundongos infectados com P. brasiliensis
tratados com CFA ou não tratados
As amostras de DNA, extraídas como descrito acima, foram submetidas a qPCR a fim
de se examinar se havia correlação entre a contagem de UFC do pulmão de animais e os valores
de Cq da corrida. Todos os ensaios de qPCR descritos a seguir foram realizados com SYBR
Green e TaqMan. Inicialmente foi feita uma curva padrão para correlacionar o Cqs com a
quantidade de DNA plasmidial (pET28a-GP43) levada para a reação. Tal curva foi constituída
de oito pontos, os quais compunham um intervalo entre 20ng até 0,2fg. Na tentativa de aumentar
a precisão da quantificação foi calculado o número de cópias de cada ponto da curva padrão. O
coeficiente de determinação para curva padrão baseada na utilização de SYBR resultou num
R²= 0,9975 (Figura 14A), enquanto a curva gerada através de reações utilizando TaqMan,
mostrou um R²=0,9946 (Figura 14B). As duas curvas mostraram um bom ajustamento do
modelo de regressão linear
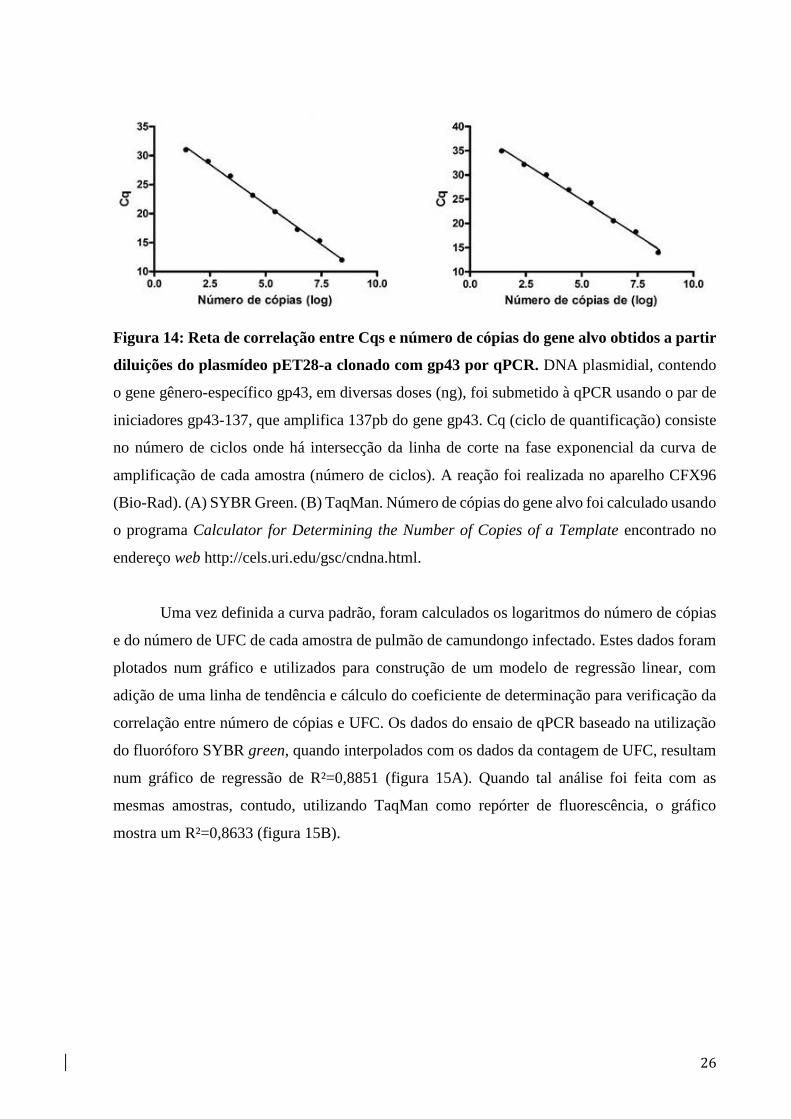
26
Figura 14: Reta de correlação entre Cqs e número de cópias do gene alvo obtidos a partir
diluições do plasmídeo pET28-a clonado com gp43 por qPCR. DNA plasmidial, contendo
o gene gênero-específico gp43, em diversas doses (ng), foi submetido à qPCR usando o par de
iniciadores gp43-137, que amplifica 137pb do gene gp43. Cq (ciclo de quantificação) consiste
no número de ciclos onde há intersecção da linha de corte na fase exponencial da curva de
amplificação de cada amostra (número de ciclos). A reação foi realizada no aparelho CFX96
(Bio-Rad). (A) SYBR Green. (B) TaqMan. Número de cópias do gene alvo foi calculado usando
o programa Calculator for Determining the Number of Copies of a Template encontrado no
endereço web http://cels.uri.edu/gsc/cndna.html.
Uma vez definida a curva padrão, foram calculados os logaritmos do número de cópias
e do número de UFC de cada amostra de pulmão de camundongo infectado. Estes dados foram
plotados num gráfico e utilizados para construção de um modelo de regressão linear, com
adição de uma linha de tendência e cálculo do coeficiente de determinação para verificação da
correlação entre número de cópias e UFC. Os dados do ensaio de qPCR baseado na utilização
do fluoróforo SYBR green, quando interpolados com os dados da contagem de UFC, resultam
num gráfico de regressão de R²=0,8851 (figura 15A). Quando tal análise foi feita com as
mesmas amostras, contudo, utilizando TaqMan como repórter de fluorescência, o gráfico
mostra um R²=0,8633 (figura 15B).

27
Figura 15: Correlação entre UFC e número de cópias do gene alvo obtidos a partir de
leveduras isoladas de pulmão de camundongos infectados com P. brasiliensis. DNA
extraído de leveduras isoladas do macerado de pulmão de camundongos infectados com P.
brasiliensis foi submetido à qPCR usando o par de iniciadores gp43-137, que amplifica 137pb
do gene gp43. O número de cópias do gene alvo foi calculado a partir das equações da reta das
regressões lineares descrito na figura 14, sendo o Cq (ciclo de quantificação) foi determinado
pela intersecção entre a linha de corte e a curva de amplificação de cada amostra. A reação foi
realizada no aparelho CFX96 (Bio-Rad). (A) SYBR Green. (B) TaqMan.
4.9. qPCR de amostras de pulmão de camundongos infectados com P. brasiliensis
tratados com CFA, MPLA ou não tratados
Assim como descrito acima, esse ensaio foi baseado na análise da correlação entre
número de cópias e contagem de UFC, embora num número maior de condições teste. A curva
padrão utilizada para calcular o número de cópias do gene gp43 foi a mesma representada no
último ensaio. Quando colocados no gráfico, os dados da qPCR confrontados com a contagem
de UFCs geraram R²= 0,9599 para a corrida com SYBR green (Figura 16A), enquanto a na
corrida com TaqMan, obteve-se um R²= 0,9007 (Figura 16B). Esse conjunto de resultados
mostram que essa abordagem se mostrou apropriada para detecção de carga fúngica em animais
infectados com P. brasiliensis.
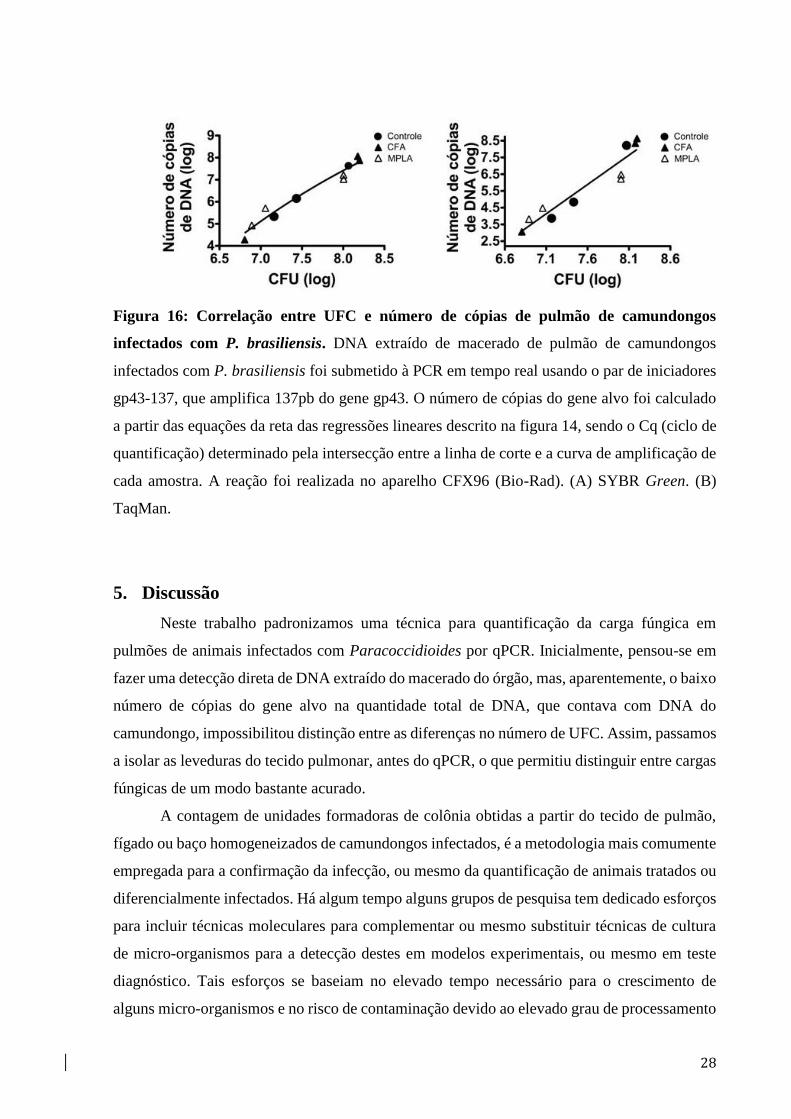
28
Figura 16: Correlação entre UFC e número de cópias de pulmão de camundongos
infectados com P. brasiliensis. DNA extraído de macerado de pulmão de camundongos
infectados com P. brasiliensis foi submetido à PCR em tempo real usando o par de iniciadores
gp43-137, que amplifica 137pb do gene gp43. O número de cópias do gene alvo foi calculado
a partir das equações da reta das regressões lineares descrito na figura 14, sendo o Cq (ciclo de
quantificação) determinado pela intersecção entre a linha de corte e a curva de amplificação de
cada amostra. A reação foi realizada no aparelho CFX96 (Bio-Rad). (A) SYBR Green. (B)
TaqMan.
5. Discussão
Neste trabalho padronizamos uma técnica para quantificação da carga fúngica em
pulmões de animais infectados com Paracoccidioides por qPCR. Inicialmente, pensou-se em
fazer uma detecção direta de DNA extraído do macerado do órgão, mas, aparentemente, o baixo
número de cópias do gene alvo na quantidade total de DNA, que contava com DNA do
camundongo, impossibilitou distinção entre as diferenças no número de UFC. Assim, passamos
a isolar as leveduras do tecido pulmonar, antes do qPCR, o que permitiu distinguir entre cargas
fúngicas de um modo bastante acurado.
A contagem de unidades formadoras de colônia obtidas a partir do tecido de pulmão,
fígado ou baço homogeneizados de camundongos infectados, é a metodologia mais comumente
empregada para a confirmação da infecção, ou mesmo da quantificação de animais tratados ou
diferencialmente infectados. Há algum tempo alguns grupos de pesquisa tem dedicado esforços
para incluir técnicas moleculares para complementar ou mesmo substituir técnicas de cultura
de micro-organismos para a detecção destes em modelos experimentais, ou mesmo em teste
diagnóstico. Tais esforços se baseiam no elevado tempo necessário para o crescimento de
alguns micro-organismos e no risco de contaminação devido ao elevado grau de processamento

29
e de passos intercorrentes até o efetivo plaqueamento da amostra. A amplificação in vitro de
sequências de DNA específicas por PCR é um método sensível que pode ser utilizado para
diagnóstico de PCM (Gomes et al., 2000). Goldani et al. (1998) buscaram comparar a
sensibilidade de detecção de P. brasiliensis através do DNA do fungo, presente no soro, com
cultura de sangue e contagem de UFC. Embora todos camundongos tivessem sido infectados
com mesma carga fúngica, algumas das amostras de cultura de sangue não mostraram
positividade à contagem de UFC, mostrando-se a variação da técnica entre as amostras. Já a
PCR, mostrou positividade para todas amostras do soro de camundongos infectados.
Uma condição experimental questionada por nós, durante esse estudo, foi a utilização
de amostras de DNA extraídas diretamente do pulmão de camundongos infectados com P.
brasiliensis. Embora tenha-se verificado amplificações razoáveis com todos os pares de
iniciadores testados com DNA genômico extraído de P. brasiliensis de cultura, de oito amostras
de camundongos infectados que foram submetidas à PCR para confirmação da ausência de
bandas inespecíficas no gel de agarose, apenas uma banda de aproximadamente 148pb foi
visualizada em todas amostras. O gráfico de regressão mostrou que a técnica de qPCR tinha
potencial para detecção de gp43 quando submetidas amostras de DNA genômico de fungos de
cultura diluído na base 10. Mas quando amostras de camundongos infectados e não infectados
foram submetidas a qPCR, observou-se que as curvas de amplificação não se encontravam no
limite de detecção da curva padrão. Amostras de camundongos infectados com diferentes
concentrações de leveduras apresentaram um intervalo de detecção muito restrito para tais
diferenças de infecção. Quando os iniciadores foram substituídos para -actina de
Paracoccidioides, não houve melhora no ensaio. Tal condição resulta na presença de grande
quantidade de DNA genômico oriundo do tecido de camundongo e uma menor fração de DNA
proveniente das leveduras infectantes do pulmão. Charbel et al. (2006) enfrentaram um
problema semelhante ao tentar amplificar amostras de DNA extraídas do soro de pacientes
infectados com P. brasiliensis. Babady et al. (2011) fizeram menção à possibilidade de inibição
da reação de qPCR mediada por matriz extracelular. Costa et al. (2000) afirmaram que a
sensibilidade verificada pelos ensaios de construção da curva padrão com diluições seriadas do
DNA de Aspergillus fumigatus, provavelmente é maior do que a sensibilidade de ensaios onde
se tem adição de amostras biológicas. Tal afirmação é suportada pela presença de DNA
genômico humano e outros possíveis inibidores.
Uma das possibilidades aventadas para a baixa detecção de cópias do gene alvo na
amostra foi buscar amplificar o gene por técnica de PCR convencional, e fazer a qPCR usando
um iniciador interno do amplicon gerado pela PCR convencional (nested-PCR). O qPCR dessas

30
e de outras amostras revelaram um problema na reação, o que nos levou a descartar os testes
com nested-PCR. A tentativa de incluir as amostras no limite de detecção da técnica de qPCR
não foi bem-sucedida devido a verificação de amplificação em amostras não infectadas.
Segundo Costa et al. (2000), além das variações biológicas, a principal razão de discrepâncias
entre diferentes estudos é a falta de padronização das técnicas utilizadas. É notório que o
aumento do número de passos aumenta as chances de contaminação das reações de nested-
qPCR.
O protocolo descrito por Marty et al. (2013), o qual foi utilizado para separação de
Blastomyces dermatites de pulmão de camundongos, mostrou-se eficiente para tal finalidade
com P. brasiliensis. A utilização de um processo de três passos que se baseia na lise de células
do hospedeiro com água, filtragem e, posterior, lavagem da amostra de pulmão infectado é
excelente para se isolar as leveduras e, desse modo, para se obter DNA de levedura com alto
grau de pureza. A eliminação de material contaminante nas reações de PCR tem sido buscada
por outros autores. O trabalho de Amicosante et al. (1995) mostrou que o uso de uma resina de
ligação aniônica para liberação do DNA de células de Mycobacterium tuberculosis resultou
num aumento da sensibilidade de detecção do DNA da bactéria por qPCR. A presença de
inibidores endógenos oriundos do escarro de pacientes pode limitar a performance da reação de
qPCR. Segundo o autor, estima-se que tal condição possa gerar mais de 20% de falsos
negativos. Bialek et al. (2000) também verificaram a presença de inibidores ao submeter
amostras de DNA de macerado de pulmão de camundongos infectados com P. brasiliensis à
nested-PCR.
Parte dos resultados que obtivemos se tornou mais interessante por conseguirmos
correlacionar UFC com número de cópias do gene alvo. Isso foi possível estabelecendo-se uma
curva a partir de resultados de qPCR com DNA plasmidial contendo o inserto de gp43.
Verificou-se um intervalo de detecção de 8 logs, no qual obtivemos razoável limite de detecção,
tanto no uso de SYBR green como de TaqMan. O coeficiente de determinação e a larga
extensão de detecção asseguram sensibilidade na detecção do gene gp43. A utilização de
PET28a clonado com gp43 como DNA molde para construção da curva padrão eleva a acurácia
ao determinar o número de cópias através do Cq obtido no ensaio. Semighini et al. (2002)
conseguiram bons resultados na construção da curva padrão utilizando iniciadores específicos
para gp43, com ótimo coeficiente de determinação (0,9952), uma extensão de detecção de 7
logs e limite de detecção de 10 cópias de gp43. Essas curvas foram interessantes para
determinação de número de cópias do gene alvo presente nas amostras, o que foi usado nos
experimentos de qPCR com DNA de leveduras isoladas de amostras de camundongos

31
infectados tratados ou não tratados com adjuvantes. Os resultados com esse protocolo
apresentaram bons coeficientes de determinação na comparação do número de cópias de gp43
e da contagem de UFC, com o uso de fluoróforo intercalante ou Taqman. Embora as amostras
tratadas nem sempre mostrem menores níveis de infecção, a correlação entre UFC e número de
cópias se mostra bastante razoável na análise de regressão linear, todas com R2 por volta de 0,9.
Brinkman et al. (2002), num estudo que avaliou o uso de qPCR para enumeração de células de
Candida patogênica em água de praia, também interpolou os dados do logaritmo de contagem
de UFC com o logaritmo da contagem de células por qPCR (TaqMan). Segundo o autor, foi
verificada correlação positiva entre tais variáveis interpoladas (R= 0,91). Pathak et al. (2012)
analisaram dados de logaritmo de UFC e quantidade de moléculas alvo da reação de qPCR
provenientes de órgãos de camundongos infectados com M. avium avium. Foi verificado
correlação linear entre as variáveis para a fase inicial da infecção (R= 0,874). Posto isso,
mostramos nesse trabalho que nossos coeficientes de correlação estão dentro do esperado.
O protocolo que desenvolvemos nesse trabalho mostrou-se muito bom. De fato, a
correlação maior que 0,90 entre número de cópias do gene alvo e UFC é muito difícil de ser
obtida devido a variáveis experimentais, das quais destacamos: a pequena, mas existente,
degradação do DNA extraído, próprio da técnica; fungos mortos que não crescem em cultura
para formar UFC, mas que podem conter ainda DNA e amplificar; perda de fungos no processo
de extração; e imprecisão na técnica de contagem por UFC. Apesar disso, com base na literatura
que mostra coeficientes de determinação de 0,9 como de boa qualidade do ensaio, acreditamos
que nossos dados de qPCR com amostras de DNA obtidas de leveduras isoladas do pulmão são
fidedignos, mostrando-se um teste robusto e reproduzível, por confirmar boa correlação em
dois experimentos com animais infectados.
6. Conclusão
Nesse estudo, demonstramos que o isolamento de leveduras oriundos do pulmão de
animais infectados com P. brasiliensis atrelado à técnica de qPCR constituem uma saída
promissora a fim de melhorar a sensibilidade dos ensaios para quantificação da carga fúngica
em animais infectados. Demostramos que existe uma forte correlação entre os dados obtidos de
qPCR e de contagem de UFC, o que constitui uma evidência de que a detecção molecular é
uma alternativa robusta para o uso em modelo experimental. Tal ferramenta pode ser de grande
valia, uma vez que, torna a quantificação mais rápida, diminui as chances de contaminação e

32
exibe sensibilidade considerável. A implementação na quantificação microbiológica através do
uso de qPCR pode significar um avanço considerável para estudos da PCM experimental.
7. Referências
Amicosante, M., Richeldi, L., et al. Inactivation of polymerase inhibitors for Mycobacterium
tuberculosis DNA amplification in sputum by using capture resin. J Clin Microbiol,, v.33,
p.629–630, 1995.
Arya, M., Shergill, I. S., et al. Basic principles of real-time quantitative PCR. Expert Rev Mol
Diagn, v.5, p.209-19, 2005.
Bagagli, E., Theodoro, R. C., Bosco, S. M., McEwen, J. G. Paracoccidioides brasiliensis:
phylogenetic and ecological aspects. Mycopathol, v.165, p.197-207, 2008.
Benard, G. An overview of the immunopathology of human paracoccidioidomycosis.
Mycopathol, v.165, p.209-21, 2008.
Benard, G., Romano, C. C., et al. Imbalance of IL-2, IFN-gamma and IL-10 secretion in the
immunosuppression associated with human paracoccidioidomycosis. Cytokine, v.13,
p.248-52, 2001.
Bialek, R., Ibricevic, A., et al. Detection of Paracoccidioides brasiliensis in tissue samples by
a nested PCR assay. J Clin Microbiol, v. 38, p.2940-2, 2000.
Blum, H., Beier, H.; Gross, J. H. Improved silver staining of plant proteins, RNA and DNA in
polyacrilamide gels. Electrophoresis, v.8, p. 93-9, 1987.
Borges-Walmsley, M. I., Chen, D., et al. The pathobiology of Paracoccidioides brasiliensis.
Trends Microbiol, v.10, p.80-7, 2002.
Brinkman, N. E., Haugland, R. A., et al. Evaluation of a rapid, quantitative real-time pcr method
for enumeration of pathogenic Candida cells in water. Appl Environ Microbiol, v.69,
p.1775–82, 2002.
Brummer, E., Castaneda E., et al. Paracoccidioidomycosis: an update. Clin Microbiol Rev,
v.6, p.89-117, 1993.
Bustin, S. A. The PCR revolution: basic technologies and applications. Cambridge, New
York: Cambridge University Press. 2010. 307p.
Bustin S. A, et al. Minimum information for publication of quantitative real-time PCR
experiments Clin Chem, v.55, p.611-22, 2009.
Calich, V. L., Kashino, S. S. Cytokines produced by susceptible and resistant mice in the course
of Paracoccidioides brasiliensis infection. Braz J Med Biol Res, v.31, p.615-23, 1998.
Calvi, S. A., Soares A. M., et al. Study of bronchoalveolar lavage fluid in
paracoccidioidomycosis: cytopathology and alveolar macrophage function in response to
gamma interferon; comparison with blood monocytes. Microbes Infect, v.5, p.1373-9,
2003.

33
Cano, L. E., Kashino S. S., et al. Protective role of gamma interferon in experimental pulmonary
paracoccidioidomycosis. Infect Immun, v.66, p.800-6, 1998.
Carrero, L. L., Nino-Vega G., et al. New Paracoccidioides brasiliensis isolate reveals
unexpected genomic variability in this human pathogen. Fungal Genet Biol, v.45, p.605-
12, 2008.
Castro, L. F., Ferreira, M. C., et al. Characterization of the immune response in human
paracoccidioidomycosis. J. Infect, v.67, p.470-85, 2013.
Charbel, C. E., Levi, J. E., Martins J. E. C. Evaluation of polymerase chain reaction for the
detection of Paracoccidioides brasiliensis DNA on serum samples from patients with
paracoccidioidomycosis. Mem Inst Oswaldo Cruz, v.101, 219-21, 2006.
Clemons, K. V., Espiritu, M., et al. Comparative efficacies of conventional amphotericin b,
liposomal amphotericin B (AmBisome), caspofungin, micafungin, and voriconazole alone
and in combination against experimental murine central nervous system aspergillosis.
Antimicrob Agents Chemother, v.49, p.4867-75, 2005.
Costa, C., Vidaud, D., Olivi, M., Bart-Delabesse, E., Vidaud, M., Stéphane, B. Development of
two real-time quantitative Taqman PCR assays to detect circulating Aspergilus fumigatus
DNA in serum. Journal Microbiological Methods, v.44, p.263-269, 2000.
Coutinho, Z. F., Silva, D., et al. Paracoccidioidomycosis mortality in Brazil (1980-1995). Cad
Saúde Pública, v.18, p.1441-54, 2002.
De Brito, T., Franco, M. F. Granulomatous inflammation. Rev Inst Med Trop São Paulo, v.36,
p.185-92, 1994.
De Mattos Grosso, D., De Almeida, S. R. et al. Characterization of gp70 and anti-gp70
monoclonal antibodies in Paracoccidioides brasiliensis pathogenesis. Infect Immun, v.71,
p.6534-42, 2003.
De Oliveira, L. L., Coltri, K. C., et al. T helper 1-inducing adjuvant protects against
experimental paracoccidioidomycosis. PLoS Negl Trop Dis, v.2, n.3, p.e183, 2008.
Diniz, S. N., Cisalpino, P. S., et al. In vitro granuloma formation, NO production and cytokines
profile from human mononuclear cells induced by fractionated antigens of Paracoccidioides
brasiliensis. Hum Immunol, v.62, p.799-808, 2001.
Diniz, S. N., Cisalpino, P. S., et al. In vitro human immune reactivity of fast protein liquid
chromatography fractionated Paracoccidioides brasiliensis soluble antigens. Microbes
Infect, v.1, p.353-60, 1999.
Fortes, M. R., Miot, H. A., et al. Immunology of paracoccidioidomycosis. An Bras Dermatol,
v.86, p.516-24, 2011.
Franco, M. Host-parasite relationships in paracoccidioidomycosis. J Med Vet Mycol, v.25,
p.5-18, 1987.
Goldani, L. Z., Sugar, A. M. Short report: Use of the polymerase chain reaction to detect
Paracoccidioides brasiliensis in murine paracoccidioidomicosis. Am. J. Trop. Med. Hyg,
58, p. 152-53, 1998.
Gomes GM, Cisalpino PS, Taborda CP, de Camargo ZP. PCR for diagnosis of
paracoccidioidomycosis. J Clin Microbiol, v.38, p. 3478-80, 2000.

34
Kashino, S. S., Fazioli, R. A., et al. Resistance to Paracoccidioides brasiliensis infection is
linked to a preferential Th1 immune response, whereas susceptibility is associated with
absence of IFN-gamma production. J Interferon Cytokine Res, v.20, p.89-97, 2000.
Koishi A. C., Vituri D. F., et al. A semi-nested PCR assay for molecular detection of
Paracoccidioides brasiliensis in tissue samples. Rev Soc Bras Med Trop, v. 43, p.728-30,
2010.
Kubista, M., Andrade J. M., et al. The real-time polymerase chain reaction. Mol Aspetcs Med,
v.27, p.95-125, 2006.
Mamoni, R. L., Blotta, M. H. Kinetics of cytokines and chemokines gene expression
distinguishes Paracoccidioides brasiliensis infection from disease. Cytokine, v.32, p.20-9,
2005.
Matute, D. R., Mcewen, J. G., et al. Cryptic speciation and recombination in the fungus
Paracoccidioides brasiliensis as revealed by gene genealogies. Mol Biol Evol, v.23, p.65-
73, 2006.
Marty, A. J., Wuthrich, M., et al. Isolation of Blastomyces dermatitidis yeast from lung tissue
during murine infection for in vivo transcriptional profiling. Fungal Genet Biol, v.56, p.1-
8, 2013.
Ministério da Saúde. Guia de vigilância epidemiológica. Brasília: Secretaria de Vigilancia em
Saude, Caderno 7, 2009. 29-38p.
Nascimento, F. R., Calich, V. L., et al. Dual role for nitric oxide in paracoccidioidomycosis:
essential for resistance, but overproduction associated with susceptibility. J Immunol,
v.168, p.4593-600, 2002.
Neworal, E. P., Altemani, A., et al. Immunocytochemical localization of cytokines and
inducible nitric oxide synthase (iNOS) in oral mucosa and lymph nodes of patients with
paracoccidioidomycosis. Cytokine, v.21, p.234-41, 2003.
Pathak, S., Awuh, J. A., Leversen, N. A., Flo, T. G., Asjo, B. Counting mycobacteria in infected
human cells and mouse tissue: A comparison between qPCR and CFU. Plos One, v.7, p.1-
11, 2012.
Pina, A., Valente-Ferreira, R. C. et al. Absence of interleukin-4 determines less severe
pulmonary paracoccidioidomycosis associated with impaired Th2 response. Infect Immun,
v.72, p.2369-78, 2004.
Popi, A. F., Lopes, J. D., et al. GP43 from Paracoccidioides brasiliensis inhibits macrophage
functions. An evasion mechanism of the fungus. Cell Immunol, v.218, p.87-94, 2002.
Restrepo, A., Mcewen, J. G., et al. The habitat of Paracoccidioides brasiliensis: how far from
solving the riddle? Med Mycol, v.39, p.233-41, 2001.
Restrepo, A., Salazar, M. E., et al. Estrogens inhibit mycelium-to-yeast transformation in the
fungus Paracoccidioides brasiliensis: implications for resistance of females to
paracoccidioidomycosis. Infect Immun, v.46, p.346-53, 1984.
Richini-Pereira, V. B., Bosco Sde, M., et al. Molecular detection of Paracoccidioides
brasiliensis in road-killed wild animals. Med Mycol, v.46, p.35-40, 2008.

35
San-Blas, G., San-Blas, F. Paracoccidioides brasiliensis: cell wall structure and virulence. A
review. Mycopathologia, v.62, p.77-86, 1977.
Saraiva E. C., Altemani, A., Franco, M. F., Unterkircher, C. S., Camargo, Z. P.
Paracoccidioides brasiliensis-gp43 used as paracoccidioidin. J Med Vet Mycol, v. 34,
p.155-61, 1996.
Semiguini, C. P., De Camargo, Z. P., Puccia, R., Goldman, M. H. S., Goldman, G. G. Molecular
identification of Paracoccidioides brasiliensis by 5’ nuclease assay. Diagnostic
Microbiology and Infectious Disease, v.44, p.383-6, 2002.
Shikanai-Yasuda, M. A., Telles Filho Fde, Q., et al. [Guidelines in paracoccidioidomycosis].
Rev Soc Bras Med Trop, v.39, p.297-310, 2006.
Sifuentes-Osornio, J., Corzo-Leon, D. E. et al. Epidemiology of Invasive Fungal Infections in
Latin America. Curr Fungal Infect Rep, v.6, p.23-34, 2012.
Singer-Vermes, L. M., Caldeira, C. B., et al. Experimental murine paracoccidioidomycosis:
relationship among the dissemination of the infection, humoral and cellular immune
responses. Clin Exp Immunol, v.94, p.75-9, 1993.
Souto, J. T., Figueiredo, F., et al. Interferon-gamma and tumor necrosis factor-alpha determine
resistance to Paracoccidioides brasiliensis infection in mice. Am J Pathol, v.156, p.1811-
20, 2000.
Teixeira, M. M., Theodoro, R. C., et al. Phylogenetic analysis reveals a high level of speciation
in the Paracoccidioides genus. Mol Phylogenet Evol, v.52, p.273-83, 2009.
Theodoro, R. C., Bagagli, E., et al. Phylogenetic analysis of PRP8 intein in Paracoccidioides
brasiliensis species complex. Fungal Genet Biol, v.45, p.1284-91, 2008.
Travassos, L. R., Puccia, R., et al. Biochemistry and molecular biology of the main diagnostic
antigen of Paracoccidioides brasiliensis. Arch Med Res, v. 26, n. 3, p. 297-304, 1995.