comunidade de mamÍferos terrestres em um mosaico de...
TRANSCRIPT

i
UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE BIOLOGIA
Camila Paula de Castilho
“COMUNIDADE DE MAMÍFEROS TERRESTRES EM UM MOSAICO DE RESTAURAÇÃO FLORESTAL: AVALIAÇÃO DOS DESCRITORES
AMBIENTAIS SOBRE A RIQUEZA, A ABUNDÂNCIA E A COMPOSIÇÃO DAS ESPÉCIES.”
Campinas
2015

ii

iii

iv

v

vi

vii
RESUMO GERAL
A ecologia da restauração tem como principal objetivo a recuperação de ambientes
degradados visando à criação de ambientes autossustentáveis. Dessa maneira, diferentes
são os modelos de restauração adotados, mas poucos são os trabalhos que avaliam a
resposta da comunidade de fauna em relação a essas restaurações. Este estudo tem como
objetivo avaliar a resposta da comunidade de mamíferos terrestres em relação aos
tratamentos de restauração presentes na fazenda Intermontes (337ha), na cidade de
Ribeirão Grande, SP. Foram avaliadas a riqueza, a abundância e a composição da
comunidade de pequenos, médios e grandes mamíferos em um mosaico de restauração em
Mata Atlântica, incluindo áreas de pasto com regeneração arbórea e ou arbustiva na qual o
plantio de restauração foi implantado, áreas de pasto com plantio, área de regeneração
natural e remanescentes florestais. Tendo em vista o modelo “trait-filter”, procuramos
evidenciar como alguns descritores ambientais influenciaram a ocorrência das espécies,
considerando como filtros a estrutura da vegetação e a disponibilidade de alimentos
(frutos, artrópodes e pequenos e médios mamíferos de acordo com a espécie avaliada).
Tanto para a assembleia de pequenos mamíferos, quanto médios e grandes, espécies
generalistas predominaram na comunidade, ocorrendo nas diferentes unidades amostrais.
Espécies especialistas foram menos abundantes e mais restritas em sua ocorrência. Para
pequenos mamíferos, estas espécies estiveram restritas aos remanescentes florestais ou às
unidades com maior complexidade de vegetação. Para médios e grandes mamíferos,
espécies especialistas estiveram presentes em diferentes tratamentos, mas foram
ocasionais. O papel dos descritores ambientais sobre cada espécie diferiu de acordo com
suas necessidades ecológicas, revelando a sua importância para a ocorrência das diferentes
espécies. Avaliar a resposta desta comunidade em função de medidas de restauração
revelou a importância dos descritores ambientais que precisam estar presentes e ser
mantidos para garantir a colonização e a permanência da comunidade de mamíferos
terrestres.

viii

ix
ABSTRACT
Restoration ecology aims to recover degraded areas creating self-sustaining
ecosystems. Different restoration techniques are being implemented nowadays but few
studies are evaluating the response of the fauna community in relation to those restored
areas. This study intended to evaluate the response of a terrestrial mammal community in
a restored area located at Intermontes farm (337ha), in the city of Ribeirão Grande, SP. We
investigate richness, abundance and composition of small, medium and large mammals in a
restoration mosaic of Atlantic forest, including pasture areas with arboreal or arbustive
regeneration where restoration where implemented, pasture areas with restoration, areas
with natural regeneration and forest remnants. Once considering the trait-filter model, we
intended to elucidate how habitat filters influenced species occurrence, considering
vegetation complexity and food resources (fruits, arthropods, small and medium mammals
depending on the species) as filters. To the small mammals assembly, as to the medium and
large ones, generalist species predominated in the community, occurring in all different
sample units. Specialized species were less abundant and were restricted in their
occurrence. Small mammals were limited to the forest remnants or to those sample units
with more complex vegetation. Specialist medium and large mammal species were present
in different sample units, but were occasional. The habitat filter role for each species
differed according to the ecological necessity of the species, revealing the importance of
those aspects to species occurrence. The evaluation of the response of this mammal
community to restoration showed that some habitat filters need to be present and
maintained in order to guarantee the colonization and the permanence of terrestrial
mammals.

x

xi
RESUMO GERAL ............................................................................................................................................ vii
ABSTRACT ....................................................................................................................................................... ix
CAPÍTULO GERAL .......................................................................................................................................... 1
OBJETIVOS GERAIS ....................................................................................................................................... 6
MATERIAL E MÉTODOS .............................................................................................................................. 7
Área de Estudo ................................................................................................................................................ 7
Delineamento amostral ............................................................................................................................... 9
Coleta de Dados ............................................................................................................................................ 13
Pequenos mamíferos .................................................................................................................................. 13
Mamíferos terrestres de médio e grande porte ............................................................................... 13
Caracterização da estrutura dos habitats ........................................................................................... 14
Disponibilidade de recursos alimentares ........................................................................................... 15
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................................................... 17
CAPÍTULO 1: “Efeito de filtros ambientais sobre a colonização de pequenos mamíferos em um mosaico de restauração de Mata Atlântica” ............................................................................... 31
RESUMO ........................................................................................................................................................... 31
ABSTRACT ...................................................................................................................................................... 32
INTRODUÇÃO ................................................................................................................................................ 33
MATERIAL E MÉTODOS ............................................................................................................................ 35
RESULTADOS ................................................................................................................................................. 37
DISCUSSÃO ..................................................................................................................................................... 54
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................................................... 59
CAPÍTULO 2: “Mamíferos terrestres de médio e grande porte em um mosaico de restauração de Mata Atlântica: avaliação dos descritores ambientais sobre a ocorrência das espécies” .......................................................................................................................................................... 71
RESUMO ........................................................................................................................................................... 71
ABSTRACT ...................................................................................................................................................... 72
INTRODUÇÃO ................................................................................................................................................ 73
MATERIAL E MÉTODOS ............................................................................................................................ 75
RESULTADOS ................................................................................................................................................. 78
DISCUSSÃO ..................................................................................................................................................... 89
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................................................... 95

xii
CONCLUSÃO GERAL ................................................................................................................................. 105
REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................................................ 107

xiii
AGRADECIMENTOS
Gostaria de agradecer ao meu querido orientador, Dr. Wesley Rodrigues Silva,
primeiro por aceitar em me orientar, mesmo não sendo um mastozoologo. Agradeço por
todo apoio nos mais diferentes aspectos para o desenvolvimento deste projeto, pelas
conversas e discussões sobre o estudo, pelas risadas no laboratório, pela confiança e
incentivo e por retornar 150 km de volta para Ribeirão Grande e me socorrer no Posto da
Polícia Rodoviária de Capão Bonito depois de seu longo dia de campo! Agradeço por além
de ser um grande orientador, por ter se tornado um grande amigo e colega de trabalho.
Agradeço também a minha co-orientadora, Dra. Natália Oliveira Leiner, que apesar
da distância sempre se colocou disponível para me auxiliar no projeto, com as sugestões e
discussões para o enriquecimento do estudo.
Agradeço a Universidade Estadual de Campinas, ao Instituto de Biologia e a
secretaria de Pós Graduação em Ecologia pela infraestrutura, atenção e auxílio.
Agradeço aos membros da pré-banca e da banca pelas sugestões e comentários em
minha tese, contribuindo para o seu aprimoramento.
Sou muito grata a todos que me ajudaram no campo, nas indas e vindas ao longo
desses quatro anos. Ao Paulo César, querido PC, sempre me auxiliando sob chuva e sob sol,
tirando os não tão fofos sapos e serpentes que igualmente caiam nos baldes. Ao Noel e ao
Emerson por me ajudarem com a fenologia. A Mari Canhoto, a Karen Santos, a Mari Reis, a
Karina Sakane, a Cristiane, ao Rodrigo Prado, ao Edi, a Emilly Sunter e a Christini por me
ajudarem igualmente nas coletas de campo.
Agradeço a Companhia de Cimento Votorantim por permitir o desenvolvimento
deste estudo em sua área de restauração e igualmente disponibilizar auxiliares para as
montagens das linhas de balde. Agradeço também toda a simpatia e ajuda do seu Zé Paraná
e de toda sua família, sempre nos recebendo muito bem e nos ajudando em tudo que era

xiv
preciso na fazenda. E agradeço ao Renato e a Solange pela simpatia e por sempre nos
disponibilizarem o trator para o uso na fazenda, assim como o Dorival e o Brás, motoristas
que nos acompanhavam o dia todo.
Sou grata igualmente ao Dr. Percequillo por gentilmente confirmar a identificação
dos roedores coletados.
Agradeço a Capes pelo fornecimento da bolsa de doutorado no primeiro ano e pela
Fapesp pelo fornecimento da bolsa nos quatro anos que se seguiram e todo o
financiamento via reserva técnica. E sou igualmente grata a IdeaWild pelo fornecimento de
equipamentos para o trabalho de campo e ao Instituto pelo fornecimento de recursos para
a aquisição de parte das armadilhas fotográficas.
Agradeço aos queridos Toyoyo, Paulo Henrique, Sebá, Martin, Léo, Christini, Jana,
Mari Reis e Carla por me auxiliarem nas análises estatísticas. E a Cristiane por enfrentar
junto todo este momento de análises no R!
Agradeço e muito meus queridos amigos do Livep, a Cris, a Jana, a Veri, a Jéssica, o
Marcelo, a Bruna, a Ileyne, a Iara, a Carol e a Natália que sempre tornaram os dias na
Unicamp mais gostosos e divertidos!
As minhas amigas mais que queridas que me ajudaram nos momentos de tristezas e
que compartilharam momentos de alegria que tive nestes anos: mais uma vez a Cris, a Jana,
a Christini, a Mari Reis, a Carla, a Ju Padilha, a Karina, a Cau, a Thata, a Michelle e a Milena
Corbo, e ao meu amigo Elson. Vocês moram no meu coração!
Aos meus amados pais, Dioraci Castilho e Maria Rita, sou eternamente grata por
proporcionarem todas as condições necessárias para que eu chegasse até aqui e por
sempre me incentivarem! As minhas irmãs também agradeço do fundo do coração por
sempre me apoiaram durante esses anos apesarem de não compreenderem a minha paixão
por ratos! E a toda a minha amada família.

xv
E gostaria de agradecer, ao meu amor, Wilson Miguel, que me apoiou e me ajudou
durante esse período com indas e vindas ao campo, ouvindo e compartilhando das minhas
preocupações e das minhas alegrias com o projeto. Agradeço pelo seu companheirismo, por
sua paciência, por seu amor e seu carinho para eu cumprir mais essa etapa em minha
carreira profissional. E igualmente aos meus filhotes de quatro patas que sempre tornaram
as minhas chegadas em casa mais alegres e divertidas: Branca, Bluma (in memorian),
Emma, Peppa, George e Vitório! Amo vocês!
E agradeço a todos aqueles que possivelmente não tenha citado nomes dado o
esquecimento, mas que foram importantes para essa etapa se concluir.

xvi

1
CAPÍTULO GERAL
A fragmentação e a perda de habitat são os principais fatores responsáveis pela
perda da biodiversidade, tanto nas regiões temperadas quanto nas tropicais (Terborgh
1988, Turner e Corlett 1996, Fahrig 2002, Fahrig 2003). No Brasil, um dos principais
biomas que vem sofrendo essa perda é a Mata Atlântica, que hoje se resume a somente
11,73% de sua cobertura vegetal original na forma de pequenos fragmentos isolados
(Gascon et al. 2000, Ribeiro et al. 2009).
Embora pequenos em área, ainda assim tais fragmentos podem atuar como os
últimos refúgios para a biota, representando uma oportunidade para a conservação das
espécies pelo menos em curto prazo (Turner e Corlett 1996, Lugo e Helmer 2004). Em
longo prazo, no entanto, somente a sua restauração pode auxiliar a reverter este cenário ao
garantir novas extensões de habitat que facilitem a dispersão das espécies e forneçam seus
recursos ecológicos (Palmer et al. 1997, Kanowski et al. 2003, Pardini et al. 2005).
Embora a prática da restauração tenha se estabelecido como uma ferramenta de
recuperação de ecossistemas desde o século XIX (Engel e Parrota 2008), a chamada
ecologia da restauração apenas se consolidou como disciplina científica recentemente, a
partir da década de 80 (Young 2000, Palmer et al. 2006). Segundo a Society for Ecological
Restoration (SER, 2004), a restauração é um processo de assistência à recuperação de um
ecossistema degradado, danificado ou destruído, com o intuito de recuperar a integridade
ecológica do ecossistema, a biodiversidade e a variabilidade estrutural e funcional dos
processos ecológicos (SER, 2004).
Com este intuito, diversas técnicas e procedimentos para a recuperação de áreas
degradadas têm sido desenvolvidos e aprimorados, tanto em áreas temperadas (Fiedler et
al. 2010, Quine e Humphrey 2010) quanto em áreas tropicais (Rodrigues et al. 2009, Jong
2010, Kanowski 2010), na busca de uma metodologia que garanta o desenvolvimento de
um ecossistema auto-sustentável (Clewell et al. 2005).
No Brasil, a própria ecologia da restauração foi marcada por uma evolução nos
métodos de recuperação que hoje são focados na diversidade florística e genética como

2
elementos-chave para a evolução e manutenção de um sistema florestal, principalmente da
Mata Atlântica (Ribeiro et al. 2009). Assim, ultimamente, todo o esforço e pesquisa em
restauração tem se centrado sobre a comunidade de plantas para garantir a recuperação do
habitat (Kanowski 2010, Kanowski et al. 2003, Rodrigues et al. 2009). Avaliações da
manutenção de outros processos ecológicos, como a ocupação e a recolonização pela fauna,
no entanto, são ainda muito pouco estudados (Patten 1997, Nicolas et al. 2009).
Os diferentes modelos de restauração podem resultar em diferentes tipos de
complexidade estrutural da vegetação, bem como proporcionar diferentes tipos de
recursos alimentares e abrigos, fatores estes determinantes para garantir a manutenção
tanto da fauna como de toda a biota (Kanowski et al. 2003). A estrutura da vegetação, a
disponibilidade de alimentos e de abrigos podem atuar como filtros (“filters”) na paisagem,
selecionando espécies de fauna colonizadoras cujas características (“traits”) de uso de
espaço, de dieta, de forrageio, por exemplo, são favorecidas pelas tais condições locais (Poff
1997).
O chamado modelo “trait-filters” tem sido utilizado pela ecologia da restauração,
mas centrado sobre a comunidade de plantas principalmente (Hough-Snee et al. 2011,
Summerville et al. 2006, Craig et al. 2012), e tem auxiliado a evidenciar quais condições
ambientais de habitat estão interferindo na composição, distribuição e abundância das
espécies locais (Poff 1997, Weiher e Keddy 2001, Lichti e Murphy 2010).
Nesse sentido, pesquisas que procurem avaliar que filtros ou descritores ambientais
influenciam a composição e a colonização da comunidade de mamíferos terrestres em
áreas restauradas são de elevada importância. Pequenos mamíferos (inferiores a 1 kg),
assim como médios e grandes (superiores a 1 kg) (Nowak 1999), destacam-se por sua
biodiversidade, colocando o Brasil como o maior país em diversidade mundial deste grupo
(Costa et al. 2005). Suas funções ecológicas são igualmente relevantes para a estruturação
das comunidades ecológicas nas florestas tropicais (Cuarón 2000), influenciando a
regeneração e a própria restauração de florestas ao atuarem como dispersores e ou
predadores de sementes e plantas (Terborgh 1988, Silva 2008). Da mesma forma, grandes
predadores de topo desempenham papel-chave no controle demográfico de herbívoros e
mesopredadores, garantindo o equilíbrio da comunidade (Crooks e Soulé 1999). Portanto,

3
garantir a presença e a ocupação desta comunidade é de grande relevância para a
manutenção de um ecossistema sustentável.
Em ambientes naturais, evidências têm demonstrado o efeito positivo da
complexidade da estrutura da vegetação sobre a ocorrência da comunidade de mamíferos
(August 1983, Robinson e Redford 1986). Médios e grandes mamíferos, por exemplo,
podem apresentar preferências e seleções de mesohabitat dentro de sua escala de habitat
ecológica (Dotta e Verdade 2007, Srbek-Araujo e Chiarello 2007, Lyra-Jorge et al. 2008,
Goulart et al. 2009). Enquanto espécies generalistas aparentemente são capazes de utilizar
diferentes ambientes, como por exemplo, o cachorro-do-mato (Cerdocyon thous) (Dotta e
Verdade 2007), espécies especialistas, como a frugívora paca (Cuniculus paca), selecionam
áreas de vegetação densa e fechada (Goulart et al. 2009).
A composição e a distribuição de pequenos mamíferos igualmente são influenciadas
pela estrutura da vegetação, tanto em uma escala de macrohabitat (August 1983, Pardini et
al. 2005, Leiner 2009), quanto em uma escala mais fina de microhabitat (Freitas et al. 1997,
Cunha e Vieira 2002, Naxara et al. 2009, Pinoti et al. 2012), evidenciando a importância
deste filtro para a comunidade como um todo. A estrutura vertical da vegetação, a abertura
de dossel, a disponibilidade de cobertura vegetal e de serapilheira sobre o solo, por
exemplo, atuam como importantes descritores ambientais (ou filtros ambientais) sobre
este grupo (Bergallo e Magnusson 1999, Gentile e Fernandez 1999, Püttker et al. 2008,
Naxara et al. 2009, Pinotti et al. 2012).
Além da estrutura da vegetação, a própria disponibilidade de recursos alimentares e
a disponibilidade de abrigos também atuam como importantes filtros para a comunidade
(Robinson e Redford 1986, Freitas et al. 1997, Munro et al. 2007, Naxara et al. 2009, Pinoti
et al. 2012). Compreender o papel e o efeito de cada um deles sobre as espécies pode ser
um primeiro passo para averiguar que condições são necessárias para a colonização dessas
espécies em ambientes restaurados.
Estudos em áreas de restauração fora do Brasil têm demonstrado que a
complexidade estrutural das áreas restauradas por plantio é determinante na colonização
destas áreas, de tal forma que somente espécies generalistas têm sido capazes de utilizar
tais habitats, diferentemente das espécies especialistas (Cunningham et al. 2007, Muron et
al. 2007).

4
Ao avaliar a ocorrência de grandes mamíferos em restaurações por plantio em
comparação a remanescentes florestais no continente australiano, por exemplo, Munro et
al. (2009) observaram que tanto as espécies arborícolas quanto as terrestres utilizaram os
dois tipos de habitat. Contudo, entre as áreas restauradas, as espécies apresentaram uma
preferência pelas unidades de maior complexidade florística e estrutural, demonstrando
indiretamente a importância deste descritor para sua ocorrência.
Igualmente, ao comparar a comunidade de pequenos mamíferos entre
remanescentes florestais e áreas de restauração por plantio na Fazenda Intermontes, São
Paulo, Leiner (2009) verificou uma maior diversidade e uma distribuição de abundância
mais uniforme nos remanescentes de mata do que nas áreas restauradas por plantio. A
composição também diferiu de tal modo que marsupiais florestais e escansoriais do gênero
Marmosops não foram verificados nas áreas restauradas aparentemente devido à baixa
complexidade estrutural e à ausência de subosque destas localidades (Malcolm 1995,
Munro et al. 2007).
Nichols e Grant (2007), por sua vez, avaliaram a colonização de espécies de
mamíferos em uma área mineradora de bauxita restaurada na Austrália após 30 anos. Os
autores verificaram que a colonização ocorreu principalmente em função da
disponibilidade de recursos alimentares, assim como de abrigos, em função das
abundâncias das espécies nas áreas florestais circundantes, demonstrando assim que
diferentes descritores ambientais podem agir conjuntamente sobre a seleção de habitat por
mamíferos terrestres, além da própria estrutura da vegetação.
Tais estudos de avaliação sobre a colonização da comunidade de mamíferos em
áreas restauradas, no entanto, não evidenciam diretamente os possíveis descritores
ambientais que podem estar influenciando a colonização das espécies em uma escala mais
fina, principalmente quando em referência a mamíferos terrestres de médio e grande porte
(Hobbs et al. 2003, Vesk et al. 2008). Para este grupo, grande parte dos estudos o fazem
numa análise de seleção de habitats dentro de uma escala regional e de paisagem (Munro et
al. 2007, Mazzolli 2010, Lantschner et al. 2012).
A avaliação na menor escala na qual um organismo responde e percebe a
variabilidade de descritores ambientais do ambiente determina o limite inferior no qual o
animal responde ao mesmo (George e Zack 2001). Identificar tais descritores neste nível de

5
escala é de grande relevância para a ecologia da restauração a fim de garantir a presença e
a manutenção destes filtros nas áreas restauradas que serão necessários para a própria
manutenção da comunidade de fauna (George e Zack 2001, Hobbs et al. 2003, Vesk et al.
2008).
Estudos que avaliem este aspecto da ecologia da restauração são escassos
mundialmente (Hobbs et al. 2003, Vesk et al. 2008) e ainda não foram realizados no Brasil.
Elucidar o efeito de filtros ambientais em áreas restauradas por plantio em comparação a
um gradiente de fisionomias distintas, como a regeneração natural, o plantio e
remanescentes florestais pode auxiliar na compreensão do papel desses descritores
ambientais sobre a ocorrência e a abundância das espécies de mamíferos terrestres.
Avaliar a resposta desta comunidade em função de medidas de restauração é
imprescindível para verificar se os modelos de restauração por plantio implantados no país
estão realmente viabilizando a colonização por este grupo. E ao mesmo tempo pode
permitir a elaboração de propostas de manejo que favoreçam a utilização dessas áreas
pelos mamíferos, como bem sua conservação em longo prazo.

6
OBJETIVOS GERAIS
Os principais objetivos deste estudo foram:
- Comparar a comunidade de pequenos, médios e grandes mamíferos terrestres em um
mosaico fisionômico de restauração, procurando avaliar:
a) se a composição, a riqueza e a abundância desta comunidade diferem ou não entre as
fisionomias deste mosaico
b) se a estrutura da vegetação, como filtro ambiental, influencia na preferência ou seleção
de habitat das espécies
c) se a disponibilidade de recursos alimentares, como filtro ambiental, influencia a
ocorrência das espécies nas fisionomias, segundo sua dieta.
Tendo em vista estes objetivos, o primeiro capítulo procurou avaliar tais aspectos
para a assembleia de pequenos mamíferos. Considerando a área de vida dessas espécies, o
estudo procurou avaliar o efeito destes filtros ambientais dentro de sua escala de micro-
habitat. Já o segundo capítulo centrou-se sobre a assembleia de mamíferos de médio e
grande porte, considerando a ocorrência e uso de espaço dentro de uma escala de
mesohabitat.

7
MATERIAL E MÉTODOS
Área de Estudo
O presente estudo foi realizado na Fazenda Intermontes, propriedade particular da
Votorantim Cimentos (337ha), localizada na cidade de Ribeirão Grande, no sudeste do
estado de São Paulo e a oeste da cadeia de montanhas que forma a Serra de Paranapiacaba
(CCRG 2003). Próximo à fazenda está presente outro contínuo florestal, o Parque Estadual
de Intervales, que se interliga a outras unidades de conservação, como o Parque Estadual
Turístico do Alto do Ribeira (PETAR) e o Parque Estadual de Carlos Botelho. Seu relevo
local é caracterizado por “montanhas com vales profundos”, cujos vales ultrapassam 800m
de altitude e os picos ultrapassam 1000m de altitude (Companhia de Cimento Ribeirão
Grande 2003) (Figura 01).
Figura 01: Localização da Fazenda Intermontes no estado de São Paulo e sua localização da na paisagem de entorno (Google Earth).
A região apresenta clima temperado úmido seco (Cfb), segundo Koeppen (1948),
com temperatura média de 18°C nos meses mais frios e 22°C nos meses mais quentes

8
(Nimer 1986). A precipitação média, por sua vez, é de aproximadamente 1300 mm por ano,
com maior concentração no verão.
A fazenda foi adquirida pela Companhia de Cimento Ribeirão Grande (CCRG),
recentemente incorporada pela Votoratim Cimento para a realização de uma medida
compensatória à supressão de vegetação nativa que a empresa empreendeu para
ampliação de sua atividade mineradora na região. Anteriormente à sua aquisição no final
de 1999, a fazenda apresentava como principal atividade produtiva a criação de gado há
pelo menos 15 a 20 anos. Após sua aquisição, um projeto de restauração foi iniciado em
agosto de 2001 com o suporte do Laboratório de Ecologia e Restauração Florestal da
Universidade de São Paulo – ESALQ (Busato et al. 2006).
O projeto de restauração da Fazenda Intermontes foi baseado em um manejo
adaptativo através de um projeto de avaliação contínua, com práticas de monitoramento
(Busato et al. 2006). O reflorestamento foi realizado por meio do plantio de espécies
nativas em áreas de pastagem, a partir de um banco de sementes alóctones formado
durante o projeto. Ao todo foram utilizadas 63 espécies de plantas pertencentes a 27
famílias, das quais 31,7% correspondiam ao grupo de plantas pioneiras e 68,3% ao grupo
de plantas de sucessão tardia e de clímax, e cerca de 50% do total de espécies
correspospondiam a zoocóricas (Nave 2005).
O processo de restauração seguiu três etapas principais e iniciou-se em 2001. A
primeira etapa foi caracterizada pelas medidas pré-plantio com a remoção de fatores
degradativos. A segunda etapa correspondeu a plantação ou a indução de auto-restauração
com o cultivo propriamente dito seguindo o modelo de plantio de mudas 3 x 2 m,
intercalando linhas de ‘grupo de preenchimento’ (com função de rápido cobrimento e
sombreamento da área) com linhas de ‘grupo de diversidade’ (caracterizado
principalmente por um alto número de espécies de sucessão tardia). E em terceiro,
medidas de pós-plantio com o controle e manutenção dos locais restaurados pelo replantio
de plântulas, fertilização superficial, controle de infestação de formigas e da espécie
herbácea exótica Urochloa decumbens (braquiária) para não prejudicar o estabelecimento
de espécies nativas (Nave 2005, Busato et al. 2006). Tais medidas de pós-plantio, no
entanto, foram mantidas somente até 2003.

9
Atualmente, após 10 anos do início da restauração, a fazenda apresenta um mosaico
de fisionomias com fragmentos florestais já presentes antes do reflorestamento, áreas de
pasto que anteriormente apresentavam ou não processos de regeneração e nos quais a
restauração arbórea foi implementada, áreas de pasto que foram mantidas somente com
regeneração natural sem intervenção da restauração por plantio, e capoeiras resultantes
igualmente da regeneração natural (Anexo I). Os fragmentos florestais, em si, possuem uma
vegetação secundária, com árvores de grande porte e estratos verticais definidos, cuja
fitofisionomia típica é Floresta Estacional Semidecídua (altura do dossel de 18 a 20m
aproximadamente) com algumas espécies típicas de Floresta Ombrófila Densa dada à
proximidade com a Serra de Paranapiacaba (CCRG 2003). As áreas de restauração
apresentam um dossel descontínuo na maioria das áreas e com uma fisionomia florestal
baixa, inferior aos fragmentos florestais (de 5 a 14m aproximadamente), e em muitos
pontos, a cobertura herbácea é dominada por braquiária com ausência de subosque
desenvolvido (Leiner 2009, Zaniratto obs. pessoal) (Anexo I).
Delineamento amostral
Para avaliar a comunidade de mamíferos terrestres, 20 parcelas de amostragem
foram sorteadas de um grid de 84 quadrantes, separados por uma distância mínima de
200m entre si e distribuídas ao longo do mosaico de fisionomias presentes na fazenda
Intermontes (Figura 02).
Os pontos amostrais representam um mosaico fisionômico que reúne fisionomias
que se separam abruptamente na paisagem, com características estruturais da vegetação
distintas entre si. Estão presentes: áreas de remanescentes florestais (03 pontos
amostrais), áreas com regeneração natural arbustiva e ou capoeira (07 pontos amostrais) e
áreas com regeneração natural arbórea (05 pontos amostrais), nas quais o plantio foi
realizado, e áreas sem regeneração (04) nas quais o plantio foi introduzido e área na qual o
processo de regeneração natural foi mantido por si só (01 ponto amostral) (Tabela 01,
Anexo I).

10
Figura 02: Localização dos 20 pontos amostrais sorteados na Fazenda Intermontes, na cidade de
Ribeirão Grande, São Paulo.

11
Tabela 01. Pontos amostrais na Fazenda Intermontes e os respectivos tratamentos
originais de restauração onde estão inseridos (siglas de referência ao tratamento em
parênteses).
Cada parcela de amostragem (30 x 30m) foi considerada como uma unidade
amostral para coleta de dados. No interior de cada parcela (30 x 30m), quatro parcelas
menores (ou sub-parcelas de 10 x 10m) foram estabelecidas para a coleta da estrutura da
vegetação, como dos demais descritores ambientais avaliados neste estudo (Figura 03).
Ponto amostral Tipo de tratamento implementado em 2001-2002
1 Remanescente florestal (RF)
2 Remanescente florestal (RF)
3 Remanescente florestal (RF)
4 Pasto com regeneração arbórea + plantio (RARBOP)
5 Pasto com regeneração arbórea + plantio (RARBOP)
6 Pasto com regeneração arbórea + plantio (RARBOP)
7 Pasto com regeneração arbórea + plantio (RARBOP)
8 Pasto com regeneração arbórea + plantio (RARBOP)
9 Pasto com regeneração arbustiva+plantio (RARBUP)
10 Pasto com regeneração arbustiva + plantio (RARBUP)
11 Pasto com regeneração arbustiva + plantio(RARBUP)
12 Pasto com regeneração arbustiva + plantio (RARBUP)
13 Pasto com regeneração arbustiva + plantio (RARBUP)
14 Pasto com regeneração arbustiva + plantio (RARBUP)
15 Capoeira + plantio (RARBUP)
16 Pasto sem regeneração + plantio (PL)
17 Pasto sem regeneração + plantio (PL)
18 Pasto sem regeneração + plantio (PL)
19 Pasto sem regeneração + plantio (PL)
20 Regeneração natural (REG)

12
Figura 03: Configuração da parcela de amostragem e das sub-parcelas presentes em cada ponto
amostral avaliado no interior da Fazenda Intermontes, na cidade de Ribeirão Grande, São Paulo.
Para determinar a posição das parcelas foi considerado um critério mínimo de
distância: cada parcela deveria estar a no mínimo 150m das demais parcelas presentes em
outras fisionomias. Tal distância mínima foi baseada em estudos de frequência de
movimento de pequenos mamíferos que indicam que tais animais podem se locomover por
cerca de 100m por noite (Gentile e Cerqueira 1995, Mendel e Vieira 2003, Leiner 2005,
Mendonça et al. 2010), a fim de garantir a independência espacial entre as unidades
amostrais com relação a assembleia de pequenos mamíferos. Para médios e grandes
mamíferos, contudo, essa independência não é garantida, uma vez que estes animais
podem se deslocar por distâncias maiores do que 150m (Emmons e Feer 1997, Reis et al.
2006). Desta forma, as unidades amostradas tiveram como objetivo representar
mesohabitat distintos que possam ser utilizados pelo animal de médio e grande porte
dentro de sua área de vida, com maior ou menor frequência.
1m
10 x 10m
Unidade amostral do estudo (parcela 30x30m)
30 x 30m
Parcela de vegetação fisionômica (parcela 10x10m)

13
Coleta de Dados
Pequenos mamíferos
Para amostrar a comunidade de pequenos mamíferos foram utilizadas armadilhas
de intercepção e queda (AIQ), método este menos seletivo em comparação às armadilhas
Sherman, por exemplo, por não utilizarem isca para a atração do animal. Comparada a esta
segunda metodologia, as AIQs capturam um maior número de espécies e de indivíduos,
provavelmente refletindo a abundância das espécies com maior precisão, além de permitir
a captura de espécies de hábitos semi-fossoriais ou que se orientam por outros sentidos
que não os visuais, raramente amostradas por outros métodos (Umetsu et al. 2006).
Desta forma, em cada unidade amostral, uma linha de 25m foi instalada com uma
sequência de cinco baldes (60 l) dispostos a cada 5 m entre si e interligados por uma cerca
guia de nylon com 50 cm de altura sustentada por estacas de madeira no centro da parcela
(Anexo II). Sessões mensais de captura foram realizadas de agosto de 2011 a dezembro de
2013, totalizando 24 meses de coletas, haja vista que o alto índice pluviométrico na região
prejudicou a realização das capturas em alguns meses. Cada sessão foi composta por
quatro a cinco dias de coletas de acordo com a intensidade de chuva durante a mesma.
Os animais capturados foram marcados com brincos numerados de cobre e níquel
resistentes a corrosão (modelo N° 01) e soltos no local da captura após registrar os dados
de espécie, de massa corpórea, de sexo, de condição reprodutiva, de medidas biométricas
(medida de comprimento, de cauda, de pata traseira e de orelha), o ponto de captura e a
data de captura. Para fins de identificação e testemunho, alguns espécimes foram coletados,
eutanasiados, taxidermizados e depositados no Museu de Zoologia da Universidade
Estadual de Campinas.
Mamíferos terrestres de médio e grande porte
Os médios e grandes mamíferos foram amostrados através de armadilhas
fotográficas, métodos não invasivos e de boa precisão que permitem estimar a riqueza, a
abundância ou a frequência de ocorrência, como o padrão de atividade de animais
solitários, de baixa densidade populacional e de grandes áreas de vida (Srbek-Araújo e
Chiarello 2007, Lyra-Jorge et al. 2008).

14
Em cada unidade amostral, uma armadilha fotográfica foi colocada na trilha de
acesso de cada unidade. No primeiro ano de coleta, 20 armadilhas da marca Wildview
(modelo Extreme 5) foram dispostas em cada unidade amostral e foram monitoradas
mensalmente a partir de junho de 2012. Dado a problemas de mau funcionamento, no
segundo ano de coleta de dados, a partir de janeiro de 2013, tais câmeras foram
substituídas por 10 armadilhas fotográficas da marca Bushnell (modelo Trophy Cam) e
através de um sistema de rodízio as armadilhas permaneceram nas unidades amostrais a
cada dois meses, e tal processo seguiu até abril de 2014. Nenhum tipo de isca foi utilizado,
garantindo que o registro da distribuição e uso do espaço por parte das diferentes espécies
fosse feito sem interferências, pois a utilização de isca pode tornar a amostra seletiva
(Srbek-Araújo e Chiarello 2007).
Tendo em vista que no segundo ano de coleta uma armadilha foi furtada em um dos
pontos (Ponto 11, RARBUP), descartou-se esse ponto amostral e foram considerados
somente os demais pontos nas análises dessa assembleia.
Caracterização da estrutura dos habitats
A caracterização dos habitats foi realizada através da coleta de dados de estrutura
da vegetação. Tais variáveis foram coletadas no centro de cada uma das quatro sub-
parcelas (10 x 10m) presentes nas parcelas maiores (30 x 30m) de cada unidade amostral
(Anexo II).
Para tal caracterização foram avaliadas sete variáveis da estrutura do habitat,
segundo Freitas et al. (2002) com alguma modificação: cobertura de dossel, cobertura
vegetal (herbácea e lenhosa), cobertura de folhiço, cobertura de solo exposto e obstrução
foliar vertical a três alturas (0 a 0,5 m; 0,5 a 1,0 m e 1,0 a 1,5 m). Para medir tais variáveis
foi utilizado um quadrado de madeira de 0,25 m² dividido em 100 quadrados abertos por
fio de nylon, de tal modo que o número de quadrados obstruídos visualmente pelo
observador (quadrados com obstrução superior a 50%) foram utilizados como medida da
determinada variável. Tal metodologia é simples, rápida e vem sendo utilizada como
método padrão para descrever a estrutura de habitat em Mata Atlântica (Freitas et al. 2002,
Anexo II).

15
As variáveis foram medidas em cinco pontos diferentes: no centro da sub-parcela
(ponto central) e a cada 3,0 m do centro da sub-parcela nas direções norte, sul, leste e oeste
que foram marcados com estacas para facilitar sua visualização (pontos limites para a
tomada das variáveis). Para verificar a medida de obstrução vertical o observador
permaneceu no ponto central e avaliou a porcentagem de quadrados obstruídos
posicionando o quadrado na posição vertical em direção a cada uma das estacas. A
cobertura de dossel foi medida como a porcentagem de quadrados obstruídos, estando o
quadrado posicionado horizontalmente sobre a cabeça do observador em cada um dos
cinco pontos. E a cobertura vegetal, a cobertura de folhiço e a cobertura de solo exposto,
que são variáveis complementares cuja soma deve ser 100%, foram medidas em cada
ponto como a porcentagem de quadrados obstruídos estando o quadrado posicionado
horizontalmente e paralelamente a cerca de 60 cm do solo (Anexo II).
Além das sete variáveis propostas pela metodologia, foi considerada como
importante a coleta de uma oitava variável: a profundidade de cobertura vegetal
(constituída predominantemente por herbácea), haja vista que a profundidade de folhiço,
serapilheira e cobertura herbácea podem influenciar a presença de espécies de pequenos
mamíferos (Naxara et al. 2009, Pinotti et al. 2012). Desta forma, com o auxílio de uma
régua, a profundidade de cobertura vegetal foi medida em cada um dos quatro pontos
cardeais nos quais as medidas de estrutura de habitat foram coletadas pela metodologia de
Freitas et al. (2002), registrando a altura que essa cobertura apresentava em relação ao
solo. Para isso foi utilizada uma régua de 30cm.
As variáveis foram medidas uma única vez na estação chuvosa no ano de 2013, dado
que as variações mensais das variáveis de habitats são desprezíveis (Leiner 2009). A média
das variáveis coletadas para cada unidade amostral foi considerada para as análises dos
dados.
Disponibilidade de recursos alimentares
A disponibilidade de recursos foi avaliada através da disponibilidade de frutos e de
artrópodes, itens estes considerados como recursos alimentares importantes para a
assembleia de pequenos, médios e grandes mamíferos (Stallings 1984, Bodmer 1990,
Paschoal e Galetti 1995, Fonseca et al. 1996, Galetti et al. 2001, Richard e Juliá 2001, Vieira

16
et al. 2003, Gayot et al. 2004, Miranda 2005, Vieira et al. 2006, Graipel et al. 2006, Rocha et
al. 2008, Aguiar et al. 2011, Pinotti et al. 2011). Dado que pequenos mamíferos são recurso
chave importante para espécies da assembleia de médios e grandes mamíferos, assim como
médios mamíferos são recurso para grandes predadores (Bordignon e Monetiro-Filho
1999, Talamoni e Assis 1999, Rau e Jimenéz 2002, Wang 2002, Bueno e Motta-Júnior 2004,
Gatti et al. 2006, Bezerra et al. 2009), essas duas categorias foram inclusas como
descritores ambientais de acordo com a espécie de mamífero avaliada.
A disponibilidade de frutos importantes para essa comunidade foi obtida através do
acompanhamento mensal de todas as espécies com frutos zoocóricos. Para a assembleia de
pequenos mamíferos, frutos com até 2,0m de altura foram considerados, por ser uma fonte
de recursos para essas espécies, tanto de solo como de subosque (Pinotti et al. 2011). Para
médios e grandes mamíferos, todos os frutos, independentes da altura, foram considerados.
O acompanhamento mensal foi realizado em duas sub-parcelas (10 x 10m) sorteadas das
quatro presentes em cada unidade amostral. A produção de frutos de cada indivíduo foi
estimada através da contagem de frutos verdes e maduros, embora somente os frutos
maduros tenham sido posteriormente considerados nas análises por se tratarem de uma
fonte imediata de recurso. A coleta ocorreu de março de 2012 a agosto de 2013 e a média
do número de frutos maduros foi utilizada para análises futuras.
A disponibilidade de artrópodes foi avaliada através de 10 coletores de artrópodes
em cada unidade amostral, constituídos de copos plásticos de 300 ml, enterrados até o
nível do solo e preenchidos com água e gotas de detergente (Almeida et al. 2003, Aquino et
al. 2006). Os coletores foram posicionados paralelamente às armadilhas de interceptação e
queda (AIQs) utilizadas para a coleta de pequenos mamíferos. Dois coletores foram
colocado a uma distância de 1,0 m de cada AIQ, resultando em dez coletores por unidade
amostral (Anexo II). A coleta de artrópodes foi realizada durante dois dias concomitante às
coletas de pequenos mamíferos realizadas mensalmente. Os artrópodes capturados
maiores de 1 mm, selecionados pelo uso de uma peneira deste calibre, foram secados em
estufa (60°C) e pesados em balança de precisão de 0,1 mg para estimar biomassa
disponível (Naxara et al. 2009). A média do peso seco para cada unidade amostral foi
utilizada para as posteriores análises. A coleta desta variável estendeu-se de agosto de
2011 a agosto de 2013.

17
A disponibilidade de pequenos mamíferos foi obtida através de coletas mensais
realizadas de agosto de 2011 a dezembro de 2013, realizada para verificar a assembleia de
pequenos mamíferos em cada ponto amostral (Capítulo 1). Desta forma, foi considerada a
abundância total de todas as espécies, como um dos recusos para algumas das espécies
médios mamíferos registrados. Já os dados de disponibilidade de médios mamíferos,
fonte de recurso considerada para onça-parda (Puma concolor) (Rau e Jimenéz 2002,
Novack et al. 2005, Martins et al. 2008, Thompson et al. 2009), foram determinados pelos
próprios registros obtidos por meio das armadilhas fotográficas durante o estudo (Capítulo
2). As espécies de presa de médio porte consideradas para este predador foi baseada na
literatura, incluindo assim o número total de registro do cachorro-do-mato, dos pequenos
felinos, dos tatus, do tamanduá, dos veados, dos esquilos e da irara (Rau e Jimenéz 2002,
Wang 2002, Novack et al. 2005, Martins et al. 2008, Thompson et al. 2009, Bianchi et al.
2011).
REFERÊNCIAS BIBLIOGRÁFICAS
Aguiar, L. M., Moro-Rios, R. F., Silvestre, T., Silva-Pereira, J. E., Bilski, D. R., Passos, F. C.,
Sekiama, M. L., Rocha, V.J. 2011. Diet of brown-nosed coatis and crab-eating raccons
from a mosaic landscape with exotic plantations in southern Brazil. Studies of
Neotropical Fauna and Environment 46(3): 153-161.
Almeida, L. M., Ribeiro-Costa, C. S. R., Marinori, L. 2003. Manual de coleta, conservação,
montagem e identificação de insetos. Holos, Ribeirão Preto.
Aquino, A. M., Aguiar-Menezes, E. L., de Queiroz, J. M. 2006. Recomendações para coleta de
artrópodes terrestres por armadilhas de queda (pitfall-traps). Circular Técnica, n.
18, Embrapa. Seropédia, Rio de Janeiro.
August, P. 1983. The role of habitat complexity and heterogeneity in structuring tropical
mammal communities. Ecology 64(6): 1495-1507.
Bergallo, H. G., Magnusson, W. E. 1999. Effects of climate and food availability on four
rodents species in southeastern Brazil. Journal of Mammalogy 80(2): 472-486.
Bezerra, B. M., Barnett, A. A., Souto, A., Jones, G. 2009. Predation by tayra on the common
marmoset and the pale-throated three-toed sloth. Journal of Ethology 27: 91-96.

18
Bianchi, R. C., Rosa, A. F., Gatti, A., Mendes, S. L. 2011. Diet of margay, Leopardus wiedii, and
jaguarundi, Puma yagouaroundi, (Carnivora: Felidae) in Atlantic Rainforest, Brazil.
Zoologia 28(1): 127-132.
Bodmer, R. E. 1990. Fruit patch size and frugivory in the lowland tapir (Tapirus terrestris).
Journal of Zoology 222:121-128.
Bordignon, M., Monteiro-Filho, E. L. A. 1999. Seasonal food resources of the squirrel Sciurus
ingrami (Thomas, 1901) in a secondary araucaria forest in southern Brazil. Studies
on Neotropical Fauna and Environment 34: 137-140.
Bueno, A. A, Motta-Junior, C. J. 2004. Food habits of two syntopic canids, tha maned wolf
(Chrysocyon brachyurus) and the crab-eating fox (Cerdocyon thous), in southeastern
Brazil. Revista Chilena de Historia Natural 77: 5-14.
Busato, L. C., Gobbo, P. R., Nave, A. G., Rodrigues, R. R. 2006. Intermontes Project in the
context of Brazilian Field works and researches on restoration. In: High diversity
forest restoration in degraded areas. Nova Science Publishers 222-244.
Clewell, A., Rieger, J., Munro, J. 2005. Guidelines for developing and managing ecological
restoration projects. Second edition. Society for Ecological Restoration International
(Disponível: www.ser.org)
Companhia de Cimento Ribeirão Grande (CCRG) 2003. Ampliação da Mina Limeira: estudo
de impacto ambiental. São Paulo.
Costa, L. P., Leite, Y. L. R., Mendes, S. L., Ditchfield, A. D. 2005. Mammal conservation in
Brazil. Conservation Biology 19(3): 672-679.
Craig, M. D., Hardy, G. E. St. J., Fontaine, J. B., Garkakalis, M. J., Grigg, A. H., Grant, C. D.,
Fleming, P. A., Hobbs, R. J. 2012. Identifying unidirectional and dynamic habitat
filters to faunal recolonisation in restored mine-pits. Journal of Applied Ecology 49:
919-928.
Crooks, K. R., Soulé, M. E. 1999. Mesopredator release and avifaunal extinctions in a
fragmented system. Nature 400:563-566.
Cuarón, A. D. 2000. A global perspective on habitat disturbance and tropical rainforet
mammals. Conservation Biology 14(6): 1574-1579.

19
Cunha, A. A., Vieira, M. V. 2002. Support diameter, incline, and vertical movements of four
didelphid marsupials in the Atlantic forest of Brazil. Journal of Zoology 258: 419-
426.
Cunningham, R. B., Lindenmayer, D. B., Crane, M. et al. 2007. Reptile and arboreal
marsupials response to replanted vegetation in agricultural landscapes. Ecological
Applications 17(2): 609-619.
Dotta, G., Verdade, L. M. 2007. Trophic categories in a mammal assemblage: diversity in an
agricultural landscape. Biota Neotropica 7(2): 287-292.
Emmons, L. H., Feer, F. 1997. Neotropical rainforest mammals – a field guide. The
University of Chicago Press. 2ª edição.
Engel, V. L., Parrotta, J. A. 2008. Definindo a restuarção ecológica: tendências e perspectivas
mundiais. In: Kageyama, P. Y., Oliveira, R. E., Moraes, L. F. D., Engel, V. L., Gandara, F.
B. (eds). Restauração ecológica de ecossistemas naturais. Pp: 1-26.
Fahrig, L. 2002. Effect of habitat fragmentation on the extinction threshold: a synthesis.
Ecological Applications 12(2): 346-353.
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology,
Evolution and Systematics 34: 487-515.
Fiedler, C. E., Metlen, K. L., Dodson, E. K. 2010. Restoration treatment effect on stand
structure, tree growth, and fire harzard in a Ponderosa Pine Douglas-Fir forest
Montana. Forest Science 56(1): 18-31.
Fonseca, G. A. B., Hermann, G., Leite, Y. L. R., Mittermeier, R. A., Rylands, A. B., Patton, J. L.
1996. Lista anotada dos mamíferos do Brasil. Occasional Papers in Conservation
Biology 4: 1-38.
Freitas, S. R., de Moraes, D. A., Santori, R. T., Cerqueira, R. 1997. Habitat preference and food
use by Metachirus nudicaudatus and Didelphis aurita (Didelphimorphia,
Didelphidae) in a restinga forest at Rio de Janeiro. Revista Brasileira de Biologia
57(1): 93-98.
Freitas, S. R., Cerqueira, R., Vieira, M. V. 2002. A device and standard variables to describe
microhabitat structure of small mammals based on plant cover. Brazilian Journal of
Biology 62(4b): 795-800.

20
Galetti, M., Keuroghlian, A., Hanada, L., Morato, M. I. 2001. Frugivory and seed dispersal by
lowland tapir (Tapirus terrestris) in Southeast Brazil. Biotropica 33(4): 723-726.
Gascon, C., Williansom, G. B., Fonseca, G. A. B. 2000. Receding forest edges and vanishing
reserves. Science 288: 1356-1358.
Gatti, A., Bianchi, R., Rosa, C. R. X., Mendes, S. L. 2006. Diet of two sympatric carnivores,
Cerdocyon thous ans Procyon cancrivorus, in a resting area of Espirito Santo State,
Brazil. Journal of Tropical Ecology 22: 227-230.
Gayot, M., Henry, O., Dubost, G., Sabatier, D. 2004. Comparative diet of the two forest
cervids of the genus Mazama in French Guiana. Journal of Tropical Ecology 20(1):
31-43.
Gentile, R., Cerqueira, R. 1995. Movement patterns of five species of small mammals in a
Brazilian restinga. Journal of Tropical Ecology 11(4): 671-677.
Gentile, R., Fernandez, F. A. S. 1999. Influence of habitat structure on a streamside small
mammal community in a Brazilian rural area. Mammalia 63(1): 29-40.
George, T. L., Zack, S. 2001. Spatial and temporal considerations in restoring habitats for
wildlife. Restoration Ecology 9(3): 272-279.
Goulart, F. V. B., Cáceres, N. C., Graipel, M. E., Tortato, M. A., Ghizoni Jr, I. R., Oliveira-Santos,
L. G. R. 2009. Habitat selection by large mammals in a southern Brazilian Atlantic
forest. Mammalian Biology 74: 182-190.
Graipel, M. E., Cherem, J. J., Monteiro-Filho, E. L. A., Glock, L. 2006. Population dynamics of
marsupials and rodents in Parque Municipal da Lagoa do Peri, Santa Catarina Island,
southern Brazil. Mastozoologia Neotropical 13(1): 31-49.
Hobbs, R., Catling, P. C., Wombey, J. C., Clayton, M., Atkins, L., Reid, A. 2003. Faunal use of
bluegum (Eucalyptus globulus) plantations in southwestern Australia. Agroforestry
Systems 58: 195-212.
Hough-Snee, N., Long, A. L., Jeroue, L., Ewing, K. 2011. Mounding alters environmental
filters that drive plant community development in a novel grassland. Ecological
Engineering 37: 1932–1936.
Jong, W. 2010. Forest Rehabilitation and its implications for forest transition theory.
Biotropica 42(1): 3-9.

21
Kanowski, J, Catterall, C. P., Wardell-Johnson, G. W., Prostor, H., Reis, T. 2003. Development
of forest structure on cleared rainforest land in Australia under different styles of
reforestation. Forest Ecology and Management 183: 265-280.
Kanowski, J. 2010. What have we learnt about rainforest restoration in the past two
decades? Ecological Managment and Restoration 11(1): 2-3.
Köeppen, W. 1948. Climatologia. México-Buenos Aires. Ed. Fundo de Cultura Econômica.
Lantschner, M. V., Rusch, V., Hayes, J. P. 2012. Habitat use by carnivores at different spatial
scales in a plantation forest landscape in Patagonia, Argentina. Forest Ecology and
Management 269: 271-278.
Leiner, N. O. 2005. Ecologia alimentar e reprodutiva de Marmosops paulensis
(Didelphimorphia: Didelphidae) em uma área de Mata Atlântica no sudeste de São
Paulo. Dissertação de Mestrado, Universidade Estadual de Campinas.
Leiner, N. O. 2009. Padrões de uso do espaço em múltiplas escalas por roedores e marsupiais
de Mata Atlântica. Tese de Doutorado, Universidade Estadual de Campinas.
Lichti, N. I., Murphy, M. T. 2001. Selection ratios on community aggregated traits estimate
ecological filters imposed on species by sites. Ecology 91(2): 347-354.
Lyra-Jorge, M. C., Ciocheti, G., Pivello, V. R. 2008. Carnivore mammals in a fragmented
landscape in northeast os São Paulo State, Brazil. Biodiversity and Conservation
17(7): 1573-1580.
Lugo, A. E., Helmer, E. 2004. Emerging forests on abandoned land: Porto Rico’s new forests.
Forest Ecology and Management 190: 145-161.
Malcom, J. R. 1995. Forest structure and abundance and diversity of small mammals. In:
Forest canopies. Academic Press, San Diego, California. Pp:179-197.
Martins, R., Quadros, J., Mazzolli, M. 2008. Hábito alimentar e interferência antrópica na
atividade de marcação territorial do Puma concolor e Leopardus pardalis (Carnivora:
Felidae) e outros carnívoros na Estação Ecológica da Juréia-Itatins, São Paulo, Brasil.
Revista Brasileira de Zoologia 25(3): 427-435.
Mazzolli, M. 2010. Mosaics of exotic forest plantations and native forests as habitat of
pumas. Environmental Management 46: 237-253.
Mendel, S. M., Vieira, M. V. 2003. Moviment distances and density estimation of small
mammals using the spool-and-line technique. Acta Theriologica 48: 289-300.

22
Mendonça, A. F., Bocchiglieri, A., Vieira, M. V. 2010. Spool-and-line in a backpack: a new
technique for studying movement of small mammals. Mammalia 74: 209-211.
Miranda, J. M. D. 2005. Dieta de Sciurus ingrami Thomas (Rodentia, Sciuridae) em um
remanescente de floresta com araucária, Paraná, Brasil. Revista Brasileira de
Zoologia 22(4): 1141-1145.
Munro, N. T., Lindenmayer, D. B., Fischer, J. 2007. Faunal response to revegetation in
agricultural areas of Australia: a review. Ecological Managmente and Restoration
8(3): 199-207.
Munro, N. T., Fischer, J., Wood, J., Lindenmayer, D.B. 2009. Revegetation in agricultural
areas: the development of structural complexity and floristic diversity. Ecological
Applications 19(5): 1197-1210.
Naxara, L., Pinotti, B. T., Pardini, R. 2009. Seasonal microhabitat selection by terrestrial
rodents in an old-growth Atlantic forest. Journal of Mammalogy 90: 404-415.
Nave, A. G. 2005. Banco de sementes autóctone e alóctone, resgate de plantas e plantio de
vegetação nativa na Fazenda Intermontes, município de Ribeirão Grande, SP. Tese de
Doutorado, Universidade de São Paulo.
Nichols, O. G., Grant, C. D. 2007. Vertebrate fauna recolonization of restored bauxite mines –
key findings from almost 30 years of monitoring and research. Restoration Ecology
15(4): S116-S126.
Nicolas, V., Barrière, P., Tapiero, A., Colyn, M. 2009. Shrew species diversity and abundance
in Ziama Biosphere Reserve, Guinea: comparison among primary forest, degraded
forest and restoration plots. Biodiversity and Conservation 18: 2043-2061.
Nimer, E. 1986. Climatologia do Brasil. IBGE, Rio de Janeiro.
Nowak, R. M. 1999. Walker’s mammals of the world. Volume 2. Sixth edition. John Hopkins
University Press, Baltimore, USA.
Novack, A. J., Main, M. B., Sunquist, M. E., Labisky, R. F. 2005. Foraging ecology of jaguar
(Panthera onca) and puma (Puma concolor) in hunted and non-hunted sites within
the Maya Biosphere Reserve, Guatemala. Journal of Zoology 267: 167-178.
Palmer, M. A., Ambrose, R. F., Poff, N. L. 1997. Ecological theory and community restoration
ecology. Restoration Ecology 5(4): 291-300.

23
Palmer, M. A., Falk, D. A., Zedler, J. B., 2006. Ecological theory and restoration ecology. In:
Falk, D. A., Palmer, M. A., Zedler, J. B. (Eds.), Foundations of Restoration Ecology.
Island Press, Washington, pp. 1–10.
Pardini, R. De Sousa, S. M., Braga-Neto, R., Metzger, J. P. 2005. The role of forest structure,
fragment size and corridors in maintaining small mammal abundance and diversity
in an Atlantic forest landscape. Biological Conservation 124: 253-266.
Paschoal, M., Galetti, M. 1995. Seasonal food use by Neotropical squirrel Sciurus ingrami in
Southeastern Brazil. Biotropica, 27(2): 268-273.
Patten, M. A. 1997. Reestablishment of a rodent community in restored desert scrub.
Restoration Ecology 5(2): 156-161.
Pinotti, B. T., Naxara, L., Pardini, R. 2011. Diet and food selection by small mammals in an
old-growth Atlantic Forest. Studies on Neotropical Fauna and Environment 46: 1-9.
Pinotti, B. T., Pagotto, C. P., Pardini, R. 2012. Habitat structure and food resources for
wildlife across sucessional stages in a tropical forest. Forest Ecology and
Management 283:119-127.
Poff, N. L. 1997. Landscape filters and species traits: towards mechanistic understanding
and prediction in stream ecology. Journal of the North American Benthological
Society 16(2): 391-409.
Püttker, T., Pardini, R., Meyer-Lucht, Y. e Sommer, S. 2008. Responses of five small mammal
species to micro-scale variations in vegetation structure in secondary Atlantic
Forest remnants, Brazil. BMC Ecology 8(9): 1-10.
Quine, C. P., Humphrey, J. W. 2010. Plantantions of exotic tree species in Britain: irrelevant
for biodiversity or novel habitat for native species? Biodiversity and Conservation
19(5): 1503-1512.
Rau, J. R., Jiménez, J. E. 2002. Diet of puma (Puma concolor, Carnivora:Felidae) in Coastal
and Andean ranges of southern Chile. Studies of Neotropical Fauna and Environment
37(3): 201-205.
Reis, N. R., Ortêncio Filho, H., Silveira, G. 2006. Mamíferos do Brasil. Eds: Reis, N. R.; Shibatta,
O. A.; Peracchi, A. L.; Pedro, W. A.; Lima, I. P. Londrina.

24
Ribeiro, M. C., Metzger, J. P., Martensen, A. C., Ponzoni, F. J., Hirota, M. M. 2009. The Brazilian
Atlantic Forest: How much is left, and how is the remaining forest distributed?
Implications for conservation. Biological Conservation 142: 1141-1153.
Richard, E., Juliá, J. P. 2001. Dieta de Mazama gouazoubira (Mammalia, Cervidae) en un
ambiente secundário de Yungas, Argentina. Série Zool. 90: 147-156.
Robinson, J. G., Redford, K. H. 1986. Body size, diet, and population density of Neotropical
forest mammal. American Naturalist 128(5): 665-680.
Rocha, V. J., Aguiar, L. M., Silva-Pereira, J. E., Moro-Rios, R. F., Passos, F. C. 2008. Feeding
habitats of the crab-eating fox, Cerdocyon thous (Carnivora: Canidae), in a mosaic
area with native and exotic vegetation in Southern Brazil. Revista Brasileira de
Zoologia 25(4): 594-600.
Rodrigues, R. R., Lima, R. A., Gandolfi, S., Nave, A. G. 2009. On the restoration of high
diversity forests: 30 years of experience in the Brazilian Atlantic forest. Biological
Conservation 142: 1242-1251.
Sbrek-Araujo, A. C., Chiarello, A. G. 2007. Armadilhas fotográficas na amostragem de
mamíferos: considerações metodológicas e comparação de equipamentos. Revista
Brasileira de Metodologia 24(3): 647-656.
Silva, W. R. 2008. A importância das interações planta-animal nos processos de restauração.
In: Kageyama, P. Y., Oliveira, R. E., Moraes, L. F. D., Engel, V. L., Gandara, F. B. (eds).
Restauração ecológica de ecossistemas naturais. Pp: 77-90.
Society for Ecological Restoration. 2004. SER International primer on ecological
restoration. Versão 2. (Disponível: http://www.ser.org/resources/)
Stallings, J. R. 1984. Notes of feeding habits of Mazama gouazoubira in the Chaco Boreal of
Paraguay. Biotropica 16(2): 155-157.
Summerville, K. S., Conoan, C. J., Steinchen, R. M. 2006. Species trait as predictors of
Lepidopteran composition in restored and remnant tallgrass prairies. Ecological
Applications 16(3): 891-900.
Talamoni, S. A., Assis, M. A. C. 2009. Feeding habit of the Brazilian tapir, Tapirus terrestris
(Perissodactyla: Tapiridae) in a vegetation transition zone in south-eastern Brazil.
Zoologia 26(2):251-254.

25
Terborgh, J. 1988. The big things that run the world – A sequel to E. O. Wilson. Conservation
Biology 2(4):402-403.
Thompson, D. J., Fecske, D. M., Jenks, J. A., Jarding, A. R. 2009. Food habits of recolonizing
cougars in the Dakotas: prey obtained from prairie and agricultural habitats.
American Midlist Naturalist 161: 69-75.
Turner, I. M., Corlett, R. T. 1996. The conservation value of small, isolated fragments of
lowland tropical rain forest. Trends in Ecology and Evolution 11(8): 330-333.
Umetsu, F., Naxara, L., Pardini, R. 2006. Evaluating the efficiency of pitfall traps for
sampling small mammals in the Neotropics. Journal of Mammalogy, 87(4): 757-765.
Vesk, P. A., Nolan, R., Thomson, J. R., Dorrough, J. W., Mac Nally, R. 2008. Time lags in
provision of habitat resources through revegetation. Biological Conservation 141:
174-186.
Vieira, E. M., Pizo, M. A., Izar, P. 2003. Fruit and seed exploitation by small rodents of the
Brazilian Atlantic forest. Mammalia. 67(4): 533-539.
Vieira, E. M., Paise, G., Machado, P. H. D. 2006. Feeding of small rodents on seeds and fruits:
a comparative analysis of three species of rodents of the Araucaria forest, southern
Brazil. Acta Theriologica 51 (3): 311-318.
Wang, E. 2002. Diets of ocelots (Leoparuds pardalis), margays (L. wiedii), and oncillas (L.
tigrinus) in the Atlantic rainforest in southeast Brazil. Studies on Neotropical Fauna
and Environment 37(3): 207-212.
Weiher, E., Keddy, P. A. 2001. Ecological assembly rules: perspectives, advances, retreats.
Cambridge University Press.
Young, T. P. 2000. Restoration ecology and conservation biology. Biological Conservation
92:73-83.

26
Anexo I. Imagens fotográficas das fisionomias características de cada ponto amostral estudado na
Fazenda Intermontes, na cidade de Ribeirão Grande, SP.
a b
c d
e f

27
g h
i j
k l

28
m n
o p
q r

29
Legenda: Pontos amostrais: Ponto 01 (a), Ponto 02 (b), Ponto 03 (c), tratamentos tipo remanescente
florestal (borda verde); Ponto 04 (d), Ponto 05 (e), Ponto 06 (f), Ponto 07 (g), Ponto 08 (h), tratamentos
tipo pasto com regeneração arbórea + plantio (borda amarela); Ponto 09 (i), Ponto 10 (j), Ponto 11 (k),
Ponto 12 (l), Ponto 13 (m), Ponto 14 (n), Ponto 15 (o), tratamentos tipo pasto com regeneração arbustiva
+ plantio (borda laranja); Ponto 16 (p), Ponto 17 (q), Ponto 18 (r), Ponto 19 (s), tratamentos tipo pasto
plantio (borda azul); Ponto 20 (t), regeneração natural (borda vermelha).
s t

30
Anexo II. Ilustração: a) da disposição das linhas de interceptação e queda em cada ponto amostral presente no
interior da Fazenda Intermontes; b) da metodologia utilizada para a coleta das variáveis estruturais da vegetação,
de acordo com Freitas et al. 2002.
a)
b)
1m5m
1m
10 x 10m
Armadilhas de interceptação e queda Parcela de vegetação fisionômica (parcela 10x10m)
Unidade amostral do estudo (parcela 30x30m) Coletores de artrópodes
Cerca guia
10 x 10m

31
CAPÍTULO 1: “Efeito de filtros ambientais sobre a colonização de pequenos
mamíferos em um mosaico de restauração de Mata Atlântica”
RESUMO
Processos de restauração têm como objetivo recuperar áreas degradadas, visando à
recomposição da vegetação, como a colonização de espécies de fauna. Todavia, estudos que
avaliem este aspecto da comunidade de fauna ainda são escassos e até o momento
nenhuma avaliação foi feita quanto aos projetos de restauração no Brasil. Compreender
quais são os descritores ou filtros ambientais que influenciam positiva ou negativamente a
presença e a colonização dessas espécies é imprescindível para que estratégias de
restauração sejam implantadas. Desta forma, este estudo teve como objetivo investigar a
resposta da comunidade de pequenos mamíferos em um mosaico de restauração de
floresta Mata Atlântica, incluindo como tratamentos, remanescentes florestais, áreas de
plantio implantadas em locais que inicialmente apresentavam regeneração arbustiva ou
arbórea e pasto em início de regeneração. Tendo em vista o modelo “trait-filter”, a riqueza,
a abundância e a composição da comunidade foram avaliadas quanto ao efeito de dois
descritores ambientais: a estrutura da vegetação e a disponibilidade de recursos
alimentares (frutos maduros e artrópodes). A riqueza, a composição e a abundância não
evidenciaram um padrão comum quanto aos tipos de tratamento. Espécies generalistas,
representadas principalmente por Oligoryzomys nigripes e Akodon montensis,
predominaram na comunidade de pequenos mamíferos, estando presente em todas as
unidades fisionômicas, mas principalmente nas áreas de plantio e de início de regeneração,
sendo favorecidas pela presença de intensa cobertura vegetal sob o solo. Espécies
especialistas estiveram restritas às áreas de remanescentes florestais e, em contrapartida,
estiveram negativamente relacionadas à presença da cobertura vegetal sob o solo. De
acordo com a dieta das espécies, artrópodes e frutos apresentaram efeitos positivos e
negativos sobre as espécies. O conjunto de variáveis da estrutura da vegetação como da
disponibilidade de recursos alimentares geralmente agiram em conjunto sobre a

32
abundância das espécies, demonstrando o importante papel sobre a distribuição das
espécies.
ABSTRACT
Restoration projects intend to recover degraded areas, promoting the restoration of
vegetation structure and also fauna recolonization. However, studies that evaluate this
fauna recolonization are still scarce and so far no study has investigated this aspect in
Brazilian restoration projects. To understand which habitat-filters may limit positively or
negatively the presence and recolonization of fauna species is paramount for the
development of new managed strategies for restoration processes. This study aimed to
investigate the small mammal community in a mosaic of Atlantic forest restored site,
including forest remnants, plantings and pasture regeneration. Considering the habitat
trait-filter model, we evaluated the richness, the abundance and the composition of the
small mammal community in relation to two kinds of habitat-filters: vegetation complexity
and food resource (mature fruits and arthropods). Richness, abundance and composition
did not present any clear pattern in relation to the vegetation treatment considered (forest
remnants, plantings and pasture). Generalized species, mainly represented by Oligoryzomys
nigripes and Akodon montensis, were the most abundant species in the community and
were registered in all vegetation treatments, especially in pasture, being favored by
vegetation cover over the ground. Specialized species were restricted to forest remnants
and, on the other hand, were negatively affected by vegetation cover over the ground.
According to species diet, arthropods and mature fruits showed positive and negative effect
over small mammal species. Vegetation structure and food resources act together over the
small mammal abundance, showing the main role of those habitat filters in species
distribution.

33
INTRODUÇÃO
A ecologia da restauração atua recentemente como um importante processo de
recuperação de áreas degradadas, visando não somente a recuperação estrutural como
funcional do habitat (Palmer et al. 1997, Young 2000). Estudos que avaliem a efetividade
desta recuperação, no entanto, têm se centrado sobre a comunidade de plantas, avaliando
questões fisionômicas, florísticas e genéticas (Kanowski 2010, Kanowski et al. 2003,
Rodrigues et al. 2009). Pouca atenção tem sido dada sobre a comunidade da fauna que
recoloniza essas áreas (Patten 1997, Nicolas et al. 2009), ainda que o sucesso da
restauração possa ser dado somente com o estabelecimento desta comunidade na nova
área recuperada (Patten 1997).
Neste sentido, pequenos mamíferos representam um grupo importante para avaliar
a efetividade desses programas de restauração, pois seu pequeno tamanho e sua alta
capacidade reprodutiva permitem que suas populações geralmente respondam rápido às
mudanças e aos distúrbios do habitat (Brown e Ernest 2000, Stone 2007). Em florestas
tropicais, este grupo é considerado um importante bioindicador dos efeitos e alterações
antrópicas ao responder à perda e à mudança de habitat, à estrutura e heterogeneidade da
vegetação (Pardini et al. 2005, Lambert et al. 2006, Umetsu e Pardini 2007).
Ainda assim, pequenos mamíferos possuem papéis ecológicos importantes ao
influenciarem a regeneração e a estruturação de florestas tropicais por atuarem como
dispersores e predadores de sementes e plantas (Fleming 1975, Terborgh 1988, Cáceres e
Lessa 2012), ao intervirem na reciclagem de nutrientes, por escavarem o solo para
remoção de sementes, de materiais vegetais, de fungos e de invertebrados, inclusive
influenciando a hidrologia do solo (Boal e Mannan 1994, Martin 2003), e por serem presas
importantes para diferentes predadores neotropicais (Núñez et al. 2000, Facure et al. 2003,
Vieira e Port 2007, Martins et al. 2008).
Algumas propostas de restauração da vegetação assumem que a colonização das
espécies de fauna irá ocorrer naturalmente, como prevê a Hipótese do “Field of Dreams”
(Palmer et al. 1997, Kurz et al. 2013). Contudo, diferentes estudos demonstram que nem
todas as espécies são capazes de colonizar as áreas recuperadas, principalmente as
espécies especialistas (Nichols e Nichols 2003, Cunningham et al. 2007, Munro et al. 2007).

34
Compreender quais são os fatores que podem influenciar positiva ou negativamente a
presença e a colonização dessas espécies são imprescindíveis para que estratégias de
restauração sejam implementadas. O chamado modelo “trait-filters”, que busca entender
quais são os filtros ou descritores ambientais que limitam e interferem na presença, na
abundância e na composição das espécies em uma dada escala de habitat (Summerville et
al. 2006, Poff 1997, Weiher e Keddy 2001, Lichti e Murphy 2010), pode representar uma
ferramenta importante para evidenciar tais fatores e permitir a aplicação dessas medidas
estratégicas para que as restaurações sejam efetivas.
Estudos em floresta de Mata Atlântica demonstram que a estrutura vertical da
vegetação, o dossel, a disponibilidade de cobertura vegetal e de serapilheira sobre o solo,
bem como a disponibilidade de artrópodes e frutos, por exemplo, atuam como importantes
descritores ambientais sobre a composição e abundância de pequenos mamíferos em
remanescentes e áreas de florestas secundárias, como áreas de florestas primárias
(Bergallo e Magnusson 1999, Gentile e Fernandez 1999, Püttker et al. 2008, Naxara et al.
2009, Pinotti 2010). Até o momento, contudo, nenhum estudo avaliou se tais descritores
ambientais também agem ou estão presentes em áreas de restauração de Mata Atlântica e
se os projetos de restauração propostos e implementados são efetivos em garantir a
recolonização das espécies de pequenos mamíferos.
Tendo isso em vista, este estudo teve como objetivo avaliar a comunidade de
pequenos mamíferos em um mosaico de restauração de Mata Atlântica (incluindo
remanescentes florestais, áreas de plantio e áreas de regeneração natural) no sudeste do
Brasil, a fim de verificar se a complexidade da vegetação e a disponibilidade de recursos
atuam como filtros ambientais sobre a riqueza, a abundância e a composição das espécies
de pequenos mamíferos, avaliando a efetividade do processo de restauração, e com base
nisso propor medidas de manejo que favoreçam a recolonização dessas espécies.
Assim como verificado em outros estudos em áreas de restauração (Nichols e
Nichols 2003, Cunningham et al. 2007, Munro et al. 2007), nossa hipótese é que a riqueza, a
abundância e a composição das espécies nas áreas de plantio e de regeneração somente
serão semelhantes aos remanescentes florestais se a complexidade estrutural da vegetação
presente nesses tratamentos atingir o mesmo padrão de complexidade dos remanescentes.
No entanto, acreditamos que não somente a estrutura da vegetação, como também a

35
disponibilidade de recursos, devem agir como filtros ambientais importantes na presença e
na abundância das espécies. Espécies especialistas devem responder a variáveis estruturais
diferentes das espécies generalistas, haja vista que as necessidades biológicas dessas
espécies não são as mesmas. E quanto maior for a disponibilidade de recursos, maior será a
abundância das espécies que os utilizam.
MATERIAL E MÉTODOS
Para Material e Métodos vide seção anterior (a partir da página 07).
Análise dos dados
Caracterização da estrutura do habitat
A coleta das variáveis estruturais teve como objetivo caracterizar cada unidade
fisionômica com relação a sua complexidade estrutural. Tendo em vista que a estrutura foi
avaliada por oito variáveis da vegetação e tais variáveis poderiam estar correlacionadas
entre si em função do método complementar das medidas estruturais, seguiu-se
primeiramente com a investigação desta possível relação. Uma matriz de correlação
múltipla foi construída e somente as variáveis não correlacionadas ou com o menor
número e intensidade de correlação foram consideradas para futuras análises. Para as
variáveis representadas por dados de porcentagem, seguiu-se a transformação das mesmas
pelo arcoseno da raiz quadrada (Zar 1999) antes de serem utilizadas em outras análises.
A fim de investigar mais especificadamente o padrão dessas variáveis estruturais
com relação aos diferentes tratamentos, uma Análise de Componentes Principais (PCA) por
matriz correlacionada e com os dados padronizados foi realizada (Quinn e Keough 2002,
Borcard et al. 2011).
Todas as análises estatísticas foram realizadas através do Programa R Studio
(RStudio: Integrated development environment for R, versão 0.98.501 2013).

36
Riqueza, composição e abundância da comunidade de pequenos mamíferos
Para cada unidade amostral, calculamos a riqueza (número de espécies) e o índice
de abundância relativa (número de indivíduos capturados, segundo Naxara et al. 2009,
Slade e Blair 2000).
Para avaliar a riqueza observada entre as unidades amostrais fisionômicas, curvas
de acúmulo médio de espécies baseadas no número de indivíduos foram construídas
utilizando a riqueza observada (Mao Tao) (Santos 2003). A eficiência do método de captura
também foi avaliada comparando-se as curvas de acúmulo baseadas no esforço amostral da
riqueza observada (Mao Tao) com a riqueza estimada (estimador não paramétrico Chao 2)
para cada unidade amostral fisionômica. Para isso foi utilizado o programa EstimateS 8.0
(Colwell 2005).
A similaridade na composição da comunidade de mamíferos entre as unidades de
amostragem foi avaliada através de uma análise de agrupamento, utilizando as médias das
distâncias euclidianas entre os grupos. Tal análise foi realizada através do Programa R
Studio (RStudio: Integrated development environment for R, versão 0.98.501 2013).
Investigação dos descritores ambientais sobre a comunidade de pequenos
mamíferos
Para avaliar o papel dos descritores ambientais (estrutura da vegetação,
disponibilidade de recursos) na composição das espécies nas unidades amostrais
fisionômicas foi realizada uma Análise de Redundância (RDA). Tal análise é um método
multivariado de ordenação constrangida, que combina uma análise de regressão com uma
PCA, permitindo associar os parâmetros ambientais com a comunidade de espécies de
pequenos mamíferos (Borcard et al. 2011).
Para o uso dos dados de espécies de pequenos mamíferos na análise de ordenação
constrangida, os mesmos foram transformados pelo método de Hellinger, uma medida
recomendada para a ordenação de dados de abundância de espécies (Legendre e Gallagher
2001). Para tal análise somente as espécies mais abundantes foram consideradas (N>10).
Embora a RDA permita um análise direta da abundância das espécies em relação às
variáveis ambientais, também investigamos a influência das variáveis de estrutura da

37
vegetação e da disponibilidade de recursos sobre abundância de cada espécie da
assembleia de pequenos mamíferos, através da construção de modelos lineares
generalizados com distribuição binomial negativa e função de ligação log, considerados
adequados para dados de contagem que possam apresentar alta dispersão (Quinn e Keough
2002, Bolker 2007). A comparação entre os modelos foi realizada pelo Critério de
Informação de Akaike modificado para amostras pequenas (AICc) (Burnham e Anderson
2002, Quinn e Keough 2002). Para cada modelo calculamos o AICc, a diferença entre o AICc
de cada modelo com aquele do melhor modelo (ΔAICc) e os pesos de evidência que indicam
que o modelo é verdadeiro (wi) (Burnham e Anderson 2002). Como critério, foram
considerados plausíveis todos os modelos com o ΔAICc < 2.
A análise foi realizada tanto para a assembleia como um todo, como separadamente
para cada uma das espécies de pequenos mamíferos que foram mais abundantes (N>10).
Em relação às variáveis ambientais de estrutura da vegetação, foram utilizados os dois
primeiros eixos da PCA da caracterização da estrutura do habitat das unidades amostrais
como representantes da variável independente para diminuir o número e complexidade
dos modelos competidores (Burnham e Anderson 2002). Com este mesmo objetivo, os
modelos múltiplos, que englobaram mais de uma variável ambiental, não consideraram a
interação entre as variáveis, até para permitir a verificação do efeito de cada filtro
ambiental exclusivamente. Desta forma, foram criados: um modelo nulo, em que não há a
influência de nenhuma variável independente; modelos simples, nos quais foi considerada
apenas uma única variável independente; e modelos múltiplos, nos quais foram
consideradas as combinações entre as variáveis independentes.
Todas as análises estatísticas foram realizadas através do Programa R Studio
(RStudio: Integrated development environment for R, versão 0.98.501 2013).
RESULTADOS
Caracterização dos habitats
A matriz de correlação múltipla construída para avaliar a correlação entre as
variáveis estruturais demonstrou haver correlação entre as seguintes variáveis: a

38
porcentagem de cobertura vegetal, a porcentagem de folhiço, profundidade de cobertura
vegetal, e entre as variáveis de obstrução vertical (p < 0,05; Tabela 01).
Tabela 01. Resultados da Análise de Correlação Múltipla realizado para as variáveis
ambientais medidas nos 20 pontos amostrais localizados na Fazenda Intermontes, na
cidade de Ribeirão Grande, SP. Legenda: Cob – quantidade de cobertura vegetal (herbácea
ou lenhosa), Folh –quantidade de cobertura de folhiço, Dos – fechamento de dossel, ObstI –
obstrução a 0,5m do solo, ObstII – obstrução a 1,0m do solo, ObstIII – obstrução a 1,5m do
solo, CobVeg – profundidade de cobertura vegetal, Solo – quantidade de solo exposto.
Variáveis R² p Variáveis R² p
Cob x Folh 0,876 < 0,0001
Solo x ObstI 0,106 0,1602
Cob x Solo 0,008 0,6993
Solo x ObstII 0,306 0,0112
Cob x Dos 0,201 0,0469
Solo x ObstIII 0,194 0,0519
Cob x ObstI 0,479 0,0007
Solo x CobVeg 0,027 0,4891
Cob x ObstII 0,136 0,1091
Dos x ObstI 0,248 0,0251
Cob x ObstIII 0,011 0,6492
Dos x ObstII 0,263 0,0206
Cob x CobVeg 0,9618 0,0001
Dos x ObstIII 0,076 0,2382
Folh x Solo 0,190 0,0542 Dos x CobVeg 0,165 0,0753
Folh x Dos 0,443 0,0013 ObstI x ObstII 0,615 < 0,0001
Folh x ObstI 0,549 0,0002
ObstI x ObstIII 0,160 0,0801
Folh x ObstII 0,280 0,0163
ObstI x CobVeg 0,006 0,7379
Folh x ObstIII 0,064 0,2806
ObstII x ObstIII 0,689 < 0,0001
Folh x CobVeg 0,002 0,8418
ObstII x CobVeg 0,002 0,8312
Solo x Dos 0,542 0,0002
ObstIII x CobVeg 0,005 0,7681

39
Desta forma, as variáveis com maiores índices de correlação foram desconsideradas
das futuras análises e foram mantidas as seguintes variáveis estruturais: solo exposto
(Solo), dossel (Dos), obstrução a 0,5m do solo (ObstI), obstrução a 1,5m do solo (a partir de
agora denominado ObstII) e profundidade de cobertura vegetal (CobVeg). Ainda que
algumas dessas variáveis apresentassem correlação, sua intensidade foi baixa e dado sua
importância biológica, elas ainda assim foram mantidas nas análises.
Com relação à Análise de Componentes Principais, o primeiro eixo da PCA explicou
48% da variação da estrutura da vegetação. Este eixo representou um gradiente de pontos
amostrais que seguem com uma maior intensidade de obstrução da vegetação a 0,5m e a
1,5m do solo, uma maior exposição do solo, e um menor fechamento de dossel (porção
esquerda do gráfico) para pontos amostrais com uma menor intensidade de obstrução da
vegetação a 0,5m e a 1,5m do solo, uma menor exposição do solo, e um maior fechamento
de dossel (porção direita do gráfico). O segundo eixo da PCA explicou 21% da variação da
estrutura da vegetação e esteve relacionado à variável de cobertura vegetal, de tal forma a
representar um gradiente de pontos amostrais com maior profundidade de cobertura
vegetal (porção inferior do gráfico) para menor profundidade de cobertura vegetal (porção
superior do gráfico, Figura 01).

40
Figura 01: Biplot do primeiro e segundo eixo da Análise de Componentes Principais realizada para
as variáveis de estrutura da vegetação dos pontos amostrais da Fazenda Intermontes, na cidade de
Ribeirão Grande, SP. Legenda: círculo vermelho – plantio (PL), círculo verde – regeneração natural
arbórea com plantio (RARBOP), círculo azul – regeneração natural arbustiva com plantio
(RARBUP), círculo amarelo – regeneração natural sem plantio (REG), círculo preto – remanescente
florestal (RF). Variáveis estruturais: Dos – dossel, Obst I – obstrução a 0,5m do solo, Obst II –
obstrução a 1,5m do solo, CobVeg – profundidade de cobertura vegetal.
A PCA demonstrou que pontos amostrais que haviam sido submetidos inicialmente
ao mesmo tipo de tratamento apresentaram um resultado distinto com relação à sua
complexidade estrutural. Os remanescentes florestais foram mantidos no mesmo
quadrante do gráfico, sendo caracterizados por uma menor obstrução da estrutura vertical
da vegetação a 0,5m e a 1,5m do solo, bem como uma maior quantidade de dossel fechado.
Os demais pontos, que representam tratamentos distintos de plantio, estiveram
dispersos sem apresentar uma ordenação clara. De modo geral, as áreas nas quais o plantio

41
foi estabelecido apresentaram árvores com baixa altura, com DAP (diâmetro a altura do
peito) pequeno e ramificadas desde sua porção mais inferior, ocasionando copas baixas e
dosséis abertos, o que foi ocasionado provavelmente pela falta de manejo no local. O tipo de
plantio 3 x 2 m também contribuiu para um padrão espacial da vegetação que não se
assemelha aos remanescentes florestais. Este espaçamento e a falta de manejo
provavelmente favoreceram o desenvolvimento de uma intensa cobertura vegetal,
dominada pela gramínea exótica braquiária (Urochloa sp.).
Os pontos de REG e um dos pontos de RARBOP estiveram reunidos por
caracteristicamente apresentarem um alto nível de obstrução vertical da vegetação nos
estratos mais inferiores e apresentarem uma profunda cobertura vegetal do solo.
Riqueza, abundância e composição da comunidade de pequenos mamíferos
Ao todo 541 indivíduos de pequenos mamíferos foram capturados ao longo de um
esforço-amostral de 9400 baldes-noite para o conjunto de 20 pontos amostrais. Ao todo,
foram capturadas 20 espécies de pequenos mamíferos (14 espécies de roedores e cinco
espécies de marsupiais). De acordo com a literatura (Tabela 02, vide Anexo I), 14 são
espécies florestais endêmicas da Mata Atlântica e do total das espécies capturadas, 35% (7)
são terrestres, 35% (7) são escansoriais ou arborícolas e 30% (6) são fossoriais ou semi-
fossoriais. Quanto à dieta, 50% (10) são insetívoras ou onívoras, seguidas de 30% (6) de
frugívoras-granívoras e as demais (20%) são herbívoras ou com dietas desconhecidas
(Anexo I).
Dado que as espécies Monodelphis cf. americana e Monodelphis cf. inheringi são de
difícil distinção em campo, as mesmas foram reunidas numa única categoria (Monodelphis
sp.).
Desta forma, ao avaliar a riqueza pelo tratamento, observou-seque a riqueza foi
similar entre RF, RARBOP e RARBUP, sendo as menores observadas no PL e na REG. As
espécies Akodon cf. montensis, Bibimys labiosus, Necromys lasiurus, Oligoryzomys nigripes e
Gracilinanus microtarsus estiveram presentes em todos os tratamentos, já as demais
variaram sua ocorrência (Tabela 02).
Já a riqueza variou de três a 11 espécies entre as unidades amostrais. Unidades que
receberam o mesmo tratamento original de plantio apresentaram valores distintos no

42
número de espécies, assim como os próprios remanescentes florestais. A riqueza média
observada apresentou valores semelhantes àqueles estimados pelo estimador não
paramétrico (Chao 2) para cada uma das unidades amostrais (Tabela 03).
Tabela 02. Distribuição geográfica (R – restrito à Mata Atlântica, N – não restrito a Mata
Atlântica); locomoção (Terr – terrestre, Semf – Semi-fossorial, Foss – fossorial, Arbo-
arbórea, Esca – escansorial), número de indivíduos capturados e número de espécies
capturadas por tipo de tratamento (RF – remanescente florestal, RARBOP – pasto com
regeneração arbórea e plantio, RARBUP – pasto com regeneração arbustiva e plantio, PL –
plantio, e REG – regeneração natural) na Fazenda Intermontes, estado de São Paulo.
Distribuição Locomoção RF RARBOP RARBUP PL REG Total
Akodon cf. montensis N Terr 6 17 27 15 2 67
Bibimys labiosus N Semf 2 3 1 2 2 10
Brucepattersonius soricinus R Semf 4 2 7 1 0 14
Delomys cf. sublineatus R Terr 2 8 11 4 0 25
Euryzygomatomys spinosus R Foss 0 1 1 0 0 2
Euryorzyomys russatus R Terr 4 7 2 0 0 13
Necromys lasiurus N Terr 1 4 12 5 3 25
Oligoryzomys nigripes N Esca 11 64 139 65 53 332
Oxymycterus cf. dasytrichus R Semf 0 2 1 1 0 4
Sooretamys angouya R Terr 2 0 5 0 0 7
Juliomys sp. R Arbo 0 0 2 2 0 4
Calomys tener N Terr 1 0 1 0 0 2
Cavia sp. N Terr 0 1 0 0 0 1
Drymoreomys albimaculatus R Terr 0 0 1 0 0 1
Gracilinanus microtarsus R Arbo 1 1 7 3 2 14
Marmosops paulensis R Esca 1 2 1 0 0 4
Marmosops incanus R Esca 2 0 0 0 0 2
Monodelphis sp. R Semf 5 3 1 1 1 11
Didelphis aurita R Arbo 0 2 0 0 1 3
Número de indivíduos
42 117 219 99 64 541
Número de espécies 13 14 16 10 7 20

43
Tabela 03. Número de espécies de pequenos mamíferos observados e estimados através de
dois estimadores de riqueza para as 20 parcelas amostrais. Desvio padrão em parênteses.
Tratamento Riqueza
Observada Estimador
Chao 2
Riqueza por Tratamento
(RF) 11 (2,55) 16,03 (5,17) (RF) 6 (2,03) 6,00 (0,10) 6-11
(RF) 6 (2,07) 6,96 (1,75)
(RARBOP) 10 (2,26) 10,96 (1,53)
(RARBOP) 9 (2,40) 12,19 (3,97)
(RARBOP) 3 (1,59) 3,00 (0,39) 3-10
(RARBOP) 6 (2,26) 8,88 (4,02)
(RARBOP) 6 (2,05) 6,32 (0,90)
(RARBUP) 9 (2,44) 11,88 (4,02)
(RARBUP) 5 (1,94) 5,32 (0,90)
(RARBUP) 7 (2,21) 8,44 (2,50)
(RARBUP) 5 (1,94) 5,32 (0,90) 3-9
(RARBUP) 8 (2,50) 15,19 (7,86)
(RARBUP) 3 (1,59) 3,00 (0,10) (RARBUP) 9 (2,63) 29,13 (16,52)
(PL) 4 (1,80) 4,48 (1,25) (PL) 7 (2,30) 16,58 (9,69) (PL) 4 (1,78) 4,00 (0,24) 4-7 (PL) 7 (2,24) 12,75 (6,79)
(REG) 8 (2,21) 8,72 (1,38) 8
A curva média de acúmulo de espécies para a assembleia como um todo (Figura 02),
bem como para cada uma das unidades amostrais fisionômicas (Figura 03), apresentaram
um comportamento de ascensão, sugerindo que novas espécies podem ser acrescentadas
com o aumento do esforço amostral.
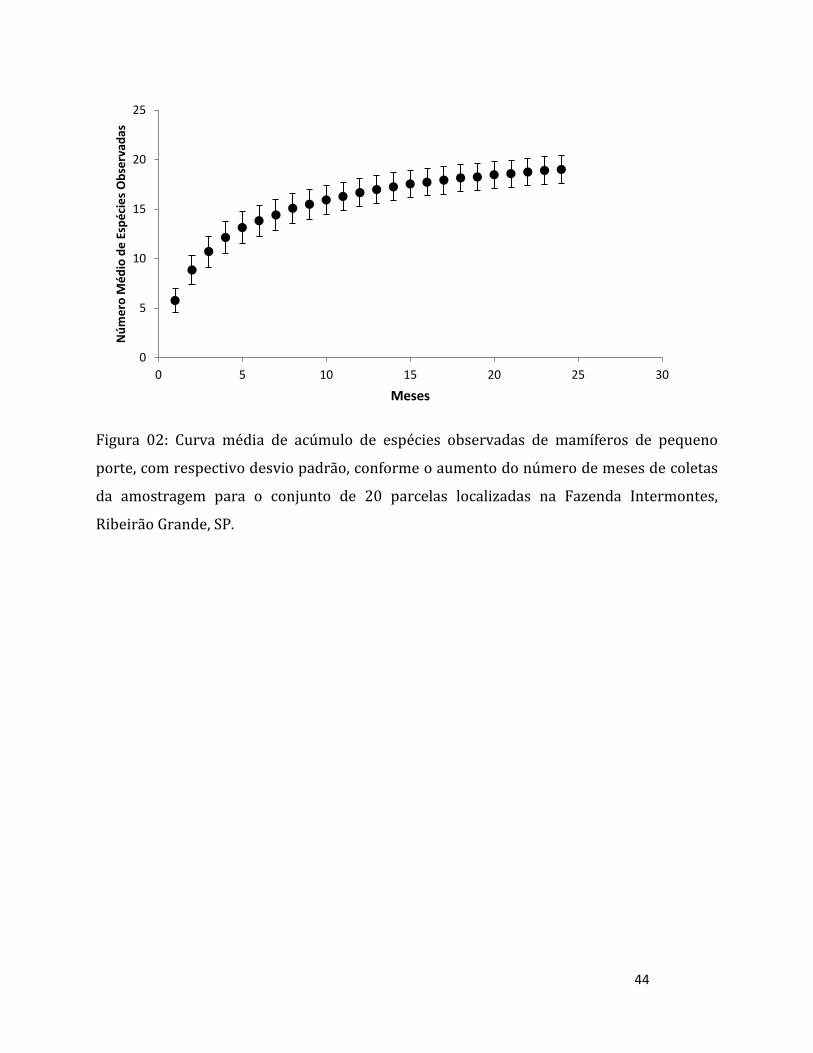
44
Figura 02: Curva média de acúmulo de espécies observadas de mamíferos de pequeno
porte, com respectivo desvio padrão, conforme o aumento do número de meses de coletas
da amostragem para o conjunto de 20 parcelas localizadas na Fazenda Intermontes,
Ribeirão Grande, SP.
0
5
10
15
20
25
0 5 10 15 20 25 30
Nú
me
ro M
éd
io d
e E
spé
cie
s O
bse
rvad
as
Meses
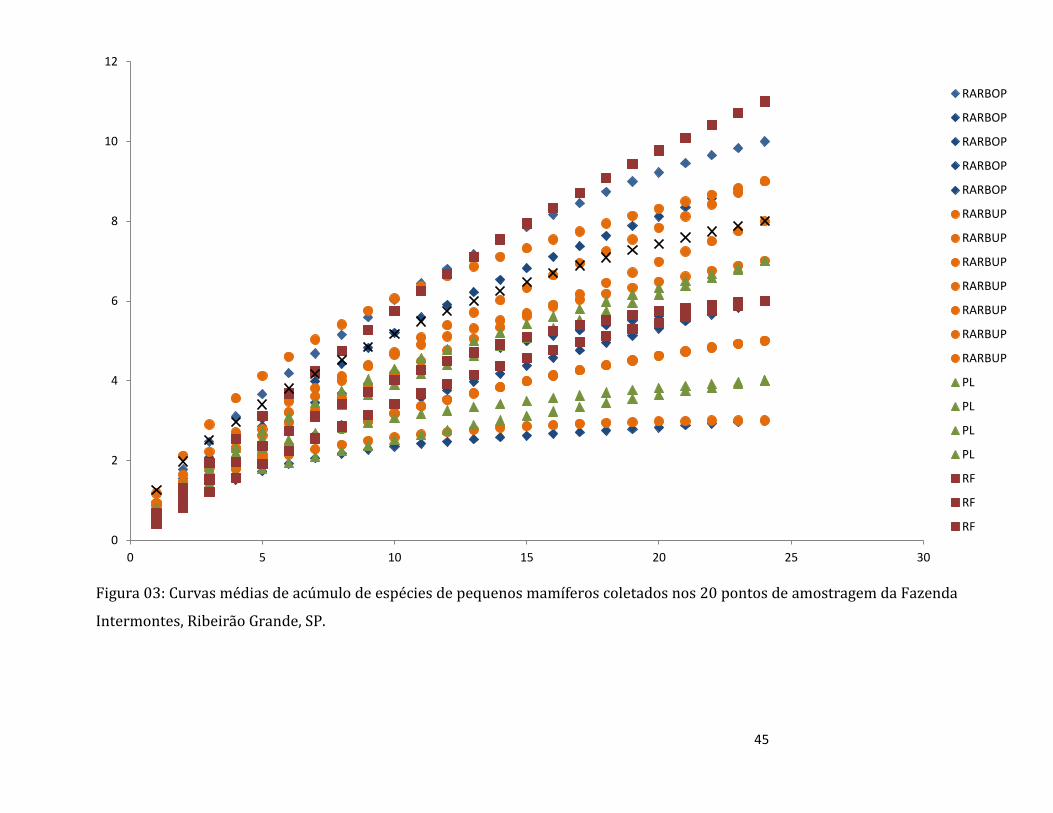
45
Figura 03: Curvas médias de acúmulo de espécies de pequenos mamíferos coletados nos 20 pontos de amostragem da Fazenda
Intermontes, Ribeirão Grande, SP.
0
2
4
6
8
10
12
0 5 10 15 20 25 30
RARBOP
RARBOP
RARBOP
RARBOP
RARBOP
RARBUP
RARBUP
RARBUP
RARBUP
RARBUP
RARBUP
RARBUP
PL
PL
PL
PL
RF
RF
RF

46
Com relação ao índice de abundância relativa, a espécie mais abundante da
assembleia de pequenos mamíferos foi Oligoryzomys nigripes (N= 332, 61%), sendo
capturada em todas as unidades amostrais, seguida de Akodon montensis (N= 67, 12%),
presente em praticamente todos os locais, mas estando presentes nos cinco tratamentos.
Tais espécies, no entanto, embora tenham sido abundantes, apresentaram uma menor
quantidade de indivíduos nas áreas de remanescentes florestais em relação aos demais
pontos amostrais (Tabela 02, Figura 04a). As espécies Bibimys labiosus, Brucepattersonius
igniventris, Delomys cf. sublineatus, Euryorzyomys russatus, Necromys lasiurus, Gracilinanus
microtarsus, Monodelphis sp., por sua vez, seguiram com N>10 (Tabela 02).
Enquanto o menor número de indivíduos foi encontrado nos remanescentes
florestais (N=42), o maior número foi encontrado na área em início de regeneração (N=64,
REG), a qual se caracteriza por um campo de gramíneas aberto, com algumas arvoretas
presentes ao longo de sua extensão (Tabela 02). Este maior índice de abundância relativa
foi devido à intensa captura de O. nigripes. Os menores índices desta espécie, por outro
lado, foram observados nos remanescentes florestais (Tabela 02, Figura 04b).
Espécies endêmicas da floresta Atlântica estiveram presentes nos diferentes
tratamentos, mas proporcionalmente sua frequência de captura foi maior nos
remanescentes florestais. Euryoryzomys russatus, por sua vez, foi a única espécie endêmica
que esteve presente somente nos remanescentes florestais e nos tratamentos de plantio em
áreas de regeneração arbórea e arbustiva (Tabela 02, Figura 04a e 04b).

47
Figura 04: Proporção do número de indivíduos capturados (a) e Índice de Abundância Relativa (b)
para cada espécie em cada uma das unidades amostrais fisionômicas. Legenda: Ako – Akodon
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
RF
RF
RARBOP
RARBOP
RARBUP
RARBUP
RARBUP
RARBUP
PL
PL
Proporção de n° de indivíduos capturados
Tra
tam
en
to
Ako
Bib
Bru*
Del*
Eru*
Nec
Oli
Gra*
Mon*
Demais*
0
10
20
30
40
50
60
70
RF
RF
RF
RA
RB
OP
RA
RB
OP
RA
RB
OP
RA
RB
OP
RA
RB
OP
RA
RB
UP
RA
RB
UP
RA
RB
UP
RA
RB
UP
RA
RB
UP
RA
RB
UP
RA
RB
UP PL
PL
PL
PL
REG
Índ
ice
de
Ab
un
dân
cia
Re
lati
va
Tratamento
Demais*
Mon*
Gra*
Oli
Nec
Eru*
Del*
Bru*
Bib
Ako
a)
b)

48
montensis, Bib - Bibimys labiosus, Bru - Brucepattersonius igniventris, Del - Delomys cf. sublineatus,
Eru -Euryoryzomys russatus, Nec - Necromys lasiurus, Oli – Oligoryzomys nigripes, Gra - Gracilinanus
microtarsus, Mon - Monodelphis sp., e Demais – demais espécies com N<10. (*) indica espécies
endêmicas da floresta Mata Atlântica (vide Anexo I).
A composição das espécies não demonstrou nenhuma tendência clara entre os
diferentes tipos de tratamentos. Unidades fisionômicas distintas foram reunidas quanto à
sua composição (Figura 05). Dois remanescentes (Ponto 02 e Ponto 03) foram reunidos
com uma área de pasto com regeneração arbórea restaurada por plantio (Ponto 05), o qual
apresentou uma abundância reduzida de Oligoryzomys nigripes e uma proporção maior de
espécies endêmicas, assim como as outras duas unidades.

49
Figura 05: Dendrograma representativo da similaridade da composição de pequenos
mamíferos entre as 20 unidades amostrais distribuídas pela Fazenda Intermontes, na
cidade de Ribeirão Grande, São Paulo (coeficiente de correlação cofonética= 0,86).
Efeito dos descritores ambientais sobre a comunidade de mamíferos
A RDA explicou 49% da variação dos dados analisados (R²ajustado=0,207) e
produziu seis eixos de ordenação, de tal forma que os dois primeiros explicaram 79,92%. O
primeiro eixo da ordenação (RDA1) representou positivamente a obstrução vertical da
vegetação a 0,5m do solo e a disponibilidade de frutos, e negativamente a abertura de
RF
0
1
RA
RB
OP
0
7
PL
1
9
RA
RB
OP
0
8
RA
RB
UP
1
1
RA
RB
UP
1
5
RA
RB
UP
0
9
PL
1
7
RA
RB
UP
1
2
RA
RB
UP
1
3
RA
RB
OP
0
6
PL
1
8
RA
RB
UP
1
4
RA
RB
OP
0
4
RA
RB
UP
1
0
PL
1
6
RE
G
20
RF
0
2
RF
0
3
RA
RB
OP
0
5
Alt
ura

50
dossel, a profundidade de cobertura vegetal e a disponibilidade de artrópodes, enquanto o
segundo eixo da ordenação (RDA2) representou a obstrução vertical da vegetação a 1,5m
do solo (Figura 06). Entre as variáveis, a obstrução a 0,5m do solo e a disponibilidade de
frutos foram as únicas variáveis a explicarem a composição das espécies da assembleia de
pequenos mamíferos (Tabela 04).
Figura 06: Diagrama de ordenação da Análise de Redundância das espécies de pequenos
mamíferos (Ako – Akodon cf. montesis; Bib - Bibimys labiosus; Bru - Brucepattersonius
soricinus; Del- Delomys cf. sublineatus; Eru - Euryorzyomys russatus; Gra – Gracilinanus
microtarsus; Mon – Monodelphis sp.; Nec - Necromys lasiurus; Oli - Oligoryzomys nigripes)
em relação a variáveis ambientais (Artro – disponibilidade de artrópodes; CobVeg –
profundidade de cobertura vegetal; Dos – quantidade de dossel fechado; Fru –
disponibilidade de frutos; ObstI – obstrução da estrutura vertical da vegetação a 0,5m do
solo; ObstII – obstrução da estrutura vertical da vegetação a 1,5m do solo; Solo –
quantidade de solo exposto). Pontos de 1-20 correspondem as áreas amostradas
(correspondência na Tabela 01, Seção Materiais e Métodos).

51
Tabela 04. Resultados da Análise de Redundância para avaliar a influência das variáveis
ambientais na composição das espécies capturadas nos 20 pontos amostrais localizados na
Fazenda Intermontes, estado de São Paulo. Número de permutações: 99. Legenda: Solo –
quantia de solo exposto, Doss – quantia de dossel fechado, ObstI – obstrução a 0,5m do
solo, ObstII – obstrução a 1,5m do solo, CobVeg – profunidade de cobertura vegetal, Artro –
peso seco médio de artrópodes, Frutos – quantia de frutos zoocóriocos, * p<0.05.
Variáveis DP Variação Permutação(>F)
Solo 1 0,0111 0,44 Doss 1 0,009 0,48 ObstI 1 0,021 0,06* ObstIII 1 0,017 0,12 CobVeg 1 0,016 0,14 Artro 1 0,013 0,28 Frutos 1 0,023 0,04*
Ao se avaliar, no entanto, a porcentagem de variação apresentada pelas variáveis
significativas, observou-se que o valor foi extremamente baixo e, portanto, pouco
explicativo (Obstrução a 0,5m do solo = 2,1% e Frutos = 2,3%, Tabela 04). Entre as espécies
consideradas, grande parte delas não apresentou uma associação clara com as variáveis,
apenas leves tendências de associação. Exceção a este padrão foi observado somente com
Monodelphis sp. que esteve fortemente relacionado ao dossel fechado e à disponibilidade
de artrópodes.
Ao analisar a influência das variáveis de estrutura da vegetação e da disponibilidade
de recursos sobre abundância de cada espécie da assembleia de pequenos mamíferos
através da construção de modelos lineares generalizados, observou-se que para a
assembleia de pequenos mamíferos terrestres, dois modelos explicaram melhor o padrão
de variação da abundância das espécies, de tal forma que a abundância esteve relacionada
negativamente à obstrução vertical da vegetação e à disponibilidade de artrópodes (Tabela
05).

52
Tabela 05. Resultados da seleção dos modelos generalizados com erros binomiais negativos
e função de ligação log para a abundância de pequenos mamíferos da Fazenda Intermontes,
na cidade de Ribeirão Grande, SP. Modelo – modelos selecionados (ΔAICc < 2); k – número
de parâmetros; AICc - Critério de Informação de Akaike modificado para amostras
pequenas; ΔAICc – diferença entre o melhor modelo e o modelo considerado; wi – peso de
evidência. Para os modelos: (+) relação positiva, (-) relação negativa, PC1 e PC2 – escores
dos pontos dos dois primeiros eixos da PCA da estrutura da vegetação, Artro –
disponibilidade de artrópodes, Frutos – disponibilidade de frutos.
Modelo K AICc ΔAICc wi
assembleia
(-)PC1 3 151,18 0,0 0,298
(-)PC1 + (-)Artro 4 151,19 0,4 0,248
Akodon cf. montensis
Nulo 2 95,35 0,0 0,207
(+)PC2 3 95,28 0,2 0,188
(+)Frutos 3 95,82 0,7 0,144
(+)PC2 + (+)Frutos 4 96,38 1,6 0,091
Bibimys labiosus
(-)PC2 + (-)Frutos 4 36,2 0,0 0,262
(-)Frutos 3 17,19 0,6 0,190
Brucepattersonius soricinus
Nulo 2 49,87 0,0 0,244
(+)Artro 3 50,26 0,6 0,177
(-)Frutos 3 51,12 1,5 0,115
Delomys sublineatus
(+)PC2 3 64,31 0,0 0,177
(-)PC1 + (+)PC2 + (-)Frutos 5 64,12 0,6 0,13
(-)PC1 + (+)PC2 4 64,75 0,8 0,119
Nulo 2 65,49 0,9 0,111
(+)PC2 + (-)Frutos 4 65,38 1,4 0,087
(+)PC2 + (-)Artro 4 65,39 1,4 0,086
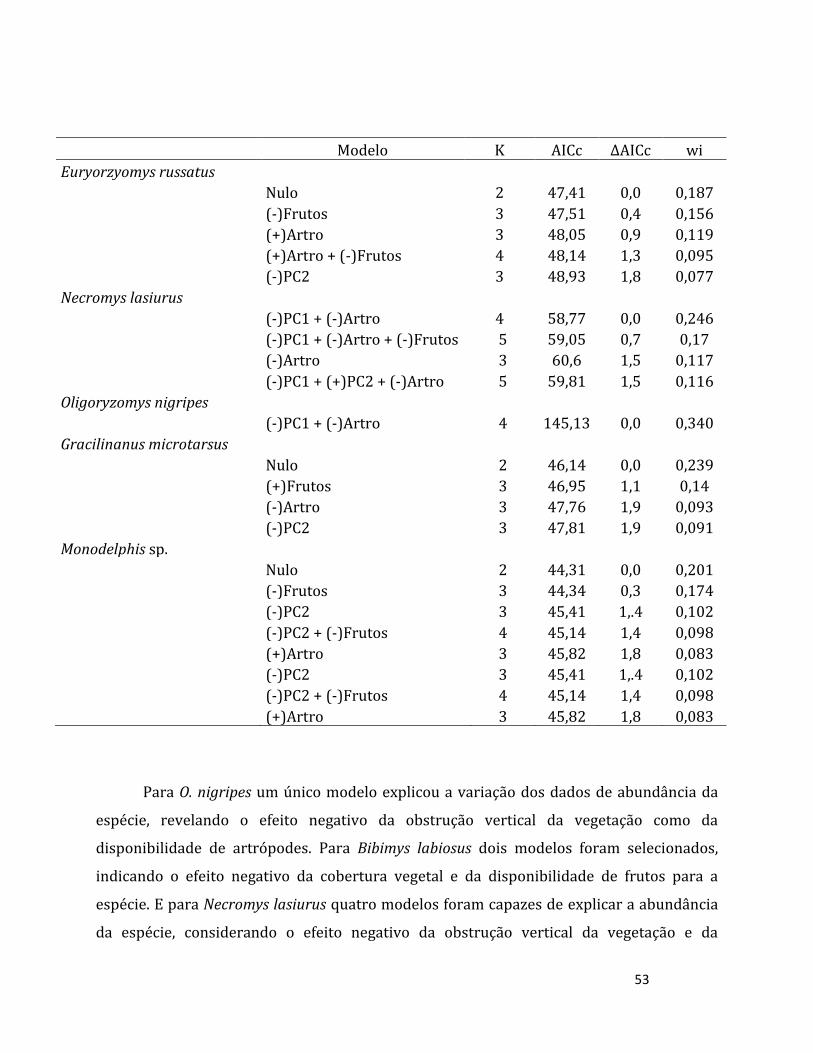
53
Modelo K AICc ΔAICc wi
Euryorzyomys russatus
Nulo 2 47,41 0,0 0,187
(-)Frutos 3 47,51 0,4 0,156
(+)Artro 3 48,05 0,9 0,119
(+)Artro + (-)Frutos 4 48,14 1,3 0,095
(-)PC2 3 48,93 1,8 0,077
Necromys lasiurus
(-)PC1 + (-)Artro 4 58,77 0,0 0,246
(-)PC1 + (-)Artro + (-)Frutos 5 59,05 0,7 0,17
(-)Artro 3 60,6 1,5 0,117
(-)PC1 + (+)PC2 + (-)Artro 5 59,81 1,5 0,116
Oligoryzomys nigripes
(-)PC1 + (-)Artro 4 145,13 0,0 0,340
Gracilinanus microtarsus
Nulo 2 46,14 0,0 0,239
(+)Frutos 3 46,95 1,1 0,14
(-)Artro 3 47,76 1,9 0,093
(-)PC2 3 47,81 1,9 0,091
Monodelphis sp.
Nulo 2 44,31 0,0 0,201
(-)Frutos 3 44,34 0,3 0,174
(-)PC2 3 45,41 1,.4 0,102
(-)PC2 + (-)Frutos 4 45,14 1,4 0,098
(+)Artro 3 45,82 1,8 0,083
(-)PC2 3 45,41 1,.4 0,102
(-)PC2 + (-)Frutos 4 45,14 1,4 0,098
(+)Artro 3 45,82 1,8 0,083
Para O. nigripes um único modelo explicou a variação dos dados de abundância da
espécie, revelando o efeito negativo da obstrução vertical da vegetação como da
disponibilidade de artrópodes. Para Bibimys labiosus dois modelos foram selecionados,
indicando o efeito negativo da cobertura vegetal e da disponibilidade de frutos para a
espécie. E para Necromys lasiurus quatro modelos foram capazes de explicar a abundância
da espécie, considerando o efeito negativo da obstrução vertical da vegetação e da

54
disponibilidade de artrópodes e de frutos, e o efeito positivo da cobertura vegetal. (Tabela
05).
Para as espécies de pequenos mamíferos analisadas individualmente, as espécies
Akodon montensis, Brucepattersonius soricinus, Delomys cf. sublineatus, Euryoryzomys
russatus, Gracilinanus microtarsus e Monodelphis sp. tiveram o modelo nulo igualmente
selecionado como aquele capaz de explicar a distribuição dos dados de abundância (Tabela
05).
DISCUSSÃO
A restauração de habitat visa a recuperação estrutural e funcional de ambientes
degradados (Palmer et al. 1997, Young 2000). Com relação a este aspecto estrutural,
diferentes tipos de modelos de restauração podem, no entanto, resultar em diferentes tipos
de complexidade estrutural da vegetação (Kanowski et al. 2003, Ribeiro et al. 2009). Em
nossa área de estudo, o modelo de restauração implementado não foi capaz de recuperar a
complexidade da vegetação encontrada nos remanescentes florestais, assim como
evidenciado pela Análise de Componentes Principais. Independente do tipo de matriz no
qual o plantio foi realizado (áreas de regeneração arbórea, arbustiva ou capoeiras, e áreas
sem regeneração), as restaurações resultaram em estruturas distintas da vegetação,
geralmente caracterizadas por ambientes com intensa obstrução vegetal até 1,0m do solo,
com dossel baixo e aberto e com a presença de maior cobertura vegetal por gramíneas.
A ausência de uma complexidade estrutural da vegetação que se assemelhe as áreas
naturais é posta como fator responsável pela falha da recolonização de espécies de
pequenos mamíferos em áreas restauradas, principalmente com relação às espécies
especialistas (Nichols e Nichols 2003, Cunningham et al. 2007, Munro et al. 2007). Em
nossa hipótese inicial acreditávamos que a riqueza, a abundância e a composição das
espécies de plantio e de regeneração somente seriam iguais aos remanescentes, desde que
tais unidades apresentassem a mesma complexidade estrutural que os fragmentos, o que
foi confirmado pelos nossos resultados.

55
Embora a riqueza dos tratamentos tenha sido semelhantes aos remanescentes
florestais para algumas unidades fisionômicas, a análise isolada desta variável não é capaz
de refletir padrões ecológicos importantes em comunidades. Junto com a abundância e a
composição das espécies foi possível observar que as diferentes áreas de restauração
foram dominadas por espécies generalistas, principalmente pela presença de Oligoryzomys
nigripes e Akodon montensis. Tais espécies não são endêmicas à floresta Atlântica e são
tipicamente encontradas em ambientes alterados, em florestas mais jovens ou perturbadas,
inclusive em áreas mais abertas (Alho et al. 1986, Carmignotto 2004, Umetsu e Pardini
2007, Naxara 2008, Putker et al. 2008, Pardini et al. 2009, Rossi 2011), como foi o
observado no local em início de regeneração, o qual apresentou o maior índice de
abundância relativa da espécie de Oligoryzomys nigripes em relação às demais localidades.
Embora algumas espécies especialistas, como Brucepattersonius soricinus, Delomys
cf. sublineatus, Euryorzyomys russatus, Gracilinanus microtarsus e Monodelphis sp., dentre
outras, tenham sido registradas nas áreas restauradas, proporcionalmente sua abundância
relativa foi menor que aquela encontrada nos remanescentes florestais, nos quais tais
espécies predominaram. A presença de espécies especialistas parece obedecer mais a
trajetória da restauração do que a condição inicial do habitat restaurado. Por exemplo, uma
área restaurada a partir de pasto com regeneração arbórea apresentou composição em
espécies similar a dois dos remanescentes florestais, mesmo que a estrutura da vegetação
tenha se mostrado distinta aos remanescentes em alguns aspectos. No entanto, o que
aproximou esta unidade amostral fisionômica aos remanescentes foi o índice de
fechamento de dossel e a altura das árvores resultantes da regeneração natural,
apresentando uma vegetação mais fechada em relação aos demais pontos, o que pode ter
favorecido a presença de espécies mais especialistas no local.
Ainda que algumas evidências demonstrem ser possível a recolonização de espécies
de pequenos mamíferos em áreas restauradas, mesmo em florestas tropicais as
comunidades de pequenos mamíferos das áreas são constituídas de espécies generalistas e
tolerantes ao efeito de perturbação de habitat (Perry e Thill 2005, Nicolas et al. 2009, Kurz
et al. 2013). Grande parte dos estudos evidencia, contudo, que a recuperação da assembleia
de espécies de pequenos mamíferos somente irá ocorrer quando as condições estruturais
do habitat necessárias à sua sobrevivência estiverem garantidas (Carey e Johson 1995,

56
Nichols e Nichols 2003, Cunningham et al. 2007, Moro e Gadal 2007, Munro et al. 2007,
Kalies et al. 2012), assim como observado em nosso estudo.
Não somente a estrutura da vegetação, mas a disponibilidade de artrópodes e de
frutos, que são recursos alimentares para estes animais, são descritores ambientais
importantes capazes de influenciar a assembleia de pequenos mamíferos (Suarez 1994,
Vieira et al. 2006, Leiner e Silva 2007, Talamoni et al. 2008, Bocchiglieri et al.2010, Lessa e
Costa 2010, Pinotti et al. 2011), podendo atuar isoladamente ou em conjunto com a
estrutura da vegetação na seleção das espécies de pequenos mamíferos. Nossa análise de
redundância como a seleção de modelos lineares generalizados evidenciaram o efeito
comum da obstrução vertical da vegetação sobre a assembleia de pequenos mamíferos,
mas não demonstraram um padrão claro quanto ao efeito dos recursos alimentares. No
entanto, em nível específico o efeito dos descritores ambientais foi mais claro para as
espécies analisadas, e a resposta de cada uma delas foi igualmente específica.
Ainda que para algumas espécies o modelo nulo tenha sido igualmente selecionado,
a cobertura vegetal foi a principal variável selecionada nos modelos das espécies
consideradas, exceto para Brucepattersonius soricinus e Oligoryzomys nigripes. O efeito
positivo desta variável para algumas espécies pode estar associado à maior proteção que
tal cobertura pode oferecer contra predadores, bem como contra variações de temperatura
e umidade que são fatores microclimáticos importantes para a sobrevivência desses
organismos (Harmon et al. 1986, Brannon 2002).
Em nosso estudo, a cobertura vegetal atual das áreas de plantio esteve associada
àquela do início da regeneração, com o predomínio de gramíneas, principalmente
braquiária. Esta situação favoreceu para duas espécies generalistas e terrestres, Akodon
montensis e Necromys lasiurus, que se encontram presentes em diferentes fisionomias no
país, desde cerrado até Mata Atlântica, e que se beneficiam de áreas alteradas, como as
áreas restauradas desde estudo (Lessa et al. 1999, Bonvicino et al. 2002, Becker et al. 2007,
Umetsu e Pardini 2007, Umetsu et al. 2008). Outros estudos em regiões temperadas
reportaram igualmente a importância desta cobertura vegetal na ocorrência de espécies
generalistas em áreas restauradas (Carey e Johnson 1995, Perry e Thill 2005, Kurz et al.
2013).

57
A única espécie especialista para a qual o efeito da cobertura vegetal também foi
positivo foi para outra espécie igualmente terrestre, Delomys cf. sublineatus. Naxara et al.
(2009) observaram a associação desta espécie com locais com grande quantidade de galhos
no chão e maior umidade de folhiço em mata contínua no Planalto do estado de São Paulo,
assim como Pinotti (2010), que inclusive registrou o mesmo efeito negativo da
disponibilidade de frutos e artrópodes sobre essa espécie. Aparentemente, algum tipo de
cobertura do solo que garanta a proteção e abrigo para essa espécie parece atuar como um
importante filtro para a sua distribuição para tais espécies terrestres.
A alta produtividade primária nos estágios iniciais da regeneração e nos plantios, em
função do predomínio de gramíneas, pode levar ao aumento de recursos alimentares, como
os frutos e os artrópodes (Malcolm 1997, DeWalt et al. 2003). Ainda que para Oligoryzomys
nigripes essa variável não tenha sido selecionada, esta produtividade pode ser
extremamente favorável, tendo em vista sua dieta frugívoro-granívora (Vieira et al. 2006,
Cademartori et al. 2008, Camargo et al. 2008, Talamoni et al. 2008, Pinotti et al. 2011),
favorecendo sua extensa ocupação nas unidades amostrais fisionômicas, em contrapartida
ao efeito negativo que a estrutura e obstrução vertical da vegetação apresentaram sobre
esta espécie.
A disponibilidade de recursos, tanto artrópodes como frutos, foram descritores
igualmente selecionados pelas diferentes espécies. Mas, seu efeito foi tanto positivo quanto
negativo sobre a abundância das espécies, diferentemente do esperado. A interferência
desses descritores também sempre esteve relacionada ao efeito da estrutura do habitat,
exceto para Brucepattersonius soricinus. Para as demais espécies, o efeito dos recursos
alimentares sempre esteve relacionado positivamente à dieta das mesmas.
Para Brucepattersonius soricinus, além do modelo nulo, outro modelo selecionou um
efeito positivo da disponibilidade de artrópodes sobre a abundância dessa espécie, e um
terceiro modelo selecionou o efeito negativo da disponibilidade de frutos, revelando a
importância deste recurso para essa espécie que é insetívora (Umetsu et al. 2006, Pinotti et
al. 2011). Tais espécies já foram capturadas em matas maduras e secundárias como
também em áreas de agricultura e áreas regeneradas de vegetação arbustiva (Naxara
2008), evidenciando, portanto, a complexidade do habitat como um fator secundário para a
ocorrência da espécie.

58
Estudos que investigam a capacidade de colonização da fauna em áreas restauradas
focalizam apenas a avaliação do efeito da estrutura e complexidade do habitat sobre a
ocorrência das espécies (Nichols e Nichols 2003, Cunningham et al. 2007, Munro et al.
2007). Considerar o efeito da disponibilidade de recursos, no entanto, é imprescindível haja
vista sua importância para a sobrevivência das espécies. A própria complexidade estrutural
gerada pelos diferentes processos de restauração pode gerar disponibilidades de recursos
insuficientes ou não à ocorrência das espécies, mas somente sua avaliação direta poderá
evidenciar qualquer padrão. Conforme evidenciado neste estudo, tanto a estrutura da
vegetação quanto a disponibilidade de recursos aparentam atuar como descritores
ambientais para a presença e abundância das espécies de pequenos mamíferos, seja pela
ação conjunta ou isolada de um deles. Portanto, estudos futuros em áreas de restauração
devem incluir estes filtros ambientais em suas investigações.
Implicações de manejo
A hipótese do “Field of Dreams” considera que a recolonização de áreas restauradas
irá ocorrer naturalmente pelas espécies de fauna, sem que medidas de manejo sejam
necessárias (Palmer et al. 1997, Kurz et al. 2013). Essa recolonização natural, no entanto,
somente deve ocorrer quando tais áreas restauradas recuperarem totalmente as
características e aspectos funcionais dos habitats originais, uma vez existindo uma área
fonte para a migração e colonização das espécies. Em nosso estudo ficou claro que, caso
esta recuperação original não seja atingida, a comunidade de pequenos mamíferos irá ser
dominada por espécies generalistas, capazes de habitar ambientes alterados. Portanto,
medidas de manejo se fazem necessárias para permitir e favorecer a colonização de novas
espécies.
Em nossa área de estudo, espécies generalistas foram favorecidas pela presença de
uma densa cobertura vegetal composta predominantemente por gramíneas, característica
esta que parece garantir proteção e recursos alimentares para essas espécies. A
predominância de gramíneas, no entanto, deve ser favorecida pelo padrão espacial das
árvores cultivadas (3 x 2 m) e pela falta de condução e manejo das mudas plantadas,
ocasionando a formação de um dossel baixo e aberto, já que as árvores ramificam-se muito
precocemente em seu crescimento. A entrada de luz, por consequência, favorece a

59
manutenção desta cobertura vegetal, enquanto deve prejudicar o desenvolvimento de
espécies secundárias que dependem de sombra. Medidas de manejo em restaurações que
permitam a formação de um dossel mais alto e fechado deve favorecer a recuperação de
áreas com aspectos mais naturais e permitir mais rapidamente a recuperação da
comunidade de pequenos mamíferos.
Por outro lado, estudos têm evidenciado a importância de galhadas no solo para a
existência de espécies mais especialistas. Embora não tenhamos medido diretamente esta
variável, este recurso não esteve presente com frequência nas áreas de plantio, mas
somente nos remanescentes florestais (obs. pessoal). Nally e Horrocks (2002)
demonstraram a efetividade da recuperação de espécies de pequenos mamíferos com
medidas de manejo que incrementaram a complexidade das galhadas no solo. Portanto,
sugerimos que a adição de galhadas pode ser um experimento válido na restauração de
ambientes muito degradados de Mata Atlântica, cuja fauna de pequenos mamíferos se
queira recuperar.
REFERÊNCIAS BIBLIOGRÁFICAS
Alho, C. J. R., Pereira, L. A., Paula, A. C. 1986. Patterns of habitat utilization by small mammal
populations in cerrado biome of central Brazil. Mammalia 50(4): 447-460.
Antunes, P. C., Arruda Campos, M. A., Oliveira-Santos, L. G. R., Graipel, M. E. 2009.
Population dynamics of Euryoryzomys russatus and Oligoryzomys nigripes (Rodentia,
Cricetidae) in an Atlantic forest area, Santa Catarina Island, Southern Brazil.
Biotemas 22 (2): 143-151.
Becker, R. G., Paise, G., Baumgarten, L. C., Vieira, E. M. 2007. Small mammal’s community
structure and density of Necromys lasiurus (Rodentia, Sigmodontinae) in open areas
of the Cerrado in central Brazil. Journal of Mastozoologia Neotropical 14(2): 157-
168.
Bergallo, H. G., Magnusson, W. E. 1999. Effects of climate and food availability on four
rodent species in southeastern Brazil. Journal of Mammalogy 80(2): 472-486.

60
Boal, C. W., Mannan, R. W. 1994. Northern goshawk diets in ponderosa pine forests on the
Kaibab Plateau. Northern Goshawk: Ecology and Management, Studies in Avian
Biology 16: 97–102.
Bocchiglieri, A. Mendonça, A. F., Campos, J. B. 2010. Diet composition of Gracilinanus agilis
(Didelphimorphia, Didelphidae) in dry woodland areas of Cerrado in central Brazil.
Mammalia 74: 225-227.
Bolker, B. 2007. Ecological models and data in R. Princeton University Press, Princeton.
Bonvicino, C. R., Lindbergh, S. M., Maroja, L. S. 2002. Small non-flying mammals from
conserved and altered areas of Atlantic forest and Cerrado: comments on their
potential use for monitoring environment. Brazilian Journal of Biology 62(4B): 765-
774.
Borcard, D., Gillet, F., Legendre, P. 2011. Numerical Ecology with R. Springer, New York,
USA.
Brannon, M. P. 2002. Distribution of Sorex cinereus and S. fumeus on north- and south-
facing slopes in the southern Appalachian Mountains. Southeastern Naturalist 1:
299-306.
Brown, J. H., Ernest, K. M. 2002. Rain and rodents: complex dynamics of desert consumers.
BioScience 52: 979-987.
Burnham, K. P., Anderson, D. R. 2002. Model selection and multimodel inference: a practical
information-theoretic approach. 2ed. Springer Verlag, New York, USA.
Cáceres, N. C., Monteiro-Filho, E. L. A. 2001. Food habits, home range and activity of
Didelphis aurita (Mammalia, Marsupialia) in a forest fragment of southern Brazil.
Studies on Neotropical Fauna and Environment 36(2): 85-92.
Cáceres, N. C. 2004. Diet of three didelphid marsupials (Mammalia, Didelphimorphia) in
southern Brazil. Mammalian Biology 69(6): 430-433.
Cáceres, N.C., Lessa, L.G. 2012. O papel dos marsupiais na dispersão de sementes. In: Os
marsupiais do Brasil – Biologia, Ecologia e Conservação. Editora Universidade
Federal do Mato Grosso 407-423.
Cademartori, C. V., Marques, R. V., Pacheco, S. M. 2008. Vertical stratification in the use of
space by small mammals (Rodentia, Sigmodontinae) in an area of mixed

61
ombrophilous forest, southern Brazil. Revista Brasileira de Zoociências 10(3): 189-
196.
Caldara, V. J., Leite, Y. L. R. 2007. Habitat use by small mammals at Parque Estadual da
Fonte Grande, Vitoria, Espirito Santo, Brazil. Boletim do Museu de Biologia Mello
Leitão 21: 57-77.
Camargo, N. F., Gurgel-Gonçalves, R., Palma, A. R. 2008. Variação morfológica de pegadas de
roedores arborícolas e cursoriais do Cerrado. Revista Brasileira de Zoologia 25(4):
696- 704.
Carmignotto, A. P. 2004. Pequenos mamíferos terrestres do Cerrado: padrões faunísticos
locais e regionais. Tese de doutorado. Universidade de São Paulo.
Carmignotto, A. P. 2006. Plano de Manejo – Parque Estadual da Serra do Mar. São Paulo.
Carey, A. B., Johnson, M. 1995. Small mammals in managed, naturally young, and old-
growth forests. Ecological Applications 5(2): 336-352.
Ceotto, P., Finotti, R., Santori, R., Cerqueira, R. 2009. Diet variation of the marsupials
Didelphis aurita and Philander frenatus (Didelphimorphia, Didelphidae) in a rural
area of Rio de Janeiro State, Brazil. Mastozoologia Neotropical 16(1): 49-58.
Cowell, R. K. 2009. Estimate S: Statistical estimation of species richness and shared species
from samples. Versão 8.2.
Cunningham, R. B., Lindenmayer, D. B., Crane, M., Michael D., MacGregor, C. 2007. Reptile
and arboreal marsupials response to replanted vegetation in agricultural
landscapes. Ecological Applications 17(2): 609-619.
Delciellos, A. C., Vieira, M. V. 2006. Arboreal walking performance in seven didelphid
marsupials as an aspect of their fundamental niche. Austral Ecology 31(4): 449-457.
Delciellos, A. C., Vieira, M. V. 2009. Allometric, phylogenetic, and adaptive components of
climbing performance in seven species of Didelphid marsupials. Journal of
Mammalogy 90(1): 104-113.
De Walt, S. J., Maliakal, S. K., Denslow, J. S. 2003. Changes in vegetation structure and
composition along a tropical forest chronosequence: implications for wildlife.
Forest Ecology and Management 182: 139-151.

62
Facure, K. G., Giaretta, A. A., Monteiro-Filho, E. L. A. 2003. Food habits of the crab-eating fox,
Cerdocyon thous, in an altitudinal forest of the Mantiqueira Range, southeastern
Brazil. Mammalia 67(4): 503-511.
Fleming, T. H. 1975. The role of small mammals in tropical ecosystems. In: F.B. Golley, K.
Petrusewicz e L. Riszkowisk (eds). Small mammals: their productivity and
population dynamics. Cambridge University Press, UK.
Fonseca, G. A. B., Kierulff, M. C. M. 1989. Biology and natural history of Brazilian Atlantic
forest small mammals. Bulletin Florida State Museum, Biological Science 34(3): 99-
152.
Fonseca, G. A. B., Hermann, G., Leite, Y. L. R., Mittermeier, R. A., Rylands, A. B., Patton, J. L.
1996. Lista anotada dos mamíferos do Brasil. Occasional Papers in Conservation
Biology 4: 1-38.
Francisco, A. D., Magnusson, W. E., Sanaiotti, T. M. 1995. Variation in growth and
reproduction of Bolomys lasiurus (Rodentia, Muridae) in an Amazonian Savanna.
Journal of Tropical Ecology 11 419-428.
Freitas, S. R., Astua de Moraes, D., Santori, R. T., Cerqueira, R. 1997. Habitat preference and
food use by Metachirus nudicaudatus and Didelphis aurita (Didelphimorphia,
Didelphidae) in a restinga forest at Rio de Janeiro. Revista Brasileira de Biologia
57(1): 93-98.
Gaspar, D. A. 2005. Comunidade de mamíferos não voadores de um fragmento de Floresta
Atlântica semidecídua do município de Campinas, SP. Tese de doutorado,
Universidade Estadual de Campinas.
Geise, L., Paresque, R., Sebastião, H., Shirai, L. T., Astúa, D., Marroig, G. 2010. Non-volant
mammals, Parque Nacional do Catimbau, Vale do Catimbau, Buíque, state of
Pernambuco, Brazil, with karyologic data. Check List 6(1): 180-186.
Gentile, R., Fernandez, F. A. S. 1999. Influence of habitat structure on a streamside small
mammal community in a Brazilian rural area. Mammalia 63(1): 29-40.
Graipel, M. E., Cherem, J. J., Monteiro-Filho, E. L. A., Glock, L. 2006. Population dynamics of
marsupials and rodents in Parque Municipal da Lagoa do Peri, Santa Catarina Island,
southern Brazil. Mastozoologia Neotropical 13(1): 31-49.

63
Harmon, M. E., Franklin, J. F., Swanson, J. F. 1986. Ecology of coarse woody debrisn
temperate ecosystems. Advances in Ecology Research 15: 133-302.
Hinojosa, F. P., Anderson, S., Patton, J. 1987. Two new species of Oxymycterus (Rodentia)
from Peru and Bolivia. American Museum Novitates 2898: 1-17.
Horn, G. B., Kindel, A., Hartz, S. M. 2008. Akodon montensis (Thomas, 1913) (Muridae) as a
disperser of endozoochoric seeds in a coastal swamp forest of southern Brazil.
Mammalian Biology 73 (4): 325-329.
Kalies, E. L., Dickson, B. G., Chambers, C. L., Covington, W. W. 2012. Community occupance
responses of small mammals to restoration treatments in ponderosa pine forests,
northern Arizona, USA. Ecological Applications 22(1): 204-217.
Kanowski, J., Catterall, C. P., Wardell-Johnson, G. W., Prostor, H., Reis, T. 2003. Development
of forest structure on cleared rainforest land in Australia under different styles of
reforestation. Forest Ecology and Management 183: 265-280.
Kanowski, J. 2010. What have we learnt about rainforest restoration in the past two
decades? Ecological Managment and Restoration 11(1): 2-3.
Kurz, D., McGinty, N. A., Stankavich, S. A. 2013. Restored wetlands can support mammalian
assemblages comparable to those in nonmitigated reference wetlands. American
Naturalist 170: 260-273.
Lambert, T. D., Malcolm, J. R., Zimmerman, B. L. 2006. Amazonian small mammal
abundances in relation to habitat structure and resource abundance. Journal of
Mammalogy 87(4): 766-776.
Legendre, P., Gallager, E. D. 2001. Ecologically meaningful transformations for ordination of
species data. Oecologia 129: 271-280.
Leite, Y. L. R., Costa, L. P., Stallings, J. R. 1996. Diet and vertical space use of three sympatric
opossums in a Brazilian Atlantic forest reserve. Journal of Tropical Ecology 12: 435-
440.
Leiner, N. O., Silva, W. R. 2007. Effects of resource availability on the use of space by the
mouse opossum Marmosops paulensis (Didelphidae) in a montane Atlantic forest
area in southeastern Brazil. Acta Theriologica 52(2): 197-204.

64
Lessa, G., Gonçalves, P. R., Morais, M. M. J., Costa, F. M., Pereira, R. F., Paglia, A. P. 1999.
Characterization and accompanying of the terrestrial small mammal fauna in a
secondary forest fragment in Viçosa, Minas Gerais. Journal of Bios 7(7): 41-49.
Lessa, L. G., Costa, F. N. 2010. Diet and seed dispersal by five marsupials (Didelphimorphia:
Didelphidae) in a Brazilian cerrado reserve. Mammalian Biology 75: 10-16.
Lichti, N. I., Murphy, M. T. 2001. Selection ratios on community aggregated traits estimate
ecological filters imposed on species by sites. Ecology 91(2): 347-354.
Malcom, J. R. 1997. Insect biomass in Amazonian forest fragments. In: Stork, N. E., Adis, J.,
Didham, R. K. (eds). Canopy Arthropods. Chapman & Hall, London.
Martins, E. G. 2004. Ecologia populacional e área de vida da cuíca Gracilinanus microtarsus
(Marsupialia: Didelphidae) em um Cerradão de Américo Brasiliense, São Paulo.
Dissertação de Mestrado, Universidade Estadual de Campinas.
Martin, G. 2003. The role of small ground-foraging mammals in topsoil health and
biodiversity: implications to management and restoration. Ecological Management
and Restoration 4(2): 114-119.
Martins, R., Quadros, J., Mazzolli, M. 2008. Hábito alimentar e interferência antrópica na
atividade de marcação territorial do Puma concolor e Leopardus pardalis (Carnivora:
Felidae) e outros carnívoros na Estação Ecológica da Juréia-Itatins, São Paulo, Brasil.
Revista Brasileira de Zoologia 25(3): 427-435.
Moro, D., Gadal, S. 2007. Benefits of habitat restoration to small mammal diversity and
abundance in a pastoral agricultural landscape in mid-Wales. Biodiversity
Conservation 16: 3543-3557.
Munro, N. T., Lindenmayer, D. B., Fischer, J. 2007. Faunal response to revegetation in
agricultural areas of Australia: a review. Ecological Managmente and Restoration
8(3): 199-207.
Nally, R. M., Horrocks, G. 2002. Habitat change and restoration: responses of forest-floor
mammal species to manipulation of fallen timber in floodplain forests. Animal
Biodiversity and Conservation 25(1): 41-52.
Naxara, L. 2008. Importância dos corredores ripários para a fauna – pequenos mamíferos em
manchas de floresta, matriz do entorno e elementos lineares em uma paisagem

65
fragmentada de Mata Atlântica. Dissertação de mestrado, Universidade de São
Paulo.
Naxara, L., Pinotti, B. T., Pardini, R. 2009. Seasonal microhabitat selection by terrestrial
rodents in an Old-Growth Atlantic Forest. Journal of Mammalogy 90(2): 404-415.
Nichols, O. G., Nichols, F. M. 2003. Long-term trends in faunal recolonization after bauxite
mining in the Jarrah forest of southwestern Australia. Restoration Ecology 11(3):
261-272.
Nicolas, V., Barrière, P., Tapiero, A., Colyn, M. 2009. Shrew species diversity and abundance
in Ziama Biosphere Reserve, Guinea: comparison among primary forest, degraded
forest and restoration plots. Biodiversity and Conservation 18: 2043-2061.
Nitikman, L. Z., Mares, M. A. 1987. Ecology of small mammals in a gallery forest of Central
Brazil. Annals of Carnegie Museum 56 (1-2): 75-95.
Núñez, R., Miller, B., Lindzey, F. 2000. Food habitat of jaguars and pumas in Jalisco, Mexico.
Journal of Zoology 252: 373-379.
Oliveira, F. F. R., Nessim, R., Costa, L. P., Leite, Y. L. R. 2007. Small mammal ecology in an
urban Atlantic Forest fragment in southeastern Brazil. Lundiana 8(1): 27-34.
Owen, R. D., Goodin, D. G., Koch, D. E., Chu, Y. K., Jonsson, C. B. 2010. Spatiotemporal
variation in Akodon montensis (Cricetidae: Sigmodontinae) and hantaviral
seroprevalence in a subtropical forest ecosystem. Journal of Mammalogy 91 (2):
467-481.
Paglia, A. P.; Fonseca, G. A. B.; Rylands, A. B.; Herrmamm, G.; Aguiar, L. M. S.; Chiarello, A. G.;
Leite, L. R. R.; Costa, L. P.; Sicilianos, S.; Kierulff, M. C. M.; Mendes, S. L.; Tavares, V. C.;
Mittermeier, R. A.;.; Patton, J. L. 2012. Lista anotada dos mamíferos do Brasil. 2°
edição. Occasional papers in Conservation Biology 6: 1-76.
Palmer, M. A., Ambrose, R. F., Poff, N. L. 1997. Ecological theory and community restoration
ecology. Restoration Ecology 5(4): 291-300.
Pardini, R., de Sousa, S. M., Braga-Neto, R., Metzger, J. P. 2005. The role of forest structure,
fragment size and corridors in maintaining small mammal abundance and diversity
in an Atlantic forest landscape. Biological Conservation 124: 253-266.
Pardini, R., Faria, D., Accacio, G. M., Laps, R. R., Mariano-Neto, E., Paciencia, M. L. B., Dixo, M.
e Baumgarten, J. 2009. The challenge of maintaining Atlantic forest biodiversity: A

66
multi-taxa conservation assessment of specialist and generalist species in an agro-
forestry mosaic in southern Bahia. Biological Conservation 142: 1178-1190.
Patten, M. A. 1997. Reestablishment of a rodent community in restored desert scrub.
Restoration Ecology 5(2): 156-161.
Percequillo, A.R., Weskler, M. e Costa, L.P. 2011. A new genus and species of rodent from
the Brazilian Atlantic Forest (Rodentia: Cricetidae: Sigmodontinae: Oryzomyini),
with comments on oryzomyine biogeography. Zoological Journal of Linean Society
161: 357-390.
Perry, R. W., Thill, R. E. 2005. Small-mammal responses to pine regeneration treatments in
the Ouachita Mountains of Arkansas and Oklahoma, USA. Forest Ecology and
Management 219: 81-94.
Pinotti, B. T. 2010. Pequenos mamíferos terrestres e a regeneração da Mata Atlântica:
influência da estrutura do habitat e da disponibilidade de alimento na recuperação da
fauna. Dissertação de Mestrado, Universidade de São Paulo.
Pinotti, B. T., Naxara, L., Pardini, R. 2011. Diet and food selection by small mammals in an
old-growth Atlantic Forest. Studies on Neotropical Fauna and Environment 46: 1-9.
Poff, N. L. 1997. Landscape filters and species traits: towards mechanistic understanding
and prediction in stream ecology. Journal of the North American Benthological
Society 16(2): 391-409.
Püttker, T., Pardini, R., Meyer-Lucht, Y. e Sommer, S. 2008. Responses of five small mammal
species to micro-scale variations in vegetation structure in secondary Atlantic
Forest remnants, Brazil. BMC Ecology 8(9): 1-10.
Quinn, G. P., Keough, M. J. 2002. Experimental design and data analysis for biologists.
Cambridge University Press, UK.
R Core Team. 2013. R: A language and environment for statistical computing. R Foundation
for Statistical Computing, Viena, Austria.
Rodrigues, R. R., Lima, R. A., Gandolfi, S., Nave, A.G. 2009. On the restoration of high
diversity forests: 30 years of experience in the Brazilian Atlantic forest. Biological
Conservation 142: 1242-1251.

67
Rossi, N. F. 2011. Pequenos mamíferos não-voadores do Planalto Atlântico de São Paulo:
Identificação, história natural e ameaças. Dissertação de mestrado, Universidade de
São Paulo.
Santos, A. J. 2003. Estimativas de riqueza em espécies. In: Cullen Jr., L.; Rudran, R. e
Valladares-Padua, C. (eds). Métodos de Estudo em Biologia da Conservação &
Manejo da Vida Silvestre. Editora UFPR. Pp: 19-39.
Slade, N. A., Blair, S. M. 2000. An empirical test of using counts of individuals as indices of
population size. Journal of Mammalogy 81:1035–1045.
Stone, E. R. 2007. Measuring impacts of restoration on small mammals in a mixed-grass
Colorado prairie. Ecological Restoration 25:183–190.
Suarez, O. V. 1994. Diet and habitat selection of Oxymycterus rutilans (Rodentia, Critecidae).
Mammalia 58(2): 225-234.
Summerville, K. S., Conoan, C. J., Steinchen, R. M. 2006. Species trait as predictors of
Lepidopteran composition in restored and remnant tallgrass prairies. Ecological
Applications 16(3): 891-900.
Talamoni, S. A., Dias, M. M. 1999. Population and community ecology of small mammal of
southeastern Brazil. Mammalia 63 (2): 167-181.
Talamoni, S. A., Couto, D., Cordeiro, D. A., Diniz, F. M. 2008. Diet of some species of
Neotropical small mammals. Mammalian Biology 73(5): 337-341.
Terborgh, J. 1988. The big things that run the world – A sequel to E. O. Wilson. Conservation
Biology 2(4):402-403.
Umetsu, F., Naxara, L., Pardini, R. 2006. Evaluating the efficiency of pitfall traps for
sampling small mammals in the Neotropics. Journal of Mammalogy, 87(4): 757-765.
Umetsu, F., Pardini, R. 2007. Small mammals in a mosaic of forest remnants and
athropogenic habitats-evaluating matrix quality in an Atlantic forest landscape.
Landscape Ecology 22(4): 517-530.
Umetsu, F., Meztger, J. P., Pardini, R. 2008. Importance of matrix quality for modeling
species distribution in complex tropical landscapes: a test with Atlantic forest small
mammals. Ecography 31: 359-370.
Vieira, E. M., Izar, P. 1999. Interactions between aroids and arboreal mammals in the
Brazilian Atlantic rainforest. Plant Ecology 145(1): 75-82.

68
Vieira, M. V. 2003. Movement patterns and the ecology of an assemblage of didelphid
marsupials in the Coastal Atlantic Forest. VI Congresso de Ecologia do Brasil,
Fortaleza: 305-307.
Vieira, E. M., Monteiro-Filho, E. L. A. 2003. Vertical stratification of small mammals in the
Atlantic rain forest of south-eastern Brazil. Journal of Tropical Ecology 19: 501-507.
Vieira, E. M., Pizo, M. A., Izar, P. 2003. Fruit and seed exploitation by small rodents of the
Brazilian Atlantic forest. Mammalia. 67(4): 533-539.
Vieira, E. M., Iob, G., Briani, D. C., Palma, A. R. T. 2005. Microhabitat selection and daily
movements of two rodents (Necromys lasiurus and Oryzomys scotti) in Brazilian
Cerrado, as revealed by a spool-and-line device. Mammalian Biology 70 (6): 359-
365.
Vieira, E. M., Paise, G., Machado, P. H. D. 2006. Feeding of small rodents on seeds and fruits:
a comparative analysis of three species of rodents of the Araucaria forest, southern
Brazil. Acta Theriologica 51 (3): 311-318.
Vieira, E. M., Port, D. 2007. Niche overlap and resource partitioning between two sympatric
fox species in southern Brazil. Journal of Zoology 272: 57-63.
Weiher, E., Keddy, P. A. 2001. Ecological assembly rules: perspectives, advances, retreats.
Cambridge University Press, UK.
Young, T. P. 2000. Restoration ecology and conservation biology. Biological Conservation
92: 73-83.
Zar, J. H. 1999. Biostatistical analysis. Prentice Hall, Jersey, 4ª edição.

69
Anexo I. Lista de pequenos mamíferos registrados na Fazenda Intermontes, SP, entre agosto de 2011 e agosto de 2013. Caracterização hábito e
da dieta para cada espécie de acordo com referências bibliográficas discriminadas.
Família/Tribo Nome científico Hábito Dieta Referências bibliográficas
Sigmodontinae
Akodontini
Akodon cf. montensis
(Thomas, 1913)
terrestre*** onívora
Talamoni e Dias 1999, Vieira et al. 2006, Talamoni
et al. 2008, Horn et al 2008, Vieira e Monteiro-Filho
2003, Graipel et al. 2006, Oliveira et al. 2007, Owen
et al. 2010,
Bibimys labiosus
(Winge, 1887)
semi-fossorial*** *
Fonseca et al 1996
Brucepattersonius soricinus
(Hershkovitz, 1998)
semi-fossorial** insetívora
Umestu et al 2006, Pinotti et al. 2011
Necromys lasiurus
(Lund, 1841)
terrestre*** onívora
Nitikman e Mares 1987, Francisco et al. 1995,
Vieira2003, Viera et al. 2005, Talamoni et al.2008,
Geise et al.2010,
Oxymycterus cf. dasytrichus
(Linnaeus, 1758)
semi-fossorial** insetívora
Hinojosa et al 1987, Vieira e Monteiro-Filho 2003,
Pinotti et al 2011
Cavidae Cavia sp. terrestre*** herbívora Paglia et al. 2012
Echimyidae Euryzygomatomys spinosus
(Fischer, 1814)
fossorial** herbívora
Fonseca et al. 1996
Oryzomyini Euryorzyomys russatus
(Wagner, 1848)
terrestre** frugívoro-
onívora
Vieira e Monteiro-Filho 2003, Graipel et al. 2006,
Antunes et al. 2009, Pinotti et al 2011
Oligoryzomys nigripes
(Olfers, 1818)
escansorial*** frugívoro-
granívora
Nitikman e Mares 1987, Fonseca e Kierulff 1989,
Fonseca et al. 1996, Vieira e Monteiro-Filho 2003, ,
Gaspar 2005, Graipel et al. 2006, Vieira et al. 2006,
Cademartori et al. 2008, Camargo et al. 2008,
Talamoni et al. 2008, Pinotti et al. 2011
Sooretamys angouya (Fischer,
1914)
terrestre** frugívora
Fonseca et al. 1996, Graipel et al. 2006, Cademartori
et al. 2008
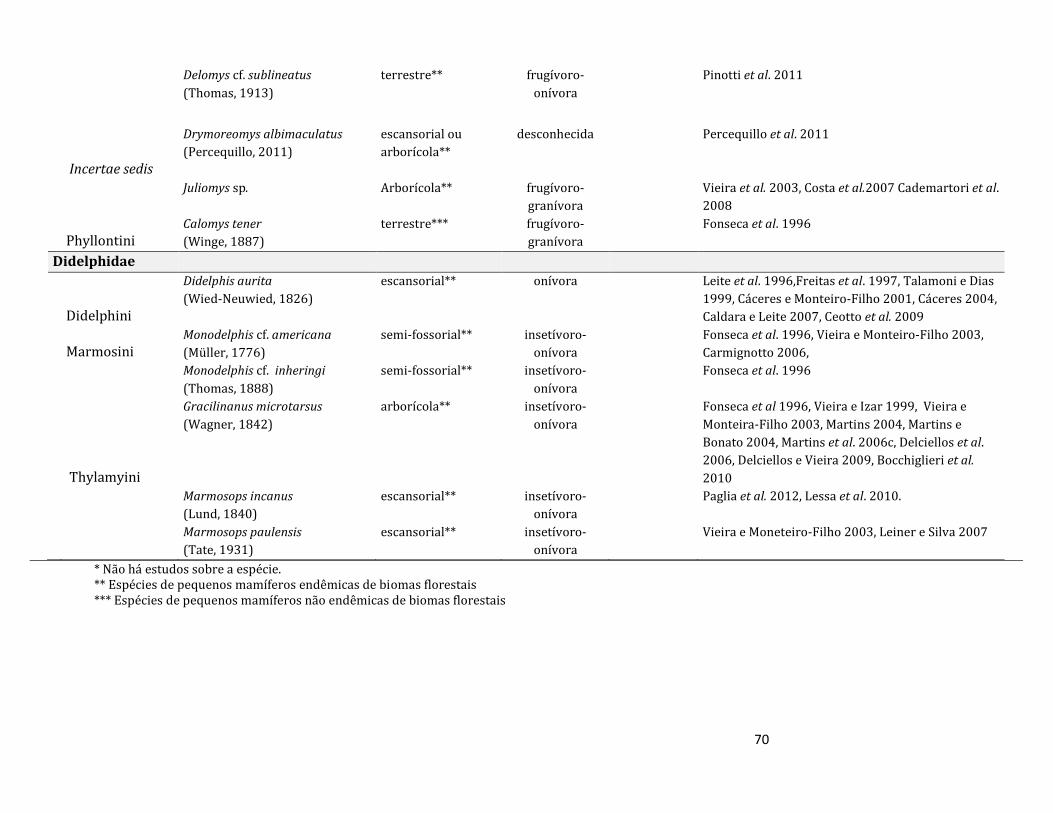
70
* Não há estudos sobre a espécie. ** Espécies de pequenos mamíferos endêmicas de biomas florestais *** Espécies de pequenos mamíferos não endêmicas de biomas florestais
Delomys cf. sublineatus
(Thomas, 1913)
terrestre**
frugívoro-
onívora
Pinotti et al. 2011
Incertae sedis
Drymoreomys albimaculatus
(Percequillo, 2011)
escansorial ou
arborícola**
desconhecida
Percequillo et al. 2011
Juliomys sp. Arborícola** frugívoro-
granívora
Vieira et al. 2003, Costa et al.2007 Cademartori et al.
2008
Phyllontini Calomys tener
(Winge, 1887)
terrestre*** frugívoro-
granívora
Fonseca et al. 1996
Didelphidae
Didelphini
Didelphis aurita
(Wied-Neuwied, 1826)
escansorial** onívora
Leite et al. 1996,Freitas et al. 1997, Talamoni e Dias
1999, Cáceres e Monteiro-Filho 2001, Cáceres 2004,
Caldara e Leite 2007, Ceotto et al. 2009
Marmosini Monodelphis cf. americana
(Müller, 1776)
semi-fossorial** insetívoro-
onívora
Fonseca et al. 1996, Vieira e Monteiro-Filho 2003,
Carmignotto 2006,
Monodelphis cf. inheringi
(Thomas, 1888)
semi-fossorial** insetívoro-
onívora
Fonseca et al. 1996
Thylamyini
Gracilinanus microtarsus
(Wagner, 1842)
arborícola** insetívoro-
onívora
Fonseca et al 1996, Vieira e Izar 1999, Vieira e
Monteira-Filho 2003, Martins 2004, Martins e
Bonato 2004, Martins et al. 2006c, Delciellos et al.
2006, Delciellos e Vieira 2009, Bocchiglieri et al.
2010
Marmosops incanus
(Lund, 1840)
escansorial** insetívoro-
onívora
Paglia et al. 2012, Lessa et al. 2010.
Marmosops paulensis
(Tate, 1931)
escansorial** insetívoro-
onívora
Vieira e Moneteiro-Filho 2003, Leiner e Silva 2007

71
CAPÍTULO 2: “Mamíferos terrestres de médio e grande porte em um mosaico de
restauração de Mata Atlântica: avaliação dos descritores ambientais sobre a
ocorrência das espécies”
RESUMO
A fragmentação e a perda de habitat são grandes ameaças à sobrevivência da
comunidade de médios e grandes mamíferos por dependerem de grandes extensões de
área de vida. Neste cenário, a restauração surge como uma importante ferramenta para
recuperar habitats que sejam favoráveis para a sobrevivência dessas espécies. Todavia,
estudos que avaliem se as restaurações florestais implantadas estão sendo ou não
adequadas para a colonização dessas espécies ainda são escassos e ainda não foram
realizados no Brasil. Com este objetivo, investigamos a presença e a influência de
descritores ambientais (estrutura da vegetação e disponibilidade de recursos alimentares)
sobre a riqueza, a composição e a frequência de ocorrência de mamíferos terrestres de
médio e grande porte em um mosaico de restauração de Mata Atlântica, incluindo
remanescentes florestais, áreas de pasto arbóreo e ou arbustivo com plantio, áreas de pasto
com plantio e pasto com regeneração natural. Com o uso de armadilhas fotográficas, 20
pontos foram avaliados quanto à presença e ocorrência dessas espécies, identificando um
total de 21 espécies, incluindo um predador de topo (Puma concolor) e um grande
herbívoro (Tapirus terrestres). Espécies generalistas, no entanto, foram mais frequentes e
foram registradas nos diferentes tipos de tratamento, já espécies especialistas foram
menos frequentes e registradas em parte dos tratamentos. O efeito dos descritores
ambientais sobre cada espécie diferiu dado suas necessidades ecológicas, revelando a sua
importância para a ocorrência das diferentes espécies. Medidas de manejo devem ser
consideradas em processos futuros de regeneração a fim de favorecer maior ocupação
pelas espécies especialistas.

72
ABSTRACT
Fragmentation and habitat loss are the major threats to the survival of the
community of medium and large mammal once they have large home ranges. In this
scenario, restoration appears as an important strategy to recover degraded habitats to
favor the survival of those species. However, studies evaluating whether forest restorations
that are being implemented are suitable or not for the colonization of those species are
scarce and have not been done in Brazil. With this goal, we investigated the presence and
the influence of habitat filters (vegetation structure and availability of food resources) on
the richness, composition and frequency of occurrence of terrestrial mammals in a mosaic
restoration of Atlantic Forest including forest remnants, pasture with arboreal or shrub
regeneration where planting were implemented, pasture areas with planting and pasture
with natural regeneration. We conducted camera-trap surveys in 20 points and identified a
total of 21 species, including a large top predator (Puma concolor) and the largest
herbivore in the biome (Tapirus terrestres). Generalist species, however, were more
frequent and were recorded in all the different types of treatment, while specialist species
were less frequent and recorded in part of them. The effect of environmental descriptors
for each species differed according to their ecological requirements, revealing its
importance for the occurrence of different species. Management measures should be
considered in future regeneration processes in order to guarantee greater occupancy by
specialist species.

73
INTRODUÇÃO
A fragmentação e a perda de habitat surgem como grandes ameaçadas para
mamíferos terrestres de médio e grande porte. Aparentemente, as primeiras espécies a
serem afetadas são os carnívoros predadores de topo, como a onça parda (Puma concolor)
e os grandes frugívoros, como a anta (Tapirus terrestris), pois suas grandes áreas de vida, a
baixa densidade e a baixa taxa de crescimento populacional os tornam mais vulneráveis à
redução de habitat (Terborgh 1974, Henle et al. 2004a), principalmente quando dependem
de recursos que se encontram agrupados espacial e temporalmente, como no caso dos
frugívoros (Chiarello 2000). Embora, em um primeiro momento, algumas espécies mais
generalistas pareçam se favorecer com esses distúrbios, dada sua plasticidade
comportamental, como a liberação de sua predação pelos animais de topo da cadeia
(Fonseca e Robinson 1990, Laurance 1994, Crooks e Soulé 1999, Michalski e Peres 2007),
em longo prazo a falta de recursos adequados ocasionará a extinção de grande parte das
espécies (Laurance et al. 2008).
Medidas que visem à recuperação e o incremento da disponibilidade de habitat
adequados são, dessa forma, uma das ferramentas que irão garantir a manutenção desta
biodiversidade no futuro. Com este objetivo, surgem os processos de restauração que
buscam recuperar áreas degradadas, recompondo as características estruturais e
funcionais do habitat (Palmer et al. 1997, Young 2000), garantindo as necessidades
ecológicas para a sobrevivência das espécies de animais.
Tais processos de restauração, no entanto, são implementados com uma série de
métodos de plantio que podem não ser capazes de garantir de maneira substancial a
complexidade estrutural da vegetação, bem como a riqueza florística dos habitats originais,
por exemplo, em virtude do conjunto de espécies de plantas e da forma que tais espécies
são implantadas durante a restauração (Kanowski et al. 2003, Munro et al. 2007, Munro et
al. 2009). Avaliar os filtros ambientais que estejam favorecendo ou prejudicando a
presença e a ocupação de mamíferos terrestres nessas áreas de restauração é, portanto, o
primeiro passo para que novas estratégias possam ser propostas a fim de incrementar a
recolonização dessas espécies.

74
Grande parte dos estudos em comunidade de médios e grandes mamíferos
geralmente avalia filtros ambientais na escala de paisagem para tentar compreender o
efeito dos tamanhos do fragmento, do grau de isolamento ou da matriz, por exemplo, no
padrão de distribuição e ocorrência dessas espécies nas paisagens modificadas (Chiarello
1999, Laurance et al. 2002, Michalski e Peres 2007, Lyra-Jorge et al. 2008, Desbiez et al.
2009, Vieira et al. 2009). Embora tais estudos sejam importantes para determinar áreas
prioritárias para restauração, é imprescindível avaliar se unidades restauradas estão sendo
efetivas e estão cumprindo o seu papel. Dessa forma, compreender o efeito de filtros
ambientais em uma escala menor é que permitirá verificar quais os aspectos dos processos
de restauração que precisam ser modificados para favorecer a ocupação dessas espécies,
aspecto este muito pouco estudado mundialmente (Hobbs et al. 2003, Vesk et al. 2008) e
ainda não realizado em áreas de restauração de um país megadiverso como o Brasil.
Segundo o modelo continuum, espécies de fauna devem responder individualmente
às mudanças gradativas do meio ambiente, de acordo com suas necessidades biológicas.
Dentro deste gradiente, não só a mudança da heterogeneidade da paisagem, mas da
complexidade estrutural, da disponibilidade de recurso, de abrigo, dos aspectos espaciais
relacionados à área de vida da espécie e do próprio clima devem atuar sobre a presença e
distribuição das espécies (Fischer e Lindernmayer 2006).
Tendo em vista a importância da variação dos filtros ambientais dentro da área de
vida do animal e de sua influência sobre as espécies numa escala menor, este estudo teve
como objetivo avaliar o papel de descritores ambientais sobre a riqueza, a composição e a
frequência de ocorrência de mamíferos de médio e grande porte em um mosaico de
restauração de Mata Atlântica, incluindo remanescentes florestais, áreas de plantio e área
de regeneração natural. Entre os descritores foram avaliados o efeito tanto da estrutura da
vegetação quanto da disponibilidade de recursos alimentares sobre as espécies de médios e
grandes mamíferos.
Baseado no modelo continuum nossa hipótese é que espécies de mamíferos de
médio e grande porte utilizem as diferentes unidades fisionômicas do mosaico de
restauração de acordo com suas necessidades biológicas de proteção, oferecida pela
estrutura da vegetação, e de recurso alimentar, tendo em mente que tais unidades devem
atuar como um mesohabitat dentro de sua área de vida de maior extensão.

75
MATERIAL E MÉTODOS
Para Material e Métodos vide seção anterior, em Capítulo Geral (a partir da página
07).
Análise dos dados
Caracterização da estrutura do habitat
A coleta das variáveis estruturais teve como objetivo caracterizar cada unidade
amostral com relação a sua complexidade estrutural. Como algumas das variáveis
poderiam estar correlacionadas entre si em função do método complementar das medidas
estruturais, foi construída uma matriz de correlação múltipla para excluí-las, utilizando-se
nas análises somente as variáveis não correlacionadas. Para as variáveis representadas por
dados de porcentagem, seguiu-se a transformação das mesmas pelo arcoseno da raiz
quadrada (Zar 1999) antes de serem utilizadas em outras análises.
A fim de investigar mais especificadamente o padrão dessas variáveis estruturais
com relação aos diferentes tratamentos, uma Análise de Componentes Principais (PCA) por
matriz correlacionada e com os dados padronizados foi realizada (Quinn e Keough 2002,
Borcard et al. 2011).
Todas as análises estatísticas foram realizadas através do Programa R Studio
(RStudio: Integrated development environment for R, versão 0.98.501 2013).
Riqueza, composição e frequência de ocorrência da comunidade de médios e grandes mamíferos
Para cada unidade fisionômica, calculamos a riqueza (número de espécies) através
do número de registros fotográficos obtidos para cada unidade amostral. A frequência de
registros das espécies pode ser considerada proporcional à sua abundância (Caughley
1977), este parâmetro (número de fotos de cada espécie) foi utilizado para estimar a
abundância (número de indivíduos). Registros de indivíduos de uma mesma espécie
obtidos em uma mesma parcela, no mesmo dia e durante o intervalo de uma hora, foram
considerados como uma captura única para evitar duplicações de registro e a consequente
superestimação da população (Goulart et al. 2009).

76
Para avaliar a eficiência do esforço amostral para a comunidade de médios e
grandes mamíferos, uma curva de acúmulo médio das espécies observadas (Mao Tao) foi
construída. A mesma foi comparada com a riqueza estimada (estimador não paramétrico
Chao 2). Para isso foi utilizado o programa EstimateS 8.0 (Colwell 2005).
A similaridade na composição da comunidade de mamíferos entre as unidades de
amostragem foi avaliada através de uma análise de agrupamento, utilizando as médias das
distâncias euclidianas entre os grupos (UPMGA, Unweighted Pair Group Method). Tal
análise foi realizada através do Programa R Studio (RStudio: Integrated development
environment for R, versão 0.98.501 2013).
Investigação dos descritores ambientais sobre a comunidade de mamíferos
Para avaliar o papel dos descritores ambientais (estrutura da vegetação,
disponibilidade de recursos) na composição das espécies nas unidades amostrais, foi
realizada uma Análise de Redundância (RDA), mas a dependência espacial foi levada em
conta por meio de uma RDA Parcial, que considera o efeito do espaço na presença dessas
espécies nos locais (Borcard et al. 2011).
Para o uso dos dados de espécies de médios e grandes mamíferos na análise de
ordenação constrangida, os mesmos foram transformados pelo método de Hellinger, uma
medida recomendada para a ordenação de dados de abundância e de frequência de
ocorrência de espécies (Legendre e Gallagher 2001). Para tal análise somente as espécies
mais frequentes foram consideradas (N>10).
Para verificar, no entanto, o quanto a variação da frequência de ocorrência (ou
registros) foi explicada unicamente pelas variáveis ambientais e pela posição espacial, ou
pelos dois conjuntos simultaneamente, uma análise de partição de variância foi realizada
(Borcard et al. 1992, Liu 1997). Para isso três modelos foram construídos: o primeiro, já
previamente analisado, no qual a frequência foi analisada somente em relação às variáveis
ambientais, ao se condicionar o espaço geográfico; outro modelo no qual o espaço
geográfico explica a frequência, quando as variáveis ambientais são condicionadas; e o
terceiro, um modelo completo com todas as variáveis ambientais e geográficas como
variáveis explanatórias.

77
A influência das variáveis de estrutura da vegetação e da disponibilidade de
recursos sobre frequência de ocorrência de cada espécie da assembleia de médios e
grandes mamíferos foi avaliada através da construção de modelos lineares generalizados
com distribuição binomial negativa e função de ligação log, considerados adequados para
dados de contagem que possam apresentar alta dispersão (Quinn e Keough 2002, Bolker
2007).
A comparação entre os modelos construídos foi realizada pelo Critério de
Informação de Akaike modificado para amostras pequenas (AICc) (Burnham e Anderson
2002, Quinn e Keough 2002). Para cada modelo calculamos o AICc, a diferença entre o AICc
de cada modelo com aquele do melhor modelo (ΔAICc) e os pesos de evidência que indicam
que o modelo é verdadeiro (wi) (Burnham e Anderson 2002). Como critério, foram
considerados plausíveis todos os modelos com o ΔAICc < 2.
A análise foi realizada tanto para a assembleia como um todo, como separadamente
para cada uma das espécies de médios e grandes mamíferos que foram mais abundantes
(N>10). Em relação às variáveis ambientais de estrutura da vegetação, foram utilizados os
dois primeiros eixos da PCA da caracterização da estrutura do habitat das unidades
amostrais como representantes da variável independente para diminuir o número e
complexidade dos modelos competidores (Burnham e Anderson 2002). Com este mesmo
objetivo, os modelos múltiplos, que englobaram mais de uma variável ambiental, não
consideraram a interação entre as variáveis, até para permitir a verificação do efeito de
cada filtro ambiental exclusivamente. Desta forma, foram criados: um modelo nulo, em que
não há a influência de nenhuma variável independente; modelos simples, nos quais foi
considerada apenas uma única variável independente; e modelos múltiplos, nos quais
foram consideradas as combinações entre as variáveis independentes.
Todas as análises estatísticas foram realizadas através do Programa R Studio
(RStudio: Integrated development environment for R, versão 0.98.501 2013).

78
RESULTADOS
Caracterização dos habitats
Para resultados em caracterização dos habitats, vide Capítulo 1 (a partir da página
37).
Riqueza, composição e frequência de ocorrência da comunidade de médios e grandes
mamíferos
Ao todo 656 registros de médios e grandes mamíferos foram obtidos ao longo de um
esforço-amostral de 6089 armadilhas-dia (310 a 320 armadilhas-dia entre os pontos
amostrais). A riqueza total incluiu 21 espécies de mamíferos terrestres de médio e grande
porte, de 11 famílias distintas. Entre elas, seis espécies estão inclusas nas listas de espécies
ameaçadas de extinção em nível estadual e nacional, entre elas o maior predador de topo
registrado, a onça-parda (Puma concolor) e o maior herbívoro brasileiro, a anta (Tapirus
terrestris). Uma espécie exótica, o cachorro-doméstico (Canis familiaris), também foi
registrada. Todas as espécies foram identificadas pelo registro fotográfico, exceto o lobo-
guará (Chrysocyon brachyurus) e o tapiti (Sylvilagus brasiliensis), os quais foram
identificados por pegada e visualização, respectivamente, sendo, portanto, utilizados
somente como dados qualitativos no estudo (Anexo I).
As espécies do gênero Mazama sp e Leopardus sp. foram reunidas em uma única
categoria para evitar suas subestimações ou superestimações, dado que sua identificação
em nível de espécie não foi possível para todos os registros fotográficos.
A riqueza média observada para a assembleia de médios e grandes mamíferos foi
igual à riqueza estimada pelo estimador não paramétrico Chao 2 (19.85, SD=2.08). E o
esforço amostral obtido não apresentou um padrão de estabilização, demonstrando que
novas espécies podem ser inseridas à assembleia caso o esforço amostral seja aumentado
(Figura 01).

79
Figura
Figura 01: Curva média de acúmulo de espécies observadas de mamíferos de médio e
grande porte, com respectivo desvio padrão, para o conjunto de 19 unidades amostrais
localizadas na Fazenda Intermontes, Ribeirão Grande, SP.
O táxon mais frequentemente registrado pelas armadilhas fotográficas foi a
categoria de pequenos felídeos (Leopardus sp.) (N=201, 30%), a qual pode representar
mais de uma espécie, seguida do cachorro-do-mato (Cerdocyon thous) (N=185, 28%).
Pequenos felídeos foram registrados em todas as unidades, exceto em uma unidade de
regeneração arbórea na qual o plantio foi implantado (Ponto 04) e em duas áreas de pasto
sem regeneração com plantio (Ponto 16 e Ponto 19). Já o cachorro-do-mato foi registrado
em todas as unidades, exceto em uma unidade de regeneração arbustiva com plantio
(Ponto 15) e na área de regeneração natural (Ponto 20), sendo bastante frequente nas
áreas que receberam tratamentos de plantio. A categoria veado (Mazama sp.), o tatu-
galinha (Dasypus novemcinctus), o tamanduá-bandeira (Myrmecophaga tridactyla) e o
esquilo (Guerlinguetus aestuans) foram as espécies que se seguiram como mais frequentes,
com 11% (N=75), 9% (N=65), 5% (N=35) e 4% (N=31), respectivamente. Tais espécies
estiveram presentes em diferentes tipos de tratamentos (Tabela 01, Figura 02).
0
2
4
6
8
10
12
14
16
18
20
0 5 10 15 20 25
Nú
me
ro M
éd
io d
e E
spé
cie
s O
bse
rva
da
s
Meses

80
Com relação à onça-parda, o maior carnívoro observado, foram obtidos 13 registros
ao longo do período do estudo, estando presentes tanto em remanescentes florestais
(Ponto 02 e Ponto 03) quanto nas áreas de regeneração arbórea com plantio (Ponto 05,
Ponto 07, Ponto 08), na área de regeneração arbustiva com plantio (Ponto 15) e nos pastos
com regeneração (Ponto 17, Ponto 18, Ponto 19). Já para a anta, somente quatro registros
foram obtidos, três deles no mês de agosto de 2012 e um deles em dezembro de 2012. Tais
registros da anta estiveram restritos às áreas de pasto com plantio (Ponto 16, Ponto 17,
Ponto 18) e a uma única área de regeneração arbustiva com plantio (Ponto 15) (Tabela 01,
Figura 02).
Já as outras espécies menos frequentes, como a irara, Eira barbara (N=12, 2%), e o
guaxinim, Procyon cancrivorus (N=9, 1%), igualmente estiveram presentes nos tratamentos
de regeneração arbórea e ou arbustiva com plantio (Ponto 05, Ponto 07, Ponto 10, Ponto
12, Ponto 14, Ponto 18) como nos remanescentes (Ponto 01, Ponto 02, Ponto 03), mas
estiveram ausentes nas unidades de pasto com plantio e na área de regeneração (Tabela
01, Figura 02).
Com relação aos tratamentos em si, os registros de médios e grandes mamíferos
foram mais frequentes na RARBUP, na RARBOP, no PL, seguido dos RF e em maior
intensidade na REG. Já comparando-se os pontos amostrais entre si, as menores
frequências e os menores números de espécies foram registrados nos pontos de
regeneração arbórea e ou arbustiva com plantio (Ponto 04, Ponto 09), dois plantios (Ponto
16 e Ponto 19), remanescente remanescente florestal (Ponto 03) e na regeneração natural
(Ponto 20). Os pontos com as maiores ocorrências igualmente estiveram presentes em
distintos tratamentos: regeneração arbórea e ou arbustiva com plantio (Ponto 05, Ponto
07, Ponto 13) e pasto sem regeneração com plantio (Ponto 18) (Tabela 01, Figura 02).

81
Tabela 01. Locomoção (Terr – terrestre, Semf – Semi-fossorial, Foss – fossorial, Arbo-
arbórea, Esca – escansorial) e número de indivíduos registrados por tipo de tratamento (RF
– remanescente florestal, RARBOP – pasto com regeneração arbórea e plantio, RARBUP –
pasto com regeneração arbustiva e plantio, PL – plantio, e REG – regeneração natural) na
Fazenda Intermontes, estado de São Paulo.
Espécies Locomoção RF RARBOP RARBUP PL REG Total
Cerdocyon thous Terr 14 73 68 30 0 185
Leopardus sp. Terr 33 71 75 21 1 201 Myrmecophaga tridactyla Terr 3 11 4 14 3 35
Dasypus novemcinctus Terr 8 16 23 17 0 64
Euphractus sexcinctus Terr 0 1 2 0 0 3
Mazama sp. Terr 4 19 19 25 8 75
Puma concolor Terr 4 3 2 4 0 13
Tapirus terrestres Terr 0 0 1 3 0 4
Guerlinguetus aestuans Arbo 2 3 26 0 0 31
Puma yagouaroundi Terr 0 0 2 0 0 2
Procyon cancrivorus Terr 2 4 2 1 0 9
Eira barbara Esca 3 4 4 1 0 12
Nasua nasua Esca 3 3 2 0 0 8
Galictis cuja Terr 0 1 1 1 0 3
Coendu spinosus Arbo 0 1 0 0 0 1
Didelphis sp. Esca 3 0 6 0 0 9
Leopardus pardalis Terr 0 0 1 0 0 1
Número de registros 79 210 238 117 12 656

82
Figura 02: Frequência de registros para as espécies de médios e grandes mamíferos nas
diferentes unidades fisionômicas estudadas na Fazenda Intermontes, na cidade de Ribeirão
Grande, São Paulo. Legenda: Cac – cachorro-do-mato (Cerdocyon thous), Fel – categoria de
pequenos felinos (Leopardus sp.), Tam – tamanduá-bandeira (Myrmecophaga tridactyla),
Tga – tatu-galinha (Dasypus novemcinctus), Vea – categoria veados (Mazama sp.), Onc –
onça-parda (Puma concolor), Esq – esquilo (Guerlinguetus aestuans), Gua – guaxinim
(Procyon cancrivorus), Ira – irara (Eira barbara) e Demais – incluem anta (Tapirus
terretres), gambá (Didelphis aurita), gato-mourisco (Puma yagouaroundi), furão (Galictis
cuja), jaguatirica (Leopardus pardalis), quati (Nasua nasua),ouriço-cacheiro (Coendu
spinosus) e tatu-peba (Euphractus sexcinctus).
0
10
20
30
40
50
60
70
80
90
100
Demais
Ira
Gua
Esq
Onc
Vea
Tga
Tam
Fel
Cac

83
A composição das espécies com relação à frequência de ocorrência revelou o
agrupamento de algumas unidades fisionômicas semelhantes entre si quanto à sua
complexidade estrutural, pelo menos em alguns dos parâmetros medidos, tais como Ponto
04 (RABOP) e Ponto 20 (REG), Ponto 05 (RABOP) e Ponto 07 (RARBOP) e Ponto 02 (RF),
Ponto 15 (RARBUP), Ponto 03 (RF). Já os demais agrupamentos não revelaram padrão
claro de similaridade, com relação à estrutura da vegetação (Figura 03).
Figura 03: Dendrograma de similaridade quanto à frequência de registros obtidos em
relação a 19 pontos amostrais coletados na Fazenda Intermontes, Ribeirão Grande, SP
(Coeficiente cofonético= 0.709).
RF
0
1
RA
RB
OP
0
5
RA
RB
OP
0
8
RF
0
2
RA
RB
OP
0
6
RA
RB
UP
1
3
RA
RB
UP
1
2
RA
RB
UP
1
4
RA
RB
UP
0
9
RA
RB
UP
1
0
RA
RB
OP
0
4
RA
RB
OP
0
7
PL
1
9
PL
1
7
PL
1
8
PL
1
6
RE
G
20
RA
RB
UP
1
5
RF
0
3
Alt
ura

84
Efeito dos descritores ambientais sobre a comunidade de mamíferos
A RDA parcial explicou apenas 50,54% da variação dos dados analisados
(R²ajustado=0,10) depois de controlar a variável espacial e produziu sete eixos de
ordenação, de tal forma que os dois primeiros explicaram 71,91%. O primeiro eixo da
ordenação (RDA1) representou positivamente e principalmente o dossel, mas também a
disponibilidade de artrópodes e de médios mamíferos, e negativamente as demais
variáveis. A profundidade de cobertura vegetal e a obstrução a 1,5m do solo foram
melhores explicadas pelo segundo eixo da ordenação (RDA2) de forma negativa, e de forma
positiva a disponibilidade de frutos. A quantidade de solo exposto e a quantidade de dossel
fechado foram as variáveis que explicaram a variação das espécies desta assembleia
(Figura 04, Tabela 02), mas a porcentagem de explicação de suas variâncias foi bastante
baixa (3,5% e 4,6%; respectivamente).
Figura 04: Diagrama de ordenação da Análise de Redundância Parcial das espécies de
médios e grandes mamíferos (Cach – Cerdocyon thous; Esq - Guerlinguetus aestuans; Feli –
pequenos felinos; Ira- Eira barbara; Onç – Puma concolor; Tam – Myrmecophaga tridactyla;
Tgal – Dasypus novemcinctus; Vea – Mazama sp.) em relação a variáveis ambientais (Artro
– disponibilidade de artrópodes; CobVeg – profundidade de cobertura vegetal; Dos –

85
quantidade de dossel fechado; Fru – disponibilidade de frutos; Peq – disponibilidade de
pequenos mamíferos; Med – disponibilidade de médios mamíferos; ObstI – obstrução da
estrutura vertical da vegetação a 0,5m do solo; ObstII – obstrução da estrutura vertical da
vegetação a 1,5m do solo; Solo – quantidade de solo exposto). Pontos de 1-20
correspondem as áreas amostradas (correspondência na Tabela 01, Seção Materiais e
Métodos).
Tabela 02. Resultados da Análise de Redundância Parcial para avaliar a influência das
variáveis ambientais na composição das espécies de médios e grandes mamíferos
registrados nos 19 pontos amostrais, localizados na Fazenda Intermontes, Ribeirão Grande,
SP. Número de permutações: 99. Legenda: Solo – quantidade de solo exposto, Doss –
quantidade de dossel fechado, ObstI – obstrução a 0,5m do solo, ObstII – obstrução a 1,5m
do solo, CobVeg – profundidade de cobertura vegetal, Artro – peso seco médio de
artrópodes, Frutos – quantidade de frutos zoocóriocos, * p<0,05.
Variáveis DP Variação Permutação(>F)
Solo 1 0,035 0,06* Doss 1 0,046 0,02* ObstI 1 0,010 0,73 ObstIII 1 0,020 0,32 CobVeg 1 0,015 0,41 Artro 1 0,003 0,97 Frutos 1 0,006 0,82
Peq 1 0,008 0,72 Med 1 0,028 0,15
Quanto às espécies, o tamanduá-bandeira esteve levemente associado à obstrução
da estrutura vertical da vegetação a 0,5m do solo, enquanto o tatu-galinha estive associado
positivamente ao dossel e a disponibilidade de médios mamíferos. No entanto, tal
associação aparenta ser reflexo de sua elevada abundância dentro da categoria de médios
mamíferos. Para as demais espécies nenhum padrão ficou claro quanto à influência das
variáveis ambientais.

86
A análise de partição demonstrou que o efeito do espaço geográfico sobre a
frequência de ocorrência das espécies de médios e grandes mamíferos foi menor (8,30%)
do que o efeito das variáveis ambientais (74,20%), logo o padrão de distribuição é
influenciado predominantemente pelas variáveis ambientais (Tabela 03). Desta forma, as
análises de seleção de modelos lineares generalizados não levaram em conta o efeito do
espaço na frequência de registros das espécies (Tabela 03).
Tabela 03. Resultados da Análise de Partição de Variância para analisar o efeito exclusivo
das variáveis ambientais e do efeito do espaço sobre a abundância de médios e grandes
mamíferos registrados nos 19 pontos amostrais da Fazenda Intermontes, Ribeirão Grande,
SP.
Variação Proporção
Explicação total 0,2365 Variáveis ambientais 0,1755 0,7420
Variáveis geográficas 0,0197 0,0830
Ambientais + geográficas 0,0413 0,1746
Ao analisar a influência das variáveis de estrutura da vegetação e da disponibilidade
de recursos sobre abundância de cada espécie da assembleia de médios e grandes
mamíferos através da construção de modelos lineares generalizados, observou-se que oito
modelos foram selecionados, entre eles o modelo nulo. Ainda assim, os demais modelos
consideraram um efeito positivo tanto da obstrução vertical da vegetação e do dossel,
quanto da disponibilidade de frutos sobre a frequência de ocorrência da assembleia, e um
efeito negativo da disponibilidade de artrópodes como de pequenos mamíferos (Tabela
03).

87
Tabela 03. Resultados da seleção dos modelos generalizados com erros binomiais negativos
e função de ligação log para a abundância de pequenos mamíferos da Fazenda Intermontes,
na cidade de Ribeirão Grande, SP. Modelo – modelos selecionados (ΔAICc < 2); k – número
de parâmetros; AICc - Critério de Informação de Akaike modificado para amostras
pequenas; ΔAICc – diferença entre o melhor modelo e o modelo considerado; wi – peso de
evidência. Para os modelos: (+) relação positiva, (-) relação negativa, PC1 e PC2 – escores
dos pontos dos dois primeiros eixos da PCA da estrutura da vegetação, Artro –
disponibilidade de artrópodes, Frutos – disponibilidade de frutos, Peq – disponibilidade de
pequenos mamíferos, Med – disponibilidade total de médios mamíferos, Medfel –
disponibilidade parcial de médios mamíferos somente para pequenos felídeos.
Modelo K AICc ΔAICc wi
Assembleia
(+)PC2 + (+)Frutos 4 170,43 0,0 0,1165
(+)PC2 3 170,84 0,4 0,0951
(+)Frutos 3 171,14 0,7 0,0818
(+)PC2 + (+)Frutos + (-)Peq 5 171,66 1,2 0,0632
Nulo 2 172,13 1,7 0,0498
(+)PC1 + (+)PC2 + (+)Frutos 5 172,17 1,7 0,0489
(+)PC2 + (-)Artro 5 172,21 1,8 0,048
(+)Frutos + (-)Peq 4 172,27 1,8 0,0464
Cerdocyon thous
(+)PC2 3 123,42 0,0 0,221
(+)PC1 + (+)PC2 4 124,78 1,4 0,112
(+)PC2 + (-)Artro 4 125,17 1,7 0,092
(+)PC2 + (+)Frutos 4 125,25 1,8 0,088
(+)PC2 + (+)Peq 4 125,4 2,0 0,082
Dasypus novemcinctus
Nulo 2 80,59 0,0 0,168
(+)PC2 3 80,9 0,3 0,143
(+)Frutos 3 81,03 0,3 0,135
(+)PC2 + (+)Frutos 4 81,95 1,4 0,085
(+)Artro 3 81,96 1,4 0,085
(-)PC1 3 82,29 1,7 0,072
Eira barbara
(-)Peq 3 40,93 0,0 0,159
(-)PC2 + (-)Artro + (-)Peq 5 41,63 0,7 0,112
(-)PC2 + (-)Peq 4 41,72 0,8 0,107
(-)Artro + (-)Peq 4 42,34 1,4 0,078
(+)Frutos + (-)Peq 4 42,56 1,6 0,070

88
Para D. novemcinctus, M. tridactyla e para G. ingrami, o modelo nulo esteve entre os
selecionados. Ainda assim, os demais modelos concorrentes revelaram o efeito positivo da
cobertura vegetal, da disponibilidade de frutos e artrópodes e um efeito negativo da
obstrução vertical da vegetação para D. novemcinctus, um efeito negativo tanto das
variáveis estruturais da vegetação quando da disponibilidade de artrópodes para M.
tridactyla, e uma relação positiva com a disponibilidade de frutos, e negativa com as
Modelo K AICc ΔAICc wi Guerlinguetus ingrami
Nulo 2 48,67 0,0 0,384
(+)Frutos 3 50,62 1,9 0,145
(-)PC2 3 50,67 2,0 0,145
(-)PC1 3 50,63 2,0 0,145
Mazama sp.
(-)PC1 + (+)Frutos 4 92,53 0,0 0,390
(-)PC1 + (-)PC2 + (+)Frutos 5 93,04 0,5 0,303
M. tridactyla Nulo 2 74,03 0,0 0,271 (-)PC1 3 74,14 0,1 0,252 (-)Artro 3 75,91 1,9 0,106 (-)PC2 3 76,01 2,0 0,101 (-)PC1 + (-)Artro 4 76,04 2,0 0,099 Leopardus sp. Nulo 2 133,09 0,0 0,229 (-)Peq 3 133,86 0,8 0,155
(+)PC2 3 134,11 1,0 0,137 (+)PC1 + (+)PC2 4 134,17 1,1 0,133 (+)PC2 + (-)Peq 4 134,28 1,2 0,126 (+) PC1 3 134,61 1,5 0,107 Puma concolor
(-)Peq 3 39,63 0,0 0,375
(-)PC1 + (-)Peq 4 40,99 1,4 0,191
(-)Peq + (+)Med 4 41,47 1,8 0,149
(+)PC2 + (-)Peq 4 41,58 1,9 0,142

89
características da vegetação para G. ingrami. Para Leopardus sp., o modelo nulo esteve
selecionado junto àqueles que revelaram um efeito negativo da disponibilidade de
pequenos mamíferos e de um efeito positivo da estrutura da vegetação.
Para o C. thous, os modelos selecionados consideraram o efeito positivo com as
variáveis da estrutura do habitat, com a disponibilidade de frutos e pequenos mamíferos, e
um efeito negativo com a disponibilidade de artrópodes.
Para a irara, Eira barbara, cinco modelos selecionados evidenciaram a relação
positiva com relação a cobertura vegetal e com a disponibilidade de frutos, e uma relação
negativa com a disponibilidade de artrópodes e de pequenos mamíferos.
Para Mazama sp. somente dois modelos foram selecionados e os mesmos
demonstraram uma relação negativa com a estrutura do habitat, mas positiva com a
disponibilidade de frutos. E por fim, para P. concolor, quatro modelos foram capazes de
explicar a variação dos dados de frequência de ocorrência da espécie. Tais modelos
evidenciaram uma relação positiva com a cobertura vegetal e com a disponibilidade de
médios mamíferos, e uma relação negativa com as variáveis de obstrução vertical da
vegetação e do fechamento do dossel e com a disponibilidade de pequenos mamíferos.
DISCUSSÃO
O grande desafio dos processos de restauração é reproduzir a mesma a
complexidade estrutural da vegetação encontrada em ambientes originais. Em nossa área
de estudo, no entanto, os diferentes tipos de tratamentos não foram capazes de recuperar a
mesma complexidade da vegetação encontrada nos remanescentes florestais amostrados
até o momento, assim como verificado em outros estudos (Kanowski et al. 2003, Munro et
al. 2007, Munro et al. 2009). Todavia, diferentes espécies de mamíferos de médio e grande
porte foram registradas nas unidades fisionômicas, mas a frequência de ocorrência entre
elas foi bastante distinta. Ainda assim, a menor ocorrência foi observada na área mais
aberta.
As espécies mais frequentemente registradas, C. thous, Leopardus sp., Mazama sp., D.
novemcinctus, M. tridactyla e G. ingrami estiveram distribuídas entre os diferentes tipos de

90
tratamentos de restauração, além dos remanescentes florestais. Tais espécies naturalmente
ocorrem em diferentes biomas do país, não demonstrando preferência de habitat (Reis et
al. 2006, Gammons et al. 2009) e vários estudos têm evidenciado a alta abundância de
cachorros-do-mato e de tatu-galinha em áreas alteradas (Naughton-Treves et al. 2003,
Harvey et al. 2006, Dotta e Verdade 2007, Michalski e Peres 2007, Gamonn et al. 2009).
Para C. thous as variáveis da estrutura do habitat, como a disponibilidade de frutos e
pequenos mamíferos apresentaram um efeito positivo em sua frequência de ocorrência.
Cerdocyon thous naturalmente faz uso de diferentes formações vegetais (florestas, campos,
savanas, restingas e bordas florestais a áreas alteradas) (Jácomo et al. 2004, Maffei e Taber
2003, Vieira e Port 2007,Di Bitetti et al. 2009, Goulart et al. 2009, Ferraz et al. 2010) e o seu
padrão de uso de habitat aparenta estar relacionado mais diretamente com o seu padrão de
forrageio (Facure e Monteiro-Filho 1996, Rocha et al. 2004) regido por uma dieta onívora e
oportunista, que apresenta composições similares em diferentes proporções em função da
localidade da espécie (MacDonald & Courtnay 1996, Juarez e Marinho-Filho 2002, Facure et
al. 2003, Bueno e Motta-Junior 2004, Jácomo et al. 2004, Rocha et al. 2004, Gatti et al. 2006,
Pedó et al. 2006, Rocha et al. 2008). Em nosso estudo, essa espécie esteve presente em
todas as unidades (exceto a regeneração natural), confirmando a falta de seletividade do
habitat, embora a presença da vegetação pareça ser relevante para lhe garantir proteção,
haja vista que tais variáveis da estrutura foram selecionadas. Mas, aparentemente a
disponibilidade de recursos do local tende a ser mais importante no seu padrão de
atividade.
A categoria Leopardus sp. esteve associada às características da estrutura do habitat.
Esta categoria igualmente está presente em diferentes biomas e fisionomias (Reis et al.
2006) e a vegetação deve lhe conferir proteção e camuflagem para a predação de suas
presas, mas a presença em todos os tratamentos demonstra não haver uma variável
restrita atuante no seu padrão de ocorrência. Embora pequenos mamíferos também
estejam entre os itens predados (Wang 2002, Bianchi et al. 2011), os mesmos não
aparentaram efeito sobre sua ocorrência.
A frequência de ocorrência dos veados (Mazama sp.) esteve relacionada
positivamente à disponibilidade de frutos, e negativamente à estrutura do habitat. Para o
gênero, tanto M. americana quanto M. gouazoubira encontram-se presentes nas diferentes

91
regiões do país vivendo em simpatria, embora M. gouazoubira esteja mais associado às
áreas mais abertas de campos e cerrados e M. americana às áreas mais florestais (Reis et al.
2006). O registro deste gênero esteve presente tanto em formações mais abertas de
regeneração, quanto em áreas de plantio e remanescentes florestais, mas as diferenças nas
preferências de habitat dessas espécies provavelmente não foram verificadas dado o nível
genérico da análise. Já a seleção positiva da disponibilidade de frutos para o grupo
representa uma importante necessidade ecológica dada sua dieta frugívoro-herbívora
composta por frutos, flores, gramíneas, leguminosas e por fungos em menor proporção
(Richard e Julia 2001, Gayot et al. 2004, Reis et al. 2006).
Para D. novemcinctus, M. tridactyla e G. ingrami a seleção do modelo nulo entre os
selecionados é evidência de que para essas espécies os descritores ambientais avaliados
podem não ser os mais importantes para a sua ocorrência. A falta de seletividade de
habitats estruturalmente distintos (Taulman e Robbins 1996, Reis et al. 2006, Gammons et
al. 2009) e a dieta onívora geralmente predominada por invertebrados, mas também
composta por material vegetal (Sikes et al. 1990, Whitaker et al. 2012), explicam este
resultado para D. novemcinctus.
Myrmecophaga tridactyla igualmente ocorre em habitats com diferentes
complexidades estruturais, desde campos a ambientes florestais (Fonseca et al. 1996,
Desbiez e Médri 2010) e para essa espécie estudos prévios têm demonstrando que a
temperatura é o fator mais importante na determinação e modificação de seu padrão de
atividade e de sua seleção de habitats, ao intervir na regulação de sua baixa taxa
metabólica, ocasionada em função de seu tamanho corpóreo (Medri e Mourão 2005a,
Camilo-Alves e Mourão 2006). Assim como observado por tais pesquisadores (Medri e
Mourão 2005a, Camilo-Alves e Mourão 2006), em nossa área de estudo, todos os registros
foram observados no período noturno, exceto por dois registros observados por volta
13h00 em remanescente florestal e por volta das 17h00 em área de regeneração arbórea
com plantio.
Já a pequena área de vida, a alta fecundidade, a rápida dispersão e a dieta
generalista são aspectos que devem favorecer a presença de G. ingrami em áreas
perturbadas (Paschoal & Galetti 1995, Cosson et al. 1999, Lugo 2001). Concorrente ao
modelo nulo, a disponibilidade de frutos demonstrou um efeito positivo sobre a ocorrência

92
dessa espécie, em contrapartida ao efeito negativo da estrutura de habitat. A presença do
recurso alimentar no entanto, deve apresentar papel essencial para garantir a existência
da espécie, uma vez presente.
Puma concolor e E. barbara foram as espécies com menores frequências. Para E.
barbara as variáveis de cobertura vegetal e a disponibilidade de frutos demonstraram um
efeito positivo sobre a ocorrência da espécie. A irara encontra-se associada às formações
florestais (Eisenberg e Redford 1999, Presley 2000, Desbiez et al. 2009). O seu próprio
hábito arborícola deve atuar na seleção desses ambientes, inclusive para permitir a captura
de suas presas, muitas vezes igualmente arborícolas (Presley 2000), mas variáveis da
estrutura vertical não demonstraram efeito, somente a cobertura vegetal. A disponibilidade
de frutos, por sua vez, confirma a importância desses descritores alimentares para a dieta
onívora (Calouro 2000, Presley 2000, Bezerra et al. 2009).
Para P. concolor a profundidade de cobertura vegetal e a disponibilidade de de
médios mamíferos demonstraram um efeito positivo sobre a ocorrência da espécie,
diferentemente das variáveis de obstrução vertical da vegetação, de fechamento do dossel e
da disponibilidade de pequenos mamíferos. No Brasil, este predador de topo está presente
em todos os biomas (Fonseca et al. 1996, Reis et al. 2006) e é considerado um generalista
quanto à seleção de habitat (Kertson et al. 2011, Kelly et al. 2008, Negrões et al. 2010). No
entanto, a presença de áreas com cobertura estrutural da vegetação capaz de lhe garantir
segurança no seu descanso e de favorecer sua camuflagem necessária para a aproximação
de suas presas são fatores considerados importantes na ocupação da espécie (Dickson e
Beier 2002, Kertson et al. 2011). A relação desta espécie com a cobertura vegetal e com a
obstrução vertical de vegetação não deixou claro o papel desses descritores sobre a
ocorrência da espécie, principalmente ao se considerar as discrepâncias estruturais da
vegetação com relação às unidades fisionômicas por onde ela foi registrada. Já sua relação
positiva com a disponibilidade de médios mamíferos e negativa com pequenos mamíferos
pode ser reflexo do papel desses grupos na dieta deste predador oportunista (Rau e
Jimenéz 2002, Novack et al. 2005, Martins et al. 2008, Thompson et al. 2009, Azevedo
2008).
Desta forma, assim como previsto em nossa hipótese inicial, as espécies de médios e
grandes mamíferos foram capazes de utilizar as diferentes unidades fisionômicas uma vez

93
que tais unidades estivessem de acordo com suas necessidades ecológicas, confirmando o
modelo continuum. O papel de cada descritor ambiental para cada espécie apresentou
efeitos distintos em vista dessas necessidades e ainda para M. tridactyla não apresentou
efeito positivo algum, evidenciando que outros filtros não estudados podem ser
importantes, como o próprio modelo considera (Fischer e Lindernmayer 2006).
Contudo, embora diferentes espécies de médios e grandes mamíferos tenham sido
registradas neste mosaico de restauração, este resultado tem que ser avaliado com grande
cautela. As espécies mais frequentemente registradas na assembleia foram representadas
por espécies generalistas que não dependem diretamente de uma estrutura complexa da
vegetação, assim como verificado em outros estudos (Nichols e Nichols 2003, Nichols e
Grant 2007, Munro et al. 2007). Espécies mais especialistas e mais suscetíveis a ambientes
alterados, como T. terrestris e L. pardalis, foram muito pouco frequentes, indicando que
suas necessidades ecológicas ainda não são garantidas pela restauração presente. Portanto,
a existência do habitat em si não significa que necessariamente todas as necessidades
ecológicas estão sendo garantidas a todas as espécies (Nichols e Nichols 2003).
Espécies mais sensíveis como a onça-pintada (Panthera onca), outro grande
predador de topo, grandes herbívoros como o queixada (Tayassu pecari), a paca (Cuniculus
paca), ou o tamanduá-mirim (Tamandua tetradactyla) encontram-se presentes no Parque
Estadual de Intervales, área contínua de Mata Atlântica localizada próxima ao local da
restauração (Vivo e Gregorin 2001), e ainda assim não foram registradas em Intermontes.
Sua ausência é, portanto, indicativo de que o processo de restauração implantado ainda não
é capaz de garantir os recursos necessários para o seu uso por toda a comunidade original
de médios e grandes mamíferos.
Embora este estudo tenha se centrado na ocorrência de mamíferos terrestres,
espécies de primatas igualmente não estão presentes na área estudada. A presença de uma
estrutura da vegetação constituída por árvores de baixo porte, ramificadas desde sua
porção mais inferior produzindo uma copa aberta e consequentemente um dossel de mata
mais aberto, são fatores que devem impedir a presença e ocupação dessas espécies que
dependem de dosséis fechados e de árvores de grande porte com diferentes estratos
arbóreos, árvores de dormir protegidas e até mesmo condição climática adequada para sua
termorregulação (Anderson 2000, Warner 2000, Gestich et al. 2014), principalmente ao se

94
considerar espécies de grandes e médios primatas presentes na região, como o mono-
cavoeiro (Brachyteles arachnoides), o bugio (Allouata fusca) e o macaco-prego (Sapajus
nigritus) (Vivo e Gregorin 2001).
Portanto, para as espécies mais generalistas a restauração pode favorecer o uso
frequente deste mesohabitat dentro de sua área de vida, mas para aquelas espécies mais
especialistas, sensíveis a ambientes alterados e com maiores áreas de vida, os ambientes
restaurados em Intermontes devem atuar como ‘trampolins ecológicos’ ou corredores
ecológicos dentro da paisagem local fragmentada (Metzger 2001. Haddad et al. 2003),
como no caso de T. terrestris e P. concolor, sendo utilizados temporariamente (Munro et al.
2007), ou ainda não devem ser capazes de suprir suas necessidades. Logo, medidas de
manejo devem ser consideradas em processos de restauração futuros, para que a
colonização abranja toda a comunidade de mamíferos terrestres de médio e grande porte,
principalmente as espécies mais especialistas que normalmente são o foco da cosnervação.
Implicações de Manejo
O presente estudo demonstrou que espécies de mamíferos terrestres de médio e
grande porte foram capazes de ocupar o mosaico de restauração de Mata Atlântica
implantada na Fazenda Intermontes. Todavia, esta ocupação ocorreu distintamente entre
as espécies, de acordo com suas necessidades ecológicas. Espécies generalistas de habitat,
que não dependem diretamente de uma estrutura mais complexa da vegetação, foram as
mais favorecidas. Espécies mais sensíveis já estiveram presentes em baixa frequência.
Não só o contexto de paisagem, mas a configuração espacial da restauração, as
espécies de plantas arbóreas e sua densidade de plantio, como as próprias espécies de sub-
bosque utilizadas podem resultar em destinos diferentes da recuperação da vegetação,
criando habitats pobres e que não garantem as condições necessárias para suportar
populações de fauna (Vesk et al. 2008).
Nosso estudo indicou que a trajetória de restauração em Intermontes não foi capaz
de igualar as características estruturais dos remanescentes florestais após 13 anos de
plantio. Unidades que se iniciaram com os mesmos tratamentos resultaram em
complexidades estruturais distintas. A estratégia de restauração adotada e ou a falta de sua
manutenção ao longo deste tempo pode ter sido responsável pelo seu destino atual na

95
estruturação da vegetação, favorecendo o uso e ocupação por espécies mais generalistas e
oportunistas.
Novas estratégias de restauração que levem em conta este aspecto podem gerar
melhores recursos às diferentes espécies de mamíferos, principalmente às especialistas.
Ainda assim, estudos que avaliem e esclareçam quais são os recursos mais importantes
para cada espécie de mamífero terrestre devem ser priorizados, pois podem determinar os
parâmetros necessários para tais estratégias, já que tais espécies respondem mais
diretamente aos gradientes de variação das variáveis ambientais em primeiro lugar
(Fischer e Lindernmayer 2006), como verificado em nosso estudo. Medidas de manejo que
sejam capazes de restaurar estes recursos espécie-específicos numa escala mais fina serão
capazes de garantir a colonização das diferentes espécies de fauna.
REFERÊNCIAS BIBLIOGRÁFICAS
Anderson, J. R. 2000. Sleep-related behavioural adaptations in free-ranging anthropoid
primates. Sleep Medicine Reviews 4(4): 355-373.
Azevedo, F. C. C. 2008. Food habitats and livestock depredation of sympatric jaguars and
pumas in the Iguaçu National Park Area, south Brazil. Biotropica 40(4): 494-500.
Bezerra, B. M., Barnett, A. A., Souto, A., Jones, G. 2009. Predation by tayra on the common
marmoset and the pale-throated three-toed sloth. Journal of Ethology 27: 91-96.
Bianchi, R. C., Rosa, A. F., Gatti, A., Mendes, S. L. 2011. Diet of margay, Leopardus wiedii, and
jaguarundi, Puma yagouaroundi, (Carnivora: Felidae) in Atlantic Rainforest, Brazil.
Zoologia 28(1): 127-132.
Bolker, B. 2007. Ecological models and data in R. Princeton University Press, Princeton.
Borcard, D., Legendre, P., Drapeau, P. 1992. Partialling out the spatial component of
ecological variation. Ecology 73: 1045-1055.
Borcard, D., Gillet, F., Legendre, P. 2011. Numerical Ecology with R. Springer, New York,
USA.

96
Bueno, A. A, Motta-Junior, C. J. 2004. Food habits of two syntopic canids, tha maned wolf
(Chrysocyon brachyurus) and the crab-eating fox (Cerdocyon thous), in southeastern
Brazil. Revista Chilena de Historia Natural 77: 5-14.
Burnham, K. P., Anderson, D. R. 2002. Model selection and multimodel inference: a practical
information-theoretic approach. 2ed. Springer Verlag, New York, USA.
Camilo-Alves, C. S. P., Mourão, G. 2006. Responses of a specialized insectivorous mammal
(Myrmecophaga tridactyla) to variation in ambient temperature. Biotropica 38(1):
52-56.
Caughley, G. 1977. Analysis of vertebrate populations. Caldwell: The Blackburn Press.
Chiarello, A. G. 1999. Effects of fragmentation of the Atlantic forest on mammal
communities in south-eastern Brazil. Biological Conservation 89: 71-82.
Chiarello, A. G. 2000. Density and population size of mammals in remnants of Brazilian
Atlantic forest. Conservation Biology 14(6): 1649-1657.
Cosson, J. F., Ringuet, S., Claessens, O., Massary, J. C. D. E., Dalecky, A., Villiers, J. F., Grajon, L.,
Pons, J. N. 1999. Ecological changes in recent land-bridge islands in French Guiana,
with emphasis on vertebrate communities. Biological Conservation 91: 213-222.
Cowell, R. K. 2009. Estimate S: Statistical estimation of species richness and shared species
from samples. Versão 8.2.
Crooks, K. R., Soulé, M. E. 1999. Mesopredator release and avifaunal extinctions in a
fragmented system. Nature 400:563-566.
Desbiez, A. L. J., Bodmer, R. E., Santos, S. A. 2009. Wildlife habitat selection and sustainable
resources management in a Neotropical wetland. International Journal of
Biodiversity and Conservation 1(1): 11-20.
Desbiez, A. L. J., Medri, I. M. 2010. Density and habitat use by giant anteaters
(Myrmecophaga tridactyla) and southern tamandúas (Tamandua tetradactyla) in the
Pantanal wetland, Brazil. Edentata 11(1): 4-10.
Di Bitetti, M. S., Blanco, Y. E., Pereira, J. A., Paviolo, A., Pérez, I. J. 2009. Time partitioning
favors the coexistence of sympatric crab-eating foxes (Cerdocyon thous) and pampas
foxes (Lycalopex gymnocercus). Journal of Mammalogy 90(2): 479-490.
Dickson, B. G., Beier, P., 2002. Home range and habitat selection by adult cougars in
southern California. Journal of Wildlife Management 66: 1235–1245.

97
Dotta, G., Verdade, L. M. 2007. Trophic categories in a mammal assemblage: diversity in an
agricultural landscape. Biota Neotropica 7(2): 287-292.
Eisenberg, J. F., Redford, K. H. 1999. Mammals of the Neotropics – The Central Neotropics
(Ecuador, Peru, Bolivia, Brazil). Chicago: University of Chicago Press.
Facure, K. G., Monteiro-Filho, E. L. A. 1996. Feeding habits of the crab-eating fox, Cerdocyon
thous (Carnivora, Canidae), in a suburban area of southeastern Brazil. Mammalia
60(1): 147-149.
Facure, K. G., Giaretta, A. A., Monteiro-Filho, E. L. A. 2003. Food habitats of the crab-eating-
fox, Cerdocyon thous, in an altitudinal forest of the Mantiqueira range, southeastern
Brazil. Mammalia 67(4): 503-511.
Ferraz, K. M. P. M. B., Siqueira, M. F., Martin, P. S., Esteves, C. F., Couto, H. T. Z. 2010.
Assessment of Cerdocyon thous distribution in an agricultural mosaic, southeastern
Brazil. Mammalia 74: 275-280.
Fischer, J., Lindenmayer, D. B. 2006. Beyond fragmentation: the continuum model for fauna
research and conservation in human-modified landscapes. Oikos 112(2): 473-480.
Fonseca, G. A. B., Robinson, J. G. 1990. Forest size and structure: competitive and predatory
effects on small mammal communities. Biological Conservation 53: 265-294.
Fonseca, G. A. B., Hermann, G., Leite, Y. L. R., Mittermeier, R. A., Rylands, A. B., Patton, J. L.
1996. Lista anotada dos mamíferos do Brasil. Occasional Papers in Conservation
Biology 4: 1-38.
Gammons, D. J., Mengak, M. T., Conner, L. M. 2009. Armadillo habitat selection in
Southwestern Georgia. Journal of Mammalogy 90(2): 356-362.
Gatti, A., Bianchi, R., Rosa, C. R. X., Mendes, S. L. 2006. Diet of two sympatric carnivores,
Cerdocyon thous ans Procyon cancrivorus, in a resting area of Espirito Santo State,
Brazil. Journal of Tropical Ecology 22: 227-230.
Gayot, M., Henry, O., Dubost, G., Sabatier, D. 2004. Comparative diet of the two forest
cervids of the genus Mazama in French Guiana. Journal of Tropical Ecology 20(1):
31-43.
Gestich, C. C., Caselli, C. B., Setz, E. Z. F. 2014. Behavioural thermorregulation in a small
neotropical primate. International Journal of Behaviour Biology 120: 331-339.

98
Goulart, F. V. B., Cáceres, N. C., Graipel, M. E., Tortato, M. A., Ghizoni Jr, I. R., Oliveira-Santos,
L. G. R. 2009. Habitat selection by large mammals in a southern Brazilian Atlantic
forest. Mammalian Biology 74: 182-190.
Haddad, N. M., Bowne, D. R., Cunningham, A., Danielson, B. J., Levey, D. J., Sargent, S., Spira,
T. 2003. Corridor use by diverse taxa. Ecology 84 (3): 609-615.
Harvey, C. A., Gonzalez, J., Somarriba, E. 2006. Dung beetle and terrestrial mammal
diversity in forests, indigenous agroforestry systems and plantain monocultures in
Talamanca, Costa Rica. Biodiversity and Conservation 15: 555-585.
Henle, K., Lindenmayer, D. B., Margules, C. R., Saunders, D. A., Wissel, C. 2004a. Species
survival in fragmented landscape: where are we now? Biodiversity and Conservation
13: 1-8.
Hobbs, R., Catling, P. C., Wombey, J. C., Clayton, M., Atkins, L., Reid, A. 2003. Faunal use of
bluegum (Eucalyptus globulus) plantations in southwestern Australia. Agroforestry
Systems 58: 195-212.
Jácomo, A. T. A, Silveira, L., Diniz-Filho, J. A. F. 2004. Niche separation between the maned
Wolf (Chrysocyon brachyurus), the crab-eating fox (Dusyon ventulus) in Central
Brazil. Journal of Zoology 262(1): 99-106.
Juarez, K. M., Marinho-Filho, J. 2002. Diet, habitat use, and home ranges of sympatric canids
in central Brazil. Journal of Mammalogy 83 (4): 925-933.
Kanowski, J., Catterall, C. P., Wardell-Johnson, G. W., Prostor, H., Reis, T. 2003. Development
of forest structure on cleared rainforest land in Australia under different styles of
reforestation. Forest Ecology and Management 183: 265-280.
Kelly, M. J., Noss, A. J., Di Bitetti, M. S., Maffei, L., Arispe, R. L. 2008. Estimating puma density
from camera trapping across three study sites: Bolivia, Argentina, and Belize.
Journal of Mammalogy 89: 408-418.
Kertson, B. N., Spencer, R. D., Marzluff, J. N., Hepinstall-Cymerman, J., Grue, C. E. 2011.
Cougar space use and movements in the wildland-urban landscape of western
Washington. Ecological Applications 21(8): 2866-2881.
Laurance, W. F. 1994. Rainforest fragmentation and the structure of small mammal
communities in Tropical Queensland. Biological Conservation 69: 23-32.

99
Laurance, W. F., Lovejoy, T. E., Vasconcellos, H. L., Bruna, E. M., Didham, R. K., Stouffer, P. C.,
Gascon, C., Bierregaard, R. O., Laurance, S. G., Sampaio, E. 2002. Ecosystem decay of
Amazonian Forest fragments: a 22-year investigation. Conservation Biology 16(3):
605-618.
Laurance, W. F., Laurance, S. G., Hilbert, D. W. 2008. Long-term dynamics of a fragmented
rainforest mammal assemblage. Conservation Biology 22(5): 1154-1164.
Legendre, P., Gallager, E. D. 2001. Ecologically meaningful transformations for ordination of
species data. Oecologia 129: 271-280.
Liu, Q. 1997. Variation partitioning by redundancy analysis (RDA). Environmentrics 8:75-
85.
Lyra-Jorge, M. C., Ciocheti, G., Pivello, V. R. 2008. Carnivore mammals in a fragmented
landscape in northeast of São Paulo State, Brazil. Biodiversity and Conservation
17(7): 1573-1580.
Lugo, A. E. 2001. El manejo de la biodiversidade en el siglo XXI. Interciencia 26(10): 484-
490.
MacDonald, D. W., Courtenay. 1996. Enduring social relationships in a population of crab-
eating zorros, Cerdocyon thous, in Amazonian Brazil (Carnivora, Canidae). Journal of
Zoology 239: 329-355.
Maffei, L., Taber, B. A. 2003. Área de accón, actividad y uso de hábitat del zorro patas
negras, Cerdoyon thous, en un bosque seco. Mastozoologia Neotropical 10: 154-160.
Martins, R., Quadros, J., Mazzolli, M. 2008. Hábito alimentar e interferência antrópica na
atividade de marcação territorial do Puma concolor e Leopardus pardalis (Carnivora:
Felidae) e outros carnívoros na Estação Ecológica da Juréia-Itatins, São Paulo, Brasil.
Revista Brasileira de Zoologia 25(3): 427-435.
Medri, I. M., Mourão, G. 2005a. Home range of giant anteaters (Myrmecophaga tridactlyla)
in the Pantanal wetland, Brazil. Journal of Zoology 266: 365-375.
Metzger, J. P. 2001. O que é ecologia de paisagens? Revista Biota Neotropica 1(1):1-9.
Michalski, F., Peres, C. A. 2007. Disturbance-mediated mammal persistence and abundance-
area relationships in Amazonian forest fragments. Conservation Biology 21(6):
1626-1640.

100
Munro, N. T., Lindenmayer, D. B., Fischer, J. 2007. Faunal response to revegetation in
agricultural areas of Australia: a review. Ecological Managmente and Restoration
8(3): 199-207.
Munro, N. T., Fischer, J., Wood, J., Lindenmayer, D.B. 2009. Revegetation in agricultural
areas: the development of structural complexity and floristic diversity. Ecological
Applications 19(5): 1197-1210.
Naughton-Treves, L., Mena, J. L., Treves, A., Alvarez, N., Radeloff, V. C. 2003. Wildlife survival
beyond park boundaries: the impact slash-and-burn agriculture and hunting on
mammals in Tambopata, Peru. Conservation Biology 17: 1106-1117.
Nichols, O. G., Nichols, F. M. 2003. Long-term trends in faunal recolonization after bauxite
mining in the Jarrah forest of southwestern Australia. Restoration Ecology 11(3):
261-272.
Nichols, O. G., Grant, C. D. 2007. Vertebrate fauna recolonization of restored bauxite mines –
key findings from almost 30 years of monitoring and research. Restoration Ecology
15(4): S116-S126.
Negrões, N., Sarmento, P., Crus, J., Eira, C., Revilla, E., Fonseca, C., Sollman, R., Tôrres, N.M.,
Furtado, M.M., Jácomo, A.T.A., Silveira, L. 2010. Use of camera-trapping to estimate
puma density and influencing factors in Central Brazil. Journal of Wildlife
Management 74(6): 1195-1203.
Novack, A. J., Main, M. B., Sunquist, M. E., Labisky, R. F. 2005. Foraging ecology of jaguar
(Panthera onca) and puma (Puma concolor) in hunted and non-hunted sites within
the Maya Biosphere Reserve, Guatemala. Journal of Zoology 267: 167-178.
Paglia, A. P., Fonseca, G. A. B., Rylands, A. B., Herrman, G., Aguiar, L. M. S., Chiarello, A. G.,
Leite, Y. L. R., Costa, L. P., Siciliano, S., Kierulff, M. C. M., Mendes, S. L., Tavares, V. C.,
Mittermeier, R. A., Patton, J. L. 2012. Lista Anotada dos Mamíferos do Brasil. 2.ed.
Occasional Papers in Conservation biology. Arlington: Conservation International
Palmer, M. A., Ambrose, R. F., Poff, N. L. 1997. Ecological theory and community restoration
ecology. Restoration Ecology 5(4): 291-300.
Paschoal, M., Galetti, M. 1995. Seasonal food use by Neotropical squirrel Sciurus ingrami in
Southeastern Brazil. Biotropica, 27(2): 268-273.

101
Pedó, E., Tomazzoni, A. C., Hartz, S. M., Christoff, A. U. 2006. Diet of crab-eating fox,
Cerdocyon thous (Linnaeus) (Carnivora, Canidae), in a suburban area of southern
Brazil. Revista Brasileira de Zoologia 23(3): 637-641.
Presley, S. J. 2000. Eira barbara. Mammalian Species 636: 1-6.
Quinn, G. P., Keough, M. J. 2002. Experimental design and data analysis for biologists.
Cambridge University Press, UK.
R Core Team. 2013. R: A language and environment for statistical computing. R Foundation
for Statistical Computing, Viena, Austria.
Rau, J. R., Jiménez, J. E. 2002. Diet of puma (Puma concolor, Carnivora:Felidae) in Coastal
and Andean ranges of southern Chile. Studies of Neotropical Fauna and Environment
37(3): 201-205.
Reis, N. R., Ortêncio Filho, H., Silveira, G. 2006. Mamíferos do Brasil. Eds: Reis, N. R.; Shibatta,
O. A.; Peracchi, A. L.; Pedro, W. A.; Lima, I. P. Londrina.
Richard, E., Juliá, J. P. 2001. Dieta de Mazama gouazoubira (Mammalia, Cervidae) en un
ambiente secundário de Yungas, Argentina. Série Zool. 90: 147-156.
Rocha, V. J., Reis, N. R., Sekiama, M. L. 2004. Dieta e dispersão de sementes por Cerdocyon
thous (Linnaeus) (Carnivora, Canidae) em um fragmento florestal no Paraná, Brasil.
Revista Brasileira de Zoologia 21(4): 871-876.
Rocha, V. J., Aguiar, L. M., Silva-Pereira, J. E., Moro-Rios, R. F., Passos, F. C. 2008. Feeding
habitats of the crab-eating fox, Cerdocyon thous (Carnivora: Canidae), in a mosaic
area with native and exotic vegetation in Southern Brazil. Revista Brasileira de
Zoologia 25(4): 594-600.
Sikes, R. S., Heidt, G. A., Elrod, D. A. 1990. Seasonal diets of the nine-banded (Dasypus
novencinctus) in a northern part of its range. American Midland Naturalist 123: 383-
389.
Taulman, J. F., Robbins, L. W. 1996. Recent range expansion and distributional limits of
nine-banded armadillo (Dasypus novemcinctus) in the United States. Journal of
Biogeography 23: 635-648.
Terborgh, J. 1974. Preservation of natural diversity: the problem of extinction of prone
species. Bioscience 24(12): 715-722.

102
Thompson, D. J., Fecske, D. M., Jenks, J. A., Jarding, A. R. 2009. Food habits of recolonizing
cougars in the Dakotas: prey obtained from prairie and agricultural habitats.
American Midlist Naturalist 161: 69-75.
Vesk, P. A., Nolan, R., Thomson, J. R., Dorrough, J. W., Mac Nally, R. 2008. Time lags in
provision of habitat resources through revegetation. Biological Conservation 141:
174-186.
Vieira, E. M., Port, D. 2007. Niche overlap and resource partitioning between two sympatric
fox species in southern Brazil. Journal of Zoology 272: 57-63.
Vieira, M. V., Olifiers, N.m Delciellos, A. C.m Antunes, V. Z.m Bernardo, L. R.; Grelle, C. E. V.m
Cerqueira, R. 2009. Land use vs. fragment size and isolation as determinants of small
mammal composition and richness in Atlantic forest remnants. Biological
Conservation 142: 1191-1200.
Vivo, M., Gregorin, R. 2001. Mamíferos. In Intervales (C. Leonel, coord.). Fundação para a
Conservação e a Produção Florestal do Estado de São Paulo p. 116-123.
Young, T. P. 2000. Restoration ecology and conservation biology. Biological Conservation
92: 73-83.
Wang, E. 2002. Diets of ocelots (Leoparuds pardalis), margays (L. wiedii), and oncillas (L.
tigrinus) in the Atlantic rainforest in southeast Brazil. Studies on Neotropical Fauna
and Environment 37(3): 207-212.
Warner, M. D. 2002. Assessing habitat utilization by Neotropical primates: a new
approcoah. Primates 43(1): 59-71.
Whitaker, J. O., Ruckdeschel, C., Bakken, L. 2012. Food of the armadilho Dasypus
novencinctus L. from Cumberland Island, GA. Southeastern Naturalist 11(3): 487-206
Zar, J. H. 1999. Biostatistical analysis. Prentice Hall, Jersey, 4ª edição.

103
Anexo I. Lista de mamíferos terrestres registrados na Fazenda Intermontes, SP, entre agosto de 2011 e março de 2014. Tipo de dieta: Car-carnívoro ; Fo-folívoro; Fru-frugívoro ; Herb-herbívoro; Ins-insetívoro; Myr-mirmecófago; Oni-onívoro. Nível de ameaça: VU- vulnerável. ¹Pode corresponder a L. tigrinus ou L. wiedii. Para todas as espécies a nomenclatura taxonômica e a dieta foram seguidos de Paglia et al. (2012).
Ordem/Família Nome científico Nome popular Tipo de
dieta
Nível de ameaça
Nacional Estadual
Artiodactyla
Cervidae
Mazama gouazoubira
(Fischer, 1814)
veado-
catingueiro Fru/Herb
Mazama americana
(Erxleben, 1777) veado-mateiro Fru/Herb VU VU
Didelphimorphia
Didelphidae
Didelphis aurita
(Wied-Neuwied, 1826)
gambá-de-
orelha-preta Oni
Carnivora
Canidae
Canis lupus familiaris
(Linnaeus, 1758)
cachorro-
doméstico Car/Oni
Cerdocyon thous
(Linnaeus, 1766)
cachorro-do-
mato Ins/Oni
Chrysocyon brachyurus
(Illiger, 1815) lobo-guará Car/Oni VU VU
Felidae
Leopardus pardalis
(Linnaeus, 1758) jaguatirica Car VU VU
Leopardus sp.¹ gato-do-mato Car VU VU
Puma yagouaroundi
(É. Geoffroy, 1803) gato-mourisco Car
Puma concolor
(Linnaeus, 1777) onça-parda Car VU VU
Mustelidae
Eira barbara
(Linnaeus,1758) irara Fru/Oni
Galictis cuja
(Molina, 1782) furão Car
Procyonidae
Procyon cancrivorus
(G. Cuvier, 1798) mão-pelada Fru/Oni
Nasua nasua
(Linnaeus, 1766) quati Fru/Oni

104
Cingulata
Dasypodidae
Dasypus novemcinctus
(Linnaeus,1758) tatu-galinha Ins/Oni
Pilosa
Euphractus sexcinctus
(Linnaeus,1758) tatu-peba Ins/Oni
Myrmecophagidae
Myrmecophaga tridactyla
(Linnaeus,1758)
tamanduá-
bandeira Myr
Lagomorpha
Leporidae
Sylvilagus brasiliensis
(Linnaeus,1758) tapiti Herb
Perissodactyla
Tapiriidae
Tapirus terrestres
(Linnaeus, 1758) anta
Herb/Fru VU VU
Rodentia
Erethizonthidae
Coendu spinosus
(F. Cuvier, 1823) ouriço-cacheiro
Fru/Fo
Sciuridae
Guerlinguetus aestuans
(Linnaeus, 1766) esquilo
Fru/Oni

105
CONCLUSÃO GERAL
A restauração tem o intuito de recuperar áreas degradadas a fim de manter
ecossistemas sustentáveis, com a recuperação de sua variabilidade estrutural e funcional
(SER, 2004). Avaliações a respeito de sua efetividade têm se centrado sobre a comunidade
de plantas (Kanowski 2010, Kanowski et al. 2003, Rodrigues et al. 2009), sendo
mundialmente escassos estudos sobre a comunidade de fauna que coloniza essas áreas
(Patten 1997, Nicolas et al. 2009). Para o Brasil, este é o primeiro trabalho que avalia a
colonização da comunidade de mamíferos em uma área de restauração de Mata Atlântica.
Estruturalmente o processo de restauração implementado na Fazenda Intermontes
demonstrou que, embora o mesmo tipo de tratamento tenha sido realizado em pontos
amostrais distintos, no seu destino atual eles apresentaram uma estrutura da vegetação
diferente entre si, após 13 anos da sua implantação.
As características estruturais da vegetação não foram capazes de recuperar a mesma
complexidade vegetacional dos remanescentes florestais analisados durante este período.
Os tratamentos de plantio geralmente foram caracterizados por uma intensa obstrução
vegetal até 1,0m do solo, com dossel baixo e aberto e com a presença de maior cobertura
vegetal por gramíneas e não por serapilheira como nos remanescentes. Logo, medidas de
manejo e intervenção ainda mostram-se necessárias para o melhor desenvolvimento da
estrutura da vegetação.
A ausência de uma complexidade estrutural da vegetação é considerada como fator
responsável pela falha da recolonização de espécies de fauna, principalmente com relação
às espécies especialistas (Nichols e Nichols 2003, Cunningham et al. 2007, Munro et al.
2007). Em nosso estudo, embora diferentes espécies tenham sido registradas, verificamos
o predomínio da comunidade por espécies generalistas, tanto para a assembleia de
pequenos mamíferos, dominada pelas espécies Oligoryzomys nigripes e Akodon montesis,
quanto para a assembleia de mamíferos de médio e grande porte, dominada pelas espécies
Cerdocyon thous e Dasypus novemcinctus (Alho et al. 1986, Naughton-Treves et al. 2003,
Carmignotto 2004, Harvey et al. 2006, Dotta e Verdade 2007, Michalski e Peres 2007,

106
Umetsu e Pardini 2007, Naxara 2008, Putker et al. 2008, Pardini et al. 2009, Gammons et al.
2009, Rossi 2011).
Para a assembleia de pequenos mamíferos, as espécies especialistas estiveram
restritas aos remanescentes florestais e ou àqueles pontos amostrais cuja estrutura da
vegetação foi mais complexa. Ainda assim, foram pouco abundantes e ocasionais ao longo
do estudo. Desta forma, a ocorrência e a abudância das espécies especialistas aumentou da
regeneração (área de menor complexidade estrutural) em direção ao remanescentes
florestais (áreas de maior complexidade estrutural), em contrapartida às espécies
generalistas (Anexo I). Para a assembleia de mamíferos de médio e grande porte, espécies
especialistas, como a onça-parda (Puma concolor) e a anta (Tapirus terrestres)
apresentaram baixa frequência de ocorrência, com poucos registros durante o estudo e a
ocorrência das espécies especialistas e ou generalistas variou de acordo com a espécie, mas
foram mais frequentes nas áreas de complexidade estrutural intermediária (Anexo I).
Tais resultados, portanto, demonstram que a restauração da Fazenda Intermontes
ainda não foi capaz de garantir a recolonização e a manutenção de toda a comunidade de
mamíferos terrestres de forma equilibrada, por não suprir as necessidades ecológicas de
todas as espécies, principalmente para as espécies especialistas. Para as espécies de
grandes áreas de vida a fazenda pode representar importantes trampolim-ecológico dentro
da paisagem local (Metzger 2001, Haddad et al. 2003), principalmente se a área oferecer
seus recursos temporariamente ou proteção para seu deslocamento. Ainda que somente 13
anos tenham se passado, caso não haja nenhum tipo de manejo que permita a evolução da
estrutura da vegetação, essa recolonização e manutenção não irá ocorrer nem em longo
prazo.
A análise dos diferentes descritores ambientais (estrutura da vegetação e
disponibilidade de recursos alimentares), por sua vez, evidenciou uma resposta espécie-
específica. Para pequenos mamíferos variáveis da estrutura da vegetação foram bastante
importantes para a ocorrência das espécies, destacando-se a intensidade de cobertura
vegetal sobre solo que aparentou favorecer as espécies generalistas. Para médios e grandes
mamíferos igualmente nenhum padrão foi observado e o efeito dos descritores revelou as
necessidades ecológicas das espécies, embora para M. tridactyla outros filtros devem atuar
sobre a espécie.

107
A identificação desses descritores ambientais em nível específico pode ser utilizada
como base para a elaboração de medidas que permitam o manejo da restauração em
favorecimento das espécies de mamíferos. Dado que as espécies respondem mais
diretamente aos gradientes de variação das variáveis ambientais em uma escala mais fina
(George e Zack 2001, Fischer e Lindernmayer 2006), a reprodução destes aspectos pode
ser capaz de garantir a permanência e a ocorrência das espécies, principalmente as
especialistas. Embora grande parte dos estudos não avalie este nível de escala da variação
dos descritores ambientais em ecologia de restauração, sugerimos que estudos futuros
avaliem este aspecto para auxiliar na compreensão da efetividade dos processos de
restauração no país, favorecendo a elaboração de medidas de manejo.
REFERÊNCIAS BIBLIOGRÁFICAS
Alho, C. J. R., Pereira, L. A., Paula, A. C. 1986. Patterns of habitat utilization by small mammal
populations in cerrado biome of central Brazil. Mammalia 50(4): 447-460.
Carmignotto, A. P. 2004. Pequenos mamíferos terrestres do Cerrado: padrões faunísticos
locais e regionais. Tese de doutorado, Universidade de São Paulo.
Cunningham, R. B., Lindenmayer, D. B., Crane, M., Michael D., MacGregor, C. 2007. Reptile
and arboreal marsupials response to replanted vegetation in agricultural
landscapes. Ecological Applications 17(2): 609-619.
Dotta, G., Verdade, L. M. 2007. Trophic categories in a mammal assemblage: diversity in an
agricultural landscape. Biota Neotropica 7(2): 287-292.
Fischer, J., Lindenmayer, D. B. 2006. Beyond fragmentation: the continuum model for fauna
research and conservation in human-modified landscapes. Oikos 112(2): 473-480.
Gammons, D. J., Mengak, M. T., Conner, L. M. 2009. Armadillo hábitat selection in
Southwestern Georgia. Journal of Mammalogy 90(2): 356-362.
George, T. L., Zack, S. 2001. Spatial and temporal considerations in restoring habitats for
wildlife. Restoration Ecology 9(3): 272-279.
Haddad, N. M., Bowne, D. R., Cunningham, A., Danielson, B. J., Levey, D. J., Sargent, S., Spira,
T. 2003. Corridor use by diverse taxa. Ecology 84 (3): 609-615.

108
Harvey, C. A., Gonzalez, J., Somarriba, E. 2006. Dung beetle and terrestrial mammal
diversity in forests, indigenous agroforestry systems and plantain monocultures in
Talamanca, Costa Rica. Biodiversity and Conservation 15: 555-585.
Kanowski, J, Catterall, C.P., Wardell-Johnson, G.W., Prostor, H., Reis, T. 2003. Development
of forest structure on cleared rainforest land in Australia under different styles of
reforestation. Forest Ecology and Management 183: 265-280.
Kanowski, J. 2010. What have we learnt about rainforest restoration in the past two
decades? Ecological Managment and Restoration 11(1): 2-3.
Metzger, J. P. 2001. O que é ecologia de paisagens? Revista Biota Neotropica 1(1):1-9.
Michalski, F., Peres, C. A. 2007. Disturbance-mediated mammal persistence and abundance-
area relationships in Amazonian forest fragments. Conservation Biology 21(6):
1626-1640.
Munro, N. T., Lindenmayer, D. B., Fischer, J. 2007. Faunal response to revegetation in
agricultural areas of Australia: a review. Ecological Management and Restoration
8(3): 199-207.
Naughton-Treves, L.; Mena, J. L.; Treves, A.; Alvarez, N.; Radeloff, V. C. 2003. Wildlife
survival beyond park boundaries: the impact slash-and-burn agriculture and
hunting on mammals in Tambopata, Peru. Conservation Biology 17: 1106-1117.
Naxara, L. 2008. Importância dos corredores ripários para a fauna – pequenos mamíferos em
manchas de floresta, matriz do entorno e elementos lineares em uma paisagem
fragmentada de Mata Atlântica. Dissertação de mestrado, Universidade de São
Paulo.
Nichols, O. G., Nichols, F. M. 2003. Long-term trends in faunal recolonization after bauxite
mining in the Jarrah forest of southwestern Australia. Restoration Ecology 11(3):
261-272.
Nicolas, V., Barrière, P., Tapiero, A., Colyn, M. 2009. Shrew species diversity and abundance
in Ziama Biosphere Reserve, Guinea: comparison among primary forest, degraded
forest and restoration plots. Biodiversity and Conservation 18: 2043-2061.
Pardini, R., Faria, D., Accacio, G. M., Laps, R. R., Mariano-Neto, E., Paciencia, M. L. B., Dixo, M.,
Baumgarten, J. 2009. The challenge of maintaining Atlantic forest biodiversity: A

109
multi-taxa conservation assessment of specialist and generalist species in an agro-
forestry mosaic in southern Bahia. Biological Conservation 142: 1178-1190.
Patten, M. A. 1997. Reestablishment of a rodent community in restored desert scrub.
Restoration Ecology 5(2): 156-161.
Püttker, T., Pardini, R., Meyer-Lucht, Y., Sommer, S. 2008. Responses of five small mammal
species to micro-scale variations in vegetation structure in secondary Atlantic
Forest remnants, Brazil. BMC Ecology 8(9): 1-10.
Rodrigues, R. R., Lima,R. A., Gandolfi, S., Nave, A. G. 2009. On the restoration of high
diversity forests: 30 years of experience in the Brazilian Atlantic forest. Biological
Conservation 142: 1242-1251.
Rossi, N. F. 2011. Pequenos mamíferos não-voadores do Planalto Atlântico de São Paulo:
Identificação, história natural e ameaças. Dissertação de mestrado, Universidade de
São Paulo.
Umetsu, F., Pardini, R. 2007. Small mammals in a mosaic of forest remnants and
athropogenic habitats-evaluating matrix quality in an Atlantic forest landscape.
Landscape Ecology 22(4): 517-530.

110
Anexo I. Esquema ilustrativo da ocorrência das espécies de pequenos, médios e grandes
mamíferos com relação aos diferentes tipos de tratamentos avaliados na fazenda
Intermontes, na cidade de Ribeirão Grande, SP.
RF RARBOP RARBUP PL REG
Espécies Especialistas de Pequenos Mamíferos
Akodon e Oligoryzomys
Dossel fechado< cobertura vegetalMais árvores (com fuste alto e poucas ramificações)
Dossel + aberto> cobertura vegetalMenos árvores (com fuste baixo e muitas ramificações)
Espécies de Médios e Grandes Mamíferos