composição e distribuição espaço-temporal da fauna bêntica
TRANSCRIPT

Relat. téc. insto oceanogr., (41):29-46, 1997
Composição e distribuição espaço-temporal da fauna bêntica no Canal de São Sebastião
Ana Maria Setubal Pires Vanin; Thais Navajas Corbisier; Emília Arasaki & Adriana Maria MõeUmann
Instituto Oceanográfico da Universidade de São Paulo (Caixa Postal 66149, 05315-970, São Paulo, SP, Brasil)
Introdução
Dentro do objetivo de descrever a organização estrutural dos componentes vivos do sistema, o subprograma Bentos se propôs a fazer o levantamento das espécies presentes dentro do Canal de São Sebastião e da plataforma interna adjacente, estudar a distribuição espaço-temporal da fauna bêntica e os fatores responsáveis pelos padrões observados, avaliar a biomassa, identificar as espécies dominantes nos dois subsistemas, identificar e quantificar a dieta alimentar das espécies mais abundantes de Brachyura e Asteroidea, visando o conhecimento das interrelações tróficas entre os vários componentes do sistema marinho costeiro de São Sebastião.
O presente relatório trata dos primeiros resultados obtidos para o Canal de São Sebastião, no que diz respeito à composição específica e distribuição espaço-temporal da mega, macro e meiofauna bênticas.
Material e métodos
Foram realizadas 05 viagens para amostragem do Canal, sendo a primeira de caráter preliminar, e as outras, uma em cada estação do ano.
Nos dois primeiros dias de cada etapa foram coletados invertebrados pertencentes à megafauna, utilizando-se rede de arrasto de portas, tipo "otter trawl". Em cada estação de coleta foram feitos 2 arrastos de 15 minutos cada. A área estimada para cada arrasto é de 5.559 m2. As coletas da megafauna ocorreram em 5 estações, escolhidas previamente em viagem piloto realizada em 6 e 7 de novembro de 1993 (estações 4,5, 7, 10 e 12) (Fig.3.1). Esses 5 locais são os únicos dentro do Canal em que é possível haver pesca. Fora deles existem fundos com forte
Contr. nº 807 do Inst. oceanogr. da Usp.
declividade, sendo q1!'~ ao norte, há numerosos e grandes blocos de pedra, extremamente danosos para a malhagem da rede.
Cada estação de coleta foi posicionada com GPS (Global Positioning System). A seguir a profundidade foi medida com ecobatímetro, no início e no fmal de cada arrasto. Os animais coletados foram triados, separados por sexo e pesados. Os pesos foram utilizados nos cálculos de biomassa (grama de peso úmido/ m2).
A macrofauna e a meiofauna bêntica foram amostradas nos 3 dias restantes. A primeira foi coletada com pegador tipo vanVeen de 0,1 m2, em duplicata, e com draga
retangular; a segunda, só com o pegador, em amostragem estratificada. A macrofauna foi obtida sazonalmente, em 15 estações situadas em 5 radiais perpendiculares à costa; a meiofauna foi coletada somente no verão e inverno, em 10 das estações (estações 1-3, 7-9 e 12-15). Foram realizadas também coletas de sedimento em cada estação para análises sedimentológicas (aproximadamente 100 g), segundo metodologia descrita em Suguio (1973). O teor de carbono orgânico foi determinado por oxidação com K2Cr07 e a titulação do dicromato restante feita com ácido clorídrico (Gaudette et ai., 1974). Os carbonatos foram determinados pelo método de dissolução ácida.
A macrofauna coletada com o pegador foi lavada e peneirada em peneiras de malhagem 2,0, 1,0 e 0,5 mm. O material obtido foi fixado e conservado em álcool 70%. Do material proveniente da draga, foram lavados 10 I de sedimento, que a seguir foram peneirados na série de peneiras acima mencionadas e fixados. Os animais foram triados e identificados sob estereomicroscópio.
As subamostras da meiofauna foram obtidas com tubos de PVC de 2,5 cm de diâmetro (5 cm2 de área) e 10 em de altura, sendo retiradas 4 amostras de cada pegador; a seguir foram fixadas com formol salino a 4% neutralizado e coradas com Rosa de Bengala. Em laboratório o material de cada subamostra foi lavado e peneirado através de

30
quatro peneiras de telas de malhagens diferentes (500, 250, 125 e 62 .um) a fim de separar a fauna nas diversas categorias de tamanho (Pfannkuche & Thiel, 1988). O material retido foi novamente fixado para posterior triagem e identificação em nível de grupo. A biomassa foi estimada para cada grupo taxonômico encontrado, separado em categoria de tamanho a partir dos pesos médios individuais, segundo Widbom (1984), Rudnick et aI. (1985) e Bodin et ai. (1989). O peso seco foi transformado em peso úmido multiplicando-se os valores pelo fator 4,70, o que equivale a considerar o peso seco sendo 21,25 % do peso úmido (LeGueUec & Bodin, 1992).
Em cada radial procurou-se amostrar a profundidade de 10 m, tanto do lado do continente quanto do lado da Ilha Bela, e o eixo longitudinal do Canal, que possui profundidades variáveis. Neste, a profundidade aumenta das bocas para o centro, indo de 23 a 25 m nas bocas sul e norte, respectivamente, e chegando a 40 m próximo ao terminal petroüfero DTCS (Dutos e Terminais Centro Sul), antigo TEBAR (Terminal Almirante Barroso), na região central do Canal. A Fígura 3.1 apresenta a localização das estações de coleta na área do Canal de São Sebastião.
Resultados e discussão
1. Dados Ambientais
A Tabela 3.1 apresenta os dados obtidos para as coletas sazonais na região do Canal, e referem-se ao nível mais profundo realizado em cada estação. Na primavera temos a presença da Água Central do Atlântico Sul (ACAS) na região mais funda do Canal, ou seja, a 40 m de profundidade. Essa massa de água caracteriza-se por sua baixa temperatura (entre 9 e 18°C) e alta salinidade (entre 34,50 e 36,(0) (Aidar et ai, 1993). O restante da área é preenchido pela Água Costeira (AC), de altas temperaturas e baixas salinidades (temperatura > 20°C e salinidade entre 32,20 e 35,40 UPS). Nos períodos do verão, outono e inverno, verificou-se a ausência de estratificação termo-salina e presença exclusiva da AC, mesmo nas estações mais profundas. No outono entretanto, as temperaturas e salinidades apresentaram valores mais elevados e variáveis em relação ao inverno. A temperatura esteve entre 24,11 e 25,82°C no outono e entre 20,2 e 2O,7"C no inverno; a salinidade situou-se entre 33,73 e 35,78 UEPS no outono e entre 33,15 e 34,21 UEPS no inverno.
Quanto às porcentagens de saturaçao do Oxigênio, os valores são superiores ou próximos de 100% na superfície, sofrendo uma diminuição de até 30% na estação mais profunda (c.a. 40 m) (est. 8). Nas estações mais rasas,
Relat. téc. insto oceanogr., (41), 1997
situadas entre 8 e 10 m de profundidade, a redução no teor de saturação é da ordem de, no máximo, 10%, mas várias estações apresentaram saturação de Oxigênio dissolvido superior a 100% (p.ex., ests 4 e 5 no outono e ests 13 e 15 no inverno). Essas porcentagens alteram-se um pouco em função das temperaturas e salinidades locais, sendo ligeiramente mais baixas nas águas mais frias.
As amostras de sedimento do Canal foram analisadas dentro do subprograma Geologia Marinha, e os dados serão apresentados no relatório do referido subprograma. Contudo, para comparações quando da distribuição da fauna bêntica, adiantamos que a região do Canal de São Sebastião é um mosaico de sedimentos variáveis quanto à composição granulométrica (porcentagens de areia, silte e argila), conteúdo de carbonatos biodetríticos e de matéria orgânica. Os sedimentos predominantes são as areias, e de um modo geral, as mais grosseiras estão distribuídas ao norte do Canal (ests. 11 a 15), e as mais finas ou süticas, ao sul e no centro, especialmente no eixo do Canal (ests 2 e 8) e na região da Ponta do Araçá (est.7).
2. Megafauna
Foram coletadas 92 espécies de invertebrados bênticos para a área do Canal. As espécies mais freqüentes e numerosas são em número de 25, que representam 95% do número total de indivíduos amostrados. Assim, as espécies dominantes foram: Callinectes ornatus, Hepatus pudibundus, Persephona punctata, Persephona lichtensteinii, Dardanus a"osor insignis, Paguristes erythrops, Penaeus brasiliensis, Penaeus schimitti, Xiphopenaeus kroyeri, P/eoticus mil/eri, Trachypenaeus constrictus, Cymothoa excisa, Siratus senega/ensis, Strombus pugi/is, Cymathium parthenopeum, Anadara brasiliensis, Lytechinus variegatus, Encope emarginata, Luidia c/athrata, Luidia senega/ensis, Luidia alternata, Astropecten marginatus, Astropecten brasiliensis, Echinaster brasiliensis, Ophiodenna januarii.
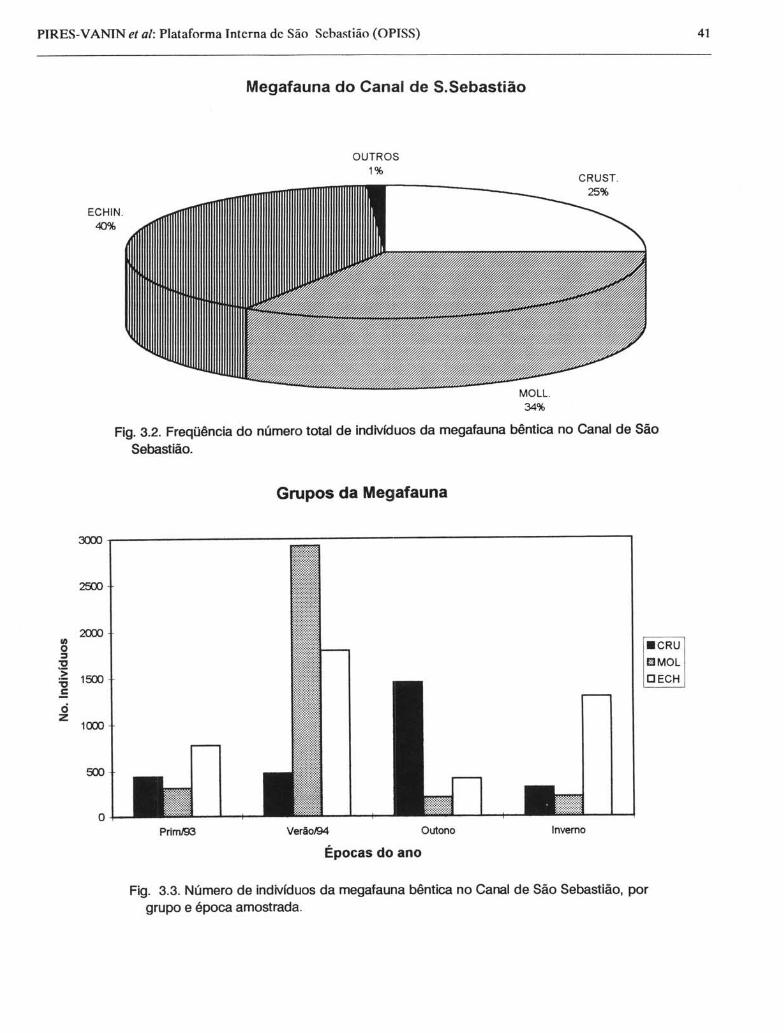
O número de indivíduos e seu peso úmido foram padronizados para 30 minutos de arrasto e uma área de 16.677 m2, e podem ser vistos nas Figuras 3.2 e 3.3. Echinodermata e Mollusca foram os grupos dominantes no Canal, seguidos por Crustacea. Os Echinodermata foram mais abundantes no verão e no inverno, enquanto que Mollusca o foi no verão e Crustacea no outono. Na primavera dominou numericamente o ouriço Lytechinus variegatus (477 ind.), o gastrópode Strombus pugilis (198 ind.) e o siri Ca/linectes ornatus (108 ind.). Já no verão, o número de indivíduos obtidos para a megafauna foi cerca de 3,6 vezes maior, em relação à primavera. Nessa época, Strombus pugilis foi extremamente abundante (2.853 ind.),

PIRES-V ANIN et ai: Plataforma Interna de São Sebastião (OPISS) 31
seguido por Encope emarginata (912 ind.), Lytechinus variegatus (480 ind.), Penaeus brasiliensis e Ca/linectes omatus (195 indo de cada espécie). Essas espécies são coletadas entre 8 e 24 m de profundidade e temperaturas entre 20 e 28°C, ou seja, na Água Costeira.
No outono predominou Callinectes omatus (531 ind) e no inverno, Encope emarginata (912 ind.) Strombus pugilis foi muito abundante nas estações 10 e 12 na primavera e 10 e 4 no verão. São estações de baixa profundidade (8 a 10 m), e de fundos com diferentes composições granulométricas. A estação 10, onde a espécie apresentou sempre a maior abundância, caracteriza-se por ter sedimento de predominância síltica (silte médio), com boa quantidade de argila (17%) e carbonato biodetrítico (20,04%). Já as estações 4 e 12 têm predominância de areias muito fmas, com quantidades variáveis de silte e de argila. Na estação 12 ocorreu também, em grande abundância, Lytechinus variegatus.
As estrelas do mar foram freqüentes e abundantes na área, localizando-se especialmente nas estações 4 e 5. Estes locais situados entre 10 e 24 m de profundidade, apresentam sedimento bem selecionado, onde predomina areia muito fma. Na estação 5 (23-24 m de prof.), com porcentagens de silte e argila mais elevadas que na estação 4 (17,14% contra 4,59%), dominaram Luidia c/athrata eAstropecten brasiliensis. Já na estação 4 (10 m de prof.) foram mais numerosas Luidia senegalensis e Astropecten marginatus.
Callinectes omatus esteve presente em toda a área amostrada e em todas as épocas. Foi mais abundante nas estações 4, 7 e 10.
No verão apareceu Penaeus brasiliensis, o camarão rosa, de valor comercial, em toda a área, mas em maior quantidade na estação 5 (132 ind.). Esta é uma espécie que prefere temperaturas altas (> 23°C) e fundos arenosos (Williams, 1984), estando presente no litoral norte do Estado de São Paulo, na Água Costeira (Pires, 1992). No outono a dominância dos Crustacea deveu-se ao camarão branco Penaeus schimitti e ao camarão sete-barbas Xiphopenaeus kroyeri. A presença de temperaturas altas nessa época do ano, favoreceu a presença de várias espécies de camarões peneídeos no Canal.
A Figura 3.3 apresenta os valores de biomassa para cada época e estação de coleta do Canal de São Sebastião, considerando-se os valores calculados em glm2. Os altos valores do outono são devidos ao siri Ca/linectes omatus, muito abundante em toda a área nessa ocasião, e ao camarão Penaeus schimitti. No inverno a maior biomassa deveu-se ao equinóide Encope emarginata, seguindo assim a tendência observada para o número de indivíduos. O inverno foi a época dos menores valores de biomassa, exceto para a estação 4. As estações que apresentaram
maiores valores de biomassa foram a 10 no verão e a 4 no inverno (7,60 e 5,86 glm2, respectivamente), enquanto que o menor valor foi encontrado na estação 7, no inverno (0,12 glm2) .
3. Macrofauna
Amostragem quantitativa
Foram identificadas 336 espécies coletadas com o pegador de fundo tipo van Veen de 0,1 m2 (Tabela 3.2), sendo Polychaeta o grupo que apresentou maior número, com 128. Mollusca foi o segundo grupo com 100 espécies, seguido de Crustacea com 75 e Echinodermata com 27. As 8 espécies restantes ficaram distribuídas em 3 grupos.
Observando-se a Figura 3.5, verificou-se que Polychaeta foi o grupo taxonômico mais abundante durante todo o estudo, totalizando 51%; Nematoda e Mollusca representaram respectivamente 15% e 14%, seguidos de Crustacea com 7%, Echinodermata com 4%, Cnidaria e Sipuncula, ambos com 3% cada. Os demais grupos estiveram presentes em apenas 2% do total. Dentre os fIlos estudados, Polychaeta ocorreu em todas as épocas e estações de coleta, principalmente nas estações 8, 9,13,14 e 15 (zonas central e norte do canal), com exceção da estação 15, no verão. Na estação 8 a alta densidade foi devida principalmente a uma espécie, o depositívoro Ci"ophorns americanus. Na estação 9 ocorreram em altas densidades, o silídeo Exogone arenosa e o sabelídeo Chone insularis. Nas estações 13, 14 e 15 (zona norte do canal) várias espécies contribuíram para os altos valores observados.
Para os grupos Crustacea, Mollusca e Echinodermata, os maiores números também foram encontrados nas estações situadas no eixo do canal e próximas à Ilha de São Sebastião, a partir da área central em direção à porção norte. Várias espécies desses grupos, nessas estações, elevaram os valores de densidade. A hipótese levantada para tentar explicar tais ocorrências, é que nesses locais, devido a uma heterogeneidade sedimentar, haveria um aumento no número de nichos ambientais e conseqüentemente, aumentaria também a possibilidade de um maior número de larvas e jovens encontrarem condições favoráveis para se estabelecerem nessas áreas.
Em termos de abundância relativa por época de ano, a Figura 3.6 mostra que o grupo Polychaeta foi mais abundante no inverno, com 4.718 indivíduos. J á para Mollusca, a primavera foi a época em que ocorreu maior número de indivíduos (1.572), principalmente de Caecum sp, C. pulchellum e C. striatum (Gastropoda). Estas espécies dominaram na estação 9, região com predomínio

32
de areia média. Para Crustacea, o inverno apresentou um total de 635 exemplares, sendo Apseudes paulensis (Tanaidacea) dominante nas estações 5 e 8, regiões com sedimentos arenosos (areia muito fma e areia síltica, respectivamente). Também Echinodermata foi mais abundante no inverno (533 exemplares), principalmente as espécies de Ophiuroidea Amphiodia atra e Hemipholis elongata, presentes em grande número na estação 8, a mais profunda do canal (40 m). Os maiores valores de abundância relativa encontrados durante o inverno, parecem estar relacionados com as mudanças no hidrodinamismo da região, seja pela ação de ventos ou de sistemas frontais, mais atuantes nessa época. Esses sistemas afetam os sedimentos superficiais, promovendo uma maior heterogeneidade granulométrica e dessa forma, influenciando na distribuição das espécies.
Já as menores densidades observadas nas estações 2 (Ponta da Sela), 7 (emissário submarino) e 10 (próximo ao Dutos e Terminais Centro Sul-DTCS) ao longo das quatro épocas estudadas, estariam relacionadas ao enriquecimento orgânico promovido pela contribuição antrópica, tal como o efluente do emissário do Araçá e a proximidade do porto comercial e do DTCS. Quanto à estação 2, a possível contribuição seria ocasionada pelas descargas ilegais, provenientes de lavagem de tanques de óleo dos petroleiros e de resíduos da casa de máquinas, quando do fundeio desses navios na entrada sul do canal.
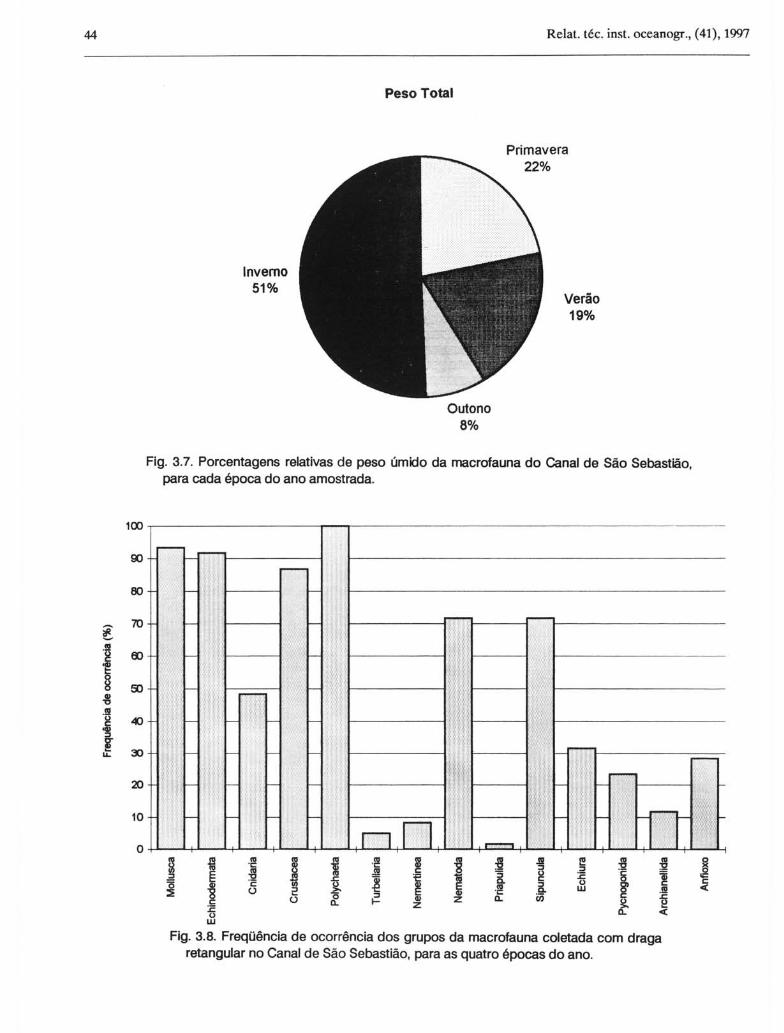
A Figura 3.7 apresenta porcentagens relativas de peso úmido por época do ano, destacando-se novamente o inverno como o período com maior valor (51%). As espécies de Ophiuroidea Amphiodia atra, Hemipholis
. e/ongata e Ophioderma januarii apresentaram diâmetros do disco e braços robustos que pesaram cerca de 170 g. Já a espécie Spirographis spallanzani (Polychaeta) foi uma das maiores encontradas no canal, principalmente na estação 8, no inverno. Apesar de ocorrerem em peque!:') número, cada exemplar pesou aproximadamente 50 g.
A primavera mostrou o segundo maior valor, com 22%, sendo Mollusca o grupo com maior peso úmido. Neste grupo, a presença de espécies de Bivalvia com conchas caIcáreas espessas, como Corbu/a caribaea e C. patagonica, contribuiu significativamente para o aumento da biomassa. Mollusca também esteve presente com mais da metade do peso úmido encontrado para o verão, sendo este resultado devido à presença de grandes exemplares de Chicoreus senega/ensis (Gastropoda), embora com poucos indivíduos (3). No outono, Mollusca contribuiu com a metade do peso total encontrado para a macrofauna, sendo que a espécie
Relat. téc. inst. oceanogr., (41), 1997
Cycline/la tenuis (Bivalvia) apresentou poucos, mas grandes exemplares.
Amostragem qualitativa
Foram identificadas 323 espécies coletadas com draga retangular, sendo novamente Polychaeta o grupo que apresentou maior número de espécies (108), vindo a seguir Mollusca com 105, Crustacea (75) e Echinodermata com 25. Foram ainda identificadas 8 espécies de Sipuncula, uma de Cnidaria e outra de Anfioxo (Tabela 3.3).
As freqüências de ocorrência calculadas para os grupos nas quatro épocas, mostradas em forma de histograma na Figura 8, indicam que Polychaeta foi também o grupo que ocorreu em todas as épocas do ano (FO = 100%), seguido de Mollusca (93,3%), Echinodermata (91,7%) e Crustacea (86,7%). Nematoda e Sipuncula tiveram a mesma freqüência (71,7%) e os demais grupos apresentaram freqüência abaixo de 50%.
Para Polychaeta, oito espécies foram as mais freqüentes: Spiochaetopterus costarum, Neanthes bruaca, Exogone arenosa, Parandalia americana, Owenia fusiformis, Eunice rubra, Lumbrineris tetraura e Aricidea (Acmira) tay/ori. Todas ocorreram em estações com fundo areno-Iodosos, exceto E. arenosa que esteve ausente nas estações 2, 7 e 10, locais com teores de silte, argila e carbono orgânico maiores do que os encontrados nos demais pontos de coleta. Eunice rubra também não ocorreu nas estações 2 e 10, devido à preferência desta espécie por fundos com alta porcentagem de cascalho e elevados teores de matéria orgânica (E. F. Nonato, IOUSP, 1996, com. pess).
Para Mollusca, houve o domínio da espécie Corbu/a caribaea (Bivalvia) em todas as épocas, em locais onde predominaram sedimento are no-lodoso. Analisando-se separadamente cada época, a primavera apresentou além de C. caribaea, a espécie Corbu/a patagonica. No verão e outono, novamente C. caribaea foi a única com alta ocorrência e no inverno, além da citada espécie, foram muito freqüentes os bivalves Nucu/a semiomata, Abra sp Govens), Abra lioica, Corbula sp Govens), c. patagonica e Corbu/a cf. cyme/la. Já para as espécies de Gastropoda, as mais freqüentes (por época do ano) foram: Calyptraea centralis e Ithycythara lanceo/ata na primavera; Natica sp no verão e outono; Caecum striatum, Macrompha/ina argentina, Natica sp e l.lanceo/ata no inverno. Todas essas espécies estiveram presentes em fundos arenosos. Quanto a Scaphopoda, somente Dentalium americanum ocorreu na primavera e D. gouldii no inverno, ambos em estações com fundos areno-Iodosos.
J á para Echinodermata, algumas espécies de Ophiuroidea foram mais freqüentes nas quatro estações do ano: Ophiactis /ymani na primavera; Amphiodia atra no

PIRES-V ANIN et ai: Plataforma Interna de São Sebastião (O PISS) 33
verão e outono;A. atra, O./ymani e O. savignyi no inverno. Todas ocorreram em diferentes tipos de sedimento.
Em relação à Crustacea, geralmente os Decapoda apresentaram um maior número de espécies, o que pode ser explicado pelo uso da draga. Como esta é um equipamento seletivo, amostra os organismos maiores da epifauna. Assim, o braquiúro Pinnixa sayana esteve presente em todas as épocas do ano e em fundos arenolodosos que, conforme consta da literatura, é o sedimento preferido 'por essa espécie (Williams, 1984). No inverno além de P. sayana, outro braquiúro, Hepatus pudibundus, foi freqüente nas estações mais rasas do canal.
Algumas considerações podem ser feitas no momento a respeito da macrofauna do Canal de São Sebastião. Polychaeta foi o grupo dominante nessa área, tanto na infauna quanto na epifauna. Algumas espécies foram abundantes em determinadas estações do centro do canal como Ci"ophorus americanus, Exogone arenosa e Chone insu/aris. Já Spiochaetopterus costarum, Neanthes bruaca, Exogone arenosa, Paranda/ia americana, Owenia fusifonnis, Eunice rubra, Lumbrineris tetraura e Aricidea (Acmira) tay/ori foram as espécies que ocorreram com as maiores freqüências nas quatro épocas estudadas. Quanto às demais espécies da macrofauna, algumas foram muito abundantes ou tiveram alta freqüência, especialmente nas estações do centro e norte do canal, como os gastrópodes Caecum pu/chellum e C. striatum, os ofiuróides Hemipholis e/ongata e Ophiactis /ymani e o tanaidáceo Apseudes pau/ensis.
As maiores abundâncias foram observadas para as estações que apresentaram sedimentos arenosos, configurando áreas com heterogeneidade granulométrica. J á os menores valores observados nas estações 7 e 10, são provavelmente devidos às ações antrópicas do emissário submarino do Araçá e do porto comercial! DTCS, respectivamente. A baixa abundância da estação 2 aparentemente não é explicada por nenhuma das variáveis ambientais coletadas. O sedimento desta estação, composto por frações peüticas, propicia a retenção de matéria orgânica e de outros compostos como hidrocarbonetos, que poderiam influenciar na densidade da macrofauna local.
4. Meiofauna
Foram analisadas 39 amostras da coleta de verão e 40 amostras da coleta de inverno, relativas à meiofauna. Observaram-se 16 grupos taxonômicos no canal no verão e 12 no inverno. No verão, os N ematoda constituíram 79,0% do número total, e nquanto que os Copepoda Harpacticoida constituíram 11,4%, as larvas de Crustacea
3,9%, os Polychaeta 2,4% e os Kinorhyncha 2,3% . Cada um dos demais grupos correspondeu a menos de 1 % da meiofauna. No inverno, os Nematoda equivaleram a 90,4% do total, os Copepoda Harpacticoida a 5,3% e os Polychaeta a 2,1 %. O restante dos grupos constituiu menos que 1% cada.
No verão foi observada uma média de 605,4 ± 442,1 indivíduos por 5 cm2, sendo destes 478,4 ± 385,7 Nematoda (Fig. 3. 9). No inverno, a média observada foi de 213,4 ± 180,8 indivíduos por 5 cm2, sendo desta, 192,0 ± 171,5 Nematoda.
No canal, durante o verão, as estações com maior densidade de meiofauna foram as de número 8 (1.310,5) e 1 (944,2). As densidades mais baixas foram observadas nas estações 2 (149,0) e 7 (163,5) (Fig. 3.10). A análise de variância sobre as amostras do canal indicou uma diferença significativa quanto à densidade entre essas estações (F =
5,13, p<O,OOl). Durante o inverno, os locais com maior densidade foram o 3 ( 496,0) e o 7 (409,5) e com menor densidade a estação 12 ( 12,7) (F = 7,33, p<O,OOl) (Fig. 10).
Em termos de biomassa, estimada em peso seco, os grupos mais importantes no verão foram Nematoda, constituindo 58,5% da biomassa total, Polychaeta, 19,6%, Copepoda, 15,4%, Oligochaeta,3,3%, Kinorhyncha,2,4%, além de Bivalvia, 1,2% . No inverno, os Nematoda constituíram 68,5% do total da biomassa, Polychaeta, 18,1 % do tota~ Copepoda, 6,5%, Oligochaeta, 4,7%, além dos Ostracoda 1,1%.
No canal a biomassa variou de 39,1 ,ug.5 cm -2 (estação 2) a 4%,1 ,ug.5 cm-2 (estação 8), com média de 223,4 ± 19O,7,ug.5 cm-2, no verão (Figs 3.11 e 3.12). No inverno, a biomassa variou de 4,5 (estação 12) a 171,1 ,ug.5 em-2
(estação 3), com média de 81,8 ± 59,5 ,ug.5 em-2.
Em termos de peso úmido, a biomassa estimada da meiofauna variou de 183,8 a 2.331,7 ,ug.5 cm-2, com média de 1.050,0 ± 896,3 ,ug.5 cm-2, no verão; no inverno, os valores variaram de 21,1 a 804,2 ,ug_5 cm-2, com média de 384,5 ± 279,6 ,ug_5 cm-2.
Comparando-se os resultados da densidade média da meiofauna com os obtidos na plataforma interna ao largo de Ubatuba (Corbisier, 1993), verifica-se um maior valor para Canal de São Sebastião no verão de 1994 (1.210,9 indo 10 cm-2) em relação à Ubatuba no verão de 1989 (723 indo 10 cm-2 ) . Esses valores estão dentro do intervalo de variação registrado em plataforma continental de diversas localidades, seja em região subtropical ou temperada, e que é da ordem de 1.000 indo 10 cm-2 (Coull, 1988; Corbisier,op. cit.) .
Analisando-se os dados de biomassa, em termos de peso seco, nota- se que o valor estimado na plataforma de Ubatuba (x = 414,ug. 10 cm-2) é semelhante ao do canal,

34
no verão (x = 432flg. 10 cm- 2). Assim como a densidade, esses dados acham-se também dentro da média geral encontrada em sistemas de plataforma de diversas localidades, principalmente de regiões temperadas (Corbisier, 1993).
Embora os valores de densidade e biomassa obtidos no presente trabalho estejam dentro de intervalos conhecidos, podem estar subestimados em função do amostrador utilizado. Há diversas críticas com relação à amostras provenientes de pegadores de fundo (Elmgren, 1973; Heip et ai., 1977).
Comparando-se os dados de verão e inverno do canal, nota-se que a densidade e a biomassa da meiofauna foram duas vezes menores no inverno do que no verão. A ordem das estações de maior ou menor densidade e biomassa também alterou-se: maiores valores foram observados na estação 8, no verão, e na estação 3, no inverno; os menores valores foram verificados na estação 7, no verão, e na estação 12, no inverno. Estes resultados mostram não só uma variação temporal na densidade e biomassa totais, como também uma variação espacial das mesmas ao longo do Canal de São Sebastião.
Os dados de biomassa da meiofauna, quando transformados para gramas por metro quadrado, mostram a importância dessa categoria do bentos para o sistema: no verão equivalem a 0,45 g de peso seco.m -2 e no inverno, a 0,17 g de peso seco.m-2, no Canal de São Sebastião; já para a plataforma interna ao largo de Ubatuba, equivale a 0,41 g de peso seco.m-2• Em termos de peso úmido, esses valores significam 2,11 e 0,80 g de peso úmido. m-2, no Canal, e 1,95 g de peso úmido. m -2 na plataforma de Ubatuba.
A dominância dos N ematoda em todas as estações do canal (exceto a estação 3 do verão) é uma característica da meiofauna, já que esses organismos geralmente são dominantes, perfazendo mais de 50% do número total de indivíduos, principalmente em sedimentos finos do sublitoral (CouB, 1988). O grupo predominante numericamente ao largo de Ubatuba também foi Nematoda, mas correspondendo a uma porcentagem do total um pouco mais baixa (78%). Esta relativamente menor dominância dos Nematoda pode ser atribuída à maior variedade de sedimentos amostrados ao largo de Ubatuba.
A grande diferença de densidade entre as estações e principalmente, entre amostras de uma mesma estação, aponta para uma distribuição agregada da meiofauna, o que concorda com os resultados da maioria dos trabalhos sobre ecologia desse grupo (Coul~ op. cit.). Os fatores que levam à tal agregação ainda não estão bem defInidos, mas é sugerido que tal fato esteja relacionado à variação da granulometria e à concentração de oxigênio dissolvido no
Relat. téc. insto oceanogr., (41), 1997
sedimento (em larga escala), e com os processos animal-habitat (em micro-escala) tais como micro topografia dos sedimentos, concentração de alimento e presença de estruturas biogênicas, entre outros (Coull,op. cit.).
Considerações gerais
A variação temporal da densidade e biomassa bênticas pode estar associada a diversos fatores, entre os quais incluem-se o aporte de nutrientes e/ou detritos provenientes de sistemas frontais ou de origem continental (Alongi, 1989; 1990), e o aporte de sedimentação pelágica, que geralmente ocorre na primavera-verão (Rudnick et ai., 1985). Ambos os fatores aumentam a disponibilidade de recursos alimentares para o bentos. Outro aspecto não menos importante e que merece ser considerado, é o papel exercido pelo hidrodinamismo local como agente causador de distúrbios físicos nos sedimentos (Aller & ABer, 1986) e que pode inclusive, determinar a ressuspensão e remoção da meiofauna (Palmer, 1984). Na plataforma interna da costa sudeste brasileira todos esses fatores parecem estar relacionados às diferenças observadas na densidade da fauna, especialmente no verão e no inverno. A entrada de frentes frias no inverno parece ser uma causa importante da ressuspensão dos sedimentos de fundo, com conseqüências diretas para as comunidades bênticas, tais como modificação da composição e dominância específica e redução de sua abundância (Paiva, 1993a, b). Além disso, a entrada da ACAS na plataforma interna durante a primavera-verão promove um aumento da produtividade primária fitoplanctônica e conseqüentemente, uma maior quantidade de recurso alimentar pode ser canalizada para o bentos (Pires, 1992).
A variação espacial da densidade ao longo do Canal de São Sebastião entre verão e inverno pode estar relacionada à entrada de frentes frias durante este último período, que alteram o regime de correntes, e ressuspendem e redistribuem os sedimentos modificando a sua distribuição, com consequências na comunidade de fundo. A fauna bêntica como um todo tem sua densidade e biomassa também muito influenciadas pela distribuição granulométrica do sedimento em plataforma continental (Hanson et ai., 1981; Pires-Vanin, 1993). A investigação sobre as causas responsáveis pelas distribuições observadas será a próxima etapa do presente projeto.

PIRES-VANTN cf ai: Plat"f0rm<t Interna de Sá0 Sehastiáo (OPISS) 35
Tabela 3.1. Dados abi6ticos do Canal de São Sebastião
ESTAÇAO DATA LAT. LONG. PROF. TEMP. SALIN. OXIGEN (S) (N) (m) co C) (UPS) (% Sat)
1 03.11.93 2350,4' 4529,7' 15 20.71 33.59 74.9 2 03.11.93 2352,7' 4527,5' 26 20.42 34.18 72.2 3 03.11 .93 2353' 4527,1' 10 21.64 32.53 92.4 4 02.11.93 2350' 4527' 10 23.61 31.45 97.6 5 02.11.93 2351,8' 4523,4' 23 19.69 34.79 59.2 6 02.11.93 23 51,8' 4526' 10 23.51 31.40 101.2 7 02.11.93 2349' 4524' 10 23.10 31.60 93.4 8 02.11 .93 2349,1' 4523,7' 40 16.81 35.57 70.2 9 02.11.93 2349,6' 4523,4' 10 22.70 32.01 98.1 10 03.11.93 2346,1' 4522,3' 10 22.39 32.14 99.2 11 04.11.93 2346,1' 4521,9' 28 20.21 33.86 74.7 12 04.11.93 2346,1' 4521,4' 9 22.36 31.42 100.7 13 03.11.93 2344,3' 4520,g 11 21.29 33.00 85.6 14 03.11.93 2343,8' 4521,2' 26 19.69 34.19 75 .7 15 03.11.93 2343,4' 4520,7' 8 22.00 32.55 88.5 1 03.02.94 23 50,5' 4529,7' 15 29.85 34.97 101.1 2 03.02.94 23 52,7' 4527,5' 25 27.50 34.87 98.7 3 03.02.94 2553' 4527,1' 10 26.83 34.95 -4 03.02.94 2550' 4527' 10 27.35 35.01 -5 03.02.94 2351,8' 4526,4' 24 24.58 35 .34 -6 04.02.94 2351,8' 4526' 10 27 .33 34.88 99.4 7 05.02.94 23 49' 4524' 8 27 .90 34.76 103.1 8 05.02.94 2349,1' 4523,6' 40 22.86 35.31 80.7 9 05 .02.94 2349,5' 4523,4' 10 27 .97 34.76 104.6 10 05.02.94 2346,1' 4522,3' 10 28.27 35.07 108.9 11 02.02.94 2346,1' 4522,3' 28 23 .21 35.15 87.3 12 05.02.94 2346,1' 4521,9' 8 26.89 34.84 106.7 13 04.02.94 23 43,S' 4521,7' 9 26.95 34.98 101.4 14 04.02.94 2343,7' 4521,2 24 25 .74 35.15 87.7 15 04.02.94 2343,7' 4520,6' 11 26.29 35.35 94.5 1 27.04.94 2350,5' 4529,6' 14 24.49 34.79 69.3 2 27.04.94 2352,7' 4527,5' 26 24.11 35.78 77.9 3 27.04.94 2353' 4527,1' 9 26.01 33.91 93.1 4 27 .04.94 2350' 4527' 10 25.06 33.73 107.3 5 27 .04.94 2351,8' 4526,4' 24 24.16 35.29 72.9 6 27.04.94 2351,8' 4526' 9 25.82 34.02 86.2 7 28.04.94 2348,9' 4524' 8 25.21 33.95 95.4 8 28.04.94 2349,1' 4523,6' 40 23.16 35.50 83 9 28.04.94 2349,5' 4523,4' 10 25.26 34.16 90.3 10 28.04.94 2346,1' 4522,3' 10 24.95 34.25 92.6 11 28.04.94 2346,1' 4521,9' 25 24.30 34.92 82.5 12 28.04.94 2346,1' 4521,4' 9 24.25 34.56 90 13 29.04.94 2343,5' 4521 ,7' 9 25.02 34.35 87.9 14 29.04.94 2343,7' 4521,1' 23 24.31 34.76 99.8 15 29.04.94 2343,6' 4520,6' 10 24.70 34.71 86.2 1 06.08.94 2350.54' 4529,73' 15 20.40 33.16 80 2 06 .08.94 2352,72' 4527,57' 26 20.20 33.76 92 3 06.08.94 2352,90' 4527,14' 12 20.40 33.24 95.1 4 06.08.94 2350,00' 4527,Og 11 20.40 33.19 83.8 5 06.08.94 2351,85' 4526,41' 24 20.40 33.16 93.8 6 06.08.94 2351 ,83' 4526,04' 10 20.60 33.25 92.7 7 10.08.94 2349,00' 4524,00' 10 20.70 33.60 95.9 8 10.08.94 2349,14' 4523,65' 41 20.70 33 .84 99.7 9 10.08.94 2349,59' 4523,41' 10 20.70 33.39 98.3 10 07.08.94 2346,11' 4522,36' 10 20.50 33.29 99.4 11 07.08.94 2346,15' 4521,95' 28 20.70 34.22 90.8 12 07.08.94 2346,18' 4521,41' 10 20.50 33.78 91.9 13 07 .08.94 2343,52' 4521,77' 10 20.60 33.74 100.2 14 07.08.94 2343,67' 4521,21' 25 20.60 34.10 96.1 15 07.08.94 2343,74' 4520,62' 10 20.60 34.05 100.5

Ta
be
la 3
.2.
Lis
tag
em
da
s e
spé
cie
s co
leta
da
s co
m
o p
eg
ad
or
de
fu
nd
o t
ipo
va
n V
ee
n n
o C
an
al
de
Sã
o S
eb
ast
ião
DE
CA
PO
DA
PE
NA
EID
EA
A
MP
HIP
OD
A (
cont
.)
GA
ST
RO
PO
DA
B
IVA
LVIA
B
IVA
LVIA
(co
nt.)
S
icyo
nia
typi
ca
Am
pelis
ciph
otis
po
do
ph
tha
lma
C
aecu
m s
p N
ucul
a se
mio
rna
ta
Pita
r fu
lmin
atus
S
icyo
nia
dors
alis
A
mph
iloch
us n
eapo
litan
us
Cae
cum
sp1
A
rca
sp
Cal
lista
mac
ulat
a T
rach
ypan
aeus
con
stri
ctus
C
apre
lla d
ilata
ta
Cae
cum
sp
2
Uo
be
rus
cast
an
eu
s D
osin
ia c
on
cen
tric
a
DE
CA
PO
DA
CA
RID
EA
C
apre
lla s
caur
a C
aecu
m p
ulch
ellu
m
Mus
culu
s la
tera
/is
Cyc
linel
la t
enui
s A
lphe
us f
lorid
anus
C
apre
lla s
p C
aecu
m a
chir
onum
U
ma
sp
C
oope
rella
atla
ntic
a A
lphe
us s
p C
heiri
phot
is m
egac
hele
s C
aecu
m s
tria
tum
C
ten
a p
ectin
ella
C
orbu
la s
p A
uto
ma
te r
ectif
rons
G
amm
arop
sis
? T
eino
stom
a sp
D
iplo
dont
a sp
C
orbu
la s
p1
Au
tom
ate
sp
Gib
bero
sus
mye
rsi
Sol
ario
rbis
sp
Dip
lodo
nta
pu
nct
ata
C
orbu
la s
p2
La
treu
tes
sp
Het
erop
hoxu
s vi
dens
M
acro
mph
alin
a a
rge
ntin
a
Fel
anie
lla v
ilard
eboa
na
Cor
bula
car
ibae
a Le
ptoc
he/a
ser
rato
rbita
Ip
anem
a ta
Ipa
Cre
pidu
la s
p P
hlyc
tider
ma
sem
iasp
era
Cor
bula
/yo
ni
Lept
oche
/a s
p U
ijebo
rgia
dub
ia
Cre
pidu
la a
cule
ata
Ch
am
a m
acer
ophy
lla
Cor
bula
pa
tag
on
ica
P
roce
ssa
hem
phill
i U
strie
lla t
iting
a C
alyp
trae
a ce
ntra
lis
Arc
ine/
la b
rasi
liana
C
orbu
la
cf c
ymel
la
Pro
cess
a sp
M
aera
hir
onde
llei
Nat
ica
sp
Cra
ssin
ella
lunu
lata
M
arl
esi
a s
p (j
ovem
) D
EC
AP
OD
A B
RA
CH
YU
RA
M
etha
rpin
ia lo
ngir
ostr
is
Nat
ica
pusi
lla
Ma
ctra
sp
T
hrac
ia s
imili
s C
alap
pida
e U
ovem
) M
icro
phox
us c
orn
utu
s P
olin
ices
lact
eus
Ma
ctra
jan
eir
oe
nsi
s C
yath
od
on
ta s
em
iru
go
sa
Cal
linec
tes
dana
e O
edic
erot
idae
P
olin
ices
ube
rinu
s A
na
tina
ana
tina
Per
iplo
ma
sp
Cal
linec
tes
sp
Par
dalis
cida
e?
Sin
um p
ars
pe
ctiv
um
S
olen
teh
ue/c
hus
En
tod
esm
a s
p D
isso
dact
ylus
crin
itich
elis
P
hotis
bre
vipe
s E
pito
nium
sp
Tel
lina
sp
En
tod
esm
a p
ata
go
nic
a
Fab
ia in
sula
ris
Pho
tis lo
ngic
auda
ta
Epi
toni
um n
ovan
glia
e T
ellin
a g
ibb
er
Car
diom
ya c
/ery
ana
Het
eroc
ript
a gr
anul
ata
Pho
xoce
phal
opsi
s cf z
imm
eri
E
ulim
a bi
fasc
iata
T
ellin
a ve
rsic
olor
S
CA
PH
OP
OD
A
Hex
apan
opeu
s pa
ulen
sis
Pho
xoce
phal
opsi
s zi
mm
eri
E
ulim
a m
ulat
a T
ellin
a n
iten
s D
enta
lium
gou
ldii
Hex
apan
opeu
s sc
hmitt
i P
htis
ica
vera
e M
e/an
ella
arc
ua
ta
Tel
lina
pu
nic
ea
P
OL
YP
LAC
OP
HO
RA
Li
thad
ia c
ario
sa
Pse
udoh
arpi
nia
de
nta
ta
Chi
core
us s
eneg
alen
sis
Tel
lina
aequ
istr
iata
C
ha
eto
ple
ura
an
gu
lata
P
erse
phon
a p
un
cta
ta
Tib
uron
ella
vis
cana
A
na
chis
sp
(jov
em)
Ma
com
a s
p O
PH
IUR
OID
EA
P
inni
xa c
haet
opte
rana
IS
OP
OD
A
Oliv
ella
sp
Ma
com
a t
en
ta
Am
phio
dia
atr
a
Pin
nixa
ra
pa
x A
nant
hura
sp1
O
livel
la f
lora
l ia
Ma
com
a b
revi
fro
ns
Am
phio
dia
pulc
hella
P
inni
xa s
ayan
a A
ncin
us b
rasi
liens
is
Nan
nodi
ella
ves
puci
na
Ma
com
a u
rug
ua
yen
sis
Am
phio
dia
riise
i P
inni
xa s
p A
pant
hura
sp
Cry
otur
ris
ad
am
si
Sem
eIe
sp
Am
phio
dia
sp
Pin
noth
eres
ost
reum
E
dote
a sp
T
ereb
ra g
em
mu
lata
S
emeI
e p
rofí
cua
A
mp
hio
plu
s lu
cya
e
Pin
noth
eres
sp
Exc
oral
lana
co
sta
ta
Ter
ebra
rio
si
Ab
ra s
p A
mph
ipho
lis ja
nu
ari
i P
inno
ther
idae
H
yssu
ridae
sp1
T
urbo
nilla
sp
Ab
ra li
oica
A
mph
ipho
lis s
p P
ortu
nida
e P
olito
lana
sp
2
Act
eo
n p
e/ec
ais
Erv
ilia
con
cen
tric
a
Am
phip
holis
sq
ua
ma
ta
AN
OM
UR
A P
AG
UR
IDE
A
Pol
itola
na s
p3
A
cte
oci
na
sp
S
o/ec
urlu
s sp
A
mph
ipho
lis s
ubtil
is
Pag
uroi
dea
Uov
em)
Roc
inel
a si
gnat
a A
cte
oci
na
cf b
ide
nta
ta
So
lecu
rlu
s cu
min
gian
us?
Am
ph
iura
fle
xuos
a A
MP
HIP
OD
A
Syn
odot
ea m
arpl
aten
sis
Act
eo
cin
a c
an
de
i G
ould
ia c
erin
a A
mp
hiu
ra jo
ub
ini
Am
pelis
ca b
revi
sim
ulat
a T
AN
AID
AC
EA
P
hilin
e m
era
C
hion
e sp
A
mp
hiu
ra s
p (j
ovem
) A
mpe
lisca
cri
stat
a A
pseu
des
paul
ensi
s V
olvu
lella
per
sim
ilis
Chi
one
inta
pu
rpu
rea
H
emip
holis
elo
ngat
a A
mpe
lisca
par
ia
Aps
eudi
dae
sp1
Vol
vule
lla t
exas
iana
C
hion
e p
ub
era
O
phia
ctis
bra
silie
nsis
A
mpe
lisca
pug
etic
a A
pseu
dida
e sp
2 C
hion
e p
ap
hia
O
phia
ctis
Iym
ani
Am
pelis
ca s
p Le
ptoc
helia
sav
igny
i T
rans
enel
la s
timp
son
i O
phia
ctis
sa
vig
nyi
A
mpe
lisci
dae
Pita
r sp
(jo
vem
) O
ph
iod
erm
a ja
nu
ari
i
~ :xl
2- a ("),
~ :r
~
:l
"'l
I") ::.> ~
:l
~
.- :'!
'-' .... §

Tab
ela
3.2.
Con
tinua
ção
OP
HIU
RO
IDE
A (
cont
.)
PO
LY
CH
AE
TA
(co
nt.)
O
phio
phra
gmus
lutk
eni
Cab
ira in
cerl
a
Oph
ioph
ragm
us p
ulc
he
r S
igam
bra
grub
ii E
CH
INO
IDE
A
Exo
gone
are
nosa
C
lype
aste
roid
ea
Odo
ntos
yl/is
het
ero-
falc
haet
a E
chin
idae
T
ypos
yl/is
hya
lina
Enc
ope
emar
gina
ta
Typ
osy/
lis s
p A
ST
ER
OID
EA
N
eant
hes
cf
succ
inea
A
stro
pect
en b
rasi
liens
is
Nea
nthe
s br
uaca
A
stro
pect
en m
argi
natu
s N
erei
s br
oa
HO
LOT
HU
RO
IDE
A
Ner
eis
sp
Cuc
umar
ia s
p N
ephy
tys
squa
mos
a P
rota
nkyr
a be
nede
ni
Gly
cera
am
eric
ana
Thy
one
paw
soni
G
lyce
ra o
xice
phal
a A
NT
HO
ZO
A
Hem
ipod
us o
Iivie
ri E
dwar
dsia
sp
Hem
ipod
us r
otun
dus
SIP
UN
CU
LA
Gon
iada
bru
nnea
A
spid
osip
hon
albu
s G
onia
da m
acul
ata
Asp
idos
ipho
n go
snol
di
Gon
iada
litto
rea
Nep
haso
ma
conf
usum
G
onia
da c
f em
erita
P
hasc
o/io
n he
drae
um
Gon
iadi
des
caro
linae
S
ipun
culu
s nu
dus
Gly
cend
e m
ultid
ens
Thy
sano
card
ia c
atha
rina
e M
oore
onup
his
linea
ta
AN
FIO
XO
M
oore
onup
his
inte
rmed
ia
Bra
nchi
osto
ma
plat
ae
Dio
patr
a cu
prea
P
OLY
CH
AE
TA
D
íopa
tra
trid
enta
ta
Eun
õe s
erra
ta
Ram
phob
rach
ium
sp
Eun
õe p
apil/
osa
Kin
berg
onup
his
ore
nsa
nzi
H
arm
otho
e lu
nula
ta
Kin
berg
onup
his
cf.
fauc
hald
i P
holo
e m
inut
a O
nuph
is e
rem
ita o
cula
ta
Sth
enel
ais
limic
ola
Eun
ice
rubr
a S
then
elan
el/a
aty
pica
E
unic
e vi
ttata
S
igal
ion
taqu
ari
Eun
ice
(Nic
idio
n) c
arib
oea
Sig
alio
nida
e N
emat
oner
eis
schm
arda
e B
haw
ania
sp
Lysi
dice
nin
etta
U
noph
erus
am
bigü
a M
arph
ysa
sang
uine
a A
naiti
des
tam
oya
Mar
phys
a sp
A c
f La
na,
1981
P
hyl/o
doce
muc
osa
Mar
phys
a sp
B c
f La
na.
1981
G
yptis
cal
lithr
ix
Mar
phys
a sp
P
erib
oea
sp
Lum
brin
eris
tet
raur
a P
aran
dalia
am
eric
ana
Lum
brin
eris
jane
iren
sis
Anc
ystr
osyl
/is jo
ne
si
Lum
brin
eris
jan
ua
rii
PO
LY
CH
AE
TA
(co
nt.)
Lu
mbr
iner
is a
ng
rae
nse
Lu
mbr
iner
is la
trei
l/i
Lum
brin
eris
sp
Lum
brin
erio
psis
mu
cro
na
ta
Nin
õe b
rasi
liens
is
Lysa
rete
bra
silie
nsis
S
colo
plos
(S
colo
plos
) tr
eadw
el/i
Sco
lopl
os (
Sco
lopl
os)
mad
agas
kare
nsis
S
colo
plos
(Le
odam
as)
rubr
a S
colo
plos
(Le
odam
as)
vera
x Le
itosc
olop
los
rob
ust
us
Leito
scol
oplo
s ke
rgue
/ens
is
Orb
inia
cf.
latr
eil/i
O
rbin
ia s
p B
cf
Bol
ivar
, 19
86
Phy
lo f
elix
S
cola
ricia
pap
il/ife
ra
Nai
nere
is s
etos
a Le
vins
enia
gra
cilis
A
ricid
ea (
Aric
idea
) cf
frag
ilis
Ari
cide
a (A
cmira
) ta
ylor
i A
rici
dea
(Acm
ira)
cf.
sim
plex
A
ricid
ea (
Acm
ira)
sp
A c
f. B
oliv
ar,
1986
A
ricid
ea (
Alli
a) c
f. al
batr
ossa
e C
irro
phor
us a
mer
ican
us
Cir
roph
orus
bra
nchi
atus
D
ispi
o re
ma
ne
i La
onic
e b
ran
chia
ta
Laon
ice
cirr
ata
Par
aprio
nosp
io p
inna
ta
Prio
nosp
io d
ayi
P
riono
spio
str
ee
nst
rup
i P
olyd
ora
soci
alis
S
piop
hane
s m
issi
onen
sis
Spi
opha
nes
bo
mb
ix
Sco
le/e
pis
squa
mat
a M
icro
spio
pig
men
tata
M
agel
ona
po
ste
relo
ng
ata
M
agel
ona
vario
lam
el/a
ta
Mag
elon
a pa
pil/i
corn
is
Mag
elon
a cr
enul
ata
Mag
elon
a rio
jai
PO
L Y
CH
AE
T A
(co
nt.
) M
agel
ona
no
na
toi
Poe
cilo
chae
tus
aust
ralis
C
ha
eto
pte
rus
vari
oped
atus
S
pioc
haet
opte
rus
cost
aru
m
Tha
ryx
sp
Oph
elin
a sp
Le
ioca
pite
l/a s
p A
cf.
Bo/
ivar
, 1
98
6
No
tom
ast
us
loba
tus
No
tom
ast
us
hem
ipod
us
Da
syb
ran
chu
s sp
A c
f. B
oliv
ar,
19
86
D
asy
bra
nch
us
sp B
cf
Bol
ivar
, 1
98
6
Axi
othe
l/a b
rasi
liens
is
Cly
men
el/a
dal
esi
Euc
lym
ene
sp A
cf
Bol
ivar
, 1
98
6
Rho
dine
love
ni
Asc
hys
sp
A c
f B
oIiv
ar,
1986
N
otop
roct
us s
p A
cf.
Bol
ivar
, 1
98
6
Ow
enia
fus
iform
is
Sa
be
llari
da
e
Pec
tinar
ia (
Pec
tinar
ia)
lael
ia
Pec
tinar
ia (
Am
phic
tene
) ca
thar
inen
sis
Isol
da p
ulch
efla
A
mph
icte
is s
p T
ereb
ellid
es a
ngui
com
us
Cho
ne in
sula
ris
Spi
rogr
aphi
s sp
alla
nzan
ii S
abel
/a s
p H
ydro
ides
pla
ten
i H
ydro
ides
unc
inat
us
Hyd
roid
es s
p
! I
'"ti ~
[Tl
VI <: » ~ z ~ ~
::2 .., C; 5'
.., 3 .., -::s n .., ::s .., a.
n :.n
~l
o VI g. ~ '" .... ;; o ...-.
O
'"ti -VI VI
'-' ~

Tab
ela
3.3
. Li
stag
em d
as
espé
cies
col
etad
as c
om
a d
rag
a r
etan
gula
r n
o C
anal
de
São
Seb
astiã
o.
DE
CA
PO
DA
PE
NA
EID
EA
A
NO
MU
RA
G
AL
AT
HE
IDE
A
GA
ST
RO
PO
DA
B
IVA
LV
IA (
cont
.)
Ace
tes
amer
ican
us
Min
yoce
rus
sp
Cal
liost
oma
ad
spe
rsu
m
Arc
a im
bric
ata
Hip
poly
tidae
P
orce
llani
dae
Cae
cum
pul
chel
lum
B
arba
tia c
andi
da
Sic
yoni
a ty
pica
A
NO
MU
RA
PA
GU
RID
EA
C
aecu
m s
tria
tum
A
na
da
ra s
p S
icyo
nia
dors
alis
D
ioge
nida
e T
eino
stom
a sp
A
na
da
ra b
rasi
liana
T
rach
ypen
aeus
con
stri
ctus
Is
oche
les
sp
Sol
ario
rbis
sp
An
ad
ara
not
abili
s X
ipho
pena
eus
kroy
eri
Loxo
pagu
rus
loxo
chel
is
Mac
rom
phal
ina
arge
ntin
a U
ob
eru
s ca
sta
ne
us
DE
CA
PO
DA
CA
RID
EA
P
agur
oide
a (m
uito
jove
m)
Str
ombu
s pu
gilis
M
uscu
lus
late
ralis
A
lphe
us s
p A
MP
HIP
OD
A
Cre
pidu
la s
p M
uscu
lus
viat
or
Latr
eute
s pa
rvul
us
Am
pelis
ca c
rist
ata
Cre
pidu
la a
cule
ata
Cte
na
pec
tinel
la
Aut
omat
e sp
A
mpe
lisca
par
ia
Cal
yptr
aea
cent
ralis
D
ivar
icel
la q
uadr
isul
cata
Le
ptoc
hela
ser
rato
rbita
A
mpe
lisca
pu
ge
tica
N
atic
a sp
D
iplo
dont
a sp
Le
ptoc
hela
sp
Am
pelis
ca s
p N
atic
a pu
silla
F
elan
iella
sp
úove
m)
Ogy
rides
alp
haer
ostr
is
Am
pelis
cida
e S
inum
pe
rsp
ect
ivu
m
Fel
anie
lla v
ilard
eboa
na
Ogy
ride
s sp
A
mpe
lisci
phot
is p
od
op
hth
alm
a
Cym
atiu
m
p. p
arl
he
no
pe
um
C
ardi
tam
era
mic
ella
P
roce
ssa
hem
phill
i A
mph
iloch
us s
p E
ulim
a au
rici
ncta
C
ha
ma
sp
Pro
cess
a sp
C
apre
lla d
ilata
ta
Eul
ima
bifa
scia
ta
Arc
inel
/a s
pú
ove
m)
DE
CA
PO
DA
BR
AC
HY
UR
A
Che
iriph
otis
meg
ache
les
Eul
ima
mu
lata
C
rass
inel
la s
p úo
vem
) C
al/in
ecte
s da
nae
Dex
amin
idae
M
elan
el/a
arc
ua
ta
Cra
ssin
el/a
lunu
lata
C
allin
ecte
s or
natu
s D
ulic
hiel
/a a
ppen
dicu
lata
C
hico
reus
sen
egal
ensi
s C
rass
inel
/a m
arlin
icen
sis
Cal
linec
tes
sp
Ga
mm
ari
da
e
An
ach
is s
p úo
vem
) T
rach
ycar
dium
mu
rica
tum
C
ol/o
des
tris
pino
sus
Gam
mar
opsi
s sp
A
na
chis
isab
el/e
i S
olen
sp
Hep
atus
pud
ibun
dus
Het
erop
hoxu
s vi
dens
A
na
chis
ob
esa
S
olen
teh
uelc
hus
Het
eroc
ript
a la
pide
a Li
ljebo
rgiid
ae
Oliv
anci
l/aria
sp
Tel
/ina
sp
Hex
apan
opeu
s pa
ulen
sis
Ust
riel/a
titi
nga
Oliv
el/a
flo
ralia
T
ellin
a g
ibb
er
Hex
apan
opeu
s sp
M
aera
gro
ssim
ana
Nan
nodi
el/a
ves
puci
na
Tel
lina
vers
icol
or
Mic
ropa
nope
sp
Mic
roph
oxus
co
rnu
tus
Cry
otur
ris
ad
am
si
Tel
/ina
pu
nic
ea
M
icro
pano
pe u
rina
tor
Mic
roph
oxus
sp
Ith
ycyt
ha
ra la
nceo
/ata
T
el/in
a ae
quis
tria
ta
Pel
ia r
otu
nd
a
Mon
ocul
odes
sp
Ter
ebra
sp
M
aco
ma
bre
vifr
on
s P
erse
phon
a m
edite
rran
ea
Oed
icer
otid
ae
Ter
ebra
rio
si
Sem
eIe
sp
Per
seph
ona
pu
nct
ata
P
hotis
bre
vipe
s T
urbo
nil/a
sp
Ab
ra s
p P
erse
phon
a sp
P
hotis
long
icau
data
T
urbo
nil/a
sp1
A
bra
lioi
ca
Pin
nixa
cha
etop
tera
na
Pho
tis s
pini
carp
a A
cte
on
pel
ecai
s E
rvili
a co
nce
ntr
ica
P
inni
xa s
ayan
a P
hoxo
ceph
alop
sis
zjm
mer
i A
cte
oci
na
sp
Gou
ldia
cer
ina
Pin
nixa
sp
Pht
isic
a ?
Act
eo
cin
a c
f bid
en
tata
C
hion
e sp
P
inno
ther
es e
mili
ai
Syn
chel
idiu
m a
mer
ican
um
Act
eo
cin
a c
an
de
i C
hion
e in
tap
urp
ure
a
Pin
noth
eres
ost
reum
T
ibur
onel
/a v
isca
na
Bul
/a s
tria
ta
Chi
one
pu
be
ra
Pin
noth
erid
ae
TA
NA
IDA
CE
A
Vol
vule
l/a p
ersi
mili
s C
hion
e p
ap
hia
P
ortu
nida
e A
pseu
des
paul
ensi
s V
olvu
lel/a
tex
asia
na
Pro
toth
aca
sp
úove
m)
Pyr
omai
a tu
berc
ulat
a A
pseu
dida
e sp
1 B
IVA
LV
IA
Tiv
ela
sp lj
ove
m)
Tet
raxa
nthu
s ra
thbu
nae
Lept
oche
lia s
avig
nyi
Nuc
ula
sem
iorn
ata
T
rans
enel
la s
timp
son
i A
dra
na
ele
cta
P
itar
sp ú
ove
m)
BIV
ALV
IA (
cont
.)
Pita
r fu
lmin
atus
P
itar
rost
ratu
s C
a/lis
ta m
acul
ata
Dos
inia
con
cent
rica
C
yclin
ella
ten
uis
Coo
pere
lla a
tlant
ica
Cor
bula
spú
oven
s)
Cor
bula
sp1
C
orbu
la s
p2
C
orbu
la c
arib
aea
Cor
bula
/yo
ni
Cor
bula
pa
tag
on
ica
C
orbu
la c
f cym
ella
T
hrac
ia d
isto
rla
T
hrac
ia s
imili
s P
erip
lom
a sp
P
erip
lom
a co
mp
ress
a
Per
iplo
ma
ova
ta
En
tod
esm
a s
p P
an
do
ra b
ushi
ana
Ca
rdio
mya
cle
ryan
a S
CA
PH
OP
OD
A
Den
taliu
m a
mer
ican
um
Den
taliu
m g
ould
ii P
OL
YP
LA
CO
PH
OR
A
Ch
ae
top
leu
ra a
ngul
ata
OP
HIU
RO
IDE
A
Am
ph
iod
ia a
tra
Am
phio
dia
pulc
hel/a
A
mp
hio
dia
riis
ei
Am
phio
dia
sp
Am
ph
iop
lus
lucy
ae
A
mph
ipho
lis s
p A
mph
ipho
lis s
qu
am
ata
A
mph
ipho
lis s
ubtil
is
Am
ph
iura
fle
xuos
a A
mp
hiu
ra jo
ub
ini
Am
ph
iura
sp
H
emip
holis
elo
ng
ata
O
ph
iact
is Iy
man
i O
ph
iact
is s
avi
gn
yi
!
. .;)
:lO
;:d
2- =. l?- r> :::l ~
:l
n t?
:-.>
:::l
:l
~
,...... ~
':-"
.... §

Tab
ela
3.3.
Con
tinua
ção
OP
HIU
RO
IDE
A (
cont
.)
PO
L YC
HA
ET
A (
cont
.)
Oph
iode
rma
jan
ua
rii
Ner
eis
bro
a
Oph
ioth
rix
rath
bu
ni
Ner
eis
sp
EC
HIN
OID
EA
N
epht
ys s
quam
osa
Ech
inid
ae
Gly
cera
am
eric
ana
Enc
ope
emar
gina
ta
Gly
cera
bru
nn
ea
A
ST
ER
OID
EA
G
onia
da m
acul
ata
Ast
rop
ect
en
bra
siH
ensi
s G
onia
da li
ttor
ea
Ast
rop
ect
en
mar
gina
tus
Gon
iada
sp
A
stro
pect
en s
p G
onia
da c
f. em
erita
E
chin
aste
r bra
silie
nsis
G
onia
dide
s ca
rolin
ae
Luid
ia s
eneg
alen
sis
Gly
cend
e m
u/tid
ens
HO
LOT
HU
RO
IDE
A
Moo
reon
uphi
s lin
eata
C
ucum
aria
sp
Moo
reon
uphi
s in
term
edia
C
RIN
OID
EA
D
iopa
tra
cupr
ea
Tro
piom
etra
c.
carin
ata
Dio
patr
a tr
iden
tata
A
NT
HO
ZO
A
Ram
phob
rach
ium
sp
Edw
ards
ia s
p K
inbe
rgon
uphi
s di
ffici
lis
PO
LYC
HA
ET
A
Kin
berg
onup
his
ore
nsa
nzi
E
unÕ
8 pa
pillo
sa
Kin
berg
onup
his
fauc
ha/d
i E
unõe
ser
rata
O
nuph
is e
rem
ita o
cu/a
ta
Har
mot
hoe
/unu
lata
E
unic
e ru
bra
Ph
%e
min
uta
Eun
ice
vitta
ta
Sth
enel
ais
limic
o/a
Eun
ice
(Nic
idio
n) c
arib
oea
Sth
ene/
aneJ
/a a
typi
ca
Nem
aton
erei
s sc
hmar
dae
Sig
a/io
n ta
qu
ari
Ly
sidi
ce n
inef
ta
Sig
alio
nida
e nã
o id
entif
. M
arp
hys
a s
angu
inea
P
sam
mo/
yce
cate
nu/a
ta
Lum
brin
eris
tet
raur
a U
no
ph
eru
s am
bigü
a Lu
mbr
iner
is ja
neir
ensi
s A
naiti
des
tam
oya
Lum
brin
eris
ang
raen
se
Phy
f/odo
ce (
A)
cf
muc
osa
Lum
brin
eris
/at
reil/
i G
yptis
ca/
lithr
ix
Nin
oe b
rasi
/iens
is
Par
anda
/ia a
mer
ican
a L
ysa
rete
bra
si/ie
nsis
A
ncys
tros
y/lis
jon
esi
S
c%p
/os
(Sc%
p/o
s) t
rea
dw
C
abira
ince
rta
Sco
/opf
os (
Leod
amas
) ru
bra
S
igam
bra
grub
ii S
c%p
los
(Leo
dam
as)
vera
x E
xogo
ne a
ren
osa
L
eit
osc
%p
/os
robu
stus
O
dont
osyl
lis h
eter
ofa/
chae
ta
Le
ito
sc%
p/o
s ke
rgue
/ens
is
Typ
osyl
lis h
yalin
a O
rbin
ia /a
teril
li T
ypos
y/lis
sp
Phy
/o f
e/ix
N
eant
hes
bru
aca
N
aine
reis
set
osa
Nea
nthe
s su
ccin
ea
Levi
nsen
ia g
raci
/is
PO
LYC
HA
ET
A (
cont
.)
Ari
cide
a (A
cmira
) ta
yfor
i A
rici
dea
(Acm
ira)
cf
sim
plex
A
rici
dea
(Acm
ira)
sp A
A
rici
dea
(Acm
ira)
cath
erin
ae
Ari
cide
a (A
li ia)
cf
alba
tros
sae
Cir
roph
orus
am
eric
anus
C
irro
phor
us
bran
chia
tus
Laon
ice
bran
chia
ta
Laon
ice
cirr
ata
Par
apri
onos
pio
pinn
ata
Pri
onos
pio
da
yi
Pol
ydor
a so
cia/
is
Prio
nosp
io s
tre
en
stru
pi
Spi
opha
nes
mis
sion
ensi
s S
piop
hane
s bo
mbi
x S
cole
lepi
s sq
uam
ata
Mic
rosp
io p
igm
enta
ta
Mag
e/on
a po
ster
e/on
gata
M
age/
ona
vario
/am
eI/a
ta
Mag
e/on
a pa
pil/i
com
is
Mag
e/on
a cr
enu/
ata
Mag
e/on
a n
on
ato
i P
oeci
/och
aetu
s au
stra
/is
Ch
ae
top
teru
s va
riop
edat
us
Spi
ocha
etop
teru
s co
sta
rum
C
irra
tulid
ae
Oph
el/in
a sp
N
oto
ma
stu
s /o
batu
s N
otom
astu
s he
mip
odus
Le
ioca
pite
l/a s
p A
D
asy
bra
nch
us
sp B
A
xiot
hella
bra
silie
nsis
C
lym
ene/
la d
ales
i E
uc/y
men
e sp
A
Rho
dine
/ov
eni
No
top
rocf
us
sp A
A
schy
s sp
A
Ow
enia
fus
iform
is
Pec
tinar
ia (
Pec
tinar
ia)
/ael
ia
Iso/
da p
u/ch
ella
A
mph
icte
is s
p
PO
LY
CH
AE
TA
(co
nt.)
S
piro
grap
his
spal
/anz
anii
Sab
ella
sp
C
ho
ne
insu
laris
S
IPU
NC
ULA
si
punc
ula
sp1
Asp
idos
ipho
n al
bus
Asp
idos
ipho
n go
snol
di
Go
lfin
gia
?
Ne
ph
aso
ma
con
fusu
m
Pha
scol
ion
he
dra
eu
m
Sip
uncu
lus
nu
du
s T
hysa
noca
rdia
cat
hari
nae
AN
FIO
XO
B
ran
chio
sto
ma
pla
tae
~ ~-
I
""é .....
;:o
['Tl
:.n <
>
~
Z
~
~ :-:- ::2
~
::.> 5'
.., 3 :lO .....
:l
rb .., :l
:lO o r.> :.n
~,
o :.n
~
cr" ~
ti>
:::to
~,
o ""' O
::g :.n
:.n
'-'
VJ
10

40 Relat. téc. inst. occanogr., (41), 1997
<4-5' J1'W <4-5' 21'W
23" «'S
N \ I I, , , \ , , , ,
,
W E , , , , , , ,
, I I ,
I \ \
S
o IIho do Toque - Toque
ILHA DE SÃO SEBASnÃO
• ESTACÕES
Pto. do Selo
23' 54'S I
FONTE: DHN (Corte Neutlco 1614)
Fig. 3.1. Mapa da área de estudo, com a localização das estações de coleta.
PIRES-V ANIN et ai: Plataforma Interna de São Sebastião (OPISS)
I/)
o :;, :2 > :s .: o z
ECHIN . 40%
Megafauna do Canal de S.Sebastião
OUTROS 1%
MOLL. 34%
Fig. 3.2. Freqüência do número total de indivfduos da megafauna bêntica no Canal de São Sebastião.
2500
2CXX)
1500
1000
500
o
Grupos da Megafauna
PrimJ93 Verãol94 Outono
Épocas do ano
Inverno
.CRU DMOL
OECH
Fig. 3.3. Número de indivfduos da megafauna bêntica no Canal de São Sebastião, por grupo e época amostrada.
41

42
180
100
140
120
o 8 100 ... ui C'G E 80 C'G ... C)
00
40
20
o
Relat. téc. insto oceanogr., (41),1997
Biomassa da Megafauna
Prim..93 Ver40194 Outono Inverno
Épocas do ano
Fig. 3.4. Peso úmido (gramas por m2 x 103) da megafauna bêntica no Canal de São Sebastião, por grupo e época amostrada.
Abundância Total da Macrofauna
Sipuncula 3%
Nematoda 16%
Polychaeta 51%
Outros 2%
Mollusca 14%
Echinodermata 4%
Cnidaria 3%
Fig. 3.5. Abundância relativa total dos grupos da macrofauna, coletados com o pegador van Veen no Canal de São Sebastião.
l -CRU
I-Mol LCECH I

PIRES-V ANTN cf 01: Plataforma Interna de São Sebastião (OPISS)
OI 1000 o ::J "g
'5 '6
500 .5 ~ . z
I o
i Primavera
CRUSTACEA
................................. ................................. •••••••• u ...................... . ::::::::::::::::m:::::::::::::: ~~~~~ ~~~~~~~~~~~~~g~~~m~;~m :
Verão Outono Inverno 1------MOllUSCA 2000 1
10o: r+~I ________ L-+-~ ______ ~~ __ L-______ ~~~ ________ ~ Primavera Verão Outono Inverno
ECHINODERMATA 111 600 o :J :2 > :o 300 .5 CP
'tJ o O z
Primavera Verão Outono Inverno
POlYCHAETA
111 5000 o :J :2 .~
2500 'tJ .5 CP 'tJ o Z O ~ I
Primavera Verão Outono Inverno L-____________________________________________________________________________ ~
Fig. 3.6. Distribuição do número de indivíduos de Crustacea, Mollusca, Echinodermata e Polychaeta, nas quatro épocas amostradas no Canal de São Sebastião.
43
44
100
5l)
80
~ 70
.lI
j €O
50 ~ .!lI
~ 40
W 30 u.
20
10
O
Inverno 51%
Peso Total
Outono 8%
Relat. téc. iost. oceaoogr., (41), 1997
Primavera 22%
Verão 19%
Fig. 3.7. Porcentagens relativas de peso úmido da macrofauna do Canal de São Sebastião, para cada época do ano amostrada.
I o I
~ w
i z I z
I!! ::J :c u W
Fig. 3.8. Freqüência de ocorrência dos grupos da macrofauna coletada com draga retangular no Canal de São Sebastião, para as quatro épocas do ano.

PIRF.S-VANTN ('( ai: Platarnrmll Interna de São Scha~tião (OPTSS)
700
100
o ~
(11
t m (11 (11 (11
~ ãí .c ãí t.I (11 t.I RI to- ~ .c ~ .c E §. 2 ~ -E 8 CII o 8 .21 z () () a.. Õ
~ ~ RI -l
Fig. 3.9. Densidade média da meiofauna total e dos principais grupos da
N
fi lO o-
8. UI
~ !! e t.I
E
meiofauna no Canal de São Sebastião, no verão e inverno de 1994.
250
200
150
fti (11 (11 RI RI RI RI Õ "8 I
CII ãí .c ãí t.I t.I to- 1V (11 RI ~
RI 1;; .c .c
E % 8 CII o 2 -E z () () o .21 a.. c
Õ RI ~ i:! RI -l
Fig. 3.10. Biomassa média da meiofauna total e dos principais grupos da meiofauna do Canal de São Sebastião, no verão e inverno de 1994.
45
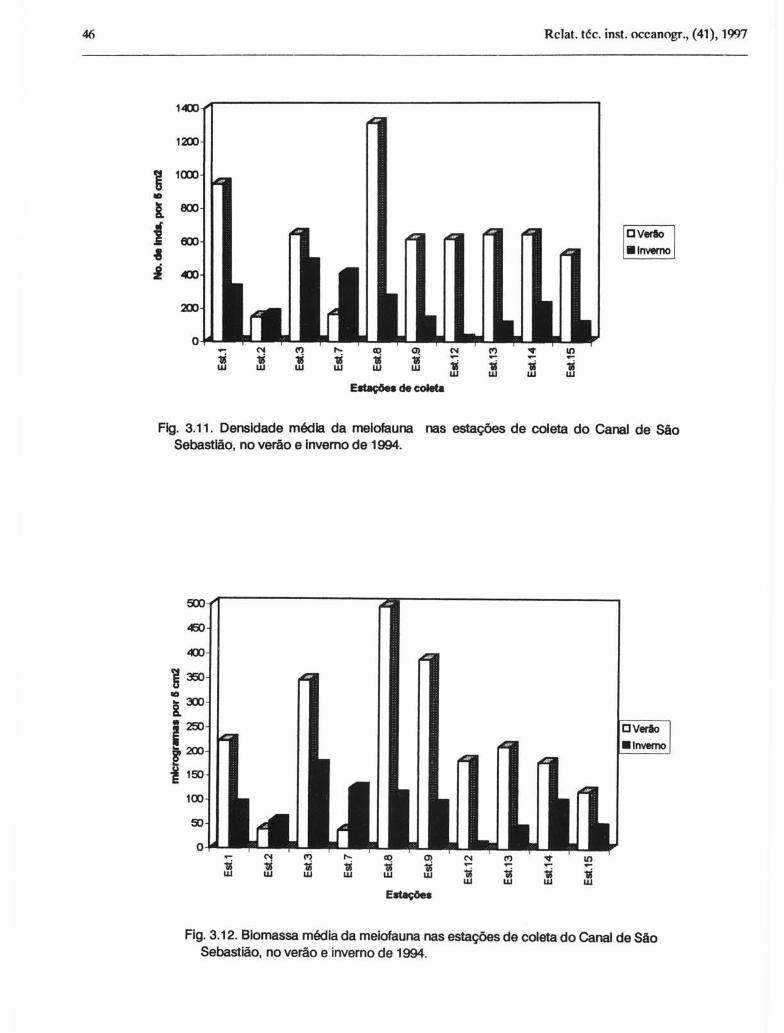
46 Rc1at. téc. inst. occanogr., (41), 1997
1cx)
1200
1 1CXXl
10
I 800
f .. Icvno I .Invemo
i
Estaç6ea de coleta
Flg. 3.11. Densidade média da melofauna nas estações de coleta do Canal de São Sebastião, no verão e inverno de 1994.
500
N ... 11i w
I/) ... ;fi
CVerlo
.Invemo
Fig. 3.12. Biomassa média da meiofauna nas estações de coleta do Canal de São Sebastião, no verão e inverno de 1994.